
トロンボスポンジン−1由来ペプチド及び治療方法
マトリックス細胞のタンパク質であるトロンボスポンジン-1(TSP-1)を利用する治療及び関連する組成物を、アテローム性動脈硬化症のプラークを安定化し、例えば、心筋梗塞、脳卒中、及び急性肢虚血を引き起こすプラーク破裂の発生を低減するために開示する。

【発明の詳細な説明】
【技術分野】
【0001】
本願は、2005年6月24日に出願の米国仮特許出願第60/693,574号の優先権の利益を享受する。
【0002】
本発明は、アテローム性動脈硬化症のプラークを安定化し、それにより、例えば、心筋梗塞、脳卒中、及び急性肢虚血を引き起こすプラーク破裂の発生を低減する、マトリックス細胞のタンパク質であるトロンボスポンジン-1(TSP-1)を利用する治療及び関連する組成物に関する。さらに、本発明は、上述の疾患を含む各種の背景下におけるプラークの安定化に使用するための一定の合成ペプチドを含む、各種のペプチドにも関連する。
【背景技術】
【0003】
アテローム性動脈硬化症の致命的な合併症、例えば、急性心筋梗塞及び脳卒中は、局所的な血管の炎症を引き起こすコレステロール及びその酸化したリン脂質の慢性的な沈着に由来するものである(1)。アテローム性動脈硬化症の病変は、動脈壁の細胞と細胞外マトリックス(ECM)構成成分との間の相互作用を部分的に介して、動脈壁において形成され、進行する(1,2)。アテローム性動脈硬化症の発症と共に、血管壁の細胞構成成分とECM構成成分の双方に変化が起こる。ECMは、アテローム性動脈硬化症の病変内における細胞の振る舞いに作用する細胞結合タンパク質及び増殖因子の貯蔵所として認識されている(3,4)。言い換えると、これらのプロセスの制御は、細胞-細胞及び細胞-ECM相互作用に直接的な効果を有する可能性があり、言い換えると、破裂する傾向があり、それに続く下流の事象、例えば心筋梗塞及び脳卒中に関連し得るアテローム性動脈硬化症のプラークのフェノタイプに直接影響を与える可能性がある。
【0004】
その環境について特異的な背景の情報を細胞に与えるECMの微量なタンパク質成分は、「マトリックス細胞の」タンパク質と称されている(4)。マトリック細胞タンパク質は、他のECM構成成分、多特異性細胞表面受容体、並びに増殖因子と相互作用して、細胞-マトリックス相互作用を調節する。これらのタンパク質は細胞機能を調節するが、原線維、コラーゲン、及びエラスチンのような他のECM成分がするように構造の物理的特性又は組織に直接寄与することはないようである(5)。これらのタンパク質は、細胞外マトリックスと細胞表面との間の接触面で働き、細胞の振る舞いを制御する。さらに、構造タンパク質は正常な血管の発生に重要な役割を有するが、マトリックス細胞のタンパク質は創傷時にのみ成人の血管において発現する(6)。具体的には、これらのタンパク質は、組織修復において主な機能を有しており、その背景において作用することで、細胞-マトリックス相互作用によって細胞機能に影響を与えるようである。その様なものとして、生殖系列における構造ECMタンパク質の標的破壊は重度な又は致死性のフェノタイプを生じるが、マトリックス細胞のタンパク質をコードする遺伝子のノックアウトは、見かけ上正常であるか又は僅かなフェノタイプを生じる(7-11)。
【0005】
TSP-1は、多数の結合ドメインを有する原始型のマトリックスタンパク質である(3,12)。TSP-1は創傷時に誘導される。当該タンパク質の無発現変異について同型接合のマウスは顕著な発生の欠陥を有さず、これらのマウスにおける主要なフェノタイプは、急性の斑であり、肺において肺炎を構成する。しかしながら、各種の疾患モデルにおいて、これらのマウスの明確なフェノタイプが存在する(10)。例えば、創傷修復モデルでは、TSP-1欠損は、炎症性細胞応答の変化及び創傷治癒の遅延に関連する。他の研究では、TSP-1は血管新生を阻害することが示されている(13-19)。その様なものとして、新生細胞におけるTSP-1の標的発現は臨床において関心がもたれており、その修飾物は臨床試験のフェーズIIにおいて現在評価されている(20)。
【非特許文献1】Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
【非特許文献2】Ross, R., Atherosclerosis~an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
【非特許文献3】Bornstein, P., Thrombospondins as matricellular modulators of cell function. J Clin Invest, 2001. 107(8): p. 929-34.
【非特許文献4】Sage, E.H., Regulation of interactions between cells and extracellular matrix: a command performance on several stages. J Clin Invest, 2001. 107(7): p. 781-3.
【非特許文献5】Murphy-Ullrich, J.E., The de-adhesive activity of matricellular proteins: is intermediate cell adhesion an adaptive state? J Clin Invest, 2001. 107(7): p. 785- 90.
【非特許文献6】Elzie, CA. and J.E. Murphy-Ullrich, The N-terminus of thrombospondin: the domain stands apart, lht J Biochem Cell Biol, 2004. 36(6): p. 1090-101.
【非特許文献7】Li, D.Y., G. Faury, D.G. Taylor, E.G. Davis, W.A. Boyle, R.P. Mecham, P. Stenzel, B. Bo ak, and M.T. Keating, Novel arterial pathology in mice and humans hemizygous for elastin. J Clin Invest, 1998. 102(10): p. 1783-7.
【非特許文献8】Aszodi, A., D. Chan, E. Hunziker, J.F. Bateman, and R. Fassler, Collagen π is essential for the removal of the notochord and the formation of intervertebral discs. J Cell Biol, 1998. 143(5): p. 1399-412.
【非特許文献9】Forsberg, E., E. Hirsch, L. Frohlich, M. Meyer, P. Ekblom, A. Aszodi, S. Werner, and R. Fassler, Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc Natl Acad Sci U S A, 1996. 93(13): p. 6594-9.
【非特許文献10】Lawler, J., M. Sunday, V. Thibert, M. Duquette, E.L. George, H. Rayburn, and R.O. Hynes, Thrombospondin- 1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J Clin Invest, 1998. 101(5): p. 982-92.
【非特許文献11】Kyriakides, T.R., J. W. Tarn, and P. Bornstein, Accelerated wound healing in mice with a disruption of the thrombospondin 2 gene. J Invest Dermatol, 1999. 113(5): p. 782-7.
【非特許文献12】Adams, J.C. and J. Lawler, The thrombospondins. Lit J Biochem Cell Biol, 2004. 36(6): p. 961-8.
【非特許文献13】Crawford, S.E., V. Stellmach, J.E. Murphy-Ullrich, S.M. Ribeiro, J. Lawler, R.O. Hynes, G.P. Boivin, and N. Bouck, Thrombospondin- 1 is a major activator of TGF-betal in vivo. Cell, 1998. 93(7): p. 1159-70.
【非特許文献14】Agah, A., T.R. Kyriakides, J. Lawler, and P. Bornstein, The lack of thrombospondin- 1 (TSPl) dictates the course of wound healing in double- TSPl/TSP2-null mice. Am J Pathol, 2002. 161(3): p. 831-9.
【非特許文献15】Yee, K.O., M. Streit, T. Hawighorst, M. Detmar, and J. Lawler, Expression of the type-1 repeats of thrombospondin- 1 inhibits tumor growth through activation of transforming growth factor-beta. Am J Pathol, 2004. 165(2): p. 541-52.
【非特許文献16】Miao, W.M., W.L. Seng, M. Duquette, P. Lawler, C. Laus, and J. Lawle r, Thrombospondin- 1 type 1 repeat recombinant proteins inhibit tumor growth through transforming growth factor-beta-dependent and -independent mechanisms. Cancer Res, 2001. 61(21): p. 7830-9.
【非特許文献17】Yano, K., H. Oura, and M. Detmar, Targeted overexpression of the angiogenesis inhibitor thrombospondin- 1 in the epidermis of transgenic mice prevents ultraviolet-B-induced angiogenesis and cutaneous photo-damage. J Invest Dermatol, 2002. 118(5): p. 800-5.
【非特許文献18】Simantov, R., M. Febbraio, R. Crombie, A.S. Asch, R.L. Nachman, and R.L. Silverstein, Histidine-rich glycoprotein inhibits the antiangiogenic effect of thrombospondin- 1. J Clin Invest, 2001. 107(1): p. 45-52.
【非特許文献19】Bornstein, P., A. Agah, and T.R. Kyriakides, The role of thrombospondins 1 and 2 in the regulation of cell-matrix interactions, collagen fibril formation, and the response to injury. Int J Biochem Cell Biol, 2004. 36(6): p. 1115-25.
【非特許文献20】Lawler, J. and M. Detmar, Tumor progression: the effects of thrombospondin- 1 and -2. Int J Biochem Cell Biol, 2004. 36(6): p. 1038-45.
【非特許文献21】Topol, EJ., J. McCarthy, S. Gabriel, DJ. Moliterno, WJ. Rogers, L.K. Newby, M. Freedman, J. Metivier, R. Cannata, CJ. O'Donnell, K. Kottke-Marchant, G. Murugesan, E.F. Plow, O. Stenina, and G.Q. Daley, Single nucleotide polymorphisms in multiple novel thrombospondin genes may be associated with familial premature myocardial infarction. Circulation, 2001. 104(22): p. 2641-4.
【非特許文献22】Stenina, O.I., T.V. Byzova, J.C. Adams, JJ. McCarthy, EJ. Topol, and E.F. Plow, Coronary artery disease and the thrombospondin single nucleotide polymorphisms. Int J Biochem Cell Biol, 2004. 36(6): p. 1013-30.
【非特許文献23】Raines, E.W., The extracellular matrix can regulate vascular cell migration, proliferation, and survival: relationships to vascular disease. Int J Exp Pathol, 2000. 81(3): p. 173-82.
【非特許文献24】Roth, JJ., V. Gahtan, J.L. Brown, C. Gerhard, V.K. Swami, V.L. Rothman, T.N. Tulenko, and G.P. Tuszynski, Thrombospondin- 1 is elevated with both intimal hyperplasia and hypercholesterolemia. J Surg Res, 1998. 74(1): p. 11-6.
【非特許文献25】Lawler, J., The functions of thrombospondin- 1 and-2. Curr Opin Cell Biol, 2000. 12(5): p. 634-40.
【非特許文献26】Chen, H., M.E. Herndon, and J. Lawler, The cell biology of thrombospondin- 1. Matrix Biol, 2000. 19(7): p. 597-614.
【非特許文献27】Majack, R.A., S.C. Cook, and P. Bornstein, Control of smooth muscle cell growth by components of the extracellular matrix: autocrine role for thrombospondin. Proc Natl Acad Sci U S A, 1986. 83(23): p. 9050-4.
【非特許文献28】Schultz-Cherry, S., S. Ribeiro, L. Gentry, and J.E. Murphy-Ullrich, Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. J Biol Chem, 1994. 269(43): p. 26775-82.
【非特許文献29】Fischer, J.W., M. Stoll, A. W. Hahn, and T. Unger, Differential regulation of thrombospondin- 1 and fibronectin by angiotensin JI receptor subtypes in cultured endothelial cells. Cardiovasc Res, 2001. 51(4): p. 784-91.
【非特許文献30】Lymn, J.S., M.K. Patel, G.F. Clunn, SJ. Rao, KX. Gallagher, and A.D. Hughes, Thrombospondin-1 differentially induces chemotaxis and DNA synthesis of human venous smooth muscle cells at the receptor-binding level. J Cell Sci, 2002. 115(Pt 22): p. 4353-60.
【非特許文献31】Patel, M.K., J.S. Lymn, G.F. Clunn, and A.D. Hughes, Thrombospondin-1 is a potent mitogen and chemoattractant for human vascular smooth muscle cells. Arterioscler Thromb Vase Biol, 1997. 17(10): p. 2107-14.
【非特許文献32】Rodriguez-Manzaneque, J.C., T.F. Lane, M.A. Ortega, R.O. Hynes, J. Lawler, and M.L. Iruela-Arispe, Thrombospondin-1 suppresses spontaneous tumor growth and inhibits activation of matrix metalloproteinase-9 and mobilization of vascular endothelial growth factor. Proc Natl Acad Sci U S A, 2001. 98(22): p. 12485-90.
【非特許文献33】Bein, K. and M. Simons, Thrombospondin type 1 repeats interact with matrix metalloproteinase 2. Regulation of metalloproteinase activity. J Biol Chem, 2000. 275(41): p. 32167-73.
【非特許文献34】Lutgens, E., M. Gijbels, M. Smook, P. Heeringa, P. Gotwals, V.E. Koteliansky, and MJ. Daemen, Transforming growth factor-beta mediates balance between inflammation and fibrosis during plaque progression. Arterioscler Thromb Vase Biol, 2002. 22(6): p. 975-82.
【非特許文献35】Mallat, Z., A. Gojova, C. Marchiol-Fournigault, B. Esposito, C. Kamate, R. Merval, D. Fradelizi, and A. Tedgui, Inhibition of transforming growth factorbeta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ Res, 2001. 89(10): p. 930-4.
【非特許文献36】Luttun, A., E. Lutgens, A. Manderveld, K. Maris, D. Collen, P. Carmeliet, and L. Moons, Loss of matrix metalloproteinase-9 or matrix metalloproteinase- 12 protects apolipoprotein E- deficient mice against atherosclerotic media destruction but differentially affects plaque growth. Circulation, 2004. 109(11): p. 1408-14.
【非特許文献37】Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
【非特許文献38】Stefanidakis, M., T. Ruohtula, N. Borregaard, CG. Gahniberg, and E. Koivunen, Intracellular and cell surface localization of a complex between alphaMbeta2 integrin and promatrix metalloproteinase-9 progelatinase in neutrophils. J Immunol, 2004. 172(11): p. 7060-8.
【非特許文献39】Wrana, J.L., L. Attisano, J. Carcamo, A. Zentella, J. Doody, M. Laiho, X.F. Wang, and J. Massague, TGF beta signals through a heteromeric protein kinase receptor complex. Cell, 1992. 71(6): p. 1003-14.
【非特許文献40】Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
【非特許文献41】Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
【非特許文献42】Ross, R., Atherosclerosis an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
【非特許文献43】FaIk, E., P.K. Shah, and V. Fuster, Coronary plaque disruption. Circulation, 1995. 92(3): p. 657-71.
【発明の開示】
【発明が解決しようとする課題】
【0006】
冠状動脈性心臓病に関して、最近の相関的及び記述的研究は、TSP-1がアテローム性動脈硬化症及び心筋梗塞において、不明確であり、且つ、未知ではあるが、役割を有している可能性があることを示唆している(21,22)。まず、調節された遺伝的変異研究は、この遺伝子のミスセンス「変異体」が、アテローム性動脈硬化症及び心筋梗塞の双方のリスクの増大に関連することを示唆している(21)。次に、ヒトのサンプルを用いた病理学的研究は、TSP-1がアテローム性動脈硬化症の病変に存在しているが、天然の血管壁には存在しないことを示している(23,24)。しかしながら、現在において、アテローム性動脈硬化症のフェノタイプに対する、プラーク内におけるTSP-1発現の意味は未知である。
【課題を解決するための手段】
【0007】
本発明は、アテローム性動脈硬化症のプラークを安定化し、任意の血管床(例えば、冠状動脈、大脳、又は肢)におけるアテローム性動脈硬化症の病変の破裂する傾向を低減するTSP-1を利用する方法及び関連する組成物を提供する。本発明は、TSP-1及びTSP-1に関連する一定の合成ペプチドを含む関連ペプチドを利用する組成物も提供する。その様なペプチドは、心筋梗塞、脳卒中、及び急性肢虚血のような疾患の状態を含む、各種の背景におけるプラークの安定化において有用性がある。これらのペプチドのいくつかは、TGF-β
1及びMMP-9活性を調節するためのTSP-1の活性部位に関連する1つ以上の配列を含む。
【0008】
ある実施態様では、本発明は、MMP-9活性化に作用することが認められた(TSP-1)に基づくペプチド配列を提供する。本発明者は、この活性についての機構は、アテローム性動脈硬化症の病変内においてMMP-9の機能と競合して白血球(主に単球)に結合する、TSP-1に特有のβ2-インテグリン結合部位によるものであると主張する。さらに、ある実施態様では、本発明は、TGF-β1活性を制御することが知られているTSP-1内の配列に基づくペプチド配列を提供する。その様な配列は、プラークの安定性に利用し得る。最後に、本発明者は、TSP-1のTGF-β1及びMMP-9活性並びにそのin vivoにおけるプラーク安定化効果の双方を摸倣する、上述の活性部位に基づく各種の特異的な合成ペプチドの配列を提供する。
【0009】
本発明の各種の実施態様の説明を容易にして、本発明を為して使用する際に用いる各種の要素及び成分の理解を提供するために、本発明において使用する以下の用語は、以下の意味を有する。
【発明を実施するための最良の形態】
【0010】
定義
各種の感染症の治療方法は、「予防上の」処置又は「治療上の」処置を含む意味を有する。「予防上の」処置は、疾患に関連する病理を発症するリスクを低減するために疾患の兆候を示していないか又は疾患の初期の兆候を示す対象に施す処置である。
【0011】
本明細書で使用する用語「治療上の」は、病理学的な兆候を減少させるか又は除くための、病理学的な兆候を示す対象に施す処置を意味する。
【0012】
本明細書で使用する用語「治療上の有効量」は、化合物を投与する対象に対して有益な効果を提供するために十分な化合物の量を意味する。有益な効果は、アテローム性動脈硬化症のプラークの安定化を意味する。
【0013】
本明細書で使用する用語「ペプチド」、「ポリペプチド」、及び「タンパク質」は、互換的に使用され、アミド結合で共有結合している少なくとも2つのアミノ酸のポリマー配列を示す。
【0014】
本明細書で使用する用語「相同の」は、2つのペプチド間で類似するアミノ酸配列を示す。双方のペプチドのアミノ酸の位置が同一のアミノ酸で占められている際に、それらは該位置で相同である。かくして、「実質的に相同」は、大部分で相同あるが、全てが相同ではなく、相同配列として大部分又は全ての活性を保持するアミノ酸配列を意味する。本明細書で使用する「実質的に相同」は、ある配列が、少なくとも80%同一であり、好ましくは少なくとも90%、更に好ましくは98%の参照ペプチドに対する相同性を有する事を意味する。
【0015】
ポリペプチドは、アミノ酸残基の配列として本明細書で開示している。それらの配列は、左から右へとアミノ末端からカルボキシル末端の方向で記載している。標準的な用語に従えば、アミノ酸残基配列は、下記の一文字又は三文字のコードのいずれかで表記される。
【0016】
【表1】
【0017】
本発明のポリペプチドの構造に修飾及び変更が為されて良く、それでもTSP-1ペプチド様特性を有する分子が得られる。例えば、ペプチドの活性の相当の損失無く、配列中のあるアミノ酸が他のアミノ酸に置換されても良い。ポリペプチドの相互作用能及び性質が、ポリペプチドの生物学的機能の活性を規定するため、あるアミノ酸配列の置換がポリペプチド配列(言うまでも無く、その基礎を成すDNAコード配列)に為されて良く、それにもかかわらず同様の特性を有するポリペプチドが得られる。
【0018】
その様な変更を為す場合には、アミノ酸のヒドロパシーが考慮されて良い。ペプチドの相互作用的生物学的機能を与えるアミノ酸のヒドロパシー指数の重要性は、当該技術分野において一般的に理解されている(Kyte, J. and R. F. Doolittle 1982)。あるアミノ酸が同様なヒドロパシー指数(スコア)を有する他のアミノ酸に置換されて良く、それでも同様の生物学的活性を有するポリペプチドを生じさせることが知られている。各アミノ酸は、その疎水性度及び電荷特性に基づいてヒドロパシーが定められている。それらのヒドロパシーは、イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(-0.4);スレオニン(-0.7);セリン(-0.8);トリプトファン(-0.9);チロシン(-1.3);プロリン(-1.6);ヒスチジン(-3.2);グルタミン(-3.5);グルタミン(-3.5);アスパルテート(-3.5);アスパラギン(-3.5);リジン(-3.9);及びアルギニン(-4.5)である。
【0019】
アミノ酸の相対的なヒドロパシーの特性が、結果として生じるポリペプチドの二次構造を決定し、言い換えると、酵素、基質、受容体、抗体、及び抗原などのような他の分子とのポリペプチドの相互作用を規定する。アミノ酸は、同様のヒドロパシー指数を有する他のアミノ酸によって置換されて良く、それでも機能的に等価のペプチドが得られることが当該技術分野において既知である。その様な変更において、ヒドロパシー指数が±2以内のアミノ酸置換が好ましく、±1以内のアミノ酸置換がより好ましく、±0.5以内のアミノ酸置換が更に好ましい。
【0020】
同様のアミノ酸の置換は、親水性度に基づいて為されても良く、特にそれによって生じる生物学的機能が等価なポリペプチド又はペプチドは、免疫学的な実施態様における使用が意図される。参照によって本明細書に取り込む米国特許第4,554,101号は、隣接するアミノ酸の親水性度によって決定されるポリペプチドの最大の局所的な平均親水性度は、その免疫原性及び抗原性、すなわちポリペプチドの生物学的特性と関連すると記述している。米国特許第4,554,101号に詳述しているように、以下の親水性度の値が、アミノ酸残基:アルギニン(+3.0);リジン(+3.0);アスパルテート(+3.0±1);グルタメート(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);プロリン(-0.5±1);スレオニン(-0.4);アラニン(-0.5);ヒスチジン(-0.5);システイン(-1.0);メチオニン(-1.3);バリン(-1.5);ロイシン(-1.8);イソロイシン(-1.8);チロシン(-2.3);フェニルアラニン(-2.5);トリプトファン(-3.4)に定められている。アミノ酸は、同様の親水性度を有する他のアミノ酸に置換されて良く、それでも生物学的に等価な、特に免疫学的に等価なポリペプチドが得られることが理解されている。その様な変更は、親水性度が±2以内のアミノ酸置換が好ましく、±1以内のアミノ酸置換が特に好ましく、±0.5以内のアミノ酸置換が更に好ましい。
【0021】
したがって上述したように、アミノ酸置換は、一般的には、アミノ酸側鎖の置換基、例えば、それらの疎水性、親水性、電荷、及び大きさなどの相対的な類似性に基づく。上述の各種の特性を考慮に入れた置換の例は当業者に既知であり、アルギニンとリジン;グルタメートとアスパルテート;セリンとスレオニン;グルタミンとアスパラギン;及びバリンと、ロイシンと、イソロイシンを含む(以下の表2参照)。本発明は、上記のペプチドの機能的又は生物学的等価物を意図する。
【0022】
【表2】
【0023】
ポリペプチドの生物学的又は機能的等価物は、部位特異的突然変異導入法を用いて調製しても良い。部位特異的突然変異導入法は、基礎とするDNAの特異的な突然変異による、それらの配列に由来する第二世代のポリペプチド、又は生物学的機能の等価なポリペプチド若しくはペプチドの調製に有用な技術である。上述のように、その様な変更は、アミノ酸置換が望まれる場合に望ましい。前記技術は、DNAに1つ以上のヌクレオチド配列の変更を導入することによって、例えば上述のものの一つ以上を含む配列の変異体を調製し試験をする準備能力を更に提供する。部位特異的突然変異導入法は、所望の変異体のDNA配列をコードする特異的なオリゴヌクレオチド配列、並びに欠失接合部の両端に安定な二本鎖を形成するために十分な大きさ及び配列の複雑性を有する一次配列を提供するのに十分な数の隣接するヌクレオチドの使用によって、変異体の製造を可能にする。
【0024】
アミノ酸残基が、ペプチドの機能を変えること無く、標準的な分子生物学的技術を用いることによって、本発明のTSP-1ペプチドに付加されるか又は欠失されて良い。例えば、TSP-1ペプチドの末端部位が除去されて、切断型のペプチドが作製されて良いが、前記切断型のペプチドは、タンパク質タンパク質相互作用を介するTGF-β1又はMMP-9の調節、並びにアテローム性動脈硬化症のプラークの安定化を含む本発明のTSP-1ペプチドの機能的な活性を保持する。
【0025】
本発明のポリペプチドは、当業者によく知られた標準的な技術によって調製される。その様な技術は、ポリペプチドを含有することが既知の組織からの単離及び精製、そのようなポリペプチドをコードするクローニングしたDNAからの形質転換細胞を用いる発現、合成ペプチド製造システムの使用を含むが、それらに限らない。
【実施例】
【0026】
TSP-1構造ドメイン及びそのマトリックス結合パートナー
TSP-1は、150kDaの分子量の鎖を有する三量体である。TSP-1の各サブユニットは、幾つかの構造ドメインからなる(12)。これらのドメインは、アミノ末端ドメイン、プロコラーゲンドメイン、タイプ1、タイプ2、及びタイプ3反復配列、並びにカルボキシル末端ドメインとして広範に定義されている(25,26)。各ドメインは、図1に示すように多数の細胞機能の調節において重要である事が示されている。さらに、特異的なリガンド結合部位が、TSP-1について定義されている(図1)。これらは、リポタンパク質受容体関連タンパク質(LRP)、CD36、β3インテグリン、及びインテグリン関連タンパク質(IAP)、並びにTGF-β1及びPDGFを含むサイトカインを含む(3,12,27,28)。その多数の結合部位を介して、TSP-1は膜タンパク質及びサイトカインを有する細胞表面で機能しているようである。この様に、TSP-1は、シグナル伝達及び転写を含む細胞のフェノタイプに影響を与える。各細胞が異なる種類の受容体を発現するため、TSP-1によって形成される複合体の組成及び個々の細胞応答は、各細胞種によって変化する。例えば、TSP-1は、平滑筋細胞移動を促進するが、内皮細胞移動を阻害する(29-31)。
【0027】
現在研究されている主な器官系において、TSP-1の生物学の大部分が、TGF-β1の細胞外活性化に寄与している(10,13-15)。具体的には、TSP-1の欠損は、TGF-β1の活性を低減し、個々の器官におけるTGF-β1欠損のフェノタイプに似ていることが示されている。例えば、TSP-1ノックアウトマウスの研究では、TSP-1の欠損が、TGF-β1ノックアウトマウスにおける知見と同様に、局所的な歯槽炎及び慢性的な肺炎を引き起こす。さらに、TGF-β1活性化について十分に認められるTSP-1の短いペプチド断片(図2に示す430から433のアミノ酸残基に対応するKRFK)による生物学的な相補性が、このフェノタイプを救い、この背景におけるフェノタイプの変化を表わす重要な細胞プロセスとしてのTGF-β1の活性化に影響を及ぼす。
【0028】
TSP-1は、マトリックスメタロプロテイナーゼ(MMP)を含む細胞外プロテアーゼの活性も制御する(MMP)(32,33)。癌の血管新生では、TSP-1の標的発現が、抗血管新生特性と一致するMMP-9活性の低減と関連する。MMP-2及びMMP-9は、TSP-1に結合することが示されている(33)。しかしながら、TSP-1に対するMMPの結合部位は明らかにされておらず、これらのプロテアーゼの活性を制御する機構も明らかにされていない。
【0029】
マトリックス代謝、白血球の補充、及び平滑筋細胞移動を含むが、それらに限らないアテローム性動脈硬化症のプラークのフェノタイプにおけるTGF-β1及びMMP-9の双方の役割を示す実験データが存在する。さらに、上述のように、TSP-1は、組織損傷に応答して、これらのリガンドの各々の活性を制御することが示されている。
【0030】
本発明者は、TSP-1が、マウスのアテローム性動脈硬化症のプラークにおいて、プラークを安定化する役割を有することを発見した。さらに、本発明者は、TGF-β1及びMMP-9に関連して、これらの活性を制御するTSP-1の活性ペプチドドメインを同定した。最後に、本発明者は、これらの活性ドメインに基づくペプチド配列を含有するペプチドを設計し、代表的なペプチドが、アテローム性動脈硬化症のマウスモデルにおいてin vivoのTSP-1の効果を刺激することを示すことができた。
【0031】
TSP-1の欠失はアテローム性動脈硬化症のプラークを変化させる
本発明者は、TSP-1マトリックス細胞タンパク質がプラーク安定化効果を有することを示すことができた。本発明者は、TSP-/-ApoE-/-ダブルノックアウトマウスを作製して、病変の形成及び組成に対するTSP-1欠失の役割を試験した。食事(3%脂肪)の24週後に、TSP-/-ApoE-/-マウス及びApoE-/-対照マウスを屠殺した。TSPノックアウトマウスと対象マウスとの間には、体重、総コレステロール、HDLコレステロール、及びトリグリセリドに優位な差はなかった。
【0032】
H&E染色を用いるアテローム性動脈硬化症の病変の程度についての大動脈洞の特性決定は、双方の群における同様な病変を明らかにした(ノックアウトマウスについては701,000μm2±95,000μm2であるのに対し、対照マウスについては724,000μm2±103,000μm2)。本発明者は、病変の組成について大動脈弁尖の詳細な形態評価を実施した(図3)。TSP-1の欠損は、内膜及び中膜の双方において病変の形態に顕著な影響を有していた。具体的には、TSPノックアウトマウスでは、(1)内膜の病変における炎症(マクロファージ)の増大、(2)内膜の病変内における脂質沈着の増大、(3)病変内におけるマトリックス(コラーゲン及びエラスチン)沈着の低減、(4)これらのマウスの内膜領域におけるコラーゲン及びエラスチン含量の低減によって示されるような、内膜の損失を伴う内膜拡張、(5)内膜内における総平滑筋細胞含量の無変化が存在した。まとめると、これらの結果は、TSP-1の欠失は、プラーク内の炎症と線維症との間のバランスを変化させることを示す。これが、細胞外マトリックス含量の低減を伴う炎症性プラークフェノタイプを引き起こす。
【0033】
本発明者は、この背景におけるTSP-1による利点の機構は、主に2つの事柄(これらに限らない):(a)TSP-1が存在しないアテローム性動脈硬化症の病変内のTGF-β1の活性化の低減、(b)TSP-1の不在下におけるMMP-9の活性化の増大である。
【0034】
TSP-/-ApoE-/-マウスにおけるアテローム性動脈硬化症の病変内でTGF-β1の活性化が低減する
上述のように、in vivoの組織創傷におけるTSP-1の他の研究において、大部分のフェノタイプの発見は、創傷部位において局所的にTGF-β1を活性化するTSP-1の機能に帰する。さらに、TGF-β1活性の中和は、TSP-/-ApoE-/-マウスにおける上述の本発明者による発見と同様に、炎症を増大し、アテローム性動脈硬化症の病変内のマトリックスを低減させることが示されている(34,35)。そのため、本発明者は、TSP-/-ApoE-/-動物由来の組織における全TGF-β1及び活性型TGF-β1の双方のレベルを測定した。本発明者は、アテローム性動脈硬化症が形成される傾向がある既知の領域である大動脈弓由来のサンプルを均質化して、ELISAアッセイにおいて、その可溶化液を使用した。これらのマウスにおける総(潜在型及び活性型)TGF-β1レベルには変化が無かったが、TSP-/-ApoE-/-マウスにおいて活性型TGF-β1レベルにおける低減が存在した(図4)。アテローム性動脈硬化症の病変の可溶化物を用いた結果と比較すると、Tsp-/-ApoE-/-マウスとApoE-/-マウスとの間の活性型TGF-β1の全身(血清)レベルにおいて差は無かった(225±87pg/mlに対して186±138pg/ml)。これらのデータは、TSP-1の欠損が、血管壁内で局所的にTGF-β1の活性化の低減を引き起こすことを示す。
【0035】
TSP-/-ApoE-/-マウスにおけるMMP-9活性化の増大
免疫沈降実験によって、ゼラチナーゼ-B(MMP-9)はTSP-1に結合することが示されている(33)。さらに、TSP-1は、プロMMP-9の活性型への変換を調節し得る(32)。しかしながら、MMP-9の修飾語翻訳のTSP-1による制御についての機構は未知である。TSP-/-ApoE-/-マウスの病変内にマトリックスの顕著な損失が存在したため(図3e)、本発明者は、ウエスタンブロットにおいて大動脈弓均質化物由来の組織可溶化物を用いてMMP-9レベルを試験した。図5に示すように、これらのマウスのアテローム性動脈硬化症の病変において、総MMP-9及び活性型MMP-9のレベルの双方の増大が存在した。
【0036】
これらの結果は、TSP-1の欠損が、アテローム性動脈硬化症の病変内で局所的にMMP-9の活性化の増大を引き起こすことを示し、これはMMP-9活性に対するTSP-1の潜在的な阻害効果と一致する。apoEノックアウトマウスにおけるMMP-9欠失の研究は、MMP-9の欠損がマクロファージ数の低減、及びコラーゲン合成の増大に関連することを示している(36)。
【0037】
本発明者の発見を更に洗練するため、本発明者は、TGF-β1とMMP-9活性の双方に対するTSP-1の「活性」部位を確立することを開始した。TSP-1によるこれらのリガンドの制御についての細胞外における機構は、プラーク内の細胞表面相互作用によって調整されていると本発明者は仮説を立てたため、本発明者は、本発明者のin vivoアテローム性動脈硬化症プラークモデルにおける主要な細胞種であるマウスのマクロファージ(RAW細胞)を用いる細胞培養系を利用した。本発明者は、TGF-β1活性のTSP-1による制御のための活性部位(過去に報告されている)及びMMP-9活性に作用する部位(本発明者が同定)を同定した。
【0038】
TSP-1は、推定のβ2インテグリン結合モチーフD(D/E)(G/L)Wによってマクロファージに結合する
TSP-1は、CD44、LAP、及びCD36を含むマクロファージに結合するための多数の潜在的な結合部位を有する(図1)。しかしながら、TSP-1には過去に確立されているβ2-インテグリン結合モチーフが存在しない。ファージ配列アッセイを用いた過去の研究は、ペプチドモチーフD(D/E)(G/L)Wが、β2インテグリンに結合するプロ-MMP-9と競合することを報告している(40)。完全なTSP-1ペプチド配列の配列探索によって、本発明者は、図2に示すように、TSP-1は2つのその様な推定のβ2インテグリン結合部位を有することを決定した:1つがTSR-1(DDGW、完全に一致)であり、他方がTSR-2(DGGW、2位の置換を有し、ほぼ一致している)。本発明者は、TSP-1のマクロファージへの結合を評価するための細胞接着アッセイを使用した(図6)。このアッセイを使用して、本発明者はマクロファージへのTSP-1の接着におけるD-Xaa-G-Wの作用を示すことができた。具体的には、本発明者は、DDGWが、TSP-1への細胞接着を十分に阻害することを示すことができた。この競合の特異性が、ペプチド配列DDGAによるTSP-1に結合するマクロファージの緩衝の損失、及び配列ADGWによる部分的な干渉によって示された。
【0039】
TSP-1の存在下におけるマクロファージによるプロMMP-9の活性化の低減は、TSR-1中のDDGWモチーフによって媒介される
TSP-1によるMMP-9の制御の機構は、ほとんど分かっていない。しかしながら、プロMMP-9は、そのD(D/E)(L/G)Wモチーフによって細胞表面aMβ2及びaLβ2インテグリンに結合し、その活性化を引き起こすことが示されている(37,38)。
【0040】
本発明者は、TSP-1が同じモチーフでマクロファージに結合することを示し、本発明者は、TSP-1によるMMP-9の阻害の1つの潜在的な機構は、結合の競合による細胞表面からのプロMMP-9の置換であるという仮説を立てた(図7)。この仮説を評価するために、本発明者は、TSP-1のβ2-インテグリン結合モチーフのいずれか又は双方を含むTSP-1の各種の変異体を作製した(図2)。RAW細胞(マクロファージ細胞株)のTSP-1を用いた一過的なトランスフェクションは、プロMMP-9の活性型への変換を低減した。この効果は、TSR-1モチーフの変異によって逆行したが、TSR-2モチーフの変異ではより少なく逆行した。これらの結果は、おそらくβ2-インテグリンに結合するプロMMP-9と競合することによる、TSP-1によるMMP-9活性化の阻害におけるDxGWモチーフ、特にTSR-1のこのモチーフの重要性を示す。
【0041】
TSP-1によるTGF-β1の活性化は、TSR-2におけるWSPWモチーフの変異によって阻害される
TSP-1におけるWSxWモチーフは、TGF-β1活性化に重要である事が過去に示されている。この仮説を評価するために、本発明者は、組織培養実験を実施して、TSP-1が実際に、活性化マクロファージの存在下において、TGR-β1を活性化することを実証した。さらに、本発明者は変異体を使用して、TSR-2のWSPWモチーフが、TSP-1によるTGF-β1の活性化に主要な役割を担っていることを観察した。
【0042】
これらのデータに基づいて、本発明者は、TSR-1内のDDGWモチーフが、白血球上のaMβ2-インテグリンに結合し、この結合によって細胞表面に媒介されるMMP-9活性に作用するという仮説を立てた。さらに、TSR-2におけるWSPWモチーフはTGF-β1活性化に重要である。
【0043】
その様なものとして、本発明者は、(1)MMP-9活性の低減及び(2)TGF-β1の活性化に重要なTSP-1における2つの活性部位を同定した。この知識を用いて、プラーク安定化特性を有する多数のペプチドを設計し、合成し得る。本発明者は、これら2つの活性部位を組み合わせた代表的な合成配列:KRFKQDDGWSPWSEWを設計して、合成した。本発明者は、これら2つのTSP-1の機能に重要な活性部位を組み合わせたペプチド断片は、in vivoにおけるプラーク安定性に対して、タンパク質全体と同じ有利な効果を奏する可能性があると仮説を立てた。これを評価するために、まず、本発明者は、組織培養物において(1)TGR-β1を活性化し、(2)MMP-9の活性化を阻害するという、このペプチドの機能を試験し、確認した。
【0044】
次に、本発明者は、5週に亘って一日当たり8-25μgの濃度でalzet輸液ポンプを使用して、このペプチドをTSP-/-マウスに注射し、このペプチドが、TSP-/-Apo-/-マウスのフェノタイプを十分に救うこと示すことができた(マクロファージを低減し、病変内のマトリックスを増大した)。さらに、apoE-/-マウスでは、前記ペプチドの輸液が有益な効果を有しており、内因性TSP-1の存在下であってもプラークを安定化するという化合物の機能を立証した。換言すると、未処理のTSP-/-マウスと比較して、(1)内膜の病変における炎症(マクロファージ)の低減、(2)内膜病変内の脂質沈着の低減、(3)病変内のマトリックス(コラーゲン及びエラスチンによって測定)沈着の増大が存在した。
【0045】
結論として、本発明者の結果は、TSP-1が、(1)アテローム性動脈硬化症のプラークの安定化に対して有益な効果を有しており、且つ、(2)MMP-9及びTGF-β1の制御に重要なTSP-1の活性部位を組み合わせた少なくとも1つの合成ペプチドが、前記疾患のマウスモデルにおいてアテローム性動脈硬化症のプラークを安定化し得ることを示す。
【0046】
一般的には、「安定化プラーク」は、プラーク内の病変部位における炎症の制限及び/又は顕著な線維症によって証明される。これらの特性は、上述のTSP-/-マウスのようなTSP-1欠失対照系との比較によって観察され得る。一般的には、安定化プラークは、処置された又はされていない動物における線維症及び炎症の比較によって同定されて良い。プラーク内の線維症と炎症との間のバランスが、多数の異なる動物モデルにおけるプラークが破裂する傾向についての良好に許容されている代理マーカーとなっている(41-43)。
【0047】
追加の実施態様
ペプチド配列
本発明は、各種の新規なペプチドを提供する。これらは、各種の結合ドメイン及び活性化配列、並びにそれらのホモログの1つ以上を含んで良い。
【0048】
以下(のドメイン)有するペプチドは、TGF-β1及び/又はMMP-9に結合するか又は活性化する。
【0049】
分類1.β2-インテグリン、特にaMβ2及びaLβ2-インテグリンに結合するためのTSP-1及び/又はMMP-9に認められるドメインに相当する配列。その様な配列は、MMP-9が通常結合するβ2-インテグリン上の部位に結合するためのドメインを提供する。代替的には、前記ドメインは、β2-インテグリン上の幾つかの他の位置に結合するための部位を提供しても良いが、そのようにする場合においても、天然のMMP-9がβ2-インテグリンに結合することを阻害する。
【0050】
各種の配列が、その様なドメインを含んで良い。幾つかのその様な配列は、天然のMMP-9上で認められる。他のその様な配列は、TSP-1上、特にタイプ1反復ドメインに認められる。1つの例では、そのドメインは、D-Xaa-(L/G)-W又は(D/E)-(D/E)-(G/L)-W又はD-(D/E)-(L/G)-Wと表わされる4つのアミノ酸配列を含む。その様な配列の具体的な例は、DDGW、DGGW、及びDELWを含む。
【0051】
分類2.本発明の新規なペプチド配列において提供されて良い他のドメインは、天然のTGF-βと結合する。これらの配列の幾つかは、TSP-1のタイプ1反復ドメインに認められる。
【0052】
具体的な例では、その様な配列は、W-S-Xaa1-W-S-Xaa2-Wを含む。具体的な例では、Xaa1はP又はHであり、Xaa2はE又はPである。その様なペプチドのより具体的な例は、WSPWSEWである。
【0053】
分類3.本発明の新規なペプチドに利用されて良い更なる他の配列は、潜在型のTGF-βを活性化するアミノ酸配列を含む。その様な配列の幾つかは、TSP-1のタイプ1反復ドメインに認められる。その様な配列の例は、KRFK及びKRFKQを含む。
【0054】
上述の配列/ドメインの3つの分類の各種の組み合わせが、本発明の実施態様に係る新規ペプチド又はペプチドの組み合わせにおいて利用されて良い。
【0055】
第一の例では、ペプチド(又はペプチドの組み合わせを含む組成物)は、上述の分類1からの配列;すなわちβ2-インテグリン結合ドメインに相当する配列のみを含む。その様なペプチドは、上述の分類2及び/又は3からの追加の配列を含まないであろう。
【0056】
第二の例では、ペプチド(又はペプチドの組み合わせを含む組成物)は、上述の分類1及び2からの配列を含む。換言すると、単独のペプチド配列(又はペプチドの組み合わせを含む組成物)が、β2-インテグリン結合ドメインに相当する配列及びTGF-β結合する異なる配列の双方を含む。
【0057】
本発明のペプチド(又はペプチドの組み合わせを含む組成物)の第3の種類が、上述の分類1及び3の配列の双方を含む。換言すると、その様なペプチド(又はペプチドの組み合わせを含む組成物) は、β2インテグリン結合ドメインに相当する配列及びTGF-βを活性化する他の配列の双方を含む。
【0058】
そして、本発明のペプチド(又はペプチドの組み合わせを含む組成物)の更に他の種類は、上述の分類1、2、及び3の各々の組み合わせを含む。その様なペプチド(又はペプチドの組み合わせを含む組成物)は、β2-インテグリン結合ドメイン、TGF-βのための結合部位、潜在型TGF-βを活性型TGF-βに変換するための活性化配列を含むであろう。
【0059】
2つ以上の結合ドメイン又は活性化ドメインを利用する任意のペプチドにおいて、個々の配列又はドメインの幾つかの重なりが許容されることに注意すべきである。例えば、ペプチド中のアミノ酸の1つ以上が、結合部位又は活性化部位の2つ以上に共有されて良い。β-インテグリン結合ドメイン及びTGF-β結合ドメインの双方を含む具体的な例では、本発明のペプチドはDGGWSPWSEWという配列を有して良く、最初のW(トリプトファン)は2つの配列に共有されている。
【0060】
上述のように、プラークの安定化において活性を有することを確認している本発明の具体的なペプチドは、KRFKQDDGWSPWSEWという配列である。配列の残基の幾つかは、活性を維持するために他のものよりも重要である。かくして、この配列の変形例が、活性を保持するであろう。配列が比較的短い場合に、各種の挿入、欠失、置換、及び伸長が、過度の実験無しで当業者に容易に識別されるであろう。
【0061】
例えば、各種の置換が、前記配列内における個々のアミノ酸について為されて良い。好ましくは、その様な置換は、通常、基礎的な配列であるKRFKQDDGWSPWSEWにおけるアミノ酸と同じ大きさ及び/又は化学特性を有するアミノ酸を利用する。
【0062】
一般的な意味では、前記ペプチド配列は、Xaa1- Xaa2- Xaa3- Xaa4- Xaa5- Xaa6- Xaa7- Xaa8- Xaa9- Xaa10- Xaa11- Xaa12- Xaa13- Xaa14- Xaa15の形態を有する。
【0063】
Xaa1、Xaa2、及びXaa4は、好ましくは塩基性の非環式アミノ酸である。例えば、R及びKを含む。
【0064】
Xaa3、Xaa9、Xaa11、Xaa12、及びXaa15は、好ましくは、中性の大きな非極性環式アミノ酸である。これらは、F、W、及び好ましくはPを含む。
【0065】
Xaa5は、好ましくは、中性の大きな極性非環式アミノ酸である。例えば、T、N、及びQが含まれる。
【0066】
Xaa6、Xaa7、及びXaa14は、好ましくは、酸性アミノ酸である。この種類の天然アミノ酸の具体的な例は、D及びEである。
【0067】
最後に、Xaa8、Xaa10、及びXaa13は、好ましくは、中性の小さな極性アミノ酸である。その様なアミノ酸の天然のものの例は、G、S、及びCである。
【0068】
1つの具体的な実施態様では、Xaa2はRである。他の具体的な実施態様では、Xaa6はDである。他の具体的な実施態様では、Xaa7はDである。他の具体的な実施態様では、Xaa9はWである。他の具体的な実施態様では、Xaa12はWである。更に他の具体的な実施態様では、Xaa15はWである。
【0069】
更に他の具体的な実施態様では、Xaa2はRであり、Xaa7はDであり、Xaa9はWである。
【0070】
本発明のペプチドは、20の天然アミノ酸を含む残基に限られないことを注意すべきである。一定の一般的に遭遇する非天然アミノ酸、例えば、脱アミノチロシン(des Tyr)、アグマチン(Agm)、n-ホルミルトリプトファン(f-Trp)、α-アミノイソ酪酸(Aib)、及びサルコシン(Sar)、スタチン、オルニチン(Orn)、ホモリジン、ホモセリン、ホモアルギニン、ノルロイシン(Nle)、ノルバリンが、本発明の化合物に組み込まれても良い。
【0071】
本発明の範囲内のペプチドは、他に開示されている配列を修飾することによって得られて良いが、かくして得られるペプチドの活性を保持する。例えば、これらの化合物のアミノ酸は、通常、天然のL光学異性体型であるが、1つ以上がD光学異性体型に置き換わってよく、又はD,L-ラセミ混合物がペプチドを含む分子において提供されて良い。さらに、ジスルフィド結合が、活性が維持される限り、本発明の化合物に存在するか又はしなくて良い。
【0072】
前記ペプチドに含有されるアミノ酸残基、特にカルボキシ又はアミノ末端のアミノ酸残基は、メチル化、アミド化、アセチル化、又はそれらの活性に悪影響を及ぼすこと無く、循環半減期、プロテアーゼに対する耐性、及び化合物の溶解度を変化し得る他の化学基による置換によって修飾されても良い。
【0073】
上述のペプチドのいずれかは、好ましくは、少なくとも7アミノ酸残基を含む。他の実施態様では、前記ペプチドは、約7から25残基の間、約7から20残基の間、約7から14残基の間、約10から12残基の間、約10から25残基の間、約15から25残基の間、及び約15から20残基の間に含まれる。本発明のペプチドは、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、又は25アミノ酸残基を含む。本発明の基礎的なスカフォールドを規定するペプチドは、好ましくは、上述の結合及び活性化配列(分類1、2、及び3)の1つ以上が、β2-インテグリン及び/又はTGF-βと相互作用するために曝露されるように適当に設計される。
【0074】
本発明の化合物は、それら自体に対してホモポリマー(すなわち、(ペプチド)n)化されるか、又は他のものに対してヘテロポリマー(すなわち、ペプチド1-ペプチド2)化されて良い。前記化合物は、生体適合性分子、例えば、脂肪酸及び/又はポリエチレングリコール(PEG)に接合されて良い。
【0075】
本発明の範囲内のペプチドは、例えば固相ペプチド合成のような当該技術分野でよく知られている手段によって化学的に合成して良い。他の実施態様では、本発明の選択される化合物が、既知の方法に従って調製される組換えDNA構築物の発現によって生産されて良い。
【0076】
簡便には、化合物は、手作業の技術、又は製造業者によって提供される説明書に与えられた指示に従って自動の技術、例えばApplied Biosystems社(Foster City, Calif.)製の各種の製品を用いて合成して良い。本発明の化合物を合成する過程の間に本明細書に従って作製される中間体自体が有用な化合物であり、本発明の範囲内であることが、ペプチド合成の分野の当業者に容易に理解されるであろう。
【0077】
治療方法
上述のように、本発明の実施態様は、TSP-1、その一部、又は関連するペプチド、例えば、上述のように同定したものを用いる治療方法に関連する。一般的には、TSP-1及び本発明のペプチドを投与して、動脈壁上のプラークを安定化する。かくして、それらは、疾患を有する動物においてプラークを安定化する方法で利用されて良い。これは、アテローム性動脈硬化症の治療方法とは区別されるべきである。
【0078】
ある実施態様では、本発明の新規ペプチドを投与して、プラーク安定化以外の結果に作用して良い。例えば、上記の分類1に区別されるモチーフ(例えば、D-Xaa-(L/G)-W含有ペプチド、例えば、DDGW、DGGW、及びDELW)を有する合成ペプチドを利用して、in vivoでMMP-9に作用させて良い。かくして、その様なペプチドを利用して、例えば、癌及び血管新生を治療して良い。ある実施態様では、D-Xaa-(L/G)-Wモチーフを有し、in vivo治療で使用する合成ペプチドは、少なくとも7アミノ酸残基、より好ましくは約7から20残基の間を含み、具体的な実施態様では約10から15残基の間を含む。
【0079】
本発明は、上述のプラーク安定化という利点を提供するために単独で役立つであろう、TSP-1及び本発明のペプチド(それらの非毒性の付加塩、アミド、及びエステルを含む)の有効量を含有する組成物を提供する。その様な組成物は、生理学的に許容される液体、ゲル、又は固体の希釈剤、アジュバント、並びに賦形剤と共に提供されても良い。
【0080】
これらの組成物は、獣医学的な使用のために家畜のような動物に投与されても良く、他の治療剤と同様にヒトに臨床的に使用されても良い。一般的には、治療上の有効性のために必要とされる用量は、対象者の体重当たり、約1μgから300mg/kg、更に通常は10μgから30mg/kgの範囲であろう。代替的には、これらの範囲内の用量が、通常は24時間を超えて、所望の治療上の利益が得られるまで経時的に一定の輸液によって投与されて良い。
【0081】
典型的には、その様な組成物は、溶液又は懸濁物のいずれかの注射可能な物として調製される;注射前の液体中の溶液又は懸濁物のために適切な固体の形状が調製されても良い。前記調製物は乳化されても良い。活性成分は、多くの場合には、生理学的に許容され、活性成分に適合する希釈剤又は賦形剤と混合する。適切な希釈剤又は賦形剤は、例えば、水、生理食塩水、デキストロース、又はグリセロールなど、並びにそれらの組み合わせである。さらに、望まれる場合には、前記組成物は、少量の付加的な物質、例えば、湿潤剤又は乳化剤、及び安定剤又はpH緩衝剤などを含有して良い。
【0082】
前記組成物は、注射、例えば、皮下又は静脈内注射によって、従来のように非経口的に投与される。他の様式の投与に適切な更に別の製剤は、坐剤、鼻内エアロゾル、を含み、ある場合には経口製剤を含む。坐剤については、従来のバインダー及び賦形剤は、例えば、ポリアルキレングリコール又はトリグリセリドを含んで良く;その様な坐剤は、0.5%から10%、好ましくは1%から2%の範囲の活性成分を含有する混合物から形成されて良い。経口製剤は、そのように通常利用される賦形剤、例えば、製薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウムなどを含む。これらの組成物は、溶液剤、懸濁剤、錠剤、丸剤、カプセル剤、持続放出型製剤、又は粉末剤の形態をとり、10%から95%、好ましくは25%から70%の活性成分を含有する。これらの経口製剤は、吸収されるまでペプチドを保護することを意図する製剤を含む。
【0083】
前記ペプチド化合物は、中性又は塩の形態として組成物に製剤化されて良い。製薬学的に許容される非毒性の塩は、無機酸、例えば塩酸又はリン酸、あるいは有機酸、例えば、酢酸、シュウ酸、酒石酸、及びマンデル酸などと形成される酸付加塩(遊離のアミノ基と形成される)を含む。遊離のカルボキシル基と形成される塩は、無機塩基、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、又は水酸化鉄、並びに有機塩基、例えば、イソプロピルアミン、トリメチルアミン、2-エチルアミノエタノール、ヒスチジン、及びプロカイン等に由来するものであって良い。
【0084】
結論
上述の記載は説明を意図するものであって、これに制限するもので無いことが理解されるべきである。各種の実施態様及び変形が、本発明の範囲及び精神を逸脱すること無く、本願に開示されている発明に為されて良いことが当業者には明らかであるはずである。したがって、本発明の範囲は、上述の記載に表わされる具体的な実施態様を参照するのではなく、その様な実施態様と均等の全範囲を参照して決定されるべきである。本明細書に挙げた全ての文献は、本発明との関係において使用され得る生物学的機構、治療、方法論、及び概念を説明するために参照される。これらの参照文献が本明細書に記載の発明との関係において従来技術であると認められるべきではない。他に示さない限り、本明細書に挙げた各参照文献は、参照することによって、全ての目的のためにその全体を本明細書に組み込む。
【0085】
参考文献
1. Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
2. Ross, R., Atherosclerosis~an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
3. Bornstein, P., Thrombospondins as matricellular modulators of cell function. J Clin Invest, 2001. 107(8): p. 929-34.
4. Sage, E.H., Regulation of interactions between cells and extracellular matrix: a command performance on several stages. J Clin Invest, 2001. 107(7): p. 781-3.
5. Murphy-Ullrich, J.E., The de-adhesive activity of matricellular proteins: is intermediate cell adhesion an adaptive state? J Clin Invest, 2001. 107(7): p. 785- 90.
6. Elzie, CA. and J.E. Murphy-Ullrich, The N-terminus of thrombospondin: the domain stands apart, lht J Biochem Cell Biol, 2004. 36(6): p. 1090-101.
7. Li, D.Y., G. Faury, D.G. Taylor, E.G. Davis, W.A. Boyle, R.P. Mecham, P. Stenzel, B. Bo ak, and M.T. Keating, Novel arterial pathology in mice and humans hemizygous for elastin. J Clin Invest, 1998. 102(10): p. 1783-7.
8. Aszodi, A., D. Chan, E. Hunziker, J.F. Bateman, and R. Fassler, Collagen π is essential for the removal of the notochord and the formation of intervertebral discs. J Cell Biol, 1998. 143(5): p. 1399-412.
9. Forsberg, E., E. Hirsch, L. Frohlich, M. Meyer, P. Ekblom, A. Aszodi, S. Werner, and R. Fassler, Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc Natl Acad Sci U S A, 1996. 93(13): p. 6594-9.
10. Lawler, J., M. Sunday, V. Thibert, M. Duquette, E.L. George, H. Rayburn, and R.O. Hynes, Thrombospondin- 1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J Clin Invest, 1998. 101(5): p. 982-92.
11. Kyriakides, T.R., J. W. Tarn, and P. Bornstein, Accelerated wound healing in mice with a disruption of the thrombospondin 2 gene. J Invest Dermatol, 1999. 113(5): p. 782-7.
12. Adams, J.C. and J. Lawler, The thrombospondins. Lit J Biochem Cell Biol, 2004. 36(6): p. 961-8.
13. Crawford, S.E., V. Stellmach, J.E. Murphy-Ullrich, S.M. Ribeiro, J. Lawler, R.O. Hynes, G.P. Boivin, and N. Bouck, Thrombospondin- 1 is a major activator of TGF-betal in vivo. Cell, 1998. 93(7): p. 1159-70.
14. Agah, A., T.R. Kyriakides, J. Lawler, and P. Bornstein, The lack of thrombospondin- 1 (TSPl) dictates the course of wound healing in double- TSPl/TSP2-null mice. Am J Pathol, 2002. 161(3): p. 831-9.
15. Yee, K.O., M. Streit, T. Hawighorst, M. Detmar, and J. Lawler, Expression of the type-1 repeats of thrombospondin- 1 inhibits tumor growth through activation of transforming growth factor-beta. Am J Pathol, 2004. 165(2): p. 541-52.
16. Miao, W.M., W.L. Seng, M. Duquette, P. Lawler, C. Laus, and J. Lawle r, Thrombospondin- 1 type 1 repeat recombinant proteins inhibit tumor growth through transforming growth factor-beta-dependent and -independent mechanisms. Cancer Res, 2001. 61(21): p. 7830-9.
17. Yano, K., H. Oura, and M. Detmar, Targeted overexpression of the angiogenesis inhibitor thrombospondin- 1 in the epidermis of transgenic mice prevents ultraviolet-B-induced angiogenesis and cutaneous photo-damage. J Invest Dermatol, 2002. 118(5): p. 800-5.
18. Simantov, R., M. Febbraio, R. Crombie, A.S. Asch, R.L. Nachman, and R.L. Silverstein, Histidine-rich glycoprotein inhibits the antiangiogenic effect of thrombospondin- 1. J Clin Invest, 2001. 107(1): p. 45-52.
19. Bornstein, P., A. Agah, and T.R. Kyriakides, The role of thrombospondins 1 and 2 in the regulation of cell-matrix interactions, collagen fibril formation, and the response to injury. Int J Biochem Cell Biol, 2004. 36(6): p. 1115-25.
20. Lawler, J. and M. Detmar, Tumor progression: the effects of thrombospondin- 1 and -2. Int J Biochem Cell Biol, 2004. 36(6): p. 1038-45.
21. Topol, EJ., J. McCarthy, S. Gabriel, DJ. Moliterno, WJ. Rogers, L.K. Newby, M. Freedman, J. Metivier, R. Cannata, CJ. O'Donnell, K. Kottke-Marchant, G. Murugesan, E.F. Plow, O. Stenina, and G.Q. Daley, Single nucleotide polymorphisms in multiple novel thrombospondin genes may be associated with familial premature myocardial infarction. Circulation, 2001. 104(22): p. 2641-4.
22. Stenina, O.I., T.V. Byzova, J.C. Adams, JJ. McCarthy, EJ. Topol, and E.F. Plow, Coronary artery disease and the thrombospondin single nucleotide polymorphisms. Int J Biochem Cell Biol, 2004. 36(6): p. 1013-30.
23. Raines, E.W., The extracellular matrix can regulate vascular cell migration, proliferation, and survival: relationships to vascular disease. Int J Exp Pathol, 2000. 81(3): p. 173-82.
24. Roth, JJ., V. Gahtan, J.L. Brown, C. Gerhard, V.K. Swami, V.L. Rothman, T.N. Tulenko, and G.P. Tuszynski, Thrombospondin- 1 is elevated with both intimal hyperplasia and hypercholesterolemia. J Surg Res, 1998. 74(1): p. 11-6.
25. Lawler, J., The functions of thrombospondin- 1 and-2. Curr Opin Cell Biol, 2000. 12(5): p. 634-40.
26. Chen, H., M.E. Herndon, and J. Lawler, The cell biology of thrombospondin- 1. Matrix Biol, 2000. 19(7): p. 597-614.
27. Majack, R.A., S.C. Cook, and P. Bornstein, Control of smooth muscle cell growth by components of the extracellular matrix: autocrine role for thrombospondin. Proc Natl Acad Sci U S A, 1986. 83(23): p. 9050-4.
28. Schultz-Cherry, S., S. Ribeiro, L. Gentry, and J.E. Murphy-Ullrich, Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. J Biol Chem, 1994. 269(43): p. 26775-82.
29. Fischer, J.W., M. Stoll, A. W. Hahn, and T. Unger, Differential regulation of thrombospondin- 1 and fibronectin by angiotensin JI receptor subtypes in cultured endothelial cells. Cardiovasc Res, 2001. 51(4): p. 784-91.
30. Lymn, J.S., M.K. Patel, G.F. Clunn, SJ. Rao, KX. Gallagher, and A.D. Hughes, Thrombospondin-1 differentially induces chemotaxis and DNA synthesis of human venous smooth muscle cells at the receptor-binding level. J Cell Sci, 2002. 115(Pt 22): p. 4353-60.
31. Patel, M.K., J.S. Lymn, G.F. Clunn, and A.D. Hughes, Thrombospondin-1 is a potent mitogen and chemoattractant for human vascular smooth muscle cells. Arterioscler Thromb Vase Biol, 1997. 17(10): p. 2107-14.
32. Rodriguez-Manzaneque, J.C., T.F. Lane, M.A. Ortega, R.O. Hynes, J. Lawler, and M.L. Iruela-Arispe, Thrombospondin-1 suppresses spontaneous tumor growth and inhibits activation of matrix metalloproteinase-9 and mobilization of vascular endothelial growth factor. Proc Natl Acad Sci U S A, 2001. 98(22): p. 12485-90.
33. Bein, K. and M. Simons, Thrombospondin type 1 repeats interact with matrix metalloproteinase 2. Regulation of metalloproteinase activity. J Biol Chem, 2000. 275(41): p. 32167-73.
34. Lutgens, E., M. Gijbels, M. Smook, P. Heeringa, P. Gotwals, V.E. Koteliansky, and MJ. Daemen, Transforming growth factor-beta mediates balance between inflammation and fibrosis during plaque progression. Arterioscler Thromb Vase Biol, 2002. 22(6): p. 975-82.
35. Mallat, Z., A. Gojova, C. Marchiol-Fournigault, B. Esposito, C. Kamate, R. Merval, D. Fradelizi, and A. Tedgui, Inhibition of transforming growth factorbeta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ Res, 2001. 89(10): p. 930-4.
36. Luttun, A., E. Lutgens, A. Manderveld, K. Maris, D. Collen, P. Carmeliet, and L. Moons, Loss of matrix metalloproteinase-9 or matrix metalloproteinase- 12 protects apolipoprotein E- deficient mice against atherosclerotic media destruction but differentially affects plaque growth. Circulation, 2004. 109(11): p. 1408-14.
37. Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
38. Stefanidakis, M., T. Ruohtula, N. Borregaard, CG. Gahniberg, and E. Koivunen, Intracellular and cell surface localization of a complex between alphaMbeta2 integrin and promatrix metalloproteinase-9 progelatinase in neutrophils. J Immunol, 2004. 172(11): p. 7060-8.
39. Wrana, J.L., L. Attisano, J. Carcamo, A. Zentella, J. Doody, M. Laiho, X.F. Wang, and J. Massague, TGF beta signals through a heteromeric protein kinase receptor complex. Cell, 1992. 71(6): p. 1003-14.
40. Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
41. Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
42. Ross, R., Atherosclerosis an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
43. FaIk, E., P.K. Shah, and V. Fuster, Coronary plaque disruption. Circulation, 1995. 92(3): p. 657-71.
【図面の簡単な説明】
【0086】
【図1】図1は、アテローム性動脈硬化症の病変において直接的な関係性を有する可能性がある、各種のTSP-1細胞及びサイトカイン結合ドメインを示す。
【図2A】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(A)TSR-1及びTSR-2には2つの推定上のβ2インテグリン結合ドメインが存在する。これら2つの結合ドメインは、TGF-β1についての2つの結合ドメインが隣接(及び1つのアミノ酸が重複)している。本発明者は、box中に示す2つのDxGWモチーフを変異させた。
【図2B】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(B)マウスマクロファージ細胞株(Raw)の一過的なトランスフェクションを使用して、本発明者は、TSP-1がMMP-9活性を低減すること(p<0.005(活性型MMP-9/総MMP-9 ELISA))を認めている。この効果は、TSP変異体:DM及びより少なくはMut1によって無効にされる。
【図2C】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(C)同じ細胞系を使用して、本発明者は、TSP-1がTGF-β1活性を増大することを認めている(p<0.005(細胞溶解物を使用して、TGF-β1プロモーター応答エレメントを介するルシフェラーゼ誘導を測定する(39)))。この作用は、TSP-1変異体Mut1及びMut2の双方によって無効にされる。まとめると、TSR-1及びTSR-2におけるDxGWモチーフは、この細胞アッセイ系においてTGF-β1活性化及びMMP-9阻害に対するTSP-1の作用を媒介するようである。
【図3A−C】図3は、アテローム性動脈硬化症病変に対するTSP-1欠失の組織学的特性を示す。(A)ヘマトキシリン及びエオシン染色大動脈弁尖。右側の棒グラフは、病変領域(内膜)を表わす。病変領域には変化が認められなかった。(B)Moma-2抗体染色(PEが接合され、赤色蛍光を表わす)は、TSP-1欠失マウスにおいて大動脈弁尖内の病変にマクロファージの増大が存在することを明らかにする(p<0.01)。(C)マクロファージの増大と一致して、Red-O染色は、TSP-1欠失マウスにおいて病変内の脂質沈着の増大が存在することを明らかにする(すなわち、泡沫細胞、p<0.05)。
【図3D−F】図3は、アテローム性動脈硬化症病変に対するTSP-1欠失の組織学的特性を示す。(D)平滑筋細胞アクチン抗体染色(FITCが接合され、緑色蛍光を表わす)。内膜病変内には平滑筋細胞(smc)の染色は存在せず、培地中の総smc含量は2つの群の間で変化がない。(E)コラーゲンについてのpicosirius red polarization染色が、大動脈弁尖における病変内のコラーゲンの損失を明らかにする(黄色-赤色染色の低減)。両種のマウスにおける弁尖及び弁の弁構造の周囲に強い(黄色-赤色)染色が存在することに注意すべきである。しかし、(矢印で示す)病変内において、TSPノックアウトマウスでは染色の低減が存在する。(F)エラスチンについてのVerhoeff's染色は、TSP-1欠失を有する病変の発生領域における中膜及び内膜のエラスチンの損失の増大を明らかにする(p<0.05)。
【図4】図4は、ApoE-/-TSP-/-(n=3マウス)及びApoE-/-マウス(n=3マウス)由来の大動脈弓の組織可溶化物からのTGF-βELISAの結果を示す。総TGF-β1のレベルについて2つの群の間に差は無いが、ApoE-/-TSP-/-マウスの大動脈内の活性型TGF-β1レベルでは有意な差(p<0.05)が存在する。
【図5】図5は、ApoE-/-TSP-/-マウス(n=3マウス)及びApoE-/-マウス(n=3マウス)由来の大動脈弓の組織可溶化物からのMMP-9ウエスタンブロットを示す。各レーンは、個々のマウスを特定する。TSP-/-マウスにおけるプロMMP-9(105kb)及び活性型MMP-9(95kb)のバンドの双方の増大が存在する。
【図6】図6は、TSP-1(40μg/ml)、BSA(5%)、又はフィブロネクチン(0.1μg/ml)の各々を事前にウェルにプレーティングした接着アッセイの結果を示す。ペプチドの実験については、TSP-1のウェルに導入される前に、細胞を、個々のペプチド(0.01M)と懸濁物中で30分間混合する。
【図7】図7は、マクロファージ上のaMβ2インテグリン部位への結合についてのTSP-1及びプロMMP-9による競合に関する機構を示す(A)。TSP-1の増大の存在下では、MMP-9活性化の低減及びTGF-β1活性化の増大と共にTSP-1結合の増大が存在する(B)。プロMMP-9の増大の背景においては、TSP-1媒介TGF-β1活性化の低減及びMMP-9活性化の増大と共にMMP-9結合の増大が存在する(C)。
【技術分野】
【0001】
本願は、2005年6月24日に出願の米国仮特許出願第60/693,574号の優先権の利益を享受する。
【0002】
本発明は、アテローム性動脈硬化症のプラークを安定化し、それにより、例えば、心筋梗塞、脳卒中、及び急性肢虚血を引き起こすプラーク破裂の発生を低減する、マトリックス細胞のタンパク質であるトロンボスポンジン-1(TSP-1)を利用する治療及び関連する組成物に関する。さらに、本発明は、上述の疾患を含む各種の背景下におけるプラークの安定化に使用するための一定の合成ペプチドを含む、各種のペプチドにも関連する。
【背景技術】
【0003】
アテローム性動脈硬化症の致命的な合併症、例えば、急性心筋梗塞及び脳卒中は、局所的な血管の炎症を引き起こすコレステロール及びその酸化したリン脂質の慢性的な沈着に由来するものである(1)。アテローム性動脈硬化症の病変は、動脈壁の細胞と細胞外マトリックス(ECM)構成成分との間の相互作用を部分的に介して、動脈壁において形成され、進行する(1,2)。アテローム性動脈硬化症の発症と共に、血管壁の細胞構成成分とECM構成成分の双方に変化が起こる。ECMは、アテローム性動脈硬化症の病変内における細胞の振る舞いに作用する細胞結合タンパク質及び増殖因子の貯蔵所として認識されている(3,4)。言い換えると、これらのプロセスの制御は、細胞-細胞及び細胞-ECM相互作用に直接的な効果を有する可能性があり、言い換えると、破裂する傾向があり、それに続く下流の事象、例えば心筋梗塞及び脳卒中に関連し得るアテローム性動脈硬化症のプラークのフェノタイプに直接影響を与える可能性がある。
【0004】
その環境について特異的な背景の情報を細胞に与えるECMの微量なタンパク質成分は、「マトリックス細胞の」タンパク質と称されている(4)。マトリック細胞タンパク質は、他のECM構成成分、多特異性細胞表面受容体、並びに増殖因子と相互作用して、細胞-マトリックス相互作用を調節する。これらのタンパク質は細胞機能を調節するが、原線維、コラーゲン、及びエラスチンのような他のECM成分がするように構造の物理的特性又は組織に直接寄与することはないようである(5)。これらのタンパク質は、細胞外マトリックスと細胞表面との間の接触面で働き、細胞の振る舞いを制御する。さらに、構造タンパク質は正常な血管の発生に重要な役割を有するが、マトリックス細胞のタンパク質は創傷時にのみ成人の血管において発現する(6)。具体的には、これらのタンパク質は、組織修復において主な機能を有しており、その背景において作用することで、細胞-マトリックス相互作用によって細胞機能に影響を与えるようである。その様なものとして、生殖系列における構造ECMタンパク質の標的破壊は重度な又は致死性のフェノタイプを生じるが、マトリックス細胞のタンパク質をコードする遺伝子のノックアウトは、見かけ上正常であるか又は僅かなフェノタイプを生じる(7-11)。
【0005】
TSP-1は、多数の結合ドメインを有する原始型のマトリックスタンパク質である(3,12)。TSP-1は創傷時に誘導される。当該タンパク質の無発現変異について同型接合のマウスは顕著な発生の欠陥を有さず、これらのマウスにおける主要なフェノタイプは、急性の斑であり、肺において肺炎を構成する。しかしながら、各種の疾患モデルにおいて、これらのマウスの明確なフェノタイプが存在する(10)。例えば、創傷修復モデルでは、TSP-1欠損は、炎症性細胞応答の変化及び創傷治癒の遅延に関連する。他の研究では、TSP-1は血管新生を阻害することが示されている(13-19)。その様なものとして、新生細胞におけるTSP-1の標的発現は臨床において関心がもたれており、その修飾物は臨床試験のフェーズIIにおいて現在評価されている(20)。
【非特許文献1】Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
【非特許文献2】Ross, R., Atherosclerosis~an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
【非特許文献3】Bornstein, P., Thrombospondins as matricellular modulators of cell function. J Clin Invest, 2001. 107(8): p. 929-34.
【非特許文献4】Sage, E.H., Regulation of interactions between cells and extracellular matrix: a command performance on several stages. J Clin Invest, 2001. 107(7): p. 781-3.
【非特許文献5】Murphy-Ullrich, J.E., The de-adhesive activity of matricellular proteins: is intermediate cell adhesion an adaptive state? J Clin Invest, 2001. 107(7): p. 785- 90.
【非特許文献6】Elzie, CA. and J.E. Murphy-Ullrich, The N-terminus of thrombospondin: the domain stands apart, lht J Biochem Cell Biol, 2004. 36(6): p. 1090-101.
【非特許文献7】Li, D.Y., G. Faury, D.G. Taylor, E.G. Davis, W.A. Boyle, R.P. Mecham, P. Stenzel, B. Bo ak, and M.T. Keating, Novel arterial pathology in mice and humans hemizygous for elastin. J Clin Invest, 1998. 102(10): p. 1783-7.
【非特許文献8】Aszodi, A., D. Chan, E. Hunziker, J.F. Bateman, and R. Fassler, Collagen π is essential for the removal of the notochord and the formation of intervertebral discs. J Cell Biol, 1998. 143(5): p. 1399-412.
【非特許文献9】Forsberg, E., E. Hirsch, L. Frohlich, M. Meyer, P. Ekblom, A. Aszodi, S. Werner, and R. Fassler, Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc Natl Acad Sci U S A, 1996. 93(13): p. 6594-9.
【非特許文献10】Lawler, J., M. Sunday, V. Thibert, M. Duquette, E.L. George, H. Rayburn, and R.O. Hynes, Thrombospondin- 1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J Clin Invest, 1998. 101(5): p. 982-92.
【非特許文献11】Kyriakides, T.R., J. W. Tarn, and P. Bornstein, Accelerated wound healing in mice with a disruption of the thrombospondin 2 gene. J Invest Dermatol, 1999. 113(5): p. 782-7.
【非特許文献12】Adams, J.C. and J. Lawler, The thrombospondins. Lit J Biochem Cell Biol, 2004. 36(6): p. 961-8.
【非特許文献13】Crawford, S.E., V. Stellmach, J.E. Murphy-Ullrich, S.M. Ribeiro, J. Lawler, R.O. Hynes, G.P. Boivin, and N. Bouck, Thrombospondin- 1 is a major activator of TGF-betal in vivo. Cell, 1998. 93(7): p. 1159-70.
【非特許文献14】Agah, A., T.R. Kyriakides, J. Lawler, and P. Bornstein, The lack of thrombospondin- 1 (TSPl) dictates the course of wound healing in double- TSPl/TSP2-null mice. Am J Pathol, 2002. 161(3): p. 831-9.
【非特許文献15】Yee, K.O., M. Streit, T. Hawighorst, M. Detmar, and J. Lawler, Expression of the type-1 repeats of thrombospondin- 1 inhibits tumor growth through activation of transforming growth factor-beta. Am J Pathol, 2004. 165(2): p. 541-52.
【非特許文献16】Miao, W.M., W.L. Seng, M. Duquette, P. Lawler, C. Laus, and J. Lawle r, Thrombospondin- 1 type 1 repeat recombinant proteins inhibit tumor growth through transforming growth factor-beta-dependent and -independent mechanisms. Cancer Res, 2001. 61(21): p. 7830-9.
【非特許文献17】Yano, K., H. Oura, and M. Detmar, Targeted overexpression of the angiogenesis inhibitor thrombospondin- 1 in the epidermis of transgenic mice prevents ultraviolet-B-induced angiogenesis and cutaneous photo-damage. J Invest Dermatol, 2002. 118(5): p. 800-5.
【非特許文献18】Simantov, R., M. Febbraio, R. Crombie, A.S. Asch, R.L. Nachman, and R.L. Silverstein, Histidine-rich glycoprotein inhibits the antiangiogenic effect of thrombospondin- 1. J Clin Invest, 2001. 107(1): p. 45-52.
【非特許文献19】Bornstein, P., A. Agah, and T.R. Kyriakides, The role of thrombospondins 1 and 2 in the regulation of cell-matrix interactions, collagen fibril formation, and the response to injury. Int J Biochem Cell Biol, 2004. 36(6): p. 1115-25.
【非特許文献20】Lawler, J. and M. Detmar, Tumor progression: the effects of thrombospondin- 1 and -2. Int J Biochem Cell Biol, 2004. 36(6): p. 1038-45.
【非特許文献21】Topol, EJ., J. McCarthy, S. Gabriel, DJ. Moliterno, WJ. Rogers, L.K. Newby, M. Freedman, J. Metivier, R. Cannata, CJ. O'Donnell, K. Kottke-Marchant, G. Murugesan, E.F. Plow, O. Stenina, and G.Q. Daley, Single nucleotide polymorphisms in multiple novel thrombospondin genes may be associated with familial premature myocardial infarction. Circulation, 2001. 104(22): p. 2641-4.
【非特許文献22】Stenina, O.I., T.V. Byzova, J.C. Adams, JJ. McCarthy, EJ. Topol, and E.F. Plow, Coronary artery disease and the thrombospondin single nucleotide polymorphisms. Int J Biochem Cell Biol, 2004. 36(6): p. 1013-30.
【非特許文献23】Raines, E.W., The extracellular matrix can regulate vascular cell migration, proliferation, and survival: relationships to vascular disease. Int J Exp Pathol, 2000. 81(3): p. 173-82.
【非特許文献24】Roth, JJ., V. Gahtan, J.L. Brown, C. Gerhard, V.K. Swami, V.L. Rothman, T.N. Tulenko, and G.P. Tuszynski, Thrombospondin- 1 is elevated with both intimal hyperplasia and hypercholesterolemia. J Surg Res, 1998. 74(1): p. 11-6.
【非特許文献25】Lawler, J., The functions of thrombospondin- 1 and-2. Curr Opin Cell Biol, 2000. 12(5): p. 634-40.
【非特許文献26】Chen, H., M.E. Herndon, and J. Lawler, The cell biology of thrombospondin- 1. Matrix Biol, 2000. 19(7): p. 597-614.
【非特許文献27】Majack, R.A., S.C. Cook, and P. Bornstein, Control of smooth muscle cell growth by components of the extracellular matrix: autocrine role for thrombospondin. Proc Natl Acad Sci U S A, 1986. 83(23): p. 9050-4.
【非特許文献28】Schultz-Cherry, S., S. Ribeiro, L. Gentry, and J.E. Murphy-Ullrich, Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. J Biol Chem, 1994. 269(43): p. 26775-82.
【非特許文献29】Fischer, J.W., M. Stoll, A. W. Hahn, and T. Unger, Differential regulation of thrombospondin- 1 and fibronectin by angiotensin JI receptor subtypes in cultured endothelial cells. Cardiovasc Res, 2001. 51(4): p. 784-91.
【非特許文献30】Lymn, J.S., M.K. Patel, G.F. Clunn, SJ. Rao, KX. Gallagher, and A.D. Hughes, Thrombospondin-1 differentially induces chemotaxis and DNA synthesis of human venous smooth muscle cells at the receptor-binding level. J Cell Sci, 2002. 115(Pt 22): p. 4353-60.
【非特許文献31】Patel, M.K., J.S. Lymn, G.F. Clunn, and A.D. Hughes, Thrombospondin-1 is a potent mitogen and chemoattractant for human vascular smooth muscle cells. Arterioscler Thromb Vase Biol, 1997. 17(10): p. 2107-14.
【非特許文献32】Rodriguez-Manzaneque, J.C., T.F. Lane, M.A. Ortega, R.O. Hynes, J. Lawler, and M.L. Iruela-Arispe, Thrombospondin-1 suppresses spontaneous tumor growth and inhibits activation of matrix metalloproteinase-9 and mobilization of vascular endothelial growth factor. Proc Natl Acad Sci U S A, 2001. 98(22): p. 12485-90.
【非特許文献33】Bein, K. and M. Simons, Thrombospondin type 1 repeats interact with matrix metalloproteinase 2. Regulation of metalloproteinase activity. J Biol Chem, 2000. 275(41): p. 32167-73.
【非特許文献34】Lutgens, E., M. Gijbels, M. Smook, P. Heeringa, P. Gotwals, V.E. Koteliansky, and MJ. Daemen, Transforming growth factor-beta mediates balance between inflammation and fibrosis during plaque progression. Arterioscler Thromb Vase Biol, 2002. 22(6): p. 975-82.
【非特許文献35】Mallat, Z., A. Gojova, C. Marchiol-Fournigault, B. Esposito, C. Kamate, R. Merval, D. Fradelizi, and A. Tedgui, Inhibition of transforming growth factorbeta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ Res, 2001. 89(10): p. 930-4.
【非特許文献36】Luttun, A., E. Lutgens, A. Manderveld, K. Maris, D. Collen, P. Carmeliet, and L. Moons, Loss of matrix metalloproteinase-9 or matrix metalloproteinase- 12 protects apolipoprotein E- deficient mice against atherosclerotic media destruction but differentially affects plaque growth. Circulation, 2004. 109(11): p. 1408-14.
【非特許文献37】Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
【非特許文献38】Stefanidakis, M., T. Ruohtula, N. Borregaard, CG. Gahniberg, and E. Koivunen, Intracellular and cell surface localization of a complex between alphaMbeta2 integrin and promatrix metalloproteinase-9 progelatinase in neutrophils. J Immunol, 2004. 172(11): p. 7060-8.
【非特許文献39】Wrana, J.L., L. Attisano, J. Carcamo, A. Zentella, J. Doody, M. Laiho, X.F. Wang, and J. Massague, TGF beta signals through a heteromeric protein kinase receptor complex. Cell, 1992. 71(6): p. 1003-14.
【非特許文献40】Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
【非特許文献41】Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
【非特許文献42】Ross, R., Atherosclerosis an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
【非特許文献43】FaIk, E., P.K. Shah, and V. Fuster, Coronary plaque disruption. Circulation, 1995. 92(3): p. 657-71.
【発明の開示】
【発明が解決しようとする課題】
【0006】
冠状動脈性心臓病に関して、最近の相関的及び記述的研究は、TSP-1がアテローム性動脈硬化症及び心筋梗塞において、不明確であり、且つ、未知ではあるが、役割を有している可能性があることを示唆している(21,22)。まず、調節された遺伝的変異研究は、この遺伝子のミスセンス「変異体」が、アテローム性動脈硬化症及び心筋梗塞の双方のリスクの増大に関連することを示唆している(21)。次に、ヒトのサンプルを用いた病理学的研究は、TSP-1がアテローム性動脈硬化症の病変に存在しているが、天然の血管壁には存在しないことを示している(23,24)。しかしながら、現在において、アテローム性動脈硬化症のフェノタイプに対する、プラーク内におけるTSP-1発現の意味は未知である。
【課題を解決するための手段】
【0007】
本発明は、アテローム性動脈硬化症のプラークを安定化し、任意の血管床(例えば、冠状動脈、大脳、又は肢)におけるアテローム性動脈硬化症の病変の破裂する傾向を低減するTSP-1を利用する方法及び関連する組成物を提供する。本発明は、TSP-1及びTSP-1に関連する一定の合成ペプチドを含む関連ペプチドを利用する組成物も提供する。その様なペプチドは、心筋梗塞、脳卒中、及び急性肢虚血のような疾患の状態を含む、各種の背景におけるプラークの安定化において有用性がある。これらのペプチドのいくつかは、TGF-β
1及びMMP-9活性を調節するためのTSP-1の活性部位に関連する1つ以上の配列を含む。
【0008】
ある実施態様では、本発明は、MMP-9活性化に作用することが認められた(TSP-1)に基づくペプチド配列を提供する。本発明者は、この活性についての機構は、アテローム性動脈硬化症の病変内においてMMP-9の機能と競合して白血球(主に単球)に結合する、TSP-1に特有のβ2-インテグリン結合部位によるものであると主張する。さらに、ある実施態様では、本発明は、TGF-β1活性を制御することが知られているTSP-1内の配列に基づくペプチド配列を提供する。その様な配列は、プラークの安定性に利用し得る。最後に、本発明者は、TSP-1のTGF-β1及びMMP-9活性並びにそのin vivoにおけるプラーク安定化効果の双方を摸倣する、上述の活性部位に基づく各種の特異的な合成ペプチドの配列を提供する。
【0009】
本発明の各種の実施態様の説明を容易にして、本発明を為して使用する際に用いる各種の要素及び成分の理解を提供するために、本発明において使用する以下の用語は、以下の意味を有する。
【発明を実施するための最良の形態】
【0010】
定義
各種の感染症の治療方法は、「予防上の」処置又は「治療上の」処置を含む意味を有する。「予防上の」処置は、疾患に関連する病理を発症するリスクを低減するために疾患の兆候を示していないか又は疾患の初期の兆候を示す対象に施す処置である。
【0011】
本明細書で使用する用語「治療上の」は、病理学的な兆候を減少させるか又は除くための、病理学的な兆候を示す対象に施す処置を意味する。
【0012】
本明細書で使用する用語「治療上の有効量」は、化合物を投与する対象に対して有益な効果を提供するために十分な化合物の量を意味する。有益な効果は、アテローム性動脈硬化症のプラークの安定化を意味する。
【0013】
本明細書で使用する用語「ペプチド」、「ポリペプチド」、及び「タンパク質」は、互換的に使用され、アミド結合で共有結合している少なくとも2つのアミノ酸のポリマー配列を示す。
【0014】
本明細書で使用する用語「相同の」は、2つのペプチド間で類似するアミノ酸配列を示す。双方のペプチドのアミノ酸の位置が同一のアミノ酸で占められている際に、それらは該位置で相同である。かくして、「実質的に相同」は、大部分で相同あるが、全てが相同ではなく、相同配列として大部分又は全ての活性を保持するアミノ酸配列を意味する。本明細書で使用する「実質的に相同」は、ある配列が、少なくとも80%同一であり、好ましくは少なくとも90%、更に好ましくは98%の参照ペプチドに対する相同性を有する事を意味する。
【0015】
ポリペプチドは、アミノ酸残基の配列として本明細書で開示している。それらの配列は、左から右へとアミノ末端からカルボキシル末端の方向で記載している。標準的な用語に従えば、アミノ酸残基配列は、下記の一文字又は三文字のコードのいずれかで表記される。
【0016】
【表1】
【0017】
本発明のポリペプチドの構造に修飾及び変更が為されて良く、それでもTSP-1ペプチド様特性を有する分子が得られる。例えば、ペプチドの活性の相当の損失無く、配列中のあるアミノ酸が他のアミノ酸に置換されても良い。ポリペプチドの相互作用能及び性質が、ポリペプチドの生物学的機能の活性を規定するため、あるアミノ酸配列の置換がポリペプチド配列(言うまでも無く、その基礎を成すDNAコード配列)に為されて良く、それにもかかわらず同様の特性を有するポリペプチドが得られる。
【0018】
その様な変更を為す場合には、アミノ酸のヒドロパシーが考慮されて良い。ペプチドの相互作用的生物学的機能を与えるアミノ酸のヒドロパシー指数の重要性は、当該技術分野において一般的に理解されている(Kyte, J. and R. F. Doolittle 1982)。あるアミノ酸が同様なヒドロパシー指数(スコア)を有する他のアミノ酸に置換されて良く、それでも同様の生物学的活性を有するポリペプチドを生じさせることが知られている。各アミノ酸は、その疎水性度及び電荷特性に基づいてヒドロパシーが定められている。それらのヒドロパシーは、イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(-0.4);スレオニン(-0.7);セリン(-0.8);トリプトファン(-0.9);チロシン(-1.3);プロリン(-1.6);ヒスチジン(-3.2);グルタミン(-3.5);グルタミン(-3.5);アスパルテート(-3.5);アスパラギン(-3.5);リジン(-3.9);及びアルギニン(-4.5)である。
【0019】
アミノ酸の相対的なヒドロパシーの特性が、結果として生じるポリペプチドの二次構造を決定し、言い換えると、酵素、基質、受容体、抗体、及び抗原などのような他の分子とのポリペプチドの相互作用を規定する。アミノ酸は、同様のヒドロパシー指数を有する他のアミノ酸によって置換されて良く、それでも機能的に等価のペプチドが得られることが当該技術分野において既知である。その様な変更において、ヒドロパシー指数が±2以内のアミノ酸置換が好ましく、±1以内のアミノ酸置換がより好ましく、±0.5以内のアミノ酸置換が更に好ましい。
【0020】
同様のアミノ酸の置換は、親水性度に基づいて為されても良く、特にそれによって生じる生物学的機能が等価なポリペプチド又はペプチドは、免疫学的な実施態様における使用が意図される。参照によって本明細書に取り込む米国特許第4,554,101号は、隣接するアミノ酸の親水性度によって決定されるポリペプチドの最大の局所的な平均親水性度は、その免疫原性及び抗原性、すなわちポリペプチドの生物学的特性と関連すると記述している。米国特許第4,554,101号に詳述しているように、以下の親水性度の値が、アミノ酸残基:アルギニン(+3.0);リジン(+3.0);アスパルテート(+3.0±1);グルタメート(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);プロリン(-0.5±1);スレオニン(-0.4);アラニン(-0.5);ヒスチジン(-0.5);システイン(-1.0);メチオニン(-1.3);バリン(-1.5);ロイシン(-1.8);イソロイシン(-1.8);チロシン(-2.3);フェニルアラニン(-2.5);トリプトファン(-3.4)に定められている。アミノ酸は、同様の親水性度を有する他のアミノ酸に置換されて良く、それでも生物学的に等価な、特に免疫学的に等価なポリペプチドが得られることが理解されている。その様な変更は、親水性度が±2以内のアミノ酸置換が好ましく、±1以内のアミノ酸置換が特に好ましく、±0.5以内のアミノ酸置換が更に好ましい。
【0021】
したがって上述したように、アミノ酸置換は、一般的には、アミノ酸側鎖の置換基、例えば、それらの疎水性、親水性、電荷、及び大きさなどの相対的な類似性に基づく。上述の各種の特性を考慮に入れた置換の例は当業者に既知であり、アルギニンとリジン;グルタメートとアスパルテート;セリンとスレオニン;グルタミンとアスパラギン;及びバリンと、ロイシンと、イソロイシンを含む(以下の表2参照)。本発明は、上記のペプチドの機能的又は生物学的等価物を意図する。
【0022】
【表2】
【0023】
ポリペプチドの生物学的又は機能的等価物は、部位特異的突然変異導入法を用いて調製しても良い。部位特異的突然変異導入法は、基礎とするDNAの特異的な突然変異による、それらの配列に由来する第二世代のポリペプチド、又は生物学的機能の等価なポリペプチド若しくはペプチドの調製に有用な技術である。上述のように、その様な変更は、アミノ酸置換が望まれる場合に望ましい。前記技術は、DNAに1つ以上のヌクレオチド配列の変更を導入することによって、例えば上述のものの一つ以上を含む配列の変異体を調製し試験をする準備能力を更に提供する。部位特異的突然変異導入法は、所望の変異体のDNA配列をコードする特異的なオリゴヌクレオチド配列、並びに欠失接合部の両端に安定な二本鎖を形成するために十分な大きさ及び配列の複雑性を有する一次配列を提供するのに十分な数の隣接するヌクレオチドの使用によって、変異体の製造を可能にする。
【0024】
アミノ酸残基が、ペプチドの機能を変えること無く、標準的な分子生物学的技術を用いることによって、本発明のTSP-1ペプチドに付加されるか又は欠失されて良い。例えば、TSP-1ペプチドの末端部位が除去されて、切断型のペプチドが作製されて良いが、前記切断型のペプチドは、タンパク質タンパク質相互作用を介するTGF-β1又はMMP-9の調節、並びにアテローム性動脈硬化症のプラークの安定化を含む本発明のTSP-1ペプチドの機能的な活性を保持する。
【0025】
本発明のポリペプチドは、当業者によく知られた標準的な技術によって調製される。その様な技術は、ポリペプチドを含有することが既知の組織からの単離及び精製、そのようなポリペプチドをコードするクローニングしたDNAからの形質転換細胞を用いる発現、合成ペプチド製造システムの使用を含むが、それらに限らない。
【実施例】
【0026】
TSP-1構造ドメイン及びそのマトリックス結合パートナー
TSP-1は、150kDaの分子量の鎖を有する三量体である。TSP-1の各サブユニットは、幾つかの構造ドメインからなる(12)。これらのドメインは、アミノ末端ドメイン、プロコラーゲンドメイン、タイプ1、タイプ2、及びタイプ3反復配列、並びにカルボキシル末端ドメインとして広範に定義されている(25,26)。各ドメインは、図1に示すように多数の細胞機能の調節において重要である事が示されている。さらに、特異的なリガンド結合部位が、TSP-1について定義されている(図1)。これらは、リポタンパク質受容体関連タンパク質(LRP)、CD36、β3インテグリン、及びインテグリン関連タンパク質(IAP)、並びにTGF-β1及びPDGFを含むサイトカインを含む(3,12,27,28)。その多数の結合部位を介して、TSP-1は膜タンパク質及びサイトカインを有する細胞表面で機能しているようである。この様に、TSP-1は、シグナル伝達及び転写を含む細胞のフェノタイプに影響を与える。各細胞が異なる種類の受容体を発現するため、TSP-1によって形成される複合体の組成及び個々の細胞応答は、各細胞種によって変化する。例えば、TSP-1は、平滑筋細胞移動を促進するが、内皮細胞移動を阻害する(29-31)。
【0027】
現在研究されている主な器官系において、TSP-1の生物学の大部分が、TGF-β1の細胞外活性化に寄与している(10,13-15)。具体的には、TSP-1の欠損は、TGF-β1の活性を低減し、個々の器官におけるTGF-β1欠損のフェノタイプに似ていることが示されている。例えば、TSP-1ノックアウトマウスの研究では、TSP-1の欠損が、TGF-β1ノックアウトマウスにおける知見と同様に、局所的な歯槽炎及び慢性的な肺炎を引き起こす。さらに、TGF-β1活性化について十分に認められるTSP-1の短いペプチド断片(図2に示す430から433のアミノ酸残基に対応するKRFK)による生物学的な相補性が、このフェノタイプを救い、この背景におけるフェノタイプの変化を表わす重要な細胞プロセスとしてのTGF-β1の活性化に影響を及ぼす。
【0028】
TSP-1は、マトリックスメタロプロテイナーゼ(MMP)を含む細胞外プロテアーゼの活性も制御する(MMP)(32,33)。癌の血管新生では、TSP-1の標的発現が、抗血管新生特性と一致するMMP-9活性の低減と関連する。MMP-2及びMMP-9は、TSP-1に結合することが示されている(33)。しかしながら、TSP-1に対するMMPの結合部位は明らかにされておらず、これらのプロテアーゼの活性を制御する機構も明らかにされていない。
【0029】
マトリックス代謝、白血球の補充、及び平滑筋細胞移動を含むが、それらに限らないアテローム性動脈硬化症のプラークのフェノタイプにおけるTGF-β1及びMMP-9の双方の役割を示す実験データが存在する。さらに、上述のように、TSP-1は、組織損傷に応答して、これらのリガンドの各々の活性を制御することが示されている。
【0030】
本発明者は、TSP-1が、マウスのアテローム性動脈硬化症のプラークにおいて、プラークを安定化する役割を有することを発見した。さらに、本発明者は、TGF-β1及びMMP-9に関連して、これらの活性を制御するTSP-1の活性ペプチドドメインを同定した。最後に、本発明者は、これらの活性ドメインに基づくペプチド配列を含有するペプチドを設計し、代表的なペプチドが、アテローム性動脈硬化症のマウスモデルにおいてin vivoのTSP-1の効果を刺激することを示すことができた。
【0031】
TSP-1の欠失はアテローム性動脈硬化症のプラークを変化させる
本発明者は、TSP-1マトリックス細胞タンパク質がプラーク安定化効果を有することを示すことができた。本発明者は、TSP-/-ApoE-/-ダブルノックアウトマウスを作製して、病変の形成及び組成に対するTSP-1欠失の役割を試験した。食事(3%脂肪)の24週後に、TSP-/-ApoE-/-マウス及びApoE-/-対照マウスを屠殺した。TSPノックアウトマウスと対象マウスとの間には、体重、総コレステロール、HDLコレステロール、及びトリグリセリドに優位な差はなかった。
【0032】
H&E染色を用いるアテローム性動脈硬化症の病変の程度についての大動脈洞の特性決定は、双方の群における同様な病変を明らかにした(ノックアウトマウスについては701,000μm2±95,000μm2であるのに対し、対照マウスについては724,000μm2±103,000μm2)。本発明者は、病変の組成について大動脈弁尖の詳細な形態評価を実施した(図3)。TSP-1の欠損は、内膜及び中膜の双方において病変の形態に顕著な影響を有していた。具体的には、TSPノックアウトマウスでは、(1)内膜の病変における炎症(マクロファージ)の増大、(2)内膜の病変内における脂質沈着の増大、(3)病変内におけるマトリックス(コラーゲン及びエラスチン)沈着の低減、(4)これらのマウスの内膜領域におけるコラーゲン及びエラスチン含量の低減によって示されるような、内膜の損失を伴う内膜拡張、(5)内膜内における総平滑筋細胞含量の無変化が存在した。まとめると、これらの結果は、TSP-1の欠失は、プラーク内の炎症と線維症との間のバランスを変化させることを示す。これが、細胞外マトリックス含量の低減を伴う炎症性プラークフェノタイプを引き起こす。
【0033】
本発明者は、この背景におけるTSP-1による利点の機構は、主に2つの事柄(これらに限らない):(a)TSP-1が存在しないアテローム性動脈硬化症の病変内のTGF-β1の活性化の低減、(b)TSP-1の不在下におけるMMP-9の活性化の増大である。
【0034】
TSP-/-ApoE-/-マウスにおけるアテローム性動脈硬化症の病変内でTGF-β1の活性化が低減する
上述のように、in vivoの組織創傷におけるTSP-1の他の研究において、大部分のフェノタイプの発見は、創傷部位において局所的にTGF-β1を活性化するTSP-1の機能に帰する。さらに、TGF-β1活性の中和は、TSP-/-ApoE-/-マウスにおける上述の本発明者による発見と同様に、炎症を増大し、アテローム性動脈硬化症の病変内のマトリックスを低減させることが示されている(34,35)。そのため、本発明者は、TSP-/-ApoE-/-動物由来の組織における全TGF-β1及び活性型TGF-β1の双方のレベルを測定した。本発明者は、アテローム性動脈硬化症が形成される傾向がある既知の領域である大動脈弓由来のサンプルを均質化して、ELISAアッセイにおいて、その可溶化液を使用した。これらのマウスにおける総(潜在型及び活性型)TGF-β1レベルには変化が無かったが、TSP-/-ApoE-/-マウスにおいて活性型TGF-β1レベルにおける低減が存在した(図4)。アテローム性動脈硬化症の病変の可溶化物を用いた結果と比較すると、Tsp-/-ApoE-/-マウスとApoE-/-マウスとの間の活性型TGF-β1の全身(血清)レベルにおいて差は無かった(225±87pg/mlに対して186±138pg/ml)。これらのデータは、TSP-1の欠損が、血管壁内で局所的にTGF-β1の活性化の低減を引き起こすことを示す。
【0035】
TSP-/-ApoE-/-マウスにおけるMMP-9活性化の増大
免疫沈降実験によって、ゼラチナーゼ-B(MMP-9)はTSP-1に結合することが示されている(33)。さらに、TSP-1は、プロMMP-9の活性型への変換を調節し得る(32)。しかしながら、MMP-9の修飾語翻訳のTSP-1による制御についての機構は未知である。TSP-/-ApoE-/-マウスの病変内にマトリックスの顕著な損失が存在したため(図3e)、本発明者は、ウエスタンブロットにおいて大動脈弓均質化物由来の組織可溶化物を用いてMMP-9レベルを試験した。図5に示すように、これらのマウスのアテローム性動脈硬化症の病変において、総MMP-9及び活性型MMP-9のレベルの双方の増大が存在した。
【0036】
これらの結果は、TSP-1の欠損が、アテローム性動脈硬化症の病変内で局所的にMMP-9の活性化の増大を引き起こすことを示し、これはMMP-9活性に対するTSP-1の潜在的な阻害効果と一致する。apoEノックアウトマウスにおけるMMP-9欠失の研究は、MMP-9の欠損がマクロファージ数の低減、及びコラーゲン合成の増大に関連することを示している(36)。
【0037】
本発明者の発見を更に洗練するため、本発明者は、TGF-β1とMMP-9活性の双方に対するTSP-1の「活性」部位を確立することを開始した。TSP-1によるこれらのリガンドの制御についての細胞外における機構は、プラーク内の細胞表面相互作用によって調整されていると本発明者は仮説を立てたため、本発明者は、本発明者のin vivoアテローム性動脈硬化症プラークモデルにおける主要な細胞種であるマウスのマクロファージ(RAW細胞)を用いる細胞培養系を利用した。本発明者は、TGF-β1活性のTSP-1による制御のための活性部位(過去に報告されている)及びMMP-9活性に作用する部位(本発明者が同定)を同定した。
【0038】
TSP-1は、推定のβ2インテグリン結合モチーフD(D/E)(G/L)Wによってマクロファージに結合する
TSP-1は、CD44、LAP、及びCD36を含むマクロファージに結合するための多数の潜在的な結合部位を有する(図1)。しかしながら、TSP-1には過去に確立されているβ2-インテグリン結合モチーフが存在しない。ファージ配列アッセイを用いた過去の研究は、ペプチドモチーフD(D/E)(G/L)Wが、β2インテグリンに結合するプロ-MMP-9と競合することを報告している(40)。完全なTSP-1ペプチド配列の配列探索によって、本発明者は、図2に示すように、TSP-1は2つのその様な推定のβ2インテグリン結合部位を有することを決定した:1つがTSR-1(DDGW、完全に一致)であり、他方がTSR-2(DGGW、2位の置換を有し、ほぼ一致している)。本発明者は、TSP-1のマクロファージへの結合を評価するための細胞接着アッセイを使用した(図6)。このアッセイを使用して、本発明者はマクロファージへのTSP-1の接着におけるD-Xaa-G-Wの作用を示すことができた。具体的には、本発明者は、DDGWが、TSP-1への細胞接着を十分に阻害することを示すことができた。この競合の特異性が、ペプチド配列DDGAによるTSP-1に結合するマクロファージの緩衝の損失、及び配列ADGWによる部分的な干渉によって示された。
【0039】
TSP-1の存在下におけるマクロファージによるプロMMP-9の活性化の低減は、TSR-1中のDDGWモチーフによって媒介される
TSP-1によるMMP-9の制御の機構は、ほとんど分かっていない。しかしながら、プロMMP-9は、そのD(D/E)(L/G)Wモチーフによって細胞表面aMβ2及びaLβ2インテグリンに結合し、その活性化を引き起こすことが示されている(37,38)。
【0040】
本発明者は、TSP-1が同じモチーフでマクロファージに結合することを示し、本発明者は、TSP-1によるMMP-9の阻害の1つの潜在的な機構は、結合の競合による細胞表面からのプロMMP-9の置換であるという仮説を立てた(図7)。この仮説を評価するために、本発明者は、TSP-1のβ2-インテグリン結合モチーフのいずれか又は双方を含むTSP-1の各種の変異体を作製した(図2)。RAW細胞(マクロファージ細胞株)のTSP-1を用いた一過的なトランスフェクションは、プロMMP-9の活性型への変換を低減した。この効果は、TSR-1モチーフの変異によって逆行したが、TSR-2モチーフの変異ではより少なく逆行した。これらの結果は、おそらくβ2-インテグリンに結合するプロMMP-9と競合することによる、TSP-1によるMMP-9活性化の阻害におけるDxGWモチーフ、特にTSR-1のこのモチーフの重要性を示す。
【0041】
TSP-1によるTGF-β1の活性化は、TSR-2におけるWSPWモチーフの変異によって阻害される
TSP-1におけるWSxWモチーフは、TGF-β1活性化に重要である事が過去に示されている。この仮説を評価するために、本発明者は、組織培養実験を実施して、TSP-1が実際に、活性化マクロファージの存在下において、TGR-β1を活性化することを実証した。さらに、本発明者は変異体を使用して、TSR-2のWSPWモチーフが、TSP-1によるTGF-β1の活性化に主要な役割を担っていることを観察した。
【0042】
これらのデータに基づいて、本発明者は、TSR-1内のDDGWモチーフが、白血球上のaMβ2-インテグリンに結合し、この結合によって細胞表面に媒介されるMMP-9活性に作用するという仮説を立てた。さらに、TSR-2におけるWSPWモチーフはTGF-β1活性化に重要である。
【0043】
その様なものとして、本発明者は、(1)MMP-9活性の低減及び(2)TGF-β1の活性化に重要なTSP-1における2つの活性部位を同定した。この知識を用いて、プラーク安定化特性を有する多数のペプチドを設計し、合成し得る。本発明者は、これら2つの活性部位を組み合わせた代表的な合成配列:KRFKQDDGWSPWSEWを設計して、合成した。本発明者は、これら2つのTSP-1の機能に重要な活性部位を組み合わせたペプチド断片は、in vivoにおけるプラーク安定性に対して、タンパク質全体と同じ有利な効果を奏する可能性があると仮説を立てた。これを評価するために、まず、本発明者は、組織培養物において(1)TGR-β1を活性化し、(2)MMP-9の活性化を阻害するという、このペプチドの機能を試験し、確認した。
【0044】
次に、本発明者は、5週に亘って一日当たり8-25μgの濃度でalzet輸液ポンプを使用して、このペプチドをTSP-/-マウスに注射し、このペプチドが、TSP-/-Apo-/-マウスのフェノタイプを十分に救うこと示すことができた(マクロファージを低減し、病変内のマトリックスを増大した)。さらに、apoE-/-マウスでは、前記ペプチドの輸液が有益な効果を有しており、内因性TSP-1の存在下であってもプラークを安定化するという化合物の機能を立証した。換言すると、未処理のTSP-/-マウスと比較して、(1)内膜の病変における炎症(マクロファージ)の低減、(2)内膜病変内の脂質沈着の低減、(3)病変内のマトリックス(コラーゲン及びエラスチンによって測定)沈着の増大が存在した。
【0045】
結論として、本発明者の結果は、TSP-1が、(1)アテローム性動脈硬化症のプラークの安定化に対して有益な効果を有しており、且つ、(2)MMP-9及びTGF-β1の制御に重要なTSP-1の活性部位を組み合わせた少なくとも1つの合成ペプチドが、前記疾患のマウスモデルにおいてアテローム性動脈硬化症のプラークを安定化し得ることを示す。
【0046】
一般的には、「安定化プラーク」は、プラーク内の病変部位における炎症の制限及び/又は顕著な線維症によって証明される。これらの特性は、上述のTSP-/-マウスのようなTSP-1欠失対照系との比較によって観察され得る。一般的には、安定化プラークは、処置された又はされていない動物における線維症及び炎症の比較によって同定されて良い。プラーク内の線維症と炎症との間のバランスが、多数の異なる動物モデルにおけるプラークが破裂する傾向についての良好に許容されている代理マーカーとなっている(41-43)。
【0047】
追加の実施態様
ペプチド配列
本発明は、各種の新規なペプチドを提供する。これらは、各種の結合ドメイン及び活性化配列、並びにそれらのホモログの1つ以上を含んで良い。
【0048】
以下(のドメイン)有するペプチドは、TGF-β1及び/又はMMP-9に結合するか又は活性化する。
【0049】
分類1.β2-インテグリン、特にaMβ2及びaLβ2-インテグリンに結合するためのTSP-1及び/又はMMP-9に認められるドメインに相当する配列。その様な配列は、MMP-9が通常結合するβ2-インテグリン上の部位に結合するためのドメインを提供する。代替的には、前記ドメインは、β2-インテグリン上の幾つかの他の位置に結合するための部位を提供しても良いが、そのようにする場合においても、天然のMMP-9がβ2-インテグリンに結合することを阻害する。
【0050】
各種の配列が、その様なドメインを含んで良い。幾つかのその様な配列は、天然のMMP-9上で認められる。他のその様な配列は、TSP-1上、特にタイプ1反復ドメインに認められる。1つの例では、そのドメインは、D-Xaa-(L/G)-W又は(D/E)-(D/E)-(G/L)-W又はD-(D/E)-(L/G)-Wと表わされる4つのアミノ酸配列を含む。その様な配列の具体的な例は、DDGW、DGGW、及びDELWを含む。
【0051】
分類2.本発明の新規なペプチド配列において提供されて良い他のドメインは、天然のTGF-βと結合する。これらの配列の幾つかは、TSP-1のタイプ1反復ドメインに認められる。
【0052】
具体的な例では、その様な配列は、W-S-Xaa1-W-S-Xaa2-Wを含む。具体的な例では、Xaa1はP又はHであり、Xaa2はE又はPである。その様なペプチドのより具体的な例は、WSPWSEWである。
【0053】
分類3.本発明の新規なペプチドに利用されて良い更なる他の配列は、潜在型のTGF-βを活性化するアミノ酸配列を含む。その様な配列の幾つかは、TSP-1のタイプ1反復ドメインに認められる。その様な配列の例は、KRFK及びKRFKQを含む。
【0054】
上述の配列/ドメインの3つの分類の各種の組み合わせが、本発明の実施態様に係る新規ペプチド又はペプチドの組み合わせにおいて利用されて良い。
【0055】
第一の例では、ペプチド(又はペプチドの組み合わせを含む組成物)は、上述の分類1からの配列;すなわちβ2-インテグリン結合ドメインに相当する配列のみを含む。その様なペプチドは、上述の分類2及び/又は3からの追加の配列を含まないであろう。
【0056】
第二の例では、ペプチド(又はペプチドの組み合わせを含む組成物)は、上述の分類1及び2からの配列を含む。換言すると、単独のペプチド配列(又はペプチドの組み合わせを含む組成物)が、β2-インテグリン結合ドメインに相当する配列及びTGF-β結合する異なる配列の双方を含む。
【0057】
本発明のペプチド(又はペプチドの組み合わせを含む組成物)の第3の種類が、上述の分類1及び3の配列の双方を含む。換言すると、その様なペプチド(又はペプチドの組み合わせを含む組成物) は、β2インテグリン結合ドメインに相当する配列及びTGF-βを活性化する他の配列の双方を含む。
【0058】
そして、本発明のペプチド(又はペプチドの組み合わせを含む組成物)の更に他の種類は、上述の分類1、2、及び3の各々の組み合わせを含む。その様なペプチド(又はペプチドの組み合わせを含む組成物)は、β2-インテグリン結合ドメイン、TGF-βのための結合部位、潜在型TGF-βを活性型TGF-βに変換するための活性化配列を含むであろう。
【0059】
2つ以上の結合ドメイン又は活性化ドメインを利用する任意のペプチドにおいて、個々の配列又はドメインの幾つかの重なりが許容されることに注意すべきである。例えば、ペプチド中のアミノ酸の1つ以上が、結合部位又は活性化部位の2つ以上に共有されて良い。β-インテグリン結合ドメイン及びTGF-β結合ドメインの双方を含む具体的な例では、本発明のペプチドはDGGWSPWSEWという配列を有して良く、最初のW(トリプトファン)は2つの配列に共有されている。
【0060】
上述のように、プラークの安定化において活性を有することを確認している本発明の具体的なペプチドは、KRFKQDDGWSPWSEWという配列である。配列の残基の幾つかは、活性を維持するために他のものよりも重要である。かくして、この配列の変形例が、活性を保持するであろう。配列が比較的短い場合に、各種の挿入、欠失、置換、及び伸長が、過度の実験無しで当業者に容易に識別されるであろう。
【0061】
例えば、各種の置換が、前記配列内における個々のアミノ酸について為されて良い。好ましくは、その様な置換は、通常、基礎的な配列であるKRFKQDDGWSPWSEWにおけるアミノ酸と同じ大きさ及び/又は化学特性を有するアミノ酸を利用する。
【0062】
一般的な意味では、前記ペプチド配列は、Xaa1- Xaa2- Xaa3- Xaa4- Xaa5- Xaa6- Xaa7- Xaa8- Xaa9- Xaa10- Xaa11- Xaa12- Xaa13- Xaa14- Xaa15の形態を有する。
【0063】
Xaa1、Xaa2、及びXaa4は、好ましくは塩基性の非環式アミノ酸である。例えば、R及びKを含む。
【0064】
Xaa3、Xaa9、Xaa11、Xaa12、及びXaa15は、好ましくは、中性の大きな非極性環式アミノ酸である。これらは、F、W、及び好ましくはPを含む。
【0065】
Xaa5は、好ましくは、中性の大きな極性非環式アミノ酸である。例えば、T、N、及びQが含まれる。
【0066】
Xaa6、Xaa7、及びXaa14は、好ましくは、酸性アミノ酸である。この種類の天然アミノ酸の具体的な例は、D及びEである。
【0067】
最後に、Xaa8、Xaa10、及びXaa13は、好ましくは、中性の小さな極性アミノ酸である。その様なアミノ酸の天然のものの例は、G、S、及びCである。
【0068】
1つの具体的な実施態様では、Xaa2はRである。他の具体的な実施態様では、Xaa6はDである。他の具体的な実施態様では、Xaa7はDである。他の具体的な実施態様では、Xaa9はWである。他の具体的な実施態様では、Xaa12はWである。更に他の具体的な実施態様では、Xaa15はWである。
【0069】
更に他の具体的な実施態様では、Xaa2はRであり、Xaa7はDであり、Xaa9はWである。
【0070】
本発明のペプチドは、20の天然アミノ酸を含む残基に限られないことを注意すべきである。一定の一般的に遭遇する非天然アミノ酸、例えば、脱アミノチロシン(des Tyr)、アグマチン(Agm)、n-ホルミルトリプトファン(f-Trp)、α-アミノイソ酪酸(Aib)、及びサルコシン(Sar)、スタチン、オルニチン(Orn)、ホモリジン、ホモセリン、ホモアルギニン、ノルロイシン(Nle)、ノルバリンが、本発明の化合物に組み込まれても良い。
【0071】
本発明の範囲内のペプチドは、他に開示されている配列を修飾することによって得られて良いが、かくして得られるペプチドの活性を保持する。例えば、これらの化合物のアミノ酸は、通常、天然のL光学異性体型であるが、1つ以上がD光学異性体型に置き換わってよく、又はD,L-ラセミ混合物がペプチドを含む分子において提供されて良い。さらに、ジスルフィド結合が、活性が維持される限り、本発明の化合物に存在するか又はしなくて良い。
【0072】
前記ペプチドに含有されるアミノ酸残基、特にカルボキシ又はアミノ末端のアミノ酸残基は、メチル化、アミド化、アセチル化、又はそれらの活性に悪影響を及ぼすこと無く、循環半減期、プロテアーゼに対する耐性、及び化合物の溶解度を変化し得る他の化学基による置換によって修飾されても良い。
【0073】
上述のペプチドのいずれかは、好ましくは、少なくとも7アミノ酸残基を含む。他の実施態様では、前記ペプチドは、約7から25残基の間、約7から20残基の間、約7から14残基の間、約10から12残基の間、約10から25残基の間、約15から25残基の間、及び約15から20残基の間に含まれる。本発明のペプチドは、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、又は25アミノ酸残基を含む。本発明の基礎的なスカフォールドを規定するペプチドは、好ましくは、上述の結合及び活性化配列(分類1、2、及び3)の1つ以上が、β2-インテグリン及び/又はTGF-βと相互作用するために曝露されるように適当に設計される。
【0074】
本発明の化合物は、それら自体に対してホモポリマー(すなわち、(ペプチド)n)化されるか、又は他のものに対してヘテロポリマー(すなわち、ペプチド1-ペプチド2)化されて良い。前記化合物は、生体適合性分子、例えば、脂肪酸及び/又はポリエチレングリコール(PEG)に接合されて良い。
【0075】
本発明の範囲内のペプチドは、例えば固相ペプチド合成のような当該技術分野でよく知られている手段によって化学的に合成して良い。他の実施態様では、本発明の選択される化合物が、既知の方法に従って調製される組換えDNA構築物の発現によって生産されて良い。
【0076】
簡便には、化合物は、手作業の技術、又は製造業者によって提供される説明書に与えられた指示に従って自動の技術、例えばApplied Biosystems社(Foster City, Calif.)製の各種の製品を用いて合成して良い。本発明の化合物を合成する過程の間に本明細書に従って作製される中間体自体が有用な化合物であり、本発明の範囲内であることが、ペプチド合成の分野の当業者に容易に理解されるであろう。
【0077】
治療方法
上述のように、本発明の実施態様は、TSP-1、その一部、又は関連するペプチド、例えば、上述のように同定したものを用いる治療方法に関連する。一般的には、TSP-1及び本発明のペプチドを投与して、動脈壁上のプラークを安定化する。かくして、それらは、疾患を有する動物においてプラークを安定化する方法で利用されて良い。これは、アテローム性動脈硬化症の治療方法とは区別されるべきである。
【0078】
ある実施態様では、本発明の新規ペプチドを投与して、プラーク安定化以外の結果に作用して良い。例えば、上記の分類1に区別されるモチーフ(例えば、D-Xaa-(L/G)-W含有ペプチド、例えば、DDGW、DGGW、及びDELW)を有する合成ペプチドを利用して、in vivoでMMP-9に作用させて良い。かくして、その様なペプチドを利用して、例えば、癌及び血管新生を治療して良い。ある実施態様では、D-Xaa-(L/G)-Wモチーフを有し、in vivo治療で使用する合成ペプチドは、少なくとも7アミノ酸残基、より好ましくは約7から20残基の間を含み、具体的な実施態様では約10から15残基の間を含む。
【0079】
本発明は、上述のプラーク安定化という利点を提供するために単独で役立つであろう、TSP-1及び本発明のペプチド(それらの非毒性の付加塩、アミド、及びエステルを含む)の有効量を含有する組成物を提供する。その様な組成物は、生理学的に許容される液体、ゲル、又は固体の希釈剤、アジュバント、並びに賦形剤と共に提供されても良い。
【0080】
これらの組成物は、獣医学的な使用のために家畜のような動物に投与されても良く、他の治療剤と同様にヒトに臨床的に使用されても良い。一般的には、治療上の有効性のために必要とされる用量は、対象者の体重当たり、約1μgから300mg/kg、更に通常は10μgから30mg/kgの範囲であろう。代替的には、これらの範囲内の用量が、通常は24時間を超えて、所望の治療上の利益が得られるまで経時的に一定の輸液によって投与されて良い。
【0081】
典型的には、その様な組成物は、溶液又は懸濁物のいずれかの注射可能な物として調製される;注射前の液体中の溶液又は懸濁物のために適切な固体の形状が調製されても良い。前記調製物は乳化されても良い。活性成分は、多くの場合には、生理学的に許容され、活性成分に適合する希釈剤又は賦形剤と混合する。適切な希釈剤又は賦形剤は、例えば、水、生理食塩水、デキストロース、又はグリセロールなど、並びにそれらの組み合わせである。さらに、望まれる場合には、前記組成物は、少量の付加的な物質、例えば、湿潤剤又は乳化剤、及び安定剤又はpH緩衝剤などを含有して良い。
【0082】
前記組成物は、注射、例えば、皮下又は静脈内注射によって、従来のように非経口的に投与される。他の様式の投与に適切な更に別の製剤は、坐剤、鼻内エアロゾル、を含み、ある場合には経口製剤を含む。坐剤については、従来のバインダー及び賦形剤は、例えば、ポリアルキレングリコール又はトリグリセリドを含んで良く;その様な坐剤は、0.5%から10%、好ましくは1%から2%の範囲の活性成分を含有する混合物から形成されて良い。経口製剤は、そのように通常利用される賦形剤、例えば、製薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウムなどを含む。これらの組成物は、溶液剤、懸濁剤、錠剤、丸剤、カプセル剤、持続放出型製剤、又は粉末剤の形態をとり、10%から95%、好ましくは25%から70%の活性成分を含有する。これらの経口製剤は、吸収されるまでペプチドを保護することを意図する製剤を含む。
【0083】
前記ペプチド化合物は、中性又は塩の形態として組成物に製剤化されて良い。製薬学的に許容される非毒性の塩は、無機酸、例えば塩酸又はリン酸、あるいは有機酸、例えば、酢酸、シュウ酸、酒石酸、及びマンデル酸などと形成される酸付加塩(遊離のアミノ基と形成される)を含む。遊離のカルボキシル基と形成される塩は、無機塩基、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、又は水酸化鉄、並びに有機塩基、例えば、イソプロピルアミン、トリメチルアミン、2-エチルアミノエタノール、ヒスチジン、及びプロカイン等に由来するものであって良い。
【0084】
結論
上述の記載は説明を意図するものであって、これに制限するもので無いことが理解されるべきである。各種の実施態様及び変形が、本発明の範囲及び精神を逸脱すること無く、本願に開示されている発明に為されて良いことが当業者には明らかであるはずである。したがって、本発明の範囲は、上述の記載に表わされる具体的な実施態様を参照するのではなく、その様な実施態様と均等の全範囲を参照して決定されるべきである。本明細書に挙げた全ての文献は、本発明との関係において使用され得る生物学的機構、治療、方法論、及び概念を説明するために参照される。これらの参照文献が本明細書に記載の発明との関係において従来技術であると認められるべきではない。他に示さない限り、本明細書に挙げた各参照文献は、参照することによって、全ての目的のためにその全体を本明細書に組み込む。
【0085】
参考文献
1. Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
2. Ross, R., Atherosclerosis~an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
3. Bornstein, P., Thrombospondins as matricellular modulators of cell function. J Clin Invest, 2001. 107(8): p. 929-34.
4. Sage, E.H., Regulation of interactions between cells and extracellular matrix: a command performance on several stages. J Clin Invest, 2001. 107(7): p. 781-3.
5. Murphy-Ullrich, J.E., The de-adhesive activity of matricellular proteins: is intermediate cell adhesion an adaptive state? J Clin Invest, 2001. 107(7): p. 785- 90.
6. Elzie, CA. and J.E. Murphy-Ullrich, The N-terminus of thrombospondin: the domain stands apart, lht J Biochem Cell Biol, 2004. 36(6): p. 1090-101.
7. Li, D.Y., G. Faury, D.G. Taylor, E.G. Davis, W.A. Boyle, R.P. Mecham, P. Stenzel, B. Bo ak, and M.T. Keating, Novel arterial pathology in mice and humans hemizygous for elastin. J Clin Invest, 1998. 102(10): p. 1783-7.
8. Aszodi, A., D. Chan, E. Hunziker, J.F. Bateman, and R. Fassler, Collagen π is essential for the removal of the notochord and the formation of intervertebral discs. J Cell Biol, 1998. 143(5): p. 1399-412.
9. Forsberg, E., E. Hirsch, L. Frohlich, M. Meyer, P. Ekblom, A. Aszodi, S. Werner, and R. Fassler, Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc Natl Acad Sci U S A, 1996. 93(13): p. 6594-9.
10. Lawler, J., M. Sunday, V. Thibert, M. Duquette, E.L. George, H. Rayburn, and R.O. Hynes, Thrombospondin- 1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J Clin Invest, 1998. 101(5): p. 982-92.
11. Kyriakides, T.R., J. W. Tarn, and P. Bornstein, Accelerated wound healing in mice with a disruption of the thrombospondin 2 gene. J Invest Dermatol, 1999. 113(5): p. 782-7.
12. Adams, J.C. and J. Lawler, The thrombospondins. Lit J Biochem Cell Biol, 2004. 36(6): p. 961-8.
13. Crawford, S.E., V. Stellmach, J.E. Murphy-Ullrich, S.M. Ribeiro, J. Lawler, R.O. Hynes, G.P. Boivin, and N. Bouck, Thrombospondin- 1 is a major activator of TGF-betal in vivo. Cell, 1998. 93(7): p. 1159-70.
14. Agah, A., T.R. Kyriakides, J. Lawler, and P. Bornstein, The lack of thrombospondin- 1 (TSPl) dictates the course of wound healing in double- TSPl/TSP2-null mice. Am J Pathol, 2002. 161(3): p. 831-9.
15. Yee, K.O., M. Streit, T. Hawighorst, M. Detmar, and J. Lawler, Expression of the type-1 repeats of thrombospondin- 1 inhibits tumor growth through activation of transforming growth factor-beta. Am J Pathol, 2004. 165(2): p. 541-52.
16. Miao, W.M., W.L. Seng, M. Duquette, P. Lawler, C. Laus, and J. Lawle r, Thrombospondin- 1 type 1 repeat recombinant proteins inhibit tumor growth through transforming growth factor-beta-dependent and -independent mechanisms. Cancer Res, 2001. 61(21): p. 7830-9.
17. Yano, K., H. Oura, and M. Detmar, Targeted overexpression of the angiogenesis inhibitor thrombospondin- 1 in the epidermis of transgenic mice prevents ultraviolet-B-induced angiogenesis and cutaneous photo-damage. J Invest Dermatol, 2002. 118(5): p. 800-5.
18. Simantov, R., M. Febbraio, R. Crombie, A.S. Asch, R.L. Nachman, and R.L. Silverstein, Histidine-rich glycoprotein inhibits the antiangiogenic effect of thrombospondin- 1. J Clin Invest, 2001. 107(1): p. 45-52.
19. Bornstein, P., A. Agah, and T.R. Kyriakides, The role of thrombospondins 1 and 2 in the regulation of cell-matrix interactions, collagen fibril formation, and the response to injury. Int J Biochem Cell Biol, 2004. 36(6): p. 1115-25.
20. Lawler, J. and M. Detmar, Tumor progression: the effects of thrombospondin- 1 and -2. Int J Biochem Cell Biol, 2004. 36(6): p. 1038-45.
21. Topol, EJ., J. McCarthy, S. Gabriel, DJ. Moliterno, WJ. Rogers, L.K. Newby, M. Freedman, J. Metivier, R. Cannata, CJ. O'Donnell, K. Kottke-Marchant, G. Murugesan, E.F. Plow, O. Stenina, and G.Q. Daley, Single nucleotide polymorphisms in multiple novel thrombospondin genes may be associated with familial premature myocardial infarction. Circulation, 2001. 104(22): p. 2641-4.
22. Stenina, O.I., T.V. Byzova, J.C. Adams, JJ. McCarthy, EJ. Topol, and E.F. Plow, Coronary artery disease and the thrombospondin single nucleotide polymorphisms. Int J Biochem Cell Biol, 2004. 36(6): p. 1013-30.
23. Raines, E.W., The extracellular matrix can regulate vascular cell migration, proliferation, and survival: relationships to vascular disease. Int J Exp Pathol, 2000. 81(3): p. 173-82.
24. Roth, JJ., V. Gahtan, J.L. Brown, C. Gerhard, V.K. Swami, V.L. Rothman, T.N. Tulenko, and G.P. Tuszynski, Thrombospondin- 1 is elevated with both intimal hyperplasia and hypercholesterolemia. J Surg Res, 1998. 74(1): p. 11-6.
25. Lawler, J., The functions of thrombospondin- 1 and-2. Curr Opin Cell Biol, 2000. 12(5): p. 634-40.
26. Chen, H., M.E. Herndon, and J. Lawler, The cell biology of thrombospondin- 1. Matrix Biol, 2000. 19(7): p. 597-614.
27. Majack, R.A., S.C. Cook, and P. Bornstein, Control of smooth muscle cell growth by components of the extracellular matrix: autocrine role for thrombospondin. Proc Natl Acad Sci U S A, 1986. 83(23): p. 9050-4.
28. Schultz-Cherry, S., S. Ribeiro, L. Gentry, and J.E. Murphy-Ullrich, Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. J Biol Chem, 1994. 269(43): p. 26775-82.
29. Fischer, J.W., M. Stoll, A. W. Hahn, and T. Unger, Differential regulation of thrombospondin- 1 and fibronectin by angiotensin JI receptor subtypes in cultured endothelial cells. Cardiovasc Res, 2001. 51(4): p. 784-91.
30. Lymn, J.S., M.K. Patel, G.F. Clunn, SJ. Rao, KX. Gallagher, and A.D. Hughes, Thrombospondin-1 differentially induces chemotaxis and DNA synthesis of human venous smooth muscle cells at the receptor-binding level. J Cell Sci, 2002. 115(Pt 22): p. 4353-60.
31. Patel, M.K., J.S. Lymn, G.F. Clunn, and A.D. Hughes, Thrombospondin-1 is a potent mitogen and chemoattractant for human vascular smooth muscle cells. Arterioscler Thromb Vase Biol, 1997. 17(10): p. 2107-14.
32. Rodriguez-Manzaneque, J.C., T.F. Lane, M.A. Ortega, R.O. Hynes, J. Lawler, and M.L. Iruela-Arispe, Thrombospondin-1 suppresses spontaneous tumor growth and inhibits activation of matrix metalloproteinase-9 and mobilization of vascular endothelial growth factor. Proc Natl Acad Sci U S A, 2001. 98(22): p. 12485-90.
33. Bein, K. and M. Simons, Thrombospondin type 1 repeats interact with matrix metalloproteinase 2. Regulation of metalloproteinase activity. J Biol Chem, 2000. 275(41): p. 32167-73.
34. Lutgens, E., M. Gijbels, M. Smook, P. Heeringa, P. Gotwals, V.E. Koteliansky, and MJ. Daemen, Transforming growth factor-beta mediates balance between inflammation and fibrosis during plaque progression. Arterioscler Thromb Vase Biol, 2002. 22(6): p. 975-82.
35. Mallat, Z., A. Gojova, C. Marchiol-Fournigault, B. Esposito, C. Kamate, R. Merval, D. Fradelizi, and A. Tedgui, Inhibition of transforming growth factorbeta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ Res, 2001. 89(10): p. 930-4.
36. Luttun, A., E. Lutgens, A. Manderveld, K. Maris, D. Collen, P. Carmeliet, and L. Moons, Loss of matrix metalloproteinase-9 or matrix metalloproteinase- 12 protects apolipoprotein E- deficient mice against atherosclerotic media destruction but differentially affects plaque growth. Circulation, 2004. 109(11): p. 1408-14.
37. Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
38. Stefanidakis, M., T. Ruohtula, N. Borregaard, CG. Gahniberg, and E. Koivunen, Intracellular and cell surface localization of a complex between alphaMbeta2 integrin and promatrix metalloproteinase-9 progelatinase in neutrophils. J Immunol, 2004. 172(11): p. 7060-8.
39. Wrana, J.L., L. Attisano, J. Carcamo, A. Zentella, J. Doody, M. Laiho, X.F. Wang, and J. Massague, TGF beta signals through a heteromeric protein kinase receptor complex. Cell, 1992. 71(6): p. 1003-14.
40. Stefanidakis, M., M. Bjorklund, E. Hianus, CG. Gahmberg, and E. Koivunen, Identification of a negatively charged peptide motif within the catalytic domain of progelatinases that mediates binding to leukocyte beta 2 integrins. J Biol Chem, 2003. 278(36): p. 34674-84.
41. Libby, P., Atheroma: more than mush. Lancet, 1996. 348 Suppl 1: p. s4-7.
42. Ross, R., Atherosclerosis an inflammatory disease. N Engl J Med, 1999. 340(2): p. 115- 26.
43. FaIk, E., P.K. Shah, and V. Fuster, Coronary plaque disruption. Circulation, 1995. 92(3): p. 657-71.
【図面の簡単な説明】
【0086】
【図1】図1は、アテローム性動脈硬化症の病変において直接的な関係性を有する可能性がある、各種のTSP-1細胞及びサイトカイン結合ドメインを示す。
【図2A】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(A)TSR-1及びTSR-2には2つの推定上のβ2インテグリン結合ドメインが存在する。これら2つの結合ドメインは、TGF-β1についての2つの結合ドメインが隣接(及び1つのアミノ酸が重複)している。本発明者は、box中に示す2つのDxGWモチーフを変異させた。
【図2B】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(B)マウスマクロファージ細胞株(Raw)の一過的なトランスフェクションを使用して、本発明者は、TSP-1がMMP-9活性を低減すること(p<0.005(活性型MMP-9/総MMP-9 ELISA))を認めている。この効果は、TSP変異体:DM及びより少なくはMut1によって無効にされる。
【図2C】図2は、TSP-1中のTSR-1及び2結合ドメイン及びこれら2つのドメイン内で作製した関連する変異体を示す。(C)同じ細胞系を使用して、本発明者は、TSP-1がTGF-β1活性を増大することを認めている(p<0.005(細胞溶解物を使用して、TGF-β1プロモーター応答エレメントを介するルシフェラーゼ誘導を測定する(39)))。この作用は、TSP-1変異体Mut1及びMut2の双方によって無効にされる。まとめると、TSR-1及びTSR-2におけるDxGWモチーフは、この細胞アッセイ系においてTGF-β1活性化及びMMP-9阻害に対するTSP-1の作用を媒介するようである。
【図3A−C】図3は、アテローム性動脈硬化症病変に対するTSP-1欠失の組織学的特性を示す。(A)ヘマトキシリン及びエオシン染色大動脈弁尖。右側の棒グラフは、病変領域(内膜)を表わす。病変領域には変化が認められなかった。(B)Moma-2抗体染色(PEが接合され、赤色蛍光を表わす)は、TSP-1欠失マウスにおいて大動脈弁尖内の病変にマクロファージの増大が存在することを明らかにする(p<0.01)。(C)マクロファージの増大と一致して、Red-O染色は、TSP-1欠失マウスにおいて病変内の脂質沈着の増大が存在することを明らかにする(すなわち、泡沫細胞、p<0.05)。
【図3D−F】図3は、アテローム性動脈硬化症病変に対するTSP-1欠失の組織学的特性を示す。(D)平滑筋細胞アクチン抗体染色(FITCが接合され、緑色蛍光を表わす)。内膜病変内には平滑筋細胞(smc)の染色は存在せず、培地中の総smc含量は2つの群の間で変化がない。(E)コラーゲンについてのpicosirius red polarization染色が、大動脈弁尖における病変内のコラーゲンの損失を明らかにする(黄色-赤色染色の低減)。両種のマウスにおける弁尖及び弁の弁構造の周囲に強い(黄色-赤色)染色が存在することに注意すべきである。しかし、(矢印で示す)病変内において、TSPノックアウトマウスでは染色の低減が存在する。(F)エラスチンについてのVerhoeff's染色は、TSP-1欠失を有する病変の発生領域における中膜及び内膜のエラスチンの損失の増大を明らかにする(p<0.05)。
【図4】図4は、ApoE-/-TSP-/-(n=3マウス)及びApoE-/-マウス(n=3マウス)由来の大動脈弓の組織可溶化物からのTGF-βELISAの結果を示す。総TGF-β1のレベルについて2つの群の間に差は無いが、ApoE-/-TSP-/-マウスの大動脈内の活性型TGF-β1レベルでは有意な差(p<0.05)が存在する。
【図5】図5は、ApoE-/-TSP-/-マウス(n=3マウス)及びApoE-/-マウス(n=3マウス)由来の大動脈弓の組織可溶化物からのMMP-9ウエスタンブロットを示す。各レーンは、個々のマウスを特定する。TSP-/-マウスにおけるプロMMP-9(105kb)及び活性型MMP-9(95kb)のバンドの双方の増大が存在する。
【図6】図6は、TSP-1(40μg/ml)、BSA(5%)、又はフィブロネクチン(0.1μg/ml)の各々を事前にウェルにプレーティングした接着アッセイの結果を示す。ペプチドの実験については、TSP-1のウェルに導入される前に、細胞を、個々のペプチド(0.01M)と懸濁物中で30分間混合する。
【図7】図7は、マクロファージ上のaMβ2インテグリン部位への結合についてのTSP-1及びプロMMP-9による競合に関する機構を示す(A)。TSP-1の増大の存在下では、MMP-9活性化の低減及びTGF-β1活性化の増大と共にTSP-1結合の増大が存在する(B)。プロMMP-9の増大の背景においては、TSP-1媒介TGF-β1活性化の低減及びMMP-9活性化の増大と共にMMP-9結合の増大が存在する(C)。
【特許請求の範囲】
【請求項1】
配列Xaa1-Xaa2-Xaa3-Xaa4-Xaa5-Xaa6-Xaa7-Xaa8-Xaa9-Xaa10-Xaa11-Xaa12-Xaa13-Xaa14-Xaa15を含むペプチドであって、Xaa1、Xaa2、及びXaa4が、Arg及びLysを含む塩基性の非環式アミノ酸からなる群から選択され;Xaa3、Xaa9、Xaa11、Xaa12、及びXaa15が、Phe、Trp、及びProを含む中性で大きな非極性環式アミノ酸からなる群から選択され;Xaa5が、Thr、Asn、及びGlnを含む中性で大きな極性非環式アミノ酸からなる群から選択され;Xaa6、Xaa7、及びXaa14が、Asp及びGlnを含む酸性アミノ酸からなる群から選択され;並びにXaa8、Xaa10、及びXaa13が、Gly、Ser、及びCysを含む中性で小さな極性アミノ酸からなる群から選択されるペプチド、又は7から14のアミノ酸を有する前記配列の切断型のセグメント。
【請求項2】
前記ペプチドが15から25アミノ酸からなる、請求項1に記載のペプチド。
【請求項3】
前記ペプチドが15から20アミノ酸からなる、請求項1に記載のペプチド。
【請求項4】
Xaa2がArgである、請求項1に記載のペプチド。
【請求項5】
Xaa6がAspである、請求項1に記載のペプチド。
【請求項6】
Xaa7がAspである、請求項1に記載のペプチド。
【請求項7】
Xaa9がTrpである、請求項1に記載のペプチド。
【請求項8】
Xaa12がTrpである、請求項1に記載のペプチド。
【請求項9】
Xaa15がTrpである、請求項1に記載のペプチド。
【請求項10】
Xaa2がArgであり、Xaa7がAspであり、Xaa9がTrpである、請求項1に記載のペプチド。
【請求項11】
配列Lys-Arg-Phe-Lys-Gln-Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む、15から25のアミノ酸からなるペプチド。
【請求項12】
配列Lys-Arg-Phe-Lys-Gln-Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpからなる、請求項11に記載のペプチド。
【請求項13】
8から14のアミノ酸残基からなる、請求項12に記載のペプチドのアミノ及び/又はカルボキシル切断型のセグメント。
【請求項14】
配列Asp-Xaa-Gly-Trp-Ser-Pro/His-Trp-Ser-Glu/Pro-Trpを含み、Xaaが任意のアミノ酸である、10から20のアミノ酸からなるペプチド。
【請求項15】
配列Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む、10から20のアミノ酸からなるペプチド。
【請求項16】
配列Asp-Xaa-Gly/Leu-Trp又はAsp/Glu-Asp/Glu-Gly/Leu-Trp又はAsp-Asp/Glu-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、7から20のアミノ酸からなるペプチド。
【請求項17】
配列Lys-Arg-Phe-Lys-Gln-Asp/Glu-Xaa-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、9から20のアミノ酸からなるペプチド。
【請求項18】
配列Lys-Arg-Phe-Lys-Asp/Glu-Xaa-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、8から20のアミノ酸からなるペプチド。
【請求項19】
配列Asp/Glu-Xaa1-Gly/Leu-Trp-Ser-Xaa2-Trp-Ser-Xaa3-Trpを含み、Xaa1、Xaa2、及びXaa3が任意のアミノ酸である、10から20のアミノ酸をからなるペプチド。
【請求項20】
Xaa1がAsp又はGluであり、Xaa2がPro又はHisであり、Xaa3がGlu又はProである、請求項19に記載のペプチド。
【請求項21】
配列Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む10から20のアミノ酸からなるペプチド。
【請求項22】
アテローム性動脈硬化症のプラークを安定化する活性がある、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項23】
1つ以上の保存的アミノ酸置換を含む、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項24】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドと95%の相同性を有するペプチド。
【請求項25】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドをコードするヌクレオチド配列。
【請求項26】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドをコードするヌクレオチド配列を含む発現ベクター。
【請求項27】
請求項26に記載の発現ベクターを含む宿主細胞。
【請求項28】
タンパク質-タンパク質相互作用によってTGF-β1及び/又はMMP-9活性を調節する、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項29】
非天然アミノ酸、D-異性体又はD,L-ラセミ混合異性体の形態、ジスルフィド結合、カルボキシ又はアミノ末端修飾、脂肪酸及びPEGを含む生体適合性分子への接合、並びにアミノ酸の化学的置換からなる群から選択される1つ以上の修飾を含む、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項30】
ホモポリマー化又はヘテロポリマー化されている、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項31】
C末端、N末端、又はそれらの側鎖に結合している請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む融合タンパク質。
【請求項32】
請求項1、11,14から19、及び21のいずれか一項に記載のペプチドを含む、純粋な医薬製剤。
【請求項33】
動物に投与するため、又はヒトに投与するための医薬製剤として調製される、請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物。
【請求項34】
液体として製剤化されている、請求項33に記載の組成物。
【請求項35】
輸液又は注射に適切なものである、請求項34に記載の液体。
【請求項36】
請求項1、11、14から19、又は21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、アテローム性動脈硬化症のプラークを安定化し、プラークが破裂する事象の発生を低減するための方法。
【請求項37】
前記組成物又は医薬製剤が液体である、請求項36に記載の方法。
【請求項38】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項36に記載の方法。
【請求項39】
前記動物が、アテローム性動脈硬化症のプラークの存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項36に記載の方法。
【請求項40】
前記疾患が、以下の特徴:心筋梗塞、脳卒中、及び急性肢虚血、又はその様な疾患に対する素因の1つ以上を含む、請求項36に記載の方法。
【請求項41】
前記動物への投与が、輸液又は注射による投与を含む、請求項36に記載の方法。
【請求項42】
請求項1、11、14から19、又は21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、内膜病変内の炎症及び脂質沈着を低減し、且つ、コラーゲン及びエラスチン沈着を増大させるための方法。
【請求項43】
前記組成物又は医薬製剤が液体である、請求項42に記載の方法。
【請求項44】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項42に記載の方法。
【請求項45】
前記動物が、内膜病変内の炎症、脂質沈着、又はコラーゲン及びエラスチン沈着の減少の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項42に記載の方法。
【請求項46】
前記疾患が、心筋梗塞、脳卒中、及び急性肢虚血、又はその様な疾患に対する素因を含む、請求項45に記載の方法。
【請求項47】
前記動物への投与が、輸液又は注射によるものである、請求項42に記載の方法。
【請求項48】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、大動脈壁でTGF-β1を活性化し、且つ、MMP-9を不活性化するための方法。
【請求項49】
前記組成物又は医薬製剤が液体である、請求項48に記載の方法。
【請求項50】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項48に記載の方法。
【請求項51】
前記動物への投与が、輸液又は注射によるものである、請求項48に記載の方法。
【請求項52】
前記動物が、大動脈壁におけるTGF-β1の不活性化及び/又はMMP-9の活性化の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項48に記載の方法。
【請求項53】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物または医薬製剤を動物に投与する工程を含む、TGF-β1の活性化及び/又はMMP-9の不活性化のための方法。
【請求項54】
前記組成物又は医薬製剤が液体である、請求項53に記載の方法。
【請求項55】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項53に記載の方法。
【請求項56】
前記動物への投与が、輸液又は注射によるものである、請求項53に記載の方法。
【請求項57】
動物が、癌及び/又は血管新生、あるいはその様な疾患に対する素因の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項53に記載の方法。
【請求項58】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤と動物を接触させる工程を含む、TGF-β1及び/又はMMP-9活性を調節するための方法。
【請求項59】
前記動物が、癌及び/又は血管新生、あるいはその様な疾患に対する素因の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項58に記載の方法。
【請求項60】
前記動物が、内膜病変内の炎症、脂質沈着、又はコラーゲン及びエラスチン沈着の減少の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項58に記載の方法。
【請求項61】
前記疾患が、心筋梗塞、脳卒中、及び急性肢虚血、あるいはその様な疾患に対する素因の1つ以上を含む、請求項60に記載の方法。
【請求項1】
配列Xaa1-Xaa2-Xaa3-Xaa4-Xaa5-Xaa6-Xaa7-Xaa8-Xaa9-Xaa10-Xaa11-Xaa12-Xaa13-Xaa14-Xaa15を含むペプチドであって、Xaa1、Xaa2、及びXaa4が、Arg及びLysを含む塩基性の非環式アミノ酸からなる群から選択され;Xaa3、Xaa9、Xaa11、Xaa12、及びXaa15が、Phe、Trp、及びProを含む中性で大きな非極性環式アミノ酸からなる群から選択され;Xaa5が、Thr、Asn、及びGlnを含む中性で大きな極性非環式アミノ酸からなる群から選択され;Xaa6、Xaa7、及びXaa14が、Asp及びGlnを含む酸性アミノ酸からなる群から選択され;並びにXaa8、Xaa10、及びXaa13が、Gly、Ser、及びCysを含む中性で小さな極性アミノ酸からなる群から選択されるペプチド、又は7から14のアミノ酸を有する前記配列の切断型のセグメント。
【請求項2】
前記ペプチドが15から25アミノ酸からなる、請求項1に記載のペプチド。
【請求項3】
前記ペプチドが15から20アミノ酸からなる、請求項1に記載のペプチド。
【請求項4】
Xaa2がArgである、請求項1に記載のペプチド。
【請求項5】
Xaa6がAspである、請求項1に記載のペプチド。
【請求項6】
Xaa7がAspである、請求項1に記載のペプチド。
【請求項7】
Xaa9がTrpである、請求項1に記載のペプチド。
【請求項8】
Xaa12がTrpである、請求項1に記載のペプチド。
【請求項9】
Xaa15がTrpである、請求項1に記載のペプチド。
【請求項10】
Xaa2がArgであり、Xaa7がAspであり、Xaa9がTrpである、請求項1に記載のペプチド。
【請求項11】
配列Lys-Arg-Phe-Lys-Gln-Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む、15から25のアミノ酸からなるペプチド。
【請求項12】
配列Lys-Arg-Phe-Lys-Gln-Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpからなる、請求項11に記載のペプチド。
【請求項13】
8から14のアミノ酸残基からなる、請求項12に記載のペプチドのアミノ及び/又はカルボキシル切断型のセグメント。
【請求項14】
配列Asp-Xaa-Gly-Trp-Ser-Pro/His-Trp-Ser-Glu/Pro-Trpを含み、Xaaが任意のアミノ酸である、10から20のアミノ酸からなるペプチド。
【請求項15】
配列Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む、10から20のアミノ酸からなるペプチド。
【請求項16】
配列Asp-Xaa-Gly/Leu-Trp又はAsp/Glu-Asp/Glu-Gly/Leu-Trp又はAsp-Asp/Glu-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、7から20のアミノ酸からなるペプチド。
【請求項17】
配列Lys-Arg-Phe-Lys-Gln-Asp/Glu-Xaa-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、9から20のアミノ酸からなるペプチド。
【請求項18】
配列Lys-Arg-Phe-Lys-Asp/Glu-Xaa-Gly/Leu-Trpを含み、Xaaが任意のアミノ酸である、8から20のアミノ酸からなるペプチド。
【請求項19】
配列Asp/Glu-Xaa1-Gly/Leu-Trp-Ser-Xaa2-Trp-Ser-Xaa3-Trpを含み、Xaa1、Xaa2、及びXaa3が任意のアミノ酸である、10から20のアミノ酸をからなるペプチド。
【請求項20】
Xaa1がAsp又はGluであり、Xaa2がPro又はHisであり、Xaa3がGlu又はProである、請求項19に記載のペプチド。
【請求項21】
配列Asp-Asp-Gly-Trp-Ser-Pro-Trp-Ser-Glu-Trpを含む10から20のアミノ酸からなるペプチド。
【請求項22】
アテローム性動脈硬化症のプラークを安定化する活性がある、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項23】
1つ以上の保存的アミノ酸置換を含む、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項24】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドと95%の相同性を有するペプチド。
【請求項25】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドをコードするヌクレオチド配列。
【請求項26】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドをコードするヌクレオチド配列を含む発現ベクター。
【請求項27】
請求項26に記載の発現ベクターを含む宿主細胞。
【請求項28】
タンパク質-タンパク質相互作用によってTGF-β1及び/又はMMP-9活性を調節する、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項29】
非天然アミノ酸、D-異性体又はD,L-ラセミ混合異性体の形態、ジスルフィド結合、カルボキシ又はアミノ末端修飾、脂肪酸及びPEGを含む生体適合性分子への接合、並びにアミノ酸の化学的置換からなる群から選択される1つ以上の修飾を含む、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項30】
ホモポリマー化又はヘテロポリマー化されている、請求項1、11、14から19、及び21のいずれか一項に記載のペプチド。
【請求項31】
C末端、N末端、又はそれらの側鎖に結合している請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む融合タンパク質。
【請求項32】
請求項1、11,14から19、及び21のいずれか一項に記載のペプチドを含む、純粋な医薬製剤。
【請求項33】
動物に投与するため、又はヒトに投与するための医薬製剤として調製される、請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物。
【請求項34】
液体として製剤化されている、請求項33に記載の組成物。
【請求項35】
輸液又は注射に適切なものである、請求項34に記載の液体。
【請求項36】
請求項1、11、14から19、又は21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、アテローム性動脈硬化症のプラークを安定化し、プラークが破裂する事象の発生を低減するための方法。
【請求項37】
前記組成物又は医薬製剤が液体である、請求項36に記載の方法。
【請求項38】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項36に記載の方法。
【請求項39】
前記動物が、アテローム性動脈硬化症のプラークの存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項36に記載の方法。
【請求項40】
前記疾患が、以下の特徴:心筋梗塞、脳卒中、及び急性肢虚血、又はその様な疾患に対する素因の1つ以上を含む、請求項36に記載の方法。
【請求項41】
前記動物への投与が、輸液又は注射による投与を含む、請求項36に記載の方法。
【請求項42】
請求項1、11、14から19、又は21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、内膜病変内の炎症及び脂質沈着を低減し、且つ、コラーゲン及びエラスチン沈着を増大させるための方法。
【請求項43】
前記組成物又は医薬製剤が液体である、請求項42に記載の方法。
【請求項44】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項42に記載の方法。
【請求項45】
前記動物が、内膜病変内の炎症、脂質沈着、又はコラーゲン及びエラスチン沈着の減少の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項42に記載の方法。
【請求項46】
前記疾患が、心筋梗塞、脳卒中、及び急性肢虚血、又はその様な疾患に対する素因を含む、請求項45に記載の方法。
【請求項47】
前記動物への投与が、輸液又は注射によるものである、請求項42に記載の方法。
【請求項48】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤を動物に投与する工程を含む、大動脈壁でTGF-β1を活性化し、且つ、MMP-9を不活性化するための方法。
【請求項49】
前記組成物又は医薬製剤が液体である、請求項48に記載の方法。
【請求項50】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項48に記載の方法。
【請求項51】
前記動物への投与が、輸液又は注射によるものである、請求項48に記載の方法。
【請求項52】
前記動物が、大動脈壁におけるTGF-β1の不活性化及び/又はMMP-9の活性化の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項48に記載の方法。
【請求項53】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物または医薬製剤を動物に投与する工程を含む、TGF-β1の活性化及び/又はMMP-9の不活性化のための方法。
【請求項54】
前記組成物又は医薬製剤が液体である、請求項53に記載の方法。
【請求項55】
前記組成物又は医薬製剤が、輸液又は注射に適切な液体である、請求項53に記載の方法。
【請求項56】
前記動物への投与が、輸液又は注射によるものである、請求項53に記載の方法。
【請求項57】
動物が、癌及び/又は血管新生、あるいはその様な疾患に対する素因の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項53に記載の方法。
【請求項58】
請求項1、11、14から19、及び21のいずれか一項に記載のペプチドを含む組成物又は医薬製剤と動物を接触させる工程を含む、TGF-β1及び/又はMMP-9活性を調節するための方法。
【請求項59】
前記動物が、癌及び/又は血管新生、あるいはその様な疾患に対する素因の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項58に記載の方法。
【請求項60】
前記動物が、内膜病変内の炎症、脂質沈着、又はコラーゲン及びエラスチン沈着の減少の存在によって、少なくとも部分的に特徴付けられる疾患を有する、請求項58に記載の方法。
【請求項61】
前記疾患が、心筋梗塞、脳卒中、及び急性肢虚血、あるいはその様な疾患に対する素因の1つ以上を含む、請求項60に記載の方法。
【図7】

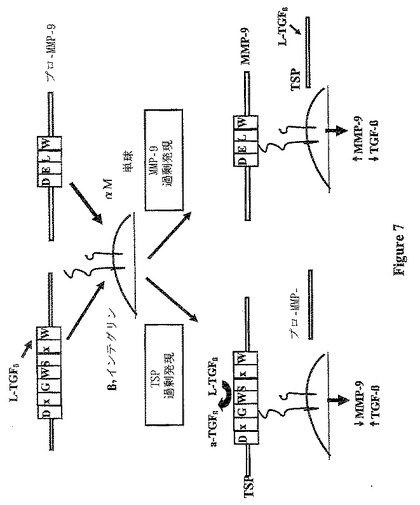
【図1】
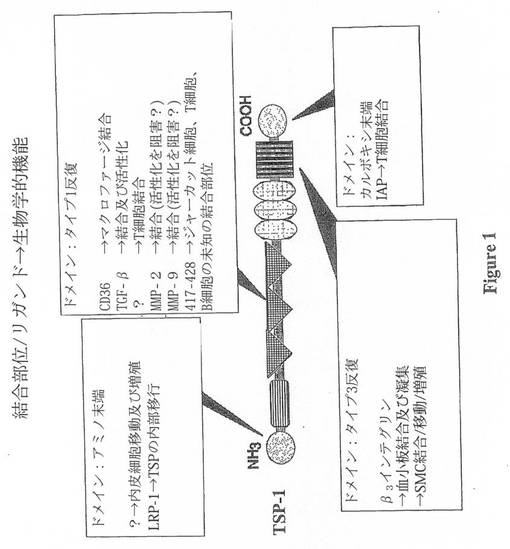
【図2A】
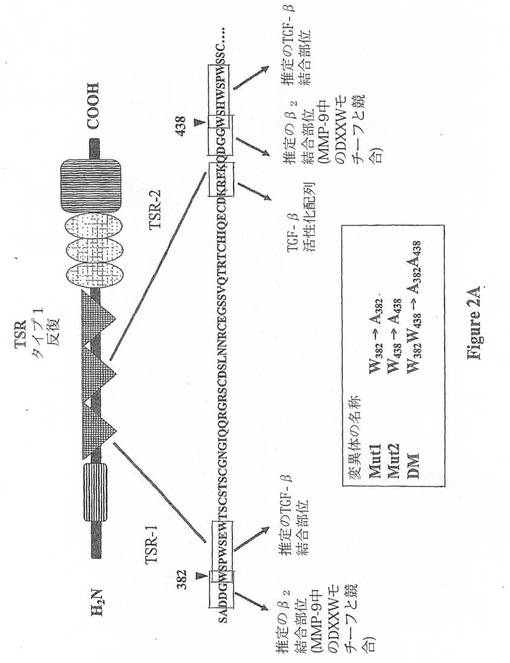
【図2B】
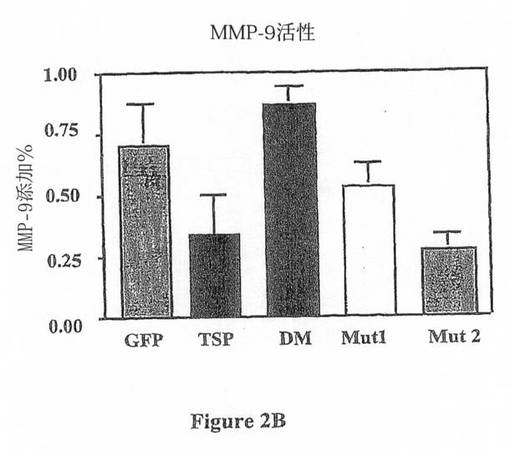
【図2C】
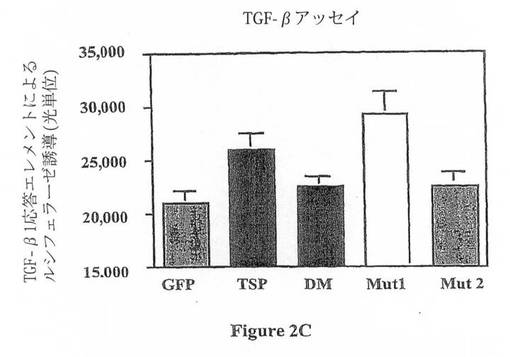
【図3A−C】
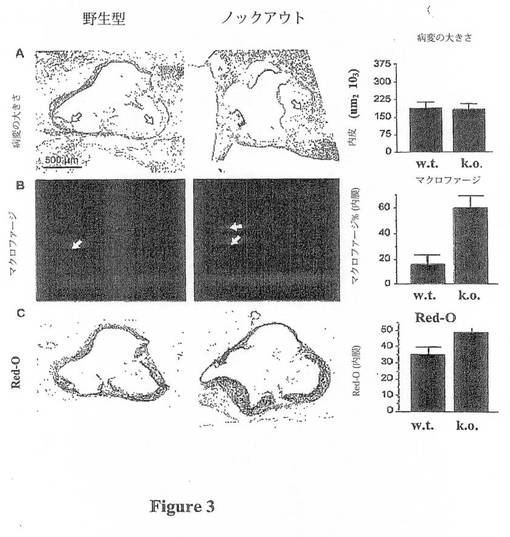
【図3D−F】
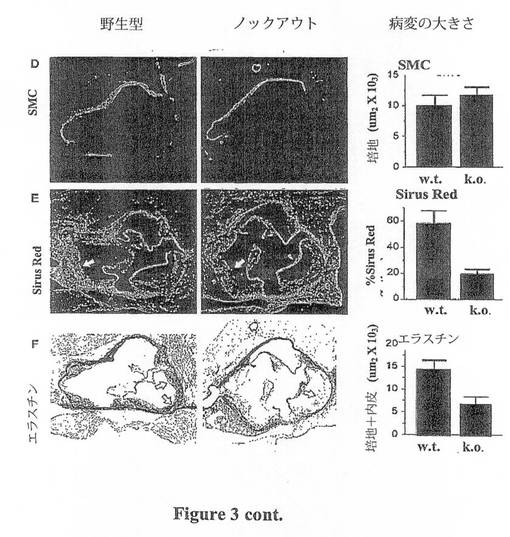
【図4】
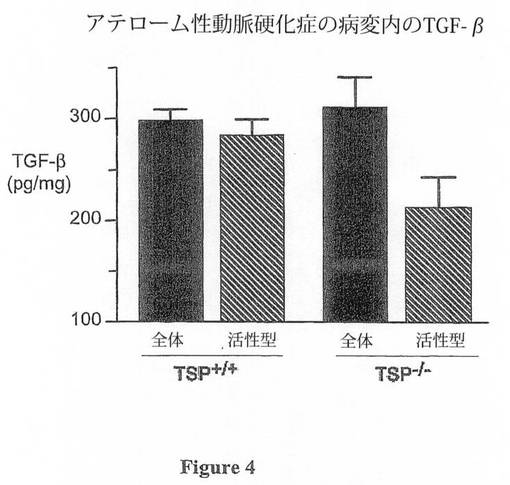
【図5】
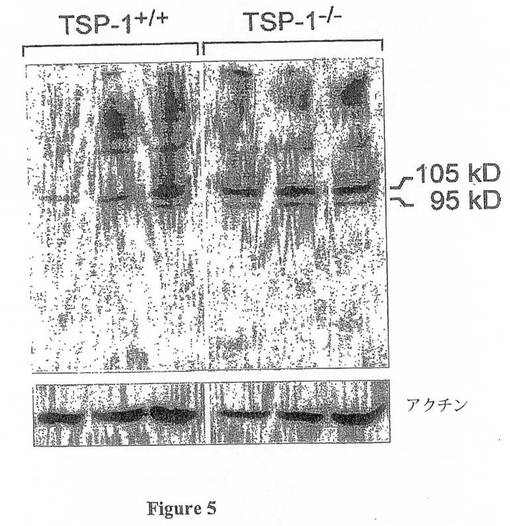
【図6】
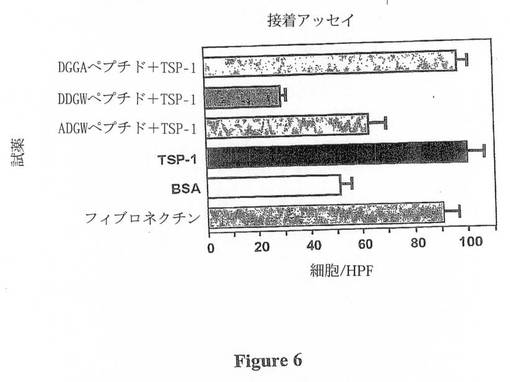
【図1】
【図2A】
【図2B】
【図2C】
【図3A−C】
【図3D−F】
【図4】
【図5】
【図6】
【公表番号】特表2008−543345(P2008−543345A)
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願番号】特願2008−518477(P2008−518477)
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024692
【国際公開番号】WO2007/002534
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(507418027)
【Fターム(参考)】
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/US2006/024692
【国際公開番号】WO2007/002534
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(507418027)
【Fターム(参考)】
[ Back to top ]