
ドナー特異的抗体ライブラリー
本発明は、本明細書で議論される1種以上の疾患に罹患した、もしくは現在罹患している患者ドナーから得られたドナー特異的抗体ライブラリーに関する。本発明はまた、ドナー特異的抗体を作製および使用するための方法に関する。本発明はさらに、上記ドナー特異的抗体ライブラリーから得られる中和抗体およびヒト疾患の予防/処置のためにこれら抗体を使用するための方法に関する。一実施形態において、本発明は、標的抗原に対する抗体生成を引き起こす疾患に罹患したか、もしくは該疾患に現在罹患しているヒト患者ドナーから得られる、抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子のレパートリーを含むベクター集団を提供し、ここで該集団は、独特のバーコードで同定される。

【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、ドナー特異的抗体ライブラリー、ならびにこれを作製および使用するための方法に関する。本発明はまた、このようなドナー特異的抗体ライブラリーから得られる中和抗体、および種々のヒト疾患および状態の予防および/もしくは処置のために得られた抗体を使用するための方法に関する。
【背景技術】
【0002】
(発明の背景)
ヒト疾患の診断、予防および処置に対して特異的な薬剤の生成および同定は、有用な化学的性質の巨大な集団の利用を要する。抗体ライブラリーの作出およびスクリーニングのための種々の技術の到来かつ迅速な開発に関して、疾患標的に対するモノクローナル抗体は、新たな薬物候補の主要なカテゴリーのうちの1つになった。ヒト使用のために、特異性および有効性に加えて、安全性は、主要な懸念であるので、ヒトモノクローナル抗体のライブラリーは、特に重要になった。
【0003】
現在では、ヒト抗体ベースの薬物候補の開発は、代表的には、代表的には、それらの意図された適用に関連しないヒトレパートリー(repertoire)に由来する抗体配列の無作為集団を含むヒト抗体ライブラリーをスクリーニングすることによって、同定される。特異的ヒトドナーから作出された各抗体ライブラリーは、上記ドナーがその個々の寿命の過程にわたって遭遇し、挑戦し、かつ乗り越えた全ての特有の挑戦(challenge)に由来するかもしくは作り出す、全ての成分、生理学、および代謝的変化に対する抗体を潜在的に含む。現在のアプローチに伴って作製された代表的なヒト抗体ライブラリーは、ドナーの健康上の履歴の理解なしに構築されるので、得られた免疫グロブリンレパートリーにおいて予測されるものはほとんど未知である。
【0004】
従って、特定の疾患との遭遇に首尾よく耐えたかもしくは現在耐えている個体から抗体ライブラリーを作り出すことは、特に重要である。なぜなら、彼らの得られたレパートリーは、関連する疾患を特異的に防御するためにドナーが使用した抗体を含むからである。ネガティブ要素もしくはポジティブ要素を除去もしくは単離し、望ましくない内容を排除し、そして改善された特性を有するヒト抗体を生成する能力を含む、このようなライブラリーの効率的なスクリーニングおよび取り扱いのための方法を提供することもまた、重要である。
【0005】
本発明は、特定の標的疾患との遭遇に曝され、これに耐えたもしくは現在耐えている個体に由来するドナー特異的抗体ライブラリーの作出、スクリーニングおよび取り扱いのための方法および手段を提供することによって、これらの必要性に対処する。
【発明の概要】
【課題を解決するための手段】
【0006】
一局面において、本発明は、標的抗原に対する抗体生成を引き起こす疾患に罹患したもしくは現在罹患しているヒト患者ドナーから得られた、抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子のレパートリーを含むベクター集団(vector collection)に関し、ここで上記集団は、特有のバーコードで同定される。
【0007】
一実施形態において、上記ベクター集団は、抗体軽鎖もしくはそのフラグメントをコードする核酸分子(例えば、抗体λ軽鎖、もしくは抗体κ軽鎖、またはこれらのフラグメント)のレパートリーを含む。
【0008】
別の実施形態において、上記ベクター集団は、抗体重鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む。
【0009】
さらに別の実施形態において、上記バーコードは、上記集団に存在するベクターに連結されるかもしくは組み込まれ、そして/または上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、上記バーコードは、上記核酸分子の発現を妨害しない。
【0010】
従って、上記バーコードは、例えば、上記核酸分子の3’非コード領域もしくは5’非コード領域に連結され得る、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列であり得る。
【0011】
さらなる実施形態において、上記バーコードは、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列であるヌクレオチド配列である。
【0012】
なおさらなる実施形態において、上記バーコードは、非連続ヌクレオチド配列である。上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記集団に存在するベクターに連結され得るかもしくは組み込まれ得る。あるいは、上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ得、その結果、上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記核酸分子の発現を妨害しない。
【0013】
別の実施形態において、上記バーコードは、ペプチドもしくはポリペプチド配列である。
【0014】
異なる実施形態において、上記ベクター集団に存在するベクターは、ファージミドベクターであり、このファージミドは、例えば、バクテリオファージ遺伝子III、および上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と上記バクテリオファージIII遺伝子との間に終止コドンを含み、そしてバーコード(例えば、上記終止コドンの後ろの非翻訳領域に挿入される連続する非コードヌクレオチド配列)を有し得る。
【0015】
別の局面において、本発明は、本発明のベクター集団を含む宿主細胞に関する。上記宿主細胞は、真核生物宿主細胞もしくは原核生物宿主細胞(例えば、E.coli脂肪)であり得る。
【0016】
さらなる局面において、本発明は、標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するライブラリーメンバーを含むドナー特異的抗体ライブラリーに関し、ここで上記抗体もしくは抗体フラグメントは、上記標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒトドナーから得られ、ここで上記抗体ライブラリーは、少なくとも1個の特有のバーコードで同定される。
【0017】
一実施形態において、上記抗体重鎖および軽鎖は、これらが得られる上記ヒトドナーに特有のバーコードで各々別個に同定される。
【0018】
別の実施形態において、上記ドナー特異的抗体ライブラリーは、1個の特有のバーコードで同定される。
【0019】
さらに別の実施形態において、上記抗体もしくは抗体フラグメントは、ベクターに存在する核酸分子によってコードされる抗体重鎖および軽鎖もしくはそのフラグメントから構成される。
【0020】
さらなる実施形態において、上記バーコードは、上記ライブラリーに存在するベクターに連結されるかもしくは組み込まれ、そして/または上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、上記バーコードは、上記核酸分子の発現を妨害しない。
【0021】
なおさらなる実施形態において、上記バーコードは、1〜約24ヌクレオチドの連続非コードヌクレオチド配列であり、これらは、例えば、上記核酸分子の3’非コード領域もしくは5’非コード領域であり得る。
【0022】
異なる実施形態において、上記バーコードは、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列によってコードされる。
【0023】
別の実施形態において、上記バーコードは、非連続ヌクレオチド配列によってコードされる。
【0024】
さらなる実施形態において、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、上記ライブラリーに存在するベクターに連結されるかもしくは組み込まれる。
【0025】
なおさらなる実施形態において、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ、その結果、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、このような核酸分子の発現を妨害しない。
【0026】
異なる実施形態において、上記バーコードは、ペプチドもしくはポリペプチド配列である。
【0027】
別の実施形態において、ベクターは、ファージミドベクターであり、例えば、バクテリオファージ遺伝子III、および上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と上記バクテリオファージIII遺伝子との間に終止コドンを含み得る。
【0028】
さらに別の実施形態において上記ヒト患者ドナーの病歴は、上記ドナーが上記疾患に罹患したかもしくは現在罹患していることを示す。いくつかの実施形態において、上記ヒトドナーが、上記疾患に罹患したかもしくは現在罹患していることは、独立して確認される。
【0029】
さらなる実施形態において、上記ドナー特異的抗体ライブラリーは、上記標的抗原とは異なる抗原を特異的に結合する抗体および抗体フラグメントを実質的に欠いている。
【0030】
一実施形態において、上記標的抗原は、インフルエンザAウイルス(例えば、インフルエンザAウイルス H1、H2、H3、H5、H7、もしくはH9サブタイプの単離株)である。
【0031】
別の実施形態において、上記ライブラリーは、1つより多くのインフルエンザAウイルスサブタイプに特異的に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0032】
さらに別の実施形態において、上記ライブラリーは、上記インフルエンザウイルスAのH5N1サブタイプに結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0033】
さらなる実施形態において、上記ヒトドナーは、以下の表1に列挙される疾患からなる群より選択される疾患に罹患したかもしくは現在罹患している。
【0034】
なおさらなる実施形態において、上記抗体ライブラリーは、上記標的疾患と関連する抗原に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0035】
さらなる実施形態において、上記抗体ライブラリーは、上記標的疾患と関連する抗原に結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0036】
上記ドナー特異的抗体ライブラリーは、例えば、ファージライブラリーであり得、一実施形態において、これは、抗体もしくは抗体フラグメントのうちの106個より多くの異なるメンバー、もしくは抗体もしくは抗体フラグメントのうちの109個より多くの異なるメンバーをコードする配列を含み得る。
【0037】
他の実施形態において、上記ドナー特異的抗体ライブラリーは、限定されることなく、芽胞−ディスプレイライブラリー、リボソームディスプレイライブラリー、mRNAディスプレイライブラリー、微生物細胞ディスプレイライブラリー、酵母ディスプレイライブラリー、もしくは哺乳動物ディスプレイライブラリーである。
【0038】
一実施形態において、上記ベクターに存在する抗体もしくは抗体フラグメントをコードする核酸は、上記ヒト患者ドナーのリンパ球から抽出されるmRNAから逆転写され、ここで上記リンパ球は、例えば、骨髄、血液、脾臓、もしくはリンパ節から発生し得る。
【0039】
望ましい場合、上記ヒトドナーの血清学的プロフィールは、上記mRNAを抽出する前に生成され得る。
【0040】
代わりに、もしくはさらに、上記ヒトドナー、および必要に応じて、上記ドナーの家族の病歴は、上記mRNAを中するする前もしくはその後に検査される。
【0041】
別の局面において、本発明は、標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するドナー特異的ライブラリーを作製するための方法に関し、上記方法は、
a)上記標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒト患者ドナーのリンパ球からmRNAを得る工程;
b)上記得られたmRNAの逆転写によって、上記患者の免疫グロブリンレパートリーをコードする配列を含む核酸の集団を生成する工程;および
c)上記核酸を標識する特有のバーコードで、上記ドナー特異的ライブラリーを同定する工程、
を包含する。
【0042】
上記方法は、上記患者の血清学的プロフィールを生成する工程、および/または工程a)の前もしくはその後に、上記患者の病歴を検査する工程をさらに包含し得る。
【0043】
上記方法は、d)上記核酸を発現ベクターに挿入する工程;e)上記免疫グロブリンレパートリーを発現させる工程;およびf)上記免疫グロブリンレパートリーをディスプレイ系においてディスプレイする工程、をさらに包含し得る。
【0044】
別の実施形態において、上記方法は、上記ライブラリーのメンバーを、上記標的抗原を中和もしくは活性化するそれらの能力に基づいて、選択する工程をさらに包含する。
【0045】
なおさらなる実施形態において、上記方法は、少なくとも1種の中和抗体を生じる。
【0046】
別の実施形態において、上記方法は、上記標的抗原を中和もしくは活性化させることが見いだされたライブラリーメンバーを含む1個以上の下位ライブラリー(sub−library)を作り出す工程をさらに包含する。
【0047】
さらに別の実施形態において、本発明は、同定された少なくとも1個のライブラリーメンバーを配列決定する工程を包含する。
【0048】
さらなる局面において、本発明は、上記方法によって選択された抗体によって中和もしくは活性化される標的抗原と関連する疾患を処置もしくは予防するための方法に関し、上記方法は、有効量の上記選択された抗体を、必要なヒト患者に投与する工程を包含する。
【0049】
上記抗体は、例えば、中和抗体(例えば、少なくとも1つのインフルエンザAウイルスサブタイプに対する中和抗体)であり得る。
【0050】
別の実施形態において、上記疾患は、以下の表1に列挙される疾患からなる群より選択される。
【図面の簡単な説明】
【0051】
【図1】図1は、本発明のヒト抗体ライブラリーの作出のための代表的な方法を模式的に例示するフローチャートである。
【図2】図2は、2つの異なる標的(AおよびB)に対する反応強度を増大させるための代表的パニング富化スキームを例示する。富化の各ラウンドは、上記個々の標的に対するプールの反応性強度を増大させる。
【図3】図3は、標的AおよびBと交叉反応性のクローンの選択のためのストラテジーを例示し、ここで各連続するラウンドは、反両方の標的に対して得られたプールの応性強度を強化する。
【図4】図4は、並行発見プール(parallel discovery pool)を組換えて、交叉反応性を生成/増大させることによって、2つの異なる標的(標的AおよびB)に対する反応性強度を増大させるためのストラテジーを例示する。上記組換え抗体ライブラリーの選択の各ラウンドは、両方の標的に対して得られたプールの反応性強度を増大させる。
【図5】図5は、標的Aに対する反応性を維持しながら、標的Bに対する交叉反応性を増大させるためのストラテジーを例示する。第1に、標的Aと反応性のクローンが選択され、次いで、上記標的Aと反応性のクローンの変異誘発ライブラリーは調製され、選択は、示されるように行われ、標的Aおよび標的Bの両方との強い反応性を示す1個以上の抗体クローンを生じる。
【図6】図6は、「指定変異誘発(destinational mutagenesis)」方法による多様な多機能抗体集団を生成するための代表的変異誘発法を例示する。
【図7A】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7B】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7C】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7D】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7E】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7F】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7G】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7H】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7I】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7J】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図8】図8は、トルコH5N1トリインフルエンザの大発生の6名のヒト生存者から得られた血液サンプルについてのH5ヘマグルチニン(HA)血清学結果を示す。上記データは、上記HA抗原に対する抗体の存在を示す。
【図9】図9は、12名のその土地のドナーの血清サンプルで得られ、H5抗原(A/ベトナム/1203/2004)およびH1N1抗原(A/ニューカレドニア/20/99)およびH3N2抗原(A/パナマ/2007/99)のウイルスに対して試験した血清学結果を示す。
【図10】図10は、抗体ファージライブラリーの構築において使用される特有のバーコード付与アプローチを例示する。
【図11】図11は、種々のインフルエンザAサブタイプに由来するヘマグルチニンに結合する抗体の分析を示す。
【図12】図12は、必要とされるH5ヘマグルチニン結合グループ1の位置およびFab 47eにおける結晶構造に対するドミナント変異を示す。
【図13】図13および図14は、トルコトリインフルエンザ生存者の血清および骨髄の分析によって同定された抗体重鎖および軽鎖配列を使用して、多様な抗体重鎖および軽鎖ライブラリーを作り出すための指定変異誘発の使用を例示する。
【図14】図13および図14は、トルコトリインフルエンザ生存者の血清および骨髄の分析によって同定された抗体重鎖および軽鎖配列を使用して、多様な抗体重鎖および軽鎖ライブラリーを作り出すための指定変異誘発の使用を例示する。
【発明を実施するための形態】
【0052】
(詳細な説明)
(A.定義)
別段定義されない限り、本明細書において使用される技術的および科学的用語は、本発明が属する分野の当業者によって一般に理解されるのと同じ意味を有する。Singletonら,Dictionary of Microbiology and Molecular Biology 第2版,J.Wiley & Sons (New York,NY 1994)は、当業者に、本願において使用される用語の多くに対する一般的ガイドを与える。
【0053】
当業者は、本発明の実施において使用され得る、本明細書に記載されるものと類似もしくは等価な多くの方法および材料を認識する。実際に、本発明は、記載される方法および材料に決して限定されない。本発明の目的に関して、以下の用語は、以下で定義される。
【0054】
語句「保存されたアミノ酸残基」とは、互いに整列させられた2種以上のアミノ酸配列の間で同一であるアミノ酸残基に言及するために使用される。
【0055】
用語「疾患」、「障害」、および「状態」は、本明細書において交換可能に使用され、通常の身体機能の混乱、もしくは任意のタイプの病状の出現に言及する。正常の生理の混乱を引き起こす原因因子は、公知であってもよいし未知であってもよい。さらに、2名の患者が同じ障害と診断され得るが、それらの個体によって示される特定の症状は、同一でってもよいし、そうでなくてもよい。
【0056】
「有効な量」とは、有益もしくは望ましい治療的(予防的を含む)結果をもたらすに十分な量である。有効な量は、1回以上の投与において投与され得る。
【0057】
「組成物」は、本明細書において使用される場合、活性成分(例えば、本発明から生成される中和抗体)、および少なくとも1種の添加剤(例えば、薬学的に受容可能なキャリア)を含むと定義され、上記薬学的に受容可能なキャリアとしては、水、ミネラル、タンパク質、および/もしくは当業者に公知の他の賦形剤が挙げられるが、これらに限定されない。
【0058】
本明細書において使用される場合、用語「処置する」もしくは「処置」とは、疾患を示す臨床的症状の改善を意味することが意図される。
【0059】
本明細書において使用される場合、用語「予防する」もしくは「予防」とは、疾患を示す臨床的症状を妨げることを意味することが意図される。
【0060】
用語「被験体」および「患者」とは、本明細書において使用される場合、交換可能に使用され、検査、処置、分析、試験もしくは診断の被験体である、動物、および好ましくは、哺乳動物のいずれかに言及し得る。従って、被験体もしくは患者は、ヒト、非ヒト霊長類、および他の哺乳動物を含み、彼らは、疾患もしくは他の病的状態を有してもよいし有さなくてもよい。
【0061】
用語「アミノ酸」もしくは「アミノ酸残基」とは、代表的には、その分野で認識される定義を有するアミノ酸(例えば、アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)からなる群より選択されるアミノ酸)をいうが、改変アミノ酸、合成アミノ酸、もしくは稀なアミノ酸が、望ましい場合には使用され得る。従って、37 CFR 1.822(b)(4)に列挙される改変アミノ酸および通常でないアミノ酸は、この定義内に含まれ、明示的に本明細書に参考として援用される。アミノ酸は、種々の亜群へと下位分類され得る。従って、アミノ酸は、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val);負に荷電した側鎖(例えば、Asp、Glu);正に荷電した側鎖(例えば、Arg、His、Lys);もしくは非荷電極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、Trp、およびTyr)を有するとして分類され得る。アミノ酸はまた、低分子アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)、および塩基性アミノ酸(Lys、Arg)として分類され得る。
【0062】
用語「改変体」とは、参照ポリペプチドに関して、ネイティブポリペプチドと比較して、少なくとも1個のアミノ酸変異もしくは改変(すなわち、変化)を有するポリペプチドをいう。「アミノ酸改変」によって生成される改変体は、上記ネイティブアミノ酸配列において、例えば、少なくとも1個のアミノ酸を置換、欠失、挿入および/もしくは化学的に改変することによって生成され得る。
【0063】
「アミノ酸改変」とは、所定のアミノ酸配列のアミノ酸配列における変化をいう。例示的な改変としては、アミノ酸置換、挿入および/もしくは欠失を含む。
【0064】
特定の位置における「アミノ酸改変」とは、上記特定の残基の置換もしくは欠失、または上記特定の位置に隣接する少なくとも1個のアミノ酸残基の挿入をいう。特定の残基に「隣接する」挿入とは、その1〜2個の残基内での挿入を意味する。上記挿入は、上記特定の残基に対してN末端側もしくはC末端側に存在し得る。
【0065】
「アミノ酸置換」とは、所定のアミノ酸配列中の少なくとも1個の既存のアミノ酸残基を、別の異なる「置換」アミノ酸残基で置換することをいう。上記置換残基は、「天然に存在するアミノ酸残基」(すなわち、遺伝コードによってコードされる)であり得、以下からなる群より選択され得る:アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)。1個以上の天然に存在しないアミノ酸残基での置換はまた、本明細書におけるアミノ酸置換の定義によって包含される。
【0066】
「天然に存在しないアミノ酸残基」とは、上記の天然に存在するアミノ酸残基以外の、ポリペプチド鎖において隣接するアミノ酸残基を共有結合することができる残基をいう。天然に存在しないアミノ酸残基の例としては、ノルロイシン、オルニチン、ノルバリン、ホモセリンおよび他のアミノ酸残基残ログ(例えば、Ellmanら Meth.Enzym.202:301 336(1991)に記載されるもの)が挙げられる。このような天然に存在しないアミノ酸残基を生成するために、Norenら Science 244:182(1989)およびEllmanら(前出)の手順が、使用され得る。簡潔には、これら手順は、天然に存在しないアミノ酸残基でサプレッサtRNAを化学的に活性化すること、続いて、上記RNAのインビトロ転写および翻訳を伴う。
【0067】
「アミノ酸挿入」とは、所定のアミノ酸配列への少なくとも1個のアミノ酸の組み込みをいう。上記挿入は、通常、1個もしくは2個のアミノ酸残基の挿入からなる一方で、本願は、より大きな「ペプチド挿入」(例えば、約3〜約5個もしくはさらに約10個までのアミノ酸残基の挿入)を企図する。上記挿入された残基は、上記で議論されるように、天然に存在していてもよいし、天然に存在していなくてもよい。
【0068】
「アミノ酸欠失」とは、少なくとも1個のアミノ酸残基を、所定のアミノ酸配列から除去することをいう。
【0069】
用語「ポリヌクレオチド」とは、核酸(例えば、DNA分子およびRNA分子ならびにこれらのアナログ(例えば、ヌクレオチドアナログを使用して、もしくは核酸化学を使用して生成されるDNAもしくはRNA)をいう。望ましい場合、上記ポリヌクレオチドは、合成的に、例えば、当該分野で認識される核酸化学を使用して、例えば、ポリメラーゼを使用して、作製され得、望ましい場合には、改変され得る。代表的な改変としては、メチル化、ビオチン化、および他の当該分野で公知の改変が挙げられる。さらに、上記核酸分子は、一本鎖もしくは二本鎖であり得、そして望ましい場合、検出可能な部分に連結され得る。
【0070】
用語「変異誘発」とは、別段特定されない限り、ポリヌクレオチドもしくはポリペプチド配列を改変するために任意の当該分野で認識されている技術をいう。変異誘発の好ましいタイプとしては、エラープローンPCR変異誘発、飽和変異誘発(saturation mutagenesis)、もしくは他の部位指向性変異誘発が挙げられる。
【0071】
「部位指向性変異誘発」とは、当該分野で標準的な技術であり、望ましい変異を表す制限されたミスマッチを除いて変異誘発されるように、一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを使用して行われる。簡潔には、上記合成オリゴヌクレオチドは、上記一本鎖ファージDNAに対して相補的な鎖の合成を指向するためにプライマーとして使用され、その得られた二本鎖DNAは、ファージ支援宿主細菌(phage−supporting host bacterium)に形質転換される。上記形質転換細菌の培養は、上部の寒天にプレートされ、上記ファージを内部にもつ単一の細胞からのプラーク形成を可能にし、上記新たなプラークのうちの50%は、一本鎖として、変異した形態を有するファージを含み;50%は、元の配列を有する。目的のプラークは、正確なマッチのハイブリダイゼーションを可能にする温度でキナーゼ処理(kinased)合成プライマーとハイブリダイズすることによって選択されるが、その温度で、上記元の鎖とのミスマッチは、ハイブリダイゼーションを妨げるのに十分である。次いで、上記プローブとハイブリダイズするプラークが選択され、配列決定され、培養され、そのDNAが回収される。
【0072】
用語「ベクター」とは、細胞における自律的複製が可能でかつDNAセグメント(例えば、遺伝子もしくはポリヌクレオチド)が、上記結合されるセグメントの複製をもたらすように作動可能に連結されるrDNA分子をいうために使用される。1種以上のポリペプチドをコードする遺伝子の発現を指向し得るベクターは、本明細書において「発現ベクター」といわれる。用語「制御配列」とは、特定の宿主生物における作動可能に連結されたコード配列の発現に必須のDNA配列をいう。原核生物に適した上記制御配列としては、例えば、プロモーター、必要に応じて、オペレーター配列、およびリボソーム結合部位が挙げられる。真核生物細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが公知である。
【0073】
核酸は、別の核酸配列と機能的関係に置かれている場合に「作動可能に連結されている」。例えば、プレ配列もしくは分泌リーダーのDNAは、上記ポリペプチドの分泌に関与するプレタンパク質として発現される場合にポリペプチドのDNAに作動可能に連結されている;プロモーターもしくはエンハンサーは、上記配列の転写に影響を及ぼす場合、コード配列に作動可能に連結されている;またはリボソーム結合部位は、翻訳を促進するように位置している場合、コード配列に作動可能に連結されている。一般に、「作動可能に連結」とは、連結されている上記DNA配列が連続しており、そして分泌リーダーの場合には、連続しておりかつリーディング相中にあることを意味する。しかし、エンハンサーは、連続している必要はない。連結は、都合のよい制限部位においてライブラリーゲーションによって達成される。このような部位が存在しない場合、上記合成オリゴヌクレオチドアダプターもしくはリンカーが、従来の経験に従って使用される。
【0074】
パーセント(%)アミノ酸配列同一性は、配列比較プログラムNCBI−BLAST2(Altschulら,Nucleic Acids Res.25:3389−3402(1997))を使用して決定され得る。上記NCBI−BLAST2配列比較プログラムは、http://www.ncbi.nlm.nih.govからダウンロードされるか、そうでなければ、National Institute of Health,Bethesda,MDから得られ得る。NCBI−BLAST2は、いくつかの検索パラメーターを使用し、ここでそれらパラメーターの全ては、以下を含むデフォルト値に設定されている:例えば、unmask=yes、strand=all、expected occurrences=10、minimum low complexity length=15/5、multi−pass e−value=0.01、constant for multi−pass=25、dropoff for final gapped alignment=25およびscoring matrix=BLOSUM62。
【0075】
「抗体依存性細胞媒介性細胞傷害性」および「ADCC」とは、FcRを発現する非特異的細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、およびマクロファージ)が、標的細胞上に結合抗体を認識し、その後、上記標的細胞の融解を引き起こす細胞媒介性反応をいうために、本明細書において使用される。種々の免疫細胞は、異なるFcレセプター(FcR)を発現する。従って、ADCCを媒介するための主な細胞(NK細胞)は、FcgRIIIのみを発現するのに対して、単球は、FcγRI、FcγRIIおよびFcγRIIIを発現する。
【0076】
用語「インフルエンザAサブタイプ」もしくは「インフルエンザAウイルスサブタイプ」とは、交換可能に使用され、ヘマグルチニン(H)およびノイラミニダーゼ(N)ウイルス表面タンパク質の種々の組み合わせによって特徴付けられ、従って、H番号およびN番号の組み合わせ(例えば、H1N1およびH3N2)によって表示されるインフルエンザAウイルス改変体をいう。上記用語は、具体的には、通常は、変異から生じ、異なる病理的側面を示す、各サブタイプ内の全ての株(絶滅株を含む)を含む。このような株はまた、ウイルスサブタイプの種々の「単離株」として言及され、全ての過去の、現在の、および将来的な単離株を含む。従って、この状況において、用語「株」および「単離株」は、交換可能に使用される。
【0077】
用語「インフルエンザ」とは、インフルエンザウイルスによって引き起こされる感染性疾患をいうために使用される。
【0078】
(B.一般的技術)
本発明の方法を行うための技術は、当該分野で周知であり、標準的な実験教科書に記載されており、これら教科書としては、例えば、Ausubelら,Current Protocols of Molecular Biology,John Wiley and Sons(1997);Molecular Cloning:A Laboratory Manual,第3版,J.Sambrook and D.W.Russell,編,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;O’Brianら,Analytical Chemistry of Bacillus Thuringiensis,Hickle and Fitch,編,Am.Chem.Soc.,1990;Bacillus thuringiensis:biology,ecology and safety,T.R.Glare and M. O’Callaghan,編,John Wiley,2000;Antibody Phage Display,Methods and Protocols,Humana Press,2001;およびAntibodies,G.Subramanian,編,Kluwer Academic,2004が挙げられる。変異誘発は、例えば、部位指向性変異誘発(Kunkelら,Proc.Natl.Acad.Sci USA 82:488−492(1985))を使用して行われ得る。PCR増幅法は、米国特許第4,683,192号、同第4,683,202号、同第4,800,159号、および同第4,965,188号に、およびいくつかの教科書(including 「PCR Technology:Principles and Applications for DNA Amplification」,H.Erlich,編,Stockton Press,New York(1989);およびPCR Protocols:A Guide to Methods and Applications,Innisら,編,Academic Press,San Diego,Calif.(1990)に記載されている。
【0079】
本発明の方法は、抗体のディスプレイのために使用されるいかなる特定の技術によっても限定されない。本発明は、ファージディスプレイを参照しながら例示されるが、本発明の抗体は、他のディスプレイおよび富化技術(例えば、リボソームもしくはmRNAディスプレイ(Mattheakisら,Proc.Natl.Acad.Sci.USA 91:9022−9026(1994);Hanes and Pluckthun,Proc.Natl.Acad.Sci.USA 94:4937−4942(1997))、微生物細胞ディスプレイ(例えば、細菌ディスプレイ(Georgiouら,Nature Biotech.15:29−34(1997))、もしくは酵母細胞ディスプレイ(Kiekeら,Protein Eng.10:1303−1310(1997))、哺乳動物細胞上でのディスプレイ、芽胞ディスプレイ(Isticatoら,J.Bacteriol.183:6294−6301(2001);Chengら,Appl.Environ.Microbiol.71:3337−3341(2005)および同時係属中の仮特許出願番号第60/955,592号(2007年8月13日出願)、ウイルスディスプレイ(例えば、レトロ・ウイルスディスプレイ(Urbanら,Nucleic Acids Res.33:e35(2005)、タンパク質−DNA連結に基づくディスプレイ(Odegripら,Proc.Acad.Natl.Sci.USA 101:2806−2810(2004);Reiersenら,Nucleic Acids Res.33:e10(2005))、および、マイクロビーズディスプレイ(Seppら,FEBS Lett.532:455−458(2002))によっても同定され得る。
【0080】
リボソームディスプレイにおいて、上記抗体およびコードmRNAは、リボソームによって連結され、上記リボソームディスプレイは、翻訳の最後に、上記mRNAが上記ポリペプチドを放出することなくとどまるようにする。選択は、全体として三成分複合体(ternary complex)に基づく。
【0081】
mRNAディスプレイライブラリーにおいて、抗体とそのコードmRNAとの間の共有結合は、アダプター分子として使用さらえるピューロマイシンを介して確立される(Wilsonら,Proc.Natl.Acad.Sci.USA 98:3750−3755(2001))。抗体をディスプレイするためのこの技術の使用に関しては、例えば、Lipovsek and Pluckthun,J.Immunol.Methods.290:51−67(2004)を参照のこと。
【0082】
微生物細胞ディスプレイ技術は、酵母(例えば、Saccharomyces cerevisiae(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997)))上での表面ディスプレイを含む。従って、例えば、抗体は、S.cerevisiaeの表面に、酵母細胞壁上に位置するα−アグルチニン酵母接着レセプターへの融合を介してディスプレイされ得る。この方法は、フローサイトメトリーによってレパートリーを選択する可能性を提供する。蛍光標識された抗原および抗エピトープタグ試薬によって上記細胞を染色することによって、上記酵母細胞は、抗原結合のレベルおよび細胞表面での抗体発現に従って、ソートされ得る。酵母ディスプレイプラットフォームはまた、ファージと組み合わされ得る(例えば、Van den Beuckenら,FEBS Lett.546:288−294(2003)を参照のこと)。
【0083】
抗体ライブラリーを選択し、スクリーニングするための技術の総説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照のこと。
【0084】
(C.好ましい実施形態の詳細な説明)
(I.ドナー特異的抗体ライブラリーの調製)
本発明は、特定の疾患との遭遇に首尾よく耐えたかもしくは現在耐えている個体に由来するドナー特異的抗体ライブラリーに関する。その得られた抗体レパートリーは、上記ドナーが関連する疾患を特異的に防御するために使用した抗体を含み、従って、例えば、標的疾患の予防および/もしくは処置のための中和抗体を開発するために、重要なツールである。
【0085】
本発明は、ヒト被験体における抗体生成を引き起こす任意の標的疾患に適用可能であるが、このような疾患の代表的な(限定ではない)例は、表1に列挙される。
【0086】
【表1】
本発明のドナー特異的ライブラリーを作り出すための方法は、図1に模式的に例示される。第1の工程として、潜在的なドナーが同定される。上記患者ドナーは、標的疾患に現在罹患していてもよいし、上記標的疾患から回復していてもよいし、上記疾患に耐えていてもよい。従って、例えば、実施例に例示されるように、上記ドナー特異的ライブラリーは、本明細書において、以前のインフルエンザ感染(季節性インフルエンザ大発生、流行、および世界的流行が挙げられる)の回復期患者の骨髄から作り出され得る。
【0087】
患者ドナーを選択もしくは同定する際に、上記患者は、実際に上記標的疾患を有したかまたは現在有していることを確認することが重要である。上記確認の一部は、上記患者ドナーの病歴の検査である。上記病歴に加えて、種々の他の要因(例えば、上記患者の家族の病歴、上記患者の性別、体重、健康状態など)が、考慮に入れられ得る。上記患者の病歴が入手可能でないもしくはあてにならない、または任意の他の理由(例えば、さらなる他の確認手段)がある場合には、上記患者の血清学的側面が、決定され得る。血清学的アッセイは、当該分野で周知であり、種々の形式において(例えば、種々のELISAアッセイフォーマットの形態において)行われ得る。従って、例えば、インフルエンザウイルスに対する抗体の存在は、周知のヘマグルチニン阻害(HAI)アッセイ(Kendal,A.P.,M.S.Pereira,and J.J.Skehel.1982.Concepts and procedures for laboratory−based influenza surveillance. U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta, Georgia)、もしくはミクロ中和アッセイ(Harmonら,J.Clin.Microbiol.26:333−337(1988))によって検出され得る。この工程は、上記血清サンプルがインフルエンザ 中和抗体を含むことが既に確認された場合には必要ではないかもしれない。
【0088】
ドナー特異的ヒト抗体ライブラリーを調製するために、リンパ球を含むサンプルを、標的疾患(例えば、表1に列挙されるものからの少なくとも1つの疾患)を発症させたことが既知の個体(患者ドナー)から集められる。上記サンプルは、例えば、骨髄、血液、脾臓、リンパ節、扁桃、胸腺などから得られ得る。骨髄は、本明細書において上記抗体ライブラリーの好ましい供給源である。なぜなら、個々のドナーの成熟抗体レパートリーの完全な「過去のアーカイブ(fossil archive)」を表すからである。
【0089】
リンパ球を含むサンプルは、種々の時点において上記患者ドナーから集められ得る。一実施形態において、リンパ球は、上記標的とした疾患から回復した患者から、少なくとも1日間、5日間、10日間、15日間、20日間、25日間、少なくとも1ヶ月間、2ヶ月間、3ヶ月間、4ヶ月間、5ヶ月間、6ヶ月間、7ヶ月間、8ヶ月間、9ヶ月間、10ヶ月間、11ヶ月間、または少なくとも1年間、2年間、3年間、4年間、5年間、6年間、7年間、8年間、9年間、10年間にわたって、集められる。別の実施形態において、リンパ球は、回収の時点で上記標的とした疾患を有している、および上記疾患を有していると診断された患者から、少なくとも回収前の1日間、5日間、10日間、15日間、20日間、25日間、もしくは少なくとも1ヶ月間、2ヶ月間、3ヶ月間、4ヶ月間、5ヶ月間、6ヶ月間、8ヶ月間、9ヶ月間、10ヶ月間、もしくは1年間、2年間、3年間、4年間、もしくは5年間にわたって集められる。
【0090】
末梢血サンプル(特に、地理的に異なる供給源に由来する)は、輸送および使用の前に安定化される必要があり得る。この目的のためのキットは周知であり、市販の(例えば、BD Vacutainer(登録商標) CPTTM 細胞調製チューブ)は、リンパ球の遠心分離精製のために使用され得、およびグアニジウム、Trizol、もしくはRNAlaterが、上記サンプルを安定化するために使用され得る。リンパ球を他の供給源(例えば、リンパ系器官)から単離するための方法およびキットはまた、周知であり、市販されている。
【0091】
上記安定化したリンパ球もしくは全骨髄を受け取ったら、RNAが抽出され、RT−PCRは、当該分野で公知の免疫グロブリンオリゴプライマーを使用して、抗体重鎖および軽鎖レパートリーをレスキューするために行われる。
【0092】
骨髄リンパ球、もしくは任意の他の供給源のリンパ球に由来するRNAの調製のための方法は、当該分野で周知である。mRNA抽出の一般的方法は、分子生物学の標準的教科書で開示されており、上記標準的教科書としては、Ausubelら,Current Protocols of Molecular Biology,John Wiley and Sons(1997)が挙げられる。RNA精製キットは、商業的製造業者(例えば、Qiagen)から市販されており、上記製造業者の説明書に従って使用され得る。
【0093】
RNAは、PCRのためのテンプレートとして働き得ないので、RNAは、cDNAへと逆転写され、これは、PCR増幅に供される。2つの最も一般的に使用される逆転写酵素は、トリ骨髄芽球症ウイルス(avilo myeloblastosis virus)逆転写酵素(AMV−RT)およびモロニーマウス白血病ウイルス逆転写酵素(MMLV−RT)である。逆転写工程は、代表的には、環境および発現プロフィールの目的に依存して、特異的プライマー、ランダムヘキサマー、もしくはオリゴ−dTプライマーを用いて始められる。例えば、抽出されたRNAは、GeneAmp RNA PCRキット(Perkin Elmer,CA,USA)を使用して、製造業者の説明書に従って逆転写され得る。次いで、得られたcDNAは、その語のPCR反応においてテンプレートとして使用され得る。
【0094】
ファージディスプレイライブラリーを作り出すために、上記PCRレパートリー生成物は、scFvライブラリーを精製するためにリンカーオリゴと合わされて、当該分野で公知の手順に従って、m13 pIIIタンパク質とインフレームで直接クリーニングされ得る。他のディスプレイ技術(例えば、上記で議論されるもの)を使用するライブラリーは、当該分野で周知の方法によって調製され得る。
【0095】
代表的プロトコルにおいて、総RNAは、Tri BD試薬(Sigma)によって、新鮮なもしくはRNAlater安定化組織から抽出される。続いて、上記単離されたドナー総RNAは、Oligotex精製(Qiagen)を使用して、mRNAへとさらに精製される。次に、第1鎖cDNA合成は、AccuScript逆転写酵素(Stratagene)のプロトコルに従って、ランダムナノマーオリゴヌクレオチドおよび/もしくはオリゴ(dT)18プライマーを使用することによって、生成される。簡潔には、100ng mRNA、0.5mM dNTPおよび300ng ランダムナノマーならびに/または500ngオリゴ(dT)18プライマーを、Accuscript RT緩衝液(Stratagene)中で、65℃で5分間にわたってインキュベートし、続いて、4℃へと迅速に冷却した。次いで、100mM DTT、Accuscript RT、およびRNAse Blockを各反応系に添加し、42℃において1時間にわたってインキュベートし、上記逆転写酵素は、70℃において15分間にわたって加熱することによって不活性化される。上記得られたcDNAは、上記抗体重鎖および軽鎖V遺伝子のRT−PCR増幅のためのテンプレートとして使用され得、これは、次いで、ベクターへと、もしくはファージディスプレイライブラリーが意図される場合には、ファージミドベクターへとクローニングされ得る。この手順は、抗体重鎖および軽鎖可変領域クローン(VHおよびVLライブラリー)のレパートリーを生成し、それは、スクリーニング目的で、別個にもしくは合わせて保持され得る。上記ベクター(例えば、ファージミドベクター)は、次いで、宿主細胞(例えば、E coli宿主)へと導入されて、抗体軽鎖もしくは重鎖またはそれらのフラグメントをコードする核酸分子のレパートリーを含むベクター集団を生成し得る。各場合において、上記ベクター集団は、1個以上の抗体軽鎖もしくは抗体重鎖サブタイプを含み得る。従って、上記ベクター集団は、抗体κ軽鎖および/もしくはλ軽鎖をコードする配列を含み得る。
【0096】
本発明の方法において、代表的には、抗体軽鎖および抗体重鎖は、上記で議論されるように最初に、別個にクローニングされ、また、上記κおよびλ軽鎖ライブラリーを分離する。上記ライブラリーはアーカイブに入れられ(archived)得、そして必要とされる場合、重鎖ライブラリーを、分離されたκ鎖ライブラリーおよびλ鎖ライブラリーと合わされ得、重鎖および軽鎖対形成が、例えば、ファージディスプレイの場合においては、パニングによって同定され得る。達成されるべき目的に依存して、これら工程は、複数回、種々のライブラリーもしくは下位ライブラリーと反復することが可能である。本発明の方法は、成功を最大化するために、パニングの間に必要とされる場合、ライブラリー、下位ライブラリーもしくはクローンを含めるもしくは排除することにおいて、優れた可撓性を提供する。
【0097】
特に、上記ベクター集団に存在する配列は、上記抗体重鎖および軽鎖(もしくはフラグメント)のコード配列をそれぞれ保有するので、上記配列は切り出され得、そして上記抗体重鎖および軽鎖、もしくはこれらのフラグメントの発現のために、1個以上の発現ベクターに挿入され得る。好ましくは、上記抗体重鎖および軽鎖もしくはそれらのフラグメントのコード配列は、上記抗体もしくは抗体フラグメントのライブラリーを生成するために重鎖および軽鎖の同時発現のために、同じ発現ベクターへと導入される。
【0098】
本発明の発現ベクターは、上記ベクターを1個以上の選択された宿主細胞において複製することを可能にする核酸配列を含む。このような配列は、種々の細菌、酵母、およびウイルスに対して周知である。上記プラスミドpBR322に由来する複製起点は、大部分のグラム陰性細菌に適しており、上記2μプラスミド起点は、酵母に適しており、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVもしくはBPV)は、哺乳動物再紡においてベクターをクローニングするために有用である。
【0099】
発現ベクターは、代表的には、選択遺伝子(選択マーカーともいわれる)を含む。代表的な選択遺伝子は、(a)抗生物質もしくた他の毒素(例えば、アンピシリン、ネオマイシン、メトトレキサート、もしくはテトラサイクリン)に対する耐性を付与するか、(b)栄養要求性欠損を補完するか、もしくは(c)複雑な培地から利用可能でない重要な栄養要素を補う(例えば、BacilliのD−アラニンラセマーゼをコードする遺伝子)、タンパク質をコードする。
【0100】
哺乳動物再紡に対する適切な選択マーカーの例は、上記再紡成分の同定が、上記抗体コード核酸を取り込みことを可能にするものである(例えば、DHFRもしくはチミジンキナーゼ)。野生型DHFRが使用される場合に適切な宿主細胞は、Urlaubら,Proc.Natl.Acad.Sci.USA,77:4216(1980)によって記載されるように調製および増殖される、DHFR活性が欠損しているCHO細胞株である。酵母において使用するための適切な選択遺伝子は、上記酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら,Nature,282:39(1979);Kingsmanら,Gene,7:141(1979);Tschemperら,Gene,10:157(1980)]。上記trp1遺伝子は、トリプトファン中で増殖する能力を欠いている酵母の変異株(例えば、ATCC番号44076もしくはPEP4−1)についての選択マーカーを提供する[Jones,Genetics,85:12(1977)]。
【0101】
発現ベクターおよびクローニングベクターは、通常、mRNA合成を指向するために、上記抗体コード核酸配列に作動可能に連結されているプロモーターを含む。種々の潜在的な宿主細胞によって認識されるプロモーターは、周知である。原核生物宿主とともに使用するために適したプロモーターとしては、β−ラクタマーゼおよびラクトースプロモーター系[Changら,Nature,275:615(1978);Goeddelら,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーター系[Goeddel,Nucleic Acids Res.,8:4057(1980);EP 36,776]、およびハイブリッドプロモーター(例えば、tacプロモーター)[deBoerら,Proc.Natl.Acad.Sci.USA,80:21−25(1983)]が挙げられる。細菌系において使用するためのプロモーターはまた、上記抗体をコードするDNAに作動可能に連結されるシャイン−ダルガーノ(S.D.)配列を含む。
【0102】
酵母宿主とともに使用するのに適したプロモーター配列(promoting sequence)の例としては、3−ホスホグリセレートキナーゼに対するプロモーター[Hitzemanら,J.Biol.Chem.,255:2073(1980)]もしくは他の糖分解性酵素[Hessら,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)](例えば、エノラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ヘキソキナーゼ、ピルベートデカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−ホスフェートイソメラーゼ、3−ホスホグリセレートムターゼ、ピルベートキナーゼ、トリオースホスフェートイソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼ)に対するプロモーターが挙げられる。
【0103】
増殖条件によって制御される転写のさらなる利点を有する誘導性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロームC、酸ホスファターゼ、窒素代謝と関連する分解酵素、メタロチオネイン、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ならびにマルトースおよびガラクトース利用を担う酵素に対するプロモーター領域である。酵母発現において使用するための適切なベクターおよびプロモーターは、EP 73,657においてさらに記載されている。
【0104】
哺乳動物宿主細胞における上記発現ベクター中の重鎖もしくは軽鎖遺伝子の転写は、例えば、ウイルス(例えば、ポリオーマウイルス、鶏痘ウイルス(UK 2,211,504(1989年7月5日公開))、アデノウイルス(例えば、アデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびシミアンウイルス40(SV40)のゲノムから得られるプロモーターによって、異種哺乳動物プロモーター(例えば、上記アクチンプロモーターもしくは免疫グロブリンプロモーター)から得られるプロモーターによって、および熱ショックプロモーターから、このようなプロモーターが上記宿主細胞系と適合性であれば、制御される。
【0105】
高等真核生物による、上記抗体遺伝子をコードするDNAの転写は、エンハンサー配列を上記ベクターに挿入することによって、増大させられ得る。エンハンサーは、その転写を増大させるようにプロモーターに対して作用する、通常、約10〜300bpのDNAのシス作用性要素である。多くのエンハンサー配列は、いまや哺乳動物遺伝子から公知である(グロビン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインスリン)。しかし、代表的には、真核生物細胞ウイルスに由来するエンハンサーを使用する。例としては、複製起点(bp100〜270)の後ろ側にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後ろ側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。上記エンハンサーは、上記抗体コード配列に対して5’側もしくは3’側にある位置において、上記ベクターにスプライスされ得るが、好ましくは、上記プロモーターから5’側の位置に位置する。
【0106】
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、もしくは他の多細胞生物に由来する核化細胞)において使用される発現ベクターはまた、転写の終結に、および上記mRNAを安定化させるために必須の配列を含む。このような配列は、真核生物のもしくはウイルスのDNAもしくはcDNAの5’非翻訳領域、およびときおり3’ 非翻訳領域から一般に入手可能である。これら領域は、上記抗体重鎖および軽鎖をコードするmRNAの非翻訳部分においてポリアデニル化フラグメントとして転写されるヌクレオチドセグメントを含む。
【0107】
組換え脊椎動物細胞培養に従ってポリペプチドの合成に適合させるために適したさらに他の方法、ベクター、および宿主細胞は、Gethingら,Nature,293:620−625(1981);Manteiら,Nature,281:40−46(1979);EP 117,060;およびEP 117,058に記載されている。
【0108】
特定の実施形態において、上記抗体ライブラリーは、ファージライブラリーの形態において生成され、ここで上記抗体重鎖および軽鎖もしくはそれらのフラグメントのコード配列は、ファージミドベクター(例えば、上記バクテリオファージ遺伝子IIIを含むベクター)にクローニングされる。ファージミドベクターは、周知であり、市販されている。これらのベクターとしては、例えば、pBluescriptベクターSKII+(Stratagene,Genbank Accession X52328)、および他のpBluscriptベクターが挙げられる。ファージディスプレイ技術は、ヒト抗体の大きなレパートリーの生成を可能にし、生体パニング(biopanning)手順は、任意の望ましい特異性もしくは他の特性を有する個々の抗体の選択を可能にする。
【0109】
例えば、初期の流行および大流行(例えば、1918年のスペイン風邪)からの生存者の末梢リンパ球に由来する免疫学的レパートリーは、上記記載されるものに類似の様式において復活させられ得、安定化され得、レスキューされ得、そして発現させられ得る。さらなるH1およびH3ライブラリーについて、レパートリーは、適切にタイミングを計ってワクチン接種したその土地の出身のドナーから回収され得る。さらなる選択肢として、市販の骨髄総RNAもしくはmRNAは、H1およびH3に適した、また、ドナーの背景に依存して、H2抗体スクリーニングに適したライブラリーを生成するための市販の供給源から購入され得る。一般に、ワクチン接種が利用可能な処置選択しである標的疾患に関しては、抗体は、免疫したヒトドナーから得られる生物学的サンプルからも単離され得る。上記特定の抗原を認識する抗体の力価を発揮した、免疫した患者から、骨髄、血液、もしくは別のリンパ球供給源が集められ、生成された抗体が単離され、上記のように増幅され発現される。
【0110】
上記で議論されるように、各ドナーについて、抗体軽鎖および重鎖ライブラリーは、別個にクローニングされ得る。従って、各ドナーについて、種々のκ軽鎖およびλ軽鎖ファミリーは、別個にプールされ得、かつ等モル量でクローニングされ得る。同様に、各ドナーについて、種々の重鎖ファミリーは、プールされ得、等モル量でクローニングされ得る。抗体の遺伝子ファミリー特異的レスキューを可能にすることによって、本発明の方法は、量が少ない抗体を含む、プールされた抗体ライブラリーの場合には、任意のおよび全ての個体からの免疫グロブリン寄与を保証する上記ドナーの抗体レパートリーをより完全に表すライブラリーを得る。例えば、実施例の実施例1に例示されるように、本明細書における
上記インフルエンザ重鎖および軽鎖ライブラリーを調製することにおいて、6個のκ軽鎖ファミリー、11個のλ軽鎖ファミリーおよび4個の重鎖ファミリーをレスキューした。
【0111】
代表的なスクリーンは、ゼロ、1個、もしくは1個より多い標的特異的ポジティブクローンを生じ得る。特定のコンビナトリアル抗体ライブラリーが、もっぱらスクリーニングされ、さらなる解決策がさらには獲得されそうにない場合、これは、上記重鎖および軽鎖レパートリーが標的に結合する能力がないことではないかもしれないが、逆に、上記集団は、標的を結合するために必要とされる上記必要な重鎖および軽鎖対を一緒にすることができなかった可能性がある。任意の個体からの代表的なレスキューされた軽鎖のレパートリーは、約105〜106個の特有の軽鎖、および約106〜108個の特有の重鎖を含み得る。このような対形成の考えられるコンビナトリアル生成物は、1011〜1014の範囲に及ぶ。このような集団は、数桁の規模で、大部分のディスプレイ系(例えば、ファージディスプレイ)の実際の限界を超える。結論として、現在のディスプレイ技術(例えば、ファージディスプレイ)を使用すると、ごく一部の上記コンビナトリアルの可能性が、任意の単一のファージ抗体ライブラリーにおいて捕獲および評価される。従って、重鎖の本来のセットと、軽鎖の本来の集団との再クローニングは、特定の標的に対するおそらく新規な抗体を有する、シャッフルされた重鎖および軽鎖の組み合わせの完全に新たなセットを生成する。このような新たな再シャッフルされた集団は、以前に存在した機能が乏しい(poorly performing)ドナー特異的ライブラリーを再現性が高い集団へと形質転換することが見いだされた。具体的には、単一ドナーに由来する集団に関して、以前に、3ラウンドの選択後のバックグラウンドと比較すると、0.3倍の富化が達成されたに過ぎなかった。しかし、この集団が再クローニングされ、再シャッフルされた場合、3ラウンドのパニングの後に、15倍の富化が可能になり、92個の選択されたクローンから55個の新規な配列が生じる。
【0112】
以前に言及されたように、代表的なスクリーニングは、標的特異的ポジティブクローンの任意の数を生じ得る。本発明は、それらの埋め込まれたバーコードによって任意のクローンの起源の同定を可能にする。代表的な抗体のスクリーンは、多くのドナー特異的ライブラリーに由来するファージ抗体を併せ得るので、上記ライブラリーのうちのいくらかおよびそれらのコンビナトリアルクローンが、抗体保有ファージ粒子として完全には表されないことが考えられる。この場合において、ポジティブクローンは、スクリーニングされる下位ライブラリーファージ集団において存在する可能なクローニングされた解決策全ての制限された物理的セットのみから生じた可能性がある。このような場合において、ドナー特異的ファージの集団をより完全に質問することはかなり重要である。この場合、ポジティブクローンからの上記バーコードは、上記クローンを担う上記特異的ライブラリーに導き、もっぱらおよびより深く上記目的の集団を深くスクリーニングすることを可能にする。
【0113】
望ましい抗体が、最初のライブラリー構築物のための基礎として最初に使用されるものを超えた機能的能力(例えば、中和もしくは活性化)を有しなければならない場合において、本発明者らは、将来を見越して、このような活性の証拠のために個々のドナー血清の輪郭を描き得る。上記望ましい活性が、任意の特定のドナー血清において合理的な力価で存在する場合、上記目的の標的に対してスクリーニングするために選択されるライブラリーに対応するものを選択し得る。他の場合において、上記関連する選択基準は、血清学に関連しない可能性があるが、ドナー特徴(例えば、年齢、性別もしくは病歴)に関連する可能性がある。いずれにしても、ドナープロフィールは、ライブラリー選択についての論理的なガイドでありかつドナー特異的かつ分離された抗体ライブラリーにおいてのみ可能ある。
【0114】
ファージパニングスクリーンを完了し、免疫優性クローンの存在を発見することは、異常ではない。さらに、反復パニングスクリーニングレジメンの際に、このようなクローンを再発見することも、異常ではない。優性クローンの場合において、より多くのもしくは異なるクローンのいずれかが望ましい場合、このクローンの存在を担うライブラリー物質を回避することが重要である。代表的なファージ抗体ライブラリーにおいて、上記特異的ライブラリーもしくは上記クローン起源の原因である物質は、上記集団から分離可能ではないが、ドナー特異的ライブラリーにおいて、上記ライブラリーを再スクリーニングし、望ましくないドナー下位ライブラリーを単純に省くことは可能であり、それによって、以前に同定された優性クローンからポジティブな選択に焦点を当てる。
【0115】
単純さのために、上記ライブラリーは、重鎖もしくは軽鎖ライブラリーとして記載されるが、同じ記載が抗体フラグメントのライブラリー、抗体重鎖および/もしくは軽鎖の抗体フラグメント、および抗体様分子のライブラリーに適用されることは、当業者にとって明らかである。
【0116】
特定の実施形態において、二重特異性(例えば、2つの異なるインフルエンザAサブタイプと、および/もしくは同じサブタイプの2つの株(単離株)と、および/もしくはヒトおよび非ヒト分離物との反応性を示す)を有する抗体は、制御される交叉反応性選択および/もしくは指向性のコンビナトリアル操作および/もしくは変異誘発操作を介して発見および最適化され得る。
【0117】
図2に例示される代表的な富化スキームにおいて、2個の標的(標的AおよびBと示される)に対する交叉反応性を示す抗体を含むライブラリーは、複数ラウンドの富化に供される。富化が標的Aとの反応性に基づいている場合、各ラウンドの富化は、標的Aに対する、上記プールの反応性強度を増大させる。同様に、富化が、標的Bとの反応性に基づく場合、各ラウンドの富化は、標的Bに対する、上記プールの反応性強度を増大させる。図2は、ファージディスプレイライブラリーをスクリーニングする場合に使用される選択方法である(以下を参照のこと)パニングに言及するが、上記アプローチは、上記で議論されるライブラリーのうちのいずれかのタイプおよびディスプレイ技術のいずれかのタイプに等しく適用可能であるか、そうでなければ当該分野で公知である。標的AおよびBは、抗体が結合する任意の標的を包含し、上記標的としては、インフルエンザウイルスの種々の分離物、タイプおよびサブタイプが挙げられるが、これらに限定されない。
【0118】
上記目的が、複数の特異性を有する中和抗体を同定することである場合、交叉反応性発見選択スキームが、使用され得る。単純性という重要性において、このスキームは、二重特異性を有する抗体の選択性を示す図3において例示される。この場合、2個の標的(標的AおよびB)との反応性を示す抗体を含む抗体ライブラリーは、最初に、上記標的のうちの一方(例えば、標的A)との反応性について、続いて、他方の標的(例えば、標的B)との反応性について選択される。各連続選択ラウンドは、両方の標的に対して生じたプールの反応性強度を強化する。従って、この方法は、二重特異性を有する抗体を同定するために特に有用である。当然のことながら、上記方法は、さらなる標的に対する反応性を示す抗体を、上記さらなる標的に対する富化のさらなるラウンドを含めることによって、同定するために拡張され得る。繰り返すと、上記スクリーニングされるライブラリーが、ファージディスプレイライブラリーである場合、選択は、交叉反応性パニングによって行われるが、他のライブラリーおよび他の選択法もまた、使用され得る。
【0119】
上記で議論した2つの方法の組み合わせは、標的Aおよび標的B、それぞれに対する反応性に対する2回の別個の富化ラウンド、得られた2個のプールを組換え、その後、上記のように、交叉反応性選択ラウンドを含む。このアプローチは、図4に例示される。ちょうど純粋な交叉反応性選択におけるように、上記組換えライブラリーの各ラウンドの選択は、両方の標的に対して得られたプールの反応性強度を増大させる。
【0120】
図5に例示されるさらなる実施形態において、標的Aとの強力な反応性を示しかつ検出可能な標的Bとの検出可能な交叉反応性を有する第1のクローンが、同定される。このクローンに基づいて、変異誘発性ライブラリーが調製され、これは、次いで交互のラウンドにおいて、それぞれ、標的Bおよび標的Aとの反応性について選択される。このスキームは、標的Aとの強力な反応性を維持し、標的Bとの増大した反応性を有する抗体を生じる。直前に、選択は、上記スクリーニングされたライブラリーが、ファージディスプレイライブラリーである場合に、パニングによって行われるが、他のライブラリー、他のディスプレイ技術、よび他の選択法もまた、同じストラテジーに従って使用され得る。
【0121】
上記で議論されるように、標的AおよびBは、例えば、上記インフルエンザAウイルスの2個の異なるサブタイプ、同じインフルエンザAウイルスの2個の異なる株(単離株)、2つの異なる種からのサブタイプもしくは単離株であり得、ここで1種は、好ましくは、ヒトである。従って、例えば、標的Aは、上記H5N1ウイルスの2004年のベトナム単離株であり得、そして標的Bは、上記H5N1ウイルスの1997年の香港単離株であり得る。これら例は、単なる例示であり、任意の2個もしくは複数の標的に対して二重および複数の特異性を有する抗体は、同定され得、選択され得、類似の様式において最適化され得ることが強調される。
【0122】
一旦所望の特性を有する中和抗体が同定されると、このような抗体の大部分によって認識される優勢なエピトープを同定することが所望である得る。エピトープマッピングのための方法は、当該分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.編,Humana Press,1996;およびEpitope Mapping:A Practical Approach,Westwood and Hay,編,Oxford University Press,2001において開示されている。
【0123】
(II.特融のバーコード付与でのドナー特異的抗体ライブラリーの同定)
本発明によれば、上記抗体重鎖および軽鎖レパートリーを、上記のように調製されたcDNA(例えば、骨髄cDNA)から増幅した後、好ましくは、抗体重鎖および軽鎖ライブラリーは、各患者ドナーに関して別個にクローニングされ、ここで上記個々のライブラリーは、特融のバーコードを使用して区別され得る。
【0124】
上記バーコードは、好ましくは、所望の抗体鎖もしくはそのフラグメントの発現を妨害することなく、標識されたクローンとともに、増殖することができるように、選択される。本発明の例示的実施形態において、上記バーコードは、上記発現ベクターの配列(好ましくは、ファージミドベクターが使用される場合には、末端pIII終止コドンの後に、3’非翻訳領域)に挿入され得る。クローン選択の際に、ベクターの特融の配列が決定され、単一の規定されたライブラリーの専用のものになる。この規定されたライブラリーは、単一のドナーのみならず、ドナーの別個のプール、または合成レパートリーもしくは半合成集団にも由来し得る。別の実施形態において、上記バーコードは、上記抗体重鎖および/もしくは軽鎖またはそれらのフラグメントのコード配列に、上記それぞれの鎖の発現を妨害しない位置もしくは形態において、挿入される。
【0125】
従って、上記バーコードは、配列決定もしくは特異的PCRプライマーによってデコンボルーションされ得る、長さが約1〜24個の連続する非コードヌクレオチドの配列であり得る。この方法、核酸の集団(例えば、抗体レパートリー)は、上記クローニング工程において連結され得る。本発明の例示的実施形態において、上記バーコードは、無作為に作製された配列の3塩基もしくは5塩基である。
【0126】
別の例において、上記バーコードは、サイレント変異のコード配列である。上記ライブラリーが、中断パリンドローム(interrupted palidrome)(例えば、Sfi GGCCNNNNNGGCC)を認識する制限酵素を利用する場合、別個のヌクレオチドが、クローンの種々の集団(例えば、抗体ライブラリー)を区別するために上記「N」の代わりに組み込まれ得る。このバーコード付与アプローチは、上記増幅工程において上記レパートリーが連結されるという利点を有する。
【0127】
さらなる実施形態において、上記バーコードは、非連続ヌクレオチド配列であり、この配列は、上記ベクター配列および/もしくは上記望ましい抗体鎖のコード配列中に存在し得る。従って、非連続配列を有するバーコードは、上記種々の個々の配列の起源を同定し、それらのその後の取り扱いをモニターすることにおいて、非常に大きな融通性を提供する。
【0128】
この例において、上記バーコードは、ファージ粒子に融合された、免疫学的に別個のペプチドもしくはタンパク質配列をコードするコード配列である。例としては、例えば、pIII、pVIII、pVII、もしくはpIXファージへのエピトープ(例えば、Myc、HA、FLAG)融合物が挙げられる。上記エピトープは、単純に、もしくは種々の組み合わせにおいて提供され得、そしてシス(ライブラリーコードプラスミドにおいて)もしくはトランス(特異的に改変されたヘルパーファージ)構成において提供され得る。
【0129】
考えられるバーコードの他の例としては、ハプテンもしくは蛍光性発色団を用いた、化学的および酵素的ファージ改変(ファージライブラリーに関して)が挙げられるが、これらに限定されない。このようなタグは、単一のラウンドの選択のために好ましい。
【0130】
個体ドナーから得られた上記個々の重鎖および軽鎖ライブラリー、もしくは他のバーコード付与クローンもしくは集団が、個々の配列の供給源を区別する能力を失うことなく、プールされ得る。
【0131】
(III.ドナー特異的抗体ライブラリー由来の中和抗体の最適化)
望ましい場合、二重特異性もしくは複数の特異性を有する上記中和抗体の交叉反応性は、米国特許出願公開第20050136428号(2005年6月23日公開)(この開示は、本明細書に参考として明示的に援用される)に記載されるように、当該分野で公知の方法(例えば、ルックスルー(Look Through)変異誘発(LTM))によってさらに改善され得る。
【0132】
ルックスルー変異誘発(LTM)は、選択されたアミノ酸のコンビナトリアル変位を同時に評価しかつ最適化する多次元変異誘発方法である。上記プロセスは、1個以上の相補性決定領域(CDR)ドメイン内の正確な分布に焦点を当て、アミノ酸側鎖化学の相乗効果的寄与を調査する。LTMは、CDR内の位置的な一連の単一の変異を生成し、ここで各野生型残基は、多くの選択されたアミノ酸のうちの1個によって、体系的に置換される。変異したCDRは、全ての改変体の定量的ディスプレイを妨げることなく、増大している複雑性およびサイズのコンビナトリアル一本鎖可変フラグメント(scFv)ライブラリーを生成するために合わされる。ポジティブ選択の後、改善された特性を有するクローンは配列決定され、それらの有益な変異がマッピングされる。改善された結合特異性のための相乗効果的変異を同定するために、全ての有益な交換(permutation)を発現するコンビナトリアルライブラリー(コンビナトリアル有益変異(combinatorial beneficial mutation)、CBM)は、混合されたDNAプローブによって生成され、ポジティブ選択され、分析されて、最適化されたscFv候補のパネルを同定し得る。上記手順は、Fvおよび他の抗体ライブラリーで、類似の様式において行われ得る。
【0133】
変異誘発はまた、上記のように、ウォークスルー(walk−through)変異誘発(WTM)によって行われ得る。
【0134】
1個より多くのインフルエンザAサブタイプおよび/もしくは同じサブタイプの1個より多くの分離株で、本明細書に記載される抗体の交叉反応性を意図的に設計するための別の有用な変異誘発方法は、「指定」変異誘発として本明細書において言及される。指定変異誘発は、好ましくは、異なる反応性の1個以上の抗体クローンに基づいて、抗体の集団を合理的に操作するために使用され得る。本発明の状況において、指定変異誘発は、抗体の個々のCDRにおけるもののような類似の配列上の類似の位置によって規定される単一もしくは複数の残基をコードするために使用され得る。この場合において、これら集団は、匹敵する位置において見いだされる残基の範囲を捕捉するために、オリゴ縮重を使用して生成される。この集団内で、特異性の連続体(continuum of specificities)は、親クローンの中にもしくは親クローンを超えたところに存在することが予測される。指定変異誘発の目的は、多様な多機能性抗体集団、ライブラリーを、2個以上の別個の実体もしくは集団の間に生成することである。指定変異誘発ライブラリーを作り出すために、両方の抗体についての上記CDR配列が最初に得られ、整列される。次に、保存された同一性の全ての位置が、最初に、単一のコドンで、マッチングされた残基に固定される。保存されていない位置において、縮重コドンが組み込まれて、両方の残基をコードする。いくつかの場合において、上記縮重コドンは、この位置において2個の親残基のみをコードする。しかし、いくつかの場合において、さらなる副産物が生成される。副産物生成のレベルは測定されて、サイズ制限もしくは目的に依存して、副産物生成を強化するかもしくはこの生成を排除し得る。
【0135】
従って、例えば、2個の抗体それぞれの最初の位置がスレオニンおよびアラニンである場合、上記縮重コドン(上記最初の2個の位置においてA/G−C−)は、第3の位置における塩基に関わらず、スレオニンもしくはアラニンのみをコードする。例えば、次の位置の残基がリジンおよびアルギニンである場合、その縮重コドンA−A/G−A/Gは、リジンもしくはアルギニンをコードするのみである。しかし、その縮重コドンA/C−A/G−A/G/C/Tが使用されれば、アスパラギン、ヒスチジン、およびセリン副産物も生成される。
【0136】
便宜上、マッチングされたCDR長を有する抗体のみを使用することは、より単純である。これを推し進めるための1つの方法は、上記CDR長およびはじめに発見された抗体によって付与される潜在的に均一なフレームワーク制限に基づいて、上記第2の抗原に対するサイズ制限されたライブラリーをスクリーニングすることである。しかし、等しい長さのCDRを使用することは、便宜のためにすぎず、必要条件ではないことに注意する。この方法は、インフルエンザAウイルス中和抗体の大きな機能的に多様なライブラリーを作り出すために有用であると同時に、その適用性は、遙かに広いことを理解することは容易である。この変異誘発技術は、任意の抗体の機能的に多様なライブラリーもしくは集団を作製するために使用され得る。従って、図6は、TNF−α抗体およびCD11a抗体のCDRを、変異誘発された親配列として使用して、指定変異誘発方法の使用を例示するために本明細書に含められる。
【0137】
他の例示的変異誘発方法は、飽和変異誘発およびエラープローンPCRを含む。
【0138】
飽和変異誘発(Hayashiら,Biotechniques 17:310−315(1994))は、全ての20アミノ酸が、タンパク質中の特定の位置、および特定の表現系についてアッセイされる各改変体に対応するクローンにおいて置換される技術である(米国特許第6,171,820号;同第6,358,709号および同第6,361,974号もまた参照のこと)。
【0139】
エラープローンPCR(Leungら,Technique 1:11−15(1989);Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、ランダム点変異をクローニングされた遺伝子に導入する改変ポリメラーゼ連鎖反応(PCR)技術である。得られたPCR生成物は、無作為変異ライブラリーを生成するためにクローニングされ得るか、またはT7プロモーターが適切なPCRプライマー内に組み込まれる場合に、直接転写され得る。
【0140】
他の変異誘発技術はまた、周知であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001に記載される。
【0141】
最適化は、単独で、もしくは任意の組み合わせにおいて、上記で議論されるライブラリーのうちのいずれか、もしくは当該分野で公知のライブラリーの任意の他のタイプに基づき得る。
【0142】
(IV.中和抗体の生成)
一旦所望の中和特性を有する抗体が同定されると、このような抗体(抗体フラグメントを含む)は、当該分野で周知の方法(例えば、ハイブリドーマ技術もしくは組換えDNA技術を含む)によって、生成され得る。
【0143】
上記ハイブリドーマ方法において、マウスもしくは他の適切な宿主動物(例えば。ハムスター)が免疫されて、免疫に使用されるタンに特異的に結合する抗体を生成するもしくは生成し得るリンパ球を顕在化させる。あるいは、リンパ球は、インビトロで免疫され得る。次いで、リンパ球は、適切な融解剤(例えば、ポリエチレングリコール)を使用して骨髄腫細胞と融合されて、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0144】
このように調製されたハイブリドーマ細胞は播種され、適切な培養培地(好ましくは、融合していない、親骨髄細胞の増殖もしくは生存を阻害する1種以上の物質を含む)中で増殖させられる。例えば、上記親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTもしくはHPRT)を欠いている場合、上記ハイブリドーマ用の培養培地は、代表的には、ヒポキサンチン、アミノプテリン、およびチミジンを含み(HAT配置)、これら物質は、HGPRT欠損細胞の増殖を妨げる。
【0145】
好ましい骨髄腫細胞は、効率的に融合し、上記選択された抗体生成細胞による抗体の安定な高レベルの生成を支援し、そして培地(例えば、HAT培地)に対して感受性であるものである。これらの中でも、好ましい骨髄腫細胞株は、マウス骨髄腫株(例えば、Salk Institute Cell Distribution Center,San Diego,California USAから市販されるMOPC−21およびMPC−11マウス腫瘍から得られるもの、ならびにAmerican Type Culture Collection,Rockville,Maryland USA.から入手可能なSP−2もしくはX63−Ag8−653細胞から得られるもの)である。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株はまた、ヒトモノクローナル抗体の生成に関して記載されてきた(Kozbor,J.Immunol.133:3001(1984);およびBrodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0146】
ハイブリドーマ細胞が増殖している培養培地は、上記抗原に対して指向されるモノクローナル抗体の生成についてアッセイされる。好ましくは、ハイブリドーマ細胞によって生成されるモノクローナル抗体の結合特異性は、免疫沈降によって、またはインビトロ結合アッセイ(例えば、ラジオイムノアッセイ(RIA)もしくは酵素結合イムノソルベントアッセイ(ELISA))によって、決定される。
【0147】
組換えモノクローナル抗体は、例えば、上記必要とされる抗体鎖をコードする上記DNAを単離し、周知の組換え発現ベクターを使用して、同時発現のためのコード配列で組換え宿主細胞を同時トランスフェクトすることによって、生成され得る。組換え宿主細胞は、原核生物細胞および真核生物細胞(例えば、上記のもの)であり得る。
【0148】
上記ヒト化抗体の作製において使用されるべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性を軽減するために非常に重要である。いわゆる「ベストフィット」方法によれば、齧歯類抗体の可変ドメインの配列は、公知のヒト改変ドメイン配列のライブラリー全体に対してスクリーニングされる。上記齧歯類のものに最も近い上記ヒト配列は、次いで、上記ヒト化抗体についてのヒトフレームワーク領域(FR)として受け入れられている(Simsら,J.Immunol.151:2296(1993);Chothiaら,J.Mol.Biol.196:901(1987))。抗体は、上記抗原および他の都合のよい生物学的特性に対して高い親和性の保持とともにヒト化されることは、重要である。この目的を達成するために、好ましい方法によれば、ヒト化抗体は、上記親配列およびヒト化配列の三次元モデルを使用して、上記親配列および種々の概念的ヒト化抗体生成物の分析プロセスによって調製される。
【0149】
さらに、ヒト抗体は、当該分野で公知の方法に従って生成され得る。例えば、トランスジェニック動物(例えば、マウス)が作製され得、上記動物は、免疫の際に、内因性免疫グロブリン生成の非存在下で、ヒト抗体の完全なレパートリーを生成し得る。例えば、Jakobovitsら,Proc.Natl.Acad.Sci.USA 90:2551(1993);Jakobovitsら,Nature 362:255−258(1993);Bruggermannら,Year in Immuno.7:33(1993);および米国特許第5,591,669号、同第5,589,369号および同第5,545,807号を参照のこと。
【0150】
(V.中和抗体の使用)
本発明の中和抗体は、標的とした疾患の予防および/もしくは処置のために使用され得る。治療的適用に関しては、上記抗体もしくは他の分子(その送達は、上記抗体もしくは抗体ベースの輸送配列を使用することによって促進される)は、通常、薬学的組成物の形態において使用される。技術および処方は、一般に、Remington’s Pharmaceutical Sciences,第18版,Mack Publishing Co.(Easton,Pa.1990)において見いだされ得る。Wang and Hanson 「Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers」,Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)もまた参照のこと。
【0151】
抗体は、代表的には、凍結乾燥処方物もしくは水溶液の形態で処方される。受容可能なキャリア、賦形剤、もしくは安定化剤は、使用される投与量および濃度においてレシピエントに対して非毒性であり、緩衝化剤(例えば、リン酸塩、クエン酸塩、および他の有機酸);抗酸化剤(例えば、アスコルビン酸およびメチオニンが挙げられる);保存剤(例えば、オクタデシルジメチルベンジルアンモニウムクロリド);ヘキサメトニウムクロリド;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;アルキルパラベン(例えば、メチルパラベンもしくはプロピルパラベン);カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾール);低分子量(約10残基未満)ポリペプチド;タンパク質(例えば、ヒト血清アルブミン、ゼラチン、もしくは免疫グロブリン);親水性ポリマー(例えば、ポリビニルピロリドン);アミノ酸(例えば、グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリジン);単糖類、二糖類および他の炭水化物(グルコース、マンノース、もしくはデキストリンを含む);キレート化剤(例えば、EDTA);糖(例えば、スクロース、マンニトール、トレハロースもしくはソルビトール);塩形成対イオン(例えば、ナトリウム);金属錯体(例えば、Zn−タンパク質錯体);および/または非イオン性界面活性剤(例えば、TWEENTM、PLURONICSTMもしくはポリエチレングリコール(PEG))が挙げられる。
【0152】
上記抗体はまた、例えば、コアセルベーション技術によって、もしくは界面重合化によって調製されるマイクロカプセル(例えば、それぞれ、ヒドロキシメチルセルロースもしくはゼラチンマイクロカプセル、およびポリ−(メチルメタクリレート)マイクロカプセル)中に、コロイド性薬物送達系(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)中に、またはマクロエマルジョン中に、捕捉され得る。このような技術は、Remington’s Pharmaceutical Sciences 第16版,Osol,A.編(1980)で開示されている。
【0153】
本明細書で開示される中和抗体はまた、イムノリポソームとして処方され得る。上記抗体を含むリポソームは、当該分野で公知の方法(例えば、Epsteinら(1985)Proc.Natl.Acad.Sci.USA 82:3688;Hwangら(1980)Proc.Natl Acad.Sci.USA 77:4030;米国特許第4,485,045号;同第4,544,545号;およびWO 97/38731(1997年10月23日公開)に記載される)によって調製される。増強された循環時間を有するリポソームは、米国特許第5,013,556号に開示されている。
【0154】
特に有用なリポソームは、ホスファチジルコリン、コレステロールおよびPEG誘導体化ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物との逆相エバポレーション法によって生成され得る。リポソームは、規定の孔サイズのフィルタを通して押し出されて、所望の直径を有するリポソームを生じ得る。本発明の抗体のFab’フラグメントは、ジスルフィド交換反応を介して、Martinら(1982)J.Biol.Chem.257:286−288に記載されるように、リポソームに結合体化され得る。化学療法剤は、必要に応じて、上記リポソーム内に含まれる。Gabizonら(1989)J.National Cancer Inst.81(19):1484を参照のこと。
【0155】
疾患の予防もしくは処置については、抗体の適切な投与量は、処置されるべき感染のタイプ、上記疾患の重篤度および経過、上記抗体が予防目的で投与されるのか、治療目的で投与されるのか、に依存する。上記抗体は、上記患者に一度に、もしくは一連の処置にわたって、適切に投与される。上記疾患のタイプおよび重篤度に依存して、約1μg/kg〜15mg/kgの抗体が、例えば、1回以上の別個の投与によろうが、または連続注入によろうが、上記患者への投与のための最初の候補投与量である。
【0156】
本発明のさらなる詳細は、以下の非限定的実施例によって例示される。
【実施例】
【0157】
(実施例1:以前のトリインフルエンザ大発生の生存者からの抗体ライブラリーおよび中和抗体の調製)
(材料および骨髄プロトコルおよび血清調製)
血液を、標準的な静脈穿刺(venopuncture)によって得、凝固させ、処理して血清を回収した。上記血清を、ドライアイス上で輸送されるまで、−20℃で3〜4日間保存した。ドナーを局所麻酔の注射で麻酔し、5mlの骨髄を、各H5N1生存者の骨盤から回収した。次に、上記5mlの骨髄を、45ml RNAlater(Ambion)を含む滅菌50mlチューブに入れた。上記混合物を、眼に見える凝集がなくなり、かつ上記骨髄およびRNAlaterが十分に混合されるまで、約8〜20回穏やかに転倒混和した。次に、上記標本を、2〜10℃の間で一晩冷蔵した。一晩冷蔵した後、上記標本を、ドライアイス上で輸送されるまで、−20℃において3〜4日間保存した。上記RNAlater/marrowを受け取ったら、上記血清含有チューブを、処理するまで−80℃で保存した。
【0158】
(血清学:HA ELISA)
ELISAプレート(Thermo,Immulon 4HBX 96W)を、1×ELISAプレートコーティング溶液(BioFX)中の100μlの100ng/mL H5ヘマグルチニン(Protein Sciences,A/ベトナム/1203/2004)で、室温において一晩インキュベーションすることによって、コーティングした。翌日、プレートを300μl PBS/0.05% Tween−20(PBST)で3回洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05% Tween−20中の4% 無脂肪乾燥乳)を添加し、室温において1時間インキュベートした。ブロッキング工程の後、上記プレートを、300μl PBS/0.05% Tween−20で3回洗浄した。次に、PBS/0.05% Tweenにおいて1:20,000希釈した100μl 血清サンプルを、室温において1=2時間インキュベートし、次いで、300μl PBS/0.05% Tween−20で3回洗浄した。PBS/0.05% Tweenにおいて1:5,000希釈した100μlの抗ヒトFc−HRP結合体を、室温において1〜2時間インキュベートし、次いで、300μl PBS/0.05% Tween−20で3回洗浄した。この最後の洗浄の後、100μlの色素生成基質溶液を添加し(TMB1基質,BioFx)、十分な時間の後、100μlの停止溶液(BioFx)の添加によって終了させた。450nmの吸光度をプレートリーダー(Softmax Proソフトウェアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、その後、Excel(Microsoft)を使用してプロットした。
【0159】
(骨髄:RNA抽出およびmRNA精製)
予め−80℃で保存しておいた骨髄(20ml RNAlater中約2.5ml)を遠心分離によって回収して、RNAlaterを除去し、次いで、300μl 酢酸を含む11.25ml TRI BD試薬(Sigma)中に再懸濁した。次いで、上記ペレットを、激しくボルテックスした。次に、1.5ml BCP(1−ブロモ−3−クロロプロパン,Sigma)を添加し、ボルテックスによって混合し、室温において5分間インキュベートし、次いで、12000×gにおいて15分間4℃で遠心分離した。その界面を廃棄しないように、上記水相を注意深く取り出した。水相からの総RNAを、次に、25ml イソプロパノールを添加することによって沈澱させ、室温において10分間インキュベートし、12000×gにおいて10分間4℃で遠心分離した。イソプロパノールを添加した後、残りのRNAlaterに起因して、2相を形成し、界面に位置した沈澱したRNAを生じた。上記残りのRNAlaterを排除し、RNAの最大の回収を可能にするために、H2O中の50% イソプロパノールの5mlのアリコートを添加し、相分離が認められなくなるまで混合し、その時点で、上記RNAを、12000×gにおいて10分間にわたって4℃で遠心分離によってペレット化した。上記RNAペレットを、75% EtOHで洗浄し、RNAseなしの1.6ml 微小遠心チューブに移し、遠心分離によって再び回収した。最後に、上記RNAペレットを、100μlの1mM リン酸Na,pH8.2中に再懸濁し、A260およびA280を読み取って、RNA純度を評価した。
【0160】
逆転写の前に、mRNAを、Qiagen Oligotex mRNA精製キットに従って、総RNAから精製した。簡潔には、50〜200μg 骨髄RNAを、RNase非含有水と一緒にして250μlにし、250μlのOBB緩衝液およびOligotex懸濁物と混合、続いて、3分間にわたって70℃においてインキュベートした。上記Oligotex粒子の上記オリゴdT30と上記mRNA ポリA−テールとの間のハイブリダイゼーションを室温において10分間行った。次いで、上記ハイブリダイズした懸濁物を、スピンカラムに移し、1分間遠心分離した。上記スピンカラムを400μl 緩衝液OW2で2回洗浄した。次いで、精製したmRNAを、20μlの熱い(70℃)緩衝液OEBと遠心分離することによって、2回溶出した。代表的な収量は、1.5μg 総RNAに対して500ngであった。
【0161】
(骨髄mRNAに対してN9およびオリゴdTを使用する逆転写)
逆転写(RT)反応を、75〜100ng mRNAと、2μl 10×Accuscript RT緩衝液(Stratagene)、0.8μl 100mM dNTP、およびeither N9(300ng)もしくはオリゴdTプライマー(100ng)を一緒に混合することによって達成し、次いで、水で最終容積17μlにした。上記混合物を、65℃において5分間加熱し、次いで、室温へと冷却した。次に、2μl DTT、0.5μl RNase Block(Stratagene)、0.5μl AccuScript RT(Stratagene)を、各反応に添加した。次に、上記N9刺激した反応系を、10分間にわたって室温でインキュベートし、上記オリゴ−dT刺激した反応系を、氷上で10分間にわたってインキュベートした。最後に、両方の反応系を、42℃において60分間、続いて、70℃において15分間インキュベートして、上記酵素を失活させた。
【0162】
(骨髄由来cDNAからのPCR)
抗体重鎖および軽鎖レパートリーを、ヒト生殖系列VおよびJ呂言う基に基づいて、以前に記載された方法および縮重プライマーを本質的に使用して(O’Brien,P.M.,Aitken R. Standard protocols for the construction of scFv Libraries. Antibody Phage Display-Methods and Protocols,vol.178,59−71,2001,Humana Press)、骨髄cDNAから増幅した。
【0163】
簡潔には、オリゴdT刺激cDNA(λ軽鎖に対して75ng mRNAから)およびN9刺激cDNA(κ軽鎖に対して75ng mRNAから、重鎖に関して100ng mRNAから)を使用して、PCR反応を、5μl 10×増幅緩衝液(Invitrogen)、1.5μl dNTP(10mM)、1μl MgSO4(50mM)、2.5μl V領域プライマー(10μM)および2.5μl J領域プライマー(10μM) VHに関して−10μM、0.5μl Platinum Pfxポリメラーゼ(Invitrogen)、および滅菌dH2Oと一緒に混合して、最終容積50μlにした。PCRパラメーターは、以下のとおりであった:工程1−95℃、5分間、工程2−95℃、30秒、工程3−58℃、30秒、工程4−68℃、1分間、工程5−工程2〜4を40回サイクルさせる、工程6−68℃、5分間。軽鎖PCR生成物を、Qiagen PCR Cleanupキットを使用してきれいにした。重鎖PCR生成物を、Qiagen Gel Extraction Kitを使用して、1.5% アガロースゲルから精製し、次いで、再増幅した。重鎖再増幅を以下のようにおこなった:10μl 10×増幅緩衝液(Invitrogen)、3μl dNTP(10mM)、2μl MgSO4(50mM)、5μl 各VHプライマー(10μM)およびJHプライマー(10μM)、5μl 重鎖プライマーPCR生成物、1μl Platinum Pfxを混合し、水で容積を100μlに調節した。サイクリングパラメーターは、以下のとおりであった:工程1−95℃、5分間、工程2−95℃、30秒、工程3−58℃、30秒、工程4−68℃、1分間、工程5−工程2〜4を20回サイクルさせる、工程6−68℃、5分間。再増幅した重鎖PCR生成物を、Qiagen Extraction Kitを使用して、1.5% アガロース−TAEゲルからきれいにした。
【0164】
(抗体ファージライブラリー構築)
各個々のトリインフルエンザ生存者についての別個の抗体ライブラリーを、繊維状ファージのpIII遺伝子の終止コドンの後ろの非翻訳領域において挿入した特融識別3−ヌクレオチドバーコードを使用して構築した。
【0165】
軽鎖クローニング:
ドナー1名につきプールしたκ軽鎖およびプールしたλ軽鎖の各1μgを、NotIおよびBamHIで消化し、Qiagen Gel Extraction Kitを用いて1.5% アガロース−TAEゲルからゲル精製した。5μgの各ベクター(pAMPFab)を、NotIおよびBamHIで消化し、Qiagen Gel Extraction Kitを使用して、1% アガロース−TAEゲルからゲル精製した。ライブラリーライゲーションを、室温において60μl中で1時間にわたって、もしくは14℃において一晩にわたって、200ngのゲル精製したκ挿入物もしくはλ挿入物および1μgのゲル精製したベクターで行った。ライゲーションを、Edge BioSystem Perfromaスピンカラムを使用して脱塩した。上記ライブラリーを、80μl TG−1もしくはXL−1 Blueアリコート中の5個のエレクトロポレーションに移し、各々、1ml SOC中に回収し、プールし、1時間にわたって37℃において増殖させた。形質転換体の総数を、上記形質転換体の各々からのアリコートをプレートすることによって、この増殖の後に決定した。その残りのエレクトロポレーションを、200ml 2YT+50μg/ml アンピシリン+2% グルコース中で、37℃において一晩増殖させることによって増幅させた。その後の軽鎖ライブラリーを、これらの一晩培養物から、Qiagen High Speed Maxiprep Kitを使用して、プラスミド精製によって回収した。
【0166】
(重鎖クローニング):
上記ドナー特異的重鎖(VH1、VH2、5、6プール、VH3、およびVH4)の1.5〜2μgの各々を、40ユニット過剰/μg DNAと、SfiIおよびXhoIで消化し、Qiagen Gel Extraction Kitを使用して、1.5% アガロース−TAEゲルからゲル精製した。15μgの各軽鎖ライブラリーベクターを、40ユニット/μg DNAと、SfiIおよびXhoIで消化し、Qiagen Gel Extraction Kitを使用して、1% アガロース−TAEゲルからゲル精製した。ライブラリーライゲーションを、消化した1.2μg SfiI/XhoI、ゲル精製重鎖ドナー集団および5μgの各軽鎖ライブラリー(κおよびλ)を一晩4℃において合わせることによって設定した。次いで、上記ライブラリーライゲーションを、Edge BioSystem Pefromaスピンカラムで脱塩し、次いで、80μl TG−1アリコート中、20エレクトロポレーション/ライブラリーを介して形質転換し、各々、1ml SOC中に回収し、プールし、37℃において1時間増殖させた。再び、この増殖の後、各々のアリコートを、1L 2YT+50μg/ml アンピシリン+2% グルコースに移した残りと一緒に、形質転換体の総数を決定するために使用し、約0.3のOD600になるように、激しく曝気して37℃において増殖させた。次に、M13K07ヘルパーファージを、感染多重度(MOI)5:1で添加し、攪拌せずに、37℃において1時間インキュベートした。次に、上記細胞を、遠心分離によって採取し、1L 2YT+50μg/ml アンピシリン、70μg/ml カナマイシン中で再懸濁し、激しく曝気しながら37℃において一晩増殖させて、scFvファージミド生成を可能にした。翌朝、上記細胞を遠心分離によって集め、ファージミドを含む上清を集めた。上記ファージミドを、0.2容積の20% PEG/5M NaCl溶液を添加し、1時間氷上でインキュベート刷ることによって、上記上清から沈澱させた。次いで、上記ファージミドライブラリーストックを、遠心分離によって回収し、20ml 滅菌PBS中に再懸濁した。残りの細菌を、さらなる遠心分離によって回収し。最終ファージミドライブラリーを、−20℃において、PBS+50% グリセロール中で保存した。
【0167】
(ファージミドパニングおよび増幅)
ELISAプレート(Immulon 4HBX平底,Nunc)を、室温において一晩のインキュベーションによって、ELISAコーティング溶液(BioFX)中、100μlの100ng/mL H5 ヘマグルチニンタンパク質(Protein Sciences,A/ベトナム/1203/2004)でコーティングした。翌日、プレートを、300μl PBSTで3回洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05% Tween−20中4% 無脂肪乾燥乳)を添加し、30分間、氷上でインキュベートした。上記ブロッキング工程の後、上記プレートを、300μl PBSTで3回洗浄した。ファージパニングの直前に、凍結したファージミドストックから、Millipore Amicon Ultraカラムを使用して、上記グリセロールを除去し、次いで、4% 無脂肪乾燥乳中、15分間にわたってブロッキングした。次に、ファージミドの100μl アリコートを、8ウェル(総ファージ約1×1012CFU)へと分配し、4℃において2時間にわたってインキュベートし、続いて、300μl PBSTで6〜8回洗浄した。ファージミドを、100μl/ウェル中、室温において10分間後にElution buffer (0.2M グリシン−HCl,pH2.2、1mg/ml BSA)中に回収した。次いで、上記溶出物を、溶出物1mlあたり56.25μl 2M トリス塩基を添加することによって中和した。中和後、5ml TG1細胞(OD600 約0.3)に、0.5ml 中和ファージを、37℃において30分間にわたって、2−YT中、振盪することなく感染させた。この工程の後、いくつかの細胞を、LB AMPグルコースプレート上にプレートして、全てのファージミド回収を決定した。残りの接種物を、10ml 2−YTAG(最終濃度2% グルコースおよび50ug/ml アンピシリン)へ入れ、OD600が約0.3になるまで37℃において激しく曝気しながら増殖させた。次に、上記培養物に、5:1のMOIでM13K07ヘルパーファージを感染させ、37℃において30〜60分間にわたって、振盪することなくインキュベートした。上記細胞を遠心分離によって集め、25ml 2−YTAK(アンピシリン 50μg/ml、カナマイシン 70μg/ml)中に再懸濁し、新しい培養フラスコに移し、振盪しながら37℃において増殖させ続けた。その後のラウンドは、同様に回収し、増幅させた。
【0168】
(scFv ELISA)
生体パニングしたファージからのE. coli HB2151形質転換細胞の個々のコロニーを、1mlの2YT+100μg/ml AMP中、37℃において一晩増殖させた。翌朝、上記細胞を遠心分離によって回収し、1.5ml ペリムラズム溶解緩衝液(1ml BBS(Teknova)+0.5ml 10mg/ml リゾチーム+10mM 終濃度になるまでEDTA)中に再懸濁した。上記細胞を、遠心分離によって再びペレット化し、上記scFvを含むペリムラズム溶解物を回収した。上記scFv溶解物を、希釈緩衝液(PBS/0.05% BSA)と1:1で合わせ、100μlを、予め抗原コーティングし、希釈緩衝液でブロッキングしたウェルに添加した。上記サンプルを、2時間にわたって室温においてインキュベートし、次いで、PBS/0.05% Tweenで3回洗浄した。次に、100μlの希釈緩衝液中で1:5000希釈したビオチン抗ヒスチジンマウス(Serotec)を各ウェルに添加し、室温において1時間インキュベートした。このインキュベーションの後、上記ウェルを、PBS/0.05% Tweenで3回洗浄し、次いで、各ウェルに、100μlの1:2500 ストレプトアビジン:HRP(Serotec)を添加し、室温において1時間インキュベートし、次いで、PBS/0.05% Tweenで3回洗浄した。この最後の洗浄の後、100μlの色素生成性基質溶液を添加し(TMB1基質,BioFx)、十分な時間の後、100μlの停止緩衝液(BioFx)を添加することによって集結させた。450nmの吸光度を、プレートリーダー(Softmax Proソフトウェアを装備したMolecular Devices Thermomaxマイクロプレートリーダー)で読み取って、データを記録し、その後、Excel(Microsoft)を使用してプロットした。
【0169】
(配列決定)
上記重鎖および軽鎖配列を推測するために、個々のクローンを増殖させ、プラスミドDNA抽出した(Qiagen)。上記プラスミドDNAを標準的DNA配列決定に供した。
【0170】
(ヘマグルチニン阻害(HAI)アッセイ)
ヘマグルチニン阻害を、丸底マイクロタイタープレート(Corning)において、4 HAU(赤血球凝集単位)のウイルスもしくはタンパク質/ウェルを使用して、Rogersら,Virology 131:394−408(1983)の方法に本質的に従って行った。HAI決定のために、精製した一本鎖可変フラグメント(scFv)の25μl サンプルを、各マイクロタイターウェルにおいて、上記試験ウイルスの4 HAUを含む25μlのPBSと混合した。室温において15分間のプレインキュベーションの後、25μlの0.75% ヒト赤血球を添加し、混合した。HAI抗体活性を、室温での60分間のインキュベーション後の視覚的検査によって決定した。
【0171】
特に、以下のプロトコルを使用した:
抗体、タンパク質およびウイルス
IgG1タンパク質を、pCIベースの(Promega)全長哺乳動物タンパク質発現ベクターへとそれらのコード領域をサブクローニングすることによってscFvもしくはFabのいずれかから生成し、次いで、293 Freestyle細胞(Invitrogen)へと、製造業者のガイドラインに従ってトランスフェクトした。簡潔には、20μgの軽鎖および10μgの重鎖をコードするプラスミドを、1.0ml 293fectinと合わせ、60分間インキュベートした。このプレインキュベーションの後、上記DNA混合物を、30ml 培地中で、3×107細胞と合わせ、その得られた細胞懸濁物を、7日間にわたって、製造業者の示唆に従って増殖させた。7日後に、上記分泌免疫グロブリンを、プロテインAクロマトグラフィー(Calbiochem)を使用して、培養上清から精製した。その得られた精製抗体を、遠心分離サイズ分画(Centricon Plus−20)を使用して滅菌PBSへと緩衝液交換し、それらのタンパク質濃度を、比色BCAアッセイ(Pierce)によって決定した。
【0172】
(交叉反応IgG ELISA)
マイクロタイタープレートを、コーティング緩衝液中で希釈した0.1mlの以下の抗原でコーティングし、室温において一晩インキュベートした:100ng/ml H5N1 ベトナム 1203/04、250ng/ml H5N1 トルコ/65596/06、1μg/ml H5N1 インドネシア/5/05、700ng/ml H1N1 ニューカレドニア/20/99、1μg/ml H1N1 ノースキャロライナ/1/18、100ng/ml and H3N2 ウィスコンシン/67/05。ブロッキングを、0.3mlのブロッキング緩衝液(PBS/0.05% Tween−20中4% 無脂肪乾燥乳)で行った。ブロッキングの後、2% 無脂肪乾燥乳ブロッキング緩衝液中、0.5μg/mlに希釈した抗体を、4℃において2時間にわたってインキュベートし、洗浄し、後に、2% 無脂肪乾燥乳ブロッキング緩衝液中1:3000希釈のペルオキシダーゼ結合体化抗ヒトFc抗体(Jackson ImmunoResearch)および標準的TMB基質検出(BioFX)を使用して検出した。450nmの吸光度を読み取り、データを記録し、本明細書において報告した。
【0173】
(ウイルス微小中和(microneutralization)
インドネシアおよびトルコのヘマグルチニン遺伝子はを、ヒトコドン最適化配列(DNA 2.0)を使用して合成的に組み立て、次いで、これを使用して、組換え操作ウイルスを生成した。組換えインフルエンザウイルスを、以前に記載されるように(Fodor,E.ら J Virol.1999;73(11):9679−82)、逆遺伝子学を使用して生成した。簡潔には、10個のプラスミドの1μg 各々を、単層において293 T細胞へとトランスフェクトした。各トランスフェクションは、vRNA(pPOL1タイプ)およびA/ベトナム/1203/04 HAおよびNA(pCAGGS発現プラスミドは、J.Miyazaki,Osaka University,Osaka,Japan)(Miyazaki,J.ら Gene 1989;79(2):269−77)によって供与)に関しては、タンパク質発現プラスミド(pCAGGS type)に加えて、A/プエルトリコ/8/34/PA、PB1、PB2、NP、M、およびNSセグメントに関して、両センス(ambisense)プラスミド(vRNAおよびmRNA両方の発現に関して)を含んでいた。トランスフェクションの20時間後、293T細胞を、細胞培養上清中に再懸濁し、これを使用して、10日齢受精卵に接種した。
【0174】
抗体を、以下のように、ウイルスに対する中和活性についてスクリーニングした。各Mabの2倍連続希釈物を、PBS中のウイルスの100 TCID50とともに、37℃において1時間インキュベートした。24ウェルプレート中のMadin−Darbyイヌ腎臓細胞単層をPBSで1回洗浄し、ウイルス−抗体混合物とともにインキュベートした。5% CO2中で37℃において1時間インキュベートした後、上記接種物を除去し、単層を、PBSでもう1回洗浄した。0.3% BSA、0.01% FBSおよび1μg/ml TPCK処理トリプシンを補充したOpti−MEMを添加し、細胞を、37℃において72時間にわたってインキュベートした。細胞培養上清中のウイルスの存在は、0.5% ニワトリ赤血球を使用して、HAアッセイによって評価した。
【0175】
(結果)
骨髄および血液サンプルを、2006年1月にトルコで起こったH5N1トリインフルエンザ大流行(大流行後約4ヶ月)の6名の生存者から集めた。6名全ての生存者について、トリインフルエンザの初期診断は、物理的検査、臨床実験室的試験、および分子診断測定によってなされ、Turkish Ministry of Healthによって確認された。これら生存者のうちの4名は、世界保健機関(WHO)によってさらに確認された。血清サンプルを分析して、上記の血清学プロトコルを使用して、H5 ヘマグルチニン(A/ベトナム/1203/2004)に対する抗体の存在を確認した。図8に示されるように、6名の患者全ての上記血液サンプル(それぞれ、SLB H1〜H6と称した)は、上記H5抗原に対する抗体の存在を示した。この確認後に、RNAを、これら個体の骨髄サンプルから抽出し、骨髄mRNAを精製し、上記のプロトコルを使用して逆転写した。次いで、上記抗体重鎖および軽鎖レパートリーを、上記のように、上記骨髄cDNAから増幅して、個々の抗体重鎖および軽鎖ファージライブラリーを、上記の3ヌクレオチドバーコード付与を使用して、各生存者について別個にクローニングして、個々のライブラリーを区別した。
【0176】
このベクターとそのコード付与系とを使用して、本発明者らは、一本鎖(scFv)およびFabファージミド両方の形式において6名の生存者のうちの5名の骨髄に由来するレパートリーを首尾よくクローニングした。個々の生存者に由来する各集団は、1.0×108メンバーより大きい多様性を有する。さらに、本発明者らは、混合した生存者軽鎖および重鎖から構成される、採取的な多様性が1.1×109のさらなるバーコード付与ライブラリーを作り出した。まとめると、上記5個のドナー特異的集団および全てのドナーからの上記プールしたライブラリーは、scFv集団として1.0×109およびFabディスプレイ集団として4.2×109の全体多様性を有する(表4)。
【0177】
表2は、scFvおよびFab両方の形式における軽鎖および全ライブラリー総形質転換体を示す。全ライブラリーによって表される全体多様性は、5.6×109である。
【0178】
【表2】
骨髄および血液サンプルをまた、2006年のインフルエンザ症状に対して処置した12名のその土地のドナーから集めた。上記のように血清学を行って、H1、H3およびH5 ヘマグルチニン、それぞれに対する抗体の存在を確認した。図8に示されるように、全ての血清サンプルを試験したところ、H1および/もしくはH3ヘマグルチニンに対する抗体について陽性であった。ここで特定のサブタイプの優性は、インフルエンザAウイルスサブタイプに依存し、このサブタイプに対して、上記特定のドナーは、彼もしくは彼女のほぼ人生全体を通して曝された。興味深いことに、血清がH5ヘマグルチニンの抗体の顕著なレベルも含んでいたドナー(図9においてドナーSLB1およびSLB5)が存在した。この確認後、RNAを、上記ドナーの骨髄サンプルから抽出し、骨髄mRNAを精製し、上記のプロトコルを使用して逆転写した。上記抗体重鎖および軽鎖レパートリーを、次いで、上記骨髄cDNAから上記のように増幅し、個々の抗体重鎖および軽鎖ファージライブラリーを、上記の3ヌクレオチドバーコード付与を使用して各ドナーについて別個にクローニングして、個々のライブラリーを区別した。
【0179】
(結合抗体の選択)
上記H5N1生存者ライブラリーを、表2にまとめ、ベトナム/1203/04ウイルスHAおよびNAタンパク質もしくは組換え精製ヘマグルチニン(Barbas,C.ら(2001)Phage Display,A Laboratory Manual(Cold Spring Harbor Laboratory Press))を含む不活性化ウイルスに対してパニングした。3〜4ラウンドのファージパニングの後、富化ファージプールからの個々のコロニーを、H5N1ウイルスもしくは精製ヘマグルチニンに対するELISAによって分析し、上記ポジティブクローンを配列決定して、それらの重鎖および軽鎖軽鎖配列を決定し、それらの生存者バーコードを読み取った(D.W.Coomber,Methods Mol Biol 178,133(2002))。このことから、本発明者らは、上記個々の生存者ライブラリーのうちの5個全てから、特定のH5ヘマグルチニン結合クローンを単離した。合計して、300,000個より多い種々の抗ウイルス抗体が回収され、そのうちの146個が、上記H5ヘマグルチニンタンパク質を特異的に結合する。
【0180】
(選択したクローンの一般的特徴)
全体として、上記個々のドナーは、重鎖および軽鎖両方に関して異なる生殖系列を利用し、このことは、個々の患者が、同じ潜在的に致死的な免疫学的攻撃に対して異なる解決策を見いだしたことを示している。得られたレパートリーの質に関する信頼を与えかつ抗体結合および/もしくは中和の化学に関する情報を与えるために使用され得るコンビナトリアル抗体ライブラリーの主要な特徴は、これらクローンにおいて認められる。これらクローンは、親和性成熟の天然の発達において、同様に選択された合成抗体ライブラリーにおいても見いだされる抗原結合に対する以前に記載された反復クローン(「ジャックポットソリューション(jackpot solution)」)の顕著な特徴を全て含む(Lerner,R.A.Angew Chem Int Ed Engl 45,8106(Dec 11,2006);A.Rajpalら,Proc Natl Acad Sci USA 102,8466(Jun 14,2005))。これら大きな集団における「ジャックポット」の存在は、上記スクリーニング手順を確認する。なぜなら、上記ファージが活性に基づいて選択されなければ、同じくローンを複数回得るという機会は、起こり得る可能性が非常に少ないからである。さらに、群内の上記重鎖の差異を分析する場合、上記アミノ酸置換の多くが、化学的にかつ構造的に保存的であることが観察された(表4)。反復クローンのように、化学的に妥当な複数のアミノ酸置換の出現は、無作為事象である可能性は低い。
【0181】
(回収された抗体の結合特異性)
6個のクローンを、全IgG1タンパク質への変換に対して異なる2個のエピトープを認識した3名の生存者から選択した。これら抗体のうちの4つの結合を、上記ヘマグルチニンタンパク質のHA1サブユニットにマッピングしたが、その残りの2つは、マッピングしなかった。上記HA1サブユニットは、感染において非常に関連しているので、本発明者らは、それら4個の抗体の活性を、以下に記載されるようにさらに分析した。
【0182】
これら研究の1つの目的は、基準から外れた(divergent)ウイルス株を広く中和する稀な抗体を回収することであった。本発明者らの抗体のうちのいくつかは、広く反応性であり得ることが示唆された。なぜなら、上記ドナー由来の血清は、彼らが感染したウイルスを超えて拡がって、基準から外れたサブファミリーのH5N1ウイルスに対して高い力価の抗体を有したからである。個々の抗体のレベルにおける交叉反応性の程度を決定するために、本発明者らは、異なるインフルエンザヘマグルチニン抗原に対する本発明者らのクローンの結合を最初に分析した(図11)。驚くべきことではく、これら抗体は、トルコ/65596/06株に感染する対応するものに由来するヘマグルチニンを認識し、そしてさらに、選択のために使用されたベトナム/1203/04株に由来する異種ヘマグルチニンを認識する。さらに、それらは、抗原的に基準から外れたインドネシア/5/05 H5ヘマグルチニンを認識する。さらに、本発明者らは、4個のプロトタイプ抗体が、密接に関連したサブタイプH1N1最新の基準株ニューカレドニア/20/99に由来するヘマグルチニンを結合しることを見いだした。顕著なことには、生存者5に属する上記3個の中和抗体はまた、1918年のスペイン風邪世界的流行の間に現れたH1N1 サウスキャロライナ/1/18単離株に由来するヘマグルチニンを結合した。逆に言えば、これら4個の抗体のうちのいずれも、最新のH3N2 ウィスコンシン/67/05基準株に由来するヘマグルチニンを結合せず、このことは、上記抗体ディスプレイが、インフルエンザサブタイプの間で広いスペクトルで結合するとしても、上記反応性は、全てのインフルエンザサブタイプに拡がらなかったことを示す。図11は、2名の生存者に由来するH5N1抗体とH1N1ウイルス由来のヘマグルチニンとの交叉反応性を示す。(A)棒は、H5N1 ベトナム 1203/04(暗い灰色)、H5N1 トルコ/65596/06(白)、H5N1 インドネシア/5/05(斜線)、H1N1 ニューカレドニア/20/99(縦縞)、H1N1 サウスキャロライナ/1/18(クロスハッチストライプ)、およびH3N2 ウィスコンシン/67/05(明るい灰色)である。
【0183】
(中和研究)
最初に、上記抗体を、H5 HA(ベトナム/1203/04)含有インフルエンザウイルスを中和するそれらの能力について評価した。共通するエピトープ(エピトープ「A」)を認識した生存者2および3、生存者5に由来する1つの抗体は、全て中和したのに対して、第2のエピトープ(エピトープ[B])を認識した生存者1に由来する2つの抗体は、中和しなかった。
【0184】
H5N1もしくはH1N1 ヘマグルチニンいずれかに対する生存者5から別個に単離されたクローンの顕著な配列類似性に基づいて、本発明者らは、それらの交叉反応性は、単純な結合を超えて拡がり、それらがまた、H5N1ウイルスおよびH1N1ウイルスの両方を中和するという非常にまれな特性を有すると推測した。上記IgGの交叉中和活性を試験するために、本発明者らは、中和アッセイにおけるH5N1スクリーニングからの代表的な抗体を試験して、抗体がまた、H1N1ウイルスもしくはH3N2ウイルスを中和するか否かを調べた(表3)。本発明者らは、H1保有ウイルスA/ニューカレドニア/20/99およびH3保有ウイルスA/香港/68を研究した。H5サブタイプヘマグルチニンを保有するウイルスの集団はまた、以下を試験した:(A/ベトナム/1203/04;A/インドネシア/5/05;A/トルコ/65596/06;A/エジプト/06)。上記抗体は、H3サブタイプインフルエンザに対して何ら活性を示さなかった。しかし、H5含有ウイルスを中和した上記モノクローナル抗体(1〜3)のうちの3つはまた、サブタイプH1からHAを保有するウイルス全てを強く中和した(表5)。
【0185】
【表3】
(中和の免疫化学的基準)
抗体ライブラリーの1つの利点は、多数の抗体を得る場合に、これらがそれらの関連性に関してグループ分けされ得ることである。従って、上記集団における所定の抗体についての機能が観察される場合、所定の抗体が属する上記群の他のメンバーが類似の活性を有することを推定し得る。
【0186】
表4は、H5N1 ベトナムパニングの後の、生存者5ライブラリーから発見された中和の免疫化学的基礎を示す例示的配列を示す。上記61個の特融の重鎖配列を、114個の特融の重鎖および軽鎖組み合わせに由来するそれらの生殖系列可変領域と整列させた。必須の変異は、太字、下線を付したテキスト(列5-PIからGMおよびAからT;列6-KSからELもしくはEMもしくはXL)で示され、優勢な変異は、斜体、下線を付したテキスト(列2-AからT;列3-ISからVT;列5-GからA;列8-KからQもしくはR)で示される。H1N1 ニューカレドニアパニングにおいて発見された重鎖配列はまた、灰色で強調される。抗体領域およびKabat番号付け範囲は、各配列の列の上に列挙される。
【0187】
【化1】
【0188】
【表4−1】
【0189】
【表4−2】
生存者5に由来するエピトープ「A」に対する上記中和抗体集団を含んだ上記群のメンバーは、表4に示される。上記群は、上記VH1e生殖系列重鎖に最も近く似ている61個の特有のメンバーから構成される。いくつかの重鎖は、1個より多くの軽鎖と対形成される。合計で、これら重鎖は、κおよびλ両方の軽鎖に対する114個の特有の対形成を有する。これら重鎖と、上記非常に関連するVH1e生殖系列とを比較すると、本発明者らは、点置換の3つのタイプを観察する。いくつかの変化は、必要とされるようであり、他は優性であり、そしていくつかの残基は、散発的にのみ変化した。必要とされる変化は、位置52A(Pro>Gly)、53(Ile>Met)、および57(Ala>Thr)においてCDR2内の群において、ならびに位置73(Lys>Glu)および74(Ser>LeuもしくはMet)においてフレームワーク3領域において、あらゆるクローンで生じ、このうちの全てが、生殖系列側鎖化学から変化し、このことは、上記変異が抗原結合および中和に必須であることを示唆する。変異の上記第2のセットは優性であり、大部分のクローンにおいて見いだされる。最初は、位置24(Ala>Thr)においてフレームワーク1において、顕著な化学的変化を表す。次の3個は、位置34(Ile>Val)および位置35(Ser>Thr)においてCDR中に、ならびに位置50(Gly>Ala)においてCDR2中にも保存的変化がある。しかし、これら優性な置換のうちの4個全ては、なくてもよい。このことは、上記置換が、有益である一方で、必須ではないことを示唆する。フレームワーク領域1、3、および4、ならびにCDR3の全体を通じて見いだされる上記散発性の変化は、全て保存的であり、おそらく小さな最適化事象を表す。
【0190】
図12は、47eといわれる非常に関連する抗HIV Fab(1rzi.pdb)(Huang,C.C.ら(2004)Proc.Nat.Acad.Sci.101,2706−2711)の結晶構造に対して重ね合わせられた抗体の構造における必要とされる変異の位置を示す。図12は、必要とされるH5 ヘマグルチニン結合群1の位置およびFab 47eの結晶構造上の優性な変異を示す。上記必要とされる変異は、G52(52A(Pro>Gly))、M53(Ile>Met)、T57(Ala>Thr)、E73(Lys>Glu)およびLM74(Ser>LeuもしくはMet)として示される。上記優性な変異は、T24(Ala>Thr)、V34(Ile>Val)、T35(Ser>Thr)、およびA50(Gly>Ala)として示される。H5 ベトナム/1203/2004 HA生体パニングにおいて同定された、上記必要とされかつ優性な群1の重鎖配列を、上記非常に関連する抗HIV Fab 47eの結晶構造に重ね合わせる。変異を、骨格モデル(上)および空間充填モデル(下)の両方において示す。密なクラスターが、CDR2中およびCDR2のみ隣接する必要とされる変異のうちの4個によって形成される。上記必要とされる変異52A(Pro>Gly)、53(Ile>Met)、73(Lys>Glu)および74(Ser>LeuもしくはMet)は、上記重鎖可変ドメインの露出された表面に顕著に密なクラスターを形成し、ここでこれらは、上記タンパク質表面から顕著に突出する隆起を形成する(図12)。その残りの必要とされる変異57(Ala>Thr)は、部分的には、上記CDR2ループの基部に埋め込まれる。CDR2およびフレームワーク3における表面露出変化は、おそらく抗原結合において直接的な役割を有することである一方で、上記あまり露出されていない必要土佐折れる変異および本質的でない優性変異は、上記CDR2ループを安定化し、そして/もしくは位置づけることを介して間接的な効果を有し得る。
【0191】
生存者2に由来する抗体は、上記VH4−4b生殖系列重鎖に最もよく似た2個の特有の重鎖からなる(表5)。上記第1の重鎖は、5個の特有のλ軽鎖と対形成していることが見いだされ、そのうちの4個は、滅多に使用されないλの6個の軽鎖ファミリーに由来し、他方は、単一のκ軽鎖と対形成される。中和プロフィールがより制限された抗体4は、この群に由来する。
【0192】
【表5】
所定の変異が、抗体の活性に重要であるという可能性は、独立して選択される時間数の関数として増大する。上記必要とされる変異が、独立したクローンから体細胞変異の間に選択されるか、またはその後の複製の間にさらに変異される単一のクローンの子孫に由来するか否かを決定するために、上記優性変異のコドン使用法を分析した(表6A〜6B)。上記データは、異なるコドンが使用されたが、上記コドンは、同じアミノ酸変化を生じたことを明らかにし、このことは、これら変異が、このなるクローンにおいて独立して発生したこと、従って、複数回選択されたことを実証する。独立して選択された事象のこの収束性の結果は、これら優性変異が、上記ウイルスへの結合および/もしくはまたはその中和における重要な役割を果たすという強力な証拠である。
【0193】
表6A〜6Bに例示されるように、個々のクローンのコドン使用法は、選択されたH5 HA結合クローンの独立した起源を示す。DNAアラインメントおよびコードされるアミノ酸は、VH1−e 生殖系列に対する6個の代表的群1の抗体である。同じアミノ酸についての異なるコドンの使用は、各特有の配列が、別個の起源のものであることを示す。表6AはCDR2に対応し、表6Bは、フレームワーク3に対応する。生殖系列コドンは、太字を付したコドンとして示される。生殖系列コドンから同じアミノ酸への変化は、通常のテキストのコドンとして示される。生殖系列アミノ酸からの第1の変化は、太字の下線を付したコドンとして示される。生殖系列アミノ酸からの第2の変化は、斜体の下線を付したコドンとして示される。生殖系列アミノ酸からの第3の変化は、下線の灰色で強調したコドンとして示される。
【0194】
【表6A】
【0195】
【表6B】
(実施例2:HIV感染患者についてのドナー特異的抗体ライブラリーの構築)
(骨髄プロトコルおよび血清調製)
血液を、標準的静脈穿刺によって得、凝固させ、処理して、血清を回収する。上記血清を、氷上で輸送されるまで、−20℃において3〜4日間保存する。ドナーを局所麻酔剤の注射で麻酔し、5mlの骨髄を、各患者ドナーの骨盤から回収する。次に、上記5mlの骨髄を、45ml RNAlater(Ambion)を含む滅菌50mlチューブに移す。上記混合物を、眼に見える塊が存在せずかつ上記骨髄およびRNAlaterが十分に混合されるまで、約8〜20回穏やかに転倒混和する。次に、上記標本を、2〜10℃の間で一晩冷蔵する。一晩冷蔵した後、上記標本を、氷上で輸送されるまで、−20℃において3〜4日間貯蔵する。受け取ったら、上記RNAlater/骨髄および血清を含むチューブを、処理するまで、−80℃において貯蔵する。候補患者は、ドナーとして選択される前に、HIVポジティブについて試験されるべきである。
【0196】
骨髄抽出およびmRNA精製、逆転写、PCR、抗体軽鎖および重鎖構築、ファージミドパニングなrばいに増幅、ELISAおよび配列決定を、実施例1に本質的に記載されるように行う。
【0197】
前述の説明において、本発明は、特定の実施形態を参照して例示されるが、そのように限定されない。実際に、本発明の種々の改変は、本明細書において示され、記載されるものに加えて、前述の説明から当業者に明らかになり、添付の特許請求の範囲内に入る。
【0198】
本明細書全体を通じて引用される全ての参考文献は、本明細書に参考として援用される。
【技術分野】
【0001】
(発明の分野)
本発明は、ドナー特異的抗体ライブラリー、ならびにこれを作製および使用するための方法に関する。本発明はまた、このようなドナー特異的抗体ライブラリーから得られる中和抗体、および種々のヒト疾患および状態の予防および/もしくは処置のために得られた抗体を使用するための方法に関する。
【背景技術】
【0002】
(発明の背景)
ヒト疾患の診断、予防および処置に対して特異的な薬剤の生成および同定は、有用な化学的性質の巨大な集団の利用を要する。抗体ライブラリーの作出およびスクリーニングのための種々の技術の到来かつ迅速な開発に関して、疾患標的に対するモノクローナル抗体は、新たな薬物候補の主要なカテゴリーのうちの1つになった。ヒト使用のために、特異性および有効性に加えて、安全性は、主要な懸念であるので、ヒトモノクローナル抗体のライブラリーは、特に重要になった。
【0003】
現在では、ヒト抗体ベースの薬物候補の開発は、代表的には、代表的には、それらの意図された適用に関連しないヒトレパートリー(repertoire)に由来する抗体配列の無作為集団を含むヒト抗体ライブラリーをスクリーニングすることによって、同定される。特異的ヒトドナーから作出された各抗体ライブラリーは、上記ドナーがその個々の寿命の過程にわたって遭遇し、挑戦し、かつ乗り越えた全ての特有の挑戦(challenge)に由来するかもしくは作り出す、全ての成分、生理学、および代謝的変化に対する抗体を潜在的に含む。現在のアプローチに伴って作製された代表的なヒト抗体ライブラリーは、ドナーの健康上の履歴の理解なしに構築されるので、得られた免疫グロブリンレパートリーにおいて予測されるものはほとんど未知である。
【0004】
従って、特定の疾患との遭遇に首尾よく耐えたかもしくは現在耐えている個体から抗体ライブラリーを作り出すことは、特に重要である。なぜなら、彼らの得られたレパートリーは、関連する疾患を特異的に防御するためにドナーが使用した抗体を含むからである。ネガティブ要素もしくはポジティブ要素を除去もしくは単離し、望ましくない内容を排除し、そして改善された特性を有するヒト抗体を生成する能力を含む、このようなライブラリーの効率的なスクリーニングおよび取り扱いのための方法を提供することもまた、重要である。
【0005】
本発明は、特定の標的疾患との遭遇に曝され、これに耐えたもしくは現在耐えている個体に由来するドナー特異的抗体ライブラリーの作出、スクリーニングおよび取り扱いのための方法および手段を提供することによって、これらの必要性に対処する。
【発明の概要】
【課題を解決するための手段】
【0006】
一局面において、本発明は、標的抗原に対する抗体生成を引き起こす疾患に罹患したもしくは現在罹患しているヒト患者ドナーから得られた、抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子のレパートリーを含むベクター集団(vector collection)に関し、ここで上記集団は、特有のバーコードで同定される。
【0007】
一実施形態において、上記ベクター集団は、抗体軽鎖もしくはそのフラグメントをコードする核酸分子(例えば、抗体λ軽鎖、もしくは抗体κ軽鎖、またはこれらのフラグメント)のレパートリーを含む。
【0008】
別の実施形態において、上記ベクター集団は、抗体重鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む。
【0009】
さらに別の実施形態において、上記バーコードは、上記集団に存在するベクターに連結されるかもしくは組み込まれ、そして/または上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、上記バーコードは、上記核酸分子の発現を妨害しない。
【0010】
従って、上記バーコードは、例えば、上記核酸分子の3’非コード領域もしくは5’非コード領域に連結され得る、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列であり得る。
【0011】
さらなる実施形態において、上記バーコードは、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列であるヌクレオチド配列である。
【0012】
なおさらなる実施形態において、上記バーコードは、非連続ヌクレオチド配列である。上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記集団に存在するベクターに連結され得るかもしくは組み込まれ得る。あるいは、上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ得、その結果、上記非連続ヌクレオチド配列のうちの少なくとも一部は、上記核酸分子の発現を妨害しない。
【0013】
別の実施形態において、上記バーコードは、ペプチドもしくはポリペプチド配列である。
【0014】
異なる実施形態において、上記ベクター集団に存在するベクターは、ファージミドベクターであり、このファージミドは、例えば、バクテリオファージ遺伝子III、および上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と上記バクテリオファージIII遺伝子との間に終止コドンを含み、そしてバーコード(例えば、上記終止コドンの後ろの非翻訳領域に挿入される連続する非コードヌクレオチド配列)を有し得る。
【0015】
別の局面において、本発明は、本発明のベクター集団を含む宿主細胞に関する。上記宿主細胞は、真核生物宿主細胞もしくは原核生物宿主細胞(例えば、E.coli脂肪)であり得る。
【0016】
さらなる局面において、本発明は、標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するライブラリーメンバーを含むドナー特異的抗体ライブラリーに関し、ここで上記抗体もしくは抗体フラグメントは、上記標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒトドナーから得られ、ここで上記抗体ライブラリーは、少なくとも1個の特有のバーコードで同定される。
【0017】
一実施形態において、上記抗体重鎖および軽鎖は、これらが得られる上記ヒトドナーに特有のバーコードで各々別個に同定される。
【0018】
別の実施形態において、上記ドナー特異的抗体ライブラリーは、1個の特有のバーコードで同定される。
【0019】
さらに別の実施形態において、上記抗体もしくは抗体フラグメントは、ベクターに存在する核酸分子によってコードされる抗体重鎖および軽鎖もしくはそのフラグメントから構成される。
【0020】
さらなる実施形態において、上記バーコードは、上記ライブラリーに存在するベクターに連結されるかもしくは組み込まれ、そして/または上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、上記バーコードは、上記核酸分子の発現を妨害しない。
【0021】
なおさらなる実施形態において、上記バーコードは、1〜約24ヌクレオチドの連続非コードヌクレオチド配列であり、これらは、例えば、上記核酸分子の3’非コード領域もしくは5’非コード領域であり得る。
【0022】
異なる実施形態において、上記バーコードは、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列によってコードされる。
【0023】
別の実施形態において、上記バーコードは、非連続ヌクレオチド配列によってコードされる。
【0024】
さらなる実施形態において、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、上記ライブラリーに存在するベクターに連結されるかもしくは組み込まれる。
【0025】
なおさらなる実施形態において、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ、その結果、上記バーコードをコードする上記非連続配列のうちの少なくとも一部は、このような核酸分子の発現を妨害しない。
【0026】
異なる実施形態において、上記バーコードは、ペプチドもしくはポリペプチド配列である。
【0027】
別の実施形態において、ベクターは、ファージミドベクターであり、例えば、バクテリオファージ遺伝子III、および上記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と上記バクテリオファージIII遺伝子との間に終止コドンを含み得る。
【0028】
さらに別の実施形態において上記ヒト患者ドナーの病歴は、上記ドナーが上記疾患に罹患したかもしくは現在罹患していることを示す。いくつかの実施形態において、上記ヒトドナーが、上記疾患に罹患したかもしくは現在罹患していることは、独立して確認される。
【0029】
さらなる実施形態において、上記ドナー特異的抗体ライブラリーは、上記標的抗原とは異なる抗原を特異的に結合する抗体および抗体フラグメントを実質的に欠いている。
【0030】
一実施形態において、上記標的抗原は、インフルエンザAウイルス(例えば、インフルエンザAウイルス H1、H2、H3、H5、H7、もしくはH9サブタイプの単離株)である。
【0031】
別の実施形態において、上記ライブラリーは、1つより多くのインフルエンザAウイルスサブタイプに特異的に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0032】
さらに別の実施形態において、上記ライブラリーは、上記インフルエンザウイルスAのH5N1サブタイプに結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0033】
さらなる実施形態において、上記ヒトドナーは、以下の表1に列挙される疾患からなる群より選択される疾患に罹患したかもしくは現在罹患している。
【0034】
なおさらなる実施形態において、上記抗体ライブラリーは、上記標的疾患と関連する抗原に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0035】
さらなる実施形態において、上記抗体ライブラリーは、上記標的疾患と関連する抗原に結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する。
【0036】
上記ドナー特異的抗体ライブラリーは、例えば、ファージライブラリーであり得、一実施形態において、これは、抗体もしくは抗体フラグメントのうちの106個より多くの異なるメンバー、もしくは抗体もしくは抗体フラグメントのうちの109個より多くの異なるメンバーをコードする配列を含み得る。
【0037】
他の実施形態において、上記ドナー特異的抗体ライブラリーは、限定されることなく、芽胞−ディスプレイライブラリー、リボソームディスプレイライブラリー、mRNAディスプレイライブラリー、微生物細胞ディスプレイライブラリー、酵母ディスプレイライブラリー、もしくは哺乳動物ディスプレイライブラリーである。
【0038】
一実施形態において、上記ベクターに存在する抗体もしくは抗体フラグメントをコードする核酸は、上記ヒト患者ドナーのリンパ球から抽出されるmRNAから逆転写され、ここで上記リンパ球は、例えば、骨髄、血液、脾臓、もしくはリンパ節から発生し得る。
【0039】
望ましい場合、上記ヒトドナーの血清学的プロフィールは、上記mRNAを抽出する前に生成され得る。
【0040】
代わりに、もしくはさらに、上記ヒトドナー、および必要に応じて、上記ドナーの家族の病歴は、上記mRNAを中するする前もしくはその後に検査される。
【0041】
別の局面において、本発明は、標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するドナー特異的ライブラリーを作製するための方法に関し、上記方法は、
a)上記標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒト患者ドナーのリンパ球からmRNAを得る工程;
b)上記得られたmRNAの逆転写によって、上記患者の免疫グロブリンレパートリーをコードする配列を含む核酸の集団を生成する工程;および
c)上記核酸を標識する特有のバーコードで、上記ドナー特異的ライブラリーを同定する工程、
を包含する。
【0042】
上記方法は、上記患者の血清学的プロフィールを生成する工程、および/または工程a)の前もしくはその後に、上記患者の病歴を検査する工程をさらに包含し得る。
【0043】
上記方法は、d)上記核酸を発現ベクターに挿入する工程;e)上記免疫グロブリンレパートリーを発現させる工程;およびf)上記免疫グロブリンレパートリーをディスプレイ系においてディスプレイする工程、をさらに包含し得る。
【0044】
別の実施形態において、上記方法は、上記ライブラリーのメンバーを、上記標的抗原を中和もしくは活性化するそれらの能力に基づいて、選択する工程をさらに包含する。
【0045】
なおさらなる実施形態において、上記方法は、少なくとも1種の中和抗体を生じる。
【0046】
別の実施形態において、上記方法は、上記標的抗原を中和もしくは活性化させることが見いだされたライブラリーメンバーを含む1個以上の下位ライブラリー(sub−library)を作り出す工程をさらに包含する。
【0047】
さらに別の実施形態において、本発明は、同定された少なくとも1個のライブラリーメンバーを配列決定する工程を包含する。
【0048】
さらなる局面において、本発明は、上記方法によって選択された抗体によって中和もしくは活性化される標的抗原と関連する疾患を処置もしくは予防するための方法に関し、上記方法は、有効量の上記選択された抗体を、必要なヒト患者に投与する工程を包含する。
【0049】
上記抗体は、例えば、中和抗体(例えば、少なくとも1つのインフルエンザAウイルスサブタイプに対する中和抗体)であり得る。
【0050】
別の実施形態において、上記疾患は、以下の表1に列挙される疾患からなる群より選択される。
【図面の簡単な説明】
【0051】
【図1】図1は、本発明のヒト抗体ライブラリーの作出のための代表的な方法を模式的に例示するフローチャートである。
【図2】図2は、2つの異なる標的(AおよびB)に対する反応強度を増大させるための代表的パニング富化スキームを例示する。富化の各ラウンドは、上記個々の標的に対するプールの反応性強度を増大させる。
【図3】図3は、標的AおよびBと交叉反応性のクローンの選択のためのストラテジーを例示し、ここで各連続するラウンドは、反両方の標的に対して得られたプールの応性強度を強化する。
【図4】図4は、並行発見プール(parallel discovery pool)を組換えて、交叉反応性を生成/増大させることによって、2つの異なる標的(標的AおよびB)に対する反応性強度を増大させるためのストラテジーを例示する。上記組換え抗体ライブラリーの選択の各ラウンドは、両方の標的に対して得られたプールの反応性強度を増大させる。
【図5】図5は、標的Aに対する反応性を維持しながら、標的Bに対する交叉反応性を増大させるためのストラテジーを例示する。第1に、標的Aと反応性のクローンが選択され、次いで、上記標的Aと反応性のクローンの変異誘発ライブラリーは調製され、選択は、示されるように行われ、標的Aおよび標的Bの両方との強い反応性を示す1個以上の抗体クローンを生じる。
【図6】図6は、「指定変異誘発(destinational mutagenesis)」方法による多様な多機能抗体集団を生成するための代表的変異誘発法を例示する。
【図7A】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7B】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7C】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7D】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7E】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7F】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7G】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7H】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7I】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図7J】図7は、15個の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示す。
【図8】図8は、トルコH5N1トリインフルエンザの大発生の6名のヒト生存者から得られた血液サンプルについてのH5ヘマグルチニン(HA)血清学結果を示す。上記データは、上記HA抗原に対する抗体の存在を示す。
【図9】図9は、12名のその土地のドナーの血清サンプルで得られ、H5抗原(A/ベトナム/1203/2004)およびH1N1抗原(A/ニューカレドニア/20/99)およびH3N2抗原(A/パナマ/2007/99)のウイルスに対して試験した血清学結果を示す。
【図10】図10は、抗体ファージライブラリーの構築において使用される特有のバーコード付与アプローチを例示する。
【図11】図11は、種々のインフルエンザAサブタイプに由来するヘマグルチニンに結合する抗体の分析を示す。
【図12】図12は、必要とされるH5ヘマグルチニン結合グループ1の位置およびFab 47eにおける結晶構造に対するドミナント変異を示す。
【図13】図13および図14は、トルコトリインフルエンザ生存者の血清および骨髄の分析によって同定された抗体重鎖および軽鎖配列を使用して、多様な抗体重鎖および軽鎖ライブラリーを作り出すための指定変異誘発の使用を例示する。
【図14】図13および図14は、トルコトリインフルエンザ生存者の血清および骨髄の分析によって同定された抗体重鎖および軽鎖配列を使用して、多様な抗体重鎖および軽鎖ライブラリーを作り出すための指定変異誘発の使用を例示する。
【発明を実施するための形態】
【0052】
(詳細な説明)
(A.定義)
別段定義されない限り、本明細書において使用される技術的および科学的用語は、本発明が属する分野の当業者によって一般に理解されるのと同じ意味を有する。Singletonら,Dictionary of Microbiology and Molecular Biology 第2版,J.Wiley & Sons (New York,NY 1994)は、当業者に、本願において使用される用語の多くに対する一般的ガイドを与える。
【0053】
当業者は、本発明の実施において使用され得る、本明細書に記載されるものと類似もしくは等価な多くの方法および材料を認識する。実際に、本発明は、記載される方法および材料に決して限定されない。本発明の目的に関して、以下の用語は、以下で定義される。
【0054】
語句「保存されたアミノ酸残基」とは、互いに整列させられた2種以上のアミノ酸配列の間で同一であるアミノ酸残基に言及するために使用される。
【0055】
用語「疾患」、「障害」、および「状態」は、本明細書において交換可能に使用され、通常の身体機能の混乱、もしくは任意のタイプの病状の出現に言及する。正常の生理の混乱を引き起こす原因因子は、公知であってもよいし未知であってもよい。さらに、2名の患者が同じ障害と診断され得るが、それらの個体によって示される特定の症状は、同一でってもよいし、そうでなくてもよい。
【0056】
「有効な量」とは、有益もしくは望ましい治療的(予防的を含む)結果をもたらすに十分な量である。有効な量は、1回以上の投与において投与され得る。
【0057】
「組成物」は、本明細書において使用される場合、活性成分(例えば、本発明から生成される中和抗体)、および少なくとも1種の添加剤(例えば、薬学的に受容可能なキャリア)を含むと定義され、上記薬学的に受容可能なキャリアとしては、水、ミネラル、タンパク質、および/もしくは当業者に公知の他の賦形剤が挙げられるが、これらに限定されない。
【0058】
本明細書において使用される場合、用語「処置する」もしくは「処置」とは、疾患を示す臨床的症状の改善を意味することが意図される。
【0059】
本明細書において使用される場合、用語「予防する」もしくは「予防」とは、疾患を示す臨床的症状を妨げることを意味することが意図される。
【0060】
用語「被験体」および「患者」とは、本明細書において使用される場合、交換可能に使用され、検査、処置、分析、試験もしくは診断の被験体である、動物、および好ましくは、哺乳動物のいずれかに言及し得る。従って、被験体もしくは患者は、ヒト、非ヒト霊長類、および他の哺乳動物を含み、彼らは、疾患もしくは他の病的状態を有してもよいし有さなくてもよい。
【0061】
用語「アミノ酸」もしくは「アミノ酸残基」とは、代表的には、その分野で認識される定義を有するアミノ酸(例えば、アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)からなる群より選択されるアミノ酸)をいうが、改変アミノ酸、合成アミノ酸、もしくは稀なアミノ酸が、望ましい場合には使用され得る。従って、37 CFR 1.822(b)(4)に列挙される改変アミノ酸および通常でないアミノ酸は、この定義内に含まれ、明示的に本明細書に参考として援用される。アミノ酸は、種々の亜群へと下位分類され得る。従って、アミノ酸は、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val);負に荷電した側鎖(例えば、Asp、Glu);正に荷電した側鎖(例えば、Arg、His、Lys);もしくは非荷電極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、Trp、およびTyr)を有するとして分類され得る。アミノ酸はまた、低分子アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)、および塩基性アミノ酸(Lys、Arg)として分類され得る。
【0062】
用語「改変体」とは、参照ポリペプチドに関して、ネイティブポリペプチドと比較して、少なくとも1個のアミノ酸変異もしくは改変(すなわち、変化)を有するポリペプチドをいう。「アミノ酸改変」によって生成される改変体は、上記ネイティブアミノ酸配列において、例えば、少なくとも1個のアミノ酸を置換、欠失、挿入および/もしくは化学的に改変することによって生成され得る。
【0063】
「アミノ酸改変」とは、所定のアミノ酸配列のアミノ酸配列における変化をいう。例示的な改変としては、アミノ酸置換、挿入および/もしくは欠失を含む。
【0064】
特定の位置における「アミノ酸改変」とは、上記特定の残基の置換もしくは欠失、または上記特定の位置に隣接する少なくとも1個のアミノ酸残基の挿入をいう。特定の残基に「隣接する」挿入とは、その1〜2個の残基内での挿入を意味する。上記挿入は、上記特定の残基に対してN末端側もしくはC末端側に存在し得る。
【0065】
「アミノ酸置換」とは、所定のアミノ酸配列中の少なくとも1個の既存のアミノ酸残基を、別の異なる「置換」アミノ酸残基で置換することをいう。上記置換残基は、「天然に存在するアミノ酸残基」(すなわち、遺伝コードによってコードされる)であり得、以下からなる群より選択され得る:アラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)。1個以上の天然に存在しないアミノ酸残基での置換はまた、本明細書におけるアミノ酸置換の定義によって包含される。
【0066】
「天然に存在しないアミノ酸残基」とは、上記の天然に存在するアミノ酸残基以外の、ポリペプチド鎖において隣接するアミノ酸残基を共有結合することができる残基をいう。天然に存在しないアミノ酸残基の例としては、ノルロイシン、オルニチン、ノルバリン、ホモセリンおよび他のアミノ酸残基残ログ(例えば、Ellmanら Meth.Enzym.202:301 336(1991)に記載されるもの)が挙げられる。このような天然に存在しないアミノ酸残基を生成するために、Norenら Science 244:182(1989)およびEllmanら(前出)の手順が、使用され得る。簡潔には、これら手順は、天然に存在しないアミノ酸残基でサプレッサtRNAを化学的に活性化すること、続いて、上記RNAのインビトロ転写および翻訳を伴う。
【0067】
「アミノ酸挿入」とは、所定のアミノ酸配列への少なくとも1個のアミノ酸の組み込みをいう。上記挿入は、通常、1個もしくは2個のアミノ酸残基の挿入からなる一方で、本願は、より大きな「ペプチド挿入」(例えば、約3〜約5個もしくはさらに約10個までのアミノ酸残基の挿入)を企図する。上記挿入された残基は、上記で議論されるように、天然に存在していてもよいし、天然に存在していなくてもよい。
【0068】
「アミノ酸欠失」とは、少なくとも1個のアミノ酸残基を、所定のアミノ酸配列から除去することをいう。
【0069】
用語「ポリヌクレオチド」とは、核酸(例えば、DNA分子およびRNA分子ならびにこれらのアナログ(例えば、ヌクレオチドアナログを使用して、もしくは核酸化学を使用して生成されるDNAもしくはRNA)をいう。望ましい場合、上記ポリヌクレオチドは、合成的に、例えば、当該分野で認識される核酸化学を使用して、例えば、ポリメラーゼを使用して、作製され得、望ましい場合には、改変され得る。代表的な改変としては、メチル化、ビオチン化、および他の当該分野で公知の改変が挙げられる。さらに、上記核酸分子は、一本鎖もしくは二本鎖であり得、そして望ましい場合、検出可能な部分に連結され得る。
【0070】
用語「変異誘発」とは、別段特定されない限り、ポリヌクレオチドもしくはポリペプチド配列を改変するために任意の当該分野で認識されている技術をいう。変異誘発の好ましいタイプとしては、エラープローンPCR変異誘発、飽和変異誘発(saturation mutagenesis)、もしくは他の部位指向性変異誘発が挙げられる。
【0071】
「部位指向性変異誘発」とは、当該分野で標準的な技術であり、望ましい変異を表す制限されたミスマッチを除いて変異誘発されるように、一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを使用して行われる。簡潔には、上記合成オリゴヌクレオチドは、上記一本鎖ファージDNAに対して相補的な鎖の合成を指向するためにプライマーとして使用され、その得られた二本鎖DNAは、ファージ支援宿主細菌(phage−supporting host bacterium)に形質転換される。上記形質転換細菌の培養は、上部の寒天にプレートされ、上記ファージを内部にもつ単一の細胞からのプラーク形成を可能にし、上記新たなプラークのうちの50%は、一本鎖として、変異した形態を有するファージを含み;50%は、元の配列を有する。目的のプラークは、正確なマッチのハイブリダイゼーションを可能にする温度でキナーゼ処理(kinased)合成プライマーとハイブリダイズすることによって選択されるが、その温度で、上記元の鎖とのミスマッチは、ハイブリダイゼーションを妨げるのに十分である。次いで、上記プローブとハイブリダイズするプラークが選択され、配列決定され、培養され、そのDNAが回収される。
【0072】
用語「ベクター」とは、細胞における自律的複製が可能でかつDNAセグメント(例えば、遺伝子もしくはポリヌクレオチド)が、上記結合されるセグメントの複製をもたらすように作動可能に連結されるrDNA分子をいうために使用される。1種以上のポリペプチドをコードする遺伝子の発現を指向し得るベクターは、本明細書において「発現ベクター」といわれる。用語「制御配列」とは、特定の宿主生物における作動可能に連結されたコード配列の発現に必須のDNA配列をいう。原核生物に適した上記制御配列としては、例えば、プロモーター、必要に応じて、オペレーター配列、およびリボソーム結合部位が挙げられる。真核生物細胞は、プロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが公知である。
【0073】
核酸は、別の核酸配列と機能的関係に置かれている場合に「作動可能に連結されている」。例えば、プレ配列もしくは分泌リーダーのDNAは、上記ポリペプチドの分泌に関与するプレタンパク質として発現される場合にポリペプチドのDNAに作動可能に連結されている;プロモーターもしくはエンハンサーは、上記配列の転写に影響を及ぼす場合、コード配列に作動可能に連結されている;またはリボソーム結合部位は、翻訳を促進するように位置している場合、コード配列に作動可能に連結されている。一般に、「作動可能に連結」とは、連結されている上記DNA配列が連続しており、そして分泌リーダーの場合には、連続しておりかつリーディング相中にあることを意味する。しかし、エンハンサーは、連続している必要はない。連結は、都合のよい制限部位においてライブラリーゲーションによって達成される。このような部位が存在しない場合、上記合成オリゴヌクレオチドアダプターもしくはリンカーが、従来の経験に従って使用される。
【0074】
パーセント(%)アミノ酸配列同一性は、配列比較プログラムNCBI−BLAST2(Altschulら,Nucleic Acids Res.25:3389−3402(1997))を使用して決定され得る。上記NCBI−BLAST2配列比較プログラムは、http://www.ncbi.nlm.nih.govからダウンロードされるか、そうでなければ、National Institute of Health,Bethesda,MDから得られ得る。NCBI−BLAST2は、いくつかの検索パラメーターを使用し、ここでそれらパラメーターの全ては、以下を含むデフォルト値に設定されている:例えば、unmask=yes、strand=all、expected occurrences=10、minimum low complexity length=15/5、multi−pass e−value=0.01、constant for multi−pass=25、dropoff for final gapped alignment=25およびscoring matrix=BLOSUM62。
【0075】
「抗体依存性細胞媒介性細胞傷害性」および「ADCC」とは、FcRを発現する非特異的細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、およびマクロファージ)が、標的細胞上に結合抗体を認識し、その後、上記標的細胞の融解を引き起こす細胞媒介性反応をいうために、本明細書において使用される。種々の免疫細胞は、異なるFcレセプター(FcR)を発現する。従って、ADCCを媒介するための主な細胞(NK細胞)は、FcgRIIIのみを発現するのに対して、単球は、FcγRI、FcγRIIおよびFcγRIIIを発現する。
【0076】
用語「インフルエンザAサブタイプ」もしくは「インフルエンザAウイルスサブタイプ」とは、交換可能に使用され、ヘマグルチニン(H)およびノイラミニダーゼ(N)ウイルス表面タンパク質の種々の組み合わせによって特徴付けられ、従って、H番号およびN番号の組み合わせ(例えば、H1N1およびH3N2)によって表示されるインフルエンザAウイルス改変体をいう。上記用語は、具体的には、通常は、変異から生じ、異なる病理的側面を示す、各サブタイプ内の全ての株(絶滅株を含む)を含む。このような株はまた、ウイルスサブタイプの種々の「単離株」として言及され、全ての過去の、現在の、および将来的な単離株を含む。従って、この状況において、用語「株」および「単離株」は、交換可能に使用される。
【0077】
用語「インフルエンザ」とは、インフルエンザウイルスによって引き起こされる感染性疾患をいうために使用される。
【0078】
(B.一般的技術)
本発明の方法を行うための技術は、当該分野で周知であり、標準的な実験教科書に記載されており、これら教科書としては、例えば、Ausubelら,Current Protocols of Molecular Biology,John Wiley and Sons(1997);Molecular Cloning:A Laboratory Manual,第3版,J.Sambrook and D.W.Russell,編,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;O’Brianら,Analytical Chemistry of Bacillus Thuringiensis,Hickle and Fitch,編,Am.Chem.Soc.,1990;Bacillus thuringiensis:biology,ecology and safety,T.R.Glare and M. O’Callaghan,編,John Wiley,2000;Antibody Phage Display,Methods and Protocols,Humana Press,2001;およびAntibodies,G.Subramanian,編,Kluwer Academic,2004が挙げられる。変異誘発は、例えば、部位指向性変異誘発(Kunkelら,Proc.Natl.Acad.Sci USA 82:488−492(1985))を使用して行われ得る。PCR増幅法は、米国特許第4,683,192号、同第4,683,202号、同第4,800,159号、および同第4,965,188号に、およびいくつかの教科書(including 「PCR Technology:Principles and Applications for DNA Amplification」,H.Erlich,編,Stockton Press,New York(1989);およびPCR Protocols:A Guide to Methods and Applications,Innisら,編,Academic Press,San Diego,Calif.(1990)に記載されている。
【0079】
本発明の方法は、抗体のディスプレイのために使用されるいかなる特定の技術によっても限定されない。本発明は、ファージディスプレイを参照しながら例示されるが、本発明の抗体は、他のディスプレイおよび富化技術(例えば、リボソームもしくはmRNAディスプレイ(Mattheakisら,Proc.Natl.Acad.Sci.USA 91:9022−9026(1994);Hanes and Pluckthun,Proc.Natl.Acad.Sci.USA 94:4937−4942(1997))、微生物細胞ディスプレイ(例えば、細菌ディスプレイ(Georgiouら,Nature Biotech.15:29−34(1997))、もしくは酵母細胞ディスプレイ(Kiekeら,Protein Eng.10:1303−1310(1997))、哺乳動物細胞上でのディスプレイ、芽胞ディスプレイ(Isticatoら,J.Bacteriol.183:6294−6301(2001);Chengら,Appl.Environ.Microbiol.71:3337−3341(2005)および同時係属中の仮特許出願番号第60/955,592号(2007年8月13日出願)、ウイルスディスプレイ(例えば、レトロ・ウイルスディスプレイ(Urbanら,Nucleic Acids Res.33:e35(2005)、タンパク質−DNA連結に基づくディスプレイ(Odegripら,Proc.Acad.Natl.Sci.USA 101:2806−2810(2004);Reiersenら,Nucleic Acids Res.33:e10(2005))、および、マイクロビーズディスプレイ(Seppら,FEBS Lett.532:455−458(2002))によっても同定され得る。
【0080】
リボソームディスプレイにおいて、上記抗体およびコードmRNAは、リボソームによって連結され、上記リボソームディスプレイは、翻訳の最後に、上記mRNAが上記ポリペプチドを放出することなくとどまるようにする。選択は、全体として三成分複合体(ternary complex)に基づく。
【0081】
mRNAディスプレイライブラリーにおいて、抗体とそのコードmRNAとの間の共有結合は、アダプター分子として使用さらえるピューロマイシンを介して確立される(Wilsonら,Proc.Natl.Acad.Sci.USA 98:3750−3755(2001))。抗体をディスプレイするためのこの技術の使用に関しては、例えば、Lipovsek and Pluckthun,J.Immunol.Methods.290:51−67(2004)を参照のこと。
【0082】
微生物細胞ディスプレイ技術は、酵母(例えば、Saccharomyces cerevisiae(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997)))上での表面ディスプレイを含む。従って、例えば、抗体は、S.cerevisiaeの表面に、酵母細胞壁上に位置するα−アグルチニン酵母接着レセプターへの融合を介してディスプレイされ得る。この方法は、フローサイトメトリーによってレパートリーを選択する可能性を提供する。蛍光標識された抗原および抗エピトープタグ試薬によって上記細胞を染色することによって、上記酵母細胞は、抗原結合のレベルおよび細胞表面での抗体発現に従って、ソートされ得る。酵母ディスプレイプラットフォームはまた、ファージと組み合わされ得る(例えば、Van den Beuckenら,FEBS Lett.546:288−294(2003)を参照のこと)。
【0083】
抗体ライブラリーを選択し、スクリーニングするための技術の総説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照のこと。
【0084】
(C.好ましい実施形態の詳細な説明)
(I.ドナー特異的抗体ライブラリーの調製)
本発明は、特定の疾患との遭遇に首尾よく耐えたかもしくは現在耐えている個体に由来するドナー特異的抗体ライブラリーに関する。その得られた抗体レパートリーは、上記ドナーが関連する疾患を特異的に防御するために使用した抗体を含み、従って、例えば、標的疾患の予防および/もしくは処置のための中和抗体を開発するために、重要なツールである。
【0085】
本発明は、ヒト被験体における抗体生成を引き起こす任意の標的疾患に適用可能であるが、このような疾患の代表的な(限定ではない)例は、表1に列挙される。
【0086】
【表1】
本発明のドナー特異的ライブラリーを作り出すための方法は、図1に模式的に例示される。第1の工程として、潜在的なドナーが同定される。上記患者ドナーは、標的疾患に現在罹患していてもよいし、上記標的疾患から回復していてもよいし、上記疾患に耐えていてもよい。従って、例えば、実施例に例示されるように、上記ドナー特異的ライブラリーは、本明細書において、以前のインフルエンザ感染(季節性インフルエンザ大発生、流行、および世界的流行が挙げられる)の回復期患者の骨髄から作り出され得る。
【0087】
患者ドナーを選択もしくは同定する際に、上記患者は、実際に上記標的疾患を有したかまたは現在有していることを確認することが重要である。上記確認の一部は、上記患者ドナーの病歴の検査である。上記病歴に加えて、種々の他の要因(例えば、上記患者の家族の病歴、上記患者の性別、体重、健康状態など)が、考慮に入れられ得る。上記患者の病歴が入手可能でないもしくはあてにならない、または任意の他の理由(例えば、さらなる他の確認手段)がある場合には、上記患者の血清学的側面が、決定され得る。血清学的アッセイは、当該分野で周知であり、種々の形式において(例えば、種々のELISAアッセイフォーマットの形態において)行われ得る。従って、例えば、インフルエンザウイルスに対する抗体の存在は、周知のヘマグルチニン阻害(HAI)アッセイ(Kendal,A.P.,M.S.Pereira,and J.J.Skehel.1982.Concepts and procedures for laboratory−based influenza surveillance. U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta, Georgia)、もしくはミクロ中和アッセイ(Harmonら,J.Clin.Microbiol.26:333−337(1988))によって検出され得る。この工程は、上記血清サンプルがインフルエンザ 中和抗体を含むことが既に確認された場合には必要ではないかもしれない。
【0088】
ドナー特異的ヒト抗体ライブラリーを調製するために、リンパ球を含むサンプルを、標的疾患(例えば、表1に列挙されるものからの少なくとも1つの疾患)を発症させたことが既知の個体(患者ドナー)から集められる。上記サンプルは、例えば、骨髄、血液、脾臓、リンパ節、扁桃、胸腺などから得られ得る。骨髄は、本明細書において上記抗体ライブラリーの好ましい供給源である。なぜなら、個々のドナーの成熟抗体レパートリーの完全な「過去のアーカイブ(fossil archive)」を表すからである。
【0089】
リンパ球を含むサンプルは、種々の時点において上記患者ドナーから集められ得る。一実施形態において、リンパ球は、上記標的とした疾患から回復した患者から、少なくとも1日間、5日間、10日間、15日間、20日間、25日間、少なくとも1ヶ月間、2ヶ月間、3ヶ月間、4ヶ月間、5ヶ月間、6ヶ月間、7ヶ月間、8ヶ月間、9ヶ月間、10ヶ月間、11ヶ月間、または少なくとも1年間、2年間、3年間、4年間、5年間、6年間、7年間、8年間、9年間、10年間にわたって、集められる。別の実施形態において、リンパ球は、回収の時点で上記標的とした疾患を有している、および上記疾患を有していると診断された患者から、少なくとも回収前の1日間、5日間、10日間、15日間、20日間、25日間、もしくは少なくとも1ヶ月間、2ヶ月間、3ヶ月間、4ヶ月間、5ヶ月間、6ヶ月間、8ヶ月間、9ヶ月間、10ヶ月間、もしくは1年間、2年間、3年間、4年間、もしくは5年間にわたって集められる。
【0090】
末梢血サンプル(特に、地理的に異なる供給源に由来する)は、輸送および使用の前に安定化される必要があり得る。この目的のためのキットは周知であり、市販の(例えば、BD Vacutainer(登録商標) CPTTM 細胞調製チューブ)は、リンパ球の遠心分離精製のために使用され得、およびグアニジウム、Trizol、もしくはRNAlaterが、上記サンプルを安定化するために使用され得る。リンパ球を他の供給源(例えば、リンパ系器官)から単離するための方法およびキットはまた、周知であり、市販されている。
【0091】
上記安定化したリンパ球もしくは全骨髄を受け取ったら、RNAが抽出され、RT−PCRは、当該分野で公知の免疫グロブリンオリゴプライマーを使用して、抗体重鎖および軽鎖レパートリーをレスキューするために行われる。
【0092】
骨髄リンパ球、もしくは任意の他の供給源のリンパ球に由来するRNAの調製のための方法は、当該分野で周知である。mRNA抽出の一般的方法は、分子生物学の標準的教科書で開示されており、上記標準的教科書としては、Ausubelら,Current Protocols of Molecular Biology,John Wiley and Sons(1997)が挙げられる。RNA精製キットは、商業的製造業者(例えば、Qiagen)から市販されており、上記製造業者の説明書に従って使用され得る。
【0093】
RNAは、PCRのためのテンプレートとして働き得ないので、RNAは、cDNAへと逆転写され、これは、PCR増幅に供される。2つの最も一般的に使用される逆転写酵素は、トリ骨髄芽球症ウイルス(avilo myeloblastosis virus)逆転写酵素(AMV−RT)およびモロニーマウス白血病ウイルス逆転写酵素(MMLV−RT)である。逆転写工程は、代表的には、環境および発現プロフィールの目的に依存して、特異的プライマー、ランダムヘキサマー、もしくはオリゴ−dTプライマーを用いて始められる。例えば、抽出されたRNAは、GeneAmp RNA PCRキット(Perkin Elmer,CA,USA)を使用して、製造業者の説明書に従って逆転写され得る。次いで、得られたcDNAは、その語のPCR反応においてテンプレートとして使用され得る。
【0094】
ファージディスプレイライブラリーを作り出すために、上記PCRレパートリー生成物は、scFvライブラリーを精製するためにリンカーオリゴと合わされて、当該分野で公知の手順に従って、m13 pIIIタンパク質とインフレームで直接クリーニングされ得る。他のディスプレイ技術(例えば、上記で議論されるもの)を使用するライブラリーは、当該分野で周知の方法によって調製され得る。
【0095】
代表的プロトコルにおいて、総RNAは、Tri BD試薬(Sigma)によって、新鮮なもしくはRNAlater安定化組織から抽出される。続いて、上記単離されたドナー総RNAは、Oligotex精製(Qiagen)を使用して、mRNAへとさらに精製される。次に、第1鎖cDNA合成は、AccuScript逆転写酵素(Stratagene)のプロトコルに従って、ランダムナノマーオリゴヌクレオチドおよび/もしくはオリゴ(dT)18プライマーを使用することによって、生成される。簡潔には、100ng mRNA、0.5mM dNTPおよび300ng ランダムナノマーならびに/または500ngオリゴ(dT)18プライマーを、Accuscript RT緩衝液(Stratagene)中で、65℃で5分間にわたってインキュベートし、続いて、4℃へと迅速に冷却した。次いで、100mM DTT、Accuscript RT、およびRNAse Blockを各反応系に添加し、42℃において1時間にわたってインキュベートし、上記逆転写酵素は、70℃において15分間にわたって加熱することによって不活性化される。上記得られたcDNAは、上記抗体重鎖および軽鎖V遺伝子のRT−PCR増幅のためのテンプレートとして使用され得、これは、次いで、ベクターへと、もしくはファージディスプレイライブラリーが意図される場合には、ファージミドベクターへとクローニングされ得る。この手順は、抗体重鎖および軽鎖可変領域クローン(VHおよびVLライブラリー)のレパートリーを生成し、それは、スクリーニング目的で、別個にもしくは合わせて保持され得る。上記ベクター(例えば、ファージミドベクター)は、次いで、宿主細胞(例えば、E coli宿主)へと導入されて、抗体軽鎖もしくは重鎖またはそれらのフラグメントをコードする核酸分子のレパートリーを含むベクター集団を生成し得る。各場合において、上記ベクター集団は、1個以上の抗体軽鎖もしくは抗体重鎖サブタイプを含み得る。従って、上記ベクター集団は、抗体κ軽鎖および/もしくはλ軽鎖をコードする配列を含み得る。
【0096】
本発明の方法において、代表的には、抗体軽鎖および抗体重鎖は、上記で議論されるように最初に、別個にクローニングされ、また、上記κおよびλ軽鎖ライブラリーを分離する。上記ライブラリーはアーカイブに入れられ(archived)得、そして必要とされる場合、重鎖ライブラリーを、分離されたκ鎖ライブラリーおよびλ鎖ライブラリーと合わされ得、重鎖および軽鎖対形成が、例えば、ファージディスプレイの場合においては、パニングによって同定され得る。達成されるべき目的に依存して、これら工程は、複数回、種々のライブラリーもしくは下位ライブラリーと反復することが可能である。本発明の方法は、成功を最大化するために、パニングの間に必要とされる場合、ライブラリー、下位ライブラリーもしくはクローンを含めるもしくは排除することにおいて、優れた可撓性を提供する。
【0097】
特に、上記ベクター集団に存在する配列は、上記抗体重鎖および軽鎖(もしくはフラグメント)のコード配列をそれぞれ保有するので、上記配列は切り出され得、そして上記抗体重鎖および軽鎖、もしくはこれらのフラグメントの発現のために、1個以上の発現ベクターに挿入され得る。好ましくは、上記抗体重鎖および軽鎖もしくはそれらのフラグメントのコード配列は、上記抗体もしくは抗体フラグメントのライブラリーを生成するために重鎖および軽鎖の同時発現のために、同じ発現ベクターへと導入される。
【0098】
本発明の発現ベクターは、上記ベクターを1個以上の選択された宿主細胞において複製することを可能にする核酸配列を含む。このような配列は、種々の細菌、酵母、およびウイルスに対して周知である。上記プラスミドpBR322に由来する複製起点は、大部分のグラム陰性細菌に適しており、上記2μプラスミド起点は、酵母に適しており、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVもしくはBPV)は、哺乳動物再紡においてベクターをクローニングするために有用である。
【0099】
発現ベクターは、代表的には、選択遺伝子(選択マーカーともいわれる)を含む。代表的な選択遺伝子は、(a)抗生物質もしくた他の毒素(例えば、アンピシリン、ネオマイシン、メトトレキサート、もしくはテトラサイクリン)に対する耐性を付与するか、(b)栄養要求性欠損を補完するか、もしくは(c)複雑な培地から利用可能でない重要な栄養要素を補う(例えば、BacilliのD−アラニンラセマーゼをコードする遺伝子)、タンパク質をコードする。
【0100】
哺乳動物再紡に対する適切な選択マーカーの例は、上記再紡成分の同定が、上記抗体コード核酸を取り込みことを可能にするものである(例えば、DHFRもしくはチミジンキナーゼ)。野生型DHFRが使用される場合に適切な宿主細胞は、Urlaubら,Proc.Natl.Acad.Sci.USA,77:4216(1980)によって記載されるように調製および増殖される、DHFR活性が欠損しているCHO細胞株である。酵母において使用するための適切な選択遺伝子は、上記酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら,Nature,282:39(1979);Kingsmanら,Gene,7:141(1979);Tschemperら,Gene,10:157(1980)]。上記trp1遺伝子は、トリプトファン中で増殖する能力を欠いている酵母の変異株(例えば、ATCC番号44076もしくはPEP4−1)についての選択マーカーを提供する[Jones,Genetics,85:12(1977)]。
【0101】
発現ベクターおよびクローニングベクターは、通常、mRNA合成を指向するために、上記抗体コード核酸配列に作動可能に連結されているプロモーターを含む。種々の潜在的な宿主細胞によって認識されるプロモーターは、周知である。原核生物宿主とともに使用するために適したプロモーターとしては、β−ラクタマーゼおよびラクトースプロモーター系[Changら,Nature,275:615(1978);Goeddelら,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーター系[Goeddel,Nucleic Acids Res.,8:4057(1980);EP 36,776]、およびハイブリッドプロモーター(例えば、tacプロモーター)[deBoerら,Proc.Natl.Acad.Sci.USA,80:21−25(1983)]が挙げられる。細菌系において使用するためのプロモーターはまた、上記抗体をコードするDNAに作動可能に連結されるシャイン−ダルガーノ(S.D.)配列を含む。
【0102】
酵母宿主とともに使用するのに適したプロモーター配列(promoting sequence)の例としては、3−ホスホグリセレートキナーゼに対するプロモーター[Hitzemanら,J.Biol.Chem.,255:2073(1980)]もしくは他の糖分解性酵素[Hessら,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)](例えば、エノラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ヘキソキナーゼ、ピルベートデカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−ホスフェートイソメラーゼ、3−ホスホグリセレートムターゼ、ピルベートキナーゼ、トリオースホスフェートイソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼ)に対するプロモーターが挙げられる。
【0103】
増殖条件によって制御される転写のさらなる利点を有する誘導性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロームC、酸ホスファターゼ、窒素代謝と関連する分解酵素、メタロチオネイン、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ならびにマルトースおよびガラクトース利用を担う酵素に対するプロモーター領域である。酵母発現において使用するための適切なベクターおよびプロモーターは、EP 73,657においてさらに記載されている。
【0104】
哺乳動物宿主細胞における上記発現ベクター中の重鎖もしくは軽鎖遺伝子の転写は、例えば、ウイルス(例えば、ポリオーマウイルス、鶏痘ウイルス(UK 2,211,504(1989年7月5日公開))、アデノウイルス(例えば、アデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびシミアンウイルス40(SV40)のゲノムから得られるプロモーターによって、異種哺乳動物プロモーター(例えば、上記アクチンプロモーターもしくは免疫グロブリンプロモーター)から得られるプロモーターによって、および熱ショックプロモーターから、このようなプロモーターが上記宿主細胞系と適合性であれば、制御される。
【0105】
高等真核生物による、上記抗体遺伝子をコードするDNAの転写は、エンハンサー配列を上記ベクターに挿入することによって、増大させられ得る。エンハンサーは、その転写を増大させるようにプロモーターに対して作用する、通常、約10〜300bpのDNAのシス作用性要素である。多くのエンハンサー配列は、いまや哺乳動物遺伝子から公知である(グロビン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインスリン)。しかし、代表的には、真核生物細胞ウイルスに由来するエンハンサーを使用する。例としては、複製起点(bp100〜270)の後ろ側にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後ろ側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。上記エンハンサーは、上記抗体コード配列に対して5’側もしくは3’側にある位置において、上記ベクターにスプライスされ得るが、好ましくは、上記プロモーターから5’側の位置に位置する。
【0106】
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、もしくは他の多細胞生物に由来する核化細胞)において使用される発現ベクターはまた、転写の終結に、および上記mRNAを安定化させるために必須の配列を含む。このような配列は、真核生物のもしくはウイルスのDNAもしくはcDNAの5’非翻訳領域、およびときおり3’ 非翻訳領域から一般に入手可能である。これら領域は、上記抗体重鎖および軽鎖をコードするmRNAの非翻訳部分においてポリアデニル化フラグメントとして転写されるヌクレオチドセグメントを含む。
【0107】
組換え脊椎動物細胞培養に従ってポリペプチドの合成に適合させるために適したさらに他の方法、ベクター、および宿主細胞は、Gethingら,Nature,293:620−625(1981);Manteiら,Nature,281:40−46(1979);EP 117,060;およびEP 117,058に記載されている。
【0108】
特定の実施形態において、上記抗体ライブラリーは、ファージライブラリーの形態において生成され、ここで上記抗体重鎖および軽鎖もしくはそれらのフラグメントのコード配列は、ファージミドベクター(例えば、上記バクテリオファージ遺伝子IIIを含むベクター)にクローニングされる。ファージミドベクターは、周知であり、市販されている。これらのベクターとしては、例えば、pBluescriptベクターSKII+(Stratagene,Genbank Accession X52328)、および他のpBluscriptベクターが挙げられる。ファージディスプレイ技術は、ヒト抗体の大きなレパートリーの生成を可能にし、生体パニング(biopanning)手順は、任意の望ましい特異性もしくは他の特性を有する個々の抗体の選択を可能にする。
【0109】
例えば、初期の流行および大流行(例えば、1918年のスペイン風邪)からの生存者の末梢リンパ球に由来する免疫学的レパートリーは、上記記載されるものに類似の様式において復活させられ得、安定化され得、レスキューされ得、そして発現させられ得る。さらなるH1およびH3ライブラリーについて、レパートリーは、適切にタイミングを計ってワクチン接種したその土地の出身のドナーから回収され得る。さらなる選択肢として、市販の骨髄総RNAもしくはmRNAは、H1およびH3に適した、また、ドナーの背景に依存して、H2抗体スクリーニングに適したライブラリーを生成するための市販の供給源から購入され得る。一般に、ワクチン接種が利用可能な処置選択しである標的疾患に関しては、抗体は、免疫したヒトドナーから得られる生物学的サンプルからも単離され得る。上記特定の抗原を認識する抗体の力価を発揮した、免疫した患者から、骨髄、血液、もしくは別のリンパ球供給源が集められ、生成された抗体が単離され、上記のように増幅され発現される。
【0110】
上記で議論されるように、各ドナーについて、抗体軽鎖および重鎖ライブラリーは、別個にクローニングされ得る。従って、各ドナーについて、種々のκ軽鎖およびλ軽鎖ファミリーは、別個にプールされ得、かつ等モル量でクローニングされ得る。同様に、各ドナーについて、種々の重鎖ファミリーは、プールされ得、等モル量でクローニングされ得る。抗体の遺伝子ファミリー特異的レスキューを可能にすることによって、本発明の方法は、量が少ない抗体を含む、プールされた抗体ライブラリーの場合には、任意のおよび全ての個体からの免疫グロブリン寄与を保証する上記ドナーの抗体レパートリーをより完全に表すライブラリーを得る。例えば、実施例の実施例1に例示されるように、本明細書における
上記インフルエンザ重鎖および軽鎖ライブラリーを調製することにおいて、6個のκ軽鎖ファミリー、11個のλ軽鎖ファミリーおよび4個の重鎖ファミリーをレスキューした。
【0111】
代表的なスクリーンは、ゼロ、1個、もしくは1個より多い標的特異的ポジティブクローンを生じ得る。特定のコンビナトリアル抗体ライブラリーが、もっぱらスクリーニングされ、さらなる解決策がさらには獲得されそうにない場合、これは、上記重鎖および軽鎖レパートリーが標的に結合する能力がないことではないかもしれないが、逆に、上記集団は、標的を結合するために必要とされる上記必要な重鎖および軽鎖対を一緒にすることができなかった可能性がある。任意の個体からの代表的なレスキューされた軽鎖のレパートリーは、約105〜106個の特有の軽鎖、および約106〜108個の特有の重鎖を含み得る。このような対形成の考えられるコンビナトリアル生成物は、1011〜1014の範囲に及ぶ。このような集団は、数桁の規模で、大部分のディスプレイ系(例えば、ファージディスプレイ)の実際の限界を超える。結論として、現在のディスプレイ技術(例えば、ファージディスプレイ)を使用すると、ごく一部の上記コンビナトリアルの可能性が、任意の単一のファージ抗体ライブラリーにおいて捕獲および評価される。従って、重鎖の本来のセットと、軽鎖の本来の集団との再クローニングは、特定の標的に対するおそらく新規な抗体を有する、シャッフルされた重鎖および軽鎖の組み合わせの完全に新たなセットを生成する。このような新たな再シャッフルされた集団は、以前に存在した機能が乏しい(poorly performing)ドナー特異的ライブラリーを再現性が高い集団へと形質転換することが見いだされた。具体的には、単一ドナーに由来する集団に関して、以前に、3ラウンドの選択後のバックグラウンドと比較すると、0.3倍の富化が達成されたに過ぎなかった。しかし、この集団が再クローニングされ、再シャッフルされた場合、3ラウンドのパニングの後に、15倍の富化が可能になり、92個の選択されたクローンから55個の新規な配列が生じる。
【0112】
以前に言及されたように、代表的なスクリーニングは、標的特異的ポジティブクローンの任意の数を生じ得る。本発明は、それらの埋め込まれたバーコードによって任意のクローンの起源の同定を可能にする。代表的な抗体のスクリーンは、多くのドナー特異的ライブラリーに由来するファージ抗体を併せ得るので、上記ライブラリーのうちのいくらかおよびそれらのコンビナトリアルクローンが、抗体保有ファージ粒子として完全には表されないことが考えられる。この場合において、ポジティブクローンは、スクリーニングされる下位ライブラリーファージ集団において存在する可能なクローニングされた解決策全ての制限された物理的セットのみから生じた可能性がある。このような場合において、ドナー特異的ファージの集団をより完全に質問することはかなり重要である。この場合、ポジティブクローンからの上記バーコードは、上記クローンを担う上記特異的ライブラリーに導き、もっぱらおよびより深く上記目的の集団を深くスクリーニングすることを可能にする。
【0113】
望ましい抗体が、最初のライブラリー構築物のための基礎として最初に使用されるものを超えた機能的能力(例えば、中和もしくは活性化)を有しなければならない場合において、本発明者らは、将来を見越して、このような活性の証拠のために個々のドナー血清の輪郭を描き得る。上記望ましい活性が、任意の特定のドナー血清において合理的な力価で存在する場合、上記目的の標的に対してスクリーニングするために選択されるライブラリーに対応するものを選択し得る。他の場合において、上記関連する選択基準は、血清学に関連しない可能性があるが、ドナー特徴(例えば、年齢、性別もしくは病歴)に関連する可能性がある。いずれにしても、ドナープロフィールは、ライブラリー選択についての論理的なガイドでありかつドナー特異的かつ分離された抗体ライブラリーにおいてのみ可能ある。
【0114】
ファージパニングスクリーンを完了し、免疫優性クローンの存在を発見することは、異常ではない。さらに、反復パニングスクリーニングレジメンの際に、このようなクローンを再発見することも、異常ではない。優性クローンの場合において、より多くのもしくは異なるクローンのいずれかが望ましい場合、このクローンの存在を担うライブラリー物質を回避することが重要である。代表的なファージ抗体ライブラリーにおいて、上記特異的ライブラリーもしくは上記クローン起源の原因である物質は、上記集団から分離可能ではないが、ドナー特異的ライブラリーにおいて、上記ライブラリーを再スクリーニングし、望ましくないドナー下位ライブラリーを単純に省くことは可能であり、それによって、以前に同定された優性クローンからポジティブな選択に焦点を当てる。
【0115】
単純さのために、上記ライブラリーは、重鎖もしくは軽鎖ライブラリーとして記載されるが、同じ記載が抗体フラグメントのライブラリー、抗体重鎖および/もしくは軽鎖の抗体フラグメント、および抗体様分子のライブラリーに適用されることは、当業者にとって明らかである。
【0116】
特定の実施形態において、二重特異性(例えば、2つの異なるインフルエンザAサブタイプと、および/もしくは同じサブタイプの2つの株(単離株)と、および/もしくはヒトおよび非ヒト分離物との反応性を示す)を有する抗体は、制御される交叉反応性選択および/もしくは指向性のコンビナトリアル操作および/もしくは変異誘発操作を介して発見および最適化され得る。
【0117】
図2に例示される代表的な富化スキームにおいて、2個の標的(標的AおよびBと示される)に対する交叉反応性を示す抗体を含むライブラリーは、複数ラウンドの富化に供される。富化が標的Aとの反応性に基づいている場合、各ラウンドの富化は、標的Aに対する、上記プールの反応性強度を増大させる。同様に、富化が、標的Bとの反応性に基づく場合、各ラウンドの富化は、標的Bに対する、上記プールの反応性強度を増大させる。図2は、ファージディスプレイライブラリーをスクリーニングする場合に使用される選択方法である(以下を参照のこと)パニングに言及するが、上記アプローチは、上記で議論されるライブラリーのうちのいずれかのタイプおよびディスプレイ技術のいずれかのタイプに等しく適用可能であるか、そうでなければ当該分野で公知である。標的AおよびBは、抗体が結合する任意の標的を包含し、上記標的としては、インフルエンザウイルスの種々の分離物、タイプおよびサブタイプが挙げられるが、これらに限定されない。
【0118】
上記目的が、複数の特異性を有する中和抗体を同定することである場合、交叉反応性発見選択スキームが、使用され得る。単純性という重要性において、このスキームは、二重特異性を有する抗体の選択性を示す図3において例示される。この場合、2個の標的(標的AおよびB)との反応性を示す抗体を含む抗体ライブラリーは、最初に、上記標的のうちの一方(例えば、標的A)との反応性について、続いて、他方の標的(例えば、標的B)との反応性について選択される。各連続選択ラウンドは、両方の標的に対して生じたプールの反応性強度を強化する。従って、この方法は、二重特異性を有する抗体を同定するために特に有用である。当然のことながら、上記方法は、さらなる標的に対する反応性を示す抗体を、上記さらなる標的に対する富化のさらなるラウンドを含めることによって、同定するために拡張され得る。繰り返すと、上記スクリーニングされるライブラリーが、ファージディスプレイライブラリーである場合、選択は、交叉反応性パニングによって行われるが、他のライブラリーおよび他の選択法もまた、使用され得る。
【0119】
上記で議論した2つの方法の組み合わせは、標的Aおよび標的B、それぞれに対する反応性に対する2回の別個の富化ラウンド、得られた2個のプールを組換え、その後、上記のように、交叉反応性選択ラウンドを含む。このアプローチは、図4に例示される。ちょうど純粋な交叉反応性選択におけるように、上記組換えライブラリーの各ラウンドの選択は、両方の標的に対して得られたプールの反応性強度を増大させる。
【0120】
図5に例示されるさらなる実施形態において、標的Aとの強力な反応性を示しかつ検出可能な標的Bとの検出可能な交叉反応性を有する第1のクローンが、同定される。このクローンに基づいて、変異誘発性ライブラリーが調製され、これは、次いで交互のラウンドにおいて、それぞれ、標的Bおよび標的Aとの反応性について選択される。このスキームは、標的Aとの強力な反応性を維持し、標的Bとの増大した反応性を有する抗体を生じる。直前に、選択は、上記スクリーニングされたライブラリーが、ファージディスプレイライブラリーである場合に、パニングによって行われるが、他のライブラリー、他のディスプレイ技術、よび他の選択法もまた、同じストラテジーに従って使用され得る。
【0121】
上記で議論されるように、標的AおよびBは、例えば、上記インフルエンザAウイルスの2個の異なるサブタイプ、同じインフルエンザAウイルスの2個の異なる株(単離株)、2つの異なる種からのサブタイプもしくは単離株であり得、ここで1種は、好ましくは、ヒトである。従って、例えば、標的Aは、上記H5N1ウイルスの2004年のベトナム単離株であり得、そして標的Bは、上記H5N1ウイルスの1997年の香港単離株であり得る。これら例は、単なる例示であり、任意の2個もしくは複数の標的に対して二重および複数の特異性を有する抗体は、同定され得、選択され得、類似の様式において最適化され得ることが強調される。
【0122】
一旦所望の特性を有する中和抗体が同定されると、このような抗体の大部分によって認識される優勢なエピトープを同定することが所望である得る。エピトープマッピングのための方法は、当該分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.編,Humana Press,1996;およびEpitope Mapping:A Practical Approach,Westwood and Hay,編,Oxford University Press,2001において開示されている。
【0123】
(II.特融のバーコード付与でのドナー特異的抗体ライブラリーの同定)
本発明によれば、上記抗体重鎖および軽鎖レパートリーを、上記のように調製されたcDNA(例えば、骨髄cDNA)から増幅した後、好ましくは、抗体重鎖および軽鎖ライブラリーは、各患者ドナーに関して別個にクローニングされ、ここで上記個々のライブラリーは、特融のバーコードを使用して区別され得る。
【0124】
上記バーコードは、好ましくは、所望の抗体鎖もしくはそのフラグメントの発現を妨害することなく、標識されたクローンとともに、増殖することができるように、選択される。本発明の例示的実施形態において、上記バーコードは、上記発現ベクターの配列(好ましくは、ファージミドベクターが使用される場合には、末端pIII終止コドンの後に、3’非翻訳領域)に挿入され得る。クローン選択の際に、ベクターの特融の配列が決定され、単一の規定されたライブラリーの専用のものになる。この規定されたライブラリーは、単一のドナーのみならず、ドナーの別個のプール、または合成レパートリーもしくは半合成集団にも由来し得る。別の実施形態において、上記バーコードは、上記抗体重鎖および/もしくは軽鎖またはそれらのフラグメントのコード配列に、上記それぞれの鎖の発現を妨害しない位置もしくは形態において、挿入される。
【0125】
従って、上記バーコードは、配列決定もしくは特異的PCRプライマーによってデコンボルーションされ得る、長さが約1〜24個の連続する非コードヌクレオチドの配列であり得る。この方法、核酸の集団(例えば、抗体レパートリー)は、上記クローニング工程において連結され得る。本発明の例示的実施形態において、上記バーコードは、無作為に作製された配列の3塩基もしくは5塩基である。
【0126】
別の例において、上記バーコードは、サイレント変異のコード配列である。上記ライブラリーが、中断パリンドローム(interrupted palidrome)(例えば、Sfi GGCCNNNNNGGCC)を認識する制限酵素を利用する場合、別個のヌクレオチドが、クローンの種々の集団(例えば、抗体ライブラリー)を区別するために上記「N」の代わりに組み込まれ得る。このバーコード付与アプローチは、上記増幅工程において上記レパートリーが連結されるという利点を有する。
【0127】
さらなる実施形態において、上記バーコードは、非連続ヌクレオチド配列であり、この配列は、上記ベクター配列および/もしくは上記望ましい抗体鎖のコード配列中に存在し得る。従って、非連続配列を有するバーコードは、上記種々の個々の配列の起源を同定し、それらのその後の取り扱いをモニターすることにおいて、非常に大きな融通性を提供する。
【0128】
この例において、上記バーコードは、ファージ粒子に融合された、免疫学的に別個のペプチドもしくはタンパク質配列をコードするコード配列である。例としては、例えば、pIII、pVIII、pVII、もしくはpIXファージへのエピトープ(例えば、Myc、HA、FLAG)融合物が挙げられる。上記エピトープは、単純に、もしくは種々の組み合わせにおいて提供され得、そしてシス(ライブラリーコードプラスミドにおいて)もしくはトランス(特異的に改変されたヘルパーファージ)構成において提供され得る。
【0129】
考えられるバーコードの他の例としては、ハプテンもしくは蛍光性発色団を用いた、化学的および酵素的ファージ改変(ファージライブラリーに関して)が挙げられるが、これらに限定されない。このようなタグは、単一のラウンドの選択のために好ましい。
【0130】
個体ドナーから得られた上記個々の重鎖および軽鎖ライブラリー、もしくは他のバーコード付与クローンもしくは集団が、個々の配列の供給源を区別する能力を失うことなく、プールされ得る。
【0131】
(III.ドナー特異的抗体ライブラリー由来の中和抗体の最適化)
望ましい場合、二重特異性もしくは複数の特異性を有する上記中和抗体の交叉反応性は、米国特許出願公開第20050136428号(2005年6月23日公開)(この開示は、本明細書に参考として明示的に援用される)に記載されるように、当該分野で公知の方法(例えば、ルックスルー(Look Through)変異誘発(LTM))によってさらに改善され得る。
【0132】
ルックスルー変異誘発(LTM)は、選択されたアミノ酸のコンビナトリアル変位を同時に評価しかつ最適化する多次元変異誘発方法である。上記プロセスは、1個以上の相補性決定領域(CDR)ドメイン内の正確な分布に焦点を当て、アミノ酸側鎖化学の相乗効果的寄与を調査する。LTMは、CDR内の位置的な一連の単一の変異を生成し、ここで各野生型残基は、多くの選択されたアミノ酸のうちの1個によって、体系的に置換される。変異したCDRは、全ての改変体の定量的ディスプレイを妨げることなく、増大している複雑性およびサイズのコンビナトリアル一本鎖可変フラグメント(scFv)ライブラリーを生成するために合わされる。ポジティブ選択の後、改善された特性を有するクローンは配列決定され、それらの有益な変異がマッピングされる。改善された結合特異性のための相乗効果的変異を同定するために、全ての有益な交換(permutation)を発現するコンビナトリアルライブラリー(コンビナトリアル有益変異(combinatorial beneficial mutation)、CBM)は、混合されたDNAプローブによって生成され、ポジティブ選択され、分析されて、最適化されたscFv候補のパネルを同定し得る。上記手順は、Fvおよび他の抗体ライブラリーで、類似の様式において行われ得る。
【0133】
変異誘発はまた、上記のように、ウォークスルー(walk−through)変異誘発(WTM)によって行われ得る。
【0134】
1個より多くのインフルエンザAサブタイプおよび/もしくは同じサブタイプの1個より多くの分離株で、本明細書に記載される抗体の交叉反応性を意図的に設計するための別の有用な変異誘発方法は、「指定」変異誘発として本明細書において言及される。指定変異誘発は、好ましくは、異なる反応性の1個以上の抗体クローンに基づいて、抗体の集団を合理的に操作するために使用され得る。本発明の状況において、指定変異誘発は、抗体の個々のCDRにおけるもののような類似の配列上の類似の位置によって規定される単一もしくは複数の残基をコードするために使用され得る。この場合において、これら集団は、匹敵する位置において見いだされる残基の範囲を捕捉するために、オリゴ縮重を使用して生成される。この集団内で、特異性の連続体(continuum of specificities)は、親クローンの中にもしくは親クローンを超えたところに存在することが予測される。指定変異誘発の目的は、多様な多機能性抗体集団、ライブラリーを、2個以上の別個の実体もしくは集団の間に生成することである。指定変異誘発ライブラリーを作り出すために、両方の抗体についての上記CDR配列が最初に得られ、整列される。次に、保存された同一性の全ての位置が、最初に、単一のコドンで、マッチングされた残基に固定される。保存されていない位置において、縮重コドンが組み込まれて、両方の残基をコードする。いくつかの場合において、上記縮重コドンは、この位置において2個の親残基のみをコードする。しかし、いくつかの場合において、さらなる副産物が生成される。副産物生成のレベルは測定されて、サイズ制限もしくは目的に依存して、副産物生成を強化するかもしくはこの生成を排除し得る。
【0135】
従って、例えば、2個の抗体それぞれの最初の位置がスレオニンおよびアラニンである場合、上記縮重コドン(上記最初の2個の位置においてA/G−C−)は、第3の位置における塩基に関わらず、スレオニンもしくはアラニンのみをコードする。例えば、次の位置の残基がリジンおよびアルギニンである場合、その縮重コドンA−A/G−A/Gは、リジンもしくはアルギニンをコードするのみである。しかし、その縮重コドンA/C−A/G−A/G/C/Tが使用されれば、アスパラギン、ヒスチジン、およびセリン副産物も生成される。
【0136】
便宜上、マッチングされたCDR長を有する抗体のみを使用することは、より単純である。これを推し進めるための1つの方法は、上記CDR長およびはじめに発見された抗体によって付与される潜在的に均一なフレームワーク制限に基づいて、上記第2の抗原に対するサイズ制限されたライブラリーをスクリーニングすることである。しかし、等しい長さのCDRを使用することは、便宜のためにすぎず、必要条件ではないことに注意する。この方法は、インフルエンザAウイルス中和抗体の大きな機能的に多様なライブラリーを作り出すために有用であると同時に、その適用性は、遙かに広いことを理解することは容易である。この変異誘発技術は、任意の抗体の機能的に多様なライブラリーもしくは集団を作製するために使用され得る。従って、図6は、TNF−α抗体およびCD11a抗体のCDRを、変異誘発された親配列として使用して、指定変異誘発方法の使用を例示するために本明細書に含められる。
【0137】
他の例示的変異誘発方法は、飽和変異誘発およびエラープローンPCRを含む。
【0138】
飽和変異誘発(Hayashiら,Biotechniques 17:310−315(1994))は、全ての20アミノ酸が、タンパク質中の特定の位置、および特定の表現系についてアッセイされる各改変体に対応するクローンにおいて置換される技術である(米国特許第6,171,820号;同第6,358,709号および同第6,361,974号もまた参照のこと)。
【0139】
エラープローンPCR(Leungら,Technique 1:11−15(1989);Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、ランダム点変異をクローニングされた遺伝子に導入する改変ポリメラーゼ連鎖反応(PCR)技術である。得られたPCR生成物は、無作為変異ライブラリーを生成するためにクローニングされ得るか、またはT7プロモーターが適切なPCRプライマー内に組み込まれる場合に、直接転写され得る。
【0140】
他の変異誘発技術はまた、周知であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001に記載される。
【0141】
最適化は、単独で、もしくは任意の組み合わせにおいて、上記で議論されるライブラリーのうちのいずれか、もしくは当該分野で公知のライブラリーの任意の他のタイプに基づき得る。
【0142】
(IV.中和抗体の生成)
一旦所望の中和特性を有する抗体が同定されると、このような抗体(抗体フラグメントを含む)は、当該分野で周知の方法(例えば、ハイブリドーマ技術もしくは組換えDNA技術を含む)によって、生成され得る。
【0143】
上記ハイブリドーマ方法において、マウスもしくは他の適切な宿主動物(例えば。ハムスター)が免疫されて、免疫に使用されるタンに特異的に結合する抗体を生成するもしくは生成し得るリンパ球を顕在化させる。あるいは、リンパ球は、インビトロで免疫され得る。次いで、リンパ球は、適切な融解剤(例えば、ポリエチレングリコール)を使用して骨髄腫細胞と融合されて、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0144】
このように調製されたハイブリドーマ細胞は播種され、適切な培養培地(好ましくは、融合していない、親骨髄細胞の増殖もしくは生存を阻害する1種以上の物質を含む)中で増殖させられる。例えば、上記親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTもしくはHPRT)を欠いている場合、上記ハイブリドーマ用の培養培地は、代表的には、ヒポキサンチン、アミノプテリン、およびチミジンを含み(HAT配置)、これら物質は、HGPRT欠損細胞の増殖を妨げる。
【0145】
好ましい骨髄腫細胞は、効率的に融合し、上記選択された抗体生成細胞による抗体の安定な高レベルの生成を支援し、そして培地(例えば、HAT培地)に対して感受性であるものである。これらの中でも、好ましい骨髄腫細胞株は、マウス骨髄腫株(例えば、Salk Institute Cell Distribution Center,San Diego,California USAから市販されるMOPC−21およびMPC−11マウス腫瘍から得られるもの、ならびにAmerican Type Culture Collection,Rockville,Maryland USA.から入手可能なSP−2もしくはX63−Ag8−653細胞から得られるもの)である。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株はまた、ヒトモノクローナル抗体の生成に関して記載されてきた(Kozbor,J.Immunol.133:3001(1984);およびBrodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0146】
ハイブリドーマ細胞が増殖している培養培地は、上記抗原に対して指向されるモノクローナル抗体の生成についてアッセイされる。好ましくは、ハイブリドーマ細胞によって生成されるモノクローナル抗体の結合特異性は、免疫沈降によって、またはインビトロ結合アッセイ(例えば、ラジオイムノアッセイ(RIA)もしくは酵素結合イムノソルベントアッセイ(ELISA))によって、決定される。
【0147】
組換えモノクローナル抗体は、例えば、上記必要とされる抗体鎖をコードする上記DNAを単離し、周知の組換え発現ベクターを使用して、同時発現のためのコード配列で組換え宿主細胞を同時トランスフェクトすることによって、生成され得る。組換え宿主細胞は、原核生物細胞および真核生物細胞(例えば、上記のもの)であり得る。
【0148】
上記ヒト化抗体の作製において使用されるべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性を軽減するために非常に重要である。いわゆる「ベストフィット」方法によれば、齧歯類抗体の可変ドメインの配列は、公知のヒト改変ドメイン配列のライブラリー全体に対してスクリーニングされる。上記齧歯類のものに最も近い上記ヒト配列は、次いで、上記ヒト化抗体についてのヒトフレームワーク領域(FR)として受け入れられている(Simsら,J.Immunol.151:2296(1993);Chothiaら,J.Mol.Biol.196:901(1987))。抗体は、上記抗原および他の都合のよい生物学的特性に対して高い親和性の保持とともにヒト化されることは、重要である。この目的を達成するために、好ましい方法によれば、ヒト化抗体は、上記親配列およびヒト化配列の三次元モデルを使用して、上記親配列および種々の概念的ヒト化抗体生成物の分析プロセスによって調製される。
【0149】
さらに、ヒト抗体は、当該分野で公知の方法に従って生成され得る。例えば、トランスジェニック動物(例えば、マウス)が作製され得、上記動物は、免疫の際に、内因性免疫グロブリン生成の非存在下で、ヒト抗体の完全なレパートリーを生成し得る。例えば、Jakobovitsら,Proc.Natl.Acad.Sci.USA 90:2551(1993);Jakobovitsら,Nature 362:255−258(1993);Bruggermannら,Year in Immuno.7:33(1993);および米国特許第5,591,669号、同第5,589,369号および同第5,545,807号を参照のこと。
【0150】
(V.中和抗体の使用)
本発明の中和抗体は、標的とした疾患の予防および/もしくは処置のために使用され得る。治療的適用に関しては、上記抗体もしくは他の分子(その送達は、上記抗体もしくは抗体ベースの輸送配列を使用することによって促進される)は、通常、薬学的組成物の形態において使用される。技術および処方は、一般に、Remington’s Pharmaceutical Sciences,第18版,Mack Publishing Co.(Easton,Pa.1990)において見いだされ得る。Wang and Hanson 「Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers」,Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)もまた参照のこと。
【0151】
抗体は、代表的には、凍結乾燥処方物もしくは水溶液の形態で処方される。受容可能なキャリア、賦形剤、もしくは安定化剤は、使用される投与量および濃度においてレシピエントに対して非毒性であり、緩衝化剤(例えば、リン酸塩、クエン酸塩、および他の有機酸);抗酸化剤(例えば、アスコルビン酸およびメチオニンが挙げられる);保存剤(例えば、オクタデシルジメチルベンジルアンモニウムクロリド);ヘキサメトニウムクロリド;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;アルキルパラベン(例えば、メチルパラベンもしくはプロピルパラベン);カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾール);低分子量(約10残基未満)ポリペプチド;タンパク質(例えば、ヒト血清アルブミン、ゼラチン、もしくは免疫グロブリン);親水性ポリマー(例えば、ポリビニルピロリドン);アミノ酸(例えば、グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、もしくはリジン);単糖類、二糖類および他の炭水化物(グルコース、マンノース、もしくはデキストリンを含む);キレート化剤(例えば、EDTA);糖(例えば、スクロース、マンニトール、トレハロースもしくはソルビトール);塩形成対イオン(例えば、ナトリウム);金属錯体(例えば、Zn−タンパク質錯体);および/または非イオン性界面活性剤(例えば、TWEENTM、PLURONICSTMもしくはポリエチレングリコール(PEG))が挙げられる。
【0152】
上記抗体はまた、例えば、コアセルベーション技術によって、もしくは界面重合化によって調製されるマイクロカプセル(例えば、それぞれ、ヒドロキシメチルセルロースもしくはゼラチンマイクロカプセル、およびポリ−(メチルメタクリレート)マイクロカプセル)中に、コロイド性薬物送達系(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)中に、またはマクロエマルジョン中に、捕捉され得る。このような技術は、Remington’s Pharmaceutical Sciences 第16版,Osol,A.編(1980)で開示されている。
【0153】
本明細書で開示される中和抗体はまた、イムノリポソームとして処方され得る。上記抗体を含むリポソームは、当該分野で公知の方法(例えば、Epsteinら(1985)Proc.Natl.Acad.Sci.USA 82:3688;Hwangら(1980)Proc.Natl Acad.Sci.USA 77:4030;米国特許第4,485,045号;同第4,544,545号;およびWO 97/38731(1997年10月23日公開)に記載される)によって調製される。増強された循環時間を有するリポソームは、米国特許第5,013,556号に開示されている。
【0154】
特に有用なリポソームは、ホスファチジルコリン、コレステロールおよびPEG誘導体化ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物との逆相エバポレーション法によって生成され得る。リポソームは、規定の孔サイズのフィルタを通して押し出されて、所望の直径を有するリポソームを生じ得る。本発明の抗体のFab’フラグメントは、ジスルフィド交換反応を介して、Martinら(1982)J.Biol.Chem.257:286−288に記載されるように、リポソームに結合体化され得る。化学療法剤は、必要に応じて、上記リポソーム内に含まれる。Gabizonら(1989)J.National Cancer Inst.81(19):1484を参照のこと。
【0155】
疾患の予防もしくは処置については、抗体の適切な投与量は、処置されるべき感染のタイプ、上記疾患の重篤度および経過、上記抗体が予防目的で投与されるのか、治療目的で投与されるのか、に依存する。上記抗体は、上記患者に一度に、もしくは一連の処置にわたって、適切に投与される。上記疾患のタイプおよび重篤度に依存して、約1μg/kg〜15mg/kgの抗体が、例えば、1回以上の別個の投与によろうが、または連続注入によろうが、上記患者への投与のための最初の候補投与量である。
【0156】
本発明のさらなる詳細は、以下の非限定的実施例によって例示される。
【実施例】
【0157】
(実施例1:以前のトリインフルエンザ大発生の生存者からの抗体ライブラリーおよび中和抗体の調製)
(材料および骨髄プロトコルおよび血清調製)
血液を、標準的な静脈穿刺(venopuncture)によって得、凝固させ、処理して血清を回収した。上記血清を、ドライアイス上で輸送されるまで、−20℃で3〜4日間保存した。ドナーを局所麻酔の注射で麻酔し、5mlの骨髄を、各H5N1生存者の骨盤から回収した。次に、上記5mlの骨髄を、45ml RNAlater(Ambion)を含む滅菌50mlチューブに入れた。上記混合物を、眼に見える凝集がなくなり、かつ上記骨髄およびRNAlaterが十分に混合されるまで、約8〜20回穏やかに転倒混和した。次に、上記標本を、2〜10℃の間で一晩冷蔵した。一晩冷蔵した後、上記標本を、ドライアイス上で輸送されるまで、−20℃において3〜4日間保存した。上記RNAlater/marrowを受け取ったら、上記血清含有チューブを、処理するまで−80℃で保存した。
【0158】
(血清学:HA ELISA)
ELISAプレート(Thermo,Immulon 4HBX 96W)を、1×ELISAプレートコーティング溶液(BioFX)中の100μlの100ng/mL H5ヘマグルチニン(Protein Sciences,A/ベトナム/1203/2004)で、室温において一晩インキュベーションすることによって、コーティングした。翌日、プレートを300μl PBS/0.05% Tween−20(PBST)で3回洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05% Tween−20中の4% 無脂肪乾燥乳)を添加し、室温において1時間インキュベートした。ブロッキング工程の後、上記プレートを、300μl PBS/0.05% Tween−20で3回洗浄した。次に、PBS/0.05% Tweenにおいて1:20,000希釈した100μl 血清サンプルを、室温において1=2時間インキュベートし、次いで、300μl PBS/0.05% Tween−20で3回洗浄した。PBS/0.05% Tweenにおいて1:5,000希釈した100μlの抗ヒトFc−HRP結合体を、室温において1〜2時間インキュベートし、次いで、300μl PBS/0.05% Tween−20で3回洗浄した。この最後の洗浄の後、100μlの色素生成基質溶液を添加し(TMB1基質,BioFx)、十分な時間の後、100μlの停止溶液(BioFx)の添加によって終了させた。450nmの吸光度をプレートリーダー(Softmax Proソフトウェアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、その後、Excel(Microsoft)を使用してプロットした。
【0159】
(骨髄:RNA抽出およびmRNA精製)
予め−80℃で保存しておいた骨髄(20ml RNAlater中約2.5ml)を遠心分離によって回収して、RNAlaterを除去し、次いで、300μl 酢酸を含む11.25ml TRI BD試薬(Sigma)中に再懸濁した。次いで、上記ペレットを、激しくボルテックスした。次に、1.5ml BCP(1−ブロモ−3−クロロプロパン,Sigma)を添加し、ボルテックスによって混合し、室温において5分間インキュベートし、次いで、12000×gにおいて15分間4℃で遠心分離した。その界面を廃棄しないように、上記水相を注意深く取り出した。水相からの総RNAを、次に、25ml イソプロパノールを添加することによって沈澱させ、室温において10分間インキュベートし、12000×gにおいて10分間4℃で遠心分離した。イソプロパノールを添加した後、残りのRNAlaterに起因して、2相を形成し、界面に位置した沈澱したRNAを生じた。上記残りのRNAlaterを排除し、RNAの最大の回収を可能にするために、H2O中の50% イソプロパノールの5mlのアリコートを添加し、相分離が認められなくなるまで混合し、その時点で、上記RNAを、12000×gにおいて10分間にわたって4℃で遠心分離によってペレット化した。上記RNAペレットを、75% EtOHで洗浄し、RNAseなしの1.6ml 微小遠心チューブに移し、遠心分離によって再び回収した。最後に、上記RNAペレットを、100μlの1mM リン酸Na,pH8.2中に再懸濁し、A260およびA280を読み取って、RNA純度を評価した。
【0160】
逆転写の前に、mRNAを、Qiagen Oligotex mRNA精製キットに従って、総RNAから精製した。簡潔には、50〜200μg 骨髄RNAを、RNase非含有水と一緒にして250μlにし、250μlのOBB緩衝液およびOligotex懸濁物と混合、続いて、3分間にわたって70℃においてインキュベートした。上記Oligotex粒子の上記オリゴdT30と上記mRNA ポリA−テールとの間のハイブリダイゼーションを室温において10分間行った。次いで、上記ハイブリダイズした懸濁物を、スピンカラムに移し、1分間遠心分離した。上記スピンカラムを400μl 緩衝液OW2で2回洗浄した。次いで、精製したmRNAを、20μlの熱い(70℃)緩衝液OEBと遠心分離することによって、2回溶出した。代表的な収量は、1.5μg 総RNAに対して500ngであった。
【0161】
(骨髄mRNAに対してN9およびオリゴdTを使用する逆転写)
逆転写(RT)反応を、75〜100ng mRNAと、2μl 10×Accuscript RT緩衝液(Stratagene)、0.8μl 100mM dNTP、およびeither N9(300ng)もしくはオリゴdTプライマー(100ng)を一緒に混合することによって達成し、次いで、水で最終容積17μlにした。上記混合物を、65℃において5分間加熱し、次いで、室温へと冷却した。次に、2μl DTT、0.5μl RNase Block(Stratagene)、0.5μl AccuScript RT(Stratagene)を、各反応に添加した。次に、上記N9刺激した反応系を、10分間にわたって室温でインキュベートし、上記オリゴ−dT刺激した反応系を、氷上で10分間にわたってインキュベートした。最後に、両方の反応系を、42℃において60分間、続いて、70℃において15分間インキュベートして、上記酵素を失活させた。
【0162】
(骨髄由来cDNAからのPCR)
抗体重鎖および軽鎖レパートリーを、ヒト生殖系列VおよびJ呂言う基に基づいて、以前に記載された方法および縮重プライマーを本質的に使用して(O’Brien,P.M.,Aitken R. Standard protocols for the construction of scFv Libraries. Antibody Phage Display-Methods and Protocols,vol.178,59−71,2001,Humana Press)、骨髄cDNAから増幅した。
【0163】
簡潔には、オリゴdT刺激cDNA(λ軽鎖に対して75ng mRNAから)およびN9刺激cDNA(κ軽鎖に対して75ng mRNAから、重鎖に関して100ng mRNAから)を使用して、PCR反応を、5μl 10×増幅緩衝液(Invitrogen)、1.5μl dNTP(10mM)、1μl MgSO4(50mM)、2.5μl V領域プライマー(10μM)および2.5μl J領域プライマー(10μM) VHに関して−10μM、0.5μl Platinum Pfxポリメラーゼ(Invitrogen)、および滅菌dH2Oと一緒に混合して、最終容積50μlにした。PCRパラメーターは、以下のとおりであった:工程1−95℃、5分間、工程2−95℃、30秒、工程3−58℃、30秒、工程4−68℃、1分間、工程5−工程2〜4を40回サイクルさせる、工程6−68℃、5分間。軽鎖PCR生成物を、Qiagen PCR Cleanupキットを使用してきれいにした。重鎖PCR生成物を、Qiagen Gel Extraction Kitを使用して、1.5% アガロースゲルから精製し、次いで、再増幅した。重鎖再増幅を以下のようにおこなった:10μl 10×増幅緩衝液(Invitrogen)、3μl dNTP(10mM)、2μl MgSO4(50mM)、5μl 各VHプライマー(10μM)およびJHプライマー(10μM)、5μl 重鎖プライマーPCR生成物、1μl Platinum Pfxを混合し、水で容積を100μlに調節した。サイクリングパラメーターは、以下のとおりであった:工程1−95℃、5分間、工程2−95℃、30秒、工程3−58℃、30秒、工程4−68℃、1分間、工程5−工程2〜4を20回サイクルさせる、工程6−68℃、5分間。再増幅した重鎖PCR生成物を、Qiagen Extraction Kitを使用して、1.5% アガロース−TAEゲルからきれいにした。
【0164】
(抗体ファージライブラリー構築)
各個々のトリインフルエンザ生存者についての別個の抗体ライブラリーを、繊維状ファージのpIII遺伝子の終止コドンの後ろの非翻訳領域において挿入した特融識別3−ヌクレオチドバーコードを使用して構築した。
【0165】
軽鎖クローニング:
ドナー1名につきプールしたκ軽鎖およびプールしたλ軽鎖の各1μgを、NotIおよびBamHIで消化し、Qiagen Gel Extraction Kitを用いて1.5% アガロース−TAEゲルからゲル精製した。5μgの各ベクター(pAMPFab)を、NotIおよびBamHIで消化し、Qiagen Gel Extraction Kitを使用して、1% アガロース−TAEゲルからゲル精製した。ライブラリーライゲーションを、室温において60μl中で1時間にわたって、もしくは14℃において一晩にわたって、200ngのゲル精製したκ挿入物もしくはλ挿入物および1μgのゲル精製したベクターで行った。ライゲーションを、Edge BioSystem Perfromaスピンカラムを使用して脱塩した。上記ライブラリーを、80μl TG−1もしくはXL−1 Blueアリコート中の5個のエレクトロポレーションに移し、各々、1ml SOC中に回収し、プールし、1時間にわたって37℃において増殖させた。形質転換体の総数を、上記形質転換体の各々からのアリコートをプレートすることによって、この増殖の後に決定した。その残りのエレクトロポレーションを、200ml 2YT+50μg/ml アンピシリン+2% グルコース中で、37℃において一晩増殖させることによって増幅させた。その後の軽鎖ライブラリーを、これらの一晩培養物から、Qiagen High Speed Maxiprep Kitを使用して、プラスミド精製によって回収した。
【0166】
(重鎖クローニング):
上記ドナー特異的重鎖(VH1、VH2、5、6プール、VH3、およびVH4)の1.5〜2μgの各々を、40ユニット過剰/μg DNAと、SfiIおよびXhoIで消化し、Qiagen Gel Extraction Kitを使用して、1.5% アガロース−TAEゲルからゲル精製した。15μgの各軽鎖ライブラリーベクターを、40ユニット/μg DNAと、SfiIおよびXhoIで消化し、Qiagen Gel Extraction Kitを使用して、1% アガロース−TAEゲルからゲル精製した。ライブラリーライゲーションを、消化した1.2μg SfiI/XhoI、ゲル精製重鎖ドナー集団および5μgの各軽鎖ライブラリー(κおよびλ)を一晩4℃において合わせることによって設定した。次いで、上記ライブラリーライゲーションを、Edge BioSystem Pefromaスピンカラムで脱塩し、次いで、80μl TG−1アリコート中、20エレクトロポレーション/ライブラリーを介して形質転換し、各々、1ml SOC中に回収し、プールし、37℃において1時間増殖させた。再び、この増殖の後、各々のアリコートを、1L 2YT+50μg/ml アンピシリン+2% グルコースに移した残りと一緒に、形質転換体の総数を決定するために使用し、約0.3のOD600になるように、激しく曝気して37℃において増殖させた。次に、M13K07ヘルパーファージを、感染多重度(MOI)5:1で添加し、攪拌せずに、37℃において1時間インキュベートした。次に、上記細胞を、遠心分離によって採取し、1L 2YT+50μg/ml アンピシリン、70μg/ml カナマイシン中で再懸濁し、激しく曝気しながら37℃において一晩増殖させて、scFvファージミド生成を可能にした。翌朝、上記細胞を遠心分離によって集め、ファージミドを含む上清を集めた。上記ファージミドを、0.2容積の20% PEG/5M NaCl溶液を添加し、1時間氷上でインキュベート刷ることによって、上記上清から沈澱させた。次いで、上記ファージミドライブラリーストックを、遠心分離によって回収し、20ml 滅菌PBS中に再懸濁した。残りの細菌を、さらなる遠心分離によって回収し。最終ファージミドライブラリーを、−20℃において、PBS+50% グリセロール中で保存した。
【0167】
(ファージミドパニングおよび増幅)
ELISAプレート(Immulon 4HBX平底,Nunc)を、室温において一晩のインキュベーションによって、ELISAコーティング溶液(BioFX)中、100μlの100ng/mL H5 ヘマグルチニンタンパク質(Protein Sciences,A/ベトナム/1203/2004)でコーティングした。翌日、プレートを、300μl PBSTで3回洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05% Tween−20中4% 無脂肪乾燥乳)を添加し、30分間、氷上でインキュベートした。上記ブロッキング工程の後、上記プレートを、300μl PBSTで3回洗浄した。ファージパニングの直前に、凍結したファージミドストックから、Millipore Amicon Ultraカラムを使用して、上記グリセロールを除去し、次いで、4% 無脂肪乾燥乳中、15分間にわたってブロッキングした。次に、ファージミドの100μl アリコートを、8ウェル(総ファージ約1×1012CFU)へと分配し、4℃において2時間にわたってインキュベートし、続いて、300μl PBSTで6〜8回洗浄した。ファージミドを、100μl/ウェル中、室温において10分間後にElution buffer (0.2M グリシン−HCl,pH2.2、1mg/ml BSA)中に回収した。次いで、上記溶出物を、溶出物1mlあたり56.25μl 2M トリス塩基を添加することによって中和した。中和後、5ml TG1細胞(OD600 約0.3)に、0.5ml 中和ファージを、37℃において30分間にわたって、2−YT中、振盪することなく感染させた。この工程の後、いくつかの細胞を、LB AMPグルコースプレート上にプレートして、全てのファージミド回収を決定した。残りの接種物を、10ml 2−YTAG(最終濃度2% グルコースおよび50ug/ml アンピシリン)へ入れ、OD600が約0.3になるまで37℃において激しく曝気しながら増殖させた。次に、上記培養物に、5:1のMOIでM13K07ヘルパーファージを感染させ、37℃において30〜60分間にわたって、振盪することなくインキュベートした。上記細胞を遠心分離によって集め、25ml 2−YTAK(アンピシリン 50μg/ml、カナマイシン 70μg/ml)中に再懸濁し、新しい培養フラスコに移し、振盪しながら37℃において増殖させ続けた。その後のラウンドは、同様に回収し、増幅させた。
【0168】
(scFv ELISA)
生体パニングしたファージからのE. coli HB2151形質転換細胞の個々のコロニーを、1mlの2YT+100μg/ml AMP中、37℃において一晩増殖させた。翌朝、上記細胞を遠心分離によって回収し、1.5ml ペリムラズム溶解緩衝液(1ml BBS(Teknova)+0.5ml 10mg/ml リゾチーム+10mM 終濃度になるまでEDTA)中に再懸濁した。上記細胞を、遠心分離によって再びペレット化し、上記scFvを含むペリムラズム溶解物を回収した。上記scFv溶解物を、希釈緩衝液(PBS/0.05% BSA)と1:1で合わせ、100μlを、予め抗原コーティングし、希釈緩衝液でブロッキングしたウェルに添加した。上記サンプルを、2時間にわたって室温においてインキュベートし、次いで、PBS/0.05% Tweenで3回洗浄した。次に、100μlの希釈緩衝液中で1:5000希釈したビオチン抗ヒスチジンマウス(Serotec)を各ウェルに添加し、室温において1時間インキュベートした。このインキュベーションの後、上記ウェルを、PBS/0.05% Tweenで3回洗浄し、次いで、各ウェルに、100μlの1:2500 ストレプトアビジン:HRP(Serotec)を添加し、室温において1時間インキュベートし、次いで、PBS/0.05% Tweenで3回洗浄した。この最後の洗浄の後、100μlの色素生成性基質溶液を添加し(TMB1基質,BioFx)、十分な時間の後、100μlの停止緩衝液(BioFx)を添加することによって集結させた。450nmの吸光度を、プレートリーダー(Softmax Proソフトウェアを装備したMolecular Devices Thermomaxマイクロプレートリーダー)で読み取って、データを記録し、その後、Excel(Microsoft)を使用してプロットした。
【0169】
(配列決定)
上記重鎖および軽鎖配列を推測するために、個々のクローンを増殖させ、プラスミドDNA抽出した(Qiagen)。上記プラスミドDNAを標準的DNA配列決定に供した。
【0170】
(ヘマグルチニン阻害(HAI)アッセイ)
ヘマグルチニン阻害を、丸底マイクロタイタープレート(Corning)において、4 HAU(赤血球凝集単位)のウイルスもしくはタンパク質/ウェルを使用して、Rogersら,Virology 131:394−408(1983)の方法に本質的に従って行った。HAI決定のために、精製した一本鎖可変フラグメント(scFv)の25μl サンプルを、各マイクロタイターウェルにおいて、上記試験ウイルスの4 HAUを含む25μlのPBSと混合した。室温において15分間のプレインキュベーションの後、25μlの0.75% ヒト赤血球を添加し、混合した。HAI抗体活性を、室温での60分間のインキュベーション後の視覚的検査によって決定した。
【0171】
特に、以下のプロトコルを使用した:
抗体、タンパク質およびウイルス
IgG1タンパク質を、pCIベースの(Promega)全長哺乳動物タンパク質発現ベクターへとそれらのコード領域をサブクローニングすることによってscFvもしくはFabのいずれかから生成し、次いで、293 Freestyle細胞(Invitrogen)へと、製造業者のガイドラインに従ってトランスフェクトした。簡潔には、20μgの軽鎖および10μgの重鎖をコードするプラスミドを、1.0ml 293fectinと合わせ、60分間インキュベートした。このプレインキュベーションの後、上記DNA混合物を、30ml 培地中で、3×107細胞と合わせ、その得られた細胞懸濁物を、7日間にわたって、製造業者の示唆に従って増殖させた。7日後に、上記分泌免疫グロブリンを、プロテインAクロマトグラフィー(Calbiochem)を使用して、培養上清から精製した。その得られた精製抗体を、遠心分離サイズ分画(Centricon Plus−20)を使用して滅菌PBSへと緩衝液交換し、それらのタンパク質濃度を、比色BCAアッセイ(Pierce)によって決定した。
【0172】
(交叉反応IgG ELISA)
マイクロタイタープレートを、コーティング緩衝液中で希釈した0.1mlの以下の抗原でコーティングし、室温において一晩インキュベートした:100ng/ml H5N1 ベトナム 1203/04、250ng/ml H5N1 トルコ/65596/06、1μg/ml H5N1 インドネシア/5/05、700ng/ml H1N1 ニューカレドニア/20/99、1μg/ml H1N1 ノースキャロライナ/1/18、100ng/ml and H3N2 ウィスコンシン/67/05。ブロッキングを、0.3mlのブロッキング緩衝液(PBS/0.05% Tween−20中4% 無脂肪乾燥乳)で行った。ブロッキングの後、2% 無脂肪乾燥乳ブロッキング緩衝液中、0.5μg/mlに希釈した抗体を、4℃において2時間にわたってインキュベートし、洗浄し、後に、2% 無脂肪乾燥乳ブロッキング緩衝液中1:3000希釈のペルオキシダーゼ結合体化抗ヒトFc抗体(Jackson ImmunoResearch)および標準的TMB基質検出(BioFX)を使用して検出した。450nmの吸光度を読み取り、データを記録し、本明細書において報告した。
【0173】
(ウイルス微小中和(microneutralization)
インドネシアおよびトルコのヘマグルチニン遺伝子はを、ヒトコドン最適化配列(DNA 2.0)を使用して合成的に組み立て、次いで、これを使用して、組換え操作ウイルスを生成した。組換えインフルエンザウイルスを、以前に記載されるように(Fodor,E.ら J Virol.1999;73(11):9679−82)、逆遺伝子学を使用して生成した。簡潔には、10個のプラスミドの1μg 各々を、単層において293 T細胞へとトランスフェクトした。各トランスフェクションは、vRNA(pPOL1タイプ)およびA/ベトナム/1203/04 HAおよびNA(pCAGGS発現プラスミドは、J.Miyazaki,Osaka University,Osaka,Japan)(Miyazaki,J.ら Gene 1989;79(2):269−77)によって供与)に関しては、タンパク質発現プラスミド(pCAGGS type)に加えて、A/プエルトリコ/8/34/PA、PB1、PB2、NP、M、およびNSセグメントに関して、両センス(ambisense)プラスミド(vRNAおよびmRNA両方の発現に関して)を含んでいた。トランスフェクションの20時間後、293T細胞を、細胞培養上清中に再懸濁し、これを使用して、10日齢受精卵に接種した。
【0174】
抗体を、以下のように、ウイルスに対する中和活性についてスクリーニングした。各Mabの2倍連続希釈物を、PBS中のウイルスの100 TCID50とともに、37℃において1時間インキュベートした。24ウェルプレート中のMadin−Darbyイヌ腎臓細胞単層をPBSで1回洗浄し、ウイルス−抗体混合物とともにインキュベートした。5% CO2中で37℃において1時間インキュベートした後、上記接種物を除去し、単層を、PBSでもう1回洗浄した。0.3% BSA、0.01% FBSおよび1μg/ml TPCK処理トリプシンを補充したOpti−MEMを添加し、細胞を、37℃において72時間にわたってインキュベートした。細胞培養上清中のウイルスの存在は、0.5% ニワトリ赤血球を使用して、HAアッセイによって評価した。
【0175】
(結果)
骨髄および血液サンプルを、2006年1月にトルコで起こったH5N1トリインフルエンザ大流行(大流行後約4ヶ月)の6名の生存者から集めた。6名全ての生存者について、トリインフルエンザの初期診断は、物理的検査、臨床実験室的試験、および分子診断測定によってなされ、Turkish Ministry of Healthによって確認された。これら生存者のうちの4名は、世界保健機関(WHO)によってさらに確認された。血清サンプルを分析して、上記の血清学プロトコルを使用して、H5 ヘマグルチニン(A/ベトナム/1203/2004)に対する抗体の存在を確認した。図8に示されるように、6名の患者全ての上記血液サンプル(それぞれ、SLB H1〜H6と称した)は、上記H5抗原に対する抗体の存在を示した。この確認後に、RNAを、これら個体の骨髄サンプルから抽出し、骨髄mRNAを精製し、上記のプロトコルを使用して逆転写した。次いで、上記抗体重鎖および軽鎖レパートリーを、上記のように、上記骨髄cDNAから増幅して、個々の抗体重鎖および軽鎖ファージライブラリーを、上記の3ヌクレオチドバーコード付与を使用して、各生存者について別個にクローニングして、個々のライブラリーを区別した。
【0176】
このベクターとそのコード付与系とを使用して、本発明者らは、一本鎖(scFv)およびFabファージミド両方の形式において6名の生存者のうちの5名の骨髄に由来するレパートリーを首尾よくクローニングした。個々の生存者に由来する各集団は、1.0×108メンバーより大きい多様性を有する。さらに、本発明者らは、混合した生存者軽鎖および重鎖から構成される、採取的な多様性が1.1×109のさらなるバーコード付与ライブラリーを作り出した。まとめると、上記5個のドナー特異的集団および全てのドナーからの上記プールしたライブラリーは、scFv集団として1.0×109およびFabディスプレイ集団として4.2×109の全体多様性を有する(表4)。
【0177】
表2は、scFvおよびFab両方の形式における軽鎖および全ライブラリー総形質転換体を示す。全ライブラリーによって表される全体多様性は、5.6×109である。
【0178】
【表2】
骨髄および血液サンプルをまた、2006年のインフルエンザ症状に対して処置した12名のその土地のドナーから集めた。上記のように血清学を行って、H1、H3およびH5 ヘマグルチニン、それぞれに対する抗体の存在を確認した。図8に示されるように、全ての血清サンプルを試験したところ、H1および/もしくはH3ヘマグルチニンに対する抗体について陽性であった。ここで特定のサブタイプの優性は、インフルエンザAウイルスサブタイプに依存し、このサブタイプに対して、上記特定のドナーは、彼もしくは彼女のほぼ人生全体を通して曝された。興味深いことに、血清がH5ヘマグルチニンの抗体の顕著なレベルも含んでいたドナー(図9においてドナーSLB1およびSLB5)が存在した。この確認後、RNAを、上記ドナーの骨髄サンプルから抽出し、骨髄mRNAを精製し、上記のプロトコルを使用して逆転写した。上記抗体重鎖および軽鎖レパートリーを、次いで、上記骨髄cDNAから上記のように増幅し、個々の抗体重鎖および軽鎖ファージライブラリーを、上記の3ヌクレオチドバーコード付与を使用して各ドナーについて別個にクローニングして、個々のライブラリーを区別した。
【0179】
(結合抗体の選択)
上記H5N1生存者ライブラリーを、表2にまとめ、ベトナム/1203/04ウイルスHAおよびNAタンパク質もしくは組換え精製ヘマグルチニン(Barbas,C.ら(2001)Phage Display,A Laboratory Manual(Cold Spring Harbor Laboratory Press))を含む不活性化ウイルスに対してパニングした。3〜4ラウンドのファージパニングの後、富化ファージプールからの個々のコロニーを、H5N1ウイルスもしくは精製ヘマグルチニンに対するELISAによって分析し、上記ポジティブクローンを配列決定して、それらの重鎖および軽鎖軽鎖配列を決定し、それらの生存者バーコードを読み取った(D.W.Coomber,Methods Mol Biol 178,133(2002))。このことから、本発明者らは、上記個々の生存者ライブラリーのうちの5個全てから、特定のH5ヘマグルチニン結合クローンを単離した。合計して、300,000個より多い種々の抗ウイルス抗体が回収され、そのうちの146個が、上記H5ヘマグルチニンタンパク質を特異的に結合する。
【0180】
(選択したクローンの一般的特徴)
全体として、上記個々のドナーは、重鎖および軽鎖両方に関して異なる生殖系列を利用し、このことは、個々の患者が、同じ潜在的に致死的な免疫学的攻撃に対して異なる解決策を見いだしたことを示している。得られたレパートリーの質に関する信頼を与えかつ抗体結合および/もしくは中和の化学に関する情報を与えるために使用され得るコンビナトリアル抗体ライブラリーの主要な特徴は、これらクローンにおいて認められる。これらクローンは、親和性成熟の天然の発達において、同様に選択された合成抗体ライブラリーにおいても見いだされる抗原結合に対する以前に記載された反復クローン(「ジャックポットソリューション(jackpot solution)」)の顕著な特徴を全て含む(Lerner,R.A.Angew Chem Int Ed Engl 45,8106(Dec 11,2006);A.Rajpalら,Proc Natl Acad Sci USA 102,8466(Jun 14,2005))。これら大きな集団における「ジャックポット」の存在は、上記スクリーニング手順を確認する。なぜなら、上記ファージが活性に基づいて選択されなければ、同じくローンを複数回得るという機会は、起こり得る可能性が非常に少ないからである。さらに、群内の上記重鎖の差異を分析する場合、上記アミノ酸置換の多くが、化学的にかつ構造的に保存的であることが観察された(表4)。反復クローンのように、化学的に妥当な複数のアミノ酸置換の出現は、無作為事象である可能性は低い。
【0181】
(回収された抗体の結合特異性)
6個のクローンを、全IgG1タンパク質への変換に対して異なる2個のエピトープを認識した3名の生存者から選択した。これら抗体のうちの4つの結合を、上記ヘマグルチニンタンパク質のHA1サブユニットにマッピングしたが、その残りの2つは、マッピングしなかった。上記HA1サブユニットは、感染において非常に関連しているので、本発明者らは、それら4個の抗体の活性を、以下に記載されるようにさらに分析した。
【0182】
これら研究の1つの目的は、基準から外れた(divergent)ウイルス株を広く中和する稀な抗体を回収することであった。本発明者らの抗体のうちのいくつかは、広く反応性であり得ることが示唆された。なぜなら、上記ドナー由来の血清は、彼らが感染したウイルスを超えて拡がって、基準から外れたサブファミリーのH5N1ウイルスに対して高い力価の抗体を有したからである。個々の抗体のレベルにおける交叉反応性の程度を決定するために、本発明者らは、異なるインフルエンザヘマグルチニン抗原に対する本発明者らのクローンの結合を最初に分析した(図11)。驚くべきことではく、これら抗体は、トルコ/65596/06株に感染する対応するものに由来するヘマグルチニンを認識し、そしてさらに、選択のために使用されたベトナム/1203/04株に由来する異種ヘマグルチニンを認識する。さらに、それらは、抗原的に基準から外れたインドネシア/5/05 H5ヘマグルチニンを認識する。さらに、本発明者らは、4個のプロトタイプ抗体が、密接に関連したサブタイプH1N1最新の基準株ニューカレドニア/20/99に由来するヘマグルチニンを結合しることを見いだした。顕著なことには、生存者5に属する上記3個の中和抗体はまた、1918年のスペイン風邪世界的流行の間に現れたH1N1 サウスキャロライナ/1/18単離株に由来するヘマグルチニンを結合した。逆に言えば、これら4個の抗体のうちのいずれも、最新のH3N2 ウィスコンシン/67/05基準株に由来するヘマグルチニンを結合せず、このことは、上記抗体ディスプレイが、インフルエンザサブタイプの間で広いスペクトルで結合するとしても、上記反応性は、全てのインフルエンザサブタイプに拡がらなかったことを示す。図11は、2名の生存者に由来するH5N1抗体とH1N1ウイルス由来のヘマグルチニンとの交叉反応性を示す。(A)棒は、H5N1 ベトナム 1203/04(暗い灰色)、H5N1 トルコ/65596/06(白)、H5N1 インドネシア/5/05(斜線)、H1N1 ニューカレドニア/20/99(縦縞)、H1N1 サウスキャロライナ/1/18(クロスハッチストライプ)、およびH3N2 ウィスコンシン/67/05(明るい灰色)である。
【0183】
(中和研究)
最初に、上記抗体を、H5 HA(ベトナム/1203/04)含有インフルエンザウイルスを中和するそれらの能力について評価した。共通するエピトープ(エピトープ「A」)を認識した生存者2および3、生存者5に由来する1つの抗体は、全て中和したのに対して、第2のエピトープ(エピトープ[B])を認識した生存者1に由来する2つの抗体は、中和しなかった。
【0184】
H5N1もしくはH1N1 ヘマグルチニンいずれかに対する生存者5から別個に単離されたクローンの顕著な配列類似性に基づいて、本発明者らは、それらの交叉反応性は、単純な結合を超えて拡がり、それらがまた、H5N1ウイルスおよびH1N1ウイルスの両方を中和するという非常にまれな特性を有すると推測した。上記IgGの交叉中和活性を試験するために、本発明者らは、中和アッセイにおけるH5N1スクリーニングからの代表的な抗体を試験して、抗体がまた、H1N1ウイルスもしくはH3N2ウイルスを中和するか否かを調べた(表3)。本発明者らは、H1保有ウイルスA/ニューカレドニア/20/99およびH3保有ウイルスA/香港/68を研究した。H5サブタイプヘマグルチニンを保有するウイルスの集団はまた、以下を試験した:(A/ベトナム/1203/04;A/インドネシア/5/05;A/トルコ/65596/06;A/エジプト/06)。上記抗体は、H3サブタイプインフルエンザに対して何ら活性を示さなかった。しかし、H5含有ウイルスを中和した上記モノクローナル抗体(1〜3)のうちの3つはまた、サブタイプH1からHAを保有するウイルス全てを強く中和した(表5)。
【0185】
【表3】
(中和の免疫化学的基準)
抗体ライブラリーの1つの利点は、多数の抗体を得る場合に、これらがそれらの関連性に関してグループ分けされ得ることである。従って、上記集団における所定の抗体についての機能が観察される場合、所定の抗体が属する上記群の他のメンバーが類似の活性を有することを推定し得る。
【0186】
表4は、H5N1 ベトナムパニングの後の、生存者5ライブラリーから発見された中和の免疫化学的基礎を示す例示的配列を示す。上記61個の特融の重鎖配列を、114個の特融の重鎖および軽鎖組み合わせに由来するそれらの生殖系列可変領域と整列させた。必須の変異は、太字、下線を付したテキスト(列5-PIからGMおよびAからT;列6-KSからELもしくはEMもしくはXL)で示され、優勢な変異は、斜体、下線を付したテキスト(列2-AからT;列3-ISからVT;列5-GからA;列8-KからQもしくはR)で示される。H1N1 ニューカレドニアパニングにおいて発見された重鎖配列はまた、灰色で強調される。抗体領域およびKabat番号付け範囲は、各配列の列の上に列挙される。
【0187】
【化1】
【0188】
【表4−1】
【0189】
【表4−2】
生存者5に由来するエピトープ「A」に対する上記中和抗体集団を含んだ上記群のメンバーは、表4に示される。上記群は、上記VH1e生殖系列重鎖に最も近く似ている61個の特有のメンバーから構成される。いくつかの重鎖は、1個より多くの軽鎖と対形成される。合計で、これら重鎖は、κおよびλ両方の軽鎖に対する114個の特有の対形成を有する。これら重鎖と、上記非常に関連するVH1e生殖系列とを比較すると、本発明者らは、点置換の3つのタイプを観察する。いくつかの変化は、必要とされるようであり、他は優性であり、そしていくつかの残基は、散発的にのみ変化した。必要とされる変化は、位置52A(Pro>Gly)、53(Ile>Met)、および57(Ala>Thr)においてCDR2内の群において、ならびに位置73(Lys>Glu)および74(Ser>LeuもしくはMet)においてフレームワーク3領域において、あらゆるクローンで生じ、このうちの全てが、生殖系列側鎖化学から変化し、このことは、上記変異が抗原結合および中和に必須であることを示唆する。変異の上記第2のセットは優性であり、大部分のクローンにおいて見いだされる。最初は、位置24(Ala>Thr)においてフレームワーク1において、顕著な化学的変化を表す。次の3個は、位置34(Ile>Val)および位置35(Ser>Thr)においてCDR中に、ならびに位置50(Gly>Ala)においてCDR2中にも保存的変化がある。しかし、これら優性な置換のうちの4個全ては、なくてもよい。このことは、上記置換が、有益である一方で、必須ではないことを示唆する。フレームワーク領域1、3、および4、ならびにCDR3の全体を通じて見いだされる上記散発性の変化は、全て保存的であり、おそらく小さな最適化事象を表す。
【0190】
図12は、47eといわれる非常に関連する抗HIV Fab(1rzi.pdb)(Huang,C.C.ら(2004)Proc.Nat.Acad.Sci.101,2706−2711)の結晶構造に対して重ね合わせられた抗体の構造における必要とされる変異の位置を示す。図12は、必要とされるH5 ヘマグルチニン結合群1の位置およびFab 47eの結晶構造上の優性な変異を示す。上記必要とされる変異は、G52(52A(Pro>Gly))、M53(Ile>Met)、T57(Ala>Thr)、E73(Lys>Glu)およびLM74(Ser>LeuもしくはMet)として示される。上記優性な変異は、T24(Ala>Thr)、V34(Ile>Val)、T35(Ser>Thr)、およびA50(Gly>Ala)として示される。H5 ベトナム/1203/2004 HA生体パニングにおいて同定された、上記必要とされかつ優性な群1の重鎖配列を、上記非常に関連する抗HIV Fab 47eの結晶構造に重ね合わせる。変異を、骨格モデル(上)および空間充填モデル(下)の両方において示す。密なクラスターが、CDR2中およびCDR2のみ隣接する必要とされる変異のうちの4個によって形成される。上記必要とされる変異52A(Pro>Gly)、53(Ile>Met)、73(Lys>Glu)および74(Ser>LeuもしくはMet)は、上記重鎖可変ドメインの露出された表面に顕著に密なクラスターを形成し、ここでこれらは、上記タンパク質表面から顕著に突出する隆起を形成する(図12)。その残りの必要とされる変異57(Ala>Thr)は、部分的には、上記CDR2ループの基部に埋め込まれる。CDR2およびフレームワーク3における表面露出変化は、おそらく抗原結合において直接的な役割を有することである一方で、上記あまり露出されていない必要土佐折れる変異および本質的でない優性変異は、上記CDR2ループを安定化し、そして/もしくは位置づけることを介して間接的な効果を有し得る。
【0191】
生存者2に由来する抗体は、上記VH4−4b生殖系列重鎖に最もよく似た2個の特有の重鎖からなる(表5)。上記第1の重鎖は、5個の特有のλ軽鎖と対形成していることが見いだされ、そのうちの4個は、滅多に使用されないλの6個の軽鎖ファミリーに由来し、他方は、単一のκ軽鎖と対形成される。中和プロフィールがより制限された抗体4は、この群に由来する。
【0192】
【表5】
所定の変異が、抗体の活性に重要であるという可能性は、独立して選択される時間数の関数として増大する。上記必要とされる変異が、独立したクローンから体細胞変異の間に選択されるか、またはその後の複製の間にさらに変異される単一のクローンの子孫に由来するか否かを決定するために、上記優性変異のコドン使用法を分析した(表6A〜6B)。上記データは、異なるコドンが使用されたが、上記コドンは、同じアミノ酸変化を生じたことを明らかにし、このことは、これら変異が、このなるクローンにおいて独立して発生したこと、従って、複数回選択されたことを実証する。独立して選択された事象のこの収束性の結果は、これら優性変異が、上記ウイルスへの結合および/もしくはまたはその中和における重要な役割を果たすという強力な証拠である。
【0193】
表6A〜6Bに例示されるように、個々のクローンのコドン使用法は、選択されたH5 HA結合クローンの独立した起源を示す。DNAアラインメントおよびコードされるアミノ酸は、VH1−e 生殖系列に対する6個の代表的群1の抗体である。同じアミノ酸についての異なるコドンの使用は、各特有の配列が、別個の起源のものであることを示す。表6AはCDR2に対応し、表6Bは、フレームワーク3に対応する。生殖系列コドンは、太字を付したコドンとして示される。生殖系列コドンから同じアミノ酸への変化は、通常のテキストのコドンとして示される。生殖系列アミノ酸からの第1の変化は、太字の下線を付したコドンとして示される。生殖系列アミノ酸からの第2の変化は、斜体の下線を付したコドンとして示される。生殖系列アミノ酸からの第3の変化は、下線の灰色で強調したコドンとして示される。
【0194】
【表6A】
【0195】
【表6B】
(実施例2:HIV感染患者についてのドナー特異的抗体ライブラリーの構築)
(骨髄プロトコルおよび血清調製)
血液を、標準的静脈穿刺によって得、凝固させ、処理して、血清を回収する。上記血清を、氷上で輸送されるまで、−20℃において3〜4日間保存する。ドナーを局所麻酔剤の注射で麻酔し、5mlの骨髄を、各患者ドナーの骨盤から回収する。次に、上記5mlの骨髄を、45ml RNAlater(Ambion)を含む滅菌50mlチューブに移す。上記混合物を、眼に見える塊が存在せずかつ上記骨髄およびRNAlaterが十分に混合されるまで、約8〜20回穏やかに転倒混和する。次に、上記標本を、2〜10℃の間で一晩冷蔵する。一晩冷蔵した後、上記標本を、氷上で輸送されるまで、−20℃において3〜4日間貯蔵する。受け取ったら、上記RNAlater/骨髄および血清を含むチューブを、処理するまで、−80℃において貯蔵する。候補患者は、ドナーとして選択される前に、HIVポジティブについて試験されるべきである。
【0196】
骨髄抽出およびmRNA精製、逆転写、PCR、抗体軽鎖および重鎖構築、ファージミドパニングなrばいに増幅、ELISAおよび配列決定を、実施例1に本質的に記載されるように行う。
【0197】
前述の説明において、本発明は、特定の実施形態を参照して例示されるが、そのように限定されない。実際に、本発明の種々の改変は、本明細書において示され、記載されるものに加えて、前述の説明から当業者に明らかになり、添付の特許請求の範囲内に入る。
【0198】
本明細書全体を通じて引用される全ての参考文献は、本明細書に参考として援用される。
【特許請求の範囲】
【請求項1】
標的抗原に対する抗体生成を引き起こす疾患に罹患したか、もしくは該疾患に現在罹患しているヒト患者ドナーから得られる、抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子のレパートリーを含むベクター集団であって、ここで該集団は、独特のバーコードで同定される、ベクター集団。
【請求項2】
抗体軽鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む、請求項1に記載のベクター集団。
【請求項3】
前記抗体軽鎖は、λ鎖である、請求項2に記載のベクター集団。
【請求項4】
前記抗体軽鎖がκ鎖である、請求項2に記載のベクター集団。
【請求項5】
抗体重鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む、請求項1に記載のベクター集団。
【請求項6】
前記バーコードは、前記集団中に存在するベクターに連結されるかもしくは組み込まれ、そして/または前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、該バーコードは、該核酸分子の発現を妨害しない、請求項1に記載のベクター集団。
【請求項7】
前記バーコードは、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列である、請求項6に記載のベクター集団。
【請求項8】
前記ヌクレオチド配列は、前記核酸分子の3’非コード領域もしくは5’非コード領域に連結される、請求項7に記載のベクター集団。
【請求項9】
前記ヌクレオチド配列は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列である、請求項6に記載のベクター集団。
【請求項10】
前記ヌクレオチド配列は、非連続である、請求項6に記載のベクター集団。
【請求項11】
前記非連続ヌクレオチド配列のうちの少なくとも一部は、前記集団に存在するベクターに連結されるかもしくは組み込まれる、請求項10に記載のベクター集団。
【請求項12】
前記非連続配列の少なくとも一部は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ、その結果、該非連続配列の少なくとも一部は、該核酸分子の発現を妨害しない、請求項10に記載のベクター集団。
【請求項13】
前記バーコードは、ペプチドもしくはポリペプチド配列である、請求項1に記載のベクター集団。
【請求項14】
前記ベクターは、ファージミドベクターである、請求項1に記載のベクター集団。
【請求項15】
前記ファージミドベクターは、バクテリオファージ遺伝子III、および前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と該バクテリオファージIII遺伝子との間に終止コドンを含む、請求項15に記載のベクター集団。
【請求項16】
前記バーコードは、前記終止コドンの後ろの非翻訳領域に挿入される、連続する非コードヌクレオチド配列である、請求項15に記載のベクター集団。
【請求項17】
請求項1に記載のベクター集団を含む、宿主細胞。
【請求項18】
E.coli宿主細胞である、請求項16に記載の宿主細胞。
【請求項19】
標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するライブラリーメンバーを含む、ドナー特異的抗体ライブラリーであって、ここで該抗体もしくは抗体フラグメントは、該標的抗原に対する抗体生成を引き起こす疾患に罹患したか、もしくは該疾患に現在罹患しているヒトドナーから得られ、ここで該抗体ライブラリーは、少なくとも1個の特有のバーコードで同定される、ドナー特異的抗体ライブラリー。
【請求項20】
前記抗体重鎖および軽鎖は、該抗体重鎖および軽鎖が得られる前記ヒトドナーに対して特有のバーコードで各々別個に同定される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項21】
1個の特有のバーコードで同定される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項22】
前記抗体もしくは抗体フラグメントは、ベクター中に存在する核酸分子によってコードされる抗体重鎖および軽鎖もしくはそのフラグメントから構成される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項23】
前記バーコードは、前記ライブラリーに存在するベクターに連結されるかもしくは組み込まれ、そして/または前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれる、ヌクレオチド配列であり、その結果、該バーコードは、該核酸分子の発現を妨害しない、請求項22に記載のドナー特異的抗体ライブラリー。
【請求項24】
前記バーコードは、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項25】
前記ヌクレオチド配列は、前記核酸分子の3’非コード領域もしくは5’非コード領域に連結される、請求項24に記載のドナー特異的抗体ライブラリー。
【請求項26】
前記ヌクレオチド配列は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項27】
前記ヌクレオチド配列は、非連続である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項28】
前記非連続配列のうちの少なくとも一部は、前記ライブラリーに存在するベクターに連結されているかもしくは組み込まれている、請求項27に記載のドナー特異的抗体ライブラリー。
【請求項29】
前記非連続配列のうちの少なくとも一部は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれており、その結果、該非連続配列のうちの少なくとも一部は、該核酸分子の発現を妨害しない、請求項27に記載のドナー特異的抗体ライブラリー。
【請求項30】
前記バーコードは、ペプチドもしくはポリペプチド配列である、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項31】
前記ベクターは、ファージミドベクターである、請求項22に記載のドナー特異的抗体ライブラリー。
【請求項32】
前記ファージミドベクターは、バクテリオファージ遺伝子III、および前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と該バクテリオファージIII遺伝子との間に終止コドンを含む、請求項31に記載のドナー特異的抗体ライブラリー。
【請求項33】
抗体重鎖もしくはそのフラグメントの集団を発現する、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項34】
1個より多くの抗体重鎖ファミリーを含む、請求項33に記載のドナー特異的抗体ライブラリー。
【請求項35】
前記ヒトドナーの病歴は、前記患者が前記疾患に罹患したか、もしくは該疾患に現在罹患していることを示す、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項36】
前記ヒトドナーが、前記疾患に罹患したか、もしくは該疾患に現在罹患していることが独立して確認される、請求項35に記載のドナー特異的抗体ライブラリー。
【請求項37】
前記標的抗原とは異なる抗原を特異的に結合する抗体および抗体フラグメントを実質的に欠いている、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項38】
前記標的抗原は、インフルエンザAウイルスである、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項39】
前記標的抗原は、インフルエンザAウイルスH1、H2もしくはH3サブタイプの単離株である、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項40】
前記標的抗原は、H5、H7およびH9インフルエンザAウイルスサブタイプを含む群から選択される、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項41】
1個より多くのインフルエンザAウイルスサブタイプに特異的に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項42】
インフルエンザウイルスAのH5N1サブタイプに結合しかつ中和する少なくとも1個の抗体もしくは抗体フラグメントを含む、請求項39に記載のドナー特異的抗体ライブラリー。
【請求項43】
前記ヒトドナーは、表1に列挙される疾患からなる群より選択される疾患に罹患したかもしくは現在罹患している、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項44】
前記標的疾患と関連する抗原に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項43に記載のドナー特異的抗体ライブラリー。
【請求項45】
前記標的疾患と関連する抗原に結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項43に記載のドナー特異的抗体ライブラリー。
【請求項46】
少なくとも1種の中和抗体を発現する、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項47】
前記ドナー特異的抗体ライブラリーは、ファージライブラリーである、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項48】
前記集団は、抗体もしくは抗体フラグメントの106個より多くの異なるメンバーをコードする配列を含む、請求項47に記載のドナー特異的抗体ライブラリー。
【請求項49】
前記集団は、抗体もしくは抗体フラグメントの109個より多くの異なるメンバーをコードする配列を含む、請求項47に記載のドナー特異的抗体ライブラリー。
【請求項50】
芽胞ディスプレイライブラリー(spore−display library)、リボソームディスプレイライブラリー、mRNAディスプレイライブラリー、微生物細胞ディスプレイライブラリー、酵母ディスプレイライブラリー、および哺乳動物ディスプレイライブラリーからなる群より選択される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項51】
前記核酸は、前記ヒトドナーのリンパ球から抽出されたmRNAから逆転写される、請求項50に記載のドナー特異的抗体ライブラリー。
【請求項52】
前記リンパ球は、骨髄、血液、脾臓、もしくはリンパ節から得られる、請求項51に記載のドナー特異的抗体ライブラリー。
【請求項53】
前記ヒトドナーの血清学的プロフィールは、前記mRNAを抽出する前に生成される、請求項52に記載のドナー特異的抗体ライブラリー。
【請求項54】
前記ヒトドナーの病歴は、前記mRNAを抽出する前もしくはその後に検査される、請求項53に記載のドナー特異的抗体ライブラリー。
【請求項55】
標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するドナー特異的ライブラリーを作製するための方法であって、該方法は、
a)該標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒト患者ドナーのリンパ球から、mRNAを得る工程;
b)該得られたmRNAの逆転写によって、該患者の免疫グロブリンレパートリーをコードする配列を含む核酸の集団を生成する工程;および
c)該ドナー特異的ライブラリーを、該核酸を標識する特有のバーコードで同定する工程、
を包含する、方法。
【請求項56】
工程a)の前もしくは後に、前記患者の血清学的プロフィールを生成する工程および/または該患者の病歴を検査する工程をさらに包含する、請求項55に記載の方法。
【請求項57】
d)前記核酸を発現ベクターに挿入する工程;
e)前記免疫グロブリンレパートリーを発現する工程;および
f)ディスプレイ系において、該免疫グロブリンレパートリーをディスプレイする工程、
をさらに包含する、請求項56に記載の方法。
【請求項58】
前記ディスプレイ系は、ファージミドである、請求項57に記載の方法。
【請求項59】
前記ライブラリーのメンバーを、前記標的抗原を中和もしくは活性化するそれらの能力に基づいて選択する工程をさらに包含する、請求項58に記載の方法。
【請求項60】
少なくとも1種の中和抗体を生じる、請求項59に記載の方法。
【請求項61】
1種より多くの中和抗体を生じる、請求項59に記載の方法。
【請求項62】
前記標的抗原を中和もしくは活性化することが見いだされたライブラリーメンバーを含む1種以上の下位ライブラリーを作り出す工程をさらに包含する、請求項61に記載の方法。
【請求項63】
同定された少なくとも1種のライブラリーメンバーを配列決定する工程を包含する、請求項59に記載の方法。
【請求項64】
請求項59に記載の方法によって選択される抗体によって中和もしくは活性化される標的抗原と関連する疾患を処置もしくは予防するための方法であって、該方法は、該選択される抗体の有効量を、必要とする該ヒト患者に投与する工程を包含する、方法。
【請求項65】
前記抗体は、中和抗体である、請求項64に記載の方法。
【請求項66】
前記疾患は、インフルエンザウイルスA感染である、請求項65に記載の方法。
【請求項67】
前記疾患は、表1に列挙される疾患からなる群より選択される、請求項64に記載の方法。
【請求項1】
標的抗原に対する抗体生成を引き起こす疾患に罹患したか、もしくは該疾患に現在罹患しているヒト患者ドナーから得られる、抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子のレパートリーを含むベクター集団であって、ここで該集団は、独特のバーコードで同定される、ベクター集団。
【請求項2】
抗体軽鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む、請求項1に記載のベクター集団。
【請求項3】
前記抗体軽鎖は、λ鎖である、請求項2に記載のベクター集団。
【請求項4】
前記抗体軽鎖がκ鎖である、請求項2に記載のベクター集団。
【請求項5】
抗体重鎖もしくはそのフラグメントをコードする核酸分子のレパートリーを含む、請求項1に記載のベクター集団。
【請求項6】
前記バーコードは、前記集団中に存在するベクターに連結されるかもしくは組み込まれ、そして/または前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれるヌクレオチド配列であり、その結果、該バーコードは、該核酸分子の発現を妨害しない、請求項1に記載のベクター集団。
【請求項7】
前記バーコードは、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列である、請求項6に記載のベクター集団。
【請求項8】
前記ヌクレオチド配列は、前記核酸分子の3’非コード領域もしくは5’非コード領域に連結される、請求項7に記載のベクター集団。
【請求項9】
前記ヌクレオチド配列は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列である、請求項6に記載のベクター集団。
【請求項10】
前記ヌクレオチド配列は、非連続である、請求項6に記載のベクター集団。
【請求項11】
前記非連続ヌクレオチド配列のうちの少なくとも一部は、前記集団に存在するベクターに連結されるかもしくは組み込まれる、請求項10に記載のベクター集団。
【請求項12】
前記非連続配列の少なくとも一部は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれ、その結果、該非連続配列の少なくとも一部は、該核酸分子の発現を妨害しない、請求項10に記載のベクター集団。
【請求項13】
前記バーコードは、ペプチドもしくはポリペプチド配列である、請求項1に記載のベクター集団。
【請求項14】
前記ベクターは、ファージミドベクターである、請求項1に記載のベクター集団。
【請求項15】
前記ファージミドベクターは、バクテリオファージ遺伝子III、および前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と該バクテリオファージIII遺伝子との間に終止コドンを含む、請求項15に記載のベクター集団。
【請求項16】
前記バーコードは、前記終止コドンの後ろの非翻訳領域に挿入される、連続する非コードヌクレオチド配列である、請求項15に記載のベクター集団。
【請求項17】
請求項1に記載のベクター集団を含む、宿主細胞。
【請求項18】
E.coli宿主細胞である、請求項16に記載の宿主細胞。
【請求項19】
標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するライブラリーメンバーを含む、ドナー特異的抗体ライブラリーであって、ここで該抗体もしくは抗体フラグメントは、該標的抗原に対する抗体生成を引き起こす疾患に罹患したか、もしくは該疾患に現在罹患しているヒトドナーから得られ、ここで該抗体ライブラリーは、少なくとも1個の特有のバーコードで同定される、ドナー特異的抗体ライブラリー。
【請求項20】
前記抗体重鎖および軽鎖は、該抗体重鎖および軽鎖が得られる前記ヒトドナーに対して特有のバーコードで各々別個に同定される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項21】
1個の特有のバーコードで同定される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項22】
前記抗体もしくは抗体フラグメントは、ベクター中に存在する核酸分子によってコードされる抗体重鎖および軽鎖もしくはそのフラグメントから構成される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項23】
前記バーコードは、前記ライブラリーに存在するベクターに連結されるかもしくは組み込まれ、そして/または前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に連結されるかもしくは組み込まれる、ヌクレオチド配列であり、その結果、該バーコードは、該核酸分子の発現を妨害しない、請求項22に記載のドナー特異的抗体ライブラリー。
【請求項24】
前記バーコードは、1〜約24ヌクレオチドの連続する非コードヌクレオチド配列である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項25】
前記ヌクレオチド配列は、前記核酸分子の3’非コード領域もしくは5’非コード領域に連結される、請求項24に記載のドナー特異的抗体ライブラリー。
【請求項26】
前記ヌクレオチド配列は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれる1個以上のサイレント変異のコード配列である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項27】
前記ヌクレオチド配列は、非連続である、請求項23に記載のドナー特異的抗体ライブラリー。
【請求項28】
前記非連続配列のうちの少なくとも一部は、前記ライブラリーに存在するベクターに連結されているかもしくは組み込まれている、請求項27に記載のドナー特異的抗体ライブラリー。
【請求項29】
前記非連続配列のうちの少なくとも一部は、前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子に組み込まれており、その結果、該非連続配列のうちの少なくとも一部は、該核酸分子の発現を妨害しない、請求項27に記載のドナー特異的抗体ライブラリー。
【請求項30】
前記バーコードは、ペプチドもしくはポリペプチド配列である、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項31】
前記ベクターは、ファージミドベクターである、請求項22に記載のドナー特異的抗体ライブラリー。
【請求項32】
前記ファージミドベクターは、バクテリオファージ遺伝子III、および前記抗体軽鎖もしくは重鎖またはそのフラグメントをコードする核酸分子と該バクテリオファージIII遺伝子との間に終止コドンを含む、請求項31に記載のドナー特異的抗体ライブラリー。
【請求項33】
抗体重鎖もしくはそのフラグメントの集団を発現する、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項34】
1個より多くの抗体重鎖ファミリーを含む、請求項33に記載のドナー特異的抗体ライブラリー。
【請求項35】
前記ヒトドナーの病歴は、前記患者が前記疾患に罹患したか、もしくは該疾患に現在罹患していることを示す、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項36】
前記ヒトドナーが、前記疾患に罹患したか、もしくは該疾患に現在罹患していることが独立して確認される、請求項35に記載のドナー特異的抗体ライブラリー。
【請求項37】
前記標的抗原とは異なる抗原を特異的に結合する抗体および抗体フラグメントを実質的に欠いている、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項38】
前記標的抗原は、インフルエンザAウイルスである、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項39】
前記標的抗原は、インフルエンザAウイルスH1、H2もしくはH3サブタイプの単離株である、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項40】
前記標的抗原は、H5、H7およびH9インフルエンザAウイルスサブタイプを含む群から選択される、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項41】
1個より多くのインフルエンザAウイルスサブタイプに特異的に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項38に記載のドナー特異的抗体ライブラリー。
【請求項42】
インフルエンザウイルスAのH5N1サブタイプに結合しかつ中和する少なくとも1個の抗体もしくは抗体フラグメントを含む、請求項39に記載のドナー特異的抗体ライブラリー。
【請求項43】
前記ヒトドナーは、表1に列挙される疾患からなる群より選択される疾患に罹患したかもしくは現在罹患している、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項44】
前記標的疾患と関連する抗原に結合する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項43に記載のドナー特異的抗体ライブラリー。
【請求項45】
前記標的疾患と関連する抗原に結合しかつ中和する少なくとも1種の抗体もしくは抗体フラグメントを発現する、請求項43に記載のドナー特異的抗体ライブラリー。
【請求項46】
少なくとも1種の中和抗体を発現する、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項47】
前記ドナー特異的抗体ライブラリーは、ファージライブラリーである、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項48】
前記集団は、抗体もしくは抗体フラグメントの106個より多くの異なるメンバーをコードする配列を含む、請求項47に記載のドナー特異的抗体ライブラリー。
【請求項49】
前記集団は、抗体もしくは抗体フラグメントの109個より多くの異なるメンバーをコードする配列を含む、請求項47に記載のドナー特異的抗体ライブラリー。
【請求項50】
芽胞ディスプレイライブラリー(spore−display library)、リボソームディスプレイライブラリー、mRNAディスプレイライブラリー、微生物細胞ディスプレイライブラリー、酵母ディスプレイライブラリー、および哺乳動物ディスプレイライブラリーからなる群より選択される、請求項19に記載のドナー特異的抗体ライブラリー。
【請求項51】
前記核酸は、前記ヒトドナーのリンパ球から抽出されたmRNAから逆転写される、請求項50に記載のドナー特異的抗体ライブラリー。
【請求項52】
前記リンパ球は、骨髄、血液、脾臓、もしくはリンパ節から得られる、請求項51に記載のドナー特異的抗体ライブラリー。
【請求項53】
前記ヒトドナーの血清学的プロフィールは、前記mRNAを抽出する前に生成される、請求項52に記載のドナー特異的抗体ライブラリー。
【請求項54】
前記ヒトドナーの病歴は、前記mRNAを抽出する前もしくはその後に検査される、請求項53に記載のドナー特異的抗体ライブラリー。
【請求項55】
標的抗原に対する抗体もしくは抗体フラグメントの集団を発現するドナー特異的ライブラリーを作製するための方法であって、該方法は、
a)該標的抗原に対する抗体生成を引き起こす疾患に罹患したかもしくは現在罹患しているヒト患者ドナーのリンパ球から、mRNAを得る工程;
b)該得られたmRNAの逆転写によって、該患者の免疫グロブリンレパートリーをコードする配列を含む核酸の集団を生成する工程;および
c)該ドナー特異的ライブラリーを、該核酸を標識する特有のバーコードで同定する工程、
を包含する、方法。
【請求項56】
工程a)の前もしくは後に、前記患者の血清学的プロフィールを生成する工程および/または該患者の病歴を検査する工程をさらに包含する、請求項55に記載の方法。
【請求項57】
d)前記核酸を発現ベクターに挿入する工程;
e)前記免疫グロブリンレパートリーを発現する工程;および
f)ディスプレイ系において、該免疫グロブリンレパートリーをディスプレイする工程、
をさらに包含する、請求項56に記載の方法。
【請求項58】
前記ディスプレイ系は、ファージミドである、請求項57に記載の方法。
【請求項59】
前記ライブラリーのメンバーを、前記標的抗原を中和もしくは活性化するそれらの能力に基づいて選択する工程をさらに包含する、請求項58に記載の方法。
【請求項60】
少なくとも1種の中和抗体を生じる、請求項59に記載の方法。
【請求項61】
1種より多くの中和抗体を生じる、請求項59に記載の方法。
【請求項62】
前記標的抗原を中和もしくは活性化することが見いだされたライブラリーメンバーを含む1種以上の下位ライブラリーを作り出す工程をさらに包含する、請求項61に記載の方法。
【請求項63】
同定された少なくとも1種のライブラリーメンバーを配列決定する工程を包含する、請求項59に記載の方法。
【請求項64】
請求項59に記載の方法によって選択される抗体によって中和もしくは活性化される標的抗原と関連する疾患を処置もしくは予防するための方法であって、該方法は、該選択される抗体の有効量を、必要とする該ヒト患者に投与する工程を包含する、方法。
【請求項65】
前記抗体は、中和抗体である、請求項64に記載の方法。
【請求項66】
前記疾患は、インフルエンザウイルスA感染である、請求項65に記載の方法。
【請求項67】
前記疾患は、表1に列挙される疾患からなる群より選択される、請求項64に記載の方法。
【図1】

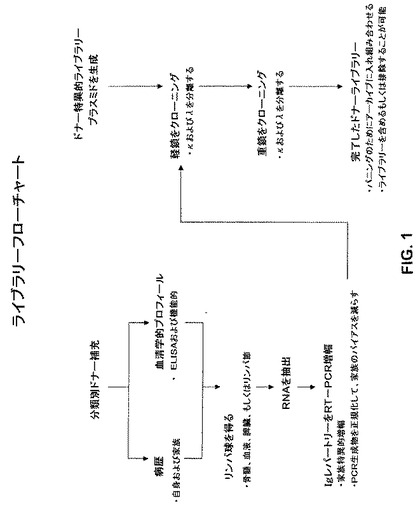
【図2】
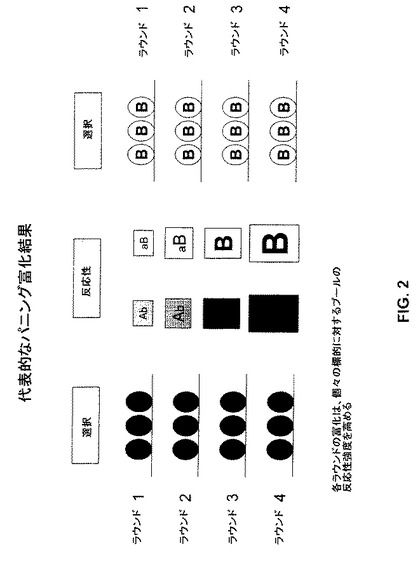
【図3】
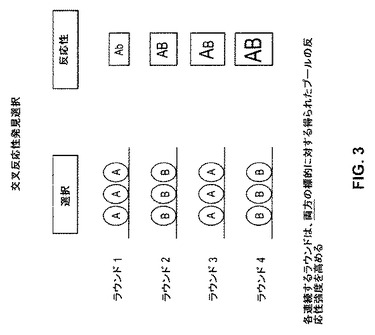
【図4】
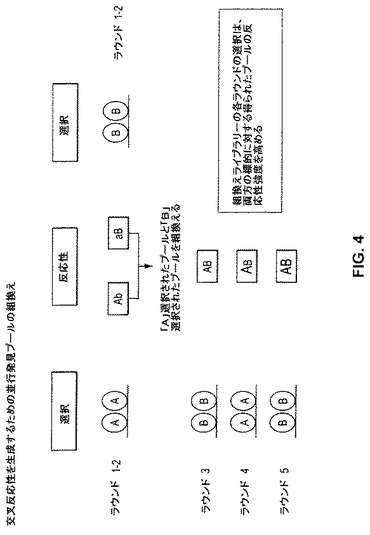
【図5】
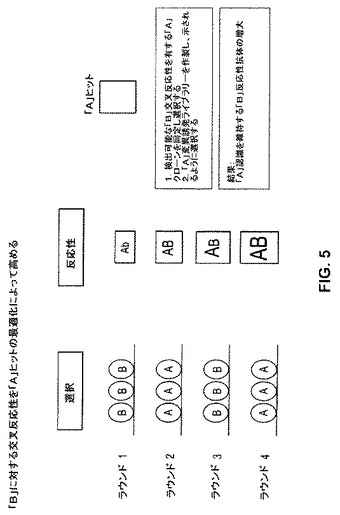
【図6】
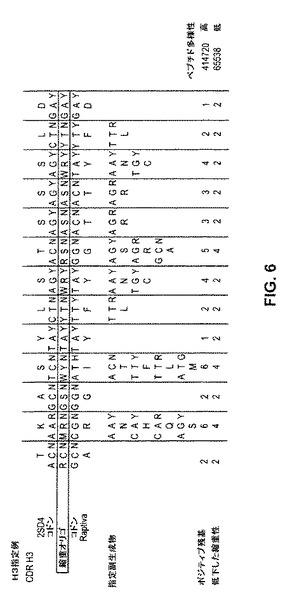
【図7A】

【図7B】

【図7C】

【図7D】

【図7E】

【図7F】

【図7G】

【図7H】

【図7I】

【図7J】

【図8】
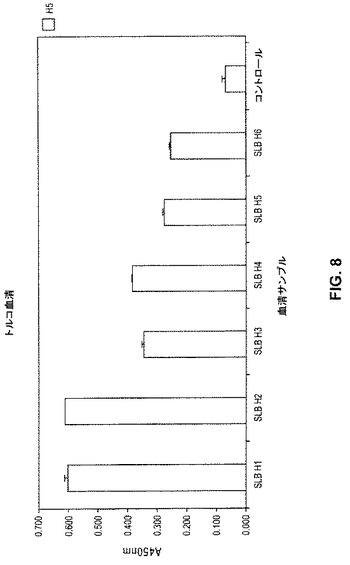
【図9】
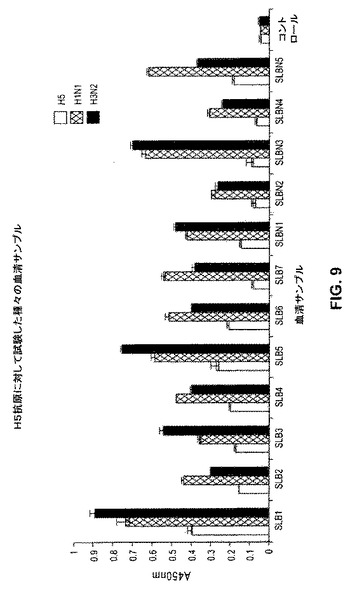
【図10】
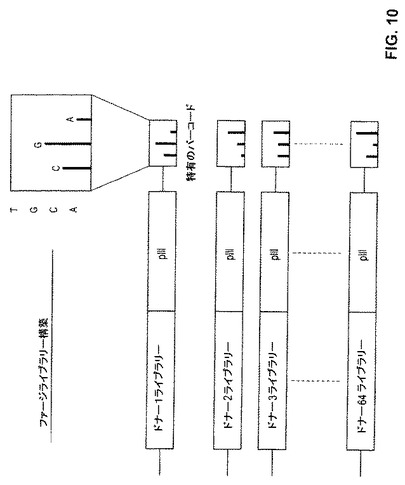
【図11】
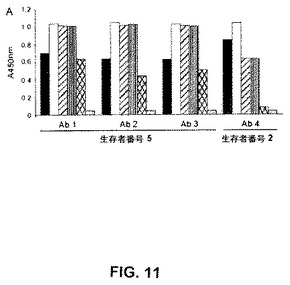
【図12】

【図13】
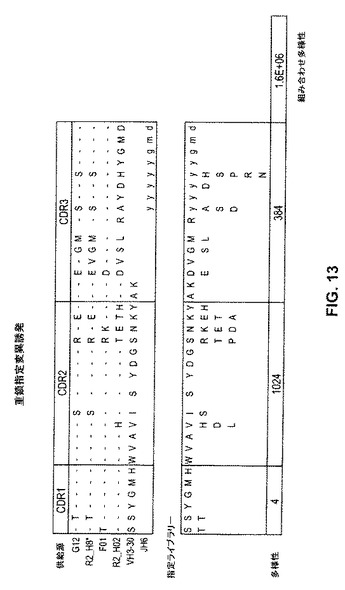
【図14】
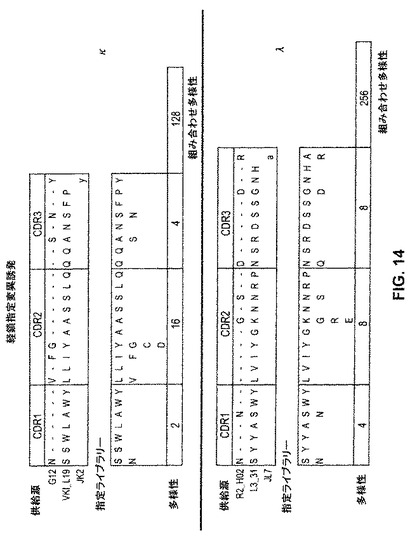
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2010−538618(P2010−538618A)
【公表日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願番号】特願2010−524259(P2010−524259)
【出願日】平成20年9月11日(2008.9.11)
【国際出願番号】PCT/US2008/075998
【国際公開番号】WO2009/036157
【国際公開日】平成21年3月19日(2009.3.19)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
【公表日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願日】平成20年9月11日(2008.9.11)
【国際出願番号】PCT/US2008/075998
【国際公開番号】WO2009/036157
【国際公開日】平成21年3月19日(2009.3.19)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
[ Back to top ]