
ニューロトロフィンの精製

【課題】治療用途に適した、成熟NGFを含むニューロトロフィンの大規模な精製方法が提供される。
【解決手段】ニューロトロフィンを、様々なあまり望ましくない誤って加工され、誤って折り畳まれた、サイズ、グリコシル化もしくは荷電型から、逆相液体クロマトグラフィー、カチオン交換クロマトグラフィーカラム等によって分離する手段を使用する。また、これらの変種が実質的に含まれない成熟NGFを含むニューロトロフィンの組成物も製造することができる。
【解決手段】ニューロトロフィンを、様々なあまり望ましくない誤って加工され、誤って折り畳まれた、サイズ、グリコシル化もしくは荷電型から、逆相液体クロマトグラフィー、カチオン交換クロマトグラフィーカラム等によって分離する手段を使用する。また、これらの変種が実質的に含まれない成熟NGFを含むニューロトロフィンの組成物も製造することができる。
【発明の詳細な説明】
【技術分野】
【0001】
この発明は、ニューロトロフィン、特にNGFファミリーのニューロトロフィン、より詳細には神経成長因子(NGF)及びニューロトロフィン−4/5(NT−4/5)、及びニューロトロフィン−3(NT−3)を、特に細菌もしくは哺乳動物細胞発酵により産生される場合に、これらに付随する変種、不純物、及び汚染物質から精製するための改良方法に関する。
【背景技術】
【0002】
比較的純粋で生物学的に活性なポリペプチド及びタンパク質を大量に生産することは、ヒトと動物の医薬製剤、酵素及びその他の特殊化学製品の製造に対して経済的に重要である。多数のタンパク質の生産に対して、多量の外来性タンパク質を哺乳動物の宿主細胞及び細菌及びその他の宿主細胞中に発現しうることから、組換えDNA技術が選択方法になった。
【0003】
哺乳動物NGF(マウスNGF)の一次構造は、非特許文献1によって最初に解明された。その前駆物質であるプレプロNGFの一次構造は、マウスNGFのcDNAのヌクレオチド配列から推論された(非特許文献2;非特許文献3)。
【0004】
相同的ヒトNGF(hNGF)遺伝子もまた同定された(非特許文献4;特許文献1)。マウスNGFに対するその相同性は、アミノ酸とヌクレオチド配列レベルでそれぞれ約90%と87%である。天然に生じるヒトNGFは希少であるため、ヒトGNFは生化学的に細かく詳細に特徴付けるのに充分な量では天然源から調製されていない。
【0005】
NGFに関連する更なる神経栄養因子がそれ以降に同定されている。これらには、脳由来神経栄養因子(BDNF)(非特許文献5)、ニューロトロフィン−3(NT−3)(非特許文献6;非特許文献7;非特許文献8)、及びニューロトロフィン4/5(NT−4/5)(非特許文献9)が含まれる。TGF−βスーパーファミリーの遠いメンバーであるGDNFとニュールツリン(neurturin:「NTN」)は、交感感覚神経細胞及び中枢神経系神経細胞に対する二つの最近同定された構造的に関連している強力な生存因子である(非特許文献10;非特許文献11;非特許文献12;非特許文献13)。
【0006】
組換えタンパク質を生産するには、そのタンパク質をコードするDNAを宿主細胞に形質移入し、組換えタンパク質の発現に好ましい条件下で細胞を成長させる。原核生物の大腸菌が、低コスト高収量で組換えタンパク質を生産することを可能にすることから、好まれる宿主であった。タンパク質をコードするDNAの一般的な細菌での発現に関しては、細胞外又は周辺質担体タンパク質に対する細菌遺伝子と非細菌遺伝子を含んでなる組換えDNA分子に関する特許文献2、凝集体形成ポリペプチドとの外来性ポリペプチドの同時生産に関する特許文献3、IGF−IのようなポリペプチドとのtrpLE融合とtrpプロモーター/オペレーターを持つ発現ベクターに関する特許文献4、外来性タンパク質と共に含む発現調節配列に関する特許文献5及びIGF−Iをコードしているもののような特異的環状DNAプラスミドに関する特許文献6を含む多くの米国特許が存在する。
【0007】
遺伝子操作により作成されるバイオ医薬品は、典型的には種々の多様な宿主細胞汚染物質を含んでいる上清から精製される。特にNGFは、多数の異なった方法を使用し、様々な度合いの努力と成功度をもって様々な度合いに精製されたことは報告されている。例えば、非特許文献14、NGFのCHO細胞による産生を開示している特許文献7、非特許文献15;非特許文献16;非特許文献17を参照せよ。これらの努力は主として研究室規模のものであった。
【0008】
【特許文献1】米国特許第5288622号
【特許文献2】米国特許第4565785号
【特許文献3】米国特許第4673641号
【特許文献4】米国特許第4738921号
【特許文献5】米国特許第4795706号
【特許文献6】米国特許第4710473号
【特許文献7】米国特許第5082774号
【非特許文献1】AngelettiとBradshaw(Proc.Natl.Acad.Aci. USA 68:2417(1971))
【非特許文献2】Scottら Nature 302:538(1983)
【非特許文献3】Ullrichら Nature 303:821(1983)
【非特許文献4】Ullrich, Symp. on Quan.Biol., Cold Spring Harbor 48:435(1983)
【非特許文献5】Leibrockら, Nature, 341:149-152(1989)
【非特許文献6】Kaishoら, FEBS Lett., 266:187(1990)
【非特許文献7】Maisonpierreら, Science, 247:1446(1990)
【非特許文献8】Rosenthalら, Neuron, 4:767(1990)
【非特許文献9】Berkmeierら, Neuron, 7:857-866(1991)
【非特許文献10】Linら, Science 260:1130-1132(1993)
【非特許文献11】Hendersonら, Science 266:1062-1064(1994)
【非特許文献12】Buj-Belloら, Neuron 15:821-828(1995)
【非特許文献13】Kotzbauerら Nature 384:467-470(1996)
【非特許文献14】Longoら(IBRO Handbook,vol.12,pp3-30(1989))
【非特許文献15】BruceとHeinrich(Neurobio. Aging 10:89-94(1989))
【非特許文献16】Schmelzerら(J.Neurochem. 59:1675-1683(1992))
【非特許文献17】Burtonら(J.Neurochem. 59:1937-1945(1992))
【発明の開示】
【発明が解決しようとする課題】
【0009】
しかし、変種が本質的に含まれない製薬的純度と高収量を達成できるような組換えヒトNGFの調製的単離は、従来技術が解決できなかった課題であった。
【0010】
従って、ニューロトロフィン、特にNGF及びNGFファミリーのニューロトロフィンを、その変種とその他の分子から、また高いpIを持つ他のポリペプチドから選択的に分離するための効率的なプロトコールに対する必要性が従来から存在している。大きな規模でニューロトロフィンを精製する方法は、治療上の必要性を満たすために、発酵ブロス、溶解細菌もしくは哺乳動物細胞を含む様々な供給源からの出発原料に適用できなければならない。
【課題を解決するための手段】
【0011】
更に、本発明者らは、過去には知られておらず分離が困難なニューロトロフィン変種、例えばNGF変種を発見したので、ここに提供する方法は、望ましくない変種が実質的に含まれない、ヒトNGF(rhNGF)、rhNT−3及びrhNT−4/5及びその所望の遺伝子操作突然変異体を含む組換えニューロトロフィンを、商業的に有用な量で提供するために特に有用である。本発明のかかる目的とその他の目的は当業者には明らかであろう。
【0012】
本発明の一実施態様において、ニューロトロフィン、特に高度に相同的なレセプターファミリー(trks)、好ましくはrhNGF、rNT−3、rhNT−4/5、rhBDNFもしくはその所望の遺伝子操作形態による認識を分かつNGF、NT−3、NT−4/5、及びBDNFを含む、特にNGFファミリーのものを、疎水的相互作用クロマトグラフィー(HIC)を使用して精製する方法が提供される。
【0013】
ここに報告したように、ニューロトロフィンの組換え生産から生じるある種の望ましくないニューロトロフィン変種を本発明者らが発見したことに鑑みると、HICの使用は、ニューロトロフィンの化学的に異なる形態ともしくは誤って折り畳まれた形態でさえも、所望の正しく折り畳まれた無傷のニューロトロフィンから分離することを可能にする。取り除くことができる変種は、部分的にプロセシングされた前駆配列、グリコシル化された成熟及び前駆物質含有形態(真核生物細胞培養にある場合)、及び誤って折り畳まれ部分的に折り畳まれた変種(一般にインビトロ折り畳み工程が使用される場合、細菌細胞培養からのもの)を含む、疎水性が成熟した正しく折り畳まれた無傷のニューロトロフィンとは異なるものである。例えば、HICは、NGFの部分的にプロセシングされた前駆配列、NGFのグリコシル化された種及び前駆体(真核細胞培養に存在する場合)、及び誤って折り畳まれ部分的に折り畳まれた変種(一般に細菌細胞培養からでインビトロ折り畳み工程からのもの)を成熟NGFの混合物から除去するのに特に有用である。NGFは一つのN結合グリコシル化部位をAsn45に有している。細菌中に発現された再折り畳みrhNT−4/5の場合は、HICは、正しく折り畳まれたNT−4/5を間違って折り畳まれた形態から分離する。ここに記載した方法によれば、ニューロトロフィンは本質的にこれらの変種を含まない。ニューロトロフィンの精製に対して、HIC樹脂官能基は、オクチル及びブチル基も有用であるが、好ましくはフェニル基である。特に好ましい実施態様は、フェニル・トーヨーパール(Phenyl Toyopearl)、フェニル・セファロース・ファースト・フロー・ロー・サブスティチューション(Phenyl Sepharose Fast Flow Low Substitution)、TSK−フェニル5PW(TSK−Phenyl 5PW)等々のHIC樹脂である。
【0014】
他の実施態様では、ニューロトロフィン、特にNGFファミリーのもの、好ましくrhNGF、rNT−3、rhNT−4/5もしくはその所望の遺伝子操作形態を、負荷修飾変種、例えば成熟ニューロトロフィン由来の酸化、isoasp(イソアスパラギン酸)及び脱アミド形態を分離する調製的カチオン交換クロマトグラフィーの使用によって精製する方法が提供される。特に好適な実施態様は、SP−セファロース・ハイ・パーフォーマンス、フラクトゲル(Fractogel)EMD SO3、又はポリアスパラギン酸樹脂を用い、なかでもポリキャット(PolyCAT)Aが特に好適である。最も好適には、大規模のSP−セファロース・ハイ・パーフォーマンス樹脂又はフラクトゲルEMD SO3樹脂が使用される。
【0015】
本発明の更に他の実施態様では、HICとカチオン交換クロマトグラフィーの両方が、所望のニューロトロフィン、例えば実質的に均質な、すなわち例えば誤って折り畳まれた変種及び化学変種のような加工及び負荷変種の双方を実質的に含まず、タンパク含量に関してもまた実質的に純粋である組換え成熟NGF、好ましくはヒトNGFの組成物を調製するために使用される。
【0016】
この発明の一実施態様では、ニューロトロフィン、特にNGFファミリーのもの、好ましく組換えヒトNGF、NT−3、NT−4/5及びその所望の遺伝子操作形態を、関連する所望されない変種、例えば発酵、プロテアーゼ切断変種、グリコシル化変種、誤って折り畳まれた変種を、逆相液体クロマトグラフィーによって精製する改良方法が提供される。より好ましくは、NGFは120/120又は118/118ホモ二量体形である。ここに記載した方法によれば、ニューロトロフィンは、最も好ましくは変種を本質的に含まない。
【0017】
他の実施態様においては、ニューロトロフィン、特にNGFファミリーのものを、生理的pHを含む溶出条件を使用して関連する変種から精製する方法が提供される。
更に他の実施態様では、均一性がかなり改善される結果が得られるニューロトロフィンの精製方法が提供される。
【0018】
他の実施態様において、本発明は、NGFファミリーのニューロトロフィンをその変種から分離する方法であって、
a)ニューロトロフィンとその変種を含む緩衝液を約5から8のpHで疎水的相互作用クロマトグラフィーカラムに装填し;
b)カラムを洗浄し、
c)約5から8のpHの緩衝液でニューロトロフィンを溶出させ、
d)約5から8のpHでカチオン交換クロマトグラフィーカラムにニューロトロフィン含有溶出物を装填し、
e)約5から8のpH、好ましくはpH6で塩勾配の緩衝液でカラムからニューロトロフィンを溶出させる、
ことを含んでなる方法を提供する。ニューロトロフィンは最も好ましくはrhNGFである。
【0019】
本発明の一実施態様では、好ましくはNGF分画であるニューロトロフィン分画から宿主細胞タンパク質を効果的に除去するシリカゲルクロマトグラフィー工程が提供される。
【0020】
本発明の一実施態様では、120アミノ酸NGFがトリプシン様プロテアーゼ処理を受けて、VRRAのC末端から末端RAジペプチドを選択的に除去して、118種を生成する方法の工程が提供される。固定化トリプシンカラムが好適である。
【0021】
本発明はまた本発明の方法により調製されたニューロトロフィン組成物と製剤及び組成物と製剤への用途にも関する。提供されるものは、実質的に均一な、すなわち例えば誤って折り畳まれた変種と化学変種のような加工及び負荷変種の双方とも実質的に含まず、またタンパク質含量に関して実質的に純粋であるニューロトロフィンの組成物である。好ましくは、成熟ヒトNGF、成熟ヒトもしくはラットNT−3、及び成熟ヒトNT−4/5がこの形で提供される。好適な実施態様では、NGFは120種で、より好ましくは118型であり、最も好ましくはホモ二量体、例えば118/118である。
【発明を実施するための最良の形態】
【0022】
<定義>
本明細書において、「ニューロトロフィン」とは、マウス、ウシ、ヒツジ、ブタ、ウマ、トリ、そして好ましくはヒトを含む任意の種から、また天然であろうと、合成であろうと組換え生産されたものであろうと、任意のソース由来の、未変性配列もしくは遺伝子操作形態のニューロトロフィン、好ましくはNGF、NT−3、NT−4/5、及びBDNFを含むNGFファミリーのニューロトロフィンをいう。例えば、「NGF」は、マウス、ウシ、ヒツジ、ブタ、ウマ、トリ、そして好ましくはヒトを含む任意の種から、また天然であろうと、合成であろうと組換え生産されたものであろうと、任意のソース由来の神経成長因子をいう。好ましくは、ニューロトロフィンは組換え的に生産される。好適な方法では、ニューロトロフィンはクローン化され、そのDNAは例えば哺乳動物細胞、細菌細胞中で発現される。ここに教示したプロセスと方法はニューロトロフィン、GDNF及びニュールツリンにも適用し得る。
【0023】
ヒトの用途に対して好ましいものは、ヒト未変性配列、成熟NGFであり、より好ましくは120アミノ酸配列、更により好ましくは118アミノ酸配列である。より好適には、この未変性配列NGFは組替え的に生産される。ヒトプレプロNGFとヒト成熟NGFに対する好適なアミノ酸配列は、出典明示によりここに特に取込む米国特許第5288622号によって提供される。更なる翻訳後の修飾のない120アミノ酸型はホモ二量体形(つまり、120/120)における好適な型である。更により好適なものは、更なる翻訳後の修飾のない、特にホモ二量体のような118型(つまり、118/118)である。
【0024】
「実質的に純粋な」が意味するところは、少なくとも70%のニューロトロフィン、より好ましくは少なくとも80%、更により好ましくは少なくとも90%、95%もしくは99%と増大する、全タンパク質に対する全ニューロトロフィン、例えばNGFの純度の度合いである。特に好適な純度は少なくとも95%である。「本質的に純粋な」が意味するところは、組成物が、所望のニューロトロフィンについて少なくとも90%もしくはそれ以上純粋であることである。
【0025】
「ニューロトロフィン変種が実質的に含まれない」が意味するところは、(望ましさが劣るニューロトロフィン種を含む)全ニューロトロフィンに対する所望のニューロトロフィンのパーセントが少なくとも70%の所望のニューロトロフィン種、より好ましくは少なくとも80%、更により好ましくは少なくとも90%、93%、95%もしくは99%と増大する組成物である。「本質的に含まれない」が意味するところは、組成物が所望のニューロトロフィンを少なくとも90%あるいはそれ以上含むことである。特に好適な量は、少なくとも95%の所望のニューロトロフィン、例えば正しく折り畳まれた無傷の118/118rhNGF種である。その他の望まれない種もしくは型は、発酵もしくは精製プロセスから生じる、例えば変更された負荷変種のような、誤ってプロセシングされた型又は化学変種、又は好ましくはここに開示した前記のものの全てである。例えば、NGFが細菌中での合成後にインビトロで折り畳まれる場合、「種」又は「変種」は誤って折り畳まれた型あるいは部分的に折り畳まれた型を含み得る。
【0026】
「誤って折り畳まれた」変種が意味するところは、そのシステイン残基対合により、もしくは遊離しているかブロックされている特定のシステイン残基により、ニューロトロフィンとは異なるニューロトロフィンの変種である。誤って折り畳まれた変種もまたニューロトロフィンと同じシステイン対合を有し得るが、誤った折り畳みから生じる異なった三次元高次構造を有する。
【0027】
「化学」変種が意味するところは、例えば変更された電荷を有することにより、カルバミル化、脱アミド、酸化、グリコシル化、又はタンパク分解性切断により、ニューロトロフィンとは化学的に異なる変種である。
【0028】
この発明のカラムの観点において、緩衝液は、一般に、約5から8の範囲のpHを有する。この範囲内にpHを調節する緩衝液には、例えば、クエン酸塩、コハク酸塩、リン酸塩、MES、ADA、BIS−TRISプロパン、PIPES、ACES、イミダゾール、ジメチルマロン酸、MOPS、MOPSO、TES、トリスバッファー、例えばトリス−HCl、HEPES、HEPPS、TRICINE、グリシンアミド、BICINE、グリシルグリシン、及びホウ酸塩バッファーが含まれる。好適な緩衝液はMOPSOバッファーである。
【0029】
ここにおいて使用するところでは、「アルコール」と「アルコール溶媒」とは、アルコールに対して通常使用されている技術用語の意味を持つものであり、好ましくは、1から10の炭素原子を持つアルコール、より好ましくは、メタノール、エタノール、イソプロパノール、n−プロパノ−ル、もしくはt−ブタノール、並びにグリセロール、プロピレングリコール、エチレングリコール、ヘキシレングリコール、ポリプロピレングリコール、及びポリエチレングリコール、最も好ましくはエタノールもしくはイソプロパノールである。このようなアルコールは、水溶液に添加したときに、溶液極性を減少させることにより溶液の疎水性を増大させる溶媒である。
【0030】
MOPSOは3−(N−モルホリノ)−2−ヒドロキシプロパンスルホン酸である。HEPESはN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸である。試薬アルコールは95容量部(特別に変成したアルコール・フォームラ3A及び5容量部のイソプロピルアルコール)である。MESは2−(N−モルホリノ)エタンスルホン酸である。UF/DFは限外濾過/ダイアフィルトレーションを意味する。TMACはテトラメチルアンモニウムクロリドである。TEACはテトラエチルアンモニウムクロリドである。NGF−120は完全長120/120神経成長因子を意味する。NGF−118は118残基のホモ二量体成熟NGFを意味する。酸化NGFは、成熟未変性NGFの約80%の生物学的活性があるとここに報告されているNGF変種分子であるメツルフォキシド(Metsulfoxide)37を意味する。IsoaspNGFはNGF異性体化変種分子、Asp93を意味する。脱アミドNGFは、Asp45に転換されたAsn45を有するNGFを意味する。RNGFはそのN末端に余分なアルギニン残基を持つNGF分子を意味する。CHOはチャイニーズハムスター卵巣細胞を意味する。
【0031】
ここに記載された樹脂は、マクロプレップ・ハイ・Sカチオン交換(バイオラッド研究所、強いカチオン交換、SO3官能基、名目粒径50ミクロン、名目孔径1000A)、シリカゲル(未誘導体化)、フェニル・セファロース・ファースト・フロー・ロー・サブスティチューション(ファーマシア;高度に架橋した6%アガロース;粒径45−165ミクロン)、SP−セファロースHP(ファーマシア;高度に架橋した6%アガロース;粒径34ミクロン)、フェニル・トーヨーパール650M(TosoHaas;粒径40−90ミクロン)、及びフラクトゲルEMD SO3−650S(E.Merck(ドイツ)の米国関連会社であるEM Separations;粒径25−40ミクロン)を含む。
【0032】
<発明を実施するための形態の説明>
ニューロトロフィンは、神経系の発達と維持に重要な役割を果たす小さい塩基性タンパク質のファミリーに属する。このファミリーの最初に同定され恐らくは最もよく理解されているメンバーは神経成長因子(NGF)である。1992年12月8日発行の米国特許第5169762号を参照せよ。
【0033】
最近、NGFと類似した機能を有している配列的に関連しているが別個のポリペプチドが同定された。例えば、ニューロトロフィン−2(NT2)ともいわれる脳由来神経栄養因子(BDNF)が、Leibrockら(Nature, 341:49-159[1989])によりクローン化され配列決定された。
【0034】
幾つかの研究グループが、ニューロン因子(NF)と当初は呼ばれ、今はニューロトロフィン−3(NT−3)といわれる神経栄養因子を同定した(Ernforsら, Proc.Natl.Acad.Sci. USA, 87:5454-5458[1990]; Hoehnら, Nature, 344:339[1990]; Maisonpierreら, Science, 247:1446[1990]; Rosenthalら, Neuron, 4:767[1990]; JonesとReichardt, Proc.Natl.Acad.Sci. USA, 87:8060-8064[1990]; Kaishoら, FEBS Lett., 266:187[1990])。
【0035】
(NT4又はNT5といわれる)ニューロトロフィン−4/5が同定された(Hallbookら, Neuron, 6:844-858[1991]; Berkmeierら, Neuron, 7:857-866[1991]; Ipら, Proc.Natl.Acad.Sci, USA, 89:3060-3064[1992])。1994年11月15日に発行された米国特許第5364769号は、ヒトNT−4/5及びその組換え発現方法を開示しており、出典明示によりここに取り込まれる。
【0036】
また報告されているのは、キメラ及びパントロピックニューロトロフィン、例えば1996年1月30日に発行された米国特許第5488099号、Urferら(EMBO J. 13(24):5896-909(1994))及び、ニューロトロフィンが、一以上のレセプターに結合するように修飾されたか、未変性ニューロトロフィンに有意な程度では通常はないレセプター結合活性を含む(ここに出典明示により取込まれる)1995年12月14日公開のWO95/33829に報告されたようなものである。
【0037】
特に興味深いものはMNTS−1及びD15A NT3と命名されたニューロトロフィンである。また特に興味深いものは、NGFアミノ酸バックボーンを有するがtrkA以外のレセプター、例えばtrkB又はtrkCに結合するように修飾されたニューロトロフィンである。
【0038】
NT−3のtrkレセプターへの結合の原因であるNT−3の対応する位置からのアミノ酸を持つNGFにアミノ酸置換がなされたものが好適である。このようなNGF突然変異体は、NGF薬物動態学的及び精製挙動を保持しながらNT−3様レセプター結合活性を有する(Urferら, Biochemistry 36(16):4775-4781(1997))。これらのNGF突然変異体はまたtrkA結合活性を欠き得る(Shihら, J.Biol.Chem. 269(44):27679-86(1994))。このようなNGF突然変異体がここに記載する本発明の用途に対して特に好適なニューロトロフィンである。
【0039】
組換えヒトニューロトロフィン、例えばrhNGFの単離は、種々の多様な宿主細胞汚染物質からのタンパク質の分離を含む。各工程は、十分な分離が生じることを可能にする特別な緩衝液を含む。ニューロトロフィンに対する最後もしくは最後から二番目のプロセシング工程は、通常のクロマトグラフィー媒体を使用して同時精製する幾つかのニューロトロフィン変種の存在により複雑化する。再折り畳み工程が回収・精製工程に含まれる場合、変種はニューロトロフィンの誤って折り畳まれた型を含む。変種はまたニューロトロフィンとは化学的に異なるもの、例えばカルバミル化、脱アミド、脱アミドもしくはタンパク分解性に切断された形態をも含みうる。NGFの場合、これらの種は主に二量体型−ホモ二量体、例えば118/118が所望の場合、120/120又は117/117、又はヘテロ二量体、例えば120/118、117/118−、化学的に修飾された変種−イソアスパラギン酸、モノ酸化、グリコシル化変種、N末端及びC末端切断型、及びその二量体からなる。
【0040】
本発明は、ニューロトロフィン、特にrhNGFを、例えばアルツハイマー病、糖尿病及びエイズ関連神経障害を含む抹消神経障害等々の治療のような治療用途に十分な量で大規模に生産することを可能にする。
【0041】
NGFとその他のニューロトロフィン、好ましくはNGFファミリーのものとの間の配列と高次構造の類似性に鑑みると、本発明の方法は、誤ってプロセシングされた変種、誤って折り畳まれもしくは部分的に折り畳まれた変種、グリコシル化変種及び/又は負荷変種を実質的に含まないこれらのニューロトロフィンを調製するために適用しうる。本発明において、これらとその他の密接に関連した変種からニューロトロフィンを選択的に分離するのに望ましいカラム樹脂と条件が特定される。
【0042】
ニューロトロフィンには、NT−3、NT−4/5、NT−6、BDNF、及びそのヘテロ二量体、キメラもしくはパントロピック形を含む操作形態が含まれる。好ましくは、ニューロトロフィンはヒトあるいはヒトアミノ酸配列と高度に相同性を有するもので、ヒト配列に対して好ましくは80%より多く、より好ましくは90%より多く、最も好ましくは95%より多い相同性を有する。操作されたニューロトロフィンは、それが模倣する未変性ニューロトロフィンのtrkレセプター結合機能の少なくとも50%を保持し、好ましくは少なくとも75%、より好ましくは少なくとも80%を保持する。これらの操作された型は、ここに記載する方法において類似の性能を保持する未変性ニューロトロフィンの疎水的性質又は十分に高いpIを保持するものである。
【0043】
以下に記載するように、ここに記載する方法は、rhNGF、rhNT−3及びrhNT−5に首尾よく適用された。例えば、大腸菌において作られたrhNT−4/5が、封入体に単離され、還元され、封入体から可溶化された。還元されたNT−4/5はDEセファロース・ファースト・フロー及びS−セファロース・ファースト・フロー・クロマトグラフィーによって部分的に精製された。S−セファロース・ファースト・フロー・プールはグアニジン含有緩衝液中で24時間の間再び折り畳まれた。NT−4/5の誤って折り畳まれた型はここに大規模なもので開示したクロマトグラフィーによって除去した。NT−4/5のカルバミル化及び切詰め型(誤ってプロセシングされた形態)が、大規模のカラム形式での高速カチオン交換クロマトグラフィーにより、ポリキャットA HPLC樹脂もしくはSPセファロースHP樹脂で除去された。精製rhNT−4/5は製剤用に酸性緩衝液中に限外濾過されダイアフィルトレーションにかけられた。
【0044】
本発明の一実施態様は、通常はニューロトロフィンを大抵の他の不純物から既に精製した後、典型的には製剤化の前の脱塩もしくはダイアフィルトレーションの前の最終工程又は最終に近い工程において、ニューロトロフィンをその関連する変種から精製することを含む。混合物中の関連した変種は、発酵の結果生じる変種残渣ばかりでなく、ニューロトロフィンが貯蔵もしくはプロセシング中に分解した場合に生成された変種も含みうる。
【0045】
本発明の実施態様において使用するのに適したニューロトロフィンは、任意の手段により調製することができるが、好ましくは組替え的に調製される。ここで考察したニューロトロフィンをコードしている核酸分子は幾つかのソースから入手可能であり、例えば既知のDNA配列を用いて化学合成により、又は当業者に知られている標準的なクローニング法を使用することにより入手できる。例えばhGNFコード配列のようなニューロトロフィンを保有しているcDNAクローンは、ニューロトロフィンの既知の配列に基づいて特に設計されたオリゴヌクレオチドハイブリダイゼーションプローブを使用して同定することができる。
【0046】
ニューロトロフィンコード配列を有する分子を得る際には、分子を、選ばれた宿主細胞中に発現させるために適したクローニングベクター内に挿入する。クローニングベクターは、コード配列の効率的な転写、翻訳及びプロセシングに必要とされる好適な調節機能をもたらすように作成される。
【0047】
ニューロトロフィンが組替え的に調製される場合、ニューロトロフィンをコードしているDNAを発現するために好適な宿主細胞は、原核生物、酵母もしくはより高等な真核生物細胞である。この目的に対して適した原核生物は、古細菌及び真正細菌のような細菌を含む。好適な細菌は真正細菌、例えばグラム陰性もしくはグラム陽性生物体、例えばエシェリチアのような腸内細菌科、例えば大腸菌、エンテロバクター、エルウィニア(Erwinia)、クレブシエラ、プロテウス、サルモネラ、例えばネズミチフス菌、セラチア属、例えばセラチア・マルセスキャンス及び赤痢菌属;桿菌、例えば枯草菌及びバシリ・リチェニフォルミス(licheniformis)(例えば1989年4月12日に公開されたDD266710に開示されたバシリ・リチェニフォルミス41P);シュードモナス属、例えば緑膿菌;ストレプトマイセス属;アゾトバクタ;根粒菌;ビトレオシラ(Vitreoscilla);及びパラコッカス(Paracoccus)である。好適な大腸菌宿主には、大腸菌W3110(ATCC27325)、大腸菌94(ATCC31446)、大腸菌B及び大腸菌X1776(ATCC31537)が含まれる。これらの例は限定するものではなくむしろ例証のためのものである。
【0048】
1995年11月16日に公開されたPCT刊行物WO95/30686の全体を出典明示によりここに取込む。この刊行物は、そのNGFの細菌合成とインビトロ折り畳みの記載が特に関連する。その方法から得られた生成物を本発明の精製方法により精製することができる。
【0049】
上述の細菌の任意のものの突然変異体細胞を使用することもできる。細菌の細胞中におけるレプリコンの複製性を考慮に入れて好適な細菌を選択することは勿論必要である。例えば、大腸菌、セラチア、又はサルモネラ種が、例えばpBR322、pBR325、pACYA177、もしくはpKN410のようなよく知られたプラスミドがレプリコンを供給するために使用される場合、宿主として好適に使用することができる。
【0050】
大腸菌株W3110は組換えDNA産物発酵に対するよく知られた宿主株であるので、好適な宿主もしくは親宿主である。好適には、宿主細胞は最小量のタンパク分解酵素を分泌する。例えば、株W3110は、宿主に対して内在性のタンパク質をコードしている遺伝子で遺伝的突然変異を行うように修飾することができ、そのような宿主の例としては、完全遺伝子型tonAΔを持つ大腸菌W3110株1A2、完全遺伝子型tonAΔptr3を持つ大腸菌W3110株9E4、完全遺伝子型tonAptr3phoAΔE15Δ(argF−lac)169ΔdegPΔompTkan<r>を持つ大腸菌W3110株27C7(ATCC55244)、完全遺伝子型tonAptr3phoAΔE15Δ(argF−lac)169ΔdegPΔompTΔrbs7ilvGkanrを持つ大腸菌W3110株37D6、非カナマイシン耐性degP欠失型変異体を持つ株37D6である大腸菌W3110株40B4、及び1990年8月7日に発行された米国特許第4946783号に開示された変異体周辺質プロテアーゼを有する大腸菌株がある。
【0051】
ヒトNGFは大腸菌中に発現された。hNGFのβサブユニットをコードしている遺伝子の単離と配列及び大腸菌中での異種性タンパク質としてのその発現は米国特許第5288622号に記載された。そこでの教唆もまた哺乳動物細胞から生成した成熟ヒトNGFを提供するのに好適である。組換え技術を使用して、ヒトβ−NGFはその他の哺乳動物のタンパク質を含まないように発現された。変化したアミノ末端を含む2つの遺伝子を用いて大腸菌中にhNGFを発現させた結果、融合タンパク質が発現したが、これはIwaiら(Chem.Pharm.Bull.34:4724(1986))によって記載された。
【0052】
原核生物に加えて、糸状菌又は酵母菌のような真核微生物は、ニューロトロフィンをコードしているベクターのための適切な発現宿主である。サッカロミセス・セレヴィシア、又は一般的なパン酵母は下等真核生物宿主微生物のなかで最も一般的に用いられる。しかしながら、例えばシゾサッカロミセス・ポンベ(Beach及びNurse, Nature, 290:140(1981);1985年5月2日発行のEP139383);クルイヴェロミセス宿主[米国特許第4943529号; FleerらBio/ Technology, 9:968-975(1991)]、例えばK.ラクティス[MW98-8C, CBS683, CBS4574; Louvencourtら, J. Bacteriol.,737(1983)]、K.フラギリス(ATCC 12424), K.ブルガリカス (ATCC 16045)、K.ウィッケラミイ(ATCC 24178)、K.ワルティイ (ATCC 56,500), K. ドゥロソフィラルム[ATCC 36,906; Van den Bergら Bio/Technology,8:135(1990)]、K.サーモトレランス、及びK. マルクシアヌス;ヤロウィア[EP 402,226];ピチア・パストリス[EP183070; Sreekrishnaら, J. Basic Microbiol.,28:265-278(1988)];カンディダ;トリコデルマ・リーシア (EP 244234); ニューロスポラ・クラッサ [Caseら, Proc. Natl. Acad. Sci. USA, 76:5259-5263 (1979)]; シュワニオミセス・オクシデンタリスのようなシュワニオミセス[1990年10月31日公開のEP394538); 及び糸状菌、例えばニューロスポラ、ペニシリウム、トリポクラディウム[WO91/00357 1991年1月10日発行]、及びアスペルギルス宿主、例えばA.ニデュランス[Balanceら, Biochem Biophys. Res. Commun.,112;284-289(1983); Tilburnら,Gene,26:205-221(1983); Yeltonら, Proc. Natl. Acad. Sci. USA, 81:1470-1474 (1984)]及びクロカビ[Kellyら,EMBO J.,4:475-479(1985)]のような、多数の他の属、種、及び菌株が一般に利用でき、ここで有用である。
【0053】
ニューロトロフィンをコードしているDNAの発現に適切な宿主細胞は、多細胞生物からもまた誘導できる。このような宿主細胞は、複雑なプロセシング及びグリコシル化活動が可能である。原則的には、脊椎動物培養由来であろうと無脊椎動物培養由来であろうと、任意の更に高等の真核生物細胞培養が適している。無脊椎動物細胞の例としては植物及び昆虫細胞が含まれる。多数のバキュロウィルス株及び変異体及び対応する許容可能な昆虫宿主細胞、例えばスポドプテラ・フルギペルダ(毛虫)、アエデス・アエジプティ(蚊)、アエデス・アルボピクトゥス(蚊)、ドゥロソフィラ・メラノガスター(ショウジョウバエ)、ボンビクス・モリのような宿主が特定されている。例えば、Luckowら, Bio/Technology, 6:47-55 (1988); Millerら, Genetic Engineering, Setlowら編 Vol. 8 (Plenum Publishing, 1986), pp.277-279; 及びMaedaら, Nature,315:592-594(1985)を参照のこと。形質移入には種々のウィルス株が公に利用できる。例えば、オートグラファ・カリフォルニカNPVのL−1変異体とボンビクス・モリNPVのBm−5株があり、このようなウィルスは、特にスポドプテラ・フルギペルダ細胞の形質移入にここで使用することができる。ヒトNGFは1993年12月21日に発行された米国特許第5272063号に報告されているように、昆虫細胞において生産されている。
【0054】
綿花、コーン、ジャガイモ、大豆、ペチュニア、トマト、及びタバコのような植物細胞培養を宿主として用いることができる。典型的には、ニューロトロフィンをコードしているDNAを含むように前もって操作しておいた細菌アグロバクテリウム・トゥメファシエンスのある菌株と共にインキュベートすることによって植物細胞は形質移入される。A.トゥメファシエンスと共に植物細胞培養をインキュベートする間に、ニューロトロフィンをコードしているDNAが、植物細胞宿主が形質移入されるようにその植物細胞宿主に移され、適切な条件下でニューロトロフィンをコードしているDNAを発現する。加えて、例えば、ノパリンシンターゼプロモーター及びポリアデニル化シグナル配列のような、植物細胞と適合しうる調節及びシグナル配列が利用できる(Depickerら,J.Mol.Appl.Gen., 1:561(1982))。また、T−DNA780遺伝子の上流領域から分離されるDNAセグメントは、組換えDNAを含む植物細胞中の植物発現遺伝子の転写レベルを活性化又は増強しうる(1989年6月21日公開のEP321196)。
【0055】
有用な哺乳動物宿主細胞株の例は、SV40によって形質転換されたサル腎臓CV1株(COS-7, ATCC CRL 1651);ヒト胚腎臓株[293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamら, J. Gen Virol., 36:59 (1977)];ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/−DHFR[CHO, UrlaubとChasin, Proc.Natl.Acad.Sci.USA,77:4216 (1980)];マウスのセルトリ細胞[TM4, Mather,Biol.Reprod., 23:243-251 (1980)];サルの腎細胞 (CVI ATCC CCL 70); アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587); ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34); バッファローラット肝臓細胞 (BRL 3A, ATCC CRL 1442); ヒト肺細胞 (W138, ATCC CCL 75); ヒト肝細胞 (Hep G2, HB 8065); マウス乳房腫瘍細胞 (MMT 060562, ATTC CCL51); TRI細胞[Motherら, Annals N.Y. Acad. Sci., 383:44-68 (1982)]; MRC5細胞; FS4細胞; 及びヒト肝癌ライン(Hep G2)である。好適な方法はCHO細胞中の発現である。プレプロNGFを含むヒトNGFのエキソンを、適切なプロモーター及びベクターを使用して分泌された成熟NGF(118及び120型を含む)の発現を達成するために使用することができる(米国特許第5288622号)。安定して形質移入された安定なCHO細胞の培養及びNGFの分泌成熟型は本明細書の実施例において検討されるように、本発明において有用である。
【0056】
宿主細胞に形質移入し、そして上述の発現又はクローニングベクターで好ましくは形質転換し、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードしている遺伝子を増幅するために適当に修飾された常套的栄養培地で培養する。形質移入は、如何なるコード配列が実際に発現されるか否かにかかわらず、宿主細胞による発現ベクターの取り上げを意味する。多数の形質移入法が当業者に知られており、例えば、CaPO4及びエレクトロポレーションである。このベクターの操作のあらゆる徴候が宿主細胞内で生じたときに一般に形質移入の成功が認められる。
【0057】
形質転換は、染色体外のエレメントとしてであろうと染色体成分によってであろうと、DNAが複製可能であるように、生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。SambrookらのMolecular Cloning:A Laboratory Manual[New York: Cold Spring Harbor Laboratory Press,1989]の1.82節に記載された塩化カルシウムを用いるカルシウム処理又はエレクトロポレーションは、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して一般に用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawら, 23:315 (1983)及び1989年6月29日公開のWO89/05859に記載されたように、ある種の植物細胞の形質転換に用いられる。加えて、1991年1月10日に公開されたWO91/00358に記載されているように、超音波処理を用いて植物をトランスフェクトすることもできる。
【0058】
このような細胞壁のない哺乳動物の細胞に対しては、Graham及びvan der Eb(Virology, 52:456-457(1978))のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞の宿主系形質転換の一般的な側面は1983年8月16日に発行された米国特許第4399216号に記載されている。酵母中の形質転換は、典型的には、Van Solingenら(J.Bact., 130:946(1977))及びHsiaoら(Proc.Natl.Acad.Sci. USA,76:3829(1979))の方法によって実施する。しかしながら、DNAを細胞中に導入する他の方法、例えば、核マイクロインジェクション、エレクトロポレーション、無傷の細胞、又はポリカチオン、例えばポリブレン、ポリオルニチン等々を用いる細菌プロトプラスト融合もまた用いることができる。哺乳動物細胞を形質転換するための種々の方法については、Keownら(Methods in Enzymology(1990),Vol185:pp527-537)及び Mansourら(Nature,336:348-352(1988))を参照のこと。
【0059】
好適には、hNGFの遺伝子が、メチオニン開始コドン、好ましくはUllrichら(Nature,303:821-825(1983))により同定されたような二つのメチオニン開始コドンの一方を利用できるようにするためにベクター内に挿入される。hNGF遺伝子(Ullrichら, Cold Spring Harbor Symposia on Quant. Biol. XLVIII, p.435(1983);米国特許第5288622号)は、転写開始コドンとして使用されそうな2つの殆ど隣接するメチオニンを有している(位置1は成熟hNGFのN末端セリン残基をいう)。これに対して、NGFに対するマウス顎下腺cDNAは、神経成長因子の最も完全に研究されているものであるが、121と119の位置のメチオニンに加えて187の位置にもメチオニンを有している。哺乳動物細胞中の発現に対する好適な実施態様では、プレプロNGF配列が存在する。
【0060】
原核生物細胞がニューロトロフィンを生産するために使用される場合には、例えばSambrookらのMolecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory Press, New York 1989)において一般に記載されているようにしてプロモーターを構成的に又は人為的に誘導することができる適切な培地中で培養される。炭素、窒素、及び無機リン酸塩源以外に、あらゆる必要な補充物質を、単独もしくは複合窒素源のような他の補充物質又は培地との混合物として導入される適当な濃度で含めることもできる。
【0061】
ニューロトロフィンを生産するために哺乳動物の宿主細胞が用いられる場合、これは種々の培地において培養することができる。ハム(Ham)のF10(Sigma)、最小必須培地([MEM] Sigma)、RPMI−1640(Sigma)及びダルベッコの改良イーグル培地([DMEM], Sigma)のような市販培地が当該宿主細胞の培養に適している。また、HamとWallace, Meth. Enz., 58:44(1979), Barnes及びSato, Anal. Biochem., 102:255 (1980), 米国特許第4767704号;4657866号;4927762号; 5122469号;4560655号;WO90/03430;WO87/00195;米国再発行特許第30985号(これらの全ての開示を出典明示によりここに取込む)に記載された任意の培地を宿主細胞の培養培地として用いることができる。これらの培地はいずれも、ホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン、又は表皮成長因子)、塩類(例えば、塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、緩衝液(例えばHEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えば、ゲンタマイシン(商標)薬)、微量元素(最終濃度がマイクロモル範囲で通常存在する無機化合物と定義される)及びグルコース又は同等のエネルギー源を必要に応じて補充することができる。任意の他の必要な補充物質もまた当業者に知られている適当な濃度で含むことができる。培養条件、例えば温度、pH等々は、発現のために選ばれた宿主細胞について従来用いられているものであり、当業者には明らかであろう。
【0062】
一般に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、及び実用技術は、Mammalian Cell Biotechnology: A Practical Approach, M. Butler編 (IRL Press at Oxford University Press,Oxford,1991)に見出すことができる。上記の方法は、ニューロトロフィンが細胞内に生産されようが、細胞膜周辺膣に生産されようが、あるいは培地に直接分泌されようが、使用することができる。
【0063】
典型的には、培養液は適当な時間後に収集され、標準的な同定アッセイ、例えばELISA及びウェスタンブロット分析のようなイムノアッセイ、又は生物学的アッセイ、例えばPC12細胞分化(Greene,L.A., Trends Neurosci. 7:91 (1986))が実施される。ここに開示された変種の種類と程度を決定するためのアッセイは従来から知られているか、あるいは実施例において提供されもしくは引用される(例えば、出典明示によりここに取込むSchmelzerらJ.Neurochem.59 (5):1675-83(1992)及びBurtonら,J.Neurochem. 59(5):1937-45(1992)を参照)。
【0064】
細胞から調製されるニューロトロフィン組成物は、好ましくはHICに先立って少なくとも一つの精製工程に通される。好適な精製工程の例には、回収すべきニューロトロフィン及び使用する出発培養に依存して、アフィニティクロマトグラフィー、シリカクロマトグラフィー、ヘパリンセファロースクロマトグラフィー、アニオンもしくはカチオン交換樹脂クロマトグラフィー(例えばポリアスパラギン酸カラム)、クロマトフォーカシング、及び調製的SDS−PAGEのようなタンパク質精製の他の方法を含むここに記載したものを含む。
【0065】
ニューロトロフィンが培地中に直接分泌される一実施態様においては、培地を遠心分離により細胞片から分離し、清澄化された発酵ブロス又は培地を次にシリカゲルでの精製に使用する。シリカクロマトグラフィーに対しては、典型的にはニューロトロフィンポリペプチドがシリカ粒子に付着するように誘導体化されていないシリカ粒子にブロスを通す;シリカ粒子を洗浄して汚染物質を除去する;そしてアルコール性もしくは極性非プロトン溶媒とアルカリ土類、アルカリ金属もしくは無機アンモニウム塩を含む緩衝液でシリカ粒子からポリペプチドを溶出させる。
【0066】
本発明の好適な実施態様では、ニューロトロフィンをその変種から分離し、またバルク汚染物質を減少させるためにマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーを用いる。この樹脂は、哺乳動物細胞培養からのニューロトロフィンの精製の初期の工程として、好ましくは収集細胞培地の分画に対して使用することができる。他の実施態様では、マクロプレップ・ハイ・Sカチオン交換クロマトグラフィーは、最も好ましくはタンパク質再折り畳み工程の直後に使用される。このカチオン交換カラムの非常に高い流れ性により、大量の希薄な再折り畳みタンパク質又は収集細胞培地ニューロトロフィンを、例えばHIC又はSP−セファロースのような次のクロマトグラフィー工程の前に、ニューロトロフィンがカラムに結合する条件を調節することにより、容易く濃縮することが可能になる。更に、樹脂のカチオン交換性により、非結合バルクタンパク質及びある種の誤ってプロセシングされた変種及び化学的に修飾された変種(例えば変更負荷、MET37−酸化変種)の除去が可能になる。最も重要なことは、未変性のニューロトロフィンと疎水性が異なる誤ってプロセシングされた変種もしくは化学的に修飾された変種(例えば(哺乳動物細胞培養生産からの)誤ってプロセシングされた変種、グリコシル化変種)が存在する場合、マクロプレップ樹脂により、これらの疎水性変種の部分的な除去が可能になり、ここで測定されるように、未変性のニューロトロフィンを実質的に富化することになることである。限定することを意図するものではないが、樹脂支持体のバックボーンはニューロトロフィンと樹脂の間の非特異的相互作用を促進する疎水性含量を含むものと信じられ、これがここに教示されるようにして利用される。典型的には、約5から8、より好ましくは6から8のpHの溶出緩衝液に対しては、0から3MのTMAC濃度が有用である。酢酸ナトリウムが、溶出緩衝液のイオン強度を増大させるために存在する場合、より低いTMAC濃度を使用することができる。塩化物は酢酸塩イオンに対する好適な代替物である。
【0067】
ニューロトロフィンが細胞膜周辺膣において生産される実施態様の一例では、培地又は可溶化物が遠心分離されて粒子状の細胞片を除去する。ついで、必要に応じて膜と可溶性タンパク質分画を分離することができる。ついで、ニューロトロフィンは、ニューロトロフィンが膜結合性か、可溶性か、凝集化形態で存在しているかに応じて、可溶性タンパク質分画から、そして培養可溶化物の膜分画から精製することができる。その後、ニューロトロフィンは可溶化され、ついで適切な緩衝液を使用して再び折り畳まれる。再折り畳みタンパク質を生産するための周辺質からのこの単離方法の詳細については以下に記載する。
【0068】
不溶性の未変性ではないニューロトロフィンは、適切な単離緩衝液中の原核生物宿主細胞から、任意の適当な技術、例えば細胞を適切なイオン強度の緩衝液に曝露して殆どの宿主タンパク質を可溶化させ(しかし凝集ニューロトロフィンは実質的に不溶性である)、封入体を放出し、例えば遠心分離により回収に利用できるようにするように細胞を破壊することにより、単離される。この技術はよく知られており、例えば米国特許第4511503号に記載されている。
【0069】
簡単には、細胞は(典型的には、約0.01から2M、好ましくは0.1から0.2Mのイオン強度を使用して、pH5から9、好ましくはpH約6から8の)緩衝液中に懸濁される。塩化ナトリウムを含む任意の適切な塩が十分なイオン強度値を維持するために有用である。細胞は、この緩衝液中に懸濁され、ついで、例えば機械的方法、例えばManton−Gaulinプレスマイクロフルイダイザー、フレンチプレスもしくは音波オシレーター、あるいは化学的又は酵素的方法のような通常用いられている技術を使用して溶解により破壊する。
【0070】
細胞破壊の化学的もしくは酵素的方法の例には、細菌壁を溶解するリゾチームの使用を伴うスフェロプラスティング(Neuら, Biochem.Biophys.Res.Comm.,17:215(1964))及びポリペプチドの放出のために生細胞を高緊張度の溶液と低緊張度の冷水洗浄で処理することを含む浸透圧ショック(Neuら, J.Biol.Chem.,240:3685-3692(1965))が含まれる。米国特許第4680262号に記載されている第3の方法は、形質転換された細菌細胞を、細胞を死滅させ溶解するのに充分な時間と温度で、有効量の2から4の炭素原子を持つ低級アルカノールに接触させるものである。
【0071】
細胞を破壊した後、懸濁液は典型的には遠心分離されて封入体をペレット化する。一実施態様では、この工程は、標準的な遠心機において、容量と遠心設計に依存する充分な時間、通常は約10分から0.5時間、約500から15000xg、好ましくは約12000xgで実施される。得られたペレットは実質的に全ての不溶性のポリペプチド分画を含むが、細胞破壊プロセスが完全でないならば、無傷の細胞もしくは破壊された細胞フラグメントも含有し得る。細胞破壊の完全性は、ペレットを少量の同じ緩衝液中に懸濁させ、懸濁液を位相差顕微鏡で調べることにより検査することができる。破壊細胞フラグメントもしくはホールセルの存在は、フラグメントもしくは細胞及び随伴する非屈折性(non−refractile)ポリペプチドを除去するために更なる破壊が必要であることを示している。そのような更なる破壊後に、必要ならば、懸濁液を再び遠心分離し、ペレットを回収し、再懸濁し、分析する。このプロセスを、視覚検査でペレット化材料中に破壊細胞フラグメントが無いことが明らかになるか、更なる処理でも得られるペレットの寸法を低減することができなくなるまで繰り返す。
【0072】
別の実施態様では、ニューロトロフィンは、適切な緩衝液中での可溶化により細胞膜周辺膣から単離される。この方法は、ニューロトロフィンが組換え的に生産された後に発酵ベッセルに試薬を直接添加して、収集、ホモジェナイゼーション、及び遠心分離という余分な工程を避けてニューロトロフィンを得るインサイツ溶解化とできる。残りの粒子は遠心分離か濾過もしくはその組合せにより除去することができる。
【0073】
ニューロトロフィンがアンフォールディングされているならば、アンフォールディングの度合いは、RP−HPLCを含む非天然ニューロトロフィンのクロマトグラフィーによって適切に決定される。非天然材料に対して増加するピーク領域は、どのくらいの非天然ニューロトロフィンが存在しているかを示す。
【0074】
可溶化された封入体から又は後の精製の段階で一度得られると、ニューロトロフィンは以下に記載されるように活性な高次構造に適切に再折り畳みされる。
【0075】
ニューロトロフィンが、再折り畳みされる前に既に可溶形態ではないならば、ニューロトロフィンを実質的に溶解させるのに必要な量のカオトロピック剤と還元剤を含有するアルカリ緩衝液におけるインキュベーションにより可溶化することができる。
【0076】
ニューロトロフィンの緩衝液中における可溶化の度合いの測定は、濁度決定法により、還元されたSDSゲル上の遠心分離後に上清とペレット間のニューロトロフィン分画を分析することにより、プロテインアッセイ(例えば、バイオ-ラッドタンパク質アッセイキット)により、又はHPLCにより好適に実施される。
【0077】
可溶化に対するアルカリ緩衝液のpH範囲は典型的には少なくとも約7.5であり、好適な範囲は約8−11である。この後の範囲のpHをもたらす適切な緩衝液の例としては、グリシン、CAPSO(3-[シクロヘキシルアミノ]-2-ヒドロキシ-1-プロパンスルホン酸)、AMP(2-アミノ-2-メチル-1-プロパノール)、CAPS(3-[シクロヘキシルアミノ]-1-プロパンスルホン酸)、CHES(2-[N-シクロヘキシルアミノ]エタンスルホン酸)、及びトリスHCl(トリス[ヒドロキシメチル]アミノメタンヒドロクロリド)が含まれる。ここで好適な緩衝液は、好ましくは約20mMの濃度、約8.5から11のpH、好ましくは約10−11のpHのグリシン又はCAPSOである。
【0078】
可溶化のための緩衝溶液中のニューロトロフィンの濃度は、ニューロトロフィンが実質的に溶解化され、部分的に又は充分に還元し、変性するようなものでなければならない。あるいは、ニューロトロフィンは最初は不溶性であってもよい。使用する正確な量は、例えば緩衝溶液中の他の成分の濃度とタイプ、特に還元剤のタイプと量、カオトロピック剤のタイプと量、および緩衝液のpHに依存する。例えば、ニューロトロフィンの濃度は、還元剤、例えばDTTの濃度が、約3:1から10:1のDDT:ニューロトロフィンの比を維持するために、同時に増加させられるならば、少なくとも3倍増加させることができる。希釈再折り畳みの前により濃縮された可溶化タンパク質溶液をつくることが望ましい。従って、ニューロトロフィンの好適な濃度は少なくとも約30mg/mLで、より好ましい範囲は30−50mg/mLである。例えば、ニューロトロフィンは、5Mから7Mの尿素中に約30−50mg/mLの濃度まで可溶化し、例えば約1mg/mLまで折り畳みのために希釈することができる。
【0079】
ニューロトロフィンを可溶化した後、これを、5−40%(v/v)のアルコール性又は非プロトン溶媒、カオトロピック剤、及びアルカリ金属、アルカリ土類、もしくはアンモニウム塩を含む再折り畳み緩衝液中に入れるか希釈する。緩衝液は、最初の緩衝溶液に対しては任意の緩衝液とすることができ、CAPSO、グリシン、及びCAPSがpH8.5−11、特に約20mMの濃度で好ましく、CAPSOとグリシンが最も好ましい。ニューロトロフィンは、好ましくは少なくとも5倍、より好ましくは少なくとも約10倍に、再折り畳み緩衝液で希釈することができる。あるいは、ニューロトロフィンは、再折り畳み緩衝液に対して透析することができる。再折り畳みは約2℃−45℃、最も好ましくは2℃−8℃で少なくとも約1時間の間、実施することができる。溶液は、場合によっては還元剤と浸透質(osmolyte)をまた含有してもよい。
【0080】
還元剤は与えられた濃度範囲における可溶化工程に対して上述したものから好適に選択される。その濃度は、特に、アルカリ土類、アルカリ金属、又はアンモニウム塩、ニューロトロフィン、及び溶媒の濃度に依存する。好ましくは、還元剤の濃度は、約0.5から8mM、より好ましくは約0.5−5mM、更により好ましくは約0.5−2mMである。好適な還元剤はDTTとシステインである。
【0081】
再折り畳み溶液中の酸素は必要に応じて不活性ガス、例えばヘリウム又はアルゴンの添加により除去して酸素を置換することもできる。
【0082】
任意の浸透質は好ましくはスクロース(約0.25−1Mの濃度)又はグリセロール(約1−4の濃度)である。より好ましくは、スクロース濃度は約1Mであり、グリセロール濃度は約4Mである。
【0083】
折り畳み緩衝液中のニューロトロフィンの初期濃度は、回収された誤って折り畳まれた配座異性体に対する正しく折り畳まれたものの比が、HPLC、RIA、もしくはバイオアッセイにより決定したとき、最大になるようなものである。(最大収量の正しく折り畳まれた配座異性体が得られる)ニューロトロフィンの好適な濃度は約0.1から15mg/mL、より好ましくは約0.1から6mg/mL、最も好ましくは約0.2から5mg/mLの範囲である。
【0084】
このインキュベーション時に生じる再折り畳みの度合いは、ニューロトロフィンのRIA力価又はHPLC分析により適切に決定され、RIA力価又は正しく折り畳まれたニューロトロフィンピーク寸法の増加は、緩衝液中に存在する正しく折り畳まれた生物学的に活性なニューロトロフィン配座異性体の量の増大に相関する。インキュベーションは、RIAもしくはHPLCにより決定される、正しく折り畳まれたニューロトロフィン配座異性体の収量と回収された誤って折り畳まれたニューロトロフィン配座異性体に対する正しく折り畳まれたニューロトロフィン配座異性体の比を最大にし、質量バランスにより決定される多量体関連ニューロトロフィンの収量を最小にするために実施される。あるいは、以下と実施例に記載する方法によって種を決定することもできる。グアニジンが再折り畳みに対する好適な変性剤である。
【0085】
ニューロトロフィンが再折り畳みされた後、ここに教示されたような次の方法が、個々にあるいは組み合わせて、より高い純度と相同性を得るための好適な精製法である。すなわち、カチオン交換カラムでの分画、疎水的相互作用クロマトグラフィー(HIC)、そしてシリカクロマトグラフィーである。
【0086】
再折り畳み工程がプロセスの一部であろうとなかろうと、ニューロトロフィンをその変種から分離するための好適な工程は疎水的相互作用クロマトグラフィー樹脂上での分離である。発酵、精製又はインビトロタンパク質再折り畳みの間、あるタンパク質は化学的に修飾され、誤ってプロセシングされ得、あるいはその未変性の三次元構造ではなくむしろその安定性、溶解性、免疫原性、もしくは生物活性について異なる他の構造に再折り畳みされうる。これらの変種は、抗原性又は作用強度の喪失のような望ましくない副作用を避けるために回収中に除去されなければならない。変種が不溶性であれば、遠心分離と濾過のような固体/液体分離法により容易く除去することができる。しかしながら、変種が可溶性であれば、クロマトグラフィーのようなより高度の分離吸着法がそれらを除去するために必要とされる。原核生物細胞中に生産されるとき、あるいはインビトロで再折り畳みされるとき、ニューロトロフィンは可溶性の安定した誤って折り畳まれた変種を生成する。誤って折り畳まれたニューロトロフィンは、未変性のニューロトロフィンに対して変更されたジスルフィド対合パターンと三次元構造を有しており、天然の薬理学的活性を欠く。真核生物細胞培養、例えば哺乳動物細胞培養中に生産されるとき、変種型は典型的には誤ってプロセシングされた型である。これらはまた典型的に天然の薬理学的活性を欠き、除去されるべきである。ここではHICが、未変性のニューロトロフィンからこれらの変種を分離するために適したものであることが見出された。
【0087】
HICは非共有性疎水的結合によって媒介された固体マトリックスに対するタンパク質の順次の吸着と脱着を含む。一般に、高塩緩衝液中の試料分子がHICカラムに装填される。緩衝液中の塩は水分子と相互作用をして溶液中の分子の溶媒和を低減させ、よってHICカラムにより結果的に吸着される試料分子の疎水性領域を暴露する。分子がより疎水性であればある程、結合を促進するために必要とされる塩は少なくなる。通常、塩勾配の低下を使用してカラムから試料を溶出させる。イオン強度が減少すると、分子の親水性領域の暴露が増加し、疎水性を増加させるために分子がカラムから溶出する。試料の溶出はまた溶出緩衝液に穏やかな有機修飾因子又は洗浄剤を添加することによっても達成される。HICは、Protein Purification2版(Springer-Verlag, New York, pp.176-179(1988))においてレビューされている。
【0088】
タンパク質とマトリックスの間の会合の強さは、固定化された官能基の大きさと疎水的性質、周囲の溶媒の極性と表面張力、及びタンパク質の疎水性を含む幾つかの要因に依存する。HICマトリックスの結合能は、固定化された疎水性リガンドが広く間隔をあけられるべきという必要性のために低くなる傾向がある。更に、あるタンパク質に対する培地の能力は試料調製における疎水性不純物の量に対して逆に変化する。所望のタンパク質を、能力を同時に最大化しながら、変種と他の不純物から分離するためには、好適なHIC固相媒体並びに装填、洗浄及び溶出のための好適な移動相を特定する必要がある。
【0089】
ここにおいて決定したところでは、無傷の正しく折り畳まれた型から正しく折り畳まれたニューロトロフィン及び誤って折り畳まれたニューロトロフィンもしくは誤ってプロセシングされた型を分離するための最も好適な媒体は、固定化されたフェニル官能基を有するものであった。異なった販売者からのフェニルベースHIC媒体はこれらのニューロトロフィン型を分離するための異なった効率を示した。最良の結果はTosoHaasによるフェニル・トーヨーパール媒体及びフェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)で達成された。TSKフェニル5PWもまた適していた。他のHIC固定化官能基も機能してこれらの型を分離しうる。例としては、オクチル基、例えばファーマシアからのオクチルセファロースCL4B媒体、及びプロピル基、たとえばBakerからのハイ・プロピル媒体であった。好ましさが劣ったものはアルコキシ、ブチル、及びイソアミル官能基樹脂である。
【0090】
HICはニューロトロフィンを哺乳動物細胞培養中のその変種から分離するのに有用である。例えば、ここで測定したところでは、rhNGF発現CHO細胞培養は不適切にタンパク分解性にプロセシングされた変種、たとえば前駆NGF、ハイブリッド前駆NGF及び切詰め前駆NGF配列のような部分的配列が存在するものを含んでいた。また哺乳動物細胞培地において見出されるものはグリコシル化NGF及び不適切にタンパク分解性にプロセシングされた変種のグリコシル化形態である。望ましくないグリコシル化型は、NGFの場合大きい分子量の種(+2000kD)として見られるが、患者に望ましくない抗原応答を発生させ、劣った製品品質又は活性をもたらす。HICは、rhNGFから疎水性変種、主にグリコシル化型を含むN末端がタンパク分解性に誤ってプロセシングされた変種を分離した。実施例に示されるように、前駆配列含有及び切詰め前駆配列NGFと、NGFと前駆配列含有NGFの両方のグリコシル化型がフェニルHICの間NGFピークの先端に溶出した。従って、これらの種が実質的に含まれておらず、高速カチオン交換クロマトグラフィーのような次の工程に特に適したrhNGF組成物を得ることができた。HICは、ソースに拘わらず、他のニューロトロフィン、並びにNGFにも適用できる。例えば、HICは、存在する単量体形に依存してホモ二量体かヘテロ二量体の何れでも、二量体からNGF単量体を分離し、並びにインビトロの再折り畳みの後、又は哺乳動物細胞から生産され分泌されたときに得られる、疎水性がまた異なる二量体を区別するのに有用である。HICに使用するニューロトロフィン混合物の好適なソースは哺乳動物細胞培養、より好ましくはCHO細胞培養である。該培養は好ましくはここで検討されたような少なくとも1回の前精製工程にかけられる。HICは、未変性の組換えニューロトロフィンから誤ってプロセシングされたグリコシル化変種(群)を分離するのに特に有効である。rhNGFの場合、グリコシル化及びプレプロNGFは未変性NGFより疎水性が低く、従って未変性NGFの前に溶出する。(細菌的に生産される場合)ニューロトロフィンの誤って折り畳まれた型もまたより疎水性で、未変性のニューロトロフィンより早く溶出する。
【0091】
ニューロトロフィン型を分離するために最も好適なHIC樹脂は固定化されたフェニル官能基を有しているものであった。異なった販売者からのフェニルベースHIC媒体はこれらのNGF型を分離するための異なった効率を示した。フェニルHIC樹脂の中で、TosoHaasによるフェニル・トーヨーパール媒体が最も好ましく、フェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)とTSKフェニル5PWが好ましい。好ましいHIC官能基としてはアルコキシ、ブチル、及びイソアミル部分が含まれる。
HICを使用して、ニューロトロフィン型を洗浄し差次的に溶出させるために様々な移動相条件を使用することができる。これらの移動相はニューロトロフィンと固定相の間の会合に異なった方法で影響を及ぼす幾つかの異なった化学種を含みうる。正しく折り畳まれ、また誤って折り畳まれたニューロトロフィン、例えばNT−4/5が、例えば硫酸アンモニウム、NaCl濃度、酢酸塩濃度のような移動相塩の段階的減少又は減少塩勾配によりHICカラム上で分離できる。塩は移動相の表面張力を調節することにより樹脂に対するニューロトロフィンの結合に影響を及ぼしうる。表面張力に影響を及ぼした他の薬剤は、実施例で検討したようなクエン酸ナトリウムとテトラメチルアンモニウムクロリドであった。変種はまた、相対的に極性の有機溶媒の濃度を増加勾配で又は段階的に増加させて結合タンパク質を溶出させることにより、カラムクロマトグラフィー中に分離することができる。好適な溶媒の例としてはエタノール、アセトニトリル、及びプロパノールがある。ニューロトロフィン型とHIC樹脂の間の会合の強さは移動相のpHにもまた依存し、中性条件が好ましい。正しく折り畳まれ、誤って折り畳まれたニューロトロフィンの相対的な疎水性はまた溶液のpHに依存する。未変性のニューロトロフィンからの変種の分離はまた、勾配もしくは段階的溶出の間、移動相のいくつかの性質を同時に変化させることにより得ることもできる。例えば、溶出時に塩濃度と非極性溶媒濃度を同時に変化させた移動相は塩のみを変化させた場合よりもより良好な分離をもたらした。
【0092】
HICに対して、硫酸アンモニウム、クエン酸塩、酢酸塩、及び塩化カリウムを含むここで検討した塩が使用できる。使用する塩に依存して、樹脂へのニューロトロフィンの結合を達成する塩の濃度は典型的に0.5Mから3M、より好ましくは0.5から2.5Mである。例えば、0.8から1.5Mの塩の結合緩衝液がNGFに対して好適であり、より高い塩濃度では樹脂上へのNGFの沈殿が生じ、回収率が低下する。NT−3に対しては、1.0から2.5の塩濃度のpH7の結合緩衝液が好ましく、1.25から1.75Mの塩が更に好ましく、1.5Mが最も好ましい。NT−4/5に対しては、1から3Mの塩のpH7の結合緩衝液が好ましく、2から2.75Mが更に好ましく、2.5MのNaClが最も好ましい。NT−4/5の場合、2.5MのNaClが装填には好ましく、2MのNaClが有機溶媒の存在する(例えば10%アルコール、pH7)溶出には好適であった。好ましくは、塩濃度の低下を使用してニューロトロフィンとその変種を溶出させ分離する。溶出を達成するには、溶出緩衝液中の塩濃度は、典型的には装填緩衝液中のものよりも低いが、有機溶媒で補償するときは同じ濃度にすることができる。
【0093】
加えて、ここで見出されたところでは、有機溶媒の使用には、有機溶媒の添加がより狭いピークプロファイルをもたらすことにより溶出パターンを改善するというもう一つの利点がある。エタノールに加えて、プロパノール、イソプロパノール、及び低級アルキレングリコール、例えばプロピレングリコール、エチレングルコール及びヘキシレングリコールを含むここで検討した他の有機溶媒を使用することができる。5から25%(v/v)、より好ましくは5から20(v/v)の有機溶媒が正しく折り畳まれたニューロトロフィンを典型的に溶出させる。有機溶媒での溶出は勾配をもたせても段階的でもよい。pH範囲は好ましくは殆ど中性から僅かに酸性で、5から8、より好ましくは6から8、6.5から7.5、最も好ましくは7である。MOPSO、MOPS、HEPES、リン酸塩、クエン酸塩、アンモニウム、酢酸塩を含むここで検討した任意の緩衝液を、所望のpHで緩衝作用を呈する限り、使用することができる。
【0094】
ここに報告したように、ニューロトロフィンの組換え生産から生じるある望まれないニューロトロフィン変種の本発明者らによる発見に鑑みると、好ましくは調製的態様で、高速カチオン交換クロマトグラフィーを使用することにより未変性ニューロトロフィンからの例えばカルバミル化、酸化、isoasp、脱アミド、およびある種の切詰め型(例えばNGFのC末端切断型)のような荷電修飾変種の分離が可能になる。例えば、NT−4/5とNT−3の場合のように細菌発酵中に生じうる負荷変更の結果として生じるN末端クリップ型(例えば2から4のN末端アミノ酸欠失)を除去することができる。哺乳動物細胞培養において生産されたニューロトロフィンの場合、C末端切断が高度に荷電された末端領域に生じ得る。例えば、NGFの118型はC末端で誤ってプロセシングされ又は切断されて117、114及び115型になる。これらは高速カチオン交換クロマトグラフィーにより未変性118NGFから分離することができる。特に好適な実施態様はSP−セファロース・ハイ・パーフォーマンス、フラクトゲルEMD SO3、またはポリアスパラギン酸樹脂を使用するもので、中でもポリキャットAが特に好ましい。最も好ましくは、大規模では、SP−セファロース・ハイ・パーフォーマンス、又はフラクトゲルEMD SO3樹脂が使用される。
【0095】
ここに記載された方法により得られる組成物は、実質的に純粋なニューロトロフィン、更に通常的には好ましくは本質的に純粋で、実質的にニューロトロフィン変種を含まず、より好ましくは本質的にニューロトロフィン変種を含まない。例えば、CHO細胞培養からのNGFの精製後の典型的なSP−セファロースプールは、約92%の118、4.6%の120、1%の脱アミド化NGF、1%の酸化NGF、及び1%のisoaspNGFを含む。各々の量は、常に、118に対しては約85から93%、(用いられる内因性及び/又は外因性タンパク分解度合いに大きく依存して)120に対しては0から5%、117に対しては0から5%、脱アミド型に対しては0から3%、isoasp型に対しては0−2%、及び酸化型に対しては0から2%の範囲である。NGF(全ての種)の純度は常に99.5%を越える。
【0096】
ニューロトロフィンは、カラムから溶出された後、担体と共に好適に組成物、好ましくは生理的に許容できる担体と共に製薬組成物に製剤化される。ニューロトロフィン組成物は好ましくは無菌である。本発明のニューロトロフィン組成物は、例えば培養中において神経細胞の成長と生存を促進するためにインビトロでの用途が見出される。
【0097】
組換えヒト神経成長因子(NGF)の水溶液中における化学的及び物理的安定性を、4.2から5.8のpH範囲で5℃と37℃の間で調べた。NGFの化学安定性はpHの増加で増大した。pH5.8のコハク酸塩緩衝液中では、NGFの物理的安定性はタンパク質凝集化のために減少した。5℃の安定性データと37℃での加速分解研究の両方に基づいて、最適な製剤はpH5.5の酢酸塩緩衝液であることが分った(ここに出典明示により取込まれるWO97/17087を参照のこと)。逆相HPLCは第1の安定性を示す方法であり、iso−AspへのAsn−93の転換が5℃での第1の分解経路であることを示した。カチオン交換クロマトグラフィーによるNGF分解の定量化はNGF単量体変種の様々な混合二量体への経時的な再構成(二量体交換)により複雑化した。希酸での試料と参照の処理は二量体中の単量体分布を平衡化させ、二量体交換の不存在下で定量化された。ベンジルアルコールとフェノールが、多用途目的に対する2つの液体製剤中へのそのrhNGFとの相容性と安定性について評価された。これらの二つの製剤は、界面活性剤として0.01%のプルロン酸(F68)を含む状態と含まない状態で、pH5.5の20mMの酢酸ナトリウムと136mMの塩化ナトリウム中の0.1mg/mLのタンパク質からなる。これらの二つの製剤の各々におけるベンジルアルコールとフェノールの最終濃度はそれぞれ0.9%と0.25%である。12ヶ月の安定性データに基づくと、rhNGFはこれらの製剤においてフェノールよりもベンジルアルコールでより安定である。界面活性剤の存在下でベンジルアルコールで保存したrhNGF製剤は界面活性剤を添加しない製剤と同じほど安定であり、rhNGF多投薬製剤へのF68の添加は安定性目的では必要とされないことを示している。従って、20mMの酢酸塩、136mMのNaCl、0.9%のベンジルアルコール、pH5.5中に0.1mg/mLのタンパク質を含んでなる製剤がフェーズIIIの臨床試験において複数回の投薬に使用されるrhNGFに対して推奨される。このrhNGF多投薬製剤は、5℃で6ヶ月後にUSP及びEP保存剤効能試験をパスしており、2mg/mLの現在の液体製剤と同じほど安定している。しかし、製剤は、光感受性がある保存剤としてベンジルアルコールを含むので、強い光への曝露は避けるべきである。
【0098】
一般に、組成物は、好ましくは安定な液体又は凍結乾燥可能形態での調製を不可能にしない量であって、効果的で安全な製薬的投与に適した量で、他の成分を含むことができる。
【0099】
ニューロトロフィンは、製薬的に許容できる担体、すなわち使用される用量と濃度でレシピエントに対して無毒性であり、製剤の他の成分と相容性があるものと共に製剤化される。例えば、製剤は、ポリペプチドに対して有害であることが知られている酸化剤及びその他の化合物を含有しない。この製剤工程は、標準的な技術を使用する脱塩又はダイアフィルトレーションにより達成される。
【0100】
一般に、製剤は、液体担体又は細かく分割された固体担体又は両方とニューロトロフィンを均一かつ密に接触させることにより調製される。ついで、必要ならば、製品は所望の製剤に成形される。好ましくは、担体は非経口担体であり、より好ましくはレシピエントの血液と等張である溶液である。そのような担体ビヒクルの例としては、水、生理食塩水、リンガー液及びブドウ糖溶液が含まれる。固定化オイル及びオレイン酸エチルのような非水性ビヒクル、並びにリポソームがここでは有用である。
【0101】
担体は、好適には、等張性及び化学安定性を高める物質のような添加物を少量含有する。このような材料は使用される用量と濃度でレシピエントに対して非毒性であり、例えば、リン酸塩、クエン酸塩、コハク酸塩、酢酸、及び他の有機酸又はその塩のような緩衝液;アスコルビン酸のような抗酸化剤;低分子量(約10残基未満)ポリペプチド、例えばポリアルギニン又はトリペプチド;タンパク質、例えば血清アルブミン、ゼラチン、又は免疫グロブリン;例えばポリビニルピロリドンのような親水性ポリマー;アミノ酸、例えばグリシン、グルタミン酸、アスパラギン酸、又はアルギニン;セルロース又はその誘導体、トレハロース、グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物;EDTAのようなキレート剤;マンニトール又はソルビトールのような糖アルコール;ナトリウムのような対イオン;及び/又はポリソルベート、ポロキサマー、又はPEGのような非イオン性界面活性剤を含む。最終調製物は液体又は凍結乾燥固体とできる。
【0102】
治療投与に対して使用されるニューロトロフィンは無菌でなければならない。無菌性は無菌濾過膜(例えば0.2ミクロン膜)を通す濾過によって直ぐに達成される。一般に、治療用ニューロトロフィン組成物は、殺菌アクセス口、例えば皮下注射針で穿孔可能なストッパーを持つ静脈内溶液袋又はバイアルを有する容器に入れられる。上記の製剤はまたインビトロ用途に適している。
【0103】
通常、ニューロトロフィンは、ユニット又は多投薬容器、例えば密封アンプル又はバイアルに、水溶液として、あるいは再構成のために凍結乾燥製剤として保存される。凍結乾燥製剤の例としては、10mLのバイアルに5mLの無菌濾過1%(w/v)ニューロトロフィン水溶液が満たされ、得られた混合物が凍結乾燥される。注入溶液は凍結乾燥ニューロトロフィンを注射用静菌水を使用して再構成することにより調製される。
【0104】
治療的に有効な用量のニューロトロフィン製剤が患者に投与される。ここにおける「治療的に有効な用量」は、投与される効果をつくりだす用量を意味する。正確な用量は治療すべき疾患に依存し、既知の技術を使用して当業者により決定される。一般に、本発明のニューロトロフィン製剤は、1日当り約0.01μg/kgから約100mg/kg投与される。好ましくは0.1から0.3μg/kgである。また、従来から知られているように、年齢並びに体重、一般的な健康状態、性別、食事、投与時間、薬物相互作用および疾患の重症度に対する調節が必要となり、当業者によりルーチン的な実験により決定される。典型的には、臨床医は、神経細胞の機能を修復し、維持し、最適には再構築する用量に達するまで本発明のニューロトロフィン製剤を投与する。この療法の進行は通常のアッセイで容易にモニターされる。
【0105】
ニューロトロフィンは、NGF、NT−4/5、NT−3、及び/又はBDNFを含む他の神経栄養因子と組み合わされ、あるいは一緒に投与され、神経疾患に対する他の通常の療法と共に使用される。
【0106】
NGFの場合、好ましくは組成物は、製薬的に有効量の神経成長因子と製薬的に許容できる酢酸塩含有緩衝液を含有する。組成物は5から6のpHを有し得る。緩衝液は好ましくは酢酸ナトリウムである。酢酸塩の濃度は好ましくは0.1から200mMである。組成物は好ましくは0.07から20mg/mlのNGF濃度を有する。そして、組成物は更に製薬的に許容できる保存剤、例えばベンジルアルコール、フェノール、m−クレゾール、メチルパラベン、又はプロピルパラベンを任意に含有する。好ましくは保存剤はベンジルアルコールである。ベンジルアルコールの濃度は好ましくは0.1から2.0%である。組成物は製薬的に許容できる界面活性剤を任意に含有し得る。そして、組成物は、生理的に許容できる濃度の塩化ナトリウムを任意であるが好ましく含有することができる。より好ましい組成物は少なくとも約0.1mg/mlの濃度で神経成長因子を、10mMから50mMの濃度の酢酸塩イオンを含有する。更により好ましくは、組成物は少なくとも約0.1から約2.0mg/mlの濃度で神経成長因子を、10mMから50mMの濃度の酢酸塩イオンを含有する。最も好ましい組成物は、0.1mg/mlの濃度のNGF、20mMの酢酸ナトリウム濃度(pH5.5)、136mMの塩化ナトリウム濃度及び0.9%(v/v)のベンジルアルコールを含有する。
【0107】
他の実施態様は、2.0mg/mlのNGF濃度、10mMの酢酸ナトリウム濃度、pH5.5、及び142mMの塩化ナトリウム濃度を含む。0.1mg/mlのNGFを20mMの酢酸ナトリウム、136mMの塩化ナトリウム、0.9%(v/v)のベンジルアルコールを5.5のpHで製剤化することが好ましい。ここで検討したところ、118/118ホモ二量体がNGFの好適な型である。NGFは正常な二量体形態を維持するために約6から8のpHで精製される。しかし、(タンパク質分解性に)切詰められた型のパーセントは単量体形態を表し、これは逆相HPLCによって明らかになり測定される。NGFの異なった二量体型−120/120、120/118、118/118等々の存在は公表されている(主に本研究に使用された分析的及びバイオアッセイについて、並びにその一般的な教示についてここに出典明示により取込まれるSchmelzerら J.Neurochem. 59:1675-1683(1992))。この刊行物は、各二量体型についてインビトロ活性が同じであったことを報告している。しかし、対照的に、本明細書における研究では、120/120二量体は活性が少なく、118/118種と比較して約80−90%の活性であることをラジオレセプターベースアッセイを使用して初めて証明している。アッセイの一形態では、ラットPC−12細胞膜を単離し、NGF標準物質と種々の被験種との間の競合結合に対して使用する。RRAはP75とtrkAレセプターを両方有している。117/117種が118/118種と同じ程活性であることもここでまた見出された。更に、PC−12ベースアッセイの使用によりレセプターベースアッセイの発見を確認し、120/120型が118/118型に対して約60%の活性であることを示す。主に本研究に使用された分析的及びバイオアッセイについて、並びにその一般的な教示についてここにその全体を出典明示により取込むのはBurtonら(J.Neurochem. 59:1937-1945(1992))である。
【0108】
118/118型は120/120型よりもヒトの患者において生物利用能が高いと信じられている。生物利用能の増加は少なくとも4から5倍である。この差は顕著で、意外であり、従来では予想できなかったことである。
【実施例】
【0109】
次の実施例は例証のために提供されるものであって限定のためではない。本明細書中の全ての引用文献の開示が出典明示により明示的にここに取込まれる。
【0110】
<実施例I.118/118NGFホモ二量体の精製>
この実施例はNGFの精製と各工程に対する原理を例証する。実施例の各々において、当業者であれば、従来からよく知られているようにして初期培養容積とタンパク質濃度を補償するようにカラム寸法と流量を容易に決定して調節することができる。
収集細胞培養液
【0111】
DNA配列をコードしている120アミノ酸ヒトNGFを含む発現ベクターを組換えCHO細胞に形質移入した。分泌とプロセシングを促進するためにNGFプレプロ配列もまた存在していた。組換えCHO細胞の培養後、細胞培地を収集した。収集細胞培地液(HCCF)はNGF種120、118及び117を含んでいた。NGFの約40−70%は典型的には118/118ホモ二量体で、残りはヘテロ二量体120/118、120/120及び少量の118/117であった。ここに教示したように、これらの種はSPセファロースHPカラムにより分離することができる。
【0112】
収穫細胞培地は、ミリポア10Kdカットオフ膜を使用して、およそ20倍に濃縮された(セルロース、複合体又はポリスルホンの何れかが交換可能に使用された)。濃縮物に0.1容量の1.0M、pH8.2のトリスを添加した。希釈材料が0.22umフィルターを使用して精密濾過され、保持タンクに移され2から18時間の間37℃で保持された。120/120型の118/118型へ転換が、保持の間、内在性プロテアーゼにより触媒された。
【0113】
シリカゲルクロマトグラフィー
精密濾過物を1MのNaClに調節し、1MのNaCl、25mMのMOPSO(pH7)で平衡化されたシリカゲルカラムに適用した。カラムを1MのNaCl、25mMのMOPSO(pH7)で洗浄した。適切なpH範囲は約6から8で、好適なpHは7である。ついで、カラムを25mMのMOPSO(pH7)で洗浄した。低伝導率洗浄により宿主細胞タンパク質を除去する。結合したNGFを50mMのMOPSO、0.5MのTMAC、20%の試薬無水等級アルコール(94−96%の特別に変性されたアルコール・フォーミュラ3A(5容量のメタノールと100容量の200プルーフ・エタノール)と4−6%のイソプロパノール)で溶出させた。例えば20%プロパのール、20%イソプロパノール及び20%メタノールのような他のアルコールを使用することもできる。ここで、「アルコール」及び「アルコール性溶媒」はアルコールについてよく用いられているところの意味を有するもので、好ましくは1から10の炭素原子のアルコール、より好ましくはメタノール、エタノール、イソプロパノール、n−プロパノール、又はt−ブタノールで、最も好ましくはエタノール又はイソプロパノールである。このようなアルコールは、水溶液に添加した場合、溶液の極性を減少させることにより溶液の疎水性を増大させる溶媒である。エタノールが最も好ましい。アルコールの下限は溶出する如何なるパーセントでもよく、上限はタンパク質の変性を避ける必要から決められる。溶媒は好ましくは5%から25%、より好ましくは5から20%、更により好ましくは5から15%である。TMACはテトラメチルアンモニウムクロリドであり、これはNGFを溶出させるために存在する。TMACは0.1から1Mの範囲とできる。0.3から0.7Mの範囲が更に好ましい。NGFを溶出するために使用されるTMACの量はpHとアルコール濃度の関数である。pHがより低く、アルコールの量がより少なくなると、TMACが必要となる。pHは約4から8の間とできる。この実施例では、好適なpHは7であり、これは、次のカラムへの装填の前のプール分画の調整を非常に少ないものにする。pHの上限は次のカラムに装填するのに必要なpHにより決まり、下限はNGFを効率よく溶出させるのに有用なものとされる。
【0114】
S−セファロース・ファースト・フロー・クロマトグラフィー
NGFを含んでいる溶出液をプールし、精製水で15.5ms/cm未満の伝導率まで希釈し、pHを7.0に調整した。幾つかのプロテアーゼがなお存在していたので、材料を8時間を越えないで保持した。しかし、120アミノ酸を118型に転換する内在性プロテアーゼの活性は観測されなかった。材料を25mMのMOPSO(pH7)で平衡化されたS−セファロース・ファースト・フロー・クロマトグラフィーカラム(カチオン交換樹脂S−セファロース TMアガロース・ファースト・フローTM(ファーマシア))に適用した。カラムは25mMのMOPSO(pH7)で洗浄される。適切なpH範囲は約6から8であり、好ましくはpH7である。ついで、カラムを0.16MのNaCl(pH7)で洗浄した。結合したNGFを0.5MのNaCl(pH7)で溶出させた。溶出液の塩モル濃度は0.3から1.0M、好ましくは0.4から0.6Mの範囲とできる。下限はNGFを全て溶出させるための有用性により決められ、上限は汚染物質を除去し、NGFの溶出に干渉しうるカラムの疎水的相互作用を生じさせることを避ける必要性により決められる。他の塩を使用することもでき、KClが好適な代替物である。0.5MのNaCl(pH7)での溶出が、小容量のプールを得るためには好適である。より高い塩濃度、例えば1Mを越えると、強固に結合した汚染物質が溶出し得る。
【0115】
フェニル・トーヨーパール650Mクロマトグラフィー
NGFを含んでいるSSFFカラム分画をプールし、1MのNaClに調節し、フェニル・トーヨーパール650Mカラムに適用した。カラムを25mMのMOPSO(pH7)で洗浄した。適切なpHは約5から8の範囲である。結合したNGFを、勾配緩衝液A(25mMのMOPSO、pH7、1.0MのNaCl)で始めて勾配緩衝液B(80%の25mMのMOPSO中の20%アルコール、pH7)で終了する10CV(カラム容量)の線形勾配で溶出させた。NGFを含む分画をSDS−PAGEポリアクリルアミドゲル電気泳動法により分析して、どの分画に前駆NGF種が溜まったかを決定した。NGFを含む分画を選択しプールして、例えば部分的な前駆配列が存在するもの、例えば前駆NGF、ハイブリッド前駆NGF、及びクリップ前駆NGF配列のような不適切にプロセシングされた変種を主に取り除いて、如何なるNGF前駆配列も実質的に含まれないNGF組成物を得た。フェニルカラムはまた少量のグリコシル化NGFとグリコシル化NGF前駆配列を除去した。前駆及び切詰め前駆NGF配列がNGFと前駆NGFの双方のグリコシル化形態と共にNGFピークの先端に溶出した。従って、このカラムは様々なNGF種からNGFを即座に分離し、これらの種が実質的に含まれないNGF組成物を得た。この工程において、タンパク分解性及びグリコシル化変種を含む、変種NGF疎水性変種、主に誤ってプロセシングされた変種がHICを使用して分離された。
【0116】
NGF型を分離するために最も好適な媒体は固定化したフェニル官能基を有するものであった。異なった販売者からのフェニルベースHIC媒体はこれらのNGF型を分離するための異なった効率を示した。最良の結果がTosoHaasのフェニル・トーヨ・パール媒体で達成された。HIC樹脂フェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)とTSKフェニル5PWはよく作用した。アルコキシ、ブチル及びイソアミル部分を含む他のHIC官能基はこれらの条件下では、適正が少なく、効果が少なかった。
【0117】
フェニル・トーヨーパール・プールは、75%の118、10%の120、7%の117、1.8%の脱アミドNGF、1.4%の酸化NGF、及び2.0%のisoaspNGFを含有し、残りの2.8%はその他の未同定NGF種であった。
【0118】
任意であるが、プールされた分画を酸処理して、最小15分の間、3.95未満のpHでウィルス不活性化を達成した。
【0119】
SP−セファロースHPクロマトグラフィー
HICプールを0.5から1容量の水で希釈し、希釈したプールをpH6に調節した。このプールを、(シリカゲルカラム工程におけるように)0.2Mの塩化ナトリウム、5%の試薬アルコールを含む20mMのコハク酸塩(pH6)で平衡化されたSP−セファロースHPカラムに装填した。カラムを0.2MのNaCl、5%の試薬アルコール(フォーミュラSDA−3Aアルコール;アルコールは任意に存在する)を含む20mMのコハク酸塩(pH6)で洗浄した。アルコールは、NGFの樹脂バックボーンとの非特異的(大抵は疎水的)相互作用を低減させる。適切なアルコール範囲は約0から10%である。負荷pHは約5から8であり、これはNGFの最大の安定性とNGF変種の分離を達成し維持するように選択される。カラムを2カラム容量の平衡緩衝液で洗浄した。結合したNGFを、11カラム容量の勾配緩衝液A(5%のアルコールを含む、0.25MのNaCl、0.02Mのコハク酸塩、pH6)と11カラム容量の勾配緩衝液B(0.5MのNaCl、pH6)を混合して線形の22カラム容量の勾配によって溶出させ、変種から分離した。アルコールは任意である。0.35Mから0.40MのNaCl濃度で118/118NGFが典型的には溶出した。
【0120】
カラム分画をNGF及び変種NGF含量について分析した。分画は好ましくは上掲のSchmelzerら(1992)及び上掲のBurtonらにより記述されたC4RP−HPLCによって分析される。分画を選択しプールして、修飾NGF変種、例えば酸化、脱アミド、isoaspNGF種が実質的に含まれないNGFの組成物を得た。先のHICカラムは酸化、isoaspNGFのようなNGFの他の変種型を効果的には除去できない。HICカラムは誤って折り畳まれたタンパク質とグリコシル化物を効果的に除去し、これは、より疎水性になる傾向があるので、正しく折り畳まれたNGFよりもHIC樹脂により強固に結合する。従って、カチオン交換樹脂、例えばSP−セファロースHPを、HIC樹脂によって除かれなかった変化した負荷変種を除去するために使用した。
【0121】
SP−セファロースプールは、典型的には約92%の118、4.6%の120、1.8%の脱アミドNGF、1%の酸化NGF、及び1%のisoaspNGFを含有していた。通常の通り、各々の種の量は、118に対しては約85から93%、120に対しては0から5%、117に対しては0から5%、脱アミド化型に対しては0から3%、isoasp型に対しては0−2%、及び酸化型に対しては0から2%の範囲である。NGF(全ての種)の純度は通常のように99.5%よりも大きい。
【0122】
製剤
SP−セファロースHPプールを製剤緩衝液中への限外濾過/ダイアフィルトレーションにより製剤のために調製した。酸性緩衝液が好ましくは用いられ、好ましくは上述のようにpH5の酢酸塩が使用される。118/118NGF組成物は実質的にNGFを含んでおらず、実質的に純粋なNGFである。処方された材料は神経疾患、特に糖尿病に伴う末梢神経障害及びエイズに伴う末梢交感神経障害を治療するのに好適である。
【0123】
<実施例II.120/120NGFホモ二量体の大規模な精製>
収集細胞培養液
一般に実施例Iに記載されているようにして12000リットルのCHO細胞培養からHCCFを得た。HCCF中のNGF種分布は、約40−65%の120/120ホモ二量体と、120/118ヘテロ二量体と残りが118/118ホモ二量体であった。典型的には培地を素早くプロセシングして120から118のタンパク分解性転換を最小化した。
【0124】
マクロプレップ・ハイ・Sカチオン交換クロマトグラフィー
HCCFをマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーカラムに装填し、1.5Mの酢酸ナトリウム、50mMのHEPES(pH7)で洗浄した。結合したNGFを、1.5MのNaCl、0.25MのTMAC、0.2%のチオジグリコール(pH7)で溶出した。マクロプレップカラムは、酢酸塩濃度を調節してpH5から8で操作することができる。塩化物は酢酸塩イオンの好ましい代替物である。NGFがTMAC勾配により溶出した。TMACはイオン性及び疎水性の双方の性質を持つ塩であり、この性質は、ある種の樹脂のバックボーン支持体がNGFと樹脂の間の非特異的相互作用を促進する疎水的含量を含むので有用である。典型的には、約6から8のpHの溶出緩衝液に対しては、0−3MのTMAC濃度が有用である。NGFを含む分画をプールした。
【0125】
シリカゲルクロマトグラフィー
プールを直接シリカゲルクロマトグラフィーカラムに適用した。シリカは、タンパク結合特性においてある役割を果たすイオン性、極性、及び疎水的相互作用を有する混合態様のクロマトグラフィー樹脂を提供する。カラムを1MのNaCl、25mMのMOPSO(pH7)で平衡化させた。カラムを1MのNaCl、25mMのMOPSO(pH7)(好ましくはpHが約5.0から8.5、より好ましくは6から8、最も好ましくは7である)で洗浄した。結合したNGFを25mMのコハク酸、pH3.9、50mMのTEAC(テトラエチルアンモニウムクロリド)で溶出させた。TEACはTMACよりも更に強力な溶出剤である。pHは好ましくは3.5から8の範囲である。しかし、長い時間の間7.5を越えるpHにするのは、脱アミドNGF種の生成を防止又は低減するために避けるべきである。一般に、緩衝液のpHが低くなればなる程、シリカカラムからNGFを溶出させるのに必要とされる例えばTMAC又はTEACのような混合特性の塩の濃度を低くする。pH4又は5の近くで良好な緩衝能力を有する緩衝液が使用に適している。溶出緩衝液中の塩の有無は任意であり、そのためカラムは、溶出緩衝液の適用に先立って塩なしにMOPSO緩衝液で洗浄することができる。
【0126】
フェニルセファロース・ファースト・フロー・クロマトグラフィー
NGFを含んでいる分画を同定しプールした。そのプールを、0.7Mの酢酸塩、pH7,25mMのMOPSOに調節した。調節したプールを、勾配緩衝液A(0.7Mの酢酸塩、25mMのMOPSO、pH7)で平衡化したフェニル・セファロース・ファースト・フロー・クロマトグラフィーカラムに装填した。カラムを、90%の勾配緩衝液A(0.7Mの酢酸塩、25mMのMOPSO、pH7)から10%の勾配緩衝液B(25mMのMOPSO、pH7、20%のプロピレングリコール)の勾配で洗浄した。例えばヘキシレングリコールのような他のグリコールに置換することもできる。典型的には、洗浄は約2から3CV、あるいは安定なベースラインODが達成されるまでとした。洗浄によりいくらかの宿主細胞タンパク質が除去された。結合したNGFを、90%の勾配緩衝液Aと10%の勾配緩衝液Bの混合物から10%の勾配緩衝液Aと90%の勾配緩衝液Bの混合物までの線形の10CV勾配で溶出させた。NaClもしくは硫酸ナトリウムをHIC緩衝液中の酢酸塩と置換することができる。pHは好ましくは約5から8、より好ましくは約5.5から7.5であり、pH6から8が許容でき、約7が最も好ましい。該カラムは、成熟NGF単量体及び存在する前駆配列の一部をなおも有しているNGF単量体のホモ二量体又はヘテロ二量体として存在する、全ての残っている前駆配列、部分的前駆配列、もしくはグリコシル化形態を分離した。NGFの前駆及びグリコシル化形態は溶出ピークの先行端に存在しており、NGF含有分画がこれらの種を実質的に排除するようにプールされた。
【0127】
分析用HPLCシステムで分離し検出したところ、HICプールは、約72%の120単量体、17%の118単量体、2.8%の117単量体、3.6%のR120単量体、0.8%のisoasp型、1.3%の酸化型、及び1%の脱アミド型を含有していた。
【0128】
SP−セファロースHPクロマトグラフィー
HIC工程からのNGFを含有する分画をプールした。大規模では、これは、カラム溶出液をプールタンク(保持タンク)に適当な時間に流すことにより達成された。プールのpHはpH6に調節し、SP−セファロースHPクロマトグラフィーカラムに適用した。カラムを20mMのコハク酸塩、0.2MのNaCl、pH6(勾配緩衝液A)で洗浄した。結合したNGFを、70%の勾配緩衝液Aと30%の勾配緩衝液Bの混合物で開始し、80%の勾配緩衝液B(0.7MのNaCl/pH6)と20%の勾配緩衝液Aの最終混合物までの22CVの勾配で溶出させた。pHは好ましくは5から8、より好ましくは5.7から6.5、最も好ましくは6である。代表的なクロマトグラムを図1に示す。
【0129】
SPセファロースHPプールはルーチン的に約95%の120型、3%のR120型、0.65%のisoasp型、0.6%の酸化型、0.6%の脱アミド型を含んでいた。NGFの他の未同定の形態は約0.6%であり、二酸化型NGF(Met37とMet92)からなり、脱アミドAsn45が存在していた。SP−セファロースクロマトグラフィー後に得られたプールと(SP−セファロース樹脂に装填された)代表的なHICプールを比較するHPIEX分析を図2に示す。NGFの3つの主要なクリップ型、120、118、及び117の各々が変種を有しているようであるが、精製中、主な切詰め型(この例では120)の変種、例えば酸化及びisoasp型が、主要ではない形態(この例では118と117)からの変種の分析を遮蔽しているようである。代表的な試行からの分画のHPLC分析を図3に示す。
【0130】
SP−セファロースHPはHICプール中に存在する変種を効果的に除去した。R120型はNGFのN末端に更なるアルギニン残基を有している;通常、rhNGFのN末端アミノ酸配列はSSSHPであるが、R120はRSSSHPのN末端配列を有している。従って、R120型は成熟NGFよりより塩基性で、SP−SHPにより分離された。これはまた、NGFのN末端がレセプター(trkA)結合に必要であるという事実に恐らくは関連して、低い生物活性を有している。酸化NGF型は酸化された37位置にメチオニンを有するモノ酸化型であり、NGFピークの先行端に溶出するより酸性の形態を生じる。isoasp型はアミノ酸93にアスパラギン酸の修飾を含む。isoasp型は僅かにより塩基性であり、従ってSP−セファロースHP樹脂にややより強固に結合する。isoAsp93を含んでいるNGF種は溶出ピークの終端に溶出した。脱アミド化がアスパラギン酸残基、典型的には45位のアスパラギン酸の位置で生じた。脱アミド化Asnを含んでいるNGFは、45位にAspを生じるが、僅かにより酸性であり、溶出ピークの先行端に現われる。
【0131】
フラクトゲルEMD SO3はNGF種の荷電変種を分離するためのSP−セファロースHP樹脂に対してあまり好適な代替樹脂ではない。このあまり好ましくない樹脂を使用する場合は、NGFを溶出させるためにより高い濃度のNaClが必要となる。
【0132】
製剤
バルク材料を、先の実施例におけるように、製剤緩衝液中への限外濾過/ダイアフィルトレーションにより製剤化した。最終のバルク生成物において、120型はルーチン的に約92から97%、R120は約1から4%、isoasp型は約0.2から1.5%、酸化型は約0.2から2%、脱アミド型は約0.2から2%の範囲であった。117及び118型はルーチン的に約2%未満であった。最終のバルク生成物はルーチン的に少なくとも99.5%の純NGF(全ての種を含む)であった。
【0133】
<実施例III.118/118の単離>
NGF変種が実質的に含まれていない実質的に純粋な118/118NGF組成物を得る好適な一実施態様においては、実施例IIの方法を、次の変更をして実施した。マクロプレップ・ハイ・Sカラムとシリカカラムの間に固定化トリプシンカラムを使用する。必要に応じてpHを約5から8.5、最も典型的には6.5から7.5に調節した後に、マクロプレッププールを固定化トリプシンカラムに直接装填した。プールをカラムに通過させ、その間に殆どのNGFが118型に転換された。プロテアーゼ消化が、C末端のVRRAをVRに切断することにより120型を118型に転換する。制限された選択的な切断を達成するために、トリプシン又はトリプシン様プロテアーゼ、好ましくはトリプシン、より好ましくは直ぐに入手できるブタトリプシン、あるいは代わりにウシトリプシン又は組換えトリプシンが使用される。制限された選択的な切断を実質的にもたらす任意のタンパク分解性方法を使用することができるが、NGF調製の汚染を最小化するために好ましいものは固定化トリプシンカラムである。カラムはプロテアーゼ活性を促すpH、好ましくは5.5から8.5のpH、より好ましくは6.0から8.0、最も好ましくは6.5から7.5のpHで操作される。
【0134】
この実施例において、グリコシル化NGFはここで検討したようにHICにより除去された。実施例IIにおいて検討したようなSP−セファロースHP工程に続いて、しかし好ましくは22カラム容量の0.3Mから0.55Mの塩勾配を用いて、治療用途に好適なNGF組成物を得た。RP−HPLCアッセイにより決定したところ、70%を超える118単量体、10%未満の120単量体及び15%未満の117単量体の組成物を得ることができる。典型的には、90%以上の118/118rhNGF、より通常的には93%以上の118/118rhGNFで、約7%以下の脱アミド、isoasp型及び酸化変種の組成物が得られる。より高い純度を達成する一つの手段は、例えば118/118rhNGFピークのような主要ニューロトロフィンピークの先端又は後行端に見出されるような有意の量の変種を伴う分画を選択することを避けることである。
【0135】
<実施例IV.細菌封入体からのrhNT−4/5の部分的精製と再折り畳み>
この実施例において、10もしくは60リットルの発酵で出発して、rhNT−4/5を精製した。この実施例において記載した発酵において組換えヒトNT−4/5を生産するために使用された宿主は27C7/pmNT5DTと命名された大腸菌株であった。但し、他の菌株と生物体から生産されるNT−4/5もまたここに記載した精製方法に適している。この実施例において使用された発現プラスミドは、大腸菌中のNT−4/5遺伝子の発現に必要とされる転写及び翻訳調節配列下に成熟NT−4/5コード配列を含んでいた。NT−4/5発現プラスミドにおいて、大腸菌中での遺伝子の発現に必要とされる転写配列はアルカリホスファターゼプロモータ配列によって提供された。転写ターミネーターに対するラムダはNT−4/5終止コドンに隣接して位置していた。細胞質からのタンパク質の分泌はSTIIシグナル配列により指示された。rhNT−4/5の大部分は屈折性体のような細胞膜周辺膣に見出された。プラスミドは、形質転換された宿主に対してテトラサイクリン耐性をもたらした。発酵プロセスは35℃から39℃、pH7.0−7.8で実施された。発酵は25−40時間続けられ、その後培養は収集前に冷却された。培養は、60℃で連続流れの装置を使用する加熱処理又はその温度で5−15分の間インタンク熱不活性化を使用して不活性化した。熱不活性化された培養は、AXアルファ−ラバル遠心機又は等価物を使用して遠心分離された。大腸菌はペレットに回収された。
【0136】
封入体中に組換えヒトNT−4/5を発現している大腸菌細胞は、封入体中にNT−4/5を含むペーストを調製するために標準的な手段により溶解させた。緩衝液にはプロテアーゼ阻害薬は含まれていなかった。
【0137】
細胞片から封入体を単離するために、大腸菌NT−5ペーストを、回転式機械的分散装置、例えばトゥラックス(Turrax)を使用して0.02Mのトリス、pH8、5mMのEDTA中に再懸濁させた。細胞懸濁液を6000psiで3回マイクロフルイダイザーを通過させた。得られたホモジェネートを約45分間、5000rpmでSorvallRC−3B遠心機で遠心分離した。上清を破棄し、ペレットを、トゥラックスを使用して2から3分間、中程度の速度で20mMのトリス、pH8、5mMのEDTA(抽出緩衝液)中に再懸濁させた。ホモジェネートを上述のようにして遠心分離した。ペレットを抽出緩衝液に再懸濁させ上述のように遠心分離した。得られたペレット(ペレット群)(NT−4/5封入体又は屈折体という)を−70℃で保存した。
【0138】
NT−4/5は次のようにして封入体から単離した。封入体ペレットを20mMのトリス、pH8、6Mの尿素、25mMのDTT(10ml緩衝液/封入体グラム)中にトゥラックスを使用して中程度の速度で約10分間懸濁した。懸濁液を2−8℃で40分間撹拌し、約45分間5000rpmでSorvallRC3Bで遠心分離した。PEI(ポリ-エチレン-イミン)を0.1%まで上清に添加し、これを2−8℃で30分間撹拌した。PEIは核酸とその他の酸性負荷分子を沈殿させる。混合物を約45分間5000rpmでSorvallRC3Bで遠心分離した。PEI上清を、0.02Mのトリス、6Mの尿素、10mMのDDT、pH8で平衡化されたDEFFセファロース・ファースト・フロー・カラム(10cmx14cm;DEFFはジエチルアミノエチル樹脂である)に装填した。1kgの可溶化屈折体の等価物がDEFFカラムに装填された。還元された変性NT−4/5はDEFF樹脂に結合しないので、NT−4/5と6Mの尿素を含む素通りプールが収集され(図6)、プールのpHが酢酸で5.0に下げられた。pH調節DEFF素通りプールを、NT−4/5が樹脂に結合する条件である、6Mの尿素を含む20mMの酢酸塩(pH5)で平衡化したセファロース・ファースト・フロー・カラム(Sは樹脂上のSO3官能基をいう)に装填した。装填後、S−セファロース・ファースト・フロー・カラムを数カラム容量の平衡緩衝液で洗浄した。結合したNT−4/5を、0.5MのNaCl、20mMの酢酸ナトリウム、6Mの尿素、pH5で溶出させた(図7)。0.5MのNaClのSSFFプールを20mMのトリス、0.14MのNaCl、pH8に対して一晩かけて透析した。この条件ではNT−4/5が不適切にではあるが再折り畳みする。誤って折り畳まれたrhNT−4/5分子は凝集して沈殿物を形成した。
【0139】
凝集した誤って折り畳まれたrhNT−4/5をプロセシングして正しく折り畳まれたNT−4/5を得た。凝集した誤って折り畳まれたrhNT−4/5を遠心分離によりペレットとして収集した。ペレットを、0.2Mのトリス、pH8、4Mの尿素、5mMのDTTに再懸濁させ、約1から2時間もしくはペレットが溶解するまで2−8℃で撹拌した。最終のタンパク質濃度を、280nmでの吸光係数1.8に基づいて約10mg/mlタンパクに調節した。酸化されたグルタチオンを可溶化されたペレット溶液に添加して20mMの最終濃度とし、2−8℃で15から30分間穏やかに撹拌した。酸化グルタチオンはNT−4/5スルフヒドリル基と反応してNT−4/5−S−グルタチオン混合ジスルフィドを生成する。NT−4/5−SG混合ジスルフィドを、100mMのトリス、20mMのグリシン、15%のPEG−300、1MのグアニジンHCl、pH8.3中に0.1から0.5mg/mlタンパクの最終濃度まで希釈した。NT−4/5の正しい再折り畳みを開始するために、再折り畳み混合物に2から4mMの最終濃度でシステインを添加し、続いて酸素を排除するために容器を密封する前に5から60分間窒素又はヘリウムで溶液をエアレーション(泡立てによる)した。NT−4/5の再折り畳みは2から8℃で18から24時間進められた。
【0140】
別法として、rhNT−4/5が次のような亜硫酸分解を使用して再折り畳みされた。封入体ペレット(110g)を1.1リットルの20mMトリス、7M尿素、10mMグリシン、100mM亜硫酸ナトリウム、10mMテトラチオン酸ナトリウムに懸濁させ、中程度の速度で10分間トゥラックスを使用して溶解させた。混合物(1260mL)をついで2から8℃で45分間撹拌した。PEIを0.1%PEIの最終濃度になるまで添加した。混合物を4℃で更に30分間撹拌し、RC3B遠心機中で5500rpmで45分間遠心分離した。上清を、20mMのトリス、6Mの尿素、pH8で平衡化したDEFFカラム(4.4cmx25cm)に装填した。DEFF素通り分を酢酸でpH5に調節して、20mMの酢酸、6Mの尿素、pH5で平衡化したS−セファロース・ファースト・フロー・カラム(4.4cmx25cm)に装填した。NT−4/5を25mMのMOPSO、0.5MのNaCl、pH7で溶出させた。
【0141】
スルホニル化rhNT−4/5を含むSSFF0.5M塩化ナトリウムプールを約0.1mg/mlタンパクに希釈し、1Mのグアニジンヒドロクロリド、100mMのトリス、20mMのグリシン、15%のPEG−300、pH8.3に調節した。2から4mMのシステインの添加によりNT−4/5の再折り畳みを開始した。再折り畳み反応は本質的に24時間以内に完了した。溶液からの酸素を置換するための例えばヘリウム又は窒素のような不活性ガスによるエアレーションを必要に応じて実施することもできる。
【0142】
<実施例V.高次構造的(誤って折り畳まれた)変種からの正しく折り畳まれたrhNT−4/5の単離>
実施例IVのrhNT−4/5の再折り畳み混合物を一晩かけてpH4から5の溶液に対して透析してグアニジンとその他の試薬を除去した。溶液を清澄にするために、溶液を5000rpmで45分間遠心分離するか、0.2umフィルターを通過させた。
【0143】
0.5から5グラムのタンパク質を含む清澄化した上清を、氷酢酸を添加してpH3から5に調節し、C4RP−HPLCカラムに装填するか、精製の準備ができるまで−20℃で凍らせて保存した。この実施例では、酸性化され清澄化された溶液を、折り畳まれたrhNT−4/5が結合した樹脂のC4RP−HPLC(3cmx50cm)カラムに装填した。正しく折り畳まれたNT−4/5を、0.05トリフルオロ酢酸(TFA)溶媒系中のアセトニトリル勾配を用いて(25ml/分の流速での0.05%TFA中の26から40%のアセトニトリル勾配(95分間を越える)からなる)溶出させた。分画を1から1.5分間隔で収集した(図8)。分析的C4HPLC Vydac(0.21x15cm)カラム上の溶出時間を正しく折り畳まれたNT−4/5標準物質の時間と比較することにより、正しく折り畳まれたNT−4/5について分画を分析した(図9)。標準的な正しく折り畳まれた無傷のNT−4/5は典型的には0.5%TFA/アセトニトリル緩衝系で毎分2.5mlの流量で19分で溶出した。正しく折り畳まれたrhNT−4/5を含む分画をプールしpHを5から7に調節した。正しく折り畳まれたrhNT−4/5のプールもまたNT−4/5のカルバミル化及びN末端切詰め型を含んでいた。
【0144】
調製的逆相液体クロマトグラフィー樹脂は好ましくは約10−40ミクロンの粒径、約200−400オングストロームの孔径、及びC4、C8、又はC18アルキル基を有する媒体である。より好ましくは、樹脂は約15−40ミクロンの粒径と約300オングストロームの孔径を有し、C4シリカ媒体である。
【0145】
<実施例VI.高次構造的(誤って折り畳まれた)変種からの正しく折り畳まれたrhNT−4/5の別の単離>
実施例IVの再折り畳みrhNT−4/5混合物を、10kDの分子量のカットオフを持つ20平方フィートのセルロース(又はポリスルホンもしくは等価物)膜を備えたミリポア−ペリコン限外濾過系を使用して約10倍に濃縮した。0.2ミクロン膜を通す濾過の前に、濃縮した混合物を一晩かけて50Lの50mM酢酸塩、pH5.5、50mMのNaClに対して透析するか、50mMの酢酸塩、50mMのNaCl、pH5.5中でダイアフィルトレーションを施した。
【0146】
濾過した再折り畳み混合物を2.5MのNaCl、20mMのMOPSO、pH7に調節し、前もって2.5MのNaCl、20mMのMOPSO、pH7で平衡化したHICカラムであるフェニル・トーヨーパール650Mカラム(10cmx19cm)に装填した。ついで、平衡緩衝液でカラムを洗浄した。rhNT−4/5分子の分子の幾つかの誤って折り畳まれた型が素通り分画に溶出し、その他の誤って折り畳まれた型が20から40%の試薬アルコールのような高濃度の有機溶媒で溶出した。正しく折り畳まれたrhNT−4/5は2Mの塩化ナトリウム、10%の試薬アルコール、pH7を使用してフェニルカラムから溶出させた(図10)。フェニルセファロースのような他のフェニル樹脂をトーヨーパールのバックボーンの代わりに使用することができる。硫酸アンモニウム、クエン酸塩、酢酸塩、及び塩化カリウムを含むここで検討した塩を使用することができる。使用する塩に依存して、塩濃度は典型的には1Mから3Mであり、2.5MのNaClが装填には好適で、2MのNaClが有機溶媒の存在する場合の溶出に好適である。好ましくは、塩濃度の低下がニューロトロフィンとその変種を溶出させ分離するために使用される。溶出を達成するには、溶出緩衝液中の塩濃度は典型的には負荷緩衝液中のものよりも低いが、有機溶媒で補償するときは同じ濃度とすることができる。また、有機溶媒の使用は、ここで見出されたところでは、有機溶媒を添加するとより狭いピークプロファイルが得られて溶出パターンが改善されるという他の利点がある。エタノールに加えて、プロパノール、イソプロパノール、及び低級アルキレングリコール、例えばプロピレングリコール、エチレングリコール及びヘキシレングリコールを含むここで検討するその他の有機溶媒を使用することもできる。5から25%(v/v)、より好ましくは5から20%(v/v)、更に好ましくは5から15%(v/v)の有機溶媒が典型的には正しく折り畳まれたニューロトロフィンを溶出する。有機溶媒での溶出は勾配によるか、段階的に行なう。pH範囲は、好ましくは中性近くから僅かに酸性であり、5から8、より好ましくは5.5から7.5、最も好ましくは7である。MOPSO、MOPS、HEPES、リン酸塩、クエン酸塩、アンモニウム、酢酸塩を含むここで検討した任意の緩衝液を、所望のpHで緩衝作用を示す限り使用することができる。
【0147】
<実施例VII.正しく折り畳まれたrhNT−4/5の化学変種からの精製>
rhNT−4/5のカルバミル化及びN末端切詰め型を含む化学変種からの正しく折り畳まれた無傷のrhNT−4/5の分離は、SP−セファロースHP樹脂又はポリキャットA HPLC樹脂を使用する高速カチオン交換クロマトグラフィーにより達成することができた。
【0148】
C4RP−HPLCカラムを誤って折り畳まれた変種を取り除くために使用した場合、C4HPLCプールをpH5から7に調節し、20mMのコハク酸塩、pH6、5%の試薬アルコール、0.2MのNaClで平衡化した7cmx19cmのSP−セファロースHPカラムに装填した。結合したNT−4/5を伴う樹脂を平衡緩衝液で洗浄した。結合したrhNT−4/5を、pH6で0.2MのNaClから0.4MのNaClまでの22カラム容量(CV)の勾配(すなわち、平衡緩衝液中の塩勾配)を使用して溶出させ、カルバミル化及びN末端切詰め型から分離した(図11)。NT−4/5を含む分画をプールし、0.05Mの酢酸塩、pH4から5に製剤化した。無傷のrhNT−4/5を同定し、好ましくはここで検討したように分析的RP−HPLC又はSDS−PAGEにより分画内の変種から区別し、標準と比較した。
【0149】
別法として、NT−4/5の変種型をポリアスパラギン酸カラムでの高速カチオン交換HPLC(ポリキャットa、ポリLC、Columbia,Md.)(9.4x200mm)により除去した(図12)。C4HPLCプールをpH5から6に調節し、ポリキャットaカラムに装填した。クロマトグラフィー条件は:緩衝液Aが20mMのリン酸塩、5%のアセトニトリル、pH6であった;緩衝液Bは20mMのリン酸塩、5%のアセトニトリル、0.8MのKCl、pH6であった。rhNT−4/5を、65分にわたって25から60%の緩衝液Bの勾配を使用して溶出させた(図12)。分画を1分間隔で収集し、上述のように分析的C4HPLCにより分析した。
【0150】
HICを用いて誤って折り畳まれた変種を除いた場合(上記の実施例VI)は、正しく折り畳まれたNT−4/5プールを、20mMのコハク酸塩、0.1MのNaCl、5%の試薬アルコール、pH6中で一晩かけて透析するか、20mMのコハク酸塩緩衝液中で限外濾過/ダイアフィルトレーションした。透析もしくはUF/DFプールをついで上述のようなSP−セファロースHP又はポリキャットaカラムに装填した。
【0151】
製剤
(SP−セファロースHP又はポリキャットaHPLC工程から得た)正しく折り畳まれた無傷のNT−4/5を含む分画をプールし、20mMの酢酸塩、pH4から5の製剤緩衝液中に1から5mg/mlまで濃縮した。別法として、NT−4/5を限外濾過/ダイアフィルトレーションを使用して製剤化した。
【0152】
最終のバルク溶液を、アミノ酸分析、N末端配列分析、質量分析、SDS−PAGE(図13)及び生物学的アッセイにより分析した。キナーゼレセプター活性化(KIRA)アッセイは、細胞膜に位置するそのチロシンキナーゼレセプター(trkB)の自己リン酸化のNT−4/5活性化を検出するものであるが、精製したrhNT−4/5を特徴付けるために使用した。gDタグを付したtrkBを発現するCHO細胞を使用した。1995年6月1日に公開されたWO95/14930はKIRAを記述しており、ここに出典明示により取込む。rhNT−4/5はこのアッセイにおいて12.6ng/mlのEC50を有していた。典型的には、ここに記載されたようにして精製される正しく折り畳まれた無傷のNT−4/5のEC50は、5から30、より好ましくは10から20である。
【0153】
非NT−4/5タンパク質に対するrhNT−4/5の純度は典型的には90から99パーセントであった。カルバミル化及びN末端切詰め変種に対してのNT−4/5の相同性は90から99パーセントであった。最も典型的かつ好ましくは、純度と相同性は99%又はそれ以上である。
【0154】
<実施例VIII.細菌封入体からのrhNT−3の初期精製、再折り畳み及び最終精製>
細胞片から封入体を単離するために、大腸菌NT−3ペースト(1kg)を、10mLの100mM酢酸ナトリウム(pH5)中に、例えばトゥラックスのような回転機械式分散装置を使用して再懸濁させた。細胞懸濁液を6000psiでマイクロフルイダイザーに3回通した。得られたホモジェネートを5000rpmで30分の間SorvallRC−3B遠心機で遠心分離した。
【0155】
NT−3を次のようにして封入体から単離した。封入体ペレットを、約10分間中程度の速度でトゥラックスを使用して、100mMのトリス、100mMのNaCl、5mMのEDTA、100mMの亜硫酸ナトリウム、10mMのテトラチオン酸ナトリウム、7.5Mの尿素、pH8.3(10ml/封入体グラム)に懸濁させた。懸濁液を2−8℃で約1時間撹拌した。PEI(ポリエチレンイミン)を約0.15%(最終濃度)になるまで添加し、2−8℃で30分間撹拌した。混合物を5000rpmで約30分の間SorvallRC3Bにおいて遠心分離した。上清をゲルマン・プレフロー(Gelman Preflow)カートリッジで濾過した。濾過した上清を3容量のS−セファロース・ファースト・フロー平衡緩衝液(50mMの酢酸ナトリウム、5Mの尿素、pH5)で希釈した。希釈した濾過上清(7mS未満の導電率を、50mMの酢酸ナトリウム、5Mの尿素、pH5.0で平衡化したS−セファロースFFカラムに装填する。カラムを最初に50mMの酢酸ナトリウム、5Mの尿素、pH5で洗浄し、ついで50mMのMOPS,5Mの尿素、10mMのグリシン、pH7.0で洗浄した。亜硫酸分解されたNT−3を、50mMのMOPS、5Mの尿素、10mMのグリシン、pH7中の0−0.6MのNaClの10カラム容量の勾配を使用してカラムから溶出させた。
【0156】
部分的に精製されたNT−3を、0.1Mのトリス、2Mの尿素、0.1MのNaCl、15%のPEG300、10mMのグリシン、25mMのエタノールアミン(pH9.1)を含む再折り畳み緩衝液中でS−セファロースFFプールをおよそ0.1mg/mlタンパクまで希釈することにより再び折り畳んだ。再折り畳みは、およそ5mMまでシステインを添加し、2−8℃で2−5日間撹拌することにより開始した。必要に応じて、再折り畳み溶液中の酸素濃度を低減させるために、再折り畳み緩衝液をHe又はアルゴンで分散させても良い。
【0157】
再折り畳みされたプールのpHは7に調節し、濾過し、50mMのHEPES(pH7)で平衡化したマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーカラムに装填した。マクロプレップカラムにpH調節再折り畳みプールを装填した後、カラムを50mMのMOPS(pH7)で最初に洗浄し、続いて50mMのMOPS、0.1MのTMAC、0.3MのNaCl(pH7)により洗浄した。NT−3を50mMのMOPS、0.25MのTMAC、1.5MのNaCl(pH7)で溶出させた。
【0158】
次に、マクロプレッププールをフェニルセファロース・ファースト・フロー・ハイ・サブスティチューションカラムで精製した。フェニルカラムを50mMのHEPES、1.5MのNaCl(pH7)で平衡化し、マクロプレッププールをフェニルカラムに直接装填した。カラムを平衡緩衝液で洗浄し、正しく再折り畳みされたNT−3を、50mMのHEPES、1.5MのNaCl(pH7)から50mMのHEPES、10%の試薬アルコール(pH7)に変わる15カラム容量の勾配を使用して溶出させた。分画をC4HPLCかSDS−PAGEによって分析し、正しく折り畳まれたNT−3を含む分画をプールした。
【0159】
フェニルプールを25mS未満に希釈(典型的には水で約2倍容量)し、25mMのMOPSO(pH7)で前もって平衡化したSP−セファロースHPカラムに装填した。カラムを最初に平衡緩衝液で洗浄し、NT−3を、25mMのMOPSO(pH7)中に0.35MのTMACから0.65MのTMACまでの20カラム容量の勾配を使用してカラムから溶出させた。(C4HPLCアッセイにより判断して)rhNT−3を含む分画をプールした。
【0160】
SP−セファロースHPプールを10000分子量の膜上で約1mg/mlまで濃縮し、ついで6容量の10mMの酢酸塩、140mMのNaCl(pH5.0)でダイアフィルトレーションを行った。
【図面の簡単な説明】
【0161】
【図1】図1はSP−セファロースHPクロマトグラムを示す。HICクロマトグラフィー後に12kLの発酵からのNGF含有混合物を、25全体適合(omnifit)SP−セファロース・ハイ・パーフォーマンス樹脂(カラム寸法1.0x35cm;総容積27.5ml)の一つのSPHP樹脂上に装填した。緩衝液Aは、0.2MのNaCl、20mMのコハク酸塩、pH6.0(23ms)であった。緩衝液Bは0.7MのNaCl、20mMのコハク酸塩、pH6.0(63ms)であった。カラムは最初に緩衝液Aで平衡にした。0.24mg/mlで345mlのHICプールが25mMのコハク酸塩、pH6.0(17ms)に調整され、その362mlを装填して、3mg/ml樹脂の負荷とした;82.5mgのNGFを装填した。装填速度は1時間当り40cmとした。NGFを、30%から80%までの緩衝液B(0.35から0.60MまでのNaCl;37から57ms)の22カラム容量の勾配を使用して溶出させた。溶出速度は1時間当り60cmとした。吸光度単位(280nm)とmS/cmを分画数と溶出容積mlに対してプロットした。NGFを含有する分画を決定し、プールした。この場合、分画43から60をプールして、0.38mg/mlNGFの99mlのサンプルを得た。これは約46%の回収率になる。
【図2】図2は、フェニル・セファロース・ファースト・フロー・プールとSP−セファロースHPプールのSP−NPR HPLCカチオン交換(HPIEX)分析を示す。それぞれに対するクロマトグラムに標しをする。
【図3】図3は、SP−セファロースHPクロマトグラフィー工程からの選択した分画(主プール、主ピークプールの先端と後端)のC4 RP−HPLC分析を示す。3つのシグナルが重ねられて描かれている。分かるように、(成熟NGFを含む)主ピークは変種NGF群を含む幾つかのより小さいピークとは分離している。
【図4】図4は、ヒトプレプロNGFの配列(配列番号:1)を示す。この図上に示されているのは、成熟NGFの最初のアミノ酸(位置1)と120NGFの最後のアミノ酸(位置120)である。好適なNGFアミノ酸配列は成熟118型のもの(位置1から位置118)である。また示されているのは変種形態:成熟120(位置1から位置120);成熟117(位置1から位置117);R120(位置1から120);及び成熟114(アミノ酸1から114)、115(アミノ酸1から115)及び117(アミノ酸1から117)変種を含む、N末端とC末端に生じるその他の誤ってプロセシングされた部位である。タンパク分解性に誤ってプロセシングされた主要なミスプロセシング変種は、NGFのプロ配列のアミノ酸R(−39)とS(−38)の間にN末端切断を有している。可能な開始メチオニンには下線が付される。他のN末端変種には、NGFの切断型が含まれ、最もありふれたものはアミノ酸H8とH9及びR9とG10の間に生じる切断を有している。
【図5】図5は、ヒトNGF(配列番号:2)、マウスNGF(配列番号:3)、BDNF(配列番号:4)、NT−3(配列番号:5)及びNT−4/5(配列番号:6)のニューロトロフィンのアミノ酸配列を示している。ボックスで囲った領域はシステインノットモチーフに関与する相同的システイン含有領域を示している図である(De Youngら, Protein Sci. 5(8):1554-66(1996))。
【図6】図6は、DEAE−セファロース・ファースト・フロー(DEFF)樹脂カラムでのrhNT−4/5のクロマトグラフィーパターンを示す。「NT45DE1:1_UV」と標されているクロマトグラムは280nmでのUV吸光度測定値である。「NT45DE1:1_Cond1」と標されているクロマトグラムは溶出する分画の伝導率測定値である。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図7】図7は、SP−セファロース・ファースト・フロー樹脂カラムでのrhNT−4/5のクロマトグラフィーパターンを示す。「NT45SFF1:1_UV」と標されているクロマトグラムは280nmでのUV吸光度測定値である。「NT45SFF1:1_Cond1」と標されているクロマトグラムは溶出する分画の伝導率測定値である。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図8】図8は、明細書に記載された条件下でのrhNT−4/5の典型的な調製用C4−RP−HPLCクロマトグラフィーパターンを示す。280nmでの吸光度をモニターした。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図9】図9は、示した時間での再折り畳みの間モニターされたNT4/5試料の分析用HPLCクロマトグラフィーパターンを提供する。カラム条件は明細書中に記載した。「SSFF(0.5m)」と標されたパターンは、再折り畳みの前にS−セファロース・ファースト・フロー・カラムから0.5MのNaClで溶出させたNT−4/5の分析を示す。NT−4/5が再折り畳みされるにつれて、溶出パターンは標準のパターンに近づく。
【図10】図10は、疎水的相互作用クロマトグラフィーカラムであるフェニル・トーヨーパール650Mカラムでの誤って折り畳まれた変種からの無傷の正しく折り畳まれたNT−4/5の分離を示すクロマトグラムを示す。正しく折り畳まれたNT−4/5よりも疎水性が低い誤って折り畳まれた変種は素通り分内に溶出する一方、より疎水性である誤って折り畳まれた変種(ピークB)は、正しく折り畳まれたNT−4/5(ピークA)を溶出するのに必要とされるものよりも高い有機溶媒濃度で溶出する。
【図11】図11は、カチオン交換樹脂であるSP−セファロースからのNT−4/5と変種の溶出パターンを示す。280nmでの吸光度がモニターされた。ピークAがカルバミル化され切詰められた(clipped)変種を含む一方、ピークBは無傷の正しく折り畳まれたNT−4/5を含む。プールされたNT−4/5含有分画は「プール」と標された水平な矢印により示されている。
【図12】図12は、明細書に記載された条件下でのrhNT−4/5の典型的な調製用ポリキャットA樹脂クロマトグラフィーパターンを示す。
【図13】図13は、本明細書に記載したrhNT4/5精製法の示された工程から採られた試料の純度と均一性を評価するための還元条件下での16%SDS−PAGE(サンジエゴのNovex,Inc.からの、予め注がれたトリス−グリシン系)分析を示す。ゲルは、タンパク質を検出するために、クーマジーR250で染色された。「DE負荷」と標されたレーンはDE−セファロース・ファースト・フロー・カラム上に負荷されたPEI混合物の試料を含み、「DE FT」と標されたレーンはDE−セファロース・ファースト・フロー・カラムの素通り分の試料を含み、レーン「Sプール」は、再折り畳み前に、S−セファロース・ファースト・フロー樹脂カラムから溶出したNT−4/5を含むプールされた分画からの試料を含み、レーン「再折り畳みプール」は再折り畳みが完了した後のプールの試料を含み、レーン「C4プール」は調製的C4RP−HPLC後のプール分画の試料を含み、レーン「ポリキャットAプール」はポリキャットA HPLCカラムからのプール分画の試料を含む。
【図14】図14は、細菌的に調製されたスルホニル化rhNT−3を含む混合物のS−セファロース・ファースト・フロー・クロマトグラフィーからの分画のUV吸光度パターンを示す。
【図15】図15は、rhNT−3のインビトロ再折り畳み型を含む混合物のマクロプレップ・ハイ・S(Macroprep High S)カチオン交換クロマトグラフィーを示す。樹脂は、バイオラッド(Biorad)から購入した。カラム寸法は9x9cmであった。pH6.8の再折り畳みされた(36時間の再折り畳み後)を含む700mlSSFFプールが約310ml/分の流量でマクロプレップ・カラム上に負荷された。条件は明細書に記載する。
【図16】図16は、誤って折り畳まれた変種を除去するための、カチオン交換精製再折り畳みrhNT−3のフェニル・セファロース・ファースト・フロー・ハイ・サブスティチューション(疎水的相互作用クロマトグラフィー)クロマトグラフィーを示す。
【図17】図17は、HIC−rhNT−3プールのSP−セファロース高速クロマトグラフィーを示す。
【技術分野】
【0001】
この発明は、ニューロトロフィン、特にNGFファミリーのニューロトロフィン、より詳細には神経成長因子(NGF)及びニューロトロフィン−4/5(NT−4/5)、及びニューロトロフィン−3(NT−3)を、特に細菌もしくは哺乳動物細胞発酵により産生される場合に、これらに付随する変種、不純物、及び汚染物質から精製するための改良方法に関する。
【背景技術】
【0002】
比較的純粋で生物学的に活性なポリペプチド及びタンパク質を大量に生産することは、ヒトと動物の医薬製剤、酵素及びその他の特殊化学製品の製造に対して経済的に重要である。多数のタンパク質の生産に対して、多量の外来性タンパク質を哺乳動物の宿主細胞及び細菌及びその他の宿主細胞中に発現しうることから、組換えDNA技術が選択方法になった。
【0003】
哺乳動物NGF(マウスNGF)の一次構造は、非特許文献1によって最初に解明された。その前駆物質であるプレプロNGFの一次構造は、マウスNGFのcDNAのヌクレオチド配列から推論された(非特許文献2;非特許文献3)。
【0004】
相同的ヒトNGF(hNGF)遺伝子もまた同定された(非特許文献4;特許文献1)。マウスNGFに対するその相同性は、アミノ酸とヌクレオチド配列レベルでそれぞれ約90%と87%である。天然に生じるヒトNGFは希少であるため、ヒトGNFは生化学的に細かく詳細に特徴付けるのに充分な量では天然源から調製されていない。
【0005】
NGFに関連する更なる神経栄養因子がそれ以降に同定されている。これらには、脳由来神経栄養因子(BDNF)(非特許文献5)、ニューロトロフィン−3(NT−3)(非特許文献6;非特許文献7;非特許文献8)、及びニューロトロフィン4/5(NT−4/5)(非特許文献9)が含まれる。TGF−βスーパーファミリーの遠いメンバーであるGDNFとニュールツリン(neurturin:「NTN」)は、交感感覚神経細胞及び中枢神経系神経細胞に対する二つの最近同定された構造的に関連している強力な生存因子である(非特許文献10;非特許文献11;非特許文献12;非特許文献13)。
【0006】
組換えタンパク質を生産するには、そのタンパク質をコードするDNAを宿主細胞に形質移入し、組換えタンパク質の発現に好ましい条件下で細胞を成長させる。原核生物の大腸菌が、低コスト高収量で組換えタンパク質を生産することを可能にすることから、好まれる宿主であった。タンパク質をコードするDNAの一般的な細菌での発現に関しては、細胞外又は周辺質担体タンパク質に対する細菌遺伝子と非細菌遺伝子を含んでなる組換えDNA分子に関する特許文献2、凝集体形成ポリペプチドとの外来性ポリペプチドの同時生産に関する特許文献3、IGF−IのようなポリペプチドとのtrpLE融合とtrpプロモーター/オペレーターを持つ発現ベクターに関する特許文献4、外来性タンパク質と共に含む発現調節配列に関する特許文献5及びIGF−Iをコードしているもののような特異的環状DNAプラスミドに関する特許文献6を含む多くの米国特許が存在する。
【0007】
遺伝子操作により作成されるバイオ医薬品は、典型的には種々の多様な宿主細胞汚染物質を含んでいる上清から精製される。特にNGFは、多数の異なった方法を使用し、様々な度合いの努力と成功度をもって様々な度合いに精製されたことは報告されている。例えば、非特許文献14、NGFのCHO細胞による産生を開示している特許文献7、非特許文献15;非特許文献16;非特許文献17を参照せよ。これらの努力は主として研究室規模のものであった。
【0008】
【特許文献1】米国特許第5288622号
【特許文献2】米国特許第4565785号
【特許文献3】米国特許第4673641号
【特許文献4】米国特許第4738921号
【特許文献5】米国特許第4795706号
【特許文献6】米国特許第4710473号
【特許文献7】米国特許第5082774号
【非特許文献1】AngelettiとBradshaw(Proc.Natl.Acad.Aci. USA 68:2417(1971))
【非特許文献2】Scottら Nature 302:538(1983)
【非特許文献3】Ullrichら Nature 303:821(1983)
【非特許文献4】Ullrich, Symp. on Quan.Biol., Cold Spring Harbor 48:435(1983)
【非特許文献5】Leibrockら, Nature, 341:149-152(1989)
【非特許文献6】Kaishoら, FEBS Lett., 266:187(1990)
【非特許文献7】Maisonpierreら, Science, 247:1446(1990)
【非特許文献8】Rosenthalら, Neuron, 4:767(1990)
【非特許文献9】Berkmeierら, Neuron, 7:857-866(1991)
【非特許文献10】Linら, Science 260:1130-1132(1993)
【非特許文献11】Hendersonら, Science 266:1062-1064(1994)
【非特許文献12】Buj-Belloら, Neuron 15:821-828(1995)
【非特許文献13】Kotzbauerら Nature 384:467-470(1996)
【非特許文献14】Longoら(IBRO Handbook,vol.12,pp3-30(1989))
【非特許文献15】BruceとHeinrich(Neurobio. Aging 10:89-94(1989))
【非特許文献16】Schmelzerら(J.Neurochem. 59:1675-1683(1992))
【非特許文献17】Burtonら(J.Neurochem. 59:1937-1945(1992))
【発明の開示】
【発明が解決しようとする課題】
【0009】
しかし、変種が本質的に含まれない製薬的純度と高収量を達成できるような組換えヒトNGFの調製的単離は、従来技術が解決できなかった課題であった。
【0010】
従って、ニューロトロフィン、特にNGF及びNGFファミリーのニューロトロフィンを、その変種とその他の分子から、また高いpIを持つ他のポリペプチドから選択的に分離するための効率的なプロトコールに対する必要性が従来から存在している。大きな規模でニューロトロフィンを精製する方法は、治療上の必要性を満たすために、発酵ブロス、溶解細菌もしくは哺乳動物細胞を含む様々な供給源からの出発原料に適用できなければならない。
【課題を解決するための手段】
【0011】
更に、本発明者らは、過去には知られておらず分離が困難なニューロトロフィン変種、例えばNGF変種を発見したので、ここに提供する方法は、望ましくない変種が実質的に含まれない、ヒトNGF(rhNGF)、rhNT−3及びrhNT−4/5及びその所望の遺伝子操作突然変異体を含む組換えニューロトロフィンを、商業的に有用な量で提供するために特に有用である。本発明のかかる目的とその他の目的は当業者には明らかであろう。
【0012】
本発明の一実施態様において、ニューロトロフィン、特に高度に相同的なレセプターファミリー(trks)、好ましくはrhNGF、rNT−3、rhNT−4/5、rhBDNFもしくはその所望の遺伝子操作形態による認識を分かつNGF、NT−3、NT−4/5、及びBDNFを含む、特にNGFファミリーのものを、疎水的相互作用クロマトグラフィー(HIC)を使用して精製する方法が提供される。
【0013】
ここに報告したように、ニューロトロフィンの組換え生産から生じるある種の望ましくないニューロトロフィン変種を本発明者らが発見したことに鑑みると、HICの使用は、ニューロトロフィンの化学的に異なる形態ともしくは誤って折り畳まれた形態でさえも、所望の正しく折り畳まれた無傷のニューロトロフィンから分離することを可能にする。取り除くことができる変種は、部分的にプロセシングされた前駆配列、グリコシル化された成熟及び前駆物質含有形態(真核生物細胞培養にある場合)、及び誤って折り畳まれ部分的に折り畳まれた変種(一般にインビトロ折り畳み工程が使用される場合、細菌細胞培養からのもの)を含む、疎水性が成熟した正しく折り畳まれた無傷のニューロトロフィンとは異なるものである。例えば、HICは、NGFの部分的にプロセシングされた前駆配列、NGFのグリコシル化された種及び前駆体(真核細胞培養に存在する場合)、及び誤って折り畳まれ部分的に折り畳まれた変種(一般に細菌細胞培養からでインビトロ折り畳み工程からのもの)を成熟NGFの混合物から除去するのに特に有用である。NGFは一つのN結合グリコシル化部位をAsn45に有している。細菌中に発現された再折り畳みrhNT−4/5の場合は、HICは、正しく折り畳まれたNT−4/5を間違って折り畳まれた形態から分離する。ここに記載した方法によれば、ニューロトロフィンは本質的にこれらの変種を含まない。ニューロトロフィンの精製に対して、HIC樹脂官能基は、オクチル及びブチル基も有用であるが、好ましくはフェニル基である。特に好ましい実施態様は、フェニル・トーヨーパール(Phenyl Toyopearl)、フェニル・セファロース・ファースト・フロー・ロー・サブスティチューション(Phenyl Sepharose Fast Flow Low Substitution)、TSK−フェニル5PW(TSK−Phenyl 5PW)等々のHIC樹脂である。
【0014】
他の実施態様では、ニューロトロフィン、特にNGFファミリーのもの、好ましくrhNGF、rNT−3、rhNT−4/5もしくはその所望の遺伝子操作形態を、負荷修飾変種、例えば成熟ニューロトロフィン由来の酸化、isoasp(イソアスパラギン酸)及び脱アミド形態を分離する調製的カチオン交換クロマトグラフィーの使用によって精製する方法が提供される。特に好適な実施態様は、SP−セファロース・ハイ・パーフォーマンス、フラクトゲル(Fractogel)EMD SO3、又はポリアスパラギン酸樹脂を用い、なかでもポリキャット(PolyCAT)Aが特に好適である。最も好適には、大規模のSP−セファロース・ハイ・パーフォーマンス樹脂又はフラクトゲルEMD SO3樹脂が使用される。
【0015】
本発明の更に他の実施態様では、HICとカチオン交換クロマトグラフィーの両方が、所望のニューロトロフィン、例えば実質的に均質な、すなわち例えば誤って折り畳まれた変種及び化学変種のような加工及び負荷変種の双方を実質的に含まず、タンパク含量に関してもまた実質的に純粋である組換え成熟NGF、好ましくはヒトNGFの組成物を調製するために使用される。
【0016】
この発明の一実施態様では、ニューロトロフィン、特にNGFファミリーのもの、好ましく組換えヒトNGF、NT−3、NT−4/5及びその所望の遺伝子操作形態を、関連する所望されない変種、例えば発酵、プロテアーゼ切断変種、グリコシル化変種、誤って折り畳まれた変種を、逆相液体クロマトグラフィーによって精製する改良方法が提供される。より好ましくは、NGFは120/120又は118/118ホモ二量体形である。ここに記載した方法によれば、ニューロトロフィンは、最も好ましくは変種を本質的に含まない。
【0017】
他の実施態様においては、ニューロトロフィン、特にNGFファミリーのものを、生理的pHを含む溶出条件を使用して関連する変種から精製する方法が提供される。
更に他の実施態様では、均一性がかなり改善される結果が得られるニューロトロフィンの精製方法が提供される。
【0018】
他の実施態様において、本発明は、NGFファミリーのニューロトロフィンをその変種から分離する方法であって、
a)ニューロトロフィンとその変種を含む緩衝液を約5から8のpHで疎水的相互作用クロマトグラフィーカラムに装填し;
b)カラムを洗浄し、
c)約5から8のpHの緩衝液でニューロトロフィンを溶出させ、
d)約5から8のpHでカチオン交換クロマトグラフィーカラムにニューロトロフィン含有溶出物を装填し、
e)約5から8のpH、好ましくはpH6で塩勾配の緩衝液でカラムからニューロトロフィンを溶出させる、
ことを含んでなる方法を提供する。ニューロトロフィンは最も好ましくはrhNGFである。
【0019】
本発明の一実施態様では、好ましくはNGF分画であるニューロトロフィン分画から宿主細胞タンパク質を効果的に除去するシリカゲルクロマトグラフィー工程が提供される。
【0020】
本発明の一実施態様では、120アミノ酸NGFがトリプシン様プロテアーゼ処理を受けて、VRRAのC末端から末端RAジペプチドを選択的に除去して、118種を生成する方法の工程が提供される。固定化トリプシンカラムが好適である。
【0021】
本発明はまた本発明の方法により調製されたニューロトロフィン組成物と製剤及び組成物と製剤への用途にも関する。提供されるものは、実質的に均一な、すなわち例えば誤って折り畳まれた変種と化学変種のような加工及び負荷変種の双方とも実質的に含まず、またタンパク質含量に関して実質的に純粋であるニューロトロフィンの組成物である。好ましくは、成熟ヒトNGF、成熟ヒトもしくはラットNT−3、及び成熟ヒトNT−4/5がこの形で提供される。好適な実施態様では、NGFは120種で、より好ましくは118型であり、最も好ましくはホモ二量体、例えば118/118である。
【発明を実施するための最良の形態】
【0022】
<定義>
本明細書において、「ニューロトロフィン」とは、マウス、ウシ、ヒツジ、ブタ、ウマ、トリ、そして好ましくはヒトを含む任意の種から、また天然であろうと、合成であろうと組換え生産されたものであろうと、任意のソース由来の、未変性配列もしくは遺伝子操作形態のニューロトロフィン、好ましくはNGF、NT−3、NT−4/5、及びBDNFを含むNGFファミリーのニューロトロフィンをいう。例えば、「NGF」は、マウス、ウシ、ヒツジ、ブタ、ウマ、トリ、そして好ましくはヒトを含む任意の種から、また天然であろうと、合成であろうと組換え生産されたものであろうと、任意のソース由来の神経成長因子をいう。好ましくは、ニューロトロフィンは組換え的に生産される。好適な方法では、ニューロトロフィンはクローン化され、そのDNAは例えば哺乳動物細胞、細菌細胞中で発現される。ここに教示したプロセスと方法はニューロトロフィン、GDNF及びニュールツリンにも適用し得る。
【0023】
ヒトの用途に対して好ましいものは、ヒト未変性配列、成熟NGFであり、より好ましくは120アミノ酸配列、更により好ましくは118アミノ酸配列である。より好適には、この未変性配列NGFは組替え的に生産される。ヒトプレプロNGFとヒト成熟NGFに対する好適なアミノ酸配列は、出典明示によりここに特に取込む米国特許第5288622号によって提供される。更なる翻訳後の修飾のない120アミノ酸型はホモ二量体形(つまり、120/120)における好適な型である。更により好適なものは、更なる翻訳後の修飾のない、特にホモ二量体のような118型(つまり、118/118)である。
【0024】
「実質的に純粋な」が意味するところは、少なくとも70%のニューロトロフィン、より好ましくは少なくとも80%、更により好ましくは少なくとも90%、95%もしくは99%と増大する、全タンパク質に対する全ニューロトロフィン、例えばNGFの純度の度合いである。特に好適な純度は少なくとも95%である。「本質的に純粋な」が意味するところは、組成物が、所望のニューロトロフィンについて少なくとも90%もしくはそれ以上純粋であることである。
【0025】
「ニューロトロフィン変種が実質的に含まれない」が意味するところは、(望ましさが劣るニューロトロフィン種を含む)全ニューロトロフィンに対する所望のニューロトロフィンのパーセントが少なくとも70%の所望のニューロトロフィン種、より好ましくは少なくとも80%、更により好ましくは少なくとも90%、93%、95%もしくは99%と増大する組成物である。「本質的に含まれない」が意味するところは、組成物が所望のニューロトロフィンを少なくとも90%あるいはそれ以上含むことである。特に好適な量は、少なくとも95%の所望のニューロトロフィン、例えば正しく折り畳まれた無傷の118/118rhNGF種である。その他の望まれない種もしくは型は、発酵もしくは精製プロセスから生じる、例えば変更された負荷変種のような、誤ってプロセシングされた型又は化学変種、又は好ましくはここに開示した前記のものの全てである。例えば、NGFが細菌中での合成後にインビトロで折り畳まれる場合、「種」又は「変種」は誤って折り畳まれた型あるいは部分的に折り畳まれた型を含み得る。
【0026】
「誤って折り畳まれた」変種が意味するところは、そのシステイン残基対合により、もしくは遊離しているかブロックされている特定のシステイン残基により、ニューロトロフィンとは異なるニューロトロフィンの変種である。誤って折り畳まれた変種もまたニューロトロフィンと同じシステイン対合を有し得るが、誤った折り畳みから生じる異なった三次元高次構造を有する。
【0027】
「化学」変種が意味するところは、例えば変更された電荷を有することにより、カルバミル化、脱アミド、酸化、グリコシル化、又はタンパク分解性切断により、ニューロトロフィンとは化学的に異なる変種である。
【0028】
この発明のカラムの観点において、緩衝液は、一般に、約5から8の範囲のpHを有する。この範囲内にpHを調節する緩衝液には、例えば、クエン酸塩、コハク酸塩、リン酸塩、MES、ADA、BIS−TRISプロパン、PIPES、ACES、イミダゾール、ジメチルマロン酸、MOPS、MOPSO、TES、トリスバッファー、例えばトリス−HCl、HEPES、HEPPS、TRICINE、グリシンアミド、BICINE、グリシルグリシン、及びホウ酸塩バッファーが含まれる。好適な緩衝液はMOPSOバッファーである。
【0029】
ここにおいて使用するところでは、「アルコール」と「アルコール溶媒」とは、アルコールに対して通常使用されている技術用語の意味を持つものであり、好ましくは、1から10の炭素原子を持つアルコール、より好ましくは、メタノール、エタノール、イソプロパノール、n−プロパノ−ル、もしくはt−ブタノール、並びにグリセロール、プロピレングリコール、エチレングリコール、ヘキシレングリコール、ポリプロピレングリコール、及びポリエチレングリコール、最も好ましくはエタノールもしくはイソプロパノールである。このようなアルコールは、水溶液に添加したときに、溶液極性を減少させることにより溶液の疎水性を増大させる溶媒である。
【0030】
MOPSOは3−(N−モルホリノ)−2−ヒドロキシプロパンスルホン酸である。HEPESはN−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸である。試薬アルコールは95容量部(特別に変成したアルコール・フォームラ3A及び5容量部のイソプロピルアルコール)である。MESは2−(N−モルホリノ)エタンスルホン酸である。UF/DFは限外濾過/ダイアフィルトレーションを意味する。TMACはテトラメチルアンモニウムクロリドである。TEACはテトラエチルアンモニウムクロリドである。NGF−120は完全長120/120神経成長因子を意味する。NGF−118は118残基のホモ二量体成熟NGFを意味する。酸化NGFは、成熟未変性NGFの約80%の生物学的活性があるとここに報告されているNGF変種分子であるメツルフォキシド(Metsulfoxide)37を意味する。IsoaspNGFはNGF異性体化変種分子、Asp93を意味する。脱アミドNGFは、Asp45に転換されたAsn45を有するNGFを意味する。RNGFはそのN末端に余分なアルギニン残基を持つNGF分子を意味する。CHOはチャイニーズハムスター卵巣細胞を意味する。
【0031】
ここに記載された樹脂は、マクロプレップ・ハイ・Sカチオン交換(バイオラッド研究所、強いカチオン交換、SO3官能基、名目粒径50ミクロン、名目孔径1000A)、シリカゲル(未誘導体化)、フェニル・セファロース・ファースト・フロー・ロー・サブスティチューション(ファーマシア;高度に架橋した6%アガロース;粒径45−165ミクロン)、SP−セファロースHP(ファーマシア;高度に架橋した6%アガロース;粒径34ミクロン)、フェニル・トーヨーパール650M(TosoHaas;粒径40−90ミクロン)、及びフラクトゲルEMD SO3−650S(E.Merck(ドイツ)の米国関連会社であるEM Separations;粒径25−40ミクロン)を含む。
【0032】
<発明を実施するための形態の説明>
ニューロトロフィンは、神経系の発達と維持に重要な役割を果たす小さい塩基性タンパク質のファミリーに属する。このファミリーの最初に同定され恐らくは最もよく理解されているメンバーは神経成長因子(NGF)である。1992年12月8日発行の米国特許第5169762号を参照せよ。
【0033】
最近、NGFと類似した機能を有している配列的に関連しているが別個のポリペプチドが同定された。例えば、ニューロトロフィン−2(NT2)ともいわれる脳由来神経栄養因子(BDNF)が、Leibrockら(Nature, 341:49-159[1989])によりクローン化され配列決定された。
【0034】
幾つかの研究グループが、ニューロン因子(NF)と当初は呼ばれ、今はニューロトロフィン−3(NT−3)といわれる神経栄養因子を同定した(Ernforsら, Proc.Natl.Acad.Sci. USA, 87:5454-5458[1990]; Hoehnら, Nature, 344:339[1990]; Maisonpierreら, Science, 247:1446[1990]; Rosenthalら, Neuron, 4:767[1990]; JonesとReichardt, Proc.Natl.Acad.Sci. USA, 87:8060-8064[1990]; Kaishoら, FEBS Lett., 266:187[1990])。
【0035】
(NT4又はNT5といわれる)ニューロトロフィン−4/5が同定された(Hallbookら, Neuron, 6:844-858[1991]; Berkmeierら, Neuron, 7:857-866[1991]; Ipら, Proc.Natl.Acad.Sci, USA, 89:3060-3064[1992])。1994年11月15日に発行された米国特許第5364769号は、ヒトNT−4/5及びその組換え発現方法を開示しており、出典明示によりここに取り込まれる。
【0036】
また報告されているのは、キメラ及びパントロピックニューロトロフィン、例えば1996年1月30日に発行された米国特許第5488099号、Urferら(EMBO J. 13(24):5896-909(1994))及び、ニューロトロフィンが、一以上のレセプターに結合するように修飾されたか、未変性ニューロトロフィンに有意な程度では通常はないレセプター結合活性を含む(ここに出典明示により取込まれる)1995年12月14日公開のWO95/33829に報告されたようなものである。
【0037】
特に興味深いものはMNTS−1及びD15A NT3と命名されたニューロトロフィンである。また特に興味深いものは、NGFアミノ酸バックボーンを有するがtrkA以外のレセプター、例えばtrkB又はtrkCに結合するように修飾されたニューロトロフィンである。
【0038】
NT−3のtrkレセプターへの結合の原因であるNT−3の対応する位置からのアミノ酸を持つNGFにアミノ酸置換がなされたものが好適である。このようなNGF突然変異体は、NGF薬物動態学的及び精製挙動を保持しながらNT−3様レセプター結合活性を有する(Urferら, Biochemistry 36(16):4775-4781(1997))。これらのNGF突然変異体はまたtrkA結合活性を欠き得る(Shihら, J.Biol.Chem. 269(44):27679-86(1994))。このようなNGF突然変異体がここに記載する本発明の用途に対して特に好適なニューロトロフィンである。
【0039】
組換えヒトニューロトロフィン、例えばrhNGFの単離は、種々の多様な宿主細胞汚染物質からのタンパク質の分離を含む。各工程は、十分な分離が生じることを可能にする特別な緩衝液を含む。ニューロトロフィンに対する最後もしくは最後から二番目のプロセシング工程は、通常のクロマトグラフィー媒体を使用して同時精製する幾つかのニューロトロフィン変種の存在により複雑化する。再折り畳み工程が回収・精製工程に含まれる場合、変種はニューロトロフィンの誤って折り畳まれた型を含む。変種はまたニューロトロフィンとは化学的に異なるもの、例えばカルバミル化、脱アミド、脱アミドもしくはタンパク分解性に切断された形態をも含みうる。NGFの場合、これらの種は主に二量体型−ホモ二量体、例えば118/118が所望の場合、120/120又は117/117、又はヘテロ二量体、例えば120/118、117/118−、化学的に修飾された変種−イソアスパラギン酸、モノ酸化、グリコシル化変種、N末端及びC末端切断型、及びその二量体からなる。
【0040】
本発明は、ニューロトロフィン、特にrhNGFを、例えばアルツハイマー病、糖尿病及びエイズ関連神経障害を含む抹消神経障害等々の治療のような治療用途に十分な量で大規模に生産することを可能にする。
【0041】
NGFとその他のニューロトロフィン、好ましくはNGFファミリーのものとの間の配列と高次構造の類似性に鑑みると、本発明の方法は、誤ってプロセシングされた変種、誤って折り畳まれもしくは部分的に折り畳まれた変種、グリコシル化変種及び/又は負荷変種を実質的に含まないこれらのニューロトロフィンを調製するために適用しうる。本発明において、これらとその他の密接に関連した変種からニューロトロフィンを選択的に分離するのに望ましいカラム樹脂と条件が特定される。
【0042】
ニューロトロフィンには、NT−3、NT−4/5、NT−6、BDNF、及びそのヘテロ二量体、キメラもしくはパントロピック形を含む操作形態が含まれる。好ましくは、ニューロトロフィンはヒトあるいはヒトアミノ酸配列と高度に相同性を有するもので、ヒト配列に対して好ましくは80%より多く、より好ましくは90%より多く、最も好ましくは95%より多い相同性を有する。操作されたニューロトロフィンは、それが模倣する未変性ニューロトロフィンのtrkレセプター結合機能の少なくとも50%を保持し、好ましくは少なくとも75%、より好ましくは少なくとも80%を保持する。これらの操作された型は、ここに記載する方法において類似の性能を保持する未変性ニューロトロフィンの疎水的性質又は十分に高いpIを保持するものである。
【0043】
以下に記載するように、ここに記載する方法は、rhNGF、rhNT−3及びrhNT−5に首尾よく適用された。例えば、大腸菌において作られたrhNT−4/5が、封入体に単離され、還元され、封入体から可溶化された。還元されたNT−4/5はDEセファロース・ファースト・フロー及びS−セファロース・ファースト・フロー・クロマトグラフィーによって部分的に精製された。S−セファロース・ファースト・フロー・プールはグアニジン含有緩衝液中で24時間の間再び折り畳まれた。NT−4/5の誤って折り畳まれた型はここに大規模なもので開示したクロマトグラフィーによって除去した。NT−4/5のカルバミル化及び切詰め型(誤ってプロセシングされた形態)が、大規模のカラム形式での高速カチオン交換クロマトグラフィーにより、ポリキャットA HPLC樹脂もしくはSPセファロースHP樹脂で除去された。精製rhNT−4/5は製剤用に酸性緩衝液中に限外濾過されダイアフィルトレーションにかけられた。
【0044】
本発明の一実施態様は、通常はニューロトロフィンを大抵の他の不純物から既に精製した後、典型的には製剤化の前の脱塩もしくはダイアフィルトレーションの前の最終工程又は最終に近い工程において、ニューロトロフィンをその関連する変種から精製することを含む。混合物中の関連した変種は、発酵の結果生じる変種残渣ばかりでなく、ニューロトロフィンが貯蔵もしくはプロセシング中に分解した場合に生成された変種も含みうる。
【0045】
本発明の実施態様において使用するのに適したニューロトロフィンは、任意の手段により調製することができるが、好ましくは組替え的に調製される。ここで考察したニューロトロフィンをコードしている核酸分子は幾つかのソースから入手可能であり、例えば既知のDNA配列を用いて化学合成により、又は当業者に知られている標準的なクローニング法を使用することにより入手できる。例えばhGNFコード配列のようなニューロトロフィンを保有しているcDNAクローンは、ニューロトロフィンの既知の配列に基づいて特に設計されたオリゴヌクレオチドハイブリダイゼーションプローブを使用して同定することができる。
【0046】
ニューロトロフィンコード配列を有する分子を得る際には、分子を、選ばれた宿主細胞中に発現させるために適したクローニングベクター内に挿入する。クローニングベクターは、コード配列の効率的な転写、翻訳及びプロセシングに必要とされる好適な調節機能をもたらすように作成される。
【0047】
ニューロトロフィンが組替え的に調製される場合、ニューロトロフィンをコードしているDNAを発現するために好適な宿主細胞は、原核生物、酵母もしくはより高等な真核生物細胞である。この目的に対して適した原核生物は、古細菌及び真正細菌のような細菌を含む。好適な細菌は真正細菌、例えばグラム陰性もしくはグラム陽性生物体、例えばエシェリチアのような腸内細菌科、例えば大腸菌、エンテロバクター、エルウィニア(Erwinia)、クレブシエラ、プロテウス、サルモネラ、例えばネズミチフス菌、セラチア属、例えばセラチア・マルセスキャンス及び赤痢菌属;桿菌、例えば枯草菌及びバシリ・リチェニフォルミス(licheniformis)(例えば1989年4月12日に公開されたDD266710に開示されたバシリ・リチェニフォルミス41P);シュードモナス属、例えば緑膿菌;ストレプトマイセス属;アゾトバクタ;根粒菌;ビトレオシラ(Vitreoscilla);及びパラコッカス(Paracoccus)である。好適な大腸菌宿主には、大腸菌W3110(ATCC27325)、大腸菌94(ATCC31446)、大腸菌B及び大腸菌X1776(ATCC31537)が含まれる。これらの例は限定するものではなくむしろ例証のためのものである。
【0048】
1995年11月16日に公開されたPCT刊行物WO95/30686の全体を出典明示によりここに取込む。この刊行物は、そのNGFの細菌合成とインビトロ折り畳みの記載が特に関連する。その方法から得られた生成物を本発明の精製方法により精製することができる。
【0049】
上述の細菌の任意のものの突然変異体細胞を使用することもできる。細菌の細胞中におけるレプリコンの複製性を考慮に入れて好適な細菌を選択することは勿論必要である。例えば、大腸菌、セラチア、又はサルモネラ種が、例えばpBR322、pBR325、pACYA177、もしくはpKN410のようなよく知られたプラスミドがレプリコンを供給するために使用される場合、宿主として好適に使用することができる。
【0050】
大腸菌株W3110は組換えDNA産物発酵に対するよく知られた宿主株であるので、好適な宿主もしくは親宿主である。好適には、宿主細胞は最小量のタンパク分解酵素を分泌する。例えば、株W3110は、宿主に対して内在性のタンパク質をコードしている遺伝子で遺伝的突然変異を行うように修飾することができ、そのような宿主の例としては、完全遺伝子型tonAΔを持つ大腸菌W3110株1A2、完全遺伝子型tonAΔptr3を持つ大腸菌W3110株9E4、完全遺伝子型tonAptr3phoAΔE15Δ(argF−lac)169ΔdegPΔompTkan<r>を持つ大腸菌W3110株27C7(ATCC55244)、完全遺伝子型tonAptr3phoAΔE15Δ(argF−lac)169ΔdegPΔompTΔrbs7ilvGkanrを持つ大腸菌W3110株37D6、非カナマイシン耐性degP欠失型変異体を持つ株37D6である大腸菌W3110株40B4、及び1990年8月7日に発行された米国特許第4946783号に開示された変異体周辺質プロテアーゼを有する大腸菌株がある。
【0051】
ヒトNGFは大腸菌中に発現された。hNGFのβサブユニットをコードしている遺伝子の単離と配列及び大腸菌中での異種性タンパク質としてのその発現は米国特許第5288622号に記載された。そこでの教唆もまた哺乳動物細胞から生成した成熟ヒトNGFを提供するのに好適である。組換え技術を使用して、ヒトβ−NGFはその他の哺乳動物のタンパク質を含まないように発現された。変化したアミノ末端を含む2つの遺伝子を用いて大腸菌中にhNGFを発現させた結果、融合タンパク質が発現したが、これはIwaiら(Chem.Pharm.Bull.34:4724(1986))によって記載された。
【0052】
原核生物に加えて、糸状菌又は酵母菌のような真核微生物は、ニューロトロフィンをコードしているベクターのための適切な発現宿主である。サッカロミセス・セレヴィシア、又は一般的なパン酵母は下等真核生物宿主微生物のなかで最も一般的に用いられる。しかしながら、例えばシゾサッカロミセス・ポンベ(Beach及びNurse, Nature, 290:140(1981);1985年5月2日発行のEP139383);クルイヴェロミセス宿主[米国特許第4943529号; FleerらBio/ Technology, 9:968-975(1991)]、例えばK.ラクティス[MW98-8C, CBS683, CBS4574; Louvencourtら, J. Bacteriol.,737(1983)]、K.フラギリス(ATCC 12424), K.ブルガリカス (ATCC 16045)、K.ウィッケラミイ(ATCC 24178)、K.ワルティイ (ATCC 56,500), K. ドゥロソフィラルム[ATCC 36,906; Van den Bergら Bio/Technology,8:135(1990)]、K.サーモトレランス、及びK. マルクシアヌス;ヤロウィア[EP 402,226];ピチア・パストリス[EP183070; Sreekrishnaら, J. Basic Microbiol.,28:265-278(1988)];カンディダ;トリコデルマ・リーシア (EP 244234); ニューロスポラ・クラッサ [Caseら, Proc. Natl. Acad. Sci. USA, 76:5259-5263 (1979)]; シュワニオミセス・オクシデンタリスのようなシュワニオミセス[1990年10月31日公開のEP394538); 及び糸状菌、例えばニューロスポラ、ペニシリウム、トリポクラディウム[WO91/00357 1991年1月10日発行]、及びアスペルギルス宿主、例えばA.ニデュランス[Balanceら, Biochem Biophys. Res. Commun.,112;284-289(1983); Tilburnら,Gene,26:205-221(1983); Yeltonら, Proc. Natl. Acad. Sci. USA, 81:1470-1474 (1984)]及びクロカビ[Kellyら,EMBO J.,4:475-479(1985)]のような、多数の他の属、種、及び菌株が一般に利用でき、ここで有用である。
【0053】
ニューロトロフィンをコードしているDNAの発現に適切な宿主細胞は、多細胞生物からもまた誘導できる。このような宿主細胞は、複雑なプロセシング及びグリコシル化活動が可能である。原則的には、脊椎動物培養由来であろうと無脊椎動物培養由来であろうと、任意の更に高等の真核生物細胞培養が適している。無脊椎動物細胞の例としては植物及び昆虫細胞が含まれる。多数のバキュロウィルス株及び変異体及び対応する許容可能な昆虫宿主細胞、例えばスポドプテラ・フルギペルダ(毛虫)、アエデス・アエジプティ(蚊)、アエデス・アルボピクトゥス(蚊)、ドゥロソフィラ・メラノガスター(ショウジョウバエ)、ボンビクス・モリのような宿主が特定されている。例えば、Luckowら, Bio/Technology, 6:47-55 (1988); Millerら, Genetic Engineering, Setlowら編 Vol. 8 (Plenum Publishing, 1986), pp.277-279; 及びMaedaら, Nature,315:592-594(1985)を参照のこと。形質移入には種々のウィルス株が公に利用できる。例えば、オートグラファ・カリフォルニカNPVのL−1変異体とボンビクス・モリNPVのBm−5株があり、このようなウィルスは、特にスポドプテラ・フルギペルダ細胞の形質移入にここで使用することができる。ヒトNGFは1993年12月21日に発行された米国特許第5272063号に報告されているように、昆虫細胞において生産されている。
【0054】
綿花、コーン、ジャガイモ、大豆、ペチュニア、トマト、及びタバコのような植物細胞培養を宿主として用いることができる。典型的には、ニューロトロフィンをコードしているDNAを含むように前もって操作しておいた細菌アグロバクテリウム・トゥメファシエンスのある菌株と共にインキュベートすることによって植物細胞は形質移入される。A.トゥメファシエンスと共に植物細胞培養をインキュベートする間に、ニューロトロフィンをコードしているDNAが、植物細胞宿主が形質移入されるようにその植物細胞宿主に移され、適切な条件下でニューロトロフィンをコードしているDNAを発現する。加えて、例えば、ノパリンシンターゼプロモーター及びポリアデニル化シグナル配列のような、植物細胞と適合しうる調節及びシグナル配列が利用できる(Depickerら,J.Mol.Appl.Gen., 1:561(1982))。また、T−DNA780遺伝子の上流領域から分離されるDNAセグメントは、組換えDNAを含む植物細胞中の植物発現遺伝子の転写レベルを活性化又は増強しうる(1989年6月21日公開のEP321196)。
【0055】
有用な哺乳動物宿主細胞株の例は、SV40によって形質転換されたサル腎臓CV1株(COS-7, ATCC CRL 1651);ヒト胚腎臓株[293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamら, J. Gen Virol., 36:59 (1977)];ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/−DHFR[CHO, UrlaubとChasin, Proc.Natl.Acad.Sci.USA,77:4216 (1980)];マウスのセルトリ細胞[TM4, Mather,Biol.Reprod., 23:243-251 (1980)];サルの腎細胞 (CVI ATCC CCL 70); アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587); ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34); バッファローラット肝臓細胞 (BRL 3A, ATCC CRL 1442); ヒト肺細胞 (W138, ATCC CCL 75); ヒト肝細胞 (Hep G2, HB 8065); マウス乳房腫瘍細胞 (MMT 060562, ATTC CCL51); TRI細胞[Motherら, Annals N.Y. Acad. Sci., 383:44-68 (1982)]; MRC5細胞; FS4細胞; 及びヒト肝癌ライン(Hep G2)である。好適な方法はCHO細胞中の発現である。プレプロNGFを含むヒトNGFのエキソンを、適切なプロモーター及びベクターを使用して分泌された成熟NGF(118及び120型を含む)の発現を達成するために使用することができる(米国特許第5288622号)。安定して形質移入された安定なCHO細胞の培養及びNGFの分泌成熟型は本明細書の実施例において検討されるように、本発明において有用である。
【0056】
宿主細胞に形質移入し、そして上述の発現又はクローニングベクターで好ましくは形質転換し、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードしている遺伝子を増幅するために適当に修飾された常套的栄養培地で培養する。形質移入は、如何なるコード配列が実際に発現されるか否かにかかわらず、宿主細胞による発現ベクターの取り上げを意味する。多数の形質移入法が当業者に知られており、例えば、CaPO4及びエレクトロポレーションである。このベクターの操作のあらゆる徴候が宿主細胞内で生じたときに一般に形質移入の成功が認められる。
【0057】
形質転換は、染色体外のエレメントとしてであろうと染色体成分によってであろうと、DNAが複製可能であるように、生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。SambrookらのMolecular Cloning:A Laboratory Manual[New York: Cold Spring Harbor Laboratory Press,1989]の1.82節に記載された塩化カルシウムを用いるカルシウム処理又はエレクトロポレーションは、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して一般に用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawら, 23:315 (1983)及び1989年6月29日公開のWO89/05859に記載されたように、ある種の植物細胞の形質転換に用いられる。加えて、1991年1月10日に公開されたWO91/00358に記載されているように、超音波処理を用いて植物をトランスフェクトすることもできる。
【0058】
このような細胞壁のない哺乳動物の細胞に対しては、Graham及びvan der Eb(Virology, 52:456-457(1978))のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞の宿主系形質転換の一般的な側面は1983年8月16日に発行された米国特許第4399216号に記載されている。酵母中の形質転換は、典型的には、Van Solingenら(J.Bact., 130:946(1977))及びHsiaoら(Proc.Natl.Acad.Sci. USA,76:3829(1979))の方法によって実施する。しかしながら、DNAを細胞中に導入する他の方法、例えば、核マイクロインジェクション、エレクトロポレーション、無傷の細胞、又はポリカチオン、例えばポリブレン、ポリオルニチン等々を用いる細菌プロトプラスト融合もまた用いることができる。哺乳動物細胞を形質転換するための種々の方法については、Keownら(Methods in Enzymology(1990),Vol185:pp527-537)及び Mansourら(Nature,336:348-352(1988))を参照のこと。
【0059】
好適には、hNGFの遺伝子が、メチオニン開始コドン、好ましくはUllrichら(Nature,303:821-825(1983))により同定されたような二つのメチオニン開始コドンの一方を利用できるようにするためにベクター内に挿入される。hNGF遺伝子(Ullrichら, Cold Spring Harbor Symposia on Quant. Biol. XLVIII, p.435(1983);米国特許第5288622号)は、転写開始コドンとして使用されそうな2つの殆ど隣接するメチオニンを有している(位置1は成熟hNGFのN末端セリン残基をいう)。これに対して、NGFに対するマウス顎下腺cDNAは、神経成長因子の最も完全に研究されているものであるが、121と119の位置のメチオニンに加えて187の位置にもメチオニンを有している。哺乳動物細胞中の発現に対する好適な実施態様では、プレプロNGF配列が存在する。
【0060】
原核生物細胞がニューロトロフィンを生産するために使用される場合には、例えばSambrookらのMolecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory Press, New York 1989)において一般に記載されているようにしてプロモーターを構成的に又は人為的に誘導することができる適切な培地中で培養される。炭素、窒素、及び無機リン酸塩源以外に、あらゆる必要な補充物質を、単独もしくは複合窒素源のような他の補充物質又は培地との混合物として導入される適当な濃度で含めることもできる。
【0061】
ニューロトロフィンを生産するために哺乳動物の宿主細胞が用いられる場合、これは種々の培地において培養することができる。ハム(Ham)のF10(Sigma)、最小必須培地([MEM] Sigma)、RPMI−1640(Sigma)及びダルベッコの改良イーグル培地([DMEM], Sigma)のような市販培地が当該宿主細胞の培養に適している。また、HamとWallace, Meth. Enz., 58:44(1979), Barnes及びSato, Anal. Biochem., 102:255 (1980), 米国特許第4767704号;4657866号;4927762号; 5122469号;4560655号;WO90/03430;WO87/00195;米国再発行特許第30985号(これらの全ての開示を出典明示によりここに取込む)に記載された任意の培地を宿主細胞の培養培地として用いることができる。これらの培地はいずれも、ホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン、又は表皮成長因子)、塩類(例えば、塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、緩衝液(例えばHEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えば、ゲンタマイシン(商標)薬)、微量元素(最終濃度がマイクロモル範囲で通常存在する無機化合物と定義される)及びグルコース又は同等のエネルギー源を必要に応じて補充することができる。任意の他の必要な補充物質もまた当業者に知られている適当な濃度で含むことができる。培養条件、例えば温度、pH等々は、発現のために選ばれた宿主細胞について従来用いられているものであり、当業者には明らかであろう。
【0062】
一般に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、及び実用技術は、Mammalian Cell Biotechnology: A Practical Approach, M. Butler編 (IRL Press at Oxford University Press,Oxford,1991)に見出すことができる。上記の方法は、ニューロトロフィンが細胞内に生産されようが、細胞膜周辺膣に生産されようが、あるいは培地に直接分泌されようが、使用することができる。
【0063】
典型的には、培養液は適当な時間後に収集され、標準的な同定アッセイ、例えばELISA及びウェスタンブロット分析のようなイムノアッセイ、又は生物学的アッセイ、例えばPC12細胞分化(Greene,L.A., Trends Neurosci. 7:91 (1986))が実施される。ここに開示された変種の種類と程度を決定するためのアッセイは従来から知られているか、あるいは実施例において提供されもしくは引用される(例えば、出典明示によりここに取込むSchmelzerらJ.Neurochem.59 (5):1675-83(1992)及びBurtonら,J.Neurochem. 59(5):1937-45(1992)を参照)。
【0064】
細胞から調製されるニューロトロフィン組成物は、好ましくはHICに先立って少なくとも一つの精製工程に通される。好適な精製工程の例には、回収すべきニューロトロフィン及び使用する出発培養に依存して、アフィニティクロマトグラフィー、シリカクロマトグラフィー、ヘパリンセファロースクロマトグラフィー、アニオンもしくはカチオン交換樹脂クロマトグラフィー(例えばポリアスパラギン酸カラム)、クロマトフォーカシング、及び調製的SDS−PAGEのようなタンパク質精製の他の方法を含むここに記載したものを含む。
【0065】
ニューロトロフィンが培地中に直接分泌される一実施態様においては、培地を遠心分離により細胞片から分離し、清澄化された発酵ブロス又は培地を次にシリカゲルでの精製に使用する。シリカクロマトグラフィーに対しては、典型的にはニューロトロフィンポリペプチドがシリカ粒子に付着するように誘導体化されていないシリカ粒子にブロスを通す;シリカ粒子を洗浄して汚染物質を除去する;そしてアルコール性もしくは極性非プロトン溶媒とアルカリ土類、アルカリ金属もしくは無機アンモニウム塩を含む緩衝液でシリカ粒子からポリペプチドを溶出させる。
【0066】
本発明の好適な実施態様では、ニューロトロフィンをその変種から分離し、またバルク汚染物質を減少させるためにマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーを用いる。この樹脂は、哺乳動物細胞培養からのニューロトロフィンの精製の初期の工程として、好ましくは収集細胞培地の分画に対して使用することができる。他の実施態様では、マクロプレップ・ハイ・Sカチオン交換クロマトグラフィーは、最も好ましくはタンパク質再折り畳み工程の直後に使用される。このカチオン交換カラムの非常に高い流れ性により、大量の希薄な再折り畳みタンパク質又は収集細胞培地ニューロトロフィンを、例えばHIC又はSP−セファロースのような次のクロマトグラフィー工程の前に、ニューロトロフィンがカラムに結合する条件を調節することにより、容易く濃縮することが可能になる。更に、樹脂のカチオン交換性により、非結合バルクタンパク質及びある種の誤ってプロセシングされた変種及び化学的に修飾された変種(例えば変更負荷、MET37−酸化変種)の除去が可能になる。最も重要なことは、未変性のニューロトロフィンと疎水性が異なる誤ってプロセシングされた変種もしくは化学的に修飾された変種(例えば(哺乳動物細胞培養生産からの)誤ってプロセシングされた変種、グリコシル化変種)が存在する場合、マクロプレップ樹脂により、これらの疎水性変種の部分的な除去が可能になり、ここで測定されるように、未変性のニューロトロフィンを実質的に富化することになることである。限定することを意図するものではないが、樹脂支持体のバックボーンはニューロトロフィンと樹脂の間の非特異的相互作用を促進する疎水性含量を含むものと信じられ、これがここに教示されるようにして利用される。典型的には、約5から8、より好ましくは6から8のpHの溶出緩衝液に対しては、0から3MのTMAC濃度が有用である。酢酸ナトリウムが、溶出緩衝液のイオン強度を増大させるために存在する場合、より低いTMAC濃度を使用することができる。塩化物は酢酸塩イオンに対する好適な代替物である。
【0067】
ニューロトロフィンが細胞膜周辺膣において生産される実施態様の一例では、培地又は可溶化物が遠心分離されて粒子状の細胞片を除去する。ついで、必要に応じて膜と可溶性タンパク質分画を分離することができる。ついで、ニューロトロフィンは、ニューロトロフィンが膜結合性か、可溶性か、凝集化形態で存在しているかに応じて、可溶性タンパク質分画から、そして培養可溶化物の膜分画から精製することができる。その後、ニューロトロフィンは可溶化され、ついで適切な緩衝液を使用して再び折り畳まれる。再折り畳みタンパク質を生産するための周辺質からのこの単離方法の詳細については以下に記載する。
【0068】
不溶性の未変性ではないニューロトロフィンは、適切な単離緩衝液中の原核生物宿主細胞から、任意の適当な技術、例えば細胞を適切なイオン強度の緩衝液に曝露して殆どの宿主タンパク質を可溶化させ(しかし凝集ニューロトロフィンは実質的に不溶性である)、封入体を放出し、例えば遠心分離により回収に利用できるようにするように細胞を破壊することにより、単離される。この技術はよく知られており、例えば米国特許第4511503号に記載されている。
【0069】
簡単には、細胞は(典型的には、約0.01から2M、好ましくは0.1から0.2Mのイオン強度を使用して、pH5から9、好ましくはpH約6から8の)緩衝液中に懸濁される。塩化ナトリウムを含む任意の適切な塩が十分なイオン強度値を維持するために有用である。細胞は、この緩衝液中に懸濁され、ついで、例えば機械的方法、例えばManton−Gaulinプレスマイクロフルイダイザー、フレンチプレスもしくは音波オシレーター、あるいは化学的又は酵素的方法のような通常用いられている技術を使用して溶解により破壊する。
【0070】
細胞破壊の化学的もしくは酵素的方法の例には、細菌壁を溶解するリゾチームの使用を伴うスフェロプラスティング(Neuら, Biochem.Biophys.Res.Comm.,17:215(1964))及びポリペプチドの放出のために生細胞を高緊張度の溶液と低緊張度の冷水洗浄で処理することを含む浸透圧ショック(Neuら, J.Biol.Chem.,240:3685-3692(1965))が含まれる。米国特許第4680262号に記載されている第3の方法は、形質転換された細菌細胞を、細胞を死滅させ溶解するのに充分な時間と温度で、有効量の2から4の炭素原子を持つ低級アルカノールに接触させるものである。
【0071】
細胞を破壊した後、懸濁液は典型的には遠心分離されて封入体をペレット化する。一実施態様では、この工程は、標準的な遠心機において、容量と遠心設計に依存する充分な時間、通常は約10分から0.5時間、約500から15000xg、好ましくは約12000xgで実施される。得られたペレットは実質的に全ての不溶性のポリペプチド分画を含むが、細胞破壊プロセスが完全でないならば、無傷の細胞もしくは破壊された細胞フラグメントも含有し得る。細胞破壊の完全性は、ペレットを少量の同じ緩衝液中に懸濁させ、懸濁液を位相差顕微鏡で調べることにより検査することができる。破壊細胞フラグメントもしくはホールセルの存在は、フラグメントもしくは細胞及び随伴する非屈折性(non−refractile)ポリペプチドを除去するために更なる破壊が必要であることを示している。そのような更なる破壊後に、必要ならば、懸濁液を再び遠心分離し、ペレットを回収し、再懸濁し、分析する。このプロセスを、視覚検査でペレット化材料中に破壊細胞フラグメントが無いことが明らかになるか、更なる処理でも得られるペレットの寸法を低減することができなくなるまで繰り返す。
【0072】
別の実施態様では、ニューロトロフィンは、適切な緩衝液中での可溶化により細胞膜周辺膣から単離される。この方法は、ニューロトロフィンが組換え的に生産された後に発酵ベッセルに試薬を直接添加して、収集、ホモジェナイゼーション、及び遠心分離という余分な工程を避けてニューロトロフィンを得るインサイツ溶解化とできる。残りの粒子は遠心分離か濾過もしくはその組合せにより除去することができる。
【0073】
ニューロトロフィンがアンフォールディングされているならば、アンフォールディングの度合いは、RP−HPLCを含む非天然ニューロトロフィンのクロマトグラフィーによって適切に決定される。非天然材料に対して増加するピーク領域は、どのくらいの非天然ニューロトロフィンが存在しているかを示す。
【0074】
可溶化された封入体から又は後の精製の段階で一度得られると、ニューロトロフィンは以下に記載されるように活性な高次構造に適切に再折り畳みされる。
【0075】
ニューロトロフィンが、再折り畳みされる前に既に可溶形態ではないならば、ニューロトロフィンを実質的に溶解させるのに必要な量のカオトロピック剤と還元剤を含有するアルカリ緩衝液におけるインキュベーションにより可溶化することができる。
【0076】
ニューロトロフィンの緩衝液中における可溶化の度合いの測定は、濁度決定法により、還元されたSDSゲル上の遠心分離後に上清とペレット間のニューロトロフィン分画を分析することにより、プロテインアッセイ(例えば、バイオ-ラッドタンパク質アッセイキット)により、又はHPLCにより好適に実施される。
【0077】
可溶化に対するアルカリ緩衝液のpH範囲は典型的には少なくとも約7.5であり、好適な範囲は約8−11である。この後の範囲のpHをもたらす適切な緩衝液の例としては、グリシン、CAPSO(3-[シクロヘキシルアミノ]-2-ヒドロキシ-1-プロパンスルホン酸)、AMP(2-アミノ-2-メチル-1-プロパノール)、CAPS(3-[シクロヘキシルアミノ]-1-プロパンスルホン酸)、CHES(2-[N-シクロヘキシルアミノ]エタンスルホン酸)、及びトリスHCl(トリス[ヒドロキシメチル]アミノメタンヒドロクロリド)が含まれる。ここで好適な緩衝液は、好ましくは約20mMの濃度、約8.5から11のpH、好ましくは約10−11のpHのグリシン又はCAPSOである。
【0078】
可溶化のための緩衝溶液中のニューロトロフィンの濃度は、ニューロトロフィンが実質的に溶解化され、部分的に又は充分に還元し、変性するようなものでなければならない。あるいは、ニューロトロフィンは最初は不溶性であってもよい。使用する正確な量は、例えば緩衝溶液中の他の成分の濃度とタイプ、特に還元剤のタイプと量、カオトロピック剤のタイプと量、および緩衝液のpHに依存する。例えば、ニューロトロフィンの濃度は、還元剤、例えばDTTの濃度が、約3:1から10:1のDDT:ニューロトロフィンの比を維持するために、同時に増加させられるならば、少なくとも3倍増加させることができる。希釈再折り畳みの前により濃縮された可溶化タンパク質溶液をつくることが望ましい。従って、ニューロトロフィンの好適な濃度は少なくとも約30mg/mLで、より好ましい範囲は30−50mg/mLである。例えば、ニューロトロフィンは、5Mから7Mの尿素中に約30−50mg/mLの濃度まで可溶化し、例えば約1mg/mLまで折り畳みのために希釈することができる。
【0079】
ニューロトロフィンを可溶化した後、これを、5−40%(v/v)のアルコール性又は非プロトン溶媒、カオトロピック剤、及びアルカリ金属、アルカリ土類、もしくはアンモニウム塩を含む再折り畳み緩衝液中に入れるか希釈する。緩衝液は、最初の緩衝溶液に対しては任意の緩衝液とすることができ、CAPSO、グリシン、及びCAPSがpH8.5−11、特に約20mMの濃度で好ましく、CAPSOとグリシンが最も好ましい。ニューロトロフィンは、好ましくは少なくとも5倍、より好ましくは少なくとも約10倍に、再折り畳み緩衝液で希釈することができる。あるいは、ニューロトロフィンは、再折り畳み緩衝液に対して透析することができる。再折り畳みは約2℃−45℃、最も好ましくは2℃−8℃で少なくとも約1時間の間、実施することができる。溶液は、場合によっては還元剤と浸透質(osmolyte)をまた含有してもよい。
【0080】
還元剤は与えられた濃度範囲における可溶化工程に対して上述したものから好適に選択される。その濃度は、特に、アルカリ土類、アルカリ金属、又はアンモニウム塩、ニューロトロフィン、及び溶媒の濃度に依存する。好ましくは、還元剤の濃度は、約0.5から8mM、より好ましくは約0.5−5mM、更により好ましくは約0.5−2mMである。好適な還元剤はDTTとシステインである。
【0081】
再折り畳み溶液中の酸素は必要に応じて不活性ガス、例えばヘリウム又はアルゴンの添加により除去して酸素を置換することもできる。
【0082】
任意の浸透質は好ましくはスクロース(約0.25−1Mの濃度)又はグリセロール(約1−4の濃度)である。より好ましくは、スクロース濃度は約1Mであり、グリセロール濃度は約4Mである。
【0083】
折り畳み緩衝液中のニューロトロフィンの初期濃度は、回収された誤って折り畳まれた配座異性体に対する正しく折り畳まれたものの比が、HPLC、RIA、もしくはバイオアッセイにより決定したとき、最大になるようなものである。(最大収量の正しく折り畳まれた配座異性体が得られる)ニューロトロフィンの好適な濃度は約0.1から15mg/mL、より好ましくは約0.1から6mg/mL、最も好ましくは約0.2から5mg/mLの範囲である。
【0084】
このインキュベーション時に生じる再折り畳みの度合いは、ニューロトロフィンのRIA力価又はHPLC分析により適切に決定され、RIA力価又は正しく折り畳まれたニューロトロフィンピーク寸法の増加は、緩衝液中に存在する正しく折り畳まれた生物学的に活性なニューロトロフィン配座異性体の量の増大に相関する。インキュベーションは、RIAもしくはHPLCにより決定される、正しく折り畳まれたニューロトロフィン配座異性体の収量と回収された誤って折り畳まれたニューロトロフィン配座異性体に対する正しく折り畳まれたニューロトロフィン配座異性体の比を最大にし、質量バランスにより決定される多量体関連ニューロトロフィンの収量を最小にするために実施される。あるいは、以下と実施例に記載する方法によって種を決定することもできる。グアニジンが再折り畳みに対する好適な変性剤である。
【0085】
ニューロトロフィンが再折り畳みされた後、ここに教示されたような次の方法が、個々にあるいは組み合わせて、より高い純度と相同性を得るための好適な精製法である。すなわち、カチオン交換カラムでの分画、疎水的相互作用クロマトグラフィー(HIC)、そしてシリカクロマトグラフィーである。
【0086】
再折り畳み工程がプロセスの一部であろうとなかろうと、ニューロトロフィンをその変種から分離するための好適な工程は疎水的相互作用クロマトグラフィー樹脂上での分離である。発酵、精製又はインビトロタンパク質再折り畳みの間、あるタンパク質は化学的に修飾され、誤ってプロセシングされ得、あるいはその未変性の三次元構造ではなくむしろその安定性、溶解性、免疫原性、もしくは生物活性について異なる他の構造に再折り畳みされうる。これらの変種は、抗原性又は作用強度の喪失のような望ましくない副作用を避けるために回収中に除去されなければならない。変種が不溶性であれば、遠心分離と濾過のような固体/液体分離法により容易く除去することができる。しかしながら、変種が可溶性であれば、クロマトグラフィーのようなより高度の分離吸着法がそれらを除去するために必要とされる。原核生物細胞中に生産されるとき、あるいはインビトロで再折り畳みされるとき、ニューロトロフィンは可溶性の安定した誤って折り畳まれた変種を生成する。誤って折り畳まれたニューロトロフィンは、未変性のニューロトロフィンに対して変更されたジスルフィド対合パターンと三次元構造を有しており、天然の薬理学的活性を欠く。真核生物細胞培養、例えば哺乳動物細胞培養中に生産されるとき、変種型は典型的には誤ってプロセシングされた型である。これらはまた典型的に天然の薬理学的活性を欠き、除去されるべきである。ここではHICが、未変性のニューロトロフィンからこれらの変種を分離するために適したものであることが見出された。
【0087】
HICは非共有性疎水的結合によって媒介された固体マトリックスに対するタンパク質の順次の吸着と脱着を含む。一般に、高塩緩衝液中の試料分子がHICカラムに装填される。緩衝液中の塩は水分子と相互作用をして溶液中の分子の溶媒和を低減させ、よってHICカラムにより結果的に吸着される試料分子の疎水性領域を暴露する。分子がより疎水性であればある程、結合を促進するために必要とされる塩は少なくなる。通常、塩勾配の低下を使用してカラムから試料を溶出させる。イオン強度が減少すると、分子の親水性領域の暴露が増加し、疎水性を増加させるために分子がカラムから溶出する。試料の溶出はまた溶出緩衝液に穏やかな有機修飾因子又は洗浄剤を添加することによっても達成される。HICは、Protein Purification2版(Springer-Verlag, New York, pp.176-179(1988))においてレビューされている。
【0088】
タンパク質とマトリックスの間の会合の強さは、固定化された官能基の大きさと疎水的性質、周囲の溶媒の極性と表面張力、及びタンパク質の疎水性を含む幾つかの要因に依存する。HICマトリックスの結合能は、固定化された疎水性リガンドが広く間隔をあけられるべきという必要性のために低くなる傾向がある。更に、あるタンパク質に対する培地の能力は試料調製における疎水性不純物の量に対して逆に変化する。所望のタンパク質を、能力を同時に最大化しながら、変種と他の不純物から分離するためには、好適なHIC固相媒体並びに装填、洗浄及び溶出のための好適な移動相を特定する必要がある。
【0089】
ここにおいて決定したところでは、無傷の正しく折り畳まれた型から正しく折り畳まれたニューロトロフィン及び誤って折り畳まれたニューロトロフィンもしくは誤ってプロセシングされた型を分離するための最も好適な媒体は、固定化されたフェニル官能基を有するものであった。異なった販売者からのフェニルベースHIC媒体はこれらのニューロトロフィン型を分離するための異なった効率を示した。最良の結果はTosoHaasによるフェニル・トーヨーパール媒体及びフェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)で達成された。TSKフェニル5PWもまた適していた。他のHIC固定化官能基も機能してこれらの型を分離しうる。例としては、オクチル基、例えばファーマシアからのオクチルセファロースCL4B媒体、及びプロピル基、たとえばBakerからのハイ・プロピル媒体であった。好ましさが劣ったものはアルコキシ、ブチル、及びイソアミル官能基樹脂である。
【0090】
HICはニューロトロフィンを哺乳動物細胞培養中のその変種から分離するのに有用である。例えば、ここで測定したところでは、rhNGF発現CHO細胞培養は不適切にタンパク分解性にプロセシングされた変種、たとえば前駆NGF、ハイブリッド前駆NGF及び切詰め前駆NGF配列のような部分的配列が存在するものを含んでいた。また哺乳動物細胞培地において見出されるものはグリコシル化NGF及び不適切にタンパク分解性にプロセシングされた変種のグリコシル化形態である。望ましくないグリコシル化型は、NGFの場合大きい分子量の種(+2000kD)として見られるが、患者に望ましくない抗原応答を発生させ、劣った製品品質又は活性をもたらす。HICは、rhNGFから疎水性変種、主にグリコシル化型を含むN末端がタンパク分解性に誤ってプロセシングされた変種を分離した。実施例に示されるように、前駆配列含有及び切詰め前駆配列NGFと、NGFと前駆配列含有NGFの両方のグリコシル化型がフェニルHICの間NGFピークの先端に溶出した。従って、これらの種が実質的に含まれておらず、高速カチオン交換クロマトグラフィーのような次の工程に特に適したrhNGF組成物を得ることができた。HICは、ソースに拘わらず、他のニューロトロフィン、並びにNGFにも適用できる。例えば、HICは、存在する単量体形に依存してホモ二量体かヘテロ二量体の何れでも、二量体からNGF単量体を分離し、並びにインビトロの再折り畳みの後、又は哺乳動物細胞から生産され分泌されたときに得られる、疎水性がまた異なる二量体を区別するのに有用である。HICに使用するニューロトロフィン混合物の好適なソースは哺乳動物細胞培養、より好ましくはCHO細胞培養である。該培養は好ましくはここで検討されたような少なくとも1回の前精製工程にかけられる。HICは、未変性の組換えニューロトロフィンから誤ってプロセシングされたグリコシル化変種(群)を分離するのに特に有効である。rhNGFの場合、グリコシル化及びプレプロNGFは未変性NGFより疎水性が低く、従って未変性NGFの前に溶出する。(細菌的に生産される場合)ニューロトロフィンの誤って折り畳まれた型もまたより疎水性で、未変性のニューロトロフィンより早く溶出する。
【0091】
ニューロトロフィン型を分離するために最も好適なHIC樹脂は固定化されたフェニル官能基を有しているものであった。異なった販売者からのフェニルベースHIC媒体はこれらのNGF型を分離するための異なった効率を示した。フェニルHIC樹脂の中で、TosoHaasによるフェニル・トーヨーパール媒体が最も好ましく、フェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)とTSKフェニル5PWが好ましい。好ましいHIC官能基としてはアルコキシ、ブチル、及びイソアミル部分が含まれる。
HICを使用して、ニューロトロフィン型を洗浄し差次的に溶出させるために様々な移動相条件を使用することができる。これらの移動相はニューロトロフィンと固定相の間の会合に異なった方法で影響を及ぼす幾つかの異なった化学種を含みうる。正しく折り畳まれ、また誤って折り畳まれたニューロトロフィン、例えばNT−4/5が、例えば硫酸アンモニウム、NaCl濃度、酢酸塩濃度のような移動相塩の段階的減少又は減少塩勾配によりHICカラム上で分離できる。塩は移動相の表面張力を調節することにより樹脂に対するニューロトロフィンの結合に影響を及ぼしうる。表面張力に影響を及ぼした他の薬剤は、実施例で検討したようなクエン酸ナトリウムとテトラメチルアンモニウムクロリドであった。変種はまた、相対的に極性の有機溶媒の濃度を増加勾配で又は段階的に増加させて結合タンパク質を溶出させることにより、カラムクロマトグラフィー中に分離することができる。好適な溶媒の例としてはエタノール、アセトニトリル、及びプロパノールがある。ニューロトロフィン型とHIC樹脂の間の会合の強さは移動相のpHにもまた依存し、中性条件が好ましい。正しく折り畳まれ、誤って折り畳まれたニューロトロフィンの相対的な疎水性はまた溶液のpHに依存する。未変性のニューロトロフィンからの変種の分離はまた、勾配もしくは段階的溶出の間、移動相のいくつかの性質を同時に変化させることにより得ることもできる。例えば、溶出時に塩濃度と非極性溶媒濃度を同時に変化させた移動相は塩のみを変化させた場合よりもより良好な分離をもたらした。
【0092】
HICに対して、硫酸アンモニウム、クエン酸塩、酢酸塩、及び塩化カリウムを含むここで検討した塩が使用できる。使用する塩に依存して、樹脂へのニューロトロフィンの結合を達成する塩の濃度は典型的に0.5Mから3M、より好ましくは0.5から2.5Mである。例えば、0.8から1.5Mの塩の結合緩衝液がNGFに対して好適であり、より高い塩濃度では樹脂上へのNGFの沈殿が生じ、回収率が低下する。NT−3に対しては、1.0から2.5の塩濃度のpH7の結合緩衝液が好ましく、1.25から1.75Mの塩が更に好ましく、1.5Mが最も好ましい。NT−4/5に対しては、1から3Mの塩のpH7の結合緩衝液が好ましく、2から2.75Mが更に好ましく、2.5MのNaClが最も好ましい。NT−4/5の場合、2.5MのNaClが装填には好ましく、2MのNaClが有機溶媒の存在する(例えば10%アルコール、pH7)溶出には好適であった。好ましくは、塩濃度の低下を使用してニューロトロフィンとその変種を溶出させ分離する。溶出を達成するには、溶出緩衝液中の塩濃度は、典型的には装填緩衝液中のものよりも低いが、有機溶媒で補償するときは同じ濃度にすることができる。
【0093】
加えて、ここで見出されたところでは、有機溶媒の使用には、有機溶媒の添加がより狭いピークプロファイルをもたらすことにより溶出パターンを改善するというもう一つの利点がある。エタノールに加えて、プロパノール、イソプロパノール、及び低級アルキレングリコール、例えばプロピレングリコール、エチレングルコール及びヘキシレングリコールを含むここで検討した他の有機溶媒を使用することができる。5から25%(v/v)、より好ましくは5から20(v/v)の有機溶媒が正しく折り畳まれたニューロトロフィンを典型的に溶出させる。有機溶媒での溶出は勾配をもたせても段階的でもよい。pH範囲は好ましくは殆ど中性から僅かに酸性で、5から8、より好ましくは6から8、6.5から7.5、最も好ましくは7である。MOPSO、MOPS、HEPES、リン酸塩、クエン酸塩、アンモニウム、酢酸塩を含むここで検討した任意の緩衝液を、所望のpHで緩衝作用を呈する限り、使用することができる。
【0094】
ここに報告したように、ニューロトロフィンの組換え生産から生じるある望まれないニューロトロフィン変種の本発明者らによる発見に鑑みると、好ましくは調製的態様で、高速カチオン交換クロマトグラフィーを使用することにより未変性ニューロトロフィンからの例えばカルバミル化、酸化、isoasp、脱アミド、およびある種の切詰め型(例えばNGFのC末端切断型)のような荷電修飾変種の分離が可能になる。例えば、NT−4/5とNT−3の場合のように細菌発酵中に生じうる負荷変更の結果として生じるN末端クリップ型(例えば2から4のN末端アミノ酸欠失)を除去することができる。哺乳動物細胞培養において生産されたニューロトロフィンの場合、C末端切断が高度に荷電された末端領域に生じ得る。例えば、NGFの118型はC末端で誤ってプロセシングされ又は切断されて117、114及び115型になる。これらは高速カチオン交換クロマトグラフィーにより未変性118NGFから分離することができる。特に好適な実施態様はSP−セファロース・ハイ・パーフォーマンス、フラクトゲルEMD SO3、またはポリアスパラギン酸樹脂を使用するもので、中でもポリキャットAが特に好ましい。最も好ましくは、大規模では、SP−セファロース・ハイ・パーフォーマンス、又はフラクトゲルEMD SO3樹脂が使用される。
【0095】
ここに記載された方法により得られる組成物は、実質的に純粋なニューロトロフィン、更に通常的には好ましくは本質的に純粋で、実質的にニューロトロフィン変種を含まず、より好ましくは本質的にニューロトロフィン変種を含まない。例えば、CHO細胞培養からのNGFの精製後の典型的なSP−セファロースプールは、約92%の118、4.6%の120、1%の脱アミド化NGF、1%の酸化NGF、及び1%のisoaspNGFを含む。各々の量は、常に、118に対しては約85から93%、(用いられる内因性及び/又は外因性タンパク分解度合いに大きく依存して)120に対しては0から5%、117に対しては0から5%、脱アミド型に対しては0から3%、isoasp型に対しては0−2%、及び酸化型に対しては0から2%の範囲である。NGF(全ての種)の純度は常に99.5%を越える。
【0096】
ニューロトロフィンは、カラムから溶出された後、担体と共に好適に組成物、好ましくは生理的に許容できる担体と共に製薬組成物に製剤化される。ニューロトロフィン組成物は好ましくは無菌である。本発明のニューロトロフィン組成物は、例えば培養中において神経細胞の成長と生存を促進するためにインビトロでの用途が見出される。
【0097】
組換えヒト神経成長因子(NGF)の水溶液中における化学的及び物理的安定性を、4.2から5.8のpH範囲で5℃と37℃の間で調べた。NGFの化学安定性はpHの増加で増大した。pH5.8のコハク酸塩緩衝液中では、NGFの物理的安定性はタンパク質凝集化のために減少した。5℃の安定性データと37℃での加速分解研究の両方に基づいて、最適な製剤はpH5.5の酢酸塩緩衝液であることが分った(ここに出典明示により取込まれるWO97/17087を参照のこと)。逆相HPLCは第1の安定性を示す方法であり、iso−AspへのAsn−93の転換が5℃での第1の分解経路であることを示した。カチオン交換クロマトグラフィーによるNGF分解の定量化はNGF単量体変種の様々な混合二量体への経時的な再構成(二量体交換)により複雑化した。希酸での試料と参照の処理は二量体中の単量体分布を平衡化させ、二量体交換の不存在下で定量化された。ベンジルアルコールとフェノールが、多用途目的に対する2つの液体製剤中へのそのrhNGFとの相容性と安定性について評価された。これらの二つの製剤は、界面活性剤として0.01%のプルロン酸(F68)を含む状態と含まない状態で、pH5.5の20mMの酢酸ナトリウムと136mMの塩化ナトリウム中の0.1mg/mLのタンパク質からなる。これらの二つの製剤の各々におけるベンジルアルコールとフェノールの最終濃度はそれぞれ0.9%と0.25%である。12ヶ月の安定性データに基づくと、rhNGFはこれらの製剤においてフェノールよりもベンジルアルコールでより安定である。界面活性剤の存在下でベンジルアルコールで保存したrhNGF製剤は界面活性剤を添加しない製剤と同じほど安定であり、rhNGF多投薬製剤へのF68の添加は安定性目的では必要とされないことを示している。従って、20mMの酢酸塩、136mMのNaCl、0.9%のベンジルアルコール、pH5.5中に0.1mg/mLのタンパク質を含んでなる製剤がフェーズIIIの臨床試験において複数回の投薬に使用されるrhNGFに対して推奨される。このrhNGF多投薬製剤は、5℃で6ヶ月後にUSP及びEP保存剤効能試験をパスしており、2mg/mLの現在の液体製剤と同じほど安定している。しかし、製剤は、光感受性がある保存剤としてベンジルアルコールを含むので、強い光への曝露は避けるべきである。
【0098】
一般に、組成物は、好ましくは安定な液体又は凍結乾燥可能形態での調製を不可能にしない量であって、効果的で安全な製薬的投与に適した量で、他の成分を含むことができる。
【0099】
ニューロトロフィンは、製薬的に許容できる担体、すなわち使用される用量と濃度でレシピエントに対して無毒性であり、製剤の他の成分と相容性があるものと共に製剤化される。例えば、製剤は、ポリペプチドに対して有害であることが知られている酸化剤及びその他の化合物を含有しない。この製剤工程は、標準的な技術を使用する脱塩又はダイアフィルトレーションにより達成される。
【0100】
一般に、製剤は、液体担体又は細かく分割された固体担体又は両方とニューロトロフィンを均一かつ密に接触させることにより調製される。ついで、必要ならば、製品は所望の製剤に成形される。好ましくは、担体は非経口担体であり、より好ましくはレシピエントの血液と等張である溶液である。そのような担体ビヒクルの例としては、水、生理食塩水、リンガー液及びブドウ糖溶液が含まれる。固定化オイル及びオレイン酸エチルのような非水性ビヒクル、並びにリポソームがここでは有用である。
【0101】
担体は、好適には、等張性及び化学安定性を高める物質のような添加物を少量含有する。このような材料は使用される用量と濃度でレシピエントに対して非毒性であり、例えば、リン酸塩、クエン酸塩、コハク酸塩、酢酸、及び他の有機酸又はその塩のような緩衝液;アスコルビン酸のような抗酸化剤;低分子量(約10残基未満)ポリペプチド、例えばポリアルギニン又はトリペプチド;タンパク質、例えば血清アルブミン、ゼラチン、又は免疫グロブリン;例えばポリビニルピロリドンのような親水性ポリマー;アミノ酸、例えばグリシン、グルタミン酸、アスパラギン酸、又はアルギニン;セルロース又はその誘導体、トレハロース、グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物;EDTAのようなキレート剤;マンニトール又はソルビトールのような糖アルコール;ナトリウムのような対イオン;及び/又はポリソルベート、ポロキサマー、又はPEGのような非イオン性界面活性剤を含む。最終調製物は液体又は凍結乾燥固体とできる。
【0102】
治療投与に対して使用されるニューロトロフィンは無菌でなければならない。無菌性は無菌濾過膜(例えば0.2ミクロン膜)を通す濾過によって直ぐに達成される。一般に、治療用ニューロトロフィン組成物は、殺菌アクセス口、例えば皮下注射針で穿孔可能なストッパーを持つ静脈内溶液袋又はバイアルを有する容器に入れられる。上記の製剤はまたインビトロ用途に適している。
【0103】
通常、ニューロトロフィンは、ユニット又は多投薬容器、例えば密封アンプル又はバイアルに、水溶液として、あるいは再構成のために凍結乾燥製剤として保存される。凍結乾燥製剤の例としては、10mLのバイアルに5mLの無菌濾過1%(w/v)ニューロトロフィン水溶液が満たされ、得られた混合物が凍結乾燥される。注入溶液は凍結乾燥ニューロトロフィンを注射用静菌水を使用して再構成することにより調製される。
【0104】
治療的に有効な用量のニューロトロフィン製剤が患者に投与される。ここにおける「治療的に有効な用量」は、投与される効果をつくりだす用量を意味する。正確な用量は治療すべき疾患に依存し、既知の技術を使用して当業者により決定される。一般に、本発明のニューロトロフィン製剤は、1日当り約0.01μg/kgから約100mg/kg投与される。好ましくは0.1から0.3μg/kgである。また、従来から知られているように、年齢並びに体重、一般的な健康状態、性別、食事、投与時間、薬物相互作用および疾患の重症度に対する調節が必要となり、当業者によりルーチン的な実験により決定される。典型的には、臨床医は、神経細胞の機能を修復し、維持し、最適には再構築する用量に達するまで本発明のニューロトロフィン製剤を投与する。この療法の進行は通常のアッセイで容易にモニターされる。
【0105】
ニューロトロフィンは、NGF、NT−4/5、NT−3、及び/又はBDNFを含む他の神経栄養因子と組み合わされ、あるいは一緒に投与され、神経疾患に対する他の通常の療法と共に使用される。
【0106】
NGFの場合、好ましくは組成物は、製薬的に有効量の神経成長因子と製薬的に許容できる酢酸塩含有緩衝液を含有する。組成物は5から6のpHを有し得る。緩衝液は好ましくは酢酸ナトリウムである。酢酸塩の濃度は好ましくは0.1から200mMである。組成物は好ましくは0.07から20mg/mlのNGF濃度を有する。そして、組成物は更に製薬的に許容できる保存剤、例えばベンジルアルコール、フェノール、m−クレゾール、メチルパラベン、又はプロピルパラベンを任意に含有する。好ましくは保存剤はベンジルアルコールである。ベンジルアルコールの濃度は好ましくは0.1から2.0%である。組成物は製薬的に許容できる界面活性剤を任意に含有し得る。そして、組成物は、生理的に許容できる濃度の塩化ナトリウムを任意であるが好ましく含有することができる。より好ましい組成物は少なくとも約0.1mg/mlの濃度で神経成長因子を、10mMから50mMの濃度の酢酸塩イオンを含有する。更により好ましくは、組成物は少なくとも約0.1から約2.0mg/mlの濃度で神経成長因子を、10mMから50mMの濃度の酢酸塩イオンを含有する。最も好ましい組成物は、0.1mg/mlの濃度のNGF、20mMの酢酸ナトリウム濃度(pH5.5)、136mMの塩化ナトリウム濃度及び0.9%(v/v)のベンジルアルコールを含有する。
【0107】
他の実施態様は、2.0mg/mlのNGF濃度、10mMの酢酸ナトリウム濃度、pH5.5、及び142mMの塩化ナトリウム濃度を含む。0.1mg/mlのNGFを20mMの酢酸ナトリウム、136mMの塩化ナトリウム、0.9%(v/v)のベンジルアルコールを5.5のpHで製剤化することが好ましい。ここで検討したところ、118/118ホモ二量体がNGFの好適な型である。NGFは正常な二量体形態を維持するために約6から8のpHで精製される。しかし、(タンパク質分解性に)切詰められた型のパーセントは単量体形態を表し、これは逆相HPLCによって明らかになり測定される。NGFの異なった二量体型−120/120、120/118、118/118等々の存在は公表されている(主に本研究に使用された分析的及びバイオアッセイについて、並びにその一般的な教示についてここに出典明示により取込まれるSchmelzerら J.Neurochem. 59:1675-1683(1992))。この刊行物は、各二量体型についてインビトロ活性が同じであったことを報告している。しかし、対照的に、本明細書における研究では、120/120二量体は活性が少なく、118/118種と比較して約80−90%の活性であることをラジオレセプターベースアッセイを使用して初めて証明している。アッセイの一形態では、ラットPC−12細胞膜を単離し、NGF標準物質と種々の被験種との間の競合結合に対して使用する。RRAはP75とtrkAレセプターを両方有している。117/117種が118/118種と同じ程活性であることもここでまた見出された。更に、PC−12ベースアッセイの使用によりレセプターベースアッセイの発見を確認し、120/120型が118/118型に対して約60%の活性であることを示す。主に本研究に使用された分析的及びバイオアッセイについて、並びにその一般的な教示についてここにその全体を出典明示により取込むのはBurtonら(J.Neurochem. 59:1937-1945(1992))である。
【0108】
118/118型は120/120型よりもヒトの患者において生物利用能が高いと信じられている。生物利用能の増加は少なくとも4から5倍である。この差は顕著で、意外であり、従来では予想できなかったことである。
【実施例】
【0109】
次の実施例は例証のために提供されるものであって限定のためではない。本明細書中の全ての引用文献の開示が出典明示により明示的にここに取込まれる。
【0110】
<実施例I.118/118NGFホモ二量体の精製>
この実施例はNGFの精製と各工程に対する原理を例証する。実施例の各々において、当業者であれば、従来からよく知られているようにして初期培養容積とタンパク質濃度を補償するようにカラム寸法と流量を容易に決定して調節することができる。
収集細胞培養液
【0111】
DNA配列をコードしている120アミノ酸ヒトNGFを含む発現ベクターを組換えCHO細胞に形質移入した。分泌とプロセシングを促進するためにNGFプレプロ配列もまた存在していた。組換えCHO細胞の培養後、細胞培地を収集した。収集細胞培地液(HCCF)はNGF種120、118及び117を含んでいた。NGFの約40−70%は典型的には118/118ホモ二量体で、残りはヘテロ二量体120/118、120/120及び少量の118/117であった。ここに教示したように、これらの種はSPセファロースHPカラムにより分離することができる。
【0112】
収穫細胞培地は、ミリポア10Kdカットオフ膜を使用して、およそ20倍に濃縮された(セルロース、複合体又はポリスルホンの何れかが交換可能に使用された)。濃縮物に0.1容量の1.0M、pH8.2のトリスを添加した。希釈材料が0.22umフィルターを使用して精密濾過され、保持タンクに移され2から18時間の間37℃で保持された。120/120型の118/118型へ転換が、保持の間、内在性プロテアーゼにより触媒された。
【0113】
シリカゲルクロマトグラフィー
精密濾過物を1MのNaClに調節し、1MのNaCl、25mMのMOPSO(pH7)で平衡化されたシリカゲルカラムに適用した。カラムを1MのNaCl、25mMのMOPSO(pH7)で洗浄した。適切なpH範囲は約6から8で、好適なpHは7である。ついで、カラムを25mMのMOPSO(pH7)で洗浄した。低伝導率洗浄により宿主細胞タンパク質を除去する。結合したNGFを50mMのMOPSO、0.5MのTMAC、20%の試薬無水等級アルコール(94−96%の特別に変性されたアルコール・フォーミュラ3A(5容量のメタノールと100容量の200プルーフ・エタノール)と4−6%のイソプロパノール)で溶出させた。例えば20%プロパのール、20%イソプロパノール及び20%メタノールのような他のアルコールを使用することもできる。ここで、「アルコール」及び「アルコール性溶媒」はアルコールについてよく用いられているところの意味を有するもので、好ましくは1から10の炭素原子のアルコール、より好ましくはメタノール、エタノール、イソプロパノール、n−プロパノール、又はt−ブタノールで、最も好ましくはエタノール又はイソプロパノールである。このようなアルコールは、水溶液に添加した場合、溶液の極性を減少させることにより溶液の疎水性を増大させる溶媒である。エタノールが最も好ましい。アルコールの下限は溶出する如何なるパーセントでもよく、上限はタンパク質の変性を避ける必要から決められる。溶媒は好ましくは5%から25%、より好ましくは5から20%、更により好ましくは5から15%である。TMACはテトラメチルアンモニウムクロリドであり、これはNGFを溶出させるために存在する。TMACは0.1から1Mの範囲とできる。0.3から0.7Mの範囲が更に好ましい。NGFを溶出するために使用されるTMACの量はpHとアルコール濃度の関数である。pHがより低く、アルコールの量がより少なくなると、TMACが必要となる。pHは約4から8の間とできる。この実施例では、好適なpHは7であり、これは、次のカラムへの装填の前のプール分画の調整を非常に少ないものにする。pHの上限は次のカラムに装填するのに必要なpHにより決まり、下限はNGFを効率よく溶出させるのに有用なものとされる。
【0114】
S−セファロース・ファースト・フロー・クロマトグラフィー
NGFを含んでいる溶出液をプールし、精製水で15.5ms/cm未満の伝導率まで希釈し、pHを7.0に調整した。幾つかのプロテアーゼがなお存在していたので、材料を8時間を越えないで保持した。しかし、120アミノ酸を118型に転換する内在性プロテアーゼの活性は観測されなかった。材料を25mMのMOPSO(pH7)で平衡化されたS−セファロース・ファースト・フロー・クロマトグラフィーカラム(カチオン交換樹脂S−セファロース TMアガロース・ファースト・フローTM(ファーマシア))に適用した。カラムは25mMのMOPSO(pH7)で洗浄される。適切なpH範囲は約6から8であり、好ましくはpH7である。ついで、カラムを0.16MのNaCl(pH7)で洗浄した。結合したNGFを0.5MのNaCl(pH7)で溶出させた。溶出液の塩モル濃度は0.3から1.0M、好ましくは0.4から0.6Mの範囲とできる。下限はNGFを全て溶出させるための有用性により決められ、上限は汚染物質を除去し、NGFの溶出に干渉しうるカラムの疎水的相互作用を生じさせることを避ける必要性により決められる。他の塩を使用することもでき、KClが好適な代替物である。0.5MのNaCl(pH7)での溶出が、小容量のプールを得るためには好適である。より高い塩濃度、例えば1Mを越えると、強固に結合した汚染物質が溶出し得る。
【0115】
フェニル・トーヨーパール650Mクロマトグラフィー
NGFを含んでいるSSFFカラム分画をプールし、1MのNaClに調節し、フェニル・トーヨーパール650Mカラムに適用した。カラムを25mMのMOPSO(pH7)で洗浄した。適切なpHは約5から8の範囲である。結合したNGFを、勾配緩衝液A(25mMのMOPSO、pH7、1.0MのNaCl)で始めて勾配緩衝液B(80%の25mMのMOPSO中の20%アルコール、pH7)で終了する10CV(カラム容量)の線形勾配で溶出させた。NGFを含む分画をSDS−PAGEポリアクリルアミドゲル電気泳動法により分析して、どの分画に前駆NGF種が溜まったかを決定した。NGFを含む分画を選択しプールして、例えば部分的な前駆配列が存在するもの、例えば前駆NGF、ハイブリッド前駆NGF、及びクリップ前駆NGF配列のような不適切にプロセシングされた変種を主に取り除いて、如何なるNGF前駆配列も実質的に含まれないNGF組成物を得た。フェニルカラムはまた少量のグリコシル化NGFとグリコシル化NGF前駆配列を除去した。前駆及び切詰め前駆NGF配列がNGFと前駆NGFの双方のグリコシル化形態と共にNGFピークの先端に溶出した。従って、このカラムは様々なNGF種からNGFを即座に分離し、これらの種が実質的に含まれないNGF組成物を得た。この工程において、タンパク分解性及びグリコシル化変種を含む、変種NGF疎水性変種、主に誤ってプロセシングされた変種がHICを使用して分離された。
【0116】
NGF型を分離するために最も好適な媒体は固定化したフェニル官能基を有するものであった。異なった販売者からのフェニルベースHIC媒体はこれらのNGF型を分離するための異なった効率を示した。最良の結果がTosoHaasのフェニル・トーヨ・パール媒体で達成された。HIC樹脂フェニル・セファロース・ファースト・フロー・ロー・サブ(低置換)とTSKフェニル5PWはよく作用した。アルコキシ、ブチル及びイソアミル部分を含む他のHIC官能基はこれらの条件下では、適正が少なく、効果が少なかった。
【0117】
フェニル・トーヨーパール・プールは、75%の118、10%の120、7%の117、1.8%の脱アミドNGF、1.4%の酸化NGF、及び2.0%のisoaspNGFを含有し、残りの2.8%はその他の未同定NGF種であった。
【0118】
任意であるが、プールされた分画を酸処理して、最小15分の間、3.95未満のpHでウィルス不活性化を達成した。
【0119】
SP−セファロースHPクロマトグラフィー
HICプールを0.5から1容量の水で希釈し、希釈したプールをpH6に調節した。このプールを、(シリカゲルカラム工程におけるように)0.2Mの塩化ナトリウム、5%の試薬アルコールを含む20mMのコハク酸塩(pH6)で平衡化されたSP−セファロースHPカラムに装填した。カラムを0.2MのNaCl、5%の試薬アルコール(フォーミュラSDA−3Aアルコール;アルコールは任意に存在する)を含む20mMのコハク酸塩(pH6)で洗浄した。アルコールは、NGFの樹脂バックボーンとの非特異的(大抵は疎水的)相互作用を低減させる。適切なアルコール範囲は約0から10%である。負荷pHは約5から8であり、これはNGFの最大の安定性とNGF変種の分離を達成し維持するように選択される。カラムを2カラム容量の平衡緩衝液で洗浄した。結合したNGFを、11カラム容量の勾配緩衝液A(5%のアルコールを含む、0.25MのNaCl、0.02Mのコハク酸塩、pH6)と11カラム容量の勾配緩衝液B(0.5MのNaCl、pH6)を混合して線形の22カラム容量の勾配によって溶出させ、変種から分離した。アルコールは任意である。0.35Mから0.40MのNaCl濃度で118/118NGFが典型的には溶出した。
【0120】
カラム分画をNGF及び変種NGF含量について分析した。分画は好ましくは上掲のSchmelzerら(1992)及び上掲のBurtonらにより記述されたC4RP−HPLCによって分析される。分画を選択しプールして、修飾NGF変種、例えば酸化、脱アミド、isoaspNGF種が実質的に含まれないNGFの組成物を得た。先のHICカラムは酸化、isoaspNGFのようなNGFの他の変種型を効果的には除去できない。HICカラムは誤って折り畳まれたタンパク質とグリコシル化物を効果的に除去し、これは、より疎水性になる傾向があるので、正しく折り畳まれたNGFよりもHIC樹脂により強固に結合する。従って、カチオン交換樹脂、例えばSP−セファロースHPを、HIC樹脂によって除かれなかった変化した負荷変種を除去するために使用した。
【0121】
SP−セファロースプールは、典型的には約92%の118、4.6%の120、1.8%の脱アミドNGF、1%の酸化NGF、及び1%のisoaspNGFを含有していた。通常の通り、各々の種の量は、118に対しては約85から93%、120に対しては0から5%、117に対しては0から5%、脱アミド化型に対しては0から3%、isoasp型に対しては0−2%、及び酸化型に対しては0から2%の範囲である。NGF(全ての種)の純度は通常のように99.5%よりも大きい。
【0122】
製剤
SP−セファロースHPプールを製剤緩衝液中への限外濾過/ダイアフィルトレーションにより製剤のために調製した。酸性緩衝液が好ましくは用いられ、好ましくは上述のようにpH5の酢酸塩が使用される。118/118NGF組成物は実質的にNGFを含んでおらず、実質的に純粋なNGFである。処方された材料は神経疾患、特に糖尿病に伴う末梢神経障害及びエイズに伴う末梢交感神経障害を治療するのに好適である。
【0123】
<実施例II.120/120NGFホモ二量体の大規模な精製>
収集細胞培養液
一般に実施例Iに記載されているようにして12000リットルのCHO細胞培養からHCCFを得た。HCCF中のNGF種分布は、約40−65%の120/120ホモ二量体と、120/118ヘテロ二量体と残りが118/118ホモ二量体であった。典型的には培地を素早くプロセシングして120から118のタンパク分解性転換を最小化した。
【0124】
マクロプレップ・ハイ・Sカチオン交換クロマトグラフィー
HCCFをマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーカラムに装填し、1.5Mの酢酸ナトリウム、50mMのHEPES(pH7)で洗浄した。結合したNGFを、1.5MのNaCl、0.25MのTMAC、0.2%のチオジグリコール(pH7)で溶出した。マクロプレップカラムは、酢酸塩濃度を調節してpH5から8で操作することができる。塩化物は酢酸塩イオンの好ましい代替物である。NGFがTMAC勾配により溶出した。TMACはイオン性及び疎水性の双方の性質を持つ塩であり、この性質は、ある種の樹脂のバックボーン支持体がNGFと樹脂の間の非特異的相互作用を促進する疎水的含量を含むので有用である。典型的には、約6から8のpHの溶出緩衝液に対しては、0−3MのTMAC濃度が有用である。NGFを含む分画をプールした。
【0125】
シリカゲルクロマトグラフィー
プールを直接シリカゲルクロマトグラフィーカラムに適用した。シリカは、タンパク結合特性においてある役割を果たすイオン性、極性、及び疎水的相互作用を有する混合態様のクロマトグラフィー樹脂を提供する。カラムを1MのNaCl、25mMのMOPSO(pH7)で平衡化させた。カラムを1MのNaCl、25mMのMOPSO(pH7)(好ましくはpHが約5.0から8.5、より好ましくは6から8、最も好ましくは7である)で洗浄した。結合したNGFを25mMのコハク酸、pH3.9、50mMのTEAC(テトラエチルアンモニウムクロリド)で溶出させた。TEACはTMACよりも更に強力な溶出剤である。pHは好ましくは3.5から8の範囲である。しかし、長い時間の間7.5を越えるpHにするのは、脱アミドNGF種の生成を防止又は低減するために避けるべきである。一般に、緩衝液のpHが低くなればなる程、シリカカラムからNGFを溶出させるのに必要とされる例えばTMAC又はTEACのような混合特性の塩の濃度を低くする。pH4又は5の近くで良好な緩衝能力を有する緩衝液が使用に適している。溶出緩衝液中の塩の有無は任意であり、そのためカラムは、溶出緩衝液の適用に先立って塩なしにMOPSO緩衝液で洗浄することができる。
【0126】
フェニルセファロース・ファースト・フロー・クロマトグラフィー
NGFを含んでいる分画を同定しプールした。そのプールを、0.7Mの酢酸塩、pH7,25mMのMOPSOに調節した。調節したプールを、勾配緩衝液A(0.7Mの酢酸塩、25mMのMOPSO、pH7)で平衡化したフェニル・セファロース・ファースト・フロー・クロマトグラフィーカラムに装填した。カラムを、90%の勾配緩衝液A(0.7Mの酢酸塩、25mMのMOPSO、pH7)から10%の勾配緩衝液B(25mMのMOPSO、pH7、20%のプロピレングリコール)の勾配で洗浄した。例えばヘキシレングリコールのような他のグリコールに置換することもできる。典型的には、洗浄は約2から3CV、あるいは安定なベースラインODが達成されるまでとした。洗浄によりいくらかの宿主細胞タンパク質が除去された。結合したNGFを、90%の勾配緩衝液Aと10%の勾配緩衝液Bの混合物から10%の勾配緩衝液Aと90%の勾配緩衝液Bの混合物までの線形の10CV勾配で溶出させた。NaClもしくは硫酸ナトリウムをHIC緩衝液中の酢酸塩と置換することができる。pHは好ましくは約5から8、より好ましくは約5.5から7.5であり、pH6から8が許容でき、約7が最も好ましい。該カラムは、成熟NGF単量体及び存在する前駆配列の一部をなおも有しているNGF単量体のホモ二量体又はヘテロ二量体として存在する、全ての残っている前駆配列、部分的前駆配列、もしくはグリコシル化形態を分離した。NGFの前駆及びグリコシル化形態は溶出ピークの先行端に存在しており、NGF含有分画がこれらの種を実質的に排除するようにプールされた。
【0127】
分析用HPLCシステムで分離し検出したところ、HICプールは、約72%の120単量体、17%の118単量体、2.8%の117単量体、3.6%のR120単量体、0.8%のisoasp型、1.3%の酸化型、及び1%の脱アミド型を含有していた。
【0128】
SP−セファロースHPクロマトグラフィー
HIC工程からのNGFを含有する分画をプールした。大規模では、これは、カラム溶出液をプールタンク(保持タンク)に適当な時間に流すことにより達成された。プールのpHはpH6に調節し、SP−セファロースHPクロマトグラフィーカラムに適用した。カラムを20mMのコハク酸塩、0.2MのNaCl、pH6(勾配緩衝液A)で洗浄した。結合したNGFを、70%の勾配緩衝液Aと30%の勾配緩衝液Bの混合物で開始し、80%の勾配緩衝液B(0.7MのNaCl/pH6)と20%の勾配緩衝液Aの最終混合物までの22CVの勾配で溶出させた。pHは好ましくは5から8、より好ましくは5.7から6.5、最も好ましくは6である。代表的なクロマトグラムを図1に示す。
【0129】
SPセファロースHPプールはルーチン的に約95%の120型、3%のR120型、0.65%のisoasp型、0.6%の酸化型、0.6%の脱アミド型を含んでいた。NGFの他の未同定の形態は約0.6%であり、二酸化型NGF(Met37とMet92)からなり、脱アミドAsn45が存在していた。SP−セファロースクロマトグラフィー後に得られたプールと(SP−セファロース樹脂に装填された)代表的なHICプールを比較するHPIEX分析を図2に示す。NGFの3つの主要なクリップ型、120、118、及び117の各々が変種を有しているようであるが、精製中、主な切詰め型(この例では120)の変種、例えば酸化及びisoasp型が、主要ではない形態(この例では118と117)からの変種の分析を遮蔽しているようである。代表的な試行からの分画のHPLC分析を図3に示す。
【0130】
SP−セファロースHPはHICプール中に存在する変種を効果的に除去した。R120型はNGFのN末端に更なるアルギニン残基を有している;通常、rhNGFのN末端アミノ酸配列はSSSHPであるが、R120はRSSSHPのN末端配列を有している。従って、R120型は成熟NGFよりより塩基性で、SP−SHPにより分離された。これはまた、NGFのN末端がレセプター(trkA)結合に必要であるという事実に恐らくは関連して、低い生物活性を有している。酸化NGF型は酸化された37位置にメチオニンを有するモノ酸化型であり、NGFピークの先行端に溶出するより酸性の形態を生じる。isoasp型はアミノ酸93にアスパラギン酸の修飾を含む。isoasp型は僅かにより塩基性であり、従ってSP−セファロースHP樹脂にややより強固に結合する。isoAsp93を含んでいるNGF種は溶出ピークの終端に溶出した。脱アミド化がアスパラギン酸残基、典型的には45位のアスパラギン酸の位置で生じた。脱アミド化Asnを含んでいるNGFは、45位にAspを生じるが、僅かにより酸性であり、溶出ピークの先行端に現われる。
【0131】
フラクトゲルEMD SO3はNGF種の荷電変種を分離するためのSP−セファロースHP樹脂に対してあまり好適な代替樹脂ではない。このあまり好ましくない樹脂を使用する場合は、NGFを溶出させるためにより高い濃度のNaClが必要となる。
【0132】
製剤
バルク材料を、先の実施例におけるように、製剤緩衝液中への限外濾過/ダイアフィルトレーションにより製剤化した。最終のバルク生成物において、120型はルーチン的に約92から97%、R120は約1から4%、isoasp型は約0.2から1.5%、酸化型は約0.2から2%、脱アミド型は約0.2から2%の範囲であった。117及び118型はルーチン的に約2%未満であった。最終のバルク生成物はルーチン的に少なくとも99.5%の純NGF(全ての種を含む)であった。
【0133】
<実施例III.118/118の単離>
NGF変種が実質的に含まれていない実質的に純粋な118/118NGF組成物を得る好適な一実施態様においては、実施例IIの方法を、次の変更をして実施した。マクロプレップ・ハイ・Sカラムとシリカカラムの間に固定化トリプシンカラムを使用する。必要に応じてpHを約5から8.5、最も典型的には6.5から7.5に調節した後に、マクロプレッププールを固定化トリプシンカラムに直接装填した。プールをカラムに通過させ、その間に殆どのNGFが118型に転換された。プロテアーゼ消化が、C末端のVRRAをVRに切断することにより120型を118型に転換する。制限された選択的な切断を達成するために、トリプシン又はトリプシン様プロテアーゼ、好ましくはトリプシン、より好ましくは直ぐに入手できるブタトリプシン、あるいは代わりにウシトリプシン又は組換えトリプシンが使用される。制限された選択的な切断を実質的にもたらす任意のタンパク分解性方法を使用することができるが、NGF調製の汚染を最小化するために好ましいものは固定化トリプシンカラムである。カラムはプロテアーゼ活性を促すpH、好ましくは5.5から8.5のpH、より好ましくは6.0から8.0、最も好ましくは6.5から7.5のpHで操作される。
【0134】
この実施例において、グリコシル化NGFはここで検討したようにHICにより除去された。実施例IIにおいて検討したようなSP−セファロースHP工程に続いて、しかし好ましくは22カラム容量の0.3Mから0.55Mの塩勾配を用いて、治療用途に好適なNGF組成物を得た。RP−HPLCアッセイにより決定したところ、70%を超える118単量体、10%未満の120単量体及び15%未満の117単量体の組成物を得ることができる。典型的には、90%以上の118/118rhNGF、より通常的には93%以上の118/118rhGNFで、約7%以下の脱アミド、isoasp型及び酸化変種の組成物が得られる。より高い純度を達成する一つの手段は、例えば118/118rhNGFピークのような主要ニューロトロフィンピークの先端又は後行端に見出されるような有意の量の変種を伴う分画を選択することを避けることである。
【0135】
<実施例IV.細菌封入体からのrhNT−4/5の部分的精製と再折り畳み>
この実施例において、10もしくは60リットルの発酵で出発して、rhNT−4/5を精製した。この実施例において記載した発酵において組換えヒトNT−4/5を生産するために使用された宿主は27C7/pmNT5DTと命名された大腸菌株であった。但し、他の菌株と生物体から生産されるNT−4/5もまたここに記載した精製方法に適している。この実施例において使用された発現プラスミドは、大腸菌中のNT−4/5遺伝子の発現に必要とされる転写及び翻訳調節配列下に成熟NT−4/5コード配列を含んでいた。NT−4/5発現プラスミドにおいて、大腸菌中での遺伝子の発現に必要とされる転写配列はアルカリホスファターゼプロモータ配列によって提供された。転写ターミネーターに対するラムダはNT−4/5終止コドンに隣接して位置していた。細胞質からのタンパク質の分泌はSTIIシグナル配列により指示された。rhNT−4/5の大部分は屈折性体のような細胞膜周辺膣に見出された。プラスミドは、形質転換された宿主に対してテトラサイクリン耐性をもたらした。発酵プロセスは35℃から39℃、pH7.0−7.8で実施された。発酵は25−40時間続けられ、その後培養は収集前に冷却された。培養は、60℃で連続流れの装置を使用する加熱処理又はその温度で5−15分の間インタンク熱不活性化を使用して不活性化した。熱不活性化された培養は、AXアルファ−ラバル遠心機又は等価物を使用して遠心分離された。大腸菌はペレットに回収された。
【0136】
封入体中に組換えヒトNT−4/5を発現している大腸菌細胞は、封入体中にNT−4/5を含むペーストを調製するために標準的な手段により溶解させた。緩衝液にはプロテアーゼ阻害薬は含まれていなかった。
【0137】
細胞片から封入体を単離するために、大腸菌NT−5ペーストを、回転式機械的分散装置、例えばトゥラックス(Turrax)を使用して0.02Mのトリス、pH8、5mMのEDTA中に再懸濁させた。細胞懸濁液を6000psiで3回マイクロフルイダイザーを通過させた。得られたホモジェネートを約45分間、5000rpmでSorvallRC−3B遠心機で遠心分離した。上清を破棄し、ペレットを、トゥラックスを使用して2から3分間、中程度の速度で20mMのトリス、pH8、5mMのEDTA(抽出緩衝液)中に再懸濁させた。ホモジェネートを上述のようにして遠心分離した。ペレットを抽出緩衝液に再懸濁させ上述のように遠心分離した。得られたペレット(ペレット群)(NT−4/5封入体又は屈折体という)を−70℃で保存した。
【0138】
NT−4/5は次のようにして封入体から単離した。封入体ペレットを20mMのトリス、pH8、6Mの尿素、25mMのDTT(10ml緩衝液/封入体グラム)中にトゥラックスを使用して中程度の速度で約10分間懸濁した。懸濁液を2−8℃で40分間撹拌し、約45分間5000rpmでSorvallRC3Bで遠心分離した。PEI(ポリ-エチレン-イミン)を0.1%まで上清に添加し、これを2−8℃で30分間撹拌した。PEIは核酸とその他の酸性負荷分子を沈殿させる。混合物を約45分間5000rpmでSorvallRC3Bで遠心分離した。PEI上清を、0.02Mのトリス、6Mの尿素、10mMのDDT、pH8で平衡化されたDEFFセファロース・ファースト・フロー・カラム(10cmx14cm;DEFFはジエチルアミノエチル樹脂である)に装填した。1kgの可溶化屈折体の等価物がDEFFカラムに装填された。還元された変性NT−4/5はDEFF樹脂に結合しないので、NT−4/5と6Mの尿素を含む素通りプールが収集され(図6)、プールのpHが酢酸で5.0に下げられた。pH調節DEFF素通りプールを、NT−4/5が樹脂に結合する条件である、6Mの尿素を含む20mMの酢酸塩(pH5)で平衡化したセファロース・ファースト・フロー・カラム(Sは樹脂上のSO3官能基をいう)に装填した。装填後、S−セファロース・ファースト・フロー・カラムを数カラム容量の平衡緩衝液で洗浄した。結合したNT−4/5を、0.5MのNaCl、20mMの酢酸ナトリウム、6Mの尿素、pH5で溶出させた(図7)。0.5MのNaClのSSFFプールを20mMのトリス、0.14MのNaCl、pH8に対して一晩かけて透析した。この条件ではNT−4/5が不適切にではあるが再折り畳みする。誤って折り畳まれたrhNT−4/5分子は凝集して沈殿物を形成した。
【0139】
凝集した誤って折り畳まれたrhNT−4/5をプロセシングして正しく折り畳まれたNT−4/5を得た。凝集した誤って折り畳まれたrhNT−4/5を遠心分離によりペレットとして収集した。ペレットを、0.2Mのトリス、pH8、4Mの尿素、5mMのDTTに再懸濁させ、約1から2時間もしくはペレットが溶解するまで2−8℃で撹拌した。最終のタンパク質濃度を、280nmでの吸光係数1.8に基づいて約10mg/mlタンパクに調節した。酸化されたグルタチオンを可溶化されたペレット溶液に添加して20mMの最終濃度とし、2−8℃で15から30分間穏やかに撹拌した。酸化グルタチオンはNT−4/5スルフヒドリル基と反応してNT−4/5−S−グルタチオン混合ジスルフィドを生成する。NT−4/5−SG混合ジスルフィドを、100mMのトリス、20mMのグリシン、15%のPEG−300、1MのグアニジンHCl、pH8.3中に0.1から0.5mg/mlタンパクの最終濃度まで希釈した。NT−4/5の正しい再折り畳みを開始するために、再折り畳み混合物に2から4mMの最終濃度でシステインを添加し、続いて酸素を排除するために容器を密封する前に5から60分間窒素又はヘリウムで溶液をエアレーション(泡立てによる)した。NT−4/5の再折り畳みは2から8℃で18から24時間進められた。
【0140】
別法として、rhNT−4/5が次のような亜硫酸分解を使用して再折り畳みされた。封入体ペレット(110g)を1.1リットルの20mMトリス、7M尿素、10mMグリシン、100mM亜硫酸ナトリウム、10mMテトラチオン酸ナトリウムに懸濁させ、中程度の速度で10分間トゥラックスを使用して溶解させた。混合物(1260mL)をついで2から8℃で45分間撹拌した。PEIを0.1%PEIの最終濃度になるまで添加した。混合物を4℃で更に30分間撹拌し、RC3B遠心機中で5500rpmで45分間遠心分離した。上清を、20mMのトリス、6Mの尿素、pH8で平衡化したDEFFカラム(4.4cmx25cm)に装填した。DEFF素通り分を酢酸でpH5に調節して、20mMの酢酸、6Mの尿素、pH5で平衡化したS−セファロース・ファースト・フロー・カラム(4.4cmx25cm)に装填した。NT−4/5を25mMのMOPSO、0.5MのNaCl、pH7で溶出させた。
【0141】
スルホニル化rhNT−4/5を含むSSFF0.5M塩化ナトリウムプールを約0.1mg/mlタンパクに希釈し、1Mのグアニジンヒドロクロリド、100mMのトリス、20mMのグリシン、15%のPEG−300、pH8.3に調節した。2から4mMのシステインの添加によりNT−4/5の再折り畳みを開始した。再折り畳み反応は本質的に24時間以内に完了した。溶液からの酸素を置換するための例えばヘリウム又は窒素のような不活性ガスによるエアレーションを必要に応じて実施することもできる。
【0142】
<実施例V.高次構造的(誤って折り畳まれた)変種からの正しく折り畳まれたrhNT−4/5の単離>
実施例IVのrhNT−4/5の再折り畳み混合物を一晩かけてpH4から5の溶液に対して透析してグアニジンとその他の試薬を除去した。溶液を清澄にするために、溶液を5000rpmで45分間遠心分離するか、0.2umフィルターを通過させた。
【0143】
0.5から5グラムのタンパク質を含む清澄化した上清を、氷酢酸を添加してpH3から5に調節し、C4RP−HPLCカラムに装填するか、精製の準備ができるまで−20℃で凍らせて保存した。この実施例では、酸性化され清澄化された溶液を、折り畳まれたrhNT−4/5が結合した樹脂のC4RP−HPLC(3cmx50cm)カラムに装填した。正しく折り畳まれたNT−4/5を、0.05トリフルオロ酢酸(TFA)溶媒系中のアセトニトリル勾配を用いて(25ml/分の流速での0.05%TFA中の26から40%のアセトニトリル勾配(95分間を越える)からなる)溶出させた。分画を1から1.5分間隔で収集した(図8)。分析的C4HPLC Vydac(0.21x15cm)カラム上の溶出時間を正しく折り畳まれたNT−4/5標準物質の時間と比較することにより、正しく折り畳まれたNT−4/5について分画を分析した(図9)。標準的な正しく折り畳まれた無傷のNT−4/5は典型的には0.5%TFA/アセトニトリル緩衝系で毎分2.5mlの流量で19分で溶出した。正しく折り畳まれたrhNT−4/5を含む分画をプールしpHを5から7に調節した。正しく折り畳まれたrhNT−4/5のプールもまたNT−4/5のカルバミル化及びN末端切詰め型を含んでいた。
【0144】
調製的逆相液体クロマトグラフィー樹脂は好ましくは約10−40ミクロンの粒径、約200−400オングストロームの孔径、及びC4、C8、又はC18アルキル基を有する媒体である。より好ましくは、樹脂は約15−40ミクロンの粒径と約300オングストロームの孔径を有し、C4シリカ媒体である。
【0145】
<実施例VI.高次構造的(誤って折り畳まれた)変種からの正しく折り畳まれたrhNT−4/5の別の単離>
実施例IVの再折り畳みrhNT−4/5混合物を、10kDの分子量のカットオフを持つ20平方フィートのセルロース(又はポリスルホンもしくは等価物)膜を備えたミリポア−ペリコン限外濾過系を使用して約10倍に濃縮した。0.2ミクロン膜を通す濾過の前に、濃縮した混合物を一晩かけて50Lの50mM酢酸塩、pH5.5、50mMのNaClに対して透析するか、50mMの酢酸塩、50mMのNaCl、pH5.5中でダイアフィルトレーションを施した。
【0146】
濾過した再折り畳み混合物を2.5MのNaCl、20mMのMOPSO、pH7に調節し、前もって2.5MのNaCl、20mMのMOPSO、pH7で平衡化したHICカラムであるフェニル・トーヨーパール650Mカラム(10cmx19cm)に装填した。ついで、平衡緩衝液でカラムを洗浄した。rhNT−4/5分子の分子の幾つかの誤って折り畳まれた型が素通り分画に溶出し、その他の誤って折り畳まれた型が20から40%の試薬アルコールのような高濃度の有機溶媒で溶出した。正しく折り畳まれたrhNT−4/5は2Mの塩化ナトリウム、10%の試薬アルコール、pH7を使用してフェニルカラムから溶出させた(図10)。フェニルセファロースのような他のフェニル樹脂をトーヨーパールのバックボーンの代わりに使用することができる。硫酸アンモニウム、クエン酸塩、酢酸塩、及び塩化カリウムを含むここで検討した塩を使用することができる。使用する塩に依存して、塩濃度は典型的には1Mから3Mであり、2.5MのNaClが装填には好適で、2MのNaClが有機溶媒の存在する場合の溶出に好適である。好ましくは、塩濃度の低下がニューロトロフィンとその変種を溶出させ分離するために使用される。溶出を達成するには、溶出緩衝液中の塩濃度は典型的には負荷緩衝液中のものよりも低いが、有機溶媒で補償するときは同じ濃度とすることができる。また、有機溶媒の使用は、ここで見出されたところでは、有機溶媒を添加するとより狭いピークプロファイルが得られて溶出パターンが改善されるという他の利点がある。エタノールに加えて、プロパノール、イソプロパノール、及び低級アルキレングリコール、例えばプロピレングリコール、エチレングリコール及びヘキシレングリコールを含むここで検討するその他の有機溶媒を使用することもできる。5から25%(v/v)、より好ましくは5から20%(v/v)、更に好ましくは5から15%(v/v)の有機溶媒が典型的には正しく折り畳まれたニューロトロフィンを溶出する。有機溶媒での溶出は勾配によるか、段階的に行なう。pH範囲は、好ましくは中性近くから僅かに酸性であり、5から8、より好ましくは5.5から7.5、最も好ましくは7である。MOPSO、MOPS、HEPES、リン酸塩、クエン酸塩、アンモニウム、酢酸塩を含むここで検討した任意の緩衝液を、所望のpHで緩衝作用を示す限り使用することができる。
【0147】
<実施例VII.正しく折り畳まれたrhNT−4/5の化学変種からの精製>
rhNT−4/5のカルバミル化及びN末端切詰め型を含む化学変種からの正しく折り畳まれた無傷のrhNT−4/5の分離は、SP−セファロースHP樹脂又はポリキャットA HPLC樹脂を使用する高速カチオン交換クロマトグラフィーにより達成することができた。
【0148】
C4RP−HPLCカラムを誤って折り畳まれた変種を取り除くために使用した場合、C4HPLCプールをpH5から7に調節し、20mMのコハク酸塩、pH6、5%の試薬アルコール、0.2MのNaClで平衡化した7cmx19cmのSP−セファロースHPカラムに装填した。結合したNT−4/5を伴う樹脂を平衡緩衝液で洗浄した。結合したrhNT−4/5を、pH6で0.2MのNaClから0.4MのNaClまでの22カラム容量(CV)の勾配(すなわち、平衡緩衝液中の塩勾配)を使用して溶出させ、カルバミル化及びN末端切詰め型から分離した(図11)。NT−4/5を含む分画をプールし、0.05Mの酢酸塩、pH4から5に製剤化した。無傷のrhNT−4/5を同定し、好ましくはここで検討したように分析的RP−HPLC又はSDS−PAGEにより分画内の変種から区別し、標準と比較した。
【0149】
別法として、NT−4/5の変種型をポリアスパラギン酸カラムでの高速カチオン交換HPLC(ポリキャットa、ポリLC、Columbia,Md.)(9.4x200mm)により除去した(図12)。C4HPLCプールをpH5から6に調節し、ポリキャットaカラムに装填した。クロマトグラフィー条件は:緩衝液Aが20mMのリン酸塩、5%のアセトニトリル、pH6であった;緩衝液Bは20mMのリン酸塩、5%のアセトニトリル、0.8MのKCl、pH6であった。rhNT−4/5を、65分にわたって25から60%の緩衝液Bの勾配を使用して溶出させた(図12)。分画を1分間隔で収集し、上述のように分析的C4HPLCにより分析した。
【0150】
HICを用いて誤って折り畳まれた変種を除いた場合(上記の実施例VI)は、正しく折り畳まれたNT−4/5プールを、20mMのコハク酸塩、0.1MのNaCl、5%の試薬アルコール、pH6中で一晩かけて透析するか、20mMのコハク酸塩緩衝液中で限外濾過/ダイアフィルトレーションした。透析もしくはUF/DFプールをついで上述のようなSP−セファロースHP又はポリキャットaカラムに装填した。
【0151】
製剤
(SP−セファロースHP又はポリキャットaHPLC工程から得た)正しく折り畳まれた無傷のNT−4/5を含む分画をプールし、20mMの酢酸塩、pH4から5の製剤緩衝液中に1から5mg/mlまで濃縮した。別法として、NT−4/5を限外濾過/ダイアフィルトレーションを使用して製剤化した。
【0152】
最終のバルク溶液を、アミノ酸分析、N末端配列分析、質量分析、SDS−PAGE(図13)及び生物学的アッセイにより分析した。キナーゼレセプター活性化(KIRA)アッセイは、細胞膜に位置するそのチロシンキナーゼレセプター(trkB)の自己リン酸化のNT−4/5活性化を検出するものであるが、精製したrhNT−4/5を特徴付けるために使用した。gDタグを付したtrkBを発現するCHO細胞を使用した。1995年6月1日に公開されたWO95/14930はKIRAを記述しており、ここに出典明示により取込む。rhNT−4/5はこのアッセイにおいて12.6ng/mlのEC50を有していた。典型的には、ここに記載されたようにして精製される正しく折り畳まれた無傷のNT−4/5のEC50は、5から30、より好ましくは10から20である。
【0153】
非NT−4/5タンパク質に対するrhNT−4/5の純度は典型的には90から99パーセントであった。カルバミル化及びN末端切詰め変種に対してのNT−4/5の相同性は90から99パーセントであった。最も典型的かつ好ましくは、純度と相同性は99%又はそれ以上である。
【0154】
<実施例VIII.細菌封入体からのrhNT−3の初期精製、再折り畳み及び最終精製>
細胞片から封入体を単離するために、大腸菌NT−3ペースト(1kg)を、10mLの100mM酢酸ナトリウム(pH5)中に、例えばトゥラックスのような回転機械式分散装置を使用して再懸濁させた。細胞懸濁液を6000psiでマイクロフルイダイザーに3回通した。得られたホモジェネートを5000rpmで30分の間SorvallRC−3B遠心機で遠心分離した。
【0155】
NT−3を次のようにして封入体から単離した。封入体ペレットを、約10分間中程度の速度でトゥラックスを使用して、100mMのトリス、100mMのNaCl、5mMのEDTA、100mMの亜硫酸ナトリウム、10mMのテトラチオン酸ナトリウム、7.5Mの尿素、pH8.3(10ml/封入体グラム)に懸濁させた。懸濁液を2−8℃で約1時間撹拌した。PEI(ポリエチレンイミン)を約0.15%(最終濃度)になるまで添加し、2−8℃で30分間撹拌した。混合物を5000rpmで約30分の間SorvallRC3Bにおいて遠心分離した。上清をゲルマン・プレフロー(Gelman Preflow)カートリッジで濾過した。濾過した上清を3容量のS−セファロース・ファースト・フロー平衡緩衝液(50mMの酢酸ナトリウム、5Mの尿素、pH5)で希釈した。希釈した濾過上清(7mS未満の導電率を、50mMの酢酸ナトリウム、5Mの尿素、pH5.0で平衡化したS−セファロースFFカラムに装填する。カラムを最初に50mMの酢酸ナトリウム、5Mの尿素、pH5で洗浄し、ついで50mMのMOPS,5Mの尿素、10mMのグリシン、pH7.0で洗浄した。亜硫酸分解されたNT−3を、50mMのMOPS、5Mの尿素、10mMのグリシン、pH7中の0−0.6MのNaClの10カラム容量の勾配を使用してカラムから溶出させた。
【0156】
部分的に精製されたNT−3を、0.1Mのトリス、2Mの尿素、0.1MのNaCl、15%のPEG300、10mMのグリシン、25mMのエタノールアミン(pH9.1)を含む再折り畳み緩衝液中でS−セファロースFFプールをおよそ0.1mg/mlタンパクまで希釈することにより再び折り畳んだ。再折り畳みは、およそ5mMまでシステインを添加し、2−8℃で2−5日間撹拌することにより開始した。必要に応じて、再折り畳み溶液中の酸素濃度を低減させるために、再折り畳み緩衝液をHe又はアルゴンで分散させても良い。
【0157】
再折り畳みされたプールのpHは7に調節し、濾過し、50mMのHEPES(pH7)で平衡化したマクロプレップ・ハイ・Sカチオン交換クロマトグラフィーカラムに装填した。マクロプレップカラムにpH調節再折り畳みプールを装填した後、カラムを50mMのMOPS(pH7)で最初に洗浄し、続いて50mMのMOPS、0.1MのTMAC、0.3MのNaCl(pH7)により洗浄した。NT−3を50mMのMOPS、0.25MのTMAC、1.5MのNaCl(pH7)で溶出させた。
【0158】
次に、マクロプレッププールをフェニルセファロース・ファースト・フロー・ハイ・サブスティチューションカラムで精製した。フェニルカラムを50mMのHEPES、1.5MのNaCl(pH7)で平衡化し、マクロプレッププールをフェニルカラムに直接装填した。カラムを平衡緩衝液で洗浄し、正しく再折り畳みされたNT−3を、50mMのHEPES、1.5MのNaCl(pH7)から50mMのHEPES、10%の試薬アルコール(pH7)に変わる15カラム容量の勾配を使用して溶出させた。分画をC4HPLCかSDS−PAGEによって分析し、正しく折り畳まれたNT−3を含む分画をプールした。
【0159】
フェニルプールを25mS未満に希釈(典型的には水で約2倍容量)し、25mMのMOPSO(pH7)で前もって平衡化したSP−セファロースHPカラムに装填した。カラムを最初に平衡緩衝液で洗浄し、NT−3を、25mMのMOPSO(pH7)中に0.35MのTMACから0.65MのTMACまでの20カラム容量の勾配を使用してカラムから溶出させた。(C4HPLCアッセイにより判断して)rhNT−3を含む分画をプールした。
【0160】
SP−セファロースHPプールを10000分子量の膜上で約1mg/mlまで濃縮し、ついで6容量の10mMの酢酸塩、140mMのNaCl(pH5.0)でダイアフィルトレーションを行った。
【図面の簡単な説明】
【0161】
【図1】図1はSP−セファロースHPクロマトグラムを示す。HICクロマトグラフィー後に12kLの発酵からのNGF含有混合物を、25全体適合(omnifit)SP−セファロース・ハイ・パーフォーマンス樹脂(カラム寸法1.0x35cm;総容積27.5ml)の一つのSPHP樹脂上に装填した。緩衝液Aは、0.2MのNaCl、20mMのコハク酸塩、pH6.0(23ms)であった。緩衝液Bは0.7MのNaCl、20mMのコハク酸塩、pH6.0(63ms)であった。カラムは最初に緩衝液Aで平衡にした。0.24mg/mlで345mlのHICプールが25mMのコハク酸塩、pH6.0(17ms)に調整され、その362mlを装填して、3mg/ml樹脂の負荷とした;82.5mgのNGFを装填した。装填速度は1時間当り40cmとした。NGFを、30%から80%までの緩衝液B(0.35から0.60MまでのNaCl;37から57ms)の22カラム容量の勾配を使用して溶出させた。溶出速度は1時間当り60cmとした。吸光度単位(280nm)とmS/cmを分画数と溶出容積mlに対してプロットした。NGFを含有する分画を決定し、プールした。この場合、分画43から60をプールして、0.38mg/mlNGFの99mlのサンプルを得た。これは約46%の回収率になる。
【図2】図2は、フェニル・セファロース・ファースト・フロー・プールとSP−セファロースHPプールのSP−NPR HPLCカチオン交換(HPIEX)分析を示す。それぞれに対するクロマトグラムに標しをする。
【図3】図3は、SP−セファロースHPクロマトグラフィー工程からの選択した分画(主プール、主ピークプールの先端と後端)のC4 RP−HPLC分析を示す。3つのシグナルが重ねられて描かれている。分かるように、(成熟NGFを含む)主ピークは変種NGF群を含む幾つかのより小さいピークとは分離している。
【図4】図4は、ヒトプレプロNGFの配列(配列番号:1)を示す。この図上に示されているのは、成熟NGFの最初のアミノ酸(位置1)と120NGFの最後のアミノ酸(位置120)である。好適なNGFアミノ酸配列は成熟118型のもの(位置1から位置118)である。また示されているのは変種形態:成熟120(位置1から位置120);成熟117(位置1から位置117);R120(位置1から120);及び成熟114(アミノ酸1から114)、115(アミノ酸1から115)及び117(アミノ酸1から117)変種を含む、N末端とC末端に生じるその他の誤ってプロセシングされた部位である。タンパク分解性に誤ってプロセシングされた主要なミスプロセシング変種は、NGFのプロ配列のアミノ酸R(−39)とS(−38)の間にN末端切断を有している。可能な開始メチオニンには下線が付される。他のN末端変種には、NGFの切断型が含まれ、最もありふれたものはアミノ酸H8とH9及びR9とG10の間に生じる切断を有している。
【図5】図5は、ヒトNGF(配列番号:2)、マウスNGF(配列番号:3)、BDNF(配列番号:4)、NT−3(配列番号:5)及びNT−4/5(配列番号:6)のニューロトロフィンのアミノ酸配列を示している。ボックスで囲った領域はシステインノットモチーフに関与する相同的システイン含有領域を示している図である(De Youngら, Protein Sci. 5(8):1554-66(1996))。
【図6】図6は、DEAE−セファロース・ファースト・フロー(DEFF)樹脂カラムでのrhNT−4/5のクロマトグラフィーパターンを示す。「NT45DE1:1_UV」と標されているクロマトグラムは280nmでのUV吸光度測定値である。「NT45DE1:1_Cond1」と標されているクロマトグラムは溶出する分画の伝導率測定値である。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図7】図7は、SP−セファロース・ファースト・フロー樹脂カラムでのrhNT−4/5のクロマトグラフィーパターンを示す。「NT45SFF1:1_UV」と標されているクロマトグラムは280nmでのUV吸光度測定値である。「NT45SFF1:1_Cond1」と標されているクロマトグラムは溶出する分画の伝導率測定値である。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図8】図8は、明細書に記載された条件下でのrhNT−4/5の典型的な調製用C4−RP−HPLCクロマトグラフィーパターンを示す。280nmでの吸光度をモニターした。プールされたニューロトロフィン含有分画は「プール」と標された水平な矢印により示されている。
【図9】図9は、示した時間での再折り畳みの間モニターされたNT4/5試料の分析用HPLCクロマトグラフィーパターンを提供する。カラム条件は明細書中に記載した。「SSFF(0.5m)」と標されたパターンは、再折り畳みの前にS−セファロース・ファースト・フロー・カラムから0.5MのNaClで溶出させたNT−4/5の分析を示す。NT−4/5が再折り畳みされるにつれて、溶出パターンは標準のパターンに近づく。
【図10】図10は、疎水的相互作用クロマトグラフィーカラムであるフェニル・トーヨーパール650Mカラムでの誤って折り畳まれた変種からの無傷の正しく折り畳まれたNT−4/5の分離を示すクロマトグラムを示す。正しく折り畳まれたNT−4/5よりも疎水性が低い誤って折り畳まれた変種は素通り分内に溶出する一方、より疎水性である誤って折り畳まれた変種(ピークB)は、正しく折り畳まれたNT−4/5(ピークA)を溶出するのに必要とされるものよりも高い有機溶媒濃度で溶出する。
【図11】図11は、カチオン交換樹脂であるSP−セファロースからのNT−4/5と変種の溶出パターンを示す。280nmでの吸光度がモニターされた。ピークAがカルバミル化され切詰められた(clipped)変種を含む一方、ピークBは無傷の正しく折り畳まれたNT−4/5を含む。プールされたNT−4/5含有分画は「プール」と標された水平な矢印により示されている。
【図12】図12は、明細書に記載された条件下でのrhNT−4/5の典型的な調製用ポリキャットA樹脂クロマトグラフィーパターンを示す。
【図13】図13は、本明細書に記載したrhNT4/5精製法の示された工程から採られた試料の純度と均一性を評価するための還元条件下での16%SDS−PAGE(サンジエゴのNovex,Inc.からの、予め注がれたトリス−グリシン系)分析を示す。ゲルは、タンパク質を検出するために、クーマジーR250で染色された。「DE負荷」と標されたレーンはDE−セファロース・ファースト・フロー・カラム上に負荷されたPEI混合物の試料を含み、「DE FT」と標されたレーンはDE−セファロース・ファースト・フロー・カラムの素通り分の試料を含み、レーン「Sプール」は、再折り畳み前に、S−セファロース・ファースト・フロー樹脂カラムから溶出したNT−4/5を含むプールされた分画からの試料を含み、レーン「再折り畳みプール」は再折り畳みが完了した後のプールの試料を含み、レーン「C4プール」は調製的C4RP−HPLC後のプール分画の試料を含み、レーン「ポリキャットAプール」はポリキャットA HPLCカラムからのプール分画の試料を含む。
【図14】図14は、細菌的に調製されたスルホニル化rhNT−3を含む混合物のS−セファロース・ファースト・フロー・クロマトグラフィーからの分画のUV吸光度パターンを示す。
【図15】図15は、rhNT−3のインビトロ再折り畳み型を含む混合物のマクロプレップ・ハイ・S(Macroprep High S)カチオン交換クロマトグラフィーを示す。樹脂は、バイオラッド(Biorad)から購入した。カラム寸法は9x9cmであった。pH6.8の再折り畳みされた(36時間の再折り畳み後)を含む700mlSSFFプールが約310ml/分の流量でマクロプレップ・カラム上に負荷された。条件は明細書に記載する。
【図16】図16は、誤って折り畳まれた変種を除去するための、カチオン交換精製再折り畳みrhNT−3のフェニル・セファロース・ファースト・フロー・ハイ・サブスティチューション(疎水的相互作用クロマトグラフィー)クロマトグラフィーを示す。
【図17】図17は、HIC−rhNT−3プールのSP−セファロース高速クロマトグラフィーを示す。
【特許請求の範囲】
【請求項1】
ニューロトロフィンを他のタンパク質を含む混合物から分離する方法において、疎水的相互作用クロマトグラフィー樹脂を使用して、混合物中の他のタンパク質からニューロトロフィンを分離することを含んでなる方法。
【請求項2】
混合物が、誤って折り畳まれた変種、不適切にタンパク分解性にプロセシングされた変種、又はそのニューロトロフィンのグリコシル化変種を含む請求項1記載の方法。
【請求項3】
分離が、ニューロトロフィンと、誤って折り畳まれた変種、不適切にタンパク分解性にプロセシングされた変種、又はそのニューロトロフィンのグリコシル化変種を含む混合物を、疎水的相互作用クロマトグラフィー樹脂上に負荷し、ニューロトロフィンが変種もしくは変種群から分離する条件下で樹脂からニューロトロフィンを溶出させることを含んでなる請求項1記載の方法。
【請求項4】
樹脂がフェニル官能基を含む請求項1記載の方法。
【請求項5】
疎水的相互作用クロマトグラフィー樹脂に装填される混合物が5から8のpHを有する請求項1記載の方法。
【請求項6】
樹脂上に装填される混合物が0.5Mから3Mの塩濃度を有する請求項1記載の方法。
【請求項7】
樹脂上に装填される混合物が0.5Mないし2.5Mの塩濃度を有する請求項6記載の方法。
【請求項8】
樹脂上に装填される混合物が0.7M酢酸塩又は1.0Mから2.5MのNaClを有する請求項7記載の方法。
【請求項9】
溶出緩衝液が有機溶媒を含有する請求項3記載の方法。
【請求項10】
有機溶媒が5容量%から5容量%から20容量%である請求項9記載の方法。
【請求項11】
溶出緩衝液のpHがpH5からpH8である請求項3記載の方法。
【請求項12】
溶出が減少する塩勾配からなる請求項3記載の方法。
【請求項13】
ニューロトロフィンがNGFスーパーファミリーのものである請求項1記載の方法。
【請求項14】
ニューロトロフィンがNGF、NT−4/5又はNT−3である請求項12記載の方法。
【請求項15】
ニューロトロフィンが細菌培養から調製され、疎水的相互作用樹脂を用いる前にインビトロで再び折り畳まれる請求項1記載の方法。
【請求項16】
ニューロトロフィンが哺乳動物細胞培養から単離される請求項1記載の方法。
【請求項17】
ニューロトロフィンを、高速カチオン交換クロマトグラフィー樹脂を使用して、その化学変種から分離することを更に含んでなる請求項1記載の方法。
【請求項18】
高速カチオン交換クロマトグラフィーによる分離工程が、ニューロトロフィンと該ニューロトロフィンの化学変種を含む混合物を高速カチオン交換クロマトグラフィー樹脂に装填し、ニューロトロフィンが化学変種から分離する条件下で樹脂からニューロトロフィンを溶出させることを含む請求項17記載の方法。
【請求項19】
カチオン交換樹脂が、SP−セファロースHP、ポリアスパラギン酸、ポリスルホエチルカチオン交換、もしくはフラクトゲルEMD SO3樹脂である請求項18記載の方法。
【請求項20】
カチオン交換樹脂からのニューロトロフィン含有溶出液を脱塩するか、ダイアフィルトレーションにかけ、ついで担体と製剤化する請求項18記載の方法。
【請求項21】
ニューロトロフィンを他のタンパク質からシリカゲル樹脂を使用して分離することを更に含む請求項1記載の方法。
【請求項22】
ニューロトロフィンをタンパク質の混合物から分離する方法であって、ニューロトロフィンを他のタンパク質からシリカゲル樹脂を使用して分離する方法。
【請求項23】
ニューロトロフィンを該ニューロトロフィンの化学変種から分離する方法であって、高速カチオン交換クロマトグラフィーを使用して、ニューロトロフィンをその化学変種から分離する方法。
【請求項24】
ニューロトロフィンとその変種が、分離前に本質的に純粋である請求項23記載の方法。
【請求項25】
樹脂が、SP−セファロースHP、ポリアスパラギン酸樹脂、ポリスルホエチルカチオン交換樹脂、もしくはフラクトゲルEMD SO3樹脂である請求項23記載の方法。
【請求項26】
調製的逆相液体クロマトグラフィー樹脂を使用してニューロトロフィンを該ニューロトロフィンの誤って折り畳まれた変種から分離する工程を更に含んでいてもよい請求項23記載の方法。
【請求項27】
樹脂がC4官能基を含む請求項26記載の方法。
【請求項28】
請求項1、22又は23に記載の方法により調製されるニューロトロフィン組成物。
【請求項29】
担体と、変種を本質的に含まないことにより本質的に純粋で均一なニューロトロフィンとを含有する組成物。
【請求項30】
無菌である請求項29記載の組成物。
【請求項31】
ニューロトロフィンを精製する方法であって、
(a) ニューロトロフィンとその変種を含む混合物を、pH5から8で疎水的相互作用クロマトグラフィー樹脂に装填し、
(b) pH5から8で緩衝液により樹脂を洗浄し、
(c) 約5から25%(v/v)の濃度でアルコール性又は極性非プロトン溶媒を含むpH5から8の緩衝液でニューロトロフィンを溶出させ、
(d) pH5から6の高速カチオン交換クロマトグラフィー樹脂にニューロトロフィン含有溶出物を装填し、
(e) 0.2から0.5Mの濃度でカチオンを含むpH5から6の緩衝液で、高速カチオン交換クロマトグラフィー樹脂からのニューロトロフィンを溶出させる方法。
【請求項1】
ニューロトロフィンを他のタンパク質を含む混合物から分離する方法において、疎水的相互作用クロマトグラフィー樹脂を使用して、混合物中の他のタンパク質からニューロトロフィンを分離することを含んでなる方法。
【請求項2】
混合物が、誤って折り畳まれた変種、不適切にタンパク分解性にプロセシングされた変種、又はそのニューロトロフィンのグリコシル化変種を含む請求項1記載の方法。
【請求項3】
分離が、ニューロトロフィンと、誤って折り畳まれた変種、不適切にタンパク分解性にプロセシングされた変種、又はそのニューロトロフィンのグリコシル化変種を含む混合物を、疎水的相互作用クロマトグラフィー樹脂上に負荷し、ニューロトロフィンが変種もしくは変種群から分離する条件下で樹脂からニューロトロフィンを溶出させることを含んでなる請求項1記載の方法。
【請求項4】
樹脂がフェニル官能基を含む請求項1記載の方法。
【請求項5】
疎水的相互作用クロマトグラフィー樹脂に装填される混合物が5から8のpHを有する請求項1記載の方法。
【請求項6】
樹脂上に装填される混合物が0.5Mから3Mの塩濃度を有する請求項1記載の方法。
【請求項7】
樹脂上に装填される混合物が0.5Mないし2.5Mの塩濃度を有する請求項6記載の方法。
【請求項8】
樹脂上に装填される混合物が0.7M酢酸塩又は1.0Mから2.5MのNaClを有する請求項7記載の方法。
【請求項9】
溶出緩衝液が有機溶媒を含有する請求項3記載の方法。
【請求項10】
有機溶媒が5容量%から5容量%から20容量%である請求項9記載の方法。
【請求項11】
溶出緩衝液のpHがpH5からpH8である請求項3記載の方法。
【請求項12】
溶出が減少する塩勾配からなる請求項3記載の方法。
【請求項13】
ニューロトロフィンがNGFスーパーファミリーのものである請求項1記載の方法。
【請求項14】
ニューロトロフィンがNGF、NT−4/5又はNT−3である請求項12記載の方法。
【請求項15】
ニューロトロフィンが細菌培養から調製され、疎水的相互作用樹脂を用いる前にインビトロで再び折り畳まれる請求項1記載の方法。
【請求項16】
ニューロトロフィンが哺乳動物細胞培養から単離される請求項1記載の方法。
【請求項17】
ニューロトロフィンを、高速カチオン交換クロマトグラフィー樹脂を使用して、その化学変種から分離することを更に含んでなる請求項1記載の方法。
【請求項18】
高速カチオン交換クロマトグラフィーによる分離工程が、ニューロトロフィンと該ニューロトロフィンの化学変種を含む混合物を高速カチオン交換クロマトグラフィー樹脂に装填し、ニューロトロフィンが化学変種から分離する条件下で樹脂からニューロトロフィンを溶出させることを含む請求項17記載の方法。
【請求項19】
カチオン交換樹脂が、SP−セファロースHP、ポリアスパラギン酸、ポリスルホエチルカチオン交換、もしくはフラクトゲルEMD SO3樹脂である請求項18記載の方法。
【請求項20】
カチオン交換樹脂からのニューロトロフィン含有溶出液を脱塩するか、ダイアフィルトレーションにかけ、ついで担体と製剤化する請求項18記載の方法。
【請求項21】
ニューロトロフィンを他のタンパク質からシリカゲル樹脂を使用して分離することを更に含む請求項1記載の方法。
【請求項22】
ニューロトロフィンをタンパク質の混合物から分離する方法であって、ニューロトロフィンを他のタンパク質からシリカゲル樹脂を使用して分離する方法。
【請求項23】
ニューロトロフィンを該ニューロトロフィンの化学変種から分離する方法であって、高速カチオン交換クロマトグラフィーを使用して、ニューロトロフィンをその化学変種から分離する方法。
【請求項24】
ニューロトロフィンとその変種が、分離前に本質的に純粋である請求項23記載の方法。
【請求項25】
樹脂が、SP−セファロースHP、ポリアスパラギン酸樹脂、ポリスルホエチルカチオン交換樹脂、もしくはフラクトゲルEMD SO3樹脂である請求項23記載の方法。
【請求項26】
調製的逆相液体クロマトグラフィー樹脂を使用してニューロトロフィンを該ニューロトロフィンの誤って折り畳まれた変種から分離する工程を更に含んでいてもよい請求項23記載の方法。
【請求項27】
樹脂がC4官能基を含む請求項26記載の方法。
【請求項28】
請求項1、22又は23に記載の方法により調製されるニューロトロフィン組成物。
【請求項29】
担体と、変種を本質的に含まないことにより本質的に純粋で均一なニューロトロフィンとを含有する組成物。
【請求項30】
無菌である請求項29記載の組成物。
【請求項31】
ニューロトロフィンを精製する方法であって、
(a) ニューロトロフィンとその変種を含む混合物を、pH5から8で疎水的相互作用クロマトグラフィー樹脂に装填し、
(b) pH5から8で緩衝液により樹脂を洗浄し、
(c) 約5から25%(v/v)の濃度でアルコール性又は極性非プロトン溶媒を含むpH5から8の緩衝液でニューロトロフィンを溶出させ、
(d) pH5から6の高速カチオン交換クロマトグラフィー樹脂にニューロトロフィン含有溶出物を装填し、
(e) 0.2から0.5Mの濃度でカチオンを含むpH5から6の緩衝液で、高速カチオン交換クロマトグラフィー樹脂からのニューロトロフィンを溶出させる方法。
【図1】



【図2】
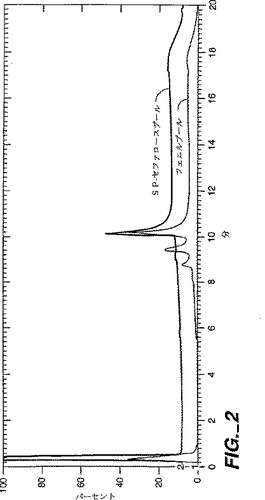
【図3】

【図4】
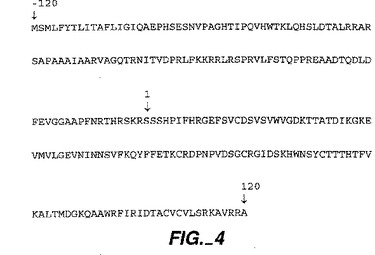
【図5】

【図6】
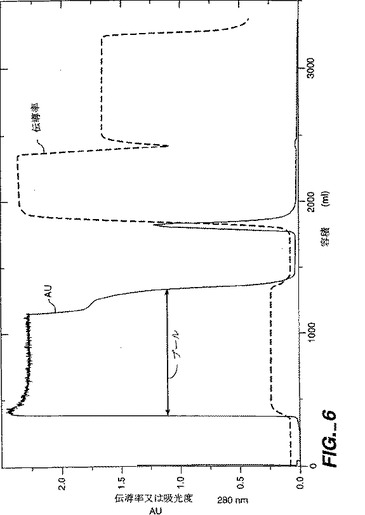
【図7】
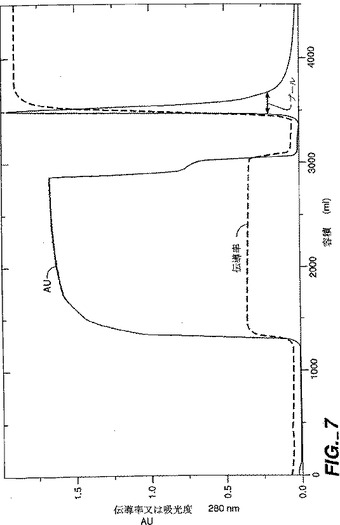
【図8】

【図9】
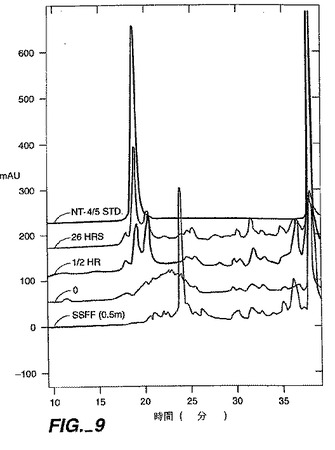
【図10】
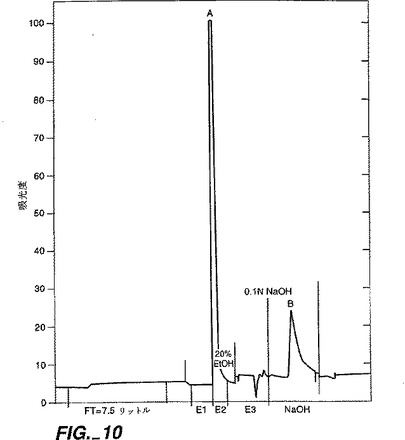
【図11】

【図12】

【図13】
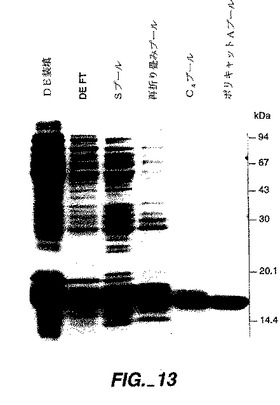
【図14】

【図15】
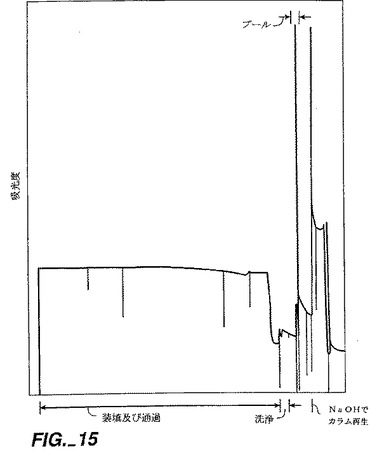
【図16】

【図17】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2009−40786(P2009−40786A)
【公開日】平成21年2月26日(2009.2.26)
【国際特許分類】
【外国語出願】
【出願番号】特願2008−208434(P2008−208434)
【出願日】平成20年8月13日(2008.8.13)
【分割の表示】特願2007−168104(P2007−168104)の分割
【原出願日】平成9年11月14日(1997.11.14)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成21年2月26日(2009.2.26)
【国際特許分類】
【出願番号】特願2008−208434(P2008−208434)
【出願日】平成20年8月13日(2008.8.13)
【分割の表示】特願2007−168104(P2007−168104)の分割
【原出願日】平成9年11月14日(1997.11.14)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]