
ニューロンの発生、再生および保護
アポトーシス性の狂犬病ウイルスG蛋白質とは反対に、特定の非アポトーシス性の狂犬病ウイルスG蛋白質(例えば、CVS-NIV株のG蛋白質)が神経突起成長促進効果を有することを本発明は実証する。この神経突起成長促進効果がアポトーシス性の狂犬病ウイルスG蛋白質の一つと比較して単一の点変異を示す前記非アポトーシス性の狂犬病ウイルスG蛋白質の細胞質尾部(より具体的には、そのPDZ-BS)によることを本発明は更に実証する。本発明は、神経突起の成長を誘導および/または刺激するための手段を提供し、これらは、ニューロンの分化の誘導のために、例えば、神経系の新生物の治療のために、同様に、障害されたニューロンの再生において、例えば、神経変性疾患、障害または状態の治療のために、または微生物感染の治療において、またはニューロンを神経毒性因子または酸化ストレスから保護することにおいて有用である。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、ニューロンの発生、再生および保護、より具体的には神経突起成長(neurite outgrowth)に関する。
【0002】
本発明は、神経突起生成作用を誘導および/または刺激する手段を提供し、これらの手段は医学の分野(より具体的には、神経系の障害の治療 および/または 緩和および/または予防)に有用である。特に、本発明は、神経突起の成長を誘導および/または刺激するための手段を提供し、これは、ニューロンの分化を誘導するために、例えば、神経系の新生物の治療のため、同様に、障害されたニューロンの再生における、例えば、神経変性疾患、障害または状態の治療のため、または微生物感染の治療における、またはニューロンを神経毒性剤または酸化ストレスからの保護において有用である。
【0003】
本発明の手段は、特定の非アポトーシス性(non-apoptotic)の狂犬病ウイルス G 蛋白質蛋白質(特に、その蛋白質の細胞質尾部)に基づく。
【発明の背景】
【0004】
神経系の発生の間に、ニューロンは適切な様式で標的に神経を分布(innervate)させるためかなりの距離で伸びる。この現象には細胞において成長円錐の活性を刺激できる特異的なシグナル伝達経路が刺激されることが必要である。
【0005】
神経系の発生(特に、中枢神経系の発生)は柔軟性(plastic)が高いが、成体の神経系(特に、成体の脳)の修復の潜在力は限定的なものである。従って、神経突起軸索の成長および変性に対する保護は、神経変性疾患、障害または状態(例えば、神経系の急性傷害または慢性の神経変性疾患)の転帰を改善すると考えられる重要な事項である。係るニューロン細胞からの神経突起成長を誘導する能力があるであろう産物は、係る疾患、障害または状態(conditions)の非常に有用な治療上および/または予防上および/または対症療法上(palliative)の解決策をもたらすであろう。
【0006】
ニューロンの発生のプロセスの別の側面として、成熟したニューロンの構造に分化しないニューロン前駆体(neuronal progenitors)の増殖によって神経系の新生物が誘発される。係る前駆体細胞から神経突起成長を誘導する能力があるであろう産物は、係る新生物の治療上および/または予防上および/または対症療法上の解決策をもたらすであろう。
【0007】
神経向性ウイルスによるニューロン細胞の感染における一般的な概念は、ニューロンの形態(特に、神経突起成長)に肯定的な影響(positive impact)を有さないとのことである。
【0008】
実際、神経向性ウイルスがアポトーシスによるニューロン細胞死を生じることを示している多数の例が存在する。この事項はDNAウイルス(例えば、疱疹ウイルス)、およびRNAウイルスの双方に関し、エンベロープウイルス、例えば、アルファウイルス、ブニヤウイルスおよびパラミクソウイルス、または非エンベロープウイルス、例えば、ピコルナウイルスおよびレオウイルスに関する。
【0009】
また、狂犬病ウイルス(特に、弱毒化狂犬病ウイルス株)は、ニューロンのアポトーシスを誘導することが記載されている。
【0010】
例えば、WO 03/048198は、ニューロン細胞の統合性の崩壊やアポトーシス小体の形成を誘導する少なくとも 100 アミノ酸の狂犬病ウイルス G 蛋白質及びその断片に関する。これらのアポトーシス小体は、体液性免疫応答、好ましくはB-依存的な体液性の免疫応答を刺激する能力がある。
【0011】
WO 03/048198は以下を示す:
・ 弱毒化した狂犬病ウイルス株(例えば、弱毒化したERA株)は、そのウイルスが感染した細胞のアポトーシスによる破裂を誘導する,
・ それによって産生されたアポトーシス小体は、体液性免疫応答、特にB-依存的な体液性免疫応答を刺激する;
・ 狂犬病ウイルス株によるアポトーシスの誘導は、そのG蛋白質の性質により決定される;
・ 弱毒化した狂犬病ウイルス株(例えば、弱毒化したERA株)からのG蛋白質を含んでいる狂犬病ウイルスはヒト細胞のアポトーシスを惹起することが可能であるが、他方で病原性の狂犬病ウイルス(例えば、Challenge Virus Standard -CVS-株)からのG蛋白質を発現させることでは惹起することは不可能である(特に、WO 03/048198の例 5 および 図 19 および 20を参照されたい)。
【0012】
また、Lay等 (Lay et al. 2003)および Prehaud等 (Prehaud et al. 2003)を参照されたい。
【0013】
以上のように、アポトーシス性の狂犬病ウイルス株のG 蛋白質(例えば、弱毒化したERA株のG蛋白質)は、体液性応答(特に、B依存的な体液性免疫応答)の刺激に有用であることが知られている。
【0014】
これらの特定の G 蛋白質はそれらが感染した細胞のアポトーシスを誘導するので、これらの蛋白質も標的細胞のアポトーシスにより望ましくない細胞を破裂させることで排除する候補因子として提案されてきた。
【0015】
狂犬病ウイルス株の病原性は、アポトーシスを誘導する能力とは逆に相関することも記載されている(WO 03/048198; Ugolini 1995; Sarmento et al. 2005; Ugolini 2008; Jackson et al. 2008を参照されたい)。
【0016】
以上のように、病原性の強い狂犬病ウイルス株は、アポトーシス性が低い。
【0017】
病原性の狂犬病ウイルス株(例えば、CVS 株)がニューロンのアポトーシスを誘導せず、これにより体液で検出(humoral detection)されることから免れるとの所見によって、なぜ病原性の狂犬病ウイルス株が疾患の徴候や症状の出現前にCNS内に非常に広範に伝播できるかが説明される。
【0018】
神経向性ウイルス〔例えば、狂犬病ウイルスまたは単純ヘルペス1型 (HSV-1)〕による感染に際して誘導される遺伝子発現パターンにおける変化を分析するためさらなる研究が実施された。
【0019】
これらの研究は、グリアの非存在下で分裂後(post-mitotic)のヒトニューロンがウイルス感染を感知する内部機構を有すること、また神経向性ウイルス(病原性の狂犬病ウイルスまたはHSV-1)が分裂後のヒトニューロンからのサイトカインの放出を誘導することを実証した(Prehaud et al. 2005)。このサイトカイン放出は、さらにニューロンがアポトーシスを回避することに寄与し、係る神経向性ウイルスを結果的に伝播させると信じられている。
【0020】
このようなことから、ニューロンの細胞死の機構(同様に、ウイルス感染に際する免疫応答を上昇させるニューロンの能力)が徹底的に調査された。
【0021】
しかしながら、神経発生(neurogeneration), 神経再生および神経保護、特に分裂前(pre-mitotic)のニューロン(例えば、ニューロン前駆体又は新生物ニューロン)からの又は変性ニューロン(degenerative neurons)からの神経突起成長に関与するプロセスについては僅かなことしか知られていない。
【0022】
狂犬病ウイルスに焦点を合わせると、現在の知識の概要は、弱毒化した狂犬病ウイルス株が該ウイルスにより誘導されるアポトーシスによる医学的な適用を有することが知られていること、また病原性の狂犬病ウイルス株がアポトーシスを誘導しないが、しかし、反対にそれらの伝播を支援するニューロンネットワークが保存されることが知られていることであろう。
【0023】
病原性の狂犬病ウイルス株がニューロンネットワークの統合性を保存することが記載されたが、それらはニューロン形態(特に、神経突起成長)に否定的な影響(negative impact)を有している(又は、最低限肯定的な影響はない)とも報告された。
【0024】
例えば、文献(Guigoni and Coulon 2002)は、病原性の 狂犬病ウイルス株 CVS-Gif-sur-Yvetteがラットの運動ニューロンの神経突起成長を誘導しないことを記載する(例えば、この文献の図 5A および 5Bを参照されたい)。
【0025】
神経突起成長における否定的な影響も報告された。例えば、文献(Scott et al., 2008)は、病原性の CVS-11 狂犬病ウイルス株が樹状突起や軸索のビーディング(beading)を誘導(即ち、ネガティブなストレス影響の特徴である小胞の形成)することを報告している。
【0026】
本発明は、特定の病原性の狂犬病ウイルス株から由来し且つ驚くべき予想外の特性を示すニューロンの発生、再生および保護のための手段を提供する。
【発明の概要】
【0027】
本発明は、幾つかの病原性(非アポトーシス性)の狂犬病ウイルス G 蛋白質が、神経突起成長促進効果を有すること〔即ち、幾つかの非アポトーシス性の狂犬病ウイルスG蛋白質が神経突起生成(neuritogenesis)を誘導および/または刺激すること〕を示している。この効果は、アポトーシス性の狂犬病ウイルス G 蛋白質により示されない。この効果は、全ての非アポトーシス性の狂犬病ウイルス G 蛋白質によっては示されない。
【0028】
発明者は、非アポトーシス性(病原性)の狂犬病ウイルス株の亜群であって、そのG蛋白質が神経突起成長に有意に陽性の効果を有する亜群を同定した。以下に記載され、説明されるように、この亜群の代表的な株は、CVS-NIV 株である。この非アポトーシス性(病原性)株のG蛋白質は、アポトーシス性(弱毒化) ERA株 (CNCM I-2760)のG蛋白質と僅か6 アミノ酸のみが異なる。
【0029】
さらに、この神経突起成長促進効果が、前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部によること、特にそれらのPDZ 結合サイト(PDZ-BS)に及び/又はCVS-NIV 株のG蛋白質の配列中のポジション 491にあるアミノ酸による、特に、それらのPDZ-BSによることを本発明は実証する。
【0030】
本発明は、特定の非アポトーシス性の狂犬病ウイルス G 蛋白質から(より具体的には、それらの細胞質尾部から)由来するポリペプチドに関する、同様に核酸、ベクター、細胞および薬学的組成物または薬物に関する。
【0031】
本発明の手段は、神経突起成長(neurite outgrowth)を刺激する及び/又は誘導する薬剤として特に有用である。それらは、ニューロンの分化を誘導するため特に使用でき、例えば、神経系の新生物の治療に、同様に、障害されたニューロンの再生のため、例えば、神経変性疾患, 障害またはコンディションの治療に、または微生物感染の治療に、またはニューロンを神経毒性剤または酸化ストレスから保護するため使用できる。
【0032】
本出願が参照する図の幾つかは、カラー印刷である。本出願は図面のカラー印刷を含んでおり、特許庁で出願ファイルを閲覧することによりこの図面にアクセスすることができる。
【図面の簡単な説明】
【0033】
【図1】神経発生、神経再生および神経保護は、神経毒性、発作(seizure)、脳卒中(stroke)、外傷、加齢、神経変性疾患、脳症(encephalopathy)を含む様々なニューロンの障害、状態および疾患に関係する。
【図2】発明者により作出され、以下の例に使用された組換え型の狂犬病ウイルス(rRABV)の構造の概要。CVS-NIV 株 (非アポトーシス性の株)のG蛋白質は、ERA株 (アポトーシス性の株)とは僅か6 aaのみが異なる。G-survival = CVS-NIV 株のG蛋白質。G-death = ERA株のG蛋白質。G-cyto death = CVS-NIV Gの遺伝背景にG-deathの細胞質尾部。G-cyto survival = ERA Gの遺伝背景にG-survivalの細胞質尾部。SP = シグナルペプチド。EC: 細胞外ドメイン。TM: 膜貫通ドメイン。Cyto: 細胞質ドメイン。
【図3A】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS または G-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルス G蛋白質。
【図3B】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERA および rRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。
【図3C】図 3A, 3B, 3C, 3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルスG蛋白質。
【図3D】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIVおよびrRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIVおよびrRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルスG蛋白質。
【図4A】図 4A、4B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、cAMP*と相乗的に作用する神経突起生成を促進する内因性エフェクターである。図 4A: db-cAMPなし。図 4B: db-cAMPなし(最初の二つのヒストグラム)、cAMP有り(最後の二つのヒストグラム)。黒星は、有意差を意味する(スチューデントt-検定、p=0.0067)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株の蛋白質G。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図4B】図 4A、4B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、cAMP*と相乗的に作用する神経突起生成を促進する内因性エフェクターである。図 4A: db-cAMPなし。図 4B: db-cAMPなし(最初の二つのヒストグラム)、cAMP有り(最後の二つのヒストグラム)。平均は有意差あり(スチューデントt-検定、p=0.0067)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図5】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部の神経突起生成効果は、分子署名に依存し、発現されたG蛋白質の量に依存しない。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図6A】図 6A、6B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、成長円錐虚脱薬(LPA)に対する神経保護を提供する。図6A: db-cAMPあり。図6B: db-cAMPなし。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図6B】図 6A、6B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、成長円錐虚脱薬(LPA)に対する神経保護を提供する。図6A: db-cAMPあり。図6B: db-cAMPなし。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図7】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部により誘導される成長円錐虚脱薬LPAに対する神経保護は強いものである。黒星は、有意差を意味する(ANOVA 検定)。黒三角は、有意差を意味する(ANOVA 検定)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株の蛋白質G。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。LPA = リゾホスファチジン酸。
【図8】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、酸化ストレス(H2O2)に対し神経保護を提供する。これらの実験は、db-cAMPなしで行われた。黒星は、有意差を意味する(スチューデントのt-検定)。黒三角は、有意差を意味する(スチューデントのt-検定)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9A】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9B】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9C】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図10A】図 10A、10B: オール-トランス-レチノイン酸 (ATRA:all-trans-retinoic acid) 分化誘導薬(pro-differentiative drug)で処理したヒト神経芽細胞腫の細胞の増殖(図10A: フローサイトメトリー; 図10B: MTTアッセイ)。N.I. = 非感染。
【図10B】図 10A、10B: オール-トランス-レチノイン酸 (ATRA:all-trans-retinoic acid) 分化誘導薬(pro-differentiative drug)で処理したヒト神経芽細胞腫の細胞の増殖(図10A: フローサイトメトリー; 図10B: MTTアッセイ)。N.I. = 非感染。
【図11A】図 11A、11B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS-NIVまたはrRABV G-ERAで処理したヒト神経芽細胞腫細胞の細胞増殖(図 11A) および 神経突起長(図 11B)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株のG蛋白質。G-ERA = ERA株のG蛋白質。
【図11B】図 11A、11B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS-NIVまたはrRABV G-ERAで処理したヒト神経芽細胞腫細胞の細胞増殖(図 11A)および神経突起長(図 11B)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株のG蛋白質。G-ERA = ERA株のG蛋白質。
【図12A】図 12A、12B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS、rRABV G-ERA、rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survivalで処理したヒト神経芽細胞腫細胞の細胞増殖(図 12A: フローサイトメトリー; 図 12B: MTT アッセイ)(N.I. = 非感染)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図12B】図 12A、12B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS、rRABV G-ERA、rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survivalで処理したヒト神経芽細胞腫細胞の細胞増殖(図 12A: フローサイトメトリー; 図 12B: MTT アッセイ)(N.I. = 非感染)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図13A】図 13A、13B: CVS-NIV 株(図 13A)の及びERA株(図 13B)のG蛋白質の核酸および蛋白質の配列。図 13A中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図13B】図 13A、13B: CVS-NIV 株(図 13A)の及びERA株(図 13B)のG蛋白質の核酸および蛋白質の配列。図 13A中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図14】CVS-NIV および ERA株のG 蛋白質の配列アラインメント; 僅かに6 アミノ酸のみが異なるG 蛋白質(図 14に太字で示す)。
【表1】

図 14に示されるアラインメントは、次のパラメータを用いて行われた:
比較マトリックス: BLOSUM62
計算したアラインメントの数: 20
ギャップオープンペナルティー: 12
ギャップエクステンションペナルティー: 4
このアラインメントの結果は:
524 残基のオーバーラップにおいて98.9%同一性;
スコア: 2787.0;
Gap 頻度: 0.0%。
【図15】CVS-NIV 株の及びERA株のG蛋白質の細胞質ドメインの核酸および蛋白質の配列。図 15中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図16】CVS-NIV 株の及びERA株のG蛋白質のPDZ-BSの核酸および蛋白質の配列。aa = アミノ酸。
【図17−1】CVS-NIV 株のG蛋白質(配列番号 2)およびCVS-Gif-sur-Yvette 株のG蛋白質(配列番号 15)のアラインメント。
【図17−2】CVS-NIV 株のG蛋白質(配列番号 2)およびCVS-Gif-sur-Yvette 株のG蛋白質(配列番号 15)のアラインメント。
【図18−1】CVS-NIV 株のG蛋白質(配列番号 2)および三つのCVS-11株のG蛋白質(配列番号 16; 配列番号17; 配列番号18)のアラインメント。
【図18−2】CVS-NIV 株のG蛋白質(配列番号 2)および三つのCVS-11株のG蛋白質(配列番号 16; 配列番号17; 配列番号18)のアラインメント。
【図19】CVS-NIV 株のG蛋白質の細胞質断片(配列番号6)及びそれから派生する二つの保存されたバリアント(配列番号19のバリアント配列 A; 配列番号20のバリアント配列 B)の配列。
【図20】PDZ-BSの欠失は、神経突起成長表現型に影響する(db-cAMPの存在下、感染後8hのμmでの平均神経突起長)。N.I. = 非感染;G-CVS-NIV = CVS-NIV 株の蛋白質G;G-CVS-NIV-DeltaPDZ-BS = PDZ-BSが欠失したCVS-NIV 株の蛋白質G。
【図21】本発明の単一の点変異体の構造の概要(バリアント B および A)。本発明の変異体 ウイルス CVS HE (I-4143) および CVS LQ (I-4142)はCVS-NIV (I-4140)とG 蛋白質において異なり、G 蛋白質中に単一の点変異が導入されている。CVS HE (バリアント B) = G蛋白質 CVS-NIV 株の第一のPDZ-BS アミノ酸の単一の点変異であり、CVS HE においてQの代わりにE(ポジション 521)である。CVS LQ (バリアント A) = ポジション 491でのアミノ酸の単一の点変異(この変異はQの代わりにEである)。SP: シグナルペプチド。EC: 細胞外ドメイン。TM: 膜貫通ドメイン。Cyto: 細胞質ドメイン。
【図22】非アポトーシス性のウイルス株の及び構築物インサートのG蛋白質のドメインのアミノ酸配列。 ・ G-CVS ドメインの配列: ・ G 全長 (配列番号 2): アポトーシス性の狂犬病ウイルス株(CVS-NIV)の完全長 G蛋白質; ・ シグナルペプチド (SP; 配列番号 21): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)のシグナルペプチド; ・ 外部ドメイン (EC; 配列番号 22): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の外部ドメイン; ・ 膜貫通ドメイン(TM. 配列番号 23): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の膜貫通ドメイン; ・ 細胞質ドメイン(Cyto; 配列番号 6): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の細胞質ドメイン; ・ 構築物インサートによりコードされたアミノ酸配列: ・ アミノ酸 M + 配列番号 6 = 配列番号 24 (構築物 G-Cyto); ・ 配列番号 23の膜貫通ドメイン + 配列番号 6の細胞質ドメイン = 配列番号 25; ・ G-CVS-NIV の外部ドメインのC端からの二つのアミノ酸(アミノ酸 GK) + 配列番号 23の膜貫通ドメイン + 配列番号 6の細胞質ドメイン = 配列番号 26; ・ 配列番号21のシグナルペプチド + G-CVS-NIVの外部ドメインのC端からの二つのアミノ酸(アミノ酸 GK) + 配列番号23の膜貫通ドメイン + 配列番号6の細胞質ドメイン = 配列番号27 (構築物 G-(SP-[2a]-TM-Cyto)。
【図23】三つの構築物〔G-full構築物によりコードされた配列番号2; G-(SP)-[2a]-TM-Cyto (=GSP2aaTMCyto) 構築物によりコードされた配列番号27; G-Cyto 構築物によりコードされた配列番号24〕のインサートによりコードされたアミノ酸配列のアラインメント。
【図24】三つのRABV G 遺伝子構築物 〔配列番号2のG-full(= G survival); 配列番号24のG-Cyto(= G survival-Cyto); 配列番号27のG-(SP)-[2a]-TM-Cyto(= G survival-ΔEC)〕の略図。
【図25】SH-SY5Y 細胞株 (ヒトニューロブラストーマ細胞株)における対照プラスミド、G-Full 構築物(配列番号2)、G-(SP-[2a]-TM-Cyto) 構築物 (配列番号27)および G-Cyto構築物 (配列番号24)に関するRABV G 発現(任意単位)。
【図26】図 26 および 27: 以下のものを発現させた際の神経突起成長における影響であって: ・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質(配列番号 2; G-Full 構築物)、 ・ 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメイン(配列番号 27; G-(SP-[2a]-TM-Cyto) 構築物)、または ・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 24; G-Cyto 構築物)、の発現をヒト神経芽細胞腫細胞(db c-AMPの存在下のSH-SY5Y 細胞株)において対照(DNAなし)および対照プラスミドと比較した影響。図 26: 一過性発現。図 27: 安定発現。
【図27】図 26 および 27: 以下のものを発現させた際の神経突起成長における影響であって: ・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質(配列番号 2; G-Full 構築物), ・ 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメイン(配列番号 27; G-(SP-[2a]-TM-Cyto) 構築物)、または ・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 24; G-Cyto 構築物)、の発現をヒト神経芽細胞腫細胞(db c-AMPの存在下のSH-SY5Y 細胞株)において対照(DNAなし)および対照プラスミドと比較した影響。図 26: 一過性発現。図 27: 安定発現。
【図28】非アポトーシス性の狂犬病ウイルス株 (配列番号 27; 構築物 G-(SP-[2a]-TM-Cyto)の膜貫通および細胞質ドメインの発現は、ヒト神経芽細胞腫の細胞 (db c-AMP存在下のSH-SY5Y 細胞株)の神経突起成長を誘導し、成長した神経突起への成長円錐虚脱薬 LPAに対する保護を与える。
【図29】図 29 および 30: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激する〔分化処置後5 日: 対照プラスミドと比較した、G-Full構築物(配列番号2のインサート)の効果、G-(SP-[2a]-TM-Cyto)構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。図 29: カラー化したイメージ。図 30: 神経突起の先端(tips)。
【図30】図 29 および 30: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激する〔分化処置後5 日: 対照プラスミドと比較した、G-Full 構築物(配列番号2のインサート)の効果、G-(SP-[2a]-TM-Cyto) 構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。図 29: カラー化したイメージ。図 30: 神経突起の先端(tips)。
【図31】非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激し、長い軸索のニューロンネットワークの組織化を誘導および/または刺激する〔生きたニューロンのカラー化されたイメージ、分化処置後50 日: 対照プラスミドと比較した、G-Full 構築物(配列番号2のインサート)の効果、 G-(SP-[2a]-TM-Cyto) 構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。
【図32】図 32 および 33: 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメインの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する〔PDL-ラミニンプラスチック器に播種したニューロン, 注射針(26GX1/2", 12-4.5)で引っ掻いた3 日後; 対照プラスミド(神経突起変性)と比べた配列番号 27 (構築物 G-(SP-[2a]-TM-Cyto)(神経突起再生)のポリペプチドの効果〕。図 32: カラー化したイメージ。図 33: 神経突起再生のパーセンテージ
【図33】図 32 および 33: 非アポトーシス性の 狂犬病ウイルス株の膜貫通および細胞質ドメインの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する〔PDL-ラミニンプラスチック器に播種したニューロン, 注射針(26GX1/2", 12-4.5)で引っ掻いた3 日後; 対照プラスミド(神経突起変性)と比べた配列番号 27 (構築物 G-(SP-[2a]-TM-Cyto)(神経突起再生)のポリペプチドの効果〕。図 32: カラー化したイメージ。図 33: 神経突起再生のパーセンテージ
【発明の詳細な記述】
【0034】
本出願は、提出され、本明細書等に記載され、説明される請求項に規定の主題に関する。
【0035】
狂犬病ウイルス(大抵の哺乳類種において致死的な脳炎を引き起こす神経向性ウイルス)の病原性は、感染したニューロンが生存する能力と相関する。生ワクチンの候補の探索で得られた研究株の弱毒化は、それらのワクチン株が細胞死を惹起する能力と常に関連する。
【0036】
所与のウイルスによる感染細胞の破壊は、特異的なシグナル伝達経路の撹乱(perturbation)があることを示唆している。
【0037】
狂犬病ウイルスは、エンベロープ型のRhabdoviridae 科、Lyssavirus属に属している弾丸型のウイルスである。ウイルス粒子は、宿主の脂質から構成される膜およびラセン状のヌクレオカプシド(NC)を囲む二つのウイルス蛋白質G および M 蛋白質からなる。NCは、N 蛋白質, P 蛋白質, および RNA-依存性RNAポリメラーゼ, L 蛋白質により保護されるウイルスのネガティブ鎖RNA分子から構成される。狂犬病ウイルス 蛋白質は、感染細胞中で等しい量で合成されない、またウイルス粒子中で同じ比で存在しない。実際、N, G および M 蛋白質は、この順序でビリオン中で最も顕著な種である。
【0038】
ニューロンネットワークの統合性 (狂犬病ウイルス伝播に有利) および ニューロンの細胞死(免疫原性に有利)の保存の間の戦略的な決定は、必然的に少なくとも二つのシグナル伝達のネットワーク間の選択を伴う。
【0039】
狂犬病ウイルス株のG蛋白質の細胞質尾部が、生存の支持(即ち、ニューロンネットワーク統合性の保存であり、狂犬病ウイルス伝播を支持する) 対 アポトーシスの支持(即ち、細胞死であり、免疫原性を支持する)の決定プロセスの指令に重大な役割を有することの実証を本発明は提供する。
【0040】
さらに本発明は、生存支持戦略(pro-survival strategy)の予想外の効果を明らかとする。
【0041】
実際, 本発明は、幾つかの非アポトーシス性(且つ病原性)の狂犬病ウイルス株(より具体的には、CVS-NIV株)のG 蛋白質が神経突起成長促進効果を有すること、即ち、これらの非アポトーシス性の狂犬病ウイルス G 蛋白質が神経突起生成を誘導および/または刺激することを示す。
【0042】
この効果は、アポトーシス性の 狂犬病ウイルス G 蛋白質(例えば、ERA および 他の弱毒化した狂犬病ウイルス株のG 蛋白質)により示されない。
【0043】
さらにまた、この効果は、全てのアポトーシス性および/または病原性の狂犬病ウイルス G 蛋白質により示されない。より具体的には、この効果はCVS-Gif-sur-Yvette 株のG蛋白質では示されないにもかかわらず(Prehaud et al. 1988)、このG蛋白質はCVS-NIV 株のG蛋白質と高い同一性 スコアを有する(図 17を参照されたい)。前記効果は、CVS-11 株(例えば、ACA57830, AAC34683, ABV24348)の G 蛋白質では示されないにもかかわらず、これらの G 蛋白質もCVS-NIV 株のG蛋白質と高い同一性スコアを有する(図 18を参照されたい)。N2CまたはCVS-24 株のG 蛋白質のどちらでも前記効果は示されない。
【0044】
従って、病原性の狂犬病ウイルス株の全てとは限られない株が、神経突起成長において有意に陽性の効果を示すG蛋白質を有する。
【0045】
このようなことから、発明者は、非アポトーシス性(且つ病原性)の狂犬病ウイルス株の亜群を同定した、この株のG蛋白質は神経突起成長に有意に陽性の効果を有する。この亜群の代表的な株は、CVS-NIV 株である。これらの G 蛋白質は、本明細書等においてCVS-NIV タイプのG 蛋白質と参照されえる。
【0046】
CVS-NIV 株は、2009年4月1日にCNCMに受託番号I-4140で寄託された。
【0047】
CVS-NIV 株のG蛋白質を発現しているプラスミドは、2001年11月30日にCNCMに受託番号I-2578で寄託された。
【0048】
CVS-NIV 株のG蛋白質は、Prehaud 等2003に記載されている。
【0049】
CVS-NIV 株のG蛋白質の配列は、アクセッション番号AF 406694で利用可能である。
【0050】
CVS-NIV 株のG蛋白質の配列は、図 13Aに示される配列番号 2の配列である。
【0051】
狂犬病ウイルス G蛋白質は、ウイルスエンベロープの三量体スパイクを形成し、感染細胞の膜に見出されるI型 膜貫通糖蛋白質である。狂犬病ウイルス G蛋白質は、外部ドメイン, 膜貫通セグメント および 44アミノ酸の長い細胞質ドメインからなる524アミノ酸長の蛋白質である。
【0052】
このC末端ドメインのまさに最後の 4 アミノ酸が、PDZ 結合サイト (PDZ-BS)を形成する。
【0053】
PDZ (PSD-95, Discs Large, ZO-1)ドメインは、パートナー蛋白質のC 末端配列が挿入されえるソケットを作る六つのベータ鎖および二つのアルファヘリックスに組織化される80-100 aaの球状の構造を形成する。
【0054】
PDZ 結合サイト (PDZ-BS)は4 アミノ酸配列であり; PDZ-BS 配列は配列番号 13の配列である、すなわち:
x1 - x2 - x3 - x4,であり、式中の:
x1 は、任意のアミノ酸であり、
x2 は、TまたはSまたはIであり、
x3 は、任意のアミノ酸であり、
x4 は、LまたはVである。
【0055】
特に、本発明は、この神経突起成長効果を担うのがCVS-NIV タイプの前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部であることを実証する。
【0056】
さらに、本発明は、この効果が特にPDZ-BSが原因であることを実証する。PDZ-BSは、CVS-NIV タイプの前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部に含まれる。
【0057】
CVS-NIV タイプの非アポトーシス性の狂犬病ウイルス G 蛋白質のPDZ-BSモチーフは、アポトーシス性の狂犬病ウイルス G 蛋白質の一つと比べて単一の点変異を示す。この単一の点変異は最初のPDZ-BS アミノ酸に関し、これは非アポトーシス性の 狂犬病ウイルス G 蛋白質においてEではない。
【0058】
さらに、本発明は、前記アミノ酸がCVS-NIV 株の全長の非アポトーシス性の狂犬病ウイルス G蛋白質配列においてポジション 491に存在し、これが神経突起成長効果に寄与することを実証する。このアミノ酸は、CVS-NIV 株のG蛋白質中でHである(配列番号 2を参照されたい)。全長 G蛋白質におけるポジション 491は、この蛋白質の細胞質断片におけるポジション 11に対応する(図 15の配列番号 6)。
【0059】
CVS-NIV 株の全長の非アポトーシス性の狂犬病ウイルス G蛋白質配列(配列番号 2)中のポジション 491 および 521のアミノ酸は、双方が本発明の神経突起成長効果に重要であることを発明者は実証する。より具体的には、発明者は、ポジション 491でのH (しかし、Lではない)およびポジション 521でのQ (しかし、Eではない)は、双方が本発明の神経突起成長効果を支持することを実証する。
G 蛋白質がH491Lの単一の点変異を生じる又はQ521Eの単一の点変異を生じるCVS-NIV 株と異なる変異体ウイルスが、発明者により構築され、作出された(CNCM I-4142 および I-4143)。
【0060】
H491L 変異体 G蛋白質は、なおもポジション 521にQを有する。
【0061】
Q521E 変異体 G蛋白質は、なおもポジション 491にHを有する。
【0062】
双方の変異体蛋白質は、双方がH をポジション 491にQ をポジション 521に有し、CVS-NIV G蛋白質よりも小さい程度ではあるが、有意に陽性の神経突起成長効果を誘導する。
【0063】
これらの変異体 G 蛋白質の細胞質断片は、図 19 (配列番号 19 および 配列番号 20)に示される。
【0064】
本発明のポリペプチドの規定:
本発明は、以下のポリペプチドに関する:
・ 以下を含むポリペプチド:
・ 配列番号 6の配列, 株 I-4140のG 蛋白質の細胞質断片, プラスミド I-2578により産生されたG 蛋白質の細胞質断片, 配列 AF406694のG 蛋白質の細胞質断片, 配列番号 2のG 蛋白質の細胞質断片から選択される親配列,
又は
・ 前記親配列の保存されたバリアント配列,
又は
・ 前記親配列の又は前記保存されたバリアント配列の保存された断片。
【0065】
前記親配列, 保存されたバリアント配列および保存された断片配列は、有利に神経突起成長を誘導する及び/又は刺激する機能を有し、例えば、以下に記載されるように、より具体的には、ヒトの分裂前のニューロンからの〔例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの〕神経突起成長を誘導する及び/又は刺激する機能を有する。
【0066】
より具体的には、本発明は、神経突起成長の刺激および/または誘導に使用するための〔より具体的には、不十分な又は障害性の神経突起成長、特に不十分な又は障害性の神経突起生成に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドに関する。
【0067】
好ましくは、前記ポリペプチドのアミノ酸長は、100 アミノ酸未満, より好ましくは90 アミノ酸未満である。
【0068】
前述のバリアント蛋白質の配列は、任意の既知の又は同定可能な狂犬病ウイルス G蛋白質に対応しない配列であってもよい。
【0069】
実際、通常の能力を発揮する当業者は、所与の狂犬病ウイルス G蛋白質の配列から出発して、一つ又は幾つかのアミノ酸の置換および/または付加および/または欠失をなすことができ、なおも神経突起成長を誘導する及び/又は刺激する能力を保持していることを認識する。かかる保存されたアミノ酸の置換および/または付加および/または欠失は、本発明に包含される。
【0070】
理論に縛られることなく、次の八つのグループの各々は、通常互いに保存された置換であるアミノ酸を含んでいると考えられる:
1) アラニン (A), グリシン (G);
2) アスパラギン酸 (D), グルタミン酸 (E);
3) アスパラギン (N), グルタミン (Q);
4) アルギニン (R), リジン (K);
5) イソロイシン (I), ロイシン (L), メチオニン (M), バリン (V);
6) フェニルアラニン (F), チロシン (Y), トリプトファン (W);
7) セリン (S), スレオニン (T); および
8) システイン (C), メチオニン (M)。
【0071】
以上のように、前述のバリアント蛋白質は、特に、人間により操作され、一もしくは数個のアミノ酸の置換および/または付加および/または欠失により前記狂犬病ウイルス G 蛋白質と異なる蛋白質を包含する〔但し、生じるバリアント蛋白質は、神経突起成長、より具体的には、ヒトの分裂前のニューロンから、例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長を誘導する及び/又は刺激する能力をなおも有する〕。
【0072】
好ましくは, 前記保存されたバリアント配列は以下の配列である:
・ 前記親配列のバリアント配列, この配列は:
・ 34〜54アミノ酸の配列であり,
・ 親およびバリアントの配列のうちの最も短い配列の全長(entire length)に対し前記親配列と少なくとも 94% 同一であり,
・ PDZ-BS配列 (好ましくは、最後の四つのC末端のアミノ酸)を含み, 前記PDZ-BS配列は以下のものであり:
x1 は、E以外の任意のアミノ酸(好ましくは、Q)であり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)であり
(配列番号14),
前記バリアント配列はバリアント 配列 Aと称される,
又は
・ 前記親配列のバリアント配列, この配列は:
・ 44 アミノ酸の配列であり,
・ 前記親配列と少なくとも 94% 同一であり,
・ PDZ-BS配列 (好ましくは、最後の四つのC末端のアミノ酸)を含み, 前記PDZ-BS配列は以下のものであり:
x1 は、Eであり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)であり
(x1 = Eの配列番号 13),
並びに
・ アミノ酸 L をポジション 11に含まなく(好ましくは、アミノ酸 H をこのポジションに含む),
前記バリアント配列はバリアント 配列 Bと称される。
【0073】
好ましくは、前記配列同一性は、少なくとも 95% (好ましくは、少なくとも 96%, より好ましくは、少なくとも 97%, なおより好ましくは、少なくとも 97.5%, さらになおより好ましくは、少なくとも 98%, 最も好ましくは、少なくとも 98.5%, なお最も好ましくは、少なくとも 99%)である。
【0074】
より好ましくは, 前記保存されたバリアント配列は、バリアント 配列 Aである。
【0075】
好ましくは, 前記保存された断片は、それぞれ前記親配列の又は前記バリアント配列 Aの又は前記バリアント配列 Bの保持されたPDZ-BS 配列を有する少なくとも 34 アミノ酸断片である。
【0076】
簡潔さのため、「ポリペプチド」の用語は、本明細書等において蛋白質が包含されることを意図しており、また逆に。
【0077】
この用語は、転写後の修飾及び/又は合成化学〔例えば、非ペプチド性の化学基の付加により及び/又はポリペプチドの三次構造の修飾により、例えば、アセチル化, アシル化, ヒドロキシル化, 環化, ラセミ化, リン酸化, など〕により修飾され、生じる修飾されたポリペプチドが神経突起成長〔より具体的には、ヒトの分裂前のニューロンからの、例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する能力を保持するポリペプチド(または蛋白質)も包含する。
【0078】
有利なことに、本発明のポリペプチドは、それ自身で神経向性(neurotropic)である。
【0079】
それでも、必要とされる又は所望される場合、本発明のポリペプチドは、神経向性(より具体的には、脳のニューロンへの向性)を改善する因子〔例えば、Kumar等(Kumar et al. 2007)により記載されたRVG-9R〕と連結できる又は融合できる。
【0080】
アミノ酸配列およびヌクレオチド配列は、標準の配向(即ち、アミノ酸配列に関してN-末端からC-末端、またヌクレオチド配列に関して5'-末端から3'-末端)にしたがって本明細書等に提供される。
【0081】
本発明の態様によると、前記バリアント配列 AのPDZ-BS 配列は、QTRL (配列番号 10)である。
【0082】
本発明の態様によると、前記バリアント 配列 Aは、44 アミノ酸の配列であり、アミノ酸 HまたはL, 好ましくは Hをポジション 11に有する。
【0083】
例示のバリアント 配列 Aは、44アミノ酸の配列であり、アミノ酸 L をポジション 11に有する配列番号 19の配列である(図 19を参照されたい)。
【0084】
本発明の態様によると、前記バリアント配列 BのPDZ-BS 配列は、ETRL (配列番号 12)である。
【0085】
本発明の態様によると、前記バリアント 配列 Bは、アミノ酸 H をポジション 11に有する。
【0086】
例示のバリアント 配列 Bは、44アミノ酸の配列であり、アミノ酸 H をポジション 11に有する配列番号 20の配列である(図 19を参照されたい)。
【0087】
本発明の態様によると、前記断片は、配列番号 6の配列の断片 11〜44である。
【0088】
本発明のポリペプチドは、神経突起成長支持配列およびG 狂犬病ウイルス 蛋白質の膜貫通部分または当業者が細胞(例えば、以下に規定される細胞)の膜上又は膜中に前記ポリペプチドをアンカーするため適切なものを見出しえる任意の他のアンカーする手段を含んでもよい又は該手段からなってもよい。
【0089】
本発明のポリペプチドは、操作された抗体(例えば、単鎖抗体)の構造中に挿入できる。
【0090】
本発明の態様によると、前記ポリペプチドは、狂犬病ウイルス株のG蛋白質, 又はその細胞質断片(例えば、配列番号 6), 又は係る細胞質断片の小断片(例えば、配列番号 6の断片11〜54)である。前記狂犬病ウイルス G蛋白質は、より好ましくは非アポトーシス性の狂犬病ウイルス G蛋白質である。例えば、前記 狂犬病ウイルス株は、CNCMにI-4140で寄託された株である。
【0091】
本発明の態様によると、前記ポリペプチドは、係る細胞質断片または小断片のバリアント 配列 A またはB (上記規定のとおり)を含む又はそれからなる。例えば、前記 ポリペプチドは、G 蛋白質がI-4140 株のG 蛋白質の単一の点変異体蛋白質であるCNCMにI-4142又はI-4143で寄託された狂犬病ウイルス株のG 蛋白質の細胞質断片(図 21を参照されたい)を含む又は該断片からなる(それぞれ配列番号 19 および 20の細胞質断片; 図19を参照されたい)。
【0092】
CNCMは、Collection Nationale de Cultures de Microorganismes(Collection Nationale de Cultures de Microorganismes ; Institut Pasteur ; 28, rue du Docteur Roux ; F-75724 Paris Cedex 15 ; France)である。前記寄託は、ブタペスト条約の条件下でなされた。
【0093】
バリアント配列 A:
本発明のバリアント 配列 Aは配列番号 14の配列、即ち、配列 x1 - x2 - x3 - x4,を含み、式中の:
x1 は、E以外の任意のアミノ酸であり、
x2 は、TまたはSまたはIであり、
x3 は、任意のアミノ酸であり、
x4 は、LまたはVである。
【0094】
この配列は、特定の PDZ-BS モチーフである。この配列は通常は前記ポリペプチドのC末端に局在する。通常、この配列は前記ポリペプチドの最後の四つのC末端アミノ酸の配列である。
【0095】
好ましくは, 前記アミノ酸 x1 は、Qである。
【0096】
好ましくは, 前記アミノ酸 x2 は、Iではなく、好ましくは, Tである。
【0097】
好ましくは, 前記アミノ酸 x3 は、Rである。
【0098】
好ましくは, 前記アミノ酸 x3 は、Lである。
【0099】
好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、TまたはSまたはIであり、
x3 は、Rであり、
x4 は、LまたはVである。
【0100】
最も好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、Tであり、
x3 は、Rであり、
x4 は、LまたはVである。
【0101】
最も好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、TまたはSまたはIであり、
x3 は、Rであり、
x4 は、Lである。
【0102】
より好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、Tであり、
x3 は、Rであり、
x4 は、Lである、
即ち、これは配列番号 10の配列である(図 16を参照されたい)。
【0103】
このQTRL配列は、CVS-NIV 株(前記株は受託番号I-4140でCNCMから利用可能である)のG蛋白質のPDZ-BS モチーフの, および/または受託番号 I-2758でCNCMから利用可能であるプラスミド(組換え型の 大腸菌含んでいる 前記 プラスミド)によりコードされた蛋白質の, および/または配列番号 2の蛋白質(図 13Aを参照されたい)の配列である。
【0104】
CVS-NIVのG蛋白質をコードしているプラスミド CNCM I-2758を含んでいる組換え型の 大腸菌株を培養するための適切な条件には、標準のLB-TYM 成長培地(アンピシリンの存在下)で前記組換え型の 大腸菌株を37°Cでインキュベーションすることを含む(WO 03/048198を参照されたい)。
【0105】
ウイルス I-4140 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0106】
好ましくは、本発明のバリアント 配列 Aは、44 アミノ酸の配列であり、アミノ酸HまたはL(好ましくは H)を44aa-配列のポジション 11に有する。
【0107】
例示のバリアント A ポリペプチドは、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株(その株のG 蛋白質はI-4140のG 蛋白質の単一の点変異体蛋白質である; 図 21を参照されたい)のG 蛋白質, その細胞質断片およびその保存された細胞質小断片を含む。
【0108】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0109】
ウイルス I-4142 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0110】
例示のバリアント A ポリペプチドは、特に配列番号 19の細胞質断片又は少なくとも一つのその保存された小断片を含んでいるポリペプチドを含む。
【0111】
配列番号 19の配列は、配列番号 6の配列の例示の保存されたバリアントである(図 19を参照されたい)。配列番号 19の配列は配列番号 6の配列よりも低いレベルにもかかわらず、陽性の神経突起成長効果をなおも有する(図 3D および例のセクションにおける関連するコメントを参照されたい)。
【0112】
バリアント 配列 B:
本発明のバリアント 配列 Bは、44 アミノ酸の配列である。
【0113】
この配列は、配列番号 13にx1 = Eを有する配列である、即ち、配列 x1 - x2 - x3 - x4を含み、配列中の:
x1 は、Eであり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)である。
【0114】
この配列は、特定の PDZ-BS モチーフである。この配列は、通常は前記ポリペプチドのC末端に局在する。通常、この配列はポリペプチドの最後の四つのC末端アミノ酸の配列である。
【0115】
好ましくは, 配列番号 13の配列は、ETRLである(配列番号 12)。
【0116】
本発明のバリアント 配列 Bにおいて、ポジション 11にあるアミノ酸は、Lではなく、最も好ましくはHである。
【0117】
例示のバリアント B ポリペプチドは、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株(そのG 蛋白質はI-4143のG 蛋白質の単一の点変異体蛋白質である; 図 21を参照されたい)のG 蛋白質, その細胞質断片およびその保存された細胞質小断片を含む。
【0118】
I-4143 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 E をポジション 521 (全長のG蛋白質に関して計算したポジション)に Q の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0119】
ウイルス I-4143 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0120】
例示のバリアント B ポリペプチドは、特に配列番号 20の細胞質断片又は少なくとも一つのその保存された小断片を含んでいるポリペプチドを含む。
【0121】
配列番号 20の配列は、例示の配列番号 6の配列の保存されたバリアントである(図 19を参照されたい)。配列番号 20の配列は配列番号 6の配列よりも低いレベルにもかかわらず、陽性の神経突起成長効果をなおも有する(図 3D および例のセクションにおける関連するコメントを参照されたい)。
【0122】
I-4142 および I-4143 株のG 蛋白質は、CVS-NIV 株 (I-4140)のG蛋白質の一つよりも有意に神経突起成長効果が低いが、有意に陽性の神経突起成長効果を誘導する(例 1 および 図 3Dを参照されたい)。
【0123】
従って、二つのアミノ酸ポジションの各々は、神経突起成長効果に寄与し、そのポジションは、即ち:
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、ポジション 491 (即ち、CVS-NIV 株のG蛋白質におけるH)であり、
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、CVS-NIV 株のG 蛋白質におけるポジション 521であり、即ち、G 蛋白質のPDZ-BSの最初のアミノ酸 (即ち、CVS-NIV 株のG蛋白質におけるアミノ酸 Q)である。
【0124】
本発明のポリペプチドの代替の又は相補的な規定:
本発明のポリペプチドは、神経突起成長支持ポリペプチド(pro-neurite outgrowth polypeptide)である。
【0125】
本発明のポリペプチドは、上記規定の親配列および/または保存されたバリアント配列(バリアント配列AまたはB)および/または保存された断片配列を含む。
【0126】
択一的に又は相補的に(Alternatively or complementarily)、本発明のポリペプチドは、狂犬病ウイルス株のG蛋白質, 又はそのバリアント配列のG蛋白質(このバリアント配列は前記G蛋白質から一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来する),または係るG蛋白質またはバリアントG蛋白質の断片配列, より具体的には、これらの細胞質断片または小断片であると規定できる。但し、前記ポリペプチドは、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を有する。
【0127】
より具体的には、本発明のポリペプチドは、択一的に又は相補的に、非アポトーシス性の狂犬病ウイルス株のG蛋白質, 又はそのバリアント配列の蛋白質(このバリアント配列は前記G蛋白質から一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来する),または係るG蛋白質またはバリアント蛋白質の断片配列, より具体的には、これらの細胞質断片または小断片であると規定できる。但し、前記バリアント配列または前記断片は、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を保持する。
【0128】
CVS-NIV株のG 蛋白質ではない及び/又はCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質以外である, および/または配列番号 2の蛋白質以外であるが、しかし、なおも適切な神経突起成長支持ポリペプチドである例示の神経突起成長支持蛋白質は、特に2009年4月1日にCNCMに受託番号I-4142 および I-4143で寄託された組換え型の狂犬病ウイルス株のG 蛋白質を含む。
【0129】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0130】
I-4143 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 E をポジション 521 (全長のG蛋白質に関して計算したポジション)に Q の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0131】
I-4142 および I-4143 株のG 蛋白質は、CVS-NIV 株 (I-4140)のG蛋白質の一つよりも有意に神経突起成長効果が低いが、有意に陽性の神経突起成長効果を誘導する。
【0132】
従って、二つのアミノ酸ポジションの各々は、神経突起成長効果に寄与し、そのポジションは、即ち:
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、ポジション 491 (即ち、CVS-NIV 株のG蛋白質におけるH)であり、
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、CVS-NIV 株のG 蛋白質におけるポジション 521であり、即ち、G 蛋白質のPDZ-BSの最初のアミノ酸(即ち、CVS-NIV 株のG蛋白質におけるアミノ酸 Q)である。
【0133】
それ故, 例示の神経突起成長支持 ポリペプチドは、I-4142 株のG蛋白質およびI-4143 株のG蛋白質、同様に、これらの蛋白質の断片、より具体的には、これらの細胞質断片(図 19における配列番号 19および配列番号 20)及びこれらの細胞質小断片を含む。但し、これらの小断片は、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を保持する。
【0134】
より具体的には、本発明は、神経突起成長を刺激および/または誘導することに使用するための〔より具体的には、不十分な又は障害性の神経突起成長、特に不十分な又は障害性の神経突起生成に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドに関する。
【0135】
好ましくは、前記ポリペプチドのアミノ酸長は、100 アミノ酸未満, より好ましくは90 アミノ酸未満である。
【0136】
「非アポトーシス性の狂犬病ウイルスG蛋白質(non-apoptotic rabies virus G protein)」の表現は、本明細書等において当該分野の通常の意味を意図している。
【0137】
非アポトーシス性の狂犬病ウイルスG蛋白質のPDZ-BSは、配列番号 14のものである。
【0138】
非アポトーシス性の狂犬病ウイルスG蛋白質は、その配列が狂犬病ウイルス株のG蛋白質の配列であること、ヒトニューロンのアポトーシスを惹起しないことの事実により付加的に又は択一的に特徴付けられることができる。
【0139】
候補の狂犬病ウイルス G蛋白質が非アポトーシス発生性(non-apoptogenic)のものであることを検査する例示の手段は、当業者に知られている。
【0140】
その手段の一つには、この G蛋白質を含み、ニューロン, より具体的には、 ヒトニューロン, 例えば、SK-N-SH 神経芽細胞腫 細胞株 (ATCC HTB11)またはSH-SY5Y 神経芽細胞腫 細胞株 (ATCC CRL-2266), 好ましくは SK-N-SH 神経芽細胞腫細胞株(ATCC HTB11)に感染する際に非アポトーシス性である狂犬病ウイルス株を検査することが含まれる。かかる手段は、候補G蛋白質を含んでいる天然の狂犬病ウイルス株が分析のため利用可能である場合に特に有用である。
【0141】
他の手段には、遺伝的に細胞を操作して候補 G蛋白質を発現させること、ニューロン細胞を前記の発現された候補G蛋白質で感染させること, およびアポトーシスが前記感染により誘導されないことを決定することが含まれる。
【0142】
かかる手段の例示は、Prehaud等(Prehaud et al. 2003)に記載される。
【0143】
例示の遺伝的に操作した細胞には、Prehaud et al.2003, より具体的には、10538頁(「誘導性のトランスジェニック細胞株」の部分を参照されたい)に記載されるトランスジェニックJurkat細胞系統が含まれる。
【0144】
例示のニューロン細胞には、SK-N-SH 神経芽細胞腫細胞株 (ATCC HTB11)またはSH-SY5Y 神経芽細胞腫細胞株 (ATCC CRL-2266), 好ましくはSK-N-SH 神経芽細胞腫細胞株 (ATCC HTB11)が含まれる。
【0145】
ニューロン細胞を感染させる遺伝的に操作した細胞を作るための例示の実験条件は、Prehaud等 (Prehaud et al.2003), より具体的には、10538頁(「誘導性のトランスジェニック細胞株」の部分を参照されたい)に記載されるものを含む。
【0146】
狂犬病ウイルス株または遺伝的に操作された細胞がニューロン細胞のアポトーシスを誘導しないことの検出は、当業者の通常の技術の範囲内である。
【0147】
例示の手段は、Prehaud等 (Prehaud et al. 2003)およびPrehaud等 (Prehaud et al. 2005)に記載のものを含む。
【0148】
例示の手段は、例えば、ヘキスト染色 (Prehaud et al. 2003, より具体的には、10538頁, § 「Detection of nuclear fragmentation by Hoechst staining」を参照されたい), および/またはTUNEL 法(Prehaud et al. 2003, より具体的には、10538頁を参照されたい(§ 「Detection of nuclear fragmentation by the TUNEL method」を参照されたい), および/またはDNA電気泳動法 (Prehaud et al. 2005を参照されたい)によりDNA 断片化が誘導されないことを検出することを含む。
【0149】
有意なDNA 断片化が誘導されないことを検出する付加的な又は代替の例示の手段は、例えば、Prehaud et al. 2003(より具体的には、10538頁, 「Detection of caspase activation」)に記載の手順にしたがってカスパーゼ 8が活性化されないことの検出を含む(但し、使用されたニューロン細胞がカスパーゼ 8を含むべきであり、ここではSK-N-SH 細胞株の場合である)。
【0150】
非有意なアポトーシスの例示の値は、Prehaud等 2003(「JrtTA-G-CVS」の表題の最後から二番目の欄を参照されたい)の図 5Dに示される以下の事項である:
・ ヘキスト染色により測定される僅か19%のアポトーシス細胞;
・ カスパーゼ 8の活性化の評価により測定される僅か4.7%のアポトーシス細胞。
【0151】
弱毒化した株(即ち、非病原性、例えば、免疫応答性マウスの筋肉内に注射された際に非病原性)である狂犬病ウイルス株、例えば、弱毒化したERA株, RV194-2 株, AVO-1 株, SN10 株, SN-10-SAD 株, SAG2 株の狂犬病ウイルス株は、アポトーシス発生性の株である。
【0152】
病原性(即ち、インビボ神経毒性)の狂犬病ウイルス株(例えば、CVS-NIV 株)は、非アポトーシス性(non-apoptotic)である。
【0153】
従って、非アポトーシス発生性の狂犬病ウイルス株は、病原性の(神経毒性の)株である(例えば、免疫応答性マウスの筋肉内に注射された際に病原性である)。
【0154】
現在の知識は、以下のものである:
・ 非アポトーシス発生性の狂犬病ウイルス株がヒトニューロン細胞に感染する際に、それがコードするG蛋白質は、核周囲に球状構造の形態で前記ニューロン細胞の細胞質に蓄積し、前記ニューロン細胞の細胞質に拡散して分布するのではなく;
・ それと反対に、アポトーシス発生性の狂犬病ウイルス株がヒトニューロン細胞に感染する際に、それがコードするG蛋白質は、核周囲に球状構造の形態で前記ニューロン細胞の細胞質に蓄積しないが、しかし、前記ニューロン細胞の細胞質に拡散して分布する。
【0155】
上述し且つ以下に説明されるとおり、本発明のポリペプチドは、狂犬病ウイルス株のG蛋白質であってもよい。
【0156】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CVS-NIV 株のG蛋白質〔前記株は、CNCMから受託番号I-4140 (寄託日は2009年4月1日である)で利用可能である〕, および/またはCNCMから受託番号I-2758(寄託日は2001年11月30日である)で利用可能なプラスミドによりコードされた蛋白質および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり、前記バリアント狂犬病ウイルス G 蛋白質のPDZ-BS モチーフの配列はなおも配列番号 14の配列である蛋白質。
【0157】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって:
・ 前記バリアント蛋白質は、i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも95%同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列からなり、
・ 前記バリアント 狂犬病ウイルス G蛋白質のPDZ-BS モチーフの配列は、配列番号 14の配列である蛋白質。
【0158】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり:
・ 前記バリアント蛋白質は、i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも95%同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列からなり、
・ 前記バリアント 狂犬病ウイルス G蛋白質のPDZ-BS モチーフの配列は、配列番号 14の配列である蛋白質。
【0159】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり、前記バリアント蛋白質は以下の配列からなるバリアント蛋白質:
・ i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも 95% 同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列, および
・ i.の前記狂犬病ウイルス G蛋白質の配列番号 14のPDZ-BS モチーフを保持する及び/又はQTRLの配列のPDZ-BS モチーフ(x1=Q; x2=T; x3=R; x4=Lを有する配列番号 14)を含む配列。
【0160】
好ましくは、ii.の前記バリアント狂犬病ウイルス G蛋白質のアミノ酸長は、i.の前記狂犬病ウイルス G蛋白質の長さ50 アミノ酸以上超えない。
【0161】
好ましくは、ii.の前記バリアント狂犬病ウイルス G蛋白質のアミノ酸長は、i.の前記狂犬病ウイルス G蛋白質の長さより50 アミノ酸以上短くない(例えば、iの前記 狂犬病ウイルス G蛋白質の配列と同じ長さである)。
【0162】
より好ましくは、ii.の前記バリアント狂犬病ウイルス G 蛋白質が含むアミノ酸の長さは、少なくともi.の前記狂犬病ウイルス G 蛋白質の長さマイナス50 アミノ酸であり、多くともi.の前記狂犬病ウイルス G 蛋白質の長さプラス 50 アミノ酸である(例えば、iの前記狂犬病ウイルス G 蛋白質の配列と同じ長さである)。
【0163】
CVS-NIV株(I-4140)のG 蛋白質ではない及び/又はCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質以外である, および/または配列番号 2の蛋白質以外であるが、しかし、なおも適切な神経突起成長支持狂犬病ウイルス G 蛋白質である例示の神経突起成長支持狂犬病ウイルス G 蛋白質は、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株のG 蛋白質を含む。
【0164】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0165】
配列番号 19の配列の例示の断片(図 19を参照されたい)。
【0166】
付加的に存在しえる他の配列:
上述し且つ以下に説明されるとおり、本発明のポリペプチドは、神経突起成長支持〔および/または神経突起の発芽(spouting)および/または軸索の成長および/または樹状突起樹の伸展(dendritic tree extension)〕の効果を有するアミノ酸配列を含む。
【0167】
上述し且つ以下に説明されるとおり、前記神経突起成長支持配列は、上記規定のとおり、親配列 および/または 保存されたバリアント配列(バリアント配列 AまたはB)および/または 保存された断片配列と規定できる。
【0168】
上述し且つ以下に説明されるとおり、前記神経突起成長支持配列は、択一的に又は相補的に(alternatively or complementarily)、(非アポトーシス性の) 狂犬病ウイルス株 (例えば、CNCM I-4140, I-4142またはI-4143のG 蛋白質)のG 蛋白質の配列と,または一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来するそのバリアント配列,または係る G 蛋白質またはバリアント G 蛋白質の断片配列, より具体的には、その細胞質断片または小断片と規定できる。
【0169】
G 蛋白質またはバリアント G 蛋白質の係る断片の例示は、特に前記 G 蛋白質またはバリアント G 蛋白質の細胞質ドメイン および 膜貫通ドメインを保持するその断片を含む。
【0170】
係る膜貫通ドメインは、特に前記細胞の細胞内区画において(より具体的には、前記細胞の小胞体および/またはゴルジ膜における)細胞質断片をアンカーして、前記細胞質断片が神経発生, 神経再生 および 神経保護の刺激および/または誘導する効果をより効率的に発揮させるとの利点を有する。以下の例4 および 5を参照されたい。
【0171】
上述のことから、本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜において前記ポリペプチドをアンカーする配列を含んでもよく、前記アンカーする配列は好ましくは前記神経突起成長支持配列のN末端にあり, 最も好ましくは前記神経突起成長支持配列のN末端での第一のアミノ酸に直接的に連結される。
それ故, 本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株、例えば、例4)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜(好ましくは、小胞体膜及びゴルジ膜)において前記ポリペプチドをアンカーするアミノ酸配列を含んでもよい。
【0172】
前記アンカーする配列は、好ましくは配列番号 6の配列またはバリアント 配列 Aまたはバリアント 配列 BのN末端に存在する。最も好ましくは, 前記アンカーする配列は、直接的もしくは間接的に(好ましくは、直接的に)配列番号 6の配列の又は前記バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結される。
【0173】
係るアンカーする配列は、例えば、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例、例えば、例4に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜において配列番号 6の細胞質ドメインをアンカーする能力を有するアミノ酸配列であってもよい。
【0174】
係るアンカーする配列の例示は、狂犬病ウイルス株(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメイン配列、好ましくは狂犬病ウイルス CVS-NIVのG蛋白質の膜貫通ドメイン配列(配列番号 23)である。
【0175】
上述のことから、前記ポリペプチドは、配列番号 6のまたは前記バリアント配列 Aの又は前記バリアント配列 Bの配列に加えて、さらに好ましくは以下の配列を、配列番号6の配列またはバリアント配列 Aまたはバリアント配列 BのN末端に含んでもよく, 最も好ましくは直接的に配列番号6の配列または前記 バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結されてもよい:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメインの配列, または
・ 狂犬病ウイルス G の膜貫通ドメインの配列と少なくとも一つのアミノ酸の置換および/または欠失および/または付加, 好ましくはアミノ酸の置換および/または欠失, より好ましくはアミノ酸置換により異なるが、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例、例えば、例4に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株)の小胞体膜および/またはゴルジ膜において、前記 ポリペプチドをアンカーする能力(より具体的には、前記アンカーする配列が配列番号 6-の前記配列のN末端で第一のアミノ酸に直接的に連結される場合、配列番号 6 の細胞質ドメインをアンカーする能力)を保持する配列。
【0176】
それ故, 本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに好ましくは以下の配列を、配列番号6の配列またはバリアント配列 Aまたはバリアント配列 BのN末端に含んでもよく, 最も好ましくは直接的に配列番号6の配列または前記 バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結されてもよい:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメインの配列, または
・ 狂犬病ウイルス G 膜貫通ドメインの配列と少なくとも一つのアミノ酸の置換および/または欠失および/または付加, 好ましくはアミノ酸の置換および/または欠失, より好ましくはアミノ酸置換により異なるが、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)の小胞体膜および/またはゴルジ膜において、(前記アンカーする配列が配列番号 6の配列のN末端で最初のアミノ酸に直接的に連結される場合に)配列番号 6のポリペプチドをアンカーする能力を保持する配列。
【0177】
係るアンカーする配列の好適な例は、特に以下を含む:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株、なおより具体的にはCVS-NIV 株)のG蛋白質の膜貫通ドメインの配列(例えば、配列番号 23の配列), または
・ そのバリアント配列であって、18〜26 アミノ酸, 好ましくは18 〜22 アミノ酸, より好ましくは22 アミノ酸の配列であり、二配列のうちの最短の配列にわたって(即ち、配列番号 23及び前記バリアント配列のうちの最短の配列にわたって)配列番号 23の配列と少なくとも 94% 同一である配列。
【0178】
係るアンカーする配列は、配列番号 6の配列または前記バリアント配列 A(より具体的には、配列番号 6の配列)である神経突起成長支持配列に連結される場合に特に有用である。
【0179】
それ故, 本発明の態様によると、本発明のポリペプチドのアミノ酸配列は、配列番号23の配列にN末端からC末端に配列番号6の配列が続く配列を含むか,または該配列からなり、1〜4アミノ酸, 好ましくは1アミノ酸(例えば、M)が任意で配列番号23の配列と配列番号6の配列との間に存在する。
【0180】
係るポリペプチドの例示は、配列番号 25のポリペプチドである。
【0181】
本発明のポリペプチドは、神経突起成長支持配列に加えて、さらに以下を含んでもよい:
・ 狂犬病ウイルス株(より具体的には非アポトーシス性の狂犬病ウイルス株, なおより具体的にはCVS-NIV 株)のG蛋白質の外部ドメインの断片, および/または
・ シグナルペプチド〔このシグナルペプチドは、好ましくは狂犬病ウイルス株(より具体的には非アポトーシス性の狂犬病ウイルス株, なおより具体的にはCVS-NIV 株)のG蛋白質のシグナルペプチドである〕。
【0182】
前記外部ドメイン断片および/またはシグナルペプチドは、前記アンカーする配列に加えて本発明のポリペプチドに存在してもよい。
【0183】
好ましくは, シグナルペプチドが前記ポリペプチドに含まれる場合、前記シグナルペプチドは前記ポリペプチドのN末端の最初の構成要素である。
【0184】
好ましくは, シグナルペプチドおよび外部ドメイン断片が前記ポリペプチドに含まれる場合、前記外部ドメイン断片は、本発明のポリペプチドに前記シグナルペプチドおよび前記神経突起成長支持配列の間(好ましくは、前記シグナルペプチドおよび前記ポリペプチドに含まれえる任意の可能なアンカーする配列の間)で含まれる。
【0185】
本発明の態様によると、本発明のポリペプチドは、N末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列を含む,またはN末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列からなる。
【0186】
本発明の別の態様によると、本発明のポリペプチドは、N末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記外部ドメイン断片(例えば、二つのアミノ酸 GK), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列を含む,または本発明のポリペプチドのN末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記外部ドメイン断片(例えば、二つのアミノ酸 GK), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列からなる。
【0187】
好ましくは, 前記外部ドメイン断片は、1〜4アミノ酸, より好ましくは2 アミノ酸である。
【0188】
好ましくは, 前記外部ドメイン断片は、前記外部ドメインのC末端の断片である, より好ましくは最後の1〜4である, より具体的には前記外部ドメインのC末端の最後の二つのアミノ酸である。
【0189】
それ故, 本発明のポリペプチドのアミノ酸配列は、さらに前記アンカーする配列のN末端での最初の一アミノ酸に(直接的にまたは間接的に, 好ましくは直接的に)以下のものが連結されたものを含んでもよい:
狂犬病ウイルス (例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の外部ドメインのC末端から1〜4アミノ酸, 好ましくは2アミノ酸, より好ましくは1〜4アミノ酸, なおより好ましくは狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の外部ドメインのC末端の最後の二つのアミノ酸、例えば、アミノ酸 GK。
【0190】
係るポリペプチドの例は、特に配列番号 26のポリペプチドおよび配列番号 26の配列を含むポリペプチドを含む。
【0191】
本発明のポリペプチドは、さらにシグナルペプチド, 好ましくは狂犬病ウイルス株(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質のシグナルペプチドを含んでもよく、前記ペプチド配列は好ましくは前記ポリペプチドのN末端(最も好ましくは前記ポリペプチドのまさにN末端、即ち、前記ポリペプチドに含まれえる任意のアンカーする配列のN-termにある)に存在する。
【0192】
係るポリペプチドの例は、特に配列番号 27のポリペプチドおよび配列番号 27の配列を含むポリペプチドを含む。
【0193】
好適なポリペプチドの例:
本発明の好適なポリペプチドは、100 アミノ酸未満のポリペプチド(より好ましくは、可能な限り少数のアミノ酸のものであるが神経突起成長効果をなおも保持しているもの, より好ましくは90 アミノ酸未満のもの)を含む。
【0194】
本発明の好適なポリペプチドは、前記 G 蛋白質の細胞質断片〔最も好ましくは、配列番号 6, 配列番号 19または配列番号 20の細胞質断片〕を含む又はからなるポリペプチドを含む(図 19を参照されたい)。
【0195】
また、本発明の好適なポリペプチドは、これらの細胞質断片の11〜44小断片を含む又はからなるポリペプチドを含む。
【0196】
本発明の好適なポリペプチドは、前記神経突起成長支持配列に加えて、上記規定の少なくとも一つのアンカーする配列(例えば、配列番号 23のアンカリング配列; 以下の例4-6を参照されたい)を含むポリペプチドを含む。
【0197】
核酸, ベクターおよび細胞:
また、本発明は、縮重を考慮した普遍的な遺伝コードに基づき本発明のポリペプチドをコードする任意の核酸(より具体的には、任意のDNAまたはRNA)に関する。
【0198】
図 15に示されるとおり、配列番号 6 の細胞質断片(即ち、CVS-NIV 株の細胞質断片)をコードする例示の核酸は、配列番号 5の核酸である。
【0199】
図 16に示されるとおり、配列番号 10 のPDZ-BS(即ち、CVS-NIV 株のPDZ-BS)をコードする例示の核酸は、配列番号 9の核酸である。
【0200】
また、本発明は、本発明のポリペプチドをコードする少なくとも一つの 核酸を含む 任意の核酸ベクターに関する。前記ベクターは、トランスフェクションベクターおよび/または 発現ベクターであってもよい。
【0201】
前記発現ベクターは、さらに前記核酸の上流および/または下流の少なくとも一つの発現シグナルおよび/または制御配列、例えば、少なくとも一つのプロモーター,または少なくとも一つのエンハンサーや少なくとも一つのプロモーター(前記核酸の上流)を含んでもよい。前記発現ベクターは、例えば、プラスミドであってもよい。
【0202】
係るベクターの例示は、CNCMから受託番号 I-2758で利用可能なプラスミドである。
【0203】
興味深いことに, レンチウイルス ベクター, AAV ベクター, アデノ-ウイルスベクターおよび単純ヘルペス無効化ウイルスベクターを使用できる。
【0204】
択一的にまたは相補的に、本発明の核酸は、デンドリマー、例えば、リポソーム,または陽イオン性ポリマー、例えば、DEAE-デキストランまたはポリエチレンイミン,または不活性な固形のナノ粒子、例えば、金粒子,または当業者が適切であることを見出しえる任意の他のトランスフェクション手段と組み合わせることができる又は会合(associated)させることができる。
【0205】
また、本発明は、少なくとも一つの本発明のポリペプチド, および/または少なくとも本発明の核酸および/または少なくとも一つの本発明のベクターを含む任意の細胞に関する。
【0206】
前記細胞は、好ましくは遺伝的に操作されて前記ポリペプチドおよび/または核酸および/またはベクターを含む細胞である。
【0207】
前記細胞は、真核生物細胞, 好ましくは哺乳類細胞、例えば、ヒト細胞または非-ヒト細胞, 最も好ましくはヒト細胞であってもよい。
【0208】
前記細胞は、原核細胞(好ましくは細菌、例えば、大腸菌)であってもよい。
【0209】
前記細胞はビリオンであってもよいが、前記ビリオンはレシピエントの対象(より具体的には、ヒト患者)に有意に有害なまたは望ましくない効果を誘発しない。
【0210】
好ましくは, 前記細胞は、ヒト胚性細胞ではない。
【0211】
使用:
本発明は、より具体的には、神経突起成長を刺激および/または誘導することに使用するための〔より具体的には、不十分な又は障害性の神経突起生成(より具体的には、不十分な又は障害性の神経突起成長)に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドおよび/または核酸および/またはベクターおよび/または細胞に関する。
【0212】
非免疫原性の使用における前記使用。
【0213】
実際, 本発明に適切である狂犬病ウイルス G 蛋白質は、非アポトーシス性の G 蛋白質と規定できる。
【0214】
従って、本発明のポリペプチドは、以前に公開されたPCT国際出願WO 03/048198に記載されたものものなどのアポトーシス小体の形成を誘導しない。
【0215】
本発明と一致して、本発明のポリペプチドは、例えば、ニューロン前駆体または新生物ニューロンからのニューロン分化のため, および/または障害性のニューロンのニューロン再生のための神経突起成長 (および/または軸索および/または樹状突起発生)のエフェクターとして意図される(双方のエフェクター効果は、神経突起成長の刺激を介して得られる)。
【0216】
上述のことから、本発明のポリペプチドは、免疫原性の薬剤(immunogenic agent)またはアジュバントではない,または最低限でも免疫原性の薬剤またはアジュバントとして使用されない、また前記ポリペプチドを免疫原性の薬剤またはアジュバントとして作用することを可能にするだろう条件下で使用されない。
【0217】
より具体的には、アポトーシス性の 狂犬病ウイルス株のG 蛋白質(例えば、弱毒化したERA株の及び他の弱毒化した狂犬病ウイルス株のG 蛋白質)とは反対に、本発明のポリペプチドは、ニューロンに投与された場合に検出可能な体液性の免疫応答を上昇させない。
【0218】
本発明のポリペプチドは、免疫原性の組成物またはワクチンであろう薬学的組成物または薬に使用されない。
【0219】
従って, 本発明のポリペプチドは、好ましくは何らかの抗原(より具体的には、過剰発現または変化により神経病理学などの病状を誘発するだろうウイルス抗原, 腫瘍抗原, 細胞抗原であろう任意の抗原)と連結されない、融合されない、および会合しない。
【0220】
疾患, 障害およびコンディション:
本発明は、ポリペプチド, 核酸, ベクターおよび細胞を提供する。これらの産物は、神経突起生成(より具体的には神経突起成長, なおより具体的にはヒト神経突起成長)を誘導および/または刺激する。
【0221】
本発明の産物は、好ましくは本発明の核酸ベクター(より具体的には、本発明のプラスミド)である。
【0222】
より具体的には、本発明の産物は、神経突起生成(分裂前のニューロン, 新生物ニューロン, ニューロン前駆体, 同様に障害があるニューロンからの神経突起成長)を誘導および/または刺激する。
【0223】
従って、本発明は、神経発生および/または神経再生および/または神経保護剤として使用するための前記産物に関する。
【0224】
本発明の産物は、神経突起の出芽および/または軸索の成長および/または樹状突起樹の伸展を刺激および/または誘導する。
【0225】
本発明の産物は、成長円錐の活性を刺激する。さらにまた、本発明の産物は、成長円錐が成長虚脱剤(例えば、LPAまたは酸化ストレス)との接触に際して虚脱(collapsing)することを予防する(以下の例 1を参照されたい)。
【0226】
本発明の産物は、結果的にシナプス形成および/または神経伝達を刺激および/または誘導する。
【0227】
本発明の産物は、新生物ニューロンの増殖を〔より具体的には、ニューロ分化剤(neuro-differentiating agent)として〕阻害する薬剤である。
【0228】
本発明の産物は、ニューロンの発生および/またはニューロンの再生および/または軸索の成長および/または樹状突起の発生および/または樹状突起樹の伸展および/またはニューロンの可塑性および/またはシナプス形成および/または神経伝達を刺激する薬剤である。
【0229】
本発明の産物は、神経突起の退縮および/または成長円錐の崩壊を誘発するであろう如何なる種類の神経毒性も予防および/または阻害および/またはブロックする薬剤である。
【0230】
本発明の産物は、神経突起 および/または錐状体が神経毒性剤と接触した後に神経突起成長および/または成長円錐活性を刺激および/または誘導する薬剤である。
【0231】
本発明の産物は、成長円錐の崩壊および/または神経突起の退縮および/または軸索樹状突起間のダメージまたは傷害および/またはシナプス統合性の崩壊および/またはニューロン結合(neuron connectivity)の欠損および/または神経終末(nerve endings)へのダメージおよび/または神経伝達の損傷を予防および/または阻害および/またはブロックする薬剤である。
【0232】
本発明の産物は神経突起成長を誘導および/または刺激する手段であり、以下の事項に特に有用である:
・ ニューロン分化を誘導すること(例えば、神経系の新生物の治療および/または緩和および/または予防)に有用である, 同様に
・ 障害されたニューロン(より具体的には、障害された神経突起)を再生することに有用である(例えば、神経変性疾患, 障害またはコンディションの治療および/または緩和および/または予防に有用である, ニューロンの微生物感染の治療および/または緩和および/または予防に有用である,または神経毒性剤または酸化ストレスからニューロンを保護すること有用である)。
【0233】
従って、本発明は、不十分なまたは障害性の神経突起生成(より具体的には、不十分なまたは障害性の神経突起成長)を伴う任意の疾患, 障害またはコンディションの治療および/または緩和および/または予防における使用のための前記産物に関する。
【0234】
前記疾患, 障害またはコンディションは、バランスを失ったニューロン細胞周期が関係する任意の疾患, 障害またはコンディションとして択一的にまたは相補的に規定され、前記ニューロン細胞周期は以下によりバランスを失う、即ち:
・ 分裂前のニューロンの過剰なまたは望まれない存在により(より具体的には、新生物が神経系において発生する場合のように、不十分なニューロンの分化および/または過剰なまたは望まれない分裂後ニューロンのニューロン細胞周期へのリエントリーにより)バランスを失う、或いは
・ 過剰なまたは望まれないニューロン変性、より具体的には過剰なまたは望まれない神経突起変性によりバランスを失う(神経変性の疾患, 障害またはコンディションやニューロンの特定の微生物感染の場合のように)。
【0235】
本発明の産物は、神経回復(neurorestorative)の治療および/または予防および/または緩和などの中枢神経系(CNS)および/または末梢神経系(PNS)を変化させる疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用できる。
【0236】
「中枢神経系」または「CNS」の表現は、本明細書等において脳や(脊椎動物の場合において)脊髄を意味することを意図している。
【0237】
末梢神経系(PNS)は、体を脳や脊髄と連結する脊髄や頭蓋の神経の広いネットワークである。PNSは、自律神経系(交感のNS および 副交感のNS)と体神経系とに細分される。PNSは、刺激レセプターからCNSに走る感覚ニューロンおよびCNSから筋肉や腺に走る運動ニューロンからなる。
【0238】
本発明の態様によると、前記疾患, 障害またはコンディションは、神経系の微生物感染(例えば、細菌および/またはウイルスの感染, より具体的にはウイルス感染)であるか又はこれらを伴う。
【0239】
前記ウイルス感染は、例えば、単純ヘルペスウイルス(HSV)感染, より具体的にはHSV 1型感染(ウイルス脳症を誘発する)であってもよい。
【0240】
前記微生物感染は、ニューロンのアポトーシスを誘導しないウイルス感染(例えば、狂犬病ウイルス感染)であってもよい。
【0241】
好ましくは, 前記微生物感染は、ニューロンのアポトーシスを誘発する微生物感染(例えば、灰白髄炎)である(Blondel et al., 2005を参照されたい)。
【0242】
本発明の別の態様によると、前記疾患, 障害またはコンディションは、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性や非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションであるか又はこれらを伴うものである。
【0243】
本発明の態様によると、前記疾患または障害は、神経変性疾患または障害(例えば、慢性の神経変性疾患または障害)、例えば、非-ウイルス性の脳症, アルツハイマー型の疾患, パーキンソン病, ALS, ハンチントン病, 多発性硬化症(MS)または稀な遺伝疾患(rare genetic disease)であるか又はこれらを伴うものである。
【0244】
好ましくは, 前記神経変性疾患または障害は、非-ウイルス性の疾患または障害, より好ましくは非-細菌性および非-ウイルス性の疾患または障害, なおより好ましくは非-微生物性の障害である。
【0245】
本発明の態様によると、前記コンディションは、神経変性コンディション(例えば、加齢)であるか又はこれを伴うものである。
【0246】
好ましくは, 前記神経変性コンディションは、非-ウイルス性のコンディション, より好ましくは非-細菌性および非-ウイルス性のコンディション, なおより好ましくは非-微生物性のコンディションである。
【0247】
本発明の態様によると、前記疾患, 障害またはコンディションは、神経系の物理的(physical)な又は虚血性の傷害、例えば、発作, 脳卒中, 外傷, 癲癇であるか又はこれらを伴うものである。
【0248】
好ましくは, 前記身体性の又は虚血性の傷害は、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性および非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションである。
【0249】
本発明の態様によると、前記疾患, 障害またはコンディションは、化学的な神経毒性剤および/または酸化ストレスの存在を伴うものである。
【0250】
好ましくは, 前記疾患, 障害またはコンディションは、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性および非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションである。
【0251】
本発明の態様によると、前記疾患は、新生物である、より具体的には新生物ニューロン(neoplastic neurons)を含む新生物である。
【0252】
「新生物(neoplasm)」の用語は、本明細書等においてより具体的には悪性新生物を意図し, より具体的には癌, なおより具体的には腫瘍または白血病, なおさらにより具体的には腫瘍を意図する。
【0253】
本発明の産物は、免疫原性剤(immunogenic agent)として作用しない。より具体的には、本発明の産物は、腫瘍抗原に対し体液性応答を上昇させるだろう免疫原性剤として作用しない。
【0254】
好ましくは、本発明の産物は、それ自身で免疫原性剤として作用できない。より具体的には、本発明の産物は、それ自身で腫瘍抗原に対し体液性応答を上昇させるだろう免疫原性剤として作用できない。
【0255】
本発明の産物は、抗増殖剤(antiproliferative agent)として作用する。
【0256】
本発明の産物は、新生物ニューロンからの神経突起成長を誘導および/または刺激して、新生物ニューロンから成熟ニューロンへの分化を誘導する及び/又は刺激する。
【0257】
前記新生物は、CNSおよび/またはPNSの新生物, 好ましくは神経節膠腫, 脳腫瘍, 中心神経細胞腫(central neurocytoma), 髄芽腫, 脳室上衣腫, 奇形腫, 神経芽細胞腫の新生物であってもよい。
【0258】
好ましくは, 前記新生物は、非-ウイルス性の新生物, より好ましくは非-細菌性および非-ウイルス性の新生物, なおより好ましくは非-微生物性の新生物である。
【0259】
当業者が適切であることを見出す任意の投与様式が本発明に包含される。
【0260】
如何に本発明の産物を製剤化するかに応じて、非経口または腸内(例えば、経口)の投与, 好ましくは非経口投与, より好ましくは非経口の注射により投与できる。
【0261】
薬学的組成物または薬; 治療の方法:
また、本発明は、少なくとも一つの本発明のポリペプチド, および/または少なくとも本発明の核酸および/または少なくとも一つの本発明の細胞を含む任意の薬学的組成物または薬に関する。
【0262】
本発明の薬学的組成物または薬は、上記により詳細に記載されとおり、不十分なまたは障害性の神経突起成長を伴う疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用できる。
【0263】
本発明の薬学的組成物または薬は、免疫原性の組成物ではなく、ワクチンでもない。
【0264】
本発明の薬学的組成物または薬は、さらに少なくとも一つの薬学的および/または生理的に許容されるビヒクル(希釈剤, 賦形剤, 添加物, pH 調整剤, 乳化剤または分散剤, 保存剤, 界面活性剤, ゲル化剤, 同様に緩衝化剤や他の安定化剤や可溶化剤, など)を含んでもよい。
【0265】
本発明の薬学的組成物または薬は、好ましくは何らかの免疫アジュバントを含まない。
【0266】
最も好ましくは、本発明の薬学的組成物または薬は、任意の抗原, より具体的には過剰な発現または変化が病状〔例えば、神経病理(neuropathology)〕を誘発するだろう任意の抗原(ウイルス抗原, 腫瘍抗原, 細胞抗原である可能性がある)を含まない。
【0267】
本発明の薬学的組成物または薬は、例えば、液体溶液剤(liquid solution), 懸濁剤, エマルジョン剤, 錠剤, ピル, カプセル剤, 徐放性製剤,または粉末剤であってもよい。好ましくは、非経口投与に適切な形態で製剤化される。
【0268】
また、本発明は、上記に記載されるとおり、対象(より具体的には、治療が必要なヒト)を治療する方法に関し、該方法は前記対象またはヒトに少なくとも一つの本発明のポリペプチドおよび/または少なくとも本発明の核酸および/または少なくとも一つの本発明のベクターおよび/または少なくとも一つの本発明の細胞を投与することを含む。
【0269】
本出願において、「含む(including)」または「含む(containing)」と同義である「含む(comprising)」の用語は、オープンエンド(open-ended)であり、付加的な未記載の要素, 構成成分または方法の工程を除外しない。他方、「からなる(consisting of)」の用語は、クローズドターム(closed term)であり、明示的に記載されない任意の付加的な要素, 工程,または構成成分を除外する。
【0270】
「から本質的になる(essentially consisting of)」の用語は、部分的なオープンタームであり、付加的な要素, 工程または構成成分が本発明の基礎および新規の特性に実質的(materially)に影響しない限り、これらの付加的な未記載の要素, 工程,または構成成分を除外しない。
【0271】
それ故、「含む」の用語は、「からなる」の用語, 同様に「から本質的になる」の用語を含む。従って、「含む」の用語は、本出願においてより具体的には「からなる」の用語および「から本質的になる」の用語を包含することを意味する。
【0272】
本明細書等において引用される全ての参照文献の関連する開示の各々は、参照によって特異的に援用される。以下の例は、例示の目的で提供され、限定の目的ではない。
【0273】
[例]
例 1: 非アポトーシス性のCVS-NIV 狂犬病ウイルス株の蛋白質Gの細胞質ドメイン(G-CVS-NIVまたは「G-survival」の細胞質ドメイン)は神経突起成長を誘導し、誘導された神経突起成長は成長円錐虚脱剤(即ち、LPA)および酸化ストレス(H2O2)に高度に耐性である。
【0274】
神経の再生や神経の保護は、広く神経毒性, 神経変性疾患, 外傷-発作-脳卒中, 脳症またはさらに加齢などの広範な分野において共通の一里塚である(図 1を参照されたい)。
【0275】
未成熟のヒトニューロン(例えば、神経芽細胞腫細胞)は、さらに適切なシグナル伝達分子で処理した場合に分化できる。このケースは、SH-SY5Y 細胞が細胞浸透性のdb-cAMPで処理された場合である。
【0276】
発明者は、ヒト神経芽細胞腫の細胞株(例えば、神経芽細胞腫細胞株 SH-SY5Y)の使用が含まれる機能的な神経保護検査を開発した。この系において、細胞はさらに分化でき、また神経突起の成長をモニターできることから成長円錐活性刺激性のエフェクターに関して試験することが可能である。そのうえ、退縮プロセス(retraction processes)に抵抗する伸長(elongated)した神経突起の能力を薬処理〔例えば、リゾホスファチジン酸 (LPA)または過酸化水素 (H2O2)〕後に評価できる。このような系により集められたデータは、インビボでなされた神経毒性分析と相関することが証明されている。
【0277】
以下の例は、特に以下の事項を実証する:
・ 非アポトーシス性の(即ち、病原性の)狂犬病ウイルス株(即ち、CVS-NIV 株)のG蛋白質は神経突起成長促進効果を有し、これは即ち、ある種の非アポトーシス性の狂犬病ウイルスG蛋白質が神経突起生成を誘発および/または刺激することである;
・ この神経突起成長効果は、ニューロンを成長円錐虚脱薬(LPA)や酸化ストレス(H2O2)から防御するために十分強いものである;
・ この神経突起成長効果を担うのは前記非アポトーシス性のCVS-NIV 狂犬病ウイルス株のG蛋白質の細胞質尾部である;
・ アポトーシス性の 狂犬病ウイルス G 蛋白質は、この神経突起成長効果を示さない; より具体的には、CVS-NIV 株のG蛋白質と僅か六アミノ酸 (また細胞質尾部においては僅か二つのアミノ酸)異なるERA株のG蛋白質は、この神経突起成長効果を示さない;
・ この効果は、特に前記 非アポトーシス性の狂犬病ウイルスG蛋白質の細胞質尾部に含まれるPDZ-BS モチーフが前記 アポトーシス性の 狂犬病ウイルス G 蛋白質の一つと比べて単一の点変異を示すとの事実によるものであり、即ち: 非アポトーシス性の CVS-NIV 株の蛋白質G(protein G)は、アミノ酸 Qをポジション521 (CVS-NIV 株の全長 G 蛋白質に対して計算されたポジション、前記 G 蛋白質の細胞質断片におけるポジション 41に対応する)に有し, 他方でG-ERAはEを同じポジションに有する;
・ CVS-NIV 株 の全長G蛋白質中のポジション491(前記 G蛋白質の細胞質断片におけるポジション 11)にあるアミノ酸もこの神経突起成長効果に寄与する (アミノ酸 H)。
【0278】
材料および方法
細胞, ウイルスおよび分子クローン
SH-SY5Yは、アメリカン・タイプ・カルチャー・コレクション(ATCC; 10801 University Blvd.; Manassas, Virginia 20110-2209; U.S.A.)から受託番号 CRL-2266)で利用可能な神経芽細胞腫の細胞株である。
【0279】
狂犬病ウイルス (RABV)のオリジナルのERA および CVS 株は、それぞれATCC から受託番号 vr332 および vr959で利用可能である。本研究において使用されたERA および CVS 株は、BSR 細胞 (乳児 ハムスター 腎臓細胞-クローン 21のクローン, ATCC 受託番号 BHK-21)においてこれまで二十年間発明者の研究室において継代されてきた。これらの継代株は、それぞれERA-NIV 株およびCVS-NIV 株である。
【0280】
本出願の例および図の全体をとおして、他で特定しない限り、CVSはCVS-NIVを意味し、ERAはERA-NIV (G蛋白質 CNCM I-2760; 配列番号 4)を意味する。
【0281】
CVS-NIV 株 (rRABV CVS HQ)は、ブタペスト条約の条件下で2009年4月1日にCollection Nationale de Cultures de Microorganismes (CNCM)に寄託された(CNCM; Institut Pasteur; 25, rue du Docteur Roux; F-75724 PARIS CEDEX 15; FRANCE)。CNCM受託番号は、I-4140である。
【0282】
ERA-NIV 株(即ち、G-ERA-NIV)のG蛋白質, およびCVS-NIV 株(即ち、G-CVS-NIV)のG蛋白質の代表的な分子クローンの取得や特徴に関する事項は、Prehaud等(Prehaud et al. 2003)に記載されている
これらの G 蛋白質の配列もアクセッション番号AF 406693 (G-ERA-NIVに関して) および AF 406694 (G-CVS-NIVに関して)で利用可能である;(図 13A および 13Bも参照されたい)。
【0283】
CVS-NIV 株のG蛋白質も2001年11月30日にブタペスト条約の条件下でCNCMに寄託された組換え型の大腸菌株から利用可能である。CNCM受託番号は、I-2758である。この組換え型の大腸菌は、CVS-NIV 株のG蛋白質を誘導性に発現するプラスミド (プラスミド pRev-TRE-G-CVS; WO 03/048198を参照されたい)を含む。
【0284】
ERA株のG蛋白質も2001年11月30日にブタペスト条約の条件下でCNCMに寄託された組換え型の大腸菌株から利用可能である。CNCM受託番号は、I-2760である。この組換え型の大腸菌は、ERA株のG蛋白質を誘導性に発現するプラスミド (プラスミド pRev-TRE-G-ERA; WO 03/048198を参照されたい)を含む。
【0285】
狂犬病ウイルス CVS-NIV 株のG蛋白質をコードしているプラスミド CNCM I-2758を含んでいる組換え型の 大腸菌株の培養のための適切な条件や狂犬病ウイルス ERA株のG蛋白質をコードしているプラスミド CNCM I-2760を含んでいる組換え型の大腸菌株の培養のための適切な条件には、標準 LB-TYM 成長培地(アンピシリンの存在下)で37°Cで前記組換え型の大腸菌株をインキュベーションすることが含まれる。
【0286】
前記組換え型の狂犬病ウイルス (rRABV)は、Faul等2008の記載の処理にしたがって産生され、単離された。rRABVsは、野生型「G survival」または「G death」配列 (それぞれCVS-NIV 株およびERA株のG 配列),またはERAの遺伝的な G 遺伝背景におけるG survivalの細胞質尾部,またはCVS-NIVの遺伝的な G 遺伝背景におけるG deathの細胞質尾部の何れかを保持するよう構築された(図 2を参照されたい)。
【0287】
組換え型の狂犬病ウイルスも産生され、このウイルスはGのコード配列中の変異によりCVS-NIV 株から由来する。これらの組換え型の狂犬病ウイルスはG蛋白質を有し、このG蛋白質はCVS-NIV 株のG蛋白質と一または二アミノ酸異なるものである、すなわち:
・ CVS-NIV 株の全長 G蛋白質 配列において、ポジション 491 (その細胞質断片中のポジション 11)のアミノ酸で異なる;および/または
・ CVS-NIV 株の全長 G蛋白質 配列において、ポジション 521 (その細胞質断片中のポジション 41)のアミノ酸で異なる。
【0288】
より具体的には、以下の組換え型の狂犬病ウイルス株が作出された:
【0289】
【表2】
【0290】
rRABV CVS LE, rRABV CVS LQ および rRABV CVS HEは、ブタペスト条約の条件下でCNCMに2009年4月1日に寄託された。CNCMの受託番号は、それぞれI-4141, I-4142 および I-4143である。
【0291】
ウイルス I-4141またはI-4142またはI-4143 (組換え型の 狂犬病 ウイルス)を増殖させる適切な条件には、グルコース (例えば、4.5 g/L), ピルビン酸ナトリウムおよび glutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地で前記 ウイルス を37°CでBHK-21 細胞 (サブ-クローン BSR)と 5% CO2の存在下でインキュベーションすることが含まれる。
【0292】
神経突起の成長および退縮のハイスループットなアッセイ
SH-SY5Y ヒト 神経芽細胞腫細胞は、プレート (Cell Bind plastic ware, Corning, USA)に40,000 細胞/ウェルの密度で非分化培地[DMEMF12 (Invitrogen, U.K.) に20% ウシ胎児血清 プラス 1%Pen:Strep および1% グルタミンを添加したもの]に播種され、一晩、37°Cで培養された。接種の24h後に、非分化培地を分化培地[Neurobasal medium (Invitrogen, U.K.) にB27 添加物(Invitrogen, U.K.), 1% P/S, 1% グルタミン および1mM db-cAMP (dibutyril c-AMPは膜浸透性, Sigma)]を添加したもので置き換え、細胞を6hインキュベーションした。それから, 細胞は、mock(MOI 3で分化培地でrRABV を感染させた)で感染させた。インキュベーションの1h後、細胞を、分化培地で一回洗浄し、分化培地を加えた後に24h、37°Cでインキュベーションした。
【0293】
自然の分化に関して、同じ手順を使用した(但し、db-cAMPを省略した)。
【0294】
分化の30h後、細胞を、3% パラホルムアルデヒドをリン酸緩衝塩類溶液 (PBS)に含むもので20 min、室温 (RT)で固定し、5分間0.1% Triton-X-100で処理し、50%の正常ヤギ血清 (NGS: normal goat serum)をPBS中に含むもので1h、RTで処理した。ニューロン特異的な抗βIII チューブリン Ab (Promega, France) および 抗-RABV ヌクレオカプシド Abを使用して軸索の突起を染色し、それぞれでのRABV感染を明らかにした。或いは、細胞は。クリスタルバイオレットで染色され、これにより軸索突起が保たれる。
【0295】
退縮アッセイは、10μM-30μMまたは50μM LPA (Sigma, USA)を分化培地に添加すること又は75μM H2O2 (Sigma, USA)をB27 マイナス 抗酸化剤含有分化培地(B27 minus anti oxidant containing differentiating medium)に添加することを除いて上記の成長アッセイと同一である。
【0296】
細胞は、Leica DM 5000B UV 顕微鏡にDC 300FX カメラ (x40またはx20 対物)を備えたものを用いて視覚化し、ImageJ 1.38X ソフトウェア (Wayne Rasband, NIH, USA, http://rsb.info.nih.gov/ij/)とそのplug-inのNeuronJ(Meijering et al. 2004; http://www.imagescience.org/meijering/software/neuronj/)を用いて分析した。ニューロン毎の平均の神経突起長は、トリプリケート実験で決定された。
【0297】
フローサイトメトリーによるRABV抗原の検出
SH-SY5Y 細胞をdb c-AMPでの処理により分化させ、分化6 時間後にrRABVsで感染させた。感染24 時間後に細胞を採取し、細胞質膜に達したRABV 糖蛋白質または感染細胞において発現したG蛋白質の総量の何れかのフローサイトメトリーによる検出のため処理した。処理は、Prehaud等(Prehaud et al. 2003)の記載に基づき行った。
【0298】
結果および考察
G-CVS-NIV 細胞質尾部(即ち、CVS-NIV 株のG蛋白質の細胞質尾部)は、神経突起成長を促進する分子署名を含む。
【0299】
感染が非存在の条件下で、db-cAMPでの処理は、神経突起の伸長を惹起する(図 3A, 左パネル二つ)。細胞がdb-cAMPで処理され、rRABV G-CVS-NIVで感染された際に、長い神経突起を示す(191%, 図 3A, 中央右パネル)。この結果によって、次の事項が確立された。即ち、成長円錐活性がcAMPシグナル伝達により刺激される場合、rRABV G-CVS-NIVが神経突起成長を促進することである。
【0300】
対照的に, 神経突起の長さは、双方がdb-cAMPで処理された非感染細胞およびrRABV G-ERA 感染細胞の間で類似し(図 3B)、rRABV ERAを発現しているG deathは神経突起成長 アッセイに関してサイレント表現型を有することを示す。実験の時間経過(即ち、分化後30hおよび感染後24 時間)は、アポトーシスの最大 レベルがrRABV ERA感染神経芽細胞腫 細胞において検出される時点(即ち、感染後48h)よりも早期の時点であることが注目される。
【0301】
図 3Cに記載のとおり、G蛋白質のエンドスワップ(end swap)を保持しているrRABVs を用いることにより(図 2を参照されたい)、この表現型が細胞質尾部のみにより完全に移されることを発明者は示す。この結果は、伸長プロセスがcAMP シグナル伝達経路により開始される場合に、G-CVS-NIV 細胞質尾部が神経突起成長を促進する分子署名を含むことを確かに確立する結果である。
【0302】
さらにまた、神経突起成長アッセイを、CVS-NIV 株 (rRABV G-CVS-NIV)とそれぞれの G 蛋白質と一つ又は二つのアミノ酸で異なる組換え型の 狂犬病ウイルスで行った。
【0303】
より具体的には、アッセイをrRABV CVS LE, rRABV CVS LQ および rRABV CVS HEで行った、これらはG-CVS-NIVとアミノ酸 491 および/または アミノ酸 521 で異なるG蛋白質を発現する(表 2 上記を参照されたい)。これらのアッセイの結果は、図 3Dで説明される。図 3Dにおいて、対照との全ての差異は、統計的に有意である(p<0.005 スチューデントのt-検定):
・ rRABV G-CVS HQ 対 N.I.: p<0.0001;
・ rRABV G-CVS LQ 対 N.I.: p=0.0002;
・ rRABV G-CVS HE 対 N.I.: p=0.0111。
【0304】
rRABV G-CVS-NIVは、それぞれG蛋白質のポジション491 および 521にH および Qを有する(CVS-NIVの全長G蛋白質に関して計算したポジション, これらのポジションはそれぞれこのG蛋白質の細胞質断片におけるポジション 11 および 41と対応する)。
【0305】
rRABV CVS LQは、L および Qをこれらのポジションに有する。
【0306】
rRABV CVS HEは、HおよびEをこれらのポジションに有する。
【0307】
rRABV CVS LEは、LおよびEをこれらのポジションに有する。
【0308】
図 3Dにより説明されるとおり、CVS-NIV (rRABV G-CVS-NIV)よりも有意に低い程度であるが、rRABV CVS LQ および rRABV CVS HEは陽性の神経突起成長効果をなおも示す[スチューデントのt-検定分析: rRABV CVS HQ 対 rRABV CVS LQ: p = 0.0156; rRABV CVS HQ 対 rRABV CVS HE: p = 0.0009]。
【0309】
以上より、CVS 株のG蛋白質のポジション 491 および ポジション 521(細胞質断片におけるポジション 11 および 41)は、双方が神経突起成長効果に密接に関係しており、ポジション 521の寄与が優勢である。
【0310】
rRABV CVS LEは、対照と比べて何らかの有意に陽性の神経突起成長効果を誘導しなかった(図 3Cを参照されたい, rRABV G-CVS-Cyto Death = rRABV CVS LE)。
【0311】
G-CVS-NIV 細胞質尾部は、cAMPと相乗的に働く神経突起生成を促進する内因性のエフェクターである。
【0312】
発明者は、G-CVS-NIV 細胞質尾部の発現がそれ自身で神経突起生成を刺激できるかどうかを調査した。SH-SY5Y 細胞を異なる rRABVで感染させ(図 2を参照されたい)、神経突起の伸長をモニターした。
【0313】
図 4Aにおいて観察されたとおり、G-CVS-NIVまたはG-ERA-Cyto Survivalの発現は、神経突起成長を促進するため十分である。これらのデータによって、G-CVS-NIVの細胞質ドメインが未知の機構により神経突起伸長プロセスにシグナルを送る内因性の分子署名を含むことが確立された。同じ実験がdb-cAMPの存在下または非存在下でなされた場合、G-CVS-NIV および cAMPの双方が媒介する神経突起成長に相乗的に働くことが観察された(図 4Bを参照されたい)。
【0314】
G-CVS-NIVの細胞質ドメインに基づく神経突起生成は分子署名に依存し、発現したG蛋白質の量には依存しない。
【0315】
産生された狂犬病ウイルス 糖蛋白質の量を、感染させdb-cAMP処理したSH-SY5Y 細胞において産生されたRABV 抗原の総量,または完全に細胞質膜へとプロセスされたG-蛋白質の量の何れかををモニターすることにより非常に慎重に分析した。G-CVS-NIVまたはG-ERAを発現している二つのrRABVsは、全体として細胞質膜に発現されたG-蛋白質の量は類似する(図 5を参照されたい)。従って、生存の表現型(即ち、神経突起成長)は、利用可能な蛋白質の量の表現型とは関連しないが、むしろ細胞質の生存ペプチドの配列に存在する特定の決定要因と関連する。
【0316】
そのうえ、rRABV G-ERA-Cyto Survivalが糖蛋白質を若干低く発現するが、このウイルスがなおも強い生存表現型を示すことは興味深い(図 3C, 4Aを参照されたい)。この結果は、ペプチドの量は主要な注意事項(caveat)ではなく、 生存決定要因が感染細胞の内側に存在する限り神経突起成長が刺激されることを意味している。
【0317】
G-CVS-NIV 細胞質尾部は、成長円錐虚脱薬(growth cone collapsing drug)に対し神経保護を与える
LPAは、細胞成長, 運動性, 形態および運命に影響する多様な細胞機能を発揮することが示された成長因子様のリン脂質として作用する生理活性脂質である。ニューロン細胞において、LPAは、アクトミオシン細胞骨格の再組織化をおこすRho および Rho-キナーゼ活性化をともなう成長円錐虚脱や神経突起退縮を媒介する。
【0318】
G蛋白質細胞質尾部の10μM LPA 誘導性の神経突起退縮に対して防御する能力が調査された。db-cAMPで分化させたSH-SY5Y 細胞がLPAで処理される場合、神経突起の退縮が観察される(図 6A, 左を参照されたい)。また、同じ効果は、細胞がrRABV G-ERAで感染される場合に観察される(図 6A, 右を参照されたい)。反対に、db-cAMP 処理し、rRABV G-CVS-NIVで感染させた細胞において検出された長い神経突起は、成長円錐虚脱剤での処理に対して完全に保存される(図 6A, 中央を参照されたい)。この結果によって、G-CVS-NIVがある種の内因性の神経保護特性を有することが確立された。
【0319】
この保護は、rRABV G-ERA-Cyto Survivalでの感染に続いても認められるが、rRABV G-CVS-Cyto Deathでの感染では認められない(図 6Bを参照されたい)。従って、エンドスワップ変異体の使用によって、神経保護表現型がG-CVS 細胞質ドメインの発現と関連することが確かに確立された。
【0320】
LPAに対するG-CVS-NIV 細胞質尾部の神経保護は強いものである
LPAの生理的な範囲は、通常1μM付近である。発明者は、高用量のLPA(即ち、10μM)を使用してG-CVS-NIV 細胞質尾部により与えられた神経保護の強さ(robustness)を確認した。
【0321】
50μMまでのLPAの亜致死量を、この神経保護の効率をモニターするため試験した。SH-SY5Y 細胞をrRABV G-CVS-NIVまたはrRABV G-ERA-Cyto Survivalで感染させた又は感染させなかった。分化後30 時間に細胞を異なる用量のLPAで処理した(図 7を参照されたい)。LPAの量を増加させて非感染細胞を処理することによって、神経突起の長さの直線的な退縮を生じた(図 7, 左を参照されたい)。反対に、rRABV G-CVS-NIVで感染された細胞は、LPAの用量がどのような用量であってもそれらの神経突起に何らかの明らかな虚脱を示さない(図 7, 中央を参照されたい)。同じデータは、30μMまでのLPAの用量に関してrRABV G-ERA-Cyto Survivalで感染された細胞に関して観察された(図 7, 右を参照されたい)。LPAの最高用量(50μM)に関して観察された軽微な減少は、このrRABVにより発現されたG蛋白質の最低量と比例する可能性がある。この場合(および成長円錐虚脱薬の非常に高濃度に関して)、G蛋白質の相対量が限定要因となる可能性がある。何れの場合ケースでも、これらのデータによって、非常に高い効率のG-CVS-NIV 細胞質尾部により与えられた神経保護が証明された。
G-CVS-NIV 細胞質尾部によって、酸化ストレスに対する神経保護が与えられる
LPAによる神経突起退縮へのG-CVS-NIV 細胞質ドメインにより提供された神経保護について上記でなされた観察に基づいて、発明者は、この表現型が他のよりユビキタスな因子に一般化できるかどうかを試験した。
【0322】
酸化ストレスは、ニューロン変性を誘導する重要な経路を代表する。酸化ストレスは、多くの神経変性の罹患者に関係するが、脳への急性ダメージ(例えば、外傷, 脳卒中および癲癇)のケースにおいても関係する。このようなことから発明者は過酸化水素処理後の神経突起の退縮表現型の研究を試みた。分化した細胞が75μM H2O2に供試される場合、神経突起は短縮する(図 8, 棒グラフの左の最初のペアを参照されたい)。同じ観察は、前記細胞が以前にrRABV G-ERAで感染された場合になされた(図 8, 棒グラフの第三のペアを参照されたい)。これに反して、神経突起の退縮は、前記細胞がrRABV G-CVS-NIVで感染された場合に観察されない(図 8, 棒グラフの第二のペアを参照されたい)。以上のとおり、G-CVS-NIV 発現は、酸化ストレスに対し神経保護を提供する。
【0323】
エンドスワップ変異体を用いることにより、我々は、この神経保護特性が図 8に記載のとおりG-CVS-NIV 細胞質尾部の発現に関連することも示した(それぞれrRABV G-CVS-Cyto Death および rRABV G-ERA-Cyto Survivalに関して、棒グラフの第四および第五のペアを参照されたい; これらのエンドスワップ変異体の構造に関して図 2を参照されたい)。
【0324】
狂犬病ウイルス G蛋白質の細胞質ドメインのCOOH 末端のPDZ-BSは、感染したニューロン細胞の生存表現型の運命に決定的な影響を与える。
【0325】
G-Cyto survival蛋白質の最後の 4aa 残基の欠失(survival G-蛋白質-delta)は、神経突起成長における影響により測定されるとおり、rRABVの生存表現型を有意に減少させるのに十分であった。
【0326】
例えば、db-cAMPの存在下で感染後8 時間で測定された神経突起成長を示す図 20を参照されたい。この図では、rRABV CVS-NIV Delta-PDZ-BS 〔アミノ酸 H をポジション 491 に有し、PDZ-BSが欠失したCVS-NIV〕により誘導されたものと、対照(N.I.)およびG-CVS-NIV 〔アミノ酸 H をポジション 491 に有し、そのPDZ-BS(即ち、ポジション 521〜524におけるQTRL)を有するCVS-NIV〕と比較して: rRABV CVS-NIV Delta-PDZ-BSにより誘導される神経突起成長効果が、対照により誘導されるものと有意に異なり(p = 0.0002 スチューデントのt-検定)、rRABV CVS-NIVにより誘導されるものと有意に異なる(p = 0.0003 スチューデントのt-検定)ことを示している。
【0327】
発明者は、特に以下の事項を実証している:
・ CVS-NIVの蛋白質G(G-CVS-NIV)は、成長円錐活性がc-AMP シグナル伝達経路(外部のエフェクターとして伝達された)により刺激される系において神経突起成長(神経突起生成)を促進することができる;
・ G-CVS-NIVの細胞質ドメインは、この表現型の原因である;
・ G-CVS-NIVの細胞質尾部の発現は、c-AMPの非存在下での神経突起成長の刺激を担うものでもある;
・ G-CVS-NIV および c-AMPの双方は、相乗的なエフェクターとして機能する;
・ 前記表現型は、G-CVS-NIVの細胞質尾部により、より具体的には全長 G-CVS-NIV 蛋白質においてポジション 491 および 521(その細胞質断片においてポジション 11 および 41)にあるアミノ酸により, なおより具体的には全長 G-CVS-NIV 蛋白質においてポジション 521(その細胞質断片においてポジション 41)にあり、PDZ-BSの部分であるアミノ酸により伝えられる分子署名と関連する;
・ G-CVS-NIV 細胞質尾部により提供される生存表現型は、成長円錐虚脱剤(即ち、LPA)による処理に高度に耐性である;
・ G-CVS-NIV 細胞質尾部により提供される生存表現型は、酸化ストレス (H2O2)にも耐性である。
【0328】
例 2: G-CVS-NIVの細胞質ドメインはHSV-1 細胞変性効果に対する神経保護を与える
単純ヘルペスウイルス タイプ I 脳炎 (HSVE)は、西洋諸国において最も一般的な散発性で致死的な中枢神経系の感染症であり、全ての年齢層の患者において年間を通じて出現する。単純ヘルペスウイルス1型が脳組織で感染する場合、HSVEは溶解性(即ち、壊死性)の状態で進行する。
【0329】
G蛋白質細胞質尾部の細胞におけるHSV-1の細胞変性効果に対して防御する能力が調査された。
【0330】
材料および方法
神経突起成長
上記の例 1に記載されているとおりである。
【0331】
HSV-1 細胞変性効果の決定
rRABVは、上記の例 1に記載されているとおり産生された。
【0332】
rRABVをBSR細胞中で増殖させた(Faul et al. 2008を参照されたい)。
【0333】
HSV-1 株 KOSをU373MG細胞(ATCC HTB 17)で増殖させた(Skare et al. 1975を参照されたい)。
【0334】
SH-SY5Y 細胞は、分化培地 マイナス db-c-AMPの条件で6 時間mockで感染させるか又はrRABVで感染させた。それから, HSV-1を感染多重度3(MOI 3)で加え、神経突起成長の表現型を24 時間後に決定した。単独(sole)の HSV-1 感染の神経突起成長の効果を確立するために、SH-SY5Y 細胞を分化培地中にdb-c-AMPを含むもので6 時間分化させ、次にHSV-1 感染(MOI 3)を行った。
【0335】
神経突起成長の表現型を同様に24 時間後に決定した。
【0336】
結果および考察
db c-AMPでの処理により分化したSH-SY5Y 神経芽細胞腫細胞は、HSV-1 感染後に神経突起の激しい退縮(〜79 %, 図 9Aを参照されたい)を示し、最終的に細胞が死に至る可能性がある。
【0337】
これに反して、前に細胞がrRABV G-CVS-NIVまたはrRABV G-ERA-Cyto Survivalで感染された場合 (上記の例 1を参照されたい)、細胞は神経突起成長を示し(図 9Bを参照されたい)、広範な神経突起退縮に対して保護される (図 9Cを参照されたい)。
【0338】
例 3: G-CVS-NIVの細胞質ドメインは神経芽細胞腫細胞に対して抗-増殖性を示す
神経芽細胞腫は、幼児期の固形腫瘍のうち二番目に一般的なものであり、米国の小児の癌による死亡の13%以上と計算される。予後は、小児の障害の臨床段階や年齢に応じて変化する。一般に, 神経芽細胞腫の大抵の患者は、外科, 放射線および細胞毒の化学療法を含む治療アプローチで治療される。
【0339】
ヒトの悪性神経芽細胞腫は未成熟の神経芽細胞の分化が不十分であることや増殖が非制御的であることにより特徴付けられるので、高用量のオールトランスレチノイン酸(ATRA)などの分化誘導薬が使用されてきた。RABV G-CVS-NIVの細胞質尾部がSH-SY5Y ヒト神経芽細胞腫細胞 (上記の例を参照されたい)の神経突起成長を促進できること(即ち、これらの細胞の分化を誘導できること)を発明者は実証した。それ故、発明者は、この分化特性が培養において係る細胞の増殖を制御するため十分に効率的であるかどうかとの疑問を解決することを望んだ。
【0340】
材料および方法
MTTアッセイ
前記アッセイは、Sargent 2003により記載された。簡単に説明すると、db c-AMPなしの分化培地で48hでmockで感染させたか又はrRABVで感染させた細胞は、MTT溶液で3.5 時間、37°C、5% CO2で染色された。次に培地および染色溶液の混合物が除去され、MTT結晶は一時間、室温で持続的に撹拌して溶媒で溶解された。前記アッセイは、550nmのODで測定した。各条件は、8のnを代表する。統計分析をANOVA 検定で行った。
【0341】
フローサイトメトリー
細胞計数および形態学的な変化を、Prehaud等(Prehaud et al. 2003)に記載のとおり、側方および前方散乱(それぞれ、SSC および FSC)により評価した。各点は、1 分間、低速で集めた細胞を代表する。フローサイトメトリー分析により、48hの時点での細胞集団の1%未満を代表する死細胞または壊死細胞を排除するために生存可能な集団をゲートした。
【0342】
神経突起成長
上記の例 1に記載されているとおりである。
【0343】
ATRA処理
神経芽細胞腫細胞のオールトランスレチノイン酸 (ATRA: All trans retinoic acid)処理をPrehaud等(Prehaud et al. 2005)により既に記載されたとおり行った。
【0344】
結果および考察
最初の例において、SH-SY5Y 細胞を無処置か又はATRAの5μMもしくは10μMを培養培地中に含むもので処置した。細胞増殖を生細胞に関してフローサイトメトリーで又はMTTアッセイを介した細胞計数でアッセイした。データを、それぞれ図 10A および 10Bに示す。
【0345】
各ケースで、細胞のATRA処理により、細胞成長をモニターするために使用された検査に応じて27%〜55%の間で変動する増殖の減少を伴う神経芽細胞の成長の遅延が生じた。このように、SH-SY5Y 細胞は、抗-増殖薬の効果に効率的に応答する。
【0346】
それゆえ、SH-SY5Y 細胞をrRABV G-CVS-NIVまたはrRABV G-ERAの何れかで感染させ、細胞の増殖(MTTアッセイにより確立された)および神経突起の成長を測定した(図 11A および 11Bを参照されたい)。我々は、細胞増殖(cell growth)が神経突起の成長と逆に相関することを見出し、G-CVS-NIVの神経生存特性が神経芽細胞における内因性の抗-増殖性の表現型と関連することを確立した。
【0347】
第二シリーズの実験において、神経芽細胞腫細胞を親のrRABV G-CVS-NIVまたはrRABV G-ERA ウイルスの何れか及びエンドスワップ変異体で感染させた(rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survival; 上記の例 1および 図 2を参照されたい)。細胞増殖をフローサイトメトリー および MTT アッセイ (それぞれ、図 12A および 12Bを参照されたい)により測定した。各ケースで、前記データは、細胞が神経突起成長促進G-cyto survival尾部ウイルスで感染された場合に神経芽細胞の増殖が影響されて18.3% 〜23%の間で変動することを示した。従って、G-Cyto survivalは、神経芽細胞の細胞増殖を緩徐にする特性を内在する。
【0348】
例 4: Cyto-Gの生存支持特性が発現ベクターに保存されることの実験による実証
ベクターがウイルス感染の非存在下で細胞で発現される場合(最初に発現ベクターとして配送されたポリペプチド)、非アポトーシス性の狂犬病ウイルス株(Cyto-G)のG蛋白質のRABV 細胞質ドメインが生存支持特性を保存する(神経突起生成および退縮に対する保護)ことを本例は実証する。
【0349】
この例は、さらに前記生存特性がサイズや細胞質または小胞体膜(endoplasmic membrane)への足場に依存するかどうかを調査する。
【0350】
突然変異誘発を例 1のrRABVs を作出するため使用されたオリゴヌクレオチド-PCR に基づく突然変異誘発処置を用いて行った(QuickChange Lightning 部位特異的変異誘発キット, Agilent Technologies; Stratagene product division; カタログ番号 210518-12; 製造者の推奨にしたがって使用されるキット; 米国特許第7,176,004; 7,132,265; 7,045,328; 6,734,293; 6,489,150; 6,444,428; 6,391,548; 6,183,997; 5,948,663; 5,932,419; 5,866,395; 5,789,166; 5,545,552, および係属している特許)
構築物の末端にそれぞれ NheI および XmaI 部位を導入するためオリゴヌクレオチドを設計した。
【0351】
突然変異誘発のプライマーは以下のものであった:
GFullATG:
GGCCGCTAGCATGGTTCCTCAGGCTCTCCTGTTT (配列番号 28)
GCytoATG:
GGCCGCTAGCATGAGAAGAGTCAATCGATCAGAACCT (配列番号 29)
GendSTOP:
GGCCCCCGGGTCACAGTCTGGTCTGACCCCCACT (配列番号 30)
mutTM.Cyto: CCCCTTCTGGTTTTTCCATTGTGTTTTGGGGGGAAGTATGTATTACTGAGT (配列番号 31)
前記変異体をシークエンシングでインサートを選別した。この方法により、特に次の三つの構築物(G-Full; G-[SP-(2aa)-TM-Cyto]; G-Cyto)を得た:
G-Full: アクセッション番号 AF 406694 (CVS-NIVのG蛋白質)
MVPQALLFVPLLVFPLCFGKFPIYTIPDKLGPWSPIDIHHLSCPNNLVVEDEGCTNLSGFSYMELKVGYILAIKMNGFTCTGVVTEAETYTNFVGYVTTTFKRKHFRPTPDACRAAYNWKMAGDPRYEESLHNPYPDYHWLRTVKTTKESLVIISPSVADLDPYDRSLHSRVFPSGKCPGVAVSSTYCSTNHDYTIWMPENPRLGMSCDIFTNSRGKRASKGSETCGFVDERGLYKSLKGACKLKLCGVLGLRLMDGTWVAMQTSNETKWCPPDQLVNLHDFRSDEIEHLVVEELVRKREECLDALESIMTTKSVSFRRLSHLRKLVPGFGKAYTIFNKTLMEADAHYKSVRTWNEILPSKGCLRVGGRCHPHVNGVFFNGIILGPDGNVLIPEMQSSLLQQHMELLESSVIPLVHPLADPSTVFKDGDEAEDFVEVHLPDVHNQVSGVDLGLPNWGKYVLLSAGALTALMLIIFLMTCCRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 2)
G-[SP-(2aa)-TM-Cyto]:
G-[SP-(2aa)-TM-Cyto]は、(N-端からC-端に)以下のものを含んでいる構築物である:
・ G蛋白質のシグナルペプチド(例えば、CVS-NIVのシグナルペプチド; 配列番号 21);
・ 2アミノ酸(例えば、G蛋白質の外部ドメインからアミノ酸、例えば、G-CVS-NIVからアミノ酸);
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通ドメイン(例えば、G-CVS-NIVの膜貫通ドメイン; 配列番号 23);
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメイン(例えば、G-CVS-NIVの細胞質ドメイン; 配列番号 6)。
【0352】
本例に使用されたG-[SP-(2aa)-TM-Cyto]の配列は、以下のものである:
MVPQALLFVPLLVFPLCFGGKYVLLSAGALTALMLIIFLMTCCRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 27)。
【0353】
G-Cyto:
G-Cytoは(N-からC-端に)次のものを含んでいる構築物である: アミノ酸(例えば、M)および非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメイン(例えば、G-CVS-NIVの細胞質ドメイン; 配列番号 6)。
【0354】
本例に使用されたG-Cytoの配列は、以下のものである:
MRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 23)。
【0355】
配列は、図 22に示される。
【0356】
配列のアラインメントは、図 23に示される。
【0357】
三つの構築物の略図は、図 24に示される。
【0358】
インサートをNheIおよびXmaI消化により除去し、標準のクローニング処置によりSAP 脱リン酸化pCI-Neo 派生物(Promega, France)にクローン化した。大腸菌XL1-blue 細胞 (Stratagene, USA)をプラスミド構築物で形質転換した。プラスミド DNAをPurelinkカラム(Invitrogen, U.K.)で大規模に精製し、インサートをプラスミドに挿入された配列の統合性を検証するためシークエンシングした。プラスミド クローンを使用してAmaxa GmbH エレクトロポレーション技術〔Amaxa(登録商標) nucleofector(登録商標) kit V Catalog # VCA 1003 Lonza Germanyを製造者の推奨にしたがって使用した〕を用いてヒト 神経芽細胞腫 細胞 [SH-SY5Y 細胞 (ATCC CRL-2266),またはNtera cl2D1 細胞 (ATCC CRL-1973)]をヌクレオフェクト(nucleofect)した。G 蛋白質の発現を、例 1に記載のとおりフローサイトメトリーでアッセイした。
【0359】
一過性発現のため、生物アッセイをヌクレオフェクションの24h後に行った。
【0360】
安定発現に関して、双方のヌクレオフェクトした細胞株 [SH-SY5YまたはNtera cl2D1]を800μg/mlのG418で三週間処理した。G418に耐性の細胞クローンを単離し、凍結した。対照の細胞株は、インサートなしのpCI-Neo単独でヌクレオフェクトした細胞からなるものである。
【0361】
神経突起の成長アッセイおよび退縮アッセイを例 1に記載のとおり行った。
【0362】
例示の結果を図 25-28に示す。
【0363】
図 25は、非アポトーシス性の狂犬病ウイルス株の細胞質ドメインを含んでいるポリペプチドの発現(より具体的には、以下のものの発現)を説明する、即ち:
・ 前記狂犬病ウイルス株の全長G蛋白質の発現,
・ 前記G蛋白質の膜貫通および細胞質ドメインを含むが、前記G蛋白質の外部ドメインを含まないポリペプチドの発現, および
・ 前記G蛋白質の細胞質ドメインを含むが、前記G蛋白質の外部ドメインおよび膜貫通ドメインを含まないポリペプチドの発現,
を対照プラスミドで測定された発現と比較して説明する。
【0364】
既に説明されたとおり、Gの発現レベルは異なり、生物学的な表現型は発現のレベルと直接的に相関しない(以下および 図 5を参照されたい)。
【0365】
ウイルス感染の非存在下で、非アポトーシス性の 狂犬病ウイルス株のG蛋白質の細胞質ドメインの一過性発現によって、ヒト神経芽細胞腫細胞からの神経突起成長が誘導および/または刺激される(図 26)。前記G蛋白質の細胞質ドメインに加えて、膜貫通ドメインを含んでいる(しかし、外部ドメインを含まない)一過性に発現されたポリペプチドは、一過性に発現された 全長 G蛋白質よりも又は前記G蛋白質の一過性に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも神経突起成長の誘導及び/又は刺激に効率的である(図 26)。
【0366】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインの,または前記ドメインを含んでいるポリペプチドまたは蛋白質の安定な発現は、ヒト 神経芽細胞腫細胞において可能である(図 27)。前記G蛋白質の細胞質ドメインに加えて、膜貫通ドメインを含んでいる(しかし、外部ドメインを含まない)安定に発現されたポリペプチドは、安定に発現された 全長 G蛋白質よりも又は前記G蛋白質の安定に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも神経突起成長の誘導及び/又は刺激に効率的である(図 27)。前記 G蛋白質の安定に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)は、神経突起成長の誘導及び/又は刺激において、安定に発現された全長G蛋白質と同等に効率的である(図 27)。
【0367】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株の細胞質ドメインを含んでいるポリペプチドの発現は、神経毒性剤〔例えば、成長円錐虚脱薬 LPA; (図 28, G-(SP-[2a]-TM-Cytoで得られた結果を示している) 構築物を参照されたい)〕への抵抗性を提供する。
【0368】
同じ実験条件において、このLPAのバッチは、図 6A, 6B, 7に説明されるとおり、成長円錐の虚脱を誘導する。
【0369】
G-(SP-[2a]-TM-Cyto)の膜貫通ドメインによって、特に本構築物により発現されるポリペプチドが細胞のゴルジ膜 および/または 小胞体膜にアンカーされることが可能となり、これにより神経発生, 神経再生および神経保護の誘導および/または刺激において発現されたポリペプチドの効率が増加する。
【0370】
それ故, 非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを係る膜-アンカリング配列とカップリングすることは、前記細胞質ドメインにより誘導および/または刺激された神経発生, 神経再生 および 神経保護効果を増加させる。
【0371】
例 5: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインがニューロン前駆体(EC 細胞)に作用して成熟ニューロンに分化させる特性(コミットメント)を内在することの実験による実証
Cyto-G (上記例 4; 例えば、配列番号 6または24を参照されたい)などの非アポトーシス性の 狂犬病ウイルス株の細胞質ドメインは、このように神経芽細胞腫細胞株 SH-SY5Yの分化を促進することにより神経突起の成長を促進する(例 4 上記を参照されたい)。
【0372】
この特性が前記細胞質ドメインの細胞株をニューロン分化へコミットすることを促進し、方向づける広範囲の能力を示すことの実験による実証を例 5は提供する。
【0373】
胚性癌腫細胞系統 NTera 2cl.-D1 (ATCC CRL-1973)からの純粋な分裂後のヒトニューロンの産生は、当該技術(例えば、Prehaud et al. 2005)において記載されている。
【0374】
上記の例 4に記載のとおり、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを発現している構築物が使用された, 例えば:
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質(配列番号 2のG蛋白質)をインサートとして含んでいるG-full 構築物,
・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 6の細胞質ドメイン)をインサートとして含んでいるG-Cyto 構築物,および
・ 非アポトーシス性の狂犬病ウイルス株のシグナルペプチド(配列番号 21のシグナルペプチド), 二つのアミノ酸(非アポトーシス性の狂犬病ウイルス株のG蛋白質の外部ドメインのC末端からのアミノ酸、例えば、GK), 非アポトーシス性のウイルス株のG蛋白質の膜貫通ドメイン(配列番号 23の膜貫通ドメイン)および非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 6の細胞質ドメイン)を含んでいるG-(SP)-[2aa]-TM-Cyto 構築物。
【0375】
NTera 2-D1 細胞のニューロンへの分化を誘導するG-発現構築物の能力を調査した。
【0376】
安定なG-発現NTera 2-D1 細胞を、Prehaud等(Prehaud et al. 2005)に記載のプロトコールのようなATRA-ニューロスフェア(ATRA-neurosphere)にしたがって処理した。
【0377】
最後の抗分裂処理および二回の複製の後、細胞を神経突起の先端(最後の複製の5 日後)またはニューロンのネットワーク構築(最後の複製の50 日後)のいずれかを計算するため播種した。
【0378】
例示の結果を図 29-31に示す。
【0379】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを含んでいるポリペプチドの発現は、神経芽細胞腫細胞から分化することにより成熟した分裂後のヒトニューロンの産生を誘導および/または刺激する: 以下のものの発現で得られた結果を説明する図 29を参照されたい:
・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質の発現,
・ 前記G蛋白質の膜貫通ドメインおよび細胞質ドメインを含んでいる(しかし、外部ドメインを含まない)ポリペプチドの発現,
・ 前記G蛋白質の細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)の発現,
を対照プラスミドと比較した発現。
【0380】
膜貫通ドメインを非アポトーシス性の狂犬病ウイルス株のG 蛋白質の細胞質ドメインに加えて含む(しかし、前記G蛋白質の外部ドメインを含まない)ポリペプチドは、全長 G蛋白質よりも又は前記G蛋白質の細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも分裂後のヒトニューロンの分化の誘導及び/又は刺激にさらに効率的である(図 30を参照されたい)。
【0381】
神経芽細胞腫細胞からの分化により成熟した分裂後のヒトニューロンの産生を誘導および/または刺激することにくわえ、ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを含んでいるポリペプチドの発現は、長い軸索を有する成熟した分裂後のヒトニューロンのネットワークの組織化を誘導および/または刺激する(図 31を参照されたい)。
【0382】
例 6: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインが傷害されたニューロンの再生を促進できることの実験による実証
傷害されたニューロンは、これらの起源や局所の環境に応じて再生する可能性がある。例えば、哺乳類の中枢神経系からのニューロンは、末梢の突起(peripheral process)への傷害が後根神経節ニューロンの再生の増加を生じるとしても、非常に限定的な再生能力を有する。内因性の再生シグナルが再生に影響して適切な再生が成功することが明らかに確立され、それらの幾つかには特異的なキナーゼ経路が関係する(Hammarlund et al. 2009)。
【0383】
Cyto-G 構築物に含まれる配列番号 6の細胞質ドメインなどの非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(上記例 4 を参照されたい)は、再生医療に関連して少なくとも二つの興味深い特性を呈する。
【0384】
第一に、前記細胞質ドメインは神経突起成長を促進し、このことは前記細胞質ドメインがニューロンの成長円錐を刺激できることを意味する。
【0385】
第二に、前記細胞質ドメインは、LPAまたはH2O2での処理後の退縮から神経突起を保存し、このことは前記細胞質ドメインが成長円錐の虚脱を避ける分子ロック(molecular locks)を刺激または強化できることを意味する。
【0386】
これらの特性が傷害されたニューロンの再生を生じることを実験により実証するため、その現象を細胞モデルで調査した。例 4のG-[SP-(2aa)-TM-Cyto] 構築物(配列番号 27をインサートとして含んでいる構築物)を安定に発現しているNT2-N 細胞株(ヒトニューロン細胞)を選択した。pCI-Neo-NT2-Nを対照として使用した。
【0387】
NT2-N 細胞株は、文献(Prehaud et al. 2005, Cheung et al. 1999 および Paquet-Durand et al. 2003)に記載の分化型の Ntera-2cl.D1 (NT2/D1; ATCC CRL-1973) 細胞から由来するものである。
【0388】
スクラッチ誘導アッセイ(scratch-induced assays)のため、細胞をポリ-D-リシン-ラミニンコートセル+(Sarstedt, Germany)12 ウェルプラスチック器に播種し、トリプシン処理後に完全に回復させるため二 日間成長させた。培地を引っ掻く10h前に変更した。個々の傷を注射針 (26GX1/2", 12-4.5)で作った。少なくとも 10の引っかき傷を個々のウェルの各々に作った。細胞をPFA (4%)で傷をつけた3 日後に固定し、クリスタルバイオレット溶液で染色した。細胞は、Leica DM 5000B顕微鏡にDC 300FX カメラ (x20 対物レンズ)を備えたものを用いて視覚化され、ImageJ 1.38X ソフトウェア (Wayne Rasband, NIH, USA, http://rsb.info.nih.gov/ij/)とそのplug-inのNeuronJを用いて分析した。再生中のニューロンの平均のパーセンテージは、8回の実験から決定された。
【0389】
例示の結果を図 32-33に示す。
【0390】
非アポトーシス性の 狂犬病ウイルス株のG蛋白質の細胞質ドメインを含むポリペプチドの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する(非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通および細胞質ドメインを含んでいるポリペプチドの発現で得られた結果を説明する図 32を参照されたい)。
【0391】
傷をつけた後にかように誘導された神経突起の再生はドラスティックである。より具体的には、前記ポリペプチドは、前記 G蛋白質の膜貫通および細胞質ドメインを含むが、前記G蛋白質の外部ドメインを含まない(非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通および細胞質ドメインを含んでいるポリペプチドの発現で得られた結果を説明する図 33を参照されたい)。
【0392】
例7:
感染性の rRABV cDNAは、固定された病原性で非アポトーシス性のRABVであるCVS-N2c 狂犬病ウイルス株から作出された(Morimoto et al. 1999)。
【0393】
このように作出された感染性の rRABV cDNA(cN2C)は、Schnell等(Schnell et al. 2010)に記載される。
【0394】
G-vir(非アポトーシス性の 狂犬病ウイルス株のG蛋白質、例えば、配列番号 2のG蛋白質)をコードしている遺伝子は、この RABV cDNAに統合された。感染性のウイルスを例 1に記載されているとおり回収した。
【0395】
組換え型のウイルスを、8個体の六週齡の雌性C57Bl6 マウスの群に筋肉内経路でマウスの80%に致死的な脳炎を生じさせるrRABV G-virのウイルス粒子数を注射した。マウスおよび実験の数を慎重に評価して、動物の負担を最小化した。
【0396】
疾患の進行を臨床徴候および死亡率をモニターすることで評価した (Camelo et al. 2001に記載のとおり)。ウイルスの神経侵襲性を測定するため、RNA抽出(脳の半分)および免疫組織化学または複数のアレイ(脳の第二の半分)のため処理する前に、感染の過程で二または三マウスのグループを灌流し、脳を除去して急速凍結し、-80°Cで貯蔵した。
【0397】
このモデルによって、さらにrRABV 株により惹起される脳の先天性の免疫応答の異なるパラメータを分析することが可能である。
【0398】
[参照文献]
Blondel et al., Poliovirus, pathogenesis of poliomyelitis, and apoptosis, CTMI, 2005, 289, 25-56.
Camelo et al. 2001 "Selective role for the p55 Kd TNF-a receptor in immune unresponsiveness induced by an acute viral encephalitis"; J. Neuroimmunol. 113: 95-108.
Cheung et al. 1999 ; BioTechniques 26: 946-954.
Faul et al. 2008; Virology 382: 226-238.
Guigoni and Coulon 2002, Journal of NeuroVirology 8: 306-317.
Hammarlund et al. 2009 "Axon regeneration requires a conserved MAP kinase pathway" Science 323: 802-806.
Kumar et al. 2007; Nature 448: 39-43 and associated Supplementary Information ("Methods").
Lay et al. 2003; Ann. N. Y. Acad. Sci. 1010: 577-581.
Loh et al. 2008; Cell Death and Differentiation 15: 283-298.
Meijering et al. 2004; Cytometry Part A 58A: 167-176; and http://www.imagescience.org/meijering/software/neuronj/
Morimoto et al. 1999, Journal of Virology 73(1): 510-518.
Paquet-Durand et al. 2003 Brain Res. Dev. Brain Res. 142: 161-167.
Prehaud et al. 1988; Journal of Virology 62(1): 1-7.
Prehaud et al. 2003; Journal of Virology 77(19): 10537-10547.
Prehaud et al. 2005; Journal of Virology 79(20): 12893-12904.
Sargent 2003; Recent Results in Cancer Research, vol. 161, Springer-Verlag Berlin Heidelberg, pages 13-25
Sarmento et al. 2005; Journal of NeuroVirology 11: 571-581.
Schnell et al. 2010; Nature Reviews Microbiology 8(1): 51-65.
Skare et al. 1975; J. Virol. 15:726-732 (Structure and function of herpesvirus genomes. I. Comparison of five HSV-1 and two HSV-2 strains by cleavage of their DNA with Eco R I restriction endonuclease).
Ugolini 1995; The Journal of Comparative Neurology 356: 457-480.
Ugolini 2008; Dodet B, Fooks AR, Muller T, Tordo N, and the Scientific & Technical Department of the OIE (eds): Towards the Elimination of Rabies in Eurasia. Dev. Biol. Basel, Karger, vol. 131, pp. 493-506.
WO 03/048198; PCT international application in the name of Institut Pasteur, Inv. Lafon et al., published on 12 June 2003; as well as its national/regional counterpart applications.
Each of the relevant disclosures of all references cited herein is specifically incorporated by reference.
【発明の分野】
【0001】
本発明は、ニューロンの発生、再生および保護、より具体的には神経突起成長(neurite outgrowth)に関する。
【0002】
本発明は、神経突起生成作用を誘導および/または刺激する手段を提供し、これらの手段は医学の分野(より具体的には、神経系の障害の治療 および/または 緩和および/または予防)に有用である。特に、本発明は、神経突起の成長を誘導および/または刺激するための手段を提供し、これは、ニューロンの分化を誘導するために、例えば、神経系の新生物の治療のため、同様に、障害されたニューロンの再生における、例えば、神経変性疾患、障害または状態の治療のため、または微生物感染の治療における、またはニューロンを神経毒性剤または酸化ストレスからの保護において有用である。
【0003】
本発明の手段は、特定の非アポトーシス性(non-apoptotic)の狂犬病ウイルス G 蛋白質蛋白質(特に、その蛋白質の細胞質尾部)に基づく。
【発明の背景】
【0004】
神経系の発生の間に、ニューロンは適切な様式で標的に神経を分布(innervate)させるためかなりの距離で伸びる。この現象には細胞において成長円錐の活性を刺激できる特異的なシグナル伝達経路が刺激されることが必要である。
【0005】
神経系の発生(特に、中枢神経系の発生)は柔軟性(plastic)が高いが、成体の神経系(特に、成体の脳)の修復の潜在力は限定的なものである。従って、神経突起軸索の成長および変性に対する保護は、神経変性疾患、障害または状態(例えば、神経系の急性傷害または慢性の神経変性疾患)の転帰を改善すると考えられる重要な事項である。係るニューロン細胞からの神経突起成長を誘導する能力があるであろう産物は、係る疾患、障害または状態(conditions)の非常に有用な治療上および/または予防上および/または対症療法上(palliative)の解決策をもたらすであろう。
【0006】
ニューロンの発生のプロセスの別の側面として、成熟したニューロンの構造に分化しないニューロン前駆体(neuronal progenitors)の増殖によって神経系の新生物が誘発される。係る前駆体細胞から神経突起成長を誘導する能力があるであろう産物は、係る新生物の治療上および/または予防上および/または対症療法上の解決策をもたらすであろう。
【0007】
神経向性ウイルスによるニューロン細胞の感染における一般的な概念は、ニューロンの形態(特に、神経突起成長)に肯定的な影響(positive impact)を有さないとのことである。
【0008】
実際、神経向性ウイルスがアポトーシスによるニューロン細胞死を生じることを示している多数の例が存在する。この事項はDNAウイルス(例えば、疱疹ウイルス)、およびRNAウイルスの双方に関し、エンベロープウイルス、例えば、アルファウイルス、ブニヤウイルスおよびパラミクソウイルス、または非エンベロープウイルス、例えば、ピコルナウイルスおよびレオウイルスに関する。
【0009】
また、狂犬病ウイルス(特に、弱毒化狂犬病ウイルス株)は、ニューロンのアポトーシスを誘導することが記載されている。
【0010】
例えば、WO 03/048198は、ニューロン細胞の統合性の崩壊やアポトーシス小体の形成を誘導する少なくとも 100 アミノ酸の狂犬病ウイルス G 蛋白質及びその断片に関する。これらのアポトーシス小体は、体液性免疫応答、好ましくはB-依存的な体液性の免疫応答を刺激する能力がある。
【0011】
WO 03/048198は以下を示す:
・ 弱毒化した狂犬病ウイルス株(例えば、弱毒化したERA株)は、そのウイルスが感染した細胞のアポトーシスによる破裂を誘導する,
・ それによって産生されたアポトーシス小体は、体液性免疫応答、特にB-依存的な体液性免疫応答を刺激する;
・ 狂犬病ウイルス株によるアポトーシスの誘導は、そのG蛋白質の性質により決定される;
・ 弱毒化した狂犬病ウイルス株(例えば、弱毒化したERA株)からのG蛋白質を含んでいる狂犬病ウイルスはヒト細胞のアポトーシスを惹起することが可能であるが、他方で病原性の狂犬病ウイルス(例えば、Challenge Virus Standard -CVS-株)からのG蛋白質を発現させることでは惹起することは不可能である(特に、WO 03/048198の例 5 および 図 19 および 20を参照されたい)。
【0012】
また、Lay等 (Lay et al. 2003)および Prehaud等 (Prehaud et al. 2003)を参照されたい。
【0013】
以上のように、アポトーシス性の狂犬病ウイルス株のG 蛋白質(例えば、弱毒化したERA株のG蛋白質)は、体液性応答(特に、B依存的な体液性免疫応答)の刺激に有用であることが知られている。
【0014】
これらの特定の G 蛋白質はそれらが感染した細胞のアポトーシスを誘導するので、これらの蛋白質も標的細胞のアポトーシスにより望ましくない細胞を破裂させることで排除する候補因子として提案されてきた。
【0015】
狂犬病ウイルス株の病原性は、アポトーシスを誘導する能力とは逆に相関することも記載されている(WO 03/048198; Ugolini 1995; Sarmento et al. 2005; Ugolini 2008; Jackson et al. 2008を参照されたい)。
【0016】
以上のように、病原性の強い狂犬病ウイルス株は、アポトーシス性が低い。
【0017】
病原性の狂犬病ウイルス株(例えば、CVS 株)がニューロンのアポトーシスを誘導せず、これにより体液で検出(humoral detection)されることから免れるとの所見によって、なぜ病原性の狂犬病ウイルス株が疾患の徴候や症状の出現前にCNS内に非常に広範に伝播できるかが説明される。
【0018】
神経向性ウイルス〔例えば、狂犬病ウイルスまたは単純ヘルペス1型 (HSV-1)〕による感染に際して誘導される遺伝子発現パターンにおける変化を分析するためさらなる研究が実施された。
【0019】
これらの研究は、グリアの非存在下で分裂後(post-mitotic)のヒトニューロンがウイルス感染を感知する内部機構を有すること、また神経向性ウイルス(病原性の狂犬病ウイルスまたはHSV-1)が分裂後のヒトニューロンからのサイトカインの放出を誘導することを実証した(Prehaud et al. 2005)。このサイトカイン放出は、さらにニューロンがアポトーシスを回避することに寄与し、係る神経向性ウイルスを結果的に伝播させると信じられている。
【0020】
このようなことから、ニューロンの細胞死の機構(同様に、ウイルス感染に際する免疫応答を上昇させるニューロンの能力)が徹底的に調査された。
【0021】
しかしながら、神経発生(neurogeneration), 神経再生および神経保護、特に分裂前(pre-mitotic)のニューロン(例えば、ニューロン前駆体又は新生物ニューロン)からの又は変性ニューロン(degenerative neurons)からの神経突起成長に関与するプロセスについては僅かなことしか知られていない。
【0022】
狂犬病ウイルスに焦点を合わせると、現在の知識の概要は、弱毒化した狂犬病ウイルス株が該ウイルスにより誘導されるアポトーシスによる医学的な適用を有することが知られていること、また病原性の狂犬病ウイルス株がアポトーシスを誘導しないが、しかし、反対にそれらの伝播を支援するニューロンネットワークが保存されることが知られていることであろう。
【0023】
病原性の狂犬病ウイルス株がニューロンネットワークの統合性を保存することが記載されたが、それらはニューロン形態(特に、神経突起成長)に否定的な影響(negative impact)を有している(又は、最低限肯定的な影響はない)とも報告された。
【0024】
例えば、文献(Guigoni and Coulon 2002)は、病原性の 狂犬病ウイルス株 CVS-Gif-sur-Yvetteがラットの運動ニューロンの神経突起成長を誘導しないことを記載する(例えば、この文献の図 5A および 5Bを参照されたい)。
【0025】
神経突起成長における否定的な影響も報告された。例えば、文献(Scott et al., 2008)は、病原性の CVS-11 狂犬病ウイルス株が樹状突起や軸索のビーディング(beading)を誘導(即ち、ネガティブなストレス影響の特徴である小胞の形成)することを報告している。
【0026】
本発明は、特定の病原性の狂犬病ウイルス株から由来し且つ驚くべき予想外の特性を示すニューロンの発生、再生および保護のための手段を提供する。
【発明の概要】
【0027】
本発明は、幾つかの病原性(非アポトーシス性)の狂犬病ウイルス G 蛋白質が、神経突起成長促進効果を有すること〔即ち、幾つかの非アポトーシス性の狂犬病ウイルスG蛋白質が神経突起生成(neuritogenesis)を誘導および/または刺激すること〕を示している。この効果は、アポトーシス性の狂犬病ウイルス G 蛋白質により示されない。この効果は、全ての非アポトーシス性の狂犬病ウイルス G 蛋白質によっては示されない。
【0028】
発明者は、非アポトーシス性(病原性)の狂犬病ウイルス株の亜群であって、そのG蛋白質が神経突起成長に有意に陽性の効果を有する亜群を同定した。以下に記載され、説明されるように、この亜群の代表的な株は、CVS-NIV 株である。この非アポトーシス性(病原性)株のG蛋白質は、アポトーシス性(弱毒化) ERA株 (CNCM I-2760)のG蛋白質と僅か6 アミノ酸のみが異なる。
【0029】
さらに、この神経突起成長促進効果が、前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部によること、特にそれらのPDZ 結合サイト(PDZ-BS)に及び/又はCVS-NIV 株のG蛋白質の配列中のポジション 491にあるアミノ酸による、特に、それらのPDZ-BSによることを本発明は実証する。
【0030】
本発明は、特定の非アポトーシス性の狂犬病ウイルス G 蛋白質から(より具体的には、それらの細胞質尾部から)由来するポリペプチドに関する、同様に核酸、ベクター、細胞および薬学的組成物または薬物に関する。
【0031】
本発明の手段は、神経突起成長(neurite outgrowth)を刺激する及び/又は誘導する薬剤として特に有用である。それらは、ニューロンの分化を誘導するため特に使用でき、例えば、神経系の新生物の治療に、同様に、障害されたニューロンの再生のため、例えば、神経変性疾患, 障害またはコンディションの治療に、または微生物感染の治療に、またはニューロンを神経毒性剤または酸化ストレスから保護するため使用できる。
【0032】
本出願が参照する図の幾つかは、カラー印刷である。本出願は図面のカラー印刷を含んでおり、特許庁で出願ファイルを閲覧することによりこの図面にアクセスすることができる。
【図面の簡単な説明】
【0033】
【図1】神経発生、神経再生および神経保護は、神経毒性、発作(seizure)、脳卒中(stroke)、外傷、加齢、神経変性疾患、脳症(encephalopathy)を含む様々なニューロンの障害、状態および疾患に関係する。
【図2】発明者により作出され、以下の例に使用された組換え型の狂犬病ウイルス(rRABV)の構造の概要。CVS-NIV 株 (非アポトーシス性の株)のG蛋白質は、ERA株 (アポトーシス性の株)とは僅か6 aaのみが異なる。G-survival = CVS-NIV 株のG蛋白質。G-death = ERA株のG蛋白質。G-cyto death = CVS-NIV Gの遺伝背景にG-deathの細胞質尾部。G-cyto survival = ERA Gの遺伝背景にG-survivalの細胞質尾部。SP = シグナルペプチド。EC: 細胞外ドメイン。TM: 膜貫通ドメイン。Cyto: 細胞質ドメイン。
【図3A】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS または G-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルス G蛋白質。
【図3B】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERA および rRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。
【図3C】図 3A, 3B, 3C, 3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIV および rRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIV および rRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルスG蛋白質。
【図3D】図 3A、3B、3C、3D: CVS-NIV 株(非アポトーシス性の株)のG蛋白質の細胞質尾部は、神経突起成長を促進する分子署名(molecular signature)を含む。図 3A: rRABV G-CVS-NIVおよびrRABV G-ERAでの神経突起成長アッセイ。図 3B: rRABV G-CVS-NIVおよびrRABV G-ERAでのdb-cAMP有りでの神経突起成長アッセイの結果。図 3C: rRABV G-CVS-NIV、rRABV G-CVS-Cyto Death、rRABV G-ERAおよびrRABV G-ERA-Cyto Survivalでのdb-cAMPなしの神経突起成長アッセイの結果。図 3D: rRABV-G-CVS-NIV、rRABV G-CVS (LQ)およびrRABV G-CVS (HE)での神経突起成長アッセイの結果。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。rRABV G-CVS (LQ) = CVS-NIV 株の全長 G蛋白質中のポジション 491にあるアミノ酸 Hがアミノ酸 Lで置換されたCVS-NIV 株の組換え型の 狂犬病ウイルス G蛋白質。rRABV G-CVS (HE) = CVS-NIV 株の全長 G蛋白質中のポジション 521にあるアミノ酸 Qがアミノ酸 Eで置換されたCVS-NIV 株の組換え型の狂犬病ウイルスG蛋白質。
【図4A】図 4A、4B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、cAMP*と相乗的に作用する神経突起生成を促進する内因性エフェクターである。図 4A: db-cAMPなし。図 4B: db-cAMPなし(最初の二つのヒストグラム)、cAMP有り(最後の二つのヒストグラム)。黒星は、有意差を意味する(スチューデントt-検定、p=0.0067)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株の蛋白質G。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図4B】図 4A、4B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、cAMP*と相乗的に作用する神経突起生成を促進する内因性エフェクターである。図 4A: db-cAMPなし。図 4B: db-cAMPなし(最初の二つのヒストグラム)、cAMP有り(最後の二つのヒストグラム)。平均は有意差あり(スチューデントt-検定、p=0.0067)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図5】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部の神経突起生成効果は、分子署名に依存し、発現されたG蛋白質の量に依存しない。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図6A】図 6A、6B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、成長円錐虚脱薬(LPA)に対する神経保護を提供する。図6A: db-cAMPあり。図6B: db-cAMPなし。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図6B】図 6A、6B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、成長円錐虚脱薬(LPA)に対する神経保護を提供する。図6A: db-cAMPあり。図6B: db-cAMPなし。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図7】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部により誘導される成長円錐虚脱薬LPAに対する神経保護は強いものである。黒星は、有意差を意味する(ANOVA 検定)。黒三角は、有意差を意味する(ANOVA 検定)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株の蛋白質G。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株の蛋白質G。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。LPA = リゾホスファチジン酸。
【図8】CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、酸化ストレス(H2O2)に対し神経保護を提供する。これらの実験は、db-cAMPなしで行われた。黒星は、有意差を意味する(スチューデントのt-検定)。黒三角は、有意差を意味する(スチューデントのt-検定)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9A】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9B】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図9C】図 9A、9B、9C: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は、単純ヘルペスウイルス1型 (HSV-1)の細胞変性効果に対する保護を提供する。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図10A】図 10A、10B: オール-トランス-レチノイン酸 (ATRA:all-trans-retinoic acid) 分化誘導薬(pro-differentiative drug)で処理したヒト神経芽細胞腫の細胞の増殖(図10A: フローサイトメトリー; 図10B: MTTアッセイ)。N.I. = 非感染。
【図10B】図 10A、10B: オール-トランス-レチノイン酸 (ATRA:all-trans-retinoic acid) 分化誘導薬(pro-differentiative drug)で処理したヒト神経芽細胞腫の細胞の増殖(図10A: フローサイトメトリー; 図10B: MTTアッセイ)。N.I. = 非感染。
【図11A】図 11A、11B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS-NIVまたはrRABV G-ERAで処理したヒト神経芽細胞腫細胞の細胞増殖(図 11A) および 神経突起長(図 11B)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株のG蛋白質。G-ERA = ERA株のG蛋白質。
【図11B】図 11A、11B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS-NIVまたはrRABV G-ERAで処理したヒト神経芽細胞腫細胞の細胞増殖(図 11A)および神経突起長(図 11B)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVS-NIV = CVS-NIV 株のG蛋白質。G-ERA = ERA株のG蛋白質。
【図12A】図 12A、12B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS、rRABV G-ERA、rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survivalで処理したヒト神経芽細胞腫細胞の細胞増殖(図 12A: フローサイトメトリー; 図 12B: MTT アッセイ)(N.I. = 非感染)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図12B】図 12A、12B: CVS-NIV 株 (非アポトーシス性の株)のG蛋白質の細胞質尾部は抗増殖性を与える: rRABV G-CVS、rRABV G-ERA、rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survivalで処理したヒト神経芽細胞腫細胞の細胞増殖(図 12A: フローサイトメトリー; 図 12B: MTT アッセイ)(N.I. = 非感染)。N.I. = 非感染。rRABV = 組換え型の狂犬病ウイルス。G-CVSまたはG-CVS-NIV = CVS-NIV 株のG蛋白質。G-CVS-Cyto Death = CVS-NIV Gの遺伝背景にERA株のG蛋白質の細胞質尾部。G-ERA = ERA株のG蛋白質。G-ERA-Cyto Survival = ERA Gの遺伝背景にCVS-NIV 株のG蛋白質の細胞質尾部。
【図13A】図 13A、13B: CVS-NIV 株(図 13A)の及びERA株(図 13B)のG蛋白質の核酸および蛋白質の配列。図 13A中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図13B】図 13A、13B: CVS-NIV 株(図 13A)の及びERA株(図 13B)のG蛋白質の核酸および蛋白質の配列。図 13A中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図14】CVS-NIV および ERA株のG 蛋白質の配列アラインメント; 僅かに6 アミノ酸のみが異なるG 蛋白質(図 14に太字で示す)。
【表1】
図 14に示されるアラインメントは、次のパラメータを用いて行われた:
比較マトリックス: BLOSUM62
計算したアラインメントの数: 20
ギャップオープンペナルティー: 12
ギャップエクステンションペナルティー: 4
このアラインメントの結果は:
524 残基のオーバーラップにおいて98.9%同一性;
スコア: 2787.0;
Gap 頻度: 0.0%。
【図15】CVS-NIV 株の及びERA株のG蛋白質の細胞質ドメインの核酸および蛋白質の配列。図 15中のCVS-NIV 株のG蛋白質のPDZ-BS モチーフを下線で示した(QTRL)。aa = アミノ酸。
【図16】CVS-NIV 株の及びERA株のG蛋白質のPDZ-BSの核酸および蛋白質の配列。aa = アミノ酸。
【図17−1】CVS-NIV 株のG蛋白質(配列番号 2)およびCVS-Gif-sur-Yvette 株のG蛋白質(配列番号 15)のアラインメント。
【図17−2】CVS-NIV 株のG蛋白質(配列番号 2)およびCVS-Gif-sur-Yvette 株のG蛋白質(配列番号 15)のアラインメント。
【図18−1】CVS-NIV 株のG蛋白質(配列番号 2)および三つのCVS-11株のG蛋白質(配列番号 16; 配列番号17; 配列番号18)のアラインメント。
【図18−2】CVS-NIV 株のG蛋白質(配列番号 2)および三つのCVS-11株のG蛋白質(配列番号 16; 配列番号17; 配列番号18)のアラインメント。
【図19】CVS-NIV 株のG蛋白質の細胞質断片(配列番号6)及びそれから派生する二つの保存されたバリアント(配列番号19のバリアント配列 A; 配列番号20のバリアント配列 B)の配列。
【図20】PDZ-BSの欠失は、神経突起成長表現型に影響する(db-cAMPの存在下、感染後8hのμmでの平均神経突起長)。N.I. = 非感染;G-CVS-NIV = CVS-NIV 株の蛋白質G;G-CVS-NIV-DeltaPDZ-BS = PDZ-BSが欠失したCVS-NIV 株の蛋白質G。
【図21】本発明の単一の点変異体の構造の概要(バリアント B および A)。本発明の変異体 ウイルス CVS HE (I-4143) および CVS LQ (I-4142)はCVS-NIV (I-4140)とG 蛋白質において異なり、G 蛋白質中に単一の点変異が導入されている。CVS HE (バリアント B) = G蛋白質 CVS-NIV 株の第一のPDZ-BS アミノ酸の単一の点変異であり、CVS HE においてQの代わりにE(ポジション 521)である。CVS LQ (バリアント A) = ポジション 491でのアミノ酸の単一の点変異(この変異はQの代わりにEである)。SP: シグナルペプチド。EC: 細胞外ドメイン。TM: 膜貫通ドメイン。Cyto: 細胞質ドメイン。
【図22】非アポトーシス性のウイルス株の及び構築物インサートのG蛋白質のドメインのアミノ酸配列。 ・ G-CVS ドメインの配列: ・ G 全長 (配列番号 2): アポトーシス性の狂犬病ウイルス株(CVS-NIV)の完全長 G蛋白質; ・ シグナルペプチド (SP; 配列番号 21): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)のシグナルペプチド; ・ 外部ドメイン (EC; 配列番号 22): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の外部ドメイン; ・ 膜貫通ドメイン(TM. 配列番号 23): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の膜貫通ドメイン; ・ 細胞質ドメイン(Cyto; 配列番号 6): アポトーシス性の狂犬病ウイルス株 (CVS-NIV)の細胞質ドメイン; ・ 構築物インサートによりコードされたアミノ酸配列: ・ アミノ酸 M + 配列番号 6 = 配列番号 24 (構築物 G-Cyto); ・ 配列番号 23の膜貫通ドメイン + 配列番号 6の細胞質ドメイン = 配列番号 25; ・ G-CVS-NIV の外部ドメインのC端からの二つのアミノ酸(アミノ酸 GK) + 配列番号 23の膜貫通ドメイン + 配列番号 6の細胞質ドメイン = 配列番号 26; ・ 配列番号21のシグナルペプチド + G-CVS-NIVの外部ドメインのC端からの二つのアミノ酸(アミノ酸 GK) + 配列番号23の膜貫通ドメイン + 配列番号6の細胞質ドメイン = 配列番号27 (構築物 G-(SP-[2a]-TM-Cyto)。
【図23】三つの構築物〔G-full構築物によりコードされた配列番号2; G-(SP)-[2a]-TM-Cyto (=GSP2aaTMCyto) 構築物によりコードされた配列番号27; G-Cyto 構築物によりコードされた配列番号24〕のインサートによりコードされたアミノ酸配列のアラインメント。
【図24】三つのRABV G 遺伝子構築物 〔配列番号2のG-full(= G survival); 配列番号24のG-Cyto(= G survival-Cyto); 配列番号27のG-(SP)-[2a]-TM-Cyto(= G survival-ΔEC)〕の略図。
【図25】SH-SY5Y 細胞株 (ヒトニューロブラストーマ細胞株)における対照プラスミド、G-Full 構築物(配列番号2)、G-(SP-[2a]-TM-Cyto) 構築物 (配列番号27)および G-Cyto構築物 (配列番号24)に関するRABV G 発現(任意単位)。
【図26】図 26 および 27: 以下のものを発現させた際の神経突起成長における影響であって: ・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質(配列番号 2; G-Full 構築物)、 ・ 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメイン(配列番号 27; G-(SP-[2a]-TM-Cyto) 構築物)、または ・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 24; G-Cyto 構築物)、の発現をヒト神経芽細胞腫細胞(db c-AMPの存在下のSH-SY5Y 細胞株)において対照(DNAなし)および対照プラスミドと比較した影響。図 26: 一過性発現。図 27: 安定発現。
【図27】図 26 および 27: 以下のものを発現させた際の神経突起成長における影響であって: ・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質(配列番号 2; G-Full 構築物), ・ 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメイン(配列番号 27; G-(SP-[2a]-TM-Cyto) 構築物)、または ・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 24; G-Cyto 構築物)、の発現をヒト神経芽細胞腫細胞(db c-AMPの存在下のSH-SY5Y 細胞株)において対照(DNAなし)および対照プラスミドと比較した影響。図 26: 一過性発現。図 27: 安定発現。
【図28】非アポトーシス性の狂犬病ウイルス株 (配列番号 27; 構築物 G-(SP-[2a]-TM-Cyto)の膜貫通および細胞質ドメインの発現は、ヒト神経芽細胞腫の細胞 (db c-AMP存在下のSH-SY5Y 細胞株)の神経突起成長を誘導し、成長した神経突起への成長円錐虚脱薬 LPAに対する保護を与える。
【図29】図 29 および 30: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激する〔分化処置後5 日: 対照プラスミドと比較した、G-Full構築物(配列番号2のインサート)の効果、G-(SP-[2a]-TM-Cyto)構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。図 29: カラー化したイメージ。図 30: 神経突起の先端(tips)。
【図30】図 29 および 30: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激する〔分化処置後5 日: 対照プラスミドと比較した、G-Full 構築物(配列番号2のインサート)の効果、G-(SP-[2a]-TM-Cyto) 構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。図 29: カラー化したイメージ。図 30: 神経突起の先端(tips)。
【図31】非アポトーシス性の狂犬病ウイルス株の細胞質ドメインの(安定な)発現は、胚性癌腫細胞系統 (NTera 2cl.-D1; ATCC CRL-1973)の成熟した分裂後のヒトニューロンへの分化を誘導および/または刺激し、長い軸索のニューロンネットワークの組織化を誘導および/または刺激する〔生きたニューロンのカラー化されたイメージ、分化処置後50 日: 対照プラスミドと比較した、G-Full 構築物(配列番号2のインサート)の効果、 G-(SP-[2a]-TM-Cyto) 構築物(配列番号27のインサート)の効果、またはG-Cyto構築物(配列番号24のインサート)の効果〕。
【図32】図 32 および 33: 非アポトーシス性の狂犬病ウイルス株の膜貫通および細胞質ドメインの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する〔PDL-ラミニンプラスチック器に播種したニューロン, 注射針(26GX1/2", 12-4.5)で引っ掻いた3 日後; 対照プラスミド(神経突起変性)と比べた配列番号 27 (構築物 G-(SP-[2a]-TM-Cyto)(神経突起再生)のポリペプチドの効果〕。図 32: カラー化したイメージ。図 33: 神経突起再生のパーセンテージ
【図33】図 32 および 33: 非アポトーシス性の 狂犬病ウイルス株の膜貫通および細胞質ドメインの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する〔PDL-ラミニンプラスチック器に播種したニューロン, 注射針(26GX1/2", 12-4.5)で引っ掻いた3 日後; 対照プラスミド(神経突起変性)と比べた配列番号 27 (構築物 G-(SP-[2a]-TM-Cyto)(神経突起再生)のポリペプチドの効果〕。図 32: カラー化したイメージ。図 33: 神経突起再生のパーセンテージ
【発明の詳細な記述】
【0034】
本出願は、提出され、本明細書等に記載され、説明される請求項に規定の主題に関する。
【0035】
狂犬病ウイルス(大抵の哺乳類種において致死的な脳炎を引き起こす神経向性ウイルス)の病原性は、感染したニューロンが生存する能力と相関する。生ワクチンの候補の探索で得られた研究株の弱毒化は、それらのワクチン株が細胞死を惹起する能力と常に関連する。
【0036】
所与のウイルスによる感染細胞の破壊は、特異的なシグナル伝達経路の撹乱(perturbation)があることを示唆している。
【0037】
狂犬病ウイルスは、エンベロープ型のRhabdoviridae 科、Lyssavirus属に属している弾丸型のウイルスである。ウイルス粒子は、宿主の脂質から構成される膜およびラセン状のヌクレオカプシド(NC)を囲む二つのウイルス蛋白質G および M 蛋白質からなる。NCは、N 蛋白質, P 蛋白質, および RNA-依存性RNAポリメラーゼ, L 蛋白質により保護されるウイルスのネガティブ鎖RNA分子から構成される。狂犬病ウイルス 蛋白質は、感染細胞中で等しい量で合成されない、またウイルス粒子中で同じ比で存在しない。実際、N, G および M 蛋白質は、この順序でビリオン中で最も顕著な種である。
【0038】
ニューロンネットワークの統合性 (狂犬病ウイルス伝播に有利) および ニューロンの細胞死(免疫原性に有利)の保存の間の戦略的な決定は、必然的に少なくとも二つのシグナル伝達のネットワーク間の選択を伴う。
【0039】
狂犬病ウイルス株のG蛋白質の細胞質尾部が、生存の支持(即ち、ニューロンネットワーク統合性の保存であり、狂犬病ウイルス伝播を支持する) 対 アポトーシスの支持(即ち、細胞死であり、免疫原性を支持する)の決定プロセスの指令に重大な役割を有することの実証を本発明は提供する。
【0040】
さらに本発明は、生存支持戦略(pro-survival strategy)の予想外の効果を明らかとする。
【0041】
実際, 本発明は、幾つかの非アポトーシス性(且つ病原性)の狂犬病ウイルス株(より具体的には、CVS-NIV株)のG 蛋白質が神経突起成長促進効果を有すること、即ち、これらの非アポトーシス性の狂犬病ウイルス G 蛋白質が神経突起生成を誘導および/または刺激することを示す。
【0042】
この効果は、アポトーシス性の 狂犬病ウイルス G 蛋白質(例えば、ERA および 他の弱毒化した狂犬病ウイルス株のG 蛋白質)により示されない。
【0043】
さらにまた、この効果は、全てのアポトーシス性および/または病原性の狂犬病ウイルス G 蛋白質により示されない。より具体的には、この効果はCVS-Gif-sur-Yvette 株のG蛋白質では示されないにもかかわらず(Prehaud et al. 1988)、このG蛋白質はCVS-NIV 株のG蛋白質と高い同一性 スコアを有する(図 17を参照されたい)。前記効果は、CVS-11 株(例えば、ACA57830, AAC34683, ABV24348)の G 蛋白質では示されないにもかかわらず、これらの G 蛋白質もCVS-NIV 株のG蛋白質と高い同一性スコアを有する(図 18を参照されたい)。N2CまたはCVS-24 株のG 蛋白質のどちらでも前記効果は示されない。
【0044】
従って、病原性の狂犬病ウイルス株の全てとは限られない株が、神経突起成長において有意に陽性の効果を示すG蛋白質を有する。
【0045】
このようなことから、発明者は、非アポトーシス性(且つ病原性)の狂犬病ウイルス株の亜群を同定した、この株のG蛋白質は神経突起成長に有意に陽性の効果を有する。この亜群の代表的な株は、CVS-NIV 株である。これらの G 蛋白質は、本明細書等においてCVS-NIV タイプのG 蛋白質と参照されえる。
【0046】
CVS-NIV 株は、2009年4月1日にCNCMに受託番号I-4140で寄託された。
【0047】
CVS-NIV 株のG蛋白質を発現しているプラスミドは、2001年11月30日にCNCMに受託番号I-2578で寄託された。
【0048】
CVS-NIV 株のG蛋白質は、Prehaud 等2003に記載されている。
【0049】
CVS-NIV 株のG蛋白質の配列は、アクセッション番号AF 406694で利用可能である。
【0050】
CVS-NIV 株のG蛋白質の配列は、図 13Aに示される配列番号 2の配列である。
【0051】
狂犬病ウイルス G蛋白質は、ウイルスエンベロープの三量体スパイクを形成し、感染細胞の膜に見出されるI型 膜貫通糖蛋白質である。狂犬病ウイルス G蛋白質は、外部ドメイン, 膜貫通セグメント および 44アミノ酸の長い細胞質ドメインからなる524アミノ酸長の蛋白質である。
【0052】
このC末端ドメインのまさに最後の 4 アミノ酸が、PDZ 結合サイト (PDZ-BS)を形成する。
【0053】
PDZ (PSD-95, Discs Large, ZO-1)ドメインは、パートナー蛋白質のC 末端配列が挿入されえるソケットを作る六つのベータ鎖および二つのアルファヘリックスに組織化される80-100 aaの球状の構造を形成する。
【0054】
PDZ 結合サイト (PDZ-BS)は4 アミノ酸配列であり; PDZ-BS 配列は配列番号 13の配列である、すなわち:
x1 - x2 - x3 - x4,であり、式中の:
x1 は、任意のアミノ酸であり、
x2 は、TまたはSまたはIであり、
x3 は、任意のアミノ酸であり、
x4 は、LまたはVである。
【0055】
特に、本発明は、この神経突起成長効果を担うのがCVS-NIV タイプの前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部であることを実証する。
【0056】
さらに、本発明は、この効果が特にPDZ-BSが原因であることを実証する。PDZ-BSは、CVS-NIV タイプの前記非アポトーシス性の狂犬病ウイルス G 蛋白質の細胞質尾部に含まれる。
【0057】
CVS-NIV タイプの非アポトーシス性の狂犬病ウイルス G 蛋白質のPDZ-BSモチーフは、アポトーシス性の狂犬病ウイルス G 蛋白質の一つと比べて単一の点変異を示す。この単一の点変異は最初のPDZ-BS アミノ酸に関し、これは非アポトーシス性の 狂犬病ウイルス G 蛋白質においてEではない。
【0058】
さらに、本発明は、前記アミノ酸がCVS-NIV 株の全長の非アポトーシス性の狂犬病ウイルス G蛋白質配列においてポジション 491に存在し、これが神経突起成長効果に寄与することを実証する。このアミノ酸は、CVS-NIV 株のG蛋白質中でHである(配列番号 2を参照されたい)。全長 G蛋白質におけるポジション 491は、この蛋白質の細胞質断片におけるポジション 11に対応する(図 15の配列番号 6)。
【0059】
CVS-NIV 株の全長の非アポトーシス性の狂犬病ウイルス G蛋白質配列(配列番号 2)中のポジション 491 および 521のアミノ酸は、双方が本発明の神経突起成長効果に重要であることを発明者は実証する。より具体的には、発明者は、ポジション 491でのH (しかし、Lではない)およびポジション 521でのQ (しかし、Eではない)は、双方が本発明の神経突起成長効果を支持することを実証する。
G 蛋白質がH491Lの単一の点変異を生じる又はQ521Eの単一の点変異を生じるCVS-NIV 株と異なる変異体ウイルスが、発明者により構築され、作出された(CNCM I-4142 および I-4143)。
【0060】
H491L 変異体 G蛋白質は、なおもポジション 521にQを有する。
【0061】
Q521E 変異体 G蛋白質は、なおもポジション 491にHを有する。
【0062】
双方の変異体蛋白質は、双方がH をポジション 491にQ をポジション 521に有し、CVS-NIV G蛋白質よりも小さい程度ではあるが、有意に陽性の神経突起成長効果を誘導する。
【0063】
これらの変異体 G 蛋白質の細胞質断片は、図 19 (配列番号 19 および 配列番号 20)に示される。
【0064】
本発明のポリペプチドの規定:
本発明は、以下のポリペプチドに関する:
・ 以下を含むポリペプチド:
・ 配列番号 6の配列, 株 I-4140のG 蛋白質の細胞質断片, プラスミド I-2578により産生されたG 蛋白質の細胞質断片, 配列 AF406694のG 蛋白質の細胞質断片, 配列番号 2のG 蛋白質の細胞質断片から選択される親配列,
又は
・ 前記親配列の保存されたバリアント配列,
又は
・ 前記親配列の又は前記保存されたバリアント配列の保存された断片。
【0065】
前記親配列, 保存されたバリアント配列および保存された断片配列は、有利に神経突起成長を誘導する及び/又は刺激する機能を有し、例えば、以下に記載されるように、より具体的には、ヒトの分裂前のニューロンからの〔例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの〕神経突起成長を誘導する及び/又は刺激する機能を有する。
【0066】
より具体的には、本発明は、神経突起成長の刺激および/または誘導に使用するための〔より具体的には、不十分な又は障害性の神経突起成長、特に不十分な又は障害性の神経突起生成に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドに関する。
【0067】
好ましくは、前記ポリペプチドのアミノ酸長は、100 アミノ酸未満, より好ましくは90 アミノ酸未満である。
【0068】
前述のバリアント蛋白質の配列は、任意の既知の又は同定可能な狂犬病ウイルス G蛋白質に対応しない配列であってもよい。
【0069】
実際、通常の能力を発揮する当業者は、所与の狂犬病ウイルス G蛋白質の配列から出発して、一つ又は幾つかのアミノ酸の置換および/または付加および/または欠失をなすことができ、なおも神経突起成長を誘導する及び/又は刺激する能力を保持していることを認識する。かかる保存されたアミノ酸の置換および/または付加および/または欠失は、本発明に包含される。
【0070】
理論に縛られることなく、次の八つのグループの各々は、通常互いに保存された置換であるアミノ酸を含んでいると考えられる:
1) アラニン (A), グリシン (G);
2) アスパラギン酸 (D), グルタミン酸 (E);
3) アスパラギン (N), グルタミン (Q);
4) アルギニン (R), リジン (K);
5) イソロイシン (I), ロイシン (L), メチオニン (M), バリン (V);
6) フェニルアラニン (F), チロシン (Y), トリプトファン (W);
7) セリン (S), スレオニン (T); および
8) システイン (C), メチオニン (M)。
【0071】
以上のように、前述のバリアント蛋白質は、特に、人間により操作され、一もしくは数個のアミノ酸の置換および/または付加および/または欠失により前記狂犬病ウイルス G 蛋白質と異なる蛋白質を包含する〔但し、生じるバリアント蛋白質は、神経突起成長、より具体的には、ヒトの分裂前のニューロンから、例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長を誘導する及び/又は刺激する能力をなおも有する〕。
【0072】
好ましくは, 前記保存されたバリアント配列は以下の配列である:
・ 前記親配列のバリアント配列, この配列は:
・ 34〜54アミノ酸の配列であり,
・ 親およびバリアントの配列のうちの最も短い配列の全長(entire length)に対し前記親配列と少なくとも 94% 同一であり,
・ PDZ-BS配列 (好ましくは、最後の四つのC末端のアミノ酸)を含み, 前記PDZ-BS配列は以下のものであり:
x1 は、E以外の任意のアミノ酸(好ましくは、Q)であり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)であり
(配列番号14),
前記バリアント配列はバリアント 配列 Aと称される,
又は
・ 前記親配列のバリアント配列, この配列は:
・ 44 アミノ酸の配列であり,
・ 前記親配列と少なくとも 94% 同一であり,
・ PDZ-BS配列 (好ましくは、最後の四つのC末端のアミノ酸)を含み, 前記PDZ-BS配列は以下のものであり:
x1 は、Eであり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)であり
(x1 = Eの配列番号 13),
並びに
・ アミノ酸 L をポジション 11に含まなく(好ましくは、アミノ酸 H をこのポジションに含む),
前記バリアント配列はバリアント 配列 Bと称される。
【0073】
好ましくは、前記配列同一性は、少なくとも 95% (好ましくは、少なくとも 96%, より好ましくは、少なくとも 97%, なおより好ましくは、少なくとも 97.5%, さらになおより好ましくは、少なくとも 98%, 最も好ましくは、少なくとも 98.5%, なお最も好ましくは、少なくとも 99%)である。
【0074】
より好ましくは, 前記保存されたバリアント配列は、バリアント 配列 Aである。
【0075】
好ましくは, 前記保存された断片は、それぞれ前記親配列の又は前記バリアント配列 Aの又は前記バリアント配列 Bの保持されたPDZ-BS 配列を有する少なくとも 34 アミノ酸断片である。
【0076】
簡潔さのため、「ポリペプチド」の用語は、本明細書等において蛋白質が包含されることを意図しており、また逆に。
【0077】
この用語は、転写後の修飾及び/又は合成化学〔例えば、非ペプチド性の化学基の付加により及び/又はポリペプチドの三次構造の修飾により、例えば、アセチル化, アシル化, ヒドロキシル化, 環化, ラセミ化, リン酸化, など〕により修飾され、生じる修飾されたポリペプチドが神経突起成長〔より具体的には、ヒトの分裂前のニューロンからの、例えば、ヒト神経芽細胞腫細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する能力を保持するポリペプチド(または蛋白質)も包含する。
【0078】
有利なことに、本発明のポリペプチドは、それ自身で神経向性(neurotropic)である。
【0079】
それでも、必要とされる又は所望される場合、本発明のポリペプチドは、神経向性(より具体的には、脳のニューロンへの向性)を改善する因子〔例えば、Kumar等(Kumar et al. 2007)により記載されたRVG-9R〕と連結できる又は融合できる。
【0080】
アミノ酸配列およびヌクレオチド配列は、標準の配向(即ち、アミノ酸配列に関してN-末端からC-末端、またヌクレオチド配列に関して5'-末端から3'-末端)にしたがって本明細書等に提供される。
【0081】
本発明の態様によると、前記バリアント配列 AのPDZ-BS 配列は、QTRL (配列番号 10)である。
【0082】
本発明の態様によると、前記バリアント 配列 Aは、44 アミノ酸の配列であり、アミノ酸 HまたはL, 好ましくは Hをポジション 11に有する。
【0083】
例示のバリアント 配列 Aは、44アミノ酸の配列であり、アミノ酸 L をポジション 11に有する配列番号 19の配列である(図 19を参照されたい)。
【0084】
本発明の態様によると、前記バリアント配列 BのPDZ-BS 配列は、ETRL (配列番号 12)である。
【0085】
本発明の態様によると、前記バリアント 配列 Bは、アミノ酸 H をポジション 11に有する。
【0086】
例示のバリアント 配列 Bは、44アミノ酸の配列であり、アミノ酸 H をポジション 11に有する配列番号 20の配列である(図 19を参照されたい)。
【0087】
本発明の態様によると、前記断片は、配列番号 6の配列の断片 11〜44である。
【0088】
本発明のポリペプチドは、神経突起成長支持配列およびG 狂犬病ウイルス 蛋白質の膜貫通部分または当業者が細胞(例えば、以下に規定される細胞)の膜上又は膜中に前記ポリペプチドをアンカーするため適切なものを見出しえる任意の他のアンカーする手段を含んでもよい又は該手段からなってもよい。
【0089】
本発明のポリペプチドは、操作された抗体(例えば、単鎖抗体)の構造中に挿入できる。
【0090】
本発明の態様によると、前記ポリペプチドは、狂犬病ウイルス株のG蛋白質, 又はその細胞質断片(例えば、配列番号 6), 又は係る細胞質断片の小断片(例えば、配列番号 6の断片11〜54)である。前記狂犬病ウイルス G蛋白質は、より好ましくは非アポトーシス性の狂犬病ウイルス G蛋白質である。例えば、前記 狂犬病ウイルス株は、CNCMにI-4140で寄託された株である。
【0091】
本発明の態様によると、前記ポリペプチドは、係る細胞質断片または小断片のバリアント 配列 A またはB (上記規定のとおり)を含む又はそれからなる。例えば、前記 ポリペプチドは、G 蛋白質がI-4140 株のG 蛋白質の単一の点変異体蛋白質であるCNCMにI-4142又はI-4143で寄託された狂犬病ウイルス株のG 蛋白質の細胞質断片(図 21を参照されたい)を含む又は該断片からなる(それぞれ配列番号 19 および 20の細胞質断片; 図19を参照されたい)。
【0092】
CNCMは、Collection Nationale de Cultures de Microorganismes(Collection Nationale de Cultures de Microorganismes ; Institut Pasteur ; 28, rue du Docteur Roux ; F-75724 Paris Cedex 15 ; France)である。前記寄託は、ブタペスト条約の条件下でなされた。
【0093】
バリアント配列 A:
本発明のバリアント 配列 Aは配列番号 14の配列、即ち、配列 x1 - x2 - x3 - x4,を含み、式中の:
x1 は、E以外の任意のアミノ酸であり、
x2 は、TまたはSまたはIであり、
x3 は、任意のアミノ酸であり、
x4 は、LまたはVである。
【0094】
この配列は、特定の PDZ-BS モチーフである。この配列は通常は前記ポリペプチドのC末端に局在する。通常、この配列は前記ポリペプチドの最後の四つのC末端アミノ酸の配列である。
【0095】
好ましくは, 前記アミノ酸 x1 は、Qである。
【0096】
好ましくは, 前記アミノ酸 x2 は、Iではなく、好ましくは, Tである。
【0097】
好ましくは, 前記アミノ酸 x3 は、Rである。
【0098】
好ましくは, 前記アミノ酸 x3 は、Lである。
【0099】
好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、TまたはSまたはIであり、
x3 は、Rであり、
x4 は、LまたはVである。
【0100】
最も好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、Tであり、
x3 は、Rであり、
x4 は、LまたはVである。
【0101】
最も好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、TまたはSまたはIであり、
x3 は、Rであり、
x4 は、Lである。
【0102】
より好ましくは, 前記PDZ-BSモチーフの配列は配列番号 14の配列であり、配列中の:
x1 は、Qであり,
x2 は、Tであり、
x3 は、Rであり、
x4 は、Lである、
即ち、これは配列番号 10の配列である(図 16を参照されたい)。
【0103】
このQTRL配列は、CVS-NIV 株(前記株は受託番号I-4140でCNCMから利用可能である)のG蛋白質のPDZ-BS モチーフの, および/または受託番号 I-2758でCNCMから利用可能であるプラスミド(組換え型の 大腸菌含んでいる 前記 プラスミド)によりコードされた蛋白質の, および/または配列番号 2の蛋白質(図 13Aを参照されたい)の配列である。
【0104】
CVS-NIVのG蛋白質をコードしているプラスミド CNCM I-2758を含んでいる組換え型の 大腸菌株を培養するための適切な条件には、標準のLB-TYM 成長培地(アンピシリンの存在下)で前記組換え型の 大腸菌株を37°Cでインキュベーションすることを含む(WO 03/048198を参照されたい)。
【0105】
ウイルス I-4140 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0106】
好ましくは、本発明のバリアント 配列 Aは、44 アミノ酸の配列であり、アミノ酸HまたはL(好ましくは H)を44aa-配列のポジション 11に有する。
【0107】
例示のバリアント A ポリペプチドは、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株(その株のG 蛋白質はI-4140のG 蛋白質の単一の点変異体蛋白質である; 図 21を参照されたい)のG 蛋白質, その細胞質断片およびその保存された細胞質小断片を含む。
【0108】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0109】
ウイルス I-4142 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0110】
例示のバリアント A ポリペプチドは、特に配列番号 19の細胞質断片又は少なくとも一つのその保存された小断片を含んでいるポリペプチドを含む。
【0111】
配列番号 19の配列は、配列番号 6の配列の例示の保存されたバリアントである(図 19を参照されたい)。配列番号 19の配列は配列番号 6の配列よりも低いレベルにもかかわらず、陽性の神経突起成長効果をなおも有する(図 3D および例のセクションにおける関連するコメントを参照されたい)。
【0112】
バリアント 配列 B:
本発明のバリアント 配列 Bは、44 アミノ酸の配列である。
【0113】
この配列は、配列番号 13にx1 = Eを有する配列である、即ち、配列 x1 - x2 - x3 - x4を含み、配列中の:
x1 は、Eであり,
x2 は、TまたはSまたはI (好ましくはIではなく, より好ましくは T)であり,
x3 は、任意のアミノ酸(好ましくは、R)であり,
x4 は、LまたはV(好ましくは、L)である。
【0114】
この配列は、特定の PDZ-BS モチーフである。この配列は、通常は前記ポリペプチドのC末端に局在する。通常、この配列はポリペプチドの最後の四つのC末端アミノ酸の配列である。
【0115】
好ましくは, 配列番号 13の配列は、ETRLである(配列番号 12)。
【0116】
本発明のバリアント 配列 Bにおいて、ポジション 11にあるアミノ酸は、Lではなく、最も好ましくはHである。
【0117】
例示のバリアント B ポリペプチドは、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株(そのG 蛋白質はI-4143のG 蛋白質の単一の点変異体蛋白質である; 図 21を参照されたい)のG 蛋白質, その細胞質断片およびその保存された細胞質小断片を含む。
【0118】
I-4143 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 E をポジション 521 (全長のG蛋白質に関して計算したポジション)に Q の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0119】
ウイルス I-4143 (組換え型の 狂犬病ウイルス)の繁殖のための適切な条件は、グルコース (例えば、 4.5 g/L), ピルビン酸ナトリウムおよびglutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地において前記ウイルスを37°Cで5% CO2存在下でBHK-21 細胞 (sub-clone BSR)とインキュベーションすることを含む(以下の例 1 を参照されたい)。
【0120】
例示のバリアント B ポリペプチドは、特に配列番号 20の細胞質断片又は少なくとも一つのその保存された小断片を含んでいるポリペプチドを含む。
【0121】
配列番号 20の配列は、例示の配列番号 6の配列の保存されたバリアントである(図 19を参照されたい)。配列番号 20の配列は配列番号 6の配列よりも低いレベルにもかかわらず、陽性の神経突起成長効果をなおも有する(図 3D および例のセクションにおける関連するコメントを参照されたい)。
【0122】
I-4142 および I-4143 株のG 蛋白質は、CVS-NIV 株 (I-4140)のG蛋白質の一つよりも有意に神経突起成長効果が低いが、有意に陽性の神経突起成長効果を誘導する(例 1 および 図 3Dを参照されたい)。
【0123】
従って、二つのアミノ酸ポジションの各々は、神経突起成長効果に寄与し、そのポジションは、即ち:
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、ポジション 491 (即ち、CVS-NIV 株のG蛋白質におけるH)であり、
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、CVS-NIV 株のG 蛋白質におけるポジション 521であり、即ち、G 蛋白質のPDZ-BSの最初のアミノ酸 (即ち、CVS-NIV 株のG蛋白質におけるアミノ酸 Q)である。
【0124】
本発明のポリペプチドの代替の又は相補的な規定:
本発明のポリペプチドは、神経突起成長支持ポリペプチド(pro-neurite outgrowth polypeptide)である。
【0125】
本発明のポリペプチドは、上記規定の親配列および/または保存されたバリアント配列(バリアント配列AまたはB)および/または保存された断片配列を含む。
【0126】
択一的に又は相補的に(Alternatively or complementarily)、本発明のポリペプチドは、狂犬病ウイルス株のG蛋白質, 又はそのバリアント配列のG蛋白質(このバリアント配列は前記G蛋白質から一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来する),または係るG蛋白質またはバリアントG蛋白質の断片配列, より具体的には、これらの細胞質断片または小断片であると規定できる。但し、前記ポリペプチドは、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を有する。
【0127】
より具体的には、本発明のポリペプチドは、択一的に又は相補的に、非アポトーシス性の狂犬病ウイルス株のG蛋白質, 又はそのバリアント配列の蛋白質(このバリアント配列は前記G蛋白質から一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来する),または係るG蛋白質またはバリアント蛋白質の断片配列, より具体的には、これらの細胞質断片または小断片であると規定できる。但し、前記バリアント配列または前記断片は、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を保持する。
【0128】
CVS-NIV株のG 蛋白質ではない及び/又はCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質以外である, および/または配列番号 2の蛋白質以外であるが、しかし、なおも適切な神経突起成長支持ポリペプチドである例示の神経突起成長支持蛋白質は、特に2009年4月1日にCNCMに受託番号I-4142 および I-4143で寄託された組換え型の狂犬病ウイルス株のG 蛋白質を含む。
【0129】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0130】
I-4143 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 E をポジション 521 (全長のG蛋白質に関して計算したポジション)に Q の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0131】
I-4142 および I-4143 株のG 蛋白質は、CVS-NIV 株 (I-4140)のG蛋白質の一つよりも有意に神経突起成長効果が低いが、有意に陽性の神経突起成長効果を誘導する。
【0132】
従って、二つのアミノ酸ポジションの各々は、神経突起成長効果に寄与し、そのポジションは、即ち:
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、ポジション 491 (即ち、CVS-NIV 株のG蛋白質におけるH)であり、
・ CVS-NIV 株のG蛋白質におけるアミノ酸ポジションが、CVS-NIV 株のG 蛋白質におけるポジション 521であり、即ち、G 蛋白質のPDZ-BSの最初のアミノ酸(即ち、CVS-NIV 株のG蛋白質におけるアミノ酸 Q)である。
【0133】
それ故, 例示の神経突起成長支持 ポリペプチドは、I-4142 株のG蛋白質およびI-4143 株のG蛋白質、同様に、これらの蛋白質の断片、より具体的には、これらの細胞質断片(図 19における配列番号 19および配列番号 20)及びこれらの細胞質小断片を含む。但し、これらの小断片は、神経突起成長〔例えば、以下に説明されるように、より具体的には、ヒトの分裂前のニューロンからの神経突起成長、例えば、ヒト神経芽細胞腫の細胞株 SH-SY5Y (ATCC CRL-2266; 例証の実験条件および材料に関して以下の例 1を参照されたい)からの神経突起成長〕を誘導する及び/又は刺激する機能を保持する。
【0134】
より具体的には、本発明は、神経突起成長を刺激および/または誘導することに使用するための〔より具体的には、不十分な又は障害性の神経突起成長、特に不十分な又は障害性の神経突起生成に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドに関する。
【0135】
好ましくは、前記ポリペプチドのアミノ酸長は、100 アミノ酸未満, より好ましくは90 アミノ酸未満である。
【0136】
「非アポトーシス性の狂犬病ウイルスG蛋白質(non-apoptotic rabies virus G protein)」の表現は、本明細書等において当該分野の通常の意味を意図している。
【0137】
非アポトーシス性の狂犬病ウイルスG蛋白質のPDZ-BSは、配列番号 14のものである。
【0138】
非アポトーシス性の狂犬病ウイルスG蛋白質は、その配列が狂犬病ウイルス株のG蛋白質の配列であること、ヒトニューロンのアポトーシスを惹起しないことの事実により付加的に又は択一的に特徴付けられることができる。
【0139】
候補の狂犬病ウイルス G蛋白質が非アポトーシス発生性(non-apoptogenic)のものであることを検査する例示の手段は、当業者に知られている。
【0140】
その手段の一つには、この G蛋白質を含み、ニューロン, より具体的には、 ヒトニューロン, 例えば、SK-N-SH 神経芽細胞腫 細胞株 (ATCC HTB11)またはSH-SY5Y 神経芽細胞腫 細胞株 (ATCC CRL-2266), 好ましくは SK-N-SH 神経芽細胞腫細胞株(ATCC HTB11)に感染する際に非アポトーシス性である狂犬病ウイルス株を検査することが含まれる。かかる手段は、候補G蛋白質を含んでいる天然の狂犬病ウイルス株が分析のため利用可能である場合に特に有用である。
【0141】
他の手段には、遺伝的に細胞を操作して候補 G蛋白質を発現させること、ニューロン細胞を前記の発現された候補G蛋白質で感染させること, およびアポトーシスが前記感染により誘導されないことを決定することが含まれる。
【0142】
かかる手段の例示は、Prehaud等(Prehaud et al. 2003)に記載される。
【0143】
例示の遺伝的に操作した細胞には、Prehaud et al.2003, より具体的には、10538頁(「誘導性のトランスジェニック細胞株」の部分を参照されたい)に記載されるトランスジェニックJurkat細胞系統が含まれる。
【0144】
例示のニューロン細胞には、SK-N-SH 神経芽細胞腫細胞株 (ATCC HTB11)またはSH-SY5Y 神経芽細胞腫細胞株 (ATCC CRL-2266), 好ましくはSK-N-SH 神経芽細胞腫細胞株 (ATCC HTB11)が含まれる。
【0145】
ニューロン細胞を感染させる遺伝的に操作した細胞を作るための例示の実験条件は、Prehaud等 (Prehaud et al.2003), より具体的には、10538頁(「誘導性のトランスジェニック細胞株」の部分を参照されたい)に記載されるものを含む。
【0146】
狂犬病ウイルス株または遺伝的に操作された細胞がニューロン細胞のアポトーシスを誘導しないことの検出は、当業者の通常の技術の範囲内である。
【0147】
例示の手段は、Prehaud等 (Prehaud et al. 2003)およびPrehaud等 (Prehaud et al. 2005)に記載のものを含む。
【0148】
例示の手段は、例えば、ヘキスト染色 (Prehaud et al. 2003, より具体的には、10538頁, § 「Detection of nuclear fragmentation by Hoechst staining」を参照されたい), および/またはTUNEL 法(Prehaud et al. 2003, より具体的には、10538頁を参照されたい(§ 「Detection of nuclear fragmentation by the TUNEL method」を参照されたい), および/またはDNA電気泳動法 (Prehaud et al. 2005を参照されたい)によりDNA 断片化が誘導されないことを検出することを含む。
【0149】
有意なDNA 断片化が誘導されないことを検出する付加的な又は代替の例示の手段は、例えば、Prehaud et al. 2003(より具体的には、10538頁, 「Detection of caspase activation」)に記載の手順にしたがってカスパーゼ 8が活性化されないことの検出を含む(但し、使用されたニューロン細胞がカスパーゼ 8を含むべきであり、ここではSK-N-SH 細胞株の場合である)。
【0150】
非有意なアポトーシスの例示の値は、Prehaud等 2003(「JrtTA-G-CVS」の表題の最後から二番目の欄を参照されたい)の図 5Dに示される以下の事項である:
・ ヘキスト染色により測定される僅か19%のアポトーシス細胞;
・ カスパーゼ 8の活性化の評価により測定される僅か4.7%のアポトーシス細胞。
【0151】
弱毒化した株(即ち、非病原性、例えば、免疫応答性マウスの筋肉内に注射された際に非病原性)である狂犬病ウイルス株、例えば、弱毒化したERA株, RV194-2 株, AVO-1 株, SN10 株, SN-10-SAD 株, SAG2 株の狂犬病ウイルス株は、アポトーシス発生性の株である。
【0152】
病原性(即ち、インビボ神経毒性)の狂犬病ウイルス株(例えば、CVS-NIV 株)は、非アポトーシス性(non-apoptotic)である。
【0153】
従って、非アポトーシス発生性の狂犬病ウイルス株は、病原性の(神経毒性の)株である(例えば、免疫応答性マウスの筋肉内に注射された際に病原性である)。
【0154】
現在の知識は、以下のものである:
・ 非アポトーシス発生性の狂犬病ウイルス株がヒトニューロン細胞に感染する際に、それがコードするG蛋白質は、核周囲に球状構造の形態で前記ニューロン細胞の細胞質に蓄積し、前記ニューロン細胞の細胞質に拡散して分布するのではなく;
・ それと反対に、アポトーシス発生性の狂犬病ウイルス株がヒトニューロン細胞に感染する際に、それがコードするG蛋白質は、核周囲に球状構造の形態で前記ニューロン細胞の細胞質に蓄積しないが、しかし、前記ニューロン細胞の細胞質に拡散して分布する。
【0155】
上述し且つ以下に説明されるとおり、本発明のポリペプチドは、狂犬病ウイルス株のG蛋白質であってもよい。
【0156】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CVS-NIV 株のG蛋白質〔前記株は、CNCMから受託番号I-4140 (寄託日は2009年4月1日である)で利用可能である〕, および/またはCNCMから受託番号I-2758(寄託日は2001年11月30日である)で利用可能なプラスミドによりコードされた蛋白質および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり、前記バリアント狂犬病ウイルス G 蛋白質のPDZ-BS モチーフの配列はなおも配列番号 14の配列である蛋白質。
【0157】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって:
・ 前記バリアント蛋白質は、i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも95%同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列からなり、
・ 前記バリアント 狂犬病ウイルス G蛋白質のPDZ-BS モチーフの配列は、配列番号 14の配列である蛋白質。
【0158】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり:
・ 前記バリアント蛋白質は、i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも95%同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列からなり、
・ 前記バリアント 狂犬病ウイルス G蛋白質のPDZ-BS モチーフの配列は、配列番号 14の配列である蛋白質。
【0159】
好ましくは, 前記狂犬病ウイルスG蛋白質は、以下のものである:
i. CNCMから受託番号 I-4140で利用可能なCVS-NIV 株のG蛋白質, および/またはCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質, および/または配列番号 2の蛋白質;または
ii. i.の前記狂犬病ウイルス G 蛋白質のバリアント蛋白質であって、前記バリアント蛋白質はなおも狂犬病ウイルス G 蛋白質であり、前記バリアント蛋白質は以下の配列からなるバリアント蛋白質:
・ i.の前記狂犬病ウイルス G蛋白質の配列の全長にわたりi.の前記狂犬病ウイルス G蛋白質の配列と少なくとも 95% 同一(好ましくは少なくとも 96%, より好ましくは少なくとも 97%, なおより好ましくは少なくとも 97.5%, さらになおより好ましくは少なくとも 98%, 最も好ましくは少なくとも 98.5%, なお最も好ましくは少なくとも 99%)である配列, および
・ i.の前記狂犬病ウイルス G蛋白質の配列番号 14のPDZ-BS モチーフを保持する及び/又はQTRLの配列のPDZ-BS モチーフ(x1=Q; x2=T; x3=R; x4=Lを有する配列番号 14)を含む配列。
【0160】
好ましくは、ii.の前記バリアント狂犬病ウイルス G蛋白質のアミノ酸長は、i.の前記狂犬病ウイルス G蛋白質の長さ50 アミノ酸以上超えない。
【0161】
好ましくは、ii.の前記バリアント狂犬病ウイルス G蛋白質のアミノ酸長は、i.の前記狂犬病ウイルス G蛋白質の長さより50 アミノ酸以上短くない(例えば、iの前記 狂犬病ウイルス G蛋白質の配列と同じ長さである)。
【0162】
より好ましくは、ii.の前記バリアント狂犬病ウイルス G 蛋白質が含むアミノ酸の長さは、少なくともi.の前記狂犬病ウイルス G 蛋白質の長さマイナス50 アミノ酸であり、多くともi.の前記狂犬病ウイルス G 蛋白質の長さプラス 50 アミノ酸である(例えば、iの前記狂犬病ウイルス G 蛋白質の配列と同じ長さである)。
【0163】
CVS-NIV株(I-4140)のG 蛋白質ではない及び/又はCNCMから受託番号I-2758で利用可能なプラスミドによりコードされた蛋白質以外である, および/または配列番号 2の蛋白質以外であるが、しかし、なおも適切な神経突起成長支持狂犬病ウイルス G 蛋白質である例示の神経突起成長支持狂犬病ウイルス G 蛋白質は、特に2009年4月1日にCNCMに受託番号I-4142 で寄託された組換え型の狂犬病ウイルス株のG 蛋白質を含む。
【0164】
I-4142 株はCVS-NIV株(CNCM I-4140)と異なり、その異なる点はG蛋白質がアミノ酸 L をポジション 491 (全長のG蛋白質に関して計算したポジション)に H の 代わりに有することである(以下の表 2 および 図 3Dを参照されたい)。
【0165】
配列番号 19の配列の例示の断片(図 19を参照されたい)。
【0166】
付加的に存在しえる他の配列:
上述し且つ以下に説明されるとおり、本発明のポリペプチドは、神経突起成長支持〔および/または神経突起の発芽(spouting)および/または軸索の成長および/または樹状突起樹の伸展(dendritic tree extension)〕の効果を有するアミノ酸配列を含む。
【0167】
上述し且つ以下に説明されるとおり、前記神経突起成長支持配列は、上記規定のとおり、親配列 および/または 保存されたバリアント配列(バリアント配列 AまたはB)および/または 保存された断片配列と規定できる。
【0168】
上述し且つ以下に説明されるとおり、前記神経突起成長支持配列は、択一的に又は相補的に(alternatively or complementarily)、(非アポトーシス性の) 狂犬病ウイルス株 (例えば、CNCM I-4140, I-4142またはI-4143のG 蛋白質)のG 蛋白質の配列と,または一もしくは数個のアミノ酸の置換および/または付加および/または欠失に由来するそのバリアント配列,または係る G 蛋白質またはバリアント G 蛋白質の断片配列, より具体的には、その細胞質断片または小断片と規定できる。
【0169】
G 蛋白質またはバリアント G 蛋白質の係る断片の例示は、特に前記 G 蛋白質またはバリアント G 蛋白質の細胞質ドメイン および 膜貫通ドメインを保持するその断片を含む。
【0170】
係る膜貫通ドメインは、特に前記細胞の細胞内区画において(より具体的には、前記細胞の小胞体および/またはゴルジ膜における)細胞質断片をアンカーして、前記細胞質断片が神経発生, 神経再生 および 神経保護の刺激および/または誘導する効果をより効率的に発揮させるとの利点を有する。以下の例4 および 5を参照されたい。
【0171】
上述のことから、本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜において前記ポリペプチドをアンカーする配列を含んでもよく、前記アンカーする配列は好ましくは前記神経突起成長支持配列のN末端にあり, 最も好ましくは前記神経突起成長支持配列のN末端での第一のアミノ酸に直接的に連結される。
それ故, 本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株、例えば、例4)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜(好ましくは、小胞体膜及びゴルジ膜)において前記ポリペプチドをアンカーするアミノ酸配列を含んでもよい。
【0172】
前記アンカーする配列は、好ましくは配列番号 6の配列またはバリアント 配列 Aまたはバリアント 配列 BのN末端に存在する。最も好ましくは, 前記アンカーする配列は、直接的もしくは間接的に(好ましくは、直接的に)配列番号 6の配列の又は前記バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結される。
【0173】
係るアンカーする配列は、例えば、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例、例えば、例4に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株)の小胞体膜(endoreticulum membrane)および/またはゴルジ膜において配列番号 6の細胞質ドメインをアンカーする能力を有するアミノ酸配列であってもよい。
【0174】
係るアンカーする配列の例示は、狂犬病ウイルス株(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメイン配列、好ましくは狂犬病ウイルス CVS-NIVのG蛋白質の膜貫通ドメイン配列(配列番号 23)である。
【0175】
上述のことから、前記ポリペプチドは、配列番号 6のまたは前記バリアント配列 Aの又は前記バリアント配列 Bの配列に加えて、さらに好ましくは以下の配列を、配列番号6の配列またはバリアント配列 Aまたはバリアント配列 BのN末端に含んでもよく, 最も好ましくは直接的に配列番号6の配列または前記 バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結されてもよい:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメインの配列, または
・ 狂犬病ウイルス G の膜貫通ドメインの配列と少なくとも一つのアミノ酸の置換および/または欠失および/または付加, 好ましくはアミノ酸の置換および/または欠失, より好ましくはアミノ酸置換により異なるが、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)(例えば、以下の例、例えば、例4に記載のヒト神経芽細胞腫細胞株 SH-SY5Y 細胞株)の小胞体膜および/またはゴルジ膜において、前記 ポリペプチドをアンカーする能力(より具体的には、前記アンカーする配列が配列番号 6-の前記配列のN末端で第一のアミノ酸に直接的に連結される場合、配列番号 6 の細胞質ドメインをアンカーする能力)を保持する配列。
【0176】
それ故, 本発明のポリペプチドは、前記神経突起成長支持配列に加えて、さらに好ましくは以下の配列を、配列番号6の配列またはバリアント配列 Aまたはバリアント配列 BのN末端に含んでもよく, 最も好ましくは直接的に配列番号6の配列または前記 バリアント配列 Aの又は前記バリアント配列 BのN末端での最初のアミノ酸に連結されてもよい:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の膜貫通ドメインの配列, または
・ 狂犬病ウイルス G 膜貫通ドメインの配列と少なくとも一つのアミノ酸の置換および/または欠失および/または付加, 好ましくはアミノ酸の置換および/または欠失, より好ましくはアミノ酸置換により異なるが、細胞(より具体的にはニューロン細胞, より具体的にはヒト ニューロン細胞)の小胞体膜および/またはゴルジ膜において、(前記アンカーする配列が配列番号 6の配列のN末端で最初のアミノ酸に直接的に連結される場合に)配列番号 6のポリペプチドをアンカーする能力を保持する配列。
【0177】
係るアンカーする配列の好適な例は、特に以下を含む:
・ 狂犬病ウイルス株(より具体的には、非アポトーシス性の狂犬病ウイルス株、なおより具体的にはCVS-NIV 株)のG蛋白質の膜貫通ドメインの配列(例えば、配列番号 23の配列), または
・ そのバリアント配列であって、18〜26 アミノ酸, 好ましくは18 〜22 アミノ酸, より好ましくは22 アミノ酸の配列であり、二配列のうちの最短の配列にわたって(即ち、配列番号 23及び前記バリアント配列のうちの最短の配列にわたって)配列番号 23の配列と少なくとも 94% 同一である配列。
【0178】
係るアンカーする配列は、配列番号 6の配列または前記バリアント配列 A(より具体的には、配列番号 6の配列)である神経突起成長支持配列に連結される場合に特に有用である。
【0179】
それ故, 本発明の態様によると、本発明のポリペプチドのアミノ酸配列は、配列番号23の配列にN末端からC末端に配列番号6の配列が続く配列を含むか,または該配列からなり、1〜4アミノ酸, 好ましくは1アミノ酸(例えば、M)が任意で配列番号23の配列と配列番号6の配列との間に存在する。
【0180】
係るポリペプチドの例示は、配列番号 25のポリペプチドである。
【0181】
本発明のポリペプチドは、神経突起成長支持配列に加えて、さらに以下を含んでもよい:
・ 狂犬病ウイルス株(より具体的には非アポトーシス性の狂犬病ウイルス株, なおより具体的にはCVS-NIV 株)のG蛋白質の外部ドメインの断片, および/または
・ シグナルペプチド〔このシグナルペプチドは、好ましくは狂犬病ウイルス株(より具体的には非アポトーシス性の狂犬病ウイルス株, なおより具体的にはCVS-NIV 株)のG蛋白質のシグナルペプチドである〕。
【0182】
前記外部ドメイン断片および/またはシグナルペプチドは、前記アンカーする配列に加えて本発明のポリペプチドに存在してもよい。
【0183】
好ましくは, シグナルペプチドが前記ポリペプチドに含まれる場合、前記シグナルペプチドは前記ポリペプチドのN末端の最初の構成要素である。
【0184】
好ましくは, シグナルペプチドおよび外部ドメイン断片が前記ポリペプチドに含まれる場合、前記外部ドメイン断片は、本発明のポリペプチドに前記シグナルペプチドおよび前記神経突起成長支持配列の間(好ましくは、前記シグナルペプチドおよび前記ポリペプチドに含まれえる任意の可能なアンカーする配列の間)で含まれる。
【0185】
本発明の態様によると、本発明のポリペプチドは、N末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列を含む,またはN末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列からなる。
【0186】
本発明の別の態様によると、本発明のポリペプチドは、N末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記外部ドメイン断片(例えば、二つのアミノ酸 GK), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列を含む,または本発明のポリペプチドのN末端からC末端に前記シグナルペプチド(例えば、配列番号 21), 前記外部ドメイン断片(例えば、二つのアミノ酸 GK), 前記アンカーする配列(例えば、配列番号 23)および前記神経突起成長支持配列からなる。
【0187】
好ましくは, 前記外部ドメイン断片は、1〜4アミノ酸, より好ましくは2 アミノ酸である。
【0188】
好ましくは, 前記外部ドメイン断片は、前記外部ドメインのC末端の断片である, より好ましくは最後の1〜4である, より具体的には前記外部ドメインのC末端の最後の二つのアミノ酸である。
【0189】
それ故, 本発明のポリペプチドのアミノ酸配列は、さらに前記アンカーする配列のN末端での最初の一アミノ酸に(直接的にまたは間接的に, 好ましくは直接的に)以下のものが連結されたものを含んでもよい:
狂犬病ウイルス (例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の外部ドメインのC末端から1〜4アミノ酸, 好ましくは2アミノ酸, より好ましくは1〜4アミノ酸, なおより好ましくは狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質の外部ドメインのC末端の最後の二つのアミノ酸、例えば、アミノ酸 GK。
【0190】
係るポリペプチドの例は、特に配列番号 26のポリペプチドおよび配列番号 26の配列を含むポリペプチドを含む。
【0191】
本発明のポリペプチドは、さらにシグナルペプチド, 好ましくは狂犬病ウイルス株(例えば、非アポトーシス性の狂犬病ウイルス株)のG蛋白質のシグナルペプチドを含んでもよく、前記ペプチド配列は好ましくは前記ポリペプチドのN末端(最も好ましくは前記ポリペプチドのまさにN末端、即ち、前記ポリペプチドに含まれえる任意のアンカーする配列のN-termにある)に存在する。
【0192】
係るポリペプチドの例は、特に配列番号 27のポリペプチドおよび配列番号 27の配列を含むポリペプチドを含む。
【0193】
好適なポリペプチドの例:
本発明の好適なポリペプチドは、100 アミノ酸未満のポリペプチド(より好ましくは、可能な限り少数のアミノ酸のものであるが神経突起成長効果をなおも保持しているもの, より好ましくは90 アミノ酸未満のもの)を含む。
【0194】
本発明の好適なポリペプチドは、前記 G 蛋白質の細胞質断片〔最も好ましくは、配列番号 6, 配列番号 19または配列番号 20の細胞質断片〕を含む又はからなるポリペプチドを含む(図 19を参照されたい)。
【0195】
また、本発明の好適なポリペプチドは、これらの細胞質断片の11〜44小断片を含む又はからなるポリペプチドを含む。
【0196】
本発明の好適なポリペプチドは、前記神経突起成長支持配列に加えて、上記規定の少なくとも一つのアンカーする配列(例えば、配列番号 23のアンカリング配列; 以下の例4-6を参照されたい)を含むポリペプチドを含む。
【0197】
核酸, ベクターおよび細胞:
また、本発明は、縮重を考慮した普遍的な遺伝コードに基づき本発明のポリペプチドをコードする任意の核酸(より具体的には、任意のDNAまたはRNA)に関する。
【0198】
図 15に示されるとおり、配列番号 6 の細胞質断片(即ち、CVS-NIV 株の細胞質断片)をコードする例示の核酸は、配列番号 5の核酸である。
【0199】
図 16に示されるとおり、配列番号 10 のPDZ-BS(即ち、CVS-NIV 株のPDZ-BS)をコードする例示の核酸は、配列番号 9の核酸である。
【0200】
また、本発明は、本発明のポリペプチドをコードする少なくとも一つの 核酸を含む 任意の核酸ベクターに関する。前記ベクターは、トランスフェクションベクターおよび/または 発現ベクターであってもよい。
【0201】
前記発現ベクターは、さらに前記核酸の上流および/または下流の少なくとも一つの発現シグナルおよび/または制御配列、例えば、少なくとも一つのプロモーター,または少なくとも一つのエンハンサーや少なくとも一つのプロモーター(前記核酸の上流)を含んでもよい。前記発現ベクターは、例えば、プラスミドであってもよい。
【0202】
係るベクターの例示は、CNCMから受託番号 I-2758で利用可能なプラスミドである。
【0203】
興味深いことに, レンチウイルス ベクター, AAV ベクター, アデノ-ウイルスベクターおよび単純ヘルペス無効化ウイルスベクターを使用できる。
【0204】
択一的にまたは相補的に、本発明の核酸は、デンドリマー、例えば、リポソーム,または陽イオン性ポリマー、例えば、DEAE-デキストランまたはポリエチレンイミン,または不活性な固形のナノ粒子、例えば、金粒子,または当業者が適切であることを見出しえる任意の他のトランスフェクション手段と組み合わせることができる又は会合(associated)させることができる。
【0205】
また、本発明は、少なくとも一つの本発明のポリペプチド, および/または少なくとも本発明の核酸および/または少なくとも一つの本発明のベクターを含む任意の細胞に関する。
【0206】
前記細胞は、好ましくは遺伝的に操作されて前記ポリペプチドおよび/または核酸および/またはベクターを含む細胞である。
【0207】
前記細胞は、真核生物細胞, 好ましくは哺乳類細胞、例えば、ヒト細胞または非-ヒト細胞, 最も好ましくはヒト細胞であってもよい。
【0208】
前記細胞は、原核細胞(好ましくは細菌、例えば、大腸菌)であってもよい。
【0209】
前記細胞はビリオンであってもよいが、前記ビリオンはレシピエントの対象(より具体的には、ヒト患者)に有意に有害なまたは望ましくない効果を誘発しない。
【0210】
好ましくは, 前記細胞は、ヒト胚性細胞ではない。
【0211】
使用:
本発明は、より具体的には、神経突起成長を刺激および/または誘導することに使用するための〔より具体的には、不十分な又は障害性の神経突起生成(より具体的には、不十分な又は障害性の神経突起成長)に関連している疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用するための〕前記ポリペプチドおよび/または核酸および/またはベクターおよび/または細胞に関する。
【0212】
非免疫原性の使用における前記使用。
【0213】
実際, 本発明に適切である狂犬病ウイルス G 蛋白質は、非アポトーシス性の G 蛋白質と規定できる。
【0214】
従って、本発明のポリペプチドは、以前に公開されたPCT国際出願WO 03/048198に記載されたものものなどのアポトーシス小体の形成を誘導しない。
【0215】
本発明と一致して、本発明のポリペプチドは、例えば、ニューロン前駆体または新生物ニューロンからのニューロン分化のため, および/または障害性のニューロンのニューロン再生のための神経突起成長 (および/または軸索および/または樹状突起発生)のエフェクターとして意図される(双方のエフェクター効果は、神経突起成長の刺激を介して得られる)。
【0216】
上述のことから、本発明のポリペプチドは、免疫原性の薬剤(immunogenic agent)またはアジュバントではない,または最低限でも免疫原性の薬剤またはアジュバントとして使用されない、また前記ポリペプチドを免疫原性の薬剤またはアジュバントとして作用することを可能にするだろう条件下で使用されない。
【0217】
より具体的には、アポトーシス性の 狂犬病ウイルス株のG 蛋白質(例えば、弱毒化したERA株の及び他の弱毒化した狂犬病ウイルス株のG 蛋白質)とは反対に、本発明のポリペプチドは、ニューロンに投与された場合に検出可能な体液性の免疫応答を上昇させない。
【0218】
本発明のポリペプチドは、免疫原性の組成物またはワクチンであろう薬学的組成物または薬に使用されない。
【0219】
従って, 本発明のポリペプチドは、好ましくは何らかの抗原(より具体的には、過剰発現または変化により神経病理学などの病状を誘発するだろうウイルス抗原, 腫瘍抗原, 細胞抗原であろう任意の抗原)と連結されない、融合されない、および会合しない。
【0220】
疾患, 障害およびコンディション:
本発明は、ポリペプチド, 核酸, ベクターおよび細胞を提供する。これらの産物は、神経突起生成(より具体的には神経突起成長, なおより具体的にはヒト神経突起成長)を誘導および/または刺激する。
【0221】
本発明の産物は、好ましくは本発明の核酸ベクター(より具体的には、本発明のプラスミド)である。
【0222】
より具体的には、本発明の産物は、神経突起生成(分裂前のニューロン, 新生物ニューロン, ニューロン前駆体, 同様に障害があるニューロンからの神経突起成長)を誘導および/または刺激する。
【0223】
従って、本発明は、神経発生および/または神経再生および/または神経保護剤として使用するための前記産物に関する。
【0224】
本発明の産物は、神経突起の出芽および/または軸索の成長および/または樹状突起樹の伸展を刺激および/または誘導する。
【0225】
本発明の産物は、成長円錐の活性を刺激する。さらにまた、本発明の産物は、成長円錐が成長虚脱剤(例えば、LPAまたは酸化ストレス)との接触に際して虚脱(collapsing)することを予防する(以下の例 1を参照されたい)。
【0226】
本発明の産物は、結果的にシナプス形成および/または神経伝達を刺激および/または誘導する。
【0227】
本発明の産物は、新生物ニューロンの増殖を〔より具体的には、ニューロ分化剤(neuro-differentiating agent)として〕阻害する薬剤である。
【0228】
本発明の産物は、ニューロンの発生および/またはニューロンの再生および/または軸索の成長および/または樹状突起の発生および/または樹状突起樹の伸展および/またはニューロンの可塑性および/またはシナプス形成および/または神経伝達を刺激する薬剤である。
【0229】
本発明の産物は、神経突起の退縮および/または成長円錐の崩壊を誘発するであろう如何なる種類の神経毒性も予防および/または阻害および/またはブロックする薬剤である。
【0230】
本発明の産物は、神経突起 および/または錐状体が神経毒性剤と接触した後に神経突起成長および/または成長円錐活性を刺激および/または誘導する薬剤である。
【0231】
本発明の産物は、成長円錐の崩壊および/または神経突起の退縮および/または軸索樹状突起間のダメージまたは傷害および/またはシナプス統合性の崩壊および/またはニューロン結合(neuron connectivity)の欠損および/または神経終末(nerve endings)へのダメージおよび/または神経伝達の損傷を予防および/または阻害および/またはブロックする薬剤である。
【0232】
本発明の産物は神経突起成長を誘導および/または刺激する手段であり、以下の事項に特に有用である:
・ ニューロン分化を誘導すること(例えば、神経系の新生物の治療および/または緩和および/または予防)に有用である, 同様に
・ 障害されたニューロン(より具体的には、障害された神経突起)を再生することに有用である(例えば、神経変性疾患, 障害またはコンディションの治療および/または緩和および/または予防に有用である, ニューロンの微生物感染の治療および/または緩和および/または予防に有用である,または神経毒性剤または酸化ストレスからニューロンを保護すること有用である)。
【0233】
従って、本発明は、不十分なまたは障害性の神経突起生成(より具体的には、不十分なまたは障害性の神経突起成長)を伴う任意の疾患, 障害またはコンディションの治療および/または緩和および/または予防における使用のための前記産物に関する。
【0234】
前記疾患, 障害またはコンディションは、バランスを失ったニューロン細胞周期が関係する任意の疾患, 障害またはコンディションとして択一的にまたは相補的に規定され、前記ニューロン細胞周期は以下によりバランスを失う、即ち:
・ 分裂前のニューロンの過剰なまたは望まれない存在により(より具体的には、新生物が神経系において発生する場合のように、不十分なニューロンの分化および/または過剰なまたは望まれない分裂後ニューロンのニューロン細胞周期へのリエントリーにより)バランスを失う、或いは
・ 過剰なまたは望まれないニューロン変性、より具体的には過剰なまたは望まれない神経突起変性によりバランスを失う(神経変性の疾患, 障害またはコンディションやニューロンの特定の微生物感染の場合のように)。
【0235】
本発明の産物は、神経回復(neurorestorative)の治療および/または予防および/または緩和などの中枢神経系(CNS)および/または末梢神経系(PNS)を変化させる疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用できる。
【0236】
「中枢神経系」または「CNS」の表現は、本明細書等において脳や(脊椎動物の場合において)脊髄を意味することを意図している。
【0237】
末梢神経系(PNS)は、体を脳や脊髄と連結する脊髄や頭蓋の神経の広いネットワークである。PNSは、自律神経系(交感のNS および 副交感のNS)と体神経系とに細分される。PNSは、刺激レセプターからCNSに走る感覚ニューロンおよびCNSから筋肉や腺に走る運動ニューロンからなる。
【0238】
本発明の態様によると、前記疾患, 障害またはコンディションは、神経系の微生物感染(例えば、細菌および/またはウイルスの感染, より具体的にはウイルス感染)であるか又はこれらを伴う。
【0239】
前記ウイルス感染は、例えば、単純ヘルペスウイルス(HSV)感染, より具体的にはHSV 1型感染(ウイルス脳症を誘発する)であってもよい。
【0240】
前記微生物感染は、ニューロンのアポトーシスを誘導しないウイルス感染(例えば、狂犬病ウイルス感染)であってもよい。
【0241】
好ましくは, 前記微生物感染は、ニューロンのアポトーシスを誘発する微生物感染(例えば、灰白髄炎)である(Blondel et al., 2005を参照されたい)。
【0242】
本発明の別の態様によると、前記疾患, 障害またはコンディションは、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性や非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションであるか又はこれらを伴うものである。
【0243】
本発明の態様によると、前記疾患または障害は、神経変性疾患または障害(例えば、慢性の神経変性疾患または障害)、例えば、非-ウイルス性の脳症, アルツハイマー型の疾患, パーキンソン病, ALS, ハンチントン病, 多発性硬化症(MS)または稀な遺伝疾患(rare genetic disease)であるか又はこれらを伴うものである。
【0244】
好ましくは, 前記神経変性疾患または障害は、非-ウイルス性の疾患または障害, より好ましくは非-細菌性および非-ウイルス性の疾患または障害, なおより好ましくは非-微生物性の障害である。
【0245】
本発明の態様によると、前記コンディションは、神経変性コンディション(例えば、加齢)であるか又はこれを伴うものである。
【0246】
好ましくは, 前記神経変性コンディションは、非-ウイルス性のコンディション, より好ましくは非-細菌性および非-ウイルス性のコンディション, なおより好ましくは非-微生物性のコンディションである。
【0247】
本発明の態様によると、前記疾患, 障害またはコンディションは、神経系の物理的(physical)な又は虚血性の傷害、例えば、発作, 脳卒中, 外傷, 癲癇であるか又はこれらを伴うものである。
【0248】
好ましくは, 前記身体性の又は虚血性の傷害は、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性および非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションである。
【0249】
本発明の態様によると、前記疾患, 障害またはコンディションは、化学的な神経毒性剤および/または酸化ストレスの存在を伴うものである。
【0250】
好ましくは, 前記疾患, 障害またはコンディションは、非-ウイルス性の疾患, 障害またはコンディション, より好ましくは非-細菌性および非-ウイルス性の疾患, 障害またはコンディション, なおより好ましくは非-微生物性の疾患, 障害またはコンディションである。
【0251】
本発明の態様によると、前記疾患は、新生物である、より具体的には新生物ニューロン(neoplastic neurons)を含む新生物である。
【0252】
「新生物(neoplasm)」の用語は、本明細書等においてより具体的には悪性新生物を意図し, より具体的には癌, なおより具体的には腫瘍または白血病, なおさらにより具体的には腫瘍を意図する。
【0253】
本発明の産物は、免疫原性剤(immunogenic agent)として作用しない。より具体的には、本発明の産物は、腫瘍抗原に対し体液性応答を上昇させるだろう免疫原性剤として作用しない。
【0254】
好ましくは、本発明の産物は、それ自身で免疫原性剤として作用できない。より具体的には、本発明の産物は、それ自身で腫瘍抗原に対し体液性応答を上昇させるだろう免疫原性剤として作用できない。
【0255】
本発明の産物は、抗増殖剤(antiproliferative agent)として作用する。
【0256】
本発明の産物は、新生物ニューロンからの神経突起成長を誘導および/または刺激して、新生物ニューロンから成熟ニューロンへの分化を誘導する及び/又は刺激する。
【0257】
前記新生物は、CNSおよび/またはPNSの新生物, 好ましくは神経節膠腫, 脳腫瘍, 中心神経細胞腫(central neurocytoma), 髄芽腫, 脳室上衣腫, 奇形腫, 神経芽細胞腫の新生物であってもよい。
【0258】
好ましくは, 前記新生物は、非-ウイルス性の新生物, より好ましくは非-細菌性および非-ウイルス性の新生物, なおより好ましくは非-微生物性の新生物である。
【0259】
当業者が適切であることを見出す任意の投与様式が本発明に包含される。
【0260】
如何に本発明の産物を製剤化するかに応じて、非経口または腸内(例えば、経口)の投与, 好ましくは非経口投与, より好ましくは非経口の注射により投与できる。
【0261】
薬学的組成物または薬; 治療の方法:
また、本発明は、少なくとも一つの本発明のポリペプチド, および/または少なくとも本発明の核酸および/または少なくとも一つの本発明の細胞を含む任意の薬学的組成物または薬に関する。
【0262】
本発明の薬学的組成物または薬は、上記により詳細に記載されとおり、不十分なまたは障害性の神経突起成長を伴う疾患, 障害またはコンディションの治療および/または緩和および/または予防に使用できる。
【0263】
本発明の薬学的組成物または薬は、免疫原性の組成物ではなく、ワクチンでもない。
【0264】
本発明の薬学的組成物または薬は、さらに少なくとも一つの薬学的および/または生理的に許容されるビヒクル(希釈剤, 賦形剤, 添加物, pH 調整剤, 乳化剤または分散剤, 保存剤, 界面活性剤, ゲル化剤, 同様に緩衝化剤や他の安定化剤や可溶化剤, など)を含んでもよい。
【0265】
本発明の薬学的組成物または薬は、好ましくは何らかの免疫アジュバントを含まない。
【0266】
最も好ましくは、本発明の薬学的組成物または薬は、任意の抗原, より具体的には過剰な発現または変化が病状〔例えば、神経病理(neuropathology)〕を誘発するだろう任意の抗原(ウイルス抗原, 腫瘍抗原, 細胞抗原である可能性がある)を含まない。
【0267】
本発明の薬学的組成物または薬は、例えば、液体溶液剤(liquid solution), 懸濁剤, エマルジョン剤, 錠剤, ピル, カプセル剤, 徐放性製剤,または粉末剤であってもよい。好ましくは、非経口投与に適切な形態で製剤化される。
【0268】
また、本発明は、上記に記載されるとおり、対象(より具体的には、治療が必要なヒト)を治療する方法に関し、該方法は前記対象またはヒトに少なくとも一つの本発明のポリペプチドおよび/または少なくとも本発明の核酸および/または少なくとも一つの本発明のベクターおよび/または少なくとも一つの本発明の細胞を投与することを含む。
【0269】
本出願において、「含む(including)」または「含む(containing)」と同義である「含む(comprising)」の用語は、オープンエンド(open-ended)であり、付加的な未記載の要素, 構成成分または方法の工程を除外しない。他方、「からなる(consisting of)」の用語は、クローズドターム(closed term)であり、明示的に記載されない任意の付加的な要素, 工程,または構成成分を除外する。
【0270】
「から本質的になる(essentially consisting of)」の用語は、部分的なオープンタームであり、付加的な要素, 工程または構成成分が本発明の基礎および新規の特性に実質的(materially)に影響しない限り、これらの付加的な未記載の要素, 工程,または構成成分を除外しない。
【0271】
それ故、「含む」の用語は、「からなる」の用語, 同様に「から本質的になる」の用語を含む。従って、「含む」の用語は、本出願においてより具体的には「からなる」の用語および「から本質的になる」の用語を包含することを意味する。
【0272】
本明細書等において引用される全ての参照文献の関連する開示の各々は、参照によって特異的に援用される。以下の例は、例示の目的で提供され、限定の目的ではない。
【0273】
[例]
例 1: 非アポトーシス性のCVS-NIV 狂犬病ウイルス株の蛋白質Gの細胞質ドメイン(G-CVS-NIVまたは「G-survival」の細胞質ドメイン)は神経突起成長を誘導し、誘導された神経突起成長は成長円錐虚脱剤(即ち、LPA)および酸化ストレス(H2O2)に高度に耐性である。
【0274】
神経の再生や神経の保護は、広く神経毒性, 神経変性疾患, 外傷-発作-脳卒中, 脳症またはさらに加齢などの広範な分野において共通の一里塚である(図 1を参照されたい)。
【0275】
未成熟のヒトニューロン(例えば、神経芽細胞腫細胞)は、さらに適切なシグナル伝達分子で処理した場合に分化できる。このケースは、SH-SY5Y 細胞が細胞浸透性のdb-cAMPで処理された場合である。
【0276】
発明者は、ヒト神経芽細胞腫の細胞株(例えば、神経芽細胞腫細胞株 SH-SY5Y)の使用が含まれる機能的な神経保護検査を開発した。この系において、細胞はさらに分化でき、また神経突起の成長をモニターできることから成長円錐活性刺激性のエフェクターに関して試験することが可能である。そのうえ、退縮プロセス(retraction processes)に抵抗する伸長(elongated)した神経突起の能力を薬処理〔例えば、リゾホスファチジン酸 (LPA)または過酸化水素 (H2O2)〕後に評価できる。このような系により集められたデータは、インビボでなされた神経毒性分析と相関することが証明されている。
【0277】
以下の例は、特に以下の事項を実証する:
・ 非アポトーシス性の(即ち、病原性の)狂犬病ウイルス株(即ち、CVS-NIV 株)のG蛋白質は神経突起成長促進効果を有し、これは即ち、ある種の非アポトーシス性の狂犬病ウイルスG蛋白質が神経突起生成を誘発および/または刺激することである;
・ この神経突起成長効果は、ニューロンを成長円錐虚脱薬(LPA)や酸化ストレス(H2O2)から防御するために十分強いものである;
・ この神経突起成長効果を担うのは前記非アポトーシス性のCVS-NIV 狂犬病ウイルス株のG蛋白質の細胞質尾部である;
・ アポトーシス性の 狂犬病ウイルス G 蛋白質は、この神経突起成長効果を示さない; より具体的には、CVS-NIV 株のG蛋白質と僅か六アミノ酸 (また細胞質尾部においては僅か二つのアミノ酸)異なるERA株のG蛋白質は、この神経突起成長効果を示さない;
・ この効果は、特に前記 非アポトーシス性の狂犬病ウイルスG蛋白質の細胞質尾部に含まれるPDZ-BS モチーフが前記 アポトーシス性の 狂犬病ウイルス G 蛋白質の一つと比べて単一の点変異を示すとの事実によるものであり、即ち: 非アポトーシス性の CVS-NIV 株の蛋白質G(protein G)は、アミノ酸 Qをポジション521 (CVS-NIV 株の全長 G 蛋白質に対して計算されたポジション、前記 G 蛋白質の細胞質断片におけるポジション 41に対応する)に有し, 他方でG-ERAはEを同じポジションに有する;
・ CVS-NIV 株 の全長G蛋白質中のポジション491(前記 G蛋白質の細胞質断片におけるポジション 11)にあるアミノ酸もこの神経突起成長効果に寄与する (アミノ酸 H)。
【0278】
材料および方法
細胞, ウイルスおよび分子クローン
SH-SY5Yは、アメリカン・タイプ・カルチャー・コレクション(ATCC; 10801 University Blvd.; Manassas, Virginia 20110-2209; U.S.A.)から受託番号 CRL-2266)で利用可能な神経芽細胞腫の細胞株である。
【0279】
狂犬病ウイルス (RABV)のオリジナルのERA および CVS 株は、それぞれATCC から受託番号 vr332 および vr959で利用可能である。本研究において使用されたERA および CVS 株は、BSR 細胞 (乳児 ハムスター 腎臓細胞-クローン 21のクローン, ATCC 受託番号 BHK-21)においてこれまで二十年間発明者の研究室において継代されてきた。これらの継代株は、それぞれERA-NIV 株およびCVS-NIV 株である。
【0280】
本出願の例および図の全体をとおして、他で特定しない限り、CVSはCVS-NIVを意味し、ERAはERA-NIV (G蛋白質 CNCM I-2760; 配列番号 4)を意味する。
【0281】
CVS-NIV 株 (rRABV CVS HQ)は、ブタペスト条約の条件下で2009年4月1日にCollection Nationale de Cultures de Microorganismes (CNCM)に寄託された(CNCM; Institut Pasteur; 25, rue du Docteur Roux; F-75724 PARIS CEDEX 15; FRANCE)。CNCM受託番号は、I-4140である。
【0282】
ERA-NIV 株(即ち、G-ERA-NIV)のG蛋白質, およびCVS-NIV 株(即ち、G-CVS-NIV)のG蛋白質の代表的な分子クローンの取得や特徴に関する事項は、Prehaud等(Prehaud et al. 2003)に記載されている
これらの G 蛋白質の配列もアクセッション番号AF 406693 (G-ERA-NIVに関して) および AF 406694 (G-CVS-NIVに関して)で利用可能である;(図 13A および 13Bも参照されたい)。
【0283】
CVS-NIV 株のG蛋白質も2001年11月30日にブタペスト条約の条件下でCNCMに寄託された組換え型の大腸菌株から利用可能である。CNCM受託番号は、I-2758である。この組換え型の大腸菌は、CVS-NIV 株のG蛋白質を誘導性に発現するプラスミド (プラスミド pRev-TRE-G-CVS; WO 03/048198を参照されたい)を含む。
【0284】
ERA株のG蛋白質も2001年11月30日にブタペスト条約の条件下でCNCMに寄託された組換え型の大腸菌株から利用可能である。CNCM受託番号は、I-2760である。この組換え型の大腸菌は、ERA株のG蛋白質を誘導性に発現するプラスミド (プラスミド pRev-TRE-G-ERA; WO 03/048198を参照されたい)を含む。
【0285】
狂犬病ウイルス CVS-NIV 株のG蛋白質をコードしているプラスミド CNCM I-2758を含んでいる組換え型の 大腸菌株の培養のための適切な条件や狂犬病ウイルス ERA株のG蛋白質をコードしているプラスミド CNCM I-2760を含んでいる組換え型の大腸菌株の培養のための適切な条件には、標準 LB-TYM 成長培地(アンピシリンの存在下)で37°Cで前記組換え型の大腸菌株をインキュベーションすることが含まれる。
【0286】
前記組換え型の狂犬病ウイルス (rRABV)は、Faul等2008の記載の処理にしたがって産生され、単離された。rRABVsは、野生型「G survival」または「G death」配列 (それぞれCVS-NIV 株およびERA株のG 配列),またはERAの遺伝的な G 遺伝背景におけるG survivalの細胞質尾部,またはCVS-NIVの遺伝的な G 遺伝背景におけるG deathの細胞質尾部の何れかを保持するよう構築された(図 2を参照されたい)。
【0287】
組換え型の狂犬病ウイルスも産生され、このウイルスはGのコード配列中の変異によりCVS-NIV 株から由来する。これらの組換え型の狂犬病ウイルスはG蛋白質を有し、このG蛋白質はCVS-NIV 株のG蛋白質と一または二アミノ酸異なるものである、すなわち:
・ CVS-NIV 株の全長 G蛋白質 配列において、ポジション 491 (その細胞質断片中のポジション 11)のアミノ酸で異なる;および/または
・ CVS-NIV 株の全長 G蛋白質 配列において、ポジション 521 (その細胞質断片中のポジション 41)のアミノ酸で異なる。
【0288】
より具体的には、以下の組換え型の狂犬病ウイルス株が作出された:
【0289】
【表2】
【0290】
rRABV CVS LE, rRABV CVS LQ および rRABV CVS HEは、ブタペスト条約の条件下でCNCMに2009年4月1日に寄託された。CNCMの受託番号は、それぞれI-4141, I-4142 および I-4143である。
【0291】
ウイルス I-4141またはI-4142またはI-4143 (組換え型の 狂犬病 ウイルス)を増殖させる適切な条件には、グルコース (例えば、4.5 g/L), ピルビン酸ナトリウムおよび glutamax (Invitrogen 31966047) および 5% FBSを含んでいるDMEM 成長培地で前記 ウイルス を37°CでBHK-21 細胞 (サブ-クローン BSR)と 5% CO2の存在下でインキュベーションすることが含まれる。
【0292】
神経突起の成長および退縮のハイスループットなアッセイ
SH-SY5Y ヒト 神経芽細胞腫細胞は、プレート (Cell Bind plastic ware, Corning, USA)に40,000 細胞/ウェルの密度で非分化培地[DMEMF12 (Invitrogen, U.K.) に20% ウシ胎児血清 プラス 1%Pen:Strep および1% グルタミンを添加したもの]に播種され、一晩、37°Cで培養された。接種の24h後に、非分化培地を分化培地[Neurobasal medium (Invitrogen, U.K.) にB27 添加物(Invitrogen, U.K.), 1% P/S, 1% グルタミン および1mM db-cAMP (dibutyril c-AMPは膜浸透性, Sigma)]を添加したもので置き換え、細胞を6hインキュベーションした。それから, 細胞は、mock(MOI 3で分化培地でrRABV を感染させた)で感染させた。インキュベーションの1h後、細胞を、分化培地で一回洗浄し、分化培地を加えた後に24h、37°Cでインキュベーションした。
【0293】
自然の分化に関して、同じ手順を使用した(但し、db-cAMPを省略した)。
【0294】
分化の30h後、細胞を、3% パラホルムアルデヒドをリン酸緩衝塩類溶液 (PBS)に含むもので20 min、室温 (RT)で固定し、5分間0.1% Triton-X-100で処理し、50%の正常ヤギ血清 (NGS: normal goat serum)をPBS中に含むもので1h、RTで処理した。ニューロン特異的な抗βIII チューブリン Ab (Promega, France) および 抗-RABV ヌクレオカプシド Abを使用して軸索の突起を染色し、それぞれでのRABV感染を明らかにした。或いは、細胞は。クリスタルバイオレットで染色され、これにより軸索突起が保たれる。
【0295】
退縮アッセイは、10μM-30μMまたは50μM LPA (Sigma, USA)を分化培地に添加すること又は75μM H2O2 (Sigma, USA)をB27 マイナス 抗酸化剤含有分化培地(B27 minus anti oxidant containing differentiating medium)に添加することを除いて上記の成長アッセイと同一である。
【0296】
細胞は、Leica DM 5000B UV 顕微鏡にDC 300FX カメラ (x40またはx20 対物)を備えたものを用いて視覚化し、ImageJ 1.38X ソフトウェア (Wayne Rasband, NIH, USA, http://rsb.info.nih.gov/ij/)とそのplug-inのNeuronJ(Meijering et al. 2004; http://www.imagescience.org/meijering/software/neuronj/)を用いて分析した。ニューロン毎の平均の神経突起長は、トリプリケート実験で決定された。
【0297】
フローサイトメトリーによるRABV抗原の検出
SH-SY5Y 細胞をdb c-AMPでの処理により分化させ、分化6 時間後にrRABVsで感染させた。感染24 時間後に細胞を採取し、細胞質膜に達したRABV 糖蛋白質または感染細胞において発現したG蛋白質の総量の何れかのフローサイトメトリーによる検出のため処理した。処理は、Prehaud等(Prehaud et al. 2003)の記載に基づき行った。
【0298】
結果および考察
G-CVS-NIV 細胞質尾部(即ち、CVS-NIV 株のG蛋白質の細胞質尾部)は、神経突起成長を促進する分子署名を含む。
【0299】
感染が非存在の条件下で、db-cAMPでの処理は、神経突起の伸長を惹起する(図 3A, 左パネル二つ)。細胞がdb-cAMPで処理され、rRABV G-CVS-NIVで感染された際に、長い神経突起を示す(191%, 図 3A, 中央右パネル)。この結果によって、次の事項が確立された。即ち、成長円錐活性がcAMPシグナル伝達により刺激される場合、rRABV G-CVS-NIVが神経突起成長を促進することである。
【0300】
対照的に, 神経突起の長さは、双方がdb-cAMPで処理された非感染細胞およびrRABV G-ERA 感染細胞の間で類似し(図 3B)、rRABV ERAを発現しているG deathは神経突起成長 アッセイに関してサイレント表現型を有することを示す。実験の時間経過(即ち、分化後30hおよび感染後24 時間)は、アポトーシスの最大 レベルがrRABV ERA感染神経芽細胞腫 細胞において検出される時点(即ち、感染後48h)よりも早期の時点であることが注目される。
【0301】
図 3Cに記載のとおり、G蛋白質のエンドスワップ(end swap)を保持しているrRABVs を用いることにより(図 2を参照されたい)、この表現型が細胞質尾部のみにより完全に移されることを発明者は示す。この結果は、伸長プロセスがcAMP シグナル伝達経路により開始される場合に、G-CVS-NIV 細胞質尾部が神経突起成長を促進する分子署名を含むことを確かに確立する結果である。
【0302】
さらにまた、神経突起成長アッセイを、CVS-NIV 株 (rRABV G-CVS-NIV)とそれぞれの G 蛋白質と一つ又は二つのアミノ酸で異なる組換え型の 狂犬病ウイルスで行った。
【0303】
より具体的には、アッセイをrRABV CVS LE, rRABV CVS LQ および rRABV CVS HEで行った、これらはG-CVS-NIVとアミノ酸 491 および/または アミノ酸 521 で異なるG蛋白質を発現する(表 2 上記を参照されたい)。これらのアッセイの結果は、図 3Dで説明される。図 3Dにおいて、対照との全ての差異は、統計的に有意である(p<0.005 スチューデントのt-検定):
・ rRABV G-CVS HQ 対 N.I.: p<0.0001;
・ rRABV G-CVS LQ 対 N.I.: p=0.0002;
・ rRABV G-CVS HE 対 N.I.: p=0.0111。
【0304】
rRABV G-CVS-NIVは、それぞれG蛋白質のポジション491 および 521にH および Qを有する(CVS-NIVの全長G蛋白質に関して計算したポジション, これらのポジションはそれぞれこのG蛋白質の細胞質断片におけるポジション 11 および 41と対応する)。
【0305】
rRABV CVS LQは、L および Qをこれらのポジションに有する。
【0306】
rRABV CVS HEは、HおよびEをこれらのポジションに有する。
【0307】
rRABV CVS LEは、LおよびEをこれらのポジションに有する。
【0308】
図 3Dにより説明されるとおり、CVS-NIV (rRABV G-CVS-NIV)よりも有意に低い程度であるが、rRABV CVS LQ および rRABV CVS HEは陽性の神経突起成長効果をなおも示す[スチューデントのt-検定分析: rRABV CVS HQ 対 rRABV CVS LQ: p = 0.0156; rRABV CVS HQ 対 rRABV CVS HE: p = 0.0009]。
【0309】
以上より、CVS 株のG蛋白質のポジション 491 および ポジション 521(細胞質断片におけるポジション 11 および 41)は、双方が神経突起成長効果に密接に関係しており、ポジション 521の寄与が優勢である。
【0310】
rRABV CVS LEは、対照と比べて何らかの有意に陽性の神経突起成長効果を誘導しなかった(図 3Cを参照されたい, rRABV G-CVS-Cyto Death = rRABV CVS LE)。
【0311】
G-CVS-NIV 細胞質尾部は、cAMPと相乗的に働く神経突起生成を促進する内因性のエフェクターである。
【0312】
発明者は、G-CVS-NIV 細胞質尾部の発現がそれ自身で神経突起生成を刺激できるかどうかを調査した。SH-SY5Y 細胞を異なる rRABVで感染させ(図 2を参照されたい)、神経突起の伸長をモニターした。
【0313】
図 4Aにおいて観察されたとおり、G-CVS-NIVまたはG-ERA-Cyto Survivalの発現は、神経突起成長を促進するため十分である。これらのデータによって、G-CVS-NIVの細胞質ドメインが未知の機構により神経突起伸長プロセスにシグナルを送る内因性の分子署名を含むことが確立された。同じ実験がdb-cAMPの存在下または非存在下でなされた場合、G-CVS-NIV および cAMPの双方が媒介する神経突起成長に相乗的に働くことが観察された(図 4Bを参照されたい)。
【0314】
G-CVS-NIVの細胞質ドメインに基づく神経突起生成は分子署名に依存し、発現したG蛋白質の量には依存しない。
【0315】
産生された狂犬病ウイルス 糖蛋白質の量を、感染させdb-cAMP処理したSH-SY5Y 細胞において産生されたRABV 抗原の総量,または完全に細胞質膜へとプロセスされたG-蛋白質の量の何れかををモニターすることにより非常に慎重に分析した。G-CVS-NIVまたはG-ERAを発現している二つのrRABVsは、全体として細胞質膜に発現されたG-蛋白質の量は類似する(図 5を参照されたい)。従って、生存の表現型(即ち、神経突起成長)は、利用可能な蛋白質の量の表現型とは関連しないが、むしろ細胞質の生存ペプチドの配列に存在する特定の決定要因と関連する。
【0316】
そのうえ、rRABV G-ERA-Cyto Survivalが糖蛋白質を若干低く発現するが、このウイルスがなおも強い生存表現型を示すことは興味深い(図 3C, 4Aを参照されたい)。この結果は、ペプチドの量は主要な注意事項(caveat)ではなく、 生存決定要因が感染細胞の内側に存在する限り神経突起成長が刺激されることを意味している。
【0317】
G-CVS-NIV 細胞質尾部は、成長円錐虚脱薬(growth cone collapsing drug)に対し神経保護を与える
LPAは、細胞成長, 運動性, 形態および運命に影響する多様な細胞機能を発揮することが示された成長因子様のリン脂質として作用する生理活性脂質である。ニューロン細胞において、LPAは、アクトミオシン細胞骨格の再組織化をおこすRho および Rho-キナーゼ活性化をともなう成長円錐虚脱や神経突起退縮を媒介する。
【0318】
G蛋白質細胞質尾部の10μM LPA 誘導性の神経突起退縮に対して防御する能力が調査された。db-cAMPで分化させたSH-SY5Y 細胞がLPAで処理される場合、神経突起の退縮が観察される(図 6A, 左を参照されたい)。また、同じ効果は、細胞がrRABV G-ERAで感染される場合に観察される(図 6A, 右を参照されたい)。反対に、db-cAMP 処理し、rRABV G-CVS-NIVで感染させた細胞において検出された長い神経突起は、成長円錐虚脱剤での処理に対して完全に保存される(図 6A, 中央を参照されたい)。この結果によって、G-CVS-NIVがある種の内因性の神経保護特性を有することが確立された。
【0319】
この保護は、rRABV G-ERA-Cyto Survivalでの感染に続いても認められるが、rRABV G-CVS-Cyto Deathでの感染では認められない(図 6Bを参照されたい)。従って、エンドスワップ変異体の使用によって、神経保護表現型がG-CVS 細胞質ドメインの発現と関連することが確かに確立された。
【0320】
LPAに対するG-CVS-NIV 細胞質尾部の神経保護は強いものである
LPAの生理的な範囲は、通常1μM付近である。発明者は、高用量のLPA(即ち、10μM)を使用してG-CVS-NIV 細胞質尾部により与えられた神経保護の強さ(robustness)を確認した。
【0321】
50μMまでのLPAの亜致死量を、この神経保護の効率をモニターするため試験した。SH-SY5Y 細胞をrRABV G-CVS-NIVまたはrRABV G-ERA-Cyto Survivalで感染させた又は感染させなかった。分化後30 時間に細胞を異なる用量のLPAで処理した(図 7を参照されたい)。LPAの量を増加させて非感染細胞を処理することによって、神経突起の長さの直線的な退縮を生じた(図 7, 左を参照されたい)。反対に、rRABV G-CVS-NIVで感染された細胞は、LPAの用量がどのような用量であってもそれらの神経突起に何らかの明らかな虚脱を示さない(図 7, 中央を参照されたい)。同じデータは、30μMまでのLPAの用量に関してrRABV G-ERA-Cyto Survivalで感染された細胞に関して観察された(図 7, 右を参照されたい)。LPAの最高用量(50μM)に関して観察された軽微な減少は、このrRABVにより発現されたG蛋白質の最低量と比例する可能性がある。この場合(および成長円錐虚脱薬の非常に高濃度に関して)、G蛋白質の相対量が限定要因となる可能性がある。何れの場合ケースでも、これらのデータによって、非常に高い効率のG-CVS-NIV 細胞質尾部により与えられた神経保護が証明された。
G-CVS-NIV 細胞質尾部によって、酸化ストレスに対する神経保護が与えられる
LPAによる神経突起退縮へのG-CVS-NIV 細胞質ドメインにより提供された神経保護について上記でなされた観察に基づいて、発明者は、この表現型が他のよりユビキタスな因子に一般化できるかどうかを試験した。
【0322】
酸化ストレスは、ニューロン変性を誘導する重要な経路を代表する。酸化ストレスは、多くの神経変性の罹患者に関係するが、脳への急性ダメージ(例えば、外傷, 脳卒中および癲癇)のケースにおいても関係する。このようなことから発明者は過酸化水素処理後の神経突起の退縮表現型の研究を試みた。分化した細胞が75μM H2O2に供試される場合、神経突起は短縮する(図 8, 棒グラフの左の最初のペアを参照されたい)。同じ観察は、前記細胞が以前にrRABV G-ERAで感染された場合になされた(図 8, 棒グラフの第三のペアを参照されたい)。これに反して、神経突起の退縮は、前記細胞がrRABV G-CVS-NIVで感染された場合に観察されない(図 8, 棒グラフの第二のペアを参照されたい)。以上のとおり、G-CVS-NIV 発現は、酸化ストレスに対し神経保護を提供する。
【0323】
エンドスワップ変異体を用いることにより、我々は、この神経保護特性が図 8に記載のとおりG-CVS-NIV 細胞質尾部の発現に関連することも示した(それぞれrRABV G-CVS-Cyto Death および rRABV G-ERA-Cyto Survivalに関して、棒グラフの第四および第五のペアを参照されたい; これらのエンドスワップ変異体の構造に関して図 2を参照されたい)。
【0324】
狂犬病ウイルス G蛋白質の細胞質ドメインのCOOH 末端のPDZ-BSは、感染したニューロン細胞の生存表現型の運命に決定的な影響を与える。
【0325】
G-Cyto survival蛋白質の最後の 4aa 残基の欠失(survival G-蛋白質-delta)は、神経突起成長における影響により測定されるとおり、rRABVの生存表現型を有意に減少させるのに十分であった。
【0326】
例えば、db-cAMPの存在下で感染後8 時間で測定された神経突起成長を示す図 20を参照されたい。この図では、rRABV CVS-NIV Delta-PDZ-BS 〔アミノ酸 H をポジション 491 に有し、PDZ-BSが欠失したCVS-NIV〕により誘導されたものと、対照(N.I.)およびG-CVS-NIV 〔アミノ酸 H をポジション 491 に有し、そのPDZ-BS(即ち、ポジション 521〜524におけるQTRL)を有するCVS-NIV〕と比較して: rRABV CVS-NIV Delta-PDZ-BSにより誘導される神経突起成長効果が、対照により誘導されるものと有意に異なり(p = 0.0002 スチューデントのt-検定)、rRABV CVS-NIVにより誘導されるものと有意に異なる(p = 0.0003 スチューデントのt-検定)ことを示している。
【0327】
発明者は、特に以下の事項を実証している:
・ CVS-NIVの蛋白質G(G-CVS-NIV)は、成長円錐活性がc-AMP シグナル伝達経路(外部のエフェクターとして伝達された)により刺激される系において神経突起成長(神経突起生成)を促進することができる;
・ G-CVS-NIVの細胞質ドメインは、この表現型の原因である;
・ G-CVS-NIVの細胞質尾部の発現は、c-AMPの非存在下での神経突起成長の刺激を担うものでもある;
・ G-CVS-NIV および c-AMPの双方は、相乗的なエフェクターとして機能する;
・ 前記表現型は、G-CVS-NIVの細胞質尾部により、より具体的には全長 G-CVS-NIV 蛋白質においてポジション 491 および 521(その細胞質断片においてポジション 11 および 41)にあるアミノ酸により, なおより具体的には全長 G-CVS-NIV 蛋白質においてポジション 521(その細胞質断片においてポジション 41)にあり、PDZ-BSの部分であるアミノ酸により伝えられる分子署名と関連する;
・ G-CVS-NIV 細胞質尾部により提供される生存表現型は、成長円錐虚脱剤(即ち、LPA)による処理に高度に耐性である;
・ G-CVS-NIV 細胞質尾部により提供される生存表現型は、酸化ストレス (H2O2)にも耐性である。
【0328】
例 2: G-CVS-NIVの細胞質ドメインはHSV-1 細胞変性効果に対する神経保護を与える
単純ヘルペスウイルス タイプ I 脳炎 (HSVE)は、西洋諸国において最も一般的な散発性で致死的な中枢神経系の感染症であり、全ての年齢層の患者において年間を通じて出現する。単純ヘルペスウイルス1型が脳組織で感染する場合、HSVEは溶解性(即ち、壊死性)の状態で進行する。
【0329】
G蛋白質細胞質尾部の細胞におけるHSV-1の細胞変性効果に対して防御する能力が調査された。
【0330】
材料および方法
神経突起成長
上記の例 1に記載されているとおりである。
【0331】
HSV-1 細胞変性効果の決定
rRABVは、上記の例 1に記載されているとおり産生された。
【0332】
rRABVをBSR細胞中で増殖させた(Faul et al. 2008を参照されたい)。
【0333】
HSV-1 株 KOSをU373MG細胞(ATCC HTB 17)で増殖させた(Skare et al. 1975を参照されたい)。
【0334】
SH-SY5Y 細胞は、分化培地 マイナス db-c-AMPの条件で6 時間mockで感染させるか又はrRABVで感染させた。それから, HSV-1を感染多重度3(MOI 3)で加え、神経突起成長の表現型を24 時間後に決定した。単独(sole)の HSV-1 感染の神経突起成長の効果を確立するために、SH-SY5Y 細胞を分化培地中にdb-c-AMPを含むもので6 時間分化させ、次にHSV-1 感染(MOI 3)を行った。
【0335】
神経突起成長の表現型を同様に24 時間後に決定した。
【0336】
結果および考察
db c-AMPでの処理により分化したSH-SY5Y 神経芽細胞腫細胞は、HSV-1 感染後に神経突起の激しい退縮(〜79 %, 図 9Aを参照されたい)を示し、最終的に細胞が死に至る可能性がある。
【0337】
これに反して、前に細胞がrRABV G-CVS-NIVまたはrRABV G-ERA-Cyto Survivalで感染された場合 (上記の例 1を参照されたい)、細胞は神経突起成長を示し(図 9Bを参照されたい)、広範な神経突起退縮に対して保護される (図 9Cを参照されたい)。
【0338】
例 3: G-CVS-NIVの細胞質ドメインは神経芽細胞腫細胞に対して抗-増殖性を示す
神経芽細胞腫は、幼児期の固形腫瘍のうち二番目に一般的なものであり、米国の小児の癌による死亡の13%以上と計算される。予後は、小児の障害の臨床段階や年齢に応じて変化する。一般に, 神経芽細胞腫の大抵の患者は、外科, 放射線および細胞毒の化学療法を含む治療アプローチで治療される。
【0339】
ヒトの悪性神経芽細胞腫は未成熟の神経芽細胞の分化が不十分であることや増殖が非制御的であることにより特徴付けられるので、高用量のオールトランスレチノイン酸(ATRA)などの分化誘導薬が使用されてきた。RABV G-CVS-NIVの細胞質尾部がSH-SY5Y ヒト神経芽細胞腫細胞 (上記の例を参照されたい)の神経突起成長を促進できること(即ち、これらの細胞の分化を誘導できること)を発明者は実証した。それ故、発明者は、この分化特性が培養において係る細胞の増殖を制御するため十分に効率的であるかどうかとの疑問を解決することを望んだ。
【0340】
材料および方法
MTTアッセイ
前記アッセイは、Sargent 2003により記載された。簡単に説明すると、db c-AMPなしの分化培地で48hでmockで感染させたか又はrRABVで感染させた細胞は、MTT溶液で3.5 時間、37°C、5% CO2で染色された。次に培地および染色溶液の混合物が除去され、MTT結晶は一時間、室温で持続的に撹拌して溶媒で溶解された。前記アッセイは、550nmのODで測定した。各条件は、8のnを代表する。統計分析をANOVA 検定で行った。
【0341】
フローサイトメトリー
細胞計数および形態学的な変化を、Prehaud等(Prehaud et al. 2003)に記載のとおり、側方および前方散乱(それぞれ、SSC および FSC)により評価した。各点は、1 分間、低速で集めた細胞を代表する。フローサイトメトリー分析により、48hの時点での細胞集団の1%未満を代表する死細胞または壊死細胞を排除するために生存可能な集団をゲートした。
【0342】
神経突起成長
上記の例 1に記載されているとおりである。
【0343】
ATRA処理
神経芽細胞腫細胞のオールトランスレチノイン酸 (ATRA: All trans retinoic acid)処理をPrehaud等(Prehaud et al. 2005)により既に記載されたとおり行った。
【0344】
結果および考察
最初の例において、SH-SY5Y 細胞を無処置か又はATRAの5μMもしくは10μMを培養培地中に含むもので処置した。細胞増殖を生細胞に関してフローサイトメトリーで又はMTTアッセイを介した細胞計数でアッセイした。データを、それぞれ図 10A および 10Bに示す。
【0345】
各ケースで、細胞のATRA処理により、細胞成長をモニターするために使用された検査に応じて27%〜55%の間で変動する増殖の減少を伴う神経芽細胞の成長の遅延が生じた。このように、SH-SY5Y 細胞は、抗-増殖薬の効果に効率的に応答する。
【0346】
それゆえ、SH-SY5Y 細胞をrRABV G-CVS-NIVまたはrRABV G-ERAの何れかで感染させ、細胞の増殖(MTTアッセイにより確立された)および神経突起の成長を測定した(図 11A および 11Bを参照されたい)。我々は、細胞増殖(cell growth)が神経突起の成長と逆に相関することを見出し、G-CVS-NIVの神経生存特性が神経芽細胞における内因性の抗-増殖性の表現型と関連することを確立した。
【0347】
第二シリーズの実験において、神経芽細胞腫細胞を親のrRABV G-CVS-NIVまたはrRABV G-ERA ウイルスの何れか及びエンドスワップ変異体で感染させた(rRABV G-CVS-Cyto DeathまたはrRABV G-ERA-Cyto Survival; 上記の例 1および 図 2を参照されたい)。細胞増殖をフローサイトメトリー および MTT アッセイ (それぞれ、図 12A および 12Bを参照されたい)により測定した。各ケースで、前記データは、細胞が神経突起成長促進G-cyto survival尾部ウイルスで感染された場合に神経芽細胞の増殖が影響されて18.3% 〜23%の間で変動することを示した。従って、G-Cyto survivalは、神経芽細胞の細胞増殖を緩徐にする特性を内在する。
【0348】
例 4: Cyto-Gの生存支持特性が発現ベクターに保存されることの実験による実証
ベクターがウイルス感染の非存在下で細胞で発現される場合(最初に発現ベクターとして配送されたポリペプチド)、非アポトーシス性の狂犬病ウイルス株(Cyto-G)のG蛋白質のRABV 細胞質ドメインが生存支持特性を保存する(神経突起生成および退縮に対する保護)ことを本例は実証する。
【0349】
この例は、さらに前記生存特性がサイズや細胞質または小胞体膜(endoplasmic membrane)への足場に依存するかどうかを調査する。
【0350】
突然変異誘発を例 1のrRABVs を作出するため使用されたオリゴヌクレオチド-PCR に基づく突然変異誘発処置を用いて行った(QuickChange Lightning 部位特異的変異誘発キット, Agilent Technologies; Stratagene product division; カタログ番号 210518-12; 製造者の推奨にしたがって使用されるキット; 米国特許第7,176,004; 7,132,265; 7,045,328; 6,734,293; 6,489,150; 6,444,428; 6,391,548; 6,183,997; 5,948,663; 5,932,419; 5,866,395; 5,789,166; 5,545,552, および係属している特許)
構築物の末端にそれぞれ NheI および XmaI 部位を導入するためオリゴヌクレオチドを設計した。
【0351】
突然変異誘発のプライマーは以下のものであった:
GFullATG:
GGCCGCTAGCATGGTTCCTCAGGCTCTCCTGTTT (配列番号 28)
GCytoATG:
GGCCGCTAGCATGAGAAGAGTCAATCGATCAGAACCT (配列番号 29)
GendSTOP:
GGCCCCCGGGTCACAGTCTGGTCTGACCCCCACT (配列番号 30)
mutTM.Cyto: CCCCTTCTGGTTTTTCCATTGTGTTTTGGGGGGAAGTATGTATTACTGAGT (配列番号 31)
前記変異体をシークエンシングでインサートを選別した。この方法により、特に次の三つの構築物(G-Full; G-[SP-(2aa)-TM-Cyto]; G-Cyto)を得た:
G-Full: アクセッション番号 AF 406694 (CVS-NIVのG蛋白質)
MVPQALLFVPLLVFPLCFGKFPIYTIPDKLGPWSPIDIHHLSCPNNLVVEDEGCTNLSGFSYMELKVGYILAIKMNGFTCTGVVTEAETYTNFVGYVTTTFKRKHFRPTPDACRAAYNWKMAGDPRYEESLHNPYPDYHWLRTVKTTKESLVIISPSVADLDPYDRSLHSRVFPSGKCPGVAVSSTYCSTNHDYTIWMPENPRLGMSCDIFTNSRGKRASKGSETCGFVDERGLYKSLKGACKLKLCGVLGLRLMDGTWVAMQTSNETKWCPPDQLVNLHDFRSDEIEHLVVEELVRKREECLDALESIMTTKSVSFRRLSHLRKLVPGFGKAYTIFNKTLMEADAHYKSVRTWNEILPSKGCLRVGGRCHPHVNGVFFNGIILGPDGNVLIPEMQSSLLQQHMELLESSVIPLVHPLADPSTVFKDGDEAEDFVEVHLPDVHNQVSGVDLGLPNWGKYVLLSAGALTALMLIIFLMTCCRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 2)
G-[SP-(2aa)-TM-Cyto]:
G-[SP-(2aa)-TM-Cyto]は、(N-端からC-端に)以下のものを含んでいる構築物である:
・ G蛋白質のシグナルペプチド(例えば、CVS-NIVのシグナルペプチド; 配列番号 21);
・ 2アミノ酸(例えば、G蛋白質の外部ドメインからアミノ酸、例えば、G-CVS-NIVからアミノ酸);
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通ドメイン(例えば、G-CVS-NIVの膜貫通ドメイン; 配列番号 23);
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメイン(例えば、G-CVS-NIVの細胞質ドメイン; 配列番号 6)。
【0352】
本例に使用されたG-[SP-(2aa)-TM-Cyto]の配列は、以下のものである:
MVPQALLFVPLLVFPLCFGGKYVLLSAGALTALMLIIFLMTCCRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 27)。
【0353】
G-Cyto:
G-Cytoは(N-からC-端に)次のものを含んでいる構築物である: アミノ酸(例えば、M)および非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメイン(例えば、G-CVS-NIVの細胞質ドメイン; 配列番号 6)。
【0354】
本例に使用されたG-Cytoの配列は、以下のものである:
MRRVNRSEPTQHNLRGTGREVSVTPQSGKIISSWESHKSGGQTRL (配列番号 23)。
【0355】
配列は、図 22に示される。
【0356】
配列のアラインメントは、図 23に示される。
【0357】
三つの構築物の略図は、図 24に示される。
【0358】
インサートをNheIおよびXmaI消化により除去し、標準のクローニング処置によりSAP 脱リン酸化pCI-Neo 派生物(Promega, France)にクローン化した。大腸菌XL1-blue 細胞 (Stratagene, USA)をプラスミド構築物で形質転換した。プラスミド DNAをPurelinkカラム(Invitrogen, U.K.)で大規模に精製し、インサートをプラスミドに挿入された配列の統合性を検証するためシークエンシングした。プラスミド クローンを使用してAmaxa GmbH エレクトロポレーション技術〔Amaxa(登録商標) nucleofector(登録商標) kit V Catalog # VCA 1003 Lonza Germanyを製造者の推奨にしたがって使用した〕を用いてヒト 神経芽細胞腫 細胞 [SH-SY5Y 細胞 (ATCC CRL-2266),またはNtera cl2D1 細胞 (ATCC CRL-1973)]をヌクレオフェクト(nucleofect)した。G 蛋白質の発現を、例 1に記載のとおりフローサイトメトリーでアッセイした。
【0359】
一過性発現のため、生物アッセイをヌクレオフェクションの24h後に行った。
【0360】
安定発現に関して、双方のヌクレオフェクトした細胞株 [SH-SY5YまたはNtera cl2D1]を800μg/mlのG418で三週間処理した。G418に耐性の細胞クローンを単離し、凍結した。対照の細胞株は、インサートなしのpCI-Neo単独でヌクレオフェクトした細胞からなるものである。
【0361】
神経突起の成長アッセイおよび退縮アッセイを例 1に記載のとおり行った。
【0362】
例示の結果を図 25-28に示す。
【0363】
図 25は、非アポトーシス性の狂犬病ウイルス株の細胞質ドメインを含んでいるポリペプチドの発現(より具体的には、以下のものの発現)を説明する、即ち:
・ 前記狂犬病ウイルス株の全長G蛋白質の発現,
・ 前記G蛋白質の膜貫通および細胞質ドメインを含むが、前記G蛋白質の外部ドメインを含まないポリペプチドの発現, および
・ 前記G蛋白質の細胞質ドメインを含むが、前記G蛋白質の外部ドメインおよび膜貫通ドメインを含まないポリペプチドの発現,
を対照プラスミドで測定された発現と比較して説明する。
【0364】
既に説明されたとおり、Gの発現レベルは異なり、生物学的な表現型は発現のレベルと直接的に相関しない(以下および 図 5を参照されたい)。
【0365】
ウイルス感染の非存在下で、非アポトーシス性の 狂犬病ウイルス株のG蛋白質の細胞質ドメインの一過性発現によって、ヒト神経芽細胞腫細胞からの神経突起成長が誘導および/または刺激される(図 26)。前記G蛋白質の細胞質ドメインに加えて、膜貫通ドメインを含んでいる(しかし、外部ドメインを含まない)一過性に発現されたポリペプチドは、一過性に発現された 全長 G蛋白質よりも又は前記G蛋白質の一過性に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも神経突起成長の誘導及び/又は刺激に効率的である(図 26)。
【0366】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインの,または前記ドメインを含んでいるポリペプチドまたは蛋白質の安定な発現は、ヒト 神経芽細胞腫細胞において可能である(図 27)。前記G蛋白質の細胞質ドメインに加えて、膜貫通ドメインを含んでいる(しかし、外部ドメインを含まない)安定に発現されたポリペプチドは、安定に発現された 全長 G蛋白質よりも又は前記G蛋白質の安定に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも神経突起成長の誘導及び/又は刺激に効率的である(図 27)。前記 G蛋白質の安定に発現された細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)は、神経突起成長の誘導及び/又は刺激において、安定に発現された全長G蛋白質と同等に効率的である(図 27)。
【0367】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株の細胞質ドメインを含んでいるポリペプチドの発現は、神経毒性剤〔例えば、成長円錐虚脱薬 LPA; (図 28, G-(SP-[2a]-TM-Cytoで得られた結果を示している) 構築物を参照されたい)〕への抵抗性を提供する。
【0368】
同じ実験条件において、このLPAのバッチは、図 6A, 6B, 7に説明されるとおり、成長円錐の虚脱を誘導する。
【0369】
G-(SP-[2a]-TM-Cyto)の膜貫通ドメインによって、特に本構築物により発現されるポリペプチドが細胞のゴルジ膜 および/または 小胞体膜にアンカーされることが可能となり、これにより神経発生, 神経再生および神経保護の誘導および/または刺激において発現されたポリペプチドの効率が増加する。
【0370】
それ故, 非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを係る膜-アンカリング配列とカップリングすることは、前記細胞質ドメインにより誘導および/または刺激された神経発生, 神経再生 および 神経保護効果を増加させる。
【0371】
例 5: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインがニューロン前駆体(EC 細胞)に作用して成熟ニューロンに分化させる特性(コミットメント)を内在することの実験による実証
Cyto-G (上記例 4; 例えば、配列番号 6または24を参照されたい)などの非アポトーシス性の 狂犬病ウイルス株の細胞質ドメインは、このように神経芽細胞腫細胞株 SH-SY5Yの分化を促進することにより神経突起の成長を促進する(例 4 上記を参照されたい)。
【0372】
この特性が前記細胞質ドメインの細胞株をニューロン分化へコミットすることを促進し、方向づける広範囲の能力を示すことの実験による実証を例 5は提供する。
【0373】
胚性癌腫細胞系統 NTera 2cl.-D1 (ATCC CRL-1973)からの純粋な分裂後のヒトニューロンの産生は、当該技術(例えば、Prehaud et al. 2005)において記載されている。
【0374】
上記の例 4に記載のとおり、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを発現している構築物が使用された, 例えば:
・ 非アポトーシス性の狂犬病ウイルス株のG蛋白質(配列番号 2のG蛋白質)をインサートとして含んでいるG-full 構築物,
・ 非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 6の細胞質ドメイン)をインサートとして含んでいるG-Cyto 構築物,および
・ 非アポトーシス性の狂犬病ウイルス株のシグナルペプチド(配列番号 21のシグナルペプチド), 二つのアミノ酸(非アポトーシス性の狂犬病ウイルス株のG蛋白質の外部ドメインのC末端からのアミノ酸、例えば、GK), 非アポトーシス性のウイルス株のG蛋白質の膜貫通ドメイン(配列番号 23の膜貫通ドメイン)および非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(配列番号 6の細胞質ドメイン)を含んでいるG-(SP)-[2aa]-TM-Cyto 構築物。
【0375】
NTera 2-D1 細胞のニューロンへの分化を誘導するG-発現構築物の能力を調査した。
【0376】
安定なG-発現NTera 2-D1 細胞を、Prehaud等(Prehaud et al. 2005)に記載のプロトコールのようなATRA-ニューロスフェア(ATRA-neurosphere)にしたがって処理した。
【0377】
最後の抗分裂処理および二回の複製の後、細胞を神経突起の先端(最後の複製の5 日後)またはニューロンのネットワーク構築(最後の複製の50 日後)のいずれかを計算するため播種した。
【0378】
例示の結果を図 29-31に示す。
【0379】
ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを含んでいるポリペプチドの発現は、神経芽細胞腫細胞から分化することにより成熟した分裂後のヒトニューロンの産生を誘導および/または刺激する: 以下のものの発現で得られた結果を説明する図 29を参照されたい:
・ 非アポトーシス性の狂犬病ウイルス株の全長G蛋白質の発現,
・ 前記G蛋白質の膜貫通ドメインおよび細胞質ドメインを含んでいる(しかし、外部ドメインを含まない)ポリペプチドの発現,
・ 前記G蛋白質の細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)の発現,
を対照プラスミドと比較した発現。
【0380】
膜貫通ドメインを非アポトーシス性の狂犬病ウイルス株のG 蛋白質の細胞質ドメインに加えて含む(しかし、前記G蛋白質の外部ドメインを含まない)ポリペプチドは、全長 G蛋白質よりも又は前記G蛋白質の細胞質ドメイン(外部ドメインなし且つ膜貫通ドメインなしの細胞質ドメイン)よりも分裂後のヒトニューロンの分化の誘導及び/又は刺激にさらに効率的である(図 30を参照されたい)。
【0381】
神経芽細胞腫細胞からの分化により成熟した分裂後のヒトニューロンの産生を誘導および/または刺激することにくわえ、ウイルス感染の非存在下で、非アポトーシス性の狂犬病ウイルス株のG蛋白質の細胞質ドメインを含んでいるポリペプチドの発現は、長い軸索を有する成熟した分裂後のヒトニューロンのネットワークの組織化を誘導および/または刺激する(図 31を参照されたい)。
【0382】
例 6: 非アポトーシス性の狂犬病ウイルス株の細胞質ドメインが傷害されたニューロンの再生を促進できることの実験による実証
傷害されたニューロンは、これらの起源や局所の環境に応じて再生する可能性がある。例えば、哺乳類の中枢神経系からのニューロンは、末梢の突起(peripheral process)への傷害が後根神経節ニューロンの再生の増加を生じるとしても、非常に限定的な再生能力を有する。内因性の再生シグナルが再生に影響して適切な再生が成功することが明らかに確立され、それらの幾つかには特異的なキナーゼ経路が関係する(Hammarlund et al. 2009)。
【0383】
Cyto-G 構築物に含まれる配列番号 6の細胞質ドメインなどの非アポトーシス性の狂犬病ウイルス株の細胞質ドメイン(上記例 4 を参照されたい)は、再生医療に関連して少なくとも二つの興味深い特性を呈する。
【0384】
第一に、前記細胞質ドメインは神経突起成長を促進し、このことは前記細胞質ドメインがニューロンの成長円錐を刺激できることを意味する。
【0385】
第二に、前記細胞質ドメインは、LPAまたはH2O2での処理後の退縮から神経突起を保存し、このことは前記細胞質ドメインが成長円錐の虚脱を避ける分子ロック(molecular locks)を刺激または強化できることを意味する。
【0386】
これらの特性が傷害されたニューロンの再生を生じることを実験により実証するため、その現象を細胞モデルで調査した。例 4のG-[SP-(2aa)-TM-Cyto] 構築物(配列番号 27をインサートとして含んでいる構築物)を安定に発現しているNT2-N 細胞株(ヒトニューロン細胞)を選択した。pCI-Neo-NT2-Nを対照として使用した。
【0387】
NT2-N 細胞株は、文献(Prehaud et al. 2005, Cheung et al. 1999 および Paquet-Durand et al. 2003)に記載の分化型の Ntera-2cl.D1 (NT2/D1; ATCC CRL-1973) 細胞から由来するものである。
【0388】
スクラッチ誘導アッセイ(scratch-induced assays)のため、細胞をポリ-D-リシン-ラミニンコートセル+(Sarstedt, Germany)12 ウェルプラスチック器に播種し、トリプシン処理後に完全に回復させるため二 日間成長させた。培地を引っ掻く10h前に変更した。個々の傷を注射針 (26GX1/2", 12-4.5)で作った。少なくとも 10の引っかき傷を個々のウェルの各々に作った。細胞をPFA (4%)で傷をつけた3 日後に固定し、クリスタルバイオレット溶液で染色した。細胞は、Leica DM 5000B顕微鏡にDC 300FX カメラ (x20 対物レンズ)を備えたものを用いて視覚化され、ImageJ 1.38X ソフトウェア (Wayne Rasband, NIH, USA, http://rsb.info.nih.gov/ij/)とそのplug-inのNeuronJを用いて分析した。再生中のニューロンの平均のパーセンテージは、8回の実験から決定された。
【0389】
例示の結果を図 32-33に示す。
【0390】
非アポトーシス性の 狂犬病ウイルス株のG蛋白質の細胞質ドメインを含むポリペプチドの発現は、傷ついた成熟した分裂後のヒトニューロンの再生を誘導および/または刺激する(非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通および細胞質ドメインを含んでいるポリペプチドの発現で得られた結果を説明する図 32を参照されたい)。
【0391】
傷をつけた後にかように誘導された神経突起の再生はドラスティックである。より具体的には、前記ポリペプチドは、前記 G蛋白質の膜貫通および細胞質ドメインを含むが、前記G蛋白質の外部ドメインを含まない(非アポトーシス性の狂犬病ウイルス株のG蛋白質の膜貫通および細胞質ドメインを含んでいるポリペプチドの発現で得られた結果を説明する図 33を参照されたい)。
【0392】
例7:
感染性の rRABV cDNAは、固定された病原性で非アポトーシス性のRABVであるCVS-N2c 狂犬病ウイルス株から作出された(Morimoto et al. 1999)。
【0393】
このように作出された感染性の rRABV cDNA(cN2C)は、Schnell等(Schnell et al. 2010)に記載される。
【0394】
G-vir(非アポトーシス性の 狂犬病ウイルス株のG蛋白質、例えば、配列番号 2のG蛋白質)をコードしている遺伝子は、この RABV cDNAに統合された。感染性のウイルスを例 1に記載されているとおり回収した。
【0395】
組換え型のウイルスを、8個体の六週齡の雌性C57Bl6 マウスの群に筋肉内経路でマウスの80%に致死的な脳炎を生じさせるrRABV G-virのウイルス粒子数を注射した。マウスおよび実験の数を慎重に評価して、動物の負担を最小化した。
【0396】
疾患の進行を臨床徴候および死亡率をモニターすることで評価した (Camelo et al. 2001に記載のとおり)。ウイルスの神経侵襲性を測定するため、RNA抽出(脳の半分)および免疫組織化学または複数のアレイ(脳の第二の半分)のため処理する前に、感染の過程で二または三マウスのグループを灌流し、脳を除去して急速凍結し、-80°Cで貯蔵した。
【0397】
このモデルによって、さらにrRABV 株により惹起される脳の先天性の免疫応答の異なるパラメータを分析することが可能である。
【0398】
[参照文献]
Blondel et al., Poliovirus, pathogenesis of poliomyelitis, and apoptosis, CTMI, 2005, 289, 25-56.
Camelo et al. 2001 "Selective role for the p55 Kd TNF-a receptor in immune unresponsiveness induced by an acute viral encephalitis"; J. Neuroimmunol. 113: 95-108.
Cheung et al. 1999 ; BioTechniques 26: 946-954.
Faul et al. 2008; Virology 382: 226-238.
Guigoni and Coulon 2002, Journal of NeuroVirology 8: 306-317.
Hammarlund et al. 2009 "Axon regeneration requires a conserved MAP kinase pathway" Science 323: 802-806.
Kumar et al. 2007; Nature 448: 39-43 and associated Supplementary Information ("Methods").
Lay et al. 2003; Ann. N. Y. Acad. Sci. 1010: 577-581.
Loh et al. 2008; Cell Death and Differentiation 15: 283-298.
Meijering et al. 2004; Cytometry Part A 58A: 167-176; and http://www.imagescience.org/meijering/software/neuronj/
Morimoto et al. 1999, Journal of Virology 73(1): 510-518.
Paquet-Durand et al. 2003 Brain Res. Dev. Brain Res. 142: 161-167.
Prehaud et al. 1988; Journal of Virology 62(1): 1-7.
Prehaud et al. 2003; Journal of Virology 77(19): 10537-10547.
Prehaud et al. 2005; Journal of Virology 79(20): 12893-12904.
Sargent 2003; Recent Results in Cancer Research, vol. 161, Springer-Verlag Berlin Heidelberg, pages 13-25
Sarmento et al. 2005; Journal of NeuroVirology 11: 571-581.
Schnell et al. 2010; Nature Reviews Microbiology 8(1): 51-65.
Skare et al. 1975; J. Virol. 15:726-732 (Structure and function of herpesvirus genomes. I. Comparison of five HSV-1 and two HSV-2 strains by cleavage of their DNA with Eco R I restriction endonuclease).
Ugolini 1995; The Journal of Comparative Neurology 356: 457-480.
Ugolini 2008; Dodet B, Fooks AR, Muller T, Tordo N, and the Scientific & Technical Department of the OIE (eds): Towards the Elimination of Rabies in Eurasia. Dev. Biol. Basel, Karger, vol. 131, pp. 493-506.
WO 03/048198; PCT international application in the name of Institut Pasteur, Inv. Lafon et al., published on 12 June 2003; as well as its national/regional counterpart applications.
Each of the relevant disclosures of all references cited herein is specifically incorporated by reference.
【特許請求の範囲】
【請求項1】
以下のものから選択される産物であって:
・以下を含むポリペプチド:
・ 配列番号6の配列、
又は
・ 以下の配列であって:
・ 34〜54アミノ酸であり、
・ 二配列のうちの短い配列の全長にわたり、配列番号6に対して少なくとも94%同一であり、
・ PDZ-BS配列を含み、前記PDZ-BS配列が:
x1は、E以外の任意のアミノ酸(好ましくは、Q)であり、および
x2は、TまたはSまたはI(好ましくはIではなく、より好ましくはT)であり、および
x3は、任意のアミノ酸(好ましくは、R)であり、および
x4は、LまたはV(好ましくは、L)であり、
(配列番号14)、
バリアント配列Aと称される配列、
又は
・ 以下の配列であって:
・ 44アミノ酸であり、
・ 配列番号6の配列に対して少なくとも94%同一であり、
・ PDZ-BS配列を含み、前記PDZ-BS配列が:
x1は、Eであり、および
x2は、TまたはSまたはI(好ましくはIではなく、より好ましくはT)であり、および
x3は、任意のアミノ酸(好ましくは、R)であり、および
x4は、LまたはV(好ましくは、L)であり
(x1=Eの配列番号13)、
および
・ アミノ酸Lをポジション11に含まない、
バリアント配列Bと称される配列、
又は
・配列番号6の配列の断片又は前記バリアント配列Aの断片又は前記バリアント配列Bの配列の断片であって、少なくとも34アミノ酸であり、且つ配列番号6の配列の保持された当該PDZ-BS配列又は前記バリアント配列Aの保持された当該PDZ-BS配列又は前記バリアント配列Bの保持された当該PDZ-BS配列をそれぞれ有する断片、
又は
・前記ポリペプチドをコードする核酸、
又は
・前記核酸を含むベクター、
又は
・前記ポリペプチドおよび/または前記核酸および/または前記ベクターを含む細胞、
又は
・少なくとも一つの前記ポリペプチド、核酸、ベクターおよび細胞を含む薬学的組成物または薬剤、
前記産物は、神経系のニューロンからの神経突起成長および/または神経突起の発芽および/または軸索の成長および/または樹状突起樹の伸展を刺激および/または誘導することによる前記神経系に影響する疾患または障害の治療および/または緩和および/または予防における使用のためのものであり、前記刺激および/または誘導のための有効成分として使用される産物。
【請求項2】
シナプス形成および/または神経伝達および/またはニューロンの可塑性を刺激および/または誘導するために使用される、請求項1に記載の産物。
【請求項3】
請求項1〜2の何れか一項に記載の産物であって、神経発生剤および/または神経再生剤および/または神経保護剤として使用される産物。
【請求項4】
請求項1〜3の何れか一項に記載の産物であって、前記使用が非免疫原性の使用であり、及び/又は前記産物が免疫原性剤またはアジュバントではない産物。
【請求項5】
請求項1〜4の何れか一項に記載の産物であって、前記バリアント配列Aの当該PDZ-BS配列は、QTRL(配列番号10)である産物。
【請求項6】
請求項1〜5の何れか一項に記載の産物であって、前記バリアント配列Aは、44アミノ酸の配列であり、アミノ酸HまたはL、好ましくはHをポジション11に有する産物。
【請求項7】
請求項1〜6の何れか一項に記載の産物であって、前記バリアント配列Bの当該PDZ-BS配列は、ETRL(配列番号12)である産物。
【請求項8】
請求項7に記載の産物であって、前記バリアント配列Bは、アミノ酸Hをポジション11に有する産物。
【請求項9】
請求項1〜8の何れか一項に記載の産物であって、前記断片は、配列番号6の配列からの断片11〜44である産物。
【請求項10】
請求項1〜9の何れか一項に記載の産物であって、前記ポリペプチドは、狂犬病ウイルス株の当該G蛋白質の細胞質断片もしくは係る細胞質断片の小断片である、又は狂犬病ウイルス株の当該G蛋白質の細胞質断片もしくは係る細胞質断片の小断片を含む産物。
【請求項11】
請求項10に記載の産物であって、前記狂犬病ウイルス株は、CNCMにI-4140,I-4142またはI-4143で寄託された株である産物。
【請求項12】
請求項1〜11の何れか一項に記載の産物であって、前記ポリペプチドは、さらに細胞、より具体的にはニューロン細胞、より具体的にはヒトニューロン細胞の小胞体膜および/またはゴルジ膜において前記ポリペプチドをアンカーするアミノ酸配列を含み、前記アンカーする配列は、配列番号6の配列またはバリアント配列Aまたはバリアント配列Bの好ましくはN末端にあり、最も好ましくは配列番号6の配列の又は前記バリアント配列Aの又は前記バリアント配列BのN末端の最初のアミノ酸に対して直接的に連結される産物。
【請求項13】
請求項1〜12の何れか一項に記載の産物であって、前記ポリペプチドは、さらに、好ましくは配列番号6の配列又はバリアント配列A又はバリアント配列BのN末端に、最も好ましくは配列番号6の配列又は前記バリアント配列A又は前記バリアント配列BのN末端の最初のアミノ酸に対して直接的に連結された以下の配列を含む産物:
・ 狂犬病ウイルス株、より具体的には、非アポトーシス性の狂犬病ウイルス株の当該G蛋白質の膜貫通ドメインの配列、または
・ 狂犬病ウイルスGの膜貫通ドメインの配列とは、少なくとも一つのアミノ酸の置換および/または欠失および/または付加、好ましくはアミノ酸の置換および/または欠失、より好ましくはアミノ酸置換により異なるが、細胞、より具体的にはニューロン細胞、より具体的にはヒトニューロン細胞の当該小胞体膜および/または当該ゴルジ膜において、(前記アンカーする配列が配列番号6の配列のN末端の最初のアミノ酸に対して直接的に連結される場合に)配列番号6のポリペプチドをアンカーする能力を保持する配列。
【請求項14】
請求項12または13に記載の産物であって、前記アンカーする配列は以下の配列である産物:
・ 配列番号23の配列、または
・ 配列番号23のバリアント配列であって、18〜26アミノ酸、好ましくは18〜22アミノ酸、より好ましくは22アミノ酸の配列であり、二つの配列のうちの最短の配列にわたって(即ち、配列番号23及び前記バリアント配列のうちの最短の配列にわたって)、配列番号23の配列と少なくとも94%同一である配列。
【請求項15】
請求項12〜14の何れか一項に記載の産物であって、前記ポリペプチドは、配列番号6の配列または前記バリアント配列A、より具体的には配列番号6の配列を含む産物。
【請求項16】
請求項15に記載の産物であって、前記ポリペプチドのアミノ酸配列は、(N末端からC末端に)配列番号6の配列が続く配列番号23の配列からなり、1〜4アミノ酸、好ましくは1アミノ酸(例えば、M)が任意に配列番号23の配列と配列番号6の配列との間に存在する産物。
【請求項17】
請求項12〜16の何れか一項に記載の産物であって、前記ポリペプチドは、配列番号25の配列であるか、又は配列番号25の配列を含む産物。
【請求項18】
請求項12〜17の何れか一項に記載の産物であって、前記ポリペプチドのアミノ酸配列は、さらに、前記アンカーする配列のN末端の最初のアミノ酸に対して(直接的にまたは間接的に、好ましくは直接的に)連結された以下の配列を含む産物:
狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質の外部ドメインのC末端からの1〜4アミノ酸、好ましくは2アミノ酸、より好ましくは1〜4アミノ酸、さらにより好ましくは狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質の外部ドメインのC末端の最後の2アミノ酸、例えば、アミノ酸GK。
【請求項19】
請求項18に記載の産物であって、前記ポリペプチドは、配列番号26の配列であるか、又は配列番号26の配列を含む産物。
【請求項20】
請求項12〜19の何れか一項に記載の産物であって、前記ポリペプチドは、さらに、シグナルペプチド、好ましくは狂犬病ウイルス株蛋白質(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質のシグナルペプチドを含み、前記ペプチド配列は好ましくは前記ポリペプチドのN末端(最も好ましくは前記ポリペプチドのまさにN末端、即ち、前記ポリペプチドに含まれ得る任意のアンカーする配列からのN末端)に存在する産物。
【請求項21】
請求項20に記載の産物であって、前記ポリペプチドは、配列番号27の配列であるか、又は配列番号27の配列を含む産物。
【請求項22】
請求項1〜21の何れか一項に記載の産物であって、前記ポリペプチドは、100アミノ酸未満からなる産物。
【請求項23】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、神経変性により障害されたニューロンである産物。
【請求項24】
請求項1〜23の何れか一項に記載の産物であって、前記疾患または障害は、神経変性疾患または障害、例えば、非ウイルス性の脳症、アルツハイマー病、パーキンソン病、ALS、ハンチントン病、加齢性の神経変性疾患または障害、MSまたは稀な遺伝疾患である産物。
【請求項25】
請求項23または24に記載の産物であって、前記疾患または障害はMSである産物。
【請求項26】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、当該神経系の微生物感染により障害されたニューロン、及び/又は当該神経系の物理的又は虚血性の傷害により障害されたニューロン、及び/又は化学的な神経毒性剤により障害されたニューロン、及び/又は酸化ストレスにより障害されたニューロンである産物。
【請求項27】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、神経系の微生物感染、例えば、灰白髄炎に関係する産物。
【請求項28】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、当該神経系の物理的又は虚血性の傷害、例えば、発作、脳卒中、外傷、癲癇である産物。
【請求項29】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、化学的な神経毒性剤および/または酸化ストレスの存在に関係する産物。
【請求項30】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、分裂前のニューロンまたはニューロン前駆体である産物。
【請求項31】
請求項1〜22および30の何れか一項に記載の産物であって、ニューロ分化剤として使用される産物。
【請求項32】
請求項1〜22、30および31の何れか一項に記載の産物であって、前記疾患は、CNSおよび/またはPNSの腫瘍、例えば、神経節膠腫、脳腫瘍、中枢神経細胞腫、髄芽腫、脳室上衣腫、奇形腫、神経芽細胞腫である産物。
【請求項1】
以下のものから選択される産物であって:
・以下を含むポリペプチド:
・ 配列番号6の配列、
又は
・ 以下の配列であって:
・ 34〜54アミノ酸であり、
・ 二配列のうちの短い配列の全長にわたり、配列番号6に対して少なくとも94%同一であり、
・ PDZ-BS配列を含み、前記PDZ-BS配列が:
x1は、E以外の任意のアミノ酸(好ましくは、Q)であり、および
x2は、TまたはSまたはI(好ましくはIではなく、より好ましくはT)であり、および
x3は、任意のアミノ酸(好ましくは、R)であり、および
x4は、LまたはV(好ましくは、L)であり、
(配列番号14)、
バリアント配列Aと称される配列、
又は
・ 以下の配列であって:
・ 44アミノ酸であり、
・ 配列番号6の配列に対して少なくとも94%同一であり、
・ PDZ-BS配列を含み、前記PDZ-BS配列が:
x1は、Eであり、および
x2は、TまたはSまたはI(好ましくはIではなく、より好ましくはT)であり、および
x3は、任意のアミノ酸(好ましくは、R)であり、および
x4は、LまたはV(好ましくは、L)であり
(x1=Eの配列番号13)、
および
・ アミノ酸Lをポジション11に含まない、
バリアント配列Bと称される配列、
又は
・配列番号6の配列の断片又は前記バリアント配列Aの断片又は前記バリアント配列Bの配列の断片であって、少なくとも34アミノ酸であり、且つ配列番号6の配列の保持された当該PDZ-BS配列又は前記バリアント配列Aの保持された当該PDZ-BS配列又は前記バリアント配列Bの保持された当該PDZ-BS配列をそれぞれ有する断片、
又は
・前記ポリペプチドをコードする核酸、
又は
・前記核酸を含むベクター、
又は
・前記ポリペプチドおよび/または前記核酸および/または前記ベクターを含む細胞、
又は
・少なくとも一つの前記ポリペプチド、核酸、ベクターおよび細胞を含む薬学的組成物または薬剤、
前記産物は、神経系のニューロンからの神経突起成長および/または神経突起の発芽および/または軸索の成長および/または樹状突起樹の伸展を刺激および/または誘導することによる前記神経系に影響する疾患または障害の治療および/または緩和および/または予防における使用のためのものであり、前記刺激および/または誘導のための有効成分として使用される産物。
【請求項2】
シナプス形成および/または神経伝達および/またはニューロンの可塑性を刺激および/または誘導するために使用される、請求項1に記載の産物。
【請求項3】
請求項1〜2の何れか一項に記載の産物であって、神経発生剤および/または神経再生剤および/または神経保護剤として使用される産物。
【請求項4】
請求項1〜3の何れか一項に記載の産物であって、前記使用が非免疫原性の使用であり、及び/又は前記産物が免疫原性剤またはアジュバントではない産物。
【請求項5】
請求項1〜4の何れか一項に記載の産物であって、前記バリアント配列Aの当該PDZ-BS配列は、QTRL(配列番号10)である産物。
【請求項6】
請求項1〜5の何れか一項に記載の産物であって、前記バリアント配列Aは、44アミノ酸の配列であり、アミノ酸HまたはL、好ましくはHをポジション11に有する産物。
【請求項7】
請求項1〜6の何れか一項に記載の産物であって、前記バリアント配列Bの当該PDZ-BS配列は、ETRL(配列番号12)である産物。
【請求項8】
請求項7に記載の産物であって、前記バリアント配列Bは、アミノ酸Hをポジション11に有する産物。
【請求項9】
請求項1〜8の何れか一項に記載の産物であって、前記断片は、配列番号6の配列からの断片11〜44である産物。
【請求項10】
請求項1〜9の何れか一項に記載の産物であって、前記ポリペプチドは、狂犬病ウイルス株の当該G蛋白質の細胞質断片もしくは係る細胞質断片の小断片である、又は狂犬病ウイルス株の当該G蛋白質の細胞質断片もしくは係る細胞質断片の小断片を含む産物。
【請求項11】
請求項10に記載の産物であって、前記狂犬病ウイルス株は、CNCMにI-4140,I-4142またはI-4143で寄託された株である産物。
【請求項12】
請求項1〜11の何れか一項に記載の産物であって、前記ポリペプチドは、さらに細胞、より具体的にはニューロン細胞、より具体的にはヒトニューロン細胞の小胞体膜および/またはゴルジ膜において前記ポリペプチドをアンカーするアミノ酸配列を含み、前記アンカーする配列は、配列番号6の配列またはバリアント配列Aまたはバリアント配列Bの好ましくはN末端にあり、最も好ましくは配列番号6の配列の又は前記バリアント配列Aの又は前記バリアント配列BのN末端の最初のアミノ酸に対して直接的に連結される産物。
【請求項13】
請求項1〜12の何れか一項に記載の産物であって、前記ポリペプチドは、さらに、好ましくは配列番号6の配列又はバリアント配列A又はバリアント配列BのN末端に、最も好ましくは配列番号6の配列又は前記バリアント配列A又は前記バリアント配列BのN末端の最初のアミノ酸に対して直接的に連結された以下の配列を含む産物:
・ 狂犬病ウイルス株、より具体的には、非アポトーシス性の狂犬病ウイルス株の当該G蛋白質の膜貫通ドメインの配列、または
・ 狂犬病ウイルスGの膜貫通ドメインの配列とは、少なくとも一つのアミノ酸の置換および/または欠失および/または付加、好ましくはアミノ酸の置換および/または欠失、より好ましくはアミノ酸置換により異なるが、細胞、より具体的にはニューロン細胞、より具体的にはヒトニューロン細胞の当該小胞体膜および/または当該ゴルジ膜において、(前記アンカーする配列が配列番号6の配列のN末端の最初のアミノ酸に対して直接的に連結される場合に)配列番号6のポリペプチドをアンカーする能力を保持する配列。
【請求項14】
請求項12または13に記載の産物であって、前記アンカーする配列は以下の配列である産物:
・ 配列番号23の配列、または
・ 配列番号23のバリアント配列であって、18〜26アミノ酸、好ましくは18〜22アミノ酸、より好ましくは22アミノ酸の配列であり、二つの配列のうちの最短の配列にわたって(即ち、配列番号23及び前記バリアント配列のうちの最短の配列にわたって)、配列番号23の配列と少なくとも94%同一である配列。
【請求項15】
請求項12〜14の何れか一項に記載の産物であって、前記ポリペプチドは、配列番号6の配列または前記バリアント配列A、より具体的には配列番号6の配列を含む産物。
【請求項16】
請求項15に記載の産物であって、前記ポリペプチドのアミノ酸配列は、(N末端からC末端に)配列番号6の配列が続く配列番号23の配列からなり、1〜4アミノ酸、好ましくは1アミノ酸(例えば、M)が任意に配列番号23の配列と配列番号6の配列との間に存在する産物。
【請求項17】
請求項12〜16の何れか一項に記載の産物であって、前記ポリペプチドは、配列番号25の配列であるか、又は配列番号25の配列を含む産物。
【請求項18】
請求項12〜17の何れか一項に記載の産物であって、前記ポリペプチドのアミノ酸配列は、さらに、前記アンカーする配列のN末端の最初のアミノ酸に対して(直接的にまたは間接的に、好ましくは直接的に)連結された以下の配列を含む産物:
狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質の外部ドメインのC末端からの1〜4アミノ酸、好ましくは2アミノ酸、より好ましくは1〜4アミノ酸、さらにより好ましくは狂犬病ウイルス(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質の外部ドメインのC末端の最後の2アミノ酸、例えば、アミノ酸GK。
【請求項19】
請求項18に記載の産物であって、前記ポリペプチドは、配列番号26の配列であるか、又は配列番号26の配列を含む産物。
【請求項20】
請求項12〜19の何れか一項に記載の産物であって、前記ポリペプチドは、さらに、シグナルペプチド、好ましくは狂犬病ウイルス株蛋白質(例えば、非アポトーシス性の狂犬病ウイルス株)の当該G蛋白質のシグナルペプチドを含み、前記ペプチド配列は好ましくは前記ポリペプチドのN末端(最も好ましくは前記ポリペプチドのまさにN末端、即ち、前記ポリペプチドに含まれ得る任意のアンカーする配列からのN末端)に存在する産物。
【請求項21】
請求項20に記載の産物であって、前記ポリペプチドは、配列番号27の配列であるか、又は配列番号27の配列を含む産物。
【請求項22】
請求項1〜21の何れか一項に記載の産物であって、前記ポリペプチドは、100アミノ酸未満からなる産物。
【請求項23】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、神経変性により障害されたニューロンである産物。
【請求項24】
請求項1〜23の何れか一項に記載の産物であって、前記疾患または障害は、神経変性疾患または障害、例えば、非ウイルス性の脳症、アルツハイマー病、パーキンソン病、ALS、ハンチントン病、加齢性の神経変性疾患または障害、MSまたは稀な遺伝疾患である産物。
【請求項25】
請求項23または24に記載の産物であって、前記疾患または障害はMSである産物。
【請求項26】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、当該神経系の微生物感染により障害されたニューロン、及び/又は当該神経系の物理的又は虚血性の傷害により障害されたニューロン、及び/又は化学的な神経毒性剤により障害されたニューロン、及び/又は酸化ストレスにより障害されたニューロンである産物。
【請求項27】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、神経系の微生物感染、例えば、灰白髄炎に関係する産物。
【請求項28】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、当該神経系の物理的又は虚血性の傷害、例えば、発作、脳卒中、外傷、癲癇である産物。
【請求項29】
請求項1〜22および26の何れか一項に記載の産物であって、前記疾患または障害は、化学的な神経毒性剤および/または酸化ストレスの存在に関係する産物。
【請求項30】
請求項1〜22の何れか一項に記載の産物であって、前記ニューロンは、分裂前のニューロンまたはニューロン前駆体である産物。
【請求項31】
請求項1〜22および30の何れか一項に記載の産物であって、ニューロ分化剤として使用される産物。
【請求項32】
請求項1〜22、30および31の何れか一項に記載の産物であって、前記疾患は、CNSおよび/またはPNSの腫瘍、例えば、神経節膠腫、脳腫瘍、中枢神経細胞腫、髄芽腫、脳室上衣腫、奇形腫、神経芽細胞腫である産物。
【図1】

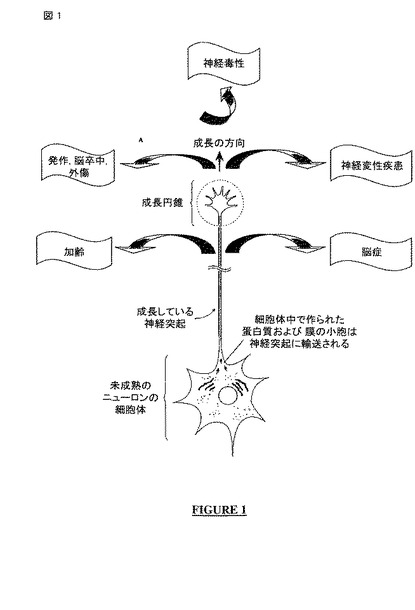
【図2】
【図3A】
【図3B】
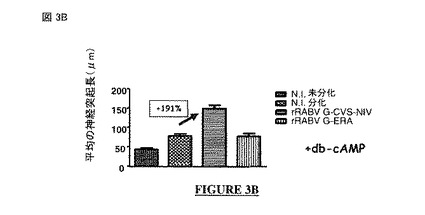
【図3C】
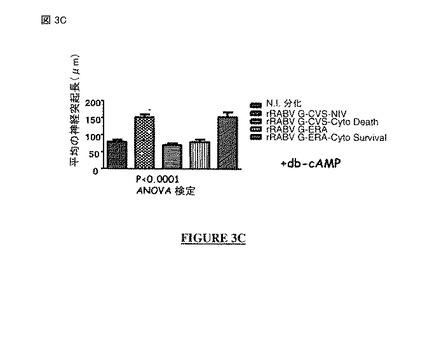
【図3D】
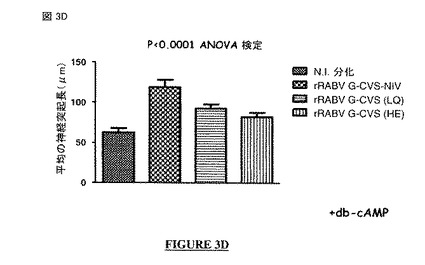
【図4A】
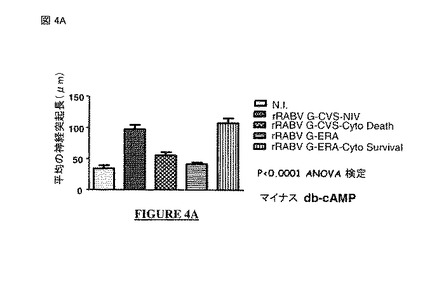
【図4B】
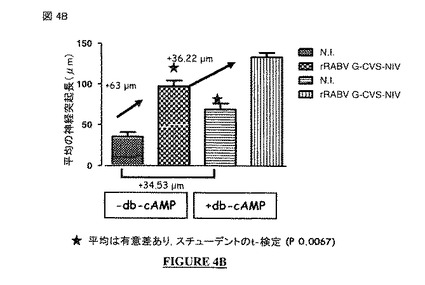
【図5】
【図6A】
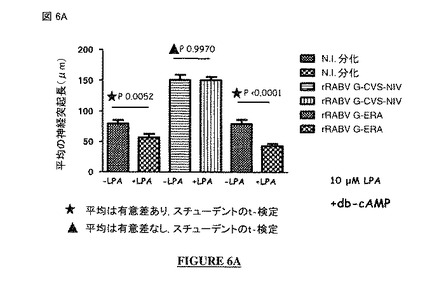
【図6B】
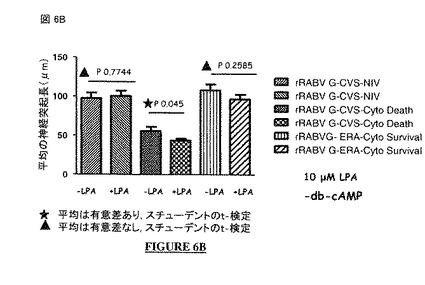
【図7】
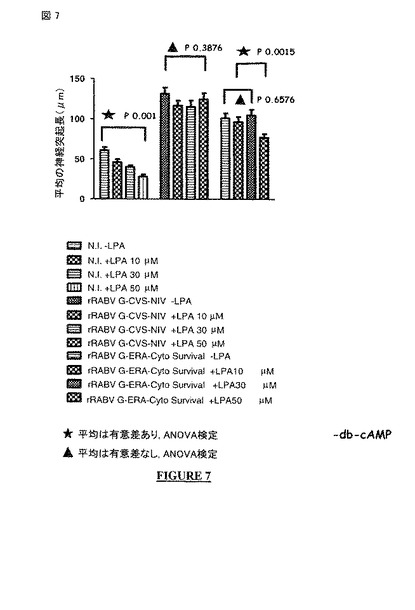
【図8】
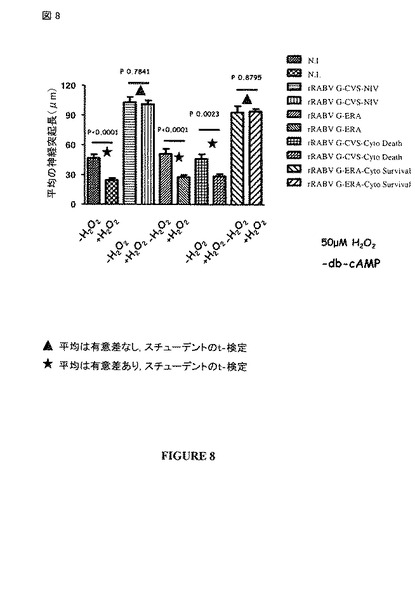
【図9A】
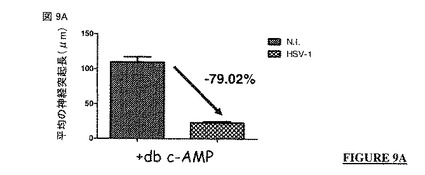
【図9B】
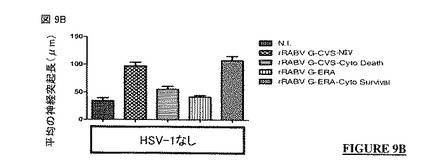
【図9C】
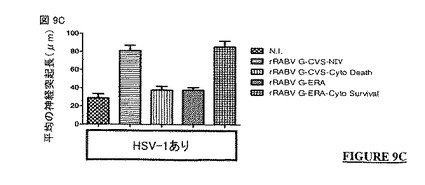
【図10A】
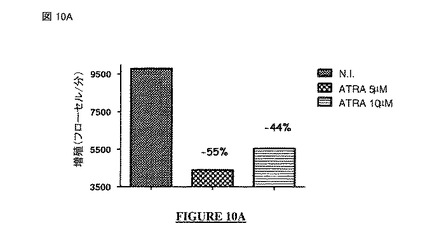
【図10B】
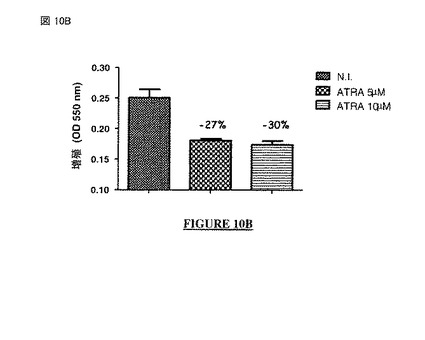
【図11A】
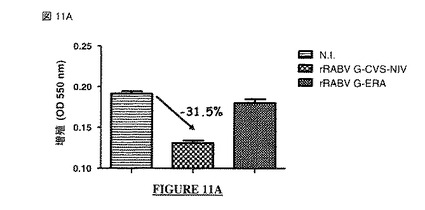
【図11B】
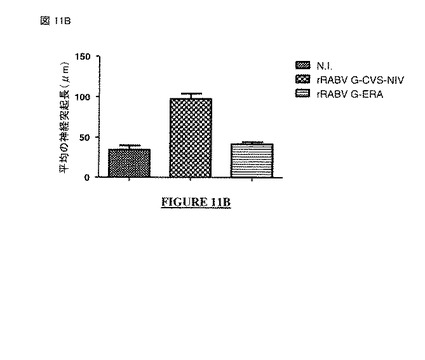
【図12A】
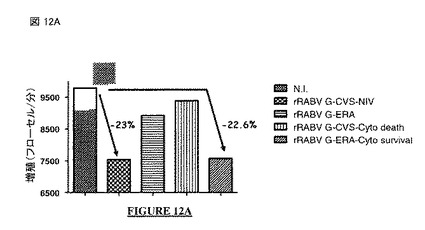
【図12B】
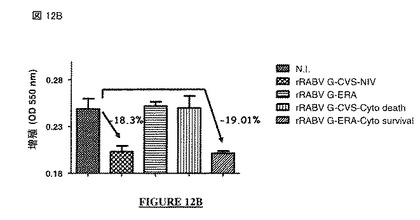
【図13A】

【図13B】

【図14】

【図15】

【図16】

【図17−1】

【図17−2】

【図18−1】

【図18−2】

【図19】

【図20】
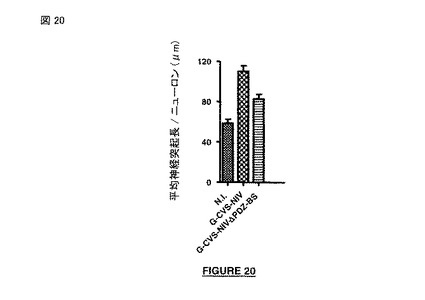
【図21】
【図22】

【図23】

【図24】
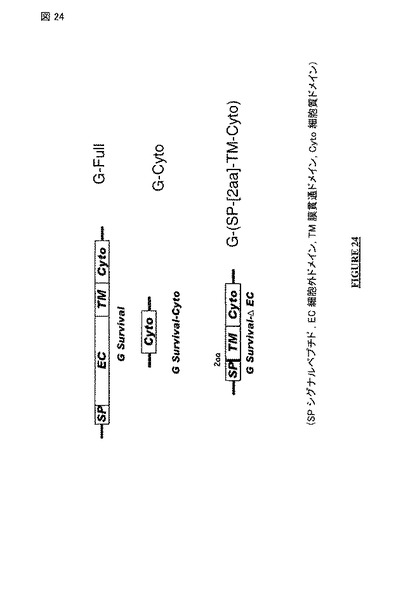
【図25】
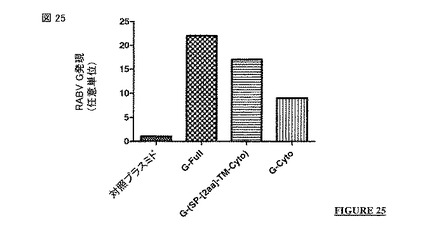
【図26】
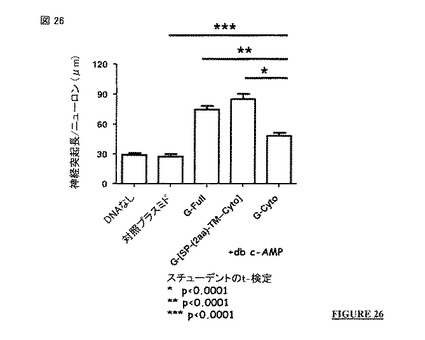
【図27】
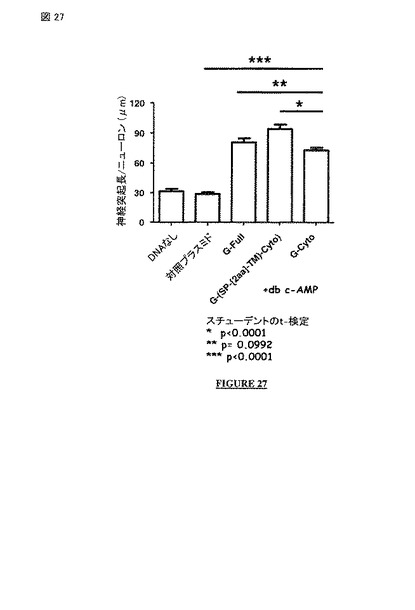
【図28】
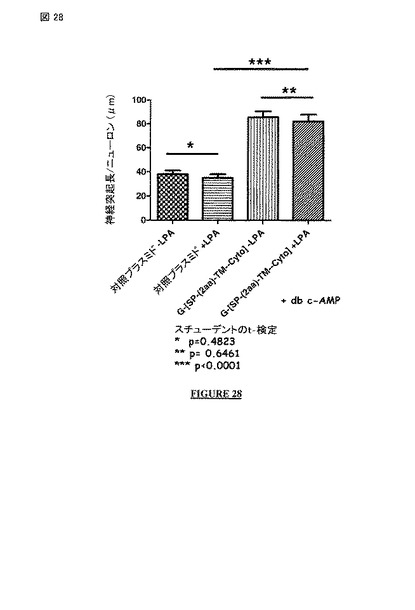
【図29】

【図30】

【図31】
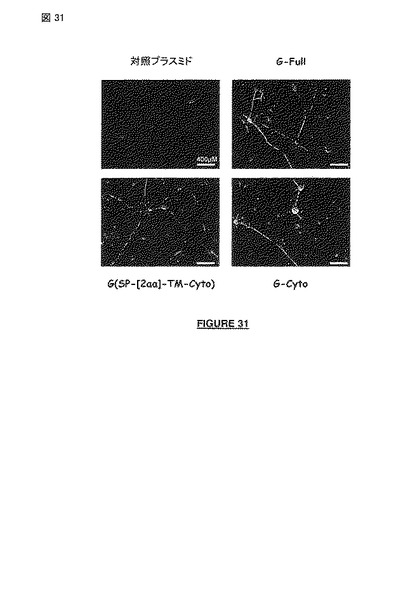
【図32】

【図33】
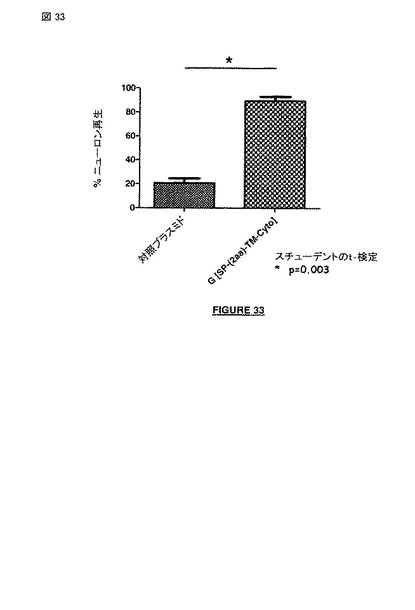
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図10A】
【図10B】
【図11A】
【図11B】
【図12A】
【図12B】
【図13A】
【図13B】
【図14】
【図15】
【図16】
【図17−1】
【図17−2】
【図18−1】
【図18−2】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【公表番号】特表2012−522831(P2012−522831A)
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−504095(P2012−504095)
【出願日】平成22年4月2日(2010.4.2)
【国際出願番号】PCT/IB2010/000967
【国際公開番号】WO2010/116258
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(399029547)トーマス・ジェファーソン・ユニバーシティー (1)
【Fターム(参考)】
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成22年4月2日(2010.4.2)
【国際出願番号】PCT/IB2010/000967
【国際公開番号】WO2010/116258
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(399029547)トーマス・ジェファーソン・ユニバーシティー (1)
【Fターム(参考)】
[ Back to top ]