
ニワトリにおける抗体の組織特異的発現
外因性抗体をコードする導入遺伝子が、ドナー細胞中に安定に組み込まれ、そしてキメラトリの体細胞組織中に存在する。これらの導入遺伝子は外因性抗体をコードし、そして好ましくは、卵における回収のために、輸卵管中で発現される。組織特異性は、導入遺伝子の内容をそれに応じて選択することによって提供される。導入遺伝子に由来する外因性抗体をコードするDNAからゲノムが構成されているトリは、細菌発現系に由来する抗体と比べて、治療的有用性の高い望ましい化学的諸特性を有する外因性抗体を発現する。

【発明の詳細な説明】
【技術分野】
【0001】
本明細書において記載される発明は、USDA SBIR 2003−33610およびNIH 2 R44 GM64261−01および2R44 HD 039583−02の下で政府により支援されている。米国政府は、本発明において特定の権利を有し得る。
【背景技術】
【0002】
(発明の背景)
遺伝的に改変された動物は、抗体など有益な薬学的製品の持続可能な製造を大きく前進させる可能性を与える。しかし、遺伝的に改変された動物の作製は、著しい技術的障害を伴い、これらの障害は少数の種でしか克服されていない。特異的発現のために、タンパク質をコードする遺伝的改変をある種のDNA中に組み入れられるようにするには、各遺伝的改変のために開発されなければならないいくつかの独特な技術が必要となる。ある動物の遺伝的特徴および肉体的特徴を改変するための1つの手法は、その動物のレシピエント胚中に細胞を導入することである。これらの細胞は、レシピエント胚から生まれる動物の組織に寄与し、結果として生じる遺伝的に改変された動物の子孫のゲノに寄与する能力を有する。
【0003】
細胞株の研究および開発、細胞ゲノムの操作、およびそのような操作された細胞が培養状態で維持されることを可能にする細胞培養技術に、かなりの時間および資源が費やされてきた。多くの試みがなされてきたが、操作された細胞の多能性を培養状態で維持する能力は、ごく少数の種、特にマウスでしか実現されていない。
【0004】
持続可能な培養物が容易に入手可能であり、そして多能性を維持しつつ遺伝子工学を受け入れることが可能である場合には、新しい技術の幅広い応用が利用可能になると思われる。培養細胞はキメラ動物の組織に寄与することができるため、胚状態のレシピエント動物中にこれらの細胞を組み入れることによって胚性幹細胞の由来元である動物の肉体的特徴をレシピエント胚に導入することができる。これは、2つの主要な利点を与える。第1に、胚性幹細胞の由来元である動物の表現型をレシピエント胚に選択的に導入することができる。第2に、上述したように、細胞培養物が特に安定である場合、これらの細胞のゲノムを遺伝的に改変して、細胞が導入されるレシピエント胚中に遺伝的改変を導入することができる。
【0005】
いくつかの場合において、抗体をコードする導入遺伝子を用いてこれらの細胞を操作することができる。導入遺伝子は、抗体を作製するための青写真の役割を果たすDNAを含み、そしてレシピエント胚中への細胞の挿入によって作り出される動物の組織における抗体の発現を可能にするのに十分なコードエレメントおよび調節エレメントを含む、遺伝子構築物である。しかし、動物の組織から有益な抗体を回収するには、通常、発現されたタンパク質の回収を容易にし、かつ望ましい化学的諸特性を伝達するいくつかの特定の組織型に発現が限定されている必要がある。例えば、雌ウシにおいては、乳汁におけるタンパク質発現により、単に雌ウシの乳汁を回収し、そして外因性タンパク質を分離することによる、タンパク質の容易な回収が可能になる。ニワトリでは、卵白における抗体の活発な産生もまた、抗体の発現および回収のための魅力的な媒介物になる。さらに、組織特異的な発現がニワトリの輸卵管に特異的である場合、この発現により、ヒト患者の治療において使用された場合に抗体の治療的有用性を高めるいくつかの特定の望ましい化学的諸特性を有する抗体が生じる。したがって、研究および商業的開発の特に魅力的な1つの分野は、卵中で抗体を選択的に発現して、望ましい化学的諸特性を有するタンパク質の単離および回収を容易にする、遺伝子操作されたニワトリである。組織特異性がない場合、動物の全組織において発現される抗体を生じるだけであるため、ある動物の特異的に選択された細胞において外因性抗体を選択的に作製する能力は、特に有益である。このような状況下では、有意義な量の抗体が動物から分離され得る見込みは少なく、外因性抗体の遍在的な発現は通常、動物の全般的健康および幸福に大きな損害を与え、また、ニワトリ輸卵管において提示される望ましい化学的諸特性は存在していない。
【0006】
細胞培養が十分に安定で、細胞のゲノム中に導入遺伝子が組み込まれるのが可能となる場合、標的細胞および導入遺伝子として使用される特定の構築物に応じたいくつかの異なる技術によって新しいキメラ生物またはトランスジェニック生物に、抗体の組織特異的発現をコードする導入遺伝子を受け渡すことができる。全ゲノムは細胞ハイブリダイゼーションによって、インタクトな染色体は微小核によって、染色体部分のセグメント(subchromosomal segment)は染色体を介した遺伝子導入によって、およびキロベース範囲のDNA断片はDNAを介した遺伝子導入によって、導入することができる(非特許文献1:Klobutcher, L.A.およびF.H.Ruddle、Annu. Rev. Biochem.、50巻:533〜554頁、1981年)。インタクトな染色体は、微小核を介した染色体導入(MMCT)によって導入することができる(非特許文献2:Fournier,R. E.およびF. H. Ruddle、Proc. Natl. Acad. Sci. U.S.A.、74巻:319〜323頁、1977年)。導入遺伝子を具体的に設計するには、抗体をコードするDNA配列の内容、標的細胞株、発現が標的される場となる特異的組織、発現が起こる場となる宿主生物、および発現される抗体を検討しなければならない。組織特異的発現のために設計される導入遺伝子は、細胞のゲノム中への成功な組み込みを可能にし、そして宿主生物の選択された組織における成功な発現を確実にするために、いくつかの条件を満たさなければならない。
【0007】
上記のように、トランスジェニック動物を作製するための遺伝的改変の導入は、ごく少数の種においてしか実証されていない。マウスの場合、キメラ子孫およびトランスジェニック子孫を作製するための、相同組換えとそれに続く胚性幹(ES)細胞への染色体導入の別々の使用が周知である。部位特異的な相同組換えまたは遺伝子ターゲティングの強力な技術が開発された(非特許文献3および4:Thomas, K. R.およびM.R. Capecchi、Cell 51巻:503〜512頁、1987年;Waldman, A.S.、Crit.Rev. Oncol. Hematol.12巻:49〜64頁、1992年による総説を参照されたい)。クローン化DNAの挿入(非特許文献5:Jakobovits,A.、Curr. Biol.4巻:761〜763頁、1994年)、ならびにCre−loxP系技術による染色体フラグメントの操作および選択(非特許文献6および7ならびに特許文献1〜4:Smith,A. J.ら、Nat. Genet.9巻:376〜385頁、1995年;Ramirez-Solis, R.ら、Nature 378巻:720〜724頁、1995年;米国特許第4,959,317号;第6,130,364号;第6,091,001号;第5,985,614号を参照されたい)は、安定な遺伝子キメラを作製するための操作およびマウスES細胞への遺伝子導入のために利用可能である。哺乳動物系において有用であることが判明したこのような多くの技術は、必要な長期の細胞培養物が入手可能である場合、ならびに、外因性タンパク質の単離および回収を容易にする、特定の組織における組織特異的発現をもたらす導入遺伝子が設計され得る場合には、非哺乳動物系に適用するために有効であると考えられる。
【0008】
組織特異的発現を可能にする導入遺伝子は複雑であり、また、レシピエント細胞株中に導入遺伝子を組み入れるのに必要である遺伝子操作は、培養条件が最適化されなければ細胞の多能性を脅かす可能性がある大規模な操作を必要とする。したがって、遺伝子導入において使用するのに適した細胞株は、培養状態で安定でなければならず、かつ組織特異的発現に必要なエレメントをすべて含むのに十分な大きさおよび複雑さを有する遺伝子構築物で細胞がトランスフェクトされた場合に、多能性を維持しなければならない。結果として生じる動物において、導入遺伝子は、導入遺伝子が発現されるように設計されている特定の個々の組織型において効果的に発現されなければならず、そして他の組織で発現されて、その結果、動物の生存能力または結果として生じるタンパク質の有利な化学的性質が損なわれるべきではない。
【0009】
外因性抗体の作製に対して、トリの生物学的系は、飼育場での効率的な飼育、迅速な成長、および経済的な生産を含めて、多くの利点を与える。さらに、トリ卵は、抗体の大量合成ならびに生成物の単離および回収の平易性の双方に関して理想的な生物学的設計を実現する。さらに、本明細書において後述するように、脊椎動物、植物、または細菌の細胞系と比較したニワトリをベースとする発現系の利点は、大量の抗体生成物に対して得ることができる。
【特許文献1】米国特許第4,959,317号明細書
【特許文献2】米国特許第6,130,364号明細書
【特許文献3】米国特許第6,091,001号明細書
【特許文献4】米国特許第5,985,614号明細書
【非特許文献1】Klobutcher, L.A.およびF.H.Ruddle、Annu. Rev. Biochem.、50巻:533〜554頁、1981年
【非特許文献2】Fournier, R. E.およびF. H. Ruddle、Proc. Natl. Acad. Sci. U.S.A.、74巻:319〜323頁、1977年
【非特許文献3】Thomas, K. R.およびM.R. Capecchi、Cell 51巻:503〜512頁、1987年
【非特許文献4】Waldman, A.S.、Crit. Rev. Oncol. Hematol.12巻:49〜64頁、1992年
【非特許文献5】Jakobovits, A.、Curr. Biol.4巻:761〜763頁、1994年
【非特許文献6】Smith, A. J.ら、Nat. Genet.9巻:376〜385頁、1995年
【非特許文献7】Ramirez-Solis, R.ら、Nature 378巻:720〜724頁、1995年
【発明の開示】
【課題を解決するための手段】
【0010】
(発明の要旨)
本発明は、ニワトリにおけるタンパク質発現、ならびにキメラニワトリおよびトランスジェニックニワトリを作製するために使用される、遺伝子工学、遺伝子導入、および細胞の長期培養などの実現技術を含む。本発明はまた、ニワトリにおいて産生される抗体およびそれらの独特な化学的性質にも関する。具体的には、これらの抗体は、ある種の用途におけるそれらの治療的有用性を高める有利な化学的諸特性を有する。ニワトリにおいて産生される抗体は、脊椎動物、植物、または細菌の細胞系において産生される抗体と比べて独特なパターンの化学的改変を有し、その結果、腫瘍などの標的組織に毒素を結合させる目的で患者に投与された場合に、標的組織は、高い治療有効性で治療される。一実施形態では、胚性幹細胞の長期培養物は、外因性抗体の組織特異的発現をもたらす導入遺伝子の挿入物を含む、キメラトリに遺伝的改変を導入するために特別に設計された遺伝子構築物を用いて作り出される。本発明のトランスジェニックトリは、導入遺伝子に由来する抗体を輸卵管中で発現し、また、抗体は、卵中に大量に蓄積する。好ましい実施形態では、天然のヒト抗体がニワトリの輸卵管において発現され、そして卵から回収することができるように、外因性抗体タンパク質は、ヒトDNA配列によってコードされる。
【0011】
本発明は、抗体の組織特異的発現を示すトリの集団、外因性抗体の発現を可能にする導入遺伝子構築物、ニワトリにおいて産生され、そして特別に定められた化学的諸特性を有する抗体の単離組成物、ならびにこれらのトリの作製、抗体の作製、およびヒトにおけるそれらの治療的使用のための関連した方法を含む。本発明は、長期細胞培養物、および持続的な細胞培養物に由来するキメラトリまたはトランスジェニックトリを作製するための特別な技術であって、ES細胞のゲノムが外因性タンパク質を発現する安定に組み込まれた導入遺伝子を有し、その結果、培養細胞の子孫が、安定に組み込まれた導入遺伝子を含む技術を使用する。後述する手順によって宿主トリの胚に導入された場合、それらの改変されたドナー細胞は、結果として生じる動物の選択された特定の体細胞組織中に導入遺伝子を発現するトリを生じる。これらのトリは、ドナー細胞に由来する表現型を提示し、また、輸卵管中で抗体を発現して、卵白における抗体の濃縮および回収を容易にする。
【0012】
本発明はまた、ニワトリ発現系において発現され、そして脊椎動物、植物、または細菌細胞の系と比べて望ましいいくつかの化学的諸特性を有する抗体の組成物も含む。具体的には、これらの抗体は、濃度の低下したフコース、ガラクトース、N−アセチルノイラミン酸、N−グリコリルノイラミン酸、および濃度の上昇したマンノースを有する。これらの諸特性のうちのいくつかまたはすべてを有する抗体は、ヒトに投与された場合に、高い治療的有用性を示す。具体的には、これらの抗体組成物は、増強された抗体依存性細胞傷害活性(ADCC)を示す。したがって、本発明の方法は、抗体の組成物をニワトリにおいて発現させることにより、ADCC作用に基づいて、それらの治療的有用性を高めることを含む。実際の応用では、これらの方法は、本明細書において説明される抗体組成物を投与するステップと、患者における細胞傷害活性を検出するステップとを含む。本発明はまた、所定の量の外因性抗体が卵白中に濃縮されるように、本明細書において定義される有利な化学的性質を有する外因性抗体を輸卵管組織中で発現するニワトリも含む。好ましい一実施形態では、外因性タンパク質は、ドナー細胞株および子孫のゲノム中に組み入れられた導入遺伝子構築物によってコードされたヒト配列モノクローナル抗体である。ヒトモノクローナル抗体をコードするポリヌクレオチド配列は、輸卵管中での発現のために特異的に構築されており、そして組織特異的発現を容易にするための適切なプロモーターおよび調節配列を含む導入遺伝子の内部に含まれている。外因性タンパク質を発現するトランスジェニックトリまたはキメラトリの実施形態では、本発明は、その動物およびタンパク質、例えば本明細書において説明される外因性抗体を含む卵白アルブミンに特異的な組成物を含む。
【発明を実施するための最良の形態】
【0013】
(発明の詳細な説明)
本発明によれば、ニワトリES細胞系は大型の核を有し、かつ顕著な核小体を含むステージX胚に由来する(図1)。これらの細胞は、長期の培養後に形態学的にニワトリ胚性幹(cES)細胞であることが確認され、また、レシピエント胚中に注入された場合にキメラを生じることが確認されている。さらに、ES細胞は、広範囲の羽毛キメラ現象によって判定されるように、体細胞組織に対する高度の寄与を可能にする。さらにまた、これらの胚性幹細胞は、外因性タンパク質をコードするDNAを有する導入遺伝子でトランスフェクトされることが実証されている。ES細胞は、導入遺伝子を安定に組み込み、そして導入遺伝子を発現して、形質転換細胞の選択を可能にする。これらの形質転換細胞はキメラを形成することができ、その際、導入遺伝子によってコードされた外因性タンパク質は、このキメラの選択された組織中に存在する。このキメラに由来する細胞は、この導入遺伝子によってコードされる外因性タンパク質を発現する。特に好ましい実施形態では、この導入遺伝子によってコードされる外因性抗体は、この導入遺伝子によってコードされる抗体に応じて、特定の組織または組織型において発現される。胚性幹細胞の子孫は、レシピエント胚へのES細胞の導入およびキメラの形成後に非ES細胞表現型に分化する、ES細胞の派生物である。
【0014】
体細胞組織における導入遺伝子の広範な発現は、胚体外組織および体細胞組織における発現によって実証される。トランスジェニック動物の卵のタンパク質内容物を解析することにより、導入遺伝子から、そして本発明の技術を用いて得られる、導入遺伝子によってコードされた外因性タンパク質の卵白における選択的発現が実証される。組織特異的発現は、1つの器官、組織、または細胞型に実質的に限局されている発現によって実証される。
【実施例】
【0015】
(実施例1)
ニワトリ胚性幹細胞(cES細胞)の誘導
ニワトリES細胞を、2種の交雑、すなわち、横斑ロック×横斑ロック、または横斑ロック×ロードアイランドレッドのうちの1つから誘導した。cES細胞の発育能を試験する場合、羽毛マーカーを得るためにこれらの品種を選択した。これらcES細胞を、優性の白色遺伝子座においてホモ接合優性である白色レグホーン胚中に注入する。これらのES細胞の注入の結果として生じるキメラニワトリは、cES細胞由来の黒色の羽毛およびレシピエント胚由来の白色の羽毛を提示する。
【0016】
cES細胞培養物の最初の樹立を、USP5,565,479に記載されているプロトコールに従って開始した。ステージXでは、胚は、約40,000〜60,000個の細胞からなり、卵黄の表面に位置する、小さな円盤状物体である。この胚を採取するために、紙でできた輪を卵黄膜上に置き、中央に胚を露出させる。この輪の外周に沿って卵黄膜を切断し、次いで卵黄からそれを持ち上げる。輪の内側に付着した胚を顕微鏡下に置き、また、細いループを用いて暗域から明域を単離する。
【0017】
表1:CES−80培地中のSTOフィーダー細胞またはポリエステル挿入物のいずれかの上で誘導されたcES細胞株。培養は、単一胚およびプールされた胚の双方から開始した。
【0018】
【表1】
胚を機械的に分散させて単細胞懸濁物とし、有糸分裂が不活化されたSTO細胞のコンフルエントなフィーダー層上に3×104細胞/cm2の濃度で播種する。cES培養培地は、10%FCS、1%ペニシリン/ストレプトマイシン、2mMグルタミン、1mMピルバート、1×ヌクレオシド、1×非必須アミノ酸、および0.1mM β−メルカプトエタノールを添加したDMEM(20%新鮮培地および80%馴化培地)からなる。使用前に、DMEM培地をバッファローラット肝臓(BRL)細胞で馴化させる。手短に言えば、BRL細胞をコンフルエントになるまで増殖させた後、5%血清含有DMEMを添加し、3日間馴化させる。この培地を除去し、培地の新しいバッチを3日間馴化させ、これを繰り返す。3回分のバッチを合わせ、cES培地を作製するために使用した。ニワトリES細胞は、胚盤葉細胞の接種後3〜7日目に可視になる。これらのcES細胞は、形態学的にmES細胞に類似していた。すなわち、これらの細胞は小型であり、大型の核および顕著な核小体を有していた(図1を参照されたい)。
【0019】
cES細胞の増殖特徴は、縁が滑らかであり、かつ区別が困難な個々の細胞を有する密な円形コロニー中で増殖するmES細胞とは異なる。ニワトリES細胞は、個々の細胞がはっきりと目に見える単層コロニー中で増殖する。密なコロニーは、cES培養物における分化の最初の徴候であることが多い。
【0020】
培養で誘導された細胞の多能性のマーカーについて試験するために、これらの細胞を固定し、そしてマウスおよびニワトリの始原生殖細胞上のエピトープを認識する、SSEA−11(Solter,D.およびB.B.Knowles、Proc.Natl.Acad.Sci、U.S.A.75巻:5565〜5569頁、1978年)、EMA−1(Hahnel,A.C.およびE.M.Eddy、GameteResearch 15巻:25〜34頁、1986年)、および同様に多能性細胞によって発現されるアルカリ性ホスファターゼ(AP)で染色した。図2に示すこれらの試験の結果により、ニワトリES細胞が、アルカリ性ホスファターゼ、ならびにSSEA−1およびEMA−1によって認識される抗原を発現することが実証される。
【0021】
上記のプロトコールを用いた後のcES細胞は可視であるが、このような培養物は、数週間より長く維持することができない。以下で考察するように、培養条件にいくつかの変更を起こすことにより、19種の細胞株(表1)の派生物を生じさせ、このうち14種をレシピエント胚中に注入することによってそれらの発育能を試験した。羽毛の色素沈着によって判定されるように、14種の細胞株のうちの11種が、レシピエント胚に寄与した(下記表2を参照されたい)。このプロトコールは、胚性幹細胞表現型を発現する多能性細胞の持続的培養物をもたらす。任意の時点で、これらの細胞を凍結保存することができ、また、これらの細胞は、損傷を与えられたレシピエント胚中に注入された場合に、体細胞組織に実質的に寄与する潜在能力を有する(以下の実施例3および実施例5を参照されたい)。
【0022】
表2:単一胚またはプールされた胚に由来する胚性幹細胞株の継代数および培養時間。ステージXレシピエント中へのこれらのES細胞の注入後の体細胞キメラ現象の出現率および程度。
【0023】
【表2】
マウスと同様に、トリ胚性幹細胞は、STO、STO−sn1、および容易に入手可能である他のものを含めて、様々なフィーダー層上で誘導される。これらのフィーダーによって産生される白血病抑制因子(LIF)、およびウシ胎児血清の添加は、ES細胞を未分化状態で維持するのに寄与する。本発明の好ましい実施形態では、ニワトリES細胞の培養は、STOフィーダー層上で開始する。STO細胞をコンフルエントになるまで増殖させ、10μg/mlマイトマイシンで3〜4時間処理し、洗浄し、トリプシン処理し、そしてゼラチンコート皿上に4×104細胞/cm2の濃度で播種する。cES細胞は、STO細胞からなるフィーダーが薄い場合に、より迅速に増殖するようである。STOフィーダー細胞濃度を103〜105細胞/cm2まで、好ましくは104細胞/cm2未満に低下させると、cES細胞の誘導および増殖が促進される。しかし、ニワトリ胚線維芽細胞およびマウス初代線維芽細胞をフィーダーとして用いた場合、cES細胞は誘導されなかった。また、予め樹立されたcES細胞をこれらのフィーダー上に播種した場合、これらはすべて1週間以内に分化した。
【0024】
フィーダーの不在下、ポリマー膜(Costar社、Transwell型)などの合成挿入物上でcES細胞を増殖させると、ES細胞を注入する際の、フィーダー細胞によるレシピエント胚の汚染が回避される。表3および表4が示すように、STOフィーダーの代わりに挿入物上で培養することにより、cES細胞の誘導が促進され、また、挿入物は、最初の誘導のために使用され得る。しかし、最初に挿入物上で迅速に増殖した後、ES細胞の有糸分裂活性は4〜6週間の培養後に低下する。培養を延長するためには、これらの細胞をSTO細胞からなるフィーダーに移さなければならない。
【0025】
表3.挿入物またはSTO細胞のフィーダー(104細胞/cm2)のいずれかの上での単一胚からのcES細胞の樹立
【0026】
【表3】
表4.STOフィーダーまたは合成挿入物のいずれかの上でのプールされた胚からのcES細胞の樹立
【0027】
【表4】
表3および表4のデータは、ニワトリの胚性フィーダー細胞およびマウス初代胎児線維芽細胞が、cES細胞の誘導も培養も支援しないことを示す。STO細胞のフィーダーは、誘導および増殖を支援するが、103〜105細胞/cm2の限定された濃度、好ましくは、本発明の実施形態では、104細胞/cm2未満または約104細胞/cm2の濃度でSTO細胞が存在する場合にのみ、支援する。濃密なSTOフィーダー層は、cES細胞の増殖を減退させるが、指定された濃度のSTO細胞は、ES細胞の増殖に必要な因子を提供する。これらの細胞が長期の培養期間に渡って維持され、かつ継続して胚性幹細胞表現型を発現し、かつin vivoで非胚性幹細胞表現型に分化する場合、これらの細胞は、「ES細胞子孫」と呼ばれる。
【0028】
cES細胞培養培地は、80%の馴化培地からなり、好ましくは、cES細胞の誘導および増殖に必要な因子を含むある種のBRL馴化培地を含む。50%の濃度では、cES細胞の増殖は、80%の馴化培地における場合ほど信頼性が高くない。馴化培地の比率が50%未満まで減少した場合、分化細胞の増加によって立証されるように、cES細胞の増殖は影響を受け、また、30%以下の濃度では、cES細胞は、1週間以内に分化する。cES細胞の誘導および維持に必要であることが判明したこの馴化培地は、mESを維持しないが、それらの分化を引き起こす。
【0029】
ウシ胎児血清は、本発明によるES細胞培地の好ましい成分であり、cES細胞を未分化状態に保つ因子を含む。しかし、血清は、分化を誘導する因子を含むことも公知である。市販の血清ロットは、ES細胞を未分化状態に保つ潜在能力について、使用者によってルーチン的に試験される。cES細胞培養物に対して使用される血清は、マウスES細胞培養物に使用される血清とは異なることが公知である。例えば、細胞毒およびヘモグロビンの濃度が低く、マウスES細胞を未分化状態で維持することが公知である、マウスES細胞の培養のために用いられる血清は、ニワトリES細胞の持続的な増殖を支援しない。
【0030】
したがって、ニワトリES細胞に対して使用される血清は、マウスES細胞において試験して培地成分としての適性を判定するべきではなく、その代わりに、ニワトリES細胞において評価すべきである。そのために、ニワトリES細胞培養物を2つに分け、血清の新しいバッチをそれぞれ試験するために使用する。試験される新しいバッチは、実験的に試験された場合に、ニワトリES細胞の増殖を明らかに支援しなければならない。
【0031】
ニワトリ染色体のスプレッドは、10本の大染色体、66本の小染色体、および1対の性染色体(雄におけるZZおよび雌におけるZW)からなる複雑な核型が原因となって、マウスとは異なる特別な評価技術を必要とする。図3に示す本発明の長期cES細胞を、培養から189日後、そして2回凍結保存した後に解析した。図3を参照すると、これらは、10本の大染色体、2本のZ染色体、および66本の小染色体を有する正常な核型を示した。
【0032】
ニワトリES細胞を、培地中10%DMSO中で凍結保存する。いくつかの細胞株を解凍し、レシピエント胚に注入した後、体細胞キメラが得られる。このことは、これらのcES細胞が、凍結保存プロセスの間、それらの発育能を維持していることを示唆する。
【0033】
(実施例2)
レシピエント胚へのニワトリ胚性幹細胞の注入
新しく産卵された卵中の胚への到達を可能にするためには、殻を割らなければならず、これは、21日のインキュベーション期間の最終時点のふ化率の低下を不可避的にもたらす。慣例としては、卵の側面を切断して小さな穴(直径10mm未満)をあけ、この穴を通して胚を操作し、そしてテープ、カバーガラス、卵殻膜、または一片の殻で再び封じた。実施するのは比較的簡単であるが、この「窓開け」法は、70%〜100%の胎児死亡率を引き起こした。胚への到達の改善、および生存およびふ化能力の増大は、2種の異なる殻および(Callebautから適応させた)方法(参照により本明細書に具体的に組み入れられる、Callebaut、Poult. Sci 60巻:723〜725頁、1981年、ならびにRowlett,K.およびK.Simkiss,J.Exp.Biol.143巻:529〜536頁、1989年)を用いて、ふ化までのインキュベーションのために代理卵殻に胚を移入した場合に実現することができ、この技術を用いた場合、平均ふ化率は、約41%(23〜70%の範囲)であり、469個のcES細胞注入胚から191匹のヒヨコがふ化した。
【0034】
レシピエント胚中へのドナーES細胞の注入後の胚のインキュベーションは、以下に説明するような系Aおよび系Bを含む、2つの部分に分けることができる。
【0035】
系Aは、産卵後発育の最初の3日間を対象として含む。レシピエント胚を含む受精卵は、3〜5グラム重い卵と一致する。鋭端(pointed pole)を切断して直径32mmの窓を開け、内容物を除去し、そして卵黄上のレシピエント胚を、周囲の卵白と共に代理殻中に注意深く移す。
【0036】
細胞を、2ミクロンフィルターを取り付けられた口式吸引器に連結された、徐々に先細になる滅菌ガラスピペット中に吸い上げる。ピペットの穴は、直径が50〜120ミクロンでよく、また、30°の尖った斜面を有する。胚は、低倍率下で、かつ青色光を用いて可視化する。ニワトリES細胞をトリプシン処理して単細胞懸濁液にし、そして約2,000〜26,000個の細胞、好ましくは約20,000個の細胞を胚中に注入する。これらの細胞は、胚の下または上のいずれかの空間中に、すなわち、胚下腔中に、または明域の先端表面と卵黄周囲の層(卵黄膜)の間に穏やかに押し出す。新鮮な受精卵から採取した余分な卵白を添加し、プラスチックフィルムのサランラップで殻を密閉する。
【0037】
系Bは、3日目からふ化までの期間を対象として含む。インキュベーション3日目に、胚は、ステージ17前後に到達した(H&H)。卵白から胚下腔に水が輸送され、それによって卵黄が拡大し、非常に脆弱になる。系Aの殻の内容物を、元の卵より30〜35グラム重い第2の代理殻(通常、シチメンチョウの卵)に、極めて慎重に移入する。細菌汚染を防止するためにペニシリンおよびストレプトマイシンを添加し、また、鈍端(blunt pole)の38〜42mmの窓をプラスチックフィルムで封じる。このより大きな殻により、人工的な気室が実現される。インキュベーションの18日目〜19日目に、これらの胚培養物を、綿密な観察のために卓上ふ卵器に移す。肺換気が確立されるとき、プラスチックフィルム中に周期的に穴が形成され、周囲空気が気室中に入ることが可能になる。ふ化の約6〜12時間前に、フィルムを、ふ化の進行中にヒヨコが容易に脇へ押しのけることができる小型ペトリ皿に交換する。
【0038】
インキュベーションの間、代理殻中の胚のために、通常の飼育温度(37.5℃〜38℃)および相対湿度(50%〜60%)を維持するが、普通は1時間毎であり、そして90度までである周期的な卵の揺り動かしは、良好な生存を確実にするように変更しなければならない。系Aでは、揺り動かしは、4〜5分間毎にに90°までであり、系Bでは、40〜45分毎に40°〜60°までである。いずれの系においても、揺り動かしの速度は、1分間当たり15°〜20°で維持される。
【0039】
cES細胞の注入の前に、レシピエント胚を(1)660ラドのガンマ線照射への曝露によって照射することによって(2)胚の中央から約1000個の細胞を機械的に除去することにより改変することによって、または(1)および(2)を組み合わせることによって調製した場合には、キメラに対するcES細胞の寄与が改善される。表5を参照すると、体細胞組織に対するcES細胞の寄与は、レシピエント胚の中央から細胞を除去することまたは放射線照射への曝露のいずれかによってレシピエント胚に損傷を与えた場合に、実質的に増加した。放射線照射および細胞の機械的除去の組合せによってレシピエント胚に損傷を与えた場合、cES細胞がより長期間培養状態にあったにもかかわらず、ES細胞の寄与はさらに増加した。結果として生じるキメラヒヨコの一部は、純粋な横斑ロックニワトリヒヨコと区別することができない(図4)。表5中のデータが示すように、キメラ現象の割合ならびに胚1つ当たりのキメラ現象の程度は、レシピエント胚に損傷を与えた後に増加する。
【0040】
表5:異なる方法によって損傷を与えられたレシピエント胚中へのcES細胞注入後の体細胞キメラ現象の頻度
【0041】
【表5】
ステージXより実質的に若いレシピエント胚もまた、ES細胞をドナーとして用いてキメラを作製するために使用され得る。初期のレシピエント胚は、オキシトシンを雌鶏に注射して、早期産卵を誘導することによって採取し、また、受精卵は、ステージVII〜IXに採取する。
【0042】
あるいは、輸卵管の膨大部領域からの胚の採取により、約4〜250個の細胞からなるステージI〜VIの胚を入手する機会が提供され、また、潜在的なレシピエント胚としてのすべての胚ステージからキメラを開発することが可能になる。
【0043】
(実施例3)
ニワトリ胚性幹細胞(cES)に由来する体細胞キメラ
ニワトリES細胞を白色レグホーンレシピエント胚中に注入する。第1期の実験では、28回の実験において合計14種の細胞株を、ステージXのレシピエント胚に注入する(表2を参照されたい)。これらのcES細胞は、4〜106日間培養して増殖させたものであり、一部の株は、凍結保存したものであった。ニワトリES細胞を軽くトリプシン処理して、cES細胞の小さな塊を得、25mM HEPES+10%ウシ胎児血清を添加したDMEM中に再懸濁する。2000〜5000個の細胞を含む3μl〜5μlの細胞懸濁液を、レシピエント胚の胚盤下腔中に注入する。羽毛を発達させた胚をすべて分析し、また、羽毛の色によって判定したところ、24パーセントの胚(83/347)が、キメラである。羽毛キメラは、14種の細胞株中11種から得られる。キメラ現象の程度は、1%〜95%まで様々であり、程度の平均は、25.9%(SD=20.4)であった。
【0044】
表2は、細胞株内および細胞株間で実施した実験間の体細胞キメラ現象の差異を示す。キメラに対するES細胞の寄与の例を、図4および図5に示す。図4では、2匹のヒヨコがキメラであり、2匹が横斑ロックである。これらのヒヨコの間に表現型の差異はなく、キメラに対するES細胞の寄与が、特に外胚葉由来の系統において多大であることが示唆されることは明らかである。図5では、左側のキメラは、ES細胞からの比較的低いレベルの寄与を有するのに対し、右のキメラは、中程度の寄与を有する。
【0045】
(実施例4)
リポフェクションおよびエレクトロポレーションによるcES細胞のトランスフェクション
表6に言及すると、トランスフェクトされるウェルのサイズと適合した適切な量のDNAを、血清も抗生物質も含まない培地中に希釈する。適切な体積のSuperfect(Stratagene社)を添加し、DNAと混合し、そして5〜10分間、反応を生じさせる。この培地を除去し、そしてトランスフェクトしようとするウェルを、CaおよびMgを含まない塩溶液で洗浄する。血清および抗生物質を含み得る適切な体積の培地を、DNA/Superfect混合物に添加する。これらのプレートを37℃で2〜3時間インキュベートする。インキュベーションが完了したら、細胞を1〜2回洗浄することによってSuperfectを除去し、そして新鮮な培養培地を添加する。
【0046】
表6:Superfectを用いたニワトリES細胞のトランスフェクションのための条件
【0047】
【表6】
ペトリ皿用パルサーを用いて、プレートの直径35mmのウェル中に付着しているcES細胞をエレクトロポレーションする。この培地を除去し、そしてCa++およびMg++を含まない塩溶液でウェルを洗浄する。1mlのエレクトロポレーション溶液をこのウェルに添加する。DNAを添加し、培地を穏やかに混合する。このペトリ皿用パルサーをウェルの底部まで下げ、電流を送達する。(電圧は好ましくは100〜500V/cmであり、また、パルス長は12〜16m秒でよい)。ペトリ皿用パルサーを取り出し、エレクトロポレーションしたウェルを室温で10分間静置する。10分後、2mlの培地を添加し、インキュベーターに皿を戻す。
【0048】
懸濁状態の細胞をトランスフェクトするために、培地を除去し、そしてCaおよびMgを含まない塩溶液で細胞を洗浄する。EDTAと共にトリプシンを添加して、単細胞懸濁液を得る。細胞を洗浄し、遠心分離し、そしてPBSなどの補正用エレクトロポレーション緩衝液中に再懸濁する。このES細胞懸濁液を無菌のキュベット中に入れ、DNA(最低濃度1mg/ml)をこの細胞懸濁液に添加し、そして上下にピペッティングすることによって混合する。これらの細胞をエレクトロポレーションし、室温で10分間放置した。細胞をキュベットから取り出し、予め準備しておいたウェルまたはディッシュに分配する。細胞をインキュベーター中に入れ、また、エレクトロポレーション後24〜48時間目の一過性トランスフェクションについて評価する。抗生物質耐性細胞の選択もまた、ピューロマイシンなどの抗生物質を培養培地中に含めることによって開始することができる。
【0049】
好ましい実施形態では、トランスフェクトされた細胞を選択するために必要とされるピューロマイシンの濃度は、死滅滴定曲線として算出される。ニワトリ胚性幹細胞に対する死滅滴定曲線は、培養中の細胞を、0.0〜1.0μg/mlの範囲のピューロマイシン濃度に10日間(表7)、および0.0〜200μg/mlの範囲のネオマイシン濃度(表8)に曝露することによって作成する。2日毎に培地を交換し、そして新鮮なピューロマイシンまたはネオマイシンを添加する。濃度0.3μg/mlのピューロマイシンに曝露した場合、ES細胞は、6日間の期間に渡って新鮮なピューロマイシンを含む培地を3回交換した後のすべてのウェルから消失していた(表7を参照されたい)。トランスフェクトされた培養物を選択するために、0.3〜1.0μg/mlのピューロマイシン濃度を使用する。40μg/mlを超えるネオマイシン濃度では、7日以内にすべてのcES細胞が消失した(表8)。
【0050】
10日間の選択後、cES細胞コロニーは目に見え、さらなる増殖のために採取することができる。
【0051】
表7:様々な濃度のピューロマイシンに様々な長さの期間(ピューロマイシン添加後の日数)曝露した後のcES細胞の形態
【0052】
【表7】
表8:様々な濃度のネオマイシンに様々な長さの期間(ネオマイシン添加後の日数)曝露した後のcES細胞の形態
【0053】
【表8】
(実施例5)
組織特異的な抗体発現
導入遺伝子の設計に応じて、外因性抗体をコードするDNAは、選択された組織おいてのみ有意なレベルの抗体を発現するように設計される。この導入遺伝子構築物は、宿主生物のゲノムに由来する遺伝子エレメントを含み得、また、選択された組織における抗体の公知の発現または発現パターンに基づいて選択され得る。特定の組織における発現のためには、選択される組織において通常発現され、また通常高度に発現される、タンパク質をコードする遺伝子を選択し、そしてこの外因性抗体の発現を駆動するために、この遺伝子に由来する調節エレメントを選択する。この外因性抗体のDNAコード配列と組み合わせる場合、オボアルブミン、オボトランスフェリン、オボムコイド、オボムチン、リゾチーム、オボグロブリン、オボインヒビター、シスタチン、オボグリコプロテイン、オボフラボプロテイン、オボマクログロブリン、またはアビジンのプロモーターなど他の調節エレメントを、選択マーカーをコードする遺伝子と組み合わせて、および/またはIRESと組み合わせて、使用してよい。この導入遺伝子は、選択された組織において、好ましくは卵白において優先的な抗体発現をもたらす。特定の組織型における優先的発現は、選択されていない組織と比較して規模が3〜4桁大きな、選択された組織における発現と定義することができる。
【0054】
トランスジェニックトリにおける組織特異的抗体発現の場合、組織特異的発現は、好ましくは、膨大部、峡部、卵殻腺、または卵管漏斗を含めて、輸卵管のある領域を対象としている。膨大部は、卵白の主なタンパク質を発現する管状腺細胞を含み、一方、峡部は、卵殻膜を発現する細胞を含む。可溶性タンパク質の発現は、好ましくは、輸卵管の膨大部の管状腺細胞に指向され、これは、好ましくはオボアルブミン、ただし、オボトランスフェリン、オボムコイド、リゾチーム、オボグロブリンG2、オボグロブリンG3、オボインヒビター、シスタチン、オボグリコプロテイン、オボフラボプロテイン、オボマクログロブリン、およびアビジンを含む、卵白タンパク質を発現する遺伝子に由来する、プロモーターを通常含む調節配列を選択することによる。上皮細胞における発現を除く、管状腺細胞における選択的発現を以下に実証し、また、2種の細胞型間の選択性を区別するために、優先的発現と命名する。
【0055】
以下の実施例では、外因性免疫グロブリン遺伝子座と共に、プロモーターを含む内因性卵白調節配列を含む導入遺伝子を構築して、輸卵管の管状腺細胞における組織特異的抗体発現を生じさせる。その場合、このようにして発現された抗体分子は、トランスジェニックニワトリの卵白中に蓄積される。この実施形態では、再配列された任意の免疫グロブリン遺伝子によってコードされる抗体は、輸卵管の膨大部領域の管状腺細胞を含む組織において特異的に発現され、そして卵白から単離することができる。モノクローナル抗体をコードする再配列された免疫グロブリン遺伝子は、他の組織における発現が検出可能なレベルより上で存在し得るものの、他の組織における発現は実質的になく、輸卵管において優先的に発現される。
【0056】
この実施形態では、オボアルブミン調節配列の制御下のモノクローナル抗体カセットは、少なくとも3.4Kb、好ましくは少なくとも約7.5Kbの5’調節配列から構成され、かつ15Kb以上の3’調節配列も含んでよい。好ましくは、この構築物は、外因性抗体コード領域の5’末端および3’末端の双方に隣接するオボアルブミン遺伝子の領域を含むが、5’隣接領域の内因性プロモーター配列のセグメントが十分に大きな場合は、3’隣接領域の必要性を回避し得る。抗体の重鎖および軽鎖の双方のコード領域は、導入遺伝子中で提供され、かつ選択されたアイソタイプの可変領域、多様性領域、連結領域、および定常領域を含む。好ましい実施形態では、この抗体は、特徴的にはヒトであり、かつ少なくともヒト重鎖を含む、免疫グロブリン遺伝子によってコードされる。また、このアイソタイプは好ましくはIgGであり、最も好ましくはIgG1である。
【0057】
オボアルブミンに由来するモノクローナル抗体構築物のために好ましい導入遺伝子構築物を、図6Aに提供する。この導入遺伝子構築物をOv7.5と呼び、これは、MAbコードに5’末端配列で隣接しているプロモーター(この特定の実施形態ではオボアルブミンプロモーター)を含む約7.5Kbの卵白調節配列、およびコード領域の3’側の15Kbのプロモーター配列を有する。コード領域は、軽鎖および重鎖の双方の可変領域、J−Cイントロン配列、κ軽鎖定常領域、IRES配列、ならびにγ1アイソタイプ重鎖定常領域を含む。この構築物の3’末端は、GFP遺伝子および選択マーカー、すなわち、この場合、本明細書において説明されるCXプロモーターによって駆動されるピューロマイシン耐性遺伝子を含む。モノクローナル抗体コード領域の3’側および5’側の双方のオボアルブミンプロモーター配列の長さは例にすぎず、類似した構築物は、25〜100Kb以上の5’配列、ならびに様々な長さの3’配列を含み得る。当業者には、GFPマーカーが、生理学的標本における検出のためにだけ存在し、導入遺伝子の有用性から逸脱することなく除去できることが理解されよう。ピューロマイシン耐性マーカーは、この導入遺伝子で成功裡に形質転換された胚性幹細胞を選択する能力を与える任意のマーカーで置換することができる。いくつかのタイプの類似した選択マーカーが当技術分野において周知であり、また、本実施形態のピューロマイシン耐性遺伝子と本質的に交換可能に使用され得る。
【0058】
上記のように、このモノクローナル抗体は、本発明の導入遺伝子構築物を用いて発現させることができるいくつかのタイプのモノクローナル抗体生成物のうちの一例にすぎない。さらに、タンパク質の1つのクラスとしてのモノクローナル抗体は、本明細書において説明される方法および技術に従って、組織特異的な様式で発現され得る多くのクラスのタンパク質生成物の一例にすぎない。
【0059】
図6Bを参照すると、エストロゲン注射によってOv7.5導入遺伝子の発現が誘導された2週齢のキメラの膨大部の切片は、形質転換された胚性幹細胞に由来する抗ダンシルモノクローナル抗体産生細胞の組織特異的発現によりGFPが発現していることを示し(図6Bの左上のパネルにおいて緑色として示される)、これにより、Ov7.5導入遺伝子で形質転換された胚性幹細胞による寄与が裏付けられる。図6Bの左下のパネルを参照すると、モノクローナル抗体は、管状腺細胞において赤色に染色されるのに対し、同じくドナー胚性幹細胞に由来する上皮細胞は緑色に染色され、赤色には染色されない。染色におけるこの差異により、構築物の発現が組織特異的であり、かつ特定の組織型に対する導入遺伝子の内容によって選択されることが実証される。以下の実施例は輸卵管の管状腺細胞における組織特異的発現を実証するが、あらゆる細胞型および組織型にわたる発現の実証により、導入遺伝子構築物の構成要素、および例えば、プロモーターまたは他の調節エレメントの対応した選択によって組織特異的発現または細胞特異的発現のために各組織型または任意の組織型を選択し得ることが実証される。
【0060】
図6Bの右上のパネルでは、細胞型のすべてがDAPI染色によって示される。右下のパネルでは、染色を重ねて、ドナー由来の管状腺細胞のみがモノクローナル抗体を発現し、一方、レシピエント由来の細胞およびドナー由来の上皮細胞がモノクローナル抗体を発現しないことを実証する。図7Cおよび図7Dは、抗ダンシルモノクローナル抗体の重鎖および軽鎖が、それぞれ、3または5キメラの輸卵管組織においてのみ選択的に発現され、また、これらのキメラの脳、腸、膵臓、または筋肉においてはRT−PCRによって検出されるレベルを上回っては発現されないことを示すRT−PCR解析である。
【0061】
ヒトIgGアイソタイプモノクローナル抗体が選択的に発現され得、そして蓄積されるタンパク質が卵白であることを実証するために、Ov7.5構築物を有するcES細胞を注入することによって合計18匹のキメラ雌を作製した。この群から6匹のキメラを用いて、初期のエストロゲン誘導による導入遺伝子発現を試験した。残りのキメラ雌12匹は、卵を回収するために性的に成熟するまで飼育した。9匹のキメラ雌が、17〜22週齢で産卵を開始し、また、これらのキメラのうちの1匹は、散発的に産卵した。キメラ雌のうちの3匹は、35週齢の時点で卵を産卵しておらず、剖検時に、それらの性腺が、雄性ES細胞に由来する組織の存在によって雄性化していることが明らかになった。
【0062】
卵を産卵する9匹のキメラに由来する卵を回収し、硫酸アンモニウム沈殿法によって代表的な卵白サンプルを調製し、そしてELISAによって解析した。ヤギ抗ヒトIgG抗体でマイクロタイタープレートをコーティングし、重鎖に対する標識されたヤギ抗ヒトIgG(γ鎖特異的)抗体および/または軽鎖に対する標識されたヤギ抗ヒトκ(κ鎖特異的)抗体のいずれかによって卵白サンプル中のヒトIgG MAbの存在を明らかにした。精製ヒトIgγ1、κタンパク質を用いて標準曲線を作成した。ELISAの感度は、0.8ng/mlであった。非トランスジェニック白色レグホーン雌鶏由来の卵白サンプルを、ネガティブコントロールとして使用した。ヒトIgG MAb蓄積は、非トランスジェニック白色レグホーン雌鶏に由来する卵(卵4個)においても、6匹のキメラ雌鶏に由来する卵(雌鶏OV11−17番由来の卵8個、雌鶏OV11−53番由来の卵8個、雌鶏OV11−73番由来の卵6個、雌鶏OV11−88番由来の卵6個、雌鶏OV12−97番由来の卵4個、および雌鶏OV13−13番由来の卵5個)においても、検出不可能であった。
【0063】
ヒトIgG MAb蓄積が、3種の異なるキメラ雌鶏に由来する卵において検出された(ELISAによってIgHを決定した場合、雌鶏OV11−13由来の卵の場合に約1.4〜6.3ng/ml、雌鶏OV11−37番由来の卵の場合に約2.0〜2.9ng/ml、および雌鶏OV11−43番由来の卵の場合に約2.9〜10.8ng/ml)。代表的な卵中のヒトIgG MAbの濃度を、表9にまとめて示す。IgLに関してELISAによって決定された卵中のヒトIgG MAbの濃度は、IgHに関してELISAによって決定された濃度より、一貫して低かった。一般に、IgLによって決定された濃度は、IgHによって決定された濃度の60%であった(表9の3列目および5列目を比較されたい)。この差異は、精製したヒトIgγ1、κタンパク質を用いて作製されたスパイクサンプルにおいても存在した。
【0064】
表9 キメラに由来する卵中のヒトIgG MAbの蓄積
【0065】
【表9】
雌鶏OV11−13番、OV11−37番(これらは、卵中にヒトIgを蓄積した)および白色レグホーンに由来する血液サンプル中のヒトIgG MAbタンパク質の濃度は、アッセイ感度(0.8ng/ml)より低かった。これらのデータは、エストロゲンによって誘導したキメラヒヨコの腸、脳、膵臓、および筋肉におけるヒトIg転写物の存在を評価するためにRT−PCRを用いて観察された、キメラニワトリにおけるヒトIgの異所発現の不在と一致している(図7Cおよび7D)。したがって、Ov7.5構築物は、組織特異的であり、ホルモンによって誘導され、そして発生に関連して調節される遺伝子発現を、トランスジェニックキメラ雌鶏に送達すると思われる。さらに、タンパク質は、膨大部中の管状腺細胞から搬出され、卵白中に蓄積されるようである。
【0066】
オボアルブミン由来の組織特異的タンパク質発現導入遺伝子を、以下のようにして構築した。
【0067】
ニワトリゲノムBACライブラリー(Crooijmans,R.P.ら、Mamm.Genome 11巻:360〜363頁、2000年)(Texas A&M BAC Center)をスクリーニングして、オボアルブミン遺伝子座中の46Kbの領域を単離する。MAbコード領域の5’側に位置するオボアルブミンプロモーターの異なる断片を有する、以下の2種の異なるベクターを構築した:(1)Ov7.5MAb−dns:42Kbの発現ベクターであり、オボアルブミン遺伝子に由来する9.2Kbの5’配列(7.5Kbプロモーターを含む)および15.5Kbの3’隣接配列を含む(図10A)。この42Kbの発現ベクターは、オボアルブミン遺伝子に由来する9.2Kbの5’配列(7.5Kbのプロモーターを含む)および15.5Kbの3’隣接配列を含む。バイシストロン性のモノクローナル抗体カセットは、抗ダンシル抗体の軽鎖、IRES、および重鎖をコードする。(2)Ov15MAb−dns:49Kbの発現ベクターであり、オボアルブミン遺伝子に由来する16.8Kbの5’配列(15Kbプロモーターを含む)および15.5Kbの3’隣接配列を含む(図示せず)。この49Kbの発現ベクターは、オボアルブミン遺伝子に由来する16.8Kbの5’配列(15Kbのプロモーターを含む)および15.5Kbの3’隣接配列を含む。このモノクローナル抗体カセットは、双方の構築物において同一である。
【0068】
上記のように、双方のベクターにおいて発現され得る遺伝子の一例は、マウス−ヒトハイブリッドの抗ダンシルモノクローナル抗体(MAbdns)である。CxEGFP/CxPuroカセットを3’側の最末端にクローニングして、cES細胞における安定なトランスフェクションをピューロマイシンを用いて選択すること、およびキメラ中のトランスフェクトされた細胞を容易に同定することを可能にする。どちらの構築物も、cES細胞中にトランスフェクションする前に直線状にし、また精製する。cES細胞のトランスフェクションは、SuperFect(Stratagene社)またはペトリ皿用パルサーによるエレクトロポレーションのいずれかを使用して、Ov7.5MAbdnsおよびOvl5MAbを用いて実施する。ピューロマイシンによる選択後、耐性クローン6つを、分子解析のために採取する。MAbdnsカセット中、GFP遺伝子中、およびPuro遺伝子中に配置されたプライマーを用いるPCRによって導入遺伝子の存在を確認する。
【0069】
発現され得る抗体の第2の例は、完全なヒト抗ヒトPSMAモノクローナル抗体(MAbF1)である。MAbF1に対応するV遺伝子をPCRによって獲得し、また、Ov15MAb−dns中のV遺伝子と入れ替えるために使用した。
【0070】
この導入遺伝子は、雌性ES細胞中に安定に組み入れられ、また、これらの安定にトランスフェクトされた細胞を、ステージXのレシピエント胚中に注入した。結果として生じるキメラを性的に成熟するまで飼育し、そして卵中のヒトモノクローナル抗体の濃度をELISAによって評価した。30匹の雌鶏のうち4匹は、1mg/卵を上回る濃度で卵中に抗体を蓄積した(表9A)。
【0071】
表9A Ov15MAbF1キメラに由来する卵中のヒトIgG MAbの濃度
【0072】
【表9A】
(実施例6)
抗体生成物の特性決定
ニワトリにおいて産生されたMAbF1を、従来のCHO細胞培養において産生されたMAbF1といくつかのアッセイにおいて比較した。
【0073】
1.抗体のSEC−HPLC解析
4.6×300mmのBioSep SEC S3000カラム(Phenomenex社)を用いて、Waters 2795 HPLCによって約10μγのIgGサンプルを解析した。0.1Mリン酸ナトリウム、0.15M NaCl、および0.1M硫酸ナトリウム、pH7.2において、流速0.4ml/分で20分間、クロマトグラフィーを実施した。A280で分離をモニターした。分子量標準物質(Bio−Rad社)を用いて、概算の分子量を決定した。SEC−HPLC解析により、双方とも90%を超えるIgGモノマーであることが示された(表10)。
【0074】
表10 MAbF1のSEC−HPLC解析
【0075】
【表10】
2.タンパク質のNano LC ESI MS/MS解析
双方のMAb調製物を還元およびアルキル化し、一晩透析し、そして25mM重炭酸アンモニウム(PH8)中、37℃で4時間、トリプシンで消化した。双方のトリプシン消化物のタンデム型質量分析を、陽イオンモードで、そしてタンデムスペクトル(tandem spectra)を得るために衝突誘起解離(CID)を用いるQSTAR pulsar質量分析計(MDS Sciex社、コンコード、オンタリオ州、カナダ)工程に連結されたNano HPLCシステム(Dionex社、サニーベール、カリフォルニア州)によって実施した。各サンプルをμl 1mlずつ注入し、75mm直径μm Pepmap C18カラムを用いて流速250nl/分で分離した。移動相は溶媒A(95%水、5%アセトニトリル、0.5%ギ酸)および溶媒B(20%水、80%アセトニトリル、0.5%ギ酸)であった。機器は、2秒間フルスペクトルを記録し、次いで、最も強度の強いイオンを選択して次の5秒間CIDスペクトルを記録する7秒間の運転サイクルを用いる情報依存的取得(information dependent acquisition)モードで操作した。スペクトルは、Mascot(Matrix Science社、ロンドン、イギリス)を用いて解析した。説明した方法による双方のMAb調製物の解析では、配列の差異は示されなかった。
【0076】
表11.ニワトリに産生されたMAbF1およびCHOに産生されたMAbF1の質量分析による配列解析
【0077】
【表11−1】
【0078】
【表11−2】
3.LC−MS解析
4MグアニジンHCl中IgGサンプル(50μγ)を、サンプルを60℃で90分間インキュベートすることによって25mM DTT中で還元した。次いで、暗所、室温で15分間、45mMヨード酢酸中でサンプルをアルキル化し、そして22mM DTTで反応を停止させた。LC−MSに先立って、サンプルを25mM重炭酸アンモニウム1Lに対して透析し、そして各サンプル50μlを、Micromass ZQ質量分析計を装備されたWaters2795 HPLCを使用するPoros R1/10 2.1×100mmカラム(Applied Biosystems社)に注入し、陽イオンモードで解析した。移動相は、(A)0.1%ギ酸および0.01%トリフルオロ酢酸(TFA)ならびに(B)0.1%ギ酸および0.01%TFAを含む100%CH3CNであった。溶出(0.25ml/分)は、100分に渡って作られる(A)中(B)の10〜60%の直線勾配によって実施した。
【0079】
L鎖の解析により、双方のMAb調製物が同一の質量(±3Da)を有することが示された(表12)。
【0080】
表12 MAb軽鎖のLC−MS解析
【0081】
【表12】
4.示差走査熱量測定による熱安定性
ニワトリ発現系およびCHO発現系において作製されたMAbF1の熱安定性を示差走査熱量測定(DSC)を用いて測定し、そしてFcドメインにおいて脱グリコシル化されたそれらの対応する型と比較した。融解温度(Tm)の熱量測定は、オートサンプラーと組み合わされたVP−Capillary DSC示差走査マイクロカロリメータープラットホーム(MicroCal LLC、ノーサンプトン、マサチューセッツ州、米国)を用いて実施した。サンプルセルの容積は0.144mLである。これらの抗体のグリコシル化型および脱グリコシル化型に関する変性データは、濃度2.3μMのこれらのサンプルを1℃/分の速度で30℃から95℃まで加熱することによって得た。これらのタンパク質サンプルは、pH7.4のリン酸緩衝化生理食塩水(PBS)中に存在した。比較してモル熱容量を得るために、同じ緩衝液を参照セルにおいて使用した。観測された温度記録図をベースライン補正し、また、標準化したデータをOrigin v7.0を用いて解析した。
【0082】
CHOおよびニワトリに由来する抗体の変性プロファイルは極めて異なるのに対し、Fcを脱グリコシル化した抗体の変性プロファイルはほぼ同一である(図8)。これらのデータから、ニワトリの管状腺細胞において産生されたMAbF1のグリコシル化の方が、CHO細胞において産生されたMAbF1に結合される糖残基より高い熱安定性を与えることが示唆される。
【0083】
5.CE−LIFによるMAbのオリゴ糖の特性決定
サンプルを12.5mUのPNGaseF(Prozyme社)と共に40℃で一晩インキュベーションすることによってIgGサンプル(100μγ)からオリゴ糖を遊離させた。使用した条件下で、CHO細胞およびニワトリにおいて発現されたHuMAbF1のFc部分に由来するN結合型グリカンが遊離された。MAbタンパク質を除去するためにエタノール沈殿を実施した後、これらのグリカンを含む上清を減圧遠心分離によって乾燥させ、そして15%酢酸中19mM APTS(Beckman社)およびTHF(Sigma社)中1Mシアノ水素化ホウ素ナトリウム中に再懸濁した。グリカン標識反応を40℃で一晩継続させ、続いて、サンプルを水で25倍に希釈した。APTSで標識したグリカンを、有効長50cm、内径50μmのN−CHOコーティングキャピラリー(Beckman社)を使用する、逆極性を有するP/ACE MDQ CEシステム(Beckman社)による、レーザー誘起蛍光法を用いたキャピラリー電気泳動に適用した。サンプルは圧力(8秒)注入し、また、分離は、Carbohydrate Separation Gel Buffer(Beckman社)を用いて、25kV、20℃で20分間実施した。これらの分離物を、3mWアルゴンイオンレーザーならびに488nmの励起波長および520nmの発光波長を用いるレーザー誘起蛍光検出システム(Beckman社)を用いて観察した。図9に示すように、ニワトリにおいて産生されたMAbF1のオリゴ糖プロファイルは、CHO細胞において産生されたMAbF1のプロファイルと大きく異なることが判明した。
【0084】
6.MALDI Q−TOF MSによるMAbのオリゴ糖特性決定
精製した抗体(100μg、各1.0mg/ml)を4μlのPNGaseF、すなわちN結合型糖質を切断するエンドグリコシダーゼ(Prozyme社)で処理した。これらのサンプルを37℃で一晩インキュベートし、次いで、50mM重炭酸アンモニウム(pH8)に対して一晩透析した。タンパク質は、エタノール沈殿によって除去した。遊離された糖質を、本質的には供給業者のプロトコールに従って、ただし溶出体積を5μlに減らして、Glygen Corp.(コロンビア、メリーランド州)製のマイクロカーボンカラムを用いて脱塩した。各グリカンサンプルの1μl分取物を、マトリックス溶液(α−シアノ−4−ヒドロキシケイ皮酸または2,5−ジヒドロキシ安息香酸(Applied Biosystems社)と1:1(体積比)で混合し、MALDIターゲットプレート上にスポットし、自然乾燥させる。MALDI−Q−TOFタンデム型解析を用いて、完全な複合糖質の解析を実施した。すべての質量スペクトルは、糖質のクラスの質量(組成)を提供する、o−MALDIソース2(MDS Sciex社、コンコード、オンタリオ州、カナダ)を装備したQSTAR pulsar i質量分析計を用いて記録した。双方のMAbのグリカンをプロファイリングした後、各グリカン中で起こり得るグリコシド結合を調査した。高CIDによる質量スペクトルを、TOF/TOFオプティクスを装備した4700Proteomics分析計(Applied Biosystems社、フォスターシティ、カリフォルニア州)を用いて記録した。高CIDのMS/MS実験の場合、衝突エネルギーは1kVに設定した。衝突セルの内部で、選択したオリゴ糖イオンを2×10−6トルの圧力でアルゴンと衝突させた。MALDI TOF質量分析法によって解析した、ニワトリに由来するMAbF1中の11種の主要なグリカンの糖質組成および起こり得るグリコシド結合を表13に要約する。これらのオリゴ糖構造体は、高マンノース型、複合型、およびハイブリッド型のNグリカンを含むことが判明した。
【0085】
表13 質量分析法による、ニワトリにおいて産生されたMAbF1の糖質解析
【0086】
【表13】
7.HPAE−PADを用いたHPLCによる単糖解析
表14 ニワトリに産生されたMAbF1およびCHOに産生されたMAbF1の単糖解析
【0087】
【表14】
IgGサンプル(200μg)を、100℃で4時間、2N TFA(中性糖を推定する場合)または6N HCl(アミノ糖を推定する場合)のいずれかを用いる酸加水分解に供した。サンプルを周囲温度で減圧遠心分離によって乾燥させ、そしてHPAE−PAD(Dionex社)による解析の前に200μlの水中に溶解した。単糖は、プレカラムのAmino TrapおよびBorate Trap(Dionex社)付きのCarboPac PA10 4×250mmカラムを用いて分離した。手順は、Dionex社の技術注記53に従った。単糖のピーク一致および相対存在量は、単糖標準物質(Dionex社)を用いて決定した。表14に要約したようなニワトリおよびCHOにおいて産生されたMAbF1の単糖解析により、糖質組成の差異が明らかになり、また、ニワトリにおいて産生されたMAbF1中にN−アセチルグルコサミン残基、マンノース残基、および極めて含有量の少ないガラクトース残基が存在することが示された。
【0088】
8.化学的諸特性の要約
要約すれば、N結合型オリゴ糖プロファイルにおける最も目を引く差異は、ニワトリにおいて産生された抗体中に高マンノース型のN−グリカンが存在し、フルコースは存在せず、またガラクトース残基の含有量が極めて少ないことであった。これらの特性はいくつかの理由から重要である。第1に、抗原性であることが公知であるα1−3 Gal結合の証拠がない。典型的には約2%未満のレベルまでガラクトース濃度が低下すると、ガラクトースを含む結合に起因する抗原性が実質的に減少する。第2に、同じく抗原性であることが公知であるN−グリコリルノイラミン酸残基の証拠がない。第3に、ニワトリ管状腺細胞において産生された抗体には、抗体のADCC活性を高めるフコシル残基が実質的にない。この文脈において、実質的にないとは、0.1%未満と定義される。第4に、ニワトリに産生された抗体は、典型的には40%を上回る高いマンノース含有率を有し、これは、CHO細胞において産生された抗体を標準物質として用いてBalb/cマウスにおいてクリアランスを評価した場合のこの抗体のクリアランス速度を速める。これらの有利な化学的諸特性に加えて、これらの抗体は、ニワトリゲノム中にランダムに組み込まれるか、または組織特異的な様式で発現されない導入遺伝子を用いた場合には観察されない濃度で卵白中に存在する。好ましい濃度は卵1個当たり抗体1mgより高く、卵1個当たり2mgより高く、卵1個当たり3mgより高く、また、卵1個当たり6mgもの高さである。各卵は約25mlの卵白を含むため、好ましい濃度は40μg/mlより高く、80μg/mlより高く、120μg/mlより高く、また、240μg/mlもの高さである。
【0089】
(実施例7)
卵白からの抗体の抽出および精製
最初に、室温で30分間、卵白を低剪断速度で混合し、次いで、以前に説明されている改良法によってオボムチンを沈殿させた。1体積のホモジナイズした卵白懸濁液を3倍体積の逆浸透水に添加し、30分間攪拌した。希釈した懸濁液を、0.5Mリン酸を用いてpH6.0に調整し、次いで12,100gで20分間遠心分離した。この方法によって主としてオボムチンを含有する卵白タンパク質の約3%を回収した。0.5Mリン酸水素二ナトリウムを用いて上清をpH7.4に調整し、そして結晶塩を用いて塩化ナトリウム濃度を150mMに調整した。Protein A−Sepharose FFカラム(Amersham Biosciences社)を用いて、線流速120cm/時でヒトIgGを精製した。吸着されたヒトIgGを5カラム容量のローディング緩衝液(PBS、pH7.4)で洗浄し、次いで、3mMリン酸を用いて溶出させた。230mM NaClを含有する60mMリン酸ナトリウム(pH7.5)を用いて、溶出されたヒトIgG画分をpH7.5に調整して、最終濃度を40mMリン酸ナトリウムおよび150mM NaClとした。次いで、サンプルを0.2mmシリンジフィルター(Pall社)に通してろ過した。図10に示すように、精製した物質の大半は、90%を超える純度を有する(ELISAおよびA280によって決定、データは示さず)、完全に構築されたH2L2であった。
【0090】
(実施例8)
結合親和性に関するアッセイ
LNCaP細胞(ATCC)上のPSMAを、結合を分析するための抗原として使用した。ウェル当たり20万個の細胞を、指定した濃度の50μlずつの抗体と共に、デュプリケートで30分間インキュベートした。細胞を2回洗浄した後、希釈度1:200のヤギ抗ヒトIgG PE標識抗体(Jackson ImmunoResearch社)をウェル当たり50μl、4℃で30分間、添加した。1%BSAを含むPBS中で細胞を2回洗浄し、FACSによって分析した。LNCaP細胞上のPSMAに対するMAb結合のEC50値は、GraphPad Prism 3.0(GraphPad Software社)を利用して、結合曲線から決定した。細胞は、10%FBS、10mM HEPES、2mM L−グルタミン、および1mMピルビン酸ナトリウムを添加したRPMI1640培地中で増殖させた。ニワトリ管状腺細胞において産生されたMAbF1の抗原結合特性を、CHO細胞において産生されたMAbF1の抗原結合特性と比較した。どちらの抗体調製物も、LNCaP細胞上で発現されたPSMAに対してほぼ同一の結合曲線を描き、EC50値は類似していた(図11)。このデータにより、ニワトリに由来する抗体およびCHOに由来する抗体はグリコシル化の様式は異なるが、同等に抗原を認識し、結合することが実証される。
【0091】
(実施例9)
抗体内部移行のアッセイ
PSMAへのMAbF1の結合は、抗体の内部移行を招く。潜在的な1つの応用において、PSMAを発現する腫瘍細胞を標的とし、そして破壊するために、MAbを細胞毒と結合させることができる。LNCaP細胞上のPSMAに対する抗体結合の内部移行は、MAbおよびHum−Zap(Advanced Targeting Systems社)と共に細胞をインキュベートすることによって判定した。HumZapは、リボソーム不活性化タンパク質であるサポリンに結合されたヤギ抗ヒトIgG抗体である。MAbF1/Hum−Zap複合体が細胞表面上のPSMAに結合し、そして内部移行された場合に細胞は死滅するが、抗体またはHum−Zapは単独ではLNCaP細胞に対して毒性ではない。300ngのHum−Zap、および300ngのF1MAbまたは対照MAbを含む培養培地150μl中で、37℃で48時間、トリプリケートでLNCaP細胞(10,000/ウェル)をインキュベートした。細胞の増殖および生存は、CellTiter−Glo Luminescent Cell Viability Assay(Promega社)を用いて決定した。ウェル当たり10,000個のLNCaP付着細胞と共に、細胞培養培地中の抗体希釈物を4℃で2時間インキュベートすることによって内部移行アッセイも実施した。抗体溶液を穏やかに除去し、HumZap 200ngを含有する培地150μlに交換した。37℃で48時間インキュベーションした後、細胞生存率を決定した。抗体内部移行に対するEC50値は、Prism3.0(GraphPad Software社)を用いてグラフから決定した。図12に示すように、双方の抗体調製物が同様の効率で内部移行された。一連の抗体濃度に渡って試験した場合、ニワトリに由来するMAbF1およびCHOに由来するMAbF1の双方の内部移行に対するEC50値は0.49nMであった。
【0092】
(実施例10)
BALB/cマウスにおけるMAbのクリアランス
放射標識した抗体の静脈注射により、BALB/cマウスにおいて、ニワトリに産生されたMAbF1のin vivo半減期を、CHOに産生された抗体と並行して解析した。MAbタンパク質10μgを、Iodobead法(Pierce社)を用いて、125Iで少しヨウ素標識した(抗体当たり1個未満のI)。6週齢の雌BALB/cマウス(Taconic Farms社、ジャーマンタウン、ニューヨーク州)に、実験前の1週間、飲料水に溶かした0.1mg/mlヨウ化カリウムを与えた。タンパク質当たり4匹のマウスの尾静脈に約600,000cpmの標識MAbを静脈注射し、そして全身ガンマカウンター(Wm.B.Johnson社、Ludlum社製計数器付きNaI結晶検出器)を用いて、選択された時間に全身の放射能を測定した。半減期は、指数関数的回帰分析または残留放射能に基づいて算出した。図13に示したように、ニワトリ管状腺細胞によって産生されたMAbF1は、102.4±0.9時間の半減期(t1/2)で浄化されたのに対し、CHO細胞によって産生されたMAbF1は、より緩徐に浄化され、半減期は207.5±18.3時間であった。
【0093】
(実施例11)
ADCCのアッセイ
改良された51CrADCCアッセイにおいてLNCaP−C42B細胞を試験した。標準的なFicoll−Paque分離法によってヘパリン処置した全血からヒト末梢血単核細胞を精製した。これらの細胞を、10%FBSおよび10U/mlヒトIL−2を含有するRPMI1640培地中に(1×10E6細胞/mLで)再懸濁し、そして37℃で一晩インキュベートした。翌日、これらの細胞を回収し、培養培地中で1回洗浄し、そして2×107細胞/mlの濃度で再懸濁した。200万個のLNCaP−C42b標的細胞を、総体積1ml中の200uCi 51Crと共に37℃で1時間インキュベートする。標的細胞を1回洗浄し、培地1ml中に再懸濁し、37℃でさらに30分間インキュベートする。最終のインキュベーションの後、標的細胞を1回洗浄し、そして最終体積を1×105細胞/mLにする。最終のADCCアッセイのために、標識LNCaP細胞100μlをエフェクター細胞50μlおよび抗体50μlと共にインキュベートする。標的とエフェクターの最終比率は1:100を選択した。すべての研究において、対照のヒトIgG1アイソタイプを試験し、CHOに由来する抗PSMA MAbF1抗体と比較した。含まれる他の対照は、a)標的細胞およびエフェクター細胞、ただし抗体はなし、b)エフェクター細胞を伴わない標的細胞、ならびにc)3%TritonX−100の存在下の標的細胞およびエフェクター細胞である。37℃で4時間インキュベーションした後、上清を回収し、240〜400keVの読取領域を有するガンマカウンター(Packard Instruments社製のCobra IIオートガンマ)を用いて計数した。抗体濃度の関数としてカウント毎分をプロットし、そしてこのデータを、Prismソフトウェア(サンディエゴ、カリフォルニア州)を用いた非線形回帰、すなわちS字形用量反応(可変勾配)によって解析した。溶解率は以下の式、すなわち、溶解率(%)=(サンプルCPM−抗体なしのCPM)/(TritonX CPM−抗体なしのCPM)×100によって決定した。本発明者らは、すべての研究においてEC50値および溶解率の双方がモニターされることが重要であることを発見した。例えば、2種の抗体を比較する場合に、EC50値もしくは溶解率または双方が変化している可能性がある。
【0094】
抗CD16抗体によるADCCの阻害を、以下の改変法を用いて実施した。5□g/mlの抗CD16抗体3G8または対照のアイソタイプ抗体の不在下または存在下で、1□g/mlまたは0.01□g/mlの、CHOに産生されたMAbF1抗体またはニワトリに産生されたMAbF1抗体と共に、細胞をインキュベートした。
【0095】
図14は、CHOに由来するMAbが用量依存的な細胞溶解を誘導することを示し、IL−2で刺激されたエフェクター細胞の場合、38%の溶解で頭打ちになり、EC50は0.11μg/mlである。一方、ニワトリの卵に由来するMAbは、より強力かつより効果的であった。ニワトリの卵に由来するMAbの最大溶解率は、抗体の2種の異なる調製物において60%であった。この物質のEC50は0.018μg/mlであったため、CHOに由来するMAbを上回る高い力価も実証された。最後に、予想されるように、対照のアイソタイプ抗体は細胞溶解を誘導しなかった。刺激されていないエフェクター細胞(新鮮なPBMC)のADCCの方が、EC50値の差はより大きいが、全体的な細胞死滅のレベルは劣る(図15)。
【0096】
CD16(FCgRIII)は、ADCCを媒介する重要なレセプターである。ADCC応答の特異性は、CD16を対象とするモノクローナル抗体を用いて標的細胞とエフェクター細胞の相互作用を妨害することによって示した。本研究においては、2種の用量、すなわち飽和用量(1μg/ml)および最適以下の用量(0.01μg/ml)のMAbF1抗体を使用した。1μg/mlのMAbF1抗体は、抗CD16抗体の不在下で、CHOに由来する抗体およびニワトリに由来する抗体の場合、それぞれ約15%および38%の溶解を誘導した。この溶解率は、抗CD16抗体の存在下では約4%に減少したが、対照のアイソタイプ抗体は影響を及ぼさなかった(図16)。
【0097】
(実施例12)
CD16結合
CHOおよびニワトリに由来するMAbF1を、Biacore社によって提供されるアミンカップリングキットを用いて、一級アミンを介してBiacoreセンサーチップ(CM5)のカルボキシメチルデキストランマトリックス表面に固定化した。どちらの抗体も、約10,000RUの密度まで塗った。固定化した抗体表面上にいくつかの濃度のタンパク質を流しかけることによってこれら2種の抗体とCD16−PheおよびCD16−Valとの結合を起こさせた。空白の表面および純粋な緩衝液の結合サイクルを考慮することによって非特異的結合の効果を説明した。HBS−EP緩衝液を希釈用および泳動用緩衝液として使用した。実験は、Biacore−3000機器を用いて25℃で実施した。データはGraphPad Prismソフトウェアによって解析し、また、平衡解離定数を推定するために、単一結合部位モデルにデータを適合させた。
【0098】
高速動態が、抗体へのFcR結合の特徴的な特色であるため、解離定数は、速度定数ではなく平衡結合実験に基づいて推定した。KDの推定のために使用した曲線適合を図17に示す。ニワトリに由来する抗体の解離定数KDは、CHO由来の対応する抗体と比べて、どちらのFcRに対しても約10分の1の小ささである。ニワトリに由来する抗体のより高い親和性は、Fc領域のグリコシル化の差異に起因している可能性があり、特に、CHOに由来する抗体中には存在するフコースがないことが寄与している可能性がある。
【0099】
(実施例13)
治療的有用性
本発明は、具体的に定められたグリコシル化パターンおよび他の化学的諸特性を有し、そして前述の遺伝的に改変されたニワトリを用いて作製された抗体を提供する。これらの特性により、標的組織中の抗原特異的な標的に結合させる目的で患者に投与した場合の治療特性が改善される。具体的には、上記のように、いくつかの臨床適応症に対して、これらの抗体は、増強された抗体依存性細胞傷害活性(ADCC)を示し、また、この効果はいくつかの臨床適応症において重要な利点を与える。
【0100】
いくつかのタイプの癌の治療に対する非結合型モノクローナル抗体(mAb)の臨床試験では、有望な結果が得られた。Dillman、1997年、Cancer Biother. & Radiopharm.12巻:223〜225頁;Deoら、1997年、ImmunologyToday 18巻:127頁。キメラの非結合型IgG1は、軽度または濾胞性のB細胞性非ホジキンリンパ腫(Dillman、1997年、前記)用に認可されており、さらに、別の非結合型mAb、すなわち充実性乳房腫瘍を標的とするヒト化IgG1もまた、臨床試験の第III相において期待できる結果を示した。Deoら、1997年、前記。これら2種のMAbの抗原は、それぞれの標的組織において高発現される。このような応用の場合、特に、抗体がADCCによって強力な腫瘍破壊を媒介する腫瘍細胞において、本発明の抗体は、患者に投与した際の治療的利点を与える。
【0101】
治療的用途のために、本発明の抗体を、(例えば、二重特異性抗体もしくは多重特異性抗体を作製するために)別の抗体、(例えば、免疫毒素などの免疫複合体を作製するために)細胞毒、細胞性リガンド、または抗原など1種または複数種の分子単位に、(例えば化学的結合、遺伝子融合、非共有結合性会合、もしくは別の方法によって)機能的に連結することもできる。本発明の抗体は、他の治療的部分、例えば放射性同位体、低分子抗癌薬、抗炎症剤、細胞毒、または免疫抑制剤に連結することができる。したがって、本発明は、ニワトリ発現系によって可能にされる化学的諸特性を有し、そして治療的用途のために、本質的にすべて公知の抗体結合、連結、および関連した技術と組み合わされた、抗体組成物を包含する。
【0102】
したがって、本発明の抗体は、特にADCC機序が標的組織において提示される場合、治療への感受性が高い標的組織において抗原を発現する細胞を伴う様々な疾患を治療および/または予防するために使用することができる。治療(例えば寛解)または予防することができる例示的な疾患には、それだけには限らないが、充実性腫瘍、リンパ腫、びまん性腫瘍、およびすべてのタイプの癌組織が含まれる。
【0103】
本発明の治療的実施形態では、ADCCの特性を提示する様式による、治療されると思われる状態の診断に具体的に従って、本発明の抗体を患者に投与する。このような臨床現場では、本発明の抗体を投与し、そして治療後に治療の細胞傷害効果を決定して、標的組織におけるADCC効果を決定する。本発明の治療用組成物に加えて、化学療法剤、放射線、またはサイトカインなど、Fc受容体の発現もしくは活性を調節、例えば増大もしくは阻害する作用物質で、患者をさらに治療してよい。治療期間中に投与するための典型的なサイトカインとしては、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターフェロン−γ(IFN−γ)、および腫瘍壊死因子(TNF)が挙げられる。典型的な治療用物質としては、とりわけ、ドキソルビシン、シスプラスチン、ブレオマイシン、カルムスチン、クロラムブシル、およびシクロホスファミドなどの抗悪性腫瘍剤が挙げられる。
【0104】
別の態様では、本発明は、組成物、例えば、ニワトリにおいて発現された本発明の抗体の1つまたは組合せを含む薬剤組成物を提供する。本発明の組成物は、当技術分野において公知の様々な方法によって投与することができる。当業者には理解されるように、投与の経路および/または様式は、所望の結果に応じて異なると考えられる。製薬上許容される担体としては、無菌の水性溶液または水性分散液が挙げられ、薬剤として有効な物質に対するこのような媒体および作用物質の使用は当技術分野において公知である。無菌の注射液剤は、適切な溶媒中の必要とされる量の活性化合物を1つの成分または成分の組合せと混合し、続いて滅菌および/または精密ろ過することによって調製することができる。一般に、分散液は、基本となる分散媒および他の任意の成分を含む無菌ビヒクル中に活性化合物を混合することによって調製される。
【0105】
投与計画は、最適な所望のADCC効果を提供するように調整する。例えば、単回ボーラスを投与してもよく、いくつかに分割した用量を時間をかけて投与してもよく、または治療状況の緊急性によって示されるように、比例的に用量を減少もしくは増加させてもよい。単位剤形の非経口組成物を調製することは、投与の容易さおよび投薬量の均一性に対して特に有利である。本明細書において使用される単位剤形とは、治療しようとする対象者に対する単位投薬量として適した物理的に個別の単位を意味し、各単位は、必要な薬剤用担体と共同して所望の治療効果を生じるように算出された所定の量の活性化合物を含む。本発明の単位剤形の仕様は、(a)活性化合物の独特な特徴および達成すべき個々の治療効果、ならびに(b)個体の感受性の治療のためにこのような活性化合物を配合する技術分野につきものの制約によって必然的に決められ、そして、直接的に依存している。
【0106】
本発明の薬剤組成物中の有効成分の実際の投薬量レベルは、個々の患者、組成物、および投与様式に対する所望の治療応答を実現するのに効果的であり、患者に対する毒性はない有効成分の量を得るために、変更してよい。選択される投薬量レベルは、使用される本発明の個々の組成物の活性、投与経路、投与時間、使用される個々の化合物の排出速度、治療の持続期間、使用される個々の組成物と組み合わせて使用される他の薬物、化合物、および/または材料、治療される患者の年齢、性別、体重、状態、全体的健康状態、および以前の病歴、ならびに医薬分野において周知である同様の因子を含めて、様々な薬物動態学的因子に依存すると考えられる。
【0107】
本発明の抗体の投与効果は、腫瘍などの標的組織において客観的に観察可能であるため、本発明の治療的方法は、具体的にはADCCを使用する治療法を含む治療法を必要とする患者を診断するステップと、効果が望まれる標的組織を同定するステップと、本発明の組成物を、それを必要とする患者に投与するステップと、患者の標的組織におけるADCCの有効性を判定することなどによって患者における治療効果を測定するステップとを含む。治療効果の判定は、細胞死、標的組織の収縮、腫瘍サイズの縮小など標的組織の特性の経時的な変化を分析することによっておよび医薬分野において公知である他の任意の診断技術によって実現することができる。
【0108】
本発明の抗体はまた、当業者が利用可能ないくつかの公知のADCCモデルのうちのいずれかにおいてADCC活性を試験することもできる。治療的用途または診断的用途に対する本発明の抗体の有用性を判定するために、ADCCの測定を独立に実施してもよく、または他の哺乳動物、非哺乳動物、植物、もしくは細菌細胞の発現系と比較してもよい。したがって、本発明の方法は、前述の系において産生された別の抗体と直接または間接的に比較して本発明の抗体を使用することによってADCCをもたらすための有用性の差異を判定するステップを含む。具体的には、この方法論は、前述のニワトリ発現系において産生された抗体のADCC効果を比較して、増強されたADCCを同定し、ニワトリ発現系のための理想的な抗体候補を同定するステップを含む。
【0109】
上記のように、治療的有用性を向上させるために、本発明の抗体は、1種または複数種の他の治療物質、例えば、細胞障害性物質、放射毒性物質、または免疫抑制剤と同時投与することができる。抗体は、(免疫複合体として)作用物質に連結されてもよく、または作用物質と別々に投与されてもよい。後者の場合(個別投与)、抗体は、作用物質の前、後、もしくは同時に投与してよく、または他の公知の治療法、例えば抗癌療法、例えば放射線療法と同時に投与してもよい。このような治療用物質としては、とりわけ、ドキソルビシン、シスプラスチン、ブレオマイシン、カルムスチン、クロラムブシル、およびシクロホスファミドなどの抗悪性腫瘍剤が挙げられる。本発明の抗体と化学療法剤との同時投与は、ヒト腫瘍細胞に対する細胞障害効果をもたらす、異なる機序を介して機能する2種の抗癌物質を提供する。このような同時投与により、薬物に対する耐性の発達、または抗体との反応性を低下させると考えられる腫瘍細胞の抗原性の変化に起因する問題が解決され得る。
【0110】
当業者には明らかであろう、開示した本発明の様々な修正、改良、および応用があると考えられ、かつ本出願は、法律によって許容される範囲まで、そのような実施形態を包含する。いくつかの好ましい実施形態において本発明を説明したが、本発明の完全な範囲はそのようには限定されず、以下の特許請求の範囲に従う。すべての参考文献、特許、または他の刊行物は、参照により本明細書に具体的に組み入れられる。
【図面の簡単な説明】
【0111】
【図1】図1は、ニワトリES細胞の特徴的な形態を示す。細胞は単層で増殖し、細胞質はほとんどなく、核小体が目立っている。
【図2】図2は、ニワトリES細胞のin vitroでの特性、具体的には、抗体SSEA−1およびEMA−1との反応、ならびにアルカリ性ホスファターゼの発現を示す。
【図3】図3は、189日間培養されたニワトリES細胞の核型である。これらの細胞は2倍体であり、38対の常染色体および1対のZ染色体を有する。
【図4】図4は、横斑ロックヒヨコ2匹、および横斑ロックES細胞を白色レグホーンのレシピエント胚中に注入することによって形成されたキメラ2匹である。これらのキメラおよび横斑ロックは区別不能であることから、メラノサイト系列へのES細胞の寄与が多大であることが示される。
【図5】図5は、横斑ロックES細胞を白色レグホーンレシピエント中に注入することによって作製されたキメラである。左のパネル中のキメラのペアは、メラノサイト系列に対して少しの寄与を示しているのに対し、左のパネル中のペアは、より大きな寄与を示している。
【図6A】図6Aは、ニワトリの輸卵管の管状腺細胞における組織特異的発現を実現する導入遺伝子構築物、および組織特異的発現を裏付ける生理学的証拠である。図6Aは、指定されたOv7.5MAbdnsを含む導入遺伝子の図である。
【図6B】図6Bは、ニワトリの輸卵管の管状腺細胞における組織特異的発現を実現する導入遺伝子構築物、および組織特異的発現を裏付ける生理学的証拠である。図6Bは、他の細胞型を除いて、選択的に管状腺細胞においてモノクローナル抗体の発現を示す、組織特異的発現導入遺伝子を含むキメラニワトリの膨大部の切片である。
【図7A】図7Aは、キメラの脳、腸、膵臓、および筋肉組織を除いて、輸卵管におけるモノクローナル抗体の軽鎖および重鎖の双方の発現をそれぞれ示すRT−PCR解析である。
【図7B】図7Bは、キメラの脳、腸、膵臓、および筋肉組織を除いて、輸卵管におけるモノクローナル抗体の軽鎖および重鎖の双方の発現をそれぞれ示すRT−PCR解析である。
【図8】図8は、ニワトリおよびCHO細胞に由来するMAbF1の熱安定性を示す。CHO細胞に由来するMAbF1(左上)およびニワトリ管状腺細胞に由来するMAbF1(右上)のDSCデータである。どちらの場合も、3つのピークを有する二状態遷移モデルに当てはまるようにデータをデコンボリュートする(赤色で示す)。CHO細胞において発現された抗体に関しては62.7℃、69.4℃、および83.4℃のTm値が得られ、ニワトリに由来する抗体に関しては63.8℃、68.5℃、および83.1℃のTm値が得られた。それぞれのFc脱グリコシル化抗体(それぞれ左下および右下)のDSCデータも同様に二状態遷移モデルに当てはめたが、いずれの場合もピークは2つであり、CHOにおいて発現された抗体に関しては61.2℃および84.0℃のTm値が得られ、ニワトリに由来する抗体に関しては62.6℃および84.0℃のTm値が得られた。
【図9】図9は、ニワトリ管状腺細胞およびCHO細胞において産生されたMAbF1のオリゴ糖プロファイルを示す。CHOに産生されたMAbの公知のグリカン構造が示されている。マンノースは黒の円として示され、ガラクトースは白の円として示され、N−アセチルグルコサミンは黒の四角形と示され、フコースは黒の三角形として示される。
【図10】図10は、卵白からのMAbF1の精製を示す。1:出発原料、2:プロテインA添加、3:プロテインA通過、4:プロテインA溶出。クーマシー染色およびウェスタンブロットの双方とも、非還元条件下である。HRP標識ヤギ抗ヒトIgG(Southern Biotechnology社)を、ウェスタンブロットにおける検出のために使用した。
【図11】図11は、ニワトリまたはCHO細胞に産生されたMAbF1の、LNCaP細胞上で発現されるPSMAに対する結合を示す。324−Eggおよび741−Eggは、卵から単離されたMAbF1の2種の異なる調製物である。F1−CHOは、CHO細胞によって産生されたMAbF1である。抗CTLA4は、LNCaP細胞上の抗原を認識しないヒトIgG1の対照MAbである。
【図12】図12は、ニワトリ管状腺細胞およびCHO細胞において産生されたMAbF1の内部移行アッセイである。570−Eggおよび741−Eggは、卵から単離されたMAbF1の2種の調製物であった。各300ng/ウェルのMAbを、Hum−Zap(抗ヒトIgG、サポリン結合体)または細胞培養培地のいずれかと共にLNCaP細胞に添加した。MAbおよびHumZapの添加後48時間目に細胞の生存率を決定した。Y軸は、相対発光量(RLU)である。
【図13】図13は、ニワトリ管状腺細胞(F1−ニワトリ)およびCHO細胞(F1−CHO)において産生されたMAbF1のin vivoクリアランスを示す。BALB/cマウス4匹に、放射標識した各抗体調製物を注射した。全身カウンターを用いて、残留放射能を350時間に渡って測定した。データは、残存している注射用量のパーセンテージとして表される。
【図14】図14は、ヒトIL−2の存在下でニワトリ管状腺細胞(F1−ニワトリ調製物IおよびII)ならびにCHO細胞(F1)において産生されたMAbF1のADCCアッセイを示す。LNCaP細胞上の抗原を認識しないヒトIgG1をアイソタイプ対照として使用した。
【図15】図15は、ヒトIL−2の不在下でニワトリ管状腺細胞(F1−ニワトリ調製物IおよびII)ならびにCHO細胞(F1)において産生されたMAbF1のADCCアッセイを示す。LNCaP細胞上の抗原を認識しないヒトIgG1をアイソタイプ対照として使用した。
【図16】図16は、抗CD16抗体を用いたADCCの妨害を示す。
【図17】図17は、ニワトリに由来するMAbF1(青色)およびCHOに由来するMAbF1(赤色)の、CD16−Phe(左のパネル)およびCD16−Val(右のパネル)との平衡結合を示す。解離定数(KD)の推定値を対応する曲線の近くに示す。
【技術分野】
【0001】
本明細書において記載される発明は、USDA SBIR 2003−33610およびNIH 2 R44 GM64261−01および2R44 HD 039583−02の下で政府により支援されている。米国政府は、本発明において特定の権利を有し得る。
【背景技術】
【0002】
(発明の背景)
遺伝的に改変された動物は、抗体など有益な薬学的製品の持続可能な製造を大きく前進させる可能性を与える。しかし、遺伝的に改変された動物の作製は、著しい技術的障害を伴い、これらの障害は少数の種でしか克服されていない。特異的発現のために、タンパク質をコードする遺伝的改変をある種のDNA中に組み入れられるようにするには、各遺伝的改変のために開発されなければならないいくつかの独特な技術が必要となる。ある動物の遺伝的特徴および肉体的特徴を改変するための1つの手法は、その動物のレシピエント胚中に細胞を導入することである。これらの細胞は、レシピエント胚から生まれる動物の組織に寄与し、結果として生じる遺伝的に改変された動物の子孫のゲノに寄与する能力を有する。
【0003】
細胞株の研究および開発、細胞ゲノムの操作、およびそのような操作された細胞が培養状態で維持されることを可能にする細胞培養技術に、かなりの時間および資源が費やされてきた。多くの試みがなされてきたが、操作された細胞の多能性を培養状態で維持する能力は、ごく少数の種、特にマウスでしか実現されていない。
【0004】
持続可能な培養物が容易に入手可能であり、そして多能性を維持しつつ遺伝子工学を受け入れることが可能である場合には、新しい技術の幅広い応用が利用可能になると思われる。培養細胞はキメラ動物の組織に寄与することができるため、胚状態のレシピエント動物中にこれらの細胞を組み入れることによって胚性幹細胞の由来元である動物の肉体的特徴をレシピエント胚に導入することができる。これは、2つの主要な利点を与える。第1に、胚性幹細胞の由来元である動物の表現型をレシピエント胚に選択的に導入することができる。第2に、上述したように、細胞培養物が特に安定である場合、これらの細胞のゲノムを遺伝的に改変して、細胞が導入されるレシピエント胚中に遺伝的改変を導入することができる。
【0005】
いくつかの場合において、抗体をコードする導入遺伝子を用いてこれらの細胞を操作することができる。導入遺伝子は、抗体を作製するための青写真の役割を果たすDNAを含み、そしてレシピエント胚中への細胞の挿入によって作り出される動物の組織における抗体の発現を可能にするのに十分なコードエレメントおよび調節エレメントを含む、遺伝子構築物である。しかし、動物の組織から有益な抗体を回収するには、通常、発現されたタンパク質の回収を容易にし、かつ望ましい化学的諸特性を伝達するいくつかの特定の組織型に発現が限定されている必要がある。例えば、雌ウシにおいては、乳汁におけるタンパク質発現により、単に雌ウシの乳汁を回収し、そして外因性タンパク質を分離することによる、タンパク質の容易な回収が可能になる。ニワトリでは、卵白における抗体の活発な産生もまた、抗体の発現および回収のための魅力的な媒介物になる。さらに、組織特異的な発現がニワトリの輸卵管に特異的である場合、この発現により、ヒト患者の治療において使用された場合に抗体の治療的有用性を高めるいくつかの特定の望ましい化学的諸特性を有する抗体が生じる。したがって、研究および商業的開発の特に魅力的な1つの分野は、卵中で抗体を選択的に発現して、望ましい化学的諸特性を有するタンパク質の単離および回収を容易にする、遺伝子操作されたニワトリである。組織特異性がない場合、動物の全組織において発現される抗体を生じるだけであるため、ある動物の特異的に選択された細胞において外因性抗体を選択的に作製する能力は、特に有益である。このような状況下では、有意義な量の抗体が動物から分離され得る見込みは少なく、外因性抗体の遍在的な発現は通常、動物の全般的健康および幸福に大きな損害を与え、また、ニワトリ輸卵管において提示される望ましい化学的諸特性は存在していない。
【0006】
細胞培養が十分に安定で、細胞のゲノム中に導入遺伝子が組み込まれるのが可能となる場合、標的細胞および導入遺伝子として使用される特定の構築物に応じたいくつかの異なる技術によって新しいキメラ生物またはトランスジェニック生物に、抗体の組織特異的発現をコードする導入遺伝子を受け渡すことができる。全ゲノムは細胞ハイブリダイゼーションによって、インタクトな染色体は微小核によって、染色体部分のセグメント(subchromosomal segment)は染色体を介した遺伝子導入によって、およびキロベース範囲のDNA断片はDNAを介した遺伝子導入によって、導入することができる(非特許文献1:Klobutcher, L.A.およびF.H.Ruddle、Annu. Rev. Biochem.、50巻:533〜554頁、1981年)。インタクトな染色体は、微小核を介した染色体導入(MMCT)によって導入することができる(非特許文献2:Fournier,R. E.およびF. H. Ruddle、Proc. Natl. Acad. Sci. U.S.A.、74巻:319〜323頁、1977年)。導入遺伝子を具体的に設計するには、抗体をコードするDNA配列の内容、標的細胞株、発現が標的される場となる特異的組織、発現が起こる場となる宿主生物、および発現される抗体を検討しなければならない。組織特異的発現のために設計される導入遺伝子は、細胞のゲノム中への成功な組み込みを可能にし、そして宿主生物の選択された組織における成功な発現を確実にするために、いくつかの条件を満たさなければならない。
【0007】
上記のように、トランスジェニック動物を作製するための遺伝的改変の導入は、ごく少数の種においてしか実証されていない。マウスの場合、キメラ子孫およびトランスジェニック子孫を作製するための、相同組換えとそれに続く胚性幹(ES)細胞への染色体導入の別々の使用が周知である。部位特異的な相同組換えまたは遺伝子ターゲティングの強力な技術が開発された(非特許文献3および4:Thomas, K. R.およびM.R. Capecchi、Cell 51巻:503〜512頁、1987年;Waldman, A.S.、Crit.Rev. Oncol. Hematol.12巻:49〜64頁、1992年による総説を参照されたい)。クローン化DNAの挿入(非特許文献5:Jakobovits,A.、Curr. Biol.4巻:761〜763頁、1994年)、ならびにCre−loxP系技術による染色体フラグメントの操作および選択(非特許文献6および7ならびに特許文献1〜4:Smith,A. J.ら、Nat. Genet.9巻:376〜385頁、1995年;Ramirez-Solis, R.ら、Nature 378巻:720〜724頁、1995年;米国特許第4,959,317号;第6,130,364号;第6,091,001号;第5,985,614号を参照されたい)は、安定な遺伝子キメラを作製するための操作およびマウスES細胞への遺伝子導入のために利用可能である。哺乳動物系において有用であることが判明したこのような多くの技術は、必要な長期の細胞培養物が入手可能である場合、ならびに、外因性タンパク質の単離および回収を容易にする、特定の組織における組織特異的発現をもたらす導入遺伝子が設計され得る場合には、非哺乳動物系に適用するために有効であると考えられる。
【0008】
組織特異的発現を可能にする導入遺伝子は複雑であり、また、レシピエント細胞株中に導入遺伝子を組み入れるのに必要である遺伝子操作は、培養条件が最適化されなければ細胞の多能性を脅かす可能性がある大規模な操作を必要とする。したがって、遺伝子導入において使用するのに適した細胞株は、培養状態で安定でなければならず、かつ組織特異的発現に必要なエレメントをすべて含むのに十分な大きさおよび複雑さを有する遺伝子構築物で細胞がトランスフェクトされた場合に、多能性を維持しなければならない。結果として生じる動物において、導入遺伝子は、導入遺伝子が発現されるように設計されている特定の個々の組織型において効果的に発現されなければならず、そして他の組織で発現されて、その結果、動物の生存能力または結果として生じるタンパク質の有利な化学的性質が損なわれるべきではない。
【0009】
外因性抗体の作製に対して、トリの生物学的系は、飼育場での効率的な飼育、迅速な成長、および経済的な生産を含めて、多くの利点を与える。さらに、トリ卵は、抗体の大量合成ならびに生成物の単離および回収の平易性の双方に関して理想的な生物学的設計を実現する。さらに、本明細書において後述するように、脊椎動物、植物、または細菌の細胞系と比較したニワトリをベースとする発現系の利点は、大量の抗体生成物に対して得ることができる。
【特許文献1】米国特許第4,959,317号明細書
【特許文献2】米国特許第6,130,364号明細書
【特許文献3】米国特許第6,091,001号明細書
【特許文献4】米国特許第5,985,614号明細書
【非特許文献1】Klobutcher, L.A.およびF.H.Ruddle、Annu. Rev. Biochem.、50巻:533〜554頁、1981年
【非特許文献2】Fournier, R. E.およびF. H. Ruddle、Proc. Natl. Acad. Sci. U.S.A.、74巻:319〜323頁、1977年
【非特許文献3】Thomas, K. R.およびM.R. Capecchi、Cell 51巻:503〜512頁、1987年
【非特許文献4】Waldman, A.S.、Crit. Rev. Oncol. Hematol.12巻:49〜64頁、1992年
【非特許文献5】Jakobovits, A.、Curr. Biol.4巻:761〜763頁、1994年
【非特許文献6】Smith, A. J.ら、Nat. Genet.9巻:376〜385頁、1995年
【非特許文献7】Ramirez-Solis, R.ら、Nature 378巻:720〜724頁、1995年
【発明の開示】
【課題を解決するための手段】
【0010】
(発明の要旨)
本発明は、ニワトリにおけるタンパク質発現、ならびにキメラニワトリおよびトランスジェニックニワトリを作製するために使用される、遺伝子工学、遺伝子導入、および細胞の長期培養などの実現技術を含む。本発明はまた、ニワトリにおいて産生される抗体およびそれらの独特な化学的性質にも関する。具体的には、これらの抗体は、ある種の用途におけるそれらの治療的有用性を高める有利な化学的諸特性を有する。ニワトリにおいて産生される抗体は、脊椎動物、植物、または細菌の細胞系において産生される抗体と比べて独特なパターンの化学的改変を有し、その結果、腫瘍などの標的組織に毒素を結合させる目的で患者に投与された場合に、標的組織は、高い治療有効性で治療される。一実施形態では、胚性幹細胞の長期培養物は、外因性抗体の組織特異的発現をもたらす導入遺伝子の挿入物を含む、キメラトリに遺伝的改変を導入するために特別に設計された遺伝子構築物を用いて作り出される。本発明のトランスジェニックトリは、導入遺伝子に由来する抗体を輸卵管中で発現し、また、抗体は、卵中に大量に蓄積する。好ましい実施形態では、天然のヒト抗体がニワトリの輸卵管において発現され、そして卵から回収することができるように、外因性抗体タンパク質は、ヒトDNA配列によってコードされる。
【0011】
本発明は、抗体の組織特異的発現を示すトリの集団、外因性抗体の発現を可能にする導入遺伝子構築物、ニワトリにおいて産生され、そして特別に定められた化学的諸特性を有する抗体の単離組成物、ならびにこれらのトリの作製、抗体の作製、およびヒトにおけるそれらの治療的使用のための関連した方法を含む。本発明は、長期細胞培養物、および持続的な細胞培養物に由来するキメラトリまたはトランスジェニックトリを作製するための特別な技術であって、ES細胞のゲノムが外因性タンパク質を発現する安定に組み込まれた導入遺伝子を有し、その結果、培養細胞の子孫が、安定に組み込まれた導入遺伝子を含む技術を使用する。後述する手順によって宿主トリの胚に導入された場合、それらの改変されたドナー細胞は、結果として生じる動物の選択された特定の体細胞組織中に導入遺伝子を発現するトリを生じる。これらのトリは、ドナー細胞に由来する表現型を提示し、また、輸卵管中で抗体を発現して、卵白における抗体の濃縮および回収を容易にする。
【0012】
本発明はまた、ニワトリ発現系において発現され、そして脊椎動物、植物、または細菌細胞の系と比べて望ましいいくつかの化学的諸特性を有する抗体の組成物も含む。具体的には、これらの抗体は、濃度の低下したフコース、ガラクトース、N−アセチルノイラミン酸、N−グリコリルノイラミン酸、および濃度の上昇したマンノースを有する。これらの諸特性のうちのいくつかまたはすべてを有する抗体は、ヒトに投与された場合に、高い治療的有用性を示す。具体的には、これらの抗体組成物は、増強された抗体依存性細胞傷害活性(ADCC)を示す。したがって、本発明の方法は、抗体の組成物をニワトリにおいて発現させることにより、ADCC作用に基づいて、それらの治療的有用性を高めることを含む。実際の応用では、これらの方法は、本明細書において説明される抗体組成物を投与するステップと、患者における細胞傷害活性を検出するステップとを含む。本発明はまた、所定の量の外因性抗体が卵白中に濃縮されるように、本明細書において定義される有利な化学的性質を有する外因性抗体を輸卵管組織中で発現するニワトリも含む。好ましい一実施形態では、外因性タンパク質は、ドナー細胞株および子孫のゲノム中に組み入れられた導入遺伝子構築物によってコードされたヒト配列モノクローナル抗体である。ヒトモノクローナル抗体をコードするポリヌクレオチド配列は、輸卵管中での発現のために特異的に構築されており、そして組織特異的発現を容易にするための適切なプロモーターおよび調節配列を含む導入遺伝子の内部に含まれている。外因性タンパク質を発現するトランスジェニックトリまたはキメラトリの実施形態では、本発明は、その動物およびタンパク質、例えば本明細書において説明される外因性抗体を含む卵白アルブミンに特異的な組成物を含む。
【発明を実施するための最良の形態】
【0013】
(発明の詳細な説明)
本発明によれば、ニワトリES細胞系は大型の核を有し、かつ顕著な核小体を含むステージX胚に由来する(図1)。これらの細胞は、長期の培養後に形態学的にニワトリ胚性幹(cES)細胞であることが確認され、また、レシピエント胚中に注入された場合にキメラを生じることが確認されている。さらに、ES細胞は、広範囲の羽毛キメラ現象によって判定されるように、体細胞組織に対する高度の寄与を可能にする。さらにまた、これらの胚性幹細胞は、外因性タンパク質をコードするDNAを有する導入遺伝子でトランスフェクトされることが実証されている。ES細胞は、導入遺伝子を安定に組み込み、そして導入遺伝子を発現して、形質転換細胞の選択を可能にする。これらの形質転換細胞はキメラを形成することができ、その際、導入遺伝子によってコードされた外因性タンパク質は、このキメラの選択された組織中に存在する。このキメラに由来する細胞は、この導入遺伝子によってコードされる外因性タンパク質を発現する。特に好ましい実施形態では、この導入遺伝子によってコードされる外因性抗体は、この導入遺伝子によってコードされる抗体に応じて、特定の組織または組織型において発現される。胚性幹細胞の子孫は、レシピエント胚へのES細胞の導入およびキメラの形成後に非ES細胞表現型に分化する、ES細胞の派生物である。
【0014】
体細胞組織における導入遺伝子の広範な発現は、胚体外組織および体細胞組織における発現によって実証される。トランスジェニック動物の卵のタンパク質内容物を解析することにより、導入遺伝子から、そして本発明の技術を用いて得られる、導入遺伝子によってコードされた外因性タンパク質の卵白における選択的発現が実証される。組織特異的発現は、1つの器官、組織、または細胞型に実質的に限局されている発現によって実証される。
【実施例】
【0015】
(実施例1)
ニワトリ胚性幹細胞(cES細胞)の誘導
ニワトリES細胞を、2種の交雑、すなわち、横斑ロック×横斑ロック、または横斑ロック×ロードアイランドレッドのうちの1つから誘導した。cES細胞の発育能を試験する場合、羽毛マーカーを得るためにこれらの品種を選択した。これらcES細胞を、優性の白色遺伝子座においてホモ接合優性である白色レグホーン胚中に注入する。これらのES細胞の注入の結果として生じるキメラニワトリは、cES細胞由来の黒色の羽毛およびレシピエント胚由来の白色の羽毛を提示する。
【0016】
cES細胞培養物の最初の樹立を、USP5,565,479に記載されているプロトコールに従って開始した。ステージXでは、胚は、約40,000〜60,000個の細胞からなり、卵黄の表面に位置する、小さな円盤状物体である。この胚を採取するために、紙でできた輪を卵黄膜上に置き、中央に胚を露出させる。この輪の外周に沿って卵黄膜を切断し、次いで卵黄からそれを持ち上げる。輪の内側に付着した胚を顕微鏡下に置き、また、細いループを用いて暗域から明域を単離する。
【0017】
表1:CES−80培地中のSTOフィーダー細胞またはポリエステル挿入物のいずれかの上で誘導されたcES細胞株。培養は、単一胚およびプールされた胚の双方から開始した。
【0018】
【表1】
胚を機械的に分散させて単細胞懸濁物とし、有糸分裂が不活化されたSTO細胞のコンフルエントなフィーダー層上に3×104細胞/cm2の濃度で播種する。cES培養培地は、10%FCS、1%ペニシリン/ストレプトマイシン、2mMグルタミン、1mMピルバート、1×ヌクレオシド、1×非必須アミノ酸、および0.1mM β−メルカプトエタノールを添加したDMEM(20%新鮮培地および80%馴化培地)からなる。使用前に、DMEM培地をバッファローラット肝臓(BRL)細胞で馴化させる。手短に言えば、BRL細胞をコンフルエントになるまで増殖させた後、5%血清含有DMEMを添加し、3日間馴化させる。この培地を除去し、培地の新しいバッチを3日間馴化させ、これを繰り返す。3回分のバッチを合わせ、cES培地を作製するために使用した。ニワトリES細胞は、胚盤葉細胞の接種後3〜7日目に可視になる。これらのcES細胞は、形態学的にmES細胞に類似していた。すなわち、これらの細胞は小型であり、大型の核および顕著な核小体を有していた(図1を参照されたい)。
【0019】
cES細胞の増殖特徴は、縁が滑らかであり、かつ区別が困難な個々の細胞を有する密な円形コロニー中で増殖するmES細胞とは異なる。ニワトリES細胞は、個々の細胞がはっきりと目に見える単層コロニー中で増殖する。密なコロニーは、cES培養物における分化の最初の徴候であることが多い。
【0020】
培養で誘導された細胞の多能性のマーカーについて試験するために、これらの細胞を固定し、そしてマウスおよびニワトリの始原生殖細胞上のエピトープを認識する、SSEA−11(Solter,D.およびB.B.Knowles、Proc.Natl.Acad.Sci、U.S.A.75巻:5565〜5569頁、1978年)、EMA−1(Hahnel,A.C.およびE.M.Eddy、GameteResearch 15巻:25〜34頁、1986年)、および同様に多能性細胞によって発現されるアルカリ性ホスファターゼ(AP)で染色した。図2に示すこれらの試験の結果により、ニワトリES細胞が、アルカリ性ホスファターゼ、ならびにSSEA−1およびEMA−1によって認識される抗原を発現することが実証される。
【0021】
上記のプロトコールを用いた後のcES細胞は可視であるが、このような培養物は、数週間より長く維持することができない。以下で考察するように、培養条件にいくつかの変更を起こすことにより、19種の細胞株(表1)の派生物を生じさせ、このうち14種をレシピエント胚中に注入することによってそれらの発育能を試験した。羽毛の色素沈着によって判定されるように、14種の細胞株のうちの11種が、レシピエント胚に寄与した(下記表2を参照されたい)。このプロトコールは、胚性幹細胞表現型を発現する多能性細胞の持続的培養物をもたらす。任意の時点で、これらの細胞を凍結保存することができ、また、これらの細胞は、損傷を与えられたレシピエント胚中に注入された場合に、体細胞組織に実質的に寄与する潜在能力を有する(以下の実施例3および実施例5を参照されたい)。
【0022】
表2:単一胚またはプールされた胚に由来する胚性幹細胞株の継代数および培養時間。ステージXレシピエント中へのこれらのES細胞の注入後の体細胞キメラ現象の出現率および程度。
【0023】
【表2】
マウスと同様に、トリ胚性幹細胞は、STO、STO−sn1、および容易に入手可能である他のものを含めて、様々なフィーダー層上で誘導される。これらのフィーダーによって産生される白血病抑制因子(LIF)、およびウシ胎児血清の添加は、ES細胞を未分化状態で維持するのに寄与する。本発明の好ましい実施形態では、ニワトリES細胞の培養は、STOフィーダー層上で開始する。STO細胞をコンフルエントになるまで増殖させ、10μg/mlマイトマイシンで3〜4時間処理し、洗浄し、トリプシン処理し、そしてゼラチンコート皿上に4×104細胞/cm2の濃度で播種する。cES細胞は、STO細胞からなるフィーダーが薄い場合に、より迅速に増殖するようである。STOフィーダー細胞濃度を103〜105細胞/cm2まで、好ましくは104細胞/cm2未満に低下させると、cES細胞の誘導および増殖が促進される。しかし、ニワトリ胚線維芽細胞およびマウス初代線維芽細胞をフィーダーとして用いた場合、cES細胞は誘導されなかった。また、予め樹立されたcES細胞をこれらのフィーダー上に播種した場合、これらはすべて1週間以内に分化した。
【0024】
フィーダーの不在下、ポリマー膜(Costar社、Transwell型)などの合成挿入物上でcES細胞を増殖させると、ES細胞を注入する際の、フィーダー細胞によるレシピエント胚の汚染が回避される。表3および表4が示すように、STOフィーダーの代わりに挿入物上で培養することにより、cES細胞の誘導が促進され、また、挿入物は、最初の誘導のために使用され得る。しかし、最初に挿入物上で迅速に増殖した後、ES細胞の有糸分裂活性は4〜6週間の培養後に低下する。培養を延長するためには、これらの細胞をSTO細胞からなるフィーダーに移さなければならない。
【0025】
表3.挿入物またはSTO細胞のフィーダー(104細胞/cm2)のいずれかの上での単一胚からのcES細胞の樹立
【0026】
【表3】
表4.STOフィーダーまたは合成挿入物のいずれかの上でのプールされた胚からのcES細胞の樹立
【0027】
【表4】
表3および表4のデータは、ニワトリの胚性フィーダー細胞およびマウス初代胎児線維芽細胞が、cES細胞の誘導も培養も支援しないことを示す。STO細胞のフィーダーは、誘導および増殖を支援するが、103〜105細胞/cm2の限定された濃度、好ましくは、本発明の実施形態では、104細胞/cm2未満または約104細胞/cm2の濃度でSTO細胞が存在する場合にのみ、支援する。濃密なSTOフィーダー層は、cES細胞の増殖を減退させるが、指定された濃度のSTO細胞は、ES細胞の増殖に必要な因子を提供する。これらの細胞が長期の培養期間に渡って維持され、かつ継続して胚性幹細胞表現型を発現し、かつin vivoで非胚性幹細胞表現型に分化する場合、これらの細胞は、「ES細胞子孫」と呼ばれる。
【0028】
cES細胞培養培地は、80%の馴化培地からなり、好ましくは、cES細胞の誘導および増殖に必要な因子を含むある種のBRL馴化培地を含む。50%の濃度では、cES細胞の増殖は、80%の馴化培地における場合ほど信頼性が高くない。馴化培地の比率が50%未満まで減少した場合、分化細胞の増加によって立証されるように、cES細胞の増殖は影響を受け、また、30%以下の濃度では、cES細胞は、1週間以内に分化する。cES細胞の誘導および維持に必要であることが判明したこの馴化培地は、mESを維持しないが、それらの分化を引き起こす。
【0029】
ウシ胎児血清は、本発明によるES細胞培地の好ましい成分であり、cES細胞を未分化状態に保つ因子を含む。しかし、血清は、分化を誘導する因子を含むことも公知である。市販の血清ロットは、ES細胞を未分化状態に保つ潜在能力について、使用者によってルーチン的に試験される。cES細胞培養物に対して使用される血清は、マウスES細胞培養物に使用される血清とは異なることが公知である。例えば、細胞毒およびヘモグロビンの濃度が低く、マウスES細胞を未分化状態で維持することが公知である、マウスES細胞の培養のために用いられる血清は、ニワトリES細胞の持続的な増殖を支援しない。
【0030】
したがって、ニワトリES細胞に対して使用される血清は、マウスES細胞において試験して培地成分としての適性を判定するべきではなく、その代わりに、ニワトリES細胞において評価すべきである。そのために、ニワトリES細胞培養物を2つに分け、血清の新しいバッチをそれぞれ試験するために使用する。試験される新しいバッチは、実験的に試験された場合に、ニワトリES細胞の増殖を明らかに支援しなければならない。
【0031】
ニワトリ染色体のスプレッドは、10本の大染色体、66本の小染色体、および1対の性染色体(雄におけるZZおよび雌におけるZW)からなる複雑な核型が原因となって、マウスとは異なる特別な評価技術を必要とする。図3に示す本発明の長期cES細胞を、培養から189日後、そして2回凍結保存した後に解析した。図3を参照すると、これらは、10本の大染色体、2本のZ染色体、および66本の小染色体を有する正常な核型を示した。
【0032】
ニワトリES細胞を、培地中10%DMSO中で凍結保存する。いくつかの細胞株を解凍し、レシピエント胚に注入した後、体細胞キメラが得られる。このことは、これらのcES細胞が、凍結保存プロセスの間、それらの発育能を維持していることを示唆する。
【0033】
(実施例2)
レシピエント胚へのニワトリ胚性幹細胞の注入
新しく産卵された卵中の胚への到達を可能にするためには、殻を割らなければならず、これは、21日のインキュベーション期間の最終時点のふ化率の低下を不可避的にもたらす。慣例としては、卵の側面を切断して小さな穴(直径10mm未満)をあけ、この穴を通して胚を操作し、そしてテープ、カバーガラス、卵殻膜、または一片の殻で再び封じた。実施するのは比較的簡単であるが、この「窓開け」法は、70%〜100%の胎児死亡率を引き起こした。胚への到達の改善、および生存およびふ化能力の増大は、2種の異なる殻および(Callebautから適応させた)方法(参照により本明細書に具体的に組み入れられる、Callebaut、Poult. Sci 60巻:723〜725頁、1981年、ならびにRowlett,K.およびK.Simkiss,J.Exp.Biol.143巻:529〜536頁、1989年)を用いて、ふ化までのインキュベーションのために代理卵殻に胚を移入した場合に実現することができ、この技術を用いた場合、平均ふ化率は、約41%(23〜70%の範囲)であり、469個のcES細胞注入胚から191匹のヒヨコがふ化した。
【0034】
レシピエント胚中へのドナーES細胞の注入後の胚のインキュベーションは、以下に説明するような系Aおよび系Bを含む、2つの部分に分けることができる。
【0035】
系Aは、産卵後発育の最初の3日間を対象として含む。レシピエント胚を含む受精卵は、3〜5グラム重い卵と一致する。鋭端(pointed pole)を切断して直径32mmの窓を開け、内容物を除去し、そして卵黄上のレシピエント胚を、周囲の卵白と共に代理殻中に注意深く移す。
【0036】
細胞を、2ミクロンフィルターを取り付けられた口式吸引器に連結された、徐々に先細になる滅菌ガラスピペット中に吸い上げる。ピペットの穴は、直径が50〜120ミクロンでよく、また、30°の尖った斜面を有する。胚は、低倍率下で、かつ青色光を用いて可視化する。ニワトリES細胞をトリプシン処理して単細胞懸濁液にし、そして約2,000〜26,000個の細胞、好ましくは約20,000個の細胞を胚中に注入する。これらの細胞は、胚の下または上のいずれかの空間中に、すなわち、胚下腔中に、または明域の先端表面と卵黄周囲の層(卵黄膜)の間に穏やかに押し出す。新鮮な受精卵から採取した余分な卵白を添加し、プラスチックフィルムのサランラップで殻を密閉する。
【0037】
系Bは、3日目からふ化までの期間を対象として含む。インキュベーション3日目に、胚は、ステージ17前後に到達した(H&H)。卵白から胚下腔に水が輸送され、それによって卵黄が拡大し、非常に脆弱になる。系Aの殻の内容物を、元の卵より30〜35グラム重い第2の代理殻(通常、シチメンチョウの卵)に、極めて慎重に移入する。細菌汚染を防止するためにペニシリンおよびストレプトマイシンを添加し、また、鈍端(blunt pole)の38〜42mmの窓をプラスチックフィルムで封じる。このより大きな殻により、人工的な気室が実現される。インキュベーションの18日目〜19日目に、これらの胚培養物を、綿密な観察のために卓上ふ卵器に移す。肺換気が確立されるとき、プラスチックフィルム中に周期的に穴が形成され、周囲空気が気室中に入ることが可能になる。ふ化の約6〜12時間前に、フィルムを、ふ化の進行中にヒヨコが容易に脇へ押しのけることができる小型ペトリ皿に交換する。
【0038】
インキュベーションの間、代理殻中の胚のために、通常の飼育温度(37.5℃〜38℃)および相対湿度(50%〜60%)を維持するが、普通は1時間毎であり、そして90度までである周期的な卵の揺り動かしは、良好な生存を確実にするように変更しなければならない。系Aでは、揺り動かしは、4〜5分間毎にに90°までであり、系Bでは、40〜45分毎に40°〜60°までである。いずれの系においても、揺り動かしの速度は、1分間当たり15°〜20°で維持される。
【0039】
cES細胞の注入の前に、レシピエント胚を(1)660ラドのガンマ線照射への曝露によって照射することによって(2)胚の中央から約1000個の細胞を機械的に除去することにより改変することによって、または(1)および(2)を組み合わせることによって調製した場合には、キメラに対するcES細胞の寄与が改善される。表5を参照すると、体細胞組織に対するcES細胞の寄与は、レシピエント胚の中央から細胞を除去することまたは放射線照射への曝露のいずれかによってレシピエント胚に損傷を与えた場合に、実質的に増加した。放射線照射および細胞の機械的除去の組合せによってレシピエント胚に損傷を与えた場合、cES細胞がより長期間培養状態にあったにもかかわらず、ES細胞の寄与はさらに増加した。結果として生じるキメラヒヨコの一部は、純粋な横斑ロックニワトリヒヨコと区別することができない(図4)。表5中のデータが示すように、キメラ現象の割合ならびに胚1つ当たりのキメラ現象の程度は、レシピエント胚に損傷を与えた後に増加する。
【0040】
表5:異なる方法によって損傷を与えられたレシピエント胚中へのcES細胞注入後の体細胞キメラ現象の頻度
【0041】
【表5】
ステージXより実質的に若いレシピエント胚もまた、ES細胞をドナーとして用いてキメラを作製するために使用され得る。初期のレシピエント胚は、オキシトシンを雌鶏に注射して、早期産卵を誘導することによって採取し、また、受精卵は、ステージVII〜IXに採取する。
【0042】
あるいは、輸卵管の膨大部領域からの胚の採取により、約4〜250個の細胞からなるステージI〜VIの胚を入手する機会が提供され、また、潜在的なレシピエント胚としてのすべての胚ステージからキメラを開発することが可能になる。
【0043】
(実施例3)
ニワトリ胚性幹細胞(cES)に由来する体細胞キメラ
ニワトリES細胞を白色レグホーンレシピエント胚中に注入する。第1期の実験では、28回の実験において合計14種の細胞株を、ステージXのレシピエント胚に注入する(表2を参照されたい)。これらのcES細胞は、4〜106日間培養して増殖させたものであり、一部の株は、凍結保存したものであった。ニワトリES細胞を軽くトリプシン処理して、cES細胞の小さな塊を得、25mM HEPES+10%ウシ胎児血清を添加したDMEM中に再懸濁する。2000〜5000個の細胞を含む3μl〜5μlの細胞懸濁液を、レシピエント胚の胚盤下腔中に注入する。羽毛を発達させた胚をすべて分析し、また、羽毛の色によって判定したところ、24パーセントの胚(83/347)が、キメラである。羽毛キメラは、14種の細胞株中11種から得られる。キメラ現象の程度は、1%〜95%まで様々であり、程度の平均は、25.9%(SD=20.4)であった。
【0044】
表2は、細胞株内および細胞株間で実施した実験間の体細胞キメラ現象の差異を示す。キメラに対するES細胞の寄与の例を、図4および図5に示す。図4では、2匹のヒヨコがキメラであり、2匹が横斑ロックである。これらのヒヨコの間に表現型の差異はなく、キメラに対するES細胞の寄与が、特に外胚葉由来の系統において多大であることが示唆されることは明らかである。図5では、左側のキメラは、ES細胞からの比較的低いレベルの寄与を有するのに対し、右のキメラは、中程度の寄与を有する。
【0045】
(実施例4)
リポフェクションおよびエレクトロポレーションによるcES細胞のトランスフェクション
表6に言及すると、トランスフェクトされるウェルのサイズと適合した適切な量のDNAを、血清も抗生物質も含まない培地中に希釈する。適切な体積のSuperfect(Stratagene社)を添加し、DNAと混合し、そして5〜10分間、反応を生じさせる。この培地を除去し、そしてトランスフェクトしようとするウェルを、CaおよびMgを含まない塩溶液で洗浄する。血清および抗生物質を含み得る適切な体積の培地を、DNA/Superfect混合物に添加する。これらのプレートを37℃で2〜3時間インキュベートする。インキュベーションが完了したら、細胞を1〜2回洗浄することによってSuperfectを除去し、そして新鮮な培養培地を添加する。
【0046】
表6:Superfectを用いたニワトリES細胞のトランスフェクションのための条件
【0047】
【表6】
ペトリ皿用パルサーを用いて、プレートの直径35mmのウェル中に付着しているcES細胞をエレクトロポレーションする。この培地を除去し、そしてCa++およびMg++を含まない塩溶液でウェルを洗浄する。1mlのエレクトロポレーション溶液をこのウェルに添加する。DNAを添加し、培地を穏やかに混合する。このペトリ皿用パルサーをウェルの底部まで下げ、電流を送達する。(電圧は好ましくは100〜500V/cmであり、また、パルス長は12〜16m秒でよい)。ペトリ皿用パルサーを取り出し、エレクトロポレーションしたウェルを室温で10分間静置する。10分後、2mlの培地を添加し、インキュベーターに皿を戻す。
【0048】
懸濁状態の細胞をトランスフェクトするために、培地を除去し、そしてCaおよびMgを含まない塩溶液で細胞を洗浄する。EDTAと共にトリプシンを添加して、単細胞懸濁液を得る。細胞を洗浄し、遠心分離し、そしてPBSなどの補正用エレクトロポレーション緩衝液中に再懸濁する。このES細胞懸濁液を無菌のキュベット中に入れ、DNA(最低濃度1mg/ml)をこの細胞懸濁液に添加し、そして上下にピペッティングすることによって混合する。これらの細胞をエレクトロポレーションし、室温で10分間放置した。細胞をキュベットから取り出し、予め準備しておいたウェルまたはディッシュに分配する。細胞をインキュベーター中に入れ、また、エレクトロポレーション後24〜48時間目の一過性トランスフェクションについて評価する。抗生物質耐性細胞の選択もまた、ピューロマイシンなどの抗生物質を培養培地中に含めることによって開始することができる。
【0049】
好ましい実施形態では、トランスフェクトされた細胞を選択するために必要とされるピューロマイシンの濃度は、死滅滴定曲線として算出される。ニワトリ胚性幹細胞に対する死滅滴定曲線は、培養中の細胞を、0.0〜1.0μg/mlの範囲のピューロマイシン濃度に10日間(表7)、および0.0〜200μg/mlの範囲のネオマイシン濃度(表8)に曝露することによって作成する。2日毎に培地を交換し、そして新鮮なピューロマイシンまたはネオマイシンを添加する。濃度0.3μg/mlのピューロマイシンに曝露した場合、ES細胞は、6日間の期間に渡って新鮮なピューロマイシンを含む培地を3回交換した後のすべてのウェルから消失していた(表7を参照されたい)。トランスフェクトされた培養物を選択するために、0.3〜1.0μg/mlのピューロマイシン濃度を使用する。40μg/mlを超えるネオマイシン濃度では、7日以内にすべてのcES細胞が消失した(表8)。
【0050】
10日間の選択後、cES細胞コロニーは目に見え、さらなる増殖のために採取することができる。
【0051】
表7:様々な濃度のピューロマイシンに様々な長さの期間(ピューロマイシン添加後の日数)曝露した後のcES細胞の形態
【0052】
【表7】
表8:様々な濃度のネオマイシンに様々な長さの期間(ネオマイシン添加後の日数)曝露した後のcES細胞の形態
【0053】
【表8】
(実施例5)
組織特異的な抗体発現
導入遺伝子の設計に応じて、外因性抗体をコードするDNAは、選択された組織おいてのみ有意なレベルの抗体を発現するように設計される。この導入遺伝子構築物は、宿主生物のゲノムに由来する遺伝子エレメントを含み得、また、選択された組織における抗体の公知の発現または発現パターンに基づいて選択され得る。特定の組織における発現のためには、選択される組織において通常発現され、また通常高度に発現される、タンパク質をコードする遺伝子を選択し、そしてこの外因性抗体の発現を駆動するために、この遺伝子に由来する調節エレメントを選択する。この外因性抗体のDNAコード配列と組み合わせる場合、オボアルブミン、オボトランスフェリン、オボムコイド、オボムチン、リゾチーム、オボグロブリン、オボインヒビター、シスタチン、オボグリコプロテイン、オボフラボプロテイン、オボマクログロブリン、またはアビジンのプロモーターなど他の調節エレメントを、選択マーカーをコードする遺伝子と組み合わせて、および/またはIRESと組み合わせて、使用してよい。この導入遺伝子は、選択された組織において、好ましくは卵白において優先的な抗体発現をもたらす。特定の組織型における優先的発現は、選択されていない組織と比較して規模が3〜4桁大きな、選択された組織における発現と定義することができる。
【0054】
トランスジェニックトリにおける組織特異的抗体発現の場合、組織特異的発現は、好ましくは、膨大部、峡部、卵殻腺、または卵管漏斗を含めて、輸卵管のある領域を対象としている。膨大部は、卵白の主なタンパク質を発現する管状腺細胞を含み、一方、峡部は、卵殻膜を発現する細胞を含む。可溶性タンパク質の発現は、好ましくは、輸卵管の膨大部の管状腺細胞に指向され、これは、好ましくはオボアルブミン、ただし、オボトランスフェリン、オボムコイド、リゾチーム、オボグロブリンG2、オボグロブリンG3、オボインヒビター、シスタチン、オボグリコプロテイン、オボフラボプロテイン、オボマクログロブリン、およびアビジンを含む、卵白タンパク質を発現する遺伝子に由来する、プロモーターを通常含む調節配列を選択することによる。上皮細胞における発現を除く、管状腺細胞における選択的発現を以下に実証し、また、2種の細胞型間の選択性を区別するために、優先的発現と命名する。
【0055】
以下の実施例では、外因性免疫グロブリン遺伝子座と共に、プロモーターを含む内因性卵白調節配列を含む導入遺伝子を構築して、輸卵管の管状腺細胞における組織特異的抗体発現を生じさせる。その場合、このようにして発現された抗体分子は、トランスジェニックニワトリの卵白中に蓄積される。この実施形態では、再配列された任意の免疫グロブリン遺伝子によってコードされる抗体は、輸卵管の膨大部領域の管状腺細胞を含む組織において特異的に発現され、そして卵白から単離することができる。モノクローナル抗体をコードする再配列された免疫グロブリン遺伝子は、他の組織における発現が検出可能なレベルより上で存在し得るものの、他の組織における発現は実質的になく、輸卵管において優先的に発現される。
【0056】
この実施形態では、オボアルブミン調節配列の制御下のモノクローナル抗体カセットは、少なくとも3.4Kb、好ましくは少なくとも約7.5Kbの5’調節配列から構成され、かつ15Kb以上の3’調節配列も含んでよい。好ましくは、この構築物は、外因性抗体コード領域の5’末端および3’末端の双方に隣接するオボアルブミン遺伝子の領域を含むが、5’隣接領域の内因性プロモーター配列のセグメントが十分に大きな場合は、3’隣接領域の必要性を回避し得る。抗体の重鎖および軽鎖の双方のコード領域は、導入遺伝子中で提供され、かつ選択されたアイソタイプの可変領域、多様性領域、連結領域、および定常領域を含む。好ましい実施形態では、この抗体は、特徴的にはヒトであり、かつ少なくともヒト重鎖を含む、免疫グロブリン遺伝子によってコードされる。また、このアイソタイプは好ましくはIgGであり、最も好ましくはIgG1である。
【0057】
オボアルブミンに由来するモノクローナル抗体構築物のために好ましい導入遺伝子構築物を、図6Aに提供する。この導入遺伝子構築物をOv7.5と呼び、これは、MAbコードに5’末端配列で隣接しているプロモーター(この特定の実施形態ではオボアルブミンプロモーター)を含む約7.5Kbの卵白調節配列、およびコード領域の3’側の15Kbのプロモーター配列を有する。コード領域は、軽鎖および重鎖の双方の可変領域、J−Cイントロン配列、κ軽鎖定常領域、IRES配列、ならびにγ1アイソタイプ重鎖定常領域を含む。この構築物の3’末端は、GFP遺伝子および選択マーカー、すなわち、この場合、本明細書において説明されるCXプロモーターによって駆動されるピューロマイシン耐性遺伝子を含む。モノクローナル抗体コード領域の3’側および5’側の双方のオボアルブミンプロモーター配列の長さは例にすぎず、類似した構築物は、25〜100Kb以上の5’配列、ならびに様々な長さの3’配列を含み得る。当業者には、GFPマーカーが、生理学的標本における検出のためにだけ存在し、導入遺伝子の有用性から逸脱することなく除去できることが理解されよう。ピューロマイシン耐性マーカーは、この導入遺伝子で成功裡に形質転換された胚性幹細胞を選択する能力を与える任意のマーカーで置換することができる。いくつかのタイプの類似した選択マーカーが当技術分野において周知であり、また、本実施形態のピューロマイシン耐性遺伝子と本質的に交換可能に使用され得る。
【0058】
上記のように、このモノクローナル抗体は、本発明の導入遺伝子構築物を用いて発現させることができるいくつかのタイプのモノクローナル抗体生成物のうちの一例にすぎない。さらに、タンパク質の1つのクラスとしてのモノクローナル抗体は、本明細書において説明される方法および技術に従って、組織特異的な様式で発現され得る多くのクラスのタンパク質生成物の一例にすぎない。
【0059】
図6Bを参照すると、エストロゲン注射によってOv7.5導入遺伝子の発現が誘導された2週齢のキメラの膨大部の切片は、形質転換された胚性幹細胞に由来する抗ダンシルモノクローナル抗体産生細胞の組織特異的発現によりGFPが発現していることを示し(図6Bの左上のパネルにおいて緑色として示される)、これにより、Ov7.5導入遺伝子で形質転換された胚性幹細胞による寄与が裏付けられる。図6Bの左下のパネルを参照すると、モノクローナル抗体は、管状腺細胞において赤色に染色されるのに対し、同じくドナー胚性幹細胞に由来する上皮細胞は緑色に染色され、赤色には染色されない。染色におけるこの差異により、構築物の発現が組織特異的であり、かつ特定の組織型に対する導入遺伝子の内容によって選択されることが実証される。以下の実施例は輸卵管の管状腺細胞における組織特異的発現を実証するが、あらゆる細胞型および組織型にわたる発現の実証により、導入遺伝子構築物の構成要素、および例えば、プロモーターまたは他の調節エレメントの対応した選択によって組織特異的発現または細胞特異的発現のために各組織型または任意の組織型を選択し得ることが実証される。
【0060】
図6Bの右上のパネルでは、細胞型のすべてがDAPI染色によって示される。右下のパネルでは、染色を重ねて、ドナー由来の管状腺細胞のみがモノクローナル抗体を発現し、一方、レシピエント由来の細胞およびドナー由来の上皮細胞がモノクローナル抗体を発現しないことを実証する。図7Cおよび図7Dは、抗ダンシルモノクローナル抗体の重鎖および軽鎖が、それぞれ、3または5キメラの輸卵管組織においてのみ選択的に発現され、また、これらのキメラの脳、腸、膵臓、または筋肉においてはRT−PCRによって検出されるレベルを上回っては発現されないことを示すRT−PCR解析である。
【0061】
ヒトIgGアイソタイプモノクローナル抗体が選択的に発現され得、そして蓄積されるタンパク質が卵白であることを実証するために、Ov7.5構築物を有するcES細胞を注入することによって合計18匹のキメラ雌を作製した。この群から6匹のキメラを用いて、初期のエストロゲン誘導による導入遺伝子発現を試験した。残りのキメラ雌12匹は、卵を回収するために性的に成熟するまで飼育した。9匹のキメラ雌が、17〜22週齢で産卵を開始し、また、これらのキメラのうちの1匹は、散発的に産卵した。キメラ雌のうちの3匹は、35週齢の時点で卵を産卵しておらず、剖検時に、それらの性腺が、雄性ES細胞に由来する組織の存在によって雄性化していることが明らかになった。
【0062】
卵を産卵する9匹のキメラに由来する卵を回収し、硫酸アンモニウム沈殿法によって代表的な卵白サンプルを調製し、そしてELISAによって解析した。ヤギ抗ヒトIgG抗体でマイクロタイタープレートをコーティングし、重鎖に対する標識されたヤギ抗ヒトIgG(γ鎖特異的)抗体および/または軽鎖に対する標識されたヤギ抗ヒトκ(κ鎖特異的)抗体のいずれかによって卵白サンプル中のヒトIgG MAbの存在を明らかにした。精製ヒトIgγ1、κタンパク質を用いて標準曲線を作成した。ELISAの感度は、0.8ng/mlであった。非トランスジェニック白色レグホーン雌鶏由来の卵白サンプルを、ネガティブコントロールとして使用した。ヒトIgG MAb蓄積は、非トランスジェニック白色レグホーン雌鶏に由来する卵(卵4個)においても、6匹のキメラ雌鶏に由来する卵(雌鶏OV11−17番由来の卵8個、雌鶏OV11−53番由来の卵8個、雌鶏OV11−73番由来の卵6個、雌鶏OV11−88番由来の卵6個、雌鶏OV12−97番由来の卵4個、および雌鶏OV13−13番由来の卵5個)においても、検出不可能であった。
【0063】
ヒトIgG MAb蓄積が、3種の異なるキメラ雌鶏に由来する卵において検出された(ELISAによってIgHを決定した場合、雌鶏OV11−13由来の卵の場合に約1.4〜6.3ng/ml、雌鶏OV11−37番由来の卵の場合に約2.0〜2.9ng/ml、および雌鶏OV11−43番由来の卵の場合に約2.9〜10.8ng/ml)。代表的な卵中のヒトIgG MAbの濃度を、表9にまとめて示す。IgLに関してELISAによって決定された卵中のヒトIgG MAbの濃度は、IgHに関してELISAによって決定された濃度より、一貫して低かった。一般に、IgLによって決定された濃度は、IgHによって決定された濃度の60%であった(表9の3列目および5列目を比較されたい)。この差異は、精製したヒトIgγ1、κタンパク質を用いて作製されたスパイクサンプルにおいても存在した。
【0064】
表9 キメラに由来する卵中のヒトIgG MAbの蓄積
【0065】
【表9】
雌鶏OV11−13番、OV11−37番(これらは、卵中にヒトIgを蓄積した)および白色レグホーンに由来する血液サンプル中のヒトIgG MAbタンパク質の濃度は、アッセイ感度(0.8ng/ml)より低かった。これらのデータは、エストロゲンによって誘導したキメラヒヨコの腸、脳、膵臓、および筋肉におけるヒトIg転写物の存在を評価するためにRT−PCRを用いて観察された、キメラニワトリにおけるヒトIgの異所発現の不在と一致している(図7Cおよび7D)。したがって、Ov7.5構築物は、組織特異的であり、ホルモンによって誘導され、そして発生に関連して調節される遺伝子発現を、トランスジェニックキメラ雌鶏に送達すると思われる。さらに、タンパク質は、膨大部中の管状腺細胞から搬出され、卵白中に蓄積されるようである。
【0066】
オボアルブミン由来の組織特異的タンパク質発現導入遺伝子を、以下のようにして構築した。
【0067】
ニワトリゲノムBACライブラリー(Crooijmans,R.P.ら、Mamm.Genome 11巻:360〜363頁、2000年)(Texas A&M BAC Center)をスクリーニングして、オボアルブミン遺伝子座中の46Kbの領域を単離する。MAbコード領域の5’側に位置するオボアルブミンプロモーターの異なる断片を有する、以下の2種の異なるベクターを構築した:(1)Ov7.5MAb−dns:42Kbの発現ベクターであり、オボアルブミン遺伝子に由来する9.2Kbの5’配列(7.5Kbプロモーターを含む)および15.5Kbの3’隣接配列を含む(図10A)。この42Kbの発現ベクターは、オボアルブミン遺伝子に由来する9.2Kbの5’配列(7.5Kbのプロモーターを含む)および15.5Kbの3’隣接配列を含む。バイシストロン性のモノクローナル抗体カセットは、抗ダンシル抗体の軽鎖、IRES、および重鎖をコードする。(2)Ov15MAb−dns:49Kbの発現ベクターであり、オボアルブミン遺伝子に由来する16.8Kbの5’配列(15Kbプロモーターを含む)および15.5Kbの3’隣接配列を含む(図示せず)。この49Kbの発現ベクターは、オボアルブミン遺伝子に由来する16.8Kbの5’配列(15Kbのプロモーターを含む)および15.5Kbの3’隣接配列を含む。このモノクローナル抗体カセットは、双方の構築物において同一である。
【0068】
上記のように、双方のベクターにおいて発現され得る遺伝子の一例は、マウス−ヒトハイブリッドの抗ダンシルモノクローナル抗体(MAbdns)である。CxEGFP/CxPuroカセットを3’側の最末端にクローニングして、cES細胞における安定なトランスフェクションをピューロマイシンを用いて選択すること、およびキメラ中のトランスフェクトされた細胞を容易に同定することを可能にする。どちらの構築物も、cES細胞中にトランスフェクションする前に直線状にし、また精製する。cES細胞のトランスフェクションは、SuperFect(Stratagene社)またはペトリ皿用パルサーによるエレクトロポレーションのいずれかを使用して、Ov7.5MAbdnsおよびOvl5MAbを用いて実施する。ピューロマイシンによる選択後、耐性クローン6つを、分子解析のために採取する。MAbdnsカセット中、GFP遺伝子中、およびPuro遺伝子中に配置されたプライマーを用いるPCRによって導入遺伝子の存在を確認する。
【0069】
発現され得る抗体の第2の例は、完全なヒト抗ヒトPSMAモノクローナル抗体(MAbF1)である。MAbF1に対応するV遺伝子をPCRによって獲得し、また、Ov15MAb−dns中のV遺伝子と入れ替えるために使用した。
【0070】
この導入遺伝子は、雌性ES細胞中に安定に組み入れられ、また、これらの安定にトランスフェクトされた細胞を、ステージXのレシピエント胚中に注入した。結果として生じるキメラを性的に成熟するまで飼育し、そして卵中のヒトモノクローナル抗体の濃度をELISAによって評価した。30匹の雌鶏のうち4匹は、1mg/卵を上回る濃度で卵中に抗体を蓄積した(表9A)。
【0071】
表9A Ov15MAbF1キメラに由来する卵中のヒトIgG MAbの濃度
【0072】
【表9A】
(実施例6)
抗体生成物の特性決定
ニワトリにおいて産生されたMAbF1を、従来のCHO細胞培養において産生されたMAbF1といくつかのアッセイにおいて比較した。
【0073】
1.抗体のSEC−HPLC解析
4.6×300mmのBioSep SEC S3000カラム(Phenomenex社)を用いて、Waters 2795 HPLCによって約10μγのIgGサンプルを解析した。0.1Mリン酸ナトリウム、0.15M NaCl、および0.1M硫酸ナトリウム、pH7.2において、流速0.4ml/分で20分間、クロマトグラフィーを実施した。A280で分離をモニターした。分子量標準物質(Bio−Rad社)を用いて、概算の分子量を決定した。SEC−HPLC解析により、双方とも90%を超えるIgGモノマーであることが示された(表10)。
【0074】
表10 MAbF1のSEC−HPLC解析
【0075】
【表10】
2.タンパク質のNano LC ESI MS/MS解析
双方のMAb調製物を還元およびアルキル化し、一晩透析し、そして25mM重炭酸アンモニウム(PH8)中、37℃で4時間、トリプシンで消化した。双方のトリプシン消化物のタンデム型質量分析を、陽イオンモードで、そしてタンデムスペクトル(tandem spectra)を得るために衝突誘起解離(CID)を用いるQSTAR pulsar質量分析計(MDS Sciex社、コンコード、オンタリオ州、カナダ)工程に連結されたNano HPLCシステム(Dionex社、サニーベール、カリフォルニア州)によって実施した。各サンプルをμl 1mlずつ注入し、75mm直径μm Pepmap C18カラムを用いて流速250nl/分で分離した。移動相は溶媒A(95%水、5%アセトニトリル、0.5%ギ酸)および溶媒B(20%水、80%アセトニトリル、0.5%ギ酸)であった。機器は、2秒間フルスペクトルを記録し、次いで、最も強度の強いイオンを選択して次の5秒間CIDスペクトルを記録する7秒間の運転サイクルを用いる情報依存的取得(information dependent acquisition)モードで操作した。スペクトルは、Mascot(Matrix Science社、ロンドン、イギリス)を用いて解析した。説明した方法による双方のMAb調製物の解析では、配列の差異は示されなかった。
【0076】
表11.ニワトリに産生されたMAbF1およびCHOに産生されたMAbF1の質量分析による配列解析
【0077】
【表11−1】
【0078】
【表11−2】
3.LC−MS解析
4MグアニジンHCl中IgGサンプル(50μγ)を、サンプルを60℃で90分間インキュベートすることによって25mM DTT中で還元した。次いで、暗所、室温で15分間、45mMヨード酢酸中でサンプルをアルキル化し、そして22mM DTTで反応を停止させた。LC−MSに先立って、サンプルを25mM重炭酸アンモニウム1Lに対して透析し、そして各サンプル50μlを、Micromass ZQ質量分析計を装備されたWaters2795 HPLCを使用するPoros R1/10 2.1×100mmカラム(Applied Biosystems社)に注入し、陽イオンモードで解析した。移動相は、(A)0.1%ギ酸および0.01%トリフルオロ酢酸(TFA)ならびに(B)0.1%ギ酸および0.01%TFAを含む100%CH3CNであった。溶出(0.25ml/分)は、100分に渡って作られる(A)中(B)の10〜60%の直線勾配によって実施した。
【0079】
L鎖の解析により、双方のMAb調製物が同一の質量(±3Da)を有することが示された(表12)。
【0080】
表12 MAb軽鎖のLC−MS解析
【0081】
【表12】
4.示差走査熱量測定による熱安定性
ニワトリ発現系およびCHO発現系において作製されたMAbF1の熱安定性を示差走査熱量測定(DSC)を用いて測定し、そしてFcドメインにおいて脱グリコシル化されたそれらの対応する型と比較した。融解温度(Tm)の熱量測定は、オートサンプラーと組み合わされたVP−Capillary DSC示差走査マイクロカロリメータープラットホーム(MicroCal LLC、ノーサンプトン、マサチューセッツ州、米国)を用いて実施した。サンプルセルの容積は0.144mLである。これらの抗体のグリコシル化型および脱グリコシル化型に関する変性データは、濃度2.3μMのこれらのサンプルを1℃/分の速度で30℃から95℃まで加熱することによって得た。これらのタンパク質サンプルは、pH7.4のリン酸緩衝化生理食塩水(PBS)中に存在した。比較してモル熱容量を得るために、同じ緩衝液を参照セルにおいて使用した。観測された温度記録図をベースライン補正し、また、標準化したデータをOrigin v7.0を用いて解析した。
【0082】
CHOおよびニワトリに由来する抗体の変性プロファイルは極めて異なるのに対し、Fcを脱グリコシル化した抗体の変性プロファイルはほぼ同一である(図8)。これらのデータから、ニワトリの管状腺細胞において産生されたMAbF1のグリコシル化の方が、CHO細胞において産生されたMAbF1に結合される糖残基より高い熱安定性を与えることが示唆される。
【0083】
5.CE−LIFによるMAbのオリゴ糖の特性決定
サンプルを12.5mUのPNGaseF(Prozyme社)と共に40℃で一晩インキュベーションすることによってIgGサンプル(100μγ)からオリゴ糖を遊離させた。使用した条件下で、CHO細胞およびニワトリにおいて発現されたHuMAbF1のFc部分に由来するN結合型グリカンが遊離された。MAbタンパク質を除去するためにエタノール沈殿を実施した後、これらのグリカンを含む上清を減圧遠心分離によって乾燥させ、そして15%酢酸中19mM APTS(Beckman社)およびTHF(Sigma社)中1Mシアノ水素化ホウ素ナトリウム中に再懸濁した。グリカン標識反応を40℃で一晩継続させ、続いて、サンプルを水で25倍に希釈した。APTSで標識したグリカンを、有効長50cm、内径50μmのN−CHOコーティングキャピラリー(Beckman社)を使用する、逆極性を有するP/ACE MDQ CEシステム(Beckman社)による、レーザー誘起蛍光法を用いたキャピラリー電気泳動に適用した。サンプルは圧力(8秒)注入し、また、分離は、Carbohydrate Separation Gel Buffer(Beckman社)を用いて、25kV、20℃で20分間実施した。これらの分離物を、3mWアルゴンイオンレーザーならびに488nmの励起波長および520nmの発光波長を用いるレーザー誘起蛍光検出システム(Beckman社)を用いて観察した。図9に示すように、ニワトリにおいて産生されたMAbF1のオリゴ糖プロファイルは、CHO細胞において産生されたMAbF1のプロファイルと大きく異なることが判明した。
【0084】
6.MALDI Q−TOF MSによるMAbのオリゴ糖特性決定
精製した抗体(100μg、各1.0mg/ml)を4μlのPNGaseF、すなわちN結合型糖質を切断するエンドグリコシダーゼ(Prozyme社)で処理した。これらのサンプルを37℃で一晩インキュベートし、次いで、50mM重炭酸アンモニウム(pH8)に対して一晩透析した。タンパク質は、エタノール沈殿によって除去した。遊離された糖質を、本質的には供給業者のプロトコールに従って、ただし溶出体積を5μlに減らして、Glygen Corp.(コロンビア、メリーランド州)製のマイクロカーボンカラムを用いて脱塩した。各グリカンサンプルの1μl分取物を、マトリックス溶液(α−シアノ−4−ヒドロキシケイ皮酸または2,5−ジヒドロキシ安息香酸(Applied Biosystems社)と1:1(体積比)で混合し、MALDIターゲットプレート上にスポットし、自然乾燥させる。MALDI−Q−TOFタンデム型解析を用いて、完全な複合糖質の解析を実施した。すべての質量スペクトルは、糖質のクラスの質量(組成)を提供する、o−MALDIソース2(MDS Sciex社、コンコード、オンタリオ州、カナダ)を装備したQSTAR pulsar i質量分析計を用いて記録した。双方のMAbのグリカンをプロファイリングした後、各グリカン中で起こり得るグリコシド結合を調査した。高CIDによる質量スペクトルを、TOF/TOFオプティクスを装備した4700Proteomics分析計(Applied Biosystems社、フォスターシティ、カリフォルニア州)を用いて記録した。高CIDのMS/MS実験の場合、衝突エネルギーは1kVに設定した。衝突セルの内部で、選択したオリゴ糖イオンを2×10−6トルの圧力でアルゴンと衝突させた。MALDI TOF質量分析法によって解析した、ニワトリに由来するMAbF1中の11種の主要なグリカンの糖質組成および起こり得るグリコシド結合を表13に要約する。これらのオリゴ糖構造体は、高マンノース型、複合型、およびハイブリッド型のNグリカンを含むことが判明した。
【0085】
表13 質量分析法による、ニワトリにおいて産生されたMAbF1の糖質解析
【0086】
【表13】
7.HPAE−PADを用いたHPLCによる単糖解析
表14 ニワトリに産生されたMAbF1およびCHOに産生されたMAbF1の単糖解析
【0087】
【表14】
IgGサンプル(200μg)を、100℃で4時間、2N TFA(中性糖を推定する場合)または6N HCl(アミノ糖を推定する場合)のいずれかを用いる酸加水分解に供した。サンプルを周囲温度で減圧遠心分離によって乾燥させ、そしてHPAE−PAD(Dionex社)による解析の前に200μlの水中に溶解した。単糖は、プレカラムのAmino TrapおよびBorate Trap(Dionex社)付きのCarboPac PA10 4×250mmカラムを用いて分離した。手順は、Dionex社の技術注記53に従った。単糖のピーク一致および相対存在量は、単糖標準物質(Dionex社)を用いて決定した。表14に要約したようなニワトリおよびCHOにおいて産生されたMAbF1の単糖解析により、糖質組成の差異が明らかになり、また、ニワトリにおいて産生されたMAbF1中にN−アセチルグルコサミン残基、マンノース残基、および極めて含有量の少ないガラクトース残基が存在することが示された。
【0088】
8.化学的諸特性の要約
要約すれば、N結合型オリゴ糖プロファイルにおける最も目を引く差異は、ニワトリにおいて産生された抗体中に高マンノース型のN−グリカンが存在し、フルコースは存在せず、またガラクトース残基の含有量が極めて少ないことであった。これらの特性はいくつかの理由から重要である。第1に、抗原性であることが公知であるα1−3 Gal結合の証拠がない。典型的には約2%未満のレベルまでガラクトース濃度が低下すると、ガラクトースを含む結合に起因する抗原性が実質的に減少する。第2に、同じく抗原性であることが公知であるN−グリコリルノイラミン酸残基の証拠がない。第3に、ニワトリ管状腺細胞において産生された抗体には、抗体のADCC活性を高めるフコシル残基が実質的にない。この文脈において、実質的にないとは、0.1%未満と定義される。第4に、ニワトリに産生された抗体は、典型的には40%を上回る高いマンノース含有率を有し、これは、CHO細胞において産生された抗体を標準物質として用いてBalb/cマウスにおいてクリアランスを評価した場合のこの抗体のクリアランス速度を速める。これらの有利な化学的諸特性に加えて、これらの抗体は、ニワトリゲノム中にランダムに組み込まれるか、または組織特異的な様式で発現されない導入遺伝子を用いた場合には観察されない濃度で卵白中に存在する。好ましい濃度は卵1個当たり抗体1mgより高く、卵1個当たり2mgより高く、卵1個当たり3mgより高く、また、卵1個当たり6mgもの高さである。各卵は約25mlの卵白を含むため、好ましい濃度は40μg/mlより高く、80μg/mlより高く、120μg/mlより高く、また、240μg/mlもの高さである。
【0089】
(実施例7)
卵白からの抗体の抽出および精製
最初に、室温で30分間、卵白を低剪断速度で混合し、次いで、以前に説明されている改良法によってオボムチンを沈殿させた。1体積のホモジナイズした卵白懸濁液を3倍体積の逆浸透水に添加し、30分間攪拌した。希釈した懸濁液を、0.5Mリン酸を用いてpH6.0に調整し、次いで12,100gで20分間遠心分離した。この方法によって主としてオボムチンを含有する卵白タンパク質の約3%を回収した。0.5Mリン酸水素二ナトリウムを用いて上清をpH7.4に調整し、そして結晶塩を用いて塩化ナトリウム濃度を150mMに調整した。Protein A−Sepharose FFカラム(Amersham Biosciences社)を用いて、線流速120cm/時でヒトIgGを精製した。吸着されたヒトIgGを5カラム容量のローディング緩衝液(PBS、pH7.4)で洗浄し、次いで、3mMリン酸を用いて溶出させた。230mM NaClを含有する60mMリン酸ナトリウム(pH7.5)を用いて、溶出されたヒトIgG画分をpH7.5に調整して、最終濃度を40mMリン酸ナトリウムおよび150mM NaClとした。次いで、サンプルを0.2mmシリンジフィルター(Pall社)に通してろ過した。図10に示すように、精製した物質の大半は、90%を超える純度を有する(ELISAおよびA280によって決定、データは示さず)、完全に構築されたH2L2であった。
【0090】
(実施例8)
結合親和性に関するアッセイ
LNCaP細胞(ATCC)上のPSMAを、結合を分析するための抗原として使用した。ウェル当たり20万個の細胞を、指定した濃度の50μlずつの抗体と共に、デュプリケートで30分間インキュベートした。細胞を2回洗浄した後、希釈度1:200のヤギ抗ヒトIgG PE標識抗体(Jackson ImmunoResearch社)をウェル当たり50μl、4℃で30分間、添加した。1%BSAを含むPBS中で細胞を2回洗浄し、FACSによって分析した。LNCaP細胞上のPSMAに対するMAb結合のEC50値は、GraphPad Prism 3.0(GraphPad Software社)を利用して、結合曲線から決定した。細胞は、10%FBS、10mM HEPES、2mM L−グルタミン、および1mMピルビン酸ナトリウムを添加したRPMI1640培地中で増殖させた。ニワトリ管状腺細胞において産生されたMAbF1の抗原結合特性を、CHO細胞において産生されたMAbF1の抗原結合特性と比較した。どちらの抗体調製物も、LNCaP細胞上で発現されたPSMAに対してほぼ同一の結合曲線を描き、EC50値は類似していた(図11)。このデータにより、ニワトリに由来する抗体およびCHOに由来する抗体はグリコシル化の様式は異なるが、同等に抗原を認識し、結合することが実証される。
【0091】
(実施例9)
抗体内部移行のアッセイ
PSMAへのMAbF1の結合は、抗体の内部移行を招く。潜在的な1つの応用において、PSMAを発現する腫瘍細胞を標的とし、そして破壊するために、MAbを細胞毒と結合させることができる。LNCaP細胞上のPSMAに対する抗体結合の内部移行は、MAbおよびHum−Zap(Advanced Targeting Systems社)と共に細胞をインキュベートすることによって判定した。HumZapは、リボソーム不活性化タンパク質であるサポリンに結合されたヤギ抗ヒトIgG抗体である。MAbF1/Hum−Zap複合体が細胞表面上のPSMAに結合し、そして内部移行された場合に細胞は死滅するが、抗体またはHum−Zapは単独ではLNCaP細胞に対して毒性ではない。300ngのHum−Zap、および300ngのF1MAbまたは対照MAbを含む培養培地150μl中で、37℃で48時間、トリプリケートでLNCaP細胞(10,000/ウェル)をインキュベートした。細胞の増殖および生存は、CellTiter−Glo Luminescent Cell Viability Assay(Promega社)を用いて決定した。ウェル当たり10,000個のLNCaP付着細胞と共に、細胞培養培地中の抗体希釈物を4℃で2時間インキュベートすることによって内部移行アッセイも実施した。抗体溶液を穏やかに除去し、HumZap 200ngを含有する培地150μlに交換した。37℃で48時間インキュベーションした後、細胞生存率を決定した。抗体内部移行に対するEC50値は、Prism3.0(GraphPad Software社)を用いてグラフから決定した。図12に示すように、双方の抗体調製物が同様の効率で内部移行された。一連の抗体濃度に渡って試験した場合、ニワトリに由来するMAbF1およびCHOに由来するMAbF1の双方の内部移行に対するEC50値は0.49nMであった。
【0092】
(実施例10)
BALB/cマウスにおけるMAbのクリアランス
放射標識した抗体の静脈注射により、BALB/cマウスにおいて、ニワトリに産生されたMAbF1のin vivo半減期を、CHOに産生された抗体と並行して解析した。MAbタンパク質10μgを、Iodobead法(Pierce社)を用いて、125Iで少しヨウ素標識した(抗体当たり1個未満のI)。6週齢の雌BALB/cマウス(Taconic Farms社、ジャーマンタウン、ニューヨーク州)に、実験前の1週間、飲料水に溶かした0.1mg/mlヨウ化カリウムを与えた。タンパク質当たり4匹のマウスの尾静脈に約600,000cpmの標識MAbを静脈注射し、そして全身ガンマカウンター(Wm.B.Johnson社、Ludlum社製計数器付きNaI結晶検出器)を用いて、選択された時間に全身の放射能を測定した。半減期は、指数関数的回帰分析または残留放射能に基づいて算出した。図13に示したように、ニワトリ管状腺細胞によって産生されたMAbF1は、102.4±0.9時間の半減期(t1/2)で浄化されたのに対し、CHO細胞によって産生されたMAbF1は、より緩徐に浄化され、半減期は207.5±18.3時間であった。
【0093】
(実施例11)
ADCCのアッセイ
改良された51CrADCCアッセイにおいてLNCaP−C42B細胞を試験した。標準的なFicoll−Paque分離法によってヘパリン処置した全血からヒト末梢血単核細胞を精製した。これらの細胞を、10%FBSおよび10U/mlヒトIL−2を含有するRPMI1640培地中に(1×10E6細胞/mLで)再懸濁し、そして37℃で一晩インキュベートした。翌日、これらの細胞を回収し、培養培地中で1回洗浄し、そして2×107細胞/mlの濃度で再懸濁した。200万個のLNCaP−C42b標的細胞を、総体積1ml中の200uCi 51Crと共に37℃で1時間インキュベートする。標的細胞を1回洗浄し、培地1ml中に再懸濁し、37℃でさらに30分間インキュベートする。最終のインキュベーションの後、標的細胞を1回洗浄し、そして最終体積を1×105細胞/mLにする。最終のADCCアッセイのために、標識LNCaP細胞100μlをエフェクター細胞50μlおよび抗体50μlと共にインキュベートする。標的とエフェクターの最終比率は1:100を選択した。すべての研究において、対照のヒトIgG1アイソタイプを試験し、CHOに由来する抗PSMA MAbF1抗体と比較した。含まれる他の対照は、a)標的細胞およびエフェクター細胞、ただし抗体はなし、b)エフェクター細胞を伴わない標的細胞、ならびにc)3%TritonX−100の存在下の標的細胞およびエフェクター細胞である。37℃で4時間インキュベーションした後、上清を回収し、240〜400keVの読取領域を有するガンマカウンター(Packard Instruments社製のCobra IIオートガンマ)を用いて計数した。抗体濃度の関数としてカウント毎分をプロットし、そしてこのデータを、Prismソフトウェア(サンディエゴ、カリフォルニア州)を用いた非線形回帰、すなわちS字形用量反応(可変勾配)によって解析した。溶解率は以下の式、すなわち、溶解率(%)=(サンプルCPM−抗体なしのCPM)/(TritonX CPM−抗体なしのCPM)×100によって決定した。本発明者らは、すべての研究においてEC50値および溶解率の双方がモニターされることが重要であることを発見した。例えば、2種の抗体を比較する場合に、EC50値もしくは溶解率または双方が変化している可能性がある。
【0094】
抗CD16抗体によるADCCの阻害を、以下の改変法を用いて実施した。5□g/mlの抗CD16抗体3G8または対照のアイソタイプ抗体の不在下または存在下で、1□g/mlまたは0.01□g/mlの、CHOに産生されたMAbF1抗体またはニワトリに産生されたMAbF1抗体と共に、細胞をインキュベートした。
【0095】
図14は、CHOに由来するMAbが用量依存的な細胞溶解を誘導することを示し、IL−2で刺激されたエフェクター細胞の場合、38%の溶解で頭打ちになり、EC50は0.11μg/mlである。一方、ニワトリの卵に由来するMAbは、より強力かつより効果的であった。ニワトリの卵に由来するMAbの最大溶解率は、抗体の2種の異なる調製物において60%であった。この物質のEC50は0.018μg/mlであったため、CHOに由来するMAbを上回る高い力価も実証された。最後に、予想されるように、対照のアイソタイプ抗体は細胞溶解を誘導しなかった。刺激されていないエフェクター細胞(新鮮なPBMC)のADCCの方が、EC50値の差はより大きいが、全体的な細胞死滅のレベルは劣る(図15)。
【0096】
CD16(FCgRIII)は、ADCCを媒介する重要なレセプターである。ADCC応答の特異性は、CD16を対象とするモノクローナル抗体を用いて標的細胞とエフェクター細胞の相互作用を妨害することによって示した。本研究においては、2種の用量、すなわち飽和用量(1μg/ml)および最適以下の用量(0.01μg/ml)のMAbF1抗体を使用した。1μg/mlのMAbF1抗体は、抗CD16抗体の不在下で、CHOに由来する抗体およびニワトリに由来する抗体の場合、それぞれ約15%および38%の溶解を誘導した。この溶解率は、抗CD16抗体の存在下では約4%に減少したが、対照のアイソタイプ抗体は影響を及ぼさなかった(図16)。
【0097】
(実施例12)
CD16結合
CHOおよびニワトリに由来するMAbF1を、Biacore社によって提供されるアミンカップリングキットを用いて、一級アミンを介してBiacoreセンサーチップ(CM5)のカルボキシメチルデキストランマトリックス表面に固定化した。どちらの抗体も、約10,000RUの密度まで塗った。固定化した抗体表面上にいくつかの濃度のタンパク質を流しかけることによってこれら2種の抗体とCD16−PheおよびCD16−Valとの結合を起こさせた。空白の表面および純粋な緩衝液の結合サイクルを考慮することによって非特異的結合の効果を説明した。HBS−EP緩衝液を希釈用および泳動用緩衝液として使用した。実験は、Biacore−3000機器を用いて25℃で実施した。データはGraphPad Prismソフトウェアによって解析し、また、平衡解離定数を推定するために、単一結合部位モデルにデータを適合させた。
【0098】
高速動態が、抗体へのFcR結合の特徴的な特色であるため、解離定数は、速度定数ではなく平衡結合実験に基づいて推定した。KDの推定のために使用した曲線適合を図17に示す。ニワトリに由来する抗体の解離定数KDは、CHO由来の対応する抗体と比べて、どちらのFcRに対しても約10分の1の小ささである。ニワトリに由来する抗体のより高い親和性は、Fc領域のグリコシル化の差異に起因している可能性があり、特に、CHOに由来する抗体中には存在するフコースがないことが寄与している可能性がある。
【0099】
(実施例13)
治療的有用性
本発明は、具体的に定められたグリコシル化パターンおよび他の化学的諸特性を有し、そして前述の遺伝的に改変されたニワトリを用いて作製された抗体を提供する。これらの特性により、標的組織中の抗原特異的な標的に結合させる目的で患者に投与した場合の治療特性が改善される。具体的には、上記のように、いくつかの臨床適応症に対して、これらの抗体は、増強された抗体依存性細胞傷害活性(ADCC)を示し、また、この効果はいくつかの臨床適応症において重要な利点を与える。
【0100】
いくつかのタイプの癌の治療に対する非結合型モノクローナル抗体(mAb)の臨床試験では、有望な結果が得られた。Dillman、1997年、Cancer Biother. & Radiopharm.12巻:223〜225頁;Deoら、1997年、ImmunologyToday 18巻:127頁。キメラの非結合型IgG1は、軽度または濾胞性のB細胞性非ホジキンリンパ腫(Dillman、1997年、前記)用に認可されており、さらに、別の非結合型mAb、すなわち充実性乳房腫瘍を標的とするヒト化IgG1もまた、臨床試験の第III相において期待できる結果を示した。Deoら、1997年、前記。これら2種のMAbの抗原は、それぞれの標的組織において高発現される。このような応用の場合、特に、抗体がADCCによって強力な腫瘍破壊を媒介する腫瘍細胞において、本発明の抗体は、患者に投与した際の治療的利点を与える。
【0101】
治療的用途のために、本発明の抗体を、(例えば、二重特異性抗体もしくは多重特異性抗体を作製するために)別の抗体、(例えば、免疫毒素などの免疫複合体を作製するために)細胞毒、細胞性リガンド、または抗原など1種または複数種の分子単位に、(例えば化学的結合、遺伝子融合、非共有結合性会合、もしくは別の方法によって)機能的に連結することもできる。本発明の抗体は、他の治療的部分、例えば放射性同位体、低分子抗癌薬、抗炎症剤、細胞毒、または免疫抑制剤に連結することができる。したがって、本発明は、ニワトリ発現系によって可能にされる化学的諸特性を有し、そして治療的用途のために、本質的にすべて公知の抗体結合、連結、および関連した技術と組み合わされた、抗体組成物を包含する。
【0102】
したがって、本発明の抗体は、特にADCC機序が標的組織において提示される場合、治療への感受性が高い標的組織において抗原を発現する細胞を伴う様々な疾患を治療および/または予防するために使用することができる。治療(例えば寛解)または予防することができる例示的な疾患には、それだけには限らないが、充実性腫瘍、リンパ腫、びまん性腫瘍、およびすべてのタイプの癌組織が含まれる。
【0103】
本発明の治療的実施形態では、ADCCの特性を提示する様式による、治療されると思われる状態の診断に具体的に従って、本発明の抗体を患者に投与する。このような臨床現場では、本発明の抗体を投与し、そして治療後に治療の細胞傷害効果を決定して、標的組織におけるADCC効果を決定する。本発明の治療用組成物に加えて、化学療法剤、放射線、またはサイトカインなど、Fc受容体の発現もしくは活性を調節、例えば増大もしくは阻害する作用物質で、患者をさらに治療してよい。治療期間中に投与するための典型的なサイトカインとしては、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターフェロン−γ(IFN−γ)、および腫瘍壊死因子(TNF)が挙げられる。典型的な治療用物質としては、とりわけ、ドキソルビシン、シスプラスチン、ブレオマイシン、カルムスチン、クロラムブシル、およびシクロホスファミドなどの抗悪性腫瘍剤が挙げられる。
【0104】
別の態様では、本発明は、組成物、例えば、ニワトリにおいて発現された本発明の抗体の1つまたは組合せを含む薬剤組成物を提供する。本発明の組成物は、当技術分野において公知の様々な方法によって投与することができる。当業者には理解されるように、投与の経路および/または様式は、所望の結果に応じて異なると考えられる。製薬上許容される担体としては、無菌の水性溶液または水性分散液が挙げられ、薬剤として有効な物質に対するこのような媒体および作用物質の使用は当技術分野において公知である。無菌の注射液剤は、適切な溶媒中の必要とされる量の活性化合物を1つの成分または成分の組合せと混合し、続いて滅菌および/または精密ろ過することによって調製することができる。一般に、分散液は、基本となる分散媒および他の任意の成分を含む無菌ビヒクル中に活性化合物を混合することによって調製される。
【0105】
投与計画は、最適な所望のADCC効果を提供するように調整する。例えば、単回ボーラスを投与してもよく、いくつかに分割した用量を時間をかけて投与してもよく、または治療状況の緊急性によって示されるように、比例的に用量を減少もしくは増加させてもよい。単位剤形の非経口組成物を調製することは、投与の容易さおよび投薬量の均一性に対して特に有利である。本明細書において使用される単位剤形とは、治療しようとする対象者に対する単位投薬量として適した物理的に個別の単位を意味し、各単位は、必要な薬剤用担体と共同して所望の治療効果を生じるように算出された所定の量の活性化合物を含む。本発明の単位剤形の仕様は、(a)活性化合物の独特な特徴および達成すべき個々の治療効果、ならびに(b)個体の感受性の治療のためにこのような活性化合物を配合する技術分野につきものの制約によって必然的に決められ、そして、直接的に依存している。
【0106】
本発明の薬剤組成物中の有効成分の実際の投薬量レベルは、個々の患者、組成物、および投与様式に対する所望の治療応答を実現するのに効果的であり、患者に対する毒性はない有効成分の量を得るために、変更してよい。選択される投薬量レベルは、使用される本発明の個々の組成物の活性、投与経路、投与時間、使用される個々の化合物の排出速度、治療の持続期間、使用される個々の組成物と組み合わせて使用される他の薬物、化合物、および/または材料、治療される患者の年齢、性別、体重、状態、全体的健康状態、および以前の病歴、ならびに医薬分野において周知である同様の因子を含めて、様々な薬物動態学的因子に依存すると考えられる。
【0107】
本発明の抗体の投与効果は、腫瘍などの標的組織において客観的に観察可能であるため、本発明の治療的方法は、具体的にはADCCを使用する治療法を含む治療法を必要とする患者を診断するステップと、効果が望まれる標的組織を同定するステップと、本発明の組成物を、それを必要とする患者に投与するステップと、患者の標的組織におけるADCCの有効性を判定することなどによって患者における治療効果を測定するステップとを含む。治療効果の判定は、細胞死、標的組織の収縮、腫瘍サイズの縮小など標的組織の特性の経時的な変化を分析することによっておよび医薬分野において公知である他の任意の診断技術によって実現することができる。
【0108】
本発明の抗体はまた、当業者が利用可能ないくつかの公知のADCCモデルのうちのいずれかにおいてADCC活性を試験することもできる。治療的用途または診断的用途に対する本発明の抗体の有用性を判定するために、ADCCの測定を独立に実施してもよく、または他の哺乳動物、非哺乳動物、植物、もしくは細菌細胞の発現系と比較してもよい。したがって、本発明の方法は、前述の系において産生された別の抗体と直接または間接的に比較して本発明の抗体を使用することによってADCCをもたらすための有用性の差異を判定するステップを含む。具体的には、この方法論は、前述のニワトリ発現系において産生された抗体のADCC効果を比較して、増強されたADCCを同定し、ニワトリ発現系のための理想的な抗体候補を同定するステップを含む。
【0109】
上記のように、治療的有用性を向上させるために、本発明の抗体は、1種または複数種の他の治療物質、例えば、細胞障害性物質、放射毒性物質、または免疫抑制剤と同時投与することができる。抗体は、(免疫複合体として)作用物質に連結されてもよく、または作用物質と別々に投与されてもよい。後者の場合(個別投与)、抗体は、作用物質の前、後、もしくは同時に投与してよく、または他の公知の治療法、例えば抗癌療法、例えば放射線療法と同時に投与してもよい。このような治療用物質としては、とりわけ、ドキソルビシン、シスプラスチン、ブレオマイシン、カルムスチン、クロラムブシル、およびシクロホスファミドなどの抗悪性腫瘍剤が挙げられる。本発明の抗体と化学療法剤との同時投与は、ヒト腫瘍細胞に対する細胞障害効果をもたらす、異なる機序を介して機能する2種の抗癌物質を提供する。このような同時投与により、薬物に対する耐性の発達、または抗体との反応性を低下させると考えられる腫瘍細胞の抗原性の変化に起因する問題が解決され得る。
【0110】
当業者には明らかであろう、開示した本発明の様々な修正、改良、および応用があると考えられ、かつ本出願は、法律によって許容される範囲まで、そのような実施形態を包含する。いくつかの好ましい実施形態において本発明を説明したが、本発明の完全な範囲はそのようには限定されず、以下の特許請求の範囲に従う。すべての参考文献、特許、または他の刊行物は、参照により本明細書に具体的に組み入れられる。
【図面の簡単な説明】
【0111】
【図1】図1は、ニワトリES細胞の特徴的な形態を示す。細胞は単層で増殖し、細胞質はほとんどなく、核小体が目立っている。
【図2】図2は、ニワトリES細胞のin vitroでの特性、具体的には、抗体SSEA−1およびEMA−1との反応、ならびにアルカリ性ホスファターゼの発現を示す。
【図3】図3は、189日間培養されたニワトリES細胞の核型である。これらの細胞は2倍体であり、38対の常染色体および1対のZ染色体を有する。
【図4】図4は、横斑ロックヒヨコ2匹、および横斑ロックES細胞を白色レグホーンのレシピエント胚中に注入することによって形成されたキメラ2匹である。これらのキメラおよび横斑ロックは区別不能であることから、メラノサイト系列へのES細胞の寄与が多大であることが示される。
【図5】図5は、横斑ロックES細胞を白色レグホーンレシピエント中に注入することによって作製されたキメラである。左のパネル中のキメラのペアは、メラノサイト系列に対して少しの寄与を示しているのに対し、左のパネル中のペアは、より大きな寄与を示している。
【図6A】図6Aは、ニワトリの輸卵管の管状腺細胞における組織特異的発現を実現する導入遺伝子構築物、および組織特異的発現を裏付ける生理学的証拠である。図6Aは、指定されたOv7.5MAbdnsを含む導入遺伝子の図である。
【図6B】図6Bは、ニワトリの輸卵管の管状腺細胞における組織特異的発現を実現する導入遺伝子構築物、および組織特異的発現を裏付ける生理学的証拠である。図6Bは、他の細胞型を除いて、選択的に管状腺細胞においてモノクローナル抗体の発現を示す、組織特異的発現導入遺伝子を含むキメラニワトリの膨大部の切片である。
【図7A】図7Aは、キメラの脳、腸、膵臓、および筋肉組織を除いて、輸卵管におけるモノクローナル抗体の軽鎖および重鎖の双方の発現をそれぞれ示すRT−PCR解析である。
【図7B】図7Bは、キメラの脳、腸、膵臓、および筋肉組織を除いて、輸卵管におけるモノクローナル抗体の軽鎖および重鎖の双方の発現をそれぞれ示すRT−PCR解析である。
【図8】図8は、ニワトリおよびCHO細胞に由来するMAbF1の熱安定性を示す。CHO細胞に由来するMAbF1(左上)およびニワトリ管状腺細胞に由来するMAbF1(右上)のDSCデータである。どちらの場合も、3つのピークを有する二状態遷移モデルに当てはまるようにデータをデコンボリュートする(赤色で示す)。CHO細胞において発現された抗体に関しては62.7℃、69.4℃、および83.4℃のTm値が得られ、ニワトリに由来する抗体に関しては63.8℃、68.5℃、および83.1℃のTm値が得られた。それぞれのFc脱グリコシル化抗体(それぞれ左下および右下)のDSCデータも同様に二状態遷移モデルに当てはめたが、いずれの場合もピークは2つであり、CHOにおいて発現された抗体に関しては61.2℃および84.0℃のTm値が得られ、ニワトリに由来する抗体に関しては62.6℃および84.0℃のTm値が得られた。
【図9】図9は、ニワトリ管状腺細胞およびCHO細胞において産生されたMAbF1のオリゴ糖プロファイルを示す。CHOに産生されたMAbの公知のグリカン構造が示されている。マンノースは黒の円として示され、ガラクトースは白の円として示され、N−アセチルグルコサミンは黒の四角形と示され、フコースは黒の三角形として示される。
【図10】図10は、卵白からのMAbF1の精製を示す。1:出発原料、2:プロテインA添加、3:プロテインA通過、4:プロテインA溶出。クーマシー染色およびウェスタンブロットの双方とも、非還元条件下である。HRP標識ヤギ抗ヒトIgG(Southern Biotechnology社)を、ウェスタンブロットにおける検出のために使用した。
【図11】図11は、ニワトリまたはCHO細胞に産生されたMAbF1の、LNCaP細胞上で発現されるPSMAに対する結合を示す。324−Eggおよび741−Eggは、卵から単離されたMAbF1の2種の異なる調製物である。F1−CHOは、CHO細胞によって産生されたMAbF1である。抗CTLA4は、LNCaP細胞上の抗原を認識しないヒトIgG1の対照MAbである。
【図12】図12は、ニワトリ管状腺細胞およびCHO細胞において産生されたMAbF1の内部移行アッセイである。570−Eggおよび741−Eggは、卵から単離されたMAbF1の2種の調製物であった。各300ng/ウェルのMAbを、Hum−Zap(抗ヒトIgG、サポリン結合体)または細胞培養培地のいずれかと共にLNCaP細胞に添加した。MAbおよびHumZapの添加後48時間目に細胞の生存率を決定した。Y軸は、相対発光量(RLU)である。
【図13】図13は、ニワトリ管状腺細胞(F1−ニワトリ)およびCHO細胞(F1−CHO)において産生されたMAbF1のin vivoクリアランスを示す。BALB/cマウス4匹に、放射標識した各抗体調製物を注射した。全身カウンターを用いて、残留放射能を350時間に渡って測定した。データは、残存している注射用量のパーセンテージとして表される。
【図14】図14は、ヒトIL−2の存在下でニワトリ管状腺細胞(F1−ニワトリ調製物IおよびII)ならびにCHO細胞(F1)において産生されたMAbF1のADCCアッセイを示す。LNCaP細胞上の抗原を認識しないヒトIgG1をアイソタイプ対照として使用した。
【図15】図15は、ヒトIL−2の不在下でニワトリ管状腺細胞(F1−ニワトリ調製物IおよびII)ならびにCHO細胞(F1)において産生されたMAbF1のADCCアッセイを示す。LNCaP細胞上の抗原を認識しないヒトIgG1をアイソタイプ対照として使用した。
【図16】図16は、抗CD16抗体を用いたADCCの妨害を示す。
【図17】図17は、ニワトリに由来するMAbF1(青色)およびCHOに由来するMAbF1(赤色)の、CD16−Phe(左のパネル)およびCD16−Val(右のパネル)との平衡結合を示す。解離定数(KD)の推定値を対応する曲線の近くに示す。
【特許請求の範囲】
【請求項1】
外因性ポリヌクレオチドにコードされ、そして遺伝的に改変されたニワトリの管状腺細胞において選択的に発現され、そして該ニワトリによって産卵された卵中で回収されるモノクローナル抗体を含む組成物であって、該抗体が、少なくとも40μg/mlの濃度で卵白中に存在する組成物。
【請求項2】
卵中に含まれる前記モノクローナル抗体がフコシル残基を実質的に含まない、請求項1に記載の組成物。
【請求項3】
マンノース濃度が約40%より高い、請求項1に記載の組成物。
【請求項4】
ガラクトース濃度が約2%より低い、請求項1に記載の組成物。
【請求項5】
フコシル残基を実質的に含まないモノクローナル抗体を作製するための方法であって、
ニワトリのゲノム中に組み入れられた外因性ポリヌクレオチドを有する遺伝的に改変されたニワトリの管状腺細胞において該抗体を発現させるステップと、
該ニワトリによって産卵された卵から、少なくとも40μg/mlの濃度で卵白中に存在する該抗体を回収するステップと
を含む方法。
【請求項6】
前記抗体のマンノース濃度が約40%より高い、請求項5に記載の方法。
【請求項7】
ガラクトース濃度が約2%より低い、請求項5に記載の方法。
【請求項8】
モノクローナル抗体が、少なくとも40μg/mlの濃度で卵白中に存在する、外因性ポリヌクレオチドにコードされたモノクローナル抗体を管状腺細胞において発現する遺伝的に改変されたニワトリ。
【請求項9】
前記抗体がフコシル残基を実質的に含まない、請求項8に記載のニワトリ。
【請求項10】
マンノース濃度が約40%より高い、請求項8に記載のニワトリ。
【請求項11】
ガラクトース濃度が約2%より低い、請求項8に記載のニワトリ。
【請求項1】
外因性ポリヌクレオチドにコードされ、そして遺伝的に改変されたニワトリの管状腺細胞において選択的に発現され、そして該ニワトリによって産卵された卵中で回収されるモノクローナル抗体を含む組成物であって、該抗体が、少なくとも40μg/mlの濃度で卵白中に存在する組成物。
【請求項2】
卵中に含まれる前記モノクローナル抗体がフコシル残基を実質的に含まない、請求項1に記載の組成物。
【請求項3】
マンノース濃度が約40%より高い、請求項1に記載の組成物。
【請求項4】
ガラクトース濃度が約2%より低い、請求項1に記載の組成物。
【請求項5】
フコシル残基を実質的に含まないモノクローナル抗体を作製するための方法であって、
ニワトリのゲノム中に組み入れられた外因性ポリヌクレオチドを有する遺伝的に改変されたニワトリの管状腺細胞において該抗体を発現させるステップと、
該ニワトリによって産卵された卵から、少なくとも40μg/mlの濃度で卵白中に存在する該抗体を回収するステップと
を含む方法。
【請求項6】
前記抗体のマンノース濃度が約40%より高い、請求項5に記載の方法。
【請求項7】
ガラクトース濃度が約2%より低い、請求項5に記載の方法。
【請求項8】
モノクローナル抗体が、少なくとも40μg/mlの濃度で卵白中に存在する、外因性ポリヌクレオチドにコードされたモノクローナル抗体を管状腺細胞において発現する遺伝的に改変されたニワトリ。
【請求項9】
前記抗体がフコシル残基を実質的に含まない、請求項8に記載のニワトリ。
【請求項10】
マンノース濃度が約40%より高い、請求項8に記載のニワトリ。
【請求項11】
ガラクトース濃度が約2%より低い、請求項8に記載のニワトリ。
【図1】


【図2】
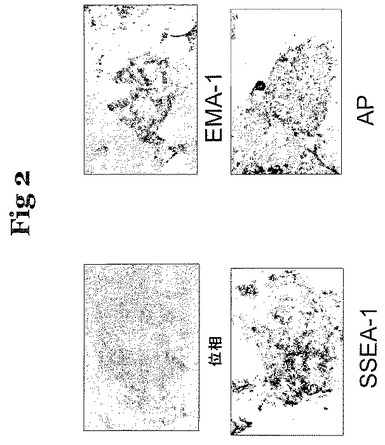
【図3】

【図4】

【図5】
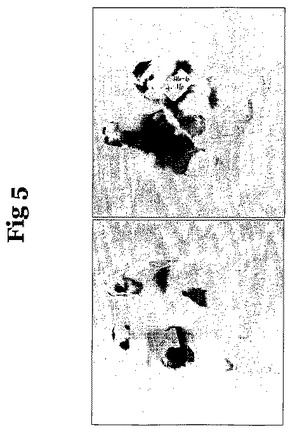
【図6A】

【図6B】
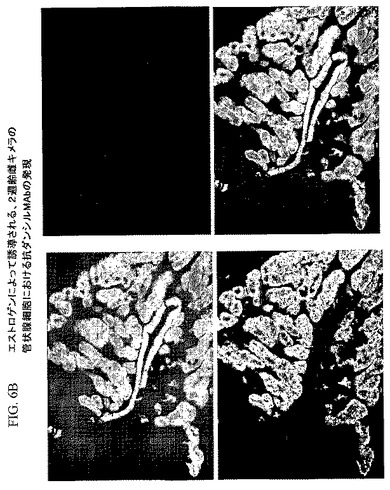
【図7A】

【図7B】
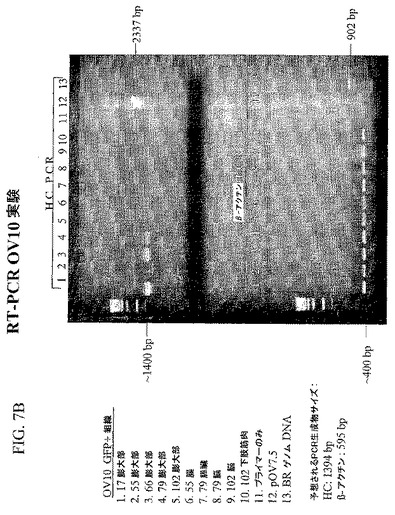
【図8】
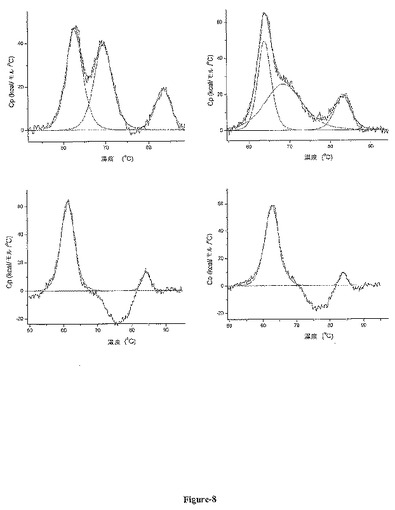
【図9】
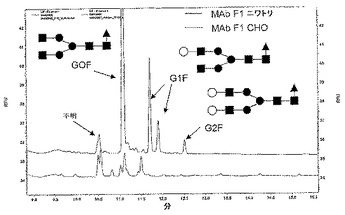
【図10】

【図11】
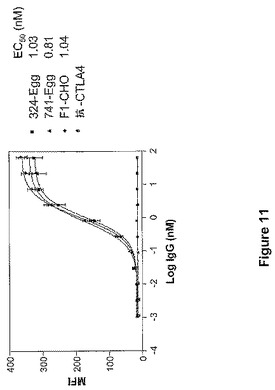
【図12】

【図13】
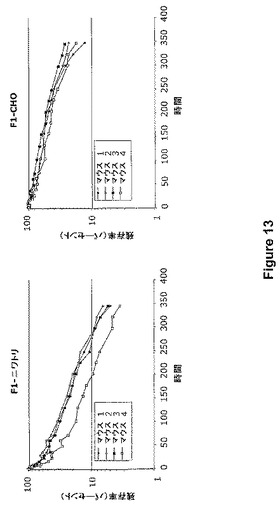
【図14】
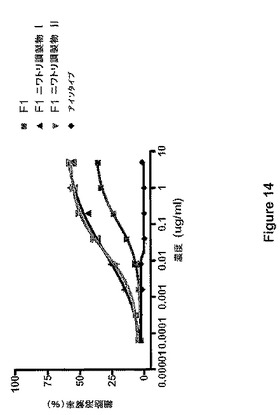
【図15】
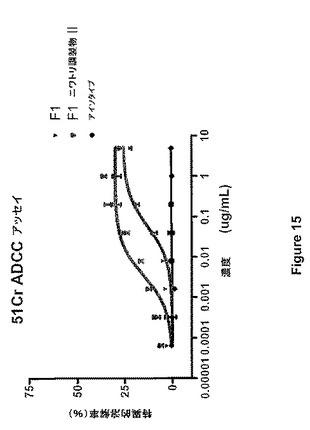
【図16】
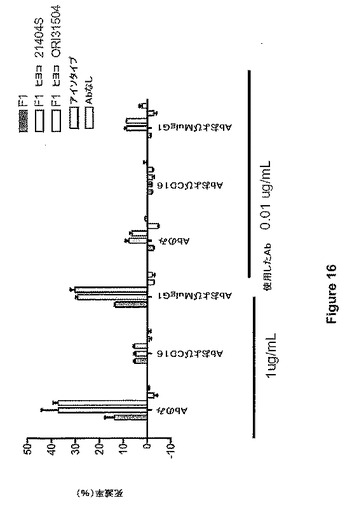
【図17】
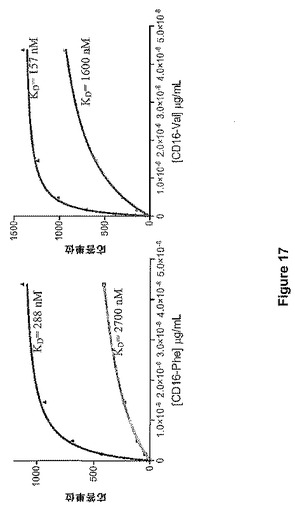
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2008−532939(P2008−532939A)
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願番号】特願2007−556416(P2007−556416)
【出願日】平成18年2月21日(2006.2.21)
【国際出願番号】PCT/US2006/006227
【国際公開番号】WO2006/089294
【国際公開日】平成18年8月24日(2006.8.24)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(507257219)オリジェン セラピューティクス, インコーポレイテッド (2)
【Fターム(参考)】
【公表日】平成20年8月21日(2008.8.21)
【国際特許分類】
【出願日】平成18年2月21日(2006.2.21)
【国際出願番号】PCT/US2006/006227
【国際公開番号】WO2006/089294
【国際公開日】平成18年8月24日(2006.8.24)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(507257219)オリジェン セラピューティクス, インコーポレイテッド (2)
【Fターム(参考)】
[ Back to top ]