
ネコ伝染性腹膜炎ウイルス(FIPV)の予防及び治療剤
【課題】ネコ伝染性腹膜炎ウイルス(FIPV)の増殖を特異的に抑制する二本鎖RNA
(dsRNA)分子およびそれを用いたFIPVの増殖を阻害するための組成物の提供。
【解決手段】FIPVのプロテアーゼ(3CLpro)のmRNAを標的とするdsRNA分子であって、該dsRNA分子の一方の鎖はFIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有するセンス鎖であり、他方の鎖は該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、上記dsRNA分子およびそれを用いたFIPVの増殖を阻害するための組成物。
(dsRNA)分子およびそれを用いたFIPVの増殖を阻害するための組成物の提供。
【解決手段】FIPVのプロテアーゼ(3CLpro)のmRNAを標的とするdsRNA分子であって、該dsRNA分子の一方の鎖はFIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有するセンス鎖であり、他方の鎖は該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、上記dsRNA分子およびそれを用いたFIPVの増殖を阻害するための組成物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子に関するものである。また本発明は、当該dsRNA分子
を含む、FIPVの複製を阻害するための組成物に関する。
【背景技術】
【0002】
ネコ伝染性腹膜炎(Feline Infectious Peritonitis:FIP)は、コロナウイルスがネコ体内で突然変異したネコ伝染性腹膜炎ウイルス(FIPV)によって起こる病気で、発症すると非治療時の致死率はほぼ100%と非常に高い恐ろしい疾患である。
【0003】
FIPの特徴的な症状には、「ウエットタイプ」と「ドライタイプ」という二つの型があ
る。症例の多い「ウエットタイプ」は、発熱や下痢、貧血などとともに腹部や胸部にひどい血管炎を起こし、腹水や胸水がたまって腹部が膨れたり、呼吸困難がみられる。「ドライタイプ」は、腹部にリンパ腫のような、大きなしこりができる。さらにこのタイプは脳内に炎症を起こして、麻痺や痙攣などの神経症状を引き起こすこともある。
【0004】
現在のところ、FIPの根治療法は存在せず、いったんFIPが発症すると、インターフェロンやステロイド剤を投与して症状を緩和する対症療法しか方法はなく、ほとんどの場合助からない。また、有効なワクチンも存在しないため、十分な治療効果を得ることは難しい(非特許文献1)。
【0005】
【非特許文献1】Niels C. Pedersen,Feline Infectious Disease, American Veterinary Publications, Inc.
【発明の開示】
【発明が解決しようとする課題】
【0006】
従来法では、FIPを十分に予防および/または治療することができなかったため、FIPを有効に治療することができる新たな方法が望まれていた。そこで、本発明では、FIPVの増殖を阻害する、FIPVの3CLproのmRNAを標的とするdsRNA分子を提供する。また、当該dsRNA分子を含む、FIPVの複製を阻害するための組成物も提供する。
【課題を解決するための手段】
【0007】
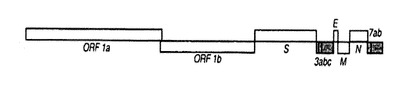
FIPVは、図1に示す構造を有するポジティブセンスの一本鎖RNAウイルスである。ゲノ
ムRNAの複製と転写に関与するするレプリカーゼは、重複を持つ2つのポリプロテインpp1a(ORF1a)とpp1ab(ORF1b)としてコードされている。ポリプロテイン中には幾つかの異なる
タンパク質が連結されており、この前駆体から個々のタンパク質がプロテアーゼによって部位特異的に切り出されて機能を発揮する。このタンパク質の切り出しに関与するプロテアーゼが、3CLproである。すなわち、3CLproは、FIPVのカプシド形成、さらには感染性のある成熟ウイルス粒子の形成に直接関与する(Journal of General Virology, 2002 vol.83 595-599, Journal of General Virology, 2002, vol. 83 581-593.)。
【0008】
そこで、本願発明者らは、上記課題を解決すべく鋭意検討した結果、FIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有するセンス鎖と該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖からなるdsRNA分子が、3CLpro遺伝子の発現を特異
的に抑制することを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明は以下のとおりである。
[1] ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子であって、一方の鎖が配列番号1で表される3CLpro遺伝子配列中の少なくとも19個以上の連続した塩基を有する配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、dsRNA分子。
【0010】
[2] dsRNA分子が、一方の鎖が配列番号1で表される3CLpro遺伝子配列の138〜1
60番目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列、および815〜837番目の塩基を有する配列からなる群から選択されるいずれかの塩基配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、[1]のdsRNA分子。
【0011】
[3] dsRNA分子が、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセ
ンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなる、[2]のdsRNA分子。
【0012】
[4] センス鎖とアンチセンス鎖がリンカー分子を介して連結されている、[1]〜[3]のいずれかのdsRNA分子。
[5] [4]のdsRNA分子の鋳型DNAを含み該dsRNA分子を発現するベクター。
[6] FIPVの複製を阻害するための、[1]〜[4]のいずれかのdsRNA分子を1つま
たは複数含む組成物。
[7] FIPVの複製を阻害するための、[5]のベクターを含む組成物。
【発明の効果】
【0013】
本発明によるdsRNA分子を用いて、FIPVの3CLpro遺伝子の発現を特異的に抑制すること
ができ、それによってFIPVの複製を阻害することができる。本発明のdsRNAは、ネコFIPの予防または治療剤として用いることができる。
【発明を実施するための最良の形態】
【0014】
一実施形態において、本発明はFIPVの3CLproのmRNAを標的とするdsRNA分子であって、
該dsRNA分子の一方の鎖はFIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有
するセンス鎖であり、他方の鎖は該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、上記dsRNA分子に関する。
【0015】
本発明のdsRNA分子は、配列特異的にFIPVの3CLproのmRNAを切断し、その結果3CLpro遺
伝子の発現を抑制するRNA干渉(RNAi)を引き起こすことができる。
【0016】
本発明のdsRNA分子は、30塩基以上の長さで提供される場合、ダイサー(Dicer)と呼ばれるRNaseIII様酵素の作用によってプロセッシングされ、3’末端に2塩基のオーバーハ
ングを有する21〜27塩基のsiRNA(small interfering RNA)分子を生じ得る。このsiRNA
分子が、RISC(RNA-induced silencing complex)と呼ばれるタンパク質複合体に取り込ま
れ、3CLpro mRNAをsiRNA分子との相同性により認識し分解し、発現抑制を行う。
【0017】
本発明においてdsRNA分子は、siRNA分子を含む。
本発明のdsRNA分子の塩基数は、特に限定されないが、15〜50塩基、好ましくは19〜35
塩基、特に好ましくは21塩基である。
【0018】
本発明において、3CLproの遺伝子配列に同一なセンス鎖とは、配列番号1で表される3CLpro遺伝子配列中のチミンがウラシルに置換された塩基配列を有するRNA配列を意味する。本発明のdsRNA分子を構成するセンス鎖は、3CLproの遺伝子配列と同一であることが望ましいが、実質的に同一、すなわち相同な配列であってもよい。すなわち、dsRNA分子のセンス鎖と実際に標的となる3CLproのmRNA配列がハイブリダイズする限り1または複数、すなわち、1〜10個、好ましくは1〜5個、さらに好ましくは、1〜3個、2個もしくは1個のミスマッチがあってもよい。したがって、本発明のdsRNA分子のセンス鎖配列と3CLproの遺伝子配列は、90%以上、好ましくは95%以上、さらに好ましくは95、96、97、98もしくは99%以上の配列相同性を有する。
【0019】
本発明において、好ましくはセンス鎖が、配列番号1で表される3CLpro遺伝子配列の138〜160番目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列および815〜837番目の塩基を有する配列からなる群から選択される塩基配列と同一であることが好ましく、特に138〜160番目の塩基を有する配列および815〜837番目の塩基を有する配列と同一であることが好ましい。
【0020】
dsRNA分子を構成するセンス鎖およびアンチセンス鎖は、公知の方法に基づいて、化学
合成や、プロモーターおよびRNAポリメラーゼを用いた転写系によってin vitroで合成す
ることができる。また、適当な発現ベクターに当該アンチセンス鎖およびセンス鎖の鋳型DNAを導入し、当該ベクターを適当な宿主細胞内に投与することによってin vivoで合成することも可能である。合成されたセンス鎖およびアンチセンス鎖のアニーリングは、当業者に公知である一般的な方法によって行うことができる。合成したセンス鎖およびアンチセンス鎖をそれぞれdsRNA用アニーリングバッファーに溶解し、等量(等モル数)を混合
し、二本鎖が解離するまで温度を加熱し、その後徐々に冷却してインキュベートすることによって行う。アニーリング条件は、例えば、90℃にて1分間、続いて37℃にて1時間静置することによって行う。その後、フェノール/クロロホルム抽出・エタノール沈殿を行うことによって、dsRNA分子を得ることができる。
【0021】
一実施形態において、本願発明のdsRNA分子は、配列番号2で表されるセンス鎖と配列
番号3で表されるアンチセンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなり、特に配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖または配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなるdsRNA分子が好ましい。
【0022】
また、本発明のdsRNA分子は、上に定義されたセンス鎖とアンチセンス鎖がリンカー分
子を介して連結され、そのリンカー部でループ構造を形成し折りたたまれたshRNA(short hairpin RNA)分子として提供され得る。shRNA分子もまた、ダイサーによってプロセッシングされ、siRNA分子を生じ得る。
【0023】
shRNA分子に含まれるリンカー分子は、センス鎖とアンチセンス鎖を連結しステムルー
プ構造を形成し得る限り、ポリヌクレオチドリンカーであっても、非ポリヌクレオチドリンカーであってもよく、特に限定しないが、当業者に公知であるポリヌクレオチドリンカーが好ましい。
【0024】
shRNA分子も、上記したように公知の方法に基づいて、in vitroまたはin vivoにて合成することが可能である。合成に際しては、センス鎖とアンチセンス鎖を逆方向配列として含む1本のRNA鎖を合成し、その後この1本のRNA鎖を自己相補的結合によって二本鎖構造を形成させshRNA分子を得ることができる。
【0025】
dsRNA分子を構成するセンス鎖またはアンチセンス鎖は必要に応じて、3’末端にオーバ
ーハングを有していてもよく、該オーバーハングの塩基の種類、数は限定されず、例えば、1〜5、好ましくは1〜3、さらに好ましくは1もしくは2塩基からなる配列が挙げられ、例えば、TTT、UUやTTが挙げられる。本発明において、オーバーハングとは、dsRNA分子の一方の鎖の末端に付加された塩基であって、もう一方の鎖の対応する位置に相補的に結合し得る塩基が存在しない塩基をいう。オーバーハングはDNAを構成する塩基であってもよい。さらに、dsRNA分子を構成するセンス鎖またはアンチセンス鎖は、遺伝子の配列決定など種々の実験操作を円滑に行うために、siRNA活性に対して影響を与えない範囲で必要に応じて1〜3塩基、さらに好ましくは1もしくは2塩基の置換、付加、欠失をさらに含んでも良い。
【0026】
また、センス鎖またはアンチセンス鎖は必要に応じて、5’末端がリン酸化されていて
もよく、shRNA分子については5’末端に三リン酸(ppp)が結合していても良い。
また、一実施形態において、本発明は、上記shRNA分子を発現するベクターに関する。
【0027】
本願発明に用いることができるベクターとしては、プラスミドベクター、ウイルスベクター、非ウイルスベクターなどを用いることができる。プラスミドベクターとしては、pBAsiベクター、pSUPERベクター等を用いることができる。ウイルスベクターとしては、アデノウイルスベクター、レトロウイルスベクター、レンチウイルスベクター等を用いることができる。非ウイルスベクターとしては、リポソームベクター等を用いることができ、特にセンダイウイルス−リポソームベクターが好ましい。
【0028】
ベクターには、プロモーターおよび/またはその他の制御配列を機能し得るかたちで連結して挿入しても良い。「機能し得るかたちで連結して挿入する」とは、当該ベクターが導入された細胞において、プロモーターおよび/またはその他の制御配列の制御の下、上記shRNA分子が発現され標的となる3CLproのmRNAが分解されるように、プロモーターおよ
び/またはその他の制御配列を連結してベクターに組み込むことを意味する。ベクターに組み込むことができるプロモーターおよび/またはその他の制御配列は特に限定されないが、構成的プロモーター、組織特異的プロモーター、時期特異的プロモーター、その他の調節エレメントなど、当該分野で公知のプロモーターおよび/またはその他の制御配列を適宜選択することが可能である。
【0029】
このように作製された上記shRNA分子を発現するベクターは、導入された細胞内でshRNA分子を発現し、3CLproのmRNAを特異的に分解する。
【0030】
また、一実施形態において、本発明は、FIPVの複製を阻害するための、本発明のdsRNA
分子を含む組成物および本発明のベクターを含む組成物に関する。本発明の組成物は、本発明のdsRNA分子またはベクターを1種または複数種を組み合わせて含んでも良い。例え
ば、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、または配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなるdsRNA分子の1種、2種、3種、または4種
を含んでも良い。本発明の組成物はまた、本発明のdsRNA分子またはベクターと共に、適
当なキャリア、希釈剤、エマルジョンなどを含んでも良い。例えば、生理的食塩水、リン酸緩衝生理食塩水、リン酸緩衝生理食塩水グルコース液、および緩衝生理食塩水などを組成物中に含めることができる。本発明の組成物が、医薬的組成物として用いられる場合には、組成物は、経口ルート、ならびに静脈内、筋肉内、皮下および腹腔内の注射または非経腸的ルートで投与することに適した形態で調製することが可能である。
【0031】
本発明は、上記dsRNA分子または発現ベクターを有効成分として含むFIPの予防または治
療剤を提供し得る。該予防または治療剤は正常ネコ、FIPVに感染しているネコまたはFIP
を発症しているネコに投与することにより、FIPVの感染を予防し、感染したFIPVの増殖を抑制しFIPVの発症を予防し、またはFIPV感染症を治療することができる。該予防または治療剤の投与量は限定されないが、例えば、腸上皮細胞やマクロファージなどの場合、一細胞あたり少なくとも1コピーのdsRNA分子が導入されるように投与することができ、例え
ば1投与単位あたり、dsRNA分子量で1nM〜100μM、好ましくは10nM〜50μM、より好まし
くは100nM〜20μMである。投与量は、患者ネコの状態、年齢、性別、重篤度などに応じて変化させることができ、専門獣医の判断により決定される。さらに、本発明は上記dsRNA
分子または発現ベクターをネコに投与してネコFIPVを予防または治療する方法を提供し得る。
【0032】
本発明の組成物は、その成分として含まれるdsRNA分子またはベクターの働きにより、FIPVの3CLproの発現を抑制し、その結果FIPVの複製を阻害し得る。
【0033】
また、本発明は、上記組成物を用いた、FIPVの複製阻害方法を提供し得る。本方法において、上記組成物をFIPVに感染した細胞、組織または個体に投与し、組成物中に含まれる本発明のdsRNA分子および発現ベクターを送達する。送達されたdsRNA分子および発現ベクターは、FIPVの3CLproの発現を抑制し、その結果FIPVの複製を阻害し得る。
【0034】
dsRNA分子および発現ベクターは、当業者に公知である核酸導入法、例えば、これらに
限定さないがHydrodynamic法、カルシウムイオンを用いる方法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法、リン酸カルシウム法、リポフェクション法、マイクロインジェクション法、遺伝子銃を用いた方法などを用いて細胞や組織に導入することが可能である。被験体が動物個体である場合には、dsRNA分子および発現ベクターは
、適当なキャリア、希釈剤、エマルジョンなどと共に含む組成物の形態で、経口ルート、ならびに静脈内、筋肉内、皮下および腹腔内の注射または非経腸的ルートで投与することが可能である。
【0035】
本発明のdsRNA分子の効果は、dsRNA分子を導入された細胞や組織、および個体における3CLproのmRNAまたはタンパク質の発現量が、当該dsRNA分子を導入していない(または導
入前の)細胞や組織、および個体における3CLproのmRNAまたはタンパク質の発現量と比較して低下していることを指標にして評価することが可能である。ここで測定される3CLproは、予めおよび/または同時に導入された3CLproを発現する発現ベクターより発現されたものであっても、FIPV感染により検出されるものであってもよい。発現量が低下しているとは、対照と比較して9割以上発現量が減少している場合のみならず、5割以上あるいは1
割以上発現量が減少している場合も含まれる。測定対象がmRNAである場合には、ノザンハイブリダイゼーション、RT-PCR、in situ hybridizationなどによって測定することがで
きる。また、測定対象がタンパク質である場合には、ウエスタンブロッティング、ELISA
、抗体を結合させたプロテインチップを用いた測定、タンパク質の活性測定などによって測定することができる。検出される3CLproが、予めおよび/または同時に導入したベクターに由来するものである場合には、当該ベクターに3CLproと共に検出可能な標識(例えば、EGFPなど)遺伝子を組み込み、3CLproと該標識からなる発現された融合タンパク質を適当な手段で検出して、比較することができる。
【実施例】
【0036】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
実施例1.3CLpro−EGFP発現ベクターの作製
FIPVは、腸コロナウイルスの変異株でありその多くは公知である。このうち、FIPV 79-
1146系統は感染性が強く、日本国内でいちばん発症しているFIPの原因ウイルスと同じ血
清型であると考えられている。そこで本実施例では、感染性が強く、日本国内で多発するFIPの原因ウイルスと同血清型のFIPV79-1146株の遺伝子を用いて、実験を行った。FIPV79-1146株の3CLpro遺伝子(配列番号1:AY994055)の配列を基に、FLAG tag 配列および制限酵素(NheIおよびBamHI)認識配列を含む以下のプライマーを作製し:
FIP 3CLpro Bam-V2: 5’-GCGGGATCCCTGAAGATTAACACCATACAT-3’(配列番号10)
FIP 3CLpro FW Nhe: 5’-CCCGCTAGCATGGACTACAAGGACGACGATGACAAGTCCGGATTGAGAAAAATGGC-3’(配列番号11)
これらプライマーと鋳型DNAとしてpMal-FIPV-3CLpro(University of WurzburgのJohn Ziebuh博士より分与されたもの)を用いてPCRを行い、3CLpro遺伝子クローニングした。クローニングした3CLpro遺伝子を、NheIおよびBamHIで制限酵素処理し、断片を精製した後、pEGFP-N3ベクター(タカラバイオ株式会社製)のNheI- BamHI 部位にクローニングした。当該ベクターより発現された3CLproタンパク質は、EGFPとの融合体であるため、蛍光顕微鏡下で可視化できる。
【0037】
実施例2.siRNAの選択および合成
FIPV79-1146株の3CLpro遺伝子配列(配列番号1:AY994055)を基に、3CLpro遺伝子を
特異的標的とする以下のセンス鎖およびアンチセンス鎖からなる4種類のsiRNAをコンピ
ュータープログラムにより選定し(RNAi社に委託)、それぞれ合成した(PROLIGO社に委
託)。
siRNA1:標的遺伝子領域138〜160塩基
センス鎖 :CAUCGCGAGUGAUCAAUUAUG (配列番号2)
アンチセンス鎖:UAAUUGAUCACUCGCGAUGUA (配列番号3)
siRNA2:標的遺伝子領域171〜193塩基
センス鎖 :CUAGUGUGCGUUUACAUAACU (配列番号4)
アンチセンス鎖:UUAUGUAAACGCACACUAGAC (配列番号5)
siRNA3:標的遺伝子領域363〜385塩基
センス鎖 :GUAGUGUCUACGGUGUUAACA (配列番号6)
アンチセンス鎖:UUAACACCGUAGACACUACCG (配列番号7)
siRNA4:標的遺伝子領域815〜837塩基
センス鎖 :GGAGGACGUACUAUACUGUCU (配列番号8)
アンチセンス鎖:ACAGUAUAGUACGUCCUCCAA (配列番号9)
ネガティブコントロール(scrambled siRNA):標的遺伝子領域はFIP 7911-46株のN遺伝
子の3’側より438〜460塩基
センス鎖 :AGUGAAGUUCGACUUUGACAC (配列番号12)
アンチセンス鎖:UCACUUCAAGCUGAAACUGUG (配列番号13)
【0038】
実施例3.siRNA分子による3CLpro発現抑制効果の解析
上記4種類の各siRNA分子を3CLpro−EGFP発現ベクターとともに、培養細胞にトランス
フェクションし、各siRNA分子による3CLpro発現抑制効果を以下の解析系を用いて調べた
。
【0039】
(I)3CLproの共焦点レーザー顕微鏡による解析
AD293細胞株を、10%ウシ胎児血清(FCS)含有Dulbecco’s modified Eagle’s medium
(D-MEM)中で37℃,5% CO2環境下にて培養及び維持した。当該細胞株を、12ウェルプレ
ートに5×105/wellの密度で播き、20時間後細胞が80〜90%コンフルエントの状態になったところで、培地を1 mlの5% FCS 含有D-MEMに置換し、上記3CLpro−EGFP発現ベクターと各種siRNAをトランスフェクトした。トランスフェクションは以下の通りに行った:3CLpro−EGFP発現ベクターを1.0μgと各種siRNA 1.0μl (終濃度50 nM) およびDsRed発現ベクター(pDsRed2-N1ベクター,Clontech)0.5μgを0pti-MEM 50μlに混合し、そこにLipofectamine 2000(Invitrogen) 4μlと50μlのOpti-MEM 培地を混合して5 分間室温で静置した溶液を添加して混合し、20分間室温でインキュベートした。この混合液を各ウェルに添加し、37℃にて24時間培養した。
【0040】
トランスフェクションから24時間後に、共焦点レーザー顕微鏡によって3CLpro−EGFPタンパク質発現の確認をおこなった。DsRedは、トランスフェクション効率を各siRNA導入細胞で比較するために用いた。
【0041】
(II)ウエスタンブロッティング解析
上述のように共焦点レーザー顕微鏡による解析を行った細胞を回収し、コンプリートミニ(ロシュ)を含む100μlの細胞溶解バッファー(10 mM Tris, pH 8.0, 1 mM EDTA, 0.15
M NaCl, 1% NP40)に溶解した。DC protein assay kit(Bio Rad)にてタンパク質の定量
を行った後、各タンパク質を6μg使用して10% SDS-PAGE還元状態で泳動した。続いてイ
モビロンP(ミリポア)に120V定電圧下、2時間、4℃にて転写した後、1%BSA, 5% 脱脂粉乳(DIFCO)を含むTBST (20 mM, 137 mM NaCl, 0.05% Tween20)で4℃にて一晩ブロッキ
ングした。その後、1次抗体として抗GFP抗体(Clontech)をTBSTで1000倍に希釈した溶液で室温にて1時間反応した後、TBSTで3回洗浄した。次いでHRP結合抗マウスIgG抗体(Promega)をTBSTで5000倍に希釈した溶液で室温にて1時間反応した後、TBSTで3回洗浄した。
【0042】
シグナルの検出にはSuperSignal West Femto Maximum Sensitivity Substrate (Pierce)を用いた。シグナル検出器はVersa Doc 5000 (Bio Rad)を使用し、定量プログラムであるQuantity One (Bio Rad)にて定量解析を行った。
【0043】
続いて、タンパク質の泳動量を補正するために、当該細胞におけるβアクチンの発現量を解析した。上記転写膜より抗体を除き、再度ブロッキングした後、1次抗体として500倍に希釈した抗βアクチン抗体(Sigma-Aldrich)、2次抗体として5000倍に希釈したHRP結合抗マウスIgG抗体(Promega)を用いて、上記同様にβアクチンを検出・定量した。
【0044】
(III)解析結果
共焦点レーザー顕微鏡およびウエスタンブロット法による解析結果
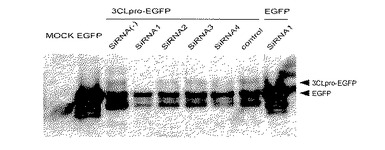
a.共焦点レーザー顕微鏡による3CLproタンパク質の発現量低下の確認
3CLpro−EGFP融合タンパク質の発現量を、EGFPの蛍光強度を共焦点レーザー顕微鏡によって解析することによって調べた結果を図2に示す。対照siRNA分子を導入した系の蛍光強度に比べて、各種siRNA分子を導入した系では蛍光強度が低下していることが観察された。特に、siRNA1分子を導入した系では顕著に蛍光強度が低下した。一方、EGFP単独の発現に対しては合成した各種siRNA分子は発現抑制を示さなかった。このことから、siRNA分子が3CLpro遺伝子の発現を特異的に抑制していることが確認された。
【0045】
b.ウエスタンブロット法による3CLproタンパク質の発現量低下の確認
3CLpro−EGFP融合タンパク質およびβアクチンの発現量を、ウエスタンブロット法によって定量解析した結果を、以下の表1ならびに図3、図4および図5に示す。
【0046】
【表1】

【0047】
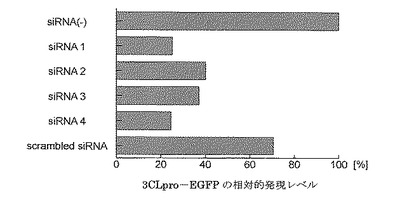
表1は、図3および4の各バンドを数値化し、各系における3CLpro−EGFP融合タンパク
質の発現量を比較した結果を示す。表1より導き出された各系における3CLpro−EGFP融合タンパク質の発現量をグラフ化したものが図5である。これらの定量解析から、対照siRNA分子を導入した系と比較して、各種siRNA分子を導入した系では検出される3CLpro−EGFP融合タンパク質量が減少していることが示された。特にsiRNA1およびsiRNA4を導入した系では3CLpro−EGFP融合タンパク質の量が著しく減少し、siRNA分子を導入していない系と比べて3CLpro−EGFP融合タンパク質の発現量が約75%抑制されることが示された。
【産業上の利用可能性】
【0048】
本発明のFIPVの3CLpro mRNAを標的とするdsRNA分子は、3CLproのタンパク質の発現量を有効に抑制することが可能であり、それによってFIPVの複製を阻害し得る。また、本発明のdsRNA分子を用いてFIPVの複製を阻害することによって、ネコ伝染性腹膜炎を有効に予
防および/または治療することができる。
【図面の簡単な説明】
【0049】
【図1】FIPVの一本鎖RNAを示す模式図である。
【図2】共焦点レーザー顕微鏡による3CLpro−EGFP融合タンパク質の解析結果を示す図である。図中a-dは、それぞれ以下の導入系を示すa: 3CLpro−EGFP+siRNA1、b:3CLpro−EGFP+siRNA2、c:3CLpro−EGFP+siRNA3、d:3CLpro−EGFP+siRNA4。それぞれ左の写真はEGFP、右の写真はDsRedの蛍光像を示す。
【図3】ウエスタンブロット法による3CLpro−EGFP融合タンパク質の発現解析結果を示す図である。
【図4】ウエスタンブロット法によるβアクチンの発現解析結果を示す図である。
【図5】各siRNA分子による3CLpro−EGFP融合タンパク質の発現抑制効果を示す図である。3CLpro−EGFP発現ベクターを単独で導入した際の3CLpro−EGFP融合タンパク質の発現量を100%として、各系における3CLpro−EGFP融合タンパク質の発現量を相対的に示す。
【配列表フリーテキスト】
【0050】
配列番号2〜13は、合成オリゴヌクレオチド配列を示す。
【技術分野】
【0001】
本発明は、ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子に関するものである。また本発明は、当該dsRNA分子
を含む、FIPVの複製を阻害するための組成物に関する。
【背景技術】
【0002】
ネコ伝染性腹膜炎(Feline Infectious Peritonitis:FIP)は、コロナウイルスがネコ体内で突然変異したネコ伝染性腹膜炎ウイルス(FIPV)によって起こる病気で、発症すると非治療時の致死率はほぼ100%と非常に高い恐ろしい疾患である。
【0003】
FIPの特徴的な症状には、「ウエットタイプ」と「ドライタイプ」という二つの型があ
る。症例の多い「ウエットタイプ」は、発熱や下痢、貧血などとともに腹部や胸部にひどい血管炎を起こし、腹水や胸水がたまって腹部が膨れたり、呼吸困難がみられる。「ドライタイプ」は、腹部にリンパ腫のような、大きなしこりができる。さらにこのタイプは脳内に炎症を起こして、麻痺や痙攣などの神経症状を引き起こすこともある。
【0004】
現在のところ、FIPの根治療法は存在せず、いったんFIPが発症すると、インターフェロンやステロイド剤を投与して症状を緩和する対症療法しか方法はなく、ほとんどの場合助からない。また、有効なワクチンも存在しないため、十分な治療効果を得ることは難しい(非特許文献1)。
【0005】
【非特許文献1】Niels C. Pedersen,Feline Infectious Disease, American Veterinary Publications, Inc.
【発明の開示】
【発明が解決しようとする課題】
【0006】
従来法では、FIPを十分に予防および/または治療することができなかったため、FIPを有効に治療することができる新たな方法が望まれていた。そこで、本発明では、FIPVの増殖を阻害する、FIPVの3CLproのmRNAを標的とするdsRNA分子を提供する。また、当該dsRNA分子を含む、FIPVの複製を阻害するための組成物も提供する。
【課題を解決するための手段】
【0007】
FIPVは、図1に示す構造を有するポジティブセンスの一本鎖RNAウイルスである。ゲノ
ムRNAの複製と転写に関与するするレプリカーゼは、重複を持つ2つのポリプロテインpp1a(ORF1a)とpp1ab(ORF1b)としてコードされている。ポリプロテイン中には幾つかの異なる
タンパク質が連結されており、この前駆体から個々のタンパク質がプロテアーゼによって部位特異的に切り出されて機能を発揮する。このタンパク質の切り出しに関与するプロテアーゼが、3CLproである。すなわち、3CLproは、FIPVのカプシド形成、さらには感染性のある成熟ウイルス粒子の形成に直接関与する(Journal of General Virology, 2002 vol.83 595-599, Journal of General Virology, 2002, vol. 83 581-593.)。
【0008】
そこで、本願発明者らは、上記課題を解決すべく鋭意検討した結果、FIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有するセンス鎖と該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖からなるdsRNA分子が、3CLpro遺伝子の発現を特異
的に抑制することを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明は以下のとおりである。
[1] ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子であって、一方の鎖が配列番号1で表される3CLpro遺伝子配列中の少なくとも19個以上の連続した塩基を有する配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、dsRNA分子。
【0010】
[2] dsRNA分子が、一方の鎖が配列番号1で表される3CLpro遺伝子配列の138〜1
60番目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列、および815〜837番目の塩基を有する配列からなる群から選択されるいずれかの塩基配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、[1]のdsRNA分子。
【0011】
[3] dsRNA分子が、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセ
ンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなる、[2]のdsRNA分子。
【0012】
[4] センス鎖とアンチセンス鎖がリンカー分子を介して連結されている、[1]〜[3]のいずれかのdsRNA分子。
[5] [4]のdsRNA分子の鋳型DNAを含み該dsRNA分子を発現するベクター。
[6] FIPVの複製を阻害するための、[1]〜[4]のいずれかのdsRNA分子を1つま
たは複数含む組成物。
[7] FIPVの複製を阻害するための、[5]のベクターを含む組成物。
【発明の効果】
【0013】
本発明によるdsRNA分子を用いて、FIPVの3CLpro遺伝子の発現を特異的に抑制すること
ができ、それによってFIPVの複製を阻害することができる。本発明のdsRNAは、ネコFIPの予防または治療剤として用いることができる。
【発明を実施するための最良の形態】
【0014】
一実施形態において、本発明はFIPVの3CLproのmRNAを標的とするdsRNA分子であって、
該dsRNA分子の一方の鎖はFIPVの3CLpro遺伝子配列またはその一部に同一な塩基配列を有
するセンス鎖であり、他方の鎖は該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、上記dsRNA分子に関する。
【0015】
本発明のdsRNA分子は、配列特異的にFIPVの3CLproのmRNAを切断し、その結果3CLpro遺
伝子の発現を抑制するRNA干渉(RNAi)を引き起こすことができる。
【0016】
本発明のdsRNA分子は、30塩基以上の長さで提供される場合、ダイサー(Dicer)と呼ばれるRNaseIII様酵素の作用によってプロセッシングされ、3’末端に2塩基のオーバーハ
ングを有する21〜27塩基のsiRNA(small interfering RNA)分子を生じ得る。このsiRNA
分子が、RISC(RNA-induced silencing complex)と呼ばれるタンパク質複合体に取り込ま
れ、3CLpro mRNAをsiRNA分子との相同性により認識し分解し、発現抑制を行う。
【0017】
本発明においてdsRNA分子は、siRNA分子を含む。
本発明のdsRNA分子の塩基数は、特に限定されないが、15〜50塩基、好ましくは19〜35
塩基、特に好ましくは21塩基である。
【0018】
本発明において、3CLproの遺伝子配列に同一なセンス鎖とは、配列番号1で表される3CLpro遺伝子配列中のチミンがウラシルに置換された塩基配列を有するRNA配列を意味する。本発明のdsRNA分子を構成するセンス鎖は、3CLproの遺伝子配列と同一であることが望ましいが、実質的に同一、すなわち相同な配列であってもよい。すなわち、dsRNA分子のセンス鎖と実際に標的となる3CLproのmRNA配列がハイブリダイズする限り1または複数、すなわち、1〜10個、好ましくは1〜5個、さらに好ましくは、1〜3個、2個もしくは1個のミスマッチがあってもよい。したがって、本発明のdsRNA分子のセンス鎖配列と3CLproの遺伝子配列は、90%以上、好ましくは95%以上、さらに好ましくは95、96、97、98もしくは99%以上の配列相同性を有する。
【0019】
本発明において、好ましくはセンス鎖が、配列番号1で表される3CLpro遺伝子配列の138〜160番目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列および815〜837番目の塩基を有する配列からなる群から選択される塩基配列と同一であることが好ましく、特に138〜160番目の塩基を有する配列および815〜837番目の塩基を有する配列と同一であることが好ましい。
【0020】
dsRNA分子を構成するセンス鎖およびアンチセンス鎖は、公知の方法に基づいて、化学
合成や、プロモーターおよびRNAポリメラーゼを用いた転写系によってin vitroで合成す
ることができる。また、適当な発現ベクターに当該アンチセンス鎖およびセンス鎖の鋳型DNAを導入し、当該ベクターを適当な宿主細胞内に投与することによってin vivoで合成することも可能である。合成されたセンス鎖およびアンチセンス鎖のアニーリングは、当業者に公知である一般的な方法によって行うことができる。合成したセンス鎖およびアンチセンス鎖をそれぞれdsRNA用アニーリングバッファーに溶解し、等量(等モル数)を混合
し、二本鎖が解離するまで温度を加熱し、その後徐々に冷却してインキュベートすることによって行う。アニーリング条件は、例えば、90℃にて1分間、続いて37℃にて1時間静置することによって行う。その後、フェノール/クロロホルム抽出・エタノール沈殿を行うことによって、dsRNA分子を得ることができる。
【0021】
一実施形態において、本願発明のdsRNA分子は、配列番号2で表されるセンス鎖と配列
番号3で表されるアンチセンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなり、特に配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖または配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなるdsRNA分子が好ましい。
【0022】
また、本発明のdsRNA分子は、上に定義されたセンス鎖とアンチセンス鎖がリンカー分
子を介して連結され、そのリンカー部でループ構造を形成し折りたたまれたshRNA(short hairpin RNA)分子として提供され得る。shRNA分子もまた、ダイサーによってプロセッシングされ、siRNA分子を生じ得る。
【0023】
shRNA分子に含まれるリンカー分子は、センス鎖とアンチセンス鎖を連結しステムルー
プ構造を形成し得る限り、ポリヌクレオチドリンカーであっても、非ポリヌクレオチドリンカーであってもよく、特に限定しないが、当業者に公知であるポリヌクレオチドリンカーが好ましい。
【0024】
shRNA分子も、上記したように公知の方法に基づいて、in vitroまたはin vivoにて合成することが可能である。合成に際しては、センス鎖とアンチセンス鎖を逆方向配列として含む1本のRNA鎖を合成し、その後この1本のRNA鎖を自己相補的結合によって二本鎖構造を形成させshRNA分子を得ることができる。
【0025】
dsRNA分子を構成するセンス鎖またはアンチセンス鎖は必要に応じて、3’末端にオーバ
ーハングを有していてもよく、該オーバーハングの塩基の種類、数は限定されず、例えば、1〜5、好ましくは1〜3、さらに好ましくは1もしくは2塩基からなる配列が挙げられ、例えば、TTT、UUやTTが挙げられる。本発明において、オーバーハングとは、dsRNA分子の一方の鎖の末端に付加された塩基であって、もう一方の鎖の対応する位置に相補的に結合し得る塩基が存在しない塩基をいう。オーバーハングはDNAを構成する塩基であってもよい。さらに、dsRNA分子を構成するセンス鎖またはアンチセンス鎖は、遺伝子の配列決定など種々の実験操作を円滑に行うために、siRNA活性に対して影響を与えない範囲で必要に応じて1〜3塩基、さらに好ましくは1もしくは2塩基の置換、付加、欠失をさらに含んでも良い。
【0026】
また、センス鎖またはアンチセンス鎖は必要に応じて、5’末端がリン酸化されていて
もよく、shRNA分子については5’末端に三リン酸(ppp)が結合していても良い。
また、一実施形態において、本発明は、上記shRNA分子を発現するベクターに関する。
【0027】
本願発明に用いることができるベクターとしては、プラスミドベクター、ウイルスベクター、非ウイルスベクターなどを用いることができる。プラスミドベクターとしては、pBAsiベクター、pSUPERベクター等を用いることができる。ウイルスベクターとしては、アデノウイルスベクター、レトロウイルスベクター、レンチウイルスベクター等を用いることができる。非ウイルスベクターとしては、リポソームベクター等を用いることができ、特にセンダイウイルス−リポソームベクターが好ましい。
【0028】
ベクターには、プロモーターおよび/またはその他の制御配列を機能し得るかたちで連結して挿入しても良い。「機能し得るかたちで連結して挿入する」とは、当該ベクターが導入された細胞において、プロモーターおよび/またはその他の制御配列の制御の下、上記shRNA分子が発現され標的となる3CLproのmRNAが分解されるように、プロモーターおよ
び/またはその他の制御配列を連結してベクターに組み込むことを意味する。ベクターに組み込むことができるプロモーターおよび/またはその他の制御配列は特に限定されないが、構成的プロモーター、組織特異的プロモーター、時期特異的プロモーター、その他の調節エレメントなど、当該分野で公知のプロモーターおよび/またはその他の制御配列を適宜選択することが可能である。
【0029】
このように作製された上記shRNA分子を発現するベクターは、導入された細胞内でshRNA分子を発現し、3CLproのmRNAを特異的に分解する。
【0030】
また、一実施形態において、本発明は、FIPVの複製を阻害するための、本発明のdsRNA
分子を含む組成物および本発明のベクターを含む組成物に関する。本発明の組成物は、本発明のdsRNA分子またはベクターを1種または複数種を組み合わせて含んでも良い。例え
ば、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、または配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなるdsRNA分子の1種、2種、3種、または4種
を含んでも良い。本発明の組成物はまた、本発明のdsRNA分子またはベクターと共に、適
当なキャリア、希釈剤、エマルジョンなどを含んでも良い。例えば、生理的食塩水、リン酸緩衝生理食塩水、リン酸緩衝生理食塩水グルコース液、および緩衝生理食塩水などを組成物中に含めることができる。本発明の組成物が、医薬的組成物として用いられる場合には、組成物は、経口ルート、ならびに静脈内、筋肉内、皮下および腹腔内の注射または非経腸的ルートで投与することに適した形態で調製することが可能である。
【0031】
本発明は、上記dsRNA分子または発現ベクターを有効成分として含むFIPの予防または治
療剤を提供し得る。該予防または治療剤は正常ネコ、FIPVに感染しているネコまたはFIP
を発症しているネコに投与することにより、FIPVの感染を予防し、感染したFIPVの増殖を抑制しFIPVの発症を予防し、またはFIPV感染症を治療することができる。該予防または治療剤の投与量は限定されないが、例えば、腸上皮細胞やマクロファージなどの場合、一細胞あたり少なくとも1コピーのdsRNA分子が導入されるように投与することができ、例え
ば1投与単位あたり、dsRNA分子量で1nM〜100μM、好ましくは10nM〜50μM、より好まし
くは100nM〜20μMである。投与量は、患者ネコの状態、年齢、性別、重篤度などに応じて変化させることができ、専門獣医の判断により決定される。さらに、本発明は上記dsRNA
分子または発現ベクターをネコに投与してネコFIPVを予防または治療する方法を提供し得る。
【0032】
本発明の組成物は、その成分として含まれるdsRNA分子またはベクターの働きにより、FIPVの3CLproの発現を抑制し、その結果FIPVの複製を阻害し得る。
【0033】
また、本発明は、上記組成物を用いた、FIPVの複製阻害方法を提供し得る。本方法において、上記組成物をFIPVに感染した細胞、組織または個体に投与し、組成物中に含まれる本発明のdsRNA分子および発現ベクターを送達する。送達されたdsRNA分子および発現ベクターは、FIPVの3CLproの発現を抑制し、その結果FIPVの複製を阻害し得る。
【0034】
dsRNA分子および発現ベクターは、当業者に公知である核酸導入法、例えば、これらに
限定さないがHydrodynamic法、カルシウムイオンを用いる方法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法、リン酸カルシウム法、リポフェクション法、マイクロインジェクション法、遺伝子銃を用いた方法などを用いて細胞や組織に導入することが可能である。被験体が動物個体である場合には、dsRNA分子および発現ベクターは
、適当なキャリア、希釈剤、エマルジョンなどと共に含む組成物の形態で、経口ルート、ならびに静脈内、筋肉内、皮下および腹腔内の注射または非経腸的ルートで投与することが可能である。
【0035】
本発明のdsRNA分子の効果は、dsRNA分子を導入された細胞や組織、および個体における3CLproのmRNAまたはタンパク質の発現量が、当該dsRNA分子を導入していない(または導
入前の)細胞や組織、および個体における3CLproのmRNAまたはタンパク質の発現量と比較して低下していることを指標にして評価することが可能である。ここで測定される3CLproは、予めおよび/または同時に導入された3CLproを発現する発現ベクターより発現されたものであっても、FIPV感染により検出されるものであってもよい。発現量が低下しているとは、対照と比較して9割以上発現量が減少している場合のみならず、5割以上あるいは1
割以上発現量が減少している場合も含まれる。測定対象がmRNAである場合には、ノザンハイブリダイゼーション、RT-PCR、in situ hybridizationなどによって測定することがで
きる。また、測定対象がタンパク質である場合には、ウエスタンブロッティング、ELISA
、抗体を結合させたプロテインチップを用いた測定、タンパク質の活性測定などによって測定することができる。検出される3CLproが、予めおよび/または同時に導入したベクターに由来するものである場合には、当該ベクターに3CLproと共に検出可能な標識(例えば、EGFPなど)遺伝子を組み込み、3CLproと該標識からなる発現された融合タンパク質を適当な手段で検出して、比較することができる。
【実施例】
【0036】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
実施例1.3CLpro−EGFP発現ベクターの作製
FIPVは、腸コロナウイルスの変異株でありその多くは公知である。このうち、FIPV 79-
1146系統は感染性が強く、日本国内でいちばん発症しているFIPの原因ウイルスと同じ血
清型であると考えられている。そこで本実施例では、感染性が強く、日本国内で多発するFIPの原因ウイルスと同血清型のFIPV79-1146株の遺伝子を用いて、実験を行った。FIPV79-1146株の3CLpro遺伝子(配列番号1:AY994055)の配列を基に、FLAG tag 配列および制限酵素(NheIおよびBamHI)認識配列を含む以下のプライマーを作製し:
FIP 3CLpro Bam-V2: 5’-GCGGGATCCCTGAAGATTAACACCATACAT-3’(配列番号10)
FIP 3CLpro FW Nhe: 5’-CCCGCTAGCATGGACTACAAGGACGACGATGACAAGTCCGGATTGAGAAAAATGGC-3’(配列番号11)
これらプライマーと鋳型DNAとしてpMal-FIPV-3CLpro(University of WurzburgのJohn Ziebuh博士より分与されたもの)を用いてPCRを行い、3CLpro遺伝子クローニングした。クローニングした3CLpro遺伝子を、NheIおよびBamHIで制限酵素処理し、断片を精製した後、pEGFP-N3ベクター(タカラバイオ株式会社製)のNheI- BamHI 部位にクローニングした。当該ベクターより発現された3CLproタンパク質は、EGFPとの融合体であるため、蛍光顕微鏡下で可視化できる。
【0037】
実施例2.siRNAの選択および合成
FIPV79-1146株の3CLpro遺伝子配列(配列番号1:AY994055)を基に、3CLpro遺伝子を
特異的標的とする以下のセンス鎖およびアンチセンス鎖からなる4種類のsiRNAをコンピ
ュータープログラムにより選定し(RNAi社に委託)、それぞれ合成した(PROLIGO社に委
託)。
siRNA1:標的遺伝子領域138〜160塩基
センス鎖 :CAUCGCGAGUGAUCAAUUAUG (配列番号2)
アンチセンス鎖:UAAUUGAUCACUCGCGAUGUA (配列番号3)
siRNA2:標的遺伝子領域171〜193塩基
センス鎖 :CUAGUGUGCGUUUACAUAACU (配列番号4)
アンチセンス鎖:UUAUGUAAACGCACACUAGAC (配列番号5)
siRNA3:標的遺伝子領域363〜385塩基
センス鎖 :GUAGUGUCUACGGUGUUAACA (配列番号6)
アンチセンス鎖:UUAACACCGUAGACACUACCG (配列番号7)
siRNA4:標的遺伝子領域815〜837塩基
センス鎖 :GGAGGACGUACUAUACUGUCU (配列番号8)
アンチセンス鎖:ACAGUAUAGUACGUCCUCCAA (配列番号9)
ネガティブコントロール(scrambled siRNA):標的遺伝子領域はFIP 7911-46株のN遺伝
子の3’側より438〜460塩基
センス鎖 :AGUGAAGUUCGACUUUGACAC (配列番号12)
アンチセンス鎖:UCACUUCAAGCUGAAACUGUG (配列番号13)
【0038】
実施例3.siRNA分子による3CLpro発現抑制効果の解析
上記4種類の各siRNA分子を3CLpro−EGFP発現ベクターとともに、培養細胞にトランス
フェクションし、各siRNA分子による3CLpro発現抑制効果を以下の解析系を用いて調べた
。
【0039】
(I)3CLproの共焦点レーザー顕微鏡による解析
AD293細胞株を、10%ウシ胎児血清(FCS)含有Dulbecco’s modified Eagle’s medium
(D-MEM)中で37℃,5% CO2環境下にて培養及び維持した。当該細胞株を、12ウェルプレ
ートに5×105/wellの密度で播き、20時間後細胞が80〜90%コンフルエントの状態になったところで、培地を1 mlの5% FCS 含有D-MEMに置換し、上記3CLpro−EGFP発現ベクターと各種siRNAをトランスフェクトした。トランスフェクションは以下の通りに行った:3CLpro−EGFP発現ベクターを1.0μgと各種siRNA 1.0μl (終濃度50 nM) およびDsRed発現ベクター(pDsRed2-N1ベクター,Clontech)0.5μgを0pti-MEM 50μlに混合し、そこにLipofectamine 2000(Invitrogen) 4μlと50μlのOpti-MEM 培地を混合して5 分間室温で静置した溶液を添加して混合し、20分間室温でインキュベートした。この混合液を各ウェルに添加し、37℃にて24時間培養した。
【0040】
トランスフェクションから24時間後に、共焦点レーザー顕微鏡によって3CLpro−EGFPタンパク質発現の確認をおこなった。DsRedは、トランスフェクション効率を各siRNA導入細胞で比較するために用いた。
【0041】
(II)ウエスタンブロッティング解析
上述のように共焦点レーザー顕微鏡による解析を行った細胞を回収し、コンプリートミニ(ロシュ)を含む100μlの細胞溶解バッファー(10 mM Tris, pH 8.0, 1 mM EDTA, 0.15
M NaCl, 1% NP40)に溶解した。DC protein assay kit(Bio Rad)にてタンパク質の定量
を行った後、各タンパク質を6μg使用して10% SDS-PAGE還元状態で泳動した。続いてイ
モビロンP(ミリポア)に120V定電圧下、2時間、4℃にて転写した後、1%BSA, 5% 脱脂粉乳(DIFCO)を含むTBST (20 mM, 137 mM NaCl, 0.05% Tween20)で4℃にて一晩ブロッキ
ングした。その後、1次抗体として抗GFP抗体(Clontech)をTBSTで1000倍に希釈した溶液で室温にて1時間反応した後、TBSTで3回洗浄した。次いでHRP結合抗マウスIgG抗体(Promega)をTBSTで5000倍に希釈した溶液で室温にて1時間反応した後、TBSTで3回洗浄した。
【0042】
シグナルの検出にはSuperSignal West Femto Maximum Sensitivity Substrate (Pierce)を用いた。シグナル検出器はVersa Doc 5000 (Bio Rad)を使用し、定量プログラムであるQuantity One (Bio Rad)にて定量解析を行った。
【0043】
続いて、タンパク質の泳動量を補正するために、当該細胞におけるβアクチンの発現量を解析した。上記転写膜より抗体を除き、再度ブロッキングした後、1次抗体として500倍に希釈した抗βアクチン抗体(Sigma-Aldrich)、2次抗体として5000倍に希釈したHRP結合抗マウスIgG抗体(Promega)を用いて、上記同様にβアクチンを検出・定量した。
【0044】
(III)解析結果
共焦点レーザー顕微鏡およびウエスタンブロット法による解析結果
a.共焦点レーザー顕微鏡による3CLproタンパク質の発現量低下の確認
3CLpro−EGFP融合タンパク質の発現量を、EGFPの蛍光強度を共焦点レーザー顕微鏡によって解析することによって調べた結果を図2に示す。対照siRNA分子を導入した系の蛍光強度に比べて、各種siRNA分子を導入した系では蛍光強度が低下していることが観察された。特に、siRNA1分子を導入した系では顕著に蛍光強度が低下した。一方、EGFP単独の発現に対しては合成した各種siRNA分子は発現抑制を示さなかった。このことから、siRNA分子が3CLpro遺伝子の発現を特異的に抑制していることが確認された。
【0045】
b.ウエスタンブロット法による3CLproタンパク質の発現量低下の確認
3CLpro−EGFP融合タンパク質およびβアクチンの発現量を、ウエスタンブロット法によって定量解析した結果を、以下の表1ならびに図3、図4および図5に示す。
【0046】
【表1】
【0047】
表1は、図3および4の各バンドを数値化し、各系における3CLpro−EGFP融合タンパク
質の発現量を比較した結果を示す。表1より導き出された各系における3CLpro−EGFP融合タンパク質の発現量をグラフ化したものが図5である。これらの定量解析から、対照siRNA分子を導入した系と比較して、各種siRNA分子を導入した系では検出される3CLpro−EGFP融合タンパク質量が減少していることが示された。特にsiRNA1およびsiRNA4を導入した系では3CLpro−EGFP融合タンパク質の量が著しく減少し、siRNA分子を導入していない系と比べて3CLpro−EGFP融合タンパク質の発現量が約75%抑制されることが示された。
【産業上の利用可能性】
【0048】
本発明のFIPVの3CLpro mRNAを標的とするdsRNA分子は、3CLproのタンパク質の発現量を有効に抑制することが可能であり、それによってFIPVの複製を阻害し得る。また、本発明のdsRNA分子を用いてFIPVの複製を阻害することによって、ネコ伝染性腹膜炎を有効に予
防および/または治療することができる。
【図面の簡単な説明】
【0049】
【図1】FIPVの一本鎖RNAを示す模式図である。
【図2】共焦点レーザー顕微鏡による3CLpro−EGFP融合タンパク質の解析結果を示す図である。図中a-dは、それぞれ以下の導入系を示すa: 3CLpro−EGFP+siRNA1、b:3CLpro−EGFP+siRNA2、c:3CLpro−EGFP+siRNA3、d:3CLpro−EGFP+siRNA4。それぞれ左の写真はEGFP、右の写真はDsRedの蛍光像を示す。
【図3】ウエスタンブロット法による3CLpro−EGFP融合タンパク質の発現解析結果を示す図である。
【図4】ウエスタンブロット法によるβアクチンの発現解析結果を示す図である。
【図5】各siRNA分子による3CLpro−EGFP融合タンパク質の発現抑制効果を示す図である。3CLpro−EGFP発現ベクターを単独で導入した際の3CLpro−EGFP融合タンパク質の発現量を100%として、各系における3CLpro−EGFP融合タンパク質の発現量を相対的に示す。
【配列表フリーテキスト】
【0050】
配列番号2〜13は、合成オリゴヌクレオチド配列を示す。
【特許請求の範囲】
【請求項1】
ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子であって、一方の鎖が配列番号1で表される3CLpro遺伝子配列中の少なくとも19個以上の連続した塩基を有する配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、dsRNA分子
。
【請求項2】
dsRNA分子が、一方の鎖が配列番号1で表される3CLpro遺伝子配列の138〜160番
目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列、および815〜837番目の塩基を有する配列からなる群から選択されるいずれかの塩基配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、請求項1記載のdsRNA分子。
【請求項3】
dsRNA分子が、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖
、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなる、請求項2記載のdsRNA分子。
【請求項4】
センス鎖とアンチセンス鎖がリンカー分子を介して連結されている、請求項1〜3のいずれか1項に記載のdsRNA分子。
【請求項5】
請求項4に記載のdsRNA分子の鋳型DNAを含み該dsRNA分子を発現するベクター。
【請求項6】
FIPVの複製を阻害するための、請求項1〜4のいずれか1項に記載されるdsRNA分子を
1つまたは複数含む組成物。
【請求項7】
FIPVの複製を阻害するための、請求項5に記載されるベクターを含む組成物。
【請求項1】
ネコ伝染性腹膜炎ウイルス(FIPV)の3C様プロテアーゼ(3CLpro)のmRNAを標的とする二本鎖RNA(dsRNA)分子であって、一方の鎖が配列番号1で表される3CLpro遺伝子配列中の少なくとも19個以上の連続した塩基を有する配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、dsRNA分子
。
【請求項2】
dsRNA分子が、一方の鎖が配列番号1で表される3CLpro遺伝子配列の138〜160番
目の塩基を有する配列、171〜193番目の塩基を有する配列、363〜385番目の塩基を有する配列、および815〜837番目の塩基を有する配列からなる群から選択されるいずれかの塩基配列に同一なセンス鎖であり、他方の鎖が該センス鎖の塩基配列に相補的な塩基配列を有するアンチセンス鎖である、請求項1記載のdsRNA分子。
【請求項3】
dsRNA分子が、配列番号2で表されるセンス鎖と配列番号3で表されるアンチセンス鎖
、配列番号4で表されるセンス鎖と配列番号5で表されるアンチセンス鎖、配列番号6で表されるセンス鎖と配列番号7で表されるアンチセンス鎖、および配列番号8で表されるセンス鎖と配列番号9で表されるアンチセンス鎖からなる群より選択されるいずれかの塩基対からなる、請求項2記載のdsRNA分子。
【請求項4】
センス鎖とアンチセンス鎖がリンカー分子を介して連結されている、請求項1〜3のいずれか1項に記載のdsRNA分子。
【請求項5】
請求項4に記載のdsRNA分子の鋳型DNAを含み該dsRNA分子を発現するベクター。
【請求項6】
FIPVの複製を阻害するための、請求項1〜4のいずれか1項に記載されるdsRNA分子を
1つまたは複数含む組成物。
【請求項7】
FIPVの複製を阻害するための、請求項5に記載されるベクターを含む組成物。
【図1】

【図2】

【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2009−165357(P2009−165357A)
【公開日】平成21年7月30日(2009.7.30)
【国際特許分類】
【出願番号】特願2008−1419(P2008−1419)
【出願日】平成20年1月8日(2008.1.8)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成19年8月27日 第144回日本獣医学会学術集会発行の「第144回日本獣医学会学術集会講演要旨集」に発表
【出願人】(803000034)学校法人日本医科大学 (37)
【Fターム(参考)】
【公開日】平成21年7月30日(2009.7.30)
【国際特許分類】
【出願日】平成20年1月8日(2008.1.8)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成19年8月27日 第144回日本獣医学会学術集会発行の「第144回日本獣医学会学術集会講演要旨集」に発表
【出願人】(803000034)学校法人日本医科大学 (37)
【Fターム(参考)】
[ Back to top ]