
ノッチをベースとした、γ−セクレターゼの可溶性基質と、その利用法およびそれを利用した組成物
本発明は、γ-セクレターゼの新規な可溶性基質に関する。より特に、本発明は、NusAタンパク質に融合したγ-セクレターゼ依存開裂部位(γとε)含有ノッチ区画を有する可溶性融合タンパク質を提供する。上述の融合タンパク質の製造法と利用法、ならびに上述の融合タンパク質を用いた組成物を開示する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、γ-セクレターゼの新規な可溶性基質に関する。さらに詳細には、本発明により、NusAタンパク質に融合したγ-セクレターゼ依存開裂部位(γとε)含有ノッチ区画を有する可溶性融合タンパク質が提供される。このような融合タンパク質の製造法と利用法、ならびにこのような融合タンパク質を利用した組成物も開示されている。
【背景技術】
【0002】
アルツハイマー病(AD)によってアミロイド斑の形成、神経原線維の変化、グリオーシス、ニューロンの破壊が起こり、進行性の痴呆が引き起こされる。この疾患には遺伝性と散発性の両方の形態があるが、臨床経過と病理学的特徴は両者で極めて似通っている。突然変異したときに常染色体優性形態のADを引き起こす3つの遺伝子がこれまでに発見されている。これら遺伝子はアミロイド・タンパク質前駆体(APP)と2つのタンパク質(構造と機能が互いに関連したプレセニリン-1(PS1)とプレセニリン-2(PS2))をコードしている。これら3つのタンパク質のどれかに突然変異があると、アミロイドβペプチド(アミノ酸が40〜42個のペプチドで、ADにおけるアミロイド斑の主要な成分であり、Aβペプチド、Aベータと呼ばれることもある)を産生する細胞内経路を通じてAPPのタンパク質分解処理が促進されることが観察されている(Younkin、Brain Pathol、第1巻(4)、253〜262ページ、1991年;Haass、J. Neurosci.、第11巻(12)、3783〜3793ページ、1991年)。
【0003】
タンパク質分解処理のための細胞内経路がうまく機能しなくなることがADの生理病理学にとって最も重要であろう。アミロイド斑が形成されるときには、APP、PS1、PS2のいずれかにおける突然変異が常にAPPのタンパク質分解処理を変化させるため、Aβ1〜42の形成が促進される。このAβ1〜42は、特にアミロイドを生成させるように見えるため、ADにおいて非常に重要なAβペプチドの一形態である。APPは、細胞表面などの分泌膜構造に局在しており、C末端の近くに膜貫通ドメインを有する(図1)。ヒトに存在することが現在知られているAPPの特別なアイソタイプの具体例としては、Kangら(Nature、第325巻、733〜736ページ、1987年)によって記載されているアミノ酸が695個のポリペプチド(“正常”APPと呼ばれる);Ponteら(Nature、第331巻、525〜527ページ、1988年)とTanziら(Nature、第331巻、528〜530ページ、1988年)によって記載されているアミノ酸が751個のポリペプチド;Kitaguchiら(Nature、第331巻、530〜532ページ、1988年)によって記載されているアミノ酸が770個のポリペプチドがある。
【0004】
Aβペプチドは、APPの領域のうちで膜貫通ドメインに隣接していてその膜貫通ドメインの一部を含む領域に由来する(図1)。通常は、α-セクレターゼ部位でAPPのプロセシングを行なうと、Aβ配列のうちで膜に隣接した中間領域が開裂し、細胞表面から可溶性細胞外ドメインが放出される。このα-セクレターゼによるAPPのプロセシングにより可溶性APP-αが生じるが、これはADの一因にならないと考えられている。しかしβ-セクレターゼ部位(α-セクレターゼ部位に対してN末端側に位置する)とγ-セクレターゼ部位(α-セクレターゼ部位に対してC末端側に位置する)でAPPの病的なプロセシングが起こるとAβペプチドが放出される。β-セクレターゼ開裂部位は、細胞膜管腔表面から28残基のところに位置し、APPγ-セクレターゼ開裂部位は膜貫通ドメインに位置する。(すべての細胞内の)細胞表面APPが再度内部化された後に、β-セクレターゼ部位とγ-セクレターゼ部位でのプロセシングが、(ニューロン内の)小胞体とエンドソーム/リソソーム経路の両方において起こる可能性がある。
【0005】
したがってβ-セクレターゼとγ-セクレターゼの酵素活性は、薬を発見するための標的である(Olson他、Curr. Opin. Drug Discovery & Develop.、第4巻、390〜401ページ、2001年)。これら2つの酵素は協力してAPPを開裂させる。APPはまず最初にβ-セクレターゼによって開裂されて、CT-100と呼ばれる膜結合C末端(CT)断片を生み出す。すると今度はこのCT-100が膜関連γ-セクレターゼの基質となる。プレセニリン1(PS1)依存γ-セクレターゼによるCT-100の膜内開裂により、Aβ1〜40とAβ1〜42が産生される。さらに、(γ様開裂またはε様開裂と名づけられた)別の開裂によって細胞膜の境界の内側約2〜5残基の位置でAPPの残基721の付近が開裂し、一連の安定なC末端APP断片が生じる(図1)。
【0006】
ノッチ-1は、細胞の運命を決める上で幅広い役割を果たしている細胞表面受容体のノッチ・ファミリーに属する。最近、APPのプロセシングが、細胞表面受容体であるノッチ-1のプロセシングに似ていることが主張されている(Wolf他、J. Biol. Chem.、第276巻、5413〜5416ページ、2001年)。実際、APPとノッチ-1は、内在性と考えられるγ-セクレターゼの競合する基質であることがわかっている。ノッチ-1は内在性膜タンパク質であり、リガンドを媒介として活性化されたときにその表面領域内でタンパク質分解処理される。ノッチ-1は、リガンドと結合した後、プレセニリンに依存した膜内γ-セクレターゼ開裂を起こす(Okochi、EMBO J.、第21巻、5408〜5416ページ、2002年)。第1の開裂は1731/1732部位(これはAβ様γ-セクレターゼ開裂に近い)であり、第2の開裂は1743/1744部位(これはAPPが生じるときのε開裂に近く、ノッチのS3開裂と呼ばれることもある;図1には“ε”開裂として示してある)である。膜貫通ドメインの端部近くで起こってノッチ-1細胞内ドメイン(NICD)を放出するのは、ノッチの1743/1744接合部におけるε開裂である。放出されたNICDは核に移動し、そこでCSL(この略号は、異なる系でこのタンパク質に与えられた3つの名前を表わしている。その系とは、哺乳動物におけるCBF1/RBP-J;ショウジョウバエとゼノプスにおける毛なしのサプレッサ[Su(H)];線虫のLag-1である)と名づけられたDNA結合タンパク質と相互作用する。NICDとCSLの間に形成される複合体は標的遺伝子の転写を変化させる。NICDは、胚発生において極めて重要なシグナル伝達経路で必要とされる(Schroeter、1998年;Hupert、2000年)。
【0007】
プレセニリンに依存したγ-セクレターゼ活性は、ノッチ受容体のプロセシングによってNICDを放出させるのに必要とされる(De Stropper他、Nature、第398巻、518〜522ページ、1999年)。細胞内では、ノッチ1とAPPが、PS1に依存したγ-セクレターゼ開裂の対象として競合する基質であることが報告されている(Berezovska他、J. Biol. Chem.、第276巻、30018〜30023ページ、2001年)。そのため、病原性Aβの産生を抑制するように設計されたγ-セクレターゼ阻害剤は、ノッチのシグナル伝達も抑制する。これは、γ-セクレターゼ阻害剤をADの治療薬として使用できる可能性があるという大きな意味を持つ。造血においてノッチ-1は重要な機能を担っているため、成人でノッチ-1のプロセシングを抑制すると、免疫不全と貧血になる可能性がある。したがってAPPの開裂によるCT-100の産生は特異的に抑制するが、ノッチの開裂は抑制しない化合物を同定する必要がある。そのような化合物は、NICDの産生を抑制するという悪い効果を生み出すことなくADを治療するための治療薬として役立つ可能性がある。
【発明の開示】
【0008】
本発明の概要
本発明により、γ-セクレターゼによるノッチの開裂を抑制しない化合物を同定する方法と組成物が提供される。この方法と組成物は、γ-セクレターゼを非特異的に抑制することに伴うNICDの産生抑制という問題を回避する。したがって本発明により、NICDの産生を抑制するという悪い効果を生み出すことなくADを治療するための治療薬を同定することが可能になる。
【0009】
本発明の第1の特徴により、NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質が提供される。このノッチ・タンパク質は、ノッチ・タンパク質の膜貫通ドメインを含んでいることが好ましい。しかしこのノッチ・タンパク質は、ノッチ・タンパク質がノッチのS3開裂部位を含んでいさえすれば、膜貫通ドメインではなく、ほぼ完全長のノッチ・タンパク質を含んでいてもよい。特別な一実施態様のノッチ・タンパク質は、配列番号4の配列を含んでおり、配列番号3の核酸配列によってコードされている。組み換えノッチ・タンパク質は、ノッチのS3開裂部位(すなわちε開裂部位)を含んでいる必要がある。組み換えノッチ・タンパク質は、脊椎動物のノッチ・タンパク質または無脊椎動物のノッチ・タンパク質が可能である。いくつかの実施態様では、組み換えノッチ・タンパク質は、配列番号5の配列[GenBank登録番号Z11886]を有するマウスのノッチ・タンパク質に由来する組み換えノッチ・タンパク質である。より特殊な実施態様では、マウスのノッチ・タンパク質のアミノ酸1703〜1860を含む組み換えノッチ・タンパク質を利用することを考える。本発明による可溶性融合タンパク質のどれも、C末端ヒスチジン・タグおよび/またはC末端フラグ・タグをさらに備えることができる。もちろん、ヒト・ノッチ配列の全体または一部(例えば配列番号6の配列[GenBank登録番号M73980])も用いることができよう。
【0010】
本発明ではさらに、この明細書に記載した融合タンパク質をコードするヌクレオチド配列を含むポリヌクレオチドを考える。そのような融合タンパク質をコードするポリヌクレオチド配列の一例は、配列番号1に示した配列を含んでいるものである。このポリヌクレオチド配列は、配列番号2の融合タンパク質をコードしている。この明細書では、本発明のポリヌクレオチドを含む発現ベクターも考える。この発現ベクターは、その内部に含まれるポリヌクレオチドがプロモータと機能上リンクしていて、そのポリヌクレオチドによってコードされるタンパク質の発現を宿主細胞の中で促進するような発現ベクターであることが好ましい。
【0011】
この明細書に記載したポリヌクレオチドまたは発現ベクターを用いた形質転換またはトランスフェクションを受けた組み換え宿主細胞も本発明の範囲に含まれる。本発明では、γ-セクレターゼ・アッセイのための基質を作る方法であって、そのような組み換え宿主細胞を増殖させて融合タンパク質の発現を可能にする操作を含む方法を考える。この方法は、その融合タンパク質を精製する操作をさらに含むことができる。このような方法では、宿主細胞は、組み換えタンパク質を産生できる任意の宿主細胞でよく、例えば哺乳動物宿主細胞、細菌宿主細胞、酵母宿主細胞などが可能である。具体的な実施態様では、宿主細胞は、Hela細胞、ヒト胚性腎臓細胞系の293細胞、線維芽細胞、CHO細胞のいずれかである。
【0012】
本発明の別の特徴により、可溶化ノッチ・タンパク質の製造方法であって、ノッチ・タンパク質をNusAタンパク質のC末端に融合させた融合タンパク質を調製する操作を含む方法が提供される。さらに詳細には、この方法は、融合タンパク質を組み換えによって産生させるため、ノッチのS3開裂部位のアミノ酸を含むノッチ・タンパク質をNusAタンパク質のC末端にリンクさせた融合タンパク質をコードしている核酸を含む発現構造体を調製し;その融合タンパク質の発現が容易になる条件下で、この発現構造体を用いて宿主細胞を形質転換し;その形質転換された宿主細胞を培地の中で増殖させる操作を含んでいる。この方法はさらに、培地内の形質転換された宿主細胞から融合タンパク質を単離する操作を含むことができる。いくつかの実施態様では、この方法は、化学的タンパク質合成を通じて融合タンパク質を製造する操作を含んでいる。そのような方法では、ノッチ・タンパク質は、マウスのノッチ・タンパク質のアミノ酸1703〜1860を含んでいることが好ましい。
【0013】
この明細書では、γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)を調べるインビトロ法であって、哺乳動物のγ-セクレターゼ複合体またはその生物学的に活性な断片を含む第1の組成物を、本発明の融合タンパク質を含む第2の組成物と接触させ;その融合タンパク質の開裂を調べる操作を含む方法も考える。
【0014】
上記方法のγ-セクレターゼ複合体は、哺乳動物の細胞から精製し単離した膜分画、あるいはこのγ-セクレターゼ複合体をコードしているヌクレオチド配列を含む発現構造体を用いた形質転換またはトランスフェクションを受けた細胞から精製し単離した膜分画を含むことができる。上記の方法では、融合タンパク質は、この明細書に記載した方法に従って調製した可溶化ノッチ・タンパク質である。
【0015】
本発明では、この明細書に記載した方法で同定されたγ-セクレターゼ・モジュレータを含む組成物を特に考える。本発明には、生体内でγ-セクレターゼの活性を変化させる方法であって、哺乳動物に対し、この明細書に記載したスクリーニング法で同定されたモジュレータを、その哺乳動物の細胞内でγ-セクレターゼの活性を変化させるのに有効な量投与するステップを含む方法も含まれる。このモジュレータは、γ-セクレターゼによるノッチ・タンパク質の開裂ではなく、γ-セクレターゼによるAPPの開裂を選択的に抑制するγ-セクレターゼ・モジュレータである。
【0016】
本発明は、本発明によって同定された1つ以上のモジュレータと、薬理学的に許容可能な基剤とを含む医薬組成物にも関する。本発明では、γ-セクレターゼの異常な活性を特徴とする疾患や症状を治療する方法であって、治療を必要としている対象に本発明の医薬組成物を投与する操作を含む方法も考える。この明細書で同定されたモジュレータを利用してアルツハイマー病の治療薬を製造することも特に考える。
【0017】
本発明の別の特徴ならびに利点は、以下の詳細な説明から明らかになろう。しかしこの詳細な説明と具体的な実施例は、本発明の好ましい実施態様を示しているとはいえ、単なる例示であることを理解する必要がある。なぜなら、当業者には、本発明の範囲内でのさまざまな変化や変更がこの詳細な説明から明らかになるだろうからである。
【0018】
図面は本発明の明細書の一部であり、本発明の特徴を詳しく説明するためのものである。本発明は、図面と、この明細書に提示した具体的な実施態様の詳細な説明とを組み合わせることによってさらによく理解できよう。
【発明を実施するための最良の形態】
【0019】
ADは進行性の痴呆と症状を伴う重要な加齢関連疾患であり、皮質の萎縮と老人斑の堆積を特徴とする。老人斑の主成分は、アミノ酸40〜42個の長さがあるペプチドAβであり、完全長APPの膜貫通ドメインの一部に隣接した領域、またはその部分を含む領域に由来する。この病原性ペプチドは、β-セクレターゼとγ-セクレターゼの活性に起因する一連のプロセシングの結果として生成される。したがってこれら2つのセクレターゼの酵素活性が、薬を発見するための標的となる(Olson他、Curr. Opin. Drug Discovery & Develop.、第4巻、390〜401ページ、2001年)。
【0020】
細胞表面受容体であるノッチのプロセシングはAPPのプロセシングと似ていることがわかっている(Wolf他、J. Biol. Chem.、第276巻、5413〜5416ページ、2001年)。図1には、APPとノッチにおけるγ-セクレターゼ様開裂部位を比較して示してある。この図からわかるように、APPの開裂部位は膜貫通ドメインの中央部にあるのに対し、ノッチは膜貫通ドメインのC末端に非常に近い位置が開裂する。似たようなε開裂部位が、カドヘリンEにおいて最近報告された(Marambaud他、EMBO J.、第21巻、1948〜1956ページ、2002年)(図1)。図2は、ノッチ・タンパク質のアミノ酸配列でノッチ内のS3開裂部位を含む1703〜1860を示している(Steiner他、J. Mol. Neurosci.、第17巻、193〜198ページ、2001年)。1743/1744接合部における特異的開裂により、V1744〜D1860ノッチ細胞内ドメイン(NICD)が生じる可能性がある。これは、S1開裂部位とS2開裂部位が欠如したmΔEノッチ構造体(1704〜2183)をトランスフェクトされた細胞で観察されるNICDの断片である(Kopan他、Proc. Natl. Acad. Sci.、第93巻、1683〜1688ページ、1996年)。ノッチ構造体(バリン1711〜グルタミン酸1809)が報告されている(Esler他、Proc. Natl. Acad. Sci.、第99巻、2720〜2725ページ、2002年)とはいえ、試験管内でのγ-セクレターゼによる1743/1744接合部の特異的開裂は記載されていない。1743/1744接合部における特異的開裂は、バリン1744抗体(セル・シグナリング・テクノロジー社)を用いて容易に明らかにすることができる。この抗体は開裂したノッチに対して特異的であり、開裂していないノッチ・タンパク質と交差反応することはない。
【0021】
したがって細胞では、PS1依存性γ-セクレターゼは、β-セクレターゼの作用によって生じるCT-100断片を開裂させることに加え、ε部位1743/1744でも開裂させてNICDを生成させる。開裂したこのNICDは核に移動し、シグナル伝達に関与する。ADを緩和するためにγ-セクレターゼの活性を抑制すると、NICDの生成も抑制される。本発明により、初めて、ノッチ-1からのNICDの生成を抑制しない治療薬を同定する方法と組成物が提供される。
【0022】
本発明の方法により、ノッチの開裂を調べるインビトロ・アッセイと、そのようなアッセイを利用してγ-セクレターゼの阻害剤を二次評価する方法が提供される。このアッセイでは、γ-セクレターゼの可溶性ノッチ基質を利用する。この基質は、NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質を含んでいる。本発明のアッセイは、γ-セクレターゼによるノッチ・タンパク質の開裂を調べる直接インビトロELISAアッセイおよび/またはウエスタン・ブロットとして実施することができる。本発明の方法により、適切な細胞系(例えば大腸菌)の中でNusAをノッチ・タンパク質配列に融合させた融合タンパク質として発現する可溶性組み換えノッチ・タンパク質(アスパラギン1703〜アスパラギン酸1860)基質の産生と精製がなされる(図6を参照のこと)。
【0023】
発明者は、本発明による精製された融合タンパク質基質を利用し、HeLa細胞からの可溶化γ-セクレターゼによってノッチの1743/1744部位が特異的に開裂することを証明した。実施例に詳しく説明したように、開裂したノッチ・タンパク質はバリン1744に対して特異的な抗体を用いて検出することができる。発明者は、このアッセイが有効であることを確認し、ノッチ・タンパク質の開裂が、γ-セクレターゼの強力な阻害剤としてよく知られているDAPTによって投与量に依存した形で抑制されることを示した。抗バリン1744抗体に基づいたELISAを開発した。さらに、開裂がγ-セクレターゼ阻害剤によって抑制されることでインビトロ・ノッチ・アッセイが有効であることを示した。阻害剤またはモジュレータは、機能的には、γ-セクレターゼを通じてAβを減少させる化合物と定義される。
【0024】
I.インビトロ・ノッチ・アッセイのためのノッチ基質
【0025】
本発明のアッセイで使用するノッチ基質は、ノッチ・ポリペプチドをNusタンパク質に融合させた可溶性融合タンパク質である。この融合タンパク質に標識すること、あるいはそれ以外の修飾を行なうことが可能である。それは、ペプチドを精製するため、あるいはノッチ融合タンパク質そのものを検出するため、あるいはγ-セクレターゼが作用したときにノッチ融合タンパク質の開裂産物の検出を容易にするためである。融合タンパク質の生成法と具体的な修飾法について以下にさらに詳しく説明する。
【0026】
本発明の好ましい1つの特徴によれば、マウスのノッチ-1タンパク質(DNA配列登録番号Z11886;図2を参照のこと)のアミノ酸N1703〜D1860を含むノッチ基質のN末端をNusAのC末端に接合した。この融合ポリペプチドを組み換えタンパク質の産生によって、あるいは自動化したペプチド合成によって得ることを考える。後者についてはこの明細書の別の箇所で説明してある。図2の膜貫通ドメインは、アミノ酸1723〜1744にまたがっている(図1を参照のこと)。γ-セクレターゼは、ノッチのアミノ酸1743と1744の間にあるε開裂部位を開裂させる。この開裂部位はS3開裂部位とも呼ばれており、NICD(すなわちアミノ酸V1744〜D1860の範囲のペプチド)を発生させる。
【0027】
本発明では、この新規な融合タンパク質に加え、上記のノッチ/NusA融合タンパク質基質、または本発明に従って同定したノッチ/NusA融合タンパク質基質の末端部付加物(融合タンパク質または融合ポリペプチドとも呼ばれる)を作ることも考える。さらに、本発明の好ましい実施態様にはマウスのノッチ-1のN1703〜D1860を含むノッチ/NusA融合ペプチドを示してあるが、ノッチ・タンパク質は任意の供給源からのものが可能であることを理解する必要がある。そのような供給源としては哺乳動物または非哺乳動物が可能である。したがってノッチ-1はヒト、マウス、ラット、あるいはそれ以外の哺乳動物に由来することが好ましいとはいえ、例えば線虫、ゼノプス、ショウジョウバエや、他の無脊椎動物からのものも可能である。
【0028】
さらに、γ-セクレターゼ開裂部位1731/1732(γ部位)と1743/1744(ε開裂)を含むあらゆるノッチ-1誘導体が本発明による融合タンパク質基質において有用であることも考慮する。ノッチ-1では、NICDを放出するγ-セクレターゼ開裂部位は、ノッチ内の1743/1744部位に位置する。この開裂部位とNusAの開始位置の距離を約500アミノ酸にして、ノッチのγ-セクレターゼ開裂ドメインの立体特性に似せることを考える。この距離は、NusAタンパク質から生じさせるか、あるいは異種ペプチド・リンカーによって作り出すことができる。この領域はNusAタンパク質からのものであることが好ましい。ノッチ/NusA融合ポリペプチドのNusAタンパク質ドメイン部分としては、そのノッチ/NusA融合ポリペプチドが可溶性を維持していて、したがってインビトロ・アッセイを実施できるようなNusAタンパク質の任意の部分が可能である。
【0029】
これまでNusAは大腸菌で可溶性タンパク質を発現させるのに用いられてきた(例えばWilkinson他、Bio/Technology、第9巻、443〜448ページ、1991年;Davis他、Biotechnol. Bioeng.、第65巻(4)、382〜388ページ、1999年を参照のこと)。この明細書では、(配列番号16の核酸配列によってコードされる)配列番号17の配列を含むNusAタンパク質をノッチと融合させる。しかし当業者であれば、配列番号17に示した配列以外のNusA配列を使用しても本発明の可溶性ノッチ/NusA融合タンパク質を作り出すことができよう。当業者は、例えば配列番号17の配列と80%、85%、90%、95%、96%、97%、98%またはそれ以上が相同であるNusAタンパク質を使用することができる。あるいは当業者は、配列番号17に由来するNusAのより小さな連続断片を用いることもできる。例えばその断片は、配列番号17の50個、100個、150個、200個、250個、300個、350個、400個、425個、450個、475個、500個、510個、520個、530個、540個、550個またはそれ以上の連続したアミノ酸にすることができる。
【0030】
融合タンパク質を設計して作り出すための一般的な原理は当業者には周知である。例えば融合体には他の種からのリーダー配列が一般に用いられているため、異種宿主の中で組み換えタンパク質または組み換えペプチドを発現させることができる。別の有用な融合体では、免疫活性のあるドメイン(例えば抗体のエピトープ)を付加して融合ポリペプチドの精製を容易にする。開裂部位を融合が起こる位置またはその近傍に組み込むと、精製後に外来性ポリペプチドを除去することが容易になる。このような融合体を組み換えによって産生させることについてはこの明細書の別の箇所でさらに詳しく説明する。他の有用な融合体としては、機能ドメイン(例えば酵素からの活性部位、グリコシル化ドメイン、細胞標的シグナル、膜貫通領域)がリンクしたものが挙げられる。
【0031】
より詳細には、本発明において、1731/1732(γ開裂)開裂部位と1743/1744(ε開裂)開裂部位を有するノッチ・タンパク質を含む第1の成分が、NusAタンパク質の全体または一部を含む第2の成分に結合した融合ポリペプチドを考える。さらに別の実施態様では、融合ポリペプチドは、レポータ遺伝子産物を含む第3の成分をさらに含むことができる。さらに別の実施態様では、融合ポリペプチドは、タグの付いた配列成分をさらに含むことができる。考慮する特別な融合ポリペプチドは、NusA部の一方の側にあるレポータ遺伝子産物と、γ-セクレターゼ開裂部位を有するノッチ・タンパク質を含む配列と、ノッチ・タンパク質の他方の側にあるタグ付き配列とを備えるものである。本発明の融合ポリペプチドで使用するレポータ遺伝子産物は、当業者が一般に使用している任意のレポータ・タンパク質でよい。具体的なレポータ・タンパク質としては、ルシフェラーゼ;分泌アルカリホスファターゼ(SEAP);β-ガラクトシダーゼ;β-グルクロニダーゼ;グリーン蛍光タンパク質;クロラムフェニコールアセチルトランスフェラーゼなどが挙げられる。
【0032】
別の特別な実施態様では、本発明の融合ポリペプチドの第4の成分としてタグ付き配列をさらに考える。市販されているさまざまな融合タンパク質発現系があり、それを本発明のこの文脈でタグ付き配列として使用することができる。特に有用な系としては、グルタチオン-S-トランスフェラーゼ(GST)系(ファルマシア社、ピスカタウェイ、ニュージャージー州)、マルトース結合タンパク質系(NEB社、ビヴァリー、マサチューセッツ州)、フラグ系(IBI社、ニュー・ヘイヴン、コネチカット州)、6×ヒスチジン系(キアジェン社、チャッツワース、カリフォルニア州)などが挙げられる。これらの系は、組み換え融合タンパク質が関係する生物学的活性に影響を与えそうにない少数の付加アミノ酸だけを有する組み換えポリペプチドを産生させることができる。例えばフラグ系と6×ヒスチジン系の両方とも、短い配列だけを付加する。両方とも抗原性が弱いことが知られており、ポリペプチドが折り畳まれて本来の立体配座になるのを妨げない。有用であると考えられる別のN末端融合体は、タンパク質またはペプチドのN末端領域にメチオニン-リシン・ジペプチドが融合したものである。このような融合体によってタンパク質の発現および/または活性が好ましく増大する可能性がある。本発明で使用することが考えられる特別なタグ付き配列としては、C末端フラグ・タグ配列DYKDDDDK(配列番号14)がある。さらに、このタグ付き配列は8ヒスチジン・タグも含んでいてよく、その場合にはフラグ/8ヒスチジン・タグ(DYKDDDDKHHHHHHHH、配列番号15)が融合タンパク質に結合する。
【0033】
本発明の好ましい融合タンパク質の一例は、γ-セクレターゼ開裂部位1731/1732と1743/1744に加えて短いC末端フラグ/8ヒスチジン・タグ尾部を含むマウスの完全長ノッチ-1タンパク質またはその一部にNusAが融合しているものである。ある融合ポリペプチドの配列は、NusAタンパク質に融合したマウスのノッチのアミノ酸1703〜1860(すなわち配列番号13に示したもの)を含んでいる。この融合ポリペプチドは配列番号2の配列を有する。γ-セクレターゼによるこのキメラ構造体の開裂をモニターするため、バリン1744残基を含むノッチ開裂産物に対する抗バリン1744抗体を用い、ノッチが開裂する結果として生成されるバリン1744断片の存在と濃度を明らかにする。キメラ構造体のC末端を検出するのに抗フラグ抗体を用いることができる。しかしキメラ構造体では、フラグ・タグの代わりにC末端にレポータ・タンパク質(例えばアルカリホスファターゼ)も用いることができる。このようなレポータ・タンパク質はノッチが開裂する結果として放出される可能性があるため、当業者に周知のアッセイを利用して検出できよう。
【0034】
本発明により、すでに説明した融合ポリペプチドが提供されることに加え、例えば標識またはそれ以外の検出可能な部分を組み込む修飾をさらに行なったノッチ融合タンパク質が提供される。
【0035】
例えばノッチ/NusA融合タンパク質基質は内部に消光状態の標識を備えており、それが、ノッチ基質が開裂した後の検出可能性を高める。ノッチ/NusA融合タンパク質基質を修飾して蛍光体(例えば7-アミノ,4-メチルクマリンと消光物質(例えば2,4-ジニトロフェノール)のペアを結合させると、γ-セクレターゼによってペプチドが開裂したとき、γ-セクレターゼによって切断されやすい結合の両側に結合させた蛍光体と消光物質が物理的に分離するために蛍光が増大する。ペアにする別の蛍光体と消光物質としては、ボディピー-テトラメチルローダミンとQSY-5(モレキュラー・プローブズ社)がある。このアッセイの別の態様では、ビオチンまたは他の適切なタグをペプチドの一端に配置してそのペプチドを基質アッセイ用プレートに固定した後、蛍光体をそのペプチドの他端に配置することができる。有用な蛍光体としては、上記のもののほか、W8044(EG&gウォラック社)などのユーロピウム標識がある。使用可能な別の好ましい標識は、システイン残基に結合させることのできるオレゴン・グリーンである。γ-セクレターゼによる融合タンパク質の開裂によってプレートから蛍光体または他のタグが放出されるため、保持された蛍光の増大から、タンパク質分解による開裂がいろいろな化合物について抑制されたかどうかを知ることができる。γ-セクレターゼのタンパク質分解活性に関する好ましい比色アッセイでは、開裂するγ-セクレターゼ認識部位を有するアミノ酸配列がアミド結合を通じてo-ニトロフェノールに結合した別のノッチ/NusA融合タンパク質を用いる。そのためγ-セクレターゼによって融合タンパク質が開裂すると、アッセイ用緩衝液がアルカリpHに変化した後に光学密度が増大する。
【0036】
さらに、ノッチ/NusA融合タンパク質には当業者に周知の標識を付けることができる。例えばビオチン標識が特に考えられる。このような標識を使用することは当業者には周知であり、例えばアメリカ合衆国特許第3,817,837号、第3,850,752号、第3,996,345号、第4,277,437号に記載されている。有用な他の標識としては、放射性標識、蛍光標識、化学的発光標識などがある。このような標識の利用に関するアメリカ合衆国特許としては、例えばアメリカ合衆国特許第3,817,837号、第3,850,752号、第3,939,350号、第3,996,345号がある。本発明のどのノッチ/NusA融合タンパク質組成物も、これら標識のうちの1つ、2つ、またはそれ以上を備えることができる。
【0037】
II.γ-セクレターゼ組成物
【0038】
本発明は、新規なノッチ融合タンパク質に加え、そのようなノッチ融合タンパク質をさまざまなγ-セクレターゼ・アッセイで利用する方法にも関する。このセクションでは、γ-セクレターゼ活性を有するタンパク質を含む分画の生成について説明する。
【0039】
γ-セクレターゼの正確な同定はまだなされていないが、γ-セクレターゼがプレセニリン1であるらしいという強い証拠がある。γ-セクレターゼ・タンパク質の配列はまだ明らかになっていないが、当業者は、γ-セクレターゼを含む細胞分画を単離するための方法と組成物は知っている。例えばLiら(Proc. Natl. Acad. Sci.、第97巻、6138〜6143ページ、2000年)は、活性のある可溶化γ-セクレターゼを含む膜調製物を得るための方法と組成物を報告している。このような方法は、本発明においてγ-セクレターゼを含む精製された分画を得るのに役立つ。そのような分画が得られると、それを本発明のアッセイで利用できる。さらに、本発明の新規な融合タンパク質も利用し、例えばアフィニティ・クロマトグラフィ法でそのような膜分画からγ-セクレターゼをさらに単離して精製することができる。
【0040】
γ-セクレターゼ分画は、一般に、γ-セクレターゼの活性を示す細胞から単離する。例えばHeLa3細胞を用いることができる。この細胞を例えばフレンチ・プレスまたは他の細胞破壊技術を利用して破壊する。細胞破壊技術としては、例えば凍結解凍法(例えば細胞をドライアイスと37℃の水浴を往復させる);ヒューズ・プレスまたはフレンチ・プレスを用いた固体剪断法;洗剤による溶解(例えばトゥイーン、トリトン、NP-40などのオン-イオン性洗剤溶液);低張液による溶解(例えば水、クエン酸緩衝液);液体剪断法(ホモジナイザー、衝突ジェット微小流体化装置);超音波処理(超音波)などがある。細胞を溶解させた後、遠心分離を利用した沈降によって細胞の残滓と核を除去することができる。膜分画は、例えば100,000×gで60分間にわたって沈殿させ、さらに洗剤を用いて可溶化させることができる。例えばLiら(上記文献)の可溶化法では、100,000×gでの遠心分離ステップからの膜分画ペレットを緩衝液の中に再び懸濁させ、1%CHAPSOを用いて4℃にて60分間にわたって処理し、60分間にわたって遠心分離する。洗剤を用いたこの可溶化法では、100,000×gで得られた分画の上澄に可溶化γ-セクレターゼが含まれている。
【0041】
この明細書に記載したすべてのγ-セクレターゼ・アッセイで上記の可溶化分画を用いる。
【0042】
III.タンパク質またはペプチドの製造と精製
【0043】
本発明により、γ-セクレターゼのモジュレータのうち、APPの開裂に特異的だがノッチの開裂は抑制しないものを同定するのに用いる可溶性ノッチ・タンパク質基質が提供される。この基質は、従来からある自動化ペプチド合成法、あるいは組み換え発現法によって得ることができる。
【0044】
A.合成ペプチドの製造
【0045】
本発明のペプチドは、あるいは本発明の完全長融合ポリペプチドでさえ、従来法に従って溶液中で、あるいは固体支持体上で合成することができる。さまざまな自動化合成装置が市販されており、公知のプロトコルに従って使用することができる。例えばStewartとYoung、『固相ペプチド合成』、第2版、ピアース・ケミカル社、1984年;Tam他、J. Am. Chem. Soc.、第105巻、6442ページ、1983年;Merrifield、Science、第232巻、341〜347ページ、1986年;BaranyとMerrifield、『ペプチド』、GrossとMeienhofer編、アカデミック出版、ニューヨーク、1〜284ページ、1979年を参照のこと。なお各文献の内容は参考としてこの明細書に組み込まれているものとする。γ-セクレターゼによって開裂するγ-セクレターゼ開裂部位1731/1732と1743/1744を含む本発明によるノッチ融合タンパク質の新規な基質は容易に合成することができ、その後γ-セクレターゼ・スクリーニング・アッセイでスクリーニングすることができる。
【0046】
特に好ましい方法では、本発明の融合タンパク質を、アプライド・バイオシステムズ社のモデル433Aを用いて固相法で合成した。自動化ペプチド合成法または組み換え法で得られたどのノッチ融合タンパク質の純度も、逆相HPLC分析を利用して測定することができる。それぞれのペプチドが化学的に本物であることは、当業者に周知の任意の方法で確認することができる。好ましい実施態様では、本物であることは質量分析によって確認される。
【0047】
さらに、融合タンパク質は、アミノ酸分析においてマイクロ波加水分解を実施することよって定量することができる。このような分析では、電子レンジ、例えばCEM社のMDS 2000電子レンジを用いるとよい。ペプチド(約2mg)を、6NのHCl(ピアース・コンスタント・ボイリング社、例えば約4ml)および約0.5%(v/v)のフェノール(マリンクロット社)と接触させる。加水分解の前にサンプルの排気とN2を用いたパージを交互に行なう。タンパク質の加水分解は、2段階プロセスで実施する。第1段階では、融合タンパク質を約100℃の反応温度にし、この温度を1分間維持する。このステップの直後に温度を150℃に上昇させ、この温度を約25分間維持する。冷却後、サンプルを乾燥させ、加水分解した融合タンパク質サンプルからのアミノ酸を6-アミノキノリル-N-ヒドロキシスクシンイミジルカルバメートを用いて誘導体化し、安定な尿素を得る。この尿素は、395nmで蛍光を出す(ウォーターズAccQタグ・ケミストリー・パッケージ)。サンプルを逆相HPLCで分析し、強力インテグレータを用いて定量する。
【0048】
B.組み換えペプチドの産生
【0049】
自動化ペプチド合成の代わりに組み換えDNA技術を利用することができる。その場合、以下に説明するように、本発明のペプチドをコードしているヌクレオチド配列を発現ベクターに組み込み、その発現ベクターを用いて適切な宿主細胞の形質転換またはトランスフェクションを行ない、発現に適した条件下でその宿主細胞を培養する。組み換え法は、本発明のペプチド配列を含む長いポリペプチドを産生させるのに特に好ましい。
【0050】
本発明による新規なノッチ融合タンパク質基質に関する説明から、組み換え技術によって融合ポリペプチドを産生させることができる。ペプチドまたは融合ポリペプチドをコードしている配列を含んでいてそのペプチドまたは融合ポリペプチドを発現させる多彩な発現ベクター/宿主系を利用することができる。そのような系としては、微生物(例えば組み換えバクテリオファージで形質転換した細菌、プラスミドDNA発現ベクター、コスミドDNA発現ベクター);酵母発現ベクターで形質転換した酵母;ウイルス発現ベクター(例えばバキュロウイルス)を感染させた昆虫細胞系;ウイルス発現ベクター(例えばカリフラワー・モザイク・ウイルス(CaMV)、タバコ・モザイク・ウイルス(TMV))をトランスフェクトした植物細胞系、または細菌発現ベクター(例えばTiプラスミドまたはpBR322プラスミド)で形質転換した植物細胞系;動物細胞系などが挙げられる。組み換えタンパク質の産生で役立つ哺乳動物の細胞としては、VERO細胞、HeLa細胞、チャイニーズ・ハムスター卵巣(CHO)細胞系、COS細胞(例えばCOS-7)、W138、BHK、HepG2、3T3、RIN、MDCK、A549、PC12、K562、293細胞などが挙げられる。ノッチ融合タンパク質を細菌、酵母、あるいは他の無脊椎動物で組み換え発現させるための具体的なプロトコルはあとで説明する。
【0051】
原核生物宿主で用いる発現ベクターは、一般に、1種類以上の表現型選択マーカー遺伝子を含んでいる。そのような遺伝子は、一般に、抗生物質に対する耐性を与えるタンパク質、または栄養要求性条件を与えるタンパク質をコードしている。多彩なそのようなベクターを市場から容易に入手することができる。具体的には、pSPORTベクター、pGEMベクター(プロメガ社)、pPROEXベクター(LTI社、ベセスダ、メリーランド州)、ブルースクリプト・ベクター(ストラタジーン社)、pETベクター(ノヴァジェン社)、pQEベクター(キアジェン社)などがある。所定のノッチ融合タンパク質をコードしているDNA配列をPCRによって増幅し、クローニングしてそのようなベクター(例えばpGEX-3X(ファルマシア社、ピスカタウェイ、ニュージャージー州))に導入する。そのベクターは、そのベクターによってコードされているグルタチオン-S-トランスフェラーゼ(GST)を含む融合タンパク質、またはそのベクターのクローニング部位に挿入されたDNA断片がコードしているタンパク質を産生するように設計されている。PCRのプライマーとしては、例えば適切な開裂部位を含むものを作ることができる。組み換え融合タンパク質をトロンビンまたはXa因子(ファルマシア社、ピスカタウェイ、ニュージャージー州)で処理すると融合タンパク質が開裂し、基質または基質含有ポリペプチドがGSTの部分から放出されることが期待される。pGEX-3X/融合タンパク質構造体を導入して大腸菌XL-1ブルー細胞(ストラタジーン社、ラ・ジョラ、カリフォルニア州)を形質転換し、個々の形質転換体を単離して増殖させる。個々の形質転換体からのプラスミドDNAを精製し、自動化シークエンサを用いて一部のシークエンシングを行ない、適切な方向の核酸挿入体をコードしている望むペプチドまたはポリペプチドの存在を確認する。
【0052】
GST/基質融合タンパク質の誘導は、形質転換したXL-1ブルー培養物を37℃にてLB培地(カルベニシリンを補足)の中で波長600nmにおける光学密度が0.4になるまで増殖させた後、0.5mMのイソプロピル*-D-チオガラクトピラノシド(シグマ・ケミカル社、セントルイス、ミズーリ州)の存在下でさらに4時間にわたってインキュベートすることによって実現される。
【0053】
細菌の中に不溶性封入体として産生されることが予想されるGST融合タンパク質は、以下のようにして精製することができる。細胞を遠心分離によって回収し;0.15MのNaCl、10mMのトリス(pH8.1)、1mMのEDTAの中で洗浄し;0.1mg/mlのリゾチーム(シグマ・ケミカル社)で室温にて15分間にわたって処理する。ライセートを超音波処理によって透明にし、細胞の残滓を10分間にわたって12,000×gで遠心分離することによってペレット化する。融合タンパク質を含むこのペレットを50mMのトリス(pH8)と10mMのEDTAの中に再び懸濁させ、50%グリセロールの上で層にし、30分間にわたって6000×gで遠心分離する。ペレットを、Mg2+とCa2+を含まない標準的なリン酸緩衝溶液(PBS)の中に再び懸濁させる。再び懸濁させたペレットを変性用SDSポリアクリルアミド・ゲルの中で分画することによって融合タンパク質をさらに精製する(Sambrook他編、コールド・スプリング・ハーバー出版、コールド・スプリング・ハーバー、ニューヨーク州、1989年)。ゲルを0.4MのKClの中に浸してタンパク質を可視化する。このタンパク質を切断し、SDSが欠けたゲル-ランニング緩衝液の中で電気溶離させる。GST/ノッチ融合タンパク質を細菌の中で可溶性タンパク質として産生させる場合には、GST精製モジュール(ファルマシア・バイオテック社)を用いて精製するとよい。
【0054】
融合タンパク質をトロンビンで消化させて成熟ノッチ融合ポリペプチドからGSTを開裂させることができる。消化反応物(20〜40μgの融合タンパク質と、20〜30単位のヒト・トロンビン(4000U/mg(シグマ社)を含む0.5mlのPBS))を室温にて16〜48時間にわたってインキュベートし、変性用SDS-PAGEゲルに充填して反応生成物を分画する。ゲルを0.4MのKClに浸してタンパク質のバンドを可視化する。融合ポリペプチドの予想分子量に対応するタンパク質のバンドは、自動化シークエンサ(アプライド・バイオシステムズ社のモデル473A、フォスター・シティ、カリフォルニア州)で一部のアミノ酸配列を分析することによって確認できる。
【0055】
融合ポリペプチドを含む予想される基質をコードしているDNA配列をクローニングし、望むプロモータを含んでいて、必要に応じてさらにリーダー配列を含むプラスミドに組み込むこともできる(例えばBetter他、Science、第240巻、1041〜1043ページ、1988年を参照のこと)。この構造体の配列は、自動化したシークエンシングによって確認できる。次にこのプラスミドを用い、CaCl2を用いたインキュベーションと熱ショック処理を行なうという標準的な方法で大腸菌を形質転換する(Sambrook他、上記文献)。形質転換した大腸菌を、カルベニシリンを補足したLB培地の中で増殖させ、適切な培地の中で増殖させることによって発現タンパク質の産生を誘導する。リーダー配列が存在している場合には、そのリーダー配列がノッチをベースとした成熟融合タンパク質の分泌に影響を与え、そのタンパク質が分泌の間に開裂する。
【0056】
この明細書に記載した方法により、分泌された組み換えタンパク質を細菌培地から精製する。
【0057】
同様に、サッカロミセス、ピキア、クルベロミセスなどの属に由来する酵母宿主細胞を用いて組み換えペプチドを産生させることができる。好ましい酵母宿主細胞は、サッカロミセス・セレビジエとピキア・パストリスである。酵母ベクターは、2T酵母プラスミドからの複製起点配列と、自律複製配列(ARS)と、プロモータ領域と、ポリアデニル化のための配列と、転写終結のための配列と、選択マーカー遺伝子とを含んでいることがしばしばある。酵母と大腸菌の両方の中で複製可能なベクター(シャトル・ベクターと呼ばれる)も使用することができる。シャトル・ベクターは、酵母ベクターの上記特徴に加え、大腸菌の中で複製と選択を行なうための配列も含んでいる。酵母宿主細胞の中で発現するポリペプチドの直接的な分泌は、基質をコードしているヌクレオチド配列の5'末端に酵母I因子リーダー配列をコードしている配列を含めることによって実現される。
【0058】
一般に、所定の基質は、市販されている発現系(例えばピキア発現系(インヴィトロジェン社、サン・ディエゴ、カリフォルニア州))を製造者の指示に従って使用して酵母の中で組み換え発現させることができる。この系は、分泌を指示するプレ-プロ-α配列にも依存しているが、挿入体の転写は、メタノールによって誘導されたときにアルコールオキシダーゼ(AOX1)プロモータによって起こる。
【0059】
分泌された組み換え基質は酵母増殖培地から精製する。そのとき、例えば細菌や哺乳動物の細胞の上澄から基質を精製するのに用いる方法を利用する。
【0060】
本発明の新規な基質をコードしている合成DNAをクローニングしてバキュロウイルス発現ベクターpVL1393(ファーミンジェン社、サン・ディエゴ、カリフォルニア州;LuckowとSummers、Bio/Technology、第6巻、47ページ、1988年)に入れることもできる。次に、基質を含むこのベクターを製造者(ファーミンジェン社)の指示に従って用い、sF9タンパク質を含まない培地の中でスポドプテラ・フルギペルダ細胞に感染させ、組み換えタンパク質を産生させる。このタンパク質またはペプチドを、ヘパリン-セファロース・カラム(ファルマシア社、ピスカタウェイ、ニュージャージー州)と直列分子サイズ分類カラム群(アミコン社、ビヴァリー、マサチューセッツ州)を用いて培地から精製し、濃縮し、PBSの中に再び懸濁させる。SDS-PAGE分析により単一のバンドが現われ、タンパク質のサイズが確認される。ポートン2090ペプチド・シークエンサでエドマン・シークエンシングを行なうことによってそのタンパク質のN末端配列が確認される。
【0061】
ノッチ融合タンパク質基質は、昆虫系で発現させることもできる。タンパク質を発現させるための昆虫系は当業者によく知られている。そのような1つの系では、オートグラファ・カリフォルニア核多角体病ウイルス(AcNPV)が、スポドプテラ・フルギペルダ細胞またはトリコプルシアの幼虫の中で外来遺伝子を発現させるためのベクターとして用いられる。基質をコードしている配列をクローニングしてウイルスの不可欠ではない領域(例えばポリヘドリン遺伝子)に挿入し、ポリヘドリン・プロモータの制御下に置く。基質がうまく挿入されるとポリヘドリン遺伝子が不活性になり、コート・タンパク質が欠けた組み換えウイルスが産生される。次に組み換えウイルスをスポドプテラ・フルギペルダ細胞またはトリコプルシアの幼虫に感染させ、その中で基質を発現させる(Smith他、J. Virol.、第46巻、584ページ、1983年;Engelhard E.K.他、Proc. Natl. Acad. Sci.、第91巻、3224〜3227ページ、1994年)。
【0062】
組み換えタンパク質を発現させるための哺乳動物宿主系も当業者にはよく知られている。宿主細胞株は、発現したタンパク質を処理したり、タンパク質に活性を与えるのに役立つある種の翻訳後修飾をしたりする特別な能力を持つものを選択することができる。ポリペプチドのこのような修飾としては、アセチル化、カルボキシル化、グリコシル化、リン酸化、脂質化、アシル化などがある。正確な挿入、折り畳み、機能のためには、タンパク質の“プレプロ”形態を開裂させる翻訳後プロセシングも重要である可能性がある。さまざまな宿主細胞(例えばCHO、HeLa、MDCK、293、WI38など)が、そのような翻訳後活性に関する特別な細胞機構と特徴的メカニズムを備えている。このような宿主細胞を選択することで、導入された外来タンパク質の正確な修飾とプロセシングを確実に行なえる。
【0063】
形質転換された細胞を長期にわたる高収率のタンパク質産生に利用できること、そしてその形質転換細胞がそのままで安定に発現することが好ましい。このような細胞を、選択マーカーと望む発現カセットとを含むベクターを用いて形質転換したときには、その細胞を1〜2日間栄養強化培地の中で増殖させた後、選択培地に切り換えるとよい。選択マーカーは選択に対する耐性を有するように設計する。したがってその選択マーカーが存在していると、導入配列をうまく発現する細胞の増殖と回収が可能になる。耐性を有する安定に形質転換された細胞の塊は、その細胞に適した組織培養技術を利用して増殖させることができる。
【0064】
多数の選択系を用い、組み換えタンパク質を産生するように形質転換された細胞を回収することができる。そのような選択系としては、tk細胞のHSVチミジンキナーゼ遺伝子、hgprt細胞のヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ遺伝子、aprt細胞のアデニンホスホリボシルトランスフェラーゼ遺伝子などが挙げられる。抗代謝物耐性を選択の基礎として利用し、メトトレキセートに対する耐性を与えるdhfr;ミコフェノール酸に対する耐性を与えるgpt;アミノグリコシドG418に対する耐性を与えるネオ;クロルスルフロンに対する耐性を与えるALS;ハイグロマイシンに対する耐性を与えるハイグロに関する選択を行なうこともできる。役に立つ可能性のある別の選択用遺伝子としては、細胞がトリプトファンの代わりにインドールを利用できるようにするtrpBと、細胞がヒスチジンの代わりにヒスチノールを利用できるようにするhisDが挙げられる。形質転換体を目で見て同定するための指標となるマーカーとしては、アントシアニン、b-グルクロニダーゼとその基質、GUS、ルシフェラーゼとその基質、ルシフェリンなどが挙げられる。
【0065】
C.部位特異的突然変異誘発
【0066】
部位特異的突然変異誘発は、γ-セクレターゼの基質となる個々のペプチドの調製、さらに特定するならばγ-セクレターゼの基質となる本発明の融合タンパク質のうちの1つを構成要素として含む融合ポリペプチドの調製に役立つ別の方法である。この方法では、(修飾の標的となるアミノ酸配列をコードしている)DNAの特異的突然変異誘発を利用する。この方法では、DNAにヌクレオチド配列の変化を1個以上導入することにより、上記の1つ以上の考慮事項を組み込んだ配列変異体を調製してテストすることが容易にできる。部位特異的突然変異誘発により、望む突然変異を有するDNA配列をコードしている特別なオリゴヌクレオチド配列と、隣接する十分な数のヌクレオチドとを利用して突然変異体を取得し、欠失接合部の両側にまたがる安定な二本鎖を形成するための、十分なサイズで配列が十分に複雑なプライマー配列を提供することができる。一般に、長さがヌクレオチド17〜25個で、変化させる配列の接合部の両側に約5〜10残基あるプライマーが好ましい。
【0067】
この方法では、一般に、一本鎖と二本鎖の両方の形態で存在するバクテリオファージ・ベクターを用いる。部位指定突然変異誘発で一般に用いられるベクターは、M13ファージなどのベクターである。ファージ・ベクターは市販されており、その使用法は当業者に一般によく知られている。二本鎖プラスミドも部位指定突然変異誘発で日常的に用いられている。この場合には、興味の対象である遺伝子をファージからプラスミドに移すステップがない。
【0068】
一般に、部位指定突然変異誘発を実施するには、まず最初に、配列内に望むタンパク質をコードしているDNA配列を有する一本鎖ベクターを取得するか、配列内に望むタンパク質をコードしているDNA配列を有する二本鎖ベクターの二本の鎖を溶かす。望む突然変異配列を有するオリゴヌクレオチド・プライマーを合成によって用意する。次に、ハイブリダイゼーション(アニーリング)条件を選択するときにミスマッチの程度を考慮してこのプライマーを一本鎖DNA調製物とともにアニールした後、DNA重合酵素(例えば大腸菌ポリメラーゼIのクレノウ・フラグメント)を作用させて突然変異を有する鎖の合成を完了させる。したがって、第1の鎖が突然変異していない元の配列をコードし、第2の鎖が望む突然変異を持つヘテロ鎖が形成される。次に、このヘテロ鎖ベクターを用いて適切な細胞(例えば大腸菌細胞)を形質転換し、突然変異した配列を有する組み換えベクターを含むクローンを選択する。
【0069】
もちろん、部位指定突然変異誘発を起こさせる上記の方法は、役に立つ可能性のある突然変異ペプチド種を作り出す唯一の方法ではなく、本発明がこの方法に限定されることはない。本発明では、突然変異誘発を実現する別の方法も考える。例えば、興味の対象である遺伝子を有する組み換えベクターを突然変異誘発剤(例えばヒドロキシルアミン)で処理して配列変異体を得る。
【0070】
D.タンパク質の精製
【0071】
本発明の融合タンパク質は精製することが望ましかろう。タンパク質精製法は当業者によく知られている。その方法には、1つの段階として、細胞培地を粗分画してポリペプチド分画と非ポリペプチド分画に分ける操作が含まれる。本発明のペプチドまたはポリペプチドが他のタンパク質から分離されると、興味の対象である融合ポリペプチドまたは融合ペプチドをクロマトグラフィ法や電気泳動法を利用してさらに精製し、部分的精製または完全な精製(または精製による均一化)を実現する。純粋なペプチドの調製に特に適した分析法は、イオン交換クロマトグラフィ、サイズ排除クロマトグラフィ;ポリアクリルアミド・ゲル電気泳動;等電点電気泳動である。融合タンパク質の精製に特に有効な方法は、高速タンパク質液体クロマトグラフィ(FPLC)または高性能液体クロマトグラフィ(HPLC)である。特に好ましい実施態様では、NusA-ノッチ融合タンパク質を、固定化金属アフィニティ・クロマトグラフィ(IMAC)を利用して分離した。
【0072】
ポリヒスチジン・タグを有する組み換えタンパク質の精製にはIMACが主に利用されている。本発明では、融合タンパク質がC末端にポリヒスチジン・タグを備えているため、この強力な方法で精製を行なうことができる。この精製は、ヒスチジンが二価の金属と自然にキレートを形成する傾向を利用している。金属イオンをクロマトグラフィの支持体上に置くと、ポリヒスチジン・タグを有するタンパク質を精製することができる。これは、当業者がこれまで利用してきた多彩なタンパク質の精製法の中でも非常に効率的な方法である。本発明のタンパク質を単離する好ましい一実施態様の具体的な条件は、実施例1に記載してある。しかし当業者であれば、その条件と媒体を変更しても本発明に従って精製を行なえることを理解されたい。その目的で、当業者は、金属アフィニティ・クロマトグラフィによって混合物から生物学的基質または関連する基質を分離する方法が詳細に記載されているアメリカ合衆国特許第4,431,546号を参考にしている。この特許文献に記載されているクロマトグラフィ媒体は、Ca2+、St2+、Ba2+、Al3+、Co2+、Ni2+、Cu2+、Zn2+からなるグループの中から選択した少なくとも1つの金属イオンの存在下で、アントラキノン、フタロシアニン、芳香族アゾからなるグループの中の少なくとも1つを含むリガンドを有する結合材料を備えている。別の媒体と条件は、例えばhttp://www.affiland.com/imac/nta.htmと、http://www.affiland.com/imac/pdc.htmに記載されている。
【0073】
本発明のいくつかの特徴は精製に関係しており、特別な実施態様では、コードされたポリペプチド、タンパク質、ペプチドいずれかの実質的な精製を行なう。この明細書では、“精製されたポリペプチド、タンパク質、ペプチド”という表現は、他の成分から分離された組成物を意味し、その組成物の中では、ポリペプチド、タンパク質、ペプチドが、そのポリペプチド、タンパク質、ペプチドを生成させるのに用いた細胞成分や合成成分よりもわずかであれ多く精製されている。したがって精製されたポリペプチド、タンパク質、ペプチドは、それが天然に発生する環境から離れたポリペプチド、タンパク質、ペプチドも意味する。
【0074】
一般に、“精製された”とは、ポリペプチド組成物、タンパク質組成物、ペプチド組成物において分画により他のさまざまな成分が除去されていて、発現した生物活性を実質的に維持していることを意味する。“実質的に精製された”という表現を用いる場合には、ポリペプチド、タンパク質、ペプチドが組成物の主要成分となっていて、組成物中のタンパク質の約50%、約60%、約70%、約80%、約90%、約95%またはそれ以上を占めていることを意味する。
【0075】
当業者は、本発明の開示内容を参考にして、ポリペプチド、タンパク質、ペプチドの精製度を定量するさまざまな方法を思い浮かべることができよう。そのような方法としては、例えば、活性分画の比活性の測定、SDS/PAGE分析による分画内のポリペプチドの量の評価などがある。分画の純度を評価する好ましい1つの方法は、分画の比活性を計算し、それを最初の抽出液の比活性と比較し、純度の大きさを計算して“〜倍純度数”として評価するというものである。活性の大きさを表わすのに用いる実際の単位は、もちろん、純度を調べるのに選択した具体的なアッセイ法と、発現したポリペプチド、タンパク質、ペプチドが検出可能な活性を示すかどうかに依存する。
【0076】
タンパク質の精製で利用するのに適したさまざまな方法が当業者にはよく知られている。例えば、硫酸アンモニウム、PEG、抗体などを用いた沈降、または熱変性を行なった後の遠心分離;クロマトグラフィ・ステップ(例えばイオン交換クロマトグラフィ、ゲル濾過クロマトグラフィ、逆相クロマトグラフィ、ヒドロキシルアパタイト・クロマトグラフィ、アフィニティ・クロマトグラフィ);等電点電気泳動;ゲル電気泳動;これらの方法と他の方法の組み合わせなどの方法がある。従来から一般に知られているように、さまざまな精製ステップを実施する順番を変えても、あるいはいくつかのステップを省略しても、実質的に精製されたポリペプチド、タンパク質、ペプチドを調製するのに適した方法になると考えられている。
【0077】
ポリペプチド、タンパク質、ペプチドが常に最高純度の状態で提供されることは一般には必要でない。実際、実質的により低い純度の生成物がいくつかの実施態様で役に立つことが考えられる。より少数の精製ステップを組み合わせて部分的精製を実現すること、あるいは同じ一般的精製スキームを異なった形で利用して部分的精製を実現することができる。例えばHPLC装置を用いて実施する陽イオン交換カラム・クロマトグラフィにより、一般に、低圧クロマトグラフィ・システムを用いた同じ方法よりも大きな“倍数”の純度が得られる。精製度が相対的に低い方法は、タンパク質生成物の全回収量や、発現するタンパク質の活性維持に関して利点を有する可能性がある。
【0078】
ポリペプチドの移動は、SDS/PAGEの条件が異なると、ときに顕著に異なる可能性があることが知られている(Capaldi他、Biochem. Biophys. Res. Comm.、第76巻、425ページ、1977年)。したがって電気泳動の条件が異なると、精製された、あるいは一部が精製された発現生成物の見かけの分子量が異なる可能性があることを理解されたい。
【0079】
IV.本発明の基質を得るのに用いる発現構造体
【0080】
本発明では、本発明のノッチ融合タンパク質を発現させる必要があろう。この発現を実現するため、本発明では、本発明のノッチ融合タンパク質をコードするためのポリヌクレオチド分子を含むベクターと、そのベクターを用いて形質転換した宿主細胞を利用する。このようなポリヌクレオチド分子は、一般に選択マーカーと複製起点を含むベクターと結合させて宿主の中に運び込む。本発明で利用する発現構造体のこれらエレメントについては、以下に詳しく説明する。
【0081】
発現ベクターは、γ-セクレターゼの基質であるこの明細書に記載したあらゆるペプチドまたはポリペプチドをコードしていて適切な転写調節配列または翻訳調節配列と機能上リンクしたDNAを含んでいる。その調節配列は、例えば哺乳動物、細菌、ウイルス、昆虫の遺伝子に由来する。調節配列の具体例としては、転写プロモータ、オペレータ、エンハンサー、mRNAリボソーム結合部位、転写と翻訳を制御する適切な配列などが挙げられる。
【0082】
“発現ベクター”、“発現構造体”、“発現カセット”という用語は、この明細書全体を通じてどれも同じ意味で使用し、1つの遺伝子産物をコードしている核酸を含んでいて、そのコードしている核酸配列の一部または全部を転写できるあらゆるタイプの遺伝子構造体を意味する。
【0083】
本発明の融合ポリペプチドを発現させるのに適した発現ベクターの選択は、もちろん使用する具体的な宿主細胞に依存するが、当業者はその選択方法を知っている。適切な発現ベクターの具体例として、pcDNA3(インヴィトロジェン社)とpSVL(ファルマシア・バイオテック社)がある。本発明における好ましい1つの発現ベクターはpcDNA3.1-ハイグロ(インヴィトロジェン社)である。哺乳動物宿主細胞で使用する発現ベクターは、ウイルスのゲノムに由来する転写制御配列と翻訳制御配列を含むことができる。本発明で用いることのできる一般に使用されているプロモータ配列とエンハンサー配列としては、ヒト・サイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマ・ウイルス、サル・ウイルス40(SV40)などがある。哺乳動物発現ベクターを構成する方法は、例えば、OkayamaとBerg(Mol. Cell. Biol.、第3巻、280ページ、1983年);Cosman他(Mol. Immunol.、第23巻、935ページ、1986年);Cosman他(Nature、第312巻、768ページ、1984年);EP-A-0367566;WO 91/18982に開示されている。
【0084】
発現構造体は、本発明による特定のノッチ融合タンパク質をコードしている核酸領域を含むことになる。上記のような発現ベクターを構成するのに用いるコード領域は、少なくともγ-セクレターゼによるこの明細書に記載した融合タンパク質の開裂をコードしている必要がある。しかし生成するペプチドの1つが、γ-セクレターゼによって開裂するγ-セクレターゼ開裂部位1731/1732と1743/1744を含んでいるのであれば、より大きなポリペプチドがコードされていてもよい。
【0085】
本発明のいくつかの特徴によると、発現構造体は、ペプチドまたはポリペプチドの発現を検出できるようにする選択マーカーをさらに含むことができる。通常は、薬剤選択マーカーを含めることによって転写体のクローニングと選択がしやすくなる。薬剤選択マーカーとしては、例えばネオマイシン、ピューロマイシン、ハイグロマイシン、DHFR、ゼオシン、ヒスチジノールがある。あるいは酵素を使用することもできる。酵素としては、例えば、単純ヘルペスウイルスのチミジンキナーゼ(tk)(真核生物)、b-ガラクトシダーゼ、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)(原核生物)がある。免疫マーカーも使用することができる。例えばエピトープ・タグとして、フラグ系(IBI、ニュー・ヘイヴン、コネチカット州)、HA、6×ヒスチジン系(キアジェン社、チャッツワース、カリフォルニア州)が使用できる。さらに、グルタチオン-S-トランスフェラーゼ(GST)系(ファルマシア社、ピスカタウェイ、ニュージャージー州)またはマルトース結合タンパク質系(NEB、ビヴァリー、マサチューセッツ州)も用いることができる。用いる選択マーカーは、遺伝子産物をコードしている核酸と同時に発現させることができるのであれば、重要であるとは考えられない。選択マーカーのさらに別の具体例は当業者には周知である。本発明で使用できる特に好ましい選択マーカーは、ネオマイシン耐性マーカーとGFPマーカーである。
【0086】
発現するには、適切なシグナルがベクターに供給される必要がある。このセクションには、さまざまな調節エレメント(例えば宿主細胞の中で興味の対象である核酸を発現させるのに用いることのできるウイルスと哺乳動物両方からのエンハンサー/プロモータ)に関する説明が含まれる。宿主細胞の中でメッセンジャーRNAの安定性と翻訳可能性を最適化するように設計されたエレメントも定義する。産物を発現する永久的に安定な細胞クローンを確立するために多数の優性薬剤選択マーカーを使用する条件と、薬剤選択マーカーの発現を突然変異表現型の発現と結びつけるエレメントも提示する。
【0087】
好ましい実施態様では、所定のペプチドをコードしている核酸、または選択マーカーをコードしている核酸は、プロモータの転写制御下にある。“プロモータ”とは、遺伝子の具体的な転写を開始させるのに必要で、細胞の合成機構または導入された合成機構によって認識されるDNA配列を意味する。
【0088】
核酸配列は、調節配列がノッチ融合タンパク質をコードしているDNAと機能の上で関係を持っている場合に、機能上リンクしている。したがってプロモータ・ヌクレオチド配列は、そのプロモータ・ヌクレオチド配列が所定のDNA配列の転写を指示するときにそのDNA配列と機能上リンクしている。同様に、“転写制御下にある”という表現は、プロモータが核酸との関係で正確な位置と向きになっていて、RNAポリメラーゼの開始と遺伝子の発現を制御することを意味する。
【0089】
プロモータという用語は、この明細書では、RNAポリメラーゼIIの開始部位の周囲に集まった一群の転写制御モジュールを意味する。プロモータをどのように構成するかについて考える時間の大部分は、いくつかのウイルスのプロモータ(例えばHSVチミジンキナーゼ(tk)やSV40初期転写ユニット)を分析することに費やされる。これらの研究にさらに最近の研究が加わることで、プロモータは離散した複数の機能モジュールで構成されており、それぞれの機能モジュールは約7〜20bpのDNAからなること、そして転写のアクチベータ・タンパク質またはリプレッサー・タンパク質のための1つ以上の認識部位を含むことが明らかになった。
【0090】
各プロモータの少なくとも1つのモジュールは、RNA合成の出発部位の位置を決める機能を有する。その最もよく知られている例はTATAボックスであるが、TATAボックスが欠けているいくつかのプロモータ(例えば、哺乳動物のターミナルデオキシヌクレオチジルトランスフェラーゼ遺伝子のためのプロモータや、SV40後期遺伝子のためのプロモータなど)では、出発部位そのものと重なっている離散エレメントが開始の位置を決めるのに役立つ。
【0091】
別のプロモータ・エレメントが転写開始の頻度を調節する。一般に、そのエレメントは出発部位の30〜110bp上流の領域に位置している。しかし出発部位の下流に機能エレメントを含むプロモータが多数あることも最近明らかになった。プロモータ・エレメント相互間のスペースはしばしば自由であるため、エレメントが互いに逆転したり移動したりしてもプロモータの機能は保持される。tkプロモータでは、プロモータ・エレメント相互間のスペースを50bpまで離したときにようやく活性が低下し始める。プロモータが何であるかに応じ、個々のエレメントは、共同して、あるいは独立に機能して転写を活性化させるように見える。
【0092】
興味の対象である核酸配列の発現制御に用いる具体的なプロモータが何であるかは、標的細胞の中で核酸の発現を指示できるのであれば重要ではないと考えられている。したがってヒト細胞を標的とする場合には、ヒト細胞の中で発現させることのできるプロモータの近くに、またはそのプロモータの制御下に核酸コード領域を置くことが好ましい。一般に、そのようなプロモータとしては、ヒトのプロモータまたはウイルスのプロモータが挙げられる。
【0093】
いくつかの実施態様では、ヒト・サイトメガロウイルス(CMV)前初期遺伝子プロモータ、SV40初期プロモータ、ラウス肉腫ウイルスの長い末端反復配列、β-アクチン、ラットのインスリン・プロモータ、ホスホグリセロールキナーゼ・プロモータ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ・プロモータを用いて興味の対象であるコード配列を高レベルで発現させることができる。これらはすべて、よく知られていて容易に入手できるプロモータである。興味の対象であるコード配列の発現を実現できることが従来技術でよく知られている他のウイルスまたは哺乳動物の細胞プロモータまたはバクテリオファージ・プロモータを用いることも、発現レベルが目的にとって十分である場合には考えられる。よく知られた特性を有するプロモータを用いることにより、トランスフェクションと形質転換を行なった後の興味の対象であるタンパク質の発現レベルと発現パターンを最適化することができる。
【0094】
特定の生理学的シグナルまたは合成シグナルに応答して調節されるプロモータを選択することにより、遺伝子産物の発現を誘導することができる。ウイルス・ベクターを作るためのいくつかの誘導プロモータ系が入手可能である。そのような1つの系はエクジソン系(インヴィトロジェン社、カールズバッド、カリフォルニア州)である。これは、哺乳動物の細胞における興味の対象である遺伝子の発現を調節できるように設計されている。これは、導入遺伝子のベースレベルの発現は実質的にないが200倍を超える誘導性を可能にする厳しく制御された発現メカニズムを備えている。
【0095】
別の有用な誘導系は、Tet-Off(登録商標)系またはTet-On(登録商標)系(クロンテック社、パロ・アルト、カリフォルニア州)であり、元々はGossenとBujardによって開発されたものである(GossenとBujard、Proc. Natl. Acad. Sci. USA、第89巻(12)、5547〜5551ページ、1992年;Gossen他、Science、第268巻(5218)、1766〜1769ページ、1995年)。
【0096】
哺乳動物の細胞では、強い転写活性を与えるのにCMV前初期プロモータがしばしば用いられている。導入遺伝子の発現レベルを小さくすることが望ましい場合には、CMVプロモータを改変して転写活性を弱めたものも用いられてきた。レトロウイルスのプロモータ(例えばMLVやMMTVからのLTR)が本発明で役に立つと考えられる。使用可能な他のウイルスのプロモータとしては、SV40、RSV LTR、HIV-1 LTR、HIV-2 LTRのプロモータ、アデノウイルスのプロモータ(例えばE1A領域、E2A領域、MLP領域からのもの)、AAV LTR、カリフラワー・モザイク・ウイルス、HSV-TK、トリ肉腫ウイルスのプロモータなどがある。
【0097】
いくつかの実施態様では、制御可能なプロモータが役に立つことがわかるであろう。そのようなプロモータとしては、ホルモンまたはサイトカインを制御できるプロモータがある。ホルモンを制御できるプロモータとしては、MMTV、MT-1、エクジソン、ルビスコのほか、ホルモンによって調節される他のプロモータ(例えば甲状腺ホルモン、下垂体ホルモン、副腎ホルモンに応答するプロモータ)がある。
【0098】
本発明で使用することが考えられる別の調節エレメントはエンハンサーである。それは、同じDNA分子上の離れた位置にあるプロモータからの転写を増大させる遺伝子エレメントである。エンハンサーはプロモータとほぼ同じよう構成されている。すなわちエンハンサーは、多数の個々のエレメントからなり、それぞれが1つ以上の転写タンパク質と結合している。エンハンサーとプロモータの基本的な違いは作用である。エンハンサー領域は全体として転写を離れた位置から刺激できる必要があるのに対し、プロモータ領域またはその成分エレメントではその必要がない。その一方で、プロモータは、特定の部位と特定の方向でRNA合成の開始を指示する1つ以上のエレメントを持っている必要があるのに対し、エンハンサーはこの特徴を欠いている。プロモータとエンハンサーはしばしば重複かつ連続していて、非常に似た構成を有するように見える。本発明で有用なエンハンサーは当業者には周知であり、使用する具体的な発現系が何であるかによって異なる(Scharf D.他、Results Probl. Cell Differ.、第20巻、125〜162ページ、1994年;Bittner他、Methods in Enzymol.、第153巻、516〜544ページ、1987年)。
【0099】
発現構造体がcDNA挿入体を利用している場合には、遺伝子転写物を適切にポリアデニル化するためのポリアデニル化シグナル配列が含まれていることが一般に望ましい。選択したトランスジェニック動物種の細胞が認識する任意のポリアデニル化シグナル配列が、本発明を実施するのに適している。それは例えば、ヒトまたはウシの成長ホルモンや、SV40ポリアデニル化シグナルである。
【0100】
発現カセットのエレメントとしてターミネータも考えられる。このエレメントは、mRNAのレベルを増大させ、カセットが読み飛ばされて他の配列になることを最少にできる。主に利用されるターミネータ領域は便利であるという理由で選ばれる。なぜなら、本発明で考えられるような用途のターミネータ領域は比較的交換可能であるように見えるからである。ターミネータ領域は、転写開始領域に元々備わっているもの、あるいは興味の対象であるDNA配列に元々備わっているもの、あるいは他の供給源に由来するものが可能である。
【0101】
本発明のいくつかの実施態様では、複数遺伝子のmRNAまたはポリシストロン性mRNAを作り出すのに内部リボソーム侵入部位(IRES)エレメントを用いることが考えられる。IRESエレメントは、5'メチル化キャップに依存した翻訳というリボソーム走査モデルを無視して内部部位で翻訳を開始できる(PelletierとSonenberg、Nature、第334巻、320〜325ページ、1988年)。ピコルナウイルス・ファミリーの2つのメンバー(ポリオウイルスと脳心筋炎)からのIRESエレメントが報告されており(PelletierとSonenberg、上記文献)、哺乳動物のmRNAからのIRESも報告されている(MacejakとSarnow、Nature、第353巻、90〜94ページ、1991年)。IRESエレメントは、異種オープン・リーディング・フレームとリンクさせることができる。IRESによって隔てられた多数のオープン・リーディング・フレームを同時に転写してポリシストロン性mRNAを作り出すことができる。IRESエレメントのおかげでそれぞれのオープン・リーディング・フレームがリボソームにアクセスして効果的な翻訳を行なうことができる。単一のプロモータ/エンハンサーを用いて単一のmRNAを転写し、複数の遺伝子を効果的に発現させることができる。
【0102】
任意の異種オープン・リーディング・フレームをIRESエレメントにリンクさせることができる。この異種オープン・リーディング・フレームには、分泌されたタンパク質、独立な遺伝子によってコードされているサブユニットを多数有するタンパク質、細胞内タンパク質または膜結合タンパク質、選択マーカーのための遺伝子が含まれている。このように単一の構造体と単一の選択マーカーを用いることで、いくつかのタンパク質の発現を1つの細胞内で同時に実現することができる。
【0103】
V.γ-セクレターゼ・アッセイにおける基質の使用
【0104】
本発明の特別な実施態様は、γ-セクレターゼの活性および/または機能をモニターするアッセイ、より詳細には、γ-セクレターゼの活性および/またはγ-セクレターゼの機能を調べるアッセイに関する。このアッセイは、溶液中でγ-セクレターゼ複合体(または精製したポリペプチド)を本発明の適切な基質とともにインキュベートし、ノッチ融合タンパク質の開裂をγ-セクレターゼのタンパク質分解活性の指標として利用する操作を含んでいる。
【0105】
A.アッセイの形式
【0106】
本発明の特別な実施態様は、ヒトγ-セクレターゼの活性を変化させる薬を同定する方法に関する。この方法の1つの特徴は、ε開裂部位1743/1744におけるγ-セクレターゼのノッチ開裂活性を、モジュレータと考えられる化合物または薬の存在下と不在下でモニターすることである。例えばノッチの開裂を調べるこのような方法は、一般に、
(a)本発明による任意のノッチ/NusA融合タンパク質を、活性なγ-セクレターゼを含む組成物と試験管内で接触させるステップと;
(b)そのノッチ/NusA融合タンパク質がγ-セクレターゼによってノッチのγ-セクレターゼ開裂部位で開裂したことを調べるステップを含んでいる。
【0107】
活性なγ-セクレターゼを含む組成物は、一般に、活性なγ-セクレターゼを含む単離された任意の組成物である。この組成物は、そのままで細胞(γ-セクレターゼ活性を有する天然の細胞、またはそのような活性を発現するようにした組み換え宿主細胞)から単離された膜分画になることができる。組成物としては、他のタンパク質を実質的に含まない単離または精製されたγ-セクレターゼも可能である。
【0108】
γ-セクレターゼのノッチ開裂活性を変化させるモジュレータを同定するため、モジュレータ候補の存在下と不在下で上記のステップ(a)と(b)を実施し、モジュレータ候補の存在下でのγ-セクレターゼのノッチ開裂活性をモジュレータ候補の不在下での活性と比較することによってそのモジュレータ候補の変更能力を評価することで、γ-セクレターゼによるこのような開裂を変化させる化合物を同定する。モジュレータ候補の存在下でノッチ開裂の量または程度がいくらかでも変化するというのは、モジュレータ候補の存在下でγ-セクレターゼの活性が変化したことの指標であり、γ-セクレターゼの活性のモジュレータとなる化合物が同定されたことになる。
【0109】
対照(試験化合物ではない)と比べて開裂を増加させる化合物は、γ-セクレターゼのタンパク質分解活性のアゴニストまたは促進剤であると評価されるのに対し、Aβ1〜40および/またはAβ1〜42において開裂を減らす化合物は阻害剤であると評価される。γ-セクレターゼの阻害剤は特に興味深い。なぜなら、γ-セクレターゼ活性の阻害剤は、ADの症状またはその進行を治療または予防する薬になる可能性があることを示しているからである。
【0110】
γ-セクレターゼによるAPPまたはCT-100の開裂を本発明の融合タンパク質の開裂と比べて選択的に抑制する試験化合物を見いだすことが望ましいため、本発明のアッセイを利用するのと並行して、あるいはその前に、試験化合物がAPPの開裂を変化させる能力を調べるとよい。γ-セクレターゼによるAPPまたはCT-100の開裂を測定する方法は従来技術において周知である。一例として、WO 01/83811には、APPまたはCT-100の開裂を測定するのに役立つ基質とインビトロ・アッセイが記載されている。WO 01/83811には、M末端メチオニン(M)と、APP597〜695と、フラグ・タグとを含むγ-セクレターゼの基質、ならびにその基質を開裂させる方法と条件、開裂産物(M-Aβ40とM-Aβ42)を検出する方法と条件が記載されている。
【0111】
γ-セクレターゼによるAPP/CT-100開裂活性をAβ1〜40および/またはAβ1〜42において抑制するが、γ-セクレターゼによる測定されたノッチ開裂活性は抑制しない阻害剤が最も好ましい。
【0112】
γ-セクレターゼは、精製されたγ-セクレターゼ・ポリペプチドまたは複合体、その生物学的に活性な断片、そのアナログ、その変異体のいずれでもよい。好ましい実施態様では、γ-セクレターゼは、γ-セクレターゼ活性を示す細胞からの膜分画に由来する。このような膜分画は、γ-セクレターゼ複合体にとって必要なすべての要素を含んでいることが好ましい。ヒト・γ-セクレターゼの非ヒト・オルトログもアッセイで使用することができる。
【0113】
本発明のアッセイは、細胞を含まない系でγ-セクレターゼ・ポリペプチドを用いて実施できるように設計する。例えば細胞を含まない系では、試験化合物の存在下と不在下で、γ-セクレターゼまたはγ-セクレターゼ含有膜分画を、本発明のペプチド基質またはタンパク質基質と混合することによって接触ステップを行なうことができる。最適な結果を得るためには、γ-セクレターゼとγ-セクレターゼの基質を実質的に精製し、制御された所定量を混合した後、酵素活性を最適にした、および/または生理学的条件を真似た適切な緩衝液の中で混合する。
【0114】
開裂を調べるステップは、N末端断片とC末端断片の一方または両方か、開裂を示す他のパラメータを測定する操作を含んでいる。例えばノッチをベースとした基質またはγ-セクレターゼの他の基質は、消光状態の標識を含むことができる。この標識は、開裂して消光部から分離したときだけより検出しやすくなる。ノッチをベースとした基質またはγ-セクレターゼの他の基質をN末端またはC末端に固定して固体支持体に取り付けることもできる。この構成では、開裂は、固体支持体から開裂断片が放出されることによって判定できる。この放出は、媒体中の標識が増加すること、あるいは固体支持体に付着した標識が減少することによって判定できる。放出は、(例えば抗体とともに)放出されたペプチドを定量的に捕捉することによっても判定できる。
【0115】
本発明の好ましい一実施態様では、バリン1744に対して特異的な抗体を用いて開裂したノッチ・タンパク質を検出する。さらに好ましい実施態様では、抗バリン1744抗体と抗フラグ抗体に基づいて開発したELISAを利用して開裂したノッチ・タンパク質を検出する。抗バリン1744抗体は開裂の1つの断片を検出するのに用いられるのに対し、抗フラグ抗体は、γ-セクレターゼの作用によって産生されたノッチ・タンパク質の断片(C末端)を検出する。このアッセイについては、実施例でさらに詳しく説明する。
【0116】
もちろん、上記のアッセイは単なる例示であり、他のアッセイも利用することができる。例えば上記のアッセイを以下のようにすることができる。384ウエルのマイクロタイター・プレートをBSAでブロックし、酵素であるγ-セクレターゼとテストするγ-セクレターゼ阻害化合物50μlを1時間にわたってインキュベートし、ノッチ/NusA融合タンパク質基質を添加することによって反応を開始させる。最終的なアッセイ条件では、体積がウエル1つにつき30μl;化合物が50μM;酵素がウエル1つにつき15ng;250nMの基質;5%DMSOと0.001%のトゥイーン20である。室温にて一晩にわたってインキュベートし、トリス-HCl(pH8.3)を添加して反応を停止させる。基質を6.25ピコモル含むアリコートを取り出し、開裂した基質および/または開裂していない基質をストレプトアビジンでコーティングしたプレートに捕捉する。このプレートを3回洗浄し、緩衝液を添加する。捕捉アッセイは、LJLアナリスト(Ex485/Em530)上でオレゴン・グリーンの蛍光放出を読み取ることによってモニターする。
【0117】
この明細書で利用できる別のアッセイは、蛍光偏光アッセイである。蛍光偏光は、感光性があって容易で非破壊的なアッセイであり、薬剤候補が本発明のγ-セクレターゼ複合体に及ぼす効果をモニターするのに用いることができる。蛍光偏光は、その基質がγ-セクレターゼと相互作用するのをモニターするのにも利用できる。制御された条件下での蛍光偏光の測定により、溶液中での蛍光分子の“分子揺動”の程度を明らかにすることができる。例えば体積が小さい小分子は、素早く揺動することが予測される。偏光した光を照射された場合には、溶液中の分子の素早い運動によって光の偏光が広範にわたって消え、“小さな”偏光値が読み取られることになろう。同じ条件下において、体積がより大きい大分子または大きな複合体は分子回転(揺動)プロセスがゆっくりしたものになろう。その結果、平面偏光した入射光の偏光度をより小さくすると、より大きな偏光値が測定されよう。
【0118】
小さなリガンドに蛍光プローブを標識することにより、そのリガンドが系の別の要素と相互作用して生じる蛍光偏光の変化を測定することができる。このような方法は、酵素(γ-セクレターゼ)と酵素の蛍光性基質の間の相互作用を測定するのに用いることもできる。
【0119】
蛍光偏光アッセイの一例では、あらかじめブロックした低アフィニティの黒いプレートの中で、酵素と抑制性/変更性化合物を30分間にわたってインキュベートし、150nMの基質(例えば蛍光標識した本発明のノッチ/NusA融合タンパク質)を最終溶液がウエル1つにつき30μlになるように添加して反応を開始させる。次にプレートを室温にて15分間にわたってインキュベートし、蛍光偏光をLJLアナリスト(Ex485/Em530)上で測定する。
【0120】
本発明では、γ-セクレターゼを単離してキャラクテリゼーションを行なうのに役立つ可能性のある1つの特徴を考える。この特徴では、酵素の活性部位またはアロステリック部位に結合する化合物を検出する結合アッセイを考える。その検出を行なうには、本発明によるノッチ基質の加水分解しない誘導体を用いるとよい。例えば開裂する結合の接合部にスタチン誘導体が存在していると、本発明のノッチが開裂部位で加水分解しなくなる。基質に対しては、検出を容易にするための適切な蛍光タグ(例えばBODIPY FL)を付加するという修飾をさらに行なうことができる。
【0121】
本発明の基質に蛍光標識を付け、γ-セクレターゼのための蛍光偏光結合アッセイを開発するのに利用できる。酵素と基質の間の相互作用に関する平衡解離定数(KD)は、酵素を用いて基質を滴定した結果として得られる蛍光偏光の変化を測定することによって決定される。
【0122】
本発明の基質とγ-セクレターゼの相互作用に関するKDを明らかにするため、さまざまな量のγ-セクレターゼを3.1nMの蛍光基質と組み合わせ、室温にて3時間にわたってインキュベートする。インキュベーションの後、LJLアナリスト(96ウエルの形式)またはパンヴェラ・ビーコン(単一のキュベットの形式)を用いて蛍光偏光を測定する。アッセイは、例えば、25mM酢酸ナトリウム、20%グリセロール、pH4.75の中で行なう。偏光の値を与える取得データを縦軸に、酵素の濃度を横軸にプロットすると、酵素と基質の相互作用に関するKDを決定するための結合等温線が得られる。次に、関係式Px=PF+(PB-PF)×[E]/(KD+[E])(ただしP=偏光値、x=サンプル、F=遊離している阻害剤、B=結合している阻害剤、E=γ-セクレターゼ)(蛍光偏光アプリケーション・ガイド、1998年;パンヴェラ社、マディソン、ウィスコンシン州)を用いてこのデータを分析し、KDを得る。このアッセイを利用し、酵素の活性部位またはアロステリック部位に結合する化合物をスクリーニングすることができる。
【0123】
試験化合物の存在下と不在下での活性測定は、並列に、あるいはいずれかの順番で実施できることに注意されたい。さらに、特定の反応条件について酵素活性の信頼できるベースラインが得られたのであれば、すべての試験化合物と並列して対照の測定(すなわち試験化合物が不在下での開裂の測定)を繰り返す必要はなかろう。特定の基質(例えば本発明のノッチ組成物またはAPPに由来する組成物)に対するγ-セクレターゼの酵素活性について阻害剤の不在下で得られた知見を、比較ステップを実施するための基礎にすることができる。
【0124】
B.基質候補
【0125】
この明細書では、“基質候補”または“試験基質”という用語は、γ-セクレターゼの活性、好ましくはヒトγ-セクレターゼの活性を変化させることのできるあらゆる分子を意味する。基質候補としては、タンパク質またはその断片、小分子阻害剤が可能であり、核酸分子さえ可能である。スクリーニング・アッセイを適用して同定される最も有用な薬理化合物は、大化合物ライブラリのスクリーニングを通じて同定される化合物、またはAPPのプロセシングとノッチのプロセシングの一方または両方に関する公知の他のモジュレータと構造的に関係のある化合物であろう。例えばアメリカ合衆国特許第6,448,229号(その内容が参考としてこの明細書に組み込まれているものとする)には、ノッチのシグナル伝達には影響を与えずにγ-セクレターゼを抑制するためADの治療または予防に使用できる特別なクラスの化合物が記載されている。このアメリカ合衆国特許第6,448,229号に記載されているような化合物は、本発明のアッセイを利用して確認することができる。さらに、そのような化合物を合理的な薬剤設計における出発材料として用い、本発明で使用する別のモジュレータ候補を同定できる可能性がある。このように合理的な薬剤設計に使用できる可能性のある他の阻害剤としては、DAPT(PHA-568638)やフェンキルアミン(PHA-512088)などが挙げられる。
【0126】
基質候補は、天然に存在する化合物の断片または一部でもよいし、組み合わせないときには不活性なのに組み合わせたときだけ活性になることがわかった既知の化合物の組み合わせでもよい。しかしそのような化合物をヒトまたはモデル動物でテストする前に、どの候補が潜在能力を持っているかを多彩な候補についてテストする必要があろう。
【0127】
したがって基質候補は、天然に存在する化合物の断片または一部でもよいし、組み合わせないときには不活性なのに組み合わせたときだけ活性になることがわかった既知の化合物の組み合わせでもよい。したがって本発明により、γ-セクレターゼによるノッチのプロセシングの促進または抑制よりも、γ-セクレターゼによる細胞APPプロセシングを促進または抑制する化合物を同定するスクリーニング・アッセイが提供される。天然の供給源(例えば動物、細菌、真菌、植物(葉や樹皮も含む)、海洋サンプル)から単離した化合物を候補として調べ、潜在的に存在している有用な医薬を探すことが提案される。
【0128】
スクリーニングする医薬は、化学的合成物または人工化合物に由来するもの、あるいは化学的合成物または人工化合物から合成することも可能であることが理解できよう。したがって、本発明によって同定されるモジュレータとしては、γ-セクレターゼの活性および/またはAPPのプロセシングを促進または阻害する公知の薬剤から出発し、合理的な薬剤設計を通じて設計することのできるポリペプチド、ポリヌクレオチド、小分子阻害剤、他の任意の無機または有機の化合物が可能であることがわかる。
【0129】
候補スクリーニング・アッセイは準備と実施が簡単である。したがってモジュレータのアッセイでは、機能的γ-セクレターゼを含む細胞膜分画(例えば可溶化γ-セクレターゼ複合体を含む細胞膜分画)を得た後、本発明による新規な基質の存在下で候補物質をそのようなγ-セクレターゼ組成物と混合する。そのとき、γ-セクレターゼの活性をその基質の開裂を通じて測定できるようにする。このようにして、候補物質の存在下でその候補物質がγ-セクレターゼを活性化する能力を測定することができる。同様に、阻害剤を探すアッセイでは、機能的γ-セクレターゼを発現する細胞膜分画を得た後、候補物質をその分画と混合する。このようにして、抑制候補物質が、γ-セクレターゼによる生物学的効果を低下、消失、減少させる能力を検出することができる。
【0130】
所定の条件下における基質の“有効な量”は、γ-セクレターゼが所定のCT-100を開裂させる活性またはAPPのプロセシングを再現性よく変化させるのに有効な量のことである。この有効量は、候補物質の不在下で見られる開裂と比べた場合に、APPが開裂して得られるCT-100の程度または量を変化させるが、本発明のノッチ融合タンパク質のγ-セクレターゼ部位における開裂は変化させない量である。治療薬として役立つであろう化合物、および/またはさらにキャラクテリゼーションを行なうのに特に役立つであろう化合物は、γ-セクレターゼによるノッチの開裂ではなくAPPの開裂を選択的に抑制する化合物である。“選択的に抑制する”とは、γ-セクレターゼによるノッチの開裂よりもAPPの開裂に対してより抑制効果を有する化合物を意味する。ノッチの開裂を抑制しない化合物が好ましいとはいえ、ノッチの開裂が幾分か抑制されることは、野生型γ-セクレターゼによるAPPの開裂よりも少ないのであれば許容できる。
【0131】
γ-セクレターゼの本発明による新規な基質を用いた上記のアッセイは、多数の高スループット・スクリーニング(HTS)法と相性がよい(概説に関してはJayawickremeとKost、Curr. Opin. Biotechnol.、第8巻、629〜634ページ、1997年を参照のこと)。例えばHoustonとBanks、Curr. Opin. Biotechnol.、第8巻、734〜740ページ、1997年に記載されているように、自動化したHTSアッセイと小型化したHTSアッセイも考えることができる。
【0132】
小分子モジュレータの同定に用いられる多数の異なったライブラリが存在している。例えば、化合物ライブラリ、天然生成物ライブラリ、コンビナトリアル・ライブラリ(ランダムに得れらた、または設計して得たペプチド、オリゴヌクレオチド、有機分子からなる)などがある。化合物ライブラリは、公知の化合物の構造的アナログからなるか、天然生成物のスクリーニングまたは潜在的治療標的に対するスクリーニングを通じてヒット化合物またはリード化合物と同定された化合物からなる。天然生成物ライブラリは、微生物、動物、植物、昆虫、海洋生物からの生成物のコレクションである。これらは、例えば土壌生物、植物、海洋生物からのブロスを発酵させた後に抽出することによってスクリーニング用混合物を作るのに用いられる。天然生成物ライブラリには、ポリペプチド、非リボソーム・ペプチド、その非天然変異体が含まれる。概説に関してはScience、第282巻、63〜68ページ、1998年を参照のこと。
【0133】
コンビナトリアル・ライブラリは、多数のペプチド、オリゴヌクレオチド、有機化合物で構成されている。コンビナトリアル・ライブラリは、伝統的な自動化合成法、PCRクローニング、あるいは他の合成法によって比較的簡単に用意できる。特に興味深いのは、ペプチド・ライブラリ、タンパク質ライブラリ、ペプチドミメティック・ライブラリ、多重並列合成コレクション・ライブラリ、リコンビナトリアル・ライブラリ、ポリペプチド・ライブラリなどのライブラリである。コンビナトリアル・ライブラリと、そこから作られるライブラリの概説に関しては、Myers、Curr. Opin. Biotechnol.、第8巻、701〜707ページ、1997年を参照のこと。次に、ここに記載したさまざまなライブラリを用いて同定したモジュレータを最適化し、例えば合理的薬剤設計を通じてγ-セクレターゼの活性を変化させる。
【0134】
もちろん、有効な候補が見つからないとしても本発明のすべてのスクリーニング法はそれ自体有効であることが理解できよう。本発明により、そのような候補を見つける方法だけでなく、そのような候補をスクリーニングする方法も提供される。
【0135】
C.インビボ・アッセイ
【0136】
本発明には、さまざまなモデル動物の利用も含まれる。すでに説明したようにインビトロ環境でモジュレータがスクリーニングされると、APPのプロセシングおよび/またはADの非ヒト・モデルを利用してそのモジュレータの生体内における効果を明らかにすることができる。これは、γ-セクレターゼが正常に発現する1つの動物系全体の中でγ-セクレターゼの機能を調べる素晴らしい機会になろう。
【0137】
γ-セクレターゼ活性のモジュレータであることが判明した試験化合物を用いた動物の治療は、その化合物を適切な形態で動物に投与する操作を含むことになる。投与は、臨床目的または非臨床目的で利用できる任意の経路(例えば経口、鼻腔、口内、直腸、膣、局所)を通じて行なうことになろう。投与は、気管内点滴、気管支点滴、皮膚内注射、皮下注射、筋肉内注射、腹腔内注射、静脈内注射によって行なうことも可能である。全身性静脈内注射、血液を通じた局所投与、脳脊髄液(CSF)への供給、リンパへの供給、腫瘍内注射などが特に考えられる。
【0138】
生体内での化合物の有効性を調べるにはさまざまな基準がある。そのような基準としては、生存率、活性レベルの上昇、食物摂取の改善などがある。他の評価法としては、γ-セクレターゼの活性が変化したことの徴候(例えばアミロイドβまたはアミロイドβ斑の産生低下、脳の萎縮減少)を探す病理学的検査(特に脳組織)がある。
【0139】
D.薬の製造
【0140】
本発明のアッセイにより、γ-セクレターゼの活性が異常なレベルであることを特徴とする疾患(例えばAD)を治療するための治療薬候補となるγ-セクレターゼのモジュレータを同定する。そこで本発明の方法は、モジュレータを同定した後、必要に応じた追加ステップとして、薬剤を製造/合成するステップと、薬理学的に許容可能な希釈剤、アジュバント、基剤を用いてその薬剤を組成物にするステップを含んでいる。医薬組成物に関しては以下に詳しく説明する。
【0141】
VI.医薬組成物
【0142】
ノッチのプロセシング、APPのプロセシング、γ-セクレターゼの開裂を変化させることが確認された本発明のモジュレータは、最終的に医薬組成物(すなわち生体内で利用するのに適した形態)にすることができる。一般に、発熱物質や、ヒトまたは動物にとって有害である可能性のある他の不純物を実質的に含まない組成物を調製する。
【0143】
一般に、同定されたモジュレータ組成物を適切な塩や緩衝液を用いて安定にし、標的細胞による取り込みが可能になるようにすることが望ましい。“薬理学的または薬物学的に許容可能な”という表現は、動物またはヒトに投与したときに好ましくない反応、アレルギー反応、不都合な反応を起こさない分子全体または組成物を意味する。この明細書では、“薬理学的に許容可能な基剤”には、あらゆる溶媒、分散媒体、コーティング、抗菌剤、抗真菌剤、等張剤、吸収遅延剤などが含まれる。薬理学的に活性な物質のためのこのような媒体や薬剤の使用法は従来技術において周知である。従来の媒体や薬剤のどれであれ、本発明によって同定されたモジュレータと適合しない場合を除き、治療用組成物で使用することが考えられる。追加活性成分も組成物に組み込むことができる。
【0144】
本発明のモジュレータ組成物には古典的な医薬調製物が含まれる。本発明の組成物は、一般的な経路で標的組織に到達させうるのであれば、その一般的な経路のうちの任意の経路を通じて投与されることになる。医薬組成物は、従来からある任意の方法で対象に導入することができる。例えば、静脈内、皮膚内、筋肉内、乳房内、腹腔内、気管内、眼内、眼球後、肺内(例えば期間限定放出)への投与;経口、舌下、鼻、肛門、膣、経皮での投与、特定部位への外科的インプランテーション(例えば脾膜や脳の下、角膜内への埋め込み)などの方法がある。治療は、1回の投与、または所定の期間にわたる複数回の投与にすることができる。
【0145】
本発明によって同定されたモジュレータ化合物は、投与するため、塩基を含まない溶液にしたり、薬理学的に許容可能な塩を含む水に適切な界面活性剤(例えばヒドロキシプロピルセルロース)を混合したものにしたりすることができる。グリセロール、液体ポリエチレングリコール、これらの混合物への分散液や、油の中への分散液も調製することができる。通常の保管条件および使用条件では、微生物の増殖を防ぐために保存剤を調製物に入れる。
【0146】
注射に適した医薬形態としては、減菌水溶液または減菌分散液や、その場で減菌注射溶液または減菌分散液を調製するための減菌粉末などがある。いずれの場合にも、減菌形態でなくてはならず、容易に注射器に入れられるよう流体形態になっている必要がある。製造条件と保管条件のもとで安定であり、微生物(例えば細菌や真菌)の汚染作用から保護されている必要がある。基剤としては、例えば水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、液体ポリエチレングリコールなど)、これらの適切な混合物、植物油などを含む溶媒または分散媒体が可能である。適切な流動性を維持するには、例えばコーティング(例えばレシチン)を使用したり、必要な粒径を維持したり(分散液の場合)、界面活性剤を用いたりする。微生物の作用は、さまざまな抗菌剤や抗真菌剤(例えばパラベン、クロロブタノール、フェノール、ソルビン酸、チメロサールなど)を用いて防止することができる。多くの場合、等張剤(例えば糖類、塩化ナトリウム)を含めることが好ましい。注射組成物の吸収時間を延ばすには、組成物の中で吸収遅延剤(例えばモノステアリン酸アルミニウム、ゼラチン)を用いるとよい。
【0147】
減菌注射溶液は、必要量の活性化合物と他のさまざまな上記成分を適切な溶媒の中に組み込んだ後、必要に応じて濾過殺菌することによって調製する。一般に、分散液は、殺菌したさまざまな活性成分を、基本的分散媒体と必要な他の上記成分とを含む減菌ビヒクルの中に組み込むことによって調製する。減菌注射溶液を調製するための減菌粉末の場合には、好ましい調製方法は真空乾燥法と凍結乾燥法であり、活性成分と望む追加成分からなる粉末を、それら成分を含むあらかじめ濾過殺菌した溶液から作る。
【0148】
この明細書では、“薬理学的に許容可能な基剤”に、あらゆる溶媒、分散媒体、コーティング、抗菌剤、抗真菌剤、等張剤、吸収遅延剤などが含まれる。このような媒体や薬剤を薬理学的に活性な物質に用いる方法は従来技術において周知である。従来の媒体や薬剤のどれであれ、活性成分と適合しない場合を除き、治療用組成物で使用することが考えられる。追加活性成分も組成物に組み込むことができる。
【0149】
経口投与するため、本発明によって同定されたモジュレータは、賦形剤に組み込み、非消化性口内洗浄剤や歯磨き剤の形態で使用することができる。口内洗浄剤は、必要量の活性成分を適切な溶媒(例えばホウ酸ナトリウム溶液(ドーベル液))の中に組み込むことによって調製できる。活性成分は、ホウ酸ナトリウム、グリセリン、炭酸水素カリウムを含む減菌口内洗浄剤に組み込むこともできる。活性成分は、歯磨き剤の中に分散させることもできる。歯磨き剤は、ゲル、ペースト、粉末、スラリーの形態がある。治療に有効な量の活性成分をペースト状歯磨き剤に添加することができる。この歯磨き剤は、水、結合剤、研磨剤、香味剤、発泡剤、湿潤剤を含むことができる。
【0150】
本発明の組成物は、中性形態または塩の形態にすることができる。薬理学的に許容可能な塩としては、(タンパク質の遊離アミノ基とで形成された)酸添加塩がある。酸添加塩は、無機酸(例えば塩酸、リン酸)または有機酸(例えば酢酸、シュウ酸、酒石酸、マンデル酸など)とで形成される。遊離カルボキシル基とで形成される塩は、無機塩基(例えば水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、水酸化第二鉄)や有機塩基(例えばイソプロピルアミン、トリメチルアミン、ヒスチジン、プロカインなど)から得られる。
【0151】
本発明の組成物は、中性形態または塩の形態にすることができる。薬理学的に許容可能な塩としては、(タンパク質の遊離アミノ基とで形成された)酸添加塩がある。酸添加塩は、無機酸(例えば塩酸、リン酸)または有機酸(例えば酢酸、シュウ酸、酒石酸、マンデル酸など)とで形成される。遊離カルボキシル基とで形成される塩は、無機塩基(例えば水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、水酸化第二鉄)や有機塩基(例えばイソプロピルアミン、トリメチルアミン、ヒスチジン、プロカインなど)から得られる。
【0152】
製剤ができると、その製剤に適合した方法で、治療に有効な量の溶液を投与する。製剤は、多彩な投与形態(例えば注射溶液、薬剤放出カプセルなど)で容易に投与できる。水溶液の形態で非経口投与するには、その水溶液を必要に応じて適切に緩衝し、希釈されたその液体をまず十分な量の生理食塩水またはグルコースで等張にする必要がある。この特別な水溶液は、静脈内、筋肉内、皮下、腹腔内に投与するのに特に適している。
【0153】
“単位用量”は、適切な基剤に分散させた離散量の治療用組成物として定義される。例えば非経口投与は、まず最初にボーラスとして実施し、その後、連続輸液を行なって治療に必要な薬剤循環レベルを維持するとよい。当業者であれば、優れた医療経験と個々の患者の臨床状態により、効果的な投与量と投与計画を容易に最適化することができよう。より詳細には、投与量は、ADになっている対象の脳内でのAβ-ペプチドの形成、さらに特定するならば斑の形成の減少、抑制、低下、阻止をもたらすように選択する必要がある。そのためには、当業者は、(例えばアメリカ合衆国特許第5,877,399号、第5,387,742号、第5,811,633号に開示されているような)ADのモデル動物を利用して投与プロトコルを最適化し、ヒト患者でADを治療するのに必要な医薬の量を予測することができよう。
【0154】
投与の頻度は、薬剤の薬物動態パラメータと投与経路によって異なる。医薬製剤の最適な形態は、当業者が、投与経路と望ましい投与量から決定する。例えば『レミントンの薬理化学』、第18版(1990年、マック出版社、イーストン、フィラデルフィア州18042)、1435〜1712ページを参照のこと。なおその内容は参考としてこの明細書に組み込まれているものとする。製剤の形態は、投与した薬剤の物理的状態、安定性、生体内の放出速度、生体内のクリアランス速度に影響を与える可能性がある。投与経路に応じ、体重、体表面積、臓器のサイズをもとにして適切な投与量を計算することができる。治療に適した投与量を決めるのに必要な計算をさらに洗練させることは、当業者であれば、特別な実験を行なうことなく、この明細書に開示した投与量情報およびアッセイと、動物またはヒトの臨床試験で観察された薬物動態データに特に基づいて日常的に行なうことができる。
【0155】
適切な投与量は、血中レベルを測定するための確立されたアッセイと、関連する投与量-応答データを利用して確認することができる。最終的な投与計画は、参加している医者が、薬の作用を変化させる因子(例えば薬の特異的活性、損傷の程度、患者の応答性、年齢、状態、体重、性別、食事、何らかの感染の程度、投与時間、他の臨床因子)を考慮して決めることになろう。研究は進行中であるため、適切な投与量のレベル、特定の疾患や症状の治療期間に関するさらなる情報が今後得られるであろう。
【0156】
本発明の医薬組成物と治療法は、ヒトの医学と動物の医学の分野で役に立つ可能性があることが理解されよう。したがって治療する対象として、哺乳動物(ヒトであることが好ましい)または他の動物が可能である。獣医学の目的では、対象として、例えば家畜(乳牛、ヒツジ、ブタ、ウマ、ヤギなど)、ペット(イヌ、ネコなど)、外国産および/または動物園の動物、実験室の動物(マウス、ラット、ウサギ、モルモット、ハムスターなど)、家禽類(ニワトリ、シチメンチョウ、アヒル、ガチョウなど)などが挙げられる。
【実施例】
【0157】
以下の実施例は、本発明の好ましい実施態様を示すために掲載してある。しかし当業者は、この明細書をもとにして、開示されている具体的な実施態様に対して本発明の精神と範囲を逸脱することなく多くの変更を施してもやはり同様の結果を得られることを理解する必要がある。
実施例1
【0158】
材料と方法
この実施例では、この明細書で説明した結果を得るのに利用する一般的な方法、試薬、アッセイについて説明する。一般的な実験室用化学薬品は、シグマ・ケミカルズ社(セントルイス、ミズーリ州)から購入した。pET43.1aベクターは、ノヴァジェン社(マディソン、ウィスコンシン州)からのものであり、制限酵素はインヴィトロジェン社(カールズバッド、カリフォルニア州)からのものである。オリゴヌクレオチドはシグマ・ジェノシス社(ザ・ウッドランズ、マサチューセッツ州)からのものである。バリン1744抗体は、セル・シグナリング・テクノロジー社(ビヴァリー、マサチューセッツ州)からのものである。PHA/PNU化合物は、ファルマシア社の化合物コレクション(ニュー・ジャージー、アメリカ合衆国)から入手した。
【0159】
ノッチ基質のクローニング:マウスのノッチ-1タンパク質(DNA配列登録番号Z11886)のアミノ酸N1703〜D1860を含むノッチ基質をクローニングし、NusAをコードしていた核酸と同じフレームのEcoRI/HindIII挿入体としてpET43.1aベクターに入れた。NusA/ノッチ融合タンパク質をコードする発現構造体は、配列番号1の配列を持っていた。この構造体はPCRによって作ったC末端フラグと8ヒスチジン・タグを含んでおり、ノッチのアミノ酸1860の後に続く延長部(DYKDDDDKHHHHHHHH、配列番号15)となっていた。このDNAでBL21(DE3)コンピテント大腸菌(ストラタジーン社)を形質転換した後、DNAコンサート・ミニプレップ・キット(ギブコ/BRL社)を用いてクローンをスクリーニングし、正しい挿入体が存在しているかどうか調べた。ノッチ-F6というクローンは、配列番号1の正しいDNA配列を含んでいることがわかった。
【0160】
ノッチ/NusA融合タンパク質の発現と精製。大腸菌をノッチ-F6というクローンを用いて形質転換してからLB/Ampに接種し、振盪インキュベータの中で37℃にて一晩にわたって増殖させた。翌日、一晩にわたる培養物35mlを2リットルのLB/Ampに接種した。大腸菌を振盪インキュベータの中で37℃にてA600が0.4になるまで増殖させた後、1mMのIPTGを用いて3時間にわたって誘導し、遠心分離した。2リットルの培養物からのペレットを、50mMのトリス(pH8.0)、100mMのNaCl、プロテアーゼ阻害剤を含む緩衝液(ペレット1gにつき5ml)の中に再び懸濁させ、フレンチ・プレスを用いて3回処理して粗抽出液を得た。抽出液のpHを2Mのトリスを用いて8.0に調節した後、その抽出液を11,000×gで45分間にわたって遠心分離した。上澄4mlを、50mMのトリス(pH8.0)、100mMのNaCl、プロテアーゼ阻害剤を含む緩衝液の中で平衡させたニッケルIMACクロマトグラフィ・カラムに充填した。このカラムを同じ緩衝液で洗浄した後、50mMのイミダゾールを含む緩衝液で洗浄した。300mMのイミダゾールを含む緩衝液を用いてNusA-ノッチ融合タンパク質を溶離させ、分画0.8mlを回収し、A280とSDS-PAGEで分析した。NusA-ノッチを含む分画をプールし、50mMのピペス(pH7.0)、100mMのNaClの中に透析した。
【0161】
ウエスタン・ブロット分析によって調べるノッチの開裂。NusA-ノッチ融合タンパク質(1.7μM)を、可溶化γ-セクレターゼを70μg/mlの割合で含む合計体積50μlの緩衝液(50mMのピペス(pH7.0)、0.25%のCHAPSO)とともに37℃にて一晩にわたってインキュベートした。DAPT(PHA-568638)をさまざまな濃度で添加した。5×レムリ緩衝液(Laemmli、1970年)12.5μlを添加して反応を停止させ、混合物30μlの電気泳動を15%SDS-PAGE上で実施した。半乾燥ブロット装置(ミリポア社)を用いてタンパク質をニトロセルロースに移し、4%BSAを含むPBS/0.5%トゥイーン20を用いて2時間にわたってブロックした。バリン1744抗体を1000倍に希釈してブロッキング溶液に添加し、1時間にわたってインキュベートした。膜をPBS/0.5%トゥイーン20で3回洗浄した後、抗ウサギIgG-HRP(4%BSAを含むPBS/0.5%トゥイーン20に1:5000の割合で希釈)とともにインキュベートした。膜を再び3回洗浄し、ECL試薬(アマーシャム社、ピスカタウェイ、ニュージャージー州)を用いて現像した。
【0162】
インビトロ・ノッチ開裂ELISA。ELISA法を利用してノッチの開裂も調べた。反応を起こさせる前に、96ウエル半面積プレート(コスター社)の中の必要数のウエルを、0.1MのNaHCO3(pH8.2)の中で200倍に希釈したバリン1744抗体50μlでコーティングした。プレートを4℃にて一晩にわたってインキュベートした。ノッチ開裂反応は以下のようにして行なわせた。NusA-ノッチ(0.9μM)を、可溶化γ-セクレターゼを70μg/mlの割合で含む合計体積が25μlの緩衝液(50mMのピペス(pH7.0)、0.25%のCHAPSO)とともに37℃にて一晩にわたってインキュベートした。翌日、バリン1744抗体でコーティングしたプレートをPBS/0.5%トゥイーン20を用いて3回洗浄し、4%BSAを含むPBS/0.5%トゥイーン20を用いて1時間にわたってブロックした。4%BSAを含むPBS/0.5%トゥイーン20を用いて開裂反応混合物を14倍に希釈し、50μlを採取したものを3つ用意し、室温にて3〜4時間にわたってインキュベートした。プレートをPBS/0.5%トゥイーン20を用いて3回洗浄し、4%BSA/PBS/0.5%トゥイーン20の中で60000倍に希釈した抗FLAG-HRP抗体(シグマ社、セントルイス、ミズーリ州)50μlを添加した。この抗体を45分間にわたってインキュベートし、プレートをPBS/0.5%トゥイーン20を用いて3回洗浄した。TMB試薬(キルケガード&ペリー社)を1:1の割合で混合し、50μlを採取してウエルに添加した。1時間にわたって放置して色を変化させ、1MのH3PO4を50μl添加し、プレートをスペクトラマックス・プラス・プレート読み取り機で450nmにて読み取った。ノッチ基質の濃度を変化させたときには、0.11〜3.6μMのNusA-ノッチを使用した。酵素の濃度を変化させたときには、2.1〜68μg/mlの可溶化γ-セクレターゼを使用した。
【0163】
ノッチの開裂抑制。50%DMSOの中で25倍の濃度にした阻害剤(1μl)を開裂反応混合物に添加した後、酵素を添加した。ブランクの対照と阻害剤なしの対照も同じ最終濃度のDMSOを含むように調節した。阻害剤のIC50は、グラフィット4.0プログラムの4パラメータ・ロジスティック・モデルを利用して計算した。
実施例2
【0164】
結果と考察
可溶性ノッチ基質を産生させるため、発明者は、大腸菌の中に多数の構造体を用意した(図3A)。カスパーゼ・リーダー、ユビキチン、N末端タウ、Nugタグを利用した。可溶性ノッチ基質の発現は、Nus融合体でだけ観察された(Wilkinson他、Bio/Technology、第9巻、443〜448ページ、1991年)。図3Bには、この構造体のクローニングに関する詳細が示してある。精製とアッセイの開発を容易にするため、ヒスチジン・タグとフラグ・タグをノッチのC末端に付着させた。
【0165】
Nusタグの付いたノッチ融合タンパク質が大腸菌の中で発現するとき、90kDaのバンドに対応する融合タンパク質の高レベルの発現がSDS-PAGEで観察された(図4、レーン1)。このサイズは、この融合タンパク質から予想される値である。全ライセートを遠心分離すると、可溶性分画の中に融合タンパク質が残った(図4、レーン2)。固定化金属イオンとしてニッケルを用いたIMACにより、ノッチを含む融合タンパク質を可溶性分画から精製した。図4(レーン5〜9)には、300mMのイミダゾールによってIMACカラムから溶離したさまざまな分画が示してある。これら分画をプールし、透析した後、γ-セクレターゼによる開裂のための基質供給源として利用した。
【0166】
ノッチ・タンパク質の特異的な開裂を調べる方法は、バリン1744抗体(セル・シグナリング・テクノロジー社)の特異性に基づいている。この抗体は開裂したノッチに対して特異的であり、開裂していないノッチ・タンパク質と交差反応することはない。図5(レーン3)に示したように、開裂したノッチ・タンパク質は、バリン1744抗体に対して特異的な抗体を用いてウエスタン・ブロット上で検出された。レーン1からわかるように、開裂していないノッチ融合タンパク質との交差反応は観察されなかった。さらに、ノッチ・タンパク質におけるこの特異的開裂は、γ-セクレターゼの強力な阻害剤であることがよく知られているDAPTの投与量に依存して抑制された(レーン4〜8、図5)(Dovey他、J. Neurochem.、第76巻、173〜181ページ、2001年)。
【0167】
ノッチ・タンパク質に別の開裂があるかどうかを調べるため、C末端フラグ抗体を用いたウエスタン・ブロットによる開裂反応のモニターも行なった。3つの免疫活性なバンドのうち、1つの開裂産物だけがDAPTによって抑制可能であった。これらの結果を合わせると、γ-セクレターゼによるインビトロでの開裂により、1743/1744に特異的に開裂が起こることが示唆される。これは、細胞に基づく研究(Kopan他、Proc. Natl. Acad. Sci.、第93巻、1683〜1688ページ、1996年;Schroeter他、Nature、第393巻、382〜386ページ、1998年)においてノッチからNICDが産生されることが示されたという結果と一致している。
【0168】
上記の結果は、ノッチ融合タンパク質がγ-セクレターゼによる特異的開裂を起こしやすく、その開裂を、非常に特異的なバリン1744モノクローナル抗体によってウエスタン・ブロット上で検出できることを示している。ウエスタン・ブロットは定量的ではないため、ノッチを抑制する化合物を評価して抑制能力をAβ ELISAと比較することは難しい。
【0169】
図6には、ノッチの開裂を定量的に調べるELISAを開発し、CT-100に基づいたELISAでさまざまな化合物の抑制能力を比較する方法が示してある。この方法は、γ-セクレターゼの存在下におけるノッチ融合タンパク質からのNICD断片のN末端とC末端を高い特異性で捕捉する抗バリン1744抗体と抗フラグ抗体を用いたサンドイッチELISAに基づいている。このアッセイは、実施例1の“インビトロ・ノッチ開裂ELISA”に記載したようにして実施する。このサンドイッチELISAでは、マイクロタイター・プレートをバリン1744抗体でコーティングすることが重要である。ところでNusA-ノッチ開裂反応は、NusA-ノッチをγ-セクレターゼとともにインキュベートすることによって実施した。バリン1744抗体でコーティングしたプレートを緩衝液で洗浄し、BSAでブロックした。次に開裂反応混合物をマイクロタイター・プレートに添加し、室温にて適切な時間にわたってインキュベートした。プレートを洗浄して抗FLAG-HRP抗体(シグマ社、セントルイス、ミズーリ州)を添加し、適切な時間にわたって再びインキュベートした。HRPのための発色基質であるTMB試薬(キルケガード&ペリー社)を添加し、変化した色をスペクトラマックス・プラス・プレート読み取り機で450nmにて読み取った。図7には、ELISAで検出したNusA-ノッチ融合タンパク質の開裂が示してある。ノイズの5倍ある信号がELISAで観察された。
【0170】
ノッチを開裂させる可溶化γ-セクレターゼの酵素活性を、所定の実験条件下でELISAによってさらに調べた。図8Aには、生成物の形成が基質に依存する様子が示してある。反応はミカエリス-メンテンの式に従う。可溶化γ-セクレターゼによるノッチ融合タンパク質基質の加水分解に関する見かけのKmは、所定の条件下で約0.7μMである。観察されたノッチ開裂活性は、γ-セクレターゼ活性を有する可溶化膜調製物に含まれるタンパク質の濃度にも比例していた(図8B)。
【0171】
文献に報告されているγ-セクレターゼの特異的阻害剤を用いた抑制の研究を行なうことにより、この酵素によるノッチの開裂をELISAでさらに確認した。DAPTと呼ばれるγ-セクレターゼの強力な阻害剤が報告されている(Dovey他、J. Neurochem.、第76巻、173〜181ページ、2001年)。図9に示したように、DAPT(PHA-568638)はノッチの開裂を投与量に依存した形で抑制し、IC50=2.4nMである。その一方で、フェンキルアミンスルホンアミド(PHA-512088)もCT-100インビトロ・アッセイにおいて小さなμMの範囲でγ-セクレターゼの活性を抑制することが報告されている(Rishton他、J. Med. Chem.、第43巻、2297〜2299ページ、2000年)。図9からわかるように、PHA-512088はγ-セクレターゼによるノッチの開裂を抑制し、所定の条件下におけるELISAでIC50は0.7μMであった。この図からわかるように、阻害剤(PHA-568638、PHA-512088)は、アッセイにおいて小さな値のnM〜小さな値のμMの範囲で非常によく機能した。これらの結果を合わせると、ノッチ融合タンパク質基質からNICD断片を生成させるインビトロ酵素活性はγ-セクレターゼによるものであることがわかる。
【0172】
最近、APPとノッチの開裂を並行して調べる細胞ベースのアッセイが報告された(Karlstrom他、J. Biol. Chem.、第277巻、6763〜6766ページ、2002年)。しかしγ-セクレターゼによるノッチの特異的な開裂(G1743-V1744)を調べるためのインビトロ定量ELISAを初めて明らかにしたのは本発明である。本発明は、HeLa細胞からのγ-セクレターゼの活性によってノッチ・タンパク質が1743-1744接合部で特異的に開裂することを初めて証明した。
【0173】
この明細書に開示したり主張したりしたすべての組成物および/または方法は、この明細書の開示内容に照らして特別な実験なしに実現することができる。本発明の組成物と方法を好ましい実施態様に関して説明してきたが、当業者にとって、本発明の考え方、精神、範囲を逸脱することなく、この明細書に記載した組成物および/または方法と、その方法における一連のステップに対してさまざまな変更を施せることは明らかであろう。さらに詳細には、この明細書に記載した薬剤を化学的に関連した薬剤や生理学的に関連した薬剤で置き換えても同じ結果または似たような結果が得られることは明らかであろう。当業者にとって明らかなそのような同様の代替物や変更は、添付の請求項に規定した本発明の精神、範囲、考え方に含まれるものと考えられる。
【0174】
この明細書全体で引用した参考文献は、この明細書に記載した具体的な手続きやそれ以外の補足的な詳細を提供している場合には、すべて参考として具体的にこの明細書に組み込まれているものとする。
【図面の簡単な説明】
【0175】
【図1】γ-セクレターゼ様開裂部位の比較図である。膜貫通ドメインを取り囲む配列を、APP(配列番号10)、ノッチ-1(配列番号12)、E-カドヘリン(配列番号11)について示してある。APPにおいてAβ1〜40とAβ1〜42を産生させるγ-セクレターゼ開裂部位と、3つの基質におけるε開裂部位(ε)またはγ-セクレターゼ様開裂部位も示してある。
【図2】ノッチ-1でγ-セクレターゼ様開裂部位を取り囲んでいる部分のアミノ酸配列である。この配列(1703〜1860;配列番号13)を選択して大腸菌で発現させた。膜貫通ドメインは下線で示してあり、1743/1744開裂部位は太字と下線で示してある。このノッチ配列の開裂により、アミノ酸1744〜1860を含むNICD断片が得られることになる。
【図3A】大腸菌においてノッチを発現させるためにクローニングした構造体。4つの構造体を示してあり、そのすべてにノッチ-1のアミノ酸配列1703〜1860が含まれている。カスパーゼ・リーダー配列、ユビキチン、タウのN末端ドメインのいずれかをN末端タグとして用いて発現を促進した場合には発現は見られなかった。ノッチ(1703〜1860)をNusAタンパク質と融合させた場合には、大腸菌において可溶性発現が見られた。
【図3B】ノッチF構造体をクローニングしてpET43.1aに組み込んだ概略図である。この構造体は、C末端にフラグと8ヒスチジン・タグも備えている。
【図4】NusA-ノッチ融合体のIMAC単離。この融合タンパク質の概略図を最上部に示してある。NusA-ノッチ融合体の発現と精製に関する詳細は「材料と方法」に見ることができる。矢印は、10%SDS-PAGEゲル上でこの融合タンパク質が発現したバンドを示している。レーン1、大腸菌の粗ライセート;レーン2、大腸菌の上澄;レーン3、IMACのフロースルー;レーン4、50mMのイミダゾールによる洗浄;レーン5〜9、300mMのイミダゾールによる溶離分画;レーン10、ベンチマーク分子量マーカー。
【図5】可溶化γ-セクレターゼを用いたNusA-ノッチ融合体の開裂を示すウエスタン・ブロット。この実験の詳細は実施例1に記載してある。要するに、さまざまな濃度のDAPT(PHA-568638)の存在下と不在下で、NusA-ノッチ融合体を可溶化γ-セクレターゼとともに一晩にわたってインキュベートした。サンプルを電気泳動にかけ、ブロットし、バリン1744での開裂に特異的なバリン1744抗体で調べた。
【図6】ノッチの特異的開裂のELISAによる検出。可溶化γ-セクレターゼによるNusA-ノッチ融合体の開裂を検出するのに用いたサンドイッチELISAの概略を示してある。特異的開裂は、バリン1744抗体を用いて検出する。
【図7】γ-セクレターゼによって開裂したNusA-ノッチ融合体のELISAによる検出。68μg/mlの可溶化γ-セクレターゼ(酵素)の不在下と存在下でNusA-ノッチ(0.9μM)基質をインキュベートし、「材料と方法」に従ってELISAを行なった。データは6回の実験の平均値を表わしている。
【図8】ELISAによる、ノッチ開裂のキャラクテリゼーション。ELISAは「材料と方法」に記載したようにして実施した。ノッチ基質の濃度を変化させたときには、0.11〜3.6μMのNusA-ノッチを68μg/mlの可溶化γ-セクレターゼとともにインキュベートし、プロットしたデータをミカエリス-メンテン曲線にフィットさせた。酵素の濃度を変化させたときには、2.1〜68μg/mlの可溶化γ-セクレターゼを0.9μMのノッチ基質とともにインキュベートした。データは3回の実験の平均値を表わしており、GraFit4.0を用いてプロットした。バックグラウンドと比べて約5倍の信号が存在していた(酵素だけを用いたバックグラウンドは無視できる)。
【図9】γ-セクレターゼによるCT-100の開裂を試験管内で抑制することが知られている化合物によるノッチ開裂の抑制。ノッチ開裂ELISAを利用して測定したDAPT(PHA-568638)とフェンキルアミン(PHA-512088)の抑制プロファイルを示してある。相対的IC50の値も示してある。
【技術分野】
【0001】
本発明は、γ-セクレターゼの新規な可溶性基質に関する。さらに詳細には、本発明により、NusAタンパク質に融合したγ-セクレターゼ依存開裂部位(γとε)含有ノッチ区画を有する可溶性融合タンパク質が提供される。このような融合タンパク質の製造法と利用法、ならびにこのような融合タンパク質を利用した組成物も開示されている。
【背景技術】
【0002】
アルツハイマー病(AD)によってアミロイド斑の形成、神経原線維の変化、グリオーシス、ニューロンの破壊が起こり、進行性の痴呆が引き起こされる。この疾患には遺伝性と散発性の両方の形態があるが、臨床経過と病理学的特徴は両者で極めて似通っている。突然変異したときに常染色体優性形態のADを引き起こす3つの遺伝子がこれまでに発見されている。これら遺伝子はアミロイド・タンパク質前駆体(APP)と2つのタンパク質(構造と機能が互いに関連したプレセニリン-1(PS1)とプレセニリン-2(PS2))をコードしている。これら3つのタンパク質のどれかに突然変異があると、アミロイドβペプチド(アミノ酸が40〜42個のペプチドで、ADにおけるアミロイド斑の主要な成分であり、Aβペプチド、Aベータと呼ばれることもある)を産生する細胞内経路を通じてAPPのタンパク質分解処理が促進されることが観察されている(Younkin、Brain Pathol、第1巻(4)、253〜262ページ、1991年;Haass、J. Neurosci.、第11巻(12)、3783〜3793ページ、1991年)。
【0003】
タンパク質分解処理のための細胞内経路がうまく機能しなくなることがADの生理病理学にとって最も重要であろう。アミロイド斑が形成されるときには、APP、PS1、PS2のいずれかにおける突然変異が常にAPPのタンパク質分解処理を変化させるため、Aβ1〜42の形成が促進される。このAβ1〜42は、特にアミロイドを生成させるように見えるため、ADにおいて非常に重要なAβペプチドの一形態である。APPは、細胞表面などの分泌膜構造に局在しており、C末端の近くに膜貫通ドメインを有する(図1)。ヒトに存在することが現在知られているAPPの特別なアイソタイプの具体例としては、Kangら(Nature、第325巻、733〜736ページ、1987年)によって記載されているアミノ酸が695個のポリペプチド(“正常”APPと呼ばれる);Ponteら(Nature、第331巻、525〜527ページ、1988年)とTanziら(Nature、第331巻、528〜530ページ、1988年)によって記載されているアミノ酸が751個のポリペプチド;Kitaguchiら(Nature、第331巻、530〜532ページ、1988年)によって記載されているアミノ酸が770個のポリペプチドがある。
【0004】
Aβペプチドは、APPの領域のうちで膜貫通ドメインに隣接していてその膜貫通ドメインの一部を含む領域に由来する(図1)。通常は、α-セクレターゼ部位でAPPのプロセシングを行なうと、Aβ配列のうちで膜に隣接した中間領域が開裂し、細胞表面から可溶性細胞外ドメインが放出される。このα-セクレターゼによるAPPのプロセシングにより可溶性APP-αが生じるが、これはADの一因にならないと考えられている。しかしβ-セクレターゼ部位(α-セクレターゼ部位に対してN末端側に位置する)とγ-セクレターゼ部位(α-セクレターゼ部位に対してC末端側に位置する)でAPPの病的なプロセシングが起こるとAβペプチドが放出される。β-セクレターゼ開裂部位は、細胞膜管腔表面から28残基のところに位置し、APPγ-セクレターゼ開裂部位は膜貫通ドメインに位置する。(すべての細胞内の)細胞表面APPが再度内部化された後に、β-セクレターゼ部位とγ-セクレターゼ部位でのプロセシングが、(ニューロン内の)小胞体とエンドソーム/リソソーム経路の両方において起こる可能性がある。
【0005】
したがってβ-セクレターゼとγ-セクレターゼの酵素活性は、薬を発見するための標的である(Olson他、Curr. Opin. Drug Discovery & Develop.、第4巻、390〜401ページ、2001年)。これら2つの酵素は協力してAPPを開裂させる。APPはまず最初にβ-セクレターゼによって開裂されて、CT-100と呼ばれる膜結合C末端(CT)断片を生み出す。すると今度はこのCT-100が膜関連γ-セクレターゼの基質となる。プレセニリン1(PS1)依存γ-セクレターゼによるCT-100の膜内開裂により、Aβ1〜40とAβ1〜42が産生される。さらに、(γ様開裂またはε様開裂と名づけられた)別の開裂によって細胞膜の境界の内側約2〜5残基の位置でAPPの残基721の付近が開裂し、一連の安定なC末端APP断片が生じる(図1)。
【0006】
ノッチ-1は、細胞の運命を決める上で幅広い役割を果たしている細胞表面受容体のノッチ・ファミリーに属する。最近、APPのプロセシングが、細胞表面受容体であるノッチ-1のプロセシングに似ていることが主張されている(Wolf他、J. Biol. Chem.、第276巻、5413〜5416ページ、2001年)。実際、APPとノッチ-1は、内在性と考えられるγ-セクレターゼの競合する基質であることがわかっている。ノッチ-1は内在性膜タンパク質であり、リガンドを媒介として活性化されたときにその表面領域内でタンパク質分解処理される。ノッチ-1は、リガンドと結合した後、プレセニリンに依存した膜内γ-セクレターゼ開裂を起こす(Okochi、EMBO J.、第21巻、5408〜5416ページ、2002年)。第1の開裂は1731/1732部位(これはAβ様γ-セクレターゼ開裂に近い)であり、第2の開裂は1743/1744部位(これはAPPが生じるときのε開裂に近く、ノッチのS3開裂と呼ばれることもある;図1には“ε”開裂として示してある)である。膜貫通ドメインの端部近くで起こってノッチ-1細胞内ドメイン(NICD)を放出するのは、ノッチの1743/1744接合部におけるε開裂である。放出されたNICDは核に移動し、そこでCSL(この略号は、異なる系でこのタンパク質に与えられた3つの名前を表わしている。その系とは、哺乳動物におけるCBF1/RBP-J;ショウジョウバエとゼノプスにおける毛なしのサプレッサ[Su(H)];線虫のLag-1である)と名づけられたDNA結合タンパク質と相互作用する。NICDとCSLの間に形成される複合体は標的遺伝子の転写を変化させる。NICDは、胚発生において極めて重要なシグナル伝達経路で必要とされる(Schroeter、1998年;Hupert、2000年)。
【0007】
プレセニリンに依存したγ-セクレターゼ活性は、ノッチ受容体のプロセシングによってNICDを放出させるのに必要とされる(De Stropper他、Nature、第398巻、518〜522ページ、1999年)。細胞内では、ノッチ1とAPPが、PS1に依存したγ-セクレターゼ開裂の対象として競合する基質であることが報告されている(Berezovska他、J. Biol. Chem.、第276巻、30018〜30023ページ、2001年)。そのため、病原性Aβの産生を抑制するように設計されたγ-セクレターゼ阻害剤は、ノッチのシグナル伝達も抑制する。これは、γ-セクレターゼ阻害剤をADの治療薬として使用できる可能性があるという大きな意味を持つ。造血においてノッチ-1は重要な機能を担っているため、成人でノッチ-1のプロセシングを抑制すると、免疫不全と貧血になる可能性がある。したがってAPPの開裂によるCT-100の産生は特異的に抑制するが、ノッチの開裂は抑制しない化合物を同定する必要がある。そのような化合物は、NICDの産生を抑制するという悪い効果を生み出すことなくADを治療するための治療薬として役立つ可能性がある。
【発明の開示】
【0008】
本発明の概要
本発明により、γ-セクレターゼによるノッチの開裂を抑制しない化合物を同定する方法と組成物が提供される。この方法と組成物は、γ-セクレターゼを非特異的に抑制することに伴うNICDの産生抑制という問題を回避する。したがって本発明により、NICDの産生を抑制するという悪い効果を生み出すことなくADを治療するための治療薬を同定することが可能になる。
【0009】
本発明の第1の特徴により、NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質が提供される。このノッチ・タンパク質は、ノッチ・タンパク質の膜貫通ドメインを含んでいることが好ましい。しかしこのノッチ・タンパク質は、ノッチ・タンパク質がノッチのS3開裂部位を含んでいさえすれば、膜貫通ドメインではなく、ほぼ完全長のノッチ・タンパク質を含んでいてもよい。特別な一実施態様のノッチ・タンパク質は、配列番号4の配列を含んでおり、配列番号3の核酸配列によってコードされている。組み換えノッチ・タンパク質は、ノッチのS3開裂部位(すなわちε開裂部位)を含んでいる必要がある。組み換えノッチ・タンパク質は、脊椎動物のノッチ・タンパク質または無脊椎動物のノッチ・タンパク質が可能である。いくつかの実施態様では、組み換えノッチ・タンパク質は、配列番号5の配列[GenBank登録番号Z11886]を有するマウスのノッチ・タンパク質に由来する組み換えノッチ・タンパク質である。より特殊な実施態様では、マウスのノッチ・タンパク質のアミノ酸1703〜1860を含む組み換えノッチ・タンパク質を利用することを考える。本発明による可溶性融合タンパク質のどれも、C末端ヒスチジン・タグおよび/またはC末端フラグ・タグをさらに備えることができる。もちろん、ヒト・ノッチ配列の全体または一部(例えば配列番号6の配列[GenBank登録番号M73980])も用いることができよう。
【0010】
本発明ではさらに、この明細書に記載した融合タンパク質をコードするヌクレオチド配列を含むポリヌクレオチドを考える。そのような融合タンパク質をコードするポリヌクレオチド配列の一例は、配列番号1に示した配列を含んでいるものである。このポリヌクレオチド配列は、配列番号2の融合タンパク質をコードしている。この明細書では、本発明のポリヌクレオチドを含む発現ベクターも考える。この発現ベクターは、その内部に含まれるポリヌクレオチドがプロモータと機能上リンクしていて、そのポリヌクレオチドによってコードされるタンパク質の発現を宿主細胞の中で促進するような発現ベクターであることが好ましい。
【0011】
この明細書に記載したポリヌクレオチドまたは発現ベクターを用いた形質転換またはトランスフェクションを受けた組み換え宿主細胞も本発明の範囲に含まれる。本発明では、γ-セクレターゼ・アッセイのための基質を作る方法であって、そのような組み換え宿主細胞を増殖させて融合タンパク質の発現を可能にする操作を含む方法を考える。この方法は、その融合タンパク質を精製する操作をさらに含むことができる。このような方法では、宿主細胞は、組み換えタンパク質を産生できる任意の宿主細胞でよく、例えば哺乳動物宿主細胞、細菌宿主細胞、酵母宿主細胞などが可能である。具体的な実施態様では、宿主細胞は、Hela細胞、ヒト胚性腎臓細胞系の293細胞、線維芽細胞、CHO細胞のいずれかである。
【0012】
本発明の別の特徴により、可溶化ノッチ・タンパク質の製造方法であって、ノッチ・タンパク質をNusAタンパク質のC末端に融合させた融合タンパク質を調製する操作を含む方法が提供される。さらに詳細には、この方法は、融合タンパク質を組み換えによって産生させるため、ノッチのS3開裂部位のアミノ酸を含むノッチ・タンパク質をNusAタンパク質のC末端にリンクさせた融合タンパク質をコードしている核酸を含む発現構造体を調製し;その融合タンパク質の発現が容易になる条件下で、この発現構造体を用いて宿主細胞を形質転換し;その形質転換された宿主細胞を培地の中で増殖させる操作を含んでいる。この方法はさらに、培地内の形質転換された宿主細胞から融合タンパク質を単離する操作を含むことができる。いくつかの実施態様では、この方法は、化学的タンパク質合成を通じて融合タンパク質を製造する操作を含んでいる。そのような方法では、ノッチ・タンパク質は、マウスのノッチ・タンパク質のアミノ酸1703〜1860を含んでいることが好ましい。
【0013】
この明細書では、γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)を調べるインビトロ法であって、哺乳動物のγ-セクレターゼ複合体またはその生物学的に活性な断片を含む第1の組成物を、本発明の融合タンパク質を含む第2の組成物と接触させ;その融合タンパク質の開裂を調べる操作を含む方法も考える。
【0014】
上記方法のγ-セクレターゼ複合体は、哺乳動物の細胞から精製し単離した膜分画、あるいはこのγ-セクレターゼ複合体をコードしているヌクレオチド配列を含む発現構造体を用いた形質転換またはトランスフェクションを受けた細胞から精製し単離した膜分画を含むことができる。上記の方法では、融合タンパク質は、この明細書に記載した方法に従って調製した可溶化ノッチ・タンパク質である。
【0015】
本発明では、この明細書に記載した方法で同定されたγ-セクレターゼ・モジュレータを含む組成物を特に考える。本発明には、生体内でγ-セクレターゼの活性を変化させる方法であって、哺乳動物に対し、この明細書に記載したスクリーニング法で同定されたモジュレータを、その哺乳動物の細胞内でγ-セクレターゼの活性を変化させるのに有効な量投与するステップを含む方法も含まれる。このモジュレータは、γ-セクレターゼによるノッチ・タンパク質の開裂ではなく、γ-セクレターゼによるAPPの開裂を選択的に抑制するγ-セクレターゼ・モジュレータである。
【0016】
本発明は、本発明によって同定された1つ以上のモジュレータと、薬理学的に許容可能な基剤とを含む医薬組成物にも関する。本発明では、γ-セクレターゼの異常な活性を特徴とする疾患や症状を治療する方法であって、治療を必要としている対象に本発明の医薬組成物を投与する操作を含む方法も考える。この明細書で同定されたモジュレータを利用してアルツハイマー病の治療薬を製造することも特に考える。
【0017】
本発明の別の特徴ならびに利点は、以下の詳細な説明から明らかになろう。しかしこの詳細な説明と具体的な実施例は、本発明の好ましい実施態様を示しているとはいえ、単なる例示であることを理解する必要がある。なぜなら、当業者には、本発明の範囲内でのさまざまな変化や変更がこの詳細な説明から明らかになるだろうからである。
【0018】
図面は本発明の明細書の一部であり、本発明の特徴を詳しく説明するためのものである。本発明は、図面と、この明細書に提示した具体的な実施態様の詳細な説明とを組み合わせることによってさらによく理解できよう。
【発明を実施するための最良の形態】
【0019】
ADは進行性の痴呆と症状を伴う重要な加齢関連疾患であり、皮質の萎縮と老人斑の堆積を特徴とする。老人斑の主成分は、アミノ酸40〜42個の長さがあるペプチドAβであり、完全長APPの膜貫通ドメインの一部に隣接した領域、またはその部分を含む領域に由来する。この病原性ペプチドは、β-セクレターゼとγ-セクレターゼの活性に起因する一連のプロセシングの結果として生成される。したがってこれら2つのセクレターゼの酵素活性が、薬を発見するための標的となる(Olson他、Curr. Opin. Drug Discovery & Develop.、第4巻、390〜401ページ、2001年)。
【0020】
細胞表面受容体であるノッチのプロセシングはAPPのプロセシングと似ていることがわかっている(Wolf他、J. Biol. Chem.、第276巻、5413〜5416ページ、2001年)。図1には、APPとノッチにおけるγ-セクレターゼ様開裂部位を比較して示してある。この図からわかるように、APPの開裂部位は膜貫通ドメインの中央部にあるのに対し、ノッチは膜貫通ドメインのC末端に非常に近い位置が開裂する。似たようなε開裂部位が、カドヘリンEにおいて最近報告された(Marambaud他、EMBO J.、第21巻、1948〜1956ページ、2002年)(図1)。図2は、ノッチ・タンパク質のアミノ酸配列でノッチ内のS3開裂部位を含む1703〜1860を示している(Steiner他、J. Mol. Neurosci.、第17巻、193〜198ページ、2001年)。1743/1744接合部における特異的開裂により、V1744〜D1860ノッチ細胞内ドメイン(NICD)が生じる可能性がある。これは、S1開裂部位とS2開裂部位が欠如したmΔEノッチ構造体(1704〜2183)をトランスフェクトされた細胞で観察されるNICDの断片である(Kopan他、Proc. Natl. Acad. Sci.、第93巻、1683〜1688ページ、1996年)。ノッチ構造体(バリン1711〜グルタミン酸1809)が報告されている(Esler他、Proc. Natl. Acad. Sci.、第99巻、2720〜2725ページ、2002年)とはいえ、試験管内でのγ-セクレターゼによる1743/1744接合部の特異的開裂は記載されていない。1743/1744接合部における特異的開裂は、バリン1744抗体(セル・シグナリング・テクノロジー社)を用いて容易に明らかにすることができる。この抗体は開裂したノッチに対して特異的であり、開裂していないノッチ・タンパク質と交差反応することはない。
【0021】
したがって細胞では、PS1依存性γ-セクレターゼは、β-セクレターゼの作用によって生じるCT-100断片を開裂させることに加え、ε部位1743/1744でも開裂させてNICDを生成させる。開裂したこのNICDは核に移動し、シグナル伝達に関与する。ADを緩和するためにγ-セクレターゼの活性を抑制すると、NICDの生成も抑制される。本発明により、初めて、ノッチ-1からのNICDの生成を抑制しない治療薬を同定する方法と組成物が提供される。
【0022】
本発明の方法により、ノッチの開裂を調べるインビトロ・アッセイと、そのようなアッセイを利用してγ-セクレターゼの阻害剤を二次評価する方法が提供される。このアッセイでは、γ-セクレターゼの可溶性ノッチ基質を利用する。この基質は、NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質を含んでいる。本発明のアッセイは、γ-セクレターゼによるノッチ・タンパク質の開裂を調べる直接インビトロELISAアッセイおよび/またはウエスタン・ブロットとして実施することができる。本発明の方法により、適切な細胞系(例えば大腸菌)の中でNusAをノッチ・タンパク質配列に融合させた融合タンパク質として発現する可溶性組み換えノッチ・タンパク質(アスパラギン1703〜アスパラギン酸1860)基質の産生と精製がなされる(図6を参照のこと)。
【0023】
発明者は、本発明による精製された融合タンパク質基質を利用し、HeLa細胞からの可溶化γ-セクレターゼによってノッチの1743/1744部位が特異的に開裂することを証明した。実施例に詳しく説明したように、開裂したノッチ・タンパク質はバリン1744に対して特異的な抗体を用いて検出することができる。発明者は、このアッセイが有効であることを確認し、ノッチ・タンパク質の開裂が、γ-セクレターゼの強力な阻害剤としてよく知られているDAPTによって投与量に依存した形で抑制されることを示した。抗バリン1744抗体に基づいたELISAを開発した。さらに、開裂がγ-セクレターゼ阻害剤によって抑制されることでインビトロ・ノッチ・アッセイが有効であることを示した。阻害剤またはモジュレータは、機能的には、γ-セクレターゼを通じてAβを減少させる化合物と定義される。
【0024】
I.インビトロ・ノッチ・アッセイのためのノッチ基質
【0025】
本発明のアッセイで使用するノッチ基質は、ノッチ・ポリペプチドをNusタンパク質に融合させた可溶性融合タンパク質である。この融合タンパク質に標識すること、あるいはそれ以外の修飾を行なうことが可能である。それは、ペプチドを精製するため、あるいはノッチ融合タンパク質そのものを検出するため、あるいはγ-セクレターゼが作用したときにノッチ融合タンパク質の開裂産物の検出を容易にするためである。融合タンパク質の生成法と具体的な修飾法について以下にさらに詳しく説明する。
【0026】
本発明の好ましい1つの特徴によれば、マウスのノッチ-1タンパク質(DNA配列登録番号Z11886;図2を参照のこと)のアミノ酸N1703〜D1860を含むノッチ基質のN末端をNusAのC末端に接合した。この融合ポリペプチドを組み換えタンパク質の産生によって、あるいは自動化したペプチド合成によって得ることを考える。後者についてはこの明細書の別の箇所で説明してある。図2の膜貫通ドメインは、アミノ酸1723〜1744にまたがっている(図1を参照のこと)。γ-セクレターゼは、ノッチのアミノ酸1743と1744の間にあるε開裂部位を開裂させる。この開裂部位はS3開裂部位とも呼ばれており、NICD(すなわちアミノ酸V1744〜D1860の範囲のペプチド)を発生させる。
【0027】
本発明では、この新規な融合タンパク質に加え、上記のノッチ/NusA融合タンパク質基質、または本発明に従って同定したノッチ/NusA融合タンパク質基質の末端部付加物(融合タンパク質または融合ポリペプチドとも呼ばれる)を作ることも考える。さらに、本発明の好ましい実施態様にはマウスのノッチ-1のN1703〜D1860を含むノッチ/NusA融合ペプチドを示してあるが、ノッチ・タンパク質は任意の供給源からのものが可能であることを理解する必要がある。そのような供給源としては哺乳動物または非哺乳動物が可能である。したがってノッチ-1はヒト、マウス、ラット、あるいはそれ以外の哺乳動物に由来することが好ましいとはいえ、例えば線虫、ゼノプス、ショウジョウバエや、他の無脊椎動物からのものも可能である。
【0028】
さらに、γ-セクレターゼ開裂部位1731/1732(γ部位)と1743/1744(ε開裂)を含むあらゆるノッチ-1誘導体が本発明による融合タンパク質基質において有用であることも考慮する。ノッチ-1では、NICDを放出するγ-セクレターゼ開裂部位は、ノッチ内の1743/1744部位に位置する。この開裂部位とNusAの開始位置の距離を約500アミノ酸にして、ノッチのγ-セクレターゼ開裂ドメインの立体特性に似せることを考える。この距離は、NusAタンパク質から生じさせるか、あるいは異種ペプチド・リンカーによって作り出すことができる。この領域はNusAタンパク質からのものであることが好ましい。ノッチ/NusA融合ポリペプチドのNusAタンパク質ドメイン部分としては、そのノッチ/NusA融合ポリペプチドが可溶性を維持していて、したがってインビトロ・アッセイを実施できるようなNusAタンパク質の任意の部分が可能である。
【0029】
これまでNusAは大腸菌で可溶性タンパク質を発現させるのに用いられてきた(例えばWilkinson他、Bio/Technology、第9巻、443〜448ページ、1991年;Davis他、Biotechnol. Bioeng.、第65巻(4)、382〜388ページ、1999年を参照のこと)。この明細書では、(配列番号16の核酸配列によってコードされる)配列番号17の配列を含むNusAタンパク質をノッチと融合させる。しかし当業者であれば、配列番号17に示した配列以外のNusA配列を使用しても本発明の可溶性ノッチ/NusA融合タンパク質を作り出すことができよう。当業者は、例えば配列番号17の配列と80%、85%、90%、95%、96%、97%、98%またはそれ以上が相同であるNusAタンパク質を使用することができる。あるいは当業者は、配列番号17に由来するNusAのより小さな連続断片を用いることもできる。例えばその断片は、配列番号17の50個、100個、150個、200個、250個、300個、350個、400個、425個、450個、475個、500個、510個、520個、530個、540個、550個またはそれ以上の連続したアミノ酸にすることができる。
【0030】
融合タンパク質を設計して作り出すための一般的な原理は当業者には周知である。例えば融合体には他の種からのリーダー配列が一般に用いられているため、異種宿主の中で組み換えタンパク質または組み換えペプチドを発現させることができる。別の有用な融合体では、免疫活性のあるドメイン(例えば抗体のエピトープ)を付加して融合ポリペプチドの精製を容易にする。開裂部位を融合が起こる位置またはその近傍に組み込むと、精製後に外来性ポリペプチドを除去することが容易になる。このような融合体を組み換えによって産生させることについてはこの明細書の別の箇所でさらに詳しく説明する。他の有用な融合体としては、機能ドメイン(例えば酵素からの活性部位、グリコシル化ドメイン、細胞標的シグナル、膜貫通領域)がリンクしたものが挙げられる。
【0031】
より詳細には、本発明において、1731/1732(γ開裂)開裂部位と1743/1744(ε開裂)開裂部位を有するノッチ・タンパク質を含む第1の成分が、NusAタンパク質の全体または一部を含む第2の成分に結合した融合ポリペプチドを考える。さらに別の実施態様では、融合ポリペプチドは、レポータ遺伝子産物を含む第3の成分をさらに含むことができる。さらに別の実施態様では、融合ポリペプチドは、タグの付いた配列成分をさらに含むことができる。考慮する特別な融合ポリペプチドは、NusA部の一方の側にあるレポータ遺伝子産物と、γ-セクレターゼ開裂部位を有するノッチ・タンパク質を含む配列と、ノッチ・タンパク質の他方の側にあるタグ付き配列とを備えるものである。本発明の融合ポリペプチドで使用するレポータ遺伝子産物は、当業者が一般に使用している任意のレポータ・タンパク質でよい。具体的なレポータ・タンパク質としては、ルシフェラーゼ;分泌アルカリホスファターゼ(SEAP);β-ガラクトシダーゼ;β-グルクロニダーゼ;グリーン蛍光タンパク質;クロラムフェニコールアセチルトランスフェラーゼなどが挙げられる。
【0032】
別の特別な実施態様では、本発明の融合ポリペプチドの第4の成分としてタグ付き配列をさらに考える。市販されているさまざまな融合タンパク質発現系があり、それを本発明のこの文脈でタグ付き配列として使用することができる。特に有用な系としては、グルタチオン-S-トランスフェラーゼ(GST)系(ファルマシア社、ピスカタウェイ、ニュージャージー州)、マルトース結合タンパク質系(NEB社、ビヴァリー、マサチューセッツ州)、フラグ系(IBI社、ニュー・ヘイヴン、コネチカット州)、6×ヒスチジン系(キアジェン社、チャッツワース、カリフォルニア州)などが挙げられる。これらの系は、組み換え融合タンパク質が関係する生物学的活性に影響を与えそうにない少数の付加アミノ酸だけを有する組み換えポリペプチドを産生させることができる。例えばフラグ系と6×ヒスチジン系の両方とも、短い配列だけを付加する。両方とも抗原性が弱いことが知られており、ポリペプチドが折り畳まれて本来の立体配座になるのを妨げない。有用であると考えられる別のN末端融合体は、タンパク質またはペプチドのN末端領域にメチオニン-リシン・ジペプチドが融合したものである。このような融合体によってタンパク質の発現および/または活性が好ましく増大する可能性がある。本発明で使用することが考えられる特別なタグ付き配列としては、C末端フラグ・タグ配列DYKDDDDK(配列番号14)がある。さらに、このタグ付き配列は8ヒスチジン・タグも含んでいてよく、その場合にはフラグ/8ヒスチジン・タグ(DYKDDDDKHHHHHHHH、配列番号15)が融合タンパク質に結合する。
【0033】
本発明の好ましい融合タンパク質の一例は、γ-セクレターゼ開裂部位1731/1732と1743/1744に加えて短いC末端フラグ/8ヒスチジン・タグ尾部を含むマウスの完全長ノッチ-1タンパク質またはその一部にNusAが融合しているものである。ある融合ポリペプチドの配列は、NusAタンパク質に融合したマウスのノッチのアミノ酸1703〜1860(すなわち配列番号13に示したもの)を含んでいる。この融合ポリペプチドは配列番号2の配列を有する。γ-セクレターゼによるこのキメラ構造体の開裂をモニターするため、バリン1744残基を含むノッチ開裂産物に対する抗バリン1744抗体を用い、ノッチが開裂する結果として生成されるバリン1744断片の存在と濃度を明らかにする。キメラ構造体のC末端を検出するのに抗フラグ抗体を用いることができる。しかしキメラ構造体では、フラグ・タグの代わりにC末端にレポータ・タンパク質(例えばアルカリホスファターゼ)も用いることができる。このようなレポータ・タンパク質はノッチが開裂する結果として放出される可能性があるため、当業者に周知のアッセイを利用して検出できよう。
【0034】
本発明により、すでに説明した融合ポリペプチドが提供されることに加え、例えば標識またはそれ以外の検出可能な部分を組み込む修飾をさらに行なったノッチ融合タンパク質が提供される。
【0035】
例えばノッチ/NusA融合タンパク質基質は内部に消光状態の標識を備えており、それが、ノッチ基質が開裂した後の検出可能性を高める。ノッチ/NusA融合タンパク質基質を修飾して蛍光体(例えば7-アミノ,4-メチルクマリンと消光物質(例えば2,4-ジニトロフェノール)のペアを結合させると、γ-セクレターゼによってペプチドが開裂したとき、γ-セクレターゼによって切断されやすい結合の両側に結合させた蛍光体と消光物質が物理的に分離するために蛍光が増大する。ペアにする別の蛍光体と消光物質としては、ボディピー-テトラメチルローダミンとQSY-5(モレキュラー・プローブズ社)がある。このアッセイの別の態様では、ビオチンまたは他の適切なタグをペプチドの一端に配置してそのペプチドを基質アッセイ用プレートに固定した後、蛍光体をそのペプチドの他端に配置することができる。有用な蛍光体としては、上記のもののほか、W8044(EG&gウォラック社)などのユーロピウム標識がある。使用可能な別の好ましい標識は、システイン残基に結合させることのできるオレゴン・グリーンである。γ-セクレターゼによる融合タンパク質の開裂によってプレートから蛍光体または他のタグが放出されるため、保持された蛍光の増大から、タンパク質分解による開裂がいろいろな化合物について抑制されたかどうかを知ることができる。γ-セクレターゼのタンパク質分解活性に関する好ましい比色アッセイでは、開裂するγ-セクレターゼ認識部位を有するアミノ酸配列がアミド結合を通じてo-ニトロフェノールに結合した別のノッチ/NusA融合タンパク質を用いる。そのためγ-セクレターゼによって融合タンパク質が開裂すると、アッセイ用緩衝液がアルカリpHに変化した後に光学密度が増大する。
【0036】
さらに、ノッチ/NusA融合タンパク質には当業者に周知の標識を付けることができる。例えばビオチン標識が特に考えられる。このような標識を使用することは当業者には周知であり、例えばアメリカ合衆国特許第3,817,837号、第3,850,752号、第3,996,345号、第4,277,437号に記載されている。有用な他の標識としては、放射性標識、蛍光標識、化学的発光標識などがある。このような標識の利用に関するアメリカ合衆国特許としては、例えばアメリカ合衆国特許第3,817,837号、第3,850,752号、第3,939,350号、第3,996,345号がある。本発明のどのノッチ/NusA融合タンパク質組成物も、これら標識のうちの1つ、2つ、またはそれ以上を備えることができる。
【0037】
II.γ-セクレターゼ組成物
【0038】
本発明は、新規なノッチ融合タンパク質に加え、そのようなノッチ融合タンパク質をさまざまなγ-セクレターゼ・アッセイで利用する方法にも関する。このセクションでは、γ-セクレターゼ活性を有するタンパク質を含む分画の生成について説明する。
【0039】
γ-セクレターゼの正確な同定はまだなされていないが、γ-セクレターゼがプレセニリン1であるらしいという強い証拠がある。γ-セクレターゼ・タンパク質の配列はまだ明らかになっていないが、当業者は、γ-セクレターゼを含む細胞分画を単離するための方法と組成物は知っている。例えばLiら(Proc. Natl. Acad. Sci.、第97巻、6138〜6143ページ、2000年)は、活性のある可溶化γ-セクレターゼを含む膜調製物を得るための方法と組成物を報告している。このような方法は、本発明においてγ-セクレターゼを含む精製された分画を得るのに役立つ。そのような分画が得られると、それを本発明のアッセイで利用できる。さらに、本発明の新規な融合タンパク質も利用し、例えばアフィニティ・クロマトグラフィ法でそのような膜分画からγ-セクレターゼをさらに単離して精製することができる。
【0040】
γ-セクレターゼ分画は、一般に、γ-セクレターゼの活性を示す細胞から単離する。例えばHeLa3細胞を用いることができる。この細胞を例えばフレンチ・プレスまたは他の細胞破壊技術を利用して破壊する。細胞破壊技術としては、例えば凍結解凍法(例えば細胞をドライアイスと37℃の水浴を往復させる);ヒューズ・プレスまたはフレンチ・プレスを用いた固体剪断法;洗剤による溶解(例えばトゥイーン、トリトン、NP-40などのオン-イオン性洗剤溶液);低張液による溶解(例えば水、クエン酸緩衝液);液体剪断法(ホモジナイザー、衝突ジェット微小流体化装置);超音波処理(超音波)などがある。細胞を溶解させた後、遠心分離を利用した沈降によって細胞の残滓と核を除去することができる。膜分画は、例えば100,000×gで60分間にわたって沈殿させ、さらに洗剤を用いて可溶化させることができる。例えばLiら(上記文献)の可溶化法では、100,000×gでの遠心分離ステップからの膜分画ペレットを緩衝液の中に再び懸濁させ、1%CHAPSOを用いて4℃にて60分間にわたって処理し、60分間にわたって遠心分離する。洗剤を用いたこの可溶化法では、100,000×gで得られた分画の上澄に可溶化γ-セクレターゼが含まれている。
【0041】
この明細書に記載したすべてのγ-セクレターゼ・アッセイで上記の可溶化分画を用いる。
【0042】
III.タンパク質またはペプチドの製造と精製
【0043】
本発明により、γ-セクレターゼのモジュレータのうち、APPの開裂に特異的だがノッチの開裂は抑制しないものを同定するのに用いる可溶性ノッチ・タンパク質基質が提供される。この基質は、従来からある自動化ペプチド合成法、あるいは組み換え発現法によって得ることができる。
【0044】
A.合成ペプチドの製造
【0045】
本発明のペプチドは、あるいは本発明の完全長融合ポリペプチドでさえ、従来法に従って溶液中で、あるいは固体支持体上で合成することができる。さまざまな自動化合成装置が市販されており、公知のプロトコルに従って使用することができる。例えばStewartとYoung、『固相ペプチド合成』、第2版、ピアース・ケミカル社、1984年;Tam他、J. Am. Chem. Soc.、第105巻、6442ページ、1983年;Merrifield、Science、第232巻、341〜347ページ、1986年;BaranyとMerrifield、『ペプチド』、GrossとMeienhofer編、アカデミック出版、ニューヨーク、1〜284ページ、1979年を参照のこと。なお各文献の内容は参考としてこの明細書に組み込まれているものとする。γ-セクレターゼによって開裂するγ-セクレターゼ開裂部位1731/1732と1743/1744を含む本発明によるノッチ融合タンパク質の新規な基質は容易に合成することができ、その後γ-セクレターゼ・スクリーニング・アッセイでスクリーニングすることができる。
【0046】
特に好ましい方法では、本発明の融合タンパク質を、アプライド・バイオシステムズ社のモデル433Aを用いて固相法で合成した。自動化ペプチド合成法または組み換え法で得られたどのノッチ融合タンパク質の純度も、逆相HPLC分析を利用して測定することができる。それぞれのペプチドが化学的に本物であることは、当業者に周知の任意の方法で確認することができる。好ましい実施態様では、本物であることは質量分析によって確認される。
【0047】
さらに、融合タンパク質は、アミノ酸分析においてマイクロ波加水分解を実施することよって定量することができる。このような分析では、電子レンジ、例えばCEM社のMDS 2000電子レンジを用いるとよい。ペプチド(約2mg)を、6NのHCl(ピアース・コンスタント・ボイリング社、例えば約4ml)および約0.5%(v/v)のフェノール(マリンクロット社)と接触させる。加水分解の前にサンプルの排気とN2を用いたパージを交互に行なう。タンパク質の加水分解は、2段階プロセスで実施する。第1段階では、融合タンパク質を約100℃の反応温度にし、この温度を1分間維持する。このステップの直後に温度を150℃に上昇させ、この温度を約25分間維持する。冷却後、サンプルを乾燥させ、加水分解した融合タンパク質サンプルからのアミノ酸を6-アミノキノリル-N-ヒドロキシスクシンイミジルカルバメートを用いて誘導体化し、安定な尿素を得る。この尿素は、395nmで蛍光を出す(ウォーターズAccQタグ・ケミストリー・パッケージ)。サンプルを逆相HPLCで分析し、強力インテグレータを用いて定量する。
【0048】
B.組み換えペプチドの産生
【0049】
自動化ペプチド合成の代わりに組み換えDNA技術を利用することができる。その場合、以下に説明するように、本発明のペプチドをコードしているヌクレオチド配列を発現ベクターに組み込み、その発現ベクターを用いて適切な宿主細胞の形質転換またはトランスフェクションを行ない、発現に適した条件下でその宿主細胞を培養する。組み換え法は、本発明のペプチド配列を含む長いポリペプチドを産生させるのに特に好ましい。
【0050】
本発明による新規なノッチ融合タンパク質基質に関する説明から、組み換え技術によって融合ポリペプチドを産生させることができる。ペプチドまたは融合ポリペプチドをコードしている配列を含んでいてそのペプチドまたは融合ポリペプチドを発現させる多彩な発現ベクター/宿主系を利用することができる。そのような系としては、微生物(例えば組み換えバクテリオファージで形質転換した細菌、プラスミドDNA発現ベクター、コスミドDNA発現ベクター);酵母発現ベクターで形質転換した酵母;ウイルス発現ベクター(例えばバキュロウイルス)を感染させた昆虫細胞系;ウイルス発現ベクター(例えばカリフラワー・モザイク・ウイルス(CaMV)、タバコ・モザイク・ウイルス(TMV))をトランスフェクトした植物細胞系、または細菌発現ベクター(例えばTiプラスミドまたはpBR322プラスミド)で形質転換した植物細胞系;動物細胞系などが挙げられる。組み換えタンパク質の産生で役立つ哺乳動物の細胞としては、VERO細胞、HeLa細胞、チャイニーズ・ハムスター卵巣(CHO)細胞系、COS細胞(例えばCOS-7)、W138、BHK、HepG2、3T3、RIN、MDCK、A549、PC12、K562、293細胞などが挙げられる。ノッチ融合タンパク質を細菌、酵母、あるいは他の無脊椎動物で組み換え発現させるための具体的なプロトコルはあとで説明する。
【0051】
原核生物宿主で用いる発現ベクターは、一般に、1種類以上の表現型選択マーカー遺伝子を含んでいる。そのような遺伝子は、一般に、抗生物質に対する耐性を与えるタンパク質、または栄養要求性条件を与えるタンパク質をコードしている。多彩なそのようなベクターを市場から容易に入手することができる。具体的には、pSPORTベクター、pGEMベクター(プロメガ社)、pPROEXベクター(LTI社、ベセスダ、メリーランド州)、ブルースクリプト・ベクター(ストラタジーン社)、pETベクター(ノヴァジェン社)、pQEベクター(キアジェン社)などがある。所定のノッチ融合タンパク質をコードしているDNA配列をPCRによって増幅し、クローニングしてそのようなベクター(例えばpGEX-3X(ファルマシア社、ピスカタウェイ、ニュージャージー州))に導入する。そのベクターは、そのベクターによってコードされているグルタチオン-S-トランスフェラーゼ(GST)を含む融合タンパク質、またはそのベクターのクローニング部位に挿入されたDNA断片がコードしているタンパク質を産生するように設計されている。PCRのプライマーとしては、例えば適切な開裂部位を含むものを作ることができる。組み換え融合タンパク質をトロンビンまたはXa因子(ファルマシア社、ピスカタウェイ、ニュージャージー州)で処理すると融合タンパク質が開裂し、基質または基質含有ポリペプチドがGSTの部分から放出されることが期待される。pGEX-3X/融合タンパク質構造体を導入して大腸菌XL-1ブルー細胞(ストラタジーン社、ラ・ジョラ、カリフォルニア州)を形質転換し、個々の形質転換体を単離して増殖させる。個々の形質転換体からのプラスミドDNAを精製し、自動化シークエンサを用いて一部のシークエンシングを行ない、適切な方向の核酸挿入体をコードしている望むペプチドまたはポリペプチドの存在を確認する。
【0052】
GST/基質融合タンパク質の誘導は、形質転換したXL-1ブルー培養物を37℃にてLB培地(カルベニシリンを補足)の中で波長600nmにおける光学密度が0.4になるまで増殖させた後、0.5mMのイソプロピル*-D-チオガラクトピラノシド(シグマ・ケミカル社、セントルイス、ミズーリ州)の存在下でさらに4時間にわたってインキュベートすることによって実現される。
【0053】
細菌の中に不溶性封入体として産生されることが予想されるGST融合タンパク質は、以下のようにして精製することができる。細胞を遠心分離によって回収し;0.15MのNaCl、10mMのトリス(pH8.1)、1mMのEDTAの中で洗浄し;0.1mg/mlのリゾチーム(シグマ・ケミカル社)で室温にて15分間にわたって処理する。ライセートを超音波処理によって透明にし、細胞の残滓を10分間にわたって12,000×gで遠心分離することによってペレット化する。融合タンパク質を含むこのペレットを50mMのトリス(pH8)と10mMのEDTAの中に再び懸濁させ、50%グリセロールの上で層にし、30分間にわたって6000×gで遠心分離する。ペレットを、Mg2+とCa2+を含まない標準的なリン酸緩衝溶液(PBS)の中に再び懸濁させる。再び懸濁させたペレットを変性用SDSポリアクリルアミド・ゲルの中で分画することによって融合タンパク質をさらに精製する(Sambrook他編、コールド・スプリング・ハーバー出版、コールド・スプリング・ハーバー、ニューヨーク州、1989年)。ゲルを0.4MのKClの中に浸してタンパク質を可視化する。このタンパク質を切断し、SDSが欠けたゲル-ランニング緩衝液の中で電気溶離させる。GST/ノッチ融合タンパク質を細菌の中で可溶性タンパク質として産生させる場合には、GST精製モジュール(ファルマシア・バイオテック社)を用いて精製するとよい。
【0054】
融合タンパク質をトロンビンで消化させて成熟ノッチ融合ポリペプチドからGSTを開裂させることができる。消化反応物(20〜40μgの融合タンパク質と、20〜30単位のヒト・トロンビン(4000U/mg(シグマ社)を含む0.5mlのPBS))を室温にて16〜48時間にわたってインキュベートし、変性用SDS-PAGEゲルに充填して反応生成物を分画する。ゲルを0.4MのKClに浸してタンパク質のバンドを可視化する。融合ポリペプチドの予想分子量に対応するタンパク質のバンドは、自動化シークエンサ(アプライド・バイオシステムズ社のモデル473A、フォスター・シティ、カリフォルニア州)で一部のアミノ酸配列を分析することによって確認できる。
【0055】
融合ポリペプチドを含む予想される基質をコードしているDNA配列をクローニングし、望むプロモータを含んでいて、必要に応じてさらにリーダー配列を含むプラスミドに組み込むこともできる(例えばBetter他、Science、第240巻、1041〜1043ページ、1988年を参照のこと)。この構造体の配列は、自動化したシークエンシングによって確認できる。次にこのプラスミドを用い、CaCl2を用いたインキュベーションと熱ショック処理を行なうという標準的な方法で大腸菌を形質転換する(Sambrook他、上記文献)。形質転換した大腸菌を、カルベニシリンを補足したLB培地の中で増殖させ、適切な培地の中で増殖させることによって発現タンパク質の産生を誘導する。リーダー配列が存在している場合には、そのリーダー配列がノッチをベースとした成熟融合タンパク質の分泌に影響を与え、そのタンパク質が分泌の間に開裂する。
【0056】
この明細書に記載した方法により、分泌された組み換えタンパク質を細菌培地から精製する。
【0057】
同様に、サッカロミセス、ピキア、クルベロミセスなどの属に由来する酵母宿主細胞を用いて組み換えペプチドを産生させることができる。好ましい酵母宿主細胞は、サッカロミセス・セレビジエとピキア・パストリスである。酵母ベクターは、2T酵母プラスミドからの複製起点配列と、自律複製配列(ARS)と、プロモータ領域と、ポリアデニル化のための配列と、転写終結のための配列と、選択マーカー遺伝子とを含んでいることがしばしばある。酵母と大腸菌の両方の中で複製可能なベクター(シャトル・ベクターと呼ばれる)も使用することができる。シャトル・ベクターは、酵母ベクターの上記特徴に加え、大腸菌の中で複製と選択を行なうための配列も含んでいる。酵母宿主細胞の中で発現するポリペプチドの直接的な分泌は、基質をコードしているヌクレオチド配列の5'末端に酵母I因子リーダー配列をコードしている配列を含めることによって実現される。
【0058】
一般に、所定の基質は、市販されている発現系(例えばピキア発現系(インヴィトロジェン社、サン・ディエゴ、カリフォルニア州))を製造者の指示に従って使用して酵母の中で組み換え発現させることができる。この系は、分泌を指示するプレ-プロ-α配列にも依存しているが、挿入体の転写は、メタノールによって誘導されたときにアルコールオキシダーゼ(AOX1)プロモータによって起こる。
【0059】
分泌された組み換え基質は酵母増殖培地から精製する。そのとき、例えば細菌や哺乳動物の細胞の上澄から基質を精製するのに用いる方法を利用する。
【0060】
本発明の新規な基質をコードしている合成DNAをクローニングしてバキュロウイルス発現ベクターpVL1393(ファーミンジェン社、サン・ディエゴ、カリフォルニア州;LuckowとSummers、Bio/Technology、第6巻、47ページ、1988年)に入れることもできる。次に、基質を含むこのベクターを製造者(ファーミンジェン社)の指示に従って用い、sF9タンパク質を含まない培地の中でスポドプテラ・フルギペルダ細胞に感染させ、組み換えタンパク質を産生させる。このタンパク質またはペプチドを、ヘパリン-セファロース・カラム(ファルマシア社、ピスカタウェイ、ニュージャージー州)と直列分子サイズ分類カラム群(アミコン社、ビヴァリー、マサチューセッツ州)を用いて培地から精製し、濃縮し、PBSの中に再び懸濁させる。SDS-PAGE分析により単一のバンドが現われ、タンパク質のサイズが確認される。ポートン2090ペプチド・シークエンサでエドマン・シークエンシングを行なうことによってそのタンパク質のN末端配列が確認される。
【0061】
ノッチ融合タンパク質基質は、昆虫系で発現させることもできる。タンパク質を発現させるための昆虫系は当業者によく知られている。そのような1つの系では、オートグラファ・カリフォルニア核多角体病ウイルス(AcNPV)が、スポドプテラ・フルギペルダ細胞またはトリコプルシアの幼虫の中で外来遺伝子を発現させるためのベクターとして用いられる。基質をコードしている配列をクローニングしてウイルスの不可欠ではない領域(例えばポリヘドリン遺伝子)に挿入し、ポリヘドリン・プロモータの制御下に置く。基質がうまく挿入されるとポリヘドリン遺伝子が不活性になり、コート・タンパク質が欠けた組み換えウイルスが産生される。次に組み換えウイルスをスポドプテラ・フルギペルダ細胞またはトリコプルシアの幼虫に感染させ、その中で基質を発現させる(Smith他、J. Virol.、第46巻、584ページ、1983年;Engelhard E.K.他、Proc. Natl. Acad. Sci.、第91巻、3224〜3227ページ、1994年)。
【0062】
組み換えタンパク質を発現させるための哺乳動物宿主系も当業者にはよく知られている。宿主細胞株は、発現したタンパク質を処理したり、タンパク質に活性を与えるのに役立つある種の翻訳後修飾をしたりする特別な能力を持つものを選択することができる。ポリペプチドのこのような修飾としては、アセチル化、カルボキシル化、グリコシル化、リン酸化、脂質化、アシル化などがある。正確な挿入、折り畳み、機能のためには、タンパク質の“プレプロ”形態を開裂させる翻訳後プロセシングも重要である可能性がある。さまざまな宿主細胞(例えばCHO、HeLa、MDCK、293、WI38など)が、そのような翻訳後活性に関する特別な細胞機構と特徴的メカニズムを備えている。このような宿主細胞を選択することで、導入された外来タンパク質の正確な修飾とプロセシングを確実に行なえる。
【0063】
形質転換された細胞を長期にわたる高収率のタンパク質産生に利用できること、そしてその形質転換細胞がそのままで安定に発現することが好ましい。このような細胞を、選択マーカーと望む発現カセットとを含むベクターを用いて形質転換したときには、その細胞を1〜2日間栄養強化培地の中で増殖させた後、選択培地に切り換えるとよい。選択マーカーは選択に対する耐性を有するように設計する。したがってその選択マーカーが存在していると、導入配列をうまく発現する細胞の増殖と回収が可能になる。耐性を有する安定に形質転換された細胞の塊は、その細胞に適した組織培養技術を利用して増殖させることができる。
【0064】
多数の選択系を用い、組み換えタンパク質を産生するように形質転換された細胞を回収することができる。そのような選択系としては、tk細胞のHSVチミジンキナーゼ遺伝子、hgprt細胞のヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ遺伝子、aprt細胞のアデニンホスホリボシルトランスフェラーゼ遺伝子などが挙げられる。抗代謝物耐性を選択の基礎として利用し、メトトレキセートに対する耐性を与えるdhfr;ミコフェノール酸に対する耐性を与えるgpt;アミノグリコシドG418に対する耐性を与えるネオ;クロルスルフロンに対する耐性を与えるALS;ハイグロマイシンに対する耐性を与えるハイグロに関する選択を行なうこともできる。役に立つ可能性のある別の選択用遺伝子としては、細胞がトリプトファンの代わりにインドールを利用できるようにするtrpBと、細胞がヒスチジンの代わりにヒスチノールを利用できるようにするhisDが挙げられる。形質転換体を目で見て同定するための指標となるマーカーとしては、アントシアニン、b-グルクロニダーゼとその基質、GUS、ルシフェラーゼとその基質、ルシフェリンなどが挙げられる。
【0065】
C.部位特異的突然変異誘発
【0066】
部位特異的突然変異誘発は、γ-セクレターゼの基質となる個々のペプチドの調製、さらに特定するならばγ-セクレターゼの基質となる本発明の融合タンパク質のうちの1つを構成要素として含む融合ポリペプチドの調製に役立つ別の方法である。この方法では、(修飾の標的となるアミノ酸配列をコードしている)DNAの特異的突然変異誘発を利用する。この方法では、DNAにヌクレオチド配列の変化を1個以上導入することにより、上記の1つ以上の考慮事項を組み込んだ配列変異体を調製してテストすることが容易にできる。部位特異的突然変異誘発により、望む突然変異を有するDNA配列をコードしている特別なオリゴヌクレオチド配列と、隣接する十分な数のヌクレオチドとを利用して突然変異体を取得し、欠失接合部の両側にまたがる安定な二本鎖を形成するための、十分なサイズで配列が十分に複雑なプライマー配列を提供することができる。一般に、長さがヌクレオチド17〜25個で、変化させる配列の接合部の両側に約5〜10残基あるプライマーが好ましい。
【0067】
この方法では、一般に、一本鎖と二本鎖の両方の形態で存在するバクテリオファージ・ベクターを用いる。部位指定突然変異誘発で一般に用いられるベクターは、M13ファージなどのベクターである。ファージ・ベクターは市販されており、その使用法は当業者に一般によく知られている。二本鎖プラスミドも部位指定突然変異誘発で日常的に用いられている。この場合には、興味の対象である遺伝子をファージからプラスミドに移すステップがない。
【0068】
一般に、部位指定突然変異誘発を実施するには、まず最初に、配列内に望むタンパク質をコードしているDNA配列を有する一本鎖ベクターを取得するか、配列内に望むタンパク質をコードしているDNA配列を有する二本鎖ベクターの二本の鎖を溶かす。望む突然変異配列を有するオリゴヌクレオチド・プライマーを合成によって用意する。次に、ハイブリダイゼーション(アニーリング)条件を選択するときにミスマッチの程度を考慮してこのプライマーを一本鎖DNA調製物とともにアニールした後、DNA重合酵素(例えば大腸菌ポリメラーゼIのクレノウ・フラグメント)を作用させて突然変異を有する鎖の合成を完了させる。したがって、第1の鎖が突然変異していない元の配列をコードし、第2の鎖が望む突然変異を持つヘテロ鎖が形成される。次に、このヘテロ鎖ベクターを用いて適切な細胞(例えば大腸菌細胞)を形質転換し、突然変異した配列を有する組み換えベクターを含むクローンを選択する。
【0069】
もちろん、部位指定突然変異誘発を起こさせる上記の方法は、役に立つ可能性のある突然変異ペプチド種を作り出す唯一の方法ではなく、本発明がこの方法に限定されることはない。本発明では、突然変異誘発を実現する別の方法も考える。例えば、興味の対象である遺伝子を有する組み換えベクターを突然変異誘発剤(例えばヒドロキシルアミン)で処理して配列変異体を得る。
【0070】
D.タンパク質の精製
【0071】
本発明の融合タンパク質は精製することが望ましかろう。タンパク質精製法は当業者によく知られている。その方法には、1つの段階として、細胞培地を粗分画してポリペプチド分画と非ポリペプチド分画に分ける操作が含まれる。本発明のペプチドまたはポリペプチドが他のタンパク質から分離されると、興味の対象である融合ポリペプチドまたは融合ペプチドをクロマトグラフィ法や電気泳動法を利用してさらに精製し、部分的精製または完全な精製(または精製による均一化)を実現する。純粋なペプチドの調製に特に適した分析法は、イオン交換クロマトグラフィ、サイズ排除クロマトグラフィ;ポリアクリルアミド・ゲル電気泳動;等電点電気泳動である。融合タンパク質の精製に特に有効な方法は、高速タンパク質液体クロマトグラフィ(FPLC)または高性能液体クロマトグラフィ(HPLC)である。特に好ましい実施態様では、NusA-ノッチ融合タンパク質を、固定化金属アフィニティ・クロマトグラフィ(IMAC)を利用して分離した。
【0072】
ポリヒスチジン・タグを有する組み換えタンパク質の精製にはIMACが主に利用されている。本発明では、融合タンパク質がC末端にポリヒスチジン・タグを備えているため、この強力な方法で精製を行なうことができる。この精製は、ヒスチジンが二価の金属と自然にキレートを形成する傾向を利用している。金属イオンをクロマトグラフィの支持体上に置くと、ポリヒスチジン・タグを有するタンパク質を精製することができる。これは、当業者がこれまで利用してきた多彩なタンパク質の精製法の中でも非常に効率的な方法である。本発明のタンパク質を単離する好ましい一実施態様の具体的な条件は、実施例1に記載してある。しかし当業者であれば、その条件と媒体を変更しても本発明に従って精製を行なえることを理解されたい。その目的で、当業者は、金属アフィニティ・クロマトグラフィによって混合物から生物学的基質または関連する基質を分離する方法が詳細に記載されているアメリカ合衆国特許第4,431,546号を参考にしている。この特許文献に記載されているクロマトグラフィ媒体は、Ca2+、St2+、Ba2+、Al3+、Co2+、Ni2+、Cu2+、Zn2+からなるグループの中から選択した少なくとも1つの金属イオンの存在下で、アントラキノン、フタロシアニン、芳香族アゾからなるグループの中の少なくとも1つを含むリガンドを有する結合材料を備えている。別の媒体と条件は、例えばhttp://www.affiland.com/imac/nta.htmと、http://www.affiland.com/imac/pdc.htmに記載されている。
【0073】
本発明のいくつかの特徴は精製に関係しており、特別な実施態様では、コードされたポリペプチド、タンパク質、ペプチドいずれかの実質的な精製を行なう。この明細書では、“精製されたポリペプチド、タンパク質、ペプチド”という表現は、他の成分から分離された組成物を意味し、その組成物の中では、ポリペプチド、タンパク質、ペプチドが、そのポリペプチド、タンパク質、ペプチドを生成させるのに用いた細胞成分や合成成分よりもわずかであれ多く精製されている。したがって精製されたポリペプチド、タンパク質、ペプチドは、それが天然に発生する環境から離れたポリペプチド、タンパク質、ペプチドも意味する。
【0074】
一般に、“精製された”とは、ポリペプチド組成物、タンパク質組成物、ペプチド組成物において分画により他のさまざまな成分が除去されていて、発現した生物活性を実質的に維持していることを意味する。“実質的に精製された”という表現を用いる場合には、ポリペプチド、タンパク質、ペプチドが組成物の主要成分となっていて、組成物中のタンパク質の約50%、約60%、約70%、約80%、約90%、約95%またはそれ以上を占めていることを意味する。
【0075】
当業者は、本発明の開示内容を参考にして、ポリペプチド、タンパク質、ペプチドの精製度を定量するさまざまな方法を思い浮かべることができよう。そのような方法としては、例えば、活性分画の比活性の測定、SDS/PAGE分析による分画内のポリペプチドの量の評価などがある。分画の純度を評価する好ましい1つの方法は、分画の比活性を計算し、それを最初の抽出液の比活性と比較し、純度の大きさを計算して“〜倍純度数”として評価するというものである。活性の大きさを表わすのに用いる実際の単位は、もちろん、純度を調べるのに選択した具体的なアッセイ法と、発現したポリペプチド、タンパク質、ペプチドが検出可能な活性を示すかどうかに依存する。
【0076】
タンパク質の精製で利用するのに適したさまざまな方法が当業者にはよく知られている。例えば、硫酸アンモニウム、PEG、抗体などを用いた沈降、または熱変性を行なった後の遠心分離;クロマトグラフィ・ステップ(例えばイオン交換クロマトグラフィ、ゲル濾過クロマトグラフィ、逆相クロマトグラフィ、ヒドロキシルアパタイト・クロマトグラフィ、アフィニティ・クロマトグラフィ);等電点電気泳動;ゲル電気泳動;これらの方法と他の方法の組み合わせなどの方法がある。従来から一般に知られているように、さまざまな精製ステップを実施する順番を変えても、あるいはいくつかのステップを省略しても、実質的に精製されたポリペプチド、タンパク質、ペプチドを調製するのに適した方法になると考えられている。
【0077】
ポリペプチド、タンパク質、ペプチドが常に最高純度の状態で提供されることは一般には必要でない。実際、実質的により低い純度の生成物がいくつかの実施態様で役に立つことが考えられる。より少数の精製ステップを組み合わせて部分的精製を実現すること、あるいは同じ一般的精製スキームを異なった形で利用して部分的精製を実現することができる。例えばHPLC装置を用いて実施する陽イオン交換カラム・クロマトグラフィにより、一般に、低圧クロマトグラフィ・システムを用いた同じ方法よりも大きな“倍数”の純度が得られる。精製度が相対的に低い方法は、タンパク質生成物の全回収量や、発現するタンパク質の活性維持に関して利点を有する可能性がある。
【0078】
ポリペプチドの移動は、SDS/PAGEの条件が異なると、ときに顕著に異なる可能性があることが知られている(Capaldi他、Biochem. Biophys. Res. Comm.、第76巻、425ページ、1977年)。したがって電気泳動の条件が異なると、精製された、あるいは一部が精製された発現生成物の見かけの分子量が異なる可能性があることを理解されたい。
【0079】
IV.本発明の基質を得るのに用いる発現構造体
【0080】
本発明では、本発明のノッチ融合タンパク質を発現させる必要があろう。この発現を実現するため、本発明では、本発明のノッチ融合タンパク質をコードするためのポリヌクレオチド分子を含むベクターと、そのベクターを用いて形質転換した宿主細胞を利用する。このようなポリヌクレオチド分子は、一般に選択マーカーと複製起点を含むベクターと結合させて宿主の中に運び込む。本発明で利用する発現構造体のこれらエレメントについては、以下に詳しく説明する。
【0081】
発現ベクターは、γ-セクレターゼの基質であるこの明細書に記載したあらゆるペプチドまたはポリペプチドをコードしていて適切な転写調節配列または翻訳調節配列と機能上リンクしたDNAを含んでいる。その調節配列は、例えば哺乳動物、細菌、ウイルス、昆虫の遺伝子に由来する。調節配列の具体例としては、転写プロモータ、オペレータ、エンハンサー、mRNAリボソーム結合部位、転写と翻訳を制御する適切な配列などが挙げられる。
【0082】
“発現ベクター”、“発現構造体”、“発現カセット”という用語は、この明細書全体を通じてどれも同じ意味で使用し、1つの遺伝子産物をコードしている核酸を含んでいて、そのコードしている核酸配列の一部または全部を転写できるあらゆるタイプの遺伝子構造体を意味する。
【0083】
本発明の融合ポリペプチドを発現させるのに適した発現ベクターの選択は、もちろん使用する具体的な宿主細胞に依存するが、当業者はその選択方法を知っている。適切な発現ベクターの具体例として、pcDNA3(インヴィトロジェン社)とpSVL(ファルマシア・バイオテック社)がある。本発明における好ましい1つの発現ベクターはpcDNA3.1-ハイグロ(インヴィトロジェン社)である。哺乳動物宿主細胞で使用する発現ベクターは、ウイルスのゲノムに由来する転写制御配列と翻訳制御配列を含むことができる。本発明で用いることのできる一般に使用されているプロモータ配列とエンハンサー配列としては、ヒト・サイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマ・ウイルス、サル・ウイルス40(SV40)などがある。哺乳動物発現ベクターを構成する方法は、例えば、OkayamaとBerg(Mol. Cell. Biol.、第3巻、280ページ、1983年);Cosman他(Mol. Immunol.、第23巻、935ページ、1986年);Cosman他(Nature、第312巻、768ページ、1984年);EP-A-0367566;WO 91/18982に開示されている。
【0084】
発現構造体は、本発明による特定のノッチ融合タンパク質をコードしている核酸領域を含むことになる。上記のような発現ベクターを構成するのに用いるコード領域は、少なくともγ-セクレターゼによるこの明細書に記載した融合タンパク質の開裂をコードしている必要がある。しかし生成するペプチドの1つが、γ-セクレターゼによって開裂するγ-セクレターゼ開裂部位1731/1732と1743/1744を含んでいるのであれば、より大きなポリペプチドがコードされていてもよい。
【0085】
本発明のいくつかの特徴によると、発現構造体は、ペプチドまたはポリペプチドの発現を検出できるようにする選択マーカーをさらに含むことができる。通常は、薬剤選択マーカーを含めることによって転写体のクローニングと選択がしやすくなる。薬剤選択マーカーとしては、例えばネオマイシン、ピューロマイシン、ハイグロマイシン、DHFR、ゼオシン、ヒスチジノールがある。あるいは酵素を使用することもできる。酵素としては、例えば、単純ヘルペスウイルスのチミジンキナーゼ(tk)(真核生物)、b-ガラクトシダーゼ、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)(原核生物)がある。免疫マーカーも使用することができる。例えばエピトープ・タグとして、フラグ系(IBI、ニュー・ヘイヴン、コネチカット州)、HA、6×ヒスチジン系(キアジェン社、チャッツワース、カリフォルニア州)が使用できる。さらに、グルタチオン-S-トランスフェラーゼ(GST)系(ファルマシア社、ピスカタウェイ、ニュージャージー州)またはマルトース結合タンパク質系(NEB、ビヴァリー、マサチューセッツ州)も用いることができる。用いる選択マーカーは、遺伝子産物をコードしている核酸と同時に発現させることができるのであれば、重要であるとは考えられない。選択マーカーのさらに別の具体例は当業者には周知である。本発明で使用できる特に好ましい選択マーカーは、ネオマイシン耐性マーカーとGFPマーカーである。
【0086】
発現するには、適切なシグナルがベクターに供給される必要がある。このセクションには、さまざまな調節エレメント(例えば宿主細胞の中で興味の対象である核酸を発現させるのに用いることのできるウイルスと哺乳動物両方からのエンハンサー/プロモータ)に関する説明が含まれる。宿主細胞の中でメッセンジャーRNAの安定性と翻訳可能性を最適化するように設計されたエレメントも定義する。産物を発現する永久的に安定な細胞クローンを確立するために多数の優性薬剤選択マーカーを使用する条件と、薬剤選択マーカーの発現を突然変異表現型の発現と結びつけるエレメントも提示する。
【0087】
好ましい実施態様では、所定のペプチドをコードしている核酸、または選択マーカーをコードしている核酸は、プロモータの転写制御下にある。“プロモータ”とは、遺伝子の具体的な転写を開始させるのに必要で、細胞の合成機構または導入された合成機構によって認識されるDNA配列を意味する。
【0088】
核酸配列は、調節配列がノッチ融合タンパク質をコードしているDNAと機能の上で関係を持っている場合に、機能上リンクしている。したがってプロモータ・ヌクレオチド配列は、そのプロモータ・ヌクレオチド配列が所定のDNA配列の転写を指示するときにそのDNA配列と機能上リンクしている。同様に、“転写制御下にある”という表現は、プロモータが核酸との関係で正確な位置と向きになっていて、RNAポリメラーゼの開始と遺伝子の発現を制御することを意味する。
【0089】
プロモータという用語は、この明細書では、RNAポリメラーゼIIの開始部位の周囲に集まった一群の転写制御モジュールを意味する。プロモータをどのように構成するかについて考える時間の大部分は、いくつかのウイルスのプロモータ(例えばHSVチミジンキナーゼ(tk)やSV40初期転写ユニット)を分析することに費やされる。これらの研究にさらに最近の研究が加わることで、プロモータは離散した複数の機能モジュールで構成されており、それぞれの機能モジュールは約7〜20bpのDNAからなること、そして転写のアクチベータ・タンパク質またはリプレッサー・タンパク質のための1つ以上の認識部位を含むことが明らかになった。
【0090】
各プロモータの少なくとも1つのモジュールは、RNA合成の出発部位の位置を決める機能を有する。その最もよく知られている例はTATAボックスであるが、TATAボックスが欠けているいくつかのプロモータ(例えば、哺乳動物のターミナルデオキシヌクレオチジルトランスフェラーゼ遺伝子のためのプロモータや、SV40後期遺伝子のためのプロモータなど)では、出発部位そのものと重なっている離散エレメントが開始の位置を決めるのに役立つ。
【0091】
別のプロモータ・エレメントが転写開始の頻度を調節する。一般に、そのエレメントは出発部位の30〜110bp上流の領域に位置している。しかし出発部位の下流に機能エレメントを含むプロモータが多数あることも最近明らかになった。プロモータ・エレメント相互間のスペースはしばしば自由であるため、エレメントが互いに逆転したり移動したりしてもプロモータの機能は保持される。tkプロモータでは、プロモータ・エレメント相互間のスペースを50bpまで離したときにようやく活性が低下し始める。プロモータが何であるかに応じ、個々のエレメントは、共同して、あるいは独立に機能して転写を活性化させるように見える。
【0092】
興味の対象である核酸配列の発現制御に用いる具体的なプロモータが何であるかは、標的細胞の中で核酸の発現を指示できるのであれば重要ではないと考えられている。したがってヒト細胞を標的とする場合には、ヒト細胞の中で発現させることのできるプロモータの近くに、またはそのプロモータの制御下に核酸コード領域を置くことが好ましい。一般に、そのようなプロモータとしては、ヒトのプロモータまたはウイルスのプロモータが挙げられる。
【0093】
いくつかの実施態様では、ヒト・サイトメガロウイルス(CMV)前初期遺伝子プロモータ、SV40初期プロモータ、ラウス肉腫ウイルスの長い末端反復配列、β-アクチン、ラットのインスリン・プロモータ、ホスホグリセロールキナーゼ・プロモータ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ・プロモータを用いて興味の対象であるコード配列を高レベルで発現させることができる。これらはすべて、よく知られていて容易に入手できるプロモータである。興味の対象であるコード配列の発現を実現できることが従来技術でよく知られている他のウイルスまたは哺乳動物の細胞プロモータまたはバクテリオファージ・プロモータを用いることも、発現レベルが目的にとって十分である場合には考えられる。よく知られた特性を有するプロモータを用いることにより、トランスフェクションと形質転換を行なった後の興味の対象であるタンパク質の発現レベルと発現パターンを最適化することができる。
【0094】
特定の生理学的シグナルまたは合成シグナルに応答して調節されるプロモータを選択することにより、遺伝子産物の発現を誘導することができる。ウイルス・ベクターを作るためのいくつかの誘導プロモータ系が入手可能である。そのような1つの系はエクジソン系(インヴィトロジェン社、カールズバッド、カリフォルニア州)である。これは、哺乳動物の細胞における興味の対象である遺伝子の発現を調節できるように設計されている。これは、導入遺伝子のベースレベルの発現は実質的にないが200倍を超える誘導性を可能にする厳しく制御された発現メカニズムを備えている。
【0095】
別の有用な誘導系は、Tet-Off(登録商標)系またはTet-On(登録商標)系(クロンテック社、パロ・アルト、カリフォルニア州)であり、元々はGossenとBujardによって開発されたものである(GossenとBujard、Proc. Natl. Acad. Sci. USA、第89巻(12)、5547〜5551ページ、1992年;Gossen他、Science、第268巻(5218)、1766〜1769ページ、1995年)。
【0096】
哺乳動物の細胞では、強い転写活性を与えるのにCMV前初期プロモータがしばしば用いられている。導入遺伝子の発現レベルを小さくすることが望ましい場合には、CMVプロモータを改変して転写活性を弱めたものも用いられてきた。レトロウイルスのプロモータ(例えばMLVやMMTVからのLTR)が本発明で役に立つと考えられる。使用可能な他のウイルスのプロモータとしては、SV40、RSV LTR、HIV-1 LTR、HIV-2 LTRのプロモータ、アデノウイルスのプロモータ(例えばE1A領域、E2A領域、MLP領域からのもの)、AAV LTR、カリフラワー・モザイク・ウイルス、HSV-TK、トリ肉腫ウイルスのプロモータなどがある。
【0097】
いくつかの実施態様では、制御可能なプロモータが役に立つことがわかるであろう。そのようなプロモータとしては、ホルモンまたはサイトカインを制御できるプロモータがある。ホルモンを制御できるプロモータとしては、MMTV、MT-1、エクジソン、ルビスコのほか、ホルモンによって調節される他のプロモータ(例えば甲状腺ホルモン、下垂体ホルモン、副腎ホルモンに応答するプロモータ)がある。
【0098】
本発明で使用することが考えられる別の調節エレメントはエンハンサーである。それは、同じDNA分子上の離れた位置にあるプロモータからの転写を増大させる遺伝子エレメントである。エンハンサーはプロモータとほぼ同じよう構成されている。すなわちエンハンサーは、多数の個々のエレメントからなり、それぞれが1つ以上の転写タンパク質と結合している。エンハンサーとプロモータの基本的な違いは作用である。エンハンサー領域は全体として転写を離れた位置から刺激できる必要があるのに対し、プロモータ領域またはその成分エレメントではその必要がない。その一方で、プロモータは、特定の部位と特定の方向でRNA合成の開始を指示する1つ以上のエレメントを持っている必要があるのに対し、エンハンサーはこの特徴を欠いている。プロモータとエンハンサーはしばしば重複かつ連続していて、非常に似た構成を有するように見える。本発明で有用なエンハンサーは当業者には周知であり、使用する具体的な発現系が何であるかによって異なる(Scharf D.他、Results Probl. Cell Differ.、第20巻、125〜162ページ、1994年;Bittner他、Methods in Enzymol.、第153巻、516〜544ページ、1987年)。
【0099】
発現構造体がcDNA挿入体を利用している場合には、遺伝子転写物を適切にポリアデニル化するためのポリアデニル化シグナル配列が含まれていることが一般に望ましい。選択したトランスジェニック動物種の細胞が認識する任意のポリアデニル化シグナル配列が、本発明を実施するのに適している。それは例えば、ヒトまたはウシの成長ホルモンや、SV40ポリアデニル化シグナルである。
【0100】
発現カセットのエレメントとしてターミネータも考えられる。このエレメントは、mRNAのレベルを増大させ、カセットが読み飛ばされて他の配列になることを最少にできる。主に利用されるターミネータ領域は便利であるという理由で選ばれる。なぜなら、本発明で考えられるような用途のターミネータ領域は比較的交換可能であるように見えるからである。ターミネータ領域は、転写開始領域に元々備わっているもの、あるいは興味の対象であるDNA配列に元々備わっているもの、あるいは他の供給源に由来するものが可能である。
【0101】
本発明のいくつかの実施態様では、複数遺伝子のmRNAまたはポリシストロン性mRNAを作り出すのに内部リボソーム侵入部位(IRES)エレメントを用いることが考えられる。IRESエレメントは、5'メチル化キャップに依存した翻訳というリボソーム走査モデルを無視して内部部位で翻訳を開始できる(PelletierとSonenberg、Nature、第334巻、320〜325ページ、1988年)。ピコルナウイルス・ファミリーの2つのメンバー(ポリオウイルスと脳心筋炎)からのIRESエレメントが報告されており(PelletierとSonenberg、上記文献)、哺乳動物のmRNAからのIRESも報告されている(MacejakとSarnow、Nature、第353巻、90〜94ページ、1991年)。IRESエレメントは、異種オープン・リーディング・フレームとリンクさせることができる。IRESによって隔てられた多数のオープン・リーディング・フレームを同時に転写してポリシストロン性mRNAを作り出すことができる。IRESエレメントのおかげでそれぞれのオープン・リーディング・フレームがリボソームにアクセスして効果的な翻訳を行なうことができる。単一のプロモータ/エンハンサーを用いて単一のmRNAを転写し、複数の遺伝子を効果的に発現させることができる。
【0102】
任意の異種オープン・リーディング・フレームをIRESエレメントにリンクさせることができる。この異種オープン・リーディング・フレームには、分泌されたタンパク質、独立な遺伝子によってコードされているサブユニットを多数有するタンパク質、細胞内タンパク質または膜結合タンパク質、選択マーカーのための遺伝子が含まれている。このように単一の構造体と単一の選択マーカーを用いることで、いくつかのタンパク質の発現を1つの細胞内で同時に実現することができる。
【0103】
V.γ-セクレターゼ・アッセイにおける基質の使用
【0104】
本発明の特別な実施態様は、γ-セクレターゼの活性および/または機能をモニターするアッセイ、より詳細には、γ-セクレターゼの活性および/またはγ-セクレターゼの機能を調べるアッセイに関する。このアッセイは、溶液中でγ-セクレターゼ複合体(または精製したポリペプチド)を本発明の適切な基質とともにインキュベートし、ノッチ融合タンパク質の開裂をγ-セクレターゼのタンパク質分解活性の指標として利用する操作を含んでいる。
【0105】
A.アッセイの形式
【0106】
本発明の特別な実施態様は、ヒトγ-セクレターゼの活性を変化させる薬を同定する方法に関する。この方法の1つの特徴は、ε開裂部位1743/1744におけるγ-セクレターゼのノッチ開裂活性を、モジュレータと考えられる化合物または薬の存在下と不在下でモニターすることである。例えばノッチの開裂を調べるこのような方法は、一般に、
(a)本発明による任意のノッチ/NusA融合タンパク質を、活性なγ-セクレターゼを含む組成物と試験管内で接触させるステップと;
(b)そのノッチ/NusA融合タンパク質がγ-セクレターゼによってノッチのγ-セクレターゼ開裂部位で開裂したことを調べるステップを含んでいる。
【0107】
活性なγ-セクレターゼを含む組成物は、一般に、活性なγ-セクレターゼを含む単離された任意の組成物である。この組成物は、そのままで細胞(γ-セクレターゼ活性を有する天然の細胞、またはそのような活性を発現するようにした組み換え宿主細胞)から単離された膜分画になることができる。組成物としては、他のタンパク質を実質的に含まない単離または精製されたγ-セクレターゼも可能である。
【0108】
γ-セクレターゼのノッチ開裂活性を変化させるモジュレータを同定するため、モジュレータ候補の存在下と不在下で上記のステップ(a)と(b)を実施し、モジュレータ候補の存在下でのγ-セクレターゼのノッチ開裂活性をモジュレータ候補の不在下での活性と比較することによってそのモジュレータ候補の変更能力を評価することで、γ-セクレターゼによるこのような開裂を変化させる化合物を同定する。モジュレータ候補の存在下でノッチ開裂の量または程度がいくらかでも変化するというのは、モジュレータ候補の存在下でγ-セクレターゼの活性が変化したことの指標であり、γ-セクレターゼの活性のモジュレータとなる化合物が同定されたことになる。
【0109】
対照(試験化合物ではない)と比べて開裂を増加させる化合物は、γ-セクレターゼのタンパク質分解活性のアゴニストまたは促進剤であると評価されるのに対し、Aβ1〜40および/またはAβ1〜42において開裂を減らす化合物は阻害剤であると評価される。γ-セクレターゼの阻害剤は特に興味深い。なぜなら、γ-セクレターゼ活性の阻害剤は、ADの症状またはその進行を治療または予防する薬になる可能性があることを示しているからである。
【0110】
γ-セクレターゼによるAPPまたはCT-100の開裂を本発明の融合タンパク質の開裂と比べて選択的に抑制する試験化合物を見いだすことが望ましいため、本発明のアッセイを利用するのと並行して、あるいはその前に、試験化合物がAPPの開裂を変化させる能力を調べるとよい。γ-セクレターゼによるAPPまたはCT-100の開裂を測定する方法は従来技術において周知である。一例として、WO 01/83811には、APPまたはCT-100の開裂を測定するのに役立つ基質とインビトロ・アッセイが記載されている。WO 01/83811には、M末端メチオニン(M)と、APP597〜695と、フラグ・タグとを含むγ-セクレターゼの基質、ならびにその基質を開裂させる方法と条件、開裂産物(M-Aβ40とM-Aβ42)を検出する方法と条件が記載されている。
【0111】
γ-セクレターゼによるAPP/CT-100開裂活性をAβ1〜40および/またはAβ1〜42において抑制するが、γ-セクレターゼによる測定されたノッチ開裂活性は抑制しない阻害剤が最も好ましい。
【0112】
γ-セクレターゼは、精製されたγ-セクレターゼ・ポリペプチドまたは複合体、その生物学的に活性な断片、そのアナログ、その変異体のいずれでもよい。好ましい実施態様では、γ-セクレターゼは、γ-セクレターゼ活性を示す細胞からの膜分画に由来する。このような膜分画は、γ-セクレターゼ複合体にとって必要なすべての要素を含んでいることが好ましい。ヒト・γ-セクレターゼの非ヒト・オルトログもアッセイで使用することができる。
【0113】
本発明のアッセイは、細胞を含まない系でγ-セクレターゼ・ポリペプチドを用いて実施できるように設計する。例えば細胞を含まない系では、試験化合物の存在下と不在下で、γ-セクレターゼまたはγ-セクレターゼ含有膜分画を、本発明のペプチド基質またはタンパク質基質と混合することによって接触ステップを行なうことができる。最適な結果を得るためには、γ-セクレターゼとγ-セクレターゼの基質を実質的に精製し、制御された所定量を混合した後、酵素活性を最適にした、および/または生理学的条件を真似た適切な緩衝液の中で混合する。
【0114】
開裂を調べるステップは、N末端断片とC末端断片の一方または両方か、開裂を示す他のパラメータを測定する操作を含んでいる。例えばノッチをベースとした基質またはγ-セクレターゼの他の基質は、消光状態の標識を含むことができる。この標識は、開裂して消光部から分離したときだけより検出しやすくなる。ノッチをベースとした基質またはγ-セクレターゼの他の基質をN末端またはC末端に固定して固体支持体に取り付けることもできる。この構成では、開裂は、固体支持体から開裂断片が放出されることによって判定できる。この放出は、媒体中の標識が増加すること、あるいは固体支持体に付着した標識が減少することによって判定できる。放出は、(例えば抗体とともに)放出されたペプチドを定量的に捕捉することによっても判定できる。
【0115】
本発明の好ましい一実施態様では、バリン1744に対して特異的な抗体を用いて開裂したノッチ・タンパク質を検出する。さらに好ましい実施態様では、抗バリン1744抗体と抗フラグ抗体に基づいて開発したELISAを利用して開裂したノッチ・タンパク質を検出する。抗バリン1744抗体は開裂の1つの断片を検出するのに用いられるのに対し、抗フラグ抗体は、γ-セクレターゼの作用によって産生されたノッチ・タンパク質の断片(C末端)を検出する。このアッセイについては、実施例でさらに詳しく説明する。
【0116】
もちろん、上記のアッセイは単なる例示であり、他のアッセイも利用することができる。例えば上記のアッセイを以下のようにすることができる。384ウエルのマイクロタイター・プレートをBSAでブロックし、酵素であるγ-セクレターゼとテストするγ-セクレターゼ阻害化合物50μlを1時間にわたってインキュベートし、ノッチ/NusA融合タンパク質基質を添加することによって反応を開始させる。最終的なアッセイ条件では、体積がウエル1つにつき30μl;化合物が50μM;酵素がウエル1つにつき15ng;250nMの基質;5%DMSOと0.001%のトゥイーン20である。室温にて一晩にわたってインキュベートし、トリス-HCl(pH8.3)を添加して反応を停止させる。基質を6.25ピコモル含むアリコートを取り出し、開裂した基質および/または開裂していない基質をストレプトアビジンでコーティングしたプレートに捕捉する。このプレートを3回洗浄し、緩衝液を添加する。捕捉アッセイは、LJLアナリスト(Ex485/Em530)上でオレゴン・グリーンの蛍光放出を読み取ることによってモニターする。
【0117】
この明細書で利用できる別のアッセイは、蛍光偏光アッセイである。蛍光偏光は、感光性があって容易で非破壊的なアッセイであり、薬剤候補が本発明のγ-セクレターゼ複合体に及ぼす効果をモニターするのに用いることができる。蛍光偏光は、その基質がγ-セクレターゼと相互作用するのをモニターするのにも利用できる。制御された条件下での蛍光偏光の測定により、溶液中での蛍光分子の“分子揺動”の程度を明らかにすることができる。例えば体積が小さい小分子は、素早く揺動することが予測される。偏光した光を照射された場合には、溶液中の分子の素早い運動によって光の偏光が広範にわたって消え、“小さな”偏光値が読み取られることになろう。同じ条件下において、体積がより大きい大分子または大きな複合体は分子回転(揺動)プロセスがゆっくりしたものになろう。その結果、平面偏光した入射光の偏光度をより小さくすると、より大きな偏光値が測定されよう。
【0118】
小さなリガンドに蛍光プローブを標識することにより、そのリガンドが系の別の要素と相互作用して生じる蛍光偏光の変化を測定することができる。このような方法は、酵素(γ-セクレターゼ)と酵素の蛍光性基質の間の相互作用を測定するのに用いることもできる。
【0119】
蛍光偏光アッセイの一例では、あらかじめブロックした低アフィニティの黒いプレートの中で、酵素と抑制性/変更性化合物を30分間にわたってインキュベートし、150nMの基質(例えば蛍光標識した本発明のノッチ/NusA融合タンパク質)を最終溶液がウエル1つにつき30μlになるように添加して反応を開始させる。次にプレートを室温にて15分間にわたってインキュベートし、蛍光偏光をLJLアナリスト(Ex485/Em530)上で測定する。
【0120】
本発明では、γ-セクレターゼを単離してキャラクテリゼーションを行なうのに役立つ可能性のある1つの特徴を考える。この特徴では、酵素の活性部位またはアロステリック部位に結合する化合物を検出する結合アッセイを考える。その検出を行なうには、本発明によるノッチ基質の加水分解しない誘導体を用いるとよい。例えば開裂する結合の接合部にスタチン誘導体が存在していると、本発明のノッチが開裂部位で加水分解しなくなる。基質に対しては、検出を容易にするための適切な蛍光タグ(例えばBODIPY FL)を付加するという修飾をさらに行なうことができる。
【0121】
本発明の基質に蛍光標識を付け、γ-セクレターゼのための蛍光偏光結合アッセイを開発するのに利用できる。酵素と基質の間の相互作用に関する平衡解離定数(KD)は、酵素を用いて基質を滴定した結果として得られる蛍光偏光の変化を測定することによって決定される。
【0122】
本発明の基質とγ-セクレターゼの相互作用に関するKDを明らかにするため、さまざまな量のγ-セクレターゼを3.1nMの蛍光基質と組み合わせ、室温にて3時間にわたってインキュベートする。インキュベーションの後、LJLアナリスト(96ウエルの形式)またはパンヴェラ・ビーコン(単一のキュベットの形式)を用いて蛍光偏光を測定する。アッセイは、例えば、25mM酢酸ナトリウム、20%グリセロール、pH4.75の中で行なう。偏光の値を与える取得データを縦軸に、酵素の濃度を横軸にプロットすると、酵素と基質の相互作用に関するKDを決定するための結合等温線が得られる。次に、関係式Px=PF+(PB-PF)×[E]/(KD+[E])(ただしP=偏光値、x=サンプル、F=遊離している阻害剤、B=結合している阻害剤、E=γ-セクレターゼ)(蛍光偏光アプリケーション・ガイド、1998年;パンヴェラ社、マディソン、ウィスコンシン州)を用いてこのデータを分析し、KDを得る。このアッセイを利用し、酵素の活性部位またはアロステリック部位に結合する化合物をスクリーニングすることができる。
【0123】
試験化合物の存在下と不在下での活性測定は、並列に、あるいはいずれかの順番で実施できることに注意されたい。さらに、特定の反応条件について酵素活性の信頼できるベースラインが得られたのであれば、すべての試験化合物と並列して対照の測定(すなわち試験化合物が不在下での開裂の測定)を繰り返す必要はなかろう。特定の基質(例えば本発明のノッチ組成物またはAPPに由来する組成物)に対するγ-セクレターゼの酵素活性について阻害剤の不在下で得られた知見を、比較ステップを実施するための基礎にすることができる。
【0124】
B.基質候補
【0125】
この明細書では、“基質候補”または“試験基質”という用語は、γ-セクレターゼの活性、好ましくはヒトγ-セクレターゼの活性を変化させることのできるあらゆる分子を意味する。基質候補としては、タンパク質またはその断片、小分子阻害剤が可能であり、核酸分子さえ可能である。スクリーニング・アッセイを適用して同定される最も有用な薬理化合物は、大化合物ライブラリのスクリーニングを通じて同定される化合物、またはAPPのプロセシングとノッチのプロセシングの一方または両方に関する公知の他のモジュレータと構造的に関係のある化合物であろう。例えばアメリカ合衆国特許第6,448,229号(その内容が参考としてこの明細書に組み込まれているものとする)には、ノッチのシグナル伝達には影響を与えずにγ-セクレターゼを抑制するためADの治療または予防に使用できる特別なクラスの化合物が記載されている。このアメリカ合衆国特許第6,448,229号に記載されているような化合物は、本発明のアッセイを利用して確認することができる。さらに、そのような化合物を合理的な薬剤設計における出発材料として用い、本発明で使用する別のモジュレータ候補を同定できる可能性がある。このように合理的な薬剤設計に使用できる可能性のある他の阻害剤としては、DAPT(PHA-568638)やフェンキルアミン(PHA-512088)などが挙げられる。
【0126】
基質候補は、天然に存在する化合物の断片または一部でもよいし、組み合わせないときには不活性なのに組み合わせたときだけ活性になることがわかった既知の化合物の組み合わせでもよい。しかしそのような化合物をヒトまたはモデル動物でテストする前に、どの候補が潜在能力を持っているかを多彩な候補についてテストする必要があろう。
【0127】
したがって基質候補は、天然に存在する化合物の断片または一部でもよいし、組み合わせないときには不活性なのに組み合わせたときだけ活性になることがわかった既知の化合物の組み合わせでもよい。したがって本発明により、γ-セクレターゼによるノッチのプロセシングの促進または抑制よりも、γ-セクレターゼによる細胞APPプロセシングを促進または抑制する化合物を同定するスクリーニング・アッセイが提供される。天然の供給源(例えば動物、細菌、真菌、植物(葉や樹皮も含む)、海洋サンプル)から単離した化合物を候補として調べ、潜在的に存在している有用な医薬を探すことが提案される。
【0128】
スクリーニングする医薬は、化学的合成物または人工化合物に由来するもの、あるいは化学的合成物または人工化合物から合成することも可能であることが理解できよう。したがって、本発明によって同定されるモジュレータとしては、γ-セクレターゼの活性および/またはAPPのプロセシングを促進または阻害する公知の薬剤から出発し、合理的な薬剤設計を通じて設計することのできるポリペプチド、ポリヌクレオチド、小分子阻害剤、他の任意の無機または有機の化合物が可能であることがわかる。
【0129】
候補スクリーニング・アッセイは準備と実施が簡単である。したがってモジュレータのアッセイでは、機能的γ-セクレターゼを含む細胞膜分画(例えば可溶化γ-セクレターゼ複合体を含む細胞膜分画)を得た後、本発明による新規な基質の存在下で候補物質をそのようなγ-セクレターゼ組成物と混合する。そのとき、γ-セクレターゼの活性をその基質の開裂を通じて測定できるようにする。このようにして、候補物質の存在下でその候補物質がγ-セクレターゼを活性化する能力を測定することができる。同様に、阻害剤を探すアッセイでは、機能的γ-セクレターゼを発現する細胞膜分画を得た後、候補物質をその分画と混合する。このようにして、抑制候補物質が、γ-セクレターゼによる生物学的効果を低下、消失、減少させる能力を検出することができる。
【0130】
所定の条件下における基質の“有効な量”は、γ-セクレターゼが所定のCT-100を開裂させる活性またはAPPのプロセシングを再現性よく変化させるのに有効な量のことである。この有効量は、候補物質の不在下で見られる開裂と比べた場合に、APPが開裂して得られるCT-100の程度または量を変化させるが、本発明のノッチ融合タンパク質のγ-セクレターゼ部位における開裂は変化させない量である。治療薬として役立つであろう化合物、および/またはさらにキャラクテリゼーションを行なうのに特に役立つであろう化合物は、γ-セクレターゼによるノッチの開裂ではなくAPPの開裂を選択的に抑制する化合物である。“選択的に抑制する”とは、γ-セクレターゼによるノッチの開裂よりもAPPの開裂に対してより抑制効果を有する化合物を意味する。ノッチの開裂を抑制しない化合物が好ましいとはいえ、ノッチの開裂が幾分か抑制されることは、野生型γ-セクレターゼによるAPPの開裂よりも少ないのであれば許容できる。
【0131】
γ-セクレターゼの本発明による新規な基質を用いた上記のアッセイは、多数の高スループット・スクリーニング(HTS)法と相性がよい(概説に関してはJayawickremeとKost、Curr. Opin. Biotechnol.、第8巻、629〜634ページ、1997年を参照のこと)。例えばHoustonとBanks、Curr. Opin. Biotechnol.、第8巻、734〜740ページ、1997年に記載されているように、自動化したHTSアッセイと小型化したHTSアッセイも考えることができる。
【0132】
小分子モジュレータの同定に用いられる多数の異なったライブラリが存在している。例えば、化合物ライブラリ、天然生成物ライブラリ、コンビナトリアル・ライブラリ(ランダムに得れらた、または設計して得たペプチド、オリゴヌクレオチド、有機分子からなる)などがある。化合物ライブラリは、公知の化合物の構造的アナログからなるか、天然生成物のスクリーニングまたは潜在的治療標的に対するスクリーニングを通じてヒット化合物またはリード化合物と同定された化合物からなる。天然生成物ライブラリは、微生物、動物、植物、昆虫、海洋生物からの生成物のコレクションである。これらは、例えば土壌生物、植物、海洋生物からのブロスを発酵させた後に抽出することによってスクリーニング用混合物を作るのに用いられる。天然生成物ライブラリには、ポリペプチド、非リボソーム・ペプチド、その非天然変異体が含まれる。概説に関してはScience、第282巻、63〜68ページ、1998年を参照のこと。
【0133】
コンビナトリアル・ライブラリは、多数のペプチド、オリゴヌクレオチド、有機化合物で構成されている。コンビナトリアル・ライブラリは、伝統的な自動化合成法、PCRクローニング、あるいは他の合成法によって比較的簡単に用意できる。特に興味深いのは、ペプチド・ライブラリ、タンパク質ライブラリ、ペプチドミメティック・ライブラリ、多重並列合成コレクション・ライブラリ、リコンビナトリアル・ライブラリ、ポリペプチド・ライブラリなどのライブラリである。コンビナトリアル・ライブラリと、そこから作られるライブラリの概説に関しては、Myers、Curr. Opin. Biotechnol.、第8巻、701〜707ページ、1997年を参照のこと。次に、ここに記載したさまざまなライブラリを用いて同定したモジュレータを最適化し、例えば合理的薬剤設計を通じてγ-セクレターゼの活性を変化させる。
【0134】
もちろん、有効な候補が見つからないとしても本発明のすべてのスクリーニング法はそれ自体有効であることが理解できよう。本発明により、そのような候補を見つける方法だけでなく、そのような候補をスクリーニングする方法も提供される。
【0135】
C.インビボ・アッセイ
【0136】
本発明には、さまざまなモデル動物の利用も含まれる。すでに説明したようにインビトロ環境でモジュレータがスクリーニングされると、APPのプロセシングおよび/またはADの非ヒト・モデルを利用してそのモジュレータの生体内における効果を明らかにすることができる。これは、γ-セクレターゼが正常に発現する1つの動物系全体の中でγ-セクレターゼの機能を調べる素晴らしい機会になろう。
【0137】
γ-セクレターゼ活性のモジュレータであることが判明した試験化合物を用いた動物の治療は、その化合物を適切な形態で動物に投与する操作を含むことになる。投与は、臨床目的または非臨床目的で利用できる任意の経路(例えば経口、鼻腔、口内、直腸、膣、局所)を通じて行なうことになろう。投与は、気管内点滴、気管支点滴、皮膚内注射、皮下注射、筋肉内注射、腹腔内注射、静脈内注射によって行なうことも可能である。全身性静脈内注射、血液を通じた局所投与、脳脊髄液(CSF)への供給、リンパへの供給、腫瘍内注射などが特に考えられる。
【0138】
生体内での化合物の有効性を調べるにはさまざまな基準がある。そのような基準としては、生存率、活性レベルの上昇、食物摂取の改善などがある。他の評価法としては、γ-セクレターゼの活性が変化したことの徴候(例えばアミロイドβまたはアミロイドβ斑の産生低下、脳の萎縮減少)を探す病理学的検査(特に脳組織)がある。
【0139】
D.薬の製造
【0140】
本発明のアッセイにより、γ-セクレターゼの活性が異常なレベルであることを特徴とする疾患(例えばAD)を治療するための治療薬候補となるγ-セクレターゼのモジュレータを同定する。そこで本発明の方法は、モジュレータを同定した後、必要に応じた追加ステップとして、薬剤を製造/合成するステップと、薬理学的に許容可能な希釈剤、アジュバント、基剤を用いてその薬剤を組成物にするステップを含んでいる。医薬組成物に関しては以下に詳しく説明する。
【0141】
VI.医薬組成物
【0142】
ノッチのプロセシング、APPのプロセシング、γ-セクレターゼの開裂を変化させることが確認された本発明のモジュレータは、最終的に医薬組成物(すなわち生体内で利用するのに適した形態)にすることができる。一般に、発熱物質や、ヒトまたは動物にとって有害である可能性のある他の不純物を実質的に含まない組成物を調製する。
【0143】
一般に、同定されたモジュレータ組成物を適切な塩や緩衝液を用いて安定にし、標的細胞による取り込みが可能になるようにすることが望ましい。“薬理学的または薬物学的に許容可能な”という表現は、動物またはヒトに投与したときに好ましくない反応、アレルギー反応、不都合な反応を起こさない分子全体または組成物を意味する。この明細書では、“薬理学的に許容可能な基剤”には、あらゆる溶媒、分散媒体、コーティング、抗菌剤、抗真菌剤、等張剤、吸収遅延剤などが含まれる。薬理学的に活性な物質のためのこのような媒体や薬剤の使用法は従来技術において周知である。従来の媒体や薬剤のどれであれ、本発明によって同定されたモジュレータと適合しない場合を除き、治療用組成物で使用することが考えられる。追加活性成分も組成物に組み込むことができる。
【0144】
本発明のモジュレータ組成物には古典的な医薬調製物が含まれる。本発明の組成物は、一般的な経路で標的組織に到達させうるのであれば、その一般的な経路のうちの任意の経路を通じて投与されることになる。医薬組成物は、従来からある任意の方法で対象に導入することができる。例えば、静脈内、皮膚内、筋肉内、乳房内、腹腔内、気管内、眼内、眼球後、肺内(例えば期間限定放出)への投与;経口、舌下、鼻、肛門、膣、経皮での投与、特定部位への外科的インプランテーション(例えば脾膜や脳の下、角膜内への埋め込み)などの方法がある。治療は、1回の投与、または所定の期間にわたる複数回の投与にすることができる。
【0145】
本発明によって同定されたモジュレータ化合物は、投与するため、塩基を含まない溶液にしたり、薬理学的に許容可能な塩を含む水に適切な界面活性剤(例えばヒドロキシプロピルセルロース)を混合したものにしたりすることができる。グリセロール、液体ポリエチレングリコール、これらの混合物への分散液や、油の中への分散液も調製することができる。通常の保管条件および使用条件では、微生物の増殖を防ぐために保存剤を調製物に入れる。
【0146】
注射に適した医薬形態としては、減菌水溶液または減菌分散液や、その場で減菌注射溶液または減菌分散液を調製するための減菌粉末などがある。いずれの場合にも、減菌形態でなくてはならず、容易に注射器に入れられるよう流体形態になっている必要がある。製造条件と保管条件のもとで安定であり、微生物(例えば細菌や真菌)の汚染作用から保護されている必要がある。基剤としては、例えば水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、液体ポリエチレングリコールなど)、これらの適切な混合物、植物油などを含む溶媒または分散媒体が可能である。適切な流動性を維持するには、例えばコーティング(例えばレシチン)を使用したり、必要な粒径を維持したり(分散液の場合)、界面活性剤を用いたりする。微生物の作用は、さまざまな抗菌剤や抗真菌剤(例えばパラベン、クロロブタノール、フェノール、ソルビン酸、チメロサールなど)を用いて防止することができる。多くの場合、等張剤(例えば糖類、塩化ナトリウム)を含めることが好ましい。注射組成物の吸収時間を延ばすには、組成物の中で吸収遅延剤(例えばモノステアリン酸アルミニウム、ゼラチン)を用いるとよい。
【0147】
減菌注射溶液は、必要量の活性化合物と他のさまざまな上記成分を適切な溶媒の中に組み込んだ後、必要に応じて濾過殺菌することによって調製する。一般に、分散液は、殺菌したさまざまな活性成分を、基本的分散媒体と必要な他の上記成分とを含む減菌ビヒクルの中に組み込むことによって調製する。減菌注射溶液を調製するための減菌粉末の場合には、好ましい調製方法は真空乾燥法と凍結乾燥法であり、活性成分と望む追加成分からなる粉末を、それら成分を含むあらかじめ濾過殺菌した溶液から作る。
【0148】
この明細書では、“薬理学的に許容可能な基剤”に、あらゆる溶媒、分散媒体、コーティング、抗菌剤、抗真菌剤、等張剤、吸収遅延剤などが含まれる。このような媒体や薬剤を薬理学的に活性な物質に用いる方法は従来技術において周知である。従来の媒体や薬剤のどれであれ、活性成分と適合しない場合を除き、治療用組成物で使用することが考えられる。追加活性成分も組成物に組み込むことができる。
【0149】
経口投与するため、本発明によって同定されたモジュレータは、賦形剤に組み込み、非消化性口内洗浄剤や歯磨き剤の形態で使用することができる。口内洗浄剤は、必要量の活性成分を適切な溶媒(例えばホウ酸ナトリウム溶液(ドーベル液))の中に組み込むことによって調製できる。活性成分は、ホウ酸ナトリウム、グリセリン、炭酸水素カリウムを含む減菌口内洗浄剤に組み込むこともできる。活性成分は、歯磨き剤の中に分散させることもできる。歯磨き剤は、ゲル、ペースト、粉末、スラリーの形態がある。治療に有効な量の活性成分をペースト状歯磨き剤に添加することができる。この歯磨き剤は、水、結合剤、研磨剤、香味剤、発泡剤、湿潤剤を含むことができる。
【0150】
本発明の組成物は、中性形態または塩の形態にすることができる。薬理学的に許容可能な塩としては、(タンパク質の遊離アミノ基とで形成された)酸添加塩がある。酸添加塩は、無機酸(例えば塩酸、リン酸)または有機酸(例えば酢酸、シュウ酸、酒石酸、マンデル酸など)とで形成される。遊離カルボキシル基とで形成される塩は、無機塩基(例えば水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、水酸化第二鉄)や有機塩基(例えばイソプロピルアミン、トリメチルアミン、ヒスチジン、プロカインなど)から得られる。
【0151】
本発明の組成物は、中性形態または塩の形態にすることができる。薬理学的に許容可能な塩としては、(タンパク質の遊離アミノ基とで形成された)酸添加塩がある。酸添加塩は、無機酸(例えば塩酸、リン酸)または有機酸(例えば酢酸、シュウ酸、酒石酸、マンデル酸など)とで形成される。遊離カルボキシル基とで形成される塩は、無機塩基(例えば水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、水酸化第二鉄)や有機塩基(例えばイソプロピルアミン、トリメチルアミン、ヒスチジン、プロカインなど)から得られる。
【0152】
製剤ができると、その製剤に適合した方法で、治療に有効な量の溶液を投与する。製剤は、多彩な投与形態(例えば注射溶液、薬剤放出カプセルなど)で容易に投与できる。水溶液の形態で非経口投与するには、その水溶液を必要に応じて適切に緩衝し、希釈されたその液体をまず十分な量の生理食塩水またはグルコースで等張にする必要がある。この特別な水溶液は、静脈内、筋肉内、皮下、腹腔内に投与するのに特に適している。
【0153】
“単位用量”は、適切な基剤に分散させた離散量の治療用組成物として定義される。例えば非経口投与は、まず最初にボーラスとして実施し、その後、連続輸液を行なって治療に必要な薬剤循環レベルを維持するとよい。当業者であれば、優れた医療経験と個々の患者の臨床状態により、効果的な投与量と投与計画を容易に最適化することができよう。より詳細には、投与量は、ADになっている対象の脳内でのAβ-ペプチドの形成、さらに特定するならば斑の形成の減少、抑制、低下、阻止をもたらすように選択する必要がある。そのためには、当業者は、(例えばアメリカ合衆国特許第5,877,399号、第5,387,742号、第5,811,633号に開示されているような)ADのモデル動物を利用して投与プロトコルを最適化し、ヒト患者でADを治療するのに必要な医薬の量を予測することができよう。
【0154】
投与の頻度は、薬剤の薬物動態パラメータと投与経路によって異なる。医薬製剤の最適な形態は、当業者が、投与経路と望ましい投与量から決定する。例えば『レミントンの薬理化学』、第18版(1990年、マック出版社、イーストン、フィラデルフィア州18042)、1435〜1712ページを参照のこと。なおその内容は参考としてこの明細書に組み込まれているものとする。製剤の形態は、投与した薬剤の物理的状態、安定性、生体内の放出速度、生体内のクリアランス速度に影響を与える可能性がある。投与経路に応じ、体重、体表面積、臓器のサイズをもとにして適切な投与量を計算することができる。治療に適した投与量を決めるのに必要な計算をさらに洗練させることは、当業者であれば、特別な実験を行なうことなく、この明細書に開示した投与量情報およびアッセイと、動物またはヒトの臨床試験で観察された薬物動態データに特に基づいて日常的に行なうことができる。
【0155】
適切な投与量は、血中レベルを測定するための確立されたアッセイと、関連する投与量-応答データを利用して確認することができる。最終的な投与計画は、参加している医者が、薬の作用を変化させる因子(例えば薬の特異的活性、損傷の程度、患者の応答性、年齢、状態、体重、性別、食事、何らかの感染の程度、投与時間、他の臨床因子)を考慮して決めることになろう。研究は進行中であるため、適切な投与量のレベル、特定の疾患や症状の治療期間に関するさらなる情報が今後得られるであろう。
【0156】
本発明の医薬組成物と治療法は、ヒトの医学と動物の医学の分野で役に立つ可能性があることが理解されよう。したがって治療する対象として、哺乳動物(ヒトであることが好ましい)または他の動物が可能である。獣医学の目的では、対象として、例えば家畜(乳牛、ヒツジ、ブタ、ウマ、ヤギなど)、ペット(イヌ、ネコなど)、外国産および/または動物園の動物、実験室の動物(マウス、ラット、ウサギ、モルモット、ハムスターなど)、家禽類(ニワトリ、シチメンチョウ、アヒル、ガチョウなど)などが挙げられる。
【実施例】
【0157】
以下の実施例は、本発明の好ましい実施態様を示すために掲載してある。しかし当業者は、この明細書をもとにして、開示されている具体的な実施態様に対して本発明の精神と範囲を逸脱することなく多くの変更を施してもやはり同様の結果を得られることを理解する必要がある。
実施例1
【0158】
材料と方法
この実施例では、この明細書で説明した結果を得るのに利用する一般的な方法、試薬、アッセイについて説明する。一般的な実験室用化学薬品は、シグマ・ケミカルズ社(セントルイス、ミズーリ州)から購入した。pET43.1aベクターは、ノヴァジェン社(マディソン、ウィスコンシン州)からのものであり、制限酵素はインヴィトロジェン社(カールズバッド、カリフォルニア州)からのものである。オリゴヌクレオチドはシグマ・ジェノシス社(ザ・ウッドランズ、マサチューセッツ州)からのものである。バリン1744抗体は、セル・シグナリング・テクノロジー社(ビヴァリー、マサチューセッツ州)からのものである。PHA/PNU化合物は、ファルマシア社の化合物コレクション(ニュー・ジャージー、アメリカ合衆国)から入手した。
【0159】
ノッチ基質のクローニング:マウスのノッチ-1タンパク質(DNA配列登録番号Z11886)のアミノ酸N1703〜D1860を含むノッチ基質をクローニングし、NusAをコードしていた核酸と同じフレームのEcoRI/HindIII挿入体としてpET43.1aベクターに入れた。NusA/ノッチ融合タンパク質をコードする発現構造体は、配列番号1の配列を持っていた。この構造体はPCRによって作ったC末端フラグと8ヒスチジン・タグを含んでおり、ノッチのアミノ酸1860の後に続く延長部(DYKDDDDKHHHHHHHH、配列番号15)となっていた。このDNAでBL21(DE3)コンピテント大腸菌(ストラタジーン社)を形質転換した後、DNAコンサート・ミニプレップ・キット(ギブコ/BRL社)を用いてクローンをスクリーニングし、正しい挿入体が存在しているかどうか調べた。ノッチ-F6というクローンは、配列番号1の正しいDNA配列を含んでいることがわかった。
【0160】
ノッチ/NusA融合タンパク質の発現と精製。大腸菌をノッチ-F6というクローンを用いて形質転換してからLB/Ampに接種し、振盪インキュベータの中で37℃にて一晩にわたって増殖させた。翌日、一晩にわたる培養物35mlを2リットルのLB/Ampに接種した。大腸菌を振盪インキュベータの中で37℃にてA600が0.4になるまで増殖させた後、1mMのIPTGを用いて3時間にわたって誘導し、遠心分離した。2リットルの培養物からのペレットを、50mMのトリス(pH8.0)、100mMのNaCl、プロテアーゼ阻害剤を含む緩衝液(ペレット1gにつき5ml)の中に再び懸濁させ、フレンチ・プレスを用いて3回処理して粗抽出液を得た。抽出液のpHを2Mのトリスを用いて8.0に調節した後、その抽出液を11,000×gで45分間にわたって遠心分離した。上澄4mlを、50mMのトリス(pH8.0)、100mMのNaCl、プロテアーゼ阻害剤を含む緩衝液の中で平衡させたニッケルIMACクロマトグラフィ・カラムに充填した。このカラムを同じ緩衝液で洗浄した後、50mMのイミダゾールを含む緩衝液で洗浄した。300mMのイミダゾールを含む緩衝液を用いてNusA-ノッチ融合タンパク質を溶離させ、分画0.8mlを回収し、A280とSDS-PAGEで分析した。NusA-ノッチを含む分画をプールし、50mMのピペス(pH7.0)、100mMのNaClの中に透析した。
【0161】
ウエスタン・ブロット分析によって調べるノッチの開裂。NusA-ノッチ融合タンパク質(1.7μM)を、可溶化γ-セクレターゼを70μg/mlの割合で含む合計体積50μlの緩衝液(50mMのピペス(pH7.0)、0.25%のCHAPSO)とともに37℃にて一晩にわたってインキュベートした。DAPT(PHA-568638)をさまざまな濃度で添加した。5×レムリ緩衝液(Laemmli、1970年)12.5μlを添加して反応を停止させ、混合物30μlの電気泳動を15%SDS-PAGE上で実施した。半乾燥ブロット装置(ミリポア社)を用いてタンパク質をニトロセルロースに移し、4%BSAを含むPBS/0.5%トゥイーン20を用いて2時間にわたってブロックした。バリン1744抗体を1000倍に希釈してブロッキング溶液に添加し、1時間にわたってインキュベートした。膜をPBS/0.5%トゥイーン20で3回洗浄した後、抗ウサギIgG-HRP(4%BSAを含むPBS/0.5%トゥイーン20に1:5000の割合で希釈)とともにインキュベートした。膜を再び3回洗浄し、ECL試薬(アマーシャム社、ピスカタウェイ、ニュージャージー州)を用いて現像した。
【0162】
インビトロ・ノッチ開裂ELISA。ELISA法を利用してノッチの開裂も調べた。反応を起こさせる前に、96ウエル半面積プレート(コスター社)の中の必要数のウエルを、0.1MのNaHCO3(pH8.2)の中で200倍に希釈したバリン1744抗体50μlでコーティングした。プレートを4℃にて一晩にわたってインキュベートした。ノッチ開裂反応は以下のようにして行なわせた。NusA-ノッチ(0.9μM)を、可溶化γ-セクレターゼを70μg/mlの割合で含む合計体積が25μlの緩衝液(50mMのピペス(pH7.0)、0.25%のCHAPSO)とともに37℃にて一晩にわたってインキュベートした。翌日、バリン1744抗体でコーティングしたプレートをPBS/0.5%トゥイーン20を用いて3回洗浄し、4%BSAを含むPBS/0.5%トゥイーン20を用いて1時間にわたってブロックした。4%BSAを含むPBS/0.5%トゥイーン20を用いて開裂反応混合物を14倍に希釈し、50μlを採取したものを3つ用意し、室温にて3〜4時間にわたってインキュベートした。プレートをPBS/0.5%トゥイーン20を用いて3回洗浄し、4%BSA/PBS/0.5%トゥイーン20の中で60000倍に希釈した抗FLAG-HRP抗体(シグマ社、セントルイス、ミズーリ州)50μlを添加した。この抗体を45分間にわたってインキュベートし、プレートをPBS/0.5%トゥイーン20を用いて3回洗浄した。TMB試薬(キルケガード&ペリー社)を1:1の割合で混合し、50μlを採取してウエルに添加した。1時間にわたって放置して色を変化させ、1MのH3PO4を50μl添加し、プレートをスペクトラマックス・プラス・プレート読み取り機で450nmにて読み取った。ノッチ基質の濃度を変化させたときには、0.11〜3.6μMのNusA-ノッチを使用した。酵素の濃度を変化させたときには、2.1〜68μg/mlの可溶化γ-セクレターゼを使用した。
【0163】
ノッチの開裂抑制。50%DMSOの中で25倍の濃度にした阻害剤(1μl)を開裂反応混合物に添加した後、酵素を添加した。ブランクの対照と阻害剤なしの対照も同じ最終濃度のDMSOを含むように調節した。阻害剤のIC50は、グラフィット4.0プログラムの4パラメータ・ロジスティック・モデルを利用して計算した。
実施例2
【0164】
結果と考察
可溶性ノッチ基質を産生させるため、発明者は、大腸菌の中に多数の構造体を用意した(図3A)。カスパーゼ・リーダー、ユビキチン、N末端タウ、Nugタグを利用した。可溶性ノッチ基質の発現は、Nus融合体でだけ観察された(Wilkinson他、Bio/Technology、第9巻、443〜448ページ、1991年)。図3Bには、この構造体のクローニングに関する詳細が示してある。精製とアッセイの開発を容易にするため、ヒスチジン・タグとフラグ・タグをノッチのC末端に付着させた。
【0165】
Nusタグの付いたノッチ融合タンパク質が大腸菌の中で発現するとき、90kDaのバンドに対応する融合タンパク質の高レベルの発現がSDS-PAGEで観察された(図4、レーン1)。このサイズは、この融合タンパク質から予想される値である。全ライセートを遠心分離すると、可溶性分画の中に融合タンパク質が残った(図4、レーン2)。固定化金属イオンとしてニッケルを用いたIMACにより、ノッチを含む融合タンパク質を可溶性分画から精製した。図4(レーン5〜9)には、300mMのイミダゾールによってIMACカラムから溶離したさまざまな分画が示してある。これら分画をプールし、透析した後、γ-セクレターゼによる開裂のための基質供給源として利用した。
【0166】
ノッチ・タンパク質の特異的な開裂を調べる方法は、バリン1744抗体(セル・シグナリング・テクノロジー社)の特異性に基づいている。この抗体は開裂したノッチに対して特異的であり、開裂していないノッチ・タンパク質と交差反応することはない。図5(レーン3)に示したように、開裂したノッチ・タンパク質は、バリン1744抗体に対して特異的な抗体を用いてウエスタン・ブロット上で検出された。レーン1からわかるように、開裂していないノッチ融合タンパク質との交差反応は観察されなかった。さらに、ノッチ・タンパク質におけるこの特異的開裂は、γ-セクレターゼの強力な阻害剤であることがよく知られているDAPTの投与量に依存して抑制された(レーン4〜8、図5)(Dovey他、J. Neurochem.、第76巻、173〜181ページ、2001年)。
【0167】
ノッチ・タンパク質に別の開裂があるかどうかを調べるため、C末端フラグ抗体を用いたウエスタン・ブロットによる開裂反応のモニターも行なった。3つの免疫活性なバンドのうち、1つの開裂産物だけがDAPTによって抑制可能であった。これらの結果を合わせると、γ-セクレターゼによるインビトロでの開裂により、1743/1744に特異的に開裂が起こることが示唆される。これは、細胞に基づく研究(Kopan他、Proc. Natl. Acad. Sci.、第93巻、1683〜1688ページ、1996年;Schroeter他、Nature、第393巻、382〜386ページ、1998年)においてノッチからNICDが産生されることが示されたという結果と一致している。
【0168】
上記の結果は、ノッチ融合タンパク質がγ-セクレターゼによる特異的開裂を起こしやすく、その開裂を、非常に特異的なバリン1744モノクローナル抗体によってウエスタン・ブロット上で検出できることを示している。ウエスタン・ブロットは定量的ではないため、ノッチを抑制する化合物を評価して抑制能力をAβ ELISAと比較することは難しい。
【0169】
図6には、ノッチの開裂を定量的に調べるELISAを開発し、CT-100に基づいたELISAでさまざまな化合物の抑制能力を比較する方法が示してある。この方法は、γ-セクレターゼの存在下におけるノッチ融合タンパク質からのNICD断片のN末端とC末端を高い特異性で捕捉する抗バリン1744抗体と抗フラグ抗体を用いたサンドイッチELISAに基づいている。このアッセイは、実施例1の“インビトロ・ノッチ開裂ELISA”に記載したようにして実施する。このサンドイッチELISAでは、マイクロタイター・プレートをバリン1744抗体でコーティングすることが重要である。ところでNusA-ノッチ開裂反応は、NusA-ノッチをγ-セクレターゼとともにインキュベートすることによって実施した。バリン1744抗体でコーティングしたプレートを緩衝液で洗浄し、BSAでブロックした。次に開裂反応混合物をマイクロタイター・プレートに添加し、室温にて適切な時間にわたってインキュベートした。プレートを洗浄して抗FLAG-HRP抗体(シグマ社、セントルイス、ミズーリ州)を添加し、適切な時間にわたって再びインキュベートした。HRPのための発色基質であるTMB試薬(キルケガード&ペリー社)を添加し、変化した色をスペクトラマックス・プラス・プレート読み取り機で450nmにて読み取った。図7には、ELISAで検出したNusA-ノッチ融合タンパク質の開裂が示してある。ノイズの5倍ある信号がELISAで観察された。
【0170】
ノッチを開裂させる可溶化γ-セクレターゼの酵素活性を、所定の実験条件下でELISAによってさらに調べた。図8Aには、生成物の形成が基質に依存する様子が示してある。反応はミカエリス-メンテンの式に従う。可溶化γ-セクレターゼによるノッチ融合タンパク質基質の加水分解に関する見かけのKmは、所定の条件下で約0.7μMである。観察されたノッチ開裂活性は、γ-セクレターゼ活性を有する可溶化膜調製物に含まれるタンパク質の濃度にも比例していた(図8B)。
【0171】
文献に報告されているγ-セクレターゼの特異的阻害剤を用いた抑制の研究を行なうことにより、この酵素によるノッチの開裂をELISAでさらに確認した。DAPTと呼ばれるγ-セクレターゼの強力な阻害剤が報告されている(Dovey他、J. Neurochem.、第76巻、173〜181ページ、2001年)。図9に示したように、DAPT(PHA-568638)はノッチの開裂を投与量に依存した形で抑制し、IC50=2.4nMである。その一方で、フェンキルアミンスルホンアミド(PHA-512088)もCT-100インビトロ・アッセイにおいて小さなμMの範囲でγ-セクレターゼの活性を抑制することが報告されている(Rishton他、J. Med. Chem.、第43巻、2297〜2299ページ、2000年)。図9からわかるように、PHA-512088はγ-セクレターゼによるノッチの開裂を抑制し、所定の条件下におけるELISAでIC50は0.7μMであった。この図からわかるように、阻害剤(PHA-568638、PHA-512088)は、アッセイにおいて小さな値のnM〜小さな値のμMの範囲で非常によく機能した。これらの結果を合わせると、ノッチ融合タンパク質基質からNICD断片を生成させるインビトロ酵素活性はγ-セクレターゼによるものであることがわかる。
【0172】
最近、APPとノッチの開裂を並行して調べる細胞ベースのアッセイが報告された(Karlstrom他、J. Biol. Chem.、第277巻、6763〜6766ページ、2002年)。しかしγ-セクレターゼによるノッチの特異的な開裂(G1743-V1744)を調べるためのインビトロ定量ELISAを初めて明らかにしたのは本発明である。本発明は、HeLa細胞からのγ-セクレターゼの活性によってノッチ・タンパク質が1743-1744接合部で特異的に開裂することを初めて証明した。
【0173】
この明細書に開示したり主張したりしたすべての組成物および/または方法は、この明細書の開示内容に照らして特別な実験なしに実現することができる。本発明の組成物と方法を好ましい実施態様に関して説明してきたが、当業者にとって、本発明の考え方、精神、範囲を逸脱することなく、この明細書に記載した組成物および/または方法と、その方法における一連のステップに対してさまざまな変更を施せることは明らかであろう。さらに詳細には、この明細書に記載した薬剤を化学的に関連した薬剤や生理学的に関連した薬剤で置き換えても同じ結果または似たような結果が得られることは明らかであろう。当業者にとって明らかなそのような同様の代替物や変更は、添付の請求項に規定した本発明の精神、範囲、考え方に含まれるものと考えられる。
【0174】
この明細書全体で引用した参考文献は、この明細書に記載した具体的な手続きやそれ以外の補足的な詳細を提供している場合には、すべて参考として具体的にこの明細書に組み込まれているものとする。
【図面の簡単な説明】
【0175】
【図1】γ-セクレターゼ様開裂部位の比較図である。膜貫通ドメインを取り囲む配列を、APP(配列番号10)、ノッチ-1(配列番号12)、E-カドヘリン(配列番号11)について示してある。APPにおいてAβ1〜40とAβ1〜42を産生させるγ-セクレターゼ開裂部位と、3つの基質におけるε開裂部位(ε)またはγ-セクレターゼ様開裂部位も示してある。
【図2】ノッチ-1でγ-セクレターゼ様開裂部位を取り囲んでいる部分のアミノ酸配列である。この配列(1703〜1860;配列番号13)を選択して大腸菌で発現させた。膜貫通ドメインは下線で示してあり、1743/1744開裂部位は太字と下線で示してある。このノッチ配列の開裂により、アミノ酸1744〜1860を含むNICD断片が得られることになる。
【図3A】大腸菌においてノッチを発現させるためにクローニングした構造体。4つの構造体を示してあり、そのすべてにノッチ-1のアミノ酸配列1703〜1860が含まれている。カスパーゼ・リーダー配列、ユビキチン、タウのN末端ドメインのいずれかをN末端タグとして用いて発現を促進した場合には発現は見られなかった。ノッチ(1703〜1860)をNusAタンパク質と融合させた場合には、大腸菌において可溶性発現が見られた。
【図3B】ノッチF構造体をクローニングしてpET43.1aに組み込んだ概略図である。この構造体は、C末端にフラグと8ヒスチジン・タグも備えている。
【図4】NusA-ノッチ融合体のIMAC単離。この融合タンパク質の概略図を最上部に示してある。NusA-ノッチ融合体の発現と精製に関する詳細は「材料と方法」に見ることができる。矢印は、10%SDS-PAGEゲル上でこの融合タンパク質が発現したバンドを示している。レーン1、大腸菌の粗ライセート;レーン2、大腸菌の上澄;レーン3、IMACのフロースルー;レーン4、50mMのイミダゾールによる洗浄;レーン5〜9、300mMのイミダゾールによる溶離分画;レーン10、ベンチマーク分子量マーカー。
【図5】可溶化γ-セクレターゼを用いたNusA-ノッチ融合体の開裂を示すウエスタン・ブロット。この実験の詳細は実施例1に記載してある。要するに、さまざまな濃度のDAPT(PHA-568638)の存在下と不在下で、NusA-ノッチ融合体を可溶化γ-セクレターゼとともに一晩にわたってインキュベートした。サンプルを電気泳動にかけ、ブロットし、バリン1744での開裂に特異的なバリン1744抗体で調べた。
【図6】ノッチの特異的開裂のELISAによる検出。可溶化γ-セクレターゼによるNusA-ノッチ融合体の開裂を検出するのに用いたサンドイッチELISAの概略を示してある。特異的開裂は、バリン1744抗体を用いて検出する。
【図7】γ-セクレターゼによって開裂したNusA-ノッチ融合体のELISAによる検出。68μg/mlの可溶化γ-セクレターゼ(酵素)の不在下と存在下でNusA-ノッチ(0.9μM)基質をインキュベートし、「材料と方法」に従ってELISAを行なった。データは6回の実験の平均値を表わしている。
【図8】ELISAによる、ノッチ開裂のキャラクテリゼーション。ELISAは「材料と方法」に記載したようにして実施した。ノッチ基質の濃度を変化させたときには、0.11〜3.6μMのNusA-ノッチを68μg/mlの可溶化γ-セクレターゼとともにインキュベートし、プロットしたデータをミカエリス-メンテン曲線にフィットさせた。酵素の濃度を変化させたときには、2.1〜68μg/mlの可溶化γ-セクレターゼを0.9μMのノッチ基質とともにインキュベートした。データは3回の実験の平均値を表わしており、GraFit4.0を用いてプロットした。バックグラウンドと比べて約5倍の信号が存在していた(酵素だけを用いたバックグラウンドは無視できる)。
【図9】γ-セクレターゼによるCT-100の開裂を試験管内で抑制することが知られている化合物によるノッチ開裂の抑制。ノッチ開裂ELISAを利用して測定したDAPT(PHA-568638)とフェンキルアミン(PHA-512088)の抑制プロファイルを示してある。相対的IC50の値も示してある。
【特許請求の範囲】
【請求項1】
NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質。
【請求項2】
前記組み換えノッチ・タンパク質がノッチのS3開裂部位を含む、請求項1に記載の可溶性融合タンパク質。
【請求項3】
前記組み換えノッチ・タンパク質が脊椎動物のノッチ・タンパク質である、請求項1に記載の可溶性融合タンパク質。
【請求項4】
前記組み換えノッチ・タンパク質が無脊椎動物のノッチ・タンパク質である、請求項1に記載の可溶性融合タンパク質。
【請求項5】
前記組み換えノッチ・タンパク質が、配列番号5の配列を持つマウス・ノッチ・タンパク質に由来する、請求項1に記載の可溶性融合タンパク質。
【請求項6】
前記組み換えノッチ・タンパク質が、マウス・ノッチ・タンパク質の第1703アミノ酸〜第1860アミノ酸を含む、請求項1に記載の可溶性融合タンパク質。
【請求項7】
C末端にヒスチジン・タグをさらに含む、請求項1〜6のいずれか1項に記載の可溶性融合タンパク質。
【請求項8】
C末端にフラグ・タグをさらに含む、請求項1〜6のいずれか1項に記載の可溶性融合タンパク質。
【請求項9】
請求項1〜8のいずれか1項に記載の融合タンパク質をコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項10】
配列番号1に示した配列を含む、請求項1に記載の融合タンパク質をコードするポリヌクレオチド配列。
【請求項11】
γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)のインビトロでのアッセイ方法であって、以下のステップ:
a.哺乳動物のγ-セクレターゼ複合体または生物学的に活性なその断片を含む第1の組成物を、請求項1〜8のいずれか1項に記載の融合タンパク質を含む第2の組成物と接触させ;そして
b.その融合タンパク質の開裂を計測する、
を含む前記方法。
【請求項12】
γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)のモジュレータのインビトロでのスクリーニング方法であって、以下のステップ:
(a)推定上のモジュレータ化合物の存在下および不存在下で、哺乳動物のγ-セクレターゼ複合体または生物学的に活性なその断片を含む第1の組成物を、請求項1〜8のいずれか1項に記載の融合タンパク質を含む第2の組成物と接触させ;
(b)推定上のモジュレータ化合物の存在下と不存在下で、上記融合タンパク質の開裂を計測し;
(c)γ-セクレターゼによる上記融合タンパク質の開裂を変化させるモジュレータを同定する、
を含み、推定上のモジュレータ化合物がステップ(b)においてγ-セクレターゼによる開裂に差異を生じさせる前記方法。
【請求項13】
請求項1〜8のいずれか1項に記載の融合タンパク質を含むγ-セクレターゼの基質を含む、γ-セクレターゼ・アッセイ用キット。
【請求項14】
配列番号17のNusA配列と90〜95%一致した配列を含むノッチ・ポリペプチドに融合したNusAポリペプチドを含む融合タンパク質であって、上記ノッチ・ポリペプチドがノッチの膜貫通ドメインを含み、かつ、上記融合タンパク質が水性溶液に溶解する前記融合タンパク質。
【請求項15】
γ-セクレターゼによるアミロイド前駆体タンパク質(APP)のプロセシングの選択的阻害剤のスクリーニング方法であって、以下のステップ:
a)APPγ-セクレターゼ部位を含むポリペプチドのγ-セクレターゼによる開裂を阻害する試験化合物を準備し;そして
b)上記試験化合物の存在下および不在下で、γ-セクレターゼによる請求項1に記載の融合タンパク質の開裂を計測する、
を含み、γ-セクレターゼによる上記融合タンパク質の開裂と比較して、γ-セクレターゼによる上記ポリペプチドの開裂を選択的に阻害する試験化合物が、γ-セクレターゼによるAPPのプロセシングの選択的阻害剤である前記方法。
【請求項1】
NusAタンパク質配列のC末端に融合した組み換えノッチ・タンパク質を含む可溶性融合タンパク質。
【請求項2】
前記組み換えノッチ・タンパク質がノッチのS3開裂部位を含む、請求項1に記載の可溶性融合タンパク質。
【請求項3】
前記組み換えノッチ・タンパク質が脊椎動物のノッチ・タンパク質である、請求項1に記載の可溶性融合タンパク質。
【請求項4】
前記組み換えノッチ・タンパク質が無脊椎動物のノッチ・タンパク質である、請求項1に記載の可溶性融合タンパク質。
【請求項5】
前記組み換えノッチ・タンパク質が、配列番号5の配列を持つマウス・ノッチ・タンパク質に由来する、請求項1に記載の可溶性融合タンパク質。
【請求項6】
前記組み換えノッチ・タンパク質が、マウス・ノッチ・タンパク質の第1703アミノ酸〜第1860アミノ酸を含む、請求項1に記載の可溶性融合タンパク質。
【請求項7】
C末端にヒスチジン・タグをさらに含む、請求項1〜6のいずれか1項に記載の可溶性融合タンパク質。
【請求項8】
C末端にフラグ・タグをさらに含む、請求項1〜6のいずれか1項に記載の可溶性融合タンパク質。
【請求項9】
請求項1〜8のいずれか1項に記載の融合タンパク質をコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項10】
配列番号1に示した配列を含む、請求項1に記載の融合タンパク質をコードするポリヌクレオチド配列。
【請求項11】
γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)のインビトロでのアッセイ方法であって、以下のステップ:
a.哺乳動物のγ-セクレターゼ複合体または生物学的に活性なその断片を含む第1の組成物を、請求項1〜8のいずれか1項に記載の融合タンパク質を含む第2の組成物と接触させ;そして
b.その融合タンパク質の開裂を計測する、
を含む前記方法。
【請求項12】
γ-セクレターゼによるノッチ・タンパク質のε開裂(1743/1744)のモジュレータのインビトロでのスクリーニング方法であって、以下のステップ:
(a)推定上のモジュレータ化合物の存在下および不存在下で、哺乳動物のγ-セクレターゼ複合体または生物学的に活性なその断片を含む第1の組成物を、請求項1〜8のいずれか1項に記載の融合タンパク質を含む第2の組成物と接触させ;
(b)推定上のモジュレータ化合物の存在下と不存在下で、上記融合タンパク質の開裂を計測し;
(c)γ-セクレターゼによる上記融合タンパク質の開裂を変化させるモジュレータを同定する、
を含み、推定上のモジュレータ化合物がステップ(b)においてγ-セクレターゼによる開裂に差異を生じさせる前記方法。
【請求項13】
請求項1〜8のいずれか1項に記載の融合タンパク質を含むγ-セクレターゼの基質を含む、γ-セクレターゼ・アッセイ用キット。
【請求項14】
配列番号17のNusA配列と90〜95%一致した配列を含むノッチ・ポリペプチドに融合したNusAポリペプチドを含む融合タンパク質であって、上記ノッチ・ポリペプチドがノッチの膜貫通ドメインを含み、かつ、上記融合タンパク質が水性溶液に溶解する前記融合タンパク質。
【請求項15】
γ-セクレターゼによるアミロイド前駆体タンパク質(APP)のプロセシングの選択的阻害剤のスクリーニング方法であって、以下のステップ:
a)APPγ-セクレターゼ部位を含むポリペプチドのγ-セクレターゼによる開裂を阻害する試験化合物を準備し;そして
b)上記試験化合物の存在下および不在下で、γ-セクレターゼによる請求項1に記載の融合タンパク質の開裂を計測する、
を含み、γ-セクレターゼによる上記融合タンパク質の開裂と比較して、γ-セクレターゼによる上記ポリペプチドの開裂を選択的に阻害する試験化合物が、γ-セクレターゼによるAPPのプロセシングの選択的阻害剤である前記方法。
【図1】

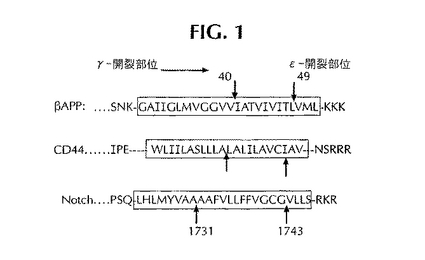
【図2】

【図3A】
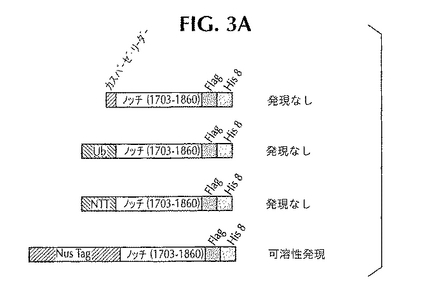
【図3B】
【図4】
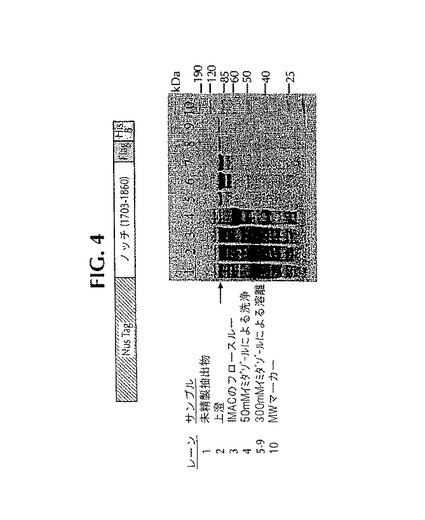
【図5】

【図6】
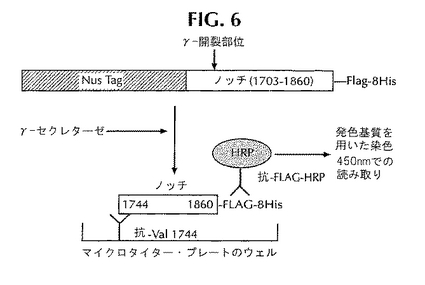
【図7】
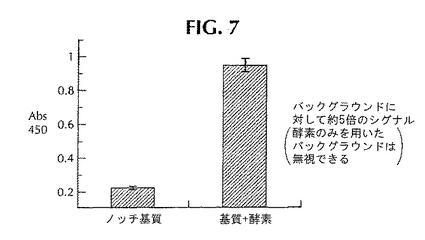
【図8】
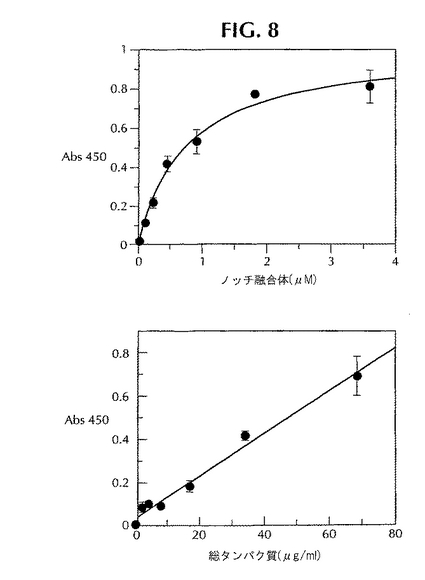
【図9】
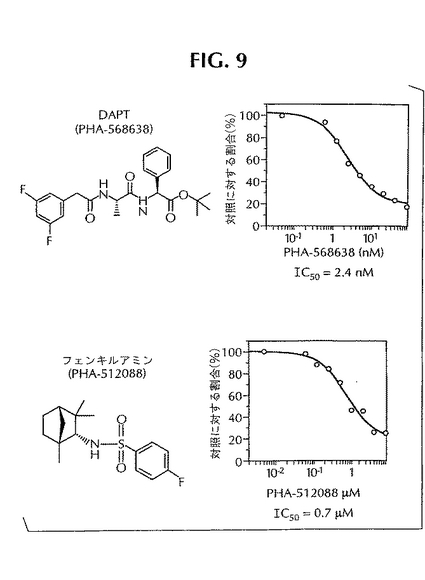
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2006−508653(P2006−508653A)
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願番号】特願2004−554808(P2004−554808)
【出願日】平成15年11月17日(2003.11.17)
【国際出願番号】PCT/IB2003/005233
【国際公開番号】WO2004/048578
【国際公開日】平成16年6月10日(2004.6.10)
【出願人】(504396379)ファルマシア・アンド・アップジョン・カンパニー・エルエルシー (130)
【Fターム(参考)】
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願日】平成15年11月17日(2003.11.17)
【国際出願番号】PCT/IB2003/005233
【国際公開番号】WO2004/048578
【国際公開日】平成16年6月10日(2004.6.10)
【出願人】(504396379)ファルマシア・アンド・アップジョン・カンパニー・エルエルシー (130)
【Fターム(参考)】
[ Back to top ]