
ノッチスペアリングガンマセクレターゼ阻害剤としてのシクロブチルスルホン
本発明はノッチシグナル伝達経路を保持しながら推定γセクレターゼによるAPPのプロセシングを阻害し、従って、ノッチ阻害による胃腸障害を併発せずにアルツハイマー病の治療又は予防に有用な新規類のシクロブチルスルホン誘導体を包含する。医薬組成物と使用方法も包含する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は新規類の化合物、その塩、前記化合物等を含有する医薬組成物及び人体の治療におけるその使用に関する。特に、本発明はγセクレターゼによるAPPのプロセシングを阻害し、従って、アルツハイマー病の治療又は予防に有用な新規シクロブチルスルホン誘導体に関する。本発明の化合物はノッチシグナル伝達経路を保持する。従って、本発明の化合物はノッチ阻害による毒性を生じずにアルツハイマー病の進行を停止又は潜在的に回復させると考えられる。
【背景技術】
【0002】
アルツハイマー病(AD)は認知症の最も一般的な形態である。主に高齢者の疾患であり、罹患率は65歳以上の人口の10%に及ぶが、遺伝的素因をもつ相当数の若年患者もADに罹患している。これは神経変性疾患であり、記憶及び認知機能の進行性低下を臨床的特徴とし、患者の皮質及び関連脳領域における細胞外蛋白プラークの沈着を病理的特徴とする。これらのプラークは主にβアミロイドペプチド(Aβ)のフィブリル凝集物から構成される(Glenner G G and Wong C W(1984)Alzheimer’s disease:Initial report of the purification and characterization of a novel cerebrovascular amyloid protein.Biochemical and Biophysical research Communications.120(3);885−890)。アミロイド前駆体蛋白質(APP)のプロセシングによりAβを形成する過程におけるγセクレターゼの役割を含むセクレターゼの役割については文献に詳細に記載されている。Aβはβアミロイド切断酵素(BACE)とγセクレターゼの2種類の酵素によるAPPの蛋白分解プロセシングにより生成される(図1;Selkoe D J(2001)Alzheimer’s disease:genes,proteins,and therapy.Physiological Review.81(2):741−766)。γセクレターゼはプレセニリン(プレセニリン1又は2)、ニカストリン、APH−1及びPEN−2の4種類の蛋白質から構成される複合体である(Takasugi N,Tomita T,Hayashi I,Tsuruoka M,Niimura M,Takahashi Y,Thinakaran G,Iwatsubo T(2003)The role of presenilin cofactors in the gamma−secretase complex.Nature.422(6930):438−441;Kimberly W T,LaVoie M J,Ostaszewski B L,Ye W,Wolfe M S,Selkoe D J(2003)Gamma−secretase is a membrane protein complex comprised of presenilin,nicastrin,Aph−1,and Pen−2.Proceedings of the National Academy of Sciences.100(11):6382−6387;Edbauer D,Winkler E,Regula J T,Pesold B,Steiner H,Haass C(2003)Reconstitution of gamma−secretase activity.Nature Cell Biology.5(5):486−488.)。プレセニリン1及び2は複合体の触媒活性に不可欠であることが分かっている膜貫通アスパラギン酸残基を含む(Wolfe M S,Xia W,Ostaszewski B L,Diehl T S,Kimberly W T,Selkoe D J(1999)Two transmembrane aspartates in presenilin−1 required for presenilin endoproteolysis and gamma−secretase activity.Nature.1999 398(6727):513−517)。早発家族性AD(FAD)に関連する突然変異の大半はPS−1又はPS−2に結び付けられる(Scheuner D,Eckman C,Jensen M,Song X,Citron M,Suzuki N,Bird T D,Hardy J,Hutton M,Kukull W,Larson E,Levy−Lahad E,Viitanen M,Peskind E,Poorkaj P,Schellenberg G,Tanzi R,Wasco W,Lannfelt L,Selkoe D,Younkin S(1996)Secreted amyloid beta−protein similar to that in the senile plaques of Alzheimer’s disease is increased in vivo by the presenilin 1 and 2 and APP mutations linked to familial Alzheimer’s disease.Nature Medicine.2(8):864−870;Duff K,Eckman C,Zehr C,Yu X,Prada C M,Perez−tur J,Hutton M,Buee L,Harigaya Y,Yager D,Morgan D,Gordon M N,Holcomb L,Refolo L,Zenk B,Hardy J,Younkin S(1996)Increased amyloid−beta42(43)in brains of mice expressing mutant presenilin 1.Nature.383(6602):710−713;Lemere CA,Lopera F,Kosik K S,Lendon C L,Ossa J,Saido T C,Yamaguchi H,Ruiz A,Martinez A,Madrigal L,Hincapie L,Arango J C,Anthony D C,Koo E H,Goate A M,Selkoe D J,Arango J C(1996)The E280A presenilin 1 Alzheimer mutation produces increased A beta 42 deposition and severe cerebellar pathology.Nature Medicine.2(10):1146−1150;Citron M,Westaway D,Xia W,Carlson G,Diehl T,Levesque G,Johnson−Wood K,Lee M,Seubert P,Davis A,Kholodenko D,Motter R,Sherrington R,Perry B,Yao H,Strome R,Lieberburg I,Rommens J,Kim S,Schenk D,Fraser P,St George Hyslop P,Selkoe D J(1997)Mutant presenilins of Alzheimer’s disease increase production of 42−residue amyloid beta−protein in both transfected cells and transgenic mice.Nature Medicine.3(1):67−72)。γセクレターゼは前提条件としてエクトドメインシェディングを受けた多数の他のI型膜蛋白質をプロセシングする(Lleo A(2008)Activity of gamma−secretase on substrates other than APP.Current Topics in Medicinal Chemistry.8(1):9−16)。
【0003】
APPのプロセシングに加え、γセクレターゼはノッチファミリー受容体を切断する。ノッチシグナル伝達及び機能にγセクレターゼ活性が不可欠であることは遺伝的に立証されている(Shen J,Bronson R T,Chen D F,Xia W,Selkoe D J,Tonegawa S(1997)Skeletal and CNS defects in Presenilin−1−deficient mice.Cell.89(4):629−639;Wong P C,Zheng H,Chen H,Becher M W,Sirinathsinghji D J,Trumbauer M E,Chen H Y,Price D L,Van der Ploeg L H,Sisodia S S(1997)Presenilin 1 is required for Notch1 and DII1 expression in the paraxial mesoderm.Nature.387(6630):288−292)。ノッチは進化的に保存されており、発生中の胚で細胞運命決定の調節に重要な役割を果たす1回貫通型I型膜貫通受容体で広く発現される(Artavanis−Tsakonas S,Rand M D,Lake R J(1999)Notch signaling:cell fate control and signal integration in development.Science.284(5415):770−776)。成人におけるノッチの役割はまだ不明であるが、ノッチ蛋白質は各種成人組織で発現され、幹細胞分化に役割を果たすと考えられる。4種類のノッチ遺伝子が哺乳動物で同定されており(ノッチ1〜4)、全4種類のノッチ蛋白質はγセクレターゼにより切断される(Saxena M T,Schroeter E H,Mumm J S,Kopan R(2001)Murine notch homologs(N1−4)undergo presenilin−dependent proteolysis.Journal of Biological Chemistry.276(43):40268−40273)。ノッチ活性化はDelta/Serrate/LAG2ファミリーの膜貫通リガンドとのトランス結合により誘導される。ノッチシグナル伝達は(a)細胞外ドメインのサイト1の切断;(b)リガンド結合後の細胞外/膜貫通ドメイン境界のちょうどN末端のサイト2の切断;及び(c)膜貫通/細胞質ドメイン境界付近の膜貫通ドメイン内のサイト3(S3)の切断の3つの切断イベントに介在される。サイト3切断はノッチ細胞内ドメイン(NICD)の遊離に必要であり、γセクレターゼに介在される(Schroeter E H,Kisslinger J A,Kopan R(1998)Notch−1 signalling requires ligand−induced proteolytic release of intracellular domain.Nature.393(6683):382−386)。NICDは(CSL)CBF1/Serrate/LAG−1ファミリーのDNA結合蛋白質に介在される転写を活性化し、各種遺伝子の発現を誘導する。NICDに調節される転写はノッチに介在されるシグナル伝達の主要な要素であると考えられる。
【0004】
APP切断とAβ生成を阻止するためのγセクレターゼ阻害剤の開発はノッチプロセシングの阻害によるメカニズム関連毒性の可能性により阻まれている。動物にγセクレターゼ阻害剤を亜慢性投与した試験ではノッチ関連毒性が観測されている。3日間以上投与後に常に腸杯細胞化生が認められる(Searfoss G H,Jordan W H,Calligaro D O,Galbreath E J,Schirtzinger L M,Berridge B R,Gao H,Higgins M A,May P C,Ryan T P(2003)Adipsin,a biomarker of gastrointestinal toxicity mediated by a functional gamma−secretase inhibitor.Journal of Biological Chemistry.278(46):46107−46116;Wong G T,Manfra D,Poulet F M,Zhang Q,Josien H,Bara T,Engstrom L,Pinzon−Ortiz M,Fine J S,Lee H J,Zhang L,Higgins G A,Parker E M(2004)Chronic treatment with the gamma−secretase inhibitor LY−411,575 inhibits beta−amyloid peptide production and alters lymphopoiesis and intestinal cell differentiation.Journal of Biological Chemistry.279(13):12876−12882;Milano J,McKay J,Dagenais C,Foster−Brown L,Pognan F,Gadient R,Jacobs R T,Zacco A,Greenberg B,Ciaccio P J(2004)Modulation of notch processing by gamma−secretase inhibitors causes intestinal goblet cell metaplasia and induction of genes known to specify gut secretory lineage differentiation.Toxicological Sciences.82(1):341−358;van Es J H,van Gijn M E,Riccio O,van den Born M,Vooijs M,Begthel H,Cozijnsen M,Robine S,Winton D J,Radtke F,Clevers H(2005)Notch/gamma−secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells.Nature.435(7044):959−963)。更に、ノッチ機能はT及びBリンパ球の適正な分化に不可欠であると思われる(Hadland B K,Manley N R,Su D,Longmore G D,Moore C L,Wolfe M S,Schroeter E H,Kopan R(2001)Gamma−secretase inhibitors repress thymocyte development.Proceedings of the National Academy of Sciences.98(13):7487−7491;Doerfler P,Shearman M S,Perlmutter R M(2001)Presenilin−dependent gamma−secretase activity modulates thymocyte development.Proceedings of the National Academy of Sciences.98(16):9312−9317)。従って、γセクレターゼ活性を薬理標的とするには、ノッチに対する活性の阻害を最小限にしながらAβを選択的に阻止する物質が必要である。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Glenner G G and Wong C W(1984)Alzheimer’s disease:Initial report of the purification and characterization of a novel cerebrovascular amyloid protein.Biochemical and Biophysical research Communications.120(3);885−890
【非特許文献2】Selkoe D J(2001)Alzheimer’s disease:genes,proteins,and therapy.Physiological Review.81(2):741−766
【非特許文献3】Takasugi N,Tomita T,Hayashi I,Tsuruoka M,Niimura M,Takahashi Y,Thinakaran G,Iwatsubo T(2003)The role of presenilin cofactors in the gamma−secretase complex.Nature.422(6930):438−441
【非特許文献4】Kimberly W T,LaVoie M J,Ostaszewski B L,Ye W,Wolfe M S,Selkoe D J(2003)Gamma−secretase is a membrane protein complex comprised of presenilin,nicastrin,Aph−1,and Pen−2.Proceedings of the National Academy of Sciences.100(11):6382−6387
【非特許文献5】Edbauer D,Winkler E,Regula J T,Pesold B,Steiner H,Haass C(2003)Reconstitution of gamma−secretase activity.Nature Cell Biology.5(5):486−488
【非特許文献6】Wolfe M S,Xia W,Ostaszewski B L,Diehl T S,Kimberly W T,Selkoe D J(1999)Two transmembrane aspartates in presenilin−1 required for presenilin endoproteolysis and gamma−secretase activity.Nature.1999 398(6727):513−517
【非特許文献7】Scheuner D,Eckman C,Jensen M,Song X,Citron M,Suzuki N,Bird T D,Hardy J,Hutton M,Kukull W,Larson E,Levy−Lahad E,Viitanen M,Peskind E,Poorkaj P,Schellenberg G,Tanzi R,Wasco W,Lannfelt L,Selkoe D,Younkin S(1996)Secreted amyloid beta−protein similar to that in the senile plaques of Alzheimer’s disease is increased in vivo by the presenilin 1 and 2 and APP mutations linked to familial Alzheimer’s disease.Nature Medicine.2(8):864−870
【非特許文献8】Duff K,Eckman C,Zehr C,Yu X,Prada C M,Perez−tur J,Hutton M,Buee L,Harigaya Y,Yager D,Morgan D,Gordon M N,Holcomb L,Refolo L,Zenk B,Hardy J,Younkin S(1996)Increased amyloid−beta42(43)in brains of mice expressing mutant presenilin 1.Nature.383(6602):710−713
【非特許文献9】Lemere CA,Lopera F,Kosik K S,Lendon C L,Ossa J,Saido T C,Yamaguchi H,Ruiz A,Martinez A,Madrigal L,Hincapie L,Arango J C,Anthony D C,Koo E H,Goate A M,Selkoe D J,Arango J C(1996)The E280A presenilin 1 Alzheimer mutation produces increased A beta 42 deposition and severe cerebellar pathology.Nature Medicine.2(10):1146−1150
【非特許文献10】Citron M,Westaway D,Xia W,Carlson G,Diehl T,Levesque G,Johnson−Wood K,Lee M,Seubert P,Davis A,Kholodenko D,Motter R,Sherrington R,Perry B,Yao H,Strome R,Lieberburg I,Rommens J,Kim S,Schenk D,Fraser P,St George Hyslop P,Selkoe D J(1997)Mutant presenilins of Alzheimer’s disease increase production of 42−residue amyloid beta−protein in both transfected cells and transgenic mice.Nature Medicine.3(1):67−72
【非特許文献11】Lleo A(2008)Activity of gamma−secretase on substrates other than APP.Current Topics in Medicinal Chemistry.8(1):9−16
【非特許文献12】Shen J,Bronson R T,Chen D F,Xia W,Selkoe D J,Tonegawa S(1997)Skeletal and CNS defects in Presenilin−1−deficient mice.Cell.89(4):629−639
【非特許文献13】Wong P C,Zheng H,Chen H,Becher M W,Sirinathsinghji D J,Trumbauer M E,Chen H Y,Price D L,Van der Ploeg L H,Sisodia S S(1997)Presenilin 1 is required for Notch1 and DII1 expression in the paraxial mesoderm.Nature.387(6630):288−292
【非特許文献14】Artavanis−Tsakonas S,Rand M D,Lake R J(1999)Notch signaling:cell fate control and signal integration in development.Science.284(5415):770−776
【非特許文献15】Saxena M T,Schroeter E H,Mumm J S,Kopan R(2001)Murine notch homologs(N1−4)undergo presenilin−dependent proteolysis.Journal of Biological Chemistry.276(43):40268−40273
【非特許文献16】Schroeter E H,Kisslinger J A,Kopan R(1998)Notch−1 signalling requires ligand−induced proteolytic release of intracellular domain.Nature.393(6683):382−386
【非特許文献17】Searfoss G H,Jordan W H,Calligaro D O,Galbreath E J,Schirtzinger L M,Berridge B R,Gao H,Higgins M A,May P C,Ryan T P(2003)Adipsin,a biomarker of gastrointestinal toxicity mediated by a functional gamma−secretase inhibitor.Journal of Biological Chemistry.278(46):46107−46116
【非特許文献18】Wong G T,Manfra D,Poulet F M,Zhang Q,Josien H,Bara T,Engstrom L,Pinzon−Ortiz M,Fine J S,Lee H J,Zhang L,Higgins G A,Parker E M(2004)Chronic treatment with the gamma−secretase inhibitor LY−411,575 inhibits beta−amyloid peptide production and alters lymphopoiesis and intestinal cell differentiation.Journal of Biological Chemistry.279(13):12876−12882
【非特許文献19】Milano J,McKay J,Dagenais C,Foster−Brown L,Pognan F,Gadient R,Jacobs R T,Zacco A,Greenberg B,Ciaccio P J(2004)Modulation of notch processing by gamma−secretase inhibitors causes intestinal goblet cell metaplasia and induction of genes known to specify gut secretory lineage differentiation.Toxicological Sciences.82(1):341−358
【非特許文献20】van Es J H,van Gijn M E,Riccio O,van den Born M,Vooijs M,Begthel H,Cozijnsen M,Robine S,Winton D J,Radtke F,Clevers H(2005)Notch/gamma−secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells.Nature.435(7044):959−963
【非特許文献21】Hadland B K,Manley N R,Su D,Longmore G D,Moore C L,Wolfe M S,Schroeter E H,Kopan R(2001)Gamma−secretase inhibitors repress thymocyte development.Proceedings of the National Academy of Sciences.98(13):7487−7491
【非特許文献22】Doerfler P,Shearman M S,Perlmutter R M(2001)Presenilin−dependent gamma−secretase activity modulates thymocyte development.Proceedings of the National Academy of Sciences.98(16):9312−9317
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明はノッチシグナル伝達経路を保持しながら推定γセクレターゼによるAPPのプロセシングを阻害し、従って、ADの治療又は予防に有用である新規類のシクロブチルスルホン誘導体を提供する。
【課題を解決するための手段】
【0007】
本発明はノッチシグナル伝達経路を保持しながら推定γセクレターゼによるAPPのプロセシングを阻害し、従って、ノッチ阻害による胃腸障害を併発せずにアルツハイマー病の治療又は予防に有用な新規類のシクロブチルスルホン誘導体を包含する。医薬組成物と使用方法も包含する。
【図面の簡単な説明】
【0008】
【図1】γセクレターゼはほぼ膜の中間(γ/S4切断)と膜のサイトゾル側(ε/S3切断)の主に2カ所でAPPとノッチを切断する。ε/S3切断は膜から基質の細胞内ドメイン(ICD)である夫々AICD及びNICDを遊離するので、重要なプロセシングイベントである。この段階はICD核転座とその後の転写モジュレーターとしての機能の前提条件である。他方、γ/S4切断は夫々APPとノッチからAβペプチドとAβ様ペプチド(Nβ)の遊離をもたらす。後者の切断はADの疾患原因物質であると考えられるAβペプチドのC末端を生じるので、特に注目されている。
【図2】他のγセクレターゼ基質の初期切断に与える影響を試験するために本願に記載するトランス活性化アッセイで試験したMRK−560と実施例2の結果を示す。UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼと共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした後、各化合物1μMで48時間処理した。MRK−560は試験した全基質のICD遊離を阻害したが、実施例2はICD遊離とその後のICD−GVP構築物の転座を維持し、レポーター活性化が可能であった。
【図3】各基質、ノッチ△E、E−カドヘリン、SCN2b及びCD43を安定的に過剰発現するHEK293細胞をTPAと共に滴定濃度のMRK−560又は実施例2で処理し、シェディングを誘導した。細胞溶解液の免疫ブロット分析の結果、MRK−560は従来のγセクレターゼ阻害剤であるDAPTと同様に、NICD生成を阻害し、γセクレターゼの直接基質であるCTFの蓄積を生じることが判明した。これに対して、実施例2はNICD生成が可能であり、CTF蓄積は全く認められないか又はMRK−560よりも少なかった。
【図4A】本願に記載する化合物結合アッセイで試験したMRK−560と実施例2の結果を示す。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。A−MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。B−実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。
【図4B】本願に記載する化合物結合アッセイで試験したMRK−560と実施例2の結果を示す。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。A−MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。B−実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。
【発明を実施するための形態】
【0009】
本発明は式I:
【0010】
【化1】
の属の化合物又はその医薬的に許容可能な塩を包含し、
式中、X1はF及びCNから構成される群から選択され;
X2はF、Cl及びCNから構成される群から選択され;
X3はF、Br、Cl、CN、CF3、OCF3、C(O)−OCH3及びS−CH3から構成される群から選択され;
X4はH、F及びClから構成される群から選択され;
R1は
(a)H、
(b)CH3、
(c)−(CH2)n−OR3、
(d)−(CH2)n−C(O)−OR4、及び
(e)−SO2−CF3
から構成される群から選択され;
式Iの化合物がシス配置であるとき、R2はH又はCH3であり、それ以外の場合にはR2はHであり;
R3は1個の酸素ヘテロ原子をもつ5又は6員非芳香族複素環であり;
R4はH又はCH3であり;
nは1〜4である。
【0011】
この属の範囲内で本発明は式Ia:
【0012】
【化2】
の第1の亜属の化合物又はその医薬的に許容可能な塩を包含する。
【0013】
第1の亜属の範囲内で本発明は、
X1がFであり、
X4がHである
式Iaの第1の分類の化合物を包含する。
【0014】
第1の分類の範囲内で本発明は、
X2がFであり、
X3がClである
式Iaの亜分類の化合物を包含する。
【0015】
この亜分類の範囲内で本発明は、R2がHである式Iaの群の化合物を包含する。
【0016】
更にこの群の範囲内で本発明は、R1が−(CH2)n−C(O)−OR4である式Iaの第2の亜群の化合物を包含する。
【0017】
この属の範囲内で本発明は式Ib:
【0018】
【化3】
の第2の亜属の化合物又はその医薬的に許容可能な塩を包含する。
【0019】
第2の亜属の範囲内で本発明は、
X1及びX2がFであり;
X3がClであり;
X4がHである
式Ibの第1の分類の化合物を包含する。
【0020】
本発明は更に下記実施例のいずれかを包含する。
【0021】
本発明は更に式Iの化合物又はその医薬的に許容可能な塩と、医薬的に許容可能なキャリヤーを含有する医薬組成物を包含する。
【0022】
本発明は更にβアミロイドの沈着を伴う病態に罹患している対象又は前記病態に罹患し易い対象の治療方法として、有効量の式Iの化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む方法を包含する。
【0023】
本発明は更にアルツハイマー病の治療又は予防用医薬の製造における式Iの化合物又はその医薬的に許容可能な塩の使用を包含する。
【0024】
式I又はその置換基に変項が2回以上出現する場合には、特に指定しない限り、前記変項の個々の出現は相互に独立している。
【0025】
医薬用として、式Iの化合物は医薬的に許容可能な塩の形態でもよい。しかし、他の塩が式Iの化合物又はその医薬的に許容可能な塩の製造に有用な場合もある。本発明の化合物の適切な医薬的に許容可能な塩としては、例えば本発明の化合物の溶液を塩酸、硫酸、メタンスルホン酸、ベンゼンスルホン酸、フマル酸、マレイン酸、琥珀酸、酢酸、安息香酸、蓚酸、クエン酸、酒石酸、炭酸又はリン酸等の医薬的に許容可能な酸の溶液と混合することにより形成することができる酸付加塩が挙げられる。あるいは、本発明の化合物が酸性部分をもつ場合には、前記酸性部分を適切な塩基で中和することにより医薬的に許容可能な塩を形成することができる。こうして形成される医薬的に許容可能な塩の例としては、ナトリウム塩やカリウム塩等のアルカリ金属塩;カルシウム塩やマグネシウム塩等のアルカリ土類金属塩;及びアミン塩(ピリジニウム塩を含む)や第四級アンモニウム塩等の適切な有機塩基と形成される塩が挙げられる。
【0026】
なお、本発明は式Iの各化合物について、ホモキラル化合物又は任意割合のエナンチオマー混合物としての両者のエナンチオマー形態を包含する。本発明の1態様において、式Iの化合物は式Iaもしくは式Ibのホモキラル化合物、又はその医薬的に許容可能な塩である。本発明の化合物が2個以上の不斉中心をもつ場合には、更にジアステレオ異性体として存在することもできる。当然のことながら、このような全異性体と任意割合のその混合物が本発明の範囲に含まれる。
【0027】
同様に当然のことながら、1つの反応から2種類以上の異性体が得られる場合には、得られた異性体の混合物を従来手段により分離することができる。本発明の化合物の製造方法により立体異性体の混合物が得られる場合には、分取クロマトグラフィー等の従来技術によりこれらの異性体を分離することができる。新規化合物をラセミ体として製造することもできるし、エナンチオ特異的合成又は分割により個々のエナンチオマーを製造することもできる。新規化合物を例えば分取HPLC等の標準技術によりその成分エナンチオマーに分割してもよいし、(−)−ジ−p−トルオイル−d−酒石酸及び/又は(+)−ジ−p−トルオイル−1−酒石酸等の光学活性酸との塩形成によるジアステレオマー対の形成後に分別結晶と遊離塩基の再生を実施してもよい。更に、ジアステレオマーエステル又はアミドの形成後にクロマトグラフィー分離とキラル助剤の除去により新規化合物を分割してもよい。あるいは、該当化合物のラセミ合成前駆体にこのような技術を実施してもよい。
【0028】
本発明の化合物はγセクレターゼの阻害剤としての活性をもつ。
【0029】
本発明は更に本発明の1種以上の化合物と医薬的に許容可能なキャリヤーを含有する医薬組成物も提供する。これらの組成物は経口、非経口、鼻孔内、舌下もしくは直腸投与用、又は吸入もしくは吹送投与用として、錠剤、ピル、カプセル剤、散剤、顆粒剤、滅菌非経口溶液剤もしくは懸濁液剤、定量式エアゾールもしくは液体スプレー、滴剤、アンプル剤、経皮パッチ、オートインジェクター装置又は坐剤等の単位剤形が好ましい。一般に主活性成分を医薬キャリヤー(例えばコーンスターチ、ラクトース、スクロース、ソルビトール、タルク、ステアリン酸、ステアリン酸マグネシウム及びリン酸二カルシウム等の従来の錠剤化成分、ガム類、分散剤、懸濁剤、又はモノオレイン酸ソルビタンやポリエチレングリコール等の界面活性剤)及び他の医薬希釈剤(例えば水)と混合し、本発明の化合物又はその医薬的に許容可能な塩を含有する均質な前製剤組成物を形成する。これらの前製剤組成物について均質と言う場合には、組成物を錠剤、ピル及びカプセル剤等の均等に有効な単位剤形に容易に細分できるように活性成分が組成物全体に均等に分散されていることを意味する。その後、この前製剤組成物を細分し、本発明の活性成分0.1〜約500mgを含有する上記型の単位剤形とする。典型的な単位剤形は活性成分1〜100mg、例えば1、2、5、10、25、50又は100mgを含有する。長期作用の利点を付与する剤形を提供するように、新規組成物の錠剤又はピルにコーティング又は他の方法で加工してもよい。例えば、錠剤又はピルを内側製剤成分と外側製剤成分から構成し、外側成分が内側成分のエンベロープを構成するようにしてもよい。胃で崩壊しないように機能すると共に内側成分を無傷のまま十二指腸に導入又は放出遅延させる腸溶層により2成分を分離することができる。このような腸溶層又はコーティングには各種材料を使用することができ、このような材料としては、多数のポリマー酸や、ポリマー酸とシェラック、セチルアルコール及び酢酸セルロース等の材料の混合物が挙げられる。
【0030】
本発明の新規組成物を経口又は注射投与用に配合することができる液体形態としては、水溶液、液体又はゲル充填カプセル剤、適切なフレーバー入りシロップ、水性又は油性懸濁液、及び綿実油、胡麻油又は椰子油等の食用油を添加したフレーバー入りエマルション、並びにエリキシル剤及び同様の医薬ビヒクルが挙げられる。水性懸濁液に適した分散剤又は懸濁剤としては、トラガカントガム、アラビアガム、アルギン酸塩、デキストラン、カルボキシメチルセルロースナトリウム、メチルセルロース、ポリ(エチレングリコール)、ポリ(ビニルピロリドン)又はゼラチン等の合成及び天然ガムが挙げられる。
【0031】
本発明は更に人体の治療方法で使用するための本発明の化合物又はその医薬的に許容可能な塩を提供する。βアミロイドの沈着を伴う病態の治療が好ましい。病態としてはアルツハイマー病等のβアミロイドの沈着を伴う神経疾患が好ましい。
【0032】
本発明は更にアルツハイマー病の治療又は予防用医薬の製造における本発明の化合物又はその医薬的に許容可能な塩の使用も提供する。
【0033】
アルツハイマー病に罹患している対象又は罹患し易い対象の治療方法として、有効量の本発明の化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む方法も開示する。
【0034】
アルツハイマー病を治療又は予防するために、適切な用量レベルは約0.01〜250mg/kg/日、好ましくは約0.01〜100mg/kg/日、より好ましくは約0.05〜50mg/kg体重/日であり、最も好ましい化合物では、約0.1〜10mg/kg体重/日である。1日1〜4回のレジメンで化合物を投与することができる。しかし、場合によってはこれらの範囲外の用量を使用してもよい。
【0035】
以下、実施例により本発明を例証する。合成スキームで使用する出発材料及び試薬が市販されていない場合には、従来手段により製造することができる。本発明は以下の実施例から選択される化合物も包含する。
【0036】
合成シーケンスのいずれかの間に、該当分子のいずれかの感受性又は反応性基を保護することが必要及び/又は望ましい場合がある。これはProtective Groups in Organic Chemistry,ed.J.F.W.McOmie,Plenum Press,1973;及びT.W.Greene & P.G.M.Wuts,Protective Groups in Organic Synthesis,John Wiley & Sons,1991に記載されているもの等の従来の保護基により実施することができる。保護基は適切な後続段階で当分野で公知の方法を使用して除去することができる。
【0037】
(実施例)
中間体A
【0038】
【化4】
2−[2−[(4−クロロフェニル)スルホニル]−2−(2,5−ジフルオロフェニル)エチル]オキシラン
WO02/081435(中間体1)に記載されているように4−クロロチオフェノールと臭化2,5−ジフルオロベンジルから2段階で4−クロロフェニル−2,5−ジフルオロベンジルスルホンを製造した。
【0039】
THF(99ml)中の4−クロロフェニル−2,5−ジフルオロベンジルスルホン(12g,39.6mmol)をnBuLi(19ml,ヘキサン中2.5M,47.6mmol)で0℃にて10分間処理後、エピクロロヒドリン(3.73ml,47.6mmol)を加えた。反応混合物を室温まで14時間かけてゆっくりと昇温し、水(100ml)でクエンチし、EtOAc(300ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中10−45%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物9.8gをオフホワイト固体として得た。1H NMR(600MHZ,CDCl3)2種類のジアステレオマー(〜1/1)δ7.52(m,2H/2H),7.38(m,2H/2H),7.30−7.22(m,1H/1H),6.99(m,1H/1H),6.86−6.80(m,1H/1H),4.76−4.70(m,1H/1H),3.03−2.48(m,4H/4H),2.37(m,1H),2.20(m,1H)。MS計算値359.0(MH+),実験値358.9(MH+)。
【0040】
中間体B
【0041】
【化5】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール
THF(2.7ml)中の2−[2−[(4−クロロフェニル)スルホニル]−2−(2,5−ジフルオロフェニル)エチル]オキシラン(100mg,0.279mmol)にMeMgBr(279μL,エーテル中3M,0.836mmol)を−78℃で加えた。反応混合物を室温まで1時間かけて昇温後、飽和NH4Cl(3ml)でクエンチし、EtOAc(10ml)で希釈した。有機相をブライン(10ml)で洗浄し、乾燥し(Na2SO4)、蒸発乾涸し、目的生成物を得た(100mg)、1H NMR(600MHZ,CDCl3)δ7.66(d,J=8.4Hz,2H),7.57(d,J=8.4Hz,2H),6.99(m,1H),6.82(m,1H),6.74(m,1H),4.28(m,1H),3.47(d,J=11.4Hz,1H,OH),3.13(m,4H)。MS計算値422.0(MNa++CH3CN),実験値421.9(MNa++CH3CN)。
【0042】
中間体C
【0043】
【化6】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール
THF(100ml)中のPPh3(8.19g,31.2mmol)にDIAD(6.31g,31.2mmol)を加え、得られた混合物を室温で0.5時間撹拌した。混合物を−50℃まで冷却し、THF(10ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(8g,22.3mmol)を加えた。反応混合物を20分間撹拌後、固体4−ニトロ安息香酸(5.22g,31.2mmol)を加えた。得られた混合物を室温まで昇温し、室温で20時間撹拌した。次に反応混合物を0℃まで冷却し、NaOMe(134ml,MeOH中0.5M,66.9mmol)を加えた。40分後に反応混合物を飽和NH4Cl(100ml)でクエンチし、EtOAc(300ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中エーテルを溶離液としてこの物質をシリカクロマトグラフィーに付し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(出発材料)〜15%を含有する白色固体として標記生成物8gを得た。1H NMR(600MHZ,CDCl3)主生成物,δ7.35(m,4H),6.96(m,1H),6.80−6.72(m,2H),4.84(m,1H),3.49(m,2H),2.59(m,2H)。MS計算値422.0(MNa++CH3CN),実験値421.9(MNa++CH3CN)。
【0044】
中間体D
【0045】
【化7】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート
DCM(27.9ml)中のトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(2g,5mmol)にトリフルオロメタンスルホン酸無水物(1.13ml,6.69mmol)とピリジン(0.902ml,11.15mmol)を0℃にて加えた。反応混合物を30分間撹拌し、飽和NH4Cl(30ml)でクエンチし、EtOAc(150ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−25%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物2.7gを白色固体として得た。1H NMR(600MHZ,CDCl3)δ7.36(m,4H),7.02(m,1H),6.79(m,2H),5.72(m,1H),3.66(broad s,2H),3.02(dd,J=14.4,6.6Hz,2H)。MS計算値554.0(MNa++CH3CN),実験値553.8(MNa++CH3CN)。
【0046】
中間体E
【0047】
【化8】
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート(3.9g,7.95mmol)とナトリウムアジド(5.17g,79mmol)をエタノール(39.7ml)と水(39.7ml)に加えた混合物を85℃に2時間加熱した。混合物を室温まで冷却し、水(100ml)とEtOAc(150ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−30%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物2.3gを白色固体として得た。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.02(m,1H),6.91(m,1H),6.80(m,1H),3.81(m,1H),3.30(m,2H),3.02(m,2H)。
【0048】
中間体F
【0049】
【化9】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン(2.07g,5.39mmol)とパラジウム(0.861g,炭素担持10%,0.809mmol)をMeOH(27ml)中でH2バルーン下に4時間撹拌した。粗製混合物をシリカゲルパックで濾過し、5:1 DCM/MeOH(100ml)で洗浄してパラジウム残渣を除去した。溶媒を除去して生成物を得、それ以上精製せずに次の変換でそのまま使用した。1H NMR(600MHZ,CDCl3)δ7.34(m,4H),6.97(m,1H),6.84(m,1H),6.77(m,1H),3.45(m,1H),3.05−2.94(m,4H)。MS計算値358.0(MH+),実験値358.0(MH+)。
【0050】
方法(b)
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン(9.5g,24.75mmol)のエタノール/THF溶液に室温で撹拌下に亜鉛(3.24g,49.5mmol)を加えた後、ギ酸アンモニウム(3.12g,49.5mmol)を加えた。反応混合物を室温で1時間撹拌した。混合物をセライトで濾過後、溶媒を除去した。得られた残渣に飽和NaHCO3溶液100mlを加え、生成物をEtOAc(2×100ml)で抽出した。有機相を合わせてブラインで洗浄し、無水硫酸ナトリウムで乾燥し、セライトで濾過した。濾液を濃縮し、目的生成物を得た。
【0051】
中間体G
【0052】
【化10】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノールを使用した以外はトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナートと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.04(m,1H),6.88(m,1H),6.81(m,1H),5.08(m,1H),3.60(m,2H),3.23(m,2H)。MS計算値554.0(MNa++CH3CN),実験値553.8(MNa++CH3CN)。
【0053】
中間体H
【0054】
【化11】
トランス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナートを使用した以外はシス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホンと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),6.98(m,1H),6.76(m,2H),4.56(m,1H),3.49(m,2H),2.66(m,2H)。
【0055】
中間体I
【0056】
【化12】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン
トランス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホンを使用した以外はシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンと同様に製造した。1H NMR(600MHZ,CD3OD)δ7.48−7.41(m,4H),7.07(m,1H),6.88(m,1H),6.79(m,1H),3.88(m,1H),3.41(m,2H),2.50(m,2H)。MS計算値358.0(MH+),実験値358.0(MH+)。
【0057】
中間体J
【0058】
【化13】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート(4g,8.15mmol)とシアン化テトラブチルアンモニウム(5.47g,20.37mmol)をDMSO(54ml)中で80℃に45分間加熱した。得られた混合物を室温まで冷却し、水(200ml)とEtOAc(250ml)で希釈した。有機相を水、ブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−50%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物(2.9g)をオフホワイト固体として得た。1H NMR(600MHZ,CDCl3)δ7.38−7.33(m,4H),7.02(m,1H),6.80−6.74(m,2H),3.74(m,1H),3.51(m,2H),3.02(m,2H)。MS計算値431.0(MNa++CH3CN),実験値431.0(MNa++CH3CN)。
【0059】
中間体K
【0060】
【化14】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリル
THF(8mL)中のトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル(600mg,1.63mmol)にLiHMDS(2.45ml,THF中1M,2.45mmol)を−78℃で加えた。10分後にMeI(306μl,4.89mmol)を反応混合物に導入した。温度をゆっくりと0℃まで上げながら反応混合物を2時間撹拌した。次に反応混合物を水でクエンチし、EtOAcで抽出した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−50%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、生成物(260mg)を単一ジアステレオマー生成物として得た。1H NMR(600MHZ,CD3OD)δ7.36(s,4H),7.02(m,1H),6.85−6.78(m,2H),3.79(d,J=14.4Hz,2H),2.72(d,J=14.4Hz,2H),1.44(s,3H)。MS計算値785.1(2M+Na+),実験値785.0(2M+Na+)。
【0061】
中間体L
【0062】
【化15】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド
DMSO(6.5ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリル(500mg,1.31mmol)とK2CO3(362mg,2.62mmol)にH2O2(1.15ml,水中35%,13.1mmol)を滴下し、反応混合物を2時間激しく撹拌した。混合物を水(50ml)とEtOAc(50ml)で希釈した。有機相を水、ブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド(500mg)をオフホワイト固体として得、それ以上精製せずに次の反応で使用した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.02(m,1H),6.87−6.79(m,2H),6.54(broad s,1H),6.40(broad s,1H),3.67(d,J=14.4Hz,2H),2.62(d,J=14.4Hz,2H),1.28(s,3H)。MS計算値400.0(MH+),実験値400.0(MH+)。
【0063】
中間体M
【0064】
【化16】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミン
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド(400mg,1mmol)とPIFA(473mg,1.1mmol)をアセトニトリル(2.5ml)と水(2.5ml)に加え、0℃で撹拌し、混合物を27時間かけて室温までゆっくりと昇温した。次に反応混合物を飽和NaHCO3でクエンチし、EtOAcで抽出した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。DCM中0−40% MeOHを溶離液としてこの物質をシリカクロマトグラフィーに付し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミン(380mg)を生成物として得た。1H NMR(600MHZ,CD3OD)δ7.41−7.36(m,4H),7.02(m,1H),6.81(m,1H),6.74(m,1H),3.58(d,J=15.6Hz,2H),2.89(d,J=15.6Hz,2H),1.58(s,3H)。MS計算値372.1(MH+),実験値372.0(MH+)。
【0065】
中間体N
【0066】
【化17】
シス−1−アリル−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリルと臭化アリルを使用した以外はシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリルと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.36(s,4H),7.03(m,1H),6.83−6.78(m,2H),5.70(m,1H),5.18(dd,J=10.2,1.2Hz,1H),5.07(dd,1H,J=16.8,1.2Hz,1H),3.70(d,J=15.0Hz,2H),2.77(d,J=15.0Hz,2H),2.32(d,J=6.6Hz,2H)。MS計算値837.1(2M+Na+),実験値837.0(2M+Na+)。
【0067】
中間体O
【0068】
【化18】
4−{[シス−3−アミノ−1−(2,5−ジフルオロフェニル)シクロブチル]スルホニル}ベンゾニトリル
シアン化亜鉛(0.295g,2.52mmol)と、Pd2(dba)3(0.384g,0.419mmol)と、亜鉛(0.030g,0.461mmol)と、DPPF(0.465g,0.838mmol)と、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン(1.5g,4.19mmol)をDMAに溶解し、25mLシュレンク管でアルゴン雰囲気下に135℃にて16時間撹拌した。次に水を加え、混合物をEtOAcで抽出した。有機層を飽和NaHCO3溶液とブラインで洗浄後、MgSO4で乾燥し、濾過し、濃縮した。残渣をシリカカラムクロマトグラフィー(0→8% MeOH/DCM)により精製し、標記化合物を得た。MS:計算値349(MH+),実験値349(MH+)。
【0069】
(実施例1)
【0070】
【化19】
N−[トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
DCM(27.9ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン(1.5g,4.19mmol)にトリエチルアミン(1.169ml,8.38mmol)とトリフルオロメタンスルホン酸無水物(0.850ml,5.03mmol)を0℃で加え、混合物を2時間撹拌した。
【0071】
混合物を水(50ml)とEtOAc(100ml)で希釈した。有機相をブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物(白色固体として1.45g)を得た。1H NMR(600MHZ,CDCl3)δ7.39(d,J=9.0Hz,2H),7.34(d,J=9.0Hz,2H),7.02(m,1H),6.77(m,2H),4.22(m,1H),3.23(m,4H)。MS計算値553.0(MNa++CH3CN),実験値553.0(MNa++CH3CN)。
【0072】
(実施例2)
【0073】
【化20】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンを使用した以外は実施例1と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.33(m,4H),6.98(m,1H),6.74(m,2H),5.35(d,J=7.8Hz,1H,NH),4.67(m,1H),3.59(m,2H),2.68(m,2H)。MS計算値553.0(MNa++CH3CN),実験値552.8(MNa++CH3CN)。
【0074】
同様の手順により以下の化合物を製造した。
【0075】
【表1】
【0076】
(実施例6)
【0077】
【化21】
N−{シス−3−(5−クロロ−2−フルオロフェニル)−3−[(4−クロロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:MS計算値506(MNa+),実験値529;1H NMR(400MHz,CDCl3)δ7.41〜7.43(d,J=8.8Hz,2H),7.34〜7.36(d,J=8.4Hz,2H),7.28〜7.33(m,1H),6.98〜7.01(m,1H),6.75〜6.83(m,2H),4.20〜4.29(m,1H),3.20〜3.32(m,4H)。
【0078】
(実施例7)
【0079】
【化22】
N−{シス−3−(2,5−ジフルオロフェニル)−3−[(4−フルオロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:1H NMR(400MHz,CDCl3):δ7.45〜7.49(m,2H),7.12〜7.29(m,2H),7.04〜7.10(m,1H),6.80〜6.85(m,3H),4.24〜4.30(m,1H),3.24〜3.35(m,4H)。
【0080】
(実施例8)
【0081】
【化23】
N−{シス−3−(2,5−ジフルオロフェニル)−3−[(3,4−ジフルオロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:MS 計算値514(MH+),実験値514;1H NMR(400MHz,CDCl3):δ7.15〜7.19(m,3H),6.97〜7.04(m,1H),6.72〜6.80(m,2H),6.63〜6.65(d,J=10.4Hz,1H),4.13〜4.23(m,1H),3.14〜3.26(m,4H)。
【0082】
(実施例9)
【0083】
【化24】
N−[シス−3−[(4−シアノフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
MS:計算値502(MNa+),実験値502(MNa+)。1H NMR(CDCl3 600MHz)7.67(d,J=4.1Hz,2H),7.58(d,J=4.1,2H),7.08(bm,1H),6.84(m,1H),6.68(m,1H),6.60(d,1H),4.24(bm,1H),3.4−3.2(bm,4H)。
【0084】
中間体P
【0085】
【化25】
4−{[3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸tert−ブチル
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(190mg,0.388mmol)をDMF(1.1mL)に加え、炭酸カリウム(59mg,0.427mmol)と4−ブロモブタン酸tert−ブチル(95mg,0.427mmol)で処理した。混合物を80℃まで加熱し、16時間撹拌した。反応混合物を周囲温度まで冷却し、酢酸エチルで希釈し、1/2飽和ブライン溶液で2回洗浄した。有機層を無水硫酸マグネシウムで乾燥し、濾過後、減圧濃縮した。残渣をMPLC(0−30% EtOAc:ヘプタン)により精製し、標記化合物を得た。MS:計算値654(M Na+),実験値654(M Na+)。
【0086】
(実施例10)
【0087】
【化26】
4−{[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸
4−{[3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸tert−ブチル(144mg,0.228mmol)を1:1 DCM:TFA(1.1mL)に加え、周囲温度で40分間撹拌した。反応混合物を減圧濃縮した。ヘプタンを加えてトリチュレーション後に標記化合物を白色固体として単離した。1H NMR(DMSO D6,600MHz)δ12.25(s,1H),7.58(d,J=8.2Hz,2H),7.43(d,J=8.2Hz,2H),7.25−7.31(m,1H),7.07−7.16(m,2H),4.18−4.28(m,1H),3.53(s br,2H),3.30−3.42(m,2H),3.08(s br,2H),2.26−2.38(m,2H),1.80−1.90(m,2H)。MS:計算値598(M Na+),実験値598(M Na+)。
【0088】
(実施例11)
【0089】
【化27】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロ−N−[2−(テトラヒドロ−2−ピラン−2−イルオキシ)エチル]メタンスルホンアミド
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(50mg,0.102mmol)を無水DMF(0.3mL)に溶解し、この溶液に撹拌下に炭酸カリウム(49mg,0.357mmol)を加えた後、2−(2−ブロモエトキシ)テトラヒドロ−2H−ピラン(53mg,0.255mmol)を加えた。得られた混合物を80℃で16時間撹拌した。混合物を周囲温度まで冷却し、水で希釈し、EtOAcで抽出した。有機層を再び飽和重炭酸塩水溶液で洗浄後、無水硫酸マグネシウムで乾燥し、減圧濃縮した。抽出層をMPLC(0−45% EtOAc/DCM)により精製し、標記化合物を得た。40% EtOAc/DCM中Rf=0.71。1H NMR(CDCl3,600MHz)δ7.31−7.38(m,4H),7.00−7.07(m,2H),6.88−6.93(m,1H),6.78−6.84(m,1H),4.65−4.68(m,1H),4.06−4.08(m,1H),3.50−4.00(m,7H),2.80−3.10(m,2H),1.20−1.90(m,7H)。MS:計算値640(M Na+),実験値640(M Na+)。
【0090】
(実施例12)
【0091】
【化28】
{[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル))スルホニル]アミノ}酢酸メチル
無水THF(0.8mL)をN−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(200mg,0.408mmol)に加えた後、反応混合物を0℃まで冷却した。水素化ナトリウム(49mg,1.225mmol)を一度に加え、混合物を0℃で15分間撹拌した。混合物は発泡し、オフホワイト懸濁液の状態で撹拌した。15分後にブロモ酢酸メチル(187mg,1.225mmol)を加えると、混合物は黄変しながら再び発泡した。混合物を16時間撹拌後、飽和塩化アンモニウム水溶液でクエンチし、EtOAcで抽出した。有機層を再び飽和塩化アンモニウム水溶液で洗浄後、無水硫酸マグネシウムで乾燥し、減圧濃縮した。抽出層をMPLC(0−45% EtOAc:ヘプタン)により精製し、標記化合物を得た。40% EtOAc:ヘプタン中Rf=0.6。1H NMR(CDCl3,600MHz)δ7.36(d,J=7.0Hz,2H),7.43(d,J=7.0Hz,2H),7.02−7.08(m,1H),6.86−6.90(m,1H),6.78−6.84(m,1H),4.30−4.60(m,3H),3.85(s,3H),3.38−3.34(m,2H),3.04(s br,2H)。MS:計算値584(M Na+),実験値584(M Na+)。
【0092】
同様の手順により以下の化合物を製造した。
【0093】
【表2】
【0094】
(実施例17)
【0095】
【化29】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミンを使用した以外は実施例1と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.33(m,4H),7.00(m,1H),6.90(s,1H,NH),6.82−6.75(m,2H),3.54(d,J=14.4Hz,2H),2.87(d,J=14.4Hz,2H),1.44(s,3H)。MS計算値526.0(MNa+),実験値525.9(MNa+)。
【0096】
(実施例18)
【0097】
【化30】
N−(シス−3−(2,5−ジフルオロフェニル)−1−メチル−3−{[4−(トリフルオロ)フェニル]スルホニル}シクロブチル)−1,1,1−トリフルオロメタンスルホンアミド
実施例17と同様の手順を使用して製造した。計算値(2M+Na)+:1097.0,実測値:1096.5。
【0098】
(実施例19)
【0099】
【化31】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロ−N−[(トリフルオロメチル)スルホニル]メタンスルホンアミド
実施例1の合成で副生物として製造した。1H NMR(600MHZ,CDCl3)δ7.38−7.33(m,4H),7.07(m,1H),6.95(m,1H),6.84(m,1H),4.44(m,1H),3.82(m,2H),3.15(m,2H)。MS計算値684.9(MNa++CH3CN),実験値684.9(MNa++CH3CN)。
【0100】
(実施例20)
【0101】
【化32】
ナトリウム[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アザニド
水素化ナトリウムをヘキサンに懸濁し、0℃まで冷却した。THF(1mL)中のN−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(200mg,0.408mmol)を水素化ナトリウム懸濁液に滴下した。得られた混合物を0℃で15分間、次いで周囲温度で30分間撹拌した。この時点で反応混合物を減圧濃縮した。乾燥白色粉末をフラスコから掻き取り、ガラスフリット付き漏斗に入れ、氷冷ペンタン(45mL)で洗浄した。次に粉末を高減圧下に16時間置いた。1H NMR(DMSO D6,600MHz)δ7.56(d,J=8.5Hz,2H),7.33(d,J=8.5Hz,2H),7.20−7.26(m,1H),7.00−7.12(m,2H),3.42−3.52(m,1H),2.66−2.80(m,4H)。
【0102】
(実施例21)
【0103】
【化33】
カリウム[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アザニド
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(1.2g,2.55mmol)を無水THF(25.5mL)中で0℃にて撹拌後、カリウムtert−ブトキシド(0.29g,2.55mmol)で処理した。反応混合物を0℃で15分間撹拌後、周囲温度まで昇温し、更に45分間撹拌した。反応混合物を減圧濃縮後、得られた白色粉末を100℃で撹拌下に最少量の3:1 IPA:トルエン(400mL)から再結晶させた。溶解後、混合物を濾紙で濾過し、4℃で20時間静置した。ガラスフリットで濾過して結晶を採取し、冷ペンタンで3回洗浄した。残留溶媒を減圧除去した。1H NMR(DMSO D6,600MHz)δ7.56(d,J=8.5Hz,2H),7.33(d,J=8.5Hz,2H),7.20−7.26(m,1H),7.00−7.12(m,2H),3.42−3.52(m,1H),2.66−2.80(m,4H)。
【0104】
(実施例22)
【0105】
【化34】
N−[シス−3−[(4−トリフルオロメトキシフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
シス−3−[(4−トリフルオロメトキシフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンを使用した以外は実施例2と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.56(m,2H),7.27(m,2H)7.088(m,1H),6.78(m,2H),6.68(d,J=10.6Hz,1H,NH),4.30(m,1H),3.31(m,2H),3.18(m,2H)。MS計算値603.46(MNa++CH3CN),実験値603.0(MNa++CH3CN)。
【0106】
生物活性
以下、本発明の化合物の生物活性を測定するためのアッセイについて記載する。
【0107】
APPプロセシング(細胞株から分泌されるAβ検体を定量するアッセイ):
アミロイドβ蛋白質を発現するSH−SY5Y細胞(SP4CT細胞)から産生されるAβ40及びAβ42ペプチドの量に与える化合物の影響をAlphaLisa(登録商標)アッセイにより判定した。ELISAアッセイと同様に、このAlphaLisa(登録商標)アッセイでシグナルを発生するには、Aβ40又はAβ42ペプチドの特異的抗体認識により「ドナー」ビーズと「アクセプター」ビーズを密接に近接させる必要がある。化合物で処理したSP4CT細胞から培地を別個のマイクロプレート2枚に移した後に、ビオチン化抗アミロイドβモノクローナル抗体(クローン4G8)と結合するストレプトアビジンを固定化したドナービーズを加えることによりアッセイを実施した。抗Aβ40モノクローナル抗体(G210)を直接固定化したアクセプタービーズを一方のマイクロプレートに加え、抗Aβ42モノクローナル抗体(12F4)アクセプタービーズを他方に加えた。Aβ40とAβ4の量はレーザー光によるドナービーズの励起後に発生した発光シグナルに正比例した。
【0108】
ノッチプロセシング:(細胞株へのノッチ細胞内ドメイン遊離量を定量するアッセイ):
ノッチ蛋白質のγセクレターゼ依存的切断の阻害を定量するために「スプリットルシフェラーゼ」アッセイを使用する。このアッセイでは、ルシフェラーゼのN末端フラグメントに融合したその細胞外ドメイン(ノッチ△E)をもたないノッチ蛋白質をHeLa細胞に発現させた。同一細胞は免疫グロブリンJκ組換えシグナル配列結合蛋白質(RBP)と融合したルシフェラーゼのC末端フラグメントも発現した。ノッチ△Eがγセクレターゼにより切断されると、ノッチ細胞内ドメイン(NICD)−N末端ルシフェラーゼ蛋白質が生成され、核転座してRBP−C末端ルシフェラーゼ融合体と結合し、ルシフェラーゼの独立して非機能的な半分ずつを一緒にして機能的ルシフェラーゼ酵素を形成する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるノッチの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。
【0109】
PXRアッセイ解説
CYP3A4−SEAPトランス活性化アッセイ(PCSTA)はチトクロームP450 CYP3A(ヒトCYP3A4又はラットCYP3A1)を誘導する可能性について化合物を有効且つ迅速に評価する。レポーター構築物は分泌型アルカリホスファターゼ(SEAP)遺伝子のすぐ上流に配置されたCYP3A4遺伝子に由来する調節配列を含む。メチオニンが野生型配列に存在するロイシンに置換する開始アミノ酸となるように、ヒトPXR核内受容体の5’末端を修飾した。HEP G2細胞にPXRプラスミド(ヒト又はラット)とレポータープラスミドをトランスフェクトする。CYP3A4の誘導の読取りはパラニトロフェニルリン酸(pNPP)を基質とするSEAP比色アッセイから構成される。各点がSEAPによるpNPPからpNPへの変換率に対応するように5点用量応答曲線を2本ずつ作成する。ヒトPCSTAでは、リファンピシンを陽性対照として使用し、100%誘導は10μMリファンピシンにより生じる最大誘導に基づく。
【0110】
ICDトランス活性化(細胞株へのγセクレターゼ基質群の細胞内ドメイン遊離量を定量するアッセイ)
ホタルルシフェラーゼによるトランス活性化アッセイを使用してγセクレターゼ基質のε/S3サイト切断の阻害を測定する。このアッセイは細胞内ドメイン(ICD)に融合したGAL4/VP16(GVP)トランス活性化ドメインを有するキメラ基質であるAPP−GVP、ノッチ△E−GVP、E−カドヘリン−GVP及びCD44−GVPを使用する。ICDが切断され、遊離されると、GVPドメインはUASプロモーターの制御下でルシフェラーゼ遺伝子の発現を誘導する。このアッセイでは、UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼ(トランスフェクション対照)と共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした。γセクレターゼにより切断されると、遊離されるICD−GVPは核転座し、UAS−ルシフェラーゼ遺伝子の発現を誘導する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるICDの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。更に、トランスフェクション効率の差を考慮するために、吸光度によるβ−ガラクトシダーゼ酵素アッセイを実施し、発光量読取を正規化する。
【0111】
全長γセクレターゼ基質の評価(γセクレターゼ基質群のプロセシングを定量的に評価するアッセイ)
他の基質に対するγセクレターゼ活性に与える化合物の影響を試験するために、C末端V5タグを付けた以下のI型膜蛋白質:CD43、CD44、E−カドヘリン及びSCN2bのうちの1種を過剰発現する4種類のHEK293安定細胞株を構築した。細胞を撒き、滴定化合物とホルボールエステルTPAで一晩処理する。全蛋白質は初期エクトドメインシェディングイベントとそれに続くγセクレターゼによるC末端フラグメント(CTF)の膜貫通切断を特徴とする調節下の膜蛋白質分解を受けるので、TPAはγセクレターゼの基質を生成する初期切断イベントを誘導する。V5タグを付けたCTFのプロセシングをウェスタンブロット分析により追跡することにより、これらの基質に対するγセクレターゼ活性に与える化合物の影響を測定する。CTFの蓄積はγセクレターゼ活性の阻害を意味する。
【0112】
ICDトランス活性化(細胞株へのγセクレターゼ基質群の細胞内ドメイン遊離量を定量するアッセイ)
ホタルルシフェラーゼによるトランス活性化アッセイを使用してγセクレターゼ基質のε/S3サイト切断の阻害を測定する。このアッセイは細胞内ドメイン(ICD)に融合したGAL4/VP16(GVP)トランス活性化ドメインを有するキメラ基質であるAPP−GVP、ノッチ△E−GVP、E−カドヘリン−GVP及びCD44−GVPを使用する。ICDが切断され、遊離されると、GVPドメインはUASプロモーターの制御下でルシフェラーゼ遺伝子の発現を誘導する。このアッセイでは、UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼ(トランスフェクション対照)と共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした。γセクレターゼにより切断されると、遊離されるICD−GVPは核転座し、UAS−ルシフェラーゼ遺伝子の発現を誘導する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるICDの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。更に、トランスフェクション効率の差を考慮するために、吸光度によるβ−ガラクトシダーゼ酵素アッセイを実施し、発光量読取を正規化する。
【0113】
インビトロAPPプロセシング(半精製γセクレターゼと共にインキュベートした組換えAPPC100Flag基質から生成されるAβ検体を定量するアッセイ)
半精製γセクレターゼにより外来C100Flag基質から生成されるAβ40及びAβ42ペプチドの量に与える化合物の影響をMESO Scale ELISAにより判定した。このMESO Scaleアッセイでシグナルを発生するには、ストレプトアビジンを固定化した抗アミロイドモノクローナル抗体(クローン4G8)がビオチンを被覆したプレートと結合する必要がある。その後、Aβ40(G210)又はAβ42(12F4)に特異的な[Ru(bpy)3]2+標識モノクローナル抗体は電気化学刺激を受けると電気化学発光シグナルを発生する。化合物と、C100Flag基質と、HeLa細胞又はマウス、ラットもしくはイヌの脳に由来するP2膜をCHAPSOで可溶化したものをインキュベートすることによりアッセイを実施した。次に反応混合物をAβ40又はAβ42検出用の別個のビオチン化マイクロプレート2枚に移した。
【0114】
インビトロノッチプロセシング(半精製γセクレターゼと共にインキュベートした組換えノッチ△E100Flag基質からのノッチ細胞内ドメイン生成を定量的に評価するアッセイ)
同様に、基質をN100Flagに代えることにより、C100Flagインビトロアッセイと同一方法を使用してノッチプロセシングをモニターすることができる。ポリクローナルビオチン標識抗DYKDDDDK抗体を捕捉用抗体として使用し、切断されたノッチ1抗体をポリクローナル[Ru(bpy)3]2+で標識して使用し、NICDを検出した。
【0115】
化合物結合:(結合したγセクレターゼ阻害剤トレーサーのインビトロ置換を定量するアッセイ)
組換えγセクレターゼを安定的に過剰発現するHEK293(γNRCF8)P2膜をCHAPSOで可溶化したものを使用して全放射性リガンド結合実験を実施する。放射性リガンド結合には、トリチウム化した阻害剤の存在下で可溶化酵素をインキュベートする。過剰の未標識阻害剤を反応に加えることにより非特異的結合を測定し、トリチウム化したリガンドの系列希釈を使用して飽和結合等温線を得る。ポリエチレンイミンを被覆したガラス繊維フィルタープレートに酵素複合体を吸着させ、セルハーベスターで迅速に濾過後に洗浄することにより、結合したリガンドを遊離リガンから分離する。プレートを乾燥後、シンチラントを加え、マイクロプレートシンチレーションカウンターでプレートを読取る。1nM 3H標識化合物として参考例L−458(遷移状態阻害剤)又は4nM 3H標識化合物として参考例L−881(非遷移状態阻害剤)の存在下で各種阻害剤の系列希釈をインキュベートすることにより結合競合アッセイを実施する。各種阻害剤のアンタゴニスト競合性を判定するために、種々の濃度のこれらの化合物の存在下で夫々の3Hトレーサー用量応答曲線を分析する。
【0116】
参考例L−458:
【0117】
【化35】
【0118】
参考例L−881:
【0119】
【化36】
【0120】
結果
上記APP及びノッチプロセシング細胞機能アッセイで本明細書の実施例を試験した。試験した化合物は下表に示すようにノッチシグナル伝達経路を保持しながらAPPプロセシングのインビトロ阻害を示した。データは少なくとも>4回の反復の平均に基づく。
【0121】
【表3】
【0122】
WO02/081435A1(公開日2002年10月17日)はγセクレターゼの活性を調節するスルホン誘導体を開示している。WO2004/031139(公開日2004年4月15日)はγセクレターゼ阻害剤としてのシクロヘキシルスルホン誘導体を開示している。WO2004/031139の実施例47は下記構造をもつ。
【0123】
【化37】
上記化合物はMRK−560とも呼ばれ、Best et al.,J Pharmacol Exp,Ther.,317:786−790,2006及びBest et al,J.Pharmacol.Exp.Ther.,320:552−558,2007に開示されている。文献はMRK−560におけるAPPのインビトロ阻害とノッチプロセシング経路の間の分離について殆ど又は全く報告していないが、この化合物はTg2576マウスでノッチシグナル伝達経路の変化に関連する毒性の不在下でアミロイドプラーク沈着に有益なインビボ効果を示した。
【0124】
薬剤毒性を予測する共有蛋白質結合アッセイでMRK−560と実施例2を試験した。該当化合物の放射性標識物を肝ミクロソームと共にインキュベートすることにより、薬剤候補が蛋白質と共有結合を生じる可能性を評価する。次にBrandel Harvester技術に基づく半自動法を使用して試験化合物と肝蛋白質の共有結合性付加物の形成を測定する(Ref.Day et.al,J Pharmacol Toxicol Mehtods.52,278−85,2005参照)。結果を下表に示す。
【0125】
【表4】
【0126】
薬剤間相互作用を予測するプレグナンX受容体(PXR)と結合及び/又はこれを活性化する能力についてMRK−560と本明細書に開示した実施例を試験した。結果を下表に示す。
【0127】
【表5】
【0128】
他のγセクレターゼ基質の初期切断に与える影響を試験するために、MRK−560と実施例2を上記トランス活性化アッセイで試験した。MRK−560は試験した全基質のICD遊離を阻害したが、ICD遊離とそれに続くICD−GVP構築物の転座によりレポーター活性化が可能であったことから明らかなように、実施例2は初期ε/S3切断を維持した。図2参照。
【0129】
MRK−560によるε/S3及びγ切断の完全な阻害の結果、SCN2b−、ECAD−及びCD43−CTF蓄積を生じることが多基質細胞アッセイにより確認された。これに対して、実施例2で処理すると、CTF蓄積は低下又はゼロになり、NS−GSIはγセクレターゼ基質の初期切断を維持すると判断された。図3参照。
【0130】
MRK−560と実施例2を上記化合物結合アッセイでも試験した。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。図4参照。
【技術分野】
【0001】
本発明は新規類の化合物、その塩、前記化合物等を含有する医薬組成物及び人体の治療におけるその使用に関する。特に、本発明はγセクレターゼによるAPPのプロセシングを阻害し、従って、アルツハイマー病の治療又は予防に有用な新規シクロブチルスルホン誘導体に関する。本発明の化合物はノッチシグナル伝達経路を保持する。従って、本発明の化合物はノッチ阻害による毒性を生じずにアルツハイマー病の進行を停止又は潜在的に回復させると考えられる。
【背景技術】
【0002】
アルツハイマー病(AD)は認知症の最も一般的な形態である。主に高齢者の疾患であり、罹患率は65歳以上の人口の10%に及ぶが、遺伝的素因をもつ相当数の若年患者もADに罹患している。これは神経変性疾患であり、記憶及び認知機能の進行性低下を臨床的特徴とし、患者の皮質及び関連脳領域における細胞外蛋白プラークの沈着を病理的特徴とする。これらのプラークは主にβアミロイドペプチド(Aβ)のフィブリル凝集物から構成される(Glenner G G and Wong C W(1984)Alzheimer’s disease:Initial report of the purification and characterization of a novel cerebrovascular amyloid protein.Biochemical and Biophysical research Communications.120(3);885−890)。アミロイド前駆体蛋白質(APP)のプロセシングによりAβを形成する過程におけるγセクレターゼの役割を含むセクレターゼの役割については文献に詳細に記載されている。Aβはβアミロイド切断酵素(BACE)とγセクレターゼの2種類の酵素によるAPPの蛋白分解プロセシングにより生成される(図1;Selkoe D J(2001)Alzheimer’s disease:genes,proteins,and therapy.Physiological Review.81(2):741−766)。γセクレターゼはプレセニリン(プレセニリン1又は2)、ニカストリン、APH−1及びPEN−2の4種類の蛋白質から構成される複合体である(Takasugi N,Tomita T,Hayashi I,Tsuruoka M,Niimura M,Takahashi Y,Thinakaran G,Iwatsubo T(2003)The role of presenilin cofactors in the gamma−secretase complex.Nature.422(6930):438−441;Kimberly W T,LaVoie M J,Ostaszewski B L,Ye W,Wolfe M S,Selkoe D J(2003)Gamma−secretase is a membrane protein complex comprised of presenilin,nicastrin,Aph−1,and Pen−2.Proceedings of the National Academy of Sciences.100(11):6382−6387;Edbauer D,Winkler E,Regula J T,Pesold B,Steiner H,Haass C(2003)Reconstitution of gamma−secretase activity.Nature Cell Biology.5(5):486−488.)。プレセニリン1及び2は複合体の触媒活性に不可欠であることが分かっている膜貫通アスパラギン酸残基を含む(Wolfe M S,Xia W,Ostaszewski B L,Diehl T S,Kimberly W T,Selkoe D J(1999)Two transmembrane aspartates in presenilin−1 required for presenilin endoproteolysis and gamma−secretase activity.Nature.1999 398(6727):513−517)。早発家族性AD(FAD)に関連する突然変異の大半はPS−1又はPS−2に結び付けられる(Scheuner D,Eckman C,Jensen M,Song X,Citron M,Suzuki N,Bird T D,Hardy J,Hutton M,Kukull W,Larson E,Levy−Lahad E,Viitanen M,Peskind E,Poorkaj P,Schellenberg G,Tanzi R,Wasco W,Lannfelt L,Selkoe D,Younkin S(1996)Secreted amyloid beta−protein similar to that in the senile plaques of Alzheimer’s disease is increased in vivo by the presenilin 1 and 2 and APP mutations linked to familial Alzheimer’s disease.Nature Medicine.2(8):864−870;Duff K,Eckman C,Zehr C,Yu X,Prada C M,Perez−tur J,Hutton M,Buee L,Harigaya Y,Yager D,Morgan D,Gordon M N,Holcomb L,Refolo L,Zenk B,Hardy J,Younkin S(1996)Increased amyloid−beta42(43)in brains of mice expressing mutant presenilin 1.Nature.383(6602):710−713;Lemere CA,Lopera F,Kosik K S,Lendon C L,Ossa J,Saido T C,Yamaguchi H,Ruiz A,Martinez A,Madrigal L,Hincapie L,Arango J C,Anthony D C,Koo E H,Goate A M,Selkoe D J,Arango J C(1996)The E280A presenilin 1 Alzheimer mutation produces increased A beta 42 deposition and severe cerebellar pathology.Nature Medicine.2(10):1146−1150;Citron M,Westaway D,Xia W,Carlson G,Diehl T,Levesque G,Johnson−Wood K,Lee M,Seubert P,Davis A,Kholodenko D,Motter R,Sherrington R,Perry B,Yao H,Strome R,Lieberburg I,Rommens J,Kim S,Schenk D,Fraser P,St George Hyslop P,Selkoe D J(1997)Mutant presenilins of Alzheimer’s disease increase production of 42−residue amyloid beta−protein in both transfected cells and transgenic mice.Nature Medicine.3(1):67−72)。γセクレターゼは前提条件としてエクトドメインシェディングを受けた多数の他のI型膜蛋白質をプロセシングする(Lleo A(2008)Activity of gamma−secretase on substrates other than APP.Current Topics in Medicinal Chemistry.8(1):9−16)。
【0003】
APPのプロセシングに加え、γセクレターゼはノッチファミリー受容体を切断する。ノッチシグナル伝達及び機能にγセクレターゼ活性が不可欠であることは遺伝的に立証されている(Shen J,Bronson R T,Chen D F,Xia W,Selkoe D J,Tonegawa S(1997)Skeletal and CNS defects in Presenilin−1−deficient mice.Cell.89(4):629−639;Wong P C,Zheng H,Chen H,Becher M W,Sirinathsinghji D J,Trumbauer M E,Chen H Y,Price D L,Van der Ploeg L H,Sisodia S S(1997)Presenilin 1 is required for Notch1 and DII1 expression in the paraxial mesoderm.Nature.387(6630):288−292)。ノッチは進化的に保存されており、発生中の胚で細胞運命決定の調節に重要な役割を果たす1回貫通型I型膜貫通受容体で広く発現される(Artavanis−Tsakonas S,Rand M D,Lake R J(1999)Notch signaling:cell fate control and signal integration in development.Science.284(5415):770−776)。成人におけるノッチの役割はまだ不明であるが、ノッチ蛋白質は各種成人組織で発現され、幹細胞分化に役割を果たすと考えられる。4種類のノッチ遺伝子が哺乳動物で同定されており(ノッチ1〜4)、全4種類のノッチ蛋白質はγセクレターゼにより切断される(Saxena M T,Schroeter E H,Mumm J S,Kopan R(2001)Murine notch homologs(N1−4)undergo presenilin−dependent proteolysis.Journal of Biological Chemistry.276(43):40268−40273)。ノッチ活性化はDelta/Serrate/LAG2ファミリーの膜貫通リガンドとのトランス結合により誘導される。ノッチシグナル伝達は(a)細胞外ドメインのサイト1の切断;(b)リガンド結合後の細胞外/膜貫通ドメイン境界のちょうどN末端のサイト2の切断;及び(c)膜貫通/細胞質ドメイン境界付近の膜貫通ドメイン内のサイト3(S3)の切断の3つの切断イベントに介在される。サイト3切断はノッチ細胞内ドメイン(NICD)の遊離に必要であり、γセクレターゼに介在される(Schroeter E H,Kisslinger J A,Kopan R(1998)Notch−1 signalling requires ligand−induced proteolytic release of intracellular domain.Nature.393(6683):382−386)。NICDは(CSL)CBF1/Serrate/LAG−1ファミリーのDNA結合蛋白質に介在される転写を活性化し、各種遺伝子の発現を誘導する。NICDに調節される転写はノッチに介在されるシグナル伝達の主要な要素であると考えられる。
【0004】
APP切断とAβ生成を阻止するためのγセクレターゼ阻害剤の開発はノッチプロセシングの阻害によるメカニズム関連毒性の可能性により阻まれている。動物にγセクレターゼ阻害剤を亜慢性投与した試験ではノッチ関連毒性が観測されている。3日間以上投与後に常に腸杯細胞化生が認められる(Searfoss G H,Jordan W H,Calligaro D O,Galbreath E J,Schirtzinger L M,Berridge B R,Gao H,Higgins M A,May P C,Ryan T P(2003)Adipsin,a biomarker of gastrointestinal toxicity mediated by a functional gamma−secretase inhibitor.Journal of Biological Chemistry.278(46):46107−46116;Wong G T,Manfra D,Poulet F M,Zhang Q,Josien H,Bara T,Engstrom L,Pinzon−Ortiz M,Fine J S,Lee H J,Zhang L,Higgins G A,Parker E M(2004)Chronic treatment with the gamma−secretase inhibitor LY−411,575 inhibits beta−amyloid peptide production and alters lymphopoiesis and intestinal cell differentiation.Journal of Biological Chemistry.279(13):12876−12882;Milano J,McKay J,Dagenais C,Foster−Brown L,Pognan F,Gadient R,Jacobs R T,Zacco A,Greenberg B,Ciaccio P J(2004)Modulation of notch processing by gamma−secretase inhibitors causes intestinal goblet cell metaplasia and induction of genes known to specify gut secretory lineage differentiation.Toxicological Sciences.82(1):341−358;van Es J H,van Gijn M E,Riccio O,van den Born M,Vooijs M,Begthel H,Cozijnsen M,Robine S,Winton D J,Radtke F,Clevers H(2005)Notch/gamma−secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells.Nature.435(7044):959−963)。更に、ノッチ機能はT及びBリンパ球の適正な分化に不可欠であると思われる(Hadland B K,Manley N R,Su D,Longmore G D,Moore C L,Wolfe M S,Schroeter E H,Kopan R(2001)Gamma−secretase inhibitors repress thymocyte development.Proceedings of the National Academy of Sciences.98(13):7487−7491;Doerfler P,Shearman M S,Perlmutter R M(2001)Presenilin−dependent gamma−secretase activity modulates thymocyte development.Proceedings of the National Academy of Sciences.98(16):9312−9317)。従って、γセクレターゼ活性を薬理標的とするには、ノッチに対する活性の阻害を最小限にしながらAβを選択的に阻止する物質が必要である。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Glenner G G and Wong C W(1984)Alzheimer’s disease:Initial report of the purification and characterization of a novel cerebrovascular amyloid protein.Biochemical and Biophysical research Communications.120(3);885−890
【非特許文献2】Selkoe D J(2001)Alzheimer’s disease:genes,proteins,and therapy.Physiological Review.81(2):741−766
【非特許文献3】Takasugi N,Tomita T,Hayashi I,Tsuruoka M,Niimura M,Takahashi Y,Thinakaran G,Iwatsubo T(2003)The role of presenilin cofactors in the gamma−secretase complex.Nature.422(6930):438−441
【非特許文献4】Kimberly W T,LaVoie M J,Ostaszewski B L,Ye W,Wolfe M S,Selkoe D J(2003)Gamma−secretase is a membrane protein complex comprised of presenilin,nicastrin,Aph−1,and Pen−2.Proceedings of the National Academy of Sciences.100(11):6382−6387
【非特許文献5】Edbauer D,Winkler E,Regula J T,Pesold B,Steiner H,Haass C(2003)Reconstitution of gamma−secretase activity.Nature Cell Biology.5(5):486−488
【非特許文献6】Wolfe M S,Xia W,Ostaszewski B L,Diehl T S,Kimberly W T,Selkoe D J(1999)Two transmembrane aspartates in presenilin−1 required for presenilin endoproteolysis and gamma−secretase activity.Nature.1999 398(6727):513−517
【非特許文献7】Scheuner D,Eckman C,Jensen M,Song X,Citron M,Suzuki N,Bird T D,Hardy J,Hutton M,Kukull W,Larson E,Levy−Lahad E,Viitanen M,Peskind E,Poorkaj P,Schellenberg G,Tanzi R,Wasco W,Lannfelt L,Selkoe D,Younkin S(1996)Secreted amyloid beta−protein similar to that in the senile plaques of Alzheimer’s disease is increased in vivo by the presenilin 1 and 2 and APP mutations linked to familial Alzheimer’s disease.Nature Medicine.2(8):864−870
【非特許文献8】Duff K,Eckman C,Zehr C,Yu X,Prada C M,Perez−tur J,Hutton M,Buee L,Harigaya Y,Yager D,Morgan D,Gordon M N,Holcomb L,Refolo L,Zenk B,Hardy J,Younkin S(1996)Increased amyloid−beta42(43)in brains of mice expressing mutant presenilin 1.Nature.383(6602):710−713
【非特許文献9】Lemere CA,Lopera F,Kosik K S,Lendon C L,Ossa J,Saido T C,Yamaguchi H,Ruiz A,Martinez A,Madrigal L,Hincapie L,Arango J C,Anthony D C,Koo E H,Goate A M,Selkoe D J,Arango J C(1996)The E280A presenilin 1 Alzheimer mutation produces increased A beta 42 deposition and severe cerebellar pathology.Nature Medicine.2(10):1146−1150
【非特許文献10】Citron M,Westaway D,Xia W,Carlson G,Diehl T,Levesque G,Johnson−Wood K,Lee M,Seubert P,Davis A,Kholodenko D,Motter R,Sherrington R,Perry B,Yao H,Strome R,Lieberburg I,Rommens J,Kim S,Schenk D,Fraser P,St George Hyslop P,Selkoe D J(1997)Mutant presenilins of Alzheimer’s disease increase production of 42−residue amyloid beta−protein in both transfected cells and transgenic mice.Nature Medicine.3(1):67−72
【非特許文献11】Lleo A(2008)Activity of gamma−secretase on substrates other than APP.Current Topics in Medicinal Chemistry.8(1):9−16
【非特許文献12】Shen J,Bronson R T,Chen D F,Xia W,Selkoe D J,Tonegawa S(1997)Skeletal and CNS defects in Presenilin−1−deficient mice.Cell.89(4):629−639
【非特許文献13】Wong P C,Zheng H,Chen H,Becher M W,Sirinathsinghji D J,Trumbauer M E,Chen H Y,Price D L,Van der Ploeg L H,Sisodia S S(1997)Presenilin 1 is required for Notch1 and DII1 expression in the paraxial mesoderm.Nature.387(6630):288−292
【非特許文献14】Artavanis−Tsakonas S,Rand M D,Lake R J(1999)Notch signaling:cell fate control and signal integration in development.Science.284(5415):770−776
【非特許文献15】Saxena M T,Schroeter E H,Mumm J S,Kopan R(2001)Murine notch homologs(N1−4)undergo presenilin−dependent proteolysis.Journal of Biological Chemistry.276(43):40268−40273
【非特許文献16】Schroeter E H,Kisslinger J A,Kopan R(1998)Notch−1 signalling requires ligand−induced proteolytic release of intracellular domain.Nature.393(6683):382−386
【非特許文献17】Searfoss G H,Jordan W H,Calligaro D O,Galbreath E J,Schirtzinger L M,Berridge B R,Gao H,Higgins M A,May P C,Ryan T P(2003)Adipsin,a biomarker of gastrointestinal toxicity mediated by a functional gamma−secretase inhibitor.Journal of Biological Chemistry.278(46):46107−46116
【非特許文献18】Wong G T,Manfra D,Poulet F M,Zhang Q,Josien H,Bara T,Engstrom L,Pinzon−Ortiz M,Fine J S,Lee H J,Zhang L,Higgins G A,Parker E M(2004)Chronic treatment with the gamma−secretase inhibitor LY−411,575 inhibits beta−amyloid peptide production and alters lymphopoiesis and intestinal cell differentiation.Journal of Biological Chemistry.279(13):12876−12882
【非特許文献19】Milano J,McKay J,Dagenais C,Foster−Brown L,Pognan F,Gadient R,Jacobs R T,Zacco A,Greenberg B,Ciaccio P J(2004)Modulation of notch processing by gamma−secretase inhibitors causes intestinal goblet cell metaplasia and induction of genes known to specify gut secretory lineage differentiation.Toxicological Sciences.82(1):341−358
【非特許文献20】van Es J H,van Gijn M E,Riccio O,van den Born M,Vooijs M,Begthel H,Cozijnsen M,Robine S,Winton D J,Radtke F,Clevers H(2005)Notch/gamma−secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells.Nature.435(7044):959−963
【非特許文献21】Hadland B K,Manley N R,Su D,Longmore G D,Moore C L,Wolfe M S,Schroeter E H,Kopan R(2001)Gamma−secretase inhibitors repress thymocyte development.Proceedings of the National Academy of Sciences.98(13):7487−7491
【非特許文献22】Doerfler P,Shearman M S,Perlmutter R M(2001)Presenilin−dependent gamma−secretase activity modulates thymocyte development.Proceedings of the National Academy of Sciences.98(16):9312−9317
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明はノッチシグナル伝達経路を保持しながら推定γセクレターゼによるAPPのプロセシングを阻害し、従って、ADの治療又は予防に有用である新規類のシクロブチルスルホン誘導体を提供する。
【課題を解決するための手段】
【0007】
本発明はノッチシグナル伝達経路を保持しながら推定γセクレターゼによるAPPのプロセシングを阻害し、従って、ノッチ阻害による胃腸障害を併発せずにアルツハイマー病の治療又は予防に有用な新規類のシクロブチルスルホン誘導体を包含する。医薬組成物と使用方法も包含する。
【図面の簡単な説明】
【0008】
【図1】γセクレターゼはほぼ膜の中間(γ/S4切断)と膜のサイトゾル側(ε/S3切断)の主に2カ所でAPPとノッチを切断する。ε/S3切断は膜から基質の細胞内ドメイン(ICD)である夫々AICD及びNICDを遊離するので、重要なプロセシングイベントである。この段階はICD核転座とその後の転写モジュレーターとしての機能の前提条件である。他方、γ/S4切断は夫々APPとノッチからAβペプチドとAβ様ペプチド(Nβ)の遊離をもたらす。後者の切断はADの疾患原因物質であると考えられるAβペプチドのC末端を生じるので、特に注目されている。
【図2】他のγセクレターゼ基質の初期切断に与える影響を試験するために本願に記載するトランス活性化アッセイで試験したMRK−560と実施例2の結果を示す。UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼと共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした後、各化合物1μMで48時間処理した。MRK−560は試験した全基質のICD遊離を阻害したが、実施例2はICD遊離とその後のICD−GVP構築物の転座を維持し、レポーター活性化が可能であった。
【図3】各基質、ノッチ△E、E−カドヘリン、SCN2b及びCD43を安定的に過剰発現するHEK293細胞をTPAと共に滴定濃度のMRK−560又は実施例2で処理し、シェディングを誘導した。細胞溶解液の免疫ブロット分析の結果、MRK−560は従来のγセクレターゼ阻害剤であるDAPTと同様に、NICD生成を阻害し、γセクレターゼの直接基質であるCTFの蓄積を生じることが判明した。これに対して、実施例2はNICD生成が可能であり、CTF蓄積は全く認められないか又はMRK−560よりも少なかった。
【図4A】本願に記載する化合物結合アッセイで試験したMRK−560と実施例2の結果を示す。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。A−MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。B−実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。
【図4B】本願に記載する化合物結合アッセイで試験したMRK−560と実施例2の結果を示す。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。A−MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。B−実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。
【発明を実施するための形態】
【0009】
本発明は式I:
【0010】
【化1】
の属の化合物又はその医薬的に許容可能な塩を包含し、
式中、X1はF及びCNから構成される群から選択され;
X2はF、Cl及びCNから構成される群から選択され;
X3はF、Br、Cl、CN、CF3、OCF3、C(O)−OCH3及びS−CH3から構成される群から選択され;
X4はH、F及びClから構成される群から選択され;
R1は
(a)H、
(b)CH3、
(c)−(CH2)n−OR3、
(d)−(CH2)n−C(O)−OR4、及び
(e)−SO2−CF3
から構成される群から選択され;
式Iの化合物がシス配置であるとき、R2はH又はCH3であり、それ以外の場合にはR2はHであり;
R3は1個の酸素ヘテロ原子をもつ5又は6員非芳香族複素環であり;
R4はH又はCH3であり;
nは1〜4である。
【0011】
この属の範囲内で本発明は式Ia:
【0012】
【化2】
の第1の亜属の化合物又はその医薬的に許容可能な塩を包含する。
【0013】
第1の亜属の範囲内で本発明は、
X1がFであり、
X4がHである
式Iaの第1の分類の化合物を包含する。
【0014】
第1の分類の範囲内で本発明は、
X2がFであり、
X3がClである
式Iaの亜分類の化合物を包含する。
【0015】
この亜分類の範囲内で本発明は、R2がHである式Iaの群の化合物を包含する。
【0016】
更にこの群の範囲内で本発明は、R1が−(CH2)n−C(O)−OR4である式Iaの第2の亜群の化合物を包含する。
【0017】
この属の範囲内で本発明は式Ib:
【0018】
【化3】
の第2の亜属の化合物又はその医薬的に許容可能な塩を包含する。
【0019】
第2の亜属の範囲内で本発明は、
X1及びX2がFであり;
X3がClであり;
X4がHである
式Ibの第1の分類の化合物を包含する。
【0020】
本発明は更に下記実施例のいずれかを包含する。
【0021】
本発明は更に式Iの化合物又はその医薬的に許容可能な塩と、医薬的に許容可能なキャリヤーを含有する医薬組成物を包含する。
【0022】
本発明は更にβアミロイドの沈着を伴う病態に罹患している対象又は前記病態に罹患し易い対象の治療方法として、有効量の式Iの化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む方法を包含する。
【0023】
本発明は更にアルツハイマー病の治療又は予防用医薬の製造における式Iの化合物又はその医薬的に許容可能な塩の使用を包含する。
【0024】
式I又はその置換基に変項が2回以上出現する場合には、特に指定しない限り、前記変項の個々の出現は相互に独立している。
【0025】
医薬用として、式Iの化合物は医薬的に許容可能な塩の形態でもよい。しかし、他の塩が式Iの化合物又はその医薬的に許容可能な塩の製造に有用な場合もある。本発明の化合物の適切な医薬的に許容可能な塩としては、例えば本発明の化合物の溶液を塩酸、硫酸、メタンスルホン酸、ベンゼンスルホン酸、フマル酸、マレイン酸、琥珀酸、酢酸、安息香酸、蓚酸、クエン酸、酒石酸、炭酸又はリン酸等の医薬的に許容可能な酸の溶液と混合することにより形成することができる酸付加塩が挙げられる。あるいは、本発明の化合物が酸性部分をもつ場合には、前記酸性部分を適切な塩基で中和することにより医薬的に許容可能な塩を形成することができる。こうして形成される医薬的に許容可能な塩の例としては、ナトリウム塩やカリウム塩等のアルカリ金属塩;カルシウム塩やマグネシウム塩等のアルカリ土類金属塩;及びアミン塩(ピリジニウム塩を含む)や第四級アンモニウム塩等の適切な有機塩基と形成される塩が挙げられる。
【0026】
なお、本発明は式Iの各化合物について、ホモキラル化合物又は任意割合のエナンチオマー混合物としての両者のエナンチオマー形態を包含する。本発明の1態様において、式Iの化合物は式Iaもしくは式Ibのホモキラル化合物、又はその医薬的に許容可能な塩である。本発明の化合物が2個以上の不斉中心をもつ場合には、更にジアステレオ異性体として存在することもできる。当然のことながら、このような全異性体と任意割合のその混合物が本発明の範囲に含まれる。
【0027】
同様に当然のことながら、1つの反応から2種類以上の異性体が得られる場合には、得られた異性体の混合物を従来手段により分離することができる。本発明の化合物の製造方法により立体異性体の混合物が得られる場合には、分取クロマトグラフィー等の従来技術によりこれらの異性体を分離することができる。新規化合物をラセミ体として製造することもできるし、エナンチオ特異的合成又は分割により個々のエナンチオマーを製造することもできる。新規化合物を例えば分取HPLC等の標準技術によりその成分エナンチオマーに分割してもよいし、(−)−ジ−p−トルオイル−d−酒石酸及び/又は(+)−ジ−p−トルオイル−1−酒石酸等の光学活性酸との塩形成によるジアステレオマー対の形成後に分別結晶と遊離塩基の再生を実施してもよい。更に、ジアステレオマーエステル又はアミドの形成後にクロマトグラフィー分離とキラル助剤の除去により新規化合物を分割してもよい。あるいは、該当化合物のラセミ合成前駆体にこのような技術を実施してもよい。
【0028】
本発明の化合物はγセクレターゼの阻害剤としての活性をもつ。
【0029】
本発明は更に本発明の1種以上の化合物と医薬的に許容可能なキャリヤーを含有する医薬組成物も提供する。これらの組成物は経口、非経口、鼻孔内、舌下もしくは直腸投与用、又は吸入もしくは吹送投与用として、錠剤、ピル、カプセル剤、散剤、顆粒剤、滅菌非経口溶液剤もしくは懸濁液剤、定量式エアゾールもしくは液体スプレー、滴剤、アンプル剤、経皮パッチ、オートインジェクター装置又は坐剤等の単位剤形が好ましい。一般に主活性成分を医薬キャリヤー(例えばコーンスターチ、ラクトース、スクロース、ソルビトール、タルク、ステアリン酸、ステアリン酸マグネシウム及びリン酸二カルシウム等の従来の錠剤化成分、ガム類、分散剤、懸濁剤、又はモノオレイン酸ソルビタンやポリエチレングリコール等の界面活性剤)及び他の医薬希釈剤(例えば水)と混合し、本発明の化合物又はその医薬的に許容可能な塩を含有する均質な前製剤組成物を形成する。これらの前製剤組成物について均質と言う場合には、組成物を錠剤、ピル及びカプセル剤等の均等に有効な単位剤形に容易に細分できるように活性成分が組成物全体に均等に分散されていることを意味する。その後、この前製剤組成物を細分し、本発明の活性成分0.1〜約500mgを含有する上記型の単位剤形とする。典型的な単位剤形は活性成分1〜100mg、例えば1、2、5、10、25、50又は100mgを含有する。長期作用の利点を付与する剤形を提供するように、新規組成物の錠剤又はピルにコーティング又は他の方法で加工してもよい。例えば、錠剤又はピルを内側製剤成分と外側製剤成分から構成し、外側成分が内側成分のエンベロープを構成するようにしてもよい。胃で崩壊しないように機能すると共に内側成分を無傷のまま十二指腸に導入又は放出遅延させる腸溶層により2成分を分離することができる。このような腸溶層又はコーティングには各種材料を使用することができ、このような材料としては、多数のポリマー酸や、ポリマー酸とシェラック、セチルアルコール及び酢酸セルロース等の材料の混合物が挙げられる。
【0030】
本発明の新規組成物を経口又は注射投与用に配合することができる液体形態としては、水溶液、液体又はゲル充填カプセル剤、適切なフレーバー入りシロップ、水性又は油性懸濁液、及び綿実油、胡麻油又は椰子油等の食用油を添加したフレーバー入りエマルション、並びにエリキシル剤及び同様の医薬ビヒクルが挙げられる。水性懸濁液に適した分散剤又は懸濁剤としては、トラガカントガム、アラビアガム、アルギン酸塩、デキストラン、カルボキシメチルセルロースナトリウム、メチルセルロース、ポリ(エチレングリコール)、ポリ(ビニルピロリドン)又はゼラチン等の合成及び天然ガムが挙げられる。
【0031】
本発明は更に人体の治療方法で使用するための本発明の化合物又はその医薬的に許容可能な塩を提供する。βアミロイドの沈着を伴う病態の治療が好ましい。病態としてはアルツハイマー病等のβアミロイドの沈着を伴う神経疾患が好ましい。
【0032】
本発明は更にアルツハイマー病の治療又は予防用医薬の製造における本発明の化合物又はその医薬的に許容可能な塩の使用も提供する。
【0033】
アルツハイマー病に罹患している対象又は罹患し易い対象の治療方法として、有効量の本発明の化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む方法も開示する。
【0034】
アルツハイマー病を治療又は予防するために、適切な用量レベルは約0.01〜250mg/kg/日、好ましくは約0.01〜100mg/kg/日、より好ましくは約0.05〜50mg/kg体重/日であり、最も好ましい化合物では、約0.1〜10mg/kg体重/日である。1日1〜4回のレジメンで化合物を投与することができる。しかし、場合によってはこれらの範囲外の用量を使用してもよい。
【0035】
以下、実施例により本発明を例証する。合成スキームで使用する出発材料及び試薬が市販されていない場合には、従来手段により製造することができる。本発明は以下の実施例から選択される化合物も包含する。
【0036】
合成シーケンスのいずれかの間に、該当分子のいずれかの感受性又は反応性基を保護することが必要及び/又は望ましい場合がある。これはProtective Groups in Organic Chemistry,ed.J.F.W.McOmie,Plenum Press,1973;及びT.W.Greene & P.G.M.Wuts,Protective Groups in Organic Synthesis,John Wiley & Sons,1991に記載されているもの等の従来の保護基により実施することができる。保護基は適切な後続段階で当分野で公知の方法を使用して除去することができる。
【0037】
(実施例)
中間体A
【0038】
【化4】
2−[2−[(4−クロロフェニル)スルホニル]−2−(2,5−ジフルオロフェニル)エチル]オキシラン
WO02/081435(中間体1)に記載されているように4−クロロチオフェノールと臭化2,5−ジフルオロベンジルから2段階で4−クロロフェニル−2,5−ジフルオロベンジルスルホンを製造した。
【0039】
THF(99ml)中の4−クロロフェニル−2,5−ジフルオロベンジルスルホン(12g,39.6mmol)をnBuLi(19ml,ヘキサン中2.5M,47.6mmol)で0℃にて10分間処理後、エピクロロヒドリン(3.73ml,47.6mmol)を加えた。反応混合物を室温まで14時間かけてゆっくりと昇温し、水(100ml)でクエンチし、EtOAc(300ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中10−45%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物9.8gをオフホワイト固体として得た。1H NMR(600MHZ,CDCl3)2種類のジアステレオマー(〜1/1)δ7.52(m,2H/2H),7.38(m,2H/2H),7.30−7.22(m,1H/1H),6.99(m,1H/1H),6.86−6.80(m,1H/1H),4.76−4.70(m,1H/1H),3.03−2.48(m,4H/4H),2.37(m,1H),2.20(m,1H)。MS計算値359.0(MH+),実験値358.9(MH+)。
【0040】
中間体B
【0041】
【化5】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール
THF(2.7ml)中の2−[2−[(4−クロロフェニル)スルホニル]−2−(2,5−ジフルオロフェニル)エチル]オキシラン(100mg,0.279mmol)にMeMgBr(279μL,エーテル中3M,0.836mmol)を−78℃で加えた。反応混合物を室温まで1時間かけて昇温後、飽和NH4Cl(3ml)でクエンチし、EtOAc(10ml)で希釈した。有機相をブライン(10ml)で洗浄し、乾燥し(Na2SO4)、蒸発乾涸し、目的生成物を得た(100mg)、1H NMR(600MHZ,CDCl3)δ7.66(d,J=8.4Hz,2H),7.57(d,J=8.4Hz,2H),6.99(m,1H),6.82(m,1H),6.74(m,1H),4.28(m,1H),3.47(d,J=11.4Hz,1H,OH),3.13(m,4H)。MS計算値422.0(MNa++CH3CN),実験値421.9(MNa++CH3CN)。
【0042】
中間体C
【0043】
【化6】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール
THF(100ml)中のPPh3(8.19g,31.2mmol)にDIAD(6.31g,31.2mmol)を加え、得られた混合物を室温で0.5時間撹拌した。混合物を−50℃まで冷却し、THF(10ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(8g,22.3mmol)を加えた。反応混合物を20分間撹拌後、固体4−ニトロ安息香酸(5.22g,31.2mmol)を加えた。得られた混合物を室温まで昇温し、室温で20時間撹拌した。次に反応混合物を0℃まで冷却し、NaOMe(134ml,MeOH中0.5M,66.9mmol)を加えた。40分後に反応混合物を飽和NH4Cl(100ml)でクエンチし、EtOAc(300ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中エーテルを溶離液としてこの物質をシリカクロマトグラフィーに付し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(出発材料)〜15%を含有する白色固体として標記生成物8gを得た。1H NMR(600MHZ,CDCl3)主生成物,δ7.35(m,4H),6.96(m,1H),6.80−6.72(m,2H),4.84(m,1H),3.49(m,2H),2.59(m,2H)。MS計算値422.0(MNa++CH3CN),実験値421.9(MNa++CH3CN)。
【0044】
中間体D
【0045】
【化7】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート
DCM(27.9ml)中のトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノール(2g,5mmol)にトリフルオロメタンスルホン酸無水物(1.13ml,6.69mmol)とピリジン(0.902ml,11.15mmol)を0℃にて加えた。反応混合物を30分間撹拌し、飽和NH4Cl(30ml)でクエンチし、EtOAc(150ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−25%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物2.7gを白色固体として得た。1H NMR(600MHZ,CDCl3)δ7.36(m,4H),7.02(m,1H),6.79(m,2H),5.72(m,1H),3.66(broad s,2H),3.02(dd,J=14.4,6.6Hz,2H)。MS計算値554.0(MNa++CH3CN),実験値553.8(MNa++CH3CN)。
【0046】
中間体E
【0047】
【化8】
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート(3.9g,7.95mmol)とナトリウムアジド(5.17g,79mmol)をエタノール(39.7ml)と水(39.7ml)に加えた混合物を85℃に2時間加熱した。混合物を室温まで冷却し、水(100ml)とEtOAc(150ml)で希釈した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−30%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物2.3gを白色固体として得た。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.02(m,1H),6.91(m,1H),6.80(m,1H),3.81(m,1H),3.30(m,2H),3.02(m,2H)。
【0048】
中間体F
【0049】
【化9】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン(2.07g,5.39mmol)とパラジウム(0.861g,炭素担持10%,0.809mmol)をMeOH(27ml)中でH2バルーン下に4時間撹拌した。粗製混合物をシリカゲルパックで濾過し、5:1 DCM/MeOH(100ml)で洗浄してパラジウム残渣を除去した。溶媒を除去して生成物を得、それ以上精製せずに次の変換でそのまま使用した。1H NMR(600MHZ,CDCl3)δ7.34(m,4H),6.97(m,1H),6.84(m,1H),6.77(m,1H),3.45(m,1H),3.05−2.94(m,4H)。MS計算値358.0(MH+),実験値358.0(MH+)。
【0050】
方法(b)
シス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン(9.5g,24.75mmol)のエタノール/THF溶液に室温で撹拌下に亜鉛(3.24g,49.5mmol)を加えた後、ギ酸アンモニウム(3.12g,49.5mmol)を加えた。反応混合物を室温で1時間撹拌した。混合物をセライトで濾過後、溶媒を除去した。得られた残渣に飽和NaHCO3溶液100mlを加え、生成物をEtOAc(2×100ml)で抽出した。有機相を合わせてブラインで洗浄し、無水硫酸ナトリウムで乾燥し、セライトで濾過した。濾液を濃縮し、目的生成物を得た。
【0051】
中間体G
【0052】
【化10】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタノールを使用した以外はトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナートと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.04(m,1H),6.88(m,1H),6.81(m,1H),5.08(m,1H),3.60(m,2H),3.23(m,2H)。MS計算値554.0(MNa++CH3CN),実験値553.8(MNa++CH3CN)。
【0053】
中間体H
【0054】
【化11】
トランス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホン
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナートを使用した以外はシス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホンと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),6.98(m,1H),6.76(m,2H),4.56(m,1H),3.49(m,2H),2.66(m,2H)。
【0055】
中間体I
【0056】
【化12】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン
トランス−3−アジド−1−(2,5−ジフルオロフェニル)シクロブチル4−クロロフェニルスルホンを使用した以外はシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンと同様に製造した。1H NMR(600MHZ,CD3OD)δ7.48−7.41(m,4H),7.07(m,1H),6.88(m,1H),6.79(m,1H),3.88(m,1H),3.41(m,2H),2.50(m,2H)。MS計算値358.0(MH+),実験値358.0(MH+)。
【0057】
中間体J
【0058】
【化13】
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチルトリフルオロメタンスルホナート(4g,8.15mmol)とシアン化テトラブチルアンモニウム(5.47g,20.37mmol)をDMSO(54ml)中で80℃に45分間加熱した。得られた混合物を室温まで冷却し、水(200ml)とEtOAc(250ml)で希釈した。有機相を水、ブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−50%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物(2.9g)をオフホワイト固体として得た。1H NMR(600MHZ,CDCl3)δ7.38−7.33(m,4H),7.02(m,1H),6.80−6.74(m,2H),3.74(m,1H),3.51(m,2H),3.02(m,2H)。MS計算値431.0(MNa++CH3CN),実験値431.0(MNa++CH3CN)。
【0059】
中間体K
【0060】
【化14】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリル
THF(8mL)中のトランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル(600mg,1.63mmol)にLiHMDS(2.45ml,THF中1M,2.45mmol)を−78℃で加えた。10分後にMeI(306μl,4.89mmol)を反応混合物に導入した。温度をゆっくりと0℃まで上げながら反応混合物を2時間撹拌した。次に反応混合物を水でクエンチし、EtOAcで抽出した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中0−50%酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、生成物(260mg)を単一ジアステレオマー生成物として得た。1H NMR(600MHZ,CD3OD)δ7.36(s,4H),7.02(m,1H),6.85−6.78(m,2H),3.79(d,J=14.4Hz,2H),2.72(d,J=14.4Hz,2H),1.44(s,3H)。MS計算値785.1(2M+Na+),実験値785.0(2M+Na+)。
【0061】
中間体L
【0062】
【化15】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド
DMSO(6.5ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリル(500mg,1.31mmol)とK2CO3(362mg,2.62mmol)にH2O2(1.15ml,水中35%,13.1mmol)を滴下し、反応混合物を2時間激しく撹拌した。混合物を水(50ml)とEtOAc(50ml)で希釈した。有機相を水、ブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド(500mg)をオフホワイト固体として得、それ以上精製せずに次の反応で使用した。1H NMR(600MHZ,CDCl3)δ7.35(m,4H),7.02(m,1H),6.87−6.79(m,2H),6.54(broad s,1H),6.40(broad s,1H),3.67(d,J=14.4Hz,2H),2.62(d,J=14.4Hz,2H),1.28(s,3H)。MS計算値400.0(MH+),実験値400.0(MH+)。
【0063】
中間体M
【0064】
【化16】
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミン
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボキサミド(400mg,1mmol)とPIFA(473mg,1.1mmol)をアセトニトリル(2.5ml)と水(2.5ml)に加え、0℃で撹拌し、混合物を27時間かけて室温までゆっくりと昇温した。次に反応混合物を飽和NaHCO3でクエンチし、EtOAcで抽出した。有機相を分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。DCM中0−40% MeOHを溶離液としてこの物質をシリカクロマトグラフィーに付し、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミン(380mg)を生成物として得た。1H NMR(600MHZ,CD3OD)δ7.41−7.36(m,4H),7.02(m,1H),6.81(m,1H),6.74(m,1H),3.58(d,J=15.6Hz,2H),2.89(d,J=15.6Hz,2H),1.58(s,3H)。MS計算値372.1(MH+),実験値372.0(MH+)。
【0065】
中間体N
【0066】
【化17】
シス−1−アリル−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリル
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンカルボニトリルと臭化アリルを使用した以外はシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンカルボニトリルと同様に製造した。1H NMR(600MHZ,CDCl3)δ7.36(s,4H),7.03(m,1H),6.83−6.78(m,2H),5.70(m,1H),5.18(dd,J=10.2,1.2Hz,1H),5.07(dd,1H,J=16.8,1.2Hz,1H),3.70(d,J=15.0Hz,2H),2.77(d,J=15.0Hz,2H),2.32(d,J=6.6Hz,2H)。MS計算値837.1(2M+Na+),実験値837.0(2M+Na+)。
【0067】
中間体O
【0068】
【化18】
4−{[シス−3−アミノ−1−(2,5−ジフルオロフェニル)シクロブチル]スルホニル}ベンゾニトリル
シアン化亜鉛(0.295g,2.52mmol)と、Pd2(dba)3(0.384g,0.419mmol)と、亜鉛(0.030g,0.461mmol)と、DPPF(0.465g,0.838mmol)と、シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン(1.5g,4.19mmol)をDMAに溶解し、25mLシュレンク管でアルゴン雰囲気下に135℃にて16時間撹拌した。次に水を加え、混合物をEtOAcで抽出した。有機層を飽和NaHCO3溶液とブラインで洗浄後、MgSO4で乾燥し、濾過し、濃縮した。残渣をシリカカラムクロマトグラフィー(0→8% MeOH/DCM)により精製し、標記化合物を得た。MS:計算値349(MH+),実験値349(MH+)。
【0069】
(実施例1)
【0070】
【化19】
N−[トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
DCM(27.9ml)中のシス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミン(1.5g,4.19mmol)にトリエチルアミン(1.169ml,8.38mmol)とトリフルオロメタンスルホン酸無水物(0.850ml,5.03mmol)を0℃で加え、混合物を2時間撹拌した。
【0071】
混合物を水(50ml)とEtOAc(100ml)で希釈した。有機相をブラインで洗浄し、分離し、乾燥し(Na2SO4)、蒸発乾涸し、油状物を得た。ヘキサン中酢酸エチルを溶離液としてこの物質をシリカクロマトグラフィーに付し、目的生成物(白色固体として1.45g)を得た。1H NMR(600MHZ,CDCl3)δ7.39(d,J=9.0Hz,2H),7.34(d,J=9.0Hz,2H),7.02(m,1H),6.77(m,2H),4.22(m,1H),3.23(m,4H)。MS計算値553.0(MNa++CH3CN),実験値553.0(MNa++CH3CN)。
【0072】
(実施例2)
【0073】
【化20】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
トランス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンを使用した以外は実施例1と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.33(m,4H),6.98(m,1H),6.74(m,2H),5.35(d,J=7.8Hz,1H,NH),4.67(m,1H),3.59(m,2H),2.68(m,2H)。MS計算値553.0(MNa++CH3CN),実験値552.8(MNa++CH3CN)。
【0074】
同様の手順により以下の化合物を製造した。
【0075】
【表1】
【0076】
(実施例6)
【0077】
【化21】
N−{シス−3−(5−クロロ−2−フルオロフェニル)−3−[(4−クロロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:MS計算値506(MNa+),実験値529;1H NMR(400MHz,CDCl3)δ7.41〜7.43(d,J=8.8Hz,2H),7.34〜7.36(d,J=8.4Hz,2H),7.28〜7.33(m,1H),6.98〜7.01(m,1H),6.75〜6.83(m,2H),4.20〜4.29(m,1H),3.20〜3.32(m,4H)。
【0078】
(実施例7)
【0079】
【化22】
N−{シス−3−(2,5−ジフルオロフェニル)−3−[(4−フルオロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:1H NMR(400MHz,CDCl3):δ7.45〜7.49(m,2H),7.12〜7.29(m,2H),7.04〜7.10(m,1H),6.80〜6.85(m,3H),4.24〜4.30(m,1H),3.24〜3.35(m,4H)。
【0080】
(実施例8)
【0081】
【化23】
N−{シス−3−(2,5−ジフルオロフェニル)−3−[(3,4−ジフルオロフェニル)スルホニル]シクロブチル}−1,1,1−トリフルオロメタンスルホンアミド:MS 計算値514(MH+),実験値514;1H NMR(400MHz,CDCl3):δ7.15〜7.19(m,3H),6.97〜7.04(m,1H),6.72〜6.80(m,2H),6.63〜6.65(d,J=10.4Hz,1H),4.13〜4.23(m,1H),3.14〜3.26(m,4H)。
【0082】
(実施例9)
【0083】
【化24】
N−[シス−3−[(4−シアノフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
MS:計算値502(MNa+),実験値502(MNa+)。1H NMR(CDCl3 600MHz)7.67(d,J=4.1Hz,2H),7.58(d,J=4.1,2H),7.08(bm,1H),6.84(m,1H),6.68(m,1H),6.60(d,1H),4.24(bm,1H),3.4−3.2(bm,4H)。
【0084】
中間体P
【0085】
【化25】
4−{[3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸tert−ブチル
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(190mg,0.388mmol)をDMF(1.1mL)に加え、炭酸カリウム(59mg,0.427mmol)と4−ブロモブタン酸tert−ブチル(95mg,0.427mmol)で処理した。混合物を80℃まで加熱し、16時間撹拌した。反応混合物を周囲温度まで冷却し、酢酸エチルで希釈し、1/2飽和ブライン溶液で2回洗浄した。有機層を無水硫酸マグネシウムで乾燥し、濾過後、減圧濃縮した。残渣をMPLC(0−30% EtOAc:ヘプタン)により精製し、標記化合物を得た。MS:計算値654(M Na+),実験値654(M Na+)。
【0086】
(実施例10)
【0087】
【化26】
4−{[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸
4−{[3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アミノ}ブタン酸tert−ブチル(144mg,0.228mmol)を1:1 DCM:TFA(1.1mL)に加え、周囲温度で40分間撹拌した。反応混合物を減圧濃縮した。ヘプタンを加えてトリチュレーション後に標記化合物を白色固体として単離した。1H NMR(DMSO D6,600MHz)δ12.25(s,1H),7.58(d,J=8.2Hz,2H),7.43(d,J=8.2Hz,2H),7.25−7.31(m,1H),7.07−7.16(m,2H),4.18−4.28(m,1H),3.53(s br,2H),3.30−3.42(m,2H),3.08(s br,2H),2.26−2.38(m,2H),1.80−1.90(m,2H)。MS:計算値598(M Na+),実験値598(M Na+)。
【0088】
(実施例11)
【0089】
【化27】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロ−N−[2−(テトラヒドロ−2−ピラン−2−イルオキシ)エチル]メタンスルホンアミド
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(50mg,0.102mmol)を無水DMF(0.3mL)に溶解し、この溶液に撹拌下に炭酸カリウム(49mg,0.357mmol)を加えた後、2−(2−ブロモエトキシ)テトラヒドロ−2H−ピラン(53mg,0.255mmol)を加えた。得られた混合物を80℃で16時間撹拌した。混合物を周囲温度まで冷却し、水で希釈し、EtOAcで抽出した。有機層を再び飽和重炭酸塩水溶液で洗浄後、無水硫酸マグネシウムで乾燥し、減圧濃縮した。抽出層をMPLC(0−45% EtOAc/DCM)により精製し、標記化合物を得た。40% EtOAc/DCM中Rf=0.71。1H NMR(CDCl3,600MHz)δ7.31−7.38(m,4H),7.00−7.07(m,2H),6.88−6.93(m,1H),6.78−6.84(m,1H),4.65−4.68(m,1H),4.06−4.08(m,1H),3.50−4.00(m,7H),2.80−3.10(m,2H),1.20−1.90(m,7H)。MS:計算値640(M Na+),実験値640(M Na+)。
【0090】
(実施例12)
【0091】
【化28】
{[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル))スルホニル]アミノ}酢酸メチル
無水THF(0.8mL)をN−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(200mg,0.408mmol)に加えた後、反応混合物を0℃まで冷却した。水素化ナトリウム(49mg,1.225mmol)を一度に加え、混合物を0℃で15分間撹拌した。混合物は発泡し、オフホワイト懸濁液の状態で撹拌した。15分後にブロモ酢酸メチル(187mg,1.225mmol)を加えると、混合物は黄変しながら再び発泡した。混合物を16時間撹拌後、飽和塩化アンモニウム水溶液でクエンチし、EtOAcで抽出した。有機層を再び飽和塩化アンモニウム水溶液で洗浄後、無水硫酸マグネシウムで乾燥し、減圧濃縮した。抽出層をMPLC(0−45% EtOAc:ヘプタン)により精製し、標記化合物を得た。40% EtOAc:ヘプタン中Rf=0.6。1H NMR(CDCl3,600MHz)δ7.36(d,J=7.0Hz,2H),7.43(d,J=7.0Hz,2H),7.02−7.08(m,1H),6.86−6.90(m,1H),6.78−6.84(m,1H),4.30−4.60(m,3H),3.85(s,3H),3.38−3.34(m,2H),3.04(s br,2H)。MS:計算値584(M Na+),実験値584(M Na+)。
【0092】
同様の手順により以下の化合物を製造した。
【0093】
【表2】
【0094】
(実施例17)
【0095】
【化29】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)−1−メチルシクロブタンアミンを使用した以外は実施例1と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.33(m,4H),7.00(m,1H),6.90(s,1H,NH),6.82−6.75(m,2H),3.54(d,J=14.4Hz,2H),2.87(d,J=14.4Hz,2H),1.44(s,3H)。MS計算値526.0(MNa+),実験値525.9(MNa+)。
【0096】
(実施例18)
【0097】
【化30】
N−(シス−3−(2,5−ジフルオロフェニル)−1−メチル−3−{[4−(トリフルオロ)フェニル]スルホニル}シクロブチル)−1,1,1−トリフルオロメタンスルホンアミド
実施例17と同様の手順を使用して製造した。計算値(2M+Na)+:1097.0,実測値:1096.5。
【0098】
(実施例19)
【0099】
【化31】
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロ−N−[(トリフルオロメチル)スルホニル]メタンスルホンアミド
実施例1の合成で副生物として製造した。1H NMR(600MHZ,CDCl3)δ7.38−7.33(m,4H),7.07(m,1H),6.95(m,1H),6.84(m,1H),4.44(m,1H),3.82(m,2H),3.15(m,2H)。MS計算値684.9(MNa++CH3CN),実験値684.9(MNa++CH3CN)。
【0100】
(実施例20)
【0101】
【化32】
ナトリウム[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アザニド
水素化ナトリウムをヘキサンに懸濁し、0℃まで冷却した。THF(1mL)中のN−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(200mg,0.408mmol)を水素化ナトリウム懸濁液に滴下した。得られた混合物を0℃で15分間、次いで周囲温度で30分間撹拌した。この時点で反応混合物を減圧濃縮した。乾燥白色粉末をフラスコから掻き取り、ガラスフリット付き漏斗に入れ、氷冷ペンタン(45mL)で洗浄した。次に粉末を高減圧下に16時間置いた。1H NMR(DMSO D6,600MHz)δ7.56(d,J=8.5Hz,2H),7.33(d,J=8.5Hz,2H),7.20−7.26(m,1H),7.00−7.12(m,2H),3.42−3.52(m,1H),2.66−2.80(m,4H)。
【0102】
(実施例21)
【0103】
【化33】
カリウム[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル][(トリフルオロメチル)スルホニル]アザニド
N−[シス−3−[(4−クロロフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド(1.2g,2.55mmol)を無水THF(25.5mL)中で0℃にて撹拌後、カリウムtert−ブトキシド(0.29g,2.55mmol)で処理した。反応混合物を0℃で15分間撹拌後、周囲温度まで昇温し、更に45分間撹拌した。反応混合物を減圧濃縮後、得られた白色粉末を100℃で撹拌下に最少量の3:1 IPA:トルエン(400mL)から再結晶させた。溶解後、混合物を濾紙で濾過し、4℃で20時間静置した。ガラスフリットで濾過して結晶を採取し、冷ペンタンで3回洗浄した。残留溶媒を減圧除去した。1H NMR(DMSO D6,600MHz)δ7.56(d,J=8.5Hz,2H),7.33(d,J=8.5Hz,2H),7.20−7.26(m,1H),7.00−7.12(m,2H),3.42−3.52(m,1H),2.66−2.80(m,4H)。
【0104】
(実施例22)
【0105】
【化34】
N−[シス−3−[(4−トリフルオロメトキシフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブチル]−1,1,1−トリフルオロメタンスルホンアミド
シス−3−[(4−トリフルオロメトキシフェニル)スルホニル]−3−(2,5−ジフルオロフェニル)シクロブタンアミンを使用した以外は実施例2と同様に製造した。1H NMR(600MHZ,CDCl3)δ7.56(m,2H),7.27(m,2H)7.088(m,1H),6.78(m,2H),6.68(d,J=10.6Hz,1H,NH),4.30(m,1H),3.31(m,2H),3.18(m,2H)。MS計算値603.46(MNa++CH3CN),実験値603.0(MNa++CH3CN)。
【0106】
生物活性
以下、本発明の化合物の生物活性を測定するためのアッセイについて記載する。
【0107】
APPプロセシング(細胞株から分泌されるAβ検体を定量するアッセイ):
アミロイドβ蛋白質を発現するSH−SY5Y細胞(SP4CT細胞)から産生されるAβ40及びAβ42ペプチドの量に与える化合物の影響をAlphaLisa(登録商標)アッセイにより判定した。ELISAアッセイと同様に、このAlphaLisa(登録商標)アッセイでシグナルを発生するには、Aβ40又はAβ42ペプチドの特異的抗体認識により「ドナー」ビーズと「アクセプター」ビーズを密接に近接させる必要がある。化合物で処理したSP4CT細胞から培地を別個のマイクロプレート2枚に移した後に、ビオチン化抗アミロイドβモノクローナル抗体(クローン4G8)と結合するストレプトアビジンを固定化したドナービーズを加えることによりアッセイを実施した。抗Aβ40モノクローナル抗体(G210)を直接固定化したアクセプタービーズを一方のマイクロプレートに加え、抗Aβ42モノクローナル抗体(12F4)アクセプタービーズを他方に加えた。Aβ40とAβ4の量はレーザー光によるドナービーズの励起後に発生した発光シグナルに正比例した。
【0108】
ノッチプロセシング:(細胞株へのノッチ細胞内ドメイン遊離量を定量するアッセイ):
ノッチ蛋白質のγセクレターゼ依存的切断の阻害を定量するために「スプリットルシフェラーゼ」アッセイを使用する。このアッセイでは、ルシフェラーゼのN末端フラグメントに融合したその細胞外ドメイン(ノッチ△E)をもたないノッチ蛋白質をHeLa細胞に発現させた。同一細胞は免疫グロブリンJκ組換えシグナル配列結合蛋白質(RBP)と融合したルシフェラーゼのC末端フラグメントも発現した。ノッチ△Eがγセクレターゼにより切断されると、ノッチ細胞内ドメイン(NICD)−N末端ルシフェラーゼ蛋白質が生成され、核転座してRBP−C末端ルシフェラーゼ融合体と結合し、ルシフェラーゼの独立して非機能的な半分ずつを一緒にして機能的ルシフェラーゼ酵素を形成する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるノッチの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。
【0109】
PXRアッセイ解説
CYP3A4−SEAPトランス活性化アッセイ(PCSTA)はチトクロームP450 CYP3A(ヒトCYP3A4又はラットCYP3A1)を誘導する可能性について化合物を有効且つ迅速に評価する。レポーター構築物は分泌型アルカリホスファターゼ(SEAP)遺伝子のすぐ上流に配置されたCYP3A4遺伝子に由来する調節配列を含む。メチオニンが野生型配列に存在するロイシンに置換する開始アミノ酸となるように、ヒトPXR核内受容体の5’末端を修飾した。HEP G2細胞にPXRプラスミド(ヒト又はラット)とレポータープラスミドをトランスフェクトする。CYP3A4の誘導の読取りはパラニトロフェニルリン酸(pNPP)を基質とするSEAP比色アッセイから構成される。各点がSEAPによるpNPPからpNPへの変換率に対応するように5点用量応答曲線を2本ずつ作成する。ヒトPCSTAでは、リファンピシンを陽性対照として使用し、100%誘導は10μMリファンピシンにより生じる最大誘導に基づく。
【0110】
ICDトランス活性化(細胞株へのγセクレターゼ基質群の細胞内ドメイン遊離量を定量するアッセイ)
ホタルルシフェラーゼによるトランス活性化アッセイを使用してγセクレターゼ基質のε/S3サイト切断の阻害を測定する。このアッセイは細胞内ドメイン(ICD)に融合したGAL4/VP16(GVP)トランス活性化ドメインを有するキメラ基質であるAPP−GVP、ノッチ△E−GVP、E−カドヘリン−GVP及びCD44−GVPを使用する。ICDが切断され、遊離されると、GVPドメインはUASプロモーターの制御下でルシフェラーゼ遺伝子の発現を誘導する。このアッセイでは、UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼ(トランスフェクション対照)と共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした。γセクレターゼにより切断されると、遊離されるICD−GVPは核転座し、UAS−ルシフェラーゼ遺伝子の発現を誘導する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるICDの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。更に、トランスフェクション効率の差を考慮するために、吸光度によるβ−ガラクトシダーゼ酵素アッセイを実施し、発光量読取を正規化する。
【0111】
全長γセクレターゼ基質の評価(γセクレターゼ基質群のプロセシングを定量的に評価するアッセイ)
他の基質に対するγセクレターゼ活性に与える化合物の影響を試験するために、C末端V5タグを付けた以下のI型膜蛋白質:CD43、CD44、E−カドヘリン及びSCN2bのうちの1種を過剰発現する4種類のHEK293安定細胞株を構築した。細胞を撒き、滴定化合物とホルボールエステルTPAで一晩処理する。全蛋白質は初期エクトドメインシェディングイベントとそれに続くγセクレターゼによるC末端フラグメント(CTF)の膜貫通切断を特徴とする調節下の膜蛋白質分解を受けるので、TPAはγセクレターゼの基質を生成する初期切断イベントを誘導する。V5タグを付けたCTFのプロセシングをウェスタンブロット分析により追跡することにより、これらの基質に対するγセクレターゼ活性に与える化合物の影響を測定する。CTFの蓄積はγセクレターゼ活性の阻害を意味する。
【0112】
ICDトランス活性化(細胞株へのγセクレターゼ基質群の細胞内ドメイン遊離量を定量するアッセイ)
ホタルルシフェラーゼによるトランス活性化アッセイを使用してγセクレターゼ基質のε/S3サイト切断の阻害を測定する。このアッセイは細胞内ドメイン(ICD)に融合したGAL4/VP16(GVP)トランス活性化ドメインを有するキメラ基質であるAPP−GVP、ノッチ△E−GVP、E−カドヘリン−GVP及びCD44−GVPを使用する。ICDが切断され、遊離されると、GVPドメインはUASプロモーターの制御下でルシフェラーゼ遺伝子の発現を誘導する。このアッセイでは、UASプロモーターに誘導されるルシフェラーゼ及びβ−ガラクトシダーゼ(トランスフェクション対照)と共にキメラ基質をHEK細胞に一過的に同時トランスフェクトした。γセクレターゼにより切断されると、遊離されるICD−GVPは核転座し、UAS−ルシフェラーゼ遺伝子の発現を誘導する。これらの細胞におけるルシフェラーゼ活性はγセクレターゼにより切断されるICDの量に正比例する。溶解細胞へのルシフェリン添加と総発光量測定の標準技術によりルシフェラーゼ活性を測定する。更に、トランスフェクション効率の差を考慮するために、吸光度によるβ−ガラクトシダーゼ酵素アッセイを実施し、発光量読取を正規化する。
【0113】
インビトロAPPプロセシング(半精製γセクレターゼと共にインキュベートした組換えAPPC100Flag基質から生成されるAβ検体を定量するアッセイ)
半精製γセクレターゼにより外来C100Flag基質から生成されるAβ40及びAβ42ペプチドの量に与える化合物の影響をMESO Scale ELISAにより判定した。このMESO Scaleアッセイでシグナルを発生するには、ストレプトアビジンを固定化した抗アミロイドモノクローナル抗体(クローン4G8)がビオチンを被覆したプレートと結合する必要がある。その後、Aβ40(G210)又はAβ42(12F4)に特異的な[Ru(bpy)3]2+標識モノクローナル抗体は電気化学刺激を受けると電気化学発光シグナルを発生する。化合物と、C100Flag基質と、HeLa細胞又はマウス、ラットもしくはイヌの脳に由来するP2膜をCHAPSOで可溶化したものをインキュベートすることによりアッセイを実施した。次に反応混合物をAβ40又はAβ42検出用の別個のビオチン化マイクロプレート2枚に移した。
【0114】
インビトロノッチプロセシング(半精製γセクレターゼと共にインキュベートした組換えノッチ△E100Flag基質からのノッチ細胞内ドメイン生成を定量的に評価するアッセイ)
同様に、基質をN100Flagに代えることにより、C100Flagインビトロアッセイと同一方法を使用してノッチプロセシングをモニターすることができる。ポリクローナルビオチン標識抗DYKDDDDK抗体を捕捉用抗体として使用し、切断されたノッチ1抗体をポリクローナル[Ru(bpy)3]2+で標識して使用し、NICDを検出した。
【0115】
化合物結合:(結合したγセクレターゼ阻害剤トレーサーのインビトロ置換を定量するアッセイ)
組換えγセクレターゼを安定的に過剰発現するHEK293(γNRCF8)P2膜をCHAPSOで可溶化したものを使用して全放射性リガンド結合実験を実施する。放射性リガンド結合には、トリチウム化した阻害剤の存在下で可溶化酵素をインキュベートする。過剰の未標識阻害剤を反応に加えることにより非特異的結合を測定し、トリチウム化したリガンドの系列希釈を使用して飽和結合等温線を得る。ポリエチレンイミンを被覆したガラス繊維フィルタープレートに酵素複合体を吸着させ、セルハーベスターで迅速に濾過後に洗浄することにより、結合したリガンドを遊離リガンから分離する。プレートを乾燥後、シンチラントを加え、マイクロプレートシンチレーションカウンターでプレートを読取る。1nM 3H標識化合物として参考例L−458(遷移状態阻害剤)又は4nM 3H標識化合物として参考例L−881(非遷移状態阻害剤)の存在下で各種阻害剤の系列希釈をインキュベートすることにより結合競合アッセイを実施する。各種阻害剤のアンタゴニスト競合性を判定するために、種々の濃度のこれらの化合物の存在下で夫々の3Hトレーサー用量応答曲線を分析する。
【0116】
参考例L−458:
【0117】
【化35】
【0118】
参考例L−881:
【0119】
【化36】
【0120】
結果
上記APP及びノッチプロセシング細胞機能アッセイで本明細書の実施例を試験した。試験した化合物は下表に示すようにノッチシグナル伝達経路を保持しながらAPPプロセシングのインビトロ阻害を示した。データは少なくとも>4回の反復の平均に基づく。
【0121】
【表3】
【0122】
WO02/081435A1(公開日2002年10月17日)はγセクレターゼの活性を調節するスルホン誘導体を開示している。WO2004/031139(公開日2004年4月15日)はγセクレターゼ阻害剤としてのシクロヘキシルスルホン誘導体を開示している。WO2004/031139の実施例47は下記構造をもつ。
【0123】
【化37】
上記化合物はMRK−560とも呼ばれ、Best et al.,J Pharmacol Exp,Ther.,317:786−790,2006及びBest et al,J.Pharmacol.Exp.Ther.,320:552−558,2007に開示されている。文献はMRK−560におけるAPPのインビトロ阻害とノッチプロセシング経路の間の分離について殆ど又は全く報告していないが、この化合物はTg2576マウスでノッチシグナル伝達経路の変化に関連する毒性の不在下でアミロイドプラーク沈着に有益なインビボ効果を示した。
【0124】
薬剤毒性を予測する共有蛋白質結合アッセイでMRK−560と実施例2を試験した。該当化合物の放射性標識物を肝ミクロソームと共にインキュベートすることにより、薬剤候補が蛋白質と共有結合を生じる可能性を評価する。次にBrandel Harvester技術に基づく半自動法を使用して試験化合物と肝蛋白質の共有結合性付加物の形成を測定する(Ref.Day et.al,J Pharmacol Toxicol Mehtods.52,278−85,2005参照)。結果を下表に示す。
【0125】
【表4】
【0126】
薬剤間相互作用を予測するプレグナンX受容体(PXR)と結合及び/又はこれを活性化する能力についてMRK−560と本明細書に開示した実施例を試験した。結果を下表に示す。
【0127】
【表5】
【0128】
他のγセクレターゼ基質の初期切断に与える影響を試験するために、MRK−560と実施例2を上記トランス活性化アッセイで試験した。MRK−560は試験した全基質のICD遊離を阻害したが、ICD遊離とそれに続くICD−GVP構築物の転座によりレポーター活性化が可能であったことから明らかなように、実施例2は初期ε/S3切断を維持した。図2参照。
【0129】
MRK−560によるε/S3及びγ切断の完全な阻害の結果、SCN2b−、ECAD−及びCD43−CTF蓄積を生じることが多基質細胞アッセイにより確認された。これに対して、実施例2で処理すると、CTF蓄積は低下又はゼロになり、NS−GSIはγセクレターゼ基質の初期切断を維持すると判断された。図3参照。
【0130】
MRK−560と実施例2を上記化合物結合アッセイでも試験した。各化合物の濃度を増加しながら、トリチウム化GSIトレーサーL−458(遷移状態,赤)又はL−881(非遷移状態,青)を半精製γセクレターゼ複合体と共にインキュベートした。MRK−560は夫々L−881及びL−458サイトの完全及び部分的置換を示した。実施例2はL−881を完全に置換することができたが、L−458はできなかった。以上の結果から、実施例2等のノッチスペアリング化合物はMRK−560等の従来の阻害剤に比較して結合部位が移動していることが明らかである。このため、全γ切断部位を潜在的に阻害しながらε/S3(AICD/NICD遊離)を保持するように酵素切断を制御解除する。図4参照。
【特許請求の範囲】
【請求項1】
式I
【化1】
[式中、X1はF及びCNから構成される群から選択され;
X2はF、Cl及びCNから構成される群から選択され;
X3はF、Br、Cl、CN、CF3、OCF3、C(O)−OCH3及びS−CH3から構成される群から選択され;
X4はH、F及びClから構成される群から選択され;
R1は
(a)H、
(b)CH3、
(c)−(CH2)n−OR3、
(d)−(CH2)n−C(O)−OR4、及び
(e)−SO2−CF3
から構成される群から選択され;
式Iの化合物がシス配置であるとき、R2はH又はCH3であり、それ以外の場合にはR2はHであり;
R3は1個の酸素ヘテロ原子をもつ5又は6員非芳香族複素環であり;
R4はH又はCH3であり;
nは1〜4である]の化合物又はその医薬的に許容可能な塩。
【請求項2】
式Ia:
【化2】
の請求項1に記載の化合物又はその医薬的に許容可能な塩。
【請求項3】
X1がFであり、X4がHである請求項2に記載の化合物。
【請求項4】
X2がFであり、X3がClである請求項3に記載の化合物。
【請求項5】
R2がHである請求項4に記載の化合物。
【請求項6】
R1が−(CH2)n−C(O)−OR4である請求項5に記載の化合物。
【請求項7】
式Ib:
【化3】
の請求項1に記載の化合物又はその医薬的に許容可能な塩。
【請求項8】
X1及びX2がFであり;
X3がClであり;
X4がHである
請求項7に記載の化合物。
【請求項9】
以下の群:
【化4】
から選択される請求項1に記載の化合物。
【請求項10】
【化5】
である請求項9に記載の化合物又はその医薬的に許容可能な塩。
【請求項11】
【化6】
である請求項10に記載の化合物。
【請求項12】
【化7】
である請求項10に記載の化合物。
【請求項13】
請求項1に記載の化合物又はその医薬的に許容可能な塩と、医薬的に許容可能なキャリヤーを含有する医薬組成物。
【請求項14】
βアミロイドの沈着を伴う病態に罹患している対象又は罹患し易い対象の治療方法であって、有効量の請求項1に記載の化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む前記方法。
【請求項15】
アルツハイマー病の治療又は予防用医薬の製造における請求項1に記載の化合物又はその医薬的に許容可能な塩の使用。
【請求項1】
式I
【化1】
[式中、X1はF及びCNから構成される群から選択され;
X2はF、Cl及びCNから構成される群から選択され;
X3はF、Br、Cl、CN、CF3、OCF3、C(O)−OCH3及びS−CH3から構成される群から選択され;
X4はH、F及びClから構成される群から選択され;
R1は
(a)H、
(b)CH3、
(c)−(CH2)n−OR3、
(d)−(CH2)n−C(O)−OR4、及び
(e)−SO2−CF3
から構成される群から選択され;
式Iの化合物がシス配置であるとき、R2はH又はCH3であり、それ以外の場合にはR2はHであり;
R3は1個の酸素ヘテロ原子をもつ5又は6員非芳香族複素環であり;
R4はH又はCH3であり;
nは1〜4である]の化合物又はその医薬的に許容可能な塩。
【請求項2】
式Ia:
【化2】
の請求項1に記載の化合物又はその医薬的に許容可能な塩。
【請求項3】
X1がFであり、X4がHである請求項2に記載の化合物。
【請求項4】
X2がFであり、X3がClである請求項3に記載の化合物。
【請求項5】
R2がHである請求項4に記載の化合物。
【請求項6】
R1が−(CH2)n−C(O)−OR4である請求項5に記載の化合物。
【請求項7】
式Ib:
【化3】
の請求項1に記載の化合物又はその医薬的に許容可能な塩。
【請求項8】
X1及びX2がFであり;
X3がClであり;
X4がHである
請求項7に記載の化合物。
【請求項9】
以下の群:
【化4】
から選択される請求項1に記載の化合物。
【請求項10】
【化5】
である請求項9に記載の化合物又はその医薬的に許容可能な塩。
【請求項11】
【化6】
である請求項10に記載の化合物。
【請求項12】
【化7】
である請求項10に記載の化合物。
【請求項13】
請求項1に記載の化合物又はその医薬的に許容可能な塩と、医薬的に許容可能なキャリヤーを含有する医薬組成物。
【請求項14】
βアミロイドの沈着を伴う病態に罹患している対象又は罹患し易い対象の治療方法であって、有効量の請求項1に記載の化合物又はその医薬的に許容可能な塩を前記対象に投与する段階を含む前記方法。
【請求項15】
アルツハイマー病の治療又は予防用医薬の製造における請求項1に記載の化合物又はその医薬的に許容可能な塩の使用。
【図1】

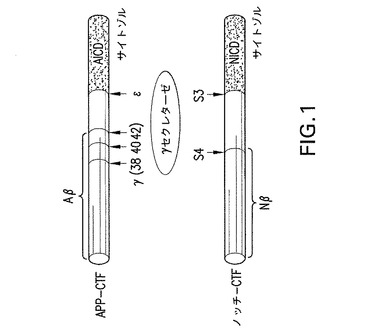
【図2】
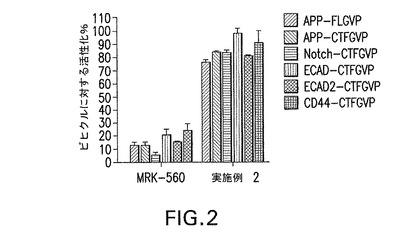
【図3】
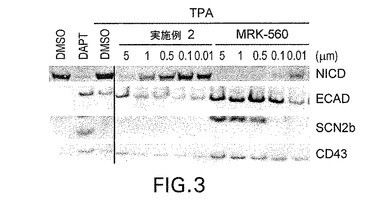
【図4A】
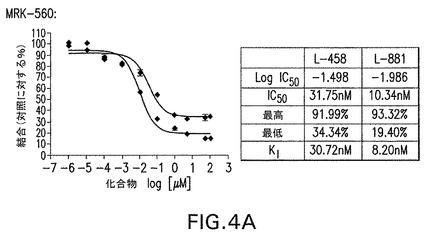
【図4B】
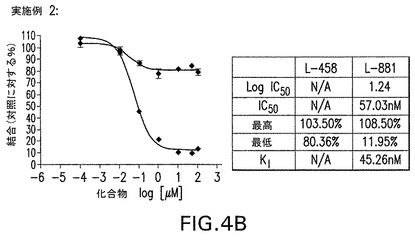
【図2】
【図3】
【図4A】
【図4B】
【公表番号】特表2011−518830(P2011−518830A)
【公表日】平成23年6月30日(2011.6.30)
【国際特許分類】
【出願番号】特願2011−506370(P2011−506370)
【出願日】平成21年4月17日(2009.4.17)
【国際出願番号】PCT/US2009/040913
【国際公開番号】WO2009/131906
【国際公開日】平成21年10月29日(2009.10.29)
【出願人】(390023526)メルク・シャープ・エンド・ドーム・コーポレイション (924)
【Fターム(参考)】
【公表日】平成23年6月30日(2011.6.30)
【国際特許分類】
【出願日】平成21年4月17日(2009.4.17)
【国際出願番号】PCT/US2009/040913
【国際公開番号】WO2009/131906
【国際公開日】平成21年10月29日(2009.10.29)
【出願人】(390023526)メルク・シャープ・エンド・ドーム・コーポレイション (924)
【Fターム(参考)】
[ Back to top ]