
ハフニア・フィターゼ
本発明は、フィターゼ活性を有する単離されたポリペプチド及び該ポリペプチドをコードする単離されたポリヌクレオチドに関する。該ポリペプチドは、ハフニア・アルベイから得られたフィターゼに関する。そのアミノ酸配列は、配列番号10として添付された配列表において示される。本発明はまた、該ポリペプチドを含む核酸構築物、ベクター及び宿主細胞、並びに特に動物飼料において該ポリペプチドを製造する方法及び用いる方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、フィターゼ活性を有する単離ポリペプチド、及び該ポリペプチドをコードする単離ポリヌクレオチドに関する。該ポリペプチドは、ハフニア・アルベイから得られたフィターゼに関し、そのアミノ酸配列は、配列番号10とし添付した配列表に示されている。本発明はまた、該ポリヌクレオチドを含む核酸構築物、ベクター及び宿主細胞、並びに該ポリペプチドを製造する方法及び特に動物飼料において該ポリペプチドを用いる方法、に関する。
【背景技術】
【0002】
配列表及び寄託微生物
配列表
本願は、配列表の書面コピー及びコンピュータ読み取り可能な形成を含む。コンピュータ読み取り可能な形態の内容は、参考によって本明細書に充分に援用される。
【0003】
生物材料の寄託
菌株を製造するフィターゼは、デンマーク土壌から単離され。該株は、酸性の最適pH及び高い熱安定性を有するフィターゼを製造することが証明された。該株は、ハフニア・アルベイ(ハフニア・アルベイ)として同定され、Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) によってブタペスト条約の下、2007年3月21日に寄託され、以下の受託番号:ハフニア・アルベイ NN020125 DSM 19197、2007年3月21日の受託番号及び受託日を与えられた。
【0004】
【表1】

【0005】
発明の背景
フィターゼは、ヒトを含む動物の飼料にそれらを添加することが有利であるので、周知の酵素である。フィターゼは、多数の真菌及び細菌株を含む、非常に多くの起源から単離されている。フィターゼ活性を有する別のポリペプチド、及び該ポリペプチドをコードするポリヌクレオチドを提供することが本発明の目的である。本発明のポリペプチドは、好ましくは、改変された、より好ましくは改善された性質、例えば、異なった基質特異性、より高い特異的活性、増加した安定性(例えば、酸-安定性、熱-安定性、及び/又はプロテアーゼ安定性、特にペプシン安定性)、変更された最適pH(例えば、より低い又はより高い最適的pH)、及び/又は 動物飼料における改善さえた能力(例えば、改善された放出及び/又はフィターゼの分解)を有する。
【発明の概要】
【0006】
発明の概要
本発明は、(a) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチドと少なくとも75%の同一性を有するアミノ酸配列を含むポリペプチド;及び/又は(b)(i)配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分の、1以上のアミノ酸の削除、挿入及び/又は保存的置換を含む変異体;及び/又は (c) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分、フラグメントからなる群より選択される、フィターゼ活性を有する単離ポリペプチドに関する。本発明はまた、(a) 配列番号10のアミノ酸1〜413と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;及び (b) 配列番号9の塩基100〜1338と少なくとも75%の同一性を有するポリヌクレオチド、からなる群より選ばれる、フィターゼ活性を有するポリペプチドをコードする単離ポリヌクレオチドに関する。本発明はまた、該ポリヌクレオチドを含む、核酸構築物、組換え発現ベクター、及び組換え宿主細胞に関する。
【0007】
本発明はまた、(a) 前記ポリペプチドの製造を誘導する条件下で、前記ポリペプチドをコードするポリヌクレオチドを含む核酸構築物を含む組換え宿主細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、フィターゼ活性を有するかかるポリペプチドを製造する方法に関する。本発明は、更に、(i) 配列番号11の塩基1〜99からなるシグナルペプチドをコードするヌクレオチド配列に作動可能に連結したタンパク質をコードする遺伝子を含む核酸構築物に関する。
【0008】
本発明はまた、動物試料において本発明のフィターゼを用いる方法、並びに前記ポリペプチドを含む動物試料及び動物試料添加組成物に関する。
【0009】
本発明はまた、発酵生成物、例えばエタノール、ビール、ワイン、の製造において本発明のフィターゼを用いる方法であって、該発酵が本発明のフィターゼの存在下で行われる、前記方法に関する。本発明は、本発明のフィターゼを用いて、植物タンパク質を含むタンパク質を処理する方法に関する。
【図面の簡単な説明】
【0010】
【図1】図1は、125〜500 FYT/kg飼料で投与された、ハフニア・アルベイ・フィターゼとシトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼとの比較におけるインビトロでのインキュベーション後の残余イノシトール-リン酸結合リン酸を示す。
【図2】図2は、250〜500 FYT/kg飼料で投与された、ハフニア・アルベイ・フィターゼとペニオホラ・リシイ(Peniophora lycii)・フィターゼとのインビトロでのインキュベーション後の残余イノシトール-リン酸結合リン酸の比較を示す。
【図3】図3は、ハフニア・アルベイ・フィターゼの解析された結晶3次元構造についての構造座標の添付を示す。
【0011】
定義
フィターゼ活性:本文脈では、フィターゼ活性を有するポリペプチド(フィターゼ)は、フィテート(myo-イノシトールイノシトール六リン酸)の、(1)myo-イノシトール及び/又は(2)そのモノ-、ジ-、トリ-、テトラ-及び/又はペンタ-リン酸エステル、及び(3)無機リン酸塩への加水分解を触媒する酵素である。
【0012】
インターネット(www.expasy.ch/enzyme/)の「酵素」サイトは、酵素の命名に対する情報の貯蔵所である。それは、主に、国際生化学分子生物学連合の命名法委員会(NC-IUBMB)の推奨に従い、EC(酵素委員会)番号は提供している特徴付けられた酵素の各種類を記載している (Bairoch A. 「酵素」データベース, 2000, Nucleic Acids Res 28: 304-305),NC-IUBMB, 1992のハンドブック酵素命名も参照)。
【0013】
酵素(ENZYME)サイトによれば、3種のフィターゼが知られている:3-フィターゼ (myo-イノシトールイノシトール六リン酸 3-ホスホヒドロラーゼ, EC 3.1.3.8)、6-フィターゼ (myo-イノシトールイノシトール六リン酸 6-ホスホヒドロラーゼ, EC 3.1.3.26)、及び5-フィターゼ (EC 3.1.3.72)。本発明の目的のためには、すべての種類がフィターゼの定義に含まれる。具体的な実施態様では、本発明のフィターゼは、エシェリヒア・コリ(Escherichia coli)pH 2.5の酸ホスファターゼ(appA遺伝子)、及び真菌フィターゼ、例えばアスペルギルス・アワモリ(Aspergillus awamorii)フィターゼA及びB(EC: 3.1.3.8)(phyA及びphyB遺伝子)を含む、酸ヒスチジンホスファターゼのファミリーに属する。ヒスチジン酸ホスファターゼは、2つの配列類似性領域を共有し、各々は、保存的ヒスチジン残基の周囲に集中している。これらの2つのヒスチジンは、酵素の触媒的メカニズムに関連していると考えられる。第1ヒスチジンは、N-末端部分に位置し、蛍光体-ヒスチジン中間体を形成するが、第2ヒスチジンは、C-末端部分に位置し、おそらくプロトン供与体として働く。
【0014】
更なる具体的な実施態様では、本発明のフィターゼは、保存的活性部位モチーフ、すなわちR-H-G-X-R-X-P、ここで、Xは、アミノ酸(配列番号10で示される成熟フィターゼのアミノ酸18〜24を参照)を有する。
【0015】
本発明の目的のために、フィターゼ活性は、FYTの単位において決定される。1つのFYTは、次の条件: pH 5.5; 温度37℃; 基質: 0.0050 mol/lの濃度中フィターゼナトリウム (C6H6O24P6Na12) の下で、1個のマイクロモル無機オルト-リン酸塩を1分当たりに放出する酵素量である。好適なフィターゼアッセイは、WO 00/20569の実施例1に記載のFYT及びFTUアアッセイである。FTUは、試料及びプレミックスにおけるフィターゼ活性を決定するためのものである。フィターゼ活性は、本明細書に記載の例のフィターゼアッセイを用いて決定してもよい。
【0016】
最適pH:本発明のポリペプチドの最適pHは、予備測定濃度での基質及び固定インキュベーション時間を用いて、様々なpH値でフィターゼを活性化することによって決定される。次いで、最適pHがフィターゼ活性-対-pHのグラフ表示決定される。具体的実施態様では、FYTアッセイが使用される。すなわち、基質は5 mMフィターゼナトリウム、反応温度37℃であり、活性は、以下の例で行われるように様々なpH-値にてFYT単位で測定される。別の特定の実施態様では、例のいずれ1つのフィターゼアッセイが使用される。比較的低い最適pHは、pH 5.0未満の最適pHを意味し、例えばpH 4.5、4.0、3.5、3.0、2.5又は更に2.0未満である。比較的高い最適pHは、pH 5.0超の最適pH、例えばpH 5.5、6.0、6.5、7.0、7.5、8.0、8.5又は更に9.0超である。
【0017】
単離ポリペプチド:本明細書で使用される用語「単離ポリペプチド」は、SDS-PAGEによって決定されるように、少なくとも20%純粋、好ましくは少なくとも40%純粋、より好ましくは少なくとも60%純粋、更により好ましくは少なくとも80%純粋、最も好ましくは少なくとも90%純粋、及び更に最も好ましくは少なくとも95%純粋であるポリペプチドを言う。
【0018】
実質的に純粋なポリペプチド:本明細書で使用される用語「実質的に純粋なポリペプチド」は、天然に関連する他のポリペプチド物質の、最大10重量%、好ましくは最大8重量%、より好ましくは最大6重量%、より好ましくは最大5重量%、より好ましくは最大4重量%、最大3重量%、更に好ましくは最大2重量%、最も好ましくは最大1重量%、及び更に最も好ましくは0.5重量%を含むポリペプチド調製物を意味する。そのため、実質的に純粋なポリペプチドが、該調製物に存在する総ポリペプチド物質の、少なくとも92重量%純粋、好ましくは少なくとも94重量%純粋、より好ましくは少なくとも95重量%純粋、より好ましくは少なくとも96重量%純粋、より好ましくは少なくとも97重量%純粋、より好ましくは少なくとも98重量%純粋、更に好ましくは少なくとも99重量%純粋、最も好ましくは少なくとも99.5重量%純粋、及び更に最も好ましくは100重量%純粋であることが好ましい。
【0019】
本発明のポリペプチドは、好ましくは、実施的に純粋な形態にある。具体的には、ポリペプチドは、「本質的に純粋な形態」、すなわち、そのポリペプチド調製物は、天然に関連する他のポリペプチド物質が本質的に含まない、ことが好ましい。このことは、例えば、周知の組換え法の手段によって又は典型的な精製法によってポリペプチドを調製することによって達成され得る。
【0020】
本明細書における用語「実質的に純粋なポリペプチド」は、用語「単離されたポリペプチド」及び「単離された形態でのポリペプチド」と同義である。
【0021】
同一性:2つのアミノ酸配列間、又は2つのヌクレオチド配列間の関係は、パラメータ「同一性」によって説明される。
【0022】
本発明の目的のために、2つのアミノ酸配列間の同一性の程度、及び2つのヌクレオチド配列間の同一性の程度は、タンパク質及びDNAアラインメントの両方に有用である、Needleman-Wunschアラインメント(すなわち、世界的なアラインメント)であるプログラム「アライン」によって決定される。デフォルトスコアリングマトリックスBLOSU M50及びデフォルト同一性マトリックスは、それぞれ、タンパク質及びDNAアラインメントのために使用される。ギャップ中の第1残基についてのペナルティは、タンパク質については-12であり、DNAについては-16である。ギャップ中の追加残基についてのペナルティは、タンパク質について-2であり、DNAについて-4である。
【0023】
「アライン」は、FASTA packageバージョンv20u6の一部である(W. R. Pearson and D. J.Lipman (1988), "生物学的配列解析のための改良されたツール", PNAS 85: 2444-2448, and W. R. Pearson (1990) "FASTP及びFASTAとの迅速かつ感度のよい配列比較," Methods in Enzymology 183: 63-98参照)。FASTAタンパク質アラインメントは、ギャップサイズに関して何の限定のないスミス・ウォーターマン(Smith-Waterman)アルゴリズムを使用する("Smith-Watermanアルゴリズム", T. F. Smith and M. S. Waterman (1981) J. Mol. Biol. 147:195-197参照)。
【0024】
ニードルマン・ウンシュ (Needleman-Wunsch)アルゴリズムは、Needleman, S. B. and Wunsch, CD., (1970), Journal of Molecular Biology, 48: 443-453、並びにMyers及びW. Millerの"線状空間での工学的アラインメント" CABIOS (バイオサイエンスにおけるコンピュータ適用)(1988)4: 11-17によるアラインプログラムに記載されている。
【0025】
標的(又は試料、又は試験)配列と特定の配列(例えば、配列番号10に示される成熟フィターゼのアミノ酸1〜413)との間の同一性の程度は、以下のように決定されてもよい:配列はプログラム「アライン」を用いて整列される。アラインメントにおける完全なマッチ(「N-完全-マッチ」)の数は、決定される(完全マッチは、通常アラインメントにおいて「I」と指定される、アラインメントの同一の位置における同一のアミノ酸残基を意味する)。特定配列の長さ(アミノ酸残基の数)は決定される(上記の例で「N-特定」は413)。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換のための)「N-完全-マッチ」と「N-特定」との比として計算される。
【0026】
別の実施態様では、標的(又は試料もしくは試験)配列と特定の配列(例えば、配列番号10mのアミノ酸1〜413)との同一性の程度は、以下のようにして決定される:該2つの配列は、プログラム「アライン」を用いて整列される。該アラインメントにおける完全なマッチ(「N-完全-マッチ」)の数が決定される(完全なマッチは、通常アラインメントにおいて「I」と指定される、該アラインメントの同一の位置における同一のアミノ酸残基を意味する)。該2つの整列された配列の共通の長さも、該アラインメントの重複部分(「N-重複」)におけるアミノ酸の総数によって決定される。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換についての)「N-完全-マッチ」と「N-重複」との比として計算される。1つの実施態様では、N-重複は、存在するならば、該アラインメントによってつくられたトレイリング及びリーディング・ギャップを含む。別の実施態様では、N-重複は、存在するならば、該アラインメントによってつくられたトレイリング及びリーディング・ギャップギャップを含まない。
【0027】
別の実施態様では、標的(又は試料又は試験)配列と特定配列(例えば、配列番号10のアミノ酸1〜413)との同一性の程度は、以下のようにして決定される:配列は、プログラム「アライン」を用いて整列される。該アラインメントにおける完全マッチ(「N-完全-マッチ」)の数が決定される(完全マッチは、通常、該アラインメントにおける「I」で設計された、アラインメントの同一位置における同一のアミノ酸残基を意味する)。標的配列の長さ(アミノ酸残基の数)が決定される)「N-標的」)。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換のための)「N-完全-マッチ」と「N-標的」との比として計算される。
【0028】
好ましくは、重複は特定配列の少なくとも20%(上で定義された「N-重複」を特定配列におけるアミノ酸の数(「N-特定」)で割って、100を乗じた)であり、より好ましくは少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、又は少なくとも95%である。このことは、試料配列が特定配列に整列される時に、特定配列のアミノ酸の少なくとも20%(好ましくは25〜95%)が該重複に結局含まれることになることを意味する。
【0029】
あるいは、前記重複が標識(又は試料、又は試験)配列の少なくとも20%(上で定義された「N-重複」を上で定義された「N-標的」で割って、100を乗じる)であり、より好ましくは少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、又は少なくとも95%である。このことは、標的配列が特定配列に整列される時に、標的配列のアミノ酸の少なくとも20%(好ましくは25〜95%)が該重複に結局含まれることになることを意味する。
【0030】
ポリペプチドフラグメント:用語「ポリペプチドフラグメント」は、本明細書では、特定配列例えば配列番号10又はその相同配列の成熟ペプチド部分のアミノ及び/又はカルボキシ末端から削除された1以上のアミノ酸を有するポリペプチドであって、該フラグメントがフィターゼ活性を有する、前記ポリペプチドとして定義される。具体的な実施態様では、該フラグメントは、少なくとも350、360、370、380、390、400、405、又は少なくとも410のアミノ酸残基を含む。
【0031】
配列:用語「配列」は、特定の配列例えば配列番号9又はその相同配列の成熟ペプチドコーディング部分の5’及び/又は3’末端から削除された1以上の塩基を有するヌクレオチド配列として本明細書で定義されている。ここで、該配列は、フィターゼ活性を有するポリペプチドフラグメントをコードする。具体的な実施態様では、該配列は、少なくとも1050、1080、1110、1140、1170、1200、1215、1230、1245、1260、1275、1290、1305、1320、又は少なくとも1335塩基を含む。
【0032】
アレル変異体:用語「アレル変異体」は、本明細書では、同一の染色体座を占める遺伝子の2以上の別形態のいずれかを意味する。アレル変異は、突然変異によって自然に起こり、集団内の多型をもたすことがある。遺伝子突然変異は、サイレントであり(コードされたポリペプチドに変化がない)、又は改変されたアミノ酸配列を有するポリペプチドをコードすることがある。ポリペプチドのアレル変異体は、遺伝子のアレル変異体によってコードされたポリペプチドである。
【0033】
実質的に純粋なポリヌクレオチド:本明細書で使用される用語「実質的に純粋なポリヌクレオチド」は、多の外因性又は望ましくない塩基がなく、そして遺伝子組換えタンパク質産生系内での使用のために好適な形態にある、ポリヌクレオチド調製物を言う。従って、実質的に純粋なポリヌクレオチドは、天然的に関連している他のポリヌクレオチド物質の、最高10重量%、好ましくは最高8重量%、より好ましくは最高6重量%、より好ましくは最高5重量%、より好ましくは最高4重量%、より好ましくは最高3重量%、更により好ましくは最高2重量%、最も好ましくは1重量%、及び更に最も好ましくは最高0.5重量%を含む。しかし、実質的に純粋なポリヌクレオチドは、天然5’及び3’の非翻訳領域、例えばプロモーター及びターミネーターを含んでもよい。実質的に純粋なポリヌクレオチドは、少なくとも90重量%の純度、少なくとも92重量%の純度、より好ましくは少なくとも94重量%の純度、より好ましくは少なくとも95重量%の純度、より好ましくは少なくとも96重量%の純度、より好ましくは少なくとも97%の純度、更により好ましくは少なくとも98重量%の純度、最も好ましくは少なくとも99重量%の純度、及び更に最も好ましくは99.5重量%の純度であることが好ましい。本発明のポリヌクレオチドは、好ましくは、実質的に純粋な形態にある。具体的には、本明細書に開示されたポリヌクレオチドが「本質的に純粋な形態」にある、すなわち、ポリククレオチド調製物が本質的に天然的に関連している他のポリヌクレオチド物質がない、ことが好ましい。本明細書では、用語「実質的に純粋なポリヌクレオチド」は、用語「単離されたポリヌクレオチド」及び「単離された形態にあるポリヌクレオチド」と同義である。ポリヌクレオチドは、ゲノム、cDNA、RNA、半合成的、合成的起源でもよく、又はこれらの任意の組合せでもよい。
【0034】
核酸構築物:本明細書で使用される用語「核酸構築物」は、天然遺伝子から単離されるか又は天然に存在しない方法で核酸の部分を含むように修飾されている、単一-又は二重鎖のいずれかの核酸分子を言う。用語「核酸構築物」は、該核酸構築物が本発明のコーディング配列の発現のために必要とされる制御配列を含む時に、用語「発現カセット」と同義である。制御配列:用語「制御配列」は、本明細書では、本発明のポリペプチドをコードするポリヌクレオチドの発現のために必要であるか又は有利である、すべての成分を含むように定義される。各制御配列は、該ポリペプチドをコードするヌクレオチド配列に対して天然型でも又は外因性でもよい。かかる制御配列は、リーダー、ポリアデニル化配列、プロペプチド配列、プロモーター、シグナルペプチド配列、及び転写終結因子Hを含むが、これらに限定されない。最低でも、制御配列は、プロモーター、及び転写及び翻訳停止シグナルを含む。制御配列は、ポリペプチドをコードするヌクレオチド配列のコーディング領域と共に、制御配列のライゲーションを助ける特定の制限部位を導入する目的のために、リンカーと共に提供され得る。
【0035】
作動可能に連結:本明細書で使用される用語「作動可能に連結された」は、制御配列がポリペプチドにコーディング配列の発現を指向するように、ポリヌクレオチド配列のコーディング配列に対して好適な位置に該制御配列が位置する配列を意味する。
【0036】
コーディング配列:本明細書で使用される用語「コーディング配列」は、そのタンパク質産物のアミノ酸配列を直接的に特定するヌクレオチド配列を意味する。
【0037】
コーディング配列の境界は、一般的に、通常、ATG開始コドン、又はGTG及びTTGのような別の開始コドンで始まる、オープンリーディングフレームによって決定される。コーディング配列は、DNA、cDNA又は組換えヌクレオチド配列でよい。
【0038】
成熟ポリペプチド部分:本明細書で使用される用語「成熟ポリペプチド部分」又は「成熟ペプチド部分」は、その遺伝子的道具の部分として、該ポリペプチドをコードするポリヌクレオチドを含む細胞によって分泌されるポリペプチドの部分を言う。言いかえれば、成熟ポリペプチド部分は、細胞の分泌経路にコードされたポリペプチドを指向するその機能を一旦満たした時に、シグナルペプチド部分が開裂される後に残るポリペプチドの部分を言う。
【0039】
配列番号10の予測されたシグナルペプチド
配列番号10は、そのアミノ酸-33〜-1である。それは、配列番号10の成熟ポリペプチド部分がそのアミノ酸1〜413に対応することを意味する。しかし、宿主細胞によってはわずかな変異が起こり得、そのため、発現成熟ポリペプチド部分は好ましい。
【0040】
成熟ポリペプチドコーディング部分:本明細書で使用される用語「成熟ポリペプチドコーディング部分」又は「成熟ポリペプチドコーディング配列」が、成熟ポリペプチド部分をコードするポリペプチドをコードするポリヌクレオチドの部分を言う。例えば、配列番号9に関して、予測された成熟ポリペプチドコーディング部分は、(配列番号10のアミノ酸1〜413をコードする)塩基100〜1338に対応する。
【0041】
発現:用語「発現」は、転写、転写後修飾、翻訳、翻訳後修飾、及び分泌を含むがこれらに限定されないポリペプチドの製造に関連する任意のステップを含む。発現ベクター:用語「発現ベクター」は、本発明のポリペプチドをコードするポリヌクレオチドを含み、その発現を提供する更なる塩基に作動可能に連結される線状又は環状DNA分子として本明細書に定義される。
【0042】
宿主細胞:本明細書で使用される用語「宿主細胞」は、本発明のポリヌクレオチドを含む核酸構築物での、形質転換、トランスフェクション、形質導入等を受けやすい任意の細胞種を含む。
【0043】
修飾:本明細書で使用する用語「修飾」は、特定のポリペプチドの任意の化学的修飾、例えば配列番号10のアミノ酸1〜413からなるポリペプチド、及び該ポリペプチドをコードするDNAの遺伝子的操作を意味する。修飾(複数)は、アミノ酸(複数)の置換(複数)、削除(複数)及び/又は挿入(複数)、並びにアミノ酸側鎖(複数)の置換(複数)でよい。人工変異体:本明細書では、用語「人工変異体」は、配列番号9の成熟フィターゼコーディング部分の修飾核酸配列を発現する生物によって産生されるフィターゼ活性を有するポリペプチドを意味する。修飾されたヌクレオチド配列は、配列番号9に開示されたヌクレオチド配列の成熟フィターゼコーディング部分の修飾によって、ヒトの介入によって得られる。
【発明を実施するための形態】
【0044】
発明の詳細な説明
フィターゼ活性を有するポリペプチド
第1の局面では、本発明は、フィターゼ活性を有し、そして配列番号10(すなわち、成熟ポリペプチド)のアミノ酸1〜143と少なくとも75%の同一性の程度を有するアミノ酸配列を有する単離ポリペプチドに関する。
【0045】
具体的な実施態様では、同一性の程度は、少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、又は少なくとも99.9%であり、これは、フィターゼ活性を有する(本明細書では、「相同ポリペプチド」)。
【0046】
他の具体的な実施態様では、相同ポリペプチドは、配列番号10のアミノ酸1〜413と20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2、又は1個のアミノ酸が異なるアミノ酸配列を有する。
【0047】
具体的な実施態様では、本発明のポリペプチドは、配列番号10のアミノ酸配列の成熟部分を含み、又はそのアレル変異体もしくはフィターゼ活性を有するそのフラグメントである。更なる具体的な実施態様では、ポリペプチドは、配列番号10のアミノ酸1〜413、又はそのアレル変異体もしくはフィターゼ活性を有するそのフラグメントを含む。
【0048】
第2の局面では、本発明は、本発明は、少なくとも媒体、好ましくは(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び (ii) のいずれか1つの相補鎖、及び/又は (iv) (i)、(ii) 又は (iii) のサブ配列(J. Sambrook, E. F. Fritsch, and T. Maniatus, 1989, Molecular Cloning, A Laboratory Manual, 2d edition, Cold Spring Harbor, New York)の緊縮条件下での媒体下でハイブリダイズするポリヌクレオチドによってコードされるフィターゼ活性を有する単離されたポリペプチドに関する。配列番号9の配列は、少なくとも100連続塩基、又は好ましくは少なくとも200連続塩基を含む。更に、配列は、フィターゼ活性を有するポリペプチドフラグメントをコードしてもよい。具体的な実施態様では、ハイブリダイゼーションは、少なくとも中程度〜高度の、少なくとも高度の、又は少なくとも非常に高い緊縮性の下で、好ましくは中程度〜高度、又は非常に高い緊縮性条件下で起こる。
【0049】
代わりの実施態様では、ハイブリダイゼーションは、非常に低い、又は低い緊縮性条件下で行われる。
【0050】
配列番号9のヌクレオチド配列又はそのサブ配列、及び配列番号10のアミノ酸配列又はそのフラグメントは、当該分野で周知の方法に従って様々な属又は種の株から、フィターゼ活性を有するポリペプチドをコードするDNAを同定し及びクローン化するための核酸プローブを設計するために使用し得る。具体的には、かかるプローブは、その中の対応する遺伝子を同定及び単離するために、標準的なサザンブロッティング手段に従って、対象の属又は種のゲノム又はcDNAとのハイブリダイゼーションのために使用され得る。かかるプローブは、配列全体よりも相当短くてよいが、少なくとも14塩基長、好ましくは少なくとも25塩基長、より好ましくは少なくとも35塩基長、及び最も好ましくは少なくとも70塩基長である。しかしながら、塩基プローブは、少なくとも100塩基長であることが好ましい。例えば、塩基プローブは、少なくとも200塩基長、好ましくは少なくとも300塩基長、より好ましくは少なくとも400塩基長、又は最も好ましくは少なくとも500塩基長でよい。更に長いプローブ、例えば、少なくとも600塩基長、少なくとも好ましくは700塩基長、より好ましくは800塩基長、又は最も好ましくは少なくとも900塩基長である核酸プローブも使用される。DNA及びRNAプローブが使用できる。プローブは、典型的に、対応する遺伝子を検出するために標識される(例えば、32P、3H、35S、ビオチン、又はアビジン)。かかるプローブはは、本発明によって包含される。
【0051】
そのため、かかる他の生物から調製されたゲノムDNAライブラリーは、上記のプローブとハイブリダイズし、そしてフィターゼ活性を有するポリペプチドをコードする、DNAについてスクリーニングされ得る。かかる他の生物由来のゲノム又は他のDNAは、アガロース又はポリアクリルアミドゲル電気泳動、又は他の分離技術によって分離され得る。該ライブラリー由来のDNA又は分離されたDNAは、ニトロセルロース又は他の好適な担体材料に移され、そしてそれに固定され得る。配列番号9又はそのサブ配列と相同であるクローン又はDNAを同定するために、担体材料はサザンブロットで使用される。
【0052】
本発明の目的のために、ハイブリダイゼーションは、ヌクレオシド配列が、非常に低い緊縮条件から常に高い緊縮条件下で、配列番号9に記載のヌクレオチド配列、その相補鎖、又はそのサブ配列に対応する標識された核酸プローブにハイブリダイズすることを示している。核酸プローブがこれらの条件下でハイブリダイズする分子は、X-線フィルムを用いて検出され得る。
【0053】
具体的な実施態様では、核酸プローブは、配列番号1〜8のいずれか1つである。別の具体的な実施態様では、核酸プローブは、配列番号9の塩基100〜450、塩基450〜900又は塩基900〜1338の相補鎖である。更なる具体的な実施態様では、核酸プローブは、配列番号10のポリヌクレオチドの成熟部分又はそのサブ配列をコードするポリヌクレオチド配列である。なお更なる具体的実施態様では、核酸プローブは、配列番号9であり、具体的には、その成熟ポリペプチドコーディング領域のいずれか1つである。
【0054】
少なくとも100塩基長の長いプローブについては、非常に低緊縮から非常に高緊縮条件は、好ましくは12〜24時間の標準的なサザンブロッティング法に従って、42℃、5X SSPE、0.3% SDS、200 マイクログラム/mlのせん断及び変性サケ精子DNA、及び非常に低い及び低い緊縮製のための25%ホルムアミド、中程度及び中〜高程度の緊縮製のための35%ホルムアミド、又は高い及び非常に高い緊縮製のため50%ホルムアミドのいずれか、でのプレハイブリダイゼーション及びハイブリダイゼーションと定義される。
【0055】
少なくとも100ヌクレオチド長の長いプローブについては、非常に低緊縮から非常に高緊縮条件は、好ましくは12〜24時間の標準的なサザンブロッティング法に従って、42℃、5X SSPE、0.3% SDS、200 マイクログラム/mlのせん断及び変性サケ精子DNA、及び非常に低い及び低い緊縮製のための25%ホルムアミド、中程度及び中〜高程度の緊縮製のための35%ホルムアミド、又は高い及び非常に高い緊縮製のため50%ホルムアミドのいずれか、でのプレハイブリダイゼーション及びハイブリダイゼーションと定義される。担体物質は、2X SSC、0.2% SDS、好ましくは少なくとも45℃ (非常に低い緊縮性)、より好ましくは少なくとも50℃ (低緊縮性)、より好ましくは少なくとも55℃ (中緊縮性)、より好ましくは少なくとも60℃ (中〜高緊縮性)、更により好ましくは少なくとも65℃ (高緊縮性)、及び最も好ましくは少なくとも70℃ (非常に高い緊縮性) を用いて、15分間3回最終的に洗浄される。
【0056】
約15ヌクレオチド長〜約70ヌクレオチド長の短いプローブについては、緊縮条件は、プレハイブリダイゼーション、ハイブリダイゼーション、並びに計算されたTm未満の約5℃〜約10℃未満で、標準的なサザンブロッティング法に従って、0.9 M NaCl、0.09 M Tris-HCl pH 7.6、6 mM EDTA、0.5% NP-40、1Xデンハート液、1 mMピロリン酸ナトリウム、1 mMりん酸二水素ナトリウム、0.1 mM ATP、及び0.2 mgの酵母RNA/ml中のBolton及びMcCarthy (1962, Proceedings of the National Academy of Sciences USA 48: 1390) に従う計算を用いる、洗浄後ハイブリダイゼーションとして定義される。
【0057】
約15ヌクレオチド長〜約70ヌクレオチド長の短いプローブについては、担体物質は、6X SCC及び0.1% SDSで1回、15分間洗浄され、計算されたTm未満の5℃〜10℃での6X SSCを用いて2回、各々15分間洗浄される。
【0058】
塩含有ハイブリダイゼーション条件下では、有効Tmは、成功的なハイブリダイゼーションのためにプローブとフィルター結合DNAとの間に要求される同一性の程度を制御するものである。有効Tmは、2つのDNAが様々な条件下でハイブリダイズするために必要とされる同一性の程度を決定するために、下記式:
【0059】
有効Tm = 81.5 + 16.6(log M[Na+]) + 0.41 (%G+C)-0.72(% ホルムアミド)
【0060】
(www.ndsu.nodak.edu/instruct/mcclean/plsc731/dna/dna6.htm参照)
を用いて決定され得る。
【0061】
「G+C」は、プローブ中のヌクレオチドG及びTの量を示す。中緊縮性については、例えば、ホルムアミドは35%であり、5X SSPE用のNa+濃度は0.75 Mである。
【0062】
1つの局面では、本発明は、フィターゼ活性及び以下の物理化学的性質(実質的に純粋なポリペプチドについて分析された)を有する単離されたポリペプチドに関する:
(i) 高い特異的活性、例えば、E. コリappA (SPTREMBLQ8GN88) の特異的活性の少なくとも50%のフィターゼでの特異的活性、該特異的活性は、好ましくは、mgフィターゼ酵素タンパク質当たりのFYTの単位で測定される;
(ii) 酸安定性、例えば
(a) HEPESバッファーpH 7.0中で37℃で終夜のインキュベーションした後の残余活性に比べて、少なくとも60%、少なくとも65%、少なくとも70%、又は少なくとも75%の、グリシン/塩酸バッファーpH 2.2中で37℃で終夜のインキュベーションした後の残余活性;
(b) HEPESバッファーpH 7.0中で37℃で終夜のインキュベーションした後の残余活性に比べて、少なくとも80%、少なくとも85%、少なくとも90%、又は少なくとも95%の、グリシン/塩酸バッファーpH 3.0中で37℃で終夜のインキュベーションした後の残余活性;及び/又は
(c) E. コリappA (SPTREMBLQ8GN88) の残余活性と比べて、少なくとも50%の、25、30、35又は37℃、好ましくは37℃の温度で、及びpH of 2.2、2.4、2.5、2.6、2.8、3.0、3.2、3.4又は3.5、好ましくはグリシン/塩酸バッファーpH 2.2又は3.0中で、2時間インキュベーション後の残余フィターゼ活性;
(iii) 熱安定性、例えば、E.コリappA (SPTREMBLQ8GN88) の残余活性と比べて少なくとも50%の、pH 5.5及び55、60、65、70、75、80、85又は95の温度、好ましくは70℃でのインキュベーションの0.5、1、1.5又は2時間、好ましくは0.5時間後の残余フィターゼ活性。
【0063】
あるいは、示差走査熱量 (DSC) 測定は、精製されたフィターゼタンパク質の変性温度Tdを決定するために使用してよい。Tdは、タンパク質の熱安定性の指標である;Tdが高くなればなるほど、熱安定性は高くなる。DSC測定は、例えばMicro CaI製のVP-DSCを用いて様々なpH値で達成され得る。スキャンは、20〜90℃で1.5℃/分の一定のスキャン速度で行われる。好ましいpH値は、4.0〜5.5、好ましくは4.0である。DSCを操作する前に、例えば好適なバッファー (例えば、25 mM酢酸ナトリウム, pH 4.0; 0.1 M酢酸ナトリウム, pH5.5) 中で平衡化されたNAP-5カラム (Pharmacia) を用いてフィターゼを脱塩する。データ操作は、MicroCal Originソフトウェア(バージョン4.10)を用いて行われ、編成温度Td(融解温度Tmとも言われる)は、サーモグラフのピークの頂点での温度として定義される。
【0064】
(iv) プロテアーゼ安定性、例えば、E. コリappA (SPTREMBLQ8GN88) の残余活性と比べて、少なくとも50%の、0.1 mg/mlのペプシンの存在下、20、25、30、35、又は37℃、好ましくは37℃の温度及びpH 5.5での0.5、1、1.5又は2時間、好ましくは1時間のインキュベーション後の残余フィターゼ活性;及び/又は
(v) pH 5.0未満の最適pH、例えば本明細書で先に記載したFYTアッセイを用いて及び/又は実施例4のアッセイを用いて決定されたpH 4.5、4.0、3.5、3.0、2.5未満又は2.0未満。
【0065】
上記の(i)の具体的実施態様では、特異的活性は、E. コリappAの特異的活性の少なくとも60、70、80、90、100、110、120、130、140、又は少なくとも150%である。上記の(ii)〜(iv)の各々の具体的実施態様では、残基活性は、E. コリappAの少なくとも60、70、80、90、100、110、120、130、140、又は少なくとも150%である。
【0066】
第5の局面では、基質pNP-ホスファターゼで測定されたpH 5.0及び37℃での本発明の酵素の活性は、基質フィターゼで測定された酵素の活性の11%未満である。好ましくは、該比は、10%、9%、8%、7%、6%未満、又は5%未満である。フィターゼ加水分解に対するpNPの比は、該酵素の真のフィターゼ特性の指標である。フィターゼについての活性に対するpNPについての活性の高い比は、問題の酵素が、比較的低い基質特異性を有するホスファターゼであることを示し、一方、低い比は、これが、基質としてより明確に受容できるフィターゼであることを示しているかもしれない。
【0067】
第6の局面では、本発明のフィターゼは、ペニオホラ・リシイ(Peniophora lycii)由来のフィターゼに比べて、好ましくはその110%、より好ましくは少なくともその120%、130%、又は少なくとも140%、インビトロのモデルにおいてより高いリン(P)放出を有する。1つの実施態様では、0.25 FYT/g飼料で投与された本発明のフィターゼは、インビトロモデルにおいて0.25 FYT/g飼料で投与されたペニオホラ・リシイ由来のフィターゼによって放出されるリンに比べて、少なくとも150%リン (P) を放出する。好ましくは、該放出は、少なくとも155%、160%、165%、170%、175%、又は少なくとも180%である。別の実施態様では、0.75 FYT/g飼料で投与された本発明のフィターゼは、インビトロモデルにおいて0.75 FYT/g飼料で投与されたペニオホラ・リシイ由来のフィターゼによって放出されるリンに比べて、少なくとも150%リン (P) を放出する。好ましくは、該放出は、少なくとも155%、160%、165%、170%、175%、180%、185%、又は少なくとも190%である。
【0068】
第7の局面では、本発明のフィターゼは、37℃、1 M グリシン/HClバッファー、pH 2.0中で、4時間のインキュベーション後に、t = 0の時点での活性と比べて少なくとも20%の残余活性を有する。該活性(及び残余活性)は、0.25 M Na-アセテートバッファーpH 5.5、ブラインドを除いたバッファーを用いて、37℃及びpH 5.5の1% (w/v) Na-フィターゼでアッセイされる。好ましい実施態様では、残余活性は、少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、又は少なくとも80%である。別の実施態様では、本発明のフィターゼは、37℃、1 M グリシン/HClバッファー、pH 2.5中で、1日(24時間)のインキュベーション後に、t = 0の時点での活性と比べて少なくとも20%の残余活性を有する。該活性(及び残余活性)は、0.25 M Na-アセテートバッファーpH 5.5、ブラインドを除いたバッファーを用いて、37℃及びpH 5.5の1% (w/v) Na-フィターゼでアッセイされる。好ましい実施態様では、残余活性は、少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、又は少なくとも80%である。
【0069】
第8の局面では、本発明は、配列番号10又はその成熟ポリペプチドの1以上のアミノ酸の保存的置換、削除及び/又は挿入を含む人工的変異体に関する。挿入は、それが伸張をも示す場合の、分子内、及び/又は分子のN-及び/又はC-末端であり得る。好ましくは、アミノ酸変化は保存的アミノ酸置換である小さな性質の変化である:典型的には、1〜約30アミノ酸の小さな削除;アミノ-末端メチオニン残基のような小さなアミノ-又はカルボキシ-末端伸張;約20〜25残基までの小さなリンカーペプチド;あるいは正味電荷又は他の機能を変化させることによって精製を助ける小さな伸張、例えばポリ-ヒスチジントラクト、抗原性エピトープ又は結合ドメイン、言い換えれば、タンパク質の折り畳み及び/又は活性にほとんど影響を与えない変化。保存的置換の例は、基本的アミノ酸(アルギニン、リジン及びヒスチジン)、酸性アミノ酸(グルタミン酸及びアスパラギン酸)、極性アミノ酸(グルタミン及びアスパラギン)、疎水性アミノ酸(ロイシン、イソロイシン及びバリン)、芳香族アミノ酸(フェニルアラニン、トリプトファン及びチロシン)、並びに小アミノ酸(グリシン、アラニン、セリン、スレオニン及びメチオニン)のグループ内の置換である。保存的置換の他の例は、非-標準的アミノ酸(例えば、4-ヒドロキシプロリン、6-N-メチルリジン、2-アミノイソ酪酸、イソバリン及びα-メチルセリン)での20個の標準的アミノ酸の置換である。保存的置換は、遺伝子コードによってコードされないアミノ酸、及び非天然アミノ酸への置換も含む。「非天然アミノ酸」は、タンパク質合成後に修飾されており、及び/又は標準的アミノ酸のそれとは異なった側鎖(複数)内の化学構造を有する。非天然アミノ酸は、化学的に合成され、好ましくは、商業的に入手可能であり、ピペコリン酸、チアゾリジンカルボン酸、デヒドロプロリン、3-及び4-メチルプロリン、及び3,3-ジメチルプロリンを含み得る。
【0070】
あるいは、アミノ酸変化は、ポリペプチドの物理化学的性質が変化しているような性質の変化である。例えば、アミノ酸変化は、ポリペプチドの熱的安定性を改善し、基質特異性を変化させ、最適pHを変化させるなどである。
【0071】
親ポリペプチドにおける必須アミノ酸は、部位指向的突然変異誘発又はアラニン・スキャニング突然変異誘発のような当該分野で公知の手段に従って同定され得る(Cunningham and Wells, 1989, Science 244: 1081-1085)。後者の技術では、単一のアラニン変異は、分子内のすべての残基に導入され、得られた変異分子は、分子の活性に重要であるアミノ酸残基を同定するための生物的活性(すなわち、フィターゼ活性)について試験される。Hilton他, 1996, J. Biol. Chem. 271: 4699-4708も参照。酵素の活性部位又は他の生物学的相互作用は、推定される接触部位アミノ酸の変異に関連して、核磁気共鳴、結晶学、電子回折、又は光親和標識のような技術によって決定されるように、構造の物理的分析によっても決定され得る。例えば、de Vos他, 1992, Science 255: 306-312; Smith他, 1992, J. Mol. Biol. 224: 899-904; Wlodaver他, 1992, FEBS Lett. 309: 59-64参照。必須アミノ酸の同一性は、本発明に従うポリペプチドに関連するポリペプチドとの同一性の分析からも推論され得る。単一又は複数のアミノ酸置換は、公知の突然変異法、組換え、及び/又はシャッフリングを用いて、次いで、Reidhaar-Olson及びSauer, 1988, Science 241: 53-57; Bowie及びSauer, 1989, Proc. Natl. Acad. Sci. USA 86: 2152-2156; WO 95/17413; 又はWO 95/22625に開示されているような関連するスクリーニング法によってなされ、そして試験され得る。使用され得るエラー-プローンPCR、ファージディスプレイ(例えば、Lowman et al., 199 , Biochem. 30: 10832-10837; 米国特許第5,223,409号明細書; WO 92/06204)、及び部位-指向的突然変異誘発(Derbyshire et al., 1986, Gene 46: 145; Ner et al., 1988, DNA 7:127)を含む。
【0072】
突然変異誘発/シャッフリング法は、宿主細胞によって発現されたクローン化された突然変異誘発化ポリペプチドの活性を検出するために、ハイスループットの自動スクリーニング法と組み合わせられ得る。活性なポリペプチドをコードする突然変異誘発DNA分子は、宿主細胞から回収され、当該分野の標準的な方法を用いて迅速にシークエンスされ得る。これらの方法は、対象の個々のポリペプチド中のアミノ酸残基の重要性の迅速決定を可能にし、未知の構造のポリペプチドに適用され得る。
【0073】
配列番号10のアミノ酸1〜413の配列中のアミノ酸置換(好ましくは保存的置換)、削除及び/又は挿入の総数は、最大10、好ましくは最大9、より好ましくは最大8、より好ましくは最大7、より好ましくは最大6、より好ましくは最大5、より好ましくは最大4、更により好ましくは最大3、より好ましくは最大2、及び更に最も好ましくは1である。
【0074】
配列番号10のアミノ酸1〜413のアミノ酸置換、削除及び/又は挿入の総数は、10,好ましくは9、より好ましくは8、より好ましくは7、より好ましくは最大6、より好ましく卯は最大5、より好ましは4、更により好ましくは3、より好ましくは2、及び更に最も好ましくは1である。代わりに、配列番号10のアミノ酸1〜413の配列中のアミノ酸置換(好ましくは保存的置換)、削除及び/又は挿入の総数は、最大50、45、40、35、30、25、20、19、18、17、16、15、14、13、12、又は最大11である。
【0075】
具体的な実施態様では、本発明のポリペプチドは、ヒトを含む動物に曝露された時に、減少された免疫応答を誘導するように設計された低-アレルギー変異体である。用語免疫応答は、ポリペプチドに曝露された動物の免疫系による任意の反応として理解されるべきである。免疫応答の1種は、曝露された動物においてIgEの増加レベルを導くアレルギー応答である。低-アレルギー変異体は、当該分野で公知の技術を用いて調製され得る。例えば、ポリペプチドは、免疫応答に関連するポリペプチドの部分又はエピトープを保護するポリマー部分と複合化され得る。ポリマーとの複合化は、例えばWO 96/17929、WO98/30682、WO98/35026、及び/又はWO99/00489に記載のポリペプチドへのポリマーのインビトロ化学結合に関連することがある。複合化は、それに加えてあるいは代わりに、ポリペプチドへのポリマーのインビボ結合に関連する。かかる複合化は、ポリペプチドをコードするヌクレオチド配列の遺伝子工学、ポリペプチド中の追加のグリコシル化部位をコードするコンセンサス配列を挿入し、ポリペプチドをグリコシル化することができる宿主中のポリペプチドを発現することによって達成され得る。例えばWO00/26354参照。低-アレルギー変異体を提供する別の方法は、ポリペプチドに自己-オリゴマー化を引き起こすような、ポリペプチドをコードするヌクレオチド配列の遺伝子工学である。これは、ポリペプチドモノマーが、他のポリペプチドモノマーのエピトープを保護し、それによってオリゴマーの抗原性を低減させることをもたらす。かかる産物及び他の調製物は、例えばWO96/16177に記載されている。免疫応答に関連するエピトープは、WO 00/26230及びWO 01/83559に記載されたファージディスプレイ法又はEP 561907に記載のランダムアプローチのような様々な方法によって同定され得る。エピトープは一旦同定されると、そのアミノ酸配列は、公知の操作方法、例えば部位指向的変異形成(例えば、WO 00/26230、WO 00/26354及び/又はWO00/22103参照)によって、ポリペプチドの改変された免疫学的性質を生成するように改変され得、及び/又はポリペプチドの複合化は、ポリマーがエピトープを遮蔽するようにエピトープの充分近くでなされ得る。
【0076】
ハフニア・アルベイ・フィターゼの3次元構造
ハフニア・アルベイ・フィターゼの3次元構造(配列番号10のアミノ酸1〜413)は、添付書類で提供される。該構造は、x-線結晶法についての原則、例えば、X-線構造決定, Stout, G. K. and Jensen, L. H., John Wiley and Sons, Inc. NY 1989で提供される、に従って解決された。ハフニア・アルベイ・フィターゼの解決された結晶構造の構造座標は、添付書類に記載の標準的なPDBフォーマットで提供される(Protein Database Bank, Brookhaven National Laboratory, Brookhaven, Conn.)。それは本願の添付形式部分と理解される、添付書類は、水素原子を除く重元素の座標を与える。酵素の第1の3つ残基は、該結晶構造及びアミノ酸180と189との間のアミノ酸では見られなかった。しかし、180と189との間の構造は、ホモロジーモデリングを組み合わせたモデリングを用いて構築された(例えば、JACKALパッケージ製のMarti-Renom et al., 2000プログラムNEST(wiki.c2b2. Columbia. edu/honiglab public/index.php/Software:Jackal)、及びCHARMmと称されるシミュレイションソフトウェア((//accelrys.com/products/scitegic/component-collections/charmm.html)参照)。
【0077】
フィターゼ活性を有するポリペプチドの起源
本発明のポリペプチドは、任意の種の微生物から得られる。本発明の目的のために、本明細書で使用される用語「から得られる」は、所定の起源と関連して、ヌクレオチド入れるによってコードされるポリペプチドは、ある起源由来のヌクレオチド配列が挿入される起源又は株によって産生されることを意味する。好ましい局面では、所定の起源から得られるポリペプチドは、細胞外に分泌される。
【0078】
本発明のポリペプチドは、細菌性ポリペプチドでよい。例えば、該ポリペプチドは、バチルス(Bacillus)ポリペプチド又はストレプトマイセス(Streptomyces)ポリペプチドのようなグラム陽性細菌性ポリペプチド、あるいはグラム陰性細菌性ポリペプチド、例えばエシェリヒア・コリ(Escherichia coli)、エルシニア(Yersinia)、クレブシエラ(Klebsiella)、サイトロバクター(Citrobacter)又はシュードモナス(Pseudomonas)ポリペプチドでよい。具体的な実施態様では、該ポリペプチドは、ガンマプロテオバクテリア(Gammaproteobacteria)のようなプロテオバクテリア(Proteobacteria)、例えばエンテロバクター目(Enterobacteriales)、例えばエンテロバクテリアセエ(Enterobacteriaceae)から得られる。
【0079】
具体的局面では、エンテロバクテリアセエ(Enterobacteriaceae)から得られるポリペプチドは、ハフニア(Hafnia)ポリペプチド、例えばハフニア・アルベイ種ポリペプチドである。
【0080】
本発明のポリペプチドは、真菌性ポリペプチド、例えば酵母ポリペプチド又は繊維状真菌性ポリペプチドでよい。
【0081】
上記の微生物株は、アメリカンタイプカルチャーコレクション (ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM)、Centraalbureau Voor Schimmelcultures (CBS)、及びアメリカ農業研究局のカルチャーコレクション、ノーザーン・リージョナル・リサーチ・センター (NRRL) のような多数のカルチャーコレクションのパブリックが容易に利用できる。
【0082】
更に、かかるポリペプチドは、上記のプローブを用いて、天然(例えば、土壌、堆肥、水等)から単離された微生物を含む他の起源から同定及び得てもよい。自然環境由来の微生物を単離するための技術は、当該分野でよく知られている。次いで、該ポリペプチドは、同様に別の微生物のゲノム又はcDNAライブラリーをスクリーニングすることによって得られる。ポリペプチドをコードするポリヌクレオチド配列がプローブ(複数)で検出されると、該ポリヌクレオチドは、当業者に周知である技術を利用することによって単離又はクローン化され得る(例えば、Sambrook et al., 1989, 前掲参照)。
【0083】
本発明のポリペプチドは、別のポリペプチドが、本発明のポリペプチドのN-末端もしくはC-末端又はそのフラグメントに融合された、融合ポリペプチド又は開裂可能な融合ポリペプチドを含むこともできる。融合ポリペプチドは、別のポリペプチドをコードするヌクレオチド配列(又はその部分)を本発明のヌクレオチド配列(又はその部分)に融合することによって製造される。融合ポリペプチドを製造するための技術は当該分野で公知であり、それらがフレーム内にあり、そして融合されたポリペプチドの発現が同一のプロモーター(複数)及びターミネーターの制御下にあるように、該ポリペプチドをコードするコーディング配列をライゲートすることを含む。
【0084】
ポリヌクレオチド
本発明はまた、本発明のポリペプチドをコードするヌクレオチド配列を有する単離されたポリヌクレオチドに関する。好ましい局面では、該ヌクレオチド配列は、配列番号9で示される。別の好ましい局面では、該ヌクレオチド配列は、配列番号9の成熟ポリペプチドコーディング領域である。本発明はまた、遺伝子コードの縮重によって、配列番号10のアミノ酸配列を有するポリペプチド又は配列番号9とは異なるその成熟ポリペプチドをコードするヌクレオチド配列を包含する。本発明はまた、配列番号10のフラグメントをコードし、フィターゼ活性を有する、配列番号10のフラグメントをコードする配列番号9のサブ配列に関する。
【0085】
本発明はまた、配列番号9のいずれか1つの成熟ポリペプチドコーディング配列における少なくとも1つの変異を含む成熟ポリヌクレオチドであって、該成熟ポリヌクレオチド配列が配列番号10のアミノ酸1〜413からなるポリペプチドをコードする、ポリヌクレオチドに関する。
【0086】
ポリペプチドをコードするポリヌクレオチドを単離又はクローン化するために使用される技術は、当該分野で知られており、ゲノムDNAからの単離、cDNAからの調製、又はその組合せを含む。かかるゲノムDNAからの本発明のポリヌクレオチドのクローニングは、例えば、周知のポリメーラーゼ連鎖反応(PCR)、又は共通の構造的特徴を有するクローン化DNAフラグメントを検出するための発現ライブラリーの抗体スクリーニングを用いて行うことができる。例えば、lnnis他, 1990, PCR:方法及び応用へのガイド, Academic Press, New York参照。リガーゼ鎖反応(LCR)、ライゲートされた活性化転写(LAT)、及びヌクレオチド配列-型増幅(NASBA)等の他の核酸増幅方法も使用され得る。該ポリヌクレオチドは、ハフニア(Hafnia)株もしくは別の又は関連する生物からクローン化され得、よって、例えば、ヌクレオチド配列のポリペプチドコーディング領域のアレル又は種変異体でもよい。
【0087】
本発明はまた、配列番号9の成熟ポリペプチドコーディング配列(すなわち、ヌクレオチド100〜1338)と少なくとも75%の同一性の程度を有し、かつフィターゼ活性を有するポリペプチドをコードする、ヌクレオチド配列を有するポリヌクレオチドに関する。具体的な実施態様では、同一性の程度は、少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、又は少なくとも99.9%である。別の実施態様では、同一性の程度は、少なくとも75%、80%、85%、90%、94、97、98、98.0、98.1、98.2、又は少なくとも98.3%である。
【0088】
本発明のポリヌクレオドをコードするヌクレオチド配列の修飾は、該ポリペプチドに実質的に類似するポリペプチドの合成のために必要であるかもしれない。用語該ポリペプチドに「実質的に類似」とは、該ポリペプチドの非-天然形態を意味する。これらのポリペプチドは、その天然源から単離されたポリペプチド由来の作製方法とは異なるかもしれない、例えば特定の活性、熱安定性、最適pH等の点で異なる人工的変異体であり得る。変異体配列は、配列番号9のポリペプチドコーディング領域として提示されるヌクレオチド配列、例えばそのサブ配列に基いて、及び/又は該ヌクレオチド配列によってコードされるポリペプチドの別のアミノ酸配列を生じないが、該ポリペプチドの製造のために意図された宿主生物のコドン使用に対応するヌクレオチド置換の導入によって、あるいは異なったアミノ酸配列を生じ得るヌクレオチド置換の導入によって、構築され得る。ヌクレオチド置換の一般的な説明については、例えばFord他, 1991, タンパク質発現及び精製 2: 95-107参照。
【0089】
かかる置換が、分子の機能に重要である領域の外側でなされ、依然として活性なポリペプチドを生じ得る、ことは当業者には明らかであろう。本発明の単離されたポリヌクレオチドによってコードされるポリペプチドの活性に必須なであり、よって好ましくは置換に供されない、アミノ酸残基は、部位指向的突然変異誘発又はアラニン・スキャニング突然変異誘発のような当該分野で公知の手段に従って同定され得る (例えば、Cunningham and Wells, 1989, Science 244: 1081-1085参照)。後者の技術では、変異は、分子のすべての正に荷電した残基において導入され、分子の活性に重要であるアミノ酸残基を同定するためにフィターゼ活性について試験される。基質-ポリペプチド相互作用の部位はまた、核磁気共鳴分析、結晶学又は光親和性標識のような技術によって決定される3次元構造の分析によって決定され得る (例えば、de Vos et al., 1992, Science 255: 306-312; Smith et al., 1992, Journal of Molecular Biology 224: 899-904; Wlodaver et al., 1992, FEBS Letters 309: 59-64参照)。
【0090】
本発明はまた、中緊縮条件下、より好ましくは中〜高緊縮条件、更により好ましくは高緊縮条件、及び最も好ましくは非常に高い緊縮条件下で、(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び/又は (ii) のいずれか1つの相補鎖とハイブリダイズする、本発明のポリペプチドをコードする単離されたポリヌクレオチド;又は本明細書で定義された、そのアレル変異体及びそのサブ配列 (Sambrook et al., 1989, 前掲) に関する。別の実施態様では、ハイブリダイゼーションは、非常に低い又は低い緊縮条件下で行われる。
【0091】
本発明はまた、(a) 非常に低い、低、中、中〜高、高又は非常に高い緊縮条件下、(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び/又は (ii) のいずれか1つの相補鎖と、DNA群をハイブリダイズし;そして、(b) フィターゼ活性を有するポリペプチドをコードする、ハイブリダイズするポリヌクレオチドを単離することによって、得られた又は得られ得る単離されたポリヌクレオチドに関する。
【0092】
核酸構築物
本発明はまた、制御配列と適合する条件下で好適な宿主細胞におけるコーディング配列の発現を指向する、1以上の制御配列に作動可能に連結した本発明の単離されたポリヌクレオチドを含む核酸構築物に関する。
【0093】
本発明のポリペプチドをコードする単離されたポリヌクレオチドは、該ポリペプチドの発現のための様々な方法において作製され得る。ベクターへのその挿入前のポリヌクレオチド配列の作製は、発現ベクターによって望ましく又は必要である。組換えDNA法を利用するポリヌクレオチド配列を改変するための技術は、当該分野でよく知られている。
【0094】
制御配列は、好適なプロモーター配列、すなわち本発明のポリペプチドをコードするポリヌクレオチドの発現用の宿主細胞によって認識されるヌクレオチド配列でよい。プロモーター配列は、ポリペプチドの発現を仲介する転写制御配列を含む。プロモーター配列は、変異、切断及びハイブリッドプロモーターを含む最適な宿主配列中において、転写活性を示す任意のヌクレオチド配列でよく、宿主細胞に相同又は非相同のいずれかである細胞外又は細胞内ポリペプチドをコードする遺伝子から得られる。
【0095】
特に細菌宿主細胞中での、本発明の核酸構築物の転写を指向するための好適なプロモーターの例は、E. コリlacオペロン、ストレプトマイセス・セリカラー(Streptomyces coelicolor)・アガラーゼ遺伝子 (dagA)、バチルス・サブティリス(Bacillus subtilis)・レバンスクラーゼ遺伝子 (sacB)、バチルス・リケニフォルミス(Bacillus licheniformis)・α-アミラーゼ遺伝子 (amyL)、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)・マルトジェニックアミラーゼ遺伝子 (amyM)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)・α-アミラーゼ遺伝子 (amyQ)、バチルス・リケニフォルミス・ペニシリナーゼ遺伝子 (penP)、バチルス・サブティリスxylA及びxylB遺伝子、及び原核生物β-ラクタマーゼ遺伝子から得られるプロモーターである (Villa-Kamaroff et al., 1978, Proceedings of the National Academy of Sciences USA 75: 3727-3731)、並びにtacプロモーター (DeBoer et al., 1983, Proceedings of the National Academy of Sciences USA 80: 21-25)。更なるプロモーターは、Scientific American, 1980, 242: 74-94; 及びSambrook他, 1989, 前掲の「組み換え細菌由来の有用タンパク質」に記載されている。
【0096】
線維状真菌宿主細胞における本発明の核酸構築物の転写を指向するための好適なプロモーターの例は、アスペルギルス・オリザエ(Aspergillus oryzae)TAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)・アスパラギンプロテイナーゼ、アスペルギルス・ニガー(Aspergillus niger)・中性α-アミラーゼ、アスペルギルス・ニガー・酸安定α-アミラーゼ、アスペルギルス・ニガー又はアスペルギルス・アワモリ(Aspergillus awamori)・グルコアミラーゼ (glaA)、リゾムコール・ミエヘイ(Rhizomucor miehei)・リパーゼ、アスペルギルス・オリザエ・アルカリプロテアーゼ、アスペルギルス・オリザエ・トリオース・ホスフェート・イソメラーゼ、アスペルギルス・ニデュランス(Aspergillus nidulans)・アセトアミダーゼ、フザリウム・ベネナツム(Fusarium venenatum)・アミログルコシダーゼ (WO 00/56900)、フザリウム・ベネナツム・ダリア (WO 00/56900)、フザリウム・ベネナツム・クイン (WO 00/56900)、フザリウム・オキシスポラム(Fusarium oxysporum)・トリプシン様プロテアーゼ (WO 96/00787)、トリコデルマ・リーゼイ(Trichoderma reesei)・β-グルコシダーゼ、トリコデルマ・リーゼイ・セロビオヒドラーゼI、トリコデルマ・リーゼイ・エンドグルカナーゼI、トリコデルマ・リーゼイ・エンドグルカナーゼII、トリコデルマ・リーゼイ・エンドグルカナーゼIII、トリコデルマ・リーゼイ・エンドグルカナーゼIV、トリコデルマ・リーゼイ・エンドグルカナーゼV、トリコデルマ・リーゼイ・キシラナーゼI、トリコデルマ・リーゼイ・キシラナーゼII、トリコデルマ・リーゼイ・β-キシロシダーゼ、並びにNA2-tpiプロモーター (アスペルギルス・ニガー・中性α-アミラーゼ及びアスペルギルス・オリザエ・トリオースリン酸イソメラーゼのための遺伝子からのプロモーターのハイブリッド) の遺伝子から得られるプロモーター、及びその変異体、切断及びハイブリッドプロモーターである。
【0097】
酵母宿主では、有用なプロモーターは、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)・エノラーゼ (ENO-1)、サッカロマイセス・セレヴィシエ・ガラクトキナーゼ (GAL1)、サッカロマイセス・セレヴィシエ・アルコール脱水素酵素/グリセルアルデヒド-3-リン酸脱水素酵素 (ADH1、ADH2/GAP)、サッカロマイセス・セレヴィシエ・トリオースリン酸イソメラーゼ (TPI)、サッカロマイセス・セレヴィシエ・メタロチオニン (CUP1)、サッカロマイセス・セレヴィシエ・3-ホスホグリセリン酸キナーゼ、及びピキア・パストリス(Pichia pastoris)・アルコール酸化酵素 (AOX1) のための遺伝子から得られる。酵母宿主細胞用の他の有用なプロモーターは、Romanos他, 1992, Yeast 8: 423-488によって記載されている。制御配列は、好適な転写終結配列、転写を終結するための宿主細胞によって認識される配列もよい。終結配列は、ポリペプチドをコードするヌクレオチド配列の3'末端に作動可能に連結される。最適な宿主細胞において機能する任意の終結因子が本発明において使用され得る。
【0098】
線維状真菌宿主細胞のための好ましい終結因子は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、アスペルギルス・ニデュランス・アントラニル酸合成酵素、アスペルギルス・ニガー・α-グルコシダーゼ、及びフザリウム・オキシスポラム・トリプシン様プロテアーゼのための遺伝子から得られる。
【0099】
酵母宿主細胞のための好ましい終結因子は、サッカロマイセス・セレヴィシエ・エノラーゼ、サッカロマイセス・セレヴィシエ・チトクローム C (CYC1)、及びサッカロマイセス・セレヴィシエ・グリセルアルデヒド-3-リン酸脱水素酵素のための遺伝子から得られる。酵母宿主細胞のための他の有用な終結因子は、Romanos他, 1992, 前掲によって記載されている。
【0100】
制御配列は、好適なリーダー配列、すなわち宿主細胞による翻訳のために重要であるmRNAの非翻訳領域でもよい。リーダー配列は、ポリペプチドをコードするヌクレオチド配列の5'末端に作動可能に連結される。最適な宿主細胞中で機能する任意のリーダー配列は、本発明において使用され得る。
【0101】
制御配列は、好適なリーダー配列、すなわち宿主細胞による翻訳のために重要であるmRNAの非翻訳領域でもよい。リーダー配列は、ポリペプチドをコードするヌクレオチド配列の5'末端に作動可能に連結される。最適な宿主細胞中で機能する任意のリーダー配列は、本発明において使用され得る。
【0102】
線維状宿主細胞のための好ましいリーダーは、アスペルギルス・オリザエTAKAアミラーゼ及びアスペルギルス・ニデュランス(Aspergillus nidulans)トリオースリン酸イソメラーゼの遺伝子から得られる。
【0103】
酵母宿主細胞のための好適なリーダーは、サッカロマイセス・セレヴィシエ・エノラーゼ (ENO-1)、サッカロマイセス・セレヴィシエ・3-ホスホグリセリン酸キナーゼ、サッカロマイセス・セレヴィシエ・α-因子、及びサッカロマイセス・セレヴィシエ・アルコール脱水素酵素/グリセルアルデヒド-3-リン酸脱水素酵素 (ADH2/GAP) のための遺伝子から得られる。
【0104】
制御配列は、ポリアデニル化配列、すなわちヌクレオチド配列の3'末端に作動可能に連結し、そして転写される時に、転写mRNAにポリアデニル化残基を付加するためのシグナルとして宿主細胞によって認識される、配列でもよい。最適な宿主細胞において機能する任意のポリアデニル化配列は、本発明において使用され得る。
【0105】
線維状真菌宿主細胞のための好ましいポリアデニル化配列は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、アスペルギルス・ニデュランス・アントラニル酸合成酵素、フザリウム・オキシスポラム・トリプシン様プロテアーゼ、及びアスペルギルス・ニガー・α-グルコシダーゼのための遺伝子から得られる。
【0106】
酵母宿主細胞のための有用ナポリアデニル化配列は、Guo及びSherman, 1995, Molecular Cellular Biology 15: 5983-5990によって記載されている。
【0107】
制御配列は、ポリペプチドのアミノ末端に連結されたアミノ酸配列をコードし、及び細胞の分泌経路にコードされたポリペプチドを指向する、シグナルペプチドコーディング領域でもよい。ヌクレオチド配列のコーディング配列の5'末端は、本来、分泌されたポリペプチドをコードするコーディング領域の断片を有する翻訳読み枠において天然に連結されたシグナルペプチドコーディング領域を含むことがある。あるいは、該コーディング配列の5'末端は、該コーディング配列には無関係であるシグナルペプチドコーディング領域を含むことがある。この無関係なシグナルペプチドコーディング領域は、該コーディング配列が天然にはシグナルペプチドコーディング領域を含まない場合に、必要とされるかもしれない。あるいは、この無権系なシグナルペプチドコーディング領域は、ポリペプチドの分泌を亢進するために、単に、天然のシグナルペプチドコーディング領域を取り替えることがある。しかしながら、最適の宿主細胞の分泌経路に発現されたポリペプチドを指向する任意のシグナルペプチドコーディング領域が、本発明において使用され得る。
【0108】
細菌宿主細胞のための効果的なシグナルペプチドコーディング領域は、バチルスNCIB 11837・マルトースアミラーゼ、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)・α-アミラーゼ、バチルス・リケニフォルミス(Bacillus licheniformis)・スブチリシン、バチルス・リケニフォルミス・β-ラクタマーゼ、バチルス・ステアロサーモフィルス・中性プロテアーゼ (nprT、nprS、nprM)、及びバチルス・バチルス・サブティリスprsAのための遺伝子から得られたシグナルペプチドコーディング領域である。更に、シグナルペプチドは、Simonen及びPalva, 1993, Microbiological Reviews 57: 109-137によって記載されている。
【0109】
線維状真菌宿主細胞のための有効なシグナルペプチドコーディング領域は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・中性アミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、リゾムコール・ミエヘイ・アスパラギンプロテイナーゼ、フミコラ・インソレンス(Humicola insolens)・セルラーゼ、及びフミコラ・ラヌギノーサ(Humicola lanuginosa)・リパーゼのための遺伝子から得られたシグナルペプチドコーディング領域である。
【0110】
酵母宿主細胞のための有用なシグナルペプチドは、サッカロマイセス・セレヴィシエ・α-因子及びサッカロマイセス・セレヴィシエ・インヴェルターゼのための遺伝子から得られる。他の有用なシグナルペプチドコーディング領域は、Romanos他, 1992, 前掲、及びXiong他のJournal of Applied Microbiology 2005, 98, 418-428に記載されている。
【0111】
好ましい局面では、シグナルペプチドコーディング領域は、配列番号10のアミノ酸1〜33をコードする配列番号9のヌクレオチド1〜99である。他の好ましい局面では、シグナルペプチドコーディング領域は、配列番号12のアミノ酸1〜27をコードする配列番号11のヌクレオチド1〜81である。
【0112】
制御配列は、ポリペプチドのアミノ末端に位置するアミノ酸をコードするプロペプチドコーディング領域でもよい。得られたポリペプチドは、プロペプチド又はプロポリペプチド(又は場合によっては酵素原)として知られている。プロポリペプチドは、一般的に、不活性であり、プロポリペプチドからのプロペプチドの触媒的又は自己触媒的開裂によって成熟活性ポリペプチドに変換され得る。プロペプチドコーディング領域は、バチルス・サブチリス・アルカリプロテアーゼ (aprE)、バチルス・サブティリス・中性プロテアーゼ (nprT)、サッカロマイセス・セレヴィシエ・α-因子、リゾムコール・ミエヘイ・アスパラギンプロテイナーゼ、及びマイセリオフトーラ・サーモフィラ(Myceliophthora thermophila)・ラクタマーゼ (WO 95/33836) のための遺伝子から得られる。
【0113】
シグナルペプチド領域及びプロペプチド領域の両方がポリペプチドのアミノ末端に存在する場合には、該プロペプチド領域は、ポリペプチドのアミノ末端に隣接して位置し、該シグナルペプチド領域は、該プロペプチド領域のアミノ末端に隣接して位置する。
【0114】
また、宿主細胞の成長に対してポリペプチドの発現のレギュレーションを可能にする調節配列を付加することが望ましいことがある。調節系の例は、調節化合物の存在を含む、遺伝子の発現を化学又は物理的刺激に反応してオン又はオフに変化させるものである。原核生物の調節系は、lac、tac及びtrpオペレーター系を含む。酵母では、ADH2系又はGAL1系が使用され得る。線維状真菌では、TAKAα-アミラーゼ・プロモーター、アスペルギルス・ニガー・グルコアミラーゼ・プロモーター及びアスペルギルス・オリザエ・グルコアミラーゼ・プロモーターが、調節配列として使用され得る。調節配列の他の例は、遺伝子増幅を可能にする配列である。
【0115】
発現ベクター
本発明はまた、本発明のポリヌクレオチド、プロモーター、及び転写及び翻訳停止シグナルを含む組換え発現ベクターに関する。上記の様々な核酸及び制御配列は、一緒に結合されて、該ポリペプチドをコードするヌクレオチド配列の挿入又は置換をかかる位置で可能にするための、1以上の簡便な制限部位を含み得る組換え発現ベクターを製造し得る。あるいは、本発明のヌクレオチド配列は、該ヌクレオチド配列又は該配列を含む核酸構築物を発現用の好適なベクターに挿入することによって発現され得る。発現ベクターの作製において、コーディング配列は、該コーディング配列が発現用の好適な制御配列と作動可能に連結されるように、該ベクターに配置される。
【0116】
組換え発現ベクターは、組換えDNA法に簡便に供され、ヌクレオチド配列の発現をもたらし得る任意ベクターでよい(例えば、プラスミド又はウイルス)。最適なベクターは、典型的には、ベクター又はベクターが導入される宿主細胞の適合性に依拠することになる。ベクターは、線状又は閉環状プラスミドでよい。
【0117】
ベクターは、自律的複製型ベクター、すなわち、染色体外物質として存在し、その複製が、染色体複製とは無関係であるベクター、例えばプラスミド、染色体外要素、人工ミニ染色体、又は人工染色体でよい。ベクターは、自己複製を確実にするための手段を含むことがある。あるいは、宿主細胞に導入される時に、ベクターはゲノムに統合され、ベクターが統合された染色体(複数)と一緒になって複製されるものでよい。更に、宿主細胞のゲノム又はトランスポソンに導入されるべき総DNAを一緒に含む、単一ベクターもしくはプラスミド又は2以上のベクターもしくはプラスミドが使用され得る。
【0118】
本発明のベクターは、好ましくは、形質転換された細胞の簡便な選択を許容する1以上の選択マーカーを含む。選択マーカーは、その産物が殺生剤又はウイルス耐性、重金属に対する耐性、栄養要求体に対する原栄養性などを与える遺伝子である。
【0119】
条件付で不可欠の遺伝子は、非-抗生物質選択的マーカーとして機能し得る。細菌性の条件付不可欠の非-抗生物質選択的マーカーの非限定例は、細菌がD-アラニンの非存在下で培養される時に不可欠であるにすぎない、バチルス・サブティリス、バチルス・リシェニフォルミス又は他のバチルス属由来のdal遺伝子である。また、UDP-がラクトースのターオーバーに関連する遺伝子コーディング酵素は、細胞がガラクトースの存在下で成長し又はガラクトースの存在を与える培地中で成長する時に、細胞中で条件付不可欠マーカーとして機能し得る。かかる遺伝子の非限定例は、UTP-依存的ホスホリラーゼ (EC 2.7.7.10)、UDP-グルコース-依存的ウリジリルトランスフェラーゼ (EC 2.7.7.12)、又はUDP-ガラクトースエピメラーゼ (EC 5.1.3.2) をコードするB. サブティリス又はB. リシェニフォルミス由来のものである。また、バチルスのxylAのようなキシロースイソメラーゼ遺伝子は、単一の炭素源としてのキシロースを含む最小培地で成長した細胞において選択マーカーとして使用され得る。グルコン酸塩、gntK及びgntPを利用するために必要な遺伝子も、単一炭素源としてのグルコン酸塩を含む最小培地で成長する細胞における選択マーカーとして使用され得る。条件付の不可欠な遺伝子の他の例は、当該分野で知られている。抗生物質選択マーカーは、アンピシリン、カナマイシン、クロラムフェニコール、エリスロマイシン、テトラサイクリン、ネオマイシン、ハイグロマイシン又はメトレキサートのような抗生物質に対する抗生物質耐性を与える。
【0120】
酵母宿主細胞のための好適なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1及びURA3である。線維状真菌宿主細胞において使用するための選択マーカーは、amdS (アセトアミダーゼ)、argB (オルニチンカルバモイル基転移酵素)、bar (ホスフィノトリシン・アセチル基転移酵素)、hph (ハイグロマイシンリン酸転移酵素)、niaD (硝酸還元酵素)、pyrG (オロチジン5'-リン酸脱炭酸酵素)、sC (サルファイト・アデニルトランスフェラーゼ)、及びtrpC (アントラニル酸合成酵素)、並びにそれらの等価体を含むが、これらに限定されない。アスペルギルス属細胞において使用するためには、アスペルギルス・ニデュランス又はアスペルギルス・オリザエのamdS及びpyrG遺伝子、及びストレプトマイセス・ハイグロスコピカス(Streptomyces hygroscopicus)のbar遺伝子が好ましい。
【0121】
本発明のベクターは、好ましくは、宿主細胞中へのベクターの統合又はゲノムに関係のない細胞中でのベクターの自律的複製を許容する、要素(複数)を含む。宿主細胞ゲノムへの統合に関して、ベクターは、ポリペプチドをコードするポリヌクレオチド配列、又は相同的もしくは非相同的な組換えによるゲノムへの統合のためのベクターの任意の他の要素に依拠し得る。あるいは、ベクターは、染色体(複数)における正確な位置(複数)での宿主細胞のゲノムへの相同組換えによる統合を指向するための、追加のヌクレオチド配列を含み得る。正確な位置での統合の可能性を増加させるために、統合要素は、相同組換えの可能性を増加させるための対応する標的配列と高い同一性の程度を有する、好ましくは、十分に多数の核酸、例えば100〜10,000塩基対、好ましくは400〜10,000塩基対、及び最も好ましくは800〜10,000塩基対を含む必要がある。統合要素は、宿主細胞のゲノム中の標的配列と相同である任意に配列でよい。更に、統合要素は、非-コーディング又はコーディングヌクレオチド配列でよい。一方、ベクターは、非-相同組換えによって宿主細胞のゲノムに統合され得る。
【0122】
自律的複製のために、ベクターは、ベクターを問題の宿主細胞において自律的に複製させる複製起源を更に含むことがある。複製起源は、細胞中で機能する自律的複製を介在する任意のプラスミド複製因子でよい。用語「複製起源」又は「プラスミド複製因子」は、インビボでプラスミド又はベクターを複製させるヌクレオチド配列として本明細書に定義される。
【0123】
細菌の複製起源の例は、E. コリ中で複製を許容するプラスミドpBR322、pUC19、pACYC177及びpACYC184、及びバチルス属中で複製を許容するpUBHO、pE194、pTA1060及びpAMβ1である。
【0124】
酵母宿主細胞中で使用するための複製起源の例は、複製の2ミクロン起源、ARS1、ARS4、ARS1とCEN3との組合せ、及びARS4とCEN6との組合せである。
【0125】
線維状真菌細胞において有用な複製起源の例は、AMA1及びANSIである (Gems et al., 1991, Gene 98:61-67; Cullen et al., 1987, Nucleic Acids Research 15: 9163-9175; WO 00/24883)。AMA1遺伝子の単離、及び該遺伝子を含むプラスミド又はベクターの構築は、WO00/24883に開示の方法に従って達成され得る。
【0126】
本発明のポリヌクレオチドの1超コピーは、遺伝子産物の産生を増加させるために宿主細胞に挿入され得る。ポリヌクレオチドのコピー数の増加は、該配列の少なくとも1つの追加のコピーを宿主細胞ゲノムに統合することによって、又は該ポリヌクレオチドと共に増幅可能な選択マーカー遺伝子を含むことによって得られ(そこでは、細胞は、選択マーカー遺伝子の増幅されたコピーを含む)、それによって、ポリヌクレオチドの追加のコピーは、好適な選択マーカーの存在下で細胞を培養することによって選択され得る。
【0127】
本発明の組換え発現ベクターを構築するための上記の要素をライゲートするために使用される方法は、当業者によく知られている (例えば、Sambrook et al., 1989, 前掲参照)。
【0128】
宿主細胞
本発明はまた、ポリペプチドの組換え産生において有利に使用される、本発明のポリヌクレオチドを含む組換え宿主細胞に関する。本発明のポリヌクレオチドを含むベクターは、該ベクターが先の記載の染色体統合因子又は自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。用語「宿主細胞」は、複製中に起こる変異に因って親細胞と同一でない親細胞の任意の子孫を包含する。最適な宿主細胞は、ポリペプチドをコードする遺伝子及びその起源にかなりの程度で依拠することになる。
【0129】
宿主細胞は、単細胞性微生物、例えば原核生物、又は非-単細胞性微生物、例えば真核生物でよい。
【0130】
有用な単細胞微生物は、細菌細胞であり、例えば、バチルス細胞、例えばバチルス・アルカロフィラス(Bacillus alkalophilus)、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)、バチルス・ブレビス(Bacillus brevis)、バチルス・サーキュランス(Bacillus circulans)、バチルス・クラウジアイ(Bacillus clausii)、バチルス・コアギュランス(Bacillus coagulans)、バチルス・ラウトゥス(Bacillus lautus)、バチルス・レンタス(Bacillus lentus)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・メガテリウム(Bacillus megaterium)、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)、バチルス・サブティリス(Bacillus subtilis)、及びバチルス・チューリンゲンシス(Bacillus thuringiensis);又はストレプトマイセス細胞、例えばストレプトマイセス・リビダンス(Streptomyces lividans)及びストレプトマイセス・ミュリナス(Streptomyces murinus)を含むがこれらに限定されないグラム陽性菌、あるいは、E. コリ及びシュードモナス属のような含むグラム陰性菌である。好ましい局面では、細菌宿主細胞は、バチルス・レンタス、バチルス・リケニフォルミス、バチルス・ステアロサーモフィルス又はバチルス・サブティリス細胞である。別の好ましい局面では、バチルス細胞は、好アルカリ性バチルスである。
【0131】
ベクターの細菌宿主細胞への導入は、例えば、コンピテント細胞を用いる (例えば、Young and Spizizin, 1961 , Journal of Bacteriology 81: 823-829、又はDubnau and Davidoff-Abelson, 1971 , Journal of Molecular Biology 56: 209-221参照) プロトラスト形質転換 (例えば、Chang and Cohen, 1979, Molecular General Genetics 168: 111-115参照)、エレクトロポレーション (例えば、Shigekawa and Dower, 1988, Biotechniques 6: 742-751参照)、又はコンジュゲーション (例えば、Koehler and Thorne, 1987, Journal of Bacteriology 169: 5771-5278参照) によって行われる。
【0132】
宿主細胞は、真核細胞、例えば哺乳動物、昆虫、植物又は真菌細胞でもよい。
【0133】
好ましい局面では、宿主細胞は真菌細胞である。本明細書で使用する「真菌」は、子嚢菌門(Ascomycota)、担子菌門(Basidiomycota)、ツボカビ門(Chytridiomycota)及び接合菌門(Zygomycota)(Hawksworth et al., In, Ainsworth and Bisby's Dictionary of The Fungi, 第8版, 1995, CAB International, University Press, Cambridge, UKによって定義された)、並びに卵菌門(Hawksworth et al., 1995, 前掲, 第171頁で引用された)及び分生子形成菌(Hawksworth et al., 1995, 前掲)を含む。
【0134】
より好ましい局面では、真菌宿主細胞は、酵母細胞である。本明細書で使用される「酵母」は、有子嚢胞子(ascosporogenous)酵母(エンドミセタレス目(Endomycetales))、担子胞子(basidiosporogenous)及び不完全菌類(Fungi lmperfecti)(不完全酵母菌綱(Blastomycetes))を含む。酵母の分類は、本発明の目的のために今後変わるかもしれないので、酵母は、「酵母の生物学及び活性」(Skinner, F.A., Passmore, S. M., and Davenport, R. R.著, Soc. App. Bacterid. Symposium Series No. 9, 1980) に記載されたように定義されべきである。
【0135】
更により好ましい局面では、酵母宿細胞は、カンジタ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロマイセス属(Kluyveromyces)、ピキア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)又はヤロウイア属(Yarrowia)細胞である。最も好ましい局面では、酵母宿主細胞は、ピキア・パストリス(Pichia pastoris)、ピキア・メタノリカ(Pichia methanolica)、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ダイアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミ・セス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)又はサッカロミセス・オビホルミス(Saccharomyces oviformis)細胞である。別の好ましい局面では、酵母宿主細胞は、クルイベロミセス・ラクチス(Kluyveromyces lactis)細胞である。別の最も好ましい局面では、酵母宿主細胞は、ヤロウイア・リポリティカ(Yarrowia lipolytica)細胞である。
【0136】
別のより好ましい局面では、真菌宿主細胞は、線維状真菌細胞である。「線維状真菌」は、(Hawksworth et al., 1995, 前掲によって定義された)サブディビジョンユーマイコタ(Eumycota)及びオオマイコタ(Oomycota)の全ての繊維型を含む。線維状真菌は、一般的に、キチン、セルロース、グルカン、キトサン、マンナン及び他の複合化多糖からなる菌糸体壁によって特徴付けられる。植物的な成長は、菌糸の伸張であり、炭素異化作用は、偏性好気性である。対照的に、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)のような酵母による植物の成長は単細胞葉状体の発芽によるものであり、炭素異化作用は発酵性でよい。
【0137】
更により好ましい局面では、線維状真菌宿主細胞は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、オーレオバシジウム(Aureobasidium)、ジェルカンデラ(Bjerkandera)、セリポリオプシス(Ceriporiopsis)、コプリナス(Coprinus)、コリオラス(Coriolus)、クリプトコッカス(Cryptococcus)、フィロバシジウム(Filobasidium)、フザリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカノマスチックス( Neocallimastix)、ネウロスポラ(Neurospora)、パエシロミセス(Paecilomyces)、ペニシリウム(Penicillium)、ファネロカエテ(Phanerochaete)、フレビア(Phlebia)、ピロミセス(Piromyces)、プレロータス(Pleurotus)、シゾフィラム(Schizophyllum)、タラロマイセス(Talaromyces)、サーモアスカス(Thermoascus)、チエラビア(Thielavia)、トイポクラディウム(Tolypocladium)、トラメテス(Trametes)、又はトリコデルマ(Trichoderma)の細胞である。
【0138】
最も好ましい局面では、線維状真菌宿主細胞は、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、アスペルギルス・フォティダス(Aspergillus foetidus)、アスペルギルス ジャポニカス(Aspergillus japonicus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ニガー(Aspergillus niger)又はアスペルギルス・オリザエ(Aspergillus oryzae)の細胞である。別の最も好ましい局面では、線維状真菌宿主細胞は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・セルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)、フザリウム・ネグンジ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レチクラツム(Fusarium reticulatum)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サムブキヌム(Fusarium sambucinum)、フザリウム・サルコクロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルフリウム(Fusarium sulphureum)、フザリウム・トルロクム(Fusarium torulosum)、フザリウム・トリコテキオイデス(Fusarium trichothecioides)、又はフザリウム・ベネナツム(Fusarium venenatum)の細胞である。別の最も好ましい局面では、線維状真菌宿主細胞は、ルカンデラ・アダスタ(Bjerkandera adusta)、セリポリオプシス・アネイニナ(Ceriporiopsis aneirina)、セリポリオプシス・カレグレア(Ceriporiopsis caregiea)、セリポリオプシス・ギルベセン.ス(Ceriporiopsis gilvescens)、セリポリオプシス・パンノシンタ(Ceriporiopsis pannocinta)、セリポリオプシス・リバロサ(Ceriporiopsis rivulosa)、セリポリオプシス・スブベルミスポラ(Ceriporiopsis subrufa)又はセリポリオプシス・サバーミスポラ(Ceriporiopsis subvermispora)、コプリヌス・シネレウス(Coprinus cinereus)、コプリヌス・ヒルスタス(Coriolus hirsutus)、ヒュミコラ・インソレンス(Humicola insolens)、ヒュミコラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヒー(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、ネウロスポラ・クラサ(Neurospora crassa)、ペニシリウム・プルプルゲヌム(Penicillium purpurogenum)、ファネロケーテ・クリプトスポリジウム(Phanerochaete chrysosporium)、フレビア・ラジアタ(Phlebia radiata)、プレオロータス・エリンジ(Pleurotus eryngii)、チエラビア・テルレストリス(Thielavia terrestris)、トラメーテス・ビロサ(Trametes villosa)、トラメテス・ベルシコロル(Trametes versicolor)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギー(Trichoderma koningii)、トリコデルマ・ロンギブラキアタム(Trichoderma longibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、又はトリコデルマ・ビリデ(Trichoderma viride)の株細胞である。
【0139】
真菌細胞は、それ自体公知の方法で、プロトプラスト形成、プロトプラストの形質転換、及び細胞壁の再生を含むプロセスによって形質転換され得る。アスペルギルス属及びトリコデルマ属宿主細胞の形質転換のための好適な方法は、EP 238 023、及びYelton他, 1984, Proceedings of the National Academy of Sciences USA 81: 1470-1474に記載されている。フザリウム属を形質転換する好適な方法は、Malardier他, 1989, Gene 78: 147-156、及びWO 96/00787に記載されている。酵母は、Becker及びGuarente, In Abelson, J.N.及びSimon, M. I.著, 酵母遺伝学及び分子生物学への指針、酵素学の方法, 第194巻, p第182-187頁, Academic Press, Inc., New York; lto他, 1983, Journal of Bacteriology 153: 163; 及びHinnen他, 1978, Proceedings of the National Academy of Sciences USA 75: 1920に記載の方法を用いて形質転換され得る。
【0140】
製造法
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、その野生型形態において、該ポリペプチドを産生することができる、細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造するための方法に関する。好ましくは、細胞h、ハフニア種の細胞であり、より好ましくはハフニア・アルベイの細胞である。
【0141】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、宿主細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造するための方法に関する。
【0142】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、宿主細胞を培養し、ここで、該宿主細胞は、配列番号9のいずれか1つの成熟ポリペプチドコーディング領域において少なくとも1つの変異を有する変異ヌクレオチド配列を含み、該変異ヌクレオチド配列は、配列番号10のアミノ酸1〜413からなるポリペプチドをコードする、及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドの製造方法に関する。
【0143】
本発明の製造方法において、細胞は、当該分野で周知の方法を用いてポリペプチドの産生のために好適な栄養培地において培養される。例えば、細胞は、好適な培地中で達成される研究室的又は工業的発酵器中で及びポリペプチドを発現させる及び/又は単離させる条件下で、振盪フラスコ培養、及び小スケール又は大スケール発光(連続的、バッチ、流加、又は固体発光を含む)によって培養され得る。培養は、当該分野で公知の方法を用いて、炭素源、窒素源及び無機塩を含む好適な栄養培地中で行う。好適な培地は、商業的供給者から入手されるか、又は(例えば、アメリカン・タイプ・カルチャー・コレクションのカタログにおいて)公表された組成に従って調製され得る。ポリペプチドが栄養培地に分泌される場合には、ポリペプチドは、培地から直接的に回収され得る。ポリペプチドが分泌されない場合には、細胞溶解物から回収され得る。
【0144】
ポリペプチドは、ポリヌクレオチドに特異的である当該分野で公知の方法を用いて検出され得る。これらの検出方法は、特異的抗体の使用、ポリペプチド産物の形成、又はポリペプチド基質の消失を含むことがある。例えば、ポリペプチドアッセイは、本明細書に記載のポリペプチドの活性を決定するために使用され得る。
【0145】
得られたポリペプチドは、当該分野で公知の方法を用いて回収され得る。例えば、ポリペプチドは、遠心、ろ過、抽出、スプレイ-乾燥、濃縮又は沈殿を含むがこれらに限定されない、慣用的な方法によって栄養培地から回収され得る。
【0146】
本発明のポリペプチドは、クロマトグラフィー(例えば、イオン交換、アフィニティー、疎水的方法、等電点電気泳動及びサイズ排除法)、電気泳動法(例えば、プレパラティブ等電点電気泳動法)、差異化された溶解度(例えば、硫酸アンモニウム沈殿)、SDS-PAGE又は抽出 (例えば、タンパク質精製, J. -C. Janson及びLars Ryden著, VCH Publishers, New York, 1989参照) を含むがこれらに限定されない当該分野で公知の様々な方法によって精製され得る。
【0147】
トランスジェニック植物
本発明はまた、本発明のフィターゼ活性を有するポリペプチドを回収可能な量で発現及び産生するように、該ポリペプチドをコードするヌクレオチド配列で形質転換された、トランスジェニック植物、植物部分又は植物細胞に関する。該ポリペプチドは、植物又は植物部分から回収され得る。あるいは、組換えポリペプチドを含む植物又は植物部分は、食品又は飼料の量を改善するためのものとして、例えば栄養価値、おいしさ及び流体力学的的性質を改善するためのものとして、あるいは抗栄養因子を破壊するために、使用され得る。
【0148】
具体的な実施態様では、該ポリペプチドは、種子中の内胚乳貯蔵小胞に標的される。これは、好適なシグナルペプチドを用いて前駆体としてそれを合成することによって得られる。Horvath他のPNAS, Feb. 15, 2000, vol. 97, no. 4, p. 1914-1919参照。
【0149】
トランスジェニック植物は、双子葉植物(子葉植物)又は単子葉植物(単子葉植物)、又はその人工変形体でよい。単子葉植物の例は、芝生、例えば牧草(ブルーグラス, Poa)、飼料草例えばウシノケグサ属、ロリウム属、温帯草、例えばアグロスチス属、及び穀物、例えば小麦、オート麦、ライ麦、大麦、米、サトウモロコシ、ライ小麦(小麦(Triticum)及びライ(Secale)及びトウモロコシ(corn)の安定化ハイブリッド)である。子葉植物の例は、タバコ、マメ科植物、例えばヒマワリ (Helianthus)、綿花 (Gossypium)、ルピナス、ジャガイモ、サトウキビ、ナシ、マメ及び大豆、及びアブラナ科植物 (Brassicaceaeファミリー)、例えば、カリフラワー、セイヨウアブラナ種子、並びに密接に関連したモデル生物体のシロイヌナズナ(Arabidopsis thaliana)である。例えば米国特許第5,689,054号明細書及び同6,111,168号明細書に記載の低-フィターゼ植物は、人工植物の例である。
【0150】
植物部分の例は、幹、カルス、葉、果実、種子及び塊茎、並びにこれらの部分を含む個々の組織、例えば表皮、葉肉、柔組織、維管束組織、分裂組織である。また、特定の植物組織区画、例えば葉緑体、アポプラスト、ミトコンドリア、小胞、ペルオキシソーム及び細胞質は植物部分と考えられる。更に、任意の植物細胞、それが組織起源であってもなくても、植物部分と考えられる。同様に、本発明の利用を助けるために単離された特定の組織及び細胞のような植物部分も、植物部分と考えられ、例えば、胚、内胚乳、澱粉(aleurone)及び種子コートである。
【0151】
かかる植物の子孫、植物部分及び植物細胞も本発明の範囲内に含まれる。本発明のポリペプチドを発現するトランスジェニック植物又は植物細胞は、当該分野で公知の方法に従って構築され得る。すなわち、該植物又は植物細胞は、本発明のポリペプチドをコードする1以上の発現構築物を植物宿主ゲノムに組み込み、そして得られた改変植物又は植物細胞をトランスジェニック植物又は植物細胞で繁殖させることによって構築される。
【0152】
好都合なことに、発現構築物は、最適な植物又は植物部分における核酸配列の発現のために必要とされる好適な調節配列と作動可能に連結した本発明のポリペプチドをコードする核酸配列を含む構築物である。更に、発現構築物は、発現構築物が統合されている宿主細胞を同定するために有用な選択マーカー、及び問題の植物への構築物の導入のために必要なDNA配列を含み得る(後者は、使用されるべきDNA導入法に依拠する)。
【0153】
最適な調節配列、例えばプロモーター及び終結配列及び場合によりシグナル又はトランジット配列が、例えば、いつ、どこで及びどのようにして本発明のポリペプチドが発現されることが望まれるかに基づいて決定される。例えば、本発明のポリペプチドをコードする遺伝子の発現は、構成的でも又は誘導可能でよく、あるいは発達特異的、段階特異的又は組織特異的でもよく、そして遺伝子産物は、特定の細胞区分、組織又は細胞部分、例えば種子又は葉に標的され得る。調節配列は、例えば、Tague他, 1988, Plant Physiology 86: 506に記載されている。
【0154】
構成的発現のために、以下のプロモーターが使用され得る: 35S-CaMVプロモーター (Franck et al., 1980, Cell 21: 285-294)、トウモロコシポリユビキチン1 (Christensen AH, Sharrock RA and Quail 1992,トウモロコシポリユビキチン遺伝子:構造、発現の熱的混乱及び転写スプライシング、及びエレクトロポレーションによるプロトプラストへの変換後のプロモーター活性)、又はコメアクチン1プロモーター (Plant Mo. Biol. 18, 675-689; Zhang W, McElroy D. and Wu R 1991, Analysis of rice Act1 5' region activity in transgenic rice plants. Plant Cell 3, 1155-1165)。生物特異的プロモーターは、例えば、種子、ジャガイモ塊茎及び果実のような貯蔵シンク組織由来のプロモーター (Edwards & Coruzzi, 1990, Ann. Rev. Genet. 24: 275-303)、又は分裂組織のような代謝性シンク組織由来のプロモーター (Ito et al., 1994, Plant Mol. Biol. 24: 863-878)、グルテリン、プロタミン、グロブリンもしくは米由来のアルブミンプロモーターのような種子特異的プロモーター (Wu et al., 1998, Plant and Cell Physiology 39: 885-889)、レグミンB4由来のヴィキア・ファバプロモーター及びヴィキア・ファバ(Vicia faba)由来の未知のタンパク質遺伝子 (Conrad et al., 1998, Journal of Plant Physiology 152: 708-711)、種子油体タンパク質由来のプロモーター (Chen et al., 1998, Plant and Cell Physiology 39: 935-941)、ブラシカ・ナパス(Brassica napus)由来の貯蔵タンパク質napAプロモーター、又は例えばWO 91/14772に記載の当該分野で公知の任意の他の種子特異的プロモーターである。更に、プロモーターは、コメ又はトマト由来のrbcsプロモーターのような葉特異的プロモーター (Kyozuka et al., 1993, Plant Physiology 102: 991-1000)、コレラウイルスアデニンメチルトランスフェラーゼ遺伝子プロモーター (Mitra and Higgins, 1994, Plant Molecular Biology 26: 85-93)、又はコメ由来のaldP遺伝子プロモーター (Kagaya et al., 1995, Molecular and General Genetics 248: 668-674)、又はジャガイモpin2プロモーターのような損傷誘導プロモーター (Xu et al., 1993, Plant Molecular Biology 22: 573-588) でよい。同様に、プロモーターは、塩分の温度、不足又は変更のような非生物的処理によって誘導され、あるいはプロモーターを活性化する外因的に適用された物質、例えばエタノール、エストロゲン、エチレンのような植物ホルモン、アブシジン酸、ジベレリン酸及び/又は重金属、によって誘導され得る。
【0155】
プロモーターエンハンサー要素は、植物におけるポリペプチドのより高い発現を達成するために使用してもよい。例えば、プロモーターエンハンサー要素は、プロモーターと本発明のポリペプチドをコードするヌクレオチド配列との間に配置されるイントロンでよい。例えば、Xu他, 1993, 前掲は、発現を亢進するためのイネアクチン1遺伝子の第1イントロンの使用を開示する。
【0156】
なお更に、コドン使用は、発現を改善するために問題の植物種について最適化され得る(上で言及したHorvath et al参照)。
【0157】
選択マーカー遺伝子及び発現構築物の任意の他の部分は、当該分野で入手可能なものから選択され得る。
【0158】
核酸構築物は、アグロバクテリウム(Agrobacterium)-介在形質転換、ウイルス-介在形質転換、マイクロインジェクション、微粒子銃、バイオリスティック形質転換、及びエレクトロポレーションを含む当該分野で公知の慣用的技術に従って、植物ゲノムに組み込まれる (Gasser et al., 1990, Science 244: 1293; Potrykus, 1990, Bio/Technology 8: 535; Shimamoto et al., 1989, Nature 338: 274)。
【0159】
現在、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)-介在遺伝子移動は、トランスジェニック双子葉植物を作製するための最適な方法であり(考察には、Hooykas and Schilperoort, 1992, Plant Molecular Biology 19: 15-38参照)、それは、単子葉植物を形質転換するためにも使用され得る。但し、他の形質転換法もこれらの植物のために通常使用される。現在、アグロバクテリウム法を補充する、トランスジェニック単子葉植物を作製するための最適な方法は、胚カルス又は発達中の胚の微粒子銃(形質転換DNAでコートされた微視的ゴールド又はタングステン粒子)(Christou, 1992, Plant Journal 2: 275-281; Shimamoto, 1994, Current Opinion Biotechnology 5: 158-162; Vasil他, 1992, Bio/Technology 10: 667-674)。単子葉植物の形質転換の別の方法は、Omirulleh他, 1993, Plant Molecular Biology 21: 415-428によって記載されたプロトプラスト形質転換に基づく。
【0160】
形質転換に続いて、発現構築物が組み込まれた形質転換体が選択され、そして当該分野で周知の方法に従って植物全体に再生される。形質転換法は、通常、例えば、2つの別個のT-DNA構築物による同時-形質転換、又は特定のリコンビナーゼによる選択遺伝子の部位特異的切除を用いて、再生の間又は次世代のいずれかにおいて選択遺伝子の選択的削除について設計されている。
【0161】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、本発明のフィターゼ活性を有するポリペプチドをコードする核酸配列を含むトランスジェニック植物又は植物細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造する方法に関する。
【0162】
トランスジェニック動物
本発明はまた、トランスジェニックな、非-ヒト動物及びその生成物又は要素に関する。その例は体液、例えばミルク及び血液、臓器、食肉、及び動物細胞である。タンパク質、例えば哺乳動物におけるタンパク質を発現するための技術は、当該分野で知られている。例えば、ハンドブック「タンパク質発現: 実際のアプローチ」Higgins and Hames (著), Oxford University Press (1999)、及び「遺伝子転写、RNAプロセッシング、及び翻訳後プロセッシン具」に関連するシリーズにおける他の3つのハンドブックを参照。一般的には、トランスジェニック動物を製造するために、選択された動物の選択された細胞は、本発明のフィターゼ活性を有するポリペプチドを発現及び産生するように、該ポリペプチドをコードする核酸配列で形質転換される。該ポリペプチドは、動物から、例えば、雌の動物のミルクから回収され、又は該ポリペプチドは、動物自体のために、例えば動物の消化を助けるために発現され得る。動物の例は、「Animal Feed」という見出しに以下のように述べられている。
【0163】
動物のミルクから該ポリペプチドを回収することを目的として、トランスジェニック動物を製造するために、該ポリペプチドをコードする遺伝子は、例えば、好適なミルクタンパク質プロモーターを含む導入遺伝子発現ベクター、及び該ポリペプチドをコードする遺伝子の使用によって、問題の動物の孵化された卵に挿入され得る。導入遺伝子発現ベクターは、孵化された卵に微量注入され、そして、好ましくは永久に染色体に統合される。該卵は成長し分割を始めると、可能性のある胚は、代理母に移植され、導入遺伝子を担持する動物が同定される。次いで、得られた動物は、慣用的な繁殖によって作製され得る。
該ポリペプチドは、動物のミルクから精製され得る、例えば、Meade, H. M. et al (1999): トランスジェニック動物のミルク中の組換えタンパク質の発現、遺伝子発現系:発現技術のための性質を使用する, J. M. Fernandez及びJ. P. Hoeffler (著), Academic Press参照。
【0164】
別の方法では、その身体細胞及び/又は細菌細胞のゲノム中にポリペプチドをコードする導入遺伝子を含む非相同導入遺伝子構築物を含む核酸配列を有するトランスジェニック非-ヒト動物を製造するために、導入遺伝子は、WO 00/064247に開示された、ポリペプチドの唾液腺特異的発現のための第1調節配列に作動可能に連結され得る。
【0165】
組成物及び使用
なお更なる局面では、本発明は、本発明のオリゴヌクレオチドを含む組成物、及びこれらの使用方法に関する。
【0166】
ポリペプチド組成物は、当該分野で公知の方法に従って調製され、液状又は乾燥組成物の形態でよい。例えば、ポリペプチド組成物は、顆粒又は微顆粒の形態でよい。ポリペプチドに含まれるポリペプチドは、当該分野で公知の方法に従って安定化され得る。
【0167】
本発明のフィターゼは、任意の産業的状況において、例えば、フィチン酸塩、フィチン酸及び/又はmyo-イノシトールのモノ-、ジ-、トリ-、テトラ-及び/又はペンタ-リン酸の分解のために使用され得る。これらの化合物のリン酸部分は、二価及び三価カチオン、例えば金属イオン、例えば栄養学的に不可欠なカルシウム、鉄、亜鉛、マグネシウムイオン及び微量元素のマンガン、銅及びモリブデンに配位する、ことが知られている。更に、フィチン酸はまた、ある程度、静電気相互作用によってタンパク質に結合する。
【0168】
従って、本発明のポリペプチドの好ましい使用は、動物飼料調製物(ヒト用食品を含む)又はかかる調製物のために添加物においてである。
【0169】
具体的態様では、本発明のポリペプチドは、動物飼料の栄養価を改善するために使用され得る。動物飼料(ヒト食品を含む)の栄養価を改善する非限定的例は、以下である:飼料消化率の改善;動物の成長の促進;飼料利用の改善;タンパク質のバイオアベイラビリティーの改善;消化できるホスフェートレベルの増加;フィターゼの放出及び/分解の改善;微量元素のバイオアベイラビリティーの改善;マクロミネラルのバイオアベイラビリティーの改善;補助的ホスフェート、微量元素及び/又はマクロミネラルを添加する必要性の削減;及び/又は卵殻品質の改善。そのため、飼料の栄養価が増加され、そして動物の成長速度及び/又は体重増加及び/又は飼料変換率(すなわち、体重増加に対する摂取飼料の重さ)が改善され得る。
【0170】
更に、本発明のポリペプチドは、肥料のフィターゼレベルを減少させるために使用され得る。
【0171】
動物、動物飼料、及び動物飼料添加物
用語動物は、ヒトを含むすべての動物を含む。動物の例は、非-反芻動物及び反芻動物である。反芻動物は、例えば、ヤギ、ヒツジ、ウマ及びウシ、例えば肉牛及び子牛、のような動物を含む。具体的な実施態様では、動物は非-反芻動物である。非-反芻動物は、単胃動物、例えばブタ(pigs)又はブタ(swine)(子豚、成長した豚及び雌豚を含むがこれらに限定されない);家禽、例えば、七面鳥、アヒル及びニワトリ(若鶏、産卵鶏を含むがこれらに限定されない);子牛;及び魚(鮭、トラウト、テラピア、ナマズ及び鯉を含むがこれらに限定されない)並びに甲殻類(エビ及び淡水エビを含むがこれらに限定されない)を含む。
【0172】
用語飼料又は飼料組成物は、動物による摂取のために好適な又はそれを意図した、任意の化合物、調製物、混合物又は組成物である。
【0173】
本発明に従う使用において、ポリペプチドは、食事の前に、後に又は同時に動物に供給され得る。後者が好ましい。
【0174】
具体的な実施態様では、飼料に添加される、又は飼料添加物として含まれる時の形態でのポリペプチドは、実質的に純粋である。具体的な実施態様ではそれは明確である。用語「明確である」は、フィターゼ調製物がサイズ排除クロマトグラフィーによって決定された少なくとも50%であることを意味する (例えば、WO 01/58275の実施例12参照)。他の具体的な実施態様では、フィターゼ調製物は、この方法によって決定される、少なくとも60%、70、80、85、88、90、92、94又は少なくとも95%純粋である。
【0175】
実質的に純粋な及び/又は明確なポリペプチド調製物は有利である。例えば、本質的に他のポリペプチドの妨害のない又は汚染のないポリペプチド飼料に正確に投薬することが非常に容易である。用語投薬は、具体的には、一貫性のある結果及び一定の結果を得る目的、及び所望の効果に基づく投薬量の最適化の可能性を言う。
【0176】
しかし、動物飼料での使用については、本発明のフィターゼポリペプチドは、純粋である必要がない;例えば他のポリペプチドを含む、それは、フィターゼ調製物と称される。
【0177】
フィターゼ調製物は、(a) 飼料に直接的に添加され(又はタンパク質の処理過程において直接的に使用される)、又は (b) 飼料に実質的に添加される(又は処理過程において使用される)飼料添加物又はプレミックスのような1以上の中間組成物の製造において使用され得る。上記の純度の程度は、上記の (a) に従って使用されるにしろ又は (b) に従って使用されるにしろ、最初のポリペプチド調製物の純度を言う。
【0178】
このようなオーダーの純度を有するポリペプチド調製物は、具体的には、組換え製造方法を用いて得られる。一方、それらはそれ程簡便に得られず、またポリペプチドは典型的な発酵法によって製造される時に非常に高いバッチごとの変化に供される。
【0179】
かかるポリペプチド調製物は、当然のことながら、他のポリペプチドと混合され得る。
【0180】
上記ポリペプチドは、相対的に純粋なポリペプチドとして任意の形態で、又は動物飼料に添加することを意図した他の成分と混合して、すなわち、いわゆる動物飼料用プレミックスのような動物飼料添加物の形態で、加えられる。
【0181】
更なる局面では、本発明は、動物飼料及び動物飼料添加物、例えばプレミックスのよな動物飼料において使用される成分に関する。
【0182】
本発明のポリペプチドとは別に、本発明の動物飼料添加物は、少なくとも1つの脂溶解ビタミン、及び/又は少なくとも1つの水溶性ビタミン、及び/又は少なくとも1つの微量元素を含む。飼料添加物は、少なくともマクロミネラルを含んでもよい。
【0183】
更に、場合により、飼料-添加物成分は、着色剤、例えばカロチノイド、例えばβ-カロチン、アスタキサンチン及びルテイン;芳香化合物;安定剤;抗菌ペプチド;ポリ不飽和脂肪酸;反応酸素生成種;及び/又はフィターゼ (EC 3.1.3.、又は3.1.3.26); キシラナーゼ (EC 3.2.1.8); ガラクタナーゼ (EC 3.2.1.89); α-ガラクトシダーゼ (EC 3.2.1.22); プロテアーゼ (EC 3.4.-.- )、ホスホリパーゼA1 (EC 3.1.1.32); ホスホリパーゼA2 (EC 3.1.1.4); リゾホスホリパーゼ (EC 3.1.1.5); ホスホリパーゼC (3.1.4.3); ホスホリパーゼD (EC 3.1.4.4); アミラーゼ、例えばα-アミラーゼ (EC 3.2.1.1); 及び/又はβ-グルカナーゼ (EC 3.2.1.4 or EC 3.2.1.6) から選ばれる少なくとも1つの他のポリペプチドである。
【0184】
具体的実施態様では、これらの他のポリペプチドは明確である(フィターゼ調製物について上で定義されたように)。
【0185】
具体的な好ましい実施態様では、比較的低い最適pHを有する本発明のフィターゼは、高い最適pHを有する少なくとも1つのフィターゼと組合わされる。高い最適pHのフィターゼの好ましい例は、バチルスフィターゼ、例えばバチルス・リケニフォルミス及びバチルス・サブティリス、並びにフィターゼ活性を有するその誘導体、変異体又はフラグメントである。
【0186】
本発明のフィターゼは、他のフィターゼ、例えば、子嚢菌フィターゼ、例えばアスペルギルス属から得られた、例えばペニオホラ・リシイ(Peniophoralycii)、アグロシベ・ペディアデス(Agrocybe pediades)、トラメテス・ピューベセンス・フィカム(Trametes pubescens ficuum)、アスペルギルス・ニガー(Aspergillus niger)、もしくはアスペルギルス・アワモリ(Aspergillus awamori)から得られたアスペルギルス・フィターゼ;あるいは担子菌フィターゼ又はパキシラス・インボルタス(Paxillus involutus)、又はフィターゼ活性を有するその誘導体、フラグメントもしくは変異体と組合わせてもよい。
【0187】
従って、本発明の動物飼料における使用の好ましい実施態様、及び本発明の動物飼料添加物及び動物飼料の好ましい実施態様では、本発明のフィターゼはかかるフィターゼと組合わされる。
【0188】
上記の子嚢菌類の植物及び担子菌のフィターゼ、特にペニオホラ・リシイ(Peniophora lycii)由来のRONOZYME P フィターゼ、及びその誘導体、変異体及びフラグメントは、動物飼料目的で、バチルス・フィターゼ、特に、B. リケニフォルミス(licheniformis)・フィターゼ及びその誘導体、フラグメント又は変異体と組み合わせてもよい。
【0189】
抗菌ペプチド(AMP)の例は、WO 03/044049及びWO 03/048148に開示された化合物及びポリペプチドを含む、CAP18、ロイコシンA、トリトリプシン、プロテグリン-1、サナチン、デフェンシン、ラクトフェリン、ラクトフェリシン及びオビスピリン、例えばノビスピリン (Robert Lehrer, 2000)、プレクタシン、及びスタチン、並びに抗菌活性を保持する上記の変異体又はフラグメントである。
【0190】
抗真菌ポリペプチド(AFP)の例は、アスペルギルス・ギガンテウス(Aspergillus giganteus)及びアスペルギルス・ニガー(Aspergillus niger)プロモーター、並びにWO 94/01459及びWO 02/090384に開示された抗真菌活性を保持するその変異体及びフラグメントである。
【0191】
ポリ不飽和脂肪酸の例は、C18、C20及びC22ポリ不飽和脂肪酸、例えばアラキドン酸、ドコサヘキサン酸、エイオコサペンタエン酸及びγ-リノール酸である。
【0192】
反応性酸素生成種の例は、化学物質、過ホウ酸塩、過硫酸塩又は過炭酸塩;及びポリペプチド、例えばオキシダーゼ、オキシゲナーゼ又はシンテターゼである。
【0193】
通常、脂溶性及び水溶性ビタミン、並びに微量元素は、飼料への添加を意図したいわゆるプレミックスの一部を形成するが、マクロミネラルは、通常、飼料に別個に添加される。これらの組成物種のいずれかは、本発明のポリペプチドが豊富に存在する場合には、本発明の動物飼料添加物である。
【0194】
具体的な実施多様では、本発明の動物飼料添加物は、0.01〜10.0%;より具体的には0.05〜5.0%;又は0.2〜1.0%(%は、添加物g/100 g飼料を意味する)のレベルで動物食品又は飼料中に含まれる(又は含まれていると指示されている)ことを意図する。このことは、具体的にはプレミックスに当てはまる。
【0195】
以下は、これらの成分の例の非限定的リストである。
【0196】
脂溶性ビタミンの例は、ビタミンA、ビタミンD3、ビタミンE及びビタミンK、例えば微ビタミンK3である。
【0197】
水溶性ビタミンの例は、ビタミンB12、ビオチン及びコリン、ビタミンB1、ビタミンB2、ビタミンB6、ナイアシン、葉酸、及びパントテン酸、例えばCa-D-パントテン酸である。微量元素の例は、マンガン、亜鉛、鉄、銅、ヨウ素、セレニウム、及びコバルトである。
【0198】
マクロミネラルの例は、カルシウム、リン及びナトリウムである。
【0199】
これらの成分の栄養的要件(家禽及び子豚/豚で代表される)は、WO 01/58275の表Aに記載されている。栄養的要件は、これらの成分が示された濃度で食品中に提供されるべきであることを意味する。
【0200】
あるいは、本発明の動物飼料添加物は、WO 01/58275の表Aで特定された個々の成分の少なくとも1つを含む。少なくとも1つは、13個の成分の内の1個以上、1個又は2個又は3個又は4個、最大13個全て、あるいは最大15個の個々の成分すべてを意味する。より具体的には、この少なくとも1つの個々の成分は、表Aのカラム4又はカラム5又はカラム6に示された範囲内の飼料中の濃度を提供するような量で、本発明の添加物中に含まれる。
【0201】
本発明は、動物飼料組成物にも関する。動物飼料組成物又は食品は比較的高いタンパク質量を有する。家禽及び豚の食品は、WO 01/58275の表Bのカラム2〜3に示されるように特徴付けられる。魚食品は、この表Bのカラム4中に示されるように特徴付けられる。更に、かかる魚食品は、通常、200〜310 g/kgの粗脂肪量を有する。
【0202】
WO 01/58275は、本明細書に参考として援用される米国出願第09/779334号に対応する。
【0203】
本発明に従う動物飼料組成物は、50〜800 g/kgの粗タンパク質量を有し、更に本願で請求された少なくとも1つのポリペプチドを含む。
【0204】
更に又はあるいは(上記の粗タンパク質量の代わりに)、本発明の動物飼料組成物は、10〜30 MJ/kgの代謝されるエネルギー量; 及び/又は0.1〜200 g/kgのカルシウム量; 及び/又は0.1〜200 g/kgの利用できるリンの量; 及び/又は0.1〜100 g/kgのメチオニン量; 及び/又は0.1〜150 g/kgのメチオニン及びシステイン量; 及び/又は0.5〜50 g/kgのリジン量を有する。
【0205】
具体的な実施態様では、代謝性エネルギー、粗タンパク質、カルシウム、リン、メチオニン、メチオニン及びシステイン、及び/又はリジンの量は、WO 01/58275の表B内のレンジ2、3、4又は5のいずれか1つにある (R. 2-5)。
【0206】
粗タンパク質は、ファクター6.25を乗じた窒素(N)として計算される、すなわち、粗タンパク質 (g/kg)= N (g/kg) x 6.25。窒素量は、Kjeldahl法 (A.O.A.C, 1984, Official Methods of Analysis 第14th版, Association of Official Analytical Chemists, Washington DC) により決定される。
【0207】
代謝エネルギーは、NRC刊行ブタの栄養要件, 第9改定版 1988, ブタ栄養についての分科委員会, 動物栄養についての委員会, 農業委員会, 米国学術研究会議に基いて計算される。National Academy Press, Washington, D. C, 第2〜6頁、及びブタ飼料のネエルギー価についての欧州表, ブタ研究及び講座のSpelderholtセンター, 7361 DA Beekbergen, The Netherlands. Grafisch bedrijf Ponsen & looijen bv, Wageningen. ISBN 90-71463-12-5。
【0208】
完全な動物食品中のカルシウム、入手できるリン及びアミノ酸の所要量は、飼料表、例えばVeevoedertabel 1997, gegevens over chemische samenstelling, verteerbaarheid en voederwaarde van voedermiddelen, Central Veevoederbureau, Runderweg 6, 8219 pk Lelystad. ISBN 90-72839-13-7に基いて計算される。
【0209】
具体的実施態様では、本発明の動物飼料組成物は、少なくとも1つのタンパク質を含む。該タンパク質は、動物タンパク質、例えば肉及び骨食品、及び/又は魚食品でよく;あるいは、野菜タンパク質でよい。本明細書で使用される用語野菜タンパク質は、任意の化合物、組成物、調製物又は混合物を言う。修飾タンパク質及びタンパク質誘導体を含む、野菜由来の又は野菜から生じる少なくとも1つのタンパク質を含む。具体的な実施態様では、野菜タンパク質のタンパク質量は、少なくとも10、20、30、40、50又は60% (w/w) である。
【0210】
野菜タンパク質は、野菜タンパク質源、例えばマメ科植物及び穀物、例えばファバセアエ(Fabaceae (レグミノサエ(Leguminosae))、クルシフェラセアエ(Cruciferaceae)、ケノポディアセアエ(Chenopodiaceae)及びピアース(Poaceae)のファミリーの植物由来の物質、例えば、大豆食品、ルピナス(lupin)食品及びマメ科植物食品から得られる。
【0211】
具体的な実施態様では、野菜タンパク質源は、ファバセアファミリーの1以上の植物、例えば、大豆、ルピナス、ナシ又は豆由来の物質である。
【0212】
別の具体的な実施態様では、野菜タンパク質源は、ケノポディアセアエファミリーの1以上の植物、例えば、ビート、砂糖大根、ホウレンソウ又はキノア、由来の物質である。野菜タンパク質源の他の例は、菜種、ヒマワリ種、綿花種及びキャベツである。
【0213】
大豆は、好ましい野菜タンパク質源である。
【0214】
野菜タンパク質源の他の例は、穀物、例えば、大麦、小麦、ライ麦、オート麦、トウモロコシ(コーン)、コメ、ライ小麦及びサトウモロコシである。更なる具体的実施態様では、本発明の動物飼料組成物は、0〜80%のトウモロコシ; 及び/又は0〜80%のサトウモロコシ; 及び/又は0〜70%の小麦; 及び/又は0〜70%の大麦; 及び/又は0〜30%のオート麦; 及び/又は0〜40%の大豆食品; 及び/又は0〜25%の魚食品; 及び/又は0〜25%の肉及び骨食品; 及び/又は0〜20%のホエイを含む。
【0215】
動物食品は、例えば、マッシュ食品(非ペレット化)又はペレット化食品として製造される。典型的には、粉砕された飼料材料は混合され、そして、必須ビタミン及びミネラルの充分量は問題の種の仕様書に従って添加される。ポリペプチドは、固体又は液状ポリペプチド製剤として添加され得る。例えば、固体ポリペプチド製剤は、典型的には、混合ステップの前又は中に加えられ;そして、液状ポリペプチド調製物は、典型的には、ペレットステップの後に加えられる。ポリペプチドは、飼料添加物又はプレミックス中に組み込まれてもよい。
【0216】
食品中の最終ポリペプチド濃度は、0.01〜200 mgポリペプチド/kg食品の範囲内にある。例えば、0.1〜10 mg/kg動物食品の範囲(典型的な投薬量は、250〜2000 FYT/kg動物食品の範囲内である)にある。
【0217】
本発明のフィターゼは、有効量、すなわち、飼料の溶解性を改善するために及び/又は栄養価を改善するためい充分な量で適用されるべきである。
【0218】
現在では、上記ポリペプチドは、以下の量(投薬範囲)の1以上で投与されることが画される:0.01〜200; 0.01〜100; 0.5〜100; 1〜50; 5〜100; 10〜100; 0.05〜50; 又は0.10〜10、これらの範囲はすべて、mgフィターゼポリペプチド/kg飼料(ppm)である。
【0219】
mgフィターゼポリペプチド/kg飼料を決定するために、フィターゼは、飼料組成物から精製され、精製されたフィターゼの特異的活性は関連するアッセイを用いて決定される。よって、飼料組成物のフィターゼ活性は、同一のアッセイを用いて決定され、これらの2つの決定に基いて、mgフィターゼポリペプチド/kg飼料での投薬量が計算される。
【0220】
mgフィターゼポリペプチド/kg飼料添加物を決定するために同一の原則が適用される。当然のことながら、試料が飼料添加物又は飼料を調製するために使用されるフィターゼが利用できる場合には、特異的活性は、この試料から決定される(飼料組成物又は添加物からフィターゼを精製する必要性はない)。
【0221】
発酵産物の製造方法
本発明の更に別の局面は、発酵産物、例えばエタノール、ビール、ワイン、蒸留乾燥種子(DDG)、の製造方法であって、該発酵が本発明のフィターゼの存在下で行われる、前記方法に関する。発酵プロセスの例は、例えば、WO01/62947に記載のプロセスを含む。発酵は、発酵微生物、例えば酵母を用いて行われる。
【0222】
具体的な実施態様では、本発明は、発酵産物の製造方法であって、(a) 本発明のフィターゼの存在下で、糖類含有物質(例えば、スターチ)を(発酵微生物、例えば酵母を用いて)発酵し、及び (b) 発酵糖類含有物質から発酵産物を製造すること、を含む、前記方法を提供する。
【0223】
具体的な実施態様では、本発明は、エタノールを製造するための方法であって、本発明のフィターゼの存在下で、糖類含有物質(例えば、スターチ)を(発酵微生物、例えば酵母を用いて)発酵し、及び (b) 発酵糖類含有物質からエタノールを製造又は回収すること、を含む、前記方法を提供する。
【0224】
別の実施態様では、本発明は、エタノールの製造方法であって、a) 例えば、液化及び/又は糖化プロセス、スターチ原料加水分解プロセスによる、加水分解スターチ、b) 本発明のフィターゼの存在下で得られたスターチを発酵させ、及び c) エタノールを製造すること、を含む、前記方法を提供する。
【0225】
フィターゼは、単独で、又は他の酵素、例えば1以上のα-アミラーゼ、グルコアミラーゼ、プロテアーゼ及び/又はセルラーゼと組み合わせて、任意の好適な段階で及び任意の好適な組成物中に、発酵プロセスに添加され得る。
【0226】
別の実施態様では、本発明は、バイオマスを加水分解し、及び(発酵微生物、例えば酵母を用いて)、本発明のフィターゼの存在下で得られたバイオマスを発酵させることを含む、エタノール製造方法を提供する。
【0227】
シグナルペプチド
本発明はまた、配列番号9のヌクレオチド1〜99からなる第1ヌクレオチド配列に作動可能に連結したタンパク質をコードし、配列番号10のアミノ酸1〜33からなるシグナルペプチドをコードする遺伝子を含み、該遺伝子が第1ヌクレオチド配列に無関係である、核酸構築物に関する。
【0228】
本発明はまた、かかる核酸構築物を含む、組換え発現ベクター及び組換え宿主細胞に関する。
【0229】
本発明はまた、タンパク質の製造法であって、(a) 該タンパク質の製造に好適な条件下でかかる組換え宿主細胞を培養し;及び (b) 該タンパク質を回収すること、を含む前記方法に関する。
【0230】
前記第1ヌクレオチド配列は、他の制御配列と共に又は他の制御配列と組み合わせて、個々に外来遺伝子に作動可能に連結され得る。かかる他の制御配列は、前掲である。該タンパク質は、天然でも又は宿主細胞に異種でもよい。用語「タンパク質」は、本明細書では、コードされた産物の特定の長さを意味せず、そのため、ペプチド、オリゴヌクレオチド及びタンパク質を包含する。用語「タンパク質」は、コードされた産物を形成するように組合わされた2以上のポリペプチドも包含する。該タンパク質は、少なくとも2つの異なったタンパク質、ここでその1以上が宿主細胞に異種又は天然である、から得られた部分的又は完全なポリペプチド配列の組合せを含むハイブリッドポリペプチドも含む。タンパク質は、上記のタンパク質及びハイブリッドタンパク質の天然アレル変異体及び人工変異体を更に含む。
【0231】
好ましくは、前記タンパク質は、ホルモン又はその変異体、ポリペプチド、例えば酵素、受容体又はその部分、抗体又はその部分、あるいはレポーターである。より好ましい局面では、該タンパク質は、酸化還元酵素、転移酵素、加水分解酵素、リアーゼ、イソメラーゼ又はリガーゼである。より更に好ましい局面では、該タンパク質は、アミノペプチダーゼ、アミラーゼ、カルボハイドラーゼ、カルボキシペプチダーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、シクロデキストリン・グリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α-ガラクトシダーゼ、β-ガラクトシダーゼ、グルコアミラーゼ、α-グルコシダーゼ、β-グルコシダーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチン質分解ポリペプチド、ペルオキシダーゼ、フィターゼ、ポリフェノールオキシダーゼ、タンパク質分解ポリペプチド、リボヌクレアーゼ、トランスグルタミナーゼ又はキシラナーゼである。
【0232】
遺伝子は、任意の原核生物、真核生物又は他の起源から得られる。
【0233】
様々な実施態様
以下は、本発明の更なる実施態様である。また、特許請求されたすべての、核酸、核酸構築物、組換え発現ベクター、組換え宿主細胞、前記ポリペプチドの製造法、トランスジェニック植物及び動物、並びに様々な使用、使用方法及び飼料組成物/添加物に関する対応する局面が本明細書に含まれる。
【0234】
フィターゼ活性を有し、37℃、0.1 M グリシン/HClバッファーpH 2.0中で24時間のインキュベーション後に、時間t = 0での活性と比べて少なくとも20%の残余活性を有する、単離されたポリペプチドであって、該活性は、0.25 M Na-酢酸バッファーpH 5.5、ブラインドを除いたバッファーを用いて、1% (w/v) Na-フィターゼで、37℃及びpH 5.5でアッセイされ;
好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0235】
フィターゼ活性を有し、37℃、0.1 M グリシン/HClバッファーpH 2.5中で24時間のインキュベーション後に、時間t = 0での活性と比べて少なくとも20%の残余活性を有する、単離されたポリペプチドであって、該活性は、0.25 M Na-酢酸バッファーpH 5.5、ブラインドを除いたバッファーを用いて、1% (w/v) Na-フィターゼで、37℃及びpH 5.5でアッセイされ;好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0236】
フィターゼ活性を有する単離されたポリペプチドであって、基質pNP-ホスホネートで測定されたpH 5.0及び37℃での該ポリペプチドの活性が、基質フィターゼで測定されたポリペプチドの活性の11%未満である;好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0237】
フィターゼ活性を有する単離されたポリペプチドであって、該ポリペプチドは、好ましくは、インビトロモデルで測定されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べてリン(P)の高い放出を有し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0238】
フィターゼ活性を有する単離されたポリペプチドであって、0.25 FYT/g飼料で投薬された該ポリペプチドは、0.25 FYT/g飼料で投薬されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べて少なくとも150%のリン(P)を放出し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0239】
フィターゼ活性を有する単離されたポリペプチドであって、0.75 FYT/g飼料で投薬された該ポリペプチドは、0.75 FYT/g飼料で投薬されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べて少なくとも150%のリン(P)を放出し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0240】
I. (a) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチド、(b) (i) 配列番号9のヌクレオチド100〜1338、(ii)配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 又は (ii) のいずれか1つの相補鎖と、少なくとも中緊縮条件下でハイブリダイズするポリヌクレオチドによってコードされるポリペプチド; (c) 1以上のアミノ酸の保存的置換、削除及び/又は挿入を含む、(a)(i)〜(a)(ii) のポリペプチドのいずれか1つの変異体;及び (d) (a)(i)〜(a)(ii) のポリペプチドのいずれか1つのフラグメント、からなる群より選ばれる、ファターゼ活性を有する単離されたポリペプチド。
【0241】
II. セクションIのポリペプチドをコードするヌクレオチド配列を含む単離されたポリヌクレオチド。
【0242】
III. 以下:(a) 配列番号10のアミノ酸1〜413と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有する、アミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;(b) 配列番号9のヌクレオチド100〜1338と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有するポリヌクレオチド;及び (c) (i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、(iii) (i) 又は(ii) のいずれか1つの相補鎖と少なくとも中緊縮条件下でハイブリダイズするポリヌクレオチド、からなる群より選ばれるフィターゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチド。
【0243】
IV. 配列番号9の成熟ポリペプチドコーディング配列において少なくとも1つの変異を有する、セクションII及びIIIのいずれか1つの単離されたポリヌクレオチドであって、該成熟ヌクレオチド配列が配列番号10のアミノ酸1〜413を含むポリペプチドをコードする、前記ポリヌクレオチド。
【0244】
V. 発現宿主においてポリペプチドの製造を指向する1以上の制御配列に作動可能に連結されたセクションII〜IVのいずれか1つのポリペプチドを含む核酸構築物。
【0245】
VI.セクションVの核酸構築物を含む組換え発現ベクター。
【0246】
VII.セクションVの核酸構築物を含む組換え宿主細胞。
【0247】
VIII. (a) ポリペプチドの製造のための条件下で、その野生型において該ポリペプチドを製造することができる細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、セクションIのポリペプチドの製造方法。
【0248】
IX. (a)ポリペプチドの製造のための条件下で、ポリペプチドをコードする核酸配列を含む核酸構築物を含む組換え宿主細胞を培養し、及び (b) 該ポリペプチドを回収すること、を含む、セクションIのポリペプチドの製造方法。
【0249】
X. セクションIのポリペプチドをコードするポリヌクレオチドで形質転換された、トランスジェニック植物、植物部分又は植物細胞。
【0250】
XI. セクションIのポリペプチドを発現することができる、トランスジェニック非-ヒト動物、又はその産物、又はその要素。
【0251】
XII. 動物飼料におけるセクションIの少なくとも1つのポリペプチドの使用。
【0252】
XIII. 動物飼料における使用のための組成物の調製における、セクションIの少なくとも1つのポリペプチドの使用。
【0253】
XIV. セクションIの少なくとも1つのポリペプチドが飼料に添加される、動物飼料の栄養価を改善するための方法。
【0254】
XV. (a) セクションIの少なくとも1つのポリペプチド;(b) 少なくとも1つの脂溶性ビタミン;(c) 少なくとも1つの水溶性ビタミン、及び/又は少なくとも1つの微量元素、を含む動物飼料添加物。
【0255】
XVI. 少なくとも1つのアミラーゼ、少なくとも1つの更なるフィターゼ、少なくとも1つのキシラナーゼ、少なくとも1つのガラクタナーゼ、少なくとも1つのα-ガラクトシダーゼ、少なくとも1つのプロテアーゼ、少なくとも1つのホスホリパーゼ、及び/又は少なくとも1つのβ-グルカナーゼを更に含む、セクションXVの動物飼料添加物。
【0256】
XVII. 更なるフィターゼが配列番号10のアミノ酸1〜413のアミノ酸配列を有するポリペプチドの最適pHよりも高い最適pHを有する、セクションXVIの動物飼料添加物。
【0257】
NXX. 50〜800 g/kgの粗タンパク質量を有し、かつセクションIの少なくとも1つのポリペプチドを含む、動物飼料組成物。
【0258】
配列番号10のアミノ酸1〜413と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有する、アミノ酸配列を含む、好ましくはそれを有する又はそれからなる、フィターゼ活性を有するポリペプチド。
【0259】
(i) 配列番号10のアミノ酸1〜413、及び/又は
(ii) 配列番号10の成熟ポリペプチド部分; 又は
(a) 1以上のアミノ酸の削除、挿入及び/又は保存的置換を含む、(i)〜(ii)のポリペプチドのいずれか1つの変異体であるポリペプチド;又は
(b) (i)〜(ii)のポリペプチドのいずれか1つのフラグメント
の配列を含み、好ましくは有する、フィターゼ活性を有するポリペプチド。
【0260】
本発明は、本発明の範囲を限定するものと解釈してはならない、以下の実施例によって更に記載される。
【実施例】
【0261】
実施例 1. ハフニア・アルベイ・フィターゼ遺伝子のクローニング
複数の配列を以下のヒスチジン酸ホスファターゼ(HAP)から作製した:appA エシェリキア・コリ(Escherichia coli)(SPTREMBLQ8GN88)、シトロバクタ・ギレニイ(Citrobacter gillenii)DSM 13694フィターゼ (geneseqp:aehO4533)、シトロバクタ・アマロナテカス(Citrobacter amalonaticus)ATCC 25407 フィターゼ (geneseqp:aehO4535)、シトロバクタ・ブラアキィ(Citrobacter braakii)フィターゼ (geneseqp:aehO4827)、及びypo1648エルシニア・ペスティス(Yersinia pestis)CO92 (SPTREMBLQ8ZFP6)。2つの縮退オリゴヌクレオチドプライマーは、コンセンサス配列に基いてデザインした:
2123fw: 5'-CATGGTGTGCGNGCNCCNACNAA-3' (配列番号1)
2065rev: 5'-CCCACCAGGNGGNGTRTTRTCNGGYTG-3' (配列番号2)、
ここで、YはT又はCを示し、RはA又はGを示し、そしてNはA、C、G又はTを示す。
【0262】
プライマーは、40℃〜50℃のアニーリング温度で多数の細菌種のPCRスクリーニングに使用した。但し、典型的には、標準的なPCRを行う前に次の10サイクルに渡って、50℃で開始して、次いで各サイクルで1℃ずつアニーリング温度を減少させるタッチダウンプログラムによって、使用した。
【0263】
PCRフラグメントによって約950 bpの形態で部分的フィターゼ遺伝子をハフニア・アルベイ (DSM 19197) において同定した。
【0264】
PCRフラグメントをアガロースゲルから単離し、該フラグメントを、該フラグメントを作製するのに使用したものと同一のPCRプライマーを用いてシークエンスした。ヌクレオチド配列の翻訳によって、DNAフラグメントがHAPフィターゼ遺伝子の一部であることが確認された。
【0265】
上記遺伝子の完全長ヌクレオチドを得るために、未知の標的部位を獲得するようにデザインされている、DNA WALKINGSPEEDUP(登録商標)キット (DWSK-V102 from Seegene, Inc., 2nd Fl., Myungji Bldg., 142-21, Samsung-dong, Kangnam-gu, Seoul, 135-090, Korea) を使用した。この目的のために、6個の特定のオリゴヌクレオチドは該キットを用いてデザインした。
【0266】
2328 TSP1dw: 5'-ACTTGCATCGACGTTGGCTG (配列番号3)
2329 TSP2dw: 5'-ACTGAG CAG CAATG GAACTCTCTG (配列番号4)
2330 TSP3dw: 5'-ACTGGGTTCCAATATCACGAGTC (配列番号5)
2331 TSP1up: 5'-ATGGTGGATCGCTAAATCACACTG (配列番号6)
2332 TSP2up: 5'-ACGTCTGCCCAAACATACACG (配列番号7)
2333 TSP3up: 5'-ACCGCCCATCAGGCTAATC (配列番号8)
【0267】
ハフニア・アルベイ DSM 19197由来のフィターゼをコードする完全長ヌクレオチド配列は、配列番号9として表される配列に示され、対応するコードされたアミノ酸配列は、配列番号10として示される。配列番号10の最初の33個のアミノ酸(すなわち、アミノ酸-33〜-1)は、ソフトウェアSignal P V3.0 (www.cbs.dtu.dk/services/SignalP/参照) によって予測されたシグナルペプチドである。
【0268】
実施例 2. ハフニア・アルベイ・フィターゼ遺伝子の発現
バチルス・リケニフォルミス(Bacillus licheniformis)由来の天然プロテアーゼ、Savinase(登録商標)の27アミノ酸シグナルペプチドコーディングポリヌクレオチドを、ハフニア・アルベイ由来の成熟フィターゼをコードする遺伝子にインフレームでPCPによって融合した。シグナンルペプチドコーディング配列は、配列番号12のシグナルペプチドをコードする配列番号11で示される。
【0269】
融合ポリペプチドをコードするDNAを、バチルス・サブティリス宿主細胞ゲノム上で相同性組換えによって統合した。遺伝子構築物は、バチルス・リケニフォルミス(Bacillus licheniformis)・α-アミラーゼ遺伝子 (amyL)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)・α-アミラーゼ遺伝子 (amyQ)、及びmRNA安定配列を含むバチルス・チューリンゲンシス(Bacillus thuringiensis)cryIIIAプロモーターからなる、三重プロモーター系(WO 99/43835に記載)の制御下で発現した。クロラムフェニコールアセチル-トランスフェラーゼをコードする遺伝子は、例えば、Diderichsen他, バチルス・サブティリス用の有用なクローニングベクター,プラスミド, 30, 第312頁, 1993に記載されているように、マーカーとして使用された。
【0270】
30℃で250 RPMで振とうされたPS-1培地(10%スクロース、4%大豆粉、1% Na3PO4-12H2O、0.5% CaCO3及び0.01 % プルロニック酸)中で、クロラムフェニコール抵抗性形質転換体を培養した。インキュベーションの2〜5日後、上清を除き、そして、0.1 M酢酸ナトリウム(pH 4.5)及び0.1%イノシトール六リン酸を含む1 % LSBアガロースプレート中で、20ミリリットルの上清を、打ち抜かれた4 mm径の孔に適用することによって、フィターゼ活性を同定した。プレートを37℃で終夜静置し、0.25 M CaCl2及び500 mM MES (4N NaOHでpH 6.5に調整) からなるバッファーをプレートに注いだ。該プレートを室温で1時間静置し、次いで、イノシトールリン酸ホスファターゼ又はフィターゼの活性を透明ゾーンとして同定した。
【0271】
数個のフィターゼ陽性形質転換体は、DNAシークエンシングによって分析して、構築物の正確なDNA配列を確立した。1つの正確なクローンを選択した。
【0272】
実施例 3 ハフニアNN020125フィターゼ宿主の発酵
ハフニア・アルベイ フィターゼ構築物を有しそして該フィターゼ(成熟部分)を発現することができる、バチルス・サブティリスの選択されたクローンを、SK-1 M培地 (カゼイン酸ナトリウム (Arla製Miprodan 30) 40 g、マルトデキストリン01 (Roquette製Glucidex 6, カタログ番号332203) 200 g、ダイズミール50 g、Dowfax 63N10 (Dow製非イオン性界面活性剤) 0.1 mlを含み、水道水で1000 mlとした、CaCO3錠剤0.5 g/100 ml) 中で、30℃で、6日間、250rpmで攪拌しながら培養した。
【0273】
実施例 4 ハフニア・アルベイ・フィターゼの精製
該フィターゼによる発酵上清を最初に、7200 rpm及び5℃で1時間遠心し、4つのワットマンガラスマイクロチューブフィルターのサンドウィッチでろ過した(2.7、1.6、1.2及び0.7μm)。次いで、圧力を用いて、Seitz-EKSデプスフィルターによって、この溶液を殺菌ろ過した。次いで、ろ過された上清を以下のように予備処理した。
【0274】
試料溶液を水で洗浄し、1 0 kDaカットオフ限界濾過膜を備えた限界濾過ユニット (Filtron, Filtron Technology Corporation製) を用いて濃縮した。次いで、pHを10% (w/v) 酢酸で4.5に調整した。これは、少量の沈殿を生じた。該沈殿には活性は見られなかった、0.22μmカットオフを有するFast PESボトル先端部フルターで濾過することによって沈殿を除いた。
【0275】
予備処理の後、バッファーAとして50 mM酢酸ナトリウム(pH 4.5)、及びバッファーBとして50 mM酢酸ナトリウム+ 1 M NaCl(pH 4.5)を用いて、XK26カラム中の約50 mlのSセファロース上でクロマトグラフィーによってフィターゼを精製した。ホスファターゼアッセイ(以下参照)を用いてカラムからのフラクションを活性について分析し、活性のあるフラクションを回収した。
【0276】
上記溶液を最終濃度1.5 Mとなるように固体硫酸アンモニウムに加え、6 M HClを用いて6.0にpHを調整した。フィターゼ含有溶液を、バッファーAとして25 mM ビス-トリス (ビス-(2-ヒドロキシエチル)イミノ-トリス(ヒドロキシメチル)メタン)) + 1.5 M 硫酸アンモニウム(pH 6.0)、及びバッファーBとして25 mM ビス-トリス(pH 6.0)を用いて、XK26カラム中約30 mlで、ブチル-セファロースカラムに適用した。カラムからのフラクションを、ホスファターゼアッセイ(以下参照)を用いて活性について分析し、回収した。最後に、精製されたフィターゼを含む溶液を50 mM酢酸ナトリウム + 0.1 M NaCl(pH 4.5)に緩衝液チャージし、30 kDaカットオフ膜を有するアミコンウルトラ-15濾過装置を用いて濃縮した。
【0277】
SDS-PAGEから見積もった分子量は、約40 kDaであり、純度は>95%であった。
【0278】
実施例 5 活性アッセイ
ホスホターゼ活性の決定
75μlのフィターゼ含有酵素溶液をマイクロタイタープレートウェル、例えばNUNC 269620に分注し、75μlの基質を加えた(基質を調製するために、2つの5 mg p-ニトロフェニルホスフェート錠剤(Sigma, カタログ番号N-9389)を10 ml 0.1 M 酢酸Naバッファー(pH 5.5)に溶解した)。該プレートを遮光し、15分間インキュベートし、37℃で750 rpmで振とうした。インキュベーション後、75μlの停止試薬を加え(停止試薬は、0.1 M 四ホウ酸二ナトリウム水溶液)、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。
【0279】
フィターゼ活性の決定
0.25 M酢酸ナトリウム、0.005% (w/v) Tween-20(pH 5.5)でおおよそ希釈した、75μlのフィターゼ含有酵素溶液を、マイクロタイタープレートウェル、例えばNUNC 269620に分注し、75μlの基質を加えた(コメ由来の100 mgフィチン酸ナトリウム(Aldrichカタログ番号274321)の10 ml 0.25 M酢酸ナトリウムバッファー(pH 5.5)によって溶解して調整した)。該プレートを遮光し、15分間インキュベートし、37℃で750 rpmで振とうした。インキュベーション後、75μlの停止試薬を加え(停止試薬は、10 mlのモリブデン酸塩溶液(10% (w/v) モリブデン酸アンモニウムの0.25% (w/v) アンモニア溶液)、10 ml バナジン酸アンモニウム(Bie&Berntsen, カタログ番号LAB17650製の、0.24%の商業的製品)及び20 ml 21.7% (w/v) 硝酸)、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。フィターゼ活性はFYT単位で表し、1FYTは、上記の条件下で、1μmol無機オルトリン酸塩/分を遊離する酵素の量である。測定されたフィターゼ活性の絶対値は、無機リン酸塩の好適な希釈から調製された標準曲線を参考に、又は公知の活性を有するフィターゼ酵素調製物(公知の活性を有するかかる標準的な酵素調製物は、請求により、Novozymes A/S, Krogshoejvej 36, DK-2880 Bagsvaerdから入手可能である)の希釈から作成された標準曲線を参考に、得られる。
【0280】
特異的フィターゼ活性の決定
酢酸ナトリウムバッファー(pH 5.5)中でフィターゼの特異的活性を測定した。該フィターゼは上記のように高度に精製した、すなわち、1成分のみをSDSポリアクリルアミドゲルで精製した。
【0281】
アミノ酸分析によって以下のようにタンパク質濃度を測定した:試料のアリコートを、空のガラスチューブ中で、110℃で16時間、6 M HCl、0.1% フェノールで加水分解した。製造者の教示に従って操作したApplied Biosystems 420Aアミノ酸分析装置を用いて、得られたアミノ酸を定量した。アミノ酸の量から、加水分解されたアリコート中の、総質量及びタンパク質濃度を計算した。
【0282】
上記のFYT単位でフィターゼ活性を決定し、そして、mgフィターゼ酵素タンパク質当たりのFYT単位で測定されたフィターゼ活性として、特異的活性を計算した。
【0283】
得られた特異的活性は980 FYT/mgタンパク質であった。特異的活性は、pH 5.5及び37℃でフィチン酸ナトリウムで決定した。
【0284】
実施例 6 フィターゼpHプロファイルの決定
0.25 M酢酸ナトリウム(pH 5.5) バッファーの代わりにバッファーカクテル(50 mMグリシン、50 mM酢酸及び5OmM ビス-トリス)を用いる以外は、「フィターゼ活性の決定」の項で述べたようにして、2.0〜7.5 (0.5のpH-単位ステップ) のpH範囲で37℃で、pHプロファイルを決定した。結果を以下の表1に纏める。2.0〜7.5の範囲の各pHについての値は、最適な値に対して標準化された相対活性(%)である。
【0285】
【表2】
【0286】
実施例 7 フィターゼ等電点の決定
フィターゼについての等電点plは、製造者によって教示されている、等電点電気泳動ゲル(Invitrogen製Novex pH 310 IEFゲル, カタログ番号EC6655A2)ランを用いて決定した。ハフニア・アルベイ・フィターゼのplは約7.4である。
【0287】
実施例 8 フィターゼ温度プロファイル
温度プロファイル (温度指数としてのフィターゼ活性) を、上記 ("Determination of フィターゼ活性の決定") のようにして基本的に20〜90℃の温度範囲でハフニア・アルベイ フィターゼについて決定した。しかしながら、酵素反応 (100μl フィターゼ-含有酵素溶液 + 100μl基質) をマイクロタイタープレートの代わにPCRチューブで行った。所望の温度での15分の反応期間の後、該チューブを20℃に20秒間冷却し、各反応混合物の150μlをマイクロタイタープレートに移した。75μlの停止試薬を加え、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。結果を以下の表2に纏める。各温度の数字は、最適値に標準化された相対活性(%)である。
【0288】
【表3】
【0289】
実施例 9 フィターゼ熱安定性
アスペルギルス・オリザエ(Aspergillus oryzae)及びバチルス・サブティリス(Bacillus subtilis)で発現されたハフニア・アルベイ・フィターゼを、示差走査熱量 (DSC) 測定による熱安定性測定に供し、E. コリ・フィターゼ (Danisco NSからPHYZYME XPとして入手可能) と比べた。
【0290】
ハフニア・アルベイ・フィターゼ (実施例4で記載されたようにして精製した) のタンパク質試料のアリコートを、2〜3時間で、4℃で2 x 500 ml 20 mM Na-酢酸、pH 4.0で透析し、その後終夜ステップを行った。試料を0.45μm濾過し、約2 A280単位にバッファーで希釈した。測定された正確な吸光度を以下の結果表に示す。示差走査熱量 (DSC) 測定で対照として、透析バッファーを使用した。真空吸引を用いて及び約10分間攪拌しながら試料を脱気した。製品PHYZYME XP由来のE. コリ・フィターゼのアリコートを実施例4に記載の方法と同様の方法で精製した。
【0291】
20〜80℃で1.5℃/分の一定のスキャン速度でDSCスキャンを行った。濾過時間:16秒。DSCを行う前に、フィターゼを好適なバッファー (例えば、0.1 Mグリシン-HCl, pH 2.5又は3.0; 20 mM酢酸ナトリウム(pH 4.0); 0.1 M酢酸ナトリウム(pH 5.5); 0.1 M Tris-HCl, pH 7.0) で透析した。MicroCal Originソフトウェア (バージョン4.10) を用いてデータ操作を行い、変性温度Td (融解温度Tmとも称される) をサーモグラムでピークの頂点の温度として定義した。アンフォールディングプロセスの可逆性を明らかにするために、短い冷却期間の後直ちに第2スキャンを行った。第2スキャンについて、ピーク面積 (ピークとベースラインとの間の面積 = 比較されるアンフォールディングのエンタルピー) を、第1スキャンのピーク面積と比較する。大きなピーク (第1スキャンのピーク面積の75〜100 %) は、可逆的アンフォールディング/フォールディングプロセスとして解釈される。アスペルギルス・オリザエ、バチルス・サブティリス及びE. コリ・フィターゼで発現されたハフニア・アルベイ・フィターゼについてのDSCの結果を以下の表3に纏める。
【0292】
【表4】
【0293】
上記の表に示すように、ハフニア・アルベイ・フィターゼは、E. コリ・フィターゼよりも高い熱安定性を有した。ハフニア・アルベイ・フィターゼの熱安定性は、発現宿主によって影響されなかったことも明らかである。
【0294】
実施例 10 ハフニア・アルベイ・フィターゼ及びE. コリ・フィターゼの胃タンパク質分解耐性
H.アルベイ・フィターゼ及びE. コリ・フィターゼ (PHYZYME XP, Daniscoから入手可能) の試料を、ペプシン (ブタ胃粘膜由来のペプシン 1:60000, Wako 162-18721, 2900単位/mg, ロットSDK5232) の250 mMグリシンバッファー pH 3.0 (1000ペプシン単位/mg フィターゼ) で処理した。振とう (750 rpm) しながら、40℃で30分間インキュベーションした。ペプシンでのインキュベーション後、ペプシンを添加することなく、フィターゼ活性を、実施例5に記載のようにしてそして同一の方法で処理した試料の活性に比べて決定した。結果を以下の表4に纏める。
【0295】
【表5】
【0296】
従って、E. コリ・フィターゼ及びH. アルベイ・フィターゼは、非常に類似した胃タンパク質分解抵抗性を有した。
【0297】
実施例 11 ハフニア・アルベイ・フィターゼ及びシトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼ用インビトロモデルにおける動物飼料での成績
インビトロモデルでのハフニア・アルベイ・フィターゼの動物試料での成績を、シトロバクタ・ブラアキィ・フィターゼの成績と比較した。インビトロモデルは、単胃動物における胃腸状態を刺激し、インビボでの動物試験で得られた結果とよく相関する。試料中のフィターゼ活性は、「フィターゼ活性の決定」の下、実施例5で記載したようにして決定した。比較は以下のようにして行った。
【0298】
30%の大豆ミールと70%のトウモロコシミールとからなる飼料試料に5 gカルシウム/kg飼料の濃度にCaCl2を添加したものを調製し、40℃で30分間、pH 3.0で、予備インキュベートし、ペプシン(3000 U/g飼料)及びフィターゼの好適な投薬量を添加した(比較を可能にするために試験されるべきすべてのフィターゼについて同一の投薬量を用いた。例えば、0.1〜1.0フィターゼ単位FYT/g飼料)。フィターゼ活性のないブランクも対照としてインキュベートした。次いで、試料を40℃で60分間、pH 3.0でインキュベートし、次いでpH 4.0で30分間インキュベートした。
【0299】
反応を停止し、そして、最終濃度0.5 MになるようにHClを加え及び40℃で2時間インキュベーションし、次いで1回の凍結融解サイクルと40℃で1時間のインキュベーションによって、フィチン酸及びイノシトールリン酸塩を抽出した。
【0300】
Chen他のJournal of Chromatography A (2003) 第1018巻, 第41-52頁に記載の高速イオンクロマトグラフィーで、フィチン酸及びイノシトールリン酸塩を分離し、Skog-lund他のJ. Agric. Food Chem. (1997), 第45巻, 第431-436頁に記載されるようにして定量した。
【0301】
図1は、125 FYT/kg飼料、250 FYT/kg飼料及び500 FYT/kg飼料の投薬量での、シトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼと比べた、ハフニア・アルベイ フィターゼの用量反応を示す。インビトロでのフィターゼの効果は、インビトロインキュベーション後の試料に残存している残余イノシトールリン酸結合リン(IP-P)として、フィターゼナシの対照試料に残存している残余IP-Pと比較して、示される。すべての数値は、4又は5レプリカ(インビトロインキュベーションでの)平均及び標準偏差である。ハフニア・フィターゼの250 FYT/kgの投薬量は、インビトロ試料中の残余IP-Pの量を、シトロバクタ・フィターゼの125 FYT/kgとほぼ同程度に減少させた。
【0302】
従って、ハフニア・フィターゼは、残余イノシトール-ホスフェート結合リンの量におて非常に優れた減少を得ることができた。
【0303】
実施例 12 ハフニア・アルベイ・フィターゼ及びペニオホラ・リシイ(Peniophora lycii)・フィターゼ・フィターゼフィターゼ用インビトロモデルにおける動物飼料での成績
250 FYT/kg及び500 FYT/kg飼料の投薬量での、インビトロモデルにおけるハフニア・アルベイ・フィターゼの動物飼料での成績も、ペニオホラ・リシイの成績と比較した。結果は、実施例11に記載の実験プロトコルに従って得た。図2から明らかなように、ハフニア・アルベイ・フィターゼは、ペニオホラ・リシイ・フィターゼよりも高く、インビトロ試料において、残余IP-P量を減少した。
【0304】
【表6】
【技術分野】
【0001】
発明の分野
本発明は、フィターゼ活性を有する単離ポリペプチド、及び該ポリペプチドをコードする単離ポリヌクレオチドに関する。該ポリペプチドは、ハフニア・アルベイから得られたフィターゼに関し、そのアミノ酸配列は、配列番号10とし添付した配列表に示されている。本発明はまた、該ポリヌクレオチドを含む核酸構築物、ベクター及び宿主細胞、並びに該ポリペプチドを製造する方法及び特に動物飼料において該ポリペプチドを用いる方法、に関する。
【背景技術】
【0002】
配列表及び寄託微生物
配列表
本願は、配列表の書面コピー及びコンピュータ読み取り可能な形成を含む。コンピュータ読み取り可能な形態の内容は、参考によって本明細書に充分に援用される。
【0003】
生物材料の寄託
菌株を製造するフィターゼは、デンマーク土壌から単離され。該株は、酸性の最適pH及び高い熱安定性を有するフィターゼを製造することが証明された。該株は、ハフニア・アルベイ(ハフニア・アルベイ)として同定され、Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) によってブタペスト条約の下、2007年3月21日に寄託され、以下の受託番号:ハフニア・アルベイ NN020125 DSM 19197、2007年3月21日の受託番号及び受託日を与えられた。
【0004】
【表1】
【0005】
発明の背景
フィターゼは、ヒトを含む動物の飼料にそれらを添加することが有利であるので、周知の酵素である。フィターゼは、多数の真菌及び細菌株を含む、非常に多くの起源から単離されている。フィターゼ活性を有する別のポリペプチド、及び該ポリペプチドをコードするポリヌクレオチドを提供することが本発明の目的である。本発明のポリペプチドは、好ましくは、改変された、より好ましくは改善された性質、例えば、異なった基質特異性、より高い特異的活性、増加した安定性(例えば、酸-安定性、熱-安定性、及び/又はプロテアーゼ安定性、特にペプシン安定性)、変更された最適pH(例えば、より低い又はより高い最適的pH)、及び/又は 動物飼料における改善さえた能力(例えば、改善された放出及び/又はフィターゼの分解)を有する。
【発明の概要】
【0006】
発明の概要
本発明は、(a) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチドと少なくとも75%の同一性を有するアミノ酸配列を含むポリペプチド;及び/又は(b)(i)配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分の、1以上のアミノ酸の削除、挿入及び/又は保存的置換を含む変異体;及び/又は (c) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分、フラグメントからなる群より選択される、フィターゼ活性を有する単離ポリペプチドに関する。本発明はまた、(a) 配列番号10のアミノ酸1〜413と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;及び (b) 配列番号9の塩基100〜1338と少なくとも75%の同一性を有するポリヌクレオチド、からなる群より選ばれる、フィターゼ活性を有するポリペプチドをコードする単離ポリヌクレオチドに関する。本発明はまた、該ポリヌクレオチドを含む、核酸構築物、組換え発現ベクター、及び組換え宿主細胞に関する。
【0007】
本発明はまた、(a) 前記ポリペプチドの製造を誘導する条件下で、前記ポリペプチドをコードするポリヌクレオチドを含む核酸構築物を含む組換え宿主細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、フィターゼ活性を有するかかるポリペプチドを製造する方法に関する。本発明は、更に、(i) 配列番号11の塩基1〜99からなるシグナルペプチドをコードするヌクレオチド配列に作動可能に連結したタンパク質をコードする遺伝子を含む核酸構築物に関する。
【0008】
本発明はまた、動物試料において本発明のフィターゼを用いる方法、並びに前記ポリペプチドを含む動物試料及び動物試料添加組成物に関する。
【0009】
本発明はまた、発酵生成物、例えばエタノール、ビール、ワイン、の製造において本発明のフィターゼを用いる方法であって、該発酵が本発明のフィターゼの存在下で行われる、前記方法に関する。本発明は、本発明のフィターゼを用いて、植物タンパク質を含むタンパク質を処理する方法に関する。
【図面の簡単な説明】
【0010】
【図1】図1は、125〜500 FYT/kg飼料で投与された、ハフニア・アルベイ・フィターゼとシトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼとの比較におけるインビトロでのインキュベーション後の残余イノシトール-リン酸結合リン酸を示す。
【図2】図2は、250〜500 FYT/kg飼料で投与された、ハフニア・アルベイ・フィターゼとペニオホラ・リシイ(Peniophora lycii)・フィターゼとのインビトロでのインキュベーション後の残余イノシトール-リン酸結合リン酸の比較を示す。
【図3】図3は、ハフニア・アルベイ・フィターゼの解析された結晶3次元構造についての構造座標の添付を示す。
【0011】
定義
フィターゼ活性:本文脈では、フィターゼ活性を有するポリペプチド(フィターゼ)は、フィテート(myo-イノシトールイノシトール六リン酸)の、(1)myo-イノシトール及び/又は(2)そのモノ-、ジ-、トリ-、テトラ-及び/又はペンタ-リン酸エステル、及び(3)無機リン酸塩への加水分解を触媒する酵素である。
【0012】
インターネット(www.expasy.ch/enzyme/)の「酵素」サイトは、酵素の命名に対する情報の貯蔵所である。それは、主に、国際生化学分子生物学連合の命名法委員会(NC-IUBMB)の推奨に従い、EC(酵素委員会)番号は提供している特徴付けられた酵素の各種類を記載している (Bairoch A. 「酵素」データベース, 2000, Nucleic Acids Res 28: 304-305),NC-IUBMB, 1992のハンドブック酵素命名も参照)。
【0013】
酵素(ENZYME)サイトによれば、3種のフィターゼが知られている:3-フィターゼ (myo-イノシトールイノシトール六リン酸 3-ホスホヒドロラーゼ, EC 3.1.3.8)、6-フィターゼ (myo-イノシトールイノシトール六リン酸 6-ホスホヒドロラーゼ, EC 3.1.3.26)、及び5-フィターゼ (EC 3.1.3.72)。本発明の目的のためには、すべての種類がフィターゼの定義に含まれる。具体的な実施態様では、本発明のフィターゼは、エシェリヒア・コリ(Escherichia coli)pH 2.5の酸ホスファターゼ(appA遺伝子)、及び真菌フィターゼ、例えばアスペルギルス・アワモリ(Aspergillus awamorii)フィターゼA及びB(EC: 3.1.3.8)(phyA及びphyB遺伝子)を含む、酸ヒスチジンホスファターゼのファミリーに属する。ヒスチジン酸ホスファターゼは、2つの配列類似性領域を共有し、各々は、保存的ヒスチジン残基の周囲に集中している。これらの2つのヒスチジンは、酵素の触媒的メカニズムに関連していると考えられる。第1ヒスチジンは、N-末端部分に位置し、蛍光体-ヒスチジン中間体を形成するが、第2ヒスチジンは、C-末端部分に位置し、おそらくプロトン供与体として働く。
【0014】
更なる具体的な実施態様では、本発明のフィターゼは、保存的活性部位モチーフ、すなわちR-H-G-X-R-X-P、ここで、Xは、アミノ酸(配列番号10で示される成熟フィターゼのアミノ酸18〜24を参照)を有する。
【0015】
本発明の目的のために、フィターゼ活性は、FYTの単位において決定される。1つのFYTは、次の条件: pH 5.5; 温度37℃; 基質: 0.0050 mol/lの濃度中フィターゼナトリウム (C6H6O24P6Na12) の下で、1個のマイクロモル無機オルト-リン酸塩を1分当たりに放出する酵素量である。好適なフィターゼアッセイは、WO 00/20569の実施例1に記載のFYT及びFTUアアッセイである。FTUは、試料及びプレミックスにおけるフィターゼ活性を決定するためのものである。フィターゼ活性は、本明細書に記載の例のフィターゼアッセイを用いて決定してもよい。
【0016】
最適pH:本発明のポリペプチドの最適pHは、予備測定濃度での基質及び固定インキュベーション時間を用いて、様々なpH値でフィターゼを活性化することによって決定される。次いで、最適pHがフィターゼ活性-対-pHのグラフ表示決定される。具体的実施態様では、FYTアッセイが使用される。すなわち、基質は5 mMフィターゼナトリウム、反応温度37℃であり、活性は、以下の例で行われるように様々なpH-値にてFYT単位で測定される。別の特定の実施態様では、例のいずれ1つのフィターゼアッセイが使用される。比較的低い最適pHは、pH 5.0未満の最適pHを意味し、例えばpH 4.5、4.0、3.5、3.0、2.5又は更に2.0未満である。比較的高い最適pHは、pH 5.0超の最適pH、例えばpH 5.5、6.0、6.5、7.0、7.5、8.0、8.5又は更に9.0超である。
【0017】
単離ポリペプチド:本明細書で使用される用語「単離ポリペプチド」は、SDS-PAGEによって決定されるように、少なくとも20%純粋、好ましくは少なくとも40%純粋、より好ましくは少なくとも60%純粋、更により好ましくは少なくとも80%純粋、最も好ましくは少なくとも90%純粋、及び更に最も好ましくは少なくとも95%純粋であるポリペプチドを言う。
【0018】
実質的に純粋なポリペプチド:本明細書で使用される用語「実質的に純粋なポリペプチド」は、天然に関連する他のポリペプチド物質の、最大10重量%、好ましくは最大8重量%、より好ましくは最大6重量%、より好ましくは最大5重量%、より好ましくは最大4重量%、最大3重量%、更に好ましくは最大2重量%、最も好ましくは最大1重量%、及び更に最も好ましくは0.5重量%を含むポリペプチド調製物を意味する。そのため、実質的に純粋なポリペプチドが、該調製物に存在する総ポリペプチド物質の、少なくとも92重量%純粋、好ましくは少なくとも94重量%純粋、より好ましくは少なくとも95重量%純粋、より好ましくは少なくとも96重量%純粋、より好ましくは少なくとも97重量%純粋、より好ましくは少なくとも98重量%純粋、更に好ましくは少なくとも99重量%純粋、最も好ましくは少なくとも99.5重量%純粋、及び更に最も好ましくは100重量%純粋であることが好ましい。
【0019】
本発明のポリペプチドは、好ましくは、実施的に純粋な形態にある。具体的には、ポリペプチドは、「本質的に純粋な形態」、すなわち、そのポリペプチド調製物は、天然に関連する他のポリペプチド物質が本質的に含まない、ことが好ましい。このことは、例えば、周知の組換え法の手段によって又は典型的な精製法によってポリペプチドを調製することによって達成され得る。
【0020】
本明細書における用語「実質的に純粋なポリペプチド」は、用語「単離されたポリペプチド」及び「単離された形態でのポリペプチド」と同義である。
【0021】
同一性:2つのアミノ酸配列間、又は2つのヌクレオチド配列間の関係は、パラメータ「同一性」によって説明される。
【0022】
本発明の目的のために、2つのアミノ酸配列間の同一性の程度、及び2つのヌクレオチド配列間の同一性の程度は、タンパク質及びDNAアラインメントの両方に有用である、Needleman-Wunschアラインメント(すなわち、世界的なアラインメント)であるプログラム「アライン」によって決定される。デフォルトスコアリングマトリックスBLOSU M50及びデフォルト同一性マトリックスは、それぞれ、タンパク質及びDNAアラインメントのために使用される。ギャップ中の第1残基についてのペナルティは、タンパク質については-12であり、DNAについては-16である。ギャップ中の追加残基についてのペナルティは、タンパク質について-2であり、DNAについて-4である。
【0023】
「アライン」は、FASTA packageバージョンv20u6の一部である(W. R. Pearson and D. J.Lipman (1988), "生物学的配列解析のための改良されたツール", PNAS 85: 2444-2448, and W. R. Pearson (1990) "FASTP及びFASTAとの迅速かつ感度のよい配列比較," Methods in Enzymology 183: 63-98参照)。FASTAタンパク質アラインメントは、ギャップサイズに関して何の限定のないスミス・ウォーターマン(Smith-Waterman)アルゴリズムを使用する("Smith-Watermanアルゴリズム", T. F. Smith and M. S. Waterman (1981) J. Mol. Biol. 147:195-197参照)。
【0024】
ニードルマン・ウンシュ (Needleman-Wunsch)アルゴリズムは、Needleman, S. B. and Wunsch, CD., (1970), Journal of Molecular Biology, 48: 443-453、並びにMyers及びW. Millerの"線状空間での工学的アラインメント" CABIOS (バイオサイエンスにおけるコンピュータ適用)(1988)4: 11-17によるアラインプログラムに記載されている。
【0025】
標的(又は試料、又は試験)配列と特定の配列(例えば、配列番号10に示される成熟フィターゼのアミノ酸1〜413)との間の同一性の程度は、以下のように決定されてもよい:配列はプログラム「アライン」を用いて整列される。アラインメントにおける完全なマッチ(「N-完全-マッチ」)の数は、決定される(完全マッチは、通常アラインメントにおいて「I」と指定される、アラインメントの同一の位置における同一のアミノ酸残基を意味する)。特定配列の長さ(アミノ酸残基の数)は決定される(上記の例で「N-特定」は413)。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換のための)「N-完全-マッチ」と「N-特定」との比として計算される。
【0026】
別の実施態様では、標的(又は試料もしくは試験)配列と特定の配列(例えば、配列番号10mのアミノ酸1〜413)との同一性の程度は、以下のようにして決定される:該2つの配列は、プログラム「アライン」を用いて整列される。該アラインメントにおける完全なマッチ(「N-完全-マッチ」)の数が決定される(完全なマッチは、通常アラインメントにおいて「I」と指定される、該アラインメントの同一の位置における同一のアミノ酸残基を意味する)。該2つの整列された配列の共通の長さも、該アラインメントの重複部分(「N-重複」)におけるアミノ酸の総数によって決定される。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換についての)「N-完全-マッチ」と「N-重複」との比として計算される。1つの実施態様では、N-重複は、存在するならば、該アラインメントによってつくられたトレイリング及びリーディング・ギャップを含む。別の実施態様では、N-重複は、存在するならば、該アラインメントによってつくられたトレイリング及びリーディング・ギャップギャップを含まない。
【0027】
別の実施態様では、標的(又は試料又は試験)配列と特定配列(例えば、配列番号10のアミノ酸1〜413)との同一性の程度は、以下のようにして決定される:配列は、プログラム「アライン」を用いて整列される。該アラインメントにおける完全マッチ(「N-完全-マッチ」)の数が決定される(完全マッチは、通常、該アラインメントにおける「I」で設計された、アラインメントの同一位置における同一のアミノ酸残基を意味する)。標的配列の長さ(アミノ酸残基の数)が決定される)「N-標的」)。同一性の程度は、(100を乗じた、同一性パーセンテージへの変換のための)「N-完全-マッチ」と「N-標的」との比として計算される。
【0028】
好ましくは、重複は特定配列の少なくとも20%(上で定義された「N-重複」を特定配列におけるアミノ酸の数(「N-特定」)で割って、100を乗じた)であり、より好ましくは少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、又は少なくとも95%である。このことは、試料配列が特定配列に整列される時に、特定配列のアミノ酸の少なくとも20%(好ましくは25〜95%)が該重複に結局含まれることになることを意味する。
【0029】
あるいは、前記重複が標識(又は試料、又は試験)配列の少なくとも20%(上で定義された「N-重複」を上で定義された「N-標的」で割って、100を乗じる)であり、より好ましくは少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、又は少なくとも95%である。このことは、標的配列が特定配列に整列される時に、標的配列のアミノ酸の少なくとも20%(好ましくは25〜95%)が該重複に結局含まれることになることを意味する。
【0030】
ポリペプチドフラグメント:用語「ポリペプチドフラグメント」は、本明細書では、特定配列例えば配列番号10又はその相同配列の成熟ペプチド部分のアミノ及び/又はカルボキシ末端から削除された1以上のアミノ酸を有するポリペプチドであって、該フラグメントがフィターゼ活性を有する、前記ポリペプチドとして定義される。具体的な実施態様では、該フラグメントは、少なくとも350、360、370、380、390、400、405、又は少なくとも410のアミノ酸残基を含む。
【0031】
配列:用語「配列」は、特定の配列例えば配列番号9又はその相同配列の成熟ペプチドコーディング部分の5’及び/又は3’末端から削除された1以上の塩基を有するヌクレオチド配列として本明細書で定義されている。ここで、該配列は、フィターゼ活性を有するポリペプチドフラグメントをコードする。具体的な実施態様では、該配列は、少なくとも1050、1080、1110、1140、1170、1200、1215、1230、1245、1260、1275、1290、1305、1320、又は少なくとも1335塩基を含む。
【0032】
アレル変異体:用語「アレル変異体」は、本明細書では、同一の染色体座を占める遺伝子の2以上の別形態のいずれかを意味する。アレル変異は、突然変異によって自然に起こり、集団内の多型をもたすことがある。遺伝子突然変異は、サイレントであり(コードされたポリペプチドに変化がない)、又は改変されたアミノ酸配列を有するポリペプチドをコードすることがある。ポリペプチドのアレル変異体は、遺伝子のアレル変異体によってコードされたポリペプチドである。
【0033】
実質的に純粋なポリヌクレオチド:本明細書で使用される用語「実質的に純粋なポリヌクレオチド」は、多の外因性又は望ましくない塩基がなく、そして遺伝子組換えタンパク質産生系内での使用のために好適な形態にある、ポリヌクレオチド調製物を言う。従って、実質的に純粋なポリヌクレオチドは、天然的に関連している他のポリヌクレオチド物質の、最高10重量%、好ましくは最高8重量%、より好ましくは最高6重量%、より好ましくは最高5重量%、より好ましくは最高4重量%、より好ましくは最高3重量%、更により好ましくは最高2重量%、最も好ましくは1重量%、及び更に最も好ましくは最高0.5重量%を含む。しかし、実質的に純粋なポリヌクレオチドは、天然5’及び3’の非翻訳領域、例えばプロモーター及びターミネーターを含んでもよい。実質的に純粋なポリヌクレオチドは、少なくとも90重量%の純度、少なくとも92重量%の純度、より好ましくは少なくとも94重量%の純度、より好ましくは少なくとも95重量%の純度、より好ましくは少なくとも96重量%の純度、より好ましくは少なくとも97%の純度、更により好ましくは少なくとも98重量%の純度、最も好ましくは少なくとも99重量%の純度、及び更に最も好ましくは99.5重量%の純度であることが好ましい。本発明のポリヌクレオチドは、好ましくは、実質的に純粋な形態にある。具体的には、本明細書に開示されたポリヌクレオチドが「本質的に純粋な形態」にある、すなわち、ポリククレオチド調製物が本質的に天然的に関連している他のポリヌクレオチド物質がない、ことが好ましい。本明細書では、用語「実質的に純粋なポリヌクレオチド」は、用語「単離されたポリヌクレオチド」及び「単離された形態にあるポリヌクレオチド」と同義である。ポリヌクレオチドは、ゲノム、cDNA、RNA、半合成的、合成的起源でもよく、又はこれらの任意の組合せでもよい。
【0034】
核酸構築物:本明細書で使用される用語「核酸構築物」は、天然遺伝子から単離されるか又は天然に存在しない方法で核酸の部分を含むように修飾されている、単一-又は二重鎖のいずれかの核酸分子を言う。用語「核酸構築物」は、該核酸構築物が本発明のコーディング配列の発現のために必要とされる制御配列を含む時に、用語「発現カセット」と同義である。制御配列:用語「制御配列」は、本明細書では、本発明のポリペプチドをコードするポリヌクレオチドの発現のために必要であるか又は有利である、すべての成分を含むように定義される。各制御配列は、該ポリペプチドをコードするヌクレオチド配列に対して天然型でも又は外因性でもよい。かかる制御配列は、リーダー、ポリアデニル化配列、プロペプチド配列、プロモーター、シグナルペプチド配列、及び転写終結因子Hを含むが、これらに限定されない。最低でも、制御配列は、プロモーター、及び転写及び翻訳停止シグナルを含む。制御配列は、ポリペプチドをコードするヌクレオチド配列のコーディング領域と共に、制御配列のライゲーションを助ける特定の制限部位を導入する目的のために、リンカーと共に提供され得る。
【0035】
作動可能に連結:本明細書で使用される用語「作動可能に連結された」は、制御配列がポリペプチドにコーディング配列の発現を指向するように、ポリヌクレオチド配列のコーディング配列に対して好適な位置に該制御配列が位置する配列を意味する。
【0036】
コーディング配列:本明細書で使用される用語「コーディング配列」は、そのタンパク質産物のアミノ酸配列を直接的に特定するヌクレオチド配列を意味する。
【0037】
コーディング配列の境界は、一般的に、通常、ATG開始コドン、又はGTG及びTTGのような別の開始コドンで始まる、オープンリーディングフレームによって決定される。コーディング配列は、DNA、cDNA又は組換えヌクレオチド配列でよい。
【0038】
成熟ポリペプチド部分:本明細書で使用される用語「成熟ポリペプチド部分」又は「成熟ペプチド部分」は、その遺伝子的道具の部分として、該ポリペプチドをコードするポリヌクレオチドを含む細胞によって分泌されるポリペプチドの部分を言う。言いかえれば、成熟ポリペプチド部分は、細胞の分泌経路にコードされたポリペプチドを指向するその機能を一旦満たした時に、シグナルペプチド部分が開裂される後に残るポリペプチドの部分を言う。
【0039】
配列番号10の予測されたシグナルペプチド
配列番号10は、そのアミノ酸-33〜-1である。それは、配列番号10の成熟ポリペプチド部分がそのアミノ酸1〜413に対応することを意味する。しかし、宿主細胞によってはわずかな変異が起こり得、そのため、発現成熟ポリペプチド部分は好ましい。
【0040】
成熟ポリペプチドコーディング部分:本明細書で使用される用語「成熟ポリペプチドコーディング部分」又は「成熟ポリペプチドコーディング配列」が、成熟ポリペプチド部分をコードするポリペプチドをコードするポリヌクレオチドの部分を言う。例えば、配列番号9に関して、予測された成熟ポリペプチドコーディング部分は、(配列番号10のアミノ酸1〜413をコードする)塩基100〜1338に対応する。
【0041】
発現:用語「発現」は、転写、転写後修飾、翻訳、翻訳後修飾、及び分泌を含むがこれらに限定されないポリペプチドの製造に関連する任意のステップを含む。発現ベクター:用語「発現ベクター」は、本発明のポリペプチドをコードするポリヌクレオチドを含み、その発現を提供する更なる塩基に作動可能に連結される線状又は環状DNA分子として本明細書に定義される。
【0042】
宿主細胞:本明細書で使用される用語「宿主細胞」は、本発明のポリヌクレオチドを含む核酸構築物での、形質転換、トランスフェクション、形質導入等を受けやすい任意の細胞種を含む。
【0043】
修飾:本明細書で使用する用語「修飾」は、特定のポリペプチドの任意の化学的修飾、例えば配列番号10のアミノ酸1〜413からなるポリペプチド、及び該ポリペプチドをコードするDNAの遺伝子的操作を意味する。修飾(複数)は、アミノ酸(複数)の置換(複数)、削除(複数)及び/又は挿入(複数)、並びにアミノ酸側鎖(複数)の置換(複数)でよい。人工変異体:本明細書では、用語「人工変異体」は、配列番号9の成熟フィターゼコーディング部分の修飾核酸配列を発現する生物によって産生されるフィターゼ活性を有するポリペプチドを意味する。修飾されたヌクレオチド配列は、配列番号9に開示されたヌクレオチド配列の成熟フィターゼコーディング部分の修飾によって、ヒトの介入によって得られる。
【発明を実施するための形態】
【0044】
発明の詳細な説明
フィターゼ活性を有するポリペプチド
第1の局面では、本発明は、フィターゼ活性を有し、そして配列番号10(すなわち、成熟ポリペプチド)のアミノ酸1〜143と少なくとも75%の同一性の程度を有するアミノ酸配列を有する単離ポリペプチドに関する。
【0045】
具体的な実施態様では、同一性の程度は、少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、又は少なくとも99.9%であり、これは、フィターゼ活性を有する(本明細書では、「相同ポリペプチド」)。
【0046】
他の具体的な実施態様では、相同ポリペプチドは、配列番号10のアミノ酸1〜413と20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2、又は1個のアミノ酸が異なるアミノ酸配列を有する。
【0047】
具体的な実施態様では、本発明のポリペプチドは、配列番号10のアミノ酸配列の成熟部分を含み、又はそのアレル変異体もしくはフィターゼ活性を有するそのフラグメントである。更なる具体的な実施態様では、ポリペプチドは、配列番号10のアミノ酸1〜413、又はそのアレル変異体もしくはフィターゼ活性を有するそのフラグメントを含む。
【0048】
第2の局面では、本発明は、本発明は、少なくとも媒体、好ましくは(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び (ii) のいずれか1つの相補鎖、及び/又は (iv) (i)、(ii) 又は (iii) のサブ配列(J. Sambrook, E. F. Fritsch, and T. Maniatus, 1989, Molecular Cloning, A Laboratory Manual, 2d edition, Cold Spring Harbor, New York)の緊縮条件下での媒体下でハイブリダイズするポリヌクレオチドによってコードされるフィターゼ活性を有する単離されたポリペプチドに関する。配列番号9の配列は、少なくとも100連続塩基、又は好ましくは少なくとも200連続塩基を含む。更に、配列は、フィターゼ活性を有するポリペプチドフラグメントをコードしてもよい。具体的な実施態様では、ハイブリダイゼーションは、少なくとも中程度〜高度の、少なくとも高度の、又は少なくとも非常に高い緊縮性の下で、好ましくは中程度〜高度、又は非常に高い緊縮性条件下で起こる。
【0049】
代わりの実施態様では、ハイブリダイゼーションは、非常に低い、又は低い緊縮性条件下で行われる。
【0050】
配列番号9のヌクレオチド配列又はそのサブ配列、及び配列番号10のアミノ酸配列又はそのフラグメントは、当該分野で周知の方法に従って様々な属又は種の株から、フィターゼ活性を有するポリペプチドをコードするDNAを同定し及びクローン化するための核酸プローブを設計するために使用し得る。具体的には、かかるプローブは、その中の対応する遺伝子を同定及び単離するために、標準的なサザンブロッティング手段に従って、対象の属又は種のゲノム又はcDNAとのハイブリダイゼーションのために使用され得る。かかるプローブは、配列全体よりも相当短くてよいが、少なくとも14塩基長、好ましくは少なくとも25塩基長、より好ましくは少なくとも35塩基長、及び最も好ましくは少なくとも70塩基長である。しかしながら、塩基プローブは、少なくとも100塩基長であることが好ましい。例えば、塩基プローブは、少なくとも200塩基長、好ましくは少なくとも300塩基長、より好ましくは少なくとも400塩基長、又は最も好ましくは少なくとも500塩基長でよい。更に長いプローブ、例えば、少なくとも600塩基長、少なくとも好ましくは700塩基長、より好ましくは800塩基長、又は最も好ましくは少なくとも900塩基長である核酸プローブも使用される。DNA及びRNAプローブが使用できる。プローブは、典型的に、対応する遺伝子を検出するために標識される(例えば、32P、3H、35S、ビオチン、又はアビジン)。かかるプローブはは、本発明によって包含される。
【0051】
そのため、かかる他の生物から調製されたゲノムDNAライブラリーは、上記のプローブとハイブリダイズし、そしてフィターゼ活性を有するポリペプチドをコードする、DNAについてスクリーニングされ得る。かかる他の生物由来のゲノム又は他のDNAは、アガロース又はポリアクリルアミドゲル電気泳動、又は他の分離技術によって分離され得る。該ライブラリー由来のDNA又は分離されたDNAは、ニトロセルロース又は他の好適な担体材料に移され、そしてそれに固定され得る。配列番号9又はそのサブ配列と相同であるクローン又はDNAを同定するために、担体材料はサザンブロットで使用される。
【0052】
本発明の目的のために、ハイブリダイゼーションは、ヌクレオシド配列が、非常に低い緊縮条件から常に高い緊縮条件下で、配列番号9に記載のヌクレオチド配列、その相補鎖、又はそのサブ配列に対応する標識された核酸プローブにハイブリダイズすることを示している。核酸プローブがこれらの条件下でハイブリダイズする分子は、X-線フィルムを用いて検出され得る。
【0053】
具体的な実施態様では、核酸プローブは、配列番号1〜8のいずれか1つである。別の具体的な実施態様では、核酸プローブは、配列番号9の塩基100〜450、塩基450〜900又は塩基900〜1338の相補鎖である。更なる具体的な実施態様では、核酸プローブは、配列番号10のポリヌクレオチドの成熟部分又はそのサブ配列をコードするポリヌクレオチド配列である。なお更なる具体的実施態様では、核酸プローブは、配列番号9であり、具体的には、その成熟ポリペプチドコーディング領域のいずれか1つである。
【0054】
少なくとも100塩基長の長いプローブについては、非常に低緊縮から非常に高緊縮条件は、好ましくは12〜24時間の標準的なサザンブロッティング法に従って、42℃、5X SSPE、0.3% SDS、200 マイクログラム/mlのせん断及び変性サケ精子DNA、及び非常に低い及び低い緊縮製のための25%ホルムアミド、中程度及び中〜高程度の緊縮製のための35%ホルムアミド、又は高い及び非常に高い緊縮製のため50%ホルムアミドのいずれか、でのプレハイブリダイゼーション及びハイブリダイゼーションと定義される。
【0055】
少なくとも100ヌクレオチド長の長いプローブについては、非常に低緊縮から非常に高緊縮条件は、好ましくは12〜24時間の標準的なサザンブロッティング法に従って、42℃、5X SSPE、0.3% SDS、200 マイクログラム/mlのせん断及び変性サケ精子DNA、及び非常に低い及び低い緊縮製のための25%ホルムアミド、中程度及び中〜高程度の緊縮製のための35%ホルムアミド、又は高い及び非常に高い緊縮製のため50%ホルムアミドのいずれか、でのプレハイブリダイゼーション及びハイブリダイゼーションと定義される。担体物質は、2X SSC、0.2% SDS、好ましくは少なくとも45℃ (非常に低い緊縮性)、より好ましくは少なくとも50℃ (低緊縮性)、より好ましくは少なくとも55℃ (中緊縮性)、より好ましくは少なくとも60℃ (中〜高緊縮性)、更により好ましくは少なくとも65℃ (高緊縮性)、及び最も好ましくは少なくとも70℃ (非常に高い緊縮性) を用いて、15分間3回最終的に洗浄される。
【0056】
約15ヌクレオチド長〜約70ヌクレオチド長の短いプローブについては、緊縮条件は、プレハイブリダイゼーション、ハイブリダイゼーション、並びに計算されたTm未満の約5℃〜約10℃未満で、標準的なサザンブロッティング法に従って、0.9 M NaCl、0.09 M Tris-HCl pH 7.6、6 mM EDTA、0.5% NP-40、1Xデンハート液、1 mMピロリン酸ナトリウム、1 mMりん酸二水素ナトリウム、0.1 mM ATP、及び0.2 mgの酵母RNA/ml中のBolton及びMcCarthy (1962, Proceedings of the National Academy of Sciences USA 48: 1390) に従う計算を用いる、洗浄後ハイブリダイゼーションとして定義される。
【0057】
約15ヌクレオチド長〜約70ヌクレオチド長の短いプローブについては、担体物質は、6X SCC及び0.1% SDSで1回、15分間洗浄され、計算されたTm未満の5℃〜10℃での6X SSCを用いて2回、各々15分間洗浄される。
【0058】
塩含有ハイブリダイゼーション条件下では、有効Tmは、成功的なハイブリダイゼーションのためにプローブとフィルター結合DNAとの間に要求される同一性の程度を制御するものである。有効Tmは、2つのDNAが様々な条件下でハイブリダイズするために必要とされる同一性の程度を決定するために、下記式:
【0059】
有効Tm = 81.5 + 16.6(log M[Na+]) + 0.41 (%G+C)-0.72(% ホルムアミド)
【0060】
(www.ndsu.nodak.edu/instruct/mcclean/plsc731/dna/dna6.htm参照)
を用いて決定され得る。
【0061】
「G+C」は、プローブ中のヌクレオチドG及びTの量を示す。中緊縮性については、例えば、ホルムアミドは35%であり、5X SSPE用のNa+濃度は0.75 Mである。
【0062】
1つの局面では、本発明は、フィターゼ活性及び以下の物理化学的性質(実質的に純粋なポリペプチドについて分析された)を有する単離されたポリペプチドに関する:
(i) 高い特異的活性、例えば、E. コリappA (SPTREMBLQ8GN88) の特異的活性の少なくとも50%のフィターゼでの特異的活性、該特異的活性は、好ましくは、mgフィターゼ酵素タンパク質当たりのFYTの単位で測定される;
(ii) 酸安定性、例えば
(a) HEPESバッファーpH 7.0中で37℃で終夜のインキュベーションした後の残余活性に比べて、少なくとも60%、少なくとも65%、少なくとも70%、又は少なくとも75%の、グリシン/塩酸バッファーpH 2.2中で37℃で終夜のインキュベーションした後の残余活性;
(b) HEPESバッファーpH 7.0中で37℃で終夜のインキュベーションした後の残余活性に比べて、少なくとも80%、少なくとも85%、少なくとも90%、又は少なくとも95%の、グリシン/塩酸バッファーpH 3.0中で37℃で終夜のインキュベーションした後の残余活性;及び/又は
(c) E. コリappA (SPTREMBLQ8GN88) の残余活性と比べて、少なくとも50%の、25、30、35又は37℃、好ましくは37℃の温度で、及びpH of 2.2、2.4、2.5、2.6、2.8、3.0、3.2、3.4又は3.5、好ましくはグリシン/塩酸バッファーpH 2.2又は3.0中で、2時間インキュベーション後の残余フィターゼ活性;
(iii) 熱安定性、例えば、E.コリappA (SPTREMBLQ8GN88) の残余活性と比べて少なくとも50%の、pH 5.5及び55、60、65、70、75、80、85又は95の温度、好ましくは70℃でのインキュベーションの0.5、1、1.5又は2時間、好ましくは0.5時間後の残余フィターゼ活性。
【0063】
あるいは、示差走査熱量 (DSC) 測定は、精製されたフィターゼタンパク質の変性温度Tdを決定するために使用してよい。Tdは、タンパク質の熱安定性の指標である;Tdが高くなればなるほど、熱安定性は高くなる。DSC測定は、例えばMicro CaI製のVP-DSCを用いて様々なpH値で達成され得る。スキャンは、20〜90℃で1.5℃/分の一定のスキャン速度で行われる。好ましいpH値は、4.0〜5.5、好ましくは4.0である。DSCを操作する前に、例えば好適なバッファー (例えば、25 mM酢酸ナトリウム, pH 4.0; 0.1 M酢酸ナトリウム, pH5.5) 中で平衡化されたNAP-5カラム (Pharmacia) を用いてフィターゼを脱塩する。データ操作は、MicroCal Originソフトウェア(バージョン4.10)を用いて行われ、編成温度Td(融解温度Tmとも言われる)は、サーモグラフのピークの頂点での温度として定義される。
【0064】
(iv) プロテアーゼ安定性、例えば、E. コリappA (SPTREMBLQ8GN88) の残余活性と比べて、少なくとも50%の、0.1 mg/mlのペプシンの存在下、20、25、30、35、又は37℃、好ましくは37℃の温度及びpH 5.5での0.5、1、1.5又は2時間、好ましくは1時間のインキュベーション後の残余フィターゼ活性;及び/又は
(v) pH 5.0未満の最適pH、例えば本明細書で先に記載したFYTアッセイを用いて及び/又は実施例4のアッセイを用いて決定されたpH 4.5、4.0、3.5、3.0、2.5未満又は2.0未満。
【0065】
上記の(i)の具体的実施態様では、特異的活性は、E. コリappAの特異的活性の少なくとも60、70、80、90、100、110、120、130、140、又は少なくとも150%である。上記の(ii)〜(iv)の各々の具体的実施態様では、残基活性は、E. コリappAの少なくとも60、70、80、90、100、110、120、130、140、又は少なくとも150%である。
【0066】
第5の局面では、基質pNP-ホスファターゼで測定されたpH 5.0及び37℃での本発明の酵素の活性は、基質フィターゼで測定された酵素の活性の11%未満である。好ましくは、該比は、10%、9%、8%、7%、6%未満、又は5%未満である。フィターゼ加水分解に対するpNPの比は、該酵素の真のフィターゼ特性の指標である。フィターゼについての活性に対するpNPについての活性の高い比は、問題の酵素が、比較的低い基質特異性を有するホスファターゼであることを示し、一方、低い比は、これが、基質としてより明確に受容できるフィターゼであることを示しているかもしれない。
【0067】
第6の局面では、本発明のフィターゼは、ペニオホラ・リシイ(Peniophora lycii)由来のフィターゼに比べて、好ましくはその110%、より好ましくは少なくともその120%、130%、又は少なくとも140%、インビトロのモデルにおいてより高いリン(P)放出を有する。1つの実施態様では、0.25 FYT/g飼料で投与された本発明のフィターゼは、インビトロモデルにおいて0.25 FYT/g飼料で投与されたペニオホラ・リシイ由来のフィターゼによって放出されるリンに比べて、少なくとも150%リン (P) を放出する。好ましくは、該放出は、少なくとも155%、160%、165%、170%、175%、又は少なくとも180%である。別の実施態様では、0.75 FYT/g飼料で投与された本発明のフィターゼは、インビトロモデルにおいて0.75 FYT/g飼料で投与されたペニオホラ・リシイ由来のフィターゼによって放出されるリンに比べて、少なくとも150%リン (P) を放出する。好ましくは、該放出は、少なくとも155%、160%、165%、170%、175%、180%、185%、又は少なくとも190%である。
【0068】
第7の局面では、本発明のフィターゼは、37℃、1 M グリシン/HClバッファー、pH 2.0中で、4時間のインキュベーション後に、t = 0の時点での活性と比べて少なくとも20%の残余活性を有する。該活性(及び残余活性)は、0.25 M Na-アセテートバッファーpH 5.5、ブラインドを除いたバッファーを用いて、37℃及びpH 5.5の1% (w/v) Na-フィターゼでアッセイされる。好ましい実施態様では、残余活性は、少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、又は少なくとも80%である。別の実施態様では、本発明のフィターゼは、37℃、1 M グリシン/HClバッファー、pH 2.5中で、1日(24時間)のインキュベーション後に、t = 0の時点での活性と比べて少なくとも20%の残余活性を有する。該活性(及び残余活性)は、0.25 M Na-アセテートバッファーpH 5.5、ブラインドを除いたバッファーを用いて、37℃及びpH 5.5の1% (w/v) Na-フィターゼでアッセイされる。好ましい実施態様では、残余活性は、少なくとも25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、又は少なくとも80%である。
【0069】
第8の局面では、本発明は、配列番号10又はその成熟ポリペプチドの1以上のアミノ酸の保存的置換、削除及び/又は挿入を含む人工的変異体に関する。挿入は、それが伸張をも示す場合の、分子内、及び/又は分子のN-及び/又はC-末端であり得る。好ましくは、アミノ酸変化は保存的アミノ酸置換である小さな性質の変化である:典型的には、1〜約30アミノ酸の小さな削除;アミノ-末端メチオニン残基のような小さなアミノ-又はカルボキシ-末端伸張;約20〜25残基までの小さなリンカーペプチド;あるいは正味電荷又は他の機能を変化させることによって精製を助ける小さな伸張、例えばポリ-ヒスチジントラクト、抗原性エピトープ又は結合ドメイン、言い換えれば、タンパク質の折り畳み及び/又は活性にほとんど影響を与えない変化。保存的置換の例は、基本的アミノ酸(アルギニン、リジン及びヒスチジン)、酸性アミノ酸(グルタミン酸及びアスパラギン酸)、極性アミノ酸(グルタミン及びアスパラギン)、疎水性アミノ酸(ロイシン、イソロイシン及びバリン)、芳香族アミノ酸(フェニルアラニン、トリプトファン及びチロシン)、並びに小アミノ酸(グリシン、アラニン、セリン、スレオニン及びメチオニン)のグループ内の置換である。保存的置換の他の例は、非-標準的アミノ酸(例えば、4-ヒドロキシプロリン、6-N-メチルリジン、2-アミノイソ酪酸、イソバリン及びα-メチルセリン)での20個の標準的アミノ酸の置換である。保存的置換は、遺伝子コードによってコードされないアミノ酸、及び非天然アミノ酸への置換も含む。「非天然アミノ酸」は、タンパク質合成後に修飾されており、及び/又は標準的アミノ酸のそれとは異なった側鎖(複数)内の化学構造を有する。非天然アミノ酸は、化学的に合成され、好ましくは、商業的に入手可能であり、ピペコリン酸、チアゾリジンカルボン酸、デヒドロプロリン、3-及び4-メチルプロリン、及び3,3-ジメチルプロリンを含み得る。
【0070】
あるいは、アミノ酸変化は、ポリペプチドの物理化学的性質が変化しているような性質の変化である。例えば、アミノ酸変化は、ポリペプチドの熱的安定性を改善し、基質特異性を変化させ、最適pHを変化させるなどである。
【0071】
親ポリペプチドにおける必須アミノ酸は、部位指向的突然変異誘発又はアラニン・スキャニング突然変異誘発のような当該分野で公知の手段に従って同定され得る(Cunningham and Wells, 1989, Science 244: 1081-1085)。後者の技術では、単一のアラニン変異は、分子内のすべての残基に導入され、得られた変異分子は、分子の活性に重要であるアミノ酸残基を同定するための生物的活性(すなわち、フィターゼ活性)について試験される。Hilton他, 1996, J. Biol. Chem. 271: 4699-4708も参照。酵素の活性部位又は他の生物学的相互作用は、推定される接触部位アミノ酸の変異に関連して、核磁気共鳴、結晶学、電子回折、又は光親和標識のような技術によって決定されるように、構造の物理的分析によっても決定され得る。例えば、de Vos他, 1992, Science 255: 306-312; Smith他, 1992, J. Mol. Biol. 224: 899-904; Wlodaver他, 1992, FEBS Lett. 309: 59-64参照。必須アミノ酸の同一性は、本発明に従うポリペプチドに関連するポリペプチドとの同一性の分析からも推論され得る。単一又は複数のアミノ酸置換は、公知の突然変異法、組換え、及び/又はシャッフリングを用いて、次いで、Reidhaar-Olson及びSauer, 1988, Science 241: 53-57; Bowie及びSauer, 1989, Proc. Natl. Acad. Sci. USA 86: 2152-2156; WO 95/17413; 又はWO 95/22625に開示されているような関連するスクリーニング法によってなされ、そして試験され得る。使用され得るエラー-プローンPCR、ファージディスプレイ(例えば、Lowman et al., 199 , Biochem. 30: 10832-10837; 米国特許第5,223,409号明細書; WO 92/06204)、及び部位-指向的突然変異誘発(Derbyshire et al., 1986, Gene 46: 145; Ner et al., 1988, DNA 7:127)を含む。
【0072】
突然変異誘発/シャッフリング法は、宿主細胞によって発現されたクローン化された突然変異誘発化ポリペプチドの活性を検出するために、ハイスループットの自動スクリーニング法と組み合わせられ得る。活性なポリペプチドをコードする突然変異誘発DNA分子は、宿主細胞から回収され、当該分野の標準的な方法を用いて迅速にシークエンスされ得る。これらの方法は、対象の個々のポリペプチド中のアミノ酸残基の重要性の迅速決定を可能にし、未知の構造のポリペプチドに適用され得る。
【0073】
配列番号10のアミノ酸1〜413の配列中のアミノ酸置換(好ましくは保存的置換)、削除及び/又は挿入の総数は、最大10、好ましくは最大9、より好ましくは最大8、より好ましくは最大7、より好ましくは最大6、より好ましくは最大5、より好ましくは最大4、更により好ましくは最大3、より好ましくは最大2、及び更に最も好ましくは1である。
【0074】
配列番号10のアミノ酸1〜413のアミノ酸置換、削除及び/又は挿入の総数は、10,好ましくは9、より好ましくは8、より好ましくは7、より好ましくは最大6、より好ましく卯は最大5、より好ましは4、更により好ましくは3、より好ましくは2、及び更に最も好ましくは1である。代わりに、配列番号10のアミノ酸1〜413の配列中のアミノ酸置換(好ましくは保存的置換)、削除及び/又は挿入の総数は、最大50、45、40、35、30、25、20、19、18、17、16、15、14、13、12、又は最大11である。
【0075】
具体的な実施態様では、本発明のポリペプチドは、ヒトを含む動物に曝露された時に、減少された免疫応答を誘導するように設計された低-アレルギー変異体である。用語免疫応答は、ポリペプチドに曝露された動物の免疫系による任意の反応として理解されるべきである。免疫応答の1種は、曝露された動物においてIgEの増加レベルを導くアレルギー応答である。低-アレルギー変異体は、当該分野で公知の技術を用いて調製され得る。例えば、ポリペプチドは、免疫応答に関連するポリペプチドの部分又はエピトープを保護するポリマー部分と複合化され得る。ポリマーとの複合化は、例えばWO 96/17929、WO98/30682、WO98/35026、及び/又はWO99/00489に記載のポリペプチドへのポリマーのインビトロ化学結合に関連することがある。複合化は、それに加えてあるいは代わりに、ポリペプチドへのポリマーのインビボ結合に関連する。かかる複合化は、ポリペプチドをコードするヌクレオチド配列の遺伝子工学、ポリペプチド中の追加のグリコシル化部位をコードするコンセンサス配列を挿入し、ポリペプチドをグリコシル化することができる宿主中のポリペプチドを発現することによって達成され得る。例えばWO00/26354参照。低-アレルギー変異体を提供する別の方法は、ポリペプチドに自己-オリゴマー化を引き起こすような、ポリペプチドをコードするヌクレオチド配列の遺伝子工学である。これは、ポリペプチドモノマーが、他のポリペプチドモノマーのエピトープを保護し、それによってオリゴマーの抗原性を低減させることをもたらす。かかる産物及び他の調製物は、例えばWO96/16177に記載されている。免疫応答に関連するエピトープは、WO 00/26230及びWO 01/83559に記載されたファージディスプレイ法又はEP 561907に記載のランダムアプローチのような様々な方法によって同定され得る。エピトープは一旦同定されると、そのアミノ酸配列は、公知の操作方法、例えば部位指向的変異形成(例えば、WO 00/26230、WO 00/26354及び/又はWO00/22103参照)によって、ポリペプチドの改変された免疫学的性質を生成するように改変され得、及び/又はポリペプチドの複合化は、ポリマーがエピトープを遮蔽するようにエピトープの充分近くでなされ得る。
【0076】
ハフニア・アルベイ・フィターゼの3次元構造
ハフニア・アルベイ・フィターゼの3次元構造(配列番号10のアミノ酸1〜413)は、添付書類で提供される。該構造は、x-線結晶法についての原則、例えば、X-線構造決定, Stout, G. K. and Jensen, L. H., John Wiley and Sons, Inc. NY 1989で提供される、に従って解決された。ハフニア・アルベイ・フィターゼの解決された結晶構造の構造座標は、添付書類に記載の標準的なPDBフォーマットで提供される(Protein Database Bank, Brookhaven National Laboratory, Brookhaven, Conn.)。それは本願の添付形式部分と理解される、添付書類は、水素原子を除く重元素の座標を与える。酵素の第1の3つ残基は、該結晶構造及びアミノ酸180と189との間のアミノ酸では見られなかった。しかし、180と189との間の構造は、ホモロジーモデリングを組み合わせたモデリングを用いて構築された(例えば、JACKALパッケージ製のMarti-Renom et al., 2000プログラムNEST(wiki.c2b2. Columbia. edu/honiglab public/index.php/Software:Jackal)、及びCHARMmと称されるシミュレイションソフトウェア((//accelrys.com/products/scitegic/component-collections/charmm.html)参照)。
【0077】
フィターゼ活性を有するポリペプチドの起源
本発明のポリペプチドは、任意の種の微生物から得られる。本発明の目的のために、本明細書で使用される用語「から得られる」は、所定の起源と関連して、ヌクレオチド入れるによってコードされるポリペプチドは、ある起源由来のヌクレオチド配列が挿入される起源又は株によって産生されることを意味する。好ましい局面では、所定の起源から得られるポリペプチドは、細胞外に分泌される。
【0078】
本発明のポリペプチドは、細菌性ポリペプチドでよい。例えば、該ポリペプチドは、バチルス(Bacillus)ポリペプチド又はストレプトマイセス(Streptomyces)ポリペプチドのようなグラム陽性細菌性ポリペプチド、あるいはグラム陰性細菌性ポリペプチド、例えばエシェリヒア・コリ(Escherichia coli)、エルシニア(Yersinia)、クレブシエラ(Klebsiella)、サイトロバクター(Citrobacter)又はシュードモナス(Pseudomonas)ポリペプチドでよい。具体的な実施態様では、該ポリペプチドは、ガンマプロテオバクテリア(Gammaproteobacteria)のようなプロテオバクテリア(Proteobacteria)、例えばエンテロバクター目(Enterobacteriales)、例えばエンテロバクテリアセエ(Enterobacteriaceae)から得られる。
【0079】
具体的局面では、エンテロバクテリアセエ(Enterobacteriaceae)から得られるポリペプチドは、ハフニア(Hafnia)ポリペプチド、例えばハフニア・アルベイ種ポリペプチドである。
【0080】
本発明のポリペプチドは、真菌性ポリペプチド、例えば酵母ポリペプチド又は繊維状真菌性ポリペプチドでよい。
【0081】
上記の微生物株は、アメリカンタイプカルチャーコレクション (ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM)、Centraalbureau Voor Schimmelcultures (CBS)、及びアメリカ農業研究局のカルチャーコレクション、ノーザーン・リージョナル・リサーチ・センター (NRRL) のような多数のカルチャーコレクションのパブリックが容易に利用できる。
【0082】
更に、かかるポリペプチドは、上記のプローブを用いて、天然(例えば、土壌、堆肥、水等)から単離された微生物を含む他の起源から同定及び得てもよい。自然環境由来の微生物を単離するための技術は、当該分野でよく知られている。次いで、該ポリペプチドは、同様に別の微生物のゲノム又はcDNAライブラリーをスクリーニングすることによって得られる。ポリペプチドをコードするポリヌクレオチド配列がプローブ(複数)で検出されると、該ポリヌクレオチドは、当業者に周知である技術を利用することによって単離又はクローン化され得る(例えば、Sambrook et al., 1989, 前掲参照)。
【0083】
本発明のポリペプチドは、別のポリペプチドが、本発明のポリペプチドのN-末端もしくはC-末端又はそのフラグメントに融合された、融合ポリペプチド又は開裂可能な融合ポリペプチドを含むこともできる。融合ポリペプチドは、別のポリペプチドをコードするヌクレオチド配列(又はその部分)を本発明のヌクレオチド配列(又はその部分)に融合することによって製造される。融合ポリペプチドを製造するための技術は当該分野で公知であり、それらがフレーム内にあり、そして融合されたポリペプチドの発現が同一のプロモーター(複数)及びターミネーターの制御下にあるように、該ポリペプチドをコードするコーディング配列をライゲートすることを含む。
【0084】
ポリヌクレオチド
本発明はまた、本発明のポリペプチドをコードするヌクレオチド配列を有する単離されたポリヌクレオチドに関する。好ましい局面では、該ヌクレオチド配列は、配列番号9で示される。別の好ましい局面では、該ヌクレオチド配列は、配列番号9の成熟ポリペプチドコーディング領域である。本発明はまた、遺伝子コードの縮重によって、配列番号10のアミノ酸配列を有するポリペプチド又は配列番号9とは異なるその成熟ポリペプチドをコードするヌクレオチド配列を包含する。本発明はまた、配列番号10のフラグメントをコードし、フィターゼ活性を有する、配列番号10のフラグメントをコードする配列番号9のサブ配列に関する。
【0085】
本発明はまた、配列番号9のいずれか1つの成熟ポリペプチドコーディング配列における少なくとも1つの変異を含む成熟ポリヌクレオチドであって、該成熟ポリヌクレオチド配列が配列番号10のアミノ酸1〜413からなるポリペプチドをコードする、ポリヌクレオチドに関する。
【0086】
ポリペプチドをコードするポリヌクレオチドを単離又はクローン化するために使用される技術は、当該分野で知られており、ゲノムDNAからの単離、cDNAからの調製、又はその組合せを含む。かかるゲノムDNAからの本発明のポリヌクレオチドのクローニングは、例えば、周知のポリメーラーゼ連鎖反応(PCR)、又は共通の構造的特徴を有するクローン化DNAフラグメントを検出するための発現ライブラリーの抗体スクリーニングを用いて行うことができる。例えば、lnnis他, 1990, PCR:方法及び応用へのガイド, Academic Press, New York参照。リガーゼ鎖反応(LCR)、ライゲートされた活性化転写(LAT)、及びヌクレオチド配列-型増幅(NASBA)等の他の核酸増幅方法も使用され得る。該ポリヌクレオチドは、ハフニア(Hafnia)株もしくは別の又は関連する生物からクローン化され得、よって、例えば、ヌクレオチド配列のポリペプチドコーディング領域のアレル又は種変異体でもよい。
【0087】
本発明はまた、配列番号9の成熟ポリペプチドコーディング配列(すなわち、ヌクレオチド100〜1338)と少なくとも75%の同一性の程度を有し、かつフィターゼ活性を有するポリペプチドをコードする、ヌクレオチド配列を有するポリヌクレオチドに関する。具体的な実施態様では、同一性の程度は、少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%、又は少なくとも99.9%である。別の実施態様では、同一性の程度は、少なくとも75%、80%、85%、90%、94、97、98、98.0、98.1、98.2、又は少なくとも98.3%である。
【0088】
本発明のポリヌクレオドをコードするヌクレオチド配列の修飾は、該ポリペプチドに実質的に類似するポリペプチドの合成のために必要であるかもしれない。用語該ポリペプチドに「実質的に類似」とは、該ポリペプチドの非-天然形態を意味する。これらのポリペプチドは、その天然源から単離されたポリペプチド由来の作製方法とは異なるかもしれない、例えば特定の活性、熱安定性、最適pH等の点で異なる人工的変異体であり得る。変異体配列は、配列番号9のポリペプチドコーディング領域として提示されるヌクレオチド配列、例えばそのサブ配列に基いて、及び/又は該ヌクレオチド配列によってコードされるポリペプチドの別のアミノ酸配列を生じないが、該ポリペプチドの製造のために意図された宿主生物のコドン使用に対応するヌクレオチド置換の導入によって、あるいは異なったアミノ酸配列を生じ得るヌクレオチド置換の導入によって、構築され得る。ヌクレオチド置換の一般的な説明については、例えばFord他, 1991, タンパク質発現及び精製 2: 95-107参照。
【0089】
かかる置換が、分子の機能に重要である領域の外側でなされ、依然として活性なポリペプチドを生じ得る、ことは当業者には明らかであろう。本発明の単離されたポリヌクレオチドによってコードされるポリペプチドの活性に必須なであり、よって好ましくは置換に供されない、アミノ酸残基は、部位指向的突然変異誘発又はアラニン・スキャニング突然変異誘発のような当該分野で公知の手段に従って同定され得る (例えば、Cunningham and Wells, 1989, Science 244: 1081-1085参照)。後者の技術では、変異は、分子のすべての正に荷電した残基において導入され、分子の活性に重要であるアミノ酸残基を同定するためにフィターゼ活性について試験される。基質-ポリペプチド相互作用の部位はまた、核磁気共鳴分析、結晶学又は光親和性標識のような技術によって決定される3次元構造の分析によって決定され得る (例えば、de Vos et al., 1992, Science 255: 306-312; Smith et al., 1992, Journal of Molecular Biology 224: 899-904; Wlodaver et al., 1992, FEBS Letters 309: 59-64参照)。
【0090】
本発明はまた、中緊縮条件下、より好ましくは中〜高緊縮条件、更により好ましくは高緊縮条件、及び最も好ましくは非常に高い緊縮条件下で、(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び/又は (ii) のいずれか1つの相補鎖とハイブリダイズする、本発明のポリペプチドをコードする単離されたポリヌクレオチド;又は本明細書で定義された、そのアレル変異体及びそのサブ配列 (Sambrook et al., 1989, 前掲) に関する。別の実施態様では、ハイブリダイゼーションは、非常に低い又は低い緊縮条件下で行われる。
【0091】
本発明はまた、(a) 非常に低い、低、中、中〜高、高又は非常に高い緊縮条件下、(i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 及び/又は (ii) のいずれか1つの相補鎖と、DNA群をハイブリダイズし;そして、(b) フィターゼ活性を有するポリペプチドをコードする、ハイブリダイズするポリヌクレオチドを単離することによって、得られた又は得られ得る単離されたポリヌクレオチドに関する。
【0092】
核酸構築物
本発明はまた、制御配列と適合する条件下で好適な宿主細胞におけるコーディング配列の発現を指向する、1以上の制御配列に作動可能に連結した本発明の単離されたポリヌクレオチドを含む核酸構築物に関する。
【0093】
本発明のポリペプチドをコードする単離されたポリヌクレオチドは、該ポリペプチドの発現のための様々な方法において作製され得る。ベクターへのその挿入前のポリヌクレオチド配列の作製は、発現ベクターによって望ましく又は必要である。組換えDNA法を利用するポリヌクレオチド配列を改変するための技術は、当該分野でよく知られている。
【0094】
制御配列は、好適なプロモーター配列、すなわち本発明のポリペプチドをコードするポリヌクレオチドの発現用の宿主細胞によって認識されるヌクレオチド配列でよい。プロモーター配列は、ポリペプチドの発現を仲介する転写制御配列を含む。プロモーター配列は、変異、切断及びハイブリッドプロモーターを含む最適な宿主配列中において、転写活性を示す任意のヌクレオチド配列でよく、宿主細胞に相同又は非相同のいずれかである細胞外又は細胞内ポリペプチドをコードする遺伝子から得られる。
【0095】
特に細菌宿主細胞中での、本発明の核酸構築物の転写を指向するための好適なプロモーターの例は、E. コリlacオペロン、ストレプトマイセス・セリカラー(Streptomyces coelicolor)・アガラーゼ遺伝子 (dagA)、バチルス・サブティリス(Bacillus subtilis)・レバンスクラーゼ遺伝子 (sacB)、バチルス・リケニフォルミス(Bacillus licheniformis)・α-アミラーゼ遺伝子 (amyL)、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)・マルトジェニックアミラーゼ遺伝子 (amyM)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)・α-アミラーゼ遺伝子 (amyQ)、バチルス・リケニフォルミス・ペニシリナーゼ遺伝子 (penP)、バチルス・サブティリスxylA及びxylB遺伝子、及び原核生物β-ラクタマーゼ遺伝子から得られるプロモーターである (Villa-Kamaroff et al., 1978, Proceedings of the National Academy of Sciences USA 75: 3727-3731)、並びにtacプロモーター (DeBoer et al., 1983, Proceedings of the National Academy of Sciences USA 80: 21-25)。更なるプロモーターは、Scientific American, 1980, 242: 74-94; 及びSambrook他, 1989, 前掲の「組み換え細菌由来の有用タンパク質」に記載されている。
【0096】
線維状真菌宿主細胞における本発明の核酸構築物の転写を指向するための好適なプロモーターの例は、アスペルギルス・オリザエ(Aspergillus oryzae)TAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)・アスパラギンプロテイナーゼ、アスペルギルス・ニガー(Aspergillus niger)・中性α-アミラーゼ、アスペルギルス・ニガー・酸安定α-アミラーゼ、アスペルギルス・ニガー又はアスペルギルス・アワモリ(Aspergillus awamori)・グルコアミラーゼ (glaA)、リゾムコール・ミエヘイ(Rhizomucor miehei)・リパーゼ、アスペルギルス・オリザエ・アルカリプロテアーゼ、アスペルギルス・オリザエ・トリオース・ホスフェート・イソメラーゼ、アスペルギルス・ニデュランス(Aspergillus nidulans)・アセトアミダーゼ、フザリウム・ベネナツム(Fusarium venenatum)・アミログルコシダーゼ (WO 00/56900)、フザリウム・ベネナツム・ダリア (WO 00/56900)、フザリウム・ベネナツム・クイン (WO 00/56900)、フザリウム・オキシスポラム(Fusarium oxysporum)・トリプシン様プロテアーゼ (WO 96/00787)、トリコデルマ・リーゼイ(Trichoderma reesei)・β-グルコシダーゼ、トリコデルマ・リーゼイ・セロビオヒドラーゼI、トリコデルマ・リーゼイ・エンドグルカナーゼI、トリコデルマ・リーゼイ・エンドグルカナーゼII、トリコデルマ・リーゼイ・エンドグルカナーゼIII、トリコデルマ・リーゼイ・エンドグルカナーゼIV、トリコデルマ・リーゼイ・エンドグルカナーゼV、トリコデルマ・リーゼイ・キシラナーゼI、トリコデルマ・リーゼイ・キシラナーゼII、トリコデルマ・リーゼイ・β-キシロシダーゼ、並びにNA2-tpiプロモーター (アスペルギルス・ニガー・中性α-アミラーゼ及びアスペルギルス・オリザエ・トリオースリン酸イソメラーゼのための遺伝子からのプロモーターのハイブリッド) の遺伝子から得られるプロモーター、及びその変異体、切断及びハイブリッドプロモーターである。
【0097】
酵母宿主では、有用なプロモーターは、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)・エノラーゼ (ENO-1)、サッカロマイセス・セレヴィシエ・ガラクトキナーゼ (GAL1)、サッカロマイセス・セレヴィシエ・アルコール脱水素酵素/グリセルアルデヒド-3-リン酸脱水素酵素 (ADH1、ADH2/GAP)、サッカロマイセス・セレヴィシエ・トリオースリン酸イソメラーゼ (TPI)、サッカロマイセス・セレヴィシエ・メタロチオニン (CUP1)、サッカロマイセス・セレヴィシエ・3-ホスホグリセリン酸キナーゼ、及びピキア・パストリス(Pichia pastoris)・アルコール酸化酵素 (AOX1) のための遺伝子から得られる。酵母宿主細胞用の他の有用なプロモーターは、Romanos他, 1992, Yeast 8: 423-488によって記載されている。制御配列は、好適な転写終結配列、転写を終結するための宿主細胞によって認識される配列もよい。終結配列は、ポリペプチドをコードするヌクレオチド配列の3'末端に作動可能に連結される。最適な宿主細胞において機能する任意の終結因子が本発明において使用され得る。
【0098】
線維状真菌宿主細胞のための好ましい終結因子は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、アスペルギルス・ニデュランス・アントラニル酸合成酵素、アスペルギルス・ニガー・α-グルコシダーゼ、及びフザリウム・オキシスポラム・トリプシン様プロテアーゼのための遺伝子から得られる。
【0099】
酵母宿主細胞のための好ましい終結因子は、サッカロマイセス・セレヴィシエ・エノラーゼ、サッカロマイセス・セレヴィシエ・チトクローム C (CYC1)、及びサッカロマイセス・セレヴィシエ・グリセルアルデヒド-3-リン酸脱水素酵素のための遺伝子から得られる。酵母宿主細胞のための他の有用な終結因子は、Romanos他, 1992, 前掲によって記載されている。
【0100】
制御配列は、好適なリーダー配列、すなわち宿主細胞による翻訳のために重要であるmRNAの非翻訳領域でもよい。リーダー配列は、ポリペプチドをコードするヌクレオチド配列の5'末端に作動可能に連結される。最適な宿主細胞中で機能する任意のリーダー配列は、本発明において使用され得る。
【0101】
制御配列は、好適なリーダー配列、すなわち宿主細胞による翻訳のために重要であるmRNAの非翻訳領域でもよい。リーダー配列は、ポリペプチドをコードするヌクレオチド配列の5'末端に作動可能に連結される。最適な宿主細胞中で機能する任意のリーダー配列は、本発明において使用され得る。
【0102】
線維状宿主細胞のための好ましいリーダーは、アスペルギルス・オリザエTAKAアミラーゼ及びアスペルギルス・ニデュランス(Aspergillus nidulans)トリオースリン酸イソメラーゼの遺伝子から得られる。
【0103】
酵母宿主細胞のための好適なリーダーは、サッカロマイセス・セレヴィシエ・エノラーゼ (ENO-1)、サッカロマイセス・セレヴィシエ・3-ホスホグリセリン酸キナーゼ、サッカロマイセス・セレヴィシエ・α-因子、及びサッカロマイセス・セレヴィシエ・アルコール脱水素酵素/グリセルアルデヒド-3-リン酸脱水素酵素 (ADH2/GAP) のための遺伝子から得られる。
【0104】
制御配列は、ポリアデニル化配列、すなわちヌクレオチド配列の3'末端に作動可能に連結し、そして転写される時に、転写mRNAにポリアデニル化残基を付加するためのシグナルとして宿主細胞によって認識される、配列でもよい。最適な宿主細胞において機能する任意のポリアデニル化配列は、本発明において使用され得る。
【0105】
線維状真菌宿主細胞のための好ましいポリアデニル化配列は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、アスペルギルス・ニデュランス・アントラニル酸合成酵素、フザリウム・オキシスポラム・トリプシン様プロテアーゼ、及びアスペルギルス・ニガー・α-グルコシダーゼのための遺伝子から得られる。
【0106】
酵母宿主細胞のための有用ナポリアデニル化配列は、Guo及びSherman, 1995, Molecular Cellular Biology 15: 5983-5990によって記載されている。
【0107】
制御配列は、ポリペプチドのアミノ末端に連結されたアミノ酸配列をコードし、及び細胞の分泌経路にコードされたポリペプチドを指向する、シグナルペプチドコーディング領域でもよい。ヌクレオチド配列のコーディング配列の5'末端は、本来、分泌されたポリペプチドをコードするコーディング領域の断片を有する翻訳読み枠において天然に連結されたシグナルペプチドコーディング領域を含むことがある。あるいは、該コーディング配列の5'末端は、該コーディング配列には無関係であるシグナルペプチドコーディング領域を含むことがある。この無関係なシグナルペプチドコーディング領域は、該コーディング配列が天然にはシグナルペプチドコーディング領域を含まない場合に、必要とされるかもしれない。あるいは、この無権系なシグナルペプチドコーディング領域は、ポリペプチドの分泌を亢進するために、単に、天然のシグナルペプチドコーディング領域を取り替えることがある。しかしながら、最適の宿主細胞の分泌経路に発現されたポリペプチドを指向する任意のシグナルペプチドコーディング領域が、本発明において使用され得る。
【0108】
細菌宿主細胞のための効果的なシグナルペプチドコーディング領域は、バチルスNCIB 11837・マルトースアミラーゼ、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)・α-アミラーゼ、バチルス・リケニフォルミス(Bacillus licheniformis)・スブチリシン、バチルス・リケニフォルミス・β-ラクタマーゼ、バチルス・ステアロサーモフィルス・中性プロテアーゼ (nprT、nprS、nprM)、及びバチルス・バチルス・サブティリスprsAのための遺伝子から得られたシグナルペプチドコーディング領域である。更に、シグナルペプチドは、Simonen及びPalva, 1993, Microbiological Reviews 57: 109-137によって記載されている。
【0109】
線維状真菌宿主細胞のための有効なシグナルペプチドコーディング領域は、アスペルギルス・オリザエ・TAKAアミラーゼ、アスペルギルス・ニガー・中性アミラーゼ、アスペルギルス・ニガー・グルコアミラーゼ、リゾムコール・ミエヘイ・アスパラギンプロテイナーゼ、フミコラ・インソレンス(Humicola insolens)・セルラーゼ、及びフミコラ・ラヌギノーサ(Humicola lanuginosa)・リパーゼのための遺伝子から得られたシグナルペプチドコーディング領域である。
【0110】
酵母宿主細胞のための有用なシグナルペプチドは、サッカロマイセス・セレヴィシエ・α-因子及びサッカロマイセス・セレヴィシエ・インヴェルターゼのための遺伝子から得られる。他の有用なシグナルペプチドコーディング領域は、Romanos他, 1992, 前掲、及びXiong他のJournal of Applied Microbiology 2005, 98, 418-428に記載されている。
【0111】
好ましい局面では、シグナルペプチドコーディング領域は、配列番号10のアミノ酸1〜33をコードする配列番号9のヌクレオチド1〜99である。他の好ましい局面では、シグナルペプチドコーディング領域は、配列番号12のアミノ酸1〜27をコードする配列番号11のヌクレオチド1〜81である。
【0112】
制御配列は、ポリペプチドのアミノ末端に位置するアミノ酸をコードするプロペプチドコーディング領域でもよい。得られたポリペプチドは、プロペプチド又はプロポリペプチド(又は場合によっては酵素原)として知られている。プロポリペプチドは、一般的に、不活性であり、プロポリペプチドからのプロペプチドの触媒的又は自己触媒的開裂によって成熟活性ポリペプチドに変換され得る。プロペプチドコーディング領域は、バチルス・サブチリス・アルカリプロテアーゼ (aprE)、バチルス・サブティリス・中性プロテアーゼ (nprT)、サッカロマイセス・セレヴィシエ・α-因子、リゾムコール・ミエヘイ・アスパラギンプロテイナーゼ、及びマイセリオフトーラ・サーモフィラ(Myceliophthora thermophila)・ラクタマーゼ (WO 95/33836) のための遺伝子から得られる。
【0113】
シグナルペプチド領域及びプロペプチド領域の両方がポリペプチドのアミノ末端に存在する場合には、該プロペプチド領域は、ポリペプチドのアミノ末端に隣接して位置し、該シグナルペプチド領域は、該プロペプチド領域のアミノ末端に隣接して位置する。
【0114】
また、宿主細胞の成長に対してポリペプチドの発現のレギュレーションを可能にする調節配列を付加することが望ましいことがある。調節系の例は、調節化合物の存在を含む、遺伝子の発現を化学又は物理的刺激に反応してオン又はオフに変化させるものである。原核生物の調節系は、lac、tac及びtrpオペレーター系を含む。酵母では、ADH2系又はGAL1系が使用され得る。線維状真菌では、TAKAα-アミラーゼ・プロモーター、アスペルギルス・ニガー・グルコアミラーゼ・プロモーター及びアスペルギルス・オリザエ・グルコアミラーゼ・プロモーターが、調節配列として使用され得る。調節配列の他の例は、遺伝子増幅を可能にする配列である。
【0115】
発現ベクター
本発明はまた、本発明のポリヌクレオチド、プロモーター、及び転写及び翻訳停止シグナルを含む組換え発現ベクターに関する。上記の様々な核酸及び制御配列は、一緒に結合されて、該ポリペプチドをコードするヌクレオチド配列の挿入又は置換をかかる位置で可能にするための、1以上の簡便な制限部位を含み得る組換え発現ベクターを製造し得る。あるいは、本発明のヌクレオチド配列は、該ヌクレオチド配列又は該配列を含む核酸構築物を発現用の好適なベクターに挿入することによって発現され得る。発現ベクターの作製において、コーディング配列は、該コーディング配列が発現用の好適な制御配列と作動可能に連結されるように、該ベクターに配置される。
【0116】
組換え発現ベクターは、組換えDNA法に簡便に供され、ヌクレオチド配列の発現をもたらし得る任意ベクターでよい(例えば、プラスミド又はウイルス)。最適なベクターは、典型的には、ベクター又はベクターが導入される宿主細胞の適合性に依拠することになる。ベクターは、線状又は閉環状プラスミドでよい。
【0117】
ベクターは、自律的複製型ベクター、すなわち、染色体外物質として存在し、その複製が、染色体複製とは無関係であるベクター、例えばプラスミド、染色体外要素、人工ミニ染色体、又は人工染色体でよい。ベクターは、自己複製を確実にするための手段を含むことがある。あるいは、宿主細胞に導入される時に、ベクターはゲノムに統合され、ベクターが統合された染色体(複数)と一緒になって複製されるものでよい。更に、宿主細胞のゲノム又はトランスポソンに導入されるべき総DNAを一緒に含む、単一ベクターもしくはプラスミド又は2以上のベクターもしくはプラスミドが使用され得る。
【0118】
本発明のベクターは、好ましくは、形質転換された細胞の簡便な選択を許容する1以上の選択マーカーを含む。選択マーカーは、その産物が殺生剤又はウイルス耐性、重金属に対する耐性、栄養要求体に対する原栄養性などを与える遺伝子である。
【0119】
条件付で不可欠の遺伝子は、非-抗生物質選択的マーカーとして機能し得る。細菌性の条件付不可欠の非-抗生物質選択的マーカーの非限定例は、細菌がD-アラニンの非存在下で培養される時に不可欠であるにすぎない、バチルス・サブティリス、バチルス・リシェニフォルミス又は他のバチルス属由来のdal遺伝子である。また、UDP-がラクトースのターオーバーに関連する遺伝子コーディング酵素は、細胞がガラクトースの存在下で成長し又はガラクトースの存在を与える培地中で成長する時に、細胞中で条件付不可欠マーカーとして機能し得る。かかる遺伝子の非限定例は、UTP-依存的ホスホリラーゼ (EC 2.7.7.10)、UDP-グルコース-依存的ウリジリルトランスフェラーゼ (EC 2.7.7.12)、又はUDP-ガラクトースエピメラーゼ (EC 5.1.3.2) をコードするB. サブティリス又はB. リシェニフォルミス由来のものである。また、バチルスのxylAのようなキシロースイソメラーゼ遺伝子は、単一の炭素源としてのキシロースを含む最小培地で成長した細胞において選択マーカーとして使用され得る。グルコン酸塩、gntK及びgntPを利用するために必要な遺伝子も、単一炭素源としてのグルコン酸塩を含む最小培地で成長する細胞における選択マーカーとして使用され得る。条件付の不可欠な遺伝子の他の例は、当該分野で知られている。抗生物質選択マーカーは、アンピシリン、カナマイシン、クロラムフェニコール、エリスロマイシン、テトラサイクリン、ネオマイシン、ハイグロマイシン又はメトレキサートのような抗生物質に対する抗生物質耐性を与える。
【0120】
酵母宿主細胞のための好適なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1及びURA3である。線維状真菌宿主細胞において使用するための選択マーカーは、amdS (アセトアミダーゼ)、argB (オルニチンカルバモイル基転移酵素)、bar (ホスフィノトリシン・アセチル基転移酵素)、hph (ハイグロマイシンリン酸転移酵素)、niaD (硝酸還元酵素)、pyrG (オロチジン5'-リン酸脱炭酸酵素)、sC (サルファイト・アデニルトランスフェラーゼ)、及びtrpC (アントラニル酸合成酵素)、並びにそれらの等価体を含むが、これらに限定されない。アスペルギルス属細胞において使用するためには、アスペルギルス・ニデュランス又はアスペルギルス・オリザエのamdS及びpyrG遺伝子、及びストレプトマイセス・ハイグロスコピカス(Streptomyces hygroscopicus)のbar遺伝子が好ましい。
【0121】
本発明のベクターは、好ましくは、宿主細胞中へのベクターの統合又はゲノムに関係のない細胞中でのベクターの自律的複製を許容する、要素(複数)を含む。宿主細胞ゲノムへの統合に関して、ベクターは、ポリペプチドをコードするポリヌクレオチド配列、又は相同的もしくは非相同的な組換えによるゲノムへの統合のためのベクターの任意の他の要素に依拠し得る。あるいは、ベクターは、染色体(複数)における正確な位置(複数)での宿主細胞のゲノムへの相同組換えによる統合を指向するための、追加のヌクレオチド配列を含み得る。正確な位置での統合の可能性を増加させるために、統合要素は、相同組換えの可能性を増加させるための対応する標的配列と高い同一性の程度を有する、好ましくは、十分に多数の核酸、例えば100〜10,000塩基対、好ましくは400〜10,000塩基対、及び最も好ましくは800〜10,000塩基対を含む必要がある。統合要素は、宿主細胞のゲノム中の標的配列と相同である任意に配列でよい。更に、統合要素は、非-コーディング又はコーディングヌクレオチド配列でよい。一方、ベクターは、非-相同組換えによって宿主細胞のゲノムに統合され得る。
【0122】
自律的複製のために、ベクターは、ベクターを問題の宿主細胞において自律的に複製させる複製起源を更に含むことがある。複製起源は、細胞中で機能する自律的複製を介在する任意のプラスミド複製因子でよい。用語「複製起源」又は「プラスミド複製因子」は、インビボでプラスミド又はベクターを複製させるヌクレオチド配列として本明細書に定義される。
【0123】
細菌の複製起源の例は、E. コリ中で複製を許容するプラスミドpBR322、pUC19、pACYC177及びpACYC184、及びバチルス属中で複製を許容するpUBHO、pE194、pTA1060及びpAMβ1である。
【0124】
酵母宿主細胞中で使用するための複製起源の例は、複製の2ミクロン起源、ARS1、ARS4、ARS1とCEN3との組合せ、及びARS4とCEN6との組合せである。
【0125】
線維状真菌細胞において有用な複製起源の例は、AMA1及びANSIである (Gems et al., 1991, Gene 98:61-67; Cullen et al., 1987, Nucleic Acids Research 15: 9163-9175; WO 00/24883)。AMA1遺伝子の単離、及び該遺伝子を含むプラスミド又はベクターの構築は、WO00/24883に開示の方法に従って達成され得る。
【0126】
本発明のポリヌクレオチドの1超コピーは、遺伝子産物の産生を増加させるために宿主細胞に挿入され得る。ポリヌクレオチドのコピー数の増加は、該配列の少なくとも1つの追加のコピーを宿主細胞ゲノムに統合することによって、又は該ポリヌクレオチドと共に増幅可能な選択マーカー遺伝子を含むことによって得られ(そこでは、細胞は、選択マーカー遺伝子の増幅されたコピーを含む)、それによって、ポリヌクレオチドの追加のコピーは、好適な選択マーカーの存在下で細胞を培養することによって選択され得る。
【0127】
本発明の組換え発現ベクターを構築するための上記の要素をライゲートするために使用される方法は、当業者によく知られている (例えば、Sambrook et al., 1989, 前掲参照)。
【0128】
宿主細胞
本発明はまた、ポリペプチドの組換え産生において有利に使用される、本発明のポリヌクレオチドを含む組換え宿主細胞に関する。本発明のポリヌクレオチドを含むベクターは、該ベクターが先の記載の染色体統合因子又は自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。用語「宿主細胞」は、複製中に起こる変異に因って親細胞と同一でない親細胞の任意の子孫を包含する。最適な宿主細胞は、ポリペプチドをコードする遺伝子及びその起源にかなりの程度で依拠することになる。
【0129】
宿主細胞は、単細胞性微生物、例えば原核生物、又は非-単細胞性微生物、例えば真核生物でよい。
【0130】
有用な単細胞微生物は、細菌細胞であり、例えば、バチルス細胞、例えばバチルス・アルカロフィラス(Bacillus alkalophilus)、バチルス・アミロリクエファシエンス(Bacillus amyloliquefaciens)、バチルス・ブレビス(Bacillus brevis)、バチルス・サーキュランス(Bacillus circulans)、バチルス・クラウジアイ(Bacillus clausii)、バチルス・コアギュランス(Bacillus coagulans)、バチルス・ラウトゥス(Bacillus lautus)、バチルス・レンタス(Bacillus lentus)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・メガテリウム(Bacillus megaterium)、バチルス・ステアロサーモフィルス(Bacillus stearothermophilus)、バチルス・サブティリス(Bacillus subtilis)、及びバチルス・チューリンゲンシス(Bacillus thuringiensis);又はストレプトマイセス細胞、例えばストレプトマイセス・リビダンス(Streptomyces lividans)及びストレプトマイセス・ミュリナス(Streptomyces murinus)を含むがこれらに限定されないグラム陽性菌、あるいは、E. コリ及びシュードモナス属のような含むグラム陰性菌である。好ましい局面では、細菌宿主細胞は、バチルス・レンタス、バチルス・リケニフォルミス、バチルス・ステアロサーモフィルス又はバチルス・サブティリス細胞である。別の好ましい局面では、バチルス細胞は、好アルカリ性バチルスである。
【0131】
ベクターの細菌宿主細胞への導入は、例えば、コンピテント細胞を用いる (例えば、Young and Spizizin, 1961 , Journal of Bacteriology 81: 823-829、又はDubnau and Davidoff-Abelson, 1971 , Journal of Molecular Biology 56: 209-221参照) プロトラスト形質転換 (例えば、Chang and Cohen, 1979, Molecular General Genetics 168: 111-115参照)、エレクトロポレーション (例えば、Shigekawa and Dower, 1988, Biotechniques 6: 742-751参照)、又はコンジュゲーション (例えば、Koehler and Thorne, 1987, Journal of Bacteriology 169: 5771-5278参照) によって行われる。
【0132】
宿主細胞は、真核細胞、例えば哺乳動物、昆虫、植物又は真菌細胞でもよい。
【0133】
好ましい局面では、宿主細胞は真菌細胞である。本明細書で使用する「真菌」は、子嚢菌門(Ascomycota)、担子菌門(Basidiomycota)、ツボカビ門(Chytridiomycota)及び接合菌門(Zygomycota)(Hawksworth et al., In, Ainsworth and Bisby's Dictionary of The Fungi, 第8版, 1995, CAB International, University Press, Cambridge, UKによって定義された)、並びに卵菌門(Hawksworth et al., 1995, 前掲, 第171頁で引用された)及び分生子形成菌(Hawksworth et al., 1995, 前掲)を含む。
【0134】
より好ましい局面では、真菌宿主細胞は、酵母細胞である。本明細書で使用される「酵母」は、有子嚢胞子(ascosporogenous)酵母(エンドミセタレス目(Endomycetales))、担子胞子(basidiosporogenous)及び不完全菌類(Fungi lmperfecti)(不完全酵母菌綱(Blastomycetes))を含む。酵母の分類は、本発明の目的のために今後変わるかもしれないので、酵母は、「酵母の生物学及び活性」(Skinner, F.A., Passmore, S. M., and Davenport, R. R.著, Soc. App. Bacterid. Symposium Series No. 9, 1980) に記載されたように定義されべきである。
【0135】
更により好ましい局面では、酵母宿細胞は、カンジタ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロマイセス属(Kluyveromyces)、ピキア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)又はヤロウイア属(Yarrowia)細胞である。最も好ましい局面では、酵母宿主細胞は、ピキア・パストリス(Pichia pastoris)、ピキア・メタノリカ(Pichia methanolica)、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ダイアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミ・セス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)又はサッカロミセス・オビホルミス(Saccharomyces oviformis)細胞である。別の好ましい局面では、酵母宿主細胞は、クルイベロミセス・ラクチス(Kluyveromyces lactis)細胞である。別の最も好ましい局面では、酵母宿主細胞は、ヤロウイア・リポリティカ(Yarrowia lipolytica)細胞である。
【0136】
別のより好ましい局面では、真菌宿主細胞は、線維状真菌細胞である。「線維状真菌」は、(Hawksworth et al., 1995, 前掲によって定義された)サブディビジョンユーマイコタ(Eumycota)及びオオマイコタ(Oomycota)の全ての繊維型を含む。線維状真菌は、一般的に、キチン、セルロース、グルカン、キトサン、マンナン及び他の複合化多糖からなる菌糸体壁によって特徴付けられる。植物的な成長は、菌糸の伸張であり、炭素異化作用は、偏性好気性である。対照的に、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)のような酵母による植物の成長は単細胞葉状体の発芽によるものであり、炭素異化作用は発酵性でよい。
【0137】
更により好ましい局面では、線維状真菌宿主細胞は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、オーレオバシジウム(Aureobasidium)、ジェルカンデラ(Bjerkandera)、セリポリオプシス(Ceriporiopsis)、コプリナス(Coprinus)、コリオラス(Coriolus)、クリプトコッカス(Cryptococcus)、フィロバシジウム(Filobasidium)、フザリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカノマスチックス( Neocallimastix)、ネウロスポラ(Neurospora)、パエシロミセス(Paecilomyces)、ペニシリウム(Penicillium)、ファネロカエテ(Phanerochaete)、フレビア(Phlebia)、ピロミセス(Piromyces)、プレロータス(Pleurotus)、シゾフィラム(Schizophyllum)、タラロマイセス(Talaromyces)、サーモアスカス(Thermoascus)、チエラビア(Thielavia)、トイポクラディウム(Tolypocladium)、トラメテス(Trametes)、又はトリコデルマ(Trichoderma)の細胞である。
【0138】
最も好ましい局面では、線維状真菌宿主細胞は、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、アスペルギルス・フォティダス(Aspergillus foetidus)、アスペルギルス ジャポニカス(Aspergillus japonicus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ニガー(Aspergillus niger)又はアスペルギルス・オリザエ(Aspergillus oryzae)の細胞である。別の最も好ましい局面では、線維状真菌宿主細胞は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・セルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)、フザリウム・ネグンジ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レチクラツム(Fusarium reticulatum)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サムブキヌム(Fusarium sambucinum)、フザリウム・サルコクロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルフリウム(Fusarium sulphureum)、フザリウム・トルロクム(Fusarium torulosum)、フザリウム・トリコテキオイデス(Fusarium trichothecioides)、又はフザリウム・ベネナツム(Fusarium venenatum)の細胞である。別の最も好ましい局面では、線維状真菌宿主細胞は、ルカンデラ・アダスタ(Bjerkandera adusta)、セリポリオプシス・アネイニナ(Ceriporiopsis aneirina)、セリポリオプシス・カレグレア(Ceriporiopsis caregiea)、セリポリオプシス・ギルベセン.ス(Ceriporiopsis gilvescens)、セリポリオプシス・パンノシンタ(Ceriporiopsis pannocinta)、セリポリオプシス・リバロサ(Ceriporiopsis rivulosa)、セリポリオプシス・スブベルミスポラ(Ceriporiopsis subrufa)又はセリポリオプシス・サバーミスポラ(Ceriporiopsis subvermispora)、コプリヌス・シネレウス(Coprinus cinereus)、コプリヌス・ヒルスタス(Coriolus hirsutus)、ヒュミコラ・インソレンス(Humicola insolens)、ヒュミコラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヒー(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、ネウロスポラ・クラサ(Neurospora crassa)、ペニシリウム・プルプルゲヌム(Penicillium purpurogenum)、ファネロケーテ・クリプトスポリジウム(Phanerochaete chrysosporium)、フレビア・ラジアタ(Phlebia radiata)、プレオロータス・エリンジ(Pleurotus eryngii)、チエラビア・テルレストリス(Thielavia terrestris)、トラメーテス・ビロサ(Trametes villosa)、トラメテス・ベルシコロル(Trametes versicolor)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギー(Trichoderma koningii)、トリコデルマ・ロンギブラキアタム(Trichoderma longibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、又はトリコデルマ・ビリデ(Trichoderma viride)の株細胞である。
【0139】
真菌細胞は、それ自体公知の方法で、プロトプラスト形成、プロトプラストの形質転換、及び細胞壁の再生を含むプロセスによって形質転換され得る。アスペルギルス属及びトリコデルマ属宿主細胞の形質転換のための好適な方法は、EP 238 023、及びYelton他, 1984, Proceedings of the National Academy of Sciences USA 81: 1470-1474に記載されている。フザリウム属を形質転換する好適な方法は、Malardier他, 1989, Gene 78: 147-156、及びWO 96/00787に記載されている。酵母は、Becker及びGuarente, In Abelson, J.N.及びSimon, M. I.著, 酵母遺伝学及び分子生物学への指針、酵素学の方法, 第194巻, p第182-187頁, Academic Press, Inc., New York; lto他, 1983, Journal of Bacteriology 153: 163; 及びHinnen他, 1978, Proceedings of the National Academy of Sciences USA 75: 1920に記載の方法を用いて形質転換され得る。
【0140】
製造法
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、その野生型形態において、該ポリペプチドを産生することができる、細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造するための方法に関する。好ましくは、細胞h、ハフニア種の細胞であり、より好ましくはハフニア・アルベイの細胞である。
【0141】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、宿主細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造するための方法に関する。
【0142】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、宿主細胞を培養し、ここで、該宿主細胞は、配列番号9のいずれか1つの成熟ポリペプチドコーディング領域において少なくとも1つの変異を有する変異ヌクレオチド配列を含み、該変異ヌクレオチド配列は、配列番号10のアミノ酸1〜413からなるポリペプチドをコードする、及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドの製造方法に関する。
【0143】
本発明の製造方法において、細胞は、当該分野で周知の方法を用いてポリペプチドの産生のために好適な栄養培地において培養される。例えば、細胞は、好適な培地中で達成される研究室的又は工業的発酵器中で及びポリペプチドを発現させる及び/又は単離させる条件下で、振盪フラスコ培養、及び小スケール又は大スケール発光(連続的、バッチ、流加、又は固体発光を含む)によって培養され得る。培養は、当該分野で公知の方法を用いて、炭素源、窒素源及び無機塩を含む好適な栄養培地中で行う。好適な培地は、商業的供給者から入手されるか、又は(例えば、アメリカン・タイプ・カルチャー・コレクションのカタログにおいて)公表された組成に従って調製され得る。ポリペプチドが栄養培地に分泌される場合には、ポリペプチドは、培地から直接的に回収され得る。ポリペプチドが分泌されない場合には、細胞溶解物から回収され得る。
【0144】
ポリペプチドは、ポリヌクレオチドに特異的である当該分野で公知の方法を用いて検出され得る。これらの検出方法は、特異的抗体の使用、ポリペプチド産物の形成、又はポリペプチド基質の消失を含むことがある。例えば、ポリペプチドアッセイは、本明細書に記載のポリペプチドの活性を決定するために使用され得る。
【0145】
得られたポリペプチドは、当該分野で公知の方法を用いて回収され得る。例えば、ポリペプチドは、遠心、ろ過、抽出、スプレイ-乾燥、濃縮又は沈殿を含むがこれらに限定されない、慣用的な方法によって栄養培地から回収され得る。
【0146】
本発明のポリペプチドは、クロマトグラフィー(例えば、イオン交換、アフィニティー、疎水的方法、等電点電気泳動及びサイズ排除法)、電気泳動法(例えば、プレパラティブ等電点電気泳動法)、差異化された溶解度(例えば、硫酸アンモニウム沈殿)、SDS-PAGE又は抽出 (例えば、タンパク質精製, J. -C. Janson及びLars Ryden著, VCH Publishers, New York, 1989参照) を含むがこれらに限定されない当該分野で公知の様々な方法によって精製され得る。
【0147】
トランスジェニック植物
本発明はまた、本発明のフィターゼ活性を有するポリペプチドを回収可能な量で発現及び産生するように、該ポリペプチドをコードするヌクレオチド配列で形質転換された、トランスジェニック植物、植物部分又は植物細胞に関する。該ポリペプチドは、植物又は植物部分から回収され得る。あるいは、組換えポリペプチドを含む植物又は植物部分は、食品又は飼料の量を改善するためのものとして、例えば栄養価値、おいしさ及び流体力学的的性質を改善するためのものとして、あるいは抗栄養因子を破壊するために、使用され得る。
【0148】
具体的な実施態様では、該ポリペプチドは、種子中の内胚乳貯蔵小胞に標的される。これは、好適なシグナルペプチドを用いて前駆体としてそれを合成することによって得られる。Horvath他のPNAS, Feb. 15, 2000, vol. 97, no. 4, p. 1914-1919参照。
【0149】
トランスジェニック植物は、双子葉植物(子葉植物)又は単子葉植物(単子葉植物)、又はその人工変形体でよい。単子葉植物の例は、芝生、例えば牧草(ブルーグラス, Poa)、飼料草例えばウシノケグサ属、ロリウム属、温帯草、例えばアグロスチス属、及び穀物、例えば小麦、オート麦、ライ麦、大麦、米、サトウモロコシ、ライ小麦(小麦(Triticum)及びライ(Secale)及びトウモロコシ(corn)の安定化ハイブリッド)である。子葉植物の例は、タバコ、マメ科植物、例えばヒマワリ (Helianthus)、綿花 (Gossypium)、ルピナス、ジャガイモ、サトウキビ、ナシ、マメ及び大豆、及びアブラナ科植物 (Brassicaceaeファミリー)、例えば、カリフラワー、セイヨウアブラナ種子、並びに密接に関連したモデル生物体のシロイヌナズナ(Arabidopsis thaliana)である。例えば米国特許第5,689,054号明細書及び同6,111,168号明細書に記載の低-フィターゼ植物は、人工植物の例である。
【0150】
植物部分の例は、幹、カルス、葉、果実、種子及び塊茎、並びにこれらの部分を含む個々の組織、例えば表皮、葉肉、柔組織、維管束組織、分裂組織である。また、特定の植物組織区画、例えば葉緑体、アポプラスト、ミトコンドリア、小胞、ペルオキシソーム及び細胞質は植物部分と考えられる。更に、任意の植物細胞、それが組織起源であってもなくても、植物部分と考えられる。同様に、本発明の利用を助けるために単離された特定の組織及び細胞のような植物部分も、植物部分と考えられ、例えば、胚、内胚乳、澱粉(aleurone)及び種子コートである。
【0151】
かかる植物の子孫、植物部分及び植物細胞も本発明の範囲内に含まれる。本発明のポリペプチドを発現するトランスジェニック植物又は植物細胞は、当該分野で公知の方法に従って構築され得る。すなわち、該植物又は植物細胞は、本発明のポリペプチドをコードする1以上の発現構築物を植物宿主ゲノムに組み込み、そして得られた改変植物又は植物細胞をトランスジェニック植物又は植物細胞で繁殖させることによって構築される。
【0152】
好都合なことに、発現構築物は、最適な植物又は植物部分における核酸配列の発現のために必要とされる好適な調節配列と作動可能に連結した本発明のポリペプチドをコードする核酸配列を含む構築物である。更に、発現構築物は、発現構築物が統合されている宿主細胞を同定するために有用な選択マーカー、及び問題の植物への構築物の導入のために必要なDNA配列を含み得る(後者は、使用されるべきDNA導入法に依拠する)。
【0153】
最適な調節配列、例えばプロモーター及び終結配列及び場合によりシグナル又はトランジット配列が、例えば、いつ、どこで及びどのようにして本発明のポリペプチドが発現されることが望まれるかに基づいて決定される。例えば、本発明のポリペプチドをコードする遺伝子の発現は、構成的でも又は誘導可能でよく、あるいは発達特異的、段階特異的又は組織特異的でもよく、そして遺伝子産物は、特定の細胞区分、組織又は細胞部分、例えば種子又は葉に標的され得る。調節配列は、例えば、Tague他, 1988, Plant Physiology 86: 506に記載されている。
【0154】
構成的発現のために、以下のプロモーターが使用され得る: 35S-CaMVプロモーター (Franck et al., 1980, Cell 21: 285-294)、トウモロコシポリユビキチン1 (Christensen AH, Sharrock RA and Quail 1992,トウモロコシポリユビキチン遺伝子:構造、発現の熱的混乱及び転写スプライシング、及びエレクトロポレーションによるプロトプラストへの変換後のプロモーター活性)、又はコメアクチン1プロモーター (Plant Mo. Biol. 18, 675-689; Zhang W, McElroy D. and Wu R 1991, Analysis of rice Act1 5' region activity in transgenic rice plants. Plant Cell 3, 1155-1165)。生物特異的プロモーターは、例えば、種子、ジャガイモ塊茎及び果実のような貯蔵シンク組織由来のプロモーター (Edwards & Coruzzi, 1990, Ann. Rev. Genet. 24: 275-303)、又は分裂組織のような代謝性シンク組織由来のプロモーター (Ito et al., 1994, Plant Mol. Biol. 24: 863-878)、グルテリン、プロタミン、グロブリンもしくは米由来のアルブミンプロモーターのような種子特異的プロモーター (Wu et al., 1998, Plant and Cell Physiology 39: 885-889)、レグミンB4由来のヴィキア・ファバプロモーター及びヴィキア・ファバ(Vicia faba)由来の未知のタンパク質遺伝子 (Conrad et al., 1998, Journal of Plant Physiology 152: 708-711)、種子油体タンパク質由来のプロモーター (Chen et al., 1998, Plant and Cell Physiology 39: 935-941)、ブラシカ・ナパス(Brassica napus)由来の貯蔵タンパク質napAプロモーター、又は例えばWO 91/14772に記載の当該分野で公知の任意の他の種子特異的プロモーターである。更に、プロモーターは、コメ又はトマト由来のrbcsプロモーターのような葉特異的プロモーター (Kyozuka et al., 1993, Plant Physiology 102: 991-1000)、コレラウイルスアデニンメチルトランスフェラーゼ遺伝子プロモーター (Mitra and Higgins, 1994, Plant Molecular Biology 26: 85-93)、又はコメ由来のaldP遺伝子プロモーター (Kagaya et al., 1995, Molecular and General Genetics 248: 668-674)、又はジャガイモpin2プロモーターのような損傷誘導プロモーター (Xu et al., 1993, Plant Molecular Biology 22: 573-588) でよい。同様に、プロモーターは、塩分の温度、不足又は変更のような非生物的処理によって誘導され、あるいはプロモーターを活性化する外因的に適用された物質、例えばエタノール、エストロゲン、エチレンのような植物ホルモン、アブシジン酸、ジベレリン酸及び/又は重金属、によって誘導され得る。
【0155】
プロモーターエンハンサー要素は、植物におけるポリペプチドのより高い発現を達成するために使用してもよい。例えば、プロモーターエンハンサー要素は、プロモーターと本発明のポリペプチドをコードするヌクレオチド配列との間に配置されるイントロンでよい。例えば、Xu他, 1993, 前掲は、発現を亢進するためのイネアクチン1遺伝子の第1イントロンの使用を開示する。
【0156】
なお更に、コドン使用は、発現を改善するために問題の植物種について最適化され得る(上で言及したHorvath et al参照)。
【0157】
選択マーカー遺伝子及び発現構築物の任意の他の部分は、当該分野で入手可能なものから選択され得る。
【0158】
核酸構築物は、アグロバクテリウム(Agrobacterium)-介在形質転換、ウイルス-介在形質転換、マイクロインジェクション、微粒子銃、バイオリスティック形質転換、及びエレクトロポレーションを含む当該分野で公知の慣用的技術に従って、植物ゲノムに組み込まれる (Gasser et al., 1990, Science 244: 1293; Potrykus, 1990, Bio/Technology 8: 535; Shimamoto et al., 1989, Nature 338: 274)。
【0159】
現在、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)-介在遺伝子移動は、トランスジェニック双子葉植物を作製するための最適な方法であり(考察には、Hooykas and Schilperoort, 1992, Plant Molecular Biology 19: 15-38参照)、それは、単子葉植物を形質転換するためにも使用され得る。但し、他の形質転換法もこれらの植物のために通常使用される。現在、アグロバクテリウム法を補充する、トランスジェニック単子葉植物を作製するための最適な方法は、胚カルス又は発達中の胚の微粒子銃(形質転換DNAでコートされた微視的ゴールド又はタングステン粒子)(Christou, 1992, Plant Journal 2: 275-281; Shimamoto, 1994, Current Opinion Biotechnology 5: 158-162; Vasil他, 1992, Bio/Technology 10: 667-674)。単子葉植物の形質転換の別の方法は、Omirulleh他, 1993, Plant Molecular Biology 21: 415-428によって記載されたプロトプラスト形質転換に基づく。
【0160】
形質転換に続いて、発現構築物が組み込まれた形質転換体が選択され、そして当該分野で周知の方法に従って植物全体に再生される。形質転換法は、通常、例えば、2つの別個のT-DNA構築物による同時-形質転換、又は特定のリコンビナーゼによる選択遺伝子の部位特異的切除を用いて、再生の間又は次世代のいずれかにおいて選択遺伝子の選択的削除について設計されている。
【0161】
本発明はまた、(a) 該ポリペプチドの産生のための条件下で、本発明のフィターゼ活性を有するポリペプチドをコードする核酸配列を含むトランスジェニック植物又は植物細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、本発明のポリペプチドを製造する方法に関する。
【0162】
トランスジェニック動物
本発明はまた、トランスジェニックな、非-ヒト動物及びその生成物又は要素に関する。その例は体液、例えばミルク及び血液、臓器、食肉、及び動物細胞である。タンパク質、例えば哺乳動物におけるタンパク質を発現するための技術は、当該分野で知られている。例えば、ハンドブック「タンパク質発現: 実際のアプローチ」Higgins and Hames (著), Oxford University Press (1999)、及び「遺伝子転写、RNAプロセッシング、及び翻訳後プロセッシン具」に関連するシリーズにおける他の3つのハンドブックを参照。一般的には、トランスジェニック動物を製造するために、選択された動物の選択された細胞は、本発明のフィターゼ活性を有するポリペプチドを発現及び産生するように、該ポリペプチドをコードする核酸配列で形質転換される。該ポリペプチドは、動物から、例えば、雌の動物のミルクから回収され、又は該ポリペプチドは、動物自体のために、例えば動物の消化を助けるために発現され得る。動物の例は、「Animal Feed」という見出しに以下のように述べられている。
【0163】
動物のミルクから該ポリペプチドを回収することを目的として、トランスジェニック動物を製造するために、該ポリペプチドをコードする遺伝子は、例えば、好適なミルクタンパク質プロモーターを含む導入遺伝子発現ベクター、及び該ポリペプチドをコードする遺伝子の使用によって、問題の動物の孵化された卵に挿入され得る。導入遺伝子発現ベクターは、孵化された卵に微量注入され、そして、好ましくは永久に染色体に統合される。該卵は成長し分割を始めると、可能性のある胚は、代理母に移植され、導入遺伝子を担持する動物が同定される。次いで、得られた動物は、慣用的な繁殖によって作製され得る。
該ポリペプチドは、動物のミルクから精製され得る、例えば、Meade, H. M. et al (1999): トランスジェニック動物のミルク中の組換えタンパク質の発現、遺伝子発現系:発現技術のための性質を使用する, J. M. Fernandez及びJ. P. Hoeffler (著), Academic Press参照。
【0164】
別の方法では、その身体細胞及び/又は細菌細胞のゲノム中にポリペプチドをコードする導入遺伝子を含む非相同導入遺伝子構築物を含む核酸配列を有するトランスジェニック非-ヒト動物を製造するために、導入遺伝子は、WO 00/064247に開示された、ポリペプチドの唾液腺特異的発現のための第1調節配列に作動可能に連結され得る。
【0165】
組成物及び使用
なお更なる局面では、本発明は、本発明のオリゴヌクレオチドを含む組成物、及びこれらの使用方法に関する。
【0166】
ポリペプチド組成物は、当該分野で公知の方法に従って調製され、液状又は乾燥組成物の形態でよい。例えば、ポリペプチド組成物は、顆粒又は微顆粒の形態でよい。ポリペプチドに含まれるポリペプチドは、当該分野で公知の方法に従って安定化され得る。
【0167】
本発明のフィターゼは、任意の産業的状況において、例えば、フィチン酸塩、フィチン酸及び/又はmyo-イノシトールのモノ-、ジ-、トリ-、テトラ-及び/又はペンタ-リン酸の分解のために使用され得る。これらの化合物のリン酸部分は、二価及び三価カチオン、例えば金属イオン、例えば栄養学的に不可欠なカルシウム、鉄、亜鉛、マグネシウムイオン及び微量元素のマンガン、銅及びモリブデンに配位する、ことが知られている。更に、フィチン酸はまた、ある程度、静電気相互作用によってタンパク質に結合する。
【0168】
従って、本発明のポリペプチドの好ましい使用は、動物飼料調製物(ヒト用食品を含む)又はかかる調製物のために添加物においてである。
【0169】
具体的態様では、本発明のポリペプチドは、動物飼料の栄養価を改善するために使用され得る。動物飼料(ヒト食品を含む)の栄養価を改善する非限定的例は、以下である:飼料消化率の改善;動物の成長の促進;飼料利用の改善;タンパク質のバイオアベイラビリティーの改善;消化できるホスフェートレベルの増加;フィターゼの放出及び/分解の改善;微量元素のバイオアベイラビリティーの改善;マクロミネラルのバイオアベイラビリティーの改善;補助的ホスフェート、微量元素及び/又はマクロミネラルを添加する必要性の削減;及び/又は卵殻品質の改善。そのため、飼料の栄養価が増加され、そして動物の成長速度及び/又は体重増加及び/又は飼料変換率(すなわち、体重増加に対する摂取飼料の重さ)が改善され得る。
【0170】
更に、本発明のポリペプチドは、肥料のフィターゼレベルを減少させるために使用され得る。
【0171】
動物、動物飼料、及び動物飼料添加物
用語動物は、ヒトを含むすべての動物を含む。動物の例は、非-反芻動物及び反芻動物である。反芻動物は、例えば、ヤギ、ヒツジ、ウマ及びウシ、例えば肉牛及び子牛、のような動物を含む。具体的な実施態様では、動物は非-反芻動物である。非-反芻動物は、単胃動物、例えばブタ(pigs)又はブタ(swine)(子豚、成長した豚及び雌豚を含むがこれらに限定されない);家禽、例えば、七面鳥、アヒル及びニワトリ(若鶏、産卵鶏を含むがこれらに限定されない);子牛;及び魚(鮭、トラウト、テラピア、ナマズ及び鯉を含むがこれらに限定されない)並びに甲殻類(エビ及び淡水エビを含むがこれらに限定されない)を含む。
【0172】
用語飼料又は飼料組成物は、動物による摂取のために好適な又はそれを意図した、任意の化合物、調製物、混合物又は組成物である。
【0173】
本発明に従う使用において、ポリペプチドは、食事の前に、後に又は同時に動物に供給され得る。後者が好ましい。
【0174】
具体的な実施態様では、飼料に添加される、又は飼料添加物として含まれる時の形態でのポリペプチドは、実質的に純粋である。具体的な実施態様ではそれは明確である。用語「明確である」は、フィターゼ調製物がサイズ排除クロマトグラフィーによって決定された少なくとも50%であることを意味する (例えば、WO 01/58275の実施例12参照)。他の具体的な実施態様では、フィターゼ調製物は、この方法によって決定される、少なくとも60%、70、80、85、88、90、92、94又は少なくとも95%純粋である。
【0175】
実質的に純粋な及び/又は明確なポリペプチド調製物は有利である。例えば、本質的に他のポリペプチドの妨害のない又は汚染のないポリペプチド飼料に正確に投薬することが非常に容易である。用語投薬は、具体的には、一貫性のある結果及び一定の結果を得る目的、及び所望の効果に基づく投薬量の最適化の可能性を言う。
【0176】
しかし、動物飼料での使用については、本発明のフィターゼポリペプチドは、純粋である必要がない;例えば他のポリペプチドを含む、それは、フィターゼ調製物と称される。
【0177】
フィターゼ調製物は、(a) 飼料に直接的に添加され(又はタンパク質の処理過程において直接的に使用される)、又は (b) 飼料に実質的に添加される(又は処理過程において使用される)飼料添加物又はプレミックスのような1以上の中間組成物の製造において使用され得る。上記の純度の程度は、上記の (a) に従って使用されるにしろ又は (b) に従って使用されるにしろ、最初のポリペプチド調製物の純度を言う。
【0178】
このようなオーダーの純度を有するポリペプチド調製物は、具体的には、組換え製造方法を用いて得られる。一方、それらはそれ程簡便に得られず、またポリペプチドは典型的な発酵法によって製造される時に非常に高いバッチごとの変化に供される。
【0179】
かかるポリペプチド調製物は、当然のことながら、他のポリペプチドと混合され得る。
【0180】
上記ポリペプチドは、相対的に純粋なポリペプチドとして任意の形態で、又は動物飼料に添加することを意図した他の成分と混合して、すなわち、いわゆる動物飼料用プレミックスのような動物飼料添加物の形態で、加えられる。
【0181】
更なる局面では、本発明は、動物飼料及び動物飼料添加物、例えばプレミックスのよな動物飼料において使用される成分に関する。
【0182】
本発明のポリペプチドとは別に、本発明の動物飼料添加物は、少なくとも1つの脂溶解ビタミン、及び/又は少なくとも1つの水溶性ビタミン、及び/又は少なくとも1つの微量元素を含む。飼料添加物は、少なくともマクロミネラルを含んでもよい。
【0183】
更に、場合により、飼料-添加物成分は、着色剤、例えばカロチノイド、例えばβ-カロチン、アスタキサンチン及びルテイン;芳香化合物;安定剤;抗菌ペプチド;ポリ不飽和脂肪酸;反応酸素生成種;及び/又はフィターゼ (EC 3.1.3.、又は3.1.3.26); キシラナーゼ (EC 3.2.1.8); ガラクタナーゼ (EC 3.2.1.89); α-ガラクトシダーゼ (EC 3.2.1.22); プロテアーゼ (EC 3.4.-.- )、ホスホリパーゼA1 (EC 3.1.1.32); ホスホリパーゼA2 (EC 3.1.1.4); リゾホスホリパーゼ (EC 3.1.1.5); ホスホリパーゼC (3.1.4.3); ホスホリパーゼD (EC 3.1.4.4); アミラーゼ、例えばα-アミラーゼ (EC 3.2.1.1); 及び/又はβ-グルカナーゼ (EC 3.2.1.4 or EC 3.2.1.6) から選ばれる少なくとも1つの他のポリペプチドである。
【0184】
具体的実施態様では、これらの他のポリペプチドは明確である(フィターゼ調製物について上で定義されたように)。
【0185】
具体的な好ましい実施態様では、比較的低い最適pHを有する本発明のフィターゼは、高い最適pHを有する少なくとも1つのフィターゼと組合わされる。高い最適pHのフィターゼの好ましい例は、バチルスフィターゼ、例えばバチルス・リケニフォルミス及びバチルス・サブティリス、並びにフィターゼ活性を有するその誘導体、変異体又はフラグメントである。
【0186】
本発明のフィターゼは、他のフィターゼ、例えば、子嚢菌フィターゼ、例えばアスペルギルス属から得られた、例えばペニオホラ・リシイ(Peniophoralycii)、アグロシベ・ペディアデス(Agrocybe pediades)、トラメテス・ピューベセンス・フィカム(Trametes pubescens ficuum)、アスペルギルス・ニガー(Aspergillus niger)、もしくはアスペルギルス・アワモリ(Aspergillus awamori)から得られたアスペルギルス・フィターゼ;あるいは担子菌フィターゼ又はパキシラス・インボルタス(Paxillus involutus)、又はフィターゼ活性を有するその誘導体、フラグメントもしくは変異体と組合わせてもよい。
【0187】
従って、本発明の動物飼料における使用の好ましい実施態様、及び本発明の動物飼料添加物及び動物飼料の好ましい実施態様では、本発明のフィターゼはかかるフィターゼと組合わされる。
【0188】
上記の子嚢菌類の植物及び担子菌のフィターゼ、特にペニオホラ・リシイ(Peniophora lycii)由来のRONOZYME P フィターゼ、及びその誘導体、変異体及びフラグメントは、動物飼料目的で、バチルス・フィターゼ、特に、B. リケニフォルミス(licheniformis)・フィターゼ及びその誘導体、フラグメント又は変異体と組み合わせてもよい。
【0189】
抗菌ペプチド(AMP)の例は、WO 03/044049及びWO 03/048148に開示された化合物及びポリペプチドを含む、CAP18、ロイコシンA、トリトリプシン、プロテグリン-1、サナチン、デフェンシン、ラクトフェリン、ラクトフェリシン及びオビスピリン、例えばノビスピリン (Robert Lehrer, 2000)、プレクタシン、及びスタチン、並びに抗菌活性を保持する上記の変異体又はフラグメントである。
【0190】
抗真菌ポリペプチド(AFP)の例は、アスペルギルス・ギガンテウス(Aspergillus giganteus)及びアスペルギルス・ニガー(Aspergillus niger)プロモーター、並びにWO 94/01459及びWO 02/090384に開示された抗真菌活性を保持するその変異体及びフラグメントである。
【0191】
ポリ不飽和脂肪酸の例は、C18、C20及びC22ポリ不飽和脂肪酸、例えばアラキドン酸、ドコサヘキサン酸、エイオコサペンタエン酸及びγ-リノール酸である。
【0192】
反応性酸素生成種の例は、化学物質、過ホウ酸塩、過硫酸塩又は過炭酸塩;及びポリペプチド、例えばオキシダーゼ、オキシゲナーゼ又はシンテターゼである。
【0193】
通常、脂溶性及び水溶性ビタミン、並びに微量元素は、飼料への添加を意図したいわゆるプレミックスの一部を形成するが、マクロミネラルは、通常、飼料に別個に添加される。これらの組成物種のいずれかは、本発明のポリペプチドが豊富に存在する場合には、本発明の動物飼料添加物である。
【0194】
具体的な実施多様では、本発明の動物飼料添加物は、0.01〜10.0%;より具体的には0.05〜5.0%;又は0.2〜1.0%(%は、添加物g/100 g飼料を意味する)のレベルで動物食品又は飼料中に含まれる(又は含まれていると指示されている)ことを意図する。このことは、具体的にはプレミックスに当てはまる。
【0195】
以下は、これらの成分の例の非限定的リストである。
【0196】
脂溶性ビタミンの例は、ビタミンA、ビタミンD3、ビタミンE及びビタミンK、例えば微ビタミンK3である。
【0197】
水溶性ビタミンの例は、ビタミンB12、ビオチン及びコリン、ビタミンB1、ビタミンB2、ビタミンB6、ナイアシン、葉酸、及びパントテン酸、例えばCa-D-パントテン酸である。微量元素の例は、マンガン、亜鉛、鉄、銅、ヨウ素、セレニウム、及びコバルトである。
【0198】
マクロミネラルの例は、カルシウム、リン及びナトリウムである。
【0199】
これらの成分の栄養的要件(家禽及び子豚/豚で代表される)は、WO 01/58275の表Aに記載されている。栄養的要件は、これらの成分が示された濃度で食品中に提供されるべきであることを意味する。
【0200】
あるいは、本発明の動物飼料添加物は、WO 01/58275の表Aで特定された個々の成分の少なくとも1つを含む。少なくとも1つは、13個の成分の内の1個以上、1個又は2個又は3個又は4個、最大13個全て、あるいは最大15個の個々の成分すべてを意味する。より具体的には、この少なくとも1つの個々の成分は、表Aのカラム4又はカラム5又はカラム6に示された範囲内の飼料中の濃度を提供するような量で、本発明の添加物中に含まれる。
【0201】
本発明は、動物飼料組成物にも関する。動物飼料組成物又は食品は比較的高いタンパク質量を有する。家禽及び豚の食品は、WO 01/58275の表Bのカラム2〜3に示されるように特徴付けられる。魚食品は、この表Bのカラム4中に示されるように特徴付けられる。更に、かかる魚食品は、通常、200〜310 g/kgの粗脂肪量を有する。
【0202】
WO 01/58275は、本明細書に参考として援用される米国出願第09/779334号に対応する。
【0203】
本発明に従う動物飼料組成物は、50〜800 g/kgの粗タンパク質量を有し、更に本願で請求された少なくとも1つのポリペプチドを含む。
【0204】
更に又はあるいは(上記の粗タンパク質量の代わりに)、本発明の動物飼料組成物は、10〜30 MJ/kgの代謝されるエネルギー量; 及び/又は0.1〜200 g/kgのカルシウム量; 及び/又は0.1〜200 g/kgの利用できるリンの量; 及び/又は0.1〜100 g/kgのメチオニン量; 及び/又は0.1〜150 g/kgのメチオニン及びシステイン量; 及び/又は0.5〜50 g/kgのリジン量を有する。
【0205】
具体的な実施態様では、代謝性エネルギー、粗タンパク質、カルシウム、リン、メチオニン、メチオニン及びシステイン、及び/又はリジンの量は、WO 01/58275の表B内のレンジ2、3、4又は5のいずれか1つにある (R. 2-5)。
【0206】
粗タンパク質は、ファクター6.25を乗じた窒素(N)として計算される、すなわち、粗タンパク質 (g/kg)= N (g/kg) x 6.25。窒素量は、Kjeldahl法 (A.O.A.C, 1984, Official Methods of Analysis 第14th版, Association of Official Analytical Chemists, Washington DC) により決定される。
【0207】
代謝エネルギーは、NRC刊行ブタの栄養要件, 第9改定版 1988, ブタ栄養についての分科委員会, 動物栄養についての委員会, 農業委員会, 米国学術研究会議に基いて計算される。National Academy Press, Washington, D. C, 第2〜6頁、及びブタ飼料のネエルギー価についての欧州表, ブタ研究及び講座のSpelderholtセンター, 7361 DA Beekbergen, The Netherlands. Grafisch bedrijf Ponsen & looijen bv, Wageningen. ISBN 90-71463-12-5。
【0208】
完全な動物食品中のカルシウム、入手できるリン及びアミノ酸の所要量は、飼料表、例えばVeevoedertabel 1997, gegevens over chemische samenstelling, verteerbaarheid en voederwaarde van voedermiddelen, Central Veevoederbureau, Runderweg 6, 8219 pk Lelystad. ISBN 90-72839-13-7に基いて計算される。
【0209】
具体的実施態様では、本発明の動物飼料組成物は、少なくとも1つのタンパク質を含む。該タンパク質は、動物タンパク質、例えば肉及び骨食品、及び/又は魚食品でよく;あるいは、野菜タンパク質でよい。本明細書で使用される用語野菜タンパク質は、任意の化合物、組成物、調製物又は混合物を言う。修飾タンパク質及びタンパク質誘導体を含む、野菜由来の又は野菜から生じる少なくとも1つのタンパク質を含む。具体的な実施態様では、野菜タンパク質のタンパク質量は、少なくとも10、20、30、40、50又は60% (w/w) である。
【0210】
野菜タンパク質は、野菜タンパク質源、例えばマメ科植物及び穀物、例えばファバセアエ(Fabaceae (レグミノサエ(Leguminosae))、クルシフェラセアエ(Cruciferaceae)、ケノポディアセアエ(Chenopodiaceae)及びピアース(Poaceae)のファミリーの植物由来の物質、例えば、大豆食品、ルピナス(lupin)食品及びマメ科植物食品から得られる。
【0211】
具体的な実施態様では、野菜タンパク質源は、ファバセアファミリーの1以上の植物、例えば、大豆、ルピナス、ナシ又は豆由来の物質である。
【0212】
別の具体的な実施態様では、野菜タンパク質源は、ケノポディアセアエファミリーの1以上の植物、例えば、ビート、砂糖大根、ホウレンソウ又はキノア、由来の物質である。野菜タンパク質源の他の例は、菜種、ヒマワリ種、綿花種及びキャベツである。
【0213】
大豆は、好ましい野菜タンパク質源である。
【0214】
野菜タンパク質源の他の例は、穀物、例えば、大麦、小麦、ライ麦、オート麦、トウモロコシ(コーン)、コメ、ライ小麦及びサトウモロコシである。更なる具体的実施態様では、本発明の動物飼料組成物は、0〜80%のトウモロコシ; 及び/又は0〜80%のサトウモロコシ; 及び/又は0〜70%の小麦; 及び/又は0〜70%の大麦; 及び/又は0〜30%のオート麦; 及び/又は0〜40%の大豆食品; 及び/又は0〜25%の魚食品; 及び/又は0〜25%の肉及び骨食品; 及び/又は0〜20%のホエイを含む。
【0215】
動物食品は、例えば、マッシュ食品(非ペレット化)又はペレット化食品として製造される。典型的には、粉砕された飼料材料は混合され、そして、必須ビタミン及びミネラルの充分量は問題の種の仕様書に従って添加される。ポリペプチドは、固体又は液状ポリペプチド製剤として添加され得る。例えば、固体ポリペプチド製剤は、典型的には、混合ステップの前又は中に加えられ;そして、液状ポリペプチド調製物は、典型的には、ペレットステップの後に加えられる。ポリペプチドは、飼料添加物又はプレミックス中に組み込まれてもよい。
【0216】
食品中の最終ポリペプチド濃度は、0.01〜200 mgポリペプチド/kg食品の範囲内にある。例えば、0.1〜10 mg/kg動物食品の範囲(典型的な投薬量は、250〜2000 FYT/kg動物食品の範囲内である)にある。
【0217】
本発明のフィターゼは、有効量、すなわち、飼料の溶解性を改善するために及び/又は栄養価を改善するためい充分な量で適用されるべきである。
【0218】
現在では、上記ポリペプチドは、以下の量(投薬範囲)の1以上で投与されることが画される:0.01〜200; 0.01〜100; 0.5〜100; 1〜50; 5〜100; 10〜100; 0.05〜50; 又は0.10〜10、これらの範囲はすべて、mgフィターゼポリペプチド/kg飼料(ppm)である。
【0219】
mgフィターゼポリペプチド/kg飼料を決定するために、フィターゼは、飼料組成物から精製され、精製されたフィターゼの特異的活性は関連するアッセイを用いて決定される。よって、飼料組成物のフィターゼ活性は、同一のアッセイを用いて決定され、これらの2つの決定に基いて、mgフィターゼポリペプチド/kg飼料での投薬量が計算される。
【0220】
mgフィターゼポリペプチド/kg飼料添加物を決定するために同一の原則が適用される。当然のことながら、試料が飼料添加物又は飼料を調製するために使用されるフィターゼが利用できる場合には、特異的活性は、この試料から決定される(飼料組成物又は添加物からフィターゼを精製する必要性はない)。
【0221】
発酵産物の製造方法
本発明の更に別の局面は、発酵産物、例えばエタノール、ビール、ワイン、蒸留乾燥種子(DDG)、の製造方法であって、該発酵が本発明のフィターゼの存在下で行われる、前記方法に関する。発酵プロセスの例は、例えば、WO01/62947に記載のプロセスを含む。発酵は、発酵微生物、例えば酵母を用いて行われる。
【0222】
具体的な実施態様では、本発明は、発酵産物の製造方法であって、(a) 本発明のフィターゼの存在下で、糖類含有物質(例えば、スターチ)を(発酵微生物、例えば酵母を用いて)発酵し、及び (b) 発酵糖類含有物質から発酵産物を製造すること、を含む、前記方法を提供する。
【0223】
具体的な実施態様では、本発明は、エタノールを製造するための方法であって、本発明のフィターゼの存在下で、糖類含有物質(例えば、スターチ)を(発酵微生物、例えば酵母を用いて)発酵し、及び (b) 発酵糖類含有物質からエタノールを製造又は回収すること、を含む、前記方法を提供する。
【0224】
別の実施態様では、本発明は、エタノールの製造方法であって、a) 例えば、液化及び/又は糖化プロセス、スターチ原料加水分解プロセスによる、加水分解スターチ、b) 本発明のフィターゼの存在下で得られたスターチを発酵させ、及び c) エタノールを製造すること、を含む、前記方法を提供する。
【0225】
フィターゼは、単独で、又は他の酵素、例えば1以上のα-アミラーゼ、グルコアミラーゼ、プロテアーゼ及び/又はセルラーゼと組み合わせて、任意の好適な段階で及び任意の好適な組成物中に、発酵プロセスに添加され得る。
【0226】
別の実施態様では、本発明は、バイオマスを加水分解し、及び(発酵微生物、例えば酵母を用いて)、本発明のフィターゼの存在下で得られたバイオマスを発酵させることを含む、エタノール製造方法を提供する。
【0227】
シグナルペプチド
本発明はまた、配列番号9のヌクレオチド1〜99からなる第1ヌクレオチド配列に作動可能に連結したタンパク質をコードし、配列番号10のアミノ酸1〜33からなるシグナルペプチドをコードする遺伝子を含み、該遺伝子が第1ヌクレオチド配列に無関係である、核酸構築物に関する。
【0228】
本発明はまた、かかる核酸構築物を含む、組換え発現ベクター及び組換え宿主細胞に関する。
【0229】
本発明はまた、タンパク質の製造法であって、(a) 該タンパク質の製造に好適な条件下でかかる組換え宿主細胞を培養し;及び (b) 該タンパク質を回収すること、を含む前記方法に関する。
【0230】
前記第1ヌクレオチド配列は、他の制御配列と共に又は他の制御配列と組み合わせて、個々に外来遺伝子に作動可能に連結され得る。かかる他の制御配列は、前掲である。該タンパク質は、天然でも又は宿主細胞に異種でもよい。用語「タンパク質」は、本明細書では、コードされた産物の特定の長さを意味せず、そのため、ペプチド、オリゴヌクレオチド及びタンパク質を包含する。用語「タンパク質」は、コードされた産物を形成するように組合わされた2以上のポリペプチドも包含する。該タンパク質は、少なくとも2つの異なったタンパク質、ここでその1以上が宿主細胞に異種又は天然である、から得られた部分的又は完全なポリペプチド配列の組合せを含むハイブリッドポリペプチドも含む。タンパク質は、上記のタンパク質及びハイブリッドタンパク質の天然アレル変異体及び人工変異体を更に含む。
【0231】
好ましくは、前記タンパク質は、ホルモン又はその変異体、ポリペプチド、例えば酵素、受容体又はその部分、抗体又はその部分、あるいはレポーターである。より好ましい局面では、該タンパク質は、酸化還元酵素、転移酵素、加水分解酵素、リアーゼ、イソメラーゼ又はリガーゼである。より更に好ましい局面では、該タンパク質は、アミノペプチダーゼ、アミラーゼ、カルボハイドラーゼ、カルボキシペプチダーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、シクロデキストリン・グリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α-ガラクトシダーゼ、β-ガラクトシダーゼ、グルコアミラーゼ、α-グルコシダーゼ、β-グルコシダーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチン質分解ポリペプチド、ペルオキシダーゼ、フィターゼ、ポリフェノールオキシダーゼ、タンパク質分解ポリペプチド、リボヌクレアーゼ、トランスグルタミナーゼ又はキシラナーゼである。
【0232】
遺伝子は、任意の原核生物、真核生物又は他の起源から得られる。
【0233】
様々な実施態様
以下は、本発明の更なる実施態様である。また、特許請求されたすべての、核酸、核酸構築物、組換え発現ベクター、組換え宿主細胞、前記ポリペプチドの製造法、トランスジェニック植物及び動物、並びに様々な使用、使用方法及び飼料組成物/添加物に関する対応する局面が本明細書に含まれる。
【0234】
フィターゼ活性を有し、37℃、0.1 M グリシン/HClバッファーpH 2.0中で24時間のインキュベーション後に、時間t = 0での活性と比べて少なくとも20%の残余活性を有する、単離されたポリペプチドであって、該活性は、0.25 M Na-酢酸バッファーpH 5.5、ブラインドを除いたバッファーを用いて、1% (w/v) Na-フィターゼで、37℃及びpH 5.5でアッセイされ;
好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0235】
フィターゼ活性を有し、37℃、0.1 M グリシン/HClバッファーpH 2.5中で24時間のインキュベーション後に、時間t = 0での活性と比べて少なくとも20%の残余活性を有する、単離されたポリペプチドであって、該活性は、0.25 M Na-酢酸バッファーpH 5.5、ブラインドを除いたバッファーを用いて、1% (w/v) Na-フィターゼで、37℃及びpH 5.5でアッセイされ;好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0236】
フィターゼ活性を有する単離されたポリペプチドであって、基質pNP-ホスホネートで測定されたpH 5.0及び37℃での該ポリペプチドの活性が、基質フィターゼで測定されたポリペプチドの活性の11%未満である;好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0237】
フィターゼ活性を有する単離されたポリペプチドであって、該ポリペプチドは、好ましくは、インビトロモデルで測定されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べてリン(P)の高い放出を有し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0238】
フィターゼ活性を有する単離されたポリペプチドであって、0.25 FYT/g飼料で投薬された該ポリペプチドは、0.25 FYT/g飼料で投薬されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べて少なくとも150%のリン(P)を放出し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0239】
フィターゼ活性を有する単離されたポリペプチドであって、0.75 FYT/g飼料で投薬された該ポリペプチドは、0.75 FYT/g飼料で投薬されたペニオホラ・リシイ(Peniophora lycii)由来のフィターゼと比べて少なくとも150%のリン(P)を放出し;及び/又は該ポリペプチドは、好ましくは、i) 配列番号10のアミノ酸1〜413と、少なくとも75%、好ましくは少なくとも76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%又は少なくとも99.9%の同一性を有する。
【0240】
I. (a) (i) 配列番号10のアミノ酸1〜413、及び/又は (ii) 配列番号10の成熟ポリペプチド部分と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチド、(b) (i) 配列番号9のヌクレオチド100〜1338、(ii)配列番号9の成熟ポリペプチドコーディング部分、及び/又は (iii) (i) 又は (ii) のいずれか1つの相補鎖と、少なくとも中緊縮条件下でハイブリダイズするポリヌクレオチドによってコードされるポリペプチド; (c) 1以上のアミノ酸の保存的置換、削除及び/又は挿入を含む、(a)(i)〜(a)(ii) のポリペプチドのいずれか1つの変異体;及び (d) (a)(i)〜(a)(ii) のポリペプチドのいずれか1つのフラグメント、からなる群より選ばれる、ファターゼ活性を有する単離されたポリペプチド。
【0241】
II. セクションIのポリペプチドをコードするヌクレオチド配列を含む単離されたポリヌクレオチド。
【0242】
III. 以下:(a) 配列番号10のアミノ酸1〜413と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有する、アミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;(b) 配列番号9のヌクレオチド100〜1338と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有するポリヌクレオチド;及び (c) (i) 配列番号9のヌクレオチド100〜1338、(ii) 配列番号9の成熟ポリペプチドコーディング部分、(iii) (i) 又は(ii) のいずれか1つの相補鎖と少なくとも中緊縮条件下でハイブリダイズするポリヌクレオチド、からなる群より選ばれるフィターゼ活性を有するポリペプチドをコードする単離されたポリヌクレオチド。
【0243】
IV. 配列番号9の成熟ポリペプチドコーディング配列において少なくとも1つの変異を有する、セクションII及びIIIのいずれか1つの単離されたポリヌクレオチドであって、該成熟ヌクレオチド配列が配列番号10のアミノ酸1〜413を含むポリペプチドをコードする、前記ポリヌクレオチド。
【0244】
V. 発現宿主においてポリペプチドの製造を指向する1以上の制御配列に作動可能に連結されたセクションII〜IVのいずれか1つのポリペプチドを含む核酸構築物。
【0245】
VI.セクションVの核酸構築物を含む組換え発現ベクター。
【0246】
VII.セクションVの核酸構築物を含む組換え宿主細胞。
【0247】
VIII. (a) ポリペプチドの製造のための条件下で、その野生型において該ポリペプチドを製造することができる細胞を培養し;及び (b) 該ポリペプチドを回収すること、を含む、セクションIのポリペプチドの製造方法。
【0248】
IX. (a)ポリペプチドの製造のための条件下で、ポリペプチドをコードする核酸配列を含む核酸構築物を含む組換え宿主細胞を培養し、及び (b) 該ポリペプチドを回収すること、を含む、セクションIのポリペプチドの製造方法。
【0249】
X. セクションIのポリペプチドをコードするポリヌクレオチドで形質転換された、トランスジェニック植物、植物部分又は植物細胞。
【0250】
XI. セクションIのポリペプチドを発現することができる、トランスジェニック非-ヒト動物、又はその産物、又はその要素。
【0251】
XII. 動物飼料におけるセクションIの少なくとも1つのポリペプチドの使用。
【0252】
XIII. 動物飼料における使用のための組成物の調製における、セクションIの少なくとも1つのポリペプチドの使用。
【0253】
XIV. セクションIの少なくとも1つのポリペプチドが飼料に添加される、動物飼料の栄養価を改善するための方法。
【0254】
XV. (a) セクションIの少なくとも1つのポリペプチド;(b) 少なくとも1つの脂溶性ビタミン;(c) 少なくとも1つの水溶性ビタミン、及び/又は少なくとも1つの微量元素、を含む動物飼料添加物。
【0255】
XVI. 少なくとも1つのアミラーゼ、少なくとも1つの更なるフィターゼ、少なくとも1つのキシラナーゼ、少なくとも1つのガラクタナーゼ、少なくとも1つのα-ガラクトシダーゼ、少なくとも1つのプロテアーゼ、少なくとも1つのホスホリパーゼ、及び/又は少なくとも1つのβ-グルカナーゼを更に含む、セクションXVの動物飼料添加物。
【0256】
XVII. 更なるフィターゼが配列番号10のアミノ酸1〜413のアミノ酸配列を有するポリペプチドの最適pHよりも高い最適pHを有する、セクションXVIの動物飼料添加物。
【0257】
NXX. 50〜800 g/kgの粗タンパク質量を有し、かつセクションIの少なくとも1つのポリペプチドを含む、動物飼料組成物。
【0258】
配列番号10のアミノ酸1〜413と少なくとも75%の同一性、好ましくは76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、96.5%、97%、97.5%、98%、98.5%、98.7%、98.8%、98.9%、99%、99.0%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%の同一性、又は少なくとも99.9%の同一性を有する、アミノ酸配列を含む、好ましくはそれを有する又はそれからなる、フィターゼ活性を有するポリペプチド。
【0259】
(i) 配列番号10のアミノ酸1〜413、及び/又は
(ii) 配列番号10の成熟ポリペプチド部分; 又は
(a) 1以上のアミノ酸の削除、挿入及び/又は保存的置換を含む、(i)〜(ii)のポリペプチドのいずれか1つの変異体であるポリペプチド;又は
(b) (i)〜(ii)のポリペプチドのいずれか1つのフラグメント
の配列を含み、好ましくは有する、フィターゼ活性を有するポリペプチド。
【0260】
本発明は、本発明の範囲を限定するものと解釈してはならない、以下の実施例によって更に記載される。
【実施例】
【0261】
実施例 1. ハフニア・アルベイ・フィターゼ遺伝子のクローニング
複数の配列を以下のヒスチジン酸ホスファターゼ(HAP)から作製した:appA エシェリキア・コリ(Escherichia coli)(SPTREMBLQ8GN88)、シトロバクタ・ギレニイ(Citrobacter gillenii)DSM 13694フィターゼ (geneseqp:aehO4533)、シトロバクタ・アマロナテカス(Citrobacter amalonaticus)ATCC 25407 フィターゼ (geneseqp:aehO4535)、シトロバクタ・ブラアキィ(Citrobacter braakii)フィターゼ (geneseqp:aehO4827)、及びypo1648エルシニア・ペスティス(Yersinia pestis)CO92 (SPTREMBLQ8ZFP6)。2つの縮退オリゴヌクレオチドプライマーは、コンセンサス配列に基いてデザインした:
2123fw: 5'-CATGGTGTGCGNGCNCCNACNAA-3' (配列番号1)
2065rev: 5'-CCCACCAGGNGGNGTRTTRTCNGGYTG-3' (配列番号2)、
ここで、YはT又はCを示し、RはA又はGを示し、そしてNはA、C、G又はTを示す。
【0262】
プライマーは、40℃〜50℃のアニーリング温度で多数の細菌種のPCRスクリーニングに使用した。但し、典型的には、標準的なPCRを行う前に次の10サイクルに渡って、50℃で開始して、次いで各サイクルで1℃ずつアニーリング温度を減少させるタッチダウンプログラムによって、使用した。
【0263】
PCRフラグメントによって約950 bpの形態で部分的フィターゼ遺伝子をハフニア・アルベイ (DSM 19197) において同定した。
【0264】
PCRフラグメントをアガロースゲルから単離し、該フラグメントを、該フラグメントを作製するのに使用したものと同一のPCRプライマーを用いてシークエンスした。ヌクレオチド配列の翻訳によって、DNAフラグメントがHAPフィターゼ遺伝子の一部であることが確認された。
【0265】
上記遺伝子の完全長ヌクレオチドを得るために、未知の標的部位を獲得するようにデザインされている、DNA WALKINGSPEEDUP(登録商標)キット (DWSK-V102 from Seegene, Inc., 2nd Fl., Myungji Bldg., 142-21, Samsung-dong, Kangnam-gu, Seoul, 135-090, Korea) を使用した。この目的のために、6個の特定のオリゴヌクレオチドは該キットを用いてデザインした。
【0266】
2328 TSP1dw: 5'-ACTTGCATCGACGTTGGCTG (配列番号3)
2329 TSP2dw: 5'-ACTGAG CAG CAATG GAACTCTCTG (配列番号4)
2330 TSP3dw: 5'-ACTGGGTTCCAATATCACGAGTC (配列番号5)
2331 TSP1up: 5'-ATGGTGGATCGCTAAATCACACTG (配列番号6)
2332 TSP2up: 5'-ACGTCTGCCCAAACATACACG (配列番号7)
2333 TSP3up: 5'-ACCGCCCATCAGGCTAATC (配列番号8)
【0267】
ハフニア・アルベイ DSM 19197由来のフィターゼをコードする完全長ヌクレオチド配列は、配列番号9として表される配列に示され、対応するコードされたアミノ酸配列は、配列番号10として示される。配列番号10の最初の33個のアミノ酸(すなわち、アミノ酸-33〜-1)は、ソフトウェアSignal P V3.0 (www.cbs.dtu.dk/services/SignalP/参照) によって予測されたシグナルペプチドである。
【0268】
実施例 2. ハフニア・アルベイ・フィターゼ遺伝子の発現
バチルス・リケニフォルミス(Bacillus licheniformis)由来の天然プロテアーゼ、Savinase(登録商標)の27アミノ酸シグナルペプチドコーディングポリヌクレオチドを、ハフニア・アルベイ由来の成熟フィターゼをコードする遺伝子にインフレームでPCPによって融合した。シグナンルペプチドコーディング配列は、配列番号12のシグナルペプチドをコードする配列番号11で示される。
【0269】
融合ポリペプチドをコードするDNAを、バチルス・サブティリス宿主細胞ゲノム上で相同性組換えによって統合した。遺伝子構築物は、バチルス・リケニフォルミス(Bacillus licheniformis)・α-アミラーゼ遺伝子 (amyL)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)・α-アミラーゼ遺伝子 (amyQ)、及びmRNA安定配列を含むバチルス・チューリンゲンシス(Bacillus thuringiensis)cryIIIAプロモーターからなる、三重プロモーター系(WO 99/43835に記載)の制御下で発現した。クロラムフェニコールアセチル-トランスフェラーゼをコードする遺伝子は、例えば、Diderichsen他, バチルス・サブティリス用の有用なクローニングベクター,プラスミド, 30, 第312頁, 1993に記載されているように、マーカーとして使用された。
【0270】
30℃で250 RPMで振とうされたPS-1培地(10%スクロース、4%大豆粉、1% Na3PO4-12H2O、0.5% CaCO3及び0.01 % プルロニック酸)中で、クロラムフェニコール抵抗性形質転換体を培養した。インキュベーションの2〜5日後、上清を除き、そして、0.1 M酢酸ナトリウム(pH 4.5)及び0.1%イノシトール六リン酸を含む1 % LSBアガロースプレート中で、20ミリリットルの上清を、打ち抜かれた4 mm径の孔に適用することによって、フィターゼ活性を同定した。プレートを37℃で終夜静置し、0.25 M CaCl2及び500 mM MES (4N NaOHでpH 6.5に調整) からなるバッファーをプレートに注いだ。該プレートを室温で1時間静置し、次いで、イノシトールリン酸ホスファターゼ又はフィターゼの活性を透明ゾーンとして同定した。
【0271】
数個のフィターゼ陽性形質転換体は、DNAシークエンシングによって分析して、構築物の正確なDNA配列を確立した。1つの正確なクローンを選択した。
【0272】
実施例 3 ハフニアNN020125フィターゼ宿主の発酵
ハフニア・アルベイ フィターゼ構築物を有しそして該フィターゼ(成熟部分)を発現することができる、バチルス・サブティリスの選択されたクローンを、SK-1 M培地 (カゼイン酸ナトリウム (Arla製Miprodan 30) 40 g、マルトデキストリン01 (Roquette製Glucidex 6, カタログ番号332203) 200 g、ダイズミール50 g、Dowfax 63N10 (Dow製非イオン性界面活性剤) 0.1 mlを含み、水道水で1000 mlとした、CaCO3錠剤0.5 g/100 ml) 中で、30℃で、6日間、250rpmで攪拌しながら培養した。
【0273】
実施例 4 ハフニア・アルベイ・フィターゼの精製
該フィターゼによる発酵上清を最初に、7200 rpm及び5℃で1時間遠心し、4つのワットマンガラスマイクロチューブフィルターのサンドウィッチでろ過した(2.7、1.6、1.2及び0.7μm)。次いで、圧力を用いて、Seitz-EKSデプスフィルターによって、この溶液を殺菌ろ過した。次いで、ろ過された上清を以下のように予備処理した。
【0274】
試料溶液を水で洗浄し、1 0 kDaカットオフ限界濾過膜を備えた限界濾過ユニット (Filtron, Filtron Technology Corporation製) を用いて濃縮した。次いで、pHを10% (w/v) 酢酸で4.5に調整した。これは、少量の沈殿を生じた。該沈殿には活性は見られなかった、0.22μmカットオフを有するFast PESボトル先端部フルターで濾過することによって沈殿を除いた。
【0275】
予備処理の後、バッファーAとして50 mM酢酸ナトリウム(pH 4.5)、及びバッファーBとして50 mM酢酸ナトリウム+ 1 M NaCl(pH 4.5)を用いて、XK26カラム中の約50 mlのSセファロース上でクロマトグラフィーによってフィターゼを精製した。ホスファターゼアッセイ(以下参照)を用いてカラムからのフラクションを活性について分析し、活性のあるフラクションを回収した。
【0276】
上記溶液を最終濃度1.5 Mとなるように固体硫酸アンモニウムに加え、6 M HClを用いて6.0にpHを調整した。フィターゼ含有溶液を、バッファーAとして25 mM ビス-トリス (ビス-(2-ヒドロキシエチル)イミノ-トリス(ヒドロキシメチル)メタン)) + 1.5 M 硫酸アンモニウム(pH 6.0)、及びバッファーBとして25 mM ビス-トリス(pH 6.0)を用いて、XK26カラム中約30 mlで、ブチル-セファロースカラムに適用した。カラムからのフラクションを、ホスファターゼアッセイ(以下参照)を用いて活性について分析し、回収した。最後に、精製されたフィターゼを含む溶液を50 mM酢酸ナトリウム + 0.1 M NaCl(pH 4.5)に緩衝液チャージし、30 kDaカットオフ膜を有するアミコンウルトラ-15濾過装置を用いて濃縮した。
【0277】
SDS-PAGEから見積もった分子量は、約40 kDaであり、純度は>95%であった。
【0278】
実施例 5 活性アッセイ
ホスホターゼ活性の決定
75μlのフィターゼ含有酵素溶液をマイクロタイタープレートウェル、例えばNUNC 269620に分注し、75μlの基質を加えた(基質を調製するために、2つの5 mg p-ニトロフェニルホスフェート錠剤(Sigma, カタログ番号N-9389)を10 ml 0.1 M 酢酸Naバッファー(pH 5.5)に溶解した)。該プレートを遮光し、15分間インキュベートし、37℃で750 rpmで振とうした。インキュベーション後、75μlの停止試薬を加え(停止試薬は、0.1 M 四ホウ酸二ナトリウム水溶液)、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。
【0279】
フィターゼ活性の決定
0.25 M酢酸ナトリウム、0.005% (w/v) Tween-20(pH 5.5)でおおよそ希釈した、75μlのフィターゼ含有酵素溶液を、マイクロタイタープレートウェル、例えばNUNC 269620に分注し、75μlの基質を加えた(コメ由来の100 mgフィチン酸ナトリウム(Aldrichカタログ番号274321)の10 ml 0.25 M酢酸ナトリウムバッファー(pH 5.5)によって溶解して調整した)。該プレートを遮光し、15分間インキュベートし、37℃で750 rpmで振とうした。インキュベーション後、75μlの停止試薬を加え(停止試薬は、10 mlのモリブデン酸塩溶液(10% (w/v) モリブデン酸アンモニウムの0.25% (w/v) アンモニア溶液)、10 ml バナジン酸アンモニウム(Bie&Berntsen, カタログ番号LAB17650製の、0.24%の商業的製品)及び20 ml 21.7% (w/v) 硝酸)、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。フィターゼ活性はFYT単位で表し、1FYTは、上記の条件下で、1μmol無機オルトリン酸塩/分を遊離する酵素の量である。測定されたフィターゼ活性の絶対値は、無機リン酸塩の好適な希釈から調製された標準曲線を参考に、又は公知の活性を有するフィターゼ酵素調製物(公知の活性を有するかかる標準的な酵素調製物は、請求により、Novozymes A/S, Krogshoejvej 36, DK-2880 Bagsvaerdから入手可能である)の希釈から作成された標準曲線を参考に、得られる。
【0280】
特異的フィターゼ活性の決定
酢酸ナトリウムバッファー(pH 5.5)中でフィターゼの特異的活性を測定した。該フィターゼは上記のように高度に精製した、すなわち、1成分のみをSDSポリアクリルアミドゲルで精製した。
【0281】
アミノ酸分析によって以下のようにタンパク質濃度を測定した:試料のアリコートを、空のガラスチューブ中で、110℃で16時間、6 M HCl、0.1% フェノールで加水分解した。製造者の教示に従って操作したApplied Biosystems 420Aアミノ酸分析装置を用いて、得られたアミノ酸を定量した。アミノ酸の量から、加水分解されたアリコート中の、総質量及びタンパク質濃度を計算した。
【0282】
上記のFYT単位でフィターゼ活性を決定し、そして、mgフィターゼ酵素タンパク質当たりのFYT単位で測定されたフィターゼ活性として、特異的活性を計算した。
【0283】
得られた特異的活性は980 FYT/mgタンパク質であった。特異的活性は、pH 5.5及び37℃でフィチン酸ナトリウムで決定した。
【0284】
実施例 6 フィターゼpHプロファイルの決定
0.25 M酢酸ナトリウム(pH 5.5) バッファーの代わりにバッファーカクテル(50 mMグリシン、50 mM酢酸及び5OmM ビス-トリス)を用いる以外は、「フィターゼ活性の決定」の項で述べたようにして、2.0〜7.5 (0.5のpH-単位ステップ) のpH範囲で37℃で、pHプロファイルを決定した。結果を以下の表1に纏める。2.0〜7.5の範囲の各pHについての値は、最適な値に対して標準化された相対活性(%)である。
【0285】
【表2】
【0286】
実施例 7 フィターゼ等電点の決定
フィターゼについての等電点plは、製造者によって教示されている、等電点電気泳動ゲル(Invitrogen製Novex pH 310 IEFゲル, カタログ番号EC6655A2)ランを用いて決定した。ハフニア・アルベイ・フィターゼのplは約7.4である。
【0287】
実施例 8 フィターゼ温度プロファイル
温度プロファイル (温度指数としてのフィターゼ活性) を、上記 ("Determination of フィターゼ活性の決定") のようにして基本的に20〜90℃の温度範囲でハフニア・アルベイ フィターゼについて決定した。しかしながら、酵素反応 (100μl フィターゼ-含有酵素溶液 + 100μl基質) をマイクロタイタープレートの代わにPCRチューブで行った。所望の温度での15分の反応期間の後、該チューブを20℃に20秒間冷却し、各反応混合物の150μlをマイクロタイタープレートに移した。75μlの停止試薬を加え、マイクロタイタープレート分光光度計で405 nmでの吸光度を測定した。結果を以下の表2に纏める。各温度の数字は、最適値に標準化された相対活性(%)である。
【0288】
【表3】
【0289】
実施例 9 フィターゼ熱安定性
アスペルギルス・オリザエ(Aspergillus oryzae)及びバチルス・サブティリス(Bacillus subtilis)で発現されたハフニア・アルベイ・フィターゼを、示差走査熱量 (DSC) 測定による熱安定性測定に供し、E. コリ・フィターゼ (Danisco NSからPHYZYME XPとして入手可能) と比べた。
【0290】
ハフニア・アルベイ・フィターゼ (実施例4で記載されたようにして精製した) のタンパク質試料のアリコートを、2〜3時間で、4℃で2 x 500 ml 20 mM Na-酢酸、pH 4.0で透析し、その後終夜ステップを行った。試料を0.45μm濾過し、約2 A280単位にバッファーで希釈した。測定された正確な吸光度を以下の結果表に示す。示差走査熱量 (DSC) 測定で対照として、透析バッファーを使用した。真空吸引を用いて及び約10分間攪拌しながら試料を脱気した。製品PHYZYME XP由来のE. コリ・フィターゼのアリコートを実施例4に記載の方法と同様の方法で精製した。
【0291】
20〜80℃で1.5℃/分の一定のスキャン速度でDSCスキャンを行った。濾過時間:16秒。DSCを行う前に、フィターゼを好適なバッファー (例えば、0.1 Mグリシン-HCl, pH 2.5又は3.0; 20 mM酢酸ナトリウム(pH 4.0); 0.1 M酢酸ナトリウム(pH 5.5); 0.1 M Tris-HCl, pH 7.0) で透析した。MicroCal Originソフトウェア (バージョン4.10) を用いてデータ操作を行い、変性温度Td (融解温度Tmとも称される) をサーモグラムでピークの頂点の温度として定義した。アンフォールディングプロセスの可逆性を明らかにするために、短い冷却期間の後直ちに第2スキャンを行った。第2スキャンについて、ピーク面積 (ピークとベースラインとの間の面積 = 比較されるアンフォールディングのエンタルピー) を、第1スキャンのピーク面積と比較する。大きなピーク (第1スキャンのピーク面積の75〜100 %) は、可逆的アンフォールディング/フォールディングプロセスとして解釈される。アスペルギルス・オリザエ、バチルス・サブティリス及びE. コリ・フィターゼで発現されたハフニア・アルベイ・フィターゼについてのDSCの結果を以下の表3に纏める。
【0292】
【表4】
【0293】
上記の表に示すように、ハフニア・アルベイ・フィターゼは、E. コリ・フィターゼよりも高い熱安定性を有した。ハフニア・アルベイ・フィターゼの熱安定性は、発現宿主によって影響されなかったことも明らかである。
【0294】
実施例 10 ハフニア・アルベイ・フィターゼ及びE. コリ・フィターゼの胃タンパク質分解耐性
H.アルベイ・フィターゼ及びE. コリ・フィターゼ (PHYZYME XP, Daniscoから入手可能) の試料を、ペプシン (ブタ胃粘膜由来のペプシン 1:60000, Wako 162-18721, 2900単位/mg, ロットSDK5232) の250 mMグリシンバッファー pH 3.0 (1000ペプシン単位/mg フィターゼ) で処理した。振とう (750 rpm) しながら、40℃で30分間インキュベーションした。ペプシンでのインキュベーション後、ペプシンを添加することなく、フィターゼ活性を、実施例5に記載のようにしてそして同一の方法で処理した試料の活性に比べて決定した。結果を以下の表4に纏める。
【0295】
【表5】
【0296】
従って、E. コリ・フィターゼ及びH. アルベイ・フィターゼは、非常に類似した胃タンパク質分解抵抗性を有した。
【0297】
実施例 11 ハフニア・アルベイ・フィターゼ及びシトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼ用インビトロモデルにおける動物飼料での成績
インビトロモデルでのハフニア・アルベイ・フィターゼの動物試料での成績を、シトロバクタ・ブラアキィ・フィターゼの成績と比較した。インビトロモデルは、単胃動物における胃腸状態を刺激し、インビボでの動物試験で得られた結果とよく相関する。試料中のフィターゼ活性は、「フィターゼ活性の決定」の下、実施例5で記載したようにして決定した。比較は以下のようにして行った。
【0298】
30%の大豆ミールと70%のトウモロコシミールとからなる飼料試料に5 gカルシウム/kg飼料の濃度にCaCl2を添加したものを調製し、40℃で30分間、pH 3.0で、予備インキュベートし、ペプシン(3000 U/g飼料)及びフィターゼの好適な投薬量を添加した(比較を可能にするために試験されるべきすべてのフィターゼについて同一の投薬量を用いた。例えば、0.1〜1.0フィターゼ単位FYT/g飼料)。フィターゼ活性のないブランクも対照としてインキュベートした。次いで、試料を40℃で60分間、pH 3.0でインキュベートし、次いでpH 4.0で30分間インキュベートした。
【0299】
反応を停止し、そして、最終濃度0.5 MになるようにHClを加え及び40℃で2時間インキュベーションし、次いで1回の凍結融解サイクルと40℃で1時間のインキュベーションによって、フィチン酸及びイノシトールリン酸塩を抽出した。
【0300】
Chen他のJournal of Chromatography A (2003) 第1018巻, 第41-52頁に記載の高速イオンクロマトグラフィーで、フィチン酸及びイノシトールリン酸塩を分離し、Skog-lund他のJ. Agric. Food Chem. (1997), 第45巻, 第431-436頁に記載されるようにして定量した。
【0301】
図1は、125 FYT/kg飼料、250 FYT/kg飼料及び500 FYT/kg飼料の投薬量での、シトロバクタ・ブラアキィ(Citrobacter braakii)・フィターゼと比べた、ハフニア・アルベイ フィターゼの用量反応を示す。インビトロでのフィターゼの効果は、インビトロインキュベーション後の試料に残存している残余イノシトールリン酸結合リン(IP-P)として、フィターゼナシの対照試料に残存している残余IP-Pと比較して、示される。すべての数値は、4又は5レプリカ(インビトロインキュベーションでの)平均及び標準偏差である。ハフニア・フィターゼの250 FYT/kgの投薬量は、インビトロ試料中の残余IP-Pの量を、シトロバクタ・フィターゼの125 FYT/kgとほぼ同程度に減少させた。
【0302】
従って、ハフニア・フィターゼは、残余イノシトール-ホスフェート結合リンの量におて非常に優れた減少を得ることができた。
【0303】
実施例 12 ハフニア・アルベイ・フィターゼ及びペニオホラ・リシイ(Peniophora lycii)・フィターゼ・フィターゼフィターゼ用インビトロモデルにおける動物飼料での成績
250 FYT/kg及び500 FYT/kg飼料の投薬量での、インビトロモデルにおけるハフニア・アルベイ・フィターゼの動物飼料での成績も、ペニオホラ・リシイの成績と比較した。結果は、実施例11に記載の実験プロトコルに従って得た。図2から明らかなように、ハフニア・アルベイ・フィターゼは、ペニオホラ・リシイ・フィターゼよりも高く、インビトロ試料において、残余IP-P量を減少した。
【0304】
【表6】
【特許請求の範囲】
【請求項1】
以下:
(a)(i)配列番号10のアミノ酸1〜413、及び/又は(ii)配列番号10の成熟ポリペプチドと少なくとも75%の同一性を有するアミノ酸配列を含むポリペプチド;及び
(b)(a)(i)〜(a)(ii)のポリペプチドのいずれか1つのフラグメント、
からなる群より選択される、フィターゼ活性を有する単離ポリペプチド。
【請求項2】
請求項1記載のポリペプチドをコードするヌクレオチド配列を含む単離ポリヌクレチド。
【請求項3】
以下:
(a)(i)配列番号10のアミノ酸1〜413と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;及び
(b)配列番号9の塩基100〜1338と少なくとも75%の同一性を有するポリヌクレオチド、
からなる群より選択される、フィターゼ活性を有する単離ポリペプチドをコードする単離ポリヌクレオチド。
【請求項4】
発現宿主において該ポリペプチドの製造を指向する1以上の制御配列に作動可能に連結された、請求項2又は3記載のポリヌクレオチドを含む核酸構築物。
【請求項5】
請求項4記載の核酸構築物を含む組換え発現ベクター。
【請求項6】
請求項5記載の核酸構築物を含む組換え宿主細胞。
【請求項7】
以下:
(a)該ポリペプチドの製造を誘導する条件下で、その野生型において、該ポリペプチドを製造することができる細胞を培養し;及び
(b)該ポリペプチドを回収すること、
を含む、請求項1記載のポリペプチドの製造方法。
【請求項8】
以下:
(a)前記ポリペプチドの製造を誘導する条件下で該ポリペプチドをコードするヌクレオチド配列を含む核酸構築物を含む組換え宿主細胞を培養し;及び
(b)該ポリペプチドを回収すること、
を含む、請求項1記載のポリペプチドの製造方法。
【請求項9】
請求項1記載のポリペプチドをコードするポリヌクレオチドによって形質転換された、トランスジェニック植物、植物部分又は植物細胞。
【請求項10】
請求項1記載のポリペプチドを発現することができる、トランスジェニック非-ヒト動物又は産物もしくはその要素。
【請求項11】
動物飼料における請求項1記載の少なくとも1つのポリペプチドの使用。
【請求項12】
動物飼料における使用のための組成物の製造における、請求項1記載の少なくとも1つのポリペプチドの使用。
【請求項13】
動物飼料の栄養値を改善するための方法であって、請求項1記載の少なくとも1つのポリペプチドが該飼料に添加される、前記方法。
【請求項14】
以下:
(a)請求項1記載の少なくとも1つのポリペプチド;及び
(b)少なくとも1つの脂溶性ビタミン、少なくとも1つの水溶性ビタミン、少なくとも1つのミネラル及びそれらの組合せ、
を含む動物飼料添加物。
【請求項15】
少なくとも1つのアミラーゼ、少なくとも1つのフィターゼ、少なくとも1つのキシラナーゼ、少なくとも1つのガラクタナーゼ、少なくとも1つのα-ガラクトシダーゼ、少なくとも1つのプロテアーゼ、少なくとも1つのホスホリパーゼ、及び/又は少なくとも1つのβ-グルカナーゼを更に含む、請求項14記載の動物飼料添加物。
【請求項16】
50〜800 g/kgの粗タンパク質量を有し、かつ請求項1記載の少なくとも1つのポリペプチドを含む、動物飼料組成物。
【請求項17】
以下:
(a)請求項1記載のフィターゼの存在下で、発酵微生物を用いて、糖質含有物質を発酵し:及び
(b)該発酵された糖質含有物質から発酵生成物又は発酵副生成物を製造すること、
を含む、発酵生成物を製造する方法。
【請求項18】
前記発酵生成物がエタノール、ビール、ワイン、又は蒸留乾燥穀類(DDG)である、請求項1記載の方法。
【請求項19】
前記ポリペプチドが、配列番号10のアミノ酸1〜413と、少なくとも80%同一性、少なくとも85%同一性、少なくとも90%同一性、少なくとも98%同一性、又は少なくとも99%同一性を有するアミノ酸配列を含む、請求項1記載のポリペプチド。
【請求項20】
前記ポリヌクレオチドが、配列番号9の塩基100〜1338と、少なくとも80%同一性、少なくとも85%同一性、少なくとも90%同一性、少なくとも98%同一性、又は少なくとも99%同一性を有するポリヌクレオチドを含む、請求項3記載のポリペプチド。
【請求項1】
以下:
(a)(i)配列番号10のアミノ酸1〜413、及び/又は(ii)配列番号10の成熟ポリペプチドと少なくとも75%の同一性を有するアミノ酸配列を含むポリペプチド;及び
(b)(a)(i)〜(a)(ii)のポリペプチドのいずれか1つのフラグメント、
からなる群より選択される、フィターゼ活性を有する単離ポリペプチド。
【請求項2】
請求項1記載のポリペプチドをコードするヌクレオチド配列を含む単離ポリヌクレチド。
【請求項3】
以下:
(a)(i)配列番号10のアミノ酸1〜413と少なくとも75%の同一性を有するアミノ酸配列を有するポリペプチドをコードするポリヌクレオチド;及び
(b)配列番号9の塩基100〜1338と少なくとも75%の同一性を有するポリヌクレオチド、
からなる群より選択される、フィターゼ活性を有する単離ポリペプチドをコードする単離ポリヌクレオチド。
【請求項4】
発現宿主において該ポリペプチドの製造を指向する1以上の制御配列に作動可能に連結された、請求項2又は3記載のポリヌクレオチドを含む核酸構築物。
【請求項5】
請求項4記載の核酸構築物を含む組換え発現ベクター。
【請求項6】
請求項5記載の核酸構築物を含む組換え宿主細胞。
【請求項7】
以下:
(a)該ポリペプチドの製造を誘導する条件下で、その野生型において、該ポリペプチドを製造することができる細胞を培養し;及び
(b)該ポリペプチドを回収すること、
を含む、請求項1記載のポリペプチドの製造方法。
【請求項8】
以下:
(a)前記ポリペプチドの製造を誘導する条件下で該ポリペプチドをコードするヌクレオチド配列を含む核酸構築物を含む組換え宿主細胞を培養し;及び
(b)該ポリペプチドを回収すること、
を含む、請求項1記載のポリペプチドの製造方法。
【請求項9】
請求項1記載のポリペプチドをコードするポリヌクレオチドによって形質転換された、トランスジェニック植物、植物部分又は植物細胞。
【請求項10】
請求項1記載のポリペプチドを発現することができる、トランスジェニック非-ヒト動物又は産物もしくはその要素。
【請求項11】
動物飼料における請求項1記載の少なくとも1つのポリペプチドの使用。
【請求項12】
動物飼料における使用のための組成物の製造における、請求項1記載の少なくとも1つのポリペプチドの使用。
【請求項13】
動物飼料の栄養値を改善するための方法であって、請求項1記載の少なくとも1つのポリペプチドが該飼料に添加される、前記方法。
【請求項14】
以下:
(a)請求項1記載の少なくとも1つのポリペプチド;及び
(b)少なくとも1つの脂溶性ビタミン、少なくとも1つの水溶性ビタミン、少なくとも1つのミネラル及びそれらの組合せ、
を含む動物飼料添加物。
【請求項15】
少なくとも1つのアミラーゼ、少なくとも1つのフィターゼ、少なくとも1つのキシラナーゼ、少なくとも1つのガラクタナーゼ、少なくとも1つのα-ガラクトシダーゼ、少なくとも1つのプロテアーゼ、少なくとも1つのホスホリパーゼ、及び/又は少なくとも1つのβ-グルカナーゼを更に含む、請求項14記載の動物飼料添加物。
【請求項16】
50〜800 g/kgの粗タンパク質量を有し、かつ請求項1記載の少なくとも1つのポリペプチドを含む、動物飼料組成物。
【請求項17】
以下:
(a)請求項1記載のフィターゼの存在下で、発酵微生物を用いて、糖質含有物質を発酵し:及び
(b)該発酵された糖質含有物質から発酵生成物又は発酵副生成物を製造すること、
を含む、発酵生成物を製造する方法。
【請求項18】
前記発酵生成物がエタノール、ビール、ワイン、又は蒸留乾燥穀類(DDG)である、請求項1記載の方法。
【請求項19】
前記ポリペプチドが、配列番号10のアミノ酸1〜413と、少なくとも80%同一性、少なくとも85%同一性、少なくとも90%同一性、少なくとも98%同一性、又は少なくとも99%同一性を有するアミノ酸配列を含む、請求項1記載のポリペプチド。
【請求項20】
前記ポリヌクレオチドが、配列番号9の塩基100〜1338と、少なくとも80%同一性、少なくとも85%同一性、少なくとも90%同一性、少なくとも98%同一性、又は少なくとも99%同一性を有するポリヌクレオチドを含む、請求項3記載のポリペプチド。
【図1】

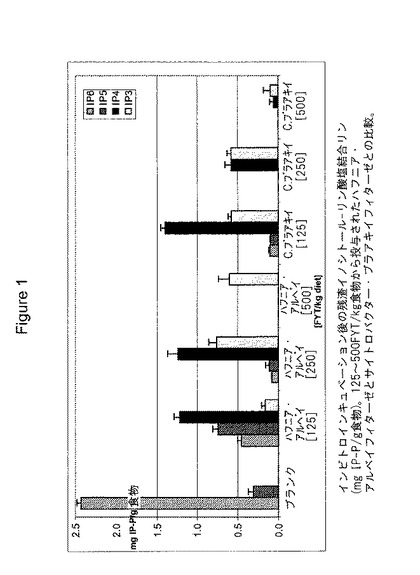
【図2】
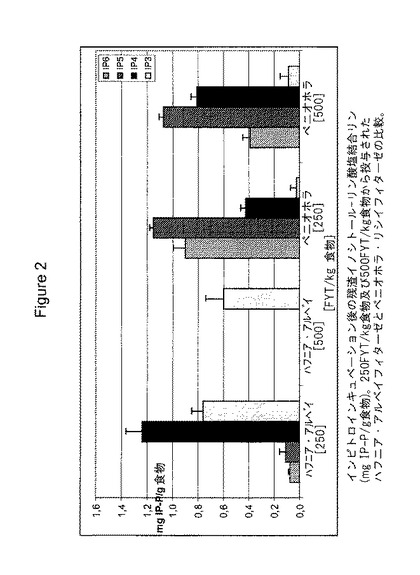
【図3−1】
【図3−2】
【図3−3】
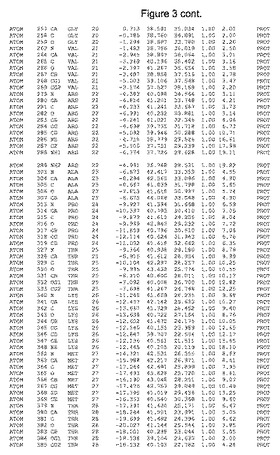
【図3−4】
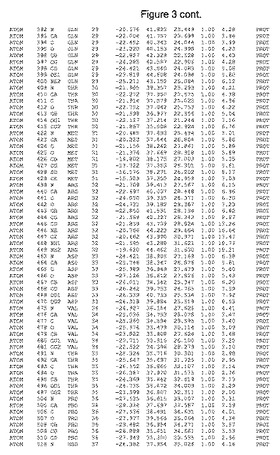
【図3−5】
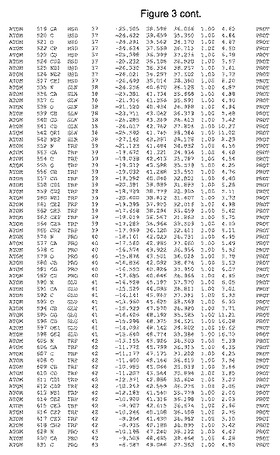
【図3−6】
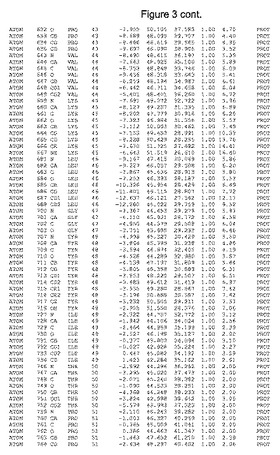
【図3−7】
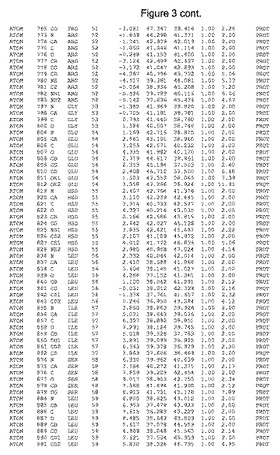
【図3−8】
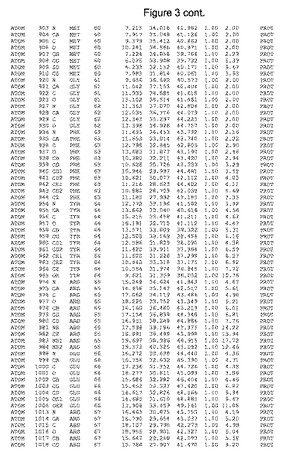
【図3−9】
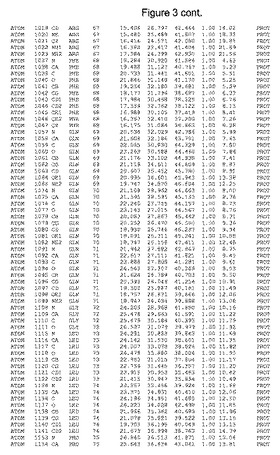
【図3−10】
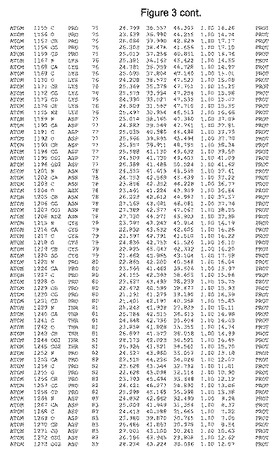
【図3−11】
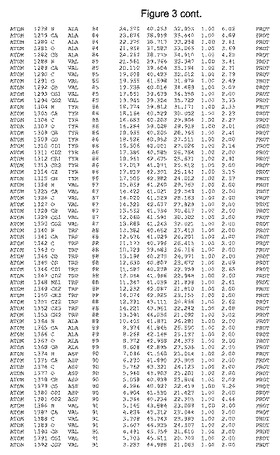
【図3−12】
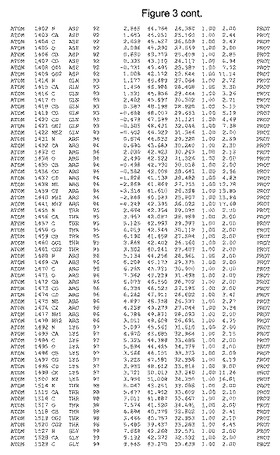
【図3−13】
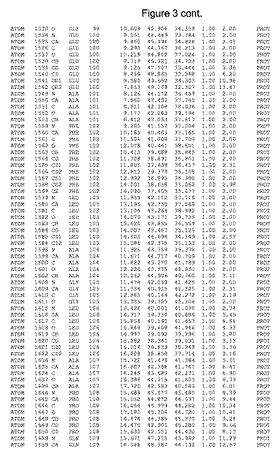
【図3−14】
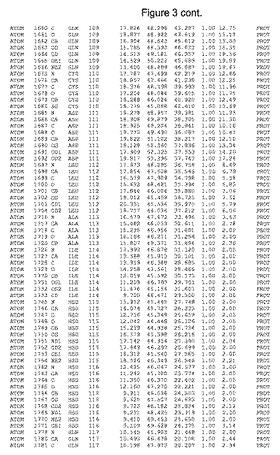
【図3−15】
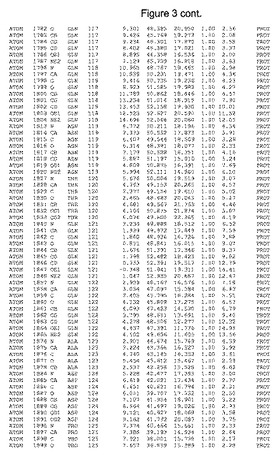
【図3−16】
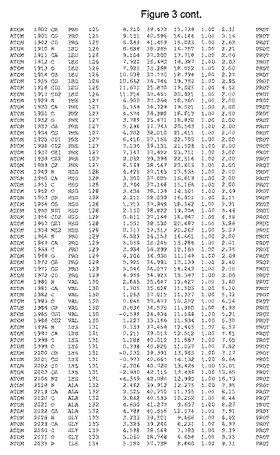
【図3−17】
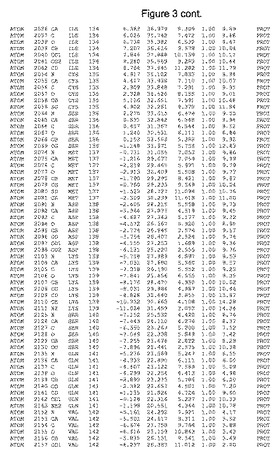
【図3−18】
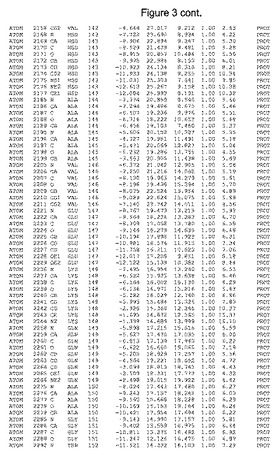
【図3−19】
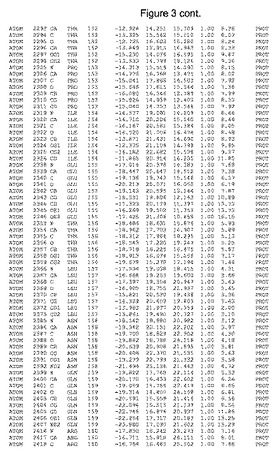
【図3−20】
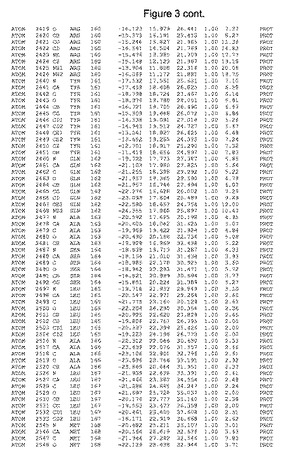
【図3−21】
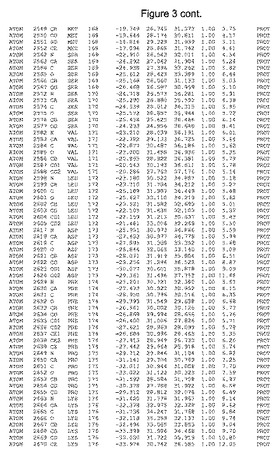
【図3−22】
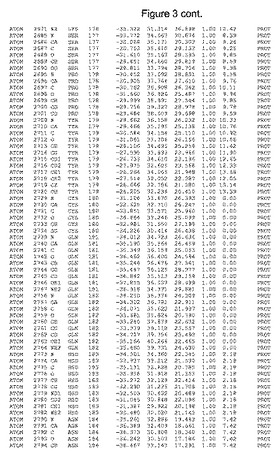
【図3−23】
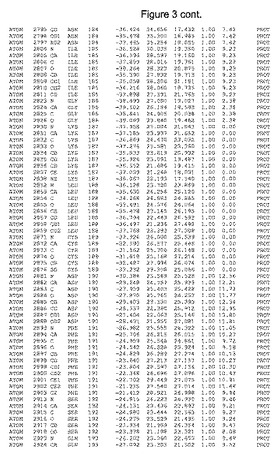
【図3−24】
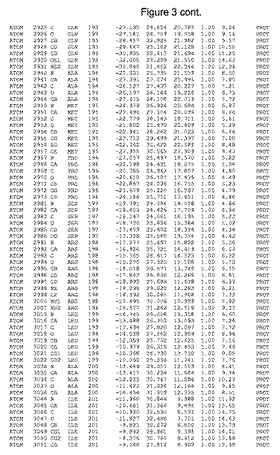
【図3−25】
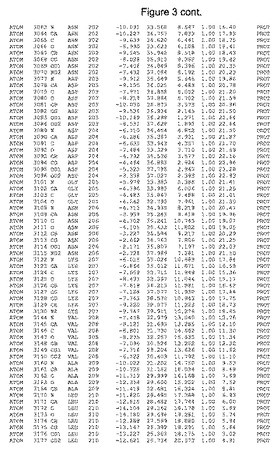
【図3−26】
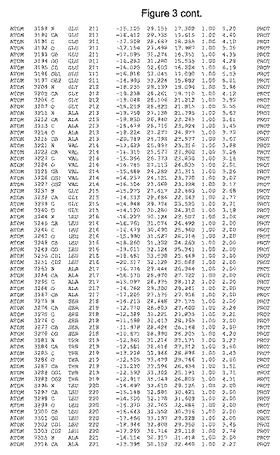
【図3−27】
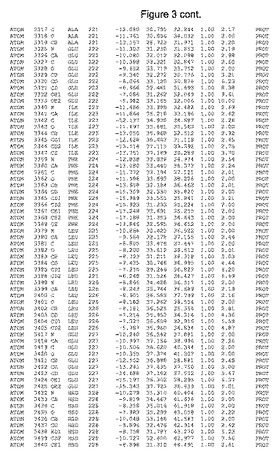
【図3−28】
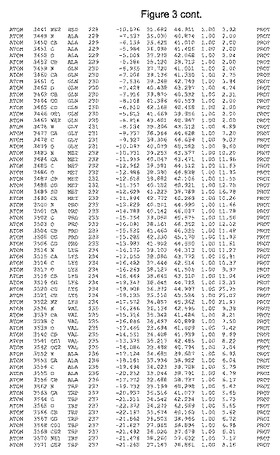
【図3−29】
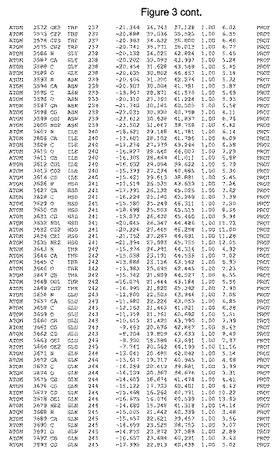
【図3−30】
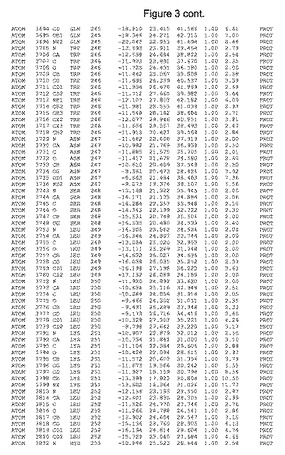
【図3−31】
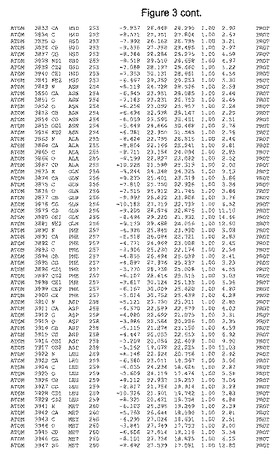
【図3−32】
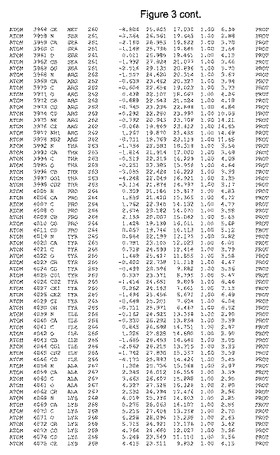
【図3−33】
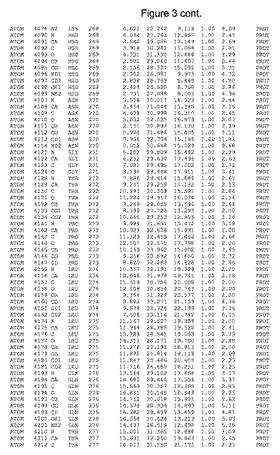
【図3−34】
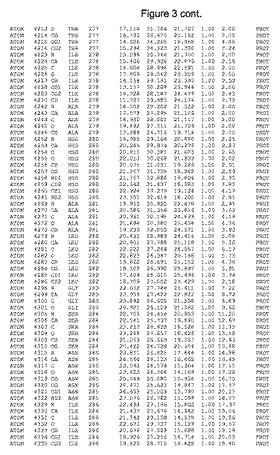
【図3−35】
【図3−36】
【図3−37】
【図3−38】
【図3−39】
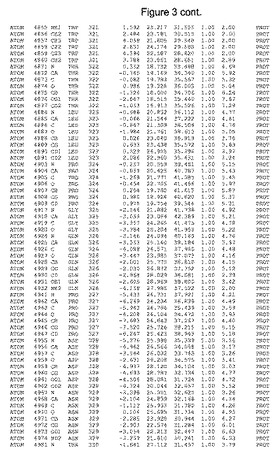
【図3−40】
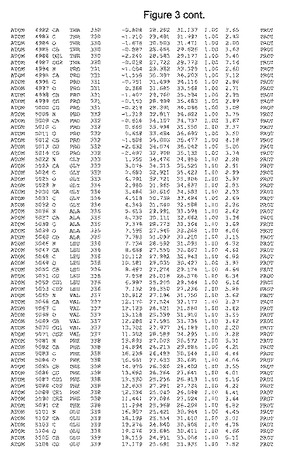
【図3−41】
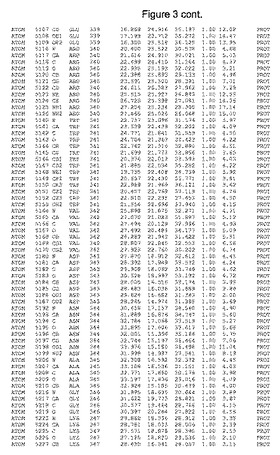
【図3−42】
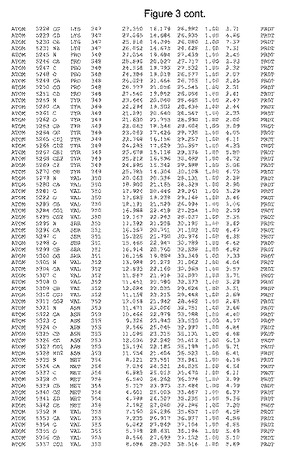
【図3−43】
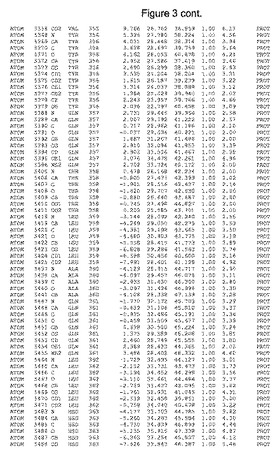
【図3−44】
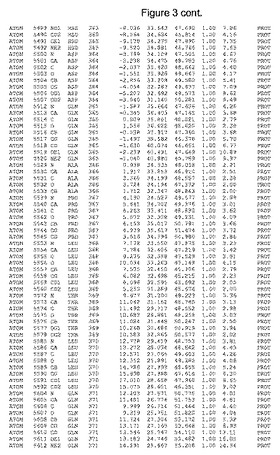
【図3−45】
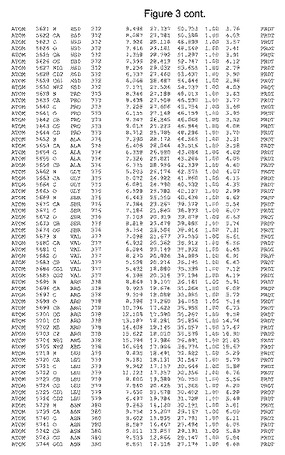
【図3−46】
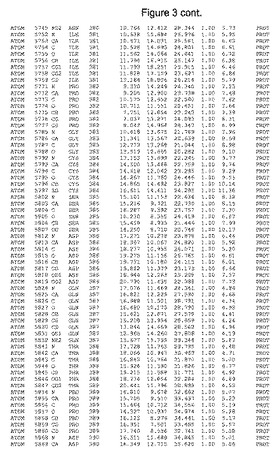
【図3−47】
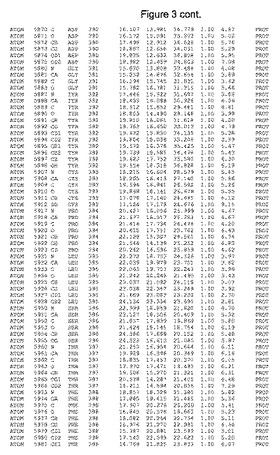
【図3−48】
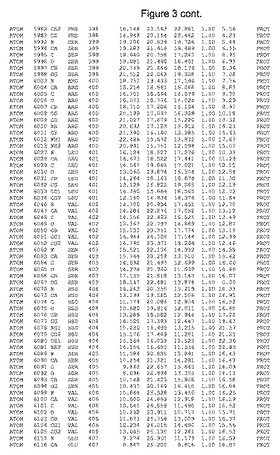
【図3−49】
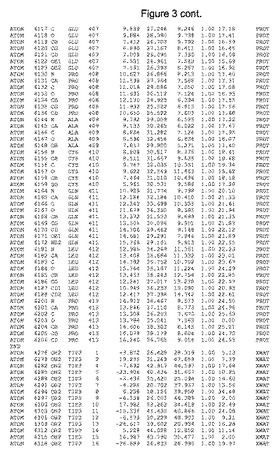
【図3−50】
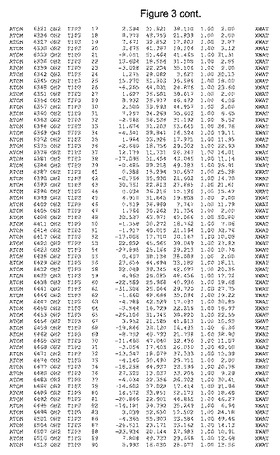
【図3−51】
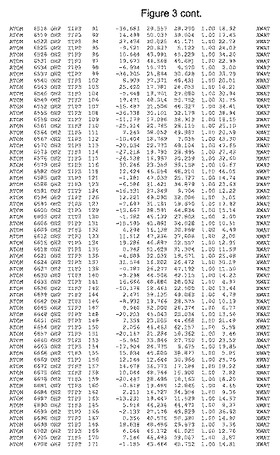
【図3−52】
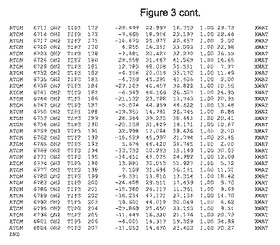
【図2】
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図3−5】
【図3−6】
【図3−7】
【図3−8】
【図3−9】
【図3−10】
【図3−11】
【図3−12】
【図3−13】
【図3−14】
【図3−15】
【図3−16】
【図3−17】
【図3−18】
【図3−19】
【図3−20】
【図3−21】
【図3−22】
【図3−23】
【図3−24】
【図3−25】
【図3−26】
【図3−27】
【図3−28】
【図3−29】
【図3−30】
【図3−31】
【図3−32】
【図3−33】
【図3−34】
【図3−35】
【図3−36】
【図3−37】
【図3−38】
【図3−39】
【図3−40】
【図3−41】
【図3−42】
【図3−43】
【図3−44】
【図3−45】
【図3−46】
【図3−47】
【図3−48】
【図3−49】
【図3−50】
【図3−51】
【図3−52】
【公表番号】特表2010−522548(P2010−522548A)
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願番号】特願2010−500256(P2010−500256)
【出願日】平成20年3月26日(2008.3.26)
【国際出願番号】PCT/EP2008/053561
【国際公開番号】WO2008/116878
【国際公開日】平成20年10月2日(2008.10.2)
【出願人】(500586299)ノボザイムス アクティーゼルスカブ (164)
【Fターム(参考)】
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願日】平成20年3月26日(2008.3.26)
【国際出願番号】PCT/EP2008/053561
【国際公開番号】WO2008/116878
【国際公開日】平成20年10月2日(2008.10.2)
【出願人】(500586299)ノボザイムス アクティーゼルスカブ (164)
【Fターム(参考)】
[ Back to top ]