
バイオセンサの細胞分析を用いたGPCRシグナリング経路の分析
本明細書に説明されているように、バイオセンサー、生細胞、及び経路活性化合物を使用したGPCRシグナリング経路の分析及び解明をなすシステムと方法を提供する。

【発明の詳細な説明】
【先願の米国出願の利益の請求】
【0001】
本出願は、2007年10月6日に出願された米国仮出願第60/997,910号、2007年11月30日に出願された米国特許出願第11/998,711号の利益を主張する。この先願の米国出願の内容及びこの明細書で引用された公報、特許、及び特許出願の全てが参考として組み込まれている。
【技術分野】
【0002】
本開示は、共鳴導波グレーティングバイオセンサンサ又は表面プラズモン共鳴(SPR)バイオセンサ等の光学バイオセンサに関し、特に、バイオセンサベースの生細胞分析を利用したGPCRシグナリング経路の分析方法に関する。
【発明の概要】
【0003】
本開示は、バイオセンサベースの生細胞分析を利用したGタンパク質共役受容体(GPCR)等受容体シグナリング経路の分析及び解明に対して直接的方法及び間接的方法を提供する。
【図面の簡単な説明】
【0004】
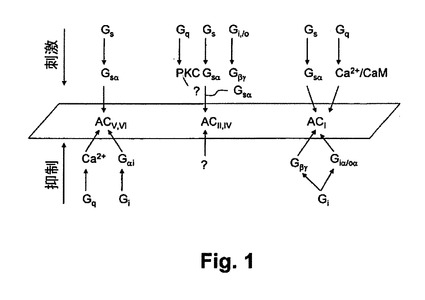
【図1】図1は、本開示の実施例におけるアデニールサイクラーゼアイソフォーム依存性細胞シグナリング経路を示す概略図である、
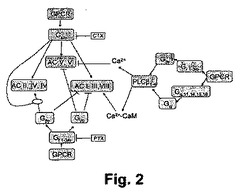
【図2】図2は、本開示の実施形態におけるGq(Gαq)、Gi(Gαi)、Gs/o(Gαs/o)、及びGβγ介在シグナリング経路から、細胞内アデニールサイクラーゼまでのクロストークと、cAMPレベルに与えるその影響と、を示す概略図である。
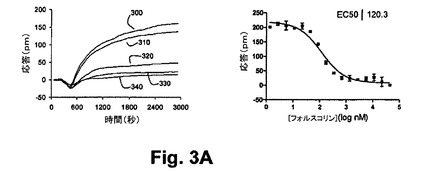
【図3A】図3Aは、本開示の実施形態における静的A431細胞におけるNECA誘起のGs応答に与えるホルスコリンの前処理の効果を示す図である。
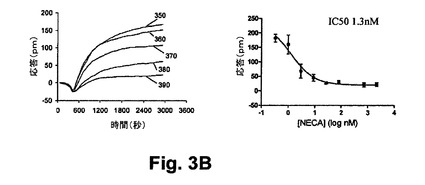
【図3B】図3Bは、本開示の実施形態における静的A431細胞におけるホルスコリン誘起のGs反応に対して与えるN−エチルアデノシン−5’−ウロン酸(NECA:N−ethyladenosine−5’−uronic acid)前処理の効果を示す図である。
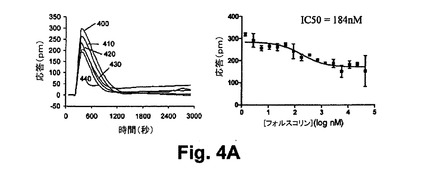
【図4A】図4Aは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のGq応答に対して与えるホルスコリン前処理の効果を示す図である。
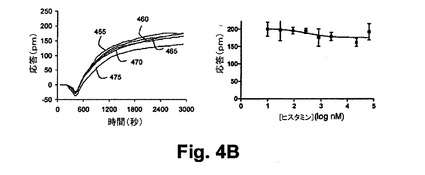
【図4B】図4Bは、本開示の実施形態における静的A431細胞におけるホルスコリン誘起のGq反応に対して与えるヒスタミン前処理の効果を示す図である。
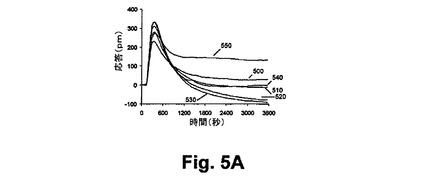
【図5A】図5Aは、本開示の実施形態における静的A431細胞におけるニコチン酸誘起のGi反応に対して与えるホルスコリン前処理の効果を示す図である。
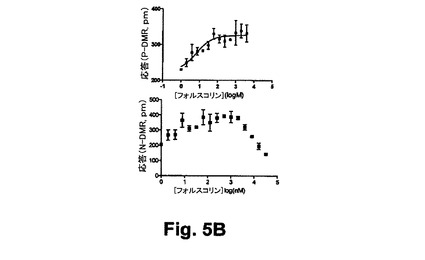
【図5B】図5Bは、本開示の実施形態における静的A431細胞におけるニコチン酸誘起のGi反応に対して与えるホルスコリン前処理の効果を示す図である。
【図6A】図6Aは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図6B】図6Bは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図6C】図6Cは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図7A】図7Aは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のDMR信号に与えるU73122の前処理の効果と、そのP−DMR振幅とをそれぞれ示す図である。
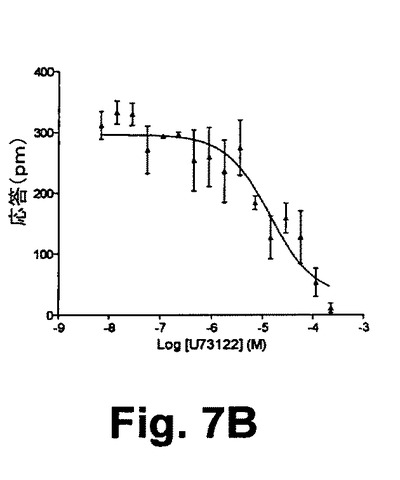
【図7B】図7Bは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のDMR信号に与えるU73122前処理の効果と、そのP−DMR振幅とをそれぞれ示す図である。
【発明を実施するための形態】
【0005】
本発明の様々な実施形態が、添付の図面を参照して詳細に説明されるであろう。様々な実施形態に対する言及は、本発明の範囲を制限するものではなく、本発明は、添付した請求の範囲によってのみ限定される。さらに、本明細書に述べられたいかなる実施例も、限定することを目的とはしていないし、特許請求の範囲に記載された発明の多くの実施形態の内、いくつかについて説明しているにすぎない。
【定義】
【0006】
「分析(Assay)」、「分析する(Assaying)」等の用語は、例えば、外因性刺激を用いた刺激に対する細胞の光学的応答又はバイオインピーダンスの応答の存在の有無、量、程度、動特性、及びタイプを決定する分析を意味する。外因性刺激は、例えば、リガンド候補化合物、アデニラート活性剤、cAMP類似体、又はGPCRリガンド若しくはリガンド候補等である。
【0007】
「付着する(Attach)」、「付着(Attachment)」、「接着(adhere)」、「接着された(adhered)」、「接着(adherent)」、「固定された(immobilized)」等の用語は、物理吸着又は化学結合等の処理若しくはそれらの組み合わせ等によって、例えば、本開示の細胞等の構成要素を、表面に、固定し、準備し、培養等することを一般に意味する。特に、「細胞の付着(cell attachment)」、「細胞の接着(cell adhesion)」等の用語は、細胞固定材料、相溶剤(例えば、フィブロネクチン、コラーゲン、ラミン、ゼラチン、ポリリジン等)、若しくはそれら双方を用いて培養し、相互作用せしめることによって、表面との細胞の相互作用又は表面との細胞の結合を意味する。実施形態において、バイオセンサ上に生細胞を固定するステップは、例えば、約1パーセントから約99パーセントのバイオセンサ表面被覆率及び同様の密集度値若しくは範囲等を有する密集度が得られるまで生細胞をバイオセンサ表面に培養するステップを含み得る。
【0008】
「付着性細胞(Adherent cells)」とは、原核生物の細胞若しくは真核細胞等の細胞、細胞株又は細胞システムを意味し、かかる細胞、細胞株又は細胞システムは、基板の外部表面に関連付けられ、基板の外部表面上に固定され、又は特定の接触を維持する。培養後のかかる細胞は、洗浄媒質交換処理、すなわち、多くの細胞ベースの分析に必須である処理に対して抵抗力を有し、耐性を有し得る。「弱付着性細胞(Weakly adherent cells)」とは、原核生物の細胞若しくは真核細胞等の細胞、細胞株又は細胞システムを意味し、かかる細胞、細胞株又は細胞システムは、細胞培養中に、基板の表面と弱く相互作用し、基板の表面と弱く関連し、基板の外部表面と弱く接触する。しかしながら、これらのタイプの細胞、例えば、人間の胎児性腎臓(HEK:human embryonic kidney)細胞は、洗浄液若しくは媒質の交換等の物理的摂動によって、基板の表面から容易に解離する傾向がある。「懸濁細胞(Suspension cells)」とは、望ましくは、媒質で培養される細胞又は細胞株を意味し、当該媒質において細胞は、培養中に、基板の表面に付着し若しくは固着しない。「細胞の培養(Cell culture)」又は「細胞培養(Cell culturing)」とは、原核生物細胞若しくは真核細胞が一定条件下で培養される処理を意味する。「細胞の培養(Cell culture)」とは、多細胞の真核細胞、特に動物細胞から得られた細胞の培養と、複合生体組織及び臓器の培養と、を意味する。
【0009】
「細胞(Cell)」等の用語は、半透明の膜によって外部的に結合された通常小なる微小質量の原形質を意味し、1つ以上の細胞核と他の多種の細胞小器官を任意で含み、かかる他の多種の細胞小器官は、生命の基本的な機能を単独で発揮し又は生命の基本的な機能を実行する他の同等の集団と相互作用することができ、独立的に機能することができる生物の最小構造ユニットを形成し、合成細胞構造体、細胞モデルシステム、人工の細胞システムを含む。
【0010】
「細胞システム(Cell system)」等の用語は、互いに相互作用して、生物学的機能、生理学的機能又は病態生理学的機能を発揮する1より多いタイプの細胞(又は単一タイプの細胞の異なる形態)の集合を意味する。かかる細胞システムは、例えば、器官、生体組織、幹細胞、分化した肝細胞等の細胞を含む。
【0011】
「マーカ(Marker)」等の用語は、少なくとも1つの細胞ターゲット(例えば、Gq−共役受容体、Gs−共役受容体、Gi−共役受容体、G12/13−共役受容体、イオンチャンネル、受容体チロシンキナーゼ、トランスポータ、ナトリウム‐プロトン交換体、細胞核受容体、セルキナーゼ、細胞性タンパク質、等)の活性を調節することができ、且つ、バイオセンサによって測定されるように、信頼性の高い検出可能な出力若しくは応答が得られる分子、生物分子、又は生物製剤を意味する。目的とする細胞ターゲットの分類及び後に生ずるセル事象に依存して、マーカは、例えば、GPCR、受容体チロシンキナーゼ(receptor tyrosine kinase)、イオンチャネル、細胞核受容体、細胞酵素アデニールサイクラーゼ等に対する、例えば、作用物質、部分的作用物質、逆作用物質等の活性剤であり得る。また、マーカは、特定の種類の細胞ターゲットに対しては抑制物質となり得るし、例えば、アクチンフィラメント若しくはマイクロチューブに対しては抑制物質又はかく乱物質となり得る。
【0012】
「検出する(Detect)」等の用語は、感知されたシグナリング経路を発見し又は感知し、かつ、感知されたシグナリング経路と経路のシグナリングがない場合とを区別する本開示の装置及び方法の能力を意味する。
【0013】
「特定する(Identify)」等の用語は、シグナリング経路を検出しかつ解明する本開示の装置及び方法の能力を意味する。
【0014】
「治療候補化合物(Therapeutic candidate compound)」、「治療候補物質(therapeutic candidate)」、「予防的候補物質(prophylactic candidate)」、「予防的物質(prophylactic agent)」、「リガンド候補(ligand candidate)」等の用語は、バイオセンサ若しくは病原体に付着した細胞と相互作用する潜在能力にとって重要であり、自然に生ずる分子若しくは材料又は合成的な分子若しくは材料を意味する。治療候補物質又は予防候補物質は、タンパク質、DNA、RNA、イオン、脂質等の細胞ターゲット若しくは病原体、又は、生細胞若しくは病原体の同様の構造体若しくは要素のうち少なくとも1つと特に結合し又は相互作用し得るし、例えば、化合物、生物学的分子、ペプチド、タンパク質、生物試料、薬物候補の小分子、薬物候補の生物学的分子、薬物候補の小分子‐生物の置換体、及び同様の材料若しくは実在の分子、又はそれら組み合わせを含む。
【0015】
「バイオセンサ(Biosensor)」等の用語は、生物学的要素を物理化学的検出構成要素に結合せしめる検体を検出するデバイスを意味する。バイオセンサは、通常3つの部分から構成される。すなわち、(生体組織、微生物、病原体、細胞、又はこれらの組合せ等の)生物学的構成部又は要素、(例えば、光学的、圧電的、熱測定、若しくは磁気的な物理化学的手法で動作する)検出要素と、双方の構成部に関連付けられたトランスデューサと、から構成され得る。生物学的構成部若しくは生物学的要素とは、例えば、生細胞であり得る。実施形態において、光学的バイオセンサは、例えば、生体細胞における分子認識事象又は分子刺激事象を定量化可能な信号に変換する光学変換装置を含み得る。
【0016】
”含む(Include)”、”含む(includes)”等は、含んで(including)を意味するがこれに限定されない。
【0017】
例えば、成分の構成要素、濃度、堆積、処理温度、処理時間、収量、流速、圧力等の値の量、及びその範囲を修飾する「約(About)」という用語は、例えば、生じうる数量における変分を意味する。かかる変分は、本開示の実施形態を説明するのに使用されており、例えば、化合物、構成、濃度、使用製剤に対して使用された通常の計測と取扱を介して、これら手法における不注意な誤動作を介して、方法を実行するのに使用される製造方法、ソース、又は出発物質若しくは成分の純度における差等を介して、例えば、生じ得る。また、「約(About)」という用語は、特定の初期濃度若しくは混合度を有する成分又は処方の経時変化に起因して異なる量、及び、特定の初期濃度若しくは混合度を有する成分又は処方の混合処理又は処理工程に起因して異なる量を包含する。“約”という用語によって修飾された本明細書に添付された特許請求の範囲は、これら量の均等物を含む。
【0018】
実施形態において「〜本質的に成る(Consisting essentially of)」とは、本明細書に定義されたGPCRシグナリング経路検出、分析、若しくはそれらの双方、バイオセンサの表面上の製剤若しくは成分、及び、本開示の物品、デバイス、若しくは装置に対する方法を例えば意味し、特許請求の範囲に記載された構成要素又はステップと、構成物、物品、若しくは装置の基本的且つ新規な特性と、特定の反応物質、特定の添加物または成分、特定の物質、特定の細胞若しくは細胞株、特定の表面修飾剤または条件、特定のリガンド候補物質、可変選択された同等の構造、材料、又は処理等の本開示の製造方法及び使用方法と、に物質的に影響を与えない他の構成要素又はステップと、を含み得る。本開示の構成要素若しくはステップの基本特性に有形的に影響し若しくは本開示に不要な特性を与え得るアイテムは、例えば、バイオセンサの表面に対する細胞の低減された親和性、リガンド候補若しくは同様の刺激に応答した異常な若しくは逆の細胞の活性化、及び同様の特性を含む。
【0019】
特に指定しない限り、本明細書で使用された不定冠詞“a”“an”、又は対応する定冠詞“the”は、少なくとも1つ、又は1つ以上を意味する。
【0020】
当業者にはよく知られている簡略が使用され得る(例えば、時間又は(複数の)時間に対して「h」若しくは「hr」、(複数の)グラムに対して「g」若しくは「gm」、ミリリットルに対して「mL」、及び室温に対して“rt”、ナノメータに対して「nm」、等の簡略)。
【0021】
要素、成分、添加剤、細胞のタイプ、抗体等、及びその範囲に対して開示された特定の値又は好ましい値は、例示する目的のためにあるに過ぎず、それらは他の定義された値又は定義された範囲内の他の値を除外するものでない。
【0022】
開示された構成、装置、及び方法は、いかなる値をも有する構成、装置、及び方法を含み、或いは、本明細書に記載された値、特定の値、より特定の値、及び好ましい値のいかなる組合せをも有する構成、装置、及び方法を含む。
【0023】
実施形態において、本開示の方法は、Gタンパク質共役受容体(GPCRs)のシグナリング経路の分析、定量、又はそれら双方を提供する。かかる分析、定量、又はそれら双方は、例えば、GPCRリガンド誘起のバイオセンサの応答に対するホルスコリン刺激された細胞の効果を測定することによって行われ、例えば、ホルスコリン誘起のバイオセンサ応答に対するGPCRリガンド刺激された細胞の効果を測定することによって行われる。
【0024】
実施形態において、方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサに生細胞を固定するステップと、当該固定された生細胞を、前記生細胞における細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に接触せしめるステップと、当該物質に接触せしめられた生細胞をGPCRリガンドに接触せしめるステップと、前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0025】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該培養された生細胞を接触せしめるステップと、これらアデニラート活性剤又はcAMP類似物質が接触した細胞を、GPCRリガンドに対して接触せしめるステップと、を含み得る。
【0026】
実施形態において、方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサ上に生細胞を固定するステップと、当該固定された生細胞をGPCRリガンドに接触せしめるステップと、前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、前記GPCRリガンドが存在する場合と存在しない場合における前記生細胞に関する前記物質誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0027】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該培養された生細胞を接触せしめるステップと、これらアデニラート活性剤又はcAMP類似物質が接触した細胞を、GPCRリガンドに接触せしめるステップを含む。
【0028】
細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質は、例えば、ホルスコリン又はその派生物質等のアデニラート活性物質を含む。その派生物質は、NKH477、1,9ジデオキシ‐ホルスコリン、7‐デアセチル‐7‐O‐へミスチニル‐ホルスコリン、又は細胞浸透性cAMP類似物質である。細胞浸透性cAMP類似物質は、(S)−アデノシン、環状3’、5’−(ハイドロゲンホスホロチオエート)トリエチルアンモニウム、8−ブロモアデノシン−3’、5’−環状リン酸(8−Br−cAMP)、8−クロロアデノシン−3’、5’−環状リン酸、又は、N6,2’−O−ジブチリルアデノシン−3’、5’−環状リン酸等である。これらcAMP類似物質は、細胞膜透過性があり、直接細胞内に取り入られ、cAMP依存性タンパク質キナーゼ(タンパク質キナーゼA、PKA)の活性化を直接的に引き起こす。
【0029】
実施形態において、代替的方法は、例えば、バイオセンサ上に生細胞を培養するステップと、当該培養された生細胞をGPCRリガンドに接触せしめるステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、を含み得る。
【0030】
実施形態において、代替的方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサ上に生細胞を固定するステップと、前記細胞内における細胞内ホスホリパーゼC(PLC)活性を抑制する物質に当該固定された生細胞を接触せしめるステップと、当該PLC抑制されたが生細胞をGPCRリガンドに接触せしめるステップと、前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0031】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、当該培養された生細胞をU73122若しくは干渉RNA等のホスポリパーゼC抑制物質若しくは抑制体に接触せしめるステップと、ホスホリパーゼCにより抑制された細胞をGPCRリガンドに接触せしめるステップと、を含み得る。
【0032】
本開示の方法は、例えば、GPCRシグナリング経路に与える周知の治療用化合物若しくは未知の治療候補化合物の陽性及び陰性の効果を特定する際に有益であり得る。したがって、前述の方法は、刺激を用いた固定生細胞の前処理、中間処理、又は後処理のさらなるステップを含み得る。
【0033】
実施形態において、本開示は、リガンド誘起バイオセンサ信号の研究において観測された詳細な生化学シグナリング経路を抽出し且つ解明する方法を提供する。本開示方法はすべての種類のGPCRシグナリングに適用可能である。本開示の方法は、異なる種類のGタンパク質シグナリングを区別し、GPCRのシグナリングと非GPCRのシグナリングとを区別し得る。本開示の方法は、アデニールサイクラーゼ(AC:adeneylate cyclase)活性物質(例えば、ホルスコリン、NKH447)を使用し得るし、又は、ホスホリパーゼC抑制物質と組み合わせて独立に使用され得る(例えば、U73122、合成アミノステロイド、すなわち、1−[6−[[17β−3−メトキシエストラ−1,3,5(10)−トリエン−17−yl]アミノ]hexyl]−1H−pyrrole−2,5−dione、又は干渉RNA)。
【0034】
本明細書に開示されたバイオセンサベースの生細胞分析を利用する経路分析若しくは経路抽出のツールとして、本開示は、バイオセンサベースの細胞分析を使用して、シグナリング経路を抽出し若しくは解明するユニバーサルな手法を提供する。
【0035】
実施形態において、本開示の方法はツーステップ分析を含み得る。また、本開示の方法は、受容体を介在する細胞シグナリングに関する有益な情報を提供し得る。
【0036】
実施形態において、本開示は、無標識バイオセンサベースの細胞分析を使用して、Gタンパク質共役受容体(GPCRs)のシグナリング経路を分析する方法を提供する。本方法は、生細胞におけるすべての種類のGPCRシグナリングに対して利用可能である。実施形態における方法は、GPCRリガンドによって誘起されたバイオセンサの応答に及ぼすcAMP増加細胞、ホスホリパーゼC抑制細胞、若しくはその双方の効果に基づいており、cAMPが生成する化学物質若しくは細胞浸透性cAMP類似物質によって誘起されたバイオセンサの応答に及ぼすGPCRリガンド刺激によって刺激された細胞の効果に基づいている。使用され得るバイオセンサは、例えば、表面プラズモン共鳴バイオセンサ、共鳴導波グレーティング(RWG)バイオセンサ等の光学バイオセンサ、バイオインピーダンスバイオセンサ、共鳴ミラー等のデバイス等の電気的バイオセンサを含む。
【0037】
Gタンパク質共役受容体(GPCRs)は細胞表面受容体の中で最も大きい群であり、細胞表面受容体は、細胞内ループ及び細胞外ループによって接合された7つの螺旋膜貫通領域の共通な構造モチーフを共有している。GPCRsは、異なる細胞システムにおいて異なる発現パターンにより、実質的に全細胞内で発現せしめられる。GPCRsの細胞外リガンドは多様的であり、生体アミン、アミノ酸、イオン、小型ペプチド、たんぱく質、及び生物活性な脂質を含む。GPCR活性物質のこのような多様性により、この受容体の種類の生理学的重要性が強調される。より詳しく述べると、GPCRsは、神経伝達、走化性、炎症、細胞の増殖等の処理を含む広範な生理学的過程を制御する。GPCRsは、例えばアストマ、癌腫、炎症、及び心臓血管疾患を含むほとんど全ての主要クラスの疾病に関わっている。
【0038】
健康及び疾病におけるそれらの役割と、小分子の薬物による治療の干渉に対するそれら潜在能力と、を想定すると、GPCRsは、ヒトゲノムにおいて最大の且つ最も成功したクラスの薬物に対して有能なターゲットに相当する。臨床的に利用可能なすべての薬物のうち約50%は直接若しくは間接的にGPCRsに対して活性的であるものの、これら薬物は、周知なすべてのGPCRsのうち数パーセントに対してのみ活性的である。ヒトゲノムは、最大1,000GPCRをエンコードしており、その内の約400が、内因性リガンドと結合すると期待された非化学的刺激受容体であると考えられている。これらの多く(約150)が、それら同族のリガンドとして分類され、若しくは生物学的機能は未だに知られていない「オーファン」GPCRsとして分類されている。これら受容体の「非−オーファン化」(すなわち、それら同族のリガンド又は生物学的機能の適用若しくは決定)は、新規治療ターゲット及び新規治療効果を提供し得る。
【0039】
GPCR薬物の発見と開発における継続的成功が、分析技術及び分析方法における発展に寄与している。GPCRスクリーニングの分析は、全体的細胞分析と無細胞分析とに広範に分類され得る。細胞ベースの分析によって、刺激に応答した細胞の振る舞いに関して高精度な指標が提供される。無細胞分析を使用して得られたデータと比較すると、細胞システムにおける化合物調節されたGPCR機能に関する直接測定によって、はるかに有益な情報が提供される。かかる情報とは、例えば、より密接に生理的環境に類似した条件下における化合物の作用、モード、及び機構である。これら利益により、薬物のスクリーニング及び試験を目的とする全体的細胞システムの使用が高められ、促進されている。
【0040】
GPCRsは、広範な一連の細胞シグナリング経路に関与している。かかる細胞シグナリング経路は、Gタンパク質依存性経路及びGタンパク質独立性経路の双方を介して、介在されており、たいては、リガンドに依存している。GPCRシグナリングは、川下のシグナリングネットワークの空間的且つ時間的フラックスによってコード化されている。かかるシグナリングネットワークは、細胞内のシグナリング及び調節機構によって厳密に制御される。GPCRシグナリングについて説明する多くのモデルにおいて、一旦、作用物質の結合によって活性構造で安定化されると、受容体が特定の細胞内ドメインを介して単一のGタンパク質と相互作用する機能的なモノメリックエンティティであることが想定されている。リガンドがGPCRに結合すると、例えば、受容体の構造に変化が生ずる。受容体の活性化は、順番に、Gαサブユニットに対するGTP−GDP交換を介して、関連Gタンパク質ヘテロトリマの活性化を引き起こす。そして、活性化されたGタンパク質は、いくつかの細胞内酵素の活性化を調節する。いくつかの細胞内酵素の活性化により、いくつかの主要な細胞内第2メッセンジャ生成が、順番に制御される。かかる第2メッセンジャは、例えば、環状AMP(cAMP)、cGMP、Ca2+、イノシトール・トリスリン酸、アラキドン酸である。そして、これら第2メッセンジャは、遺伝子転写及び細胞機能を調整するイオンチャンネル及びキナーゼを含む下流にあるいくつかのターゲットに作用する。
【0041】
合成化合物によって介在された細胞内シグナリングにおける予測不可能性が原因で、測定されたシグナリング事象に依存して単一化合物は作用物質又は拮抗物質として作用するであろう。さらに、測定された所定の事象において、効果不足により、受容体の活性化が不足するとは限らない。したがって、理想的なGPCRスクリーンは、単一の分析に依存するべきでなく、むしろ、統合された手法が、多くのシグナリング事象を測定するのに採用されるべきである。細胞シグナリング経路から独立した技術は、GPCR活性化が、多くの状況下においてGタンパク質独立シグナリングをも引き起こし得るという最近の調査結果を想定すると、それら経路バイアスの分析技術よりも優れているだろう。
【0042】
バイオセンサベースの細胞分析によって、より生理的に関連した環境下で、内因性受容体の活性化を観測することが可能となる。かかる非侵襲的能力は、リガンド薬理学に関してより魅力的な指標を与え得る。大抵のGPCR細胞分析は、細胞に関して特定の工学技術若しくは操作を利用しており、これは、確実であり且つ信頼性のある検出を得るために所定の感度を達成することを目的とする。これら人工的システムにおいて、リガンドの力価又は効果が変更され得る。これは、例えば、受容体の高い発現レベル又はターゲットの細胞生理学における特定の操作(例えば、GFP−タグ付け、又はトランスフェクション作用)の干渉に帰属する。例えば、α2A−アドレナリン作用受容体(α2AAR)は、Gαiプロテインに結合すると一般に信じられており、アデニレートサイククラーゼ活性の抑制を引き起こす。天然のHEL92.1.7細胞において、薬物のLevomedは、α2AARの逆作用物質であり、cAMP生成の増大を引き起こす。しかしながら、トランスフェクトされたPC10において、線維芽細胞Levomedは、陽性の作用物質であることが見出され、cAMP生成の抑制を引き起こす。
【0043】
さらに、バイオセンサベースの細胞分析においては、蛍光標識を必要としない。かかる蛍光標識は、多数の従来分析において検出の際に広く使用されている。かかる標識の非依存性により、GPCRスクリーンのデータ品質が大幅に改善され得る。蛍光性細胞の構成要素、標識、若しくは化合物の干渉が最小化され若しくは排除されるからである。
【0044】
バイオセンサベースの細胞分析は、積分されたバイオセンサの応答を測定している。例えば、RWGバイオセンサは、細胞内におけるリガンドによって誘起された動的質量再分布(DMR:dynamic mass redistribution)を観測する。これに反して、電気バイオセンサ、生細胞においてリガンドによって誘起されたバイオインピーダンスの変化を測定している。リガンドは細胞内の1つより多い数の受容体を活性化し得るし、受容体によって、複数のシグナリング経路が導かれる。この交絡の可能性が原因で、活性化された特定の経路を、リガンドによって誘起されたバイオセンサの応答に帰属させることは、バイオセンサ応答の形状及び動特性を測定するだけでは、困難である。
【0045】
1.バイオセンサベースの細胞分析とバイオセンサ基板
一般に、無標識細胞に基づく分析は、生体細胞におけるリガンド誘起の応答を観測することを目的としてバイオセンサを採用している。バイオセンサは、光学的、電気的、熱的、音響的、磁気的トランスデューサ等のトランスデューサを通常利用しており、分子認識事象又は細胞層におけるリガンド誘起の変化を定量化可能な信号に変換する。これら無標識バイオセンサは分子の相互作用の分析に通常使用され得る。かかる分析は、分子複合体が経時によってどのように形成し且つ分離するのかを特徴付けることを含む。本発明は、ほとんどすべてのタイプのバイオセンサ表面に利用可能であるものの、RWGバイオセンサと電気バイオセンサのみが例示されている。
【0046】
RWGバイオセンサ
RWGバイオセンサは、例えば、埋め込まれたグレーティング構造を有する基板(例えば、ガラス)と、細胞層と、から構成される。RWGバイオセンサは回折格子によって導波路に対する光の共鳴結合を利用し、界面において次々に電磁場が形成される溶液表面の界面において全内反射が生ずる。かかる電磁場はセンサの表面から指数関数的に減衰する事実上エバネッセントである。電磁場が初期値に関して1/eに減衰する距離は、侵入深さとして知られており、特定のRWGバイオセンサに関して、設計に応じた関数となるものの、通常約200nmのオーダーである。このタイプのバイオセンサは、かかるエバネセント波を利用して、センサ表面の近傍の若しくはセンサの表面における細胞層のリガンド誘起の変化を特性化している。
【0047】
電気バイオセンサ −
電気バイオセンサは、例えば、基板(例えば、プラスチック)、電極、及び細胞層から構成される。この電気的な検知手法において、細胞は基板上に並べられた小さな金の電極上に培養され、システムの電気インピーダンスが経時的に調べられる。インピーダンスは細胞層の電気伝導率の変化の指標となる。通常、固定周波数若しくは変動周波数を有する小定電圧が、電極又は一連の電極に印加され、回路を介した電流の経時的に観測される。リガンド誘起の電流変動により、細胞応答に関する指標が与えられる。細胞全体を検出するインピーダンス測定のアプリケーションは1984年に実証された。それ以来、インピーダンスベースの測定は、細胞の付着及び拡散、細胞の微小運動、細胞の構造変化、及び細胞の死を含む広い範囲の細胞の事象を研究することを目的として利用されてきた。古典的インピーダンスシステムは、小さな検出電極を使用しかつ大きな基準電極を使用しているのが原因で、分析に際して変動が大きいという欠点を有する。かかる変動性を克服するために、CellKey system(MDS Sciex、南サンフランシスコ、カリフォルニア州)やRT‐CES(ACEA Biosciences株式会社、サンディエゴ、カリフォルニア州)等の最新世代のシステムは、ミクロ電極アレイを有する集積回路を利用している。
【0048】
RWGバイオセンサを用いたGPCR活性の光信号 −
細胞は、例えば、通常数十ミクロンを有する比較的大きい寸法の動的な物体である。RWGバイオセンサは、細胞の底部におけるリガンド誘起の変化の検出を可能にし、エバネセント波の侵入度によって決定される。その上、光学的バイオセンサの空間分解能は、入射光源のスポットサイズ(約100ミクロン)によって決定される。したがって、一般に、非常に密集した細胞層が、最適な分析結果を得るのに使用され、センサの構成は、例えば基板、導波薄膜、及び細胞層から構成される3層導波複合体と見なされ得る。細胞生物物理学と組み合わせて3層導波バイオセンサ理論に従って、全体の細胞の検知に関しては、有効屈折率におけるリガンドによって誘起された変化、すなわち検出された信号ΔNが数式(1)により与えられる。
【0049】
(数式1)
【0050】
ここで、S(C)は細胞層に対するシステムの感度であり、Δncはバイオセンサによって検知された細胞層の局所的な屈折率のリガンド誘起の変化である。ΔZcは細胞層への侵入深さであり、αは特定の屈折率の増大(タンパク質に対しては約0.18/mL/g)であり、ziは、質量の再分配が生ずる距離であり、dは細胞層内の薄片の仮想的厚さである。ここで、細胞層は垂直方向において均等に離間された部分に分割される。検出された信号が細胞層の底部分の屈折率Δncの変化に一次的に正比例することが想定されている。細胞内の所定体積の屈折率は、主にタンパク質等の生物分子の濃度によって主に決定されると仮定すると、Δncは検出ボリューム内の細胞のターゲット又は分子集合の局所濃度変化に正比例する。重み付け因子exp(−zi/ΔZc)は、指数関数的に減衰するエバネッセント波の特性を考慮して、生ずる局所的タンパク質濃度の変化が考慮されている。したがって、検出された信号は、センサの表面から異なる距離で生じる質量再分布を合計したものであり、各々が全ての応答に対して不均一に貢献する。数式(1)が示していることは、RWGバイオセンサを用いて検出された信号は、所的なタンパク質濃度変化の結果として、かかる変化が生ずる場所、時間の結果として、主として垂直な集団再分布に対して敏感であることである。検出された信号は、動的集団再分布(DMR:dynamic mass redistribution)信号と呼ばれている。
【0051】
GPCRの活性化は、一連の空間的かつ経時的な事象を引き起こす。かかる事象は、リガンドの結合、受容体の活性化、タンパク質の補充、受容体の内部化及び再利用化、第2メッセンジャの代替、細胞骨格のリモデリング、遺伝子の発現、細胞付着の変化等の事象を含む。各細胞の事象は、細胞の動力学、持続時間、振幅、及び質量の移動に関してそれ自身の特性を有する。したがって、細胞事象が生ずる位置に依存して、これらの細胞事象が、DMR信号全体に対して異なる寄与をなし得ることが合理的に想定されている。さまざまなGPCRsをターゲットとする作用物質のパネルを使用して、介在されたシグナリング経路を反映する3つのクラスのDMR信号がヒト類表皮癌A431細胞において特定された(Fang, Y., et al., “Non−invasive optical biosensor for assaying endogenous G protein−coupled receptors in adherent cells,” J. Pharmacol.Toxicol.Methods, 2007, 55, 314−322)(図6を参照)。受容体が結合したGタンパク質に依存して、DMR信号の各々は特定のクラスのGPCRs活性と相関しているので、得られたDMR信号は、Gq−DMR信号、Gs−DMR信号、及びGi−DMR信号とそれぞれ称されている。各クラスのDMR信号は、異なるクラスのGPCRsを介して介在された固有のシグナリングの集合を反映した異なる運動特性及び動的特性を示す。興味深いことに、Gq‐タイプのDMR信号は急激に変化するものの、Gs‐タイプDMR信号は比較的遅い。本明細書に開示されるようにオーファンGPCRsのGタンパク質カップリング機構を特定するのにDMR信号に関して固有の特性が使用され得る。
【0052】
しかしながら、両方のGq−DMR信号とGs−DMR信号は、テストされた多数の細胞株にわたって遍在しない場合、かなり普遍的であることがわかったが、Gi−DMR信号は受容体と同様に細胞の成分に強い依存を示す。その上、同一の細胞株における異なる受容体の活性化又は異なる細胞株における同一の受容体の活性化を介して介在された同じクラスのDMR信号に対して、振幅や動力学等の微細な特徴において、有意差がしばしばある。かかる差は、各受容体固有のシグナリングと同様に、GPCRシグナリングにおける細胞の状況関して重要な役割を反映している。したがって、その動特性及び形状に基づいて、特定のシグナリング経路をGPCRリガンド誘起の光学応答に帰属せしめることは、大抵困難である。GPCRリガンドによって活性化され且つリガンドによって誘起された光学応答を支配するその動特性及び形状に基づいて、経路の抽出が、主シグナリング経路を決定するのに、大抵は必要である。
【0053】
GPCR活性に関するバイオインピーダンス信号−
典型的なインピーダンスベースの細胞分析において、細胞は搬送されて、培養ウェルの底に配置された金電極に接触せしめられる。センサーシステムの総インピーダンスはバイオセンサを囲むイオン環境により、主に決定される。電界を印加すると、イオンは電界によって指向された運動を行い、濃度勾配による拡散を受ける。細胞全体について検出する際には、全電気インピーダンスは、電界溶液の抵抗、細胞のインピーダンス、電極/溶液界面におけるインピーダンス、及び電極/細胞界面におけるインピーダンスの4つの成分を有する。さらに、細胞のインピーダンスは、抵抗とリアクタンスとの2つの成分を含む。細胞のイオン強度に関する伝導特性は抵抗成分を与えるものの、不完全なコンデンサとして作用する細胞膜は周波数に依存する応答成分に寄与する。したがって、全インピーダンスは、例えば、細胞の生存能力、細胞の密集度、細胞の数、細胞の形態、細胞の付着度、イオン環境、細胞内の水分、検出周波数等を含む多くの要因の関数となる。
【0054】
RT−CESシステムにおいては、印加された小電位の割合は細胞内部に連結されている。細胞に印加されたかかる信号は、典型的ほ乳動物細胞の静止膜電位よりもはるかに小さいと信じられている。その結果、細胞の機能に対してほとんど妨害しないか、若しくは全く妨害しない。RT−CESシステムは、インピーダンスのこれら変化を測定し、細胞インデックスと呼ばれるパラメータとしてそれを表示する。数式(2)に従って、細胞インデックスが計算される。
【0055】
(数式2)
【0056】
Nは、インピーダンスが測定された振動数ポイントの数であり(例えば、10kHz、25kHz、及び50kHzに対してN=3)、R0(f)とRcell(f)は、それぞれ、ウェルにおいて細胞が存在しない場合と存在する場合における周波数電極抵抗である。
【0057】
CellKeyシステムにおいて、センサーシステムのインピーダンス変化は、受容体刺激に応答して生ずる細胞層のインピーダンス(デルタZ又はdZ)の変化に帰属される。低周波数において、印加された小電位は、細胞外の電流(iec)を誘起する。かかる細胞外の電流(iec)は、層内の個々の細胞の周囲に流れる。しかしながら、イオンチャンネルによる細胞膜を介した伝導電流も低測定周波数において重要であり得る。高周波数において、それらは、細胞性の膜に侵入する細胞透過電流(itc)を誘起する。各ウェルに対して測定された電流に対する印加電圧の比が、オームの法則により説明されるように、インピーダンス(Z)となる。
【0058】
細胞が受容体リガンド等の刺激に曝される際に、シグナル変換事象が活性化され、アクチン細胞骨格の調節等、複雑な細胞事象が引き起こされる。かかる複雑な細胞事象は、例えば、細胞の密着性、細胞の形態、及び体積、細胞間相互作用について変化を生じせしめる。これら細胞の変化は、細胞外電流及び細胞内電流の流れに対して個別的又は集団的に影響を与え、これにより、測定されたインピーダンスの強度及び特性に影響を与える。3つのクラスのGPCRs活性化を介して介在する3つのタイプのインピーダンス信号が知られている(Leung,G.,et al.,J.Assoc.Lab.Automat.,2005,10,258)。理論によって制限されないものの、これらインピーダンス信号はアクチン細胞骨格に与える異なる効果に起因すると信じられている。かかるアクチン細胞骨格は、異なるクラスのGPCRsの活性化に応答して、インピーダンスによって測定された細胞パラメータに影響を与える。Gq−GPCRs及びGi−GPCRsの活性化によって、アクチン重合が増強されるものの、Gs−GPCRsの刺激によって、アクチン解重合が引き起こされることが示されている。
【0059】
2.GPCRシグナリングとシグナリングの積分
生体組織に共通の特性は、環境変化と共に有機体の活性化を絶えず調整する動的能力にある。環境と交信する機能は多数経路を介して達成される。かかる多数経路は、外部環境や細胞内の異なる領域等から信号を受信しかつ処理する。個々の経路は、直線的な器官の管に沿って信号を伝え、その結果、個々の細胞機能が調整される。このタイプの情報転送は、調節機構に関する細胞の能力範囲に関して重要な部分となっている。しかしながら、より多数の細胞シグナリング成分及び経路が特定されかつ研究されると、これらの直線的経路は、独立のエンティティではなく、大なるネットワークの部分であることが明らかになっている。
【0060】
異なるタイプ細胞のネットワーク及び構成に関する初期研究から得られた1つの驚くべき結果は、細胞のタイプ特有の入力(すなわち、受容体)から信号を受信し、かつ、細胞のタイプに特有の機構に係合する一般的な信号通信ネットワークがあり得るということである。シグナリング成分及びそれら相互作用パターンに関する分子の特定は、細胞のタイプに特有なものであり得るが、これらの構成の全体的な機能と化学的信号回路の論理は、細胞のタイプに応じて維持される。
【0061】
理論によって制限されないものの、ネットワークはシグナリング経路間の内部接続から生じ得ると信じられている。同一のシグナリング成分が複数入力から信号を受信することができるので、かかる内部接続が生じ得る。かかるネットワークは、同様のクラスのシグナリング経路内で生じ得る。かかる同様のクラスのシグナリング経路は、例えば、Ras経路とRho経路との間、又はGsα/cAMP及びMAPキナーゼ経路等の異なる経路間である。
【0062】
GPCRシグナリング
GPCRsは、一連の様々な知覚性刺激及び化学的刺激に応答する7つの膜貫通型スパニングタンパク質のスーパーファミリを構成している。その様々な知覚性刺激及び化学的刺激とは、例えば、光、におい、味、フェロモン、ホルモン、化学的伝達物質である。GPCRsは、これら刺激によって与えられた情報を、細胞内の第2メッセンジャ変換する。その細胞内の第2メッセンジャは、重要な信号として細胞によって解釈される。この処理過程は、ヘテロトリメリックグアニンヌクレオチド結合タンパク質(Gタンパク質)を用いて、種々のエフェクタシステムに対して、それら相互作用を介して作用物質活性のGPCRsの結合を含む。GPCRに対する作用物質の結合は、受容体の形態状態を選択する。その受容体の形態状態は、Gタンパク質サブユニット上のGTPへのGDPの交換を促進し、Gタンパク質G−サブユニット及びG−サブユニットの解離を可能にせしめるように想定されている。その後に、活性されたG−及びとG−サブユニットは、エフェクタ酵素とイオンチャンネルの活性化を、正の調整、負の調整、若しくはその双方の調整を実行する。GPCRの作用物質の活性化はエフェクタシステムのGたんぱく質依存の活性化をもたらし、1)Gタンパク質結合のフィードバック調節、2)受容体エンドサイトーシス、及び3)Gタンパク質独立性信号変換経路を介したシグナリングを可能にする一連の分子相互作用を適切に設定する。
【0063】
ヘテロ3量体Gタンパク質を介したシグナリングに加えて、GPCRsは、Gタンパク質独立性信号変換錯体の形成と区分化を促進する足場として作用することが現在認識されている。GPCRsを結合せしめ、GPCRsをGタンパク質独立性信号変換経路に結合せしめ若しくはGタンパク質の特異性及び作用物質の選択性を変更するより多くのタンパク質が特定されている。GPCR相互作用タンパク質のリストは、例えば、GRKs、アレスチン、カルモジュリン、カルシオン、Aキナーゼアンカータンパク質(AKAP)、チューブリン、受容体活性変更ヤヌスキナーゼ2、PDZドメイン含有タンパク質(例えば、NHERF、RGS12)、SH3ドメイン含有アダプター分子(例えば、Grb2、Nck、c−Src、及びエンドフィリン)、小Gタンパク質、及び同様のたんぱく質を含む。
【0064】
受容体に結合されたGタンパク質に依存して、GPCRsは少なくとも4つの主カテゴリに分類され得る。かかる受容体は、Ca2+の可動化を引き起こすGq−共役受容体、Ca2+の可動を引き起こすGq−共役受容体、cAMPを低減せしめるGi/o−共役受容体、cAMPの集積を生じせしめるGs−共役受容体、Rho等の小GTPを活性化するG12/13−共役受容体を含む。
【0065】
細胞内cAMPレベルのGPCRシグナリングと積分
アデニリルシクラーゼ(ACs)は、細胞情報伝達ネットワーク内のジャンクション若しくは信号インテグレータとして見なされてきた。異なるアデニリルシクラーゼアイソフォームには、異なる信号受信能力がある。すべてのアデニリルシクラーゼアイソフォームは、Gs共役受容体からの信号に対応してcAMPを生成することができる。しかしながら、これらアイソフォームは、種々の入力から信号を受信する能力において異なる。細胞内のcAMPレベルは多くの経路間の信号のバランスに関する指標として機能し得る。アデニリルシクラーゼの信号受信能力について詳細に説明するこの複雑な状況が、図1に示されている。図面を参照すると、図1は、アデニールサイクラーゼイソフォーム依存性細胞シグナリング経路を示す概略図である。アデニールサイクラーゼは、複数の調整力のあるシグナリング経路のターゲットであり、自身が属するシクラーゼ群に依存して、異なる応答をし得る。
【0066】
細胞において、9つのACイソフォームがあり、そのすべてがGsαによって活性化され、全てではないがいくつかかが、Gi/oαサブユニットによって抑制される。多かれ少なかれ、ACアイソフォームの各々は、1つより多いシグナリング経路から入力を受信するので、細胞応答は、自身が発現するイソ酵素によって調整される。実施例において、Ca2+/CaMII(カルモジュリン依存性プロテインキナーゼII)はACI(アデニールサイクラーゼサブタイプ1)を活性化するものの、Ca2+はACV(アデニールサイクラーゼサブタイプ5)及びACVI(ACサブタイプ6)を抑制する。別の実施例において、ACV及びACVIはGiαによって抑制されるものの、ACII(ACサブタイプ2)及びACIV(ACサブタイプ4)は、Giαに対して無反応であるように見える。
【0067】
また、細胞のcAMP反応は刺激される受容体のGタンパク質の選択度/特異性に依存している。Ca2+は、反対に、異なるアデニリルシクラーゼに影響する。ACIはCa2+/CaMIIによって刺激され、ACV及びACVIはCa2+によって抑制され、リン酸化反応は種々の形態のPKC(プロテインキナーゼC)及びPKAによって抑制され、ACII、IV、及びVIIのACは、細胞内のCa2+変化に対して鈍感であるようである。無傷細胞において、Ca2+の効果は、発現されたイソ酵素のみに依存するのではなく、Ca2+の増加が生ずるサイトにも依存し得る。ACIIを発現する細胞において、ACIV若しくはACVIIはCa2+を増加させ、ジアシルグリセロール(DAG)は、cAMPの増加を引き起こす。かかるcAMPは、Ca2+及び酵素を形成するcAMPのDAG刺激されたリン酸化反応によって調節され得る。対照的に、PKA介在リン酸化反応は、ACVとACVIの負のフィードバックレギュレータとして作用し得る。
【0068】
酵素アデニールサイクラーゼの9つの細胞膜に束縛されたアイソフォームは、細胞内cAMPレベルを調節するためにGタンパク質共役受容体を通して作用する化学的伝達物質と薬物によって大きく調整される。一般に、Gs共役受容体の急性の活性化はcAMPの蓄積を刺激するものの、Gi/o共役受容体の急性の活性化はcAMPの蓄積を通常は抑制する。Gタンパク質共役受容体の永続的な活性化によって、その後の薬物調節されたcAMPの蓄積が変更されることも確立されている。持続する受容体の活性化に続いて、これら変更は細胞適応型の応答を表していると信じられている。
【0069】
異種感作
一般的に観測された1つの現象、すなわち、アデニールサイクラーゼの異種感作は、Gi/o共役受容体持続的な活性化に続く薬物による刺激されたcAMPの累積に対する高応答性によって特徴付けられる。アデニールサイクラーゼの異種感作は、慢性の鎮静剤投与に続く耐性及び使用中止に関して説明するために当初は提案され、細胞が抑制受容体の持続された活性化に適応する機構であり得る。かかる適応型の機構は、例えば、薬物依存及び薬物乱用からの使用中止の処理において、及び、分裂病と鬱病を含む精神神経系症状において、役割を果たすように提案されてきた。また、異種感作は、複数のGi/oたんぱく質の同時活性化を必要とし得る。例えば、全内因性のGi/oプールが利用可能であった細胞の感作と比べると、選択性Go−誘起の異種感作の強さは、低減されるようである。
【0070】
異種脱感作
受容体介在の細胞応答の脱感作は、複数の経路を要する複雑な処理である。かかる複雑な処理は、a)Gタンパク質から解かれた受容体、b)原形質膜から離間した受容体の隔離、c)シグナリングタンパク質の下降調整を含む。受容体脱感作は一般的に相同である(すなわち、特有の受容体でありかつ受容体の占有に依存する)。
対照的に、活性物質によるアデニールサイクラーゼの直接活性化又は細胞透過性cAMPによるPKAの直接的な活性化は、異種脱感作と関連付けられた受容体のリン酸化反応を大部分は引き起こし得る(すなわち、非特定的の受容体であり、受容体の占有に依存しない)。
【0071】
細胞内Ca2+レベルのGPCRシグナリングと積分
同様に、また、細胞内Ca2+レベルはGPCRシグナリングにおいて重要な役割を果たす。細胞内Ca2+レベルはイオンチャンネルによって一般的に調整される。複数のGタンパク質介在経路は1つにまとまって、カルシウムチャンネルを調節することが知られている。事象の複雑なタイミングはいくつかのシグナリング経路の構成部と細胞骨格構造及び足場構造の動的ネットワーク間の相互作用によって制御される。その細胞骨格構造及び足場構造は、これら構成とカルシウムチャンネル間の相互作用のための層として機能し得る。カルシウムチャンネルの変更を誘起することが知られた受容体の多くが、百日咳毒素(PTX)を有する細胞の前処理がカルシウムチャンネルの抑制を止めるという観測によって示されているように、ヘテロトリメリックGi/oたんぱく質に結合される。カルシウムチャンネルの抑制は、細胞膜によって範囲を定められた信号若しくは第2のメッセンジャによって調節された信号を介して生じ得る。細胞膜によって範囲を定められた経路は、Gタンパク質βγーサブユニット(Gβγ)の直接結合によって調節され、Cav2.2チャンネルの電圧依存性抑制という結果となる。
【0072】
無標識バイオセンサ分析用いた経路の抽出
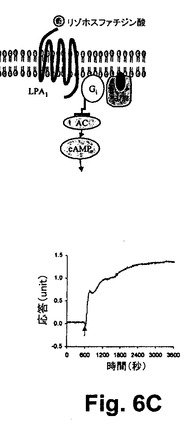
GPCRシグナリングは高度化され、非常に複雑である。受容体は、Gタンパク質依存性経路及びGタンパク質独立性経路を介して、シグナリングを介在し得るし、受容体は複数のGたんぱく質依存性経路を介して介在し得る。無標識バイオセンサ分析は、積分された細胞の応答(例えば、光学バイオセンサを用いたDMR信号、若しくは、電気バイオセンサを用いたバイオインピーダンス)を測定するので、受容体の活性化により、非定型的なバイオセンサ反応若しくは出力信号が引き起こされる。これは、例えば、リガンドと細胞のコンテクスト(又は、バックグラウンド)の特性に依存している。例えば、2つの内因性のGi−共役受容体、LPA受容体(図6C)、及びニコチン酸受容体(図5A)は、同一のA431細胞膜において2つの異なるタイプのDMR信号を引き起こす。これは、受容体によって誘起されたシグナリング処理の複雑性及び特異性を反映している。その上、A431細胞における同一のLPA受容体は、完全に静的な状態対増殖状態(図6C、データは示されていない)等の細胞の状況に依存して、異なるタイプのDMR信号を引き起こすであろう。したがって、無標識バイオセンサ分析は、リアルタイムで、受容体シグナリングに続いて、明確な動力学応答を提供するものの、動力学応答の動特性のみが、経路の特定のための予測的な指標でなくてもよい。
【0073】
バイオセンサシグナリングを統合する方法を利用することによって、少なくとも2つの異なる手法は、無標識バイオセンサ分析を使用して、GPCRシグナリングを抽出するのに使用され得る。ホルスコリン若しくはNKH447等のAC活性物質、又は細胞プリアンブルcAMP類似物質等のPKA活性物質を使用して、第1方法(方法1)は、細胞内cAMPレベルを上昇せしめる。GPCRリガンドによって誘起されたバイオセンサの出力に及ぼす高cAMPレベルの特異な効果若しくは作用に基づいて、GPCRシグナリングを抽出することができる。前記増大された細胞内cAMPレベルは、Gq若しくはG12/13シグナリングに対してほとんど抑制効果を与えないか若しくは全く与えず、異種脱感作によりGsシグナリングを完全に減衰せしめ、異種感作によりEC50濃度におけるGiシグナリングを増強せしめる。
【0074】
U73122等のPLC抑制物質若しくはPLC遺伝子抑制干渉RNAのうちの一方を使用して、第2方法(方法2)は細胞内のホスホリパーゼC(PLC)活性の活性化を抑制する。かかる手法は、無標識分析を使用して、GPCRシグナリングを抽出する補完的な方法を提供する。PLC活性の抑制はGs若しくはG12/13シグナリングに対してはほとんど効果を示さないものの、Gqシグナリングを完全に減衰せしめ、Giシグナリングを部分的に減衰せしめる。2つの手法は、無標識細胞分析を使用して、GPCRシグナリングを抽出するのに独立して使用され得るし、組み合わせても使用され得る。上記2つの方法と、示されたGPCRに与えるそれら方法のそれぞれの効果が、添付した表にまとめられている。
【0075】
【表1】
【0076】
実施例
以下の実施例は、上述した開示を使用する方法をより完全に説明し、さらに開示の種々な態様を行うように想定された最良の態様を詳しく説明するために用いている。これらの実施例は本開示の範囲を限定してはおらず、説明及び例示を目的として提示されていると理解される。
【0077】
バイオセンサベースの生細胞分析によって、細胞応答の検出結果が、ホルスコリンによるアデニールサイクラーゼの活性化に続いてGタンパク質共役受容体等の活性化によって得られる。ホルスコリンによるアデニールサイクラーゼの持続的活性化により、受容体のタイプに依存して、作用物質刺激に対してGタンパク質共役受容体が敏感になり若しくは鈍感になる。
【0078】
実験方法
材料 −
ホルスコリン及びエピネフリンは、Tocris(セントルイス、ミズーリ州)から入手した。ヒスタミン及びニコチン酸は、シグマ・ケミカル・カンパニー(SigmaChemicalCompany)(ミズリー州、セントルイス)から入手した。Epic社登録商標384ウェル細胞分析マイクロプレートはコーニング株式会社(コーニング、ニューヨーク州)から入手した。各ウェルが共鳴導波グレーティング(RWG)バイオセンサから成るマイクロプレートは、すぐに培養することができ、いかなる前処理も必要とせずに直接的に使用された。
【0079】
細胞の培養 −
人間の表皮癌A431細胞(American Type Cell Culture)が、ダルベッコの未改質Eagle媒質(DMEM:Dulbecco’s modified Eagle’s medium)内で培養された。かかる媒質には、10%の牛胎児血清(FBS)、4.5g/リットルのグルコース、2mMのグルタミン、及び抗生物質が加えられた。10%のFBSを含む50LのDMEM媒質に3〜8回浮遊された約1.8×104の細胞が384ウェルマイクロプレートの各々のウェル内に設けられ、37℃で空気/5%CO2下で1日間培養され、かかる細胞を続いて無血清DMEMにおいて継続して培養することによって、約20時間、放置した。
【0080】
光学的バイオセンサシステム及び細胞分析 −
コーニング登録商標Epic登録商標波長送受観測システムが使用された。システムは、独立型とすることができ、温度制御ユニット、光学検知ユニット、及びロボティックスを有するオンボード液体処理ユニットから構成され得る。温度制御ユニットが、必要に応じて温度変動を最小にするべく内蔵されている。このユニット内に、センサーマイクロプレートと化合物ソースプレートの双方を処理する2つの並列スタックがある。一旦、安定した温度が得られると(通常1時間以内)、センサーマイクロプレートは、ロボティクスによって、検知システムの直上のプレートホルダに自動的に搬送され、ソースプレートは適切なコンパートメントに移動されるので、オンボード液体処理ユニットによって容易に処理可能である。
【0081】
検知ユニットが、統合されたファイバオプティクスの中心に設けられ、例えば、生細胞内のリガンド誘起の動的質量再分布に起因する共鳴光の波長シフトが測定される。マイクロプレートの底部を介した垂直入射によって、光ファイバとコリメータレンズとを介して生成された830nmを中心とする広帯域白色光源が、グレーティングの小領域を照明するのに使用された。反射光を記録する検出ファイバは照光ファイバを用いて1束にされた。一連の照光/検出ヘッドが直線的に配置されるので、反射スペクトルは、384ウェルのマイクロプレートの同じカラム内にある16ウェルから一度に収集される。プレート全体が照光/検出ヘッドによってスキャンされて、各センサが複数回扱われ、各カラムは連続して扱われ、6秒若しくは13秒の時間間隔で細胞応答の動的測定が行われた。
【0082】
動的分析を目的として、細胞は、HBSS(20mMのHEPESを有するHanks Balanced Salt Solution)緩衝液を用いて洗浄された。検知システム内で1時間培養した後に、センサープレートがスキャンされ、ベースラインの応答が記録された。そして、化合溶液が、オンボード液体処理システムを使用して、センサープレートに移され、細胞応答が別の期間に記録された。同様に、第2の任意のステップは、例えば、異なる化合溶液を添加するステップを含み、かかるステップが実行されると、第2リガンド誘起の応答に対する第1の化合物の効果が決定される。化合物が導入されると、センサーマイクロプレートのリッドが、短期間(例えば、約1分)を除いて、分析中、維持された。プレートリッドは、ロボティクスによって自動的に処理された。すべての分析測定が被制御温度(28℃)にて実行された。
【0083】
統計的分析 −
明確に言及しない限り、各測定若しくは各化合物に対して、3回測定が行われた。標準偏差がこれら測定値(n=3)から得られた。分析係数の変動は、通常約10%未満であることが見出された。投与量依存性応答の全てが、(Graph Padから利用可能な)Prismソフトウェアを用いた非線形回帰法を使用して分析された。
【0084】
結果および議論
図2は、標的細胞内に出現するアデニールサイクラーゼの下位集合に依存して、cAMPレベルを増大させ又は低減せしめ得るGq(Gαq)、Gi(Gαi)、Gs/o(Gαs/o)、及びGβγ介在シグナリング経路から、細胞内アデニールサイクラーゼまでのクロストークと、を示す概略図である。
【0085】
図3Aは、ホルスコリン前処理から生じる光学応答の実施例を示しており、かかるホルスコリン前処理に続いて静的A431細胞におけるGs共役受容体(A2アデノシン受容体)の活性化処理が行われた。通常、ホルスコリンによるアデニールサイクラーゼの活性化が、Gs共役受容体の活性化によって相乗的に作用するものの、種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理は、続いて100nMのNECAの暴露処理がなされて(N−エチルアデノシンー5’−ウロン酸、A2‐特有作用物質)、異種脱感作を引き起こす、すなわち、その作用物質による刺激に対するA2受容体の応答能力が低減された。同様に、種々の濃度のNECAを用いたA431細胞の1時間の前処理は、アデニールサイクラーゼを減感せしめ、ホルスコリン誘起応答(図3B)を減衰せしめる。異種脱感作に関する観測が示していることは、クロストークがシグナリング経路間において生じ、これはアデニールサイクラーゼの活性化とA2受容体の活性化から生ずることである。理論によって制限されないものの、これら2つの経路間のクロストークに関して考えられる機構は、ホルスコリンによるアデニールサイクラーゼの活性化の際に、cAMPが生成され且つPKAを活性化することにある。PKAがGs受容体をリン酸化し得るし、Gs介在反応を完全に抑制せしめる。したがって、図3A及び3Bは、3A)NECA誘起のGs応答に対するホルスコリン前処理の効果と、3B)静的A431細胞におけるホルスコリン誘起の反応に対するNECA前処理の効果と、をそれぞれ示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(300=5nm、310=44nm、320=174nmと、330=348nm、340=690nm)が行われ、続いて100nMのNECAの暴露処理がなされた結果、NECA誘起の応答の減衰を引き起こす。ホルスコリンによる抑制効果は、シグナリング経路間のクロストークに帰属すると信じられており、ホルスコリンによるアデニールサイクラーゼの活性化とNECAによるA2アデノシン受容体の活性化から生じる。同様に、種々の濃度のNECAを用いたA431細胞の前処理(350=3nm、360=384nm、370=696nm、380=5nm、390=11nm)に続いて1Mのホルスコリンによる処理をした結果、A2受容体作用物質が1.3nMのIC50とのホルスコリンの結合を抑制することを示した。
【0086】
図4Aは、ホルスコリン前処理から生じる細胞応答の実施例を示している。かかるホルスコリン前処理に続いて静的A431細胞においてGq−共役受容体(H1ヒスタミン受容体)の活性化処理が行われた。種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理に続いて100nMのヒスタミン(H1−特有作用物質)の暴露によって処理したところ、部分的な脱感作をも引き起こした。
【0087】
ホルスコリンの前処理による投与量依存の部分的な抑制は約184nMのIC50を引き起こした。これは、ホルスコリンの効能に対応する。作用物質誘起の脱感作に関する観測は、この場合、アデニールサイクラーゼの特定のアイソフォームにおけるいくつかのクロストークがシグナリング経路間において生じ、これは、ホルスコリンによるすべての内因性のアデニールサイクラーゼの活性化とH1受容体の活性化から生ずることを再度示している。図4A及び4Bは、4A)ヒスタミン誘起のGq応答に対して与えるホルスコリン前処理の効果、及び、4B)静的A431細胞におけるホルスコリン誘起の反応に対するヒスタミン前処理の効果をそれぞれ示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(400=3nm、410=5nm、420=174nm、430=696nm、440=2,784nm)に続いて100nMのヒスタミンの暴露処理がなされた結果、ヒスタミン−誘起の応答の減衰を引き起こした。ホルスコリンによる抑制効果は、シグナリング経路間のクロストークに帰属すると信じられている。これは、ホルスコリンによるアデニールサイクラーゼの活性化とヒスタミンによるH1ヒスタミン受容体の活性化とから生じる。同様に、種々の濃度のホルスコリンを用いたA431細胞の前処理(455=30nM、460=90nM、465=9.9nM、470=270nM、475=810nM)に続いて1Mのホルスコリンによる処理をしたところ、H1受容体作用物質が、301nMのIC50とのホルスコリンの結合を抑制することを示した。
【0088】
図5は、ホルスコリン前処理から生じる細胞応答の実施例を示している。かかるホルスコリン前処理に続いて静的A431細胞においてGi‐共役受容体(ニコチン酸受容体HM74受容体)の活性化処理が行われた。Gs−共有受容体及びGq−共有受容体の活性化と対比して、種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理に続いて25Mニコチン酸(HM74−特有作用物質)の暴露によって処理したところ、H1−誘起の応答の増強を引き起こし、ホルスコリンのEC50において最大値に達する。図5Aは、静的A431細胞におけるニコチン酸誘起のGi応答に対するヒスタミン前処理の効果を示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(すなわち、500=0nm、510=4nm、520=32nm、530=125nm、540=4,000nm、550=32,000nm)に続いて25Mのニコチン酸によって処理したところ、ニコチン酸誘起の応答の増強が引き起こされた。Gi応答の増強作用は、異種感作に帰属すると信じられている。
【0089】
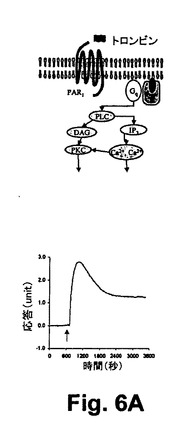
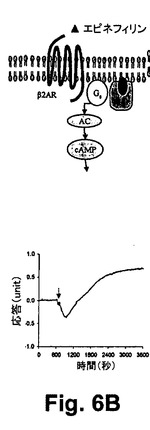
図6A乃至6Cは、RWGバイオセンサを使用した生細胞における3つのクラスのDMR信号のGPCRシグナリング(Gq‐、Gs‐、及びGi‐)を示している。すなわち、6A)GqシグナリングとそのDMR信号(チャイニーズハムスター細胞、10ユニット/mLのトロンビンにおけるプロテアーゼ活性されたレセプターサブタイプ1)と、6B)Gsシグナリング及びそのDMR信号(A431細胞、2nMエピネフリンにおける2ーアドレナリン作用受容体)と、6C)Giシグナリング及びそのDMR信号(A431,100nM LPAのリゾホスファチジン酸(LPA)受容体サブタイプ1)と、を示している。破線矢印は作用溶液が導入された時間を示している。
【0090】
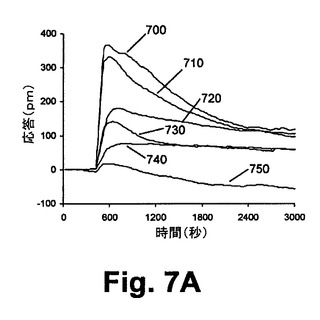
図7Aは、静的A431細胞におけるヒスタミン誘起のDMR信号に対するU73122の効果を示している。種々の濃度のU73122を用いたA431細胞の前処理(すなわち、700=0M、710=1.78M、720=7.15M、730=28.5M、740=114M、750=228M)に続いて1,000nMのヒスタミンを用いて刺激の処理を行ったところ、ヒスタミン誘起のDMR応答の投与量依存性が抑制される。ヒスタミンは、A431細胞に内因的に発現したGq−共役受容体ヒスタミン受容体サブタイプ1に対して、天然の作用物質である。図7Bに示されたヒスタミン反応のP−DMR振幅に基づいて、高投与量におけるU73122がヒスタミン反応を完全に抑制し得る。
【0091】
本開示は、種々の特定の実施形態及び技術を参照して説明されてきた。しかしながら、本開示の精神及び範囲内において多くの変更及び改良を行うことができることが理解されるはずである。
【参照】
【0092】
1.Y, Fang et al., ”Label−Free Biosensors and Cells”, PCT App. No. PCT/US2006/013539 (Pub.No. WO 2006/108183)(Attorney Docket No. SP05−037).
2.Fang, Y., et al., ”Resonant waveguide grating biosensor for living cell sensing,” Biophys. J., (2006) 91, 1925−1940.
【先願の米国出願の利益の請求】
【0001】
本出願は、2007年10月6日に出願された米国仮出願第60/997,910号、2007年11月30日に出願された米国特許出願第11/998,711号の利益を主張する。この先願の米国出願の内容及びこの明細書で引用された公報、特許、及び特許出願の全てが参考として組み込まれている。
【技術分野】
【0002】
本開示は、共鳴導波グレーティングバイオセンサンサ又は表面プラズモン共鳴(SPR)バイオセンサ等の光学バイオセンサに関し、特に、バイオセンサベースの生細胞分析を利用したGPCRシグナリング経路の分析方法に関する。
【発明の概要】
【0003】
本開示は、バイオセンサベースの生細胞分析を利用したGタンパク質共役受容体(GPCR)等受容体シグナリング経路の分析及び解明に対して直接的方法及び間接的方法を提供する。
【図面の簡単な説明】
【0004】
【図1】図1は、本開示の実施例におけるアデニールサイクラーゼアイソフォーム依存性細胞シグナリング経路を示す概略図である、
【図2】図2は、本開示の実施形態におけるGq(Gαq)、Gi(Gαi)、Gs/o(Gαs/o)、及びGβγ介在シグナリング経路から、細胞内アデニールサイクラーゼまでのクロストークと、cAMPレベルに与えるその影響と、を示す概略図である。
【図3A】図3Aは、本開示の実施形態における静的A431細胞におけるNECA誘起のGs応答に与えるホルスコリンの前処理の効果を示す図である。
【図3B】図3Bは、本開示の実施形態における静的A431細胞におけるホルスコリン誘起のGs反応に対して与えるN−エチルアデノシン−5’−ウロン酸(NECA:N−ethyladenosine−5’−uronic acid)前処理の効果を示す図である。
【図4A】図4Aは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のGq応答に対して与えるホルスコリン前処理の効果を示す図である。
【図4B】図4Bは、本開示の実施形態における静的A431細胞におけるホルスコリン誘起のGq反応に対して与えるヒスタミン前処理の効果を示す図である。
【図5A】図5Aは、本開示の実施形態における静的A431細胞におけるニコチン酸誘起のGi反応に対して与えるホルスコリン前処理の効果を示す図である。
【図5B】図5Bは、本開示の実施形態における静的A431細胞におけるニコチン酸誘起のGi反応に対して与えるホルスコリン前処理の効果を示す図である。
【図6A】図6Aは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図6B】図6Bは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図6C】図6Cは、本開示の実施形態におけるRWGバイオセンサを使用した生細胞における3つのクラスの(Gq‐、Gs‐、及びGi‐)DMR信号のGPCRシグナリングを示す図である。
【図7A】図7Aは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のDMR信号に与えるU73122の前処理の効果と、そのP−DMR振幅とをそれぞれ示す図である。
【図7B】図7Bは、本開示の実施形態における静的A431細胞におけるヒスタミン誘起のDMR信号に与えるU73122前処理の効果と、そのP−DMR振幅とをそれぞれ示す図である。
【発明を実施するための形態】
【0005】
本発明の様々な実施形態が、添付の図面を参照して詳細に説明されるであろう。様々な実施形態に対する言及は、本発明の範囲を制限するものではなく、本発明は、添付した請求の範囲によってのみ限定される。さらに、本明細書に述べられたいかなる実施例も、限定することを目的とはしていないし、特許請求の範囲に記載された発明の多くの実施形態の内、いくつかについて説明しているにすぎない。
【定義】
【0006】
「分析(Assay)」、「分析する(Assaying)」等の用語は、例えば、外因性刺激を用いた刺激に対する細胞の光学的応答又はバイオインピーダンスの応答の存在の有無、量、程度、動特性、及びタイプを決定する分析を意味する。外因性刺激は、例えば、リガンド候補化合物、アデニラート活性剤、cAMP類似体、又はGPCRリガンド若しくはリガンド候補等である。
【0007】
「付着する(Attach)」、「付着(Attachment)」、「接着(adhere)」、「接着された(adhered)」、「接着(adherent)」、「固定された(immobilized)」等の用語は、物理吸着又は化学結合等の処理若しくはそれらの組み合わせ等によって、例えば、本開示の細胞等の構成要素を、表面に、固定し、準備し、培養等することを一般に意味する。特に、「細胞の付着(cell attachment)」、「細胞の接着(cell adhesion)」等の用語は、細胞固定材料、相溶剤(例えば、フィブロネクチン、コラーゲン、ラミン、ゼラチン、ポリリジン等)、若しくはそれら双方を用いて培養し、相互作用せしめることによって、表面との細胞の相互作用又は表面との細胞の結合を意味する。実施形態において、バイオセンサ上に生細胞を固定するステップは、例えば、約1パーセントから約99パーセントのバイオセンサ表面被覆率及び同様の密集度値若しくは範囲等を有する密集度が得られるまで生細胞をバイオセンサ表面に培養するステップを含み得る。
【0008】
「付着性細胞(Adherent cells)」とは、原核生物の細胞若しくは真核細胞等の細胞、細胞株又は細胞システムを意味し、かかる細胞、細胞株又は細胞システムは、基板の外部表面に関連付けられ、基板の外部表面上に固定され、又は特定の接触を維持する。培養後のかかる細胞は、洗浄媒質交換処理、すなわち、多くの細胞ベースの分析に必須である処理に対して抵抗力を有し、耐性を有し得る。「弱付着性細胞(Weakly adherent cells)」とは、原核生物の細胞若しくは真核細胞等の細胞、細胞株又は細胞システムを意味し、かかる細胞、細胞株又は細胞システムは、細胞培養中に、基板の表面と弱く相互作用し、基板の表面と弱く関連し、基板の外部表面と弱く接触する。しかしながら、これらのタイプの細胞、例えば、人間の胎児性腎臓(HEK:human embryonic kidney)細胞は、洗浄液若しくは媒質の交換等の物理的摂動によって、基板の表面から容易に解離する傾向がある。「懸濁細胞(Suspension cells)」とは、望ましくは、媒質で培養される細胞又は細胞株を意味し、当該媒質において細胞は、培養中に、基板の表面に付着し若しくは固着しない。「細胞の培養(Cell culture)」又は「細胞培養(Cell culturing)」とは、原核生物細胞若しくは真核細胞が一定条件下で培養される処理を意味する。「細胞の培養(Cell culture)」とは、多細胞の真核細胞、特に動物細胞から得られた細胞の培養と、複合生体組織及び臓器の培養と、を意味する。
【0009】
「細胞(Cell)」等の用語は、半透明の膜によって外部的に結合された通常小なる微小質量の原形質を意味し、1つ以上の細胞核と他の多種の細胞小器官を任意で含み、かかる他の多種の細胞小器官は、生命の基本的な機能を単独で発揮し又は生命の基本的な機能を実行する他の同等の集団と相互作用することができ、独立的に機能することができる生物の最小構造ユニットを形成し、合成細胞構造体、細胞モデルシステム、人工の細胞システムを含む。
【0010】
「細胞システム(Cell system)」等の用語は、互いに相互作用して、生物学的機能、生理学的機能又は病態生理学的機能を発揮する1より多いタイプの細胞(又は単一タイプの細胞の異なる形態)の集合を意味する。かかる細胞システムは、例えば、器官、生体組織、幹細胞、分化した肝細胞等の細胞を含む。
【0011】
「マーカ(Marker)」等の用語は、少なくとも1つの細胞ターゲット(例えば、Gq−共役受容体、Gs−共役受容体、Gi−共役受容体、G12/13−共役受容体、イオンチャンネル、受容体チロシンキナーゼ、トランスポータ、ナトリウム‐プロトン交換体、細胞核受容体、セルキナーゼ、細胞性タンパク質、等)の活性を調節することができ、且つ、バイオセンサによって測定されるように、信頼性の高い検出可能な出力若しくは応答が得られる分子、生物分子、又は生物製剤を意味する。目的とする細胞ターゲットの分類及び後に生ずるセル事象に依存して、マーカは、例えば、GPCR、受容体チロシンキナーゼ(receptor tyrosine kinase)、イオンチャネル、細胞核受容体、細胞酵素アデニールサイクラーゼ等に対する、例えば、作用物質、部分的作用物質、逆作用物質等の活性剤であり得る。また、マーカは、特定の種類の細胞ターゲットに対しては抑制物質となり得るし、例えば、アクチンフィラメント若しくはマイクロチューブに対しては抑制物質又はかく乱物質となり得る。
【0012】
「検出する(Detect)」等の用語は、感知されたシグナリング経路を発見し又は感知し、かつ、感知されたシグナリング経路と経路のシグナリングがない場合とを区別する本開示の装置及び方法の能力を意味する。
【0013】
「特定する(Identify)」等の用語は、シグナリング経路を検出しかつ解明する本開示の装置及び方法の能力を意味する。
【0014】
「治療候補化合物(Therapeutic candidate compound)」、「治療候補物質(therapeutic candidate)」、「予防的候補物質(prophylactic candidate)」、「予防的物質(prophylactic agent)」、「リガンド候補(ligand candidate)」等の用語は、バイオセンサ若しくは病原体に付着した細胞と相互作用する潜在能力にとって重要であり、自然に生ずる分子若しくは材料又は合成的な分子若しくは材料を意味する。治療候補物質又は予防候補物質は、タンパク質、DNA、RNA、イオン、脂質等の細胞ターゲット若しくは病原体、又は、生細胞若しくは病原体の同様の構造体若しくは要素のうち少なくとも1つと特に結合し又は相互作用し得るし、例えば、化合物、生物学的分子、ペプチド、タンパク質、生物試料、薬物候補の小分子、薬物候補の生物学的分子、薬物候補の小分子‐生物の置換体、及び同様の材料若しくは実在の分子、又はそれら組み合わせを含む。
【0015】
「バイオセンサ(Biosensor)」等の用語は、生物学的要素を物理化学的検出構成要素に結合せしめる検体を検出するデバイスを意味する。バイオセンサは、通常3つの部分から構成される。すなわち、(生体組織、微生物、病原体、細胞、又はこれらの組合せ等の)生物学的構成部又は要素、(例えば、光学的、圧電的、熱測定、若しくは磁気的な物理化学的手法で動作する)検出要素と、双方の構成部に関連付けられたトランスデューサと、から構成され得る。生物学的構成部若しくは生物学的要素とは、例えば、生細胞であり得る。実施形態において、光学的バイオセンサは、例えば、生体細胞における分子認識事象又は分子刺激事象を定量化可能な信号に変換する光学変換装置を含み得る。
【0016】
”含む(Include)”、”含む(includes)”等は、含んで(including)を意味するがこれに限定されない。
【0017】
例えば、成分の構成要素、濃度、堆積、処理温度、処理時間、収量、流速、圧力等の値の量、及びその範囲を修飾する「約(About)」という用語は、例えば、生じうる数量における変分を意味する。かかる変分は、本開示の実施形態を説明するのに使用されており、例えば、化合物、構成、濃度、使用製剤に対して使用された通常の計測と取扱を介して、これら手法における不注意な誤動作を介して、方法を実行するのに使用される製造方法、ソース、又は出発物質若しくは成分の純度における差等を介して、例えば、生じ得る。また、「約(About)」という用語は、特定の初期濃度若しくは混合度を有する成分又は処方の経時変化に起因して異なる量、及び、特定の初期濃度若しくは混合度を有する成分又は処方の混合処理又は処理工程に起因して異なる量を包含する。“約”という用語によって修飾された本明細書に添付された特許請求の範囲は、これら量の均等物を含む。
【0018】
実施形態において「〜本質的に成る(Consisting essentially of)」とは、本明細書に定義されたGPCRシグナリング経路検出、分析、若しくはそれらの双方、バイオセンサの表面上の製剤若しくは成分、及び、本開示の物品、デバイス、若しくは装置に対する方法を例えば意味し、特許請求の範囲に記載された構成要素又はステップと、構成物、物品、若しくは装置の基本的且つ新規な特性と、特定の反応物質、特定の添加物または成分、特定の物質、特定の細胞若しくは細胞株、特定の表面修飾剤または条件、特定のリガンド候補物質、可変選択された同等の構造、材料、又は処理等の本開示の製造方法及び使用方法と、に物質的に影響を与えない他の構成要素又はステップと、を含み得る。本開示の構成要素若しくはステップの基本特性に有形的に影響し若しくは本開示に不要な特性を与え得るアイテムは、例えば、バイオセンサの表面に対する細胞の低減された親和性、リガンド候補若しくは同様の刺激に応答した異常な若しくは逆の細胞の活性化、及び同様の特性を含む。
【0019】
特に指定しない限り、本明細書で使用された不定冠詞“a”“an”、又は対応する定冠詞“the”は、少なくとも1つ、又は1つ以上を意味する。
【0020】
当業者にはよく知られている簡略が使用され得る(例えば、時間又は(複数の)時間に対して「h」若しくは「hr」、(複数の)グラムに対して「g」若しくは「gm」、ミリリットルに対して「mL」、及び室温に対して“rt”、ナノメータに対して「nm」、等の簡略)。
【0021】
要素、成分、添加剤、細胞のタイプ、抗体等、及びその範囲に対して開示された特定の値又は好ましい値は、例示する目的のためにあるに過ぎず、それらは他の定義された値又は定義された範囲内の他の値を除外するものでない。
【0022】
開示された構成、装置、及び方法は、いかなる値をも有する構成、装置、及び方法を含み、或いは、本明細書に記載された値、特定の値、より特定の値、及び好ましい値のいかなる組合せをも有する構成、装置、及び方法を含む。
【0023】
実施形態において、本開示の方法は、Gタンパク質共役受容体(GPCRs)のシグナリング経路の分析、定量、又はそれら双方を提供する。かかる分析、定量、又はそれら双方は、例えば、GPCRリガンド誘起のバイオセンサの応答に対するホルスコリン刺激された細胞の効果を測定することによって行われ、例えば、ホルスコリン誘起のバイオセンサ応答に対するGPCRリガンド刺激された細胞の効果を測定することによって行われる。
【0024】
実施形態において、方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサに生細胞を固定するステップと、当該固定された生細胞を、前記生細胞における細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に接触せしめるステップと、当該物質に接触せしめられた生細胞をGPCRリガンドに接触せしめるステップと、前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0025】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該培養された生細胞を接触せしめるステップと、これらアデニラート活性剤又はcAMP類似物質が接触した細胞を、GPCRリガンドに対して接触せしめるステップと、を含み得る。
【0026】
実施形態において、方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサ上に生細胞を固定するステップと、当該固定された生細胞をGPCRリガンドに接触せしめるステップと、前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、前記GPCRリガンドが存在する場合と存在しない場合における前記生細胞に関する前記物質誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0027】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該培養された生細胞を接触せしめるステップと、これらアデニラート活性剤又はcAMP類似物質が接触した細胞を、GPCRリガンドに接触せしめるステップを含む。
【0028】
細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質は、例えば、ホルスコリン又はその派生物質等のアデニラート活性物質を含む。その派生物質は、NKH477、1,9ジデオキシ‐ホルスコリン、7‐デアセチル‐7‐O‐へミスチニル‐ホルスコリン、又は細胞浸透性cAMP類似物質である。細胞浸透性cAMP類似物質は、(S)−アデノシン、環状3’、5’−(ハイドロゲンホスホロチオエート)トリエチルアンモニウム、8−ブロモアデノシン−3’、5’−環状リン酸(8−Br−cAMP)、8−クロロアデノシン−3’、5’−環状リン酸、又は、N6,2’−O−ジブチリルアデノシン−3’、5’−環状リン酸等である。これらcAMP類似物質は、細胞膜透過性があり、直接細胞内に取り入られ、cAMP依存性タンパク質キナーゼ(タンパク質キナーゼA、PKA)の活性化を直接的に引き起こす。
【0029】
実施形態において、代替的方法は、例えば、バイオセンサ上に生細胞を培養するステップと、当該培養された生細胞をGPCRリガンドに接触せしめるステップと、細胞内cAMPレベルの増大を生じせしめ、次にタンパク質キナーゼAの活性化を生じせしめる物質に当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、を含み得る。
【0030】
実施形態において、代替的方法は、例えば、GPCRシグナリング経路の決定方法を含み得る。かかる方法は、バイオセンサ上に生細胞を固定するステップと、前記細胞内における細胞内ホスホリパーゼC(PLC)活性を抑制する物質に当該固定された生細胞を接触せしめるステップと、当該PLC抑制されたが生細胞をGPCRリガンドに接触せしめるステップと、前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む。
【0031】
実施形態において、方法は、例えば、バイオセンサ上に生細胞を培養するステップと、当該培養された生細胞をU73122若しくは干渉RNA等のホスポリパーゼC抑制物質若しくは抑制体に接触せしめるステップと、ホスホリパーゼCにより抑制された細胞をGPCRリガンドに接触せしめるステップと、を含み得る。
【0032】
本開示の方法は、例えば、GPCRシグナリング経路に与える周知の治療用化合物若しくは未知の治療候補化合物の陽性及び陰性の効果を特定する際に有益であり得る。したがって、前述の方法は、刺激を用いた固定生細胞の前処理、中間処理、又は後処理のさらなるステップを含み得る。
【0033】
実施形態において、本開示は、リガンド誘起バイオセンサ信号の研究において観測された詳細な生化学シグナリング経路を抽出し且つ解明する方法を提供する。本開示方法はすべての種類のGPCRシグナリングに適用可能である。本開示の方法は、異なる種類のGタンパク質シグナリングを区別し、GPCRのシグナリングと非GPCRのシグナリングとを区別し得る。本開示の方法は、アデニールサイクラーゼ(AC:adeneylate cyclase)活性物質(例えば、ホルスコリン、NKH447)を使用し得るし、又は、ホスホリパーゼC抑制物質と組み合わせて独立に使用され得る(例えば、U73122、合成アミノステロイド、すなわち、1−[6−[[17β−3−メトキシエストラ−1,3,5(10)−トリエン−17−yl]アミノ]hexyl]−1H−pyrrole−2,5−dione、又は干渉RNA)。
【0034】
本明細書に開示されたバイオセンサベースの生細胞分析を利用する経路分析若しくは経路抽出のツールとして、本開示は、バイオセンサベースの細胞分析を使用して、シグナリング経路を抽出し若しくは解明するユニバーサルな手法を提供する。
【0035】
実施形態において、本開示の方法はツーステップ分析を含み得る。また、本開示の方法は、受容体を介在する細胞シグナリングに関する有益な情報を提供し得る。
【0036】
実施形態において、本開示は、無標識バイオセンサベースの細胞分析を使用して、Gタンパク質共役受容体(GPCRs)のシグナリング経路を分析する方法を提供する。本方法は、生細胞におけるすべての種類のGPCRシグナリングに対して利用可能である。実施形態における方法は、GPCRリガンドによって誘起されたバイオセンサの応答に及ぼすcAMP増加細胞、ホスホリパーゼC抑制細胞、若しくはその双方の効果に基づいており、cAMPが生成する化学物質若しくは細胞浸透性cAMP類似物質によって誘起されたバイオセンサの応答に及ぼすGPCRリガンド刺激によって刺激された細胞の効果に基づいている。使用され得るバイオセンサは、例えば、表面プラズモン共鳴バイオセンサ、共鳴導波グレーティング(RWG)バイオセンサ等の光学バイオセンサ、バイオインピーダンスバイオセンサ、共鳴ミラー等のデバイス等の電気的バイオセンサを含む。
【0037】
Gタンパク質共役受容体(GPCRs)は細胞表面受容体の中で最も大きい群であり、細胞表面受容体は、細胞内ループ及び細胞外ループによって接合された7つの螺旋膜貫通領域の共通な構造モチーフを共有している。GPCRsは、異なる細胞システムにおいて異なる発現パターンにより、実質的に全細胞内で発現せしめられる。GPCRsの細胞外リガンドは多様的であり、生体アミン、アミノ酸、イオン、小型ペプチド、たんぱく質、及び生物活性な脂質を含む。GPCR活性物質のこのような多様性により、この受容体の種類の生理学的重要性が強調される。より詳しく述べると、GPCRsは、神経伝達、走化性、炎症、細胞の増殖等の処理を含む広範な生理学的過程を制御する。GPCRsは、例えばアストマ、癌腫、炎症、及び心臓血管疾患を含むほとんど全ての主要クラスの疾病に関わっている。
【0038】
健康及び疾病におけるそれらの役割と、小分子の薬物による治療の干渉に対するそれら潜在能力と、を想定すると、GPCRsは、ヒトゲノムにおいて最大の且つ最も成功したクラスの薬物に対して有能なターゲットに相当する。臨床的に利用可能なすべての薬物のうち約50%は直接若しくは間接的にGPCRsに対して活性的であるものの、これら薬物は、周知なすべてのGPCRsのうち数パーセントに対してのみ活性的である。ヒトゲノムは、最大1,000GPCRをエンコードしており、その内の約400が、内因性リガンドと結合すると期待された非化学的刺激受容体であると考えられている。これらの多く(約150)が、それら同族のリガンドとして分類され、若しくは生物学的機能は未だに知られていない「オーファン」GPCRsとして分類されている。これら受容体の「非−オーファン化」(すなわち、それら同族のリガンド又は生物学的機能の適用若しくは決定)は、新規治療ターゲット及び新規治療効果を提供し得る。
【0039】
GPCR薬物の発見と開発における継続的成功が、分析技術及び分析方法における発展に寄与している。GPCRスクリーニングの分析は、全体的細胞分析と無細胞分析とに広範に分類され得る。細胞ベースの分析によって、刺激に応答した細胞の振る舞いに関して高精度な指標が提供される。無細胞分析を使用して得られたデータと比較すると、細胞システムにおける化合物調節されたGPCR機能に関する直接測定によって、はるかに有益な情報が提供される。かかる情報とは、例えば、より密接に生理的環境に類似した条件下における化合物の作用、モード、及び機構である。これら利益により、薬物のスクリーニング及び試験を目的とする全体的細胞システムの使用が高められ、促進されている。
【0040】
GPCRsは、広範な一連の細胞シグナリング経路に関与している。かかる細胞シグナリング経路は、Gタンパク質依存性経路及びGタンパク質独立性経路の双方を介して、介在されており、たいては、リガンドに依存している。GPCRシグナリングは、川下のシグナリングネットワークの空間的且つ時間的フラックスによってコード化されている。かかるシグナリングネットワークは、細胞内のシグナリング及び調節機構によって厳密に制御される。GPCRシグナリングについて説明する多くのモデルにおいて、一旦、作用物質の結合によって活性構造で安定化されると、受容体が特定の細胞内ドメインを介して単一のGタンパク質と相互作用する機能的なモノメリックエンティティであることが想定されている。リガンドがGPCRに結合すると、例えば、受容体の構造に変化が生ずる。受容体の活性化は、順番に、Gαサブユニットに対するGTP−GDP交換を介して、関連Gタンパク質ヘテロトリマの活性化を引き起こす。そして、活性化されたGタンパク質は、いくつかの細胞内酵素の活性化を調節する。いくつかの細胞内酵素の活性化により、いくつかの主要な細胞内第2メッセンジャ生成が、順番に制御される。かかる第2メッセンジャは、例えば、環状AMP(cAMP)、cGMP、Ca2+、イノシトール・トリスリン酸、アラキドン酸である。そして、これら第2メッセンジャは、遺伝子転写及び細胞機能を調整するイオンチャンネル及びキナーゼを含む下流にあるいくつかのターゲットに作用する。
【0041】
合成化合物によって介在された細胞内シグナリングにおける予測不可能性が原因で、測定されたシグナリング事象に依存して単一化合物は作用物質又は拮抗物質として作用するであろう。さらに、測定された所定の事象において、効果不足により、受容体の活性化が不足するとは限らない。したがって、理想的なGPCRスクリーンは、単一の分析に依存するべきでなく、むしろ、統合された手法が、多くのシグナリング事象を測定するのに採用されるべきである。細胞シグナリング経路から独立した技術は、GPCR活性化が、多くの状況下においてGタンパク質独立シグナリングをも引き起こし得るという最近の調査結果を想定すると、それら経路バイアスの分析技術よりも優れているだろう。
【0042】
バイオセンサベースの細胞分析によって、より生理的に関連した環境下で、内因性受容体の活性化を観測することが可能となる。かかる非侵襲的能力は、リガンド薬理学に関してより魅力的な指標を与え得る。大抵のGPCR細胞分析は、細胞に関して特定の工学技術若しくは操作を利用しており、これは、確実であり且つ信頼性のある検出を得るために所定の感度を達成することを目的とする。これら人工的システムにおいて、リガンドの力価又は効果が変更され得る。これは、例えば、受容体の高い発現レベル又はターゲットの細胞生理学における特定の操作(例えば、GFP−タグ付け、又はトランスフェクション作用)の干渉に帰属する。例えば、α2A−アドレナリン作用受容体(α2AAR)は、Gαiプロテインに結合すると一般に信じられており、アデニレートサイククラーゼ活性の抑制を引き起こす。天然のHEL92.1.7細胞において、薬物のLevomedは、α2AARの逆作用物質であり、cAMP生成の増大を引き起こす。しかしながら、トランスフェクトされたPC10において、線維芽細胞Levomedは、陽性の作用物質であることが見出され、cAMP生成の抑制を引き起こす。
【0043】
さらに、バイオセンサベースの細胞分析においては、蛍光標識を必要としない。かかる蛍光標識は、多数の従来分析において検出の際に広く使用されている。かかる標識の非依存性により、GPCRスクリーンのデータ品質が大幅に改善され得る。蛍光性細胞の構成要素、標識、若しくは化合物の干渉が最小化され若しくは排除されるからである。
【0044】
バイオセンサベースの細胞分析は、積分されたバイオセンサの応答を測定している。例えば、RWGバイオセンサは、細胞内におけるリガンドによって誘起された動的質量再分布(DMR:dynamic mass redistribution)を観測する。これに反して、電気バイオセンサ、生細胞においてリガンドによって誘起されたバイオインピーダンスの変化を測定している。リガンドは細胞内の1つより多い数の受容体を活性化し得るし、受容体によって、複数のシグナリング経路が導かれる。この交絡の可能性が原因で、活性化された特定の経路を、リガンドによって誘起されたバイオセンサの応答に帰属させることは、バイオセンサ応答の形状及び動特性を測定するだけでは、困難である。
【0045】
1.バイオセンサベースの細胞分析とバイオセンサ基板
一般に、無標識細胞に基づく分析は、生体細胞におけるリガンド誘起の応答を観測することを目的としてバイオセンサを採用している。バイオセンサは、光学的、電気的、熱的、音響的、磁気的トランスデューサ等のトランスデューサを通常利用しており、分子認識事象又は細胞層におけるリガンド誘起の変化を定量化可能な信号に変換する。これら無標識バイオセンサは分子の相互作用の分析に通常使用され得る。かかる分析は、分子複合体が経時によってどのように形成し且つ分離するのかを特徴付けることを含む。本発明は、ほとんどすべてのタイプのバイオセンサ表面に利用可能であるものの、RWGバイオセンサと電気バイオセンサのみが例示されている。
【0046】
RWGバイオセンサ
RWGバイオセンサは、例えば、埋め込まれたグレーティング構造を有する基板(例えば、ガラス)と、細胞層と、から構成される。RWGバイオセンサは回折格子によって導波路に対する光の共鳴結合を利用し、界面において次々に電磁場が形成される溶液表面の界面において全内反射が生ずる。かかる電磁場はセンサの表面から指数関数的に減衰する事実上エバネッセントである。電磁場が初期値に関して1/eに減衰する距離は、侵入深さとして知られており、特定のRWGバイオセンサに関して、設計に応じた関数となるものの、通常約200nmのオーダーである。このタイプのバイオセンサは、かかるエバネセント波を利用して、センサ表面の近傍の若しくはセンサの表面における細胞層のリガンド誘起の変化を特性化している。
【0047】
電気バイオセンサ −
電気バイオセンサは、例えば、基板(例えば、プラスチック)、電極、及び細胞層から構成される。この電気的な検知手法において、細胞は基板上に並べられた小さな金の電極上に培養され、システムの電気インピーダンスが経時的に調べられる。インピーダンスは細胞層の電気伝導率の変化の指標となる。通常、固定周波数若しくは変動周波数を有する小定電圧が、電極又は一連の電極に印加され、回路を介した電流の経時的に観測される。リガンド誘起の電流変動により、細胞応答に関する指標が与えられる。細胞全体を検出するインピーダンス測定のアプリケーションは1984年に実証された。それ以来、インピーダンスベースの測定は、細胞の付着及び拡散、細胞の微小運動、細胞の構造変化、及び細胞の死を含む広い範囲の細胞の事象を研究することを目的として利用されてきた。古典的インピーダンスシステムは、小さな検出電極を使用しかつ大きな基準電極を使用しているのが原因で、分析に際して変動が大きいという欠点を有する。かかる変動性を克服するために、CellKey system(MDS Sciex、南サンフランシスコ、カリフォルニア州)やRT‐CES(ACEA Biosciences株式会社、サンディエゴ、カリフォルニア州)等の最新世代のシステムは、ミクロ電極アレイを有する集積回路を利用している。
【0048】
RWGバイオセンサを用いたGPCR活性の光信号 −
細胞は、例えば、通常数十ミクロンを有する比較的大きい寸法の動的な物体である。RWGバイオセンサは、細胞の底部におけるリガンド誘起の変化の検出を可能にし、エバネセント波の侵入度によって決定される。その上、光学的バイオセンサの空間分解能は、入射光源のスポットサイズ(約100ミクロン)によって決定される。したがって、一般に、非常に密集した細胞層が、最適な分析結果を得るのに使用され、センサの構成は、例えば基板、導波薄膜、及び細胞層から構成される3層導波複合体と見なされ得る。細胞生物物理学と組み合わせて3層導波バイオセンサ理論に従って、全体の細胞の検知に関しては、有効屈折率におけるリガンドによって誘起された変化、すなわち検出された信号ΔNが数式(1)により与えられる。
【0049】
(数式1)
【0050】
ここで、S(C)は細胞層に対するシステムの感度であり、Δncはバイオセンサによって検知された細胞層の局所的な屈折率のリガンド誘起の変化である。ΔZcは細胞層への侵入深さであり、αは特定の屈折率の増大(タンパク質に対しては約0.18/mL/g)であり、ziは、質量の再分配が生ずる距離であり、dは細胞層内の薄片の仮想的厚さである。ここで、細胞層は垂直方向において均等に離間された部分に分割される。検出された信号が細胞層の底部分の屈折率Δncの変化に一次的に正比例することが想定されている。細胞内の所定体積の屈折率は、主にタンパク質等の生物分子の濃度によって主に決定されると仮定すると、Δncは検出ボリューム内の細胞のターゲット又は分子集合の局所濃度変化に正比例する。重み付け因子exp(−zi/ΔZc)は、指数関数的に減衰するエバネッセント波の特性を考慮して、生ずる局所的タンパク質濃度の変化が考慮されている。したがって、検出された信号は、センサの表面から異なる距離で生じる質量再分布を合計したものであり、各々が全ての応答に対して不均一に貢献する。数式(1)が示していることは、RWGバイオセンサを用いて検出された信号は、所的なタンパク質濃度変化の結果として、かかる変化が生ずる場所、時間の結果として、主として垂直な集団再分布に対して敏感であることである。検出された信号は、動的集団再分布(DMR:dynamic mass redistribution)信号と呼ばれている。
【0051】
GPCRの活性化は、一連の空間的かつ経時的な事象を引き起こす。かかる事象は、リガンドの結合、受容体の活性化、タンパク質の補充、受容体の内部化及び再利用化、第2メッセンジャの代替、細胞骨格のリモデリング、遺伝子の発現、細胞付着の変化等の事象を含む。各細胞の事象は、細胞の動力学、持続時間、振幅、及び質量の移動に関してそれ自身の特性を有する。したがって、細胞事象が生ずる位置に依存して、これらの細胞事象が、DMR信号全体に対して異なる寄与をなし得ることが合理的に想定されている。さまざまなGPCRsをターゲットとする作用物質のパネルを使用して、介在されたシグナリング経路を反映する3つのクラスのDMR信号がヒト類表皮癌A431細胞において特定された(Fang, Y., et al., “Non−invasive optical biosensor for assaying endogenous G protein−coupled receptors in adherent cells,” J. Pharmacol.Toxicol.Methods, 2007, 55, 314−322)(図6を参照)。受容体が結合したGタンパク質に依存して、DMR信号の各々は特定のクラスのGPCRs活性と相関しているので、得られたDMR信号は、Gq−DMR信号、Gs−DMR信号、及びGi−DMR信号とそれぞれ称されている。各クラスのDMR信号は、異なるクラスのGPCRsを介して介在された固有のシグナリングの集合を反映した異なる運動特性及び動的特性を示す。興味深いことに、Gq‐タイプのDMR信号は急激に変化するものの、Gs‐タイプDMR信号は比較的遅い。本明細書に開示されるようにオーファンGPCRsのGタンパク質カップリング機構を特定するのにDMR信号に関して固有の特性が使用され得る。
【0052】
しかしながら、両方のGq−DMR信号とGs−DMR信号は、テストされた多数の細胞株にわたって遍在しない場合、かなり普遍的であることがわかったが、Gi−DMR信号は受容体と同様に細胞の成分に強い依存を示す。その上、同一の細胞株における異なる受容体の活性化又は異なる細胞株における同一の受容体の活性化を介して介在された同じクラスのDMR信号に対して、振幅や動力学等の微細な特徴において、有意差がしばしばある。かかる差は、各受容体固有のシグナリングと同様に、GPCRシグナリングにおける細胞の状況関して重要な役割を反映している。したがって、その動特性及び形状に基づいて、特定のシグナリング経路をGPCRリガンド誘起の光学応答に帰属せしめることは、大抵困難である。GPCRリガンドによって活性化され且つリガンドによって誘起された光学応答を支配するその動特性及び形状に基づいて、経路の抽出が、主シグナリング経路を決定するのに、大抵は必要である。
【0053】
GPCR活性に関するバイオインピーダンス信号−
典型的なインピーダンスベースの細胞分析において、細胞は搬送されて、培養ウェルの底に配置された金電極に接触せしめられる。センサーシステムの総インピーダンスはバイオセンサを囲むイオン環境により、主に決定される。電界を印加すると、イオンは電界によって指向された運動を行い、濃度勾配による拡散を受ける。細胞全体について検出する際には、全電気インピーダンスは、電界溶液の抵抗、細胞のインピーダンス、電極/溶液界面におけるインピーダンス、及び電極/細胞界面におけるインピーダンスの4つの成分を有する。さらに、細胞のインピーダンスは、抵抗とリアクタンスとの2つの成分を含む。細胞のイオン強度に関する伝導特性は抵抗成分を与えるものの、不完全なコンデンサとして作用する細胞膜は周波数に依存する応答成分に寄与する。したがって、全インピーダンスは、例えば、細胞の生存能力、細胞の密集度、細胞の数、細胞の形態、細胞の付着度、イオン環境、細胞内の水分、検出周波数等を含む多くの要因の関数となる。
【0054】
RT−CESシステムにおいては、印加された小電位の割合は細胞内部に連結されている。細胞に印加されたかかる信号は、典型的ほ乳動物細胞の静止膜電位よりもはるかに小さいと信じられている。その結果、細胞の機能に対してほとんど妨害しないか、若しくは全く妨害しない。RT−CESシステムは、インピーダンスのこれら変化を測定し、細胞インデックスと呼ばれるパラメータとしてそれを表示する。数式(2)に従って、細胞インデックスが計算される。
【0055】
(数式2)
【0056】
Nは、インピーダンスが測定された振動数ポイントの数であり(例えば、10kHz、25kHz、及び50kHzに対してN=3)、R0(f)とRcell(f)は、それぞれ、ウェルにおいて細胞が存在しない場合と存在する場合における周波数電極抵抗である。
【0057】
CellKeyシステムにおいて、センサーシステムのインピーダンス変化は、受容体刺激に応答して生ずる細胞層のインピーダンス(デルタZ又はdZ)の変化に帰属される。低周波数において、印加された小電位は、細胞外の電流(iec)を誘起する。かかる細胞外の電流(iec)は、層内の個々の細胞の周囲に流れる。しかしながら、イオンチャンネルによる細胞膜を介した伝導電流も低測定周波数において重要であり得る。高周波数において、それらは、細胞性の膜に侵入する細胞透過電流(itc)を誘起する。各ウェルに対して測定された電流に対する印加電圧の比が、オームの法則により説明されるように、インピーダンス(Z)となる。
【0058】
細胞が受容体リガンド等の刺激に曝される際に、シグナル変換事象が活性化され、アクチン細胞骨格の調節等、複雑な細胞事象が引き起こされる。かかる複雑な細胞事象は、例えば、細胞の密着性、細胞の形態、及び体積、細胞間相互作用について変化を生じせしめる。これら細胞の変化は、細胞外電流及び細胞内電流の流れに対して個別的又は集団的に影響を与え、これにより、測定されたインピーダンスの強度及び特性に影響を与える。3つのクラスのGPCRs活性化を介して介在する3つのタイプのインピーダンス信号が知られている(Leung,G.,et al.,J.Assoc.Lab.Automat.,2005,10,258)。理論によって制限されないものの、これらインピーダンス信号はアクチン細胞骨格に与える異なる効果に起因すると信じられている。かかるアクチン細胞骨格は、異なるクラスのGPCRsの活性化に応答して、インピーダンスによって測定された細胞パラメータに影響を与える。Gq−GPCRs及びGi−GPCRsの活性化によって、アクチン重合が増強されるものの、Gs−GPCRsの刺激によって、アクチン解重合が引き起こされることが示されている。
【0059】
2.GPCRシグナリングとシグナリングの積分
生体組織に共通の特性は、環境変化と共に有機体の活性化を絶えず調整する動的能力にある。環境と交信する機能は多数経路を介して達成される。かかる多数経路は、外部環境や細胞内の異なる領域等から信号を受信しかつ処理する。個々の経路は、直線的な器官の管に沿って信号を伝え、その結果、個々の細胞機能が調整される。このタイプの情報転送は、調節機構に関する細胞の能力範囲に関して重要な部分となっている。しかしながら、より多数の細胞シグナリング成分及び経路が特定されかつ研究されると、これらの直線的経路は、独立のエンティティではなく、大なるネットワークの部分であることが明らかになっている。
【0060】
異なるタイプ細胞のネットワーク及び構成に関する初期研究から得られた1つの驚くべき結果は、細胞のタイプ特有の入力(すなわち、受容体)から信号を受信し、かつ、細胞のタイプに特有の機構に係合する一般的な信号通信ネットワークがあり得るということである。シグナリング成分及びそれら相互作用パターンに関する分子の特定は、細胞のタイプに特有なものであり得るが、これらの構成の全体的な機能と化学的信号回路の論理は、細胞のタイプに応じて維持される。
【0061】
理論によって制限されないものの、ネットワークはシグナリング経路間の内部接続から生じ得ると信じられている。同一のシグナリング成分が複数入力から信号を受信することができるので、かかる内部接続が生じ得る。かかるネットワークは、同様のクラスのシグナリング経路内で生じ得る。かかる同様のクラスのシグナリング経路は、例えば、Ras経路とRho経路との間、又はGsα/cAMP及びMAPキナーゼ経路等の異なる経路間である。
【0062】
GPCRシグナリング
GPCRsは、一連の様々な知覚性刺激及び化学的刺激に応答する7つの膜貫通型スパニングタンパク質のスーパーファミリを構成している。その様々な知覚性刺激及び化学的刺激とは、例えば、光、におい、味、フェロモン、ホルモン、化学的伝達物質である。GPCRsは、これら刺激によって与えられた情報を、細胞内の第2メッセンジャ変換する。その細胞内の第2メッセンジャは、重要な信号として細胞によって解釈される。この処理過程は、ヘテロトリメリックグアニンヌクレオチド結合タンパク質(Gタンパク質)を用いて、種々のエフェクタシステムに対して、それら相互作用を介して作用物質活性のGPCRsの結合を含む。GPCRに対する作用物質の結合は、受容体の形態状態を選択する。その受容体の形態状態は、Gタンパク質サブユニット上のGTPへのGDPの交換を促進し、Gタンパク質G−サブユニット及びG−サブユニットの解離を可能にせしめるように想定されている。その後に、活性されたG−及びとG−サブユニットは、エフェクタ酵素とイオンチャンネルの活性化を、正の調整、負の調整、若しくはその双方の調整を実行する。GPCRの作用物質の活性化はエフェクタシステムのGたんぱく質依存の活性化をもたらし、1)Gタンパク質結合のフィードバック調節、2)受容体エンドサイトーシス、及び3)Gタンパク質独立性信号変換経路を介したシグナリングを可能にする一連の分子相互作用を適切に設定する。
【0063】
ヘテロ3量体Gタンパク質を介したシグナリングに加えて、GPCRsは、Gタンパク質独立性信号変換錯体の形成と区分化を促進する足場として作用することが現在認識されている。GPCRsを結合せしめ、GPCRsをGタンパク質独立性信号変換経路に結合せしめ若しくはGタンパク質の特異性及び作用物質の選択性を変更するより多くのタンパク質が特定されている。GPCR相互作用タンパク質のリストは、例えば、GRKs、アレスチン、カルモジュリン、カルシオン、Aキナーゼアンカータンパク質(AKAP)、チューブリン、受容体活性変更ヤヌスキナーゼ2、PDZドメイン含有タンパク質(例えば、NHERF、RGS12)、SH3ドメイン含有アダプター分子(例えば、Grb2、Nck、c−Src、及びエンドフィリン)、小Gタンパク質、及び同様のたんぱく質を含む。
【0064】
受容体に結合されたGタンパク質に依存して、GPCRsは少なくとも4つの主カテゴリに分類され得る。かかる受容体は、Ca2+の可動化を引き起こすGq−共役受容体、Ca2+の可動を引き起こすGq−共役受容体、cAMPを低減せしめるGi/o−共役受容体、cAMPの集積を生じせしめるGs−共役受容体、Rho等の小GTPを活性化するG12/13−共役受容体を含む。
【0065】
細胞内cAMPレベルのGPCRシグナリングと積分
アデニリルシクラーゼ(ACs)は、細胞情報伝達ネットワーク内のジャンクション若しくは信号インテグレータとして見なされてきた。異なるアデニリルシクラーゼアイソフォームには、異なる信号受信能力がある。すべてのアデニリルシクラーゼアイソフォームは、Gs共役受容体からの信号に対応してcAMPを生成することができる。しかしながら、これらアイソフォームは、種々の入力から信号を受信する能力において異なる。細胞内のcAMPレベルは多くの経路間の信号のバランスに関する指標として機能し得る。アデニリルシクラーゼの信号受信能力について詳細に説明するこの複雑な状況が、図1に示されている。図面を参照すると、図1は、アデニールサイクラーゼイソフォーム依存性細胞シグナリング経路を示す概略図である。アデニールサイクラーゼは、複数の調整力のあるシグナリング経路のターゲットであり、自身が属するシクラーゼ群に依存して、異なる応答をし得る。
【0066】
細胞において、9つのACイソフォームがあり、そのすべてがGsαによって活性化され、全てではないがいくつかかが、Gi/oαサブユニットによって抑制される。多かれ少なかれ、ACアイソフォームの各々は、1つより多いシグナリング経路から入力を受信するので、細胞応答は、自身が発現するイソ酵素によって調整される。実施例において、Ca2+/CaMII(カルモジュリン依存性プロテインキナーゼII)はACI(アデニールサイクラーゼサブタイプ1)を活性化するものの、Ca2+はACV(アデニールサイクラーゼサブタイプ5)及びACVI(ACサブタイプ6)を抑制する。別の実施例において、ACV及びACVIはGiαによって抑制されるものの、ACII(ACサブタイプ2)及びACIV(ACサブタイプ4)は、Giαに対して無反応であるように見える。
【0067】
また、細胞のcAMP反応は刺激される受容体のGタンパク質の選択度/特異性に依存している。Ca2+は、反対に、異なるアデニリルシクラーゼに影響する。ACIはCa2+/CaMIIによって刺激され、ACV及びACVIはCa2+によって抑制され、リン酸化反応は種々の形態のPKC(プロテインキナーゼC)及びPKAによって抑制され、ACII、IV、及びVIIのACは、細胞内のCa2+変化に対して鈍感であるようである。無傷細胞において、Ca2+の効果は、発現されたイソ酵素のみに依存するのではなく、Ca2+の増加が生ずるサイトにも依存し得る。ACIIを発現する細胞において、ACIV若しくはACVIIはCa2+を増加させ、ジアシルグリセロール(DAG)は、cAMPの増加を引き起こす。かかるcAMPは、Ca2+及び酵素を形成するcAMPのDAG刺激されたリン酸化反応によって調節され得る。対照的に、PKA介在リン酸化反応は、ACVとACVIの負のフィードバックレギュレータとして作用し得る。
【0068】
酵素アデニールサイクラーゼの9つの細胞膜に束縛されたアイソフォームは、細胞内cAMPレベルを調節するためにGタンパク質共役受容体を通して作用する化学的伝達物質と薬物によって大きく調整される。一般に、Gs共役受容体の急性の活性化はcAMPの蓄積を刺激するものの、Gi/o共役受容体の急性の活性化はcAMPの蓄積を通常は抑制する。Gタンパク質共役受容体の永続的な活性化によって、その後の薬物調節されたcAMPの蓄積が変更されることも確立されている。持続する受容体の活性化に続いて、これら変更は細胞適応型の応答を表していると信じられている。
【0069】
異種感作
一般的に観測された1つの現象、すなわち、アデニールサイクラーゼの異種感作は、Gi/o共役受容体持続的な活性化に続く薬物による刺激されたcAMPの累積に対する高応答性によって特徴付けられる。アデニールサイクラーゼの異種感作は、慢性の鎮静剤投与に続く耐性及び使用中止に関して説明するために当初は提案され、細胞が抑制受容体の持続された活性化に適応する機構であり得る。かかる適応型の機構は、例えば、薬物依存及び薬物乱用からの使用中止の処理において、及び、分裂病と鬱病を含む精神神経系症状において、役割を果たすように提案されてきた。また、異種感作は、複数のGi/oたんぱく質の同時活性化を必要とし得る。例えば、全内因性のGi/oプールが利用可能であった細胞の感作と比べると、選択性Go−誘起の異種感作の強さは、低減されるようである。
【0070】
異種脱感作
受容体介在の細胞応答の脱感作は、複数の経路を要する複雑な処理である。かかる複雑な処理は、a)Gタンパク質から解かれた受容体、b)原形質膜から離間した受容体の隔離、c)シグナリングタンパク質の下降調整を含む。受容体脱感作は一般的に相同である(すなわち、特有の受容体でありかつ受容体の占有に依存する)。
対照的に、活性物質によるアデニールサイクラーゼの直接活性化又は細胞透過性cAMPによるPKAの直接的な活性化は、異種脱感作と関連付けられた受容体のリン酸化反応を大部分は引き起こし得る(すなわち、非特定的の受容体であり、受容体の占有に依存しない)。
【0071】
細胞内Ca2+レベルのGPCRシグナリングと積分
同様に、また、細胞内Ca2+レベルはGPCRシグナリングにおいて重要な役割を果たす。細胞内Ca2+レベルはイオンチャンネルによって一般的に調整される。複数のGタンパク質介在経路は1つにまとまって、カルシウムチャンネルを調節することが知られている。事象の複雑なタイミングはいくつかのシグナリング経路の構成部と細胞骨格構造及び足場構造の動的ネットワーク間の相互作用によって制御される。その細胞骨格構造及び足場構造は、これら構成とカルシウムチャンネル間の相互作用のための層として機能し得る。カルシウムチャンネルの変更を誘起することが知られた受容体の多くが、百日咳毒素(PTX)を有する細胞の前処理がカルシウムチャンネルの抑制を止めるという観測によって示されているように、ヘテロトリメリックGi/oたんぱく質に結合される。カルシウムチャンネルの抑制は、細胞膜によって範囲を定められた信号若しくは第2のメッセンジャによって調節された信号を介して生じ得る。細胞膜によって範囲を定められた経路は、Gタンパク質βγーサブユニット(Gβγ)の直接結合によって調節され、Cav2.2チャンネルの電圧依存性抑制という結果となる。
【0072】
無標識バイオセンサ分析用いた経路の抽出
GPCRシグナリングは高度化され、非常に複雑である。受容体は、Gタンパク質依存性経路及びGタンパク質独立性経路を介して、シグナリングを介在し得るし、受容体は複数のGたんぱく質依存性経路を介して介在し得る。無標識バイオセンサ分析は、積分された細胞の応答(例えば、光学バイオセンサを用いたDMR信号、若しくは、電気バイオセンサを用いたバイオインピーダンス)を測定するので、受容体の活性化により、非定型的なバイオセンサ反応若しくは出力信号が引き起こされる。これは、例えば、リガンドと細胞のコンテクスト(又は、バックグラウンド)の特性に依存している。例えば、2つの内因性のGi−共役受容体、LPA受容体(図6C)、及びニコチン酸受容体(図5A)は、同一のA431細胞膜において2つの異なるタイプのDMR信号を引き起こす。これは、受容体によって誘起されたシグナリング処理の複雑性及び特異性を反映している。その上、A431細胞における同一のLPA受容体は、完全に静的な状態対増殖状態(図6C、データは示されていない)等の細胞の状況に依存して、異なるタイプのDMR信号を引き起こすであろう。したがって、無標識バイオセンサ分析は、リアルタイムで、受容体シグナリングに続いて、明確な動力学応答を提供するものの、動力学応答の動特性のみが、経路の特定のための予測的な指標でなくてもよい。
【0073】
バイオセンサシグナリングを統合する方法を利用することによって、少なくとも2つの異なる手法は、無標識バイオセンサ分析を使用して、GPCRシグナリングを抽出するのに使用され得る。ホルスコリン若しくはNKH447等のAC活性物質、又は細胞プリアンブルcAMP類似物質等のPKA活性物質を使用して、第1方法(方法1)は、細胞内cAMPレベルを上昇せしめる。GPCRリガンドによって誘起されたバイオセンサの出力に及ぼす高cAMPレベルの特異な効果若しくは作用に基づいて、GPCRシグナリングを抽出することができる。前記増大された細胞内cAMPレベルは、Gq若しくはG12/13シグナリングに対してほとんど抑制効果を与えないか若しくは全く与えず、異種脱感作によりGsシグナリングを完全に減衰せしめ、異種感作によりEC50濃度におけるGiシグナリングを増強せしめる。
【0074】
U73122等のPLC抑制物質若しくはPLC遺伝子抑制干渉RNAのうちの一方を使用して、第2方法(方法2)は細胞内のホスホリパーゼC(PLC)活性の活性化を抑制する。かかる手法は、無標識分析を使用して、GPCRシグナリングを抽出する補完的な方法を提供する。PLC活性の抑制はGs若しくはG12/13シグナリングに対してはほとんど効果を示さないものの、Gqシグナリングを完全に減衰せしめ、Giシグナリングを部分的に減衰せしめる。2つの手法は、無標識細胞分析を使用して、GPCRシグナリングを抽出するのに独立して使用され得るし、組み合わせても使用され得る。上記2つの方法と、示されたGPCRに与えるそれら方法のそれぞれの効果が、添付した表にまとめられている。
【0075】
【表1】
【0076】
実施例
以下の実施例は、上述した開示を使用する方法をより完全に説明し、さらに開示の種々な態様を行うように想定された最良の態様を詳しく説明するために用いている。これらの実施例は本開示の範囲を限定してはおらず、説明及び例示を目的として提示されていると理解される。
【0077】
バイオセンサベースの生細胞分析によって、細胞応答の検出結果が、ホルスコリンによるアデニールサイクラーゼの活性化に続いてGタンパク質共役受容体等の活性化によって得られる。ホルスコリンによるアデニールサイクラーゼの持続的活性化により、受容体のタイプに依存して、作用物質刺激に対してGタンパク質共役受容体が敏感になり若しくは鈍感になる。
【0078】
実験方法
材料 −
ホルスコリン及びエピネフリンは、Tocris(セントルイス、ミズーリ州)から入手した。ヒスタミン及びニコチン酸は、シグマ・ケミカル・カンパニー(SigmaChemicalCompany)(ミズリー州、セントルイス)から入手した。Epic社登録商標384ウェル細胞分析マイクロプレートはコーニング株式会社(コーニング、ニューヨーク州)から入手した。各ウェルが共鳴導波グレーティング(RWG)バイオセンサから成るマイクロプレートは、すぐに培養することができ、いかなる前処理も必要とせずに直接的に使用された。
【0079】
細胞の培養 −
人間の表皮癌A431細胞(American Type Cell Culture)が、ダルベッコの未改質Eagle媒質(DMEM:Dulbecco’s modified Eagle’s medium)内で培養された。かかる媒質には、10%の牛胎児血清(FBS)、4.5g/リットルのグルコース、2mMのグルタミン、及び抗生物質が加えられた。10%のFBSを含む50LのDMEM媒質に3〜8回浮遊された約1.8×104の細胞が384ウェルマイクロプレートの各々のウェル内に設けられ、37℃で空気/5%CO2下で1日間培養され、かかる細胞を続いて無血清DMEMにおいて継続して培養することによって、約20時間、放置した。
【0080】
光学的バイオセンサシステム及び細胞分析 −
コーニング登録商標Epic登録商標波長送受観測システムが使用された。システムは、独立型とすることができ、温度制御ユニット、光学検知ユニット、及びロボティックスを有するオンボード液体処理ユニットから構成され得る。温度制御ユニットが、必要に応じて温度変動を最小にするべく内蔵されている。このユニット内に、センサーマイクロプレートと化合物ソースプレートの双方を処理する2つの並列スタックがある。一旦、安定した温度が得られると(通常1時間以内)、センサーマイクロプレートは、ロボティクスによって、検知システムの直上のプレートホルダに自動的に搬送され、ソースプレートは適切なコンパートメントに移動されるので、オンボード液体処理ユニットによって容易に処理可能である。
【0081】
検知ユニットが、統合されたファイバオプティクスの中心に設けられ、例えば、生細胞内のリガンド誘起の動的質量再分布に起因する共鳴光の波長シフトが測定される。マイクロプレートの底部を介した垂直入射によって、光ファイバとコリメータレンズとを介して生成された830nmを中心とする広帯域白色光源が、グレーティングの小領域を照明するのに使用された。反射光を記録する検出ファイバは照光ファイバを用いて1束にされた。一連の照光/検出ヘッドが直線的に配置されるので、反射スペクトルは、384ウェルのマイクロプレートの同じカラム内にある16ウェルから一度に収集される。プレート全体が照光/検出ヘッドによってスキャンされて、各センサが複数回扱われ、各カラムは連続して扱われ、6秒若しくは13秒の時間間隔で細胞応答の動的測定が行われた。
【0082】
動的分析を目的として、細胞は、HBSS(20mMのHEPESを有するHanks Balanced Salt Solution)緩衝液を用いて洗浄された。検知システム内で1時間培養した後に、センサープレートがスキャンされ、ベースラインの応答が記録された。そして、化合溶液が、オンボード液体処理システムを使用して、センサープレートに移され、細胞応答が別の期間に記録された。同様に、第2の任意のステップは、例えば、異なる化合溶液を添加するステップを含み、かかるステップが実行されると、第2リガンド誘起の応答に対する第1の化合物の効果が決定される。化合物が導入されると、センサーマイクロプレートのリッドが、短期間(例えば、約1分)を除いて、分析中、維持された。プレートリッドは、ロボティクスによって自動的に処理された。すべての分析測定が被制御温度(28℃)にて実行された。
【0083】
統計的分析 −
明確に言及しない限り、各測定若しくは各化合物に対して、3回測定が行われた。標準偏差がこれら測定値(n=3)から得られた。分析係数の変動は、通常約10%未満であることが見出された。投与量依存性応答の全てが、(Graph Padから利用可能な)Prismソフトウェアを用いた非線形回帰法を使用して分析された。
【0084】
結果および議論
図2は、標的細胞内に出現するアデニールサイクラーゼの下位集合に依存して、cAMPレベルを増大させ又は低減せしめ得るGq(Gαq)、Gi(Gαi)、Gs/o(Gαs/o)、及びGβγ介在シグナリング経路から、細胞内アデニールサイクラーゼまでのクロストークと、を示す概略図である。
【0085】
図3Aは、ホルスコリン前処理から生じる光学応答の実施例を示しており、かかるホルスコリン前処理に続いて静的A431細胞におけるGs共役受容体(A2アデノシン受容体)の活性化処理が行われた。通常、ホルスコリンによるアデニールサイクラーゼの活性化が、Gs共役受容体の活性化によって相乗的に作用するものの、種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理は、続いて100nMのNECAの暴露処理がなされて(N−エチルアデノシンー5’−ウロン酸、A2‐特有作用物質)、異種脱感作を引き起こす、すなわち、その作用物質による刺激に対するA2受容体の応答能力が低減された。同様に、種々の濃度のNECAを用いたA431細胞の1時間の前処理は、アデニールサイクラーゼを減感せしめ、ホルスコリン誘起応答(図3B)を減衰せしめる。異種脱感作に関する観測が示していることは、クロストークがシグナリング経路間において生じ、これはアデニールサイクラーゼの活性化とA2受容体の活性化から生ずることである。理論によって制限されないものの、これら2つの経路間のクロストークに関して考えられる機構は、ホルスコリンによるアデニールサイクラーゼの活性化の際に、cAMPが生成され且つPKAを活性化することにある。PKAがGs受容体をリン酸化し得るし、Gs介在反応を完全に抑制せしめる。したがって、図3A及び3Bは、3A)NECA誘起のGs応答に対するホルスコリン前処理の効果と、3B)静的A431細胞におけるホルスコリン誘起の反応に対するNECA前処理の効果と、をそれぞれ示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(300=5nm、310=44nm、320=174nmと、330=348nm、340=690nm)が行われ、続いて100nMのNECAの暴露処理がなされた結果、NECA誘起の応答の減衰を引き起こす。ホルスコリンによる抑制効果は、シグナリング経路間のクロストークに帰属すると信じられており、ホルスコリンによるアデニールサイクラーゼの活性化とNECAによるA2アデノシン受容体の活性化から生じる。同様に、種々の濃度のNECAを用いたA431細胞の前処理(350=3nm、360=384nm、370=696nm、380=5nm、390=11nm)に続いて1Mのホルスコリンによる処理をした結果、A2受容体作用物質が1.3nMのIC50とのホルスコリンの結合を抑制することを示した。
【0086】
図4Aは、ホルスコリン前処理から生じる細胞応答の実施例を示している。かかるホルスコリン前処理に続いて静的A431細胞においてGq−共役受容体(H1ヒスタミン受容体)の活性化処理が行われた。種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理に続いて100nMのヒスタミン(H1−特有作用物質)の暴露によって処理したところ、部分的な脱感作をも引き起こした。
【0087】
ホルスコリンの前処理による投与量依存の部分的な抑制は約184nMのIC50を引き起こした。これは、ホルスコリンの効能に対応する。作用物質誘起の脱感作に関する観測は、この場合、アデニールサイクラーゼの特定のアイソフォームにおけるいくつかのクロストークがシグナリング経路間において生じ、これは、ホルスコリンによるすべての内因性のアデニールサイクラーゼの活性化とH1受容体の活性化から生ずることを再度示している。図4A及び4Bは、4A)ヒスタミン誘起のGq応答に対して与えるホルスコリン前処理の効果、及び、4B)静的A431細胞におけるホルスコリン誘起の反応に対するヒスタミン前処理の効果をそれぞれ示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(400=3nm、410=5nm、420=174nm、430=696nm、440=2,784nm)に続いて100nMのヒスタミンの暴露処理がなされた結果、ヒスタミン−誘起の応答の減衰を引き起こした。ホルスコリンによる抑制効果は、シグナリング経路間のクロストークに帰属すると信じられている。これは、ホルスコリンによるアデニールサイクラーゼの活性化とヒスタミンによるH1ヒスタミン受容体の活性化とから生じる。同様に、種々の濃度のホルスコリンを用いたA431細胞の前処理(455=30nM、460=90nM、465=9.9nM、470=270nM、475=810nM)に続いて1Mのホルスコリンによる処理をしたところ、H1受容体作用物質が、301nMのIC50とのホルスコリンの結合を抑制することを示した。
【0088】
図5は、ホルスコリン前処理から生じる細胞応答の実施例を示している。かかるホルスコリン前処理に続いて静的A431細胞においてGi‐共役受容体(ニコチン酸受容体HM74受容体)の活性化処理が行われた。Gs−共有受容体及びGq−共有受容体の活性化と対比して、種々の濃度のホルスコリンを用いたA431細胞の1時間の前処理に続いて25Mニコチン酸(HM74−特有作用物質)の暴露によって処理したところ、H1−誘起の応答の増強を引き起こし、ホルスコリンのEC50において最大値に達する。図5Aは、静的A431細胞におけるニコチン酸誘起のGi応答に対するヒスタミン前処理の効果を示している。種々の濃度のホルスコリンを用いたA431細胞の前処理(すなわち、500=0nm、510=4nm、520=32nm、530=125nm、540=4,000nm、550=32,000nm)に続いて25Mのニコチン酸によって処理したところ、ニコチン酸誘起の応答の増強が引き起こされた。Gi応答の増強作用は、異種感作に帰属すると信じられている。
【0089】
図6A乃至6Cは、RWGバイオセンサを使用した生細胞における3つのクラスのDMR信号のGPCRシグナリング(Gq‐、Gs‐、及びGi‐)を示している。すなわち、6A)GqシグナリングとそのDMR信号(チャイニーズハムスター細胞、10ユニット/mLのトロンビンにおけるプロテアーゼ活性されたレセプターサブタイプ1)と、6B)Gsシグナリング及びそのDMR信号(A431細胞、2nMエピネフリンにおける2ーアドレナリン作用受容体)と、6C)Giシグナリング及びそのDMR信号(A431,100nM LPAのリゾホスファチジン酸(LPA)受容体サブタイプ1)と、を示している。破線矢印は作用溶液が導入された時間を示している。
【0090】
図7Aは、静的A431細胞におけるヒスタミン誘起のDMR信号に対するU73122の効果を示している。種々の濃度のU73122を用いたA431細胞の前処理(すなわち、700=0M、710=1.78M、720=7.15M、730=28.5M、740=114M、750=228M)に続いて1,000nMのヒスタミンを用いて刺激の処理を行ったところ、ヒスタミン誘起のDMR応答の投与量依存性が抑制される。ヒスタミンは、A431細胞に内因的に発現したGq−共役受容体ヒスタミン受容体サブタイプ1に対して、天然の作用物質である。図7Bに示されたヒスタミン反応のP−DMR振幅に基づいて、高投与量におけるU73122がヒスタミン反応を完全に抑制し得る。
【0091】
本開示は、種々の特定の実施形態及び技術を参照して説明されてきた。しかしながら、本開示の精神及び範囲内において多くの変更及び改良を行うことができることが理解されるはずである。
【参照】
【0092】
1.Y, Fang et al., ”Label−Free Biosensors and Cells”, PCT App. No. PCT/US2006/013539 (Pub.No. WO 2006/108183)(Attorney Docket No. SP05−037).
2.Fang, Y., et al., ”Resonant waveguide grating biosensor for living cell sensing,” Biophys. J., (2006) 91, 1925−1940.
【特許請求の範囲】
【請求項1】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に、当該固定された生細胞を接触せしめるステップと、
当該物質に接触せしめられた生細胞をGPCRリガンドに接触せしめるステップと、
前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項2】
前記物質はアデニラート活性剤を含むことを特徴とする請求項1に記載の方法。
【請求項3】
請求項2に記載の方法であって、
前記アデニラート活性剤は、ホルスコリン、NKH477、1,9ジデオキシ‐ホルスコリン、7‐デアセチル‐7‐O‐へミスチニル‐ホルスコリン、又はそれら組み合わせの内の少なくとも1つを含むことを特徴とする方法。
【請求項4】
前記物質は細胞浸透性cAMP類似物質を含むことを特徴とする請求項1に記載の方法。
【請求項5】
請求項5に記載の方法であって、
前記細胞浸透性cAMP類似物質は、(S)−アデノシン、環状3’、5’−(ハイドロゲンホスホロチオエート)トリエチルアンモニウム、8−ブロモアデノシン−3’、5’−環状リン酸、8−クロロアデノシン−3’、5’−環状リン酸、N6,2’−O−ジブチリルアデノシン−3’、5’−環状リン酸、又はそれらの組合せのうちの少なくとも1つことを特徴とする方法。
【請求項6】
請求項1に記載の方法であって、
前記GPCRリガンドは、前記生細胞内に発現した受容体を介したシグナリングを調節する作用物質、部分作用物質、逆作用物質、又はそれらの組合せ、のうちの少なくとも1つを含むことを特徴とする方法。
【請求項7】
請求項1に記載の方法であって、
前記固定された生細胞を前記物質に接触せしめる前記ステップは、第1に細胞内cAMPを増大せしめ、かつ、第2にタンパク質キナーゼAを活性せしめることを特徴とする方法。
【請求項8】
請求項1に記載の方法であって、
バイオセンサ上に生細胞を固定する前記ステップは、約1パーセントから約99パーセントのバイオセンサ表面被覆率を有する密集度が得られるまで生細胞をバイオセンサ表面に培養するステップを含むことを特徴とする方法。
【請求項9】
請求項1に記載の方法であって、
前記増大された細胞内cAMPレベルは、
Gq若しくはG12/13シグナリングに対して部分的な抑制効果を与え、
異種脱感作によりGsシグナリングを完全に減衰せしめ、
異種感作によりEC50濃度においてGiシグナリングを増強せしめ、
生細胞GPCRシグナリングの抽出及び決定を可能にせしめることを特徴とする方法。
【請求項10】
ホスホリパーゼC抑制剤をアデニールサイクラーゼ活性剤と合成するステップをさらに含む請求項1に記載の方法。
【請求項11】
請求項1に記載の方法であって、
刺激を用いて、前記固定された生細胞の前処理、中間処理、後処理、又はそれら組合せのうち少なくとも1つをさらに含む方法。
【請求項12】
請求項1に記載の方法であって、
前記GPCRリガンド誘起のバイオセンサの応答により、GPCRシグナリング経路に与える周知の治療用化合物若しくは治療候補化合物の陽性若しくは陰性の効果が特定されることを特徴とする方法。
【請求項13】
請求項1に記載の方法であって、
前記GPCRリガンド誘起のバイオセンサの応答は、異なるクラスのGタンパク質シグナリングを識別し、GPCRのシグナリングと非GPCRのシグナリングとを識別し、又は、それら双方を識別することを特徴とする方法。
【請求項14】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
当該固定された生細胞をGPCRリガンドに接触せしめるステップと、
前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に、当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、
前記GPCRリガンドが存在する場合と存在しない場合における前記生細胞に関する前記物質誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項15】
請求項14に記載の方法であって、
前記GPCRは、ヒト類表皮A431細胞に発現した内因性ニコチン酸受容体HM74aであることを特徴とする方法。
【請求項16】
請求項14に記載の方法であって、
前記GPCRリガンドは、前記生細胞内に発現した前記GPCRを介してシグナリングを調整する作用物質、部分作用物質、逆作用物質、又はそれらの組合せ、のうちの少なくとも1つを含むことを特徴する方法。
【請求項17】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
前記細胞内において細胞内ホスホリパーゼC(PLC)活性を抑制する物質に、当該固定された生細胞を接触せしめるステップと、
当該PLC抑制されたが生細胞をGPCRリガンドに接触せしめるステップと、
前記物質が存在する場合と存在しない場合における前記生細胞の前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項18】
請求項17に記載の方法であって、
前記細胞内において細胞内ホスホリパーゼC(PLC)活性を抑制する前記物質は、
PLC抑制剤、PLC遺伝子抑制干渉RNA、又はそれらの組合せ、を含むことを特徴とする方法。
【請求項19】
前記PLC抑制剤はU73122を含むことを特徴とする請求項18に記載の方法。
【請求項20】
請求項19に記載の方法であって、
前記PLC活性の前記抑制は、Gs若しくはG12/13シグナリングに対して限定的効果を与え、
Gq信号通信を完全に減衰せしめ、
Giシグナリングを部分的に減衰せしめ、
生細胞のGPCRシグナリングの抽出及び決定を可能にせしめることを特徴とする方法。
【請求項1】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に、当該固定された生細胞を接触せしめるステップと、
当該物質に接触せしめられた生細胞をGPCRリガンドに接触せしめるステップと、
前記物質が存在する場合と存在しない場合における前記生細胞に関する前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項2】
前記物質はアデニラート活性剤を含むことを特徴とする請求項1に記載の方法。
【請求項3】
請求項2に記載の方法であって、
前記アデニラート活性剤は、ホルスコリン、NKH477、1,9ジデオキシ‐ホルスコリン、7‐デアセチル‐7‐O‐へミスチニル‐ホルスコリン、又はそれら組み合わせの内の少なくとも1つを含むことを特徴とする方法。
【請求項4】
前記物質は細胞浸透性cAMP類似物質を含むことを特徴とする請求項1に記載の方法。
【請求項5】
請求項5に記載の方法であって、
前記細胞浸透性cAMP類似物質は、(S)−アデノシン、環状3’、5’−(ハイドロゲンホスホロチオエート)トリエチルアンモニウム、8−ブロモアデノシン−3’、5’−環状リン酸、8−クロロアデノシン−3’、5’−環状リン酸、N6,2’−O−ジブチリルアデノシン−3’、5’−環状リン酸、又はそれらの組合せのうちの少なくとも1つことを特徴とする方法。
【請求項6】
請求項1に記載の方法であって、
前記GPCRリガンドは、前記生細胞内に発現した受容体を介したシグナリングを調節する作用物質、部分作用物質、逆作用物質、又はそれらの組合せ、のうちの少なくとも1つを含むことを特徴とする方法。
【請求項7】
請求項1に記載の方法であって、
前記固定された生細胞を前記物質に接触せしめる前記ステップは、第1に細胞内cAMPを増大せしめ、かつ、第2にタンパク質キナーゼAを活性せしめることを特徴とする方法。
【請求項8】
請求項1に記載の方法であって、
バイオセンサ上に生細胞を固定する前記ステップは、約1パーセントから約99パーセントのバイオセンサ表面被覆率を有する密集度が得られるまで生細胞をバイオセンサ表面に培養するステップを含むことを特徴とする方法。
【請求項9】
請求項1に記載の方法であって、
前記増大された細胞内cAMPレベルは、
Gq若しくはG12/13シグナリングに対して部分的な抑制効果を与え、
異種脱感作によりGsシグナリングを完全に減衰せしめ、
異種感作によりEC50濃度においてGiシグナリングを増強せしめ、
生細胞GPCRシグナリングの抽出及び決定を可能にせしめることを特徴とする方法。
【請求項10】
ホスホリパーゼC抑制剤をアデニールサイクラーゼ活性剤と合成するステップをさらに含む請求項1に記載の方法。
【請求項11】
請求項1に記載の方法であって、
刺激を用いて、前記固定された生細胞の前処理、中間処理、後処理、又はそれら組合せのうち少なくとも1つをさらに含む方法。
【請求項12】
請求項1に記載の方法であって、
前記GPCRリガンド誘起のバイオセンサの応答により、GPCRシグナリング経路に与える周知の治療用化合物若しくは治療候補化合物の陽性若しくは陰性の効果が特定されることを特徴とする方法。
【請求項13】
請求項1に記載の方法であって、
前記GPCRリガンド誘起のバイオセンサの応答は、異なるクラスのGタンパク質シグナリングを識別し、GPCRのシグナリングと非GPCRのシグナリングとを識別し、又は、それら双方を識別することを特徴とする方法。
【請求項14】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
当該固定された生細胞をGPCRリガンドに接触せしめるステップと、
前記生細胞において細胞内cAMPを増加させ且つタンパク質キナーゼAを活性化する物質に、当該GPCRリガンドに接触せしめられた生細胞を接触せしめるステップと、
前記GPCRリガンドが存在する場合と存在しない場合における前記生細胞に関する前記物質誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項15】
請求項14に記載の方法であって、
前記GPCRは、ヒト類表皮A431細胞に発現した内因性ニコチン酸受容体HM74aであることを特徴とする方法。
【請求項16】
請求項14に記載の方法であって、
前記GPCRリガンドは、前記生細胞内に発現した前記GPCRを介してシグナリングを調整する作用物質、部分作用物質、逆作用物質、又はそれらの組合せ、のうちの少なくとも1つを含むことを特徴する方法。
【請求項17】
GPCRシグナリング経路の決定方法であって、
バイオセンサに生細胞を固定するステップと、
前記細胞内において細胞内ホスホリパーゼC(PLC)活性を抑制する物質に、当該固定された生細胞を接触せしめるステップと、
当該PLC抑制されたが生細胞をGPCRリガンドに接触せしめるステップと、
前記物質が存在する場合と存在しない場合における前記生細胞の前記GPCRリガンド誘起のバイオセンサの応答を検出し且つ比較するステップと、を含む方法。
【請求項18】
請求項17に記載の方法であって、
前記細胞内において細胞内ホスホリパーゼC(PLC)活性を抑制する前記物質は、
PLC抑制剤、PLC遺伝子抑制干渉RNA、又はそれらの組合せ、を含むことを特徴とする方法。
【請求項19】
前記PLC抑制剤はU73122を含むことを特徴とする請求項18に記載の方法。
【請求項20】
請求項19に記載の方法であって、
前記PLC活性の前記抑制は、Gs若しくはG12/13シグナリングに対して限定的効果を与え、
Gq信号通信を完全に減衰せしめ、
Giシグナリングを部分的に減衰せしめ、
生細胞のGPCRシグナリングの抽出及び決定を可能にせしめることを特徴とする方法。
【図1】

【図2】
【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図2】
【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【公表番号】特表2011−515065(P2011−515065A)
【公表日】平成23年5月19日(2011.5.19)
【国際特許分類】
【出願番号】特願2010−527984(P2010−527984)
【出願日】平成20年10月2日(2008.10.2)
【国際出願番号】PCT/US2008/011401
【国際公開番号】WO2009/045463
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(397068274)コーニング インコーポレイテッド (1,222)
【Fターム(参考)】
【公表日】平成23年5月19日(2011.5.19)
【国際特許分類】
【出願日】平成20年10月2日(2008.10.2)
【国際出願番号】PCT/US2008/011401
【国際公開番号】WO2009/045463
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(397068274)コーニング インコーポレイテッド (1,222)
【Fターム(参考)】
[ Back to top ]