
バクテリオファージPHIC31組み換え系による真核細胞におけるDNA組み換え
【課題】本発明は、核酸の真核細胞への特異的かつ安定した組み込みを得るための方法を提供する。
【解決手段】本発明は、組み換え部位間の組み換えを仲介するが、組み換えで形成されたハイブリッド組み換え部位間では仲介しないΦC31インテグラーゼなどの原核リコンビナーゼを使用した部位特異的組み換え系を応用する。それゆえ、組み換えは、追加の因子の不存在下では不可逆である。リコンビナーゼポリペプチド又はリコンビナーゼをコード化する遺伝子を含む真核細胞も提供する。
【解決手段】本発明は、組み換え部位間の組み換えを仲介するが、組み換えで形成されたハイブリッド組み換え部位間では仲介しないΦC31インテグラーゼなどの原核リコンビナーゼを使用した部位特異的組み換え系を応用する。それゆえ、組み換えは、追加の因子の不存在下では不可逆である。リコンビナーゼポリペプチド又はリコンビナーゼをコード化する遺伝子を含む真核細胞も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
連邦政府が後援する研究に関する説明 本発明は、政府の指示で米国農務省、農業リサーチサービスによって授与された許可No.5335-21000-009-06Sの下でなされた。政府は本発明において一定の権利を有する。
【0002】
関連出願への相互参照 本出願は、ここで参照として組み込む1999年7月23日に出願された米国仮出願番号60/145,469の利益をクレームする。
【0003】
本発明は、真核細胞の染色体の中への特異的かつ安定した核酸の組み込みを得るための方法の分野に関する。本発明は、ΦC31インテグラーゼなどの原核リコンビナーゼポリペプチドを使用する部位特異的組み換え系を利用する。
【背景技術】
【0004】
原核の遺伝子形質転換は、しばしば重大な短所に苦しむ。例えば、トランス遺伝子の組み込みを興味ある特定の座で再生産可能に得ることはしばしば困難である。相同組み換えは、一般に非常に低い頻度でのみ生じる。この問題を克服するために、部位特異的組み換え系を用いた。これらの方法は、より高等な原核細胞において機能することが可能な部位特異的組み換え系の使用を含む。多くのバクテリオファージ及び組み込みプラスミドは、それらのゲノムをそれらの宿主のものの中へ安定に組み込むことが可能な部位特異的組み換え系をコード化する。これらの系において、組み換え反応の最小必要条件は、組み換え現象を触媒するリコンビナーゼ酵素又はインテグラーゼ、及び2つの組み換え部位である(Sadowski(1986)J.Bacteriol.165:341-347;Sadowski(1993)FASEB J.7:760-767)。ファージ組み込み系について、これらは、付着(att)部位といい、ファージDNA由来のattP因子と、バクテリアゲノムによってコード化されるattB因子がある。2つの付着部位は、2,3の塩基対と同じくらい小さい配列を共有する。リコンビナーゼタンパク質は、両方のatt部位に結合し、宿主への環状ファージ又はプラスミドDNAの組み込みを生ずる、DNA鎖のコンサーバティブ(保存型)及び相互作用交換を触媒する。DNAベンディングタンパク質IHF、組み込み宿主因子などの追加のファージ又は宿主因子波、効率的な反応に対して要求されるかもしれない(Freidman (1988)Cell 55:545-554;Finkel & Johnson(1992)Mol.Microbiol. 6:3257-3265)。逆切り取り反応は、時折、ファージλのxis遺伝子産物などの追加のファージ因子を必要とする。(Weisberg & Landy(1983)“ファージλにおける部位特異的組み換え”In Lambda II、eds. Hendrix et al.(Cold Dpring Harbor Laboratory, Cold Spring Harbor, NY)pp.211−250;Landy(1989)Ann.Rev.Biochem. 58:913−949)。
【0005】
リコンビナーゼは、2つのグループ、λインテグラーゼ(Argos et al.(1986)EMBO J.5:433-44;Voziyanov et al(1999)Nucl. Acids Res.27:930-941)、及びリゾルバーゼ/転化酵素属(Hatfull & Grindley (1988)“Resolvases and DNA−invertase:a family of enzymes active in site-specific recombination” In Genetic Recombination, eds. Kucherlipati, R., & Smith, G.R. (Am. Soc. Microbiol., Wachington DC), pp. 357-396)に分類される。これらは、インテグラーゼ酵素の構造、及び触媒のそれらのモードの分子詳細を変える(Stark et al. (1992)Trends Genetics 8:432−439)。鋳型Streptomyces ファージΦC31は、後者の分類の68kDリコンビナーゼをコード化する。同系の付着部位を組み換える際にΦC31インテグラーゼ酵素の有効性が、試験官内及び生体内でrecA突然変異体大腸菌において最近証明された(Thorpe & Smith(1998)Proc.Nat’l. Acad.Sci.USA95:5505−5510)。ΦC31インテグラーゼ反応は、おそらく追加のファージタンパク質が切り取りに要求されないために、宿主因子を必要とせず、不可逆であるようであるという点においてシンプルである。ファージ及びバクテリアatt部位は、クロスオーバー点で相同性の3つの塩基対だけを共有する。この相同関係は、逆方向反復、おそらくインテグラーゼタンパク質の結合部位によって隣接する。両方のattB及びattPに対する最小の既知の機能的な大きさは、〜50bpである。
【0006】
バクテリオファージP1のCre-lox系、及びSaccharomyces cerevisiae のFLP−FRT系は、動物及び植物におけるトランス遺伝子及び染色体工学技術に広く使用されている(Sauer(1994)Curr.Opin.Biotechnol.5:521−527;Ow(1996)Curr.Opin.Biotechnol.7:181−186)。動物又は植物細胞において作用する他の系は、以下すなわち、1)Zygosaccharomyces rouxii 由来のR−RS系(Onouchi et al(1995)Mol. Gen.Genet.247:653−660)、2)バクテリオファージMu由来のGin−gix系(Maeser & Kahmann(1991)Mol.Gen.Genet.230:170−176)、及び3)バクテリアプラスミドpSM19035 由来のβリコンビナーゼ−six系(Diaz et al.(1999)J.Biol.Chem.274:6634−6640)、を含む。部位特異的リコンビナーゼを使用することによって、より高頻度で組み込みを得ることができる。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】国際公開第99/18222号
【発明の概要】
【発明が解決しようとする課題】
【0008】
しかしながら、これら5つの系は、重大な短所を有する。各これらの系は、単一のポリペプチドリコンビナーゼは、同一かほぼ同一の配列の2つの部位間の組み換えを触媒するという共通した特性を有する。組み換えによって生じた生産部位は、それ自体次の組み換えに対する基質である。結果として、組み換え反応は容易に可逆可能である。分子内相互作用の反応速度論は、分子間相互作用に渡って好ましいので、これら組み換え系は、DNAを組み込むよりむしろ削除するのに有効である。したがって、トランス遺伝子の安定的な部位特異的組み込みを得るための方法と系に対する必要性が存在する。本発明は、この及び他の必要性を、満たす。
【課題を解決するための手段】
【0009】
本発明は、真核細胞において、安定で、部位特異的な組み換えを得るための方法を提供する。以前に知られた部位特異的組み換え方法と違って、本発明の方法を使用して得られた組み換え体は、安定である。組み換え反応は、可逆的ではない。
【0010】
本方法は、第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として機能することが可能な第二の組み換え部位とを含む真核細胞を提供することを含む。第一及び第二の組み換え部位は、原核リコンビナーゼポリペプチドと接触し、組み換え部位間に組み換えを生じ、それによって1又は2つのハイブリッド組み換え部位を形成する。有意義なことに、リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の部位特異的組み換えを仲介することができるが、真核細胞において存在しない追加のファージ生産因子の不存在下で2つのハイブリッド組み換え部位間の組み換えを仲介することができないものである。組み換え部位のいずれか又は両方が真核細胞の染色体において存在することができる。いくつかの実施態様において、組み換え部位の1つは染色体に存在し、他は染色体の中へ組み込まれるべき核酸内に含まれる。
【0011】
本発明は、原核リコンビナーゼポリペプチド又は原核リコンビナーゼをコード化する核酸を含む真核細胞も提供する。これらの実施態様において、リコンビナーゼは、第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として作用することが可能な第二の組み込み部位との間の部位特異的組み換えを仲介をすることができるが、第一の組み換え部位と第二の組み換え部位との間の組み換えに形成された2つのハイブリッド組み換え部位間の組み換えを、真核細胞に存在しない追加の因子の不存在下では仲介することができないものである。現在好ましい実施態様において、本発明の細胞は、リコンビナーゼポリペプチドのコード化配列を有する核酸を含む。リコンビナーゼコード化配列は、真核細胞においてリコンビナーゼコード化ポリヌクレオチドの発現を仲介するプロモーターと作用可能に結合することが好ましい。本発明の真核細胞は、例えば、動物細胞、植物細胞、酵母細胞、又はカビ細胞とすることができる。
【0012】
別の実施態様において、本発明は、安定に組み込んだトランス遺伝子を有する真核細胞を得るための方法を提供する。これらの方法は、第一の組み換え部位を含む真核細胞の中へ核酸を導入することを含み、前記核酸は興味あるトランス遺伝子と前記第一の組み換え部位との組み換えの基質として作用することが可能な第二の組み換え部位とを含むことを特徴とする。第一及び第二の組み換え部位は、原核リコンビナーゼポリペプチドと接触する。リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の組み換えを触媒し、第一の組み換え部位で核酸の組み込みを生じ、それによって核酸の各末端でハイブリッド組み換え部位を形成する。かさねて、リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の部位特異的組み換えを仲介するが、真核細胞において存在しない付加的因子の不存在下では2つのハイブリッド組み換え部位間の組み換えを仲介することができないものである。
【0013】
本発明の別の実施態様において、真核細胞において機能するプロモーターと作用可能に結合するバクテリアリコンビナーゼポリペプチドをコード化するポリヌクレオチド配列を含む核酸を提供する。本発明のこれらの核酸によってコード化されるリコンビナーゼポリペプチドは、真核細胞において存在しないバクテリオファージ因子の不存在下では第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位間の組み換えを仲介することができない。いくつかの実施態様において、さらに、核酸は、リコンビナーゼポリペプチドによって認識される少なくとも1つの組み換え部位を含む。
【0014】
本発明によって提供される真核細胞は、また、真核細胞において存在しないバクテリオファージ因子の不存在下では第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位を仲介できない1又はそれ以上のバクテリオファージΦC31組み換え部位、または他のリコンビナーゼに対する組み換え部位を有するポリヌクレオチドを含む。
【図面の簡単な説明】
【0015】
【図1】S.pombe leul Locusで染色体構造の概略的な外観を示す(縮小)。PLT44の染色体への相同的挿入(図1A)は、図1Bで示されるようなleul 対立遺伝子間のΦC31 attP標的を設置する。 pLT45の染色体attP標的内へのpLT43促進部位特異的組み込みは、図1Cに示す構造を導入する。矢印は、T7プロモーター(T7)、T3プロモーター(T3)及びura4+コード化領域(U4)に対応するPCRプライマーを示す。XbalI(X)切断産物の予想される大きさを示す。
【図2】ΦC31インテグラーゼが、CHO細胞におけるグリーン蛍光タンパク質(GFP)をコード化するトランス遺伝子の部位特異的組み込みを触媒することを証明する実験の概略を示す。
【図3】ΦC31が、ハイグロマイシンホスホトランスフェラーゼ遺伝子を染色体に位置したプロモーターの下流に挿入するためにattB部位での特異的組み換えを触媒することを証明する実験の概略図を示す。成功した組み込みは、Pc-att-hptリンケージ及びハイグロマイシン耐性表現型を生産する。attP及びattB部位の異なる長さの影響を、指標したプラスミドを使用して分析した。
【図4】タバコゲノム由来のattB及びattP部位によって隣接させたDNAの切り取りを触媒することを示す実験の概略的な図表を示す。
【図5】ΦC31インテグラーゼがタバコゲノムへのトランス遺伝子の組み込みを触媒したことを示す実験の概略的な図表を示す。
【発明を実施するための形態】
【0016】
定義
ここで使用するような「外因性DNA断片」、「異質ポリヌクレオチド」、「トランス遺伝子」、「異質核酸」は、異質の起源から特定の宿主へ創造したものか、あるいは、もし同じ起源由来ならその原型由来のものから修飾したものである。したがって、宿主細胞における異質遺伝子は、特定の宿主細胞へ内生的であるが、修飾されていない遺伝子を含む。それゆえ、前記用語は、細胞に対して外来性か、異質か又は相同であるが、因子が通常見られない宿主細胞核酸内に位置するDNA断片を言う。外因性のDNA断片を発現させて外因性ポリペプチドを生産する。
【0017】
「遺伝子」の語は、生物学上の機能と関係するいずれのDNA断片を表すものとして広く使用する。したがって、遺伝子は、それらの発現に要求されるコード化配列及び/又は調節配列を含む。遺伝子は、例えば、他のタンパク質に対する認識配列を形成する非発現DNA断片も含むことができる。遺伝子は、種々の起源から得ることができ、興味ある起源由来のクローン、または既知の若しくは予想した配列情報からゴウセイしたもの、所望のパラメーターを有するように設計された配列を含むことができる。
【0018】
核酸又はタンパク質に適用するとき、「単離」の語は、核酸又はタンパク質が自然の状態に関係する他の細胞成分が本質的に存在しないことを表す。乾燥しているか水溶液のいずれかとすることができるが、均質な状態が好ましい。純度や均質性は、例えばポリアクリルアミドゲル電気泳動又は高性能液体クロマトグラフィーなどの分析化学技術を使用して典型的に決定される。調製において存在する有力種であるタンパク質は、本質的に精製される。特に、遺伝子を隣接させて、興味ある遺伝子以外のタンパク質をコード化するオープンリーディングフレームから単離した遺伝子を分離する。「精製」の語は、電気泳動ゲルにおいて核酸又はタンパク質が1つのバンドを本質的に生じることを意味する。特に、核酸又はタンパク質が少なくとも約50%純粋、より好ましくは少なくとも約85%純粋、最も好ましくは少なくとも約99%純粋であることを意味する。
【0019】
「自然に生じる」の語は、ヒトによって人工的に生産されることから区別するものとして天然に見出すことができる対象を記述するために使用する。例えば、実験室においてヒトによって意図的に修飾されていない天然の起源から単離することが可能な(ウイルスを含む)組織において存在するポリペプチド、又はポリヌクレオチド配列は、自然に生じる。
【0020】
「核酸」又は「ポリヌクレオチド」の語は、一本鎖又は二本鎖型のいずれかにおけるデオキシリボヌクレオチド又はリボヌクレオチド、及びその重合体を言う。特に限定しない場合、前記語は、引用核酸と同様の結合特性を有し、自然に生ずるヌクレオチドと同様の方法で代謝される自然のヌクレオチドの既知のアナログを含有する核酸を包含する。他に指示しない場合、特定の核酸配列は、伝統的に修飾したその変異体(例えば、縮退コドン置換)、相補的配列、及び明確に指示した配列も絶対的に包含する。具体的に、1又はそれ以上選択した(又はすべての)コドンの3つの部分を、混合した塩基及び/又はデオキシイノシン残基で置換した配列を発生させることによって縮退コドン置換を達成することができる(Batzer et al.(1991)Nucleic Acid Res. 19:5081;Ohtsuka et al.(1985)J.Biol.Chem。260:2605−2608;Cassol et al.(1992);Rossolini et al.(1994)Mol.Cell.Probes 8:91−98)。核酸の語は、遺伝子、遺伝子によってコード化されたcDNA及びmRNAと交互に使用される。
【0021】
「遺伝子から誘導される核酸」とは、遺伝子又はそのサブ配列(例えば、コード化領域)を合成し、最終的に鋳型として機能する核酸を言う。それゆえ、mRNA、mRNAから逆転写されたcDNA、そのcDNAから転写されたRNA、cDNAから増幅したDNA、増幅したDNAから転写したRNAなどは、すべて遺伝子から誘導され、そのように誘導された産物の検出は、オリジナルの存在及び/又は不存在の指示となる。
【0022】
DNA断片は、別のDNA断片と機能ある関係に置かれたとき、「作用可能に結合」する。例えば、ポリペプチドの分泌に加わるプレタンパク質として発現するなら、ポリペプチドをコード化するDNAに、単一の配列についてのDNAは作用可能に結合する。プロモーター又はエンハンサーは、配列の転写を刺激するならコード化配列に作用可能に結合する。一般に作用可能に結合するDNA配列は、隣接と読み取り段階の両方のシグナル配列の場合、隣接する。しかしながら、例えば、エンハンサーは、それらが制御する転写のコード化配列と隣接する必要がない。結合は、従来の制限酵素部位、又はその代わりに挿入したアダプター、リンカーでのライゲーションによって達成される。
【0023】
「植物」の語は、全植物、植物組織(例えば、葉、茎、根など)、種子、植物細胞、及び同様の子孫を含む。本発明の方法で使用可能な植物の分類は、形質転換技術に修正可能なより高い植物の分類と同様に一般に広く、単子葉、双子葉植物の両方を含む。
【0024】
「プロモーター」の語は、転写を開始するためにRNAポリメラーゼとの結合に関わるDNA領域を言う。「誘導プロモーター」は、発現のレベルを、例えば、温度、pH、転写因子及び化学薬品などの環境又は開発因子によって変更可能である遺伝子の発現を導くプロモーターをいう。
【0025】
細胞に関して使用するとき「組み換え」の語は、細胞が異質核酸を複製するか、又は異質核酸によってコード化されるペプチド又はタンパク質を発現することを表す。組み換え細胞は、細胞の野生型(非組み換え型)内では見出すことができないポリヌクレオチドを含むことができる。組み換え細胞は、ポリヌクレオチドが人工的な手段によって細胞の中へ修正又は再導入される細胞の野生型に見出されるポリヌクレオチドも含む。語は、細胞から核酸を除去することなしに修飾された細胞に内在する核酸を含む細胞も包含し、そのような修飾は、遺伝子置換、部位特異的突然変異体、及び関連技術等によって得られるものを含む。
【0026】
「組み換え発現カセット」又は単なる「発現カセット」は、構造遺伝子の発現をそのような配列と両立できる宿主において達成することが可能な核酸因子を持つ組み換え的、又は合成的に発生させた核酸構築物である。発現カセットは、少なくともプロモーター、及び選択的に、転写終結シグナルを含む。典型的には、組み換え発現カセットは、転写すべき核酸(例えば、所望のポリペプチドをコード化する核酸)、及びプロモーターを含む。発現を達成する際に必要な又は役立つ追加の因子をここで述べたように使用することができる。例えば、発現カセットは、宿主細胞由来の発現タンパク質の分泌を指示するシグナル配列をコード化するヌクレオチド配列も含むことができる。遺伝子発現に影響する転写終結シグナル、エンハンサー、及び他のヌクレオチド配列も、発現カセットに含まれる。
【0027】
「リコンビナーゼ」の語は、2つ又はそれ以上の組み換え部位間の組み換えを触媒する酵素をいう。本発明のおいて役立つリコンビナーゼは、特定のリコンビナーゼによって認識される特異的ポリヌクレオチド配列である特異的組み換え部位での組み換えを触媒する。「インテグラーゼ」の語は、リコンビナーゼのタイプをいう。
【0028】
「形質転換率」は、異質ポリヌクレオチドをそのゲノム及び残存物の中へ上手く組み込んだ細胞の割合をいう。
【0029】
「トランスジェニック」の語は、細胞の中へか、細胞の原型の中へ導入された特異的修飾を含む細胞をいう。そのような修飾は、一又はそれ以上の点突然変異、欠失、挿入、又はその組み合わせを含むことができる。動物に言及するとき、「トランスジェニック」の語は、トランスジェニックである細胞を含む動物を意味する。トランスジェニック及び非トランスジェニック細胞の両方からなる動物は、ここでは「キメラ」動物という。
【0030】
「ベクター」の語は、核酸(又は複数の核酸)を宿主細胞へ運搬するための組成物を言う。ベクターは、運搬すべき核酸をコード化する核酸を含み、核酸の宿主細胞内への加入及び/又はベクターの宿主細胞への複製を容易にするために、ウイルスキャプシド又は他の物質(例えば、キャプシド内、又はキャプシドの一部としてパッケージされた逆転写酵素又は他の酵素)を選択的に含む。
【0031】
「組み換え部位」はここで述べるリコンビナーゼ酵素によって認識される特異的ポリヌクレオチド配列である。典型的には、(相補部位と呼ばれる)2つの異なる部位を含み、1つは目的の核酸に存在し(例えば、真核の染色体又はエピソームなど)、別のものは、目的の組み換え部位で組み込まれるべき核酸上に存在する。特別の酵素の組み換え部位が異なる名前を持つかもしれないが、「attB」及び「attP」の語は、それぞれ、本来的にバクテリアターゲット、及びファージドナー由来の付着(又は組み換え)部位をいう。組み換え部位は、典型的に、コア又スペーサー領域によって分離された左及び右アームを含む。それゆえ、attB組み換え部位は、BOB‘からなり、B及びB’は、それぞれ左及び右アームであり、Oはコア領域である。同様に、attPは「POP‘」であり、P及びP’はアームであり、Oは重ねてコア領域である。前記attB及びattP部位との間の組み換えでは、核酸の標的での付随する組み込み、組み込んだDNAに隣接する組み換え部位は、「attL」及び「attR」という。それゆえ、上述した用語を使用してattL及びattRは、それぞれBOP’及びPOB‘からなる。ここでのいくつかの表示において、「O」を省略し、attB及びattPはそれぞれBB’及びPP’と示す。
【0032】
(好適な態様の説明)
本発明は、真核細胞において部位特異的な組み換えを得るための方法を提供するものである。部位特異的な組み換えを得るための過去に知られたシステムとは異なり、本発明の方法を用いて行われた組み換えの産物は安定である。そこで、例えば、真核細胞の染色体へトランスジーンを導入するために本方法を用いることが可能であり、そして既に知られた部位特異的な組み換えシステムを用いる場合にしばしば起こる、トランスジーンの切除を避けることができる。安定な逆位(inversion )、トランスロケーション(translocation )及び他の転位(rearrangement )もまた、得ることができる。
【0033】
本発明は、バクテリオファージのインテグラーゼの様な原核細胞のリコンビナーゼを採用し、その様な酵素は2つの相補的な組み換え部位の間の組み換えを触媒するが、この組み換えにより形成されたハイブリッド部位の間の組み換えを触媒することができない、という点において単方向性である。その様なリコンビナーゼの一つであるΦC31 インテグラーゼは、それ自身ではattB x attP 反応のみを触媒する。そのインテグラーゼは、attBとattPの間の組み換えにより形成された、attLとattR部位の間の組み換えをを仲介することはできない。ΦC31 インテグラーゼの様なリコンビナーゼは単独で逆反応を触媒することはできないために、ΦC31 のattB x attP 組み換えは安定である。この性質により、真核細胞において現在使用されている部位特異的な組み換えシステムから、本発明の方法は離れている。現在使用されているシステムにはCre-lox システム又はFLP-FRT システムの様なものがあり、そこでは組み換え反応は容易に逆転される。本発明の組み換えシステムの使用により、真核細胞において、安定なトランスジーン及び染色体転位(rearrangement )に向けての新たな機会が提供される。
【0034】
本方法は、対応するリコンビナーゼを用いて、真核細胞に存在している一組の組み換え部位(例えば、attB及びattP)に接触することを含む。そこでリコンビナーゼは組み換え部位の間の組み換えを仲介する。二つの組み換え部位の相対的な位置に応じて、組み換えの結果としての数多くのイベントのいずれか一つが起こることがある。例えば、もし二つの組み換え部位が異なった核酸分子上に存在しているならば、その組み換えは一つの核酸分子を第二の分子へと組み込む結果と成り得るであろう。そこで、対応する組み換え部位を含んでいる真核細胞染色体へ、一つの組み換え部位を含んでいるプラスミドを組み込むことができるであろう。本発明の方法において用いられたリコンビナーゼは逆反応を触媒することができないから、その組み込みは安定である。その様な方法は、例えば、プラスミド上に存在しているトランスジーンを、真核細胞の染色体へ安定に組み込むために有用である。
【0035】
二つの組み換え部位が、同一の核酸分子上に存在することもまたできる。その様な場合には、得られた産物は典型的には、その部位の相対的な方向性に依存する。例えば、直線方向にある部位の間の組み換えは一般的に、その2つの組み換え部位の間にある任意のDNA が切り出されるという結果となる。対照的に、逆方向にある部位の間の組み換えは、仲介しているDNA のインバージョンが起こるという結果となることがある。再び、付加的因子の非存在下において組み換えは不可逆的であるという点において、得られた転位した(rearranged)核酸は安定であり、付加的因子は一般的にそのリコンビナーゼが由来する特定のバクテリオファージによってコードされ、正常には真核細胞においては見られないものである。この方法が有用である適用の一つの例は、2つの組み換え部位の間のプロモーターの交換に伴って生じる。もしプロモーターが、そのプロモーターによって発現されるべきコード配列に関して最初に反対向きであって、そしてそのプロモーターをフランク(flank )する組み換え部位が逆方法であるならば、組み換え部位を接触させることはプロモーターの逆位(inversion )が起こる結果となり、従ってプロモーターを正しい方向に置いてコード配列の発現を駆り立てるであろう。同様に、もしプロモーターが最初に発現のために正しい方向性にあり、組み換え部位が同じ方向性にあるならば、組み換え部位がプロモーターと接触することによりプロモーターフ断片を切り出して、そしてコード配列の発現を停止させるという結果を得ることができる。
【0036】
本発明の方法は、例えば、染色体のトランスロケーションを得るためにもまた有用である。これらの態様において、一つの組み換え部位は一つの染色体上に置かれ、そして第一の組み換え部位と共に組み換えの基質として働くことができる第二の組み換え部位が第二の染色体上に置かれている。その二つの組み換え部位をリコンビナーゼと接触させると組み換えが起こって、2つの染色体アームが交換されるという結果となる。例えば、生物の2つの系統を構築することが可能であり、そのうちの一つの系統は第一の組み換え部位を含み、第二の系統は第二の組み換え部位を含んでいる。その2つの系統は掛け合わされ、その組み換え部位の両者を含む子孫の系統が得られる。その部位をリコンビナーゼと接触させると、染色体アームの交換が起こる。
【0037】
(リコンビナーゼと組み換え部位)
本方法は、真核細胞において、安定な核酸の組み込み又は他の転位(rearrangement )を達成するために、リコンビナーゼのシステムを使用する。リコンビナーゼのシステムは典型的には3つの要素から成り、その要素は2つの特異的なDNA 配列(「組み換え部位」)及び特異的な酵素(「リコンビナーゼ」)である。リコンビナーゼは、特異的な組み換え部位の間の組み換え反応を触媒する。
【0038】
組み換え部位は方向性を有している。言い換えれば、それらは回文(バリンドローム)ではない。相互の関連性における組み換え部位の方向性により、どの様な組み換えイベントが起こるか決定される。組み換え部位は、平行(同じ方向性)または反対という、2つの方向性をとることが可能である。組み換え部位が単一の核酸分子上に存在し、そして相互に平行な方向性であるときには、その際にリコンビナーゼにより触媒される典型的な組み換えイベントは、仲介している核酸の切り出して単一の組み換え部位を離すことである。組み換え部位が反対の方向性にあるときには、その際には任意の仲介配列が逆位(inverted)となるのが典型的である。
【0039】
本発明の方法において使用されるリコンビナーゼは、第一の組み換え部位と第二の組み換え部位の間の部位特異的な組み換えを仲介することができ、その第二の組み換え部位は第一の組み換え部位と共に組み換えのための基質として働くことができる。しかしながら、正常には真核細胞に存在しない、付加的因子の非存在下では、第一の組み換え部位と第二の組み換え部位の間の組み換えにより形成される、二つのハイブリッド組み換え部位の間の組み換えを仲介することができない。これらのリコンビナーゼの例には、例えば、バクテリオファージΦC31 インテグラーゼ(例えばThorpe & Smith (1998) Proc.Natl.Acad.Sci.USA 95:5505-5510 ; Kuhstoss & Rao (1991) J.Mol.Biol.222:897-890 ;米国特許番号5,190,871 を見よ)、ファージP4リコンビナーゼ(Ow & Ausubel (1983) J.Bacteriol.155:704-713 )、リステリアファージリコンビナーゼ、バクテリオファージR4 Sreリコンビナーゼ(Matsuura et al. (1996) J.Bacteriol.178:3374-3376)、CisAリコンビナーゼ(Sato et al.(1990) J.Bacteriol.172:1092-1098 ; Stragier et al.(1989) Science 243:507-512) 、XisFリコンビナーゼ(Carrasco et al.(1994)Genes Dev.8:74-83)、トランスポゾンTn4451 TnpX リコンビナーゼ(Bannam et al.(1995) Mol.Microbiol. 16:535-551 ; Crelin & Rood (1997) J.Bacteriol.179:5148-5156)が含まれる。
【0040】
リコンビナーゼポリペプチド、及びそのリコンビナーゼポリペプチドをコードする核酸は本技術分野において述べられており、日常的な方法を用いて得ることが可能である。例えば、ΦC31 インテグラーゼをコードする核酸フラグメントを含むベクターは米国特許番号5,190,871 の中に述べられており、B-18477 というアクセッション番号の下でノザンレジョナル リサーチラボラトリー(ぺオリア、イリノイ 61604)から入手可能である。
【0041】
任意の適切な方法により、組み換えを望む場所において、組み換え部位を含む真核細胞へリコンビナーゼを導入することができる。例えばリコンビナーゼをポリペプチドの形で、例えばマイクロインジェクション又は他の方法によって導入することができる。しかしながら、現在において好ましい態様においては、リコンビナーゼをコードする遺伝子が細胞に導入される。遺伝子の発現はリコンビナーゼが作られるという結果となり、そのリコンビナーゼは対応する組み換え部位の間の組み換えを触媒する。興味の対象である外来性のポリヌクレオチドを導入する前、後、又はそれと同時に、リコンビナーゼ遺伝子を細胞中に導入することができる。一つの態様において、リコンビナーゼ遺伝子は挿入されるべきポリヌクレオチドを運搬するベクター中に存在し、そのリコンビナーゼ遺伝子はポリヌクレオチド中に含まれることさえも可能である。他の態様においては、リコンビナーゼ遺伝子は、形質転換した植物、動物、カビ又はその類似物の様な形質転換した真核生物中へ導入され、その後にそれらは対応する組み換え部位を含む生物と掛け合わされる。
【0042】
(標的生物)
本発明の方法は、任意の型の真核細胞において、DNA の安定した組み込み及び/又は転位(rearrangement )を得るために有用である。例えば、本方法は動物、植物、カビ、細菌、及び他の微生物の細胞において有用である。いくつかの態様において、その細胞は多細胞生物、例えば形質転換植物又は動物等の一部分である。本発明の方法は、形質転換したコムギ、トウモロコシ、及び動物の様な、形質転換した材料を得ることが困難である状況において特に有用である。これらの状況において、まれにしかない単一コピー挿入物を見出すには、独立して得られた多数の形質転換したクローンを前もって得ることが必要となるが、それはそれ自身非常に多くの労力を払う必要がある。
【0043】
特に興味が持たれる植物標的には単子葉植物があり、その中には、例えば、イネ、トウモロコシ、コムギ、ライムギ、オオムギ、バナナ、ヤシ、ユリ、ラン及びスゲが含まれる。双子葉植物もまた適切な標的であり、例えば、タバコ、リンゴ、ジャガイモ、テンサイ、ニンジン、ヤナギ、ニレ、カエデ、バラ、キンポウゲ、ペチュニア、フロックス、スミレ及びヒマワリが含まれる。他の標的には、動物及びカビ細胞が含まれる。これらのリストは単に説明するためのものであり、限定するものではない。
【0044】
(標的細胞内へ外来DNA を導入するためのコンストラクト)
本発明の方法は、標的細胞内へ外来DNA を導入することをしばしば含む。例えば、一つ又はそれ以上の組み換え部位を含む核酸が細胞中へしばしば導入される。細胞内へ導入されるべきポリヌクレオチドコンストラクトは、一つ又は複数の組み換え部位に加えて、望まれる形質型を細胞に与えるであろう遺伝子又は機能的な配列を含むことができる。
【0045】
いくつかの態様において、組み換え部位に加えて、リコンビナーゼをコードするポリヌクレオチドコンストラクトが真核細胞内へ導入される。リコンビナーゼをコードしているポリペプチドは、一つ又は複数の組み換え部位として同一の核酸上に含まれることが可能であり、また別個の核酸として細胞内へ導入されることも可能である。本発明は組み換え部位を含む核酸のみならず、その核酸内においてリコンビナーゼをコードしているポリヌクレオチド配列が標的真核細胞内において機能しているプロモーターに作用可能に結合している核酸もまた提供する。
【0046】
一般的には、発現するべきポリヌクレオチド(例えばリコンビナーゼをコードしているポリヌクレオチド又は興味の対象であるトランスジーン)は発現カセットに存在しているであろうし、それはそのポリヌクレオチドが発現制御シグナルに作用可能に結合していることを意味しており、その発現制御シグナルは、例えばプロモーターやターミネーターであって興味の対象である宿主細胞において機能を有する。リコンビナーゼ及び選択可能なマーカーをコードする遺伝子もまた、宿主細胞内において機能するその様なシグナルの制御下にあるであろう。発現の制御はプロモーターの選択によって最も容易に達成される。転写ターミネーターは一般的には重要でなく、細胞によって認識される限り種々の既知の因子を使用することができる。
【0047】
研究がなされている遺伝子からプロモーターを得ることが可能であり、または異なった遺伝子または異なった種から得られた異種プロモーターであることも可能である。形質転換植物又は他の生物の全ての組織において遺伝子の直接的な発現が望まれる場合には、「構成的な」プロモーターを使用することが可能であり、「構成的な」プロモーターは多くの環境条件や発達又は細胞分化の状況において一般的に活性である。
【0048】
植物において使用するために適切な構成的なプロモーターには、例えば、カリフラワーモザイクウイルス(CaMV)35S 転写開始領域及び領域VIプロモーター、アグロバクテリウムツメファシエンスのT-DNA から得られた1'- 又は2'- プロモーター、及び本技術分野の当業者に知られている植物細胞中で活性である他のプロモーターが含まれる。他の適切なプロモーターには、フィグウオート(Figwort )モザイクウイルス由来の全長の転写プロモーター、アクチンプロモーター、ヒストンプロモーター、チュブリンプロモーター、又はマンノピン合成酵素プロモーター(MAS )が含まれる。
【0049】
他の構成的な植物プロモーターには、とりわけシロイヌナズナ(Sun and Callis, Plant J.,11(5):1017-1027(1997))より得られた、種々のユビキチン又はポリユビキチンプロモーター、マス(mas )、マック(Mac )又はダブルマック(DoubleMac )プロモーター(米国特許番号5,106,739 、及びComai et al によってPlant Mol.Biol.15:373-381 (1990)において述べられている)、及び本技術分野の当業者に知られている、種々の植物遺伝子に由来する他の転写開始領域が含まれる。その様な遺伝子には、例えば、シロイヌナズナ由来のACT11 (Huang et al.,Plant Mol.Biol.33:125-139(1996))、シロイヌナズナ由来のCat3((GenBank No.U43147,Zhong et al.,Mol.Gen.Genet.251:196-203(1996))、セイヨウアブラナ(Brassica napus)由来のステアロイルアシルキャリア蛋白質不飽和化酵素をコードしている遺伝子(GenBank No.X74782,Solocombe et al.,Plant Physiol.104:1167-1176(1997))、トウモロコシ由来のGPc1(GenBank No.X15596 Martinez et al.,J.Mol.Biol 208:551-565(1989))、及びトウモロコシ由来のGpc2(GenBank No.U45855,Manjunath et al.,Plant Mol.Biol.33:97-112(1997) )が含まれる。
【0050】
植物のための有用なプロモーターには、Ti- 又はRi- プラスミドから、植物細胞から得られたプロモーター、プロモーターが植物中において機能を有することが見出されているときには植物ウイルス又は他の宿主が含まれる。植物中で機能し、そしてそのため本発明の方法において使用するのに適切である細菌プロモーターには、オクトピン合成酵素プロモーター、ノパリン合成酵素プロモーター、マノピン(manopine)合成酵素プロモーターが含まれる。適切な内在性植物プロモーターには、リブロース-1,6- ビスホスファターゼ(RUBP)、カルボキシラーゼスモールサブユニット(ssu )プロモーター、α- コングリチニン(conglycinin )プロモーター、ファゼオリン(phaseolin )プロモーター、ADH プロモーター、及び熱ショックプロモーターが含まれる。
【0051】
大腸菌において使用するためのプロモーターには、T7、trp 、又はラムダプロモーター、リボソーム結合部位を含み、及び好ましくは転写終結シグナルである。真核細胞のためには、制御配列は典型的にはプロモーターを含み、そのプロモーターは随意にイムノグロブリン遺伝子、SV40、サイトメガロウイルス等に由来するエンハンサー及びポリアデニレーション配列を含み、そしてスプライスドナー及びアクセプター配列を含むこともできる。酵母において、便利なプロモーターにはGAL1-10 (Johnson and Davies (1984) Mol.Cell.Biol.4:1440-1448 )、ADH2(Russell et al.(1983) J.Biol.Chem.258:2674-2682)、PHO5(EMBO J.(1982) 6:675-680 )及びMFα(Herskowitz and Oshima (1982) The Molecular Biology of the Yeast Saccharomyces (酵母サッカロミセスの分子生物学)(eds.Strathem,Jones, and Broach)Cold Spring Harbor Lab.,Cold Spring Haror,N.Y.,PP181-209の中において)が含まれる。
【0052】
代わりに、興味の対象である遺伝子の特異的な組織における発現を標的としたプロモーター、又はさもなければより厳密な環境又は発達による制御の下にあるプロモーターを使用することが可能である。その様なプロモーターをここでは、「誘導可能な」又は「抑制可能な」プロモーターと称する。誘導可能なプロモーターの転写によって転写に影響するかもしれない環境条件の例には、病原体の攻撃、嫌気性の条件、エチレン、又は光の存在が含まれる。発達による制御下にあるプロモーターには、葉、根、果実、種、又は花の様な特定の組織においてのみ転写を開始するプロモーターが含まれる。プロモーターの操作はゲノム中のそれの位置によってもまた変化する可能性がある。そこで、誘導可能なプロモーターはある位置において完全に又は部分的に構成的となることがある。誘導可能なプロモーターは、リコンビナーゼ遺伝子の発現を制御するためにしばしば使用され、そこで組み換え反応のタイミングを制御することが可能となる。
【0053】
発達による制御の下にある組織特異的な植物プロモーターの例には、果実、種、花の様な特定の組織においてのみ転写を開始するプロモーターが含まれる。トマト由来の組織特異的なE8プロモーターは遺伝子発現を方向付けるために特に有用であり、そのために望まれる遺伝子発現産物は果実の中に位置する。例えば、Lincoln et al.(1988) Proc.Natl.Acad.Sci.USA 84:2793-2797; Deikman et al.(1988) EMBO J.7:3315-3320; Deikman et al.(1992) Plant Physiol.100:2013-2017 を見よ。他の適切なプロモーターには胚の貯蔵蛋白質をコードしている遺伝子が含まれる。
【0054】
誘導可能なプロモーターによる転写に影響するかもしれな環境条件の例には、嫌気的な条件、上昇した温度、又は光の存在が含まれる。更なる器官特異的、組織特異的、及び/又は誘導可能な外来のプロモーターもまた知られており(例えば、Kuhlemeier et al (1987) Ann.Rev.Plant Physiol.38:221において引用された参考文献を見よ)、それには、光誘導性であって光合成を行う組織においてのみ活性である、シロイヌナズナ(Arabidopsis thaliana)の1,5-リブロースビスホスフェイトカルボキシラーゼ スモールサブユニット遺伝子類("ssu" プロモーター)プロモーター、葯特異的なプロモーター(EP 344029 )、及び例えばシロイヌナズナ由来の(Arabidopsis thaliana)種子特異的なプロモーター(Krebbers et al.(1988) Plant Physiol.87:859)が含まれる。見本となる緑色組織特異的なプロモーターにはトウモロコシのホスフォエノールピルビン酸カルボキシラーゼ(PEPC)プロモーター、スモールサブユニットリブロース ビスカルボキシラーゼプロモーター(ssRUBISCO )及びクロロフィルa/b 結合蛋白質プロモーターが含まれる。プロモーターは、国際公開番号WO93/07278の中で述べられた通りの、植物TrpA遺伝子から単離されたプロモーターの様な樹心(pith)特異的なプロモーターであることもできる。
【0055】
他の生物のための誘導可能なプロモーターには、例えば、アラビノースプロモーアー、lacZプロモーター、メタロチオネインプロモーター、及び熱ショックプロモーターのみならず、本技術分野の当業者に知られている他の多くのものが含まれる。S.pombe の様な酵母において有用な抑制可能なプロモーターは、ビタミンB1によって抑制可能なPmntプロモーターである。
【0056】
典型的には、これらの細胞へ導入されるコンストラクトは組み換え発現技術を用いて調製される。組み換え発現技術は、組み換えた核酸の構築と感染した細胞内での遺伝子の発現を伴う。これらの末端に到達するための分子クローニング技術は本技術分野において知られている。組み換え核酸の構築に適する広範囲のクローニングとインビトロ増幅の方法は本技術部分野の当業者に良く知られている。これらの技術の例と、多くのクローニング演習を通じて当業者を教示するのに十分である指示は、Berger and Kimmel,Guide to Molecular Cloning Techniques, Methods in Enzymology(分子クローニング技術への案内 酵素学における方法論), volume 152, Academic Press, Inc.,San Diego,CA (Berger); 及びCurrent Protocols in Molecular Biology(分子生物学における現在のプロトコール),F.M. Ausubel et al., eds.,Current Protocols (現在のプロトコール),a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons,Inc.,(1998 Supplement) (Ausubel) の中に見出される。
【0057】
ポリヌクレオチドコンストラクトの構築には、細菌内で複製可能なベクターを使用することが通常必要となる。細菌からプラスミドを精製するために使用することができるキットは有り余る程市販されている。それらを適切に使用するために製造者の指示に従うこと(例えば、EasyPrepJ,FlexiPrepJ:両者はファルマシアバイオテック由来、StrataCleanJ:ストラタジーン由来、及びQIAexpress Expression System:キアゲン、を見よ)。単離されて精製されたプラスミドを更に操作して他のプラスミドを作製することも可能であり、細胞をトランスフェクトするか、又はアグロバクテリウムツメファシエンスへ導入して感染させて、植物を形質転換するのに使用される。アグロバクテリウムが形質転換の手段である場合にはシャトルベクターが構築される。ストレプトマイセス又はバシラスにおけるクローニングもまた可能である。
【0058】
標的細胞中へ取り込まれるために使用されるポリヌクレオチドコンストラクト及び/又はベクター中へ、選択可能なマーカーがしばしば導入される。これらのマーカーによって、興味の対象であるポリヌクレオチドを含んでいる細胞のコロニーを選択することができる。ベクターは、例えば大腸菌又は他の細胞中で機能する選択マーカーをしばしば有するであろうし、標的細胞へ導入される前にその細胞中でベクターは複製される。大腸菌内で選択可能なマーカーの例には、例えばアンピシリン、テトラサイクリン、カナマイシン、エリスロマイシンなどの抗生物質に対する耐性を特定している遺伝子、又はβ−ガラクトシダーゼの様な他の型の選択可能な酵素活性を与える遺伝子、又はラクトースオペロンが含まれる。哺乳細胞において使用するのに適している選択可能なマーカーには、例えば、ジヒドロ葉酸還元酵素(DHFR)、シミジンキナーゼ遺伝子(TK)、又は薬剤耐性を特定している原核細胞の遺伝子が含まれ、薬剤耐性を特定している原核細胞の遺伝子には、ミコフェノール酸(mycophenolic acid )で選択することができるgpt (キサンチン−グアニン ホスフォリボシルトランスフェラーゼ); G418、ハイグロマイシン又はピュロマイシンで選択することができるneo (ネオマイシンホスフォトランスフェラーゼ); 及びメトトレキサートで選択することができるDHFR(ジヒドロ葉酸還元酵素)(Mulligan & Berg (1981) Proc.Natl.Acad.Sci.USA 78:2072;Southern & Berg (1982) J.Mol.Appl.Genet.1:327)がある。
【0059】
植物細胞のための選択マーカーはしばしば、例えば、カナマイシン、G418、ブレオマイシン、ハイグロマイシン又はクロラムフェニコールの様な殺生剤又は抗生物質に対する耐性、又はクロロスルフロン(chlorsulfuron )又はバスタ(Basta )の様な除草剤に対する耐性を与える。選択可能なマーカーのための適切なコード配列の例には、抗生物質カナマイシンに対する耐性を与えるネオマイシンホスフォトランスフェラーゼ酵素をコードするneo 遺伝子(Beck et al (1982)Gene 19:327 )、ハイグロマイシンホスフォトランスフェラーゼ酵素をコードして抗生物質ハイグロマイシンに対する耐性を与えるhyg (hpt) 遺伝子(Gritz and Davies (1983) Gene 25:179 )、及び除草剤物質であるホスフィノスリチン(phosphinothricin)及びビアラフォス(bialaphos )に対する耐性をあたえるホスフィノスリチンアセチルトランスフェラーゼをコードするbar 遺伝子(EP 242236 )がある。
【0060】
もし一つ以上の外来遺伝子を標的とする真核細胞に導入するならば、通常は、各々の外来性の核酸について異なった選択可能なマーカーを使用することが望ましい。これにより、望まれる外来性の核酸の両者を含む細胞について同時に選択することができるようになる。
【0061】
(標的細胞へコンストラクトを導入するための方法)
組み換え部位及び/又はリコンビナーゼをコードする遺伝子を含むポリヌクレオチドコンストラクトを、標的細胞及び/又は生物へ、当業者に知られているいくつかの手段のいずれかを用いて導入することができる。例えば、植物の培養中又は器官中のいずれかで、種々の通常の技術によって、DNA コンストラクトを植物細胞中へ導入することができる。例えば、DNA パーティクルボンバートメントの様なバイオリスティックな方法を用いてDNA コンストラクトを植物細胞へ直接に導入することが可能であり、又は植物細胞プロトプラストの電気穿孔法及びマイクロインジェクションの様な技術を用いてDNA コンストラクトを導入することが可能である。パーティクルにより仲介された形質転換技術(「バイオリスティック」としても知られている)は、Klein et al.,Nature,327:70-73 (1987); Vasil,V.et al.,Bio/Technol.11:1553-1558 (1993)及びBecker,D.et al.,Plant J.,5:299-307 (1994) において述べられている。これらの方法は、微小ビーズ又はパーティクルの基質の中又はその表面上のいずれかにおいて核酸を有する小さなパーティクルにより細胞を穿孔することを伴う。
【0062】
バイオリスティックなSDS-1000遺伝子銃(バイオラッド、ヘラキュレス、CA)は、DNA を被覆した金又はタングステンの微小担体を標的細胞へ加速するためにヘリウムガスの圧力を使用する。その過程は広範囲の生物由来の組織及び細胞に適用可能であり、その生物には植物、細菌、カビ、藻、無傷動物組織、組織培養細胞及び動物胚が含まれる。電気パルス送達を用いることも可能であり、それは動物及び患者の生組織のための、本質的に緩和な電気穿孔の型である。Zhao,Advanced Drug Derivery Reviews (進歩した薬物送達の総論)17:257-262 (1995)。
【0063】
他の形質転換方法が当業者に知られている。マイクロインジェクションの技術が本技術分野において知られ、科学及び特許文献において良く述べられている。ポリエチレングリコール(PEG )沈殿を用いたDNA コンストラクトの導入は、Paszkowski et al.,EMBO J.3:2717 (1984)において述べられている。電気穿孔技術は、Fromm et al.,Proc.Natl.Acad.Sci.USA,82:5824 (1985)において述べられている。PEG により仲介された植物プロトプラストの形質転換及び電気穿孔もまた、Lazzeri,P.,Methods Mol.Biol.49:95-106 (1995)において議論されている。異種遺伝子の導入及び発現のための方法は単子葉及び双子葉植物の両者において知られている。米国特許番号5,633,446, 5,317,096, 5,689,052, 5,159,135及び5,679,558; Weising et al.(1988) Ann.Rev.Genet.22:421- 477 を見よ。単子葉植物の形質転換には特に種々の技術を使用することが可能であり、それには電気穿孔法(例えばShimamoto et al.,Nature (1992),338:274-276)、バイオリスティック(例えば欧州特許出願 270,356)、及びアグロバクテリウム(例えば、Bytebier et al.,Proc.Natl.Acad.Sci.USA (1987) 84:5345-5349)が含まれる。
【0064】
植物の形質転換のために、DNA コンストラクトを適切なT-DNA フランキング領域と結合させて、一般的なアグロバクテリウムツメファシエンス宿主ベクターへ導入することができる。アグロバクテリウムツメファシエンス宿主の毒性機能により、細胞が細菌によって感染した時に、トランスジーン及び隣接したマーカー遺伝子(もし存在しているならば)が植物細胞DNA へ挿入される。アグロバクテリウムツメファシエンスにより仲介された形質転換技術は、科学文献において良く述べられている。例えば、Horsch et al.Science,233:496 (1984), Fraley et al.,Proc.Natl.Acad.Sci.USA,80:4803 (1983)及びHooykaas,Plant Mol.Biol.,13:327-336 (1989), Bechtold et al.,Comptes Rendus De L Academie Des Sciences Serie Iii-Sciences De La Vie-Life Sciences,316:1194-1199 (1993), Valvekens et al.,Proc.Natl.Acad.SCI.USA,85:5536-5540 (1988)を見よ。植物及び細胞培養物の遺伝子移送方法の総論としては、Fisk et al.,Scientia Horticulturae 55:5-36 (1993) 及びPotrykus,CIBA Found.Symp.154:198 (1990) を見よ。
【0065】
ポリヌクレオチド配列を細胞内へ送達するための他の方法には、例えば、リポソームに基づいた遺伝子送達(Debs and Zhu (1993) WO93/24640; Mannino andGould-Fogerite (1988) BioTechniques 6(7); Rose米国特許番号5,279,833; Brigham (1991) WO91/06309; 及びFelgner et al.(1987) Proc.Natl.Acad.Sci.USA84:7413-7414)のみならず、ウイルスベクターの使用、(例えば、アデノウイルス(総論のために、Berns et al.(1995) Ann.NY Acad.Sci.772:95-104; Ali etal.(1994) Gene Ther.1:367-384;及びHaddada et al.(1995) Curr.Top.Microbiol.Immunol.199 (Pt3):297-306 を見よ)、パピローマ(papilloma) ウイルス、レトロウイルス(例えば、Buchscher et al.(1992) J.Virol.66(5) 2731-2739; Johann et al.(1992) J.Virol.66(5):1635-1640; Sommerfelt et al.,(1990) Virol.176:58-59; Wilson et al.(1989))J.Virol.63:2374-2378; Miller et al.,J.Virol.65:2220-2224 (1991); Wong-Staal et al.,PCT/US94/05700,及びRosenburg and Fauci (1993) in Fundamental Immunology(基本的免疫学),Third Edition Paul (ed) Raven Press,Ltd.,New York及びその中の引用文献、及びYu et al.,Gene Therapy (1994) supra を見よ)、及びアデノ関連ウイルスベクター(West et al.(1987) Virology 160:38-47; Canter et al.(1989) 米国特許番号4,797,368; Canter et al.WO93/24641 (1993); Kotin (1994) Human Gene Therapy 5:793-801; Muzyczka (1994) J.Clin.Invst.94:1351及びSamulski(supra) をAAVベクターの概観のために見よ。また、Lebkowski,米国特許番号5,173,414; Tratschin et al.(1985) Mol.Cell.Biol.,5(11):3251-3260; Tratschin et al.(1984) Mol.Cell.Biol.,4:2072-2081; Hermonat and Muzyczka (1984) Proc.Natl.Acad.Sci.USA,81:6466-6470; McLaughlin et al.(1988)及びSamulski et al. (1989) J.Virol.,63:03822-3828 もまた見よ)及びそれに類似なものが含まれる。
【0066】
導入された外来DNA が組み込まれたパターンを解析することができる方法は、当業者に良く知られている。例えば形質転換した細胞からDNA を抽出し、一つ又はそれ以上の制限酵素によってそのDNA を消化し、そしてポリヌクレオチドコンストラクトのラベル化された断片をハイブリダイズさせることができる。挿入された配列をポリメラーゼ連鎖反応(PCR )を用いて同定することもまた可能である。例えば、Sambrook et al.,Molecular Cloning - A Laboratory Manual (分子クローニング、実験室マニュアル), Cold Spring Harbor Laboratory, ColdSpring Harbor, New York,1989を、これら及び他の適切な方法の記載のために見よ。
【0067】
(形質転換植物及び動物の再生)
本発明の方法は、安定して組み込まれた外来性ポリヌクレオチド又は他の細胞内核酸の安定な転位(rearrangement )を有する、形質転換したキメラ型の多細胞生物を得るために特に有用である。形質転換したキメラ型の生物を得るための方法は、植物と動物の両者について当業者に良く知られている。
【0068】
上記のいずれかの形質転換技術によって得られた形質転換した植物細胞は培養され、形質転換した遺伝子型及びそのために望みの表現型を有している完全な植物が再生される。その様な再生技術は組織培養成長培地の中の特定の植物ホルモンの取り扱いに依り、典型的には望みの核酸配列と共に導入された殺生剤及び/又は除草剤マーカーに依る。培養されたプロトプラストからの植物再生は、Evans et al.,Protoplast Isolation and Culture, Handbook of Plant Cell Culture (プロトプラストの単離及び培養、植物細胞培養のハンドブック),pp.124-176, Macmillian Publishing Company, New York (1983);及びその結びつきで、Regeneration of Plants, Plant Protoplast(植物、植物プロトプラストの再生),pp.21-73,CRC Press, Boca Raton,(1985)において述べられている。植物カルス、外植片、体細胞胚(Dandekar et al.,J.Tissue Cult.Meth.,12:145 (1989); MacGranahan et al.,Plant Cell Rep.,8:512 (1990))、器官、又はその一部分からもまた再生することができる。その様な再生技術は一般的にKlee et al.,Ann.Rev.of Plant Phys.,38:467-486 (1987)において述べられている。
【0069】
その方法は大部分の脊椎動物種の形質転換したキメラ型の動物を作製するのに有用である。その様な種には、それに限定されるものではないが、ネズミ及びラットの様なゲッシ類、ウサギ、ヒツジやヤギの様なヒツジ類(ovines)、ブタの様なブタ類(porcines)、ウシ(cattle)やバッファローの様なウシ類(bovine)を含む、非ヒト哺乳類が含まれる。形質転換動物を得る為の方法は、例えば、Puhler,A.,Ed.,Genetic Engineering of Animals(動物の遺伝工学)VCH Publ.,1993; Murphy and Carter, Eds.,Transgenesis Techniques:Principles and Protocols(形質転換技術:原理とプロトコール)(Methods in Molecular Biology, Vol.18),1993;及びPinkert,CA,Ed.,Transgenic Animal Technology:A Laboratory Handbook (形質転換動物の技術工学:実験室ハンドブック),Academic Press,1994において述べられている。特異的な遺伝的修飾を有している形質転換魚類もまた、クレイムされた方法を用いて作製することができる。形質転換魚類を作製するための一般的な方法のためには、例えば、Iyengar et al.(1996) Transgenic Res.5:147-166 を見よ。
【0070】
そのゲノム中に特異的な修飾を有している、形質転換した又はキメラ型の動物を得る一つの方法は、受精した卵母細胞を興味の対象であるポリヌクレオチドを組み換え部位に隣接して含んでいるベクターと接触させることである。ネズミの様ないくつかの動物についてインビボで受精が行われ、受精した卵子は手術により移された。他の動物、特にウシおいては、生きた又は屠殺された動物から卵子を移して来て、そしてその卵子をインビトロで受精させることは好ましい。DeBoer et al.,WO91/08216を見よ。インビトロで受精させることにより、実質的に同調した細胞に導入すべき修飾を行うことができる。そして、着床前の胚が約16-150個の細胞を含むようになるまで受精した卵母細胞をインビトロで培養した。16-32 細胞期の胚は桑実胚として述べられている。32細胞以上を含んでいる着床前の胚を胞胚と称する。これらの胚は、典型的には64細胞期において胞胚腔の空洞(cavity)の発達を示した。もし望むならば、胚細胞の中における望みの外来性ポリヌクレオチドの存在を当業者に知られている方法によって検出することができる。
【0071】
受精した卵母細胞を着床前の段階まで培養するための方法は、Gordon et al.(1984) Methods Enzymol.101:414; Hogan et al. Manipulation of the Mouse Embryo:A Laboartory Manual(マウス胚の操作:実験室マニュアル)C.S.H.L. N.Y.(1986) (マウス胚); Hammer et al.(1985) Nature 315:680(ウサギ及びブタ胚);Gandolfi et al.(1987) J.Reprod.Fert.81:23-28;Rexroad et al.(1988) J.Anim.Sci.66:947-953(ヒツジ胚)及びEyestone et al.(1989) J.Reprod.Fert.85:715-720; Camous et al.(1984) J.Reprod.Fert.72:779-785及びHeyman et al.(1987) Theriogenology 27:5968(ウシ胚)において述べられている。時々、着床を待つ間の期間着床前の胚を凍結して保存した。着床前の胚を適切なメスに移植し、その結果、トランスジーンが組み込まれたときの発達の段階に依って、形質転換した又はキメラ型の動物が誕生するという結果となった。正しい生殖系列の形質転換動物を形成するために、キメラ型の哺乳動物を繁殖させることができる。
【0072】
替わりに本方法を用いて、望みの外来性ポリヌクレオチドの単一コピーを有する胚幹細胞(ES)を得ることができる。これらの細胞は、インビトロで培養された着床前の胚から得られた。例えば、Hooper,ML,Embryonal Stem Cells:Introducing Planned Changes into the Animal Germline (胚性幹細胞;計画された変化の動物生殖系列への導入)(Modern Genetics,v.1) Int'1.Pub.Distrib.,Inc.,1993; Bradley et al.(1984) Nature 309,255-258 を見よ。形質転換したES細胞を非ヒト動物由来の胞胚と結合させた。ES細胞は胚をコロナイズし、いくつかの胚においては得られたキメラ動物の生殖系列を形成する。Jaenisch,Science,240:1468-1474 (1988) を見よ。代わりに、生物を再構成することができるES細胞又は体細胞(”体細胞再集団細胞:somatic repopulating cells”)を、形質転換した哺乳動物を生み出す、除核された受精卵母細胞へ着床させるための核の起源として使用することができる。例えば、Wilmut et al.(1997) Nature 385:810-813を見よ。
【0073】
(実施例)
下記の実施例は説明の目的で与えられたものであって本発明を限定するものではない。
【0074】
(実施例1)
(シゾサッカロミセス・ボンベ(Schizosaccharomyces pombe )におけるΦC31組み換えシステム)
この実施例は、ストレプトミセスバクテリオファージΦC31 部位特異的な組み換えシステムが、真核細胞において機能することを示している。S.Pombe のleu1遺伝子座において、バクテリオファージ接着部位(attP)をシゾサッカロミセス・ボンベの染色体において導入した。続いてこの標的株を、ura4+選択可能なマーカーと連結した細菌接着部位(attB)を含むプラスミドにより形質転換された。ΦC31 インテグラーゼ遺伝子を有している第二のプラスミドによって共トランスフェクトしたとき、インテグラーゼ遺伝子が発現している条件下で、Ura+へ高い効率で形質転換していることが観察された。
【0075】
組み換えイベントのサザン解析により、leu1遺伝子座のattPへ、attB-ura4+プラスミドが挿入されていることが示された。ハイブリッドジャンクジョンのヌクレオチド配列により、attB x attP 組み換え反応は正確であることが明らかとなった。
【0076】
(材料及び方法)
(組み換えDNA )
全体として標準的な方法が用いられた。大腸菌株XL2-Blue(recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F' proAB lAcIq Z △M15Tn10(Tetr) Amy Camr、ストラタジーン]がDNA コンストラクトとして働いた。
【0077】
(培地)
分裂酵母株を、必要に応じて225mg/l のアデニン、ヒスチジン、ロイシン又はウラシルを添加した、最小培地(バイオ101 由来EMM-低グルコース)で生育させた。5-FOA (5-フロロオロチン酸、ザイモリサーチ社より)を含む最小プレートをGrimm ら((1988) Mol.Gen.Genet.215:81-86)に従って調製し、アデニン、ヒスチジン及びロイシンを添加した。使用した時、5 μg/mlとなるようにチアミンを添加した。
【0078】
(ΦC31 attP標的を有するS.prombe)
pHS282からApaI-SacI 断片として単離された(Thorpe & Smith (1998) Proc.NATL.Acad.Sci.USA 95:5505-5510 )84塩基対のΦC31 attP部位(PP' と略称)を、S.prombe組み込みベクターpJK148の同じ部位にクローン化し(Keeney & Boeke (1994) Genetics 136:849-856)、pLT44 を作製した。NdeI切断DNA を用いた酢酸リチウムにより仲介された形質転換により、このプラスミドはS.prombe leu1-32アリールへ標的化された。pLT44 による相同組み換えによってLeu+へ変換された、レシピエント宿主FY527(h-ade6-M216 his3-D1 leu1-32 ura4-D18) を、サザンブロットによって試験した。FY527attP と名付けられた一つのLeu+型は単一コピーのpLT44 を含んでいることが見出された。FY527attPx2 と名付けられたもう一つの形質転換体は直列(tandem)のプラスミド挿入を有していた。
【0079】
(ΦC31 attB部位を有する組み込み可能なura4+ベクター)
1.8kb のEcoRI-BamHI 断片上でpTZura4 (S.Forsburg)から切り出した、S.prombeのura4+遺伝子を、同じ酵素を用いて切断されたpJK へ挿入し、pLT40 を作製した。500bp のBamHI-XbaI断片としてpHS21 から単離されたΦC31 attB部位(BB'と略称)を、それらの酵素を用いて切断されたpLT40 へ連結してpLT42 を作製した。XhoI断片を削除することによりpLT42 からleu1遺伝子の大部分を除去し、pLT45 を作製した。これにより、pLT45 からleu1の全部であるが229bp が除去されて、その形質転換効率はleu1相同性を持たないプラスミドの形質転換効率まで低下した。ura4のすぐ反対側に同じ方向の第二でattB部位を有しているpLT50 が構築され、それは最初にpLT42 由来のattB BamHI-SacI 断片をpUC19 へサブクローニングし、それをEcoRI とSalII によって切り出し、続いてそれをEcoRI とXhoIによって切断したpLT45 へ挿入することにより構築された。最終構築物中の第二のattB部位を各鎖について一回配列決定し、最初のattB部位と同一であることが見出された。
【0080】
(平滑DNA 形質転換)
attB-ura4+-attB 平滑DNA は、pLT50 から精製されたattII-AlwNl 断片として、又はpLT50 を鋳型として用いたPCR 産物として調製された。T3プライマーとpJK148のプラスミド骨格に対応している第二のプライマー(5' ggc cct gaa att gtt gct tct gcc 3' )を用いて、標準的な条件によりPCR は行われた。
【0081】
(ΦC31 インテグラーゼの抑制的な合成)
ビタミンB1によって抑制される、S.prombe Pmntプロモーターは、pMO147から1.2kb のPstI-SacI 断片として切り出され、同じ酵素を用いて切断されたhis3+,ars1 ベクターpBG2(Ohi et al.(1996))Gene 174:315-318)へ挿入され、pLT41 が作製された。ΦC31 int コード領域を含んでいる2.0kb SacI断片を、pHS33(Thorpe & Smith (1998) supra )からpLT41 のSacI部位へ移した。発現がPmntの制御下にある様にint コード領域が配向されたクローンを、pLT43 と名付けた。
【0082】
(分子解析)
ベーリンガーマンハイムのGeniusTMシステムを用いてサザン解析を行った。leu1の998bp の内部EcoRV 断片、ura4の1.8kb 断片及び2.0kb のΦC31 int 遺伝子をランダムプライマー法によってジゴキシゲン標識し、プローブとして使用した。ストラタジーンTurbo PFU 酵素又はVENTポリメラーゼを用いて、パーキンエルマーCetus Gene Amp PCR 9600 で、ポリメラーゼ連鎖反応を行った。可能なところでは標準的なT3及びT7プライマーを用いた。ura4プライマー(5' gtc aaa aag ttt cgt caa tat cac 3' )(配列番号1)及びpJK148プライマーをオペロンテクノロジーから購入した。全てのPCR 反応において、51℃のアニーリニング温度と30秒の伸長時間が用いられた。
【0083】
(結果と考察)
(S.prombeゲノムへの標的部位の挿入)
ΦC31 により仲介された組み込みのための標的部位を有する宿主株を作製するために、相同組み換えによって、ΦC31 attP部位を分裂酵母ゲノムのleu1遺伝子座へ挿入し、Leu+株FY527 attPが形成された(図1 A)。過去の研究により、S.prombeのDNA をXbaIによって開裂し、leu1+遺伝子の内部の1kp 断片により検証したら、そのプローブにより14kbのバンドが検出されることが示されている(Keeney & Boeke (1994) Genetics 136:849-856)。leu+プラスミドpJK148がleu1-32 遺伝子座において挿入されていると、3 と18kbのバンドが検出されるという結果となる(図1 A)。84bpのΦC31 attP因子が封入されている点で、pLT44はpJK148から異なっているために、leu1-32 におけるpLT44 の組み込みは、FY527 attPにおいて、同様の3kb と18kbのハイブリダイゼーションパターンをもたらした。他のハイブリダイズしている断片が存在しないことは、単一に組み込まれたコピーとしてのpLT44 のDNA 残基を示している。
【0084】
ΦC31インテグラーゼ仲介形質転換 ura4+およびattB配列(BB’)を有するが、複製の原点を欠くFY527attPをpLT45で形質転換した。このコンストラクトを、それ自体により、またはΦC31インテグラーゼを産生するhis3+複製ベクターpLT43を用いて導入した。pLT43を封入することにより、Ura+形質転換体の数が平均で15倍増加した(表1)。この効果はインテグラーゼ遺伝子発現によるものなので、この増加がpLT45と複製にプロフィシエントな(replication-proficient)pLT43との間の組み換えに起因するものではありえない。インテグラーゼ遺伝子の転写は、高濃度のビタミンB1により抑制可能なプロモーターPmntの支配を受ける(Maundrell,K.(1993年)Gene 123:127〜130頁)。抑制は完全ではないが(Forsburg,S.L.(1993年)Nucleic Acids Res 21:2955〜2956頁)、インテグラーゼたん白の生成が減少する。発育培地にチアミンを加えた場合、Ura+形質転換体の数は背景レベル付近にまで減少した。培地からヒスチジンを省略することによりインテグラーゼプラスミドを共選択してもしなくても、Ura+形質転換体の頻度が顕著に変化することは無かった。pLT43またはその前駆体プラスミドpBG2を用いて得たHis+形質転換体の数から、FY527atttPの形質転換能力を推定した。両方の複製プラスミドの頻度を比較したところ、FY527attPのpLT43依存形質転換は平均で約15%であった。
【0085】
【表1】

【0086】
ΦC31インテグラーゼをプロモートしたattP x attB組み換え pLT45をコードしたΦC31attBエレメントと染色体的に位置の定まったattP配列との間の組み換えにより、図1Bに示すように、環状DNAがleu1遺伝子座に組み込まれた。この反応が起きた場合、Ura+形質転換体から得たXbaI分別ゲノムのDNAをleu1 DNAでプローブすると、3kbバンドは変わらないが、一方18kbバンドは約23kbまで増大する(図1C)。無作為に選択したUra+コロニーをハイブリダイゼーション分析により検査した。チアミンの省略によりΦC31インテグラーゼ遺伝子発現を抑制解除した場合の実験から得られた8つの分離株のうち7つが、この約23kbバンドの存在を示した。この同サイズバンドをura4プローブにハイブリダイズした。ura4−D18欠損対立遺伝子から期待されるように、これは親株にura4ハイブリダイゼーションしなかったものと対照的である。これら7つの分離株の一つが、両方のプローブにハイブリタイズする付加的なバンドを示した。部位に特異的な組み換え現象に加えて、この候補物質がleu1遺伝子座にDNA再配列を有すると思われる。leu1再配列が、有効なS.pombe相同体組み換え系によりおそらく触媒された。残る分離株は部位特異的組み換え現象を経験したことが無く、pLT45とpLT43との間の組み換えにより、ウラシル原栄養株が増大したと思われた。これら8つの分離株のうち、半分をUra+およびHis+の両方として選択したが、このグループとUra+のみで選択したグループとの間に顕著な差は見られなかった。
【0087】
ビタミンB1の存在下で行った形質転換実験から、DNAハイブリダイゼーションにより同数のUra+形質転換体を調べた。チアミン抑制可能なPnmtプロモーターがインテグラーゼ産生を制限し、したがって部位特異的組込みを制限すると考えられる。この低頻度形質転換から分離された8つのUra+候補物質のうちの2つにより、23kbバンドがleu1およびura4プローブにハイブリダイズしていることが示された。しかし、いずれのプローブも付加的バンドを検出したので、これらは正確な組込み現象を表しておらず、発明者らはこれらをクラスbインテグラントとして分類した。他の6つの分離体では、ハイブリダイゼーションのパターンを翻訳することは困難である。これらのいくつかにおいて、遺伝子座は再配列を経験しているが、leu1プローブでは3kbバンドを検出できなかった。これらの多くにおいて、ura4に対するハイブリダイゼーションが弱いことから、Ura+表現型はゲノム中のpLT45の安定な保守によるものではないであろう。
【0088】
選択しない場合にインテグラーゼプラスミドを維持する形質転換体の比率を確実にするために、インテグラーゼ遺伝子配列でブロットを再プローブした。Ura+His+として選択されたそれらはプラスミドを維持すると考えられており、かつハイブリダイゼーションが明らかになるにつれてそうなった。His表現型に関係なくUra+として選択された8つの分離株のうちの5つは、インテグラーゼプローブにハイブリダイズするバンドも与えた。intの損失が安定な組込みに影響を及ぼすことを確認するため、無作為に選択したUra+細胞の別の組を数世代にわたり非選択的に成長させ、pLT43を失ったHis−子孫に対してスクリーニングをした。8つの代表的Ura+His−クローンの分析は、全てが正確に染色体が位置するattP部位に組込まれたpLT45の単一コピーを有することを示した。これらの組込み体のDNAはインテグラーゼプローブでハイブリダイズしなかった。対照的に、pLT45単体の形質転換により誘導された背景頻度Ura+クローンにより、leu1遺伝子座にハイブリダイジングバンドの親構造が得られ、かつ5kbおよび7kbに更なる微弱バンドが得られた。これらの知見は、ゲノムのどこかのpLT45の組込みまたはS.pombe複製原点の欠損にもかかわらずいくつかの細胞中のプラスミドの維持のいずれかと一致している。
【0089】
保守的部位特異的組み換え PCRを用いて、3つの代表的Ura+候補物質からattP/attB組み換え接合部を回収した。ハイブリッド部位の一つでは、attR(PB’)がT3およびT7プロモーターに隣接していた。他の部位では、attL(BP’)がT3プロモーターおよびura4DNAに隣接していた(図1C)。いずれの場合も、プライマー対が、期待される大きさのバンドを増幅したこれらの配列に向けられ、一方、本来のattP(PP’)はもはや見つからない。attPは検出されるがattLおよびattRのいずれもが検出されない場合には、これは親株FY527aatPと対照的である。3つの代表的attLおよびattR PCR製品のヌクレオチド配列は、付随的な突然変異体が無いことを示した。したがって、バクテリアおよび哺乳動物細胞において、S.pombe中のΦC31仲介部位特異的組み換えは保守的組み換え反応である。
【0090】
ΦC31インテグラーゼは組込みした分子を除去しない。
ThorpeとSmith((1998年)Proc.Natl.Acad.Sci.USA95:5505〜5510)は、ゲル分別DNA断片の分析でΦC31インテグラーゼ反応の反転を検出しなかった。発明者らは遺伝学的選択法により反転反応の可能性を調べた。pLT45のFY527attPへの正確な組込みが、サザン分析により3つのクローンに対して確認された。次いで、これらの株をpLT43で再転換した。pLT45を切り取ることにより、ura4+マーカーの欠損を招いた。Ura−表現型を5−FOAを用いてプレート上に得点することができる(Grimmら、(1988年)Mol. Gen. Genet.215:81〜86頁)。3つのUra+His−前駆体の培養体から得たUra−分離個体の頻度は5.7×10−4、7.1×10−4および5.6×10−4であった。対照的に、3つのUra+His+誘導体から得たUra−コロニーの頻度はある程度高く、1.1×10−2、3.8×10−3および2.3×10−3であり、それぞれ19、5および4倍増加した。インテグラーゼ遺伝子を有しない制御ベクターpBG2を代わりに用いて、5−FOA耐性の速度が増大することも分かった。それぞれ1.0×10−2、1.0×10−2および8.0×10−3であった。形質転換プロセス自体は変異原性を示す。
【0091】
pLT45で形質転換された3つの培養体のそれぞれから得られた3つのUra−His+クローンをサザンブロッティングにより分析した。1つの分離体が、pLT45のFY527attPへの安定な組込みと一致するDNAパターンを有していた。したがって、このクローンにおいて、Ura−表現型は、部位特異的組み換え反応の反転によるよりも、むしろ制限パターンが感知可能なほどに変わっていない変異体により引き起こされた。第2クローンはpLT45挿入欠損FY527attPのサザンパターン特徴を示し、第3クローンはpLT45挿入の無いFY527attP等の細胞とFY527attP前駆体株FY527等の細胞の2つの型の細胞の混合物と一致するパターンを有していた。後者の構造は、pLT44の挿入を反転するleu反復の間の染色体内相同組み換えから生じる場合がある(図1A)。組込みしたプラスミドDNAの正確な切り取りが前述の2つの候補物質に現れた場合、attP部位が再生した。これはPCRで検出可能であった。PCR製品の大きさは、完全なハイブリッド部位に期待される大きさであり、ハイブリッド部位の存在はPCR製品を配列決定することにより確認された。これらの観察は、ura4遺伝子の欠損がΦC31仲介切り取り以外の機構により発生するという考えと一致している。
【0092】
(まとめ)
単一目的部位での環状分子の組込みは、ほとんど全ての形質転換体の正確な挿入を得る効果的な方法であった。発明者らが観察したわずかの異常現象は、おそらくleu1反復DNAに作用するS.pombe組み換え系に主として起因する。インテグラーゼ産生がそのプロモーターの発現により制限されていた場合、形質転換体の数を背景レベル付近にまで減らした。この条件下で、ΦC31部位特異的組み換えから誘導された修復形質転換体はほとんどなかった。真核細胞におけるΦC31部位特異的組み換え系の機能的操作により、トランス遺伝子および染色体のマニピュレーションを行う新たな機会が与えられた。ΦC31系をattBおよびattP部位の選択的配置と共に用いて、DNAの削除、反転または挿入を行うことができる。この系の重要な特徴は、attB×attP反応が切り取り特異的タンパク質の不存在下で不可逆という点である。
【0093】
(実施例2)
ΦC31インテグラーゼはCHO細胞中で安定な組込みを作る役目をする。
この実施例では、チャイニーズハムスター卵巣(CHO)細胞におけるattB組み換え部位とattP組み換え部位との間の組み換えを仲介する能力に関してΦC31インテグラーゼをテストする実験を説明する。
【0094】
(方法)
ゼオシン耐性を与える選択マーカーを含んだCHO細胞株51YT211をattP含有プラスミドpFY1で形質転換した(図2)。単一コロニーを2回精製した後、6つのゼオシン耐性細胞株を分離した。サザンDNAハイブリダイゼーションによる分析により、6つの細胞株のそれぞれが、ゲノム内に組込みされたpFY1の少なくとも一分子を有することが確認された。
【0095】
6つの細胞株のそれぞれをattB含有プラスミドpFY9およびint含有プラスミドpFY6でトランスフェクトし、pFY9上のattB部位とpFY1の染色体コピー上のattP部位との間の部位特異的組み換えに対するテストを行った。コントロールとして、同じ細胞株をpFY9で形質転換したが、int含有pFY6は用いなかった。pFY9プラスミドは、SV40初期プロモーターの制御下でネオマイシン耐性選択マーカーおよびプロモーターに結合していない緑蛍光タンパク質(GFP)をコードした配列を含んでいた。したがって、部位特異的組み換えが、pFY1に含まれるヒト・サイトメガロウイルス・プロモーターの制御下でGFPコード化配列を配置し、その結果GFPを発現することが予想された。
【0096】
(結果)
トランスフェクションの結果:ネオマイシン耐性コロニーを顕微鏡下に置き、GFP遺伝子の活性の有無を観察した。pFT9+pFY6でトランスフェクションした細胞では大部分が、しかしpFY9のみでトランスフェクションした細胞ではごく僅かが、GFP活性を示した。このことは、pFY6でトランスフェクションした場合のpFY9の部位特異的組込みおよび共トランスフェクションint遺伝子の不存在下のpFY9の無作為挿入と一致する。
【0097】
Pc(ヒト・サイトメガロウイルス・プロモーター)およびGFP(図2)に対応するプライマー対を用いてPCR分析を行った。これらのプライマーは、組込み接合に対応する約0.6kbのバンドを増幅すると予想された。部位特異的組込みと無作為組込みの両者からネオマイシン耐性コロニーが生じる場合があり、かつGFPマーカーは選択的形質を与えないので、構成クローンの純粋な培養株を得ることは困難であった。したがって、各トランスフェクション株から得たネオマイシン体制細胞のプールをPCR分析に供し、ネオマイシン耐性細胞間に組込み接合が存在するか否かを調査した。大きさが約0.6kbと予想されるバンドを2つの株から得た。これは、attB×attP組み換え接合により、PcとGFPの結合が形成されることを示している。
【0098】
(実施例3)
ΦC31インテグラーゼはCHO細胞中で部位特異的組み換えを触媒する。
本実施例では、ΦC31インテグラーゼをテストして、attP部位とattB部位との間の組み換えにより、DNA分子をチャイニーズハムスター卵巣(CHO)細胞の染色体中に組込む能力を調べた。
【0099】
(方法)
プラスミドコンストラクト染色体attB標的コンストラクトpFY12、pFY14およびpFY15 プラスミドpcDNA3.1/His/lacZ(Invitrogen)をベクターバックボーンとして用いた。長さの異なるattB部位を含有し、HindIII部位とKpnI部位に隣接した合成オリゴヌクレオチドをpcDNA3.1/His/lacZのHindIII (AAGCTT)部位とKpnI (GGTACC)部位との間に挿入した。
プラスミドpFY12は90bpのattB 配列(AAGCTT gacggtctcg aagccgcggt gcgggtgcca gggcgtgccc ttgggctccc cgggcgcgta ctccacctca cccatctggt ccatcatgat GGTACC)(SEQ ID NO:2)を含有する。
プラスミドpFY14は50 bpのattB部位(AAGCTT gcgggtgcca gggcgtgccc ttgggctccc cgggcgcgta ctccacctca TGGTACC)(SEQ ID NO:3)を含有する。
プラスミドpFY15は30 bpのattB(AAGCTT ccagggcgtg cccttgggct ccccgggcgc ATGGTACC)(SEQ ID NO:4)を含有する。
【0100】
attPプラスミドpFY17、pFY19、pFY20の組込み ハイグロマイシンに対する耐性のためにコードし、pED113から得られた1.6kbのBamHI−KpnI断片として得られたhpt遺伝子を、pBluescript II SKのBamHIとKpnIとの間に挿入してコントロールプラスミドpBSK−hptを作った。
【0101】
長さの異なるattP部位を含有する合成オリゴヌクレオチドをpBSK−hpt中のSacI(GTCGAC)部位とBamHI(GGATCC)部位との間に挿入し、 次のプラスミドを作った。
a)90bpのattP部位(GAGCTC−gaagcggttt tcgggagtagtgccccaact ggggtaacct ttgagttctc tcagttgggg gcgtagggtc gccgacatga cacaaggggt−GGATCC)(SEQ ID NO:5)を含有するプラスミドpFY17。
b)50bpのattP部位(GAGCTC−tgccccaact ggggtaacct ttgagttctc tcagttgggg gcgtagggtc−GGATCC)(SEQ ID NO:6)を含有するプラスミドpFY19。
c)32bpのattP部位(GAGCTC−actggggtaa cctttgagtt ctctcagttg ggATCC)(SEQ ID NO:7) を含有するプラスミドpFY20。
【0102】
インテグラーゼ発現構成pFY6 インテグラーゼ遺伝子のほぼ完全なオープンリーディングフレームを含有するEcoRI−BamHI断片を、pcDNA3.1/Zeo(−)(Invitrogen)のEcoRI部位とBamHI部位との間に挿入した。次いで、Kozack配列およびインテグラーゼ遺伝子の配列をコードするN末端アミノ酸を含有する合成オリゴヌクレオチド(GGGCCCGCCACGATGACACAAGGGGTTGTGACCGGGGTGGACACGTACGCGGGTGCTTACGACCGTCAGTCGCGCGAGCGCGAGAATTC)(SEQ ID NO:8)をApaI部位とEcoRI部位との間に挿入して、オープンリーディングフレームを再構成した。この方向は、pcDNA3.1/Zeo(−)中でCMV(ヒト・サイトメガロウイルス)プロモーターの制御下で、完全なインテグラーゼコード化領域を配置する。
【0103】
トランスフェクション・プロトコール CHO細胞株K−1をattB標的コンストラクトpFY12、pFY14またはpFY15でトランスフェクションした(図3)。これらのプラスミドは、ネオマイシン耐性に対する選択マーカー、およびpC(ヒト・サイトメガロウイルス・プロモーター)とlacZコード化領域との間に位置する種々の長さのattB部位を有する。プラスミドpFY12、pFY14およびpFY15は、それぞれ90、50および30bpのattB配列を含有する。ネオマイシン耐性細胞株を単一コロニーの連続精製により得た。各コンストラクトの4つの株を組込み実験に用いた。
【0104】
12株のそれぞれに、ΦC31インテグラーゼ発現プラスミドpFY6を、組込みベクターpFY17、pFY19またはpFY20と共にトランスフェクションした。プラスミドpFY17、pFY19およびpFY20は、それぞれ長さ90、50および32bpのattP配列を有する。attP配列をhptオープンリーディングフレームの上流に置いた。このattP配列は、ハイグロマイシン耐性を与えるハイグロマイシン・ホスホトランスフェラーゼをコードする。attP−hpt部分の上流にプロモーターは無く、したがってhptコード化領域にゲノムプロモーターを融合するようにプラスミドをゲノムに組込みしない限り、hptは発現しない。pBSK−hptをコントロールに用いて、hptへのプロモーター融合の頻度を監視した。attP配列が無いという点を除いて、プラスミドpBSK−hptはpFY17、pFY19およびpFY20と同一である。attP部位とattB部位との間の組み換えは、組込みベクターを染色体標的に挿入し、Pc−attL−hpt結合を発生すると予想される。hptの発現はハイグロマイシン耐性を与える。
【0105】
結果トランスフェクションの結果:12細胞株にトランスフェクションされた各組込みプラスミドに対して、ハイグロマイシン耐性コロニーを記録した(表2)。1×106細胞を塗布したものから、pBSK−hptトランスフェクションは顕著に多数の耐性コロニーの産生に失敗した。これは、ゲノムプロモーターに与えるhtpコード化領域の頻度が著しく低いことを示している。対照的に、pFY17、pFY19およびpFY20は、特定の組込みプラスミドおよび特定の細胞株によっては、最大で千倍のハイグロマイシン耐性コロニーを生じた。pFY19またはpFY17をFY12中にトランスフェクションしたものから、より多くのハイグロマイシン耐性コロニーが生じた。これは、より長いattB配列とattP配列との間の組み換えは、より短いattB部位とattP部位との間の組換えよりもより効率的であることを示している。
【0106】
PCRを用いて、代表的コロニーから予想される約0.8kb接合バンドを検出した。ヒト・サイトメラロウイルス・プロモーターおよびhptコード化領域に対応するプライマーは、予想サイズ(0.8kb)のPCR産生物を増幅した。これは、Pcがhptコード化領域に結合していることを示しており、ゲノムのattB部位とプラスミドattP配列との間の組み換えと一致する。
【0107】
【表2】
【0108】
(実施例4)
ΦC31インテグラーゼは植物染色体中でattP部位およびattB部位を組み換える働きをする。
本実施例は、ΦC31インテグラーゼをテストして、植物染色体中に存在するattP部位およびattB部位を組み換える能力を調べた実験についての説明である。本実験の作成物および方法を図4に示す。
【0109】
方法
コンストラクトpWP29は、35S−attP−npt−attB−gusからなりRBおよびLBに隣接した断片を含有する。ここにおいて、35Sはカリフラワー・モザイクウイルス・プロモーターであり、nptはネオマイシン・ホスホトランスフェラーゼをコードする領域であり、gusはグルクロニダーゼをコードする領域である。RBおよびLBは、それぞれ右および左アグロバクテリウムT−DNA境界配列である。35Sとnptとの間のattP部位は非翻訳リーダー配列としての機能を果す。35Sによるnptの転写によりカナマイシン耐性が与えられる。上流にプロモーターが無いので、gusコード化領域は転写されない。
【0110】
植物形質転換に用いる第2のコンストラクトはpWP24である。このコンストラクトはRBおよびLBに隣接する断片Pnos−npt−35S−intを含有する。ここにおいて、Pnosはノパリンシンターゼ・プロモーターであり、intはΦC31インテグラーゼコード化領域である。nptとintの両者が、それぞれ上流のプロモーターから転写される。
【0111】
この2つのコンストラクトが同じゲノム中に存在する場合、pWP24を有する染色体からのintの発現により、機能性ΦC31インテグラーゼが発生し、pWP29を有する染色体上に位置するattB部位とattP部位との間の組み換えを触媒すると考えられた。組み換え現象により、pWP29構造体からnpt遺伝子を削除すること、およびgusに35Sを与えることが予想された。得られる構造は35S−attR−gusであり、ここでattRは、attPとattBとの間の組み換えにより形成されたハイブリッド部位であり、PB’とも表される(図4)。nptの削除により35Sの転写下でgusが得られ、かつGUS酵素活性を有する植物を生じると予想された。この活性は、植物組織を組織化学的に染色することにより検出することができる。
【0112】
結果
一時発現アッセイを行って、pWP29が組み換えに対して機能的であるか否かを調べた。裸のDNAの遺伝子銃で仲介した送達により、pWP29をトウモロコシBMS細胞中にpWP8と共導入した。コンストラクトpWP8は、単子葉植物細胞中で発現するためのトウモロコシ・ユビキチン・プロモーターの後方に融合したインテグラーゼ遺伝子を有する。両方のプラスミドを共導入した場合には青色点が観察されたが、プラスミドの一方のみを用いた場合には、これは見つからなかった。このことは、トウモロコシ細胞中で部位特異的組み換えが起きたこと、およびpWP29中のattP部位とattB部位とが機能性部位であることを示している。
【0113】
カナマイシン耐性タバコ植物体を、pWP29またはpWP24を用いてアグロバクテリウムで仲介した形質転換により再生した。もう一度、一時発現アッセイを行って、pWP24株が機能性インテグラーゼを産生するか否かを調べた。コンストラクトpWP29を、裸のDNAの遺伝子銃で仲介した送達により、pWP24植物体に導入した。pWP29DNAを有する細胞は、35S−attR−gus構成の形成の結果、GUS酵素活性を発現すると予想された。実際、2つの株24.3および24.4は、attP部位とattB部位との間の機能性インテグラーゼ仲介部位特異的組み換えと一致した青色点を生じた。
【0114】
これら2つのpWP24インテグラーゼ株をpWP29テスター株と交雑して、pWP29とpWP24とを同じゲノムに保有する染色体を有する子孫を作った。インテグラーゼ(24.3、24.4)とテスター株(29.2、29.4、29.5、29.19)との間の遺伝子交雑から得られた結果を表3にまとめた。いずれの場合も、代表的子孫苗を選択せずに発芽させ、GUS酵素活性に対して組織化学的に染色した。青色に染色された子孫の数を表に示した。一次形質転換したpWP24およびpWP29株は代表的トランス遺伝子に対してヘミ接合性であるので、子孫の4分の1だけが両方のトランス遺伝子型を保有することが期待された。サンプル数が少ないので、期待頻度からの顕著な偏差は異常ではない。
【0115】
【表3】
【0116】
親株として用いた株の組合せにより、染色の強度が変わった。組織の大部分が青色に染まった子孫を有するものは、組み換え現象がより効率的であったことを示している。逆に、染色が均一でない子孫を与えるものは、組み換え現象がより非効率であったことを示している。異なる子孫プールの間のこの偏差は、恐らくトランス遺伝子の組込み位置により生じる効果によるものである。2つのインテグラーゼ株のうち、24.4は促進部位特異的組み換えにおいてより効率的に現れる。これは、恐らくint遺伝子発現のレベルが高いことによる。24.4を29.4および29.19に交雑して作った染色パターンは、attBおよびattPのint仲介部位特異的組み換えによりgus遺伝子活性の活性化を招くという、本実験の目的に一致する。
【0117】
(実施例5)
ΦC31インテグラーゼは、植物染色体中への環状プラスミドの組み込みを触媒する。
本実施例は、ΦC31インテグラーゼをテストして、attP×attB部位特異的組み換えにより植物染色体中に環状プラスミドを挿入する能力を調べた実験についての説明である。本実験を図5に図示する。
【0118】
方法
標的コンストラクトpWP6は、RBおよびLBに隣接し35S−attP−nptからなる断片を含む。35Sとnptとの間のattP部位は非翻訳リーダー配列としての役割を果す。35Sによるnptの転写によりカナマイシンに対する耐性が得られる。
【0119】
組み込んだコンストラクトpYJC43は断片attB−hptを有する。ここで、hptはハイグロマイシンに対する耐性をコードしている。インテグラーゼ発現コンストラクトはpYJC41であり、35Sはintを転写する。
【0120】
標的コンストラクトpWP6を、pWP6DNAの無作為組込みにより植物染色体中に配置した。次いで、pWP6トランス遺伝子の単一コピーを含んだカナマイシン耐性植物をpYJC43およびpYJC41で形質転換した。pYJC41から得たintの一時発現が、pYJC43のattB部位とpWP6トランス遺伝子の染色体的に定まったattP部位との間の組み換えを触媒することが期待された。attB部位とattP部位との間の特異的組み換えにより、pYJC43環状分子が染色体に挿入され、35S−attL−hpt連鎖を作ることが期待された。attP部位およびattB部位は逆方向に図示されているので、attL部位も同様に逆方向にするか、または図中のBP’を逆方向にしたものと同じP’Bで表すことに留意されたい。機能性35S−attL−hpt連鎖はハイグロマイシン耐性表現型を与えた。
【0121】
結果
pWP6を有するカナマイシン耐性タバコ植生を、アグロバクテリウム仲介形質転換により得た。サザンハイブリダイゼーション分析により、pWP6トランス遺伝子の単一コピーを有する株を検出した。この株の子孫WP6.1を無菌的に発芽させ、これらの植物からプロトプラストを作った。このプロトプラストを、pYJC43とpYJC41DNAとを組合せ、直接DNA形質転換のためのポリエチレングリコール法により形質転換した。次いで、プロトプラストをアガロース中に埋め込み、培養してハイグロマイシン存在下でカルスを形成した。ハイグロマイシンを選択しない場合には、カルス形成速度は4×10−4であった。これは通常の10分の1であるが、プロトプラスト形質転換実験で観察される偏差の範囲内である。ハイグロマイシンを選択する場合には、カルス形成速度は7×10−5であった。これは、プロトプラストから再生されたカルスの約18%が、標的部位に組込みベクターを含有することを示している。インテグラーゼコンストラクトpYJC41を形質転換から除外した場合、カルス形成速度は1×10−5未満であった。プラスミドpYJC41を発現するインテグラーゼの封入により作られたハイグロマイシン耐性カルスの頻度がより高いことは、pYJC43の染色体attP標的へのインテグラーゼ促進部位特異的組込みと一致する。
【0122】
本明細書で説明した実施例および実施態様は例示目的のためだけのものであって、当業者にとってこれらを考慮した種々の変更または修正は示唆されており、かつこれらの変更または修正は本明細書の精神および範囲、ならびに特許請求の範囲に含まれることを理解されたい。本明細書中で引用した刊行物、特許および特許出願書類の全てを、全ての目的のためにここに参照により組み入れる。
【技術分野】
【0001】
連邦政府が後援する研究に関する説明 本発明は、政府の指示で米国農務省、農業リサーチサービスによって授与された許可No.5335-21000-009-06Sの下でなされた。政府は本発明において一定の権利を有する。
【0002】
関連出願への相互参照 本出願は、ここで参照として組み込む1999年7月23日に出願された米国仮出願番号60/145,469の利益をクレームする。
【0003】
本発明は、真核細胞の染色体の中への特異的かつ安定した核酸の組み込みを得るための方法の分野に関する。本発明は、ΦC31インテグラーゼなどの原核リコンビナーゼポリペプチドを使用する部位特異的組み換え系を利用する。
【背景技術】
【0004】
原核の遺伝子形質転換は、しばしば重大な短所に苦しむ。例えば、トランス遺伝子の組み込みを興味ある特定の座で再生産可能に得ることはしばしば困難である。相同組み換えは、一般に非常に低い頻度でのみ生じる。この問題を克服するために、部位特異的組み換え系を用いた。これらの方法は、より高等な原核細胞において機能することが可能な部位特異的組み換え系の使用を含む。多くのバクテリオファージ及び組み込みプラスミドは、それらのゲノムをそれらの宿主のものの中へ安定に組み込むことが可能な部位特異的組み換え系をコード化する。これらの系において、組み換え反応の最小必要条件は、組み換え現象を触媒するリコンビナーゼ酵素又はインテグラーゼ、及び2つの組み換え部位である(Sadowski(1986)J.Bacteriol.165:341-347;Sadowski(1993)FASEB J.7:760-767)。ファージ組み込み系について、これらは、付着(att)部位といい、ファージDNA由来のattP因子と、バクテリアゲノムによってコード化されるattB因子がある。2つの付着部位は、2,3の塩基対と同じくらい小さい配列を共有する。リコンビナーゼタンパク質は、両方のatt部位に結合し、宿主への環状ファージ又はプラスミドDNAの組み込みを生ずる、DNA鎖のコンサーバティブ(保存型)及び相互作用交換を触媒する。DNAベンディングタンパク質IHF、組み込み宿主因子などの追加のファージ又は宿主因子波、効率的な反応に対して要求されるかもしれない(Freidman (1988)Cell 55:545-554;Finkel & Johnson(1992)Mol.Microbiol. 6:3257-3265)。逆切り取り反応は、時折、ファージλのxis遺伝子産物などの追加のファージ因子を必要とする。(Weisberg & Landy(1983)“ファージλにおける部位特異的組み換え”In Lambda II、eds. Hendrix et al.(Cold Dpring Harbor Laboratory, Cold Spring Harbor, NY)pp.211−250;Landy(1989)Ann.Rev.Biochem. 58:913−949)。
【0005】
リコンビナーゼは、2つのグループ、λインテグラーゼ(Argos et al.(1986)EMBO J.5:433-44;Voziyanov et al(1999)Nucl. Acids Res.27:930-941)、及びリゾルバーゼ/転化酵素属(Hatfull & Grindley (1988)“Resolvases and DNA−invertase:a family of enzymes active in site-specific recombination” In Genetic Recombination, eds. Kucherlipati, R., & Smith, G.R. (Am. Soc. Microbiol., Wachington DC), pp. 357-396)に分類される。これらは、インテグラーゼ酵素の構造、及び触媒のそれらのモードの分子詳細を変える(Stark et al. (1992)Trends Genetics 8:432−439)。鋳型Streptomyces ファージΦC31は、後者の分類の68kDリコンビナーゼをコード化する。同系の付着部位を組み換える際にΦC31インテグラーゼ酵素の有効性が、試験官内及び生体内でrecA突然変異体大腸菌において最近証明された(Thorpe & Smith(1998)Proc.Nat’l. Acad.Sci.USA95:5505−5510)。ΦC31インテグラーゼ反応は、おそらく追加のファージタンパク質が切り取りに要求されないために、宿主因子を必要とせず、不可逆であるようであるという点においてシンプルである。ファージ及びバクテリアatt部位は、クロスオーバー点で相同性の3つの塩基対だけを共有する。この相同関係は、逆方向反復、おそらくインテグラーゼタンパク質の結合部位によって隣接する。両方のattB及びattPに対する最小の既知の機能的な大きさは、〜50bpである。
【0006】
バクテリオファージP1のCre-lox系、及びSaccharomyces cerevisiae のFLP−FRT系は、動物及び植物におけるトランス遺伝子及び染色体工学技術に広く使用されている(Sauer(1994)Curr.Opin.Biotechnol.5:521−527;Ow(1996)Curr.Opin.Biotechnol.7:181−186)。動物又は植物細胞において作用する他の系は、以下すなわち、1)Zygosaccharomyces rouxii 由来のR−RS系(Onouchi et al(1995)Mol. Gen.Genet.247:653−660)、2)バクテリオファージMu由来のGin−gix系(Maeser & Kahmann(1991)Mol.Gen.Genet.230:170−176)、及び3)バクテリアプラスミドpSM19035 由来のβリコンビナーゼ−six系(Diaz et al.(1999)J.Biol.Chem.274:6634−6640)、を含む。部位特異的リコンビナーゼを使用することによって、より高頻度で組み込みを得ることができる。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】国際公開第99/18222号
【発明の概要】
【発明が解決しようとする課題】
【0008】
しかしながら、これら5つの系は、重大な短所を有する。各これらの系は、単一のポリペプチドリコンビナーゼは、同一かほぼ同一の配列の2つの部位間の組み換えを触媒するという共通した特性を有する。組み換えによって生じた生産部位は、それ自体次の組み換えに対する基質である。結果として、組み換え反応は容易に可逆可能である。分子内相互作用の反応速度論は、分子間相互作用に渡って好ましいので、これら組み換え系は、DNAを組み込むよりむしろ削除するのに有効である。したがって、トランス遺伝子の安定的な部位特異的組み込みを得るための方法と系に対する必要性が存在する。本発明は、この及び他の必要性を、満たす。
【課題を解決するための手段】
【0009】
本発明は、真核細胞において、安定で、部位特異的な組み換えを得るための方法を提供する。以前に知られた部位特異的組み換え方法と違って、本発明の方法を使用して得られた組み換え体は、安定である。組み換え反応は、可逆的ではない。
【0010】
本方法は、第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として機能することが可能な第二の組み換え部位とを含む真核細胞を提供することを含む。第一及び第二の組み換え部位は、原核リコンビナーゼポリペプチドと接触し、組み換え部位間に組み換えを生じ、それによって1又は2つのハイブリッド組み換え部位を形成する。有意義なことに、リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の部位特異的組み換えを仲介することができるが、真核細胞において存在しない追加のファージ生産因子の不存在下で2つのハイブリッド組み換え部位間の組み換えを仲介することができないものである。組み換え部位のいずれか又は両方が真核細胞の染色体において存在することができる。いくつかの実施態様において、組み換え部位の1つは染色体に存在し、他は染色体の中へ組み込まれるべき核酸内に含まれる。
【0011】
本発明は、原核リコンビナーゼポリペプチド又は原核リコンビナーゼをコード化する核酸を含む真核細胞も提供する。これらの実施態様において、リコンビナーゼは、第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として作用することが可能な第二の組み込み部位との間の部位特異的組み換えを仲介をすることができるが、第一の組み換え部位と第二の組み換え部位との間の組み換えに形成された2つのハイブリッド組み換え部位間の組み換えを、真核細胞に存在しない追加の因子の不存在下では仲介することができないものである。現在好ましい実施態様において、本発明の細胞は、リコンビナーゼポリペプチドのコード化配列を有する核酸を含む。リコンビナーゼコード化配列は、真核細胞においてリコンビナーゼコード化ポリヌクレオチドの発現を仲介するプロモーターと作用可能に結合することが好ましい。本発明の真核細胞は、例えば、動物細胞、植物細胞、酵母細胞、又はカビ細胞とすることができる。
【0012】
別の実施態様において、本発明は、安定に組み込んだトランス遺伝子を有する真核細胞を得るための方法を提供する。これらの方法は、第一の組み換え部位を含む真核細胞の中へ核酸を導入することを含み、前記核酸は興味あるトランス遺伝子と前記第一の組み換え部位との組み換えの基質として作用することが可能な第二の組み換え部位とを含むことを特徴とする。第一及び第二の組み換え部位は、原核リコンビナーゼポリペプチドと接触する。リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の組み換えを触媒し、第一の組み換え部位で核酸の組み込みを生じ、それによって核酸の各末端でハイブリッド組み換え部位を形成する。かさねて、リコンビナーゼポリペプチドは、第一及び第二の組み換え部位間の部位特異的組み換えを仲介するが、真核細胞において存在しない付加的因子の不存在下では2つのハイブリッド組み換え部位間の組み換えを仲介することができないものである。
【0013】
本発明の別の実施態様において、真核細胞において機能するプロモーターと作用可能に結合するバクテリアリコンビナーゼポリペプチドをコード化するポリヌクレオチド配列を含む核酸を提供する。本発明のこれらの核酸によってコード化されるリコンビナーゼポリペプチドは、真核細胞において存在しないバクテリオファージ因子の不存在下では第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位間の組み換えを仲介することができない。いくつかの実施態様において、さらに、核酸は、リコンビナーゼポリペプチドによって認識される少なくとも1つの組み換え部位を含む。
【0014】
本発明によって提供される真核細胞は、また、真核細胞において存在しないバクテリオファージ因子の不存在下では第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位を仲介できない1又はそれ以上のバクテリオファージΦC31組み換え部位、または他のリコンビナーゼに対する組み換え部位を有するポリヌクレオチドを含む。
【図面の簡単な説明】
【0015】
【図1】S.pombe leul Locusで染色体構造の概略的な外観を示す(縮小)。PLT44の染色体への相同的挿入(図1A)は、図1Bで示されるようなleul 対立遺伝子間のΦC31 attP標的を設置する。 pLT45の染色体attP標的内へのpLT43促進部位特異的組み込みは、図1Cに示す構造を導入する。矢印は、T7プロモーター(T7)、T3プロモーター(T3)及びura4+コード化領域(U4)に対応するPCRプライマーを示す。XbalI(X)切断産物の予想される大きさを示す。
【図2】ΦC31インテグラーゼが、CHO細胞におけるグリーン蛍光タンパク質(GFP)をコード化するトランス遺伝子の部位特異的組み込みを触媒することを証明する実験の概略を示す。
【図3】ΦC31が、ハイグロマイシンホスホトランスフェラーゼ遺伝子を染色体に位置したプロモーターの下流に挿入するためにattB部位での特異的組み換えを触媒することを証明する実験の概略図を示す。成功した組み込みは、Pc-att-hptリンケージ及びハイグロマイシン耐性表現型を生産する。attP及びattB部位の異なる長さの影響を、指標したプラスミドを使用して分析した。
【図4】タバコゲノム由来のattB及びattP部位によって隣接させたDNAの切り取りを触媒することを示す実験の概略的な図表を示す。
【図5】ΦC31インテグラーゼがタバコゲノムへのトランス遺伝子の組み込みを触媒したことを示す実験の概略的な図表を示す。
【発明を実施するための形態】
【0016】
定義
ここで使用するような「外因性DNA断片」、「異質ポリヌクレオチド」、「トランス遺伝子」、「異質核酸」は、異質の起源から特定の宿主へ創造したものか、あるいは、もし同じ起源由来ならその原型由来のものから修飾したものである。したがって、宿主細胞における異質遺伝子は、特定の宿主細胞へ内生的であるが、修飾されていない遺伝子を含む。それゆえ、前記用語は、細胞に対して外来性か、異質か又は相同であるが、因子が通常見られない宿主細胞核酸内に位置するDNA断片を言う。外因性のDNA断片を発現させて外因性ポリペプチドを生産する。
【0017】
「遺伝子」の語は、生物学上の機能と関係するいずれのDNA断片を表すものとして広く使用する。したがって、遺伝子は、それらの発現に要求されるコード化配列及び/又は調節配列を含む。遺伝子は、例えば、他のタンパク質に対する認識配列を形成する非発現DNA断片も含むことができる。遺伝子は、種々の起源から得ることができ、興味ある起源由来のクローン、または既知の若しくは予想した配列情報からゴウセイしたもの、所望のパラメーターを有するように設計された配列を含むことができる。
【0018】
核酸又はタンパク質に適用するとき、「単離」の語は、核酸又はタンパク質が自然の状態に関係する他の細胞成分が本質的に存在しないことを表す。乾燥しているか水溶液のいずれかとすることができるが、均質な状態が好ましい。純度や均質性は、例えばポリアクリルアミドゲル電気泳動又は高性能液体クロマトグラフィーなどの分析化学技術を使用して典型的に決定される。調製において存在する有力種であるタンパク質は、本質的に精製される。特に、遺伝子を隣接させて、興味ある遺伝子以外のタンパク質をコード化するオープンリーディングフレームから単離した遺伝子を分離する。「精製」の語は、電気泳動ゲルにおいて核酸又はタンパク質が1つのバンドを本質的に生じることを意味する。特に、核酸又はタンパク質が少なくとも約50%純粋、より好ましくは少なくとも約85%純粋、最も好ましくは少なくとも約99%純粋であることを意味する。
【0019】
「自然に生じる」の語は、ヒトによって人工的に生産されることから区別するものとして天然に見出すことができる対象を記述するために使用する。例えば、実験室においてヒトによって意図的に修飾されていない天然の起源から単離することが可能な(ウイルスを含む)組織において存在するポリペプチド、又はポリヌクレオチド配列は、自然に生じる。
【0020】
「核酸」又は「ポリヌクレオチド」の語は、一本鎖又は二本鎖型のいずれかにおけるデオキシリボヌクレオチド又はリボヌクレオチド、及びその重合体を言う。特に限定しない場合、前記語は、引用核酸と同様の結合特性を有し、自然に生ずるヌクレオチドと同様の方法で代謝される自然のヌクレオチドの既知のアナログを含有する核酸を包含する。他に指示しない場合、特定の核酸配列は、伝統的に修飾したその変異体(例えば、縮退コドン置換)、相補的配列、及び明確に指示した配列も絶対的に包含する。具体的に、1又はそれ以上選択した(又はすべての)コドンの3つの部分を、混合した塩基及び/又はデオキシイノシン残基で置換した配列を発生させることによって縮退コドン置換を達成することができる(Batzer et al.(1991)Nucleic Acid Res. 19:5081;Ohtsuka et al.(1985)J.Biol.Chem。260:2605−2608;Cassol et al.(1992);Rossolini et al.(1994)Mol.Cell.Probes 8:91−98)。核酸の語は、遺伝子、遺伝子によってコード化されたcDNA及びmRNAと交互に使用される。
【0021】
「遺伝子から誘導される核酸」とは、遺伝子又はそのサブ配列(例えば、コード化領域)を合成し、最終的に鋳型として機能する核酸を言う。それゆえ、mRNA、mRNAから逆転写されたcDNA、そのcDNAから転写されたRNA、cDNAから増幅したDNA、増幅したDNAから転写したRNAなどは、すべて遺伝子から誘導され、そのように誘導された産物の検出は、オリジナルの存在及び/又は不存在の指示となる。
【0022】
DNA断片は、別のDNA断片と機能ある関係に置かれたとき、「作用可能に結合」する。例えば、ポリペプチドの分泌に加わるプレタンパク質として発現するなら、ポリペプチドをコード化するDNAに、単一の配列についてのDNAは作用可能に結合する。プロモーター又はエンハンサーは、配列の転写を刺激するならコード化配列に作用可能に結合する。一般に作用可能に結合するDNA配列は、隣接と読み取り段階の両方のシグナル配列の場合、隣接する。しかしながら、例えば、エンハンサーは、それらが制御する転写のコード化配列と隣接する必要がない。結合は、従来の制限酵素部位、又はその代わりに挿入したアダプター、リンカーでのライゲーションによって達成される。
【0023】
「植物」の語は、全植物、植物組織(例えば、葉、茎、根など)、種子、植物細胞、及び同様の子孫を含む。本発明の方法で使用可能な植物の分類は、形質転換技術に修正可能なより高い植物の分類と同様に一般に広く、単子葉、双子葉植物の両方を含む。
【0024】
「プロモーター」の語は、転写を開始するためにRNAポリメラーゼとの結合に関わるDNA領域を言う。「誘導プロモーター」は、発現のレベルを、例えば、温度、pH、転写因子及び化学薬品などの環境又は開発因子によって変更可能である遺伝子の発現を導くプロモーターをいう。
【0025】
細胞に関して使用するとき「組み換え」の語は、細胞が異質核酸を複製するか、又は異質核酸によってコード化されるペプチド又はタンパク質を発現することを表す。組み換え細胞は、細胞の野生型(非組み換え型)内では見出すことができないポリヌクレオチドを含むことができる。組み換え細胞は、ポリヌクレオチドが人工的な手段によって細胞の中へ修正又は再導入される細胞の野生型に見出されるポリヌクレオチドも含む。語は、細胞から核酸を除去することなしに修飾された細胞に内在する核酸を含む細胞も包含し、そのような修飾は、遺伝子置換、部位特異的突然変異体、及び関連技術等によって得られるものを含む。
【0026】
「組み換え発現カセット」又は単なる「発現カセット」は、構造遺伝子の発現をそのような配列と両立できる宿主において達成することが可能な核酸因子を持つ組み換え的、又は合成的に発生させた核酸構築物である。発現カセットは、少なくともプロモーター、及び選択的に、転写終結シグナルを含む。典型的には、組み換え発現カセットは、転写すべき核酸(例えば、所望のポリペプチドをコード化する核酸)、及びプロモーターを含む。発現を達成する際に必要な又は役立つ追加の因子をここで述べたように使用することができる。例えば、発現カセットは、宿主細胞由来の発現タンパク質の分泌を指示するシグナル配列をコード化するヌクレオチド配列も含むことができる。遺伝子発現に影響する転写終結シグナル、エンハンサー、及び他のヌクレオチド配列も、発現カセットに含まれる。
【0027】
「リコンビナーゼ」の語は、2つ又はそれ以上の組み換え部位間の組み換えを触媒する酵素をいう。本発明のおいて役立つリコンビナーゼは、特定のリコンビナーゼによって認識される特異的ポリヌクレオチド配列である特異的組み換え部位での組み換えを触媒する。「インテグラーゼ」の語は、リコンビナーゼのタイプをいう。
【0028】
「形質転換率」は、異質ポリヌクレオチドをそのゲノム及び残存物の中へ上手く組み込んだ細胞の割合をいう。
【0029】
「トランスジェニック」の語は、細胞の中へか、細胞の原型の中へ導入された特異的修飾を含む細胞をいう。そのような修飾は、一又はそれ以上の点突然変異、欠失、挿入、又はその組み合わせを含むことができる。動物に言及するとき、「トランスジェニック」の語は、トランスジェニックである細胞を含む動物を意味する。トランスジェニック及び非トランスジェニック細胞の両方からなる動物は、ここでは「キメラ」動物という。
【0030】
「ベクター」の語は、核酸(又は複数の核酸)を宿主細胞へ運搬するための組成物を言う。ベクターは、運搬すべき核酸をコード化する核酸を含み、核酸の宿主細胞内への加入及び/又はベクターの宿主細胞への複製を容易にするために、ウイルスキャプシド又は他の物質(例えば、キャプシド内、又はキャプシドの一部としてパッケージされた逆転写酵素又は他の酵素)を選択的に含む。
【0031】
「組み換え部位」はここで述べるリコンビナーゼ酵素によって認識される特異的ポリヌクレオチド配列である。典型的には、(相補部位と呼ばれる)2つの異なる部位を含み、1つは目的の核酸に存在し(例えば、真核の染色体又はエピソームなど)、別のものは、目的の組み換え部位で組み込まれるべき核酸上に存在する。特別の酵素の組み換え部位が異なる名前を持つかもしれないが、「attB」及び「attP」の語は、それぞれ、本来的にバクテリアターゲット、及びファージドナー由来の付着(又は組み換え)部位をいう。組み換え部位は、典型的に、コア又スペーサー領域によって分離された左及び右アームを含む。それゆえ、attB組み換え部位は、BOB‘からなり、B及びB’は、それぞれ左及び右アームであり、Oはコア領域である。同様に、attPは「POP‘」であり、P及びP’はアームであり、Oは重ねてコア領域である。前記attB及びattP部位との間の組み換えでは、核酸の標的での付随する組み込み、組み込んだDNAに隣接する組み換え部位は、「attL」及び「attR」という。それゆえ、上述した用語を使用してattL及びattRは、それぞれBOP’及びPOB‘からなる。ここでのいくつかの表示において、「O」を省略し、attB及びattPはそれぞれBB’及びPP’と示す。
【0032】
(好適な態様の説明)
本発明は、真核細胞において部位特異的な組み換えを得るための方法を提供するものである。部位特異的な組み換えを得るための過去に知られたシステムとは異なり、本発明の方法を用いて行われた組み換えの産物は安定である。そこで、例えば、真核細胞の染色体へトランスジーンを導入するために本方法を用いることが可能であり、そして既に知られた部位特異的な組み換えシステムを用いる場合にしばしば起こる、トランスジーンの切除を避けることができる。安定な逆位(inversion )、トランスロケーション(translocation )及び他の転位(rearrangement )もまた、得ることができる。
【0033】
本発明は、バクテリオファージのインテグラーゼの様な原核細胞のリコンビナーゼを採用し、その様な酵素は2つの相補的な組み換え部位の間の組み換えを触媒するが、この組み換えにより形成されたハイブリッド部位の間の組み換えを触媒することができない、という点において単方向性である。その様なリコンビナーゼの一つであるΦC31 インテグラーゼは、それ自身ではattB x attP 反応のみを触媒する。そのインテグラーゼは、attBとattPの間の組み換えにより形成された、attLとattR部位の間の組み換えをを仲介することはできない。ΦC31 インテグラーゼの様なリコンビナーゼは単独で逆反応を触媒することはできないために、ΦC31 のattB x attP 組み換えは安定である。この性質により、真核細胞において現在使用されている部位特異的な組み換えシステムから、本発明の方法は離れている。現在使用されているシステムにはCre-lox システム又はFLP-FRT システムの様なものがあり、そこでは組み換え反応は容易に逆転される。本発明の組み換えシステムの使用により、真核細胞において、安定なトランスジーン及び染色体転位(rearrangement )に向けての新たな機会が提供される。
【0034】
本方法は、対応するリコンビナーゼを用いて、真核細胞に存在している一組の組み換え部位(例えば、attB及びattP)に接触することを含む。そこでリコンビナーゼは組み換え部位の間の組み換えを仲介する。二つの組み換え部位の相対的な位置に応じて、組み換えの結果としての数多くのイベントのいずれか一つが起こることがある。例えば、もし二つの組み換え部位が異なった核酸分子上に存在しているならば、その組み換えは一つの核酸分子を第二の分子へと組み込む結果と成り得るであろう。そこで、対応する組み換え部位を含んでいる真核細胞染色体へ、一つの組み換え部位を含んでいるプラスミドを組み込むことができるであろう。本発明の方法において用いられたリコンビナーゼは逆反応を触媒することができないから、その組み込みは安定である。その様な方法は、例えば、プラスミド上に存在しているトランスジーンを、真核細胞の染色体へ安定に組み込むために有用である。
【0035】
二つの組み換え部位が、同一の核酸分子上に存在することもまたできる。その様な場合には、得られた産物は典型的には、その部位の相対的な方向性に依存する。例えば、直線方向にある部位の間の組み換えは一般的に、その2つの組み換え部位の間にある任意のDNA が切り出されるという結果となる。対照的に、逆方向にある部位の間の組み換えは、仲介しているDNA のインバージョンが起こるという結果となることがある。再び、付加的因子の非存在下において組み換えは不可逆的であるという点において、得られた転位した(rearranged)核酸は安定であり、付加的因子は一般的にそのリコンビナーゼが由来する特定のバクテリオファージによってコードされ、正常には真核細胞においては見られないものである。この方法が有用である適用の一つの例は、2つの組み換え部位の間のプロモーターの交換に伴って生じる。もしプロモーターが、そのプロモーターによって発現されるべきコード配列に関して最初に反対向きであって、そしてそのプロモーターをフランク(flank )する組み換え部位が逆方法であるならば、組み換え部位を接触させることはプロモーターの逆位(inversion )が起こる結果となり、従ってプロモーターを正しい方向に置いてコード配列の発現を駆り立てるであろう。同様に、もしプロモーターが最初に発現のために正しい方向性にあり、組み換え部位が同じ方向性にあるならば、組み換え部位がプロモーターと接触することによりプロモーターフ断片を切り出して、そしてコード配列の発現を停止させるという結果を得ることができる。
【0036】
本発明の方法は、例えば、染色体のトランスロケーションを得るためにもまた有用である。これらの態様において、一つの組み換え部位は一つの染色体上に置かれ、そして第一の組み換え部位と共に組み換えの基質として働くことができる第二の組み換え部位が第二の染色体上に置かれている。その二つの組み換え部位をリコンビナーゼと接触させると組み換えが起こって、2つの染色体アームが交換されるという結果となる。例えば、生物の2つの系統を構築することが可能であり、そのうちの一つの系統は第一の組み換え部位を含み、第二の系統は第二の組み換え部位を含んでいる。その2つの系統は掛け合わされ、その組み換え部位の両者を含む子孫の系統が得られる。その部位をリコンビナーゼと接触させると、染色体アームの交換が起こる。
【0037】
(リコンビナーゼと組み換え部位)
本方法は、真核細胞において、安定な核酸の組み込み又は他の転位(rearrangement )を達成するために、リコンビナーゼのシステムを使用する。リコンビナーゼのシステムは典型的には3つの要素から成り、その要素は2つの特異的なDNA 配列(「組み換え部位」)及び特異的な酵素(「リコンビナーゼ」)である。リコンビナーゼは、特異的な組み換え部位の間の組み換え反応を触媒する。
【0038】
組み換え部位は方向性を有している。言い換えれば、それらは回文(バリンドローム)ではない。相互の関連性における組み換え部位の方向性により、どの様な組み換えイベントが起こるか決定される。組み換え部位は、平行(同じ方向性)または反対という、2つの方向性をとることが可能である。組み換え部位が単一の核酸分子上に存在し、そして相互に平行な方向性であるときには、その際にリコンビナーゼにより触媒される典型的な組み換えイベントは、仲介している核酸の切り出して単一の組み換え部位を離すことである。組み換え部位が反対の方向性にあるときには、その際には任意の仲介配列が逆位(inverted)となるのが典型的である。
【0039】
本発明の方法において使用されるリコンビナーゼは、第一の組み換え部位と第二の組み換え部位の間の部位特異的な組み換えを仲介することができ、その第二の組み換え部位は第一の組み換え部位と共に組み換えのための基質として働くことができる。しかしながら、正常には真核細胞に存在しない、付加的因子の非存在下では、第一の組み換え部位と第二の組み換え部位の間の組み換えにより形成される、二つのハイブリッド組み換え部位の間の組み換えを仲介することができない。これらのリコンビナーゼの例には、例えば、バクテリオファージΦC31 インテグラーゼ(例えばThorpe & Smith (1998) Proc.Natl.Acad.Sci.USA 95:5505-5510 ; Kuhstoss & Rao (1991) J.Mol.Biol.222:897-890 ;米国特許番号5,190,871 を見よ)、ファージP4リコンビナーゼ(Ow & Ausubel (1983) J.Bacteriol.155:704-713 )、リステリアファージリコンビナーゼ、バクテリオファージR4 Sreリコンビナーゼ(Matsuura et al. (1996) J.Bacteriol.178:3374-3376)、CisAリコンビナーゼ(Sato et al.(1990) J.Bacteriol.172:1092-1098 ; Stragier et al.(1989) Science 243:507-512) 、XisFリコンビナーゼ(Carrasco et al.(1994)Genes Dev.8:74-83)、トランスポゾンTn4451 TnpX リコンビナーゼ(Bannam et al.(1995) Mol.Microbiol. 16:535-551 ; Crelin & Rood (1997) J.Bacteriol.179:5148-5156)が含まれる。
【0040】
リコンビナーゼポリペプチド、及びそのリコンビナーゼポリペプチドをコードする核酸は本技術分野において述べられており、日常的な方法を用いて得ることが可能である。例えば、ΦC31 インテグラーゼをコードする核酸フラグメントを含むベクターは米国特許番号5,190,871 の中に述べられており、B-18477 というアクセッション番号の下でノザンレジョナル リサーチラボラトリー(ぺオリア、イリノイ 61604)から入手可能である。
【0041】
任意の適切な方法により、組み換えを望む場所において、組み換え部位を含む真核細胞へリコンビナーゼを導入することができる。例えばリコンビナーゼをポリペプチドの形で、例えばマイクロインジェクション又は他の方法によって導入することができる。しかしながら、現在において好ましい態様においては、リコンビナーゼをコードする遺伝子が細胞に導入される。遺伝子の発現はリコンビナーゼが作られるという結果となり、そのリコンビナーゼは対応する組み換え部位の間の組み換えを触媒する。興味の対象である外来性のポリヌクレオチドを導入する前、後、又はそれと同時に、リコンビナーゼ遺伝子を細胞中に導入することができる。一つの態様において、リコンビナーゼ遺伝子は挿入されるべきポリヌクレオチドを運搬するベクター中に存在し、そのリコンビナーゼ遺伝子はポリヌクレオチド中に含まれることさえも可能である。他の態様においては、リコンビナーゼ遺伝子は、形質転換した植物、動物、カビ又はその類似物の様な形質転換した真核生物中へ導入され、その後にそれらは対応する組み換え部位を含む生物と掛け合わされる。
【0042】
(標的生物)
本発明の方法は、任意の型の真核細胞において、DNA の安定した組み込み及び/又は転位(rearrangement )を得るために有用である。例えば、本方法は動物、植物、カビ、細菌、及び他の微生物の細胞において有用である。いくつかの態様において、その細胞は多細胞生物、例えば形質転換植物又は動物等の一部分である。本発明の方法は、形質転換したコムギ、トウモロコシ、及び動物の様な、形質転換した材料を得ることが困難である状況において特に有用である。これらの状況において、まれにしかない単一コピー挿入物を見出すには、独立して得られた多数の形質転換したクローンを前もって得ることが必要となるが、それはそれ自身非常に多くの労力を払う必要がある。
【0043】
特に興味が持たれる植物標的には単子葉植物があり、その中には、例えば、イネ、トウモロコシ、コムギ、ライムギ、オオムギ、バナナ、ヤシ、ユリ、ラン及びスゲが含まれる。双子葉植物もまた適切な標的であり、例えば、タバコ、リンゴ、ジャガイモ、テンサイ、ニンジン、ヤナギ、ニレ、カエデ、バラ、キンポウゲ、ペチュニア、フロックス、スミレ及びヒマワリが含まれる。他の標的には、動物及びカビ細胞が含まれる。これらのリストは単に説明するためのものであり、限定するものではない。
【0044】
(標的細胞内へ外来DNA を導入するためのコンストラクト)
本発明の方法は、標的細胞内へ外来DNA を導入することをしばしば含む。例えば、一つ又はそれ以上の組み換え部位を含む核酸が細胞中へしばしば導入される。細胞内へ導入されるべきポリヌクレオチドコンストラクトは、一つ又は複数の組み換え部位に加えて、望まれる形質型を細胞に与えるであろう遺伝子又は機能的な配列を含むことができる。
【0045】
いくつかの態様において、組み換え部位に加えて、リコンビナーゼをコードするポリヌクレオチドコンストラクトが真核細胞内へ導入される。リコンビナーゼをコードしているポリペプチドは、一つ又は複数の組み換え部位として同一の核酸上に含まれることが可能であり、また別個の核酸として細胞内へ導入されることも可能である。本発明は組み換え部位を含む核酸のみならず、その核酸内においてリコンビナーゼをコードしているポリヌクレオチド配列が標的真核細胞内において機能しているプロモーターに作用可能に結合している核酸もまた提供する。
【0046】
一般的には、発現するべきポリヌクレオチド(例えばリコンビナーゼをコードしているポリヌクレオチド又は興味の対象であるトランスジーン)は発現カセットに存在しているであろうし、それはそのポリヌクレオチドが発現制御シグナルに作用可能に結合していることを意味しており、その発現制御シグナルは、例えばプロモーターやターミネーターであって興味の対象である宿主細胞において機能を有する。リコンビナーゼ及び選択可能なマーカーをコードする遺伝子もまた、宿主細胞内において機能するその様なシグナルの制御下にあるであろう。発現の制御はプロモーターの選択によって最も容易に達成される。転写ターミネーターは一般的には重要でなく、細胞によって認識される限り種々の既知の因子を使用することができる。
【0047】
研究がなされている遺伝子からプロモーターを得ることが可能であり、または異なった遺伝子または異なった種から得られた異種プロモーターであることも可能である。形質転換植物又は他の生物の全ての組織において遺伝子の直接的な発現が望まれる場合には、「構成的な」プロモーターを使用することが可能であり、「構成的な」プロモーターは多くの環境条件や発達又は細胞分化の状況において一般的に活性である。
【0048】
植物において使用するために適切な構成的なプロモーターには、例えば、カリフラワーモザイクウイルス(CaMV)35S 転写開始領域及び領域VIプロモーター、アグロバクテリウムツメファシエンスのT-DNA から得られた1'- 又は2'- プロモーター、及び本技術分野の当業者に知られている植物細胞中で活性である他のプロモーターが含まれる。他の適切なプロモーターには、フィグウオート(Figwort )モザイクウイルス由来の全長の転写プロモーター、アクチンプロモーター、ヒストンプロモーター、チュブリンプロモーター、又はマンノピン合成酵素プロモーター(MAS )が含まれる。
【0049】
他の構成的な植物プロモーターには、とりわけシロイヌナズナ(Sun and Callis, Plant J.,11(5):1017-1027(1997))より得られた、種々のユビキチン又はポリユビキチンプロモーター、マス(mas )、マック(Mac )又はダブルマック(DoubleMac )プロモーター(米国特許番号5,106,739 、及びComai et al によってPlant Mol.Biol.15:373-381 (1990)において述べられている)、及び本技術分野の当業者に知られている、種々の植物遺伝子に由来する他の転写開始領域が含まれる。その様な遺伝子には、例えば、シロイヌナズナ由来のACT11 (Huang et al.,Plant Mol.Biol.33:125-139(1996))、シロイヌナズナ由来のCat3((GenBank No.U43147,Zhong et al.,Mol.Gen.Genet.251:196-203(1996))、セイヨウアブラナ(Brassica napus)由来のステアロイルアシルキャリア蛋白質不飽和化酵素をコードしている遺伝子(GenBank No.X74782,Solocombe et al.,Plant Physiol.104:1167-1176(1997))、トウモロコシ由来のGPc1(GenBank No.X15596 Martinez et al.,J.Mol.Biol 208:551-565(1989))、及びトウモロコシ由来のGpc2(GenBank No.U45855,Manjunath et al.,Plant Mol.Biol.33:97-112(1997) )が含まれる。
【0050】
植物のための有用なプロモーターには、Ti- 又はRi- プラスミドから、植物細胞から得られたプロモーター、プロモーターが植物中において機能を有することが見出されているときには植物ウイルス又は他の宿主が含まれる。植物中で機能し、そしてそのため本発明の方法において使用するのに適切である細菌プロモーターには、オクトピン合成酵素プロモーター、ノパリン合成酵素プロモーター、マノピン(manopine)合成酵素プロモーターが含まれる。適切な内在性植物プロモーターには、リブロース-1,6- ビスホスファターゼ(RUBP)、カルボキシラーゼスモールサブユニット(ssu )プロモーター、α- コングリチニン(conglycinin )プロモーター、ファゼオリン(phaseolin )プロモーター、ADH プロモーター、及び熱ショックプロモーターが含まれる。
【0051】
大腸菌において使用するためのプロモーターには、T7、trp 、又はラムダプロモーター、リボソーム結合部位を含み、及び好ましくは転写終結シグナルである。真核細胞のためには、制御配列は典型的にはプロモーターを含み、そのプロモーターは随意にイムノグロブリン遺伝子、SV40、サイトメガロウイルス等に由来するエンハンサー及びポリアデニレーション配列を含み、そしてスプライスドナー及びアクセプター配列を含むこともできる。酵母において、便利なプロモーターにはGAL1-10 (Johnson and Davies (1984) Mol.Cell.Biol.4:1440-1448 )、ADH2(Russell et al.(1983) J.Biol.Chem.258:2674-2682)、PHO5(EMBO J.(1982) 6:675-680 )及びMFα(Herskowitz and Oshima (1982) The Molecular Biology of the Yeast Saccharomyces (酵母サッカロミセスの分子生物学)(eds.Strathem,Jones, and Broach)Cold Spring Harbor Lab.,Cold Spring Haror,N.Y.,PP181-209の中において)が含まれる。
【0052】
代わりに、興味の対象である遺伝子の特異的な組織における発現を標的としたプロモーター、又はさもなければより厳密な環境又は発達による制御の下にあるプロモーターを使用することが可能である。その様なプロモーターをここでは、「誘導可能な」又は「抑制可能な」プロモーターと称する。誘導可能なプロモーターの転写によって転写に影響するかもしれない環境条件の例には、病原体の攻撃、嫌気性の条件、エチレン、又は光の存在が含まれる。発達による制御下にあるプロモーターには、葉、根、果実、種、又は花の様な特定の組織においてのみ転写を開始するプロモーターが含まれる。プロモーターの操作はゲノム中のそれの位置によってもまた変化する可能性がある。そこで、誘導可能なプロモーターはある位置において完全に又は部分的に構成的となることがある。誘導可能なプロモーターは、リコンビナーゼ遺伝子の発現を制御するためにしばしば使用され、そこで組み換え反応のタイミングを制御することが可能となる。
【0053】
発達による制御の下にある組織特異的な植物プロモーターの例には、果実、種、花の様な特定の組織においてのみ転写を開始するプロモーターが含まれる。トマト由来の組織特異的なE8プロモーターは遺伝子発現を方向付けるために特に有用であり、そのために望まれる遺伝子発現産物は果実の中に位置する。例えば、Lincoln et al.(1988) Proc.Natl.Acad.Sci.USA 84:2793-2797; Deikman et al.(1988) EMBO J.7:3315-3320; Deikman et al.(1992) Plant Physiol.100:2013-2017 を見よ。他の適切なプロモーターには胚の貯蔵蛋白質をコードしている遺伝子が含まれる。
【0054】
誘導可能なプロモーターによる転写に影響するかもしれな環境条件の例には、嫌気的な条件、上昇した温度、又は光の存在が含まれる。更なる器官特異的、組織特異的、及び/又は誘導可能な外来のプロモーターもまた知られており(例えば、Kuhlemeier et al (1987) Ann.Rev.Plant Physiol.38:221において引用された参考文献を見よ)、それには、光誘導性であって光合成を行う組織においてのみ活性である、シロイヌナズナ(Arabidopsis thaliana)の1,5-リブロースビスホスフェイトカルボキシラーゼ スモールサブユニット遺伝子類("ssu" プロモーター)プロモーター、葯特異的なプロモーター(EP 344029 )、及び例えばシロイヌナズナ由来の(Arabidopsis thaliana)種子特異的なプロモーター(Krebbers et al.(1988) Plant Physiol.87:859)が含まれる。見本となる緑色組織特異的なプロモーターにはトウモロコシのホスフォエノールピルビン酸カルボキシラーゼ(PEPC)プロモーター、スモールサブユニットリブロース ビスカルボキシラーゼプロモーター(ssRUBISCO )及びクロロフィルa/b 結合蛋白質プロモーターが含まれる。プロモーターは、国際公開番号WO93/07278の中で述べられた通りの、植物TrpA遺伝子から単離されたプロモーターの様な樹心(pith)特異的なプロモーターであることもできる。
【0055】
他の生物のための誘導可能なプロモーターには、例えば、アラビノースプロモーアー、lacZプロモーター、メタロチオネインプロモーター、及び熱ショックプロモーターのみならず、本技術分野の当業者に知られている他の多くのものが含まれる。S.pombe の様な酵母において有用な抑制可能なプロモーターは、ビタミンB1によって抑制可能なPmntプロモーターである。
【0056】
典型的には、これらの細胞へ導入されるコンストラクトは組み換え発現技術を用いて調製される。組み換え発現技術は、組み換えた核酸の構築と感染した細胞内での遺伝子の発現を伴う。これらの末端に到達するための分子クローニング技術は本技術分野において知られている。組み換え核酸の構築に適する広範囲のクローニングとインビトロ増幅の方法は本技術部分野の当業者に良く知られている。これらの技術の例と、多くのクローニング演習を通じて当業者を教示するのに十分である指示は、Berger and Kimmel,Guide to Molecular Cloning Techniques, Methods in Enzymology(分子クローニング技術への案内 酵素学における方法論), volume 152, Academic Press, Inc.,San Diego,CA (Berger); 及びCurrent Protocols in Molecular Biology(分子生物学における現在のプロトコール),F.M. Ausubel et al., eds.,Current Protocols (現在のプロトコール),a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons,Inc.,(1998 Supplement) (Ausubel) の中に見出される。
【0057】
ポリヌクレオチドコンストラクトの構築には、細菌内で複製可能なベクターを使用することが通常必要となる。細菌からプラスミドを精製するために使用することができるキットは有り余る程市販されている。それらを適切に使用するために製造者の指示に従うこと(例えば、EasyPrepJ,FlexiPrepJ:両者はファルマシアバイオテック由来、StrataCleanJ:ストラタジーン由来、及びQIAexpress Expression System:キアゲン、を見よ)。単離されて精製されたプラスミドを更に操作して他のプラスミドを作製することも可能であり、細胞をトランスフェクトするか、又はアグロバクテリウムツメファシエンスへ導入して感染させて、植物を形質転換するのに使用される。アグロバクテリウムが形質転換の手段である場合にはシャトルベクターが構築される。ストレプトマイセス又はバシラスにおけるクローニングもまた可能である。
【0058】
標的細胞中へ取り込まれるために使用されるポリヌクレオチドコンストラクト及び/又はベクター中へ、選択可能なマーカーがしばしば導入される。これらのマーカーによって、興味の対象であるポリヌクレオチドを含んでいる細胞のコロニーを選択することができる。ベクターは、例えば大腸菌又は他の細胞中で機能する選択マーカーをしばしば有するであろうし、標的細胞へ導入される前にその細胞中でベクターは複製される。大腸菌内で選択可能なマーカーの例には、例えばアンピシリン、テトラサイクリン、カナマイシン、エリスロマイシンなどの抗生物質に対する耐性を特定している遺伝子、又はβ−ガラクトシダーゼの様な他の型の選択可能な酵素活性を与える遺伝子、又はラクトースオペロンが含まれる。哺乳細胞において使用するのに適している選択可能なマーカーには、例えば、ジヒドロ葉酸還元酵素(DHFR)、シミジンキナーゼ遺伝子(TK)、又は薬剤耐性を特定している原核細胞の遺伝子が含まれ、薬剤耐性を特定している原核細胞の遺伝子には、ミコフェノール酸(mycophenolic acid )で選択することができるgpt (キサンチン−グアニン ホスフォリボシルトランスフェラーゼ); G418、ハイグロマイシン又はピュロマイシンで選択することができるneo (ネオマイシンホスフォトランスフェラーゼ); 及びメトトレキサートで選択することができるDHFR(ジヒドロ葉酸還元酵素)(Mulligan & Berg (1981) Proc.Natl.Acad.Sci.USA 78:2072;Southern & Berg (1982) J.Mol.Appl.Genet.1:327)がある。
【0059】
植物細胞のための選択マーカーはしばしば、例えば、カナマイシン、G418、ブレオマイシン、ハイグロマイシン又はクロラムフェニコールの様な殺生剤又は抗生物質に対する耐性、又はクロロスルフロン(chlorsulfuron )又はバスタ(Basta )の様な除草剤に対する耐性を与える。選択可能なマーカーのための適切なコード配列の例には、抗生物質カナマイシンに対する耐性を与えるネオマイシンホスフォトランスフェラーゼ酵素をコードするneo 遺伝子(Beck et al (1982)Gene 19:327 )、ハイグロマイシンホスフォトランスフェラーゼ酵素をコードして抗生物質ハイグロマイシンに対する耐性を与えるhyg (hpt) 遺伝子(Gritz and Davies (1983) Gene 25:179 )、及び除草剤物質であるホスフィノスリチン(phosphinothricin)及びビアラフォス(bialaphos )に対する耐性をあたえるホスフィノスリチンアセチルトランスフェラーゼをコードするbar 遺伝子(EP 242236 )がある。
【0060】
もし一つ以上の外来遺伝子を標的とする真核細胞に導入するならば、通常は、各々の外来性の核酸について異なった選択可能なマーカーを使用することが望ましい。これにより、望まれる外来性の核酸の両者を含む細胞について同時に選択することができるようになる。
【0061】
(標的細胞へコンストラクトを導入するための方法)
組み換え部位及び/又はリコンビナーゼをコードする遺伝子を含むポリヌクレオチドコンストラクトを、標的細胞及び/又は生物へ、当業者に知られているいくつかの手段のいずれかを用いて導入することができる。例えば、植物の培養中又は器官中のいずれかで、種々の通常の技術によって、DNA コンストラクトを植物細胞中へ導入することができる。例えば、DNA パーティクルボンバートメントの様なバイオリスティックな方法を用いてDNA コンストラクトを植物細胞へ直接に導入することが可能であり、又は植物細胞プロトプラストの電気穿孔法及びマイクロインジェクションの様な技術を用いてDNA コンストラクトを導入することが可能である。パーティクルにより仲介された形質転換技術(「バイオリスティック」としても知られている)は、Klein et al.,Nature,327:70-73 (1987); Vasil,V.et al.,Bio/Technol.11:1553-1558 (1993)及びBecker,D.et al.,Plant J.,5:299-307 (1994) において述べられている。これらの方法は、微小ビーズ又はパーティクルの基質の中又はその表面上のいずれかにおいて核酸を有する小さなパーティクルにより細胞を穿孔することを伴う。
【0062】
バイオリスティックなSDS-1000遺伝子銃(バイオラッド、ヘラキュレス、CA)は、DNA を被覆した金又はタングステンの微小担体を標的細胞へ加速するためにヘリウムガスの圧力を使用する。その過程は広範囲の生物由来の組織及び細胞に適用可能であり、その生物には植物、細菌、カビ、藻、無傷動物組織、組織培養細胞及び動物胚が含まれる。電気パルス送達を用いることも可能であり、それは動物及び患者の生組織のための、本質的に緩和な電気穿孔の型である。Zhao,Advanced Drug Derivery Reviews (進歩した薬物送達の総論)17:257-262 (1995)。
【0063】
他の形質転換方法が当業者に知られている。マイクロインジェクションの技術が本技術分野において知られ、科学及び特許文献において良く述べられている。ポリエチレングリコール(PEG )沈殿を用いたDNA コンストラクトの導入は、Paszkowski et al.,EMBO J.3:2717 (1984)において述べられている。電気穿孔技術は、Fromm et al.,Proc.Natl.Acad.Sci.USA,82:5824 (1985)において述べられている。PEG により仲介された植物プロトプラストの形質転換及び電気穿孔もまた、Lazzeri,P.,Methods Mol.Biol.49:95-106 (1995)において議論されている。異種遺伝子の導入及び発現のための方法は単子葉及び双子葉植物の両者において知られている。米国特許番号5,633,446, 5,317,096, 5,689,052, 5,159,135及び5,679,558; Weising et al.(1988) Ann.Rev.Genet.22:421- 477 を見よ。単子葉植物の形質転換には特に種々の技術を使用することが可能であり、それには電気穿孔法(例えばShimamoto et al.,Nature (1992),338:274-276)、バイオリスティック(例えば欧州特許出願 270,356)、及びアグロバクテリウム(例えば、Bytebier et al.,Proc.Natl.Acad.Sci.USA (1987) 84:5345-5349)が含まれる。
【0064】
植物の形質転換のために、DNA コンストラクトを適切なT-DNA フランキング領域と結合させて、一般的なアグロバクテリウムツメファシエンス宿主ベクターへ導入することができる。アグロバクテリウムツメファシエンス宿主の毒性機能により、細胞が細菌によって感染した時に、トランスジーン及び隣接したマーカー遺伝子(もし存在しているならば)が植物細胞DNA へ挿入される。アグロバクテリウムツメファシエンスにより仲介された形質転換技術は、科学文献において良く述べられている。例えば、Horsch et al.Science,233:496 (1984), Fraley et al.,Proc.Natl.Acad.Sci.USA,80:4803 (1983)及びHooykaas,Plant Mol.Biol.,13:327-336 (1989), Bechtold et al.,Comptes Rendus De L Academie Des Sciences Serie Iii-Sciences De La Vie-Life Sciences,316:1194-1199 (1993), Valvekens et al.,Proc.Natl.Acad.SCI.USA,85:5536-5540 (1988)を見よ。植物及び細胞培養物の遺伝子移送方法の総論としては、Fisk et al.,Scientia Horticulturae 55:5-36 (1993) 及びPotrykus,CIBA Found.Symp.154:198 (1990) を見よ。
【0065】
ポリヌクレオチド配列を細胞内へ送達するための他の方法には、例えば、リポソームに基づいた遺伝子送達(Debs and Zhu (1993) WO93/24640; Mannino andGould-Fogerite (1988) BioTechniques 6(7); Rose米国特許番号5,279,833; Brigham (1991) WO91/06309; 及びFelgner et al.(1987) Proc.Natl.Acad.Sci.USA84:7413-7414)のみならず、ウイルスベクターの使用、(例えば、アデノウイルス(総論のために、Berns et al.(1995) Ann.NY Acad.Sci.772:95-104; Ali etal.(1994) Gene Ther.1:367-384;及びHaddada et al.(1995) Curr.Top.Microbiol.Immunol.199 (Pt3):297-306 を見よ)、パピローマ(papilloma) ウイルス、レトロウイルス(例えば、Buchscher et al.(1992) J.Virol.66(5) 2731-2739; Johann et al.(1992) J.Virol.66(5):1635-1640; Sommerfelt et al.,(1990) Virol.176:58-59; Wilson et al.(1989))J.Virol.63:2374-2378; Miller et al.,J.Virol.65:2220-2224 (1991); Wong-Staal et al.,PCT/US94/05700,及びRosenburg and Fauci (1993) in Fundamental Immunology(基本的免疫学),Third Edition Paul (ed) Raven Press,Ltd.,New York及びその中の引用文献、及びYu et al.,Gene Therapy (1994) supra を見よ)、及びアデノ関連ウイルスベクター(West et al.(1987) Virology 160:38-47; Canter et al.(1989) 米国特許番号4,797,368; Canter et al.WO93/24641 (1993); Kotin (1994) Human Gene Therapy 5:793-801; Muzyczka (1994) J.Clin.Invst.94:1351及びSamulski(supra) をAAVベクターの概観のために見よ。また、Lebkowski,米国特許番号5,173,414; Tratschin et al.(1985) Mol.Cell.Biol.,5(11):3251-3260; Tratschin et al.(1984) Mol.Cell.Biol.,4:2072-2081; Hermonat and Muzyczka (1984) Proc.Natl.Acad.Sci.USA,81:6466-6470; McLaughlin et al.(1988)及びSamulski et al. (1989) J.Virol.,63:03822-3828 もまた見よ)及びそれに類似なものが含まれる。
【0066】
導入された外来DNA が組み込まれたパターンを解析することができる方法は、当業者に良く知られている。例えば形質転換した細胞からDNA を抽出し、一つ又はそれ以上の制限酵素によってそのDNA を消化し、そしてポリヌクレオチドコンストラクトのラベル化された断片をハイブリダイズさせることができる。挿入された配列をポリメラーゼ連鎖反応(PCR )を用いて同定することもまた可能である。例えば、Sambrook et al.,Molecular Cloning - A Laboratory Manual (分子クローニング、実験室マニュアル), Cold Spring Harbor Laboratory, ColdSpring Harbor, New York,1989を、これら及び他の適切な方法の記載のために見よ。
【0067】
(形質転換植物及び動物の再生)
本発明の方法は、安定して組み込まれた外来性ポリヌクレオチド又は他の細胞内核酸の安定な転位(rearrangement )を有する、形質転換したキメラ型の多細胞生物を得るために特に有用である。形質転換したキメラ型の生物を得るための方法は、植物と動物の両者について当業者に良く知られている。
【0068】
上記のいずれかの形質転換技術によって得られた形質転換した植物細胞は培養され、形質転換した遺伝子型及びそのために望みの表現型を有している完全な植物が再生される。その様な再生技術は組織培養成長培地の中の特定の植物ホルモンの取り扱いに依り、典型的には望みの核酸配列と共に導入された殺生剤及び/又は除草剤マーカーに依る。培養されたプロトプラストからの植物再生は、Evans et al.,Protoplast Isolation and Culture, Handbook of Plant Cell Culture (プロトプラストの単離及び培養、植物細胞培養のハンドブック),pp.124-176, Macmillian Publishing Company, New York (1983);及びその結びつきで、Regeneration of Plants, Plant Protoplast(植物、植物プロトプラストの再生),pp.21-73,CRC Press, Boca Raton,(1985)において述べられている。植物カルス、外植片、体細胞胚(Dandekar et al.,J.Tissue Cult.Meth.,12:145 (1989); MacGranahan et al.,Plant Cell Rep.,8:512 (1990))、器官、又はその一部分からもまた再生することができる。その様な再生技術は一般的にKlee et al.,Ann.Rev.of Plant Phys.,38:467-486 (1987)において述べられている。
【0069】
その方法は大部分の脊椎動物種の形質転換したキメラ型の動物を作製するのに有用である。その様な種には、それに限定されるものではないが、ネズミ及びラットの様なゲッシ類、ウサギ、ヒツジやヤギの様なヒツジ類(ovines)、ブタの様なブタ類(porcines)、ウシ(cattle)やバッファローの様なウシ類(bovine)を含む、非ヒト哺乳類が含まれる。形質転換動物を得る為の方法は、例えば、Puhler,A.,Ed.,Genetic Engineering of Animals(動物の遺伝工学)VCH Publ.,1993; Murphy and Carter, Eds.,Transgenesis Techniques:Principles and Protocols(形質転換技術:原理とプロトコール)(Methods in Molecular Biology, Vol.18),1993;及びPinkert,CA,Ed.,Transgenic Animal Technology:A Laboratory Handbook (形質転換動物の技術工学:実験室ハンドブック),Academic Press,1994において述べられている。特異的な遺伝的修飾を有している形質転換魚類もまた、クレイムされた方法を用いて作製することができる。形質転換魚類を作製するための一般的な方法のためには、例えば、Iyengar et al.(1996) Transgenic Res.5:147-166 を見よ。
【0070】
そのゲノム中に特異的な修飾を有している、形質転換した又はキメラ型の動物を得る一つの方法は、受精した卵母細胞を興味の対象であるポリヌクレオチドを組み換え部位に隣接して含んでいるベクターと接触させることである。ネズミの様ないくつかの動物についてインビボで受精が行われ、受精した卵子は手術により移された。他の動物、特にウシおいては、生きた又は屠殺された動物から卵子を移して来て、そしてその卵子をインビトロで受精させることは好ましい。DeBoer et al.,WO91/08216を見よ。インビトロで受精させることにより、実質的に同調した細胞に導入すべき修飾を行うことができる。そして、着床前の胚が約16-150個の細胞を含むようになるまで受精した卵母細胞をインビトロで培養した。16-32 細胞期の胚は桑実胚として述べられている。32細胞以上を含んでいる着床前の胚を胞胚と称する。これらの胚は、典型的には64細胞期において胞胚腔の空洞(cavity)の発達を示した。もし望むならば、胚細胞の中における望みの外来性ポリヌクレオチドの存在を当業者に知られている方法によって検出することができる。
【0071】
受精した卵母細胞を着床前の段階まで培養するための方法は、Gordon et al.(1984) Methods Enzymol.101:414; Hogan et al. Manipulation of the Mouse Embryo:A Laboartory Manual(マウス胚の操作:実験室マニュアル)C.S.H.L. N.Y.(1986) (マウス胚); Hammer et al.(1985) Nature 315:680(ウサギ及びブタ胚);Gandolfi et al.(1987) J.Reprod.Fert.81:23-28;Rexroad et al.(1988) J.Anim.Sci.66:947-953(ヒツジ胚)及びEyestone et al.(1989) J.Reprod.Fert.85:715-720; Camous et al.(1984) J.Reprod.Fert.72:779-785及びHeyman et al.(1987) Theriogenology 27:5968(ウシ胚)において述べられている。時々、着床を待つ間の期間着床前の胚を凍結して保存した。着床前の胚を適切なメスに移植し、その結果、トランスジーンが組み込まれたときの発達の段階に依って、形質転換した又はキメラ型の動物が誕生するという結果となった。正しい生殖系列の形質転換動物を形成するために、キメラ型の哺乳動物を繁殖させることができる。
【0072】
替わりに本方法を用いて、望みの外来性ポリヌクレオチドの単一コピーを有する胚幹細胞(ES)を得ることができる。これらの細胞は、インビトロで培養された着床前の胚から得られた。例えば、Hooper,ML,Embryonal Stem Cells:Introducing Planned Changes into the Animal Germline (胚性幹細胞;計画された変化の動物生殖系列への導入)(Modern Genetics,v.1) Int'1.Pub.Distrib.,Inc.,1993; Bradley et al.(1984) Nature 309,255-258 を見よ。形質転換したES細胞を非ヒト動物由来の胞胚と結合させた。ES細胞は胚をコロナイズし、いくつかの胚においては得られたキメラ動物の生殖系列を形成する。Jaenisch,Science,240:1468-1474 (1988) を見よ。代わりに、生物を再構成することができるES細胞又は体細胞(”体細胞再集団細胞:somatic repopulating cells”)を、形質転換した哺乳動物を生み出す、除核された受精卵母細胞へ着床させるための核の起源として使用することができる。例えば、Wilmut et al.(1997) Nature 385:810-813を見よ。
【0073】
(実施例)
下記の実施例は説明の目的で与えられたものであって本発明を限定するものではない。
【0074】
(実施例1)
(シゾサッカロミセス・ボンベ(Schizosaccharomyces pombe )におけるΦC31組み換えシステム)
この実施例は、ストレプトミセスバクテリオファージΦC31 部位特異的な組み換えシステムが、真核細胞において機能することを示している。S.Pombe のleu1遺伝子座において、バクテリオファージ接着部位(attP)をシゾサッカロミセス・ボンベの染色体において導入した。続いてこの標的株を、ura4+選択可能なマーカーと連結した細菌接着部位(attB)を含むプラスミドにより形質転換された。ΦC31 インテグラーゼ遺伝子を有している第二のプラスミドによって共トランスフェクトしたとき、インテグラーゼ遺伝子が発現している条件下で、Ura+へ高い効率で形質転換していることが観察された。
【0075】
組み換えイベントのサザン解析により、leu1遺伝子座のattPへ、attB-ura4+プラスミドが挿入されていることが示された。ハイブリッドジャンクジョンのヌクレオチド配列により、attB x attP 組み換え反応は正確であることが明らかとなった。
【0076】
(材料及び方法)
(組み換えDNA )
全体として標準的な方法が用いられた。大腸菌株XL2-Blue(recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F' proAB lAcIq Z △M15Tn10(Tetr) Amy Camr、ストラタジーン]がDNA コンストラクトとして働いた。
【0077】
(培地)
分裂酵母株を、必要に応じて225mg/l のアデニン、ヒスチジン、ロイシン又はウラシルを添加した、最小培地(バイオ101 由来EMM-低グルコース)で生育させた。5-FOA (5-フロロオロチン酸、ザイモリサーチ社より)を含む最小プレートをGrimm ら((1988) Mol.Gen.Genet.215:81-86)に従って調製し、アデニン、ヒスチジン及びロイシンを添加した。使用した時、5 μg/mlとなるようにチアミンを添加した。
【0078】
(ΦC31 attP標的を有するS.prombe)
pHS282からApaI-SacI 断片として単離された(Thorpe & Smith (1998) Proc.NATL.Acad.Sci.USA 95:5505-5510 )84塩基対のΦC31 attP部位(PP' と略称)を、S.prombe組み込みベクターpJK148の同じ部位にクローン化し(Keeney & Boeke (1994) Genetics 136:849-856)、pLT44 を作製した。NdeI切断DNA を用いた酢酸リチウムにより仲介された形質転換により、このプラスミドはS.prombe leu1-32アリールへ標的化された。pLT44 による相同組み換えによってLeu+へ変換された、レシピエント宿主FY527(h-ade6-M216 his3-D1 leu1-32 ura4-D18) を、サザンブロットによって試験した。FY527attP と名付けられた一つのLeu+型は単一コピーのpLT44 を含んでいることが見出された。FY527attPx2 と名付けられたもう一つの形質転換体は直列(tandem)のプラスミド挿入を有していた。
【0079】
(ΦC31 attB部位を有する組み込み可能なura4+ベクター)
1.8kb のEcoRI-BamHI 断片上でpTZura4 (S.Forsburg)から切り出した、S.prombeのura4+遺伝子を、同じ酵素を用いて切断されたpJK へ挿入し、pLT40 を作製した。500bp のBamHI-XbaI断片としてpHS21 から単離されたΦC31 attB部位(BB'と略称)を、それらの酵素を用いて切断されたpLT40 へ連結してpLT42 を作製した。XhoI断片を削除することによりpLT42 からleu1遺伝子の大部分を除去し、pLT45 を作製した。これにより、pLT45 からleu1の全部であるが229bp が除去されて、その形質転換効率はleu1相同性を持たないプラスミドの形質転換効率まで低下した。ura4のすぐ反対側に同じ方向の第二でattB部位を有しているpLT50 が構築され、それは最初にpLT42 由来のattB BamHI-SacI 断片をpUC19 へサブクローニングし、それをEcoRI とSalII によって切り出し、続いてそれをEcoRI とXhoIによって切断したpLT45 へ挿入することにより構築された。最終構築物中の第二のattB部位を各鎖について一回配列決定し、最初のattB部位と同一であることが見出された。
【0080】
(平滑DNA 形質転換)
attB-ura4+-attB 平滑DNA は、pLT50 から精製されたattII-AlwNl 断片として、又はpLT50 を鋳型として用いたPCR 産物として調製された。T3プライマーとpJK148のプラスミド骨格に対応している第二のプライマー(5' ggc cct gaa att gtt gct tct gcc 3' )を用いて、標準的な条件によりPCR は行われた。
【0081】
(ΦC31 インテグラーゼの抑制的な合成)
ビタミンB1によって抑制される、S.prombe Pmntプロモーターは、pMO147から1.2kb のPstI-SacI 断片として切り出され、同じ酵素を用いて切断されたhis3+,ars1 ベクターpBG2(Ohi et al.(1996))Gene 174:315-318)へ挿入され、pLT41 が作製された。ΦC31 int コード領域を含んでいる2.0kb SacI断片を、pHS33(Thorpe & Smith (1998) supra )からpLT41 のSacI部位へ移した。発現がPmntの制御下にある様にint コード領域が配向されたクローンを、pLT43 と名付けた。
【0082】
(分子解析)
ベーリンガーマンハイムのGeniusTMシステムを用いてサザン解析を行った。leu1の998bp の内部EcoRV 断片、ura4の1.8kb 断片及び2.0kb のΦC31 int 遺伝子をランダムプライマー法によってジゴキシゲン標識し、プローブとして使用した。ストラタジーンTurbo PFU 酵素又はVENTポリメラーゼを用いて、パーキンエルマーCetus Gene Amp PCR 9600 で、ポリメラーゼ連鎖反応を行った。可能なところでは標準的なT3及びT7プライマーを用いた。ura4プライマー(5' gtc aaa aag ttt cgt caa tat cac 3' )(配列番号1)及びpJK148プライマーをオペロンテクノロジーから購入した。全てのPCR 反応において、51℃のアニーリニング温度と30秒の伸長時間が用いられた。
【0083】
(結果と考察)
(S.prombeゲノムへの標的部位の挿入)
ΦC31 により仲介された組み込みのための標的部位を有する宿主株を作製するために、相同組み換えによって、ΦC31 attP部位を分裂酵母ゲノムのleu1遺伝子座へ挿入し、Leu+株FY527 attPが形成された(図1 A)。過去の研究により、S.prombeのDNA をXbaIによって開裂し、leu1+遺伝子の内部の1kp 断片により検証したら、そのプローブにより14kbのバンドが検出されることが示されている(Keeney & Boeke (1994) Genetics 136:849-856)。leu+プラスミドpJK148がleu1-32 遺伝子座において挿入されていると、3 と18kbのバンドが検出されるという結果となる(図1 A)。84bpのΦC31 attP因子が封入されている点で、pLT44はpJK148から異なっているために、leu1-32 におけるpLT44 の組み込みは、FY527 attPにおいて、同様の3kb と18kbのハイブリダイゼーションパターンをもたらした。他のハイブリダイズしている断片が存在しないことは、単一に組み込まれたコピーとしてのpLT44 のDNA 残基を示している。
【0084】
ΦC31インテグラーゼ仲介形質転換 ura4+およびattB配列(BB’)を有するが、複製の原点を欠くFY527attPをpLT45で形質転換した。このコンストラクトを、それ自体により、またはΦC31インテグラーゼを産生するhis3+複製ベクターpLT43を用いて導入した。pLT43を封入することにより、Ura+形質転換体の数が平均で15倍増加した(表1)。この効果はインテグラーゼ遺伝子発現によるものなので、この増加がpLT45と複製にプロフィシエントな(replication-proficient)pLT43との間の組み換えに起因するものではありえない。インテグラーゼ遺伝子の転写は、高濃度のビタミンB1により抑制可能なプロモーターPmntの支配を受ける(Maundrell,K.(1993年)Gene 123:127〜130頁)。抑制は完全ではないが(Forsburg,S.L.(1993年)Nucleic Acids Res 21:2955〜2956頁)、インテグラーゼたん白の生成が減少する。発育培地にチアミンを加えた場合、Ura+形質転換体の数は背景レベル付近にまで減少した。培地からヒスチジンを省略することによりインテグラーゼプラスミドを共選択してもしなくても、Ura+形質転換体の頻度が顕著に変化することは無かった。pLT43またはその前駆体プラスミドpBG2を用いて得たHis+形質転換体の数から、FY527atttPの形質転換能力を推定した。両方の複製プラスミドの頻度を比較したところ、FY527attPのpLT43依存形質転換は平均で約15%であった。
【0085】
【表1】
【0086】
ΦC31インテグラーゼをプロモートしたattP x attB組み換え pLT45をコードしたΦC31attBエレメントと染色体的に位置の定まったattP配列との間の組み換えにより、図1Bに示すように、環状DNAがleu1遺伝子座に組み込まれた。この反応が起きた場合、Ura+形質転換体から得たXbaI分別ゲノムのDNAをleu1 DNAでプローブすると、3kbバンドは変わらないが、一方18kbバンドは約23kbまで増大する(図1C)。無作為に選択したUra+コロニーをハイブリダイゼーション分析により検査した。チアミンの省略によりΦC31インテグラーゼ遺伝子発現を抑制解除した場合の実験から得られた8つの分離株のうち7つが、この約23kbバンドの存在を示した。この同サイズバンドをura4プローブにハイブリダイズした。ura4−D18欠損対立遺伝子から期待されるように、これは親株にura4ハイブリダイゼーションしなかったものと対照的である。これら7つの分離株の一つが、両方のプローブにハイブリタイズする付加的なバンドを示した。部位に特異的な組み換え現象に加えて、この候補物質がleu1遺伝子座にDNA再配列を有すると思われる。leu1再配列が、有効なS.pombe相同体組み換え系によりおそらく触媒された。残る分離株は部位特異的組み換え現象を経験したことが無く、pLT45とpLT43との間の組み換えにより、ウラシル原栄養株が増大したと思われた。これら8つの分離株のうち、半分をUra+およびHis+の両方として選択したが、このグループとUra+のみで選択したグループとの間に顕著な差は見られなかった。
【0087】
ビタミンB1の存在下で行った形質転換実験から、DNAハイブリダイゼーションにより同数のUra+形質転換体を調べた。チアミン抑制可能なPnmtプロモーターがインテグラーゼ産生を制限し、したがって部位特異的組込みを制限すると考えられる。この低頻度形質転換から分離された8つのUra+候補物質のうちの2つにより、23kbバンドがleu1およびura4プローブにハイブリダイズしていることが示された。しかし、いずれのプローブも付加的バンドを検出したので、これらは正確な組込み現象を表しておらず、発明者らはこれらをクラスbインテグラントとして分類した。他の6つの分離体では、ハイブリダイゼーションのパターンを翻訳することは困難である。これらのいくつかにおいて、遺伝子座は再配列を経験しているが、leu1プローブでは3kbバンドを検出できなかった。これらの多くにおいて、ura4に対するハイブリダイゼーションが弱いことから、Ura+表現型はゲノム中のpLT45の安定な保守によるものではないであろう。
【0088】
選択しない場合にインテグラーゼプラスミドを維持する形質転換体の比率を確実にするために、インテグラーゼ遺伝子配列でブロットを再プローブした。Ura+His+として選択されたそれらはプラスミドを維持すると考えられており、かつハイブリダイゼーションが明らかになるにつれてそうなった。His表現型に関係なくUra+として選択された8つの分離株のうちの5つは、インテグラーゼプローブにハイブリダイズするバンドも与えた。intの損失が安定な組込みに影響を及ぼすことを確認するため、無作為に選択したUra+細胞の別の組を数世代にわたり非選択的に成長させ、pLT43を失ったHis−子孫に対してスクリーニングをした。8つの代表的Ura+His−クローンの分析は、全てが正確に染色体が位置するattP部位に組込まれたpLT45の単一コピーを有することを示した。これらの組込み体のDNAはインテグラーゼプローブでハイブリダイズしなかった。対照的に、pLT45単体の形質転換により誘導された背景頻度Ura+クローンにより、leu1遺伝子座にハイブリダイジングバンドの親構造が得られ、かつ5kbおよび7kbに更なる微弱バンドが得られた。これらの知見は、ゲノムのどこかのpLT45の組込みまたはS.pombe複製原点の欠損にもかかわらずいくつかの細胞中のプラスミドの維持のいずれかと一致している。
【0089】
保守的部位特異的組み換え PCRを用いて、3つの代表的Ura+候補物質からattP/attB組み換え接合部を回収した。ハイブリッド部位の一つでは、attR(PB’)がT3およびT7プロモーターに隣接していた。他の部位では、attL(BP’)がT3プロモーターおよびura4DNAに隣接していた(図1C)。いずれの場合も、プライマー対が、期待される大きさのバンドを増幅したこれらの配列に向けられ、一方、本来のattP(PP’)はもはや見つからない。attPは検出されるがattLおよびattRのいずれもが検出されない場合には、これは親株FY527aatPと対照的である。3つの代表的attLおよびattR PCR製品のヌクレオチド配列は、付随的な突然変異体が無いことを示した。したがって、バクテリアおよび哺乳動物細胞において、S.pombe中のΦC31仲介部位特異的組み換えは保守的組み換え反応である。
【0090】
ΦC31インテグラーゼは組込みした分子を除去しない。
ThorpeとSmith((1998年)Proc.Natl.Acad.Sci.USA95:5505〜5510)は、ゲル分別DNA断片の分析でΦC31インテグラーゼ反応の反転を検出しなかった。発明者らは遺伝学的選択法により反転反応の可能性を調べた。pLT45のFY527attPへの正確な組込みが、サザン分析により3つのクローンに対して確認された。次いで、これらの株をpLT43で再転換した。pLT45を切り取ることにより、ura4+マーカーの欠損を招いた。Ura−表現型を5−FOAを用いてプレート上に得点することができる(Grimmら、(1988年)Mol. Gen. Genet.215:81〜86頁)。3つのUra+His−前駆体の培養体から得たUra−分離個体の頻度は5.7×10−4、7.1×10−4および5.6×10−4であった。対照的に、3つのUra+His+誘導体から得たUra−コロニーの頻度はある程度高く、1.1×10−2、3.8×10−3および2.3×10−3であり、それぞれ19、5および4倍増加した。インテグラーゼ遺伝子を有しない制御ベクターpBG2を代わりに用いて、5−FOA耐性の速度が増大することも分かった。それぞれ1.0×10−2、1.0×10−2および8.0×10−3であった。形質転換プロセス自体は変異原性を示す。
【0091】
pLT45で形質転換された3つの培養体のそれぞれから得られた3つのUra−His+クローンをサザンブロッティングにより分析した。1つの分離体が、pLT45のFY527attPへの安定な組込みと一致するDNAパターンを有していた。したがって、このクローンにおいて、Ura−表現型は、部位特異的組み換え反応の反転によるよりも、むしろ制限パターンが感知可能なほどに変わっていない変異体により引き起こされた。第2クローンはpLT45挿入欠損FY527attPのサザンパターン特徴を示し、第3クローンはpLT45挿入の無いFY527attP等の細胞とFY527attP前駆体株FY527等の細胞の2つの型の細胞の混合物と一致するパターンを有していた。後者の構造は、pLT44の挿入を反転するleu反復の間の染色体内相同組み換えから生じる場合がある(図1A)。組込みしたプラスミドDNAの正確な切り取りが前述の2つの候補物質に現れた場合、attP部位が再生した。これはPCRで検出可能であった。PCR製品の大きさは、完全なハイブリッド部位に期待される大きさであり、ハイブリッド部位の存在はPCR製品を配列決定することにより確認された。これらの観察は、ura4遺伝子の欠損がΦC31仲介切り取り以外の機構により発生するという考えと一致している。
【0092】
(まとめ)
単一目的部位での環状分子の組込みは、ほとんど全ての形質転換体の正確な挿入を得る効果的な方法であった。発明者らが観察したわずかの異常現象は、おそらくleu1反復DNAに作用するS.pombe組み換え系に主として起因する。インテグラーゼ産生がそのプロモーターの発現により制限されていた場合、形質転換体の数を背景レベル付近にまで減らした。この条件下で、ΦC31部位特異的組み換えから誘導された修復形質転換体はほとんどなかった。真核細胞におけるΦC31部位特異的組み換え系の機能的操作により、トランス遺伝子および染色体のマニピュレーションを行う新たな機会が与えられた。ΦC31系をattBおよびattP部位の選択的配置と共に用いて、DNAの削除、反転または挿入を行うことができる。この系の重要な特徴は、attB×attP反応が切り取り特異的タンパク質の不存在下で不可逆という点である。
【0093】
(実施例2)
ΦC31インテグラーゼはCHO細胞中で安定な組込みを作る役目をする。
この実施例では、チャイニーズハムスター卵巣(CHO)細胞におけるattB組み換え部位とattP組み換え部位との間の組み換えを仲介する能力に関してΦC31インテグラーゼをテストする実験を説明する。
【0094】
(方法)
ゼオシン耐性を与える選択マーカーを含んだCHO細胞株51YT211をattP含有プラスミドpFY1で形質転換した(図2)。単一コロニーを2回精製した後、6つのゼオシン耐性細胞株を分離した。サザンDNAハイブリダイゼーションによる分析により、6つの細胞株のそれぞれが、ゲノム内に組込みされたpFY1の少なくとも一分子を有することが確認された。
【0095】
6つの細胞株のそれぞれをattB含有プラスミドpFY9およびint含有プラスミドpFY6でトランスフェクトし、pFY9上のattB部位とpFY1の染色体コピー上のattP部位との間の部位特異的組み換えに対するテストを行った。コントロールとして、同じ細胞株をpFY9で形質転換したが、int含有pFY6は用いなかった。pFY9プラスミドは、SV40初期プロモーターの制御下でネオマイシン耐性選択マーカーおよびプロモーターに結合していない緑蛍光タンパク質(GFP)をコードした配列を含んでいた。したがって、部位特異的組み換えが、pFY1に含まれるヒト・サイトメガロウイルス・プロモーターの制御下でGFPコード化配列を配置し、その結果GFPを発現することが予想された。
【0096】
(結果)
トランスフェクションの結果:ネオマイシン耐性コロニーを顕微鏡下に置き、GFP遺伝子の活性の有無を観察した。pFT9+pFY6でトランスフェクションした細胞では大部分が、しかしpFY9のみでトランスフェクションした細胞ではごく僅かが、GFP活性を示した。このことは、pFY6でトランスフェクションした場合のpFY9の部位特異的組込みおよび共トランスフェクションint遺伝子の不存在下のpFY9の無作為挿入と一致する。
【0097】
Pc(ヒト・サイトメガロウイルス・プロモーター)およびGFP(図2)に対応するプライマー対を用いてPCR分析を行った。これらのプライマーは、組込み接合に対応する約0.6kbのバンドを増幅すると予想された。部位特異的組込みと無作為組込みの両者からネオマイシン耐性コロニーが生じる場合があり、かつGFPマーカーは選択的形質を与えないので、構成クローンの純粋な培養株を得ることは困難であった。したがって、各トランスフェクション株から得たネオマイシン体制細胞のプールをPCR分析に供し、ネオマイシン耐性細胞間に組込み接合が存在するか否かを調査した。大きさが約0.6kbと予想されるバンドを2つの株から得た。これは、attB×attP組み換え接合により、PcとGFPの結合が形成されることを示している。
【0098】
(実施例3)
ΦC31インテグラーゼはCHO細胞中で部位特異的組み換えを触媒する。
本実施例では、ΦC31インテグラーゼをテストして、attP部位とattB部位との間の組み換えにより、DNA分子をチャイニーズハムスター卵巣(CHO)細胞の染色体中に組込む能力を調べた。
【0099】
(方法)
プラスミドコンストラクト染色体attB標的コンストラクトpFY12、pFY14およびpFY15 プラスミドpcDNA3.1/His/lacZ(Invitrogen)をベクターバックボーンとして用いた。長さの異なるattB部位を含有し、HindIII部位とKpnI部位に隣接した合成オリゴヌクレオチドをpcDNA3.1/His/lacZのHindIII (AAGCTT)部位とKpnI (GGTACC)部位との間に挿入した。
プラスミドpFY12は90bpのattB 配列(AAGCTT gacggtctcg aagccgcggt gcgggtgcca gggcgtgccc ttgggctccc cgggcgcgta ctccacctca cccatctggt ccatcatgat GGTACC)(SEQ ID NO:2)を含有する。
プラスミドpFY14は50 bpのattB部位(AAGCTT gcgggtgcca gggcgtgccc ttgggctccc cgggcgcgta ctccacctca TGGTACC)(SEQ ID NO:3)を含有する。
プラスミドpFY15は30 bpのattB(AAGCTT ccagggcgtg cccttgggct ccccgggcgc ATGGTACC)(SEQ ID NO:4)を含有する。
【0100】
attPプラスミドpFY17、pFY19、pFY20の組込み ハイグロマイシンに対する耐性のためにコードし、pED113から得られた1.6kbのBamHI−KpnI断片として得られたhpt遺伝子を、pBluescript II SKのBamHIとKpnIとの間に挿入してコントロールプラスミドpBSK−hptを作った。
【0101】
長さの異なるattP部位を含有する合成オリゴヌクレオチドをpBSK−hpt中のSacI(GTCGAC)部位とBamHI(GGATCC)部位との間に挿入し、 次のプラスミドを作った。
a)90bpのattP部位(GAGCTC−gaagcggttt tcgggagtagtgccccaact ggggtaacct ttgagttctc tcagttgggg gcgtagggtc gccgacatga cacaaggggt−GGATCC)(SEQ ID NO:5)を含有するプラスミドpFY17。
b)50bpのattP部位(GAGCTC−tgccccaact ggggtaacct ttgagttctc tcagttgggg gcgtagggtc−GGATCC)(SEQ ID NO:6)を含有するプラスミドpFY19。
c)32bpのattP部位(GAGCTC−actggggtaa cctttgagtt ctctcagttg ggATCC)(SEQ ID NO:7) を含有するプラスミドpFY20。
【0102】
インテグラーゼ発現構成pFY6 インテグラーゼ遺伝子のほぼ完全なオープンリーディングフレームを含有するEcoRI−BamHI断片を、pcDNA3.1/Zeo(−)(Invitrogen)のEcoRI部位とBamHI部位との間に挿入した。次いで、Kozack配列およびインテグラーゼ遺伝子の配列をコードするN末端アミノ酸を含有する合成オリゴヌクレオチド(GGGCCCGCCACGATGACACAAGGGGTTGTGACCGGGGTGGACACGTACGCGGGTGCTTACGACCGTCAGTCGCGCGAGCGCGAGAATTC)(SEQ ID NO:8)をApaI部位とEcoRI部位との間に挿入して、オープンリーディングフレームを再構成した。この方向は、pcDNA3.1/Zeo(−)中でCMV(ヒト・サイトメガロウイルス)プロモーターの制御下で、完全なインテグラーゼコード化領域を配置する。
【0103】
トランスフェクション・プロトコール CHO細胞株K−1をattB標的コンストラクトpFY12、pFY14またはpFY15でトランスフェクションした(図3)。これらのプラスミドは、ネオマイシン耐性に対する選択マーカー、およびpC(ヒト・サイトメガロウイルス・プロモーター)とlacZコード化領域との間に位置する種々の長さのattB部位を有する。プラスミドpFY12、pFY14およびpFY15は、それぞれ90、50および30bpのattB配列を含有する。ネオマイシン耐性細胞株を単一コロニーの連続精製により得た。各コンストラクトの4つの株を組込み実験に用いた。
【0104】
12株のそれぞれに、ΦC31インテグラーゼ発現プラスミドpFY6を、組込みベクターpFY17、pFY19またはpFY20と共にトランスフェクションした。プラスミドpFY17、pFY19およびpFY20は、それぞれ長さ90、50および32bpのattP配列を有する。attP配列をhptオープンリーディングフレームの上流に置いた。このattP配列は、ハイグロマイシン耐性を与えるハイグロマイシン・ホスホトランスフェラーゼをコードする。attP−hpt部分の上流にプロモーターは無く、したがってhptコード化領域にゲノムプロモーターを融合するようにプラスミドをゲノムに組込みしない限り、hptは発現しない。pBSK−hptをコントロールに用いて、hptへのプロモーター融合の頻度を監視した。attP配列が無いという点を除いて、プラスミドpBSK−hptはpFY17、pFY19およびpFY20と同一である。attP部位とattB部位との間の組み換えは、組込みベクターを染色体標的に挿入し、Pc−attL−hpt結合を発生すると予想される。hptの発現はハイグロマイシン耐性を与える。
【0105】
結果トランスフェクションの結果:12細胞株にトランスフェクションされた各組込みプラスミドに対して、ハイグロマイシン耐性コロニーを記録した(表2)。1×106細胞を塗布したものから、pBSK−hptトランスフェクションは顕著に多数の耐性コロニーの産生に失敗した。これは、ゲノムプロモーターに与えるhtpコード化領域の頻度が著しく低いことを示している。対照的に、pFY17、pFY19およびpFY20は、特定の組込みプラスミドおよび特定の細胞株によっては、最大で千倍のハイグロマイシン耐性コロニーを生じた。pFY19またはpFY17をFY12中にトランスフェクションしたものから、より多くのハイグロマイシン耐性コロニーが生じた。これは、より長いattB配列とattP配列との間の組み換えは、より短いattB部位とattP部位との間の組換えよりもより効率的であることを示している。
【0106】
PCRを用いて、代表的コロニーから予想される約0.8kb接合バンドを検出した。ヒト・サイトメラロウイルス・プロモーターおよびhptコード化領域に対応するプライマーは、予想サイズ(0.8kb)のPCR産生物を増幅した。これは、Pcがhptコード化領域に結合していることを示しており、ゲノムのattB部位とプラスミドattP配列との間の組み換えと一致する。
【0107】
【表2】
【0108】
(実施例4)
ΦC31インテグラーゼは植物染色体中でattP部位およびattB部位を組み換える働きをする。
本実施例は、ΦC31インテグラーゼをテストして、植物染色体中に存在するattP部位およびattB部位を組み換える能力を調べた実験についての説明である。本実験の作成物および方法を図4に示す。
【0109】
方法
コンストラクトpWP29は、35S−attP−npt−attB−gusからなりRBおよびLBに隣接した断片を含有する。ここにおいて、35Sはカリフラワー・モザイクウイルス・プロモーターであり、nptはネオマイシン・ホスホトランスフェラーゼをコードする領域であり、gusはグルクロニダーゼをコードする領域である。RBおよびLBは、それぞれ右および左アグロバクテリウムT−DNA境界配列である。35Sとnptとの間のattP部位は非翻訳リーダー配列としての機能を果す。35Sによるnptの転写によりカナマイシン耐性が与えられる。上流にプロモーターが無いので、gusコード化領域は転写されない。
【0110】
植物形質転換に用いる第2のコンストラクトはpWP24である。このコンストラクトはRBおよびLBに隣接する断片Pnos−npt−35S−intを含有する。ここにおいて、Pnosはノパリンシンターゼ・プロモーターであり、intはΦC31インテグラーゼコード化領域である。nptとintの両者が、それぞれ上流のプロモーターから転写される。
【0111】
この2つのコンストラクトが同じゲノム中に存在する場合、pWP24を有する染色体からのintの発現により、機能性ΦC31インテグラーゼが発生し、pWP29を有する染色体上に位置するattB部位とattP部位との間の組み換えを触媒すると考えられた。組み換え現象により、pWP29構造体からnpt遺伝子を削除すること、およびgusに35Sを与えることが予想された。得られる構造は35S−attR−gusであり、ここでattRは、attPとattBとの間の組み換えにより形成されたハイブリッド部位であり、PB’とも表される(図4)。nptの削除により35Sの転写下でgusが得られ、かつGUS酵素活性を有する植物を生じると予想された。この活性は、植物組織を組織化学的に染色することにより検出することができる。
【0112】
結果
一時発現アッセイを行って、pWP29が組み換えに対して機能的であるか否かを調べた。裸のDNAの遺伝子銃で仲介した送達により、pWP29をトウモロコシBMS細胞中にpWP8と共導入した。コンストラクトpWP8は、単子葉植物細胞中で発現するためのトウモロコシ・ユビキチン・プロモーターの後方に融合したインテグラーゼ遺伝子を有する。両方のプラスミドを共導入した場合には青色点が観察されたが、プラスミドの一方のみを用いた場合には、これは見つからなかった。このことは、トウモロコシ細胞中で部位特異的組み換えが起きたこと、およびpWP29中のattP部位とattB部位とが機能性部位であることを示している。
【0113】
カナマイシン耐性タバコ植物体を、pWP29またはpWP24を用いてアグロバクテリウムで仲介した形質転換により再生した。もう一度、一時発現アッセイを行って、pWP24株が機能性インテグラーゼを産生するか否かを調べた。コンストラクトpWP29を、裸のDNAの遺伝子銃で仲介した送達により、pWP24植物体に導入した。pWP29DNAを有する細胞は、35S−attR−gus構成の形成の結果、GUS酵素活性を発現すると予想された。実際、2つの株24.3および24.4は、attP部位とattB部位との間の機能性インテグラーゼ仲介部位特異的組み換えと一致した青色点を生じた。
【0114】
これら2つのpWP24インテグラーゼ株をpWP29テスター株と交雑して、pWP29とpWP24とを同じゲノムに保有する染色体を有する子孫を作った。インテグラーゼ(24.3、24.4)とテスター株(29.2、29.4、29.5、29.19)との間の遺伝子交雑から得られた結果を表3にまとめた。いずれの場合も、代表的子孫苗を選択せずに発芽させ、GUS酵素活性に対して組織化学的に染色した。青色に染色された子孫の数を表に示した。一次形質転換したpWP24およびpWP29株は代表的トランス遺伝子に対してヘミ接合性であるので、子孫の4分の1だけが両方のトランス遺伝子型を保有することが期待された。サンプル数が少ないので、期待頻度からの顕著な偏差は異常ではない。
【0115】
【表3】
【0116】
親株として用いた株の組合せにより、染色の強度が変わった。組織の大部分が青色に染まった子孫を有するものは、組み換え現象がより効率的であったことを示している。逆に、染色が均一でない子孫を与えるものは、組み換え現象がより非効率であったことを示している。異なる子孫プールの間のこの偏差は、恐らくトランス遺伝子の組込み位置により生じる効果によるものである。2つのインテグラーゼ株のうち、24.4は促進部位特異的組み換えにおいてより効率的に現れる。これは、恐らくint遺伝子発現のレベルが高いことによる。24.4を29.4および29.19に交雑して作った染色パターンは、attBおよびattPのint仲介部位特異的組み換えによりgus遺伝子活性の活性化を招くという、本実験の目的に一致する。
【0117】
(実施例5)
ΦC31インテグラーゼは、植物染色体中への環状プラスミドの組み込みを触媒する。
本実施例は、ΦC31インテグラーゼをテストして、attP×attB部位特異的組み換えにより植物染色体中に環状プラスミドを挿入する能力を調べた実験についての説明である。本実験を図5に図示する。
【0118】
方法
標的コンストラクトpWP6は、RBおよびLBに隣接し35S−attP−nptからなる断片を含む。35Sとnptとの間のattP部位は非翻訳リーダー配列としての役割を果す。35Sによるnptの転写によりカナマイシンに対する耐性が得られる。
【0119】
組み込んだコンストラクトpYJC43は断片attB−hptを有する。ここで、hptはハイグロマイシンに対する耐性をコードしている。インテグラーゼ発現コンストラクトはpYJC41であり、35Sはintを転写する。
【0120】
標的コンストラクトpWP6を、pWP6DNAの無作為組込みにより植物染色体中に配置した。次いで、pWP6トランス遺伝子の単一コピーを含んだカナマイシン耐性植物をpYJC43およびpYJC41で形質転換した。pYJC41から得たintの一時発現が、pYJC43のattB部位とpWP6トランス遺伝子の染色体的に定まったattP部位との間の組み換えを触媒することが期待された。attB部位とattP部位との間の特異的組み換えにより、pYJC43環状分子が染色体に挿入され、35S−attL−hpt連鎖を作ることが期待された。attP部位およびattB部位は逆方向に図示されているので、attL部位も同様に逆方向にするか、または図中のBP’を逆方向にしたものと同じP’Bで表すことに留意されたい。機能性35S−attL−hpt連鎖はハイグロマイシン耐性表現型を与えた。
【0121】
結果
pWP6を有するカナマイシン耐性タバコ植生を、アグロバクテリウム仲介形質転換により得た。サザンハイブリダイゼーション分析により、pWP6トランス遺伝子の単一コピーを有する株を検出した。この株の子孫WP6.1を無菌的に発芽させ、これらの植物からプロトプラストを作った。このプロトプラストを、pYJC43とpYJC41DNAとを組合せ、直接DNA形質転換のためのポリエチレングリコール法により形質転換した。次いで、プロトプラストをアガロース中に埋め込み、培養してハイグロマイシン存在下でカルスを形成した。ハイグロマイシンを選択しない場合には、カルス形成速度は4×10−4であった。これは通常の10分の1であるが、プロトプラスト形質転換実験で観察される偏差の範囲内である。ハイグロマイシンを選択する場合には、カルス形成速度は7×10−5であった。これは、プロトプラストから再生されたカルスの約18%が、標的部位に組込みベクターを含有することを示している。インテグラーゼコンストラクトpYJC41を形質転換から除外した場合、カルス形成速度は1×10−5未満であった。プラスミドpYJC41を発現するインテグラーゼの封入により作られたハイグロマイシン耐性カルスの頻度がより高いことは、pYJC43の染色体attP標的へのインテグラーゼ促進部位特異的組込みと一致する。
【0122】
本明細書で説明した実施例および実施態様は例示目的のためだけのものであって、当業者にとってこれらを考慮した種々の変更または修正は示唆されており、かつこれらの変更または修正は本明細書の精神および範囲、ならびに特許請求の範囲に含まれることを理解されたい。本明細書中で引用した刊行物、特許および特許出願書類の全てを、全ての目的のためにここに参照により組み入れる。
【特許請求の範囲】
【請求項1】
原核リコンビナーゼポリペプチド又は原核リコンビナーゼをコード化する核酸を含有する真核細胞であり、リコンビナーゼは、第一の組み換え部位と前記第一の組み換え部位との組み換えの基質として作用可能な第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で第一の組み換え部位と第二の組み換え部位との間の組み換えで形成される2つのハイブリッドリコンビナーゼ組み換え部位間の組み換えを仲介することが不可能であることを特徴とする真核細胞。
【請求項2】
リコンビナーゼが、バクテリオファージΦC31インテグラーゼ、大腸菌ファージP4リコンビナーゼ、リステリアファージリコンビナーゼ、バクテリオファージR4 Sre リコンビナーゼ、CisA リコンビナーゼ、XisFリコンビナーゼ及びトランスポゾンTn4451 TnpXリコンビナーゼからなる群から選択されることを特徴とする請求項1記載の真核細胞。
【請求項3】
リコンビナーゼがバクテリオファージΦC31インテグラーゼであることを特徴とする請求項1記載の真核細胞。
【請求項4】
第一の組み換え部位がattB部位であり、第二の組み換え部位がattP部位であることを特徴とする請求項1記載の真核細胞。
【請求項5】
さらに、細胞が第一リコンビナーゼ組み換え部位を含むことを特徴とする請求項1記載の真核細胞。
【請求項6】
細胞が、真核細胞においてリコンビナーゼコード化ポリヌクレオチドの発現を仲介するプロモーターに作用可能に結合した、リコンビナーゼポリペプチドに対するコード化配列を含む核酸を含有することを特徴とする請求項1記載の真核細胞。
【請求項7】
さらに、核酸が選択マーカーを含むことを特徴とする請求項6記載の真核細胞。
【請求項8】
プロモーターが誘導又は抑制プロモーターであることを特徴とする請求項6項記載の真核細胞。
【請求項9】
核酸がプラスミドpLT43であることを特徴とする請求項8項記載の真核細胞。
【請求項10】
真核細胞が、動物細胞、植物細胞、酵母細胞、昆虫細胞、及びカビ細胞からなる群から選択されることを特徴とする請求項1記載の真核細胞。
【請求項11】
真核細胞が、哺乳類の細胞であること特徴とする請求項10記載の真核細胞。
【請求項12】
真核細胞が、多細胞生物に存在することを特徴とする請求項10記載の真核細胞。
【請求項13】
真核細胞において部位特異的組み換えを得るための方法であり、
第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として作用可能な第二の組み換え部位とを含む真核細胞を供給し、
第一及び第二の組み換え部位を原核リコンビナーゼポリペプチドと接触させて、組み換え部位間で組み換えを生じ、それによって一又は二つのハイブリッド組み換え部位を形成することからなる方法であって、
リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で2つのハイブリッド組み換え部位間の組み換えを仲介することが不可能であることを特徴とする方法。
【請求項14】
真核細胞が、酵母細胞、カビ細胞、植物細胞、昆虫細胞、及び動物細胞からなる群から選択されることを特徴賭する請求項13記載の方法。
【請求項15】
第一の組み換え部位が、真核細胞の染色体に存在することを特徴とする請求項13記載の方法。
【請求項16】
第二の組み換え部位が真核細胞の第二の染色体に存在し、第一及び第二の組み換え部位をリコンビナーゼと接触させて染色体アームのトランスロケーションを生じることを特徴とする請求項15記載の方法。
【請求項17】
第一の組み換え部位と第二の組み換え部位が単一核酸分子に存在することを特徴とする請求項13記載の方法。
【請求項18】
第一の組み換え部位及び第二の組み換え部位が直線方向(direct orientation)であることを特徴とする請求項17記載の方法。
【請求項19】
組み換えが、第一と第二の組み換え部位との間にある核酸分子の部分の切り取りを生じることを特徴とする請求項18記載の方法。
【請求項20】
第一の組み換え部位と第二の組み換え部位が逆方向(inverted orientation)であることを特徴とする請求項17記載の方法。
【請求項21】
組み換えが、第一と第二の組み換え部位との間にある核酸分子の部分の逆位を生じることを特徴とする請求項20記載の方法。
【請求項22】
真核細胞が、リコンビナーゼポリペプチドをコード化するポリヌクレオチドを含むことを特徴とする請求項13記載の方法。
【請求項23】
リコンビナーゼコード化ポリヌクレオチドは、真核細胞におけるポリヌクレオチドの発現を仲介するプロモーターに作用可能に結合することを特徴とする請求項22記載の方法。
【請求項24】
プロモーターが、誘導又は抑制プロモーターであることを特徴とする請求項23項記載の方法。
【請求項25】
プロモーターが、Pmntプロモーターであることを特徴とする請求項24記載の方法。
【請求項26】
安定的に組み込んだトランス遺伝子を有する真核細胞を得るための方法であり、
トランス遺伝子と、第一の組み換え部位との組み換えに対して基質として作用することが可能な第二の組み換え部位とを含む核酸を第一の組み換え部位を含む真核細胞の中に導入し、
第一及び第二の組み換え部位を原核リコンビナーゼポリペプチドに接触させて、リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の組み換えを触媒し、第一の組み換え部位で核酸を組み込みませて、それによって、核酸の各末端でハイブリッド組み換え部位を形成することからなる方法であって、
リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で2つのハイブリット組み換え部位間の組み換えを仲介することが不可能であることを特徴とする方法。
【請求項27】
リコンビナーゼポリペプチドが、バクテリオファージΦC31インテグラーゼ、大腸菌ファージP4リコンビナーゼ、リステリアファージリコンビナーゼ、バクテリオファージR4 Sre リコンビナーゼ、CisA リコンビナーゼ、XisFリコンビナーゼ及びトランスポゾンTn4451 TnpXリコンビナーゼからなる群から選択されることを特徴とする請求項26記載の方法。
【請求項28】
リコンビナーゼが、バクテリオファージΦC31インテグラーゼであることを特徴とする請求項27記載の方法。
【請求項29】
リコンビナーゼポリペプチドを真核細胞の中へリコンビナーゼポリペプチドをコード化するポリヌクレオチドの発現によって導入することを特徴とする請求項26記載の方法。
【請求項30】
リコンビナーゼポリペプチドをコード化するポリヌクレオチドが、真核細胞において機能するプロモーターに作用可能に結合することを特徴とする請求項29記載の方法。
【請求項31】
プロモーターが、誘導又は抑制プロモーターであることを特徴とする請求項30記載の方法。
【請求項32】
真核細胞において機能するプロモーターに作用可能に結合した細菌性リコンビナーゼポリペプチドをコード化するポリヌクレオチド配列を含む核酸であり、リコンビナーゼポリペプチドが付加的因子の不存在下において第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位間の組み換えを仲介できないことを特徴とする核酸。
【請求項33】
さらに、核酸がリコンビナーゼポリペプチドによって認識される少なくとも1つの組み換え部位を含むことを特徴とする請求項32記載の核酸。
【請求項34】
核酸がプラスミドベクターを含むことを特徴とする請求項32記載の核酸
【請求項35】
ベクターがpLT43であることを特徴とする請求項34記載の核酸。
【請求項36】
第一のバクテリオファージΦC31インテグラーゼ組み換え部位を含むポリヌクレオチドを含有することを特徴とする真核細胞。
【請求項37】
組み換え部位がattP及びattBからなる群から選択されることを特徴とする請求項36記載の真核細胞。
【請求項38】
さらに、真核細胞が、ΦC31インテグラーゼポリペプチドと接触したとき、第一のΦC31組み換え部位との組み換えを経た第二のΦC31組み換え部位を含む第二のポリヌクレオチドを含有することを特徴とする請求項36記載の真核細胞。
【請求項39】
第一の組み換え部位がattBであり、第二の組み換え部位がattPであるか、第一の組み換え部位がattPであり、第二の組み換え部位がattBであることを特徴とする請求項38記載の真核細胞。
【請求項40】
さらに、第二のポリヌクレオチドがトランス遺伝子を含むことを特徴とする請求項38記載の真核細胞。
【請求項41】
さらに、第二のポリヌクレオチドが選択マーカーを含むことを特徴とする請求項38記載の真核細胞。
【請求項42】
さらに、真核細胞がΦC31インテグラーゼポリペプチドを含むことを特徴とする請求項36記載の真核細胞。
【請求項43】
さらに、真核細胞がΦC31インテグラーゼポリペプチドをコード化するポリヌクレオチドを含む核酸を含有することを特徴とする請求項36記載の真核細胞。
【請求項44】
さらに、核酸が選択マーカーを含むことを特徴とする請求項43記載の真核細胞。
【請求項45】
さらに、核酸が、細胞においてΦC31インテグラーゼコード化ポリペプチドの発現を生じるプロモーターを含有することを特徴とする請求項43記載の真核細胞。
【請求項46】
プロモーターが誘導プロモーターであることを特徴とする請求項45記載の真核細胞。
【請求項47】
真核細胞が、酵母細胞、カビ細胞、植物細胞、及び動物細胞からなる群から選択されることを特徴とする請求項36記載の真核細胞。
【請求項1】
原核リコンビナーゼポリペプチド又は原核リコンビナーゼをコード化する核酸を含有する真核細胞であり、リコンビナーゼは、第一の組み換え部位と前記第一の組み換え部位との組み換えの基質として作用可能な第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で第一の組み換え部位と第二の組み換え部位との間の組み換えで形成される2つのハイブリッドリコンビナーゼ組み換え部位間の組み換えを仲介することが不可能であることを特徴とする真核細胞。
【請求項2】
リコンビナーゼが、バクテリオファージΦC31インテグラーゼ、大腸菌ファージP4リコンビナーゼ、リステリアファージリコンビナーゼ、バクテリオファージR4 Sre リコンビナーゼ、CisA リコンビナーゼ、XisFリコンビナーゼ及びトランスポゾンTn4451 TnpXリコンビナーゼからなる群から選択されることを特徴とする請求項1記載の真核細胞。
【請求項3】
リコンビナーゼがバクテリオファージΦC31インテグラーゼであることを特徴とする請求項1記載の真核細胞。
【請求項4】
第一の組み換え部位がattB部位であり、第二の組み換え部位がattP部位であることを特徴とする請求項1記載の真核細胞。
【請求項5】
さらに、細胞が第一リコンビナーゼ組み換え部位を含むことを特徴とする請求項1記載の真核細胞。
【請求項6】
細胞が、真核細胞においてリコンビナーゼコード化ポリヌクレオチドの発現を仲介するプロモーターに作用可能に結合した、リコンビナーゼポリペプチドに対するコード化配列を含む核酸を含有することを特徴とする請求項1記載の真核細胞。
【請求項7】
さらに、核酸が選択マーカーを含むことを特徴とする請求項6記載の真核細胞。
【請求項8】
プロモーターが誘導又は抑制プロモーターであることを特徴とする請求項6項記載の真核細胞。
【請求項9】
核酸がプラスミドpLT43であることを特徴とする請求項8項記載の真核細胞。
【請求項10】
真核細胞が、動物細胞、植物細胞、酵母細胞、昆虫細胞、及びカビ細胞からなる群から選択されることを特徴とする請求項1記載の真核細胞。
【請求項11】
真核細胞が、哺乳類の細胞であること特徴とする請求項10記載の真核細胞。
【請求項12】
真核細胞が、多細胞生物に存在することを特徴とする請求項10記載の真核細胞。
【請求項13】
真核細胞において部位特異的組み換えを得るための方法であり、
第一の組み換え部位と、前記第一の組み換え部位との組み換えの基質として作用可能な第二の組み換え部位とを含む真核細胞を供給し、
第一及び第二の組み換え部位を原核リコンビナーゼポリペプチドと接触させて、組み換え部位間で組み換えを生じ、それによって一又は二つのハイブリッド組み換え部位を形成することからなる方法であって、
リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で2つのハイブリッド組み換え部位間の組み換えを仲介することが不可能であることを特徴とする方法。
【請求項14】
真核細胞が、酵母細胞、カビ細胞、植物細胞、昆虫細胞、及び動物細胞からなる群から選択されることを特徴賭する請求項13記載の方法。
【請求項15】
第一の組み換え部位が、真核細胞の染色体に存在することを特徴とする請求項13記載の方法。
【請求項16】
第二の組み換え部位が真核細胞の第二の染色体に存在し、第一及び第二の組み換え部位をリコンビナーゼと接触させて染色体アームのトランスロケーションを生じることを特徴とする請求項15記載の方法。
【請求項17】
第一の組み換え部位と第二の組み換え部位が単一核酸分子に存在することを特徴とする請求項13記載の方法。
【請求項18】
第一の組み換え部位及び第二の組み換え部位が直線方向(direct orientation)であることを特徴とする請求項17記載の方法。
【請求項19】
組み換えが、第一と第二の組み換え部位との間にある核酸分子の部分の切り取りを生じることを特徴とする請求項18記載の方法。
【請求項20】
第一の組み換え部位と第二の組み換え部位が逆方向(inverted orientation)であることを特徴とする請求項17記載の方法。
【請求項21】
組み換えが、第一と第二の組み換え部位との間にある核酸分子の部分の逆位を生じることを特徴とする請求項20記載の方法。
【請求項22】
真核細胞が、リコンビナーゼポリペプチドをコード化するポリヌクレオチドを含むことを特徴とする請求項13記載の方法。
【請求項23】
リコンビナーゼコード化ポリヌクレオチドは、真核細胞におけるポリヌクレオチドの発現を仲介するプロモーターに作用可能に結合することを特徴とする請求項22記載の方法。
【請求項24】
プロモーターが、誘導又は抑制プロモーターであることを特徴とする請求項23項記載の方法。
【請求項25】
プロモーターが、Pmntプロモーターであることを特徴とする請求項24記載の方法。
【請求項26】
安定的に組み込んだトランス遺伝子を有する真核細胞を得るための方法であり、
トランス遺伝子と、第一の組み換え部位との組み換えに対して基質として作用することが可能な第二の組み換え部位とを含む核酸を第一の組み換え部位を含む真核細胞の中に導入し、
第一及び第二の組み換え部位を原核リコンビナーゼポリペプチドに接触させて、リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の組み換えを触媒し、第一の組み換え部位で核酸を組み込みませて、それによって、核酸の各末端でハイブリッド組み換え部位を形成することからなる方法であって、
リコンビナーゼポリペプチドが、第一と第二の組み換え部位との間の部位特異的組み換えを仲介することが可能であるが、真核細胞に存在しない付加的因子の不存在下で2つのハイブリット組み換え部位間の組み換えを仲介することが不可能であることを特徴とする方法。
【請求項27】
リコンビナーゼポリペプチドが、バクテリオファージΦC31インテグラーゼ、大腸菌ファージP4リコンビナーゼ、リステリアファージリコンビナーゼ、バクテリオファージR4 Sre リコンビナーゼ、CisA リコンビナーゼ、XisFリコンビナーゼ及びトランスポゾンTn4451 TnpXリコンビナーゼからなる群から選択されることを特徴とする請求項26記載の方法。
【請求項28】
リコンビナーゼが、バクテリオファージΦC31インテグラーゼであることを特徴とする請求項27記載の方法。
【請求項29】
リコンビナーゼポリペプチドを真核細胞の中へリコンビナーゼポリペプチドをコード化するポリヌクレオチドの発現によって導入することを特徴とする請求項26記載の方法。
【請求項30】
リコンビナーゼポリペプチドをコード化するポリヌクレオチドが、真核細胞において機能するプロモーターに作用可能に結合することを特徴とする請求項29記載の方法。
【請求項31】
プロモーターが、誘導又は抑制プロモーターであることを特徴とする請求項30記載の方法。
【請求項32】
真核細胞において機能するプロモーターに作用可能に結合した細菌性リコンビナーゼポリペプチドをコード化するポリヌクレオチド配列を含む核酸であり、リコンビナーゼポリペプチドが付加的因子の不存在下において第一の組み換え部位と第二の組み換え部位との間の組み換えで形成された2つのハイブリッド組み換え部位間の組み換えを仲介できないことを特徴とする核酸。
【請求項33】
さらに、核酸がリコンビナーゼポリペプチドによって認識される少なくとも1つの組み換え部位を含むことを特徴とする請求項32記載の核酸。
【請求項34】
核酸がプラスミドベクターを含むことを特徴とする請求項32記載の核酸
【請求項35】
ベクターがpLT43であることを特徴とする請求項34記載の核酸。
【請求項36】
第一のバクテリオファージΦC31インテグラーゼ組み換え部位を含むポリヌクレオチドを含有することを特徴とする真核細胞。
【請求項37】
組み換え部位がattP及びattBからなる群から選択されることを特徴とする請求項36記載の真核細胞。
【請求項38】
さらに、真核細胞が、ΦC31インテグラーゼポリペプチドと接触したとき、第一のΦC31組み換え部位との組み換えを経た第二のΦC31組み換え部位を含む第二のポリヌクレオチドを含有することを特徴とする請求項36記載の真核細胞。
【請求項39】
第一の組み換え部位がattBであり、第二の組み換え部位がattPであるか、第一の組み換え部位がattPであり、第二の組み換え部位がattBであることを特徴とする請求項38記載の真核細胞。
【請求項40】
さらに、第二のポリヌクレオチドがトランス遺伝子を含むことを特徴とする請求項38記載の真核細胞。
【請求項41】
さらに、第二のポリヌクレオチドが選択マーカーを含むことを特徴とする請求項38記載の真核細胞。
【請求項42】
さらに、真核細胞がΦC31インテグラーゼポリペプチドを含むことを特徴とする請求項36記載の真核細胞。
【請求項43】
さらに、真核細胞がΦC31インテグラーゼポリペプチドをコード化するポリヌクレオチドを含む核酸を含有することを特徴とする請求項36記載の真核細胞。
【請求項44】
さらに、核酸が選択マーカーを含むことを特徴とする請求項43記載の真核細胞。
【請求項45】
さらに、核酸が、細胞においてΦC31インテグラーゼコード化ポリペプチドの発現を生じるプロモーターを含有することを特徴とする請求項43記載の真核細胞。
【請求項46】
プロモーターが誘導プロモーターであることを特徴とする請求項45記載の真核細胞。
【請求項47】
真核細胞が、酵母細胞、カビ細胞、植物細胞、及び動物細胞からなる群から選択されることを特徴とする請求項36記載の真核細胞。
【図1】

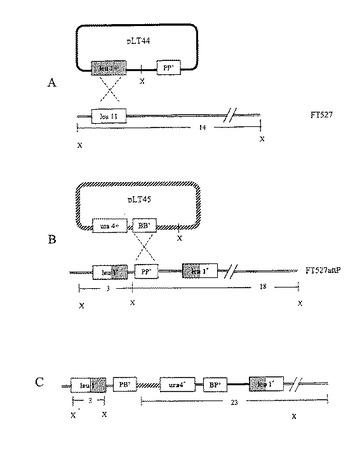
【図2】
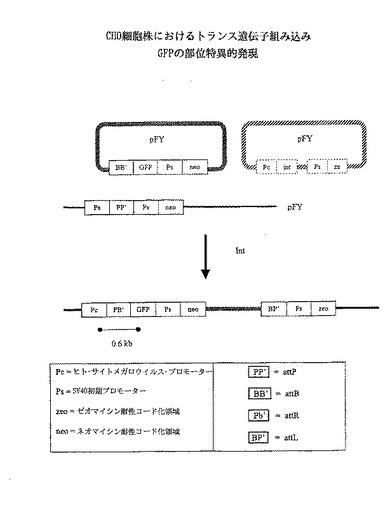
【図3】
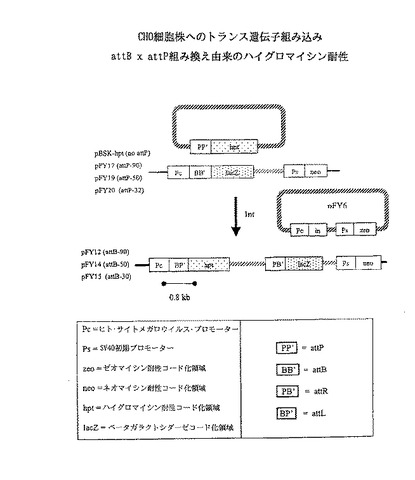
【図4】
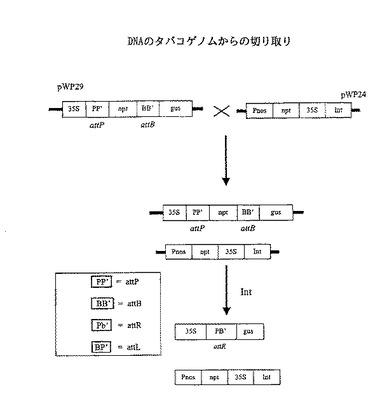
【図5】
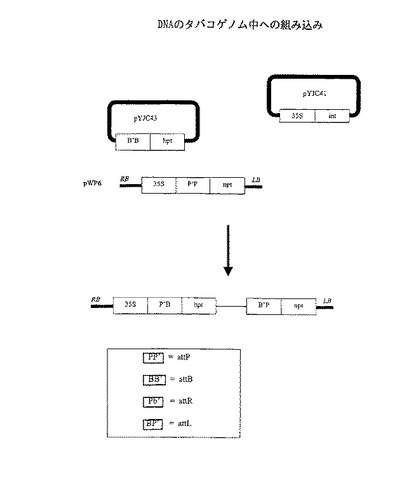
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−45381(P2011−45381A)
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−241383(P2010−241383)
【出願日】平成22年10月27日(2010.10.27)
【分割の表示】特願2001−512843(P2001−512843)の分割
【原出願日】平成12年7月21日(2000.7.21)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【出願人】(500031032)
【Fターム(参考)】
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願番号】特願2010−241383(P2010−241383)
【出願日】平成22年10月27日(2010.10.27)
【分割の表示】特願2001−512843(P2001−512843)の分割
【原出願日】平成12年7月21日(2000.7.21)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【出願人】(500031032)
【Fターム(参考)】
[ Back to top ]