
バチルスにおけるタンパク質発現の増大
【課題】発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞の提供。
【解決手段】目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失され、プロテアーゼであるズブチリシンの発現量が増大されたバチルス株。
【解決手段】目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失され、プロテアーゼであるズブチリシンの発現量が増大されたバチルス株。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。
【背景技術】
【0002】
発明の背景
遺伝子工学は工業用生物反応器、細胞工場として、及び食物発酵において用いる微生物の向上を可能としてきた。特に、バチルス種は多数の有用なタンパク質及び代謝産物を生成及び分泌する(Zukowski、“Production of commercially valuable products(商業的に有用な産物の生成)”、In:Doi and McGlouglin(編集)Biology of Bachilli:Applications to Industry,Butterworth−Heinemann,Stoneham.Mass第311−337頁[1992])。工業的に用いられる最も一般的なバチルス種はB.リケニフォルミス、B.アミロリケファシエンス及びB.ズブチリスである。これらはGRAS(通常、安全であると認識される)状態であるので、これらバチルス種株は食品及び医薬業界において利用されるタンパク質生成のもっともな候補である。重要な生成酵素はα−アミラーゼ、中性プロテアーゼ、及びアルカリ(またはセリン)プロテアーゼを含む。しかしながら、バチルス宿主細胞中におけるタンパク質生成の理解の進展にも関わらず、これらタンパク質の発現を増加させるための方法に関する必要性が以前としてある。
【発明の概要】
【0003】
発明の概要
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。いくつかの好ましい実施態様において、本発明は、バチルスにおける目的タンパク質の発現及び/または分泌を向上させる方法及び組成物を提供する。
【0004】
特に好ましい実施態様において、本発明はバチルスにおける目的タンパク質の発現及び/または分泌を向上させる手段を提供する。より具体的には、これらの実施態様において、本発明はバチルス宿主株における1以上の染色体遺伝子の不活性化に関し、ここで不活化遺伝子は株の生存に必須のものではない。1以上の染色体遺伝子を不活化した結果、変化したバチルス株が生成され、対応する変化していないバチルス宿主株よりも高いレベルの目的タンパク質を発現することができる。
【0005】
さらに、別の実施態様において、本発明はバチルス宿主株における大きな領域の染色体DNAを除去する手段を提供し、ここで欠失された固有染色体領域は株生成に必須のものではない。1以上の固有染色体領域を除去した結果、変化したバチルス株が生成され、対応する変化していないバチルス宿主株よりも高いレベルの目的タンパク質を発現することができる。いくつかの好ましい実施態様において、バチルス宿主株は目的タンパク質をエンコードするポリヌクレオチドを含む組換え宿主株である。特に好ましい実施態様において、変異バチルス株はB.ズブチリス株である。下記に詳細に説明するように、欠失固有染色体領域は、限定されないが、プロファージ領域、抗菌(例えば、抗生物質)領域、調節領域、複数隣接単一遺伝子(multi−contiguous single gene)領域及びオペロン領域を含む。
【0006】
いくつかの実施態様において、本発明はバチルス細胞由来の目的タンパク質発現を増大させる方法及び組成物を提供する。いくつかの好ましい実施態様において、本方法はバチルス宿主株におけるsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子を不活性させて変異バチルス株を生成する工程;変異バチルス株を適当な成長条件下で成長させる工程;及び目的タンパク質を変異バチルス中で発現させる工程を含み、ここでタンパク質発現は対応する非変異バチルス宿主株と比較して増大している。いくつかの実施態様において、目的タンパク質は相同タンパク質であり、一方、他の実施態様において、目的タンパク質は異種タンパク質である。いくつかの実施態様において、1以上の目的タンパク質が生成される。いくつかの好ましい実施態様において、バチルス種はB.ズブチリス株である。さらなる実施態様において、染色体遺伝子の不活性化は遺伝子の欠失を含み、変異バチルス株を生成する。さらなる実施態様において、染色体遺伝子の不活性化は挿入不活性化を含む。いくつかの好ましい実施態様において。目的タンパク質は酵素である。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0007】
さらに別の実施態様において、本発明は変異バチルス株を提供し、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDの群より選択される1以上染色体遺伝子の欠失を含む。いくつかの実施態様において、変異株はプロテアーゼ生成バチルス株である。別の実施態様において、変異バチルス株はズブチリシン生成株である。さらに別の実施態様において、変異バチルス株はさらにdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子の変異を含む。
【0008】
さらなる実施態様において、本発明は入来配列(incoming sequence)を含むDNA構築体を提供する。いくつかの実施態様において、入来配列は、選択マーカー及びsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子または遺伝子断片を含む。別の実施態様において、選択マーカーは遺伝子の2つの断片間に位置する。他の実施態様において、入来配列は選択マーカー及び相同ボックス(homology box)を含み、ここで相同ボックスはマーカーの5’及び/または3’末端の側面に位置する。別の実施態様において、宿主細胞DNA構築体を用いて形質転換される。さらなる実施態様において宿主細胞は大腸菌またはバチルス細胞である。いくつかの好ましい実施態様において、DNA構築体は宿主細胞内の染色体上に統合される。
【0009】
本発明は、目的タンパク質を発現する変異バチルス株を得る方法も提供し、本発明のDNA構築体を含むバチルス宿主細胞を形質転換する工程を含み、ここでDNA構築体はバチルス宿主細胞の染色体中に統合され;変異バチルス株を生成する工程であって、1以上の染色体遺伝子が不活性される工程;及び目的タンパク質発現のための適当な成長条件下で変異バチルス株を成長させる工程を含む。いくつかの実施態様において、目的タンパク質は、プロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。さらに別の実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ、B.メガテリウム及び好ましくはB.ズブチリスからなる群より選択される。いくつかの実施態様において、バチルス宿主株は組換え宿主である。さらに別の実施態様において、目的タンパク質を回収する。さらなる実施態様において、選択マーカーは変異バチルスから切除される。
【0010】
本発明はさらに目的タンパク質を発現する変異バチルス株を得る方法を提供する。いくつかの実施態様において、本方法は入来配列を含むDNA構築体を含むバチルス宿主細胞を形質転換する工程を含み、ここで入来配列は選択マーカー及びsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子を含み、ここでDNA構築体はバチルス宿主細胞の染色体内に統合され、1以上の遺伝子欠失を生じ;変異バチルス株を得る工程及び変異バチルス株を目的タンパク質の発現に適した成長条件下で成長させる工程を含む。
【0011】
いくつかの実施態様において、本発明は入来配列を含むDNA構築体を提供し、ここで入来配列は選択マーカー、及びcssS遺伝子、cssS遺伝子断片またはそれに相同な配列を含む。いくつかの実施態様において、選択マーカーは当該遺伝子の2つの断片間に位置する。別の実施態様において、入来配列は選択マーカー及び相同ボックスを含み、ここで相同ボックスはマーカーの5’及び/または3’末端の側面に位置する。さらに別の実施態様において、宿主細胞はDNA構築体を用いて形質転換される。別の実施態様において、宿主細胞は大腸菌またはバチルス細胞である。さらに別の実施態様において、DNA構築体は宿主細胞の染色体上に統合される。
【0012】
また、本発明はプロテアーゼ生成が増大したバチルス・ズブチリス株を得る方法を提供する。いくつかの実施態様において、本方法は、本発明に従うDNA構築体を用いてバチルス・ズブチリス宿主細胞を形質転換する工程;DNA構築体とバチルス染色体の相同領域を相同組換えさせる工程であって、少なくとも1の以下の遺伝子がバチルス染色体から欠失される工程、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD;変異バチルス・ズブチリス株を得る工程;及びプロテアーゼ発現に適した条件下で変異バチルス株を成長させる工程を含む。いくつかの実施態様において、プロテアーゼ生成バチルスはズブチリシン生成株である。別の実施態様において、プロテアーゼは異種プロテアーゼである。別の実施態様において、プロテアーゼ生成株は、degU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子の変異をさらに含む。いくつかの実施態様において、本発明は遺伝子の挿入不活性化を含む。
【0013】
本発明はさらにsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子の欠失を含む変異バチルス・ズブチリス株を提供し、ここで変異バチルス・ズブチリス株は目的タンパク質を発現することが可能である。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの別の実施態様において、目的タンパク質は異種タンパク質である。
【0014】
いくつかの実施態様において、本発明は1以上の固有染色体領域またはそれらの断片の欠失を含む変異バチルス株を提供し、固有染色体領域は約0.5〜500キロ塩基(kb)を含み、及びここで変異バチルス株は本質的に同じ成長条件下で成長させた場合、対応する非変異バチルス株と比較して増大したレベルの目的タンパク質発現を有する。
【0015】
さらに別の実施態様において、本発明は、PBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、yvfF−yveK領域、DHB領域及びそれらの断片からなる群より選択される固有染色体領域の少なくとも1の欠失を含むプロテアーゼ生成バチルス株を提供する。
【0016】
さらなる実施態様において、本発明はバチルスにおいて目的タンパク質の発現を増大させる方法を提供し、以下を含む:選択マーカーを含むDNA構築体を導入し、及びバチルス宿主株内の染色体断片を不活性化させることにより生成した変異バチルス株を得る工程あって、ここでDNA構築体はバチルス染色体内に統合され、バチルス宿主細胞由来の固有染色体領域またはそれらの断片の欠失を生じる工程;及び成長に適した条件下で変異バチルスを成長させる工程であって、ここで目的タンパク質の発現は変異バチルス株において対応する非変異バチルス宿主細胞の目的タンパク質の発現と比較して大きい。
【0017】
また、本発明はバチルス株から目的タンパク質を得る方法を提供し、以下の工程を含む:選択マーカーを含むDNA構築体を用いてバチルス宿主細胞を形質転換し、染色体断片を不活性化する工程であって、ここでDNA構築体はバチルス株の染色体内に統合され、固有染色体領域またはそれらの断片の欠失を生じて変異バチルス株を形成する工程;目的タンパク質が発現できるように適した成長条件下で変異バチルス株を培養する工程;及び目的タンパク質を回収する工程。
【0018】
また、本発明は、有益な変異をスクリーン及び/または同定するためにDNAマイクロアレイデータを使用する手段を提供する。いくつかの好ましい実施態様において、これらの変異はtrpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子に関する。いくつかの好ましい実施態様において、これらの有益な変異は所定のアミノ酸生合成経路及び生分解経路の同時発現に関するトランスクリプトーム証拠及び/または分解経路の欠失がより良い性能の株を生じるという証拠及び/または生合成経路の過剰発現がより良い性能の株を生じるという証拠に基づく。別の実施態様において、本発明は有益な変異を提供するためにDNAマイクロアレイデータを使用する手段を提供する。いくつかの好ましい実施態様において、これらの変異はtrpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子に関し、アミノ酸生合成経路を含む遺伝子由来のmRNAの発現がバランスが取れていない場合、全体的な経路の過剰発現は親(すなわち野生型及び/または最初の)株よりも優れた性能の株を提供する。さらに、本発明は糖新生遺伝子の不活性化により株生成を向上させる手段を提供する。これらの好ましい実施態様のいくつかにおいて、不活化糖新生遺伝子はpckA、gapB及びfbpからなる群より選択される。
【0019】
本発明はバチルスから目的タンパク質の発現を増大させる方法を提供し、以下の工程を含む:目的タンパク質を生成できる変異バチルス株を得る工程であって、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも2の不活化染色体遺伝子を有する工程、及び変異バチルス株を目的タンパク質が変異バチルス株により発現される条件下で成長させる工程であって、目的タンパク質の発現が非変異バチルス宿主株における目的タンパク質の発現と比較して増大している工程。いくつかの実施態様において、目的タンパク質は、相同タンパク質及び異種タンパク質からなる群より選択される。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。いくつかの特に好ましい実施態様において、目的タンパク質はプロテアーゼである。さらに別の実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子を欠失することにより得られる。
【0020】
また、本発明はここに記載する方法を用いて得られる変異バチルス株も提供する。好ましい実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の遺伝子の染色体欠失を含む。いくつかの実施態様において、これらの染色体遺伝子の1以上が欠失される。特に好ましい実施態様において、変異株はB.ズブチリス株である。別の好ましい実施態様において、変異バチルス株はプロテアーゼ生成株である。いくつかの特に好ましい実施態様において、プロテアーゼはズブチリシンである。さらに別の実施態様において、ズブチリシンはズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309及びこれらの変異体からなる群より選択される。さらに別の実施態様において、変異バチルス株はdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される少なくとも1の遺伝子において変異をさらに含む。特に好ましい実施態様において、変異バチルス株はさらに目的の異種タンパク質を含む。
【0021】
また、本発明はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD、それらの遺伝子断片及びそれらの相同配列からなる群より選択される少なくとも1の遺伝子を含むDNA構築体を提供する。いくつかの好ましい実施態様において、DNA構築体は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号29、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33からなる群より選択される少なくとも1の核酸配列を含む。いくつかの実施態様において、DNA構築体はさらに少なくとも1の目的タンパク質をエンコードする少なくとも1のポリヌクレオチド配列を含む。
【0022】
本発明はDNA構築体を含むプラスミドも提供する。さらなる実施態様において、本発明はDNA構築体を含むプラスミドを含んだ宿主細胞を提供する。いくつかの実施態様において、宿主細胞はバチルス細胞及び大腸菌細胞からなる群より選択される。いくつかの好ましい実施態様において、宿主細胞はB.ズブチリスである。特に好ましい実施態様において、DNA構築体は宿主細胞の染色体内に統合される。別の実施態様において、DNA構築体は、配列番号2、配列番号3、配列番号4、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号30、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される少なくとも1のアミノ酸配列をエンコードする少なくとも1の遺伝子を含む。別の実施態様において、DNA構築体はさらに少なくとも1の選択マーカーを含み、ここで選択マーカーは当該遺伝子配列またはそれに相同な遺伝子配列の断片の両側に位置する。
【0023】
本発明は入来配列を含むDNA構築体も提供し、ここで入来配列は目的タンパク質をエンコードする核酸及び相同ボックスを含む両側に位置する選択マーカーを含み、相同ボックスはsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の遺伝子のコード領域のすぐ隣に位置する配列に対して80〜100%配列同一性を有する核酸配列を含む。いくつかの実施態様において、DNA構築体は遺伝子のコード配列の側面に位置する少なくとも1の核酸をさらに含む。本発明はDNA構築体を含むプラスミドも提供する。さらなる実施態様において、本発明はDNA構築体を含むプラスミドを含む宿主細胞を提供する。いくつかの実施態様において、宿主細胞はバチルス細胞及び大腸菌細胞からなる群より選択される。いくつかの好ましい実施態様において、宿主細胞はB.スブチリスである。いくつかの特に好ましい実施態様において、DNA構築体は宿主細胞の染色体内に統合される。別の好ましい実施態様において、選択マーカーは宿主細胞染色体から切除される。
【0024】
本発明はプロテアーゼ生成が増大した変異バチルス株を得るための方法をさらに提供し、以下を含む:少なくとも1の本発明のDNA構築体を含むバチルス宿主細胞を形質転換する工程であって、DNA構築体中の目的タンパク質がプロテアーゼであり、DNA構築体が少なくとも1の遺伝子が変異バチルス株を生成するために不活性化される条件下でバチルス宿主細胞染色体内に統合され;及び増大したプロテアーゼ生成が得られる条件下で変異バチルス株を成長させる工程。いくつかの特に好ましい実施態様において、本方法はプロテアーゼを回収する工程をさらに含む。別の好ましい実施態様において、少なくとも1の不活化遺伝子は変異バチルス株の染色体から欠失される。本発明はここに記載される方法を用いて生成した変異バチルス株も提供する。いくつかの実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、バチルス宿主細胞はB.ズブチリスである。
【0025】
本発明は変異バチルスにおいてプロテアーゼ発現を増大させる方法も提供し、以下を含む:本発明のDNA構築体を用いてバチルス宿主細胞を形質転換させる工程;DNA構築体とバチルス宿主細胞の染色体領域を相同組換えさせる工程であって、バチルス宿主細胞の少なくとも1の染色体遺伝子が不活性化され、変異バチルス株を生成する工程;及びプロテアーゼ発現に適した条件下で変異バチルス株を成長させる工程であって、ここでプロテアーゼ生成は形質転換前のバチルス・ズブチリス宿主と比較して変異バチルス・ズブチリス株が大きい。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。別の実施態様において、プロテアーゼは組換えプロテアーゼである。さらに別の実施態様において、不活性化は少なくとも1の遺伝子の欠失により達成される。さらに別の実施態様において、不活性化は少なくとも1の遺伝子の挿入不活性化による。本発明はここに記載の方法を用いて得た変異バチルス株も提供する。いくつかの実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の不活性化遺伝子を含む。いくつかの好ましい実施態様において、不活性化遺伝子は欠失により不活性化される。別の実施態様において、変異バチルス株はdegU、degS、degQ、scoC4、spollE及びoppAからなる群より選択される遺伝子に少なくとも1の変異を含む。いくつかの好ましい実施態様において、変異はdegU(Hy)32である。さらに別の実施態様において、株は組換えプロテアーゼ生成株である。いくつかの好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。
【0026】
本発明は1以上の固有染色体領域またはその断片を含む変異バチルス株も提供し、ここで固有染色体領域は約0.5〜500kbを含み、変異バチルス株は変異及び非変異バチルス株を本質的に同じ成長条件下で成長させた場合、対応する非変異バチルス株と比較して目的タンパク質の発現が増大している。好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、変異バチルス株はB.ズブチリス、B.リケニフォルミ及びB.アミロリケファシエンスからなる群より選択される。いくつかの特に好ましい実施態様において、変異バチルス株はB.ズブチリス株である。さらに別の実施態様において、固有染色体領域はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVEF−YVEK領域、DHB領域及びそれらの断片からなる群より選択される。いくつかの好ましい実施態様において、2つの固有染色体領域またはその断片は欠失される。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。さらに別の実施態様において、目的タンパク質はプロテアーゼである。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。いくつかの特に好ましい実施態様において、ズブチリシンはズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、及びズブチリシン309及びこれらの変異体からなる群より選択される。さらに好ましい実施態様において、バチルス宿主は組換え体株である。いくつかの特に好ましい実施態様において、変異バチルス株は、degU、degQ、degS、sco4、spollE及びoppAからなる群より選択される遺伝子において少なくとも1の変異をさらに含む。いくつかの好ましい実施態様において、変異はdegU(Hy)32でる。
【0027】
本発明はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVFF−YVEK領域、DHB領域及びそれらの断片からなる群より選択される固有染色体領域の欠失を含むプロテアーゼ生成バチルス株をさらに提供する。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。いくつかの実施態様において、プロテアーゼは異種プロテアーゼである。いくつかの好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。別の実施態様において、バチルス株はB.ズブチリス株である。
【0028】
本発明はバチルスにおいて目的タンパク質の発現を増大する方法も提供し、以下を含む:選択マーカー及び不活性化染色体断片を含むDNA構築体をバチルス宿主株内に導入する工程であって、ここでDNA構築体をバチルス宿主株の染色体内に統合し、バチルス宿主細胞から固有染色体領域またはその断片の欠失を生じて変異バチルス株を生じる工程;及び適当な条件下で変異バチルス株を成長させる工程であって、ここで目的タンパク質発現は変異していないバチルス宿主細胞における目的タンパク質の発現と比較して変異バチルス株が大きい。いくつかの好ましい実施態様において、本方法は目的タンパク質を回収する工程をさらに含む。いくつかの実施態様において、本方法は変異バチルス株から選択マーカーを切除する工程をさらに含む。別の実施態様において、固有染色体領域はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVFF−YVEK領域、DHB及びそれらの断片からなる領域の群より選択される。さらに別の実施態様において、変異バチルス株は少なくとも2の固有染色体領域の欠失を含む。いくつかの好ましい実施態様において、目的タンパク質は酵素である。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。いくつかの実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。本発明はここに記載の方法を用いて生成した変異バチルス株も提供する。
【0029】
本発明はバチルス株から目的タンパク質を得る方法も提供し、以下を含む:選択マーカー及び不活性化染色体断片を含むDNA構築体を用いてバチルス宿主細胞を形質転換する工程であって、ここでDNA構築体はバチルス株の染色体内に統合され、固有染色体領域またはその断片の欠失を生じ、変異バチルス株を生成し、目的タンパク質の発現を可能にするのに適した条件下で変異バチルス株を培養する工程及び目的タンパク質を回収する工程。いくつかの好ましい実施態様において、目的タンパク質は酵素である。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、固有染色体領域はPBSX、スキン、プロファージ7、SPβ、プロファージ1、プロファージ2、プロファージ3、プロファージ4、プロファージ5、プロファージ6、PPS、PKS、YVFF−YVEK、DHB及びそれらの断片からなる群より選択される。いくつかの特に好ましい実施態様において、変異バチルス株はdegU、degQ、degS、sco4、spollE及びoppAからなる群より選択される遺伝子において少なくとも1の変異をさらに含む。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼからなる群より選択される酵素である。いくつかの特に好ましい実施態様において、当該酵素はプロテアーゼである。いくつかの好ましい実施態様において、目的タンパク質は酵素である。別の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0030】
本発明はバチルスにおいて目的タンパク質の発現を増大させる方法をさらに提供し、以下を含む:少なくとも1のバチルス細胞から核酸を得る工程;前記バチルス細胞由来核酸についてトランスクリプトームDNAアレイ分析を行い、少なくとも1の目的遺伝子を同定する工程;DNA構築体を生成するために少なくとも1の目的遺伝子を修飾する工程;DNA構築体をバチルス宿主細胞内に導入して変異バチルス株を生成する工程であって、ここで変異体は目的タンパク質発現が非変異のバチルスにおける目的タンパク質の発現と比較して増大するような条件下で目的タンパク質を生成することができる。いくつかの実施態様において、目的タンパク質はアミノ酸生合成経路及び生分解経路からなる群より選択される少なくとも1の生化学経路に関係する。いくつかの実施態様において、本方法は少なくとも1の生分解経路を無効にすることを含む。いくつかの実施態様において、生分解経路は目的遺伝子の転写により無効にされる。しかしながら、本発明はこれらの経路に限定されるものではなく、本方法は目的タンパク質の発現の増加が生じるように細胞内のその他の生化学経路の修飾においても使用できることが考えられる。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの好ましい実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0031】
本発明はバチルス中において目的タンパク質の発現を増大させる方法をさらに提供し、以下を含む:少なくとも1のバチルス細胞から少なくとも1の目的遺伝子を含む核酸を得る工程;前記核酸を断片化する工程;前記少なくとも1の目的遺伝子を含む増殖断片プールを生成するために前記断片を増殖する工程;前記増殖断片をDNA構築体を生成するために連結する工程;前記DNA構築体をバチルス宿主細胞内に直接形質転換し、変異バチルス株を生成する工程;変異されていないバチルスにおける前記目的タンパク質の発現と比較して前記目的タンパク質の発現が増大するような条件下で前記変異バチルス株を培養する工程。いくつかの好ましい実施態様において、前記増殖はポリメラーゼ連鎖反応を用いることを含む。いくつかの実施態様において、変異バチルス株はprpC、sigD及びtdh/kblからなる群より選択される修飾遺伝子を含む。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの好ましい実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0032】
本発明はさらに配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33からなる群より選択される核酸配列に示す配列を含む単離核酸をさらに提供する。
【0033】
また、本発明はアミノ酸をエンコードする単離核酸配列を提供し、ここで当該アミノ酸は配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される。
【0034】
本発明はさらに単離アミノ酸配列を提供し、当該アミノ酸は配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される。
【図面の簡単な説明】
【0035】
【図1A】図1Aは本発明により提供されるの1の方法(“方法1”実施例1参照)の一般的な説明図を示すものである。この方法において、遺伝子のフランキング領域及び/または固有染色体領域は野生型バチルス染色体から増殖され、制限酵素(少なくともBamHIを含む)で切断し、pJM102に連結する。当該構築体は大腸菌によりクローンされ、及びプラスミドを単離し、BamHIを用いて線形化し、相補的末端を含む抗菌マーカーに連結する。大腸菌で再度クローニングした後、液体培養物を成長させ、バチルス宿主株(好ましくは、コンピテントバチルス宿主株)の形質転換に用いるプラスミドDNAを単離するために用いる。
【図1B】図1Bは本発明により提供されるの1の方法(“方法1”実施例1参照)の一般的な説明図を示すものである。この方法において、遺伝子のフランキング領域及び/または固有染色体領域は野生型バチルス染色体から増殖され、制限酵素(少なくともBamHIを含む)で切断し、pJM102に連結する。当該構築体は大腸菌によりクローンされ、及びプラスミドを単離し、BamHIを用いて線形化し、相補的末端を含む抗菌マーカーに連結する。大腸菌で再度クローニングした後、液体培養物を成長させ、バチルス宿主株(好ましくは、コンピテントバチルス宿主株)の形質転換に用いるプラスミドDNAを単離するために用いる
【図2】図2は本発明のいくつかの実施態様に従うDNAカセットの構築において用いるプライマーの位置を示す。図はここで用いるプライマー命名システムの説明を提供する。プライマー1及び4は欠失の存在をチェックするために用いる。これらのプライマーは“欠失X−UF−chk”及び“欠失X−UR−chk−del”と呼ぶ。欠失X−UF−chkも、染色体中に存在するカセットの陽性チェックのための抗生物質マーカー内のリバースプライマー(プライマー11:PBSX−UR−chk−Delと呼ばれる)を用いてPCR反応において用いる。プライマー2及び6は上流フランキング領域を増殖するために用いる。これらのプライマーは“欠失X−UF”及び“欠失X−UR”と呼び、黒の縦棒のところで組換え制限部位を含む。プライマー5及び8は下流フランキング領域を増幅するために用いる。これらのプライマーは“欠失X−DF”及び“欠失X−DR”と呼ぶ。これらのプライマーは連結及びクローニングのための組換えBamHI部位またはPCR融合で用いるバチルス・ズブチリス染色体の適当な部分に相同な25塩基対末端を含むことができる。いくつかの実施態様において、プライマー3及び7はPCR融合により作成したカセットの場合、当該カセットを一緒に融合するために用いられ、一方、他の実施態様において、それらは挿入の存在をチェックするために用いられる。これらのプライマーは“欠失X−UF−ネスティド”及び“欠失X−DR−ネスティド”と呼ぶ。いくつかの実施態様において、“抗生物質マーカー”に対応する配列はSpc耐性マーカーであり、欠失する領域はCssS遺伝子である。
【図3】図3は本発明の1の方法(“方法2”;実施例2を参照)の一般的な説明図である。フランキング領域は選択マーカー配列に相補的な25bpの配列を含むように設計される。選択マーカー配列は1のフランキング領域のDNAに相補的である25bpの末端も含む。フランキング領域末端に近いプライマーは単一反応チューブにおける3つすべてのテンプレートを増幅するために用いられ、それにより融合断片を作成する。この融合断片またはDNA構築体はコンピテントバチルス宿主株に直接形質転換される。
【図4】図4はバチルスDHB欠失クローンのエレクトロフォレーシスゲルを提供する。レーン1及び2はプライマー1及び11を用いて増幅されたDHB欠失を有する2つの株を描き、不活性化染色体断片の上流からフレオマイシン・マーカー内に増幅した1.2バンドを表す。レーン3はこの反応の野生型対照を表す。非特異増幅のみが観測される。レーン4及び5はプライマー9及び12を用いて増幅されたDHB欠失株を表す。この2kbバンドは抗生物質領域から不活性化染色体断片の下流部分以下まで増幅する。レーン6はこの反応の陰性対照であり、バンドは示されていない。レーン7及び8はプライマー1及び4を用いて増幅した欠失株を表し、当該説明図によりDHB領域がないことが確認される。レーン9は野生型対照である。
【図5】図5はバチルス・ズブチリス(野生型)の生成株の2つのクローンのゲルエレクトロフォレーシスを表し、s/rはs/r遺伝子の欠失を生じるフレオマイシン(phleo)マーカーで置換される。レーン1及び2は位置1及び11でプライマーを用いて増幅したクローン表す。レーン3は同じプライマーを用いて増幅した野生型染色体DNAである。1.2kbのバンドが挿入に関して観測される。レーン4及び5は位置9及び12でプライマーを用いて増幅したクローンを表す。レーン6は同じプライマーを用いて増幅した野生型染色体DNAである。正確な形質転換体は2kbのバンドを含む。レーン7及び8は位置2及び4でプライマーを用いて増幅したクローンを表す。レーン9は同じプライマーを用いて増幅した野生型染色体DNAである。欠失株に関してバンドは観測されないが、1kb周辺のバンドが野生型において観測される。プライマー位置の説明のため、図2を参照されたい。
【図6】図6はバチルス・ズブチリス(野生型)の生成株のクローンのエレクトロフォレーシスゲルを提供し、cssSはspcマーカーを染色体内に統合することにより不活性化される。レーン1は非統合の対照であり、約1.5kb小さい。
【図7】図7はバチルス・ズブチリス野生型株(非変異)及び種々の欠失を有する対応する変異バチルス・ズブチリス株を含む振とうフラスコ培養物から測定したズブチリシン分泌の改善を示す棒グラフである。プロテアーゼ活性(g/L)は17、24及び40時間後、または24及び40時間の時点で測定した。
【図8】図8はバチルス・ズブチリス野生型株(非変異)及び対応する変異欠失株(−sbo)及び(−slr)における振とうフラスコ培養物から測定したズブチリシン分泌の改善を示す棒グラフである。プロテアーゼ活性(g/L)は17、24及び40時間後に測定した。
【発明を実施するための形態】
【0036】
発明の説明
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、または修飾される。好ましい実施態様において、1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。
【0037】
定義
ここで言及する全ての特許及び文献、並びに当該特許及び文献に開示される全ての配列はここに明示的に引用するものとする。別に定めない限り、ここで用いる全ての技術及び科学的用語は本発明が属する当業者に通常理解される意味と同じ意味を有する(例えば、Singleton et al.,Dictionary of Microbiology and Molecular Biology,第2版、John Wiley and Sons,ニューヨーク[1994];及びHale and Marham、The Harper Collins Dictionary of Biology,Harper Perennial,NY[1991]を参照、これらはここで用いる多くの用語の当業者の一般的な辞書となる)。ここに記載の方法及び材料に類似または同等のいかなる方法及び材料も本発明を実施または試験するために用いることができるが、好ましい方法及び材料を記載する。数値範囲はその範囲を定める数字を含むものとする。明細書及び請求の範囲で用いるように、文脈において明らかに示される場合を除いて、単数の記載は複数も含むものとする。従って、例えば“宿主細胞”への言及は複数の当該宿主細胞も含む。
【0038】
別に示さない限り、核酸は5’から3’の左から右方向に記載し、アミノ酸配列はアミノからカルボキシの左から右方向にそれぞれ記載する。ここに記載する見出しは本発明の種々の側面または実施態様を限定するものではなく、明細書全体に対して参照され得るものである。従って、下記に定義する用語は明細書全体を参照することによりさらに十分に定義されるものである。
【0039】
ここで用いる、“宿主細胞”とは新しく導入するDNA配列のための宿主または発現媒体として作用する能力を有する細胞をいう。本発明の好ましい実施態様において、宿主細胞はバチルス種または大腸菌細胞である。
【0040】
ここで用いる“バチルス属”とは、当業者に公知の“バチルス”属に含まれるすべての種類を含み、限定されないが、B.ズブチリス、B.リケニフォルミス、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.アミロリケファシエンス、B.クラウジイ、B.ハロデュランス、B.メガテリウム、B.コアグランス、B.サーキュランス、B.ロータス、及びB.チューリンゲンシスが挙げられる。バチルス属は分類上再編成を受け続けている。従って、当該属は再分類された種も含むものとし、限定されないが、現在は“ゲオバチルス・ステロサーモフィラス”と呼ばれるB.ステロサーモフィラスなどの微生物を含む。酸素存在下での耐性内生胞子の生成はバチルス属の重要な特徴と考えられるが、この特徴は最近ではアリシクロバチルス、アンフィバチルス(amphibacillus)、アニューリーニバチルス(aneurinibacillus)、アノキシバチルス(anoxybacillus)、ブレビバチルス、フィロバチルス、グラシリバチルス(Gracilibacillus)、ハロバチルス、ペニバチルス、サリバチルス、サーモバチルス、ウレーバチルス(Ureibacillus)及びヴァージバチルス(Virgibacillus)と称されるものにも当てはまる。
【0041】
ここで用いる、“核酸”の語はヌクレオチドまたはポリヌクレオチド配列及びそれらの断片または部分、及びセンスまたはアンチセンスストランドのいずれを表すかにかかわらず、2本鎖または1本鎖のゲノムまたは合成起源のDNA、cDNA及びRNAをいう。遺伝子コードの縮退の結果、多数のヌクレオチド配列が所定のタンパク質をエンコードすることは当然理解されるであろう。
【0042】
ここで用いる、“遺伝子”の語はポリぺプチド鎖の生成に関するDNAの染色体断片を意味し、コード領域(例えば、5’非翻訳(5’UTR)またはリーダー配列及び3’非翻訳(3’UTR)またはトレーラー配列)に先行または後に続く領域、及び個々のコード断片(エクソン)間の介在配列(イントロン)を含んでも含まなくてもよい。
【0043】
いくつかの実施態様において、遺伝子は治療的に重要なタンパク質またはぺプチド、例えば成長因子、サイトカイン、配位子、受容体及び阻害物質、並びにワクチン及び抗体などをエンコードする。遺伝子は商業的に重要な工業タンパク質またはぺプチド、例えば酵素(例えば、プロテアーゼ、アミラーゼ及びグルコアミラーゼなどのカルボヒドラーゼ、セルラーゼ、オキシダーゼ及びリパーゼ)もエンコードし得る。しかしながら、本発明は特定の酵素またはタンパク質に限定されるものではない。いくつかの実施態様において、目的遺伝子は天然遺伝子であり、一方、他の実施態様において、変異遺伝子または合成遺伝子である。
【0044】
ここで用いる“ベクター”の語は、細胞内で複製でき、新しい遺伝子またはDNA断片を細胞内に運ぶことができる核酸をいう。従って、当該用語は異なる宿主細胞間を移動するために設計された核酸構築体をいう。“発現ベクター”とは、外来細胞中に異種DNA断片を導入し、発現できるベクターをいう。多くの原核及び真核発現ベクターは市販されている。適当な発現ベクターの選択は当業者の知識の範囲内である。
【0045】
ここで用いる、“DNA構築体”、“発現カセット”及び“発現ベクター”の語は標的細胞中で特定の核酸の転写を可能とする、一連の具体的な核酸成分を用いて組換えにより、または合成的に生成した核酸構築体をいう(すなわち、これらは上述のベクターまたはベクター成分である)。組換え発現カセットはプラスミド、染色体、ミトコンドリアDNA、色素体DNA、ウイルスまたは核酸断片内に組み込むことができる。通常、発現ベクターの組換え発現カセット部分は、数ある配列の中でも、転写される核酸配列及びプロモーターを含む。いくつかの実施態様において、DNA構築体は標的細胞中で特定の核酸の転写を可能とする一連の特定核酸成分も含む。1の実施態様において、本発明のDNA構築体はここで定義する選択マーカー及び不活性化染色体断片を含む。
【0046】
ここで用いる“形質転換DNA”、“形質転換配列”及び“DNA構築体”とは、配列を宿主細胞または微生物内に導入するために用いられるDNAをいう。形質転換DNAはPCRまたはその他の適当な技術によりin vitroで生成できる。いくつかの好ましい実施態様において、形質転換DNAは入来配列を含み、一方、他の好ましい実施態様において、さらに相同ボックスの側面に位置する入来配列を含む。さらに別の実施態様において、形質転換DNAはその他の非相同配列を含み。末端に加えられる(すなわち、stuffer配列またはフランキング配列)。末端は、例えばベクター内への挿入など形質転換DNAが閉じた環を形成するように閉じることができる。
【0047】
ここで用いる、“プラスミド”の語はクローニングベクターとして用いる環状2本鎖(ds)DNA構築体をいい、多くの細菌及びいくつかの真核生物内において染色体外自己複製遺伝子要素を形成する。いくつかの実施態様において、プラスミドは宿主細胞のゲノム内に組み込まれるようになる。
【0048】
ここで用いる“単離”及び“精製”の語は、天然に結合している少なくとも1の成分から取り除いた核酸またはアミノ酸(またはその他の成分)をいう。
【0049】
ここで用いる“発現の増大”とは、目的タンパク質生成の増大を含むように広く解釈される。発現の増大とは、ここでの記載に従った変異をさせていないが、本質的に同じ成長条件下で成長させた対応宿主株における通常の発現レベル以上の発現をいう。
【0050】
いくつかの好ましい実施態様において、“増大”は望ましい特性の増加を生じる任意の修飾により達成される。例えば、いくつかの特に好ましい実施態様において、本発明はタンパク質生成を増大させる手段を提供し、増大株が親株(例えば、野生型及び/または始発株)よりも目的タンパク質を多く生成し及び/または優れた質を生じるようにする。
【0051】
ここで用いる“発現”とは、ポリぺプチドが遺伝子の核酸配列に基づいて生成されるプロセスをいう。当該プロセスは転写及び翻訳の両方を含む。
【0052】
ここで用いる、核酸配列を細胞内に導入することに関する“導入”の語は核酸配列を細胞内に移動させるのに適した任意の方法をいう。導入のための手段は、限定されないが、原形質融合、トランスフェクション、形質転換、接合及び形質導入などが挙げられる(例えば、Ferrari et al.、“Genetics(遺伝学)”、Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.,第57〜72頁、[1989]を参照)。
【0053】
ここで用いる“形質転換”及び“安定な形質転換”の語は、ゲノム内に統合された、または少なくとも2世代の間維持されたエピソームプラスミドとして非天然(異種)ポリヌクレオチド配列を有する細胞をいう。
【0054】
ここで用いる“入来配列”の語はバチルス染色体内に導入されたDNA配列をいう。いくつかの好ましい実施態様において、入来配列はDNA構築体の一部である。好ましい実施態様において、入来配列は1以上の目的タンパク質をエンコードする。いくつかの実施態様において、入来配列は形質転換される細胞のゲノム中に既に存在する、または存在しない配列であってもよい(すなわち、相同または異種配列である)。いくつかの実施態様において、入来配列は1以上の目的タンパク質、遺伝子及び/または変異または修飾遺伝子をエンコードする。別の実施態様において、入来配列は機能野生型遺伝子またはオペロン、機能変異遺伝子またはオペロン、または非機能遺伝子またはオペロンをエンコードする。いくつかの実施態様において、非機能配列は遺伝子の機能を妨害するために遺伝子内に導入できる。いくつかの実施態様において、入来配列は1以上の機能野生型遺伝子をエンコードし、一方他の実施態様において、入来配列は1以上の機能変異遺伝子をエンコードし、さらに他の実施態様において、入来配列は1以上の非機能遺伝子をエンコードする。他の実施態様において、入来配列は形質転換される宿主細胞の染色体内に既に存在する配列をエンコードする。好ましい実施態様において、入来配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD及びこれらの断片からなる群より選択される遺伝子を含む。さらに別の実施態様において、入来配列は選択マーカーを含む。さらなる実施態様において、入来配列は2つの相同ボックスを含む。
【0055】
いくつかの実施態様において、入来配列は少なくとも1の異種タンパク質をエンコードし、限定されないが、ホルモン、酵素及び成長因子が挙げられる。他の実施態様において、酵素は、限定されないが、プロテアーゼ、エステラーゼ、リパーゼ、フェノールオキシダーゼ、パーミアーゼ、アミラーゼ、プルラナーゼ、セルラーゼ、グルコースイソメラーゼ、ラッカーゼ及びタンパク質ジスルフィドイソメラーゼが挙げられる。
【0056】
ここで用いる“相同ボックス”とはバチルス染色体の配列と相同な核酸配列をいう。より具体的には、相同ボックスは、本発明に従って不活性化される遺伝子に隣接するコード領域または遺伝子の一部と約80〜100%配列同一性、約90〜100%配列同一性または約95%〜100%配列同一性を有する上流または下流領域をいう。これらの配列はバチルス染色体におけるDNA構築体の統合を目的とし、及び入来配列によりバチルス染色体の一部が置換されることを目的とする。本発明を限定するものではないが、相同ボックスは約1塩基対(bp)〜200キロ塩基(kb)を含む。好ましくは、相同ボックスは約1bp〜10.0kb;1bp〜5.0kb;1bp〜2.5kb;1bp〜1.0kb及び0.25kb〜2.5kbを含む。また、相同ボックスは約10.0kb、5.0kb、2.5kb、2.0kb、1.5kb、1.0kb、0.5kb、0.25kb及び0.1kbを含む。いくつかの実施態様において、選択マーカーの5’及び3’末端は相同ボックスに隣接し、ここで相同ボックスは当該遺伝子のコード領域にすぐ隣接した核酸配列を含む。
【0057】
ここで用いる“選択マーカー−エンコードヌクレオチド配列”の語は宿主細胞において発現できるヌクレオチド配列をいい、選択マーカーの発現により対応する選択因子の存在下、または必須栄養素の不存在下で発現遺伝子を含む細胞は成長できるようになる。
【0058】
ここで用いる“選択可能マーカー”及び“選択マーカー”の語は宿主細胞中で発現できる核酸(例えば遺伝子)をいい、ベクターを含むこれら宿主の選択が簡単にできるようにする。当該選択マーカーの例としては、限定されないが、抗微生物などがある。従って、“選択マーカー”とは、宿主細胞が目的の入来DNAを取り込んだ、またはその他の反応が起こった印を与える遺伝子をいう。通常、選択マーカーは、形質転換時に外来配列を受け取らなかった細胞と外来DNAを含む細胞との区別を可能にするために宿主細胞に抗菌耐性または代謝利点を与える遺伝子をいう。“存在(residing)選択マーカー”とは、形質転換される微生物の染色体上に位置するマーカーである。存在選択マーカーは遺伝子を形質転換DNA構築体上の選択マーカーとは異なる遺伝子をエンコードする。選択マーカーは当業者に公知である。上述の通り、好ましくは当該マーカーは抗微生物耐性マーカーである(例えば、ampR;phleoR;specR;kanR;eryR;tetR;cmpR;及びneoR;例えばGuerot−Fleury,Gene,167:335−337[1995];Palmeros et al.、Gene 247:255−264[2000];及びTrieu−Cuot et al.、Gene、23:331−341[1983]を参照)。いくつかの特に好ましい実施態様において、本発明はクロラムフェニコール耐性遺伝子を提供する(例えば、pC194上に存在する遺伝子、及びバチルス・リケニフォルミスゲノム中に存在する耐性遺伝子)。この耐性遺伝子は本発明及び染色体上統合されたカセット及び統合プラスミドの染色体増幅に関する実施態様において特に有用である(例えば、Albertini and Galizzi、Bacteriol.,162:1203−1211[1985];及びStahl and Ferrari,J.Bacteriol.158:411−418[1984]を参照)。この天然クロラムフェニコール耐性遺伝子のDNA配列を以下に示す:
【式1】
【0059】

このクロラムフェニコール耐性タンパク質の推定アミノ酸配列は以下の通り:
【式2】
【0060】
本発明に従う有用なその他のマーカーは、限定されないが、トリプトファンなどの栄養素要求性マーカー及びβ−グラクトシダーゼなどの検出マーカーである。
【0061】
ここで用いる“プロモーター”の語は下流遺伝子の転写に直接機能する核酸配列をいう。好ましい実施態様において、プロモーターは標的遺伝子が発現するように宿主細胞に適したものである。プロモーターはその他の転写及び翻訳調節核酸配列(“制御配列”とも称される)と一緒に、所定の遺伝子を発現するために必要なものである。通常、転写及び翻訳調節配列は、限定されないが、プロモーター配列、リボソーム結合部位、転写開始及び停止配列、翻訳開始及び停止配列、及びエンハンサーまたは活性化配列などが挙げられる。
【0062】
核酸は他の核酸配列と機能的に関連するように位置する場合、“作動可能に連結する”。例えば、分泌リーダー(すなわち、1のぺプチド)をエンコードするDNAはポリぺプチドの分泌に参加するプレタンパク質として発現する場合、当該ポリぺプチドに関するDNAに作動可能に連結し;プロモーターまたはエンハンサーは配列転写に影響を与える場合、コード配列に作動可能に連結し;またはリボソーム結合部位は翻訳を促進するように位置する場合、コード配列に作動可能に連結する。一般的に“作動可能に連結する”とは、連結するDNA配列が連続していることを意味し、分泌リーダーの場合、連続かつ読取り層にある。しかしながら、エンハンサーは必ずしも連続している必要はない。結合は都合のよい制限酵素認識部位で連結することにより達成される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを従来実務に従って用いる。
【0063】
“不活性化”の語は、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD染色体遺伝子のうち1以上の機能発現を妨げる方法を含み、当該遺伝子または遺伝子産物はその公知機能を発揮するための使用に適している。不活性化または増大化は任意の適した手段によって生じ、核酸遺伝子配列における欠失、置換(例えば変異)、阻害、及び/または挿入などがある。1の実施態様において、不活性化遺伝子の発現産物は対応するタンパク質の生物活性変化を有する短いタンパク質である。いくつかの実施態様において、生物活性変化は活性の増加であり、一方、好ましい実施態様において、当該変化は生物活性の損失を生じる。いくつかの実施態様において、変異バチルス株は好ましくは安定かつ不可逆的な不活性化を生じる1以上の遺伝子の不活性化を含む。
【0064】
いくつかの好ましい実施態様において、不活性化は欠失により達成される。いくつかの好ましい実施態様において、遺伝子は相同組換えにより欠失する。例えば、いくつかの実施態様において、欠失する遺伝子がsboの場合、相同ボックスの両側に隣接する選択マーカーを有する入来配列を含むDNA構築体を用いる。相同ボックスは染色体sbo遺伝子の核酸フランキング領域に相同なヌクレオチド配列を含む。DNA構築体はバチルス宿主染色体の相同配列と一致し、二重交差(double crossover)の場合、sbo遺伝子は宿主染色体から切除される。
【0065】
ここで用いる遺伝子の“欠失”の語は、コード配列全体の欠失、コード配列の一部の欠失、またはフランキング領域を含むコード配列の欠失をいう。欠失は染色体に残った配列が望ましい遺伝子の生物活性を与えるのであれば部分的であってもよい。コード配列のフランキング領域は約1bp〜約500bpを5’及び3’末端に含むことができる。フランキング領域は500bpよりも大きくてもよいが、好ましくは本発明に従って不活性化または欠失され得るその他の遺伝子を当該領域に含まない。最終的には、欠失遺伝子は事実上、非機能的である。簡単に言えば、“欠失”の語は1以上のヌクレオチドまたはアミノ酸残基がそれぞれ除去された(すなわち、存在しない)、ヌクレオチドまたはアミノ酸配列の変化として定義される。従って、“欠失変異体”はそれぞれの野生型生物よりも少ないヌクレオチドまたはアミノ酸を有する。
【0066】
本発明の他の実施態様において、DNAアレイ分析(例えば、ここに記載するトランスクリプトーム分析)により測定して不適当な時に活性な遺伝子の欠失は生成タンパク質の発現の増加をもたらす。いくつかの好ましい実施態様において、pckA,gapB、fbp及び/またはalsDからなる群より選択される1以上の遺伝子の欠失は供給物の効率が向上した改善株を提供する。ここで用いる“トランスクリプトーム分析”とは、遺伝子転写分析をいう。
【0067】
本発明の他の実施態様において、遺伝子は調節配列の欠失により“最適化”されると考えられ、この欠失により所望の生成物の発現が増加する。本発明のいくつかの好ましい実施態様において、トリプトファンオペロン(すなわち、遺伝子trpA、trpB、trpC、trpD、trpE、trpFを含む)はTRAP結合RNA配列をコードするDNA配列の欠失により最適化される(Yang et al.、J.Mol.Biol.,270:696−710[1997]を参照)。この欠失は宿主細胞由来の所望の生成物の発現を増加させると考えられる。
【0068】
他の好ましい実施態様において、不活性化は挿入による。例えば、いくつかの実施態様において、sboが不活性化される遺伝子である場合、DNA構築体は選択マーカーにより阻害されるsbo遺伝子を有する入来配列を含む。選択マーカーはsboコード配列部分の両側に隣接する。DNA構築体は宿主染色体中のsbo遺伝子の本質的に同一配列と一致し、二重交差の場合、sbo遺伝子は選択マーカーの挿入により不活性化される。簡単に言えば、“挿入”または“付加”は、それぞれ天然配列と比較して1以上のヌクレオチドまたはアミノ酸残基の付加を生じたヌクレオチドまたはアミノ酸配列の変化である。
【0069】
他の実施態様において、一重交差の場合、ベクターとしてプラスミドを用いた挿入により活性化される。例えば、sbo染色体遺伝子は遺伝子または遺伝子コード配列の一部及び選択マーカーを含むプラスミドに合わせる。いくつかの実施態様において、選択マーカーは遺伝子コード配列内に位置し、または遺伝子から分離されたプラスミドの一部に位置する。ベクターはバチルス染色体内に統合され、当該遺伝子はコード配列中のベクター挿入により不活性化される。
【0070】
別の実施態様において、不活性化は遺伝子の変異により生じる。遺伝子を変異する方法は当業者に公知であり、限定されないが、部位特異的変異、ランダム変異発生、及びgapped−duplex法が挙げられる(例えば米国特許第4,760,025号;Moring et al.、Biotech.2:646[1984];及びKramer et al.、Nucleic Acids Res.,12:9441[1984]を参照)。
【0071】
ここで用いる“置換”とは1以上のヌクレオチドまたはアミノ酸と、それぞれ異なるヌクレオチドまたはアミノ酸との置き換えにより生じる。
【0072】
ここで用いる“相同遺伝子”とは、異なるが、通常は関連種由来の遺伝子対をいい、それぞれが対応し、それぞれ同一または非常に類似しているものをいう。当該用語は種形成(すなわち、新しい種の発達)により分離した遺伝子(例えばオーソロガス遺伝子)、及び遺伝子重複により分離された遺伝子(例えばパラロガス遺伝子)を包含する。
【0073】
ここで用いる“オーソログ”及び“オーソロガス遺伝子”とは種形成により共通祖先の遺伝子(すなわち相同遺伝子)から進化した異なる種の遺伝子をいう。通常、オーソロガスは進化の過程において同じ機能を保持する。オーソロガスの同定により新しく配列決定されたゲノムの遺伝子機能の信頼性の高い推測ができる。
【0074】
ここで用いる“パラログ”及び“パラロガス遺伝子”とはゲノム内で重複していることにより関連した遺伝子をいう。オーソログは進化の過程を通して同じ機能を維持するが、パラログはいくつかの機能は最初の機能に関連することが多いとしても、新しい機能を進化させる。パラログ遺伝子の例としては、限定されないが、トリプシン、キモトリプシン、エラスターゼ及びトロンビンをエンコードする遺伝子が挙げられ、これらは全てセリンプロテイナーゼであり、同じ種内で一緒に生じる。
【0075】
ここで用いる“相同性”とは、好ましい同一性を有する配列類似性または同一性をいう。この相同性は当業者に公知の標準技術を用いて決定する(例えば、Smith and Waterman,Adv.Appl.Math.、2:482[1981];Needlman and Wunschを、J.Mol.Biol.,48:443[1970];Pearson and Lipman、Proc.Natl.Acad.Sci.USA 85:2444[1988];ウィスコンシン・ジェネティックス・ソフトウェアパッケージ(Genetics Computer Group,マディソン、ウィスコンシン州)のGAP、BESTFIT、FASTA及びTFASTAなどのプログラム;Devereux et al.、Nucl.Acid Res.,12:387−395[1984]を参照)。
【0076】
ここで用いる“類似配列”は遺伝子機能が本質的にバチルス・ズブチリス株168から示される遺伝子と同じであるものをいう。さらに、類似遺伝子はバチルス・ズブチリス株168遺伝子の配列と少なくとも60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%または100%の配列同一性を含む。もしくは、類似配列はB.ズブチリス内で見られる70〜100%の遺伝子配列を有し、及び/またはB.ズブチリス168染色体内の遺伝子と一致する領域に見られる少なくとも5〜10の遺伝子を有する。類似配列は配列決定の公知方法により決定できる。一般的に用いられる配列決定方法はBLASTであるが、上述及び下記に示す通りその他の方法も配列を決定するために用いることができる。
【0077】
有用なアルゴリズムの1の例はPILEUPである。PILEUPは連続(progressive)、対合配列を用いて関連配列グループから複数の配列決定を行うことができる。また、配列決定に用いる集積性関係を示す系図をプロットすることもできる。PILEUPはFeng及びDoolittleの連続配列法の単純化を用いる(Feng and Doolittle、J.Mol.Evol.、35:351−360[1987])。当該方法はHigginsとSharpが説明したものと類似する(Higgins and Sharp、CABIOS 5:151−153[1989])。有用なPILEUPパラメーターは初期設定ギャップウエイトが3.00、初期設定ギャップ長さウエイトが0.10であり、及びエンドギャップを用いる。
【0078】
有用なアルゴリズムの他の例としてはBLASTアルゴリズムであり、Altschul et al.により説明されている(Altschul et al.,J.Mol.Biol.,215:403−410、[1990];及びKarlin et al.、Proc.Natl.Acad.Sci.USA 90:5873−5787[1993])。特に有用なBLASTプログラムはWU−BLAST−2プログラムである(Altschul et al.,Meth.Enzymol.,266:460−480[1996]を参照)。WU−BLAST−2はいくつかのサーチパラメーターを用い、ほとんどは初期値に設定されている。調節可能なパラメーターは以下の値に設定する:オーバーラップスパン=1、オーバーラップフラクション=0.125、言語閾値(T)=11。HSP S及びHSP S2パラメーターは動的値であり、特定配列の組成物及び目的配列がサーチされる特定データベースの組成物に応じてプログラム自体により設定される。しかしながら、当該値は感度を増加させるため調節できる。A%のアミノ酸配列同一性値は適合同一残基の数を配列領域の“より長い”配列残基の合計数で割ることにより決定する。“より長い”配列は配列領域において実際に存在している残基を最も有する配列である(配列スコアを最大化するためにWU−BLAST−2により導入されたギャップは無視する)。
【0079】
従って、“核酸配列同一性比率(%)”は、図の核酸において示されるヌクレオチド配列残基と同一である、候補配列におけるヌクレオチド残基の比率として定義される。好ましい方法はオーバーラップスパン及びオーバーラップファンクションがそれぞれ1及び0.125に設定された初期設定パラメーター設定のWU−BLAST−2のBLASTNモジュールを利用する。
【0080】
配列は配列決定される配列においてギャップ導入を含むことができる。さらに、図の核酸よりも多いまたは少ないヌクレオチドを含む配列については、相同性比率はヌクレオチド合計数と比較した相同ヌクレオチドの数に基づいて決定されることが理解される。従って、例えば、ここで同定する配列よりも短い配列の相同性は下記に説明するように、短い配列中のヌクレオチドの数を用いて決定される。
【0081】
ここで用いる“ハイブリダイゼーション”の語は当業者に公知の塩基対合により、核酸鎖を相補鎖と接合させるプロセスをいう。
【0082】
核酸配列は、2つの配列が中〜高ストリンジェンシーなハイブリダイゼーション条件及び洗浄条件下でお互いに特異的にハイブリダイズする場合、対照核酸配列に “選択的にハイブリダイズ可能”であると考えられる。ハイブリダイゼーション条件は核酸結合複合体またはプローブの融点(Tm)に基づく。例えば、“最大ストリンジェンシー”は通常約Tm−5℃(プローブのTmより5°低い)で起こり、“高ストリンジェンシー”はTmより約5〜10℃低く;“中間ストリンジェンシー”はプローブのTmより約10〜20℃低く;及び“低ストリンジェンシー”はTmより約20〜25℃低い。機能上、最大ストリンジェンシー条件はハイブリダイゼーションプローブと厳密な同一性またはほぼ厳密な同一性を有する配列を同定するために用いることができ、一方、中間または低ストリンジェンシーハイブリダイゼーションはポリヌクレオチド配列相同体を同定または検出するために用いることができる。
【0083】
中〜高ストリンジェンシーハイブリダイゼーション条件は当業者に公知である。高ストリンジェンシー条件の例としては、50%ホルムアミド、5X SSC、5X Denhardt’s溶液、0.5%SDS及び100μg/ml変性キャリアDNA中、約42℃でのハイブリダイゼーションを含み、続いて2X SSC及び0.5%SDS中、室温で2回洗浄し、0.1X SSC及び0.5%SDS中、42℃でさらに2回洗浄する。中ストリンジェント条件の例としては、20%ホルムアミド、5X SSC(150mM NaCl、15mM クエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5X Denhardt’s溶液、10% 硫酸デキストラン及び20mg/ml変性せん断サケ精子DNAを含む溶液中、37℃で一晩中培養し、続いて1X SSC中、約37〜50℃でフィルターを洗浄する。プローブ長さ等の因子を適合させるために必要な温度、イオン強度等の調節の仕方は、当業者に公知である。
【0084】
ここで用いる“組換え体”は、異種核酸配列の導入により修飾された、または細胞がそのように修飾された細胞に由来する、細胞またはベクターに関するものを含む。従って、例えば組換え細胞は、意図的なヒトの介入の結果として発現または発現されない条件下で、当該細胞の天然型(非組換え体)内で同一の形態で見られない遺伝子を発現し、または異常発現される天然遺伝子を発現する。“組換え”または“組換え”核酸の生成は通常、2以上の核酸断片の組み立てであり、当該組み立てによりキメラ遺伝子が生じる。
【0085】
好ましい実施態様において、変異DNA断片は少なくとも1のコドンにおいて部位飽和性(site saturation)突然変異誘発を用いて生じる。他の好ましい実施態様において、部位飽和性突然変異誘発は2以上のコドンについて行われる。さらなる実施態様において、変異DNA配列は野生型配列と40%以上、45%以上、50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上または98%以上の相同性を有する。別の実施態様において、変異DNAは公知の変異誘発手順、例えば、放射、ニトロソグアニジン等を用いてin vivoで生じる。それから望ましいDNA配列は単離し、ここに提供する方法において用いられる。
【0086】
別の実施態様において、形質転換DNA配列は入来配列が存在しない相同ボックスを含む。この実施態様において、2つの相同ボックス間の内生DNA配列を欠失することが望ましい。さらに、いくつかの実施態様において、形質転換する配列は野生型であり、一方他の実施態様において、それらは変異または修飾配列である。さらに、いくつかの実施態様において、形質転換する配列は相同であり、一方他の実施態様において、それらは異種のものである。
【0087】
ここで用いる“標的配列”の語は、入来配列が宿主細胞ゲノム内に挿入されることが望ましい配列をエンコードする宿主細胞中のDNA配列をいう。いくつかの実施態様において、標的配列は機能野生型遺伝子またはオペロンをエンコードし、一方、他の実施態様において、標的配列は機能変異遺伝子またはオペロンまたは非機能遺伝子またはオペロンをエンコードする。
【0088】
ここで用いる“フランキング配列”とは対象の配列の上流または下流にある配列をいう(例えば、遺伝子A−B−Cは、遺伝子BはA及びC遺伝子配列にフランキング(側面に位置)している)。好ましい実施態様において、入来配列は相同ボックスの両側の側面に位置する。他の実施態様において、入来配列及び相同ボックスはstuffer配列の両側の側面に位置する単位を含む。いくつかの実施態様において、フランキング配列は一方の側(3’または5’)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。各相同ボックスの配列はバチルス染色体中の配列に相同である。これらの配列はバチルス染色体に新しい構築体が統合されること、及びバチルス染色体の一部が入来配列により置換されることを目的とする。好ましい実施態様において、選択マーカーの5’及び3’末端は不活性化染色体断片の一部を含むポリヌクレオチド配列の側面に位置する。いくつかの実施態様において、フランキング配列は一方の側(3’または5’)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。
【0089】
ここで用いる“stuffer配列”の語は相同ボックスの側面に位置する任意の余分なDNAをいう(通常ベクター配列)。しかしながら、当該用語は非相同DNA配列を包含する。いかなる理論にも限定するものではないが、stuffer配列はDNA取込みを開始するために細胞に重要でない標的を提供する。
【0090】
ここで用いる、“変異体ライブラリー”の語は、そのゲノムのほとんどは同一だが、1以上の遺伝子が異なる相同体を含む細胞集団をいう。当該ライブラリーは例えば、改善形質を有する遺伝子またはオペロンを同定するための方法において使用できる。
【0091】
ここで用いる“ハイパーコンピテント”及び“スーパーコンピテント”とは、細胞集団の1%以上が染色体DNA(例えば、バチルスDNA)と形質転換可能であることを意味する。もしくは、当該用語は細胞集団の10%以上が自己複製プラスミド(例えば、バチルスプラスミド)と形質転換可能である細胞集団に関して用いられる。好ましくは、スーパーコンピテント細胞は野生型または親細胞集団について観測されるよりも大きな割合で形質転換される。スーパーコンピテント及びハイパーコンピテントはここでは交換可能に用いられる。
【0092】
ここで用いる、“増幅”及び“遺伝子増幅”の語は特定DNAが増幅遺伝子が初期ゲノムに存在した数より多いコピー数で存在するように偏って複製されるプロセスをいう。いくつかの実施態様において、薬物存在下(例えば、阻害可能酵素のインヒビター)での増殖による細胞選択は、薬物存在下での増殖に必要な遺伝子産物をエンコードする内生遺伝子の増幅、またはこの遺伝子産物をエンコードする外来(すなわち、入来)配列の増幅または両方をもたらす。
【0093】
“増幅”はテンプレート特異性に関する核酸複製の特殊なケースである。これは非特異性テンプレート複製(すなわち、テンプレート依存性であるが特定テンプレートに依存しない複製)と対比される。テンプレート特異性は複製の忠実度(すなわち、適正なポリヌクレオチド配列の合成)及びヌクレオチド(リボ−またはデオキシリボ−)特異性とはここでは区別される。テンプレート特異性は“標的”特異性に関して説明されることが多い。標的配列は、それらがその他の核酸から選別するために探されるという意味において“標的”である。増幅技術はこの選別を第一に考えて設計されてきた。
【0094】
ここで用いる“共増幅”の語は、その他の遺伝子配列が一緒に導入される増幅マーカーの1の細胞内への導入(すなわち、発現ベクター内に含まれる遺伝子のような1以上の非選択遺伝子を含む)及び細胞が増幅マーカーとその他の非選択遺伝子配列の両方を増幅するような適当な選択圧の増幅をいう。増幅マーカーはその他の遺伝子配列に物理的に結合していてもよく、またはDNAが2つの別個の断片、すなわち一方が増幅マーカーを含み、及びもう一方が非選択マーカーを含むものが同じ細胞内に導入されてもよい。
【0095】
ここで用いる、“増幅マーカー”、“増幅遺伝子”及び“増幅ベクター”の語は適当な増殖条件下で遺伝子の増幅を可能にする遺伝子をエンコードする遺伝子またはベクターをいう。
【0096】
“テンプレート特異性”はほとんどの増幅技術において酵素選択により達成される。増幅酵素は使用される条件下において、核酸の外来混合物において核酸の特定配列のみ処理する酵素である。例えば、Qβレプリカーゼの場合、MDV−1 RNAはレプリカーゼの特異テンプレートである(例えば、Kacian et al.、Proc.Natl.Acad.Sci.USA 69:3038[1972]を参照)。その他の核酸はこの増幅酵素によっては複製されない。同様に、T7 RNAポリメラーゼの場合、この増幅酵素はそれ自身のプロモーターにストリンジェントな特異性を有する(Chamberlin et al.、Nature 228:227[1970]を参照)。T4 DNAリガーゼの場合、当該酵素は2つのオリゴヌクレオチドまたはポリヌクレオチドを連結せず、ここで連結接合部分にオリゴヌクレオチドまたはポリヌクレオチド基質とテンプレート間の不整合が存在する(Wu and Wallace,Genomics 4:560[1989]を参照)。最後に、Taq及びPfuポリメラーゼは、高温で機能する能力を有するため、配列結合の高い特異性を示し、従って、プライマーにより確定され;高温が標的配列とのプライマーハイブリダイゼーションに有利に働き、非標的配列とのハイブリダイゼーションには有利でない熱力学的条件を生じる。
【0097】
ここで用いる“増幅核酸”の語は任意の増幅方法により増幅され得る核酸をいう。“増幅核酸”は通常“サンプルテンプレート”を含むことが考えられる。
【0098】
ここで用いる“サンプルテンプレート”の語は、“標的”(下記に定義)の存在に関して分析されるサンプルから生じる核酸をいう。一方、“背景テンプレート”はサンプルテンプレート以外の核酸に関して用いられ、サンプル内に存在していてもいなくてもよい。背景テンプレートはほとんどの場合、故意のものではない。それは以前のものからの影響の結果であり、またはサンプルから精製すべき核酸汚染物質の存在によるものである。例えば、検出すべき微生物以外の生物由来の核酸は試験サンプル内で背景として存在し得る。
【0099】
ここで用いる、“プライマー”の語は、精製制限消化などにおいて自然に生じるまたは合成的に生成されるオリゴヌクレオチドをいい、核酸鎖に相補的なプライマー伸長生成物の合成が導入される条件下に置かれた場合(すなわち、ヌクレオチド及びDNAポリメラーゼなどの誘発剤の存在下、及び適した温度及びpHで)、合成開始点として作用できる。当該プライマーは好ましくは増幅効率を最大にする1本鎖であるが、あるいは2本鎖であってもよい。2本鎖の場合、プライマーは伸長生成物の準備のために用いる前にそのストランドを分離処理する。好ましくは、プライマーはオリゴデオキシリボヌクレオチドである。プライマーは誘発剤の存在下、伸長生成物の合成開始のため十分に長くなければならない。プライマーの正確な長さは、誘発温度、プライマーの供給源及び使用する方法など多くの因子に左右される。
【0100】
ここで用いる“プローブ”の語は、精製制限消化などにおいて自然に生じる、または合成、組換えまたはPCR増幅により生成されるオリゴヌクレオチド(すなわちヌクレオチド配列)をいい、別の目的オリゴヌクレオチドにハイブリダイズできる。プローブは1本鎖または2本鎖である。プローブは特定遺伝子配列の検出、同定及び単離に有用である。本発明で使用されるいかなるプローブも任意の“リポーター分子”で標識するので、任意の検出システムにおいて検出可能であり、限定されないが、酵素(例えば、ELISA及び酵素ベースの組織化学的分析)、蛍光、放射性及び発光性システムなどがある。本発明はいかなる特定の検出システムまたは標識にも限定されない。
【0101】
ここで用いる“標的”の語はポリメラーゼ連鎖反応に関して用いる場合、ポリメラーゼ連鎖反応に用いるプライマーが結合する核酸領域をいう。従って、“標的”はその他の核酸配列から選別するために探求されるものである。“断片”は標的配列内の核酸領域として定義される。
【0102】
ここで用いる“ポリメラーゼ連鎖反応”(“PCR”)の語はここに引用する米国特許第4,683,195号、第4,683,202号及び第4,965,188号の方法をいい、クローニングまたは精製することなくゲノムDNAの混合物における標的配列の断片濃度を増加させる方法を含む。標的配列を増幅するこの方法は、所望の標的配列を含むDNA混合物に過剰な2つのオリゴヌクレオチドプライマーを導入し、DNAポリメラーゼの存在下、サーモサイクルの正確な配列を行うことからなる。2つのプライマーは2本鎖標的配列のそれぞれの鎖に相補的である。増幅をもたらすために、混合物を変性させ、それからプライマーを標的分子内のそれらの相補配列にアニールする。アニーリングに続いて、プライマーは新しい相補鎖ペアを形成するためにポリメラーゼを用いて伸長する。変性、プライマーアニーリング及びポリメラーゼ伸長工程は何回も繰り返すことができ(すなわち変性、アニーリング、及び伸長は1“サイクル”を構成し、多数の“サイクル”を行うことができる)、所望の標的配列の増幅断片を高濃度で得ることができる。所望の標的配列の増幅断片の長さはお互いのプライマーの相対位置により決定され、従って、この長さは制御可能なパラメーターである。当該プロセスの繰返し側面のため、当該方法は“ポリメラーゼ連鎖反応”(以下“PCR”とする)と呼ばれる。標的配列の所望の増幅断片は混合物中で主要な配列(濃度に関して)となるので、それらは“PCR増幅された”という。
【0103】
ここで用いる“増幅試薬”の語は、プライマー、核酸テンプレート及び増幅酵素以外の増幅に必要な試薬をいう(デオキシリボヌクレオチド3リン酸、バッファー等)。通常、増幅試薬はその他の反応成分と一緒に反応容器(試験管、マイクロウェル等)内に加えて含まれる。
【0104】
PCRを用いて、ゲノムDNA中の特異標的配列の1のコピーをいくつかの異なる方法で検出可能なレベルまで増幅することが可能である(例えば、標識プローブを用いるハイブリダイゼーション;ビオチン化プライマーの混合、続いてアビジン−酵素結合検出;dCTPまたはdATPなどの32P標識デオキシヌクレオチド3リン酸の増幅断片への混合)。ゲノムDNAに加えて、オリゴヌクレオチドまたはポリヌクレオチド配列が適当なプライマー分子のセットを用いて増幅できる。特に、PCRプロセスにより作成された増幅断片はそれ自身、続くPCR増幅の効率的なテンプレートである。
【0105】
ここで用いる“PCR生成物”“PCR断片”及び“増幅生成物”の語は変性、アニーリング及び伸長が完結する2以上のサイクルのPCR工程後の化合物の最終混合物をいう。これらの語は1以上の標的配列の1以上の断片の増幅が行われる場合を含む。
【0106】
ここで用いる“RT−PCR”の語はRNA配列の複製及び増幅をいう。この方法では、ここに引用する米国特許第5,322,770号に記載されているように、ほとんどの場合熱安定性ポリメラーゼを用いる1の酵素手順を用いて、逆転写とPCRを組み合わせる。RT−PCRにおいて、RNAテンプレートはポリメラーゼの逆転写酵素活性によりcDNAに転換され、それからポリメラーゼの重合活性を用いて増幅させる(すなわち、他のPCR法と同様)。
【0107】
ここで用いる“制限エンドヌクレアーゼ”及び“制限酵素”の語は細菌性酵素をいい、それぞれ特定ヌクレオチド配列で、またはその近くで2本鎖DNAを切断する。
【0108】
“制限酵素認識部位”とは所定の制限エンドヌクレアーゼにより認識及び切断されるヌクレオチド配列をいい、DNA断片の挿入部位であることが多い。本発明の特定の実施態様において、制限酵素認識部位は選択マーカー内及びDNA構築体の5’及び3’末端内に設計される。
【0109】
ここで用いる“不活性化染色体断片”とは2つの部分を含む。各部分は、ここに定義するように固有の染色体領域にすぐ隣接するゲノム染色体DNAの上流または下流に相同なポリヌクレオチドを含む。“すぐ隣接する”とは、不活性化染色体断片を含むヌクレオチドが固有染色体領域を画定するヌクレオチドを含まないことを意味する。不活性化染色体断片はバチルス染色体中にDNA構築体を統合し、バチルス染色体の一部を置換することを目的とする。
【0110】
ここで用いる“固有染色体領域”及び“固有染色体領域の断片”とは本発明のいくつかの実施態様においてバチルス宿主細胞から欠失されるバチルス染色体の断片をいう。通常、“断片”、“領域”、“部分”及び“要素”の語はここでは交換可能に用いられる。いくつかの実施態様において、欠失断片は公知の機能を有する1以上の遺伝子を含み、一方他の実施態様において、欠失断片は未知の機能を有する1以上の遺伝子を含み、及び他の実施態様において、欠失断片は公知及び未知の機能を含む遺伝子の組み合わせを含む。いくつかの実施態様において、固有染色体領域またはそれらの断片は200程度またはそれ以上の遺伝子を含む。
【0111】
いくつかの実施態様において、固有染色体領域またはそれらの断片は特定の条件下で必要な機能を有するが、当該領域は研究室条件下では必ずしもバチルス株生存に必須のものではない。好ましい研究室条件は、限定されないが、標準温度及び大気条件(例えば好気性)での発酵槽、振とうフラスコ中、プレート培地上等での成長などの条件がある。
【0112】
固有染色体領域またはそれらの断片は約0.5kb〜500kb、約1.0kb〜500kb、約5kb〜500kb、約10kb〜500kb、約10kb〜200kb、約10kb〜100kb、約10kb〜50kb、約100kb〜500kb、約200〜500kbの範囲のバチルス染色体を含む。他の側面において、固有染色体領域またはそれらの断片が欠失された場合、変異バチルス株の染色体は99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、85%、80%、75%または70%の対応する非変異バチルス宿主染色体を含むことができる。好ましくは、本発明に従う変異バチルス株の染色体は約99〜90%、99〜92%及び98〜94%の対応する非変異バチルス宿主株染色体ゲノムを含む。
【0113】
ここで用いる“株生存性”とは繁殖可能性をいう。固有染色体領域またはそれらの断片の欠失は研究所条件下において変異バチルス株の分割及び生存に有害な影響を与えるものではない。
【0114】
ここで用いる、“変異バチルス株”とは遺伝子組換えバチルス種をいい、ここで目的タンパク質は本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現及び/または生成レベルが増大している。いくつかの実施態様において、発現レベルの増加は1以上の染色体遺伝子の不活性化により生じる。1の実施態様において、発現レベルの増加は1以上の染色体遺伝子の欠失により生じる。いくつかの実施態様において、変異バチルス株は1以上の固有染色体領域またはそれらの断片が欠失した遺伝子組換えバチルス種であり、ここで目的タンパク質は本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現または生成レベルが増大している。別の実施態様において、発現レベルの増加は1以上の染色体遺伝子の挿入不活性化により生じる。いくつかの別の実施態様において、発現レベルの増加は活性の増加または最適化遺伝子により生じる。いくつかの好ましい実施態様において、不活性化遺伝子はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される。
【0115】
特定の実施態様において、変異バチルス株は不活性化遺伝子を2つ含み、一方他の実施態様において不活性化遺伝子3つ、不活性化遺伝子4つ、不活性化遺伝子5つ、不活性化遺伝子6つまたはそれ以上存在する。従って、不活性化遺伝子の数は特定の遺伝子数に限定されない。いくつかの実施態様において、不活性化遺伝子はお互いに連続であり、一方他の実施態様において、それらはバチルス染色体の離れた領域に位置する。いくつかの実施態様において、不活性化染色体遺伝子は特定の条件下において必要な機能を有するが、当該遺伝子は研究所条件下において必ずしもバチルス株生存に必須のものではない。好ましい研究室条件は、限定されないが、微生物の成長に適した発酵槽、振とうフラスコ中、プレート培地上等での成長などの条件がある。
【0116】
ここで用いる“対応する非変異バチルス株”とは、固有染色体領域またはそれらの断片がそこから欠失または修飾される、及び変化株が生成する宿主株(例えば、最初の及び/または野生型株)である。
【0117】
ここで用いる“染色体統合”の語は入来配列が宿主細胞(例えば、バチルス)の染色体内に導入されるプロセスをいう。形質転換DNAの相同領域は染色体の相同領域と一致する。続いて、相同ボックス間の配列を二重交差の入来配列で置換する(すなわち、相同組換え)。本発明のいくつかの実施態様において、DNA構築体の不活性化染色体断片の相同部分はバチルス染色体の固有染色体領域のフランキング相同領域と一致する。続いて、固有染色体領域は二重交差でDNA構築体により欠失される(すなわち相同組換え)。
【0118】
“相同組換え”とは、同一またはほぼ同一のヌクレオチド配列部位での2つの分子間または対の染色体間のDNA断片の交換を意味する。好ましい実施態様において、染色体統合は相同組換えである。
【0119】
ここで用いる“相同配列”は比較のために最適に配列した場合、他の核酸またはポリぺプチド配列と100%、99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、88%、85%、80%、75%または70%配列同一性を有する核酸またはポリぺプチド配列を意味する。いくつかの実施態様において、相同配列は85%〜100%の配列同一性を有し、一方他の実施態様において、90%〜100%の配列同一性、及びより好ましい実施態様において、95%〜100%の配列同一性を有する。
【0120】
ここで用いる“アミノ酸”とは、ぺプチドまたはタンパク質配列またはそれらの一部をいう。“タンパク質”、“ぺプチド”及び“ポリぺプチド”の語は交換可能に用いられる。
【0121】
ここで用いる“目的タンパク質”及び“目的ポリぺプチド”とは望ましい及び/または評価対象のタンパク質/ポリぺプチドをいう。いくつかの実施態様において、目的タンパク質は細胞内にあり、一方他の実施態様において、それは分泌ポリぺプチドである。特に好ましいポリぺプチドは酵素を含み、限定されないが、でんぷん分解酵素、タンパク質分解酵素、細胞分解(cellulytic)酵素、酸化還元酵素及び植物細胞壁分解酵素から選択される酵素である。より具体的には、これらの酵素は限定されないが、アミラーゼ、プロテアーゼ、キシラナーゼ、リパーゼ、ラッカーゼ、フェノールオキシダーゼ、オキシダーゼ、クチナーゼ、セルラーゼ、ヘミセルラーゼ、エステラーゼ、ペルオキシダーゼ、カタラーゼ、グルコースオキシダーゼ、フィターゼ、ペクチナーゼ、グルコシダーゼ、イソメラーゼ、トランスフェラーゼ、ガラクトシダーゼ及びキチナーゼを含む。本発明のいくつかの特に好ましい実施態様において、目的ポリぺプチドはプロテアーゼである。いくつかの実施態様において、目的タンパク質は1のぺプチドに融合される分泌ポリぺプチドである(すなわち、分泌されるタンパク質上でのアミノ末端伸長)。ほとんど全ての分泌タンパク質は、膜透過の前駆体タンパク質のターゲッティング及び翻訳において重要な役割を果たすアミノ末端タンパク質伸長を用いる。この伸長は、膜移動の間またはそのすぐ後にシグナルぺプチダーゼによりタンパク質分解により除去される。
【0122】
本発明のいくつかの実施態様において、目的ポリぺプチドはホルモン、抗体、成長因子、受容体等から選択される。ホルモンは本発明に含まれ、限定されないが、卵胞刺激ホルモン、黄体形成ホルモン、副腎皮質刺激ホルモン放出因子、ソマトスタチン、ゴナドトロピンホルモン、バソプレシン、オキシトシン、エリスロポエチン、インスリン等が挙げられる。成長因子は限定されないが、血小板由来成長因子、インスリン様成長因子、上皮成長因子、神経成長因子、線維芽細胞成長因子、形質転換成長因子、サイトカイン、例えばインターロイキン(例えば、IL−1からIL−13)、インターフェロン、コロニー刺激因子等が挙げられる。抗体は限定されないが、抗体を生成するのが望ましい任意の種から直接得られる免疫グロブリンが挙げられる。さらに、本発明は修飾抗体を含む。ポリクローナル及びモノクローナル抗体も本発明に含まれる。特に好ましい実施態様において、抗体はヒト抗体である。
【0123】
ここで用いる“異種タンパク質”の語は宿主細胞に天然に生じないタンパク質またはポリぺプチドをいう。異種タンパク質の例としては、加水分解酵素、例えばプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ及びリパーゼなど;イソメラーゼ、例えばラセマーゼ、エピメラーゼ、トートメラーゼまたはムターゼなど;トランスフェラーゼ、キナーゼ及びホスファターゼなどの酵素を含む。いくつかの実施態様において、タンパク質は治療的に重要なタンパク質またはぺプチドであり、限定されないが、成長因子、サイトカイン、配位子、受容体及びインヒビター並びにワクチン及び抗体を含む。別の実施態様において、タンパク質は商業的に重要な工業タンパク質/ぺプチドである(例えば、プロテアーゼ、アミラーゼ及びグルコアミラーゼなどのカルボヒドラーゼ、セルラーゼ、オキシダーゼ及びリパーゼ)。いくつかの実施態様において、タンパク質をエンコードする遺伝子は天然遺伝子であり、一方他の実施態様において、変異及び/または合成遺伝子が使用される。
【0124】
ここで用いる“相同タンパク質”とは細胞内の野生型または天然タンパク質またはポリぺプチドをいう。好ましい実施態様において、当該細胞はグラム陽性細胞であり、一方特に好ましい実施態様において、当該細胞はバチルス宿主細胞である。別の実施態様において、相同タンパク質はその他の生物、限定されないが大腸菌などにより生成された天然タンパク質である。本発明は組換えDNA技術により相同タンパク質を生成する宿主細胞を包含する。
【0125】
ここで用いる“オペロン領域”は、共有プロモーターからの1の転写単位として転写され、それにより共調節される隣接遺伝子のグループを含む。いくつかの実施態様において、オペロンは調節遺伝子を含む。最も好ましい実施態様において、RNAレベルで測定して高度に発現するが未知または不必要な機能を有するオペロンが用いられる。
【0126】
ここで用いる“複数−隣接単一遺伝子領域”とは、少なくとも2つの遺伝子のコード領域が一列になって起こる領域であり、及びいくつかの実施態様において、コード領域に先行及び後に続く介在配列を含む。いくつかの実施態様において、抗菌領域を導入する。
【0127】
ここで用いる“抗菌領域”は抗菌タンパク質をエンコードする少なくとも1の遺伝子を含む領域である。
【0128】
発明の詳細な説明
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。実際、本発明は1または複数の遺伝子を欠失する方法、及び大量の染色体を欠失させる方法を提供する。好ましい実施態様において、当該欠失は目的タンパク質の生成向上などの利点を提供する。
【0129】
A.遺伝子欠失
上述の通り、本発明は1または複数の遺伝子欠失及び/または変異及び大量の染色体欠失に関する実施態様を含む。
【0130】
いくつかの好ましい実施態様において、本発明は入来配列を含むDNA構築体を含む。DNA構築体はin vitroで組み立てられ、それからコンピテントバチルス宿主内に当該構築体の直接クローニングを行い、DNA構築体がバチルス染色体内に統合されるようにする。例えば、PCR融合及び/または連結はDNA構築体をin vitroで組み立てるために用いることができる。いくつかの実施態様において、DNA構築体は非プラスミド構築体であり、一方他の実施態様において、ベクター(例えばプラスミド)内に組み込まれる。いくつかの実施態様において、環状プラスミドを使用する。好ましい実施態様において、環状プラスミドは適当な制限酵素を用いるために設計される(すなわち、DNA構築を阻害しないもの)。従って、線状プラスミドは本発明で使用できる(図1を参照)。しかしながら、その他の方法も本発明での使用に適しており、当業者に公知である(例えば、Perego,“Integration Vectors for Genetic Manipulation in Bacillus subtilis(バチルス・ズブチリスにおける遺伝子操作のための統合ベクター)”、(Sonenshein et al.(編集)、Bacillus subtilis and Other Gram−Positive Bacteria(バチルス・ズブチリス及びその他のグラム陽性細菌),American Society for Microbiology,ワシントン、DC[1993]を参照)。
【0131】
いくつかの実施態様において、入来配列は選択マーカーを含む。いくつかの好ましい実施態様において、入来配列は、sbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDまたはこれらの遺伝子のいずれかの断片(単独または組み合わせ)からなる群より選択される染色体遺伝子を含む。別の実施態様において、入来配列は相同sbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及び/またはrocD遺伝子配列を含む。相同配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD遺伝子またはそれらの遺伝子断片と少なくとも99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、88%、85%、または80%配列同一性を有する核酸配列であり、入来配列に含まれる。好ましい実施態様において、相同配列を含む入来配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocFまたはrocD遺伝子またはそれらの遺伝子断片と少なくとも95%配列同一性を有する。さらに別の実施態様において、入来配列は遺伝子配列の断片を有する5’及び3’末端の側面に位置する選択マーカーを含む。いくつかの実施態様において、DNA構築体が選択マーカー及び遺伝子を含む場合、遺伝子断片またはそれに相同な配列は宿主細胞内に形質転換され、選択マーカーの配置は当該遺伝子の本来の目的に関して非機能的なものにする。いくつかの実施態様において、入来配列は遺伝子のプロモーター領域に位置する選択マーカーを含む。他の実施態様において、入来配列は遺伝子のプロモーター領域の後に位置する選択マーカーを含む。さらに他の実施態様において、入来配列は遺伝子のコード領域に位置する選択マーカーを含む。さらに別の実施態様において、入来配列は両末端上の相同ボックスの側面に位置する選択マーカーを含む。さらに別の実施態様において、入来配列はコード配列の転写及び/または翻訳を妨害する配列を含む。さらに別の実施態様において、DNA構築体は当該構築体の上流及び下流末端に設計された制限酵素認識部位を含む。
【0132】
DNA構築体はベクター内に組み込まれ、またはプラスミドDNAの不存在下で使用されるかに関わらず、微生物を形質転換するために使用される。形質転換に適したいかなる方法も本発明で使用できる。好ましい実施態様において、少なくとも1のDNA構築体のコピーが宿主バチルス染色体内に統合される。いくつかの実施態様において、本発明の1以上のDNA構築体は宿主細胞を形質転換するために用いる。例えば、1のDNA構築体はs/r遺伝子を不活性化するために用い、他の構築体はphrC遺伝子を不活性化するために用いることができる。もちろん、別の組み合わせも考えられ、本発明により提供される。
【0133】
いくつかの好ましい実施態様において、DNA構築体は目的タンパク質をエンコードするポリヌクレオチドも含む。これらの好ましい実施態様のいくつかにおいて、DNA構築体は目的タンパク質をエンコードする配列に作動可能に連結する構成または導入プロモーターも含む。目的タンパク質がプロテアーゼであるいくつかの好ましい実施態様において、プロモーターはtacプロモーター、β−ラクタマーゼプロモーター、またはaprEプロモーター(DeBoer et al.、Proc.Natl.Acad.Sci.USA 80:21−25[1983])からなる群より選択される。しかしながら、本発明はいかなる特定のプロモーターにも限定されるものではなく、当業者に公知の任意の適したプロモーターが本発明で使用できる。とはいえ、特に好ましい実施態様において、プロモーターはB.ズブチリスaprEプロモーターである。
【0134】
バチルス種の形質転換に関する様々な方法が公知である。実際に、プラスミド構築体及び当該プラスミドを大腸菌内に形質転換することを含むバチルス染色体を改変させる方法が公知である。ほとんどの方法において、プラスミドは大腸菌からその後単離され、バチルス内に形質転換される。しかしながら、大腸菌などの介在微生物を使用することは必須ではなく、いくつかの好ましい実施態様においてDNA構築体はコンピテントバチルス宿主内に直接形質転換される。
【0135】
いくつかの実施態様において、公知のバチルスズブチリス株168は本発明で使用できる。実際にこの株のゲノムはよく特性付けられている(Kunst et al.、Nature 390:249−256[1997];及びHenner et al.、Microbiol.Rev.,44:57−82[1980]を参照)。ゲノムは4215kb染色体からなる。ここで用いる統合は168株に言及するが、本発明はバチルス株に類似の配列を包含する。
【0136】
いくつかの実施態様において、入来染色体配列は、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD、それらの遺伝子断片及びそれらに相同な配列からなる群より選択される1以上の遺伝子を含む。B.ズブチリス168由来のこれら遺伝子のDNAコード配列は配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号29、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33において提供される。
【0137】
上述の通り、いくつかの実施態様において、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh、kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD遺伝子、それらの遺伝子断片またはそれらに相同な配列を含む入来配列はコード領域を含み、すぐ隣に染色体コード領域フランキング配列をさらに含むことができる。いくつかの実施態様において、コード領域フランキング配列は約1bp〜2500bp;約1bp〜1500bp、約1bp〜1000bp、約1bp〜500bp、及び1bp〜250bpの範囲を含む。コード領域フランキング配列を含む核酸配列の数は遺伝子コード配列の各末端ごとに異なる。例えば、いくつかの実施態様において、コード配列の5’末端は25bp以下を含み、コード配列の3’末端は100bp以上を含む。これらの遺伝子の配列及び遺伝子産物を以下に示す。ここで用いる番号はsubtilistで用いられるものである(例えば、Moszer et al.、Microbiol.,141:261−268[1995]を参照)。
【0138】
B.ズブチリス168のsboコード配列を以下に示す:
【式3】
【0139】
sboの推定アミノ酸配列は:
【式4】
【0140】
1の実施態様において、B.ズブチリス168染色体の約3834868〜3835219で見られる遺伝子領域は本発明を用いて欠失させた。約3835081から3835209で見られるsboコード領域はズブチリシンA、すなわちいくつかのグラム陽性細菌に対する活性を有する抗菌薬を生成する(Zheng et al.、J.Bacteriol.,181:7346−7355[1994]を参照)。
【0141】
B.ズブチリス168のslrコード配列を以下に示す:
【式5】
【0142】
Slrの推定アミノ酸配列は:
【式6】
【0143】
1の実施態様において、B.ズブチリス168染色体の約3529014−3529603bpで見られる配列を本発明を用いて欠失させた。slrコード領域は染色体の約3529131から3529586で見られる。
【0144】
B.ズブチリス168のphrCコード配列を以下に示す:
【式7】
【0145】
phrCの推定アミノ酸配列は:
【式8】
【0146】
さらに、B.ズブチリス168染色体の約429531−429650bpで見られるコード領域をコード配列の429591で選択マーカー挿入により不活性化させた。
【0147】
B.ズブチリス168のsigBコード配列を以下に示す:
【式9】
【0148】
sigBの推定アミノ酸配列は:
【式10】
【0149】
さらに、当該コード配列はB.ズブチリス168染色体の約522417から5232085bpで見られる。
【0150】
B.ズブチリス168のspollSAコード配列を以下に示す:
【式11】
【0151】
spollSAの推定アミノ酸配列は:
【式12】
【0152】
さらに、当該コード領域はB.ズブチリス168染色体の約1347587から1348714bpで見られる。
【0153】
B.ズブチリス168のcsnコード配列を以下に示す:
【式13】
【0154】
Csnの推定アミノ酸配列は:
【式14】
【0155】
さらに、当該コード領域はB.ズブチリス168染色体の約2747213から2748043bpで見られる。
【0156】
B.ズブチリス168のybcOコード配列を以下に示す:
【式15】
【0157】
ybcOの推定アミノ酸配列は:
【式16】
【0158】
さらに、当該コード領域はB.ズブチリス168染色体の約213926から214090bpで見られる。
【0159】
B.ズブチリス168のrapAコード配列を以下に示す:
【式17】
【0160】
RapAの推定アミノ酸配列は:
【式18】
【0161】
さらに、当該コード領域はB.ズブチリス168染色体の約1315179から1316312bpで見られる。
【0162】
B.ズブチリス168のCssコード配列を以下に示す:
【式19】
【0163】
Cssの推定アミノ酸配列は(GenBankアクセッション番O32193):
【式20】
【0164】
さらに、当該コード領域はB.ズブチリス168染色体の約3384612から3386774bpで見られる。
【0165】
B.ズブチリス168のFbpタンパク質(フルクトース−1,6−ビオオスファターゼ(biophosphatase))のfbpコード配列を以下に示す:
【式21】
【0166】
Fbpタンパク質の推定アミノ酸配列は:
【式22】
【0167】
さらに、当該コード領域はB.ズブチリス168染色体の約4127053から4129065bpで見られる。
【0168】
B.ズブチリス168のalsDタンパク質(α−アセト乳酸デカルボキシラーゼ)のalsDコード配列を以下に示す:
【式23】
【0169】
alsDタンパク質配列の推定アミノ酸配列は:
【式24】
【0170】
さらに、当該コード領域はB.ズブチリス168染色体の約3707829から3708593bpで見られる。
【0171】
B.ズブチリス168のgapBタンパク質(グリセルアルデヒド−3−リン酸デヒドロゲナーゼ)のgapBコード配列を以下に示す:
【式25】
【0172】
GapBタンパク質の推定アミノ酸配列は:
【式26】
【0173】
さらに、当該コード領域はB.ズブチリス168染色体の約2966075から2967094bpで見られる。
【0174】
Kblタンパク質(2−アミノ−3−ケトブチラートCoAリガーゼ)のKblコード配列を以下に示す:
【式27】
【0175】
Kblタンパク質の推定アミノ酸配列は:
【式28】
【0176】
さらに、当該コード領域はB.ズブチリス168染色体の約1770787から1771962bpで見られる。
【0177】
B.ズブチリス168のPckA(ホスホエノールピルビン酸カルボキシキナーゼ)のPckAコード配列を以下に示す:
【式29】
【0178】
PckAタンパク質の推定アミノ酸配列は:
【式30】
【0179】
さらに、当該コード領域はB.ズブチリス168染色体の約3128579から3130159bpで見られる。
【0180】
B.ズブチリス168のprpCタンパク質(タンパク質ホスファターゼ)のprpCコード配列を以下に示す:
【式31】
【0181】
prpCタンパク質の推定アミノ酸配列は:
【式32】
【0182】
さらに、当該コード領域はB.ズブチリス168染色体の約1649684から1650445bpで見られる。
【0183】
B.ズブチリス168のrocAタンパク質(ピロリン−5−カルボン酸デヒドロゲナーゼ)のrocAコード配列を以下に示す:
【式33】
【0184】
RocAタンパク質の推定アミノ酸配列は:
【式34】
【0185】
さらに、当該コード領域はB.ズブチリス168染色体の約3877991から3879535bpで見られる。
【0186】
B.ズブチリス168のrocDタンパク質(オルニチンアミノトランスフェラーゼ)のrocDコード配列を以下に示す:
【式35】
【0187】
RocDタンパク質の推定アミノ酸配列は:
【式36】
【0188】
さらに、当該コード領域はB.ズブチリス168染色体の約4143328から4144530bpで見られる。
【0189】
B.ズブチリス168のrocFタンパク質(アルギナーゼ)のrocFコード配列を以下に示す:
【式37】
【0190】
RocFタンパク質の推定アミノ酸配列は:
【式38】
【0191】
さらに、当該コード領域はB.ズブチリス168染色体の約4140738から4141625bpで見られる。
【0192】
B.ズブチリス168のTdhタンパク質(トレオニン−3−デヒドロゲナーゼ)のTdhコード配列を以下に示す:
【式39】
【0193】
Tdhタンパク質の推定アミノ酸配列は:
【式40】
【0194】
さらに、当該コード領域はB.ズブチリス168染色体の約1769731から1770771bpで見られる。
【0195】
トリプトファンオペロン調節領域のコード配列及び遺伝子trpE(配列番号48)、trpD(配列番号46)、trpC(配列番号44)、trpF(配列番号50)、trpB(配列番号42)及びtrpA(配列番号40)は以下に示す。下線部はオペロン調節領域である。太字はtrpE開始(ATG)であり、trpD、trpC、trpF、trpB及びtrpA開始が同様に続く(示した順番に太字で表す)。
【式41】
【0196】
推定TrpAタンパク質(トリプトファンシンターゼ(αサブユニット))の配列は:
【式42】
【0197】
推定TrpBタンパク質(トリプトファンシンターゼ(βサブユニット))の配列は:
【式43】
【0198】
推定TrpCタンパク質(インドール−3−グリセロールリン酸シンターゼ)の配列は:
【式44】
【0199】
推定TrpDタンパク質(アントラニル酸ホスフォリボシルトランスフェラーゼ)の配列は:
【式45】
【0200】
推定TrpEタンパク質(アントラニル酸シンターゼ)の配列は:
【式46】
【0201】
推定TrpFタンパク質(ホスフォリボシルアントラニル酸イソメラーゼ)の配列は:
【式47】
【0202】
さらに、当該コード領域はB.ズブチリス168染色体の約2370707から2376834bp(最初のbp=2376834;最後のbp=2370707)で見られる。
【0203】
B.ズブチリス168のycgMタンパク質(プロリンオキシダーゼに類似)のycgMコード配列を以下に示す:
【式48】
【0204】
ycgMタンパク質の推定アミノ酸配列は:
【式49】
【0205】
さらに、当該コード領域はB.ズブチリス168染色体の約344111から345019bpで見られる。
【0206】
B.ズブチリス168のycgNタンパク質(1−ピロリン−5−カルボン酸デヒドロゲナーゼに類似)のycgNコード配列を以下に示す:
【式50】
【0207】
ycgNタンパク質の推定アミノ酸配列は:
【式51】
【0208】
さらに、当該コード領域はB.ズブチリス168染色体の約345039から346583bpで見られる。
【0209】
B.ズブチリス168のsigDタンパク質(RNAポリメラーゼ鞭毛、運動、走化性及び自己分解シグマ因子)のsigDコード配列を以下に示す:
【式52】
【0210】
sigDの推定アミノ酸配列は:
【式53】
【0211】
さらに、当該コード領域はB.ズブチリス168染色体の約1715786から1716547bpで見られる。
【0212】
上述の通り、その他のバチルス宿主で見られる不活性化類似遺伝子は本発明で使用できると考えられる。
【0213】
いくつかの好ましい実施態様において、宿主細胞はバチルス属のメンバーであり、一方他の実施態様において、目的バチルス株は好アルカリ性である。多数の好アルカリ性バチルス株が公知である(例えば、米国特許第5,217,878号及びAunstrup et al.、Proc IV IFS:Ferment.Technol.Today,299−305[1972]を参照)。いくつかの好ましい実施態様において、目的のバチルス株は工業バチルス株である。工業バチルス株の例としては、限定されないが、B.リケニフォルミス、B.レンタス、B.ズブチリス及びB.アミロリケファシエンスが挙げられる。別の実施態様において、バチルス宿主株は、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ及びB.メガテリウム、並びに上述のバチルス属内のその他の微生物からなる群より選択される。いくつかの特に好ましい実施態様において、B.ズブチリスを用いる。例えば、米国特許第5,264,366号及び第4,760,025号(RE34,606)は本発明で使用できる種々のバチルス宿主株について記載しているが、その他の適当な株も本発明に使用できると考えられる。
【0214】
工業株はバチルス種の非組換え体であってもよく、天然株の変異体または組換え体株であってもよい。好ましくは、宿主株は組換え宿主株であり、目的のポリぺプチドをエンコードするポリヌクレオチドが宿主内に導入されている。さらに好ましい宿主株はバチルス・ズブチリス宿主株であり、特に組換えバチルス・ズブチリス宿主株である。多数のB.ズブチリス株が公知であり、限定されないが、1A6(ATCC39085)、168(1A01)、SB19、W23、Ts85、B637、PB1753〜PB1758、PB3360、JH642、1A243(ATCC39,087)、ATCC21332、ATCC6051、MI113、DE100(ATCC39,094)、GX4931、PBT110、及びPEP211株(例えばHoch et al.、Genetics,73:215−228[1973];米国特許第4,450,235号;米国特許第4,302,544及び欧州特許0134048号を参照)が挙げられる。発現宿主としてのB.ズブチリスの使用はさらにPalva et al.などに記載されている(Palva et al.、Gene19:81−87[1982];Fahnestock and Fischer、J.Bacteriol.,165:796−804[1986];及びWang et al.,Gene 69:39−47[1988]を参照)。
【0215】
工業プロテアーゼ生成バチルス株は特に好ましい発現宿主を提供する。いくつかの好ましい実施態様において、本発明でのこれらの株の使用はさらに効率及びプロテアーゼ生成を増大させる。通常バチルス種により2つの一般的な種類のプロテアーゼが分泌され、すなわち、中性(または“メタロプロテアーゼ”)及びアルカリ性(または“セリン”)プロテアーゼである。セリンプロテアーゼはぺプチド結合の加水分解を触媒する酵素であり、活性部位に必須のセリン残基がある。セリンプロテアーゼは分子量が25,000〜30,000である(Priest、Bacteriol.Rev.,41:711−753[1977]を参照)。ズブチリシンは本発明での使用に好ましいセリンプロテアーゼである。多種多様のバチルス・ズブチリシンが同定及び配列決定されており、例えば、ズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147及びズブチリシン309である(例えば、欧州特許第414279号、WO89/06279号;及びStahl et al.、J.Bacteriol.,159:811−818[1984]を参照)。本発明のいくつかの実施態様において、バチルス宿主株は変異(例えば変異体)プロテアーゼを生成する。多数の参考文献が変異体プロテアーゼの例及び参照を提供している(例えば、WO99/20770、WO99/20726、WO99/20769、WO89/06279、RE34,606、米国特許第4,914,031号、米国特許第4,980,288号、米国特許第5,208,158号、米国特許第5,310,675号、米国特許第5,336,611号、米国特許第5,399,283号、米国特許第5,441,882号、米国特許第5,482,849号、米国特許第5,631,217号、米国特許第5,665,587号、米国特許第5,700,676号、米国特許第5,741,694号、米国特許第5,858,757号、米国特許第5,880,080号、米国特許第6,197,567号及び米国特許第6,218,165号を参照)。
【0216】
さらに他の実施態様において、好ましいバチルス宿主は、以下の遺伝子の少なくとも1つにおいて変異または欠失を有するバチルス種である:degU、degS、degR及びdegQ。好ましくは、変異はdegU遺伝子にあり、より好ましくは変異はdegU(Hy)32である。(Msadek et al.,J.Bacteriol.、172:824−834[1990];及びOlmos et al.、Mol.Gen.Genet.,253:562−567[1997]参照)。最も好ましくは宿主株はdegU32(Hy)変異を有するバチルス・ズブチリスである。さらなる実施態様において、バチルス宿主はscoC4(Caldwell et al.、J.Bacteriol.,183:7329−7340[2001]を参照);spollE(Arigoni et al.、Mol.Microbiol.,31:1407−1415[1999]を参照);oppAまたはその他のoppオペロンの遺伝子(Perego et al.、Mol.Mictobiol.,5:173−185[1991]を参照)における変異または欠失を含む。実際に、oppA遺伝子における変異と同じ表現型を生じるoppオペロンにおける変異は本発明の変異バチルス株のいくつかの実施態様において使用できると考えられる。いくつかの実施態様において、これらの変異は単独で起こり、一方他の実施態様において、変異の組み合わせが存在する。いくつかの実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異をすでに含むバチルス宿主株から得られる。別の実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異を含むようにさらに設計される。
【0217】
さらに別の実施態様において、入来配列は2つのloxP部位間に位置する選択マーカーを含み(Kuhn and Torres、Meth.Mol.Biol.,180:175−204[2002]を参照)、抗菌剤はそれからCreタンパク質の活性により欠失される。いくつかの実施態様において、これは、相同フランキングDNA及び抗菌剤含有入来DNAを構築するために用いるプライマーにより測定される1のloxP部位の挿入及び天然DNAの欠失を生じる。
【0218】
当業者はバチルス細胞内にポリヌクレオチド配列を導入する適切な方法をよく知っている(例えば、Ferrari et al.、“Genetics”、Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.[1989]、57〜72頁、また、Saunders et al.、J.Bacteriol.157:718−726[1984];Hoch et al.、J.Bacteriol.、93:1925−1937[1967];Mann et al.、Current Microbiol.,13:131−135[1986];及びHolubova、Folia Microbiol.,30:97[1985];B.ズブチリス、Chang et al.、Mol.Gen.Genet.,168:11−115[1979];B.メガテリウム、Vorobjeva et al.、FEMS Microbiol.Lett.,7:261−263[1980];B.アミロリケファシエンス、Smith et al.、Appl.Env.Microbiol.,51:634(1986);B.チューリンゲンシス、Fisher et al.、Arch.Microbiol.,139:213−217「1981」;及びB.スファエリカス、McDonald、J.Gen.Microbiol.,130:203[1984]を参照)。実際、プロトプラスト形質転換及び配向(congression)、形質導入及びプロプラスト融合などの形質転換の方法は公知であり、本発明での使用に適している。形質転換の方法は特に本発明により提供されるDNA構築体を宿主細胞に導入するために好ましい。
【0219】
一般的に使用される方法に加えて、いくつかの実施態様において、宿主細胞は直接形質転換される(すなわち、媒介細胞を増幅のために使用しない、または別の方法では、DNA構築体を宿主細胞内に前もって導入する)。宿主細胞内へのDNA構築体の導入は、プラスミドまたはベクター内への導入なしで、宿主細胞内に導入するための当業者に公知の物理的及び化学的方法を含む。当該方法は、限定されないが、塩化カルシウム沈殿法、エレクトロポレーション、裸のDNA(naked DNA)、リポソーム等が挙げられる。別の実施態様において、DNA構築体は、プラスミド内に導入されることなく、プラスミドを用いて共形質転換される。さらに別の実施態様において、選択マーカーは当業者に公知の方法により変異バチルス株から欠失される(Stahl et al.、J.Bacteriol.,158:411−418[1984];及びPalmeros et al.、Gene 247:255−264[2000]を参照)。
【0220】
いくつかの実施態様において、宿主細胞は、本発明に従う1以上のDNA構築体を用いて形質転換され、2以上の遺伝子が宿主細胞中で不活性化されている変異バチルス株を生成する。いくつかの実施態様において、2以上の遺伝子は宿主細胞染色体から欠失される。別の実施態様において、2以上の遺伝子はDNA構築体の挿入により不活性化される。いくつかの実施態様において、不活性化遺伝子は連続しており(欠失及び/または挿入による不活性化に関わらず)、一方、他の実施態様において、連続した遺伝子ではない。
【0221】
細胞内及び細胞外で発現したポリぺプチドの活性化を検出及び測定する当業者に公知の様々な分析法が存在する。特に、プロテアーゼに関しては、吸光度280nmで測定、またはFolin法を用いた比色分析により測定したカゼインまたはヘモグロビンからの酸溶性ぺプチドの放出に基づく分析法がある(例えば、Bergmeyer et al.、“Methods of Enzymatic Analysis”vol.5、Peptidases,Proteinases and their Inhibitors(ペプチダーゼ、プロテイナーゼ、及びそれらの阻害因子)、Verlag Chemie、Weinheim「1984」を参照)。その他の分析法は発色基質の可溶化を含む(例えば、Ward、“Proteinases(プロテイナーゼ)”、Fogarty(編集)、Microbial Enzymes and Biotechnology(微生物酵素及びバイオテクノロジー)、Applied Science,ロンドン[1983]、251〜317頁を参照)。その他の分析法の例としては、スクシニル−Ala−Ala−Pro−Phe−paraニトロアニリド分析(SAAPFpNA)及び2,4,6−トリニトロベンゼンスルフォン酸ナトリウム塩分析(TNBS分析)が挙げられる。当業者に公知の多数の別の文献は適切な方法を提供する(例えば、Wells et al.、Nucleic Acids Res.11:7911−7925[1983];Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal Biochem.,242:221−227[1999]を参照)。
【0222】
宿主細胞内の目的タンパク質の分泌レベルの測定方法及び発現タンパク質の検出方法はタンパク質に特異なポリクローナルまたはモノクローナル抗体を用いる免疫学的分析法の使用を含む。例としては、酵素免疫測定法(ELISA)、放射免疫分析法(RIA)、蛍光免疫分析法(FIA)、及び蛍光活性化細胞分類法(FACS)を含む。しかしながら、その他の方法も当業者に公知であり、目的タンパク質の評価に使用できる(例えば、Hampton et al.、Serological Methods,A Laboratory Manual(血清学的方法、研究室マニュアル),APS Press、St.Paul,MN[1990];及びMaddox et al.、J.Exp.Med.、158:1211[1983]を参照)。いくつかの好ましい実施態様において、目的タンパク質の分泌は、対応する非変異宿主よりも本発明を用いて得た変異株におけるほうが高い。当業界で公知の通り、本発明を用いて生成した変異バチルス細胞は発現及び細胞培養物からの目的ポリぺプチドの回収に適した条件下で維持及び成長する(例えば、Harwood and Cutting(編集)Molecular Biological Methods for Bacillus(バチルスの分子生物学的方法)、John Wiley&Sons[1990]を参照)。
【0223】
B.大量の染色体欠失
上述の通り、1及び複数の遺伝子欠失に加えて、本発明は大量の染色体欠失も提供する。本発明のいくつかの好ましい実施態様において、固有染色体領域またはそれらの断片はバチルス宿主細胞から欠失され、変異バチルス株を生成する。いくつかの実施態様において、固有染色体領域はプロファージ領域、抗菌領域(例えば、抗生物質領域)、調節領域、複数連続シングル遺伝子領域及び/またはオペロン領域を含む。ここで言及する固有染色体領域を描写する座標はバチルス・ズブチリス株168染色体地図に従って特定される。数字は通常、存在する場合は、リボソーム結合部位の開始部位に関するものであり、またはコード領域の末端に関し、及び存在するであろうターミネーターは通常含まない。株168のバチルス・ズブチリスゲノムは公知であり(Kunst et al.、Nature 390:249−256[1997];及びHenner et al.、Microbiol.Rev.、44:57−82[1980]を参照)、及び1の4215kb染色体からなる。しかしながら、本発明は任意のバチルス株由来の類似配列も含む。特に好ましくは、その他のB.ズブチリス株、B.リケニフォルミス株及びB.アミロリケファシエンス株である。
【0224】
いくつかの実施態様において、固有染色体領域はプロファージ断片及びそれらの断片を含む。“プロファージ断片”は細菌性染色体内に挿入されたウイルス性DNAであり、ウイルス性DNAは通常の細菌性遺伝子と事実上区別できない。B.ズブチリスゲノムは多数のプロファージ断片からなり、これらの断片は感染性ではない(Seaman et al.、Biochem.,3:607−613[1964];及びStickler et al.、Vitrol.,26:142−145[1965])。バチルス・ズブチリスプロファージ領域のいずれを欠失してもよいが、以下の非限定的例に関して説明する。
【0225】
本発明の実施態様において欠失される1のプロファージ領域はシグマK介在“スキン”要素である。この領域はB.ズブチリス168染色体の約2652600bp(spolVCA)〜2700579bp(yqaB)で見られる。本発明を用いて、染色体の2653562bp〜2699604bpに対応する、約46kb断片を欠失した。この要素は、SIGK ORF内、遺伝子spolVCB〜spolllC間に位置する祖先から受け継いだ溶原性ファージの残余物であると考えられる。しかしながら、本発明は欠失領域に関するいかなる特定のメカニズムまたは作用形態に限定されるものではない。当該要素は推定リボソーム結合部位を含む57のオープン読取りフレームを含むように示されている(Takemaru et al.、Microbiol.,141:323−327 [1995]を参照)。母細胞における胞子形成時に、スキン要素は削除されてsigK遺伝子の再構築を導く。
【0226】
欠失に適したその他の領域はプロファージ7領域である。この領域はB.ズブチリス168染色体の約2701208bp(yrkS)〜2749572bp(yraK)で見られる。本発明を用いて、染色体の2701087bp〜2749642bpに対応する、約48.5kb断片を欠失した。
【0227】
さらなる領域はスキン+プロファージ7領域である。この領域はB.ズブチリス168染色体の約2652151bp〜2749642bp(yraK)で見られる。本発明を用いて、約97.5kbの断片を欠失した。この領域は介在spolllC遺伝子も含む。スキン/プロファージ7領域は、限定されないが、以下の遺伝子を含む:spolVCA−DNAリコンビナーゼ、blt(多剤耐性)、cypA(シトクロムP450様酵素)、czcD(カチオン流出システム膜タンパク質)、及びrapE(応答制御因子アスパラギン酸ホスファターゼ)。
【0228】
さらに別の領域はPBSX領域である。この領域はB.ズブチリス168染色体の約1319884bp(wkdA)〜1347491bp(xlyA)で見られる。本発明を用いて、染色体の1319663bp〜1348691bpに対応する、約29kb断片を欠失した。通常の非導入状態において、このプロファージ要素は非感染性であり、殺菌性ではない(W23及びS31などの数種の敏感な株を除く)。マイトマイシンCが導入可能であり、SOS応答により活性化され、ファージ様粒子を放出する細胞溶解を生じる。ファージ粒子は細菌性染色体DNAを含み、敏感な細菌をDNAを注入することなく殺すことができる(Canosi et al.、J.Gen.Virol.39:81−90[1978])。この領域は以下の遺伝子の非限定的リストを含む:xtmA−B;xkdA−K及びM−X、xre、xtrA、xpf、xep、xhlA−B及びxlyA。
【0229】
別の領域はSPβ領域である。この領域はB.ズブチリス168染色体の約2150824bp(yodU)〜2286246bp(ypqP)で見られる。本発明を用いて、染色体の2151827bp〜2285246bpに対応する、約133.5kb断片を欠失した。この要素は機能がまだ特性付けられていない溶原性プロファージである。しかしながら、この領域の遺伝子は推定上の胞子殻タンパク質(yodU、sspC、yokH)、推定上のストレス応答性タンパク質(yorD、yppQ、ypnP)及び胞子殻における遺伝子と相同性を有するその他の遺伝子及びyomオペロンのメンバーなどストレス応答性遺伝子を含む。その他の遺伝子は以下を含む領域である:yot;yos、yoq、yop、yon、yom、yoz、yol、yok、ypo、及びypm。
【0230】
別の領域はプロファージ1領域である。この領域はB.ズブチリス168染色体の約202098bp(ybbU)〜220015bp(ybdE)で見られる。本発明を用いて、染色体の202112bp〜220141bpに対応する、約18.0kb断片を欠失した。この領域の遺伝子は、DNAアルキル化に適応反応を与えるAdaA/Bオペロン及びNADHデヒドロゲナーゼ、サブユニット5をコードするndhFを含む。
【0231】
別の領域はプロファージ2領域である。この領域はB.ズブチリス168染色体の約529069bp(ybcL)〜569493bp(ydeJ)で見られる。本発明を用いて、染色体の529067bp〜569578bpに対応する、約40.5kb断片を欠失した。この領域の遺伝子はrapl/phrlwp(応答制御因子アスパラギン酸ホスファターゼ)、sacV(レバンスクラーゼの転写調節因子)及びcspCを含む。
【0232】
別の領域はプロファージ3領域である。本発明を用いて、B.ズブチリス168染色体の652000bp〜664300bpに対応する、約50.7kb断片を欠失した。
【0233】
さらに別の領域はプロファージ4領域である。この領域はB.ズブチリス168染色体の約1263017(yjcM)〜1313627bp(yjoA)で見られる。本発明を用いて、染色体の1262987bp〜1313692bpに対応する、約2.3kb断片を欠失した。
【0234】
別の領域はプロファージ5領域である。本発明を用いて、B.ズブチリス168染色体の1879200bp〜1900000bpに対応する、約20.8kb断片を欠失した。
【0235】
別の領域はプロファージ6領域である。本発明を用いて、B.ズブチリス168染色体の2046050bp〜2078000bpに対応する、約31.9kb断片を欠失した。
【0236】
別の実施態様において、固有染色体領域は1以上のオペロン領域、複数隣接シングル遺伝子領域、及び/または抗菌領域を含む。いくつかの実施態様において、これらの領域は以下を含む:
1)PPSオペロン領域:
この領域はバチルス・ズブチリス168染色体の約1959410(ppsE)〜1997178bp(ppsA)で見られる。本発明を用いて、染色体の1960409bp〜1998026pに対応する、約38.6kb断片を欠失した。このオペロン領域は抗菌合成に関係し、プリパスタチン・シンテターゼをエンコードする;
2)PKSオペロン領域:
この領域はバチルス・ズブチリス168染色体の約1781110(pksA)〜1857712bp(pksR)で見られる。本発明を用いて、染色体の1781795bp〜1857985pに対応する、約76.2kb断片を欠失した。この領域はポリケチドシンテアーゼをエンコードし、抗菌合成に関係する(Scotti et al.、Gene、130:65−71[1993]);
3)yvfF−yveKオペロン領域:
この領域はバチルス・ズブチリス168染色体の約3513149(yvfF)〜3528184bp(yveK)で見られる。本発明を用いて、染色体の3513137bp〜3528896bpに対応する、約15.8kb断片を欠失した。この領域は推定多糖類をコードする(Dartois et al.、バチルスに関する第7回国際会議(1993)Institute Pasteur[1993]、56頁を参照)。この領域は以下の遺伝子を含む:yvfA−F、yveK−T及びslr。B.ズブチリス168染色体の約3529014〜3529603で見られるslr遺伝子領域は約589bpの断片を包含する。この領域はyvfF−yveKオペロンの調節領域である;
4)DHBオペロン領域:
この領域はバチルス・ズブチリス168染色体の約3279750(yukL)〜3293206bp(yuiH)で見られる。本発明を用いて、染色体の3279418bp〜3292920bpに対応する、約13.0kb断片を欠失した。この領域はcatecholic siderophone2,3−ジヒドロキシ安息香酸−グリシン−トレオニン三量体エステルbacilibactinの生合成テンプレートをエンコードする(May et al.、J.Biol.Chem.,276:7209−7217[2001]を参照)。この領域は以下の遺伝子を含む:yukL、yukM、dhbA−C、E及びF及びyuiI−H。
【0237】
上述の領域は欠失される好ましい固有染色体領域の例であるが、本発明のいくつかの実施態様において、当該領域の断片も欠失される。いくつかの実施態様において、当該断片は固有染色体領域の約1%〜99%を含む。他の実施態様において、断片は固有染色体領域の5%〜95%を含む。さらに別の実施態様において、断片は固有染色体領域の少なくとも99%、98%、97%、96%、95%、94%、93%、92%、90%、88%、85%、80%、75%、70%、65%、50%、40%、30%、25%、20%及び10%を含む。
【0238】
B.ズブチリス168染色体における染色体上の位置に関してさらに欠失される固有染色体領域の断片の非限定的な例は以下を含む:
a)スキン領域について:
i)約2666663〜2693807の座標位置であって、yqcC〜yqaMを含む、及び
ii)約2658440〜2659688の座標位置であって、rapE〜phrEを含む;
b)PBSXプロファージ領域について:
i)約1320043〜1345263の座標位置であって、xkdA〜xkdXを含む、及び
ii)約1326662〜1345102の座標位置であって、xkdE〜xkdWを含む;
c)SPβ領域について:
i)約2149354〜2237029の座標位置であって、xkdA〜xkdXを含む;
d)DHB領域について:
i)約3282879〜3291353の座標位置であって、dhbF〜dhbAを含む;
e)yvfF−yveK領域について
i)約3516549〜3522333の座標位置であって、yvfB〜yveQを含み、
ii)約3513181〜3528915の座標位置であって、yvfF〜yveKを含み、及び
iii)約3521233〜3528205の座標位置であって、yveQ〜yveLを含む;
f)プロファージ1領域について:
i)約213926〜220015の座標位置であって、ybcO〜ybdEを含み、及び
ii)約214146〜220015の座標位置であって、ybcP〜ybdEを含む;
g)プロファージ2領域について:
i)約546867〜559005座標位置であって、rapI〜cspCを含む;及び
h)プロファージ4領域について:
i)約1263017〜675421座標位置であって、yjcM〜ydjJを含む。
【0239】
断片は同定した固有染色体領域よりも少ない2、3のbpsのみからなることができるので、欠失に適した固有染色体領域の断片の数は多数ある。さらに、多くの同定された固有染色体領域は多数の遺伝子を包含する。当業者は特定の用途の使用のために固有染色体領域のどの断片を欠失させるのが適当かを容易に決定することができる。
【0240】
固有染色体領域の測定は多数の特定断片に隣接したヌクレオチドを排除することに関しては正確ではない。例えば、SPβ領域はB.ズブチリス168染色体の座標2150824〜2286246に位置するものとしてここで定義され、固有染色体領域はさらに10〜5000bp、さらに100〜4000bp、またはさらに100〜1000bpを当該領域の両側に含むことができる。当該領域の両側のbpの数は欠失の標的とされる固有染色体領域に含まれない他の遺伝子の存在により制限される。
【0241】
上述の通り、ここに開示する特定領域の位置はB.ズブチリス168染色体に関するものである。バチルス株由来のその他の類似領域は固有染色体領域の定義に含まれる。類似領域は任意のバチルス株内で見られるが、特に好ましい類似領域はその他のバチルス・ズブチリス株、バチルス・リケニフォルミス株及びバチルス・アミロリケファシエンス株で見られる領域である。
【0242】
特定の実施態様において、1以上の固有染色体領域またはそれらの断片がバチルス株から欠失される。しかしながら、1以上の固有染色体領域またはそれらの断片の欠失は当該欠失を含む株の再生可能性に悪影響を与えるものではない。いくつかの実施態様において、2つの固有染色体領域またはそれらの断片が欠失される。別の実施態様において、3つの固有染色体領域またはそれらの断片が欠失される。さらに別の実施態様において、4つの固有染色体領域またはそれらの断片が欠失される。さらなる実施態様において、5つの固有染色体領域またはそれらの断片が欠失される。別の実施態様において、14程度の固有染色体領域またはそれらの断片が欠失される。いくつかの実施態様において、固有染色体領域またはそれらの断片は連続しており、一方他の実施態様において、それらはバチルス染色体の別々の領域に位置する。
【0243】
欠失した固有染色体領域またはそれらの断片を含むバチルス属の任意のメンバー株は本発明で使用できる。いくつかの好ましい実施態様において、バチルス株はB.ズブチリス株、B.アミロリケファシエンス株、B.レンタス株及びB.リケニフォルミス株からなる群より選択される。いくつかの好ましい実施態様において、当該株は工業バチルス株であり、最も好ましくは工業B.ズブチリス株である。さらに好ましい実施態様において、変異バチルス株はプロテアーゼ生成株である。いくつかの特に好ましい実施態様において、前もってプロテアーゼ酵素をエンコードするポリヌクレオチドを含むように設計されたB.ズブチリス株である。
【0244】
上述の通り、固有染色体領域またはそれらの断片が欠失されたバチルス株はここで“変異バチルス株”という。本発明の特に好ましい実施態様において、変異バチルス株は目的タンパク質の発現レベルが増大している(すなわち、目的タンパク質の発現が同じ成長条件下で成長させた対応する非変異バチルス株と比較して増大している)。
【0245】
増大の1の指標は目的タンパク質の分泌である。いくつかの実施態様において、目的タンパク質の生成は対応する非変異バチルス株と比較して少なくとも0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、4.0%、5.0%、8.0%、10%、15%、20%及び25%またはそれ以上、増大している。他の実施態様において、目的タンパク質の生成は、1リットル当たりに生成したタンパク質(g)で測定すると対応する非変異バチルス株と比較して0.25%〜20%、0.5%〜15%、及び1.0%〜10%増大している。
【0246】
固有染色体領域またはそれらの断片の欠失を含む本発明によって提供される変異バチルス株は任意の適当な方法により生成され、限定されないが以下の方法を含む。1の一般的な実施態様において、DNA構築体をバチルス宿主中に導入する。DNA構築体は不活性化染色体断片を含み、いくつかの実施態様において、さらに選択マーカーを含む。好ましくは、選択マーカーは不活性化染色体断片の5’及び3’末端の一部の両方に隣接している。
【0247】
いくつかの実施態様において、不活性化染色体断片は、好ましくは欠失される固有染色体領域(または当該領域の断片)のすぐ上流及び下流ヌクレオチドと100%配列同一性を有するが、固有染色体領域の上流及び下流ヌクレオチドと約70〜100%、約80〜100%、約90〜100%、及び約95〜100%配列同一性を有する。不活性化染色体断片の各部分は、非変異宿主中で固有染色体領域と相同組換えするために、固有染色体領域の十分な5’及び3’フランキング配列を含まなければならない。
【0248】
いくつかの実施態様において、不活性化染色体断片の各部分は約50〜10,000塩基対(bp)を含む。しかしながら、より小さいまたは大きいbp部分を本発明で用いることができる。好ましくは、各部分は約50〜5000bp、約100〜5000bp、約100〜3000bp、100〜2000bp、約100〜1000bp、約200〜4000bp、約400〜3000bp、約500〜2000bp、及び約800〜1500bpである。
【0249】
いくつかの実施態様において、選択マーカー及び不活性化染色体断片を含むDNA構築体はin vitroで組み立てられ、続いてコンピテントバチルス宿主内に前記構築体を直接クローニングし、DNA構築体がバチルス染色体内に統合されるようにする。例えば、PCR融合及び/または連結はin vitroでのDNA構築体の組み立てに適している。いくつかの実施態様において、DNA構築体は非プラスミド構築体であり、一方、他の実施態様において、ベクター内に組み込まれる(すなわち、プラスミド)。いくつかの実施態様において、環状プラスミドを用い、当該環状プラスミドは適当な制限酵素(すなわち、DNA構築を阻害しないもの)を用いて切断する。従って、線形プラスミドは本発明で使用できる(例えば、図1;及びBacillus subtilis and other Gram−Positive Bacteria(バチルス・ズブチリス及びその他のグラム陽性細菌)、Sonenshein et al.、編集、Am.Soc.Microbiol.,ワシントン、DC[1993]のPerego、“Integrational Vectors for Genetic Manipulation in Bacillus subtilis(バチルス・ズブチリスにおける遺伝子操作のための統合ベクター)”、を参照)。
【0250】
いくつかの実施態様において、DNA構築体またはベクター、好ましくは不活性化染色体断片を含むプラスミドは非変異宿主中で固有染色体領域またはそれらの断片と相同組換えするために十分な量の固有染色体断片の5’及び3’フランキング配列(seq)またはそれらの断片を含む。他の実施態様において、DNA構築体は当該構築体の上流及び下流末端で設計される制限酵素認識部位を含む。本発明に従い、座標位置に従い同定される有用なDNA構築体の非限定的な例は以下を含む:
1.PBSX領域を欠失するDNA構築体:[yjqB末端及びリボソーム結合部位(RBS)を含むyjpC全体を含む5’フランキング配列1318874−1319860bp]−マーカー遺伝子−[pitのターミネーター及び上流部分を含む3’フランキング配列1348691−1349656bp]。
【0251】
2.プロファージ1領域を欠失するDNA構築体:[RBS及びターミネーターを含むglmS全体及びybbU RBSを含む5’フランキング配列201248−202112bp]−マーカー遺伝子−[RBSを含むybgd全体を含む3’フランキング配列220141−221195bp]。
【0252】
3. プロファージ2領域を欠失するDNA構築体:[ydcK末端、以下のtRNA全体:trnS−Asn、trnS−Ser、trnS−Glu、trnS−Glu、trnS−Leu1及びtrnS−leu2を含む5’フランキング配列527925−529067bp]−マーカー遺伝子−[ydeK全体及びydeLの上流部分を含む3’フランキング配列569578−571062bp]。
【0253】
4. プロファージ4領域を欠失するDNA構築体:[yjcMの一部を含む5’フランキング配列1263127−1264270bp]−マーカー遺伝子−[RBSを含むyjoBの一部を含む3’フランキング配列1313660−1314583bp]。
【0254】
5. yvfF−yveK領域を欠失するDNA構築体:[sigLの一部、yvfF全体及びyvfFの開始を含む5’フランキング配列3512061−3513161bp]−マーカー遺伝子−[slr全体及びpnbAの開始を含む3’フランキング配列3528896−3529810bp]。
【0255】
6. DHBオペロン領域を欠失するDNA構築体:[ターミネーターを含むald末端、RBSを含むyuxlF全体、RBS及びターミネーターを含むyukJ全体及びyukL全体を含む5’フランキング配列3278457−3280255bp]−マーカー遺伝子−[RBSを含むyuiH末端、RBS及びターミネーターを含むyuiG全体及びターミネーターを含むyuiE上流末端を含む3’フランキング配列3292919−3294076bp]。
【0256】
DNA構築体はベクター内に組み込まれる、またはプラスミドDNAなしで用いられるかに関わらず、微生物、好ましくは大腸菌細胞またはコンピテントバチルス細胞内に組み込まれる。
【0257】
プラスミド構築体を含むバチルス細胞内にDNAを導入する及び大腸菌内にプラスミドを形質転換する方法は公知である。プラスミドは続いて大腸菌から単離され、バチルス内に形質転換される。しかしながら、大腸菌などの介在微生物を使用することは必須ではなく、いくつかの実施態様において、DNA構築体またはベクターはバチルス宿主内に直接導入される。
【0258】
好ましい実施態様において、宿主細胞はバチルス種である(例えば、米国特許第5,264,366号、米国特許第4,760,025号及びRE34,6060を参照)。いくつかの実施態様において、目的のバチルス株はアルカロフィリック(Alkalophilic)バチルスである。多数のアルカロフィリックバチルス株が公知である(例えば、米国特許第5,217,878号;及びAunstrup et al.、Proc IV IFS:Ferment.Tech.Today,299−305[1972]を参照)。特定の目的の他の種類のバチルス株は工業バチルス株の細胞である。工業バチルス株の例としては、限定されないが、B.リケニフォルミス、B.レンタス、B.ズブチリス及びB.アミロリケファシエンスが挙げられる。別の実施態様において、バチルス宿主株は、B.リケニフォルミス、B.ズブチリス、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.アミロリケファシエンス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ及びB.メガテリウムからなる群より選択される。特に好ましい実施態様において、B.ズブチリス細胞を用いる。
【0259】
いくつかの実施態様において、工業宿主株はバチルス種の非組換え体株、対応する天然バチルス株の変異体及び組換えバチルス宿主株からなる群より選択される。好ましくは、宿主株は組換え宿主株であり、ここで目的ポリぺプチドをエンコードするポリヌクレオチドは前もって宿主内に導入されている。さらに好ましい宿主株はバチルス・ズブチリス宿主株であり、特に組換えバチルス・ズブチリス宿主株である。多数のB.ズブチリス株が公知であり、本発明の使用に適している(例えば、1A6(ATCC 39085)、168(1A01)、SB19、W23、Ts85、B637、PB1753〜PB1758、PB3360、JH642、1A243(ATCC 39,087)、ATCC 21332、ATCC 6051、MI113、DE100(ATCC 39,094)、GX4931、PBT110及びPEP211株;Hoch et al.、Genetics、73:215−228[1973];米国特許第4,450,235号;米国特許第4,302,544号;EP0134048;Palva et al.、Gene、19:81−87[1982];Fahnestock and Fischer,J.Bacteriol.,(1986)165:796−804[1986];及びWang et al.、Gene 69:39−47[1988]を参照)。発現宿主として特に興味深いのは、工業プロテアーゼ生成バチルス株である。これらの3つの株を用いることにより、プロテアーゼ生成に関する高効率は本発明の変異バチルス株によりさらに増大する。
【0260】
工業プロテアーゼ生成バチルス株は特に好ましい発現宿主を提供する。いくつかの実施態様において、本発明におけるこれらの株の使用は効率及びプロテアーゼ生成をさらに増大させる。上述の通り、2つの一般的なプロテアーゼの種類、すなわち中性(または“メタロプロテアーゼ”)及びアルカリ性(または“セリン”)プロテアーゼがあり、通常はバチルス種により分泌される。また、上述の通り、ズブチリシンは本発明の使用に好ましいセリンである。多種多様のズブチリシンが同定及び配列決定されており、例えば、ズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309である(例えば、EP414279B;WO89/06279;及びStahl et al.、J.Bacteriol.,159:811−818[1984]を参照)。本発明の1の実施態様において、バチルス宿主株は改変(例えば、変異体)プロテアーゼを生成する。多数の文献が変異体プロテアーゼの例及び引例を提供している(例えば、WO99/20770;WO/20726;WO99/20769;WO89/06279;RE34,606;米国特許第4,914,031号;米国特許第4,980,288号;米国特許第5,208,158号;米国特許第5,310,675号;米国特許第5,336,611号;米国特許第5,399,283号;米国特許第5,441,882号;米国特許第5,482,849号;米国特許第5,631,217号;米国特許第5,665,587号;米国特許第5,700,676号;米国特許第5,741,694号;米国特許第5,858,757号;米国特許第5,880,080号;米国特許第6,197,567号及び米国特許第6,218,165号を参照)。
【0261】
さらに別の実施態様において、好ましいバチルス宿主はdegU、degS、degR及びdegQの遺伝子の少なくとも1つにおいて変異または欠失を有するバチルス種である。好ましくは、変異はdegU遺伝子中にあり、より好ましくは変異はdegU(Hy)32である。(例えば、Msadek et al.、J.Bacteriol.,172:824−834[1990];及びOlmos et al.、Mol.Gen.Genet.,253:562−567[1997]を参照)。最も好ましい宿主株はdegU32(Hy)変異を有するバチルス・ズブチリスである。さらなる実施態様において、バチルス宿主はscoC4(Caldwell et al.、J.Bacteriol.,183:7329−7340[2001]を参照);spollE(Argoni et al.、Mol.Microbiol.,31:1407−1415[1999]を参照);oppAまたはその他のoppオペロン遺伝子(Perego et al.、Mol.Microbiol.,5:173−185[1991]を参照)における変異または欠失を含む。実際に、oppA遺伝子の変異と同じ表現型を生じるoppオペロンの変異が考えられ、本発明の変異バチルス株のいくつかの実施態様において使用できる。いくつかの実施態様において、これらの変異は単独で生じ、一方他の実施態様において、変異は組み合わせて存在する。いくつかの実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異をすでに含むバチルス宿主株から得られる。別の実施態様において、本発明の変異バチルスはさらに、上述の遺伝子の1以上の変異を含むように設計される。
【0262】
いくつかの実施態様において、2以上のDNA構築体はバチルス宿主細胞内に導入され、変異バチルスの2以上の固有染色体領域の欠失を生じる。いくつかの実施態様において、これらの領域は連続しており(例えば、スキンとプロファージ7領域)、一方、他の実施態様において、当該領域は分離している(例えば、PBSX領域及びPKS領域;スキン領域及びDHB領域;またはPKS領域、SPβ領域及びyvfF−yveK領域)。
【0263】
当業者はポリヌクレオチド配列を細菌(例えば、大腸菌及びバチルス)細胞内に導入する適切な方法をよく把握している(例えば、Ferrari et al.、“Genetics”、in Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.[1989]、57−72頁;例えば、Saunders et al.、J.Bacteriol.,157:718−726[1984];Hoch et al.、J.Bacteriol.,93:1925−1937[1967];Mann et al.、Current Microbiol.、13:131−135[1986];及びHolubova,Folia Microbiol.,30:97[1985];B.ズブチリスについて、Chang et al.、Mol.Gen.Genet.,168:11−115[1979];B.メガテリウムについて、Vorobjeva et al.、FEMS Microbiol.Lett.,7:261−263[1980];B.アミロリケファシエンスについて、Smith et al.、Appl.Env.Microbiol.,51:634(1986);B.チューリンゲンシスについて、Fisher et al.、Arch.Mictobiol.、139:213−217[1981];及びB.スフェリカスについて、McDonald,J.Gen.Microbiol.,130:203[1984]を参照)。実際に、プロトプラスト形質転換及び配向、形質導入及びプロプラスト融合などの形質転換の方法は公知であり、本発明での使用に適している。形質転換の方法は特に本発明により提供されるDNA構築体を宿主細胞内に導入するのに好ましい。
【0264】
一般的に用いられる方法に加えて、いくつかの実施態様において、宿主細胞は直接形質転換される(すなわち、媒介細胞を増幅のために用いず、または宿主細胞内への導入前にDNA構築のその他のプロセスを用いない)。DNA構築体の宿主細胞内への導入は、プラスミドまたはベクター内への挿入なしでDNA構築体を宿主細胞に導入するための当業者に公知の物理的及び化学的方法を含む。当該方法は、限定されないが、塩化カルシウム沈殿法、エレクトロポレーション、裸のDNA(naked DNA)、リポソーム等が挙げられる。別の実施態様において、DNA構築体は、プラスミド内に導入されることなく、プラスミドを用いて共形質転換される。さらに別の実施態様において、選択マーカーは当業者に公知の方法により変異バチルス株から欠失または実質的に切除される(Stahl et al.、J.Bacteriol.,158:411−418[1984];及びPalmeros et al.の保存部位特異的組換え[CSSR]方法、Palmeros et al.、Gene 247:255−264[2000]に記載、を参照)。いくつかの好ましい実施態様において、宿主染色体由来のベクター分解は固有染色体領域を除去しながら染色体中のフランキング領域を残す。
【0265】
いくつかの実施態様において、宿主細胞は変異バチルス株を生成するために本発明に従う1以上のDNA構築体を用いて形質転換され、ここで2以上の遺伝子が宿主細胞中で不活性化される。いくつかの実施態様において、2以上の遺伝子は宿主細胞染色体から欠失される。別の実施態様において、不活性化遺伝子は連続しており(欠失及び/または挿入による不活性化に関わらず)、一方他の実施態様において、連続した遺伝子ではない。
【0266】
上述の通り、細胞内及び細胞外発現ポリぺプチド活性を検出及び測定するための当業者に公知の種々の分析法がある。特に、プロテアーゼに関して、吸光度280nmで測定、またはFolin法を用いた比色分析により測定したカゼインまたはヘモグロビンからの酸溶性ぺプチドの放出に基づく分析法がある(例えば、Bergmeyer et al.、“Methods of Enzymatic Analysis”vol.5、Peptidases,Proteinases and their Inhibitors(ペプチダーゼ、プロテイナーゼ、及びそれらの阻害因子)、Verlag Chemie、Weinheim「1984」を参照)。その他の分析法は発色基質の可溶化を含む(例えば、Ward、“Proteinases(プロテイナーゼ)”、Fogarty(編集)、Microbial Enzymes and Biotechnology、Applied Science,ロンドン[1983]、251〜317頁を参照)。その他の分析法の例としては、スクシニル−Ala−Ala−Pro−Phe−paraニトロアニリド分析(SAAPFpNA)及び2,4,6−トリニトロベンゼンスルフォン酸ナトリウム塩分析(TNBS分析)が挙げられる。他の当業者に公知の多数の文献が適切な方法を提供する(例えば、Wells et al.、Nucleic Acids Res.11:7911−7925[1983];Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal Biochem.,242:221−227[1999]を参照)。
【0267】
上述の通り、宿主細胞中の目的タンパク質の分泌レベルの決定方法及び発現タンパク質の検出方法は当該タンパク質に特異なポリクローナルまたはモノクローナル抗体を用いる免疫学的分析法の使用を含む。例としては、酵素免疫測定法(ELISA)、放射免疫分析法(RIA)、蛍光免疫分析法(FIA)、及び蛍光活性化細胞分類法(FACS)を含む。しかしながら、その他の方法も当業者に公知であり、目的タンパク質の評価に使用できる(例えば、Hampton et al.、Serological Methods,A Laboratory Manual,APS Press、St.Paul,MN[1990];及びMaddox et al.、J.Exp.Med.、158:1211[1983]を参照)。いくつかの好ましい実施態様において、目的タンパク質の分泌は、対応する非変異宿主よりも本発明を用いて得た変異株におけるほうが高い。当業界で公知の通り、本発明を用いて生成した変異バチルス細胞は発現及び細胞培養物からの目的ポリぺプチドの回収に適した条件下で維持及び成長する(例えば、Harwood and Cutting(編集)Molecular Biological Methods for Bacillus(バチルスの分子生物学的方法)、John Wiley&Sons[1990]を参照)。
【0268】
本発明を実施する手段及び方法は以下の実施例を参照することにより、当業者により十分理解されるものであり、当該実施例は本発明の範囲またはその請求の範囲をいかなる方法によっても限定するものではない。
【0269】
実験
以下の実施例は特定の好ましい実施態様及び本発明の特徴を示し、さらに説明するためのものであり、本発明の範囲を限定するものとして解釈されるものではない。
【0270】
実験の開示において、以下の通り、以下の略語を適用する:℃(摂氏度);rpm(1分間当たりの回転数);H2O(水);dH2O(脱イオン水);HCl(塩酸);aa(アミノ酸);bp(塩基対);kb(キロ塩基対);kD(キロドルトン);gm(グラム);μg(マイクログラム);mg(ミリグラム);ng(ナノグラム);μl(マイクロリットル);ml(ミリリットル);mm(ミリメーター);nm(ナノメーター);μm(マイクロメーター);M(モル濃度);mM(ミリモル濃度);μM(マイクロモル濃度);U(単位);V(ボルト);MW(分子量);sec(秒);min(s)(分);hr(s)(時間);MgCl2(塩化マグネシウム);NaCl(塩化ナトリウム);OD280(280nmでの光学密度);OD600(600nmでの光学密度);PAGE(ポリアクリルアミドゲルエレクトロフォレーシス);PBS(リン酸緩衝生理食塩水[150mM NaCl、10mM リン酸ナトリウム緩衝液、pH7.2]);PEG(ポリエチレングリコール);PCR(ポリメラーゼ連鎖反応);RT−PCR(逆転写PCR);SDS(ドデシル硫酸ナトリウム);Tris(トリス(ヒドロキシメチル)アミノメタン);w/v(体積当たり重量);v/v(体積対体積);LA培地(リットル当たり:Difcoトリプトン・ペプトン 20g、Difco酵母抽出物 10g、EMサイエンス NaCl 1g、EMサイエンス 寒天17.5g、dH20〜1L);ATCC(American Type Culture Collection、ロックビル、メリーランド州);Clontech(CLONTECHラボラトリーズ、パロ・アルト、カリフォルニア州);Difco(Difcoラボラトリーズ、デトロイト、ミシガン州);GIBCO BRLまたはGibco BRL(ライフ・テクノロジーズ、Inc.,ゲーサーズバーグ、MD);Invitrogen(Invitrogen Corp.,サンディエゴ、カリフォルニア州);NEB(ニューイングランド Biolabs、ビバリー、マサチューセッツ州);Sigma(シグマ・ケミカル Co.,セントルイス、ミズーリ州);Takara(タカラ・バイオ Inc.Otsu,日本);Roche Diagnostics and Roche(ロシュ Diagnostics,F.Hoffmann La Roche部門、Ltd.,Basel、スイス);EMサイエンス(EMサイエンス、Gibbstown,ニュージャージー州);Qiagen(Qiagen,Inc.,バレンシア、カリフォルニア州);Stratagene(Stratagene Cloning System,ラ・ホーヤ、カリフォルニア州);Affymetrix(Affymetrix、サンタクララ、カリフォルニア州)。
【実施例】
【0271】
実施例1
欠失株の作成
この実施例は“方法1”について説明し、図1にも示される。この方法では、バチルス株内に形質転換されるDNA構築体を運ぶpJM102プラスミドベクターを生成するために大腸菌を用いた(Perego、上述を参照)。欠失部位の5’及び3’末端にすぐ隣接する領域をPCR増幅した。PCRプライマーを約37塩基対長さになるように設計し、バチルス・ズブチリス染色体に相同な31塩基対及びプライマーの5’末端から6塩基対に位置する6塩基対の制限酵素認識部位を含むようにした。プライマーを当該構築体の上流及び下流末端の固有制限酵素認識部位及びクローニング使用のための2つの断片間のBamHI部位構築のため設計した。抗菌マーカーのためのプライマーは当該断片の両側末端にBamHI部位を含んだ。可能であれば、PCRプライマーは欠失固有染色体領域のプロモーターを除去するが、隣接領域の全てのターミネーターを残すように設計した。染色体配列、遺伝子決定、及びプロモーター及びターミネーター情報の主な供給源はKunst et al.、(1997)上述から得ており、また当業者に公知のSubtiListワールドワイドウェブサーバーからも入手可能である(例えば、Moszer et al.、上述を参照)。本発明を用いて多数の欠失を作った。本発明により作成した欠失からのプライマー配列リストを表1に示す。また、プライマー命名システムの説明に関して図2も参照されたい。
【表1】
【0272】
【0273】
この方法では、100μL PCR反応を84μL水、10μL PCR緩衝液、1μLの各プライマー(すなわち、PKS−UF及びPKS−UR)、2μLのdNTPs、1μLの野生型バチルス染色体DNAテンプレート、及び1μLのポリメラーゼを含む150μLのエッペンドルフチューブ中で行った。使用したDNAポリメラーゼはTaq Plus Precision ポリメラーゼ及びHerculase(Stratagene)を含ませた。反応は以下のプログラムを用いてハイブリッドPCRエキスプレス熱サイクラー中で行った。サンプルは最初に94℃で5分間加熱し、それから冷却して50℃に保った。ポリメラーゼをこの時点で加えた。25サイクルの増幅は95℃で1分間、50℃で1分、及び72℃で1分からなる。最終的に72℃で10分間行うことにより、伸長完了を確実にした。サンプルを分析のため4℃で保存した。
【0274】
PCR完了後、10μlの各反応をInvitrogen 1.2% アガロースE−ゲル上で、60ボルト、30分間で行い、正しいサイズのバンドの存在を確認した。ここに記載する全てのゲルエレクトロフォレーシス法はこれらの条件を用いた。バンドが存在する場合、反応チューブの残余物は製造メーカーの取扱説明書に従いQiagen Qiaquick(登録商標)PCR精製キットを用いて精製し、それから、適当な制限酵素対を用いて切断した。消化を37℃、1時間、9μLの水、、2μLの10×BSA、2μLの適当なNEB制限緩衝液(2000−01 NEBカタログ及び技術文献に従う)、5μLのテンプレート、及び1μLの各制限酵素からなる20μLの反応として行った。例えば、PBSX上流断片及びCssS上流断片をNEB(ニューイングランド・バイオラボ)制限緩衝液B中でXbal及びBamHIを用いて切断した。消化断片をゲル・エレクトロフォレーシス及び製造メーカー取扱説明書に従いQiagen Qiaquickゲル抽出キットを用いた抽出により精製した。図5及び6は種々の欠失の結果を示すゲルを表す。
【0275】
プラスミドベクター内への断片の連結は2つの工程で行われ、製造メーカー取扱説明書に従ってTakara連結キットまたはT4DNAリガーゼ(反応内容:5μL 各挿入断片、1μL 切断pJM102プラスミド、3μL T4DNAリガーゼ緩衝液及び1μL T4DNAリガーゼ)を用いる。まず、切断上流及び下流断片をpJM102プラスミド・ポリリンカー中の固有制限酵素認識部位内に15℃で一晩中連結させ、共通BamHI部位で接続し、環状プラスミドを再形成させた。pJM102プラスミドを各欠失に適した固有制限酵素認識部位を用いて切断し(表2を参照;cssS、Xbal及びSaclを用いた)、連結前に上述の通り精製した。この再環状化プラスミドをInvitrogenの“Top Ten”大腸菌細胞内に形質転換し、製造メーカーのワン・ショット形質転換手順を用いた。
【0276】
形質転換体は1.5%寒天(LA)とブルーホワイトスクリーニングのためのX−galを含んだ50ppmカルバニリシン(carbanicillin)を用いて凝固したLB培地上で選択した。クローンを採取し、5mlのLB培地(Luria−Bertani)プラス50ppmカルバニシリン(carbanicillin)中、37℃で一晩中成長させ、プラスミドをQiagen’s Qiaquick ミニ−準備キットを用いて単離した。制限分析により、挿入の各末端で制限酵素認識部位を用いて切断し、プラスミドから約2kbのバンドにすることにより挿入の存在を確認した。挿入を確認したプラスミドはBamHIを用いて切断し、上述の通り消化反応において線形化し(第2の制限酵素に代えてさらに1μLの水を用いて)、再環状化を防止するために1μLの子牛の腸(calf intestinal)ホスファターゼ及びshrimpホスファターゼを用いて1時間、37℃で処理し、及び表2に記載するように抗菌剤耐性マーカーに連結させた。抗菌剤耐性マーカーはBamHIを用いて切断し、連結前に製造メーカー取扱説明書に従いQiagenゲル抽出キットを用いて洗浄した。このプラスミドを既に述べたように大腸菌内にクローンし、選択に応じて適切な、5ppmのフレオマイシン(phl)または100ppmのスペクチノマイシン(spc)を用いる。単離プラスミド中のマーカー挿入の確認は上述の通り、BamHIを用いる制限分析により行った。B.ズブチリス内への形質転換前に、プラスミドをScalを用いて線形化し、二重交差結合を確実にさせた。
【表2】
【0277】
実施例2
大腸菌をバイパスするためにPCR融合を用いるDNA構築体の作成
この実施例は“方法2”について説明し、また図3にも示される。プライマーが抗菌マーカーのプライマー配列に相補的な25bp“テイル”を用いて設計されること以外は、上流及び下流断片は方法1の通り増幅する。“テイル”とは、直接増幅される配列に相同でないが、他のDNA配列に相補的であるプライマーの5’末端上の塩基対としてここで定義する。同様に、抗菌剤を増幅するためのプライマーは断片プライマーに相補的な“テイル”を含む。任意の所定欠失について、欠失X−UFfus及び欠失X−URfusは互いに直接の相補体である。これは、DF−fusとDR−fusプライマーセットに関しても当てはまる。さらに、いくつかの実施態様において、これらのプライマーはプラスミドベクター作成で使用する方法1において用いたものに類似した制限酵素認識部位を含む(表3及び米国特許第5,023,171号を参照)。表3はPCR融合による欠失構築体の作成に有用なプライマーのリストを提供する。表4はPCR融合による欠失構築体の作成に有用な別のプライマーのリストを提供する。しかしながら、この表では、全ての欠失構築体はphleoRマーカーを含む。
【表3】
【表4】
【0278】
上述の通りゲル・エレクトロフォレーシスにより表3及び4に記載した断片の大きさを確認した。正しい場合には、1μLの各上流、下流及び抗菌剤耐性マーカー断片を記載された欠失X−UF及び欠失X−DRプライマーまたはネスティドプライマーを含む1の反応管に加えた。ネスティドプライマーは上流または下流断片の内部に相同な25塩基対のDNAであり、通常、当該断片の外側末端由来の約100塩基対である(図2を参照)。ネスティドプライマーの使用は融合成功度を高めることが多い。PCR反応成分は、82μLの水をテンプレート体積を追加して補うために用いた以外は上述のものに類似した。PCR反応条件は72℃の伸長を3分に延長した以外は、上述のものと類似した。伸長の間、抗菌剤耐性遺伝子を上流と下流断片間に融合した。この融合断片は、精製工程を用いずに、または製造メーカー取扱説明書に従って行われるQiagen Qiaquick PRC精製を用いてバチルス内に直接統合できる。
【0279】
実施例3
PCR断片の連結を用いるDNA構築体の作成及び大腸菌クローニングをバイパスするためのバチルス・ズブチリスの直接形質転換
この実施例では、PCR断片の連結を用いるDNA構築体を作成するための方法(“方法3”)及びバチルスの直接形質転換について説明する。例として、prpC、sigD及びtdh/kblの修飾を連結方法を示すために提供する。実際に、sigD及びtdh/kblを1の方法で構築し、prpCを別の方法により構築した。
【0280】
A.Tdh/Kbl及びSigD
バチルス・ズブチリス染色体のtdh/kbl領域に隣接した上流及び下流断片を、フランキングDNAの内部プライマーがII型の制限酵素認識部位を含むように設計した以外は方法1に記載した類似のPCRにより増幅した。loxP−スペクチノマイシン−loxPカセットのプライマーをフランキング及び相補的突出部と同じII型制限酵素認識部位を用いて設計した。左側面及び右側面の固有突出部は上流及び下流のフランキングDNA間の抗菌カセットの直接連結を可能にする。全てのDNA断片は適当な制限酵素により消化し、当該断片を製造メーカー取扱説明書を用いてQiagen Qiaquick PCR精製キットで精製した。この精製に続いて、BioRad P−6ゲルを含む1mLの回転カラム中で脱塩を行い、2mM Tris−HCl、pH7.5で釣り合いを取った。断片をSavant Speed Vac SC110システムを用いて124〜250ng/μLに濃縮した。0.8〜1μgの各断片の3つの断片連結を、16時間、14〜16℃で15〜25μL反応体積中で12U T4リガーゼ(ロシュ)を用いて行った。所望の連結生成物の合計収量は、アガロースゲル上の標準DNAラダー(ladder)との比較により評価して、反応当たり100ngより大きかった。当該連結混合物は形質転換反応のための精製を行わないで用いた。この構築体のためのプライマーは下記の表5に示す。
【表5】
【0281】
sigD欠失の構築はtdh/kblの構築にほとんど従う。sigD構築に用いるプライマーは表6に示す。
【表6】
【0282】
B.PrpC
バチルスの直接形質転換のためのPCR増幅DNA断片の連結によりDNA分子を作成する別の実施例は遺伝子prpCの部分フレーム内欠失を含む。prpC遺伝子を含むバチルス・ズブチリス染色体DNAの3953bp断片をp95及びp96を用いてPCRにより増幅した。当該断片を固有制限酵素認識部位PflMI及びBstXIで切断した。これにより3つの断片、上流、下流及び中心断片を生じた。後者は欠失される断片であり、内部からprpC遺伝子に位置した170bpからなる。消化混合物をQiagen Qiaquick PCR精製キットを用いて精製し、続いてBioRad P−6ゲルを含む1mLの回転カラム中、脱塩し、2mMのTris−HCl、pH7.5で釣り合いを取った。2回目のPCR反応で、抗菌カセット、すなわちloxP−スペクチノマイシン−loxPをBstXI部位を含むプライマー及びPflMI部位を含む下流プライマーであって、両方がゲノムDNA断片の部位に相補的な切断部位を含むものを用いて増幅させた。当該断片をPflMI及びBstXIを用いて消化し、上述の染色体断片について説明した通り精製した。上流、抗菌カセット及び下流断片の3つの断片連結を上述のtdh/kblに関しての通り、行った。下記にさらに詳細を説明する通り、所望の連結生成物の収量は類似しており、連結生成物をxylRcomKコンピテントバチルス・ズブチリスの形質転換に関するさらなる処理を行うことなく用いた。
【表7】
【0283】
C.PckA欠失
上述の欠失に加えて、pckAも修飾した。PCRプライマーのpckAUF、pckA−2Urfus、spc ffus、spc rfus、pckA Dffus及びpckA DRをPCR及びテンプレートとしてバチルス・ズブチリス168誘導体の染色体DNA及びpDG1726(Guerout−Fleury et al.、Gene 167(1−2):335−6[1995]を参照)を用いるPCR融合反応に関して用いた。プライマーを表8に示す。これらの欠失変異体を構築するのに使用する方法は上述の方法1と同じである。
【表8】
【0284】
D.連結DNAを用いるキシロース導入コンピテンス宿主細胞形質転換
2001年8月10日に出願され、ここに引用する米国特許出願番号第09/927,161号に記載されているように、部分遺伝子型xylRcomKを有する宿主株バチルス・ズブチリスの細胞を1%キシロースを含むLB培地中で2時間成長させることによりコンピテントにし、OD550を1とした。この培養物を6時間培養物から取り除いた。全ての培養物を37℃で、300rpmで振とうさせながら成長させた。0.3mLのアリコートを1:1の培養物と30%グリセロール混合物として丸底2mL管内に冷凍し、その後の使用のために液体窒素中で保存した。
【0285】
形質転換のために、凍結コンピテント細胞を37℃で解凍し、解凍が完了してすぐに、連結反応混合物由来のDNAを5〜15μL/管のレベルで加えた。当該管をそれから60分間、37℃で1400rpmで振とうした(Tekmar VXR S−10)。形質転換混合物を100ppmのスペクチノマイシンを含む8cmLAプレート上の100μLアリコート中で希釈しないでプレートした。一晩中成長させた後、形質転換体をLB培地(100ppmスペクチノマイシン)中に採取し、及び当業者に公知の通り実施されるゲノムDNA単離のために37℃で成長させた(例えば、Harwood and Cuttings,Molecular Biological Methods for Bacillus,John Wiley and Son,ニューヨーク、N.Y.[1990]、23頁を参照)。5μLの連結反応混合物を形質転換に用いた場合、通常、400〜1400の形質転換体が100μLの形質転換混合物から得られた。
【0286】
抗菌マーカーが入来DNA内の2つのloxP部位間に位置する場合、マーカーはCreタンパク質を発現できるcre遺伝子を含むプラスミドを含む株を形質転換することにより除去できる。細胞は37℃〜42℃で培養したpCRM−TSpleo(下記参照)を用いて形質転換し、LA上にプレートし、コロニー生成後、100ppmのスペクチノマイシンを含むLA上に継ぎ合せた。一晩中培養後に成長していなかったパッチは抗菌マーカーを失ったと考えられる。マーカーの損失は所定の遺伝子に適したプライマーを用いるPCR分析により確認した。
【0287】
pCRM−TS−pleoは以下の配列を有する(配列番号205):
【式54】
【0288】
E. トランスクリプトームDNAアレイ法
上述の方法に加えて、トランスクリプトームDNAアレイ法を本発明の変異体を発達させるために用いた。まず、標的RNAを当業者に公知のグアニジン酸(guanidinium acid)フェノール抽出によりバチルス株から取り入れ(例えば、Farrell、RNA Methodologies、(第2版).Academic Press,サンディエゴ、81頁を参照)、及び時点をdeSaizieu et al.(deSaizieu et al.、J.Bacteriol.,182:4696−4703[2003])及びここに記載する方法から適合させた方法によりビオチン標識cDNA内に逆転写した。合計RNA(25mg)を37℃、100mL反応中で一晩中培養した:1×GIBCO 一本鎖(first−strand)緩衝液(50mM Tris−HCl pH8.3、75mM KCl、3mM MgCl2);10mM DTT;40mMランダムヘキサマー;0.3mMの各dCTP、dGTP及びdTTP;0.12mM dATP;0.3mM ビオチン−dATP(NEN0;2500単位 SuperScript II 逆転写酵素(ロシュ)。RNAを除去するために、当該反応を0.25M NaOHに加え、30分間、65℃で培養した。当該反応をHCl及び2.5M 酢酸アンモニウムを含むエタノール中で−20℃で沈殿させた核酸を用いて中和させた。ペレットを洗浄、空気乾燥し、水中で再懸濁させ、UV分光法により量を測った。反応収量は約20〜25mgビオチン標識cDNAであった。
【0289】
12mgのこのcDNAを、10分間、37℃で3.75ミリ単位のDNAアーゼを含む33mL 1×One−Phor−All緩衝液(Amersham−Pharmacia番号27−0901−02)において断片化した。DNAアーゼを熱殺菌後、3%アガロースゲル上の2mgの断片化cDNAを管理することにより確認した。ビオチン含有cDNAの大きさは通常、25〜125ヌクレオチドの範囲である。残りの10mgのcDNAをAffymetrixバチルス遺伝子チップアレイにハイブリダイズした。
【0290】
ハイブリダイゼーションをAffymetrix発現分析技術マニュアル(Affymetrix)に記載されている通りに、供給元が提示する試薬を用いて行った。要するに、10mgの断片化ビオチン標識cDNAを220mLの以下を含むハイブリダイゼーション混合物に加えた:100mM MES(N−モルホリノエタンスルホン酸)、1M Na+、20mM EDTA、0.01% Tween20;5mg/mL合計酵母RNA;0.5mg/mL BSA;0.1mg/mLニシン精子DNA;50pM対照オリゴヌクレオチド(AFFX−B1)。当該混合物を5分間、95℃に加熱し、5分間40℃に冷却し、粒子を除去するために手短に遠心分離し、及び200mLをそれぞれ前もって温め、すすいでおいた(1×MES緩衝液+5mg/ml酵母RNA)遺伝子チップカートリッジ内に注入した。当該アレイを40℃で一晩中回転させた。
【0291】
当該サンプルを除去し、アレイを非ストリンジェントな洗浄緩衝液(6×SSPE、0.01%Tween20)を用いて充填し、手順Euk−GE−WS2を用いるAffymetrix 流体ステーション上で非ストリンジェント及びストリンジェント(0.1M MES,0.1M[Na+]、0.01%Tween20)洗浄緩衝液を用いて洗浄した。アレイを3つの工程で着色した:(1)ストレプトアビジン;(2)ビオチンで標識した抗ストレプトアビジン抗体;(3)ストレプトアビジン−フィコエリトリン結合。
【0292】
アレイ中のシグナルをヒューレット・パッカード遺伝子アレイスキャナーで3mmピクセル解像度の570nmレーザー光を用いて検出した。4351ORFプローブセットのシグナル強度を時間的経過実験を含む全ての時点にわたって測定及び標準化した。これらのシグナルをそれから実験中、遺伝子の相対発現レベルを推測するために比較した。トレオニン生合成系及び分解系遺伝子を同時に転写し、非効率なトレオニン利用を示した。分解系トレオニン経路の欠失は所望の生成物の発現を向上させた(図7を参照)。本発明はトレオニン生合成系及び分解系プロファイルに類似した転写プロファイルを含む経路を修飾する手段を提供する。従って、本発明はバチルス株を最適化するためにトレオニンに類似した転写プロファイルを含む経路の修飾にも使用できる。いくつかの好ましい実施態様において、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の遺伝子が欠失または修飾される。ここに記載した本発明を用いて、sigDレギュロンが転写されたという驚くべき発見をした。この遺伝子の欠失は所望の生成物のより優れた発現を生じる(図7を参照)。gapB及びpckAの転写も驚くべき発見であった。pckAの欠失は改善または損失のいずれももたらさなかった。しかしながら、本発明はpckA欠失または修飾とgapB及び/またはfbpの欠失または修飾の組み合わせにより株タンパク質の生成を向上させる手段を提供する。さらに、本発明の開発時に、トリプトファン生合成経路遺伝子は不均衡な転写を示すことが観測された。従って、本発明は、trpA、trpB、trpC、trpD、trpE、及び/またはtrpFからなる群より選択される遺伝子などの転写が増加した株を生成するのに使用でき、改善株は母体(すなわち、野生型及び/または起源株)と比較して所望の生成物の発現が向上すると考えられる。実際に、これらの遺伝子修飾の任意の組み合わせは所望の生成物の発現の向上をもたらすと考えられる。
【0293】
F.発酵
上述の構築体を用いて生成した株分析は以下の発酵を行った。大きさ14Lの培養はBiolafitte(登録商標)発酵器中で行った。7リットル当たりの培養成分を表9に記載する。
【表9】
【0294】
750rpmで槽を攪拌し、気流を11リットル/分、温度37℃に調整し、pHをNH4OHを用いて6.8に維持した。60%グルコース溶液を0.5〜2.1グラム/分で開始から発酵の終わりまで直線ランプ中で約14時間送った。炭素収支及び効率はグルコース供給、タンパク質生成物の収量、細胞集団収量、培養液中のその他の炭素及び発生CO2から計算した。変異体株を改善点を検証するために母体株と比較した。この変異pckA株はこれらの条件下では改善を示さなかったが、改善は修飾培養条件(すなわち、当業者に公知のもの)下及び/または別の遺伝子と組み合わせて生じると考えられる。いくつかの好ましい実施態様において、これらの別の遺伝子はgapB、alsD及び/またはfbpからなる群より選択される。
【0295】
実施例4
変異バチルス株を得るための宿主細胞形質転換
上述の方法1または2によりDNA構築体を作成すると、それを適当なバチルス・ズブチリス研究所株内に形質転換した(例えば、BG2036または、BG2097;任意のコンピテントバチルス直接宿主細胞は本発明の方法に使用できる)。当該細胞は0.5ppmのフレオマイシンまたは100ppmのスペクチノマイシンの選択培地上に必要に応じてプレートした(Ferrari and Miller、Gene Expression Systems:Using Nature for the Art of ExpressionのBacillus Expression:A Gram−Positive Model(バチルス発現:グラム陽性モデル)、65〜94頁[1999])。研究所株はバチルス・ズブチリス生成宿主株内に2回、またはBG3594及びそれからMDT98−113に一度形質転換した欠失を有する染色体DNAの供給源として用いた。形質転換体は1のコロニーに単離するためにストリーキングし、5mLのLBと適当な抗菌剤中で一晩中採取及び成長させた。染色体DNAを当業者に公知の通り単離した(例えば、Hardwood et al.、上述参照)。
【0296】
統合DNA構築体の存在は、上述の成分及び条件を用いて3つのPCR反応により確認した。例えば、2つの反応は欠失カセット外側由来領域を1の場合において(プライマー1及び11)抗菌遺伝子内に増幅するために設計し、他の場合において(プライマー1及び12)全体的な挿入により増幅するために設計した。第3の照合は、欠失カセット外側由来の領域を欠失領域内(プライマー1及び4))に増幅した。図4は、正しいクローンが最初の2つの場合にはバンドを示すが、3番目の場合には示さないということを示す。野生型バチルス・ズブチリス染色体DNAは全ての反応で陰性対照として用い、第3のプライマーセットのバンドのみを増幅すべきである。
【0297】
実施例5
振とうフラスコ分析−プロテアーゼ活性測定
DNA構築体を安定にコンピテントバチルス・ズブチリス株内に統合すると、ズブチリシン活性を振とうフラスコ分析により測定し、当該活性を野生型レベルと比較した。分析は当業者に公知のズブチリシン生成に適した50mLの成長培地を含む250mlのバッフルフラスコ内で行った(Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal.Biochem.242:221−227[1996]を参照)。当該培地を50μLの8時間、5mL培養物と接種し、250rpmで振とうしながら37℃、40時間成長させた。そして、1mLのサンプルをプロテアーゼ活性分析のために17、24及び40時間の時点で取った。プロテアーゼ活性をモナコ自動分析器(Monarch Automatic Analyser)を用いて405nMで測定した。2つのサンプルを緩衝液中で1:11(3.131g/L)に希釈した。正確な機械較正を保証する対照として、1のサンプルを1:6(5.585g/L)、1:12(2.793g/L)及び1:18(1.862g/L)に希釈した。図7は種々の変異バチルス・ズブチリスクローンにおけるプロテアーゼ活性を示す。図8は、バチルス・ズブチリス野生型株(非変異)及び対応する変異欠失株(−sbo)及び(−slr)の振とうフラスコ培養物から測定した改善されたプロテアーゼ分泌を示すグラフを提供する。プロテアーゼ活性(g/L)を17、24及び40時間後に測定した。
【0298】
また、細胞密度はOD600で分光光度測定を用いて測定した。測定時間でのサンプルの著しい違いは見られなかった(データは示していない)。
【0299】
上述の明細書中に言及した全ての文献及び特許はここに引用するものとする。言及した本発明の方法及びシステムの種々の修正及び変更は本発明の範囲及び精神を逸脱することなく当業者に明らかである。本発明は特定の好ましい実施態様に関連して説明したが、本発明は当該特定の実施態様に過度に限定されるものではないことは当然のことである。実際に、当業界及び関連分野の当業者に明らかである本発明を実施するための前述の方法の種々の変更は本発明の範囲内である。
【技術分野】
【0001】
発明の分野
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。
【背景技術】
【0002】
発明の背景
遺伝子工学は工業用生物反応器、細胞工場として、及び食物発酵において用いる微生物の向上を可能としてきた。特に、バチルス種は多数の有用なタンパク質及び代謝産物を生成及び分泌する(Zukowski、“Production of commercially valuable products(商業的に有用な産物の生成)”、In:Doi and McGlouglin(編集)Biology of Bachilli:Applications to Industry,Butterworth−Heinemann,Stoneham.Mass第311−337頁[1992])。工業的に用いられる最も一般的なバチルス種はB.リケニフォルミス、B.アミロリケファシエンス及びB.ズブチリスである。これらはGRAS(通常、安全であると認識される)状態であるので、これらバチルス種株は食品及び医薬業界において利用されるタンパク質生成のもっともな候補である。重要な生成酵素はα−アミラーゼ、中性プロテアーゼ、及びアルカリ(またはセリン)プロテアーゼを含む。しかしながら、バチルス宿主細胞中におけるタンパク質生成の理解の進展にも関わらず、これらタンパク質の発現を増加させるための方法に関する必要性が以前としてある。
【発明の概要】
【0003】
発明の概要
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。いくつかの好ましい実施態様において、本発明は、バチルスにおける目的タンパク質の発現及び/または分泌を向上させる方法及び組成物を提供する。
【0004】
特に好ましい実施態様において、本発明はバチルスにおける目的タンパク質の発現及び/または分泌を向上させる手段を提供する。より具体的には、これらの実施態様において、本発明はバチルス宿主株における1以上の染色体遺伝子の不活性化に関し、ここで不活化遺伝子は株の生存に必須のものではない。1以上の染色体遺伝子を不活化した結果、変化したバチルス株が生成され、対応する変化していないバチルス宿主株よりも高いレベルの目的タンパク質を発現することができる。
【0005】
さらに、別の実施態様において、本発明はバチルス宿主株における大きな領域の染色体DNAを除去する手段を提供し、ここで欠失された固有染色体領域は株生成に必須のものではない。1以上の固有染色体領域を除去した結果、変化したバチルス株が生成され、対応する変化していないバチルス宿主株よりも高いレベルの目的タンパク質を発現することができる。いくつかの好ましい実施態様において、バチルス宿主株は目的タンパク質をエンコードするポリヌクレオチドを含む組換え宿主株である。特に好ましい実施態様において、変異バチルス株はB.ズブチリス株である。下記に詳細に説明するように、欠失固有染色体領域は、限定されないが、プロファージ領域、抗菌(例えば、抗生物質)領域、調節領域、複数隣接単一遺伝子(multi−contiguous single gene)領域及びオペロン領域を含む。
【0006】
いくつかの実施態様において、本発明はバチルス細胞由来の目的タンパク質発現を増大させる方法及び組成物を提供する。いくつかの好ましい実施態様において、本方法はバチルス宿主株におけるsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子を不活性させて変異バチルス株を生成する工程;変異バチルス株を適当な成長条件下で成長させる工程;及び目的タンパク質を変異バチルス中で発現させる工程を含み、ここでタンパク質発現は対応する非変異バチルス宿主株と比較して増大している。いくつかの実施態様において、目的タンパク質は相同タンパク質であり、一方、他の実施態様において、目的タンパク質は異種タンパク質である。いくつかの実施態様において、1以上の目的タンパク質が生成される。いくつかの好ましい実施態様において、バチルス種はB.ズブチリス株である。さらなる実施態様において、染色体遺伝子の不活性化は遺伝子の欠失を含み、変異バチルス株を生成する。さらなる実施態様において、染色体遺伝子の不活性化は挿入不活性化を含む。いくつかの好ましい実施態様において。目的タンパク質は酵素である。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0007】
さらに別の実施態様において、本発明は変異バチルス株を提供し、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDの群より選択される1以上染色体遺伝子の欠失を含む。いくつかの実施態様において、変異株はプロテアーゼ生成バチルス株である。別の実施態様において、変異バチルス株はズブチリシン生成株である。さらに別の実施態様において、変異バチルス株はさらにdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子の変異を含む。
【0008】
さらなる実施態様において、本発明は入来配列(incoming sequence)を含むDNA構築体を提供する。いくつかの実施態様において、入来配列は、選択マーカー及びsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子または遺伝子断片を含む。別の実施態様において、選択マーカーは遺伝子の2つの断片間に位置する。他の実施態様において、入来配列は選択マーカー及び相同ボックス(homology box)を含み、ここで相同ボックスはマーカーの5’及び/または3’末端の側面に位置する。別の実施態様において、宿主細胞DNA構築体を用いて形質転換される。さらなる実施態様において宿主細胞は大腸菌またはバチルス細胞である。いくつかの好ましい実施態様において、DNA構築体は宿主細胞内の染色体上に統合される。
【0009】
本発明は、目的タンパク質を発現する変異バチルス株を得る方法も提供し、本発明のDNA構築体を含むバチルス宿主細胞を形質転換する工程を含み、ここでDNA構築体はバチルス宿主細胞の染色体中に統合され;変異バチルス株を生成する工程であって、1以上の染色体遺伝子が不活性される工程;及び目的タンパク質発現のための適当な成長条件下で変異バチルス株を成長させる工程を含む。いくつかの実施態様において、目的タンパク質は、プロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。さらに別の実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ、B.メガテリウム及び好ましくはB.ズブチリスからなる群より選択される。いくつかの実施態様において、バチルス宿主株は組換え宿主である。さらに別の実施態様において、目的タンパク質を回収する。さらなる実施態様において、選択マーカーは変異バチルスから切除される。
【0010】
本発明はさらに目的タンパク質を発現する変異バチルス株を得る方法を提供する。いくつかの実施態様において、本方法は入来配列を含むDNA構築体を含むバチルス宿主細胞を形質転換する工程を含み、ここで入来配列は選択マーカー及びsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子を含み、ここでDNA構築体はバチルス宿主細胞の染色体内に統合され、1以上の遺伝子欠失を生じ;変異バチルス株を得る工程及び変異バチルス株を目的タンパク質の発現に適した成長条件下で成長させる工程を含む。
【0011】
いくつかの実施態様において、本発明は入来配列を含むDNA構築体を提供し、ここで入来配列は選択マーカー、及びcssS遺伝子、cssS遺伝子断片またはそれに相同な配列を含む。いくつかの実施態様において、選択マーカーは当該遺伝子の2つの断片間に位置する。別の実施態様において、入来配列は選択マーカー及び相同ボックスを含み、ここで相同ボックスはマーカーの5’及び/または3’末端の側面に位置する。さらに別の実施態様において、宿主細胞はDNA構築体を用いて形質転換される。別の実施態様において、宿主細胞は大腸菌またはバチルス細胞である。さらに別の実施態様において、DNA構築体は宿主細胞の染色体上に統合される。
【0012】
また、本発明はプロテアーゼ生成が増大したバチルス・ズブチリス株を得る方法を提供する。いくつかの実施態様において、本方法は、本発明に従うDNA構築体を用いてバチルス・ズブチリス宿主細胞を形質転換する工程;DNA構築体とバチルス染色体の相同領域を相同組換えさせる工程であって、少なくとも1の以下の遺伝子がバチルス染色体から欠失される工程、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD;変異バチルス・ズブチリス株を得る工程;及びプロテアーゼ発現に適した条件下で変異バチルス株を成長させる工程を含む。いくつかの実施態様において、プロテアーゼ生成バチルスはズブチリシン生成株である。別の実施態様において、プロテアーゼは異種プロテアーゼである。別の実施態様において、プロテアーゼ生成株は、degU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子の変異をさらに含む。いくつかの実施態様において、本発明は遺伝子の挿入不活性化を含む。
【0013】
本発明はさらにsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子の欠失を含む変異バチルス・ズブチリス株を提供し、ここで変異バチルス・ズブチリス株は目的タンパク質を発現することが可能である。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの別の実施態様において、目的タンパク質は異種タンパク質である。
【0014】
いくつかの実施態様において、本発明は1以上の固有染色体領域またはそれらの断片の欠失を含む変異バチルス株を提供し、固有染色体領域は約0.5〜500キロ塩基(kb)を含み、及びここで変異バチルス株は本質的に同じ成長条件下で成長させた場合、対応する非変異バチルス株と比較して増大したレベルの目的タンパク質発現を有する。
【0015】
さらに別の実施態様において、本発明は、PBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、yvfF−yveK領域、DHB領域及びそれらの断片からなる群より選択される固有染色体領域の少なくとも1の欠失を含むプロテアーゼ生成バチルス株を提供する。
【0016】
さらなる実施態様において、本発明はバチルスにおいて目的タンパク質の発現を増大させる方法を提供し、以下を含む:選択マーカーを含むDNA構築体を導入し、及びバチルス宿主株内の染色体断片を不活性化させることにより生成した変異バチルス株を得る工程あって、ここでDNA構築体はバチルス染色体内に統合され、バチルス宿主細胞由来の固有染色体領域またはそれらの断片の欠失を生じる工程;及び成長に適した条件下で変異バチルスを成長させる工程であって、ここで目的タンパク質の発現は変異バチルス株において対応する非変異バチルス宿主細胞の目的タンパク質の発現と比較して大きい。
【0017】
また、本発明はバチルス株から目的タンパク質を得る方法を提供し、以下の工程を含む:選択マーカーを含むDNA構築体を用いてバチルス宿主細胞を形質転換し、染色体断片を不活性化する工程であって、ここでDNA構築体はバチルス株の染色体内に統合され、固有染色体領域またはそれらの断片の欠失を生じて変異バチルス株を形成する工程;目的タンパク質が発現できるように適した成長条件下で変異バチルス株を培養する工程;及び目的タンパク質を回収する工程。
【0018】
また、本発明は、有益な変異をスクリーン及び/または同定するためにDNAマイクロアレイデータを使用する手段を提供する。いくつかの好ましい実施態様において、これらの変異はtrpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子に関する。いくつかの好ましい実施態様において、これらの有益な変異は所定のアミノ酸生合成経路及び生分解経路の同時発現に関するトランスクリプトーム証拠及び/または分解経路の欠失がより良い性能の株を生じるという証拠及び/または生合成経路の過剰発現がより良い性能の株を生じるという証拠に基づく。別の実施態様において、本発明は有益な変異を提供するためにDNAマイクロアレイデータを使用する手段を提供する。いくつかの好ましい実施態様において、これらの変異はtrpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される遺伝子に関し、アミノ酸生合成経路を含む遺伝子由来のmRNAの発現がバランスが取れていない場合、全体的な経路の過剰発現は親(すなわち野生型及び/または最初の)株よりも優れた性能の株を提供する。さらに、本発明は糖新生遺伝子の不活性化により株生成を向上させる手段を提供する。これらの好ましい実施態様のいくつかにおいて、不活化糖新生遺伝子はpckA、gapB及びfbpからなる群より選択される。
【0019】
本発明はバチルスから目的タンパク質の発現を増大させる方法を提供し、以下の工程を含む:目的タンパク質を生成できる変異バチルス株を得る工程であって、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも2の不活化染色体遺伝子を有する工程、及び変異バチルス株を目的タンパク質が変異バチルス株により発現される条件下で成長させる工程であって、目的タンパク質の発現が非変異バチルス宿主株における目的タンパク質の発現と比較して増大している工程。いくつかの実施態様において、目的タンパク質は、相同タンパク質及び異種タンパク質からなる群より選択される。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。いくつかの特に好ましい実施態様において、目的タンパク質はプロテアーゼである。さらに別の実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の染色体遺伝子を欠失することにより得られる。
【0020】
また、本発明はここに記載する方法を用いて得られる変異バチルス株も提供する。好ましい実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の遺伝子の染色体欠失を含む。いくつかの実施態様において、これらの染色体遺伝子の1以上が欠失される。特に好ましい実施態様において、変異株はB.ズブチリス株である。別の好ましい実施態様において、変異バチルス株はプロテアーゼ生成株である。いくつかの特に好ましい実施態様において、プロテアーゼはズブチリシンである。さらに別の実施態様において、ズブチリシンはズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309及びこれらの変異体からなる群より選択される。さらに別の実施態様において、変異バチルス株はdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される少なくとも1の遺伝子において変異をさらに含む。特に好ましい実施態様において、変異バチルス株はさらに目的の異種タンパク質を含む。
【0021】
また、本発明はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD、それらの遺伝子断片及びそれらの相同配列からなる群より選択される少なくとも1の遺伝子を含むDNA構築体を提供する。いくつかの好ましい実施態様において、DNA構築体は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号29、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33からなる群より選択される少なくとも1の核酸配列を含む。いくつかの実施態様において、DNA構築体はさらに少なくとも1の目的タンパク質をエンコードする少なくとも1のポリヌクレオチド配列を含む。
【0022】
本発明はDNA構築体を含むプラスミドも提供する。さらなる実施態様において、本発明はDNA構築体を含むプラスミドを含んだ宿主細胞を提供する。いくつかの実施態様において、宿主細胞はバチルス細胞及び大腸菌細胞からなる群より選択される。いくつかの好ましい実施態様において、宿主細胞はB.ズブチリスである。特に好ましい実施態様において、DNA構築体は宿主細胞の染色体内に統合される。別の実施態様において、DNA構築体は、配列番号2、配列番号3、配列番号4、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号30、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される少なくとも1のアミノ酸配列をエンコードする少なくとも1の遺伝子を含む。別の実施態様において、DNA構築体はさらに少なくとも1の選択マーカーを含み、ここで選択マーカーは当該遺伝子配列またはそれに相同な遺伝子配列の断片の両側に位置する。
【0023】
本発明は入来配列を含むDNA構築体も提供し、ここで入来配列は目的タンパク質をエンコードする核酸及び相同ボックスを含む両側に位置する選択マーカーを含み、相同ボックスはsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の遺伝子のコード領域のすぐ隣に位置する配列に対して80〜100%配列同一性を有する核酸配列を含む。いくつかの実施態様において、DNA構築体は遺伝子のコード配列の側面に位置する少なくとも1の核酸をさらに含む。本発明はDNA構築体を含むプラスミドも提供する。さらなる実施態様において、本発明はDNA構築体を含むプラスミドを含む宿主細胞を提供する。いくつかの実施態様において、宿主細胞はバチルス細胞及び大腸菌細胞からなる群より選択される。いくつかの好ましい実施態様において、宿主細胞はB.スブチリスである。いくつかの特に好ましい実施態様において、DNA構築体は宿主細胞の染色体内に統合される。別の好ましい実施態様において、選択マーカーは宿主細胞染色体から切除される。
【0024】
本発明はプロテアーゼ生成が増大した変異バチルス株を得るための方法をさらに提供し、以下を含む:少なくとも1の本発明のDNA構築体を含むバチルス宿主細胞を形質転換する工程であって、DNA構築体中の目的タンパク質がプロテアーゼであり、DNA構築体が少なくとも1の遺伝子が変異バチルス株を生成するために不活性化される条件下でバチルス宿主細胞染色体内に統合され;及び増大したプロテアーゼ生成が得られる条件下で変異バチルス株を成長させる工程。いくつかの特に好ましい実施態様において、本方法はプロテアーゼを回収する工程をさらに含む。別の好ましい実施態様において、少なくとも1の不活化遺伝子は変異バチルス株の染色体から欠失される。本発明はここに記載される方法を用いて生成した変異バチルス株も提供する。いくつかの実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、バチルス宿主細胞はB.ズブチリスである。
【0025】
本発明は変異バチルスにおいてプロテアーゼ発現を増大させる方法も提供し、以下を含む:本発明のDNA構築体を用いてバチルス宿主細胞を形質転換させる工程;DNA構築体とバチルス宿主細胞の染色体領域を相同組換えさせる工程であって、バチルス宿主細胞の少なくとも1の染色体遺伝子が不活性化され、変異バチルス株を生成する工程;及びプロテアーゼ発現に適した条件下で変異バチルス株を成長させる工程であって、ここでプロテアーゼ生成は形質転換前のバチルス・ズブチリス宿主と比較して変異バチルス・ズブチリス株が大きい。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。別の実施態様において、プロテアーゼは組換えプロテアーゼである。さらに別の実施態様において、不活性化は少なくとも1の遺伝子の欠失により達成される。さらに別の実施態様において、不活性化は少なくとも1の遺伝子の挿入不活性化による。本発明はここに記載の方法を用いて得た変異バチルス株も提供する。いくつかの実施態様において、変異バチルス株はsbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の不活性化遺伝子を含む。いくつかの好ましい実施態様において、不活性化遺伝子は欠失により不活性化される。別の実施態様において、変異バチルス株はdegU、degS、degQ、scoC4、spollE及びoppAからなる群より選択される遺伝子に少なくとも1の変異を含む。いくつかの好ましい実施態様において、変異はdegU(Hy)32である。さらに別の実施態様において、株は組換えプロテアーゼ生成株である。いくつかの好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。
【0026】
本発明は1以上の固有染色体領域またはその断片を含む変異バチルス株も提供し、ここで固有染色体領域は約0.5〜500kbを含み、変異バチルス株は変異及び非変異バチルス株を本質的に同じ成長条件下で成長させた場合、対応する非変異バチルス株と比較して目的タンパク質の発現が増大している。好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、変異バチルス株はB.ズブチリス、B.リケニフォルミ及びB.アミロリケファシエンスからなる群より選択される。いくつかの特に好ましい実施態様において、変異バチルス株はB.ズブチリス株である。さらに別の実施態様において、固有染色体領域はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVEF−YVEK領域、DHB領域及びそれらの断片からなる群より選択される。いくつかの好ましい実施態様において、2つの固有染色体領域またはその断片は欠失される。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。さらに別の実施態様において、目的タンパク質はプロテアーゼである。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。いくつかの特に好ましい実施態様において、ズブチリシンはズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、及びズブチリシン309及びこれらの変異体からなる群より選択される。さらに好ましい実施態様において、バチルス宿主は組換え体株である。いくつかの特に好ましい実施態様において、変異バチルス株は、degU、degQ、degS、sco4、spollE及びoppAからなる群より選択される遺伝子において少なくとも1の変異をさらに含む。いくつかの好ましい実施態様において、変異はdegU(Hy)32でる。
【0027】
本発明はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVFF−YVEK領域、DHB領域及びそれらの断片からなる群より選択される固有染色体領域の欠失を含むプロテアーゼ生成バチルス株をさらに提供する。いくつかの好ましい実施態様において、プロテアーゼはズブチリシンである。いくつかの実施態様において、プロテアーゼは異種プロテアーゼである。いくつかの好ましい実施態様において、変異バチルス株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される。別の実施態様において、バチルス株はB.ズブチリス株である。
【0028】
本発明はバチルスにおいて目的タンパク質の発現を増大する方法も提供し、以下を含む:選択マーカー及び不活性化染色体断片を含むDNA構築体をバチルス宿主株内に導入する工程であって、ここでDNA構築体をバチルス宿主株の染色体内に統合し、バチルス宿主細胞から固有染色体領域またはその断片の欠失を生じて変異バチルス株を生じる工程;及び適当な条件下で変異バチルス株を成長させる工程であって、ここで目的タンパク質発現は変異していないバチルス宿主細胞における目的タンパク質の発現と比較して変異バチルス株が大きい。いくつかの好ましい実施態様において、本方法は目的タンパク質を回収する工程をさらに含む。いくつかの実施態様において、本方法は変異バチルス株から選択マーカーを切除する工程をさらに含む。別の実施態様において、固有染色体領域はPBSX領域、スキン領域、プロファージ7領域、SPβ領域、プロファージ1領域、プロファージ2領域、プロファージ3領域、プロファージ4領域、プロファージ5領域、プロファージ6領域、PPS領域、PKS領域、YVFF−YVEK領域、DHB及びそれらの断片からなる領域の群より選択される。さらに別の実施態様において、変異バチルス株は少なくとも2の固有染色体領域の欠失を含む。いくつかの好ましい実施態様において、目的タンパク質は酵素である。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。いくつかの実施態様において、バチルス宿主株はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。本発明はここに記載の方法を用いて生成した変異バチルス株も提供する。
【0029】
本発明はバチルス株から目的タンパク質を得る方法も提供し、以下を含む:選択マーカー及び不活性化染色体断片を含むDNA構築体を用いてバチルス宿主細胞を形質転換する工程であって、ここでDNA構築体はバチルス株の染色体内に統合され、固有染色体領域またはその断片の欠失を生じ、変異バチルス株を生成し、目的タンパク質の発現を可能にするのに適した条件下で変異バチルス株を培養する工程及び目的タンパク質を回収する工程。いくつかの好ましい実施態様において、目的タンパク質は酵素である。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの好ましい実施態様において、固有染色体領域はPBSX、スキン、プロファージ7、SPβ、プロファージ1、プロファージ2、プロファージ3、プロファージ4、プロファージ5、プロファージ6、PPS、PKS、YVFF−YVEK、DHB及びそれらの断片からなる群より選択される。いくつかの特に好ましい実施態様において、変異バチルス株はdegU、degQ、degS、sco4、spollE及びoppAからなる群より選択される遺伝子において少なくとも1の変異をさらに含む。いくつかの実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼからなる群より選択される酵素である。いくつかの特に好ましい実施態様において、当該酵素はプロテアーゼである。いくつかの好ましい実施態様において、目的タンパク質は酵素である。別の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0030】
本発明はバチルスにおいて目的タンパク質の発現を増大させる方法をさらに提供し、以下を含む:少なくとも1のバチルス細胞から核酸を得る工程;前記バチルス細胞由来核酸についてトランスクリプトームDNAアレイ分析を行い、少なくとも1の目的遺伝子を同定する工程;DNA構築体を生成するために少なくとも1の目的遺伝子を修飾する工程;DNA構築体をバチルス宿主細胞内に導入して変異バチルス株を生成する工程であって、ここで変異体は目的タンパク質発現が非変異のバチルスにおける目的タンパク質の発現と比較して増大するような条件下で目的タンパク質を生成することができる。いくつかの実施態様において、目的タンパク質はアミノ酸生合成経路及び生分解経路からなる群より選択される少なくとも1の生化学経路に関係する。いくつかの実施態様において、本方法は少なくとも1の生分解経路を無効にすることを含む。いくつかの実施態様において、生分解経路は目的遺伝子の転写により無効にされる。しかしながら、本発明はこれらの経路に限定されるものではなく、本方法は目的タンパク質の発現の増加が生じるように細胞内のその他の生化学経路の修飾においても使用できることが考えられる。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの好ましい実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0031】
本発明はバチルス中において目的タンパク質の発現を増大させる方法をさらに提供し、以下を含む:少なくとも1のバチルス細胞から少なくとも1の目的遺伝子を含む核酸を得る工程;前記核酸を断片化する工程;前記少なくとも1の目的遺伝子を含む増殖断片プールを生成するために前記断片を増殖する工程;前記増殖断片をDNA構築体を生成するために連結する工程;前記DNA構築体をバチルス宿主細胞内に直接形質転換し、変異バチルス株を生成する工程;変異されていないバチルスにおける前記目的タンパク質の発現と比較して前記目的タンパク質の発現が増大するような条件下で前記変異バチルス株を培養する工程。いくつかの好ましい実施態様において、前記増殖はポリメラーゼ連鎖反応を用いることを含む。いくつかの実施態様において、変異バチルス株はprpC、sigD及びtdh/kblからなる群より選択される修飾遺伝子を含む。いくつかの特に好ましい実施態様において、バチルス宿主は目的タンパク質をエンコードする異種遺伝子を含む。別の実施態様において、バチルス宿主細胞はB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.クラウジイ、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス及びB.チューリンゲンシスからなる群より選択される。いくつかの実施態様において、目的タンパク質は酵素である。いくつかの好ましい実施態様において、目的タンパク質はプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ、リパーゼ、イソメラーゼ、トランスフェラーゼ、キナーゼ及びホスファターゼから選択され、他の実施態様において、目的タンパク質は抗体、ホルモン及び成長因子からなる群より選択される。
【0032】
本発明はさらに配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33からなる群より選択される核酸配列に示す配列を含む単離核酸をさらに提供する。
【0033】
また、本発明はアミノ酸をエンコードする単離核酸配列を提供し、ここで当該アミノ酸は配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される。
【0034】
本発明はさらに単離アミノ酸配列を提供し、当該アミノ酸は配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号38、配列番号26、配列番号22、配列番号57、配列番号24、配列番号28、配列番号20、配列番号32、配列番号55、配列番号53、配列番号36及び配列番号34からなる群より選択される。
【図面の簡単な説明】
【0035】
【図1A】図1Aは本発明により提供されるの1の方法(“方法1”実施例1参照)の一般的な説明図を示すものである。この方法において、遺伝子のフランキング領域及び/または固有染色体領域は野生型バチルス染色体から増殖され、制限酵素(少なくともBamHIを含む)で切断し、pJM102に連結する。当該構築体は大腸菌によりクローンされ、及びプラスミドを単離し、BamHIを用いて線形化し、相補的末端を含む抗菌マーカーに連結する。大腸菌で再度クローニングした後、液体培養物を成長させ、バチルス宿主株(好ましくは、コンピテントバチルス宿主株)の形質転換に用いるプラスミドDNAを単離するために用いる。
【図1B】図1Bは本発明により提供されるの1の方法(“方法1”実施例1参照)の一般的な説明図を示すものである。この方法において、遺伝子のフランキング領域及び/または固有染色体領域は野生型バチルス染色体から増殖され、制限酵素(少なくともBamHIを含む)で切断し、pJM102に連結する。当該構築体は大腸菌によりクローンされ、及びプラスミドを単離し、BamHIを用いて線形化し、相補的末端を含む抗菌マーカーに連結する。大腸菌で再度クローニングした後、液体培養物を成長させ、バチルス宿主株(好ましくは、コンピテントバチルス宿主株)の形質転換に用いるプラスミドDNAを単離するために用いる
【図2】図2は本発明のいくつかの実施態様に従うDNAカセットの構築において用いるプライマーの位置を示す。図はここで用いるプライマー命名システムの説明を提供する。プライマー1及び4は欠失の存在をチェックするために用いる。これらのプライマーは“欠失X−UF−chk”及び“欠失X−UR−chk−del”と呼ぶ。欠失X−UF−chkも、染色体中に存在するカセットの陽性チェックのための抗生物質マーカー内のリバースプライマー(プライマー11:PBSX−UR−chk−Delと呼ばれる)を用いてPCR反応において用いる。プライマー2及び6は上流フランキング領域を増殖するために用いる。これらのプライマーは“欠失X−UF”及び“欠失X−UR”と呼び、黒の縦棒のところで組換え制限部位を含む。プライマー5及び8は下流フランキング領域を増幅するために用いる。これらのプライマーは“欠失X−DF”及び“欠失X−DR”と呼ぶ。これらのプライマーは連結及びクローニングのための組換えBamHI部位またはPCR融合で用いるバチルス・ズブチリス染色体の適当な部分に相同な25塩基対末端を含むことができる。いくつかの実施態様において、プライマー3及び7はPCR融合により作成したカセットの場合、当該カセットを一緒に融合するために用いられ、一方、他の実施態様において、それらは挿入の存在をチェックするために用いられる。これらのプライマーは“欠失X−UF−ネスティド”及び“欠失X−DR−ネスティド”と呼ぶ。いくつかの実施態様において、“抗生物質マーカー”に対応する配列はSpc耐性マーカーであり、欠失する領域はCssS遺伝子である。
【図3】図3は本発明の1の方法(“方法2”;実施例2を参照)の一般的な説明図である。フランキング領域は選択マーカー配列に相補的な25bpの配列を含むように設計される。選択マーカー配列は1のフランキング領域のDNAに相補的である25bpの末端も含む。フランキング領域末端に近いプライマーは単一反応チューブにおける3つすべてのテンプレートを増幅するために用いられ、それにより融合断片を作成する。この融合断片またはDNA構築体はコンピテントバチルス宿主株に直接形質転換される。
【図4】図4はバチルスDHB欠失クローンのエレクトロフォレーシスゲルを提供する。レーン1及び2はプライマー1及び11を用いて増幅されたDHB欠失を有する2つの株を描き、不活性化染色体断片の上流からフレオマイシン・マーカー内に増幅した1.2バンドを表す。レーン3はこの反応の野生型対照を表す。非特異増幅のみが観測される。レーン4及び5はプライマー9及び12を用いて増幅されたDHB欠失株を表す。この2kbバンドは抗生物質領域から不活性化染色体断片の下流部分以下まで増幅する。レーン6はこの反応の陰性対照であり、バンドは示されていない。レーン7及び8はプライマー1及び4を用いて増幅した欠失株を表し、当該説明図によりDHB領域がないことが確認される。レーン9は野生型対照である。
【図5】図5はバチルス・ズブチリス(野生型)の生成株の2つのクローンのゲルエレクトロフォレーシスを表し、s/rはs/r遺伝子の欠失を生じるフレオマイシン(phleo)マーカーで置換される。レーン1及び2は位置1及び11でプライマーを用いて増幅したクローン表す。レーン3は同じプライマーを用いて増幅した野生型染色体DNAである。1.2kbのバンドが挿入に関して観測される。レーン4及び5は位置9及び12でプライマーを用いて増幅したクローンを表す。レーン6は同じプライマーを用いて増幅した野生型染色体DNAである。正確な形質転換体は2kbのバンドを含む。レーン7及び8は位置2及び4でプライマーを用いて増幅したクローンを表す。レーン9は同じプライマーを用いて増幅した野生型染色体DNAである。欠失株に関してバンドは観測されないが、1kb周辺のバンドが野生型において観測される。プライマー位置の説明のため、図2を参照されたい。
【図6】図6はバチルス・ズブチリス(野生型)の生成株のクローンのエレクトロフォレーシスゲルを提供し、cssSはspcマーカーを染色体内に統合することにより不活性化される。レーン1は非統合の対照であり、約1.5kb小さい。
【図7】図7はバチルス・ズブチリス野生型株(非変異)及び種々の欠失を有する対応する変異バチルス・ズブチリス株を含む振とうフラスコ培養物から測定したズブチリシン分泌の改善を示す棒グラフである。プロテアーゼ活性(g/L)は17、24及び40時間後、または24及び40時間の時点で測定した。
【図8】図8はバチルス・ズブチリス野生型株(非変異)及び対応する変異欠失株(−sbo)及び(−slr)における振とうフラスコ培養物から測定したズブチリシン分泌の改善を示す棒グラフである。プロテアーゼ活性(g/L)は17、24及び40時間後に測定した。
【発明を実施するための形態】
【0036】
発明の説明
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、または修飾される。好ましい実施態様において、1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。
【0037】
定義
ここで言及する全ての特許及び文献、並びに当該特許及び文献に開示される全ての配列はここに明示的に引用するものとする。別に定めない限り、ここで用いる全ての技術及び科学的用語は本発明が属する当業者に通常理解される意味と同じ意味を有する(例えば、Singleton et al.,Dictionary of Microbiology and Molecular Biology,第2版、John Wiley and Sons,ニューヨーク[1994];及びHale and Marham、The Harper Collins Dictionary of Biology,Harper Perennial,NY[1991]を参照、これらはここで用いる多くの用語の当業者の一般的な辞書となる)。ここに記載の方法及び材料に類似または同等のいかなる方法及び材料も本発明を実施または試験するために用いることができるが、好ましい方法及び材料を記載する。数値範囲はその範囲を定める数字を含むものとする。明細書及び請求の範囲で用いるように、文脈において明らかに示される場合を除いて、単数の記載は複数も含むものとする。従って、例えば“宿主細胞”への言及は複数の当該宿主細胞も含む。
【0038】
別に示さない限り、核酸は5’から3’の左から右方向に記載し、アミノ酸配列はアミノからカルボキシの左から右方向にそれぞれ記載する。ここに記載する見出しは本発明の種々の側面または実施態様を限定するものではなく、明細書全体に対して参照され得るものである。従って、下記に定義する用語は明細書全体を参照することによりさらに十分に定義されるものである。
【0039】
ここで用いる、“宿主細胞”とは新しく導入するDNA配列のための宿主または発現媒体として作用する能力を有する細胞をいう。本発明の好ましい実施態様において、宿主細胞はバチルス種または大腸菌細胞である。
【0040】
ここで用いる“バチルス属”とは、当業者に公知の“バチルス”属に含まれるすべての種類を含み、限定されないが、B.ズブチリス、B.リケニフォルミス、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.アミロリケファシエンス、B.クラウジイ、B.ハロデュランス、B.メガテリウム、B.コアグランス、B.サーキュランス、B.ロータス、及びB.チューリンゲンシスが挙げられる。バチルス属は分類上再編成を受け続けている。従って、当該属は再分類された種も含むものとし、限定されないが、現在は“ゲオバチルス・ステロサーモフィラス”と呼ばれるB.ステロサーモフィラスなどの微生物を含む。酸素存在下での耐性内生胞子の生成はバチルス属の重要な特徴と考えられるが、この特徴は最近ではアリシクロバチルス、アンフィバチルス(amphibacillus)、アニューリーニバチルス(aneurinibacillus)、アノキシバチルス(anoxybacillus)、ブレビバチルス、フィロバチルス、グラシリバチルス(Gracilibacillus)、ハロバチルス、ペニバチルス、サリバチルス、サーモバチルス、ウレーバチルス(Ureibacillus)及びヴァージバチルス(Virgibacillus)と称されるものにも当てはまる。
【0041】
ここで用いる、“核酸”の語はヌクレオチドまたはポリヌクレオチド配列及びそれらの断片または部分、及びセンスまたはアンチセンスストランドのいずれを表すかにかかわらず、2本鎖または1本鎖のゲノムまたは合成起源のDNA、cDNA及びRNAをいう。遺伝子コードの縮退の結果、多数のヌクレオチド配列が所定のタンパク質をエンコードすることは当然理解されるであろう。
【0042】
ここで用いる、“遺伝子”の語はポリぺプチド鎖の生成に関するDNAの染色体断片を意味し、コード領域(例えば、5’非翻訳(5’UTR)またはリーダー配列及び3’非翻訳(3’UTR)またはトレーラー配列)に先行または後に続く領域、及び個々のコード断片(エクソン)間の介在配列(イントロン)を含んでも含まなくてもよい。
【0043】
いくつかの実施態様において、遺伝子は治療的に重要なタンパク質またはぺプチド、例えば成長因子、サイトカイン、配位子、受容体及び阻害物質、並びにワクチン及び抗体などをエンコードする。遺伝子は商業的に重要な工業タンパク質またはぺプチド、例えば酵素(例えば、プロテアーゼ、アミラーゼ及びグルコアミラーゼなどのカルボヒドラーゼ、セルラーゼ、オキシダーゼ及びリパーゼ)もエンコードし得る。しかしながら、本発明は特定の酵素またはタンパク質に限定されるものではない。いくつかの実施態様において、目的遺伝子は天然遺伝子であり、一方、他の実施態様において、変異遺伝子または合成遺伝子である。
【0044】
ここで用いる“ベクター”の語は、細胞内で複製でき、新しい遺伝子またはDNA断片を細胞内に運ぶことができる核酸をいう。従って、当該用語は異なる宿主細胞間を移動するために設計された核酸構築体をいう。“発現ベクター”とは、外来細胞中に異種DNA断片を導入し、発現できるベクターをいう。多くの原核及び真核発現ベクターは市販されている。適当な発現ベクターの選択は当業者の知識の範囲内である。
【0045】
ここで用いる、“DNA構築体”、“発現カセット”及び“発現ベクター”の語は標的細胞中で特定の核酸の転写を可能とする、一連の具体的な核酸成分を用いて組換えにより、または合成的に生成した核酸構築体をいう(すなわち、これらは上述のベクターまたはベクター成分である)。組換え発現カセットはプラスミド、染色体、ミトコンドリアDNA、色素体DNA、ウイルスまたは核酸断片内に組み込むことができる。通常、発現ベクターの組換え発現カセット部分は、数ある配列の中でも、転写される核酸配列及びプロモーターを含む。いくつかの実施態様において、DNA構築体は標的細胞中で特定の核酸の転写を可能とする一連の特定核酸成分も含む。1の実施態様において、本発明のDNA構築体はここで定義する選択マーカー及び不活性化染色体断片を含む。
【0046】
ここで用いる“形質転換DNA”、“形質転換配列”及び“DNA構築体”とは、配列を宿主細胞または微生物内に導入するために用いられるDNAをいう。形質転換DNAはPCRまたはその他の適当な技術によりin vitroで生成できる。いくつかの好ましい実施態様において、形質転換DNAは入来配列を含み、一方、他の好ましい実施態様において、さらに相同ボックスの側面に位置する入来配列を含む。さらに別の実施態様において、形質転換DNAはその他の非相同配列を含み。末端に加えられる(すなわち、stuffer配列またはフランキング配列)。末端は、例えばベクター内への挿入など形質転換DNAが閉じた環を形成するように閉じることができる。
【0047】
ここで用いる、“プラスミド”の語はクローニングベクターとして用いる環状2本鎖(ds)DNA構築体をいい、多くの細菌及びいくつかの真核生物内において染色体外自己複製遺伝子要素を形成する。いくつかの実施態様において、プラスミドは宿主細胞のゲノム内に組み込まれるようになる。
【0048】
ここで用いる“単離”及び“精製”の語は、天然に結合している少なくとも1の成分から取り除いた核酸またはアミノ酸(またはその他の成分)をいう。
【0049】
ここで用いる“発現の増大”とは、目的タンパク質生成の増大を含むように広く解釈される。発現の増大とは、ここでの記載に従った変異をさせていないが、本質的に同じ成長条件下で成長させた対応宿主株における通常の発現レベル以上の発現をいう。
【0050】
いくつかの好ましい実施態様において、“増大”は望ましい特性の増加を生じる任意の修飾により達成される。例えば、いくつかの特に好ましい実施態様において、本発明はタンパク質生成を増大させる手段を提供し、増大株が親株(例えば、野生型及び/または始発株)よりも目的タンパク質を多く生成し及び/または優れた質を生じるようにする。
【0051】
ここで用いる“発現”とは、ポリぺプチドが遺伝子の核酸配列に基づいて生成されるプロセスをいう。当該プロセスは転写及び翻訳の両方を含む。
【0052】
ここで用いる、核酸配列を細胞内に導入することに関する“導入”の語は核酸配列を細胞内に移動させるのに適した任意の方法をいう。導入のための手段は、限定されないが、原形質融合、トランスフェクション、形質転換、接合及び形質導入などが挙げられる(例えば、Ferrari et al.、“Genetics(遺伝学)”、Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.,第57〜72頁、[1989]を参照)。
【0053】
ここで用いる“形質転換”及び“安定な形質転換”の語は、ゲノム内に統合された、または少なくとも2世代の間維持されたエピソームプラスミドとして非天然(異種)ポリヌクレオチド配列を有する細胞をいう。
【0054】
ここで用いる“入来配列”の語はバチルス染色体内に導入されたDNA配列をいう。いくつかの好ましい実施態様において、入来配列はDNA構築体の一部である。好ましい実施態様において、入来配列は1以上の目的タンパク質をエンコードする。いくつかの実施態様において、入来配列は形質転換される細胞のゲノム中に既に存在する、または存在しない配列であってもよい(すなわち、相同または異種配列である)。いくつかの実施態様において、入来配列は1以上の目的タンパク質、遺伝子及び/または変異または修飾遺伝子をエンコードする。別の実施態様において、入来配列は機能野生型遺伝子またはオペロン、機能変異遺伝子またはオペロン、または非機能遺伝子またはオペロンをエンコードする。いくつかの実施態様において、非機能配列は遺伝子の機能を妨害するために遺伝子内に導入できる。いくつかの実施態様において、入来配列は1以上の機能野生型遺伝子をエンコードし、一方他の実施態様において、入来配列は1以上の機能変異遺伝子をエンコードし、さらに他の実施態様において、入来配列は1以上の非機能遺伝子をエンコードする。他の実施態様において、入来配列は形質転換される宿主細胞の染色体内に既に存在する配列をエンコードする。好ましい実施態様において、入来配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD及びこれらの断片からなる群より選択される遺伝子を含む。さらに別の実施態様において、入来配列は選択マーカーを含む。さらなる実施態様において、入来配列は2つの相同ボックスを含む。
【0055】
いくつかの実施態様において、入来配列は少なくとも1の異種タンパク質をエンコードし、限定されないが、ホルモン、酵素及び成長因子が挙げられる。他の実施態様において、酵素は、限定されないが、プロテアーゼ、エステラーゼ、リパーゼ、フェノールオキシダーゼ、パーミアーゼ、アミラーゼ、プルラナーゼ、セルラーゼ、グルコースイソメラーゼ、ラッカーゼ及びタンパク質ジスルフィドイソメラーゼが挙げられる。
【0056】
ここで用いる“相同ボックス”とはバチルス染色体の配列と相同な核酸配列をいう。より具体的には、相同ボックスは、本発明に従って不活性化される遺伝子に隣接するコード領域または遺伝子の一部と約80〜100%配列同一性、約90〜100%配列同一性または約95%〜100%配列同一性を有する上流または下流領域をいう。これらの配列はバチルス染色体におけるDNA構築体の統合を目的とし、及び入来配列によりバチルス染色体の一部が置換されることを目的とする。本発明を限定するものではないが、相同ボックスは約1塩基対(bp)〜200キロ塩基(kb)を含む。好ましくは、相同ボックスは約1bp〜10.0kb;1bp〜5.0kb;1bp〜2.5kb;1bp〜1.0kb及び0.25kb〜2.5kbを含む。また、相同ボックスは約10.0kb、5.0kb、2.5kb、2.0kb、1.5kb、1.0kb、0.5kb、0.25kb及び0.1kbを含む。いくつかの実施態様において、選択マーカーの5’及び3’末端は相同ボックスに隣接し、ここで相同ボックスは当該遺伝子のコード領域にすぐ隣接した核酸配列を含む。
【0057】
ここで用いる“選択マーカー−エンコードヌクレオチド配列”の語は宿主細胞において発現できるヌクレオチド配列をいい、選択マーカーの発現により対応する選択因子の存在下、または必須栄養素の不存在下で発現遺伝子を含む細胞は成長できるようになる。
【0058】
ここで用いる“選択可能マーカー”及び“選択マーカー”の語は宿主細胞中で発現できる核酸(例えば遺伝子)をいい、ベクターを含むこれら宿主の選択が簡単にできるようにする。当該選択マーカーの例としては、限定されないが、抗微生物などがある。従って、“選択マーカー”とは、宿主細胞が目的の入来DNAを取り込んだ、またはその他の反応が起こった印を与える遺伝子をいう。通常、選択マーカーは、形質転換時に外来配列を受け取らなかった細胞と外来DNAを含む細胞との区別を可能にするために宿主細胞に抗菌耐性または代謝利点を与える遺伝子をいう。“存在(residing)選択マーカー”とは、形質転換される微生物の染色体上に位置するマーカーである。存在選択マーカーは遺伝子を形質転換DNA構築体上の選択マーカーとは異なる遺伝子をエンコードする。選択マーカーは当業者に公知である。上述の通り、好ましくは当該マーカーは抗微生物耐性マーカーである(例えば、ampR;phleoR;specR;kanR;eryR;tetR;cmpR;及びneoR;例えばGuerot−Fleury,Gene,167:335−337[1995];Palmeros et al.、Gene 247:255−264[2000];及びTrieu−Cuot et al.、Gene、23:331−341[1983]を参照)。いくつかの特に好ましい実施態様において、本発明はクロラムフェニコール耐性遺伝子を提供する(例えば、pC194上に存在する遺伝子、及びバチルス・リケニフォルミスゲノム中に存在する耐性遺伝子)。この耐性遺伝子は本発明及び染色体上統合されたカセット及び統合プラスミドの染色体増幅に関する実施態様において特に有用である(例えば、Albertini and Galizzi、Bacteriol.,162:1203−1211[1985];及びStahl and Ferrari,J.Bacteriol.158:411−418[1984]を参照)。この天然クロラムフェニコール耐性遺伝子のDNA配列を以下に示す:
【式1】
【0059】
このクロラムフェニコール耐性タンパク質の推定アミノ酸配列は以下の通り:
【式2】
【0060】
本発明に従う有用なその他のマーカーは、限定されないが、トリプトファンなどの栄養素要求性マーカー及びβ−グラクトシダーゼなどの検出マーカーである。
【0061】
ここで用いる“プロモーター”の語は下流遺伝子の転写に直接機能する核酸配列をいう。好ましい実施態様において、プロモーターは標的遺伝子が発現するように宿主細胞に適したものである。プロモーターはその他の転写及び翻訳調節核酸配列(“制御配列”とも称される)と一緒に、所定の遺伝子を発現するために必要なものである。通常、転写及び翻訳調節配列は、限定されないが、プロモーター配列、リボソーム結合部位、転写開始及び停止配列、翻訳開始及び停止配列、及びエンハンサーまたは活性化配列などが挙げられる。
【0062】
核酸は他の核酸配列と機能的に関連するように位置する場合、“作動可能に連結する”。例えば、分泌リーダー(すなわち、1のぺプチド)をエンコードするDNAはポリぺプチドの分泌に参加するプレタンパク質として発現する場合、当該ポリぺプチドに関するDNAに作動可能に連結し;プロモーターまたはエンハンサーは配列転写に影響を与える場合、コード配列に作動可能に連結し;またはリボソーム結合部位は翻訳を促進するように位置する場合、コード配列に作動可能に連結する。一般的に“作動可能に連結する”とは、連結するDNA配列が連続していることを意味し、分泌リーダーの場合、連続かつ読取り層にある。しかしながら、エンハンサーは必ずしも連続している必要はない。結合は都合のよい制限酵素認識部位で連結することにより達成される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを従来実務に従って用いる。
【0063】
“不活性化”の語は、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD染色体遺伝子のうち1以上の機能発現を妨げる方法を含み、当該遺伝子または遺伝子産物はその公知機能を発揮するための使用に適している。不活性化または増大化は任意の適した手段によって生じ、核酸遺伝子配列における欠失、置換(例えば変異)、阻害、及び/または挿入などがある。1の実施態様において、不活性化遺伝子の発現産物は対応するタンパク質の生物活性変化を有する短いタンパク質である。いくつかの実施態様において、生物活性変化は活性の増加であり、一方、好ましい実施態様において、当該変化は生物活性の損失を生じる。いくつかの実施態様において、変異バチルス株は好ましくは安定かつ不可逆的な不活性化を生じる1以上の遺伝子の不活性化を含む。
【0064】
いくつかの好ましい実施態様において、不活性化は欠失により達成される。いくつかの好ましい実施態様において、遺伝子は相同組換えにより欠失する。例えば、いくつかの実施態様において、欠失する遺伝子がsboの場合、相同ボックスの両側に隣接する選択マーカーを有する入来配列を含むDNA構築体を用いる。相同ボックスは染色体sbo遺伝子の核酸フランキング領域に相同なヌクレオチド配列を含む。DNA構築体はバチルス宿主染色体の相同配列と一致し、二重交差(double crossover)の場合、sbo遺伝子は宿主染色体から切除される。
【0065】
ここで用いる遺伝子の“欠失”の語は、コード配列全体の欠失、コード配列の一部の欠失、またはフランキング領域を含むコード配列の欠失をいう。欠失は染色体に残った配列が望ましい遺伝子の生物活性を与えるのであれば部分的であってもよい。コード配列のフランキング領域は約1bp〜約500bpを5’及び3’末端に含むことができる。フランキング領域は500bpよりも大きくてもよいが、好ましくは本発明に従って不活性化または欠失され得るその他の遺伝子を当該領域に含まない。最終的には、欠失遺伝子は事実上、非機能的である。簡単に言えば、“欠失”の語は1以上のヌクレオチドまたはアミノ酸残基がそれぞれ除去された(すなわち、存在しない)、ヌクレオチドまたはアミノ酸配列の変化として定義される。従って、“欠失変異体”はそれぞれの野生型生物よりも少ないヌクレオチドまたはアミノ酸を有する。
【0066】
本発明の他の実施態様において、DNAアレイ分析(例えば、ここに記載するトランスクリプトーム分析)により測定して不適当な時に活性な遺伝子の欠失は生成タンパク質の発現の増加をもたらす。いくつかの好ましい実施態様において、pckA,gapB、fbp及び/またはalsDからなる群より選択される1以上の遺伝子の欠失は供給物の効率が向上した改善株を提供する。ここで用いる“トランスクリプトーム分析”とは、遺伝子転写分析をいう。
【0067】
本発明の他の実施態様において、遺伝子は調節配列の欠失により“最適化”されると考えられ、この欠失により所望の生成物の発現が増加する。本発明のいくつかの好ましい実施態様において、トリプトファンオペロン(すなわち、遺伝子trpA、trpB、trpC、trpD、trpE、trpFを含む)はTRAP結合RNA配列をコードするDNA配列の欠失により最適化される(Yang et al.、J.Mol.Biol.,270:696−710[1997]を参照)。この欠失は宿主細胞由来の所望の生成物の発現を増加させると考えられる。
【0068】
他の好ましい実施態様において、不活性化は挿入による。例えば、いくつかの実施態様において、sboが不活性化される遺伝子である場合、DNA構築体は選択マーカーにより阻害されるsbo遺伝子を有する入来配列を含む。選択マーカーはsboコード配列部分の両側に隣接する。DNA構築体は宿主染色体中のsbo遺伝子の本質的に同一配列と一致し、二重交差の場合、sbo遺伝子は選択マーカーの挿入により不活性化される。簡単に言えば、“挿入”または“付加”は、それぞれ天然配列と比較して1以上のヌクレオチドまたはアミノ酸残基の付加を生じたヌクレオチドまたはアミノ酸配列の変化である。
【0069】
他の実施態様において、一重交差の場合、ベクターとしてプラスミドを用いた挿入により活性化される。例えば、sbo染色体遺伝子は遺伝子または遺伝子コード配列の一部及び選択マーカーを含むプラスミドに合わせる。いくつかの実施態様において、選択マーカーは遺伝子コード配列内に位置し、または遺伝子から分離されたプラスミドの一部に位置する。ベクターはバチルス染色体内に統合され、当該遺伝子はコード配列中のベクター挿入により不活性化される。
【0070】
別の実施態様において、不活性化は遺伝子の変異により生じる。遺伝子を変異する方法は当業者に公知であり、限定されないが、部位特異的変異、ランダム変異発生、及びgapped−duplex法が挙げられる(例えば米国特許第4,760,025号;Moring et al.、Biotech.2:646[1984];及びKramer et al.、Nucleic Acids Res.,12:9441[1984]を参照)。
【0071】
ここで用いる“置換”とは1以上のヌクレオチドまたはアミノ酸と、それぞれ異なるヌクレオチドまたはアミノ酸との置き換えにより生じる。
【0072】
ここで用いる“相同遺伝子”とは、異なるが、通常は関連種由来の遺伝子対をいい、それぞれが対応し、それぞれ同一または非常に類似しているものをいう。当該用語は種形成(すなわち、新しい種の発達)により分離した遺伝子(例えばオーソロガス遺伝子)、及び遺伝子重複により分離された遺伝子(例えばパラロガス遺伝子)を包含する。
【0073】
ここで用いる“オーソログ”及び“オーソロガス遺伝子”とは種形成により共通祖先の遺伝子(すなわち相同遺伝子)から進化した異なる種の遺伝子をいう。通常、オーソロガスは進化の過程において同じ機能を保持する。オーソロガスの同定により新しく配列決定されたゲノムの遺伝子機能の信頼性の高い推測ができる。
【0074】
ここで用いる“パラログ”及び“パラロガス遺伝子”とはゲノム内で重複していることにより関連した遺伝子をいう。オーソログは進化の過程を通して同じ機能を維持するが、パラログはいくつかの機能は最初の機能に関連することが多いとしても、新しい機能を進化させる。パラログ遺伝子の例としては、限定されないが、トリプシン、キモトリプシン、エラスターゼ及びトロンビンをエンコードする遺伝子が挙げられ、これらは全てセリンプロテイナーゼであり、同じ種内で一緒に生じる。
【0075】
ここで用いる“相同性”とは、好ましい同一性を有する配列類似性または同一性をいう。この相同性は当業者に公知の標準技術を用いて決定する(例えば、Smith and Waterman,Adv.Appl.Math.、2:482[1981];Needlman and Wunschを、J.Mol.Biol.,48:443[1970];Pearson and Lipman、Proc.Natl.Acad.Sci.USA 85:2444[1988];ウィスコンシン・ジェネティックス・ソフトウェアパッケージ(Genetics Computer Group,マディソン、ウィスコンシン州)のGAP、BESTFIT、FASTA及びTFASTAなどのプログラム;Devereux et al.、Nucl.Acid Res.,12:387−395[1984]を参照)。
【0076】
ここで用いる“類似配列”は遺伝子機能が本質的にバチルス・ズブチリス株168から示される遺伝子と同じであるものをいう。さらに、類似遺伝子はバチルス・ズブチリス株168遺伝子の配列と少なくとも60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%または100%の配列同一性を含む。もしくは、類似配列はB.ズブチリス内で見られる70〜100%の遺伝子配列を有し、及び/またはB.ズブチリス168染色体内の遺伝子と一致する領域に見られる少なくとも5〜10の遺伝子を有する。類似配列は配列決定の公知方法により決定できる。一般的に用いられる配列決定方法はBLASTであるが、上述及び下記に示す通りその他の方法も配列を決定するために用いることができる。
【0077】
有用なアルゴリズムの1の例はPILEUPである。PILEUPは連続(progressive)、対合配列を用いて関連配列グループから複数の配列決定を行うことができる。また、配列決定に用いる集積性関係を示す系図をプロットすることもできる。PILEUPはFeng及びDoolittleの連続配列法の単純化を用いる(Feng and Doolittle、J.Mol.Evol.、35:351−360[1987])。当該方法はHigginsとSharpが説明したものと類似する(Higgins and Sharp、CABIOS 5:151−153[1989])。有用なPILEUPパラメーターは初期設定ギャップウエイトが3.00、初期設定ギャップ長さウエイトが0.10であり、及びエンドギャップを用いる。
【0078】
有用なアルゴリズムの他の例としてはBLASTアルゴリズムであり、Altschul et al.により説明されている(Altschul et al.,J.Mol.Biol.,215:403−410、[1990];及びKarlin et al.、Proc.Natl.Acad.Sci.USA 90:5873−5787[1993])。特に有用なBLASTプログラムはWU−BLAST−2プログラムである(Altschul et al.,Meth.Enzymol.,266:460−480[1996]を参照)。WU−BLAST−2はいくつかのサーチパラメーターを用い、ほとんどは初期値に設定されている。調節可能なパラメーターは以下の値に設定する:オーバーラップスパン=1、オーバーラップフラクション=0.125、言語閾値(T)=11。HSP S及びHSP S2パラメーターは動的値であり、特定配列の組成物及び目的配列がサーチされる特定データベースの組成物に応じてプログラム自体により設定される。しかしながら、当該値は感度を増加させるため調節できる。A%のアミノ酸配列同一性値は適合同一残基の数を配列領域の“より長い”配列残基の合計数で割ることにより決定する。“より長い”配列は配列領域において実際に存在している残基を最も有する配列である(配列スコアを最大化するためにWU−BLAST−2により導入されたギャップは無視する)。
【0079】
従って、“核酸配列同一性比率(%)”は、図の核酸において示されるヌクレオチド配列残基と同一である、候補配列におけるヌクレオチド残基の比率として定義される。好ましい方法はオーバーラップスパン及びオーバーラップファンクションがそれぞれ1及び0.125に設定された初期設定パラメーター設定のWU−BLAST−2のBLASTNモジュールを利用する。
【0080】
配列は配列決定される配列においてギャップ導入を含むことができる。さらに、図の核酸よりも多いまたは少ないヌクレオチドを含む配列については、相同性比率はヌクレオチド合計数と比較した相同ヌクレオチドの数に基づいて決定されることが理解される。従って、例えば、ここで同定する配列よりも短い配列の相同性は下記に説明するように、短い配列中のヌクレオチドの数を用いて決定される。
【0081】
ここで用いる“ハイブリダイゼーション”の語は当業者に公知の塩基対合により、核酸鎖を相補鎖と接合させるプロセスをいう。
【0082】
核酸配列は、2つの配列が中〜高ストリンジェンシーなハイブリダイゼーション条件及び洗浄条件下でお互いに特異的にハイブリダイズする場合、対照核酸配列に “選択的にハイブリダイズ可能”であると考えられる。ハイブリダイゼーション条件は核酸結合複合体またはプローブの融点(Tm)に基づく。例えば、“最大ストリンジェンシー”は通常約Tm−5℃(プローブのTmより5°低い)で起こり、“高ストリンジェンシー”はTmより約5〜10℃低く;“中間ストリンジェンシー”はプローブのTmより約10〜20℃低く;及び“低ストリンジェンシー”はTmより約20〜25℃低い。機能上、最大ストリンジェンシー条件はハイブリダイゼーションプローブと厳密な同一性またはほぼ厳密な同一性を有する配列を同定するために用いることができ、一方、中間または低ストリンジェンシーハイブリダイゼーションはポリヌクレオチド配列相同体を同定または検出するために用いることができる。
【0083】
中〜高ストリンジェンシーハイブリダイゼーション条件は当業者に公知である。高ストリンジェンシー条件の例としては、50%ホルムアミド、5X SSC、5X Denhardt’s溶液、0.5%SDS及び100μg/ml変性キャリアDNA中、約42℃でのハイブリダイゼーションを含み、続いて2X SSC及び0.5%SDS中、室温で2回洗浄し、0.1X SSC及び0.5%SDS中、42℃でさらに2回洗浄する。中ストリンジェント条件の例としては、20%ホルムアミド、5X SSC(150mM NaCl、15mM クエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5X Denhardt’s溶液、10% 硫酸デキストラン及び20mg/ml変性せん断サケ精子DNAを含む溶液中、37℃で一晩中培養し、続いて1X SSC中、約37〜50℃でフィルターを洗浄する。プローブ長さ等の因子を適合させるために必要な温度、イオン強度等の調節の仕方は、当業者に公知である。
【0084】
ここで用いる“組換え体”は、異種核酸配列の導入により修飾された、または細胞がそのように修飾された細胞に由来する、細胞またはベクターに関するものを含む。従って、例えば組換え細胞は、意図的なヒトの介入の結果として発現または発現されない条件下で、当該細胞の天然型(非組換え体)内で同一の形態で見られない遺伝子を発現し、または異常発現される天然遺伝子を発現する。“組換え”または“組換え”核酸の生成は通常、2以上の核酸断片の組み立てであり、当該組み立てによりキメラ遺伝子が生じる。
【0085】
好ましい実施態様において、変異DNA断片は少なくとも1のコドンにおいて部位飽和性(site saturation)突然変異誘発を用いて生じる。他の好ましい実施態様において、部位飽和性突然変異誘発は2以上のコドンについて行われる。さらなる実施態様において、変異DNA配列は野生型配列と40%以上、45%以上、50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上または98%以上の相同性を有する。別の実施態様において、変異DNAは公知の変異誘発手順、例えば、放射、ニトロソグアニジン等を用いてin vivoで生じる。それから望ましいDNA配列は単離し、ここに提供する方法において用いられる。
【0086】
別の実施態様において、形質転換DNA配列は入来配列が存在しない相同ボックスを含む。この実施態様において、2つの相同ボックス間の内生DNA配列を欠失することが望ましい。さらに、いくつかの実施態様において、形質転換する配列は野生型であり、一方他の実施態様において、それらは変異または修飾配列である。さらに、いくつかの実施態様において、形質転換する配列は相同であり、一方他の実施態様において、それらは異種のものである。
【0087】
ここで用いる“標的配列”の語は、入来配列が宿主細胞ゲノム内に挿入されることが望ましい配列をエンコードする宿主細胞中のDNA配列をいう。いくつかの実施態様において、標的配列は機能野生型遺伝子またはオペロンをエンコードし、一方、他の実施態様において、標的配列は機能変異遺伝子またはオペロンまたは非機能遺伝子またはオペロンをエンコードする。
【0088】
ここで用いる“フランキング配列”とは対象の配列の上流または下流にある配列をいう(例えば、遺伝子A−B−Cは、遺伝子BはA及びC遺伝子配列にフランキング(側面に位置)している)。好ましい実施態様において、入来配列は相同ボックスの両側の側面に位置する。他の実施態様において、入来配列及び相同ボックスはstuffer配列の両側の側面に位置する単位を含む。いくつかの実施態様において、フランキング配列は一方の側(3’または5’)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。各相同ボックスの配列はバチルス染色体中の配列に相同である。これらの配列はバチルス染色体に新しい構築体が統合されること、及びバチルス染色体の一部が入来配列により置換されることを目的とする。好ましい実施態様において、選択マーカーの5’及び3’末端は不活性化染色体断片の一部を含むポリヌクレオチド配列の側面に位置する。いくつかの実施態様において、フランキング配列は一方の側(3’または5’)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。
【0089】
ここで用いる“stuffer配列”の語は相同ボックスの側面に位置する任意の余分なDNAをいう(通常ベクター配列)。しかしながら、当該用語は非相同DNA配列を包含する。いかなる理論にも限定するものではないが、stuffer配列はDNA取込みを開始するために細胞に重要でない標的を提供する。
【0090】
ここで用いる、“変異体ライブラリー”の語は、そのゲノムのほとんどは同一だが、1以上の遺伝子が異なる相同体を含む細胞集団をいう。当該ライブラリーは例えば、改善形質を有する遺伝子またはオペロンを同定するための方法において使用できる。
【0091】
ここで用いる“ハイパーコンピテント”及び“スーパーコンピテント”とは、細胞集団の1%以上が染色体DNA(例えば、バチルスDNA)と形質転換可能であることを意味する。もしくは、当該用語は細胞集団の10%以上が自己複製プラスミド(例えば、バチルスプラスミド)と形質転換可能である細胞集団に関して用いられる。好ましくは、スーパーコンピテント細胞は野生型または親細胞集団について観測されるよりも大きな割合で形質転換される。スーパーコンピテント及びハイパーコンピテントはここでは交換可能に用いられる。
【0092】
ここで用いる、“増幅”及び“遺伝子増幅”の語は特定DNAが増幅遺伝子が初期ゲノムに存在した数より多いコピー数で存在するように偏って複製されるプロセスをいう。いくつかの実施態様において、薬物存在下(例えば、阻害可能酵素のインヒビター)での増殖による細胞選択は、薬物存在下での増殖に必要な遺伝子産物をエンコードする内生遺伝子の増幅、またはこの遺伝子産物をエンコードする外来(すなわち、入来)配列の増幅または両方をもたらす。
【0093】
“増幅”はテンプレート特異性に関する核酸複製の特殊なケースである。これは非特異性テンプレート複製(すなわち、テンプレート依存性であるが特定テンプレートに依存しない複製)と対比される。テンプレート特異性は複製の忠実度(すなわち、適正なポリヌクレオチド配列の合成)及びヌクレオチド(リボ−またはデオキシリボ−)特異性とはここでは区別される。テンプレート特異性は“標的”特異性に関して説明されることが多い。標的配列は、それらがその他の核酸から選別するために探されるという意味において“標的”である。増幅技術はこの選別を第一に考えて設計されてきた。
【0094】
ここで用いる“共増幅”の語は、その他の遺伝子配列が一緒に導入される増幅マーカーの1の細胞内への導入(すなわち、発現ベクター内に含まれる遺伝子のような1以上の非選択遺伝子を含む)及び細胞が増幅マーカーとその他の非選択遺伝子配列の両方を増幅するような適当な選択圧の増幅をいう。増幅マーカーはその他の遺伝子配列に物理的に結合していてもよく、またはDNAが2つの別個の断片、すなわち一方が増幅マーカーを含み、及びもう一方が非選択マーカーを含むものが同じ細胞内に導入されてもよい。
【0095】
ここで用いる、“増幅マーカー”、“増幅遺伝子”及び“増幅ベクター”の語は適当な増殖条件下で遺伝子の増幅を可能にする遺伝子をエンコードする遺伝子またはベクターをいう。
【0096】
“テンプレート特異性”はほとんどの増幅技術において酵素選択により達成される。増幅酵素は使用される条件下において、核酸の外来混合物において核酸の特定配列のみ処理する酵素である。例えば、Qβレプリカーゼの場合、MDV−1 RNAはレプリカーゼの特異テンプレートである(例えば、Kacian et al.、Proc.Natl.Acad.Sci.USA 69:3038[1972]を参照)。その他の核酸はこの増幅酵素によっては複製されない。同様に、T7 RNAポリメラーゼの場合、この増幅酵素はそれ自身のプロモーターにストリンジェントな特異性を有する(Chamberlin et al.、Nature 228:227[1970]を参照)。T4 DNAリガーゼの場合、当該酵素は2つのオリゴヌクレオチドまたはポリヌクレオチドを連結せず、ここで連結接合部分にオリゴヌクレオチドまたはポリヌクレオチド基質とテンプレート間の不整合が存在する(Wu and Wallace,Genomics 4:560[1989]を参照)。最後に、Taq及びPfuポリメラーゼは、高温で機能する能力を有するため、配列結合の高い特異性を示し、従って、プライマーにより確定され;高温が標的配列とのプライマーハイブリダイゼーションに有利に働き、非標的配列とのハイブリダイゼーションには有利でない熱力学的条件を生じる。
【0097】
ここで用いる“増幅核酸”の語は任意の増幅方法により増幅され得る核酸をいう。“増幅核酸”は通常“サンプルテンプレート”を含むことが考えられる。
【0098】
ここで用いる“サンプルテンプレート”の語は、“標的”(下記に定義)の存在に関して分析されるサンプルから生じる核酸をいう。一方、“背景テンプレート”はサンプルテンプレート以外の核酸に関して用いられ、サンプル内に存在していてもいなくてもよい。背景テンプレートはほとんどの場合、故意のものではない。それは以前のものからの影響の結果であり、またはサンプルから精製すべき核酸汚染物質の存在によるものである。例えば、検出すべき微生物以外の生物由来の核酸は試験サンプル内で背景として存在し得る。
【0099】
ここで用いる、“プライマー”の語は、精製制限消化などにおいて自然に生じるまたは合成的に生成されるオリゴヌクレオチドをいい、核酸鎖に相補的なプライマー伸長生成物の合成が導入される条件下に置かれた場合(すなわち、ヌクレオチド及びDNAポリメラーゼなどの誘発剤の存在下、及び適した温度及びpHで)、合成開始点として作用できる。当該プライマーは好ましくは増幅効率を最大にする1本鎖であるが、あるいは2本鎖であってもよい。2本鎖の場合、プライマーは伸長生成物の準備のために用いる前にそのストランドを分離処理する。好ましくは、プライマーはオリゴデオキシリボヌクレオチドである。プライマーは誘発剤の存在下、伸長生成物の合成開始のため十分に長くなければならない。プライマーの正確な長さは、誘発温度、プライマーの供給源及び使用する方法など多くの因子に左右される。
【0100】
ここで用いる“プローブ”の語は、精製制限消化などにおいて自然に生じる、または合成、組換えまたはPCR増幅により生成されるオリゴヌクレオチド(すなわちヌクレオチド配列)をいい、別の目的オリゴヌクレオチドにハイブリダイズできる。プローブは1本鎖または2本鎖である。プローブは特定遺伝子配列の検出、同定及び単離に有用である。本発明で使用されるいかなるプローブも任意の“リポーター分子”で標識するので、任意の検出システムにおいて検出可能であり、限定されないが、酵素(例えば、ELISA及び酵素ベースの組織化学的分析)、蛍光、放射性及び発光性システムなどがある。本発明はいかなる特定の検出システムまたは標識にも限定されない。
【0101】
ここで用いる“標的”の語はポリメラーゼ連鎖反応に関して用いる場合、ポリメラーゼ連鎖反応に用いるプライマーが結合する核酸領域をいう。従って、“標的”はその他の核酸配列から選別するために探求されるものである。“断片”は標的配列内の核酸領域として定義される。
【0102】
ここで用いる“ポリメラーゼ連鎖反応”(“PCR”)の語はここに引用する米国特許第4,683,195号、第4,683,202号及び第4,965,188号の方法をいい、クローニングまたは精製することなくゲノムDNAの混合物における標的配列の断片濃度を増加させる方法を含む。標的配列を増幅するこの方法は、所望の標的配列を含むDNA混合物に過剰な2つのオリゴヌクレオチドプライマーを導入し、DNAポリメラーゼの存在下、サーモサイクルの正確な配列を行うことからなる。2つのプライマーは2本鎖標的配列のそれぞれの鎖に相補的である。増幅をもたらすために、混合物を変性させ、それからプライマーを標的分子内のそれらの相補配列にアニールする。アニーリングに続いて、プライマーは新しい相補鎖ペアを形成するためにポリメラーゼを用いて伸長する。変性、プライマーアニーリング及びポリメラーゼ伸長工程は何回も繰り返すことができ(すなわち変性、アニーリング、及び伸長は1“サイクル”を構成し、多数の“サイクル”を行うことができる)、所望の標的配列の増幅断片を高濃度で得ることができる。所望の標的配列の増幅断片の長さはお互いのプライマーの相対位置により決定され、従って、この長さは制御可能なパラメーターである。当該プロセスの繰返し側面のため、当該方法は“ポリメラーゼ連鎖反応”(以下“PCR”とする)と呼ばれる。標的配列の所望の増幅断片は混合物中で主要な配列(濃度に関して)となるので、それらは“PCR増幅された”という。
【0103】
ここで用いる“増幅試薬”の語は、プライマー、核酸テンプレート及び増幅酵素以外の増幅に必要な試薬をいう(デオキシリボヌクレオチド3リン酸、バッファー等)。通常、増幅試薬はその他の反応成分と一緒に反応容器(試験管、マイクロウェル等)内に加えて含まれる。
【0104】
PCRを用いて、ゲノムDNA中の特異標的配列の1のコピーをいくつかの異なる方法で検出可能なレベルまで増幅することが可能である(例えば、標識プローブを用いるハイブリダイゼーション;ビオチン化プライマーの混合、続いてアビジン−酵素結合検出;dCTPまたはdATPなどの32P標識デオキシヌクレオチド3リン酸の増幅断片への混合)。ゲノムDNAに加えて、オリゴヌクレオチドまたはポリヌクレオチド配列が適当なプライマー分子のセットを用いて増幅できる。特に、PCRプロセスにより作成された増幅断片はそれ自身、続くPCR増幅の効率的なテンプレートである。
【0105】
ここで用いる“PCR生成物”“PCR断片”及び“増幅生成物”の語は変性、アニーリング及び伸長が完結する2以上のサイクルのPCR工程後の化合物の最終混合物をいう。これらの語は1以上の標的配列の1以上の断片の増幅が行われる場合を含む。
【0106】
ここで用いる“RT−PCR”の語はRNA配列の複製及び増幅をいう。この方法では、ここに引用する米国特許第5,322,770号に記載されているように、ほとんどの場合熱安定性ポリメラーゼを用いる1の酵素手順を用いて、逆転写とPCRを組み合わせる。RT−PCRにおいて、RNAテンプレートはポリメラーゼの逆転写酵素活性によりcDNAに転換され、それからポリメラーゼの重合活性を用いて増幅させる(すなわち、他のPCR法と同様)。
【0107】
ここで用いる“制限エンドヌクレアーゼ”及び“制限酵素”の語は細菌性酵素をいい、それぞれ特定ヌクレオチド配列で、またはその近くで2本鎖DNAを切断する。
【0108】
“制限酵素認識部位”とは所定の制限エンドヌクレアーゼにより認識及び切断されるヌクレオチド配列をいい、DNA断片の挿入部位であることが多い。本発明の特定の実施態様において、制限酵素認識部位は選択マーカー内及びDNA構築体の5’及び3’末端内に設計される。
【0109】
ここで用いる“不活性化染色体断片”とは2つの部分を含む。各部分は、ここに定義するように固有の染色体領域にすぐ隣接するゲノム染色体DNAの上流または下流に相同なポリヌクレオチドを含む。“すぐ隣接する”とは、不活性化染色体断片を含むヌクレオチドが固有染色体領域を画定するヌクレオチドを含まないことを意味する。不活性化染色体断片はバチルス染色体中にDNA構築体を統合し、バチルス染色体の一部を置換することを目的とする。
【0110】
ここで用いる“固有染色体領域”及び“固有染色体領域の断片”とは本発明のいくつかの実施態様においてバチルス宿主細胞から欠失されるバチルス染色体の断片をいう。通常、“断片”、“領域”、“部分”及び“要素”の語はここでは交換可能に用いられる。いくつかの実施態様において、欠失断片は公知の機能を有する1以上の遺伝子を含み、一方他の実施態様において、欠失断片は未知の機能を有する1以上の遺伝子を含み、及び他の実施態様において、欠失断片は公知及び未知の機能を含む遺伝子の組み合わせを含む。いくつかの実施態様において、固有染色体領域またはそれらの断片は200程度またはそれ以上の遺伝子を含む。
【0111】
いくつかの実施態様において、固有染色体領域またはそれらの断片は特定の条件下で必要な機能を有するが、当該領域は研究室条件下では必ずしもバチルス株生存に必須のものではない。好ましい研究室条件は、限定されないが、標準温度及び大気条件(例えば好気性)での発酵槽、振とうフラスコ中、プレート培地上等での成長などの条件がある。
【0112】
固有染色体領域またはそれらの断片は約0.5kb〜500kb、約1.0kb〜500kb、約5kb〜500kb、約10kb〜500kb、約10kb〜200kb、約10kb〜100kb、約10kb〜50kb、約100kb〜500kb、約200〜500kbの範囲のバチルス染色体を含む。他の側面において、固有染色体領域またはそれらの断片が欠失された場合、変異バチルス株の染色体は99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、85%、80%、75%または70%の対応する非変異バチルス宿主染色体を含むことができる。好ましくは、本発明に従う変異バチルス株の染色体は約99〜90%、99〜92%及び98〜94%の対応する非変異バチルス宿主株染色体ゲノムを含む。
【0113】
ここで用いる“株生存性”とは繁殖可能性をいう。固有染色体領域またはそれらの断片の欠失は研究所条件下において変異バチルス株の分割及び生存に有害な影響を与えるものではない。
【0114】
ここで用いる、“変異バチルス株”とは遺伝子組換えバチルス種をいい、ここで目的タンパク質は本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現及び/または生成レベルが増大している。いくつかの実施態様において、発現レベルの増加は1以上の染色体遺伝子の不活性化により生じる。1の実施態様において、発現レベルの増加は1以上の染色体遺伝子の欠失により生じる。いくつかの実施態様において、変異バチルス株は1以上の固有染色体領域またはそれらの断片が欠失した遺伝子組換えバチルス種であり、ここで目的タンパク質は本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現または生成レベルが増大している。別の実施態様において、発現レベルの増加は1以上の染色体遺伝子の挿入不活性化により生じる。いくつかの別の実施態様において、発現レベルの増加は活性の増加または最適化遺伝子により生じる。いくつかの好ましい実施態様において、不活性化遺伝子はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される。
【0115】
特定の実施態様において、変異バチルス株は不活性化遺伝子を2つ含み、一方他の実施態様において不活性化遺伝子3つ、不活性化遺伝子4つ、不活性化遺伝子5つ、不活性化遺伝子6つまたはそれ以上存在する。従って、不活性化遺伝子の数は特定の遺伝子数に限定されない。いくつかの実施態様において、不活性化遺伝子はお互いに連続であり、一方他の実施態様において、それらはバチルス染色体の離れた領域に位置する。いくつかの実施態様において、不活性化染色体遺伝子は特定の条件下において必要な機能を有するが、当該遺伝子は研究所条件下において必ずしもバチルス株生存に必須のものではない。好ましい研究室条件は、限定されないが、微生物の成長に適した発酵槽、振とうフラスコ中、プレート培地上等での成長などの条件がある。
【0116】
ここで用いる“対応する非変異バチルス株”とは、固有染色体領域またはそれらの断片がそこから欠失または修飾される、及び変化株が生成する宿主株(例えば、最初の及び/または野生型株)である。
【0117】
ここで用いる“染色体統合”の語は入来配列が宿主細胞(例えば、バチルス)の染色体内に導入されるプロセスをいう。形質転換DNAの相同領域は染色体の相同領域と一致する。続いて、相同ボックス間の配列を二重交差の入来配列で置換する(すなわち、相同組換え)。本発明のいくつかの実施態様において、DNA構築体の不活性化染色体断片の相同部分はバチルス染色体の固有染色体領域のフランキング相同領域と一致する。続いて、固有染色体領域は二重交差でDNA構築体により欠失される(すなわち相同組換え)。
【0118】
“相同組換え”とは、同一またはほぼ同一のヌクレオチド配列部位での2つの分子間または対の染色体間のDNA断片の交換を意味する。好ましい実施態様において、染色体統合は相同組換えである。
【0119】
ここで用いる“相同配列”は比較のために最適に配列した場合、他の核酸またはポリぺプチド配列と100%、99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、88%、85%、80%、75%または70%配列同一性を有する核酸またはポリぺプチド配列を意味する。いくつかの実施態様において、相同配列は85%〜100%の配列同一性を有し、一方他の実施態様において、90%〜100%の配列同一性、及びより好ましい実施態様において、95%〜100%の配列同一性を有する。
【0120】
ここで用いる“アミノ酸”とは、ぺプチドまたはタンパク質配列またはそれらの一部をいう。“タンパク質”、“ぺプチド”及び“ポリぺプチド”の語は交換可能に用いられる。
【0121】
ここで用いる“目的タンパク質”及び“目的ポリぺプチド”とは望ましい及び/または評価対象のタンパク質/ポリぺプチドをいう。いくつかの実施態様において、目的タンパク質は細胞内にあり、一方他の実施態様において、それは分泌ポリぺプチドである。特に好ましいポリぺプチドは酵素を含み、限定されないが、でんぷん分解酵素、タンパク質分解酵素、細胞分解(cellulytic)酵素、酸化還元酵素及び植物細胞壁分解酵素から選択される酵素である。より具体的には、これらの酵素は限定されないが、アミラーゼ、プロテアーゼ、キシラナーゼ、リパーゼ、ラッカーゼ、フェノールオキシダーゼ、オキシダーゼ、クチナーゼ、セルラーゼ、ヘミセルラーゼ、エステラーゼ、ペルオキシダーゼ、カタラーゼ、グルコースオキシダーゼ、フィターゼ、ペクチナーゼ、グルコシダーゼ、イソメラーゼ、トランスフェラーゼ、ガラクトシダーゼ及びキチナーゼを含む。本発明のいくつかの特に好ましい実施態様において、目的ポリぺプチドはプロテアーゼである。いくつかの実施態様において、目的タンパク質は1のぺプチドに融合される分泌ポリぺプチドである(すなわち、分泌されるタンパク質上でのアミノ末端伸長)。ほとんど全ての分泌タンパク質は、膜透過の前駆体タンパク質のターゲッティング及び翻訳において重要な役割を果たすアミノ末端タンパク質伸長を用いる。この伸長は、膜移動の間またはそのすぐ後にシグナルぺプチダーゼによりタンパク質分解により除去される。
【0122】
本発明のいくつかの実施態様において、目的ポリぺプチドはホルモン、抗体、成長因子、受容体等から選択される。ホルモンは本発明に含まれ、限定されないが、卵胞刺激ホルモン、黄体形成ホルモン、副腎皮質刺激ホルモン放出因子、ソマトスタチン、ゴナドトロピンホルモン、バソプレシン、オキシトシン、エリスロポエチン、インスリン等が挙げられる。成長因子は限定されないが、血小板由来成長因子、インスリン様成長因子、上皮成長因子、神経成長因子、線維芽細胞成長因子、形質転換成長因子、サイトカイン、例えばインターロイキン(例えば、IL−1からIL−13)、インターフェロン、コロニー刺激因子等が挙げられる。抗体は限定されないが、抗体を生成するのが望ましい任意の種から直接得られる免疫グロブリンが挙げられる。さらに、本発明は修飾抗体を含む。ポリクローナル及びモノクローナル抗体も本発明に含まれる。特に好ましい実施態様において、抗体はヒト抗体である。
【0123】
ここで用いる“異種タンパク質”の語は宿主細胞に天然に生じないタンパク質またはポリぺプチドをいう。異種タンパク質の例としては、加水分解酵素、例えばプロテアーゼ、セルラーゼ、アミラーゼ、カルボヒドラーゼ及びリパーゼなど;イソメラーゼ、例えばラセマーゼ、エピメラーゼ、トートメラーゼまたはムターゼなど;トランスフェラーゼ、キナーゼ及びホスファターゼなどの酵素を含む。いくつかの実施態様において、タンパク質は治療的に重要なタンパク質またはぺプチドであり、限定されないが、成長因子、サイトカイン、配位子、受容体及びインヒビター並びにワクチン及び抗体を含む。別の実施態様において、タンパク質は商業的に重要な工業タンパク質/ぺプチドである(例えば、プロテアーゼ、アミラーゼ及びグルコアミラーゼなどのカルボヒドラーゼ、セルラーゼ、オキシダーゼ及びリパーゼ)。いくつかの実施態様において、タンパク質をエンコードする遺伝子は天然遺伝子であり、一方他の実施態様において、変異及び/または合成遺伝子が使用される。
【0124】
ここで用いる“相同タンパク質”とは細胞内の野生型または天然タンパク質またはポリぺプチドをいう。好ましい実施態様において、当該細胞はグラム陽性細胞であり、一方特に好ましい実施態様において、当該細胞はバチルス宿主細胞である。別の実施態様において、相同タンパク質はその他の生物、限定されないが大腸菌などにより生成された天然タンパク質である。本発明は組換えDNA技術により相同タンパク質を生成する宿主細胞を包含する。
【0125】
ここで用いる“オペロン領域”は、共有プロモーターからの1の転写単位として転写され、それにより共調節される隣接遺伝子のグループを含む。いくつかの実施態様において、オペロンは調節遺伝子を含む。最も好ましい実施態様において、RNAレベルで測定して高度に発現するが未知または不必要な機能を有するオペロンが用いられる。
【0126】
ここで用いる“複数−隣接単一遺伝子領域”とは、少なくとも2つの遺伝子のコード領域が一列になって起こる領域であり、及びいくつかの実施態様において、コード領域に先行及び後に続く介在配列を含む。いくつかの実施態様において、抗菌領域を導入する。
【0127】
ここで用いる“抗菌領域”は抗菌タンパク質をエンコードする少なくとも1の遺伝子を含む領域である。
【0128】
発明の詳細な説明
本発明は発現タンパク質を生成する能力を変化させるために遺伝子組換えした細胞を提供する。特に、本発明は、目的タンパク質の発現が増大したバチルス種などのグラム陽性細菌に関し、ここで1以上の染色体遺伝子が不活性化され、好ましくは1以上の染色体遺伝子がバチルス染色体から欠失される。いくつかの実施態様において、1以上の固有の染色体領域が対応する野生型バチルス宿主染色体から欠失される。実際、本発明は1または複数の遺伝子を欠失する方法、及び大量の染色体を欠失させる方法を提供する。好ましい実施態様において、当該欠失は目的タンパク質の生成向上などの利点を提供する。
【0129】
A.遺伝子欠失
上述の通り、本発明は1または複数の遺伝子欠失及び/または変異及び大量の染色体欠失に関する実施態様を含む。
【0130】
いくつかの好ましい実施態様において、本発明は入来配列を含むDNA構築体を含む。DNA構築体はin vitroで組み立てられ、それからコンピテントバチルス宿主内に当該構築体の直接クローニングを行い、DNA構築体がバチルス染色体内に統合されるようにする。例えば、PCR融合及び/または連結はDNA構築体をin vitroで組み立てるために用いることができる。いくつかの実施態様において、DNA構築体は非プラスミド構築体であり、一方他の実施態様において、ベクター(例えばプラスミド)内に組み込まれる。いくつかの実施態様において、環状プラスミドを使用する。好ましい実施態様において、環状プラスミドは適当な制限酵素を用いるために設計される(すなわち、DNA構築を阻害しないもの)。従って、線状プラスミドは本発明で使用できる(図1を参照)。しかしながら、その他の方法も本発明での使用に適しており、当業者に公知である(例えば、Perego,“Integration Vectors for Genetic Manipulation in Bacillus subtilis(バチルス・ズブチリスにおける遺伝子操作のための統合ベクター)”、(Sonenshein et al.(編集)、Bacillus subtilis and Other Gram−Positive Bacteria(バチルス・ズブチリス及びその他のグラム陽性細菌),American Society for Microbiology,ワシントン、DC[1993]を参照)。
【0131】
いくつかの実施態様において、入来配列は選択マーカーを含む。いくつかの好ましい実施態様において、入来配列は、sbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDまたはこれらの遺伝子のいずれかの断片(単独または組み合わせ)からなる群より選択される染色体遺伝子を含む。別の実施態様において、入来配列は相同sbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及び/またはrocD遺伝子配列を含む。相同配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD遺伝子またはそれらの遺伝子断片と少なくとも99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、88%、85%、または80%配列同一性を有する核酸配列であり、入来配列に含まれる。好ましい実施態様において、相同配列を含む入来配列はsbo、slr、ybcO、csn、spollSA、phrC、sigB、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocFまたはrocD遺伝子またはそれらの遺伝子断片と少なくとも95%配列同一性を有する。さらに別の実施態様において、入来配列は遺伝子配列の断片を有する5’及び3’末端の側面に位置する選択マーカーを含む。いくつかの実施態様において、DNA構築体が選択マーカー及び遺伝子を含む場合、遺伝子断片またはそれに相同な配列は宿主細胞内に形質転換され、選択マーカーの配置は当該遺伝子の本来の目的に関して非機能的なものにする。いくつかの実施態様において、入来配列は遺伝子のプロモーター領域に位置する選択マーカーを含む。他の実施態様において、入来配列は遺伝子のプロモーター領域の後に位置する選択マーカーを含む。さらに他の実施態様において、入来配列は遺伝子のコード領域に位置する選択マーカーを含む。さらに別の実施態様において、入来配列は両末端上の相同ボックスの側面に位置する選択マーカーを含む。さらに別の実施態様において、入来配列はコード配列の転写及び/または翻訳を妨害する配列を含む。さらに別の実施態様において、DNA構築体は当該構築体の上流及び下流末端に設計された制限酵素認識部位を含む。
【0132】
DNA構築体はベクター内に組み込まれ、またはプラスミドDNAの不存在下で使用されるかに関わらず、微生物を形質転換するために使用される。形質転換に適したいかなる方法も本発明で使用できる。好ましい実施態様において、少なくとも1のDNA構築体のコピーが宿主バチルス染色体内に統合される。いくつかの実施態様において、本発明の1以上のDNA構築体は宿主細胞を形質転換するために用いる。例えば、1のDNA構築体はs/r遺伝子を不活性化するために用い、他の構築体はphrC遺伝子を不活性化するために用いることができる。もちろん、別の組み合わせも考えられ、本発明により提供される。
【0133】
いくつかの好ましい実施態様において、DNA構築体は目的タンパク質をエンコードするポリヌクレオチドも含む。これらの好ましい実施態様のいくつかにおいて、DNA構築体は目的タンパク質をエンコードする配列に作動可能に連結する構成または導入プロモーターも含む。目的タンパク質がプロテアーゼであるいくつかの好ましい実施態様において、プロモーターはtacプロモーター、β−ラクタマーゼプロモーター、またはaprEプロモーター(DeBoer et al.、Proc.Natl.Acad.Sci.USA 80:21−25[1983])からなる群より選択される。しかしながら、本発明はいかなる特定のプロモーターにも限定されるものではなく、当業者に公知の任意の適したプロモーターが本発明で使用できる。とはいえ、特に好ましい実施態様において、プロモーターはB.ズブチリスaprEプロモーターである。
【0134】
バチルス種の形質転換に関する様々な方法が公知である。実際に、プラスミド構築体及び当該プラスミドを大腸菌内に形質転換することを含むバチルス染色体を改変させる方法が公知である。ほとんどの方法において、プラスミドは大腸菌からその後単離され、バチルス内に形質転換される。しかしながら、大腸菌などの介在微生物を使用することは必須ではなく、いくつかの好ましい実施態様においてDNA構築体はコンピテントバチルス宿主内に直接形質転換される。
【0135】
いくつかの実施態様において、公知のバチルスズブチリス株168は本発明で使用できる。実際にこの株のゲノムはよく特性付けられている(Kunst et al.、Nature 390:249−256[1997];及びHenner et al.、Microbiol.Rev.,44:57−82[1980]を参照)。ゲノムは4215kb染色体からなる。ここで用いる統合は168株に言及するが、本発明はバチルス株に類似の配列を包含する。
【0136】
いくつかの実施態様において、入来染色体配列は、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD、それらの遺伝子断片及びそれらに相同な配列からなる群より選択される1以上の遺伝子を含む。B.ズブチリス168由来のこれら遺伝子のDNAコード配列は配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号39、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号37、配列番号25、配列番号21、配列番号50、配列番号29、配列番号23、配列番号27、配列番号19、配列番号31、配列番号48、配列番号46、配列番号35及び配列番号33において提供される。
【0137】
上述の通り、いくつかの実施態様において、sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh、kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocD遺伝子、それらの遺伝子断片またはそれらに相同な配列を含む入来配列はコード領域を含み、すぐ隣に染色体コード領域フランキング配列をさらに含むことができる。いくつかの実施態様において、コード領域フランキング配列は約1bp〜2500bp;約1bp〜1500bp、約1bp〜1000bp、約1bp〜500bp、及び1bp〜250bpの範囲を含む。コード領域フランキング配列を含む核酸配列の数は遺伝子コード配列の各末端ごとに異なる。例えば、いくつかの実施態様において、コード配列の5’末端は25bp以下を含み、コード配列の3’末端は100bp以上を含む。これらの遺伝子の配列及び遺伝子産物を以下に示す。ここで用いる番号はsubtilistで用いられるものである(例えば、Moszer et al.、Microbiol.,141:261−268[1995]を参照)。
【0138】
B.ズブチリス168のsboコード配列を以下に示す:
【式3】
【0139】
sboの推定アミノ酸配列は:
【式4】
【0140】
1の実施態様において、B.ズブチリス168染色体の約3834868〜3835219で見られる遺伝子領域は本発明を用いて欠失させた。約3835081から3835209で見られるsboコード領域はズブチリシンA、すなわちいくつかのグラム陽性細菌に対する活性を有する抗菌薬を生成する(Zheng et al.、J.Bacteriol.,181:7346−7355[1994]を参照)。
【0141】
B.ズブチリス168のslrコード配列を以下に示す:
【式5】
【0142】
Slrの推定アミノ酸配列は:
【式6】
【0143】
1の実施態様において、B.ズブチリス168染色体の約3529014−3529603bpで見られる配列を本発明を用いて欠失させた。slrコード領域は染色体の約3529131から3529586で見られる。
【0144】
B.ズブチリス168のphrCコード配列を以下に示す:
【式7】
【0145】
phrCの推定アミノ酸配列は:
【式8】
【0146】
さらに、B.ズブチリス168染色体の約429531−429650bpで見られるコード領域をコード配列の429591で選択マーカー挿入により不活性化させた。
【0147】
B.ズブチリス168のsigBコード配列を以下に示す:
【式9】
【0148】
sigBの推定アミノ酸配列は:
【式10】
【0149】
さらに、当該コード配列はB.ズブチリス168染色体の約522417から5232085bpで見られる。
【0150】
B.ズブチリス168のspollSAコード配列を以下に示す:
【式11】
【0151】
spollSAの推定アミノ酸配列は:
【式12】
【0152】
さらに、当該コード領域はB.ズブチリス168染色体の約1347587から1348714bpで見られる。
【0153】
B.ズブチリス168のcsnコード配列を以下に示す:
【式13】
【0154】
Csnの推定アミノ酸配列は:
【式14】
【0155】
さらに、当該コード領域はB.ズブチリス168染色体の約2747213から2748043bpで見られる。
【0156】
B.ズブチリス168のybcOコード配列を以下に示す:
【式15】
【0157】
ybcOの推定アミノ酸配列は:
【式16】
【0158】
さらに、当該コード領域はB.ズブチリス168染色体の約213926から214090bpで見られる。
【0159】
B.ズブチリス168のrapAコード配列を以下に示す:
【式17】
【0160】
RapAの推定アミノ酸配列は:
【式18】
【0161】
さらに、当該コード領域はB.ズブチリス168染色体の約1315179から1316312bpで見られる。
【0162】
B.ズブチリス168のCssコード配列を以下に示す:
【式19】
【0163】
Cssの推定アミノ酸配列は(GenBankアクセッション番O32193):
【式20】
【0164】
さらに、当該コード領域はB.ズブチリス168染色体の約3384612から3386774bpで見られる。
【0165】
B.ズブチリス168のFbpタンパク質(フルクトース−1,6−ビオオスファターゼ(biophosphatase))のfbpコード配列を以下に示す:
【式21】
【0166】
Fbpタンパク質の推定アミノ酸配列は:
【式22】
【0167】
さらに、当該コード領域はB.ズブチリス168染色体の約4127053から4129065bpで見られる。
【0168】
B.ズブチリス168のalsDタンパク質(α−アセト乳酸デカルボキシラーゼ)のalsDコード配列を以下に示す:
【式23】
【0169】
alsDタンパク質配列の推定アミノ酸配列は:
【式24】
【0170】
さらに、当該コード領域はB.ズブチリス168染色体の約3707829から3708593bpで見られる。
【0171】
B.ズブチリス168のgapBタンパク質(グリセルアルデヒド−3−リン酸デヒドロゲナーゼ)のgapBコード配列を以下に示す:
【式25】
【0172】
GapBタンパク質の推定アミノ酸配列は:
【式26】
【0173】
さらに、当該コード領域はB.ズブチリス168染色体の約2966075から2967094bpで見られる。
【0174】
Kblタンパク質(2−アミノ−3−ケトブチラートCoAリガーゼ)のKblコード配列を以下に示す:
【式27】
【0175】
Kblタンパク質の推定アミノ酸配列は:
【式28】
【0176】
さらに、当該コード領域はB.ズブチリス168染色体の約1770787から1771962bpで見られる。
【0177】
B.ズブチリス168のPckA(ホスホエノールピルビン酸カルボキシキナーゼ)のPckAコード配列を以下に示す:
【式29】
【0178】
PckAタンパク質の推定アミノ酸配列は:
【式30】
【0179】
さらに、当該コード領域はB.ズブチリス168染色体の約3128579から3130159bpで見られる。
【0180】
B.ズブチリス168のprpCタンパク質(タンパク質ホスファターゼ)のprpCコード配列を以下に示す:
【式31】
【0181】
prpCタンパク質の推定アミノ酸配列は:
【式32】
【0182】
さらに、当該コード領域はB.ズブチリス168染色体の約1649684から1650445bpで見られる。
【0183】
B.ズブチリス168のrocAタンパク質(ピロリン−5−カルボン酸デヒドロゲナーゼ)のrocAコード配列を以下に示す:
【式33】
【0184】
RocAタンパク質の推定アミノ酸配列は:
【式34】
【0185】
さらに、当該コード領域はB.ズブチリス168染色体の約3877991から3879535bpで見られる。
【0186】
B.ズブチリス168のrocDタンパク質(オルニチンアミノトランスフェラーゼ)のrocDコード配列を以下に示す:
【式35】
【0187】
RocDタンパク質の推定アミノ酸配列は:
【式36】
【0188】
さらに、当該コード領域はB.ズブチリス168染色体の約4143328から4144530bpで見られる。
【0189】
B.ズブチリス168のrocFタンパク質(アルギナーゼ)のrocFコード配列を以下に示す:
【式37】
【0190】
RocFタンパク質の推定アミノ酸配列は:
【式38】
【0191】
さらに、当該コード領域はB.ズブチリス168染色体の約4140738から4141625bpで見られる。
【0192】
B.ズブチリス168のTdhタンパク質(トレオニン−3−デヒドロゲナーゼ)のTdhコード配列を以下に示す:
【式39】
【0193】
Tdhタンパク質の推定アミノ酸配列は:
【式40】
【0194】
さらに、当該コード領域はB.ズブチリス168染色体の約1769731から1770771bpで見られる。
【0195】
トリプトファンオペロン調節領域のコード配列及び遺伝子trpE(配列番号48)、trpD(配列番号46)、trpC(配列番号44)、trpF(配列番号50)、trpB(配列番号42)及びtrpA(配列番号40)は以下に示す。下線部はオペロン調節領域である。太字はtrpE開始(ATG)であり、trpD、trpC、trpF、trpB及びtrpA開始が同様に続く(示した順番に太字で表す)。
【式41】
【0196】
推定TrpAタンパク質(トリプトファンシンターゼ(αサブユニット))の配列は:
【式42】
【0197】
推定TrpBタンパク質(トリプトファンシンターゼ(βサブユニット))の配列は:
【式43】
【0198】
推定TrpCタンパク質(インドール−3−グリセロールリン酸シンターゼ)の配列は:
【式44】
【0199】
推定TrpDタンパク質(アントラニル酸ホスフォリボシルトランスフェラーゼ)の配列は:
【式45】
【0200】
推定TrpEタンパク質(アントラニル酸シンターゼ)の配列は:
【式46】
【0201】
推定TrpFタンパク質(ホスフォリボシルアントラニル酸イソメラーゼ)の配列は:
【式47】
【0202】
さらに、当該コード領域はB.ズブチリス168染色体の約2370707から2376834bp(最初のbp=2376834;最後のbp=2370707)で見られる。
【0203】
B.ズブチリス168のycgMタンパク質(プロリンオキシダーゼに類似)のycgMコード配列を以下に示す:
【式48】
【0204】
ycgMタンパク質の推定アミノ酸配列は:
【式49】
【0205】
さらに、当該コード領域はB.ズブチリス168染色体の約344111から345019bpで見られる。
【0206】
B.ズブチリス168のycgNタンパク質(1−ピロリン−5−カルボン酸デヒドロゲナーゼに類似)のycgNコード配列を以下に示す:
【式50】
【0207】
ycgNタンパク質の推定アミノ酸配列は:
【式51】
【0208】
さらに、当該コード領域はB.ズブチリス168染色体の約345039から346583bpで見られる。
【0209】
B.ズブチリス168のsigDタンパク質(RNAポリメラーゼ鞭毛、運動、走化性及び自己分解シグマ因子)のsigDコード配列を以下に示す:
【式52】
【0210】
sigDの推定アミノ酸配列は:
【式53】
【0211】
さらに、当該コード領域はB.ズブチリス168染色体の約1715786から1716547bpで見られる。
【0212】
上述の通り、その他のバチルス宿主で見られる不活性化類似遺伝子は本発明で使用できると考えられる。
【0213】
いくつかの好ましい実施態様において、宿主細胞はバチルス属のメンバーであり、一方他の実施態様において、目的バチルス株は好アルカリ性である。多数の好アルカリ性バチルス株が公知である(例えば、米国特許第5,217,878号及びAunstrup et al.、Proc IV IFS:Ferment.Technol.Today,299−305[1972]を参照)。いくつかの好ましい実施態様において、目的のバチルス株は工業バチルス株である。工業バチルス株の例としては、限定されないが、B.リケニフォルミス、B.レンタス、B.ズブチリス及びB.アミロリケファシエンスが挙げられる。別の実施態様において、バチルス宿主株は、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ及びB.メガテリウム、並びに上述のバチルス属内のその他の微生物からなる群より選択される。いくつかの特に好ましい実施態様において、B.ズブチリスを用いる。例えば、米国特許第5,264,366号及び第4,760,025号(RE34,606)は本発明で使用できる種々のバチルス宿主株について記載しているが、その他の適当な株も本発明に使用できると考えられる。
【0214】
工業株はバチルス種の非組換え体であってもよく、天然株の変異体または組換え体株であってもよい。好ましくは、宿主株は組換え宿主株であり、目的のポリぺプチドをエンコードするポリヌクレオチドが宿主内に導入されている。さらに好ましい宿主株はバチルス・ズブチリス宿主株であり、特に組換えバチルス・ズブチリス宿主株である。多数のB.ズブチリス株が公知であり、限定されないが、1A6(ATCC39085)、168(1A01)、SB19、W23、Ts85、B637、PB1753〜PB1758、PB3360、JH642、1A243(ATCC39,087)、ATCC21332、ATCC6051、MI113、DE100(ATCC39,094)、GX4931、PBT110、及びPEP211株(例えばHoch et al.、Genetics,73:215−228[1973];米国特許第4,450,235号;米国特許第4,302,544及び欧州特許0134048号を参照)が挙げられる。発現宿主としてのB.ズブチリスの使用はさらにPalva et al.などに記載されている(Palva et al.、Gene19:81−87[1982];Fahnestock and Fischer、J.Bacteriol.,165:796−804[1986];及びWang et al.,Gene 69:39−47[1988]を参照)。
【0215】
工業プロテアーゼ生成バチルス株は特に好ましい発現宿主を提供する。いくつかの好ましい実施態様において、本発明でのこれらの株の使用はさらに効率及びプロテアーゼ生成を増大させる。通常バチルス種により2つの一般的な種類のプロテアーゼが分泌され、すなわち、中性(または“メタロプロテアーゼ”)及びアルカリ性(または“セリン”)プロテアーゼである。セリンプロテアーゼはぺプチド結合の加水分解を触媒する酵素であり、活性部位に必須のセリン残基がある。セリンプロテアーゼは分子量が25,000〜30,000である(Priest、Bacteriol.Rev.,41:711−753[1977]を参照)。ズブチリシンは本発明での使用に好ましいセリンプロテアーゼである。多種多様のバチルス・ズブチリシンが同定及び配列決定されており、例えば、ズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147及びズブチリシン309である(例えば、欧州特許第414279号、WO89/06279号;及びStahl et al.、J.Bacteriol.,159:811−818[1984]を参照)。本発明のいくつかの実施態様において、バチルス宿主株は変異(例えば変異体)プロテアーゼを生成する。多数の参考文献が変異体プロテアーゼの例及び参照を提供している(例えば、WO99/20770、WO99/20726、WO99/20769、WO89/06279、RE34,606、米国特許第4,914,031号、米国特許第4,980,288号、米国特許第5,208,158号、米国特許第5,310,675号、米国特許第5,336,611号、米国特許第5,399,283号、米国特許第5,441,882号、米国特許第5,482,849号、米国特許第5,631,217号、米国特許第5,665,587号、米国特許第5,700,676号、米国特許第5,741,694号、米国特許第5,858,757号、米国特許第5,880,080号、米国特許第6,197,567号及び米国特許第6,218,165号を参照)。
【0216】
さらに他の実施態様において、好ましいバチルス宿主は、以下の遺伝子の少なくとも1つにおいて変異または欠失を有するバチルス種である:degU、degS、degR及びdegQ。好ましくは、変異はdegU遺伝子にあり、より好ましくは変異はdegU(Hy)32である。(Msadek et al.,J.Bacteriol.、172:824−834[1990];及びOlmos et al.、Mol.Gen.Genet.,253:562−567[1997]参照)。最も好ましくは宿主株はdegU32(Hy)変異を有するバチルス・ズブチリスである。さらなる実施態様において、バチルス宿主はscoC4(Caldwell et al.、J.Bacteriol.,183:7329−7340[2001]を参照);spollE(Arigoni et al.、Mol.Microbiol.,31:1407−1415[1999]を参照);oppAまたはその他のoppオペロンの遺伝子(Perego et al.、Mol.Mictobiol.,5:173−185[1991]を参照)における変異または欠失を含む。実際に、oppA遺伝子における変異と同じ表現型を生じるoppオペロンにおける変異は本発明の変異バチルス株のいくつかの実施態様において使用できると考えられる。いくつかの実施態様において、これらの変異は単独で起こり、一方他の実施態様において、変異の組み合わせが存在する。いくつかの実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異をすでに含むバチルス宿主株から得られる。別の実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異を含むようにさらに設計される。
【0217】
さらに別の実施態様において、入来配列は2つのloxP部位間に位置する選択マーカーを含み(Kuhn and Torres、Meth.Mol.Biol.,180:175−204[2002]を参照)、抗菌剤はそれからCreタンパク質の活性により欠失される。いくつかの実施態様において、これは、相同フランキングDNA及び抗菌剤含有入来DNAを構築するために用いるプライマーにより測定される1のloxP部位の挿入及び天然DNAの欠失を生じる。
【0218】
当業者はバチルス細胞内にポリヌクレオチド配列を導入する適切な方法をよく知っている(例えば、Ferrari et al.、“Genetics”、Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.[1989]、57〜72頁、また、Saunders et al.、J.Bacteriol.157:718−726[1984];Hoch et al.、J.Bacteriol.、93:1925−1937[1967];Mann et al.、Current Microbiol.,13:131−135[1986];及びHolubova、Folia Microbiol.,30:97[1985];B.ズブチリス、Chang et al.、Mol.Gen.Genet.,168:11−115[1979];B.メガテリウム、Vorobjeva et al.、FEMS Microbiol.Lett.,7:261−263[1980];B.アミロリケファシエンス、Smith et al.、Appl.Env.Microbiol.,51:634(1986);B.チューリンゲンシス、Fisher et al.、Arch.Microbiol.,139:213−217「1981」;及びB.スファエリカス、McDonald、J.Gen.Microbiol.,130:203[1984]を参照)。実際、プロトプラスト形質転換及び配向(congression)、形質導入及びプロプラスト融合などの形質転換の方法は公知であり、本発明での使用に適している。形質転換の方法は特に本発明により提供されるDNA構築体を宿主細胞に導入するために好ましい。
【0219】
一般的に使用される方法に加えて、いくつかの実施態様において、宿主細胞は直接形質転換される(すなわち、媒介細胞を増幅のために使用しない、または別の方法では、DNA構築体を宿主細胞内に前もって導入する)。宿主細胞内へのDNA構築体の導入は、プラスミドまたはベクター内への導入なしで、宿主細胞内に導入するための当業者に公知の物理的及び化学的方法を含む。当該方法は、限定されないが、塩化カルシウム沈殿法、エレクトロポレーション、裸のDNA(naked DNA)、リポソーム等が挙げられる。別の実施態様において、DNA構築体は、プラスミド内に導入されることなく、プラスミドを用いて共形質転換される。さらに別の実施態様において、選択マーカーは当業者に公知の方法により変異バチルス株から欠失される(Stahl et al.、J.Bacteriol.,158:411−418[1984];及びPalmeros et al.、Gene 247:255−264[2000]を参照)。
【0220】
いくつかの実施態様において、宿主細胞は、本発明に従う1以上のDNA構築体を用いて形質転換され、2以上の遺伝子が宿主細胞中で不活性化されている変異バチルス株を生成する。いくつかの実施態様において、2以上の遺伝子は宿主細胞染色体から欠失される。別の実施態様において、2以上の遺伝子はDNA構築体の挿入により不活性化される。いくつかの実施態様において、不活性化遺伝子は連続しており(欠失及び/または挿入による不活性化に関わらず)、一方、他の実施態様において、連続した遺伝子ではない。
【0221】
細胞内及び細胞外で発現したポリぺプチドの活性化を検出及び測定する当業者に公知の様々な分析法が存在する。特に、プロテアーゼに関しては、吸光度280nmで測定、またはFolin法を用いた比色分析により測定したカゼインまたはヘモグロビンからの酸溶性ぺプチドの放出に基づく分析法がある(例えば、Bergmeyer et al.、“Methods of Enzymatic Analysis”vol.5、Peptidases,Proteinases and their Inhibitors(ペプチダーゼ、プロテイナーゼ、及びそれらの阻害因子)、Verlag Chemie、Weinheim「1984」を参照)。その他の分析法は発色基質の可溶化を含む(例えば、Ward、“Proteinases(プロテイナーゼ)”、Fogarty(編集)、Microbial Enzymes and Biotechnology(微生物酵素及びバイオテクノロジー)、Applied Science,ロンドン[1983]、251〜317頁を参照)。その他の分析法の例としては、スクシニル−Ala−Ala−Pro−Phe−paraニトロアニリド分析(SAAPFpNA)及び2,4,6−トリニトロベンゼンスルフォン酸ナトリウム塩分析(TNBS分析)が挙げられる。当業者に公知の多数の別の文献は適切な方法を提供する(例えば、Wells et al.、Nucleic Acids Res.11:7911−7925[1983];Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal Biochem.,242:221−227[1999]を参照)。
【0222】
宿主細胞内の目的タンパク質の分泌レベルの測定方法及び発現タンパク質の検出方法はタンパク質に特異なポリクローナルまたはモノクローナル抗体を用いる免疫学的分析法の使用を含む。例としては、酵素免疫測定法(ELISA)、放射免疫分析法(RIA)、蛍光免疫分析法(FIA)、及び蛍光活性化細胞分類法(FACS)を含む。しかしながら、その他の方法も当業者に公知であり、目的タンパク質の評価に使用できる(例えば、Hampton et al.、Serological Methods,A Laboratory Manual(血清学的方法、研究室マニュアル),APS Press、St.Paul,MN[1990];及びMaddox et al.、J.Exp.Med.、158:1211[1983]を参照)。いくつかの好ましい実施態様において、目的タンパク質の分泌は、対応する非変異宿主よりも本発明を用いて得た変異株におけるほうが高い。当業界で公知の通り、本発明を用いて生成した変異バチルス細胞は発現及び細胞培養物からの目的ポリぺプチドの回収に適した条件下で維持及び成長する(例えば、Harwood and Cutting(編集)Molecular Biological Methods for Bacillus(バチルスの分子生物学的方法)、John Wiley&Sons[1990]を参照)。
【0223】
B.大量の染色体欠失
上述の通り、1及び複数の遺伝子欠失に加えて、本発明は大量の染色体欠失も提供する。本発明のいくつかの好ましい実施態様において、固有染色体領域またはそれらの断片はバチルス宿主細胞から欠失され、変異バチルス株を生成する。いくつかの実施態様において、固有染色体領域はプロファージ領域、抗菌領域(例えば、抗生物質領域)、調節領域、複数連続シングル遺伝子領域及び/またはオペロン領域を含む。ここで言及する固有染色体領域を描写する座標はバチルス・ズブチリス株168染色体地図に従って特定される。数字は通常、存在する場合は、リボソーム結合部位の開始部位に関するものであり、またはコード領域の末端に関し、及び存在するであろうターミネーターは通常含まない。株168のバチルス・ズブチリスゲノムは公知であり(Kunst et al.、Nature 390:249−256[1997];及びHenner et al.、Microbiol.Rev.、44:57−82[1980]を参照)、及び1の4215kb染色体からなる。しかしながら、本発明は任意のバチルス株由来の類似配列も含む。特に好ましくは、その他のB.ズブチリス株、B.リケニフォルミス株及びB.アミロリケファシエンス株である。
【0224】
いくつかの実施態様において、固有染色体領域はプロファージ断片及びそれらの断片を含む。“プロファージ断片”は細菌性染色体内に挿入されたウイルス性DNAであり、ウイルス性DNAは通常の細菌性遺伝子と事実上区別できない。B.ズブチリスゲノムは多数のプロファージ断片からなり、これらの断片は感染性ではない(Seaman et al.、Biochem.,3:607−613[1964];及びStickler et al.、Vitrol.,26:142−145[1965])。バチルス・ズブチリスプロファージ領域のいずれを欠失してもよいが、以下の非限定的例に関して説明する。
【0225】
本発明の実施態様において欠失される1のプロファージ領域はシグマK介在“スキン”要素である。この領域はB.ズブチリス168染色体の約2652600bp(spolVCA)〜2700579bp(yqaB)で見られる。本発明を用いて、染色体の2653562bp〜2699604bpに対応する、約46kb断片を欠失した。この要素は、SIGK ORF内、遺伝子spolVCB〜spolllC間に位置する祖先から受け継いだ溶原性ファージの残余物であると考えられる。しかしながら、本発明は欠失領域に関するいかなる特定のメカニズムまたは作用形態に限定されるものではない。当該要素は推定リボソーム結合部位を含む57のオープン読取りフレームを含むように示されている(Takemaru et al.、Microbiol.,141:323−327 [1995]を参照)。母細胞における胞子形成時に、スキン要素は削除されてsigK遺伝子の再構築を導く。
【0226】
欠失に適したその他の領域はプロファージ7領域である。この領域はB.ズブチリス168染色体の約2701208bp(yrkS)〜2749572bp(yraK)で見られる。本発明を用いて、染色体の2701087bp〜2749642bpに対応する、約48.5kb断片を欠失した。
【0227】
さらなる領域はスキン+プロファージ7領域である。この領域はB.ズブチリス168染色体の約2652151bp〜2749642bp(yraK)で見られる。本発明を用いて、約97.5kbの断片を欠失した。この領域は介在spolllC遺伝子も含む。スキン/プロファージ7領域は、限定されないが、以下の遺伝子を含む:spolVCA−DNAリコンビナーゼ、blt(多剤耐性)、cypA(シトクロムP450様酵素)、czcD(カチオン流出システム膜タンパク質)、及びrapE(応答制御因子アスパラギン酸ホスファターゼ)。
【0228】
さらに別の領域はPBSX領域である。この領域はB.ズブチリス168染色体の約1319884bp(wkdA)〜1347491bp(xlyA)で見られる。本発明を用いて、染色体の1319663bp〜1348691bpに対応する、約29kb断片を欠失した。通常の非導入状態において、このプロファージ要素は非感染性であり、殺菌性ではない(W23及びS31などの数種の敏感な株を除く)。マイトマイシンCが導入可能であり、SOS応答により活性化され、ファージ様粒子を放出する細胞溶解を生じる。ファージ粒子は細菌性染色体DNAを含み、敏感な細菌をDNAを注入することなく殺すことができる(Canosi et al.、J.Gen.Virol.39:81−90[1978])。この領域は以下の遺伝子の非限定的リストを含む:xtmA−B;xkdA−K及びM−X、xre、xtrA、xpf、xep、xhlA−B及びxlyA。
【0229】
別の領域はSPβ領域である。この領域はB.ズブチリス168染色体の約2150824bp(yodU)〜2286246bp(ypqP)で見られる。本発明を用いて、染色体の2151827bp〜2285246bpに対応する、約133.5kb断片を欠失した。この要素は機能がまだ特性付けられていない溶原性プロファージである。しかしながら、この領域の遺伝子は推定上の胞子殻タンパク質(yodU、sspC、yokH)、推定上のストレス応答性タンパク質(yorD、yppQ、ypnP)及び胞子殻における遺伝子と相同性を有するその他の遺伝子及びyomオペロンのメンバーなどストレス応答性遺伝子を含む。その他の遺伝子は以下を含む領域である:yot;yos、yoq、yop、yon、yom、yoz、yol、yok、ypo、及びypm。
【0230】
別の領域はプロファージ1領域である。この領域はB.ズブチリス168染色体の約202098bp(ybbU)〜220015bp(ybdE)で見られる。本発明を用いて、染色体の202112bp〜220141bpに対応する、約18.0kb断片を欠失した。この領域の遺伝子は、DNAアルキル化に適応反応を与えるAdaA/Bオペロン及びNADHデヒドロゲナーゼ、サブユニット5をコードするndhFを含む。
【0231】
別の領域はプロファージ2領域である。この領域はB.ズブチリス168染色体の約529069bp(ybcL)〜569493bp(ydeJ)で見られる。本発明を用いて、染色体の529067bp〜569578bpに対応する、約40.5kb断片を欠失した。この領域の遺伝子はrapl/phrlwp(応答制御因子アスパラギン酸ホスファターゼ)、sacV(レバンスクラーゼの転写調節因子)及びcspCを含む。
【0232】
別の領域はプロファージ3領域である。本発明を用いて、B.ズブチリス168染色体の652000bp〜664300bpに対応する、約50.7kb断片を欠失した。
【0233】
さらに別の領域はプロファージ4領域である。この領域はB.ズブチリス168染色体の約1263017(yjcM)〜1313627bp(yjoA)で見られる。本発明を用いて、染色体の1262987bp〜1313692bpに対応する、約2.3kb断片を欠失した。
【0234】
別の領域はプロファージ5領域である。本発明を用いて、B.ズブチリス168染色体の1879200bp〜1900000bpに対応する、約20.8kb断片を欠失した。
【0235】
別の領域はプロファージ6領域である。本発明を用いて、B.ズブチリス168染色体の2046050bp〜2078000bpに対応する、約31.9kb断片を欠失した。
【0236】
別の実施態様において、固有染色体領域は1以上のオペロン領域、複数隣接シングル遺伝子領域、及び/または抗菌領域を含む。いくつかの実施態様において、これらの領域は以下を含む:
1)PPSオペロン領域:
この領域はバチルス・ズブチリス168染色体の約1959410(ppsE)〜1997178bp(ppsA)で見られる。本発明を用いて、染色体の1960409bp〜1998026pに対応する、約38.6kb断片を欠失した。このオペロン領域は抗菌合成に関係し、プリパスタチン・シンテターゼをエンコードする;
2)PKSオペロン領域:
この領域はバチルス・ズブチリス168染色体の約1781110(pksA)〜1857712bp(pksR)で見られる。本発明を用いて、染色体の1781795bp〜1857985pに対応する、約76.2kb断片を欠失した。この領域はポリケチドシンテアーゼをエンコードし、抗菌合成に関係する(Scotti et al.、Gene、130:65−71[1993]);
3)yvfF−yveKオペロン領域:
この領域はバチルス・ズブチリス168染色体の約3513149(yvfF)〜3528184bp(yveK)で見られる。本発明を用いて、染色体の3513137bp〜3528896bpに対応する、約15.8kb断片を欠失した。この領域は推定多糖類をコードする(Dartois et al.、バチルスに関する第7回国際会議(1993)Institute Pasteur[1993]、56頁を参照)。この領域は以下の遺伝子を含む:yvfA−F、yveK−T及びslr。B.ズブチリス168染色体の約3529014〜3529603で見られるslr遺伝子領域は約589bpの断片を包含する。この領域はyvfF−yveKオペロンの調節領域である;
4)DHBオペロン領域:
この領域はバチルス・ズブチリス168染色体の約3279750(yukL)〜3293206bp(yuiH)で見られる。本発明を用いて、染色体の3279418bp〜3292920bpに対応する、約13.0kb断片を欠失した。この領域はcatecholic siderophone2,3−ジヒドロキシ安息香酸−グリシン−トレオニン三量体エステルbacilibactinの生合成テンプレートをエンコードする(May et al.、J.Biol.Chem.,276:7209−7217[2001]を参照)。この領域は以下の遺伝子を含む:yukL、yukM、dhbA−C、E及びF及びyuiI−H。
【0237】
上述の領域は欠失される好ましい固有染色体領域の例であるが、本発明のいくつかの実施態様において、当該領域の断片も欠失される。いくつかの実施態様において、当該断片は固有染色体領域の約1%〜99%を含む。他の実施態様において、断片は固有染色体領域の5%〜95%を含む。さらに別の実施態様において、断片は固有染色体領域の少なくとも99%、98%、97%、96%、95%、94%、93%、92%、90%、88%、85%、80%、75%、70%、65%、50%、40%、30%、25%、20%及び10%を含む。
【0238】
B.ズブチリス168染色体における染色体上の位置に関してさらに欠失される固有染色体領域の断片の非限定的な例は以下を含む:
a)スキン領域について:
i)約2666663〜2693807の座標位置であって、yqcC〜yqaMを含む、及び
ii)約2658440〜2659688の座標位置であって、rapE〜phrEを含む;
b)PBSXプロファージ領域について:
i)約1320043〜1345263の座標位置であって、xkdA〜xkdXを含む、及び
ii)約1326662〜1345102の座標位置であって、xkdE〜xkdWを含む;
c)SPβ領域について:
i)約2149354〜2237029の座標位置であって、xkdA〜xkdXを含む;
d)DHB領域について:
i)約3282879〜3291353の座標位置であって、dhbF〜dhbAを含む;
e)yvfF−yveK領域について
i)約3516549〜3522333の座標位置であって、yvfB〜yveQを含み、
ii)約3513181〜3528915の座標位置であって、yvfF〜yveKを含み、及び
iii)約3521233〜3528205の座標位置であって、yveQ〜yveLを含む;
f)プロファージ1領域について:
i)約213926〜220015の座標位置であって、ybcO〜ybdEを含み、及び
ii)約214146〜220015の座標位置であって、ybcP〜ybdEを含む;
g)プロファージ2領域について:
i)約546867〜559005座標位置であって、rapI〜cspCを含む;及び
h)プロファージ4領域について:
i)約1263017〜675421座標位置であって、yjcM〜ydjJを含む。
【0239】
断片は同定した固有染色体領域よりも少ない2、3のbpsのみからなることができるので、欠失に適した固有染色体領域の断片の数は多数ある。さらに、多くの同定された固有染色体領域は多数の遺伝子を包含する。当業者は特定の用途の使用のために固有染色体領域のどの断片を欠失させるのが適当かを容易に決定することができる。
【0240】
固有染色体領域の測定は多数の特定断片に隣接したヌクレオチドを排除することに関しては正確ではない。例えば、SPβ領域はB.ズブチリス168染色体の座標2150824〜2286246に位置するものとしてここで定義され、固有染色体領域はさらに10〜5000bp、さらに100〜4000bp、またはさらに100〜1000bpを当該領域の両側に含むことができる。当該領域の両側のbpの数は欠失の標的とされる固有染色体領域に含まれない他の遺伝子の存在により制限される。
【0241】
上述の通り、ここに開示する特定領域の位置はB.ズブチリス168染色体に関するものである。バチルス株由来のその他の類似領域は固有染色体領域の定義に含まれる。類似領域は任意のバチルス株内で見られるが、特に好ましい類似領域はその他のバチルス・ズブチリス株、バチルス・リケニフォルミス株及びバチルス・アミロリケファシエンス株で見られる領域である。
【0242】
特定の実施態様において、1以上の固有染色体領域またはそれらの断片がバチルス株から欠失される。しかしながら、1以上の固有染色体領域またはそれらの断片の欠失は当該欠失を含む株の再生可能性に悪影響を与えるものではない。いくつかの実施態様において、2つの固有染色体領域またはそれらの断片が欠失される。別の実施態様において、3つの固有染色体領域またはそれらの断片が欠失される。さらに別の実施態様において、4つの固有染色体領域またはそれらの断片が欠失される。さらなる実施態様において、5つの固有染色体領域またはそれらの断片が欠失される。別の実施態様において、14程度の固有染色体領域またはそれらの断片が欠失される。いくつかの実施態様において、固有染色体領域またはそれらの断片は連続しており、一方他の実施態様において、それらはバチルス染色体の別々の領域に位置する。
【0243】
欠失した固有染色体領域またはそれらの断片を含むバチルス属の任意のメンバー株は本発明で使用できる。いくつかの好ましい実施態様において、バチルス株はB.ズブチリス株、B.アミロリケファシエンス株、B.レンタス株及びB.リケニフォルミス株からなる群より選択される。いくつかの好ましい実施態様において、当該株は工業バチルス株であり、最も好ましくは工業B.ズブチリス株である。さらに好ましい実施態様において、変異バチルス株はプロテアーゼ生成株である。いくつかの特に好ましい実施態様において、前もってプロテアーゼ酵素をエンコードするポリヌクレオチドを含むように設計されたB.ズブチリス株である。
【0244】
上述の通り、固有染色体領域またはそれらの断片が欠失されたバチルス株はここで“変異バチルス株”という。本発明の特に好ましい実施態様において、変異バチルス株は目的タンパク質の発現レベルが増大している(すなわち、目的タンパク質の発現が同じ成長条件下で成長させた対応する非変異バチルス株と比較して増大している)。
【0245】
増大の1の指標は目的タンパク質の分泌である。いくつかの実施態様において、目的タンパク質の生成は対応する非変異バチルス株と比較して少なくとも0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、4.0%、5.0%、8.0%、10%、15%、20%及び25%またはそれ以上、増大している。他の実施態様において、目的タンパク質の生成は、1リットル当たりに生成したタンパク質(g)で測定すると対応する非変異バチルス株と比較して0.25%〜20%、0.5%〜15%、及び1.0%〜10%増大している。
【0246】
固有染色体領域またはそれらの断片の欠失を含む本発明によって提供される変異バチルス株は任意の適当な方法により生成され、限定されないが以下の方法を含む。1の一般的な実施態様において、DNA構築体をバチルス宿主中に導入する。DNA構築体は不活性化染色体断片を含み、いくつかの実施態様において、さらに選択マーカーを含む。好ましくは、選択マーカーは不活性化染色体断片の5’及び3’末端の一部の両方に隣接している。
【0247】
いくつかの実施態様において、不活性化染色体断片は、好ましくは欠失される固有染色体領域(または当該領域の断片)のすぐ上流及び下流ヌクレオチドと100%配列同一性を有するが、固有染色体領域の上流及び下流ヌクレオチドと約70〜100%、約80〜100%、約90〜100%、及び約95〜100%配列同一性を有する。不活性化染色体断片の各部分は、非変異宿主中で固有染色体領域と相同組換えするために、固有染色体領域の十分な5’及び3’フランキング配列を含まなければならない。
【0248】
いくつかの実施態様において、不活性化染色体断片の各部分は約50〜10,000塩基対(bp)を含む。しかしながら、より小さいまたは大きいbp部分を本発明で用いることができる。好ましくは、各部分は約50〜5000bp、約100〜5000bp、約100〜3000bp、100〜2000bp、約100〜1000bp、約200〜4000bp、約400〜3000bp、約500〜2000bp、及び約800〜1500bpである。
【0249】
いくつかの実施態様において、選択マーカー及び不活性化染色体断片を含むDNA構築体はin vitroで組み立てられ、続いてコンピテントバチルス宿主内に前記構築体を直接クローニングし、DNA構築体がバチルス染色体内に統合されるようにする。例えば、PCR融合及び/または連結はin vitroでのDNA構築体の組み立てに適している。いくつかの実施態様において、DNA構築体は非プラスミド構築体であり、一方、他の実施態様において、ベクター内に組み込まれる(すなわち、プラスミド)。いくつかの実施態様において、環状プラスミドを用い、当該環状プラスミドは適当な制限酵素(すなわち、DNA構築を阻害しないもの)を用いて切断する。従って、線形プラスミドは本発明で使用できる(例えば、図1;及びBacillus subtilis and other Gram−Positive Bacteria(バチルス・ズブチリス及びその他のグラム陽性細菌)、Sonenshein et al.、編集、Am.Soc.Microbiol.,ワシントン、DC[1993]のPerego、“Integrational Vectors for Genetic Manipulation in Bacillus subtilis(バチルス・ズブチリスにおける遺伝子操作のための統合ベクター)”、を参照)。
【0250】
いくつかの実施態様において、DNA構築体またはベクター、好ましくは不活性化染色体断片を含むプラスミドは非変異宿主中で固有染色体領域またはそれらの断片と相同組換えするために十分な量の固有染色体断片の5’及び3’フランキング配列(seq)またはそれらの断片を含む。他の実施態様において、DNA構築体は当該構築体の上流及び下流末端で設計される制限酵素認識部位を含む。本発明に従い、座標位置に従い同定される有用なDNA構築体の非限定的な例は以下を含む:
1.PBSX領域を欠失するDNA構築体:[yjqB末端及びリボソーム結合部位(RBS)を含むyjpC全体を含む5’フランキング配列1318874−1319860bp]−マーカー遺伝子−[pitのターミネーター及び上流部分を含む3’フランキング配列1348691−1349656bp]。
【0251】
2.プロファージ1領域を欠失するDNA構築体:[RBS及びターミネーターを含むglmS全体及びybbU RBSを含む5’フランキング配列201248−202112bp]−マーカー遺伝子−[RBSを含むybgd全体を含む3’フランキング配列220141−221195bp]。
【0252】
3. プロファージ2領域を欠失するDNA構築体:[ydcK末端、以下のtRNA全体:trnS−Asn、trnS−Ser、trnS−Glu、trnS−Glu、trnS−Leu1及びtrnS−leu2を含む5’フランキング配列527925−529067bp]−マーカー遺伝子−[ydeK全体及びydeLの上流部分を含む3’フランキング配列569578−571062bp]。
【0253】
4. プロファージ4領域を欠失するDNA構築体:[yjcMの一部を含む5’フランキング配列1263127−1264270bp]−マーカー遺伝子−[RBSを含むyjoBの一部を含む3’フランキング配列1313660−1314583bp]。
【0254】
5. yvfF−yveK領域を欠失するDNA構築体:[sigLの一部、yvfF全体及びyvfFの開始を含む5’フランキング配列3512061−3513161bp]−マーカー遺伝子−[slr全体及びpnbAの開始を含む3’フランキング配列3528896−3529810bp]。
【0255】
6. DHBオペロン領域を欠失するDNA構築体:[ターミネーターを含むald末端、RBSを含むyuxlF全体、RBS及びターミネーターを含むyukJ全体及びyukL全体を含む5’フランキング配列3278457−3280255bp]−マーカー遺伝子−[RBSを含むyuiH末端、RBS及びターミネーターを含むyuiG全体及びターミネーターを含むyuiE上流末端を含む3’フランキング配列3292919−3294076bp]。
【0256】
DNA構築体はベクター内に組み込まれる、またはプラスミドDNAなしで用いられるかに関わらず、微生物、好ましくは大腸菌細胞またはコンピテントバチルス細胞内に組み込まれる。
【0257】
プラスミド構築体を含むバチルス細胞内にDNAを導入する及び大腸菌内にプラスミドを形質転換する方法は公知である。プラスミドは続いて大腸菌から単離され、バチルス内に形質転換される。しかしながら、大腸菌などの介在微生物を使用することは必須ではなく、いくつかの実施態様において、DNA構築体またはベクターはバチルス宿主内に直接導入される。
【0258】
好ましい実施態様において、宿主細胞はバチルス種である(例えば、米国特許第5,264,366号、米国特許第4,760,025号及びRE34,6060を参照)。いくつかの実施態様において、目的のバチルス株はアルカロフィリック(Alkalophilic)バチルスである。多数のアルカロフィリックバチルス株が公知である(例えば、米国特許第5,217,878号;及びAunstrup et al.、Proc IV IFS:Ferment.Tech.Today,299−305[1972]を参照)。特定の目的の他の種類のバチルス株は工業バチルス株の細胞である。工業バチルス株の例としては、限定されないが、B.リケニフォルミス、B.レンタス、B.ズブチリス及びB.アミロリケファシエンスが挙げられる。別の実施態様において、バチルス宿主株は、B.リケニフォルミス、B.ズブチリス、B.レンタス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.アミロリケファシエンス、B.コアグランス、B.サーキュランス、B.プミルス、B.チューリンゲンシス、B.クラウジイ及びB.メガテリウムからなる群より選択される。特に好ましい実施態様において、B.ズブチリス細胞を用いる。
【0259】
いくつかの実施態様において、工業宿主株はバチルス種の非組換え体株、対応する天然バチルス株の変異体及び組換えバチルス宿主株からなる群より選択される。好ましくは、宿主株は組換え宿主株であり、ここで目的ポリぺプチドをエンコードするポリヌクレオチドは前もって宿主内に導入されている。さらに好ましい宿主株はバチルス・ズブチリス宿主株であり、特に組換えバチルス・ズブチリス宿主株である。多数のB.ズブチリス株が公知であり、本発明の使用に適している(例えば、1A6(ATCC 39085)、168(1A01)、SB19、W23、Ts85、B637、PB1753〜PB1758、PB3360、JH642、1A243(ATCC 39,087)、ATCC 21332、ATCC 6051、MI113、DE100(ATCC 39,094)、GX4931、PBT110及びPEP211株;Hoch et al.、Genetics、73:215−228[1973];米国特許第4,450,235号;米国特許第4,302,544号;EP0134048;Palva et al.、Gene、19:81−87[1982];Fahnestock and Fischer,J.Bacteriol.,(1986)165:796−804[1986];及びWang et al.、Gene 69:39−47[1988]を参照)。発現宿主として特に興味深いのは、工業プロテアーゼ生成バチルス株である。これらの3つの株を用いることにより、プロテアーゼ生成に関する高効率は本発明の変異バチルス株によりさらに増大する。
【0260】
工業プロテアーゼ生成バチルス株は特に好ましい発現宿主を提供する。いくつかの実施態様において、本発明におけるこれらの株の使用は効率及びプロテアーゼ生成をさらに増大させる。上述の通り、2つの一般的なプロテアーゼの種類、すなわち中性(または“メタロプロテアーゼ”)及びアルカリ性(または“セリン”)プロテアーゼがあり、通常はバチルス種により分泌される。また、上述の通り、ズブチリシンは本発明の使用に好ましいセリンである。多種多様のズブチリシンが同定及び配列決定されており、例えば、ズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309である(例えば、EP414279B;WO89/06279;及びStahl et al.、J.Bacteriol.,159:811−818[1984]を参照)。本発明の1の実施態様において、バチルス宿主株は改変(例えば、変異体)プロテアーゼを生成する。多数の文献が変異体プロテアーゼの例及び引例を提供している(例えば、WO99/20770;WO/20726;WO99/20769;WO89/06279;RE34,606;米国特許第4,914,031号;米国特許第4,980,288号;米国特許第5,208,158号;米国特許第5,310,675号;米国特許第5,336,611号;米国特許第5,399,283号;米国特許第5,441,882号;米国特許第5,482,849号;米国特許第5,631,217号;米国特許第5,665,587号;米国特許第5,700,676号;米国特許第5,741,694号;米国特許第5,858,757号;米国特許第5,880,080号;米国特許第6,197,567号及び米国特許第6,218,165号を参照)。
【0261】
さらに別の実施態様において、好ましいバチルス宿主はdegU、degS、degR及びdegQの遺伝子の少なくとも1つにおいて変異または欠失を有するバチルス種である。好ましくは、変異はdegU遺伝子中にあり、より好ましくは変異はdegU(Hy)32である。(例えば、Msadek et al.、J.Bacteriol.,172:824−834[1990];及びOlmos et al.、Mol.Gen.Genet.,253:562−567[1997]を参照)。最も好ましい宿主株はdegU32(Hy)変異を有するバチルス・ズブチリスである。さらなる実施態様において、バチルス宿主はscoC4(Caldwell et al.、J.Bacteriol.,183:7329−7340[2001]を参照);spollE(Argoni et al.、Mol.Microbiol.,31:1407−1415[1999]を参照);oppAまたはその他のoppオペロン遺伝子(Perego et al.、Mol.Microbiol.,5:173−185[1991]を参照)における変異または欠失を含む。実際に、oppA遺伝子の変異と同じ表現型を生じるoppオペロンの変異が考えられ、本発明の変異バチルス株のいくつかの実施態様において使用できる。いくつかの実施態様において、これらの変異は単独で生じ、一方他の実施態様において、変異は組み合わせて存在する。いくつかの実施態様において、本発明の変異バチルスは上述の遺伝子の1以上の変異をすでに含むバチルス宿主株から得られる。別の実施態様において、本発明の変異バチルスはさらに、上述の遺伝子の1以上の変異を含むように設計される。
【0262】
いくつかの実施態様において、2以上のDNA構築体はバチルス宿主細胞内に導入され、変異バチルスの2以上の固有染色体領域の欠失を生じる。いくつかの実施態様において、これらの領域は連続しており(例えば、スキンとプロファージ7領域)、一方、他の実施態様において、当該領域は分離している(例えば、PBSX領域及びPKS領域;スキン領域及びDHB領域;またはPKS領域、SPβ領域及びyvfF−yveK領域)。
【0263】
当業者はポリヌクレオチド配列を細菌(例えば、大腸菌及びバチルス)細胞内に導入する適切な方法をよく把握している(例えば、Ferrari et al.、“Genetics”、in Harwood et al.(編集)、Bacillus、Plenum Publishing Corp.[1989]、57−72頁;例えば、Saunders et al.、J.Bacteriol.,157:718−726[1984];Hoch et al.、J.Bacteriol.,93:1925−1937[1967];Mann et al.、Current Microbiol.、13:131−135[1986];及びHolubova,Folia Microbiol.,30:97[1985];B.ズブチリスについて、Chang et al.、Mol.Gen.Genet.,168:11−115[1979];B.メガテリウムについて、Vorobjeva et al.、FEMS Microbiol.Lett.,7:261−263[1980];B.アミロリケファシエンスについて、Smith et al.、Appl.Env.Microbiol.,51:634(1986);B.チューリンゲンシスについて、Fisher et al.、Arch.Mictobiol.、139:213−217[1981];及びB.スフェリカスについて、McDonald,J.Gen.Microbiol.,130:203[1984]を参照)。実際に、プロトプラスト形質転換及び配向、形質導入及びプロプラスト融合などの形質転換の方法は公知であり、本発明での使用に適している。形質転換の方法は特に本発明により提供されるDNA構築体を宿主細胞内に導入するのに好ましい。
【0264】
一般的に用いられる方法に加えて、いくつかの実施態様において、宿主細胞は直接形質転換される(すなわち、媒介細胞を増幅のために用いず、または宿主細胞内への導入前にDNA構築のその他のプロセスを用いない)。DNA構築体の宿主細胞内への導入は、プラスミドまたはベクター内への挿入なしでDNA構築体を宿主細胞に導入するための当業者に公知の物理的及び化学的方法を含む。当該方法は、限定されないが、塩化カルシウム沈殿法、エレクトロポレーション、裸のDNA(naked DNA)、リポソーム等が挙げられる。別の実施態様において、DNA構築体は、プラスミド内に導入されることなく、プラスミドを用いて共形質転換される。さらに別の実施態様において、選択マーカーは当業者に公知の方法により変異バチルス株から欠失または実質的に切除される(Stahl et al.、J.Bacteriol.,158:411−418[1984];及びPalmeros et al.の保存部位特異的組換え[CSSR]方法、Palmeros et al.、Gene 247:255−264[2000]に記載、を参照)。いくつかの好ましい実施態様において、宿主染色体由来のベクター分解は固有染色体領域を除去しながら染色体中のフランキング領域を残す。
【0265】
いくつかの実施態様において、宿主細胞は変異バチルス株を生成するために本発明に従う1以上のDNA構築体を用いて形質転換され、ここで2以上の遺伝子が宿主細胞中で不活性化される。いくつかの実施態様において、2以上の遺伝子は宿主細胞染色体から欠失される。別の実施態様において、不活性化遺伝子は連続しており(欠失及び/または挿入による不活性化に関わらず)、一方他の実施態様において、連続した遺伝子ではない。
【0266】
上述の通り、細胞内及び細胞外発現ポリぺプチド活性を検出及び測定するための当業者に公知の種々の分析法がある。特に、プロテアーゼに関して、吸光度280nmで測定、またはFolin法を用いた比色分析により測定したカゼインまたはヘモグロビンからの酸溶性ぺプチドの放出に基づく分析法がある(例えば、Bergmeyer et al.、“Methods of Enzymatic Analysis”vol.5、Peptidases,Proteinases and their Inhibitors(ペプチダーゼ、プロテイナーゼ、及びそれらの阻害因子)、Verlag Chemie、Weinheim「1984」を参照)。その他の分析法は発色基質の可溶化を含む(例えば、Ward、“Proteinases(プロテイナーゼ)”、Fogarty(編集)、Microbial Enzymes and Biotechnology、Applied Science,ロンドン[1983]、251〜317頁を参照)。その他の分析法の例としては、スクシニル−Ala−Ala−Pro−Phe−paraニトロアニリド分析(SAAPFpNA)及び2,4,6−トリニトロベンゼンスルフォン酸ナトリウム塩分析(TNBS分析)が挙げられる。他の当業者に公知の多数の文献が適切な方法を提供する(例えば、Wells et al.、Nucleic Acids Res.11:7911−7925[1983];Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal Biochem.,242:221−227[1999]を参照)。
【0267】
上述の通り、宿主細胞中の目的タンパク質の分泌レベルの決定方法及び発現タンパク質の検出方法は当該タンパク質に特異なポリクローナルまたはモノクローナル抗体を用いる免疫学的分析法の使用を含む。例としては、酵素免疫測定法(ELISA)、放射免疫分析法(RIA)、蛍光免疫分析法(FIA)、及び蛍光活性化細胞分類法(FACS)を含む。しかしながら、その他の方法も当業者に公知であり、目的タンパク質の評価に使用できる(例えば、Hampton et al.、Serological Methods,A Laboratory Manual,APS Press、St.Paul,MN[1990];及びMaddox et al.、J.Exp.Med.、158:1211[1983]を参照)。いくつかの好ましい実施態様において、目的タンパク質の分泌は、対応する非変異宿主よりも本発明を用いて得た変異株におけるほうが高い。当業界で公知の通り、本発明を用いて生成した変異バチルス細胞は発現及び細胞培養物からの目的ポリぺプチドの回収に適した条件下で維持及び成長する(例えば、Harwood and Cutting(編集)Molecular Biological Methods for Bacillus(バチルスの分子生物学的方法)、John Wiley&Sons[1990]を参照)。
【0268】
本発明を実施する手段及び方法は以下の実施例を参照することにより、当業者により十分理解されるものであり、当該実施例は本発明の範囲またはその請求の範囲をいかなる方法によっても限定するものではない。
【0269】
実験
以下の実施例は特定の好ましい実施態様及び本発明の特徴を示し、さらに説明するためのものであり、本発明の範囲を限定するものとして解釈されるものではない。
【0270】
実験の開示において、以下の通り、以下の略語を適用する:℃(摂氏度);rpm(1分間当たりの回転数);H2O(水);dH2O(脱イオン水);HCl(塩酸);aa(アミノ酸);bp(塩基対);kb(キロ塩基対);kD(キロドルトン);gm(グラム);μg(マイクログラム);mg(ミリグラム);ng(ナノグラム);μl(マイクロリットル);ml(ミリリットル);mm(ミリメーター);nm(ナノメーター);μm(マイクロメーター);M(モル濃度);mM(ミリモル濃度);μM(マイクロモル濃度);U(単位);V(ボルト);MW(分子量);sec(秒);min(s)(分);hr(s)(時間);MgCl2(塩化マグネシウム);NaCl(塩化ナトリウム);OD280(280nmでの光学密度);OD600(600nmでの光学密度);PAGE(ポリアクリルアミドゲルエレクトロフォレーシス);PBS(リン酸緩衝生理食塩水[150mM NaCl、10mM リン酸ナトリウム緩衝液、pH7.2]);PEG(ポリエチレングリコール);PCR(ポリメラーゼ連鎖反応);RT−PCR(逆転写PCR);SDS(ドデシル硫酸ナトリウム);Tris(トリス(ヒドロキシメチル)アミノメタン);w/v(体積当たり重量);v/v(体積対体積);LA培地(リットル当たり:Difcoトリプトン・ペプトン 20g、Difco酵母抽出物 10g、EMサイエンス NaCl 1g、EMサイエンス 寒天17.5g、dH20〜1L);ATCC(American Type Culture Collection、ロックビル、メリーランド州);Clontech(CLONTECHラボラトリーズ、パロ・アルト、カリフォルニア州);Difco(Difcoラボラトリーズ、デトロイト、ミシガン州);GIBCO BRLまたはGibco BRL(ライフ・テクノロジーズ、Inc.,ゲーサーズバーグ、MD);Invitrogen(Invitrogen Corp.,サンディエゴ、カリフォルニア州);NEB(ニューイングランド Biolabs、ビバリー、マサチューセッツ州);Sigma(シグマ・ケミカル Co.,セントルイス、ミズーリ州);Takara(タカラ・バイオ Inc.Otsu,日本);Roche Diagnostics and Roche(ロシュ Diagnostics,F.Hoffmann La Roche部門、Ltd.,Basel、スイス);EMサイエンス(EMサイエンス、Gibbstown,ニュージャージー州);Qiagen(Qiagen,Inc.,バレンシア、カリフォルニア州);Stratagene(Stratagene Cloning System,ラ・ホーヤ、カリフォルニア州);Affymetrix(Affymetrix、サンタクララ、カリフォルニア州)。
【実施例】
【0271】
実施例1
欠失株の作成
この実施例は“方法1”について説明し、図1にも示される。この方法では、バチルス株内に形質転換されるDNA構築体を運ぶpJM102プラスミドベクターを生成するために大腸菌を用いた(Perego、上述を参照)。欠失部位の5’及び3’末端にすぐ隣接する領域をPCR増幅した。PCRプライマーを約37塩基対長さになるように設計し、バチルス・ズブチリス染色体に相同な31塩基対及びプライマーの5’末端から6塩基対に位置する6塩基対の制限酵素認識部位を含むようにした。プライマーを当該構築体の上流及び下流末端の固有制限酵素認識部位及びクローニング使用のための2つの断片間のBamHI部位構築のため設計した。抗菌マーカーのためのプライマーは当該断片の両側末端にBamHI部位を含んだ。可能であれば、PCRプライマーは欠失固有染色体領域のプロモーターを除去するが、隣接領域の全てのターミネーターを残すように設計した。染色体配列、遺伝子決定、及びプロモーター及びターミネーター情報の主な供給源はKunst et al.、(1997)上述から得ており、また当業者に公知のSubtiListワールドワイドウェブサーバーからも入手可能である(例えば、Moszer et al.、上述を参照)。本発明を用いて多数の欠失を作った。本発明により作成した欠失からのプライマー配列リストを表1に示す。また、プライマー命名システムの説明に関して図2も参照されたい。
【表1】
【0272】
【0273】
この方法では、100μL PCR反応を84μL水、10μL PCR緩衝液、1μLの各プライマー(すなわち、PKS−UF及びPKS−UR)、2μLのdNTPs、1μLの野生型バチルス染色体DNAテンプレート、及び1μLのポリメラーゼを含む150μLのエッペンドルフチューブ中で行った。使用したDNAポリメラーゼはTaq Plus Precision ポリメラーゼ及びHerculase(Stratagene)を含ませた。反応は以下のプログラムを用いてハイブリッドPCRエキスプレス熱サイクラー中で行った。サンプルは最初に94℃で5分間加熱し、それから冷却して50℃に保った。ポリメラーゼをこの時点で加えた。25サイクルの増幅は95℃で1分間、50℃で1分、及び72℃で1分からなる。最終的に72℃で10分間行うことにより、伸長完了を確実にした。サンプルを分析のため4℃で保存した。
【0274】
PCR完了後、10μlの各反応をInvitrogen 1.2% アガロースE−ゲル上で、60ボルト、30分間で行い、正しいサイズのバンドの存在を確認した。ここに記載する全てのゲルエレクトロフォレーシス法はこれらの条件を用いた。バンドが存在する場合、反応チューブの残余物は製造メーカーの取扱説明書に従いQiagen Qiaquick(登録商標)PCR精製キットを用いて精製し、それから、適当な制限酵素対を用いて切断した。消化を37℃、1時間、9μLの水、、2μLの10×BSA、2μLの適当なNEB制限緩衝液(2000−01 NEBカタログ及び技術文献に従う)、5μLのテンプレート、及び1μLの各制限酵素からなる20μLの反応として行った。例えば、PBSX上流断片及びCssS上流断片をNEB(ニューイングランド・バイオラボ)制限緩衝液B中でXbal及びBamHIを用いて切断した。消化断片をゲル・エレクトロフォレーシス及び製造メーカー取扱説明書に従いQiagen Qiaquickゲル抽出キットを用いた抽出により精製した。図5及び6は種々の欠失の結果を示すゲルを表す。
【0275】
プラスミドベクター内への断片の連結は2つの工程で行われ、製造メーカー取扱説明書に従ってTakara連結キットまたはT4DNAリガーゼ(反応内容:5μL 各挿入断片、1μL 切断pJM102プラスミド、3μL T4DNAリガーゼ緩衝液及び1μL T4DNAリガーゼ)を用いる。まず、切断上流及び下流断片をpJM102プラスミド・ポリリンカー中の固有制限酵素認識部位内に15℃で一晩中連結させ、共通BamHI部位で接続し、環状プラスミドを再形成させた。pJM102プラスミドを各欠失に適した固有制限酵素認識部位を用いて切断し(表2を参照;cssS、Xbal及びSaclを用いた)、連結前に上述の通り精製した。この再環状化プラスミドをInvitrogenの“Top Ten”大腸菌細胞内に形質転換し、製造メーカーのワン・ショット形質転換手順を用いた。
【0276】
形質転換体は1.5%寒天(LA)とブルーホワイトスクリーニングのためのX−galを含んだ50ppmカルバニリシン(carbanicillin)を用いて凝固したLB培地上で選択した。クローンを採取し、5mlのLB培地(Luria−Bertani)プラス50ppmカルバニシリン(carbanicillin)中、37℃で一晩中成長させ、プラスミドをQiagen’s Qiaquick ミニ−準備キットを用いて単離した。制限分析により、挿入の各末端で制限酵素認識部位を用いて切断し、プラスミドから約2kbのバンドにすることにより挿入の存在を確認した。挿入を確認したプラスミドはBamHIを用いて切断し、上述の通り消化反応において線形化し(第2の制限酵素に代えてさらに1μLの水を用いて)、再環状化を防止するために1μLの子牛の腸(calf intestinal)ホスファターゼ及びshrimpホスファターゼを用いて1時間、37℃で処理し、及び表2に記載するように抗菌剤耐性マーカーに連結させた。抗菌剤耐性マーカーはBamHIを用いて切断し、連結前に製造メーカー取扱説明書に従いQiagenゲル抽出キットを用いて洗浄した。このプラスミドを既に述べたように大腸菌内にクローンし、選択に応じて適切な、5ppmのフレオマイシン(phl)または100ppmのスペクチノマイシン(spc)を用いる。単離プラスミド中のマーカー挿入の確認は上述の通り、BamHIを用いる制限分析により行った。B.ズブチリス内への形質転換前に、プラスミドをScalを用いて線形化し、二重交差結合を確実にさせた。
【表2】
【0277】
実施例2
大腸菌をバイパスするためにPCR融合を用いるDNA構築体の作成
この実施例は“方法2”について説明し、また図3にも示される。プライマーが抗菌マーカーのプライマー配列に相補的な25bp“テイル”を用いて設計されること以外は、上流及び下流断片は方法1の通り増幅する。“テイル”とは、直接増幅される配列に相同でないが、他のDNA配列に相補的であるプライマーの5’末端上の塩基対としてここで定義する。同様に、抗菌剤を増幅するためのプライマーは断片プライマーに相補的な“テイル”を含む。任意の所定欠失について、欠失X−UFfus及び欠失X−URfusは互いに直接の相補体である。これは、DF−fusとDR−fusプライマーセットに関しても当てはまる。さらに、いくつかの実施態様において、これらのプライマーはプラスミドベクター作成で使用する方法1において用いたものに類似した制限酵素認識部位を含む(表3及び米国特許第5,023,171号を参照)。表3はPCR融合による欠失構築体の作成に有用なプライマーのリストを提供する。表4はPCR融合による欠失構築体の作成に有用な別のプライマーのリストを提供する。しかしながら、この表では、全ての欠失構築体はphleoRマーカーを含む。
【表3】
【表4】
【0278】
上述の通りゲル・エレクトロフォレーシスにより表3及び4に記載した断片の大きさを確認した。正しい場合には、1μLの各上流、下流及び抗菌剤耐性マーカー断片を記載された欠失X−UF及び欠失X−DRプライマーまたはネスティドプライマーを含む1の反応管に加えた。ネスティドプライマーは上流または下流断片の内部に相同な25塩基対のDNAであり、通常、当該断片の外側末端由来の約100塩基対である(図2を参照)。ネスティドプライマーの使用は融合成功度を高めることが多い。PCR反応成分は、82μLの水をテンプレート体積を追加して補うために用いた以外は上述のものに類似した。PCR反応条件は72℃の伸長を3分に延長した以外は、上述のものと類似した。伸長の間、抗菌剤耐性遺伝子を上流と下流断片間に融合した。この融合断片は、精製工程を用いずに、または製造メーカー取扱説明書に従って行われるQiagen Qiaquick PRC精製を用いてバチルス内に直接統合できる。
【0279】
実施例3
PCR断片の連結を用いるDNA構築体の作成及び大腸菌クローニングをバイパスするためのバチルス・ズブチリスの直接形質転換
この実施例では、PCR断片の連結を用いるDNA構築体を作成するための方法(“方法3”)及びバチルスの直接形質転換について説明する。例として、prpC、sigD及びtdh/kblの修飾を連結方法を示すために提供する。実際に、sigD及びtdh/kblを1の方法で構築し、prpCを別の方法により構築した。
【0280】
A.Tdh/Kbl及びSigD
バチルス・ズブチリス染色体のtdh/kbl領域に隣接した上流及び下流断片を、フランキングDNAの内部プライマーがII型の制限酵素認識部位を含むように設計した以外は方法1に記載した類似のPCRにより増幅した。loxP−スペクチノマイシン−loxPカセットのプライマーをフランキング及び相補的突出部と同じII型制限酵素認識部位を用いて設計した。左側面及び右側面の固有突出部は上流及び下流のフランキングDNA間の抗菌カセットの直接連結を可能にする。全てのDNA断片は適当な制限酵素により消化し、当該断片を製造メーカー取扱説明書を用いてQiagen Qiaquick PCR精製キットで精製した。この精製に続いて、BioRad P−6ゲルを含む1mLの回転カラム中で脱塩を行い、2mM Tris−HCl、pH7.5で釣り合いを取った。断片をSavant Speed Vac SC110システムを用いて124〜250ng/μLに濃縮した。0.8〜1μgの各断片の3つの断片連結を、16時間、14〜16℃で15〜25μL反応体積中で12U T4リガーゼ(ロシュ)を用いて行った。所望の連結生成物の合計収量は、アガロースゲル上の標準DNAラダー(ladder)との比較により評価して、反応当たり100ngより大きかった。当該連結混合物は形質転換反応のための精製を行わないで用いた。この構築体のためのプライマーは下記の表5に示す。
【表5】
【0281】
sigD欠失の構築はtdh/kblの構築にほとんど従う。sigD構築に用いるプライマーは表6に示す。
【表6】
【0282】
B.PrpC
バチルスの直接形質転換のためのPCR増幅DNA断片の連結によりDNA分子を作成する別の実施例は遺伝子prpCの部分フレーム内欠失を含む。prpC遺伝子を含むバチルス・ズブチリス染色体DNAの3953bp断片をp95及びp96を用いてPCRにより増幅した。当該断片を固有制限酵素認識部位PflMI及びBstXIで切断した。これにより3つの断片、上流、下流及び中心断片を生じた。後者は欠失される断片であり、内部からprpC遺伝子に位置した170bpからなる。消化混合物をQiagen Qiaquick PCR精製キットを用いて精製し、続いてBioRad P−6ゲルを含む1mLの回転カラム中、脱塩し、2mMのTris−HCl、pH7.5で釣り合いを取った。2回目のPCR反応で、抗菌カセット、すなわちloxP−スペクチノマイシン−loxPをBstXI部位を含むプライマー及びPflMI部位を含む下流プライマーであって、両方がゲノムDNA断片の部位に相補的な切断部位を含むものを用いて増幅させた。当該断片をPflMI及びBstXIを用いて消化し、上述の染色体断片について説明した通り精製した。上流、抗菌カセット及び下流断片の3つの断片連結を上述のtdh/kblに関しての通り、行った。下記にさらに詳細を説明する通り、所望の連結生成物の収量は類似しており、連結生成物をxylRcomKコンピテントバチルス・ズブチリスの形質転換に関するさらなる処理を行うことなく用いた。
【表7】
【0283】
C.PckA欠失
上述の欠失に加えて、pckAも修飾した。PCRプライマーのpckAUF、pckA−2Urfus、spc ffus、spc rfus、pckA Dffus及びpckA DRをPCR及びテンプレートとしてバチルス・ズブチリス168誘導体の染色体DNA及びpDG1726(Guerout−Fleury et al.、Gene 167(1−2):335−6[1995]を参照)を用いるPCR融合反応に関して用いた。プライマーを表8に示す。これらの欠失変異体を構築するのに使用する方法は上述の方法1と同じである。
【表8】
【0284】
D.連結DNAを用いるキシロース導入コンピテンス宿主細胞形質転換
2001年8月10日に出願され、ここに引用する米国特許出願番号第09/927,161号に記載されているように、部分遺伝子型xylRcomKを有する宿主株バチルス・ズブチリスの細胞を1%キシロースを含むLB培地中で2時間成長させることによりコンピテントにし、OD550を1とした。この培養物を6時間培養物から取り除いた。全ての培養物を37℃で、300rpmで振とうさせながら成長させた。0.3mLのアリコートを1:1の培養物と30%グリセロール混合物として丸底2mL管内に冷凍し、その後の使用のために液体窒素中で保存した。
【0285】
形質転換のために、凍結コンピテント細胞を37℃で解凍し、解凍が完了してすぐに、連結反応混合物由来のDNAを5〜15μL/管のレベルで加えた。当該管をそれから60分間、37℃で1400rpmで振とうした(Tekmar VXR S−10)。形質転換混合物を100ppmのスペクチノマイシンを含む8cmLAプレート上の100μLアリコート中で希釈しないでプレートした。一晩中成長させた後、形質転換体をLB培地(100ppmスペクチノマイシン)中に採取し、及び当業者に公知の通り実施されるゲノムDNA単離のために37℃で成長させた(例えば、Harwood and Cuttings,Molecular Biological Methods for Bacillus,John Wiley and Son,ニューヨーク、N.Y.[1990]、23頁を参照)。5μLの連結反応混合物を形質転換に用いた場合、通常、400〜1400の形質転換体が100μLの形質転換混合物から得られた。
【0286】
抗菌マーカーが入来DNA内の2つのloxP部位間に位置する場合、マーカーはCreタンパク質を発現できるcre遺伝子を含むプラスミドを含む株を形質転換することにより除去できる。細胞は37℃〜42℃で培養したpCRM−TSpleo(下記参照)を用いて形質転換し、LA上にプレートし、コロニー生成後、100ppmのスペクチノマイシンを含むLA上に継ぎ合せた。一晩中培養後に成長していなかったパッチは抗菌マーカーを失ったと考えられる。マーカーの損失は所定の遺伝子に適したプライマーを用いるPCR分析により確認した。
【0287】
pCRM−TS−pleoは以下の配列を有する(配列番号205):
【式54】
【0288】
E. トランスクリプトームDNAアレイ法
上述の方法に加えて、トランスクリプトームDNAアレイ法を本発明の変異体を発達させるために用いた。まず、標的RNAを当業者に公知のグアニジン酸(guanidinium acid)フェノール抽出によりバチルス株から取り入れ(例えば、Farrell、RNA Methodologies、(第2版).Academic Press,サンディエゴ、81頁を参照)、及び時点をdeSaizieu et al.(deSaizieu et al.、J.Bacteriol.,182:4696−4703[2003])及びここに記載する方法から適合させた方法によりビオチン標識cDNA内に逆転写した。合計RNA(25mg)を37℃、100mL反応中で一晩中培養した:1×GIBCO 一本鎖(first−strand)緩衝液(50mM Tris−HCl pH8.3、75mM KCl、3mM MgCl2);10mM DTT;40mMランダムヘキサマー;0.3mMの各dCTP、dGTP及びdTTP;0.12mM dATP;0.3mM ビオチン−dATP(NEN0;2500単位 SuperScript II 逆転写酵素(ロシュ)。RNAを除去するために、当該反応を0.25M NaOHに加え、30分間、65℃で培養した。当該反応をHCl及び2.5M 酢酸アンモニウムを含むエタノール中で−20℃で沈殿させた核酸を用いて中和させた。ペレットを洗浄、空気乾燥し、水中で再懸濁させ、UV分光法により量を測った。反応収量は約20〜25mgビオチン標識cDNAであった。
【0289】
12mgのこのcDNAを、10分間、37℃で3.75ミリ単位のDNAアーゼを含む33mL 1×One−Phor−All緩衝液(Amersham−Pharmacia番号27−0901−02)において断片化した。DNAアーゼを熱殺菌後、3%アガロースゲル上の2mgの断片化cDNAを管理することにより確認した。ビオチン含有cDNAの大きさは通常、25〜125ヌクレオチドの範囲である。残りの10mgのcDNAをAffymetrixバチルス遺伝子チップアレイにハイブリダイズした。
【0290】
ハイブリダイゼーションをAffymetrix発現分析技術マニュアル(Affymetrix)に記載されている通りに、供給元が提示する試薬を用いて行った。要するに、10mgの断片化ビオチン標識cDNAを220mLの以下を含むハイブリダイゼーション混合物に加えた:100mM MES(N−モルホリノエタンスルホン酸)、1M Na+、20mM EDTA、0.01% Tween20;5mg/mL合計酵母RNA;0.5mg/mL BSA;0.1mg/mLニシン精子DNA;50pM対照オリゴヌクレオチド(AFFX−B1)。当該混合物を5分間、95℃に加熱し、5分間40℃に冷却し、粒子を除去するために手短に遠心分離し、及び200mLをそれぞれ前もって温め、すすいでおいた(1×MES緩衝液+5mg/ml酵母RNA)遺伝子チップカートリッジ内に注入した。当該アレイを40℃で一晩中回転させた。
【0291】
当該サンプルを除去し、アレイを非ストリンジェントな洗浄緩衝液(6×SSPE、0.01%Tween20)を用いて充填し、手順Euk−GE−WS2を用いるAffymetrix 流体ステーション上で非ストリンジェント及びストリンジェント(0.1M MES,0.1M[Na+]、0.01%Tween20)洗浄緩衝液を用いて洗浄した。アレイを3つの工程で着色した:(1)ストレプトアビジン;(2)ビオチンで標識した抗ストレプトアビジン抗体;(3)ストレプトアビジン−フィコエリトリン結合。
【0292】
アレイ中のシグナルをヒューレット・パッカード遺伝子アレイスキャナーで3mmピクセル解像度の570nmレーザー光を用いて検出した。4351ORFプローブセットのシグナル強度を時間的経過実験を含む全ての時点にわたって測定及び標準化した。これらのシグナルをそれから実験中、遺伝子の相対発現レベルを推測するために比較した。トレオニン生合成系及び分解系遺伝子を同時に転写し、非効率なトレオニン利用を示した。分解系トレオニン経路の欠失は所望の生成物の発現を向上させた(図7を参照)。本発明はトレオニン生合成系及び分解系プロファイルに類似した転写プロファイルを含む経路を修飾する手段を提供する。従って、本発明はバチルス株を最適化するためにトレオニンに類似した転写プロファイルを含む経路の修飾にも使用できる。いくつかの好ましい実施態様において、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される少なくとも1の遺伝子が欠失または修飾される。ここに記載した本発明を用いて、sigDレギュロンが転写されたという驚くべき発見をした。この遺伝子の欠失は所望の生成物のより優れた発現を生じる(図7を参照)。gapB及びpckAの転写も驚くべき発見であった。pckAの欠失は改善または損失のいずれももたらさなかった。しかしながら、本発明はpckA欠失または修飾とgapB及び/またはfbpの欠失または修飾の組み合わせにより株タンパク質の生成を向上させる手段を提供する。さらに、本発明の開発時に、トリプトファン生合成経路遺伝子は不均衡な転写を示すことが観測された。従って、本発明は、trpA、trpB、trpC、trpD、trpE、及び/またはtrpFからなる群より選択される遺伝子などの転写が増加した株を生成するのに使用でき、改善株は母体(すなわち、野生型及び/または起源株)と比較して所望の生成物の発現が向上すると考えられる。実際に、これらの遺伝子修飾の任意の組み合わせは所望の生成物の発現の向上をもたらすと考えられる。
【0293】
F.発酵
上述の構築体を用いて生成した株分析は以下の発酵を行った。大きさ14Lの培養はBiolafitte(登録商標)発酵器中で行った。7リットル当たりの培養成分を表9に記載する。
【表9】
【0294】
750rpmで槽を攪拌し、気流を11リットル/分、温度37℃に調整し、pHをNH4OHを用いて6.8に維持した。60%グルコース溶液を0.5〜2.1グラム/分で開始から発酵の終わりまで直線ランプ中で約14時間送った。炭素収支及び効率はグルコース供給、タンパク質生成物の収量、細胞集団収量、培養液中のその他の炭素及び発生CO2から計算した。変異体株を改善点を検証するために母体株と比較した。この変異pckA株はこれらの条件下では改善を示さなかったが、改善は修飾培養条件(すなわち、当業者に公知のもの)下及び/または別の遺伝子と組み合わせて生じると考えられる。いくつかの好ましい実施態様において、これらの別の遺伝子はgapB、alsD及び/またはfbpからなる群より選択される。
【0295】
実施例4
変異バチルス株を得るための宿主細胞形質転換
上述の方法1または2によりDNA構築体を作成すると、それを適当なバチルス・ズブチリス研究所株内に形質転換した(例えば、BG2036または、BG2097;任意のコンピテントバチルス直接宿主細胞は本発明の方法に使用できる)。当該細胞は0.5ppmのフレオマイシンまたは100ppmのスペクチノマイシンの選択培地上に必要に応じてプレートした(Ferrari and Miller、Gene Expression Systems:Using Nature for the Art of ExpressionのBacillus Expression:A Gram−Positive Model(バチルス発現:グラム陽性モデル)、65〜94頁[1999])。研究所株はバチルス・ズブチリス生成宿主株内に2回、またはBG3594及びそれからMDT98−113に一度形質転換した欠失を有する染色体DNAの供給源として用いた。形質転換体は1のコロニーに単離するためにストリーキングし、5mLのLBと適当な抗菌剤中で一晩中採取及び成長させた。染色体DNAを当業者に公知の通り単離した(例えば、Hardwood et al.、上述参照)。
【0296】
統合DNA構築体の存在は、上述の成分及び条件を用いて3つのPCR反応により確認した。例えば、2つの反応は欠失カセット外側由来領域を1の場合において(プライマー1及び11)抗菌遺伝子内に増幅するために設計し、他の場合において(プライマー1及び12)全体的な挿入により増幅するために設計した。第3の照合は、欠失カセット外側由来の領域を欠失領域内(プライマー1及び4))に増幅した。図4は、正しいクローンが最初の2つの場合にはバンドを示すが、3番目の場合には示さないということを示す。野生型バチルス・ズブチリス染色体DNAは全ての反応で陰性対照として用い、第3のプライマーセットのバンドのみを増幅すべきである。
【0297】
実施例5
振とうフラスコ分析−プロテアーゼ活性測定
DNA構築体を安定にコンピテントバチルス・ズブチリス株内に統合すると、ズブチリシン活性を振とうフラスコ分析により測定し、当該活性を野生型レベルと比較した。分析は当業者に公知のズブチリシン生成に適した50mLの成長培地を含む250mlのバッフルフラスコ内で行った(Christianson et al.、Anal.Biochem.,223:119−129[1994];及びHsia et al.、Anal.Biochem.242:221−227[1996]を参照)。当該培地を50μLの8時間、5mL培養物と接種し、250rpmで振とうしながら37℃、40時間成長させた。そして、1mLのサンプルをプロテアーゼ活性分析のために17、24及び40時間の時点で取った。プロテアーゼ活性をモナコ自動分析器(Monarch Automatic Analyser)を用いて405nMで測定した。2つのサンプルを緩衝液中で1:11(3.131g/L)に希釈した。正確な機械較正を保証する対照として、1のサンプルを1:6(5.585g/L)、1:12(2.793g/L)及び1:18(1.862g/L)に希釈した。図7は種々の変異バチルス・ズブチリスクローンにおけるプロテアーゼ活性を示す。図8は、バチルス・ズブチリス野生型株(非変異)及び対応する変異欠失株(−sbo)及び(−slr)の振とうフラスコ培養物から測定した改善されたプロテアーゼ分泌を示すグラフを提供する。プロテアーゼ活性(g/L)を17、24及び40時間後に測定した。
【0298】
また、細胞密度はOD600で分光光度測定を用いて測定した。測定時間でのサンプルの著しい違いは見られなかった(データは示していない)。
【0299】
上述の明細書中に言及した全ての文献及び特許はここに引用するものとする。言及した本発明の方法及びシステムの種々の修正及び変更は本発明の範囲及び精神を逸脱することなく当業者に明らかである。本発明は特定の好ましい実施態様に関連して説明したが、本発明は当該特定の実施態様に過度に限定されるものではないことは当然のことである。実際に、当業界及び関連分野の当業者に明らかである本発明を実施するための前述の方法の種々の変更は本発明の範囲内である。
【特許請求の範囲】
【請求項1】
sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の遺伝子の染色体欠失を含む変異バチルス株であって、
プロテアーゼである目的タンパク質が本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現及び/または生成レベルが増大している、前記変異バチルス株。
【請求項2】
前記変異バチルス株がB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される、請求項1の変異バチルス株。
【請求項3】
前記変異バチルス株がB.ズブチリス株である、請求項2の変異バチルス株。
【請求項4】
前記変異バチルス株がプロテアーゼ生成株である、請求項1の変異バチルス株。
【請求項5】
前記プロテアーゼがズブチリシンである、請求項4の変異バチルス株。
【請求項6】
前記ズブチリシンがズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309及びこれらの変異体からなる群より選択される、請求項5の変異バチルス株。
【請求項7】
前記変異バチルス株がdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子における少なくとも1つの変異をさらに含む、請求項1の変異バチルス株。
【請求項8】
前記変異がdegU(Hy)32である、請求項7の変異バチルス株。
【請求項9】
前記変異バチルス株がさらに目的異種タンパク質を含む、請求項1の変異バチルス株。
【請求項1】
sbo、slr、ybcO、csn、spollSA、sigB、phrC、rapA、CssS、trpA、trpB、trpC、trpD、trpE、trpF、tdh/kbl、alsD、sigD、prpC、gapB、pckA、fbp、rocA、ycgN、ycgM、rocF及びrocDからなる群より選択される1以上の遺伝子の染色体欠失を含む変異バチルス株であって、
プロテアーゼである目的タンパク質が本質的に同じ成長条件下で成長させた対応する非変異バチルス宿主株における同じ目的タンパク質の発現及び/または生成と比較して発現及び/または生成レベルが増大している、前記変異バチルス株。
【請求項2】
前記変異バチルス株がB.リケニフォルミス、B.レンタス、B.ズブチリス、B.アミロリケファシエンス、B.ブレビス、B.ステロサーモフィラス、B.アルカロフィラス、B.コアグランス、B.サーキュランス、B.プミルス、B.ロータス、B.クラウジイ、B.メガテリウム及びB.チューリンゲンシスからなる群より選択される、請求項1の変異バチルス株。
【請求項3】
前記変異バチルス株がB.ズブチリス株である、請求項2の変異バチルス株。
【請求項4】
前記変異バチルス株がプロテアーゼ生成株である、請求項1の変異バチルス株。
【請求項5】
前記プロテアーゼがズブチリシンである、請求項4の変異バチルス株。
【請求項6】
前記ズブチリシンがズブチリシン168、ズブチリシンBPN’、ズブチリシン・カールスバーグ、ズブチリシンDY、ズブチリシン147、ズブチリシン309及びこれらの変異体からなる群より選択される、請求項5の変異バチルス株。
【請求項7】
前記変異バチルス株がdegU、degQ、degS、scoC4、spollE及びoppAからなる群より選択される遺伝子における少なくとも1つの変異をさらに含む、請求項1の変異バチルス株。
【請求項8】
前記変異がdegU(Hy)32である、請求項7の変異バチルス株。
【請求項9】
前記変異バチルス株がさらに目的異種タンパク質を含む、請求項1の変異バチルス株。
【図1A】

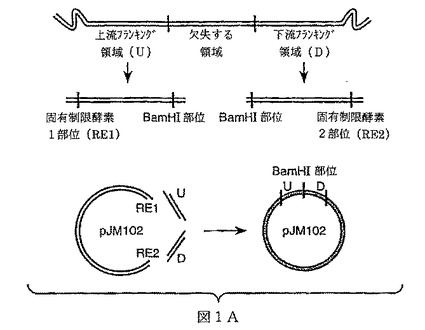
【図1B】
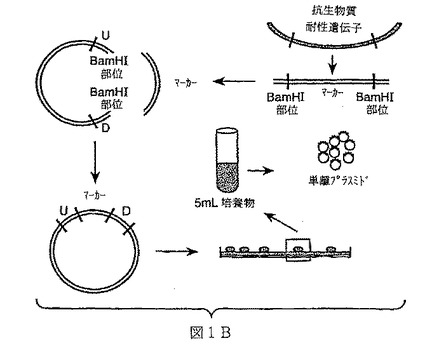
【図2】
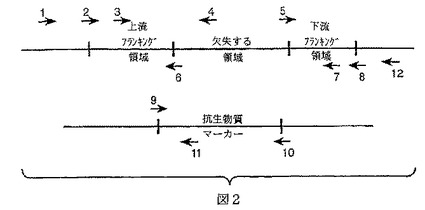
【図3】
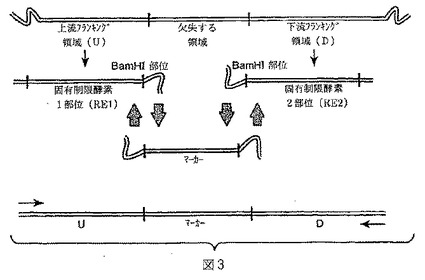
【図4】
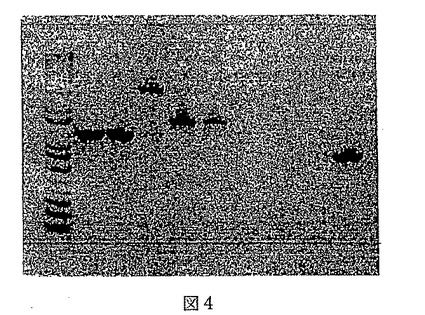
【図5】

【図6】

【図7】
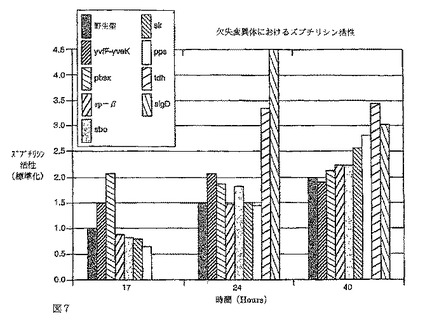
【図8】
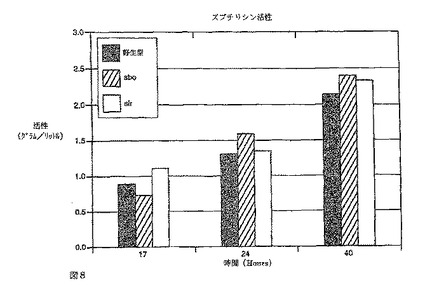
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2011−4759(P2011−4759A)
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願番号】特願2010−183904(P2010−183904)
【出願日】平成22年8月19日(2010.8.19)
【分割の表示】特願2003−580558(P2003−580558)の分割
【原出願日】平成15年3月28日(2003.3.28)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願日】平成22年8月19日(2010.8.19)
【分割の表示】特願2003−580558(P2003−580558)の分割
【原出願日】平成15年3月28日(2003.3.28)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
[ Back to top ]