
バラ科果樹の形質転換法及び形質転換体
【課題】ウメなどのバラ科果樹を容易かつ高い効率で形質転換することができる形質転換法、この方法によって形質転換された形質転換体、遺伝子組換えバラ科果樹などを提供する。
【解決手段】アグロバクテリウム法によるバラ科果樹の形質転換において、外植体の作成に使用する種子の採取時期を5月上旬から中旬の間、共存培養期間が3日間程度、除菌に使用する抗生物質をメロペンに限定することによって、高い確率で形質転換体を得ることができた。また、共存培養の前に超音波処理を施すことによって、形質転換効率を向上させることができた。さらに、この形質転換体に発根処理を施すことによって遺伝子組換えバラ科果樹を得ることができた。
【解決手段】アグロバクテリウム法によるバラ科果樹の形質転換において、外植体の作成に使用する種子の採取時期を5月上旬から中旬の間、共存培養期間が3日間程度、除菌に使用する抗生物質をメロペンに限定することによって、高い確率で形質転換体を得ることができた。また、共存培養の前に超音波処理を施すことによって、形質転換効率を向上させることができた。さらに、この形質転換体に発根処理を施すことによって遺伝子組換えバラ科果樹を得ることができた。
【発明の詳細な説明】
【技術分野】
【0001】
この発明は、バラ科果樹、特にウメの形質転換法及びこの方法により作出された形質転換体などに関するものである。
【背景技術】
【0002】
ウメ等のバラ科果樹は、その実が食用となるため、従来から耐寒性や耐病性などの優れた性質を備えた品種が求められている。
【0003】
さて、これらバラ科果樹の繁殖は、品種の形質(特長)を安定させ、収穫までの期間を短縮するため、結実した種子から行う(種子繁殖)のではなく、接ぎ木(栄養繁殖)によって行うのが一般的である。なお、接ぎ木とは、有用形質を持った親の枝(穂木)を近縁種の台木に接着・再生させて新たな個体を得る繁殖方法である。
【0004】
ところで、接ぎ木においては、使用する台木により耐干性・耐水性・耐凍性等の各種形質が左右されることが一般的に知られており、バラ科果樹においても、台木の選択によって果実の生産性やその品質が大きく変化することが知られている。なお、現在使用されているウメ台木の多くは実生台木であり、遺伝的な性質が不揃いなため、優れた台木を見分けるのは難しい。
【0005】
そこで、以前から優れた性質を備えた台木を開発することが試みられてはいる。しかし、バラ科果樹類には、台木として利用できる大きさまで成長するのに長時間かかること、種子繁殖では品種の形質が安定しにくいことなどから、一般的な交雑育種を利用することは困難であった。そこで、従来から、育種を効率化することを目的として、組織培養や遺伝子導入などのバイオテクノロジー技術を利用した研究がなされている。
【0006】
ところで、バラ科果樹は、果樹の中ではゲノムサイズが小さく、また幼木期が短いため、遺伝解析が比較的容易である。しかし、バラ科果樹類の組織培養からの再分化は困難であり、これが形質転換を行う上での障壁となっている。
【0007】
このことはウメに関しても当てはまり、ウメについてはこれまで充分効率的な再分化系は確立されておらず、形質転換の成功例も報告されていない。なお、現在までに報告されているウメの再分化系としては、ウメ「紅さし」の未熟子葉からの再分化系が挙げられるが、この再分化系では未熟子葉上にいったんカルスを誘導させ、そのカルスを組成の異なる培地に植え替えることにより、不定芽を誘導させたものである(非特許文献1を参照。)。
【0008】
しかし、上記の再分化系は、組成の異なる培地に植え替えるため、不定芽を形成させるまでに時間と手間がかかり、カルス形成を経由しているため突然変異体が生じやすいという問題点があった。また、不定芽は不定胚とは異なり増殖能力を備えておらず、台木の生産には利用できないとの問題点もあった。
【0009】
また、同じバラ科果樹であるモモでは、成熟子葉を外植体として形質転換体が得られたとの報告がある(非特許文献2を参照。)。この報告に記載の方法は、アグロバクテリウム菌を感染した子葉上にカルスを誘導させ、このカルスから形質転換不定芽を誘導させたのち、形質転換シュートを形成させるものである。しかし、この方法はウメなどのように不定芽再分化効率が低い植物には、利用できないとの問題点があった。
【0010】
さらに、バラ科果樹に含まれるセイヨウナシでは、その腋芽の直近に遺伝子導入のための切り口を設けた培養シュートの茎からなる外植体を使用する形質転換法が研究されている(特許文献1)。ただ、この方法の形質転換効率は4〜5%であり、実用化するには形質転換効率が低すぎるとの問題点があった。
【特許文献1】特開2004-283051号公報
【非特許文献1】中川文雄ら 「ウメ未熟子葉由来カルスからの不定芽形成」 園芸学会雑誌 第63巻 別冊1 1994年 p88-89
【非特許文献2】Rosa M. et al., 2004. Molecular Breeding 14:419-427
【発明の開示】
【発明が解決しようとする課題】
【0011】
この発明は、ウメなどのバラ科果樹を容易かつ高い効率で形質転換することができる形質転換法、この方法によって形質転換された形質転換体、遺伝子組換えバラ科果樹などを提供することを課題とする。
【課題を解決するための手段】
【0012】
この発明は、アグロバクテリウム法による形質転換において、外植体の作成に使用する種子の採取時期、共存培養期間、除菌に使用する抗生物質などを特定することによって、バラ科果樹からの形質転換体の作出を可能にすることを最も主要な特徴とする。また、共存培養する前に超音波処理することによって、形質転換効率を高めることをもう一つの特徴とする。
【発明の効果】
【0013】
この発明により、バラ科果樹であるウメで、初めて形質転換体を作出することができた。これにより、優良な形質を備えた形質転換体、例えば耐病性や耐寒性に優れたウメを育種することができるようになり、優れた性質を備えた台木の育種が期待できるようになった。また、ウメと同じ科に属する、スモモ、モモ、サクラ、アンズ、アーモンドについても優れた形質転換体を容易に得ることが期待できるようになった。
【発明を実施するための最良の形態】
【0014】
この発明の形質転換法は、バラ科果樹の未熟果実から種子を摘出し、この種子から未熟子葉を取出して外植体を作成する工程と、この外植体をアグロバクテリウム菌が分散している感染液に浸漬したのち、外植体とアグロバクテリウム菌とを共存培養する工程と、アグロバクテリウム菌に感染した外植体を、抗生物質メロペンを含む培地で培養することによってアグロバクテリウム菌を除菌する工程とを含むバラ科果樹の形質転換法である。また、この発明の形質転換体はこの発明の方法によって得られたものである。さらに、この発明の遺伝子組換えバラ科果樹の作出法は、前記形質転換体により得られた培養シュートに発根処理を行なうものであり、この発明の遺伝子組換えバラ科果樹は前記作出法で作出したものである。そこで、この発明の形質転換法及び作出法などについて以下に詳説する。
【0015】
(1)形質転換法の対象となる果実
対象となる果実は、主にウメを想定しているが、同じバラ科果樹の果実であるスモモ、モモ、桜桃、アンズ、アーモンド等にも適用できると考えられる。なお、ウメの品種としては、市場において取引されている品種、例えば、「紅さし」、「越の梅」など通常栽培されている品種であれば特にこだわることなく使用できるが、穂木との親和性や果実の商品力等を考えると「南高」が好ましい。
【0016】
また、不定胚、形質転換体の作出に利用する果実は未熟果実であり、具体的には、開花後4月下旬から5月下旬の間に採取された未熟果実であり、好ましくは5月上旬から5月中旬以内、さらに好ましくは5月10日前後のものである。
【0017】
(2)外植体の作成
外植体の作成は、例えば、バラ科果樹の未熟果実をエタノールで滅菌し、滅菌したメス、ピンセット等で果実から種子を摘出して種皮を取り除き、二枚の子葉を縦に切断することによって行う。作成した外植体は、適当な培地(例えば、後述する共存培地など)上に置床して乾燥による劣化を防ぐ。
【0018】
(3)アグロバクテリウム菌の感染及び共存培養
アグロバクテリウム菌の外植体への感染は、外植体をアグロバクテリウム菌が分散した感染液に10分程度浸漬して余分な感染液を除去したのち、この外植体を適当な固体培地に置床して暗黒下、常温(例えば25℃)で共存培養することによって行う。共存培養の期間は、外植体が枯死しない範囲であれば特に限定する必要はないが、共存培養を始めてから3日間程度が適当である。これは1日〜2日間程度では充分感染せず、5日間では多くの外植体が衰弱するからである。
【0019】
なお、外植体がアグロバクテリウム菌の分散した感染液に浸漬した状態で超音波処理することによって、アグロバクテリウム菌の感染効率を向上することができる。超音波処理は、例えば、外植体と感染液が入った試験管を超音波洗浄機に入れて超音波を処理することによって行う。
【0020】
ここで、超音波処理は、超音波の周波数が略28kHz、処理時間が10秒〜2分間、好ましくは10秒〜30秒間、強度が略0.23W/cm2となるように行う。
【0021】
また、使用するアグロバクテリウム菌は、予め導入を目的とする遺伝子、選択マーカー等を組込んで構築したベクターを導入したものであれば、特にその種を限定することなく使用できる。そして、感染液は、前記アグロバクテリウム菌を培地に接種して、28℃で24時間程度培養したものである。ここで導入を目的とする遺伝子、選択マーカーは、必要に応じて任意の遺伝子や選択マーカー、例えば耐寒性遺伝子、耐病性遺伝子、抗生物質耐性遺伝子等を選択することができる。
【0022】
(4)除菌培養
除菌培養は、共存培養が完了した外植体を、抗生物質メロペン(大日本住友製薬株式会社製)を含む固体培地に移植し、暗黒下、常温(例えば25℃)で1週間程度培養することによって行う。ここで、抗生物質としてメロペンを使用したのは他の抗生物質と比べてアグロバクテリウムを殺菌能力が高く、外植体への毒性が低いからである。なお、メロペンの濃度は、50mg/l程度が適当である。
【0023】
(5)形質転換体の選抜
除菌培養が完了した外植体から遺伝子が導入された不定胚の誘導および選抜は、例えば、導入したベクターに抗生物質耐性が組込まれている場合には、この外植体を前記抗生物質が添加された選抜培地に移植して、通常2月程度選抜培養行うことによって行う。この他にも、例えば、蛍光を発する遺伝子を利用して選抜することもできる。
【0024】
(6)形質転換体の作出
このようにして選抜された形質転換不定胚は、さらに前記抗生物質が添加された選抜培地で発芽させる。その後,発芽した不定胚を同抗生物質が添加されたシュート育成培地に移植し,形質転換シュートを育成する。このようにして得た形質転換シュートを発根培地に移植して、常温(例えば25℃)で2ヶ月間程度培養することによって発根処理を行ったのち、植木鉢などに収容された土壌に移植して、バラ科果樹の植物体とすることができる。
【0025】
以下にこの発明を実施例に従ってさらに詳しく説明するが、この発明の特許請求の範囲は如何なる意味においても下記の実施例により限定されるものではない。
【実施例1】
【0026】
未熟果実の採取時期及び培地が不定胚形成率に与える影響を調べた。具体的には、以下に示す方法により、「南高」の未熟子葉からの不定胚誘導率等を比較した。
【0027】
(1)外植体の採取
2005年4月7日、4月19日、4月27日、5月9日、5月19日に採取した未熟果実を採取して石鹸水でよく洗ったのち、70%エタノールに15分間浸漬して、クリーンベンチの中で滅菌蒸留水により3回すすいだ。そして、この果実をその中央線に沿って包丁により割って種子を摘出した。最後に、この種子から種皮を取り除き、2枚の子葉をそれぞれ縦に2つに切断して外植体を得た。
【0028】
(2)不定胚の誘導
このようにして得た外植体を、WP+ソルビトール培地又はMS+ショ糖培地に置床して、25℃暗所で培養し、不定胚形成率の違いを置床60日後に比較した。その結果を表1に示す。なお、かっこ内は不定胚誘導率を示す。
【0029】
なお、WP+ソルビトール培地は、WP基本培地(成分は表2を参照。)に2,4-ジクロロフェノキシ酢酸(以下、2,4-Dと省略する。)(1μM)、ベンジルアデニン(以下、BAと省略)(1μM)、寒天(0.7%)及びソルビトール3重量%を添加した培地であり、MS+ショ糖は、MS基本培地(成分は表2を参照。)に2,4-D(1μM)、BA(1μM)、寒天(0.7%)及びショ糖3重量%を添加した培地である。
【0030】
【表1】

【0031】
【表2】
【0032】
表1から明らかなように、5月上旬ごろ(5月9日)に採取した未熟果実をMS+ショ糖培地で培養したときが最も不定胚の形成率が高かった。
【実施例2】
【0033】
ウメの品種の違いが、不定胚の誘導効率に与える影響を調べた。具体的には、以下に示す方法により、異なる品種のウメ未熟子葉からの不定胚誘導率を比較した。
【0034】
(1)外植体の採取
2005年5月9日〜12日に、8品種のウメ、具体的には「南高」、「剣先」、「改良内田梅」、「八郎」、「小粒南高」、「白加賀」、「越の梅」、「紅さし」の未熟果実を採取して実施例1(1)と同様の方法により外植体を得た。
【0035】
(2)不定胚の誘導
このようにして得た外植体を不定胚誘導培地に置床し、25℃、暗黒下で50日間培養したのち、品種の違いによる不定胚形成率の違いを比較した。その結果を図1に示す。なお、不定胚誘導培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、ショ糖(3%)、寒天(0.7%)を添加して、pHをpH5.8に調整した培地である。
【0036】
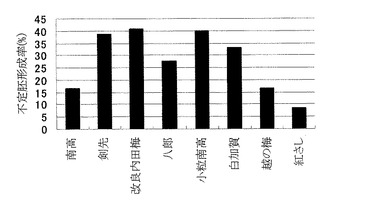
図1からも明らかなように、不定胚形成率は、品種によって違いがあり、「剣先」、「改良内田梅」、「小粒南高」、「白加賀」では高く、「南高」、「越の梅」、「紅さし」では低かった。特に、「改良内田梅」、「小粒南高」の不定胚形成率は約40%であり、従来法と比較して、高い形成率が得られた。また、目視による観察では、不定胚形成に要する時間も品種による違いがあり、「剣先」、「白加賀」、「小粒南高」では早く、「南高」や「越の梅」では遅かった。
【実施例3】
【0037】
共存培養後の除菌に使用する抗生物質の違いが、形質転換に与える影響を調べた。具体的には、「南高」の未熟種子を使用し、使用する抗生物質がメロペンである場合とセフォタキシンである場合の除菌効果の違いを以下に示す方法により比較した。
【0038】
(1)外植体の採取と共存培養
2005年4月28日に「南高」の未熟果実を採取して実施例1と同様の方法により外植体を得た。得られた外植体をアグロバクテリウム感染液に10分間浸漬した。外植体を浸漬している間に、共存培養培地に滅菌したろ紙を敷いた。感染後、外植体についた余分な感染液を滅菌ペーパータオルで吸い取り、ろ紙を敷いた共存培養培地に向軸側を下にして外植体を置床し、25℃、暗黒下で3日間共存培養を行った。なお、前記アグロバクテリウム感染液は次のようにして調製した。
【0039】
まず、感染の前日、523液体培地10mlにゲンタマイシン(20mg/l)とカナマイシン(50mg/l)を添加した液体培地に、pG-GUSプラスミドベクター(T-DNA上にイントロン−GUS(GUS-INT)遺伝子が挿入され、カナマイシン耐性遺伝子を組込んだバイナリーベクター)を保持するアグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)EHA101株を接種し、28℃で24時間振とう培養した。
【0040】
つぎに、感染当日、培養したアグロバクテリウムを4000rpmで15分間遠心して培地を除去し、沈殿した菌をMS誘導培地でOD420=0.5となるように菌濃度を調整したのち、28℃で5時間振とう培養して感染液とした。
【0041】
なお、523液体培地は、Yeast extract(4g/l)、Casein hydrolysate(8g/l)、K2HPO4(2g/l)、 MgSO4・7H20(0.31g/l)、ショ糖(10g/l)を含み、そのpHをpH7.1に調整した培地である。また、MS誘導培地は、MS基本培地にショ糖(20g/l)、プロリン(1mM)、アセトシリンゴン(0.1mM)を加えてpHをpH5.2に調製したのち、0.2μmのフィルターで濾過滅菌した培地である。
【0042】
(2)除菌培養
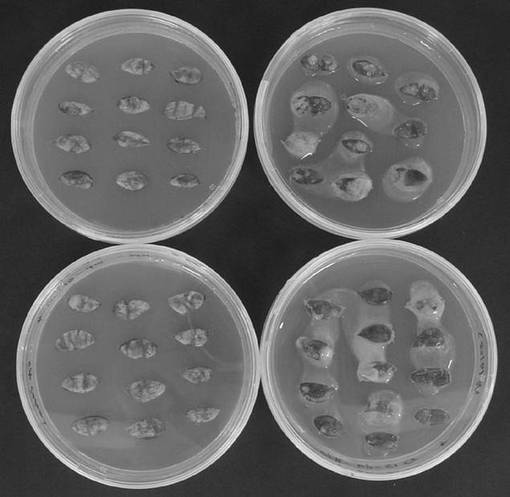
共存培養した子葉外植体を除菌培地に置床し、25℃、暗黒下で1週間(5月2日〜5月9日)1回目の除菌培養を行なったのち、選抜培地に移植して1月ほど(5月9日〜6月6日)選抜培養を行って、抗生物質の違いによる除菌効果の違いを比較した。その結果を図2に示す。
【0043】
なお、図2において、左列がメロペンを使用した区の培養写真であり、右列がセフォタキシンを利用した区の培養写真である。また、除菌培地は、MS基本培地にメロペン(50mg/l)又はセフォタキシン(300mg/l)、2,4-D(1μM)、BA(1μM)、Sucrose(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。また、選抜培地は各除菌培地にそれぞれカナマイシン(25mg/l)を添加した培地である。
【0044】
図2からも明らかなように、抗生物質としてメロペンを使用した場合には、アグロバクテリウム菌が目的どおり除菌されたのに対して、セフォタキシンを使用した場合にはアグロバクテリウム菌は除菌されていなかった。このことから、除菌の際に使用する抗生物質としてはメロペンが適切であることが確認できた。
【実施例4】
【0045】
前培養の有無、共存培養の期間が、アグロバクテリウム菌の感染率に与える影響を以下に示す方法により調べた。
【0046】
(1)形質転換体の作出
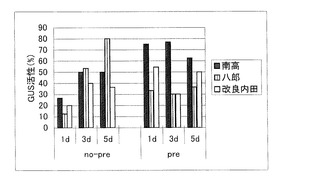
2005年5月10日に「南高」、「八郎」、「改良内田」の未熟果実を採取し、実施例1と同様の方法により外植体を得て、前培養の有無や共存培養の期間が異なる6つの区に振り分けた。前培養なし(no-pre)の区は、当日(5月10日)に、実施例3と同様の方法によりアグロバクテリウム菌を感染させ、1日〜3日間共存培養を行ったのち、7日間除菌培養を行ってGUS活性(X-Gluc法)を調べた。また、前培養あり(pre)の区は、不定胚誘導培地に置床して1週間(5月10日〜5月17日)培養したのち、同じく実施例3と同様の方法により、アグロバクテリウム菌感染させ、1日〜3日間共存培養を行ったのち、7日間除菌培養を行ってGUS活性(X-Gluc法)を調べた。具体的には、形質転換が認められた外植体の割合から形質転換効率を調べ、表3及び図3にまとめた。なお、表3中の値は活性の認められた外植体数/全外植体数×100(%)を表示してあり、図3は表3をグラフ化したものである。さらに、得られた外植体の写真を図4に示す。この写真から外植体中に変色した部分が存在し、除菌後の外植体には形質転換細胞が含まれていることが分かった。
【0047】
【表3】
【0048】
(2)発現の確認
なお、X-GLUC法による発現の確認は定法に従って次のようにして行った。まず、100μlのX-Gluc stock溶液を5mlのリン酸バーファーに加えた液体に、GUS活性を測定する植物組織を浸漬し、37℃で一晩インキュベーションした。インキュベーションののち、液体を捨て、植物組織を70%のエタノールで数回すすいだのち、目視により観察した。なお、X-Gluc stock溶液は25mgのx-gluc (5-Brome-4-chloro-3-indolyl-β-D-glucuronide Cyclohexylammonium Salt)を500μlのDMF (N, N-Dimethylformamide)に溶かしたものである。
【0049】
表3と図3から明らかなように、前培養の有無が感染率に及ぼす影響については明確な傾向を見出すことができなかった。また、共存培養期間が1日間では短すぎるようであったが、3日間と5日間では形質転換率に大きな差異がなく、5日間共存培養を行った外植体は衰弱しているものが多かった。これらのことを考慮すれば、不定胚の前培養にはあまり効果がなく、共存培養期間は3日間が適していると考えられる。
【実施例5】
【0050】
形質転換体の形成率を最大化することを目的に、形質転換に関わる各種条件を変えてその最適条件を検討した。
【0051】
(1)外植体の採取
2005年4月26日、5月10日、5月17日及び5月24日に、8品種のウメ、具体的には「南高」、「剣先」、「改良内田梅」、「八郎」、「小粒南高」、「白加賀」、「越の梅」、「紅さし」の未熟果実を採取して実施例1と同様の方法により外植体を得た。なお、得られた果実、子葉、外植体の一例を図5のそれぞれ左側、中央、右側に示す。
【0052】
(2)アグロバクテリウム菌の感染と共存培養
得られた外植体をA区、B区に分け、A区は採取日当日にアグロバクテリウム菌に感染させたのち、B区は7日間不定胚誘導培地上で培養したのちに感染させたのち、それぞれ3日間共存培養した。なお、感染に使用した菌株や感染方法、共存培養の方法は実施例3と同じである。
【0053】
(3)除菌培養
共存培養した外植体を除菌培地に置床し、25℃、暗黒下で1週間アグロバクテリウム菌の除菌を行った。なお、除菌培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【0054】
(4)不定胚の誘導
除菌された外植体を選抜培地1に置床して25℃、暗黒下で培養し、3〜4週間ごとに新しい培地に移植した。その結果、約2ヶ月後に不定胚の発生が見られた。このようにして発生した不定胚を選抜培地2に移植して、25℃、16時間日長で培養した結果、約1ヶ月後に不定胚を大量増殖できた。
【0055】
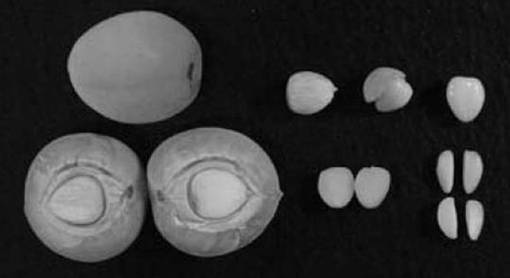
形成された不定胚を図6に示す。ここで図6の左側は「剣先」、中央は「小粒南高」、右側は「白加賀」にそれぞれ由来する不定胚の写真である。また、5月10日に採取した未熟子葉の不定胚の形成率を表4に、5月17日に採取した未熟子葉の不定胚の形成率を表5にそれぞれ示す。なお、4月26日に採取した外植体は実験期間中にすべて枯死し、5月24日に採取した外植体は不定胚を形成しなかった。そのため、これらのデータは省略した。
【0056】
【表4】
【0057】
【表5】
【0058】
表4と表5から、不定胚の形成率には品種間に差のあること、5月10日に採取した場合と5月17日に採取した場合では不定胚の形成率に大きく差があること、すなわち、果実の採取時期は不定胚の形成率に大きく影響すること、及び前培養は不定胚の形成率向上に影響しないこと、が分かった。
【0059】
なお、選抜培地1は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。また、選抜培地2は、MS基本培地にNAA(0.1μM)、BA(5μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ショ糖(3%)、Gellan Gum(0.2%)を添加し、pHをpH5.8に調整した培地である。
【0060】
(5)形質転換体の検出
選抜培地2に移植した約1ヶ月後に、外植体から発生した「白加賀」由来の4つの不定胚系統をサンプリングし、4-MUG法によるGUS活性の測定と、PCR反応による遺伝子増幅とを行なって、遺伝子の導入及び導入した遺伝子の働きを確認した。
【0061】
なお、4-MUG法によるGUS活性の測定は、定法に従って次のようにして行なった。まず、サンプルとなる不定胚と、石英砂と、200μlの抽出用緩衝液とを試験管に入れ、滅菌したガラス棒により氷中で不定胚を砕いて一様な溶液にした。つぎに、試験管を遠心分離機によって遠心し、上澄みを抽出液とした。そして、この抽出液50μlと、4-MUG(sigma)を抽出用緩衝液に4mMとなるように溶かした溶液250μlとを混ぜ合わせて混合液としたのち、37℃で20分間インキュベートした。その後、混合液にNa2CO3(0.2M)を含む抽出用緩衝液を添加して反応を停止し、その蛍光を比較した。その結果を図7に示す。ここで、図の左端のサンプルは遺伝子を導入していないネガティブコントロールの結果であり、その右側から右端にあるサンプルが「白加賀」由来するNo.1からNo.4の各形質転換系統の結果である。
【0062】
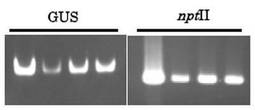
また、遺伝子導入は次のようにして確認した。まず、Nucleon extraction and purification kit(GEヘルスケアバイオサイエンス社製)よって、不定胚からゲノムDNAを抽出した。つぎに、Ex Taq(タカラバイオ株式会社製)を使用して、PCR反応を行った。最後に、PCR産物を電気泳動してPCR産物を比較した。その結果を図8に示す。なお、図の左端から右端に向かって、遺伝子導入に使用したプラスミドベクター(ポジティブコントロール)、「白加賀」由来する不定胚No.2からNo.4の各形質転換系統の結果を示している。なお、No.1は栽培中に枯死したため、遺伝子導入は確認できなかった。
【0063】
ここで、PCR反応のプライマーには、gusA Forward(配列表の配列番号1を参照)、gusA Reverse(配列表の配列番号2を参照)、nptII Forward(配列表の配列番号3を参照)、nptII Rewerse(配列表の配列番号4を参照)を使用した。また、PCR反応は、94℃、45秒の変性工程、60度、60秒のアーニリング工程、72℃、120秒の伸張工程からなる反応サイクルを30サイクル行った。
【0064】
図7及び図8からも明らかなように、カナマイシン耐性により選抜した4系統は、GUS活性が確認できるとともに、カナマイシン耐性遺伝子及びnptII遺伝子の導入が確認できた。これらの結果から、前記の4系統においては遺伝子が導入され、当該遺伝子が作動していることが確認できた。
【0065】
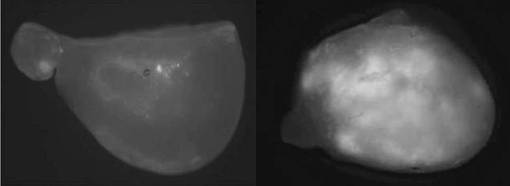
(6)形質転換体の育成
4-MUG法及びPCR法によって遺伝子の導入が確認できた不定胚を、シュート誘導培地に移植し、25℃、16時間日長で培養した。その結果、不定胚が発芽してシュートになった。その結果を図9に示す。なお、シュート誘導培地は、MS基本培地にBA(5μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ソルビトール(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【実施例6】
【0066】
超音波処理が、アグロバクテリウム菌の感染効率、形質転換効率に与える影響を以下に示すような方法により調べた。
【0067】
(1)外植体の採取
2006年5月17日に和歌山県暖地園芸センター植栽のウメ「南高」と「二青」の成木から採取した未熟果実、および同年5月31日に京都大学附属農場植栽の「小粒南高」の成木から採取した未熟果実から、実施例1と同様の方法により外植体を得た。
【0068】
(2)アグロバクテリウム菌の感染及び共存培養
得られた外植体を超音波処理区(SAAT)と対照区(control)の2つの区に分けた。そのうち、超音波処理区に属する外植体は、アグロバクテリウム感染液の入った試験管に入れ、この試験管を超音波洗浄機(柴田科学株式会社製、SU-27TH)にそれぞれ2分間(5月17日採取分)、10秒間、20秒間、30秒間(5月31日採取分)漬けることによって超音波処理し、そのままアグロバクテリウム感染液に10分間浸漬しておいた。また、対照区に属する外植体は、超音波処理せず、アグロバクテリウム感染液に10分間浸漬した。
【0069】
外植体を浸漬している間に、共存培養培地に滅菌したろ紙を敷いた。感染後、外植体についた余分な感染液を滅菌ペーパータオルで吸い取り、ろ紙を敷いた共存培養培地に向軸側を下にして外植体を置床し、25℃、暗黒下で3日間共存培養を行った。なお、前記アグロバクテリウム感染液は次のようにして調製した。
【0070】
まず、感染の前日、523液体培地10mlにゲンタマイシン(20mg/l)とカナマイシン(50mg/l)を添加した液体培地に、pDU92-sGFPプラスミドベクター(T-DNA上にGFP遺伝子が挿入され、カナマイシン耐性遺伝子を組込んだバイナリーベクター)を保持するアグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)EHA101株を接種し、28℃で24時間振とう培養した。
【0071】
つぎに、感染当日、培養したアグロバクテリウムを4000rpmで15分間遠心して培地を除去し、沈殿した菌をMS誘導培地でOD420=0.5となるように菌濃度を調整したのち、28℃で5時間振とう培養して感染液とした。
【0072】
(3)除菌培養
共存培養した外植体を除菌培地に置床し、25℃、暗黒下で1週間アグロバクテリウム菌の除菌を行った。なお、除菌培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【0073】
(4)選抜培養
また、除菌された外植体を選抜培地(Km 50)及び選抜培地(No Km)に置床して25℃、暗黒下で培養し、3〜4週間ごとに新しい培地に移植した。なお、選抜培地(Km 50)は除菌培地にカナマイシン(終濃度50mg/l)を添加した培地である。また、選抜培地(No Km)はカナマイシンを添加していない培地である。
【0074】
(5)GFP遺伝子の発現率及び不定胚再生率の調査
共存培養直後、除菌培養後、さらに不定胚誘導培養の1月後と2月後にGFP遺伝子の発現率とを調査し、不定胚誘導培養1月後と2月後には不定胚再生率を調査した。具体的には、外植体を載置した培養プレートごと、蛍光顕微鏡(オリンパス株式会社製、マクロズーム顕微鏡 MVX10)及び実体顕微鏡(オリンパス株式会社製、SZX12)により観察して、蛍光を発している外植体と不定胚を数えた。その結果を表6及び表7に示す。
【0075】
なお、表6は5月17日採取の未熟果実を使用した結果を示しており、表7は5月31日採取の未熟果実を使用した結果を示している。また、5月17日に採取した未熟子葉におけるGFP遺伝子の発現を蛍光顕微鏡で観察した結果を図10に示す。ここで、図10の左側は対照区(Control)の未熟子葉の写真であり、右側は超音波処理区(SAAT)の未熟子葉の写真である。
【0076】
【表6】
【0077】
【表7】
【0078】
表6から明らかなように、超音波処理により、除菌後の外植体においてGFP遺伝子の発現率が著しく増加している。一方で、超音波処理することにより、不定胚再生率は低下していた。このことは、2分間の超音波処理は、外植体に大きなダメージを与えていることを示唆している。
【0079】
また、表7から明らかなように、超音波処理の時間を表6の結果よりも短縮することによって、GFP遺伝子の発現率をそれほど低下させることなく、1月後及び2月後の不定胚形成率を向上できることが明らかになった。なかでも、長期的に不定胚再生率が維持されることを考慮すれば、超音波処理の時間を20秒間程度にすることが最も有効であることが分かった。
【0080】
さらに、選抜培地にカナマイシンを添加することにより、不定胚の再生率は低下したものの、GPF遺伝子の発現率が高まっていることから、形質転換効率を高めるため、選抜培地にカナマイシンを添加することが有効であることが分かった。
【図面の簡単な説明】
【0081】
【図1】ウメの品種の違いによる不定胚形成率の違いを示すグラフである。
【図2】抗生物質の違いによる除菌効果の違いを示す写真である。
【図3】前培養の有無、共存培養の期間が、アグロバクテリウムの感染率に与える影響を示すグラフである。
【図4】メロペンで除菌後にX-Gluc法によりGUSの一過性の発現を調べた結果を示す写真である。
【図5】採取された果実、子葉、外植体の一例を示す写真である。
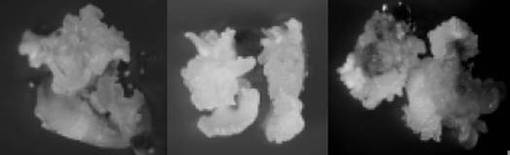
【図6】大量増殖した不定胚の写真である。左側は「剣先」、中央は「小粒南高」、右側は「白加賀」にそれぞれ由来する不定胚の写真である。
【図7】形質転換不定胚のGUS活性の測定結果である。
【図8】GUS活性を有する形質転換不定胚にGUS及びnptII遺伝子断片が含まれていることをPCR法により確認した結果を示す図である。
【図9】不定胚から発芽したシュートの写真である。
【図10】GFP遺伝子を持つアグロバクテリウムに感染させた未熟子葉におけるGFP遺伝子の発現を蛍光顕微鏡で観察した結果を示す写真である。ここで、図10の左側は対照区(Control)の未熟子葉を共存培養した後のGFP遺伝子の発現を示す写真であり、右側は超音波処理区(SAAT)の未熟子葉を共存培養した後のGFP遺伝子の発現を示す写真である。
【技術分野】
【0001】
この発明は、バラ科果樹、特にウメの形質転換法及びこの方法により作出された形質転換体などに関するものである。
【背景技術】
【0002】
ウメ等のバラ科果樹は、その実が食用となるため、従来から耐寒性や耐病性などの優れた性質を備えた品種が求められている。
【0003】
さて、これらバラ科果樹の繁殖は、品種の形質(特長)を安定させ、収穫までの期間を短縮するため、結実した種子から行う(種子繁殖)のではなく、接ぎ木(栄養繁殖)によって行うのが一般的である。なお、接ぎ木とは、有用形質を持った親の枝(穂木)を近縁種の台木に接着・再生させて新たな個体を得る繁殖方法である。
【0004】
ところで、接ぎ木においては、使用する台木により耐干性・耐水性・耐凍性等の各種形質が左右されることが一般的に知られており、バラ科果樹においても、台木の選択によって果実の生産性やその品質が大きく変化することが知られている。なお、現在使用されているウメ台木の多くは実生台木であり、遺伝的な性質が不揃いなため、優れた台木を見分けるのは難しい。
【0005】
そこで、以前から優れた性質を備えた台木を開発することが試みられてはいる。しかし、バラ科果樹類には、台木として利用できる大きさまで成長するのに長時間かかること、種子繁殖では品種の形質が安定しにくいことなどから、一般的な交雑育種を利用することは困難であった。そこで、従来から、育種を効率化することを目的として、組織培養や遺伝子導入などのバイオテクノロジー技術を利用した研究がなされている。
【0006】
ところで、バラ科果樹は、果樹の中ではゲノムサイズが小さく、また幼木期が短いため、遺伝解析が比較的容易である。しかし、バラ科果樹類の組織培養からの再分化は困難であり、これが形質転換を行う上での障壁となっている。
【0007】
このことはウメに関しても当てはまり、ウメについてはこれまで充分効率的な再分化系は確立されておらず、形質転換の成功例も報告されていない。なお、現在までに報告されているウメの再分化系としては、ウメ「紅さし」の未熟子葉からの再分化系が挙げられるが、この再分化系では未熟子葉上にいったんカルスを誘導させ、そのカルスを組成の異なる培地に植え替えることにより、不定芽を誘導させたものである(非特許文献1を参照。)。
【0008】
しかし、上記の再分化系は、組成の異なる培地に植え替えるため、不定芽を形成させるまでに時間と手間がかかり、カルス形成を経由しているため突然変異体が生じやすいという問題点があった。また、不定芽は不定胚とは異なり増殖能力を備えておらず、台木の生産には利用できないとの問題点もあった。
【0009】
また、同じバラ科果樹であるモモでは、成熟子葉を外植体として形質転換体が得られたとの報告がある(非特許文献2を参照。)。この報告に記載の方法は、アグロバクテリウム菌を感染した子葉上にカルスを誘導させ、このカルスから形質転換不定芽を誘導させたのち、形質転換シュートを形成させるものである。しかし、この方法はウメなどのように不定芽再分化効率が低い植物には、利用できないとの問題点があった。
【0010】
さらに、バラ科果樹に含まれるセイヨウナシでは、その腋芽の直近に遺伝子導入のための切り口を設けた培養シュートの茎からなる外植体を使用する形質転換法が研究されている(特許文献1)。ただ、この方法の形質転換効率は4〜5%であり、実用化するには形質転換効率が低すぎるとの問題点があった。
【特許文献1】特開2004-283051号公報
【非特許文献1】中川文雄ら 「ウメ未熟子葉由来カルスからの不定芽形成」 園芸学会雑誌 第63巻 別冊1 1994年 p88-89
【非特許文献2】Rosa M. et al., 2004. Molecular Breeding 14:419-427
【発明の開示】
【発明が解決しようとする課題】
【0011】
この発明は、ウメなどのバラ科果樹を容易かつ高い効率で形質転換することができる形質転換法、この方法によって形質転換された形質転換体、遺伝子組換えバラ科果樹などを提供することを課題とする。
【課題を解決するための手段】
【0012】
この発明は、アグロバクテリウム法による形質転換において、外植体の作成に使用する種子の採取時期、共存培養期間、除菌に使用する抗生物質などを特定することによって、バラ科果樹からの形質転換体の作出を可能にすることを最も主要な特徴とする。また、共存培養する前に超音波処理することによって、形質転換効率を高めることをもう一つの特徴とする。
【発明の効果】
【0013】
この発明により、バラ科果樹であるウメで、初めて形質転換体を作出することができた。これにより、優良な形質を備えた形質転換体、例えば耐病性や耐寒性に優れたウメを育種することができるようになり、優れた性質を備えた台木の育種が期待できるようになった。また、ウメと同じ科に属する、スモモ、モモ、サクラ、アンズ、アーモンドについても優れた形質転換体を容易に得ることが期待できるようになった。
【発明を実施するための最良の形態】
【0014】
この発明の形質転換法は、バラ科果樹の未熟果実から種子を摘出し、この種子から未熟子葉を取出して外植体を作成する工程と、この外植体をアグロバクテリウム菌が分散している感染液に浸漬したのち、外植体とアグロバクテリウム菌とを共存培養する工程と、アグロバクテリウム菌に感染した外植体を、抗生物質メロペンを含む培地で培養することによってアグロバクテリウム菌を除菌する工程とを含むバラ科果樹の形質転換法である。また、この発明の形質転換体はこの発明の方法によって得られたものである。さらに、この発明の遺伝子組換えバラ科果樹の作出法は、前記形質転換体により得られた培養シュートに発根処理を行なうものであり、この発明の遺伝子組換えバラ科果樹は前記作出法で作出したものである。そこで、この発明の形質転換法及び作出法などについて以下に詳説する。
【0015】
(1)形質転換法の対象となる果実
対象となる果実は、主にウメを想定しているが、同じバラ科果樹の果実であるスモモ、モモ、桜桃、アンズ、アーモンド等にも適用できると考えられる。なお、ウメの品種としては、市場において取引されている品種、例えば、「紅さし」、「越の梅」など通常栽培されている品種であれば特にこだわることなく使用できるが、穂木との親和性や果実の商品力等を考えると「南高」が好ましい。
【0016】
また、不定胚、形質転換体の作出に利用する果実は未熟果実であり、具体的には、開花後4月下旬から5月下旬の間に採取された未熟果実であり、好ましくは5月上旬から5月中旬以内、さらに好ましくは5月10日前後のものである。
【0017】
(2)外植体の作成
外植体の作成は、例えば、バラ科果樹の未熟果実をエタノールで滅菌し、滅菌したメス、ピンセット等で果実から種子を摘出して種皮を取り除き、二枚の子葉を縦に切断することによって行う。作成した外植体は、適当な培地(例えば、後述する共存培地など)上に置床して乾燥による劣化を防ぐ。
【0018】
(3)アグロバクテリウム菌の感染及び共存培養
アグロバクテリウム菌の外植体への感染は、外植体をアグロバクテリウム菌が分散した感染液に10分程度浸漬して余分な感染液を除去したのち、この外植体を適当な固体培地に置床して暗黒下、常温(例えば25℃)で共存培養することによって行う。共存培養の期間は、外植体が枯死しない範囲であれば特に限定する必要はないが、共存培養を始めてから3日間程度が適当である。これは1日〜2日間程度では充分感染せず、5日間では多くの外植体が衰弱するからである。
【0019】
なお、外植体がアグロバクテリウム菌の分散した感染液に浸漬した状態で超音波処理することによって、アグロバクテリウム菌の感染効率を向上することができる。超音波処理は、例えば、外植体と感染液が入った試験管を超音波洗浄機に入れて超音波を処理することによって行う。
【0020】
ここで、超音波処理は、超音波の周波数が略28kHz、処理時間が10秒〜2分間、好ましくは10秒〜30秒間、強度が略0.23W/cm2となるように行う。
【0021】
また、使用するアグロバクテリウム菌は、予め導入を目的とする遺伝子、選択マーカー等を組込んで構築したベクターを導入したものであれば、特にその種を限定することなく使用できる。そして、感染液は、前記アグロバクテリウム菌を培地に接種して、28℃で24時間程度培養したものである。ここで導入を目的とする遺伝子、選択マーカーは、必要に応じて任意の遺伝子や選択マーカー、例えば耐寒性遺伝子、耐病性遺伝子、抗生物質耐性遺伝子等を選択することができる。
【0022】
(4)除菌培養
除菌培養は、共存培養が完了した外植体を、抗生物質メロペン(大日本住友製薬株式会社製)を含む固体培地に移植し、暗黒下、常温(例えば25℃)で1週間程度培養することによって行う。ここで、抗生物質としてメロペンを使用したのは他の抗生物質と比べてアグロバクテリウムを殺菌能力が高く、外植体への毒性が低いからである。なお、メロペンの濃度は、50mg/l程度が適当である。
【0023】
(5)形質転換体の選抜
除菌培養が完了した外植体から遺伝子が導入された不定胚の誘導および選抜は、例えば、導入したベクターに抗生物質耐性が組込まれている場合には、この外植体を前記抗生物質が添加された選抜培地に移植して、通常2月程度選抜培養行うことによって行う。この他にも、例えば、蛍光を発する遺伝子を利用して選抜することもできる。
【0024】
(6)形質転換体の作出
このようにして選抜された形質転換不定胚は、さらに前記抗生物質が添加された選抜培地で発芽させる。その後,発芽した不定胚を同抗生物質が添加されたシュート育成培地に移植し,形質転換シュートを育成する。このようにして得た形質転換シュートを発根培地に移植して、常温(例えば25℃)で2ヶ月間程度培養することによって発根処理を行ったのち、植木鉢などに収容された土壌に移植して、バラ科果樹の植物体とすることができる。
【0025】
以下にこの発明を実施例に従ってさらに詳しく説明するが、この発明の特許請求の範囲は如何なる意味においても下記の実施例により限定されるものではない。
【実施例1】
【0026】
未熟果実の採取時期及び培地が不定胚形成率に与える影響を調べた。具体的には、以下に示す方法により、「南高」の未熟子葉からの不定胚誘導率等を比較した。
【0027】
(1)外植体の採取
2005年4月7日、4月19日、4月27日、5月9日、5月19日に採取した未熟果実を採取して石鹸水でよく洗ったのち、70%エタノールに15分間浸漬して、クリーンベンチの中で滅菌蒸留水により3回すすいだ。そして、この果実をその中央線に沿って包丁により割って種子を摘出した。最後に、この種子から種皮を取り除き、2枚の子葉をそれぞれ縦に2つに切断して外植体を得た。
【0028】
(2)不定胚の誘導
このようにして得た外植体を、WP+ソルビトール培地又はMS+ショ糖培地に置床して、25℃暗所で培養し、不定胚形成率の違いを置床60日後に比較した。その結果を表1に示す。なお、かっこ内は不定胚誘導率を示す。
【0029】
なお、WP+ソルビトール培地は、WP基本培地(成分は表2を参照。)に2,4-ジクロロフェノキシ酢酸(以下、2,4-Dと省略する。)(1μM)、ベンジルアデニン(以下、BAと省略)(1μM)、寒天(0.7%)及びソルビトール3重量%を添加した培地であり、MS+ショ糖は、MS基本培地(成分は表2を参照。)に2,4-D(1μM)、BA(1μM)、寒天(0.7%)及びショ糖3重量%を添加した培地である。
【0030】
【表1】
【0031】
【表2】
【0032】
表1から明らかなように、5月上旬ごろ(5月9日)に採取した未熟果実をMS+ショ糖培地で培養したときが最も不定胚の形成率が高かった。
【実施例2】
【0033】
ウメの品種の違いが、不定胚の誘導効率に与える影響を調べた。具体的には、以下に示す方法により、異なる品種のウメ未熟子葉からの不定胚誘導率を比較した。
【0034】
(1)外植体の採取
2005年5月9日〜12日に、8品種のウメ、具体的には「南高」、「剣先」、「改良内田梅」、「八郎」、「小粒南高」、「白加賀」、「越の梅」、「紅さし」の未熟果実を採取して実施例1(1)と同様の方法により外植体を得た。
【0035】
(2)不定胚の誘導
このようにして得た外植体を不定胚誘導培地に置床し、25℃、暗黒下で50日間培養したのち、品種の違いによる不定胚形成率の違いを比較した。その結果を図1に示す。なお、不定胚誘導培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、ショ糖(3%)、寒天(0.7%)を添加して、pHをpH5.8に調整した培地である。
【0036】
図1からも明らかなように、不定胚形成率は、品種によって違いがあり、「剣先」、「改良内田梅」、「小粒南高」、「白加賀」では高く、「南高」、「越の梅」、「紅さし」では低かった。特に、「改良内田梅」、「小粒南高」の不定胚形成率は約40%であり、従来法と比較して、高い形成率が得られた。また、目視による観察では、不定胚形成に要する時間も品種による違いがあり、「剣先」、「白加賀」、「小粒南高」では早く、「南高」や「越の梅」では遅かった。
【実施例3】
【0037】
共存培養後の除菌に使用する抗生物質の違いが、形質転換に与える影響を調べた。具体的には、「南高」の未熟種子を使用し、使用する抗生物質がメロペンである場合とセフォタキシンである場合の除菌効果の違いを以下に示す方法により比較した。
【0038】
(1)外植体の採取と共存培養
2005年4月28日に「南高」の未熟果実を採取して実施例1と同様の方法により外植体を得た。得られた外植体をアグロバクテリウム感染液に10分間浸漬した。外植体を浸漬している間に、共存培養培地に滅菌したろ紙を敷いた。感染後、外植体についた余分な感染液を滅菌ペーパータオルで吸い取り、ろ紙を敷いた共存培養培地に向軸側を下にして外植体を置床し、25℃、暗黒下で3日間共存培養を行った。なお、前記アグロバクテリウム感染液は次のようにして調製した。
【0039】
まず、感染の前日、523液体培地10mlにゲンタマイシン(20mg/l)とカナマイシン(50mg/l)を添加した液体培地に、pG-GUSプラスミドベクター(T-DNA上にイントロン−GUS(GUS-INT)遺伝子が挿入され、カナマイシン耐性遺伝子を組込んだバイナリーベクター)を保持するアグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)EHA101株を接種し、28℃で24時間振とう培養した。
【0040】
つぎに、感染当日、培養したアグロバクテリウムを4000rpmで15分間遠心して培地を除去し、沈殿した菌をMS誘導培地でOD420=0.5となるように菌濃度を調整したのち、28℃で5時間振とう培養して感染液とした。
【0041】
なお、523液体培地は、Yeast extract(4g/l)、Casein hydrolysate(8g/l)、K2HPO4(2g/l)、 MgSO4・7H20(0.31g/l)、ショ糖(10g/l)を含み、そのpHをpH7.1に調整した培地である。また、MS誘導培地は、MS基本培地にショ糖(20g/l)、プロリン(1mM)、アセトシリンゴン(0.1mM)を加えてpHをpH5.2に調製したのち、0.2μmのフィルターで濾過滅菌した培地である。
【0042】
(2)除菌培養
共存培養した子葉外植体を除菌培地に置床し、25℃、暗黒下で1週間(5月2日〜5月9日)1回目の除菌培養を行なったのち、選抜培地に移植して1月ほど(5月9日〜6月6日)選抜培養を行って、抗生物質の違いによる除菌効果の違いを比較した。その結果を図2に示す。
【0043】
なお、図2において、左列がメロペンを使用した区の培養写真であり、右列がセフォタキシンを利用した区の培養写真である。また、除菌培地は、MS基本培地にメロペン(50mg/l)又はセフォタキシン(300mg/l)、2,4-D(1μM)、BA(1μM)、Sucrose(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。また、選抜培地は各除菌培地にそれぞれカナマイシン(25mg/l)を添加した培地である。
【0044】
図2からも明らかなように、抗生物質としてメロペンを使用した場合には、アグロバクテリウム菌が目的どおり除菌されたのに対して、セフォタキシンを使用した場合にはアグロバクテリウム菌は除菌されていなかった。このことから、除菌の際に使用する抗生物質としてはメロペンが適切であることが確認できた。
【実施例4】
【0045】
前培養の有無、共存培養の期間が、アグロバクテリウム菌の感染率に与える影響を以下に示す方法により調べた。
【0046】
(1)形質転換体の作出
2005年5月10日に「南高」、「八郎」、「改良内田」の未熟果実を採取し、実施例1と同様の方法により外植体を得て、前培養の有無や共存培養の期間が異なる6つの区に振り分けた。前培養なし(no-pre)の区は、当日(5月10日)に、実施例3と同様の方法によりアグロバクテリウム菌を感染させ、1日〜3日間共存培養を行ったのち、7日間除菌培養を行ってGUS活性(X-Gluc法)を調べた。また、前培養あり(pre)の区は、不定胚誘導培地に置床して1週間(5月10日〜5月17日)培養したのち、同じく実施例3と同様の方法により、アグロバクテリウム菌感染させ、1日〜3日間共存培養を行ったのち、7日間除菌培養を行ってGUS活性(X-Gluc法)を調べた。具体的には、形質転換が認められた外植体の割合から形質転換効率を調べ、表3及び図3にまとめた。なお、表3中の値は活性の認められた外植体数/全外植体数×100(%)を表示してあり、図3は表3をグラフ化したものである。さらに、得られた外植体の写真を図4に示す。この写真から外植体中に変色した部分が存在し、除菌後の外植体には形質転換細胞が含まれていることが分かった。
【0047】
【表3】
【0048】
(2)発現の確認
なお、X-GLUC法による発現の確認は定法に従って次のようにして行った。まず、100μlのX-Gluc stock溶液を5mlのリン酸バーファーに加えた液体に、GUS活性を測定する植物組織を浸漬し、37℃で一晩インキュベーションした。インキュベーションののち、液体を捨て、植物組織を70%のエタノールで数回すすいだのち、目視により観察した。なお、X-Gluc stock溶液は25mgのx-gluc (5-Brome-4-chloro-3-indolyl-β-D-glucuronide Cyclohexylammonium Salt)を500μlのDMF (N, N-Dimethylformamide)に溶かしたものである。
【0049】
表3と図3から明らかなように、前培養の有無が感染率に及ぼす影響については明確な傾向を見出すことができなかった。また、共存培養期間が1日間では短すぎるようであったが、3日間と5日間では形質転換率に大きな差異がなく、5日間共存培養を行った外植体は衰弱しているものが多かった。これらのことを考慮すれば、不定胚の前培養にはあまり効果がなく、共存培養期間は3日間が適していると考えられる。
【実施例5】
【0050】
形質転換体の形成率を最大化することを目的に、形質転換に関わる各種条件を変えてその最適条件を検討した。
【0051】
(1)外植体の採取
2005年4月26日、5月10日、5月17日及び5月24日に、8品種のウメ、具体的には「南高」、「剣先」、「改良内田梅」、「八郎」、「小粒南高」、「白加賀」、「越の梅」、「紅さし」の未熟果実を採取して実施例1と同様の方法により外植体を得た。なお、得られた果実、子葉、外植体の一例を図5のそれぞれ左側、中央、右側に示す。
【0052】
(2)アグロバクテリウム菌の感染と共存培養
得られた外植体をA区、B区に分け、A区は採取日当日にアグロバクテリウム菌に感染させたのち、B区は7日間不定胚誘導培地上で培養したのちに感染させたのち、それぞれ3日間共存培養した。なお、感染に使用した菌株や感染方法、共存培養の方法は実施例3と同じである。
【0053】
(3)除菌培養
共存培養した外植体を除菌培地に置床し、25℃、暗黒下で1週間アグロバクテリウム菌の除菌を行った。なお、除菌培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【0054】
(4)不定胚の誘導
除菌された外植体を選抜培地1に置床して25℃、暗黒下で培養し、3〜4週間ごとに新しい培地に移植した。その結果、約2ヶ月後に不定胚の発生が見られた。このようにして発生した不定胚を選抜培地2に移植して、25℃、16時間日長で培養した結果、約1ヶ月後に不定胚を大量増殖できた。
【0055】
形成された不定胚を図6に示す。ここで図6の左側は「剣先」、中央は「小粒南高」、右側は「白加賀」にそれぞれ由来する不定胚の写真である。また、5月10日に採取した未熟子葉の不定胚の形成率を表4に、5月17日に採取した未熟子葉の不定胚の形成率を表5にそれぞれ示す。なお、4月26日に採取した外植体は実験期間中にすべて枯死し、5月24日に採取した外植体は不定胚を形成しなかった。そのため、これらのデータは省略した。
【0056】
【表4】
【0057】
【表5】
【0058】
表4と表5から、不定胚の形成率には品種間に差のあること、5月10日に採取した場合と5月17日に採取した場合では不定胚の形成率に大きく差があること、すなわち、果実の採取時期は不定胚の形成率に大きく影響すること、及び前培養は不定胚の形成率向上に影響しないこと、が分かった。
【0059】
なお、選抜培地1は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。また、選抜培地2は、MS基本培地にNAA(0.1μM)、BA(5μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ショ糖(3%)、Gellan Gum(0.2%)を添加し、pHをpH5.8に調整した培地である。
【0060】
(5)形質転換体の検出
選抜培地2に移植した約1ヶ月後に、外植体から発生した「白加賀」由来の4つの不定胚系統をサンプリングし、4-MUG法によるGUS活性の測定と、PCR反応による遺伝子増幅とを行なって、遺伝子の導入及び導入した遺伝子の働きを確認した。
【0061】
なお、4-MUG法によるGUS活性の測定は、定法に従って次のようにして行なった。まず、サンプルとなる不定胚と、石英砂と、200μlの抽出用緩衝液とを試験管に入れ、滅菌したガラス棒により氷中で不定胚を砕いて一様な溶液にした。つぎに、試験管を遠心分離機によって遠心し、上澄みを抽出液とした。そして、この抽出液50μlと、4-MUG(sigma)を抽出用緩衝液に4mMとなるように溶かした溶液250μlとを混ぜ合わせて混合液としたのち、37℃で20分間インキュベートした。その後、混合液にNa2CO3(0.2M)を含む抽出用緩衝液を添加して反応を停止し、その蛍光を比較した。その結果を図7に示す。ここで、図の左端のサンプルは遺伝子を導入していないネガティブコントロールの結果であり、その右側から右端にあるサンプルが「白加賀」由来するNo.1からNo.4の各形質転換系統の結果である。
【0062】
また、遺伝子導入は次のようにして確認した。まず、Nucleon extraction and purification kit(GEヘルスケアバイオサイエンス社製)よって、不定胚からゲノムDNAを抽出した。つぎに、Ex Taq(タカラバイオ株式会社製)を使用して、PCR反応を行った。最後に、PCR産物を電気泳動してPCR産物を比較した。その結果を図8に示す。なお、図の左端から右端に向かって、遺伝子導入に使用したプラスミドベクター(ポジティブコントロール)、「白加賀」由来する不定胚No.2からNo.4の各形質転換系統の結果を示している。なお、No.1は栽培中に枯死したため、遺伝子導入は確認できなかった。
【0063】
ここで、PCR反応のプライマーには、gusA Forward(配列表の配列番号1を参照)、gusA Reverse(配列表の配列番号2を参照)、nptII Forward(配列表の配列番号3を参照)、nptII Rewerse(配列表の配列番号4を参照)を使用した。また、PCR反応は、94℃、45秒の変性工程、60度、60秒のアーニリング工程、72℃、120秒の伸張工程からなる反応サイクルを30サイクル行った。
【0064】
図7及び図8からも明らかなように、カナマイシン耐性により選抜した4系統は、GUS活性が確認できるとともに、カナマイシン耐性遺伝子及びnptII遺伝子の導入が確認できた。これらの結果から、前記の4系統においては遺伝子が導入され、当該遺伝子が作動していることが確認できた。
【0065】
(6)形質転換体の育成
4-MUG法及びPCR法によって遺伝子の導入が確認できた不定胚を、シュート誘導培地に移植し、25℃、16時間日長で培養した。その結果、不定胚が発芽してシュートになった。その結果を図9に示す。なお、シュート誘導培地は、MS基本培地にBA(5μM)、メロペン(50mg/l)、カナマイシン(25mg/l)、ソルビトール(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【実施例6】
【0066】
超音波処理が、アグロバクテリウム菌の感染効率、形質転換効率に与える影響を以下に示すような方法により調べた。
【0067】
(1)外植体の採取
2006年5月17日に和歌山県暖地園芸センター植栽のウメ「南高」と「二青」の成木から採取した未熟果実、および同年5月31日に京都大学附属農場植栽の「小粒南高」の成木から採取した未熟果実から、実施例1と同様の方法により外植体を得た。
【0068】
(2)アグロバクテリウム菌の感染及び共存培養
得られた外植体を超音波処理区(SAAT)と対照区(control)の2つの区に分けた。そのうち、超音波処理区に属する外植体は、アグロバクテリウム感染液の入った試験管に入れ、この試験管を超音波洗浄機(柴田科学株式会社製、SU-27TH)にそれぞれ2分間(5月17日採取分)、10秒間、20秒間、30秒間(5月31日採取分)漬けることによって超音波処理し、そのままアグロバクテリウム感染液に10分間浸漬しておいた。また、対照区に属する外植体は、超音波処理せず、アグロバクテリウム感染液に10分間浸漬した。
【0069】
外植体を浸漬している間に、共存培養培地に滅菌したろ紙を敷いた。感染後、外植体についた余分な感染液を滅菌ペーパータオルで吸い取り、ろ紙を敷いた共存培養培地に向軸側を下にして外植体を置床し、25℃、暗黒下で3日間共存培養を行った。なお、前記アグロバクテリウム感染液は次のようにして調製した。
【0070】
まず、感染の前日、523液体培地10mlにゲンタマイシン(20mg/l)とカナマイシン(50mg/l)を添加した液体培地に、pDU92-sGFPプラスミドベクター(T-DNA上にGFP遺伝子が挿入され、カナマイシン耐性遺伝子を組込んだバイナリーベクター)を保持するアグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)EHA101株を接種し、28℃で24時間振とう培養した。
【0071】
つぎに、感染当日、培養したアグロバクテリウムを4000rpmで15分間遠心して培地を除去し、沈殿した菌をMS誘導培地でOD420=0.5となるように菌濃度を調整したのち、28℃で5時間振とう培養して感染液とした。
【0072】
(3)除菌培養
共存培養した外植体を除菌培地に置床し、25℃、暗黒下で1週間アグロバクテリウム菌の除菌を行った。なお、除菌培地は、MS基本培地に2,4-D(1μM)、BA(1μM)、メロペン(50mg/l)、ショ糖(3%)、寒天(0.7%)を添加し、pHをpH5.8に調整した培地である。
【0073】
(4)選抜培養
また、除菌された外植体を選抜培地(Km 50)及び選抜培地(No Km)に置床して25℃、暗黒下で培養し、3〜4週間ごとに新しい培地に移植した。なお、選抜培地(Km 50)は除菌培地にカナマイシン(終濃度50mg/l)を添加した培地である。また、選抜培地(No Km)はカナマイシンを添加していない培地である。
【0074】
(5)GFP遺伝子の発現率及び不定胚再生率の調査
共存培養直後、除菌培養後、さらに不定胚誘導培養の1月後と2月後にGFP遺伝子の発現率とを調査し、不定胚誘導培養1月後と2月後には不定胚再生率を調査した。具体的には、外植体を載置した培養プレートごと、蛍光顕微鏡(オリンパス株式会社製、マクロズーム顕微鏡 MVX10)及び実体顕微鏡(オリンパス株式会社製、SZX12)により観察して、蛍光を発している外植体と不定胚を数えた。その結果を表6及び表7に示す。
【0075】
なお、表6は5月17日採取の未熟果実を使用した結果を示しており、表7は5月31日採取の未熟果実を使用した結果を示している。また、5月17日に採取した未熟子葉におけるGFP遺伝子の発現を蛍光顕微鏡で観察した結果を図10に示す。ここで、図10の左側は対照区(Control)の未熟子葉の写真であり、右側は超音波処理区(SAAT)の未熟子葉の写真である。
【0076】
【表6】
【0077】
【表7】
【0078】
表6から明らかなように、超音波処理により、除菌後の外植体においてGFP遺伝子の発現率が著しく増加している。一方で、超音波処理することにより、不定胚再生率は低下していた。このことは、2分間の超音波処理は、外植体に大きなダメージを与えていることを示唆している。
【0079】
また、表7から明らかなように、超音波処理の時間を表6の結果よりも短縮することによって、GFP遺伝子の発現率をそれほど低下させることなく、1月後及び2月後の不定胚形成率を向上できることが明らかになった。なかでも、長期的に不定胚再生率が維持されることを考慮すれば、超音波処理の時間を20秒間程度にすることが最も有効であることが分かった。
【0080】
さらに、選抜培地にカナマイシンを添加することにより、不定胚の再生率は低下したものの、GPF遺伝子の発現率が高まっていることから、形質転換効率を高めるため、選抜培地にカナマイシンを添加することが有効であることが分かった。
【図面の簡単な説明】
【0081】
【図1】ウメの品種の違いによる不定胚形成率の違いを示すグラフである。
【図2】抗生物質の違いによる除菌効果の違いを示す写真である。
【図3】前培養の有無、共存培養の期間が、アグロバクテリウムの感染率に与える影響を示すグラフである。
【図4】メロペンで除菌後にX-Gluc法によりGUSの一過性の発現を調べた結果を示す写真である。
【図5】採取された果実、子葉、外植体の一例を示す写真である。
【図6】大量増殖した不定胚の写真である。左側は「剣先」、中央は「小粒南高」、右側は「白加賀」にそれぞれ由来する不定胚の写真である。
【図7】形質転換不定胚のGUS活性の測定結果である。
【図8】GUS活性を有する形質転換不定胚にGUS及びnptII遺伝子断片が含まれていることをPCR法により確認した結果を示す図である。
【図9】不定胚から発芽したシュートの写真である。
【図10】GFP遺伝子を持つアグロバクテリウムに感染させた未熟子葉におけるGFP遺伝子の発現を蛍光顕微鏡で観察した結果を示す写真である。ここで、図10の左側は対照区(Control)の未熟子葉を共存培養した後のGFP遺伝子の発現を示す写真であり、右側は超音波処理区(SAAT)の未熟子葉を共存培養した後のGFP遺伝子の発現を示す写真である。
【特許請求の範囲】
【請求項1】
バラ科果樹の未熟果実から種子を摘出し、この種子から未熟子葉を取出して外植体を作成する工程と、
この外植体をアグロバクテリウム菌が分散している感染液に浸漬したのち、外植体とアグロバクテリウム菌とを共存培養する工程と、
アグロバクテリウム菌に感染した外植体を、抗生物質メロペンを含む培地で培養することによってアグロバクテリウム菌を除菌する工程と、
を含むバラ科果樹の形質転換法。
【請求項2】
アグロバクテリウム菌が分散している感染液に外植体を浸漬した状態で超音波処理したのち、外植体とアグロバクテリウム菌とを共存培養する請求項1に記載のバラ科果樹の形質転換法。
【請求項3】
バラ科果樹がウメである請求項1に記載のバラ科果樹の形質転換法。
【請求項4】
5月初旬から中旬の間に採取された未熟果実から外植体を作成する請求項1に記載のバラ科果樹の形質転換法。
【請求項5】
共存培養期間が3日間である請求項1に記載のバラ科果樹の形質転換法。
【請求項6】
請求項1から請求項5の何れかに記載のバラ科果樹の形質転換法により形質転換されたバラ科果樹の形質転換体。
【請求項7】
請求項1から請求項5の何れかに記載のバラ科果樹の形質転換法により形質転換して得られたバラ科果樹の培養シュートに、発根処理を行なう遺伝子組換えバラ科果樹の作出法。
【請求項8】
請求項7に記載の遺伝子組換えバラ科果樹の作出法により作出された遺伝子組換えバラ科果樹。
【請求項1】
バラ科果樹の未熟果実から種子を摘出し、この種子から未熟子葉を取出して外植体を作成する工程と、
この外植体をアグロバクテリウム菌が分散している感染液に浸漬したのち、外植体とアグロバクテリウム菌とを共存培養する工程と、
アグロバクテリウム菌に感染した外植体を、抗生物質メロペンを含む培地で培養することによってアグロバクテリウム菌を除菌する工程と、
を含むバラ科果樹の形質転換法。
【請求項2】
アグロバクテリウム菌が分散している感染液に外植体を浸漬した状態で超音波処理したのち、外植体とアグロバクテリウム菌とを共存培養する請求項1に記載のバラ科果樹の形質転換法。
【請求項3】
バラ科果樹がウメである請求項1に記載のバラ科果樹の形質転換法。
【請求項4】
5月初旬から中旬の間に採取された未熟果実から外植体を作成する請求項1に記載のバラ科果樹の形質転換法。
【請求項5】
共存培養期間が3日間である請求項1に記載のバラ科果樹の形質転換法。
【請求項6】
請求項1から請求項5の何れかに記載のバラ科果樹の形質転換法により形質転換されたバラ科果樹の形質転換体。
【請求項7】
請求項1から請求項5の何れかに記載のバラ科果樹の形質転換法により形質転換して得られたバラ科果樹の培養シュートに、発根処理を行なう遺伝子組換えバラ科果樹の作出法。
【請求項8】
請求項7に記載の遺伝子組換えバラ科果樹の作出法により作出された遺伝子組換えバラ科果樹。
【図1】

【図3】
【図2】
【図4】

【図5】
【図6】
【図7】

【図8】
【図9】

【図10】
【図3】
【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2007−289150(P2007−289150A)
【公開日】平成19年11月8日(2007.11.8)
【国際特許分類】
【出願番号】特願2007−58198(P2007−58198)
【出願日】平成19年3月8日(2007.3.8)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2005年10月1日 園芸学会発行の「園芸学会雑誌 第74巻別冊2−2005−(園芸学会平成17年度秋季大会研究発表)」に発表
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成19年11月8日(2007.11.8)
【国際特許分類】
【出願日】平成19年3月8日(2007.3.8)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2005年10月1日 園芸学会発行の「園芸学会雑誌 第74巻別冊2−2005−(園芸学会平成17年度秋季大会研究発表)」に発表
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]