
パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体又は可溶性Nタンパク質の調製及びそれらのワクチンとしての使用
本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法、このように調製した複合体、及び当該複合体から単離し得る可溶性Nタンパク質に関する。本発明はさらに、このパラミクソウイルス科のNタンパク質−トランケート型Pタンパク質複合体又はNタンパク質を含むワクチン組成物に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法、こうして調製した複合体、さらにこの複合体から単離することが可能な可溶性Nタンパク質に関する。また、本発明はパラミクソウイルス科のこのNタンパク質−トランケート型Pタンパク質複合体又はNタンパク質を含むワクチン組成物に関する。好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片である。
【背景技術】
【0002】
フランスでは、大部分の国と同様、ウシの呼吸器合胞体ウイルス(RSV)が、70%以上の農場内の仔牛における、また約70%の生後一年までの仔牛における重い呼吸器疾患(細気管支炎、肺炎)の主要な原因となる因子となっている(Perrin et al., 1979、 Ames, 1993、 Elvander, 1996)。死亡率は20%に達する(Wellemans, 1992)。これは、毎年冬のこの疾患の定期的な発生を止める術がない飼育業者にとって大きな悩みの種である。したがって、有効な予防法の開発という課題に対して牧畜業者の強い要求がある。ワクチンは市販されているが、実際、効果は少ないか、または効果が見られない。
【0003】
ヒトにおいても同じ疾患が存在する。それは新生児の細気管支炎の原因となる因子である(Virologie、第7巻、2003年10月特別号、infections a virus respiratoire syncytial en pediatrie参照)。このヒトの疾患についてのデータは極めて詳細である。ウシと同様、フランスでは、ヒトにおいて、それは冬に流行する疾患である。生後一年の間に、乳児の70%がRSVに感染し、生後二年の間に100%が感染する。これは、フランスでは毎年500,000人の小児が感染していることを意味する。感染した乳児の20%で下気道の疾患(細気管支炎)が生じ、フランスにおいては入院率が2〜5%であり、それは8〜9日間にわたり、未熟児の10〜25%、肺異形成の場合では14〜45%、先天性心異常の場合では15〜25%で下気道の疾患が生じる。入院患者の約10%が集中治療室で治療されなければならない。死亡率は0.1%に近い。世界保健機関の最近の報告によれば、毎年世界で6400万人が感染し160,000人がRSVにより死亡すると推定されている。アメリカでは毎年18,000〜75,000人が入院し、2,000人近くが直接的に、17,000人が間接的に死亡している(Magon and Barik,2004)。RSVはまた、成人におけるインフルエンザ症状の原因の3分の1を占める。実際、RSVにより小児の4倍の高齢者が死亡すると推定されている。全体では、RSVにより年間300万〜500万人が死亡する。さらに、極めて若い年齢(生後4か月前)で罹患した成人において、まだ明らかでない複雑な免疫機構により、この疾患はぜんそくを起こすリスクを増加させると考えられる。
【0004】
たとえヒトRSV及びウシRSVが異なる二種のウイルスであるとしても、構造的、分子的、抗原的側面、またそれらが誘発する疾患(臨床像、感染の季節性、若年者の感染)が等しいこと、そして、非常に高価であるため稀にしか使用されないモノクローナル抗体と、効果が弱く細胞内で実際にATPを消費してしまい毒性が強い抗ウイルス剤の他には有効なワクチンも抗ウイルス剤も存在しないという事実から、それらは極めて近いものである。
【0005】
1960年代に、ホルマリンで固定したウイルスを用いた、幼い小児に対するワクチン接種の試験がアメリカで行われた。結果は悲惨なものであり、ワクチン接種を受けた小児は数ヵ月後のウイルスの自然感染の際に激しい症状を発し、しかもそのうちの数人は死亡する結果となった。
【0006】
この不幸な試験の後、この疾患に対するヒト用のワクチンは未だ存在しておらず、また、たとえフランスにおいてこの課題について本格的な研究がなされていなくても、ウシ用の市販ワクチンの効果は極めて疑わしい。しかし、複数のワクチンが、ウシにおけるワクチン接種後の事故の後に市場からなくなった。
【0007】
RSVに対するワクチンの開発の困難性は、次のような理由により説明することができる。
(1)RSVに対する局所的免疫記憶が短期的であるため、自然感染は再感染に対する防御免疫を付与しないこと、
(2)強い細胞性免疫及び液性免疫の誘導が、疾患の重篤さの増大と関連すること(ワクチン接種による感作)、
(3)経粘膜投与で効果的なワクチン接種を可能にする、アジュバント又はワクチン戦略が存在しないこと、
(4)新生児(生後2ヵ月以内)がワクチン接種の主要な対象であるが、彼らの免疫系は従来のワクチンに対する反応が悪いこと、
(5)感染を防がない移行抗体が血液中に存在すること、
(6)流行しているウイルス株が変化すること。
【0008】
サブユニットワクチンが開発されつつあり、そのいくつかはG表面タンパク質に基づいている。新しいタイプのワクチンは逆遺伝学によって(すなわち遺伝的修飾による株の弱毒化によって)得られ、BRSVと同様HRSVに対しても数年前から開発されている。目下のところ、ヒトに対する弱毒株の使用は確認されていない。これはおそらく弱毒株の免疫応答が弱いためであろう。
【0009】
仔牛に対するものも含めて、HRSV又はBRSVのF膜タンパク質及びG膜タンパク質に基づくDNAワクチンが開発中である(Taylor et al. 2005、 Martinez et al. 1999)。このタイプのワクチンはいくつかの免疫病理学的研究に関連する防御免疫を付与する。このワクチンの効果は改良の余地がある。
【0010】
RSVは、マイナス鎖の一本鎖RNAのゲノムを有するエンベロープウイルスである。このRNAは11個のタンパク質をコードし、ヌクレオカプシド(N)タンパク質によりカプシドに包まれ、L(ラージ)つまりポリメラーゼとコファクターP(リンタンパク質)の二つのタンパク質から成る、ポリメラーゼ複合体と関連する(図1)。この分子は、PによりLがゲノムRNAを認識することが可能になるという、ポリメラーゼの機能における重要な役割を有する。さらにRSVには二つのコファクターM2−1及びM2−2が存在し、これらはプロセシング(M2−1)及び転写/複製バランスの調節(M2−2)において役割を有する。
【0011】
ウイルス粒子の表面上には二つの主要なFタンパク質及びGタンパク質がある。Fタンパク質はウイルスのエンベロープと細胞膜の融合を可能にし、合胞体の形成に関与する。Gタンパク質はウイルスが細胞の表面に付着することを可能にする。Mタンパク質(マトリックス)はウイルスのエンベロープとポリメラーゼ/ゲノム複合体の仲介をする。F表面タンパク質及びG表面タンパク質の二つのタンパク質は、中和抗体の標的であるため、RSVの主要な免疫原性タンパク質である。
【0012】
しかし、マウスについてなされた研究は、Gタンパク質がTh2型の免疫病理学的ワクチン反応(IL−4とIL−5の産生及び好酸球の集合)の誘発に関与することを示した(Sparer et al. 1998)。それに対して、ウシの場合には、これら二つのタンパク質のいずれも免疫病理学的応答に関与せず、これらの二つのタンパク質は防御応答の確立を可能にする(Taylor et al. 1997)。
【0013】
現在、新規なワクチン候補の研究は、多数がFタンパク質及びGタンパク質に関するものである。
【0014】
しかし、ヒト及びウシにおいては、細胞を介する免疫、特に細胞傷害性T細胞の応答は、RSVに対する防御の不可欠な構成要素である。ウシの場合と同様ヒトの場合以外では、Nタンパク質が細胞傷害性T細胞の応答の主要な基礎である(Goulder et al., 2000)。仔牛がRSVに対するワクチン接種に適切なモデルである。RSVのNタンパク質を発現する組換えワクチンを使用すると、免疫応答のバランスの回復を可能にする細胞型(Th1)の応答が生じることが示されている(Taylor et al.,1997、Gaddum et al.,2003)。したがって、現在の研究全体が、複数のタンパク質、特にF表面タンパク質及びG表面タンパク質並びに内部のNタンパク質の組み合わせで構成される抗RSVワクチンに有利に働く。
【0015】
Nタンパク質は、感染細胞で最も多く発現するタンパク質であり、ウイルス粒子中で量的観点から最も重要なタンパク質の一つである(Collins et al., 2001)。それは、一本鎖RNAから構成されるウイルスゲノムを包囲し、大きなサイズのらせん構造を形成している。それが組換え体の形で単独で発現した時、Nタンパク質は非特異的に細胞RNA上に重合する。それらはそこで極めて巨大な、不溶性で精製不可能な、感染細胞で見られるヌクレオカプシドに似たらせん構造(RNA/N)を形成する(Meric et al., 1994、 Bhella et al., 2002)。
【0016】
このNタンパク質は、ウイルスポリメラーゼLのコファクターであるRSVのPタンパク質と相互作用することができる。ツーハイブリッド法と免疫沈降法を主として使用する相互作用ドメインの位置決定の研究が、様々なグループにより行われたが、スクリーニングの結果は否定的(相互作用は認められない)であった。ヒトウイルス又はウシウイルスのPタンパク質のC末端ドメインはNタンパク質との相互作用において重要な役割を有する疑いがあった(Garcia Barreno et al., 1996、 Mallipeddi et al., 1996、 Slack and Easton, 1998、 Khattar et al., 2001a, 2001b)。しかし、ツーハイブリッドシステムはNタンパク質とPタンパク質との間の相互作用の実際の性質を反映しないことが分かった(Khattar et al., 2001a)。その上、P−N複合体の正確な性質(各分子の数又は化学量、構造)は報告されておらず、相互作用のドメインは明示されても特徴を明らかにされてもいない。
【0017】
パラミクソウイルス科に属する近縁のウイルス(センダイウイルス、麻疹ウイルス)についてなされた研究から、Pタンパク質は、Nタンパク質と共にN°Pと呼ばれる可溶性複合体を形成し、細胞RNAに自身が非特異的に固定することを防ぐとの考えが導かれた(Kolakofsky et al., 2004)。Pタンパク質は、PがLのコファクターとして作用してLがその基質を「発見する」ことを可能にするため、Nタンパク質によりカプシドに包まれたRNAから形成されるヌクレオカプシドを認識することもできる。
【0018】
パラミクソウイルス科については、二つの相互作用ドメインがP上に発見されている。一方は、タンパク質のC末端の位置にあり、N−RNA複合体を認識するドメインを構成し、もう一方は、N末端の位置にあり、N°P複合体の形成を可能にする(Kolakowsky et al., 2004)。RSVについては、これらの複合体は明確に同定されておらず、Nと相互作用するPタンパク質のC末端ドメインの役割は明確に定義されていない。
【0019】
現在まで、Nタンパク質に基づくサブユニットワクチンの開発は、可溶性の形でNタンパク質を単離することが困難なため不可能であった。
【0020】
最近、本発明者は、細菌におけるPタンパク質及びNタンパク質の同時発現系を開発し、これをパラミクソウイルス科のRSVのモデルとして選んだ(Castagne et al., 2004)。Pタンパク質は、アンピシリン耐性を有するプラスミドにおいてグルタチオン−S−トランスフェラーゼ(GST)と融合させ、Nタンパク質はカナマイシン耐性を有するプラスミド中にクローニングした。同じ細菌中におけるこれらプラスミドの同時発現により、融合タンパク質GST−Pを精製すること及びNタンパク質を伴わせることが可能になった。
【非特許文献1】Kolakofsky D, Le Mercier P, Iseni F, Garcin D. 2004. Viral DNA polymerase scanning and the gymnastics of Sendai virus RNA synthesis. Virology.; 318(2):463−73. Review.
【非特許文献2】Castagne, N., A. Barbier, J. Bernard, H. Rezaei, J.−C. Huet, C. Henry, B. Da Costa, and J.−F. Eleouet. 2004. Biochemical characterization of the Respiratory Syncytial Virus P−P and P−N protein complexes and localization of the P protein oligomerization domain. Journal of General Virology; 85: 1643−1653
【発明の開示】
【発明が解決しようとする課題】
【0021】
しかしながら、依然として溶解性の問題が間違いなく存在するため、可溶性の形でのNタンパク質の生産量が、工業的規模におけるこの系の実施を検討するには依然として著しく不十分である。その上、こうして生産されたNタンパク質の性質も明らかにされなかった。
【課題を解決するための手段】
【0022】
本発明者は、Pタンパク質のN末端が欠失した突然変異体とRSVのNタンパク質との同時発現により、完全なPタンパク質で得られる場合と比較して格段に多い量のNタンパク質を精製することが可能になるということを明らかにした。
【0023】
定義
「パラミクソウイルス科」なる科は、パラミクソウイルス亜科とニューモウイルス亜科に再分類される。パラミクソウイルス亜科は、プロトタイプがセンダイウイルスであるレスピロウイルス属、そして、ルブラウイルス属(特に流行性耳下腺炎ウイルス)、及び麻疹ウイルスのようなモービリウイルス属を含む。レスピロウイルス属とルブラウイルス属の各属は、パラインフルエンザウイルス株を構成する。ニューモウイルス亜科は、ニューモウイルス属とメタニューモウイルス属の二つの属に再分類され、後者はヒトメタニューモウイルスを含む。ヒト呼吸器合胞体ウイルス(RSV)は、ニューモウイルス亜科に属するニューモウイルス属のプロトタイプのウイルスを構成する。ニューモウイルス属はまた、ウシRSV株及びマウスRSV株を含む。
【0024】
他に明言しない限り、「呼吸器合胞体ウイルス」は通常、考えられるあらゆる形態(ヒト、ウシ・・・)、サブグループ(例えばヒトRSVで同定されているサブグループA、B及びS)又は株であるRSVを指す。
【0025】
「Pタンパク質」という記載は、パラミクソウイルス科ウイルスのポリメラーゼ複合体から得られるリンタンパク質すなわちPタンパク質を指す。Pタンパク質はウイルスポリメラーゼ(レプリカーゼ/トランスクリプターゼ)のコファクターであり、リン酸化され得る。パラミクソウイルス科のPタンパク質の配列は当業者に知られている。例えば、ヒトRSVのLong株のPタンパク質は241個のアミノ酸からなる配列を有し、これは受託番号P12579でSwissprotデータベースに寄託されている。この配列は、配列番号1に示されている。ウシRSVのPタンパク質も同様に241個のアミノ酸からなる(配列番号23)。センダイウイルス(Harris株)、麻疹ウイルス(Edmonston B株)、流行性耳下腺炎ウイルス(SBL−1株)、及びヒトメタニューモウイルス(00−1株)のPタンパク質も同様に、受託番号P04859(配列番号2)、CAA91364(配列番号3)、P19717(配列番号4)、及びQ91KZ5(配列番号5)でそれぞれSwissprotデータベースに登録されている。「Pタンパク質」という記載は、完全なPタンパク質、トランケート型Pタンパク質、又はPタンパク質の断片を示し得る。
【0026】
パラミクソウイルス科のPタンパク質は、例えばセンダイウイルス又はRSVの場合、ホモオリゴマー特にホモテトラマーを形成する。RSVについては、オリゴマー化(P−Pオリゴマー化)することが可能なPタンパク質のドメインは、このタンパク質の120〜150番アミノ酸のところに位置決定されている(Castagne et al., 2004)。したがって、例えば、RSVのPタンパク質の161〜241番アミノ酸で構成される断片はオリゴマーを形成しない。センダイウイルスのPタンパク質のオリゴマー化ドメインは、Tarbouriechら(2000)により、Pタンパク質の320〜446番残基で構成されるものとして報告されている。他方、麻疹ウイルスのPタンパク質については、Pタンパク質のオリゴマー化領域は304〜376番アミノ酸のところに同定されている(Johansson et al., 2003)。
【0027】
「トランケート型Pタンパク質」という記載は、連続する一つ又は複数のアミノ酸配列が欠失したPタンパク質を指す。それらの欠失は、Pタンパク質の一次配列に関する、C末端配列、N末端配列、及び「内側」配列のトランケーション、又はこれらのトランケーションの組み合わせであり得る。
【0028】
本発明によるトランケート型Pタンパク質は、Pタンパク質のオリゴマー化ドメインを含まず、Nタンパク質と相互作用することができる。パラミクソウイルス科のPタンパク質とNタンパク質との相互作用ドメインがC末端のところに位置決定されたので、トランケート型Pタンパク質の例は好ましくは、Pタンパク質のC末端断片、又はPタンパク質のC末端断片(Nタンパク質と相互作用することができる)とPタンパク質の連続する少なくとも一つの他のアミノ酸配列との融合により構成された「キメラ」Pタンパク質を含む。前記C末端断片と前記Pタンパク質の他の配列は、それ自体は天然に連続しておらず、その中に配列のオーバーラップは存在しない。例えば、RSVのトランケート型Pタンパク質は天然Pタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸から構成される配列を有し得る。参照ポリペプチドの「断片」は、参照ポリペプチド配列中に見られるあらゆる連続したアミノ酸配列を指す。
【0029】
「Pタンパク質の断片」又は「PΔ」は、配列がPタンパク質のアミノ酸鎖からなり、Pタンパク質の連続する一つ又は複数のアミノ酸がN末端及び/又はC末端の末端から欠失しているポリペプチドを指す。
【0030】
「Pタンパク質のC末端断片」又は「PΔN」は、連続する一つ又は複数のアミノ酸がN末端の末端から欠失しているPタンパク質を指す。好ましくは、Pタンパク質のC末端断片はPタンパク質の一次配列のC末端側の半分にあるアミノ酸鎖を指す(配列のアミノ酸の数が奇数の場合には、N末端側の半分との兼ね合いで、一個の追加のアミノ酸をタンパク質のC末端側の半分に任意に加えることができる)。例えば、241個のアミノ酸を含むRSVのPタンパク質では、PΔ161NはPタンパク質の161〜241番アミノ酸により構成されるC末端断片を指す。同様に、例えば、507個のアミノ酸からなる麻疹ウイルス(Edmonston B株)のPタンパク質では、PΔ386NはPタンパク質の386〜507番アミノ酸により構成されるC末端断片を指す。
【0031】
「Pタンパク質のN末端断片」又は「PΔC」は、連続する一つ又は複数のアミノ酸がC末端の末端から欠失しているPタンパク質を意味する。
【0032】
「Pタンパク質の内側断片」又は「PΔNC」は、連続する一つ又は複数のアミノ酸がN末端の末端から欠失し、かつ連続する一つ又は複数のアミノ酸がC末端の末端から欠失しているPタンパク質を意味する。
【0033】
「Nタンパク質」は、ウイルスゲノムを囲むためにらせん構造を形成している、パラミクソウイルス科のヌクレオカプシドタンパク質を指す。ヒトRSVのLong株のNタンパク質は、391個のアミノ酸からなる配列を有し、これは配列番号6に示されている。ウシRSVのNタンパク質も同様に391個のアミノ酸からなる(配列番号24参照)。センダイウイルス(Hamamatsu株)、麻疹ウイルス(Edmonston B株)、流行性耳下腺炎ウイルス(SBL−1株)、及びヒトメタニューモウイルス(00−1株)のNタンパク質も同様に、受託番号Q9DUE3(配列番号7)、受託番号Q89933(配列番号8)、受託番号P21277(配列番号9)、及び受託番号Q91F57(配列番号10)でSwissprotデータベースにそれぞれ登録されている。
【0034】
上記Pタンパク質及びNタンパク質の配列は例示的な特徴を有し、これらの配列は所与のウイルスについて考えられる特定の株に応じて変化する可能性がある。このように、本出願の明細書の以下に記載されるアミノ酸の位置はこれらの参照配列について示したものである。当業者は、これらの例示以外のウイルス株における対応するドメインを完全に同定することが可能である。
【0035】
パラミクソウイルス科ウイルスのこれらのNタンパク質及びPタンパク質をコードする配列も当業者に知られている。
【0036】
「タグタンパク質」とも呼ばれる「標識タンパク質」は、関心のあるタンパク質と融合して精製を容易にするために使用されるタンパク質を指す。標識タンパク質は当業者に知られている。標識タンパク質の例としては、グルタチオン−S−トランスフェラーゼ(GST)、又は4〜10個のヒスチジン残基鎖を通常含む配列であるヒスチジンタグが挙げられる。
【0037】
本発明の内容においては、「相同」という語は、同じ進化上の起源を有するタンパク質間、例えば異なる種、又はウイルスの場合では異なるウイルス株が有する相同タンパク質間に存在する関係に関するものである。このようなタンパク質(及びそれをコードする遺伝子)は配列の相同性を示すものであるが、その相同性は、それらタンパク質の配列の類似性に、つまり、類似の割合、すなわち保存された位置における特定の残基又はモチーフの存在に関して反映される。
【0038】
「配列の類似性」という表現は、同じ進化上の起源を共有し得るか又はそうでない核酸配列間又はタンパク質のアミノ酸配列間の同一性の程度を示す。通常、相同と類似性という語は区別なく使用される。二つのアミノ酸が少なくとも80%同一又は少なくとも90%類似(すなわち機能的に同一)の際、それら二つのアミノ酸の配列は「基本的に相同」である。類似又は相同の配列は、例えばBLASTプログラム又はFASTAプログラムを使用して、アラインメントにより同定することができる。
【0039】
本発明によるタンパク質又は複合体の溶解度は、1×PBS、150mMのNaClを加えた10mMのTris緩衝液(pH7.4〜8.0)、0.2×TBE、さらに例えば、60mMのNaCl、1mMのEDTA、2mMのDTT、0.2%のTriton X−100、10mMのMgSO4、1mMのCaCl2及び1mg/mlのリゾチームを加えた50mMのTris−HCl(pH7.8)を含む細菌溶解緩衝液のような、緩衝化水性溶媒に対して定義される。
【0040】
可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法
本発明者は、Pタンパク質とGSTとの融合物及びパラミクソウイルス科のNタンパク質をそれぞれコードするプラスミドの同時発現により、Nタンパク質を伴う融合タンパク質GST−Pを得ることが可能になることを先に示した(Castagne et al., 2004)。しかし、N−P複合体の生産量は少ないので、この複合体の工業的規模における生産と相容れない。
【0041】
本発明者は、Pタンパク質の欠失突然変異体の特徴を明らかにし、Nタンパク質と相互作用するその能力を判定した。そして、この突然変異体のいくつかもまたRNA−N複合体すなわちリボヌクレオカプシド(RNP)の形のNタンパク質と相互作用することができること、また同時に、完全なPタンパク質で得られるものよりも極めて顕著に多い収量のN−P複合体の調製を可能にすることを明らかにした。この特定の突然変異体は、分子のC末端部分を含みPタンパク質のオリゴマー化ドメインを欠くPタンパク質の断片に相当する。
【0042】
したがって、これらPタンパク質の突然変異体とNタンパク質との同時発現は、大量の、可溶性RNPの形、特にNタンパク質−トランケート型Pタンパク質複合体、特にNタンパク質−Pタンパク質C末端断片複合体の形でのNタンパク質の調製を可能にする。
【0043】
したがって、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法に関するものであって、前記方法は、
a)パラミクソウイルス科ウイルスのNタンパク質を、パラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、トランケート型Pタンパク質と同時発現させること、及び
b)こうして形成された可溶性Nタンパク質−トランケート型Pタンパク質複合体を回収すること
から成る過程を含む。
【0044】
トランケート型Pタンパク質は、好ましくはPタンパク質のC末端断片からなる。パラミクソウイルス科のPタンパク質と、場合によりN−RNA複合体の形であるNタンパク質との相互作用ドメインは、実際、Pタンパク質のC末端側に位置する。
【0045】
トランケート型Pタンパク質は、Pタンパク質のC末端断片とPタンパク質の連続する少なくとも一つの他のアミノ酸配列との融合により構成された、上に定義したような「キメラ」Pタンパク質であり得る。
【0046】
好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片である。
【0047】
従って、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)を調製する方法に関するものであって、前記方法は、
a)パラミクソウイルス科ウイルスのNタンパク質とパラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、Pタンパク質のC末端断片とを同時発現させること、
b)こうして形成されたN−PΔN可溶性複合体を回収すること
から成る過程を含むものである。
【0048】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスはヒト又はウシの呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0049】
当業者は、Nタンパク質と相互作用することができる、トランケート型Pタンパク質、又はさらに具体的にはPタンパク質のC末端断片を知っているか又は決定することができる。
【0050】
例えば、RSVの場合、本発明者はPタンパク質とNタンパク質との相互作用ドメインの位置決定をするために、先に報告された、大腸菌におけるNタンパク質及びPタンパク質の同時発現手段(Castagne et al., 2004)を使用した。そのために、Nタンパク質を、GSTと融合したPタンパク質の欠失突然変異体と同時発現させた。本発明者はまた、Pタンパク質とNタンパク質との相互作用ドメインがPタンパク質のC末端の末端のところに位置していることを明らかにした(図1)。本発明者は、特に、RSVのPタンパク質のC末端の9個のアミノ酸から構成されるオリゴペプチド(233〜241番アミノ酸)までのPタンパク質のC末端断片が、Nタンパク質と相互作用することができることを示した。
【0051】
他方で、例えば、センダイウイルスのPタンパク質の、「Xドメイン」又はXDと呼ばれる、RNA−N複合体すなわちRNPの形のNタンパク質との相互作用ドメインが、473〜568番アミノ酸に該当することが報告されている(Kolakofsky et al. 2004)。
【0052】
それ以外のパラミクソウイルス科については、必要に応じて、当業者は本発明者が報告した手段を使用して、ヌクレオカプシドの形のNタンパク質と相互作用するドメインをPタンパク質に同定することができる。
【0053】
本発明者はまた、RSVのPタンパク質の特定のC末端断片、とりわけPΔ161N断片(161〜241番アミノ酸)が、実際には工業的規模に充分な収量を得ることができない完全なPタンパク質の場合と比較して、大量のNタンパク質の調製を可能にすることを明らかにした。非常に小さな、9個のアミノ酸しか有さないPΔ233N(233〜241番アミノ酸)までの欠失突然変異体が、PΔ161Nの場合の収量に匹敵する収量を得ることを可能にする。
【0054】
PΔ161Nよりも小さいこれらの断片は、Nタンパク質と相互作用することはできるがもはやオリゴマー化することができない、すなわちもはやPタンパク質のオリゴマー化ドメインを含まない、RSVタンパク質の断片に対応する。RSVのPタンパク質の最小のオリゴマー化ドメインは、実際、Pタンパク質の120〜150番アミノ酸付近で定義される。
【0055】
この同じ手段により、本発明者は、386〜507番アミノ酸残基から構成される麻疹ウイルスのPタンパク質のC末端断片(PΔ386N)が、このウイルスのNタンパク質と相互作用し、その精製を可能にするということを示すことができた。一方、Pタンパク質のN末端部分において456番残基までが欠失していると(PΔ457N断片を含む)、Nタンパク質の精製はできない。リボヌクレオカプシドと相互作用するPタンパク質のC末端領域の構造は、Johansson et al.(2003)により特定されている。Pタンパク質のオリゴマー化領域は、304〜376番アミノ酸で定義されたような推定と欠失により特定された。
【0056】
したがって、RNPの形のNタンパク質と相互作用するドメインを含むがPタンパク質のオリゴマー化ドメインが欠失している、Pタンパク質のC末端断片を使用すると、Pタンパク質断片とNタンパク質との相互作用、可溶性N−PΔN複合体の形成、及び高収量でのこの複合体の生産が同時に可能になる。特定のメカニズムに結びつけることはしないが、Pタンパク質のオリゴマー化ドメインが存在しないと、この複合体のPタンパク質同士間の相互作用に関連したN−PΔN複合体の不溶性の問題が回避されると推定される。
【0057】
さらに、一実施形態によると、N−PΔN複合体を調製する方法は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠失する、RSVのPタンパク質のC末端断片を発現させることを含む。
【0058】
さらに具体的には、本発明による方法では、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、RSVのPタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を、RSVのNタンパク質と同時発現させることができる。
【0059】
RSVのPタンパク質のC末端断片は、例えば、PΔ120N(Pタンパク質の120〜241番アミノ酸)、PΔ150N(Pタンパク質の150〜241番アミノ酸)、PΔ161N(Pタンパク質の161〜241番アミノ酸)、PΔ180N(Pタンパク質の180〜241番アミノ酸)、PΔ200N(Pタンパク質の200〜241番アミノ酸)、PΔ220N(Pタンパク質の220〜241番アミノ酸)、PΔ230N(Pタンパク質の230〜241番アミノ酸)、及びPΔ233N(Pタンパク質の233〜241番アミノ酸)から構成される群から選択され得る。
【0060】
本発明はまた、RSVのNタンパク質と、上記のようなRSVのPタンパク質のC末端断片を含み、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、トランケート型Pタンパク質とを同時発現させる方法に関するものである。
【0061】
例えば、Pタンパク質のC末端断片を含むトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合から構成され得る。それは例えば、配列番号1に示すようなヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸と161〜241番アミノ酸との連結から構成され得る。
【0062】
別の一実施形態によれば、パラミクソウイルス科は麻疹ウイルスであり、N−PΔN複合体を調製する方法は、Pタンパク質のC末端のアミノ酸を最大122個含むか、又はPタンパク質のC末端の122個のアミノ酸で構成される、麻疹ウイルスのPタンパク質のC末端断片を発現させることを含む。それらの断片は、特に、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜507番アミノ酸から構成されるC末端断片(PΔ386N)、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であり得る。
【0063】
本発明による方法を利用するに当たり、分子生物学、微生物学、又は組換えDNAにおける通常のあらゆる技術を使用することができる。このような技術は当業者に既知であり、特に次のものに記載されている。Sambrook,Fritsch&Maniatis、分子クローニング:A Laboratory Manual 第二版(1989)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(「Sambrook et al., 1989」)。DNAクローニング:A Practical Approach、第I巻及び第II巻(D.N. Glover編、1985)。オリゴヌクレオチド合成(M.J. Gait編、1984)。核酸のハイブリダイゼーション[B.D.Hames&S.J.Higgins編(1985)]。転写及び翻訳[B.D.Hames&S.J.Higgins編(1984)]。動物細胞の培養[R.I.Freshney編(1986)]。固定化細胞及び固定化酵素[IRL Press,(1986)]。B. Perbal,A Practical Guide To Molecular Cloning(1984)。F.M.Ausubel et al.(編)、Current Protocols in Molecular Biology,John Wiley&Sons,Inc.(1994)。
【0064】
「発現する」又は「発現」は、遺伝子又はDNA配列に含まれる情報が現れることを可能にすること又は現れるようにすること、例えば、対応する遺伝子配列又はDNA配列の転写と翻訳に関与する細胞の機能を活性化することによるタンパク質の産生を意味する。二つの遺伝子又はDNA配列に含まれる情報が同じ宿主細胞内で発現することを「同時発現」という。
【0065】
「コード配列」は、それが発現するとRNA、ポリペプチド、タンパク質等が生成するヌクレオチド配列を意味する。タンパク質をコードする配列は一般に開始コドン(ATG)と終止コドンを含む。
【0066】
コード配列は、RNAポリメラーゼがコード配列をRNA、特にmRNAに転写し、次いでそれがイントロンを含む場合にはスプライシングされ、コードする配列によりコードされているタンパク質に翻訳されるとき、転写及び翻訳の制御配列「の制御下にある」か又はそれら「と機能的に関連している」。
【0067】
「ベクター」、「クローニングベクター」及び「発現ベクター」という記載は、DNA配列又はRNA配列(例えば異種遺伝子)を宿主細胞に導入し、その結果、宿主細胞を形質転換し、導入された配列を発現させることができる担体を意味する。ベクターの例としては、プラスミド、ファージ、ウイルスが挙げられる。もっとも一般的なベクターは、通常は細菌に由来する、自己複製ユニットである、二本鎖DNAの形態であり得るプラスミドである。プラスミドは外来DNA配列を容易に組み込むことができ、それを適当な宿主内に容易に導入することができる。一つのプラスミドベクターは通常、一つのコードDNA配列、一つのプロモーターDNA配列を含み、外来DNAの導入が可能な一つ又は複数の制限部位を有する。プラスミドの非制限的な例としては、pKK(Clonetech)、pUC及びpET(Novagen,Inc.、マディソン、ウィスコンシン州)、pRSET及びpREP(Invitrogen、サンディエゴ、カリフォルニア州)、pMAL(New England Biolabs、ビバリー、マサチューセッツ州)、又はpGEX−4T−3(Pharmacia)のプラスミドが挙げられる。
【0068】
「宿主細胞」は、細胞による物質の生産、例えば、細胞による、遺伝子、DNA配列又はRNA配列、タンパク質又は酵素の発現のために選択され、修飾され、培養され又は操作される、あらゆる細胞又は生物を意味する。
【0069】
「発現系」は、ベクターにより担持され宿主細胞内に導入される外来DNAによってコードされるタンパク質の生産に適した条件下で使用される、宿主細胞及び適当なベクターを指す。通常の発現系は、大腸菌宿主細胞及びプラスミドベクター、昆虫細胞、及びバキュロウイルスベクター、またさらには哺乳類細胞及びベクターを含む。
【0070】
本発明の方法による発現系は、有利には、例えばpGEX−4T−3のようなベクターを有する、細菌特に大腸菌の発現系である。細菌の発現系は実際、一般にきわめて高い生産量が得られることを可能にする発現系である。
【0071】
有利には、トランケート型Pタンパク質及び特にPタンパク質のC末端断片は、Nタンパク質−トランケート型Pタンパク質複合体の精製を容易にするタンパク質と融合した形、とりわけアフィニティークロマトグラフィーに使用できるタンパク質と融合した形で発現する。それらのタンパク質は、トランケート型Pタンパク質−GST融合タンパク質が、グルタチオンを結合した固体支持体上でのクロマトグラフィーにより単離できる場合、グルタチオン−S−トランスフェラーゼ(GST)のような標識タンパク質であり得る。ポリヒスチジンすなわち「His−Tag」のような他の標識又は「タグ」も使用することができる。
【0072】
こうして、Nタンパク質−トランケート型Pタンパク質−標識タンパク質複合体(トランケート型Pタンパク質、特にPΔN断片と融合したGST又は他の標識タンパク質)が得られ、標識タンパク質は酵素切断により除去することができる。例えば、GSTはトロンビンによる切断により、又は融合体がGST以外のタンパク質を含む場合は他の全ての適当な酵素により除去することができる。
【0073】
本発明による方法の利用を可能にするベクター構築物の具体例を以下の実施例に示す。
【0074】
可溶性Nタンパク質−トランケート型Pタンパク質複合体
【0075】
上述したような、可溶性Nタンパク質−トランケート型Pタンパク質複合体、特に可溶性Nタンパク質−Pタンパク質C末端断片複合体の調製方法により、単離又は精製された形でNタンパク質−トランケート型Pタンパク質複合体を容易に得ることを可能にする。
【0076】
したがって、本発明はまた、本発明の調製方法により得ることが可能な、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体に関するものである。
【0077】
好ましくは、トランケート型Pタンパク質は、Pタンパク質のC末端断片を含むか、又はPタンパク質のC末端断片である。
【0078】
本発明は、さらに具体的には、本発明の調製方法により得ることが可能な、パラミクソウイルス科のウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)に関するものである。
【0079】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(HMPV)及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスは、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0080】
一実施形態によれば、パラミクソウイルス科のウイルスは呼吸器合胞体ウイルス(RSV)であり、前記Pタンパク質のC末端断片は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている。
【0081】
さらに具体的には、前記Pタンパク質のC末端断片は、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びているか、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片を含む
ことができる。
【0082】
それらの断片は、特に、PΔ120N、PΔ150N、PΔ161N、PΔ180N、PΔ200N、PΔ220N、PΔ230N、及びPΔ233Nから構成される群から選択されるPタンパク質のC末端断片であり得る。
【0083】
また、本発明は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、上記に記載のようなRSVのPタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質を含む、可溶性の複合体に関する。
【0084】
例えば、Pタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合により構成され得る。それは、例えば配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸との連結により構成され得る。
【0085】
別の一実施形態によれば、パラミクソウイルス科のウイルスは麻疹ウイルスであり、前記Pタンパク質の断片は、Pタンパク質のC末端のアミノ酸を最大122個含むか又はPタンパク質のC末端の122個のアミノ酸で構成されている、Pタンパク質のC末端断片である。さらに具体的には、麻疹ウイルスのPタンパク質の前記C末端断片は、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜514番アミノ酸(PΔ386N)から構成されていてもよく、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であってもよい。
【0086】
N−PΔN複合体において、タンパク質PΔNは、場合により、例えばGST、ヒスチジンタグ、又はN−PΔN複合体の精製を容易にする他のあらゆる適当なタンパク質のような標識タンパク質と融合した形で提供され得る。
【0087】
RSVについての複合体生成物の電子顕微鏡分析から、それが10分子のNタンパク質を含む環状化合物であり、この可溶性の環はまた、アガロースゲル電気泳動で確認することが可能な約70塩基の小さなRNAを含むことが明らかになった(図3)。この複合体中には同様の量のNタンパク質とPΔN−GSTタンパク質がみられる。RNAは、このタンパク質を変性させることなくNタンパク質の環から分離することができない。
【0088】
このように、本発明はまた、上記で定義したような、10分子のNタンパク質を含み、そのうちのそれぞれ又はその多数がPタンパク質のC末端断片と関連し、前記Pタンパク質のC末端断片がRSVのPタンパク質のC末端の先端の9個のアミノ酸を含みRSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、呼吸器合胞体ウイルス(RSV)の単離された可溶性Nタンパク質−Pタンパク質C末端断片複合体にも関する。RSVのこのN−PΔN複合体は、さらに約70塩基のRNAを含む。
【0089】
可溶性Nタンパク質を調製する方法
【0090】
このNタンパク質−トランケート型Pタンパク質複合体から、又はさらに具体的にはN−PΔNから、例えばサイズ排除クロマトグラフィー(ゲル濾過)により、可溶性Nタンパク質を環状でそのRNAと共に容易に単離することができる。この分離は、必要に応じて、トランケート型Pタンパク質と、場合によりトランケート型Pタンパク質が融合している標識タンパク質との、酵素切断による分離の後に行うことができる。
【0091】
したがって、本発明はさらに、
a)上記に定義したような方法により可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製すること、そして
b)可溶性Nタンパク質−トランケート型Pタンパク質複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む、パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法に関する。
【0092】
好ましくは、トランケート型Pタンパク質は、Pタンパク質のC末端断片を含むか、またはPタンパク質のC末端断片である。
【0093】
本発明は、さらに具体的には、
a)上記に定義したような方法により可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)を調製すること、そして
b)可溶性N−PΔN複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む、パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法に関する。
【0094】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス、及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスは、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0095】
上記の方法により得ることができるパラミクソウイルス科ウイルスの可溶性Nタンパク質もまた、本発明の一部をなす。
【0096】
RSVの場合、利用できる最大のPタンパク質のC末端断片(PΔ161N)が15kDaの分子量を有するのに対し、Nタンパク質は450kDaの分子量を有する。したがって、Nタンパク質の環は、例えば下記の実施例3(及び図2)に記載するようなセファデックスカラム上でのクロマトグラフィーにより、Pタンパク質のC末端断片から分離することができる。
【0097】
一実施形態によると、本発明はしたがって、可溶性のRSVのNタンパク質を提案し、この前記Nタンパク質は、約7nmの直径を有し10個のサブユニットを含む環を形成している。しかし、ある種の部分的で10個より少ないサブユニットを含む環を除外することしかできない。環はさらに約70塩基のRNAを含む。
【0098】
ワクチン組成物
【0099】
RSV、さらに一般的にはパラミクソウイルス科のNタンパク質は、ワクチン接種にとって有益な抗原であるが、現在までのところ、可溶性の形でそれを精製することに誰も成功していない。本発明による方法は、極めて純度が高くかつ極めて均質な、環状構造のNタンパク質を容易かつ大量に得ることを可能にする。
【0100】
環状Nタンパク質の免疫原性を評価するため、本発明者は、本発明によるパラミクソウイルス科のN−PΔN複合体でマウスに免疫を付与した。ワクチンとして使用するために、Nタンパク質とPΔNは場合により分離され得るが、この操作は必要なものではなく、PΔNが存在しても弊害はない。
【0101】
本発明者は、マウスにRSVのN−PΔ161N複合体でさらに特異的に免疫を付与し、RSVのPΔ161Nポリペプチドをコントロールとして使用した。
【0102】
気道に対するこのウイルスの特異性を考慮して、非経口のワクチン接種の従来の方法である皮下経路、及び呼吸粘膜とそれに関連するリンパ組織レベルでの局所的な免疫付与の誘発を可能にする鼻経路の、二つの免疫付与経路を比較した。
【0103】
ワクチンアジュバントの不存在下で可溶性の組換えタンパク質に対する免疫応答を得ることが困難なので、本発明者は、粘膜経路でのアジュバント性が報告されている(McNeal et al. 2002, Freytag et Clements 2005)、大腸菌(R192G)の無毒化リンホトキシン(LT)を共に使用した(Dr.J.D.Clements、アメリカ、から取得)。
【0104】
免疫応答のパラメーター、(i)血清及び粘液の抗体産生(気管支肺胞洗浄による)、及び(ii)脾臓又は肺から単離された記憶Tリンパ球によるIFN−γの産生を介した細胞性免疫応答について追跡した。
【0105】
アジュバントLT(R192G)の存在下で、Nタンパク質(N−PΔ161N複合体)は、投与経路のいかんにかかわらず(経鼻又は皮下)血清抗体の産生を強く誘導する(図4A)。この応答は最初の免疫付与の後(14日目)に検出可能であり、二度目の免疫付与の後(28日目)に増強される。アジュバントはまた、二度目の免疫付与の後にPΔ161Nに対する抗体の産生を誘導することを可能にする。
【0106】
大いに注目すべきことには、アジュバントの不存在下でのN−PΔ161N複合体の経鼻及び皮下投与は、最初の免疫付与からすぐに血清抗体を多く産生させる。同じ条件下でPΔ161Nは応答を誘導しない。比較した応答の特徴は、粘液の抗体、総Igの産生に見られる(図4B)。
【0107】
これに対して、LT(R192G)存在下でNタンパク質(N−PΔ161N複合体)を単独で経鼻投与すると、呼吸粘膜レベルでのIgA産生を可能にする(図4B)。
【0108】
細胞性免疫応答は、脾臓又は肺の白血球の、N−PΔ161N又はPΔ161Nのみの存在下においてIFN−γをインビトロで産生する能力により測定した。IFN−γは、CD4及びCD8Tリンパ球により産生されるサイトカインである。CD4及びCD8記憶Tリンパ球は、それらが特異的な抗原の存在下で、インビトロで反応性がある。IFN−γは、ウイルスに対する防御における主要なエフェクターであるTh1リンパ球及び細胞傷害性Tリンパ球により産生される。
【0109】
最も強い応答はN−PΔ161N複合体による再刺激後に得られる(図5)。抗体の場合のように、N−PΔ161N複合体の経鼻又は皮下投与により、Nタンパク質に特異的な末梢細胞(脾臓)の免疫性が刺激される。注目すべきことには、アジュバントとともに経鼻経路で投与したNタンパク質により局所細胞(肺)の免疫が生じる(図5)。
【0110】
結論として、環状構造のNタンパク質は局所応答(呼吸粘膜)を刺激することを含め、強い免疫原性を示す。
【0111】
したがって、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質を含むワクチン組成物であって、前記可溶性Nタンパク質が医薬的に許容される担体中に存在するワクチン組成物を提案する。この可溶性Nタンパク質は、特に、上記のような可溶性Nタンパク質−トランケート型Pタンパク質複合体、さらに具体的には可溶性N−PΔN複合体から可溶性Nタンパク質を分離する方法により得ることが可能である。
【0112】
上述したように、可溶性Nタンパク質はPタンパク質との複合体の形で、好ましくない作用を伴わずにワクチン接種に使用され得る。したがって、本発明によるワクチン組成物は、医薬的に許容される担体中にある、パラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質複合体を含むことができる。
【0113】
これら複合体の生産量の問題については、本発明は好ましくは、医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を含むワクチン組成物であって、前記トランケート型Pタンパク質がPタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができるワクチン組成物を対象とする。
【0114】
好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片を含むか、又はPタンパク質のC末端断片である。
【0115】
本発明はまた、さらに具体的には、医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体であって、前記Pタンパク質のC末端断片がPタンパク質のオリゴマー化ドメインを含まず、Nタンパク質と相互作用することができる複合体に関する。
【0116】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、パラミクソウイルス科ウイルスの可溶性Nタンパク質は、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)の可溶性Nタンパク質である。
【0117】
一実施形態によれば、パラミクソウイルス科のウイルスは呼吸器合胞体ウイルス(RSV)であり、前記Pタンパク質のC末端断片は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いているC末端断片(PΔN)である。
【0118】
さらに具体的には、前記組成物において、RSVのPタンパク質のC末端断片は、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、C末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
であり得る。
【0119】
それらの断片は、特に、PΔ120N、PΔ150N、PΔ161N、PΔ180N、PΔ200N、PΔ220N、PΔ230N、及びPΔ233Nから構成される群から選択されるRSVのPタンパク質のC末端断片であり得る。
【0120】
本発明はまた、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、上記のようなRSVのPタンパク質のC末端断片をRSVのトランケート型Pタンパク質が含んでいる組成物に関する。
【0121】
例えば、Pタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合により構成され得る。それは、例えば配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸との連結により構成され得る。
【0122】
別の一実施形態によれば、パラミクソウイルス科のウイルスは麻疹ウイルスであり、前記Pタンパク質の断片は、Pタンパク質のC末端のアミノ酸を最大122個含むか又はPタンパク質のC末端の122個のアミノ酸で構成されている、Pタンパク質のC末端断片である。さらに具体的には、麻疹ウイルスのPタンパク質の前記C末端断片は、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜514番アミノ酸(PΔ386N)から構成されていてもよく、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であってもよい。
【0123】
「医薬的に許容される担体」は、ヒト又は動物において例えばアレルギーのような副作用を生じさせない、あらゆる溶媒、分散媒、吸収遅延剤等を意味する。
【0124】
有利には、本発明によるワクチン組成物はさらにアジュバントを含み得る。「アジュバント」は、同時に投与された抗原に対する細胞性又は液性免疫反応を増大させ、刺激し、活性化し、強化し、又は調節する物質を指す。従来のアジュバントの例は、完全フロイントアジュバント、LPS及びそれらの派生物、細菌毒素(コレラ毒素及びエンテロトキシン)及びその無毒化変異体(例えばLT(R192G))のような細菌抗原、CpGモチーフを含むオリゴヌクレオチド配列、水酸化アルミニウム(Alum)、リン酸カルシウム又はリン酸カリウムのような無機アジュバント、油性エマルジョン及び乳化剤(サポニン、例えばQS21)、サイトカインを含むアジュバントを含む。
【0125】
本発明によるワクチン組成物は、パラミクソウイルス科のウイルスによる感染に対する防御を提供し、すなわちワクチン組成物により免疫を付与されていない対象と比較してこの感染の影響の程度を軽減する。
【0126】
本発明はまた、ワクチン接種方法における上記で定義したようなワクチン組成物の使用に関する。
【0127】
したがって、本発明は、少なくとも本発明によるワクチン組成物を対象に投与することを含む、ワクチン接種方法を対象とする。好ましくは、ワクチン接種方法は、対象へのワクチン組成物の最初の投与と、同じ対象に対する前記ワクチン組成物の二度目の投与を含む。二度目の投与は、患者を抗原に再びさらし、より強い二次免疫応答を誘導する。
【0128】
「対象」という記載は、パラミクソウイルス科ウイルスによる感染にさらされるか又はさらされるおそれがある、ヒト又はヒトでない動物、例えば鳥、又はウシ、げっ歯類、イヌ、ネコ、ブタ、サルといった哺乳類を指す。好ましくは、本発明の趣旨の範囲内の対象はヒト又はウシである。
【0129】
ワクチン組成物は、有利には、パラミクソウイルス科ウイルスによる感染に対する防御的又は治療的な免疫応答の誘導に有効な量を投与される。用量は当然、検討される有効成分、投与方法、対象の年齢及び状態に依存する。投与量当たりのN−P複合体、N−PΔN複合体、又はNタンパク質は0.1〜200μgの間とすることができ、好ましくはワクチン投与量当たり10〜100μgの間である。
【0130】
ワクチン組成物は、いかなる経路、特に粘膜経路(例えば眼内、鼻腔内、経口)、又は非経口経路(例えば皮下、皮内、筋肉内、静脈内、又は腹腔内)によっても投与することができる。
【0131】
診断への使用
【0132】
場合により可溶性Nタンパク質−トランケート型Pタンパク質複合体の形である、パラミクソウイルス科ウイルスの可溶性Nタンパク質は、加えて、前記パラミクソウイルス科ウイルスのNタンパク質に対する抗体を検出するための診断用途に使用することができる試薬を構成する。
【0133】
したがって、本発明はさらに、上記のような、場合によりNタンパク質−Pタンパク質複合体の形であるパラミクソウイルス科ウイルスのNタンパク質を含む診断試薬に関する。
【0134】
前記試薬と適当な検出手段を含む診断キットもまた、本発明の範囲にある。
【0135】
本発明はまた、インビトロ又はインビボで前記Nタンパク質に対する抗体を検出するための、本発明によるパラミクソウイルス科ウイルスのNタンパク質の使用を提案する。
【0136】
本発明はまた、生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に対する特異的な抗体の検出方法であって、
a)前記生物サンプルとパラミクソウイルス科ウイルスのNタンパク質とを接触させること、
b)形成されたNタンパク質/抗体複合体を検出すること
から構成される過程を含み、この複合体の存在が生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に特異的な抗体の存在の指標となる方法に関する。
【0137】
生物サンプルは、例えば筋肉、肝臓、心臓、脳などの生検により得られた組織サンプル、又は、例えば血液、血漿、又は脳脊髄液のような生体液のような液体サンプルであり得る。
【0138】
複合体の検出は、(サイズ排除クロマトグラフィー、アフィニティークロマトグラフィーなどの)クロマトグラフィー又は非変性条件下での電気泳動のような、当業者に周知の常用の手段により実施され得る。
【0139】
上記で定義したようなNタンパク質に対する特異的な抗体の検出方法において、生物サンプルと接触させるNタンパク質は、Nタンパク質−Pタンパク質複合体の形を有し得る。
【0140】
以下の実施例及び図面は本発明の範囲を限定することなく説明するものである。
【0141】
図1は、Pタンパク質に対するPタンパク質−Nタンパク質の相互作用ドメインの位置決定を示す図である。Pタンパク質をGSTに融合し、別のプラスミド上で発現したNタンパク質と大腸菌内で同時発現させた。
【0142】
図2は、サイズ排除クロマトグラフィーにおけるN−PΔ161N複合体の溶離特性を示している。図2AはTSKカラムにおける220nmでの溶離特性を示す。図2Bはクマシーブルーによる染色後の、種々の断片のアクリルアミドゲル電気泳動による分析結果を示す。17〜22の断片はRNA−環状Nタンパク質のみを含んでいる。
【0143】
図3は、RSVのNタンパク質の環状構造を示している。図3Aは、P161−241により精製した環状Nタンパク質−RNAの電子顕微鏡分析の結果を示す。図3Bは、低温顕微鏡による再構築の結果を示す。図3Cは、環中に存在するRNAのアガロースゲル上での分析結果を示す。
【0144】
図4は、N−PΔ161N複合体に対する抗体の産生を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示している。BALB/cマウスに、粘液性アジュバントLT(R192G)の存在下又は非存在下で20μgのN−PΔ161N複合体又はPΔ161Nを鼻腔内投与(i.n.)又は皮下投与(s.c.)して、免疫を付与した。二週間後に二度目の投与を行う(14日目)。マウスを二度目の投与の二週間後に安楽死させる(28日目)。血清抗体を測定するため、血清を0日目、14日目及び28日目に採取する(図4A)。粘液抗体を測定するため、気管支肺胞洗浄を28日目に行う(図4B)。N−PΔ161Nに対する抗体の割合をELISAにより測定する。データは、平均値±平均値における標準偏差(n=5)で示し、対数目盛で表示する。
【0145】
図5は、PΔ161N及びN−PΔ161Nに特異的な細胞性免疫応答を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示している。BALB/cマウスに、粘液性アジュバントLT(R192G)の存在下又は非存在下で20μgのN−PΔ161N複合体又はPΔ161Nを鼻腔内投与(i.n.)又は皮下投与(s.c.)して、免疫を付与した。二週間後に二度目の投与を行う(14日目)。脾臓と肺を摘出するため、マウスを二度目の投与の二週間後に安楽死させる(28日目)。脾臓細胞及び肺細胞を、N−PΔ161N、PΔ161Nの存在下で、又は再刺激を行わずに72時間培養する。IFN−γの分泌をELISAにより測定する。再刺激を行わない場合、IFN−γのベースレベルは15pg/ml未満である。データは、平均値±平均値における標準偏差(n=5)で示す。
【実施例1】
【0146】
RSVのリンタンパク質のC末端領域を含むプラスミドの構築
【0147】
RSVのLONG株のPタンパク質は241個のアミノ酸残基から構成される。
【0148】
RSVのPタンパク質のC末端部分を増幅するために使用したオリゴヌクレオチドプライマー配列(5’から3’)(BamHI制限部位に下線を付し、Pタンパク質遺伝子の「開始」コドンATGは太字で表す)は以下である。
【0149】

【0150】
RSVのLONG株のPタンパク質遺伝子のcDNAを、プライマーLONG−PBam+及びLONG−P−(Castagne et al., 2004)を用い、ヒトRSVのLONG株に感染させたHep−2細胞からRT−PCRにより増幅した。PCR産物を制限酵素BamHIで消化し、プラスミドpGEX−4T−3(Pharmacia)のBamHI−SmaI部位において、グルタチオン−S−トランスフェラーゼすなわちGSTをコードする遺伝子とクローニングした。このプラスミドをpGEX−Pと称する。
【0151】
・P161−241(PΔ161N)のクローニング
Pタンパク質のC末端領域(161〜241番アミノ酸)を、以下の条件でプラスミドpGEX−PからPCRにより増幅した。
PCRプライマー:P161B+及びLONG−P−を各100ng(各1μl)
pGEX−Pの鋳型DNA:10ng(1μl)
酵素:StratageneのPfu Turbo(登録商標)(μl当たりのユニット):1μl
dATP:最終濃度0.2mM
dGTP:最終濃度0.2mM
dCTP:最終濃度0.2mM
dTTP:最終濃度0.2mM
最終濃度1×Pfu緩衝液(Stratagene)
最終容量:100μl
PCRは以下の条件下で実施した。
94℃で15秒、40℃で2分、72℃で1分を5サイクル、
94℃で15秒、55℃で1分、72℃で1分を25サイクル
【0152】
増幅したDNAを1体積のフェノール/クロロホルム(1体積/1体積)(100μl)、次いで1体積のクロロホルムで抽出し、最後に10分の1体積の5MのNaCl(10μl)と2体積の100%エタノール(200μl)を加えて沈殿させた。DNAを13000gで20分間遠心し、1体積の70%エタノールで洗浄し、乾燥させ、1体積の水90μl中に再懸濁した。10×BamHI酵素緩衝液を10μl加えたのち、DNAを10ユニットのBamHI酵素の存在下で37℃で2時間消化した。消化したDNAを、エチジウムブロマイドの存在下で、1×トリス−ホウ酸−EDTA緩衝液(TBE)中の1.5%アガロースゲル上に置き、電気泳動した。P161−241のDNAに対応するバンドを切り取り、DNAを電気溶出により抽出した。それをさらに1体積のフェノール−クロロホルム、1体積のクロロホルムで抽出し、エタノールで沈殿した。それを、1%アガロースゲルにおける精製の後、BamHIとSmaIで消化したpGEX−4T−3ベクターにライゲーションした。
pGEX4T−3のDNA:100ng
P161−241のDNA:100ng
最終濃度1×リガーゼ緩衝液
リガーゼ(5U/μl):1μl
最終体積20μl
【0153】
全体を14℃で一晩インキュベートした。翌日、適当な細菌DH5−アルファTM(Life Technologies)を10μlのライゲーション産物で形質転換し、最終濃度100μg/mlのアンピシリンを補足したL−寒天培地を含むペトリ皿上に塗布した。組換え細菌のコロニーをプラスミドのミニプレップと制限酵素BamHI及びXhoIによる消化でスクリーニングした。組換えプラスミドはアガロースゲル上で、一方はベクター(4.9kb)に対応し、もう一方はPタンパク質のC末端部分(246bp)に対応する、二つのバンドを示す。組換えプラスミド全体を配列決定した。
【0154】
・P180−241、P201−241、P221−241のクローニング
180〜241、200〜241、220〜241番アミノ酸部分に対応するPタンパク質の断片を、以下のプライマーを用いてpGEX−PプラスミドからPCRにより得た。
P180−241:プライマーP180B+及びLONG−P
P200−241:プライマーP201B+及びLONG−P−
P220−241:プライマーP221B+及びLONG−P−
【0155】
これらを、P161−241(上記参照)と同様に増幅しクローニングした。
【0156】
・RSVのLong株のヌクレオカプシドタンパク質をコードする遺伝子のクローニング
ヒトRSVのLONG株のNタンパク質をコードする遺伝子を、そのウイルスに感染したHep−2細胞からRT−PCRにより得た。使用したプライマーは、
LONG−Nbam+:GAGGGATCCATGGCTCTTAGCAAAGTCAAGTTG(配列番号19)
LONG−N−:TTAACTCAAAGCTCTACATCATTATCTTTTGG(配列番号20)
であった。
【0157】
PCR産物をBamHIで消化し、プラスミドpGEX−4T−3のBamHI−SmaI部位においてクローニングした。BamHI−XhoIでpGEX−Nプラスミドを消化して、Nタンパク質(配列番号)をコードする領域をpET28+プラスミド(Novagen、配列番号及び図面参照)中にサブクローニングした。
【0158】
・P231−241のクローニング
以下のプライマーを94℃で5分間加熱して変性させ、室温で冷却した。
P231NB+:GATCCGATAGTGACAATGATCTATCACTTGAAGATTTCTGA(配列番号21)
P231N−:TCAGAAATCTTCAAGTGATAGATCATTGTCACTATCG(配列番号22)
【0159】
ハイブリダイズ後、10ngの二本鎖オリゴヌクレオチドを、BamHI酵素及びSmaI酵素で消化しアガロースゲル上での電気泳動により精製した、pGEX−4T−3プラスミドのDNA100ngとライゲーションした。組換えプラスミドは、Nタンパク質遺伝子に関して配列決定することにより確認した。
【実施例2】
【0160】
複合体の発現と精製
【0161】
適当な細菌BL21(DE3)(Novagen)をpGEX−PΔのDNA1μgとpET−NのDNA1μgで形質転換し、次いで最終濃度100μg/mlのアンピシリンと最終濃度50μg/mlのカナマイシンを補足したL−寒天培地を含むペトリ皿に塗布した。コロニーを、100μg/mlのアンピシリンと50μg/mlのカナマイシンを含む2mlのLB培地に移植し、37℃で一晩生育させた。翌朝、コンフルエントな状態の培養物1mlを、抗生物質を添加した1リットルのLB培地に播種し、夕方まで生育させた。夕方に、160μg/mlの濃度のIPTG(タンパク質の発現を誘導する)を含む1体積の新鮮なLB培地を培養物に加え、全体を28℃で一晩生育させる。翌日、細菌を5000rpmで15分間遠心し、ペレットを100mlの以下の緩衝液中に再び入れる。
50mMのトリス(PH7.8)
60mMのNaCl
2mMのDTT
1mMのEDTA
4mMのベンズアミジン
1×アンチプロテアーゼ(Complete EDTA−free protease inhibitor cocktail、Roche、参照番号11873580001)すなわち50mlの溶解緩衝液に対して一錠
0.1%Triton−X100
【0162】
10mg/ml(最終濃度1mg/ml)のリゾチームを添加した同様の緩衝液10mlを加える。細菌を氷中で1時間インキュベートする(溶解)。全体が粘性になったら、全体を、混合物中に入れたプローブを用いて氷中で1分間の超音波処理を3回行い、各超音波処理の間に5分間放置する。全体を4℃、10000gで30分間遠心し、次いで上清を回収する。上清を4℃、10000gで30分間再び遠心し、次いで新たに上清を回収する。8mlのビーズ緩衝液(体積/体積)混合物を回収して4mlのグルタチオンセファロース4Bビーズ(Amersham−Pharmacia)を溶解緩衝液で洗浄する。同量の緩衝液中にビーズを置き、澄ました細菌溶解液を加え、4℃で一晩回転させる。翌日、ビーズを2000rpmで3分間遠心し、次いで上清を除去し、ビーズを、アンチプロテアーゼを含まない溶解緩衝液で三回洗浄し、1×PBS緩衝液で三回洗浄する。
【0163】
ビーズを、ビオチン化したThrombin(Novagen)をビーズ1ml当たりトロンビン(Thrombin Cleavage Capture Kit、 No.69022−3FRZ)1μl(1U)の比率で用いて、トロンビン部位で切断する。ビーズを20℃で一晩インキュベートし、翌日、2000rpmで3分間遠心し、上清を回収するため15分間沈殿させる。等量の1×PBSをビーズに加え、混合物を攪拌し沈殿させる。上清を新たに回収し、あらかじめ採取しておいた上清に加える。回収した上清にストレプトアビジンアガロースビーズ(Novagen、参照番号69203)を樹脂16μl(すなわち樹脂/緩衝液混合物(vol/vol)32μl)の割合で加える。混合物を一時間攪拌し、次いで2000rpmで3分間遠心し、上清を回収する。タンパク質濃縮物2mg/mlを得る。
【0164】
切断産物を含む10μlの上清を、1×Laemmli緩衝液中で変性し、沸騰させ、0.1%トリス−グリシンSDS緩衝液中の12%ポリアクリルアミドゲル上に置き、次いでタンパク質を可視化するために、電気泳動後にクマシーブルーで染色する。
【実施例3】
【0165】
Nタンパク質とPΔ161N(P161−241)の分離及びNタンパク質の環の精製
【0166】
上清中に存在するタンパク質は、1×PBS中におけるサイズ排除クロマトグラフィー(ゲル濾過、図2)により分離することができる。Nタンパク質は450000Daの見かけのサイズで、PΔ161Nは15kDaの分子量で排除される。
【0167】
サイズ排除クロマトグラフィーにより得られた「Nタンパク質」画分を電子顕微鏡により観察した結果、Nタンパク質は、直径7nmを有しNタンパク質のサブユニットを10個含む(図3B)、環の形(図3A)であることが分かった。環は約70bpのRNAを含む(図3C)。
【実施例4】
【0168】
環状構造の組換えNタンパク質の免疫原性の評価
【0169】
・経鼻投与又は皮下投与によるワクチン接種を、アジュバントの存在下又は非存在下でマウスで実施する。
マウス:l’Unite Experimentale Animalerie Rongeur(INRA、Jouy−en−Josas)で育成した10〜12週齢の雌のBALB/cマウス30頭
抗原:トロンビンでの切断によりGSTを分離し、ストレプトアビジンと結合したビーズによりビオチン化トロンビンを除去した後の、濃度1mg/mlの可溶性のP161−241及び{P161−241+N}複合体
アジュバント:1mg/mlの、大腸菌のリンホトキシンLT(R192G)(Choi et al., 2004)
サンプル採取
−0日目、14日目及び28日目に、眼窩後洞から血液を採取
−28日目:
・1.5mlのHBSS培地及び1mMのEDTAによる気管支肺胞洗浄物(LBA)
・氷上での、抗生物質(100UI/mlのペニシリン及び100μg/mlのストレプトマイシン、PS)を加えたRPMI培地における脾臓と肺の解剖
【0170】
実験計画:
【表1】
【0171】
・抗Nタンパク質抗体の産生
血清を血液から採取し(4℃で一晩滲出)、次いで−20℃で凍結する。
【0172】
LBAを1700rpmで5分間遠心し、上清を集め(約1ml)、−20℃で凍結する。
【0173】
ELISAにより血清及びLBAにおける抗Nタンパク質抗体(総Ig、IgG1,IgG2a、及びIgA)を調べる。96ウェルプレート(Immulon 2HB、ThermoLabsystems)を、0.1Mの重炭酸緩衝液(pH9.5)において、P161−241+Nタンパク質複合体(ウェル当たり200ng)により4℃で一晩感作する。プレートをウェル当たり200μlの、0.05%tween20を含むPBSで5回洗浄する(自動装置Wellwash、Labsystemsを使用)。次にプレートを、37℃で1時間、ウェル当たり150μlの、0.05%tween20及び5%ウシ胎児血清を含むPBS緩衝液(PBS−T−FBS)で満たす。5回洗浄した後、滴定サンプルをPBS−T−FBSで希釈する(血清については最初の希釈度から1/30になるまで3倍の連続希釈を7回行い、LBAについては1/3になるまで行う)。プレートを37℃で2時間インキュベートする。5回洗浄した後、PBS−T−FBSで希釈した二次抗体をウェル当たり100μlの割合で添加する。使用する二次抗体は、ペルオキシダーゼと結合しており、マウスの免疫グロブリン、総IgG(1/4000、PARIS)、IgG1(1/2000、BD biosciences)、IgG2a(1/2000、BD biosciences)又はIgA(1/1000、Caltag)に対するものである。プレートを37℃で2時間インキュベートし、5回洗浄する。次にプレートをペルオキシダーゼの基質(TMB、ウェル当たり100μl)と共に10分間暗所でインキュベートする。2MのH3PO4を50μl加えて酵素反応を停止する。光学密度(OD)を450nmで読み取る(読み取り装置Dynex)。OD450=f(希釈度)の曲線を、回帰曲線y=(b+cx)/(1+ax)によりモデル化する。抗体の力価は、最大限に希釈されたコントロールサンプル(0日目)の二倍のOD450を示す希釈値として決定する。
【0174】
・P161−241及びNタンパク質に特異的なTリンパ球によるIFN−γの産生
摘出した脾臓及び肺を同じ手順で処理する。脾臓は個別に処理し、肺は実験群にグループ化する(群当たり5個の肺)。
【0175】
組織を薄切りにし、次いでRPMI培地及びPS培地中において、篩上(100μmのセルストレーナー、BD Falcon)で注意深く破砕する。細胞懸濁液を1700rpmで10分間、4℃で遠心する。細胞を1mlの赤血球溶解緩衝液(低張生理食塩緩衝液)中に懸濁し、室温で5分間インキュベートする。溶解反応は、10mlの完全RPMI(PS、2mMのL−グルタミン及び10%FBS)を添加することにより停止する。膜の破片を沈殿させ、細胞を遠心(1700rpmで10分間、4℃)により3回洗浄する。細胞懸濁液を、Malassezの計算盤を用いて計数する。
【0176】
細胞を、細胞培養用に処理された96ウェルマイクロプレート(Falcon)中で、200μlの完全RPMI培地においてウェル当たり200000個の細胞の比率で培養する。
【0177】
各細胞懸濁液に対して、四つの培養条件を三回試験する。
−10ng/mlのPMA(ホルボール12ミリステート13アセテート、Sigma)及び1μg/mlのイオノマイシン(Sigma)(ポジティブコントロール、ポリクローナル活性化)
−完全RPMI(ネガティブコントロール)
−10μg/mlのP161−241
−10μg/mlのP161−241+N
【0178】
5%CO2下において37℃で72時間培養した後、培養上清を回収し、ELISAによりIFN−γを滴定するまで−20℃で凍結する。
【0179】
IFN−γのELISA:96ウェルプレート(Immulon 2HB、 ThermoLabsystems)を、0.1Mの重炭酸緩衝液(100μl/ウェル)(pH9.5)中で、4μg/mlのマウス抗IFN−γ捕捉抗体(BD Biosciences)で4℃で一晩感作する。プレートをウェル当たり200μlの、0.05%tween20を加えたPBSで5回洗浄する(自動装置Wellwash、Labsystemsを使用)。次にプレートをウェル当たり150μlの、0.05%tween20及び2%ウシ血清アルブミン(PBS−T−BSA)を加えたPBS緩衝液で、37℃で2時間、コンフルエントな状態にする。5回洗浄した後、参照用の組換えマウスIFN−γ(R&D systems)及び滴定サンプルをPBS−T−BSAで二倍連続希釈により希釈する。IFN−γの希釈範囲は3312.5pg/mlから3.235pg/mlまでとする。四回の二倍連続希釈を純粋なサンプルに対して行う。続いてプレートを4℃で一晩インキュベートする。5回洗浄した後、ビオチン化した検出抗体(BD Biosciences)を添加し(PBS−T−BSA中に1μg/ml、100μl/ウェル)、4℃で3時間インキュベートする。5回洗浄した後、ストレプトアビジン−ペルオキシダーゼコンジュゲート(Pierce)を添加し(PBS−T−BSA中に1μg/ml、100μl/ウェル)、4℃で1時間インキュベートする。5回洗浄した後、ペルオキシダーゼの基質(ABTS+H2O2)をウェルに添加する。45分間のインキュベーションの後、光学密度を405nmで読み取る(読み取り装置ELISA Dynex)。サンプルのIFN−γ濃度の計算は、IFN−γの希釈範囲との比較から行う。
【図面の簡単な説明】
【0180】
【図1】Pタンパク質に対するPタンパク質−Nタンパク質の相互作用ドメインの位置決定を示す図である。
【図2】サイズ排除クロマトグラフィーにおけるN−PΔ161N複合体の溶離特性を示す図である。
【図3】RSVのNタンパク質の環状構造を示す図である。
【図4】N−PΔ161N複合体に対する抗体の産生を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示す図である。
【図5】PΔ161N及びN−PΔ161Nに特異的な細胞性免疫応答を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示すグラフである。
【0181】
参考文献
−Ames,T.R.1993.The epidemiology of BRSV infection.Vet.Med.881−884.
−Bhella,D.,Ralph,A.,Murphy L.B.,& Yeo,R.P.2002.Significant differences in nucleocapsid morphology within the Paramyxoviridae.Journal of General Virology; 83,1831−1839.
−Castagne,N.,A.Barbier,J.Bernard,H.Rezaei,J.−C.Huet,C.Henry,B.Da Costa,and J.−F.Eleouet.2004.Biochemical characterization of the Respiratory Syncytial Virus P−P and P−N protein complexes and localization of the P protein oligomerization domain.Journal of General Virology; 85: 1643−1653.
−Choi et al.,2004,Protein Expression and Purification; 38,pp205
−Elvander,M.1996.Severe respiratory disease in dairy cows caused by infection with bovine respiratory syncytial virus.Vet.Rec.; 138,101−105.
−Freytag,LC et Clements,JD.2005.Mucosal adjuvants.Vaccine.; 23(15): 1804−13.
−Gaddum,R.M.,R.S.Cook,J.M.Furze,S.A.Ellis & G.Taylor.2003.Recognition of bovine respiratory syncytial virus proteins by bovine CD8a T lymphocytes.Immunology; 108,220−229;
−Goulder PJ,Lechner F,Klenerman P,Mclntosh K,Walker BD.2000.Characterization of a novel respiratory syncytial virus−specific human cytotoxic T−lymphocyte epitope.J Virol.; 74(16): 7694−7.
−Johansson et al.,2003; Journal of Biological Chemistry vol.278 p 44567−44573.
−Khattar SK,Yunus AS,Samal SK.2001a.Mapping the domains on the phosphoprotein of bovine respiratory syncytial virus required for N−P and P−L interactions using a minigenome system.J Gen Virol.; 82(Pt4):775−9.
−Khattar SK,Yunus AS,Collins PL,Samal SK..2001b.Deletion and substitution analysis defines regions and residues within the phosphoprotein of bovine respiratory syncytial virus that affect transcription,RNA replication,and interaction with the nucleoprotein.Virology.; 285(2): 253−69.
−Kolakofsky D,Le Mercier P,Iseni F,Garcin D.2004.Viral DNA polymerase scanning and the gymnastics of Sendai virus RNA synthesis.Virology.; 318(2): 463−73.Review.
−Maggon K,Barik S.2004.New drugs and treatment for respiratory syncytial virus.Rev Med Virol.14(3): 149−68.Review.
−Mallipeddi SK,Lupiani B,Samal SK.1996.Mapping the domains on the phosphoprotein of bovine respiratory syncytial virus required for N−P interaction using a two−hybrid system.J Gen Virol.; 77(Pt5): 1019−23.
−Martinez X,Li X,Kovarik J,Klein M,Lambert PH,Siegrist CA.1999.Combining DNA and protein vaccines for early life immunization against respiratory syncytial virus in mice.Eur J Immunol.; 29(10): 3390−400.
−Mavrakis M,Iseni F,Mazza C,Schoehn G,Ebel C,Gentzel M,Franz T,Ruigrok RW.2003.Isolation and characterisation of the rabies virus N゜−P complex produced in insect cells.Virology.; 305(2): 406−14.
−McNeal MM,VanCott JL,Choi AH,Basu M,Flint JA,Stone SC,Clements JD,Ward RL.2002.CD4 T cells are the only lymphocytes needed to protect mice against rotavirus shedding after intranasal immunization with a chimeric VP6 protein and the adjuvant LT(R192G).J Virol.; 76(2): 560−8.
−Meric C,Spehner D,Mazarin V.1994.Respiratory syncytial virus nucleocapsid protein (N) expressed in insect cells forms nucleocapsid−like structures.Virus Res.31(2): 187−201.
−Perrin,B.,Dannacher,G.,et Solsona,M.1979.Mise en evidence des anticorps contre le virus respiratoire syncytial chez les bovins francais.Rec.Med.Vet.155,465−471.
−Samal SK,Pastey MK,McPhillips TH,Mohanty SB.1993.Bovine respiratory syncytial virus nucleocapsid protein expressed in insect cells specifically interacts with the phosphoprotein and the M2 protein.Virology.; 193(1):470−3.
−Slack MS,Easton AJ.1998.Characterization of the interaction of the human respiratory syncytial virus phosphoprotein and nucleocapsid protein using the two−hybrid system.Virus Res.; 55(2): 167−76.
−Sparer TE,Matthews S,Hussell T,Rae AJ,Garcia−Barreno B,Melero JA,Openshaw PJ.1998.Eliminating a region of respiratory syncytial virus attachment protein allows induction of protective immunity without vaccine−enhanced lung eosinophilia.J Exp Med; 187(11): 1921−6.
−Tarbouriech,N.,Curran,J.,Ruigrok,R.W.,& Burmeister,W.P.(2000).Tetrameric coiled coil domain of Sendai virus phosphoprotein.Nature Structural Biology 7,777−781.
−Taylor G,Bruce C,Barbet AF,Wyld SG,Thomas LH.2005.DNA vaccination against respiratory syncytial virus in young calves.Vaccine; 23(10): 1242−50
−Taylor,G.L.H.Thomas,J.M.Furze,R.S.Cook,S.G.Wyld,R.Lerch,R.Hardy and G.W.Wertz.1997.Recombinant vaccinia viruses expressing the F,G or N,but not the M2,protein of bovine respiratory syncytial virus (BRSV) induce resistance to BRSV challenge in the calf and protect against the development of pneumonic lesions.Journal of General Virology; 78,3195−3206.
−Thompson W.W.,D.K.Shay,E.Weintraub,L Brammer,N.Cox,L.J.Anderson,K.Fukuda.2003.Mortality associated with influenza and respiratory syncytial virus in the United States.JAMA.; 289(2): 179−86.
−Wellemans,G.,and J.Leunen.1975.Le virus respiratoire syncytial et les troubles respiratoires des bovins. Ann. Med. Vet.; 119,359−369.
【技術分野】
【0001】
本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法、こうして調製した複合体、さらにこの複合体から単離することが可能な可溶性Nタンパク質に関する。また、本発明はパラミクソウイルス科のこのNタンパク質−トランケート型Pタンパク質複合体又はNタンパク質を含むワクチン組成物に関する。好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片である。
【背景技術】
【0002】
フランスでは、大部分の国と同様、ウシの呼吸器合胞体ウイルス(RSV)が、70%以上の農場内の仔牛における、また約70%の生後一年までの仔牛における重い呼吸器疾患(細気管支炎、肺炎)の主要な原因となる因子となっている(Perrin et al., 1979、 Ames, 1993、 Elvander, 1996)。死亡率は20%に達する(Wellemans, 1992)。これは、毎年冬のこの疾患の定期的な発生を止める術がない飼育業者にとって大きな悩みの種である。したがって、有効な予防法の開発という課題に対して牧畜業者の強い要求がある。ワクチンは市販されているが、実際、効果は少ないか、または効果が見られない。
【0003】
ヒトにおいても同じ疾患が存在する。それは新生児の細気管支炎の原因となる因子である(Virologie、第7巻、2003年10月特別号、infections a virus respiratoire syncytial en pediatrie参照)。このヒトの疾患についてのデータは極めて詳細である。ウシと同様、フランスでは、ヒトにおいて、それは冬に流行する疾患である。生後一年の間に、乳児の70%がRSVに感染し、生後二年の間に100%が感染する。これは、フランスでは毎年500,000人の小児が感染していることを意味する。感染した乳児の20%で下気道の疾患(細気管支炎)が生じ、フランスにおいては入院率が2〜5%であり、それは8〜9日間にわたり、未熟児の10〜25%、肺異形成の場合では14〜45%、先天性心異常の場合では15〜25%で下気道の疾患が生じる。入院患者の約10%が集中治療室で治療されなければならない。死亡率は0.1%に近い。世界保健機関の最近の報告によれば、毎年世界で6400万人が感染し160,000人がRSVにより死亡すると推定されている。アメリカでは毎年18,000〜75,000人が入院し、2,000人近くが直接的に、17,000人が間接的に死亡している(Magon and Barik,2004)。RSVはまた、成人におけるインフルエンザ症状の原因の3分の1を占める。実際、RSVにより小児の4倍の高齢者が死亡すると推定されている。全体では、RSVにより年間300万〜500万人が死亡する。さらに、極めて若い年齢(生後4か月前)で罹患した成人において、まだ明らかでない複雑な免疫機構により、この疾患はぜんそくを起こすリスクを増加させると考えられる。
【0004】
たとえヒトRSV及びウシRSVが異なる二種のウイルスであるとしても、構造的、分子的、抗原的側面、またそれらが誘発する疾患(臨床像、感染の季節性、若年者の感染)が等しいこと、そして、非常に高価であるため稀にしか使用されないモノクローナル抗体と、効果が弱く細胞内で実際にATPを消費してしまい毒性が強い抗ウイルス剤の他には有効なワクチンも抗ウイルス剤も存在しないという事実から、それらは極めて近いものである。
【0005】
1960年代に、ホルマリンで固定したウイルスを用いた、幼い小児に対するワクチン接種の試験がアメリカで行われた。結果は悲惨なものであり、ワクチン接種を受けた小児は数ヵ月後のウイルスの自然感染の際に激しい症状を発し、しかもそのうちの数人は死亡する結果となった。
【0006】
この不幸な試験の後、この疾患に対するヒト用のワクチンは未だ存在しておらず、また、たとえフランスにおいてこの課題について本格的な研究がなされていなくても、ウシ用の市販ワクチンの効果は極めて疑わしい。しかし、複数のワクチンが、ウシにおけるワクチン接種後の事故の後に市場からなくなった。
【0007】
RSVに対するワクチンの開発の困難性は、次のような理由により説明することができる。
(1)RSVに対する局所的免疫記憶が短期的であるため、自然感染は再感染に対する防御免疫を付与しないこと、
(2)強い細胞性免疫及び液性免疫の誘導が、疾患の重篤さの増大と関連すること(ワクチン接種による感作)、
(3)経粘膜投与で効果的なワクチン接種を可能にする、アジュバント又はワクチン戦略が存在しないこと、
(4)新生児(生後2ヵ月以内)がワクチン接種の主要な対象であるが、彼らの免疫系は従来のワクチンに対する反応が悪いこと、
(5)感染を防がない移行抗体が血液中に存在すること、
(6)流行しているウイルス株が変化すること。
【0008】
サブユニットワクチンが開発されつつあり、そのいくつかはG表面タンパク質に基づいている。新しいタイプのワクチンは逆遺伝学によって(すなわち遺伝的修飾による株の弱毒化によって)得られ、BRSVと同様HRSVに対しても数年前から開発されている。目下のところ、ヒトに対する弱毒株の使用は確認されていない。これはおそらく弱毒株の免疫応答が弱いためであろう。
【0009】
仔牛に対するものも含めて、HRSV又はBRSVのF膜タンパク質及びG膜タンパク質に基づくDNAワクチンが開発中である(Taylor et al. 2005、 Martinez et al. 1999)。このタイプのワクチンはいくつかの免疫病理学的研究に関連する防御免疫を付与する。このワクチンの効果は改良の余地がある。
【0010】
RSVは、マイナス鎖の一本鎖RNAのゲノムを有するエンベロープウイルスである。このRNAは11個のタンパク質をコードし、ヌクレオカプシド(N)タンパク質によりカプシドに包まれ、L(ラージ)つまりポリメラーゼとコファクターP(リンタンパク質)の二つのタンパク質から成る、ポリメラーゼ複合体と関連する(図1)。この分子は、PによりLがゲノムRNAを認識することが可能になるという、ポリメラーゼの機能における重要な役割を有する。さらにRSVには二つのコファクターM2−1及びM2−2が存在し、これらはプロセシング(M2−1)及び転写/複製バランスの調節(M2−2)において役割を有する。
【0011】
ウイルス粒子の表面上には二つの主要なFタンパク質及びGタンパク質がある。Fタンパク質はウイルスのエンベロープと細胞膜の融合を可能にし、合胞体の形成に関与する。Gタンパク質はウイルスが細胞の表面に付着することを可能にする。Mタンパク質(マトリックス)はウイルスのエンベロープとポリメラーゼ/ゲノム複合体の仲介をする。F表面タンパク質及びG表面タンパク質の二つのタンパク質は、中和抗体の標的であるため、RSVの主要な免疫原性タンパク質である。
【0012】
しかし、マウスについてなされた研究は、Gタンパク質がTh2型の免疫病理学的ワクチン反応(IL−4とIL−5の産生及び好酸球の集合)の誘発に関与することを示した(Sparer et al. 1998)。それに対して、ウシの場合には、これら二つのタンパク質のいずれも免疫病理学的応答に関与せず、これらの二つのタンパク質は防御応答の確立を可能にする(Taylor et al. 1997)。
【0013】
現在、新規なワクチン候補の研究は、多数がFタンパク質及びGタンパク質に関するものである。
【0014】
しかし、ヒト及びウシにおいては、細胞を介する免疫、特に細胞傷害性T細胞の応答は、RSVに対する防御の不可欠な構成要素である。ウシの場合と同様ヒトの場合以外では、Nタンパク質が細胞傷害性T細胞の応答の主要な基礎である(Goulder et al., 2000)。仔牛がRSVに対するワクチン接種に適切なモデルである。RSVのNタンパク質を発現する組換えワクチンを使用すると、免疫応答のバランスの回復を可能にする細胞型(Th1)の応答が生じることが示されている(Taylor et al.,1997、Gaddum et al.,2003)。したがって、現在の研究全体が、複数のタンパク質、特にF表面タンパク質及びG表面タンパク質並びに内部のNタンパク質の組み合わせで構成される抗RSVワクチンに有利に働く。
【0015】
Nタンパク質は、感染細胞で最も多く発現するタンパク質であり、ウイルス粒子中で量的観点から最も重要なタンパク質の一つである(Collins et al., 2001)。それは、一本鎖RNAから構成されるウイルスゲノムを包囲し、大きなサイズのらせん構造を形成している。それが組換え体の形で単独で発現した時、Nタンパク質は非特異的に細胞RNA上に重合する。それらはそこで極めて巨大な、不溶性で精製不可能な、感染細胞で見られるヌクレオカプシドに似たらせん構造(RNA/N)を形成する(Meric et al., 1994、 Bhella et al., 2002)。
【0016】
このNタンパク質は、ウイルスポリメラーゼLのコファクターであるRSVのPタンパク質と相互作用することができる。ツーハイブリッド法と免疫沈降法を主として使用する相互作用ドメインの位置決定の研究が、様々なグループにより行われたが、スクリーニングの結果は否定的(相互作用は認められない)であった。ヒトウイルス又はウシウイルスのPタンパク質のC末端ドメインはNタンパク質との相互作用において重要な役割を有する疑いがあった(Garcia Barreno et al., 1996、 Mallipeddi et al., 1996、 Slack and Easton, 1998、 Khattar et al., 2001a, 2001b)。しかし、ツーハイブリッドシステムはNタンパク質とPタンパク質との間の相互作用の実際の性質を反映しないことが分かった(Khattar et al., 2001a)。その上、P−N複合体の正確な性質(各分子の数又は化学量、構造)は報告されておらず、相互作用のドメインは明示されても特徴を明らかにされてもいない。
【0017】
パラミクソウイルス科に属する近縁のウイルス(センダイウイルス、麻疹ウイルス)についてなされた研究から、Pタンパク質は、Nタンパク質と共にN°Pと呼ばれる可溶性複合体を形成し、細胞RNAに自身が非特異的に固定することを防ぐとの考えが導かれた(Kolakofsky et al., 2004)。Pタンパク質は、PがLのコファクターとして作用してLがその基質を「発見する」ことを可能にするため、Nタンパク質によりカプシドに包まれたRNAから形成されるヌクレオカプシドを認識することもできる。
【0018】
パラミクソウイルス科については、二つの相互作用ドメインがP上に発見されている。一方は、タンパク質のC末端の位置にあり、N−RNA複合体を認識するドメインを構成し、もう一方は、N末端の位置にあり、N°P複合体の形成を可能にする(Kolakowsky et al., 2004)。RSVについては、これらの複合体は明確に同定されておらず、Nと相互作用するPタンパク質のC末端ドメインの役割は明確に定義されていない。
【0019】
現在まで、Nタンパク質に基づくサブユニットワクチンの開発は、可溶性の形でNタンパク質を単離することが困難なため不可能であった。
【0020】
最近、本発明者は、細菌におけるPタンパク質及びNタンパク質の同時発現系を開発し、これをパラミクソウイルス科のRSVのモデルとして選んだ(Castagne et al., 2004)。Pタンパク質は、アンピシリン耐性を有するプラスミドにおいてグルタチオン−S−トランスフェラーゼ(GST)と融合させ、Nタンパク質はカナマイシン耐性を有するプラスミド中にクローニングした。同じ細菌中におけるこれらプラスミドの同時発現により、融合タンパク質GST−Pを精製すること及びNタンパク質を伴わせることが可能になった。
【非特許文献1】Kolakofsky D, Le Mercier P, Iseni F, Garcin D. 2004. Viral DNA polymerase scanning and the gymnastics of Sendai virus RNA synthesis. Virology.; 318(2):463−73. Review.
【非特許文献2】Castagne, N., A. Barbier, J. Bernard, H. Rezaei, J.−C. Huet, C. Henry, B. Da Costa, and J.−F. Eleouet. 2004. Biochemical characterization of the Respiratory Syncytial Virus P−P and P−N protein complexes and localization of the P protein oligomerization domain. Journal of General Virology; 85: 1643−1653
【発明の開示】
【発明が解決しようとする課題】
【0021】
しかしながら、依然として溶解性の問題が間違いなく存在するため、可溶性の形でのNタンパク質の生産量が、工業的規模におけるこの系の実施を検討するには依然として著しく不十分である。その上、こうして生産されたNタンパク質の性質も明らかにされなかった。
【課題を解決するための手段】
【0022】
本発明者は、Pタンパク質のN末端が欠失した突然変異体とRSVのNタンパク質との同時発現により、完全なPタンパク質で得られる場合と比較して格段に多い量のNタンパク質を精製することが可能になるということを明らかにした。
【0023】
定義
「パラミクソウイルス科」なる科は、パラミクソウイルス亜科とニューモウイルス亜科に再分類される。パラミクソウイルス亜科は、プロトタイプがセンダイウイルスであるレスピロウイルス属、そして、ルブラウイルス属(特に流行性耳下腺炎ウイルス)、及び麻疹ウイルスのようなモービリウイルス属を含む。レスピロウイルス属とルブラウイルス属の各属は、パラインフルエンザウイルス株を構成する。ニューモウイルス亜科は、ニューモウイルス属とメタニューモウイルス属の二つの属に再分類され、後者はヒトメタニューモウイルスを含む。ヒト呼吸器合胞体ウイルス(RSV)は、ニューモウイルス亜科に属するニューモウイルス属のプロトタイプのウイルスを構成する。ニューモウイルス属はまた、ウシRSV株及びマウスRSV株を含む。
【0024】
他に明言しない限り、「呼吸器合胞体ウイルス」は通常、考えられるあらゆる形態(ヒト、ウシ・・・)、サブグループ(例えばヒトRSVで同定されているサブグループA、B及びS)又は株であるRSVを指す。
【0025】
「Pタンパク質」という記載は、パラミクソウイルス科ウイルスのポリメラーゼ複合体から得られるリンタンパク質すなわちPタンパク質を指す。Pタンパク質はウイルスポリメラーゼ(レプリカーゼ/トランスクリプターゼ)のコファクターであり、リン酸化され得る。パラミクソウイルス科のPタンパク質の配列は当業者に知られている。例えば、ヒトRSVのLong株のPタンパク質は241個のアミノ酸からなる配列を有し、これは受託番号P12579でSwissprotデータベースに寄託されている。この配列は、配列番号1に示されている。ウシRSVのPタンパク質も同様に241個のアミノ酸からなる(配列番号23)。センダイウイルス(Harris株)、麻疹ウイルス(Edmonston B株)、流行性耳下腺炎ウイルス(SBL−1株)、及びヒトメタニューモウイルス(00−1株)のPタンパク質も同様に、受託番号P04859(配列番号2)、CAA91364(配列番号3)、P19717(配列番号4)、及びQ91KZ5(配列番号5)でそれぞれSwissprotデータベースに登録されている。「Pタンパク質」という記載は、完全なPタンパク質、トランケート型Pタンパク質、又はPタンパク質の断片を示し得る。
【0026】
パラミクソウイルス科のPタンパク質は、例えばセンダイウイルス又はRSVの場合、ホモオリゴマー特にホモテトラマーを形成する。RSVについては、オリゴマー化(P−Pオリゴマー化)することが可能なPタンパク質のドメインは、このタンパク質の120〜150番アミノ酸のところに位置決定されている(Castagne et al., 2004)。したがって、例えば、RSVのPタンパク質の161〜241番アミノ酸で構成される断片はオリゴマーを形成しない。センダイウイルスのPタンパク質のオリゴマー化ドメインは、Tarbouriechら(2000)により、Pタンパク質の320〜446番残基で構成されるものとして報告されている。他方、麻疹ウイルスのPタンパク質については、Pタンパク質のオリゴマー化領域は304〜376番アミノ酸のところに同定されている(Johansson et al., 2003)。
【0027】
「トランケート型Pタンパク質」という記載は、連続する一つ又は複数のアミノ酸配列が欠失したPタンパク質を指す。それらの欠失は、Pタンパク質の一次配列に関する、C末端配列、N末端配列、及び「内側」配列のトランケーション、又はこれらのトランケーションの組み合わせであり得る。
【0028】
本発明によるトランケート型Pタンパク質は、Pタンパク質のオリゴマー化ドメインを含まず、Nタンパク質と相互作用することができる。パラミクソウイルス科のPタンパク質とNタンパク質との相互作用ドメインがC末端のところに位置決定されたので、トランケート型Pタンパク質の例は好ましくは、Pタンパク質のC末端断片、又はPタンパク質のC末端断片(Nタンパク質と相互作用することができる)とPタンパク質の連続する少なくとも一つの他のアミノ酸配列との融合により構成された「キメラ」Pタンパク質を含む。前記C末端断片と前記Pタンパク質の他の配列は、それ自体は天然に連続しておらず、その中に配列のオーバーラップは存在しない。例えば、RSVのトランケート型Pタンパク質は天然Pタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸から構成される配列を有し得る。参照ポリペプチドの「断片」は、参照ポリペプチド配列中に見られるあらゆる連続したアミノ酸配列を指す。
【0029】
「Pタンパク質の断片」又は「PΔ」は、配列がPタンパク質のアミノ酸鎖からなり、Pタンパク質の連続する一つ又は複数のアミノ酸がN末端及び/又はC末端の末端から欠失しているポリペプチドを指す。
【0030】
「Pタンパク質のC末端断片」又は「PΔN」は、連続する一つ又は複数のアミノ酸がN末端の末端から欠失しているPタンパク質を指す。好ましくは、Pタンパク質のC末端断片はPタンパク質の一次配列のC末端側の半分にあるアミノ酸鎖を指す(配列のアミノ酸の数が奇数の場合には、N末端側の半分との兼ね合いで、一個の追加のアミノ酸をタンパク質のC末端側の半分に任意に加えることができる)。例えば、241個のアミノ酸を含むRSVのPタンパク質では、PΔ161NはPタンパク質の161〜241番アミノ酸により構成されるC末端断片を指す。同様に、例えば、507個のアミノ酸からなる麻疹ウイルス(Edmonston B株)のPタンパク質では、PΔ386NはPタンパク質の386〜507番アミノ酸により構成されるC末端断片を指す。
【0031】
「Pタンパク質のN末端断片」又は「PΔC」は、連続する一つ又は複数のアミノ酸がC末端の末端から欠失しているPタンパク質を意味する。
【0032】
「Pタンパク質の内側断片」又は「PΔNC」は、連続する一つ又は複数のアミノ酸がN末端の末端から欠失し、かつ連続する一つ又は複数のアミノ酸がC末端の末端から欠失しているPタンパク質を意味する。
【0033】
「Nタンパク質」は、ウイルスゲノムを囲むためにらせん構造を形成している、パラミクソウイルス科のヌクレオカプシドタンパク質を指す。ヒトRSVのLong株のNタンパク質は、391個のアミノ酸からなる配列を有し、これは配列番号6に示されている。ウシRSVのNタンパク質も同様に391個のアミノ酸からなる(配列番号24参照)。センダイウイルス(Hamamatsu株)、麻疹ウイルス(Edmonston B株)、流行性耳下腺炎ウイルス(SBL−1株)、及びヒトメタニューモウイルス(00−1株)のNタンパク質も同様に、受託番号Q9DUE3(配列番号7)、受託番号Q89933(配列番号8)、受託番号P21277(配列番号9)、及び受託番号Q91F57(配列番号10)でSwissprotデータベースにそれぞれ登録されている。
【0034】
上記Pタンパク質及びNタンパク質の配列は例示的な特徴を有し、これらの配列は所与のウイルスについて考えられる特定の株に応じて変化する可能性がある。このように、本出願の明細書の以下に記載されるアミノ酸の位置はこれらの参照配列について示したものである。当業者は、これらの例示以外のウイルス株における対応するドメインを完全に同定することが可能である。
【0035】
パラミクソウイルス科ウイルスのこれらのNタンパク質及びPタンパク質をコードする配列も当業者に知られている。
【0036】
「タグタンパク質」とも呼ばれる「標識タンパク質」は、関心のあるタンパク質と融合して精製を容易にするために使用されるタンパク質を指す。標識タンパク質は当業者に知られている。標識タンパク質の例としては、グルタチオン−S−トランスフェラーゼ(GST)、又は4〜10個のヒスチジン残基鎖を通常含む配列であるヒスチジンタグが挙げられる。
【0037】
本発明の内容においては、「相同」という語は、同じ進化上の起源を有するタンパク質間、例えば異なる種、又はウイルスの場合では異なるウイルス株が有する相同タンパク質間に存在する関係に関するものである。このようなタンパク質(及びそれをコードする遺伝子)は配列の相同性を示すものであるが、その相同性は、それらタンパク質の配列の類似性に、つまり、類似の割合、すなわち保存された位置における特定の残基又はモチーフの存在に関して反映される。
【0038】
「配列の類似性」という表現は、同じ進化上の起源を共有し得るか又はそうでない核酸配列間又はタンパク質のアミノ酸配列間の同一性の程度を示す。通常、相同と類似性という語は区別なく使用される。二つのアミノ酸が少なくとも80%同一又は少なくとも90%類似(すなわち機能的に同一)の際、それら二つのアミノ酸の配列は「基本的に相同」である。類似又は相同の配列は、例えばBLASTプログラム又はFASTAプログラムを使用して、アラインメントにより同定することができる。
【0039】
本発明によるタンパク質又は複合体の溶解度は、1×PBS、150mMのNaClを加えた10mMのTris緩衝液(pH7.4〜8.0)、0.2×TBE、さらに例えば、60mMのNaCl、1mMのEDTA、2mMのDTT、0.2%のTriton X−100、10mMのMgSO4、1mMのCaCl2及び1mg/mlのリゾチームを加えた50mMのTris−HCl(pH7.8)を含む細菌溶解緩衝液のような、緩衝化水性溶媒に対して定義される。
【0040】
可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法
本発明者は、Pタンパク質とGSTとの融合物及びパラミクソウイルス科のNタンパク質をそれぞれコードするプラスミドの同時発現により、Nタンパク質を伴う融合タンパク質GST−Pを得ることが可能になることを先に示した(Castagne et al., 2004)。しかし、N−P複合体の生産量は少ないので、この複合体の工業的規模における生産と相容れない。
【0041】
本発明者は、Pタンパク質の欠失突然変異体の特徴を明らかにし、Nタンパク質と相互作用するその能力を判定した。そして、この突然変異体のいくつかもまたRNA−N複合体すなわちリボヌクレオカプシド(RNP)の形のNタンパク質と相互作用することができること、また同時に、完全なPタンパク質で得られるものよりも極めて顕著に多い収量のN−P複合体の調製を可能にすることを明らかにした。この特定の突然変異体は、分子のC末端部分を含みPタンパク質のオリゴマー化ドメインを欠くPタンパク質の断片に相当する。
【0042】
したがって、これらPタンパク質の突然変異体とNタンパク質との同時発現は、大量の、可溶性RNPの形、特にNタンパク質−トランケート型Pタンパク質複合体、特にNタンパク質−Pタンパク質C末端断片複合体の形でのNタンパク質の調製を可能にする。
【0043】
したがって、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製する方法に関するものであって、前記方法は、
a)パラミクソウイルス科ウイルスのNタンパク質を、パラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、トランケート型Pタンパク質と同時発現させること、及び
b)こうして形成された可溶性Nタンパク質−トランケート型Pタンパク質複合体を回収すること
から成る過程を含む。
【0044】
トランケート型Pタンパク質は、好ましくはPタンパク質のC末端断片からなる。パラミクソウイルス科のPタンパク質と、場合によりN−RNA複合体の形であるNタンパク質との相互作用ドメインは、実際、Pタンパク質のC末端側に位置する。
【0045】
トランケート型Pタンパク質は、Pタンパク質のC末端断片とPタンパク質の連続する少なくとも一つの他のアミノ酸配列との融合により構成された、上に定義したような「キメラ」Pタンパク質であり得る。
【0046】
好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片である。
【0047】
従って、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)を調製する方法に関するものであって、前記方法は、
a)パラミクソウイルス科ウイルスのNタンパク質とパラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、Pタンパク質のC末端断片とを同時発現させること、
b)こうして形成されたN−PΔN可溶性複合体を回収すること
から成る過程を含むものである。
【0048】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスはヒト又はウシの呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0049】
当業者は、Nタンパク質と相互作用することができる、トランケート型Pタンパク質、又はさらに具体的にはPタンパク質のC末端断片を知っているか又は決定することができる。
【0050】
例えば、RSVの場合、本発明者はPタンパク質とNタンパク質との相互作用ドメインの位置決定をするために、先に報告された、大腸菌におけるNタンパク質及びPタンパク質の同時発現手段(Castagne et al., 2004)を使用した。そのために、Nタンパク質を、GSTと融合したPタンパク質の欠失突然変異体と同時発現させた。本発明者はまた、Pタンパク質とNタンパク質との相互作用ドメインがPタンパク質のC末端の末端のところに位置していることを明らかにした(図1)。本発明者は、特に、RSVのPタンパク質のC末端の9個のアミノ酸から構成されるオリゴペプチド(233〜241番アミノ酸)までのPタンパク質のC末端断片が、Nタンパク質と相互作用することができることを示した。
【0051】
他方で、例えば、センダイウイルスのPタンパク質の、「Xドメイン」又はXDと呼ばれる、RNA−N複合体すなわちRNPの形のNタンパク質との相互作用ドメインが、473〜568番アミノ酸に該当することが報告されている(Kolakofsky et al. 2004)。
【0052】
それ以外のパラミクソウイルス科については、必要に応じて、当業者は本発明者が報告した手段を使用して、ヌクレオカプシドの形のNタンパク質と相互作用するドメインをPタンパク質に同定することができる。
【0053】
本発明者はまた、RSVのPタンパク質の特定のC末端断片、とりわけPΔ161N断片(161〜241番アミノ酸)が、実際には工業的規模に充分な収量を得ることができない完全なPタンパク質の場合と比較して、大量のNタンパク質の調製を可能にすることを明らかにした。非常に小さな、9個のアミノ酸しか有さないPΔ233N(233〜241番アミノ酸)までの欠失突然変異体が、PΔ161Nの場合の収量に匹敵する収量を得ることを可能にする。
【0054】
PΔ161Nよりも小さいこれらの断片は、Nタンパク質と相互作用することはできるがもはやオリゴマー化することができない、すなわちもはやPタンパク質のオリゴマー化ドメインを含まない、RSVタンパク質の断片に対応する。RSVのPタンパク質の最小のオリゴマー化ドメインは、実際、Pタンパク質の120〜150番アミノ酸付近で定義される。
【0055】
この同じ手段により、本発明者は、386〜507番アミノ酸残基から構成される麻疹ウイルスのPタンパク質のC末端断片(PΔ386N)が、このウイルスのNタンパク質と相互作用し、その精製を可能にするということを示すことができた。一方、Pタンパク質のN末端部分において456番残基までが欠失していると(PΔ457N断片を含む)、Nタンパク質の精製はできない。リボヌクレオカプシドと相互作用するPタンパク質のC末端領域の構造は、Johansson et al.(2003)により特定されている。Pタンパク質のオリゴマー化領域は、304〜376番アミノ酸で定義されたような推定と欠失により特定された。
【0056】
したがって、RNPの形のNタンパク質と相互作用するドメインを含むがPタンパク質のオリゴマー化ドメインが欠失している、Pタンパク質のC末端断片を使用すると、Pタンパク質断片とNタンパク質との相互作用、可溶性N−PΔN複合体の形成、及び高収量でのこの複合体の生産が同時に可能になる。特定のメカニズムに結びつけることはしないが、Pタンパク質のオリゴマー化ドメインが存在しないと、この複合体のPタンパク質同士間の相互作用に関連したN−PΔN複合体の不溶性の問題が回避されると推定される。
【0057】
さらに、一実施形態によると、N−PΔN複合体を調製する方法は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠失する、RSVのPタンパク質のC末端断片を発現させることを含む。
【0058】
さらに具体的には、本発明による方法では、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、RSVのPタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を、RSVのNタンパク質と同時発現させることができる。
【0059】
RSVのPタンパク質のC末端断片は、例えば、PΔ120N(Pタンパク質の120〜241番アミノ酸)、PΔ150N(Pタンパク質の150〜241番アミノ酸)、PΔ161N(Pタンパク質の161〜241番アミノ酸)、PΔ180N(Pタンパク質の180〜241番アミノ酸)、PΔ200N(Pタンパク質の200〜241番アミノ酸)、PΔ220N(Pタンパク質の220〜241番アミノ酸)、PΔ230N(Pタンパク質の230〜241番アミノ酸)、及びPΔ233N(Pタンパク質の233〜241番アミノ酸)から構成される群から選択され得る。
【0060】
本発明はまた、RSVのNタンパク質と、上記のようなRSVのPタンパク質のC末端断片を含み、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、トランケート型Pタンパク質とを同時発現させる方法に関するものである。
【0061】
例えば、Pタンパク質のC末端断片を含むトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合から構成され得る。それは例えば、配列番号1に示すようなヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸と161〜241番アミノ酸との連結から構成され得る。
【0062】
別の一実施形態によれば、パラミクソウイルス科は麻疹ウイルスであり、N−PΔN複合体を調製する方法は、Pタンパク質のC末端のアミノ酸を最大122個含むか、又はPタンパク質のC末端の122個のアミノ酸で構成される、麻疹ウイルスのPタンパク質のC末端断片を発現させることを含む。それらの断片は、特に、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜507番アミノ酸から構成されるC末端断片(PΔ386N)、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であり得る。
【0063】
本発明による方法を利用するに当たり、分子生物学、微生物学、又は組換えDNAにおける通常のあらゆる技術を使用することができる。このような技術は当業者に既知であり、特に次のものに記載されている。Sambrook,Fritsch&Maniatis、分子クローニング:A Laboratory Manual 第二版(1989)、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York(「Sambrook et al., 1989」)。DNAクローニング:A Practical Approach、第I巻及び第II巻(D.N. Glover編、1985)。オリゴヌクレオチド合成(M.J. Gait編、1984)。核酸のハイブリダイゼーション[B.D.Hames&S.J.Higgins編(1985)]。転写及び翻訳[B.D.Hames&S.J.Higgins編(1984)]。動物細胞の培養[R.I.Freshney編(1986)]。固定化細胞及び固定化酵素[IRL Press,(1986)]。B. Perbal,A Practical Guide To Molecular Cloning(1984)。F.M.Ausubel et al.(編)、Current Protocols in Molecular Biology,John Wiley&Sons,Inc.(1994)。
【0064】
「発現する」又は「発現」は、遺伝子又はDNA配列に含まれる情報が現れることを可能にすること又は現れるようにすること、例えば、対応する遺伝子配列又はDNA配列の転写と翻訳に関与する細胞の機能を活性化することによるタンパク質の産生を意味する。二つの遺伝子又はDNA配列に含まれる情報が同じ宿主細胞内で発現することを「同時発現」という。
【0065】
「コード配列」は、それが発現するとRNA、ポリペプチド、タンパク質等が生成するヌクレオチド配列を意味する。タンパク質をコードする配列は一般に開始コドン(ATG)と終止コドンを含む。
【0066】
コード配列は、RNAポリメラーゼがコード配列をRNA、特にmRNAに転写し、次いでそれがイントロンを含む場合にはスプライシングされ、コードする配列によりコードされているタンパク質に翻訳されるとき、転写及び翻訳の制御配列「の制御下にある」か又はそれら「と機能的に関連している」。
【0067】
「ベクター」、「クローニングベクター」及び「発現ベクター」という記載は、DNA配列又はRNA配列(例えば異種遺伝子)を宿主細胞に導入し、その結果、宿主細胞を形質転換し、導入された配列を発現させることができる担体を意味する。ベクターの例としては、プラスミド、ファージ、ウイルスが挙げられる。もっとも一般的なベクターは、通常は細菌に由来する、自己複製ユニットである、二本鎖DNAの形態であり得るプラスミドである。プラスミドは外来DNA配列を容易に組み込むことができ、それを適当な宿主内に容易に導入することができる。一つのプラスミドベクターは通常、一つのコードDNA配列、一つのプロモーターDNA配列を含み、外来DNAの導入が可能な一つ又は複数の制限部位を有する。プラスミドの非制限的な例としては、pKK(Clonetech)、pUC及びpET(Novagen,Inc.、マディソン、ウィスコンシン州)、pRSET及びpREP(Invitrogen、サンディエゴ、カリフォルニア州)、pMAL(New England Biolabs、ビバリー、マサチューセッツ州)、又はpGEX−4T−3(Pharmacia)のプラスミドが挙げられる。
【0068】
「宿主細胞」は、細胞による物質の生産、例えば、細胞による、遺伝子、DNA配列又はRNA配列、タンパク質又は酵素の発現のために選択され、修飾され、培養され又は操作される、あらゆる細胞又は生物を意味する。
【0069】
「発現系」は、ベクターにより担持され宿主細胞内に導入される外来DNAによってコードされるタンパク質の生産に適した条件下で使用される、宿主細胞及び適当なベクターを指す。通常の発現系は、大腸菌宿主細胞及びプラスミドベクター、昆虫細胞、及びバキュロウイルスベクター、またさらには哺乳類細胞及びベクターを含む。
【0070】
本発明の方法による発現系は、有利には、例えばpGEX−4T−3のようなベクターを有する、細菌特に大腸菌の発現系である。細菌の発現系は実際、一般にきわめて高い生産量が得られることを可能にする発現系である。
【0071】
有利には、トランケート型Pタンパク質及び特にPタンパク質のC末端断片は、Nタンパク質−トランケート型Pタンパク質複合体の精製を容易にするタンパク質と融合した形、とりわけアフィニティークロマトグラフィーに使用できるタンパク質と融合した形で発現する。それらのタンパク質は、トランケート型Pタンパク質−GST融合タンパク質が、グルタチオンを結合した固体支持体上でのクロマトグラフィーにより単離できる場合、グルタチオン−S−トランスフェラーゼ(GST)のような標識タンパク質であり得る。ポリヒスチジンすなわち「His−Tag」のような他の標識又は「タグ」も使用することができる。
【0072】
こうして、Nタンパク質−トランケート型Pタンパク質−標識タンパク質複合体(トランケート型Pタンパク質、特にPΔN断片と融合したGST又は他の標識タンパク質)が得られ、標識タンパク質は酵素切断により除去することができる。例えば、GSTはトロンビンによる切断により、又は融合体がGST以外のタンパク質を含む場合は他の全ての適当な酵素により除去することができる。
【0073】
本発明による方法の利用を可能にするベクター構築物の具体例を以下の実施例に示す。
【0074】
可溶性Nタンパク質−トランケート型Pタンパク質複合体
【0075】
上述したような、可溶性Nタンパク質−トランケート型Pタンパク質複合体、特に可溶性Nタンパク質−Pタンパク質C末端断片複合体の調製方法により、単離又は精製された形でNタンパク質−トランケート型Pタンパク質複合体を容易に得ることを可能にする。
【0076】
したがって、本発明はまた、本発明の調製方法により得ることが可能な、パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体に関するものである。
【0077】
好ましくは、トランケート型Pタンパク質は、Pタンパク質のC末端断片を含むか、又はPタンパク質のC末端断片である。
【0078】
本発明は、さらに具体的には、本発明の調製方法により得ることが可能な、パラミクソウイルス科のウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)に関するものである。
【0079】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(HMPV)及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスは、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0080】
一実施形態によれば、パラミクソウイルス科のウイルスは呼吸器合胞体ウイルス(RSV)であり、前記Pタンパク質のC末端断片は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている。
【0081】
さらに具体的には、前記Pタンパク質のC末端断片は、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びているか、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片を含む
ことができる。
【0082】
それらの断片は、特に、PΔ120N、PΔ150N、PΔ161N、PΔ180N、PΔ200N、PΔ220N、PΔ230N、及びPΔ233Nから構成される群から選択されるPタンパク質のC末端断片であり得る。
【0083】
また、本発明は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、上記に記載のようなRSVのPタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質を含む、可溶性の複合体に関する。
【0084】
例えば、Pタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合により構成され得る。それは、例えば配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸との連結により構成され得る。
【0085】
別の一実施形態によれば、パラミクソウイルス科のウイルスは麻疹ウイルスであり、前記Pタンパク質の断片は、Pタンパク質のC末端のアミノ酸を最大122個含むか又はPタンパク質のC末端の122個のアミノ酸で構成されている、Pタンパク質のC末端断片である。さらに具体的には、麻疹ウイルスのPタンパク質の前記C末端断片は、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜514番アミノ酸(PΔ386N)から構成されていてもよく、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であってもよい。
【0086】
N−PΔN複合体において、タンパク質PΔNは、場合により、例えばGST、ヒスチジンタグ、又はN−PΔN複合体の精製を容易にする他のあらゆる適当なタンパク質のような標識タンパク質と融合した形で提供され得る。
【0087】
RSVについての複合体生成物の電子顕微鏡分析から、それが10分子のNタンパク質を含む環状化合物であり、この可溶性の環はまた、アガロースゲル電気泳動で確認することが可能な約70塩基の小さなRNAを含むことが明らかになった(図3)。この複合体中には同様の量のNタンパク質とPΔN−GSTタンパク質がみられる。RNAは、このタンパク質を変性させることなくNタンパク質の環から分離することができない。
【0088】
このように、本発明はまた、上記で定義したような、10分子のNタンパク質を含み、そのうちのそれぞれ又はその多数がPタンパク質のC末端断片と関連し、前記Pタンパク質のC末端断片がRSVのPタンパク質のC末端の先端の9個のアミノ酸を含みRSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、呼吸器合胞体ウイルス(RSV)の単離された可溶性Nタンパク質−Pタンパク質C末端断片複合体にも関する。RSVのこのN−PΔN複合体は、さらに約70塩基のRNAを含む。
【0089】
可溶性Nタンパク質を調製する方法
【0090】
このNタンパク質−トランケート型Pタンパク質複合体から、又はさらに具体的にはN−PΔNから、例えばサイズ排除クロマトグラフィー(ゲル濾過)により、可溶性Nタンパク質を環状でそのRNAと共に容易に単離することができる。この分離は、必要に応じて、トランケート型Pタンパク質と、場合によりトランケート型Pタンパク質が融合している標識タンパク質との、酵素切断による分離の後に行うことができる。
【0091】
したがって、本発明はさらに、
a)上記に定義したような方法により可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製すること、そして
b)可溶性Nタンパク質−トランケート型Pタンパク質複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む、パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法に関する。
【0092】
好ましくは、トランケート型Pタンパク質は、Pタンパク質のC末端断片を含むか、またはPタンパク質のC末端断片である。
【0093】
本発明は、さらに具体的には、
a)上記に定義したような方法により可溶性Nタンパク質−Pタンパク質C末端断片複合体(「N−PΔN複合体」)を調製すること、そして
b)可溶性N−PΔN複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む、パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法に関する。
【0094】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、ヒトメタニューモウイルス、及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、ウイルスは、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)のようなニューモウイルスである。
【0095】
上記の方法により得ることができるパラミクソウイルス科ウイルスの可溶性Nタンパク質もまた、本発明の一部をなす。
【0096】
RSVの場合、利用できる最大のPタンパク質のC末端断片(PΔ161N)が15kDaの分子量を有するのに対し、Nタンパク質は450kDaの分子量を有する。したがって、Nタンパク質の環は、例えば下記の実施例3(及び図2)に記載するようなセファデックスカラム上でのクロマトグラフィーにより、Pタンパク質のC末端断片から分離することができる。
【0097】
一実施形態によると、本発明はしたがって、可溶性のRSVのNタンパク質を提案し、この前記Nタンパク質は、約7nmの直径を有し10個のサブユニットを含む環を形成している。しかし、ある種の部分的で10個より少ないサブユニットを含む環を除外することしかできない。環はさらに約70塩基のRNAを含む。
【0098】
ワクチン組成物
【0099】
RSV、さらに一般的にはパラミクソウイルス科のNタンパク質は、ワクチン接種にとって有益な抗原であるが、現在までのところ、可溶性の形でそれを精製することに誰も成功していない。本発明による方法は、極めて純度が高くかつ極めて均質な、環状構造のNタンパク質を容易かつ大量に得ることを可能にする。
【0100】
環状Nタンパク質の免疫原性を評価するため、本発明者は、本発明によるパラミクソウイルス科のN−PΔN複合体でマウスに免疫を付与した。ワクチンとして使用するために、Nタンパク質とPΔNは場合により分離され得るが、この操作は必要なものではなく、PΔNが存在しても弊害はない。
【0101】
本発明者は、マウスにRSVのN−PΔ161N複合体でさらに特異的に免疫を付与し、RSVのPΔ161Nポリペプチドをコントロールとして使用した。
【0102】
気道に対するこのウイルスの特異性を考慮して、非経口のワクチン接種の従来の方法である皮下経路、及び呼吸粘膜とそれに関連するリンパ組織レベルでの局所的な免疫付与の誘発を可能にする鼻経路の、二つの免疫付与経路を比較した。
【0103】
ワクチンアジュバントの不存在下で可溶性の組換えタンパク質に対する免疫応答を得ることが困難なので、本発明者は、粘膜経路でのアジュバント性が報告されている(McNeal et al. 2002, Freytag et Clements 2005)、大腸菌(R192G)の無毒化リンホトキシン(LT)を共に使用した(Dr.J.D.Clements、アメリカ、から取得)。
【0104】
免疫応答のパラメーター、(i)血清及び粘液の抗体産生(気管支肺胞洗浄による)、及び(ii)脾臓又は肺から単離された記憶Tリンパ球によるIFN−γの産生を介した細胞性免疫応答について追跡した。
【0105】
アジュバントLT(R192G)の存在下で、Nタンパク質(N−PΔ161N複合体)は、投与経路のいかんにかかわらず(経鼻又は皮下)血清抗体の産生を強く誘導する(図4A)。この応答は最初の免疫付与の後(14日目)に検出可能であり、二度目の免疫付与の後(28日目)に増強される。アジュバントはまた、二度目の免疫付与の後にPΔ161Nに対する抗体の産生を誘導することを可能にする。
【0106】
大いに注目すべきことには、アジュバントの不存在下でのN−PΔ161N複合体の経鼻及び皮下投与は、最初の免疫付与からすぐに血清抗体を多く産生させる。同じ条件下でPΔ161Nは応答を誘導しない。比較した応答の特徴は、粘液の抗体、総Igの産生に見られる(図4B)。
【0107】
これに対して、LT(R192G)存在下でNタンパク質(N−PΔ161N複合体)を単独で経鼻投与すると、呼吸粘膜レベルでのIgA産生を可能にする(図4B)。
【0108】
細胞性免疫応答は、脾臓又は肺の白血球の、N−PΔ161N又はPΔ161Nのみの存在下においてIFN−γをインビトロで産生する能力により測定した。IFN−γは、CD4及びCD8Tリンパ球により産生されるサイトカインである。CD4及びCD8記憶Tリンパ球は、それらが特異的な抗原の存在下で、インビトロで反応性がある。IFN−γは、ウイルスに対する防御における主要なエフェクターであるTh1リンパ球及び細胞傷害性Tリンパ球により産生される。
【0109】
最も強い応答はN−PΔ161N複合体による再刺激後に得られる(図5)。抗体の場合のように、N−PΔ161N複合体の経鼻又は皮下投与により、Nタンパク質に特異的な末梢細胞(脾臓)の免疫性が刺激される。注目すべきことには、アジュバントとともに経鼻経路で投与したNタンパク質により局所細胞(肺)の免疫が生じる(図5)。
【0110】
結論として、環状構造のNタンパク質は局所応答(呼吸粘膜)を刺激することを含め、強い免疫原性を示す。
【0111】
したがって、本発明は、パラミクソウイルス科ウイルスの可溶性Nタンパク質を含むワクチン組成物であって、前記可溶性Nタンパク質が医薬的に許容される担体中に存在するワクチン組成物を提案する。この可溶性Nタンパク質は、特に、上記のような可溶性Nタンパク質−トランケート型Pタンパク質複合体、さらに具体的には可溶性N−PΔN複合体から可溶性Nタンパク質を分離する方法により得ることが可能である。
【0112】
上述したように、可溶性Nタンパク質はPタンパク質との複合体の形で、好ましくない作用を伴わずにワクチン接種に使用され得る。したがって、本発明によるワクチン組成物は、医薬的に許容される担体中にある、パラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質複合体を含むことができる。
【0113】
これら複合体の生産量の問題については、本発明は好ましくは、医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を含むワクチン組成物であって、前記トランケート型Pタンパク質がPタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができるワクチン組成物を対象とする。
【0114】
好ましくは、トランケート型Pタンパク質はPタンパク質のC末端断片を含むか、又はPタンパク質のC末端断片である。
【0115】
本発明はまた、さらに具体的には、医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−Pタンパク質C末端断片複合体であって、前記Pタンパク質のC末端断片がPタンパク質のオリゴマー化ドメインを含まず、Nタンパク質と相互作用することができる複合体に関する。
【0116】
前記パラミクソウイルス科のウイルスは、パラミクソウイルス亜科又はニューモウイルス亜科であり得る。特に、ウイルスは、流行性耳下腺炎ウイルス、麻疹ウイルス、及びパラインフルエンザウイルスから構成される群から選択され得る。好ましくは、パラミクソウイルス科ウイルスの可溶性Nタンパク質は、例えばヒト又はウシの、呼吸器合胞体ウイルス(RSV)の可溶性Nタンパク質である。
【0117】
一実施形態によれば、パラミクソウイルス科のウイルスは呼吸器合胞体ウイルス(RSV)であり、前記Pタンパク質のC末端断片は、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いているC末端断片(PΔN)である。
【0118】
さらに具体的には、前記組成物において、RSVのPタンパク質のC末端断片は、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から、120位、好ましくは150位、さらに好ましくは161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、C末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
であり得る。
【0119】
それらの断片は、特に、PΔ120N、PΔ150N、PΔ161N、PΔ180N、PΔ200N、PΔ220N、PΔ230N、及びPΔ233Nから構成される群から選択されるRSVのPタンパク質のC末端断片であり得る。
【0120】
本発明はまた、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも119個、好ましくは149個、さらに好ましくは160個のアミノ酸を欠いている、上記のようなRSVのPタンパク質のC末端断片をRSVのトランケート型Pタンパク質が含んでいる組成物に関する。
【0121】
例えば、Pタンパク質のC末端断片を含むRSVのトランケート型Pタンパク質は、RSVのPタンパク質のN末端の先端の122個のアミノ酸とC末端の先端の80個のアミノ酸との融合により構成され得る。それは、例えば配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の1〜121番アミノ酸及び161〜241番アミノ酸との連結により構成され得る。
【0122】
別の一実施形態によれば、パラミクソウイルス科のウイルスは麻疹ウイルスであり、前記Pタンパク質の断片は、Pタンパク質のC末端のアミノ酸を最大122個含むか又はPタンパク質のC末端の122個のアミノ酸で構成されている、Pタンパク質のC末端断片である。さらに具体的には、麻疹ウイルスのPタンパク質の前記C末端断片は、配列番号3に示すような、麻疹ウイルスのEdmonston B株のPタンパク質の386〜514番アミノ酸(PΔ386N)から構成されていてもよく、又はEdmonston株のPタンパク質について定義したものと相同な、麻疹ウイルスの他の株に由来するPタンパク質のC末端断片であってもよい。
【0123】
「医薬的に許容される担体」は、ヒト又は動物において例えばアレルギーのような副作用を生じさせない、あらゆる溶媒、分散媒、吸収遅延剤等を意味する。
【0124】
有利には、本発明によるワクチン組成物はさらにアジュバントを含み得る。「アジュバント」は、同時に投与された抗原に対する細胞性又は液性免疫反応を増大させ、刺激し、活性化し、強化し、又は調節する物質を指す。従来のアジュバントの例は、完全フロイントアジュバント、LPS及びそれらの派生物、細菌毒素(コレラ毒素及びエンテロトキシン)及びその無毒化変異体(例えばLT(R192G))のような細菌抗原、CpGモチーフを含むオリゴヌクレオチド配列、水酸化アルミニウム(Alum)、リン酸カルシウム又はリン酸カリウムのような無機アジュバント、油性エマルジョン及び乳化剤(サポニン、例えばQS21)、サイトカインを含むアジュバントを含む。
【0125】
本発明によるワクチン組成物は、パラミクソウイルス科のウイルスによる感染に対する防御を提供し、すなわちワクチン組成物により免疫を付与されていない対象と比較してこの感染の影響の程度を軽減する。
【0126】
本発明はまた、ワクチン接種方法における上記で定義したようなワクチン組成物の使用に関する。
【0127】
したがって、本発明は、少なくとも本発明によるワクチン組成物を対象に投与することを含む、ワクチン接種方法を対象とする。好ましくは、ワクチン接種方法は、対象へのワクチン組成物の最初の投与と、同じ対象に対する前記ワクチン組成物の二度目の投与を含む。二度目の投与は、患者を抗原に再びさらし、より強い二次免疫応答を誘導する。
【0128】
「対象」という記載は、パラミクソウイルス科ウイルスによる感染にさらされるか又はさらされるおそれがある、ヒト又はヒトでない動物、例えば鳥、又はウシ、げっ歯類、イヌ、ネコ、ブタ、サルといった哺乳類を指す。好ましくは、本発明の趣旨の範囲内の対象はヒト又はウシである。
【0129】
ワクチン組成物は、有利には、パラミクソウイルス科ウイルスによる感染に対する防御的又は治療的な免疫応答の誘導に有効な量を投与される。用量は当然、検討される有効成分、投与方法、対象の年齢及び状態に依存する。投与量当たりのN−P複合体、N−PΔN複合体、又はNタンパク質は0.1〜200μgの間とすることができ、好ましくはワクチン投与量当たり10〜100μgの間である。
【0130】
ワクチン組成物は、いかなる経路、特に粘膜経路(例えば眼内、鼻腔内、経口)、又は非経口経路(例えば皮下、皮内、筋肉内、静脈内、又は腹腔内)によっても投与することができる。
【0131】
診断への使用
【0132】
場合により可溶性Nタンパク質−トランケート型Pタンパク質複合体の形である、パラミクソウイルス科ウイルスの可溶性Nタンパク質は、加えて、前記パラミクソウイルス科ウイルスのNタンパク質に対する抗体を検出するための診断用途に使用することができる試薬を構成する。
【0133】
したがって、本発明はさらに、上記のような、場合によりNタンパク質−Pタンパク質複合体の形であるパラミクソウイルス科ウイルスのNタンパク質を含む診断試薬に関する。
【0134】
前記試薬と適当な検出手段を含む診断キットもまた、本発明の範囲にある。
【0135】
本発明はまた、インビトロ又はインビボで前記Nタンパク質に対する抗体を検出するための、本発明によるパラミクソウイルス科ウイルスのNタンパク質の使用を提案する。
【0136】
本発明はまた、生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に対する特異的な抗体の検出方法であって、
a)前記生物サンプルとパラミクソウイルス科ウイルスのNタンパク質とを接触させること、
b)形成されたNタンパク質/抗体複合体を検出すること
から構成される過程を含み、この複合体の存在が生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に特異的な抗体の存在の指標となる方法に関する。
【0137】
生物サンプルは、例えば筋肉、肝臓、心臓、脳などの生検により得られた組織サンプル、又は、例えば血液、血漿、又は脳脊髄液のような生体液のような液体サンプルであり得る。
【0138】
複合体の検出は、(サイズ排除クロマトグラフィー、アフィニティークロマトグラフィーなどの)クロマトグラフィー又は非変性条件下での電気泳動のような、当業者に周知の常用の手段により実施され得る。
【0139】
上記で定義したようなNタンパク質に対する特異的な抗体の検出方法において、生物サンプルと接触させるNタンパク質は、Nタンパク質−Pタンパク質複合体の形を有し得る。
【0140】
以下の実施例及び図面は本発明の範囲を限定することなく説明するものである。
【0141】
図1は、Pタンパク質に対するPタンパク質−Nタンパク質の相互作用ドメインの位置決定を示す図である。Pタンパク質をGSTに融合し、別のプラスミド上で発現したNタンパク質と大腸菌内で同時発現させた。
【0142】
図2は、サイズ排除クロマトグラフィーにおけるN−PΔ161N複合体の溶離特性を示している。図2AはTSKカラムにおける220nmでの溶離特性を示す。図2Bはクマシーブルーによる染色後の、種々の断片のアクリルアミドゲル電気泳動による分析結果を示す。17〜22の断片はRNA−環状Nタンパク質のみを含んでいる。
【0143】
図3は、RSVのNタンパク質の環状構造を示している。図3Aは、P161−241により精製した環状Nタンパク質−RNAの電子顕微鏡分析の結果を示す。図3Bは、低温顕微鏡による再構築の結果を示す。図3Cは、環中に存在するRNAのアガロースゲル上での分析結果を示す。
【0144】
図4は、N−PΔ161N複合体に対する抗体の産生を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示している。BALB/cマウスに、粘液性アジュバントLT(R192G)の存在下又は非存在下で20μgのN−PΔ161N複合体又はPΔ161Nを鼻腔内投与(i.n.)又は皮下投与(s.c.)して、免疫を付与した。二週間後に二度目の投与を行う(14日目)。マウスを二度目の投与の二週間後に安楽死させる(28日目)。血清抗体を測定するため、血清を0日目、14日目及び28日目に採取する(図4A)。粘液抗体を測定するため、気管支肺胞洗浄を28日目に行う(図4B)。N−PΔ161Nに対する抗体の割合をELISAにより測定する。データは、平均値±平均値における標準偏差(n=5)で示し、対数目盛で表示する。
【0145】
図5は、PΔ161N及びN−PΔ161Nに特異的な細胞性免疫応答を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示している。BALB/cマウスに、粘液性アジュバントLT(R192G)の存在下又は非存在下で20μgのN−PΔ161N複合体又はPΔ161Nを鼻腔内投与(i.n.)又は皮下投与(s.c.)して、免疫を付与した。二週間後に二度目の投与を行う(14日目)。脾臓と肺を摘出するため、マウスを二度目の投与の二週間後に安楽死させる(28日目)。脾臓細胞及び肺細胞を、N−PΔ161N、PΔ161Nの存在下で、又は再刺激を行わずに72時間培養する。IFN−γの分泌をELISAにより測定する。再刺激を行わない場合、IFN−γのベースレベルは15pg/ml未満である。データは、平均値±平均値における標準偏差(n=5)で示す。
【実施例1】
【0146】
RSVのリンタンパク質のC末端領域を含むプラスミドの構築
【0147】
RSVのLONG株のPタンパク質は241個のアミノ酸残基から構成される。
【0148】
RSVのPタンパク質のC末端部分を増幅するために使用したオリゴヌクレオチドプライマー配列(5’から3’)(BamHI制限部位に下線を付し、Pタンパク質遺伝子の「開始」コドンATGは太字で表す)は以下である。
【0149】
【0150】
RSVのLONG株のPタンパク質遺伝子のcDNAを、プライマーLONG−PBam+及びLONG−P−(Castagne et al., 2004)を用い、ヒトRSVのLONG株に感染させたHep−2細胞からRT−PCRにより増幅した。PCR産物を制限酵素BamHIで消化し、プラスミドpGEX−4T−3(Pharmacia)のBamHI−SmaI部位において、グルタチオン−S−トランスフェラーゼすなわちGSTをコードする遺伝子とクローニングした。このプラスミドをpGEX−Pと称する。
【0151】
・P161−241(PΔ161N)のクローニング
Pタンパク質のC末端領域(161〜241番アミノ酸)を、以下の条件でプラスミドpGEX−PからPCRにより増幅した。
PCRプライマー:P161B+及びLONG−P−を各100ng(各1μl)
pGEX−Pの鋳型DNA:10ng(1μl)
酵素:StratageneのPfu Turbo(登録商標)(μl当たりのユニット):1μl
dATP:最終濃度0.2mM
dGTP:最終濃度0.2mM
dCTP:最終濃度0.2mM
dTTP:最終濃度0.2mM
最終濃度1×Pfu緩衝液(Stratagene)
最終容量:100μl
PCRは以下の条件下で実施した。
94℃で15秒、40℃で2分、72℃で1分を5サイクル、
94℃で15秒、55℃で1分、72℃で1分を25サイクル
【0152】
増幅したDNAを1体積のフェノール/クロロホルム(1体積/1体積)(100μl)、次いで1体積のクロロホルムで抽出し、最後に10分の1体積の5MのNaCl(10μl)と2体積の100%エタノール(200μl)を加えて沈殿させた。DNAを13000gで20分間遠心し、1体積の70%エタノールで洗浄し、乾燥させ、1体積の水90μl中に再懸濁した。10×BamHI酵素緩衝液を10μl加えたのち、DNAを10ユニットのBamHI酵素の存在下で37℃で2時間消化した。消化したDNAを、エチジウムブロマイドの存在下で、1×トリス−ホウ酸−EDTA緩衝液(TBE)中の1.5%アガロースゲル上に置き、電気泳動した。P161−241のDNAに対応するバンドを切り取り、DNAを電気溶出により抽出した。それをさらに1体積のフェノール−クロロホルム、1体積のクロロホルムで抽出し、エタノールで沈殿した。それを、1%アガロースゲルにおける精製の後、BamHIとSmaIで消化したpGEX−4T−3ベクターにライゲーションした。
pGEX4T−3のDNA:100ng
P161−241のDNA:100ng
最終濃度1×リガーゼ緩衝液
リガーゼ(5U/μl):1μl
最終体積20μl
【0153】
全体を14℃で一晩インキュベートした。翌日、適当な細菌DH5−アルファTM(Life Technologies)を10μlのライゲーション産物で形質転換し、最終濃度100μg/mlのアンピシリンを補足したL−寒天培地を含むペトリ皿上に塗布した。組換え細菌のコロニーをプラスミドのミニプレップと制限酵素BamHI及びXhoIによる消化でスクリーニングした。組換えプラスミドはアガロースゲル上で、一方はベクター(4.9kb)に対応し、もう一方はPタンパク質のC末端部分(246bp)に対応する、二つのバンドを示す。組換えプラスミド全体を配列決定した。
【0154】
・P180−241、P201−241、P221−241のクローニング
180〜241、200〜241、220〜241番アミノ酸部分に対応するPタンパク質の断片を、以下のプライマーを用いてpGEX−PプラスミドからPCRにより得た。
P180−241:プライマーP180B+及びLONG−P
P200−241:プライマーP201B+及びLONG−P−
P220−241:プライマーP221B+及びLONG−P−
【0155】
これらを、P161−241(上記参照)と同様に増幅しクローニングした。
【0156】
・RSVのLong株のヌクレオカプシドタンパク質をコードする遺伝子のクローニング
ヒトRSVのLONG株のNタンパク質をコードする遺伝子を、そのウイルスに感染したHep−2細胞からRT−PCRにより得た。使用したプライマーは、
LONG−Nbam+:GAGGGATCCATGGCTCTTAGCAAAGTCAAGTTG(配列番号19)
LONG−N−:TTAACTCAAAGCTCTACATCATTATCTTTTGG(配列番号20)
であった。
【0157】
PCR産物をBamHIで消化し、プラスミドpGEX−4T−3のBamHI−SmaI部位においてクローニングした。BamHI−XhoIでpGEX−Nプラスミドを消化して、Nタンパク質(配列番号)をコードする領域をpET28+プラスミド(Novagen、配列番号及び図面参照)中にサブクローニングした。
【0158】
・P231−241のクローニング
以下のプライマーを94℃で5分間加熱して変性させ、室温で冷却した。
P231NB+:GATCCGATAGTGACAATGATCTATCACTTGAAGATTTCTGA(配列番号21)
P231N−:TCAGAAATCTTCAAGTGATAGATCATTGTCACTATCG(配列番号22)
【0159】
ハイブリダイズ後、10ngの二本鎖オリゴヌクレオチドを、BamHI酵素及びSmaI酵素で消化しアガロースゲル上での電気泳動により精製した、pGEX−4T−3プラスミドのDNA100ngとライゲーションした。組換えプラスミドは、Nタンパク質遺伝子に関して配列決定することにより確認した。
【実施例2】
【0160】
複合体の発現と精製
【0161】
適当な細菌BL21(DE3)(Novagen)をpGEX−PΔのDNA1μgとpET−NのDNA1μgで形質転換し、次いで最終濃度100μg/mlのアンピシリンと最終濃度50μg/mlのカナマイシンを補足したL−寒天培地を含むペトリ皿に塗布した。コロニーを、100μg/mlのアンピシリンと50μg/mlのカナマイシンを含む2mlのLB培地に移植し、37℃で一晩生育させた。翌朝、コンフルエントな状態の培養物1mlを、抗生物質を添加した1リットルのLB培地に播種し、夕方まで生育させた。夕方に、160μg/mlの濃度のIPTG(タンパク質の発現を誘導する)を含む1体積の新鮮なLB培地を培養物に加え、全体を28℃で一晩生育させる。翌日、細菌を5000rpmで15分間遠心し、ペレットを100mlの以下の緩衝液中に再び入れる。
50mMのトリス(PH7.8)
60mMのNaCl
2mMのDTT
1mMのEDTA
4mMのベンズアミジン
1×アンチプロテアーゼ(Complete EDTA−free protease inhibitor cocktail、Roche、参照番号11873580001)すなわち50mlの溶解緩衝液に対して一錠
0.1%Triton−X100
【0162】
10mg/ml(最終濃度1mg/ml)のリゾチームを添加した同様の緩衝液10mlを加える。細菌を氷中で1時間インキュベートする(溶解)。全体が粘性になったら、全体を、混合物中に入れたプローブを用いて氷中で1分間の超音波処理を3回行い、各超音波処理の間に5分間放置する。全体を4℃、10000gで30分間遠心し、次いで上清を回収する。上清を4℃、10000gで30分間再び遠心し、次いで新たに上清を回収する。8mlのビーズ緩衝液(体積/体積)混合物を回収して4mlのグルタチオンセファロース4Bビーズ(Amersham−Pharmacia)を溶解緩衝液で洗浄する。同量の緩衝液中にビーズを置き、澄ました細菌溶解液を加え、4℃で一晩回転させる。翌日、ビーズを2000rpmで3分間遠心し、次いで上清を除去し、ビーズを、アンチプロテアーゼを含まない溶解緩衝液で三回洗浄し、1×PBS緩衝液で三回洗浄する。
【0163】
ビーズを、ビオチン化したThrombin(Novagen)をビーズ1ml当たりトロンビン(Thrombin Cleavage Capture Kit、 No.69022−3FRZ)1μl(1U)の比率で用いて、トロンビン部位で切断する。ビーズを20℃で一晩インキュベートし、翌日、2000rpmで3分間遠心し、上清を回収するため15分間沈殿させる。等量の1×PBSをビーズに加え、混合物を攪拌し沈殿させる。上清を新たに回収し、あらかじめ採取しておいた上清に加える。回収した上清にストレプトアビジンアガロースビーズ(Novagen、参照番号69203)を樹脂16μl(すなわち樹脂/緩衝液混合物(vol/vol)32μl)の割合で加える。混合物を一時間攪拌し、次いで2000rpmで3分間遠心し、上清を回収する。タンパク質濃縮物2mg/mlを得る。
【0164】
切断産物を含む10μlの上清を、1×Laemmli緩衝液中で変性し、沸騰させ、0.1%トリス−グリシンSDS緩衝液中の12%ポリアクリルアミドゲル上に置き、次いでタンパク質を可視化するために、電気泳動後にクマシーブルーで染色する。
【実施例3】
【0165】
Nタンパク質とPΔ161N(P161−241)の分離及びNタンパク質の環の精製
【0166】
上清中に存在するタンパク質は、1×PBS中におけるサイズ排除クロマトグラフィー(ゲル濾過、図2)により分離することができる。Nタンパク質は450000Daの見かけのサイズで、PΔ161Nは15kDaの分子量で排除される。
【0167】
サイズ排除クロマトグラフィーにより得られた「Nタンパク質」画分を電子顕微鏡により観察した結果、Nタンパク質は、直径7nmを有しNタンパク質のサブユニットを10個含む(図3B)、環の形(図3A)であることが分かった。環は約70bpのRNAを含む(図3C)。
【実施例4】
【0168】
環状構造の組換えNタンパク質の免疫原性の評価
【0169】
・経鼻投与又は皮下投与によるワクチン接種を、アジュバントの存在下又は非存在下でマウスで実施する。
マウス:l’Unite Experimentale Animalerie Rongeur(INRA、Jouy−en−Josas)で育成した10〜12週齢の雌のBALB/cマウス30頭
抗原:トロンビンでの切断によりGSTを分離し、ストレプトアビジンと結合したビーズによりビオチン化トロンビンを除去した後の、濃度1mg/mlの可溶性のP161−241及び{P161−241+N}複合体
アジュバント:1mg/mlの、大腸菌のリンホトキシンLT(R192G)(Choi et al., 2004)
サンプル採取
−0日目、14日目及び28日目に、眼窩後洞から血液を採取
−28日目:
・1.5mlのHBSS培地及び1mMのEDTAによる気管支肺胞洗浄物(LBA)
・氷上での、抗生物質(100UI/mlのペニシリン及び100μg/mlのストレプトマイシン、PS)を加えたRPMI培地における脾臓と肺の解剖
【0170】
実験計画:
【表1】
【0171】
・抗Nタンパク質抗体の産生
血清を血液から採取し(4℃で一晩滲出)、次いで−20℃で凍結する。
【0172】
LBAを1700rpmで5分間遠心し、上清を集め(約1ml)、−20℃で凍結する。
【0173】
ELISAにより血清及びLBAにおける抗Nタンパク質抗体(総Ig、IgG1,IgG2a、及びIgA)を調べる。96ウェルプレート(Immulon 2HB、ThermoLabsystems)を、0.1Mの重炭酸緩衝液(pH9.5)において、P161−241+Nタンパク質複合体(ウェル当たり200ng)により4℃で一晩感作する。プレートをウェル当たり200μlの、0.05%tween20を含むPBSで5回洗浄する(自動装置Wellwash、Labsystemsを使用)。次にプレートを、37℃で1時間、ウェル当たり150μlの、0.05%tween20及び5%ウシ胎児血清を含むPBS緩衝液(PBS−T−FBS)で満たす。5回洗浄した後、滴定サンプルをPBS−T−FBSで希釈する(血清については最初の希釈度から1/30になるまで3倍の連続希釈を7回行い、LBAについては1/3になるまで行う)。プレートを37℃で2時間インキュベートする。5回洗浄した後、PBS−T−FBSで希釈した二次抗体をウェル当たり100μlの割合で添加する。使用する二次抗体は、ペルオキシダーゼと結合しており、マウスの免疫グロブリン、総IgG(1/4000、PARIS)、IgG1(1/2000、BD biosciences)、IgG2a(1/2000、BD biosciences)又はIgA(1/1000、Caltag)に対するものである。プレートを37℃で2時間インキュベートし、5回洗浄する。次にプレートをペルオキシダーゼの基質(TMB、ウェル当たり100μl)と共に10分間暗所でインキュベートする。2MのH3PO4を50μl加えて酵素反応を停止する。光学密度(OD)を450nmで読み取る(読み取り装置Dynex)。OD450=f(希釈度)の曲線を、回帰曲線y=(b+cx)/(1+ax)によりモデル化する。抗体の力価は、最大限に希釈されたコントロールサンプル(0日目)の二倍のOD450を示す希釈値として決定する。
【0174】
・P161−241及びNタンパク質に特異的なTリンパ球によるIFN−γの産生
摘出した脾臓及び肺を同じ手順で処理する。脾臓は個別に処理し、肺は実験群にグループ化する(群当たり5個の肺)。
【0175】
組織を薄切りにし、次いでRPMI培地及びPS培地中において、篩上(100μmのセルストレーナー、BD Falcon)で注意深く破砕する。細胞懸濁液を1700rpmで10分間、4℃で遠心する。細胞を1mlの赤血球溶解緩衝液(低張生理食塩緩衝液)中に懸濁し、室温で5分間インキュベートする。溶解反応は、10mlの完全RPMI(PS、2mMのL−グルタミン及び10%FBS)を添加することにより停止する。膜の破片を沈殿させ、細胞を遠心(1700rpmで10分間、4℃)により3回洗浄する。細胞懸濁液を、Malassezの計算盤を用いて計数する。
【0176】
細胞を、細胞培養用に処理された96ウェルマイクロプレート(Falcon)中で、200μlの完全RPMI培地においてウェル当たり200000個の細胞の比率で培養する。
【0177】
各細胞懸濁液に対して、四つの培養条件を三回試験する。
−10ng/mlのPMA(ホルボール12ミリステート13アセテート、Sigma)及び1μg/mlのイオノマイシン(Sigma)(ポジティブコントロール、ポリクローナル活性化)
−完全RPMI(ネガティブコントロール)
−10μg/mlのP161−241
−10μg/mlのP161−241+N
【0178】
5%CO2下において37℃で72時間培養した後、培養上清を回収し、ELISAによりIFN−γを滴定するまで−20℃で凍結する。
【0179】
IFN−γのELISA:96ウェルプレート(Immulon 2HB、 ThermoLabsystems)を、0.1Mの重炭酸緩衝液(100μl/ウェル)(pH9.5)中で、4μg/mlのマウス抗IFN−γ捕捉抗体(BD Biosciences)で4℃で一晩感作する。プレートをウェル当たり200μlの、0.05%tween20を加えたPBSで5回洗浄する(自動装置Wellwash、Labsystemsを使用)。次にプレートをウェル当たり150μlの、0.05%tween20及び2%ウシ血清アルブミン(PBS−T−BSA)を加えたPBS緩衝液で、37℃で2時間、コンフルエントな状態にする。5回洗浄した後、参照用の組換えマウスIFN−γ(R&D systems)及び滴定サンプルをPBS−T−BSAで二倍連続希釈により希釈する。IFN−γの希釈範囲は3312.5pg/mlから3.235pg/mlまでとする。四回の二倍連続希釈を純粋なサンプルに対して行う。続いてプレートを4℃で一晩インキュベートする。5回洗浄した後、ビオチン化した検出抗体(BD Biosciences)を添加し(PBS−T−BSA中に1μg/ml、100μl/ウェル)、4℃で3時間インキュベートする。5回洗浄した後、ストレプトアビジン−ペルオキシダーゼコンジュゲート(Pierce)を添加し(PBS−T−BSA中に1μg/ml、100μl/ウェル)、4℃で1時間インキュベートする。5回洗浄した後、ペルオキシダーゼの基質(ABTS+H2O2)をウェルに添加する。45分間のインキュベーションの後、光学密度を405nmで読み取る(読み取り装置ELISA Dynex)。サンプルのIFN−γ濃度の計算は、IFN−γの希釈範囲との比較から行う。
【図面の簡単な説明】
【0180】
【図1】Pタンパク質に対するPタンパク質−Nタンパク質の相互作用ドメインの位置決定を示す図である。
【図2】サイズ排除クロマトグラフィーにおけるN−PΔ161N複合体の溶離特性を示す図である。
【図3】RSVのNタンパク質の環状構造を示す図である。
【図4】N−PΔ161N複合体に対する抗体の産生を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示す図である。
【図5】PΔ161N及びN−PΔ161Nに特異的な細胞性免疫応答を測定することによる、環状構造のNタンパク質の免疫原性の分析結果を示すグラフである。
【0181】
参考文献
−Ames,T.R.1993.The epidemiology of BRSV infection.Vet.Med.881−884.
−Bhella,D.,Ralph,A.,Murphy L.B.,& Yeo,R.P.2002.Significant differences in nucleocapsid morphology within the Paramyxoviridae.Journal of General Virology; 83,1831−1839.
−Castagne,N.,A.Barbier,J.Bernard,H.Rezaei,J.−C.Huet,C.Henry,B.Da Costa,and J.−F.Eleouet.2004.Biochemical characterization of the Respiratory Syncytial Virus P−P and P−N protein complexes and localization of the P protein oligomerization domain.Journal of General Virology; 85: 1643−1653.
−Choi et al.,2004,Protein Expression and Purification; 38,pp205
−Elvander,M.1996.Severe respiratory disease in dairy cows caused by infection with bovine respiratory syncytial virus.Vet.Rec.; 138,101−105.
−Freytag,LC et Clements,JD.2005.Mucosal adjuvants.Vaccine.; 23(15): 1804−13.
−Gaddum,R.M.,R.S.Cook,J.M.Furze,S.A.Ellis & G.Taylor.2003.Recognition of bovine respiratory syncytial virus proteins by bovine CD8a T lymphocytes.Immunology; 108,220−229;
−Goulder PJ,Lechner F,Klenerman P,Mclntosh K,Walker BD.2000.Characterization of a novel respiratory syncytial virus−specific human cytotoxic T−lymphocyte epitope.J Virol.; 74(16): 7694−7.
−Johansson et al.,2003; Journal of Biological Chemistry vol.278 p 44567−44573.
−Khattar SK,Yunus AS,Samal SK.2001a.Mapping the domains on the phosphoprotein of bovine respiratory syncytial virus required for N−P and P−L interactions using a minigenome system.J Gen Virol.; 82(Pt4):775−9.
−Khattar SK,Yunus AS,Collins PL,Samal SK..2001b.Deletion and substitution analysis defines regions and residues within the phosphoprotein of bovine respiratory syncytial virus that affect transcription,RNA replication,and interaction with the nucleoprotein.Virology.; 285(2): 253−69.
−Kolakofsky D,Le Mercier P,Iseni F,Garcin D.2004.Viral DNA polymerase scanning and the gymnastics of Sendai virus RNA synthesis.Virology.; 318(2): 463−73.Review.
−Maggon K,Barik S.2004.New drugs and treatment for respiratory syncytial virus.Rev Med Virol.14(3): 149−68.Review.
−Mallipeddi SK,Lupiani B,Samal SK.1996.Mapping the domains on the phosphoprotein of bovine respiratory syncytial virus required for N−P interaction using a two−hybrid system.J Gen Virol.; 77(Pt5): 1019−23.
−Martinez X,Li X,Kovarik J,Klein M,Lambert PH,Siegrist CA.1999.Combining DNA and protein vaccines for early life immunization against respiratory syncytial virus in mice.Eur J Immunol.; 29(10): 3390−400.
−Mavrakis M,Iseni F,Mazza C,Schoehn G,Ebel C,Gentzel M,Franz T,Ruigrok RW.2003.Isolation and characterisation of the rabies virus N゜−P complex produced in insect cells.Virology.; 305(2): 406−14.
−McNeal MM,VanCott JL,Choi AH,Basu M,Flint JA,Stone SC,Clements JD,Ward RL.2002.CD4 T cells are the only lymphocytes needed to protect mice against rotavirus shedding after intranasal immunization with a chimeric VP6 protein and the adjuvant LT(R192G).J Virol.; 76(2): 560−8.
−Meric C,Spehner D,Mazarin V.1994.Respiratory syncytial virus nucleocapsid protein (N) expressed in insect cells forms nucleocapsid−like structures.Virus Res.31(2): 187−201.
−Perrin,B.,Dannacher,G.,et Solsona,M.1979.Mise en evidence des anticorps contre le virus respiratoire syncytial chez les bovins francais.Rec.Med.Vet.155,465−471.
−Samal SK,Pastey MK,McPhillips TH,Mohanty SB.1993.Bovine respiratory syncytial virus nucleocapsid protein expressed in insect cells specifically interacts with the phosphoprotein and the M2 protein.Virology.; 193(1):470−3.
−Slack MS,Easton AJ.1998.Characterization of the interaction of the human respiratory syncytial virus phosphoprotein and nucleocapsid protein using the two−hybrid system.Virus Res.; 55(2): 167−76.
−Sparer TE,Matthews S,Hussell T,Rae AJ,Garcia−Barreno B,Melero JA,Openshaw PJ.1998.Eliminating a region of respiratory syncytial virus attachment protein allows induction of protective immunity without vaccine−enhanced lung eosinophilia.J Exp Med; 187(11): 1921−6.
−Tarbouriech,N.,Curran,J.,Ruigrok,R.W.,& Burmeister,W.P.(2000).Tetrameric coiled coil domain of Sendai virus phosphoprotein.Nature Structural Biology 7,777−781.
−Taylor G,Bruce C,Barbet AF,Wyld SG,Thomas LH.2005.DNA vaccination against respiratory syncytial virus in young calves.Vaccine; 23(10): 1242−50
−Taylor,G.L.H.Thomas,J.M.Furze,R.S.Cook,S.G.Wyld,R.Lerch,R.Hardy and G.W.Wertz.1997.Recombinant vaccinia viruses expressing the F,G or N,but not the M2,protein of bovine respiratory syncytial virus (BRSV) induce resistance to BRSV challenge in the calf and protect against the development of pneumonic lesions.Journal of General Virology; 78,3195−3206.
−Thompson W.W.,D.K.Shay,E.Weintraub,L Brammer,N.Cox,L.J.Anderson,K.Fukuda.2003.Mortality associated with influenza and respiratory syncytial virus in the United States.JAMA.; 289(2): 179−86.
−Wellemans,G.,and J.Leunen.1975.Le virus respiratoire syncytial et les troubles respiratoires des bovins. Ann. Med. Vet.; 119,359−369.
【特許請求の範囲】
【請求項1】
パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質の複合体を調製する方法であって、
a)パラミクソウイルス科ウイルスのNタンパク質を、パラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、トランケート型Pタンパク質と同時発現させること、及び
b)こうして形成された可溶性Nタンパク質−トランケート型Pタンパク質複合体を回収すること
から構成される過程を含む方法。
【請求項2】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項1に記載の方法。
【請求項3】
パラミクソウイルス科ウイルスが、ヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項1又は2に記載の方法。
【請求項4】
前記トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項3に記載の方法。
【請求項5】
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位〜161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、RSVのPタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を同時発現する、請求項4に記載の方法。
【請求項6】
前記トランケート型Pタンパク質が標識タンパク質との融合タンパク質の形で発現する、請求項1〜5のいずれか一つに記載の方法。
【請求項7】
前記Nタンパク質と前記トランケート型Pタンパク質とが、細菌の発現系中で同時発現する、請求項1〜6のいずれか一つに記載の方法。
【請求項8】
請求項1〜7のいずれか一つに記載の方法により得ることができる、パラミクソウイルス科ウイルスの単離された可溶性Nタンパク質−トランケート型Pタンパク質複合体。
【請求項9】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項8に記載の単離された可溶性の複合体。
【請求項10】
パラミクソウイルス科ウイルスがヒト又はウシの呼吸器合胞体ウイルス(RSV)であり、トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項9に記載の単離された可溶性の複合体。
【請求項11】
前記Pタンパク質のC末端断片が、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、C末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
である、請求項10に記載の単離された可溶性の複合体。
【請求項12】
10分子のNタンパク質を含み、Pタンパク質のC末端断片がRSVのPタンパク質のC末端の先端の9個のアミノ酸を含みRSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、呼吸器合胞体ウイルス(RSV)の単離された可溶性Nタンパク質−Pタンパク質C末端断片複合体。
【請求項13】
パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法であって、
a)請求項1〜7のいずれか一つに記載の方法により可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製すること、及び
b)可溶性Nタンパク質−トランケート型Pタンパク質複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む方法。
【請求項14】
前記トランケート型Pタンパク質がPタンパク質のC末端断片である、請求項13に記載の可溶性Nタンパク質を調製する方法。
【請求項15】
請求項13又は14に記載の方法により得ることができる、パラミクソウイルス科ウイルスの可溶性Nタンパク質。
【請求項16】
医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質を含むワクチン組成物。
【請求項17】
パラミクソウイルス科ウイルスの可溶性Nタンパク質が請求項19又は20に記載の方法により得ることができる、請求項16に記載のワクチン組成物。
【請求項18】
パラミクソウイルス科ウイルスが、ヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項13又は14に記載のワクチン組成物。
【請求項19】
医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を含むワクチン組成物であって、前記トランケート型Pタンパク質がPタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができるワクチン組成物。
【請求項20】
前記トランケート型Pタンパク質がPタンパク質のC末端断片である、請求項19に記載のワクチン組成物。
【請求項21】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項19又は20に記載のワクチン組成物。
【請求項22】
パラミクソウイルス科ウイルスがヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項19〜21のいずれか一つに記載のワクチン組成物。
【請求項23】
前記トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項22に記載のワクチン組成物。
【請求項24】
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、Pタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を含む、請求項23に記載のワクチン組成物。
【請求項25】
請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質を含む診断試薬。
【請求項26】
前記Nタンパク質が、請求項8〜12のいずれか一つで定義したようなパラミクソウイルス科の同じウイルスのNタンパク質−トランケート型Pタンパク質複合体の形である、請求項25に記載の診断試薬。
【請求項27】
前記Nタンパク質に対する抗体をインビトロで検出するための、請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質の使用方法。
【請求項28】
生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に対する特異的な抗体の検出方法であって、
a)前記生物サンプルと請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質とを接触させること、及び
b)形成されたNタンパク質/抗体複合体を検出すること
から構成される過程を含み、この複合体の存在が生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に特異的な抗体の存在の指標となる方法。
【請求項29】
Nタンパク質を、請求項8〜12のいずれか一つに定義したようなパラミクソウイルス科の同じウイルスのNタンパク質−トランケート型Pタンパク質複合体の形で生物サンプルと接触させる、請求項28に記載の方法。
【請求項1】
パラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質の複合体を調製する方法であって、
a)パラミクソウイルス科ウイルスのNタンパク質を、パラミクソウイルス科の同じウイルスの、Pタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができる、トランケート型Pタンパク質と同時発現させること、及び
b)こうして形成された可溶性Nタンパク質−トランケート型Pタンパク質複合体を回収すること
から構成される過程を含む方法。
【請求項2】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項1に記載の方法。
【請求項3】
パラミクソウイルス科ウイルスが、ヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項1又は2に記載の方法。
【請求項4】
前記トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項3に記載の方法。
【請求項5】
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位〜161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、RSVのPタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を同時発現する、請求項4に記載の方法。
【請求項6】
前記トランケート型Pタンパク質が標識タンパク質との融合タンパク質の形で発現する、請求項1〜5のいずれか一つに記載の方法。
【請求項7】
前記Nタンパク質と前記トランケート型Pタンパク質とが、細菌の発現系中で同時発現する、請求項1〜6のいずれか一つに記載の方法。
【請求項8】
請求項1〜7のいずれか一つに記載の方法により得ることができる、パラミクソウイルス科ウイルスの単離された可溶性Nタンパク質−トランケート型Pタンパク質複合体。
【請求項9】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項8に記載の単離された可溶性の複合体。
【請求項10】
パラミクソウイルス科ウイルスがヒト又はウシの呼吸器合胞体ウイルス(RSV)であり、トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項9に記載の単離された可溶性の複合体。
【請求項11】
前記Pタンパク質のC末端断片が、
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、C末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
である、請求項10に記載の単離された可溶性の複合体。
【請求項12】
10分子のNタンパク質を含み、Pタンパク質のC末端断片がRSVのPタンパク質のC末端の先端の9個のアミノ酸を含みRSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、呼吸器合胞体ウイルス(RSV)の単離された可溶性Nタンパク質−Pタンパク質C末端断片複合体。
【請求項13】
パラミクソウイルス科ウイルスの可溶性Nタンパク質を調製する方法であって、
a)請求項1〜7のいずれか一つに記載の方法により可溶性Nタンパク質−トランケート型Pタンパク質複合体を調製すること、及び
b)可溶性Nタンパク質−トランケート型Pタンパク質複合体から可溶性Nタンパク質を分離すること
から構成される過程を含む方法。
【請求項14】
前記トランケート型Pタンパク質がPタンパク質のC末端断片である、請求項13に記載の可溶性Nタンパク質を調製する方法。
【請求項15】
請求項13又は14に記載の方法により得ることができる、パラミクソウイルス科ウイルスの可溶性Nタンパク質。
【請求項16】
医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質を含むワクチン組成物。
【請求項17】
パラミクソウイルス科ウイルスの可溶性Nタンパク質が請求項19又は20に記載の方法により得ることができる、請求項16に記載のワクチン組成物。
【請求項18】
パラミクソウイルス科ウイルスが、ヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項13又は14に記載のワクチン組成物。
【請求項19】
医薬的に許容される担体中のパラミクソウイルス科ウイルスの可溶性Nタンパク質−トランケート型Pタンパク質複合体を含むワクチン組成物であって、前記トランケート型Pタンパク質がPタンパク質のオリゴマー化ドメインを含まずNタンパク質と相互作用することができるワクチン組成物。
【請求項20】
前記トランケート型Pタンパク質がPタンパク質のC末端断片である、請求項19に記載のワクチン組成物。
【請求項21】
パラミクソウイルス科ウイルスがニューモウイルスである、請求項19又は20に記載のワクチン組成物。
【請求項22】
パラミクソウイルス科ウイルスがヒト又はウシの呼吸器合胞体ウイルス(RSV)である、請求項19〜21のいずれか一つに記載のワクチン組成物。
【請求項23】
前記トランケート型Pタンパク質が、RSVのPタンパク質のC末端の先端の9個のアミノ酸を含み、RSVのPタンパク質のN末端の少なくとも160個のアミノ酸を欠いている、Pタンパク質のC末端断片である、請求項22に記載のワクチン組成物。
【請求項24】
a)配列番号1に示すような、ヒトRSVのLONG株のPタンパク質の233〜241番のアミノ酸配列を含み、配列番号1に示すような、RSVのPタンパク質配列の233位から161位までの間に含まれるアミノ酸残基までN末端の方向に延びている、Pタンパク質のC末端断片、又は
b)他のヒトRSV株又はウシRSV株に由来するPタンパク質の、a)で定義した断片と相同であるC末端断片
を含む、請求項23に記載のワクチン組成物。
【請求項25】
請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質を含む診断試薬。
【請求項26】
前記Nタンパク質が、請求項8〜12のいずれか一つで定義したようなパラミクソウイルス科の同じウイルスのNタンパク質−トランケート型Pタンパク質複合体の形である、請求項25に記載の診断試薬。
【請求項27】
前記Nタンパク質に対する抗体をインビトロで検出するための、請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質の使用方法。
【請求項28】
生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に対する特異的な抗体の検出方法であって、
a)前記生物サンプルと請求項15に記載のパラミクソウイルス科ウイルスのNタンパク質とを接触させること、及び
b)形成されたNタンパク質/抗体複合体を検出すること
から構成される過程を含み、この複合体の存在が生物サンプルにおけるパラミクソウイルス科ウイルスのNタンパク質に特異的な抗体の存在の指標となる方法。
【請求項29】
Nタンパク質を、請求項8〜12のいずれか一つに定義したようなパラミクソウイルス科の同じウイルスのNタンパク質−トランケート型Pタンパク質複合体の形で生物サンプルと接触させる、請求項28に記載の方法。
【図1】


【図2】
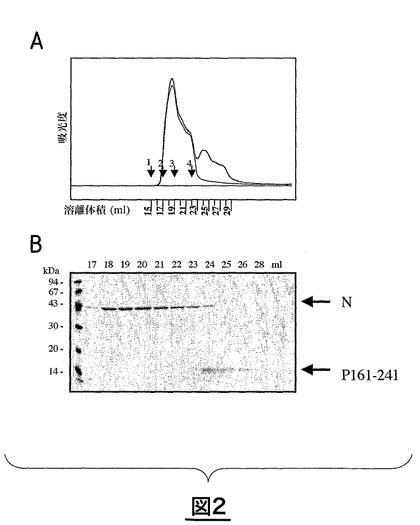
【図3】
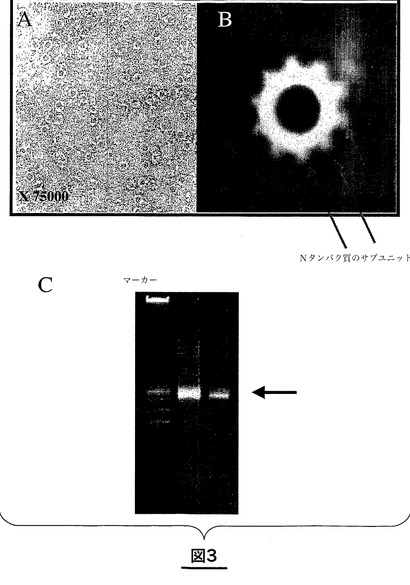
【図4】
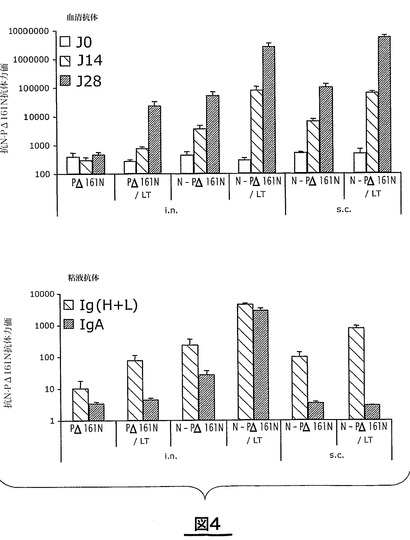
【図5】
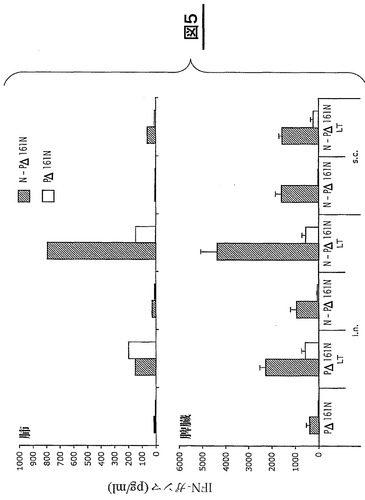
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2008−538905(P2008−538905A)
【公表日】平成20年11月13日(2008.11.13)
【国際特許分類】
【出願番号】特願2008−508259(P2008−508259)
【出願日】平成18年4月27日(2006.4.27)
【国際出願番号】PCT/FR2006/000949
【国際公開番号】WO2006/117456
【国際公開日】平成18年11月9日(2006.11.9)
【出願人】(507356305)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (インラ) (3)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE (INRA)
【Fターム(参考)】
【公表日】平成20年11月13日(2008.11.13)
【国際特許分類】
【出願日】平成18年4月27日(2006.4.27)
【国際出願番号】PCT/FR2006/000949
【国際公開番号】WO2006/117456
【国際公開日】平成18年11月9日(2006.11.9)
【出願人】(507356305)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (インラ) (3)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE (INRA)
【Fターム(参考)】
[ Back to top ]