
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質−対象タンパク質の融合タンパク質
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質-対象タンパク質の融合タンパク質。
本発明は、場合により、溶解性Nタンパク質-対象タンパク質/Pタンパク質複合体の形態の、Nタンパク質-対象タンパク質の融合タンパク質であって、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーのウイルスのタンパク質である、前記融合タンパク質に関する。対象タンパク質が抗原である時、本発明は、Nタンパク質-対象タンパク質の融合タンパク質又はNタンパク質-抗原/Pタンパク質複合体を含む、ワクチン組成物及び診断試薬にも関する。Nタンパク質-対象タンパク質の融合タンパク質は、抗ウイルス薬又は抗癌剤のような対象の治療的分子を細胞に移送するための「ベクター」としても使用されることもできる。
本発明は、場合により、溶解性Nタンパク質-対象タンパク質/Pタンパク質複合体の形態の、Nタンパク質-対象タンパク質の融合タンパク質であって、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーのウイルスのタンパク質である、前記融合タンパク質に関する。対象タンパク質が抗原である時、本発明は、Nタンパク質-対象タンパク質の融合タンパク質又はNタンパク質-抗原/Pタンパク質複合体を含む、ワクチン組成物及び診断試薬にも関する。Nタンパク質-対象タンパク質の融合タンパク質は、抗ウイルス薬又は抗癌剤のような対象の治療的分子を細胞に移送するための「ベクター」としても使用されることもできる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Nタンパク質-対象タンパク質の融合タンパク質、場合により、溶解性Nタンパク質-対象タンパク質/Pタンパク質の複合体の形態にあり、Nタンパク質及びPタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質である、に関する。対象タンパク質が抗原である時に、本発明は、Nタンパク質-抗原の融合タンパク質又はNタンパク質-抗原/Pタンパク質の複合体を含む、ワクチン組成物及び診断試薬にも関する。対象融合タンパク質のNタンパク質は、抗ウイルス薬又は抗癌剤のような関心の治療分子を細胞に移送するための「ベクター」としても使用され得る。
【背景技術】
【0002】
呼吸器合胞体ウイルス(RSV)は、幼児の気管支梢炎の原因である(500,000人の子供がフランスで毎年罹患している)。該疾患に対する真に効果的な予防処置(ワクチン)又は抗体ウイルス薬は存在しない。同一の疾患は、ウシにも存在し、最初の年に子牛の70%がかかり、致死率は20%に到達し、ウシRSVは、子牛の重大な気管支疾患を引き起こす主な薬剤である。臨床発表はヒトでのものと同一である。このウイルスは、モノネガウイルス(Mononegavirales)系列及びパラミクソウイルス科(Paramyxoviridae)ファミリーに属する。ウイルス粒子は、2つの主なタンパク質、融合タンパク質(F)及び糖タンパク質(G)、を含む脂質外皮膜によって取り囲まれている。粒子の内側には、ヌクレオカプシドタンパク質(N)と関連する、約15 kbの陰極の単一鎖RNAが存在する。そのRNA-N複合体は、RNA-依存的RNAポリメラーゼであるLタンパク質(大きい断片)及びビリオンにも存在するその補助因子P(リンタンパク質)によって構成されるポリメラーゼ複合体のマトリックスを構成する。
【0003】
溶解形態のNタンパク質を精製するのはこれまで可能でなかった。なぜなら、該タンパク質は組換え体で発現される時に、細胞RNAに自発的かつ非-特異的に結合し、非常に高分子量の不溶解性構造を形成するからである(Meric et al., 1994 Virus Res. 31(2): 187-201; Bhella et al., 2002 Journal of General Virology; 83, 1831-1839)。本発明者らは、環状様の構造を有する溶解性組み換え体として呼吸器合胞体ウイルス(RSV)のヌクレオカプシド(N)タンパク質を入手するための方法、及びワクチンとしてのその用途を開発してきた。Nタンパク質は、約7 nmの径を有する非常に規則的な環の形態にあり、Nタンパク質の10分子及び約70塩基の細菌起源のRNAを含む。Nタンパク質は、グルタチオン-S-トランスフェラーゼ(GST)に融合されたRSVのリンタンパク質(P)、より具体的にはそのC-末端部分で、E.コリ中で共発現される。それは、複合体をセファロース-グルタチオンビーズでの親和性によって精製させる。この環状様構造は、特に、鼻腔内投与によって、アジュバントの非存在下でも、マウスでの強い免疫反応を誘導する。該方法は、FR 05 04 426で2005年4月29日にフランスで出願された特許出願の対象である。
【0004】
本発明者らは、ワクチンベクターとしてその環状様構造を使用できることを示している。その技術の実現可能性は、プラスミド構築によって、Nタンパク質の他のタンパク質、GFP(緑色蛍光タンパク質)への融合によって証明されている。それは、GSTに融合されたPタンパク質(アミノ酸161〜241)のC-末端部分で、E.コリ中で共発現されてきた。電子顕微鏡を用いる精製構造体の分析は、Nタンパク質のC-末端でのGFPの接合が、Nタンパク質を、約10 nmの径を有する環の形態でなお産生させる、ことを示している。N-GFP融合タンパク質は蛍光を発する。
【0005】
N-RNA環は非常に免疫原性であるので、本発明者らは、特にGFPに対する強力な抗体反応を誘導できるかどうかをみるためにマウスにその構造体を注入した。その反応を、E.コリからの組み換え体でも精製された「通常の」GFPタンパク質に対して得られたものと比較した。得られた結果は、環状様構造体が使用される時に、GFPに対してより顕著な強い反応を示した(第1実験について約40倍超)。
【0006】
Nタンパク質への対象タンパク質の融合の可能性、並びに工業的スケール及び適当なコストでの溶解性環の形態での複合体の精製の可能性(特に、E.コリにおいて)は、それ自体に注入された時に高い免疫原性でないか又は免疫原性でない任意の種類のエピトープに対するワクチンの開発を可能にするだろう。
【発明の開示】
【0007】
本発明者らは、そのNタンパク質-GFP/Pタンパク質構造体が吸収され及び異なった細胞種によって効果的に内面化される、ことを更に示した。従ってその結果は、ワクチン化のための抗原ベクターとして又はより一般的には治療的関心の分子としてそのようなNタンパク質-対象タンパク質の融合体の価値を、場合により、Nタンパク質-対象タンパク質/Pタンパク質複合体の形態で、強調する。
【0008】
定義
パラミクソウイルス科(Paramyxoviridae)ファミリーは、パラミクソウイルス(Paramyxoviridae)及びニューモウイルス(Pneumoviridae)サブファミリーを包含する。パラミクソウイルス科は、センダイウイルスのプロトタイプであるレスピロウイルス(Respirovirus)属、ルブラウイルス(Rubulavirus)(特におたふく風邪ウイルス)、モルビリウイルス(Morbilivirus)、例えば麻疹ウイルスを含む。レスピロウイルス及びルブラウイルス属の各々は、共にパラインフルエンザウイルスの株に分類される。ニューモウイルス・サブファミリーは、2つの属、ニューモウイルスとメタニューモウイルス(Metapneumoviridae)に分類され、後者はヒトメタニューモウイルスを含む。ヒト呼吸器合胞体ウイルス(RSV)は、ニューモウイルス・サブファミリーに属するニューモウイルス属のプロトタイプウイルスを構成する。ニューモウイルスは、RSVのウシ及びネズミ株をも含む。
【0009】
他に特定しない限り、「呼吸器合胞体ウイルス」は、問題となっている、その形態(ヒト、ウシ等)、サブグループ(例えば、ヒトRSVにおいて特定されたサブグループA、B及びS)又は株がそのようなものであれ、一般的に、RSVを意味すると理解される。
【0010】
「Nタンパク質」は、ウイルスゲノムを囲むように螺旋構造を形成する、パラミクソウイルスのヌクレオカプシドタンパク質を称する。ヒトRSV長鎖のNタンパク質は、配列番号1によって記載される391アミノ酸の配列を有する。同様に、ウシRSVのNタンパク質は、391アミノ酸を含む(配列番号2)。センダイウイルス(Hamamatsu株)、パラミクソウイルス科、おたふく風ウイルス(SBL-1株)、及びヒトメタニューモウイルス(00-1株)のNタンパク質も、各々、受入番号Q9DUE3(配列番号3)、Q89933(配列番号4)、P21277(配列番号5)及びQ91F57(配列番号6)としてスイスプロットデータベースに記載されている。
【0011】
表現「Pタンパク質」は、リンタンパク質、又はパラミクソウイルス科ファミリーのウイルスのポリメラーゼ複合体のPタンパク質形成部分を称する。Pタンパク質は、ウイルスポリメラーゼ(レプリカーゼ/転写酵素)の補助因子であり、リン酸化される。パラミクソウイルスのPタンパク質の配列は、当業者に知られている。例えば、ヒトRSV長鎖株のPタンパク質は241アミノ酸の配列を有し、それは受入番号P12579としてスイスプロットデータベースに寄託されている。その配列は、配列番号7で示される。ウシRSVのPタンパク質も241アミン酸を含む(配列番号8)。センダイウイルス(Harris株)、麻疹ウイルス(Edmonston B株)、おたふく風ウイルス(SBL-1株)、及びヒトメタニューモウイルス(00-1株)のPタンパク質も、各々、受入番号P04559(配列番号9)、CAA91364(配列番号10)、P19717(配列番号11)及びQ91KZ5(配列番号12)としてスイスプロットデータベースに記載されている。表現「Pタンパク質」は、Pタンパク質全体、切断されたPタンパク質又はPタンパク質の断片を意味することがある。
【0012】
パラミクソウイルスのPタンパク質は、ホモオリゴマー特にホモテトラマーを、例えばセンダイウイルス又はRSV中で形成する。RSVの場合には、オリゴマー化(P-Pオリゴマー化)できるPタンパク質のドメインは、そのタンパク質の120〜150アミノ酸領域にマップされている(Castagne et al., 2004; Journal of General Virology; 85: 1643-1653)。従って、例えば、RSVのPタンパク質の161〜241アミノ酸によって構成される断片は、オリゴマーを形成しない。センダイウイルスのPタンパク質のオリゴマー化ドメインは、Pタンパク質の320〜446残基によって構成されるものとして、Tarbouriech et al. (2000; Nature Structural Biology; 7, 777-781)によって記載されている。一方、Pオリゴマー化領域は、麻疹ウイルスのPタンパク質については304〜376アミノ酸の領域に同定されている(Johannson et al., 2003; Journal of Biological Chemistry; 278, p.44567-44573)。
【0013】
上記のPタンパク質及びNタンパク質の配列は、本来、例証であり、所与のウイルスを考慮すると特定の株に従って変形を示す配列も可能である。従って、出願のリマインダーで述べたアミノ酸の位置は、参照配列に対して示されている。当業者は、完全に、特に例えばClustalwのようなソフトウェアを用いて実行された配列アラインメントの助けを借りて、例示された株以外のウイルス株における対応するドメインを特定することができる。
【0014】
パラミクソウイルスファミリーのウイルスのNタンパク質及びPタンパク質のコーディング配列も当業者に知られている。
【0015】
用語「タンパク質ラベル」は「タンパク質タグ」とも称され、その精製を助けるために対象タンパク質への融合のために使用されるタンパク質を言う。タンパク質タグは、当業者に知られている。タンパク質タグの例は、グルタチオン-S-トランスフェラーゼ(GST)、又は4〜10ヒスチジン残基の鎖を一般的に含む配列であるヒスチジンタグを含む。
【0016】
「対象タンパク質」は、任意のタンパク質、ポリペプチド又はペプチド(これらの用語は無差別に使用される)、例えばマーカータンパク質又は治療的もしくはワクチン的関心、を称する。
【0017】
対象タンパク質は、例えば、gfpレポーター遺伝子によってコードされる、タンパク質GFP(緑色蛍光タンパク質)でもよい。
【0018】
治療対象のタンパク質は、例えば、抗癌剤として使用され得る、血管新生抑制ポリペプチド、例えばRGD又はRGDを含む配列、エンドスタチン、又はアポトーシス促進性ポリペプチド例えばアポトーシス-誘導因子(AIF)、ウイルスタンパク質と特異的に相互作用し、ウイルス又は毒素の複製を許容するメカニズムを妨害することができるポリペプチドでよい。
【0019】
対象の異種タンパク質は、抗原、特にワクチン的関心の抗原性タンパク質でよい。「抗原」又は「Ag」は、投与される宿主内での免疫反応を誘導することができるペプチド性又は糖ペプチド性の性質の配列であると理解される。従って、抗原は、タンパク質又はタンパク質の一部(ポリペプチド)でもよく、あるいは潜在的にエピトープに対応する小ペプチドでもよい。
【0020】
「エピトープ」は、抗体によって又はリンパ球受容体によって認識される抗原の一部である。(直鎖)エピトープは、一般的に、7〜15アミノ酸の配列によって構成される。本発明の範囲内の抗原は、エピトープによって構成されてもよく、エピトープを含んでもよく、又は抗原性タンパク質でもよい。
【0021】
好ましくは、抗原は、病原性微生物、例えばウイルス、バクテリア、真菌又は寄生的な後生生物もしくは原生生物から得られる抗原である。
【0022】
ウイルスの例は、ヒト免疫不全ウイルス(HIV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、ヘルペスウイルス(Herpes simplex)、インフルエンザウイルス(インフルエンザウイルス又はトリインフルエンザウイルス)、西洋ニールウイルス、黄熱ウイルス、サイトメガロウイルス、パピローマウイルス(HPV)、エプスタイン・バール・ウイルス(EBV)、RSV、デングウイルス、及びチクングニアウイルスを含む。
【0023】
寄生動物の例は、マラリア(プラスモディウム(Plasmodium)、特にP. ファルシファルム(P. falciparum)、三日熱マラリア(P. vivax)、卵形マラリア原虫(P. ovale)、及び四日熱マラリア原虫(P. malariae))及びトリパノソミアシス(特に、睡眠病)(T. brucei)及びシャーガス病(T. cruzi))の原因となる寄生動物を含む。病原は、カンジタ症の原因となるカンジタ・アルビカンスのような真菌でもよい。
【0024】
細菌は、例えば、ヘリコバクター・ピロリ(Heliobacter pylori)、クロストリジウム・テタニ(Clostridium tetani)、結核菌(Mycobacterium tuberculosis)又はウシ型結核菌(Mycobacterium bovis)でよい。
【0025】
例えば、抗原は、GFP、ウイルス性糖タンパク質、例えばRSV融合タンパク質(F)、インフルエンザウイルス血球凝集素、又はHIVgp120タンパク質、細菌性アナトキシン、例えば破傷風抗毒素又はそのエピトープでよい。
【0026】
Nタンパク質-対象タンパク質の融合タンパク質、及びその調製
本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質、及び該Nタンパク質のC-末端のインフレームに融合された対象タンパク質を含む、Nタンパク質と対象タンパク質との融合タンパク質に関する。
【0027】
本発明の文脈において、対象タンパク質は、Nタンパク質のC-末端に特異的に融合される。本明細書で以下に記載される融合タンパク質を産生する方法が使用される時に、この構造は、Nタンパク質に、約10 nmの径を有する環状構造を取り込むことを可能にする。
【0028】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス(Paramyxoviridae)科及びニューモウイルス(Pneumoviridae)科でよい。特に、該ウイルスは、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルスであり、特にヒト又はウシ呼吸器合胞体ウイルス(RSV)である。
【0029】
本発明に従って融合タンパク質に存在するNタンパク質は、一般的に、天然Nタンパク質のアミノ酸配列を有し、すなわちパラミクソウイルス科ウイルスに天然に存在するNタンパク質である。
【0030】
しかしながら、融合タンパク質の発現の目的のために、Nタンパク質がPタンパク質と相互作用するその能力を保持することを条件に、配列修飾は、Nタンパク質のC-末端に導入され得る。特に、パラミクソウイルス科ウイルスの天然Nタンパク質は、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されているかもしれない。
【0031】
このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、換及び/又は挿入から成る。
【0032】
Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質と相互作用することができる。
【0033】
対象タンパク質は、マーカータンパク質、治療的又はワクチン的関心のタンパク質でよいが、これらに限定されない。好ましくは、対象タンパク質は、抗原性タンパク質である。
【0034】
更に、対象タンパク質は、それ自体、融合タンパク質でもよい。例えば、それは、対象タンパク質と、そのC-末端のインフレームに融合されたGFPを含む構造、Nタンパク質のC-末端のインフレームに融合されたGFP自体(すなわち、Nタンパク質-GFP-対象タンパク質の融合タンパク質)でもよい。
【0035】
好ましくは、N-GFP-対象タンパク質の構造は、NとGFPタンパク質との間に、配列KLRILQSTVPSERPQASGVYMGNLTTRGPVAT(配列番号32)を有するリンカー配列を含んでもよい。これは、N-GFP融合タンパク質の産生収量を最適化し(実施例6参照)、それによってN-GFP-対象タンパク質の融合タンパク質の産生収量を最適化する。
【0036】
対象タンパク質は、特に、GFP-抗原性タンパク質、又はGFP-治療対象タンパク質の融合タンパク質でよい。それは、対象タンパク質に融合された「リンカー配列」を含むキメラタンパク質でもよい。リンカー配列は、典型的には、最大30アミノ酸、好ましくは最大20アミノ酸、より好ましくは最大10アミノ酸を含み、Nタンパク質と対象タンパク質との間のスペーサとして働き、これらのタンパク質の各々を正確に折り畳むことができる、ポリペプチドである。
【0037】
従って、ある実施態様に従って、対象タンパク質は抗原であり、本発明に従う融合タンパク質はNタンパク質-抗原(「N-Ag」)融合体である。好ましくは、それは、ヒト又はウシRSVのNタンパク質と、抗原性タンパク質との融合タンパク質である。
【0038】
別の実施態様によれば、対象タンパク質は治療対象のタンパク質であり、本発明に従う融合タンパク質はN-タンパク質-治療対象のタンパク質との融合体である。好ましくは、それは、ヒト又はウシRSVのNタンパク質と、治療対象のタンパク質との融合タンパク質である。
【0039】
有利なことに、タグ(例えばヒスチジンタグ)は、Nタンパク質-対象タンパク質の融合タンパク質の精製を助けるために、Nタンパク質のN-末端に融合され得る。
【0040】
代わりに、Nタンパク質-対象タンパク質の融合タンパク質の精製は、本明細書で以下に詳しく説明するように、パラミクソウイルス科ウイルスのPタンパク質との融合タンパク質、特に、GSTに融合されたPタンパク質を共発現させることによって実行され得る。
【0041】
任意の慣用的な分子生物的な、微生物的な又は組換えDNA技術は、本発明に従う融合タンパク質を産生するため使用され得る。このような技術は、当業者の範囲内にあり、特にSambrook, Fritsch & Maniatis, Molecular Cloning: A Laboratory Manual, Second Edition (1989) Cold Spring Harbor laboratory Press, Cold Spring Harbor, New York ("Sambrook et al., 1989"); DNA Cloning: A Practical Approach, Volumes I and II (D.N. Glover ed. 1995); Oligonucleotide Synthesis (M.J. Gait ed. 1984); Nucleic Acid Hybridization [B.D. Hames & S.J. Higgins eds. (1985)]; Transcription and Translation [B.D. Hames & S.J. Higgins eds. (1984)]; Animal Cell Culture [R.I. Freshney, ed. (1986)]; Immobilized Cells and Enzyemes [IRL Press, (1986)]; B. Perbal, A Practica; Guide To Molecular Cloning (1984); F. M. Ausubel et al (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (1994)に記載されている。
【0042】
従って、本発明は、上記のNタンパク質-対象タンパク質の融合タンパク質をコードする核酸にも関する。
【0043】
「コーディング配列」は、ヌクレオチド配列が発現された時にRNA、ポリペプチド、タンパク質等の産生を生じるヌクレオチド配列を称する。タンパク質のコーディング配列は、一般的に、開始コドン(ATG)及び終始コドンを含む。
【0044】
「発現する」又は「発現」は、例えば、遺伝子配列又は対応するDNAの転写及び翻訳に関連する細胞機能の活性化によってタンパク質を産生することにより、遺伝子又はDNA配列に含まれる情報をそれ自体明らかにすることを許容する又は可能にすることを意味する。用語「共発現」は、2つの遺伝子又はDNA配列に含まれる情報が同一の宿主細胞で発現される時に使用される。
【0045】
従って、本発明は、Nタンパク質-対象タンパク質の融合タンパク質をコードする、核酸(cDNA、ゲノムDNA、合成DNA又はRNA)を含む。核酸は、二重鎖でも単一鎖(すなわち、センス鎖又はアンチセンス鎖)でもよい。核酸は、融合タンパク質をコードする配列に限定されず、融合タンパク質をコードする配列の上流又は下流のコーディング配列又は非-コーディング配列を含んでもよい。
【0046】
本発明に従う融合タンパク質をコードする配列は、少なくとも2つの部分、5'から3'方向に連続して、Nタンパク質をコードする部分、及び対象タンパク質をコードする3'側上の部分、を含むハイブリッド又はキメラ配列である。対象タンパク質がそれ自体、融合体である時、又はタグがNタンパク質に結合されている時には、コーディング配列は、好適な数の部分を含む。該部分の各々の間には、コドンがリンカー配列をコードすることがある。
【0047】
本発明は、Nタンパク質-対象タンパク質の融合タンパク質をコードする核酸を含む発現ベクターにも関する。かかるベクターは、DNAと機能的に連結された転写-調節要素を含むことがある。
【0048】
RNAポリメラーゼがコーディング配列をRNA、特にmRNAに転写する時に、コーディング配列は、転写及び翻訳制御配列と「機能的に連結される」。このRNAは、イントロンを含む場合にその後につなぎ合わされ、コーディング配列によってコードされるタンパク質に翻訳される。
【0049】
表現「ベクター」、「クローニングベクター」及び「発現ベクター」は、宿主を形質転換し、導入された配列の発現を促進するために、DNA又はRNA配列(例えば、非相同遺伝子)を宿主に導入できるベヒクルを称する。ベクターの例は、プラスミド、ファージ、ウイルスを含む。最も一般的なベクターは、一般的には細菌起源の自律的複製単位であり、二重鎖DNAの形態にあり得る、プラスミドである。プラスミドは、外来DNA配列を容易に結合し、これは次いで好適な宿主に容易に導入され得る。プラスミドベクターは、一般的に、コーディングDNA配列、プロモーターDNA配列を含み、外来DNAの導入を可能にする1以上の制限部位を有する。プラスミドの非-限定的例は、プラスミドpKK(Clonetech)、pUC及びpET(Novagen, Inc., Madison, WI)、pRSET又はPREP(Invitrogen, San Diego, CA)、pMAL(New England Biolabs, Beverly, MA)、又はpGEX-4T-3(Pharmacia)を含む。
【0050】
本発明は、本発明に従う発現ベクターを含む宿主にも関する。これらの宿主細胞は、前記ベクターにより「形質転換される」。
【0051】
「宿主細胞」は、細胞によって物質の産生、例えば、遺伝子の細胞、DNA又はRNA配列、タンパク質又は酵素による発現、のために選択され、修飾され、培養され又は設計される、任意の細胞又は生物を意味するものと理解される。
【0052】
「発現系」は、ベクターによって運ばれる外来DNAによってコードされそして宿主細胞内に導入されるタンパク質を産生するために、好適な条件下で使用される、宿主細胞及び対応するベクターを称する。慣用的な発現系は、E.コリ宿主細胞及びプラスミドベクター、昆虫細胞及びバキュロウイルスベクター、又は哺乳動物細胞及びウイルス性起源の強力なプロモーターを有するベクター(例えば、サイトメガロウイルス)を含む。
【0053】
本発明の方法に従う発現系は、有利なことに、例えばpGEX-4T-3をベクターとして有する細菌発現系、特にE.コリである。細菌系は、実際に、最も高い産生収量が得られる発現系である。
【0054】
本発明は、従って、上で定義されたNタンパク質-対象タンパク質の融合タンパク質を産生するための方法にも関する。本方法は、場合により、宿主細胞の形質転換、次いでNタンパク質-対象タンパク質の融合タンパク質の発現を許容する条件下で、Nタンパク質-対象タンパク質の融合タンパク質をコードする核酸を含むベクターで形質転換された宿主細胞を培養すること、及び場合により、Nタンパク質-対象タンパク質の融合タンパク質の精製を含む。培養条件は、選択された発現系(すなわち、宿主細胞及びベクター)に依拠し、これの条件の決定は、当業者の範囲内にある。
【0055】
Nタンパク質-対象タンパク質/Pタンパク質複合体の調製法
本発明者らは、パラミクソウイルス科ファミリーのウイルスのNタンパク質がパラミクソウイルス科ファミリーの同一のウイルスのPタンパク質との共発現系で産生され得る、ことを先に示している。同一の系は、Pタンパク質との複合体の形態でのNタンパク質-対象タンパク質の融合タンパク質を発現し、次いで場合によりその複合体から融合タンパク質を精製するために使用され得る。
【0056】
本発明は、Nタンパク質-対象タンパク質/Pタンパク質複合体の調製法であって、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーのウイルスのタンパク質であり、以下のステップ:
a) パラミクソウイルス科ファミリーのウイルスのNタンパク質を共発現し、該対象タンパク質は、パラミクソウイルス科ファミリーの同一のウイルスのPタンパク質で、Nタンパク質のC-末端のインフレームに融合され;
b) そのように形成されたNタンパク質と対象タンパク質/Pタンパク質との複合体を回収すること、
を含む、前記方法に関する。
【0057】
上記のように、Nタンパク質は、ヒスチジンタグのようなタグをそのN-末端で有することがある。更に、対象タンパク質は、それ自体、融合構築物でもよい。
【0058】
更に、上記のように、Nタンパク質は、天然Nタンパク質であるか、あるいは修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義された領域で修飾されてもよい。このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質と相互作用することができる。
【0059】
好ましくは、前記のPタンパク質は、オリゴマー化ドメインPを含まず、かつNタンパク質への結合ドメインを含む、切断Pタンパク質(「PΔ」)である。本発明者らは、この実施態様が溶解形態での大量のN-P複合体の発現を許容したことを先に示している(特許出願FR 05 04 426)。溶解性は、生理食塩水溶液、例えば1XPBS(NaCl 140 mM, KCl 27 mM, Na2HPO4 8mM, KH2PO4 1.5 mM, pH 7.4)又はトリス緩衝液(10 mM pH 7.4, 150 mM NaCl)のような界面活性剤を含まない水性媒体に関して、10,000×gで30分間遠心することによって決定される。
【0060】
表現「切断されたPタンパク質」は、1個以上の隣接アミノ酸配列が削除されたPタンパク質を称する。それは、C-末端配列、N-末端配列、Pタンパク質の一次構造に対して「内部の」配列の切断でも、又はこれらの切断の組合せでもよい。
【0061】
本発明に従う切断されたPタンパク質は、オリゴマー化ドメインPを含まず、Nタンパク質と相互作用することができ、すなわち、それらは、Nタンパク質に対する結合ドメインを有する。パラミクソウイルス科Pタンパク質とNタンパク質との相互作用のドメインは、C-末端の領域でマップされてきたが、切断されたPタンパク質の例は、好ましくはPタンパク質のC-末端断片、又は(Nタンパク質と相互作用できる)Pタンパク質のC-末端断片とPタンパク質の隣接アミノ酸の少なくとも1つの他の配列との融合によって構成される「キメラ」Pタンパク質を含む。前記C-末端断片及び前記Pタンパク質の他の配列は、それ自体、本来隣接してなく、配列重複を示さない。例えば、切断されたRSV Pタンパク質は、天然Pタンパク質のアミノ酸1〜121及び161〜241によって構成される配列を有することがある。参照ポリペプチドの「断片」は、参照ポリペプチドの配列に見出される隣接アミノ酸の任意の配列を称する。
【0062】
表現「Pタンパク質のC-末端断片」又は「PΔN」は、1以上の連続アミノ酸がN-末端から削除されているPタンパク質を意味するものと理解される。好ましくは、Pタンパク質のC-末端断片は、Pタンパク質の一次構造のC-末端半分に位置するアミノ酸鎖を称する(配列中のアミノ酸数が奇数の時、追加のアミノ酸は、N-末端半分に比べて、タンパク質のC-末端半分に任意に配置され得る)。例えば、241アミノ酸を含むRSV Pタンパク質の場合には、PΔ161Nは、Pタンパク質のアミノ酸161〜241によって構成されるC-末端断片を称する。例えば、507アミノ酸を含む麻疹ウイルス(Edmonston B株)のPタンパク質の場合には、PΔ386Nは、Pタンパク質のアミノ酸386〜507によって構成されるC-末端断片を称する。
【0063】
切断されたPタンパク質がPタンパク質のC-末端である時に、本発明は、パラミクソウイルス科ファミリーのウイルスの溶解性Nタンパク質-対象タンパク質/Pタンパク質のC-末端断片の複合体(「N-PI/PΔN複合体」の調製法であって、以下のステップ:
a) パラミクソウイルス科ファミリーの同一のウイルスのPタンパク質と、対象タンパク質のC-末端のインフレームに融合された、パラミクソウイルス科ファミリーのウイルスのNタンパク質を共発現し、ここで、該Pタンパク質のC-末端断片はオリゴマー化ドメインPを含まず、Nタンパク質-対象タンパク質の融合タンパク質と相互作用できる;
b) そのように形成された溶解性N-PI/PΔN複合体を回収すること、
を含む、前記方法に関する。
【0064】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス科又はニューモウイルス科でよい。特に、該ウイルスは、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルス、例えばヒト又はウシの呼吸器合胞体ウイルス(RSV)である。
【0065】
当業者は、抗原-Nタンパク質の融合タンパク質と相互作用することができる、切断されたPタンパク質、より具体的にはPタンパク質のC-末端断片を知っているか、又は決定することができる。
【0066】
例えば、Castagne et al.(2004; Journal of General Virology; 85: 1643-1653)に記載のE.コリのNタンパク質及びPタンパク質を共発現する方法は、P及びNとの相互作用ドメインをマップするために使用できる。よって、本発明者らは、Pタンパク質の9個のC-末端アミノ酸(アミノ酸233〜241)によって構成されるオリゴぺプチドを含むRSV Pタンパク質のC-末端断片は、Nタンパク質と相互作用する、ことを明らかにしている。
【0067】
更に、例えば、センダイウイルスのPタンパク質とRNA-N複合体の形態のNタンパク質又はリボヌクレオタンパク質(RNP)、すなわち「Xドメイン」又はXDとも称される、との相互作用ドメインが、アミノ酸473〜568によって定義される、ことを記載してきた(Kolakofsky et al., 2004,; Virology; 318(2): 463-73)。
【0068】
本発明者らはまた、PSV Pたんぱく質のあるC-末端断片、特に断片PΔ161N(アミノ酸161〜241)が、Pタンパク質全体と比べて、大量のNタンパク質の調製を可能にしたことを証明した。但し、実際に、十分な収量及び工業的スケールを達成することは可能でない。たった9個のアミノ酸を含むPΔ233N(アミノ酸233〜241)に至るまでのより小さな削除変異体ははPΔ161Nの変異体に匹敵する収量を達成できる。
【0069】
PΔ161Nより小さい断片は、Nタンパク質と相互作用することができ、オリゴマー化をすることができず、そのためオリゴマー化ドメインPを含まない、RSVタンパク質の断片に相当する。RSVの最小オリゴマー化ドメインPは、事実、Pタンパク質のアミノ酸120〜150によって定義される。
【0070】
この同一の方法は、本発明者らに、アミノ酸残基386〜507(PΔ386N)によって構成される麻疹ウイルスのPタンパク質のC-末端断片が、該ウイルスのNタンパク質と相互作用し、その精製を可能にした、ことを示させた。対照的に、Pタンパク質のN-末端部分の削除は、残基456(断片PΔ457Nを含む)である限り、Nタンパク質の精製を許容しない。リボヌクレオカプシドと相互作用するPタンパク質のC-末端領域の構造は、Johansson et al.によって決定された(2003 Journal of Biological Chemistry Vol.278, p.44567-44573)。オリゴマー化領域Pは、削除によって決定され、アミノ酸304〜376によって定義されるものと予測されている。
【0071】
RNPの形態でNタンパク質との相互作用ドメインを含むPタンパク質のC-末端断片であるが、オリゴマー化ドメインPが削除されていないPタンパク質のC-末端断片の使用は、Nタンパク質との、タンパク質P断片の相互作用、溶解性N-PΔN複合体の形成、及び高収率での該複合体の産生を許容する。特定のメカニズムに限定するものではないが、オリゴマー化ドメインPの非存在は、これらの複合体のPタンパク質間の相互作用に関連するN-ΔPN複合体の不溶解性の問題を避ける。
【0072】
従って、1つの実施態様によれば、N-PI-PΔN複合体の調製法は、RSV Pタンパク質の最後の9個のC-末端アミノ酸を含み、そしてRSV Pタンパク質の少なくとも119、好ましくは149、より好ましくは160個のN-末端アミノ酸が欠けている、RSV Pタンパク質のC-末端断片の発現、を含む。
【0073】
より具体的には、本発明に従う方法において、RSVのNタンパク質-対象タンパク質の融合タンパク質で共発現することが可能である。
a) 配列番号1で示されるヒトRSVロング株のPタンパク質のアミノ酸配列233〜241を含み、そして配列番号1で示されるRSV Pタンパク質の配列の位置233と120、好ましくは150、より好ましくは161との間に位置するアミノ酸残基である限り、N-末端方向に伸びる、RSV Pタンパク質のC-末端断片;又は
b) C-末端断片、すなわち、異なったヒトRSV株由来又はウシRSV株から得られたPタンパク質の、a) で定義された断片の相同体。
【0074】
RSVのNタンパク質のC-末端断片は、例えば、PΔ120N(Pのアミノ酸120〜241)、PΔ150N(Pのアミノ酸150〜241)、PΔ161N(Pのアミノ酸161〜241)、PΔ180N(Pのアミノ酸180〜241)、PΔ200N(Pのアミノ酸200〜241)、PΔ220N(Pのアミノ酸220〜241)、PΔ230N(Pのアミノ酸230〜241)及びPΔ233N(Pのアミノ酸233〜241)からなる群より選択される。
【0075】
本発明は、RSVのNタンパク質-対象タンパク質の融合タンパク質により、RSV Pタンパク質の最後の9個のC-末端アミノ酸を含み、そしてRSV Pタンパク質の少なくとも119、好ましくは149、より好ましくは160個のN-末端アミノ酸が欠けている、上記のRSV Pタンパク質のC-末端断片を含む切断されたPタンパク質が共発現される方法にも関する。
【0076】
例えば、Pタンパク質のC-末端断片を含む切断されたPタンパク質は、RSV Pタンパク質の最後の122個のN-末端アミノ酸と、最後の80個のC-末端アミノ酸との融合によって構成され得る;例えば、配列番号7で示されるヒトRSVロング株のPタンパク質のアミノ酸鎖1〜121及び161〜241によって構成され得る。
【0077】
別の実施態様によれば、パラミクソウイルス科は、麻疹ウイルスであり、N-PI-PΔN複合体の調製法は、Pタンパク質の最大122個のC-末端アミノ酸を含む麻疹ウイルスのPタンパク質のC-末端断片を発現し、それによって構成されることを含む。特に、配列番号10で示される麻疹ウイルス株Edmonston BのPタンパク質の、株Edmonston BのPタンパク質について定義されたものと相同であるC-末端断片の、又は麻疹ウイルスの異なった株から得られたPタンパク質の、アミノ酸386〜507(PΔ386N)によって構成されるC-末端断片でよい。
【0078】
本発明の文脈では、用語「相同的」は、同一の進化的起源を有するタンパク質、例えば、異なった種に属する、又はウイルス、ウイルス株の場合の、相同タンパク質、の間に存在する関係に関する。かかるタンパク質(及びそれらをコードする遺伝子)は、それらの配列類似性、それがパーセント類似性であろうと又は保存的位置での特定の残基もしくはモチーフの存在の点からであろうと、その配列類似性によって影響される、配列同一性を有する。
【0079】
表現「配列類似性」は、同一の進化的起源を有しても又は有さなくてもよいタンパク質の核酸又はアミノ酸の配列間の同一性の程度を称する。用語相同性と類似性とは、慣用的に区別なく使用される。2つのアミノ酸配列は、それらのアミノ酸が少なくとも80%の同一性又は少なくとも90%の類似性である時に(いわゆる機能的に同一である時に)、「実質的に相同」である。類似の又は相同の配列は、例えばプログラムBLAST又はFASTAを用いてアラインメントによって同定され得る。
【0080】
有利なことに、Pタンパク質、例えば切断されたPタンパク質及び特にPタンパク質のC-末端断片は、Nタンパク質-対象タンパク質/Pタンパク質の複合体の精製を助けるタンパク質、特に、アフィニティクロマトグラフィーで使用されるタンパク質、との融合体の形態で発現される。それは、タンパク質タグ、例えばグルタチオン-S-トランスフェラーゼ(GST)でよい。そこでは、Pタンパク質-融合タンパク質がグルタチオンと結合された固体支持体上のクロマトグラフィーによって分離される。他のラベル又は「タグ」、例えばポリヒスチジン又は「His-タグ」が使用できる。
【0081】
従って、Nタンパク質-対象タンパク質/Pタンパク質-タンパク質タグ(GST又はPタンパク質と融合された別のタンパク質タグ)複合体が得られる。そこではタンパク質タグは酵素的開裂によって除かれる。例えば、該融合体がGST以外のタンパク質を含む時に、GSTは、トロンビン又は任意の他の好適な酵素による開裂によって除かれる。
【0082】
本発明に従う方法を実行させるベクターの構築の具体的な例は、以下の例で記載される。
【0083】
1つの実施態様によれば、対象タンパク質は抗原であり、本発明に従う方法は、溶解性Nタンパク質-抗原/Pタンパク質複合体(N-Ag/P複合体)、好ましくはNタンパク質-抗原/切断されたPタンパク質複合体(N-Ag/PΔ複合体)及びより好ましくはNタンパク質-抗原/Pタンパク質複合体のC-末端断片(N-Ag/PΔN)の形成を生じる。
【0084】
Nタンパク質-対象タンパク質/Pタンパク質複合体からの融合タンパク質の分離
Pタンパク質が切断され、特にPタンパク質のC-末端断片で切断される、Nタンパク質-対象タンパク質/Pタンパク質複合体を調製する方法は、上記のように、Nタンパク質-対象タンパク質/Pタンパク質複合体を、単離された又は精製された形態で容易に取得させる。
【0085】
従って、本発明は、Nタンパク質-対象タンパク質/Pタンパク質複合体であって、Nタンパク質及びPタンパク質が、パラミクソウイルス科ファミリーのウイルスのタンパク質であり、上記の調製法によって得られる、前記複合体にも関する。
【0086】
本発明は、特に、溶解性Nタンパク質-対象タンパク質/切断されたPタンパク質複合体(N-PI/Δ複合体)に関し、より好ましくは溶解性Nタンパク質-対象タンパク質/Pタンパク質複合体のC-末端断片(N-PI/PΔN複合体)に関する。ここで、N及びPは、パラミクソウイルス科ファミリーのウイルスのタンパク質である。
【0087】
このNタンパク質-対象タンパク質/Pタンパク質複合体、又はより具体的には、N-PI/PΔもしくはN-PI/PΔNから、Nタンパク質-対象タンパク質の融合タンパク質は、そのRNAを有する環の形態で、例えばサイズ排除法クロマトグラフィー(ゲル濾過)によって容易に単離され得る。この分離は、好適には、Pタンパク質、及びPタンパク質が場合により融合されるタンパク質タグの酵素的開裂による分離後に、実行され得る。
【0088】
従って、本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質-対象タンパク質の融合タンパク質を調製する方法であって、以下のステップ:
a) 上で定義された方法によってNタンパク質-対象タンパク質/Pタンパク質の複合体を調製し;及び
b) Nタンパク質-対象タンパク質/Pタンパク質の複合体から、Nタンパク質-対象タンパク質の融合タンパク質を分離すること、
を含む、前記方法に更に関する。
【0089】
好ましくは、Pタンパク質は、切断タンパク質、より好ましくはPタンパク質のC-末端断片である。
【0090】
本発明は、より具体的には、溶解性Nタンパク質-パラミクソウイルス科ファミリーのウイルスの対象タンパク質の融合タンパク質を調製する方法であって、以下のステップ:
a)上で定義された方法によって溶解性Nタンパク質-対象タンパク質/P-タンパク質のC-末端の複合体(N-PI/PΔN複合体)を調製し;及び
b) 溶解性N-PI/PΔN複合体から、Nタンパク質-対象タンパク質の融合タンパク質を分離すること、を含む、前記方法に関する。
【0091】
上記のように、Nタンパク質は、天然Nタンパク質でもよく、又は修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されていてもよい。かかる修飾は典型的には、Nタンパク質のC-末端配列における、1以上のアミノ酸(1〜25、又は1〜20、1〜15、1〜10、あるいは1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾性の例は、6又は12個のC-末端アミノ酸の削除から成ってもよい。その切断されたNタンパク質(NΔ6C及びNΔ12C)は依然としてPタンパク質と相互作用することができる。
【0092】
本発明は、上記の方法によって得られるNタンパク質-対象タンパク質の融合タンパク質にも関する。
【0093】
対象タンパク質は、治療的又はワクチン的関心のタンパク質でよい。好ましくは、対象タンパク質は抗原である。
【0094】
免疫原性、ワクチン又は治療的組成物
本発明は、単離された、又はPタンパク質複合体のNタンパク質/C-末端断片(N-PΔN)の形態での、環状構造を有するRSVNタンパク質が、高度に免疫原性であり、特に例えば気管支粘膜での局所反応の刺激を許容する、ことを先に明らかにしてきた。
【0095】
本発明者らは、このような環状構造が治療的又はワクチンタンパク質のためのベクターとして使用され得ることを証明してきた。
【0096】
本発明は、従って、治療的もしくはワクチン的関心の分子又はタンパク質のためのベクターとしての、パラミクソウイルス科ファミリーのウイルスのNタンパク質の使用に関する。
【0097】
1つの実施態様によれば、治療的又はワクチンタンパク質は、上記のように、Nタンパク質に融合される。
【0098】
本発明者らは、単離された、又はNタンパク質/Pタンパク質のC-末端断片の複合体(N-PΔN)の形態での、環状構造を有するRSV Nタンパク質が、免疫原性が高く、特に例えば気管支粘膜での局所的反応の刺激を許容する、ことを先に明らかにしている。
【0099】
本発明者らは、このような環状構造が治療的又はワクチンのタンパク質のためのベクターとして使用され得ることを証明してきた。
【0100】
従って、本発明は、治療的又はワクチン的関心の分子又はタンパク質のためのベクターとしてのパラミクソウイルス科ファミリーのウイルスのNタンパク質の使用に関する。
【0101】
1つの実施態様によれば、治療的又はワクチンタンパク質は、上記のように、Nタンパク質に融合される。
【0102】
別の実施態様によれば、治療的又はワクチン分子は、Nタンパク質に化学的に結合される。Nタンパク質のC-末端へのシステインの付加によって、実際に、タンパク質性又は非-タンパク質性(有機又は無機的な)な性質の様々な分子の化学的結合を許容する部位がつくられる。ジスルフィド結合によってシステインを有するタンパク質をつなぐこともできる。よって、様々な分子に結合されたマレイミド自体は、共有結合によってこのシステインにつながれる。マレイミドは、有機分子のすべての種類、例えばデキストラン(dextrone)、ビオチン、金、又は任意のタンパク質に結合され得る。
【0103】
従って、本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質と、関心の分子、特に、治療的又はワクチン的分子、との複合体にも関する。
【0104】
本発明は、治療的又はワクチン分子又はタンパク質を対象に送達する方法にも関する。その方法は、パラミクソウイルス科ファミリーのウイルスのNタンパク質に共有結合的に結合された治療的又はワクチン分子を、該対象に送達することを含み、あるいはパラミクソウイルス科ファミリーのウイルスのNタンパク質との融合体の形態で該タンパク質を送達することを含む。該治療的又はワクチンタンパク質は、Nタンパク質のC-末端のフレームに融合されている。
【0105】
本発明は、薬学的に許容される担体と共に、場合により、Nタンパク質-対象タンパク質/Pタンパク質の複合体の形態で、Nタンパク質-対象タンパク質の融合タンパク質を含む医薬組成物をも目的とする。ここで、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーの同一のウイルスから得られる。
【0106】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス科でも又はニューモウイルス科でもよい。特に、該ウイルスは、おたふく風ウイルス、麻疹ウイルス及びパラインフルエンザウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルスであり、特に例えばヒト又はウシの呼吸器合胞体ウイルス(RSV)である。
【0107】
対象タンパク質は、治療的又はワクチン的関心のタンパク質でよい。
【0108】
1つの実施態様によれば、医薬組成物は治療的使用に好適である。この実施態様によれば、対象タンパク質は、治療対象のタンパク質である。本発明者らは、Nタンパク質-対象タンパク質の融合タンパク質が細胞を通過し、よってNタンパク質が治療対象の分子又はタンパク質、例えば血管新生抑制又はアポトーシス促進のポリペプチド、のための可能性のあるベクターを構築する、ことを証明してきた。
【0109】
1つの実施態様によれば、医薬組成物は、免疫原性又はワクチン的使用のために好適である。この実施態様によれば、対象タンパク質は、血管新生タンパク質、好ましくは病原性微生物、例えばウイルス、細菌、真菌又は寄生的な後生生物又は原生生物、から得られる抗原である。
【0110】
本発明者らは、実際に、抗原がNタンパク質との融合タンパク質の形態で提供される時に、抗原に対する宿主の反応が非常に顕著に増加する、ことを証明してきた。Nタンパク質-抗原融合タンパク質は、細胞性及び体液の経路による免疫反応を刺激することができる。
【0111】
溶解性Nタンパク質-抗原融合タンパク質は、逆作用なしに、Pタンパク質との複合体の形態でワクチン接種に使用される。従って、本発明に従う免疫原性又はワクチン組成物は、薬学的に許容される担体と共に、溶解性Nタンパク質-抗原/Pタンパク質複合体を含むことがある。
【0112】
「薬学的に許容される担体」は、副作用、例えばヒトマ又は動物におけるアレルギー性反応を生じない、任意の溶媒、分散媒体、吸収-抑制剤等を意味すると理解される。
【0113】
生理学的に許容される担体の例は、当業者に知られている。液状担体の例は、活性成分以外の物質及び水を含まない殺菌性水溶液、あるいは生理学的塩濃度を有する生理学的pH値のリン酸ナトリウムのような緩衝剤、又はその両方、例えばリン酸緩衝生理食塩水(PBS)を含む殺菌性水溶液、を含む。水性担体は、1つ超の緩衝塩、並びに塩化ナトリウム又は塩化カリウムのような塩、デキストロース、ポリエチレングリコール及び他の溶質を含むことができる。
【0114】
本組成物は、生薬製剤に匹敵する方法で、治療上有効量で投与される。投与される量は、治療される対象、活性成分を使用する対象系の能力、及び治療効果の所望の程度に依拠する。投与に必要とされる融合タンパク質の正確な量は、医師の選択及び個人の特徴に依拠する。
【0115】
有利なことに、医薬組成物が免疫原性又はワクチン組成物である時に、アジュバントを更に含んでもよい。「アジュバント」は、同時に投与される抗原に対して細胞又は体液レベルで免疫反応を増加し、刺激し、活性化し、改良し又は調節する、産物を称する。慣用的なアジュバントの例は、細菌性抗原を含むアジュバント、例えば完全フロインドアジュバント、LPS及びその誘導体、細菌性毒素(コレラ毒素及びエンテロトキシン)、及びそれらの解毒変異体(例えば、LT(R192G))、CpGモチーフを含むオリゴヌクレオチド配列、無機アジュバント、例えば水酸化アルミニウム(alum)、リン酸カルシウム又はリン酸カリウム、油状エマルション及び乳化剤(サポニン、例えばQS21)、サイトカインを含む。
【0116】
本発明に従う免疫原性組成物は、免疫反応を、ワクチン接種された対象における抗原に対して誘導する。又はより具体的には抗原が得られる病原体に対して誘導する。
【0117】
本発明に従うワクチン組成物は、抗原を含む病原体による感染に対して保護する。すなわち、ワクチン組成物で免疫されていない対象と比べて感染のような効果の重度が減少する。
【0118】
本発明は、抗原が得られる病原体に対するワクチン接種の方法において上記のワクチン組成物の使用にも関する。
【0119】
従って、本発明は、本発明に従うワクチン組成物の対象への少なくとも1つの投与を含むワクチン接種法に関する。好ましくは、該ワクチン接種法は、該ワクチン組成物の対象への第1投与、及び該ワクチン組成物の同一の対象への少なくとも1つのブースター投与を含む。ブースター投与は、抗原に患者を再曝露することによって、より強い二次免疫反応を誘導する。
【0120】
ワクチン組成物は、有利なことに、抗原が得られる病原体による感染に保護的又は治療的免疫反応を誘導することにおいて、効果的である量で投与される。投薬量は、必然的に、問題となっている活性成分、投与形式、及び対象の年齢及び症状に依拠する。1投薬量当たりのN-Ag融合タンパク質のN-Ag-P、N-Ag-PΔ、又はN-Ag-PΔN複合体の量は、ワクチン投薬量当たり、0.1〜200μg、好ましくは10〜100μgでよい。
【0121】
免疫原性、ワクチンの又は医薬組成物は、任意の経路、特に、粘膜(例えば、眼内、鼻腔内、経口的)経路によって、非経口的(例えば、皮下、皮内、筋肉内、静脈内又は腹腔内)経路によって投与される。
【0122】
表現「対象」は、パラミクソウイルス科ウイルスによる又は任意の他の病原体により感染に曝露されたことがある又は曝露される可能性がある、ヒト又は非-ヒト動物、例えば鳥又は哺乳動物、例えばウシ、ヒツジ、齧歯動物、イヌ科、特にイヌ、ネコ科、特にネコ、ブタ、モンキーを称する。本発明の範囲内の対象は、好ましくは、ヒト又はウシである。
【0123】
それらの局面の各々について、上記のように、Nタンパク質は、天然Nタンパク質でよく、あるいは修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されていてもよい。かかる修飾は、典型的には、Nタンパク質のC-末端配列における、1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、そのような切断されたNタンパク質(NΔ6C及びNΔ12C)は依然としてPタンパク質と相互作用することができる。
【0124】
診断的適用
パラミクソウイルス科ウイルスのNタンパク質と、少なくとも1つのエピトープを含む抗原との融合は、更に、融合タンパク質によって運ばれるエピトープの少なくとも1つに対する抗体を検出するための診断的適用で使用される可能性のある試薬を構築する。
【0125】
本発明は、従って、更に、上記のように、パラミクソウイルス科ファミリーのウイルスのNタンパク質のC-末端のフレームに融合された抗原を含有するNタンパク質-抗原融合タンパク質を含む診断試薬に関する。
【0126】
上記試薬を含む診断キット及び好適な検出手段は、同様に、本発明の範囲内にある。
【0127】
前記抗原は、少なくとも1つのエピトープを含み、同様に、2以上の同一又は異なったエピトープを含んでもよい。
【0128】
本発明は、融合タンパク質の前記抗原に対する抗体のin vitro又はin vivoでの検出のためのNタンパク質-抗原の融合タンパク質の使用をも目的とする。
【0129】
抗原が単一のエピトープを含む時に、検出された抗体はそのエピトープに対して特異的である。
【0130】
抗原が2以上の異なったエピトープを含む時に、検出された抗体は、前記抗原のエピトープの1つ、2つ又は3以上に特異的である。
【0131】
本発明は、生物試料中の抗原に特異的な抗体を検出する方法であって、以下のステップ:
a) 該生物試料を、パラミクソウイルス科ファミリーのウイルスのNタンパク質のC-末端のフレームに融合された抗原を含むN-抗原融合タンパク質と接触させ;
b) 得られたN-抗原融合タンパク質/抗体複合体を検出すること、
を含む、前記方法にも関する。
【0132】
生物試料は、筋肉、肝臓、心臓、脳等よって得られた組織試料、唾液又は液体試料、例えば血液、血漿又は脳脊髄液のような生物液体でよい。
【0133】
複合体の検出は、当業者によく知られている慣用的手段、例えばクロマトグラフィー(サイズ排除、アフィニティー等)、又は非-変性条件下での電気泳動によって行われる。
【0134】
N-抗原/抗体複合体の検出は、検出可能な方法でNタンパク質を標識することによって更に促進され得る。
【0135】
これらの局面の各々について、Nタンパク質は、天然Nタンパク質であるか、又は修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されてもよい。このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。
【0136】
Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質となお相互作用することができる。
【0137】
以下の実施例及び図面は、本発明の範囲を限定することなく、本発明を説明するものである。
【実施例】
【0138】
実施例1:RSVリンタンパク質及びGFPに融合されたRSV Nタンパク質のC-末端領域を含むプラスミドの構築
RSV長鎖のPタンパク質は、241アミノ酸残基からなる。
【0139】
RSV Pタンパク質のC-末端部分を増幅するために使用されるオリゴヌクレオチドプライマー(5'〜3')の配列(BamHI制限部位を強調する;P遺伝子の開始コドンATGは太字で示す)。
【0140】
【表1】

【0141】
RSVロング株のP遺伝子のcDNAは、プライマーであるロング-PBam+及びロング-P-を用いてヒトRSVのロング株によって感染したHep-2細胞からRT-PCRによって増福した(Castagne et al., 2004; Journal of General Virology; 85: 1643-1653)。PCR産物を制限酵素BamHIによって消化し、プラスミドpGEX-4X-3(Pharamacia)の、グルタチオン-S-トランスフェラーゼ又はGSTをコードする遺伝子を有するインフレームのBamHI-SamI部位にクローン化した。このプラスミドをpGEX-Pと呼ぶ。
【0142】
・P161-241のクローニング(PΔ161N)
以下の条件下で、pGEX-Pプラスミドから、PのC-末端領域(アミノ酸161〜241)を増幅した。
PCRプライマー:P16B+及びロング-P- 各100 ng(各1μl)
DNAテンプレートpGEX-P:10 ng(1μl)
酵素:Pfu Turbo(商標)Strategene(2.5 U/μl):1μl
dATP:最終0.2 mM
dGTP:最終0.2 mM
dCTP:最終0.2 mM
dTTP:最終0.2 mM
Pfu緩衝液 最終1X(Strategene)
最終容積:100μl
【0143】
以下の条件下でPCRを行った。
5サイクル:94℃で15秒間、40℃で2分間、72℃で1分間;
25サイクル:94℃で15秒間、55℃で1分間、72℃で1分間。
【0144】
増福されたDNAは、1倍容(100μl)のフェノール/クロロホルム(1vol/1vol)で抽出し、次いで1倍容のクロロホルムで抽出し、最後に10分の1容の5M NaCl(10μl)及び2倍容の100%エタノール(200μl)を加えて沈殿させた。DNAは、13,000 gで20分間遠心し、1倍容の70%エタノールで洗浄し、乾燥し、1倍容の水(90μl)に再懸濁した。10ユニットの酵素BamHIの存在下で、酵素BamHI用の10μlの10X緩衝剤を加えた後、DNAを37℃で2時間消化した。消化したDNAは、エチジウムブロミドの存在下で、1Xトリス-ホウ酸-EDTA緩衝剤(TBE)中の1.5%アガロースゲル上に充填し、電気泳動によって移動させた。P161-241のDNAに対応するバンドを切り取り、DNAを電気溶出によって抽出した。1倍容のフェノール-クロロホルム、1倍容のクロロホルムで再度抽出し、エタノールで沈殿させた。1%アガロースゲル上での精製の後、BamHI及びSmaIによって消化したベクターpGEX-4T-3にライゲートした:
pGEX-4T-3 DNA:100 ng
P161-241 DNA:100 ng
リガーゼ緩衝液 最終1X
リガーゼ(5 U/μl):1μl
最終容積20μl。
【0145】
全体を14℃で終夜インキュベートした。翌日に、コンピテントDH5-アルファTM細菌(Life Technologies)を10μlのライゲーション産物で形質転換し、最終100μg/mlのアンピシリンで補充したLB-アガー培地を含むペトリ皿上に拡げた。組換え菌コロニーをミニ-プラスミド調製物でスクリーニングし、制限酵素BamHI及びXhoIによって消化した。次いで組換えプラスミドはアガロースゲルに2つのバンドを示し、1つはベクター(4.9 kb)をもう1つはPのC-末端(246 bp)に対応した。組換えプラスミドを完全にシークエンスした。
【0146】
・P180-241、P201-241、P221-241のクローニング
アミノ酸180〜241、200〜241、220〜241の部分に対応するPの断片は、以下のプライマー:
P180-241:プライマーP180B+及びロング-P
P201-241:プライマーP201B+及びロング-P-
P220-241:プライマーP221B+及びロング-P-
【0147】
を用いて、pGEX-PプラスミドからPCRによって得た。それらはP161〜241(上記参照)と同一の方法によって増幅し、クローン化した。
【0148】
・RSVロング株のヌクレオカプシドタンパク質をコードする遺伝子のクローニング
ウイルスによって感染されたHep-2細胞からRT-PCRによってヒトRSVロング鎖のNタンパク質をコードする遺伝子を得た。使用したプライマーは以下のとおりである。
【0149】
【表2】
【0150】
PCR産物は、BamHIによって消化し、BamHI-SmaI部位でpGEX-4T-3プラスミドにクローン化した。Nをコードする領域は、BamHI-XhoIによってpGEX-Nプラスミドの消化によってサブクローン化し、pET28a+プラスミド(Novagen)にサブクローン化した。
【0151】
・pET-Nプラスミドの突然変異誘発及び停止コドンの上流の特定のSaI制限部位の生成
以下のオリゴヌクレオチドを用いて指定の突然変異誘発によってヒトRSV(長鎖)のNタンパク質の停止コドンの直ぐ前に、SaI制限部位を導入した。
【0152】
【表3】
【0153】
このことにより、pET-N-Sacプラスミドが得られた。
【0154】
・GFPに融合されたNタンパク質の入手
GFPを制限酵素EagIによる消化によってpEGFPN1プラスミドから除き、次いでDNA上に平滑末端を得るために、ヌクレオチドの存在下でクレノウ処理を行い、次いで、SacIにより消化した。プラスミドpET-N-Sacを酵素XhoIによって消化し、ヌクレオチドの存在下でクレノウ処理し、次いでSaIで消化した。従って、GFPはpET-N-Sacプラスミドに挿入し、pET-N-GFPプラスミドを得た。
【0155】
・Nタンパク質-Fエピトープの取得
RSVのFタンパク質の「ミモトープ」及び「ヘプタッド」エピトープを、二重鎖オリゴヌクレオチドの挿入により、Nタンパク質のC-末端にクローン化した。
【0156】
ミモトープ・エピトープ:
【0157】
【表4】
【0158】
これらの互いにハイブリダイズされたオリゴヌクレオチドは、pET-N-GFPプラスミドのSacI-SaII部位に挿入した。
【0159】
ヘプタッド・エピトープ:
以下のオリゴヌクレオチドを互いにハイブリダイズした。
【0160】
【表5】
【0161】
それらは、pET-N-GFPプラスミドのAgeI部位でクローン化した。
【0162】
・P231-241のクローニング
以下のプライマーを94℃で5分間加熱することによって変性し、次いで室温まで冷却した。
【0163】
【表6】
【0164】
ハイブリダイゼーション後、10 ngの二重鎖オリゴヌクレオチドを、BamHI及びSmaIによって消化された100 ngのPEX-4T-3プラスミドのDNAとライゲートし、アガロースゲル電気泳動によって精製した。組換えプラスミドは、N遺伝子の領域でシークエンシングすることによって確認した。
【0165】
実施例2:複合体の発現及び精製
コンピテントBL21(DE3)細菌(Novagen)を1μgのpGEX-PΔ DNA、及び1μgのpET-N DNAで形質転換し、次いで、100μg/ml終濃度のアンピシリン及び50μg/ml終濃度のカナマイシンで補充されたLB-アガー培地を含むペトリ皿に拡げた。コロニーを選択し、それぞれ100μg/ml及び50μg/mlの濃度のアンピシリン及びカナマイシンを含む2 mlのLB培地で、37℃で終夜培養した。翌日、抗体で補充された1リットルのLB培地を接種するために1 mlの飽和培養液を用い、夜になるまで培養した。夜に、160μg/mlの(タンパク質発現を誘導する)IPTGを含む新鮮LB培地の1容を、培養物に加え、全体を28℃で終夜培養した。翌日、細菌を5000 rpmで15分間遠心し、ペレットを100 mlの以下の緩衝剤:
50 mM トリス pH 708
60 mM NaCl
2 mM DTT
1 mM EDTA
4 mM ベンザミジン
1X 抗蛋白質分解酵素(完全なEDTA-無しのプロテアーゼ阻害剤カクテル、Roche番号11 873 580 001参照)、すなわち、50 mlの溶解緩衝剤につき、1錠
0.1% トリトン-X100
に再懸濁した。
【0166】
10 mg/ml(1 mg/mlの最終濃度)の量のリゾチームで補充した、10 mlの同一の緩衝剤を加えた。細菌を氷上で1時間インキュベートした(溶解)。全体が粘性になったら、全体を混合物に浸漬されたプローブを用いて1分間に3回、氷上で超音波処理し、各超音波処理の間に5分間の間隔を設けた。全体を4℃で30分間遠心し、次いで上清を回収した。上清を4℃で10,000 gで30分間、再度遠心し、次いで新しい上清を回収した。4 mlのセファロース4B-グルタチオンビーズ(Amersham-Pharmacia)を8 mlのビーズ/緩衝剤の混合物(vol/vol)を溶解緩衝液で除くことによって、洗浄した。ビーズを等量の緩衝剤中に放置し、洗浄した細菌溶解物に加え、4℃で終夜回転した。翌日、ビーズを2000 rpmで3分間遠心し、次いで上清を除き、抗蛋白質分解酵素を含まない溶解緩衝剤でビーズを3回、1XPBS緩衝剤で3回、洗浄した。
【0167】
1 mlのビーズについて、1μl(1U)のトロンビン量のビオチン化トロンビン(Novagen)(「トロンビン開裂捕獲キット」、Novagen参照69022-3FRZ)を用いて、トロンビン部位で、ビーズを分けた。ビーズを20℃で終夜インキュベートし、翌日、2000 rpmで3分間遠心し、上清を得るために15分間デカントした。等量の1XPBSをビーズに加え;混合物を攪拌し、デカントした。上清を再度、回収し、先に回収した上清に加えた。回収された上清に、16μlの量の樹脂(すなわち、32μlの樹脂/緩衝剤の混合物(vol/vol))中のストレプトアビジン-アガロースビーズ(Novagen参照69023)を加えた。混合物を1時間攪拌し、次いで2000 rpmで3分間遠心し、上清を回収した。2 mg/mlのタンパク質濃度を得た。
【0168】
開裂産物を含む10μlの上清を1Xラエムリ緩衝液中で変性し、次いで沸騰させ、0.1%トリス-グリシンSDS緩衝液中の12%ポリアクリルアミドゲル上に沈殿させ、次いでタンパク質を明らかにするために電気泳動後にクマシ・ブルーで染色した。
【0169】
実施例3:N-GFP及びPΔ161N(P161-241)の分離、及びN-GFP環の精製
上清に存在するタンパク質は、1XPBS中でサイズ排除クロマトグラフィー(ゲルろ過)によって分離され得る。
【0170】
実施例4:異なった細胞株によるN-PΔN複合体の捕獲:N-緑色蛍光タンパク質(GFP)融合タンパク質の補助付きの試験
I.材料及び方法
I.1 細胞株及び食作用試験:
−HEp-2:EMEM(イーグル最小必須培地)+10% FCS+L-グルタミン+PS(ペニシリン、ストレプトマイシン)中の単層で培養した、ヒト喉頭上皮株(Cancer Res 1955; 15: 598)。
−RAW:DMEM(ダルベッコ最小必須培地)+10% FCS+L-グルタミン+PS中の単層で培養した、ネズミ腹膜単球-マクロファージ(J. Immunol 1977; 119: 950)。
【0171】
細胞株をD-PBS(ダルベッコリン酸緩衝生理食塩水)+0.33 %リドカイン+5 mM EDTA中で分解した。単離された細胞を洗浄し、107細胞/mlのPBS+2% FCSの濃度で再懸濁し、
−20μg/mlのP161-241+N-GFP(N-GF-PΔN)
−6.5μg/mlのGFP
−1 mg/mlのデキストラン-FITC(DX-FITC)(参照:D-1844,Molecular Probes)
の存在下で5 mlの丸底チューブ内で、緩やかに攪拌しながらインキュベートした。
【0172】
エンドサイトーシスのメカニズムが抑制される温度である4℃、内在化が許容される温度である37℃で並行してインキュベーションを行った。1時間又は2時間のインキュベーションの後、細胞を3回洗浄し、PBS+2% FCSに再懸濁した。
【0173】
I.2 フローサイトメトリーによる吸収/内在化の分析
37℃で4℃で蛍光分子(DX-FITC又はN-GFP-PΔN)でインキュベートされた細胞(HEP-2,RAW)と関連する蛍光(FL1)は、フローサイトメトリー(FACSCalibur, Becton)によって比較した。該細胞の自己蛍光のレベルは、蛍光分子の非存在下で、37℃で2時間、インキュベートされた細胞によって得た。
【0174】
I.3 蛍光顕微鏡による吸収/内在化の分析
インキュベーションの後、105細胞をスーパーフロスト(Superfrost)付きスライド(SFPLUS-42, Milian)での細胞遠心によって沈殿させた。スライドは、15分間乾燥し、PERTEX(参照:00814,Histolab)でマウントした。
【0175】
顕微鏡(Axiovert200M, Zeiss)で蛍光レベルを観察した。Metavueソフトウェアを用いてカメラ(Coolsnap HQ)で写真を撮った。
【0176】
I.4 細胞の浸透後のNタンパク質の免疫検出
目的は、Nタンパク質の免疫検出によって、非-浸透細胞(細胞外N-GFP-PΔN複合体)で検出されたシグナルと、浸透細胞(細胞内又は細胞外N-GFP-PΔN複合体)で検出されたシグナルを比較することである。
【0177】
細胞をA緩衝液(参照:GAS-003,Caltag)中に15分間固定し、洗浄した。
【0178】
非-浸透細胞の場合には、細胞洗浄(Cell Wash)緩衝液(Becton)+5% FCSで標識を行った。標識中に細胞を浸透するために、緩衝液B(参照:RGAS-003,Caltag)を用いた。1/100に希釈した抗-CD16/CD32抗体(参照:553141,Becton)で、Fc受容体を20分間飽和した。次いで、1/1000に希釈したNタンパク質に対するラビット血清で、該細胞を30分間インキュベートした。ウサギIgの検出は、1/200に希釈したビオチン化抗体(参照:BA-1000,Vector)で30分間インキュベーションすることによって行った。1/500でのAPC-結合ストレプトアビジン(554067,Becton)の使用は、フローサイトメトリー(FACSCalibur, Becton)によるNタンパク質のFL4チャンネルでの直接検出を可能にした。
【0179】
II.結果:
II-1 N-GFP-PΔN複合体と、RAW細胞(マクロファージ)及びHEp2細胞(上皮)との相互作用の分析
RAW細胞は単球/マクロファージの株である。該細胞は、蛍光ポリマー、DX-FITC、と該細胞を接触させることによって観察され得る食作用能力を有する。DX-FITCの捕獲は、4℃と比べると、37℃では顕著に増加した(図1B)。HEp-2株は、上皮株である。それは、食作用能力を有さない。そのようなわけで、DX-FITCと該細胞との相互作用に関連した蛍光が4℃と37℃でのインキュベーションでほとんど差がない(図1A)。
【0180】
N-GFP-PΔN複合体がRAW及びHEp-2細胞と接触する時に、基本レベル(PBS)に対して蛍光レベルの相当な増加が観察された(図1C、D)。このことは、そのような細胞の膜での効果的な吸収を示した。37℃でのインキュベーション後に、N-GFP-PΔNの存在下で検出された蛍光が、両方の細胞種で減少したが、それにもかかわらず、基本レベルより優れていた(図1C、D)。細胞を組換えGFPでインキュベートする時には、蛍光シグナルは、基本レベルのままであった(データ非表示)。このことは、観察された捕獲現象がN-PΔN環に関連し、GFPには関連しないことを示した。
【0181】
RAW及びHEp-2株でのN-GFP-PΔN環の捕獲は、顕微鏡によって確認した。
【0182】
Nタンパク質に融合されたGFPに特異的な蛍光が、HEp-2又はRAW株の内側で顆粒の形態で見られた。その蛍光は、インキュベーションが組換えGFPで行った場合には見られなかった。N-GFP-PΔN複合体は、従って、RAWマクロファージ株及びHEp-2上皮株で吸収及び/又は内在化された。
【0183】
II-2 RAW細胞によるN-GFP-PΔN複合体の内在化
37℃対4℃でのRAW細胞におけるN-GFP-PΔN複合体の最終結果を理解するために、細胞を透過させ、Nタンパク質の存在を免疫標識及びフローサイトメトリーによる分析によって明らかにした(図2)。透過の非存在下では、N保護(蛍光シグナル)の量は、4℃よりも37℃において少なかった(図2A)。一方、透過後、シグナルレベルは、37℃及び4℃のインキュベーション条件で同一となった(図2B)。従って、N-GFP-PΔN複合体は、37℃でのインキュベーションで内在化された。
【0184】
結局、フローサイトメトリー及び顕微鏡によって、N-GFP-PΔN複合体を用いて行われた実験の全体性は、これらのタンパク質の構造の新規性を証明することができた。核タンパク質の環状集合体は、様々な細胞種、例えばマクロファージ、上皮細胞及び樹状細胞によって効果的に吸収され、内在化される能力を有する(データ非表示)。これらの性質は、抗原ベクター又はワクチンとしてのその構造の使用に特に有用である。
【0185】
実施例5:融合体中で発現された緑色蛍光タンパク質(GFP)と比べた環状形態のNタンパク質の抗原ベクター性の評価
I.材料及び方法
I.1 マウス:
Unite Experimentale Animalerie Rongeur(INRA, Jouy-en-Josas)で飼育された、雄性BALB/c、8〜10週齢。
【0186】
I.2 抗原:
−0.1064μg/μlのP161-241+N-GFP(N-GFP-PΔN)濃度。
−0.5μg/μlのGFP濃度。
−1μg/μlのP161-241+N-PΔN濃度。
【0187】
I.3 アジュバント:
−E.コリのLT(R192G)解毒リンホトキシン、1 mg/ml (John Clements; Choi et al ., 2004, Protein Expression and Purification 38, pp.205によって送られたバッチ)。
【0188】
I.4 免疫:
アベルチン麻酔(300 li.p.)下で、以下の混合物(60μl/マウス):
−5μgのLT(R192G)
−5μGのN-GFP-PΔN
又は、1.7μgの溶解性GFP、又は3.3μgのN-PΔN(5μgのN-GFP-PΔN中に存在するものに対応するGFP量又はN-PΔN量)
−非発熱性の生理的血清を有するgsp 60μl
の鼻腔内経路(i.n.)による投与。
【0189】
すべての溶液は、注入前に0.22μmのフィルターを通過させた。
【0190】
【表7】
【0191】
I.5 抗-N-PΔN及び抗-GFP抗体の産生
ELISAによって、血清及びBAW中の抗-N-PΔN又は抗-GFP抗体(IgH+L及びIgA)を検索した。
−血清を血液試料から集め(4℃での1晩exsudation)、次いで-20℃で冷凍した。
−BAWを1700 rpmで5分間遠心し、上清を集め(約1 ml)、-20℃で冷凍した。
【0192】
96-ウェルプレート(Immulon 2HB, ThermoLabsystems)を、0.1 M重炭酸ナトリウム緩衝液、pH 9.5中のN-PΔN複合体又は組換えGFPタンパク質(200 ng/ウェル、100μl/ウェル)で、4℃で1晩感作した。このプレートを0.05% PBS-Tween 20(Wellwash device, Labsysytems)の200μl/ウェルで5回洗浄した。次いで、プレートを、0.05% PBS-Tween 20緩衝液及び5%ウシ胎児血清(PBS-T-FCS)の150μl/ウェルで、37℃で1時間飽和した。5回洗浄後、滴定する試料をPBS-T-FCSで希釈した(血清については、最初の希釈から1/30まで3のファクターで7連続希釈、BAWについては、最初の希釈から1/3までの希釈)。このプレートを37℃で2時間インキュベートした。5回洗浄後、PBS-T-FCSで希釈した二次抗体を100μl/ウェルの量で分配した。使用した二次抗体は、ペルオキシダーゼにコンジュゲートされ、マウス免疫グロブリン:Ig(H+L)(4000th, P.A.I.S.)又はIgA(1000th, Caltag)に対するものであった。このプレートを次いで37℃で2時間インキュベートし、5回洗浄した。次いで、このプレートをペルオキシダーゼ基質(TMB, 100μl/ウェル)で10分間、暗闇でインキュベートした。酵素反応を50μlの2M H3PO4で停止した。光学密度(OD)を450 nmで読んだ(Dynexリーダー)。OD450=f(希釈)曲線をオリジンソフトウェアによって回帰曲線y=(b+cx)/(1+ax)によってモデル化した。抗体力価を、最も薄い希釈における対照試料(d0)のOD450を2回与える希釈値として決定した。
【0193】
I.6 N-PΔN又はGFPに特異的なTリンパ球によるIFN-γ/IL-5/IL-10の産生
脾臓、及び気道(顔の、首の及び縦隔の気道)をドレインするガングリオンを、同一のプロトコールに従って処理した。脾臓を個々に処理し、ガングリオンを試験群によって分類した。
【0194】
リンパ器官を薄くスライスし、次いで、RPMI培地及びPSでフィルター(100μm細胞シーブ,BD Falcon)上でそっと粉にした。細胞懸濁液を4℃で1700 rpmで10分間遠心した。細胞を1 mlの赤血球溶解緩衝液(低張生理食塩水緩衝液)に再懸濁し、室温で5分間インキュベートした。溶解反応を10 mlの完全RPMI(PS, 2 mM L-グルタミン及び10% FCS)の添加により停止した。膜の残骸をデカントし、細胞を遠心(1700 rpmで4℃で10分間)により3回洗浄した。細胞懸濁液をマラセズ細胞によってカウントした。
【0195】
該細胞を、細胞培養処理した96-ウェルマイクロプレート培養(Falcon)で、200μlの完全RPMI培地で400,000細胞/ウェルの量で培養した。4つの培養条件は、各々の懸濁液について3点で試験した:
−PMA(ホルボール 12-ミリスチン酸エステル 13-酢酸エステル,Sigma)10 ng/ml、及びイオノマイシン(Sigma)1μg/ml(陽性対照、ポリクローナル活性化)
−完全RPMI(陰性対照)
−N-PΔN 10μg/ml
-GFP 10μg/ml。
【0196】
5% CO2で37℃での48時間後(IL-5及びIL-10)又は72時間(IFN-γ)培養後、培養上清を回収し、ELISAのサイトカインの滴定まで-20℃で冷凍した。
【0197】
96-ウェルプレート(Immulon 2HB, ThermoLabsystems)を、0.1 M重炭酸ナトリウム緩衝液、pH 9.5中の、4μg/ml(IFN-γ)又は2μg/ml(IL-5/IL-10)のマウス抗-サイトカイン捕獲抗体(100μl/ウェル)で、4℃で終夜、感作した。このプレートを0.05% PBS-Tween 20(Wellwash device, Labsysytems)の200μl/ウェルで5回洗浄した。次いで、プレートを、0.05% PBS-Tween 20緩衝液及び0.2%ウシ胎児血清(PBS-T-BSA)の150μl/ウェルで、37℃で2時間飽和した。5回洗浄後、滴定される、組換えマウスサイトカイン対照及び試料を連続希釈によってPBS-T-BSAで1/2に希釈した。1/2への4つの連続希釈は、純水な試料で行った。プレートを次いで4℃で終夜インキュベートした。5回洗浄後、ビオチン化検出抗体を分配し(PBS-T-BSA中、IFN-γ/IL-10について1μg/ml、IL-5について0.5μg/ml、100μl/ウェル)、4℃で3時間インキュベートした。5回洗浄後、ストレプトアビジン-ペルオキシダーゼコンジュゲート(Pierce)を分配し(PBS-T-BSA中1μg/ml、100μl/ウェル)、4℃で1時間インキュベートした。5回洗浄後、ペルオキシダーゼ基質(ABTS+H2O)をウェルに分配した。45分間インキュベーションした後、光学密度を405 nmで読んだ(Dynex ELISAリーダー)。リーダーに結合した分析ソフトウェア「Revelation」を用いて標準値に対して試料中のIFN-γ/IL-5/IL-10の濃度を計算した。
【0198】
抗体参照(BD Bioscience):
−IFN-γ:捕獲抗体:クローンR4-6A2(BD Bioscience,551216)
−検出抗体:クローンXMG1.2(BD Bioscience,参照554410)
−組換えmIFN-γ:R&D装置485-MI
−IL-5:捕獲抗体:クローンTRFK5(BD Bioscience,554393)
−検出抗体:クローンTRFK4(BD Bioscience,参照554397
−組換えmIL-5:BD Bioscience 554581
−IL-10:捕獲抗体:クローンJES5-2A5(BD Bioscience,551215
−検出抗体:クローンSXC-1(BD Bioscience,参照554423)
−組換えmIL-10:BD Bioscience 550070
−ストレプトアビジンペルオキシダーゼImmunopure(Pierce21126)。
【0199】
I.7 統計的分析
マン-ホイットニーU試験を行うことによって群を対で比較した(http://eatworms.swmed.edu/〜leon/stats/utest.html)。p<0.05の値が有意であると考えられた。
【0200】
II.結果:
ワクチンベクターとしてN-PΔN複合体によって形成される環の効力を評価するために、RSVのN核タンパク質との融合タンパク質の形態のGFP(モデル抗原)で、BALB/cマウスを免疫した。N-GFP融合タンパク質は、先に記載の方法(特許第FR0504426号)によって精製され得る溶解性環構造を形成する。このようにしてNGFP-PΔNタンパク質複合体は得られた。ワクチン用途については、N-GFP及びPタンパク質が分離できるが、その操作は必ずしも必要でなく、N環の免疫原性についての発明者の先の結果は、PΔNの存在が負の効果を有さないことを示している。
【0201】
気道を標的とする病原体に関する研究、及び鼻腔内経路によって投与される時の、N-PΔN複合体の証明された免疫原性を考慮すると、それは、その複合体のベクター性を証明するための好ましい投与経路である。マウス群を、天然組換え体のGFPタンパク質、及びN-PΔN複合体でも免疫した。E.コリの解毒リンンフォトキシン、LT(R192G)、ここで粘膜経路によるそのアジュバント性は、十分に記載されているが(McNeal et al., 2005, Freytag and Clements 2005)。
【0202】
GFP及びN-PΔNに対する免疫反応のパラメーターは、(i)(気管支肺胞の洗浄)による血清及び粘膜抗体の産生、及び (ii) 脾臓、及び気道をドレインするガングリオンから単離された記憶Tリンパ球による、サイトカイン(IFN-γ、IL-5及びIL-10)の産生を介する細胞反応、である。
【0203】
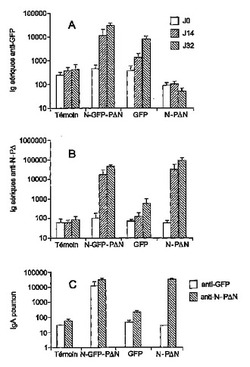
II-1 Nの環は、外来抗原、例えばGFPmに対する抗体を標的化するための効果的なベクターである
N-PΔN複合体は、全身的経路及び粘膜経路によって免疫原性が非常に高い(特許第FR0504426)。GFPとNタンパク質との融合体は、血清レベル(図3B)又は粘膜レベル(図3C)の抗-N-PΔN Ac反応の強度を変更しない:すなわち、群N-GFP-PΔNと群N-PΔNとには有意な差はない。
【0204】
鼻腔経路によって投与されたGFPは、全身性Ac反応を誘導するが(図3A;GFP対対照、p=0.003)、局所的反応を誘導しない(図3C;GFP対対照、有意でない)。
【0205】
一方、GFPとN-PΔN複合体との関係は、GFPのみと比較して、抗-GFP抗体の割合をかなり増加する。抗-GF抗体のレベルは、血清レベルで4のファクターで乗じられ(N-GFP-PΔN対GFP、p=0.039)、BAWレベルで100のファクターで乗じられる(N-GFP-PΔN対GFP、p=0.014)。
【0206】
II-2 N環でのGFPのベクター化は、特定のT反応を増加しない:
抗体反応を刺激するその強い能力に加えて、N-PΔNは、全身的レベルの抗原依存的IFN-γの合成によって証明される記憶T反応を効果的に刺激するが(特許第FR0504426及び図4、N-PΔN脾臓対対照、P=0.01)、局所レベルの抗原依存的IFN-γの合成によって証明される記憶T反応を効果的に刺激しない(図5、局所的ガングリオン)。GFPのような外来抗原のNへの融合は、脾臓レベル(N-GFP-PΔN対N-PΔN、差はない)又はガングリオンのレベルで(図4)、そのT反応の確立を妨害するものではない。
【0207】
一方、GFPの提示形式によらず(単独でも又はNと融合しても)、IFN-γ(図4)又はIL-5/IL-10(図示せず)の産生によって測定された、GFPに特異的なT反応は観察されなかった:すなわち、対照群、N-GFP-PΔN及びGFP、間で有意な差はない。
【0208】
結局、GFPの例は、N-PΔN複合体が、特に粘膜レベルで、抗体反応を刺激するための非常に極力な抗原ベクターであることを証明することができる。一方、それらは、同一の抗原に対する記憶T反応を刺激する点でほとんど効果がないことが判る。重要なことに、外来抗原の融合は、N-PΔN複合体に固有の免疫原性を変更しなかった。
【0209】
これらの結果は、強力な抗体反応を刺激す目的で抗原ベクターとしてN-PΔN複合体を用いて考えることを可能にする。RSVのワクチン化の範囲内で、得られた結果は、核タンパク質に対する細胞反応と、抗体を中和する標的であることが知られているRSVの抗原/エピトープに対する抗体反応とを利用することができることを示している。免疫防御のこれらの2つの事実は、RSVに対する殺菌免疫を得る点で重要である。
【0210】
一般的に、多くの病原体(特にウイルス)に関して、標的抗原又は中和抗体の抗原は、文献に十分に記載され、感染の効果的な制御に関連している。従って、これらの反応を効果的に刺激するための新規ベクターは、ワクチン化には非常に好ましい。N-PΔN複合体は、宿主又は環境のための危険性がない、不活性で、非-複製的構造であるという利点を有している。
【0211】
実施例6:Nタンパク質とGFPタンパク質とのリンカー配列の最適化
N-GFP融合タンパク質の精製は、GSTに融合されたタンパク質P(断片161〜241)を用いる親和性によって行った。実施例1に従って構築されたタンパク質N-GFP(配列番号33)は、タンパク質Nの収量よりも10倍低い収量で精製された。加えて、約43 kDaの見掛け質量で移動する2つのポリペプチドの存在が依然として観察された。これらのバンドは、質量分析(MALDI-TOF)によって分析した。それらは、(i) Nタンパク質のC-末端の追加の11残基を生じる、(アルギニンのレベルでの)GFPからNを分離するリンカー配列において、及び (ii) 最後の6残基を除くことによって(リジンのレベルでの)Nタンパク質のC-末端において、起こった開裂に相当する。C-末端の6、12及び27残基が削除されたNタンパク質の構築は、NΔ5C及びNΔ12Cのタンパク質が依然としてPと相互作用し、切断された形態NΔ27Cとは異なる、ことを示している。
【0212】
従って、NとGFPとの間のリンカー配列は修飾された。すなわち、その配列:
【0213】
【表8】
【0214】
(配列番号34、矢印は、E.コリでの開裂部位を示す)は、配列:
【0215】
【表9】
【0216】
(配列番号32)によって置換された。
【0217】
このことは、細菌中でもはや分解されず(SDS-PAGEゲル中の更なるバンドの非存在)、そして天然Nタンパク質単独と同一の収率で精製された、より安定なN-GFP融合タンパク質(配列番号35)をもたらした。
【図面の簡単な説明】
【0218】
【図1】図1は、HEp2又はRAW細胞によるN-GFP-PΔNの捕獲を示す。HEp-2(A;C)又はRAW(B;D)細胞を、DX-FITC(A;B)又はN-GFP-PΔN(C;D)の存在下にインキュベートした。インキュベーションを4℃で1時間(点線の曲線)、4℃で2時間(太い実線の曲線)、37℃で1時間続けた(実線の曲線)又は37℃で2時間続けた(太い点線の曲線)。100,000事象についてフローサイトメトリーで蛍光(FITC又はGFP)を分析した。
【図2】図2は、N-GFP-PΔN複合体がRAW細胞中に内在化されることを示している。RAW細胞は、4℃でN-GFP-PΔNの存在下で(太い実線の曲線)又は37℃でN-GFP-PΔNの存在下で(点線の曲線)、1時間インキュベートした。Nタンパク質を、非-浸透細胞(A)又は浸透細胞(B)についてのフローサイトメトリーによる直接的な免疫標識によって検出した。細胞の自己蛍光は、PBS中で37℃でインキュベートされた細胞のシグナルによって得られた(実線の曲線)。
【図3】図3は、N-PΔN複合体に対するGFPのベクター化による抗体反応の増加を示す。アジュバントのみ(対照)、アジュバントの存在下でのN-GFP-PΔN複合体、アジュバントの存在下でのN-PΔN又はGFPを用いて、d0日に鼻腔内経路によってBALB/cマウスを免疫した。2週間後(d14)にブースター注射を行った。動物をブースター注射後2週間に屠殺した(d28)。d0、d14及びd28の血清を回収した(A及びB)。d28に気管支肺胞洗浄を行った(C)。抗-N-PΔN抗体(B及びC)、及び抗-GFP抗体(A及びC)の力価は、ELISAによって決定した。データを平均±平均の標準誤差として表し(対照及びGFP群についてはn=8、N-GFP-PΔ群についてはn=6、N-PΔ群についてはn=4)、対数目盛に従って表した。
【図4】図4は、N-PΔNによってベクター化されたその形態を含む、GFPに対する記憶T反応の非存在を示す。アジュバントのみ(対照)、アジュバントの存在下でのN-GFP-PΔN複合体、アジュバントの存在下でのN-PΔN又はGFPを用いて、d0日に鼻腔内経路によってBALB/cマウスを免疫した。2週間後(d14)にブースター注射を行った。脾臓及び局所的ガングリオンを除くために、ブースター注射の2週間後に動物を解剖した。細胞懸濁液を脾臓については個別に(菱形)、及びガングリオンについては1プール/群で処理した(丸)。細胞をGFP(白抜き菱形及び丸)又はN-PΔ(黒色菱形及び丸)で72時間再刺激した。IFN-γの分泌をELISAで測定した。細胞を培地のみで培養した時、IFN-γの基本レベルは、15 pg/ml未満であった。
【技術分野】
【0001】
本発明は、Nタンパク質-対象タンパク質の融合タンパク質、場合により、溶解性Nタンパク質-対象タンパク質/Pタンパク質の複合体の形態にあり、Nタンパク質及びPタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質である、に関する。対象タンパク質が抗原である時に、本発明は、Nタンパク質-抗原の融合タンパク質又はNタンパク質-抗原/Pタンパク質の複合体を含む、ワクチン組成物及び診断試薬にも関する。対象融合タンパク質のNタンパク質は、抗ウイルス薬又は抗癌剤のような関心の治療分子を細胞に移送するための「ベクター」としても使用され得る。
【背景技術】
【0002】
呼吸器合胞体ウイルス(RSV)は、幼児の気管支梢炎の原因である(500,000人の子供がフランスで毎年罹患している)。該疾患に対する真に効果的な予防処置(ワクチン)又は抗体ウイルス薬は存在しない。同一の疾患は、ウシにも存在し、最初の年に子牛の70%がかかり、致死率は20%に到達し、ウシRSVは、子牛の重大な気管支疾患を引き起こす主な薬剤である。臨床発表はヒトでのものと同一である。このウイルスは、モノネガウイルス(Mononegavirales)系列及びパラミクソウイルス科(Paramyxoviridae)ファミリーに属する。ウイルス粒子は、2つの主なタンパク質、融合タンパク質(F)及び糖タンパク質(G)、を含む脂質外皮膜によって取り囲まれている。粒子の内側には、ヌクレオカプシドタンパク質(N)と関連する、約15 kbの陰極の単一鎖RNAが存在する。そのRNA-N複合体は、RNA-依存的RNAポリメラーゼであるLタンパク質(大きい断片)及びビリオンにも存在するその補助因子P(リンタンパク質)によって構成されるポリメラーゼ複合体のマトリックスを構成する。
【0003】
溶解形態のNタンパク質を精製するのはこれまで可能でなかった。なぜなら、該タンパク質は組換え体で発現される時に、細胞RNAに自発的かつ非-特異的に結合し、非常に高分子量の不溶解性構造を形成するからである(Meric et al., 1994 Virus Res. 31(2): 187-201; Bhella et al., 2002 Journal of General Virology; 83, 1831-1839)。本発明者らは、環状様の構造を有する溶解性組み換え体として呼吸器合胞体ウイルス(RSV)のヌクレオカプシド(N)タンパク質を入手するための方法、及びワクチンとしてのその用途を開発してきた。Nタンパク質は、約7 nmの径を有する非常に規則的な環の形態にあり、Nタンパク質の10分子及び約70塩基の細菌起源のRNAを含む。Nタンパク質は、グルタチオン-S-トランスフェラーゼ(GST)に融合されたRSVのリンタンパク質(P)、より具体的にはそのC-末端部分で、E.コリ中で共発現される。それは、複合体をセファロース-グルタチオンビーズでの親和性によって精製させる。この環状様構造は、特に、鼻腔内投与によって、アジュバントの非存在下でも、マウスでの強い免疫反応を誘導する。該方法は、FR 05 04 426で2005年4月29日にフランスで出願された特許出願の対象である。
【0004】
本発明者らは、ワクチンベクターとしてその環状様構造を使用できることを示している。その技術の実現可能性は、プラスミド構築によって、Nタンパク質の他のタンパク質、GFP(緑色蛍光タンパク質)への融合によって証明されている。それは、GSTに融合されたPタンパク質(アミノ酸161〜241)のC-末端部分で、E.コリ中で共発現されてきた。電子顕微鏡を用いる精製構造体の分析は、Nタンパク質のC-末端でのGFPの接合が、Nタンパク質を、約10 nmの径を有する環の形態でなお産生させる、ことを示している。N-GFP融合タンパク質は蛍光を発する。
【0005】
N-RNA環は非常に免疫原性であるので、本発明者らは、特にGFPに対する強力な抗体反応を誘導できるかどうかをみるためにマウスにその構造体を注入した。その反応を、E.コリからの組み換え体でも精製された「通常の」GFPタンパク質に対して得られたものと比較した。得られた結果は、環状様構造体が使用される時に、GFPに対してより顕著な強い反応を示した(第1実験について約40倍超)。
【0006】
Nタンパク質への対象タンパク質の融合の可能性、並びに工業的スケール及び適当なコストでの溶解性環の形態での複合体の精製の可能性(特に、E.コリにおいて)は、それ自体に注入された時に高い免疫原性でないか又は免疫原性でない任意の種類のエピトープに対するワクチンの開発を可能にするだろう。
【発明の開示】
【0007】
本発明者らは、そのNタンパク質-GFP/Pタンパク質構造体が吸収され及び異なった細胞種によって効果的に内面化される、ことを更に示した。従ってその結果は、ワクチン化のための抗原ベクターとして又はより一般的には治療的関心の分子としてそのようなNタンパク質-対象タンパク質の融合体の価値を、場合により、Nタンパク質-対象タンパク質/Pタンパク質複合体の形態で、強調する。
【0008】
定義
パラミクソウイルス科(Paramyxoviridae)ファミリーは、パラミクソウイルス(Paramyxoviridae)及びニューモウイルス(Pneumoviridae)サブファミリーを包含する。パラミクソウイルス科は、センダイウイルスのプロトタイプであるレスピロウイルス(Respirovirus)属、ルブラウイルス(Rubulavirus)(特におたふく風邪ウイルス)、モルビリウイルス(Morbilivirus)、例えば麻疹ウイルスを含む。レスピロウイルス及びルブラウイルス属の各々は、共にパラインフルエンザウイルスの株に分類される。ニューモウイルス・サブファミリーは、2つの属、ニューモウイルスとメタニューモウイルス(Metapneumoviridae)に分類され、後者はヒトメタニューモウイルスを含む。ヒト呼吸器合胞体ウイルス(RSV)は、ニューモウイルス・サブファミリーに属するニューモウイルス属のプロトタイプウイルスを構成する。ニューモウイルスは、RSVのウシ及びネズミ株をも含む。
【0009】
他に特定しない限り、「呼吸器合胞体ウイルス」は、問題となっている、その形態(ヒト、ウシ等)、サブグループ(例えば、ヒトRSVにおいて特定されたサブグループA、B及びS)又は株がそのようなものであれ、一般的に、RSVを意味すると理解される。
【0010】
「Nタンパク質」は、ウイルスゲノムを囲むように螺旋構造を形成する、パラミクソウイルスのヌクレオカプシドタンパク質を称する。ヒトRSV長鎖のNタンパク質は、配列番号1によって記載される391アミノ酸の配列を有する。同様に、ウシRSVのNタンパク質は、391アミノ酸を含む(配列番号2)。センダイウイルス(Hamamatsu株)、パラミクソウイルス科、おたふく風ウイルス(SBL-1株)、及びヒトメタニューモウイルス(00-1株)のNタンパク質も、各々、受入番号Q9DUE3(配列番号3)、Q89933(配列番号4)、P21277(配列番号5)及びQ91F57(配列番号6)としてスイスプロットデータベースに記載されている。
【0011】
表現「Pタンパク質」は、リンタンパク質、又はパラミクソウイルス科ファミリーのウイルスのポリメラーゼ複合体のPタンパク質形成部分を称する。Pタンパク質は、ウイルスポリメラーゼ(レプリカーゼ/転写酵素)の補助因子であり、リン酸化される。パラミクソウイルスのPタンパク質の配列は、当業者に知られている。例えば、ヒトRSV長鎖株のPタンパク質は241アミノ酸の配列を有し、それは受入番号P12579としてスイスプロットデータベースに寄託されている。その配列は、配列番号7で示される。ウシRSVのPタンパク質も241アミン酸を含む(配列番号8)。センダイウイルス(Harris株)、麻疹ウイルス(Edmonston B株)、おたふく風ウイルス(SBL-1株)、及びヒトメタニューモウイルス(00-1株)のPタンパク質も、各々、受入番号P04559(配列番号9)、CAA91364(配列番号10)、P19717(配列番号11)及びQ91KZ5(配列番号12)としてスイスプロットデータベースに記載されている。表現「Pタンパク質」は、Pタンパク質全体、切断されたPタンパク質又はPタンパク質の断片を意味することがある。
【0012】
パラミクソウイルスのPタンパク質は、ホモオリゴマー特にホモテトラマーを、例えばセンダイウイルス又はRSV中で形成する。RSVの場合には、オリゴマー化(P-Pオリゴマー化)できるPタンパク質のドメインは、そのタンパク質の120〜150アミノ酸領域にマップされている(Castagne et al., 2004; Journal of General Virology; 85: 1643-1653)。従って、例えば、RSVのPタンパク質の161〜241アミノ酸によって構成される断片は、オリゴマーを形成しない。センダイウイルスのPタンパク質のオリゴマー化ドメインは、Pタンパク質の320〜446残基によって構成されるものとして、Tarbouriech et al. (2000; Nature Structural Biology; 7, 777-781)によって記載されている。一方、Pオリゴマー化領域は、麻疹ウイルスのPタンパク質については304〜376アミノ酸の領域に同定されている(Johannson et al., 2003; Journal of Biological Chemistry; 278, p.44567-44573)。
【0013】
上記のPタンパク質及びNタンパク質の配列は、本来、例証であり、所与のウイルスを考慮すると特定の株に従って変形を示す配列も可能である。従って、出願のリマインダーで述べたアミノ酸の位置は、参照配列に対して示されている。当業者は、完全に、特に例えばClustalwのようなソフトウェアを用いて実行された配列アラインメントの助けを借りて、例示された株以外のウイルス株における対応するドメインを特定することができる。
【0014】
パラミクソウイルスファミリーのウイルスのNタンパク質及びPタンパク質のコーディング配列も当業者に知られている。
【0015】
用語「タンパク質ラベル」は「タンパク質タグ」とも称され、その精製を助けるために対象タンパク質への融合のために使用されるタンパク質を言う。タンパク質タグは、当業者に知られている。タンパク質タグの例は、グルタチオン-S-トランスフェラーゼ(GST)、又は4〜10ヒスチジン残基の鎖を一般的に含む配列であるヒスチジンタグを含む。
【0016】
「対象タンパク質」は、任意のタンパク質、ポリペプチド又はペプチド(これらの用語は無差別に使用される)、例えばマーカータンパク質又は治療的もしくはワクチン的関心、を称する。
【0017】
対象タンパク質は、例えば、gfpレポーター遺伝子によってコードされる、タンパク質GFP(緑色蛍光タンパク質)でもよい。
【0018】
治療対象のタンパク質は、例えば、抗癌剤として使用され得る、血管新生抑制ポリペプチド、例えばRGD又はRGDを含む配列、エンドスタチン、又はアポトーシス促進性ポリペプチド例えばアポトーシス-誘導因子(AIF)、ウイルスタンパク質と特異的に相互作用し、ウイルス又は毒素の複製を許容するメカニズムを妨害することができるポリペプチドでよい。
【0019】
対象の異種タンパク質は、抗原、特にワクチン的関心の抗原性タンパク質でよい。「抗原」又は「Ag」は、投与される宿主内での免疫反応を誘導することができるペプチド性又は糖ペプチド性の性質の配列であると理解される。従って、抗原は、タンパク質又はタンパク質の一部(ポリペプチド)でもよく、あるいは潜在的にエピトープに対応する小ペプチドでもよい。
【0020】
「エピトープ」は、抗体によって又はリンパ球受容体によって認識される抗原の一部である。(直鎖)エピトープは、一般的に、7〜15アミノ酸の配列によって構成される。本発明の範囲内の抗原は、エピトープによって構成されてもよく、エピトープを含んでもよく、又は抗原性タンパク質でもよい。
【0021】
好ましくは、抗原は、病原性微生物、例えばウイルス、バクテリア、真菌又は寄生的な後生生物もしくは原生生物から得られる抗原である。
【0022】
ウイルスの例は、ヒト免疫不全ウイルス(HIV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、ヘルペスウイルス(Herpes simplex)、インフルエンザウイルス(インフルエンザウイルス又はトリインフルエンザウイルス)、西洋ニールウイルス、黄熱ウイルス、サイトメガロウイルス、パピローマウイルス(HPV)、エプスタイン・バール・ウイルス(EBV)、RSV、デングウイルス、及びチクングニアウイルスを含む。
【0023】
寄生動物の例は、マラリア(プラスモディウム(Plasmodium)、特にP. ファルシファルム(P. falciparum)、三日熱マラリア(P. vivax)、卵形マラリア原虫(P. ovale)、及び四日熱マラリア原虫(P. malariae))及びトリパノソミアシス(特に、睡眠病)(T. brucei)及びシャーガス病(T. cruzi))の原因となる寄生動物を含む。病原は、カンジタ症の原因となるカンジタ・アルビカンスのような真菌でもよい。
【0024】
細菌は、例えば、ヘリコバクター・ピロリ(Heliobacter pylori)、クロストリジウム・テタニ(Clostridium tetani)、結核菌(Mycobacterium tuberculosis)又はウシ型結核菌(Mycobacterium bovis)でよい。
【0025】
例えば、抗原は、GFP、ウイルス性糖タンパク質、例えばRSV融合タンパク質(F)、インフルエンザウイルス血球凝集素、又はHIVgp120タンパク質、細菌性アナトキシン、例えば破傷風抗毒素又はそのエピトープでよい。
【0026】
Nタンパク質-対象タンパク質の融合タンパク質、及びその調製
本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質、及び該Nタンパク質のC-末端のインフレームに融合された対象タンパク質を含む、Nタンパク質と対象タンパク質との融合タンパク質に関する。
【0027】
本発明の文脈において、対象タンパク質は、Nタンパク質のC-末端に特異的に融合される。本明細書で以下に記載される融合タンパク質を産生する方法が使用される時に、この構造は、Nタンパク質に、約10 nmの径を有する環状構造を取り込むことを可能にする。
【0028】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス(Paramyxoviridae)科及びニューモウイルス(Pneumoviridae)科でよい。特に、該ウイルスは、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルスであり、特にヒト又はウシ呼吸器合胞体ウイルス(RSV)である。
【0029】
本発明に従って融合タンパク質に存在するNタンパク質は、一般的に、天然Nタンパク質のアミノ酸配列を有し、すなわちパラミクソウイルス科ウイルスに天然に存在するNタンパク質である。
【0030】
しかしながら、融合タンパク質の発現の目的のために、Nタンパク質がPタンパク質と相互作用するその能力を保持することを条件に、配列修飾は、Nタンパク質のC-末端に導入され得る。特に、パラミクソウイルス科ウイルスの天然Nタンパク質は、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されているかもしれない。
【0031】
このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、換及び/又は挿入から成る。
【0032】
Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質と相互作用することができる。
【0033】
対象タンパク質は、マーカータンパク質、治療的又はワクチン的関心のタンパク質でよいが、これらに限定されない。好ましくは、対象タンパク質は、抗原性タンパク質である。
【0034】
更に、対象タンパク質は、それ自体、融合タンパク質でもよい。例えば、それは、対象タンパク質と、そのC-末端のインフレームに融合されたGFPを含む構造、Nタンパク質のC-末端のインフレームに融合されたGFP自体(すなわち、Nタンパク質-GFP-対象タンパク質の融合タンパク質)でもよい。
【0035】
好ましくは、N-GFP-対象タンパク質の構造は、NとGFPタンパク質との間に、配列KLRILQSTVPSERPQASGVYMGNLTTRGPVAT(配列番号32)を有するリンカー配列を含んでもよい。これは、N-GFP融合タンパク質の産生収量を最適化し(実施例6参照)、それによってN-GFP-対象タンパク質の融合タンパク質の産生収量を最適化する。
【0036】
対象タンパク質は、特に、GFP-抗原性タンパク質、又はGFP-治療対象タンパク質の融合タンパク質でよい。それは、対象タンパク質に融合された「リンカー配列」を含むキメラタンパク質でもよい。リンカー配列は、典型的には、最大30アミノ酸、好ましくは最大20アミノ酸、より好ましくは最大10アミノ酸を含み、Nタンパク質と対象タンパク質との間のスペーサとして働き、これらのタンパク質の各々を正確に折り畳むことができる、ポリペプチドである。
【0037】
従って、ある実施態様に従って、対象タンパク質は抗原であり、本発明に従う融合タンパク質はNタンパク質-抗原(「N-Ag」)融合体である。好ましくは、それは、ヒト又はウシRSVのNタンパク質と、抗原性タンパク質との融合タンパク質である。
【0038】
別の実施態様によれば、対象タンパク質は治療対象のタンパク質であり、本発明に従う融合タンパク質はN-タンパク質-治療対象のタンパク質との融合体である。好ましくは、それは、ヒト又はウシRSVのNタンパク質と、治療対象のタンパク質との融合タンパク質である。
【0039】
有利なことに、タグ(例えばヒスチジンタグ)は、Nタンパク質-対象タンパク質の融合タンパク質の精製を助けるために、Nタンパク質のN-末端に融合され得る。
【0040】
代わりに、Nタンパク質-対象タンパク質の融合タンパク質の精製は、本明細書で以下に詳しく説明するように、パラミクソウイルス科ウイルスのPタンパク質との融合タンパク質、特に、GSTに融合されたPタンパク質を共発現させることによって実行され得る。
【0041】
任意の慣用的な分子生物的な、微生物的な又は組換えDNA技術は、本発明に従う融合タンパク質を産生するため使用され得る。このような技術は、当業者の範囲内にあり、特にSambrook, Fritsch & Maniatis, Molecular Cloning: A Laboratory Manual, Second Edition (1989) Cold Spring Harbor laboratory Press, Cold Spring Harbor, New York ("Sambrook et al., 1989"); DNA Cloning: A Practical Approach, Volumes I and II (D.N. Glover ed. 1995); Oligonucleotide Synthesis (M.J. Gait ed. 1984); Nucleic Acid Hybridization [B.D. Hames & S.J. Higgins eds. (1985)]; Transcription and Translation [B.D. Hames & S.J. Higgins eds. (1984)]; Animal Cell Culture [R.I. Freshney, ed. (1986)]; Immobilized Cells and Enzyemes [IRL Press, (1986)]; B. Perbal, A Practica; Guide To Molecular Cloning (1984); F. M. Ausubel et al (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (1994)に記載されている。
【0042】
従って、本発明は、上記のNタンパク質-対象タンパク質の融合タンパク質をコードする核酸にも関する。
【0043】
「コーディング配列」は、ヌクレオチド配列が発現された時にRNA、ポリペプチド、タンパク質等の産生を生じるヌクレオチド配列を称する。タンパク質のコーディング配列は、一般的に、開始コドン(ATG)及び終始コドンを含む。
【0044】
「発現する」又は「発現」は、例えば、遺伝子配列又は対応するDNAの転写及び翻訳に関連する細胞機能の活性化によってタンパク質を産生することにより、遺伝子又はDNA配列に含まれる情報をそれ自体明らかにすることを許容する又は可能にすることを意味する。用語「共発現」は、2つの遺伝子又はDNA配列に含まれる情報が同一の宿主細胞で発現される時に使用される。
【0045】
従って、本発明は、Nタンパク質-対象タンパク質の融合タンパク質をコードする、核酸(cDNA、ゲノムDNA、合成DNA又はRNA)を含む。核酸は、二重鎖でも単一鎖(すなわち、センス鎖又はアンチセンス鎖)でもよい。核酸は、融合タンパク質をコードする配列に限定されず、融合タンパク質をコードする配列の上流又は下流のコーディング配列又は非-コーディング配列を含んでもよい。
【0046】
本発明に従う融合タンパク質をコードする配列は、少なくとも2つの部分、5'から3'方向に連続して、Nタンパク質をコードする部分、及び対象タンパク質をコードする3'側上の部分、を含むハイブリッド又はキメラ配列である。対象タンパク質がそれ自体、融合体である時、又はタグがNタンパク質に結合されている時には、コーディング配列は、好適な数の部分を含む。該部分の各々の間には、コドンがリンカー配列をコードすることがある。
【0047】
本発明は、Nタンパク質-対象タンパク質の融合タンパク質をコードする核酸を含む発現ベクターにも関する。かかるベクターは、DNAと機能的に連結された転写-調節要素を含むことがある。
【0048】
RNAポリメラーゼがコーディング配列をRNA、特にmRNAに転写する時に、コーディング配列は、転写及び翻訳制御配列と「機能的に連結される」。このRNAは、イントロンを含む場合にその後につなぎ合わされ、コーディング配列によってコードされるタンパク質に翻訳される。
【0049】
表現「ベクター」、「クローニングベクター」及び「発現ベクター」は、宿主を形質転換し、導入された配列の発現を促進するために、DNA又はRNA配列(例えば、非相同遺伝子)を宿主に導入できるベヒクルを称する。ベクターの例は、プラスミド、ファージ、ウイルスを含む。最も一般的なベクターは、一般的には細菌起源の自律的複製単位であり、二重鎖DNAの形態にあり得る、プラスミドである。プラスミドは、外来DNA配列を容易に結合し、これは次いで好適な宿主に容易に導入され得る。プラスミドベクターは、一般的に、コーディングDNA配列、プロモーターDNA配列を含み、外来DNAの導入を可能にする1以上の制限部位を有する。プラスミドの非-限定的例は、プラスミドpKK(Clonetech)、pUC及びpET(Novagen, Inc., Madison, WI)、pRSET又はPREP(Invitrogen, San Diego, CA)、pMAL(New England Biolabs, Beverly, MA)、又はpGEX-4T-3(Pharmacia)を含む。
【0050】
本発明は、本発明に従う発現ベクターを含む宿主にも関する。これらの宿主細胞は、前記ベクターにより「形質転換される」。
【0051】
「宿主細胞」は、細胞によって物質の産生、例えば、遺伝子の細胞、DNA又はRNA配列、タンパク質又は酵素による発現、のために選択され、修飾され、培養され又は設計される、任意の細胞又は生物を意味するものと理解される。
【0052】
「発現系」は、ベクターによって運ばれる外来DNAによってコードされそして宿主細胞内に導入されるタンパク質を産生するために、好適な条件下で使用される、宿主細胞及び対応するベクターを称する。慣用的な発現系は、E.コリ宿主細胞及びプラスミドベクター、昆虫細胞及びバキュロウイルスベクター、又は哺乳動物細胞及びウイルス性起源の強力なプロモーターを有するベクター(例えば、サイトメガロウイルス)を含む。
【0053】
本発明の方法に従う発現系は、有利なことに、例えばpGEX-4T-3をベクターとして有する細菌発現系、特にE.コリである。細菌系は、実際に、最も高い産生収量が得られる発現系である。
【0054】
本発明は、従って、上で定義されたNタンパク質-対象タンパク質の融合タンパク質を産生するための方法にも関する。本方法は、場合により、宿主細胞の形質転換、次いでNタンパク質-対象タンパク質の融合タンパク質の発現を許容する条件下で、Nタンパク質-対象タンパク質の融合タンパク質をコードする核酸を含むベクターで形質転換された宿主細胞を培養すること、及び場合により、Nタンパク質-対象タンパク質の融合タンパク質の精製を含む。培養条件は、選択された発現系(すなわち、宿主細胞及びベクター)に依拠し、これの条件の決定は、当業者の範囲内にある。
【0055】
Nタンパク質-対象タンパク質/Pタンパク質複合体の調製法
本発明者らは、パラミクソウイルス科ファミリーのウイルスのNタンパク質がパラミクソウイルス科ファミリーの同一のウイルスのPタンパク質との共発現系で産生され得る、ことを先に示している。同一の系は、Pタンパク質との複合体の形態でのNタンパク質-対象タンパク質の融合タンパク質を発現し、次いで場合によりその複合体から融合タンパク質を精製するために使用され得る。
【0056】
本発明は、Nタンパク質-対象タンパク質/Pタンパク質複合体の調製法であって、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーのウイルスのタンパク質であり、以下のステップ:
a) パラミクソウイルス科ファミリーのウイルスのNタンパク質を共発現し、該対象タンパク質は、パラミクソウイルス科ファミリーの同一のウイルスのPタンパク質で、Nタンパク質のC-末端のインフレームに融合され;
b) そのように形成されたNタンパク質と対象タンパク質/Pタンパク質との複合体を回収すること、
を含む、前記方法に関する。
【0057】
上記のように、Nタンパク質は、ヒスチジンタグのようなタグをそのN-末端で有することがある。更に、対象タンパク質は、それ自体、融合構築物でもよい。
【0058】
更に、上記のように、Nタンパク質は、天然Nタンパク質であるか、あるいは修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義された領域で修飾されてもよい。このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質と相互作用することができる。
【0059】
好ましくは、前記のPタンパク質は、オリゴマー化ドメインPを含まず、かつNタンパク質への結合ドメインを含む、切断Pタンパク質(「PΔ」)である。本発明者らは、この実施態様が溶解形態での大量のN-P複合体の発現を許容したことを先に示している(特許出願FR 05 04 426)。溶解性は、生理食塩水溶液、例えば1XPBS(NaCl 140 mM, KCl 27 mM, Na2HPO4 8mM, KH2PO4 1.5 mM, pH 7.4)又はトリス緩衝液(10 mM pH 7.4, 150 mM NaCl)のような界面活性剤を含まない水性媒体に関して、10,000×gで30分間遠心することによって決定される。
【0060】
表現「切断されたPタンパク質」は、1個以上の隣接アミノ酸配列が削除されたPタンパク質を称する。それは、C-末端配列、N-末端配列、Pタンパク質の一次構造に対して「内部の」配列の切断でも、又はこれらの切断の組合せでもよい。
【0061】
本発明に従う切断されたPタンパク質は、オリゴマー化ドメインPを含まず、Nタンパク質と相互作用することができ、すなわち、それらは、Nタンパク質に対する結合ドメインを有する。パラミクソウイルス科Pタンパク質とNタンパク質との相互作用のドメインは、C-末端の領域でマップされてきたが、切断されたPタンパク質の例は、好ましくはPタンパク質のC-末端断片、又は(Nタンパク質と相互作用できる)Pタンパク質のC-末端断片とPタンパク質の隣接アミノ酸の少なくとも1つの他の配列との融合によって構成される「キメラ」Pタンパク質を含む。前記C-末端断片及び前記Pタンパク質の他の配列は、それ自体、本来隣接してなく、配列重複を示さない。例えば、切断されたRSV Pタンパク質は、天然Pタンパク質のアミノ酸1〜121及び161〜241によって構成される配列を有することがある。参照ポリペプチドの「断片」は、参照ポリペプチドの配列に見出される隣接アミノ酸の任意の配列を称する。
【0062】
表現「Pタンパク質のC-末端断片」又は「PΔN」は、1以上の連続アミノ酸がN-末端から削除されているPタンパク質を意味するものと理解される。好ましくは、Pタンパク質のC-末端断片は、Pタンパク質の一次構造のC-末端半分に位置するアミノ酸鎖を称する(配列中のアミノ酸数が奇数の時、追加のアミノ酸は、N-末端半分に比べて、タンパク質のC-末端半分に任意に配置され得る)。例えば、241アミノ酸を含むRSV Pタンパク質の場合には、PΔ161Nは、Pタンパク質のアミノ酸161〜241によって構成されるC-末端断片を称する。例えば、507アミノ酸を含む麻疹ウイルス(Edmonston B株)のPタンパク質の場合には、PΔ386Nは、Pタンパク質のアミノ酸386〜507によって構成されるC-末端断片を称する。
【0063】
切断されたPタンパク質がPタンパク質のC-末端である時に、本発明は、パラミクソウイルス科ファミリーのウイルスの溶解性Nタンパク質-対象タンパク質/Pタンパク質のC-末端断片の複合体(「N-PI/PΔN複合体」の調製法であって、以下のステップ:
a) パラミクソウイルス科ファミリーの同一のウイルスのPタンパク質と、対象タンパク質のC-末端のインフレームに融合された、パラミクソウイルス科ファミリーのウイルスのNタンパク質を共発現し、ここで、該Pタンパク質のC-末端断片はオリゴマー化ドメインPを含まず、Nタンパク質-対象タンパク質の融合タンパク質と相互作用できる;
b) そのように形成された溶解性N-PI/PΔN複合体を回収すること、
を含む、前記方法に関する。
【0064】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス科又はニューモウイルス科でよい。特に、該ウイルスは、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルス、例えばヒト又はウシの呼吸器合胞体ウイルス(RSV)である。
【0065】
当業者は、抗原-Nタンパク質の融合タンパク質と相互作用することができる、切断されたPタンパク質、より具体的にはPタンパク質のC-末端断片を知っているか、又は決定することができる。
【0066】
例えば、Castagne et al.(2004; Journal of General Virology; 85: 1643-1653)に記載のE.コリのNタンパク質及びPタンパク質を共発現する方法は、P及びNとの相互作用ドメインをマップするために使用できる。よって、本発明者らは、Pタンパク質の9個のC-末端アミノ酸(アミノ酸233〜241)によって構成されるオリゴぺプチドを含むRSV Pタンパク質のC-末端断片は、Nタンパク質と相互作用する、ことを明らかにしている。
【0067】
更に、例えば、センダイウイルスのPタンパク質とRNA-N複合体の形態のNタンパク質又はリボヌクレオタンパク質(RNP)、すなわち「Xドメイン」又はXDとも称される、との相互作用ドメインが、アミノ酸473〜568によって定義される、ことを記載してきた(Kolakofsky et al., 2004,; Virology; 318(2): 463-73)。
【0068】
本発明者らはまた、PSV Pたんぱく質のあるC-末端断片、特に断片PΔ161N(アミノ酸161〜241)が、Pタンパク質全体と比べて、大量のNタンパク質の調製を可能にしたことを証明した。但し、実際に、十分な収量及び工業的スケールを達成することは可能でない。たった9個のアミノ酸を含むPΔ233N(アミノ酸233〜241)に至るまでのより小さな削除変異体ははPΔ161Nの変異体に匹敵する収量を達成できる。
【0069】
PΔ161Nより小さい断片は、Nタンパク質と相互作用することができ、オリゴマー化をすることができず、そのためオリゴマー化ドメインPを含まない、RSVタンパク質の断片に相当する。RSVの最小オリゴマー化ドメインPは、事実、Pタンパク質のアミノ酸120〜150によって定義される。
【0070】
この同一の方法は、本発明者らに、アミノ酸残基386〜507(PΔ386N)によって構成される麻疹ウイルスのPタンパク質のC-末端断片が、該ウイルスのNタンパク質と相互作用し、その精製を可能にした、ことを示させた。対照的に、Pタンパク質のN-末端部分の削除は、残基456(断片PΔ457Nを含む)である限り、Nタンパク質の精製を許容しない。リボヌクレオカプシドと相互作用するPタンパク質のC-末端領域の構造は、Johansson et al.によって決定された(2003 Journal of Biological Chemistry Vol.278, p.44567-44573)。オリゴマー化領域Pは、削除によって決定され、アミノ酸304〜376によって定義されるものと予測されている。
【0071】
RNPの形態でNタンパク質との相互作用ドメインを含むPタンパク質のC-末端断片であるが、オリゴマー化ドメインPが削除されていないPタンパク質のC-末端断片の使用は、Nタンパク質との、タンパク質P断片の相互作用、溶解性N-PΔN複合体の形成、及び高収率での該複合体の産生を許容する。特定のメカニズムに限定するものではないが、オリゴマー化ドメインPの非存在は、これらの複合体のPタンパク質間の相互作用に関連するN-ΔPN複合体の不溶解性の問題を避ける。
【0072】
従って、1つの実施態様によれば、N-PI-PΔN複合体の調製法は、RSV Pタンパク質の最後の9個のC-末端アミノ酸を含み、そしてRSV Pタンパク質の少なくとも119、好ましくは149、より好ましくは160個のN-末端アミノ酸が欠けている、RSV Pタンパク質のC-末端断片の発現、を含む。
【0073】
より具体的には、本発明に従う方法において、RSVのNタンパク質-対象タンパク質の融合タンパク質で共発現することが可能である。
a) 配列番号1で示されるヒトRSVロング株のPタンパク質のアミノ酸配列233〜241を含み、そして配列番号1で示されるRSV Pタンパク質の配列の位置233と120、好ましくは150、より好ましくは161との間に位置するアミノ酸残基である限り、N-末端方向に伸びる、RSV Pタンパク質のC-末端断片;又は
b) C-末端断片、すなわち、異なったヒトRSV株由来又はウシRSV株から得られたPタンパク質の、a) で定義された断片の相同体。
【0074】
RSVのNタンパク質のC-末端断片は、例えば、PΔ120N(Pのアミノ酸120〜241)、PΔ150N(Pのアミノ酸150〜241)、PΔ161N(Pのアミノ酸161〜241)、PΔ180N(Pのアミノ酸180〜241)、PΔ200N(Pのアミノ酸200〜241)、PΔ220N(Pのアミノ酸220〜241)、PΔ230N(Pのアミノ酸230〜241)及びPΔ233N(Pのアミノ酸233〜241)からなる群より選択される。
【0075】
本発明は、RSVのNタンパク質-対象タンパク質の融合タンパク質により、RSV Pタンパク質の最後の9個のC-末端アミノ酸を含み、そしてRSV Pタンパク質の少なくとも119、好ましくは149、より好ましくは160個のN-末端アミノ酸が欠けている、上記のRSV Pタンパク質のC-末端断片を含む切断されたPタンパク質が共発現される方法にも関する。
【0076】
例えば、Pタンパク質のC-末端断片を含む切断されたPタンパク質は、RSV Pタンパク質の最後の122個のN-末端アミノ酸と、最後の80個のC-末端アミノ酸との融合によって構成され得る;例えば、配列番号7で示されるヒトRSVロング株のPタンパク質のアミノ酸鎖1〜121及び161〜241によって構成され得る。
【0077】
別の実施態様によれば、パラミクソウイルス科は、麻疹ウイルスであり、N-PI-PΔN複合体の調製法は、Pタンパク質の最大122個のC-末端アミノ酸を含む麻疹ウイルスのPタンパク質のC-末端断片を発現し、それによって構成されることを含む。特に、配列番号10で示される麻疹ウイルス株Edmonston BのPタンパク質の、株Edmonston BのPタンパク質について定義されたものと相同であるC-末端断片の、又は麻疹ウイルスの異なった株から得られたPタンパク質の、アミノ酸386〜507(PΔ386N)によって構成されるC-末端断片でよい。
【0078】
本発明の文脈では、用語「相同的」は、同一の進化的起源を有するタンパク質、例えば、異なった種に属する、又はウイルス、ウイルス株の場合の、相同タンパク質、の間に存在する関係に関する。かかるタンパク質(及びそれらをコードする遺伝子)は、それらの配列類似性、それがパーセント類似性であろうと又は保存的位置での特定の残基もしくはモチーフの存在の点からであろうと、その配列類似性によって影響される、配列同一性を有する。
【0079】
表現「配列類似性」は、同一の進化的起源を有しても又は有さなくてもよいタンパク質の核酸又はアミノ酸の配列間の同一性の程度を称する。用語相同性と類似性とは、慣用的に区別なく使用される。2つのアミノ酸配列は、それらのアミノ酸が少なくとも80%の同一性又は少なくとも90%の類似性である時に(いわゆる機能的に同一である時に)、「実質的に相同」である。類似の又は相同の配列は、例えばプログラムBLAST又はFASTAを用いてアラインメントによって同定され得る。
【0080】
有利なことに、Pタンパク質、例えば切断されたPタンパク質及び特にPタンパク質のC-末端断片は、Nタンパク質-対象タンパク質/Pタンパク質の複合体の精製を助けるタンパク質、特に、アフィニティクロマトグラフィーで使用されるタンパク質、との融合体の形態で発現される。それは、タンパク質タグ、例えばグルタチオン-S-トランスフェラーゼ(GST)でよい。そこでは、Pタンパク質-融合タンパク質がグルタチオンと結合された固体支持体上のクロマトグラフィーによって分離される。他のラベル又は「タグ」、例えばポリヒスチジン又は「His-タグ」が使用できる。
【0081】
従って、Nタンパク質-対象タンパク質/Pタンパク質-タンパク質タグ(GST又はPタンパク質と融合された別のタンパク質タグ)複合体が得られる。そこではタンパク質タグは酵素的開裂によって除かれる。例えば、該融合体がGST以外のタンパク質を含む時に、GSTは、トロンビン又は任意の他の好適な酵素による開裂によって除かれる。
【0082】
本発明に従う方法を実行させるベクターの構築の具体的な例は、以下の例で記載される。
【0083】
1つの実施態様によれば、対象タンパク質は抗原であり、本発明に従う方法は、溶解性Nタンパク質-抗原/Pタンパク質複合体(N-Ag/P複合体)、好ましくはNタンパク質-抗原/切断されたPタンパク質複合体(N-Ag/PΔ複合体)及びより好ましくはNタンパク質-抗原/Pタンパク質複合体のC-末端断片(N-Ag/PΔN)の形成を生じる。
【0084】
Nタンパク質-対象タンパク質/Pタンパク質複合体からの融合タンパク質の分離
Pタンパク質が切断され、特にPタンパク質のC-末端断片で切断される、Nタンパク質-対象タンパク質/Pタンパク質複合体を調製する方法は、上記のように、Nタンパク質-対象タンパク質/Pタンパク質複合体を、単離された又は精製された形態で容易に取得させる。
【0085】
従って、本発明は、Nタンパク質-対象タンパク質/Pタンパク質複合体であって、Nタンパク質及びPタンパク質が、パラミクソウイルス科ファミリーのウイルスのタンパク質であり、上記の調製法によって得られる、前記複合体にも関する。
【0086】
本発明は、特に、溶解性Nタンパク質-対象タンパク質/切断されたPタンパク質複合体(N-PI/Δ複合体)に関し、より好ましくは溶解性Nタンパク質-対象タンパク質/Pタンパク質複合体のC-末端断片(N-PI/PΔN複合体)に関する。ここで、N及びPは、パラミクソウイルス科ファミリーのウイルスのタンパク質である。
【0087】
このNタンパク質-対象タンパク質/Pタンパク質複合体、又はより具体的には、N-PI/PΔもしくはN-PI/PΔNから、Nタンパク質-対象タンパク質の融合タンパク質は、そのRNAを有する環の形態で、例えばサイズ排除法クロマトグラフィー(ゲル濾過)によって容易に単離され得る。この分離は、好適には、Pタンパク質、及びPタンパク質が場合により融合されるタンパク質タグの酵素的開裂による分離後に、実行され得る。
【0088】
従って、本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質-対象タンパク質の融合タンパク質を調製する方法であって、以下のステップ:
a) 上で定義された方法によってNタンパク質-対象タンパク質/Pタンパク質の複合体を調製し;及び
b) Nタンパク質-対象タンパク質/Pタンパク質の複合体から、Nタンパク質-対象タンパク質の融合タンパク質を分離すること、
を含む、前記方法に更に関する。
【0089】
好ましくは、Pタンパク質は、切断タンパク質、より好ましくはPタンパク質のC-末端断片である。
【0090】
本発明は、より具体的には、溶解性Nタンパク質-パラミクソウイルス科ファミリーのウイルスの対象タンパク質の融合タンパク質を調製する方法であって、以下のステップ:
a)上で定義された方法によって溶解性Nタンパク質-対象タンパク質/P-タンパク質のC-末端の複合体(N-PI/PΔN複合体)を調製し;及び
b) 溶解性N-PI/PΔN複合体から、Nタンパク質-対象タンパク質の融合タンパク質を分離すること、を含む、前記方法に関する。
【0091】
上記のように、Nタンパク質は、天然Nタンパク質でもよく、又は修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されていてもよい。かかる修飾は典型的には、Nタンパク質のC-末端配列における、1以上のアミノ酸(1〜25、又は1〜20、1〜15、1〜10、あるいは1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾性の例は、6又は12個のC-末端アミノ酸の削除から成ってもよい。その切断されたNタンパク質(NΔ6C及びNΔ12C)は依然としてPタンパク質と相互作用することができる。
【0092】
本発明は、上記の方法によって得られるNタンパク質-対象タンパク質の融合タンパク質にも関する。
【0093】
対象タンパク質は、治療的又はワクチン的関心のタンパク質でよい。好ましくは、対象タンパク質は抗原である。
【0094】
免疫原性、ワクチン又は治療的組成物
本発明は、単離された、又はPタンパク質複合体のNタンパク質/C-末端断片(N-PΔN)の形態での、環状構造を有するRSVNタンパク質が、高度に免疫原性であり、特に例えば気管支粘膜での局所反応の刺激を許容する、ことを先に明らかにしてきた。
【0095】
本発明者らは、このような環状構造が治療的又はワクチンタンパク質のためのベクターとして使用され得ることを証明してきた。
【0096】
本発明は、従って、治療的もしくはワクチン的関心の分子又はタンパク質のためのベクターとしての、パラミクソウイルス科ファミリーのウイルスのNタンパク質の使用に関する。
【0097】
1つの実施態様によれば、治療的又はワクチンタンパク質は、上記のように、Nタンパク質に融合される。
【0098】
本発明者らは、単離された、又はNタンパク質/Pタンパク質のC-末端断片の複合体(N-PΔN)の形態での、環状構造を有するRSV Nタンパク質が、免疫原性が高く、特に例えば気管支粘膜での局所的反応の刺激を許容する、ことを先に明らかにしている。
【0099】
本発明者らは、このような環状構造が治療的又はワクチンのタンパク質のためのベクターとして使用され得ることを証明してきた。
【0100】
従って、本発明は、治療的又はワクチン的関心の分子又はタンパク質のためのベクターとしてのパラミクソウイルス科ファミリーのウイルスのNタンパク質の使用に関する。
【0101】
1つの実施態様によれば、治療的又はワクチンタンパク質は、上記のように、Nタンパク質に融合される。
【0102】
別の実施態様によれば、治療的又はワクチン分子は、Nタンパク質に化学的に結合される。Nタンパク質のC-末端へのシステインの付加によって、実際に、タンパク質性又は非-タンパク質性(有機又は無機的な)な性質の様々な分子の化学的結合を許容する部位がつくられる。ジスルフィド結合によってシステインを有するタンパク質をつなぐこともできる。よって、様々な分子に結合されたマレイミド自体は、共有結合によってこのシステインにつながれる。マレイミドは、有機分子のすべての種類、例えばデキストラン(dextrone)、ビオチン、金、又は任意のタンパク質に結合され得る。
【0103】
従って、本発明は、パラミクソウイルス科ファミリーのウイルスのNタンパク質と、関心の分子、特に、治療的又はワクチン的分子、との複合体にも関する。
【0104】
本発明は、治療的又はワクチン分子又はタンパク質を対象に送達する方法にも関する。その方法は、パラミクソウイルス科ファミリーのウイルスのNタンパク質に共有結合的に結合された治療的又はワクチン分子を、該対象に送達することを含み、あるいはパラミクソウイルス科ファミリーのウイルスのNタンパク質との融合体の形態で該タンパク質を送達することを含む。該治療的又はワクチンタンパク質は、Nタンパク質のC-末端のフレームに融合されている。
【0105】
本発明は、薬学的に許容される担体と共に、場合により、Nタンパク質-対象タンパク質/Pタンパク質の複合体の形態で、Nタンパク質-対象タンパク質の融合タンパク質を含む医薬組成物をも目的とする。ここで、Nタンパク質及びPタンパク質は、パラミクソウイルス科ファミリーの同一のウイルスから得られる。
【0106】
前記のパラミクソウイルス科ファミリーのウイルスは、パラミクソウイルス科でも又はニューモウイルス科でもよい。特に、該ウイルスは、おたふく風ウイルス、麻疹ウイルス及びパラインフルエンザウイルスからなる群より選ばれる。好ましくは、該ウイルスは、ニューモウイルスであり、特に例えばヒト又はウシの呼吸器合胞体ウイルス(RSV)である。
【0107】
対象タンパク質は、治療的又はワクチン的関心のタンパク質でよい。
【0108】
1つの実施態様によれば、医薬組成物は治療的使用に好適である。この実施態様によれば、対象タンパク質は、治療対象のタンパク質である。本発明者らは、Nタンパク質-対象タンパク質の融合タンパク質が細胞を通過し、よってNタンパク質が治療対象の分子又はタンパク質、例えば血管新生抑制又はアポトーシス促進のポリペプチド、のための可能性のあるベクターを構築する、ことを証明してきた。
【0109】
1つの実施態様によれば、医薬組成物は、免疫原性又はワクチン的使用のために好適である。この実施態様によれば、対象タンパク質は、血管新生タンパク質、好ましくは病原性微生物、例えばウイルス、細菌、真菌又は寄生的な後生生物又は原生生物、から得られる抗原である。
【0110】
本発明者らは、実際に、抗原がNタンパク質との融合タンパク質の形態で提供される時に、抗原に対する宿主の反応が非常に顕著に増加する、ことを証明してきた。Nタンパク質-抗原融合タンパク質は、細胞性及び体液の経路による免疫反応を刺激することができる。
【0111】
溶解性Nタンパク質-抗原融合タンパク質は、逆作用なしに、Pタンパク質との複合体の形態でワクチン接種に使用される。従って、本発明に従う免疫原性又はワクチン組成物は、薬学的に許容される担体と共に、溶解性Nタンパク質-抗原/Pタンパク質複合体を含むことがある。
【0112】
「薬学的に許容される担体」は、副作用、例えばヒトマ又は動物におけるアレルギー性反応を生じない、任意の溶媒、分散媒体、吸収-抑制剤等を意味すると理解される。
【0113】
生理学的に許容される担体の例は、当業者に知られている。液状担体の例は、活性成分以外の物質及び水を含まない殺菌性水溶液、あるいは生理学的塩濃度を有する生理学的pH値のリン酸ナトリウムのような緩衝剤、又はその両方、例えばリン酸緩衝生理食塩水(PBS)を含む殺菌性水溶液、を含む。水性担体は、1つ超の緩衝塩、並びに塩化ナトリウム又は塩化カリウムのような塩、デキストロース、ポリエチレングリコール及び他の溶質を含むことができる。
【0114】
本組成物は、生薬製剤に匹敵する方法で、治療上有効量で投与される。投与される量は、治療される対象、活性成分を使用する対象系の能力、及び治療効果の所望の程度に依拠する。投与に必要とされる融合タンパク質の正確な量は、医師の選択及び個人の特徴に依拠する。
【0115】
有利なことに、医薬組成物が免疫原性又はワクチン組成物である時に、アジュバントを更に含んでもよい。「アジュバント」は、同時に投与される抗原に対して細胞又は体液レベルで免疫反応を増加し、刺激し、活性化し、改良し又は調節する、産物を称する。慣用的なアジュバントの例は、細菌性抗原を含むアジュバント、例えば完全フロインドアジュバント、LPS及びその誘導体、細菌性毒素(コレラ毒素及びエンテロトキシン)、及びそれらの解毒変異体(例えば、LT(R192G))、CpGモチーフを含むオリゴヌクレオチド配列、無機アジュバント、例えば水酸化アルミニウム(alum)、リン酸カルシウム又はリン酸カリウム、油状エマルション及び乳化剤(サポニン、例えばQS21)、サイトカインを含む。
【0116】
本発明に従う免疫原性組成物は、免疫反応を、ワクチン接種された対象における抗原に対して誘導する。又はより具体的には抗原が得られる病原体に対して誘導する。
【0117】
本発明に従うワクチン組成物は、抗原を含む病原体による感染に対して保護する。すなわち、ワクチン組成物で免疫されていない対象と比べて感染のような効果の重度が減少する。
【0118】
本発明は、抗原が得られる病原体に対するワクチン接種の方法において上記のワクチン組成物の使用にも関する。
【0119】
従って、本発明は、本発明に従うワクチン組成物の対象への少なくとも1つの投与を含むワクチン接種法に関する。好ましくは、該ワクチン接種法は、該ワクチン組成物の対象への第1投与、及び該ワクチン組成物の同一の対象への少なくとも1つのブースター投与を含む。ブースター投与は、抗原に患者を再曝露することによって、より強い二次免疫反応を誘導する。
【0120】
ワクチン組成物は、有利なことに、抗原が得られる病原体による感染に保護的又は治療的免疫反応を誘導することにおいて、効果的である量で投与される。投薬量は、必然的に、問題となっている活性成分、投与形式、及び対象の年齢及び症状に依拠する。1投薬量当たりのN-Ag融合タンパク質のN-Ag-P、N-Ag-PΔ、又はN-Ag-PΔN複合体の量は、ワクチン投薬量当たり、0.1〜200μg、好ましくは10〜100μgでよい。
【0121】
免疫原性、ワクチンの又は医薬組成物は、任意の経路、特に、粘膜(例えば、眼内、鼻腔内、経口的)経路によって、非経口的(例えば、皮下、皮内、筋肉内、静脈内又は腹腔内)経路によって投与される。
【0122】
表現「対象」は、パラミクソウイルス科ウイルスによる又は任意の他の病原体により感染に曝露されたことがある又は曝露される可能性がある、ヒト又は非-ヒト動物、例えば鳥又は哺乳動物、例えばウシ、ヒツジ、齧歯動物、イヌ科、特にイヌ、ネコ科、特にネコ、ブタ、モンキーを称する。本発明の範囲内の対象は、好ましくは、ヒト又はウシである。
【0123】
それらの局面の各々について、上記のように、Nタンパク質は、天然Nタンパク質でよく、あるいは修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されていてもよい。かかる修飾は、典型的には、Nタンパク質のC-末端配列における、1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、そのような切断されたNタンパク質(NΔ6C及びNΔ12C)は依然としてPタンパク質と相互作用することができる。
【0124】
診断的適用
パラミクソウイルス科ウイルスのNタンパク質と、少なくとも1つのエピトープを含む抗原との融合は、更に、融合タンパク質によって運ばれるエピトープの少なくとも1つに対する抗体を検出するための診断的適用で使用される可能性のある試薬を構築する。
【0125】
本発明は、従って、更に、上記のように、パラミクソウイルス科ファミリーのウイルスのNタンパク質のC-末端のフレームに融合された抗原を含有するNタンパク質-抗原融合タンパク質を含む診断試薬に関する。
【0126】
上記試薬を含む診断キット及び好適な検出手段は、同様に、本発明の範囲内にある。
【0127】
前記抗原は、少なくとも1つのエピトープを含み、同様に、2以上の同一又は異なったエピトープを含んでもよい。
【0128】
本発明は、融合タンパク質の前記抗原に対する抗体のin vitro又はin vivoでの検出のためのNタンパク質-抗原の融合タンパク質の使用をも目的とする。
【0129】
抗原が単一のエピトープを含む時に、検出された抗体はそのエピトープに対して特異的である。
【0130】
抗原が2以上の異なったエピトープを含む時に、検出された抗体は、前記抗原のエピトープの1つ、2つ又は3以上に特異的である。
【0131】
本発明は、生物試料中の抗原に特異的な抗体を検出する方法であって、以下のステップ:
a) 該生物試料を、パラミクソウイルス科ファミリーのウイルスのNタンパク質のC-末端のフレームに融合された抗原を含むN-抗原融合タンパク質と接触させ;
b) 得られたN-抗原融合タンパク質/抗体複合体を検出すること、
を含む、前記方法にも関する。
【0132】
生物試料は、筋肉、肝臓、心臓、脳等よって得られた組織試料、唾液又は液体試料、例えば血液、血漿又は脳脊髄液のような生物液体でよい。
【0133】
複合体の検出は、当業者によく知られている慣用的手段、例えばクロマトグラフィー(サイズ排除、アフィニティー等)、又は非-変性条件下での電気泳動によって行われる。
【0134】
N-抗原/抗体複合体の検出は、検出可能な方法でNタンパク質を標識することによって更に促進され得る。
【0135】
これらの局面の各々について、Nタンパク質は、天然Nタンパク質であるか、又は修飾されたNタンパク質がPタンパク質と相互作用する能力を保持することを条件に、最後の25、好ましくは最後の20、15、10又は5個のC-末端アミノ酸によって定義される領域で修飾されてもよい。このような修飾は、典型的には、Nタンパク質のC-末端配列の1以上のアミノ酸(例えば、1〜25、1〜20、1〜15、1〜10、又は1〜5個の隣接もしくは非-隣接アミノ酸)の削除、置換及び/又は挿入から成る。
【0136】
Nタンパク質の配列修飾の例は、6又は12個のC-末端アミノ酸の削除から成ってもよく、切断されたNタンパク質(NΔ6C及びNΔ12C)はPタンパク質となお相互作用することができる。
【0137】
以下の実施例及び図面は、本発明の範囲を限定することなく、本発明を説明するものである。
【実施例】
【0138】
実施例1:RSVリンタンパク質及びGFPに融合されたRSV Nタンパク質のC-末端領域を含むプラスミドの構築
RSV長鎖のPタンパク質は、241アミノ酸残基からなる。
【0139】
RSV Pタンパク質のC-末端部分を増幅するために使用されるオリゴヌクレオチドプライマー(5'〜3')の配列(BamHI制限部位を強調する;P遺伝子の開始コドンATGは太字で示す)。
【0140】
【表1】
【0141】
RSVロング株のP遺伝子のcDNAは、プライマーであるロング-PBam+及びロング-P-を用いてヒトRSVのロング株によって感染したHep-2細胞からRT-PCRによって増福した(Castagne et al., 2004; Journal of General Virology; 85: 1643-1653)。PCR産物を制限酵素BamHIによって消化し、プラスミドpGEX-4X-3(Pharamacia)の、グルタチオン-S-トランスフェラーゼ又はGSTをコードする遺伝子を有するインフレームのBamHI-SamI部位にクローン化した。このプラスミドをpGEX-Pと呼ぶ。
【0142】
・P161-241のクローニング(PΔ161N)
以下の条件下で、pGEX-Pプラスミドから、PのC-末端領域(アミノ酸161〜241)を増幅した。
PCRプライマー:P16B+及びロング-P- 各100 ng(各1μl)
DNAテンプレートpGEX-P:10 ng(1μl)
酵素:Pfu Turbo(商標)Strategene(2.5 U/μl):1μl
dATP:最終0.2 mM
dGTP:最終0.2 mM
dCTP:最終0.2 mM
dTTP:最終0.2 mM
Pfu緩衝液 最終1X(Strategene)
最終容積:100μl
【0143】
以下の条件下でPCRを行った。
5サイクル:94℃で15秒間、40℃で2分間、72℃で1分間;
25サイクル:94℃で15秒間、55℃で1分間、72℃で1分間。
【0144】
増福されたDNAは、1倍容(100μl)のフェノール/クロロホルム(1vol/1vol)で抽出し、次いで1倍容のクロロホルムで抽出し、最後に10分の1容の5M NaCl(10μl)及び2倍容の100%エタノール(200μl)を加えて沈殿させた。DNAは、13,000 gで20分間遠心し、1倍容の70%エタノールで洗浄し、乾燥し、1倍容の水(90μl)に再懸濁した。10ユニットの酵素BamHIの存在下で、酵素BamHI用の10μlの10X緩衝剤を加えた後、DNAを37℃で2時間消化した。消化したDNAは、エチジウムブロミドの存在下で、1Xトリス-ホウ酸-EDTA緩衝剤(TBE)中の1.5%アガロースゲル上に充填し、電気泳動によって移動させた。P161-241のDNAに対応するバンドを切り取り、DNAを電気溶出によって抽出した。1倍容のフェノール-クロロホルム、1倍容のクロロホルムで再度抽出し、エタノールで沈殿させた。1%アガロースゲル上での精製の後、BamHI及びSmaIによって消化したベクターpGEX-4T-3にライゲートした:
pGEX-4T-3 DNA:100 ng
P161-241 DNA:100 ng
リガーゼ緩衝液 最終1X
リガーゼ(5 U/μl):1μl
最終容積20μl。
【0145】
全体を14℃で終夜インキュベートした。翌日に、コンピテントDH5-アルファTM細菌(Life Technologies)を10μlのライゲーション産物で形質転換し、最終100μg/mlのアンピシリンで補充したLB-アガー培地を含むペトリ皿上に拡げた。組換え菌コロニーをミニ-プラスミド調製物でスクリーニングし、制限酵素BamHI及びXhoIによって消化した。次いで組換えプラスミドはアガロースゲルに2つのバンドを示し、1つはベクター(4.9 kb)をもう1つはPのC-末端(246 bp)に対応した。組換えプラスミドを完全にシークエンスした。
【0146】
・P180-241、P201-241、P221-241のクローニング
アミノ酸180〜241、200〜241、220〜241の部分に対応するPの断片は、以下のプライマー:
P180-241:プライマーP180B+及びロング-P
P201-241:プライマーP201B+及びロング-P-
P220-241:プライマーP221B+及びロング-P-
【0147】
を用いて、pGEX-PプラスミドからPCRによって得た。それらはP161〜241(上記参照)と同一の方法によって増幅し、クローン化した。
【0148】
・RSVロング株のヌクレオカプシドタンパク質をコードする遺伝子のクローニング
ウイルスによって感染されたHep-2細胞からRT-PCRによってヒトRSVロング鎖のNタンパク質をコードする遺伝子を得た。使用したプライマーは以下のとおりである。
【0149】
【表2】
【0150】
PCR産物は、BamHIによって消化し、BamHI-SmaI部位でpGEX-4T-3プラスミドにクローン化した。Nをコードする領域は、BamHI-XhoIによってpGEX-Nプラスミドの消化によってサブクローン化し、pET28a+プラスミド(Novagen)にサブクローン化した。
【0151】
・pET-Nプラスミドの突然変異誘発及び停止コドンの上流の特定のSaI制限部位の生成
以下のオリゴヌクレオチドを用いて指定の突然変異誘発によってヒトRSV(長鎖)のNタンパク質の停止コドンの直ぐ前に、SaI制限部位を導入した。
【0152】
【表3】
【0153】
このことにより、pET-N-Sacプラスミドが得られた。
【0154】
・GFPに融合されたNタンパク質の入手
GFPを制限酵素EagIによる消化によってpEGFPN1プラスミドから除き、次いでDNA上に平滑末端を得るために、ヌクレオチドの存在下でクレノウ処理を行い、次いで、SacIにより消化した。プラスミドpET-N-Sacを酵素XhoIによって消化し、ヌクレオチドの存在下でクレノウ処理し、次いでSaIで消化した。従って、GFPはpET-N-Sacプラスミドに挿入し、pET-N-GFPプラスミドを得た。
【0155】
・Nタンパク質-Fエピトープの取得
RSVのFタンパク質の「ミモトープ」及び「ヘプタッド」エピトープを、二重鎖オリゴヌクレオチドの挿入により、Nタンパク質のC-末端にクローン化した。
【0156】
ミモトープ・エピトープ:
【0157】
【表4】
【0158】
これらの互いにハイブリダイズされたオリゴヌクレオチドは、pET-N-GFPプラスミドのSacI-SaII部位に挿入した。
【0159】
ヘプタッド・エピトープ:
以下のオリゴヌクレオチドを互いにハイブリダイズした。
【0160】
【表5】
【0161】
それらは、pET-N-GFPプラスミドのAgeI部位でクローン化した。
【0162】
・P231-241のクローニング
以下のプライマーを94℃で5分間加熱することによって変性し、次いで室温まで冷却した。
【0163】
【表6】
【0164】
ハイブリダイゼーション後、10 ngの二重鎖オリゴヌクレオチドを、BamHI及びSmaIによって消化された100 ngのPEX-4T-3プラスミドのDNAとライゲートし、アガロースゲル電気泳動によって精製した。組換えプラスミドは、N遺伝子の領域でシークエンシングすることによって確認した。
【0165】
実施例2:複合体の発現及び精製
コンピテントBL21(DE3)細菌(Novagen)を1μgのpGEX-PΔ DNA、及び1μgのpET-N DNAで形質転換し、次いで、100μg/ml終濃度のアンピシリン及び50μg/ml終濃度のカナマイシンで補充されたLB-アガー培地を含むペトリ皿に拡げた。コロニーを選択し、それぞれ100μg/ml及び50μg/mlの濃度のアンピシリン及びカナマイシンを含む2 mlのLB培地で、37℃で終夜培養した。翌日、抗体で補充された1リットルのLB培地を接種するために1 mlの飽和培養液を用い、夜になるまで培養した。夜に、160μg/mlの(タンパク質発現を誘導する)IPTGを含む新鮮LB培地の1容を、培養物に加え、全体を28℃で終夜培養した。翌日、細菌を5000 rpmで15分間遠心し、ペレットを100 mlの以下の緩衝剤:
50 mM トリス pH 708
60 mM NaCl
2 mM DTT
1 mM EDTA
4 mM ベンザミジン
1X 抗蛋白質分解酵素(完全なEDTA-無しのプロテアーゼ阻害剤カクテル、Roche番号11 873 580 001参照)、すなわち、50 mlの溶解緩衝剤につき、1錠
0.1% トリトン-X100
に再懸濁した。
【0166】
10 mg/ml(1 mg/mlの最終濃度)の量のリゾチームで補充した、10 mlの同一の緩衝剤を加えた。細菌を氷上で1時間インキュベートした(溶解)。全体が粘性になったら、全体を混合物に浸漬されたプローブを用いて1分間に3回、氷上で超音波処理し、各超音波処理の間に5分間の間隔を設けた。全体を4℃で30分間遠心し、次いで上清を回収した。上清を4℃で10,000 gで30分間、再度遠心し、次いで新しい上清を回収した。4 mlのセファロース4B-グルタチオンビーズ(Amersham-Pharmacia)を8 mlのビーズ/緩衝剤の混合物(vol/vol)を溶解緩衝液で除くことによって、洗浄した。ビーズを等量の緩衝剤中に放置し、洗浄した細菌溶解物に加え、4℃で終夜回転した。翌日、ビーズを2000 rpmで3分間遠心し、次いで上清を除き、抗蛋白質分解酵素を含まない溶解緩衝剤でビーズを3回、1XPBS緩衝剤で3回、洗浄した。
【0167】
1 mlのビーズについて、1μl(1U)のトロンビン量のビオチン化トロンビン(Novagen)(「トロンビン開裂捕獲キット」、Novagen参照69022-3FRZ)を用いて、トロンビン部位で、ビーズを分けた。ビーズを20℃で終夜インキュベートし、翌日、2000 rpmで3分間遠心し、上清を得るために15分間デカントした。等量の1XPBSをビーズに加え;混合物を攪拌し、デカントした。上清を再度、回収し、先に回収した上清に加えた。回収された上清に、16μlの量の樹脂(すなわち、32μlの樹脂/緩衝剤の混合物(vol/vol))中のストレプトアビジン-アガロースビーズ(Novagen参照69023)を加えた。混合物を1時間攪拌し、次いで2000 rpmで3分間遠心し、上清を回収した。2 mg/mlのタンパク質濃度を得た。
【0168】
開裂産物を含む10μlの上清を1Xラエムリ緩衝液中で変性し、次いで沸騰させ、0.1%トリス-グリシンSDS緩衝液中の12%ポリアクリルアミドゲル上に沈殿させ、次いでタンパク質を明らかにするために電気泳動後にクマシ・ブルーで染色した。
【0169】
実施例3:N-GFP及びPΔ161N(P161-241)の分離、及びN-GFP環の精製
上清に存在するタンパク質は、1XPBS中でサイズ排除クロマトグラフィー(ゲルろ過)によって分離され得る。
【0170】
実施例4:異なった細胞株によるN-PΔN複合体の捕獲:N-緑色蛍光タンパク質(GFP)融合タンパク質の補助付きの試験
I.材料及び方法
I.1 細胞株及び食作用試験:
−HEp-2:EMEM(イーグル最小必須培地)+10% FCS+L-グルタミン+PS(ペニシリン、ストレプトマイシン)中の単層で培養した、ヒト喉頭上皮株(Cancer Res 1955; 15: 598)。
−RAW:DMEM(ダルベッコ最小必須培地)+10% FCS+L-グルタミン+PS中の単層で培養した、ネズミ腹膜単球-マクロファージ(J. Immunol 1977; 119: 950)。
【0171】
細胞株をD-PBS(ダルベッコリン酸緩衝生理食塩水)+0.33 %リドカイン+5 mM EDTA中で分解した。単離された細胞を洗浄し、107細胞/mlのPBS+2% FCSの濃度で再懸濁し、
−20μg/mlのP161-241+N-GFP(N-GF-PΔN)
−6.5μg/mlのGFP
−1 mg/mlのデキストラン-FITC(DX-FITC)(参照:D-1844,Molecular Probes)
の存在下で5 mlの丸底チューブ内で、緩やかに攪拌しながらインキュベートした。
【0172】
エンドサイトーシスのメカニズムが抑制される温度である4℃、内在化が許容される温度である37℃で並行してインキュベーションを行った。1時間又は2時間のインキュベーションの後、細胞を3回洗浄し、PBS+2% FCSに再懸濁した。
【0173】
I.2 フローサイトメトリーによる吸収/内在化の分析
37℃で4℃で蛍光分子(DX-FITC又はN-GFP-PΔN)でインキュベートされた細胞(HEP-2,RAW)と関連する蛍光(FL1)は、フローサイトメトリー(FACSCalibur, Becton)によって比較した。該細胞の自己蛍光のレベルは、蛍光分子の非存在下で、37℃で2時間、インキュベートされた細胞によって得た。
【0174】
I.3 蛍光顕微鏡による吸収/内在化の分析
インキュベーションの後、105細胞をスーパーフロスト(Superfrost)付きスライド(SFPLUS-42, Milian)での細胞遠心によって沈殿させた。スライドは、15分間乾燥し、PERTEX(参照:00814,Histolab)でマウントした。
【0175】
顕微鏡(Axiovert200M, Zeiss)で蛍光レベルを観察した。Metavueソフトウェアを用いてカメラ(Coolsnap HQ)で写真を撮った。
【0176】
I.4 細胞の浸透後のNタンパク質の免疫検出
目的は、Nタンパク質の免疫検出によって、非-浸透細胞(細胞外N-GFP-PΔN複合体)で検出されたシグナルと、浸透細胞(細胞内又は細胞外N-GFP-PΔN複合体)で検出されたシグナルを比較することである。
【0177】
細胞をA緩衝液(参照:GAS-003,Caltag)中に15分間固定し、洗浄した。
【0178】
非-浸透細胞の場合には、細胞洗浄(Cell Wash)緩衝液(Becton)+5% FCSで標識を行った。標識中に細胞を浸透するために、緩衝液B(参照:RGAS-003,Caltag)を用いた。1/100に希釈した抗-CD16/CD32抗体(参照:553141,Becton)で、Fc受容体を20分間飽和した。次いで、1/1000に希釈したNタンパク質に対するラビット血清で、該細胞を30分間インキュベートした。ウサギIgの検出は、1/200に希釈したビオチン化抗体(参照:BA-1000,Vector)で30分間インキュベーションすることによって行った。1/500でのAPC-結合ストレプトアビジン(554067,Becton)の使用は、フローサイトメトリー(FACSCalibur, Becton)によるNタンパク質のFL4チャンネルでの直接検出を可能にした。
【0179】
II.結果:
II-1 N-GFP-PΔN複合体と、RAW細胞(マクロファージ)及びHEp2細胞(上皮)との相互作用の分析
RAW細胞は単球/マクロファージの株である。該細胞は、蛍光ポリマー、DX-FITC、と該細胞を接触させることによって観察され得る食作用能力を有する。DX-FITCの捕獲は、4℃と比べると、37℃では顕著に増加した(図1B)。HEp-2株は、上皮株である。それは、食作用能力を有さない。そのようなわけで、DX-FITCと該細胞との相互作用に関連した蛍光が4℃と37℃でのインキュベーションでほとんど差がない(図1A)。
【0180】
N-GFP-PΔN複合体がRAW及びHEp-2細胞と接触する時に、基本レベル(PBS)に対して蛍光レベルの相当な増加が観察された(図1C、D)。このことは、そのような細胞の膜での効果的な吸収を示した。37℃でのインキュベーション後に、N-GFP-PΔNの存在下で検出された蛍光が、両方の細胞種で減少したが、それにもかかわらず、基本レベルより優れていた(図1C、D)。細胞を組換えGFPでインキュベートする時には、蛍光シグナルは、基本レベルのままであった(データ非表示)。このことは、観察された捕獲現象がN-PΔN環に関連し、GFPには関連しないことを示した。
【0181】
RAW及びHEp-2株でのN-GFP-PΔN環の捕獲は、顕微鏡によって確認した。
【0182】
Nタンパク質に融合されたGFPに特異的な蛍光が、HEp-2又はRAW株の内側で顆粒の形態で見られた。その蛍光は、インキュベーションが組換えGFPで行った場合には見られなかった。N-GFP-PΔN複合体は、従って、RAWマクロファージ株及びHEp-2上皮株で吸収及び/又は内在化された。
【0183】
II-2 RAW細胞によるN-GFP-PΔN複合体の内在化
37℃対4℃でのRAW細胞におけるN-GFP-PΔN複合体の最終結果を理解するために、細胞を透過させ、Nタンパク質の存在を免疫標識及びフローサイトメトリーによる分析によって明らかにした(図2)。透過の非存在下では、N保護(蛍光シグナル)の量は、4℃よりも37℃において少なかった(図2A)。一方、透過後、シグナルレベルは、37℃及び4℃のインキュベーション条件で同一となった(図2B)。従って、N-GFP-PΔN複合体は、37℃でのインキュベーションで内在化された。
【0184】
結局、フローサイトメトリー及び顕微鏡によって、N-GFP-PΔN複合体を用いて行われた実験の全体性は、これらのタンパク質の構造の新規性を証明することができた。核タンパク質の環状集合体は、様々な細胞種、例えばマクロファージ、上皮細胞及び樹状細胞によって効果的に吸収され、内在化される能力を有する(データ非表示)。これらの性質は、抗原ベクター又はワクチンとしてのその構造の使用に特に有用である。
【0185】
実施例5:融合体中で発現された緑色蛍光タンパク質(GFP)と比べた環状形態のNタンパク質の抗原ベクター性の評価
I.材料及び方法
I.1 マウス:
Unite Experimentale Animalerie Rongeur(INRA, Jouy-en-Josas)で飼育された、雄性BALB/c、8〜10週齢。
【0186】
I.2 抗原:
−0.1064μg/μlのP161-241+N-GFP(N-GFP-PΔN)濃度。
−0.5μg/μlのGFP濃度。
−1μg/μlのP161-241+N-PΔN濃度。
【0187】
I.3 アジュバント:
−E.コリのLT(R192G)解毒リンホトキシン、1 mg/ml (John Clements; Choi et al ., 2004, Protein Expression and Purification 38, pp.205によって送られたバッチ)。
【0188】
I.4 免疫:
アベルチン麻酔(300 li.p.)下で、以下の混合物(60μl/マウス):
−5μgのLT(R192G)
−5μGのN-GFP-PΔN
又は、1.7μgの溶解性GFP、又は3.3μgのN-PΔN(5μgのN-GFP-PΔN中に存在するものに対応するGFP量又はN-PΔN量)
−非発熱性の生理的血清を有するgsp 60μl
の鼻腔内経路(i.n.)による投与。
【0189】
すべての溶液は、注入前に0.22μmのフィルターを通過させた。
【0190】
【表7】
【0191】
I.5 抗-N-PΔN及び抗-GFP抗体の産生
ELISAによって、血清及びBAW中の抗-N-PΔN又は抗-GFP抗体(IgH+L及びIgA)を検索した。
−血清を血液試料から集め(4℃での1晩exsudation)、次いで-20℃で冷凍した。
−BAWを1700 rpmで5分間遠心し、上清を集め(約1 ml)、-20℃で冷凍した。
【0192】
96-ウェルプレート(Immulon 2HB, ThermoLabsystems)を、0.1 M重炭酸ナトリウム緩衝液、pH 9.5中のN-PΔN複合体又は組換えGFPタンパク質(200 ng/ウェル、100μl/ウェル)で、4℃で1晩感作した。このプレートを0.05% PBS-Tween 20(Wellwash device, Labsysytems)の200μl/ウェルで5回洗浄した。次いで、プレートを、0.05% PBS-Tween 20緩衝液及び5%ウシ胎児血清(PBS-T-FCS)の150μl/ウェルで、37℃で1時間飽和した。5回洗浄後、滴定する試料をPBS-T-FCSで希釈した(血清については、最初の希釈から1/30まで3のファクターで7連続希釈、BAWについては、最初の希釈から1/3までの希釈)。このプレートを37℃で2時間インキュベートした。5回洗浄後、PBS-T-FCSで希釈した二次抗体を100μl/ウェルの量で分配した。使用した二次抗体は、ペルオキシダーゼにコンジュゲートされ、マウス免疫グロブリン:Ig(H+L)(4000th, P.A.I.S.)又はIgA(1000th, Caltag)に対するものであった。このプレートを次いで37℃で2時間インキュベートし、5回洗浄した。次いで、このプレートをペルオキシダーゼ基質(TMB, 100μl/ウェル)で10分間、暗闇でインキュベートした。酵素反応を50μlの2M H3PO4で停止した。光学密度(OD)を450 nmで読んだ(Dynexリーダー)。OD450=f(希釈)曲線をオリジンソフトウェアによって回帰曲線y=(b+cx)/(1+ax)によってモデル化した。抗体力価を、最も薄い希釈における対照試料(d0)のOD450を2回与える希釈値として決定した。
【0193】
I.6 N-PΔN又はGFPに特異的なTリンパ球によるIFN-γ/IL-5/IL-10の産生
脾臓、及び気道(顔の、首の及び縦隔の気道)をドレインするガングリオンを、同一のプロトコールに従って処理した。脾臓を個々に処理し、ガングリオンを試験群によって分類した。
【0194】
リンパ器官を薄くスライスし、次いで、RPMI培地及びPSでフィルター(100μm細胞シーブ,BD Falcon)上でそっと粉にした。細胞懸濁液を4℃で1700 rpmで10分間遠心した。細胞を1 mlの赤血球溶解緩衝液(低張生理食塩水緩衝液)に再懸濁し、室温で5分間インキュベートした。溶解反応を10 mlの完全RPMI(PS, 2 mM L-グルタミン及び10% FCS)の添加により停止した。膜の残骸をデカントし、細胞を遠心(1700 rpmで4℃で10分間)により3回洗浄した。細胞懸濁液をマラセズ細胞によってカウントした。
【0195】
該細胞を、細胞培養処理した96-ウェルマイクロプレート培養(Falcon)で、200μlの完全RPMI培地で400,000細胞/ウェルの量で培養した。4つの培養条件は、各々の懸濁液について3点で試験した:
−PMA(ホルボール 12-ミリスチン酸エステル 13-酢酸エステル,Sigma)10 ng/ml、及びイオノマイシン(Sigma)1μg/ml(陽性対照、ポリクローナル活性化)
−完全RPMI(陰性対照)
−N-PΔN 10μg/ml
-GFP 10μg/ml。
【0196】
5% CO2で37℃での48時間後(IL-5及びIL-10)又は72時間(IFN-γ)培養後、培養上清を回収し、ELISAのサイトカインの滴定まで-20℃で冷凍した。
【0197】
96-ウェルプレート(Immulon 2HB, ThermoLabsystems)を、0.1 M重炭酸ナトリウム緩衝液、pH 9.5中の、4μg/ml(IFN-γ)又は2μg/ml(IL-5/IL-10)のマウス抗-サイトカイン捕獲抗体(100μl/ウェル)で、4℃で終夜、感作した。このプレートを0.05% PBS-Tween 20(Wellwash device, Labsysytems)の200μl/ウェルで5回洗浄した。次いで、プレートを、0.05% PBS-Tween 20緩衝液及び0.2%ウシ胎児血清(PBS-T-BSA)の150μl/ウェルで、37℃で2時間飽和した。5回洗浄後、滴定される、組換えマウスサイトカイン対照及び試料を連続希釈によってPBS-T-BSAで1/2に希釈した。1/2への4つの連続希釈は、純水な試料で行った。プレートを次いで4℃で終夜インキュベートした。5回洗浄後、ビオチン化検出抗体を分配し(PBS-T-BSA中、IFN-γ/IL-10について1μg/ml、IL-5について0.5μg/ml、100μl/ウェル)、4℃で3時間インキュベートした。5回洗浄後、ストレプトアビジン-ペルオキシダーゼコンジュゲート(Pierce)を分配し(PBS-T-BSA中1μg/ml、100μl/ウェル)、4℃で1時間インキュベートした。5回洗浄後、ペルオキシダーゼ基質(ABTS+H2O)をウェルに分配した。45分間インキュベーションした後、光学密度を405 nmで読んだ(Dynex ELISAリーダー)。リーダーに結合した分析ソフトウェア「Revelation」を用いて標準値に対して試料中のIFN-γ/IL-5/IL-10の濃度を計算した。
【0198】
抗体参照(BD Bioscience):
−IFN-γ:捕獲抗体:クローンR4-6A2(BD Bioscience,551216)
−検出抗体:クローンXMG1.2(BD Bioscience,参照554410)
−組換えmIFN-γ:R&D装置485-MI
−IL-5:捕獲抗体:クローンTRFK5(BD Bioscience,554393)
−検出抗体:クローンTRFK4(BD Bioscience,参照554397
−組換えmIL-5:BD Bioscience 554581
−IL-10:捕獲抗体:クローンJES5-2A5(BD Bioscience,551215
−検出抗体:クローンSXC-1(BD Bioscience,参照554423)
−組換えmIL-10:BD Bioscience 550070
−ストレプトアビジンペルオキシダーゼImmunopure(Pierce21126)。
【0199】
I.7 統計的分析
マン-ホイットニーU試験を行うことによって群を対で比較した(http://eatworms.swmed.edu/〜leon/stats/utest.html)。p<0.05の値が有意であると考えられた。
【0200】
II.結果:
ワクチンベクターとしてN-PΔN複合体によって形成される環の効力を評価するために、RSVのN核タンパク質との融合タンパク質の形態のGFP(モデル抗原)で、BALB/cマウスを免疫した。N-GFP融合タンパク質は、先に記載の方法(特許第FR0504426号)によって精製され得る溶解性環構造を形成する。このようにしてNGFP-PΔNタンパク質複合体は得られた。ワクチン用途については、N-GFP及びPタンパク質が分離できるが、その操作は必ずしも必要でなく、N環の免疫原性についての発明者の先の結果は、PΔNの存在が負の効果を有さないことを示している。
【0201】
気道を標的とする病原体に関する研究、及び鼻腔内経路によって投与される時の、N-PΔN複合体の証明された免疫原性を考慮すると、それは、その複合体のベクター性を証明するための好ましい投与経路である。マウス群を、天然組換え体のGFPタンパク質、及びN-PΔN複合体でも免疫した。E.コリの解毒リンンフォトキシン、LT(R192G)、ここで粘膜経路によるそのアジュバント性は、十分に記載されているが(McNeal et al., 2005, Freytag and Clements 2005)。
【0202】
GFP及びN-PΔNに対する免疫反応のパラメーターは、(i)(気管支肺胞の洗浄)による血清及び粘膜抗体の産生、及び (ii) 脾臓、及び気道をドレインするガングリオンから単離された記憶Tリンパ球による、サイトカイン(IFN-γ、IL-5及びIL-10)の産生を介する細胞反応、である。
【0203】
II-1 Nの環は、外来抗原、例えばGFPmに対する抗体を標的化するための効果的なベクターである
N-PΔN複合体は、全身的経路及び粘膜経路によって免疫原性が非常に高い(特許第FR0504426)。GFPとNタンパク質との融合体は、血清レベル(図3B)又は粘膜レベル(図3C)の抗-N-PΔN Ac反応の強度を変更しない:すなわち、群N-GFP-PΔNと群N-PΔNとには有意な差はない。
【0204】
鼻腔経路によって投与されたGFPは、全身性Ac反応を誘導するが(図3A;GFP対対照、p=0.003)、局所的反応を誘導しない(図3C;GFP対対照、有意でない)。
【0205】
一方、GFPとN-PΔN複合体との関係は、GFPのみと比較して、抗-GFP抗体の割合をかなり増加する。抗-GF抗体のレベルは、血清レベルで4のファクターで乗じられ(N-GFP-PΔN対GFP、p=0.039)、BAWレベルで100のファクターで乗じられる(N-GFP-PΔN対GFP、p=0.014)。
【0206】
II-2 N環でのGFPのベクター化は、特定のT反応を増加しない:
抗体反応を刺激するその強い能力に加えて、N-PΔNは、全身的レベルの抗原依存的IFN-γの合成によって証明される記憶T反応を効果的に刺激するが(特許第FR0504426及び図4、N-PΔN脾臓対対照、P=0.01)、局所レベルの抗原依存的IFN-γの合成によって証明される記憶T反応を効果的に刺激しない(図5、局所的ガングリオン)。GFPのような外来抗原のNへの融合は、脾臓レベル(N-GFP-PΔN対N-PΔN、差はない)又はガングリオンのレベルで(図4)、そのT反応の確立を妨害するものではない。
【0207】
一方、GFPの提示形式によらず(単独でも又はNと融合しても)、IFN-γ(図4)又はIL-5/IL-10(図示せず)の産生によって測定された、GFPに特異的なT反応は観察されなかった:すなわち、対照群、N-GFP-PΔN及びGFP、間で有意な差はない。
【0208】
結局、GFPの例は、N-PΔN複合体が、特に粘膜レベルで、抗体反応を刺激するための非常に極力な抗原ベクターであることを証明することができる。一方、それらは、同一の抗原に対する記憶T反応を刺激する点でほとんど効果がないことが判る。重要なことに、外来抗原の融合は、N-PΔN複合体に固有の免疫原性を変更しなかった。
【0209】
これらの結果は、強力な抗体反応を刺激す目的で抗原ベクターとしてN-PΔN複合体を用いて考えることを可能にする。RSVのワクチン化の範囲内で、得られた結果は、核タンパク質に対する細胞反応と、抗体を中和する標的であることが知られているRSVの抗原/エピトープに対する抗体反応とを利用することができることを示している。免疫防御のこれらの2つの事実は、RSVに対する殺菌免疫を得る点で重要である。
【0210】
一般的に、多くの病原体(特にウイルス)に関して、標的抗原又は中和抗体の抗原は、文献に十分に記載され、感染の効果的な制御に関連している。従って、これらの反応を効果的に刺激するための新規ベクターは、ワクチン化には非常に好ましい。N-PΔN複合体は、宿主又は環境のための危険性がない、不活性で、非-複製的構造であるという利点を有している。
【0211】
実施例6:Nタンパク質とGFPタンパク質とのリンカー配列の最適化
N-GFP融合タンパク質の精製は、GSTに融合されたタンパク質P(断片161〜241)を用いる親和性によって行った。実施例1に従って構築されたタンパク質N-GFP(配列番号33)は、タンパク質Nの収量よりも10倍低い収量で精製された。加えて、約43 kDaの見掛け質量で移動する2つのポリペプチドの存在が依然として観察された。これらのバンドは、質量分析(MALDI-TOF)によって分析した。それらは、(i) Nタンパク質のC-末端の追加の11残基を生じる、(アルギニンのレベルでの)GFPからNを分離するリンカー配列において、及び (ii) 最後の6残基を除くことによって(リジンのレベルでの)Nタンパク質のC-末端において、起こった開裂に相当する。C-末端の6、12及び27残基が削除されたNタンパク質の構築は、NΔ5C及びNΔ12Cのタンパク質が依然としてPと相互作用し、切断された形態NΔ27Cとは異なる、ことを示している。
【0212】
従って、NとGFPとの間のリンカー配列は修飾された。すなわち、その配列:
【0213】
【表8】
【0214】
(配列番号34、矢印は、E.コリでの開裂部位を示す)は、配列:
【0215】
【表9】
【0216】
(配列番号32)によって置換された。
【0217】
このことは、細菌中でもはや分解されず(SDS-PAGEゲル中の更なるバンドの非存在)、そして天然Nタンパク質単独と同一の収率で精製された、より安定なN-GFP融合タンパク質(配列番号35)をもたらした。
【図面の簡単な説明】
【0218】
【図1】図1は、HEp2又はRAW細胞によるN-GFP-PΔNの捕獲を示す。HEp-2(A;C)又はRAW(B;D)細胞を、DX-FITC(A;B)又はN-GFP-PΔN(C;D)の存在下にインキュベートした。インキュベーションを4℃で1時間(点線の曲線)、4℃で2時間(太い実線の曲線)、37℃で1時間続けた(実線の曲線)又は37℃で2時間続けた(太い点線の曲線)。100,000事象についてフローサイトメトリーで蛍光(FITC又はGFP)を分析した。
【図2】図2は、N-GFP-PΔN複合体がRAW細胞中に内在化されることを示している。RAW細胞は、4℃でN-GFP-PΔNの存在下で(太い実線の曲線)又は37℃でN-GFP-PΔNの存在下で(点線の曲線)、1時間インキュベートした。Nタンパク質を、非-浸透細胞(A)又は浸透細胞(B)についてのフローサイトメトリーによる直接的な免疫標識によって検出した。細胞の自己蛍光は、PBS中で37℃でインキュベートされた細胞のシグナルによって得られた(実線の曲線)。
【図3】図3は、N-PΔN複合体に対するGFPのベクター化による抗体反応の増加を示す。アジュバントのみ(対照)、アジュバントの存在下でのN-GFP-PΔN複合体、アジュバントの存在下でのN-PΔN又はGFPを用いて、d0日に鼻腔内経路によってBALB/cマウスを免疫した。2週間後(d14)にブースター注射を行った。動物をブースター注射後2週間に屠殺した(d28)。d0、d14及びd28の血清を回収した(A及びB)。d28に気管支肺胞洗浄を行った(C)。抗-N-PΔN抗体(B及びC)、及び抗-GFP抗体(A及びC)の力価は、ELISAによって決定した。データを平均±平均の標準誤差として表し(対照及びGFP群についてはn=8、N-GFP-PΔ群についてはn=6、N-PΔ群についてはn=4)、対数目盛に従って表した。
【図4】図4は、N-PΔNによってベクター化されたその形態を含む、GFPに対する記憶T反応の非存在を示す。アジュバントのみ(対照)、アジュバントの存在下でのN-GFP-PΔN複合体、アジュバントの存在下でのN-PΔN又はGFPを用いて、d0日に鼻腔内経路によってBALB/cマウスを免疫した。2週間後(d14)にブースター注射を行った。脾臓及び局所的ガングリオンを除くために、ブースター注射の2週間後に動物を解剖した。細胞懸濁液を脾臓については個別に(菱形)、及びガングリオンについては1プール/群で処理した(丸)。細胞をGFP(白抜き菱形及び丸)又はN-PΔ(黒色菱形及び丸)で72時間再刺激した。IFN-γの分泌をELISAで測定した。細胞を培地のみで培養した時、IFN-γの基本レベルは、15 pg/ml未満であった。
【特許請求の範囲】
【請求項1】
Nタンパク質と対象タンパク質との融合タンパク質であって、該Nタンパク質がパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質であり、該対象タンパク質がNタンパク質のC-末端のインフレームに融合される、前記融合タンパク質。
【請求項2】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる、請求項1記載の融合タンパク質。
【請求項3】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、ヒト又はウシ呼吸器合胞体ウイルスである、請求項1又は2記載の融合タンパク質。
【請求項4】
前記の対象タンパク質が、マーカータンパク質、又は治療的もしくはワクチン的関心のタンパク質である、請求項1〜3のいずれか1項記載の融合タンパク質。
【請求項5】
前記のワクチン的関心のタンパク質が、病原性微生物から得られる抗原である、請求項4記載の融合タンパク質。
【請求項6】
前記の対象タンパク質が、そのC-末端のインフレームに融合されたGFPを、治療的又はワクチンタンパク質と共に含む構造である、請求項4記載の融合タンパク質。
【請求項7】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、天然タンパク質である、請求項1〜6のいずれか1項記載の融合タンパク質。
【請求項8】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、最後の25個のC-末端アミノ酸によって定義される領域で修飾されたNタンパク質であり、該修飾されたNタンパク質は、Pタンパク質と相互作用する能力を保持する、請求項1〜6のいずれか1項記載の融合タンパク質。
【請求項9】
請求項1〜8のいずれか1項記載の融合タンパク質をコードする核酸。
【請求項10】
請求項9記載の核酸を含む発現ベクター。
【請求項11】
請求項10記載の発現ベクターを含む宿主細胞。
【請求項12】
Nタンパク質と対象タンパク質との融合タンパク質の製造法であって、該Nタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質であり、該対象タンパク質は、Nタンパク質のC-末端のインフレームに融合され、以下:
Nタンパク質と対象タンパク質との融合タンパク質の発現を許容する条件下での請求項11記載の宿主細胞の培養、及び
場合により発現されたNタンパク質と対象タンパク質との融合タンパク質の生成、
を含む、前記方法。
【請求項13】
Nタンパク質と対象タンパク質/Pタンパク質との複合体の製造法であって、該Nタンパク質及びPタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質であり、以下のステップ:
a) パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質を共発現し、該対象タンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーと同一のウイルスのPタンパク質で、Nタンパク質のC-末端のインフレームに融合され;
b) そのように形成されたNタンパク質と対象タンパク質/Pタンパク質との複合体を回収すること、
を含む、前記方法。
【請求項14】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、天然タンパク質である、請求項13記載の方法。
【請求項15】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、最後の25個のC-末端アミノ酸によって定義される領域で修飾されたNタンパク質であり、該修飾されたNタンパク質は、Pタンパク質と相互作用する能力を保持する、請求項13記載の方法。
【請求項16】
請求項13〜15のいずれか1項記載の方法によって得られる、Nタンパク質と対象タンパク質/Pタンパク質との複合体。
【請求項17】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質の製造法であって、そのC-末端のインフレームに対象タンパク質と融合され、以下:
請求項16記載のNタンパク質と対象タンパク質/Pタンパク質との複合体から、Nタンパク質と対象タンパク質との融合タンパク質の分離、を含む、前記方法。
【請求項18】
Nタンパク質と対象タンパク質との融合タンパク質であって、該Nタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質であり、該対象タンパク質は、Nタンパク質のC-末端のインフレームに融合され、請求項13〜15のいずれか1項記載の方法によって得られる、前記融合タンパク質。
【請求項19】
薬学的に許容される担体中、請求項1〜8又は18で定義された対象タンパク質と融合された、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質を含む、医薬組成物。
【請求項20】
薬学的に許容される担体中、請求項16で定義されたNタンパク質と対象タンパク質/Pタンパク質との複合体を含む、請求項19記載の医薬組成物。
【請求項21】
免疫原性の又はワクチン組成物である、請求項19又は20記載の組成物。
【請求項22】
治療的もしくはワクチン的関心の分子又はタンパク質のためのベクターの製造における、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質の使用。
【請求項23】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、おたふく風邪ウイルス、麻疹ウイルス、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる、請求項22記載の使用。
【請求項24】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質に化学的に結合された、治療的又はワクチン的関心の分子を含む複合物。
【請求項25】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質を含む診断試薬。
【請求項26】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質の、融合タンパク質の抗原に対する抗体のin vitro検出における使用。
【請求項27】
生物試料中の抗原に特異的な抗体を検出する方法であって、以下のステップ:
a) 該生物試料を、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質と接触させ、
b) 得られたN-抗原融合タンパク質/抗体複合体を検出すること、
を含み、
かかる複合体の存在は、抗原特異的抗体の生物試料中の存在を示唆する、前記方法。
【請求項1】
Nタンパク質と対象タンパク質との融合タンパク質であって、該Nタンパク質がパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質であり、該対象タンパク質がNタンパク質のC-末端のインフレームに融合される、前記融合タンパク質。
【請求項2】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、おたふく風邪ウイルス、麻疹ウイルス、ヒトメタニューモウイルス(Metapneumovirus)、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる、請求項1記載の融合タンパク質。
【請求項3】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、ヒト又はウシ呼吸器合胞体ウイルスである、請求項1又は2記載の融合タンパク質。
【請求項4】
前記の対象タンパク質が、マーカータンパク質、又は治療的もしくはワクチン的関心のタンパク質である、請求項1〜3のいずれか1項記載の融合タンパク質。
【請求項5】
前記のワクチン的関心のタンパク質が、病原性微生物から得られる抗原である、請求項4記載の融合タンパク質。
【請求項6】
前記の対象タンパク質が、そのC-末端のインフレームに融合されたGFPを、治療的又はワクチンタンパク質と共に含む構造である、請求項4記載の融合タンパク質。
【請求項7】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、天然タンパク質である、請求項1〜6のいずれか1項記載の融合タンパク質。
【請求項8】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、最後の25個のC-末端アミノ酸によって定義される領域で修飾されたNタンパク質であり、該修飾されたNタンパク質は、Pタンパク質と相互作用する能力を保持する、請求項1〜6のいずれか1項記載の融合タンパク質。
【請求項9】
請求項1〜8のいずれか1項記載の融合タンパク質をコードする核酸。
【請求項10】
請求項9記載の核酸を含む発現ベクター。
【請求項11】
請求項10記載の発現ベクターを含む宿主細胞。
【請求項12】
Nタンパク質と対象タンパク質との融合タンパク質の製造法であって、該Nタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質であり、該対象タンパク質は、Nタンパク質のC-末端のインフレームに融合され、以下:
Nタンパク質と対象タンパク質との融合タンパク質の発現を許容する条件下での請求項11記載の宿主細胞の培養、及び
場合により発現されたNタンパク質と対象タンパク質との融合タンパク質の生成、
を含む、前記方法。
【請求項13】
Nタンパク質と対象タンパク質/Pタンパク質との複合体の製造法であって、該Nタンパク質及びPタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのタンパク質であり、以下のステップ:
a) パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質を共発現し、該対象タンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーと同一のウイルスのPタンパク質で、Nタンパク質のC-末端のインフレームに融合され;
b) そのように形成されたNタンパク質と対象タンパク質/Pタンパク質との複合体を回収すること、
を含む、前記方法。
【請求項14】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、天然タンパク質である、請求項13記載の方法。
【請求項15】
前記のパラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質が、最後の25個のC-末端アミノ酸によって定義される領域で修飾されたNタンパク質であり、該修飾されたNタンパク質は、Pタンパク質と相互作用する能力を保持する、請求項13記載の方法。
【請求項16】
請求項13〜15のいずれか1項記載の方法によって得られる、Nタンパク質と対象タンパク質/Pタンパク質との複合体。
【請求項17】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質の製造法であって、そのC-末端のインフレームに対象タンパク質と融合され、以下:
請求項16記載のNタンパク質と対象タンパク質/Pタンパク質との複合体から、Nタンパク質と対象タンパク質との融合タンパク質の分離、を含む、前記方法。
【請求項18】
Nタンパク質と対象タンパク質との融合タンパク質であって、該Nタンパク質は、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質であり、該対象タンパク質は、Nタンパク質のC-末端のインフレームに融合され、請求項13〜15のいずれか1項記載の方法によって得られる、前記融合タンパク質。
【請求項19】
薬学的に許容される担体中、請求項1〜8又は18で定義された対象タンパク質と融合された、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質を含む、医薬組成物。
【請求項20】
薬学的に許容される担体中、請求項16で定義されたNタンパク質と対象タンパク質/Pタンパク質との複合体を含む、請求項19記載の医薬組成物。
【請求項21】
免疫原性の又はワクチン組成物である、請求項19又は20記載の組成物。
【請求項22】
治療的もしくはワクチン的関心の分子又はタンパク質のためのベクターの製造における、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質の使用。
【請求項23】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスが、おたふく風邪ウイルス、麻疹ウイルス、パラインフルエンザウイルス、及び呼吸器合胞体ウイルスからなる群より選ばれる、請求項22記載の使用。
【請求項24】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質に化学的に結合された、治療的又はワクチン的関心の分子を含む複合物。
【請求項25】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質を含む診断試薬。
【請求項26】
パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質の、融合タンパク質の抗原に対する抗体のin vitro検出における使用。
【請求項27】
生物試料中の抗原に特異的な抗体を検出する方法であって、以下のステップ:
a) 該生物試料を、パラミクソウイルス科(Paramyxoviridae)ファミリーのウイルスのNタンパク質のC-末端のインフレームに融合された抗原を含む融合タンパク質と接触させ、
b) 得られたN-抗原融合タンパク質/抗体複合体を検出すること、
を含み、
かかる複合体の存在は、抗原特異的抗体の生物試料中の存在を示唆する、前記方法。
【図1】

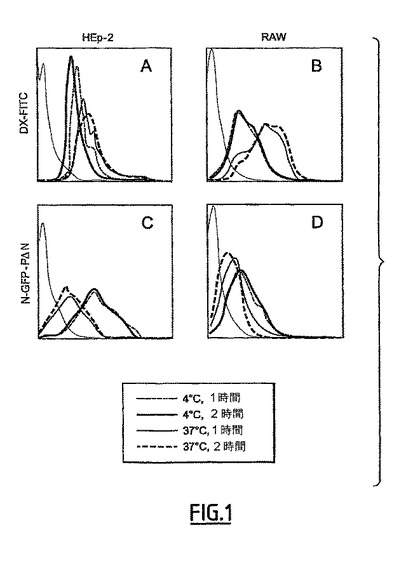
【図2】
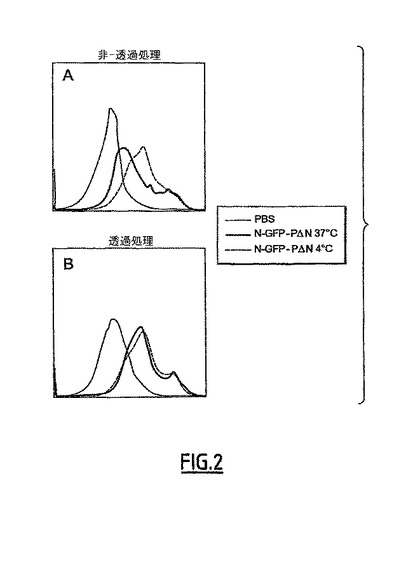
【図3】
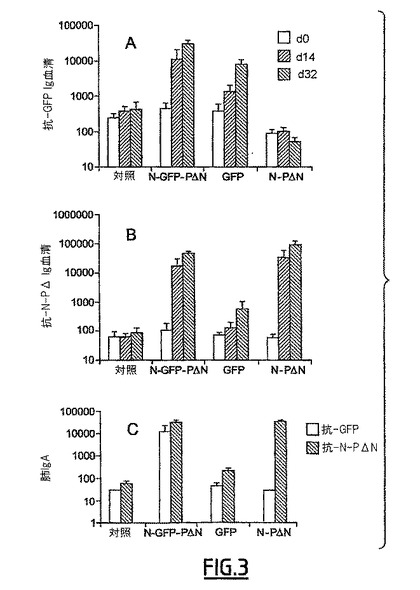
【図4】
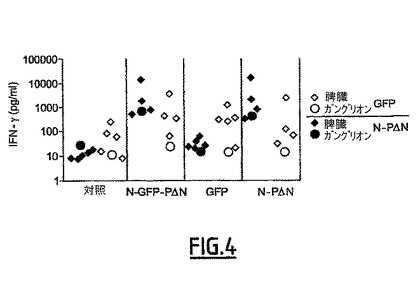
【図2】
【図3】
【図4】
【公表番号】特表2009−534024(P2009−534024A)
【公表日】平成21年9月24日(2009.9.24)
【国際特許分類】
【出願番号】特願2009−505929(P2009−505929)
【出願日】平成19年4月18日(2007.4.18)
【国際出願番号】PCT/FR2007/000651
【国際公開番号】WO2007/119011
【国際公開日】平成19年10月25日(2007.10.25)
【出願人】(594184698)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック(イエヌエールア) (3)
【Fターム(参考)】
【公表日】平成21年9月24日(2009.9.24)
【国際特許分類】
【出願日】平成19年4月18日(2007.4.18)
【国際出願番号】PCT/FR2007/000651
【国際公開番号】WO2007/119011
【国際公開日】平成19年10月25日(2007.10.25)
【出願人】(594184698)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック(イエヌエールア) (3)
【Fターム(参考)】
[ Back to top ]