
パーキン相互作用ポリペプチドおよび使用方法
本発明はパーキン結合ポリペプチドおよびそれをコードする核酸を提供する。本発明はパーキン結合ポリペプチドに特異的な抗体も提供する。本発明は、さらに、パーキン結合ポリペプチドを検出する方法およびパーキン結合ポリペプチドをコードする核酸を検出する方法を提供する。本発明はパーキン結合ポリペプチドを使用する方法を更に提供する。1つの実施態様において、本発明は、パーキン結合ポリペプチドと1つ以上の化合物とを接触させパーキン結合ポリペプチドの活性を変える化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を提供する。
【発明の詳細な説明】
【背景技術】
【0001】
本発明は、一般的に、分子生物学、細胞生物学および医薬の分野に関し、より具体的にはパーキンソン病に関する。
【0002】
パーキンソン病(PD)は筋硬直、震え、および動作緩慢により特徴付けられる主な神経変性病である(DunnettおよびBjorklund,Nature 399:A32−A39(1999))。姿勢障害(postural deficit)、歩行障害、および痴呆などの他の症状もPD患者の部分母集団に観察される。特発性PD症例の大半は散発性であり恐らく環境要因に影響されるが、症例の家族集積性およびPD特性の稀なメンデル遺伝は遺伝の重要性を明示している。
【0003】
パーキンソン症候群は、四つの主要な徴候、即ち、安静時振戦、動作緩慢、自発性および運動の減少、硬直、ならびに姿勢の不安定性により特色付けられる臨床的症候群である。顕著でない徴候は、気分および知性、自律機能ならびに感覚系に関する。発症時の平均年齢は55歳であり、60歳以上の人の約1%がこの病を患う。男性は女性より高い頻度で冒される。
【0004】
パーキンソン症候群は、四つの主要な徴候、即ち、安静時振戦、動作緩慢、自発性および運動の減少、硬直、ならびに姿勢の不安定性により特色付けられる臨床的症候群である。顕著でない徴候は、気分および知性、自律機能ならびに感覚系に関する。発症時の平均年齢は55歳であり、60歳以上の人の約1%がこの病を患う。男性は女性より高い頻度で冒される。
【0005】
安静時振戦および動作緩慢は最も典型的なパーキンソン症候群の徴候であり診断では事実上同義である。動作緩慢は、関連するパーキンソン症候群の症状および徴候、即ち運動および日常生活動作の一般的減速;無表情((hypomimia)または仮面様顔貌);まばたきの頻度の減少による凝視表情;よだれを惹起する嚥下障害;運動低下性および発声不全性の構語障害;単調な発話;小さな字体筆記(小字症);反復運動および同時運動の困難;椅子から立ち上がることが困難およびベッドで寝返りを打つのが困難;小股に引きずり歩行;腕振りおよび他の自動運動の減少;ならびに躊躇およびすくみの開始の大半を占める。突然および頻繁の予測できない運動不能により明示されるすくみは、あらゆるパーキンソン症候群の症状の最も生活に支障を来す1つである。
【0006】
人口が高齢化し60歳以上の人の数が増すにつれて、パーキンソン病を発症する個体数が増えつつあるらしい。パーキンソン病を処置するために治療が利用できるが、これらの治療の多くは望ましくない副作用を有する薬物を使用する。パーキンソン病に関連する衰弱の症状を考えると、更なる治療様式が開発できるようにパーキンソン病の原因を理解することが重要である。
【発明の開示】
【発明が解決しようとする課題】
【0007】
従って、パーキンソン病の発症に関連する遺伝子および遺伝子産物を同定し特徴決定する必要がある。本発明は、この必要性を満たし、さらに、関連する利点を提供する。
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明はパーキン結合ポリペプチドおよびそれをコードする核酸を提供する。本発明はパーキン結合ポリペプチドに特異的な抗体も提供する。本発明は、さらに、パーキン結合ポリペプチドを検出する方法およびパーキン結合ポリペプチドをコードする核酸を検出する方法を提供する。本発明はパーキン結合ポリペプチドを使用する方法を更に提供する。1つの実施態様において、本発明は、パーキン結合ポリペプチドと1つ以上の化合物とを接触させパーキン結合ポリペプチドの活性を変える化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を提供する。
【0009】
(発明の詳細な説明)
本発明はパーキン結合ポリペプチド(PBP)を提供する。本発明はさらにパーキン結合ポリペプチドを使用する方法を提供する。
【0010】
パーキンソン病(PD)において、ドーパミンのレベルは線条体で低下するが、被殻で最も高い。これは主として黒質緻密部でドーパミンを産生するニューロンの変性の結果である(Yamadaら、Brain Res.526:303−307(1990); Damierら、Brain 122:1437−1448(1999); およびNaoiら、Mech.Ageing Dev.111:175−188(1999))。NR4A2(Leら、Nat.Genet.33:85−89(2003))、α−シヌクレイン(synuclein)(Polymeropoulosら、Science 276:2045−2047(1997))およびユビキチンC末端ヒドロキシラーゼL1(UCHL1)(Wintermeyerら、Neuroreport 11:2079−2082(2000))の三遺伝子は常染色体優性PDに関連する。常染色体劣性PDに関連する2つの遺伝子はパーキン(Kitadaら、Nature 392:605−608(1998))およびDJ−1(Bonifatiら、Science 299:256−259(2003))である。パーキン遺伝子の不活性化変異はPARK2常染色体劣性若年性パーキンソン症候群(AR−JP)を惹起する。他のPD形態と同様に、PARK2は黒質のドーパミン作動性ニューロンの欠如により特徴付けられる。しかしながら、PARK2は、黒質ニューロンのレーヴィ小体がAR−JPの大半の症例において存在しないという点で独特である(IshikawaおよびTsuji,Neurology 47:160−166(1996); IshikawaおよびTakahashi,J.Neurol.245:4−9(1998);ならびにMatsumine,J.Neurol.245:10−14(1998))。パーキン遺伝子の変異はAR−JPの形態を惹起するが高齢のPD患者にも見出され、パーキンの変異が若年の発症に限定されないことを証明する(Abbasら、Hum.Mol.Genet.8:567−574(1999))。
【0011】
パーキンをコードする遺伝子の不活性化変異は、常染色体劣性若年性パーキンソン病の幾つかの形態の原因である。パーキンはプロテアソーム依存性タンパク質の分解経路で標的とされるミスフォールドしたタンパク質をユビキチン化するユビキチン・リガーゼである。パーキンの機能の手掛かりはパーキンの一次構造およびパーキン遺伝子の変異部位の局在化により示唆される。パーキンはN末端ドメインのユビキチン様ドメインおよびC末端ドメイン寄りの2つのRINGフィンガー・モチーフから構成される(Kitadaら、前出、1998; およびShimuraら、Nat.Genet.25:302−305(2000))。幾つかの不活性化変異は、RINGフィンガー・ドメインに見出され、これらのドメインが機能的に重要であることを示唆している(Shimuraら、Ann.Neurol.45:668−672(1999))。今日までに、唯1つのミスセンス(Arg42Pro)および三つのフレームシフト変異がユビキチン様ドメインで見出された。部位42のアルギニンは標的タンパク質の結合で機能するようであ。Shimuraら(前出、2000年)は、ユビキチンと共役するH7タンパク質がRINGフィンガー・ドメインに結合し且つパーキンのRINGドメインがヒトのドーパミン作動性SH−Sy5Y神経芽腫細胞のユビキチン化に必要であることを証明した。これらの観察は、ユビキチン様ドメインおよびRINGフィンガー・ドメインの異なる役割を示唆している。ユビキチン様ドメインはパーキンの安定性(Finneyら、J.Biol.Chem.278:16054−16058(2003))ならびに恐らくプロテアソームへのユビキチン化基質の標的化に重要であることが見出された。他方で、RINGフィンガーは基質およびUbcH7(E2)などのユビキチン−リガーゼ活性に必要な他のユビキチン構成要素に結合する。この観察は確認され(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000))、パーキンはセプチンと呼ばれるシナプス小胞に関連するタンパク質ファミリーの一員であるCDCrel−1に結合し且つパーキンはこのタンパク質のユビキチン化および代謝回転を刺激することが更に見出された。併せて、これらのデータはパーキンがE3ユビキチン・リガーゼとして機能するという示唆に至った。
【0012】
活性化E1およびしばしば共役するE2酵素とともに、E3ユビキチン・リガーゼは、26Sプロテアソーム複合体での分解で標的とされる細胞質タンパク質へのユビキチン鎖の共役を触媒し、細胞周期、細胞死、および細胞分化などの重要な細胞過程を調節する。ユビキチン化基質は、基質が多ユビキチン化(polyubiquitinated)(5ユビキチン単位以上の鎖を含む)される場合、プロテアソーム依存性経路(JoazeiroおよびWeissman,Cell 102:549−552(2000))を介して、または、このタンパク質が単ユビキチン化(monoubiquitinated)(5未満の短いユビキチン鎖を含む)される場合、リソソーム分解経路を介してのいずれかで分解できる。単ユビキチン化は、例えばEGF受容体などのある種の細胞表面受容体を内在化(エンドサイトース)させ得る、またはエンドソーム経路のタンパク質選別シグナルとして機能し得る(Helliwellら、J.Cell.Biol.153:649−662(2001); Hicke,Cell 106:527−530(2001); およびHicke,Nat.Rev.Mol.Cell.Biol.2:195−201(2001))、ならびにこれらの単ユビキチン化タンパク質をリソソームに導く。パーキンは、Pael−1受容体(Imaiら、Cell 105:891−902(2001));CDCrel−1(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000));グリコシル化α−シヌクレイン(Shimuraら、Science 293:263−269(2001));シンフィリン(synphilin)−1(Chungら、Nat.Med.7:1144−1150(2001));CHIP(Imaiら、Mol.Cell.10:55−67(2002));サイクリンE(Starpoliら、Neuron 37:735−749(2003));HSP70(Tsaiら、J.Biol.Chem.278:22044−22055(2003));α/β−チューブリン(Renら、J.Neurosci.23:3316−3324(2003));およびアミノアシルt−RNAシンセターゼ複合体のP38サブユニット(Cortiら、Hum.Mol.Genet.12:1427−1437(2003))を含む幾つかのタンパク質と相互作用することが見出された。パーキンが媒介するユビキチン化はプロテアソーム系によるこのタンパク質の分解につながる。Pael−1受容体のパーキン媒介性分解の欠如はPael受容体の蓄積をもたらし、細胞死を惹起する(Imaiら、前出、2001)。
【0013】
本明細書で開示するように、酵母2−ハイブリッド系はパーキン相互作用ポリペプチドを同定するために用いられた。具体的には、パーキンはシナプトタグミン1およびシナプトタグミン11(実施例I〜VIを参照)ならびにSLP(実施例VII)と相互作用することが見出された。パーキンとの相互作用はシナプトタグミンのユビキチン化を惹起しシナプトタグミンの細胞内局在化を変更する。酵母2−ハイブリッド系および共免疫沈降法は、パーキンがそれらのC2AドメインおよびC2Bドメインを通してシナプトタグミン・ファミリーのメンバーと相互作用することを同定するために用いられた。パーキンは多ユビキチン化しシナプトタグミン1およびシナプトタグミン11を分解する。パーキンおよびシナプトタグミンの共発現は、パーキンおよびシナプトタグミンの両方を含む核周辺構造への正常なシナプトタグミンの局在化を変化させる。疾患を惹起するアミノ酸置換を含有するパーキンを含む欠失パーキンおよびミスセンス・パーキンは、シナプトタグミン1およびシナプトタグミン11との相互作用ならびにそれらのユビキチン化を阻害した。変異パーキンはシナプトタグミンの細胞内局在化を変えなかった。パーキン媒介性ユビキチン化はシナプトタグミン11の代謝回転も増強する。シナプトタグミンは小胞の形成およびドッキングに対する重要性においてよく特徴付けられるため、これらの結果はシナプス小胞プールの調節および小胞の放出におけるパーキンおよびシナプトタグミンの役割を示している。従って、シナプトタグミン・ファミリーのメンバーとパーキンとの相互作用は、シナプス前末端での神経伝達物質輸送の制御に関与するタンパク質の調節にパーキンが関与することを示唆する。パーキンは、シナプス機能に関与する多数のタンパク質で見出されるカルシウム感知ドメインであるシナプトタグミンのC2ドメインに結合する。従って、パーキンの欠如は小胞のプール、ドッキングおよび放出を制御する多種のタンパク質に影響を及ぼし、パーキン変異をもつ患者に見られるドーパミン機能の欠損を説明する。
【0014】
本明細書に開示するように、パーキンと相互作用するシナプトタグミン・ファミリーの2つのメンバーが同定され特徴決定された。本明細書に開示する結果は、パーキンが以下の観察に基づいてシナプトタグミン1およびシナプトタグミン11に結合することを確認した。第一に、パーキンはGFPで標識されたシナプトタグミン1またはシナプトタグミン11とのみ共免疫沈降したがGFP標識単独とは共免疫沈降しなかった(実施例IIIおよび図2B)。第二に、内因性パーキンは内因性syt1と相互作用する(実施例IIIおよび図2C)。第三に、野生型のパーキンおよびRINGフィンガー・モチーフを含む欠失パーキンのみがシナプトタグミン1およびシナプトタグミン11に結合した(実施例IVおよび図3C)。第四に、RINGフィンガー・モチーフを欠く欠失パーキンおよびアミノ酸置換のパーキンはシナプトタグミンと相互作用できなかったまたは弱く相互作用した(実施例IVおよび図3C)。第五に、野生型のパーキンのみがシナプトタグミンをユビキチン化し分解に至った一方で、全ての欠失パーキンおよび変異パーキンはシナプトタグミンのユビキチン化の減少または欠如を示した(実施例V、図4および図6)。第六に、内因性パーキンは、NGF誘導性PC12細胞の神経突起に沿ったシナプス・ボタンでおよび核周辺の位置で、シナプトタグミン1およびシナプトタグミン11と共局在化した(実施例VIおよび図5)。最後に、パーキンおよびシナプトタグミンの共発現により、パーキン−シナプトタグミン複合体は核周辺分布の構造に補充された(実施例VIおよび図6)。
【0015】
パーキンは、シンフィリン−1(Chungら、前出(2001))、Pael−1受容体(Imaiら、前出(2001))、CDCrel−1(Zhangら、前出、2000)、およびグリコシル化シヌクレイン(Shimuraら、Science(2001))と相互作用することが見出された。これらのタンパク質の2つ、即ちCDCrel−1およびシンフィリン−1はシナプス小胞に関連するタンパク質である(Ribeiroら、J.Biol.Chem.277:23927−23933(2002); Wakabayashiら、Acta Neuropathol 103−209−214(2002);ならびにBeitesら、Nat.Neurosci.2:434−439(1999))一方、Pael−1受容体は未知の機能をもつ膜貫通タンパク質であり、グリコシル化シヌクレインはレーヴィ小体で見出される希少タンパク質である。CDCrel−1はシンタキシンと相互作用し、野生型CDCrel−1の過剰発現はHIT−T15細胞での分泌を阻害する(Ribeiroら、前出(2002); Wakabayashiら、前出(2002);およびBeitesら、前出(1999))。シンフィリン−1はα−シヌクレインと相互作用しPDの細胞質レヴィー小体の形成を刺激する(Engelenderら、Nat.Ganet.22:110−114(1999))。野生型パーキンの存在はシンフィリン−1が誘導するレーヴィ小体の形成に必須であるらしい。現在、CDCrel−1またはシンフィリン−1がどのようにシナプス前神経伝達の調節に参加するかは知られていない。ならびにCDCrel−1またはシンフィリン−1がドーパミンのシナプス前分泌の調節に関与するか否かも不明である。パーキンがシナプトタグミン・ファミリーのメンバーと相互作用し且つユビキチン化するという発見は、パーキンがシナプス小胞に関連するタンパク質の調節に重要な役割を果たすという仮説を更に支持する。
【0016】
シナプトタグミン1およびシナプトタグミン11(本明細書ではシナプトタグミンIおよびシナプトタグミンXIまたはsytIおよびsytXIとも呼ばれる)は、C2AドメインおよびC2Bドメインに高い相同性を有する(BLAST検索)約50個のカルシウム結合タンパク質から成る大ファミリーに属する。これらのタンパク質は、シナプトタグミンの1〜13、ラフィリン(raphilin)−2a、タンパク質キナーゼC、GTPアーゼ活性化タンパク質(GAP)、ラット/酵母ユビキチン・リガーゼNedd4、およびホスホリパーゼを含む。併せて、これらのタンパク質はカルシウム・センサー(sytおよびタンパク質キナーゼC)からリン酸化(GAP)およびリン脂質分解(ホスホリパーゼC)の範囲に及ぶ細胞シグナル伝達の調節物質として共通の機能を果たす。syt1は、シナプトタグミンの中でsyt2と最も高い相同性を有し、シナプス小胞および分泌顆粒で豊富に発現する(Sudhof,J.Biol.Chem.277:7629−7632(2002))。Syt1およびsyt2は、syt3、syt5−7およびsyt10と同様に、早いシナプス前神経伝達のカルシウム・センサーとして機能する(Fernandez−Chaconら、Nature 410;41−49(2001)およびGeppertら、Cell 79:717−727(1994))。シナプトタグミン11は、対照的に、C2Aドメインのセリン残基によるアスパラギン酸塩の同類置換によりsyt4と類似し、この置換はこのドメインへのCa2+結合の欠損をもたらす(von Poserら、J.Biol.Chem.272:14314−14319(1997))。Syt11の細胞局在は知られていないが、syt4はゴルジ体に局在する(Fukudaら、J.Neurochem.77:730−740(2001)およびBertonら、Eur.J.Neurosci.12:1294−1302(2000))。現在、syt4およびsyt11、syt8、syt9、syt12、およびsyt13の機能は推論的であるが、syt4は早いシナプス前神経伝達の下流調節物質として機能すると考えられる(Wangら、Science 294:1111−1115(2001))。概して、sytファミリーのメンバーはC2ドメインで高い相同性を有し、アミノ酸同一性は30%から50%の範囲に及ぶ。パーキンはsyt11のC2AドメインおよびC2Bドメインに結合するため、パーキンは他のsytとも相互作用するらしい。パーキンがまたsyt1(syt11と最も低いC2ドメインの相同性(30%同一性)を含むタンパク質)と相互作用し且つ調節するという観察は、パーキンがC2AおよびC2Bの配列と関連するドメインを含む広範囲のタンパク質と相互作用しうることを示唆する。
【0017】
シナプス前神経伝達は三つの過程を伴う:1)ドッキング、2)融合、および3)シナプス小胞の再利用である。実験的証拠により、シナプトタグミン1は三過程全てと結び付いた。ドッキングの段階では、シナプトタグミン1はt−SNAREタンパク質、シンタキシンおよびSNAP25と相互作用しシナプス小胞のドッキングを刺激する(Schiavoら、Proc.Natl.Acad.Sci.USA 94:997−1001(1997)およびLiら、Nature 375:594−599(1995))。融合の段階では、syt1は会合したSNARE複合体およびリン脂質と相互作用しシナプス小胞の融合を刺激し安定化する(Levequeら、J.Neurochem.74:367−374(2000); Geronaら、J.Biol.Chem.275:6328−6336(2000);およびDavisら、Neuron.24:363−376(1999))。再利用の段階では、syt1とクラスリン会合タンパク質複合体AP−2との相互作用はシナプス小胞の再利用に重要である(Zhangら、Cell 78:7510760(1994))。これらの相互作用に加えて、機能データは、シナプトタグミン1がシナプス小胞のドッキング(Reistら、J.Neurosci.18:7662−7673(1998))、融合(Geppertら、前出、(1994); Elferinkら、Cell 72:153−159(1993); DiAntonioら、Cell 73:1281−1290(1993); DiAntonioら、Neuron.12:909−920(1993);およびBommertら、Nature 363:163−165(1993))、および再利用(Jorgensenら、Nature 378:196−199(1995))に重要な役割を果たすことも示唆している。
【0018】
更に、syt1ノックアウト・マウスの研究は、syt1が迅速な神経伝達物質のエキソサイトーシス(Fernandez−Chaconら、前出(2001))およびCa++感受性の大きな有芯小胞のエキソサイトーシス(Voetsら、Proc.Natl.Acad.Sci.USA 98:11680−11685(20010))についての主要なCa++センサーであることを示唆する。Syt1の過剰発現は融合孔の開口時間を延長し、一方、syt4の過剰発現は融合孔の開口時間を短縮する(Wangら、前出、(2001))。これはシナプトタグミン1およびシナプトタグミン4が補完機能を有することを示唆している。Sytファミリーのメンバーがドーパミン作動性ニューロンのドーパミン分泌を調節することに関与するか否かは知られていない。変異パーキンによるsytの調節分解の欠如がシナプス形成障害をもたらし(MurpheyおよびGodenschwege,Neuron 36:5(2002))ドーパミン作動性ニューロンでドーパミン分泌の減少に至るか否かも知られていない。しかしながら、野生型のパーキンはsyt1およびsyt11と相互作用し調節するが変異パーキンまたは欠失パーキンはしないという本明細書で開示される観察により、パーキンが重要なE3ユビキチン・リガーゼであり且つシナプス前膜でシナプス小胞の機能を調節することが示される。
【0019】
SytXIは特発性PDの患者から得られる黒質ニューロンのLBの中核で見出される(図6を参照)。この分布は他のパーキン基質であるアミノアシルtRNAシンセターゼ複合体のp38サブユニットおよびシンフィリン−1についても観察される(Cortiら、Hum.Mol.Genet.12:1427−1437(2003); Wakabayashiら、Ann.Neurol.47:521−523(2000); Schlossmacherら、Am.J.Pathol.160:1655−1667(2002))。LBのsytXIの発見はPDにおいてシナプトタグミンの異常なプロセシングが関与する可能性を示唆している。LBがパーキンソン症候群のドパーミン作動性神経細胞死に役割を果たすか否かは推論の域を出ない。パーキンが関連するパーキンソン症候群のLBの欠如(Hayashiら、Mov.Discord.15:884−888(2000); Moriら、Neurology 51:890−892(1998); van de Warrenburgら、Neurology 56:555−557(2001))は、これらの封入がパーキンが関連するパーキンソン症候群におけるドパーミン作動性ニューロン変性の根本原因でないことを示している。
【0020】
パーキンは、三つの機能ドメイン、即ち、ユビキチン様、RING1およびRING2のドメインから成る(Shimuraら、Nat.Genet.25:302−305(2000))。RING2ドメインはユビキチンと共役した酵素への結合に必要であることが見出された(Shimuraら、前出、2000; Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000); Imaiら、J.Biol.Chem.275:35661−35664(2000))。ユビキチン様ドメインはパーキンの安定性に重要である(Finneyら、J.Biol.Chem.278:16054−16058(2003))。しかしながら、RINGフィンガー・モチーフは、パーキンの2つの基質、即ちCDCrel−1(Zhangら、前出、2000)およびPael−R(Imaiら、前出、2000)への結合に必須であることが後に見出された。これらの発見と一致して、本明細書に開示されるように、sytXIはアミノ酸残基の204〜293の領域に結合することが見出された(図3)。この領域はRINGモチーフを含む。更に、ユビキチン様ドメイン(p78−465)のみを欠くパーキン・ペプチドはユビキチン様ドメインを含むパーキンより弱くシナプトタグミンに結合した(図3B)。これらの観察は、ユビキチン様ドメインがシナプトタグミンの結合でRINGフィンガー・モチーフを曝すため全長パーキンの正しい折り畳みに重要であることを示唆する。本発明者らは三つのパーキン・ドメインが異なる機能を果たすことを示唆する。即ち、ユビキチン様ドメインはパーキンの正しい折り畳みおよび安定性に必要であり、RING1フィンガー・モチーフを含むp204−293ドメインはsytXIなどのC2ドメインを含むタンパク質との相互作用に必須である一方、RING2フィンガー・モチーフはE1ユビキチン活性化酵素およびE2ユビキチン共役タンパク質との複合体形成に重要である。
【0021】
パーキンは標的タンパク質のユビキチン化を触媒するE3ユビキチン・リガーゼである(Shimuraら、前出、2000; Zhangら、前出、2000)。多ユビキチン化はプロテアソームによるユビキチン共役基質の分解をもたらす。野生型パーキンは、欠失パーキン、ミスセンス変異パーキン、または負の対照と比べてsytXIの多ユビキチン化を強く触媒する(図4)。パーキン依存性ユビキチン化もsytXIの急速な代謝回転をもたらし(図5)、パーキンがsytXIのレベルを調節するという仮説を更に支持する。欠失パーキンまたはミスセンス変異パーキンを発現する細胞(C289GおよびC418R)は、GFP−sytXIのみを発現する細胞と同量のユビキチン化sytXIを産生した(図4)。これらの観察は、パーキンの欠失またはミスセンス変異がsytXIのユビキチン化を減らすまたはなくすことを示唆する。PARK2 AR−JPにおいて、パーキンの変異はおそらく特異的タンパク質のユビキチン化を減らし、sytXIおよびパーキンにより調節される他のタンパク質の細胞内レベルが増加する。パーキンが調節するタンパク質の細胞内レベルの異常な増加の最終的な効果はAR−JPの病的状態に寄与するであろう。
【0022】
SytXI mRNAは脳で豊富に発現するが、非神経組織では低レベルである(von Poserら、J.Biol.Chem.272:14314−14319(1997))。しかしながら、内因性sytXIタンパク質の特異的な細胞内分布についての情報は知られていない。PC12細胞の外因性sytXIは主にゴルジ・ネットワークに局在した(FukudaおよびMikoshiba,Biochem.J.354:249−257(2001))。トランスフェクションされていないNGF誘導性PC12細胞で、内因性のパーキンおよびsytXIはNGF誘導過程において核周辺分布および有芯小胞に共局在化することが見出された(図6)。内因性パーキンの分布様式は先の観察(Huynhら、Ann.Neurol.48:737−744(2000))と類似した。SytXIの分布はsytXIと48%の同一性を有するタンパク質であるsytIVの細胞内分布にも類似した。PC12細胞において、sytIVは主にゴルジおよび未熟小胞に局在する(Bertonら、Eur.J.Neurosci.12:1294−1302(2000); Ibataら、J.Neurochem.74:518−526(2000); Fukudaら、J.Neurochem.77:730−740(2003); Fukudaら、J.Biol.Chem.278:3220−3226(2003))。PC12細胞がNGFで処理される場合、sytIVタンパク質は成熟な有芯小胞に再分布する(Fukudaら、J.Biol.Chem.278:3220−3226(2003))。有芯小胞は、神経ペプチドまたは生体アミンを運ぶ分泌顆粒であり、カルシウム・イオンの刺激下でそれらの内容物を放出する。従って、sytXIおよびパーキンの両方が有芯小胞に共局在するという観察は、両タンパク質がおそらくカルシウム依存性エキソサイトーシスで役割を果たすことを示唆している。この仮説は、パーキンおよびsytXIの両方がヒトの黒質の神経突起およびニューロンの細胞体で類似する分布様式を有するという観察(図6)により更に支持される。
【0023】
AR−JP患者のパーキン機能の喪失は、シナプトタグミンXIの機能を変化させ、ドーパミンの放出の変化をもたらすことが予想される。これはジストニアおよびパーキンソン症候群の症状を順に惹起する。小胞機能の変化は、放出または再利用の段階であれば、細胞質ドーパミンの増加を惹起し、酸化的損傷の増加、続いて細胞死をもたらしうる。これはパーキン変異の患者に見られる神経変性を説明する。
【0024】
本明細書で開示される結果は、パーキンの喪失がドーパミンの放出を変化させ、ジストニアおよびパーキンソン症候群の初期症状をもたらしうることを示している。放出または再利用のいずれかの段階での小胞機能の変化は、細胞質ドーパミンを増加させ、酸化的損傷の増加、続いて細胞死をもたらしうる。
【0025】
本発明はシナプトタグミン1およびシナプトタグミン11およびSLPを含む典型的なパーキン結合ポリペプチドを提供する。本発明は、本明細書に開示するように、パーキン結合ポリペプチドを同定する方法も提供する。本発明は更にパーキン結合ポリペプチドを使用する方法を提供する。
【0026】
本発明はパーキン結合ポリペプチド(PBP)をコードする単離されたポリペプチドを提供する。具体的な実施態様において、本発明は配列番号2として示されるアミノ酸配列を有する単離されたポリペプチドを提供する。本発明はPBPの機能断片も提供する。
【0027】
本明細書で用いられるように、「機能断片」という用語は、パーキン結合ポリペプチド(PBP)に関して用いる場合、パーキン結合ポリペプチドの活性の一部または全てを保持するパーキン結合ポリペプチドの部分を指すことが意図される。典型的なPBPの機能断片はPBPのパーキン結合断片を含む。他の典型的なPBPの機能断片はPBPに特異的な抗体に特異的に結合する機能断片である。PBPの他の機能断片は抗原断片として機能するエピトープであるペプチド断片を含み、これは個々のPBPに特異的な抗体を作製するために使用できる。
【0028】
本明細書で用いられるように、「ポリペプチド」という用語は、パーキン結合ポリペプチド(PBP)に関して用いる場合、全長以下のパーキン結合ポリペプチドを含む、10以上のアミノ酸のペプチドまたはポリペプチドを指す。本明細書で用いられるように、「ペプチド断片」は2つ以上のアミノ酸のペプチドまたはポリペプチドを指す。パーキン結合ポリペプチドの「修飾」は、修飾がPBPの機能を保持する限り、PBPのアミノ酸配列の同義置換を含む。コードされるアミノ酸の同義置換は、例えば下記の群内に属するアミノ酸を含む。1)非極性アミノ酸(Gly,Ala,Val,Leu,およびIle);2)極性中性アミノ酸(Cys,Met,Ser,Thr,Asn,およびGln);3)極性酸性アミノ酸(AspおよびGlu);4)極性塩基性アミノ酸(Lys,ArgおよびHis);および5)芳香族アミノ酸(Phe,Typ,Tyr,およびHis)。他の微修飾は、ポリペプチドが本明細書に記載される機能の一部または全てを保持する限り、PBP内に含まれる。
【0029】
ポリペプチドの修飾は、修飾がPBPの機能を保持する限り、その誘導体、類似体および機能模倣体も含み得る。例えば、誘導体は、アルキル化、アシル化、カルバミル化、ヨード化などのポリペプチドの化学修飾、またはポリペプチドを誘導体化する任意の修飾を含み得る。類似体は、例えばヒドロキシプロリンまたはカルボキシグルタメートの修飾アミノ酸を含み得る、ならびにペプチド結合により連結していないアミノ酸を含み得る。模倣体はポリペプチドの機能を模倣する化学部分を含む化学物質を包含する。例えば、ポリペプチドが機能活性を有する2つの荷電化学部分を含む場合、模倣体は、この荷電化学機能が三次元空間で維持されるように、2つの荷電化学部分を空間的定位および制約構造に置く。従って、PBPの機能を与える官能基を配置する模倣体はPBPの誘導体の意味内に含まれる。
【0030】
本明細書で用いられるように、「実質的に」または「実質的に同じ」という用語は、ヌクレオチドまたはアミノ酸の配列に関して用いられる場合、ヌクレオチドまたはアミノ酸の配列が参照配列と比べて相当な程度、量または範囲の配列同一性を示すことを意味すると意図する。実質的に同じアミノ酸配列は参照ポリペプチドに特徴的な機能活性および/または生物活性を保持する。
【0031】
本明細書で用いられるように、「核酸」という用語は、デオキシリボ核酸(DNA)またはリボ核酸(RNA)などのポリヌクレオチドを意味し、一本鎖および二本鎖の両核酸ならびにオリゴヌクレオチドを包含する。本発明に有用な核酸は、ゲノムDNA、cDNAおよびmRNAを含み、センス鎖、アンチセンス鎖、または両方を表し得る。本発明のゲノム配列は、PBP遺伝子の発現を調節するプロモーターおよびエンハンサーなどの調節領域ならびにPBPをコードするエキソンを除いたイントロンを含むが、PBPをコードしない近接遺伝子を含まない。典型的なPBP核酸は、配列番号1、配列番号3および配列番号4として示されるヌクレオチド配列、またはそれらの断片を含む。PBP核酸分子に関して用いられる「単離された」という用語は、この分子が生来付随する構成要素からまたは他の方法で人為的に修飾された構成要素から実質的に取り出されるまたは分離されることを意味することにより、天然で存在するようなPBP核酸分子を除くことを意図する。
【0032】
本明細書で用いられるように、「オリゴヌクレオチド」という用語は、参照ヌクレオチド配列から少なくとも15個の連続したヌクレオチドを含む核酸分子を指し、少なくとも16、17、18、19、20または少なくとも25個の連続したヌクレオチドを含み得る、ならびに参照ヌクレオチド配列から少なくとも30、40、50、60、70、80、90、100、125、150、175、200、225、250、275、300、325、350までの連続したヌクレオチドをしばしば含む。参照ヌクレオチド配列はセンス鎖またはアンチセンス鎖であり得る。このオリゴヌクレオチドは化学合成され得るまたは組換え発現し得る。
【0033】
本明細書で用いられるように、核酸の「修飾」は参照配列に対して1つまたは幾つかのヌクレオチドの付加、欠失または置換を含み得る。核酸の修飾は、遺伝コードの縮重によりコードするアミノ酸配列を変更しない置換を含み得る。このような修飾は意図的に為される変異または核酸の複製中に突然変異として起きる変異に対応し得る。このようなものとして、核酸の修飾は実質的に同じ配列を含み、これは本明細書で開示されるPBPヌクレオチド配列などの親核酸分子に類縁すると認識できる。実質的に同じヌクレオチド配列は中程度にストリンジェントな条件下または極めてストリンジェントな条件下で参照ヌクレオチド配列とハイブリダイズできる。
【0034】
本明細書で開示するPBP核酸配列の典型的な修飾は、ヒト、霊長類、ラット、ウサギ、ウシ、ブタ、ヒツジ、イヌ、ネコまたは他の動物種などの他の種のホモログに対応する配列を含む。ヒト以外の種の対応するPBPの配列は、ポリメラーゼ連鎖反応(PCR)によるまたはゲノム・ライブラリー、cDNAライブラリーもしくは発現ライブラリーのスクリーニングによるなどの当該分野で公知の方法により決定できる。PBP核酸分子の他の典型的な修飾はPBPヌクレオチド配列のスプライス変異型に相当し得る。さらに、ヌクレオチド配列の修飾は、例えば塩基、糖もしくはリン酸塩部分への修飾を有する、または修飾されたホスホジエステル結合を有する1つ以上の非野生型ヌクレオチドを含み得る。このような修飾は核酸分子の安定性を増すのに有利であり得る。
【0035】
更に、ヌクレオチド配列の修飾は、例えば放射性標識、蛍光色素、強磁性体、発光標識などの検出可能な部分またはビオチンなどの検出可能な結合剤を含み得る。このような修飾はPBP核酸分子の検出が望まれる適用において好都合であり得る。
【0036】
本明細書で用いられるように、「ベクター」とはポリヌクレオチドを含む組換えのDNAもしくはRNAのプラスミドまたはウイルスを指す。ベクターは、発現エレメントがポリヌクレオチドの発現を制御するようポリヌクレオチドに機能的に連結された発現エレメントを含み得る。「発現エレメント」とは、ポリヌクレオチドの複製、転写、スプライシング、翻訳、または分解を含む、ポリヌクレオチドの機能調節に寄与する分子の相互作用に関与するヌクレオチド配列である。遺伝子の転写を制御する発現エレメントは、プロモーター、転写開始部位、または転写速度を増すDNA配列のエンハンサーであり得る。
【0037】
本明細書で用いられるように、「サンプル」という用語は、パーキンと結合するタンパク質、核酸またはポリペプチドを含むまたは潜在的に含む生体液、細胞、組織、器官またはそれらの一部を意味することが意図される。この用語は、個体に存在するサンプルならびに個体から得られるサンプルまたは個体に由来するサンプルを含む。例えば、サンプルは、生検により得られる検体の組織切片、または組織培養に置かれたまたは適応された細胞であり得る。サンプルは更に抽出物の細胞下画分または精製していないもしくは実質的に純粋な核酸またはタンパク質の調製物であり得る。サンプルは、例えば、縮重オリゴヌクレオチドを合成することにより、化学合成もされ得る。
【0038】
本明細書で用いられるように、「特異的にハイブリダイズする」という用語は、本明細書に記載するように、少なくとも中程度にストリンジェントな条件下またはよりストリンジェントな条件下で、参照PBP核酸分子にハイブリダイズする核酸分子の能力を指し、参照PBP核酸分子でない核酸分子(例えばアクチンcDNAなどの負の対照)と同条件下でハイブリダイズしない。
【0039】
本発明は単離されたパーキン結合ポリペプチド(PBP)またはその機能断片を提供する。典型的なパーキン結合ポリペプチドは本明細書に開示するシナプシン様タンパク質を含む(実施例VIIIを参照)。本発明の単離されたPBPおよびペプチドは、生化学的方法、組換え方法および合成方法を含む当該分野で既知の方法により調製できる。例えば、PBPは、PBPをコードする該当の転写産物またはPBPを発現する細胞または組織の起源から常套の生化学的方法により精製できる。本明細書に開示する方法は、どの細胞および組織、ならびにどの細胞下画分が適切な出発物質であるかを決めるために適応できる。生化学的精製は、例えば適切な組織または細胞の可溶化、所望の細胞下画分の単離、サイズ・クロマトグラフィー、イオン交換クロマトグラフィー、疎水クロマトグラフィーまたはアフィニティー・クロマトグラフィー、電気泳動、および免疫アフィニティーの手法などの段階を含み得る。本発明のポリペプチドの生化学的精製の方法および条件は当業者により選択でき、精製は、例えば、免疫アッセイまたは機能アッセイにより観察される。
【0040】
本発明はパーキン結合ポリペプチド(PBP)に特異的に結合する抗体も提供する。具体的な実施態様において、本発明は配列番号2として示されるアミノ配列を有するPBPに特異的に結合する抗体を提供する。本明細書で用いられるように、「抗体」という用語は、その最も広い意味で用いられ、ポリクローナル抗体およびモノクローナル抗体ならびにこの抗体の抗原結合断片を含む。PBPに特異的な本発明の抗体に関して、「抗原」という用語は野生型PBPもしくは合成PBPまたはそれらの断片を意味する。
【0041】
PBPに特異的な抗体、またはこの抗体の抗原結合断片は少なくとも約1x105 M−1のPBPまたはそのペプチド部分に対して特異的な結合活性を有することにより特徴付けられる。従って、PBPに特異的な抗体(PBPに対して特異的な結合活性を保持する)のFab、F(ab′)2、FdおよびFvの断片は抗体の定義内に含まれる。PBPの特異的結合活性は、例えばPBP対PBPでない対照ポリペプチドに対する抗体の結合活性を比較することにより、当業者により容易に測定できる。当業者は個々のPBPに特異的な結合活性を有する抗体の意味を容易に理解するであろう。抗体はポリクローナル抗体またはモノクローナル抗体であり得る。ポリクローナル抗体またはモノクローナル抗体を調製する方法は当業者によく知られている(例えば、HarlowおよびLane,Antibodies: A Laboratory Manual,Cold Spring Harbor Laboratory Press(1988))。ポリクローナル抗体を使用する場合、ポリクローナル血清は、バックグラウンド結合が少なく且つ抗原に特異的な抗体の割合が高い単一特異的な抗体を作製するために、抗原を用いてアフィニティー精製され得る。
【0042】
加えて、本明細書で用いられる「抗体」という用語は、天然に存在する抗体、ならびに、例えば一本鎖抗体、キメラ、二元機能およびヒト化抗体を含む天然に存在しない抗体、ならびにそれらの抗原結合断片を含む。このような天然に存在しない抗体は、固相ペプチド合成を用いて構築され得る、組換え産生され得る、または例えばHuseら、(Science 246:1275−1281(1989))により記載される可変重鎖および可変軽鎖から成るコンビナトリアル・ライブラリーをスクリーニングすることにより得られ得る。これらの方法、ならびに例えば、キメラ、ヒト化、CDR−グラフト化、一本鎖、および二元機能の抗体を作製する他の方法は当業者によく知られている(WinterおよびHarris,Immunol.Today 14:243−246(1993); Wardら、Nature 341:544−546(1989); HarlowおよびLane,前出,1988; Hilyardら、Protein Engineering: A practical approach(IRL Press 1992); Borrabeck,Antibody Engineering,2d ed.(Oxford University Press 1995))。
【0043】
PBPに特異的な抗体は、単離されたPBP、またはその断片(天然源から調製され得るまたは組換え産生され得る)、またはエピトープとして機能できるPBPのペプチド部分などの免疫原を用いて生じ得る。このようなPBPのペプチド部分は、抗原ペプチドがPBPに特異的な抗体を作製するために使用できる場合、機能的抗原断片である。非免疫原性もしくは弱い免疫原性のPBPまたはそれらの一部は、ウシ血清アルブミン(BSA)またはキーホール・リンペット・ヘモシアニン(KLH)などの担体分子にハプテンを結合することにより免疫原性にされ得る。種々の他の担体分子およびハプテンを担体分子に結合する方法は当該分野で周知である(例えば、HarlowおよびLane,前出,1988を参照)。免疫原性PBP断片は、例えばグルタチオンSトランスフェラーゼ(GST)、ポリHis等との融合タンパク質としてペプチド部分を発現することによっても作製できる。ペプチド融合を発現する方法は当業者によく知られている(Ausubelら、Current Protocols in Molecular Biology(Supplement 47),John Wiley & Sons,New York(1999))。
【0044】
本発明はPBPに特異的に結合する抗体とサンプルとを接触させ抗体の特異的結合を検出することによりPBPを検出する方法も提供する。従って、PBPに特異的な抗体は、例えばサンプル中のPBPの存在および/またはレベルを決定するために有用である。PBPに特異的な抗体は、適切な発現ライブラリー、例えばラムダgt11ライブラリー、または他の種類の発現ライブラリーから、免疫学的にPBPに関連するポリペプチドをコードする遺伝子をコードする核酸分子をクローニングするためにも有用である。PBPに特異的な抗体は、サンプルから、例えばPBPを発現する細胞もしくは組織の細胞抽出物から、または組換え核酸分子からPBPを発現する細胞から得られる細胞抽出物から、PBPを実質的に精製するためにも使用できる。
【0045】
PBPを検出するアッセイは、PBPに特異的な抗体または抗原結合断片を用いて、例えば免疫組織化学、免疫蛍光、ELISAアッセイ、放射免疫アッセイ、FACS分析、免疫沈降、免疫ブロット分析、およびフローサイトメトリーを含む(HarlowおよびLane,前出,1988; HarlowおよびLane,Using Antibodies: A Laboratory Manual,Cold Spring Harbor Press(1999))。種々の免疫アッセイは、当該分野で周知であり、要望どおり、当業者により容易に改変できる。例えば、免疫アッセイで使用する抗体は、検出可能部分の取り込みもしくは共役により、またはそれ自体検出できるまたは検出できるよう標識される二次分子に結合することにより、検出できる。
【0046】
PBPまたはPBPに特異的な抗体は当該分野で周知の方法を用いて検出できるように標識され得る(Hermanson,Bioconjugate Techniques,Academic Press,1996; HarlowおよびLane,前出,1988)。例えば、ペプチドまたは抗体は放射標識、酵素、ビオチンまたは蛍光色素を含む種々の検出可能部分で標識され得る。ペプチドまたは抗体を標識する試薬は、ペプチドまたは抗体を含むキットに包含され得るまたは商業的供給源から個別に購入できる。本発明は、PBP、PBPに特異的な抗体、または両方を含むキットを更に提供する。このようなキットは、サンプル中のPBPの発現レベルを測定するために、例えばELISAまたは他の免疫アッセイなどのアッセイを実施するのに適した条件を供する反応カクテルも含み得る、ならびに既知量のPBP、および必要ならばPBPに特異的な抗体に結合できる二次抗体を含む対照サンプルを含み得る。キットが免疫アッセイに使用される場合、抗体に結合するサンプル中のPBPの存在または量を検出する簡単な方法を含み得る。
【0047】
本発明は、本明細書に開示するPBPのアミノ酸配列、例えば配列番号2として示されるアミノ酸配列をコードする単離された核酸分子も提供する。本発明はこのような核酸分子の修飾も提供する。このような核酸分子は示されるアミノ酸配列をコードする縮重ヌクレオチド配列を含む。さらに、本発明は配列番号1、3または4として示されるヌクレオチド配列を含む単離されたPBP核酸分子ならびにそれらの修飾物を提供する。本発明は、本明細書に開示するように、PBPの機能的断片をコードするヌクレオチド配列を有する核酸分子を更に提供する。
【0048】
本発明は、少なくとも中程度にストリンジェントな条件下で、PBP核酸分子、例えば配列番号1、3または4として示される核酸分子とハイブリダイズするPBPヌクレオチド配列の修飾物も提供する。修飾物がPBPヌクレオチド配列と少なくとも60%の同一性を有する場合、PBPヌクレオチド配列の修飾物も提供される。本発明は、配列番号1、3または4として示されるものなどのPBP核酸に対して、少なくとも65%の同一性、少なくとも70%の同一性、少なくとも75%の同一性、少なくとも80%の同一性、少なくとも85%の同一性、少なくとも90%の同一性、少なくとも95%の同一性、少なくとも98%の同一性、または少なくとも99%の同一性を有するPBPのヌクレオチド配列の修飾物も提供する。
【0049】
本明細書で用いられるように、中程度にストリンジェントな条件は、核酸分子が参照配列と実質的な同一性を有する核酸に結合できるハイブリダイゼーションの条件を指す。中程度にストリンジェントな条件は、42℃で、50%ホルムアミド、5Xデンハルト溶液、5XSSPE、0.2%SDSでのフィルターに結合した核酸のハイブリダイゼーション、続いて42℃で0.2XSSPE、0.2%SDSでの洗浄に等価な条件を含む。対照的に、「極めてストリンジェントな条件」は、42℃で、50%ホルムアミド、5Xデンハルト溶液、5XSSPE、0.2%SDSでのフィルターに結合した核酸のハイブリダイゼーション、続いて65℃で0.2XSSPE、0.2%SDSでの洗浄に等価な条件を含む。デンハルト溶液は1%フィコール、1%ポリビニルピロリドン、および1%ウシ血清アルブミン(BSA)を含む。20XSSPE(塩化ナトリウム、リン酸ナトリウム、エチレンジアミンテトラ酢酸(EDTA))は、3M塩化ナトリウム、0.2Mリン酸ナトリウム、および0.025M(EDTA)を含む。他の適切な中程度にストリンジェントなおよび極めてストリンジェントなハイブリダイゼーションの緩衝液および条件は、種々の塩および温度の条件を含み、当業者に周知であり、例えばSambrookら、Molecular Cloning: A Laboratory Manual,2nd ed.,Cold Spring Harbor Press,Plainview,New York(1989);およびAusubelら、前出,1999に記載されている。
【0050】
一般的に、中程度にストリンジェントな条件下で列挙されたの配列にハイブリダイズする核酸分子は、比較される2つの配列の長さにわたって、参照配列と約70%以上の同一性または約80%以上の同一性など、約60%以上の同一性を有する。極めてストリンジェントな条件下で再言及の配列にハイブリダイズする核酸分子は、一般的に、比較される2つの配列の長さにわたって、参照配列と約95%以上の同一性を含む、約90%以上の同一性を有する。任意の2つの核酸配列の同一性は、例えばBLASTコンピュータ整列、またはデフォルト・パラメータもしくは他の望ましいパラメータを用いて、配列を整列する類似の方法に基づいて、当業者により決定できる(例えば、Tatianaら、FEMS Microbiol Lett.174:247−250(1999); Altschulら、J.Mol.Biol.215:403−410(1990); GishおよびStates,(1993) Nature Genet.3:266−272(1993); Maddenら、Meth.Enzymol.266:131−141(1996); Altschulら、Nucleic Acids Res.25:3389−3402(1997))。
【0051】
本発明の単離されたPBP核酸分子は種々の診断および治療の適用に使用できる。例えば、本発明の単離されたPBP核酸分子もしくはその断片は、本明細書に記載するように、プローブとして、またはPBP核酸分子もしくはその断片の増幅に適したプローブもしくはプライマーを得るために、パーキン結合ポリペプチドの組換え発現用の鋳型として、またはパーキンへのPBPの結合に使用されるものと類似して、PBPに結合する細胞分子を同定するために2−ハイブリッドアッセイなどのスクリーニングアッセイに、使用できる。
【0052】
本発明は、本明細書に開示するPBPヌクレオチド配列の少なくとも15個の連続したヌクレオチドを含むオリゴヌクレオチド、またはそのアンチセンス鎖も提供する。参照PBPヌクレオチド配列の少なくとも15個の連続したヌクレオチドを含む本発明のオリゴヌクレオチドは、中程度にストリンジェントなまたは極めてストリンジェントなハイブリダイゼーション条件下で、PBP核酸分子にハイブリダイズできるので、例えば、サンプル中でPBPのDNAもしくはRNAを検出するためまたはそのスプライス変異体を検出するためのプローブとして;配列決定もしくはPCRプライマーとして;本明細書で開示するように、細胞でのRNAの転写を阻害するためもしくは低分子干渉RNA(siRNA)を作製するためのアンチセンス試薬として;またはPBP核酸分子へのハイブリダイゼーションが望ましい分野の当業者に知られる他の適応で、有利に使用できる。
【0053】
本明細書で用いられるように、PBP核酸分子は、nr、dbest、dbsts、gssおよびhtgsのデータベース(これらはデータベースの検索に利用できる)などの公共データベースに寄託されている、例えば発現配列標識(EST)、配列標識部位(STS)およびゲノム断片など、本明細書に開示されるように、配列番号1、3または4などのPBPヌクレオチド配列と同一性を有するヌクレオチド配列から成る予め知られる核酸分子を特異的に排除すると理解される(Altshculら、Nucleic Acids Res.25:3389−3402(1997))。
【0054】
具体的な実施態様において、本発明は、配列番号1のヌクレオチド796〜955と100%の同一性を有する20〜200の連続したヌクレオチドを含むオリゴヌクレオチド、またはそれらのアンチセンス鎖を提供する。オリゴヌクレオチドは、配列番号1のヌクレオチド896〜955と100%の同一性を有する少なくとも25、30、40、50、60、70、80、90、100、125、150、175の連続したヌクレオチド、および200までの連続したヌクレオチドを含み得る。GenBank登録番号のBI041917;CD614598;CD614596;CD614594;CD614592;CD614590;CD614588;CD614576;CD614574;CD614570;BU542453;BF666086;AW374529;BU687172;BM975158;BM910986;BG765308;BG745915;BG745175;BG698661;BG113587;BF837913;BE909317;BE899012;AL135049;AI143229に相当するヌクレオチド配列、ならびに本発明の核酸分子と同一性を有する他の既知の配列は、本発明のオリゴヌクレオチドから特異的に排除される。
【0055】
本発明のPBP核酸分子およびオリゴヌクレオチドは、当該分野で公知の方法により産生または単離できる(例えば、Sambroolら、前出,1989; Ausubelら、前出,1999を参照)。選択される方法は、例えば所望する核酸分子の種類に応じて変わる。当業者は、本明細書に開示するヌクレオチド配列の知見に基づいて、ゲノムDNAとしてのPBP核酸分子を容易に単離できる、または当該分野で公知の方法により、それらから所望のイントロン、エキソンもしくは調節配列を;それらから全長のcDNAもしくは所望の断片として;もしくはそれらから全長のmRNAもしくは所望の断片として、容易に単離できる。
【0056】
本発明のPBP核酸分子を作製する1つの有用な方法は、PCRおよび適切なオリゴヌクレオチドを用いた核酸分子の増幅を含む。PCRまたはRT−PCRのいずれかは、所望するヌクレオチドの境界を有するPBP核酸分子を作製するために用いられ得る。核酸配列への望ましい修飾は1つ以上の付加、欠失または置換をもつ適切なオリゴヌクレオチド・プライマーを選択することによっても導入できる。このような核酸分子は、僅かな単一遺伝子またはmRNAコピーから、対象とする任意の細胞、組織または種から、出発して、指数関数的に増幅され得る。
【0057】
本発明は、PBP核酸分子に特異的にハイブリダイズできる条件下で、PBP核酸分子またはそれらに由来する1つ以上のオリゴヌクレオチドとサンプルとを接触させ、特異的なハイブリダイゼーションを検出することにより、サンプル中のPBP核酸分子を検出する方法を更に提供する。PBP核酸分子は、例えば配列番号1、3もしくは4として示されるPBPヌクレオチド配列、または配列番号1、3もしくは4などの参照PBPヌクレオチド配列の少なくとも15の連続したヌクレオチドを含み且つこの配列に由来するオリゴヌクレオチドであり得る。このようなPBP核酸分子またはそれらに由来するオリゴヌクレオチドは、所望の検出方法の必要に応じて、センスまたはアンチセンスであり得ることが理解される。
【0058】
本発明は、所望する核酸分子の増幅に適した2つ以上のオリゴヌクレオチドとサンプルとを接触させ、核酸分子を増幅し、ならびにこの増幅を検出することにより、サンプル中のPBP核酸分子を検出する方法を更に提供する。サンプル中のPBP核酸を検出する方法は、要望どおり、定性的または定量的のいずれかであり得る。例えば、PBP核酸の存在、存在量、完全性または構造は、要望どおり、アッセイ形式および選択されるプローブまたはプライマーに応じて決定できる。
【0059】
単離されたPBP核酸分子との特異的ハイブリダイゼーションに基づいてPBP核酸を検出するための有用なアッセイは、当該分野でよく知られており、例えば、用いるアッセイ形式に応じて、核酸分子の染色体位置の変化、遺伝子コピー数の変化、およびRNAの存在量を検出するために使用できるインサイツ・ハイブリダイゼーションを含む。他のハイブリダイゼーションアッセイは、例えば、異なるRNAスプライス変異体の存在量および完全性を決定するために用いられ得るノーザン・ブロットおよびRNアーゼ保護アッセイ、ならびにDNAのコピー数および完全性を決定するために用いられ得るサザン・ブロットを含む。ハイブリダイゼーション・プローブは、放射性同位体、蛍光色素、化学発光マーカー、ビオチンなどの任意の適切な検出可能部分、または分析方法により検出でき且つ当該分野で公知の他の検出可能部分を用いて標識できる。
【0060】
2つ以上のオリゴヌクレオチドを用いたPBP核酸の増幅に基づいてサンプル中のPBP核酸を検出する有用なアッセイも、当該分野でよく知られており、例えば定性的または定量的なポリメラーゼ連鎖反応(PCR);逆転写PCR(RT−PCR);一本鎖立体構造多型(SSCP)分析(これは非変性ゲル電気泳動時に電気泳動移動度の変化を生じる一本鎖DNAの二次構造の差異に基づいてDNAの単一点突然変異を容易に同定できる);ならびにタンパク質欠失試験(DNAの突然変異が電気泳動ゲルでタンパク質産物の変化により決定できる)などの共役したPCR、転写および翻訳のアッセイを含む。加えて、増幅されたPBP核酸は突然変異および突然変異多発点を検出するために配列決定でき、このような突然変異を同定するためにサンプルを大規模スクリーニングする特異的なアッセイが開発され得る。
【0061】
本発明は、PBP核酸分子、例えば配列番号1、3もしくは4として示されるPBPヌクレオチド配列または本発明のPBPオリゴヌクレオチドを含むキットを更に提供する。例えば、診断用核酸は、配列番号1、3もしくは4などのPBP核酸分子の任意部分またはそのアンチセンス鎖に由来し得る。本発明のキットは、ゲノムDNA、mRNAまたはcDNAのいずれかにおけるPBPをコードする核酸の有無をアッセイする診断システムとして有用である。適切な診断システムは、少なくとも1つの発明核酸を含み、少なくとも1つのアッセイに十分な量の個別に包装された化学試薬として2つ以上の発明核酸を含み得る。包装された試薬の使用説明書も通常包含される。当業者は、本明細書に記載する発明方法の実施に適した緩衝液および溶液と組み合わせて、発明核酸のプローブおよび/またはプライマーとして有用なオリゴヌクレオチドをキット形態に容易に組み込み得る。
【0062】
本発明のPBP核酸分子は、PBP核酸分子に関連する核酸分子をスクリーニングするために使用できる。PBP核酸分子に関連する核酸分子は、例えば、ゲノム・ライブラリー、cDNAライブラリーまたは発現ライブラリーなどのライブラリーを検出可能試薬を用いてスクリーニングすることにより同定できる。このようなライブラリーは、市販されている、または当該分野で公知の方法を用いて対象とする任意の望ましい組織、細胞もしくは種から作製できる。例えば、cDNAライブラリーまたはゲノム・ライブラリーは検出可能なように標識されたPBP核酸分子を用いたハイブリダイゼーションによりスクリーニングできる。加えて、発現ライブラリーはPBP核酸のコード配列に相当するポリペプチドに対して生じる抗体を用いてスクリーニングできる。本発明のPBP核酸分子を含むライブラリー・クローンは当該分野で公知の方法により他のクローンから単離でき、必要ならば、それらの断片は制限酵素消化およびゲル電気泳動により単離できる。
【0063】
本発明はPBP核酸分子を含むベクターも提供する。本発明のベクターは、PBP核酸分子をサブクローニングし増幅するのに有用でありならびにPBPポリペプチドを組換え発現するのに有用である。本発明のベクターは、例えばバクテリオファージ、バキュロウイルスまたはレトロウイルスなどのウイルス・ベクター;コスミドまたはプラスミド;ならびに特に大きな核酸分子をクローニングするために、細菌人工染色体ベクター(BAC)および酵母人工染色体ベクター(YAC)を含み得る。このようなベクターは市販されており、それらの使用は当該分野で周知である。
【0064】
本発明はPBP核酸分子を含むベクターを含む宿主細胞を更に提供する。組換えPBP分子を発現するために使用できる典型的な宿主細胞は、哺乳動物初代細胞;COS、CHO、HeLa、NIH3T3、HEK293およびPC12細胞などの確立された哺乳動物細胞株;アフリカツメガエルの胚および卵母細胞などの両性動物細胞;ならびに他の脊椎動物細胞を含む。典型的な宿主細胞は、ショウジョウバエなどの昆虫細胞、サッカロミセス・セレヴィシエ、サッカロミセス・ポンベ、またはピチア・パストリスなどの酵母細胞、および大腸菌などの原核細胞も含む。
【0065】
本発明はPBPの発現および/または活性を調節する分子を同定する方法も提供する。これらの分子は、例えばPBPの生物機能を促進または阻害するために治療への応用に使用できる。
【0066】
タンパク質結合ドメインと相互作用する細胞タンパク質を同定するための種々の結合アッセイが当該分野で知られており、例えば酵母2−ハイブリッド・スクリーニングアッセイ(例えば、U.S.patent Nos.5,283,173,5,468,614および5,667,973;Ausubelら、前出,1999; Lubanら、Curr.Opin.Biotechnol.6:59−64(1995))を含む。このアッセイは、本明細書に開示するように、典型的なパーキン結合ポリペプチド(PBP)を同定するために用いた。他の方法は細胞抽出物を用いるアフィニティー・カラム・クロマトグラフィー方法を含む。種々のPBP配列または欠失を含むポリペプチド断片を合成または発現することにより、PBPの結合界面は容易に同定できる。
【0067】
本発明は、PBP調節化合物など、PBPの発現および/または活性を調節する分子を同定する方法も提供する。PBP調節化合物は、PBP核酸分子またはPBPに特異的に結合し、その発現または活性を変える分子である。PBP調節化合物は、ペプチドもしくはポリペプチド、核酸、炭水化物、脂質、またはそれらの任意の組み合わせなど、天然に存在する高分子であり得る。PBP調節化合物は、部分的にもしくは完全にこの高分子の合成の誘導体、類似体もしくは模倣体、またはコンビナトリアル化学方法により部分的にもしくは完全に調製された有機もしくは無機の小分子でもあり得る。典型的なPBP調節化合物は、本明細書に開示するように、阻害剤を含む。単純もしくは複雑な有機分子、金属含有化合物、炭水化物、ペプチド、タンパク質、ペプチド模倣体、糖タンパク質、リポタンパク質、核酸、抗体等の化学分子または生体分子を含む、PBP調節化合物をスクリーニングする際に使用する複数の化合物を作製する方法は、本明細書に記載するように、当該分野でよく知られている。
【0068】
当該分野で公知の種々の低処理能および高処理能のアッセイは、PBP核酸分子またはポリペプチドと候補PBP調節化合物との特異的な結合相互作用を検出するのに適している。例えば、蛍光相関分光法(FCS)およびシンチレーション近似アッセイ(SPA)(reviewed in Major,J.Receptor Signal Transduction Res.15:595−607(1995);およびin Sterrerら、J.Receptor Signal Transduction Res.17:511−520(1997))を含む、直接アッセイおよび競合アッセイの両方が実施できる。特異的な結合相互作用を検出するアッセイは、PBPに特異的なリガンド、例えばELISAアッセイ、FACS分析またはアフィニティー分離で使用される抗体を用いるアフィニティー分離方法を含み得る。
【0069】
PBPの遺伝子発現を調節する化合物を同定するアッセイは、β−ラクタマーゼ、ルシフェラーゼ、緑色蛍光タンパク質またはβ−ガラクトシダーゼなどのタンパク質の発現の変化がPBPの発現を上流調節または下流調節するPBP調節化合物と細胞との接触に応答して検出されるように、PBPプロモーター−レポーター核酸構築物を細胞にまず形質導入する工程を含み得る。このようなアッセイおよびレポーター系は、当該分野で周知であり、例えば、Ausubelら、前出,1999に記載されている。PBPの遺伝子発現を調節する化合物を同定する他のアッセイは、ノーザン・ブロット、RNアーゼ保護アッセイ、およびRT−PCRなどのPBP転写産物のレベルを測定するアッセイを含む。PBPをコードするゲノムDNAからプロモーターおよび/またはエンハンサーを同定する方法は当該分野でよく知られている。レポーター遺伝子構築物は、PBPのプロモーター領域を用いて作製でき、PBP遺伝子のプロモーター活性を増減する化合物についてスクリーニングできる。このような化合物はPBPの発現を改変するためにも使用できる。
【0070】
PBPの発現を調節する化合物を同定するアッセイは、PBP調節化合物と細胞との接触に応答して、PBPの存在量の変化を検出する工程を含み得る。ポリペプチドの発現の変化を検出するアッセイは、例えば免疫ブロッティング、免疫蛍光、免疫組織化学および免疫沈降のアッセイなど、特異的なPBP抗体を用いた免疫アッセイを含む。
【0071】
PBP調節化合物がPBPの活性を阻害または促進するか否かを決定するのに適したアッセイは、PBPの生物活性に基づいて当業者により決定できる。例えば、PBPは、PBP核酸もしくはPBPの発現を改変するまたはPBPの生物活性を改変するPBP調節化合物を同定するために、上述するように、種々の化合物を用いてスクリーニングできる。
【0072】
本発明のポリペプチドおよび核酸分子は種々の診断または治療の適用に使用できる。診断および治療の適用はPBPの生物活性に基づき得る。例えば、PBP核酸分子は、変化したPBP活性を有する個体を処置するための治療方法で使用できる。AR−JPの患者におけるパーキン機能の喪失は、シナプトタグミン1またはシナプトタグミン11またはSLPなどのPBPなど、PBPの機能を変化させ得る。パーキンはPBPをユビキチン化するため、パーキン機能の喪失はPBPの発現を増加させるのに役立ち得る。治療方法において、正常なPBPの発現に比べて増加する変化のPBP活性は、本明細書に開示するように、PBPアンチセンス核酸またはsiRNAを投与することにより低下し得る。
【0073】
PBPの発現を阻害する核酸分子を含むベクターは、インビボまたはエクスビボの方法により個体に導入されPBPの発現を減少させ得る。このような治療方法に有用なベクターは、例えばレトロウイルス、アデノウイルス、レンチウイルス、ヘルペスウイルス、ポックスウイルスのDNAまたは対象とする異種のポリヌクレオチドを発現できる任意のウイルスDNAを含む。例えば、DNAベクター、偽型レトロウイルス・ベクター、アデノ随伴ウイルス、テナガザル(gibbon ape)白血病ベクター、水疱性口内炎ウイルス(VSL)、VL30ベクター、リポソーム媒介性ベクターなど、他のベクターも利用できる。
【0074】
PBP調節化合物は治療方法でも使用できる。例えば、PBP調節化合物は、例えばパーキン機能の喪失に起因する発現の増加など、異常に発現するPBPの発現または活性を変えるために使用できる。例えば、増加したPBPの発現はPBPの発現および/または活性を低下させるPBP調節化合物を用いて、低減できる。
【0075】
本発明は、増大したレベルのパーキン結合ポリペプチドを発現するトランスジェニック動物を作製することにより、パーキンソン病の動物モデルを作製する方法を更に提供する。パーキン結合ポリペプチドは、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択できる。本発明はこのような方法により作製されたパーキンソン病の動物モデルを更に提供する。
【0076】
本発明は、PBPをコードする外因性核酸を発現できるヒト以外のトランスジェニック哺乳動物を更に提供する。パーキンの欠如はPBPのユビキチン化の減少をもたらすため、発現を増加させるので、ヒト以外のトランスジェニック哺乳動物におけるPBPの発現はパーキンソン病の少なくとも幾つかの側面について動物として役立ち得る。PBP導入遺伝子は、本明細書に開示するように(実施例VI〜VIIIを参照)、PBPを発現することが知られる細胞または組織を標的とし得る。外因性核酸とは、宿主を原産としない核酸配列、またはその天然環境以外で、例えば遺伝操作されたDNA構築物の一部として、宿主に存在する核酸配列を指す。天然に存在するレベルのPBPに加えて、本発明のPBPは、例えば周知のノックアウト・トランスジェニックのように、トランスジェニック哺乳動物で、上述の過剰発現、あるいは過少発現し得る。
【0077】
内在性遺伝子を組換えまたは変異のPBP遺伝子で置換することによりPBPの発現または構造の調節を変えるため、トランスジェニック動物において生来の遺伝子座をもつPBP遺伝子の変異型または正常型の相同的組換えの使用も本明細書で意図される。ヒト以外の遺伝子ノックアウト哺乳動物を含む、ヒト以外のトランスジェニック哺乳動物を作製する方法は当業者によく知られている(Capecchiら、Science 244:1288(1989); Zimmerら、Nature 338:150(1989); Shastry,Experentia,51: 1028−1039(1995); Shastry,Mol.Cell.Biochem.,181:163−179(1998);および米国特許第5,616,491号,1997年4月1日公表,同第5,750,826号,1998年5月12日公表,および同第5,981,830号,1999年11月9日を参照)。
【0078】
正常に活性できないよう変異したPBPをコードする核酸を発現でき、従って天然のPBPを発現しないヒト以外のトランスジェニック哺乳動物も提供される。本発明は、PBPをコードするmRNAに相補的なアンチセンスmRNAに転写されるよう配置されて(このアンチセンスmRNAはこのmRNAにハイブリダイズすることによりその翻訳を低減する)、PBPをコードする核酸に相補的なアンチセンス核酸を含むゲノムを有するヒト以外のトランスジェニック哺乳動物も提供する。核酸は、発現が誘導され得るようにまたは特異的な細胞型に限定され得るように、誘導性プロモーターおよび/または組織特異的調節要素を更に含み得る。ヒト以外のトランスジェニック哺乳動物の例はトランスジェニック・マウスである。組織特異性を決定する要素の例はメタロチオネイン・プロモーターおよびL7プロモーターである。
【0079】
PBPの生理学的役割および挙動役割を解明する動物モデル・システムも提供され、PBPの発現が種々の技術を用いて改変されるトランスジェニック動物を創出することにより作製される。このような技術の例は、マイクロインジェクション、レトロウイルス感染または当業者に周知の他の手段により、PBPをコードする核酸の正常型または変異型を、トランスジェニック動物を作製するのに適した受精胚に挿入することを含む(例えば、Hoganら、Manipulating the Mouse Embryo: A Laboratory Manual(Cold Spring Harbor Laboratory,(1986)を参照)。
【0080】
本明細書で論じるように、パーキンはE3ユビキチン・リガーゼとして機能する。パーキンはシナプトタグミン1およびシナプトタグミン11をユビキチン化することが見出された。パーキン遺伝子の突然変異の不活性化は常染色体劣性若年性パーキンソン症候群を惹起する。従って、パーキンの不活性化は、例えばユビキチン化による、パーキン相互作用ポリペプチドのプロセシング化能力に影響を及ぼし得る。パーキンの活性の模倣、即ち、パーキン相互作用ポリペプチドの活性の低下は、パーキンソン病に関連する徴候または症状を改善するために使用できることが可能である。当業者はパーキンソン病に関連する徴候または症状の改善を容易に認識または決定できる。
【0081】
ポリペプチドの活性を低下させる方法は当業者によく知られている。ポリペプチドの活性の低下はポリペプチドの発現レベルの低下およびポリペプチドにより示される生物活性の低下の両方を含むと理解される。
【0082】
ポリペプチドの発現を減少させる方法は、例えばリボザイム、アンチセンス核酸またはRNA干渉の使用を含み得る。RNA干渉は以前に記載されている(Fireら、Nature 391:806−811(1998))。天然で起きるRNA干渉において、開始段階中、入力dsRNAは21〜23のヌクレオチド小干渉RNA(siRNA)に消化される。siRNAは、Hammondら、Nature Rev Gen 2:110−119(2001); Sharp,Genes Dev 15:485−490(2001);ならびに HutvagnerおよびZamore,Curr Opin Genetics & Development 12:225−232(2002)に記載されるように、「ガイドRNA」とも呼ばれる。siRNAは、dsRNAに特異的なリボヌクレアーゼのRNアーゼIIIファミリーに属する酵素が漸次dsRNAを切断する場合に産生される。dsRNAは直接または導入遺伝子もしくはベクターを介して導入され得る。連続切断事象はRNAを19〜21の塩基対二本鎖(siRNA)に分解し、HutvagnerおよびZamore,Curr.Opin.Genetics & Development 12: 225−232(2002); Bernsteinら、Nature 409:363−366(2001)により記載されるように、それぞれが2−ヌクレオチド3′オーバーハングを有する。エフェクター段階において、siRNA二本鎖は、ヌクレアーゼ複合体に結合して、RNA誘導性サイレンシング複合体、即ちRISCとして知られるものを形成する。次いで、活性なRISCは、塩基対の相互作用により相同な転写産物を標的にし、siRNAの3′末端から約12ヌクレオチドのmRNAを切断する(Nykanenら、Cell 107: 309−321(2001))。
【0083】
パーキン相互作用ポリペプチドの活性は阻害剤を用いても低下し得る。阻害剤は、パーキン相互作用ポリペプチドの発現、活性または細胞内シグナル伝達を低減する化合物であり得る。このような阻害剤は、例えば小分子、タンパク質、ペプチド、ペプチド模倣体、リボザイム、核酸分子もしくはオリゴヌクレオチド、オリゴ糖、またはそれらの組み合わせであり得る。このような分子を作製する方法は当業者によく知られている(Huse,U.S.Patent No.5,264,563; Francisら、Curr.Opin.Chem.Biol.2:422−428(1998); Tietzeら、Curr.Biol.,2: 363−371(1998); Sofia,Mol.Divers.3: 75−94(1998); Eichlerら、Med.Res.Rev.15: 481−496(1995); Gordonら、J.Med.Chem.37: 1233−1251(1994); Gordonら、J.Med.Chem.37:1385−1401(1994); Gordonら、Acc.Chem.Res.29: 144−154(1996); WilsonおよびCzarnik,eds.,Combinatorial Chemistry: Synthesis and Application,John Wiley & Sons,New York(1997))。多数の天然化合物および合成化合物を含むライブラリーも市販から入手できる。分子のコンビナトリアル・ライブラリーは、上記で論じたように、周知のコンビナトリアル化学方法を用いて調製できる。阻害剤は、例えば、アンタゴニスト;パーキン相互作用ポリペプチドの活性化を妨げる優性阻害分子;パーキン相互作用ポリペプチドの活性または発現を阻害する抗体、タンパク質、小分子およびオリゴヌクレオチド;リボザイム、アンチセンス核酸分子、およびパーキン相互作用ポリペプチドの発現を妨げるまたは減少させる負の調節転写因子をコードする核酸分子、ならびにこのようなリボザイムおよび核酸分子を含む細胞またはウイルスを含む。当業者は、これらの分子、ならびにパーキン相互作用ポリペプチドの発現、活性またはシグナリングを阻害する他の分子が阻害剤として使用できることを容易に理解するであろう。
【0084】
リボザイムまたはアンチセンス核酸分子などの配列特異的リボヌクレアーゼもパーキン相互作用ポリペプチドの発現を阻害するために使用できる。
配列特異的リボヌクレアーゼは、所定のリボヌクレオチド配列でRNAの切断を触媒する分子を指す。リボザイムは所定のリボヌクレオチド配列でRNAの切断を触媒するRNA分子を指す。ハンマーヘッドおよびヘアピンなどのリボザイムは常套方法により設計され調製できる。標的切断部位に対するハンマーヘッドおよびヘアピンなどのリボザイムの特異性は、リボザイムとそのRNA標的との塩基対合により決定される。リボザイムを設計する方法は、例えばHauswirthおよびLewin,Prog.Retin.Eye Res.19:689−710(2000),ならびにLewinおよびHauswirth,Trends.Mol.Med.7: 221−228(2001)に記載されるように、よく知られている。
【0085】
リボザイムおよびDNA酵素を含む配列特異的リボヌクレアーゼは、上述のように設計され、核酸分子の標準的な合成方法により調製され得る。Keら、Int.J.Oncol.12:1391−1396(1998); Dohertyら、Ann.Rev.Biophys.Biomol.Struct.30:457−475(2001); HauswirthおよびLewin,前出,2000;ならびにLewinおよびHauswirth,前出,2001も参照のこと。配列特異的リボザイムはランダム配列のプールからインビトロ選別によっても同定できる。このような方法は、例えばBartelおよびSzostak,Science 261:1411−1418(1993),Breaker,Chem.Rev.97: 371−390(1997)ならびにSantoroおよびJoyce,Proc.Natl.Acad.Sci.,USA 94:4262−4266(1997)に記載されているように、十分に確立されている。
【0086】
リボザイムがウイルス・ベクターまたは他のベクターを用いて送達されずに患者に投与されるべきである場合、必要ならば、リボザイムは安定性を向上するために修飾され得る。治療用リボザイムに有用な修飾は、この分子の3′末端およびピリミジンの2′部位の遮断を含むが、これらに限定されない。安定化されたリボザイムは、数時間の半減期を有することができ、例えば静脈内または局所の注射を用いて反復投与できる。当業者は、リボザイムがウイルス遺伝子治療ベクターでの発現によっても投与できることを理解している。リボザイムをコードするDNAオリゴヌクレオチドは、RNA polIIまたはRNA polIIIのプロモーターの下流にクローニングでき、必要ならば、tRNAval、U6 snRNAまたはアデノウイルスVA1 RNAなどの遺伝子の転写産物内に組み込める。
【0087】
アンチセンス核酸分子とは、メッセンジャーRNAまたは他の特異的RNA転写産物の分子の全てまたは一部に対して配列の相補的な核酸分子を指す。アンチセンス核酸分子は、例えばDNAまたはRNAであり得る、ならびに天然に存在するヌクレオチド、および合成ヌクレオチドまたは安定性を向上するリン酸バックボーンへの修飾などの他の天然に存在しない修飾物を含み得る。ホスホロチオエートを含むアンチセンス・オリゴヌクレオチドおよび他の修飾オリゴヌクレオチドは、本明細書で用いられるように、アンチセンス核酸分子という用語に包含される。下記に束縛されることなく、本発明に有用なアンチセンス核酸分子はmRNAの翻訳を減らせるまたはmRNAの分解を増やせることにより、標的mRNAの発現を減少させる。
【0088】
アンチセンス方法を用いて発現を減少させるための相同性の必要条件は実験的に決定できる。一般的に、少なくとも約80〜90%の核酸配列の同一性は本発明に有用なアンチセンス核酸分子に存在し、より高い核酸配列の同一性がアンチセンス・オリゴヌクレオチドでしばしば用いられ、これは患者の内因性転写産物と完全に同一であり得る。標的配列は、必要ならば、鎖の1つを置換するためにアンチセンス分子により侵入される二重鎖のらせん状に整列した幹に加えて、核形成が起きる小さな一本鎖の領域を有するよう選択され得る(MirおよびSouthern,Nature Biotech.17:788−792(1999))。アンチセンス核酸分子を選択し調製する方法は、当該分野で周知であり、コンピュータ手法を含む(Patzelら、Nucl.Acids.Res.27:4328−4334(1999);Chengら、Proc.Natl.Acad.Sci.,USA 93:8502−8507(1996);LebedevaおよびStein,Ann.Rev.Pharmacol.Toxicol.41:403−419(2001);JulianoおよびYoo,Curr.Opin.Mol.Ther.2:297−303(2000);ならびにCho−Chung,Pharmacol.Ther.82:437−449(1999))。
【0089】
当業者はパーキン結合ポリペプチドの活性または発現の減少を容易に測定できる。例えば、核酸のプローブまたはプライマーはパーキン相互作用ポリペプチドのmRNAの発現を調べるために使用でき、パーキン相互作用ポリペプチドの抗体はポリペプチドの発現レベルを調べるために使用できる。阻害剤の作用は、パーキン相互作用ポリペプチドの生物活性に及ぼすその影響をアッセイすることにより、容易に測定できる。例えば、シナプトタグミンの活性が測定できる(Sudhof,J.Biol.Chem.277:7629−7632(2002))。これらの方法および他の適切な方法は、当業者により容易に測定でき、パーキン相互作用ポリペプチドの潜在的な阻害剤として化合物の作用を試験するために用いることができる。化合物は、パーキンソン病においてパーキンのユビキチン化活性の低下を補うためにパーキン相互作用ポリペプチドのユビキチン化を増す能力についてもスクリーニングされ得る。
【0090】
本発明は、他の実施態様において、1つ以上の化合物とパーキン結合ポリペプチドとを接触させパーキン結合ポリペプチドの活性を変える化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を提供する。典型的なパーキン結合ポリペプチドは、シナプトタグミンI、シナプトタグミンXI、およびシナプシン様タンパク質を含む。この方法はパーキン結合ポリペプチドの活性を低下させる化合物についてスクリーニングするために用いられ得る。
【0091】
本発明は、パーキン結合ポリペプチドを発現する細胞と1つ以上の化合物とを接触させパーキン結合ポリペプチドの発現を減らす化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を更に提供する。他の実施態様において、本発明はパーキン結合ポリペプチドの発現または活性を低減する分子を投与することによりパーキンソン病を処置する方法を提供する。このような分子は本明細書に開示する方法により同定できる。
【0092】
本発明の様々な実施態様の活性に実質的に影響を及ぼさない修飾も本明細書で供される発明の定義内で提供されることが理解される。従って、下記の実施例は本発明を説明することを意図し限定するものではない。
【実施例】
【0093】
(実施例1)
(酵母によるパーキン相互作用ポリペプチドの同定)
本実施例は酵母2−ハイブリッド系を用いたパーキン相互作用ポリペプチドの同定を記載する。
【0094】
酵母2−ハイブリッド方法はpGBT9−パーキン(1〜465)構築物を用いてヒト胎児脳pGAD10−cDNAライブラリーをスクリーニングするために用いた(FieldsおよびSong,Nature 340:245−246(1989))。酵母2−ハイブリッドのバイトプラスミドpGBT9−パーキン(1〜465)を調製するために、アミノ酸1〜465をコードする全長のパーキンcDNAはpEGFPC1−パーキン(Huynh et al Ann.Neurol.48:737−744(2000))から切り出しpGBT9プラスミド(Clontech; Palo Alto CA)に連結した。
【0095】
パーキンと相互作用するタンパク質を同定するために、GAL4活性化ドメイン・ベクターPGAD10にクローニングされたヒト成人脳cDNAライブラリーの酵母2−ハイブリッド・スクリーニングは、バイトとして、GAL4結合ドメインと融合したパーキン・アミノ酸1〜465をコードするpGBT9−パーキン(1〜465)を用いて実施した(Clontech社のベクターおよびライブラリー)。先に記載されるように(Shibataら、Hum.Mol.Genet.9:1303−1313(2000)およびScolesら、Nat.Genet.18:354−359(1998))、バイトプラスミドは酵母株Y190に共形質転換し、ロイシン、トリプトファン、およびヒスチジンを含まず且つ25mM3−アミノ−1,2,4−トリアゾールおよび2%のグルコースを含む合成培地で増殖した。Y190株は、ヒスチジンを含まない培地で増殖でき且つβ−ガラクトシダーゼをコードするLacZ遺伝子を用いて色選別できるHIS3遺伝子の栄養選別を可能にする。β−ガラクトシダーゼ・レポーターは、37℃で15分間から8時間、Z緩衝液(60mM Na2HPO4、40mM NaH2PO4、10mM KCl、1mM MgSO4、pH 7.0、0.03mM β−メルカプトエタノール、および2.5μM X−gal)で凍結破砕されたコロニーをインキュベーションすることにより、スタンプされたニトロセルロース・フィルターでアッセイされた。陽性クローンはロイシンまたはトリプトファンを含まない合成培地で再画線され、β−ガラクトシダーゼ活性について再試験された後、pGAD10の「獲物」プラスミドが単離された。
【0096】
ヒトのシナプトタグミン11遺伝子をコードする部分配列を含むpGAD10プラスミドは精製された後、pGBT9−パーキン(1〜465)または負の対照プラスミド(pGBT9ベクター、pGBT9−NF2、pGBT9−HRS)を用いて再形質転換され、ならびにβ−ガラクトシダーゼ活性について再試験した。種々のパーキンとシナプトタグミン11タンパク質断片との相互作用強度の半定量的評価を得るために、β−ガラクトシダーゼの液体アッセイは、0.6 mg/mlのo−ニトロフェニルガラクトシドを添加したZ緩衝液および5%クロロホルムで抽出されたY190酵母細胞を2分間から1時間インキュベートすることにより実施した。標準偏差は同型培養の三回の測定から計算した。β−ガラクトシダーゼ活性はミラー単位として表した(ミラー単位=1000x[OD420/(OD600x時間x体積)](PoulletおよびTamanoi Methods Enzymol.255:488−497(1995))。
【0097】
6個の潜在クローンは2x106個の独立したヒト胎児脳pGAD10−cDNAコロニーで同定された。これらのクローンは、個々に単離され、配列決定され、更に酵母2−ハイブリッド・フィルターアッセイにかけられ相互作用を確認した。これらのクローンの内2つは長い読み取り枠を有していたため、精製し配列決定した。ヌクレオチド配列は、2つのクローンの内1つはシナプトタグミン11のC2AおよびC2Bのドメインをコードすることを示した。このクローンはhsyt11ABと呼ばれた(ヒトのシナプトタグミン11、C2AドメインおよびC2Bドメイン)。hsyt11AB断片が真のパーキン相互作用物質か否かを更に決定するために、精製したpGAD10−hsyt11ABはpGBT9−パーキン(1〜465)または2つの無関係なおとりのpGBT9−NF2(神経鞘腫抑制物質をコードする)およびpGBT9−Hrs(肝細胞増殖因子に調節されたキナーゼ基質をコードする)の構築物(Scolesら、前出 2000)、またはpGBT9ベクターを用いて、Y190酵母細胞に共トランスフェクションした。フィルター結合アッセイは、pGAD10−hystXIABクローンがpGBT9−パーキン構築物と陽性の相互作用を示すがpGBT9ベクター対照または無関係のタンパク質とは示さないことを証明した(図1A)。
【0098】
パーキンはsyt11のC2AドメインおよびC2Bドメインと相互作用することが見出された。図1Aは酵母2−ハイブリッド・フィルターアッセイを示す。pGAD10−hsyt11ABおよびpGBT9−パーキンで形質転換された酵母Y190細胞はβ−ガラクトシダーゼ基質と陽性反応を生じた(灰色の斑)一方、PGAD10−hsyt11ABおよびpGBT9ベクター対照で形質転換された酵母細胞は生じなかった(図1A)。pGAD10−hsyt11抗体ならびに神経フィブロミンおよびHrs(pGBT9−NF1、およびpGBT9−Hrs)を発現する2つの他の対照を用いて形質転換された酵母細胞は、β−ガラクトシダーゼ基質と陽性反応を生成しなかった。これらの対照タンパク質は、先に記載される(Scolesら、前出(1998))ように、酵母2−ハイブリッド系で機能的である。これらのフィルター結合アッセイは、pGAD10−hsyt11ABクローンのみがpGBT9−パーキン構築物と陽性の相互作用を示すがpGBT9ベクター対照または無関係のタンパク質とは示さないことを示した(図1A)。
【0099】
酵母2−ハイブリッド液体アッセイによりパーキンの相互作用を確認するために、単一トランスフェクションされた3通りの酵母コロニーは液体培地で増殖しβ−ガラクトシダーゼ活性について試験した。pGAD10−hsyt11ABおよびpGBT9−パーキンで共トランスフェクションされた酵母細胞は、pGAD10−hsyt11ABおよびpGBT9ベクター対照を共発現する酵母細胞(図1Bの「2」)と比べて、約20倍高いβ−ガラクトシダーゼ活性を生じた(図1Bの「1」)。図1Bに示されるそれぞれの棒グラフはn=3を表す。
【0100】
これらの結果は、パーキンがシナプトタグミン11(syt11)のC2AドメインおよびC2Bドメインと相互作用することを証明する。
【0101】
(実施例II)
(シナプトタグミン1およびシナプトタグミン11に対する抗体の作製および特異性)
本実施例はシナプトタグミン1およびシナプトタグミン11の抗体の産生および特徴決定を記載する。
【0102】
抗シナプトタグミン11抗体を作製するために、ウサギ抗syt11抗体はQCPによりsytXIAペプチド(HQQAEKKQKNPPYKF;配列番号9)に対して作製した。sytXIBペプチド(KVRRDKDGPRRESGRG;配列番号10)に対する他の抗体が作製されたが、この抗体はヒトおよびPC12細胞のタンパク質抽出物のウェスタン・ブロットで複数のバンドを認識した。抗sytXIA抗体はセファロース(商標)sytXIAカラムによりアフィニティー精製された。ウサギおよびニワトリの抗パーキンは、先に記載するペプチドParkA(Huynhら、Ann.Neurol.48:737−744(2000))に対して作製された。
【0103】
シナプトタグミン11の3ペプチドに対するウサギの抗体(抗syt11)が作製された。シナプトタグミン1に対するマウスの抗体(抗syt65)は購入した(Stressgen; Victoria,British Columbia,Canada)。β−アクチン(Sigma)およびβ−COP(Sigma)に対するマウスのモノクローナル抗体、ならびにウサギ・ユビキチン抗体(DAKO;Carpinteria CA)は購入した。sytIおよびsytIIは関連するタンパク質であり少なくとも11の他のシナプトタグミンに密接な相同性を有するため、抗syt65および抗syt11の抗体は、これらの抗体を用いる前に他の抗原との交差反応性について試験しパーキン−syt相互作用を調べた。
【0104】
タンパク質抽出のため、細胞はトランスフェクション後の所定時点でCO−IP緩衝液(20mM Hepes、pH 7.2、150mM NaCl、0.5% TritonX100)を用いて抽出した。ユビキチン化アッセイおよび抗体分析のため、細胞または組織は強力な3界面活性剤緩衝液(20mMHepes、pH 7.2、150mM NaCl、1% Triton X−100、0.1% SDS、0.5%デオキシコール酸塩)を用いて抽出した。タンパク質抽出物は卓上型Beckmann Microfuge(Beckman Coulter; Fullerton CA)を用いて30分間の遠心分離により清澄化した。ウェスタン・ブロット分析では、10μlのタンパク質抽出物は15ウェル、4〜20%勾配のミニ・ドデシル硫酸ナトリウム(SDS)−ポリアクリルアミド・ゲルの1ウェルにローディングした。タンパク質は、100Vで2時間解離し、低温室で一晩30VでAmersham社のニトロセルロース・フィルター(Amersham; Piscataway NJ)に転写した。次いで、このフィルターはウェスタン・ブロット装置から取り出し室温で1時間5%の脱脂乳でブロッキングした。次に、ブロッキング溶液は所望濃度の一次抗体を含むブロッキング溶液と取り換えられた。ウェスタン・ブロットはAmersham Chemiluminescent Western blot detection kitで可視化した。
【0105】
シナプトタグミンXIの2ペプチドに対するウサギの抗体(抗sytXIAおよび抗sytXIB)は作製され、シナプトタグミン1に対するマウスの抗体(抗syt65)は購入した。sytIおよびsytXIは関連タンパク質であり他のシナプトタグミンに密接な相同性を有するため、抗syt65および抗sytXIの抗体は、これらの抗体を用いる前に、各抗原との交差反応性について試験しパーキン−sytXI相互作用を調べた。図2Aは免疫ブロッティング(ウェスタン・ブロッティング)を用いたシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。タンパク質抽出物はGFP−syt1、GFP−syt4、GFP−syt11、またはGFPを発現するヒト胎児腎臓(HEK)293T細胞から単離された。GFPまたはGFP−syt1、4および11のプラスミドでトランスフェクションされたHEK293細胞からのタンパク質抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した(図2A)。ウェスタン・ブロットは、抗GFP、抗syt65または抗sytXIAの抗体を用いて個別に検出した。抗syt1および抗syt11の両抗体はそれらの個々のエピトープを特異的に検出したが、GFP−syt4融合タンパク質を検出しなかった。予想どおり、抗syt65および抗syt11の両抗体はそれぞれのGFP−syt1またはGFP−syt11のタンパク質のみを検出した一方で、抗GFP抗体はGFPで標識した全てのタンパク質を検出した。抗syt11抗体は、syt4がsyt11と最も近い相同性を有するにもかかわらず、GFP−syt4を検出しなかった。「nt」として図2Aに示されるレーンは非トランスフェクション細胞であった。分子量マーカーの位置は右側に示す。
【0106】
sytXIA抗体の特異性はPC12細胞から得るタンパク質抽出物のウェスタン・ブロットで更に確認された(図2B)。強力な3界面活性剤の緩衝液におけるPC12細胞のタンパク質抽出物は、抗sytXIA、sytXIAペプチドで前吸収された抗sytXIA、またはsytXIBペプチドで免疫ブロットした。抗sytXIA抗体は64kDaの単独のバンドを認識した(図2B、レーン1)。p64バンドは、抗sytXIA抗体がsytXIAペプチドで前吸収された場合、検出されなかった(図2B、レーン2)。一方、異なるペプチド(sytXIB)との前吸収は抗sytXIA免疫反応性を阻害しなかった(図2B、レーン3)。総合すれば、これらの観察は抗sytXIA抗体の特異性を更に確認する。弱い界面活性剤の緩衝液(0.2%もしくは0.5%のNP40またはTriton−X100)を用いて単離されたタンパク質抽出物のウェスタン・ブロットにより、64kDaおよび110kDaの2つのsytXIA陽性バンドを得た。これはsytXIAがホモ二量体を形成しうることを示唆する。
【0107】
これらの結果は抗syt11抗体の特異性を確認する。
【0108】
(実施例III)
(パーキンは全長のシナプトタグミン1およびシナプトタグミン11と相互作用する)
本実施例は、パーキンが全長のシナプトタグミン1およびシナプトタグミン11と相互作用することを示す共免疫沈降実験を記載する。
【0109】
GFP−hsyt11AB融合タンパク質およびヘマグルチニン−パーキン(HA−パーキン)と共発現する293T細胞の共免疫沈降は、パーキンがhsyt11AB断片と共沈することを見出した。パーキンが全長のシナプトタグミンと相互作用するか否かを決定するために、全長のシナプトタグミン1(syt1)およびsyt11のcDNAはPEGFPベクターにクローニングされた。簡潔に述べれば、ヒトのsyt1およびsyt11の全長cDNAは、全読み枠に及ぶプライマー対を用いてpGAD10発現プラスミド(Clontech)にクローニングされたヒト成人脳cDNAライブラリーからポリメラーゼ連鎖反応(PCR)により得られた。ヒトのシナプトタグミンIをクローニングするために用いたプライマーは、前プライマー:TGGTGAGCGAGAGTCACCATGA(配列番号11);逆プライマー:B1、TTTCCTTTACTTCTTGACG(配列番号12);B2、TGAAGGACTTAGGGGCTCTCT(配列番号13)であった。ヒトのシナプトタグミンXIをクローニングするために用いたプライマーは、前プライマー:GAGGGTTCCCAGAGCTGTCT(配列番号14);逆プライマー:CACATCCCTCCCCAGCTTG(配列番号15)であった。ヒトのシナプトタグミンIおよびシナプトタグミンXIのヒトcDNA配列は、GenBank登録番号のBC058917(図12Aおよび図12B)およびBC039205(図12Cおよび図12D)で表されるように、それぞれ図12Aおよび図12Cに示される。他の発現プラスミドは全て特異的なPCRプライマー対を用いて同様に構築された。突然変異体パーキンのcDNAであるパーキンG289RおよびパーキンC418RはAlexis Brice教授(INSERM U289,Hopital de la Salpetriere,Paris,France)から賜った。
【0110】
細胞のトランスフェクションのため、細胞はトランスフェクションの24時間前にプレートに播種した。翌日、cDNAプラスミドは、polyfect形質転換体試薬(Qiagen)で処理し、製造業者のプロトコルに従ってHEK293細胞にトランスフェクションした。トランスフェクション後の望ましい時点(24時間、48時間および72時間)で、細胞は免疫蛍光標識用に固定されたまたは免疫沈降およびウェスタン・ブロット用に抽出された。トランスフェクションの24時間以上後に試験された細胞については、培地を一度交換した。
【0111】
インビトロ共免疫沈降実験を実施するために、HAで標識したパーキン発現cDNA(pCMV−HA−パーキン、pCMV−HA−欠失パーキン、pCMV−HA−パーキンC418R、pCMV−HA−パーキンG289R)は、個々のGFPで標識した融合タンパク質発現ベクター(pEGFP−hsyt11AB、pEGFP−syt1、pEGFP−syt11、またはpEGFP)を用いて、100mM2皿に60〜80%の密集度で標準培地条件で増殖したHEK293細胞に共トランスフェクションした。対照はpCMV−HA−パーキンでのみトランスフェクションされた細胞または非トランスフェクション細胞であった。ある実験において、β−アクチン抗体(Sigma;St.Louis,MO)およびβ−COP抗体が用いられた。下記の試薬:マウス抗HA、抗HA−ペルオキシダーゼ、抗myc−ペルオキシダーゼ、抗HA−アガロースはRoche Diagnostics社から購入した。48時間後、タンパク質は、0.5%のNP40およびプロテアーゼ阻害剤混合物(Roche Molecular Biochemicals;Indianapolis,IN)を含む界面活性剤緩衝液を用いて、基本的に実施例IIに記載するように抽出した。タンパク質抽出物は抗GFP(Chemicon;Temecula,CA)またはラットの抗HAアガロース基質(Roche)を用いて免疫沈降(ip)した。免疫沈降産物は抗GFP抗体および抗HA共役ペルオキシダーゼを用いて免疫ブロットした。
【0112】
図2CはHA−パーキンとGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体を用いて共免疫沈降(共IP)した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)を用いて検出した。
【0113】
pEGFP−syt1およびpEGFP−syt11のベクターはpCMV−HA−パーキン・プラスミドと共に293T細胞に個別に共トランスフェクションした。pEGFPプラスミドはベクター対照として使用した。24時間後、タンパク質抽出物が得られマウス抗GFP抗体と共沈しGFP融合タンパク質を沈降させた。共IP産物はウサギ抗GFP(図2C、下側パネル)またはラット抗HA共役ペルオキシダーゼ(図2C、上側パネル)を用いて免疫ブロットした。抗HAペルオキシダーゼは、GFP−syt1およびGFP−syt11を含むサンプル中でのみHA−パーキン・バンドを検出したが、GFP対照を含むサンプル中では検出しなかった。これは、HA−パーキンがシナプトタグミン融合タンパク質と特異的に共沈したがGFP標識とは共沈しなかったことを示す。従って、syt1およびsyt11の両タンパク質はパーキンと共沈したが、GFP標識はしなかった。
【0114】
大きなGFP標識はパーキンとシナプトタグミンとの相互作用に影響を及ぼしうる可能性を明らかにするため、syt1 cDNAはpCMV−myc発現プラスミドに転移された。抗HA−アガロースとHA−パーキンおよびGFP−syt1とを共発現する293T細胞からのタンパク質抽出物を共免疫沈降した後、抗myc共役ペルオキシダーゼまたは抗HA共役ペルオキシダーゼを用いて検出し、HA−パーキンもmyc−syt1と沈降することを見出した。これらの観察は、大きなGFP標識がパーキンとシナプトタグミンとの相互作用に影響を及ぼさないことを示している。従って、GFPで標識されたシナプトタグミンはその後の分析に用いた。
【0115】
内因性のパーキンがシナプトタグミンと相互作用するか否かを決定するために、インビボ共免疫沈降実験がPC12細胞で行われた。PC12細胞は、10%CO2、37℃のインキュベーターで、10%加熱不活性化血清、5%ウシ胎児血清、およびペニシリン/ストレプトマイシンを含むDMEMで増殖した。培地は三日毎に交換した。PC12細胞は48時間増殖し、タンパク質は共ip緩衝液(50mM Tris−HCl、pH 7.5、0.2% TritonX−100、150mM NaCl、および10mlの緩衝液当たり1ペレットのプロテアーゼ阻害剤(Roche))で抽出した。タンパク質抽出物は、マウスまたはウサギのIgG共役アガロースで前清澄化(preclear)し、低温室で一晩個々の抗体とインキュベートした。一次抗体は抗マウスまたは抗ウサギのIgG共役アガロースと2時間沈降させた。最終のペレットは共ip緩衝液で7回洗浄し、共沈物はSDS−PAGE緩衝液を用いて二次抗体と共役したアガロースから溶出した。共ip産物は抗パーキン抗体または抗syt抗体のいずれかと免疫ブロットした。
【0116】
図2DはPC12細胞におけるsyt1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞のタンパク質抽出物との共ip。共IP産物はウサギ抗パーキン抗体およびマウス抗syt1抗体を同時に用いて検出した。内因性パーキンは抗syt1免疫沈降物中に検出されたが、マウスIgGを用いた対照の免疫沈降物中には検出されなかった。レーン4はPC12タンパク質溶解物のブロットを示す。
【0117】
マウス抗syt1抗体を用いた共免疫沈降はsyt1および内因性パーキンの両方を沈降させた(図2D)。マウスIgGはsyt1またはパーキンのいずれも沈降させなかった。これは内因性共ipの特異性を示唆する。抗パーキン抗体または抗syt11抗体を用いた共ipはsyt11とパーキンを共沈させなかった。いかなる内因性のパーキン−syt沈降物も見出せなかったことは、おそらく共免疫沈降で使用した緩衝液への内因性syt11の不溶性の結果であっただろう。
【0118】
パーキンが内因性sytXIと相互作用したか否かを決定するために、ヒトの大脳皮質から得るタンパク質抽出物は抗parkA抗体と免疫沈降した。ヒトの脳抽出物の共免疫沈降物については、1.4 gのヒトの大脳皮質サンプルは小片に刻まれ7mlの冷溶解緩衝液(100mM Tris−HCl、pH 7.5、150mM NaCl、1% TritonX−100、0.05% SDS、0.05% デオキシコール酸、および1プロテアーゼ阻害剤ペレット/10ml緩衝液)に再懸濁した。細胞懸濁液はガラス製のホモジナイザーによりホモジナイズした。タンパク質溶解物は1mlのアリコートに等分し低温室で最高速度で遠心分離した。タンパク質抽出物はウサギIgG共役アガロースおよびタンパク質A共役アガロースを用いて4℃で3時間前清澄化し、前清澄化した溶解物はウサギ抗parkA抗体またはマウス抗sytXI抗体を用いて低温室で一晩インキュベートした。一次抗体はタンパク質A共役アガロースと4℃で4時間沈降した。最終ペレットは共ip緩衝液で5回洗浄し、共沈物はSDS−PAGE緩衝液を用いて二次抗体と共役したアガロースから溶出した。共ip産物はニワトリ抗parkA抗体または抗sytXIA抗体のいずれかと免疫ブロットした。
【0119】
ヒトの大脳皮質抽出物の共ip後、沈降物は次いでニワトリ抗parkA抗体または抗sytXIA抗体のいずれかを用いて検出した(図2E、レーン1および2)。抗parkA抗体は抗parkA沈降物に単独のパーキン・バンドを検出した(図2E、レーン1、上側パネル)一方、抗sytXIA抗体はsytXIタンパク質バンドを検出した(図2E、レーン1、下側パネル)。同じタンパク質抽出物が抗sytXI抗体と共免疫沈降した場合、パーキンはsytXIと特異的に共沈した(図2E、レーン3)。対照反応におけるsytXIおよびパーキンの両バンドの欠如(図2E、レーン2および4)は、この細胞でのパーキン−sytXI相互作用の特異性を証明し、内因性パーキンが内因性sytXIと相互作用したことを確認した。
【0120】
これらの結果はパーキンがシナプトタグミン1およびシナプトタグミン11と相互作用することを示す。
【0121】
(実施例IV)
(RING2モチーフはシナプトタグミンの結合に必須である)
これらの実験はシナプトタグミンの結合におけるパーキンのRINGモチーフの役割の特徴決定を記載する。
【0122】
パーキンのどのドメインがシナプトタグミンに結合するかを決定するために、HAエピトープで標識された幾つかの欠失パーキンが構築された(図3A)。欠失パーキンcDNA発現プラスミドはGFP−sytとの共発現に十分な欠失パーキンを発現した(図3C)。それぞれの欠失構築物はGFP−sytXIと共発現した。24時間後、タンパク質抽出物はラット抗HAと共役したアガロースを用いて免疫沈降した後、抗GFP抗体でウェスタン・ブロット検出した。アミノ酸残基204〜293(p1〜203、p294〜385、およびp294〜465)を欠く欠失パーキンは全長のシナプトタグミンXIと相互作用できなかった。これらの欠失パーキンがsytXIと相互作用できないことは、欠失パーキンの発現レベルの低下の結果ではなかった。p1〜203およびp294〜465のパーキンの発現レベルはsytXIと相互作用した構築物よりはるかに高かった(図3C)。p204〜293ドメインの全てまたは一部(p1〜465、p1〜314、p78〜465、p78〜238、p257〜465)を含む欠失パーキンはsytXIと相互作用し、全て異なる結合親和性を有した(図3B)。これらの観察は、RING1ドメインを含むアミノ酸残基204〜293がsytXIとパーキンの相互作用に重要であることを示す。
【0123】
結合アッセイから得るデータは、パーキンに少なくとも2つのsytXI結合部位が存在することを示唆する。第一の結合部位はアミノ酸残基の204〜238に位置する。これは、RING1ドメインを含まないp78〜238のペプチドがsytXIと相互作用したがp1〜203のペプチドが結合しなかったという観察(図3)により支持された。第二の結合部位はアミノ酸残基の257〜293のRING1ドメイン内に位置する。これは、ペプチドp257〜465がsytXIと相互作用したが、ペプチドのp294〜385およびp294〜465が結合しなかった(図3)という観察により支持された。この観察はパーキンにおける病気を惹起するアミノ酸置換の作用により更に支持された。RING1フィンガー・モチーフにミスセンス変異を含むパーキン(パーキンC289G)はパーキンC418Rと比べてsytXIと弱く(2.5倍未満)相互作用した(図3B)。さらに、両変異パーキンはsytXIをユビキチン化できない(図4)が、パーキンC418RはsytXIへの結合能を喪失しない(図3B)。
【0124】
異なる境界を有する他のパーキン欠失変異体について同様な結果が見られた。パーキンの結合部位はRING2フィンガー・モチーフにマッピングされた。同様な結果はsyt1についても観察された。
【0125】
これらの結果はsyt11の結合部位がRINGフィンガー・モチーフに位置することを示す。
【0126】
(実施例V)
(パーキンによるシナプトタグミンのユビキチン化)
本実施例はパーキンによるシナプトタグミンのユビキチン化を記載する。
【0127】
パーキンはE3ユビキチン・リガーゼであり、プロテアソーム複合体またはリソソームによる分解の標的となる特異的基質のユビキチン化に必要な必須酵素である(Shimuraら、前出,2000;Zhangら、前出,2000)。シナプトタグミンがパーキンの基質であるか否かを決定するために、インビトロ・ユビキチン化アッセイは、引用文献により本明細書に組み込まれるZhangら、Proc.Natl.Acad.Sci.USA 97:13354〜13359(2000)により記載される通りに実施した。簡潔に述べれば、HEK293細胞は5μgのpCMV−Myc−ユビキチン、pEGFP−syt1または−syt11、および異なるpCMV−HA−パーキン(野生型、欠失、およびミスセンス)を用いてトランスフェクションした。24(または時折36)時間後、細胞は20μMラクタシスチン(lactacystin)または20μMプロテアソーム阻害剤Iを含む標準培地で4時間インキュベートした。細胞は冷DPBSで洗浄した。ユビキチン化アッセイで、細胞は3界面活性剤緩衝液(20mM Hepes、pH 7.2、150mM NaCl、1% Triton X−100、0.1% SDS、0.5%デオキシコール酸塩)で抽出した。タンパク質はプロテアーゼ阻害剤ペレット(Roche社、10mlの緩衝液当たり1ペレット)および脱ユビキチン化酵素を阻害するための2μM N−エチルアミド(ethylamimide)を含むRIPA緩衝液で抽出した。タンパク質抽出物はマウス抗GFP抗体を用いて免疫沈降し、IP産物は、抗HA、抗myc、およびウサギ抗GFPで検出した。
【0128】
ユビキチン化実験において、HEK293細胞は、GFP標識されたsyt1またはGFP標識されたsyt11のいずれかと、HA標識されたまたは対照のパーキンcDNAプラスミドおよびmyc標識されたユビキチンとで共トランスフェクションされた(図4)。GFPは基質特異性の負の対照として使用し、一方、HA標識した欠失パーキン・タンパク質(図4A〜4Cのp78〜465、p1〜203、p294〜385、p294〜465)(図4D〜図4Fのp1〜p314およびp77〜p465)は野生型パーキンの負の対照として使用した。HA−パーキンならびに対応するmyc−ユビキチンおよびGFPで標識されたタンパク質を過剰発現するHEK293細胞はラクタシスチンで4時間処理し、タンパク質抽出物は抗GFP抗体で免疫沈降した。syt1のユビキチン化アッセイの産物は図4Dにmyc−(上端)、GFP−(中間)、およびHA−標識(下端)に対する抗体を用いて検出した。図4Eに示されるようにsyt11のパーキン・ユビキチン化の検出では、ウェスタン・ブロットが抗myc(上側)および抗GFP(下側)の抗体を用いて検出した。両アッセイにおいて、GFP−syt1およびHA−パーキンを共発現する細胞は、HA−パーキンおよび対照を発現する細胞より多くのユビキチンと共役したsyt1複合体を形成した。HA−パーキンおよびGFPを発現する細胞ならびに他の負の対照におけるユビキチン化産物の欠如に留意せよ。
【0129】
図4に示すように、細胞がプロテアソーム複合体の阻害剤であるラクタシスチンとインキュベートした場合、野生型パーキンおよびGFP−syt11(図4A〜4Cおよび4E)またはGFP−syt1(図4D)を発現する細胞はバックグランド対照を上回るユビキチン−シナプトタグミン共役体の蓄積を示した。野生型パーキンはGFP標識の多ユビキチン化に影響を及ぼさなかった。ペプチドの1つ(p78〜465)はシナプトタグミンに結合できた(図3)が、欠失パーキンはユビキチン化されたシナプトタグミン共役体のレベルにほとんど影響を示さず、欠失パーキンの発現レベルは野生型パーキンと比べて高かった。GFP−sytおよびmyc−ユビキチンのみを共発現する細胞で見出されるユビキチンと共役するsytの存在は、HEK293細胞における内因性パーキンの存在の結果であるらしかった。ユビキチン化sytの様式は、異なる長さのユビキチン鎖を含む、多様な種類の存在を示した。
【0130】
病気に関連する点突然変異がsytをユビキチン化するパーキンの能力に影響するか否かを決定するために、ミスセンス変異パーキンC289GおよびパーキンC418Rについてユビキチン化アッセイも実施した。両変異パーキンは野生型パーキンと比べて検出不能なレベルのユビキチン化sytXIを生じた(図4)。より長い暴露下で、欠失およびミスセンス変異のパーキンでトランスフェクションした細胞は全てGFP−sytXIのみでトランスフェクションした細胞と同程度の弱いレベルのユビキチンと共役したsytXIを生じたが、ユビキチン化sytXIのレベルは野生型パーキンにより生じるレベルより実質的に低かった。このユビキチン化sytXIのバックグランド・レベルは、おそらく内因性パーキンによってまたはHEK293細胞の未同定のE3ユビキチン・リガーゼによって生じたと思われ、HEK293細胞を用いた他の研究者による観察(Renら、J.Neurosci.23:3316−3324(2003))と一致している。共ip(図3)およびユビキチン化(図4)の両実験において、大半の変異パーキンはSDS−PAGEサンプル緩衝液に溶解するペレット画分で検出できた。
【0131】
抗GFP抗体を用いた同じタンパク質サンプルのウェスタン・ブロット分析は、パーキン−sytXIを共発現するサンプルをローディングしたウェルの上端近くに高分子量のGFP−sytXIバンドを検出した(図4)。抗myc抗体もユビキチン化sytXI複合体を濃く検出したため、このバンドは不溶性の同複合体であると思われた。このバンドは対照でかすかに観察された。より小さな分子量のユビキチン化sytXI種は抗GFP抗体により検出できなかった。これらの発見はHEK293細胞におけるα−およびβ−チューブリン(Renら、前出,2003)ならびにシンフィリン−1(Chungら、Nat.Med.7:1144−1150(2001))のユビキチン化実験と一致している。これらの実験において、ユビキチン化基質は各自のタンパク質に対する抗体により検出できなかったが、ユビキチンまたはその標識に対する抗体は各自のパーキンが媒介するユビキチン化基質を濃く検出した。
【0132】
同様な結果はsyt1についても見出された。図4Fに示すように、変異パーキンのC289GおよびC418Rは、syt1およびsyt11のユビキチン化の減少を示した。パーキン変異体およびGFP−syt1またはGFP−syt11を発現する細胞は少量のユビキチンと共役したsytを生じた。変異体C289Gによるsyt11の弱いユビキチン化に留意せよ。従って、パーキンC289G変異体はsyt1のユビキチン化を阻害するが、syt11を弱くユビキチン化する。対照的に、パーキンC418R変異体はsyt1およびsyt11の両ユビキチン化を等しく阻害する。変異体パーキンおよびsytを共発現する細胞のユビキチン化様式は、野生型パーキンおよびsytを発現する細胞より弱かった(図4F、上端パネル)。パーキンおよびsytを発現しラクタシスチンで4時間処理された細胞から得るタンパク質抽出物のウェスタン・ブロットは、野生型パーキンと比べて少量の変異体を検出した(図4F)。変異体パーキンは不溶性ペレットで豊富に見出された(図4F)。
【0133】
パーキンが媒介するユビキチン化基質はプロテアソーム依存性経路による分解を受ける(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000);Imaiら、Cell 105:891−902(2001);Imaiら、Mol.Cell.10:55−67(2002);Renら、J.Neurosci.23:3316−3324(2003);Cortiら、Hum.Mol.Genet.12:1427−1437(2003);Engelenderら、Nat.Genet.22:110−114(1999))ため、HAベクターおよびHA−パーキン1−465でトランスフェクションしたHEK293細胞でのGFP−sytXIの代謝回転を測定するためにパルス・チェイス実験が実施された。パーキンがGFP−sytXIの分解を促進するか否かを決定するため、HEK293細胞はGFP−sytXIおよびHAベクターまたはGFP−sytXIおよびHA−パーキン・プラスミドで共トランスフェクションされた。24時間後、細胞は5%透析FBSを含み且つMetおよびCysのアミノ酸を含まないDMEMで一度洗浄した。細胞は、この培地で30分間インキュベートし、100μCi/mlの35S−Met/Cys(EXPRE35S35S(35S)Protein Labeling Mix,Amersham)を含む同培地で30分間増殖した。その後、細胞は表示する時点で追跡された。タンパク質抽出物はRIPA緩衝液(20mM HEPES,pH 7.5,150mM NaCl,0.1% SDS,0.5% デオキシコール酸塩、1% Triton−X100,1mM EDTA,プロテアーゼ混合ペレット)を用いて単離した。GFP−sytXIは、上述したように、ウサギ抗GFP抗体(CHEMICON;Temecula、CA)と免疫沈降した。
【0134】
パーキンはHA−パーキン1−465でトランスフェクションしたHEK293細胞においてGFP−sytXIの分解を増加させた(図5)。HA−パーキン1−465を発現する細胞で1.5時間の追跡後、新しく合成されたGFP−sytXIの約40%が分解された一方で、HAベクターでトランスフェクションした細胞では等量のGFP−sytXIを分解するのに3時間かかった。
【0135】
これらの結果はパーキンがsyt1およびsyt11の両方をユビキチン化することを証明する。
【0136】
(実施例VI)
(パーキンはシナプトタグミンと共局在化しシナプトタグミンを核周辺複合体に補充する)
本実施例はパーキンおよびシナプトタグミンの細胞位置を記載する。
【0137】
2つのタンパク質が同じ細胞内区画を占めるならば、それらの相互作用は生理的に関連すると思われる。パーキン−syt共局在化を調べるため、免疫蛍光実験が行われた。簡潔に述べれば、COS1細胞培養で、COS1細胞は5%CO2で37℃のインキュベーターにおいて10%FBSおよびペニシリン/ストレプトマイシンを補足したDMEM培地で増殖した。培地は3日毎に交換した。トランスフェクションの一日前に、予め20μg/mlのコラーゲンIVで被覆した1cmのカバースリップに50,000個の細胞が播種された。PC12細胞は実施例IIIに記載するように増殖した。
【0138】
免疫蛍光標識および共焦点レーザー顕微鏡が下記の通りに実施された。細胞は、氷上で20分間4%のパラホルムアルデヒドを含むDPBSで固定し、溶液A(DPBS,3% ヤギ血清、0.05% TritonX−1000)で30分間インキュベートした。細胞は次いで選択され且つ溶液Aで希釈されたマウスまたはウサギの一次抗体と室温で1時間インキュベートした。次に、細胞は、冷DPBSで5回洗浄し、溶液Aで希釈したフルオレセイン・イソチオシアネート(FITC)またはテトラメチルローダミン・イソチオシアネート(TRITC)のいずれかと共役した該当する二次抗体と室温で1時間インキュベートした。次に、細胞は、冷DPBSで5回再洗浄し、退色保護のために80%グリセロールおよび10mM没食子酸ナトリウム中スライドで被覆した。細胞は油浸100Xレンズを通してLeica・TCSSP(真走査型共焦点分光測光)顕微鏡を用いて観察した。TRITC蛍光とFITCまたはGFPで漂白を妨げるため画像は順次得た。
【0139】
パーキン−sytの共局在化を調べるため、PC12細胞はNGFで7日間誘導した。PC12細胞は、50ng/mlのNGFで7日間誘導し、ウサギ・パーキン(赤に染色、図6P)およびマウスsyt1(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6D)に対する抗体を用いて免疫蛍光で共標識した。画像は100x油浸レンズを用いてLeica・TCSSP顕微鏡により得た。積層画像が併合された(図6Cおよび6R)。併合画像の黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび6P〜6Rの挿入物は長い神経突起が生じる同細胞の細胞体に由来する(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。
【0140】
NGF誘導性PC12ニューロンは、ウサギ・パーキンおよびマウスsyt1(図6P〜6R)、またはニワトリ・パーキンおよびウサギsyt11(図6A〜6C)に対する抗体で共標識した。パーキンは核膜周辺の位置(図6R、下の矢印)および神経突起に沿ったボタン(図6Cおよび6R、白矢印)でsyt1およびsyt11の両方と共局在化した。細胞の他の領域に存在するパーキンまたはGFP−sytは共局在しなかった。これらの結果は、内因性のパーキンがPC12細胞で内因性のsyt1およびsyt11と共局在化することを示す。
【0141】
シナプトタグミンの分布およびレベルに及ぼすパーキンの影響を調べるため、HEK293細胞はGFP−sytおよびHA−パーキンの両ベクターで共トランスフェクションした。36時間後、細胞は抗HA抗体で標識した。
【0142】
細胞は、GFP−syt1およびHA−パーキン(図7A〜7C)、GFP−syt1およびHA−ベクター(図7D〜7F)、GFP−syt11およびHA−パーキン(図7G〜7I)、GFP−syt11およびHA−ベクター(図7J〜7L)またはGFP−ベクターおよびHA−パーキン(図7M〜7O)で共トランスフェクションした。トランスフェクションされた細胞は抗HAで標識し、画像は100x油浸レンズを用いてLeica・TCSSPにより得られた。syt1およびsyt11とパーキンとの共局在化の差異に留意せよ。
【0143】
パーキンおよびsytの両方を共発現するHEK293細胞において、HA−パーキンおよびGFP−sytは核周辺膜に近接した凝集体に共局在化した(図7A〜7C、7G〜7I)。これらの細胞において、GFP−sytのレベルは対照と比べてはるかに低かった。対照では、HEK293細胞がGFP−sytおよびHAベクターのみで共トランスフェクションされた場合、GFP−sytは細胞全体にわたって細胞質小胞に分布することが見出された。同様に、HA−パーキンがGFPと発現した場合、細胞全体にわたって拡散して分布した(図7M〜7O)。
【0144】
これらの結果は、パーキンが共トランスフェクションされたHEK293細胞でsytと共局在化し、それらの正常な細胞分布を変えることを示す。
【0145】
(実施例VII)
(SytXIはヒトの黒質ニューロンの細胞体および神経突起に局在する)
黒質ニューロンの死およびレヴィー小体の形成はPDの多くの型の特質であり、レヴィー小体の幾つかの構成タンパク質はPDの遺伝形が変異する。古典的PDの神経変性の病因におけるsytXIおよびパーキンの潜在的な関連性を確立するため、二人の標準的なPD患者および二人の散発性PD患者から得られる黒質切片の免疫組織化学標識がsytXI(抗sytXIA)、パーキンおよびユビキチンに対する抗体を用いて実施された。
【0146】
ヒトの黒質切片の免疫組織化学標識では、Huynhら、(Ann.Neurol.48:737−744(2000))に記載される免疫組織化学標識プロトコルを用いて、ヒトの脳の7μmの切片がウサギ抗sytXIA(10μg/ml)、parkA(5μg/ml)、ユビキチン(1/500)の抗体で染色された。簡潔に述べれば、脳の切片はBiomedia‘s Autozyme溶液(Fisher社)により脱パラフィン処理し(deparafinize)脱マスキングした。切片は、次いで、3%の標準ヤギ血清でブロッキングし低温室で一晩一次抗体とインキュベートした。翌日、切片はElite Vector ABCキット(Vector,San Diego,CA,USA)およびBiomedia社のジアミノベンジデン基質キット(Fisher)を用いて顕色した。ペプチド前吸収では、10μgの抗sytXIA抗体は低温室で一晩1000倍のsytXIAペプチドと前インキュベートした。翌日、前インキュベートした抗体は10分間遠心分離し1mlの染色緩衝液で希釈した。画像は20Xおよび63xの油浸レンズを用いて得られSPOTデジタル・カメラにより保存された。
【0147】
図6D〜6Oに示されるように、正常なヒトの黒質ニューロンはsytXIおよびパーキンに対する抗体により濃く染色された。抗sytXIA抗体は、抗parkA抗体(図6F、6Iおよび6L)と同様に、黒質の細胞体および神経突起伸長物の両方を標識した(図6D、6Gおよび6J)。sytXIAペプチドと前吸収した抗sytXIAは黒質ニューロンを標識しなかった(図6E、6Hおよび6K)ため、sytXI免疫反応性は特異的であった。
【0148】
図6M〜6Oは散発性PDの個体から得られる黒質ニューロンの免疫組織化学標識を表す。抗sytXIA抗体は、抗ユビキチン抗体(図6M)と同様に、細胞内レヴィー小体の核を標識した(LB;図6N、黒矢印)。sytXIのLB染色はユビキチン染色と比べて薄かった。神経網の濃い標識が注目される。sytXIAペプチドと前吸収した抗sytXIA抗体がレヴィー小体または神経網を標識しなかったため、この標識は特異的であった(図6O、黒矢印)。先に報告されたように(Huynhら、Ann.Neurol.48:737−744(2000))、これらの2つの散発性PD脳におけるレヴィー小体は抗parkA抗体に染色しなかった。
【0149】
これらの結果はsytXIがヒトの黒質ニューロンの細胞体および神経突起に局在することを示す。
【0150】
(実施例VIII)
(パーキンと相互作用するポリペプチドの同定および特徴決定)
本実施例はパーキンと相互作用するポリペプチドの同定を記載する。
【0151】
酵母2−ハイブリッド・スクリーニングは実施例Iに記載するように実施した。簡潔に述べれば、パーキンと相互作用するCNSタンパク質を同定するために、酵母2−ハイブリッドアッセイはヒトの脳cDNAライブラリーから1x106個の独立した酵母コロニーについて実施した。スクリーニングはおとりとして全長のパーキンを用いた。酵母フィルターを用いた次のスクリーニングおよび液体アッセイはクローンが陽性であることを確認した。
【0152】
ヌクレオチド配列決定は、酵母2−ハイブリッド・スクリーニングで同定された1つのクローンが新規の小さなシナプシン様タンパク質(SLP)のN末端ドメインをコードすることを示した。ポリペプチドの配列は図8に示す。
【0153】
図9Aに示すように、シナプシン様タンパク質(SLP)とも呼ばれるMP36aおよびMP23aの両型は酵母フィルターアッセイでパーキンと相互作用した。負対照のプラスミドであるpGBT9ベクター、pGBT9−NF2、およびpGBT9−Hrsでは結合は観察されなかった。パーキンとの結合は実施例Iに記載するように液体培養アッセイを用いて確認した。図9Bに示すように、MP36aおよびMP23aは、負対照のpGBT9ベクターより、パーキンの存在下でそれぞれ約20倍および30倍高いβ−ガラクトシダーゼ活性を示した。
【0154】
相互作用を更に試験するため、実施例IIIに記載するように共免疫沈降実験が実施された。共免疫沈降実験では、構築物はGFP融合として作製した。共免疫沈降実験は全長のタンパク質がパーキンと相互作用することを示した。細胞は、図9Cに示すように、個々のプラスミドでトランスフェクションした。HA−パーキンおよびMP36aまたはMP23a GFP融合を発現する細胞から得るタンパク質抽出物の共免疫沈降はHA−アガロースを用いて実施し、個々のGFP融合タンパク質の位置はGFP抗体を用いてウェスタン・ブロッティングにより決定した。図9Cに示すように、MP36aおよびMP23aは共にパーキンと共免疫沈降した。
【0155】
内因性パーキンが野生型SLPと結合しうるか否かを決定するために、共免疫沈降実験がPC12細胞で行われた。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照を用いて免疫沈降した(図9D)。IP産物はニワトリ抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキンのバンドを検出した。このバンドはニワトリIgG免疫沈降物のサンプルでは存在しなかった。抗SLP抗体は抗parkAの免疫沈降物およびPC12タンパク質抽出物において36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性パーキンが野生型SLPと共沈降したことを示す。
【0156】
ヒトの黒質のSLPの発現も試験した(図10)。黒質ち密部切片は、10μg/mlのアフィニティー精製された抗SLP(図10Aおよび10C)、抗SLP+SLPペプチド(図10B)、抗ユビキチン(図10D)の抗体を用いて免疫組織化学的に染色した。一次抗体はVector Elite Vectastain Rabbit ABCキットを用いて検出し、3,3′−ジアミノベンジジン(DAB)で可視化した。全ての切片は同一に処理し染色した。抗SLPおよび抗sytXIの両抗体は黒質ち密部におけるニューロンの神経突起を濃く標識した。SLPペプチドと前吸収した抗SLP抗体は反応しなかった。これは免疫組織化学標識の特異性を示す。細胞体で見られるこげ茶色の染色はドパーミン作動性ニューロンで見出されるニューロメラニン(neuromelanin)である。図10Cおよび図10Dは抗SLP(図10C)および抗ユビキチン(図10D)の抗体を用いたレヴィー小体(LB)の標識を示す。
【0157】
パーキンおよびSLPの共局在化は基本的に実施例VIに記載されるように決定した。細胞はパーキン抗体(緑に染色、図11A)、およびSLPを認識する抗体(赤に染色、図11B)で共標識した。積層画像はパーキンおよびSLPが共局在化することを示す(黄色に染色、図11C)。NGF誘導性PC12ニューロンの共焦点免疫蛍光研究は両SLPがシナプス・ボタンでパーキンと共局在化することを確認した。図11Dおよび図11Eはそれぞれ黒質および大脳皮質の染色を示す。ユビキチン化実験は実施例Vに記載するように実施した。
【0158】
これらの結果は、SLPがインビボでパーキンと共局在化するパーキンと結合するタンパク質であることを示す。これらの結果は、SLPが特発性PDの病理学的特質であるレヴィー小体に局在することも示し、SLPが直接的に(突然変異により)あるいは間接的に(枯渇により)PDの病因に機能し得ることを示す。
【0159】
本出願を通して、種々の刊行物が参照された。これらの刊行物の開示は、本発明に係る技術の状況をより十分に記載するために、本出願の参照文献により本明細書にそのまま組み込まれる。本発明は上記で提供した実施例に関して記載したが、種々の改変が本発明の精神を逸脱することなく為され得ることが理解されるべきである。
【図面の簡単な説明】
【0160】
【図1】図1Aは酵母2−ハイブリッド・フィルターアッセイを示す。pGAD10−hyst11ABおよびpGBT9−パーキンで形質転換した酵母細胞は、β−ガラクトシダーゼ基質と陽性反応した。図1Bは典型的な酵母2−ハイブリッド液体アッセイを示す。酵母はpGAD10−hsyt11ABおよびpGBT9−パーキンのプラスミド(図1Bの「1」)またはpGAD10−hsyt11ABおよびpGBT9ベクターの対照(図1Bの「2」)で形質転換した。
【図2−1】図2Aはシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。緑色蛍光タンパク質(GFP)およびGFP−syt1、GFP−syt4、およびGFP−syt11のプラスミドでトランスフェクションされたHEK293細胞から得られる細胞抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した。「nt」の印が付いたレーンはトランスフェクションされていない細胞をロードした。図2Bはウサギ抗sytXIA抗体で検出されたPC12細胞から得られるタンパク質抽出物のウェスタン・ブロットを示す。レーン1、1μg/mlの抗sytXIA抗体とインキュベーション;レーン2、sytXIAペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション;レーン3、sytXIBペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション。抗sytXIA抗体は64kDaの単独のバンドを検出する。このバンドはsytXIAペプチドとの前インキュベーションで前吸収したがsytXIBペプチドでは前吸収しなかった。図2Cはヘマグルチニン−パーキン(HA−パーキン)とGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体と共免疫沈降した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)で検出した。図2DはPC12細胞におけるシナプトタグミン1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞からのタンパク質抽出物との共ip。共IP産物はウサギ抗パーキンおよびマウス抗syt1抗体で同時に検出した。レーン4はPC12タンパク質の溶解物のブロットを示す。図2Eは内因性のパーキンおよびsytXIAの共免疫沈降を示す。ヒトの大脳皮質から得られるタンパク質抽出物は、ウサギ抗parkA抗体(レーン1)、または対照のウサギIgG(レーン2)を用いて共免疫沈降した。沈降物は抗parkA(上側パネル)または抗sytXIA抗体(下側パネル)を用いて検出した。抗parkA抗体はsytXIを共免疫沈降させたがウサギIgG対照はしなかった。逆共ipを行うために、タンパク質抽出物は同一の希釈でウサギ抗sytXI抗血清(レーン3)または対応する前血清(preserum)(レーン4)と共沈した。ウェスタン・ブロットはニワトリ抗parkAを用いて検出した(レーン3およびレーン4)。このニワトリ抗parkAは抗sytXI抗血清で内因性パーキンのバンドを検出した(レーン3)が、前血清の対照では検出しなかった(レーン4)。
【図2−2】図2Aはシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。緑色蛍光タンパク質(GFP)およびGFP−syt1、GFP−syt4、およびGFP−syt11のプラスミドでトランスフェクションされたHEK293細胞から得られる細胞抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した。「nt」の印が付いたレーンはトランスフェクションされていない細胞をロードした。図2Bはウサギ抗sytXIA抗体で検出されたPC12細胞から得られるタンパク質抽出物のウェスタン・ブロットを示す。レーン1、1μg/mlの抗sytXIA抗体とインキュベーション;レーン2、sytXIAペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション;レーン3、sytXIBペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション。抗sytXIA抗体は64kDaの単独のバンドを検出する。このバンドはsytXIAペプチドとの前インキュベーションで前吸収したがsytXIBペプチドでは前吸収しなかった。図2Cはヘマグルチニン−パーキン(HA−パーキン)とGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体と共免疫沈降した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)で検出した。図2DはPC12細胞におけるシナプトタグミン1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞からのタンパク質抽出物との共ip。共IP産物はウサギ抗パーキンおよびマウス抗syt1抗体で同時に検出した。レーン4はPC12タンパク質の溶解物のブロットを示す。図2Eは内因性のパーキンおよびsytXIAの共免疫沈降を示す。ヒトの大脳皮質から得られるタンパク質抽出物は、ウサギ抗parkA抗体(レーン1)、または対照のウサギIgG(レーン2)を用いて共免疫沈降した。沈降物は抗parkA(上側パネル)または抗sytXIA抗体(下側パネル)を用いて検出した。抗parkA抗体はsytXIを共免疫沈降させたがウサギIgG対照はしなかった。逆共ipを行うために、タンパク質抽出物は同一の希釈でウサギ抗sytXI抗血清(レーン3)または対応する前血清(preserum)(レーン4)と共沈した。ウェスタン・ブロットはニワトリ抗parkAを用いて検出した(レーン3およびレーン4)。このニワトリ抗parkAは抗sytXI抗血清で内因性パーキンのバンドを検出した(レーン3)が、前血清の対照では検出しなかった(レーン4)。
【図3】図3A〜3Cはパーキンのドメインに対するsytXIの結合部位の地図のマッピングを示す。図3Aはパーキンの地図を示す。全長および欠失のパーキンはPCRによって構築しHA−エピトープ標識と枠内にクローニングした。C289GおよびC418Rはそれぞれアミノ酸位置の289および418にミスセンス変異を含むパーキンを示す。図3BはsytXIの結合部位がパーキンのRING1モチーフに位置することを示す。アミノ酸残基204〜293(RING1フィンガー・モチーフを包含する)を欠く欠失パーキンはsytXIに結合できなかった。C289Gミスセンス変異パーキンはC418R変異と比べてsytXIと弱く相互作用した。図3CはHEK293細胞におけるHAで標識したパーキンの発現を示す。全長野生型、ミスセンス変異、または多様な欠失のパーキンを過剰発現するHEK293細胞のウェスタン・ブロットは抗HA−ペルオキシダーゼを用いて検出した。
【図4−1】図4A〜4CはsytXIのユビキチン化アッセイを示す。HA−パーキンまたはMyc−ユビキチンおよびGFPで標識したタンパク質の対照を過剰発現するHEK293細胞はラクタシスチン(lactacystin)で4時間処理し、タンパク質抽出物は抗GFP抗体と免疫沈降した。ユビキチン化アッセイのIP産物はmyc標識に対する抗体(図4A)および抗GFP抗体(図4B)を用いて検出した。HA−パーキンおよびGFPを発現する細胞のユビキチン化産物の欠如、ならびに他の対照におけるGFP−sytXIのユビキチン化の検出不能なレベルに留意せよ。パーキン変異体およびGFP−sytXIを発現する細胞は野生型のパーキンと比べて少ない量のユビキチンと共役したsytXIを産生する。抗GFP抗体は、GFP−sytXIを含む全サンプルにおいてほぼ等量のGFP−sytXI単量体、ならびにGFP−sytXIおよびHA−パーキンの両方と共発現したサンプルを含むウェルの近くに大きなGFP−sytXIバンドを検出した。図4Cは抗HA抗体を用いた同溶解物のウェスタン・ブロットを示す。これは欠失パーキンおよび変異パーキンが野生型パーキンより高いレベルで発現することを示している。図4DはHA−パーキンおよび対応するmyc−ユビキチンおよびGFPで標識したsyt1タンパク質を過剰発現するHEK293細胞のタンパク質抽出物の抗GFP抗体を用いた免疫沈降を示す。この細胞はプロテオソーム複合体の阻害剤であるラクタシスチンで4時間処理した。ユビキチン化アッセイの免疫沈降産物はmyc(上端パネル)、GFP(中間パネル)、およびHA−標識(下端パネル)に対する抗体を用いて検出した。図4Eは、GFP−syt11が使用されたことを除くと、図4Dに類似する。ウェスタン・ブロットは抗myc(上端パネル)および抗GFP(下端パネル)の抗体を用いて検出した。図4Fは、変異パーキン、C289GおよびC418Rを用いた共ipおよびウェスタン・ブロットを示す。
【図4−2】図4A〜4CはsytXIのユビキチン化アッセイを示す。HA−パーキンまたはMyc−ユビキチンおよびGFPで標識したタンパク質の対照を過剰発現するHEK293細胞はラクタシスチン(lactacystin)で4時間処理し、タンパク質抽出物は抗GFP抗体と免疫沈降した。ユビキチン化アッセイのIP産物はmyc標識に対する抗体(図4A)および抗GFP抗体(図4B)を用いて検出した。HA−パーキンおよびGFPを発現する細胞のユビキチン化産物の欠如、ならびに他の対照におけるGFP−sytXIのユビキチン化の検出不能なレベルに留意せよ。パーキン変異体およびGFP−sytXIを発現する細胞は野生型のパーキンと比べて少ない量のユビキチンと共役したsytXIを産生する。抗GFP抗体は、GFP−sytXIを含む全サンプルにおいてほぼ等量のGFP−sytXI単量体、ならびにGFP−sytXIおよびHA−パーキンの両方と共発現したサンプルを含むウェルの近くに大きなGFP−sytXIバンドを検出した。図4Cは抗HA抗体を用いた同溶解物のウェスタン・ブロットを示す。これは欠失パーキンおよび変異パーキンが野生型パーキンより高いレベルで発現することを示している。図4DはHA−パーキンおよび対応するmyc−ユビキチンおよびGFPで標識したsyt1タンパク質を過剰発現するHEK293細胞のタンパク質抽出物の抗GFP抗体を用いた免疫沈降を示す。この細胞はプロテオソーム複合体の阻害剤であるラクタシスチンで4時間処理した。ユビキチン化アッセイの免疫沈降産物はmyc(上端パネル)、GFP(中間パネル)、およびHA−標識(下端パネル)に対する抗体を用いて検出した。図4Eは、GFP−syt11が使用されたことを除くと、図4Dに類似する。ウェスタン・ブロットは抗myc(上端パネル)および抗GFP(下端パネル)の抗体を用いて検出した。図4Fは、変異パーキン、C289GおよびC418Rを用いた共ipおよびウェスタン・ブロットを示す。
【図5】図5Aおよび図5BはパーキンがGFP−sytXIの代謝回転を促進することを示す。HAベクターまたはHA−パーキンのいずれかを発現するHEK293細胞における0時間、1.5時間、3時間、6時間および24時間でのGFP−sytXIの分解のパルス・チェイス分析が実施され、GFP−sytXIは抗GFP抗体と免疫沈降した。免疫沈降物はゲル電気泳動により分析し(図5A)、定量化した(図5B)。データは2つの独立実験の1つから得られる。2つ目の実験はさらに強力なパーキンの効果を及ぼした。
【図6−1】図6A〜6RはトランスフェクションされていないPC12細胞およびヒトの黒質ニューロンにおける内因性のシナプトタグミンIおよびシナプトタグミンXIの細胞内分布を示す。図6A〜6Cおよび図6P〜6Rは7日間50ng/mlngFで誘導されたPC12細胞の免疫蛍光を示す。この細胞はウサギ・パーキン(赤に染色、図6P)およびマウスsytI(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6B)に対する抗体を用いて免疫蛍光で共標識された。100x油浸レンズを用いてLeica・TCSSP顕微鏡により画像が得られた。積層画像は併合した(図6Cおよび6R)。黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび図6P〜6Rの挿入物は、長い神経突起が生じる同じ細胞の細胞体から得られる(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。図6D〜6Lは正常なヒトの黒質切片におけるシナプトタグミンXIおよびパーキンの分布を示す。ヒトの黒質切片は、ウサギ抗sytXIA抗体(D,G,J)、100sytXIAペプチドで前吸収した抗sytXIA抗体(E,H,K)、またはウサギ抗parkA抗体(F,I,L)で標識した。画像は黒質におけるドパーミン作動性ニューロンの細胞体および神経突起を示す。図6M〜6Oは、ウサギ抗ユビキチン(M)、抗sytXIA(N)、および抗sytXIA p sytXIAペプチド(O)で標識された隣接のPD脳切片を示し;黒矢印はレヴィー小体を指す。前吸収したsytXIA抗体によるレヴィー小体の標識の欠如に留意せよ。画像は20xレンズ(図6D〜6F)および63x油浸レンズ(図6G〜6O)を用いて得られた。
【図6−2】図6A〜6RはトランスフェクションされていないPC12細胞およびヒトの黒質ニューロンにおける内因性のシナプトタグミンIおよびシナプトタグミンXIの細胞内分布を示す。図6A〜6Cおよび図6P〜6Rは7日間50ng/mlngFで誘導されたPC12細胞の免疫蛍光を示す。この細胞はウサギ・パーキン(赤に染色、図6P)およびマウスsytI(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6B)に対する抗体を用いて免疫蛍光で共標識された。100x油浸レンズを用いてLeica・TCSSP顕微鏡により画像が得られた。積層画像は併合した(図6Cおよび6R)。黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび図6P〜6Rの挿入物は、長い神経突起が生じる同じ細胞の細胞体から得られる(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。図6D〜6Lは正常なヒトの黒質切片におけるシナプトタグミンXIおよびパーキンの分布を示す。ヒトの黒質切片は、ウサギ抗sytXIA抗体(D,G,J)、100sytXIAペプチドで前吸収した抗sytXIA抗体(E,H,K)、またはウサギ抗parkA抗体(F,I,L)で標識した。画像は黒質におけるドパーミン作動性ニューロンの細胞体および神経突起を示す。図6M〜6Oは、ウサギ抗ユビキチン(M)、抗sytXIA(N)、および抗sytXIA p sytXIAペプチド(O)で標識された隣接のPD脳切片を示し;黒矢印はレヴィー小体を指す。前吸収したsytXIA抗体によるレヴィー小体の標識の欠如に留意せよ。画像は20xレンズ(図6D〜6F)および63x油浸レンズ(図6G〜6O)を用いて得られた。
【図7】図7A〜7OはHEK293細胞の免疫蛍光を示す。この細胞はGFP−syt1およびHA−パーキン(図7A〜7C)、GFP−syt1およびHA−ベクター(図7D〜7F)、GFP−syt11およびHA−パーキン(図7G〜7I)、GFP−syt11およびHA−ベクター(図7J〜7L)またはGFP−ベクターおよびHA−パーキン(図7M〜7O)で共トランスフェクションした。トランスフェクションされた細胞は抗HAで標識し、画像は100x油浸レンズを用いてLeica・TCSSPにより得られた。
【図8】図8は、パーキン結合ポリペプチドであり本明細書中MP23とも呼ばれるヒトのシナプシン様タンパク質(SLP)のヌクレオチド配列(配列番号1)およびアミノ酸配列(配列番号2)を示す。SLP cDNAコード領域はヌクレオチド272〜955である(配列番号3)。図8BはSLP(MP23a)の部分cDNA配列(配列番号4)を示す。小文字はpGAD10ベクターである。SLP配列の第一ヌクレオチドは図8Aに示すヌクレオチド配列のヌクレオチド535に対応する。
【図9−1】図9Aはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド・フィルター結合アッセイを示す。図9Bはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド液体アッセイを示す。図9CはHA−パーキンとMP36aおよびMP23a GFP融合の共免疫沈降を示す。免疫沈降はHA−アガロース基質を用いて実施した。MP36aおよびMP23a GFP融合はGFP抗体で検出し(上側パネル)、パーキンはParkA抗体で検出した(下側パネル)。図9DはPC12細胞におけるSLPと内因性パーキンとの共免疫沈降を示す。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照と免疫沈降した。IP産物はニワトリの抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキン・バンドを検出した。このバンドはニワトリIgGのipサンプルには存在しなかった。抗SLP抗体は抗parkA免疫沈降物およびPC12タンパク質抽出物で36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性のパーキンが野生型SLPと共沈したことを示している。
【図9−2】図9Aはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド・フィルター結合アッセイを示す。図9Bはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド液体アッセイを示す。図9CはHA−パーキンとMP36aおよびMP23a GFP融合の共免疫沈降を示す。免疫沈降はHA−アガロース基質を用いて実施した。MP36aおよびMP23a GFP融合はGFP抗体で検出し(上側パネル)、パーキンはParkA抗体で検出した(下側パネル)。図9DはPC12細胞におけるSLPと内因性パーキンとの共免疫沈降を示す。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照と免疫沈降した。IP産物はニワトリの抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキン・バンドを検出した。このバンドはニワトリIgGのipサンプルには存在しなかった。抗SLP抗体は抗parkA免疫沈降物およびPC12タンパク質抽出物で36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性のパーキンが野生型SLPと共沈したことを示している。
【図10】図10A〜10Dはヒトの黒質におけるSLPおよびユビキチンの発現を示す。黒質ち密部切片は10μg/mlのアフィニティー精製した抗SLP(図10Aおよび10C)、抗SLP+SLPペプチド(図10B)、抗ユビキチン(図10D)の抗体を用いて免疫組織化学的に染色した。一次抗体はVector Elite Vectastain Rabbit ABCキットを用いて検出し、DABで可視化した。切片は全て同一に処理し染色した。抗SLPおよび抗sytXIの両抗体は黒質ち密部のニューロンの神経突起を濃く標識した。SLPペプチドで前吸収した抗SLP抗体は反応しなかった。これは免疫組織化学標識の特異性を示す。細胞体に見られるこげ茶色の染色はドパーミン作動性ニューロンで見出されるニューロメラニンである。図10Cおよび10Dは抗SLP(図10C)および抗ユビキチン(図10D)の抗体を用いたLBの標識を示す。
【図11】図11A〜11Cは細胞の免疫蛍光を示し、パーキンおよびSLPの位置を示す。細胞はパーキン抗体(緑に染色、図11A)およびSLP(赤に染色、図11B)で共染色した。積層画像が図11Cに示され、黄色はパーキンおよびSLPの共局在化を示す。図11Dおよび11Eはそれぞれ黒質および大脳皮質の染色を示す。
【図12】図12Aおよび12BはヒトのシナプトタグミンI(syt1)cDNAのヌクレオチド(配列番号5)およびアミノ酸(配列番号6)の配列をそれぞれ示す(GenBank登録番号BC058917)。図12Cおよび12DはヒトのシナプトタグミンXI(syt11)cDNAのヌクレオチド(配列番号7)およびアミノ酸(配列番号8)の配列をそれぞれ示す(GenBank登録番号BC039205)。
【背景技術】
【0001】
本発明は、一般的に、分子生物学、細胞生物学および医薬の分野に関し、より具体的にはパーキンソン病に関する。
【0002】
パーキンソン病(PD)は筋硬直、震え、および動作緩慢により特徴付けられる主な神経変性病である(DunnettおよびBjorklund,Nature 399:A32−A39(1999))。姿勢障害(postural deficit)、歩行障害、および痴呆などの他の症状もPD患者の部分母集団に観察される。特発性PD症例の大半は散発性であり恐らく環境要因に影響されるが、症例の家族集積性およびPD特性の稀なメンデル遺伝は遺伝の重要性を明示している。
【0003】
パーキンソン症候群は、四つの主要な徴候、即ち、安静時振戦、動作緩慢、自発性および運動の減少、硬直、ならびに姿勢の不安定性により特色付けられる臨床的症候群である。顕著でない徴候は、気分および知性、自律機能ならびに感覚系に関する。発症時の平均年齢は55歳であり、60歳以上の人の約1%がこの病を患う。男性は女性より高い頻度で冒される。
【0004】
パーキンソン症候群は、四つの主要な徴候、即ち、安静時振戦、動作緩慢、自発性および運動の減少、硬直、ならびに姿勢の不安定性により特色付けられる臨床的症候群である。顕著でない徴候は、気分および知性、自律機能ならびに感覚系に関する。発症時の平均年齢は55歳であり、60歳以上の人の約1%がこの病を患う。男性は女性より高い頻度で冒される。
【0005】
安静時振戦および動作緩慢は最も典型的なパーキンソン症候群の徴候であり診断では事実上同義である。動作緩慢は、関連するパーキンソン症候群の症状および徴候、即ち運動および日常生活動作の一般的減速;無表情((hypomimia)または仮面様顔貌);まばたきの頻度の減少による凝視表情;よだれを惹起する嚥下障害;運動低下性および発声不全性の構語障害;単調な発話;小さな字体筆記(小字症);反復運動および同時運動の困難;椅子から立ち上がることが困難およびベッドで寝返りを打つのが困難;小股に引きずり歩行;腕振りおよび他の自動運動の減少;ならびに躊躇およびすくみの開始の大半を占める。突然および頻繁の予測できない運動不能により明示されるすくみは、あらゆるパーキンソン症候群の症状の最も生活に支障を来す1つである。
【0006】
人口が高齢化し60歳以上の人の数が増すにつれて、パーキンソン病を発症する個体数が増えつつあるらしい。パーキンソン病を処置するために治療が利用できるが、これらの治療の多くは望ましくない副作用を有する薬物を使用する。パーキンソン病に関連する衰弱の症状を考えると、更なる治療様式が開発できるようにパーキンソン病の原因を理解することが重要である。
【発明の開示】
【発明が解決しようとする課題】
【0007】
従って、パーキンソン病の発症に関連する遺伝子および遺伝子産物を同定し特徴決定する必要がある。本発明は、この必要性を満たし、さらに、関連する利点を提供する。
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明はパーキン結合ポリペプチドおよびそれをコードする核酸を提供する。本発明はパーキン結合ポリペプチドに特異的な抗体も提供する。本発明は、さらに、パーキン結合ポリペプチドを検出する方法およびパーキン結合ポリペプチドをコードする核酸を検出する方法を提供する。本発明はパーキン結合ポリペプチドを使用する方法を更に提供する。1つの実施態様において、本発明は、パーキン結合ポリペプチドと1つ以上の化合物とを接触させパーキン結合ポリペプチドの活性を変える化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を提供する。
【0009】
(発明の詳細な説明)
本発明はパーキン結合ポリペプチド(PBP)を提供する。本発明はさらにパーキン結合ポリペプチドを使用する方法を提供する。
【0010】
パーキンソン病(PD)において、ドーパミンのレベルは線条体で低下するが、被殻で最も高い。これは主として黒質緻密部でドーパミンを産生するニューロンの変性の結果である(Yamadaら、Brain Res.526:303−307(1990); Damierら、Brain 122:1437−1448(1999); およびNaoiら、Mech.Ageing Dev.111:175−188(1999))。NR4A2(Leら、Nat.Genet.33:85−89(2003))、α−シヌクレイン(synuclein)(Polymeropoulosら、Science 276:2045−2047(1997))およびユビキチンC末端ヒドロキシラーゼL1(UCHL1)(Wintermeyerら、Neuroreport 11:2079−2082(2000))の三遺伝子は常染色体優性PDに関連する。常染色体劣性PDに関連する2つの遺伝子はパーキン(Kitadaら、Nature 392:605−608(1998))およびDJ−1(Bonifatiら、Science 299:256−259(2003))である。パーキン遺伝子の不活性化変異はPARK2常染色体劣性若年性パーキンソン症候群(AR−JP)を惹起する。他のPD形態と同様に、PARK2は黒質のドーパミン作動性ニューロンの欠如により特徴付けられる。しかしながら、PARK2は、黒質ニューロンのレーヴィ小体がAR−JPの大半の症例において存在しないという点で独特である(IshikawaおよびTsuji,Neurology 47:160−166(1996); IshikawaおよびTakahashi,J.Neurol.245:4−9(1998);ならびにMatsumine,J.Neurol.245:10−14(1998))。パーキン遺伝子の変異はAR−JPの形態を惹起するが高齢のPD患者にも見出され、パーキンの変異が若年の発症に限定されないことを証明する(Abbasら、Hum.Mol.Genet.8:567−574(1999))。
【0011】
パーキンをコードする遺伝子の不活性化変異は、常染色体劣性若年性パーキンソン病の幾つかの形態の原因である。パーキンはプロテアソーム依存性タンパク質の分解経路で標的とされるミスフォールドしたタンパク質をユビキチン化するユビキチン・リガーゼである。パーキンの機能の手掛かりはパーキンの一次構造およびパーキン遺伝子の変異部位の局在化により示唆される。パーキンはN末端ドメインのユビキチン様ドメインおよびC末端ドメイン寄りの2つのRINGフィンガー・モチーフから構成される(Kitadaら、前出、1998; およびShimuraら、Nat.Genet.25:302−305(2000))。幾つかの不活性化変異は、RINGフィンガー・ドメインに見出され、これらのドメインが機能的に重要であることを示唆している(Shimuraら、Ann.Neurol.45:668−672(1999))。今日までに、唯1つのミスセンス(Arg42Pro)および三つのフレームシフト変異がユビキチン様ドメインで見出された。部位42のアルギニンは標的タンパク質の結合で機能するようであ。Shimuraら(前出、2000年)は、ユビキチンと共役するH7タンパク質がRINGフィンガー・ドメインに結合し且つパーキンのRINGドメインがヒトのドーパミン作動性SH−Sy5Y神経芽腫細胞のユビキチン化に必要であることを証明した。これらの観察は、ユビキチン様ドメインおよびRINGフィンガー・ドメインの異なる役割を示唆している。ユビキチン様ドメインはパーキンの安定性(Finneyら、J.Biol.Chem.278:16054−16058(2003))ならびに恐らくプロテアソームへのユビキチン化基質の標的化に重要であることが見出された。他方で、RINGフィンガーは基質およびUbcH7(E2)などのユビキチン−リガーゼ活性に必要な他のユビキチン構成要素に結合する。この観察は確認され(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000))、パーキンはセプチンと呼ばれるシナプス小胞に関連するタンパク質ファミリーの一員であるCDCrel−1に結合し且つパーキンはこのタンパク質のユビキチン化および代謝回転を刺激することが更に見出された。併せて、これらのデータはパーキンがE3ユビキチン・リガーゼとして機能するという示唆に至った。
【0012】
活性化E1およびしばしば共役するE2酵素とともに、E3ユビキチン・リガーゼは、26Sプロテアソーム複合体での分解で標的とされる細胞質タンパク質へのユビキチン鎖の共役を触媒し、細胞周期、細胞死、および細胞分化などの重要な細胞過程を調節する。ユビキチン化基質は、基質が多ユビキチン化(polyubiquitinated)(5ユビキチン単位以上の鎖を含む)される場合、プロテアソーム依存性経路(JoazeiroおよびWeissman,Cell 102:549−552(2000))を介して、または、このタンパク質が単ユビキチン化(monoubiquitinated)(5未満の短いユビキチン鎖を含む)される場合、リソソーム分解経路を介してのいずれかで分解できる。単ユビキチン化は、例えばEGF受容体などのある種の細胞表面受容体を内在化(エンドサイトース)させ得る、またはエンドソーム経路のタンパク質選別シグナルとして機能し得る(Helliwellら、J.Cell.Biol.153:649−662(2001); Hicke,Cell 106:527−530(2001); およびHicke,Nat.Rev.Mol.Cell.Biol.2:195−201(2001))、ならびにこれらの単ユビキチン化タンパク質をリソソームに導く。パーキンは、Pael−1受容体(Imaiら、Cell 105:891−902(2001));CDCrel−1(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000));グリコシル化α−シヌクレイン(Shimuraら、Science 293:263−269(2001));シンフィリン(synphilin)−1(Chungら、Nat.Med.7:1144−1150(2001));CHIP(Imaiら、Mol.Cell.10:55−67(2002));サイクリンE(Starpoliら、Neuron 37:735−749(2003));HSP70(Tsaiら、J.Biol.Chem.278:22044−22055(2003));α/β−チューブリン(Renら、J.Neurosci.23:3316−3324(2003));およびアミノアシルt−RNAシンセターゼ複合体のP38サブユニット(Cortiら、Hum.Mol.Genet.12:1427−1437(2003))を含む幾つかのタンパク質と相互作用することが見出された。パーキンが媒介するユビキチン化はプロテアソーム系によるこのタンパク質の分解につながる。Pael−1受容体のパーキン媒介性分解の欠如はPael受容体の蓄積をもたらし、細胞死を惹起する(Imaiら、前出、2001)。
【0013】
本明細書で開示するように、酵母2−ハイブリッド系はパーキン相互作用ポリペプチドを同定するために用いられた。具体的には、パーキンはシナプトタグミン1およびシナプトタグミン11(実施例I〜VIを参照)ならびにSLP(実施例VII)と相互作用することが見出された。パーキンとの相互作用はシナプトタグミンのユビキチン化を惹起しシナプトタグミンの細胞内局在化を変更する。酵母2−ハイブリッド系および共免疫沈降法は、パーキンがそれらのC2AドメインおよびC2Bドメインを通してシナプトタグミン・ファミリーのメンバーと相互作用することを同定するために用いられた。パーキンは多ユビキチン化しシナプトタグミン1およびシナプトタグミン11を分解する。パーキンおよびシナプトタグミンの共発現は、パーキンおよびシナプトタグミンの両方を含む核周辺構造への正常なシナプトタグミンの局在化を変化させる。疾患を惹起するアミノ酸置換を含有するパーキンを含む欠失パーキンおよびミスセンス・パーキンは、シナプトタグミン1およびシナプトタグミン11との相互作用ならびにそれらのユビキチン化を阻害した。変異パーキンはシナプトタグミンの細胞内局在化を変えなかった。パーキン媒介性ユビキチン化はシナプトタグミン11の代謝回転も増強する。シナプトタグミンは小胞の形成およびドッキングに対する重要性においてよく特徴付けられるため、これらの結果はシナプス小胞プールの調節および小胞の放出におけるパーキンおよびシナプトタグミンの役割を示している。従って、シナプトタグミン・ファミリーのメンバーとパーキンとの相互作用は、シナプス前末端での神経伝達物質輸送の制御に関与するタンパク質の調節にパーキンが関与することを示唆する。パーキンは、シナプス機能に関与する多数のタンパク質で見出されるカルシウム感知ドメインであるシナプトタグミンのC2ドメインに結合する。従って、パーキンの欠如は小胞のプール、ドッキングおよび放出を制御する多種のタンパク質に影響を及ぼし、パーキン変異をもつ患者に見られるドーパミン機能の欠損を説明する。
【0014】
本明細書に開示するように、パーキンと相互作用するシナプトタグミン・ファミリーの2つのメンバーが同定され特徴決定された。本明細書に開示する結果は、パーキンが以下の観察に基づいてシナプトタグミン1およびシナプトタグミン11に結合することを確認した。第一に、パーキンはGFPで標識されたシナプトタグミン1またはシナプトタグミン11とのみ共免疫沈降したがGFP標識単独とは共免疫沈降しなかった(実施例IIIおよび図2B)。第二に、内因性パーキンは内因性syt1と相互作用する(実施例IIIおよび図2C)。第三に、野生型のパーキンおよびRINGフィンガー・モチーフを含む欠失パーキンのみがシナプトタグミン1およびシナプトタグミン11に結合した(実施例IVおよび図3C)。第四に、RINGフィンガー・モチーフを欠く欠失パーキンおよびアミノ酸置換のパーキンはシナプトタグミンと相互作用できなかったまたは弱く相互作用した(実施例IVおよび図3C)。第五に、野生型のパーキンのみがシナプトタグミンをユビキチン化し分解に至った一方で、全ての欠失パーキンおよび変異パーキンはシナプトタグミンのユビキチン化の減少または欠如を示した(実施例V、図4および図6)。第六に、内因性パーキンは、NGF誘導性PC12細胞の神経突起に沿ったシナプス・ボタンでおよび核周辺の位置で、シナプトタグミン1およびシナプトタグミン11と共局在化した(実施例VIおよび図5)。最後に、パーキンおよびシナプトタグミンの共発現により、パーキン−シナプトタグミン複合体は核周辺分布の構造に補充された(実施例VIおよび図6)。
【0015】
パーキンは、シンフィリン−1(Chungら、前出(2001))、Pael−1受容体(Imaiら、前出(2001))、CDCrel−1(Zhangら、前出、2000)、およびグリコシル化シヌクレイン(Shimuraら、Science(2001))と相互作用することが見出された。これらのタンパク質の2つ、即ちCDCrel−1およびシンフィリン−1はシナプス小胞に関連するタンパク質である(Ribeiroら、J.Biol.Chem.277:23927−23933(2002); Wakabayashiら、Acta Neuropathol 103−209−214(2002);ならびにBeitesら、Nat.Neurosci.2:434−439(1999))一方、Pael−1受容体は未知の機能をもつ膜貫通タンパク質であり、グリコシル化シヌクレインはレーヴィ小体で見出される希少タンパク質である。CDCrel−1はシンタキシンと相互作用し、野生型CDCrel−1の過剰発現はHIT−T15細胞での分泌を阻害する(Ribeiroら、前出(2002); Wakabayashiら、前出(2002);およびBeitesら、前出(1999))。シンフィリン−1はα−シヌクレインと相互作用しPDの細胞質レヴィー小体の形成を刺激する(Engelenderら、Nat.Ganet.22:110−114(1999))。野生型パーキンの存在はシンフィリン−1が誘導するレーヴィ小体の形成に必須であるらしい。現在、CDCrel−1またはシンフィリン−1がどのようにシナプス前神経伝達の調節に参加するかは知られていない。ならびにCDCrel−1またはシンフィリン−1がドーパミンのシナプス前分泌の調節に関与するか否かも不明である。パーキンがシナプトタグミン・ファミリーのメンバーと相互作用し且つユビキチン化するという発見は、パーキンがシナプス小胞に関連するタンパク質の調節に重要な役割を果たすという仮説を更に支持する。
【0016】
シナプトタグミン1およびシナプトタグミン11(本明細書ではシナプトタグミンIおよびシナプトタグミンXIまたはsytIおよびsytXIとも呼ばれる)は、C2AドメインおよびC2Bドメインに高い相同性を有する(BLAST検索)約50個のカルシウム結合タンパク質から成る大ファミリーに属する。これらのタンパク質は、シナプトタグミンの1〜13、ラフィリン(raphilin)−2a、タンパク質キナーゼC、GTPアーゼ活性化タンパク質(GAP)、ラット/酵母ユビキチン・リガーゼNedd4、およびホスホリパーゼを含む。併せて、これらのタンパク質はカルシウム・センサー(sytおよびタンパク質キナーゼC)からリン酸化(GAP)およびリン脂質分解(ホスホリパーゼC)の範囲に及ぶ細胞シグナル伝達の調節物質として共通の機能を果たす。syt1は、シナプトタグミンの中でsyt2と最も高い相同性を有し、シナプス小胞および分泌顆粒で豊富に発現する(Sudhof,J.Biol.Chem.277:7629−7632(2002))。Syt1およびsyt2は、syt3、syt5−7およびsyt10と同様に、早いシナプス前神経伝達のカルシウム・センサーとして機能する(Fernandez−Chaconら、Nature 410;41−49(2001)およびGeppertら、Cell 79:717−727(1994))。シナプトタグミン11は、対照的に、C2Aドメインのセリン残基によるアスパラギン酸塩の同類置換によりsyt4と類似し、この置換はこのドメインへのCa2+結合の欠損をもたらす(von Poserら、J.Biol.Chem.272:14314−14319(1997))。Syt11の細胞局在は知られていないが、syt4はゴルジ体に局在する(Fukudaら、J.Neurochem.77:730−740(2001)およびBertonら、Eur.J.Neurosci.12:1294−1302(2000))。現在、syt4およびsyt11、syt8、syt9、syt12、およびsyt13の機能は推論的であるが、syt4は早いシナプス前神経伝達の下流調節物質として機能すると考えられる(Wangら、Science 294:1111−1115(2001))。概して、sytファミリーのメンバーはC2ドメインで高い相同性を有し、アミノ酸同一性は30%から50%の範囲に及ぶ。パーキンはsyt11のC2AドメインおよびC2Bドメインに結合するため、パーキンは他のsytとも相互作用するらしい。パーキンがまたsyt1(syt11と最も低いC2ドメインの相同性(30%同一性)を含むタンパク質)と相互作用し且つ調節するという観察は、パーキンがC2AおよびC2Bの配列と関連するドメインを含む広範囲のタンパク質と相互作用しうることを示唆する。
【0017】
シナプス前神経伝達は三つの過程を伴う:1)ドッキング、2)融合、および3)シナプス小胞の再利用である。実験的証拠により、シナプトタグミン1は三過程全てと結び付いた。ドッキングの段階では、シナプトタグミン1はt−SNAREタンパク質、シンタキシンおよびSNAP25と相互作用しシナプス小胞のドッキングを刺激する(Schiavoら、Proc.Natl.Acad.Sci.USA 94:997−1001(1997)およびLiら、Nature 375:594−599(1995))。融合の段階では、syt1は会合したSNARE複合体およびリン脂質と相互作用しシナプス小胞の融合を刺激し安定化する(Levequeら、J.Neurochem.74:367−374(2000); Geronaら、J.Biol.Chem.275:6328−6336(2000);およびDavisら、Neuron.24:363−376(1999))。再利用の段階では、syt1とクラスリン会合タンパク質複合体AP−2との相互作用はシナプス小胞の再利用に重要である(Zhangら、Cell 78:7510760(1994))。これらの相互作用に加えて、機能データは、シナプトタグミン1がシナプス小胞のドッキング(Reistら、J.Neurosci.18:7662−7673(1998))、融合(Geppertら、前出、(1994); Elferinkら、Cell 72:153−159(1993); DiAntonioら、Cell 73:1281−1290(1993); DiAntonioら、Neuron.12:909−920(1993);およびBommertら、Nature 363:163−165(1993))、および再利用(Jorgensenら、Nature 378:196−199(1995))に重要な役割を果たすことも示唆している。
【0018】
更に、syt1ノックアウト・マウスの研究は、syt1が迅速な神経伝達物質のエキソサイトーシス(Fernandez−Chaconら、前出(2001))およびCa++感受性の大きな有芯小胞のエキソサイトーシス(Voetsら、Proc.Natl.Acad.Sci.USA 98:11680−11685(20010))についての主要なCa++センサーであることを示唆する。Syt1の過剰発現は融合孔の開口時間を延長し、一方、syt4の過剰発現は融合孔の開口時間を短縮する(Wangら、前出、(2001))。これはシナプトタグミン1およびシナプトタグミン4が補完機能を有することを示唆している。Sytファミリーのメンバーがドーパミン作動性ニューロンのドーパミン分泌を調節することに関与するか否かは知られていない。変異パーキンによるsytの調節分解の欠如がシナプス形成障害をもたらし(MurpheyおよびGodenschwege,Neuron 36:5(2002))ドーパミン作動性ニューロンでドーパミン分泌の減少に至るか否かも知られていない。しかしながら、野生型のパーキンはsyt1およびsyt11と相互作用し調節するが変異パーキンまたは欠失パーキンはしないという本明細書で開示される観察により、パーキンが重要なE3ユビキチン・リガーゼであり且つシナプス前膜でシナプス小胞の機能を調節することが示される。
【0019】
SytXIは特発性PDの患者から得られる黒質ニューロンのLBの中核で見出される(図6を参照)。この分布は他のパーキン基質であるアミノアシルtRNAシンセターゼ複合体のp38サブユニットおよびシンフィリン−1についても観察される(Cortiら、Hum.Mol.Genet.12:1427−1437(2003); Wakabayashiら、Ann.Neurol.47:521−523(2000); Schlossmacherら、Am.J.Pathol.160:1655−1667(2002))。LBのsytXIの発見はPDにおいてシナプトタグミンの異常なプロセシングが関与する可能性を示唆している。LBがパーキンソン症候群のドパーミン作動性神経細胞死に役割を果たすか否かは推論の域を出ない。パーキンが関連するパーキンソン症候群のLBの欠如(Hayashiら、Mov.Discord.15:884−888(2000); Moriら、Neurology 51:890−892(1998); van de Warrenburgら、Neurology 56:555−557(2001))は、これらの封入がパーキンが関連するパーキンソン症候群におけるドパーミン作動性ニューロン変性の根本原因でないことを示している。
【0020】
パーキンは、三つの機能ドメイン、即ち、ユビキチン様、RING1およびRING2のドメインから成る(Shimuraら、Nat.Genet.25:302−305(2000))。RING2ドメインはユビキチンと共役した酵素への結合に必要であることが見出された(Shimuraら、前出、2000; Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000); Imaiら、J.Biol.Chem.275:35661−35664(2000))。ユビキチン様ドメインはパーキンの安定性に重要である(Finneyら、J.Biol.Chem.278:16054−16058(2003))。しかしながら、RINGフィンガー・モチーフは、パーキンの2つの基質、即ちCDCrel−1(Zhangら、前出、2000)およびPael−R(Imaiら、前出、2000)への結合に必須であることが後に見出された。これらの発見と一致して、本明細書に開示されるように、sytXIはアミノ酸残基の204〜293の領域に結合することが見出された(図3)。この領域はRINGモチーフを含む。更に、ユビキチン様ドメイン(p78−465)のみを欠くパーキン・ペプチドはユビキチン様ドメインを含むパーキンより弱くシナプトタグミンに結合した(図3B)。これらの観察は、ユビキチン様ドメインがシナプトタグミンの結合でRINGフィンガー・モチーフを曝すため全長パーキンの正しい折り畳みに重要であることを示唆する。本発明者らは三つのパーキン・ドメインが異なる機能を果たすことを示唆する。即ち、ユビキチン様ドメインはパーキンの正しい折り畳みおよび安定性に必要であり、RING1フィンガー・モチーフを含むp204−293ドメインはsytXIなどのC2ドメインを含むタンパク質との相互作用に必須である一方、RING2フィンガー・モチーフはE1ユビキチン活性化酵素およびE2ユビキチン共役タンパク質との複合体形成に重要である。
【0021】
パーキンは標的タンパク質のユビキチン化を触媒するE3ユビキチン・リガーゼである(Shimuraら、前出、2000; Zhangら、前出、2000)。多ユビキチン化はプロテアソームによるユビキチン共役基質の分解をもたらす。野生型パーキンは、欠失パーキン、ミスセンス変異パーキン、または負の対照と比べてsytXIの多ユビキチン化を強く触媒する(図4)。パーキン依存性ユビキチン化もsytXIの急速な代謝回転をもたらし(図5)、パーキンがsytXIのレベルを調節するという仮説を更に支持する。欠失パーキンまたはミスセンス変異パーキンを発現する細胞(C289GおよびC418R)は、GFP−sytXIのみを発現する細胞と同量のユビキチン化sytXIを産生した(図4)。これらの観察は、パーキンの欠失またはミスセンス変異がsytXIのユビキチン化を減らすまたはなくすことを示唆する。PARK2 AR−JPにおいて、パーキンの変異はおそらく特異的タンパク質のユビキチン化を減らし、sytXIおよびパーキンにより調節される他のタンパク質の細胞内レベルが増加する。パーキンが調節するタンパク質の細胞内レベルの異常な増加の最終的な効果はAR−JPの病的状態に寄与するであろう。
【0022】
SytXI mRNAは脳で豊富に発現するが、非神経組織では低レベルである(von Poserら、J.Biol.Chem.272:14314−14319(1997))。しかしながら、内因性sytXIタンパク質の特異的な細胞内分布についての情報は知られていない。PC12細胞の外因性sytXIは主にゴルジ・ネットワークに局在した(FukudaおよびMikoshiba,Biochem.J.354:249−257(2001))。トランスフェクションされていないNGF誘導性PC12細胞で、内因性のパーキンおよびsytXIはNGF誘導過程において核周辺分布および有芯小胞に共局在化することが見出された(図6)。内因性パーキンの分布様式は先の観察(Huynhら、Ann.Neurol.48:737−744(2000))と類似した。SytXIの分布はsytXIと48%の同一性を有するタンパク質であるsytIVの細胞内分布にも類似した。PC12細胞において、sytIVは主にゴルジおよび未熟小胞に局在する(Bertonら、Eur.J.Neurosci.12:1294−1302(2000); Ibataら、J.Neurochem.74:518−526(2000); Fukudaら、J.Neurochem.77:730−740(2003); Fukudaら、J.Biol.Chem.278:3220−3226(2003))。PC12細胞がNGFで処理される場合、sytIVタンパク質は成熟な有芯小胞に再分布する(Fukudaら、J.Biol.Chem.278:3220−3226(2003))。有芯小胞は、神経ペプチドまたは生体アミンを運ぶ分泌顆粒であり、カルシウム・イオンの刺激下でそれらの内容物を放出する。従って、sytXIおよびパーキンの両方が有芯小胞に共局在するという観察は、両タンパク質がおそらくカルシウム依存性エキソサイトーシスで役割を果たすことを示唆している。この仮説は、パーキンおよびsytXIの両方がヒトの黒質の神経突起およびニューロンの細胞体で類似する分布様式を有するという観察(図6)により更に支持される。
【0023】
AR−JP患者のパーキン機能の喪失は、シナプトタグミンXIの機能を変化させ、ドーパミンの放出の変化をもたらすことが予想される。これはジストニアおよびパーキンソン症候群の症状を順に惹起する。小胞機能の変化は、放出または再利用の段階であれば、細胞質ドーパミンの増加を惹起し、酸化的損傷の増加、続いて細胞死をもたらしうる。これはパーキン変異の患者に見られる神経変性を説明する。
【0024】
本明細書で開示される結果は、パーキンの喪失がドーパミンの放出を変化させ、ジストニアおよびパーキンソン症候群の初期症状をもたらしうることを示している。放出または再利用のいずれかの段階での小胞機能の変化は、細胞質ドーパミンを増加させ、酸化的損傷の増加、続いて細胞死をもたらしうる。
【0025】
本発明はシナプトタグミン1およびシナプトタグミン11およびSLPを含む典型的なパーキン結合ポリペプチドを提供する。本発明は、本明細書に開示するように、パーキン結合ポリペプチドを同定する方法も提供する。本発明は更にパーキン結合ポリペプチドを使用する方法を提供する。
【0026】
本発明はパーキン結合ポリペプチド(PBP)をコードする単離されたポリペプチドを提供する。具体的な実施態様において、本発明は配列番号2として示されるアミノ酸配列を有する単離されたポリペプチドを提供する。本発明はPBPの機能断片も提供する。
【0027】
本明細書で用いられるように、「機能断片」という用語は、パーキン結合ポリペプチド(PBP)に関して用いる場合、パーキン結合ポリペプチドの活性の一部または全てを保持するパーキン結合ポリペプチドの部分を指すことが意図される。典型的なPBPの機能断片はPBPのパーキン結合断片を含む。他の典型的なPBPの機能断片はPBPに特異的な抗体に特異的に結合する機能断片である。PBPの他の機能断片は抗原断片として機能するエピトープであるペプチド断片を含み、これは個々のPBPに特異的な抗体を作製するために使用できる。
【0028】
本明細書で用いられるように、「ポリペプチド」という用語は、パーキン結合ポリペプチド(PBP)に関して用いる場合、全長以下のパーキン結合ポリペプチドを含む、10以上のアミノ酸のペプチドまたはポリペプチドを指す。本明細書で用いられるように、「ペプチド断片」は2つ以上のアミノ酸のペプチドまたはポリペプチドを指す。パーキン結合ポリペプチドの「修飾」は、修飾がPBPの機能を保持する限り、PBPのアミノ酸配列の同義置換を含む。コードされるアミノ酸の同義置換は、例えば下記の群内に属するアミノ酸を含む。1)非極性アミノ酸(Gly,Ala,Val,Leu,およびIle);2)極性中性アミノ酸(Cys,Met,Ser,Thr,Asn,およびGln);3)極性酸性アミノ酸(AspおよびGlu);4)極性塩基性アミノ酸(Lys,ArgおよびHis);および5)芳香族アミノ酸(Phe,Typ,Tyr,およびHis)。他の微修飾は、ポリペプチドが本明細書に記載される機能の一部または全てを保持する限り、PBP内に含まれる。
【0029】
ポリペプチドの修飾は、修飾がPBPの機能を保持する限り、その誘導体、類似体および機能模倣体も含み得る。例えば、誘導体は、アルキル化、アシル化、カルバミル化、ヨード化などのポリペプチドの化学修飾、またはポリペプチドを誘導体化する任意の修飾を含み得る。類似体は、例えばヒドロキシプロリンまたはカルボキシグルタメートの修飾アミノ酸を含み得る、ならびにペプチド結合により連結していないアミノ酸を含み得る。模倣体はポリペプチドの機能を模倣する化学部分を含む化学物質を包含する。例えば、ポリペプチドが機能活性を有する2つの荷電化学部分を含む場合、模倣体は、この荷電化学機能が三次元空間で維持されるように、2つの荷電化学部分を空間的定位および制約構造に置く。従って、PBPの機能を与える官能基を配置する模倣体はPBPの誘導体の意味内に含まれる。
【0030】
本明細書で用いられるように、「実質的に」または「実質的に同じ」という用語は、ヌクレオチドまたはアミノ酸の配列に関して用いられる場合、ヌクレオチドまたはアミノ酸の配列が参照配列と比べて相当な程度、量または範囲の配列同一性を示すことを意味すると意図する。実質的に同じアミノ酸配列は参照ポリペプチドに特徴的な機能活性および/または生物活性を保持する。
【0031】
本明細書で用いられるように、「核酸」という用語は、デオキシリボ核酸(DNA)またはリボ核酸(RNA)などのポリヌクレオチドを意味し、一本鎖および二本鎖の両核酸ならびにオリゴヌクレオチドを包含する。本発明に有用な核酸は、ゲノムDNA、cDNAおよびmRNAを含み、センス鎖、アンチセンス鎖、または両方を表し得る。本発明のゲノム配列は、PBP遺伝子の発現を調節するプロモーターおよびエンハンサーなどの調節領域ならびにPBPをコードするエキソンを除いたイントロンを含むが、PBPをコードしない近接遺伝子を含まない。典型的なPBP核酸は、配列番号1、配列番号3および配列番号4として示されるヌクレオチド配列、またはそれらの断片を含む。PBP核酸分子に関して用いられる「単離された」という用語は、この分子が生来付随する構成要素からまたは他の方法で人為的に修飾された構成要素から実質的に取り出されるまたは分離されることを意味することにより、天然で存在するようなPBP核酸分子を除くことを意図する。
【0032】
本明細書で用いられるように、「オリゴヌクレオチド」という用語は、参照ヌクレオチド配列から少なくとも15個の連続したヌクレオチドを含む核酸分子を指し、少なくとも16、17、18、19、20または少なくとも25個の連続したヌクレオチドを含み得る、ならびに参照ヌクレオチド配列から少なくとも30、40、50、60、70、80、90、100、125、150、175、200、225、250、275、300、325、350までの連続したヌクレオチドをしばしば含む。参照ヌクレオチド配列はセンス鎖またはアンチセンス鎖であり得る。このオリゴヌクレオチドは化学合成され得るまたは組換え発現し得る。
【0033】
本明細書で用いられるように、核酸の「修飾」は参照配列に対して1つまたは幾つかのヌクレオチドの付加、欠失または置換を含み得る。核酸の修飾は、遺伝コードの縮重によりコードするアミノ酸配列を変更しない置換を含み得る。このような修飾は意図的に為される変異または核酸の複製中に突然変異として起きる変異に対応し得る。このようなものとして、核酸の修飾は実質的に同じ配列を含み、これは本明細書で開示されるPBPヌクレオチド配列などの親核酸分子に類縁すると認識できる。実質的に同じヌクレオチド配列は中程度にストリンジェントな条件下または極めてストリンジェントな条件下で参照ヌクレオチド配列とハイブリダイズできる。
【0034】
本明細書で開示するPBP核酸配列の典型的な修飾は、ヒト、霊長類、ラット、ウサギ、ウシ、ブタ、ヒツジ、イヌ、ネコまたは他の動物種などの他の種のホモログに対応する配列を含む。ヒト以外の種の対応するPBPの配列は、ポリメラーゼ連鎖反応(PCR)によるまたはゲノム・ライブラリー、cDNAライブラリーもしくは発現ライブラリーのスクリーニングによるなどの当該分野で公知の方法により決定できる。PBP核酸分子の他の典型的な修飾はPBPヌクレオチド配列のスプライス変異型に相当し得る。さらに、ヌクレオチド配列の修飾は、例えば塩基、糖もしくはリン酸塩部分への修飾を有する、または修飾されたホスホジエステル結合を有する1つ以上の非野生型ヌクレオチドを含み得る。このような修飾は核酸分子の安定性を増すのに有利であり得る。
【0035】
更に、ヌクレオチド配列の修飾は、例えば放射性標識、蛍光色素、強磁性体、発光標識などの検出可能な部分またはビオチンなどの検出可能な結合剤を含み得る。このような修飾はPBP核酸分子の検出が望まれる適用において好都合であり得る。
【0036】
本明細書で用いられるように、「ベクター」とはポリヌクレオチドを含む組換えのDNAもしくはRNAのプラスミドまたはウイルスを指す。ベクターは、発現エレメントがポリヌクレオチドの発現を制御するようポリヌクレオチドに機能的に連結された発現エレメントを含み得る。「発現エレメント」とは、ポリヌクレオチドの複製、転写、スプライシング、翻訳、または分解を含む、ポリヌクレオチドの機能調節に寄与する分子の相互作用に関与するヌクレオチド配列である。遺伝子の転写を制御する発現エレメントは、プロモーター、転写開始部位、または転写速度を増すDNA配列のエンハンサーであり得る。
【0037】
本明細書で用いられるように、「サンプル」という用語は、パーキンと結合するタンパク質、核酸またはポリペプチドを含むまたは潜在的に含む生体液、細胞、組織、器官またはそれらの一部を意味することが意図される。この用語は、個体に存在するサンプルならびに個体から得られるサンプルまたは個体に由来するサンプルを含む。例えば、サンプルは、生検により得られる検体の組織切片、または組織培養に置かれたまたは適応された細胞であり得る。サンプルは更に抽出物の細胞下画分または精製していないもしくは実質的に純粋な核酸またはタンパク質の調製物であり得る。サンプルは、例えば、縮重オリゴヌクレオチドを合成することにより、化学合成もされ得る。
【0038】
本明細書で用いられるように、「特異的にハイブリダイズする」という用語は、本明細書に記載するように、少なくとも中程度にストリンジェントな条件下またはよりストリンジェントな条件下で、参照PBP核酸分子にハイブリダイズする核酸分子の能力を指し、参照PBP核酸分子でない核酸分子(例えばアクチンcDNAなどの負の対照)と同条件下でハイブリダイズしない。
【0039】
本発明は単離されたパーキン結合ポリペプチド(PBP)またはその機能断片を提供する。典型的なパーキン結合ポリペプチドは本明細書に開示するシナプシン様タンパク質を含む(実施例VIIIを参照)。本発明の単離されたPBPおよびペプチドは、生化学的方法、組換え方法および合成方法を含む当該分野で既知の方法により調製できる。例えば、PBPは、PBPをコードする該当の転写産物またはPBPを発現する細胞または組織の起源から常套の生化学的方法により精製できる。本明細書に開示する方法は、どの細胞および組織、ならびにどの細胞下画分が適切な出発物質であるかを決めるために適応できる。生化学的精製は、例えば適切な組織または細胞の可溶化、所望の細胞下画分の単離、サイズ・クロマトグラフィー、イオン交換クロマトグラフィー、疎水クロマトグラフィーまたはアフィニティー・クロマトグラフィー、電気泳動、および免疫アフィニティーの手法などの段階を含み得る。本発明のポリペプチドの生化学的精製の方法および条件は当業者により選択でき、精製は、例えば、免疫アッセイまたは機能アッセイにより観察される。
【0040】
本発明はパーキン結合ポリペプチド(PBP)に特異的に結合する抗体も提供する。具体的な実施態様において、本発明は配列番号2として示されるアミノ配列を有するPBPに特異的に結合する抗体を提供する。本明細書で用いられるように、「抗体」という用語は、その最も広い意味で用いられ、ポリクローナル抗体およびモノクローナル抗体ならびにこの抗体の抗原結合断片を含む。PBPに特異的な本発明の抗体に関して、「抗原」という用語は野生型PBPもしくは合成PBPまたはそれらの断片を意味する。
【0041】
PBPに特異的な抗体、またはこの抗体の抗原結合断片は少なくとも約1x105 M−1のPBPまたはそのペプチド部分に対して特異的な結合活性を有することにより特徴付けられる。従って、PBPに特異的な抗体(PBPに対して特異的な結合活性を保持する)のFab、F(ab′)2、FdおよびFvの断片は抗体の定義内に含まれる。PBPの特異的結合活性は、例えばPBP対PBPでない対照ポリペプチドに対する抗体の結合活性を比較することにより、当業者により容易に測定できる。当業者は個々のPBPに特異的な結合活性を有する抗体の意味を容易に理解するであろう。抗体はポリクローナル抗体またはモノクローナル抗体であり得る。ポリクローナル抗体またはモノクローナル抗体を調製する方法は当業者によく知られている(例えば、HarlowおよびLane,Antibodies: A Laboratory Manual,Cold Spring Harbor Laboratory Press(1988))。ポリクローナル抗体を使用する場合、ポリクローナル血清は、バックグラウンド結合が少なく且つ抗原に特異的な抗体の割合が高い単一特異的な抗体を作製するために、抗原を用いてアフィニティー精製され得る。
【0042】
加えて、本明細書で用いられる「抗体」という用語は、天然に存在する抗体、ならびに、例えば一本鎖抗体、キメラ、二元機能およびヒト化抗体を含む天然に存在しない抗体、ならびにそれらの抗原結合断片を含む。このような天然に存在しない抗体は、固相ペプチド合成を用いて構築され得る、組換え産生され得る、または例えばHuseら、(Science 246:1275−1281(1989))により記載される可変重鎖および可変軽鎖から成るコンビナトリアル・ライブラリーをスクリーニングすることにより得られ得る。これらの方法、ならびに例えば、キメラ、ヒト化、CDR−グラフト化、一本鎖、および二元機能の抗体を作製する他の方法は当業者によく知られている(WinterおよびHarris,Immunol.Today 14:243−246(1993); Wardら、Nature 341:544−546(1989); HarlowおよびLane,前出,1988; Hilyardら、Protein Engineering: A practical approach(IRL Press 1992); Borrabeck,Antibody Engineering,2d ed.(Oxford University Press 1995))。
【0043】
PBPに特異的な抗体は、単離されたPBP、またはその断片(天然源から調製され得るまたは組換え産生され得る)、またはエピトープとして機能できるPBPのペプチド部分などの免疫原を用いて生じ得る。このようなPBPのペプチド部分は、抗原ペプチドがPBPに特異的な抗体を作製するために使用できる場合、機能的抗原断片である。非免疫原性もしくは弱い免疫原性のPBPまたはそれらの一部は、ウシ血清アルブミン(BSA)またはキーホール・リンペット・ヘモシアニン(KLH)などの担体分子にハプテンを結合することにより免疫原性にされ得る。種々の他の担体分子およびハプテンを担体分子に結合する方法は当該分野で周知である(例えば、HarlowおよびLane,前出,1988を参照)。免疫原性PBP断片は、例えばグルタチオンSトランスフェラーゼ(GST)、ポリHis等との融合タンパク質としてペプチド部分を発現することによっても作製できる。ペプチド融合を発現する方法は当業者によく知られている(Ausubelら、Current Protocols in Molecular Biology(Supplement 47),John Wiley & Sons,New York(1999))。
【0044】
本発明はPBPに特異的に結合する抗体とサンプルとを接触させ抗体の特異的結合を検出することによりPBPを検出する方法も提供する。従って、PBPに特異的な抗体は、例えばサンプル中のPBPの存在および/またはレベルを決定するために有用である。PBPに特異的な抗体は、適切な発現ライブラリー、例えばラムダgt11ライブラリー、または他の種類の発現ライブラリーから、免疫学的にPBPに関連するポリペプチドをコードする遺伝子をコードする核酸分子をクローニングするためにも有用である。PBPに特異的な抗体は、サンプルから、例えばPBPを発現する細胞もしくは組織の細胞抽出物から、または組換え核酸分子からPBPを発現する細胞から得られる細胞抽出物から、PBPを実質的に精製するためにも使用できる。
【0045】
PBPを検出するアッセイは、PBPに特異的な抗体または抗原結合断片を用いて、例えば免疫組織化学、免疫蛍光、ELISAアッセイ、放射免疫アッセイ、FACS分析、免疫沈降、免疫ブロット分析、およびフローサイトメトリーを含む(HarlowおよびLane,前出,1988; HarlowおよびLane,Using Antibodies: A Laboratory Manual,Cold Spring Harbor Press(1999))。種々の免疫アッセイは、当該分野で周知であり、要望どおり、当業者により容易に改変できる。例えば、免疫アッセイで使用する抗体は、検出可能部分の取り込みもしくは共役により、またはそれ自体検出できるまたは検出できるよう標識される二次分子に結合することにより、検出できる。
【0046】
PBPまたはPBPに特異的な抗体は当該分野で周知の方法を用いて検出できるように標識され得る(Hermanson,Bioconjugate Techniques,Academic Press,1996; HarlowおよびLane,前出,1988)。例えば、ペプチドまたは抗体は放射標識、酵素、ビオチンまたは蛍光色素を含む種々の検出可能部分で標識され得る。ペプチドまたは抗体を標識する試薬は、ペプチドまたは抗体を含むキットに包含され得るまたは商業的供給源から個別に購入できる。本発明は、PBP、PBPに特異的な抗体、または両方を含むキットを更に提供する。このようなキットは、サンプル中のPBPの発現レベルを測定するために、例えばELISAまたは他の免疫アッセイなどのアッセイを実施するのに適した条件を供する反応カクテルも含み得る、ならびに既知量のPBP、および必要ならばPBPに特異的な抗体に結合できる二次抗体を含む対照サンプルを含み得る。キットが免疫アッセイに使用される場合、抗体に結合するサンプル中のPBPの存在または量を検出する簡単な方法を含み得る。
【0047】
本発明は、本明細書に開示するPBPのアミノ酸配列、例えば配列番号2として示されるアミノ酸配列をコードする単離された核酸分子も提供する。本発明はこのような核酸分子の修飾も提供する。このような核酸分子は示されるアミノ酸配列をコードする縮重ヌクレオチド配列を含む。さらに、本発明は配列番号1、3または4として示されるヌクレオチド配列を含む単離されたPBP核酸分子ならびにそれらの修飾物を提供する。本発明は、本明細書に開示するように、PBPの機能的断片をコードするヌクレオチド配列を有する核酸分子を更に提供する。
【0048】
本発明は、少なくとも中程度にストリンジェントな条件下で、PBP核酸分子、例えば配列番号1、3または4として示される核酸分子とハイブリダイズするPBPヌクレオチド配列の修飾物も提供する。修飾物がPBPヌクレオチド配列と少なくとも60%の同一性を有する場合、PBPヌクレオチド配列の修飾物も提供される。本発明は、配列番号1、3または4として示されるものなどのPBP核酸に対して、少なくとも65%の同一性、少なくとも70%の同一性、少なくとも75%の同一性、少なくとも80%の同一性、少なくとも85%の同一性、少なくとも90%の同一性、少なくとも95%の同一性、少なくとも98%の同一性、または少なくとも99%の同一性を有するPBPのヌクレオチド配列の修飾物も提供する。
【0049】
本明細書で用いられるように、中程度にストリンジェントな条件は、核酸分子が参照配列と実質的な同一性を有する核酸に結合できるハイブリダイゼーションの条件を指す。中程度にストリンジェントな条件は、42℃で、50%ホルムアミド、5Xデンハルト溶液、5XSSPE、0.2%SDSでのフィルターに結合した核酸のハイブリダイゼーション、続いて42℃で0.2XSSPE、0.2%SDSでの洗浄に等価な条件を含む。対照的に、「極めてストリンジェントな条件」は、42℃で、50%ホルムアミド、5Xデンハルト溶液、5XSSPE、0.2%SDSでのフィルターに結合した核酸のハイブリダイゼーション、続いて65℃で0.2XSSPE、0.2%SDSでの洗浄に等価な条件を含む。デンハルト溶液は1%フィコール、1%ポリビニルピロリドン、および1%ウシ血清アルブミン(BSA)を含む。20XSSPE(塩化ナトリウム、リン酸ナトリウム、エチレンジアミンテトラ酢酸(EDTA))は、3M塩化ナトリウム、0.2Mリン酸ナトリウム、および0.025M(EDTA)を含む。他の適切な中程度にストリンジェントなおよび極めてストリンジェントなハイブリダイゼーションの緩衝液および条件は、種々の塩および温度の条件を含み、当業者に周知であり、例えばSambrookら、Molecular Cloning: A Laboratory Manual,2nd ed.,Cold Spring Harbor Press,Plainview,New York(1989);およびAusubelら、前出,1999に記載されている。
【0050】
一般的に、中程度にストリンジェントな条件下で列挙されたの配列にハイブリダイズする核酸分子は、比較される2つの配列の長さにわたって、参照配列と約70%以上の同一性または約80%以上の同一性など、約60%以上の同一性を有する。極めてストリンジェントな条件下で再言及の配列にハイブリダイズする核酸分子は、一般的に、比較される2つの配列の長さにわたって、参照配列と約95%以上の同一性を含む、約90%以上の同一性を有する。任意の2つの核酸配列の同一性は、例えばBLASTコンピュータ整列、またはデフォルト・パラメータもしくは他の望ましいパラメータを用いて、配列を整列する類似の方法に基づいて、当業者により決定できる(例えば、Tatianaら、FEMS Microbiol Lett.174:247−250(1999); Altschulら、J.Mol.Biol.215:403−410(1990); GishおよびStates,(1993) Nature Genet.3:266−272(1993); Maddenら、Meth.Enzymol.266:131−141(1996); Altschulら、Nucleic Acids Res.25:3389−3402(1997))。
【0051】
本発明の単離されたPBP核酸分子は種々の診断および治療の適用に使用できる。例えば、本発明の単離されたPBP核酸分子もしくはその断片は、本明細書に記載するように、プローブとして、またはPBP核酸分子もしくはその断片の増幅に適したプローブもしくはプライマーを得るために、パーキン結合ポリペプチドの組換え発現用の鋳型として、またはパーキンへのPBPの結合に使用されるものと類似して、PBPに結合する細胞分子を同定するために2−ハイブリッドアッセイなどのスクリーニングアッセイに、使用できる。
【0052】
本発明は、本明細書に開示するPBPヌクレオチド配列の少なくとも15個の連続したヌクレオチドを含むオリゴヌクレオチド、またはそのアンチセンス鎖も提供する。参照PBPヌクレオチド配列の少なくとも15個の連続したヌクレオチドを含む本発明のオリゴヌクレオチドは、中程度にストリンジェントなまたは極めてストリンジェントなハイブリダイゼーション条件下で、PBP核酸分子にハイブリダイズできるので、例えば、サンプル中でPBPのDNAもしくはRNAを検出するためまたはそのスプライス変異体を検出するためのプローブとして;配列決定もしくはPCRプライマーとして;本明細書で開示するように、細胞でのRNAの転写を阻害するためもしくは低分子干渉RNA(siRNA)を作製するためのアンチセンス試薬として;またはPBP核酸分子へのハイブリダイゼーションが望ましい分野の当業者に知られる他の適応で、有利に使用できる。
【0053】
本明細書で用いられるように、PBP核酸分子は、nr、dbest、dbsts、gssおよびhtgsのデータベース(これらはデータベースの検索に利用できる)などの公共データベースに寄託されている、例えば発現配列標識(EST)、配列標識部位(STS)およびゲノム断片など、本明細書に開示されるように、配列番号1、3または4などのPBPヌクレオチド配列と同一性を有するヌクレオチド配列から成る予め知られる核酸分子を特異的に排除すると理解される(Altshculら、Nucleic Acids Res.25:3389−3402(1997))。
【0054】
具体的な実施態様において、本発明は、配列番号1のヌクレオチド796〜955と100%の同一性を有する20〜200の連続したヌクレオチドを含むオリゴヌクレオチド、またはそれらのアンチセンス鎖を提供する。オリゴヌクレオチドは、配列番号1のヌクレオチド896〜955と100%の同一性を有する少なくとも25、30、40、50、60、70、80、90、100、125、150、175の連続したヌクレオチド、および200までの連続したヌクレオチドを含み得る。GenBank登録番号のBI041917;CD614598;CD614596;CD614594;CD614592;CD614590;CD614588;CD614576;CD614574;CD614570;BU542453;BF666086;AW374529;BU687172;BM975158;BM910986;BG765308;BG745915;BG745175;BG698661;BG113587;BF837913;BE909317;BE899012;AL135049;AI143229に相当するヌクレオチド配列、ならびに本発明の核酸分子と同一性を有する他の既知の配列は、本発明のオリゴヌクレオチドから特異的に排除される。
【0055】
本発明のPBP核酸分子およびオリゴヌクレオチドは、当該分野で公知の方法により産生または単離できる(例えば、Sambroolら、前出,1989; Ausubelら、前出,1999を参照)。選択される方法は、例えば所望する核酸分子の種類に応じて変わる。当業者は、本明細書に開示するヌクレオチド配列の知見に基づいて、ゲノムDNAとしてのPBP核酸分子を容易に単離できる、または当該分野で公知の方法により、それらから所望のイントロン、エキソンもしくは調節配列を;それらから全長のcDNAもしくは所望の断片として;もしくはそれらから全長のmRNAもしくは所望の断片として、容易に単離できる。
【0056】
本発明のPBP核酸分子を作製する1つの有用な方法は、PCRおよび適切なオリゴヌクレオチドを用いた核酸分子の増幅を含む。PCRまたはRT−PCRのいずれかは、所望するヌクレオチドの境界を有するPBP核酸分子を作製するために用いられ得る。核酸配列への望ましい修飾は1つ以上の付加、欠失または置換をもつ適切なオリゴヌクレオチド・プライマーを選択することによっても導入できる。このような核酸分子は、僅かな単一遺伝子またはmRNAコピーから、対象とする任意の細胞、組織または種から、出発して、指数関数的に増幅され得る。
【0057】
本発明は、PBP核酸分子に特異的にハイブリダイズできる条件下で、PBP核酸分子またはそれらに由来する1つ以上のオリゴヌクレオチドとサンプルとを接触させ、特異的なハイブリダイゼーションを検出することにより、サンプル中のPBP核酸分子を検出する方法を更に提供する。PBP核酸分子は、例えば配列番号1、3もしくは4として示されるPBPヌクレオチド配列、または配列番号1、3もしくは4などの参照PBPヌクレオチド配列の少なくとも15の連続したヌクレオチドを含み且つこの配列に由来するオリゴヌクレオチドであり得る。このようなPBP核酸分子またはそれらに由来するオリゴヌクレオチドは、所望の検出方法の必要に応じて、センスまたはアンチセンスであり得ることが理解される。
【0058】
本発明は、所望する核酸分子の増幅に適した2つ以上のオリゴヌクレオチドとサンプルとを接触させ、核酸分子を増幅し、ならびにこの増幅を検出することにより、サンプル中のPBP核酸分子を検出する方法を更に提供する。サンプル中のPBP核酸を検出する方法は、要望どおり、定性的または定量的のいずれかであり得る。例えば、PBP核酸の存在、存在量、完全性または構造は、要望どおり、アッセイ形式および選択されるプローブまたはプライマーに応じて決定できる。
【0059】
単離されたPBP核酸分子との特異的ハイブリダイゼーションに基づいてPBP核酸を検出するための有用なアッセイは、当該分野でよく知られており、例えば、用いるアッセイ形式に応じて、核酸分子の染色体位置の変化、遺伝子コピー数の変化、およびRNAの存在量を検出するために使用できるインサイツ・ハイブリダイゼーションを含む。他のハイブリダイゼーションアッセイは、例えば、異なるRNAスプライス変異体の存在量および完全性を決定するために用いられ得るノーザン・ブロットおよびRNアーゼ保護アッセイ、ならびにDNAのコピー数および完全性を決定するために用いられ得るサザン・ブロットを含む。ハイブリダイゼーション・プローブは、放射性同位体、蛍光色素、化学発光マーカー、ビオチンなどの任意の適切な検出可能部分、または分析方法により検出でき且つ当該分野で公知の他の検出可能部分を用いて標識できる。
【0060】
2つ以上のオリゴヌクレオチドを用いたPBP核酸の増幅に基づいてサンプル中のPBP核酸を検出する有用なアッセイも、当該分野でよく知られており、例えば定性的または定量的なポリメラーゼ連鎖反応(PCR);逆転写PCR(RT−PCR);一本鎖立体構造多型(SSCP)分析(これは非変性ゲル電気泳動時に電気泳動移動度の変化を生じる一本鎖DNAの二次構造の差異に基づいてDNAの単一点突然変異を容易に同定できる);ならびにタンパク質欠失試験(DNAの突然変異が電気泳動ゲルでタンパク質産物の変化により決定できる)などの共役したPCR、転写および翻訳のアッセイを含む。加えて、増幅されたPBP核酸は突然変異および突然変異多発点を検出するために配列決定でき、このような突然変異を同定するためにサンプルを大規模スクリーニングする特異的なアッセイが開発され得る。
【0061】
本発明は、PBP核酸分子、例えば配列番号1、3もしくは4として示されるPBPヌクレオチド配列または本発明のPBPオリゴヌクレオチドを含むキットを更に提供する。例えば、診断用核酸は、配列番号1、3もしくは4などのPBP核酸分子の任意部分またはそのアンチセンス鎖に由来し得る。本発明のキットは、ゲノムDNA、mRNAまたはcDNAのいずれかにおけるPBPをコードする核酸の有無をアッセイする診断システムとして有用である。適切な診断システムは、少なくとも1つの発明核酸を含み、少なくとも1つのアッセイに十分な量の個別に包装された化学試薬として2つ以上の発明核酸を含み得る。包装された試薬の使用説明書も通常包含される。当業者は、本明細書に記載する発明方法の実施に適した緩衝液および溶液と組み合わせて、発明核酸のプローブおよび/またはプライマーとして有用なオリゴヌクレオチドをキット形態に容易に組み込み得る。
【0062】
本発明のPBP核酸分子は、PBP核酸分子に関連する核酸分子をスクリーニングするために使用できる。PBP核酸分子に関連する核酸分子は、例えば、ゲノム・ライブラリー、cDNAライブラリーまたは発現ライブラリーなどのライブラリーを検出可能試薬を用いてスクリーニングすることにより同定できる。このようなライブラリーは、市販されている、または当該分野で公知の方法を用いて対象とする任意の望ましい組織、細胞もしくは種から作製できる。例えば、cDNAライブラリーまたはゲノム・ライブラリーは検出可能なように標識されたPBP核酸分子を用いたハイブリダイゼーションによりスクリーニングできる。加えて、発現ライブラリーはPBP核酸のコード配列に相当するポリペプチドに対して生じる抗体を用いてスクリーニングできる。本発明のPBP核酸分子を含むライブラリー・クローンは当該分野で公知の方法により他のクローンから単離でき、必要ならば、それらの断片は制限酵素消化およびゲル電気泳動により単離できる。
【0063】
本発明はPBP核酸分子を含むベクターも提供する。本発明のベクターは、PBP核酸分子をサブクローニングし増幅するのに有用でありならびにPBPポリペプチドを組換え発現するのに有用である。本発明のベクターは、例えばバクテリオファージ、バキュロウイルスまたはレトロウイルスなどのウイルス・ベクター;コスミドまたはプラスミド;ならびに特に大きな核酸分子をクローニングするために、細菌人工染色体ベクター(BAC)および酵母人工染色体ベクター(YAC)を含み得る。このようなベクターは市販されており、それらの使用は当該分野で周知である。
【0064】
本発明はPBP核酸分子を含むベクターを含む宿主細胞を更に提供する。組換えPBP分子を発現するために使用できる典型的な宿主細胞は、哺乳動物初代細胞;COS、CHO、HeLa、NIH3T3、HEK293およびPC12細胞などの確立された哺乳動物細胞株;アフリカツメガエルの胚および卵母細胞などの両性動物細胞;ならびに他の脊椎動物細胞を含む。典型的な宿主細胞は、ショウジョウバエなどの昆虫細胞、サッカロミセス・セレヴィシエ、サッカロミセス・ポンベ、またはピチア・パストリスなどの酵母細胞、および大腸菌などの原核細胞も含む。
【0065】
本発明はPBPの発現および/または活性を調節する分子を同定する方法も提供する。これらの分子は、例えばPBPの生物機能を促進または阻害するために治療への応用に使用できる。
【0066】
タンパク質結合ドメインと相互作用する細胞タンパク質を同定するための種々の結合アッセイが当該分野で知られており、例えば酵母2−ハイブリッド・スクリーニングアッセイ(例えば、U.S.patent Nos.5,283,173,5,468,614および5,667,973;Ausubelら、前出,1999; Lubanら、Curr.Opin.Biotechnol.6:59−64(1995))を含む。このアッセイは、本明細書に開示するように、典型的なパーキン結合ポリペプチド(PBP)を同定するために用いた。他の方法は細胞抽出物を用いるアフィニティー・カラム・クロマトグラフィー方法を含む。種々のPBP配列または欠失を含むポリペプチド断片を合成または発現することにより、PBPの結合界面は容易に同定できる。
【0067】
本発明は、PBP調節化合物など、PBPの発現および/または活性を調節する分子を同定する方法も提供する。PBP調節化合物は、PBP核酸分子またはPBPに特異的に結合し、その発現または活性を変える分子である。PBP調節化合物は、ペプチドもしくはポリペプチド、核酸、炭水化物、脂質、またはそれらの任意の組み合わせなど、天然に存在する高分子であり得る。PBP調節化合物は、部分的にもしくは完全にこの高分子の合成の誘導体、類似体もしくは模倣体、またはコンビナトリアル化学方法により部分的にもしくは完全に調製された有機もしくは無機の小分子でもあり得る。典型的なPBP調節化合物は、本明細書に開示するように、阻害剤を含む。単純もしくは複雑な有機分子、金属含有化合物、炭水化物、ペプチド、タンパク質、ペプチド模倣体、糖タンパク質、リポタンパク質、核酸、抗体等の化学分子または生体分子を含む、PBP調節化合物をスクリーニングする際に使用する複数の化合物を作製する方法は、本明細書に記載するように、当該分野でよく知られている。
【0068】
当該分野で公知の種々の低処理能および高処理能のアッセイは、PBP核酸分子またはポリペプチドと候補PBP調節化合物との特異的な結合相互作用を検出するのに適している。例えば、蛍光相関分光法(FCS)およびシンチレーション近似アッセイ(SPA)(reviewed in Major,J.Receptor Signal Transduction Res.15:595−607(1995);およびin Sterrerら、J.Receptor Signal Transduction Res.17:511−520(1997))を含む、直接アッセイおよび競合アッセイの両方が実施できる。特異的な結合相互作用を検出するアッセイは、PBPに特異的なリガンド、例えばELISAアッセイ、FACS分析またはアフィニティー分離で使用される抗体を用いるアフィニティー分離方法を含み得る。
【0069】
PBPの遺伝子発現を調節する化合物を同定するアッセイは、β−ラクタマーゼ、ルシフェラーゼ、緑色蛍光タンパク質またはβ−ガラクトシダーゼなどのタンパク質の発現の変化がPBPの発現を上流調節または下流調節するPBP調節化合物と細胞との接触に応答して検出されるように、PBPプロモーター−レポーター核酸構築物を細胞にまず形質導入する工程を含み得る。このようなアッセイおよびレポーター系は、当該分野で周知であり、例えば、Ausubelら、前出,1999に記載されている。PBPの遺伝子発現を調節する化合物を同定する他のアッセイは、ノーザン・ブロット、RNアーゼ保護アッセイ、およびRT−PCRなどのPBP転写産物のレベルを測定するアッセイを含む。PBPをコードするゲノムDNAからプロモーターおよび/またはエンハンサーを同定する方法は当該分野でよく知られている。レポーター遺伝子構築物は、PBPのプロモーター領域を用いて作製でき、PBP遺伝子のプロモーター活性を増減する化合物についてスクリーニングできる。このような化合物はPBPの発現を改変するためにも使用できる。
【0070】
PBPの発現を調節する化合物を同定するアッセイは、PBP調節化合物と細胞との接触に応答して、PBPの存在量の変化を検出する工程を含み得る。ポリペプチドの発現の変化を検出するアッセイは、例えば免疫ブロッティング、免疫蛍光、免疫組織化学および免疫沈降のアッセイなど、特異的なPBP抗体を用いた免疫アッセイを含む。
【0071】
PBP調節化合物がPBPの活性を阻害または促進するか否かを決定するのに適したアッセイは、PBPの生物活性に基づいて当業者により決定できる。例えば、PBPは、PBP核酸もしくはPBPの発現を改変するまたはPBPの生物活性を改変するPBP調節化合物を同定するために、上述するように、種々の化合物を用いてスクリーニングできる。
【0072】
本発明のポリペプチドおよび核酸分子は種々の診断または治療の適用に使用できる。診断および治療の適用はPBPの生物活性に基づき得る。例えば、PBP核酸分子は、変化したPBP活性を有する個体を処置するための治療方法で使用できる。AR−JPの患者におけるパーキン機能の喪失は、シナプトタグミン1またはシナプトタグミン11またはSLPなどのPBPなど、PBPの機能を変化させ得る。パーキンはPBPをユビキチン化するため、パーキン機能の喪失はPBPの発現を増加させるのに役立ち得る。治療方法において、正常なPBPの発現に比べて増加する変化のPBP活性は、本明細書に開示するように、PBPアンチセンス核酸またはsiRNAを投与することにより低下し得る。
【0073】
PBPの発現を阻害する核酸分子を含むベクターは、インビボまたはエクスビボの方法により個体に導入されPBPの発現を減少させ得る。このような治療方法に有用なベクターは、例えばレトロウイルス、アデノウイルス、レンチウイルス、ヘルペスウイルス、ポックスウイルスのDNAまたは対象とする異種のポリヌクレオチドを発現できる任意のウイルスDNAを含む。例えば、DNAベクター、偽型レトロウイルス・ベクター、アデノ随伴ウイルス、テナガザル(gibbon ape)白血病ベクター、水疱性口内炎ウイルス(VSL)、VL30ベクター、リポソーム媒介性ベクターなど、他のベクターも利用できる。
【0074】
PBP調節化合物は治療方法でも使用できる。例えば、PBP調節化合物は、例えばパーキン機能の喪失に起因する発現の増加など、異常に発現するPBPの発現または活性を変えるために使用できる。例えば、増加したPBPの発現はPBPの発現および/または活性を低下させるPBP調節化合物を用いて、低減できる。
【0075】
本発明は、増大したレベルのパーキン結合ポリペプチドを発現するトランスジェニック動物を作製することにより、パーキンソン病の動物モデルを作製する方法を更に提供する。パーキン結合ポリペプチドは、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択できる。本発明はこのような方法により作製されたパーキンソン病の動物モデルを更に提供する。
【0076】
本発明は、PBPをコードする外因性核酸を発現できるヒト以外のトランスジェニック哺乳動物を更に提供する。パーキンの欠如はPBPのユビキチン化の減少をもたらすため、発現を増加させるので、ヒト以外のトランスジェニック哺乳動物におけるPBPの発現はパーキンソン病の少なくとも幾つかの側面について動物として役立ち得る。PBP導入遺伝子は、本明細書に開示するように(実施例VI〜VIIIを参照)、PBPを発現することが知られる細胞または組織を標的とし得る。外因性核酸とは、宿主を原産としない核酸配列、またはその天然環境以外で、例えば遺伝操作されたDNA構築物の一部として、宿主に存在する核酸配列を指す。天然に存在するレベルのPBPに加えて、本発明のPBPは、例えば周知のノックアウト・トランスジェニックのように、トランスジェニック哺乳動物で、上述の過剰発現、あるいは過少発現し得る。
【0077】
内在性遺伝子を組換えまたは変異のPBP遺伝子で置換することによりPBPの発現または構造の調節を変えるため、トランスジェニック動物において生来の遺伝子座をもつPBP遺伝子の変異型または正常型の相同的組換えの使用も本明細書で意図される。ヒト以外の遺伝子ノックアウト哺乳動物を含む、ヒト以外のトランスジェニック哺乳動物を作製する方法は当業者によく知られている(Capecchiら、Science 244:1288(1989); Zimmerら、Nature 338:150(1989); Shastry,Experentia,51: 1028−1039(1995); Shastry,Mol.Cell.Biochem.,181:163−179(1998);および米国特許第5,616,491号,1997年4月1日公表,同第5,750,826号,1998年5月12日公表,および同第5,981,830号,1999年11月9日を参照)。
【0078】
正常に活性できないよう変異したPBPをコードする核酸を発現でき、従って天然のPBPを発現しないヒト以外のトランスジェニック哺乳動物も提供される。本発明は、PBPをコードするmRNAに相補的なアンチセンスmRNAに転写されるよう配置されて(このアンチセンスmRNAはこのmRNAにハイブリダイズすることによりその翻訳を低減する)、PBPをコードする核酸に相補的なアンチセンス核酸を含むゲノムを有するヒト以外のトランスジェニック哺乳動物も提供する。核酸は、発現が誘導され得るようにまたは特異的な細胞型に限定され得るように、誘導性プロモーターおよび/または組織特異的調節要素を更に含み得る。ヒト以外のトランスジェニック哺乳動物の例はトランスジェニック・マウスである。組織特異性を決定する要素の例はメタロチオネイン・プロモーターおよびL7プロモーターである。
【0079】
PBPの生理学的役割および挙動役割を解明する動物モデル・システムも提供され、PBPの発現が種々の技術を用いて改変されるトランスジェニック動物を創出することにより作製される。このような技術の例は、マイクロインジェクション、レトロウイルス感染または当業者に周知の他の手段により、PBPをコードする核酸の正常型または変異型を、トランスジェニック動物を作製するのに適した受精胚に挿入することを含む(例えば、Hoganら、Manipulating the Mouse Embryo: A Laboratory Manual(Cold Spring Harbor Laboratory,(1986)を参照)。
【0080】
本明細書で論じるように、パーキンはE3ユビキチン・リガーゼとして機能する。パーキンはシナプトタグミン1およびシナプトタグミン11をユビキチン化することが見出された。パーキン遺伝子の突然変異の不活性化は常染色体劣性若年性パーキンソン症候群を惹起する。従って、パーキンの不活性化は、例えばユビキチン化による、パーキン相互作用ポリペプチドのプロセシング化能力に影響を及ぼし得る。パーキンの活性の模倣、即ち、パーキン相互作用ポリペプチドの活性の低下は、パーキンソン病に関連する徴候または症状を改善するために使用できることが可能である。当業者はパーキンソン病に関連する徴候または症状の改善を容易に認識または決定できる。
【0081】
ポリペプチドの活性を低下させる方法は当業者によく知られている。ポリペプチドの活性の低下はポリペプチドの発現レベルの低下およびポリペプチドにより示される生物活性の低下の両方を含むと理解される。
【0082】
ポリペプチドの発現を減少させる方法は、例えばリボザイム、アンチセンス核酸またはRNA干渉の使用を含み得る。RNA干渉は以前に記載されている(Fireら、Nature 391:806−811(1998))。天然で起きるRNA干渉において、開始段階中、入力dsRNAは21〜23のヌクレオチド小干渉RNA(siRNA)に消化される。siRNAは、Hammondら、Nature Rev Gen 2:110−119(2001); Sharp,Genes Dev 15:485−490(2001);ならびに HutvagnerおよびZamore,Curr Opin Genetics & Development 12:225−232(2002)に記載されるように、「ガイドRNA」とも呼ばれる。siRNAは、dsRNAに特異的なリボヌクレアーゼのRNアーゼIIIファミリーに属する酵素が漸次dsRNAを切断する場合に産生される。dsRNAは直接または導入遺伝子もしくはベクターを介して導入され得る。連続切断事象はRNAを19〜21の塩基対二本鎖(siRNA)に分解し、HutvagnerおよびZamore,Curr.Opin.Genetics & Development 12: 225−232(2002); Bernsteinら、Nature 409:363−366(2001)により記載されるように、それぞれが2−ヌクレオチド3′オーバーハングを有する。エフェクター段階において、siRNA二本鎖は、ヌクレアーゼ複合体に結合して、RNA誘導性サイレンシング複合体、即ちRISCとして知られるものを形成する。次いで、活性なRISCは、塩基対の相互作用により相同な転写産物を標的にし、siRNAの3′末端から約12ヌクレオチドのmRNAを切断する(Nykanenら、Cell 107: 309−321(2001))。
【0083】
パーキン相互作用ポリペプチドの活性は阻害剤を用いても低下し得る。阻害剤は、パーキン相互作用ポリペプチドの発現、活性または細胞内シグナル伝達を低減する化合物であり得る。このような阻害剤は、例えば小分子、タンパク質、ペプチド、ペプチド模倣体、リボザイム、核酸分子もしくはオリゴヌクレオチド、オリゴ糖、またはそれらの組み合わせであり得る。このような分子を作製する方法は当業者によく知られている(Huse,U.S.Patent No.5,264,563; Francisら、Curr.Opin.Chem.Biol.2:422−428(1998); Tietzeら、Curr.Biol.,2: 363−371(1998); Sofia,Mol.Divers.3: 75−94(1998); Eichlerら、Med.Res.Rev.15: 481−496(1995); Gordonら、J.Med.Chem.37: 1233−1251(1994); Gordonら、J.Med.Chem.37:1385−1401(1994); Gordonら、Acc.Chem.Res.29: 144−154(1996); WilsonおよびCzarnik,eds.,Combinatorial Chemistry: Synthesis and Application,John Wiley & Sons,New York(1997))。多数の天然化合物および合成化合物を含むライブラリーも市販から入手できる。分子のコンビナトリアル・ライブラリーは、上記で論じたように、周知のコンビナトリアル化学方法を用いて調製できる。阻害剤は、例えば、アンタゴニスト;パーキン相互作用ポリペプチドの活性化を妨げる優性阻害分子;パーキン相互作用ポリペプチドの活性または発現を阻害する抗体、タンパク質、小分子およびオリゴヌクレオチド;リボザイム、アンチセンス核酸分子、およびパーキン相互作用ポリペプチドの発現を妨げるまたは減少させる負の調節転写因子をコードする核酸分子、ならびにこのようなリボザイムおよび核酸分子を含む細胞またはウイルスを含む。当業者は、これらの分子、ならびにパーキン相互作用ポリペプチドの発現、活性またはシグナリングを阻害する他の分子が阻害剤として使用できることを容易に理解するであろう。
【0084】
リボザイムまたはアンチセンス核酸分子などの配列特異的リボヌクレアーゼもパーキン相互作用ポリペプチドの発現を阻害するために使用できる。
配列特異的リボヌクレアーゼは、所定のリボヌクレオチド配列でRNAの切断を触媒する分子を指す。リボザイムは所定のリボヌクレオチド配列でRNAの切断を触媒するRNA分子を指す。ハンマーヘッドおよびヘアピンなどのリボザイムは常套方法により設計され調製できる。標的切断部位に対するハンマーヘッドおよびヘアピンなどのリボザイムの特異性は、リボザイムとそのRNA標的との塩基対合により決定される。リボザイムを設計する方法は、例えばHauswirthおよびLewin,Prog.Retin.Eye Res.19:689−710(2000),ならびにLewinおよびHauswirth,Trends.Mol.Med.7: 221−228(2001)に記載されるように、よく知られている。
【0085】
リボザイムおよびDNA酵素を含む配列特異的リボヌクレアーゼは、上述のように設計され、核酸分子の標準的な合成方法により調製され得る。Keら、Int.J.Oncol.12:1391−1396(1998); Dohertyら、Ann.Rev.Biophys.Biomol.Struct.30:457−475(2001); HauswirthおよびLewin,前出,2000;ならびにLewinおよびHauswirth,前出,2001も参照のこと。配列特異的リボザイムはランダム配列のプールからインビトロ選別によっても同定できる。このような方法は、例えばBartelおよびSzostak,Science 261:1411−1418(1993),Breaker,Chem.Rev.97: 371−390(1997)ならびにSantoroおよびJoyce,Proc.Natl.Acad.Sci.,USA 94:4262−4266(1997)に記載されているように、十分に確立されている。
【0086】
リボザイムがウイルス・ベクターまたは他のベクターを用いて送達されずに患者に投与されるべきである場合、必要ならば、リボザイムは安定性を向上するために修飾され得る。治療用リボザイムに有用な修飾は、この分子の3′末端およびピリミジンの2′部位の遮断を含むが、これらに限定されない。安定化されたリボザイムは、数時間の半減期を有することができ、例えば静脈内または局所の注射を用いて反復投与できる。当業者は、リボザイムがウイルス遺伝子治療ベクターでの発現によっても投与できることを理解している。リボザイムをコードするDNAオリゴヌクレオチドは、RNA polIIまたはRNA polIIIのプロモーターの下流にクローニングでき、必要ならば、tRNAval、U6 snRNAまたはアデノウイルスVA1 RNAなどの遺伝子の転写産物内に組み込める。
【0087】
アンチセンス核酸分子とは、メッセンジャーRNAまたは他の特異的RNA転写産物の分子の全てまたは一部に対して配列の相補的な核酸分子を指す。アンチセンス核酸分子は、例えばDNAまたはRNAであり得る、ならびに天然に存在するヌクレオチド、および合成ヌクレオチドまたは安定性を向上するリン酸バックボーンへの修飾などの他の天然に存在しない修飾物を含み得る。ホスホロチオエートを含むアンチセンス・オリゴヌクレオチドおよび他の修飾オリゴヌクレオチドは、本明細書で用いられるように、アンチセンス核酸分子という用語に包含される。下記に束縛されることなく、本発明に有用なアンチセンス核酸分子はmRNAの翻訳を減らせるまたはmRNAの分解を増やせることにより、標的mRNAの発現を減少させる。
【0088】
アンチセンス方法を用いて発現を減少させるための相同性の必要条件は実験的に決定できる。一般的に、少なくとも約80〜90%の核酸配列の同一性は本発明に有用なアンチセンス核酸分子に存在し、より高い核酸配列の同一性がアンチセンス・オリゴヌクレオチドでしばしば用いられ、これは患者の内因性転写産物と完全に同一であり得る。標的配列は、必要ならば、鎖の1つを置換するためにアンチセンス分子により侵入される二重鎖のらせん状に整列した幹に加えて、核形成が起きる小さな一本鎖の領域を有するよう選択され得る(MirおよびSouthern,Nature Biotech.17:788−792(1999))。アンチセンス核酸分子を選択し調製する方法は、当該分野で周知であり、コンピュータ手法を含む(Patzelら、Nucl.Acids.Res.27:4328−4334(1999);Chengら、Proc.Natl.Acad.Sci.,USA 93:8502−8507(1996);LebedevaおよびStein,Ann.Rev.Pharmacol.Toxicol.41:403−419(2001);JulianoおよびYoo,Curr.Opin.Mol.Ther.2:297−303(2000);ならびにCho−Chung,Pharmacol.Ther.82:437−449(1999))。
【0089】
当業者はパーキン結合ポリペプチドの活性または発現の減少を容易に測定できる。例えば、核酸のプローブまたはプライマーはパーキン相互作用ポリペプチドのmRNAの発現を調べるために使用でき、パーキン相互作用ポリペプチドの抗体はポリペプチドの発現レベルを調べるために使用できる。阻害剤の作用は、パーキン相互作用ポリペプチドの生物活性に及ぼすその影響をアッセイすることにより、容易に測定できる。例えば、シナプトタグミンの活性が測定できる(Sudhof,J.Biol.Chem.277:7629−7632(2002))。これらの方法および他の適切な方法は、当業者により容易に測定でき、パーキン相互作用ポリペプチドの潜在的な阻害剤として化合物の作用を試験するために用いることができる。化合物は、パーキンソン病においてパーキンのユビキチン化活性の低下を補うためにパーキン相互作用ポリペプチドのユビキチン化を増す能力についてもスクリーニングされ得る。
【0090】
本発明は、他の実施態様において、1つ以上の化合物とパーキン結合ポリペプチドとを接触させパーキン結合ポリペプチドの活性を変える化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を提供する。典型的なパーキン結合ポリペプチドは、シナプトタグミンI、シナプトタグミンXI、およびシナプシン様タンパク質を含む。この方法はパーキン結合ポリペプチドの活性を低下させる化合物についてスクリーニングするために用いられ得る。
【0091】
本発明は、パーキン結合ポリペプチドを発現する細胞と1つ以上の化合物とを接触させパーキン結合ポリペプチドの発現を減らす化合物を同定することにより、パーキンソン病を処置するための候補薬物を同定する方法を更に提供する。他の実施態様において、本発明はパーキン結合ポリペプチドの発現または活性を低減する分子を投与することによりパーキンソン病を処置する方法を提供する。このような分子は本明細書に開示する方法により同定できる。
【0092】
本発明の様々な実施態様の活性に実質的に影響を及ぼさない修飾も本明細書で供される発明の定義内で提供されることが理解される。従って、下記の実施例は本発明を説明することを意図し限定するものではない。
【実施例】
【0093】
(実施例1)
(酵母によるパーキン相互作用ポリペプチドの同定)
本実施例は酵母2−ハイブリッド系を用いたパーキン相互作用ポリペプチドの同定を記載する。
【0094】
酵母2−ハイブリッド方法はpGBT9−パーキン(1〜465)構築物を用いてヒト胎児脳pGAD10−cDNAライブラリーをスクリーニングするために用いた(FieldsおよびSong,Nature 340:245−246(1989))。酵母2−ハイブリッドのバイトプラスミドpGBT9−パーキン(1〜465)を調製するために、アミノ酸1〜465をコードする全長のパーキンcDNAはpEGFPC1−パーキン(Huynh et al Ann.Neurol.48:737−744(2000))から切り出しpGBT9プラスミド(Clontech; Palo Alto CA)に連結した。
【0095】
パーキンと相互作用するタンパク質を同定するために、GAL4活性化ドメイン・ベクターPGAD10にクローニングされたヒト成人脳cDNAライブラリーの酵母2−ハイブリッド・スクリーニングは、バイトとして、GAL4結合ドメインと融合したパーキン・アミノ酸1〜465をコードするpGBT9−パーキン(1〜465)を用いて実施した(Clontech社のベクターおよびライブラリー)。先に記載されるように(Shibataら、Hum.Mol.Genet.9:1303−1313(2000)およびScolesら、Nat.Genet.18:354−359(1998))、バイトプラスミドは酵母株Y190に共形質転換し、ロイシン、トリプトファン、およびヒスチジンを含まず且つ25mM3−アミノ−1,2,4−トリアゾールおよび2%のグルコースを含む合成培地で増殖した。Y190株は、ヒスチジンを含まない培地で増殖でき且つβ−ガラクトシダーゼをコードするLacZ遺伝子を用いて色選別できるHIS3遺伝子の栄養選別を可能にする。β−ガラクトシダーゼ・レポーターは、37℃で15分間から8時間、Z緩衝液(60mM Na2HPO4、40mM NaH2PO4、10mM KCl、1mM MgSO4、pH 7.0、0.03mM β−メルカプトエタノール、および2.5μM X−gal)で凍結破砕されたコロニーをインキュベーションすることにより、スタンプされたニトロセルロース・フィルターでアッセイされた。陽性クローンはロイシンまたはトリプトファンを含まない合成培地で再画線され、β−ガラクトシダーゼ活性について再試験された後、pGAD10の「獲物」プラスミドが単離された。
【0096】
ヒトのシナプトタグミン11遺伝子をコードする部分配列を含むpGAD10プラスミドは精製された後、pGBT9−パーキン(1〜465)または負の対照プラスミド(pGBT9ベクター、pGBT9−NF2、pGBT9−HRS)を用いて再形質転換され、ならびにβ−ガラクトシダーゼ活性について再試験した。種々のパーキンとシナプトタグミン11タンパク質断片との相互作用強度の半定量的評価を得るために、β−ガラクトシダーゼの液体アッセイは、0.6 mg/mlのo−ニトロフェニルガラクトシドを添加したZ緩衝液および5%クロロホルムで抽出されたY190酵母細胞を2分間から1時間インキュベートすることにより実施した。標準偏差は同型培養の三回の測定から計算した。β−ガラクトシダーゼ活性はミラー単位として表した(ミラー単位=1000x[OD420/(OD600x時間x体積)](PoulletおよびTamanoi Methods Enzymol.255:488−497(1995))。
【0097】
6個の潜在クローンは2x106個の独立したヒト胎児脳pGAD10−cDNAコロニーで同定された。これらのクローンは、個々に単離され、配列決定され、更に酵母2−ハイブリッド・フィルターアッセイにかけられ相互作用を確認した。これらのクローンの内2つは長い読み取り枠を有していたため、精製し配列決定した。ヌクレオチド配列は、2つのクローンの内1つはシナプトタグミン11のC2AおよびC2Bのドメインをコードすることを示した。このクローンはhsyt11ABと呼ばれた(ヒトのシナプトタグミン11、C2AドメインおよびC2Bドメイン)。hsyt11AB断片が真のパーキン相互作用物質か否かを更に決定するために、精製したpGAD10−hsyt11ABはpGBT9−パーキン(1〜465)または2つの無関係なおとりのpGBT9−NF2(神経鞘腫抑制物質をコードする)およびpGBT9−Hrs(肝細胞増殖因子に調節されたキナーゼ基質をコードする)の構築物(Scolesら、前出 2000)、またはpGBT9ベクターを用いて、Y190酵母細胞に共トランスフェクションした。フィルター結合アッセイは、pGAD10−hystXIABクローンがpGBT9−パーキン構築物と陽性の相互作用を示すがpGBT9ベクター対照または無関係のタンパク質とは示さないことを証明した(図1A)。
【0098】
パーキンはsyt11のC2AドメインおよびC2Bドメインと相互作用することが見出された。図1Aは酵母2−ハイブリッド・フィルターアッセイを示す。pGAD10−hsyt11ABおよびpGBT9−パーキンで形質転換された酵母Y190細胞はβ−ガラクトシダーゼ基質と陽性反応を生じた(灰色の斑)一方、PGAD10−hsyt11ABおよびpGBT9ベクター対照で形質転換された酵母細胞は生じなかった(図1A)。pGAD10−hsyt11抗体ならびに神経フィブロミンおよびHrs(pGBT9−NF1、およびpGBT9−Hrs)を発現する2つの他の対照を用いて形質転換された酵母細胞は、β−ガラクトシダーゼ基質と陽性反応を生成しなかった。これらの対照タンパク質は、先に記載される(Scolesら、前出(1998))ように、酵母2−ハイブリッド系で機能的である。これらのフィルター結合アッセイは、pGAD10−hsyt11ABクローンのみがpGBT9−パーキン構築物と陽性の相互作用を示すがpGBT9ベクター対照または無関係のタンパク質とは示さないことを示した(図1A)。
【0099】
酵母2−ハイブリッド液体アッセイによりパーキンの相互作用を確認するために、単一トランスフェクションされた3通りの酵母コロニーは液体培地で増殖しβ−ガラクトシダーゼ活性について試験した。pGAD10−hsyt11ABおよびpGBT9−パーキンで共トランスフェクションされた酵母細胞は、pGAD10−hsyt11ABおよびpGBT9ベクター対照を共発現する酵母細胞(図1Bの「2」)と比べて、約20倍高いβ−ガラクトシダーゼ活性を生じた(図1Bの「1」)。図1Bに示されるそれぞれの棒グラフはn=3を表す。
【0100】
これらの結果は、パーキンがシナプトタグミン11(syt11)のC2AドメインおよびC2Bドメインと相互作用することを証明する。
【0101】
(実施例II)
(シナプトタグミン1およびシナプトタグミン11に対する抗体の作製および特異性)
本実施例はシナプトタグミン1およびシナプトタグミン11の抗体の産生および特徴決定を記載する。
【0102】
抗シナプトタグミン11抗体を作製するために、ウサギ抗syt11抗体はQCPによりsytXIAペプチド(HQQAEKKQKNPPYKF;配列番号9)に対して作製した。sytXIBペプチド(KVRRDKDGPRRESGRG;配列番号10)に対する他の抗体が作製されたが、この抗体はヒトおよびPC12細胞のタンパク質抽出物のウェスタン・ブロットで複数のバンドを認識した。抗sytXIA抗体はセファロース(商標)sytXIAカラムによりアフィニティー精製された。ウサギおよびニワトリの抗パーキンは、先に記載するペプチドParkA(Huynhら、Ann.Neurol.48:737−744(2000))に対して作製された。
【0103】
シナプトタグミン11の3ペプチドに対するウサギの抗体(抗syt11)が作製された。シナプトタグミン1に対するマウスの抗体(抗syt65)は購入した(Stressgen; Victoria,British Columbia,Canada)。β−アクチン(Sigma)およびβ−COP(Sigma)に対するマウスのモノクローナル抗体、ならびにウサギ・ユビキチン抗体(DAKO;Carpinteria CA)は購入した。sytIおよびsytIIは関連するタンパク質であり少なくとも11の他のシナプトタグミンに密接な相同性を有するため、抗syt65および抗syt11の抗体は、これらの抗体を用いる前に他の抗原との交差反応性について試験しパーキン−syt相互作用を調べた。
【0104】
タンパク質抽出のため、細胞はトランスフェクション後の所定時点でCO−IP緩衝液(20mM Hepes、pH 7.2、150mM NaCl、0.5% TritonX100)を用いて抽出した。ユビキチン化アッセイおよび抗体分析のため、細胞または組織は強力な3界面活性剤緩衝液(20mMHepes、pH 7.2、150mM NaCl、1% Triton X−100、0.1% SDS、0.5%デオキシコール酸塩)を用いて抽出した。タンパク質抽出物は卓上型Beckmann Microfuge(Beckman Coulter; Fullerton CA)を用いて30分間の遠心分離により清澄化した。ウェスタン・ブロット分析では、10μlのタンパク質抽出物は15ウェル、4〜20%勾配のミニ・ドデシル硫酸ナトリウム(SDS)−ポリアクリルアミド・ゲルの1ウェルにローディングした。タンパク質は、100Vで2時間解離し、低温室で一晩30VでAmersham社のニトロセルロース・フィルター(Amersham; Piscataway NJ)に転写した。次いで、このフィルターはウェスタン・ブロット装置から取り出し室温で1時間5%の脱脂乳でブロッキングした。次に、ブロッキング溶液は所望濃度の一次抗体を含むブロッキング溶液と取り換えられた。ウェスタン・ブロットはAmersham Chemiluminescent Western blot detection kitで可視化した。
【0105】
シナプトタグミンXIの2ペプチドに対するウサギの抗体(抗sytXIAおよび抗sytXIB)は作製され、シナプトタグミン1に対するマウスの抗体(抗syt65)は購入した。sytIおよびsytXIは関連タンパク質であり他のシナプトタグミンに密接な相同性を有するため、抗syt65および抗sytXIの抗体は、これらの抗体を用いる前に、各抗原との交差反応性について試験しパーキン−sytXI相互作用を調べた。図2Aは免疫ブロッティング(ウェスタン・ブロッティング)を用いたシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。タンパク質抽出物はGFP−syt1、GFP−syt4、GFP−syt11、またはGFPを発現するヒト胎児腎臓(HEK)293T細胞から単離された。GFPまたはGFP−syt1、4および11のプラスミドでトランスフェクションされたHEK293細胞からのタンパク質抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した(図2A)。ウェスタン・ブロットは、抗GFP、抗syt65または抗sytXIAの抗体を用いて個別に検出した。抗syt1および抗syt11の両抗体はそれらの個々のエピトープを特異的に検出したが、GFP−syt4融合タンパク質を検出しなかった。予想どおり、抗syt65および抗syt11の両抗体はそれぞれのGFP−syt1またはGFP−syt11のタンパク質のみを検出した一方で、抗GFP抗体はGFPで標識した全てのタンパク質を検出した。抗syt11抗体は、syt4がsyt11と最も近い相同性を有するにもかかわらず、GFP−syt4を検出しなかった。「nt」として図2Aに示されるレーンは非トランスフェクション細胞であった。分子量マーカーの位置は右側に示す。
【0106】
sytXIA抗体の特異性はPC12細胞から得るタンパク質抽出物のウェスタン・ブロットで更に確認された(図2B)。強力な3界面活性剤の緩衝液におけるPC12細胞のタンパク質抽出物は、抗sytXIA、sytXIAペプチドで前吸収された抗sytXIA、またはsytXIBペプチドで免疫ブロットした。抗sytXIA抗体は64kDaの単独のバンドを認識した(図2B、レーン1)。p64バンドは、抗sytXIA抗体がsytXIAペプチドで前吸収された場合、検出されなかった(図2B、レーン2)。一方、異なるペプチド(sytXIB)との前吸収は抗sytXIA免疫反応性を阻害しなかった(図2B、レーン3)。総合すれば、これらの観察は抗sytXIA抗体の特異性を更に確認する。弱い界面活性剤の緩衝液(0.2%もしくは0.5%のNP40またはTriton−X100)を用いて単離されたタンパク質抽出物のウェスタン・ブロットにより、64kDaおよび110kDaの2つのsytXIA陽性バンドを得た。これはsytXIAがホモ二量体を形成しうることを示唆する。
【0107】
これらの結果は抗syt11抗体の特異性を確認する。
【0108】
(実施例III)
(パーキンは全長のシナプトタグミン1およびシナプトタグミン11と相互作用する)
本実施例は、パーキンが全長のシナプトタグミン1およびシナプトタグミン11と相互作用することを示す共免疫沈降実験を記載する。
【0109】
GFP−hsyt11AB融合タンパク質およびヘマグルチニン−パーキン(HA−パーキン)と共発現する293T細胞の共免疫沈降は、パーキンがhsyt11AB断片と共沈することを見出した。パーキンが全長のシナプトタグミンと相互作用するか否かを決定するために、全長のシナプトタグミン1(syt1)およびsyt11のcDNAはPEGFPベクターにクローニングされた。簡潔に述べれば、ヒトのsyt1およびsyt11の全長cDNAは、全読み枠に及ぶプライマー対を用いてpGAD10発現プラスミド(Clontech)にクローニングされたヒト成人脳cDNAライブラリーからポリメラーゼ連鎖反応(PCR)により得られた。ヒトのシナプトタグミンIをクローニングするために用いたプライマーは、前プライマー:TGGTGAGCGAGAGTCACCATGA(配列番号11);逆プライマー:B1、TTTCCTTTACTTCTTGACG(配列番号12);B2、TGAAGGACTTAGGGGCTCTCT(配列番号13)であった。ヒトのシナプトタグミンXIをクローニングするために用いたプライマーは、前プライマー:GAGGGTTCCCAGAGCTGTCT(配列番号14);逆プライマー:CACATCCCTCCCCAGCTTG(配列番号15)であった。ヒトのシナプトタグミンIおよびシナプトタグミンXIのヒトcDNA配列は、GenBank登録番号のBC058917(図12Aおよび図12B)およびBC039205(図12Cおよび図12D)で表されるように、それぞれ図12Aおよび図12Cに示される。他の発現プラスミドは全て特異的なPCRプライマー対を用いて同様に構築された。突然変異体パーキンのcDNAであるパーキンG289RおよびパーキンC418RはAlexis Brice教授(INSERM U289,Hopital de la Salpetriere,Paris,France)から賜った。
【0110】
細胞のトランスフェクションのため、細胞はトランスフェクションの24時間前にプレートに播種した。翌日、cDNAプラスミドは、polyfect形質転換体試薬(Qiagen)で処理し、製造業者のプロトコルに従ってHEK293細胞にトランスフェクションした。トランスフェクション後の望ましい時点(24時間、48時間および72時間)で、細胞は免疫蛍光標識用に固定されたまたは免疫沈降およびウェスタン・ブロット用に抽出された。トランスフェクションの24時間以上後に試験された細胞については、培地を一度交換した。
【0111】
インビトロ共免疫沈降実験を実施するために、HAで標識したパーキン発現cDNA(pCMV−HA−パーキン、pCMV−HA−欠失パーキン、pCMV−HA−パーキンC418R、pCMV−HA−パーキンG289R)は、個々のGFPで標識した融合タンパク質発現ベクター(pEGFP−hsyt11AB、pEGFP−syt1、pEGFP−syt11、またはpEGFP)を用いて、100mM2皿に60〜80%の密集度で標準培地条件で増殖したHEK293細胞に共トランスフェクションした。対照はpCMV−HA−パーキンでのみトランスフェクションされた細胞または非トランスフェクション細胞であった。ある実験において、β−アクチン抗体(Sigma;St.Louis,MO)およびβ−COP抗体が用いられた。下記の試薬:マウス抗HA、抗HA−ペルオキシダーゼ、抗myc−ペルオキシダーゼ、抗HA−アガロースはRoche Diagnostics社から購入した。48時間後、タンパク質は、0.5%のNP40およびプロテアーゼ阻害剤混合物(Roche Molecular Biochemicals;Indianapolis,IN)を含む界面活性剤緩衝液を用いて、基本的に実施例IIに記載するように抽出した。タンパク質抽出物は抗GFP(Chemicon;Temecula,CA)またはラットの抗HAアガロース基質(Roche)を用いて免疫沈降(ip)した。免疫沈降産物は抗GFP抗体および抗HA共役ペルオキシダーゼを用いて免疫ブロットした。
【0112】
図2CはHA−パーキンとGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体を用いて共免疫沈降(共IP)した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)を用いて検出した。
【0113】
pEGFP−syt1およびpEGFP−syt11のベクターはpCMV−HA−パーキン・プラスミドと共に293T細胞に個別に共トランスフェクションした。pEGFPプラスミドはベクター対照として使用した。24時間後、タンパク質抽出物が得られマウス抗GFP抗体と共沈しGFP融合タンパク質を沈降させた。共IP産物はウサギ抗GFP(図2C、下側パネル)またはラット抗HA共役ペルオキシダーゼ(図2C、上側パネル)を用いて免疫ブロットした。抗HAペルオキシダーゼは、GFP−syt1およびGFP−syt11を含むサンプル中でのみHA−パーキン・バンドを検出したが、GFP対照を含むサンプル中では検出しなかった。これは、HA−パーキンがシナプトタグミン融合タンパク質と特異的に共沈したがGFP標識とは共沈しなかったことを示す。従って、syt1およびsyt11の両タンパク質はパーキンと共沈したが、GFP標識はしなかった。
【0114】
大きなGFP標識はパーキンとシナプトタグミンとの相互作用に影響を及ぼしうる可能性を明らかにするため、syt1 cDNAはpCMV−myc発現プラスミドに転移された。抗HA−アガロースとHA−パーキンおよびGFP−syt1とを共発現する293T細胞からのタンパク質抽出物を共免疫沈降した後、抗myc共役ペルオキシダーゼまたは抗HA共役ペルオキシダーゼを用いて検出し、HA−パーキンもmyc−syt1と沈降することを見出した。これらの観察は、大きなGFP標識がパーキンとシナプトタグミンとの相互作用に影響を及ぼさないことを示している。従って、GFPで標識されたシナプトタグミンはその後の分析に用いた。
【0115】
内因性のパーキンがシナプトタグミンと相互作用するか否かを決定するために、インビボ共免疫沈降実験がPC12細胞で行われた。PC12細胞は、10%CO2、37℃のインキュベーターで、10%加熱不活性化血清、5%ウシ胎児血清、およびペニシリン/ストレプトマイシンを含むDMEMで増殖した。培地は三日毎に交換した。PC12細胞は48時間増殖し、タンパク質は共ip緩衝液(50mM Tris−HCl、pH 7.5、0.2% TritonX−100、150mM NaCl、および10mlの緩衝液当たり1ペレットのプロテアーゼ阻害剤(Roche))で抽出した。タンパク質抽出物は、マウスまたはウサギのIgG共役アガロースで前清澄化(preclear)し、低温室で一晩個々の抗体とインキュベートした。一次抗体は抗マウスまたは抗ウサギのIgG共役アガロースと2時間沈降させた。最終のペレットは共ip緩衝液で7回洗浄し、共沈物はSDS−PAGE緩衝液を用いて二次抗体と共役したアガロースから溶出した。共ip産物は抗パーキン抗体または抗syt抗体のいずれかと免疫ブロットした。
【0116】
図2DはPC12細胞におけるsyt1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞のタンパク質抽出物との共ip。共IP産物はウサギ抗パーキン抗体およびマウス抗syt1抗体を同時に用いて検出した。内因性パーキンは抗syt1免疫沈降物中に検出されたが、マウスIgGを用いた対照の免疫沈降物中には検出されなかった。レーン4はPC12タンパク質溶解物のブロットを示す。
【0117】
マウス抗syt1抗体を用いた共免疫沈降はsyt1および内因性パーキンの両方を沈降させた(図2D)。マウスIgGはsyt1またはパーキンのいずれも沈降させなかった。これは内因性共ipの特異性を示唆する。抗パーキン抗体または抗syt11抗体を用いた共ipはsyt11とパーキンを共沈させなかった。いかなる内因性のパーキン−syt沈降物も見出せなかったことは、おそらく共免疫沈降で使用した緩衝液への内因性syt11の不溶性の結果であっただろう。
【0118】
パーキンが内因性sytXIと相互作用したか否かを決定するために、ヒトの大脳皮質から得るタンパク質抽出物は抗parkA抗体と免疫沈降した。ヒトの脳抽出物の共免疫沈降物については、1.4 gのヒトの大脳皮質サンプルは小片に刻まれ7mlの冷溶解緩衝液(100mM Tris−HCl、pH 7.5、150mM NaCl、1% TritonX−100、0.05% SDS、0.05% デオキシコール酸、および1プロテアーゼ阻害剤ペレット/10ml緩衝液)に再懸濁した。細胞懸濁液はガラス製のホモジナイザーによりホモジナイズした。タンパク質溶解物は1mlのアリコートに等分し低温室で最高速度で遠心分離した。タンパク質抽出物はウサギIgG共役アガロースおよびタンパク質A共役アガロースを用いて4℃で3時間前清澄化し、前清澄化した溶解物はウサギ抗parkA抗体またはマウス抗sytXI抗体を用いて低温室で一晩インキュベートした。一次抗体はタンパク質A共役アガロースと4℃で4時間沈降した。最終ペレットは共ip緩衝液で5回洗浄し、共沈物はSDS−PAGE緩衝液を用いて二次抗体と共役したアガロースから溶出した。共ip産物はニワトリ抗parkA抗体または抗sytXIA抗体のいずれかと免疫ブロットした。
【0119】
ヒトの大脳皮質抽出物の共ip後、沈降物は次いでニワトリ抗parkA抗体または抗sytXIA抗体のいずれかを用いて検出した(図2E、レーン1および2)。抗parkA抗体は抗parkA沈降物に単独のパーキン・バンドを検出した(図2E、レーン1、上側パネル)一方、抗sytXIA抗体はsytXIタンパク質バンドを検出した(図2E、レーン1、下側パネル)。同じタンパク質抽出物が抗sytXI抗体と共免疫沈降した場合、パーキンはsytXIと特異的に共沈した(図2E、レーン3)。対照反応におけるsytXIおよびパーキンの両バンドの欠如(図2E、レーン2および4)は、この細胞でのパーキン−sytXI相互作用の特異性を証明し、内因性パーキンが内因性sytXIと相互作用したことを確認した。
【0120】
これらの結果はパーキンがシナプトタグミン1およびシナプトタグミン11と相互作用することを示す。
【0121】
(実施例IV)
(RING2モチーフはシナプトタグミンの結合に必須である)
これらの実験はシナプトタグミンの結合におけるパーキンのRINGモチーフの役割の特徴決定を記載する。
【0122】
パーキンのどのドメインがシナプトタグミンに結合するかを決定するために、HAエピトープで標識された幾つかの欠失パーキンが構築された(図3A)。欠失パーキンcDNA発現プラスミドはGFP−sytとの共発現に十分な欠失パーキンを発現した(図3C)。それぞれの欠失構築物はGFP−sytXIと共発現した。24時間後、タンパク質抽出物はラット抗HAと共役したアガロースを用いて免疫沈降した後、抗GFP抗体でウェスタン・ブロット検出した。アミノ酸残基204〜293(p1〜203、p294〜385、およびp294〜465)を欠く欠失パーキンは全長のシナプトタグミンXIと相互作用できなかった。これらの欠失パーキンがsytXIと相互作用できないことは、欠失パーキンの発現レベルの低下の結果ではなかった。p1〜203およびp294〜465のパーキンの発現レベルはsytXIと相互作用した構築物よりはるかに高かった(図3C)。p204〜293ドメインの全てまたは一部(p1〜465、p1〜314、p78〜465、p78〜238、p257〜465)を含む欠失パーキンはsytXIと相互作用し、全て異なる結合親和性を有した(図3B)。これらの観察は、RING1ドメインを含むアミノ酸残基204〜293がsytXIとパーキンの相互作用に重要であることを示す。
【0123】
結合アッセイから得るデータは、パーキンに少なくとも2つのsytXI結合部位が存在することを示唆する。第一の結合部位はアミノ酸残基の204〜238に位置する。これは、RING1ドメインを含まないp78〜238のペプチドがsytXIと相互作用したがp1〜203のペプチドが結合しなかったという観察(図3)により支持された。第二の結合部位はアミノ酸残基の257〜293のRING1ドメイン内に位置する。これは、ペプチドp257〜465がsytXIと相互作用したが、ペプチドのp294〜385およびp294〜465が結合しなかった(図3)という観察により支持された。この観察はパーキンにおける病気を惹起するアミノ酸置換の作用により更に支持された。RING1フィンガー・モチーフにミスセンス変異を含むパーキン(パーキンC289G)はパーキンC418Rと比べてsytXIと弱く(2.5倍未満)相互作用した(図3B)。さらに、両変異パーキンはsytXIをユビキチン化できない(図4)が、パーキンC418RはsytXIへの結合能を喪失しない(図3B)。
【0124】
異なる境界を有する他のパーキン欠失変異体について同様な結果が見られた。パーキンの結合部位はRING2フィンガー・モチーフにマッピングされた。同様な結果はsyt1についても観察された。
【0125】
これらの結果はsyt11の結合部位がRINGフィンガー・モチーフに位置することを示す。
【0126】
(実施例V)
(パーキンによるシナプトタグミンのユビキチン化)
本実施例はパーキンによるシナプトタグミンのユビキチン化を記載する。
【0127】
パーキンはE3ユビキチン・リガーゼであり、プロテアソーム複合体またはリソソームによる分解の標的となる特異的基質のユビキチン化に必要な必須酵素である(Shimuraら、前出,2000;Zhangら、前出,2000)。シナプトタグミンがパーキンの基質であるか否かを決定するために、インビトロ・ユビキチン化アッセイは、引用文献により本明細書に組み込まれるZhangら、Proc.Natl.Acad.Sci.USA 97:13354〜13359(2000)により記載される通りに実施した。簡潔に述べれば、HEK293細胞は5μgのpCMV−Myc−ユビキチン、pEGFP−syt1または−syt11、および異なるpCMV−HA−パーキン(野生型、欠失、およびミスセンス)を用いてトランスフェクションした。24(または時折36)時間後、細胞は20μMラクタシスチン(lactacystin)または20μMプロテアソーム阻害剤Iを含む標準培地で4時間インキュベートした。細胞は冷DPBSで洗浄した。ユビキチン化アッセイで、細胞は3界面活性剤緩衝液(20mM Hepes、pH 7.2、150mM NaCl、1% Triton X−100、0.1% SDS、0.5%デオキシコール酸塩)で抽出した。タンパク質はプロテアーゼ阻害剤ペレット(Roche社、10mlの緩衝液当たり1ペレット)および脱ユビキチン化酵素を阻害するための2μM N−エチルアミド(ethylamimide)を含むRIPA緩衝液で抽出した。タンパク質抽出物はマウス抗GFP抗体を用いて免疫沈降し、IP産物は、抗HA、抗myc、およびウサギ抗GFPで検出した。
【0128】
ユビキチン化実験において、HEK293細胞は、GFP標識されたsyt1またはGFP標識されたsyt11のいずれかと、HA標識されたまたは対照のパーキンcDNAプラスミドおよびmyc標識されたユビキチンとで共トランスフェクションされた(図4)。GFPは基質特異性の負の対照として使用し、一方、HA標識した欠失パーキン・タンパク質(図4A〜4Cのp78〜465、p1〜203、p294〜385、p294〜465)(図4D〜図4Fのp1〜p314およびp77〜p465)は野生型パーキンの負の対照として使用した。HA−パーキンならびに対応するmyc−ユビキチンおよびGFPで標識されたタンパク質を過剰発現するHEK293細胞はラクタシスチンで4時間処理し、タンパク質抽出物は抗GFP抗体で免疫沈降した。syt1のユビキチン化アッセイの産物は図4Dにmyc−(上端)、GFP−(中間)、およびHA−標識(下端)に対する抗体を用いて検出した。図4Eに示されるようにsyt11のパーキン・ユビキチン化の検出では、ウェスタン・ブロットが抗myc(上側)および抗GFP(下側)の抗体を用いて検出した。両アッセイにおいて、GFP−syt1およびHA−パーキンを共発現する細胞は、HA−パーキンおよび対照を発現する細胞より多くのユビキチンと共役したsyt1複合体を形成した。HA−パーキンおよびGFPを発現する細胞ならびに他の負の対照におけるユビキチン化産物の欠如に留意せよ。
【0129】
図4に示すように、細胞がプロテアソーム複合体の阻害剤であるラクタシスチンとインキュベートした場合、野生型パーキンおよびGFP−syt11(図4A〜4Cおよび4E)またはGFP−syt1(図4D)を発現する細胞はバックグランド対照を上回るユビキチン−シナプトタグミン共役体の蓄積を示した。野生型パーキンはGFP標識の多ユビキチン化に影響を及ぼさなかった。ペプチドの1つ(p78〜465)はシナプトタグミンに結合できた(図3)が、欠失パーキンはユビキチン化されたシナプトタグミン共役体のレベルにほとんど影響を示さず、欠失パーキンの発現レベルは野生型パーキンと比べて高かった。GFP−sytおよびmyc−ユビキチンのみを共発現する細胞で見出されるユビキチンと共役するsytの存在は、HEK293細胞における内因性パーキンの存在の結果であるらしかった。ユビキチン化sytの様式は、異なる長さのユビキチン鎖を含む、多様な種類の存在を示した。
【0130】
病気に関連する点突然変異がsytをユビキチン化するパーキンの能力に影響するか否かを決定するために、ミスセンス変異パーキンC289GおよびパーキンC418Rについてユビキチン化アッセイも実施した。両変異パーキンは野生型パーキンと比べて検出不能なレベルのユビキチン化sytXIを生じた(図4)。より長い暴露下で、欠失およびミスセンス変異のパーキンでトランスフェクションした細胞は全てGFP−sytXIのみでトランスフェクションした細胞と同程度の弱いレベルのユビキチンと共役したsytXIを生じたが、ユビキチン化sytXIのレベルは野生型パーキンにより生じるレベルより実質的に低かった。このユビキチン化sytXIのバックグランド・レベルは、おそらく内因性パーキンによってまたはHEK293細胞の未同定のE3ユビキチン・リガーゼによって生じたと思われ、HEK293細胞を用いた他の研究者による観察(Renら、J.Neurosci.23:3316−3324(2003))と一致している。共ip(図3)およびユビキチン化(図4)の両実験において、大半の変異パーキンはSDS−PAGEサンプル緩衝液に溶解するペレット画分で検出できた。
【0131】
抗GFP抗体を用いた同じタンパク質サンプルのウェスタン・ブロット分析は、パーキン−sytXIを共発現するサンプルをローディングしたウェルの上端近くに高分子量のGFP−sytXIバンドを検出した(図4)。抗myc抗体もユビキチン化sytXI複合体を濃く検出したため、このバンドは不溶性の同複合体であると思われた。このバンドは対照でかすかに観察された。より小さな分子量のユビキチン化sytXI種は抗GFP抗体により検出できなかった。これらの発見はHEK293細胞におけるα−およびβ−チューブリン(Renら、前出,2003)ならびにシンフィリン−1(Chungら、Nat.Med.7:1144−1150(2001))のユビキチン化実験と一致している。これらの実験において、ユビキチン化基質は各自のタンパク質に対する抗体により検出できなかったが、ユビキチンまたはその標識に対する抗体は各自のパーキンが媒介するユビキチン化基質を濃く検出した。
【0132】
同様な結果はsyt1についても見出された。図4Fに示すように、変異パーキンのC289GおよびC418Rは、syt1およびsyt11のユビキチン化の減少を示した。パーキン変異体およびGFP−syt1またはGFP−syt11を発現する細胞は少量のユビキチンと共役したsytを生じた。変異体C289Gによるsyt11の弱いユビキチン化に留意せよ。従って、パーキンC289G変異体はsyt1のユビキチン化を阻害するが、syt11を弱くユビキチン化する。対照的に、パーキンC418R変異体はsyt1およびsyt11の両ユビキチン化を等しく阻害する。変異体パーキンおよびsytを共発現する細胞のユビキチン化様式は、野生型パーキンおよびsytを発現する細胞より弱かった(図4F、上端パネル)。パーキンおよびsytを発現しラクタシスチンで4時間処理された細胞から得るタンパク質抽出物のウェスタン・ブロットは、野生型パーキンと比べて少量の変異体を検出した(図4F)。変異体パーキンは不溶性ペレットで豊富に見出された(図4F)。
【0133】
パーキンが媒介するユビキチン化基質はプロテアソーム依存性経路による分解を受ける(Zhangら、Proc.Natl.Acad.Sci.USA 97:13354−13359(2000);Imaiら、Cell 105:891−902(2001);Imaiら、Mol.Cell.10:55−67(2002);Renら、J.Neurosci.23:3316−3324(2003);Cortiら、Hum.Mol.Genet.12:1427−1437(2003);Engelenderら、Nat.Genet.22:110−114(1999))ため、HAベクターおよびHA−パーキン1−465でトランスフェクションしたHEK293細胞でのGFP−sytXIの代謝回転を測定するためにパルス・チェイス実験が実施された。パーキンがGFP−sytXIの分解を促進するか否かを決定するため、HEK293細胞はGFP−sytXIおよびHAベクターまたはGFP−sytXIおよびHA−パーキン・プラスミドで共トランスフェクションされた。24時間後、細胞は5%透析FBSを含み且つMetおよびCysのアミノ酸を含まないDMEMで一度洗浄した。細胞は、この培地で30分間インキュベートし、100μCi/mlの35S−Met/Cys(EXPRE35S35S(35S)Protein Labeling Mix,Amersham)を含む同培地で30分間増殖した。その後、細胞は表示する時点で追跡された。タンパク質抽出物はRIPA緩衝液(20mM HEPES,pH 7.5,150mM NaCl,0.1% SDS,0.5% デオキシコール酸塩、1% Triton−X100,1mM EDTA,プロテアーゼ混合ペレット)を用いて単離した。GFP−sytXIは、上述したように、ウサギ抗GFP抗体(CHEMICON;Temecula、CA)と免疫沈降した。
【0134】
パーキンはHA−パーキン1−465でトランスフェクションしたHEK293細胞においてGFP−sytXIの分解を増加させた(図5)。HA−パーキン1−465を発現する細胞で1.5時間の追跡後、新しく合成されたGFP−sytXIの約40%が分解された一方で、HAベクターでトランスフェクションした細胞では等量のGFP−sytXIを分解するのに3時間かかった。
【0135】
これらの結果はパーキンがsyt1およびsyt11の両方をユビキチン化することを証明する。
【0136】
(実施例VI)
(パーキンはシナプトタグミンと共局在化しシナプトタグミンを核周辺複合体に補充する)
本実施例はパーキンおよびシナプトタグミンの細胞位置を記載する。
【0137】
2つのタンパク質が同じ細胞内区画を占めるならば、それらの相互作用は生理的に関連すると思われる。パーキン−syt共局在化を調べるため、免疫蛍光実験が行われた。簡潔に述べれば、COS1細胞培養で、COS1細胞は5%CO2で37℃のインキュベーターにおいて10%FBSおよびペニシリン/ストレプトマイシンを補足したDMEM培地で増殖した。培地は3日毎に交換した。トランスフェクションの一日前に、予め20μg/mlのコラーゲンIVで被覆した1cmのカバースリップに50,000個の細胞が播種された。PC12細胞は実施例IIIに記載するように増殖した。
【0138】
免疫蛍光標識および共焦点レーザー顕微鏡が下記の通りに実施された。細胞は、氷上で20分間4%のパラホルムアルデヒドを含むDPBSで固定し、溶液A(DPBS,3% ヤギ血清、0.05% TritonX−1000)で30分間インキュベートした。細胞は次いで選択され且つ溶液Aで希釈されたマウスまたはウサギの一次抗体と室温で1時間インキュベートした。次に、細胞は、冷DPBSで5回洗浄し、溶液Aで希釈したフルオレセイン・イソチオシアネート(FITC)またはテトラメチルローダミン・イソチオシアネート(TRITC)のいずれかと共役した該当する二次抗体と室温で1時間インキュベートした。次に、細胞は、冷DPBSで5回再洗浄し、退色保護のために80%グリセロールおよび10mM没食子酸ナトリウム中スライドで被覆した。細胞は油浸100Xレンズを通してLeica・TCSSP(真走査型共焦点分光測光)顕微鏡を用いて観察した。TRITC蛍光とFITCまたはGFPで漂白を妨げるため画像は順次得た。
【0139】
パーキン−sytの共局在化を調べるため、PC12細胞はNGFで7日間誘導した。PC12細胞は、50ng/mlのNGFで7日間誘導し、ウサギ・パーキン(赤に染色、図6P)およびマウスsyt1(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6D)に対する抗体を用いて免疫蛍光で共標識した。画像は100x油浸レンズを用いてLeica・TCSSP顕微鏡により得た。積層画像が併合された(図6Cおよび6R)。併合画像の黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび6P〜6Rの挿入物は長い神経突起が生じる同細胞の細胞体に由来する(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。
【0140】
NGF誘導性PC12ニューロンは、ウサギ・パーキンおよびマウスsyt1(図6P〜6R)、またはニワトリ・パーキンおよびウサギsyt11(図6A〜6C)に対する抗体で共標識した。パーキンは核膜周辺の位置(図6R、下の矢印)および神経突起に沿ったボタン(図6Cおよび6R、白矢印)でsyt1およびsyt11の両方と共局在化した。細胞の他の領域に存在するパーキンまたはGFP−sytは共局在しなかった。これらの結果は、内因性のパーキンがPC12細胞で内因性のsyt1およびsyt11と共局在化することを示す。
【0141】
シナプトタグミンの分布およびレベルに及ぼすパーキンの影響を調べるため、HEK293細胞はGFP−sytおよびHA−パーキンの両ベクターで共トランスフェクションした。36時間後、細胞は抗HA抗体で標識した。
【0142】
細胞は、GFP−syt1およびHA−パーキン(図7A〜7C)、GFP−syt1およびHA−ベクター(図7D〜7F)、GFP−syt11およびHA−パーキン(図7G〜7I)、GFP−syt11およびHA−ベクター(図7J〜7L)またはGFP−ベクターおよびHA−パーキン(図7M〜7O)で共トランスフェクションした。トランスフェクションされた細胞は抗HAで標識し、画像は100x油浸レンズを用いてLeica・TCSSPにより得られた。syt1およびsyt11とパーキンとの共局在化の差異に留意せよ。
【0143】
パーキンおよびsytの両方を共発現するHEK293細胞において、HA−パーキンおよびGFP−sytは核周辺膜に近接した凝集体に共局在化した(図7A〜7C、7G〜7I)。これらの細胞において、GFP−sytのレベルは対照と比べてはるかに低かった。対照では、HEK293細胞がGFP−sytおよびHAベクターのみで共トランスフェクションされた場合、GFP−sytは細胞全体にわたって細胞質小胞に分布することが見出された。同様に、HA−パーキンがGFPと発現した場合、細胞全体にわたって拡散して分布した(図7M〜7O)。
【0144】
これらの結果は、パーキンが共トランスフェクションされたHEK293細胞でsytと共局在化し、それらの正常な細胞分布を変えることを示す。
【0145】
(実施例VII)
(SytXIはヒトの黒質ニューロンの細胞体および神経突起に局在する)
黒質ニューロンの死およびレヴィー小体の形成はPDの多くの型の特質であり、レヴィー小体の幾つかの構成タンパク質はPDの遺伝形が変異する。古典的PDの神経変性の病因におけるsytXIおよびパーキンの潜在的な関連性を確立するため、二人の標準的なPD患者および二人の散発性PD患者から得られる黒質切片の免疫組織化学標識がsytXI(抗sytXIA)、パーキンおよびユビキチンに対する抗体を用いて実施された。
【0146】
ヒトの黒質切片の免疫組織化学標識では、Huynhら、(Ann.Neurol.48:737−744(2000))に記載される免疫組織化学標識プロトコルを用いて、ヒトの脳の7μmの切片がウサギ抗sytXIA(10μg/ml)、parkA(5μg/ml)、ユビキチン(1/500)の抗体で染色された。簡潔に述べれば、脳の切片はBiomedia‘s Autozyme溶液(Fisher社)により脱パラフィン処理し(deparafinize)脱マスキングした。切片は、次いで、3%の標準ヤギ血清でブロッキングし低温室で一晩一次抗体とインキュベートした。翌日、切片はElite Vector ABCキット(Vector,San Diego,CA,USA)およびBiomedia社のジアミノベンジデン基質キット(Fisher)を用いて顕色した。ペプチド前吸収では、10μgの抗sytXIA抗体は低温室で一晩1000倍のsytXIAペプチドと前インキュベートした。翌日、前インキュベートした抗体は10分間遠心分離し1mlの染色緩衝液で希釈した。画像は20Xおよび63xの油浸レンズを用いて得られSPOTデジタル・カメラにより保存された。
【0147】
図6D〜6Oに示されるように、正常なヒトの黒質ニューロンはsytXIおよびパーキンに対する抗体により濃く染色された。抗sytXIA抗体は、抗parkA抗体(図6F、6Iおよび6L)と同様に、黒質の細胞体および神経突起伸長物の両方を標識した(図6D、6Gおよび6J)。sytXIAペプチドと前吸収した抗sytXIAは黒質ニューロンを標識しなかった(図6E、6Hおよび6K)ため、sytXI免疫反応性は特異的であった。
【0148】
図6M〜6Oは散発性PDの個体から得られる黒質ニューロンの免疫組織化学標識を表す。抗sytXIA抗体は、抗ユビキチン抗体(図6M)と同様に、細胞内レヴィー小体の核を標識した(LB;図6N、黒矢印)。sytXIのLB染色はユビキチン染色と比べて薄かった。神経網の濃い標識が注目される。sytXIAペプチドと前吸収した抗sytXIA抗体がレヴィー小体または神経網を標識しなかったため、この標識は特異的であった(図6O、黒矢印)。先に報告されたように(Huynhら、Ann.Neurol.48:737−744(2000))、これらの2つの散発性PD脳におけるレヴィー小体は抗parkA抗体に染色しなかった。
【0149】
これらの結果はsytXIがヒトの黒質ニューロンの細胞体および神経突起に局在することを示す。
【0150】
(実施例VIII)
(パーキンと相互作用するポリペプチドの同定および特徴決定)
本実施例はパーキンと相互作用するポリペプチドの同定を記載する。
【0151】
酵母2−ハイブリッド・スクリーニングは実施例Iに記載するように実施した。簡潔に述べれば、パーキンと相互作用するCNSタンパク質を同定するために、酵母2−ハイブリッドアッセイはヒトの脳cDNAライブラリーから1x106個の独立した酵母コロニーについて実施した。スクリーニングはおとりとして全長のパーキンを用いた。酵母フィルターを用いた次のスクリーニングおよび液体アッセイはクローンが陽性であることを確認した。
【0152】
ヌクレオチド配列決定は、酵母2−ハイブリッド・スクリーニングで同定された1つのクローンが新規の小さなシナプシン様タンパク質(SLP)のN末端ドメインをコードすることを示した。ポリペプチドの配列は図8に示す。
【0153】
図9Aに示すように、シナプシン様タンパク質(SLP)とも呼ばれるMP36aおよびMP23aの両型は酵母フィルターアッセイでパーキンと相互作用した。負対照のプラスミドであるpGBT9ベクター、pGBT9−NF2、およびpGBT9−Hrsでは結合は観察されなかった。パーキンとの結合は実施例Iに記載するように液体培養アッセイを用いて確認した。図9Bに示すように、MP36aおよびMP23aは、負対照のpGBT9ベクターより、パーキンの存在下でそれぞれ約20倍および30倍高いβ−ガラクトシダーゼ活性を示した。
【0154】
相互作用を更に試験するため、実施例IIIに記載するように共免疫沈降実験が実施された。共免疫沈降実験では、構築物はGFP融合として作製した。共免疫沈降実験は全長のタンパク質がパーキンと相互作用することを示した。細胞は、図9Cに示すように、個々のプラスミドでトランスフェクションした。HA−パーキンおよびMP36aまたはMP23a GFP融合を発現する細胞から得るタンパク質抽出物の共免疫沈降はHA−アガロースを用いて実施し、個々のGFP融合タンパク質の位置はGFP抗体を用いてウェスタン・ブロッティングにより決定した。図9Cに示すように、MP36aおよびMP23aは共にパーキンと共免疫沈降した。
【0155】
内因性パーキンが野生型SLPと結合しうるか否かを決定するために、共免疫沈降実験がPC12細胞で行われた。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照を用いて免疫沈降した(図9D)。IP産物はニワトリ抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキンのバンドを検出した。このバンドはニワトリIgG免疫沈降物のサンプルでは存在しなかった。抗SLP抗体は抗parkAの免疫沈降物およびPC12タンパク質抽出物において36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性パーキンが野生型SLPと共沈降したことを示す。
【0156】
ヒトの黒質のSLPの発現も試験した(図10)。黒質ち密部切片は、10μg/mlのアフィニティー精製された抗SLP(図10Aおよび10C)、抗SLP+SLPペプチド(図10B)、抗ユビキチン(図10D)の抗体を用いて免疫組織化学的に染色した。一次抗体はVector Elite Vectastain Rabbit ABCキットを用いて検出し、3,3′−ジアミノベンジジン(DAB)で可視化した。全ての切片は同一に処理し染色した。抗SLPおよび抗sytXIの両抗体は黒質ち密部におけるニューロンの神経突起を濃く標識した。SLPペプチドと前吸収した抗SLP抗体は反応しなかった。これは免疫組織化学標識の特異性を示す。細胞体で見られるこげ茶色の染色はドパーミン作動性ニューロンで見出されるニューロメラニン(neuromelanin)である。図10Cおよび図10Dは抗SLP(図10C)および抗ユビキチン(図10D)の抗体を用いたレヴィー小体(LB)の標識を示す。
【0157】
パーキンおよびSLPの共局在化は基本的に実施例VIに記載されるように決定した。細胞はパーキン抗体(緑に染色、図11A)、およびSLPを認識する抗体(赤に染色、図11B)で共標識した。積層画像はパーキンおよびSLPが共局在化することを示す(黄色に染色、図11C)。NGF誘導性PC12ニューロンの共焦点免疫蛍光研究は両SLPがシナプス・ボタンでパーキンと共局在化することを確認した。図11Dおよび図11Eはそれぞれ黒質および大脳皮質の染色を示す。ユビキチン化実験は実施例Vに記載するように実施した。
【0158】
これらの結果は、SLPがインビボでパーキンと共局在化するパーキンと結合するタンパク質であることを示す。これらの結果は、SLPが特発性PDの病理学的特質であるレヴィー小体に局在することも示し、SLPが直接的に(突然変異により)あるいは間接的に(枯渇により)PDの病因に機能し得ることを示す。
【0159】
本出願を通して、種々の刊行物が参照された。これらの刊行物の開示は、本発明に係る技術の状況をより十分に記載するために、本出願の参照文献により本明細書にそのまま組み込まれる。本発明は上記で提供した実施例に関して記載したが、種々の改変が本発明の精神を逸脱することなく為され得ることが理解されるべきである。
【図面の簡単な説明】
【0160】
【図1】図1Aは酵母2−ハイブリッド・フィルターアッセイを示す。pGAD10−hyst11ABおよびpGBT9−パーキンで形質転換した酵母細胞は、β−ガラクトシダーゼ基質と陽性反応した。図1Bは典型的な酵母2−ハイブリッド液体アッセイを示す。酵母はpGAD10−hsyt11ABおよびpGBT9−パーキンのプラスミド(図1Bの「1」)またはpGAD10−hsyt11ABおよびpGBT9ベクターの対照(図1Bの「2」)で形質転換した。
【図2−1】図2Aはシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。緑色蛍光タンパク質(GFP)およびGFP−syt1、GFP−syt4、およびGFP−syt11のプラスミドでトランスフェクションされたHEK293細胞から得られる細胞抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した。「nt」の印が付いたレーンはトランスフェクションされていない細胞をロードした。図2Bはウサギ抗sytXIA抗体で検出されたPC12細胞から得られるタンパク質抽出物のウェスタン・ブロットを示す。レーン1、1μg/mlの抗sytXIA抗体とインキュベーション;レーン2、sytXIAペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション;レーン3、sytXIBペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション。抗sytXIA抗体は64kDaの単独のバンドを検出する。このバンドはsytXIAペプチドとの前インキュベーションで前吸収したがsytXIBペプチドでは前吸収しなかった。図2Cはヘマグルチニン−パーキン(HA−パーキン)とGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体と共免疫沈降した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)で検出した。図2DはPC12細胞におけるシナプトタグミン1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞からのタンパク質抽出物との共ip。共IP産物はウサギ抗パーキンおよびマウス抗syt1抗体で同時に検出した。レーン4はPC12タンパク質の溶解物のブロットを示す。図2Eは内因性のパーキンおよびsytXIAの共免疫沈降を示す。ヒトの大脳皮質から得られるタンパク質抽出物は、ウサギ抗parkA抗体(レーン1)、または対照のウサギIgG(レーン2)を用いて共免疫沈降した。沈降物は抗parkA(上側パネル)または抗sytXIA抗体(下側パネル)を用いて検出した。抗parkA抗体はsytXIを共免疫沈降させたがウサギIgG対照はしなかった。逆共ipを行うために、タンパク質抽出物は同一の希釈でウサギ抗sytXI抗血清(レーン3)または対応する前血清(preserum)(レーン4)と共沈した。ウェスタン・ブロットはニワトリ抗parkAを用いて検出した(レーン3およびレーン4)。このニワトリ抗parkAは抗sytXI抗血清で内因性パーキンのバンドを検出した(レーン3)が、前血清の対照では検出しなかった(レーン4)。
【図2−2】図2Aはシナプトタグミン1およびシナプトタグミン11に対する抗体の特異性を示す。緑色蛍光タンパク質(GFP)およびGFP−syt1、GFP−syt4、およびGFP−syt11のプラスミドでトランスフェクションされたHEK293細胞から得られる細胞抽出物のウェスタン・ブロットは、GFP、syt1、およびsyt11に対する抗体を用いて検出した。「nt」の印が付いたレーンはトランスフェクションされていない細胞をロードした。図2Bはウサギ抗sytXIA抗体で検出されたPC12細胞から得られるタンパク質抽出物のウェスタン・ブロットを示す。レーン1、1μg/mlの抗sytXIA抗体とインキュベーション;レーン2、sytXIAペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション;レーン3、sytXIBペプチドと前インキュベーションした抗sytXIA抗体とインキュベーション。抗sytXIA抗体は64kDaの単独のバンドを検出する。このバンドはsytXIAペプチドとの前インキュベーションで前吸収したがsytXIBペプチドでは前吸収しなかった。図2Cはヘマグルチニン−パーキン(HA−パーキン)とGFP−syt1およびsyt11とのインビトロ相互作用を示す。HA−パーキンおよび対応するGFP融合タンパク質を過剰発現するHEK293細胞から得られるタンパク質抽出物はマウス抗GFP抗体と共免疫沈降した。免疫沈降産物はラット抗HA−ペルオキシダーゼ(上側パネル)およびウサギ抗GFP抗体(下側パネル)で検出した。図2DはPC12細胞におけるシナプトタグミン1と内因性パーキンとのインビボ相互作用を示す。5μl(レーン1)および1μl(レーン2)のマウス抗syt1、ならびに1μlのマウスIgG(レーン3)とPC12細胞からのタンパク質抽出物との共ip。共IP産物はウサギ抗パーキンおよびマウス抗syt1抗体で同時に検出した。レーン4はPC12タンパク質の溶解物のブロットを示す。図2Eは内因性のパーキンおよびsytXIAの共免疫沈降を示す。ヒトの大脳皮質から得られるタンパク質抽出物は、ウサギ抗parkA抗体(レーン1)、または対照のウサギIgG(レーン2)を用いて共免疫沈降した。沈降物は抗parkA(上側パネル)または抗sytXIA抗体(下側パネル)を用いて検出した。抗parkA抗体はsytXIを共免疫沈降させたがウサギIgG対照はしなかった。逆共ipを行うために、タンパク質抽出物は同一の希釈でウサギ抗sytXI抗血清(レーン3)または対応する前血清(preserum)(レーン4)と共沈した。ウェスタン・ブロットはニワトリ抗parkAを用いて検出した(レーン3およびレーン4)。このニワトリ抗parkAは抗sytXI抗血清で内因性パーキンのバンドを検出した(レーン3)が、前血清の対照では検出しなかった(レーン4)。
【図3】図3A〜3Cはパーキンのドメインに対するsytXIの結合部位の地図のマッピングを示す。図3Aはパーキンの地図を示す。全長および欠失のパーキンはPCRによって構築しHA−エピトープ標識と枠内にクローニングした。C289GおよびC418Rはそれぞれアミノ酸位置の289および418にミスセンス変異を含むパーキンを示す。図3BはsytXIの結合部位がパーキンのRING1モチーフに位置することを示す。アミノ酸残基204〜293(RING1フィンガー・モチーフを包含する)を欠く欠失パーキンはsytXIに結合できなかった。C289Gミスセンス変異パーキンはC418R変異と比べてsytXIと弱く相互作用した。図3CはHEK293細胞におけるHAで標識したパーキンの発現を示す。全長野生型、ミスセンス変異、または多様な欠失のパーキンを過剰発現するHEK293細胞のウェスタン・ブロットは抗HA−ペルオキシダーゼを用いて検出した。
【図4−1】図4A〜4CはsytXIのユビキチン化アッセイを示す。HA−パーキンまたはMyc−ユビキチンおよびGFPで標識したタンパク質の対照を過剰発現するHEK293細胞はラクタシスチン(lactacystin)で4時間処理し、タンパク質抽出物は抗GFP抗体と免疫沈降した。ユビキチン化アッセイのIP産物はmyc標識に対する抗体(図4A)および抗GFP抗体(図4B)を用いて検出した。HA−パーキンおよびGFPを発現する細胞のユビキチン化産物の欠如、ならびに他の対照におけるGFP−sytXIのユビキチン化の検出不能なレベルに留意せよ。パーキン変異体およびGFP−sytXIを発現する細胞は野生型のパーキンと比べて少ない量のユビキチンと共役したsytXIを産生する。抗GFP抗体は、GFP−sytXIを含む全サンプルにおいてほぼ等量のGFP−sytXI単量体、ならびにGFP−sytXIおよびHA−パーキンの両方と共発現したサンプルを含むウェルの近くに大きなGFP−sytXIバンドを検出した。図4Cは抗HA抗体を用いた同溶解物のウェスタン・ブロットを示す。これは欠失パーキンおよび変異パーキンが野生型パーキンより高いレベルで発現することを示している。図4DはHA−パーキンおよび対応するmyc−ユビキチンおよびGFPで標識したsyt1タンパク質を過剰発現するHEK293細胞のタンパク質抽出物の抗GFP抗体を用いた免疫沈降を示す。この細胞はプロテオソーム複合体の阻害剤であるラクタシスチンで4時間処理した。ユビキチン化アッセイの免疫沈降産物はmyc(上端パネル)、GFP(中間パネル)、およびHA−標識(下端パネル)に対する抗体を用いて検出した。図4Eは、GFP−syt11が使用されたことを除くと、図4Dに類似する。ウェスタン・ブロットは抗myc(上端パネル)および抗GFP(下端パネル)の抗体を用いて検出した。図4Fは、変異パーキン、C289GおよびC418Rを用いた共ipおよびウェスタン・ブロットを示す。
【図4−2】図4A〜4CはsytXIのユビキチン化アッセイを示す。HA−パーキンまたはMyc−ユビキチンおよびGFPで標識したタンパク質の対照を過剰発現するHEK293細胞はラクタシスチン(lactacystin)で4時間処理し、タンパク質抽出物は抗GFP抗体と免疫沈降した。ユビキチン化アッセイのIP産物はmyc標識に対する抗体(図4A)および抗GFP抗体(図4B)を用いて検出した。HA−パーキンおよびGFPを発現する細胞のユビキチン化産物の欠如、ならびに他の対照におけるGFP−sytXIのユビキチン化の検出不能なレベルに留意せよ。パーキン変異体およびGFP−sytXIを発現する細胞は野生型のパーキンと比べて少ない量のユビキチンと共役したsytXIを産生する。抗GFP抗体は、GFP−sytXIを含む全サンプルにおいてほぼ等量のGFP−sytXI単量体、ならびにGFP−sytXIおよびHA−パーキンの両方と共発現したサンプルを含むウェルの近くに大きなGFP−sytXIバンドを検出した。図4Cは抗HA抗体を用いた同溶解物のウェスタン・ブロットを示す。これは欠失パーキンおよび変異パーキンが野生型パーキンより高いレベルで発現することを示している。図4DはHA−パーキンおよび対応するmyc−ユビキチンおよびGFPで標識したsyt1タンパク質を過剰発現するHEK293細胞のタンパク質抽出物の抗GFP抗体を用いた免疫沈降を示す。この細胞はプロテオソーム複合体の阻害剤であるラクタシスチンで4時間処理した。ユビキチン化アッセイの免疫沈降産物はmyc(上端パネル)、GFP(中間パネル)、およびHA−標識(下端パネル)に対する抗体を用いて検出した。図4Eは、GFP−syt11が使用されたことを除くと、図4Dに類似する。ウェスタン・ブロットは抗myc(上端パネル)および抗GFP(下端パネル)の抗体を用いて検出した。図4Fは、変異パーキン、C289GおよびC418Rを用いた共ipおよびウェスタン・ブロットを示す。
【図5】図5Aおよび図5BはパーキンがGFP−sytXIの代謝回転を促進することを示す。HAベクターまたはHA−パーキンのいずれかを発現するHEK293細胞における0時間、1.5時間、3時間、6時間および24時間でのGFP−sytXIの分解のパルス・チェイス分析が実施され、GFP−sytXIは抗GFP抗体と免疫沈降した。免疫沈降物はゲル電気泳動により分析し(図5A)、定量化した(図5B)。データは2つの独立実験の1つから得られる。2つ目の実験はさらに強力なパーキンの効果を及ぼした。
【図6−1】図6A〜6RはトランスフェクションされていないPC12細胞およびヒトの黒質ニューロンにおける内因性のシナプトタグミンIおよびシナプトタグミンXIの細胞内分布を示す。図6A〜6Cおよび図6P〜6Rは7日間50ng/mlngFで誘導されたPC12細胞の免疫蛍光を示す。この細胞はウサギ・パーキン(赤に染色、図6P)およびマウスsytI(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6B)に対する抗体を用いて免疫蛍光で共標識された。100x油浸レンズを用いてLeica・TCSSP顕微鏡により画像が得られた。積層画像は併合した(図6Cおよび6R)。黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび図6P〜6Rの挿入物は、長い神経突起が生じる同じ細胞の細胞体から得られる(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。図6D〜6Lは正常なヒトの黒質切片におけるシナプトタグミンXIおよびパーキンの分布を示す。ヒトの黒質切片は、ウサギ抗sytXIA抗体(D,G,J)、100sytXIAペプチドで前吸収した抗sytXIA抗体(E,H,K)、またはウサギ抗parkA抗体(F,I,L)で標識した。画像は黒質におけるドパーミン作動性ニューロンの細胞体および神経突起を示す。図6M〜6Oは、ウサギ抗ユビキチン(M)、抗sytXIA(N)、および抗sytXIA p sytXIAペプチド(O)で標識された隣接のPD脳切片を示し;黒矢印はレヴィー小体を指す。前吸収したsytXIA抗体によるレヴィー小体の標識の欠如に留意せよ。画像は20xレンズ(図6D〜6F)および63x油浸レンズ(図6G〜6O)を用いて得られた。
【図6−2】図6A〜6RはトランスフェクションされていないPC12細胞およびヒトの黒質ニューロンにおける内因性のシナプトタグミンIおよびシナプトタグミンXIの細胞内分布を示す。図6A〜6Cおよび図6P〜6Rは7日間50ng/mlngFで誘導されたPC12細胞の免疫蛍光を示す。この細胞はウサギ・パーキン(赤に染色、図6P)およびマウスsytI(緑に染色、図6Q)、またはニワトリ・パーキン(緑に染色、図6A)およびウサギsyt11(赤に染色、図6B)に対する抗体を用いて免疫蛍光で共標識された。100x油浸レンズを用いてLeica・TCSSP顕微鏡により画像が得られた。積層画像は併合した(図6Cおよび6R)。黄色は2つのタンパク質の共局在化を示す。図6A〜6Cおよび図6P〜6Rの挿入物は、長い神経突起が生じる同じ細胞の細胞体から得られる(低倍率で示す)。パーキンおよびsytは核周辺領域および神経突起に沿ったボタン(矢印)に共局在化する。図6D〜6Lは正常なヒトの黒質切片におけるシナプトタグミンXIおよびパーキンの分布を示す。ヒトの黒質切片は、ウサギ抗sytXIA抗体(D,G,J)、100sytXIAペプチドで前吸収した抗sytXIA抗体(E,H,K)、またはウサギ抗parkA抗体(F,I,L)で標識した。画像は黒質におけるドパーミン作動性ニューロンの細胞体および神経突起を示す。図6M〜6Oは、ウサギ抗ユビキチン(M)、抗sytXIA(N)、および抗sytXIA p sytXIAペプチド(O)で標識された隣接のPD脳切片を示し;黒矢印はレヴィー小体を指す。前吸収したsytXIA抗体によるレヴィー小体の標識の欠如に留意せよ。画像は20xレンズ(図6D〜6F)および63x油浸レンズ(図6G〜6O)を用いて得られた。
【図7】図7A〜7OはHEK293細胞の免疫蛍光を示す。この細胞はGFP−syt1およびHA−パーキン(図7A〜7C)、GFP−syt1およびHA−ベクター(図7D〜7F)、GFP−syt11およびHA−パーキン(図7G〜7I)、GFP−syt11およびHA−ベクター(図7J〜7L)またはGFP−ベクターおよびHA−パーキン(図7M〜7O)で共トランスフェクションした。トランスフェクションされた細胞は抗HAで標識し、画像は100x油浸レンズを用いてLeica・TCSSPにより得られた。
【図8】図8は、パーキン結合ポリペプチドであり本明細書中MP23とも呼ばれるヒトのシナプシン様タンパク質(SLP)のヌクレオチド配列(配列番号1)およびアミノ酸配列(配列番号2)を示す。SLP cDNAコード領域はヌクレオチド272〜955である(配列番号3)。図8BはSLP(MP23a)の部分cDNA配列(配列番号4)を示す。小文字はpGAD10ベクターである。SLP配列の第一ヌクレオチドは図8Aに示すヌクレオチド配列のヌクレオチド535に対応する。
【図9−1】図9Aはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド・フィルター結合アッセイを示す。図9Bはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド液体アッセイを示す。図9CはHA−パーキンとMP36aおよびMP23a GFP融合の共免疫沈降を示す。免疫沈降はHA−アガロース基質を用いて実施した。MP36aおよびMP23a GFP融合はGFP抗体で検出し(上側パネル)、パーキンはParkA抗体で検出した(下側パネル)。図9DはPC12細胞におけるSLPと内因性パーキンとの共免疫沈降を示す。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照と免疫沈降した。IP産物はニワトリの抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキン・バンドを検出した。このバンドはニワトリIgGのipサンプルには存在しなかった。抗SLP抗体は抗parkA免疫沈降物およびPC12タンパク質抽出物で36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性のパーキンが野生型SLPと共沈したことを示している。
【図9−2】図9Aはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド・フィルター結合アッセイを示す。図9Bはβ−ガラクトシダーゼの検出を用いた酵母2−ハイブリッド液体アッセイを示す。図9CはHA−パーキンとMP36aおよびMP23a GFP融合の共免疫沈降を示す。免疫沈降はHA−アガロース基質を用いて実施した。MP36aおよびMP23a GFP融合はGFP抗体で検出し(上側パネル)、パーキンはParkA抗体で検出した(下側パネル)。図9DはPC12細胞におけるSLPと内因性パーキンとの共免疫沈降を示す。タンパク質抽出物はウサギ抗parkAまたはウサギIgG対照と免疫沈降した。IP産物はニワトリの抗パーキン抗体(左)、またはウサギ抗SLP(右)で免疫ブロットした。抗parkA抗体は抗parkA共ipにおいて50kDaのパーキン・バンドを検出した。このバンドはニワトリIgGのipサンプルには存在しなかった。抗SLP抗体は抗parkA免疫沈降物およびPC12タンパク質抽出物で36kDaの予測サイズのバンドを検出したが、ウサギIgGと沈降したサンプルでは検出しなかった。これは内因性のパーキンが野生型SLPと共沈したことを示している。
【図10】図10A〜10Dはヒトの黒質におけるSLPおよびユビキチンの発現を示す。黒質ち密部切片は10μg/mlのアフィニティー精製した抗SLP(図10Aおよび10C)、抗SLP+SLPペプチド(図10B)、抗ユビキチン(図10D)の抗体を用いて免疫組織化学的に染色した。一次抗体はVector Elite Vectastain Rabbit ABCキットを用いて検出し、DABで可視化した。切片は全て同一に処理し染色した。抗SLPおよび抗sytXIの両抗体は黒質ち密部のニューロンの神経突起を濃く標識した。SLPペプチドで前吸収した抗SLP抗体は反応しなかった。これは免疫組織化学標識の特異性を示す。細胞体に見られるこげ茶色の染色はドパーミン作動性ニューロンで見出されるニューロメラニンである。図10Cおよび10Dは抗SLP(図10C)および抗ユビキチン(図10D)の抗体を用いたLBの標識を示す。
【図11】図11A〜11Cは細胞の免疫蛍光を示し、パーキンおよびSLPの位置を示す。細胞はパーキン抗体(緑に染色、図11A)およびSLP(赤に染色、図11B)で共染色した。積層画像が図11Cに示され、黄色はパーキンおよびSLPの共局在化を示す。図11Dおよび11Eはそれぞれ黒質および大脳皮質の染色を示す。
【図12】図12Aおよび12BはヒトのシナプトタグミンI(syt1)cDNAのヌクレオチド(配列番号5)およびアミノ酸(配列番号6)の配列をそれぞれ示す(GenBank登録番号BC058917)。図12Cおよび12DはヒトのシナプトタグミンXI(syt11)cDNAのヌクレオチド(配列番号7)およびアミノ酸(配列番号8)の配列をそれぞれ示す(GenBank登録番号BC039205)。
【特許請求の範囲】
【請求項1】
配列番号2として示されるアミノ酸配列を有する単離されたポリペプチド。
【請求項2】
請求項1のポリペプチドに特異的に結合する抗体。
【請求項3】
前記抗体がポリクローナル抗体である、請求項2に記載の抗体。
【請求項4】
前記抗体がモノクローナル抗体である、請求項2に記載の抗体。
【請求項5】
ポリペプチドを検出する方法であって、サンプルを請求項2に記載の抗体と接触させる工程、および該抗体の特異的結合を検出する工程を包含する、方法。
【請求項6】
配列番号2として示されるポリペプチドのアミノ酸配列をコードする単離された核酸分子。
【請求項7】
配列番号3として示されるヌクレオチド配列を含む、請求項6に記載の単離された核酸分子。
【請求項8】
配列番号3に記載の15と300との間の連続したヌクレオチドを含むオリゴヌクレオチドまたはそのアンチセンス鎖。
【請求項9】
請求項6に記載のヌクレオチド配列に作動可能に連結された発現エレメントを含むベクター。
【請求項10】
請求項9に記載のベクターを含む宿主細胞。
【請求項11】
サンプル中の核酸分子を検出する方法であって、該サンプルを、該サンプル中の核酸分子に特異的にハイブリダイズする条件下で、請求項8に記載のオリゴヌクレオチドと接触させる工程、および特異的なハイブリダイゼーションを検出する工程を包含する、方法。
【請求項12】
サンプル中の核酸分子を検出する方法であって、該サンプルを請求項8に記載の2つ以上のオリゴヌクレオチドと接触させる工程、核酸分子を増殖する工程、および該増殖された核酸分子を検出する工程を包含する、方法。
【請求項13】
前記増幅がポリメラーゼ連鎖反応を用いて実施される、請求項12に記載の方法。
【請求項14】
配列番号3の15と300との間の連続したヌクレオチドを含む1つ以上のオリゴヌクレオチド、またはそのアンチセンス鎖を含む、キット。
【請求項15】
パーキンソン病を処置するための候補薬物を同定する方法であって、パーキン結合ポリペプチドを1つ以上の化合物と接触させる工程、および該パーキン結合ポリペプチドの活性を変化させる化合物を同定する工程を包含する、方法。
【請求項16】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項15に記載の方法。
【請求項17】
前記化合物は、前記パーキン結合ポリペプチドの活性を低下させる、請求項15に記載の方法。
【請求項18】
パーキンソン病を処置するための候補薬物を同定する方法であって、パーキン結合ポリペプチドを発現する細胞を1つ以上の化合物と接触させる工程、および該パーキン結合ポリペプチドの発現を減少させる化合物を同定する工程を包含する、方法。
【請求項19】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項18に記載の方法。
【請求項20】
パーキンソン病を処置する方法であって、パーキン結合ポリペプチドの発現または活性を低減する分子を投与する工程を包含する、方法。
【請求項21】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項20に記載の方法。
【請求項22】
パーキンソン病の動物モデルを発生させる方法であって、増加したレベルのパーキン結合ポリペプチドを発現するトランスジェニック動物を発生させ工程を包含する、方法。
【請求項23】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項22に記載の方法。
【請求項24】
請求項22に記載の方法により発生される、パーキンソン病の動物モデル。
【請求項1】
配列番号2として示されるアミノ酸配列を有する単離されたポリペプチド。
【請求項2】
請求項1のポリペプチドに特異的に結合する抗体。
【請求項3】
前記抗体がポリクローナル抗体である、請求項2に記載の抗体。
【請求項4】
前記抗体がモノクローナル抗体である、請求項2に記載の抗体。
【請求項5】
ポリペプチドを検出する方法であって、サンプルを請求項2に記載の抗体と接触させる工程、および該抗体の特異的結合を検出する工程を包含する、方法。
【請求項6】
配列番号2として示されるポリペプチドのアミノ酸配列をコードする単離された核酸分子。
【請求項7】
配列番号3として示されるヌクレオチド配列を含む、請求項6に記載の単離された核酸分子。
【請求項8】
配列番号3に記載の15と300との間の連続したヌクレオチドを含むオリゴヌクレオチドまたはそのアンチセンス鎖。
【請求項9】
請求項6に記載のヌクレオチド配列に作動可能に連結された発現エレメントを含むベクター。
【請求項10】
請求項9に記載のベクターを含む宿主細胞。
【請求項11】
サンプル中の核酸分子を検出する方法であって、該サンプルを、該サンプル中の核酸分子に特異的にハイブリダイズする条件下で、請求項8に記載のオリゴヌクレオチドと接触させる工程、および特異的なハイブリダイゼーションを検出する工程を包含する、方法。
【請求項12】
サンプル中の核酸分子を検出する方法であって、該サンプルを請求項8に記載の2つ以上のオリゴヌクレオチドと接触させる工程、核酸分子を増殖する工程、および該増殖された核酸分子を検出する工程を包含する、方法。
【請求項13】
前記増幅がポリメラーゼ連鎖反応を用いて実施される、請求項12に記載の方法。
【請求項14】
配列番号3の15と300との間の連続したヌクレオチドを含む1つ以上のオリゴヌクレオチド、またはそのアンチセンス鎖を含む、キット。
【請求項15】
パーキンソン病を処置するための候補薬物を同定する方法であって、パーキン結合ポリペプチドを1つ以上の化合物と接触させる工程、および該パーキン結合ポリペプチドの活性を変化させる化合物を同定する工程を包含する、方法。
【請求項16】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項15に記載の方法。
【請求項17】
前記化合物は、前記パーキン結合ポリペプチドの活性を低下させる、請求項15に記載の方法。
【請求項18】
パーキンソン病を処置するための候補薬物を同定する方法であって、パーキン結合ポリペプチドを発現する細胞を1つ以上の化合物と接触させる工程、および該パーキン結合ポリペプチドの発現を減少させる化合物を同定する工程を包含する、方法。
【請求項19】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項18に記載の方法。
【請求項20】
パーキンソン病を処置する方法であって、パーキン結合ポリペプチドの発現または活性を低減する分子を投与する工程を包含する、方法。
【請求項21】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項20に記載の方法。
【請求項22】
パーキンソン病の動物モデルを発生させる方法であって、増加したレベルのパーキン結合ポリペプチドを発現するトランスジェニック動物を発生させ工程を包含する、方法。
【請求項23】
前記パーキン結合ポリペプチドが、シナプトタグミンI、シナプトタグミンXI、またはシナプシン様タンパク質から選択される、請求項22に記載の方法。
【請求項24】
請求項22に記載の方法により発生される、パーキンソン病の動物モデル。
【図1】


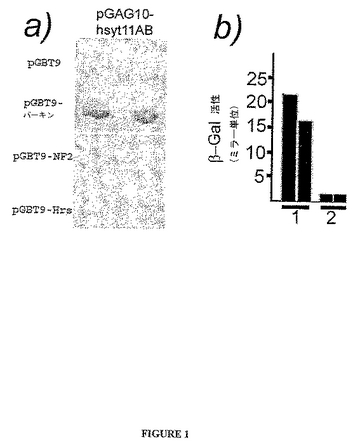
【図2−1】
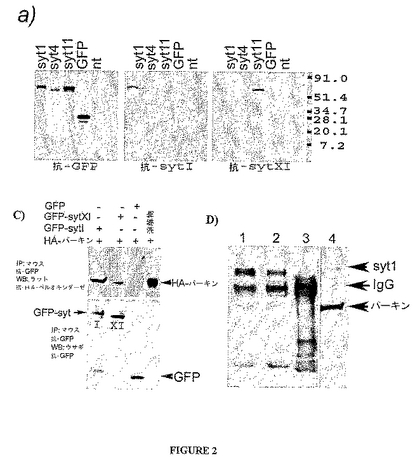
【図2−2】
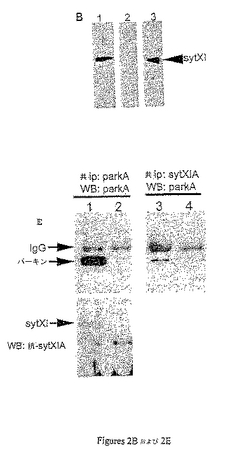
【図3】
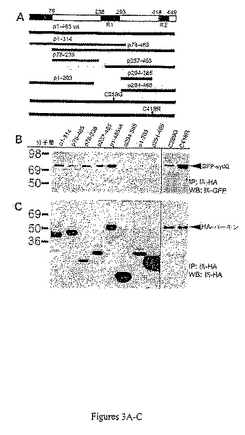
【図4−1】
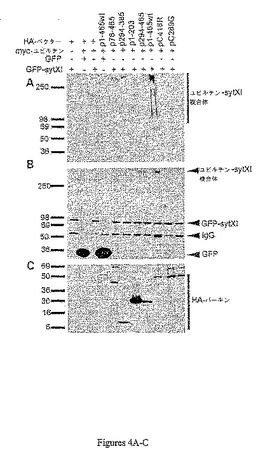
【図4−2】
【図5】
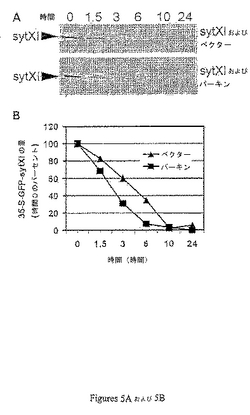
【図6−1】
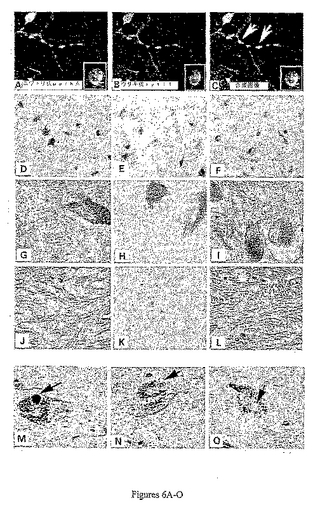
【図6−2】
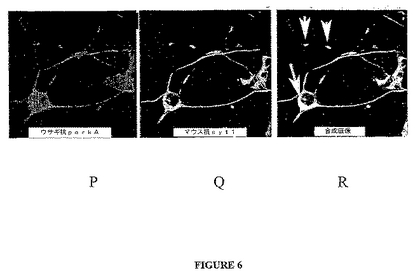
【図7】
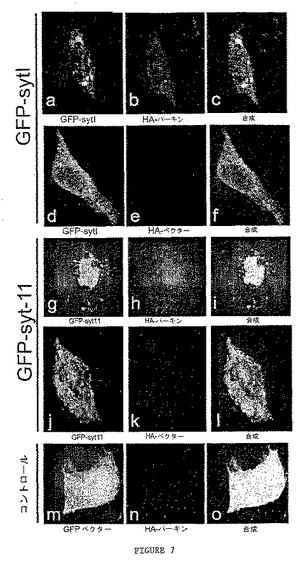




【図9−1】
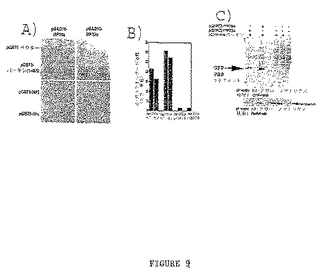
【図9−2】
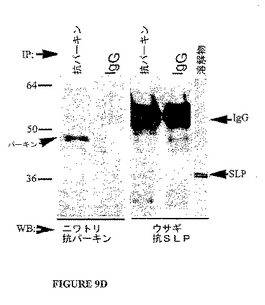
【図10】
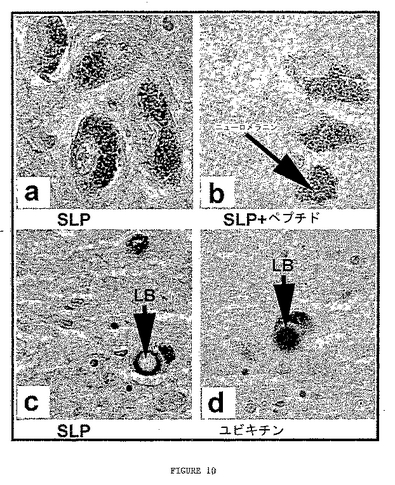
【図11】
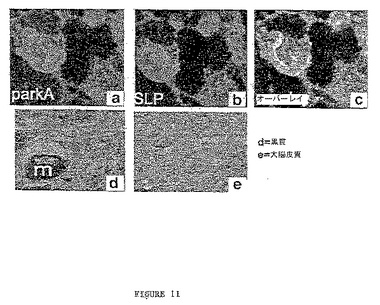








【図2−1】
【図2−2】
【図3】
【図4−1】
【図4−2】
【図5】
【図6−1】
【図6−2】
【図7】
【図9−1】
【図9−2】
【図10】
【図11】
【公表番号】特表2006−518218(P2006−518218A)
【公表日】平成18年8月10日(2006.8.10)
【国際特許分類】
【出願番号】特願2006−503681(P2006−503681)
【出願日】平成16年2月18日(2004.2.18)
【国際出願番号】PCT/US2004/004809
【国際公開番号】WO2004/074446
【国際公開日】平成16年9月2日(2004.9.2)
【出願人】(398062149)セダーズ−シナイ メディカル センター (34)
【Fターム(参考)】
【公表日】平成18年8月10日(2006.8.10)
【国際特許分類】
【出願日】平成16年2月18日(2004.2.18)
【国際出願番号】PCT/US2004/004809
【国際公開番号】WO2004/074446
【国際公開日】平成16年9月2日(2004.9.2)
【出願人】(398062149)セダーズ−シナイ メディカル センター (34)
【Fターム(参考)】
[ Back to top ]