
ヒスタミン分泌能を有する欠失型IgE依存的ヒスタミン放出因子、HRF結合ペプチドおよびその利用方法

【課題】IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor,HRF)およびそれに結合するペプチドを提供する。
【解決手段】特定のアミノ酸配列を含むダイマーを形成できる欠失型HRF、前記欠失型HRFをコードする遺伝子およびHRFの活性を抑制することができる新規なHRF結合ペプチド。ヒスタミンおよびIL-8分泌を可能にする前記ダイマーを形成できる欠失型HRFを用いた、HRFによるアレルギー発生を抑制する薬物のスクリーニング、およびアレルギー患者血清内HRF検出のためのキット。また、前記HRF結合ペプチドの、動物の喘息、鼻炎等のアレルギー疾患またはマラリアの予防および治療への使用。
【解決手段】特定のアミノ酸配列を含むダイマーを形成できる欠失型HRF、前記欠失型HRFをコードする遺伝子およびHRFの活性を抑制することができる新規なHRF結合ペプチド。ヒスタミンおよびIL-8分泌を可能にする前記ダイマーを形成できる欠失型HRFを用いた、HRFによるアレルギー発生を抑制する薬物のスクリーニング、およびアレルギー患者血清内HRF検出のためのキット。また、前記HRF結合ペプチドの、動物の喘息、鼻炎等のアレルギー疾患またはマラリアの予防および治療への使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)およびそれに結合するペプチドに関するものである。詳細には、配列番号:3で表されるアミノ酸配列を含むダイマーを形成できる欠失型HRF、HRFの活性を抑制することができる新規なHRF結合ペプチドおよびこれらの医薬分野における用途に関するものである。
【背景技術】
【0002】
喘息、鼻炎、蕁麻疹、アナフィラキーショック(anaphylaxis)、アレルギー性気管支拡張症、食べ物・薬物・植木鉢・虫によるアレルギー、枯草熱(hay fever)、寒冷じんま疹、アトピー性皮膚炎などのアレルギー(非特許文献1、非特許文献2)は、遺伝的にアレルギー抗原(allergen)に対して過敏に反応し大量生成されたIgE(Immunoglobulin E)およびIgE分泌調節に関与するサイトカイン(cytokine)間の均衡破壊から起因すると知られている。
【0003】
一旦アレルギー抗原に露出すれば、即発反応(immediate reaction)が起きて、肥満細胞(mast cell)から分泌したサイトカイン等によって炎症に関連した細胞が抗原露出部位に集まるようになる。数時間後には好塩基球(basophil)、好酸球(eosinophil)およびリンパ球(lymphocyte)から分泌するヒスタミンと様々なサイトカインなどによって遅発相反応(late-phase reaction, 以下「LPR」とする)が起きるようになる。アレルギー患者の半分程度がLPRに進行する。LPRでは好塩基球からヒスタミンが分泌され、その時には即発反応を誘発したアレルギー抗原がないので、好塩基球からヒスタミン分泌およびLPRへの進行を誘発する原因物質を糾明することが主要関心事だった。今まで、MCP-3(Monocyte Chemotatic Protein-3)、MCP-1(Monocyte Chemotatic Protein-1)またはRANTES(Regulated upon Activation Normal T-cell Expressed and Secreted)のようなサイトカイン(cytokine)がヒスタミンを分泌させることが知られていたが、IgE依存的LPRでは「HRF」と名付けられたタンパク質だけが好塩基球からヒスタミンを分泌させることができることが明らかにされた(非特許文献3)。しかし、HRFがどのような作用機序によって好塩基球からヒスタミン分泌を誘導するかに対しては報告されたことがない。
【0004】
HRFは、すべての細胞質に存在する、172個のアミノ酸で構成された公知のタンパク質である(非特許文献4)、その中の79〜123番アミノ酸は塩基性ドメイン(basic domain)を形成していて、この部位は微小管結合タンパク質である MAP-1B(microtubule-associated protein-1B)と約46%の相同性を示すため、微小管に 結合する機能を有することができると推定されている。ガチェット(Gachet)らは共焦点顕微鏡(confocal microscope)を利用してHRFの分布と細胞骨格網分布がどの程度一致するかを観察したが(非特許文献5)、これはHRFが細胞骨格と結合していることを示唆するものである。一方、サンチェスらは、HRFが一般的なカルシウム結合タンパク質ファミリーに属さないにもかかわらず、Ca2+と結合することを発表した。また、サカロマイセスセレビシエ(Saccaromyces cerevisiae)でHRF遺伝子を欠失させても酵母が生存できることを確認した(非特許文献6)。これは、HRFが重複経路(redundant pathway)を持っている遺伝子ファミリーに属するという事実を暗示するものである。
【0005】
一方、HRFは親水性で細胞質内に存在するタンパク質であるにもかかわらず、マクドナルドらが、HRFを細胞の外で見つけ、LPRアレルギー患者の血漿から多量に検出されるため、細胞死滅(apoptosis)や他の作用機序によって細胞外に分泌した後、細胞膜に存在するHRF受容体を通してヒスタミンを遊離させると考えられている(非特許文献7)。また、HRFはIgE感作された(sensitized)好塩基球を刺激してヒスタミンを分泌するようにすることが知られているが、これに対するIgEの作用機序は、まだ糾明されていない。一方、ビーカエスクラ(Bheekha-Escura)らは、IgE受容体がない細胞でもHRFが炎症反応を起こすことができることを観察することにより、HRFがIgEと結合して作用するのではなく、特異な細胞膜受容体に結合して作用することを提示した(非特許文献8)。
【0006】
本発明者らは、I)HRFが親水性にもかかわらず細胞膜を透過することができるという事実、II)酵母2-ハイブリッド分析(yeast two-hybrid)によってHRF受容体が(Na,K)ATPaseの第3細胞質ドメイン(CD3)という事実を明らかにしてきた(特許文献1、 特許文献2)。同時に、細胞外に分泌したHRFが、好塩基球細胞内でヒスタミン分泌を促進する作用機序も明らかにしてきた。
【0007】
また、I)HRFが細胞内に入って行く工程を遮断して/遮断したり、II)HRFが (Na,K)ATPaseに結合する工程を遮断して、HRFのヒスタミン分泌を阻害することができるペプチドを見つけて、アレルギー疾患を予防または治療することができることを立証してきた(特許文献1)。
【0008】
最近ブデ(Budde)らは、活性化した単核球培養上澄み液から分離した HRF(HRFmn)と組換えHRF(recombinant HRF, rHRF)は、好塩基球でのヒスタミン分泌能が同じではないことを報告したが、これはHRFmnとrHRFが、同じ作用因子(factor)ではない可能性を提示する(非特許文献9)。また、ヒトHRF特異的な酵素免疫吸着検出法(enzyme-linked immunoabsorbant assay)によってHRFmnからHRFが検出されなかったことに言及したが、これもHRFmnとrHRFとの構造的差異を示唆するものであると言える。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】大韓民国特許第457350号
【特許文献2】大韓民国特許第457351号
【非特許文献】
【0010】
【非特許文献1】Bachertら, Clinical and Experimental Allergy, 1998年,第28巻, 15-19頁
【非特許文献2】MacDonald and Lichtenstein, Springer Semin Immunopathol., 1990年,第12巻, 415-428頁
【非特許文献3】MacDonaldら., Science, 1995年,第269巻, 688-690頁
【非特許文献4】Bohmら, Biochem. Int., 1989年、第19巻, 277-286頁
【非特許文献5】Gachetら., J. Cell Sci., 1999年,第112巻, 1257-1271頁
【非特許文献6】Sanchezら, Electrophoresis, 1997年,第18巻, 150-155頁
【非特許文献7】MacDonaldら, Science, 1995年,第269巻, 688-690頁
【非特許文献8】Bheekha-Escuraら, Blood, 2000年,第96巻, 2191-2198頁
【非特許文献9】Buddeら, Ann. Allergy Asthma Immunol., 2002年、第89巻, 606-612頁
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明の目的は、細胞からヒスタミンおよびIL-8を分泌させることができるダイマーを形成できる欠失型HRF、HRFの作用を抑制することができる、新規なHRF結合ペプチドおよびこれらの用途を提供することである。
【課題を解決するための手段】
【0012】
本発明者らは、HRFmnとrHRFの活性の差は、アレルギー患者の血液の中に多量存在するタンパク質分解酵素(protease)によるものと判断して、ヒスタミン分泌能を有するHRFを捜そうと努力した結果、野生型HRFに比べ非常に高い活性を有する欠失型HRFを見つけた。これら欠失型HRFは、野生型HRFとは異なり分子内ジスルフィド結合を有していて、このジスルフィド結合による二量体化によって活性を持つようになることを確認した。また、すでに発見されているHRF結合ペプチドを変形させた後、やはりこれが欠失型HRFに結合してHRFのヒスタミンおよびIL-8分泌能を効果的に妨害することを確認して、これら欠失型HRFおよびその結合ペプチドが抗アレルギー薬物開発に有効に使用できることを確認して本発明を完成した。
【0013】
上記目的を達成するために、本発明は、野生型IgE-依存的ヒスタミン放出因子のN末端が除去されることによりダイマーを形成でき、ヒスタミンおよびIL-8分泌能が向上された組換え欠失型IgE-依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)を提供する。
【0014】
また、配列番号:3で表されるアミノ酸配列を含むダイマーを形成できる欠失型HRFを提供する。
【0015】
また、前記組換え欠失型IgE-依存的ヒスタミン放出因子の同形または異形ダイマーおよびその製造方法を提供する。
【0016】
また、前記組換え欠失型IgE依存的ヒスタミン放出因子または野生型 IgE依存的ヒスタミン放出因子間に形成される同形または異形ダイマーおよびその製造方法を提供する。
【0017】
また、本発明は、前記欠失型HRFをコードする遺伝子、前記遺伝子を含む組換え発現ベクターおよび前記発現ベクターで形質転換された形質転換体を提供する。
【0018】
また、本発明は、前記欠失型HRFを有効成分として含むヒスタミンおよびIL-8放出誘導剤を提供する。
【0019】
また、本発明は、前記欠失型HRFを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法を提供する。
【0020】
また、本発明は、前記欠失型HRFを利用したアレルギー患者血清内HRF検出方法を提供する。
【0021】
また、本発明は、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)配列を持つ新規なHRF結合ペプチドを提供する。
【0022】
また、本発明は、前記HRF結合ペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物を提供する。
【0023】
また、本発明は、前記HRF結合ペプチドおよび抗-HRFモノクローナル抗体を含むアレルギー診断キットを提供する。
【0024】
また、本発明は、前記HRF結合ペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物を提供する。
【0025】
合わせて、本発明は、タンパク質-タンパク質相互作用分析を行なう工程を含む前記欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法を提供する。
【0026】
本発明(1)は、野生型IgE依存的ヒスタミン放出因子のN末端が除去され、ダイマー形成が可能でヒスタミンおよびIL-8分泌能が向上した組換え欠失型IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)である。
本発明(2)は、HRFが、脊椎動物のHRFまたはHRFの類似体であることを特徴とする、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(3)は、HRFが、野生型HRFのN末端の少なくとも11乃至35アミノ酸が欠失したことを特徴とする、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(4)は、配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(5)は、HRFが、配列番号:2(Del-N11HRF)または配列番号:3(Del-N35HRF)で表されるアミノ酸配列を有することを特徴とする、本発明(4)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(6)は、本発明(1)の組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成されるIgE依存的ヒスタミン放出因子の同形または異形ダイマーである。
本発明(7)は、本発明(1)のHRFまたは本発明(6)のHRF同形または異形ダイマーに特異的な抗体である。
本発明(8)は、本発明(1)の欠失型HRFおよび/または野生型HRFに架橋剤を処理する工程を含む、本発明(6)の同形または異形ダイマーを製造する方法である。
本発明(9)は、架橋剤が、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン( 1,4-Di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-Bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-Bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(Bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-Bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(Bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-Hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(Dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bismaleimidyl-2,3-dihydroxybutane;BMDB)からなる群より選択されることを特徴とする、本発明(8)の欠失型HRFの同形または異形ダイマーを製造する方法である。
本発明(10)は、本発明(1)の欠失型HRFをコードする遺伝子である。
本発明(11)は、遺伝子が、配列番号:11(Del-N35HRF)で表される塩基配列を含むことを特徴とする、本発明(10)の遺伝子である。
本発明(12)は、遺伝子が、配列番号:10(Del-N11HRF)または配列番号:11(Del-N35HRF)で表される塩基配列を持つことを特徴とする、本発明(11)の遺伝子である。
本発明(13)は、本発明(10)の遺伝子を含む組換え発現ベクターである。
本発明(14)は、pRSET-A-Del-N11HRFまたはpRSET-A-Del-N35HRFであることを特徴とする、本発明(13)の組換え発現ベクターである。
本発明(15)は、本発明(13)の発現ベクターで形質転換された形質転換体である。
本発明(16)は、大腸菌であることを特徴とする、本発明(15)の形質転換体である。
本発明(17)は、BL21(DE3)-pRSET-A- Del-N11HRF、BL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A- Del-N11HRF、またはBL21(DE3)pLysS-pRSET-A-Del-N35HRFであることを特徴とする、本発明(16)の形質転換体である。
本発明(18)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子または本発明(6)の同形または異形ダイマーを有効成分として含むヒスタミン放出誘導剤である。
本発明(19)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子または本発明(6)の同形または異形ダイマーを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法である。
本発明(20)は、欠失型IgE依存的ヒスタミン放出因子が、配列番号:8のアミノ酸配列を有することを特徴とする、本発明(15)の方法である。
本発明(21)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子を利用したアレルギー患者血清内HRFの検出方法である。
本発明(22)は、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を有するHRF結合ペプチドである。
本発明(23)は、配列番号:17のアミノ酸配列を有することを特徴とする、本発明(22)のペプチドである。
本発明(24)は、L-アミノ酸、D-アミノ酸、またはL-アミノ酸およびD-アミノ酸からなる、本発明(22)のペプチドである。
本発明(25)は、一つ以上の変形アミノ酸を含む、本発明(22)のペプチドである。
本発明(26)は、変形アミノ酸が、アミノ酸誘導体またはアルキル化されたアミノ酸であることを特徴とする、本発明(25)のペプチドである。
本発明(27)は、本発明(22)のペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物である。
本発明(28)は、本発明(22)のペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物である。
本発明(29)は、本発明(22)のペプチドおよび抗HRFモノクローナル抗体を含むアレルギー診断キットである。
本発明(30)は、タンパク質-タンパク質相互作用分析を行なう工程を含む欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法である。
本発明(31)は、タンパク質-タンパク質相互作用分析が、同時精製(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)またはタンパク質チップシステムにより行なわれることを特徴とする、本発明(30)のHRF特異的受容体を同定する方法である。
本発明(32)は、同時精製が、
1)HRF-HRF受容体複合体を分離する工程;
2)前記分離された複合体を精製する工程;および、
3)前記精製された複合体からHRF受容体の正体を確認する工程;
を含むことを特徴とする、本発明(31)のHRF特異的受容体を同定する方法である。
本発明(33)は、タンパク質チップシステムが、
1)機能が明らかにされているか明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程;および、
2)前記欠失型HRFまたはHRFダイマーに、特異的な抗体がないかまたは抗体がある状態で前記言及された分析方法を利用して、タンパク質-タンパク質相互作用が起きるかどうかを確認する工程;
を含むことを特徴とする、本発明(31)のHRF特異的受容体を同定する方法である。
【発明の効果】
【0027】
本発明により、細胞からヒスタミンおよびIL-8を分泌させることができるダイマーを形成できる欠失型HRF、HRFの作用を抑制することができる、新規なHRF結合ペプチドおよびこれらの用途が提供された。
【図面の簡単な説明】
【0028】
【図1】本発明で製造した多様な欠失型HRFの一模式図である。
【図2】本発明で製造した多様な欠失型HRFの一模式図である。
【図3】図1と図2に記載した各々の組換え欠失型HRF(pRSET-A-rHRF, pRSET-A-Del-N11HRF, pRSET-A-Del-N35HRF, pRSET-A-Del-C112HRF, pRSET-A-Del-N39C110HRF, pRSET-A-Del-C38HRF, pRSET-A-Del-N111HRF, pRSET-A-Del-N84C108HRF)製造のためのpRSET-Aベクターの遺伝子地図を示した模式図である。
【図4】図3に記載したpRSET-AベクターのMCS(multiple cloning site)を示した遺伝子配列および前記遺伝子のコード部位にあたるアミノ酸配列である。
【図5】図1に記載した欠失型HRFによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図6】IgE存在下で図1に記載した欠失型HRFによるヒト好塩基球でのヒスタミン分泌率を比べた結果を示したグラフである。
【図7】図1に記載されたRrHRFの還元剤有無による移動性(mobility)の差を示したゲル電気泳動写真である。
【図8】図1に記載した欠失型HRFの還元剤の有無による移動性(mobility)の差を示したゲル電気泳動写真である。
【図9】組換え欠失型HRFであるpRSET-A-Del-N11HRFとpRSET-A-Del-N11HRFの突然変異体であるpRSET-A-Del-N11HRF-C28SとpRSET-A-Del-N11HRF-C172Sのゲル電気泳動写真である。
【図10】組換え欠失型HRFであるpRSET-A-Del-N11HRFとpRSET-A-Del-N11HRFの突然変異体であるpRSET-A-Del-N11HRF-C28SとpRSET-A-Del-N11HRF-C172SによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図11】組換え欠失型HRFであるpRSET-A-Del-N35HRFとpRSET-A-Del-N35HRFの突然変異体であるpRSET-A-Del-N35HRF-C172SのPAGE(ポリアクリルアミドゲル電気泳動)写真である。
【図12】組換え欠失型HRFであるpRSET-A-Del-N35HRFとpRSET-A-Del-N35HRFの突然変異体であるpRSET-A-Del-N35HRF-C172SによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図13】架橋剤を利用して欠失型HRFと野生型HRFのダイマー形成能を確認した結果を示したSDS-PAGE写真である。
【図14】HRF結合ペプチドを発現しているファージ(phage)であるΦ-HBP-2と図1に記載した欠失型HRFとの親和度を比べた結果を示したグラフである。
【図15】HRF結合ペプチドを発現しているファージ(phage)であるΦ-HBP-2と図2に記載した欠失型HRFとの親和度を比べた結果を示したグラフである。
【図16】HRF結合ペプチドであるΦ-HBP-2 が、BEAS-2B細胞株で欠失型HRFによるIL-8分泌を抑制することを示したグラフである。
【図17】HRF結合ペプチドであるΦ-HBP-2 が、RBL-2H3細胞株で欠失型HRFによるヒスタミン分泌を抑制することを示したグラフである。
【図18】HRF結合ペプチドであるΦ-HBP-2 、そのアラニン置換体(HBP2-M1、HBP2-M2m、HBP2-M3、HBP2-M4、HBP2-M5、HBP2-M6、HBP2-M7)および1番残基が欠失した6mer HRF結合ペプチド(HBP2-D1)によるIL-8分泌抑制程度を比較した結果を示したグラフである。
【発明を実施するための形態】
【0029】
以下、本発明を詳しく説明する。
本発明は、野生型IgE-依存的ヒスタミン放出因子のN末端が除去されることによりダイマー形成が可能で、ヒスタミンおよびIL-8分泌能が向上された組換え欠失型 IgE-依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)を提供する。
【0030】
本発明の欠失型HRFは、本発明が提供する配列によって化学的に合成することができる。また他の方法として、本発明の欠失型HRFは、これをコードする塩基配列を持つDNAが挿入された発現ベクターを利用する組換えDNA技術で製造することができる。前記ベクターは、生体内に存在するように製造され、サムブロック(Sambrook)らの方法(Sambrookら.,Molecular cloning, 1989年, Cold Spring Harbor, Cold Spring Harbor Laboratory Press)によって適当な宿主細胞で形質転換され適当な条件下で発現するように製造される。また、本発明によるアミノ酸配列を含む融合タンパク質を利用して本発明の欠失型HRFを製造することができる。
【0031】
一方、本発明による欠失型HRFのアミノ酸配列は、当技術分野の公知された通常の技術により変形され得る。例えば、本発明の欠失型HRFは、アミノ酸数を増加させたり減少させたりすることにより変形できる。また、本発明による欠失型HRFの活性を減少させない範囲内で、特定残基やその順序を変えることにより変形することができる。変形可能なアミノ酸は、自然的に存在する L-α-アミノ酸だけではなく、β、γ、δアミノ酸はもちろんD-α-アミノ酸の誘導体にも変形可能である。
【0032】
したがって、当技術分野で通常の知識を持った者は、前記欠失型HRFをそのヒスタミン分泌活性を維持乃至増加させたり損傷させたりしない範囲内で通常の技術を使って変形させることができる。これは、本発明の範疇内に属するものである。
【0033】
本発明では、配列番号:1で表されるラットの全長配列(Accession number U20525, アミノ酸1-172)、配列番号:2で表されるアミノ酸11-172切片、配列番号:3で表されるアミノ酸35-172切片、配列番号:4で表されるアミノ酸1-112切片、配列番号:5で表されるアミノ酸39-110切片、配列番号:6で表されるアミノ酸1-38切片、配列番号:7で表されるアミノ酸111-172切片、配列番号:8で表されるアミノ酸84-108切片を有する多様な欠失型HRFを製造するために、配列番号:9で表されるラットHRF全長配列(Accession number U20525)遺伝子から前記断片をコードする遺伝子をpRSET-Aベクターにクローニング(図1乃至図4参照)して大腸菌に形質転換させ、その後発現を誘導して分離、精製した。
【0034】
続いて、製造された各欠失型HRFのN末端の欠失による各欠失型HRFの BEAS-2B細胞株でのIL-8分泌能(図5参照)と好塩基球でのヒスタミン分泌能(図6参照)を比べた。その結果、配列番号:2で表される野生型HRFのN末端の10個アミノ酸残基が欠失されたタンパク質(Del-N11HRF)および配列番号:3で表される野生型HRFのN末端の34個アミノ酸残基が欠失したタンパク質(Del-N35HRF)は、BEAS-2B細胞株で野生型HRFに比べて増加したIL-8分泌能を見せた。特に、これらは野生型HRFの場合、測定可能なIL-8を分泌することができない低濃度(1μg/ml)でもIL-8分泌を誘導した。また、これら欠失型HRFは、IgEに感作されたヒト好塩基球細胞でも野生型HRFに比べて増加したヒスタミン分泌能を見せた。
【0035】
したがって、HRFはその活性に主要な役目を持つ部位がN末端によって選り分けられているものと思われ、遅発相反応(LPR)アレルギー患者の場合、ヒスタミンおよびIL-8の分泌を誘発する活性型のHRFを持っているものと推定される。N末端の一部アミノ酸残基を除去した場合(Del-N11HRFとDel-N35HRF)は、ヒスタミン分泌能が増加するが、C末端のみを除去した場合(Del-C112HRF)には野生型HRFと等しい活性を見せることも、HRFのN末端がHRF機能調節に重要な役目を持つという事実を裏付けてくれる。
【0036】
欠失型HRFタンパク質は、HRFに対して特異的結合を見せるヘプタマー(heptamer)ペプチドを発現しているファージに対する親和度の面でも差を示したが、野生型HRFとDel-C112HRFがファージに対して弱い親和度を示した一方、Del-N11HRFとDel-N35HRFはとても強い親和度を示した(図14参照)。これは、各HRFタンパク質の活性と相応するものであり、以上、言及した結果から実験的にHRFをターゲットにするアレルギー制御薬物開発において利用することができるHRFの形態がいくつかの欠失型に限られることを確認した。
【0037】
本発明によって欠失型HRFの活性型とその機序が明かされた以上、当技術分野で通常の知識を持った者なら誰でもマウス、ヒト、ウサギ、ニワトリ等で活性型HRFを容易に同定することができる。よって、マウス、ヒト、ウサギ、ニワトリの活性型HRFもまた本発明の範疇内に属するものである。
【0038】
本発明は、配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、ダイマーを形成できる欠失型HRFを提供する。
【0039】
配列番号:3のHRFは、配列番号:1で表されるラット(Rattus norvegicus)の全長HRF アミノ酸配列のN末端を34個欠失させた形態であり、前記アミノ酸配列を含む欠失型HRFがヒスタミンおよびIL-8分泌能がすぐれたものであることを確認し、この活性が分子内ジスルフィド結合による二量体化によるものであることを確認した。
【0040】
また、本発明は、前記組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成される同形または異形ダイマーおよびその製造方法を提供する。
【0041】
上記のようにHRFのN末端がHRF機能調節に重要な役割を持つとみられるため、本発明者らは、欠失型HRFと野生型HRFの間に構造的差異があると推定して、これらが還元剤であるβ-メルカプトエタノールの有無によってその移動性に変化を見せるかどうかを確認した(図7、図8参照)。非還元SDS-PAGE結果、強い活性を見せる欠失型HRFは弱い活性を見せる野生型HRFやC末端欠失型HRFとは異なり、ダイマー位置で観察された。これは、これらN末端欠失型HRFが分子内ジスルフィド結合によるダイマー構造を成していることを示唆する。HRFの場合、残基28番と172番にシスチンを含んでいるのでpRSET-A-Del-N11HRFの場合は、これらアミノ酸を各々セリンに変える突然変異を実施した。pRSET-A-Del-N35HRFの場合は、172番のシスチンのみをセリンに変える突然変異を実施した後、これらの非還元SDS-PAGEでの移動性を比較した(図9、11参照)。pRSET-A-Del-N11HRFとpRSET-A-Del-N11HRF-C28S、pRSET-A-Del-N35HRFがダイマー位置に移動するのとは異なり、pRSET-A-Del-N11HRF-C172S、pRSET-A-Del-N35HRF-C172Sは、モノマー位置に移動することが観察された。これは、172番のシスチンがHRFの二量体化に主な役目を持つことを意味する。
【0042】
本発明者らは、この二量体化がHRFの活性に影響を及ぼすか否かを調べるために、BEAS-2B細胞株でのIL-8分泌能を比較した(図10、12参照)。分泌したIL-8の量は、pRSET-A-Del-N11HRF-C172Sの場合、野生型の46%、pRSET-A-Del-N35HRF-C172Sの場合、野生型の26%に各々減少した活性が見られた。これは、HRF内で28番シスチンに比べて172番シスチンの活性に寄与するところが大きいことを示唆し、これはPAGEでの移動性結果と相応する。以上の結果は、欠失型HRFは野生型HRFとは異なり、分子間ジスルフィド結合(intermolecular disulfide bond)により、ダイマー形態で存在し、その構造的な差がこれらの間の活性差を誘発する原因であることを示している。
【0043】
本発明者らは、架橋剤(crosslinker)を利用して野生型HRFもダイマーを形成できるか否か調べた(図13参照)。その結果、欠失型HRFとは異なり野生型HRFは分子間ジスルフィド結合(intermolecular disulfide)をほとんど形成することができず、大部分が分子内ジスルフィド結合(intramolecular disulfide bond)を形成した。これは、欠失型HRFと野生型HRFの構造的な差を再び示す結果で、HRFの場合化学的交差結合よりN末端欠失がその活性においてさらに効果的であることが分かる。
【0044】
但し、前記野生型HRFのダイマー形成の失敗は、分子間ジスルフィド結合の不在に起因するものであり、ジスルフィド結合以外の他の方法による試験管内ダイマーの製造方法によるヒスタミンおよびIL-8分泌能が向上された野生型HRFの同形ダイマーの存在可能性を排除するものではない。したがって、本発明のダイマーには、前記多様な長さを有する欠失型HRF間に形成される同形または異形ダイマーの他にもジスルフィド結合以外の方法を使用して形成される野生型HRFの同形ダイマーまたは前記多様な長さを有する欠失型HRFと野生型HRF間に形成される異形ダイマーまでも含まれる。
【0045】
本発明のHRF同形または異形ダイマーの製造方法は、特別にそれに制限されるものではなく、公知のすべてのタンパク質ダイマー製造方法が使用可能であるが、欧州特許出願公開0261616A2号、日本特許出願公開第1993-032941A号、 日本特許出願公開第 1986-069759号、 国際公開第 WO05/105994A1号および国際公開92/013965A1号等に記載された方法を使用することが好ましい。前記文献は、すべて本明細書に参考文献として挿入される。
【0046】
本発明の欠失型HRFは、本発明が提供する配列により製造された後、市販の架橋剤を処理することにより化学的にダイマーを形成させられる。例えば、-SHを標的にする架橋剤等を使用してイオウを交差結合させてダイマーを製造できる。架橋剤としては、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン(1,4-di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bis-maleimidyl-2,3-dihydroxybutane;BMDB)が使用可能である。
【0047】
また、本発明は、前記欠失型HRFをコードする遺伝子を提供する。
【0048】
本発明では、配列番号:9で表されるラットHRF全長配列から下記プライマー配列を利用して配列番号:10から配列番号:16までの多様な欠失型HRF遺伝子をクローニングした。
【0049】
配列番号:9のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ATG ATT ATC TAC CGG GAC(配列番号:22)。
【0050】
配列番号:9のクローニングのためのリバースプライマー:CCG CTCGAG(XhoI)TGT CCT AAG TCC TGG TGT(配列番号:23)。
【0051】
配列番号:10のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)GAC GAG CTG TCC TCC GAC AT(配列番号:24)。
【0052】
配列番号:10のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:25)。
【0053】
配列番号:11のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)AGT GTC AGT AGA ACA GAG(配列番号:26)。
【0054】
配列番号:11のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:27)。
【0055】
配列番号:12のクローニングのためのフォワードプライマー:TAACAAATTGGATCTATCGCCCGCGGAC(配列番号:28)。
【0056】
配列番号:12のクローニングのためのリバースプライマー:CTTTACCCTTTCTGGTTTCTGTTCTTC(配列番号:29)。
【0057】
配列番号:13のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ACA GAG GGT GCC ATC GA(配列番号:30)。
【0058】
配列番号:13のクローニングのためのリバースプライマー:G GAATTC(EcoRI)CCT TTC TGG TTT CTG TT(配列番号:31)。
【0059】
配列番号:14のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ATG ATT ATC TAC CGG GAC(配列番号:32)。
【0060】
配列番号:14のクローニングのためのリバースプライマー:G GAATTC(EcoRI)TCT ACT GAC CAT CTT GC(配列番号:33)。
【0061】
配列番号:15のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)GTA AAG CCT TTT ATG ACT(配列番号:34)。
【0062】
配列番号:15のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:35)。
【0063】
配列番号:16のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ACA AAA GAG GCC TAC AAA(配列番号:36)。
【0064】
配列番号:16のクローニングのためのリバースプライマー:CG GGATCC(BamHIII)TGG TTT CTG TTC TTC AAG(配列番号:37)。
【0065】
配列番号:10は、配列番号:2で表されるアミノ酸11-172切片をコードする遺伝子の塩基配列であり、配列番号:11は、配列番号:3で表されるアミノ酸35-172切片をコードする遺伝子の塩基配列であり、配列番号:12は、配列番号:4で表されるアミノ酸1-112切片をコードする遺伝子の塩基配列であり、配列番号:13は、配列番号:5で表されるアミノ酸39-110切片をコードする遺伝子の塩基配列であり、配列番号:14は、配列番号:6で表されるアミノ酸1-38切片をコードする遺伝子の塩基配列であり、配列番号:15は、配列番号:7で表されるアミノ酸111-172切片をコードする遺伝子の塩基配列であり、配列番号:16は、配列番号:8で表されるアミノ酸84-108切片をコードする遺伝子の塩基配列を示す。
【0066】
本発明の欠失型HRFタンパク質をコードする遺伝子配列は、配列番号:10、11、12、13、14、15および16からなる群より選択される塩基配列のいずれかであり、配列番号:11で表される塩基配列を含むことが好ましく、配列番号:10または配列番号:11で表される塩基配列を有することがさらに好ましい。
【0067】
また、本発明は、前記欠失型HRFをコードする遺伝子を含む組換え発現ベクターを提供する。
【0068】
本発明の組換え発現ベクターは、特別に限定されるものではなく、欠失型HRFをコードする遺伝子を含むものなら微生物、植物または動物形質転換用組換え発現ベクターのいずれでも本発明の範疇に属する。
【0069】
本発明では、大腸菌発現ベクターpRSET-Aを利用して配列番号:9、10、11、12、13、14、15および16の塩基配列の群より選択される遺伝子をクローニングして各々の組換えベクター、pRSET-A-RrHRF、pRSET-A-Del-N11HRF、pRSET-A-Del-N35HRF、pRSET-A-Del-C112HRF、pRSET-A-Del-N39C110HRF、pRSET-A-Del-C38HRF、pRSET-A-Del-N111HRF、pRSET-A-Del-N84C108HRFを製造した。
【0070】
また、本発明は、前記組換え発現ベクターで形質転換された形質転換体を提供する。
【0071】
前記形質転換体は、特別に限定されるものではなく、本発明の欠失型HRFをコードする遺伝子を含む組換え発現ベクターで形質転換されたものなら、大腸菌、植物、動物細胞のいずれであっても本発明の範疇に含まれる。
【0072】
本発明者らは、大腸菌BL21(DE3)またはBL21(DE3)pLysS菌株に前記組換え発現ベクターを導入して形質転換体を製造した。その中で、前記組換え発現ベクターpRSET-A-Del-N11HRFをBL21(DE3)およびBL21(DE3)pLysS菌株に形質転換させたものを各々BL21(DE3)-pRSET-A-Del-N11HRF、BL21(DE3)pLysS-pRSET-A-Del-N11HRFと、組換え発現ベクターpRSET-A-Del-N35HRFをBL21(DE3)およびBL21(DE3)pLysS菌株に形質転換させたものをBL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A-Del-N35HRFと名付けた。これら大腸菌は、lacUV5プローモーター調節下でのみT7RNA重合酵素が転写されるように考案されたもので、IPTG(isopropyl-β-D-thiogalactopyranoside)添加時、T7RNA重合酵素発現が誘導されて欠失型HRFが転写されるようになる。また精製中にタンパク質を分解することができるlonプロテアーゼとompT外膜プロテアーゼが欠乏していて多量の欠失型HRFを得ることができるようにする利点がある。
【0073】
また、本発明は、前記欠失型HRFを有効成分として含むヒスタミンおよびIL-8放出誘導剤を提供する。
【0074】
配列番号:2と配列番号:3のアミノ酸配列を有する欠失型HRFタンパク質は、野生型HRFに比べてBEAS-2B細胞株で増加されたIL-8分泌能を示し、ヒト好塩基球でも同様の方式でヒスタミン放出を誘導した。よって、前記欠失型HRFは、IL-8およびヒスタミン放出誘導剤として効果的に使用できる。本発明のIL-8およびヒスタミン放出誘導剤は、アレルギー治療を目的にする薬物開発における実験的モデルを提示する。すなわち、本発明の誘導剤は、多量のアレルギー誘発物質を必要とするアレルギー制御薬物の開発および検証過程に利用できる。
【0075】
また、本発明は、欠失型HRFを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法を提供する。
【0076】
すなわち、本発明の欠失型HRFは、HRF結合ペプチドの結合部位を提供することにより、これを利用してHRF抑制を通じたアレルギー制御薬物開発に利用することができる。HBP2 ペプチド(大韓民国特許出願第2001-27896号)を利用した親和性実験では、配列番号:5および 配列番号:8のアミノ酸配列を持つ欠失型HRFが前記ペプチドへの親和度が高く現われた(図9参照)。よって、これら配列を有するHRFペプチドは、これと結合する物質や化合物の検索に利用でき、このような物質、化合物は、アレルギー制御薬物の候補になり得る。
【0077】
すなわち、競争結合分析を使って、欠失型HRFをコードする遺伝子を含む組換えベクターで形質転換された細胞から欠失型HRFを分離した後、試験薬物および前記欠失型HRFと相互作用することが知られた物質[例えば、HRF結合ペプチド(大韓民国特許出願第2001-27896号)]と一緒に反応させて、前記試験薬物の中で欠失型HRFとHRF結合ペプチドとの間の相互作用を減少させる薬物を選別する。最終的に、HRF結合ペプチドと似た構造を有し、細胞で HRFによるヒスタミン分泌を抑制することができる薬物をスクリーニングできる。
【0078】
また、本発明は、欠失型HRFを利用したアレルギー患者の血清内HRFの検出方法を提供する。
【0079】
本発明の欠失型HRFを利用して通常の方法によって抗欠失型HRFのモノクローナル抗体を製造することができ、製造された抗-欠失型HRFモノクローナル抗体も本発明の範疇に含まれる。
【0080】
これを利用して患者の血清内HRFを検出する方法を説明すると、底にHRF結合ペプチドまたは抗欠失型HRFモノクローナル抗体を付着させた後、血液サンプルと反応させて、そこに通常の標識物質(HRP(horse reddish peroxydase)、塩基性脱リン酸化酵素(alkaline phosphatase)、蛍光物質(fluorescein) または色素(dye)等)がコンジュゲートされた抗欠失型HRFモノクローナル抗体、または抗HRFモノクローナル抗体を加える。発色または蛍光を発して陽性と判定されれば、血液内活性型HRFが含まれているため、アレルゲンがなくてもアレルギーを起こし得る疾患を持っていると判断することができる。
【0081】
また、本発明は、HRFに特異的に高い親和度で結合してヒスタミン分泌を阻害する新規なペプチドを提供する。
【0082】
本発明のHRF結合ペプチドは、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を持つことができ、配列番号:17に記載したYVYPSM配列を持つことが好ましい。
【0083】
本発明のHRF結合ペプチドは、ペプチドの活性を減少させない範囲内で、結合に直接的に関与したり保存しなければならない残基を除いた特定残基成分やその順序を変えることにより変形することもできる。変形可能なアミノ酸は、自然界に存在するL-α-アミノ酸だけでなく、β、γ、δアミノ酸はもちろんD-α-アミノ酸の誘導体へも変形可能である。
【0084】
典型的に、一つのアミノ酸が置換されたペプチドを利用して静電気力や親水性が結合に及ぼす影響を調査した結果、陽電荷を帯びたアミノ酸(例:Lys、Arg)や陰電荷を帯びたアミノ酸(例:Glu)が置換されると敏感度が変化することが分かる。このように、置き換えられたり付加されたりする残基の数や形態は、必須の結合点の間に必要な空間と親水性または疎水性と共に要求される機能によって決まる。このような置換によって本発明によるペプチドの目的タンパク質に対する親和度をさらに増加させることができる。
【0085】
置換によって機能において重要な変化が現われたりする。変化する残基の選択は、目的位置に存在する分子の電気度、疎水性、側鎖の変化またはらせん構造の変化等のようにペプチドの基本骨格を維持するのに大きな影響を及ぼしたりする。一般的に、ペプチドの性質に重大な変化を起こすものは、セリンのような親水性残基が、ロイシン、イソロイシン、フェニルアラニン、バリンまたはアラニンのような疎水性残基に置換されたり、リジン、アルギニンまたはヒスチジンのように電気的に陽性である残基が、グルタミン酸やアスパラギン酸のような電気的に陰性である残基に置き換えられたり、グリシンのように側鎖を持たないアミノ酸がかさだかい側鎖を持つ残基に置き換えられる場合である。
【0086】
以上、言及した事実を考慮して、当技術分野で通常の知識を有する者は、前記HRF結合ペプチドをその欠失型HRFに対する結合力、およびそれによるヒスタミン分泌阻害活性を維持乃至増加させたり損傷させたりしない範囲内で、通常の技術を利用して変形させることができ、それは本発明の範疇内に属する。
【0087】
本発明者らは、前記ペプチドをファージディスプレーライブラリスクリーニングを通して得、合成ペプチドで再び確認した。本発明のペプチドは化学的に合成したり遺伝子組換え技術を利用して製造したりすることができる。または、本発明のペプチドを構成するドメインを血液内に存在するタンパク質やその一部から製造することができる。また他の方法として、本発明のペプチドはそれをコードする塩基配列を有するDNAが挿入された発現ベクターを利用する組換えDNA技術で製造することができる。前記ベクターは、サムブルック(Sambrook)ら(Molecular Cloning, 1989年, Cold Spring Harbor, Cold Spring Harbor Laboratory Press)の方法によって適当な宿主細胞で形質転換され適当な条件下で発現するように製造される。また、本発明のアミノ酸配列を含む融合タンパク質を利用して、本発明のペプチドを製造することもできる。
【0088】
また、本発明は、前記HRF結合ペプチドを有効成分として含むアレルギーまたはマラリア予防または治療用薬剤学的組成物を提供する。
【0089】
本発明のHRF結合ペプチドは、HRFに結合してそのヒスタミンおよびIL-8分泌能を抑制することを示したため(図16、17および図18参照)、アレルギー予防または治療に効果的に使用できることを確認した。
【0090】
一方、HRFは、翻訳によって調節される腫瘍タンパク質(translationlly controlled tumor protein)とも呼ばれる。1998年ビスチバンらによって抗マラリア剤であるアルテミシニン(Artemisinin)がマラリアタンパク質であるHRFに結合して作用することが明らかにされた。したがって、HRFに結合する活性を有する本発明のHRF結合ペプチドも前記アルテミシンと同様にマラリアの予防および治療に使用することができる。
【0091】
本発明のHRF結合ペプチドの有効量は、体重1kg当たり約30μg〜1mgである。本発明の組成物は、溶液またはミセル形態で直接注入したり製剤化して使用できる。本発明による組成物は、非経口または局所投与によって人体に適用することができ、静脈注射、皮下注射、内皮注射、筋肉注射など注射によって投与することが好ましい。このような目的のために、本発明のペプチドを薬剤学的に許容可能な担体に懸濁させたり溶解させたりする。ここで、特に水溶性担体を使うのが好ましい。
【0092】
また、本発明は、前記HRF結合ペプチドおよび抗-HRFモノクローナル抗体を含むアレルギー診断キットを提供する。
【0093】
本発明のHRF結合ペプチドと抗HRFモノクローナル抗体を含む診断キットを利用する試験で、血液反応の時に陽性と判定されれば、アレルゲンがなくてもアレルギーを持っていると判断することができる。すなわち、LPRアレルギー患者の血液にはHRFが浮遊しているため、血液検査の時に本発明のキットを利用すれば、HRFが血液に存在するか否かが分かり、それをもってLPR患者か否かを確認することができる。本発明では底に本発明のHRF結合ペプチドを付着させた後、血液と反応させて、そこに標識物質がコンジュゲートされた抗HRFモノクローナル抗体を加える方法を使用できる。
【0094】
合わせて、本発明は、タンパク質-タンパク質相互作用分析を行なう工程を含む、前記欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法を提供する。ここで、前記タンパク質-タンパク質相互作用分析は、特別にそれに制限されるものではなく、今までに公知された通常のタンパク質-タンパク質相互作用分析方法をすべて使用でき、詳細には、同時精製方法(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)、タンパク質チップ等を使用して行なうことが好ましい。
【0095】
本発明の一実施態様として、同時精製方法は、1)HRF-HRF受容体複合体を適切な方法で分離する工程、2)前記分離された複合体を精製する工程;3)前記精製された複合体からHRF受容体の正体を確認する工程を含む。本発明の好ましい実施態様で、工程1は1次元電気泳動(1D)や二次元電気泳動(2D)または液体クロマトグラフィー(ligquid chromatography)方法を含む。
【0096】
本発明による好ましい実施態様で工程2は、内在タンパク質複合体(endogenous protein complex)を直接抗体で精製したり抗原決定基標識(epitope tagging)をした後、抗体で精製する免疫沈澱法(immunoprecipitation, Barrett等, J. Lab. Clin. Med. 1960年, 第55巻: 605-15頁)、GST-沈澱法(GST-pulldown, Magnaghi-Jaulin等, Nucleic Acids Res., 1996年, 第24(6)巻: 1052-8頁)のような非免疫的親和精製法(affinity purification)、および非特異的反応抑制と回収率および再現性を高めるために2個以上の標識を利用したタンデムアフィニティー精製(tandem affinity purification)(TAP)システム(Russell等, Infect Immun., 1980年, 第29(3)巻,999-1006頁)のような多標識親和精製法(multi-tag affinity purification)からなる群より選択される親和性クロマトグラフィーを含む。
【0097】
本発明の実施態様で、工程3は基質補助レーザー脱離/イオン化飛行時間質量分光法(matrix-assisted laser desorption/ ionization time-of-flight mass spectrometry, MALDI-TOF MS, Hill 等, Rapid Commun. Mass Spectrom., 1991年, 第5(9)巻、395-9頁)のようなペプチド質量分析(peptide mass fingerprinting)またはナノエレクトロスプレーイオン化質量分光法(nanoelectrospray ionization (nanoES)-MS/MS, Shevchenko等, Proc. Natl. Acad. Sci. 米国, 1996年, 第93(25)巻, 14440-5頁)、ナノLC質量分光法(nanoLC-MS/MS, Oosterkamp等, J. Mass Spectrom. 1998年, 第33(10)巻, 976-83頁)およびLC/LC-MS/MSからなる群より選択されるタンデム質量分光法(tandem MS (MS/MS))が含まれる。
【0098】
本発明の好ましい実施態様で、本発明のHRF-特異的受容体の同定方法は、選択的に下記の工程を含む:4)工程3で確認されたHRF受容体が、実際の生体内または試験管内条件でHRFによって媒介されるヒスタミン放出過程に関与するか否かを判定する工程。ここで、前記判定工程は、今まで公知されたタンパク質の機能分析方法をすべて使用でき、詳細には、前記工程4は、前記確認されたHRF受容体をコードする遺伝子をクローニングする工程、それを哺乳動物細胞発現ベクターにクローニングする工程、前記クローニングされた発現ベクターでヒスタミン放出細胞株を形質感染させる工程;および本発明の欠失型HRFまたはHRFダイマーを処理した後モックベクター(mock vector)で形質感染された対照細胞株と比較して、ヒスタミン放出程度が有意に増加するかどうかを観察する工程を通じて達成できる。選択的には、HRF受容体遺伝子が欠損したノックアウト形質転換動物または細胞株を製造する工程、前記形質転換動物または細胞株にHRFを投与した後、野生型と比較してヒスタミン放出が有意に減少するかどうかを分析する工程を通じて達成できる。
【0099】
酵母ツーハイブリッドシステム(Fields S. and Song O., Nature., 1989年, 第340(6230)巻, 245-6頁)は、DNA結合ドメイン(DBD; DNA binding domain)と転写活性ドメイン(TAD; transcription activation domain)を遺伝子組換え方法で分離して相互作用する二タンパク質(X,Y)の遺伝子を各々DBDとTADにクローニングした後、それらを酵母内で発現されるように誘導すると、DBDとTADが物理的に分離されたタンパク質が発現されるが、X/Yタンパク質相互作用により転写因子で再構成され指示遺伝子(reporter gene)の遺伝子発現が可能になるという点を利用した分析方法である。前記システムは、遺伝子操作技術を基盤技術としてタンパク質の特性に関係なしに同一な実験方法で相互作用を検索できるので、前記システムは大規模相互作用分析が容易である。
【0100】
本発明の一実施態様で、指示遺伝子は、1)色素選別(color selection)遺伝子のβ-ガラクトシダーゼ(β-galactosidase, LacZ)、α-ガラクトシダーゼ(α-galactosidase(MEL1)、β-グルクロニダーゼ(β-glucuronidase, gusA)、緑色蛍光タンパク質(green fluorescence protein, GFP)、グルコアミラーゼ(glucoamylase)等と、2)成長選別(growth selection)遺伝子であるイミダゾールグリセロールホスファートジハイドラターゼ(imidazoleglycerol-phosphate dehydratase, HIS3)、ホスホリボシルアミノイミダゾールカルボキシラーゼ(phosphoribosylaminoimidazole-carboxylase, ADE2)、α-アミノアジパートリダクターゼ(α-aminoadipate reductase, LYS2)、β-イソプロピルマレートジヒイドロゲナーゼ(β-isopropylmalate dehydrogenase, LEU2)、オルチジン-5'-ホスファートジカルボクシラーゼ(orotidine-5'-phosphate decarboxylase, URA3)等を含む。
【0101】
タンパク質チップは、マイクロアレイによって固体基板上に点滴されたタンパク質とタンパク質、ヘキサン、炭水化物、脂質、コレステロールまたは他の小さな化合物のような異なる分子間の相互作用を分析するのに使用されるバイオチップを意味する。生体分子(biomolecule)間の相互作用を測定できるBiacoreのセンサーチップ(sensor chip)が1990年代に商用化されて以来、タンパク質チップ開発が加速化されてきた。タンパク質チップ技術は疾病診断、タンパク質活性研究、新薬候補物質検索、生体分子相互作用検索等の分野に応用できるのでタンパク質体学の核心技術として浮上してきている。
【0102】
本発明の好ましい実施態様で、タンパク質チップを利用した本発明のHRF-特異的受容体の同定方法は、表面プラスモン共鳴(surface plasmon resonance; SPR)分析、質量分析(surface enhanced laser desorption/ ionization time of flight mass spectrometry; SELDI-TOF)、蛍光分析、電気化学的分析等を通して行なえる。
【0103】
したがって、タンパク質チップを利用した本発明のHRF特異的受容体の同定は、1)機能が明らかにされたり明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程、2)前記HRFまたはHRFダイマーに特異的な抗体がなかったりあったりする状態で、前記言及された分析方法を利用してタンパク質-タンパク質相互作用が起きるかどうかを確認する工程を通して行なえる。
【0104】
上記以外にも、現在まで公知された多様なタンパク質-タンパク質相互作用分析方法が本発明のHRF受容体の同定方法に使用できる。その例として、 欧州特許第 EP1003853号、 欧州特許第 1098967号、 欧州特許出願公開第 EP1184463A1号、 欧州特許第 1224324号、 米国特許第6114111号、 米国特許第6562576号、 米国特許第6828112号、 米国特許出願公開第 US20020094519A1号、 米国特許出願公開第20020106693A1号、 米国特許出願公開第20020106698A1号、 米国特許出願公開第20020142348A1号、米国特許出願公開第20020177217A1号、米国特許出願公開第20030003439A1号、米国特許出願公開第20030040012A1号、米国特許出願公開第20030170723A1号、米国特許出願公開第20030211523A1号、米国特許出願公開第20040146931A1号、米国特許出願公開第20040157279A1号、米国特許出願公開第20050106636A1号、米国特許出願公開第20050176005A1号または米国特許出願公開第20050221280A1号に開示された方法が使用できる。前記文献は、すべて本発明に参考文献として挿入される。
【0105】
以下、本発明を実施例によって詳しく説明する。
ただ、下記の実施例は、本発明を例示するだけのものであって、本発明の内容が下記の実施例に限定されるものではない。
【実施例】
【0106】
実施例1
多様な欠失型HRFの製造
1-1:遺伝子の分離および増幅
まず、大韓民国特許出願第2001-27896号および第2001-27921号に記載した方法によってHRFをコードする遺伝子のcDNAを製造した。詳細には、ラット(Rattus norvegicus)骨格筋から総細胞質RNAを抽出した後、pJG4-5ベクター(Invitrogen, Inc., 米国)を利用して酵母2-ハイブリッド分析のためのcDNAライブラリを製造した。
【0107】
(Na,K)ATPaseのα2サブユニットを利用してCD3部位を pEG202ベクター(Brent, R., and Finley, R. L., Jr. Annu. Rev. Genet. 1997年、第31巻, 663-704頁)のLexA DNA結合ドメイン(pEG202 ベクター)に挿入した後、それをスクリーニングのためのバイト(bait)に使用した。リポーター遺伝子を活性化させた陽性クローンを選択して塩基配列分析(sequencing)、制限酵素マッピングと BLASTサーチを通して配列分析を行なった。これらの中で一つのクローンがIgE依存的ヒスタミン放出因子(HRF)と完全に一致した。
【0108】
前記rHRF cDNA遺伝子から配列番号:1で表されるラットHRF全長配列(Accession number U20525、アミノ酸1-172)をコードする遺伝子を配列番号:9、配列番号:2で表されるアミノ酸11-172切片をコードする遺伝子を配列番号:10、配列番号:3で表されるアミノ酸35-172切片をコードする遺伝子を配列番号:11、配列番号:4で表されるアミノ酸1-112切片をコードする遺伝子を配列番号:12、配列番号:5で表されるアミノ酸39-110切片をコードする遺伝子を配列番号:13、配列番号:6で表されるアミノ酸1-38切片をコードする遺伝子を配列番号:14、配列番号:7で表されるアミノ酸111-172切片をコードする遺伝子を配列番号:15、配列番号:8で表されるアミノ酸84-108切片をコードする遺伝子を配列番号:16で示した。各々の遺伝子をPCRを遂行して増幅させた(図1および図2)。PCRは前変性(pre-denaturation)工程として94℃で5分、変性(denaturation)工程として94℃で1分、アニーリング工程として45〜50℃で1分、延長(elongation)工程として72℃で1分、後延長(post-elongation)工程として72℃で7分の条件で30サイクルの間反応させた。PCR反応に使用したプライマー配列は、前記で記載した。
【0109】
1-2:組換え発現ベクターの製造
実施例1-1で増幅させた配列番号:9乃至配列番号:16の遺伝子を各々 pRSET-Aベクター(Invitrogen)にクローニングして組換え発現ベクターを製造し、それを各々pRSET-A-RrHRF、pRSET-A-Del-N11HRF、pRSET-A-Del-N35HRF、pRSET-A-Del-C112HRF、pRSET-A-Del-N39C110HRF、pRSET-A-Del-C38HRF、pRSET-A-Del-N111HRF、pRSET-A-Del-N84C108HRFと名付けた(図3および図4)。
【0110】
1-3:形質転換体の製造および各欠失型HRFの分離精製
各欠失型HRFを過発現(overexpression)させるために実施例1-2で製造した組換え発現ベクターを大腸菌BL21(DE3)(Novagen)またはBL21(DE3)pLysS(Novagen)に形質転換させた。
【0111】
前記大腸菌形質転換体は、アムピシリンまたはアムピシリンとクロラムフェニコール(chloramphenicol)を含んだLB培地で培養した後、ODが0.6に到達した時、0.4mMになるようにIPTG(isopropyl β-D-thiogalactoside)を添加して三時間さらに培養した後、5,500xgで5分間遠心分離した。回収した大腸菌をバインディングバッファー(5mMイミダゾール、500mM NaCl、20mM Tris-HCl、pH7.9)に再懸濁させた後、超音波を使って粉砕した。溶菌された大腸菌を39,000xgで遠心分離した後、上澄み液はNiカラム(Novagen)を利用して精製した。pRSET-A ベクターにクローニングした欠失型HRFは、N末端に6個のヒスチジンを持つようになるため、ニッケルで充電したHIS-結合カラムに付くようになる。これらタンパク質を高濃度のイミダゾン(imidazone)(1M イミダゾール、500mM NaCl、20mM Tris-Hcl、pH7.9)を含んだイルーションバッファーを使って抽出した後、PD-10カラムを利用して多量の塩分を除去した。 精製された欠失型HRFをモノ-Q陰イオン交換(anion exchange)カラム(Amersham Pharmacia Biotech)でNaCl濃度勾配を利用して再び精製した後、以下の実施例で BEAS-2B細胞(ATCC)とヒト好塩基球を刺激するために使用した。
【0112】
実施例2
BEAS-2B細胞でHRF形態によるIL-8分泌能の検査
各欠失型HRFの活性をBEAS-2B細胞(ATCC)でのIL-8分泌能を通して比較した。BEAS-2B細胞を48ウェルプレートで70%程度育つまで培養した後、1%ペニシリンストレプトマイシン/BEBM(Clonetics)で2回洗浄した。実施例1-3で分離した各々の組換えタンパク質(RrHRFまたは各欠失型HRF)を1μg/mlまたは10μg/mlずつ添加した。48時間または24時間後、上澄み液を取って遊離したIL-8を酵素免疫吸着検出法(PIERCE)を利用して定量した(図5)。
【0113】
実施例3
ヒト好塩基球でHRF形態によるヒスタミン分泌能の検査
各欠失型HRFの活性をヒト好塩基球でのヒスタミン分泌能を通して比較した。アトピー性皮膚炎を持っている血液提供者から40mlの静脈血を採取した後、抗凝血剤であるEDTAを10 mMになるように添加して、食塩水に6%になるように溶解したデキストラン10mlを入れて混合した。室温で90分間静置した後、白血球が集まっている上層を分離して150xgで8分間遠心分離した。回収した白血球をPAG-EDTA(4mM EDTA、25mM PIPES、110mM NaCl、5mM KCl、0.003%HSA、0.1%D-グルコース)で二度洗浄し、冷たい乳酸バッファー(13.4mM 乳酸、140mM NaCl、5mM Kcl、pH3.9)で3〜5分間静置する過程を通して細胞表面にある IgEを除去した。続いて、直ちに30mlのPAG-EDTAを加えて二回洗浄して再びPAG-EDTAに懸濁させた後、ヒトIgE(1μg/ml、Serotec)で2時間感作させた。5%FBSを添加したIMDM培地(Gibco BRL)で2回洗浄した後、実施例1-3で分離した各々の組換えタンパク質(rHRFまたは各欠失型HRF)を20μg/mlずつ添加した。15分後、ヒト抗IgE抗体を加えて4時間後、上澄み液を取って遊離したヒスタミンをヒスタミンアナライザー(Astoria Analyzer, Series 300 system)を利用して定量した(図6)。
【0114】
その結果、配列番号:2で表されるN末端の11個アミノ酸残基が欠失したタンパク質(Del-N11HRF)および配列番号:3で表されるN末端の35個アミノ酸残基が欠失したタンパク質(Del-N35HRF)は、BEAS-2B細胞のように野生型HRFに比べて効果的にヒスタミン放出を誘導することを確認した。
【0115】
実施例4
分子内ジスルフィド結合による二量体化がHRF活性に及ぼす影響比較
4-1:非還元SDS-PAGEによる移動性の差の比較
実施例1-3で製造した野生型HRFと各欠失型HRFのN末端の欠失による電気泳動での移動性(mobility)変化を確認した(図7、図8)。SDS-PAGEは、ラエムリ(Laemmli )(Laemmli U.K., Nature, 第227巻, 680-685頁)の方法を変形して使用し、各タンパク質を還元サンプルバッファー(reducing sample buffer)[0.125M Tris-HCl、pH6.8、4%(w/v)SDS、20%(v/v)グリセロール、and 2%β-メルカプトエタノール(β-ME)]または、非還元サンプルバッファー(non-reducing sample buffer)(w/o β-ME)と混合した後、15%ゲルを作って分離した。その結果、N末端を除去したタンパク質は、野生型HRFとC-末端を除去したタンパク質とは異なり、ダイマー位置に移動することが観察された。これは、N末端欠失型HRFと野生型HRFの構造的な差を裏付けし、HRFのN末端がHRF機能調節に重要な役目を果たすという事実を裏付けしてくれる。また、N末端を除去したタンパク質が分子内ジスルフィド結合を持っていることを示唆する。
【0116】
4-2:ジスルフィド結合を除去するための部位特異的突然変異の製作
N末端欠失型HRFのシスチン残基をセリンに置換して、ジスルフィド結合が形成されることを阻害した場合、HRF活性に及ぼす影響を調べるために部位特異的突然変異キット(site-directed mutagenesis kit, stratagene)を利用して突然変異を製作した。HRFの場合、28番と172番残基にシスチンを含んでいるため、プライマーにCG GAC GGG CTG TCT CTG GAG GTG GA(配列番号:18)とTC CAC CTC CAG AGA CAG CCC GTC CG(配列番号:19)を使用して、pRSET-A-Del-N11HRF-C28Sを製造した。また他のプライマーにGAG ATG GAA AAA TCT AAG CTT GAT CCG(配列番号:20)とCGG ATC AAG CTT AGA TTT TTC CAT CTC(配列番号:21)を使用して、pRSET-A-Del-N11HRF-C172SとpRSET-A-Del-N35HRF-C172Sを製造した。
【0117】
PCR増幅条件は、DNA重合酵素(Pfu DNA polymerase, Stratagene)を使用して前記に記載のプライマー対と鋳型を95℃で5分間転変性させて、95℃で1分、55℃で1分および 68℃で7分間16回反応させて、68℃で7分間後延長させた後、反応を終結した。この反応液にDpnI(Stratagene)を入れて 37℃で1時間反応させて鋳型DNAのみを選択的に切断させた後、XL1-blueに形質転換させた。
【0118】
製作したコンストラクトは、DNA配列分析を通して突然変異生成の有無を確認した後、BL21(DE3)/pLysSに形質転換させて実施例1-3により分離、精製した。これら突然変異の中で特にpRSET-A-Del-N35HRF-C172Sは、非還元的(non-reducing)SDS-PAGEを通して細胞間ジスルフィド結合(intermolecular disulfide bond)を形成しないものが現れ、BEAS-2B細胞株でIL-8を分泌させる能力も顕著に減少したことを確認できた(図11、図12)。これは、N末端欠失型HRFの活性において重要な役割を持つものが、172番残基にあるシスチンによる細胞間ジスルフィド結合であることを示唆する結果であり、HRF活性においてダイマー形成が非常に重要であることが分かる。
【0119】
4-3:化学的方法を利用したHRFの交差結合
架橋剤(crosslinker)を利用して野生型HRFと欠失型HRFがダイマーを形成する能力面でいかなる差があるか調べてみた。-SHと反応する二種類の架橋剤(BMOE, BM(PEO)4, Pierce, 米国)を使用して野生型HRFとDel-N35HRFを交差結合させた。Del-N35HRFの場合、架橋剤と反応可能な-SHを作るために前もって20mMのDTTを使用して細胞間ジスルフィド結合を還元させた後、カラム(Vivaspin column, Vivascience, 米国)を使用してDTTを除去した後、PBSで緩衝溶液組成を変えた。以後、5倍過量の架橋剤を処理した後、BMOEの場合は4℃で4時間、BM(PEO)4の場合は37℃で30分放置した後、上記カラム(vivaspin column)を使用して過量の架橋剤を除去した。これらタンパク質を定量した後、SDS-PAGEで分離した(図13)。還元されたDel-N35HRFが架橋剤により細胞間ジスルフィド結合を形成するのに対して、野生型HRFは細胞内ジスルフィド結合を形成し、架橋剤によってもダイマーを形成できなかった。
【0120】
実施例5
HRF形態によるファージディスプレーペプチドクローンの結合能力の差の比較
各々の欠失型HRFをプラスチックウェルに固定(immobilization)させた後、すでにHRFと結合することが知られているペプチドを発現しているファージ(phage)を添加して、その結合能力に差があるかどうかを調べた。
【0121】
具体的に、コーティング緩衝液(0.1M NaHCO3、pH8.6)に20μg/ml濃度で溶解させた各々の欠失型HRFを50ulずつポリスチレンマイクロタイタープレートに加えて、4℃で一晩中コーティングした後、BSAで非特異的結合を遮断した。0.1%Tween/TBS(TBST)で6回洗浄した後、ファージ溶液30ulを6%BSA/PBS30ulに希釈した溶液を加えて室温で2時間静置した。PBSTで5回洗浄した後、3%BSA/PBSに1:5000で希釈したHRP-コンジュゲートされた抗-M13抗体(Pharmacia)100ulずつを加えて1時間静置した。PBSTで6回、PBSで1回洗浄した後、ペルオキシダーゼ(peroxydase)基質溶液100μlずつを加えて発色程度をエライザ(ELISA)リーダー(Bio-Rad)を使用して405nmで測定した。
【0122】
その結果、前記HRFは、HRFに対して特異的結合を示すヘプタマー(heptamer)ペプチドを発現しているファージに対する親和度にも差を示したが、RrHRFとDel-C112HRFがファージに対してとても弱い親和度を示した一方、Del-N11HRFとDel-N35HRFはとても強い親和度を示した(図14)。このことから、各HRFタンパク質間のヒスタミン分泌能と相応するもので、以上言及した結果から実験的にHRFをターゲットにしたアレルギー制御薬物開発において利用することができるHRFの形態がいくつかの欠失型に限られることが分かる。
【0123】
また、HRF内のHRF結合ペプチドの結合部位を調べるためにHRFを三等分した後、HRF結合ペプチドとの親和度を比較した。Del-N39C110HRFの場合、前記欠失型HRFに似た程度の親和度を示し、アミノ酸をもう少し除去したDel-N84C108HRFも高い親和度を示した(図15)。これは、HRF結合ペプチドがHRFのアミノ酸84-108部位に結合することを意味し、これにより、HRF抑制を通したアレルギー制御薬物開発の検索に利用することができることを確認した。
【0124】
実施例6
欠失型HRFの活性を抑制することができるHRF結合ペプチドの作用比較
RBL-2H3(ATCC)を5×104細胞で24ウェルで増殖させた後、ラットIgE抗体(0.2μg/ml、Serotec)で60分間感作させて、5%FBS/MEM培地で組換えHRFタンパク質を1.56mMで処理した(陽性対照群)。また、上記で準備した細胞にヘプタマー(heptamer)ペプチド(大韓民国特許出願第2001-27896号)を用量依存的方法で処理した(0.0156-15.6mM)。15分後、ラット抗IgE抗体を加えて4時間後、上澄み液を取って遊離したヒスタミンをヒスタミンアナライザー(Astoria Analyzer, Series 300 system)を使用して定量した。
【0125】
HRF結合ペプチドの欠失型HRFに対する作用は、BEAS-2B細胞株でも分析された。実施例3と同じ方法でDel-N35HRF(61nM)をBEAS-2Bに処理した。ここでHRF結合ペプチドおよびそのアラニン置換体(大韓民国特許出願第2001-27896号)、1番または7番アミノ酸が欠失した本発明のHRF結合ペプチドを色々な濃度(6.1-610nM)でDel-N35HRFに混合して室温で10分静置後、添加した。24時間後、上澄み液を取って遊離したIL-8を酵素免疫吸着検出法を利用して定量した。
【0126】
その結果、図16と図17に示したように、HRF結合ペプチドは、RBL-2H3細胞で濃度依存的に欠失型HRFによるヒスタミン分泌を阻害し、BEAS-2B細胞でも濃度依存的に欠失型HRFによるIL-8分泌を抑制した。WYVYPSM配列を持つHRF結合ペプチドは、変形されたアミノ酸位置によって活性の差を示した。N末端から2および6位置のアミノ酸残基をアラニンに変えた置換体は、欠失型HRFに対する抑制能力を喪失するとみえ、HRF結合ペプチドの作用においてチロシン残基とセリン残基が重要な作用をするものと考えられる(図18)。また、前記ペプチドのN末端のトリプトファン残基を除去した本発明の配列番号:17のHRF結合ペプチド(HBP2-D1)の場合も元来の 7-merペプチドとよく似た活性を示したため、2〜6番残基だけでも活性型HRFに対する抑制能力を持つことを確認した。
【産業上の利用可能性】
【0127】
上記で詳しく見たように、本発明のダイマーを形成できる欠失型HRFは、活性型のHRFを提供することによりHRFによるアレルギー制御薬物開発およびアレルギー患者血清内のHRF量を検出するためのキット製作などにも活用することができる。また、本発明の欠失型HRF結合ペプチドは、細胞内でIL-8およびヒスタミン分泌を効果的に制御することにより、動物の喘息や鼻炎などのようなアレルギー疾患およびマラリア予防または治療用薬物開発に有効に利用できる。
【技術分野】
【0001】
本発明は、IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)およびそれに結合するペプチドに関するものである。詳細には、配列番号:3で表されるアミノ酸配列を含むダイマーを形成できる欠失型HRF、HRFの活性を抑制することができる新規なHRF結合ペプチドおよびこれらの医薬分野における用途に関するものである。
【背景技術】
【0002】
喘息、鼻炎、蕁麻疹、アナフィラキーショック(anaphylaxis)、アレルギー性気管支拡張症、食べ物・薬物・植木鉢・虫によるアレルギー、枯草熱(hay fever)、寒冷じんま疹、アトピー性皮膚炎などのアレルギー(非特許文献1、非特許文献2)は、遺伝的にアレルギー抗原(allergen)に対して過敏に反応し大量生成されたIgE(Immunoglobulin E)およびIgE分泌調節に関与するサイトカイン(cytokine)間の均衡破壊から起因すると知られている。
【0003】
一旦アレルギー抗原に露出すれば、即発反応(immediate reaction)が起きて、肥満細胞(mast cell)から分泌したサイトカイン等によって炎症に関連した細胞が抗原露出部位に集まるようになる。数時間後には好塩基球(basophil)、好酸球(eosinophil)およびリンパ球(lymphocyte)から分泌するヒスタミンと様々なサイトカインなどによって遅発相反応(late-phase reaction, 以下「LPR」とする)が起きるようになる。アレルギー患者の半分程度がLPRに進行する。LPRでは好塩基球からヒスタミンが分泌され、その時には即発反応を誘発したアレルギー抗原がないので、好塩基球からヒスタミン分泌およびLPRへの進行を誘発する原因物質を糾明することが主要関心事だった。今まで、MCP-3(Monocyte Chemotatic Protein-3)、MCP-1(Monocyte Chemotatic Protein-1)またはRANTES(Regulated upon Activation Normal T-cell Expressed and Secreted)のようなサイトカイン(cytokine)がヒスタミンを分泌させることが知られていたが、IgE依存的LPRでは「HRF」と名付けられたタンパク質だけが好塩基球からヒスタミンを分泌させることができることが明らかにされた(非特許文献3)。しかし、HRFがどのような作用機序によって好塩基球からヒスタミン分泌を誘導するかに対しては報告されたことがない。
【0004】
HRFは、すべての細胞質に存在する、172個のアミノ酸で構成された公知のタンパク質である(非特許文献4)、その中の79〜123番アミノ酸は塩基性ドメイン(basic domain)を形成していて、この部位は微小管結合タンパク質である MAP-1B(microtubule-associated protein-1B)と約46%の相同性を示すため、微小管に 結合する機能を有することができると推定されている。ガチェット(Gachet)らは共焦点顕微鏡(confocal microscope)を利用してHRFの分布と細胞骨格網分布がどの程度一致するかを観察したが(非特許文献5)、これはHRFが細胞骨格と結合していることを示唆するものである。一方、サンチェスらは、HRFが一般的なカルシウム結合タンパク質ファミリーに属さないにもかかわらず、Ca2+と結合することを発表した。また、サカロマイセスセレビシエ(Saccaromyces cerevisiae)でHRF遺伝子を欠失させても酵母が生存できることを確認した(非特許文献6)。これは、HRFが重複経路(redundant pathway)を持っている遺伝子ファミリーに属するという事実を暗示するものである。
【0005】
一方、HRFは親水性で細胞質内に存在するタンパク質であるにもかかわらず、マクドナルドらが、HRFを細胞の外で見つけ、LPRアレルギー患者の血漿から多量に検出されるため、細胞死滅(apoptosis)や他の作用機序によって細胞外に分泌した後、細胞膜に存在するHRF受容体を通してヒスタミンを遊離させると考えられている(非特許文献7)。また、HRFはIgE感作された(sensitized)好塩基球を刺激してヒスタミンを分泌するようにすることが知られているが、これに対するIgEの作用機序は、まだ糾明されていない。一方、ビーカエスクラ(Bheekha-Escura)らは、IgE受容体がない細胞でもHRFが炎症反応を起こすことができることを観察することにより、HRFがIgEと結合して作用するのではなく、特異な細胞膜受容体に結合して作用することを提示した(非特許文献8)。
【0006】
本発明者らは、I)HRFが親水性にもかかわらず細胞膜を透過することができるという事実、II)酵母2-ハイブリッド分析(yeast two-hybrid)によってHRF受容体が(Na,K)ATPaseの第3細胞質ドメイン(CD3)という事実を明らかにしてきた(特許文献1、 特許文献2)。同時に、細胞外に分泌したHRFが、好塩基球細胞内でヒスタミン分泌を促進する作用機序も明らかにしてきた。
【0007】
また、I)HRFが細胞内に入って行く工程を遮断して/遮断したり、II)HRFが (Na,K)ATPaseに結合する工程を遮断して、HRFのヒスタミン分泌を阻害することができるペプチドを見つけて、アレルギー疾患を予防または治療することができることを立証してきた(特許文献1)。
【0008】
最近ブデ(Budde)らは、活性化した単核球培養上澄み液から分離した HRF(HRFmn)と組換えHRF(recombinant HRF, rHRF)は、好塩基球でのヒスタミン分泌能が同じではないことを報告したが、これはHRFmnとrHRFが、同じ作用因子(factor)ではない可能性を提示する(非特許文献9)。また、ヒトHRF特異的な酵素免疫吸着検出法(enzyme-linked immunoabsorbant assay)によってHRFmnからHRFが検出されなかったことに言及したが、これもHRFmnとrHRFとの構造的差異を示唆するものであると言える。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】大韓民国特許第457350号
【特許文献2】大韓民国特許第457351号
【非特許文献】
【0010】
【非特許文献1】Bachertら, Clinical and Experimental Allergy, 1998年,第28巻, 15-19頁
【非特許文献2】MacDonald and Lichtenstein, Springer Semin Immunopathol., 1990年,第12巻, 415-428頁
【非特許文献3】MacDonaldら., Science, 1995年,第269巻, 688-690頁
【非特許文献4】Bohmら, Biochem. Int., 1989年、第19巻, 277-286頁
【非特許文献5】Gachetら., J. Cell Sci., 1999年,第112巻, 1257-1271頁
【非特許文献6】Sanchezら, Electrophoresis, 1997年,第18巻, 150-155頁
【非特許文献7】MacDonaldら, Science, 1995年,第269巻, 688-690頁
【非特許文献8】Bheekha-Escuraら, Blood, 2000年,第96巻, 2191-2198頁
【非特許文献9】Buddeら, Ann. Allergy Asthma Immunol., 2002年、第89巻, 606-612頁
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明の目的は、細胞からヒスタミンおよびIL-8を分泌させることができるダイマーを形成できる欠失型HRF、HRFの作用を抑制することができる、新規なHRF結合ペプチドおよびこれらの用途を提供することである。
【課題を解決するための手段】
【0012】
本発明者らは、HRFmnとrHRFの活性の差は、アレルギー患者の血液の中に多量存在するタンパク質分解酵素(protease)によるものと判断して、ヒスタミン分泌能を有するHRFを捜そうと努力した結果、野生型HRFに比べ非常に高い活性を有する欠失型HRFを見つけた。これら欠失型HRFは、野生型HRFとは異なり分子内ジスルフィド結合を有していて、このジスルフィド結合による二量体化によって活性を持つようになることを確認した。また、すでに発見されているHRF結合ペプチドを変形させた後、やはりこれが欠失型HRFに結合してHRFのヒスタミンおよびIL-8分泌能を効果的に妨害することを確認して、これら欠失型HRFおよびその結合ペプチドが抗アレルギー薬物開発に有効に使用できることを確認して本発明を完成した。
【0013】
上記目的を達成するために、本発明は、野生型IgE-依存的ヒスタミン放出因子のN末端が除去されることによりダイマーを形成でき、ヒスタミンおよびIL-8分泌能が向上された組換え欠失型IgE-依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)を提供する。
【0014】
また、配列番号:3で表されるアミノ酸配列を含むダイマーを形成できる欠失型HRFを提供する。
【0015】
また、前記組換え欠失型IgE-依存的ヒスタミン放出因子の同形または異形ダイマーおよびその製造方法を提供する。
【0016】
また、前記組換え欠失型IgE依存的ヒスタミン放出因子または野生型 IgE依存的ヒスタミン放出因子間に形成される同形または異形ダイマーおよびその製造方法を提供する。
【0017】
また、本発明は、前記欠失型HRFをコードする遺伝子、前記遺伝子を含む組換え発現ベクターおよび前記発現ベクターで形質転換された形質転換体を提供する。
【0018】
また、本発明は、前記欠失型HRFを有効成分として含むヒスタミンおよびIL-8放出誘導剤を提供する。
【0019】
また、本発明は、前記欠失型HRFを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法を提供する。
【0020】
また、本発明は、前記欠失型HRFを利用したアレルギー患者血清内HRF検出方法を提供する。
【0021】
また、本発明は、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)配列を持つ新規なHRF結合ペプチドを提供する。
【0022】
また、本発明は、前記HRF結合ペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物を提供する。
【0023】
また、本発明は、前記HRF結合ペプチドおよび抗-HRFモノクローナル抗体を含むアレルギー診断キットを提供する。
【0024】
また、本発明は、前記HRF結合ペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物を提供する。
【0025】
合わせて、本発明は、タンパク質-タンパク質相互作用分析を行なう工程を含む前記欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法を提供する。
【0026】
本発明(1)は、野生型IgE依存的ヒスタミン放出因子のN末端が除去され、ダイマー形成が可能でヒスタミンおよびIL-8分泌能が向上した組換え欠失型IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)である。
本発明(2)は、HRFが、脊椎動物のHRFまたはHRFの類似体であることを特徴とする、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(3)は、HRFが、野生型HRFのN末端の少なくとも11乃至35アミノ酸が欠失したことを特徴とする、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(4)は、配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、本発明(1)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(5)は、HRFが、配列番号:2(Del-N11HRF)または配列番号:3(Del-N35HRF)で表されるアミノ酸配列を有することを特徴とする、本発明(4)の欠失型IgE依存的ヒスタミン放出因子である。
本発明(6)は、本発明(1)の組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成されるIgE依存的ヒスタミン放出因子の同形または異形ダイマーである。
本発明(7)は、本発明(1)のHRFまたは本発明(6)のHRF同形または異形ダイマーに特異的な抗体である。
本発明(8)は、本発明(1)の欠失型HRFおよび/または野生型HRFに架橋剤を処理する工程を含む、本発明(6)の同形または異形ダイマーを製造する方法である。
本発明(9)は、架橋剤が、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン( 1,4-Di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-Bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-Bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(Bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-Bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(Bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-Hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(Dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bismaleimidyl-2,3-dihydroxybutane;BMDB)からなる群より選択されることを特徴とする、本発明(8)の欠失型HRFの同形または異形ダイマーを製造する方法である。
本発明(10)は、本発明(1)の欠失型HRFをコードする遺伝子である。
本発明(11)は、遺伝子が、配列番号:11(Del-N35HRF)で表される塩基配列を含むことを特徴とする、本発明(10)の遺伝子である。
本発明(12)は、遺伝子が、配列番号:10(Del-N11HRF)または配列番号:11(Del-N35HRF)で表される塩基配列を持つことを特徴とする、本発明(11)の遺伝子である。
本発明(13)は、本発明(10)の遺伝子を含む組換え発現ベクターである。
本発明(14)は、pRSET-A-Del-N11HRFまたはpRSET-A-Del-N35HRFであることを特徴とする、本発明(13)の組換え発現ベクターである。
本発明(15)は、本発明(13)の発現ベクターで形質転換された形質転換体である。
本発明(16)は、大腸菌であることを特徴とする、本発明(15)の形質転換体である。
本発明(17)は、BL21(DE3)-pRSET-A- Del-N11HRF、BL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A- Del-N11HRF、またはBL21(DE3)pLysS-pRSET-A-Del-N35HRFであることを特徴とする、本発明(16)の形質転換体である。
本発明(18)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子または本発明(6)の同形または異形ダイマーを有効成分として含むヒスタミン放出誘導剤である。
本発明(19)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子または本発明(6)の同形または異形ダイマーを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法である。
本発明(20)は、欠失型IgE依存的ヒスタミン放出因子が、配列番号:8のアミノ酸配列を有することを特徴とする、本発明(15)の方法である。
本発明(21)は、本発明(1)の欠失型 IgE依存的ヒスタミン放出因子を利用したアレルギー患者血清内HRFの検出方法である。
本発明(22)は、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を有するHRF結合ペプチドである。
本発明(23)は、配列番号:17のアミノ酸配列を有することを特徴とする、本発明(22)のペプチドである。
本発明(24)は、L-アミノ酸、D-アミノ酸、またはL-アミノ酸およびD-アミノ酸からなる、本発明(22)のペプチドである。
本発明(25)は、一つ以上の変形アミノ酸を含む、本発明(22)のペプチドである。
本発明(26)は、変形アミノ酸が、アミノ酸誘導体またはアルキル化されたアミノ酸であることを特徴とする、本発明(25)のペプチドである。
本発明(27)は、本発明(22)のペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物である。
本発明(28)は、本発明(22)のペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物である。
本発明(29)は、本発明(22)のペプチドおよび抗HRFモノクローナル抗体を含むアレルギー診断キットである。
本発明(30)は、タンパク質-タンパク質相互作用分析を行なう工程を含む欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法である。
本発明(31)は、タンパク質-タンパク質相互作用分析が、同時精製(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)またはタンパク質チップシステムにより行なわれることを特徴とする、本発明(30)のHRF特異的受容体を同定する方法である。
本発明(32)は、同時精製が、
1)HRF-HRF受容体複合体を分離する工程;
2)前記分離された複合体を精製する工程;および、
3)前記精製された複合体からHRF受容体の正体を確認する工程;
を含むことを特徴とする、本発明(31)のHRF特異的受容体を同定する方法である。
本発明(33)は、タンパク質チップシステムが、
1)機能が明らかにされているか明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程;および、
2)前記欠失型HRFまたはHRFダイマーに、特異的な抗体がないかまたは抗体がある状態で前記言及された分析方法を利用して、タンパク質-タンパク質相互作用が起きるかどうかを確認する工程;
を含むことを特徴とする、本発明(31)のHRF特異的受容体を同定する方法である。
【発明の効果】
【0027】
本発明により、細胞からヒスタミンおよびIL-8を分泌させることができるダイマーを形成できる欠失型HRF、HRFの作用を抑制することができる、新規なHRF結合ペプチドおよびこれらの用途が提供された。
【図面の簡単な説明】
【0028】
【図1】本発明で製造した多様な欠失型HRFの一模式図である。
【図2】本発明で製造した多様な欠失型HRFの一模式図である。
【図3】図1と図2に記載した各々の組換え欠失型HRF(pRSET-A-rHRF, pRSET-A-Del-N11HRF, pRSET-A-Del-N35HRF, pRSET-A-Del-C112HRF, pRSET-A-Del-N39C110HRF, pRSET-A-Del-C38HRF, pRSET-A-Del-N111HRF, pRSET-A-Del-N84C108HRF)製造のためのpRSET-Aベクターの遺伝子地図を示した模式図である。
【図4】図3に記載したpRSET-AベクターのMCS(multiple cloning site)を示した遺伝子配列および前記遺伝子のコード部位にあたるアミノ酸配列である。
【図5】図1に記載した欠失型HRFによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図6】IgE存在下で図1に記載した欠失型HRFによるヒト好塩基球でのヒスタミン分泌率を比べた結果を示したグラフである。
【図7】図1に記載されたRrHRFの還元剤有無による移動性(mobility)の差を示したゲル電気泳動写真である。
【図8】図1に記載した欠失型HRFの還元剤の有無による移動性(mobility)の差を示したゲル電気泳動写真である。
【図9】組換え欠失型HRFであるpRSET-A-Del-N11HRFとpRSET-A-Del-N11HRFの突然変異体であるpRSET-A-Del-N11HRF-C28SとpRSET-A-Del-N11HRF-C172Sのゲル電気泳動写真である。
【図10】組換え欠失型HRFであるpRSET-A-Del-N11HRFとpRSET-A-Del-N11HRFの突然変異体であるpRSET-A-Del-N11HRF-C28SとpRSET-A-Del-N11HRF-C172SによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図11】組換え欠失型HRFであるpRSET-A-Del-N35HRFとpRSET-A-Del-N35HRFの突然変異体であるpRSET-A-Del-N35HRF-C172SのPAGE(ポリアクリルアミドゲル電気泳動)写真である。
【図12】組換え欠失型HRFであるpRSET-A-Del-N35HRFとpRSET-A-Del-N35HRFの突然変異体であるpRSET-A-Del-N35HRF-C172SによるBEAS-2B細胞株でのIL-8分泌量を比べた結果を示したグラフである。
【図13】架橋剤を利用して欠失型HRFと野生型HRFのダイマー形成能を確認した結果を示したSDS-PAGE写真である。
【図14】HRF結合ペプチドを発現しているファージ(phage)であるΦ-HBP-2と図1に記載した欠失型HRFとの親和度を比べた結果を示したグラフである。
【図15】HRF結合ペプチドを発現しているファージ(phage)であるΦ-HBP-2と図2に記載した欠失型HRFとの親和度を比べた結果を示したグラフである。
【図16】HRF結合ペプチドであるΦ-HBP-2 が、BEAS-2B細胞株で欠失型HRFによるIL-8分泌を抑制することを示したグラフである。
【図17】HRF結合ペプチドであるΦ-HBP-2 が、RBL-2H3細胞株で欠失型HRFによるヒスタミン分泌を抑制することを示したグラフである。
【図18】HRF結合ペプチドであるΦ-HBP-2 、そのアラニン置換体(HBP2-M1、HBP2-M2m、HBP2-M3、HBP2-M4、HBP2-M5、HBP2-M6、HBP2-M7)および1番残基が欠失した6mer HRF結合ペプチド(HBP2-D1)によるIL-8分泌抑制程度を比較した結果を示したグラフである。
【発明を実施するための形態】
【0029】
以下、本発明を詳しく説明する。
本発明は、野生型IgE-依存的ヒスタミン放出因子のN末端が除去されることによりダイマー形成が可能で、ヒスタミンおよびIL-8分泌能が向上された組換え欠失型 IgE-依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)を提供する。
【0030】
本発明の欠失型HRFは、本発明が提供する配列によって化学的に合成することができる。また他の方法として、本発明の欠失型HRFは、これをコードする塩基配列を持つDNAが挿入された発現ベクターを利用する組換えDNA技術で製造することができる。前記ベクターは、生体内に存在するように製造され、サムブロック(Sambrook)らの方法(Sambrookら.,Molecular cloning, 1989年, Cold Spring Harbor, Cold Spring Harbor Laboratory Press)によって適当な宿主細胞で形質転換され適当な条件下で発現するように製造される。また、本発明によるアミノ酸配列を含む融合タンパク質を利用して本発明の欠失型HRFを製造することができる。
【0031】
一方、本発明による欠失型HRFのアミノ酸配列は、当技術分野の公知された通常の技術により変形され得る。例えば、本発明の欠失型HRFは、アミノ酸数を増加させたり減少させたりすることにより変形できる。また、本発明による欠失型HRFの活性を減少させない範囲内で、特定残基やその順序を変えることにより変形することができる。変形可能なアミノ酸は、自然的に存在する L-α-アミノ酸だけではなく、β、γ、δアミノ酸はもちろんD-α-アミノ酸の誘導体にも変形可能である。
【0032】
したがって、当技術分野で通常の知識を持った者は、前記欠失型HRFをそのヒスタミン分泌活性を維持乃至増加させたり損傷させたりしない範囲内で通常の技術を使って変形させることができる。これは、本発明の範疇内に属するものである。
【0033】
本発明では、配列番号:1で表されるラットの全長配列(Accession number U20525, アミノ酸1-172)、配列番号:2で表されるアミノ酸11-172切片、配列番号:3で表されるアミノ酸35-172切片、配列番号:4で表されるアミノ酸1-112切片、配列番号:5で表されるアミノ酸39-110切片、配列番号:6で表されるアミノ酸1-38切片、配列番号:7で表されるアミノ酸111-172切片、配列番号:8で表されるアミノ酸84-108切片を有する多様な欠失型HRFを製造するために、配列番号:9で表されるラットHRF全長配列(Accession number U20525)遺伝子から前記断片をコードする遺伝子をpRSET-Aベクターにクローニング(図1乃至図4参照)して大腸菌に形質転換させ、その後発現を誘導して分離、精製した。
【0034】
続いて、製造された各欠失型HRFのN末端の欠失による各欠失型HRFの BEAS-2B細胞株でのIL-8分泌能(図5参照)と好塩基球でのヒスタミン分泌能(図6参照)を比べた。その結果、配列番号:2で表される野生型HRFのN末端の10個アミノ酸残基が欠失されたタンパク質(Del-N11HRF)および配列番号:3で表される野生型HRFのN末端の34個アミノ酸残基が欠失したタンパク質(Del-N35HRF)は、BEAS-2B細胞株で野生型HRFに比べて増加したIL-8分泌能を見せた。特に、これらは野生型HRFの場合、測定可能なIL-8を分泌することができない低濃度(1μg/ml)でもIL-8分泌を誘導した。また、これら欠失型HRFは、IgEに感作されたヒト好塩基球細胞でも野生型HRFに比べて増加したヒスタミン分泌能を見せた。
【0035】
したがって、HRFはその活性に主要な役目を持つ部位がN末端によって選り分けられているものと思われ、遅発相反応(LPR)アレルギー患者の場合、ヒスタミンおよびIL-8の分泌を誘発する活性型のHRFを持っているものと推定される。N末端の一部アミノ酸残基を除去した場合(Del-N11HRFとDel-N35HRF)は、ヒスタミン分泌能が増加するが、C末端のみを除去した場合(Del-C112HRF)には野生型HRFと等しい活性を見せることも、HRFのN末端がHRF機能調節に重要な役目を持つという事実を裏付けてくれる。
【0036】
欠失型HRFタンパク質は、HRFに対して特異的結合を見せるヘプタマー(heptamer)ペプチドを発現しているファージに対する親和度の面でも差を示したが、野生型HRFとDel-C112HRFがファージに対して弱い親和度を示した一方、Del-N11HRFとDel-N35HRFはとても強い親和度を示した(図14参照)。これは、各HRFタンパク質の活性と相応するものであり、以上、言及した結果から実験的にHRFをターゲットにするアレルギー制御薬物開発において利用することができるHRFの形態がいくつかの欠失型に限られることを確認した。
【0037】
本発明によって欠失型HRFの活性型とその機序が明かされた以上、当技術分野で通常の知識を持った者なら誰でもマウス、ヒト、ウサギ、ニワトリ等で活性型HRFを容易に同定することができる。よって、マウス、ヒト、ウサギ、ニワトリの活性型HRFもまた本発明の範疇内に属するものである。
【0038】
本発明は、配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、ダイマーを形成できる欠失型HRFを提供する。
【0039】
配列番号:3のHRFは、配列番号:1で表されるラット(Rattus norvegicus)の全長HRF アミノ酸配列のN末端を34個欠失させた形態であり、前記アミノ酸配列を含む欠失型HRFがヒスタミンおよびIL-8分泌能がすぐれたものであることを確認し、この活性が分子内ジスルフィド結合による二量体化によるものであることを確認した。
【0040】
また、本発明は、前記組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成される同形または異形ダイマーおよびその製造方法を提供する。
【0041】
上記のようにHRFのN末端がHRF機能調節に重要な役割を持つとみられるため、本発明者らは、欠失型HRFと野生型HRFの間に構造的差異があると推定して、これらが還元剤であるβ-メルカプトエタノールの有無によってその移動性に変化を見せるかどうかを確認した(図7、図8参照)。非還元SDS-PAGE結果、強い活性を見せる欠失型HRFは弱い活性を見せる野生型HRFやC末端欠失型HRFとは異なり、ダイマー位置で観察された。これは、これらN末端欠失型HRFが分子内ジスルフィド結合によるダイマー構造を成していることを示唆する。HRFの場合、残基28番と172番にシスチンを含んでいるのでpRSET-A-Del-N11HRFの場合は、これらアミノ酸を各々セリンに変える突然変異を実施した。pRSET-A-Del-N35HRFの場合は、172番のシスチンのみをセリンに変える突然変異を実施した後、これらの非還元SDS-PAGEでの移動性を比較した(図9、11参照)。pRSET-A-Del-N11HRFとpRSET-A-Del-N11HRF-C28S、pRSET-A-Del-N35HRFがダイマー位置に移動するのとは異なり、pRSET-A-Del-N11HRF-C172S、pRSET-A-Del-N35HRF-C172Sは、モノマー位置に移動することが観察された。これは、172番のシスチンがHRFの二量体化に主な役目を持つことを意味する。
【0042】
本発明者らは、この二量体化がHRFの活性に影響を及ぼすか否かを調べるために、BEAS-2B細胞株でのIL-8分泌能を比較した(図10、12参照)。分泌したIL-8の量は、pRSET-A-Del-N11HRF-C172Sの場合、野生型の46%、pRSET-A-Del-N35HRF-C172Sの場合、野生型の26%に各々減少した活性が見られた。これは、HRF内で28番シスチンに比べて172番シスチンの活性に寄与するところが大きいことを示唆し、これはPAGEでの移動性結果と相応する。以上の結果は、欠失型HRFは野生型HRFとは異なり、分子間ジスルフィド結合(intermolecular disulfide bond)により、ダイマー形態で存在し、その構造的な差がこれらの間の活性差を誘発する原因であることを示している。
【0043】
本発明者らは、架橋剤(crosslinker)を利用して野生型HRFもダイマーを形成できるか否か調べた(図13参照)。その結果、欠失型HRFとは異なり野生型HRFは分子間ジスルフィド結合(intermolecular disulfide)をほとんど形成することができず、大部分が分子内ジスルフィド結合(intramolecular disulfide bond)を形成した。これは、欠失型HRFと野生型HRFの構造的な差を再び示す結果で、HRFの場合化学的交差結合よりN末端欠失がその活性においてさらに効果的であることが分かる。
【0044】
但し、前記野生型HRFのダイマー形成の失敗は、分子間ジスルフィド結合の不在に起因するものであり、ジスルフィド結合以外の他の方法による試験管内ダイマーの製造方法によるヒスタミンおよびIL-8分泌能が向上された野生型HRFの同形ダイマーの存在可能性を排除するものではない。したがって、本発明のダイマーには、前記多様な長さを有する欠失型HRF間に形成される同形または異形ダイマーの他にもジスルフィド結合以外の方法を使用して形成される野生型HRFの同形ダイマーまたは前記多様な長さを有する欠失型HRFと野生型HRF間に形成される異形ダイマーまでも含まれる。
【0045】
本発明のHRF同形または異形ダイマーの製造方法は、特別にそれに制限されるものではなく、公知のすべてのタンパク質ダイマー製造方法が使用可能であるが、欧州特許出願公開0261616A2号、日本特許出願公開第1993-032941A号、 日本特許出願公開第 1986-069759号、 国際公開第 WO05/105994A1号および国際公開92/013965A1号等に記載された方法を使用することが好ましい。前記文献は、すべて本明細書に参考文献として挿入される。
【0046】
本発明の欠失型HRFは、本発明が提供する配列により製造された後、市販の架橋剤を処理することにより化学的にダイマーを形成させられる。例えば、-SHを標的にする架橋剤等を使用してイオウを交差結合させてダイマーを製造できる。架橋剤としては、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン(1,4-di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bis-maleimidyl-2,3-dihydroxybutane;BMDB)が使用可能である。
【0047】
また、本発明は、前記欠失型HRFをコードする遺伝子を提供する。
【0048】
本発明では、配列番号:9で表されるラットHRF全長配列から下記プライマー配列を利用して配列番号:10から配列番号:16までの多様な欠失型HRF遺伝子をクローニングした。
【0049】
配列番号:9のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ATG ATT ATC TAC CGG GAC(配列番号:22)。
【0050】
配列番号:9のクローニングのためのリバースプライマー:CCG CTCGAG(XhoI)TGT CCT AAG TCC TGG TGT(配列番号:23)。
【0051】
配列番号:10のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)GAC GAG CTG TCC TCC GAC AT(配列番号:24)。
【0052】
配列番号:10のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:25)。
【0053】
配列番号:11のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)AGT GTC AGT AGA ACA GAG(配列番号:26)。
【0054】
配列番号:11のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:27)。
【0055】
配列番号:12のクローニングのためのフォワードプライマー:TAACAAATTGGATCTATCGCCCGCGGAC(配列番号:28)。
【0056】
配列番号:12のクローニングのためのリバースプライマー:CTTTACCCTTTCTGGTTTCTGTTCTTC(配列番号:29)。
【0057】
配列番号:13のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ACA GAG GGT GCC ATC GA(配列番号:30)。
【0058】
配列番号:13のクローニングのためのリバースプライマー:G GAATTC(EcoRI)CCT TTC TGG TTT CTG TT(配列番号:31)。
【0059】
配列番号:14のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ATG ATT ATC TAC CGG GAC(配列番号:32)。
【0060】
配列番号:14のクローニングのためのリバースプライマー:G GAATTC(EcoRI)TCT ACT GAC CAT CTT GC(配列番号:33)。
【0061】
配列番号:15のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)GTA AAG CCT TTT ATG ACT(配列番号:34)。
【0062】
配列番号:15のクローニングのためのリバースプライマー:CCC AAGCTT(HindIII)ACA TTT TTC CAT CTC TAA(配列番号:35)。
【0063】
配列番号:16のクローニングのためのフォワードプライマー:CG GGATCC(BamHI)ACA AAA GAG GCC TAC AAA(配列番号:36)。
【0064】
配列番号:16のクローニングのためのリバースプライマー:CG GGATCC(BamHIII)TGG TTT CTG TTC TTC AAG(配列番号:37)。
【0065】
配列番号:10は、配列番号:2で表されるアミノ酸11-172切片をコードする遺伝子の塩基配列であり、配列番号:11は、配列番号:3で表されるアミノ酸35-172切片をコードする遺伝子の塩基配列であり、配列番号:12は、配列番号:4で表されるアミノ酸1-112切片をコードする遺伝子の塩基配列であり、配列番号:13は、配列番号:5で表されるアミノ酸39-110切片をコードする遺伝子の塩基配列であり、配列番号:14は、配列番号:6で表されるアミノ酸1-38切片をコードする遺伝子の塩基配列であり、配列番号:15は、配列番号:7で表されるアミノ酸111-172切片をコードする遺伝子の塩基配列であり、配列番号:16は、配列番号:8で表されるアミノ酸84-108切片をコードする遺伝子の塩基配列を示す。
【0066】
本発明の欠失型HRFタンパク質をコードする遺伝子配列は、配列番号:10、11、12、13、14、15および16からなる群より選択される塩基配列のいずれかであり、配列番号:11で表される塩基配列を含むことが好ましく、配列番号:10または配列番号:11で表される塩基配列を有することがさらに好ましい。
【0067】
また、本発明は、前記欠失型HRFをコードする遺伝子を含む組換え発現ベクターを提供する。
【0068】
本発明の組換え発現ベクターは、特別に限定されるものではなく、欠失型HRFをコードする遺伝子を含むものなら微生物、植物または動物形質転換用組換え発現ベクターのいずれでも本発明の範疇に属する。
【0069】
本発明では、大腸菌発現ベクターpRSET-Aを利用して配列番号:9、10、11、12、13、14、15および16の塩基配列の群より選択される遺伝子をクローニングして各々の組換えベクター、pRSET-A-RrHRF、pRSET-A-Del-N11HRF、pRSET-A-Del-N35HRF、pRSET-A-Del-C112HRF、pRSET-A-Del-N39C110HRF、pRSET-A-Del-C38HRF、pRSET-A-Del-N111HRF、pRSET-A-Del-N84C108HRFを製造した。
【0070】
また、本発明は、前記組換え発現ベクターで形質転換された形質転換体を提供する。
【0071】
前記形質転換体は、特別に限定されるものではなく、本発明の欠失型HRFをコードする遺伝子を含む組換え発現ベクターで形質転換されたものなら、大腸菌、植物、動物細胞のいずれであっても本発明の範疇に含まれる。
【0072】
本発明者らは、大腸菌BL21(DE3)またはBL21(DE3)pLysS菌株に前記組換え発現ベクターを導入して形質転換体を製造した。その中で、前記組換え発現ベクターpRSET-A-Del-N11HRFをBL21(DE3)およびBL21(DE3)pLysS菌株に形質転換させたものを各々BL21(DE3)-pRSET-A-Del-N11HRF、BL21(DE3)pLysS-pRSET-A-Del-N11HRFと、組換え発現ベクターpRSET-A-Del-N35HRFをBL21(DE3)およびBL21(DE3)pLysS菌株に形質転換させたものをBL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A-Del-N35HRFと名付けた。これら大腸菌は、lacUV5プローモーター調節下でのみT7RNA重合酵素が転写されるように考案されたもので、IPTG(isopropyl-β-D-thiogalactopyranoside)添加時、T7RNA重合酵素発現が誘導されて欠失型HRFが転写されるようになる。また精製中にタンパク質を分解することができるlonプロテアーゼとompT外膜プロテアーゼが欠乏していて多量の欠失型HRFを得ることができるようにする利点がある。
【0073】
また、本発明は、前記欠失型HRFを有効成分として含むヒスタミンおよびIL-8放出誘導剤を提供する。
【0074】
配列番号:2と配列番号:3のアミノ酸配列を有する欠失型HRFタンパク質は、野生型HRFに比べてBEAS-2B細胞株で増加されたIL-8分泌能を示し、ヒト好塩基球でも同様の方式でヒスタミン放出を誘導した。よって、前記欠失型HRFは、IL-8およびヒスタミン放出誘導剤として効果的に使用できる。本発明のIL-8およびヒスタミン放出誘導剤は、アレルギー治療を目的にする薬物開発における実験的モデルを提示する。すなわち、本発明の誘導剤は、多量のアレルギー誘発物質を必要とするアレルギー制御薬物の開発および検証過程に利用できる。
【0075】
また、本発明は、欠失型HRFを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法を提供する。
【0076】
すなわち、本発明の欠失型HRFは、HRF結合ペプチドの結合部位を提供することにより、これを利用してHRF抑制を通じたアレルギー制御薬物開発に利用することができる。HBP2 ペプチド(大韓民国特許出願第2001-27896号)を利用した親和性実験では、配列番号:5および 配列番号:8のアミノ酸配列を持つ欠失型HRFが前記ペプチドへの親和度が高く現われた(図9参照)。よって、これら配列を有するHRFペプチドは、これと結合する物質や化合物の検索に利用でき、このような物質、化合物は、アレルギー制御薬物の候補になり得る。
【0077】
すなわち、競争結合分析を使って、欠失型HRFをコードする遺伝子を含む組換えベクターで形質転換された細胞から欠失型HRFを分離した後、試験薬物および前記欠失型HRFと相互作用することが知られた物質[例えば、HRF結合ペプチド(大韓民国特許出願第2001-27896号)]と一緒に反応させて、前記試験薬物の中で欠失型HRFとHRF結合ペプチドとの間の相互作用を減少させる薬物を選別する。最終的に、HRF結合ペプチドと似た構造を有し、細胞で HRFによるヒスタミン分泌を抑制することができる薬物をスクリーニングできる。
【0078】
また、本発明は、欠失型HRFを利用したアレルギー患者の血清内HRFの検出方法を提供する。
【0079】
本発明の欠失型HRFを利用して通常の方法によって抗欠失型HRFのモノクローナル抗体を製造することができ、製造された抗-欠失型HRFモノクローナル抗体も本発明の範疇に含まれる。
【0080】
これを利用して患者の血清内HRFを検出する方法を説明すると、底にHRF結合ペプチドまたは抗欠失型HRFモノクローナル抗体を付着させた後、血液サンプルと反応させて、そこに通常の標識物質(HRP(horse reddish peroxydase)、塩基性脱リン酸化酵素(alkaline phosphatase)、蛍光物質(fluorescein) または色素(dye)等)がコンジュゲートされた抗欠失型HRFモノクローナル抗体、または抗HRFモノクローナル抗体を加える。発色または蛍光を発して陽性と判定されれば、血液内活性型HRFが含まれているため、アレルゲンがなくてもアレルギーを起こし得る疾患を持っていると判断することができる。
【0081】
また、本発明は、HRFに特異的に高い親和度で結合してヒスタミン分泌を阻害する新規なペプチドを提供する。
【0082】
本発明のHRF結合ペプチドは、(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を持つことができ、配列番号:17に記載したYVYPSM配列を持つことが好ましい。
【0083】
本発明のHRF結合ペプチドは、ペプチドの活性を減少させない範囲内で、結合に直接的に関与したり保存しなければならない残基を除いた特定残基成分やその順序を変えることにより変形することもできる。変形可能なアミノ酸は、自然界に存在するL-α-アミノ酸だけでなく、β、γ、δアミノ酸はもちろんD-α-アミノ酸の誘導体へも変形可能である。
【0084】
典型的に、一つのアミノ酸が置換されたペプチドを利用して静電気力や親水性が結合に及ぼす影響を調査した結果、陽電荷を帯びたアミノ酸(例:Lys、Arg)や陰電荷を帯びたアミノ酸(例:Glu)が置換されると敏感度が変化することが分かる。このように、置き換えられたり付加されたりする残基の数や形態は、必須の結合点の間に必要な空間と親水性または疎水性と共に要求される機能によって決まる。このような置換によって本発明によるペプチドの目的タンパク質に対する親和度をさらに増加させることができる。
【0085】
置換によって機能において重要な変化が現われたりする。変化する残基の選択は、目的位置に存在する分子の電気度、疎水性、側鎖の変化またはらせん構造の変化等のようにペプチドの基本骨格を維持するのに大きな影響を及ぼしたりする。一般的に、ペプチドの性質に重大な変化を起こすものは、セリンのような親水性残基が、ロイシン、イソロイシン、フェニルアラニン、バリンまたはアラニンのような疎水性残基に置換されたり、リジン、アルギニンまたはヒスチジンのように電気的に陽性である残基が、グルタミン酸やアスパラギン酸のような電気的に陰性である残基に置き換えられたり、グリシンのように側鎖を持たないアミノ酸がかさだかい側鎖を持つ残基に置き換えられる場合である。
【0086】
以上、言及した事実を考慮して、当技術分野で通常の知識を有する者は、前記HRF結合ペプチドをその欠失型HRFに対する結合力、およびそれによるヒスタミン分泌阻害活性を維持乃至増加させたり損傷させたりしない範囲内で、通常の技術を利用して変形させることができ、それは本発明の範疇内に属する。
【0087】
本発明者らは、前記ペプチドをファージディスプレーライブラリスクリーニングを通して得、合成ペプチドで再び確認した。本発明のペプチドは化学的に合成したり遺伝子組換え技術を利用して製造したりすることができる。または、本発明のペプチドを構成するドメインを血液内に存在するタンパク質やその一部から製造することができる。また他の方法として、本発明のペプチドはそれをコードする塩基配列を有するDNAが挿入された発現ベクターを利用する組換えDNA技術で製造することができる。前記ベクターは、サムブルック(Sambrook)ら(Molecular Cloning, 1989年, Cold Spring Harbor, Cold Spring Harbor Laboratory Press)の方法によって適当な宿主細胞で形質転換され適当な条件下で発現するように製造される。また、本発明のアミノ酸配列を含む融合タンパク質を利用して、本発明のペプチドを製造することもできる。
【0088】
また、本発明は、前記HRF結合ペプチドを有効成分として含むアレルギーまたはマラリア予防または治療用薬剤学的組成物を提供する。
【0089】
本発明のHRF結合ペプチドは、HRFに結合してそのヒスタミンおよびIL-8分泌能を抑制することを示したため(図16、17および図18参照)、アレルギー予防または治療に効果的に使用できることを確認した。
【0090】
一方、HRFは、翻訳によって調節される腫瘍タンパク質(translationlly controlled tumor protein)とも呼ばれる。1998年ビスチバンらによって抗マラリア剤であるアルテミシニン(Artemisinin)がマラリアタンパク質であるHRFに結合して作用することが明らかにされた。したがって、HRFに結合する活性を有する本発明のHRF結合ペプチドも前記アルテミシンと同様にマラリアの予防および治療に使用することができる。
【0091】
本発明のHRF結合ペプチドの有効量は、体重1kg当たり約30μg〜1mgである。本発明の組成物は、溶液またはミセル形態で直接注入したり製剤化して使用できる。本発明による組成物は、非経口または局所投与によって人体に適用することができ、静脈注射、皮下注射、内皮注射、筋肉注射など注射によって投与することが好ましい。このような目的のために、本発明のペプチドを薬剤学的に許容可能な担体に懸濁させたり溶解させたりする。ここで、特に水溶性担体を使うのが好ましい。
【0092】
また、本発明は、前記HRF結合ペプチドおよび抗-HRFモノクローナル抗体を含むアレルギー診断キットを提供する。
【0093】
本発明のHRF結合ペプチドと抗HRFモノクローナル抗体を含む診断キットを利用する試験で、血液反応の時に陽性と判定されれば、アレルゲンがなくてもアレルギーを持っていると判断することができる。すなわち、LPRアレルギー患者の血液にはHRFが浮遊しているため、血液検査の時に本発明のキットを利用すれば、HRFが血液に存在するか否かが分かり、それをもってLPR患者か否かを確認することができる。本発明では底に本発明のHRF結合ペプチドを付着させた後、血液と反応させて、そこに標識物質がコンジュゲートされた抗HRFモノクローナル抗体を加える方法を使用できる。
【0094】
合わせて、本発明は、タンパク質-タンパク質相互作用分析を行なう工程を含む、前記欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法を提供する。ここで、前記タンパク質-タンパク質相互作用分析は、特別にそれに制限されるものではなく、今までに公知された通常のタンパク質-タンパク質相互作用分析方法をすべて使用でき、詳細には、同時精製方法(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)、タンパク質チップ等を使用して行なうことが好ましい。
【0095】
本発明の一実施態様として、同時精製方法は、1)HRF-HRF受容体複合体を適切な方法で分離する工程、2)前記分離された複合体を精製する工程;3)前記精製された複合体からHRF受容体の正体を確認する工程を含む。本発明の好ましい実施態様で、工程1は1次元電気泳動(1D)や二次元電気泳動(2D)または液体クロマトグラフィー(ligquid chromatography)方法を含む。
【0096】
本発明による好ましい実施態様で工程2は、内在タンパク質複合体(endogenous protein complex)を直接抗体で精製したり抗原決定基標識(epitope tagging)をした後、抗体で精製する免疫沈澱法(immunoprecipitation, Barrett等, J. Lab. Clin. Med. 1960年, 第55巻: 605-15頁)、GST-沈澱法(GST-pulldown, Magnaghi-Jaulin等, Nucleic Acids Res., 1996年, 第24(6)巻: 1052-8頁)のような非免疫的親和精製法(affinity purification)、および非特異的反応抑制と回収率および再現性を高めるために2個以上の標識を利用したタンデムアフィニティー精製(tandem affinity purification)(TAP)システム(Russell等, Infect Immun., 1980年, 第29(3)巻,999-1006頁)のような多標識親和精製法(multi-tag affinity purification)からなる群より選択される親和性クロマトグラフィーを含む。
【0097】
本発明の実施態様で、工程3は基質補助レーザー脱離/イオン化飛行時間質量分光法(matrix-assisted laser desorption/ ionization time-of-flight mass spectrometry, MALDI-TOF MS, Hill 等, Rapid Commun. Mass Spectrom., 1991年, 第5(9)巻、395-9頁)のようなペプチド質量分析(peptide mass fingerprinting)またはナノエレクトロスプレーイオン化質量分光法(nanoelectrospray ionization (nanoES)-MS/MS, Shevchenko等, Proc. Natl. Acad. Sci. 米国, 1996年, 第93(25)巻, 14440-5頁)、ナノLC質量分光法(nanoLC-MS/MS, Oosterkamp等, J. Mass Spectrom. 1998年, 第33(10)巻, 976-83頁)およびLC/LC-MS/MSからなる群より選択されるタンデム質量分光法(tandem MS (MS/MS))が含まれる。
【0098】
本発明の好ましい実施態様で、本発明のHRF-特異的受容体の同定方法は、選択的に下記の工程を含む:4)工程3で確認されたHRF受容体が、実際の生体内または試験管内条件でHRFによって媒介されるヒスタミン放出過程に関与するか否かを判定する工程。ここで、前記判定工程は、今まで公知されたタンパク質の機能分析方法をすべて使用でき、詳細には、前記工程4は、前記確認されたHRF受容体をコードする遺伝子をクローニングする工程、それを哺乳動物細胞発現ベクターにクローニングする工程、前記クローニングされた発現ベクターでヒスタミン放出細胞株を形質感染させる工程;および本発明の欠失型HRFまたはHRFダイマーを処理した後モックベクター(mock vector)で形質感染された対照細胞株と比較して、ヒスタミン放出程度が有意に増加するかどうかを観察する工程を通じて達成できる。選択的には、HRF受容体遺伝子が欠損したノックアウト形質転換動物または細胞株を製造する工程、前記形質転換動物または細胞株にHRFを投与した後、野生型と比較してヒスタミン放出が有意に減少するかどうかを分析する工程を通じて達成できる。
【0099】
酵母ツーハイブリッドシステム(Fields S. and Song O., Nature., 1989年, 第340(6230)巻, 245-6頁)は、DNA結合ドメイン(DBD; DNA binding domain)と転写活性ドメイン(TAD; transcription activation domain)を遺伝子組換え方法で分離して相互作用する二タンパク質(X,Y)の遺伝子を各々DBDとTADにクローニングした後、それらを酵母内で発現されるように誘導すると、DBDとTADが物理的に分離されたタンパク質が発現されるが、X/Yタンパク質相互作用により転写因子で再構成され指示遺伝子(reporter gene)の遺伝子発現が可能になるという点を利用した分析方法である。前記システムは、遺伝子操作技術を基盤技術としてタンパク質の特性に関係なしに同一な実験方法で相互作用を検索できるので、前記システムは大規模相互作用分析が容易である。
【0100】
本発明の一実施態様で、指示遺伝子は、1)色素選別(color selection)遺伝子のβ-ガラクトシダーゼ(β-galactosidase, LacZ)、α-ガラクトシダーゼ(α-galactosidase(MEL1)、β-グルクロニダーゼ(β-glucuronidase, gusA)、緑色蛍光タンパク質(green fluorescence protein, GFP)、グルコアミラーゼ(glucoamylase)等と、2)成長選別(growth selection)遺伝子であるイミダゾールグリセロールホスファートジハイドラターゼ(imidazoleglycerol-phosphate dehydratase, HIS3)、ホスホリボシルアミノイミダゾールカルボキシラーゼ(phosphoribosylaminoimidazole-carboxylase, ADE2)、α-アミノアジパートリダクターゼ(α-aminoadipate reductase, LYS2)、β-イソプロピルマレートジヒイドロゲナーゼ(β-isopropylmalate dehydrogenase, LEU2)、オルチジン-5'-ホスファートジカルボクシラーゼ(orotidine-5'-phosphate decarboxylase, URA3)等を含む。
【0101】
タンパク質チップは、マイクロアレイによって固体基板上に点滴されたタンパク質とタンパク質、ヘキサン、炭水化物、脂質、コレステロールまたは他の小さな化合物のような異なる分子間の相互作用を分析するのに使用されるバイオチップを意味する。生体分子(biomolecule)間の相互作用を測定できるBiacoreのセンサーチップ(sensor chip)が1990年代に商用化されて以来、タンパク質チップ開発が加速化されてきた。タンパク質チップ技術は疾病診断、タンパク質活性研究、新薬候補物質検索、生体分子相互作用検索等の分野に応用できるのでタンパク質体学の核心技術として浮上してきている。
【0102】
本発明の好ましい実施態様で、タンパク質チップを利用した本発明のHRF-特異的受容体の同定方法は、表面プラスモン共鳴(surface plasmon resonance; SPR)分析、質量分析(surface enhanced laser desorption/ ionization time of flight mass spectrometry; SELDI-TOF)、蛍光分析、電気化学的分析等を通して行なえる。
【0103】
したがって、タンパク質チップを利用した本発明のHRF特異的受容体の同定は、1)機能が明らかにされたり明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程、2)前記HRFまたはHRFダイマーに特異的な抗体がなかったりあったりする状態で、前記言及された分析方法を利用してタンパク質-タンパク質相互作用が起きるかどうかを確認する工程を通して行なえる。
【0104】
上記以外にも、現在まで公知された多様なタンパク質-タンパク質相互作用分析方法が本発明のHRF受容体の同定方法に使用できる。その例として、 欧州特許第 EP1003853号、 欧州特許第 1098967号、 欧州特許出願公開第 EP1184463A1号、 欧州特許第 1224324号、 米国特許第6114111号、 米国特許第6562576号、 米国特許第6828112号、 米国特許出願公開第 US20020094519A1号、 米国特許出願公開第20020106693A1号、 米国特許出願公開第20020106698A1号、 米国特許出願公開第20020142348A1号、米国特許出願公開第20020177217A1号、米国特許出願公開第20030003439A1号、米国特許出願公開第20030040012A1号、米国特許出願公開第20030170723A1号、米国特許出願公開第20030211523A1号、米国特許出願公開第20040146931A1号、米国特許出願公開第20040157279A1号、米国特許出願公開第20050106636A1号、米国特許出願公開第20050176005A1号または米国特許出願公開第20050221280A1号に開示された方法が使用できる。前記文献は、すべて本発明に参考文献として挿入される。
【0105】
以下、本発明を実施例によって詳しく説明する。
ただ、下記の実施例は、本発明を例示するだけのものであって、本発明の内容が下記の実施例に限定されるものではない。
【実施例】
【0106】
実施例1
多様な欠失型HRFの製造
1-1:遺伝子の分離および増幅
まず、大韓民国特許出願第2001-27896号および第2001-27921号に記載した方法によってHRFをコードする遺伝子のcDNAを製造した。詳細には、ラット(Rattus norvegicus)骨格筋から総細胞質RNAを抽出した後、pJG4-5ベクター(Invitrogen, Inc., 米国)を利用して酵母2-ハイブリッド分析のためのcDNAライブラリを製造した。
【0107】
(Na,K)ATPaseのα2サブユニットを利用してCD3部位を pEG202ベクター(Brent, R., and Finley, R. L., Jr. Annu. Rev. Genet. 1997年、第31巻, 663-704頁)のLexA DNA結合ドメイン(pEG202 ベクター)に挿入した後、それをスクリーニングのためのバイト(bait)に使用した。リポーター遺伝子を活性化させた陽性クローンを選択して塩基配列分析(sequencing)、制限酵素マッピングと BLASTサーチを通して配列分析を行なった。これらの中で一つのクローンがIgE依存的ヒスタミン放出因子(HRF)と完全に一致した。
【0108】
前記rHRF cDNA遺伝子から配列番号:1で表されるラットHRF全長配列(Accession number U20525、アミノ酸1-172)をコードする遺伝子を配列番号:9、配列番号:2で表されるアミノ酸11-172切片をコードする遺伝子を配列番号:10、配列番号:3で表されるアミノ酸35-172切片をコードする遺伝子を配列番号:11、配列番号:4で表されるアミノ酸1-112切片をコードする遺伝子を配列番号:12、配列番号:5で表されるアミノ酸39-110切片をコードする遺伝子を配列番号:13、配列番号:6で表されるアミノ酸1-38切片をコードする遺伝子を配列番号:14、配列番号:7で表されるアミノ酸111-172切片をコードする遺伝子を配列番号:15、配列番号:8で表されるアミノ酸84-108切片をコードする遺伝子を配列番号:16で示した。各々の遺伝子をPCRを遂行して増幅させた(図1および図2)。PCRは前変性(pre-denaturation)工程として94℃で5分、変性(denaturation)工程として94℃で1分、アニーリング工程として45〜50℃で1分、延長(elongation)工程として72℃で1分、後延長(post-elongation)工程として72℃で7分の条件で30サイクルの間反応させた。PCR反応に使用したプライマー配列は、前記で記載した。
【0109】
1-2:組換え発現ベクターの製造
実施例1-1で増幅させた配列番号:9乃至配列番号:16の遺伝子を各々 pRSET-Aベクター(Invitrogen)にクローニングして組換え発現ベクターを製造し、それを各々pRSET-A-RrHRF、pRSET-A-Del-N11HRF、pRSET-A-Del-N35HRF、pRSET-A-Del-C112HRF、pRSET-A-Del-N39C110HRF、pRSET-A-Del-C38HRF、pRSET-A-Del-N111HRF、pRSET-A-Del-N84C108HRFと名付けた(図3および図4)。
【0110】
1-3:形質転換体の製造および各欠失型HRFの分離精製
各欠失型HRFを過発現(overexpression)させるために実施例1-2で製造した組換え発現ベクターを大腸菌BL21(DE3)(Novagen)またはBL21(DE3)pLysS(Novagen)に形質転換させた。
【0111】
前記大腸菌形質転換体は、アムピシリンまたはアムピシリンとクロラムフェニコール(chloramphenicol)を含んだLB培地で培養した後、ODが0.6に到達した時、0.4mMになるようにIPTG(isopropyl β-D-thiogalactoside)を添加して三時間さらに培養した後、5,500xgで5分間遠心分離した。回収した大腸菌をバインディングバッファー(5mMイミダゾール、500mM NaCl、20mM Tris-HCl、pH7.9)に再懸濁させた後、超音波を使って粉砕した。溶菌された大腸菌を39,000xgで遠心分離した後、上澄み液はNiカラム(Novagen)を利用して精製した。pRSET-A ベクターにクローニングした欠失型HRFは、N末端に6個のヒスチジンを持つようになるため、ニッケルで充電したHIS-結合カラムに付くようになる。これらタンパク質を高濃度のイミダゾン(imidazone)(1M イミダゾール、500mM NaCl、20mM Tris-Hcl、pH7.9)を含んだイルーションバッファーを使って抽出した後、PD-10カラムを利用して多量の塩分を除去した。 精製された欠失型HRFをモノ-Q陰イオン交換(anion exchange)カラム(Amersham Pharmacia Biotech)でNaCl濃度勾配を利用して再び精製した後、以下の実施例で BEAS-2B細胞(ATCC)とヒト好塩基球を刺激するために使用した。
【0112】
実施例2
BEAS-2B細胞でHRF形態によるIL-8分泌能の検査
各欠失型HRFの活性をBEAS-2B細胞(ATCC)でのIL-8分泌能を通して比較した。BEAS-2B細胞を48ウェルプレートで70%程度育つまで培養した後、1%ペニシリンストレプトマイシン/BEBM(Clonetics)で2回洗浄した。実施例1-3で分離した各々の組換えタンパク質(RrHRFまたは各欠失型HRF)を1μg/mlまたは10μg/mlずつ添加した。48時間または24時間後、上澄み液を取って遊離したIL-8を酵素免疫吸着検出法(PIERCE)を利用して定量した(図5)。
【0113】
実施例3
ヒト好塩基球でHRF形態によるヒスタミン分泌能の検査
各欠失型HRFの活性をヒト好塩基球でのヒスタミン分泌能を通して比較した。アトピー性皮膚炎を持っている血液提供者から40mlの静脈血を採取した後、抗凝血剤であるEDTAを10 mMになるように添加して、食塩水に6%になるように溶解したデキストラン10mlを入れて混合した。室温で90分間静置した後、白血球が集まっている上層を分離して150xgで8分間遠心分離した。回収した白血球をPAG-EDTA(4mM EDTA、25mM PIPES、110mM NaCl、5mM KCl、0.003%HSA、0.1%D-グルコース)で二度洗浄し、冷たい乳酸バッファー(13.4mM 乳酸、140mM NaCl、5mM Kcl、pH3.9)で3〜5分間静置する過程を通して細胞表面にある IgEを除去した。続いて、直ちに30mlのPAG-EDTAを加えて二回洗浄して再びPAG-EDTAに懸濁させた後、ヒトIgE(1μg/ml、Serotec)で2時間感作させた。5%FBSを添加したIMDM培地(Gibco BRL)で2回洗浄した後、実施例1-3で分離した各々の組換えタンパク質(rHRFまたは各欠失型HRF)を20μg/mlずつ添加した。15分後、ヒト抗IgE抗体を加えて4時間後、上澄み液を取って遊離したヒスタミンをヒスタミンアナライザー(Astoria Analyzer, Series 300 system)を利用して定量した(図6)。
【0114】
その結果、配列番号:2で表されるN末端の11個アミノ酸残基が欠失したタンパク質(Del-N11HRF)および配列番号:3で表されるN末端の35個アミノ酸残基が欠失したタンパク質(Del-N35HRF)は、BEAS-2B細胞のように野生型HRFに比べて効果的にヒスタミン放出を誘導することを確認した。
【0115】
実施例4
分子内ジスルフィド結合による二量体化がHRF活性に及ぼす影響比較
4-1:非還元SDS-PAGEによる移動性の差の比較
実施例1-3で製造した野生型HRFと各欠失型HRFのN末端の欠失による電気泳動での移動性(mobility)変化を確認した(図7、図8)。SDS-PAGEは、ラエムリ(Laemmli )(Laemmli U.K., Nature, 第227巻, 680-685頁)の方法を変形して使用し、各タンパク質を還元サンプルバッファー(reducing sample buffer)[0.125M Tris-HCl、pH6.8、4%(w/v)SDS、20%(v/v)グリセロール、and 2%β-メルカプトエタノール(β-ME)]または、非還元サンプルバッファー(non-reducing sample buffer)(w/o β-ME)と混合した後、15%ゲルを作って分離した。その結果、N末端を除去したタンパク質は、野生型HRFとC-末端を除去したタンパク質とは異なり、ダイマー位置に移動することが観察された。これは、N末端欠失型HRFと野生型HRFの構造的な差を裏付けし、HRFのN末端がHRF機能調節に重要な役目を果たすという事実を裏付けしてくれる。また、N末端を除去したタンパク質が分子内ジスルフィド結合を持っていることを示唆する。
【0116】
4-2:ジスルフィド結合を除去するための部位特異的突然変異の製作
N末端欠失型HRFのシスチン残基をセリンに置換して、ジスルフィド結合が形成されることを阻害した場合、HRF活性に及ぼす影響を調べるために部位特異的突然変異キット(site-directed mutagenesis kit, stratagene)を利用して突然変異を製作した。HRFの場合、28番と172番残基にシスチンを含んでいるため、プライマーにCG GAC GGG CTG TCT CTG GAG GTG GA(配列番号:18)とTC CAC CTC CAG AGA CAG CCC GTC CG(配列番号:19)を使用して、pRSET-A-Del-N11HRF-C28Sを製造した。また他のプライマーにGAG ATG GAA AAA TCT AAG CTT GAT CCG(配列番号:20)とCGG ATC AAG CTT AGA TTT TTC CAT CTC(配列番号:21)を使用して、pRSET-A-Del-N11HRF-C172SとpRSET-A-Del-N35HRF-C172Sを製造した。
【0117】
PCR増幅条件は、DNA重合酵素(Pfu DNA polymerase, Stratagene)を使用して前記に記載のプライマー対と鋳型を95℃で5分間転変性させて、95℃で1分、55℃で1分および 68℃で7分間16回反応させて、68℃で7分間後延長させた後、反応を終結した。この反応液にDpnI(Stratagene)を入れて 37℃で1時間反応させて鋳型DNAのみを選択的に切断させた後、XL1-blueに形質転換させた。
【0118】
製作したコンストラクトは、DNA配列分析を通して突然変異生成の有無を確認した後、BL21(DE3)/pLysSに形質転換させて実施例1-3により分離、精製した。これら突然変異の中で特にpRSET-A-Del-N35HRF-C172Sは、非還元的(non-reducing)SDS-PAGEを通して細胞間ジスルフィド結合(intermolecular disulfide bond)を形成しないものが現れ、BEAS-2B細胞株でIL-8を分泌させる能力も顕著に減少したことを確認できた(図11、図12)。これは、N末端欠失型HRFの活性において重要な役割を持つものが、172番残基にあるシスチンによる細胞間ジスルフィド結合であることを示唆する結果であり、HRF活性においてダイマー形成が非常に重要であることが分かる。
【0119】
4-3:化学的方法を利用したHRFの交差結合
架橋剤(crosslinker)を利用して野生型HRFと欠失型HRFがダイマーを形成する能力面でいかなる差があるか調べてみた。-SHと反応する二種類の架橋剤(BMOE, BM(PEO)4, Pierce, 米国)を使用して野生型HRFとDel-N35HRFを交差結合させた。Del-N35HRFの場合、架橋剤と反応可能な-SHを作るために前もって20mMのDTTを使用して細胞間ジスルフィド結合を還元させた後、カラム(Vivaspin column, Vivascience, 米国)を使用してDTTを除去した後、PBSで緩衝溶液組成を変えた。以後、5倍過量の架橋剤を処理した後、BMOEの場合は4℃で4時間、BM(PEO)4の場合は37℃で30分放置した後、上記カラム(vivaspin column)を使用して過量の架橋剤を除去した。これらタンパク質を定量した後、SDS-PAGEで分離した(図13)。還元されたDel-N35HRFが架橋剤により細胞間ジスルフィド結合を形成するのに対して、野生型HRFは細胞内ジスルフィド結合を形成し、架橋剤によってもダイマーを形成できなかった。
【0120】
実施例5
HRF形態によるファージディスプレーペプチドクローンの結合能力の差の比較
各々の欠失型HRFをプラスチックウェルに固定(immobilization)させた後、すでにHRFと結合することが知られているペプチドを発現しているファージ(phage)を添加して、その結合能力に差があるかどうかを調べた。
【0121】
具体的に、コーティング緩衝液(0.1M NaHCO3、pH8.6)に20μg/ml濃度で溶解させた各々の欠失型HRFを50ulずつポリスチレンマイクロタイタープレートに加えて、4℃で一晩中コーティングした後、BSAで非特異的結合を遮断した。0.1%Tween/TBS(TBST)で6回洗浄した後、ファージ溶液30ulを6%BSA/PBS30ulに希釈した溶液を加えて室温で2時間静置した。PBSTで5回洗浄した後、3%BSA/PBSに1:5000で希釈したHRP-コンジュゲートされた抗-M13抗体(Pharmacia)100ulずつを加えて1時間静置した。PBSTで6回、PBSで1回洗浄した後、ペルオキシダーゼ(peroxydase)基質溶液100μlずつを加えて発色程度をエライザ(ELISA)リーダー(Bio-Rad)を使用して405nmで測定した。
【0122】
その結果、前記HRFは、HRFに対して特異的結合を示すヘプタマー(heptamer)ペプチドを発現しているファージに対する親和度にも差を示したが、RrHRFとDel-C112HRFがファージに対してとても弱い親和度を示した一方、Del-N11HRFとDel-N35HRFはとても強い親和度を示した(図14)。このことから、各HRFタンパク質間のヒスタミン分泌能と相応するもので、以上言及した結果から実験的にHRFをターゲットにしたアレルギー制御薬物開発において利用することができるHRFの形態がいくつかの欠失型に限られることが分かる。
【0123】
また、HRF内のHRF結合ペプチドの結合部位を調べるためにHRFを三等分した後、HRF結合ペプチドとの親和度を比較した。Del-N39C110HRFの場合、前記欠失型HRFに似た程度の親和度を示し、アミノ酸をもう少し除去したDel-N84C108HRFも高い親和度を示した(図15)。これは、HRF結合ペプチドがHRFのアミノ酸84-108部位に結合することを意味し、これにより、HRF抑制を通したアレルギー制御薬物開発の検索に利用することができることを確認した。
【0124】
実施例6
欠失型HRFの活性を抑制することができるHRF結合ペプチドの作用比較
RBL-2H3(ATCC)を5×104細胞で24ウェルで増殖させた後、ラットIgE抗体(0.2μg/ml、Serotec)で60分間感作させて、5%FBS/MEM培地で組換えHRFタンパク質を1.56mMで処理した(陽性対照群)。また、上記で準備した細胞にヘプタマー(heptamer)ペプチド(大韓民国特許出願第2001-27896号)を用量依存的方法で処理した(0.0156-15.6mM)。15分後、ラット抗IgE抗体を加えて4時間後、上澄み液を取って遊離したヒスタミンをヒスタミンアナライザー(Astoria Analyzer, Series 300 system)を使用して定量した。
【0125】
HRF結合ペプチドの欠失型HRFに対する作用は、BEAS-2B細胞株でも分析された。実施例3と同じ方法でDel-N35HRF(61nM)をBEAS-2Bに処理した。ここでHRF結合ペプチドおよびそのアラニン置換体(大韓民国特許出願第2001-27896号)、1番または7番アミノ酸が欠失した本発明のHRF結合ペプチドを色々な濃度(6.1-610nM)でDel-N35HRFに混合して室温で10分静置後、添加した。24時間後、上澄み液を取って遊離したIL-8を酵素免疫吸着検出法を利用して定量した。
【0126】
その結果、図16と図17に示したように、HRF結合ペプチドは、RBL-2H3細胞で濃度依存的に欠失型HRFによるヒスタミン分泌を阻害し、BEAS-2B細胞でも濃度依存的に欠失型HRFによるIL-8分泌を抑制した。WYVYPSM配列を持つHRF結合ペプチドは、変形されたアミノ酸位置によって活性の差を示した。N末端から2および6位置のアミノ酸残基をアラニンに変えた置換体は、欠失型HRFに対する抑制能力を喪失するとみえ、HRF結合ペプチドの作用においてチロシン残基とセリン残基が重要な作用をするものと考えられる(図18)。また、前記ペプチドのN末端のトリプトファン残基を除去した本発明の配列番号:17のHRF結合ペプチド(HBP2-D1)の場合も元来の 7-merペプチドとよく似た活性を示したため、2〜6番残基だけでも活性型HRFに対する抑制能力を持つことを確認した。
【産業上の利用可能性】
【0127】
上記で詳しく見たように、本発明のダイマーを形成できる欠失型HRFは、活性型のHRFを提供することによりHRFによるアレルギー制御薬物開発およびアレルギー患者血清内のHRF量を検出するためのキット製作などにも活用することができる。また、本発明の欠失型HRF結合ペプチドは、細胞内でIL-8およびヒスタミン分泌を効果的に制御することにより、動物の喘息や鼻炎などのようなアレルギー疾患およびマラリア予防または治療用薬物開発に有効に利用できる。
【特許請求の範囲】
【請求項1】
野生型IgE依存的ヒスタミン放出因子のN末端が除去され、ダイマー形成が可能でヒスタミンおよびIL-8分泌能が向上した組換え欠失型IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)。
【請求項2】
HRFが、脊椎動物のHRFまたはHRFの類似体であることを特徴とする、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項3】
HRFが、野生型HRFのN末端の少なくとも11乃至35アミノ酸が欠失したことを特徴とする、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項4】
配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項5】
HRFが、配列番号:2(Del-N11HRF)または配列番号:3(Del-N35HRF)で表されるアミノ酸配列を有することを特徴とする、請求項4記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項6】
請求項1の組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成されるIgE依存的ヒスタミン放出因子の同形または異形ダイマー。
【請求項7】
請求項1のHRFまたは請求項6のHRF同形または異形ダイマーに特異的な抗体。
【請求項8】
請求項1の欠失型HRFおよび/または野生型HRFに架橋剤を処理する工程を含む、請求項6の同形または異形ダイマーを製造する方法。
【請求項9】
架橋剤が、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン( 1,4-Di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-Bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-Bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(Bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-Bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(Bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-Hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(Dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bismaleimidyl-2,3-dihydroxybutane;BMDB)からなる群より選択されることを特徴とする、請求項8記載の欠失型HRFの同形または異形ダイマーを製造する方法。
【請求項10】
請求項1の欠失型HRFをコードする遺伝子。
【請求項11】
遺伝子が、配列番号:11(Del-N35HRF)で表される塩基配列を含むことを特徴とする、請求項10記載の遺伝子。
【請求項12】
遺伝子が、配列番号:10(Del-N11HRF)または配列番号:11(Del-N35HRF)で表される塩基配列を持つことを特徴とする、請求項11記載の遺伝子。
【請求項13】
請求項10の遺伝子を含む組換え発現ベクター。
【請求項14】
pRSET-A-Del-N11HRFまたはpRSET-A-Del-N35HRFであることを特徴とする、請求項13記載の組換え発現ベクター。
【請求項15】
請求項13の発現ベクターで形質転換された形質転換体。
【請求項16】
大腸菌であることを特徴とする、請求項15記載の形質転換体。
【請求項17】
BL21(DE3)-pRSET-A- Del-N11HRF、BL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A- Del-N11HRF、またはBL21(DE3)pLysS-pRSET-A-Del-N35HRFであることを特徴とする、請求項16記載の形質転換体 。
【請求項18】
請求項1の欠失型 IgE依存的ヒスタミン放出因子または請求項6の同形または異形ダイマーを有効成分として含むヒスタミン放出誘導剤。
【請求項19】
請求項1の欠失型 IgE依存的ヒスタミン放出因子または請求項6の同形または異形ダイマーを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法。
【請求項20】
欠失型IgE依存的ヒスタミン放出因子が、配列番号:8のアミノ酸配列を有することを特徴とする、請求項15記載の方法。
【請求項21】
請求項1の欠失型 IgE依存的ヒスタミン放出因子を利用したアレルギー患者血清内HRFの検出方法。
【請求項22】
(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を有するHRF結合ペプチド。
【請求項23】
配列番号:17のアミノ酸配列を有することを特徴とする、請求項22記載のペプチド。
【請求項24】
L-アミノ酸、D-アミノ酸、またはL-アミノ酸およびD-アミノ酸からなる、請求項22記載のペプチド。
【請求項25】
一つ以上の変形アミノ酸を含む、請求項22記載のペプチド。
【請求項26】
変形アミノ酸が、アミノ酸誘導体またはアルキル化されたアミノ酸であることを特徴とする、請求項25記載のペプチド。
【請求項27】
請求項22のペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物。
【請求項28】
請求項22のペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物。
【請求項29】
請求項22のペプチドおよび抗HRFモノクローナル抗体を含むアレルギー診断キット。
【請求項30】
タンパク質-タンパク質相互作用分析を行なう工程を含む欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法。
【請求項31】
タンパク質-タンパク質相互作用分析が、同時精製(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)またはタンパク質チップシステムにより行なわれることを特徴とする、請求項30記載のHRF特異的受容体を同定する方法。
【請求項32】
同時精製が、
1)HRF-HRF受容体複合体を分離する工程;
2)前記分離された複合体を精製する工程;および、
3)前記精製された複合体からHRF受容体の正体を確認する工程;
を含むことを特徴とする、請求項31記載のHRF特異的受容体を同定する方法。
【請求項33】
タンパク質チップシステムが、
1)機能が明らかにされているか明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程;および、
2)前記欠失型HRFまたはHRFダイマーに、特異的な抗体がないかまたは抗体がある状態で前記言及された分析方法を利用して、タンパク質-タンパク質相互作用が起きるかどうかを確認する工程;
を含むことを特徴とする、請求項31記載のHRF特異的受容体を同定する方法。
【請求項1】
野生型IgE依存的ヒスタミン放出因子のN末端が除去され、ダイマー形成が可能でヒスタミンおよびIL-8分泌能が向上した組換え欠失型IgE依存的ヒスタミン放出因子(IgE-dependent histamine releasing factor, HRF)。
【請求項2】
HRFが、脊椎動物のHRFまたはHRFの類似体であることを特徴とする、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項3】
HRFが、野生型HRFのN末端の少なくとも11乃至35アミノ酸が欠失したことを特徴とする、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項4】
配列番号:3(Del-N35HRF)で表されるアミノ酸配列を含む、請求項1記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項5】
HRFが、配列番号:2(Del-N11HRF)または配列番号:3(Del-N35HRF)で表されるアミノ酸配列を有することを特徴とする、請求項4記載の欠失型IgE依存的ヒスタミン放出因子。
【請求項6】
請求項1の組換え欠失型IgE依存的ヒスタミン放出因子または野生型IgE依存的ヒスタミン放出因子間に形成されるIgE依存的ヒスタミン放出因子の同形または異形ダイマー。
【請求項7】
請求項1のHRFまたは請求項6のHRF同形または異形ダイマーに特異的な抗体。
【請求項8】
請求項1の欠失型HRFおよび/または野生型HRFに架橋剤を処理する工程を含む、請求項6の同形または異形ダイマーを製造する方法。
【請求項9】
架橋剤が、1,4-ジ-[3'-(2'ピリジルジチオ)プロピオナミド]ブタン( 1,4-Di-[3'-(2'pyridyldithio)propionamido]butane;DPDPB)、1,8-ビス-マレイミドジエチレングリコール(1,8-Bis-maleimidodiethylene glycol;BM[PEO]2)、1,11-ビス-マレイミドトリエチレングリコール(1,11-Bis-maleimidotriethylene glycol;BM[PEO]3)、ビス-マレイミドエタン(Bis-maleimidoethane;BMOE)、1,4-ビス-マレイミドブタン(1,4-Bis-maleimidobutane;BMB)、ビス-マレイミドへキサン(Bis-maleimidohexane;BMH)、1,6-へキサン-ビス-ビニルスルホン(1,6-Hexane-bis-vinylsulfone;HBVS)、ジチオ-ビス-マレイミドエタン(Dithio-bis-maleimidoethane;DTME)および1,4-ビス-マレイミジル-2,3-ジヒドロキシブタン(1,4-bismaleimidyl-2,3-dihydroxybutane;BMDB)からなる群より選択されることを特徴とする、請求項8記載の欠失型HRFの同形または異形ダイマーを製造する方法。
【請求項10】
請求項1の欠失型HRFをコードする遺伝子。
【請求項11】
遺伝子が、配列番号:11(Del-N35HRF)で表される塩基配列を含むことを特徴とする、請求項10記載の遺伝子。
【請求項12】
遺伝子が、配列番号:10(Del-N11HRF)または配列番号:11(Del-N35HRF)で表される塩基配列を持つことを特徴とする、請求項11記載の遺伝子。
【請求項13】
請求項10の遺伝子を含む組換え発現ベクター。
【請求項14】
pRSET-A-Del-N11HRFまたはpRSET-A-Del-N35HRFであることを特徴とする、請求項13記載の組換え発現ベクター。
【請求項15】
請求項13の発現ベクターで形質転換された形質転換体。
【請求項16】
大腸菌であることを特徴とする、請求項15記載の形質転換体。
【請求項17】
BL21(DE3)-pRSET-A- Del-N11HRF、BL21(DE3)-pRSET-A-Del-N35HRF、BL21(DE3)pLysS-pRSET-A- Del-N11HRF、またはBL21(DE3)pLysS-pRSET-A-Del-N35HRFであることを特徴とする、請求項16記載の形質転換体 。
【請求項18】
請求項1の欠失型 IgE依存的ヒスタミン放出因子または請求項6の同形または異形ダイマーを有効成分として含むヒスタミン放出誘導剤。
【請求項19】
請求項1の欠失型 IgE依存的ヒスタミン放出因子または請求項6の同形または異形ダイマーを利用したHRFによるアレルギー発生を抑制することができる薬物の検出方法。
【請求項20】
欠失型IgE依存的ヒスタミン放出因子が、配列番号:8のアミノ酸配列を有することを特徴とする、請求項15記載の方法。
【請求項21】
請求項1の欠失型 IgE依存的ヒスタミン放出因子を利用したアレルギー患者血清内HRFの検出方法。
【請求項22】
(V、Y、EまたはA)-(T、V、FまたはA)-(Y、PまたはA)-(P、GまたはK)-(A、L、SまたはW)-(A、PまたはM)のアミノ酸配列を有するHRF結合ペプチド。
【請求項23】
配列番号:17のアミノ酸配列を有することを特徴とする、請求項22記載のペプチド。
【請求項24】
L-アミノ酸、D-アミノ酸、またはL-アミノ酸およびD-アミノ酸からなる、請求項22記載のペプチド。
【請求項25】
一つ以上の変形アミノ酸を含む、請求項22記載のペプチド。
【請求項26】
変形アミノ酸が、アミノ酸誘導体またはアルキル化されたアミノ酸であることを特徴とする、請求項25記載のペプチド。
【請求項27】
請求項22のペプチドを有効成分として含むアレルギー予防または治療用薬剤学的組成物。
【請求項28】
請求項22のペプチドを有効成分として含むマラリア予防または治療用薬剤学的組成物。
【請求項29】
請求項22のペプチドおよび抗HRFモノクローナル抗体を含むアレルギー診断キット。
【請求項30】
タンパク質-タンパク質相互作用分析を行なう工程を含む欠失型HRFまたは前記同形または異形HRFダイマーを利用してHRF特異的受容体を同定する方法。
【請求項31】
タンパク質-タンパク質相互作用分析が、同時精製(Co-purification)、酵母ツーハイブリッドシステム(Yeast Two-Hybrid system)またはタンパク質チップシステムにより行なわれることを特徴とする、請求項30記載のHRF特異的受容体を同定する方法。
【請求項32】
同時精製が、
1)HRF-HRF受容体複合体を分離する工程;
2)前記分離された複合体を精製する工程;および、
3)前記精製された複合体からHRF受容体の正体を確認する工程;
を含むことを特徴とする、請求項31記載のHRF特異的受容体を同定する方法。
【請求項33】
タンパク質チップシステムが、
1)機能が明らかにされているか明らかにされていない多様なタンパク質が集積されたタンパク質チップに、本発明の欠失型HRFまたは本発明の同形または異形HRFダイマーを処理する工程;および、
2)前記欠失型HRFまたはHRFダイマーに、特異的な抗体がないかまたは抗体がある状態で前記言及された分析方法を利用して、タンパク質-タンパク質相互作用が起きるかどうかを確認する工程;
を含むことを特徴とする、請求項31記載のHRF特異的受容体を同定する方法。
【図1】


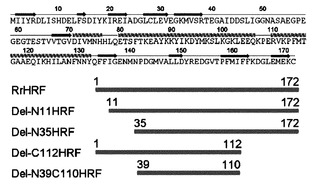
【図2】
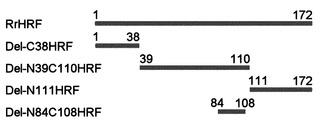
【図3】

【図4】

【図5】

【図6】
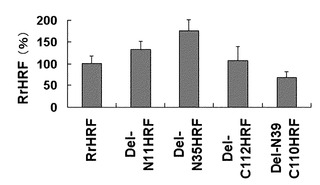
【図7】
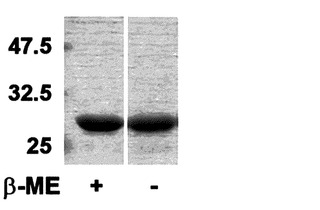
【図8】
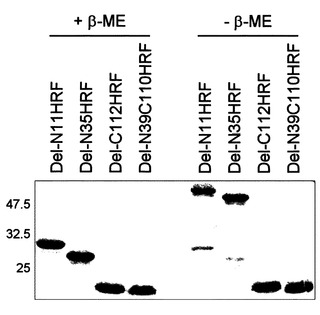
【図9】
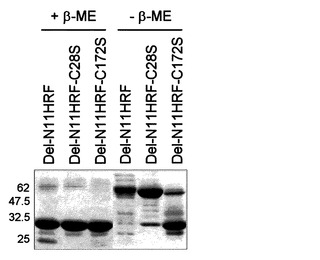
【図10】

【図11】

【図12】
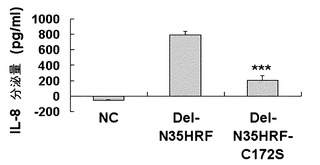
【図13】
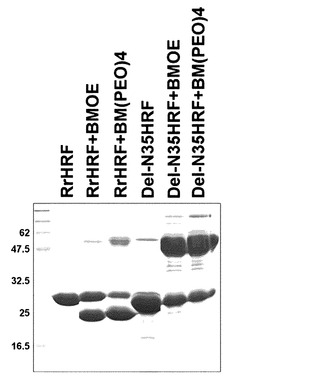
【図14】
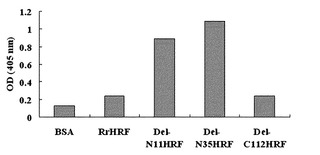
【図15】
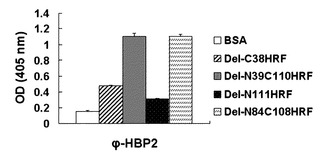
【図16】
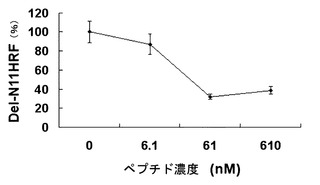
【図17】

【図18】
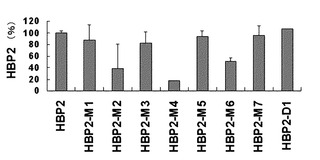
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公開番号】特開2010−158246(P2010−158246A)
【公開日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願番号】特願2010−33371(P2010−33371)
【出願日】平成22年2月18日(2010.2.18)
【分割の表示】特願2006−15614(P2006−15614)の分割
【原出願日】平成18年1月24日(2006.1.24)
【出願人】(506027387)エワ ユニバーシティ−インダストリー コラボレーション ファウンデーション (2)
【Fターム(参考)】
【公開日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願日】平成22年2月18日(2010.2.18)
【分割の表示】特願2006−15614(P2006−15614)の分割
【原出願日】平成18年1月24日(2006.1.24)
【出願人】(506027387)エワ ユニバーシティ−インダストリー コラボレーション ファウンデーション (2)
【Fターム(参考)】
[ Back to top ]