
ヒストン脱アセチル化酵素−8のタンパク質、核酸およびその使用方法
【課題】本発明は、クロマチン構造および転写に対する影響を示す新規HDACタンパク質をコードする新規核酸を指向するものである。
【解決手段】本明細書中で提供されるものは、上記核酸配列を含有するベクターおよび宿主細胞、ヘテロなポリペプチド配列と融合した本発明のポリペプチドを含むキメラポリペプチド分子、本発明のポリペプチドと結合する抗体および本発明のポリペプチドを産生する方法である。さらに、本発明によって提供されるものは、HDAC8を媒介する新規組成物を同定する方法と、疾病の診断および処置におけるそのような組成物の使用である。
【解決手段】本明細書中で提供されるものは、上記核酸配列を含有するベクターおよび宿主細胞、ヘテロなポリペプチド配列と融合した本発明のポリペプチドを含むキメラポリペプチド分子、本発明のポリペプチドと結合する抗体および本発明のポリペプチドを産生する方法である。さらに、本発明によって提供されるものは、HDAC8を媒介する新規組成物を同定する方法と、疾病の診断および処置におけるそのような組成物の使用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、クロマチン構造の制御、即ち転写制御に関与する組成物および使用方法に及ぶ。より具体的には、本発明は、転写制御に関与するヒストン脱アセチル化酵素タンパク質をコードする核酸を指向する。使用方法には、転写モデュレーターのスクリーニングアッセイにおける使用および治療薬としての使用が含まれる。
【背景技術】
【0002】
(発明の分野)
ヌクレオソームヒストンの可逆的アセチル化によるクロマチン構造の修飾は、真核細胞における転写制御において重要な役割を担っている(参照:Grunstein. 1997. Nature 389:349-352)。コアヒストンのアミノ酸末端尾内にある特定リジン残基のεアミノ基のアセチル化により局所的なクロマチン弛緩が生じる。このアセチル化は、転写中のヌクレオソームがアンフォールディング(unfolding)構造(変性構造)を維持するために必要である(Walia et al. 1998. J. Biol. Chem. 273:14516-14522)。一般的に、ヒストンアセチル化活性は転写活性と関連しており、一方脱アセチル化活性は転写抑制と関連している。遺伝子発現中の広範な変化は、細胞核内のヒストンアセチル基転移酵素(HAT)活性とヒストン脱アセチル化酵素(HDAC)活性との間の動的平行により起こると考えられている。
【0003】
HDACは、比較的大きいマルチサブユニット複合体との相互作用によって転写抑制を媒介する。例えば、HDAC1として知られる特定のHDACは、コリプレッサーSin3と結合し、HDAC1−Sin3はサイレンシング媒介物質NCoRおよびSMRTとさらに結合することが知られている(Alland, et al. 1997. Nature 387:49-55)。それから、N−CoR/SMRT-HDAC1は、核内のプロモーターエレメントと結合している特定の転写因子によって供給される。例えば、網膜芽腫(Rb) 遺伝子産物は、N-CoR/SMRT-HDAC1を供給し、転写因子E2Fと結合してE2F調節プロモーターを抑制する(Luo et al. 1998. Cell 92:463-473; Brehm et al. 1998. Nature. 391:597-600, Magnaghi-Jaulin et al. 1998. Nature 391:601-605)。HDAC1-Sin3もまた結合し、Mad/MAXリプレッサーヘテロダイマーによって抑制を媒介する (Laherty et al. 1997. Cell 89:349-356)。HDAC1のヒストン脱アセチル化活性は、この転写抑制にとって必須のものである(Hassig et al. 1998. PNAS 95:3519-3524)。
【0004】
ヒストン脱アセチル化酵素をコードする幾つかのcDNAが特性分析されている。クローンされた最初のものは酵母タンパク質RPD3であり、これは転写抑制因子に関する遺伝子的スクリーンにおいて最初に同定された(Vidal et al. 1991. Mol. Cell Biol. 11:6317-6327)。哺乳類HDAC1はTSAの標的分子として別個にクローンされた(Taunton et al. 1996. Science 272:408-411)。HDAC1は、酵母RPD3のオルトログ(ortholog)であることが観察され、また両方ともインビトロでHDAC活性を持つことが示された。RPD3に対して相同なタンパク質をコードする3つのcDNA、HDAC1、HDAC2およびHDAC3が記載されている(Yang et al. 1997. J. Biol. Chem. 272:28001-28007; Emiliani et al. 1998. PNAS 95:2795-2800)。3つ全ては、偏在組織分布を持ち、またその活性は重複するようである。現在まで、HDACをコードする5つのS. cerevisiae遺伝子が発見されており、そのファミリーは配列保存および提案された生物学的機能を基にした2つのクラスに分けられてきた。S. cerevisiaeのHos1、Hos2、Hos3およびRPD3は哺乳類HDAC1、2および3と共にクラス1に含まれる。第2のクラスは、酵母HDA1およびHDA3および最近同定されたヒトホモログHDAC4、HDAC5およびHDAC6から構成される(Grozinger et al. 1999. PNAS 96:4868-4873; Fischle et al. 1999. J. Biol. Chem. 274:11713-11720)。
【0005】
HDACはマルチサブユニット転写制御複合体の一部として機能していることが知られているので、制御HDACは、HDACを含有する制御複合体の制御下で、特定遺伝子または一組の特定遺伝子の転写を調節する、即ち増加または低下させるように使用され得る(Carmen et al. 1996. J. Biol. Chem. 271(26):15837-15844)。
【0006】
HDAC阻害活性を持つ多様な化合物はヒストンの過アセチル化をもたらすことがわかっており、この調節は、多くの細胞型で細胞周期の拘束および末端分化を伴う (参照:Proceedings of the American Association for Cancer Research 40, March, 1999)。NaBuなどの、強力で特異な可逆的阻害剤である該菌類毒素トリコスタチンA(TSA)は、多様なシステムにおける細胞周期拘束および/または分化を誘導することがわかってきた。近年、HDAC阻害剤活性を有する合成ベンザミド誘導体(MS-27275)が記載された(Saito et al. 1999. PNAS 96:4592-5497)。MS-27275は、ヒストンの過アセチル化を誘導し、腫瘍罹患ヌードマウスにおける腫瘍増殖を阻害する点で効果を示した。
【0007】
急性前骨髄性白血病(APL)では、遺伝的再配列により、PMLまたはPLZFタンパク質のいずれかのレチノイン酸受容体α(RARα)の融合が生じる。PML-RARαおよびPLZF-RARαの両方は、N-CoR/SMRT-HDACを異常に補充し、レチノイン酸制御遺伝子を抑制することによって白血病誘発発生を媒介する(Lin et al. 1998. Nature 391:811-814; Grignani et al. 1998. Nature 391:815-818)。実際、APLにおける転写を標的とする非特異的HDAC阻害剤であるブチル酸ナトリウム(NaBu)の治療的使用は、その疾患に関するレチノイン酸耐性型においてさえも成功してきた(Warrell et al. 1998. J. Natl. Cancer Inst. 90:1621-1625)。HDACの役割は、ETO(「821(eight twenty one)」 または MT68) 遺伝子産物とAML1 転写因子の融合の結果として生じる急性骨髄性白血病においてもまた実証されている(Gelmetti et al. 1998. Mol. Cell Biol. 18:7185-7191; Wang et al. 1998. PNAS 95:10860-10865)。併せて、これは、HDACの確実な白血病の治療に対する有効な方法であることを示唆している。さらに、大腸癌(Hassig et al. 1997. Chem. Biol. 4:783-789; Archer et al. 1998. PNAS 95:6791-6796)、扁平上皮細胞癌(Gillenwater et al., 1998. Int. J. Cancer 75:217-224; Saunders et al. 1999. Cancer Res. 59:399-404)、腺癌(McBain et al. 1997. Biochem. Pharmacol. 53:1357-1368)および神経芽腫(Swendeman et al. 1999. Proc. Amer. Assoc. Cancer Res. 40, Abst. #3836)を含む別の種類の癌においても、HDACの役割に関するさらなる証拠がある。HDACは、BRCA1(Yarden et al. 1999. Proc. Ameri. Assoc. Cancer Res. 40, Abstr. #3387)およびc-fos(Bakin and Curran. 1999. Science 283:387-390)に媒介される転写制御にも関わっている。
【0008】
一方でまた、制御された方法において細胞増殖を増強することが、ときに望まれる。例えば、細胞増殖は創傷治癒および組織の増殖が望ましいところで有用である。即ち、増殖の阻害を助長、増強または遅延させる調節因子を同定することが望ましい。
【発明の概要】
【発明が解決しようとする課題】
【0009】
転写成分および調節因子を同定することは望ましいが、そのような化合物の分野では十分になされていない。従って、転写の調節因子をスクリーニングするための有用な組成物および方法を提供することは利点がある。また、転写制御に関与する新規組成物を提供することも又利点がある。
【課題を解決するための手段】
【0010】
(発明の要旨)
上記目的に従って、本発明は、HDACタンパク質をコードする核酸を提供する。そのポリペプチドを本出願において「HDAC8」として示す。
【0011】
ある実施態様において、本発明はHDAC8をコードする単離された核酸分子を提供する。また、本発明はHDAC8タンパク質およびその断片を提供する。
【0012】
本発明は、HDAC8タンパク質をコードする核酸に高緊縮条件下でハイブリダイズするHDACタンパク質をコードする核酸を提供する。それに従って、本発明はHDAC8タンパク質またはその断片をコードする核酸を同定する方法を提供する。
【0013】
本発明は、HDAC8タンパク質をコードし、発現する発現ベクターをさらに提供する。
【0014】
別の態様において、本発明はHDAC8タンパク質をコードする組換え核酸および発現ベクターを含む宿主細胞を提供する。それに従って、本発明は、HDAC8タンパク質を産生する方法を提供する。
【0015】
さらなる態様において、本発明はHDAC8アミノ酸配列を含む組換えポリペプチドおよびHDAC8アミノ酸配列を含有するポリペプチドを同定する方法を提供する。
【0016】
別の態様において、本発明はHDAC8ポリペプチドと結合する抗体を提供する。
【0017】
さらに別の態様において、本発明はHDAC8タンパク質に結合するバイオ活性物質をスクリーニングする方法を提供する。
【0018】
さらに別の態様において、本発明は異常なHDAC8発現と関連のある疾患状態を同定する方法を提供する。
【0019】
本発明の別の態様は、本発明に関する以下の記載によって当業者に明らかであろう。
【図面の簡単な説明】
【0020】
【図1−1】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−2】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−3】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−4】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−5】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図2】図2は、HDAC1(配列番号:4)、HDAC2(配列番号:5)、HDAC3 (配列番号:6)およびHDAC8(配列番号:7)との間のアミノ酸の類似性を示す。同一残基は暗灰色のボックスとし、保存性のある置換基は明るい灰色で示す。推定377アミノ酸HDAC8(約42kDa)産物は、HDAC1、2および3のC末端の〜100アミノ酸に相同性のある領域を失っていることがわかる。
【発明を実施するための形態】
【0021】
(発明の詳細な説明)
本発明は、ヒストン脱アセチル化-8(HDAC8)タンパク質とそのタンパク質をコードする核酸を提供する。HDAC8タンパク質を結合し得る、また好ましくはHDAC8タンパク質の活性を調節し得るバイオ活性物質をスクリーニングする方法を提供する。その方法は、HDAC8タンパク質および候補バイオ活性物質および細胞または細胞集団を合わせ、候補物質の存在下および非存在下で細胞への効果を測定することを含む。結合アッセイを含む他のスクリーニングアッセイも、下記のように本発明で提供される。HDAC8タンパク質を制御するか、もしくは調節するための治療薬も提供され、本明細書に記載されている。以下にさらに記載するように、診断薬も提供される。
【0022】
本発明のHDAC8タンパク質は、いくつかの方法で同定され得る。この意味での「タンパク質」には、タンパク質、ポリペプチドおよびペプチドが含まれる。本発明のHDAC8タンパク質は、2つの一般的クラスに分類される:完全に新しいタンパク質、即ち、発見時点で公共のデータベースの一部分になっていないものである。但し、それらは発現配列タグ(EST)によってコードされた既知のタンパク質またはペプチドとホモロジーを持ち得る。今一つは、HDAC8タンパク質が既知のタンパク質であるが、クロマチン構造制御、ヒストンアセチル化、転写制御などに関与することが知られていないものである。即ち、本明細書中で新規生物学的機能を持つものとして同定される。従って、HDAC8タンパク質は、ヒストンアセチル化および/または転写に関与すると知られているタンパク質との関連で最初に同定され得る。HDAC8タンパク質および核酸が新規である場合には、組成物および使用の方法が本明細書で提供される。HDAC8タンパク質および核酸が既知であるが、しかし本明細書中に記載のようにクロマチン構造の制御、ヒストンアセチル化および/または転写に関与することが知られていない場合には、使用方法、即ち機能スクリーンが提供される。
【0023】
本発明で提供されるある実施態様では、HDAC8タンパク質は、本明細書で定義するように、1つまたはそれ以上の下記特性を有する:ヒストンのアセチル化;HDAC1(配列番号:4)、HDAC2(配列番号:5)、HDAC3(配列番号:6)およびHDAC8(配列番号:7)との相同性。該相同性は、下記データベース、アルゴリズムおよびパラメーターを用いて見出される。本明細書中の「相同性」とは、配列の類似性および好ましくは同一性を意味する。
【0024】
ある実施態様では、HDAC8核酸またはHDAC8タンパク質は、本発明で提供される配列に対するへの核酸および/またはアミノ酸配列の実質的同一性または類似性により最初に同定される。好ましい実施態様では、HDAC8核酸またはHDAC8タンパク質は、下記のように本発明で提供される配列に対して配列同一性または類似性を有し、1またはそれ以上のさらに後述するようなHDAC8タンパク質バイオ活性を有する。そのような配列同一性または類似性は、全体的な核酸またはアミノ酸配列に基づくことができる。
【0025】
好ましい実施態様では、あるタンパク質は、図1のアミノ酸配列に対する全体的な配列同一性が好ましくは約60%より大きく、より好ましくは約70%より大きく、さらに好ましくは約80%より大きく、最も好ましくは約90%より大きいならば、本明細書で定義する「HDAC8タンパク質」である。ある実施態様では、該配列同一性は約93ないし95または98%にも上る。
【0026】
技術上周知のように、タンパク質(または以下に考察するように核酸)が既知配列と配列同一性または類似性を有しているか否かを同定するために多数の異なるプログラムを使用することができる。配列同一性および/または類似性は技術上周知の標準的技法を用いて測定することができ、それらは、Smith & Waterman, Adv. Appl. Math., 2: 482 (1981)の局所配列同一性アルゴリズム、Needleman & Wunsch, J. Mol. Biol., 48: 443 (1970)の配列同一性アラインメント、Pearson & Lipman, PNAS U.S.A., 85: 2444 (1988)の類似性検索法、これらのアルゴリズムのコンピュータによる実行(Wisconsin Genetics Software Package、Genetics Computer Group, 575 Science Drive, Madison, WI、中のGAP、BESTFIT、FASTAおよびTFASTA)、Devereux et al., Nucl. Acid Res., 12: 387-395 (1984)に記載のBest Fit配列プログラム、を非限定的に含み、好ましくはデフォルト設定を用い、または検定により使用する。好ましくは、FstDBにより以下のパラメータに基づいてパーセント同一性を計算する:ミスマッチペナルティ(mismatch penalty) 1; ギャップペナルティ(gap penalty)1; ギャプサイズペナルティ(gap size penalty) 0.33; 接合ペナルティ(joining penalty) 30、「Current Methods in Sequence Comparison and Analysis,」Macromolecule Sequencing and Synthesis, Selected Methods and Applications, pp 127-149 (1988), Alan R. Liss, Inc.である。
【0027】
有用なアルゴリズムの一例はPILEUPである。PILEUPは、漸進的対アラインメントを用いて関連配列から多重配列アラインメントを創成する。それはまた、アラインメント創成に使用される、クラスタリングの関係を示すツリーを描くことができる。PILEUPは、Feng & Doolittle, J. Mol. Evol. 35: 351-360 (1987)の漸進的アラインメント法の簡略化したものを用いる;この方法はHiggins & Sharp CABIOS 5: 151-153 (1989)記載の方法と類似している。有用なPILEUPパラメータは、デフォルトギャップウェイト(default gap weight) 3.00, デフォルトギャップレングスウェイト(default gap length weight) 0.10, ウェイテッド・ギャップ・エンド・ギャップ(weighted end gaps)を含む。
【0028】
有用なアルゴリズムのもう1つの例は、Altschul et al., J. Mol. Biol. 215, 403-410, (1990);およびKarlin et al., PNAS U.S.A. 90: 5873-5787 (1993)に記載のBLASTアルゴリズムである。特に有用なBLASTプログラムは、Altschul et al., Methods in Enzymology, 266: 460-480 (1996); http://blast.wustl/edu/blast/ README.htmlから得られるWU−BLAST−2である。WU−BLAST−2はいくつかの検索パラメータを使用するがその殆どはデフォルト値に設定されている。調節可能なパラメータは以下の値で設定する:オーバーラップスパン(overlap span)=1, オーバーラップフラクション(overlap fraction) = 0.125, 閾値(word threshold)(T) = 11である。HSP SおよびHSP S2パラメータは動的数値であり、特定の配列の組成および興味の対象である配列を検索する特定のデータベースの組成に依存してプログラム自身により決定されるが、値は感度を上げるように調節することができる。
【0029】
これに加えて有用なアルゴリズムは、Altschul et al., Nucl. Acids Res., 25: 3389-3402に報告されているギャップドBLASTである。ギャップドBLASTはBLOSUM−62代替スコアを使用する。ここで閾値Tパラメータは9に設定し;2−ヒット法によりギャップのない伸長を開始し;ギャップ長kにコスト10+kを課し;Xuを16に設定し;Xgをデータベース検索段階では40に、そしてアルゴリズムのアウトプット段階では67に設定する。ギャップドアラインメントは約22ビットまでに相当するスコアで開始される。
【0030】
パーセントアミノ酸配列同一性の値は、マッチする同一残基の数を、アラインメントを行った領域における「より長い」配列の総残基数で割って決定される。「より長い」配列は、アラインメントを行った領域中で実際の残基を最も多く有する配列である(アラインメントスコアを最大化するためにWU−Blast−2により導入されたギャップを無視する)。
【0031】
同様にして、ここで同定されるポリペプチドのコード配列に関する「パーセント(%)核酸配列同一性」を、HDAC8タンパク質のコード配列中のヌクレオチド残基と同一な、候補配列中のヌクレオチド残基の割合として定義する。好ましい方法は、WU−BLAST−2のBLASTNモジュールをデフォルトパラメータに設定し、オーバーラップスパンおよびオーバーラップフラクションをそれぞれ1および0.125に設定して利用するものである。
【0032】
アラインメントは、アラインメントを行う配列中へのギャップ導入を含んでもよい。加えて、図中の配列でコードされるタンパク質より多いかまたは少ないアミノ酸を含有する配列については、ある実施態様では、配列同一性の割合は、アミノ酸総数に対する同一アミノ酸数に基づいて決定されると理解される。かくして、例えば、後で論じるように、ある実施態様では、図1に示す配列より短い配列の配列同一性は、より短い配列中のアミノ酸数を用いて決定される。パーセント同一性の計算においては、相対的重みは、挿入、欠失、置換その他のような種々の配列変化の表出に起因するとはされない。
【0033】
ある実施態様では、同一性のみがプラスのスコアを与えられ(+1)、ギャップを含むあらゆる形の配列変化に「0」の値が与えられる。これにより、配列類似性計算について後述するような、重みをつけた(加重値を与える)目盛りまたはパラメータの必要性がなくなる。例えば、配列同一性の割合は、アミノ酸総数に対する同一アミノ酸数に基づいて決定されると理解される。例えば、パーセントアミノ酸配列同一性は、マッチする同一残基の数を、アラインメントを行った領域における「より短い」配列の総残基数で割り、100を掛けて算出される。「より長い」配列は、アラインメントを行った領域中で実際の残基を最も多く有する配列である。
【0034】
当業者に認識されるであろうように、本発明の配列は、配列のエラーを含んでもよい。つまり、誤ったヌクレオシド、フレームシフト、未知のヌクレオシド、または他のタイプの配列エラーが、配列のどこにあってもよい。しかし、正しい配列は、本発明の相同性および緊縮の定義の範囲内にある。
【0035】
本発明のHDAC8タンパク質は、図1に示す核酸にコードされるアミノ酸配列よりも、短くても長くてもよい。ゆえに、好ましい実施態様では、本発明で提供される核酸配列にコードされるアミノ酸配列の部分または断片が、HDAC8タンパク質の定義に包含される。本発明のある実施態様では、HDAC8タンパク質の断片は、a)少なくとも1つの抗原のエピトープを共有する;b)少なくとも表示される配列同一性を有する;およびc)好ましくは本発明でさらに定義されるヒストン脱アセチル化酵素−8の生物学的活性を有する場合、HDAC8タンパク質である。該配列が診断的に用いられる場合、つまり、HDAC8タンパク質の核酸の有無が求められる場合には、表示される配列の同一性のみが要求される。本発明の核酸もまた、図1の配列よりも、短くても長くてもよい。核酸断片には、以前に正確には同定されていない配列を有する、本発明で提供される核酸のあらゆる部分が含まれる;以前に同定されていない部分に対して、表示される配列同一性がある配列を有する断片が、本発明のある実施態様で提供される。
【0036】
さらに、より詳しく後で概説するように、図1に示されるものよりも長いHDAC8タンパク質を作成し得る。例えばエピトープもしくは精製用タグの付加、他の融合配列の付加、または付加的なコードおよび非コード配列の解明による。後述のように、HDAC8ペプチドを、グリーン蛍光ペプチド(GFP)などの蛍光ペプチドと融合させることが、特に好ましい。
【0037】
HDAC8タンパク質はまた、本明細書で概説するように、図1に示す配列またはその相補鎖とハイブリダイズするHDAC8核酸にコードされるものとしても同定され得る。ハイブリダイゼーションの条件は、さらに後述する。
【0038】
好ましい実施態様では、抗体を作成するためにHDAC8タンパク質を用いようとするとき、HDAC8タンパク質は、完全長タンパク質と、少なくとも1つのエピトープまたは決定因子を共有せねばならない。本明細書では、「エピトープ」または「決定因子」は、抗体を作成および/または結合するであろうタンパク質の一部分を意味する。ゆえに、ほとんどの例で、より小さいHDAC8タンパク質に対して作られた抗体は、完全長タンパク質に結合できる。好ましい実施態様では、エピトープは独特である。つまり、独特なエピトープに対して作成された抗体は、交差反応性をほとんど、または全く示さない。「抗体」という用語には、当分野で周知のように、抗体全体を修飾して作成されたか、または組換えDNA技術を用いて新規に合成された、Fab、Fab2、1本鎖抗体(例えば、Fv)、キメラ抗体などを含む抗体断片が含まれる。
【0039】
後述するように、好ましい実施態様では、HDAC8タンパク質に結合するHDAC8タンパク質に対する抗体には、本明細書に記載のHDAC8タンパク質の生物学的機能を低減するか、または除去する能力がある。つまり、抗HDAC8タンパク質抗体(ポリクローナル、または好ましくはモノクローナルのどちらでも)をHDAC8タンパク質(またはHDAC8タンパク質を含有する細胞)に加えることで、HDAC8活性を低減するか、または除去し得る。一般的に、少なくとも25%の活性の減少が好ましく、少なくとも約50%が特に好ましく、約95−100%の減少がことさら好ましい。
【0040】
本発明のHDAC8抗体は、HDAC8タンパク質に特異的に結合する。本明細書では、「特異的に結合する」は、少なくとも10−4−10−6M−1の範囲の結合定数で、好ましくは10−7−10−9M−1の範囲で、抗体がタンパク質に結合することを意味する。抗体について、さらに後述する。
【0041】
核酸の場合、核酸配列の包括的な配列同一性は、アミノ酸の配列同一性と同一基準であるが、遺伝暗号の縮重と異なる生物間のコドンの偏りを考慮にいれる。従って、核酸配列の同一性は、タンパク質配列のそれと比べて低くも高くもあり得る。ゆえに、図の核酸配列と比較した核酸配列の配列同一性は、好ましくは75%より大きく、より好ましくは約80%より大きく、特に約85%より大きく、そして最も好ましくは90%より大きい。いくつかの実施態様では、該配列同一性は約93ないし95または98%ほどに高い。
【0042】
好ましい実施態様では、HDAC8核酸は、HDAC8タンパク質をコードする。当業者には理解されるであろうように、遺伝暗号の縮重のために、すべて本発明のHDAC8タンパク質をコードする、極めて多数の核酸を作り得る。ゆえに、特定のアミノ酸配列を同定すれば、当業者は、HDAC8タンパク質のアミノ酸配列を変えない方法で、単純に1つまたはそれ以上のコドンの配列を変更することにより、異なる核酸をいくつでも作成し得る。
【0043】
ある実施態様では、核酸はハイブリダイゼーション検査により決定される。ゆえに、例えば、図1に示す核酸配列またはその相補鎖と、高緊縮条件下のもとでハイブリダイゼーションする核酸は、HDAC8核酸と考えられる。高緊縮条件下条件は当分野で既知である;例えば、出典明示により本明細書の一部とするManiatis et al., Molecular Cloning: A Laboratory Manual, 2d Edition, 1989およびShort Protocols in Molecular Biology, ed. Ausubel, et al.を参照のこと。高緊縮条件は配列依存的であり、環境が違えば異なる。より長い配列はより高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションの発広範な手引きは、Tijssen, Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993)に記載されている。一般的に、特異的な配列について、緊縮条件は、規定されたイオン強度、pHにおいて熱融解点(Tm)より約5−10℃低いように選択される。Tmは、標的に相補的なプローブの50%が、平衡状態で標的配列に(規定のイオン強度、pHおよび核酸濃度で)ハイブリダイズする温度である(標的配列が過剰に存在するので、Tmでは、50%のプローブが平衡状態において占められる)。緊縮条件は、塩濃度が約1.0Mナトリウムイオンより小さく、典型的には0.01ないし1.0Mのナトリウムイオン(または他の塩)濃度、pH7.0ないし8.3、温度が、短いプローブ(例えば、10ないし50のヌクレオチド)には低くても約30℃、長いプローブ(例えば、50ヌクレオチド以上)には低くても約60℃、である。緊縮条件は、ホルムアミドのような不安定化させる物質の付加により達せられてもよい。
【0044】
他の実施態様では、より低い緊縮条件のハイブリダイゼーション条件を用いる;例えば、当分野で周知のように、穏やかなまたは低緊縮条件を用いてもよい;Maniatis and Ausubel, 前出, and Tijssen(前出)を参照のこと。
【0045】
本発明のHDAC8タンパク質と核酸は、好ましくは組換えである。ここで用いられる「核酸」は、さらに定義するように、DNAもしくはRNA、またはデオキシおよびリボヌクレオチドの両者を意味し得る。核酸は、ゲノムDNA、cDNAおよびオリゴヌクレオチドを含むセンスおよびアンチセンス核酸を含む。このような核酸はまた、生理的環境におけるそのような分子の安定性および半減期を増加させるために、リボース−リン酸バックボーンに修飾を含んでいてもよい。
【0046】
核酸は2本鎖、1本鎖であってよく、または2本鎖もしくは1本鎖の配列の両方の部分を含んでいてもよい。当業者により認識されるであろうように、1本鎖(「ワトソン」)を描けばもう1つの鎖(「クリック」)の配列が定義されるので、図に示す配列は、該配列の相補鎖もまた含む。ここで「組換え核酸」という用語は、一般的に、核酸をエンドヌクレアーゼによって操作して、自然界には存在しない形状で、独創的にインビトロで形成した核酸を意味する。ゆえに、線状の形状で単離されたHDAC8核酸や、通常は結合していないDNA分子を連結することによりインビトロで形成した発現ベクターは、両者とも本発明のためには組換え体と考えられる。一旦組換え核酸が作成され宿主細胞または生物に再導入されれば、それは非組換え的に、即ち、インビトロの操作ではなく宿主のインビボの細胞機構を用いて複製すると理解される;しかしながら、そのような核酸は一旦組換え的に生産されれば、以後は非組換え的に複製しても、本発明のためにはなお組換え体と考えられる。
【0047】
同様に、「組換えタンパク質」は組換え技術を用いて、即ち上述のように組換え核酸の発現を通して作成されたタンパク質である。組換えタンパク質は、少なくとも1つまたはそれ以上の特性によって、天然産生のタンパク質と区別される。例えば、このタンパク質は、野生型宿主中で通常会合しているタンパク質または化合物の一部または全てから単離または精製され、実質的に純粋であり得る。例えば、単離されたタンパク質は、自然状態では通常会合している物質の少なくとも一部を伴わないで、所定の試料中の総タンパク質重量の好ましくは少なくとも約0.5%、より好ましくは少なくとも5%を構成している。実質的に純粋なタンパク質は、総タンパク質重量の少なくとも約75%、好ましくは少なくとも約80%、そして特に好ましくは少なくとも約90%を含む。この定義には、ある生物由来のHDAC8タンパク質を異なる生物または宿主中で生産することが含まれる。あるいは、タンパク質がより増加した濃度レベルで作られるように誘導可能プロモーターもしくは高発現プロモーターを使用することにより、タンパク質を通常見られるよりも有意に高濃度で作ることができる。あるいは、以下に考察するように、該タンパク質は、エピトープタグの付加またはアミノ酸置換、挿入および欠失を含むような、自然界に通常見出されない形であり得る。
【0048】
ある実施態様では、本発明は、HDAC8タンパク質の変異体を提供する。これらの変異体は置換、挿入または欠失変異体の3つの分類の1つまたはそれ以上に相当する。これらの変異体は通常、カセット式もしくはPCR式変異誘発または技術上周知の他の技術を用いて、HDAC8タンパク質をコードするDNA中のヌクレオチドの部位特異的変異誘発により、変異体をコードするDNAを生産し、そして次いでDNAを上に略述した組換え培養細胞中で発現させることにより調製する。しかしながら、約100〜150残基までを有する変異HDAC8タンパク質断片を、確立された技術を用いてインビトロ合成により調製することができる。アミノ酸配列変異体は変化が予め決定されているという特徴を有し、この特徴によって、これらの変異体は、HDAC8タンパク質のアミノ酸配列に対する天然産生の対立遺伝子変異体または種間変異体から区別される。変異体は典型的に、天然産生の類似体と定性的に同じバイオ活性を発揮するが、ただし、以下にさらに詳しく略述するように、変更された特性を有する変異体を選択することもできる。
【0049】
アミノ酸配列変異を導入する部位または領域は予め決定されるが、変異自体は予め決定しておく必要はない。例えば、所定の部位における変異能を最適化するために、標的コドンまたは領域にランダム変異誘発を起こし、発現したHDAC8細胞周期変異体をスクリーニングして所望の活性の最適な組合わせを有するものを探してもよい。既知の配列を有するDNA中の予め定められた部位に置換変異を作成する技術は周知であり、例えば、M13プライマーによる変異誘発およびPCRによる変異誘発がある。変異体のスクリーニングは、HDAC8タンパク質活性のアッセイを用いて行われる。
【0050】
アミノ酸置換は典型的には単一の残基置換である;かなり大きな挿入も許容されるが、挿入は通常、約1〜20アミノ酸の単位で行われる。欠失は、より大きな場合もあるが、約1から約20残基の範囲である。
【0051】
最終誘導体に到達するために、置換、欠失、挿入またはそれらのいずれの組み合わせを用いてもよい。一般的に、これらの変化は、分子の変化を最小限にするために少数のアミノ酸について行われる。しかしながら、より大きな変化も一定の状況では許容される。HDAC8タンパク質の特徴について小さな変化が望まれる場合は、置換は一般的に次のチャートに従ってなされる。
(表1)
チャートI

【0052】
機能または免疫学的同一性における実質的な変化は、チャートIに示したものより保存性の低い置換を選択することによってもたらされる。例えば、より大きく影響する置換を行うことができる:それらは、変化する区域のポリペプチドバックボーンの構造、例えばアルファ−ヘリックス構造またはベータ−シート構造;標的部位での分子の電荷または疎水性;または側鎖の大きさである。一般的にポリペプチドの性質に最も大きな変化を生じると期待される置換は(a)親水性残基、例えばセリルまたはスレオニルを、疎水性残基、例えばロイシル、イソロイシル、フェニルアラニル、バリル、またはアラニルに置換する(またはその逆)、(b)システインまたはプロリンを他のいずれかの残基に置換する(またはその逆)、(c)正電荷を持つ側鎖、例えばリシル、アルギニル、またはヒスチジルを負電荷を持つ側鎖、例えばグルタミル、アスパルチルに置換する(またはその逆)、(d)大きい側鎖を持つ残基、例えばフェニルアラニンを側鎖を持たない残基、例えばグリシンに置換する(またはその逆)、ものである。
【0053】
変異体は典型的には天然産生の類似体と定性的に同じ生物活性を発揮し、同じ免疫応答を誘起するが、ただし、必要に応じてHDAC8タンパク質の特性を変更するような変異体もまた選択される。あるいは、変異体をHDAC8タンパク質の生物活性が変わるようにデザインすることができる。例えば、グリコシル化部位を加えたり、変えたりまたは除去したりすることができる。HDAC8変異体は、リン酸化部位を加え、変え、除去することが可能である。例えばcAMP仲介性リン酸化コンセンサス配列「KRAS」が見出されるアミノ酸部位36−39で変更できる。チロシンリン酸化コンセンサス配列は、Try−174で見出され、これも変更可能である。
【0054】
HDAC8ポリペプチドの共有結合による修飾は、本発明の範囲内に含まれる。共有結合による修飾の1つのタイプは、HDAC8ポリペプチドの標的アミノ酸残基を、HDAC8ポリペプチドの選択された側鎖またはN−もしくはC−末端と反応できる有機誘導体化試薬と反応させることを含む。以下に詳述するように、二官能性試薬による誘導体化は、例えば、抗HDAC8抗体の精製またはスクリーニングアッセイの方法において使用する水不溶性の支持マトリックスまたは表面に、HDAC8タンパク質を架橋するために有用である。一般的に用いられる架橋試薬としては、例えば、1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、例えば4−アジドサリチル酸とのエステルのようなN−ヒドロキシスクシンイミドエステル、3,3'−ジチオビス(スクシンイミジルプロピオン酸エステル)のようなジスクシンイミジルエステルを含むホモ二官能イミドエステル、ビス−N−マレインイミド−1,8−オクタンのような二官能マレインイミドおよびメチル‐3−[(p−アジドフェニル)ジチオ]プロピオンイミダートのような試薬が挙げられる。
【0055】
他の修飾としては、グルタミニルおよびアスパラギニル残基からそれぞれ対応するグルタミルおよびアスパルチル残基への脱アミド化、プロリンおよびリシンのヒドロキシル化、セリルまたはスレオニル残基の水酸基のリン酸化、リシン、アルギニン、およびヒスチジン側鎖のアミノ基のメチル化[T.E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)]、N末端アミンのアセチル化、およびC末端カルボキシル基のアミド化が挙げられる。
【0056】
本発明の範囲内に含まれるHDAC8ポリペプチドの別のタイプの共有結合的修飾は、ポリペプチドの天然のグリコシル化パターンを変えることより成る。「天然のグリコシル化パターンを変える」とは、ここでの目的には、HDAC8ポリペプチドの天然の配列中に見出される1個もしくはそれ以上の炭水化物部分を削除、および/またはHDAC8ポリペプチドの天然の配列に存在しない1個またはそれ以上のグリコシル化部位を付加することを意味する。
【0057】
HDAC8ポリペプチドへのグリコシル化部位の付加はそのアミノ酸配列を変化させることにより達成される。例えば、その変化は、1個またはそれ以上のセリンまたはスレオニン残基をHDAC8ポリペプチドの天然配列に添加または置換することによって行われる(O−グリコシル部位)。また、その変更は、例えば、天然配列のHDAC8ポリペプチド中で、1以上のAxn−Xaa−Ser/Thr部位(Xaa=任意のアミノ酸)の添加もしくは置換でなされ得る(N−グリコシル部位)。HDAC8アミノ酸配列は、DNAレベルでの変化を通して任意に変化させ得るが、特に、所望のアミノ酸に翻訳されるであろうコドンを作成するように予め選択した塩基において、HDAC8ポリペプチドをコードするDNAを変異させることによって変化させ得る。
【0058】
HDAC8ポリペプチド上の炭水化物部分の数を増加させるもう1つの方法は、ポリペプチドにグリコシドを化学的もしくは酵素的に結合させることによる。このような方法は、技術上で、例えば、1987年9月11日に公開されたWO 87/05330およびAplin and Wriston, CRC Crit. Rev. Biochem., pp. 259-306 (1981)に記述されている。
【0059】
HDAC8ポリペプチド上に存在する炭水化物部分の除去は、化学的にもしくは酵素的に、またはグリコシル化の標的となっているアミノ酸残基をコードするコドンを置換する変異により達成される。化学的脱グリコシル化の手法は技術上周知であり、例えば、Hakimuddin, et al., Arch. Biochem. Biophys., 259: 52 (1987)およびEdge et al., Anal. Biochem., 118: 131 (1981)に記述されている。ポリペプチド上の炭水化物部分の酵素的切断は、Thotakura et al., Meth. Enzymol., 138: 350 (1987)に記述されているように種々のエンド−およびエキソ−グリコシダーゼの使用により達成できる。
【0060】
もう1つのタイプのHDAC8ポリペプチドの共有結合的修飾は、特許第4640835号、第4496689号、第4301144号、第4670417号、第4791192号または第4179337号に示されている方法で、HDAC8ポリペプチドを種々の非タンパク質ポリマー、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンの1つに結合させることを含む。
【0061】
本発明のHDAC8ポリペプチドはまた、もう1つの、異種のポリペプチドまたはアミノ酸配列に融合したHDAC8ポリペプチドを含むキメラ分子を形成するように修飾し得る。ある実施態様では、そのようなキメラ分子は、HDAC8ポリペプチドと、抗タグ抗体が選択的に結合できるエピトープを提供するタグポリペプチドとの融合を含む。エピトープタグは一般的に、HDAC8ポリペプチドのアミノ−(N−)またはカルボキシル−(C−)末端に置かれる。HDAC8ポリペプチドにそのようなエピトープタグが付いた形のものの存在は、タグポリペプチドに対する抗体によって検出することができる。また、エピトープタグをつけることにより、HDAC8ポリペプチドが抗タグ抗体、またはエピトープタグに結合する別のタイプのアフィニティーマトリックスによる精製が容易になる。別の実施態様では、キメラ分子は、HDAC8ポリペプチドと免疫グロブリンまたは免疫グロブリンの特定の領域との融合を含んでもよい。以下でさらに論じるように、2価形態のキメラ分子について、このような融合はIgG分子のFc領域に対してなし得る。
【0062】
種々のタグポリペプチドおよびその各々の抗体は技術上周知である。例としては、ポリ−ヒスチジン(poly-his)またはポリ−ヒスチジン−グリシン(poly-his-gly)タグ、flu HAタグポリペプチドおよびその抗体12CA5[Field et al., Mol. Cell. Biol. 8: 2159-2165 (1988)];c−mycタグおよびそれに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evan et al., Molecular and Cellular Biology, 5: 3610-3616 (1985)];ならびに単純ヘルペスウイルスの糖タンパク質D(gD)タグおよびその抗体[Paborsky et al., Protein Engineering, 3(6): 547-553 (1990)]が挙げられる。その他のタグポリペプチドとしては、Flag−ペプチド[Hopp et al., BioTechnology 6: 1204-1210 (1988)];KT3エピトープペプチド[Martin et al., Science 255: 192-194 (1992)];チューブリンエピトープペプチド[Skinner et al., J. Biol. Chem. 266: 15163-15166 (1991)];T7遺伝子10タンパク質ペプチドタグ[Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. U.S.A. 87: 6393-6397 (1990)];ヒスチジンタグおよび金属結合部位[Smith, Ann. NY. Acad. Sci.,646:315-321(1991) ]が挙げられるが、Flgおよびヒスチジンタグが好ましい。
【0063】
以下に概説するように、ある実施態様では、HDACファミリーのHDAC8タンパク質および他の生物由来のHDAC8タンパク質を、クローンとして発生させ、発現させる。ゆえに、プローブまたはポリメラーゼ連鎖反応(PCR)の配列を、ヒトまたは他の生物由来の他の関連するHDAC8タンパク質を見つけ出すのに用いてもよい。当業者には理解されるであろうように、特に有用なプローブおよび/またはPCRプライマー配列は、HDAC8核酸配列の独特な領域を含む。当分野で一般的に知られるように、好ましいPCRプライマーは、約15ないし約35ヌクレオチド長であり、約20ないし約30が好ましく、必要ならイノシンを含んでもよい。PCR反応の条件は当分野で周知である。ゆえに、ここで挙げられた配列の中で配列にともなって提供されるものは、これらの配列の各部分であり、15ヌクレオチドまたはそれ以上の独特の各部分が特に好ましいこともまた理解される。当業者は、日常的にヌクレオチド配列を所望の長さに合成したり切断することができる。
【0064】
一旦、プラスミドまたは他のベクターに含めたり、そこから直線状の核酸断片として切り出すなどして、その天然の供給源から単離すると、組換えHDAC8核酸を、他のHDAC8核酸を同定および単離するためのプローブとしてさらに用いることができる。それをまた、修飾された、または変異型のHDAC8核酸およびタンパク質を作成するための「前駆」核酸として用いることができる。
【0065】
HDAC8タンパク質をコードする本発明の核酸を用いて、種々の発現ベクターが作られる。発現ベクターは、自己複製的な染色体外ベクターでも宿主ゲノムに組み込まれるベクターでもよい。一般的に、これらの発現ベクターは、HDAC8タンパク質をコードする核酸に機能し得るように結合された転写および翻訳の制御核酸を含む。「制御配列」という用語は、特定の宿主生物内で、機能し得るように結合されたコード配列が発現するのに必要な核酸配列を意味する。原核生物に適した制御配列は、例えば、プロモーター、任意にオペレーター配列、およびリボゾーム結合部位を含む。真核細胞はプロモーター、ポリアデニル化シグナル、およびエンハンサーを使用することが知られている。
【0066】
核酸は、それが他の核酸配列と機能的な関係に置かれている場合に「機能し得るように結合されて」いる。例えば、前配列もしくは分泌リーダーに対するDNAは、もしそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドに対するDNAに機能し得るように結合されている;プロモーターまたはエンハンサーは、もしそれが配列の転写に影響を与えるならば、コード配列に機能し得るように結合されている;または、リボゾーム結合部位は、もしそれが翻訳を促進するように位置しているならば、コード配列に機能し得るように結合されている。他の例として、機能し得るように結合されたとは、隣接するように、そして、分泌リーダー配列の場合は、隣接しかつ読みとり段階にあるように結合されたDNA配列を意味する。しかしながら、エンハンサーは隣接している必要はない。結合は都合のよい制限部位におけるライゲーションにより達成される。もしそのような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーを従来の実施法に従って使用する。転写および翻訳制御核酸は、HDAC8タンパク質の発現に使用する宿主細胞にとって一般的に適当であろう;例えば、Bacillus由来の転写および翻訳の制御核酸配列は、BacillusでHDAC8タンパク質を発現させるために好ましく使用される。夥しい数の適当な発現ベクターおよび適切な制御配列が、様々な宿主細胞に関して技術上公知である。
【0067】
一般的に、転写および翻訳の制御核酸は、プロモーター配列、リボゾーム結合部位、転写開始および終結配列、翻訳開始および終結配列、およびエンハンサーまたはアクチベーター配列を非限定的に含む。好ましい実施態様では、制御配列はプロモーター並びに転写の開始および終結配列を含む。
【0068】
プロモーター配列は構成または誘導可能プロモーターをコードする。該プロモーターは天然産生のプロモーターでもハイブリッドプロモーターでもよい。2以上の多くのプロモーターのエレメントを組み合わせたハイブリッドプロモーターも技術上公知であり、本発明において有用である。
【0069】
加えて、発現ベクターは付加的エレメントを含み得る。例えば、発現ベクターは、2つの複製システムを有し、これにより2種の生物で、例えば発現のために哺乳動物または昆虫の細胞で、そしてクローニングおよび増幅のために原核宿主で、維持することができる。さらに、発現ベクターを組込むために、発現ベクターは宿主細胞のゲノムと相同な配列を少なくとも1個、また好ましくは発現構築体に隣接する2個の相同配列を含む。ベクターに取り入れるのに適当な相同配列を選択することにより、組込みベクターを宿主細胞の特定の座位を目指して組込ませ得る。組込みベクターの構築は技術上周知である。
【0070】
加えて、好ましい実施態様では、該発現ベクターは形質転換した宿主細胞の選択を可能にする選択マーカー遺伝子を含む。選択遺伝子は技術上周知であり、用いる宿主により異なる。
【0071】
好ましい発現ベクター系は、PCT/US97/01019およびPCT/US97/01048に一般的に記述されているレトロウイルスベクター系であり、両者とも特に出典明示により本明細書の一部とする。
【0072】
本発明のHDAC8タンパク質は、HDAC8タンパク質をコードする核酸を含む発現ベクターで形質転換した宿主細胞を、HDAC8タンパク質の発現を誘導または誘起する適当な条件下で培養することによって生産される。HDAC8タンパク質の発現に適当な条件は、発現ベクターと宿主細胞の選択によって異なるが、当業者は日常的な実験により容易に確かめることができる。例えば、発現ベクター中に構成的プロモーターを使用している場合には、宿主細胞の生育および増殖を最適化することが要求されるだろうし、一方、誘導可能プロモーターを使用している場合には、誘導に適した生育条件が要求される。加えて、いくつかの実施態様では、収穫のタイミングが重要である。例えば、昆虫細胞での発現に用いられるバキュロウイルスシステムは、溶原性ウイルスであるので、採取時期の選択は生産物の収量にとって極めて重要である。
【0073】
適当な宿主細胞としては、酵母、細菌、古細菌、真菌、ならびに昆虫および哺乳動物細胞を含む動物細胞が含まれる。特に興味の持たれるのはDrosophila melanogaster細胞、Saccharomyces cerevisiaeおよび他の酵母、E. coli, Bacillus subtilis、SF9細胞、C129細胞、293細胞、ニューロスポラ、BHK、CHO、COS、HeLa細胞、繊維芽細胞、シュワン腫細胞株、不死骨髄およびリンパ細胞であり、HeLaおよびSF9細胞が特に好ましい。
【0074】
好ましい実施態様では、HDAC8タンパク質は哺乳動物細胞で発現される。哺乳動物の発現系もまた技術上公知であり、レトロウイルスシステムが含まれる。哺乳動物のプロモーターは、哺乳動物のRNAポリメラーゼを結合し、mRNA中のHDAC8タンパク質をコードする配列の下流(3'側)の転写を開始できる全てのDNAである。プロモーターは、通常コード配列の5'末端に近接して配置されている転写開始領域、および転写開始部位の25〜30塩基対上流に配置されているTATAボックスを有している。該TATAボックスは、RNA合成を正しい部位からRNA合成を開始するようにRNAポリメラーゼIIを誘導すると考えられている。哺乳動物のプロモーターはまた、典型的にTATAボックスの上流100〜200塩基対以内に位置している上流のプロモーターエレメント(エンハンサーエレメント)を含有している。上流のプロモーターエレメントは、転写開始速度を決定し、そしてどちら向きにも作用する。哺乳動物プロモーターで特に有用なのは哺乳動物ウイルスの遺伝子由来のプロモーターであるが、これはウイルスの遺伝子はしばしば高度に発現され、また宿主範囲が広いためである。例としては、SV40の初期プロモーター、マウス乳癌ウイルスのLTRプロモーター、アデノウイルスの主要後期プロモーター、単純ヘルペスウイルスのプロモーター、およびCMVのプロモーターが挙げられる。
【0075】
典型的には、哺乳動物細胞によって認識される転写終結配列およびポリアデニル化配列は、転写終結コドンの3'側に位置する制御領域であり、従って、プロモーターエレメントと共にコード配列に隣接している。成熟mRNAの3'末端は、部位特異的な翻訳後切断およびポリアデニル化により形成される。転写ターミネーターおよびポリアデニル化シグナルの例としてはSV40由来のものが含まれる。
【0076】
外来性核酸を哺乳動物宿主およびその他の宿主中に導入する方法は技術上周知であり、用いる宿主細胞により異なるであろう。この技術としては、デキストラン仲介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン仲介性トランスフェクション、プロトプラスト融合、電気穿孔法、ウイルス感染、ポリヌクレオチド(類)のリポソームへの封入、およびDNAの核内への直接マイクロインジェクションが挙げられる。
【0077】
好ましい実施態様では、HDAC8タンパク質は細菌のシステムで発現される。細菌の発現システムは技術上周知である。
【0078】
適当な細菌のプロモーターは、細菌のRNAポリメラーゼを結合し、HDAC8タンパク質の下流(3'側)のコード配列の転写を開始できるすべての核酸配列である。細菌のプロモーターは転写開始領域を有し、これは通常コード配列の5'末端に隣接している。この転写開始領域は典型的に、RNAポリメラーゼ結合部位と転写開始部位を含んでいる。代謝経路上の酵素をコード化する配列は、特に有用なプロモーター配列を提供する。例としては、ガラクトース、乳糖および麦芽糖のような糖代謝酵素由来のプロモーター配列、およびトリプトファンのような物質の生合成にかかわる酵素由来の配列が挙げられる。バクテリオファージ由来のプロモーターも使用でき、技術上公知である。加えて、合成プロモーターおよびハイブリッドプロモーターも有用である;例えば、tacプロモーターはtrpおよびlacプロモーター配列のハイブリッドである。さらに、細菌のプロモーターには、細菌起源でないが細菌のRNAポリメラーゼを結合し転写を開始する能力のある天然産生のプロモーターを含めることができる。
【0079】
機能的プロモーター配列に加えて、効率の良いリボゾーム結合部位が望ましい。E. coliでは、リボゾーム結合部位はシャイン・ダルガルノ(SD)配列と呼ばれ、開始コドン、および開始コドンの約3〜11ヌクレオチド上流にある約3〜9ヌクレオチド長の配列を含む。
【0080】
発現ベクターも、細菌中のHDAC8タンパク質の分泌を起こさせるシグナルペプチド配列を含有していてもよい。技術上周知のように、シグナル配列は典型的に、細胞からのタンパク質の分泌を指令する疎水性アミノ酸よりなるシグナルペプチドをコードする。タンパク質は、成育培地中(グラム陽性細菌)、もしくは細胞の内膜と外膜の間に位置する周辺腔内(グラム陰性細菌)に分泌される。
【0081】
細菌の発現ベクターはまた、形質転換した細菌株の選択を可能にするために、選択マーカー遺伝子を含んでよい。適当な選択遺伝子は、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン、ネオマイシンおよびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含む。選択マーカーはまた、ヒスチジン、トリプトファンおよびロイシンの生合成経路上の遺伝子のような生合成遺伝子を含む。
【0082】
これらの成分は発現ベクターに組み立てて入れる。細菌用の発現ベクターは技術上周知であり、特にBacillus subtilis、E. coli、Streptococcus cremoris、およびStreptococcus lividans用のベクターが含まれる。
【0083】
細菌の発現ベクターは、塩化カルシウム処理、電気穿孔法、その他のような技術上周知の技術を用いて細菌宿主細胞に形質転換される。
【0084】
ある実施態様では、HDAC8タンパク質は昆虫細胞中に生産される。昆虫細胞の形質転換用発現ベクター、特にバキュロウイルスに基づく発現ベクターは技術上周知である。
【0085】
好ましい実施態様では、HDAC8タンパク質は酵母細胞中に生産される。酵母の発現システムは技術上周知であり、Saccharomyces cerevisiae、Candida albicansおよびC. maltosa、Hansenula polymorpha、Kluyveromyces fragilisおよびK. lactis、Pichia guillerimondiiおよびP. pastoris、Schizosaccharomyces pombe、ならびにYarrowia lipolytica用の発現ベクターを含む。酵母での発現用に好ましいプロモーター配列としては、誘導性GAL1、10プロモーターおよびアルコール脱水素酵素、エノラーゼ、グルコキナーゼ、グルコース−6リン酸イソメラーゼ、グリセルアルデヒド−3リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ホスホフラクトキナーゼ、3−ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、および酸ホスファターゼ遺伝子由来のプロモーターが挙げられる。酵母の選択マーカーとしては、ツニカマイシン耐性を付与するADE2、HIS4、LEU2、TRP1、およびALG7、G418に対する耐性を付与するネオマイシンホスホトランスフェラーゼ遺伝子、および銅イオン存在下における酵母の生育を可能にするCUP1遺伝子が挙げられる。
【0086】
HDAC8タンパク質は、当分野で周知の技術を用いて、融合タンパク質としても作成され得る。ゆえに、例えば、モノクローナル抗体作成のために、もし目的のエピトープが小さければ、そのHDAC8タンパク質を、キャリアータンパク質と融合させて免疫原を形成させ得る。あるいは、HDAC8タンパク質を、発現の増加、または他の目的のために、融合タンパク質として作成し得る。例えば、HDAC8タンパク質がHDAC8ペプチドであるとき、そのペプチドをコードする核酸は、発現の目的のために、他の核酸と連結され得る。同様に、本発明のHDAC8タンパク質を、グリーン蛍光タンパク質(GFP)、レッド蛍光タンパク質(RFP)、ブルー蛍光タンパク質(BFP)、イエロー蛍光タンパク質(YFP)などのような、タンパク質の標識と連結させることができる。
【0087】
ある実施態様では、本発明のHDAC8核酸、タンパク質および抗体を標識化する。本明細書中で「標識化」とは、ある化合物が、その化合物の検出を可能にするように付加された少なくとも一成分、同位元素または化合物、を有することを意味する。一般的に、標識は次の3つの分類に分けられる:a)放射活性を有する、または重同位元素であり得る、同位元素標識;b)抗体または抗原であり得る、免疫標識;c)有色または蛍光染料。これらの標識は化合物のどの位置にでも組み込まれ得る。
【0088】
好ましい実施態様では、該HDAC8タンパク質は発現後精製または単離される。HDAC8タンパク質は、どのような他の成分が試料中に存在するかに依存して、当業者周知の種々の方法で単離または精製し得る。標準的な精製法としては、電気泳動的、分子的、免疫学的技法ならびにイオン交換、疎水、アフィニティー、および逆相HPLCクロマトグラフィーを含むクロマトグラフィー技法、ならびにクロマトフォーカシングが挙げられる。例えば、HDAC8タンパク質は、標準的な抗HDAC8抗体カラムを用いて精製することができる。タンパク質濃縮と組合わせた限外ろ過および膜分離精製法も有用である。適当な精製技法の一般的手引書としては、Scopes, R., Protein Purification, Springer-Verlag, NY (1982)を参照のこと。必要な精製の程度はHDAC8タンパク質の用途によって変わるであろう。場合によっては精製は不要である。
【0089】
一旦発現され、必要なら精製されると、HDAC8タンパク質および核酸は、多数の応用において有用である。
【0090】
HDAC8タンパク質類をコードしているこのヌクレオチド配列(またはそれらの相補鎖)には、分子生物学の分野で、ハイブリダイゼーションプローブとしての、クロモソームおよび遺伝子マッピングにおける、およびアンチ−センスRNAおよびDNAの生成における利用を含む、多様な応用がある。HDAC8タンパク質核酸は、本明細書中に記載した組換え技術によるHDAC8タンパク質類の調製にも有用であろう。
【0091】
天然の完全長配列のHDAC8タンパク質遺伝子またはその各部分は、該HDAC8タンパク質をコードしている配列との所望の配列同一性を有する他の遺伝子(例えば、天然に産生するHDAC8タンパク質の変種または他種由来のHDAC8タンパク質をコードしている遺伝子)を単離するために、cDNAライブラリーについてのハイブリダイゼーションプローブとして使用し得る。場合により、これらのプローブの長さは約20ないし約50塩基である。これらのハイブリダイゼーションプローブは、本明細書中のヌクレオチド配列から、または本明細書中に提供されたような天然配列のプロモーター、エンハンサーエレメントおよびイントロンを含むゲノム配列から誘導し得る。例をあげれば、スクリーニング方法は、既知DNA配列を使用して約40塩基の選択プローブを合成する、HDAC8タンパク質遺伝子のコード領域の単離を含む。ハイブリダイゼーションプローブは、32Pまたは35Sのような放射性ヌクレオチド類、またはアビジン/ビオチンカップリング系を介して該プローブと連結したアルカリホスファターゼのような酵素標識を含む各種標識により標識化し得る。本発明のHDAC8タンパク質遺伝子の配列と相補的な配列を有する標識化したプローブは、ヒトcDNA、ゲノムDNAまたはmRNAの各ライブラリーをスクリーニングし、該プローブがそれらのライブラリーのどのメンバーとハイブリダイズするかを測定するのに使用できる。
【0092】
HDAC8タンパク質をコードしているヌクレオチド配列は、該HDAC8タンパク質をコードしている遺伝子のマッピングのため、および遺伝的異常がある個体の遺伝子分析のためにハイブリダイゼーションプローブを構築するのにも使用し得る。本明細書中に記載したこれらのヌクレオチド配列は、既知技術、例えば、その場でのハイブリダイゼーション、既知染色体マーカーに対する連結分析、およびライブラリーについてのハイブリダイゼーションスクリーニングを使用して、染色体や染色体の特定領域に対してマッピングし得る。
【0093】
HDAC8タンパク質またはその修飾形態をコードしている核酸は、トランスジェニック動物または「ノックアウト」動物を発生させるのにも使用でき、ひいては治療的有用物質の開発およびスクリーニングにおいても有用である。トランスジェニック動物(例えば、マウスまたはラット)とは、導入遺伝子含有細胞を有する動物であり、該導入遺伝子が該動物または該動物の祖先に胎内期(prenatal)、例えば、胚期(embryonic stage)において導入されたものである。導入遺伝子とは、それからトランスジェニック動物が発育する細胞のゲノム中に集積されたDNAである。ある実施態様では、HDAC8タンパク質をコードしているcDNAは、確立された技術に従ってHDAC8タンパク質をコードしているゲノムDNAをクローン化するのに使用でき、該ゲノム配列は、所望のDNAを発現する細胞を含有するトランスジェニック動物の発生に使用される。トランスジェニック動物、殊にマウスまたはラットのような動物を発生させる方法は、当技術分野で日常化してきており、例えば、米国特許番号第4736866号および第4870009号に記載されている。典型的には、特定の細胞が、組織特異的エンハンサーによるHDAC8タンパク質導入遺伝子導入の標的になる。胚期に該動物の生殖ライン中に導入された、HDAC8タンパク質をコードしている導入遺伝子のコピーを含むトランスジェニック動物は、所望の核酸の増加した発現の効果を調べるのに使用できる。それらの動物は、例えば、その過剰発現に伴う病理的状況からの保護をもたらすと考えられる物質に対するテスター動物として使用できる。本発明のこの側面によれば、動物を該物質で処理し、該導入遺伝子を有する非処理動物と対比した病理的状況の減少効果が、該病理的状況に対する治療的干渉のポテンシャルを示す。
【0094】
あるいは、このHDAC8タンパク質の非ヒト相同体をHDAC8タンパク質「ノックアウト」動物を構築するのに使用することができる。このノックアウト動物は、HDAC8タンパク質をコードしている内因性遺伝子と、該動物の胚細胞中に導入されたHDAC8タンパク質をコードしている変化させたゲノムDNAとの間の、相同組換えの結果HDAC8タンパク質をコードする遺伝子が欠落または変化している動物である。例えば、HDAC8タンパク質をコードしているcDNAを、HDAC8タンパク質をコードしているゲノムDNAのクローン化に、確立された技術に従って使用することができる。HDAC8タンパク質をコードしているゲノムDNAの一部は、削除したり、または他の遺伝子例えば、モニター集積に使用し得る選択マーカーをコードしている遺伝子と置き換えたりすることができる。典型的には、数キロ塩基の(5'または3'末端の両方における)不変フランキング(側位)DNAを、ベクター中に含める[相同組換えベクター類については、例えば、Thomas and Capecchi, Cell, 51:503 (1987) の既述を参照]。このベクターを、胚幹細胞系中に(例えば、電気穿孔法により)導入し、そして、導入したDNAが内因性DNAと相同的に組換えられている細胞を選択する[例えば、Li et al., Cell, 69:915 (1992) 参照]。選択した細胞を次いで動物(例えば、マウスまたはラット)の胚盤胞中に注入し、キメラ集合体を形成させる[例えば、Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp.113-152参照]。キメラ胚を、次いで適切な偽妊娠雌フォスター動物中に移植し、該胚を「ノックアウト」動物作成期に導く。生殖細胞中にこの相同組換えDNAを内包する後代は、標準的技術で同定することができ、そして該動物の全細胞が相同的に組換えられたDNAを含有する動物を産出することに使用できる。ノックアウト動物には、例えば、ある種の病理学的状況に対して防御する能力があることや、HDAC8タンパク質の不存在により病理学的状況が進展することなどが特徴であると言える。
【0095】
本明細書中に記載したモデルは変更可能と理解される。例えば、「ノックイン」モデルを作成することができ、あるいは該モデルを動物モデルよりむしろ細胞ベースとすることができる。
【0096】
これらのHDAC8ポリペプチドをコードしている核酸、アンタゴニストまたはアゴニストは、遺伝子治療にも使用できる。遺伝治療への応用では、遺伝子を細胞内に導入し、例えば欠陥遺伝子置換のための、治療的に有効な遺伝子産物のインビボ合成を達成させる。「遺伝子治療」には、単回の処置により持続的効果を達成させる在来の遺伝子治療と、治療的に有効なDNAまたはmRNAの単回または反復した投与を含む、遺伝子治療的物質の投与との両方が含まれる。アンチセンスRNAおよびDNAを、インビボである種の遺伝子の発現を阻止する治療的物質として使用できる。短いアンチセンスオリゴヌクレオチドが細胞内に導入され、細胞膜による取り込みが制限されることにより細胞内濃度が低いにもかかわらず、そこで阻害物質として働き得ることが既に示されている(Zamecnik et al., Proc. Natl. Acad. Sci. USA 83, 4143-4146 [1986])。これらのオリゴヌクレオチドは、修飾、例えばそれらの負荷電ホスフォジエステル基を非荷電基と置換、してそれらの取り込みを増強することができる
【0097】
生存細胞内に核酸類を導入するには各種の技術が利用できる。これらの技術は、核酸がインビトロで培養細胞内に移入されるのか、またはインビボで意図する宿主の細胞内に移入されるのかによって変る。インビトロで哺乳動物細胞内に核酸を移入するのに適した技術には、リポソーム、電気穿孔法、マイクロインジェクション、細胞癒合、DEAE−デキストラン、リン酸カルシウム沈殿法、等の使用が含まれる。現今好適とされているインビボ遺伝子移入技術には、ウイルス(典型的にはレトロウイルス)ベクターを用いるトランスフェクション、およびウイルス被覆タンパク質-リポソーム媒介トランスフェクションが含まれる(Dzau et al., Trends in Biotechnology 11, 205-210 [1993])。ある状況では、標的細胞を標的とする物質、例えば、細胞表面膜タンパク質または標的細胞に特異的な抗体、標的細胞上のレセプターに対するリガンドを有する核酸源を供給するのが望ましい。リポソームを採用した場合は、エンドサイトーシスに伴って細胞表面膜タンパク質に結合するタンパク質、例えば特定の細胞型に親和性のカプシドタンパク質またはその断片、サイクリング中に内在化を受けるタンパク質に対する抗体、細胞間局在化を標的とし細胞内半減期を強化するタンパク質、を標的化および/または取り込み増強のために使用することができる。レセプター媒介エンドサイトーシスの技術は、例えば、Wu et al., J. Biol. Chem. 262,4429-2232 (1987): および Wagnernet al., Proc. Natl. Acad. Sci. USA 87, 3410-3414(1990) に記載されている。遺伝子マーキングおよび遺伝子治療プロトコールについての総説 Andersen et al., Science 256, 808-813 (1992)参照。
【0098】
好ましい態様では、HDAC8タンパク質、核酸、変異種、修飾タンパク質、該核酸またはタンパク質を含有する細胞および/またはトランスジェニック体をスクリーニングアッセイに使用する。本明細書中に記載したHDAC8タンパク質の同定により、HDAC8タンパク質に結合するかまたはその結合を妨げる化合物についての、およびHDAC8活性を制御する化合物についての薬剤スクリーニングアッセイの設計が可能となる。
【0099】
本明細書に記載したアッセイでは、好ましくはヒトHDAC8タンパク質を利用するが、齧歯類(マウス、ラット、ハムスター、モルモット、その他)、家畜類(牛、羊、豚、馬、その他)および霊長類を含む他の哺乳動物のタンパク質も使用できる。これらの後者の実施態様はヒトの疾患の動物モデルの開発に好適であろう。ある実施態様では、本明細書中で概説したように、変異体または誘導体HDAC8タンパク質、上記したようなHDAC8タンパク質の除去を含めて、を使用してもよい。
【0100】
好ましい態様では、本発明の方法は、HDAC8タンパク質と候補バイオ活性物質とを合わせること、および該HDAC8タンパク質への該候補物質の結合を測定することを含む。他の態様では、以下に詳論するが、結合の阻害またはバイオ活性を測定する。
【0101】
本明細書において「候補バイオ活性物質」または「外来性化合物」とは、例えば、タンパク質、小型の有機分子、炭水化物(多糖類を含む)、ポリヌクレオチド、脂質などのいずれかの分子をいう。一般に、複数のアッセイ混合物を種々の薬物濃度を用いて並行して流し、種々の濃度に対しての種々の示差応答を得る。典型的には、これらの濃度のうち1つ、即ち0または検出レベルに満たない濃度がネガティブコントロールとして用いられる。さらに、ポジティブコントロールを用いてもよい。即ち、HDAC8活性を変化させるか調節すると知られている薬剤の使用である。
【0102】
候補物質は多くの化学クラスを包含するが、典型的にはそれらは有機分子、好ましくは分子量が100ダルトンより大きく約2,500ダルトン未満の小型の有機化合物である。候補物質はタンパク質との構造相互作用、特に水素結合に必要な官能基を含み、典型的には少なくとも1つのアミン、カルボニル、ヒドロキシルまたはカルボキシル基、好ましくは少なくとも2つの官能化学基を含む。該候補物質は環式炭素または複素環式構造および/または上記の1以上の官能基で置換された芳香族もしくはポリ芳香族構造を含む。候補物質はまた、ペプチド、糖類、脂肪酸、ステロイド、プリン、ピリミジン、誘導体、構造類似体またはそれらの組合せを含む生体分子間に見出せる。特に好ましいのはペプチドである。
【0103】
候補物質は、合成または天然化合物のライブラリーを含む多様な供給源から得られる。例えば、ランダム化オリゴヌクレオチドの発現をはじめ、多様な有機化合物および生体分子のランダムな、また所定の合成には多くの手段が利用できる。あるいは、細菌、真菌、植物および動物の抽出物形態における天然化合物のライブラリーが入手可能であるか、または容易に作製できる。さらに、天然の、または合成により作製したライブラリーまたは化合物は従来の化学的、物理的、生化学的手段により容易に修飾される。公知の薬理活性物質は、アシル化、アルキル化、エステル化、アミド化などの所定のまたはランダムな化学修飾を受けて構造類似体を形成し得る。
【0104】
好ましい態様では、種々の候補バイオ活性物質のライブラリーを用いる。好ましくは、該ライブラリーはランダム化された物質の、構造的に十分多様な集団を提供して、特定の標的との結合を可能にするために十分と見込まれる範囲の多様性を達成する。従って、相互作用ライブラリーは、メンバーの少なくとも1つが、標的に対する親和性を有する構造を持つよう、十分大きくなければならない。相互作用ライブラリーに要求される絶対的なサイズを測るのは難しいが、自然が免疫応答でヒントを与えてくれる。即ち、多様な107ないし108個の異なる抗体は、生物が直面する大部分のあり得る抗原と相互作用するに十分な親和性を有する少なくとも1つの組合せを与える。また、公表されているインビトロ選択技術も、107ないし108のライブラリーサイズが、標的への親和性を有する構造を見出すに十分であることを示している。本明細書で一般に提案されるものなど7ないし20アミノ酸長のペプチドのすべての組合せに関するライブラリーは、207(109)ないし2020をコードする可能性を有する。従って、107ないし108の異なる分子のライブラリーについて、本方法では、理論的に完全な相互作用ライブラリーの「実働」サブセットをアミノ酸7個と見積り、形態サブセットを2020ライブラリーと見積もる。このように好ましい態様では、主題の方法において少なくとも106、好ましくは107、より好ましくは少なくとも108、最も好ましくは少なくとも109個の異なる配列が同時に解析される。好ましい方法はライブラリーのサイズおよび多様性を最大にする。
【0105】
好ましい実施態様では、該候補バイオ活性物質はタンパク質である。本明細書において「タンパク質」とは、共有結合した少なくとも2つのアミノ酸を意味し、タンパク質、ポリペプチド、オリゴペプチドおよびペプチドが含まれる。該タンパク質は、天然産生のアミノ酸およびペプチド結合または合成ペプチド模倣構造からなってもよい。従って本明細書において「アミノ酸」または「ペプチド残基」とは、天然産生のアミノ酸と合成アミノ酸の双方を意味する。例えば、ホモフェニルアラニン、シトルリンおよびノルロイシンは、本発明の目的のためのアミノ酸と考えられる。また「アミノ酸」には、プロリンおよびヒドロキシプロリンなどのイミノ酸残基が含まれる。その側鎖は(R)配置または(S)配置のいずれであってもよい。好ましい態様では、該アミノ酸は(S)またはL配置である。天然産生ではない側鎖を用いる場合は、例えばインビボ分解を妨げるまたは遅延させるために非アミノ酸置換基を用いてもよい。また、化学保護基またはその他の化学置換基を付加してもよい。
【0106】
好ましい態様では、候補バイオ活性物質は天然産生のタンパク質または天然産生のタンパク質の断片である。従って、例えば、タンパク質またはタンパク質性細胞抽出物のランダムなもしくは所定の消化物を含む、細胞抽出物を用いてもよい。このように、本明細書に記載の系のスクリーニングのためには、原核生物および真核生物のタンパク質ライブラリーを作製すればよい。本態様において特に好ましいのは、細菌、真菌、ウイルスおよび哺乳類タンパク質のライブラリーであり、後者が好ましく、ヒトタンパク質が特に好ましい。
【0107】
好ましい態様では、該候補バイオ活性物質は、約5ないし約30のアミノ酸からなるペプチドであり、約5ないし約20のアミノ酸からなるものが好ましく、約7ないし約15のアミノ酸からなるものが特に好ましい。該ペプチドは上記に概説したような天然産生のタンパク質の消化物、ランダムペプチド、または「偏りのある」ランダムペプチドであってもよい。本明細書において「ランダム化された」または文法的均等語は、核酸およびペプチドが、それぞれ本質的にランダムなヌクレオチドおよびアミノ酸からなることを意味する。一般に、これらのランダムペプチド(または以下に論じる核酸)は化学的に合成されるので、いかなる位置にいかなるヌクレオチドまたはアミノ酸をも組み込める。この合成法は、ランダム化されたタンパク質または核酸を作成し、配列全長にわたって、あり得る組合せのすべてまたは大部分の形成を可能にし、ランダム化されたタンパク質性候補バイオ活性物質のライブラリーを形成するよう設計することができる。
【0108】
ある実施態様では、該ライブラリーは完全にランダム化され、いかなる位置にも優先のまたは一定の配列は存在しない。好ましい態様では、該ライブラリーには偏りがある。即ち、配列内のある位置では一定であるか、または限られた数の可能性から選択される。例えば、好ましい態様では、ヌクレオチドまたはアミノ酸残基は、例えば、疎水性アミノ酸、親水性残基、立体的に偏りのある(小さいか、または大きい)残基、架橋のためのシステイン、SH−3ドメインのためのプロリン、リン酸化部位のためのセリン、トレオニン、チロシンもしくはヒスチジンの作出などに対して、またはプリンに対してといった所定のクラス内でランダム化される。
【0109】
好ましい態様では、該候補バイオ活性物質は核酸である。本明細書において「核酸」もしくは「オリゴヌクレオチド」または文法的均等語は、共有結合した少なくとも2つのヌクレオチドを意味する。本発明の核酸は一般にホスホジエステル結合を含むが、以下に概説するように、例えば、ホスホルアミド(Beaucage, et al., Tetrahedron, 49(10):1925 (1993)およびその参照文献; Letsinger, J. Org. Chem., 35:3800 (1970); Sprinzl, et al., Eur. J. Biochem., 81:579 (1977); Letsinger, et al., Nucl. Acids Res., 14:3487 (1986); Sawai, et al., Chem. Lett., 805 (1984); Letsinger , et al., J. Am. Chem. Soc., 110:4470 (1988); およびPauwels, et al., Chemica Scripta, 26:141 (1986))、ホスホロチオエート(Mag, et al., Nucleic Acids Res., 19:1437 (1991);および米国特許番号第5644048号)、ホスホロジチオエート(Briu, et al., J. Am. Chem. Soc., 111:2321 (1989))、O−メチルホスホロアミダイト結合(Eckstein , Oligonucleotides and Analogues: A Practical Approach, Oxford University Press参照)、ならびにペプチド核酸主鎖および結合(Egholm, J. Am. Chem. Sci., 114:1895 (1992); Meie, et al., Chem. Int. Ed. Engl., 31:1008 (1992); Nielsen, Nature, 365:566 (1993); Carisson, et al., Nature, 380:207 (1996)参照。なお、これらは出典明示により本明細書の一部とする)を含む、代替的な主鎖を有し得る核酸類似体が含まれる場合がある。その他の核酸類似体としては、正電荷主鎖(Denpcy, et al., Proc. Natl. Acad. Sci. USA, 92:6097 (1995));非イオン性主鎖(米国特許番号第5386023号、第5637684号;第5602240号;第5216141号;および第4469863号; Kiedroeshi, et al., Angew. Chem. Intl. Ed. English, 30;423 (1991); Letsinger, et al., J. Am. Chem. Sci., 110:4470 (1988); Letsinger, et al., Nucleoside & Nucleotide, 13:1597 (1994); Chapters 2 and 3, ASC Symposium Series 580, "Carbohydrate modifications in Antisense Research", Ed. Y.S. Sanghui and P. Dan Cook; Mesmaeker, et al., Bioorganic & Medicinal Chem. Lett., 4:395 (1994); Jeffs, et al., J. Biomolecular NMR, 34:17 (1994); Tetrahedron Lett., 37:743 (1996))、ならびに米国特許第5235033号、第5034506号、およびChapters 6 and 7, ASC Symposium Series 580, "Carbohydrate Modifications in Antisense Research", Ed. Y.S. Sanghui and P.Dan Cookに記載のものをはじめとする非リボース主鎖を有するものが挙げられる。1以上の炭素環式糖を含む核酸もまた、本核酸定義内に含まれる(Jenkins, et al., Chem. Soc. Rev., (1995) pp. 169-176参照)。いくつかの核酸類似体が、Rawls, C & E News, June 2, 1997, page 35に記載されている。これらの参照文献は、すべて出典明示により本明細書の一部とする。リボース−リン酸主鎖のこれらの修飾を行い、標識などの付加部分の付加を助けたり、あるいは生理学的環境下でかかる分子の安定性を高め、半減期を延ばしてもよい。さらに、天然産生の核酸と類似体の混合物を作成することもできる。あるいは、種々の核酸類似体の混合物や天然産生の核酸と類似体の混合物を作製してもよい。該核酸は明示したように1本鎖であっても2本鎖であってもよく、あるいは2本鎖または1本鎖配列の双方の部分を含んでもよい。該核酸は、デオキシリボヌクレオチドとリボヌクレオチドのいずれかの組合せ、およびウラシル、アデニン、チミン、シトシン、グアニン、イノシン、キサンチン、ヒポキサンチン、イソシトシン、イソグアニンなどを含む塩基のあらゆる組合せを含む、DNA(ゲノムDNAおよびcDNAの双方)、RNAまたはハイブリッドであってもよい。
【0110】
一般にタンパク質に関して上記したように、核酸候補バイオ活性物質は天然産生の核酸、ランダム核酸、または「偏りのある」ランダム核酸であってよい。例えば、タンパク質に関して先に概説したように原核生物ゲノムまたは真核生物ゲノムの消化物を用いてもよい。
【0111】
好ましい態様では、該候補バイオ活性物質は有機化学部分であり、多様なものが文献から得られる。
【0112】
好ましい態様では、候補バイオ活性物質は融合パートナーに結合している。本明細書において「融合パートナー」または「機能的グループ」とは、候補バイオ活性物質と会合し、そのクラスのライブラリーのすべてのメンバーに、共通の機能または能力を付与する配列を意味する。融合パートナーは異種(即ち、宿主にとり天然でない)、または合成のもの(いずれの細胞にとっても天然でない)であり得る。好適な融合パートナーとしては、限定されるものではないが、a)候補バイオ活性物質を立体配位上限定的なまたは安定な形で提供する提示構造、b)候補バイオ活性物質を細胞内または細胞外区画へ局在化させる標的配列、c)候補バイオ活性物質またはそれらをコードする核酸のいずれかの精製または単離を可能にするレスキュー配列、d)候補バイオ活性物質またはそれをコードする核酸に、安定性または分解からの保護、例えばタンパク質分解耐性を付与する安定化配列、e)ペプチド二量化を可能にする二量化配列、あるいはf)a)、b)、c)、d)およびe)ならびに要すればリンカー配列のいずれかの組合せ、が挙げられる。
【0113】
本明細書に記載した方法のある実施態様では、HDAC8タンパク質の各部分を使用する;好ましい態様では、HDAC8活性を有する部分を使用してHDAC8に結合する物質を同定する。加えて、本明細書中に記載したアッセイでは、単離したHDAC8タンパク質またはHDAC8タンパク質を含む細胞を利用し得る。
【0114】
該して言えば、本発明の方法の好ましい態様では、例えば結合アッセイのために、HDAC8タンパク質または候補物質を、単離試料受容領域を有する不溶性支持体(例えば、マイクロタイタープレート、アレイ、その他)に非拡散的に結合させる。これらの不溶性支持体は、本発明の組成物がそれに結合することができ、容易に可溶性材料から分離でき、そしてスクリーニングの全体方法と他の点では適合性である任意の組成で作成できる。それらの支持体の表面は固体または多孔質であり、任意の都合のよい形状であり得る。好適な不溶性支持体の例には、マイクロタイタープレート、アレイ、膜およびビーズが含まれる。これらは、典型的にはガラス、プラスチック(例えば、ポリスチレン)、ポリサッカライド、ナイロンまたはニトロセルロース、テフロン(登録商標)、その他で作成する。マイクロタイタープレートおよびアレイは、多数のアッセイを同時に実施でき、試薬や試料の使用量が少なくてすむので特に好都合である。ある場合には、磁性ビーズおよび類似物が含まれる。組成物結合の個々の様式は、本発明の試薬類および全体方法と適合性であり、組成物の活性を維持し、そして非拡散的である限り、厳密なものではない。好ましい結合方法には、抗体(それは、該タンパク質が支持体に結合したとき、リガンド結合部位または活性化配列のいずれをも立体的にブロックしないものである)の使用、「粘着性」またはイオン性支持体への直接結合、化学的架橋結合、表面上での該タンパク質または物質の合成、その他が含まれる。該タンパク質または物質の結合につづいて、過剰の未結合材料を洗浄により除去する。試料受容領域を次いで牛血清アルブミン(BSA)、カゼインまたは他の無害なタンパク質または他の部位とインキュベートしてブロックする。本発明には、固体支持体を使用しないスクリーニングアッセイも含まれる:その例は以下に述べる。
【0115】
好ましい実施態様では、HDAC8タンパク質を支持体に結合させ、それから候補バイオ活性物質をアッセイに加える。あるいは候補物質を支持体に結合させ、それからHDAC8タンパク質を加える。新規な結合物質には、特殊な抗体、化学的ライブラリーのスクリーニングで同定された非天然産結合物質、ペプチド類似体などが含まれる。特に重要であるのは、ヒト細胞に対する毒性が低い物質に対するスクリーニングアッセイである。多様なアッセイがこの目的のために使用でき、それらには標識化インビトロタンパク質―タンパク質結合アッセイ、電気泳動シフトアッセイ、タンパク質結合に対する免疫アッセイ、機能的アッセイ、好ましくは短いアセチル化ペプチドまたは標識アセチル化ペプチドの脱アセチル化が含まれる。
【0116】
候補バイオ活性物質のHDAC8タンパク質への結合の測定は、各種の方法で実施できる。好ましい態様では、候補バイオ活性物質を標識化し、結合を直接測定する。例えば、これは、HDAC8タンパク質の全部または一部を固体支持体に結合させ、標識化(例えば、蛍光標識)候補物質を加え、過剰の物質を洗い去り、そして標識が固体支持体上に存在するかどうかを測定することにより実施できる。当分野で既知の各種ブロックおよび洗浄段階を利用することができる。
【0117】
本明細書中では「標識化」とは、その化合物が、検出可能なシグナルを提供する標識、例えば放射性同位元素、蛍光剤、酵素、抗体、磁性粒子のような粒子、化学的発光剤、または特異的結合分子、その他、により、直接的かまたは間接的かのいずれかで標識されていることを意味する。特異的結合分子には、ビオチンとストレプトアビジン、ジゴキシンとアンチジゴキシンのような対、その他、が含まれる。特異的結合メンバーでは、前記概説のように、相補的メンバーは通常、既知操作により検出のための分子により標識化されている。標識は直接または間接的に検出可能なシグナルを提供する。
【0118】
ある実施態様では、成分のひとつだけを標識化する。例えば、タンパク質(またはタンパク質性候補物質)をチロシン位で、125Iまたは発蛍光団を用いて標識化することができる。あるいは、1以上の成分を異なる標識で;例えばタンパク質は125Iを用いて標識化し、候補物質は発蛍光団で標識化してもよい。
【0119】
好ましい態様では、候補バイオ活性物質の結合は、競合的結合アッセイを使用して測定できる。この実施態様では、競合物質は標的分子と結合することが既知の結合部分(即ち、HDAC8タンパク質)、例えば、抗体、結合パートナー、リガンド、その他である。ある状況では、バイオ活性物質が結合部位と置き換わる、バイオ活性物質と結合部位間でのような競合的結合があり得る。このアッセイは、HDAC8タンパク質と結合パートナー間の結合を妨害する候補物質を測定するために使用することができる。本明細書中では「結合の妨害(interference)」とは、HDAC8タンパク質の本来の結合が、候補物質の存在下に異なることを意味する。この結合は除去することができるか、またはその親和性を低減することができる。それ故に、ある実施態様では、妨害は例えば、本来の結合部位に対する直接競合というよりむしろ立体配置の変化により生じるものである。
【0120】
ある実施態様では、候補バイオ活性物質を標識化する。候補バイオ活性物質または競合物質のいずれか、または両方を、まずこのタンパク質に、それが存在するならば充分結合できるだけの時間加える。インキュベーションは、最適活性を促す任意の温度、典型的には4ないし40℃で実施できる。インキュベーションの時間は、最適活性となるように選択するが、迅速高度生産スクリーニングを促すように最適化してもよい。典型的には0.1ないし1時間で充分であろう。過剰の試薬は一般的に除去するかまたは洗い去る。次に、第2の成分を加え、結合を示す標識化成分の存在または不存在を調べる。
【0121】
好ましい態様では、競合物質を最初に加え、その後候補バイオ活性物質を加える。競合物質が置換することは、候補バイオ活性物質がHDAC8タンパク質と結合していること、従ってHDAC8タンパク質活性に対して、それと結合しそれを強力に調節する能力があることを示す指標である。この実施態様では、両成分を標識化し得る。従って、例えば、競合物質を標識化した場合、洗浄液中に標識が存在することは該物質による置換があったことを示す。あるいは、候補バイオ活性物質を標識化した場合、支持体上の標識の存在は置換があったことを示す。
【0122】
別の実施態様では、候補バイオ活性物質を最初に加え、インキュベーションし洗浄し、その後競合物質を加える。競合物質による結合の不存在は、該バイオ活性物質がHDAC8タンパク質とより高度な親和性で結合していることの証しとなる。かくて、候補バイオ活性物質を標識化した場合は、支持体上の標識の存在が、競合物質結合がないために、該候補物質がHDAC8タンパク質と結合する能力があることを示すこととなる。
【0123】
好ましい実施態様では、本発明の方法は、HDAC8タンパク質の活性を調節し得る候補バイオ活性物質を同定するための分別スクリーニングを含む。そのようなアッセイは、HDAC8タンパク質または該HDAC8タンパク質を含んでいる細胞により実施できる。ある実施態様では、本発明の方法は、あるHDAC8タンパク質と、第1試料中の競合物質とを合わせることを含む。第2試料には、候補バイオ活性物質、HDAC8タンパク質および競合物質を含める。競合物質の結合を両方の試料について測定すれば、2試料間の結合の変化または差異が、HDAC8タンパク質と結合しその活性を強力に調節する能力のある物質の存在を示す。即ち、競合物質の結合が第1試料と対比して第2試料中では異なるならば、その物質はHDAC8タンパク質と結合する能力がある。
【0124】
あるいは、好ましい実施態様は、天然産生のHDAC8タンパク質とは結合するが修飾HDAC8タンパク質とは結合できない薬物候補を同定するために分別スクリーニングを利用する。このHDAC8タンパク質の構造をモデルとし、該部位と相互作用する物質を合成するための理論的薬物設計に使用する。HDAC8のバイオ活性に作用する薬物候補も、該タンパク質の活性を増強または低減させる能力について薬物をスクリーニングすることによって同定される。
【0125】
これらのアッセイでは、ポジティブコントロールおよびネガティブコントロールを使用することができる。好ましくは、全てのコントロールおよびテスト試料は、統計的に有意な結果を得るために、少なくとも3部づつとする。全試料のインキュベーションは、その物質が該タンパク質に結合するのに充分なだけの時間行う。インキュベーションに続いて、全試料を、特異的結合をしていない材料がなくなるまで洗浄し、結合の通常標識された物質の量を測定する。例えば、放射性標識を採用した場合は、試料をシンチレーションカウンターでカウントして結合化合物の量を測定する。
【0126】
このスクリーニングアッセイには各種の他の試薬を含めることができる。それらには、塩類、アルブミンのような中性タンパク質、洗浄剤、その他のような試薬が含まれ、それらは最適のタンパク質−タンパク質結合を促す、および/または非特異的またはバックグラウンド相互作用を低減するために用いられる。このアッセイの効率を別の方法で改善する試薬、例えば、プロテアーゼインヒビター、ヌクレアーゼインヒビター、抗微生物剤なども使用できる。これらの成分の混合物は、必要な結合をもたらす任意の順序で添加できる。
【0127】
HDAC8タンパク質の活性を調節する物質についてのスクリーニングも実施できる。好ましい態様では、HDAC8の活性を調節し得るバイオ活性物質のためのスクリーニング方法は、候補バイオ活性物質をHDAC8タンパク質(またはHDAC8タンパク質を含む細胞)の試料に加える段階、および該HDAC8タンパク質の生物学的活性の変化を測定する段階を含む。「HDAC8タンパク質の活性を調節する」には、存在する活性の増加、活性の減少、または活性の型または種類の変化が含まれる。従って、この実施態様では、候補物質は、本明細書中で定義したように、HDAC8タンパク質と結合(これは多分不要であるが)し、かつ生物学的または生化学的活性を変更する。この方法は、一般的に上で説明したようなインビトロスクリーニング方法、およびHDAC8タンパク質の存在、分布、活性または量における変更についての細胞のインビボスクリーニング方法の両方を含む。
【0128】
従ってこの態様では、本発明の方法は、HDAC8試料と候補バイオ活性物質とを合わせること、およびそのHDAC8への影響を評価することを含む。本明細書中で「HDAC8タンパク質活性」またはその文法的均等語は、HDAC8タンパク質の生物学的活性の少なくともひとつを意味し、それには、クロマチン構造、ヒストンアセチル化、翻訳、細胞周期、細胞複製、細胞活性などでインビトロまたはインビボにおいて影響を与える、各能力が含まれるがそれらに限定されない。
【0129】
好ましい実施態様では、HDAC8タンパク質の活性が減少し;他の好ましい実施態様ではHDAC8タンパク質の活性が増加する。従って、ある実施態様ではアンタゴニストであるバイオ活性物質が好適であり、他の実施態様ではアゴニストであるバイオ活性物質が好適である。
【0130】
好ましい実施態様では、本発明は、HDAC8タンパク質の活性を調節する能力のあるバイオ活性物質のスクリーニング方法を提供する。この方法は、上記したように、HDAC8タンパク質を含む細胞に候補バイオ活性物質を加えることを含む。好適な細胞型には殆ど全ての細胞が含まれる。それらの細胞は、HDAC8タンパク質をコードしている組換え核酸を含有する。好ましい実施態様では、候補物質のライブラリーを複数の細胞について試験する。
【0131】
HDAC8制御の検出は、当業者には明らかなように実施できる。ある実施態様では、HDAC8活性のインディケーター、例えばリジンがアセチル化されたペプチドRTKRなどのアセチル化合成ペプチドを使用する。HDAC8制御における変更の検出を可能とするために評価またはアッセイするパラメータは多数あり、それらには、細胞生存力アッセイ、細胞が特定の細胞周期段階に留まっているかどうかを測定するアッセイ(「細胞増殖アッセイ」)、およびその細胞がどの細胞段階に留まっているかを決定するアッセイ(「細胞相(細胞期)アッセイ」)、が含まれるがそれらに限定されない。別のパラメーターとして、上記に記載のようなmRNA合成、翻訳、アセチル化ペプチドの脱アセチル化がある。これらのパラメータの1またはそれ以上をアッセイまたは測定することにより、HDAC8制御における変更のみならず、該HDAC8制御過程の異なる段階での変更をも検出することが可能となる。これは、天然産生細胞を評価するために、例えば、腫瘍細胞型の攻撃性を測るために、またはHDAC8制御に対する作用を試験中の候補薬物の作用を評価するために、実施してもよい。このようにして、候補物質の迅速で正確なスクリーニングを達成し、HDAC8制御を調節する物質を同定することができる。
【0132】
従って、本発明の組成物および方法はバイオ活性物質を明示するために有用である。この物質によって細胞または細胞集団が、そのクロマチン構造、ヒストンアセチル化、転写活性を変更するか、あるいはある増殖期から他の期(相)に移行するか、または増殖期(成育相)に留まることができる。ある実施態様では、細胞はある特定の増殖期に留まっているが、それらをその相から出て新しい相中に入れることが望まれる。あるいは、細胞をある相、例えばG1内に強制的に留まらせることが、細胞周期を経由して移行しつづけるよりもむしろ望ましいことがあろう。同様に、ある場合には、滞留してはいないが移行が緩慢な細胞集団を次の相へまたはHDAC8に忠実に沿って加速させること、または次相の開始を遅らせることが望ましいこともあろう。例えば、ある種の酵素、例えばキナーゼ、ホスファターゼ、プロテアーゼの活性を変更させること、または酵素をユビキチン化させることが可能であり、これは細胞相変化の開始に役立つ。
【0133】
好ましい態様では、本明細書中で概説した方法を、G1期に留まっていない細胞上で:即ち、迅速にまたは制御不能に成育し複製しつづけている細胞、例えば腫瘍細胞上で実施することができる。このようにして候補物質を評価し、HDAC8制御を変更し得る、即ち細胞を各HDAC8チェックポイント、例えばG1(他の相、例えばS,G2またはMに留まることも望ましいが)に留まらせ得る物質を見出す。あるいは、候補物質を評価し、細胞集団を増殖させ得る、即ち一般にG1に留まっている細胞:例えば、末梢血液細胞、最終分化細胞、培地中の幹細胞その他、を再増殖させる物質を見出す。
【0134】
従って、本発明は、細胞集団のHDAC8制御における変更についてのスクリーニング方法を提供する。「変更」または「調節」(本明細書中では互変的に使用)により、一般に、2つのことのどちらかを意味する。好ましい実施態様では、この変更の結果、例えばヒストンアセチル化において、ある細胞の細胞周期に変化が生じる。即ち、増殖細胞が各相のどれかひとつに滞留するか、または滞留細胞が滞留相外へ動き出し、その結果別の細胞と比較して、または異なる条件下の同じ細胞内で、細胞周期が開始する。あるいは、どれか特定の相を経由する細胞の進行が変更される;即ち、特定の増殖期を経由して細胞が移動するのに費やす時間の長さに加速または遅延が生じ得る。例えば、該細胞は通常G1期に数時間留まるが;ある物質の添加によりG1期の延長を来し得る。
【0135】
計測は、各計測が同じ条件にあるようにして行うことも、各種の条件下に、バイオ活性物質を用いるかまたは用いずに、または細胞周期過程の異なる段階で行うこともできる。例えば、HDAC8制御の計測は、候補バイオ活性物質が存在する、および候補バイオ活性物質が存在しない、細胞または細胞集団中で行うことができる。別の実施例では、HDAC8制御の計測を、条件または環境がそれぞれ異なる細胞または細胞集団について行う。例えば、細胞を、生理学的シグナル、例えば、ホルモン、抗体、ペプチド、抗原、サイトカイン、成長因子、アクションポテンシャル、化学療法剤を含む医薬物質、放射能、カルシノゲン物質、その他の細胞(即ち、細胞−細胞接触)への曝露、の存在下または不存在下に、またはそれに先立ちまたはその後に、評価してもよい。別の態様では、HDAC8制御の計測を、HDAC8過程の異なる段階で行う。さらに別の実施例では、HDAC8制御の計測を、条件は同じであるが、ある細胞または細胞集団と別の細胞または細胞集団との間での変更について行う。
【0136】
本明細書中で「細胞集団」または「細胞ライブラリー」とは、少なくとも2個の細胞を意味するが、少なくとも約103個であることが好ましく、少なくとも約106個であることが特に好ましく、少なくとも約108個ないし109個であることが格別に好ましい。該集団または試料は、第1または第2培養物と異なる細胞型の混合物を含有してもよいが、単一の細胞型のみを含有する試料、例えば上記したように1種の細胞株由来であり得る、特に腫瘍細胞株由来の試料が好ましい。これらの細胞は、M、G1、S、およびG2を含む、同位相(同位期)または同位相でない、任意の細胞相(細胞期)であり得る。好ましい実施態様では、複製または増殖しつつある細胞を使用するが:このようにすると候補バイオ活性物質の導入にレトロウイルスベクターの使用が可能になる。あるいは、複製中でない細胞を使用することもでき、かつ他のベクター類(例えば、アデノウイルスおよびレンチウイルスベクター)を使用することもできる。加えて、必須ではないが、これらの細胞は染料および抗体と適合性である。
【0137】
本発明での使用に好適な細胞のタイプには、動物(マウス、ラット、ハムスター、アレチネズミを含む齧歯類)、霊長類、およびヒトの細胞、殊に乳癌、皮膚癌、肺癌、頚癌、直腸癌、白血病、脳腫瘍、その他を含む全タイプの腫瘍細胞を含み、哺乳動物細胞が含まれるが、それらに限定されない。
【0138】
好ましい実施態様では、この方法は、細胞生存能力、細胞増殖および細胞相を含むが、それらに限定されない1種または数種以上の細胞パラメータをアッセイすることを含む。
【0139】
好ましい実施態様では、細胞の生存能力をアッセイし、細胞性変化のないことが実験的条件による(即ち、候補バイオ活性物質の導入)ものではなく、細胞死によるものではないことを確認する。光散乱法、生存能力染色法、排除染色法を含むが、それらに限定されない各種の適切な細胞生存能力アッセイを使用できる。
【0140】
好ましい実施態様において、光散乱測定は、当分野において周知であるように、生存能力アッセイとして使用される。例えば、FACSで観察した場合には、細胞は前方および90度(側方)光散乱特性により測定される特異的な特徴を有する。これらの散乱特性は細胞のサイズ、形態および顆粒含有量を示す。これらの特性は、生存能力の読み取り(readout)として測定されるパラメータの説明となる。簡単に言えば、瀕死細胞または死細胞のDNAは一般に凝集しており、このことは90°散乱を変える;同様に、膜の気泡は前方散乱を変える。光散乱の強度、または細胞反射値の変動は生存能力の変動を示す。
【0141】
したがって、一般的に、光散乱アッセイのため、特定の細胞タイプの生存細胞集団をその前方および側方散乱特性を測定して評価する。このことは後に使用され得る散乱の規準を与える。
【0142】
好ましい実施態様において、生存能力アッセイは生存可能な色素 (viability dye)を使用する。死細胞または瀕死細胞を染色するが、増殖中の細胞は染色しない、多くの既知の生存可能な色素が存在する。例えば、アネキシンVは2価イオンに依存した方法でリン脂質(ホスファチジルセリン)に特異的な結合を示すタンパクファミリーのメンバーである。このタンパク質は、ホスファチジルセリンの細胞表面への露出がこのプロセスの初期シグナルの特徴であるので、アポトーシス(プログラム細胞死)の測定に広く使用されている。適当な生存可能な色素は、アネキシン、エチジウムホモダイマー−1、DEAD Red、ヨウ化プロピジウム、SYTOX Greenなど、および当分野で既知の他の物質を含むが、これらに限定されない;the Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Haugland, Sixth Editionを参照。引用により本明細書に含める;特に285ページのApoptosis Assay、およびChapter 16を参照。
【0143】
細胞が生存可能な生存色素染色(viability dye staining)のプロトコールは既知である。上記のMolecular Probesの目録を参照。本実施態様において、アネキシンのような生存可能な色素を直接的または間接的に標識し、細胞集団と組み合わせる。アネキシンは商業的に、即ちPharMingen, San Diego, CaliforniaまたはCaltag Laboratories, Millbrae, Californiaから入手可能である。好ましくは、生存可能な色素は、色素の濃度が約100ng/ml〜約500ng/ml、さらに好ましくは約500ng/ml〜約1μg/ml、および最も好ましくは約1μg/ml〜約5μg/mlの溶液で提供される。好ましい実施態様において、生存可能な色素は直接的に標識される;例えば、アネキシンはフルオレセイン イソチオシアネート(FITC)、Alexa色素、TRITC、AMCA、APC、tri−color、Cy−5のような蛍光色素、および当分野で既知または商業的に入手可能なものであり得る。別の好ましい実施態様において、生存可能な色素はハプテン(例えばビオチン)のような第1の標識で標識され、第2の蛍光標識として、例えば蛍光ストレプトアビジンが使用される。当業者に分かるであろう他の第1および第2の標識の組合せが使用され得る。
【0144】
添加後、生存可能な色素は、ある時間、細胞とともにインキュベートされ、必要であれば洗浄される。次いで、以下に概説したように細胞をソートして非生存細胞を取り除く。
【0145】
好ましい実施態様において、色素排除染色は生存能力アッセイとして使用される。排除色素は生存細胞から排除されるものであり、即ちそれらは受動的に取り込まれない(それらは生存細胞の細胞膜を浸透しない)。しかしながら、死細胞または瀕死細胞の浸透性により、それらは死細胞に取り込まれる。一般的に(必ずというわけではないが)、排除色素はDNAと、例えばインターカレーションにより結合する。好ましくは、排除色素はDNAの不存在下において蛍光を発っしないか、または不十分に蛍光するを発する;このことにより洗浄の段階の必要がなくなる。代わりに、第2の標識の使用を必要とする排除色素も使用され得る。好適な排除色素は、エチジウムブロマイド;エチジウムホモダイマー−1;ヨウ化プロピジウム;SYTOX緑色核酸鎖(green nucleic acid chain);Calcein AM、BCECF AM;フルオレセイン ジアセテート;TOTO(登録商標)およびTO−PRO(商標)(Molecular Probesより;上記、chapter 16参照)および当分野で既知の他の物質を含むが、これらに限定されない。
【0146】
細胞が生存したまま色素排除染色するためのプロトコールは既知である。上記のMolecular Probes参照。一般的に、排除色素は、約100ng/ml〜約500ng/ml、さらに好ましくは約500ng/ml〜約1μg/ml、および最も好ましくは約0.1μg/ml〜約5μg/mlの濃度で細胞に加えられ、特に約0.5μg/mLが好ましい。細胞および排除色素を、ある時間インキュベートし、必要であれば洗浄し、次いで該細胞を以下に概説したようにソートして集団から非生存細胞を取り除く。
【0147】
さらに、実施可能な他の細胞生存能力アッセイが存在し、該方法は生存細胞(即ち分泌されたプロテアーゼなど)または死細胞(即ち培地中に細胞内酵素、例えばプロテアーゼ、ミトコンドリア酵素などが存在すること)の細胞外酵素活性の測定が可能である。the Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Haugland, Sixth Editionを参照。引用により本明細書に含める;特にChapter 16を参照。
【0148】
好ましい実施態様において、少なくとも1つの細胞生存能力アッセイが実施され、蛍光が互換可能な場合には2つの異なる細胞生存能力アッセイが好ましい。1つの生存能力アッセイのみを実施する場合、好ましい実施態様では光分散測定が利用され(前方および側方分散の両方)。2つの生存能力アッセイを実施する場合、好ましい実施態様では光分散と色素排除が利用され、光分散と生存可能な色素染色も利用可能であり、そして場合によっては3つすべてが利用され得る。したがって、生存能力アッセイにより非生存細胞または瀕死細胞から生存細胞を分離することが可能となる。
【0149】
細胞生存能力アッセイに加えて、好ましい実施態様では細胞増殖アッセイが利用される。本明細書において「増殖アッセイ(proliferation assay)」は、細胞集団が増殖している(即ち複製している)か、または複製していないことの測定を可能にするアッセイを意味する。
【0150】
好ましい実施態様において、増殖アッセイは色素封入アッセイである。色素封入アッセイは細胞相を区別する希釈効果によるものである。簡単に言えば、色素(一般に、以下に概説した蛍光色素)を細胞に導入し、細胞に取り込ませる。一度取り込まれると、色素は細胞中にトラップされ、拡散しない。細胞集団が分裂するにつれて、色素は比例して希釈される。即ち、封入色素の導入後、細胞を、ある時間、インキュベートするのである;時間の経過にともない蛍光を失う細胞は分裂しており、蛍光を維持している細胞は非増殖期にある。
【0151】
一般に、封入色素の導入は2つの方法のうちの一方で行われ得る。色素が細胞内に受動的には入れずに(例えば、荷電を有する)、色素が取り込まれるように、例えば電気パルスを使用して;細胞を処理しなければならない。もう1つは、色素は受動的に細胞内に入るが、取り込まれた後、細胞外に拡散できないように修飾される。例えば、封入色素の酵素学的修飾によりそれを帯電させ、したがって、細胞外に拡散できないようにし得る。例えば、Molecular Probes CellTracker(登録商標)は蛍光クロロメチル誘導体であり、自由に細胞内へと拡散し、次いでグルタチオンS−トランスフェラーゼ介在反応により膜不浸透性色素を生じる。
【0152】
適当な封入色素は、Molecular Probes CellTracker(登録商標)のMolecular Probes系を含むが、これらに限定されず、CellTracker(登録商標) Blue、CellTracker(登録商標) Yellow-Green、CellTracker(登録商標) Green、CellTracker(登録商標) Orange、PKH26(Sigma)、および当分野で既知の他の物質を含むが、これらに限定されない;上記のMolecular Probes Handbook;特にchapter 15参照。
【0153】
一般に、封入色素は約100ng/ml〜約5μg/ml、好ましくは約500ng/ml〜約1μg/mlの範囲の濃度で細胞に供与される。洗浄の段階を使用しても、使用しなくてもよい。好ましい実施態様において、候補バイオ活性物質を本明細書に記載のような細胞と組み合わせる。細胞および封入色素を、ある時間、インキュベートすると細胞分裂し、したがって色素が希釈化される。時間の長さは、細胞特有の細胞周期時間に依存する;一般に、少なくとも約2回の細胞分裂が好ましく、少なくとも約3回が特に好ましく、そして少なくとも約4回がとりわけ好ましい。次いで細胞を以下に概略するようにソートし、複製している細胞集団とそうでない集団を作成する。当業者には分かるであろうが、場合によっては、例えば抗増殖剤をスクリーニングする場合、発光(即ち蛍光)している細胞を回収する;他の実施態様において、例えば増殖剤をスクリーニングするため、蛍光の少ない細胞を回収する。変動は、異なる時点での、または異なる細胞集団における蛍光を測定することにより決定され、そして該決定を互いにまたは基準と比較することにより決定される。
【0154】
好ましい実施態様において、増殖アッセイは代謝拮抗物質アッセイである。代謝拮抗物質アッセイは、G1またはG2休止期における休止の原因となる薬品を所望の場合に最も使用される。代謝拮抗物質増殖アッセイでは、毒性があり、分裂中の細胞を死滅させる代謝拮抗物質の使用の結果、分裂していない細胞のみが生き残ることになる。適当な代謝拮抗物質は、メソトレキセート、シスプラチン、タキソール、ヒドロキシウレア、AraCのような核酸アナログ、などのような通常の化学療法剤を含むが、これらに限定されない。さらに、代謝拮抗アッセイは、発現すると細胞死の原因となる遺伝子の使用を含み得る。
【0155】
代謝拮抗物質が添加される濃度は代謝拮抗物質に特有の毒性に依存し、当分野で既知のように決定される。代謝拮抗物質が添加され、細胞は、一般に、ある時間インキュベートされる;再び、正確な時間は代謝拮抗物質の特徴および独自性、ならびに細胞集団に特有の細胞周期の時間に依存する。一般に、少なくとも1回の細胞分裂が起こるのに十分な時間である。
【0156】
好ましい実施態様において、少なくとも1つの増殖アッセイが実施され、2以上が好ましい。したがって、増殖アッセイにより、結果的に増殖中の細胞集団と休止の細胞集団になる。さらに他の増殖アッセイ、即ち当分野において既知の比色定量アッセイを使用し得る。
【0157】
好ましい実施態様において、上で概説した1以上の増殖アッセイの後または同時に、少なくとも1つの細胞相アッセイが行われる。「細胞相」アッセイは、細胞が滞留している細胞相、M、G1、S、またはG2を決定する。
【0158】
好ましい実施態様において、細胞相アッセイはDNA結合色素アッセイである。簡単に言えば、DNA結合色素は細胞に導入され、受動的に取り込まれる。一度細胞内に入ると、DNA結合色素はDNAに、一般的にはインターカレーションにより結合するが、場合によっては、それらは主溝または副溝結合性化合物であり得る。したがって、色素の量は細胞中のDNAの量に直接関連し、これは細胞の相によって変動する;G2およびM期の細胞はG1期の細胞の2倍のDNA含有量を有し、S期の細胞は中間の量を有し、その細胞がS期のどの点であるかに依存する。適当なDNA結合色素は浸透性であり、Hoechst 33342および33258、アクリジンオレンジ、7−AAD、LDS751、DAPI、SYTO16、上記のMolecular Probes Handbook;特にchapter 8および16を含むが、これらに限定されない。
【0159】
一般的に、DNA結合色素は約1μg/ml〜約5μg/mlの範囲の濃度で加えられる。該色素は細胞に加えられ、ある時間、インキュベートされる;時間の長さは選択された色素に一部依存する。1つの実施態様において、色素の添加直後に測定が行われる。次いで、以下に概説したように細胞をソートし、異なる量の色素、したがって異なる量のDNAを含む細胞集団を作成する;本方法において、複製中の細胞は、そうでないものと分離される。当業者には分かるように、場合によっては、例えば抗増殖剤をスクリーニングする場合に、最も弱い蛍光を有する細胞(したがって1セットのゲノム)を増殖中の細胞(したがって1セットのゲノムより多いDNA含む)と分離することができる。変動は、異なる時点での、または異なる細胞集団における蛍光を測定することにより決定され、そして該決定を互いにまたは基準と比較することにより決定される。
【0160】
好ましい実施態様において、細胞相アッセイはサイクリン破壊アッセイである。本実施態様において、スクリーニングの前(および一般的に、以下に概略したように、候補バイオ活性物質の導入前)に融合核酸を細胞に導入する。融合核酸はサイクリン破壊ボックスをコードする核酸および検出可能な分子をコードしている核酸を含む。「サイクリン破壊ボックス」は当分野において既知であり、特定の細胞相の間に該ボックスを含むタンパク質のユビキチン化経路を通して分解の原因となる配列である。即ち、例えば、G1サイクリンはG1期の間安定であり得るが、G1サイクリン破壊ボックスのためS期の間に分解され得る。したがって、サイクリン破壊ボックスを検出可能な分子、例えばグリーン蛍光タンパク質と結合させることにより、検出可能な分子の存在または不存在により細胞集団の細胞相の同定が可能となる。好ましい実施態様において、好ましくはそれぞれ、さまざまな蛍光とともに多数のボックスが使用され、その結果、細胞相の検出が可能となる。
【0161】
多くのサイクリン破壊ボックスが当分野において既知であり、例えば、サイクリンAは配列RTVLGVIGD(配列番号:8)を含む破壊ボックスを有する;サイクリンB1の破壊ボックスは配列RTALGDIGN(配列番号:9)を含む。Glotzer et al., Nature 349:132-138 (1991)参照。さらに他の破壊ボックスとして:
YMTVSIIDRFMQDSCVPKKMLQLVGVT(配列番号:10:ラットサイクリンB);
KFRLLQETMYMTVSIIDRFMQNSCVPKK(配列番号:11:マウスサイクリンB);
RAILIDWLIQVQMKFRLLQETMYMTVS(配列番号:12:マウスサイクリンB1);
DRFLQAQLVCRKKLQVVGITALLLASK(配列番号:13:マウスサイクリンB2);および
MSVLRGKLQLVGTAAMLL(配列番号:14:マウスサイクリンA2)
が知られている。
【0162】
サイクリン破壊ボックスをコードする核酸は検出可能な分子をコードする核酸と機能し得るように結合させる。融合タンパク質は当分野において既知の方法で作成される。例えば、サイクリン破壊ボックスをコードする核酸は検出可能な分子をコードする核酸と結合させる。本明細書の「検出可能な分子」は、検出可能な分子を含む細胞または化合物と、それを含まないものとの区別を可能にする分子を意味する。即ちエピトープ(抗原TAGと呼ばれることもある)、特定の酵素または蛍光分子である。好適な蛍光分子は、グリーン蛍光タンパク質(GFP)、ブルー蛍光タンパク質(BFP)、イエロー蛍光タンパク質(YFP)、レッド蛍光タンパク質(RFP)、およびルシフェラーゼおよびβ−ガラクトシダーゼを含む酵素を含むが、これらに限定されない。抗原TAGが使用される場合、好適な実施態様では細胞表面抗原を使用する。エピトープは、好ましくは、一般的には細胞膜上で見出されない任意の検出可能なペプチドであるが、いくつかの例において、エピトープが細胞上で普通に見出されるものである場合には、増加が検出され得る(このことは一般的には好ましくないのだが)。同様に酵素学的に検出可能な分子;例えば新規または発色性の生成物を産生する酵素も使用され得る。
【0163】
したがって、細胞相アッセイ後のソートの結果、一般的に、異なる細胞相にある少なくとも2つの細胞集団が生じる。
【0164】
ここで提供されるタンパク質および核酸を、スクリーニング目的のために使用することができ、HDAC8タンパク質のタンパク質−タンパク質相互作用を同定することができる。タンパク質−タンパク質相互作用を検出するための遺伝学的システムが記載されている。酵母のシステムにおいて最初の研究が実施された。即ち「酵母ツー−ハイブリッド」システムである。基本的なシステムは、レポーター遺伝子の転写を開始するために、タンパク質−タンパク質相互作用を必要とする。次いで、研究は哺乳類細胞において実施された。Fields et al., Nature 340: 245(1989); Vasavada et al., PNAS USA 88: 10686(1991); Fearon et al., PNAS USA 89: 7958(1992); Dang et al., Mol. Cell. Biol. 11: 954(1991); Chien et al., PNAS USA 88: 9578(1991); 並びに米国特許番号第5283173号、第5667973号、第5468614号、第5525490号、および第5637463号を参照されたい。好ましいシステムは、出願番号第09/050863号、1998年3月30日出願、および第09/359081号、1999年7月22日出願、表題「哺乳動物タンパク質のインターカレーションクローニングシステム(Mammalian Protein Interaction Cloning System)」に記載されている。これらのシステムの合同的な使用について、特に有用なシャトルベクターが、出願番号09/133944、1998年8月14日出願、表題「シャトルベクター(Shuttle Vectors)」に記載されている。
【0165】
一般に、2つの核酸を1細胞に形質転換する。1つの核酸はHDAC8タンパク質またはその一部をコードする遺伝子のような「ベイト(bait: えさ)」であり、もう1つの核酸は試験候補をコードしている。2つの発現産物が互いに結合する場合のみ、蛍光タンパク質などの指示薬は発現する。その指示薬の発現は、いつ試験候補がHDAC8タンパク質に結合するかを示し、試験候補をHDAC8タンパク質として同定することができる。同じシステムと同定されるHDAC8タンパク質を使用して、逆のことを実施することができる。即ち、本発明で提供されるHDAC8タンパク質を、新規なベイト、またはHDAC8タンパク質と相互作用する物質を同定するために、使用することができる。さらに、ベイトおよびHDAC8タンパク質をコードしている核酸のほかに試験候補を添加し、ベイトとHDAC8タンパク質との相互作用を妨害する物質を調べる場合に、ツー−ハイブリッドシステムを使用することができる。
【0166】
ある実施態様において、哺乳類ツー−ハイブリッドシステムが好ましい。哺乳類のシステムは、タンパク質のポスト翻訳修飾を提供し、その修飾は、タンパク質が相互作用する能力に有意に寄与し得る。さらに、哺乳類ツー−ハイブリッドシステムを広範な哺乳類細胞のタイプにおいて使用し、特定の細胞タイプ内の特異的タンパク質の制御、誘導、プロセッシングなどを模倣することができる。例えば、疾患状態(即ち、癌、アポトーシス関連障害)に関連するタンパク質を、関連する疾患細胞において試験し得る。同様に、ランダムタンパク質の試験のために、関連する細胞内条件でアッセイすることにより、最高の肯定的な結果が得られる。さらに、哺乳類細胞を、細胞間タンパク質−タンパク質相互作用に影響を及ぼし得る広範な実験条件下(例えば、ホルモン、薬剤、成長因子およびサイトカイン、放射線、化学療法剤、細胞内および化学的刺激などの存在)で試験することができ、このことは、タンパク質−タンパク質相互作用に影響を及ぼすことのできる条件(特に癌に関連する条件)に寄与し得る。
【0167】
ツー−ハイブリッドシステムなどの結合に関連するアッセイは、非特異的結合タンパク質(NSB)を考慮に入れ得る。
【0168】
様々な細胞タイプにおける発現、およびHDAC8活性のアッセイを、前記した。例えば、クロマチン構造、ヒストンアセチル化、転写などに対する作用についての活性のアッセイを実施し、PCNAとの配列同一性/類似性またはそれとの結合によって、既に同定されていたHDAC8タンパク質の活性、転写などに影響を示すことを確認することができる。さらに、HDAC8の調節因子として同定されるリード化合物の活性をさらに確認することができる。
【0169】
本発明で提供されるアッセイ用の、本発明で提供される構成成分を組合せて、キットを形成してもよい。該キットは、タンパク質および/またはHDAC8タンパク質をコードする核酸の使用をベースとすることができる。ある実施態様では、他の構成成分がキット中で提供される。そのような構成成分には、1つまたはそれ以上の包装、指示書、抗体、および標識が含まれる。診断に使われるようなさらなるアッセイは、さらに後述する。
【0170】
このようにして、バイオ活性物質が同定される。医薬的活性のある化合物は、HDAC8タンパク質の活性を高めたり、妨害したりできる。さらに後述するように、所望の医薬的活性を有する化合物を、医薬的に許容し得る担体中で宿主に投与してもよい。
【0171】
即ち、細胞中でのHDAC8タンパク質の役割に関する今回の開発は、細胞中で細胞増殖を誘導または防止する方法を提供する。好ましい実施態様では、HDAC8タンパク質、および特にHDAC8タンパク質断片は、HDAC8タンパク質が介在する症状の研究または処置において、即ち、HDAC8関連障害の診断、処置、または予防において、有用である。ゆえに、「HDAC8関連障害」または「疾患状態」には、細胞増殖の不足または過剰の両方に関連する症状が含まれる。細胞増殖の不足または過剰の例には、老化、アポトーシス、ネクローシス、癌などが含まれるが、癌が好ましい。癌には、腫瘍形成、癌転移および脈管形成と関連する全ての機構が含まれる。
【0172】
ゆえに、ある実施態様では、細胞または生物内でのHDAC8制御が提供される。ある実施態様では、その方法は、治療的な量でHDAC8タンパク質を、それを必要としている細胞または個体に投与することを含む。あるいは、内在性のHDAC8タンパク質の生物学的活性を低減または排除する抗HDAC8抗体を投与する。他の実施態様では、本発明で提供される方法で同定したバイオ活性物質を投与する。あるいは、その方法は、HDAC8タンパク質をコードする組換え核酸を、細胞または個体に投与することを含む。当業者に理解されるであろうように、このことはいかなる数の方法で達成されてもよい。好ましい実施態様では、例えば、内因性HDAC8タンパク質を過剰発現させるか、または、遺伝子治療技術などでHDAC8タンパク質をコードする遺伝子を投与することで、細胞中のHDAC8タンパク質の量を増やすことによって、HDAC8の活性は、増大する。好ましい実施態様では、該遺伝子治療技術は、例えば出典明示により本明細書の一部とするPCT/US93/03868に記載のように、強化相同組換え(EHR)を用いて、外来性遺伝子を組込むことを含む。
【0173】
理論によって制限を受けないが、HDAC8タンパク質は、転写およびHDAC8を制御する点でHDAC8において重要なタンパク質であることがわかった。従って、HDAC8遺伝子の突然変異または変異に基づく障害が確定され得る。ある実施態様では、本発明は、細胞中の少なくとも1つの内因性HDAC8遺伝子配列の全部または部分を解読することを含む、変異型HDAC8遺伝子を有する細胞の同定法を提供する。当業者に理解されるであろうように、このことは、配列解読技術をいくつでも用いて行われ得る。好ましい実施態様では、本発明は、個体の少なくとも1つのHDAC8遺伝子配列の全部または一部を解読することを含む、個体のHDAC8遺伝子型の同定法を提供する。このことは、一般的に、個体の少なくとも1組織について行われ、多数の組織または同じ組織の異なる試料の評価を含んでもよい。該方法は、解読されたHDAC8遺伝子の配列を、既知のHDAC8遺伝子、即ち、野生型遺伝子と比較することを含んでもよい。
【0174】
HDAC8遺伝子の全部または一部の配列を、既知のHDAC8遺伝子の配列と比較して、差異が存在するかどうかを決定することができる。このことは、Bestfitなどの周知の配列同一性プログラムをいくつでも用いて行うことができる。好ましい実施態様では、患者のHDAC8遺伝子と既知のHDAC8遺伝子との間の配列の差異が存在することは、疾患状態または疾患状態になる傾向を示している。従って、本発明は、例えば遺伝子の完全性に影響を与える変化、置換に関する欠損および挿入、染色体の配置転換、mRNAの異常型スプライシングパターンなどによって特徴付けられるHDAC8核酸における遺伝的障害の存在もしくは非存在を決定する方法を提供する。
【0175】
ある実施態様では、本発明は、個体におけるHDAC8関連症状の診断法を提供する。この方法には、個体または患者由来の組織または体液などの試料中におけるHDAC8活性の測定が含まれる。HDAC8の活性は、ある試料がHDAC8の既知の基質を脱アセチル化する能力についてのアッセイによって測定するのが好ましい。例えば、特定の抗血清を用いるラジオイムノアッセイは、アセチル化と非アセチル化ペプチドに対して行った。一方、患者由来のHDAC8試料の能力は、インビボでのラジオアセチル化された天然または合成基質を脱アセチル化するその能力が試験され得る。この活性を、罹患していない第2の個体または第1の個体の罹患していない組織のどちらかに由来するHDAC8の活性と比較する。これらの活性が異なる場合、第1の個体にはHDAC8関連異常のリスクが有り得る。例えば、癌細胞は、末端分化に関与する遺伝子の転写抑制の手段としてHDAC8をアップレギュレートし「得る」。このことは、細胞を増殖状態にさせる。従って、HDAC8の阻害により、癌細胞の細胞分化を非疾患状態へと誘導する遺伝子の転写が可能となる。このようにして、例えば、タンパク質のレベルまたはそのmRNAの発現をモニターすることによって、様々な疾患状態をモニターし得る。同様に、発現レベルは、予後に相関し得る。
【0176】
ある態様では、HDAC8タンパク質遺伝子の発現レベルを、診断または予後情報が求められる異なる患者の試料または細胞について測定する。遺伝子発現のモニターは、HDAC8タンパク質をコードする遺伝子について行われる。ある態様では、HDAC8タンパク質遺伝子の発現レベルは、正常細胞およびアポトーシスまたは形質転換をしている細胞のような、細胞の異なる状態について測定される。異なる状態の細胞においてHDAC8タンパク質遺伝子の発現レベルを比較することによって、HDAC8タンパク質遺伝子のアップレギュレーションおよびダウンレギュレーションの両者を含む情報が得られ、この情報は多数の方法で用いられる。例えば、特定の処置方式を評価し得る:ある化学療法剤が特定の患者の長期的予後を改善するのに役立っているかを評価し得る。同様に、患者の試料を比較して、診断を行うか、または確認してもよい。さらに、これらの遺伝子発現レベルによって、特定の発現レベルを模倣または変化させる目的で、薬物候補をスクリーニングすることが可能になる。このことは、このようなスクリーニングに用いることができ、本発明のもののような重要なHDAC8タンパク質遺伝子の一揃えを含むバイオチップを作成することによって行ってもよい。これらの方法は、タンパク質に基づいて行うこともできる;つまり、HDAC8タンパク質のタンパク質発現レベルを、診断の目的や候補物質のスクリーニングのために評価できる。さらに、HDAC8タンパク質の核酸配列を遺伝子治療の目的で投与することができる。このことには、アンチセンス核酸の投与、または治療薬としてのHDAC8タンパク質の投与が含まれる。
【0177】
バイオチップに結合するHDAC8タンパク質配列には、上記定義のように核酸およびアミノ酸の両者の配列が含まれる。好ましい実施態様では、HDAC8タンパク質の核酸(図に概説した配列を有する核酸配列および/またはその相補鎖の両者)に対する核酸プローブが作成される。バイオチップに取付けられる核酸プローブは、HDAC8タンパク質の核酸、即ち、標的配列(試料の標的配列、または例えばサンドイッチアッセイにおける他のプローブ配列への標的配列)に実質的に相補的に設計し、標的配列と本発明のプローブとのハイブリダイゼーションが起こるようにする。以下に概説するように、この相補性は、完全である必要は無い;標的配列と本発明の1本鎖核酸の間のハイブリダイゼーションを妨害するミスマッチ塩基対がいくつあってもよい。しかし、もし変異の数が多すぎて、緊縮性が最低のハイブリダイゼーション条件下でさえハイブリダイゼーションが起こらないならば、その配列は相補的標的配列ではない。ゆえに、本明細書で「実質的に相補的」は、本明細書で概説するように、通常の反応条件、特に高緊縮条件下な条件下でハイブリダイズするのに、プローブが標的配列に十分相補的であることを意味する。
【0178】
「核酸プローブ」は、一般的に1本鎖であるが、部分的に1本鎖、および部分的に2本鎖であり得る。プローブの鎖形成は、構造、組成、および標的配列の性質によって決定される。一般的に、核酸プローブは約8ないし約100塩基長の範囲にあり、約10ないし約80塩基が好ましく、約30ないし約50塩基が特に好ましい。ある実施態様では、数百塩基に上るさらに長い核酸(例えば、遺伝子全長)を用いることができる。
【0179】
当業者に理解されるであろうように、核酸を、幅広い方法で、固体支持体に結合または固定することができる。 本明細書での「固定された」および文法的均等語は、以下に概説するように、核酸プローブと固体支持体の会合または結合が、結合、洗浄、分析、および除去の条件下で安定であるために十分であることを意味する。結合は、共有的または非共有的であり得る。本明細書における「非共有結合」および文法的均等語は、1またはそれ以上の、電気的、親水的、および疎水的相互作用のいずれかを意味する。ストレプトアビジンのような分子の支持体への共有結合、およびビオチン化プローブのストレプトアビジンへの非共有結合は、非共有結合に含まれる。本明細書における「共有結合」および文法的均等語は、2つの部分、固体支持体とプローブが、シグマ結合、パイ結合および配位結合を含む少なくとも1つの結合によって結合していることを意味する。共有結合は、プローブと固体支持体との間に直接形成でき、あるいは、クロスリンカーによって、または固体支持体とプローブのいずれかもしくは両者に特異的反応基を含むことによって形成できる。固定化は、共有および非共有相互作用の組合せも含み得る。
【0180】
一般的に、当業者に理解されるであろうように、プローブを幅広い方法でバイオチップに結合させる。本明細書に記載のように、核酸は、最初に合成して次にバイオチップに結合するか、またはバイオチップ上で直接合成することができる。
【0181】
バイオチップは、好適な固体基板を含む。本明細書の「基板」または「固体支持体」または他の文法的均等語は、核酸プローブの結合または会合に適する独立した個々の部位を含有するように変更することができ、そして少なくとも1つの検出法で分析できる、あらゆる材料を意味する。当業者に理解されるであろうように、可能な材料の数は非常に多く、ガラスおよび修飾または機能化ガラス、プラスチック(アクリル、ポリスチレン、ならびにスチレンおよび他の材料の共重合体、ポリプロピレン、ポリエチレン、ポリブチレン、ポリウレタン、テフロンJなどを含む)、多糖類、ナイロンまたはニトロセルロース、樹脂、シリカまたはシリコンもしくは修飾シリコンを含むシリカベースの材料、炭素、金属、無機ガラス、プラスチックなどを含むが、これらに限定されるものではない。一般的に、該基板は、好都合な検出を可能にし、感知できるほどの蛍光を示さない。
【0182】
好ましい実施態様では、バイオチップとプローブの表面を、両者の結合のために、化学官能基で誘導し得る。ゆえに、例えば、バイオチップは、アミノ基、カルボキシル基、オキソ基およびチオール基を含むがこれらに限定されない化学官能基で誘導され、アミノ基が特に好ましい。これらの官能基を用いると、プローブをプローブ上の官能基を用いて結合させることができる。例えば、アミノ基を含む核酸は、例えば、当分野で周知のようにリンカーを用いて、アミノ基を含む表面に取付けることができる;例えば、ホモ―またはヘテロ―両機能性リンカーは、周知である(1994 Pierce Chemical Company catalog, technical section on cross-linkers, pages 155-200, 参照。出典明示により本明細書の一部とする)。さらに、ある場合では、アルキル基(置換およびヘテロアルキル基を含む)のような追加のリンカーを用いてもよい。
【0183】
この実施態様では、核酸プローブに対応するオリゴヌクレオチドを当分野で周知のように合成し、固体支持体の表面に結合させる。当業者には理解されるであろうように、5'または3'の末端のどちらを固体支持体に結合させてもよく、また内側のヌクレオチドを介して結合させてもよい。
【0184】
さらなる実施態様では、固体支持体への固定化は、非共有結合であっても非常に強固であり得る。例えば、ストレプトアビジンで共有結合により被膜された表面に結合するビオチン化オリゴヌクレオチドを作成することができ、結合させる。
【0185】
または、当分野で周知のように、オリゴヌクレオチドを表面上で合成できる。たとえば、光重合の化合物および技術を利用する光活性化技術を用いる。好ましい実施態様では、核酸は、WO95/25116;WO95/35505;米国特許番号第5700637号および第5445934号;およびそれらの中で引用された参照文献(それらすべてを出典明示により本明細書の一部とする)に記載のように、周知の写真平板技術を用いてin situで合成できる;これらの結合の方法は、Affimetrix GeneChip(登録商標)技術の基礎をなしている。
【0186】
本明細書で使用される「ディファレンシャル発現」または文法的均等語は、遺伝子の時間的および/または細胞の発現パターンにおける、細胞間の量的差異と質的差異の両方を表わす。ゆえに、異なって発現された遺伝子は、例えば正常細胞対アポトーシスの細胞において、活性化と不活性化を含む、定性的に変化した発現を有することができる。つまり、遺伝子は、特定の状態において、他の状態に関連して点いたり切れたりし得る。当業者には明らかなように、2種またはそれ以上の状態のどのような比較も行うことができる。そのような定性的に制御される遺伝子は、ある状態または細胞のタイプにおいて、発現パターンを示すであろう。そのパターンは、状態または細胞のタイプが1種の場合は標準的な技術で検出できるが、両方の場合は検出できない。あるいは、発現が増加または減少する場合には、測定は定量的である;つまり、遺伝子の発現はアップレギュレーションされ、転写物の量が増加するか、または遺伝子の発現はダウンレギュレーションされ、転写物の量が減少するかのどちらかである。発現が異なる程度は、以下に概説するように、Affymetrix GeneChip(登録商標)発現アレイ(Lockhart, Nature Biotechnology 14:1675-1680 (1996)、出典明示により本明細書の一部とする)のような、標準的な特性分析技術を介して定量するのに十分なほどに大きい必要があるだけである。他の技術には、定量的逆転写酵素PCR、ノーザン分析およびRNase保護が含まれるが、これらに限定されるものではない。
【0187】
当業者に理解されるであろうように、このことは遺伝子転写物、またはタンパク質レベルのいずれかを評価することによってなし得る;つまり遺伝子発現の量は、遺伝子転写物と等価のDNAまたはRNAに対する核酸プローブ、および遺伝子発現のレベルの定量を用いてモニターすることができ、または、例えばHDAC8タンパク質の抗体および標準的なイムノアッセイ(ELAISAなど)、または質量分析アッセイ、2次元ゲル電気泳動アッセイなどの他の技術を介して、遺伝子の最終産物そのもの(タンパク質)をモニターすることができる。
【0188】
他の方法では、mRNAの検出はin situで行われる。この方法では、透過性にした細胞または組織の試料を、検出可能な標識化核酸プローブに十分な時間接触させ、プローブを標的mRNAにハイブリダイズさせる。非特異的に結合したプローブを除去するために洗浄した後、標識を検出する。例えば、HDAC8タンパク質をコードするmRNAに相補的なジゴキシゲニン標識化リボプローブ(RNAプローブ)は、ジゴキシゲニンを抗ジゴキシゲニン二次抗体と結合させ、ニトロブルーテトラゾリウムおよびリン酸5−ブロモ4−クロロ−3−インドリルと反応させることで検出される。
【0189】
他の好ましい方法では、HDAC8タンパク質の発現は、HDAC8タンパク質抗体を使用するin situイメージング技術を用いて行われる。この方法では、細胞をHDAC8タンパク質(群)に対する1種ないし多種の抗体に接触させる。非特異的に結合している抗体を除去するために洗浄し、抗体または抗体の存在を検出する。ある実施態様では、抗体は、検出可能な標識を含む二次抗体とインキュベートすることにより検出される。他の方法では、HDAC8タンパク質(群)に対する一次抗体が、検出可能な標識を有する。他の好ましい実施態様では、多種類の一次抗体の各々が、別個の検出可能な標識を含む。この方法には、多数のHDAC8タンパク質の同時スクリーニングにおいて特に有用性がある。標識は、異なる波長の発光を検出し、区別できる性能を有する蛍光光度計で検出し得る。さらに、蛍光標示式細胞分取器(FACS)を、この方法で用いることができる。通常の技術のある当業者に理解されるであろうように、膨大な数の他の組織学的イメージング技術が本発明で有用であり、抗体をELISA,イムノブロット(ウエスタンブロット)、免疫沈降、BIACORE技術などに使用できる。
【0190】
ある実施態様において、HDAC8タンパク質に対するポリクローナルおよびモノクローナル抗体を生成するために、本発明のHDAC8タンパク質を使用し得る。それらの抗体は、ここで記載されるように有用である。同様に、HDAC8タンパク質を、標準的な技術を使用してアフィニティ・クロマトグラフィ・カラムに結合することができる。そして、これらのカラムをHDAC8抗体を精製するために使用し得る。好ましい実施態様において、抗体をHDAC8タンパク質に対してエピトープ特異的に生成する;即ち、抗体は、他のタンパク質とほとんどまたは全く交差反応性を示さない。これらの抗体について、多くの応用における使用が見出される。例えば、HDAC8タンパク質抗体を標準的なアフィニティ・クロマトグラフィ・カラムに結合し得る。そして、以下にさらに記載するように、HDAC8タンパク質を精製するために使用し得る。抗体を、前記で概説したように、ブロッキングポリペプチドとしても使用し得る。というのは、これらは、HDAC8タンパク質と特異的に結合するからである。
【0191】
抗HDAC8タンパク質抗体は、ポリクローナル抗体を含み得る。ポリクローナル抗体を調製する方法は、当業者にとって既知である。ポリクローナル抗体を哺乳類中で産生し得る。例えば、1以上の免疫化物質および所望によりアジュバントの注射による。典型的には、免疫化物質および/またはアジュバントを、数回の皮下または腹膜内への注射によって哺乳類に注射する。免疫化物質は、HDAC8タンパク質またはその融合タンパク質を含み得る。免疫化物質と、免疫化される哺乳類において免疫原であることが既知であるタンパク質とを結合することは、有用である。そのような免疫原タンパク質の例は、キーホール・リンペット・ヘモシアニン、血清アルブミン、ウシ・チログロブリンおよびダイズ・トリプシンインヒビターを含むが、これらに限定されない。使用され得るアジュバントの例は、フロイントの完全アジュバントおよびMPL−TDMアジュバント(モノフォスフィリル・リピッドA、合成トレハロース・ジコリノミコレート)を含む。免疫化プロトコールを当業者は過度な実験なくして選択し得る。
【0192】
あるいは、抗HDAC8タンパク質抗体はモノクローナル抗体であってよい。モノクローナル抗体は、ハイブリドーマ法を使用して調製し得る。その方法は、Kohler and Milstein, Nature, 256: 495(1975)に記載されている。ハイブリドーマ法では、マウス、ハムスターなどの適当な宿主動物を典型的には免疫化物質で免疫化する。そして、免疫化物質に特異的に結合する抗体を、産生または産生し得るリンパ球を誘発する。あるいはリンパ球をインビトロで免疫化し得る。
【0193】
免疫化物質は、典型的には、HDAC8タンパク質またはその融合タンパク質を含む。一般的には、ヒト起源の細胞が望まれるならば、末梢血液リンパ球(「PBL」)を使用し、または、非ヒト哺乳類ソースが望まれるなら、脾臓細胞またはリンパ節細胞を使用する。そして、リンパ球を不死化細胞系統と融合化する。その際に、ポリエチレングリコールのような適当な融合化物質を使用してハイブリドーマ細胞を形成させる[Goding, Monoclonal Antibodies: Pronciples and Practice, Academic Press, (1986) pp. 59-103]。不死化細胞系統として、通常、形質転換された哺乳類細胞、特に齧歯類、ウシおよびヒト起源のミエローマ細胞を使用する。通常、ラットまたはマウスミエローマ細胞系統を使用する。ハイブリドーマ細胞を適当な培地中で培養し得る。それは、好ましくは、融合されず、不死化していない細胞の成長または生存を阻害する1以上の物質を含む。例えば、親細胞が酵素ヒポキサンチン・グアニン・ホスホリボシル・トランスフェラーゼ(HGPRTまたはHPRT)を欠くならば、ハイブリドーマのための培地(「HAT培地」)は、典型的には、HGPRT欠失細胞の成長を妨げるヒポキサンチン、アミノプテリン、およびチミジンを含む。
【0194】
好ましい不死化細胞系統は効率的に融合し、選択された抗体生産細胞による抗体の安定的高水準の発現を支持し、そしてHAT培地のような培地に感受性である。さらに好ましい不死化細胞系統は、ネズミミエローマ系統であり、例えば、Salk Institute Cell Distribution Center, San Diego, California and the American Type Culture Collection, Rockville, Marylandから得ることができる。ヒトミエローマおよびマウス−ヒトへテロミエローマ細胞系統は、ヒトモノクローナル抗体の生産に関して記載されている[Kozbor, J. Immunol., 133:3001(1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63]。
【0195】
そして、ハイブリドーマ細胞を培養する培地を、HDAC8タンパク質に対するモノクローナル抗体の存在についてアッセイすることができる。好ましくは、ハイブリドーマ細胞によって産生されるモノクローナル抗体の結合特異性を、免疫沈降によって、または放射性免疫測定法(RIA)もしくは酵素結合免疫吸着アッセイ(ELISA)などのインビトロ結合アッセイによって測定する。これらの技術およびアッセイは当技術分野で既知である。例えば、モノクローナル抗体の結合親和性をMunson and Pollard, Anal. Biochem., 107:220(1980)のScatchard分析によって測定することができる。
【0196】
所望のハイブリドーマ細胞の同定の後、クローンを限界希釈法によってサブクローン化し、標準的な方法によって成長させ得る[Goding、前出]。この目的のための適当な培地は、例えば、ダルベッコ変法イーグル培地(Dulbecco's Modified Eagle's Medium)およびPPMI−1640培地を含む。あるいは、ハイブリドーマ細胞を哺乳類内の腹水のようなインビボで成長させ得る。
【0197】
サブクローン化によって分泌されたモノクローナル抗体を、通常の免疫グロブリン精製方法により培地または腹水液から単離し精製し得る。この精製方法には、例えば、タンパク質A−セファロース、ヒドロキシルアパタイト・クロマトグラフィー、ゲル電気泳動、透析、またはアフィニティー・クロマトグラフィーがある。
【0198】
モノクローナル抗体を組み換えDNA法によっても作成し得る。例えば、米国特許番号第4816567号に記載されている。本発明のモノクローナル抗体をコードするDNAを通常の方法を使用して容易に単離し、配列決定することができる(例えば、ネズミ抗体の重鎖および軽鎖をコードしている遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを使用して)。本発明のハイブリドーマ細胞を、そのようなDNAの好ましいソースとして使用することができる。単離すると、DNAを発現ベクター内に入れ、宿主細胞にトランスフェクトし得る。そのような細胞には、トランスフェクトされなければ免疫グロブリンタンパク質を産生しないシミアンCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、またはミエローマ細胞がある。次いで組換え宿主細胞においてモノクローナル抗体が合成される。例えば、ヒトの重鎖および軽鎖の定常ドメインのコード配列を、相同的なネズミ配列の場所に置換することにより[特許第4816567号;Morrison et al., 前出]、または免疫グロブリン・コード配列と、非免疫グロブリン・ポリペプチドのコード配列のすべてまたは一部とを共有結合させることなどによりDNAもまた修飾し得る。そのような非免疫グロブリン・ポリペプチドを、本発明の抗体の定常ドメインと置換することができる。あるいは、本発明の抗体のある抗原結合部位の可変ドメインと置換し、キメラ二価抗体を生成することができる。
【0199】
抗体は一価の抗体であってよい。一価の抗体を調製する方法は、当技術分野で周知である。例えば、ある方法は、免疫グロブリンの軽鎖および修飾重鎖の組換え発現を含む。重鎖は、Fc領域における任意の点において末端切除し、重鎖の交差結合(cross linking)を防止する。あるいは、関連するシステイン残基を、別のアミノ酸残基で置換するか削除して、交差結合を防ぐ。
【0200】
インビトロ法は、一価抗体を調製するためにも適当である。抗体の消化によって、その断片、特にFab断片を生成することは、当分野で既知の日常的な技術を使用し実施することができる。
【0201】
本発明の抗HDAC8タンパク質抗体は、ヒト化抗体またはヒト抗体をさらに含み得る。ヒト化型の非ヒト(例えばネズミ)抗体は、キメラ免疫グロブリン、免疫グロブリン鎖またはその断片(Fv、Fab、Fab'、F(ab')2または抗体の他の抗原結合サブシーケンスなど)である。これらは、非ヒト免疫グロブリンに由来する最小配列を含む。ヒト化抗体はヒト免疫グロブリン(レシピエント抗体)を含み、レシピエントの相補性決定領域(CDR)からの残基が、マウス、ラットまたはウサギなどの所望の特異性、親和性および能力を有する非ヒト種(ドナー抗体)のCDRからの残基で置換されている。いくつかの例では、ヒト免疫グロブリンのFvフレームワーク残基が、対応する非ヒト残基で置換されている。ヒト化抗体は、レシピエント抗体および導入されたCDRまたはフレームワーク配列のいずれにも見出されない残基も含む。一般に、ヒト化抗体は、実質的に少なくとも1つの、そして典型的には2つの可変ドメインの全部を含む。このドメインでは、すべてまたは実質的にすべてのCDR領域が、非ヒト免疫グロブリンのCDR領域に一致し、すべてまたは実質的にすべてのFR領域が、ヒト免疫グロブリン共通配列のFR領域である。ヒト化抗体は、所望により、免疫グロブリン定常領域(Fc)、通常はヒト免疫グロブリンの定常領域の少なくとも一部も含む[Jones et al., Nature, 321: 522-525(1986); Riechmann et al., Nature, 332: 323-329(1988); and Presta, Curr. Op. Struct. Biol., 2: 593-596(1992)]。
【0202】
非ヒト抗体をヒト化する方法は当技術分野で周知である。一般に、ヒト化抗体は、非ヒトのソースから抗体に導入された1以上のアミノ酸残基を有している。これらの非ヒトアミノ酸残基をしばしば「導入」残基と称する。それは、典型的には「導入」可変ドメインに由来する。ヒト化をWinterおよびその共同研究者の方法[Jones et al., Nature, 321: 522-525(1986); Riechmann et al., Nature, 332: 323-327(1988); Verhoeyen et al., Science, 239: 1534-1536(1988)]に従って、齧歯類CDR配列と、ヒト抗体の対応する配列を置換することによって、基本的に実施することができる。従って、該「ヒト化」抗体は、キメラ抗体であり(米国特許第4816567号)、無処置のヒト可変ドメインより実質的に小さいドメインが非ヒト種からの対応する配列によって置換されている。実際、ヒト化抗体は、典型的にはいくつかのCDR残基およびおそらくいくつかのFR残基が、齧歯類抗体の同種の部位からの残基によって置換されているヒト抗体である。
【0203】
ヒト抗体は、当技術分野で既知の様々な技術を使用して産生することができる。この技術はファージ・ディスプレイ・ライブラリーを含む[Hoogenboom and Winter, J. Mol. Biol., 227: 381(1991); Marks et al., J. Mol. Biol., 222: 581(1991)]。ColeらおよびBoernerらの技術は、ヒトモノクローナル抗体の調製のためにも利用可能である(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77(1985) and Boerner et al., J. Immunol., 147(1):86-95(1991)]。同様に、ヒト抗体は、内因性免疫グロブリン遺伝子を部分的にまたは完全に不活性化したマウスなどのトランスジェニック動物への、ヒト免疫グロブリン遺伝子座の導入によって作成することができる。チャレンジしたとき、ヒト抗体生産が観察され、それはすべての点でヒトに見出されるものとよく類似し、遺伝子の再配列、集合、およびレパートリーを含む。この方法は、例えば、米国特許第5545807号;第5545806号;第5569825号;第5625126号;第5633425号;第5661016号、および以下の科学的出版物: Marks et al., Bio/Technology 10, 779-783(1992); Lonberg et al., Nature 368 856-859(1994); Morrison, Nature 368, 812-13(1994); Fishwild et al., Nature Biotechnology 14, 845-51(1996); Neuberger, Nature Biotechnology 14, 826(1996); Lonberg and Huszar, Intern Rsv. Immunol. 13 65-93(1995)に記載されている。
【0204】
二重特異的抗体は、少なくとも2の異なる抗原に結合特異性を有するモノクローナル抗体、好ましくはヒトまたはヒト化抗体である。今回の場合において、結合特異性の1つはHDAC8タンパク質について、他方は任意の他の抗原について、そして好ましくは細胞表面タンパク質または受容体もしくは受容体サブユニットについての抗体である。
【0205】
二重特異的抗体を生成するための方法は、当技術分野で既知である。通常、二重特異的抗体の組換え産物は、2つの免疫グロブリン重鎖/軽鎖組の共発現に基づき、この場合2本の重鎖が異なる特異性を有する。[Milstein and Cuello, Nature, 305: 537-539(1983)]。免疫グロブリン重鎖および軽鎖のランダムな組合わせのために、これらのハイブリドーマ(クアドローマ)は、10種の異なる抗体分子があり得る混合物を産生する。このうち1種のみが適当な二重特異的構造を有する。適当な分子の精製を通常アフィニティ・クロマトグラフィ法によって実施する。類似方法は、WO93/08829、1993年5月13日公開、およびTraunecker et al., EMBO J., 10: 3655-3659(1991)に記載されている。
【0206】
所望の結合特異性を有する抗体可変ドメイン(抗体抗原結合部位)を、免疫グロブリン定常ドメイン配列に融合することができる。その融合は、好ましくは免疫グロブリン重鎖定常ドメインとのものである。そしてヒンジ、CH2、およびCH3領域の少なくとも一部を含む。第1の重鎖定常領域(CH1)を有するのが好ましい。それは、融合の少なくとも1つに存在する軽鎖結合に必要である部位を含む。免疫グロブリン重鎖融合、および所望により免疫グロブリン軽鎖をコードするDNAを別の発現ベクターに挿入し、適当な宿主生物に共トランスフェクトさせる。二重特異的抗体の生成のさらなる詳細について、例えば、Suresh et al., Methods in Enzymology, 121: 210(1986)を参照されたい。
【0207】
ヘテロ結合抗体もまた、本発明の範囲内である。ヘテロ結合抗体は、2つの共有結合した抗体から成る。そのような抗体は、例えば、好ましくない細胞に免疫系細胞の標的を定めるために[米国特許第4676980号]、HIV感染の処置のために[WO91/00360; WO92/200373; EP03089]提案されている。抗体は、合成タンパク質化学(関連交差結合物質を含む)における既知の方法を使用して、インビトロで調製し得る。例えば抗毒素を、ジスルフィド置換反応を使用して、またはチオエーテル結合の形成によって構築し得る。この目的のために適当な物質の例は、イミノチオレートおよびメチル−4−メルカプトブチルイミデートを含み、例えば、米国特許第4676980号に記載されている。
【0208】
本発明の抗HDAC8タンパク質抗体は、多様な有用性を有する。例えば、抗HDAC8タンパク質抗体は、HDAC8タンパク質についての診断アッセイに使用しうる。例えば、特定の細胞、組織、または血清におけるその発現の検出である。当技術分野で既知の多様な診断アッセイ技術を使用し得る。例えば、競合的結合アッセイ、直接的または間接的サンドイッチアッセイ、および異種性または同種性の相のいずれかにおいて実施される免疫沈降法である[Zola, Monoclonal Antibodies: A Manual of Techniques, CRC Press, Inc.(1987)pp. 147-158]。診断アッセイに使用される抗体を、検出可能部分で標識化することができる。検出可能部分は、直接的にまたは間接的に検出可能なシグナルを生産するものである。例えば、検出可能部分は、3H、14C、32P、35S、もしくは125Iのような放射性同位体、フルオレセイン・イソチオシアネート、ローダミン、もしくはルシフェリンのような蛍光もしくは化学発光化合物、またはアルカリホスファターゼ、ベータ−ガラクトシダーゼもしくはホースラディッシュ・ペルオキシダーゼなどの酵素であってよい。抗体の検出可能部分への結合について、当技術分野で既知の任意の方法を使用し得る。それらの方法は、Hunter et al., Nature, 144: 945(1962); David et al., Biochemistry, 13: 1014(1974); Pain et al., J. Immunol. Meth., 40: 219(1981); およびNygren, J. Histochem. and Cytochem., 30: 407(1982)に記載された方法を含む。
【0209】
抗HDAC8タンパク質抗体は、組換え細胞培養または天然起源からのHDAC8タンパク質の親和性精製にとっても有用である。この方法では、HDAC8タンパク質に対する抗体を、例えば、Sephadfex樹脂または濾紙などの適当な支持体に、当技術分野で周知の方法を使用して固定する。次いで、固定した抗体を、精製するHDAC8タンパク質を含む試料と接触させる。その後、該支持体を適当な溶剤で洗浄し、固定した抗体に結合している、試料中のHDAC8タンパク質以外の実質的にすべての物質を除去する。最終的に、HDAC8タンパク質を抗体から遊離する別の適当な溶剤で、支持体を洗浄する。
【0210】
抗HDAC8タンパク質抗体を医療処置にも使用し得る。ある実施態様において、抗体をコードしている遺伝子を提供する。抗体は細胞内のHDAC8タンパク質に結合し、それを調節する。
【0211】
ある実施態様において、HDAC8タンパク質、アゴニストまたはアンタゴニストの治療有効量を患者に投与する。ここで「治療有効量」は、投与の目的である効果を生じる用量を意味する。正確な用量は処置目的により異なり、公知技術を用いて当業者は確認し得る。当分野で既知の通り、HDAC8タンパク質分解、全身的対局所的送達、並びに年齢、体重、全般的な健康状態、性別、食事、投与時間、薬剤相互作用および病状の重篤度による調節が必要であり、当業者は常用の試行で確認し得る。
【0212】
本発明の目的における「患者」は、ヒトおよび他の動物の両方、特に哺乳類および生物を包含する。即ち、本発明の方法は、ヒトの治療および獣医学的利用の両方に適用され得る。好ましい実施態様において、患者は哺乳類であり、最も好ましい実施態様において患者はヒトである。
【0213】
本発明のHDAC8タンパク質、アゴニストまたはアンタゴニストの投与を、様々な経路、例えば、経口、皮下、静脈内、鼻腔内、経皮、腹腔内、筋肉内、肺内、膣、直腸または眼内経路で行い得るがこれらに限定されない。場合によっては、例えば、損傷および炎症の処置において、組成物を溶液またはスプレーとして直接適用し得る。導入の方法に応じて、化合物を様々な方法で製剤化し得る。製剤中の治療的に活性な化合物の濃度は、約0.1ないし100重量%で変化し得る。
【0214】
本発明の医薬組成物は、患者への投与に適した形態のHDAC8タンパク質、アゴニストまたはアンタゴニスト(本明細書に記載の抗体およびバイオ活性物質を含む)を含む。好ましい実施態様において、この医薬組成物は、水溶性形態、例えば、医薬的に許容し得る塩類として存在しており、これらは酸および塩基の両付加塩類を包含するものとする。「医薬的に許容し得る酸付加塩類」は、遊離塩基の生物学的有効性を保持し、かつ生物学上またはその他の点で許容できるものであり、無機酸、例えば、塩酸、臭化水素酸、硫酸、硝酸、リン酸など、および有機酸、例えば、酢酸、プロピオン酸、グリコール酸、ピルビン酸、蓚酸、マレイン酸、マロン酸、コハク酸、フマル酸、酒石酸、クエン酸、安息香酸、桂皮酸、マンデル酸、メタンスルホン酸、エタンスルホン酸、p−トルエンスルホン酸、サリチル酸などと形成する。「医薬的に許容し得る塩基付加塩類」は、無機塩基、例えば、ナトリウム、カリウム、リチウム、アンモニウム、カルシウム、マグネシウム、鉄、亜鉛、銅、マンガン、アルミニウム塩類などから誘導されたものを含む。特に好ましいのは、アンモニウム、カリウム、ナトリウム、カルシウムおよびマグネシウム塩類などである。医薬的に許容し得る有機非毒性塩基から誘導される塩類は、第1級、第2級および第3級アミン類、置換アミン類、例えば、天然産生の置換アミン類、環状アミン類および塩基性イオン交換樹脂、例えば、イロプロピルアミン、トリメチルアミン、ジエチルアミン、トリエチルアミン、トリプロピルアミンおよびエタノールアミンの塩類がある。
【0215】
これらの医薬組成物はまた次の物質;即ち担体タンパク質、例えば、血清アルブミン;緩衝液;充填剤、例えば、微晶性セルロース、ラクトース、トウモロコシおよび他の澱粉類などの増量剤;結合剤;甘味料および他の着香剤;着色剤;およびポリエチレングリコールのうちの1種以上を含み得る。添加物は当業界ではよく知られており、様々な製剤で使用される。
【0216】
組成物の組合せを投与し得る。さらに、組成物を成長因子または化学療法剤および/もしくは放射線を含む他の治療剤と組み合わせて投与してもよい。標的化物質(即ち、癌細胞の受容体のリガンド)も、本発明で提供される組成物と組合わせてよい。
【0217】
本発明で提供されるある実施態様では、抗体を免疫療法に用いる。ゆえに、免疫療法の方法が提供される。「免疫療法」は、HDAC8タンパク質に対して生成された抗体を用いて、HDAC8タンパク質に関連する疾患を処置することを意味する。本発明で使用されるように、免疫療法は、受動的または能動的であり得る。本明細書に記載のように、受動的免疫療法は、抗体を受容者(患者)に受動的に輸送することである。能動的免疫療法は、抗体および/またはT細胞反応を受容者(患者)の中で誘導することである。HDAC8タンパク質抗原(これに対して抗体を生成する)を受容者に与えると、その結果免疫反応が誘導され得る。当業者に理解されるように、HDAC8タンパク質抗原は、抗体を生成しようとしているHDAC8タンパク質を受容者に注射するか、またはHDAC8タンパク質抗原を発現する条件下で、HDAC8タンパク質抗原を発現できるHDAC8タンパク質核酸を、受容者に接触させるかして与えられる。
【0218】
好ましい実施態様では、治療的化合物が抗体、好ましくはHDAC8タンパク質抗体に結合している。治療的化合物は細胞毒性物質であってもよい。この方法では、アポトーシス細胞または癌組織もしくは細胞に細胞毒性物質の標的を定めることにより、罹患した細胞の数を低減し、その結果、アポトーシス、癌HDAC8関連障害に付随する症状を低減する結果をもたらす。細胞毒性物質は膨大かつ多様であり、細胞毒性薬もしくは毒物またはそのような毒物の活性断片を含むが、これらに限定されるものではない。好適な毒物およびその対応する断片には、ジフテリアA鎖、エクソトキシンA鎖、リシンA鎖、アブリンA鎖、カルシン(curcin)、クロチン、フェノマイシン(phenomycin)、エノマイシン(enomycin)などが含まれる。細胞毒性物質には、HDAC8タンパク質に対して生成された抗体に放射性同位元素を結合させるか、または抗体に共有結合で結合させたキレート化剤に放射性核種を結合させて作成される放射性化学物質も含まれる。
【0219】
好ましい実施態様では、HDAC8タンパク質遺伝子を、DNA免疫原として投与する。1種類の核酸またはHDAC8タンパク質遺伝子の組合せのどちらかである。むき出しのDNA免疫原は、当分野で一般的に知られている;Brower, Nature Biotechnology 16:1304-1305 (1998)を参照のこと。DNA免疫原としての核酸の使用は、当業者に周知であり、HDAC8タンパク質遺伝子またはHDAC8タンパク質核酸の部分を、患者中での発現用のプロモーターの制御下におくことを含む。DNA免疫原に使われるHDAC8タンパク質遺伝子は、HDAC8タンパク質完全長をコードできるが、より好ましくはHDAC8タンパク質から生じるペプチドを含む、HDAC8タンパク質の部分をコードする。好ましい実施態様では、患者は、HDAC8タンパク質遺伝子から生じる多数の核酸配列を含むDNA免疫原で免疫性を与えられる。同様に、本明細書に記載のように、多数のHDAC8タンパク質遺伝子またはその部分で、患者に免疫性を与えることが可能である。理論によって制限を受けないが、DNA免疫原にコードされるポリペプチドの発現に続いて、HDAC8タンパク質を発現している細胞を破壊または除去する細胞障害性T細胞、ヘルパーT細胞および抗体が誘導される。
【0220】
下記記載の実施例は、上述した本発明の使用態様をより詳しく記載するためのものであるとともに、本発明の多様な側面を実施するために企図された最良の態様を記載するものである。これらの実施例は、如何なる意味においても本発明の真の範囲を制限しようとするものではなく、例示な目的で説明するためのものである。本明細書中で参照したすべての文献は、出典明示により本明細書の一部としている。さらに、開示した全ての配列、引用文献または引例中の各個別番号は、出典明示により本明細書の一部とする。
【実施例】
【0221】
実施例1
Basic BLASTを用いて発現配列タグ(EST)に関するGenBankデータベースの探索は、デフォルトパラメーターを用いるNCBIウェブサイト(http://www.ncbi.nlm.nih.gov/blast/cgi)から利用できるが、ウテリンライブラリーからのEST(40936;受託番号AA33308)がHDAC3のアミノ末端部分に対して相同性を持つことを明らかにした。完全長のクローンを得るために、Marathon- Ready human Uterus cDNAライブラリー(Clontech, Palo Alto, CA)を3'RACEの技術を用いてスクリーンし、そこでcDNAの3'末端をPCRで増幅した。Marathon-Ready cDNAライブラリーを、アダプタープライマーによってフランキングした完全長cDNAのライブラリーとして産生した。3’RACEを行うために、PCR反応をアダプター領域および遺伝子特異的プライマーと相同性のあるプライマーを用いて行った。HDAC8特異的プライマーは:TGCGGAACGGTTTTAAGCGGAG(配列番号:15)であった。
【0222】
この技術から、おおよそ等しい発生量の2つの転写物、1つは1700塩基ともう一つは800塩基の存在が明らかとなった。両形態の配列分析から、800塩基は1700塩基の5’末端の転写物と同一であり、アミノ末端の146残基を示す。
【0223】
HDAC8タンパク質は、HDACのRPD3クラス(I)と相同である。HDAC1、HDAC2およびHDAC3タンパク質は、推定377アミノ酸のHDAC8タンパク質(図2)に関連する短いカルボキシ末端伸長物を包含する。HDAC1、2および3および8のアミノ酸同一性%を、MacVector Prigram(Version 6.5r1 for Macintosh Oxfored Molecular Group)により測定した。
結果は以下である:
HDAC8対HDAC1: 146/486同一性=30%
HDAC8対HDAC2: 151/491同一性=31%
HDAC8対HDAC3: 151/439同一性=34%
HDAC1対HDAC2: 410/488同一性=84%
HDAC2対HDAC3: 250/491同一性=51%
HDAC1対HDAC3: 252/484同一性=52%
上記に示したように、HDAC1−3は、51%から84%のアミノ酸配列の同一性を有する。対照的に、これらタンパク質がHDAC8と共有するアミノ酸同一性は、約30%〜34%であった。即ち、上記HDACタンパク質の全ては、配列同一性を有するがHDAC8はより近密に相関した基の形態を有する他のHDACタンパク質に比べて比較的距離をおいて関連している。
【技術分野】
【0001】
本発明は、クロマチン構造の制御、即ち転写制御に関与する組成物および使用方法に及ぶ。より具体的には、本発明は、転写制御に関与するヒストン脱アセチル化酵素タンパク質をコードする核酸を指向する。使用方法には、転写モデュレーターのスクリーニングアッセイにおける使用および治療薬としての使用が含まれる。
【背景技術】
【0002】
(発明の分野)
ヌクレオソームヒストンの可逆的アセチル化によるクロマチン構造の修飾は、真核細胞における転写制御において重要な役割を担っている(参照:Grunstein. 1997. Nature 389:349-352)。コアヒストンのアミノ酸末端尾内にある特定リジン残基のεアミノ基のアセチル化により局所的なクロマチン弛緩が生じる。このアセチル化は、転写中のヌクレオソームがアンフォールディング(unfolding)構造(変性構造)を維持するために必要である(Walia et al. 1998. J. Biol. Chem. 273:14516-14522)。一般的に、ヒストンアセチル化活性は転写活性と関連しており、一方脱アセチル化活性は転写抑制と関連している。遺伝子発現中の広範な変化は、細胞核内のヒストンアセチル基転移酵素(HAT)活性とヒストン脱アセチル化酵素(HDAC)活性との間の動的平行により起こると考えられている。
【0003】
HDACは、比較的大きいマルチサブユニット複合体との相互作用によって転写抑制を媒介する。例えば、HDAC1として知られる特定のHDACは、コリプレッサーSin3と結合し、HDAC1−Sin3はサイレンシング媒介物質NCoRおよびSMRTとさらに結合することが知られている(Alland, et al. 1997. Nature 387:49-55)。それから、N−CoR/SMRT-HDAC1は、核内のプロモーターエレメントと結合している特定の転写因子によって供給される。例えば、網膜芽腫(Rb) 遺伝子産物は、N-CoR/SMRT-HDAC1を供給し、転写因子E2Fと結合してE2F調節プロモーターを抑制する(Luo et al. 1998. Cell 92:463-473; Brehm et al. 1998. Nature. 391:597-600, Magnaghi-Jaulin et al. 1998. Nature 391:601-605)。HDAC1-Sin3もまた結合し、Mad/MAXリプレッサーヘテロダイマーによって抑制を媒介する (Laherty et al. 1997. Cell 89:349-356)。HDAC1のヒストン脱アセチル化活性は、この転写抑制にとって必須のものである(Hassig et al. 1998. PNAS 95:3519-3524)。
【0004】
ヒストン脱アセチル化酵素をコードする幾つかのcDNAが特性分析されている。クローンされた最初のものは酵母タンパク質RPD3であり、これは転写抑制因子に関する遺伝子的スクリーンにおいて最初に同定された(Vidal et al. 1991. Mol. Cell Biol. 11:6317-6327)。哺乳類HDAC1はTSAの標的分子として別個にクローンされた(Taunton et al. 1996. Science 272:408-411)。HDAC1は、酵母RPD3のオルトログ(ortholog)であることが観察され、また両方ともインビトロでHDAC活性を持つことが示された。RPD3に対して相同なタンパク質をコードする3つのcDNA、HDAC1、HDAC2およびHDAC3が記載されている(Yang et al. 1997. J. Biol. Chem. 272:28001-28007; Emiliani et al. 1998. PNAS 95:2795-2800)。3つ全ては、偏在組織分布を持ち、またその活性は重複するようである。現在まで、HDACをコードする5つのS. cerevisiae遺伝子が発見されており、そのファミリーは配列保存および提案された生物学的機能を基にした2つのクラスに分けられてきた。S. cerevisiaeのHos1、Hos2、Hos3およびRPD3は哺乳類HDAC1、2および3と共にクラス1に含まれる。第2のクラスは、酵母HDA1およびHDA3および最近同定されたヒトホモログHDAC4、HDAC5およびHDAC6から構成される(Grozinger et al. 1999. PNAS 96:4868-4873; Fischle et al. 1999. J. Biol. Chem. 274:11713-11720)。
【0005】
HDACはマルチサブユニット転写制御複合体の一部として機能していることが知られているので、制御HDACは、HDACを含有する制御複合体の制御下で、特定遺伝子または一組の特定遺伝子の転写を調節する、即ち増加または低下させるように使用され得る(Carmen et al. 1996. J. Biol. Chem. 271(26):15837-15844)。
【0006】
HDAC阻害活性を持つ多様な化合物はヒストンの過アセチル化をもたらすことがわかっており、この調節は、多くの細胞型で細胞周期の拘束および末端分化を伴う (参照:Proceedings of the American Association for Cancer Research 40, March, 1999)。NaBuなどの、強力で特異な可逆的阻害剤である該菌類毒素トリコスタチンA(TSA)は、多様なシステムにおける細胞周期拘束および/または分化を誘導することがわかってきた。近年、HDAC阻害剤活性を有する合成ベンザミド誘導体(MS-27275)が記載された(Saito et al. 1999. PNAS 96:4592-5497)。MS-27275は、ヒストンの過アセチル化を誘導し、腫瘍罹患ヌードマウスにおける腫瘍増殖を阻害する点で効果を示した。
【0007】
急性前骨髄性白血病(APL)では、遺伝的再配列により、PMLまたはPLZFタンパク質のいずれかのレチノイン酸受容体α(RARα)の融合が生じる。PML-RARαおよびPLZF-RARαの両方は、N-CoR/SMRT-HDACを異常に補充し、レチノイン酸制御遺伝子を抑制することによって白血病誘発発生を媒介する(Lin et al. 1998. Nature 391:811-814; Grignani et al. 1998. Nature 391:815-818)。実際、APLにおける転写を標的とする非特異的HDAC阻害剤であるブチル酸ナトリウム(NaBu)の治療的使用は、その疾患に関するレチノイン酸耐性型においてさえも成功してきた(Warrell et al. 1998. J. Natl. Cancer Inst. 90:1621-1625)。HDACの役割は、ETO(「821(eight twenty one)」 または MT68) 遺伝子産物とAML1 転写因子の融合の結果として生じる急性骨髄性白血病においてもまた実証されている(Gelmetti et al. 1998. Mol. Cell Biol. 18:7185-7191; Wang et al. 1998. PNAS 95:10860-10865)。併せて、これは、HDACの確実な白血病の治療に対する有効な方法であることを示唆している。さらに、大腸癌(Hassig et al. 1997. Chem. Biol. 4:783-789; Archer et al. 1998. PNAS 95:6791-6796)、扁平上皮細胞癌(Gillenwater et al., 1998. Int. J. Cancer 75:217-224; Saunders et al. 1999. Cancer Res. 59:399-404)、腺癌(McBain et al. 1997. Biochem. Pharmacol. 53:1357-1368)および神経芽腫(Swendeman et al. 1999. Proc. Amer. Assoc. Cancer Res. 40, Abst. #3836)を含む別の種類の癌においても、HDACの役割に関するさらなる証拠がある。HDACは、BRCA1(Yarden et al. 1999. Proc. Ameri. Assoc. Cancer Res. 40, Abstr. #3387)およびc-fos(Bakin and Curran. 1999. Science 283:387-390)に媒介される転写制御にも関わっている。
【0008】
一方でまた、制御された方法において細胞増殖を増強することが、ときに望まれる。例えば、細胞増殖は創傷治癒および組織の増殖が望ましいところで有用である。即ち、増殖の阻害を助長、増強または遅延させる調節因子を同定することが望ましい。
【発明の概要】
【発明が解決しようとする課題】
【0009】
転写成分および調節因子を同定することは望ましいが、そのような化合物の分野では十分になされていない。従って、転写の調節因子をスクリーニングするための有用な組成物および方法を提供することは利点がある。また、転写制御に関与する新規組成物を提供することも又利点がある。
【課題を解決するための手段】
【0010】
(発明の要旨)
上記目的に従って、本発明は、HDACタンパク質をコードする核酸を提供する。そのポリペプチドを本出願において「HDAC8」として示す。
【0011】
ある実施態様において、本発明はHDAC8をコードする単離された核酸分子を提供する。また、本発明はHDAC8タンパク質およびその断片を提供する。
【0012】
本発明は、HDAC8タンパク質をコードする核酸に高緊縮条件下でハイブリダイズするHDACタンパク質をコードする核酸を提供する。それに従って、本発明はHDAC8タンパク質またはその断片をコードする核酸を同定する方法を提供する。
【0013】
本発明は、HDAC8タンパク質をコードし、発現する発現ベクターをさらに提供する。
【0014】
別の態様において、本発明はHDAC8タンパク質をコードする組換え核酸および発現ベクターを含む宿主細胞を提供する。それに従って、本発明は、HDAC8タンパク質を産生する方法を提供する。
【0015】
さらなる態様において、本発明はHDAC8アミノ酸配列を含む組換えポリペプチドおよびHDAC8アミノ酸配列を含有するポリペプチドを同定する方法を提供する。
【0016】
別の態様において、本発明はHDAC8ポリペプチドと結合する抗体を提供する。
【0017】
さらに別の態様において、本発明はHDAC8タンパク質に結合するバイオ活性物質をスクリーニングする方法を提供する。
【0018】
さらに別の態様において、本発明は異常なHDAC8発現と関連のある疾患状態を同定する方法を提供する。
【0019】
本発明の別の態様は、本発明に関する以下の記載によって当業者に明らかであろう。
【図面の簡単な説明】
【0020】
【図1−1】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−2】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−3】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−4】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図1−5】図1は、HDAC8と呼ばれるcDNAクローンの完全なヌクレオチド配列を示す。その配列(配列番号:1)により、HDACのRPD3クラス(I)との相同性を有するタンパク質(配列番号2)産物を推定する。翻訳開始因子および終結因子のコドンは、下線した。
【図2】図2は、HDAC1(配列番号:4)、HDAC2(配列番号:5)、HDAC3 (配列番号:6)およびHDAC8(配列番号:7)との間のアミノ酸の類似性を示す。同一残基は暗灰色のボックスとし、保存性のある置換基は明るい灰色で示す。推定377アミノ酸HDAC8(約42kDa)産物は、HDAC1、2および3のC末端の〜100アミノ酸に相同性のある領域を失っていることがわかる。
【発明を実施するための形態】
【0021】
(発明の詳細な説明)
本発明は、ヒストン脱アセチル化-8(HDAC8)タンパク質とそのタンパク質をコードする核酸を提供する。HDAC8タンパク質を結合し得る、また好ましくはHDAC8タンパク質の活性を調節し得るバイオ活性物質をスクリーニングする方法を提供する。その方法は、HDAC8タンパク質および候補バイオ活性物質および細胞または細胞集団を合わせ、候補物質の存在下および非存在下で細胞への効果を測定することを含む。結合アッセイを含む他のスクリーニングアッセイも、下記のように本発明で提供される。HDAC8タンパク質を制御するか、もしくは調節するための治療薬も提供され、本明細書に記載されている。以下にさらに記載するように、診断薬も提供される。
【0022】
本発明のHDAC8タンパク質は、いくつかの方法で同定され得る。この意味での「タンパク質」には、タンパク質、ポリペプチドおよびペプチドが含まれる。本発明のHDAC8タンパク質は、2つの一般的クラスに分類される:完全に新しいタンパク質、即ち、発見時点で公共のデータベースの一部分になっていないものである。但し、それらは発現配列タグ(EST)によってコードされた既知のタンパク質またはペプチドとホモロジーを持ち得る。今一つは、HDAC8タンパク質が既知のタンパク質であるが、クロマチン構造制御、ヒストンアセチル化、転写制御などに関与することが知られていないものである。即ち、本明細書中で新規生物学的機能を持つものとして同定される。従って、HDAC8タンパク質は、ヒストンアセチル化および/または転写に関与すると知られているタンパク質との関連で最初に同定され得る。HDAC8タンパク質および核酸が新規である場合には、組成物および使用の方法が本明細書で提供される。HDAC8タンパク質および核酸が既知であるが、しかし本明細書中に記載のようにクロマチン構造の制御、ヒストンアセチル化および/または転写に関与することが知られていない場合には、使用方法、即ち機能スクリーンが提供される。
【0023】
本発明で提供されるある実施態様では、HDAC8タンパク質は、本明細書で定義するように、1つまたはそれ以上の下記特性を有する:ヒストンのアセチル化;HDAC1(配列番号:4)、HDAC2(配列番号:5)、HDAC3(配列番号:6)およびHDAC8(配列番号:7)との相同性。該相同性は、下記データベース、アルゴリズムおよびパラメーターを用いて見出される。本明細書中の「相同性」とは、配列の類似性および好ましくは同一性を意味する。
【0024】
ある実施態様では、HDAC8核酸またはHDAC8タンパク質は、本発明で提供される配列に対するへの核酸および/またはアミノ酸配列の実質的同一性または類似性により最初に同定される。好ましい実施態様では、HDAC8核酸またはHDAC8タンパク質は、下記のように本発明で提供される配列に対して配列同一性または類似性を有し、1またはそれ以上のさらに後述するようなHDAC8タンパク質バイオ活性を有する。そのような配列同一性または類似性は、全体的な核酸またはアミノ酸配列に基づくことができる。
【0025】
好ましい実施態様では、あるタンパク質は、図1のアミノ酸配列に対する全体的な配列同一性が好ましくは約60%より大きく、より好ましくは約70%より大きく、さらに好ましくは約80%より大きく、最も好ましくは約90%より大きいならば、本明細書で定義する「HDAC8タンパク質」である。ある実施態様では、該配列同一性は約93ないし95または98%にも上る。
【0026】
技術上周知のように、タンパク質(または以下に考察するように核酸)が既知配列と配列同一性または類似性を有しているか否かを同定するために多数の異なるプログラムを使用することができる。配列同一性および/または類似性は技術上周知の標準的技法を用いて測定することができ、それらは、Smith & Waterman, Adv. Appl. Math., 2: 482 (1981)の局所配列同一性アルゴリズム、Needleman & Wunsch, J. Mol. Biol., 48: 443 (1970)の配列同一性アラインメント、Pearson & Lipman, PNAS U.S.A., 85: 2444 (1988)の類似性検索法、これらのアルゴリズムのコンピュータによる実行(Wisconsin Genetics Software Package、Genetics Computer Group, 575 Science Drive, Madison, WI、中のGAP、BESTFIT、FASTAおよびTFASTA)、Devereux et al., Nucl. Acid Res., 12: 387-395 (1984)に記載のBest Fit配列プログラム、を非限定的に含み、好ましくはデフォルト設定を用い、または検定により使用する。好ましくは、FstDBにより以下のパラメータに基づいてパーセント同一性を計算する:ミスマッチペナルティ(mismatch penalty) 1; ギャップペナルティ(gap penalty)1; ギャプサイズペナルティ(gap size penalty) 0.33; 接合ペナルティ(joining penalty) 30、「Current Methods in Sequence Comparison and Analysis,」Macromolecule Sequencing and Synthesis, Selected Methods and Applications, pp 127-149 (1988), Alan R. Liss, Inc.である。
【0027】
有用なアルゴリズムの一例はPILEUPである。PILEUPは、漸進的対アラインメントを用いて関連配列から多重配列アラインメントを創成する。それはまた、アラインメント創成に使用される、クラスタリングの関係を示すツリーを描くことができる。PILEUPは、Feng & Doolittle, J. Mol. Evol. 35: 351-360 (1987)の漸進的アラインメント法の簡略化したものを用いる;この方法はHiggins & Sharp CABIOS 5: 151-153 (1989)記載の方法と類似している。有用なPILEUPパラメータは、デフォルトギャップウェイト(default gap weight) 3.00, デフォルトギャップレングスウェイト(default gap length weight) 0.10, ウェイテッド・ギャップ・エンド・ギャップ(weighted end gaps)を含む。
【0028】
有用なアルゴリズムのもう1つの例は、Altschul et al., J. Mol. Biol. 215, 403-410, (1990);およびKarlin et al., PNAS U.S.A. 90: 5873-5787 (1993)に記載のBLASTアルゴリズムである。特に有用なBLASTプログラムは、Altschul et al., Methods in Enzymology, 266: 460-480 (1996); http://blast.wustl/edu/blast/ README.htmlから得られるWU−BLAST−2である。WU−BLAST−2はいくつかの検索パラメータを使用するがその殆どはデフォルト値に設定されている。調節可能なパラメータは以下の値で設定する:オーバーラップスパン(overlap span)=1, オーバーラップフラクション(overlap fraction) = 0.125, 閾値(word threshold)(T) = 11である。HSP SおよびHSP S2パラメータは動的数値であり、特定の配列の組成および興味の対象である配列を検索する特定のデータベースの組成に依存してプログラム自身により決定されるが、値は感度を上げるように調節することができる。
【0029】
これに加えて有用なアルゴリズムは、Altschul et al., Nucl. Acids Res., 25: 3389-3402に報告されているギャップドBLASTである。ギャップドBLASTはBLOSUM−62代替スコアを使用する。ここで閾値Tパラメータは9に設定し;2−ヒット法によりギャップのない伸長を開始し;ギャップ長kにコスト10+kを課し;Xuを16に設定し;Xgをデータベース検索段階では40に、そしてアルゴリズムのアウトプット段階では67に設定する。ギャップドアラインメントは約22ビットまでに相当するスコアで開始される。
【0030】
パーセントアミノ酸配列同一性の値は、マッチする同一残基の数を、アラインメントを行った領域における「より長い」配列の総残基数で割って決定される。「より長い」配列は、アラインメントを行った領域中で実際の残基を最も多く有する配列である(アラインメントスコアを最大化するためにWU−Blast−2により導入されたギャップを無視する)。
【0031】
同様にして、ここで同定されるポリペプチドのコード配列に関する「パーセント(%)核酸配列同一性」を、HDAC8タンパク質のコード配列中のヌクレオチド残基と同一な、候補配列中のヌクレオチド残基の割合として定義する。好ましい方法は、WU−BLAST−2のBLASTNモジュールをデフォルトパラメータに設定し、オーバーラップスパンおよびオーバーラップフラクションをそれぞれ1および0.125に設定して利用するものである。
【0032】
アラインメントは、アラインメントを行う配列中へのギャップ導入を含んでもよい。加えて、図中の配列でコードされるタンパク質より多いかまたは少ないアミノ酸を含有する配列については、ある実施態様では、配列同一性の割合は、アミノ酸総数に対する同一アミノ酸数に基づいて決定されると理解される。かくして、例えば、後で論じるように、ある実施態様では、図1に示す配列より短い配列の配列同一性は、より短い配列中のアミノ酸数を用いて決定される。パーセント同一性の計算においては、相対的重みは、挿入、欠失、置換その他のような種々の配列変化の表出に起因するとはされない。
【0033】
ある実施態様では、同一性のみがプラスのスコアを与えられ(+1)、ギャップを含むあらゆる形の配列変化に「0」の値が与えられる。これにより、配列類似性計算について後述するような、重みをつけた(加重値を与える)目盛りまたはパラメータの必要性がなくなる。例えば、配列同一性の割合は、アミノ酸総数に対する同一アミノ酸数に基づいて決定されると理解される。例えば、パーセントアミノ酸配列同一性は、マッチする同一残基の数を、アラインメントを行った領域における「より短い」配列の総残基数で割り、100を掛けて算出される。「より長い」配列は、アラインメントを行った領域中で実際の残基を最も多く有する配列である。
【0034】
当業者に認識されるであろうように、本発明の配列は、配列のエラーを含んでもよい。つまり、誤ったヌクレオシド、フレームシフト、未知のヌクレオシド、または他のタイプの配列エラーが、配列のどこにあってもよい。しかし、正しい配列は、本発明の相同性および緊縮の定義の範囲内にある。
【0035】
本発明のHDAC8タンパク質は、図1に示す核酸にコードされるアミノ酸配列よりも、短くても長くてもよい。ゆえに、好ましい実施態様では、本発明で提供される核酸配列にコードされるアミノ酸配列の部分または断片が、HDAC8タンパク質の定義に包含される。本発明のある実施態様では、HDAC8タンパク質の断片は、a)少なくとも1つの抗原のエピトープを共有する;b)少なくとも表示される配列同一性を有する;およびc)好ましくは本発明でさらに定義されるヒストン脱アセチル化酵素−8の生物学的活性を有する場合、HDAC8タンパク質である。該配列が診断的に用いられる場合、つまり、HDAC8タンパク質の核酸の有無が求められる場合には、表示される配列の同一性のみが要求される。本発明の核酸もまた、図1の配列よりも、短くても長くてもよい。核酸断片には、以前に正確には同定されていない配列を有する、本発明で提供される核酸のあらゆる部分が含まれる;以前に同定されていない部分に対して、表示される配列同一性がある配列を有する断片が、本発明のある実施態様で提供される。
【0036】
さらに、より詳しく後で概説するように、図1に示されるものよりも長いHDAC8タンパク質を作成し得る。例えばエピトープもしくは精製用タグの付加、他の融合配列の付加、または付加的なコードおよび非コード配列の解明による。後述のように、HDAC8ペプチドを、グリーン蛍光ペプチド(GFP)などの蛍光ペプチドと融合させることが、特に好ましい。
【0037】
HDAC8タンパク質はまた、本明細書で概説するように、図1に示す配列またはその相補鎖とハイブリダイズするHDAC8核酸にコードされるものとしても同定され得る。ハイブリダイゼーションの条件は、さらに後述する。
【0038】
好ましい実施態様では、抗体を作成するためにHDAC8タンパク質を用いようとするとき、HDAC8タンパク質は、完全長タンパク質と、少なくとも1つのエピトープまたは決定因子を共有せねばならない。本明細書では、「エピトープ」または「決定因子」は、抗体を作成および/または結合するであろうタンパク質の一部分を意味する。ゆえに、ほとんどの例で、より小さいHDAC8タンパク質に対して作られた抗体は、完全長タンパク質に結合できる。好ましい実施態様では、エピトープは独特である。つまり、独特なエピトープに対して作成された抗体は、交差反応性をほとんど、または全く示さない。「抗体」という用語には、当分野で周知のように、抗体全体を修飾して作成されたか、または組換えDNA技術を用いて新規に合成された、Fab、Fab2、1本鎖抗体(例えば、Fv)、キメラ抗体などを含む抗体断片が含まれる。
【0039】
後述するように、好ましい実施態様では、HDAC8タンパク質に結合するHDAC8タンパク質に対する抗体には、本明細書に記載のHDAC8タンパク質の生物学的機能を低減するか、または除去する能力がある。つまり、抗HDAC8タンパク質抗体(ポリクローナル、または好ましくはモノクローナルのどちらでも)をHDAC8タンパク質(またはHDAC8タンパク質を含有する細胞)に加えることで、HDAC8活性を低減するか、または除去し得る。一般的に、少なくとも25%の活性の減少が好ましく、少なくとも約50%が特に好ましく、約95−100%の減少がことさら好ましい。
【0040】
本発明のHDAC8抗体は、HDAC8タンパク質に特異的に結合する。本明細書では、「特異的に結合する」は、少なくとも10−4−10−6M−1の範囲の結合定数で、好ましくは10−7−10−9M−1の範囲で、抗体がタンパク質に結合することを意味する。抗体について、さらに後述する。
【0041】
核酸の場合、核酸配列の包括的な配列同一性は、アミノ酸の配列同一性と同一基準であるが、遺伝暗号の縮重と異なる生物間のコドンの偏りを考慮にいれる。従って、核酸配列の同一性は、タンパク質配列のそれと比べて低くも高くもあり得る。ゆえに、図の核酸配列と比較した核酸配列の配列同一性は、好ましくは75%より大きく、より好ましくは約80%より大きく、特に約85%より大きく、そして最も好ましくは90%より大きい。いくつかの実施態様では、該配列同一性は約93ないし95または98%ほどに高い。
【0042】
好ましい実施態様では、HDAC8核酸は、HDAC8タンパク質をコードする。当業者には理解されるであろうように、遺伝暗号の縮重のために、すべて本発明のHDAC8タンパク質をコードする、極めて多数の核酸を作り得る。ゆえに、特定のアミノ酸配列を同定すれば、当業者は、HDAC8タンパク質のアミノ酸配列を変えない方法で、単純に1つまたはそれ以上のコドンの配列を変更することにより、異なる核酸をいくつでも作成し得る。
【0043】
ある実施態様では、核酸はハイブリダイゼーション検査により決定される。ゆえに、例えば、図1に示す核酸配列またはその相補鎖と、高緊縮条件下のもとでハイブリダイゼーションする核酸は、HDAC8核酸と考えられる。高緊縮条件下条件は当分野で既知である;例えば、出典明示により本明細書の一部とするManiatis et al., Molecular Cloning: A Laboratory Manual, 2d Edition, 1989およびShort Protocols in Molecular Biology, ed. Ausubel, et al.を参照のこと。高緊縮条件は配列依存的であり、環境が違えば異なる。より長い配列はより高い温度で特異的にハイブリダイズする。核酸のハイブリダイゼーションの発広範な手引きは、Tijssen, Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993)に記載されている。一般的に、特異的な配列について、緊縮条件は、規定されたイオン強度、pHにおいて熱融解点(Tm)より約5−10℃低いように選択される。Tmは、標的に相補的なプローブの50%が、平衡状態で標的配列に(規定のイオン強度、pHおよび核酸濃度で)ハイブリダイズする温度である(標的配列が過剰に存在するので、Tmでは、50%のプローブが平衡状態において占められる)。緊縮条件は、塩濃度が約1.0Mナトリウムイオンより小さく、典型的には0.01ないし1.0Mのナトリウムイオン(または他の塩)濃度、pH7.0ないし8.3、温度が、短いプローブ(例えば、10ないし50のヌクレオチド)には低くても約30℃、長いプローブ(例えば、50ヌクレオチド以上)には低くても約60℃、である。緊縮条件は、ホルムアミドのような不安定化させる物質の付加により達せられてもよい。
【0044】
他の実施態様では、より低い緊縮条件のハイブリダイゼーション条件を用いる;例えば、当分野で周知のように、穏やかなまたは低緊縮条件を用いてもよい;Maniatis and Ausubel, 前出, and Tijssen(前出)を参照のこと。
【0045】
本発明のHDAC8タンパク質と核酸は、好ましくは組換えである。ここで用いられる「核酸」は、さらに定義するように、DNAもしくはRNA、またはデオキシおよびリボヌクレオチドの両者を意味し得る。核酸は、ゲノムDNA、cDNAおよびオリゴヌクレオチドを含むセンスおよびアンチセンス核酸を含む。このような核酸はまた、生理的環境におけるそのような分子の安定性および半減期を増加させるために、リボース−リン酸バックボーンに修飾を含んでいてもよい。
【0046】
核酸は2本鎖、1本鎖であってよく、または2本鎖もしくは1本鎖の配列の両方の部分を含んでいてもよい。当業者により認識されるであろうように、1本鎖(「ワトソン」)を描けばもう1つの鎖(「クリック」)の配列が定義されるので、図に示す配列は、該配列の相補鎖もまた含む。ここで「組換え核酸」という用語は、一般的に、核酸をエンドヌクレアーゼによって操作して、自然界には存在しない形状で、独創的にインビトロで形成した核酸を意味する。ゆえに、線状の形状で単離されたHDAC8核酸や、通常は結合していないDNA分子を連結することによりインビトロで形成した発現ベクターは、両者とも本発明のためには組換え体と考えられる。一旦組換え核酸が作成され宿主細胞または生物に再導入されれば、それは非組換え的に、即ち、インビトロの操作ではなく宿主のインビボの細胞機構を用いて複製すると理解される;しかしながら、そのような核酸は一旦組換え的に生産されれば、以後は非組換え的に複製しても、本発明のためにはなお組換え体と考えられる。
【0047】
同様に、「組換えタンパク質」は組換え技術を用いて、即ち上述のように組換え核酸の発現を通して作成されたタンパク質である。組換えタンパク質は、少なくとも1つまたはそれ以上の特性によって、天然産生のタンパク質と区別される。例えば、このタンパク質は、野生型宿主中で通常会合しているタンパク質または化合物の一部または全てから単離または精製され、実質的に純粋であり得る。例えば、単離されたタンパク質は、自然状態では通常会合している物質の少なくとも一部を伴わないで、所定の試料中の総タンパク質重量の好ましくは少なくとも約0.5%、より好ましくは少なくとも5%を構成している。実質的に純粋なタンパク質は、総タンパク質重量の少なくとも約75%、好ましくは少なくとも約80%、そして特に好ましくは少なくとも約90%を含む。この定義には、ある生物由来のHDAC8タンパク質を異なる生物または宿主中で生産することが含まれる。あるいは、タンパク質がより増加した濃度レベルで作られるように誘導可能プロモーターもしくは高発現プロモーターを使用することにより、タンパク質を通常見られるよりも有意に高濃度で作ることができる。あるいは、以下に考察するように、該タンパク質は、エピトープタグの付加またはアミノ酸置換、挿入および欠失を含むような、自然界に通常見出されない形であり得る。
【0048】
ある実施態様では、本発明は、HDAC8タンパク質の変異体を提供する。これらの変異体は置換、挿入または欠失変異体の3つの分類の1つまたはそれ以上に相当する。これらの変異体は通常、カセット式もしくはPCR式変異誘発または技術上周知の他の技術を用いて、HDAC8タンパク質をコードするDNA中のヌクレオチドの部位特異的変異誘発により、変異体をコードするDNAを生産し、そして次いでDNAを上に略述した組換え培養細胞中で発現させることにより調製する。しかしながら、約100〜150残基までを有する変異HDAC8タンパク質断片を、確立された技術を用いてインビトロ合成により調製することができる。アミノ酸配列変異体は変化が予め決定されているという特徴を有し、この特徴によって、これらの変異体は、HDAC8タンパク質のアミノ酸配列に対する天然産生の対立遺伝子変異体または種間変異体から区別される。変異体は典型的に、天然産生の類似体と定性的に同じバイオ活性を発揮するが、ただし、以下にさらに詳しく略述するように、変更された特性を有する変異体を選択することもできる。
【0049】
アミノ酸配列変異を導入する部位または領域は予め決定されるが、変異自体は予め決定しておく必要はない。例えば、所定の部位における変異能を最適化するために、標的コドンまたは領域にランダム変異誘発を起こし、発現したHDAC8細胞周期変異体をスクリーニングして所望の活性の最適な組合わせを有するものを探してもよい。既知の配列を有するDNA中の予め定められた部位に置換変異を作成する技術は周知であり、例えば、M13プライマーによる変異誘発およびPCRによる変異誘発がある。変異体のスクリーニングは、HDAC8タンパク質活性のアッセイを用いて行われる。
【0050】
アミノ酸置換は典型的には単一の残基置換である;かなり大きな挿入も許容されるが、挿入は通常、約1〜20アミノ酸の単位で行われる。欠失は、より大きな場合もあるが、約1から約20残基の範囲である。
【0051】
最終誘導体に到達するために、置換、欠失、挿入またはそれらのいずれの組み合わせを用いてもよい。一般的に、これらの変化は、分子の変化を最小限にするために少数のアミノ酸について行われる。しかしながら、より大きな変化も一定の状況では許容される。HDAC8タンパク質の特徴について小さな変化が望まれる場合は、置換は一般的に次のチャートに従ってなされる。
(表1)
チャートI
【0052】
機能または免疫学的同一性における実質的な変化は、チャートIに示したものより保存性の低い置換を選択することによってもたらされる。例えば、より大きく影響する置換を行うことができる:それらは、変化する区域のポリペプチドバックボーンの構造、例えばアルファ−ヘリックス構造またはベータ−シート構造;標的部位での分子の電荷または疎水性;または側鎖の大きさである。一般的にポリペプチドの性質に最も大きな変化を生じると期待される置換は(a)親水性残基、例えばセリルまたはスレオニルを、疎水性残基、例えばロイシル、イソロイシル、フェニルアラニル、バリル、またはアラニルに置換する(またはその逆)、(b)システインまたはプロリンを他のいずれかの残基に置換する(またはその逆)、(c)正電荷を持つ側鎖、例えばリシル、アルギニル、またはヒスチジルを負電荷を持つ側鎖、例えばグルタミル、アスパルチルに置換する(またはその逆)、(d)大きい側鎖を持つ残基、例えばフェニルアラニンを側鎖を持たない残基、例えばグリシンに置換する(またはその逆)、ものである。
【0053】
変異体は典型的には天然産生の類似体と定性的に同じ生物活性を発揮し、同じ免疫応答を誘起するが、ただし、必要に応じてHDAC8タンパク質の特性を変更するような変異体もまた選択される。あるいは、変異体をHDAC8タンパク質の生物活性が変わるようにデザインすることができる。例えば、グリコシル化部位を加えたり、変えたりまたは除去したりすることができる。HDAC8変異体は、リン酸化部位を加え、変え、除去することが可能である。例えばcAMP仲介性リン酸化コンセンサス配列「KRAS」が見出されるアミノ酸部位36−39で変更できる。チロシンリン酸化コンセンサス配列は、Try−174で見出され、これも変更可能である。
【0054】
HDAC8ポリペプチドの共有結合による修飾は、本発明の範囲内に含まれる。共有結合による修飾の1つのタイプは、HDAC8ポリペプチドの標的アミノ酸残基を、HDAC8ポリペプチドの選択された側鎖またはN−もしくはC−末端と反応できる有機誘導体化試薬と反応させることを含む。以下に詳述するように、二官能性試薬による誘導体化は、例えば、抗HDAC8抗体の精製またはスクリーニングアッセイの方法において使用する水不溶性の支持マトリックスまたは表面に、HDAC8タンパク質を架橋するために有用である。一般的に用いられる架橋試薬としては、例えば、1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、例えば4−アジドサリチル酸とのエステルのようなN−ヒドロキシスクシンイミドエステル、3,3'−ジチオビス(スクシンイミジルプロピオン酸エステル)のようなジスクシンイミジルエステルを含むホモ二官能イミドエステル、ビス−N−マレインイミド−1,8−オクタンのような二官能マレインイミドおよびメチル‐3−[(p−アジドフェニル)ジチオ]プロピオンイミダートのような試薬が挙げられる。
【0055】
他の修飾としては、グルタミニルおよびアスパラギニル残基からそれぞれ対応するグルタミルおよびアスパルチル残基への脱アミド化、プロリンおよびリシンのヒドロキシル化、セリルまたはスレオニル残基の水酸基のリン酸化、リシン、アルギニン、およびヒスチジン側鎖のアミノ基のメチル化[T.E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)]、N末端アミンのアセチル化、およびC末端カルボキシル基のアミド化が挙げられる。
【0056】
本発明の範囲内に含まれるHDAC8ポリペプチドの別のタイプの共有結合的修飾は、ポリペプチドの天然のグリコシル化パターンを変えることより成る。「天然のグリコシル化パターンを変える」とは、ここでの目的には、HDAC8ポリペプチドの天然の配列中に見出される1個もしくはそれ以上の炭水化物部分を削除、および/またはHDAC8ポリペプチドの天然の配列に存在しない1個またはそれ以上のグリコシル化部位を付加することを意味する。
【0057】
HDAC8ポリペプチドへのグリコシル化部位の付加はそのアミノ酸配列を変化させることにより達成される。例えば、その変化は、1個またはそれ以上のセリンまたはスレオニン残基をHDAC8ポリペプチドの天然配列に添加または置換することによって行われる(O−グリコシル部位)。また、その変更は、例えば、天然配列のHDAC8ポリペプチド中で、1以上のAxn−Xaa−Ser/Thr部位(Xaa=任意のアミノ酸)の添加もしくは置換でなされ得る(N−グリコシル部位)。HDAC8アミノ酸配列は、DNAレベルでの変化を通して任意に変化させ得るが、特に、所望のアミノ酸に翻訳されるであろうコドンを作成するように予め選択した塩基において、HDAC8ポリペプチドをコードするDNAを変異させることによって変化させ得る。
【0058】
HDAC8ポリペプチド上の炭水化物部分の数を増加させるもう1つの方法は、ポリペプチドにグリコシドを化学的もしくは酵素的に結合させることによる。このような方法は、技術上で、例えば、1987年9月11日に公開されたWO 87/05330およびAplin and Wriston, CRC Crit. Rev. Biochem., pp. 259-306 (1981)に記述されている。
【0059】
HDAC8ポリペプチド上に存在する炭水化物部分の除去は、化学的にもしくは酵素的に、またはグリコシル化の標的となっているアミノ酸残基をコードするコドンを置換する変異により達成される。化学的脱グリコシル化の手法は技術上周知であり、例えば、Hakimuddin, et al., Arch. Biochem. Biophys., 259: 52 (1987)およびEdge et al., Anal. Biochem., 118: 131 (1981)に記述されている。ポリペプチド上の炭水化物部分の酵素的切断は、Thotakura et al., Meth. Enzymol., 138: 350 (1987)に記述されているように種々のエンド−およびエキソ−グリコシダーゼの使用により達成できる。
【0060】
もう1つのタイプのHDAC8ポリペプチドの共有結合的修飾は、特許第4640835号、第4496689号、第4301144号、第4670417号、第4791192号または第4179337号に示されている方法で、HDAC8ポリペプチドを種々の非タンパク質ポリマー、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンの1つに結合させることを含む。
【0061】
本発明のHDAC8ポリペプチドはまた、もう1つの、異種のポリペプチドまたはアミノ酸配列に融合したHDAC8ポリペプチドを含むキメラ分子を形成するように修飾し得る。ある実施態様では、そのようなキメラ分子は、HDAC8ポリペプチドと、抗タグ抗体が選択的に結合できるエピトープを提供するタグポリペプチドとの融合を含む。エピトープタグは一般的に、HDAC8ポリペプチドのアミノ−(N−)またはカルボキシル−(C−)末端に置かれる。HDAC8ポリペプチドにそのようなエピトープタグが付いた形のものの存在は、タグポリペプチドに対する抗体によって検出することができる。また、エピトープタグをつけることにより、HDAC8ポリペプチドが抗タグ抗体、またはエピトープタグに結合する別のタイプのアフィニティーマトリックスによる精製が容易になる。別の実施態様では、キメラ分子は、HDAC8ポリペプチドと免疫グロブリンまたは免疫グロブリンの特定の領域との融合を含んでもよい。以下でさらに論じるように、2価形態のキメラ分子について、このような融合はIgG分子のFc領域に対してなし得る。
【0062】
種々のタグポリペプチドおよびその各々の抗体は技術上周知である。例としては、ポリ−ヒスチジン(poly-his)またはポリ−ヒスチジン−グリシン(poly-his-gly)タグ、flu HAタグポリペプチドおよびその抗体12CA5[Field et al., Mol. Cell. Biol. 8: 2159-2165 (1988)];c−mycタグおよびそれに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evan et al., Molecular and Cellular Biology, 5: 3610-3616 (1985)];ならびに単純ヘルペスウイルスの糖タンパク質D(gD)タグおよびその抗体[Paborsky et al., Protein Engineering, 3(6): 547-553 (1990)]が挙げられる。その他のタグポリペプチドとしては、Flag−ペプチド[Hopp et al., BioTechnology 6: 1204-1210 (1988)];KT3エピトープペプチド[Martin et al., Science 255: 192-194 (1992)];チューブリンエピトープペプチド[Skinner et al., J. Biol. Chem. 266: 15163-15166 (1991)];T7遺伝子10タンパク質ペプチドタグ[Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. U.S.A. 87: 6393-6397 (1990)];ヒスチジンタグおよび金属結合部位[Smith, Ann. NY. Acad. Sci.,646:315-321(1991) ]が挙げられるが、Flgおよびヒスチジンタグが好ましい。
【0063】
以下に概説するように、ある実施態様では、HDACファミリーのHDAC8タンパク質および他の生物由来のHDAC8タンパク質を、クローンとして発生させ、発現させる。ゆえに、プローブまたはポリメラーゼ連鎖反応(PCR)の配列を、ヒトまたは他の生物由来の他の関連するHDAC8タンパク質を見つけ出すのに用いてもよい。当業者には理解されるであろうように、特に有用なプローブおよび/またはPCRプライマー配列は、HDAC8核酸配列の独特な領域を含む。当分野で一般的に知られるように、好ましいPCRプライマーは、約15ないし約35ヌクレオチド長であり、約20ないし約30が好ましく、必要ならイノシンを含んでもよい。PCR反応の条件は当分野で周知である。ゆえに、ここで挙げられた配列の中で配列にともなって提供されるものは、これらの配列の各部分であり、15ヌクレオチドまたはそれ以上の独特の各部分が特に好ましいこともまた理解される。当業者は、日常的にヌクレオチド配列を所望の長さに合成したり切断することができる。
【0064】
一旦、プラスミドまたは他のベクターに含めたり、そこから直線状の核酸断片として切り出すなどして、その天然の供給源から単離すると、組換えHDAC8核酸を、他のHDAC8核酸を同定および単離するためのプローブとしてさらに用いることができる。それをまた、修飾された、または変異型のHDAC8核酸およびタンパク質を作成するための「前駆」核酸として用いることができる。
【0065】
HDAC8タンパク質をコードする本発明の核酸を用いて、種々の発現ベクターが作られる。発現ベクターは、自己複製的な染色体外ベクターでも宿主ゲノムに組み込まれるベクターでもよい。一般的に、これらの発現ベクターは、HDAC8タンパク質をコードする核酸に機能し得るように結合された転写および翻訳の制御核酸を含む。「制御配列」という用語は、特定の宿主生物内で、機能し得るように結合されたコード配列が発現するのに必要な核酸配列を意味する。原核生物に適した制御配列は、例えば、プロモーター、任意にオペレーター配列、およびリボゾーム結合部位を含む。真核細胞はプロモーター、ポリアデニル化シグナル、およびエンハンサーを使用することが知られている。
【0066】
核酸は、それが他の核酸配列と機能的な関係に置かれている場合に「機能し得るように結合されて」いる。例えば、前配列もしくは分泌リーダーに対するDNAは、もしそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドに対するDNAに機能し得るように結合されている;プロモーターまたはエンハンサーは、もしそれが配列の転写に影響を与えるならば、コード配列に機能し得るように結合されている;または、リボゾーム結合部位は、もしそれが翻訳を促進するように位置しているならば、コード配列に機能し得るように結合されている。他の例として、機能し得るように結合されたとは、隣接するように、そして、分泌リーダー配列の場合は、隣接しかつ読みとり段階にあるように結合されたDNA配列を意味する。しかしながら、エンハンサーは隣接している必要はない。結合は都合のよい制限部位におけるライゲーションにより達成される。もしそのような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーを従来の実施法に従って使用する。転写および翻訳制御核酸は、HDAC8タンパク質の発現に使用する宿主細胞にとって一般的に適当であろう;例えば、Bacillus由来の転写および翻訳の制御核酸配列は、BacillusでHDAC8タンパク質を発現させるために好ましく使用される。夥しい数の適当な発現ベクターおよび適切な制御配列が、様々な宿主細胞に関して技術上公知である。
【0067】
一般的に、転写および翻訳の制御核酸は、プロモーター配列、リボゾーム結合部位、転写開始および終結配列、翻訳開始および終結配列、およびエンハンサーまたはアクチベーター配列を非限定的に含む。好ましい実施態様では、制御配列はプロモーター並びに転写の開始および終結配列を含む。
【0068】
プロモーター配列は構成または誘導可能プロモーターをコードする。該プロモーターは天然産生のプロモーターでもハイブリッドプロモーターでもよい。2以上の多くのプロモーターのエレメントを組み合わせたハイブリッドプロモーターも技術上公知であり、本発明において有用である。
【0069】
加えて、発現ベクターは付加的エレメントを含み得る。例えば、発現ベクターは、2つの複製システムを有し、これにより2種の生物で、例えば発現のために哺乳動物または昆虫の細胞で、そしてクローニングおよび増幅のために原核宿主で、維持することができる。さらに、発現ベクターを組込むために、発現ベクターは宿主細胞のゲノムと相同な配列を少なくとも1個、また好ましくは発現構築体に隣接する2個の相同配列を含む。ベクターに取り入れるのに適当な相同配列を選択することにより、組込みベクターを宿主細胞の特定の座位を目指して組込ませ得る。組込みベクターの構築は技術上周知である。
【0070】
加えて、好ましい実施態様では、該発現ベクターは形質転換した宿主細胞の選択を可能にする選択マーカー遺伝子を含む。選択遺伝子は技術上周知であり、用いる宿主により異なる。
【0071】
好ましい発現ベクター系は、PCT/US97/01019およびPCT/US97/01048に一般的に記述されているレトロウイルスベクター系であり、両者とも特に出典明示により本明細書の一部とする。
【0072】
本発明のHDAC8タンパク質は、HDAC8タンパク質をコードする核酸を含む発現ベクターで形質転換した宿主細胞を、HDAC8タンパク質の発現を誘導または誘起する適当な条件下で培養することによって生産される。HDAC8タンパク質の発現に適当な条件は、発現ベクターと宿主細胞の選択によって異なるが、当業者は日常的な実験により容易に確かめることができる。例えば、発現ベクター中に構成的プロモーターを使用している場合には、宿主細胞の生育および増殖を最適化することが要求されるだろうし、一方、誘導可能プロモーターを使用している場合には、誘導に適した生育条件が要求される。加えて、いくつかの実施態様では、収穫のタイミングが重要である。例えば、昆虫細胞での発現に用いられるバキュロウイルスシステムは、溶原性ウイルスであるので、採取時期の選択は生産物の収量にとって極めて重要である。
【0073】
適当な宿主細胞としては、酵母、細菌、古細菌、真菌、ならびに昆虫および哺乳動物細胞を含む動物細胞が含まれる。特に興味の持たれるのはDrosophila melanogaster細胞、Saccharomyces cerevisiaeおよび他の酵母、E. coli, Bacillus subtilis、SF9細胞、C129細胞、293細胞、ニューロスポラ、BHK、CHO、COS、HeLa細胞、繊維芽細胞、シュワン腫細胞株、不死骨髄およびリンパ細胞であり、HeLaおよびSF9細胞が特に好ましい。
【0074】
好ましい実施態様では、HDAC8タンパク質は哺乳動物細胞で発現される。哺乳動物の発現系もまた技術上公知であり、レトロウイルスシステムが含まれる。哺乳動物のプロモーターは、哺乳動物のRNAポリメラーゼを結合し、mRNA中のHDAC8タンパク質をコードする配列の下流(3'側)の転写を開始できる全てのDNAである。プロモーターは、通常コード配列の5'末端に近接して配置されている転写開始領域、および転写開始部位の25〜30塩基対上流に配置されているTATAボックスを有している。該TATAボックスは、RNA合成を正しい部位からRNA合成を開始するようにRNAポリメラーゼIIを誘導すると考えられている。哺乳動物のプロモーターはまた、典型的にTATAボックスの上流100〜200塩基対以内に位置している上流のプロモーターエレメント(エンハンサーエレメント)を含有している。上流のプロモーターエレメントは、転写開始速度を決定し、そしてどちら向きにも作用する。哺乳動物プロモーターで特に有用なのは哺乳動物ウイルスの遺伝子由来のプロモーターであるが、これはウイルスの遺伝子はしばしば高度に発現され、また宿主範囲が広いためである。例としては、SV40の初期プロモーター、マウス乳癌ウイルスのLTRプロモーター、アデノウイルスの主要後期プロモーター、単純ヘルペスウイルスのプロモーター、およびCMVのプロモーターが挙げられる。
【0075】
典型的には、哺乳動物細胞によって認識される転写終結配列およびポリアデニル化配列は、転写終結コドンの3'側に位置する制御領域であり、従って、プロモーターエレメントと共にコード配列に隣接している。成熟mRNAの3'末端は、部位特異的な翻訳後切断およびポリアデニル化により形成される。転写ターミネーターおよびポリアデニル化シグナルの例としてはSV40由来のものが含まれる。
【0076】
外来性核酸を哺乳動物宿主およびその他の宿主中に導入する方法は技術上周知であり、用いる宿主細胞により異なるであろう。この技術としては、デキストラン仲介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン仲介性トランスフェクション、プロトプラスト融合、電気穿孔法、ウイルス感染、ポリヌクレオチド(類)のリポソームへの封入、およびDNAの核内への直接マイクロインジェクションが挙げられる。
【0077】
好ましい実施態様では、HDAC8タンパク質は細菌のシステムで発現される。細菌の発現システムは技術上周知である。
【0078】
適当な細菌のプロモーターは、細菌のRNAポリメラーゼを結合し、HDAC8タンパク質の下流(3'側)のコード配列の転写を開始できるすべての核酸配列である。細菌のプロモーターは転写開始領域を有し、これは通常コード配列の5'末端に隣接している。この転写開始領域は典型的に、RNAポリメラーゼ結合部位と転写開始部位を含んでいる。代謝経路上の酵素をコード化する配列は、特に有用なプロモーター配列を提供する。例としては、ガラクトース、乳糖および麦芽糖のような糖代謝酵素由来のプロモーター配列、およびトリプトファンのような物質の生合成にかかわる酵素由来の配列が挙げられる。バクテリオファージ由来のプロモーターも使用でき、技術上公知である。加えて、合成プロモーターおよびハイブリッドプロモーターも有用である;例えば、tacプロモーターはtrpおよびlacプロモーター配列のハイブリッドである。さらに、細菌のプロモーターには、細菌起源でないが細菌のRNAポリメラーゼを結合し転写を開始する能力のある天然産生のプロモーターを含めることができる。
【0079】
機能的プロモーター配列に加えて、効率の良いリボゾーム結合部位が望ましい。E. coliでは、リボゾーム結合部位はシャイン・ダルガルノ(SD)配列と呼ばれ、開始コドン、および開始コドンの約3〜11ヌクレオチド上流にある約3〜9ヌクレオチド長の配列を含む。
【0080】
発現ベクターも、細菌中のHDAC8タンパク質の分泌を起こさせるシグナルペプチド配列を含有していてもよい。技術上周知のように、シグナル配列は典型的に、細胞からのタンパク質の分泌を指令する疎水性アミノ酸よりなるシグナルペプチドをコードする。タンパク質は、成育培地中(グラム陽性細菌)、もしくは細胞の内膜と外膜の間に位置する周辺腔内(グラム陰性細菌)に分泌される。
【0081】
細菌の発現ベクターはまた、形質転換した細菌株の選択を可能にするために、選択マーカー遺伝子を含んでよい。適当な選択遺伝子は、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン、ネオマイシンおよびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含む。選択マーカーはまた、ヒスチジン、トリプトファンおよびロイシンの生合成経路上の遺伝子のような生合成遺伝子を含む。
【0082】
これらの成分は発現ベクターに組み立てて入れる。細菌用の発現ベクターは技術上周知であり、特にBacillus subtilis、E. coli、Streptococcus cremoris、およびStreptococcus lividans用のベクターが含まれる。
【0083】
細菌の発現ベクターは、塩化カルシウム処理、電気穿孔法、その他のような技術上周知の技術を用いて細菌宿主細胞に形質転換される。
【0084】
ある実施態様では、HDAC8タンパク質は昆虫細胞中に生産される。昆虫細胞の形質転換用発現ベクター、特にバキュロウイルスに基づく発現ベクターは技術上周知である。
【0085】
好ましい実施態様では、HDAC8タンパク質は酵母細胞中に生産される。酵母の発現システムは技術上周知であり、Saccharomyces cerevisiae、Candida albicansおよびC. maltosa、Hansenula polymorpha、Kluyveromyces fragilisおよびK. lactis、Pichia guillerimondiiおよびP. pastoris、Schizosaccharomyces pombe、ならびにYarrowia lipolytica用の発現ベクターを含む。酵母での発現用に好ましいプロモーター配列としては、誘導性GAL1、10プロモーターおよびアルコール脱水素酵素、エノラーゼ、グルコキナーゼ、グルコース−6リン酸イソメラーゼ、グリセルアルデヒド−3リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ホスホフラクトキナーゼ、3−ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、および酸ホスファターゼ遺伝子由来のプロモーターが挙げられる。酵母の選択マーカーとしては、ツニカマイシン耐性を付与するADE2、HIS4、LEU2、TRP1、およびALG7、G418に対する耐性を付与するネオマイシンホスホトランスフェラーゼ遺伝子、および銅イオン存在下における酵母の生育を可能にするCUP1遺伝子が挙げられる。
【0086】
HDAC8タンパク質は、当分野で周知の技術を用いて、融合タンパク質としても作成され得る。ゆえに、例えば、モノクローナル抗体作成のために、もし目的のエピトープが小さければ、そのHDAC8タンパク質を、キャリアータンパク質と融合させて免疫原を形成させ得る。あるいは、HDAC8タンパク質を、発現の増加、または他の目的のために、融合タンパク質として作成し得る。例えば、HDAC8タンパク質がHDAC8ペプチドであるとき、そのペプチドをコードする核酸は、発現の目的のために、他の核酸と連結され得る。同様に、本発明のHDAC8タンパク質を、グリーン蛍光タンパク質(GFP)、レッド蛍光タンパク質(RFP)、ブルー蛍光タンパク質(BFP)、イエロー蛍光タンパク質(YFP)などのような、タンパク質の標識と連結させることができる。
【0087】
ある実施態様では、本発明のHDAC8核酸、タンパク質および抗体を標識化する。本明細書中で「標識化」とは、ある化合物が、その化合物の検出を可能にするように付加された少なくとも一成分、同位元素または化合物、を有することを意味する。一般的に、標識は次の3つの分類に分けられる:a)放射活性を有する、または重同位元素であり得る、同位元素標識;b)抗体または抗原であり得る、免疫標識;c)有色または蛍光染料。これらの標識は化合物のどの位置にでも組み込まれ得る。
【0088】
好ましい実施態様では、該HDAC8タンパク質は発現後精製または単離される。HDAC8タンパク質は、どのような他の成分が試料中に存在するかに依存して、当業者周知の種々の方法で単離または精製し得る。標準的な精製法としては、電気泳動的、分子的、免疫学的技法ならびにイオン交換、疎水、アフィニティー、および逆相HPLCクロマトグラフィーを含むクロマトグラフィー技法、ならびにクロマトフォーカシングが挙げられる。例えば、HDAC8タンパク質は、標準的な抗HDAC8抗体カラムを用いて精製することができる。タンパク質濃縮と組合わせた限外ろ過および膜分離精製法も有用である。適当な精製技法の一般的手引書としては、Scopes, R., Protein Purification, Springer-Verlag, NY (1982)を参照のこと。必要な精製の程度はHDAC8タンパク質の用途によって変わるであろう。場合によっては精製は不要である。
【0089】
一旦発現され、必要なら精製されると、HDAC8タンパク質および核酸は、多数の応用において有用である。
【0090】
HDAC8タンパク質類をコードしているこのヌクレオチド配列(またはそれらの相補鎖)には、分子生物学の分野で、ハイブリダイゼーションプローブとしての、クロモソームおよび遺伝子マッピングにおける、およびアンチ−センスRNAおよびDNAの生成における利用を含む、多様な応用がある。HDAC8タンパク質核酸は、本明細書中に記載した組換え技術によるHDAC8タンパク質類の調製にも有用であろう。
【0091】
天然の完全長配列のHDAC8タンパク質遺伝子またはその各部分は、該HDAC8タンパク質をコードしている配列との所望の配列同一性を有する他の遺伝子(例えば、天然に産生するHDAC8タンパク質の変種または他種由来のHDAC8タンパク質をコードしている遺伝子)を単離するために、cDNAライブラリーについてのハイブリダイゼーションプローブとして使用し得る。場合により、これらのプローブの長さは約20ないし約50塩基である。これらのハイブリダイゼーションプローブは、本明細書中のヌクレオチド配列から、または本明細書中に提供されたような天然配列のプロモーター、エンハンサーエレメントおよびイントロンを含むゲノム配列から誘導し得る。例をあげれば、スクリーニング方法は、既知DNA配列を使用して約40塩基の選択プローブを合成する、HDAC8タンパク質遺伝子のコード領域の単離を含む。ハイブリダイゼーションプローブは、32Pまたは35Sのような放射性ヌクレオチド類、またはアビジン/ビオチンカップリング系を介して該プローブと連結したアルカリホスファターゼのような酵素標識を含む各種標識により標識化し得る。本発明のHDAC8タンパク質遺伝子の配列と相補的な配列を有する標識化したプローブは、ヒトcDNA、ゲノムDNAまたはmRNAの各ライブラリーをスクリーニングし、該プローブがそれらのライブラリーのどのメンバーとハイブリダイズするかを測定するのに使用できる。
【0092】
HDAC8タンパク質をコードしているヌクレオチド配列は、該HDAC8タンパク質をコードしている遺伝子のマッピングのため、および遺伝的異常がある個体の遺伝子分析のためにハイブリダイゼーションプローブを構築するのにも使用し得る。本明細書中に記載したこれらのヌクレオチド配列は、既知技術、例えば、その場でのハイブリダイゼーション、既知染色体マーカーに対する連結分析、およびライブラリーについてのハイブリダイゼーションスクリーニングを使用して、染色体や染色体の特定領域に対してマッピングし得る。
【0093】
HDAC8タンパク質またはその修飾形態をコードしている核酸は、トランスジェニック動物または「ノックアウト」動物を発生させるのにも使用でき、ひいては治療的有用物質の開発およびスクリーニングにおいても有用である。トランスジェニック動物(例えば、マウスまたはラット)とは、導入遺伝子含有細胞を有する動物であり、該導入遺伝子が該動物または該動物の祖先に胎内期(prenatal)、例えば、胚期(embryonic stage)において導入されたものである。導入遺伝子とは、それからトランスジェニック動物が発育する細胞のゲノム中に集積されたDNAである。ある実施態様では、HDAC8タンパク質をコードしているcDNAは、確立された技術に従ってHDAC8タンパク質をコードしているゲノムDNAをクローン化するのに使用でき、該ゲノム配列は、所望のDNAを発現する細胞を含有するトランスジェニック動物の発生に使用される。トランスジェニック動物、殊にマウスまたはラットのような動物を発生させる方法は、当技術分野で日常化してきており、例えば、米国特許番号第4736866号および第4870009号に記載されている。典型的には、特定の細胞が、組織特異的エンハンサーによるHDAC8タンパク質導入遺伝子導入の標的になる。胚期に該動物の生殖ライン中に導入された、HDAC8タンパク質をコードしている導入遺伝子のコピーを含むトランスジェニック動物は、所望の核酸の増加した発現の効果を調べるのに使用できる。それらの動物は、例えば、その過剰発現に伴う病理的状況からの保護をもたらすと考えられる物質に対するテスター動物として使用できる。本発明のこの側面によれば、動物を該物質で処理し、該導入遺伝子を有する非処理動物と対比した病理的状況の減少効果が、該病理的状況に対する治療的干渉のポテンシャルを示す。
【0094】
あるいは、このHDAC8タンパク質の非ヒト相同体をHDAC8タンパク質「ノックアウト」動物を構築するのに使用することができる。このノックアウト動物は、HDAC8タンパク質をコードしている内因性遺伝子と、該動物の胚細胞中に導入されたHDAC8タンパク質をコードしている変化させたゲノムDNAとの間の、相同組換えの結果HDAC8タンパク質をコードする遺伝子が欠落または変化している動物である。例えば、HDAC8タンパク質をコードしているcDNAを、HDAC8タンパク質をコードしているゲノムDNAのクローン化に、確立された技術に従って使用することができる。HDAC8タンパク質をコードしているゲノムDNAの一部は、削除したり、または他の遺伝子例えば、モニター集積に使用し得る選択マーカーをコードしている遺伝子と置き換えたりすることができる。典型的には、数キロ塩基の(5'または3'末端の両方における)不変フランキング(側位)DNAを、ベクター中に含める[相同組換えベクター類については、例えば、Thomas and Capecchi, Cell, 51:503 (1987) の既述を参照]。このベクターを、胚幹細胞系中に(例えば、電気穿孔法により)導入し、そして、導入したDNAが内因性DNAと相同的に組換えられている細胞を選択する[例えば、Li et al., Cell, 69:915 (1992) 参照]。選択した細胞を次いで動物(例えば、マウスまたはラット)の胚盤胞中に注入し、キメラ集合体を形成させる[例えば、Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp.113-152参照]。キメラ胚を、次いで適切な偽妊娠雌フォスター動物中に移植し、該胚を「ノックアウト」動物作成期に導く。生殖細胞中にこの相同組換えDNAを内包する後代は、標準的技術で同定することができ、そして該動物の全細胞が相同的に組換えられたDNAを含有する動物を産出することに使用できる。ノックアウト動物には、例えば、ある種の病理学的状況に対して防御する能力があることや、HDAC8タンパク質の不存在により病理学的状況が進展することなどが特徴であると言える。
【0095】
本明細書中に記載したモデルは変更可能と理解される。例えば、「ノックイン」モデルを作成することができ、あるいは該モデルを動物モデルよりむしろ細胞ベースとすることができる。
【0096】
これらのHDAC8ポリペプチドをコードしている核酸、アンタゴニストまたはアゴニストは、遺伝子治療にも使用できる。遺伝治療への応用では、遺伝子を細胞内に導入し、例えば欠陥遺伝子置換のための、治療的に有効な遺伝子産物のインビボ合成を達成させる。「遺伝子治療」には、単回の処置により持続的効果を達成させる在来の遺伝子治療と、治療的に有効なDNAまたはmRNAの単回または反復した投与を含む、遺伝子治療的物質の投与との両方が含まれる。アンチセンスRNAおよびDNAを、インビボである種の遺伝子の発現を阻止する治療的物質として使用できる。短いアンチセンスオリゴヌクレオチドが細胞内に導入され、細胞膜による取り込みが制限されることにより細胞内濃度が低いにもかかわらず、そこで阻害物質として働き得ることが既に示されている(Zamecnik et al., Proc. Natl. Acad. Sci. USA 83, 4143-4146 [1986])。これらのオリゴヌクレオチドは、修飾、例えばそれらの負荷電ホスフォジエステル基を非荷電基と置換、してそれらの取り込みを増強することができる
【0097】
生存細胞内に核酸類を導入するには各種の技術が利用できる。これらの技術は、核酸がインビトロで培養細胞内に移入されるのか、またはインビボで意図する宿主の細胞内に移入されるのかによって変る。インビトロで哺乳動物細胞内に核酸を移入するのに適した技術には、リポソーム、電気穿孔法、マイクロインジェクション、細胞癒合、DEAE−デキストラン、リン酸カルシウム沈殿法、等の使用が含まれる。現今好適とされているインビボ遺伝子移入技術には、ウイルス(典型的にはレトロウイルス)ベクターを用いるトランスフェクション、およびウイルス被覆タンパク質-リポソーム媒介トランスフェクションが含まれる(Dzau et al., Trends in Biotechnology 11, 205-210 [1993])。ある状況では、標的細胞を標的とする物質、例えば、細胞表面膜タンパク質または標的細胞に特異的な抗体、標的細胞上のレセプターに対するリガンドを有する核酸源を供給するのが望ましい。リポソームを採用した場合は、エンドサイトーシスに伴って細胞表面膜タンパク質に結合するタンパク質、例えば特定の細胞型に親和性のカプシドタンパク質またはその断片、サイクリング中に内在化を受けるタンパク質に対する抗体、細胞間局在化を標的とし細胞内半減期を強化するタンパク質、を標的化および/または取り込み増強のために使用することができる。レセプター媒介エンドサイトーシスの技術は、例えば、Wu et al., J. Biol. Chem. 262,4429-2232 (1987): および Wagnernet al., Proc. Natl. Acad. Sci. USA 87, 3410-3414(1990) に記載されている。遺伝子マーキングおよび遺伝子治療プロトコールについての総説 Andersen et al., Science 256, 808-813 (1992)参照。
【0098】
好ましい態様では、HDAC8タンパク質、核酸、変異種、修飾タンパク質、該核酸またはタンパク質を含有する細胞および/またはトランスジェニック体をスクリーニングアッセイに使用する。本明細書中に記載したHDAC8タンパク質の同定により、HDAC8タンパク質に結合するかまたはその結合を妨げる化合物についての、およびHDAC8活性を制御する化合物についての薬剤スクリーニングアッセイの設計が可能となる。
【0099】
本明細書に記載したアッセイでは、好ましくはヒトHDAC8タンパク質を利用するが、齧歯類(マウス、ラット、ハムスター、モルモット、その他)、家畜類(牛、羊、豚、馬、その他)および霊長類を含む他の哺乳動物のタンパク質も使用できる。これらの後者の実施態様はヒトの疾患の動物モデルの開発に好適であろう。ある実施態様では、本明細書中で概説したように、変異体または誘導体HDAC8タンパク質、上記したようなHDAC8タンパク質の除去を含めて、を使用してもよい。
【0100】
好ましい態様では、本発明の方法は、HDAC8タンパク質と候補バイオ活性物質とを合わせること、および該HDAC8タンパク質への該候補物質の結合を測定することを含む。他の態様では、以下に詳論するが、結合の阻害またはバイオ活性を測定する。
【0101】
本明細書において「候補バイオ活性物質」または「外来性化合物」とは、例えば、タンパク質、小型の有機分子、炭水化物(多糖類を含む)、ポリヌクレオチド、脂質などのいずれかの分子をいう。一般に、複数のアッセイ混合物を種々の薬物濃度を用いて並行して流し、種々の濃度に対しての種々の示差応答を得る。典型的には、これらの濃度のうち1つ、即ち0または検出レベルに満たない濃度がネガティブコントロールとして用いられる。さらに、ポジティブコントロールを用いてもよい。即ち、HDAC8活性を変化させるか調節すると知られている薬剤の使用である。
【0102】
候補物質は多くの化学クラスを包含するが、典型的にはそれらは有機分子、好ましくは分子量が100ダルトンより大きく約2,500ダルトン未満の小型の有機化合物である。候補物質はタンパク質との構造相互作用、特に水素結合に必要な官能基を含み、典型的には少なくとも1つのアミン、カルボニル、ヒドロキシルまたはカルボキシル基、好ましくは少なくとも2つの官能化学基を含む。該候補物質は環式炭素または複素環式構造および/または上記の1以上の官能基で置換された芳香族もしくはポリ芳香族構造を含む。候補物質はまた、ペプチド、糖類、脂肪酸、ステロイド、プリン、ピリミジン、誘導体、構造類似体またはそれらの組合せを含む生体分子間に見出せる。特に好ましいのはペプチドである。
【0103】
候補物質は、合成または天然化合物のライブラリーを含む多様な供給源から得られる。例えば、ランダム化オリゴヌクレオチドの発現をはじめ、多様な有機化合物および生体分子のランダムな、また所定の合成には多くの手段が利用できる。あるいは、細菌、真菌、植物および動物の抽出物形態における天然化合物のライブラリーが入手可能であるか、または容易に作製できる。さらに、天然の、または合成により作製したライブラリーまたは化合物は従来の化学的、物理的、生化学的手段により容易に修飾される。公知の薬理活性物質は、アシル化、アルキル化、エステル化、アミド化などの所定のまたはランダムな化学修飾を受けて構造類似体を形成し得る。
【0104】
好ましい態様では、種々の候補バイオ活性物質のライブラリーを用いる。好ましくは、該ライブラリーはランダム化された物質の、構造的に十分多様な集団を提供して、特定の標的との結合を可能にするために十分と見込まれる範囲の多様性を達成する。従って、相互作用ライブラリーは、メンバーの少なくとも1つが、標的に対する親和性を有する構造を持つよう、十分大きくなければならない。相互作用ライブラリーに要求される絶対的なサイズを測るのは難しいが、自然が免疫応答でヒントを与えてくれる。即ち、多様な107ないし108個の異なる抗体は、生物が直面する大部分のあり得る抗原と相互作用するに十分な親和性を有する少なくとも1つの組合せを与える。また、公表されているインビトロ選択技術も、107ないし108のライブラリーサイズが、標的への親和性を有する構造を見出すに十分であることを示している。本明細書で一般に提案されるものなど7ないし20アミノ酸長のペプチドのすべての組合せに関するライブラリーは、207(109)ないし2020をコードする可能性を有する。従って、107ないし108の異なる分子のライブラリーについて、本方法では、理論的に完全な相互作用ライブラリーの「実働」サブセットをアミノ酸7個と見積り、形態サブセットを2020ライブラリーと見積もる。このように好ましい態様では、主題の方法において少なくとも106、好ましくは107、より好ましくは少なくとも108、最も好ましくは少なくとも109個の異なる配列が同時に解析される。好ましい方法はライブラリーのサイズおよび多様性を最大にする。
【0105】
好ましい実施態様では、該候補バイオ活性物質はタンパク質である。本明細書において「タンパク質」とは、共有結合した少なくとも2つのアミノ酸を意味し、タンパク質、ポリペプチド、オリゴペプチドおよびペプチドが含まれる。該タンパク質は、天然産生のアミノ酸およびペプチド結合または合成ペプチド模倣構造からなってもよい。従って本明細書において「アミノ酸」または「ペプチド残基」とは、天然産生のアミノ酸と合成アミノ酸の双方を意味する。例えば、ホモフェニルアラニン、シトルリンおよびノルロイシンは、本発明の目的のためのアミノ酸と考えられる。また「アミノ酸」には、プロリンおよびヒドロキシプロリンなどのイミノ酸残基が含まれる。その側鎖は(R)配置または(S)配置のいずれであってもよい。好ましい態様では、該アミノ酸は(S)またはL配置である。天然産生ではない側鎖を用いる場合は、例えばインビボ分解を妨げるまたは遅延させるために非アミノ酸置換基を用いてもよい。また、化学保護基またはその他の化学置換基を付加してもよい。
【0106】
好ましい態様では、候補バイオ活性物質は天然産生のタンパク質または天然産生のタンパク質の断片である。従って、例えば、タンパク質またはタンパク質性細胞抽出物のランダムなもしくは所定の消化物を含む、細胞抽出物を用いてもよい。このように、本明細書に記載の系のスクリーニングのためには、原核生物および真核生物のタンパク質ライブラリーを作製すればよい。本態様において特に好ましいのは、細菌、真菌、ウイルスおよび哺乳類タンパク質のライブラリーであり、後者が好ましく、ヒトタンパク質が特に好ましい。
【0107】
好ましい態様では、該候補バイオ活性物質は、約5ないし約30のアミノ酸からなるペプチドであり、約5ないし約20のアミノ酸からなるものが好ましく、約7ないし約15のアミノ酸からなるものが特に好ましい。該ペプチドは上記に概説したような天然産生のタンパク質の消化物、ランダムペプチド、または「偏りのある」ランダムペプチドであってもよい。本明細書において「ランダム化された」または文法的均等語は、核酸およびペプチドが、それぞれ本質的にランダムなヌクレオチドおよびアミノ酸からなることを意味する。一般に、これらのランダムペプチド(または以下に論じる核酸)は化学的に合成されるので、いかなる位置にいかなるヌクレオチドまたはアミノ酸をも組み込める。この合成法は、ランダム化されたタンパク質または核酸を作成し、配列全長にわたって、あり得る組合せのすべてまたは大部分の形成を可能にし、ランダム化されたタンパク質性候補バイオ活性物質のライブラリーを形成するよう設計することができる。
【0108】
ある実施態様では、該ライブラリーは完全にランダム化され、いかなる位置にも優先のまたは一定の配列は存在しない。好ましい態様では、該ライブラリーには偏りがある。即ち、配列内のある位置では一定であるか、または限られた数の可能性から選択される。例えば、好ましい態様では、ヌクレオチドまたはアミノ酸残基は、例えば、疎水性アミノ酸、親水性残基、立体的に偏りのある(小さいか、または大きい)残基、架橋のためのシステイン、SH−3ドメインのためのプロリン、リン酸化部位のためのセリン、トレオニン、チロシンもしくはヒスチジンの作出などに対して、またはプリンに対してといった所定のクラス内でランダム化される。
【0109】
好ましい態様では、該候補バイオ活性物質は核酸である。本明細書において「核酸」もしくは「オリゴヌクレオチド」または文法的均等語は、共有結合した少なくとも2つのヌクレオチドを意味する。本発明の核酸は一般にホスホジエステル結合を含むが、以下に概説するように、例えば、ホスホルアミド(Beaucage, et al., Tetrahedron, 49(10):1925 (1993)およびその参照文献; Letsinger, J. Org. Chem., 35:3800 (1970); Sprinzl, et al., Eur. J. Biochem., 81:579 (1977); Letsinger, et al., Nucl. Acids Res., 14:3487 (1986); Sawai, et al., Chem. Lett., 805 (1984); Letsinger , et al., J. Am. Chem. Soc., 110:4470 (1988); およびPauwels, et al., Chemica Scripta, 26:141 (1986))、ホスホロチオエート(Mag, et al., Nucleic Acids Res., 19:1437 (1991);および米国特許番号第5644048号)、ホスホロジチオエート(Briu, et al., J. Am. Chem. Soc., 111:2321 (1989))、O−メチルホスホロアミダイト結合(Eckstein , Oligonucleotides and Analogues: A Practical Approach, Oxford University Press参照)、ならびにペプチド核酸主鎖および結合(Egholm, J. Am. Chem. Sci., 114:1895 (1992); Meie, et al., Chem. Int. Ed. Engl., 31:1008 (1992); Nielsen, Nature, 365:566 (1993); Carisson, et al., Nature, 380:207 (1996)参照。なお、これらは出典明示により本明細書の一部とする)を含む、代替的な主鎖を有し得る核酸類似体が含まれる場合がある。その他の核酸類似体としては、正電荷主鎖(Denpcy, et al., Proc. Natl. Acad. Sci. USA, 92:6097 (1995));非イオン性主鎖(米国特許番号第5386023号、第5637684号;第5602240号;第5216141号;および第4469863号; Kiedroeshi, et al., Angew. Chem. Intl. Ed. English, 30;423 (1991); Letsinger, et al., J. Am. Chem. Sci., 110:4470 (1988); Letsinger, et al., Nucleoside & Nucleotide, 13:1597 (1994); Chapters 2 and 3, ASC Symposium Series 580, "Carbohydrate modifications in Antisense Research", Ed. Y.S. Sanghui and P. Dan Cook; Mesmaeker, et al., Bioorganic & Medicinal Chem. Lett., 4:395 (1994); Jeffs, et al., J. Biomolecular NMR, 34:17 (1994); Tetrahedron Lett., 37:743 (1996))、ならびに米国特許第5235033号、第5034506号、およびChapters 6 and 7, ASC Symposium Series 580, "Carbohydrate Modifications in Antisense Research", Ed. Y.S. Sanghui and P.Dan Cookに記載のものをはじめとする非リボース主鎖を有するものが挙げられる。1以上の炭素環式糖を含む核酸もまた、本核酸定義内に含まれる(Jenkins, et al., Chem. Soc. Rev., (1995) pp. 169-176参照)。いくつかの核酸類似体が、Rawls, C & E News, June 2, 1997, page 35に記載されている。これらの参照文献は、すべて出典明示により本明細書の一部とする。リボース−リン酸主鎖のこれらの修飾を行い、標識などの付加部分の付加を助けたり、あるいは生理学的環境下でかかる分子の安定性を高め、半減期を延ばしてもよい。さらに、天然産生の核酸と類似体の混合物を作成することもできる。あるいは、種々の核酸類似体の混合物や天然産生の核酸と類似体の混合物を作製してもよい。該核酸は明示したように1本鎖であっても2本鎖であってもよく、あるいは2本鎖または1本鎖配列の双方の部分を含んでもよい。該核酸は、デオキシリボヌクレオチドとリボヌクレオチドのいずれかの組合せ、およびウラシル、アデニン、チミン、シトシン、グアニン、イノシン、キサンチン、ヒポキサンチン、イソシトシン、イソグアニンなどを含む塩基のあらゆる組合せを含む、DNA(ゲノムDNAおよびcDNAの双方)、RNAまたはハイブリッドであってもよい。
【0110】
一般にタンパク質に関して上記したように、核酸候補バイオ活性物質は天然産生の核酸、ランダム核酸、または「偏りのある」ランダム核酸であってよい。例えば、タンパク質に関して先に概説したように原核生物ゲノムまたは真核生物ゲノムの消化物を用いてもよい。
【0111】
好ましい態様では、該候補バイオ活性物質は有機化学部分であり、多様なものが文献から得られる。
【0112】
好ましい態様では、候補バイオ活性物質は融合パートナーに結合している。本明細書において「融合パートナー」または「機能的グループ」とは、候補バイオ活性物質と会合し、そのクラスのライブラリーのすべてのメンバーに、共通の機能または能力を付与する配列を意味する。融合パートナーは異種(即ち、宿主にとり天然でない)、または合成のもの(いずれの細胞にとっても天然でない)であり得る。好適な融合パートナーとしては、限定されるものではないが、a)候補バイオ活性物質を立体配位上限定的なまたは安定な形で提供する提示構造、b)候補バイオ活性物質を細胞内または細胞外区画へ局在化させる標的配列、c)候補バイオ活性物質またはそれらをコードする核酸のいずれかの精製または単離を可能にするレスキュー配列、d)候補バイオ活性物質またはそれをコードする核酸に、安定性または分解からの保護、例えばタンパク質分解耐性を付与する安定化配列、e)ペプチド二量化を可能にする二量化配列、あるいはf)a)、b)、c)、d)およびe)ならびに要すればリンカー配列のいずれかの組合せ、が挙げられる。
【0113】
本明細書に記載した方法のある実施態様では、HDAC8タンパク質の各部分を使用する;好ましい態様では、HDAC8活性を有する部分を使用してHDAC8に結合する物質を同定する。加えて、本明細書中に記載したアッセイでは、単離したHDAC8タンパク質またはHDAC8タンパク質を含む細胞を利用し得る。
【0114】
該して言えば、本発明の方法の好ましい態様では、例えば結合アッセイのために、HDAC8タンパク質または候補物質を、単離試料受容領域を有する不溶性支持体(例えば、マイクロタイタープレート、アレイ、その他)に非拡散的に結合させる。これらの不溶性支持体は、本発明の組成物がそれに結合することができ、容易に可溶性材料から分離でき、そしてスクリーニングの全体方法と他の点では適合性である任意の組成で作成できる。それらの支持体の表面は固体または多孔質であり、任意の都合のよい形状であり得る。好適な不溶性支持体の例には、マイクロタイタープレート、アレイ、膜およびビーズが含まれる。これらは、典型的にはガラス、プラスチック(例えば、ポリスチレン)、ポリサッカライド、ナイロンまたはニトロセルロース、テフロン(登録商標)、その他で作成する。マイクロタイタープレートおよびアレイは、多数のアッセイを同時に実施でき、試薬や試料の使用量が少なくてすむので特に好都合である。ある場合には、磁性ビーズおよび類似物が含まれる。組成物結合の個々の様式は、本発明の試薬類および全体方法と適合性であり、組成物の活性を維持し、そして非拡散的である限り、厳密なものではない。好ましい結合方法には、抗体(それは、該タンパク質が支持体に結合したとき、リガンド結合部位または活性化配列のいずれをも立体的にブロックしないものである)の使用、「粘着性」またはイオン性支持体への直接結合、化学的架橋結合、表面上での該タンパク質または物質の合成、その他が含まれる。該タンパク質または物質の結合につづいて、過剰の未結合材料を洗浄により除去する。試料受容領域を次いで牛血清アルブミン(BSA)、カゼインまたは他の無害なタンパク質または他の部位とインキュベートしてブロックする。本発明には、固体支持体を使用しないスクリーニングアッセイも含まれる:その例は以下に述べる。
【0115】
好ましい実施態様では、HDAC8タンパク質を支持体に結合させ、それから候補バイオ活性物質をアッセイに加える。あるいは候補物質を支持体に結合させ、それからHDAC8タンパク質を加える。新規な結合物質には、特殊な抗体、化学的ライブラリーのスクリーニングで同定された非天然産結合物質、ペプチド類似体などが含まれる。特に重要であるのは、ヒト細胞に対する毒性が低い物質に対するスクリーニングアッセイである。多様なアッセイがこの目的のために使用でき、それらには標識化インビトロタンパク質―タンパク質結合アッセイ、電気泳動シフトアッセイ、タンパク質結合に対する免疫アッセイ、機能的アッセイ、好ましくは短いアセチル化ペプチドまたは標識アセチル化ペプチドの脱アセチル化が含まれる。
【0116】
候補バイオ活性物質のHDAC8タンパク質への結合の測定は、各種の方法で実施できる。好ましい態様では、候補バイオ活性物質を標識化し、結合を直接測定する。例えば、これは、HDAC8タンパク質の全部または一部を固体支持体に結合させ、標識化(例えば、蛍光標識)候補物質を加え、過剰の物質を洗い去り、そして標識が固体支持体上に存在するかどうかを測定することにより実施できる。当分野で既知の各種ブロックおよび洗浄段階を利用することができる。
【0117】
本明細書中では「標識化」とは、その化合物が、検出可能なシグナルを提供する標識、例えば放射性同位元素、蛍光剤、酵素、抗体、磁性粒子のような粒子、化学的発光剤、または特異的結合分子、その他、により、直接的かまたは間接的かのいずれかで標識されていることを意味する。特異的結合分子には、ビオチンとストレプトアビジン、ジゴキシンとアンチジゴキシンのような対、その他、が含まれる。特異的結合メンバーでは、前記概説のように、相補的メンバーは通常、既知操作により検出のための分子により標識化されている。標識は直接または間接的に検出可能なシグナルを提供する。
【0118】
ある実施態様では、成分のひとつだけを標識化する。例えば、タンパク質(またはタンパク質性候補物質)をチロシン位で、125Iまたは発蛍光団を用いて標識化することができる。あるいは、1以上の成分を異なる標識で;例えばタンパク質は125Iを用いて標識化し、候補物質は発蛍光団で標識化してもよい。
【0119】
好ましい態様では、候補バイオ活性物質の結合は、競合的結合アッセイを使用して測定できる。この実施態様では、競合物質は標的分子と結合することが既知の結合部分(即ち、HDAC8タンパク質)、例えば、抗体、結合パートナー、リガンド、その他である。ある状況では、バイオ活性物質が結合部位と置き換わる、バイオ活性物質と結合部位間でのような競合的結合があり得る。このアッセイは、HDAC8タンパク質と結合パートナー間の結合を妨害する候補物質を測定するために使用することができる。本明細書中では「結合の妨害(interference)」とは、HDAC8タンパク質の本来の結合が、候補物質の存在下に異なることを意味する。この結合は除去することができるか、またはその親和性を低減することができる。それ故に、ある実施態様では、妨害は例えば、本来の結合部位に対する直接競合というよりむしろ立体配置の変化により生じるものである。
【0120】
ある実施態様では、候補バイオ活性物質を標識化する。候補バイオ活性物質または競合物質のいずれか、または両方を、まずこのタンパク質に、それが存在するならば充分結合できるだけの時間加える。インキュベーションは、最適活性を促す任意の温度、典型的には4ないし40℃で実施できる。インキュベーションの時間は、最適活性となるように選択するが、迅速高度生産スクリーニングを促すように最適化してもよい。典型的には0.1ないし1時間で充分であろう。過剰の試薬は一般的に除去するかまたは洗い去る。次に、第2の成分を加え、結合を示す標識化成分の存在または不存在を調べる。
【0121】
好ましい態様では、競合物質を最初に加え、その後候補バイオ活性物質を加える。競合物質が置換することは、候補バイオ活性物質がHDAC8タンパク質と結合していること、従ってHDAC8タンパク質活性に対して、それと結合しそれを強力に調節する能力があることを示す指標である。この実施態様では、両成分を標識化し得る。従って、例えば、競合物質を標識化した場合、洗浄液中に標識が存在することは該物質による置換があったことを示す。あるいは、候補バイオ活性物質を標識化した場合、支持体上の標識の存在は置換があったことを示す。
【0122】
別の実施態様では、候補バイオ活性物質を最初に加え、インキュベーションし洗浄し、その後競合物質を加える。競合物質による結合の不存在は、該バイオ活性物質がHDAC8タンパク質とより高度な親和性で結合していることの証しとなる。かくて、候補バイオ活性物質を標識化した場合は、支持体上の標識の存在が、競合物質結合がないために、該候補物質がHDAC8タンパク質と結合する能力があることを示すこととなる。
【0123】
好ましい実施態様では、本発明の方法は、HDAC8タンパク質の活性を調節し得る候補バイオ活性物質を同定するための分別スクリーニングを含む。そのようなアッセイは、HDAC8タンパク質または該HDAC8タンパク質を含んでいる細胞により実施できる。ある実施態様では、本発明の方法は、あるHDAC8タンパク質と、第1試料中の競合物質とを合わせることを含む。第2試料には、候補バイオ活性物質、HDAC8タンパク質および競合物質を含める。競合物質の結合を両方の試料について測定すれば、2試料間の結合の変化または差異が、HDAC8タンパク質と結合しその活性を強力に調節する能力のある物質の存在を示す。即ち、競合物質の結合が第1試料と対比して第2試料中では異なるならば、その物質はHDAC8タンパク質と結合する能力がある。
【0124】
あるいは、好ましい実施態様は、天然産生のHDAC8タンパク質とは結合するが修飾HDAC8タンパク質とは結合できない薬物候補を同定するために分別スクリーニングを利用する。このHDAC8タンパク質の構造をモデルとし、該部位と相互作用する物質を合成するための理論的薬物設計に使用する。HDAC8のバイオ活性に作用する薬物候補も、該タンパク質の活性を増強または低減させる能力について薬物をスクリーニングすることによって同定される。
【0125】
これらのアッセイでは、ポジティブコントロールおよびネガティブコントロールを使用することができる。好ましくは、全てのコントロールおよびテスト試料は、統計的に有意な結果を得るために、少なくとも3部づつとする。全試料のインキュベーションは、その物質が該タンパク質に結合するのに充分なだけの時間行う。インキュベーションに続いて、全試料を、特異的結合をしていない材料がなくなるまで洗浄し、結合の通常標識された物質の量を測定する。例えば、放射性標識を採用した場合は、試料をシンチレーションカウンターでカウントして結合化合物の量を測定する。
【0126】
このスクリーニングアッセイには各種の他の試薬を含めることができる。それらには、塩類、アルブミンのような中性タンパク質、洗浄剤、その他のような試薬が含まれ、それらは最適のタンパク質−タンパク質結合を促す、および/または非特異的またはバックグラウンド相互作用を低減するために用いられる。このアッセイの効率を別の方法で改善する試薬、例えば、プロテアーゼインヒビター、ヌクレアーゼインヒビター、抗微生物剤なども使用できる。これらの成分の混合物は、必要な結合をもたらす任意の順序で添加できる。
【0127】
HDAC8タンパク質の活性を調節する物質についてのスクリーニングも実施できる。好ましい態様では、HDAC8の活性を調節し得るバイオ活性物質のためのスクリーニング方法は、候補バイオ活性物質をHDAC8タンパク質(またはHDAC8タンパク質を含む細胞)の試料に加える段階、および該HDAC8タンパク質の生物学的活性の変化を測定する段階を含む。「HDAC8タンパク質の活性を調節する」には、存在する活性の増加、活性の減少、または活性の型または種類の変化が含まれる。従って、この実施態様では、候補物質は、本明細書中で定義したように、HDAC8タンパク質と結合(これは多分不要であるが)し、かつ生物学的または生化学的活性を変更する。この方法は、一般的に上で説明したようなインビトロスクリーニング方法、およびHDAC8タンパク質の存在、分布、活性または量における変更についての細胞のインビボスクリーニング方法の両方を含む。
【0128】
従ってこの態様では、本発明の方法は、HDAC8試料と候補バイオ活性物質とを合わせること、およびそのHDAC8への影響を評価することを含む。本明細書中で「HDAC8タンパク質活性」またはその文法的均等語は、HDAC8タンパク質の生物学的活性の少なくともひとつを意味し、それには、クロマチン構造、ヒストンアセチル化、翻訳、細胞周期、細胞複製、細胞活性などでインビトロまたはインビボにおいて影響を与える、各能力が含まれるがそれらに限定されない。
【0129】
好ましい実施態様では、HDAC8タンパク質の活性が減少し;他の好ましい実施態様ではHDAC8タンパク質の活性が増加する。従って、ある実施態様ではアンタゴニストであるバイオ活性物質が好適であり、他の実施態様ではアゴニストであるバイオ活性物質が好適である。
【0130】
好ましい実施態様では、本発明は、HDAC8タンパク質の活性を調節する能力のあるバイオ活性物質のスクリーニング方法を提供する。この方法は、上記したように、HDAC8タンパク質を含む細胞に候補バイオ活性物質を加えることを含む。好適な細胞型には殆ど全ての細胞が含まれる。それらの細胞は、HDAC8タンパク質をコードしている組換え核酸を含有する。好ましい実施態様では、候補物質のライブラリーを複数の細胞について試験する。
【0131】
HDAC8制御の検出は、当業者には明らかなように実施できる。ある実施態様では、HDAC8活性のインディケーター、例えばリジンがアセチル化されたペプチドRTKRなどのアセチル化合成ペプチドを使用する。HDAC8制御における変更の検出を可能とするために評価またはアッセイするパラメータは多数あり、それらには、細胞生存力アッセイ、細胞が特定の細胞周期段階に留まっているかどうかを測定するアッセイ(「細胞増殖アッセイ」)、およびその細胞がどの細胞段階に留まっているかを決定するアッセイ(「細胞相(細胞期)アッセイ」)、が含まれるがそれらに限定されない。別のパラメーターとして、上記に記載のようなmRNA合成、翻訳、アセチル化ペプチドの脱アセチル化がある。これらのパラメータの1またはそれ以上をアッセイまたは測定することにより、HDAC8制御における変更のみならず、該HDAC8制御過程の異なる段階での変更をも検出することが可能となる。これは、天然産生細胞を評価するために、例えば、腫瘍細胞型の攻撃性を測るために、またはHDAC8制御に対する作用を試験中の候補薬物の作用を評価するために、実施してもよい。このようにして、候補物質の迅速で正確なスクリーニングを達成し、HDAC8制御を調節する物質を同定することができる。
【0132】
従って、本発明の組成物および方法はバイオ活性物質を明示するために有用である。この物質によって細胞または細胞集団が、そのクロマチン構造、ヒストンアセチル化、転写活性を変更するか、あるいはある増殖期から他の期(相)に移行するか、または増殖期(成育相)に留まることができる。ある実施態様では、細胞はある特定の増殖期に留まっているが、それらをその相から出て新しい相中に入れることが望まれる。あるいは、細胞をある相、例えばG1内に強制的に留まらせることが、細胞周期を経由して移行しつづけるよりもむしろ望ましいことがあろう。同様に、ある場合には、滞留してはいないが移行が緩慢な細胞集団を次の相へまたはHDAC8に忠実に沿って加速させること、または次相の開始を遅らせることが望ましいこともあろう。例えば、ある種の酵素、例えばキナーゼ、ホスファターゼ、プロテアーゼの活性を変更させること、または酵素をユビキチン化させることが可能であり、これは細胞相変化の開始に役立つ。
【0133】
好ましい態様では、本明細書中で概説した方法を、G1期に留まっていない細胞上で:即ち、迅速にまたは制御不能に成育し複製しつづけている細胞、例えば腫瘍細胞上で実施することができる。このようにして候補物質を評価し、HDAC8制御を変更し得る、即ち細胞を各HDAC8チェックポイント、例えばG1(他の相、例えばS,G2またはMに留まることも望ましいが)に留まらせ得る物質を見出す。あるいは、候補物質を評価し、細胞集団を増殖させ得る、即ち一般にG1に留まっている細胞:例えば、末梢血液細胞、最終分化細胞、培地中の幹細胞その他、を再増殖させる物質を見出す。
【0134】
従って、本発明は、細胞集団のHDAC8制御における変更についてのスクリーニング方法を提供する。「変更」または「調節」(本明細書中では互変的に使用)により、一般に、2つのことのどちらかを意味する。好ましい実施態様では、この変更の結果、例えばヒストンアセチル化において、ある細胞の細胞周期に変化が生じる。即ち、増殖細胞が各相のどれかひとつに滞留するか、または滞留細胞が滞留相外へ動き出し、その結果別の細胞と比較して、または異なる条件下の同じ細胞内で、細胞周期が開始する。あるいは、どれか特定の相を経由する細胞の進行が変更される;即ち、特定の増殖期を経由して細胞が移動するのに費やす時間の長さに加速または遅延が生じ得る。例えば、該細胞は通常G1期に数時間留まるが;ある物質の添加によりG1期の延長を来し得る。
【0135】
計測は、各計測が同じ条件にあるようにして行うことも、各種の条件下に、バイオ活性物質を用いるかまたは用いずに、または細胞周期過程の異なる段階で行うこともできる。例えば、HDAC8制御の計測は、候補バイオ活性物質が存在する、および候補バイオ活性物質が存在しない、細胞または細胞集団中で行うことができる。別の実施例では、HDAC8制御の計測を、条件または環境がそれぞれ異なる細胞または細胞集団について行う。例えば、細胞を、生理学的シグナル、例えば、ホルモン、抗体、ペプチド、抗原、サイトカイン、成長因子、アクションポテンシャル、化学療法剤を含む医薬物質、放射能、カルシノゲン物質、その他の細胞(即ち、細胞−細胞接触)への曝露、の存在下または不存在下に、またはそれに先立ちまたはその後に、評価してもよい。別の態様では、HDAC8制御の計測を、HDAC8過程の異なる段階で行う。さらに別の実施例では、HDAC8制御の計測を、条件は同じであるが、ある細胞または細胞集団と別の細胞または細胞集団との間での変更について行う。
【0136】
本明細書中で「細胞集団」または「細胞ライブラリー」とは、少なくとも2個の細胞を意味するが、少なくとも約103個であることが好ましく、少なくとも約106個であることが特に好ましく、少なくとも約108個ないし109個であることが格別に好ましい。該集団または試料は、第1または第2培養物と異なる細胞型の混合物を含有してもよいが、単一の細胞型のみを含有する試料、例えば上記したように1種の細胞株由来であり得る、特に腫瘍細胞株由来の試料が好ましい。これらの細胞は、M、G1、S、およびG2を含む、同位相(同位期)または同位相でない、任意の細胞相(細胞期)であり得る。好ましい実施態様では、複製または増殖しつつある細胞を使用するが:このようにすると候補バイオ活性物質の導入にレトロウイルスベクターの使用が可能になる。あるいは、複製中でない細胞を使用することもでき、かつ他のベクター類(例えば、アデノウイルスおよびレンチウイルスベクター)を使用することもできる。加えて、必須ではないが、これらの細胞は染料および抗体と適合性である。
【0137】
本発明での使用に好適な細胞のタイプには、動物(マウス、ラット、ハムスター、アレチネズミを含む齧歯類)、霊長類、およびヒトの細胞、殊に乳癌、皮膚癌、肺癌、頚癌、直腸癌、白血病、脳腫瘍、その他を含む全タイプの腫瘍細胞を含み、哺乳動物細胞が含まれるが、それらに限定されない。
【0138】
好ましい実施態様では、この方法は、細胞生存能力、細胞増殖および細胞相を含むが、それらに限定されない1種または数種以上の細胞パラメータをアッセイすることを含む。
【0139】
好ましい実施態様では、細胞の生存能力をアッセイし、細胞性変化のないことが実験的条件による(即ち、候補バイオ活性物質の導入)ものではなく、細胞死によるものではないことを確認する。光散乱法、生存能力染色法、排除染色法を含むが、それらに限定されない各種の適切な細胞生存能力アッセイを使用できる。
【0140】
好ましい実施態様において、光散乱測定は、当分野において周知であるように、生存能力アッセイとして使用される。例えば、FACSで観察した場合には、細胞は前方および90度(側方)光散乱特性により測定される特異的な特徴を有する。これらの散乱特性は細胞のサイズ、形態および顆粒含有量を示す。これらの特性は、生存能力の読み取り(readout)として測定されるパラメータの説明となる。簡単に言えば、瀕死細胞または死細胞のDNAは一般に凝集しており、このことは90°散乱を変える;同様に、膜の気泡は前方散乱を変える。光散乱の強度、または細胞反射値の変動は生存能力の変動を示す。
【0141】
したがって、一般的に、光散乱アッセイのため、特定の細胞タイプの生存細胞集団をその前方および側方散乱特性を測定して評価する。このことは後に使用され得る散乱の規準を与える。
【0142】
好ましい実施態様において、生存能力アッセイは生存可能な色素 (viability dye)を使用する。死細胞または瀕死細胞を染色するが、増殖中の細胞は染色しない、多くの既知の生存可能な色素が存在する。例えば、アネキシンVは2価イオンに依存した方法でリン脂質(ホスファチジルセリン)に特異的な結合を示すタンパクファミリーのメンバーである。このタンパク質は、ホスファチジルセリンの細胞表面への露出がこのプロセスの初期シグナルの特徴であるので、アポトーシス(プログラム細胞死)の測定に広く使用されている。適当な生存可能な色素は、アネキシン、エチジウムホモダイマー−1、DEAD Red、ヨウ化プロピジウム、SYTOX Greenなど、および当分野で既知の他の物質を含むが、これらに限定されない;the Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Haugland, Sixth Editionを参照。引用により本明細書に含める;特に285ページのApoptosis Assay、およびChapter 16を参照。
【0143】
細胞が生存可能な生存色素染色(viability dye staining)のプロトコールは既知である。上記のMolecular Probesの目録を参照。本実施態様において、アネキシンのような生存可能な色素を直接的または間接的に標識し、細胞集団と組み合わせる。アネキシンは商業的に、即ちPharMingen, San Diego, CaliforniaまたはCaltag Laboratories, Millbrae, Californiaから入手可能である。好ましくは、生存可能な色素は、色素の濃度が約100ng/ml〜約500ng/ml、さらに好ましくは約500ng/ml〜約1μg/ml、および最も好ましくは約1μg/ml〜約5μg/mlの溶液で提供される。好ましい実施態様において、生存可能な色素は直接的に標識される;例えば、アネキシンはフルオレセイン イソチオシアネート(FITC)、Alexa色素、TRITC、AMCA、APC、tri−color、Cy−5のような蛍光色素、および当分野で既知または商業的に入手可能なものであり得る。別の好ましい実施態様において、生存可能な色素はハプテン(例えばビオチン)のような第1の標識で標識され、第2の蛍光標識として、例えば蛍光ストレプトアビジンが使用される。当業者に分かるであろう他の第1および第2の標識の組合せが使用され得る。
【0144】
添加後、生存可能な色素は、ある時間、細胞とともにインキュベートされ、必要であれば洗浄される。次いで、以下に概説したように細胞をソートして非生存細胞を取り除く。
【0145】
好ましい実施態様において、色素排除染色は生存能力アッセイとして使用される。排除色素は生存細胞から排除されるものであり、即ちそれらは受動的に取り込まれない(それらは生存細胞の細胞膜を浸透しない)。しかしながら、死細胞または瀕死細胞の浸透性により、それらは死細胞に取り込まれる。一般的に(必ずというわけではないが)、排除色素はDNAと、例えばインターカレーションにより結合する。好ましくは、排除色素はDNAの不存在下において蛍光を発っしないか、または不十分に蛍光するを発する;このことにより洗浄の段階の必要がなくなる。代わりに、第2の標識の使用を必要とする排除色素も使用され得る。好適な排除色素は、エチジウムブロマイド;エチジウムホモダイマー−1;ヨウ化プロピジウム;SYTOX緑色核酸鎖(green nucleic acid chain);Calcein AM、BCECF AM;フルオレセイン ジアセテート;TOTO(登録商標)およびTO−PRO(商標)(Molecular Probesより;上記、chapter 16参照)および当分野で既知の他の物質を含むが、これらに限定されない。
【0146】
細胞が生存したまま色素排除染色するためのプロトコールは既知である。上記のMolecular Probes参照。一般的に、排除色素は、約100ng/ml〜約500ng/ml、さらに好ましくは約500ng/ml〜約1μg/ml、および最も好ましくは約0.1μg/ml〜約5μg/mlの濃度で細胞に加えられ、特に約0.5μg/mLが好ましい。細胞および排除色素を、ある時間インキュベートし、必要であれば洗浄し、次いで該細胞を以下に概説したようにソートして集団から非生存細胞を取り除く。
【0147】
さらに、実施可能な他の細胞生存能力アッセイが存在し、該方法は生存細胞(即ち分泌されたプロテアーゼなど)または死細胞(即ち培地中に細胞内酵素、例えばプロテアーゼ、ミトコンドリア酵素などが存在すること)の細胞外酵素活性の測定が可能である。the Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Haugland, Sixth Editionを参照。引用により本明細書に含める;特にChapter 16を参照。
【0148】
好ましい実施態様において、少なくとも1つの細胞生存能力アッセイが実施され、蛍光が互換可能な場合には2つの異なる細胞生存能力アッセイが好ましい。1つの生存能力アッセイのみを実施する場合、好ましい実施態様では光分散測定が利用され(前方および側方分散の両方)。2つの生存能力アッセイを実施する場合、好ましい実施態様では光分散と色素排除が利用され、光分散と生存可能な色素染色も利用可能であり、そして場合によっては3つすべてが利用され得る。したがって、生存能力アッセイにより非生存細胞または瀕死細胞から生存細胞を分離することが可能となる。
【0149】
細胞生存能力アッセイに加えて、好ましい実施態様では細胞増殖アッセイが利用される。本明細書において「増殖アッセイ(proliferation assay)」は、細胞集団が増殖している(即ち複製している)か、または複製していないことの測定を可能にするアッセイを意味する。
【0150】
好ましい実施態様において、増殖アッセイは色素封入アッセイである。色素封入アッセイは細胞相を区別する希釈効果によるものである。簡単に言えば、色素(一般に、以下に概説した蛍光色素)を細胞に導入し、細胞に取り込ませる。一度取り込まれると、色素は細胞中にトラップされ、拡散しない。細胞集団が分裂するにつれて、色素は比例して希釈される。即ち、封入色素の導入後、細胞を、ある時間、インキュベートするのである;時間の経過にともない蛍光を失う細胞は分裂しており、蛍光を維持している細胞は非増殖期にある。
【0151】
一般に、封入色素の導入は2つの方法のうちの一方で行われ得る。色素が細胞内に受動的には入れずに(例えば、荷電を有する)、色素が取り込まれるように、例えば電気パルスを使用して;細胞を処理しなければならない。もう1つは、色素は受動的に細胞内に入るが、取り込まれた後、細胞外に拡散できないように修飾される。例えば、封入色素の酵素学的修飾によりそれを帯電させ、したがって、細胞外に拡散できないようにし得る。例えば、Molecular Probes CellTracker(登録商標)は蛍光クロロメチル誘導体であり、自由に細胞内へと拡散し、次いでグルタチオンS−トランスフェラーゼ介在反応により膜不浸透性色素を生じる。
【0152】
適当な封入色素は、Molecular Probes CellTracker(登録商標)のMolecular Probes系を含むが、これらに限定されず、CellTracker(登録商標) Blue、CellTracker(登録商標) Yellow-Green、CellTracker(登録商標) Green、CellTracker(登録商標) Orange、PKH26(Sigma)、および当分野で既知の他の物質を含むが、これらに限定されない;上記のMolecular Probes Handbook;特にchapter 15参照。
【0153】
一般に、封入色素は約100ng/ml〜約5μg/ml、好ましくは約500ng/ml〜約1μg/mlの範囲の濃度で細胞に供与される。洗浄の段階を使用しても、使用しなくてもよい。好ましい実施態様において、候補バイオ活性物質を本明細書に記載のような細胞と組み合わせる。細胞および封入色素を、ある時間、インキュベートすると細胞分裂し、したがって色素が希釈化される。時間の長さは、細胞特有の細胞周期時間に依存する;一般に、少なくとも約2回の細胞分裂が好ましく、少なくとも約3回が特に好ましく、そして少なくとも約4回がとりわけ好ましい。次いで細胞を以下に概略するようにソートし、複製している細胞集団とそうでない集団を作成する。当業者には分かるであろうが、場合によっては、例えば抗増殖剤をスクリーニングする場合、発光(即ち蛍光)している細胞を回収する;他の実施態様において、例えば増殖剤をスクリーニングするため、蛍光の少ない細胞を回収する。変動は、異なる時点での、または異なる細胞集団における蛍光を測定することにより決定され、そして該決定を互いにまたは基準と比較することにより決定される。
【0154】
好ましい実施態様において、増殖アッセイは代謝拮抗物質アッセイである。代謝拮抗物質アッセイは、G1またはG2休止期における休止の原因となる薬品を所望の場合に最も使用される。代謝拮抗物質増殖アッセイでは、毒性があり、分裂中の細胞を死滅させる代謝拮抗物質の使用の結果、分裂していない細胞のみが生き残ることになる。適当な代謝拮抗物質は、メソトレキセート、シスプラチン、タキソール、ヒドロキシウレア、AraCのような核酸アナログ、などのような通常の化学療法剤を含むが、これらに限定されない。さらに、代謝拮抗アッセイは、発現すると細胞死の原因となる遺伝子の使用を含み得る。
【0155】
代謝拮抗物質が添加される濃度は代謝拮抗物質に特有の毒性に依存し、当分野で既知のように決定される。代謝拮抗物質が添加され、細胞は、一般に、ある時間インキュベートされる;再び、正確な時間は代謝拮抗物質の特徴および独自性、ならびに細胞集団に特有の細胞周期の時間に依存する。一般に、少なくとも1回の細胞分裂が起こるのに十分な時間である。
【0156】
好ましい実施態様において、少なくとも1つの増殖アッセイが実施され、2以上が好ましい。したがって、増殖アッセイにより、結果的に増殖中の細胞集団と休止の細胞集団になる。さらに他の増殖アッセイ、即ち当分野において既知の比色定量アッセイを使用し得る。
【0157】
好ましい実施態様において、上で概説した1以上の増殖アッセイの後または同時に、少なくとも1つの細胞相アッセイが行われる。「細胞相」アッセイは、細胞が滞留している細胞相、M、G1、S、またはG2を決定する。
【0158】
好ましい実施態様において、細胞相アッセイはDNA結合色素アッセイである。簡単に言えば、DNA結合色素は細胞に導入され、受動的に取り込まれる。一度細胞内に入ると、DNA結合色素はDNAに、一般的にはインターカレーションにより結合するが、場合によっては、それらは主溝または副溝結合性化合物であり得る。したがって、色素の量は細胞中のDNAの量に直接関連し、これは細胞の相によって変動する;G2およびM期の細胞はG1期の細胞の2倍のDNA含有量を有し、S期の細胞は中間の量を有し、その細胞がS期のどの点であるかに依存する。適当なDNA結合色素は浸透性であり、Hoechst 33342および33258、アクリジンオレンジ、7−AAD、LDS751、DAPI、SYTO16、上記のMolecular Probes Handbook;特にchapter 8および16を含むが、これらに限定されない。
【0159】
一般的に、DNA結合色素は約1μg/ml〜約5μg/mlの範囲の濃度で加えられる。該色素は細胞に加えられ、ある時間、インキュベートされる;時間の長さは選択された色素に一部依存する。1つの実施態様において、色素の添加直後に測定が行われる。次いで、以下に概説したように細胞をソートし、異なる量の色素、したがって異なる量のDNAを含む細胞集団を作成する;本方法において、複製中の細胞は、そうでないものと分離される。当業者には分かるように、場合によっては、例えば抗増殖剤をスクリーニングする場合に、最も弱い蛍光を有する細胞(したがって1セットのゲノム)を増殖中の細胞(したがって1セットのゲノムより多いDNA含む)と分離することができる。変動は、異なる時点での、または異なる細胞集団における蛍光を測定することにより決定され、そして該決定を互いにまたは基準と比較することにより決定される。
【0160】
好ましい実施態様において、細胞相アッセイはサイクリン破壊アッセイである。本実施態様において、スクリーニングの前(および一般的に、以下に概略したように、候補バイオ活性物質の導入前)に融合核酸を細胞に導入する。融合核酸はサイクリン破壊ボックスをコードする核酸および検出可能な分子をコードしている核酸を含む。「サイクリン破壊ボックス」は当分野において既知であり、特定の細胞相の間に該ボックスを含むタンパク質のユビキチン化経路を通して分解の原因となる配列である。即ち、例えば、G1サイクリンはG1期の間安定であり得るが、G1サイクリン破壊ボックスのためS期の間に分解され得る。したがって、サイクリン破壊ボックスを検出可能な分子、例えばグリーン蛍光タンパク質と結合させることにより、検出可能な分子の存在または不存在により細胞集団の細胞相の同定が可能となる。好ましい実施態様において、好ましくはそれぞれ、さまざまな蛍光とともに多数のボックスが使用され、その結果、細胞相の検出が可能となる。
【0161】
多くのサイクリン破壊ボックスが当分野において既知であり、例えば、サイクリンAは配列RTVLGVIGD(配列番号:8)を含む破壊ボックスを有する;サイクリンB1の破壊ボックスは配列RTALGDIGN(配列番号:9)を含む。Glotzer et al., Nature 349:132-138 (1991)参照。さらに他の破壊ボックスとして:
YMTVSIIDRFMQDSCVPKKMLQLVGVT(配列番号:10:ラットサイクリンB);
KFRLLQETMYMTVSIIDRFMQNSCVPKK(配列番号:11:マウスサイクリンB);
RAILIDWLIQVQMKFRLLQETMYMTVS(配列番号:12:マウスサイクリンB1);
DRFLQAQLVCRKKLQVVGITALLLASK(配列番号:13:マウスサイクリンB2);および
MSVLRGKLQLVGTAAMLL(配列番号:14:マウスサイクリンA2)
が知られている。
【0162】
サイクリン破壊ボックスをコードする核酸は検出可能な分子をコードする核酸と機能し得るように結合させる。融合タンパク質は当分野において既知の方法で作成される。例えば、サイクリン破壊ボックスをコードする核酸は検出可能な分子をコードする核酸と結合させる。本明細書の「検出可能な分子」は、検出可能な分子を含む細胞または化合物と、それを含まないものとの区別を可能にする分子を意味する。即ちエピトープ(抗原TAGと呼ばれることもある)、特定の酵素または蛍光分子である。好適な蛍光分子は、グリーン蛍光タンパク質(GFP)、ブルー蛍光タンパク質(BFP)、イエロー蛍光タンパク質(YFP)、レッド蛍光タンパク質(RFP)、およびルシフェラーゼおよびβ−ガラクトシダーゼを含む酵素を含むが、これらに限定されない。抗原TAGが使用される場合、好適な実施態様では細胞表面抗原を使用する。エピトープは、好ましくは、一般的には細胞膜上で見出されない任意の検出可能なペプチドであるが、いくつかの例において、エピトープが細胞上で普通に見出されるものである場合には、増加が検出され得る(このことは一般的には好ましくないのだが)。同様に酵素学的に検出可能な分子;例えば新規または発色性の生成物を産生する酵素も使用され得る。
【0163】
したがって、細胞相アッセイ後のソートの結果、一般的に、異なる細胞相にある少なくとも2つの細胞集団が生じる。
【0164】
ここで提供されるタンパク質および核酸を、スクリーニング目的のために使用することができ、HDAC8タンパク質のタンパク質−タンパク質相互作用を同定することができる。タンパク質−タンパク質相互作用を検出するための遺伝学的システムが記載されている。酵母のシステムにおいて最初の研究が実施された。即ち「酵母ツー−ハイブリッド」システムである。基本的なシステムは、レポーター遺伝子の転写を開始するために、タンパク質−タンパク質相互作用を必要とする。次いで、研究は哺乳類細胞において実施された。Fields et al., Nature 340: 245(1989); Vasavada et al., PNAS USA 88: 10686(1991); Fearon et al., PNAS USA 89: 7958(1992); Dang et al., Mol. Cell. Biol. 11: 954(1991); Chien et al., PNAS USA 88: 9578(1991); 並びに米国特許番号第5283173号、第5667973号、第5468614号、第5525490号、および第5637463号を参照されたい。好ましいシステムは、出願番号第09/050863号、1998年3月30日出願、および第09/359081号、1999年7月22日出願、表題「哺乳動物タンパク質のインターカレーションクローニングシステム(Mammalian Protein Interaction Cloning System)」に記載されている。これらのシステムの合同的な使用について、特に有用なシャトルベクターが、出願番号09/133944、1998年8月14日出願、表題「シャトルベクター(Shuttle Vectors)」に記載されている。
【0165】
一般に、2つの核酸を1細胞に形質転換する。1つの核酸はHDAC8タンパク質またはその一部をコードする遺伝子のような「ベイト(bait: えさ)」であり、もう1つの核酸は試験候補をコードしている。2つの発現産物が互いに結合する場合のみ、蛍光タンパク質などの指示薬は発現する。その指示薬の発現は、いつ試験候補がHDAC8タンパク質に結合するかを示し、試験候補をHDAC8タンパク質として同定することができる。同じシステムと同定されるHDAC8タンパク質を使用して、逆のことを実施することができる。即ち、本発明で提供されるHDAC8タンパク質を、新規なベイト、またはHDAC8タンパク質と相互作用する物質を同定するために、使用することができる。さらに、ベイトおよびHDAC8タンパク質をコードしている核酸のほかに試験候補を添加し、ベイトとHDAC8タンパク質との相互作用を妨害する物質を調べる場合に、ツー−ハイブリッドシステムを使用することができる。
【0166】
ある実施態様において、哺乳類ツー−ハイブリッドシステムが好ましい。哺乳類のシステムは、タンパク質のポスト翻訳修飾を提供し、その修飾は、タンパク質が相互作用する能力に有意に寄与し得る。さらに、哺乳類ツー−ハイブリッドシステムを広範な哺乳類細胞のタイプにおいて使用し、特定の細胞タイプ内の特異的タンパク質の制御、誘導、プロセッシングなどを模倣することができる。例えば、疾患状態(即ち、癌、アポトーシス関連障害)に関連するタンパク質を、関連する疾患細胞において試験し得る。同様に、ランダムタンパク質の試験のために、関連する細胞内条件でアッセイすることにより、最高の肯定的な結果が得られる。さらに、哺乳類細胞を、細胞間タンパク質−タンパク質相互作用に影響を及ぼし得る広範な実験条件下(例えば、ホルモン、薬剤、成長因子およびサイトカイン、放射線、化学療法剤、細胞内および化学的刺激などの存在)で試験することができ、このことは、タンパク質−タンパク質相互作用に影響を及ぼすことのできる条件(特に癌に関連する条件)に寄与し得る。
【0167】
ツー−ハイブリッドシステムなどの結合に関連するアッセイは、非特異的結合タンパク質(NSB)を考慮に入れ得る。
【0168】
様々な細胞タイプにおける発現、およびHDAC8活性のアッセイを、前記した。例えば、クロマチン構造、ヒストンアセチル化、転写などに対する作用についての活性のアッセイを実施し、PCNAとの配列同一性/類似性またはそれとの結合によって、既に同定されていたHDAC8タンパク質の活性、転写などに影響を示すことを確認することができる。さらに、HDAC8の調節因子として同定されるリード化合物の活性をさらに確認することができる。
【0169】
本発明で提供されるアッセイ用の、本発明で提供される構成成分を組合せて、キットを形成してもよい。該キットは、タンパク質および/またはHDAC8タンパク質をコードする核酸の使用をベースとすることができる。ある実施態様では、他の構成成分がキット中で提供される。そのような構成成分には、1つまたはそれ以上の包装、指示書、抗体、および標識が含まれる。診断に使われるようなさらなるアッセイは、さらに後述する。
【0170】
このようにして、バイオ活性物質が同定される。医薬的活性のある化合物は、HDAC8タンパク質の活性を高めたり、妨害したりできる。さらに後述するように、所望の医薬的活性を有する化合物を、医薬的に許容し得る担体中で宿主に投与してもよい。
【0171】
即ち、細胞中でのHDAC8タンパク質の役割に関する今回の開発は、細胞中で細胞増殖を誘導または防止する方法を提供する。好ましい実施態様では、HDAC8タンパク質、および特にHDAC8タンパク質断片は、HDAC8タンパク質が介在する症状の研究または処置において、即ち、HDAC8関連障害の診断、処置、または予防において、有用である。ゆえに、「HDAC8関連障害」または「疾患状態」には、細胞増殖の不足または過剰の両方に関連する症状が含まれる。細胞増殖の不足または過剰の例には、老化、アポトーシス、ネクローシス、癌などが含まれるが、癌が好ましい。癌には、腫瘍形成、癌転移および脈管形成と関連する全ての機構が含まれる。
【0172】
ゆえに、ある実施態様では、細胞または生物内でのHDAC8制御が提供される。ある実施態様では、その方法は、治療的な量でHDAC8タンパク質を、それを必要としている細胞または個体に投与することを含む。あるいは、内在性のHDAC8タンパク質の生物学的活性を低減または排除する抗HDAC8抗体を投与する。他の実施態様では、本発明で提供される方法で同定したバイオ活性物質を投与する。あるいは、その方法は、HDAC8タンパク質をコードする組換え核酸を、細胞または個体に投与することを含む。当業者に理解されるであろうように、このことはいかなる数の方法で達成されてもよい。好ましい実施態様では、例えば、内因性HDAC8タンパク質を過剰発現させるか、または、遺伝子治療技術などでHDAC8タンパク質をコードする遺伝子を投与することで、細胞中のHDAC8タンパク質の量を増やすことによって、HDAC8の活性は、増大する。好ましい実施態様では、該遺伝子治療技術は、例えば出典明示により本明細書の一部とするPCT/US93/03868に記載のように、強化相同組換え(EHR)を用いて、外来性遺伝子を組込むことを含む。
【0173】
理論によって制限を受けないが、HDAC8タンパク質は、転写およびHDAC8を制御する点でHDAC8において重要なタンパク質であることがわかった。従って、HDAC8遺伝子の突然変異または変異に基づく障害が確定され得る。ある実施態様では、本発明は、細胞中の少なくとも1つの内因性HDAC8遺伝子配列の全部または部分を解読することを含む、変異型HDAC8遺伝子を有する細胞の同定法を提供する。当業者に理解されるであろうように、このことは、配列解読技術をいくつでも用いて行われ得る。好ましい実施態様では、本発明は、個体の少なくとも1つのHDAC8遺伝子配列の全部または一部を解読することを含む、個体のHDAC8遺伝子型の同定法を提供する。このことは、一般的に、個体の少なくとも1組織について行われ、多数の組織または同じ組織の異なる試料の評価を含んでもよい。該方法は、解読されたHDAC8遺伝子の配列を、既知のHDAC8遺伝子、即ち、野生型遺伝子と比較することを含んでもよい。
【0174】
HDAC8遺伝子の全部または一部の配列を、既知のHDAC8遺伝子の配列と比較して、差異が存在するかどうかを決定することができる。このことは、Bestfitなどの周知の配列同一性プログラムをいくつでも用いて行うことができる。好ましい実施態様では、患者のHDAC8遺伝子と既知のHDAC8遺伝子との間の配列の差異が存在することは、疾患状態または疾患状態になる傾向を示している。従って、本発明は、例えば遺伝子の完全性に影響を与える変化、置換に関する欠損および挿入、染色体の配置転換、mRNAの異常型スプライシングパターンなどによって特徴付けられるHDAC8核酸における遺伝的障害の存在もしくは非存在を決定する方法を提供する。
【0175】
ある実施態様では、本発明は、個体におけるHDAC8関連症状の診断法を提供する。この方法には、個体または患者由来の組織または体液などの試料中におけるHDAC8活性の測定が含まれる。HDAC8の活性は、ある試料がHDAC8の既知の基質を脱アセチル化する能力についてのアッセイによって測定するのが好ましい。例えば、特定の抗血清を用いるラジオイムノアッセイは、アセチル化と非アセチル化ペプチドに対して行った。一方、患者由来のHDAC8試料の能力は、インビボでのラジオアセチル化された天然または合成基質を脱アセチル化するその能力が試験され得る。この活性を、罹患していない第2の個体または第1の個体の罹患していない組織のどちらかに由来するHDAC8の活性と比較する。これらの活性が異なる場合、第1の個体にはHDAC8関連異常のリスクが有り得る。例えば、癌細胞は、末端分化に関与する遺伝子の転写抑制の手段としてHDAC8をアップレギュレートし「得る」。このことは、細胞を増殖状態にさせる。従って、HDAC8の阻害により、癌細胞の細胞分化を非疾患状態へと誘導する遺伝子の転写が可能となる。このようにして、例えば、タンパク質のレベルまたはそのmRNAの発現をモニターすることによって、様々な疾患状態をモニターし得る。同様に、発現レベルは、予後に相関し得る。
【0176】
ある態様では、HDAC8タンパク質遺伝子の発現レベルを、診断または予後情報が求められる異なる患者の試料または細胞について測定する。遺伝子発現のモニターは、HDAC8タンパク質をコードする遺伝子について行われる。ある態様では、HDAC8タンパク質遺伝子の発現レベルは、正常細胞およびアポトーシスまたは形質転換をしている細胞のような、細胞の異なる状態について測定される。異なる状態の細胞においてHDAC8タンパク質遺伝子の発現レベルを比較することによって、HDAC8タンパク質遺伝子のアップレギュレーションおよびダウンレギュレーションの両者を含む情報が得られ、この情報は多数の方法で用いられる。例えば、特定の処置方式を評価し得る:ある化学療法剤が特定の患者の長期的予後を改善するのに役立っているかを評価し得る。同様に、患者の試料を比較して、診断を行うか、または確認してもよい。さらに、これらの遺伝子発現レベルによって、特定の発現レベルを模倣または変化させる目的で、薬物候補をスクリーニングすることが可能になる。このことは、このようなスクリーニングに用いることができ、本発明のもののような重要なHDAC8タンパク質遺伝子の一揃えを含むバイオチップを作成することによって行ってもよい。これらの方法は、タンパク質に基づいて行うこともできる;つまり、HDAC8タンパク質のタンパク質発現レベルを、診断の目的や候補物質のスクリーニングのために評価できる。さらに、HDAC8タンパク質の核酸配列を遺伝子治療の目的で投与することができる。このことには、アンチセンス核酸の投与、または治療薬としてのHDAC8タンパク質の投与が含まれる。
【0177】
バイオチップに結合するHDAC8タンパク質配列には、上記定義のように核酸およびアミノ酸の両者の配列が含まれる。好ましい実施態様では、HDAC8タンパク質の核酸(図に概説した配列を有する核酸配列および/またはその相補鎖の両者)に対する核酸プローブが作成される。バイオチップに取付けられる核酸プローブは、HDAC8タンパク質の核酸、即ち、標的配列(試料の標的配列、または例えばサンドイッチアッセイにおける他のプローブ配列への標的配列)に実質的に相補的に設計し、標的配列と本発明のプローブとのハイブリダイゼーションが起こるようにする。以下に概説するように、この相補性は、完全である必要は無い;標的配列と本発明の1本鎖核酸の間のハイブリダイゼーションを妨害するミスマッチ塩基対がいくつあってもよい。しかし、もし変異の数が多すぎて、緊縮性が最低のハイブリダイゼーション条件下でさえハイブリダイゼーションが起こらないならば、その配列は相補的標的配列ではない。ゆえに、本明細書で「実質的に相補的」は、本明細書で概説するように、通常の反応条件、特に高緊縮条件下な条件下でハイブリダイズするのに、プローブが標的配列に十分相補的であることを意味する。
【0178】
「核酸プローブ」は、一般的に1本鎖であるが、部分的に1本鎖、および部分的に2本鎖であり得る。プローブの鎖形成は、構造、組成、および標的配列の性質によって決定される。一般的に、核酸プローブは約8ないし約100塩基長の範囲にあり、約10ないし約80塩基が好ましく、約30ないし約50塩基が特に好ましい。ある実施態様では、数百塩基に上るさらに長い核酸(例えば、遺伝子全長)を用いることができる。
【0179】
当業者に理解されるであろうように、核酸を、幅広い方法で、固体支持体に結合または固定することができる。 本明細書での「固定された」および文法的均等語は、以下に概説するように、核酸プローブと固体支持体の会合または結合が、結合、洗浄、分析、および除去の条件下で安定であるために十分であることを意味する。結合は、共有的または非共有的であり得る。本明細書における「非共有結合」および文法的均等語は、1またはそれ以上の、電気的、親水的、および疎水的相互作用のいずれかを意味する。ストレプトアビジンのような分子の支持体への共有結合、およびビオチン化プローブのストレプトアビジンへの非共有結合は、非共有結合に含まれる。本明細書における「共有結合」および文法的均等語は、2つの部分、固体支持体とプローブが、シグマ結合、パイ結合および配位結合を含む少なくとも1つの結合によって結合していることを意味する。共有結合は、プローブと固体支持体との間に直接形成でき、あるいは、クロスリンカーによって、または固体支持体とプローブのいずれかもしくは両者に特異的反応基を含むことによって形成できる。固定化は、共有および非共有相互作用の組合せも含み得る。
【0180】
一般的に、当業者に理解されるであろうように、プローブを幅広い方法でバイオチップに結合させる。本明細書に記載のように、核酸は、最初に合成して次にバイオチップに結合するか、またはバイオチップ上で直接合成することができる。
【0181】
バイオチップは、好適な固体基板を含む。本明細書の「基板」または「固体支持体」または他の文法的均等語は、核酸プローブの結合または会合に適する独立した個々の部位を含有するように変更することができ、そして少なくとも1つの検出法で分析できる、あらゆる材料を意味する。当業者に理解されるであろうように、可能な材料の数は非常に多く、ガラスおよび修飾または機能化ガラス、プラスチック(アクリル、ポリスチレン、ならびにスチレンおよび他の材料の共重合体、ポリプロピレン、ポリエチレン、ポリブチレン、ポリウレタン、テフロンJなどを含む)、多糖類、ナイロンまたはニトロセルロース、樹脂、シリカまたはシリコンもしくは修飾シリコンを含むシリカベースの材料、炭素、金属、無機ガラス、プラスチックなどを含むが、これらに限定されるものではない。一般的に、該基板は、好都合な検出を可能にし、感知できるほどの蛍光を示さない。
【0182】
好ましい実施態様では、バイオチップとプローブの表面を、両者の結合のために、化学官能基で誘導し得る。ゆえに、例えば、バイオチップは、アミノ基、カルボキシル基、オキソ基およびチオール基を含むがこれらに限定されない化学官能基で誘導され、アミノ基が特に好ましい。これらの官能基を用いると、プローブをプローブ上の官能基を用いて結合させることができる。例えば、アミノ基を含む核酸は、例えば、当分野で周知のようにリンカーを用いて、アミノ基を含む表面に取付けることができる;例えば、ホモ―またはヘテロ―両機能性リンカーは、周知である(1994 Pierce Chemical Company catalog, technical section on cross-linkers, pages 155-200, 参照。出典明示により本明細書の一部とする)。さらに、ある場合では、アルキル基(置換およびヘテロアルキル基を含む)のような追加のリンカーを用いてもよい。
【0183】
この実施態様では、核酸プローブに対応するオリゴヌクレオチドを当分野で周知のように合成し、固体支持体の表面に結合させる。当業者には理解されるであろうように、5'または3'の末端のどちらを固体支持体に結合させてもよく、また内側のヌクレオチドを介して結合させてもよい。
【0184】
さらなる実施態様では、固体支持体への固定化は、非共有結合であっても非常に強固であり得る。例えば、ストレプトアビジンで共有結合により被膜された表面に結合するビオチン化オリゴヌクレオチドを作成することができ、結合させる。
【0185】
または、当分野で周知のように、オリゴヌクレオチドを表面上で合成できる。たとえば、光重合の化合物および技術を利用する光活性化技術を用いる。好ましい実施態様では、核酸は、WO95/25116;WO95/35505;米国特許番号第5700637号および第5445934号;およびそれらの中で引用された参照文献(それらすべてを出典明示により本明細書の一部とする)に記載のように、周知の写真平板技術を用いてin situで合成できる;これらの結合の方法は、Affimetrix GeneChip(登録商標)技術の基礎をなしている。
【0186】
本明細書で使用される「ディファレンシャル発現」または文法的均等語は、遺伝子の時間的および/または細胞の発現パターンにおける、細胞間の量的差異と質的差異の両方を表わす。ゆえに、異なって発現された遺伝子は、例えば正常細胞対アポトーシスの細胞において、活性化と不活性化を含む、定性的に変化した発現を有することができる。つまり、遺伝子は、特定の状態において、他の状態に関連して点いたり切れたりし得る。当業者には明らかなように、2種またはそれ以上の状態のどのような比較も行うことができる。そのような定性的に制御される遺伝子は、ある状態または細胞のタイプにおいて、発現パターンを示すであろう。そのパターンは、状態または細胞のタイプが1種の場合は標準的な技術で検出できるが、両方の場合は検出できない。あるいは、発現が増加または減少する場合には、測定は定量的である;つまり、遺伝子の発現はアップレギュレーションされ、転写物の量が増加するか、または遺伝子の発現はダウンレギュレーションされ、転写物の量が減少するかのどちらかである。発現が異なる程度は、以下に概説するように、Affymetrix GeneChip(登録商標)発現アレイ(Lockhart, Nature Biotechnology 14:1675-1680 (1996)、出典明示により本明細書の一部とする)のような、標準的な特性分析技術を介して定量するのに十分なほどに大きい必要があるだけである。他の技術には、定量的逆転写酵素PCR、ノーザン分析およびRNase保護が含まれるが、これらに限定されるものではない。
【0187】
当業者に理解されるであろうように、このことは遺伝子転写物、またはタンパク質レベルのいずれかを評価することによってなし得る;つまり遺伝子発現の量は、遺伝子転写物と等価のDNAまたはRNAに対する核酸プローブ、および遺伝子発現のレベルの定量を用いてモニターすることができ、または、例えばHDAC8タンパク質の抗体および標準的なイムノアッセイ(ELAISAなど)、または質量分析アッセイ、2次元ゲル電気泳動アッセイなどの他の技術を介して、遺伝子の最終産物そのもの(タンパク質)をモニターすることができる。
【0188】
他の方法では、mRNAの検出はin situで行われる。この方法では、透過性にした細胞または組織の試料を、検出可能な標識化核酸プローブに十分な時間接触させ、プローブを標的mRNAにハイブリダイズさせる。非特異的に結合したプローブを除去するために洗浄した後、標識を検出する。例えば、HDAC8タンパク質をコードするmRNAに相補的なジゴキシゲニン標識化リボプローブ(RNAプローブ)は、ジゴキシゲニンを抗ジゴキシゲニン二次抗体と結合させ、ニトロブルーテトラゾリウムおよびリン酸5−ブロモ4−クロロ−3−インドリルと反応させることで検出される。
【0189】
他の好ましい方法では、HDAC8タンパク質の発現は、HDAC8タンパク質抗体を使用するin situイメージング技術を用いて行われる。この方法では、細胞をHDAC8タンパク質(群)に対する1種ないし多種の抗体に接触させる。非特異的に結合している抗体を除去するために洗浄し、抗体または抗体の存在を検出する。ある実施態様では、抗体は、検出可能な標識を含む二次抗体とインキュベートすることにより検出される。他の方法では、HDAC8タンパク質(群)に対する一次抗体が、検出可能な標識を有する。他の好ましい実施態様では、多種類の一次抗体の各々が、別個の検出可能な標識を含む。この方法には、多数のHDAC8タンパク質の同時スクリーニングにおいて特に有用性がある。標識は、異なる波長の発光を検出し、区別できる性能を有する蛍光光度計で検出し得る。さらに、蛍光標示式細胞分取器(FACS)を、この方法で用いることができる。通常の技術のある当業者に理解されるであろうように、膨大な数の他の組織学的イメージング技術が本発明で有用であり、抗体をELISA,イムノブロット(ウエスタンブロット)、免疫沈降、BIACORE技術などに使用できる。
【0190】
ある実施態様において、HDAC8タンパク質に対するポリクローナルおよびモノクローナル抗体を生成するために、本発明のHDAC8タンパク質を使用し得る。それらの抗体は、ここで記載されるように有用である。同様に、HDAC8タンパク質を、標準的な技術を使用してアフィニティ・クロマトグラフィ・カラムに結合することができる。そして、これらのカラムをHDAC8抗体を精製するために使用し得る。好ましい実施態様において、抗体をHDAC8タンパク質に対してエピトープ特異的に生成する;即ち、抗体は、他のタンパク質とほとんどまたは全く交差反応性を示さない。これらの抗体について、多くの応用における使用が見出される。例えば、HDAC8タンパク質抗体を標準的なアフィニティ・クロマトグラフィ・カラムに結合し得る。そして、以下にさらに記載するように、HDAC8タンパク質を精製するために使用し得る。抗体を、前記で概説したように、ブロッキングポリペプチドとしても使用し得る。というのは、これらは、HDAC8タンパク質と特異的に結合するからである。
【0191】
抗HDAC8タンパク質抗体は、ポリクローナル抗体を含み得る。ポリクローナル抗体を調製する方法は、当業者にとって既知である。ポリクローナル抗体を哺乳類中で産生し得る。例えば、1以上の免疫化物質および所望によりアジュバントの注射による。典型的には、免疫化物質および/またはアジュバントを、数回の皮下または腹膜内への注射によって哺乳類に注射する。免疫化物質は、HDAC8タンパク質またはその融合タンパク質を含み得る。免疫化物質と、免疫化される哺乳類において免疫原であることが既知であるタンパク質とを結合することは、有用である。そのような免疫原タンパク質の例は、キーホール・リンペット・ヘモシアニン、血清アルブミン、ウシ・チログロブリンおよびダイズ・トリプシンインヒビターを含むが、これらに限定されない。使用され得るアジュバントの例は、フロイントの完全アジュバントおよびMPL−TDMアジュバント(モノフォスフィリル・リピッドA、合成トレハロース・ジコリノミコレート)を含む。免疫化プロトコールを当業者は過度な実験なくして選択し得る。
【0192】
あるいは、抗HDAC8タンパク質抗体はモノクローナル抗体であってよい。モノクローナル抗体は、ハイブリドーマ法を使用して調製し得る。その方法は、Kohler and Milstein, Nature, 256: 495(1975)に記載されている。ハイブリドーマ法では、マウス、ハムスターなどの適当な宿主動物を典型的には免疫化物質で免疫化する。そして、免疫化物質に特異的に結合する抗体を、産生または産生し得るリンパ球を誘発する。あるいはリンパ球をインビトロで免疫化し得る。
【0193】
免疫化物質は、典型的には、HDAC8タンパク質またはその融合タンパク質を含む。一般的には、ヒト起源の細胞が望まれるならば、末梢血液リンパ球(「PBL」)を使用し、または、非ヒト哺乳類ソースが望まれるなら、脾臓細胞またはリンパ節細胞を使用する。そして、リンパ球を不死化細胞系統と融合化する。その際に、ポリエチレングリコールのような適当な融合化物質を使用してハイブリドーマ細胞を形成させる[Goding, Monoclonal Antibodies: Pronciples and Practice, Academic Press, (1986) pp. 59-103]。不死化細胞系統として、通常、形質転換された哺乳類細胞、特に齧歯類、ウシおよびヒト起源のミエローマ細胞を使用する。通常、ラットまたはマウスミエローマ細胞系統を使用する。ハイブリドーマ細胞を適当な培地中で培養し得る。それは、好ましくは、融合されず、不死化していない細胞の成長または生存を阻害する1以上の物質を含む。例えば、親細胞が酵素ヒポキサンチン・グアニン・ホスホリボシル・トランスフェラーゼ(HGPRTまたはHPRT)を欠くならば、ハイブリドーマのための培地(「HAT培地」)は、典型的には、HGPRT欠失細胞の成長を妨げるヒポキサンチン、アミノプテリン、およびチミジンを含む。
【0194】
好ましい不死化細胞系統は効率的に融合し、選択された抗体生産細胞による抗体の安定的高水準の発現を支持し、そしてHAT培地のような培地に感受性である。さらに好ましい不死化細胞系統は、ネズミミエローマ系統であり、例えば、Salk Institute Cell Distribution Center, San Diego, California and the American Type Culture Collection, Rockville, Marylandから得ることができる。ヒトミエローマおよびマウス−ヒトへテロミエローマ細胞系統は、ヒトモノクローナル抗体の生産に関して記載されている[Kozbor, J. Immunol., 133:3001(1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63]。
【0195】
そして、ハイブリドーマ細胞を培養する培地を、HDAC8タンパク質に対するモノクローナル抗体の存在についてアッセイすることができる。好ましくは、ハイブリドーマ細胞によって産生されるモノクローナル抗体の結合特異性を、免疫沈降によって、または放射性免疫測定法(RIA)もしくは酵素結合免疫吸着アッセイ(ELISA)などのインビトロ結合アッセイによって測定する。これらの技術およびアッセイは当技術分野で既知である。例えば、モノクローナル抗体の結合親和性をMunson and Pollard, Anal. Biochem., 107:220(1980)のScatchard分析によって測定することができる。
【0196】
所望のハイブリドーマ細胞の同定の後、クローンを限界希釈法によってサブクローン化し、標準的な方法によって成長させ得る[Goding、前出]。この目的のための適当な培地は、例えば、ダルベッコ変法イーグル培地(Dulbecco's Modified Eagle's Medium)およびPPMI−1640培地を含む。あるいは、ハイブリドーマ細胞を哺乳類内の腹水のようなインビボで成長させ得る。
【0197】
サブクローン化によって分泌されたモノクローナル抗体を、通常の免疫グロブリン精製方法により培地または腹水液から単離し精製し得る。この精製方法には、例えば、タンパク質A−セファロース、ヒドロキシルアパタイト・クロマトグラフィー、ゲル電気泳動、透析、またはアフィニティー・クロマトグラフィーがある。
【0198】
モノクローナル抗体を組み換えDNA法によっても作成し得る。例えば、米国特許番号第4816567号に記載されている。本発明のモノクローナル抗体をコードするDNAを通常の方法を使用して容易に単離し、配列決定することができる(例えば、ネズミ抗体の重鎖および軽鎖をコードしている遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを使用して)。本発明のハイブリドーマ細胞を、そのようなDNAの好ましいソースとして使用することができる。単離すると、DNAを発現ベクター内に入れ、宿主細胞にトランスフェクトし得る。そのような細胞には、トランスフェクトされなければ免疫グロブリンタンパク質を産生しないシミアンCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、またはミエローマ細胞がある。次いで組換え宿主細胞においてモノクローナル抗体が合成される。例えば、ヒトの重鎖および軽鎖の定常ドメインのコード配列を、相同的なネズミ配列の場所に置換することにより[特許第4816567号;Morrison et al., 前出]、または免疫グロブリン・コード配列と、非免疫グロブリン・ポリペプチドのコード配列のすべてまたは一部とを共有結合させることなどによりDNAもまた修飾し得る。そのような非免疫グロブリン・ポリペプチドを、本発明の抗体の定常ドメインと置換することができる。あるいは、本発明の抗体のある抗原結合部位の可変ドメインと置換し、キメラ二価抗体を生成することができる。
【0199】
抗体は一価の抗体であってよい。一価の抗体を調製する方法は、当技術分野で周知である。例えば、ある方法は、免疫グロブリンの軽鎖および修飾重鎖の組換え発現を含む。重鎖は、Fc領域における任意の点において末端切除し、重鎖の交差結合(cross linking)を防止する。あるいは、関連するシステイン残基を、別のアミノ酸残基で置換するか削除して、交差結合を防ぐ。
【0200】
インビトロ法は、一価抗体を調製するためにも適当である。抗体の消化によって、その断片、特にFab断片を生成することは、当分野で既知の日常的な技術を使用し実施することができる。
【0201】
本発明の抗HDAC8タンパク質抗体は、ヒト化抗体またはヒト抗体をさらに含み得る。ヒト化型の非ヒト(例えばネズミ)抗体は、キメラ免疫グロブリン、免疫グロブリン鎖またはその断片(Fv、Fab、Fab'、F(ab')2または抗体の他の抗原結合サブシーケンスなど)である。これらは、非ヒト免疫グロブリンに由来する最小配列を含む。ヒト化抗体はヒト免疫グロブリン(レシピエント抗体)を含み、レシピエントの相補性決定領域(CDR)からの残基が、マウス、ラットまたはウサギなどの所望の特異性、親和性および能力を有する非ヒト種(ドナー抗体)のCDRからの残基で置換されている。いくつかの例では、ヒト免疫グロブリンのFvフレームワーク残基が、対応する非ヒト残基で置換されている。ヒト化抗体は、レシピエント抗体および導入されたCDRまたはフレームワーク配列のいずれにも見出されない残基も含む。一般に、ヒト化抗体は、実質的に少なくとも1つの、そして典型的には2つの可変ドメインの全部を含む。このドメインでは、すべてまたは実質的にすべてのCDR領域が、非ヒト免疫グロブリンのCDR領域に一致し、すべてまたは実質的にすべてのFR領域が、ヒト免疫グロブリン共通配列のFR領域である。ヒト化抗体は、所望により、免疫グロブリン定常領域(Fc)、通常はヒト免疫グロブリンの定常領域の少なくとも一部も含む[Jones et al., Nature, 321: 522-525(1986); Riechmann et al., Nature, 332: 323-329(1988); and Presta, Curr. Op. Struct. Biol., 2: 593-596(1992)]。
【0202】
非ヒト抗体をヒト化する方法は当技術分野で周知である。一般に、ヒト化抗体は、非ヒトのソースから抗体に導入された1以上のアミノ酸残基を有している。これらの非ヒトアミノ酸残基をしばしば「導入」残基と称する。それは、典型的には「導入」可変ドメインに由来する。ヒト化をWinterおよびその共同研究者の方法[Jones et al., Nature, 321: 522-525(1986); Riechmann et al., Nature, 332: 323-327(1988); Verhoeyen et al., Science, 239: 1534-1536(1988)]に従って、齧歯類CDR配列と、ヒト抗体の対応する配列を置換することによって、基本的に実施することができる。従って、該「ヒト化」抗体は、キメラ抗体であり(米国特許第4816567号)、無処置のヒト可変ドメインより実質的に小さいドメインが非ヒト種からの対応する配列によって置換されている。実際、ヒト化抗体は、典型的にはいくつかのCDR残基およびおそらくいくつかのFR残基が、齧歯類抗体の同種の部位からの残基によって置換されているヒト抗体である。
【0203】
ヒト抗体は、当技術分野で既知の様々な技術を使用して産生することができる。この技術はファージ・ディスプレイ・ライブラリーを含む[Hoogenboom and Winter, J. Mol. Biol., 227: 381(1991); Marks et al., J. Mol. Biol., 222: 581(1991)]。ColeらおよびBoernerらの技術は、ヒトモノクローナル抗体の調製のためにも利用可能である(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77(1985) and Boerner et al., J. Immunol., 147(1):86-95(1991)]。同様に、ヒト抗体は、内因性免疫グロブリン遺伝子を部分的にまたは完全に不活性化したマウスなどのトランスジェニック動物への、ヒト免疫グロブリン遺伝子座の導入によって作成することができる。チャレンジしたとき、ヒト抗体生産が観察され、それはすべての点でヒトに見出されるものとよく類似し、遺伝子の再配列、集合、およびレパートリーを含む。この方法は、例えば、米国特許第5545807号;第5545806号;第5569825号;第5625126号;第5633425号;第5661016号、および以下の科学的出版物: Marks et al., Bio/Technology 10, 779-783(1992); Lonberg et al., Nature 368 856-859(1994); Morrison, Nature 368, 812-13(1994); Fishwild et al., Nature Biotechnology 14, 845-51(1996); Neuberger, Nature Biotechnology 14, 826(1996); Lonberg and Huszar, Intern Rsv. Immunol. 13 65-93(1995)に記載されている。
【0204】
二重特異的抗体は、少なくとも2の異なる抗原に結合特異性を有するモノクローナル抗体、好ましくはヒトまたはヒト化抗体である。今回の場合において、結合特異性の1つはHDAC8タンパク質について、他方は任意の他の抗原について、そして好ましくは細胞表面タンパク質または受容体もしくは受容体サブユニットについての抗体である。
【0205】
二重特異的抗体を生成するための方法は、当技術分野で既知である。通常、二重特異的抗体の組換え産物は、2つの免疫グロブリン重鎖/軽鎖組の共発現に基づき、この場合2本の重鎖が異なる特異性を有する。[Milstein and Cuello, Nature, 305: 537-539(1983)]。免疫グロブリン重鎖および軽鎖のランダムな組合わせのために、これらのハイブリドーマ(クアドローマ)は、10種の異なる抗体分子があり得る混合物を産生する。このうち1種のみが適当な二重特異的構造を有する。適当な分子の精製を通常アフィニティ・クロマトグラフィ法によって実施する。類似方法は、WO93/08829、1993年5月13日公開、およびTraunecker et al., EMBO J., 10: 3655-3659(1991)に記載されている。
【0206】
所望の結合特異性を有する抗体可変ドメイン(抗体抗原結合部位)を、免疫グロブリン定常ドメイン配列に融合することができる。その融合は、好ましくは免疫グロブリン重鎖定常ドメインとのものである。そしてヒンジ、CH2、およびCH3領域の少なくとも一部を含む。第1の重鎖定常領域(CH1)を有するのが好ましい。それは、融合の少なくとも1つに存在する軽鎖結合に必要である部位を含む。免疫グロブリン重鎖融合、および所望により免疫グロブリン軽鎖をコードするDNAを別の発現ベクターに挿入し、適当な宿主生物に共トランスフェクトさせる。二重特異的抗体の生成のさらなる詳細について、例えば、Suresh et al., Methods in Enzymology, 121: 210(1986)を参照されたい。
【0207】
ヘテロ結合抗体もまた、本発明の範囲内である。ヘテロ結合抗体は、2つの共有結合した抗体から成る。そのような抗体は、例えば、好ましくない細胞に免疫系細胞の標的を定めるために[米国特許第4676980号]、HIV感染の処置のために[WO91/00360; WO92/200373; EP03089]提案されている。抗体は、合成タンパク質化学(関連交差結合物質を含む)における既知の方法を使用して、インビトロで調製し得る。例えば抗毒素を、ジスルフィド置換反応を使用して、またはチオエーテル結合の形成によって構築し得る。この目的のために適当な物質の例は、イミノチオレートおよびメチル−4−メルカプトブチルイミデートを含み、例えば、米国特許第4676980号に記載されている。
【0208】
本発明の抗HDAC8タンパク質抗体は、多様な有用性を有する。例えば、抗HDAC8タンパク質抗体は、HDAC8タンパク質についての診断アッセイに使用しうる。例えば、特定の細胞、組織、または血清におけるその発現の検出である。当技術分野で既知の多様な診断アッセイ技術を使用し得る。例えば、競合的結合アッセイ、直接的または間接的サンドイッチアッセイ、および異種性または同種性の相のいずれかにおいて実施される免疫沈降法である[Zola, Monoclonal Antibodies: A Manual of Techniques, CRC Press, Inc.(1987)pp. 147-158]。診断アッセイに使用される抗体を、検出可能部分で標識化することができる。検出可能部分は、直接的にまたは間接的に検出可能なシグナルを生産するものである。例えば、検出可能部分は、3H、14C、32P、35S、もしくは125Iのような放射性同位体、フルオレセイン・イソチオシアネート、ローダミン、もしくはルシフェリンのような蛍光もしくは化学発光化合物、またはアルカリホスファターゼ、ベータ−ガラクトシダーゼもしくはホースラディッシュ・ペルオキシダーゼなどの酵素であってよい。抗体の検出可能部分への結合について、当技術分野で既知の任意の方法を使用し得る。それらの方法は、Hunter et al., Nature, 144: 945(1962); David et al., Biochemistry, 13: 1014(1974); Pain et al., J. Immunol. Meth., 40: 219(1981); およびNygren, J. Histochem. and Cytochem., 30: 407(1982)に記載された方法を含む。
【0209】
抗HDAC8タンパク質抗体は、組換え細胞培養または天然起源からのHDAC8タンパク質の親和性精製にとっても有用である。この方法では、HDAC8タンパク質に対する抗体を、例えば、Sephadfex樹脂または濾紙などの適当な支持体に、当技術分野で周知の方法を使用して固定する。次いで、固定した抗体を、精製するHDAC8タンパク質を含む試料と接触させる。その後、該支持体を適当な溶剤で洗浄し、固定した抗体に結合している、試料中のHDAC8タンパク質以外の実質的にすべての物質を除去する。最終的に、HDAC8タンパク質を抗体から遊離する別の適当な溶剤で、支持体を洗浄する。
【0210】
抗HDAC8タンパク質抗体を医療処置にも使用し得る。ある実施態様において、抗体をコードしている遺伝子を提供する。抗体は細胞内のHDAC8タンパク質に結合し、それを調節する。
【0211】
ある実施態様において、HDAC8タンパク質、アゴニストまたはアンタゴニストの治療有効量を患者に投与する。ここで「治療有効量」は、投与の目的である効果を生じる用量を意味する。正確な用量は処置目的により異なり、公知技術を用いて当業者は確認し得る。当分野で既知の通り、HDAC8タンパク質分解、全身的対局所的送達、並びに年齢、体重、全般的な健康状態、性別、食事、投与時間、薬剤相互作用および病状の重篤度による調節が必要であり、当業者は常用の試行で確認し得る。
【0212】
本発明の目的における「患者」は、ヒトおよび他の動物の両方、特に哺乳類および生物を包含する。即ち、本発明の方法は、ヒトの治療および獣医学的利用の両方に適用され得る。好ましい実施態様において、患者は哺乳類であり、最も好ましい実施態様において患者はヒトである。
【0213】
本発明のHDAC8タンパク質、アゴニストまたはアンタゴニストの投与を、様々な経路、例えば、経口、皮下、静脈内、鼻腔内、経皮、腹腔内、筋肉内、肺内、膣、直腸または眼内経路で行い得るがこれらに限定されない。場合によっては、例えば、損傷および炎症の処置において、組成物を溶液またはスプレーとして直接適用し得る。導入の方法に応じて、化合物を様々な方法で製剤化し得る。製剤中の治療的に活性な化合物の濃度は、約0.1ないし100重量%で変化し得る。
【0214】
本発明の医薬組成物は、患者への投与に適した形態のHDAC8タンパク質、アゴニストまたはアンタゴニスト(本明細書に記載の抗体およびバイオ活性物質を含む)を含む。好ましい実施態様において、この医薬組成物は、水溶性形態、例えば、医薬的に許容し得る塩類として存在しており、これらは酸および塩基の両付加塩類を包含するものとする。「医薬的に許容し得る酸付加塩類」は、遊離塩基の生物学的有効性を保持し、かつ生物学上またはその他の点で許容できるものであり、無機酸、例えば、塩酸、臭化水素酸、硫酸、硝酸、リン酸など、および有機酸、例えば、酢酸、プロピオン酸、グリコール酸、ピルビン酸、蓚酸、マレイン酸、マロン酸、コハク酸、フマル酸、酒石酸、クエン酸、安息香酸、桂皮酸、マンデル酸、メタンスルホン酸、エタンスルホン酸、p−トルエンスルホン酸、サリチル酸などと形成する。「医薬的に許容し得る塩基付加塩類」は、無機塩基、例えば、ナトリウム、カリウム、リチウム、アンモニウム、カルシウム、マグネシウム、鉄、亜鉛、銅、マンガン、アルミニウム塩類などから誘導されたものを含む。特に好ましいのは、アンモニウム、カリウム、ナトリウム、カルシウムおよびマグネシウム塩類などである。医薬的に許容し得る有機非毒性塩基から誘導される塩類は、第1級、第2級および第3級アミン類、置換アミン類、例えば、天然産生の置換アミン類、環状アミン類および塩基性イオン交換樹脂、例えば、イロプロピルアミン、トリメチルアミン、ジエチルアミン、トリエチルアミン、トリプロピルアミンおよびエタノールアミンの塩類がある。
【0215】
これらの医薬組成物はまた次の物質;即ち担体タンパク質、例えば、血清アルブミン;緩衝液;充填剤、例えば、微晶性セルロース、ラクトース、トウモロコシおよび他の澱粉類などの増量剤;結合剤;甘味料および他の着香剤;着色剤;およびポリエチレングリコールのうちの1種以上を含み得る。添加物は当業界ではよく知られており、様々な製剤で使用される。
【0216】
組成物の組合せを投与し得る。さらに、組成物を成長因子または化学療法剤および/もしくは放射線を含む他の治療剤と組み合わせて投与してもよい。標的化物質(即ち、癌細胞の受容体のリガンド)も、本発明で提供される組成物と組合わせてよい。
【0217】
本発明で提供されるある実施態様では、抗体を免疫療法に用いる。ゆえに、免疫療法の方法が提供される。「免疫療法」は、HDAC8タンパク質に対して生成された抗体を用いて、HDAC8タンパク質に関連する疾患を処置することを意味する。本発明で使用されるように、免疫療法は、受動的または能動的であり得る。本明細書に記載のように、受動的免疫療法は、抗体を受容者(患者)に受動的に輸送することである。能動的免疫療法は、抗体および/またはT細胞反応を受容者(患者)の中で誘導することである。HDAC8タンパク質抗原(これに対して抗体を生成する)を受容者に与えると、その結果免疫反応が誘導され得る。当業者に理解されるように、HDAC8タンパク質抗原は、抗体を生成しようとしているHDAC8タンパク質を受容者に注射するか、またはHDAC8タンパク質抗原を発現する条件下で、HDAC8タンパク質抗原を発現できるHDAC8タンパク質核酸を、受容者に接触させるかして与えられる。
【0218】
好ましい実施態様では、治療的化合物が抗体、好ましくはHDAC8タンパク質抗体に結合している。治療的化合物は細胞毒性物質であってもよい。この方法では、アポトーシス細胞または癌組織もしくは細胞に細胞毒性物質の標的を定めることにより、罹患した細胞の数を低減し、その結果、アポトーシス、癌HDAC8関連障害に付随する症状を低減する結果をもたらす。細胞毒性物質は膨大かつ多様であり、細胞毒性薬もしくは毒物またはそのような毒物の活性断片を含むが、これらに限定されるものではない。好適な毒物およびその対応する断片には、ジフテリアA鎖、エクソトキシンA鎖、リシンA鎖、アブリンA鎖、カルシン(curcin)、クロチン、フェノマイシン(phenomycin)、エノマイシン(enomycin)などが含まれる。細胞毒性物質には、HDAC8タンパク質に対して生成された抗体に放射性同位元素を結合させるか、または抗体に共有結合で結合させたキレート化剤に放射性核種を結合させて作成される放射性化学物質も含まれる。
【0219】
好ましい実施態様では、HDAC8タンパク質遺伝子を、DNA免疫原として投与する。1種類の核酸またはHDAC8タンパク質遺伝子の組合せのどちらかである。むき出しのDNA免疫原は、当分野で一般的に知られている;Brower, Nature Biotechnology 16:1304-1305 (1998)を参照のこと。DNA免疫原としての核酸の使用は、当業者に周知であり、HDAC8タンパク質遺伝子またはHDAC8タンパク質核酸の部分を、患者中での発現用のプロモーターの制御下におくことを含む。DNA免疫原に使われるHDAC8タンパク質遺伝子は、HDAC8タンパク質完全長をコードできるが、より好ましくはHDAC8タンパク質から生じるペプチドを含む、HDAC8タンパク質の部分をコードする。好ましい実施態様では、患者は、HDAC8タンパク質遺伝子から生じる多数の核酸配列を含むDNA免疫原で免疫性を与えられる。同様に、本明細書に記載のように、多数のHDAC8タンパク質遺伝子またはその部分で、患者に免疫性を与えることが可能である。理論によって制限を受けないが、DNA免疫原にコードされるポリペプチドの発現に続いて、HDAC8タンパク質を発現している細胞を破壊または除去する細胞障害性T細胞、ヘルパーT細胞および抗体が誘導される。
【0220】
下記記載の実施例は、上述した本発明の使用態様をより詳しく記載するためのものであるとともに、本発明の多様な側面を実施するために企図された最良の態様を記載するものである。これらの実施例は、如何なる意味においても本発明の真の範囲を制限しようとするものではなく、例示な目的で説明するためのものである。本明細書中で参照したすべての文献は、出典明示により本明細書の一部としている。さらに、開示した全ての配列、引用文献または引例中の各個別番号は、出典明示により本明細書の一部とする。
【実施例】
【0221】
実施例1
Basic BLASTを用いて発現配列タグ(EST)に関するGenBankデータベースの探索は、デフォルトパラメーターを用いるNCBIウェブサイト(http://www.ncbi.nlm.nih.gov/blast/cgi)から利用できるが、ウテリンライブラリーからのEST(40936;受託番号AA33308)がHDAC3のアミノ末端部分に対して相同性を持つことを明らかにした。完全長のクローンを得るために、Marathon- Ready human Uterus cDNAライブラリー(Clontech, Palo Alto, CA)を3'RACEの技術を用いてスクリーンし、そこでcDNAの3'末端をPCRで増幅した。Marathon-Ready cDNAライブラリーを、アダプタープライマーによってフランキングした完全長cDNAのライブラリーとして産生した。3’RACEを行うために、PCR反応をアダプター領域および遺伝子特異的プライマーと相同性のあるプライマーを用いて行った。HDAC8特異的プライマーは:TGCGGAACGGTTTTAAGCGGAG(配列番号:15)であった。
【0222】
この技術から、おおよそ等しい発生量の2つの転写物、1つは1700塩基ともう一つは800塩基の存在が明らかとなった。両形態の配列分析から、800塩基は1700塩基の5’末端の転写物と同一であり、アミノ末端の146残基を示す。
【0223】
HDAC8タンパク質は、HDACのRPD3クラス(I)と相同である。HDAC1、HDAC2およびHDAC3タンパク質は、推定377アミノ酸のHDAC8タンパク質(図2)に関連する短いカルボキシ末端伸長物を包含する。HDAC1、2および3および8のアミノ酸同一性%を、MacVector Prigram(Version 6.5r1 for Macintosh Oxfored Molecular Group)により測定した。
結果は以下である:
HDAC8対HDAC1: 146/486同一性=30%
HDAC8対HDAC2: 151/491同一性=31%
HDAC8対HDAC3: 151/439同一性=34%
HDAC1対HDAC2: 410/488同一性=84%
HDAC2対HDAC3: 250/491同一性=51%
HDAC1対HDAC3: 252/484同一性=52%
上記に示したように、HDAC1−3は、51%から84%のアミノ酸配列の同一性を有する。対照的に、これらタンパク質がHDAC8と共有するアミノ酸同一性は、約30%〜34%であった。即ち、上記HDACタンパク質の全ては、配列同一性を有するがHDAC8はより近密に相関した基の形態を有する他のHDACタンパク質に比べて比較的距離をおいて関連している。
【特許請求の範囲】
【請求項1】
図1(配列番号:1)に記載の配列と相補的な配列に高緊縮条件下でハイブリダイズする核酸を含む、HDAC8タンパク質をコードする組換え核酸。
【請求項2】
図1(配列番号:1)に記載の核酸配列を含む、請求項1に記載の組換え核酸。
【請求項3】
図1(配列番号:1)に記載の配列と少なくとも85%の配列同一性を持つ核酸を含む、HDACタンパク質をコードする組換え核酸。
【請求項4】
図1(配列番号:2)に記載のアミノ酸配列をコードする組換え核酸。
【請求項5】
該核酸で形質転換された宿主細胞により認識される制御配列と機能的に結合する請求項1、2、3または4のいずれかに記載の組換え核酸を含む、発現ベクター。
【請求項6】
請求項1、2、3、4または5のいずれかに記載の組換え核酸を含む、宿主細胞。
【請求項7】
請求項6のベクターを含む、宿主細胞。
【請求項8】
請求項6または7に記載の宿主細胞を該HDACタンパク質の発現に適切な条件下で培養することを含む、HDACタンパク質を産生する方法。
【請求項9】
該タンパク質を回収することをさらに含む、請求項8に記載の方法。
【請求項10】
請求項1、2、3または4のいずれかに記載の核酸によりコードされる、組換えHDACタンパク質。
【請求項11】
図1(配列番号:2)に記載のアミノ酸配列と少なくとも80%の配列同一性を持つアミノ酸配列を含む、組換えポリペプチド。
【請求項12】
該配列が図2(配列番号:2)に記載されている、請求項11に記載の組換えポリペプチド。
【請求項13】
請求項12に記載のHDACタンパク質に特異的に結合する、単離されたポリペプチド。
【請求項14】
抗体である、請求項13に記載のポリペプチド。
【請求項15】
該抗体がモノクローナル抗体である、請求項14に記載のポリペプチド。
【請求項16】
該抗体が該タンパク質の生物学的機能を低下または排除する、請求項15に記載のモノクローナル抗体。
【請求項17】
HDAC8タンパク質と結合し得るバイオ活性物質をスクリーニングする方法であって、下記
a)HDAC8タンパク質と候補バイオ活性物質とを組合せること、および
b)該候補バイオ活性物質と該HDAC8タンパク質との結合を測定すること
を含む方法。
【請求項18】
HDAC8タンパク質の活性を調節し得るバイオ活性物質をスクリーニングする方法であって、下記
a)HDAC8タンパク質をコードする組換え核酸を含む細胞に候補バイオ活性物質を加えること、および
b)該細胞に対する該候補バイオ活性物質の作用を測定すること、
を含む方法。
【請求項19】
候補バイオ活性物質のライブラリーが、HDAC8タンパク質をコードする組換え核酸を含む複数の細胞に添加される、請求項20に記載の方法。
【請求項20】
該細胞を標識する標識物質を細胞周期の特定の期(相)において加えることをさらに含む、請求項21に記載の方法。
【請求項21】
該細胞が腫瘍細胞である、請求項20記載の方法。
【請求項1】
図1(配列番号:1)に記載の配列と相補的な配列に高緊縮条件下でハイブリダイズする核酸を含む、HDAC8タンパク質をコードする組換え核酸。
【請求項2】
図1(配列番号:1)に記載の核酸配列を含む、請求項1に記載の組換え核酸。
【請求項3】
図1(配列番号:1)に記載の配列と少なくとも85%の配列同一性を持つ核酸を含む、HDACタンパク質をコードする組換え核酸。
【請求項4】
図1(配列番号:2)に記載のアミノ酸配列をコードする組換え核酸。
【請求項5】
該核酸で形質転換された宿主細胞により認識される制御配列と機能的に結合する請求項1、2、3または4のいずれかに記載の組換え核酸を含む、発現ベクター。
【請求項6】
請求項1、2、3、4または5のいずれかに記載の組換え核酸を含む、宿主細胞。
【請求項7】
請求項6のベクターを含む、宿主細胞。
【請求項8】
請求項6または7に記載の宿主細胞を該HDACタンパク質の発現に適切な条件下で培養することを含む、HDACタンパク質を産生する方法。
【請求項9】
該タンパク質を回収することをさらに含む、請求項8に記載の方法。
【請求項10】
請求項1、2、3または4のいずれかに記載の核酸によりコードされる、組換えHDACタンパク質。
【請求項11】
図1(配列番号:2)に記載のアミノ酸配列と少なくとも80%の配列同一性を持つアミノ酸配列を含む、組換えポリペプチド。
【請求項12】
該配列が図2(配列番号:2)に記載されている、請求項11に記載の組換えポリペプチド。
【請求項13】
請求項12に記載のHDACタンパク質に特異的に結合する、単離されたポリペプチド。
【請求項14】
抗体である、請求項13に記載のポリペプチド。
【請求項15】
該抗体がモノクローナル抗体である、請求項14に記載のポリペプチド。
【請求項16】
該抗体が該タンパク質の生物学的機能を低下または排除する、請求項15に記載のモノクローナル抗体。
【請求項17】
HDAC8タンパク質と結合し得るバイオ活性物質をスクリーニングする方法であって、下記
a)HDAC8タンパク質と候補バイオ活性物質とを組合せること、および
b)該候補バイオ活性物質と該HDAC8タンパク質との結合を測定すること
を含む方法。
【請求項18】
HDAC8タンパク質の活性を調節し得るバイオ活性物質をスクリーニングする方法であって、下記
a)HDAC8タンパク質をコードする組換え核酸を含む細胞に候補バイオ活性物質を加えること、および
b)該細胞に対する該候補バイオ活性物質の作用を測定すること、
を含む方法。
【請求項19】
候補バイオ活性物質のライブラリーが、HDAC8タンパク質をコードする組換え核酸を含む複数の細胞に添加される、請求項20に記載の方法。
【請求項20】
該細胞を標識する標識物質を細胞周期の特定の期(相)において加えることをさらに含む、請求項21に記載の方法。
【請求項21】
該細胞が腫瘍細胞である、請求項20記載の方法。
【図1−1】

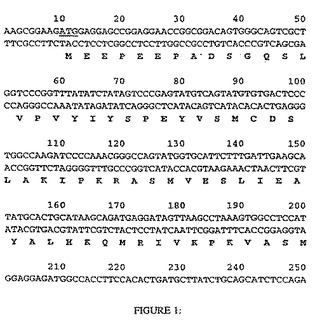
【図1−2】

【図1−3】
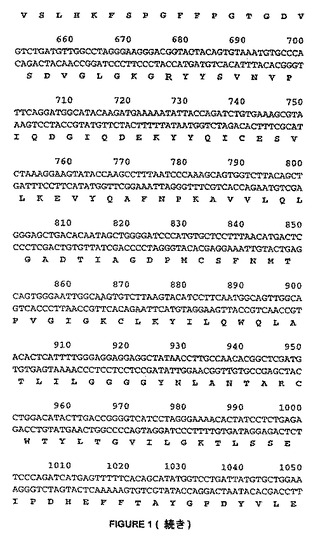
【図1−4】

【図1−5】
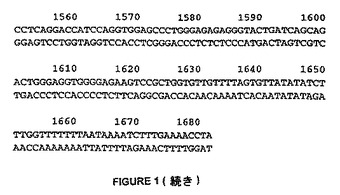
【図2】

【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図2】
【公開番号】特開2011−182800(P2011−182800A)
【公開日】平成23年9月22日(2011.9.22)
【国際特許分類】
【出願番号】特願2011−123587(P2011−123587)
【出願日】平成23年6月1日(2011.6.1)
【分割の表示】特願2001−544313(P2001−544313)の分割
【原出願日】平成12年12月8日(2000.12.8)
【出願人】(399031229)ファーマサイクリクス,インコーポレイテッド (9)
【Fターム(参考)】
【公開日】平成23年9月22日(2011.9.22)
【国際特許分類】
【出願日】平成23年6月1日(2011.6.1)
【分割の表示】特願2001−544313(P2001−544313)の分割
【原出願日】平成12年12月8日(2000.12.8)
【出願人】(399031229)ファーマサイクリクス,インコーポレイテッド (9)
【Fターム(参考)】
[ Back to top ]