
ヒトの肝臓前駆細胞およびその使用方法
【課題】機能性の肝細胞へと分化し得る肝臓前駆細胞の供給源を提供すること。
【解決手段】培養物中で増殖を可能にするCD117およびCD34に対して免疫反応性であり;そして肝細胞、胆管細胞、または類洞細胞へインビボで分化する、肝臓前駆細胞が、提供される。この培養物は、多数の継代にわたって消費され得、そして移植の後、成人の肝臓にうまく結合し得る。本発明のインビトロ細胞培養物は、ヒトの肝臓組織由来の細胞を含み、この培養物中の細胞は、CD117+、CD34+、およびLin−であり、培養物中で増殖し得、そしてインビトロで肝細胞、胆管細胞、または類洞細胞へ分化し得る細胞である。
【解決手段】培養物中で増殖を可能にするCD117およびCD34に対して免疫反応性であり;そして肝細胞、胆管細胞、または類洞細胞へインビボで分化する、肝臓前駆細胞が、提供される。この培養物は、多数の継代にわたって消費され得、そして移植の後、成人の肝臓にうまく結合し得る。本発明のインビトロ細胞培養物は、ヒトの肝臓組織由来の細胞を含み、この培養物中の細胞は、CD117+、CD34+、およびLin−であり、培養物中で増殖し得、そしてインビトロで肝細胞、胆管細胞、または類洞細胞へ分化し得る細胞である。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、前駆細胞に関する。
【背景技術】
【0002】
(発明の背景)
急性肝不全は、依然として高い死亡率を有する重大な問題である。肝機能障害および肝不全を生じる疾患の発症率が高いにもかかわらず、内科的治療における主な進歩は、現在、特定の形態のウィルス性肝炎の予防および処置に限られている。急性および慢性の肝疾患は、治療的なアプローチよりも対症的なアプローチで、依然として処置されている。同所性肝移植は、今までのところ、末期の肝疾患を有する患者のための唯一利用可能な治療である。残念なことに、ドナーの臓器のアベイラビリティは制限されているので、多くの患者が、肝臓移植を待ちながら毎年亡くなっている。最近では、正常肝細胞を罹患した肝臓へと移植することが、代替治療として用いられている。しかし、臓器ドナーの不足が、肝細胞移植の臨床適用を制限している。
【0003】
幹細胞およびそれらの子孫細胞を用いた細胞治療は、肝疾患を有する患者のための、大部分が未対処の医学的必要性に対する有望な新しいアプローチである。多くの研究が、実験的に誘導した急性肝不全において、(肝外肝臓組織および肝内肝臓組織から得られた)幹/前駆細胞の移植の可能性を調べている。成人の臓器由来の幹/前駆細胞は、機能性の肝細胞を産出し得るが、このような細胞は、稀である。遺伝性疾患および後天性疾患を回復するための肝臓前駆細胞移植に関する臨床治療プロトコルは、完全に発達した機能を依然として保持したままの細胞を大量に生じる能力に、大いに恩恵を受ける。
【発明の概要】
【発明が解決しようとする課題】
【0004】
従って、機能性の肝細胞へと分化し得る肝臓前駆細胞の供給源に対する必要性が存在する。
【課題を解決するための手段】
【0005】
(発明の要旨)
本発明は、肝臓前駆細胞の発見に基づいている。前駆細胞は、多分化能細胞である。前駆細胞は、(例えば、インビボまたはインビトロで)肝細胞(例えば、肝細胞、胆管細胞、または肝臓内皮細胞(すなわち、類洞細胞))へと分化することが可能である。従って、本発明は、肝臓前駆細胞培養物(例えば、インビトロ培養物)を特色とする。この培養物は、付着培養物である。あるいは、培養物中の細胞は、懸濁液中にある。この細胞は、肝臓組織(例えば、胎児肝臓組織)由来である。上記組織は、哺乳動物(例えば、ヒト、霊長類、マウス、ラット、イヌ、ネコ、ウシ、ウマ、ブタ)由来である。上記細胞は、CD117およびCD34に対して免疫反応性であり、Linには免疫反応しない。上記細胞は、インビトロで増殖する。上記細胞は、2回、3回、4回、5回、6回、7回、8回、9回、10回、11回、12回、15回、20回、25回またはそれ以上倍加することが可能であり、そして肝細胞、胆管細胞、または肝臓内皮細胞へと分化する能力を保持する。
【0006】
さらに提供されるものは、1つ以上の分化因子を含有する培養培地中で肝臓前駆細胞培養物を分化させることにより、肝細胞、胆管細胞、または肝臓内皮細胞を生成する方法である。分化因子としては、例えば、血管内皮成長因子(VEGF)、肝細胞成長因子(HGF)および表皮成長因子(EGF)が挙げられる。
【0007】
本発明はさらに、CD117+、CD34+、およびLin−のヒト肝臓前駆細胞を提供し、そしてその細胞を宿主へ移植することにより、(例えば、ヒト、霊長類、マウス、ラット、イヌ、ネコ、ウシ、ウマ、ブタのような哺乳動物である)宿主中に、前駆細胞または前駆細胞の子孫を移植する方法を、特徴とする。必要に応じて、肝臓の間質細胞または間葉細胞は、宿主中に共移植される。宿主は、前駆細胞の前、後、または同時に肝細胞増殖因子を投与される。宿主は、肝臓障害または肝臓組織損傷に罹患している。例えば、被験体は、肝炎、肝硬変、肝臓癌、脂肪肝疾患、ライ症候群、糖原病、肝嚢腫、またはウィルソン病を罹患している。移植は、臨床上の利益(例えば、特定の肝臓障害の1つ以上の症状を軽減すること)を与える。肝臓障害は、当該分野で公知の方法を使用して、医師によって診断される。
【0008】
肝細胞の増殖、分化、または生存をもたらす化合物は、肝臓前駆細胞培養物を試験化合物に接触させ、そして、その化合物が肝臓前駆細胞の増殖、分化、または生存に対する効果を有するかどうかを決定することによって、同定される。同様に、試験化合物の代謝産物が決定される。代謝産物は、培養物を試験化合物に接触させた後、培養培地をスクリーニングすることにより同定される。代謝産物は、当該分野で公知の方法(例えば、HPLC、質量分析、またはゲル電気泳動)によって同定される。試験化合物の抗ウィルス活性は、試験化合物の存在下または非存在下で、ウィルスを前駆細胞培養物へと導入し、その細胞の生存率を決定することによって決定される。試験化合物の非存在下と比較した、試験化合物の存在下における生存率の増加は、この試験化合物が抗ウィルス活性を有することを示す。ウィルスの感染性は、前駆細胞培養物をウィルスに接触させ、そしてこの細胞における増殖または生存の効果を決定することによって決定される。ウィルスと接触させなかった細胞培養物と比較した、生存または増殖の減少は、このウィルスが肝細胞に感染し得ることを示す。その一方、ウィルスと接触させなかった細胞培養物と比較した、生存または増殖における類似性は、このウィルスが肝細胞に感染しないことを示す。
【0009】
必要に応じて、培養物は、その培養物を試験化合物に接触させる前に、分化する。増殖および/または生存は、当該分野で公知の方法(例えば、BrdUアッセイ)によって決定される。分化は、肝細胞表面マーカーを形態学的かまたは組織学的に決定することによって決定される。ウィルスは、肝臓栄養性ウィルス(例えば、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルス)である。
本発明はまた、以下の項目を提供する。
(項目1)
ヒトの肝臓組織由来の細胞を含むインビトロ細胞培養物であって、該培養物中の該細胞は、
a.CD117+、CD34+、およびLin−であり;
b.培養物中で増殖し得;そして
c.インビトロで肝細胞、胆管細胞、または類洞細胞へ分化し得る、
細胞である、培養物。
(項目2)
項目1に記載の培養物であって、前記培養物は、少なくとも6回、倍加し得る、培養物。
(項目3)
項目1に記載の培養物であって、前記培養物は、少なくとも12回、倍加し得る、培養物。
(項目4)
項目1に記載の培養物であって、前記培養物は、付着培養物である、培養物。
(項目5)
インビトロでヒトの肝類洞細胞を生成する方法であって、該方法は、
a.CD117+、CD34+、およびLin−の細胞を含む細胞懸濁物を提供する工程;
b.該細胞懸濁液を培養する工程;ならびに
c.血管内皮成長因子を含有する培地中で該細胞の子孫を分化させる工程
を包含する、方法。
(項目6)
ヒトの肝臓の肝細胞または胆管細胞をインビトロで生成する方法であって、該方法は、
a.CD117+、CD34+、およびLin−の細胞を含む細胞懸濁物を提供する工程;
b.該細胞懸濁液を培養する工程;ならびに
c.EGFおよびHGFを含有する培地中で該細胞の子孫を分化させる工程
を包含する、方法。
(項目7)
インビボで肝細胞、または胆管細胞または類洞細胞へ分化する、ヒトの肝臓前駆細胞の集団を生成する方法であって、CD117+、CD34+、およびLin−である細胞を、ヒトの肝臓由来細胞の集団から選択する工程を包含する、方法。
(項目8)
多分化能肝臓前駆細胞の子孫を宿主へ移植する方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;ならびに
b.該細胞を該宿主中に移植する工程
を包含する、方法。
(項目9)
肝細胞の増殖、分化、または生存をもたらす化合物についてスクリーニングする方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化ための多分化能力を維持する、工程;
b.該培養物を試験化合物に接触させる工程;ならびに
c.該化合物が、肝細胞の増殖、分化、または生存をもたらすかどうかを決定する工程;
を包含する、方法。
(項目10)
項目9に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目11)
試験化合物の代謝産物を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;
b.該培養物を試験化合物に接触させる工程;ならびに
c.該培養物とインキュベーションした後に、該試験化合物由来の代謝産物を同定する工程;
を包含する、方法。
(項目12)
項目11に記載の方法であって、工程bを実施する前に、前記培養物の分化を誘導する工程を包含する、方法。
(項目13)
試験化合物の抗ウィルス活性を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させるための多分化能力を維持する、工程;
b.該培養物をウィルスおよび試験化合物に接触させる工程;ならびに
c.該培養物の生存率をコントロール細胞培養物と比較する工程;
を包含し、該コントロール培養物と比較された該培養物の生存率の増加が、該試験化合物の抗ウィルス活性を示している、方法。
(項目14)
項目13に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目15)
項目13に記載の方法であって、前記ウィルスは、肝臓栄養性ウィルスである、方法。
(項目16)
項目15に記載の方法であって、前記肝臓栄養性ウィルスは、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルスである、方法。
(項目17)
ウィルスの感染性を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;
b.該培養物をウィルスに接触させる工程;ならびに
c.該ウィルスが、該培養物中の該細胞の増殖または生存に対する効果を有するかどうかを決定する工程;
を包含する、方法。
(項目18)
項目17に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目19)
項目17に記載の方法であって、前記ウィルスは、肝臓栄養性ウィルスである、方法。
(項目20)
項目19に記載の方法であって、前記肝臓栄養性ウィルスは、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルスである、方法。
(項目21)
項目8に記載の方法であって、胎児肝臓間質細胞または胎児肝臓間葉細胞を前記宿主中に共移植する工程をさらに包含する、方法。
(項目22)
項目8に記載の方法であって、前記宿主に肝細胞成長因子を投与する工程をさらに包含する、方法。
【0010】
他に規定されない限り、本明細書中で使用されるすべての技術用語および科学用語は、本発明が属する分野の当業者に一般に理解されるような同じ意味を有する。本明細書中に記載される方法および材料と類似するかまたはそれらと同等である方法および材料が、本発明の実施または試験において使用され得るが、適切な方法および材料が、以下に記載される。本明細書中で言及された他のすべての刊行物、特許出願、特許、および参考文献は、その全体が参考して援用される。矛盾する場合は、定義を含む本明細書が統制する。さらに、材料、方法、実施例は、例示するのみであり、限定されることを意図しない。
本発明の他の特徴および利点は、以下の詳細な説明および上記特許請求の範囲から明かである。
【図面の簡単な説明】
【0011】
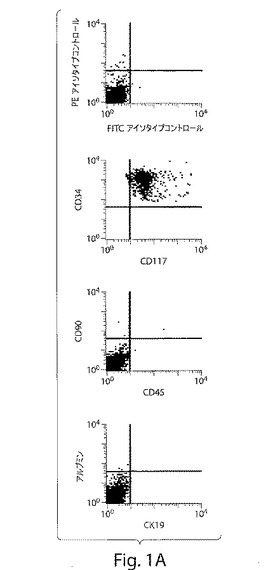
【図1A】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Aは、単離の直後に磁気によって分類したFL細胞を、CD117およびCD34については二重染色したが、CD90、CD45、アルブミン、およびサイトケラチン19(CK19)については染色しなかった。
【図1B】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Bは、CD117+/CD34+/Lin−細胞は、培養される際にコロニー内で成長し、そして発現される第1マーカーは、肝細胞成長因子レセプター(c−Met)であった(拡大率:40倍)。
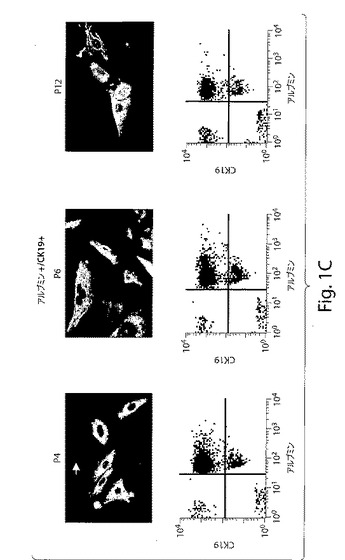
【図1C】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Cは、種々の継代において新たに単離し膨張させた細胞は、ALB−CK19−(白矢印)、ALB+(緑)、CK19+(赤)、ALB+CK19−(緑;肝細胞)およびALB−CK19+(赤;胆管細胞)を生じた(拡大率:60倍)。
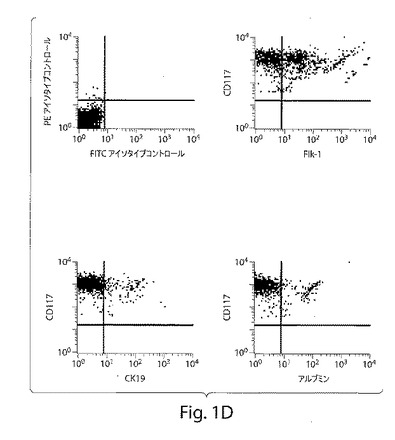
【図1D】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Dは、血管内皮成長因子を含有する培養培地中で成長させた際の、磁気によって分類したCD117+/CD34+/Lin−付着細胞は、Flk−1+内皮細胞(約50%)、CK19+胆管細胞(約13%)およびアルブミン+肝細胞(約17%)へと分化した。およそ20%の細胞は、これらのマーカーのうちのいずれも発現しなかった。
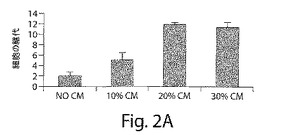
【図2A】図2Aは、肝臓前駆体およびそれらの子孫の増殖を示す棒グラフである。20%に調製された培地(CM)の使用は、肝臓前駆細胞の増殖を有意に増加し、そしてCMを使用しないものと比較して数回継代され得る(p<0.001;スチューデントt検定)。
【図2B】図2Bは、フローサイトメトリー分析の結果を示す棒グラフである。このグラフは、高い増殖度(BrdU+細胞)が、アルブミンCK19+、アルブミン+CK19−、およびアルブミン−CK19+細胞において観察されたが、その一方で、両方ともネガティブな(アルブミン−CK19−)細胞は、より静止状態であったことを示す。
【図2C】図2Cは、種々の継代における肝臓前駆細胞を示す一連の写真であり、アルブミン(緑)およびCK19(赤)については、蛍光染色され、そして、BrdUの取り込みの検出については酵素学的に染色した。これらは、種々の部分集団において増殖能力を示した(拡大率:40倍)。
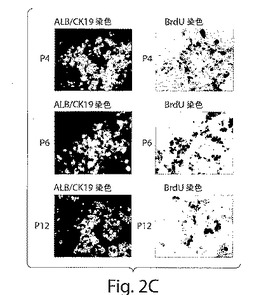
【図3】図3A−3Nは、マウス肝臓中のヒト肝臓前駆細胞の局在化の一連の写真である。a−b;ヒト肝臓(DAB)において核シグナルを示すが、マウス肝臓においては示さない、ヒト動原体プローブを用いたインサイチュハイブリダイゼーション。c−d:免疫組織化学を使用し、抗ヒト核抗体(DAB−Ni)で同様の結果を得た。e−j;新たに単離した細胞(e−g)である、第6継代および第12継代細胞が、D−ガラクトサミン処置(GalN)マウスへ移植される際に、肝細胞、胆管細胞、および内皮細胞への分化を示した(DAB−Ni;矢印の先端)。k;移植された細胞は、肝臓中で観察されたが、l−m;GalN処置マウスの脾臓、肺では、観察されなかった。n;偽移植されたマウスの肝臓を、コントロールとして用いた(拡大率:60倍)。
【図4A−J】図4A−Jは、ヒト肝臓前駆細胞のインビボでの発生運命を示す一連の写真である。a−d;GalN処置マウスへの移植の1ヶ月後のヒト肝臓前駆細胞は、グルコース−6−リン酸(茶)、グリコーゲン(ピンク)、ジペプチジルペプチダーゼIV(赤茶)、γグルタミルトランスペプチダーゼ(茶)を含んだ。抗ヒト核抗体(DAB−Ni;矢印の先端)による二重染色は、ヒト核(黒)を視覚化した。e;ヒト肝臓前駆体(核;黒矢印の先端)によるマウス組織切片全体の再生。f;サイトケラチン19(赤茶)の発現および、g−j;ヒトアルブミン(緑)は、偽移植したマウスでは観察されなかったが、新たに単離した、第6〜第12継代のヒト肝臓前駆細胞を移植した、GalN処置マウスの肝臓中で観察された(矢印の先端)。a−j;(拡大率:60倍)。f;(拡大率:200倍)。
【図4K】図4Kは、マウス肝臓におけるヒト肝臓特異的遺伝子の転写を示す写真である。ヒトサイトケラチン19、αフェトプロテイン、およびアルブミンは、ヒト肝臓前駆細胞を受容したGalN処置マウスの肝臓では検出されたが、偽移植されたマウスの肝臓では検出されなかった。しかし、α1アンチトリプシンは、コントロール中でもまたわずかに増幅した。グルコース−6−リン酸デヒドロゲナーゼを、ハウスキーピング遺伝子として使用した。
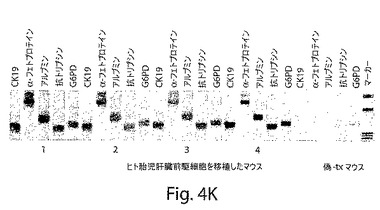
【図5】図5は、胎児肝臓間質参謀および胎児肝臓前駆細胞の共移植の効果を示す一連の写真である。A;抗ヒト核抗体で染色した正常ヒト肝臓切片は、陽性染色を示す(黒/茶)。B;正常マウス肝臓切片では、同じ抗体による陽性染色が見られなかったが、抗体の特異性を示した。C;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして肝細胞成長因子(HGF)を皮下注入して、そして胎児肝臓前駆細胞を移植した後のマウスは、高い細胞移植度をもたらさなかった。D;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして胎児肝臓から単離された間質細胞を注入した後のマウスは、高い細胞移植度をもたらさなかった。E−H;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして胎児肝臓間質細胞と前駆/幹細胞との混合物を移植した後のマウスは、高い肝細胞移植度をもたらした。I−K;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして肝細胞成長因子(HGF)を皮下注入して、そして胎児肝臓間質細胞および肝臓前駆細胞を移植した後のマウスは、高い細胞移植度をもたらした。
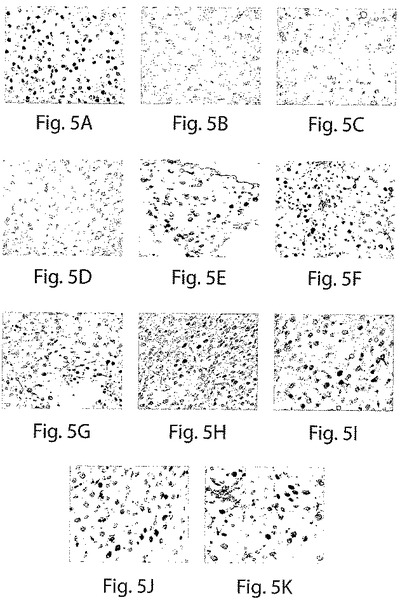
【図6】図6は、種々の方法で処置した正常ヌードC57ブラックマウスおよびC57ブラックマウスにおける、ヒト第VIII因子の検出レベルを示すチャートである。
【図7】図7は、胎児肝臓および成人肝臓に対する、種々の造血細胞、肝細胞、および脾臓細胞の表面マーカーの発現を示すチャートである。
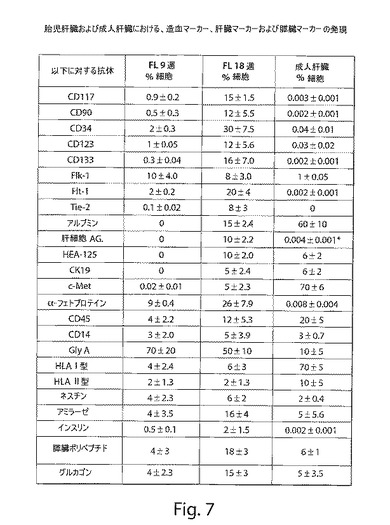
【図8】図8は、ヒトの胎児肝臓および成人肝臓における肝臓マーカーを示す一連の写真である。
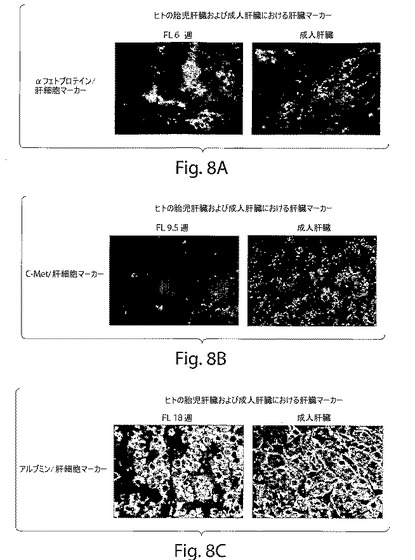
【図9A】図9Aは、ヒト肝臓前駆細胞の特徴づけを示す一連のFACS分析である。
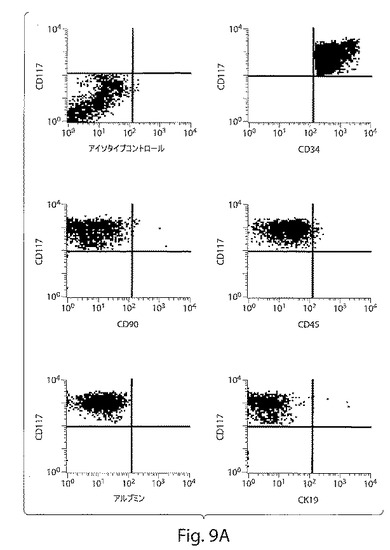
【図9B−F】図9B−Fは、異なるマトリックス上のヒト肝臓前駆細胞の形態を示す一連の写真である。
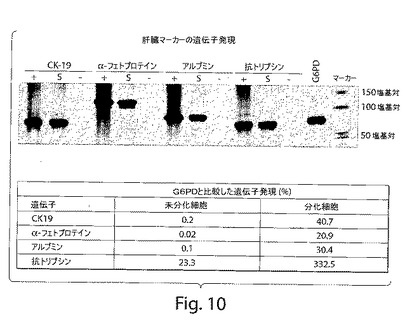
【図10】図10の上図は、ヒト肝臓前駆細胞における肝臓マーカーの遺伝子発現を示すノーザンブロットの写真である。図10の下図は、ヒト肝臓前駆細胞における肝臓マーカーの遺伝子発現を示すチャートである。
【発明を実施するための形態】
【0012】
(詳細な説明)
本発明は、数世代の間インビトロで拡張しかつ祖先の表現型を維持する、ヒト胎児肝臓内の非造血性前駆細胞の規定された集団における予想外の発見に基づいている。これらの細胞は、急性肝損傷を有する動物へと移植される際に、肝細胞、胆管細胞、および類洞細胞への機能的な分化を示した。
【0013】
肝臓疾患としては、世界的にみて顕著な罹患率および死亡率に関連する、広範囲にわたる急性的な状態および慢性的な状態の両方が挙げられる。肝細胞移植は、肝臓疾患の処置において強大な治療的潜在能力を有するが、肝細胞移植の臨床的使用は、ドナー組織の不足によって阻まれている。成人または胎児の肝細胞前駆体からのインビトロでの肝細胞の生成、または成熟肝細胞をインビボで生成し得る前駆体集団の同定は、この問題を解決する。本明細書中に記載されたデータは、ヒト胎児肝臓からの規定された細胞集団の同定を実証する。このヒト胎児肝臓は、エキソビボで数世代の間、首尾よく拡張され、そして急性肝損傷を有する動物へと移植される際に、肝細胞、胆管細胞、および類洞細胞への機能的な分化を示す。肝臓前駆細胞のインビトロで成功した拡張および分化は、肝細胞移植、候補治療薬の代謝試験および毒性試験、ならびに遺伝子治療のビヒクルに有用である。
【0014】
本発明は、インビトロまたはインビボ(すなわち、インサイチュ)で増殖させるためにヒト胎児肝臓組織からの多分化能肝臓前駆細胞を誘導して、肝細胞、胆管細胞、および類洞細胞へと分化することが可能な多分化能ヒト前駆細胞の子孫を大量に生成するための方法を提供する。ヒト肝臓前駆細胞の子孫を分化するための方法もまた、提供される。
【0015】
(ヒト肝臓前駆細胞)
本発明は、ヒト肝臓前駆細胞(本明細書中でLPCと呼ばれる)を提供する。LPC細胞は、本発明の方法を用いて増殖させるために誘導され得る未分化細胞である。LPCは、各細胞分裂において、少なくとも1つの娘細胞がまたLPC細胞であるように、自己保持することが可能である。LPCは、100倍、250倍、500倍、1000倍、2000倍、3000倍、4000倍、5000倍以上、拡張されることが可能である。
【0016】
LPCの表現型決定は、これらの細胞が、どの関係付けられた(committed)肝細胞マーカーも発現しないが、この細胞が幹細胞マーカーを発現することを明らかにする。例えば、LPCは、CD117およびCD34の両方に対して免疫反応性であり、そして系統表面抗原(Lineage surface antigen)(Lin)に対しては非免疫反応性である。LPCは、多分化能前駆細胞である。多分化能前駆細胞とは、この細胞が1つより多くの細胞型へと分化することが可能であることを意味する。例えば、その細胞は、肝細胞、胆管細胞、または類洞細胞へと分化することが可能である。
【0017】
CD117はまた、c−kit(造血幹細胞因子(steel factor)レセプターまたは幹細胞因子レセプター)としても公知である。CD117は、IIIクラスレセプターチロシンキナーゼファミリーに属する、145kDの細胞表面糖タンパク質である。これは、多分化能造血性幹細胞を含む大多数の造血前駆細胞上で発現され、同様に関係付けられた骨髄性前駆細胞、赤血球系前駆細胞、およびリンパ系前駆細胞上でも発現される。CD117はまた、わずかな成熟造血性細胞(例えば、肥満細胞)上で発現される。CD34は、ヒトのリンパ系造血性前駆細胞および骨髄性造血性前駆細胞上で発現された110kDの単鎖膜貫通糖タンパク質である。系統表面抗原とは、異なる14個の細胞表面タンパク質のうちの13個が成熟血液細胞系統のマーカーである場合の混合物である。
【0018】
LPCは、胎児の肝臓組織から得られる。肝臓組織は、肝臓組織を有する任意の動物(例えば、魚、爬虫類、鳥、両生類、および哺乳動物(例えば、好ましくは、マウスのようなげっ歯類およびヒト))から入手され得る。この組織は、少なくとも4週齢、5週齢、6週齢、7週齢、8週齢、9週齢、10週齢以上の胎児から得られる。LPCは、全体の胎児肝臓のうちのおよそ0.5〜0.7%に相当する。
【0019】
LPCは、インビトロの長期培養において維持され得る。LPCは、培養中で2回、3回、4回、5回、6回、7回、8回、9回、10回、11回、12回以上継代されることが可能である。
【0020】
移植の前には、新たに単離されたLPC(CD117+/CD34+/Lin−細胞)は、アルブミンおよびCK19を発現しなかった。しかし、移植後には、これらの細胞は、成熟肝細胞および胆管細胞へと分化された。このことは、ヒトのアルブミン、CK19、G−6−P、GGT、DPPIV、およびグリコーゲンの発現によって決定され、そして、これらの細胞は高い増殖能力を有していた。肝臓前駆細胞は、2つの造血性関連マーカー(c−kitおよびCD34)を発現したが、非造血性細胞から造血性細胞を区別するマーカーであるCD45は、発現しなかった。従って、本明細書中に記載される大規模な増殖能力を有するLPCは、造血性起源ではない。さらに、インビトロで成人造血性幹細胞を拡張する限定された能力とは異なり、胎児肝臓からの肝臓前駆体は、高い増殖能力を有する。単離されたCD117+/CD34+/lin−細胞のすべてが、インビトロでの培養の間に、培養プレートに付着し、そして肝細胞へと分化するわけではなかったという観察は、これらの細胞の部分集団のみが肝細胞の前駆体であることを示す。CD117+/CD34+/lin−細胞は、HGFおよびEGFを含有する培地中で培養した場合、4タイプの細胞(ALB−CK19−肝細胞、ALB+CK19+肝細胞、ALB+CK19−肝細胞、およびALB−CK19+胆管細胞)を生じた。ダブルネガティブ細胞を除いてすべての他の部分集団は、増殖細胞(BrdU+)を高い比率で有した。ALB−CK19−細胞の部分セットは、インビトロで非常に静止状態であったが、インビボでのP0継代において移植された細胞数の10倍の増加は、これらの細胞が、高い増殖能力を有し得ることを示す。肝臓前駆細胞の高い増殖能力にもかかわらず、腫瘍は、ヒトの細胞移植の4週間後にインビボで観察されなかった。興味深いことに、早期胎児肝臓からのCD117+/CD34+/Lin−細胞は、VEGFを含有する倍地中で培養した場合、肝細胞および胆管細胞へと分化しただけでなくまた、類洞内皮細胞へ、分化した。
【0021】
(培養条件)
LPCは、本明細書中に記載された方法を用いて増殖される。細胞は、ドナー組織の連結している細胞外マトリクスから個々の細胞を分離することによって、その組織から入手される。胎児からの組織は、滅菌手順を用いて除去され、そして細胞は、酵素(例えば、トリプシン、コラゲナーゼなど)を用いる処置を含む当該分野で公知の任意の方法を用いてか、または例えば鈍器もしくはホモジナイザーを用いる物理的な分離方法を用いて、分離される。胎児細胞の分離は、組織培養培地中で行われ得る。
【0022】
例えば、細胞の分離は、0.1%トリプシンおよび0.05%DNaseを含むDMEM中で行われ得る。分離された細胞は、200〜2000rpmの間(通常は400〜800rpmの間)の低速で遠心分離され、そして次いで、培養培地中に再懸濁される。肝細胞は、懸濁液中かまたは固定された基質上で培養され得る。分離された細胞懸濁液は、細胞を支持することが可能な任意の容器中(具体的には、培養フラスコ、培養プレート、またはローラー瓶、そしてより具代的には、25cm2培養フラスコのような小さな培養フラスコ中)に播種される。懸濁液中で培養された細胞は、およそ5×104〜2×105(細胞/ml)(例えば、1×105(細胞/ml))で再懸濁される。固定された基質上にプレートされた細胞は、およそ2〜3×103(細胞/cm3)でプレートされる。必要に応じて、培養プレートは、コラーゲンのようなマトリクスタンパク質で被覆される。分離された肝細胞は、細胞増殖を支持することが可能な任意の公知の培養培地(HEM、DMEM、PRMI、F−12などを含む)へと配置され得る。この培地は、細胞代謝に必要な補充物(例えば、グルタミンおよび他のアミノ酸、ビタミン、ミネラル、およびトランスフェリンのようなタンパク質など)を含有する。培養培地はまた、酵母、細菌、および真菌類による汚染を防ぐための抗生物質(例えば、ペニシリン、ストレプトマイシン、ゲンタマイシンなど)を含み得る。培養培地は、ウシ、ウマ、ニワトリなどに由来する血清を含み得る。
【0023】
培養条件は、生理学的条件に近いべきである。培養培地のpHは、生理学的pH(例えば、pH6〜8の間、pH約7〜7.8の間、またはpH7.4)に近いべきである。生理学的温度の範囲は、約30℃〜40℃の間である。LPCは、約32℃〜約38℃の間(例えば、約35℃〜約37℃の間)の温度にて培養される。
【0024】
必要に応じて、培養培地は、少なくとも1つの増殖誘導(「マイトジェン」)成長因子を補充される。「成長因子」とは、LPCに対する成長効果、増殖誘導効果、分化誘導効果、または栄養効果を有する、タンパク質、ペプチド、または他の分子である。「増殖誘導成長因子」は、LPCを増殖させることを可能にする栄養因子であり、細胞の表面上のレセプターと結合して、細胞に栄養効果または成長誘導効果を与える任意の分子を含む。増殖誘導成長因子としては、EGF、アンフィレグリン(amphiregulin)、酸性線維芽細胞成長因子(aFGFまたはFGF−1)、塩基性線維芽細胞成長因子(bFGFまたはFGF−2)、トランスホーミング増殖因子α(TGFα)、VEGFおよびこれらの組み合わせが挙げられる。成長因子は、通常、約1fg/ml〜1mg/mlの間の範囲の濃度で培養培地へと加えられる。約1〜100ng/mlの間の濃度で、通常は十分である。簡単な滴定アッセイは、特定の成長因子の最適濃度を決定するために容易に実施され得る。
【0025】
成長因子および栄養因子の生物学的効果は、一般に、細胞表面のレセプターとの結合を介して仲介される。これら多くの因子に対するレセプターは、同定されてきており、そして特定のレセプターに対する抗体および分子プローブが、利用可能である。LPCは、分化のすべての段階において成長因子レセプターの存在について分析され得る。多くの場合、特定のレセプターの同定は、外因性成長因子または栄養因子の付加を伴う、特定の発生経路に沿ったさらなる細胞の分化における使用のためのストラテジーについての手引きを提供する。
【0026】
一般に、インビトロで約3〜10日後に、培地を吸引し、そして培養フラスコへ新鮮な培地を加えることによって、LPCを増殖する。必要に応じて、吸引された培地は、収集され、フィルタ処理され、そして次に続くLPCの継代のための条件培地として使用される。例えば、10%、20%、30%、40%以上の条件培地が使用される。
【0027】
LPC細胞培養物は、増殖を再開するために用意に継代され得る。例えば、インビトロで約3〜7日後に、培養フラスコは、十分に振盪され、そして次いで、LPCは、50ml遠心管へと移され、低速で遠心される。この培地は吸引され、LPCは、少量の培養培地中に再懸濁される。次いで細胞は、カウントされ、そして増殖を再開するために所望の密度にて再度プレートされる。この手順は、毎週繰り返され、各継代における生存可能な細胞数の対数的な増加をもたらし得る。この手順は、所望の数のLPCが入手されるまで続けられる。
【0028】
LPCおよびLPCの子孫は、それらが必要とされるまで、当該分野における任意の公知の方法によって凍結保存され得る(例えば、米国特許出願第5,071,741号、PCT国際特許出願第WO93/14191号、同WO95/07611号、同WO96/27287号、同WO96/29862号、および同WO98/14058号、Karlssonら、65 Biophysical J.2524−2536(1993)を参照のこと)。LPCは、等張液中、好ましくは、特定の凍結保存剤(cryopreservant)を含有する細胞培養培地中に懸濁され得る。このような凍結保存剤としては、ジメチルスルホキシド(DMSO)、グリセロールなどが挙げられる。これらの凍結保存剤は、5〜5%(例えば、8〜10%)の濃度で使用される。細胞は、−10℃〜−150℃(例えば、−20℃〜−100℃、または−70℃〜−80℃)の温度まで徐々に凍結される。
(ヒト肝臓前駆細胞の分化)
培養条件に依存して、LPCは、肝細胞、胆管細胞、または類洞細胞へと分化され得る。
【0029】
LPCは、HGFおよびEGFを有する培養培地中の固定された基質上でLPCを培養することによって、肝細胞、または胆管細胞へと分化され得る。あるいは、LPCは、VEGFを有する培養培地中の固定された基質上でLPCを培養することによって、類洞細胞へと分化され得る。
【0030】
LPCの分化はまた、成長を引き起こす生物学的事象のカスケードを活性化する当該分野で公知の任意の方法によって誘導され得る。この生物学的事象としては、イノシトール三リン酸塩および細胞内のCa2+の遊離、ジアシルグリセロールの遊離、ならびにプロテインキナーゼCおよび他の細胞キナーゼの活性化などが挙げられる。ホルボールエステル、分化誘導成長因子、および他の化学シグナルによる処置は、分化を誘導し得る。LPCの増殖のための増殖誘導成長因子(上記参照)の代わりに、分化誘導成長因子は、LPCの分化に影響を与えるために培養培地に加えられ得る。成長因子を誘導する他の分化としては、血小板由来成長因子(PDGF)、チロトロピン放出ホルモン(TRH)、トランスホーミング成長因子β(TGF)、インスリン様成長因子(IGF−1)などが挙げられる。
【0031】
分化した肝細胞、胆管細胞、または類洞細胞は、当該分野で公知の免疫細胞化学的技術を用いて検出される。免疫細胞化学(例えば、二重標識免疫蛍光法および免疫ペルオキシダーゼ法)は、肝細胞の細胞特徴または表現型特性を区別するために、細胞タンパク質を検出する抗体を使用する。肝細胞および胆管細胞に対する細胞マーカーとしては、アルブミンおよびCK10が挙げられ、一方で類洞細胞に対する細胞マーカーとしては、Flkが挙げられる。他の適切なマーカーとしては、グルコース6ホスファターゼ、グリコーゲン、ジペピジルペプチダーゼIV、γグルタリルトランスペプチダーゼが挙げられる。
【0032】
免疫細胞化学はまた、例えば、アルブミン、α1−抗トリプシン、CK−19、α胎児タンパク質、またはヒト第VIII因子のような肝臓機能を担う肝臓遺伝子の発現を検出することによって、肝細胞を同定するために使用され得る。
【0033】
インサイチュハイブリダイゼーション組織化学はまた、肝臓遺伝子のmRNAに特異的なcDNAプローブまたはRNAプローブを用いて行われ得る。これらの技術は、特定の表現型の同定を増強するための免疫細胞化学的方法と組み合わされ得る。必要であれば、上で議論された抗体および分子プローブは、ウェスタンブロット手順およびノーザンブロット手順(それぞれが細胞の同定に役立つ)に適用され得る。
【0034】
(ヒト肝臓前駆細胞の移植)
新しい細胞の損傷した肝臓への移植は、損傷した肝臓組織を修復し、それによって肝臓機能を回復する可能性を有する。必要に応じて胎児間質細胞および/またはHGFは、LPCとともに共移植される。しかし、移植目的に適する細胞の欠如は、この手順が満たすべき最大の可能性を妨げている。「適切な」細胞とは、以下の基準:(1)大量に入手され得ること;(2)必要に応じて、遺伝物質の挿入を可能とするためにインビトロで増殖され得ること;(3)無制限に生存することが可能であり、肝臓における移植に対する肝臓修復を促進することが可能であること;そして(4)非免疫原性であり、好ましくは、患者自身の組織からかまたは適合し得るドナーから入手されることを、満たす細胞である。
【0035】
胎児の肝臓組織から入手可能なLPC(これは、本明細書中に記載された培養条件を用いてインビトロで維持される場合に、長期間にわたって分裂することが可能である)は、肝臓移植目的に適する細胞の望ましい必要条件のすべてを満たし、かつ不死化されていない細胞として特に適切な細胞株であり、かつ腫瘍形成の発生源ではない。肝臓障害の処置におけるLPCの使用は、動物モデルの使用によって実証され得る。
【0036】
LPCは、異常な肝臓または肝不全症状を有する任意の動物に投与される。LPCは、宿主に対して異種であるドナー組織から調製され得る。異種移植を成功させるために、移植された組織に対する免疫応答を低減させるか排除する何らかの方法が、通常、利用される。従って、LPCレシピエントは、免疫抑制薬物(例えば、シクロスポリン)の使用を介してか、または局所的に適用された免疫抑制剤を利用する局所的免疫抑制ストラテジーを介して、免疫抑制され得る。局所的免疫抑制は、Gruber、54 Transplantation 1−11(1992)によって開示される。米国特許第5,026,365号は、局所的免疫抑制に適切なカプセル化方法を開示する。
【0037】
免疫抑制技術の利用の代替として、胎児幹細胞における相同組み換えを用いる遺伝子置換法または遺伝子ノックアウト法(Smithiesら、317 Nature 230−234(1985)によって教示され、細胞株における遺伝子置換または遺伝子ノックアウトまで拡張された(Zhengら、88 Proc.Natl.Acad.Sci.8067−8071(1991))が、主要組織適合遺伝子複合体(MHC)遺伝子の切除のために、LPCに適用され得る。MHC発現を欠くLPCは、同種異系を越えて濃縮された肝細胞集団の移植を可能にし、そしておそらくレシピエントを免疫抑制する必要なく異種間の組織適合性の障壁を越えて濃縮された肝細胞集団の移植でさえも可能にする。レシピエントを免疫抑制する必要なく組織適合性の障壁をなくす。ドナー細胞の抗原性を低減させるための組み換え方法の使用についての一般的な総説および引用はまた、Gruber、54 Transplantation 1−11(1992)によって開示される。表面修飾によって移植片の免疫原性を低減する例示的なアプローチは、PCT国際特許出願第WO92/04033号および同第PCT/US99/24630号によって開示される。あるいは、移植片の免疫原性は、MHC抗原を変化させたかまたは欠失させたトランスジェニック動物からLPCを調製することによって低減され得る。
【0038】
LPCは、マイクロカプセル化(例えば、本明細書中で参考として援用される、米国特許出願第4,352,883号;同第4,353,888号;および同第5,084,350号を参照のこと)、およびマクロカプセル化(例えば、それぞれが本明細書中で参考として援用される、米国特許第5,284,761号;同第5,158,881号、同第4,976,859号、および同第4,968,733号、ならびにPCT国際特許出願第WO92/19195号および同第WO95/05452号を参照のこと)を含む、公知のカプセル化技術に従って、因子を宿主へと送達するためにカプセル化され得、かつ使用され得る。マクロカプセル化は、米国特許第5,284,761号;同第5,158,881号;同第4,976,859号;同第4,968,733号;同第5,800,828号およびPCT国際特許出願第WO95/05452号において開示され、それぞれが本明細書中で参考として援用される。デバイス中の細胞数は、変動され得る;好ましくは、各デバイスは、103〜109の間の細胞を含む(例えば、105〜107の細胞)。複数のマクロカプセル化デバイスが、宿主中に移植され得る。
【0039】
レシピエントの組織と同種異系である組織から調製されたLPCは、レシピエントの組織適合性型とほぼ一致させるように、周知の組織型決定方法によって使用のために試験される。
【0040】
LPCは時折、レシピエント自身の肝臓から調製され得る(例えば、腫瘍除去生検の場合)。このような場合、LPCは、分離された組織から生成され得、そして上述の方法を用いてインビトロで増殖され得る。適切な細胞数が増殖された際に、LPCは、収集され得、必要に応じて遺伝的に改変され得、そしてレシピエントの肝臓へと直接注入するために整えられ得る。LPCは、肝臓移植片を形成し得る肝臓領域に投与され、その結果、その細胞は、隣接する肝細胞との通常の結合を形成し、移植されたかまたは既存の肝細胞との接触を維持する。従って、移植されたLPCは、疾患および加齢が原因で損傷している肝臓組織を再構築する。
【0041】
宿主の肝臓組織への移植片の機能的な組み込みは、種々の機能を回復することに関して移植片の有効性を検査することによって評価され得る。この検査としては、アラニントランスアミナーゼ(ALT)、アスパラギン酸トランスアミナーゼ(AST)、アルカリホスフォターゼ(ALP)、アルブミン、総タンパク質、ならびに総ビリルビンおよび直接ビリルビンについての血液検査が挙げられる。
【0042】
移植における使用のためにインビトロでLPCを拡張する能力はまた、エキソビボの遺伝子治療にも有用である。従って、LPCは、エキソビボでの遺伝子治療試行におけるビヒクルとしての使用のために、肝臓細胞を回収および拡張するさらなる方法を提供する。
【0043】
(肝臓前駆細胞の遺伝子改変)
LPCは、形質転換されてない初代細胞であるが、それらは、連続的な細胞株の特徴を保有する。未分化の状態において、LPCは、連続的に分裂し、そしてそれ故に、遺伝子改変の標的となる。いくつかの実施形態において、遺伝子改変された細胞は、上述の任意の方法によって、肝細胞、胆管細胞、または類洞細胞へと分化されるように誘導される。
【0044】
用語「遺伝子改変」とは、外来DNAの意図的な導入によって、LPCの遺伝子型を、安定にかまたは一時的に変更することをいう。DNAは、合成DNAまたは天然に由来するDNAであり得、そして遺伝子、遺伝子部分、または他の有用なDNA配列を含み得る。本明細書中で使用される場合、用語「遺伝子改変」とは、天然に起こる変化(例えば、天然のウィルス活性、天然の遺伝子組み換えなどを介して起こる変化)を含むことを意図しない。
【0045】
細胞の有用な遺伝子改変のいずれもが、本発明の範囲内にある。例えば、LPCは、生物学的に活性な物質(例えば、成長因子など)を生成するか、または生成を増加させるために改変され得る。一実施形態では、生物学的に活性な物質は、転写因子(例えば、遺伝的分化を調節する転写因子)である。代替的な実施形態では、生物学的に活性な物質は、非分裂促進的な増殖因子(例えば、v−myc、SV−40ラージT、またはテロメアーゼ)である。
【0046】
遺伝的改変は、ウィルスベクター(レトロウィルス、改変されたヘルペスウィルス、ヘルペスウィルス、アデノウィルス、アデノ関連ウィルスなど)を用いる感染によってか、または、当該分野において公知の方法を使用するトランスフェクション(リポフェクション、リン酸カルシウムトランスフェクション、DEAE−デキストラン、エレクトロポレーションなど)のいずれかによって行われる(Maniatisら、Molecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory,N.Y.,1982)を参照のこと)。例えば、キメラ遺伝子構築物は、ウィルス(例えば、レトロウィルス)の末端反復配列(LTR)、サルウィルス40(SV40)、サイトメガロウイルス(CMV);または哺乳動物細胞特異的プロモーター(例えば、チロシンヒドロキシラーゼ(TH;ドーパミン細胞についてのマーカー))、DBH、フェニルエタノールアミンN−メチルトランスフェラーゼ(PNMT)、ChAT、GFAP、NSE、所望のタンパク質をコードする構造遺伝子の発現を指示するNFタンパク質(NE−L、NF−M、NF−Hなど)を含み得る。加えて、ベクターは、薬物選択マーカー(例えば、E.coliアミノグリコシドホスフォトランスフェラーゼ遺伝子)を含み得、これは、試験遺伝子とともに共感染(co−infected)される際に、タンパク質合成インヒビターであるジェネテシン(G418)に対する耐性を与える。
【0047】
LPCは、発現ベクターによるトランスフェクションを使用して遺伝子的に改変され得る。1つのプロトコルでは、遺伝子を含むベクターDNAを、0.1×TE(1mM Tris(pH8.0)、0.1mM EDTA)中で40μg/mlの濃度まで希釈する。22μlのDNAを、使い捨ての滅菌5mlプラスチックチューブ中の、250μlの2×HBS(280mM NaCl、10mM KCl、1.5mM Na2HPO4、12mMブドウ糖、50mM HEPES)に加える。31μlの2M CaCl2をゆっくりと加え、そして混合物を、室温にて30分間インキュベートする。この30分のインキュベーションの間に、細胞を、800gで5分間、4℃にて遠心分離する。細胞を、20容量の氷冷されたPBS中に再懸濁し、そして1×107の細胞のアリコートに分ける。これらを再度、遠心分離する。細胞のアリコートの各々を、1mlのDNA−CaCl2懸濁液中に再懸濁し、そして室温にて20分間インキュベートする。次いで、細胞を成長培地中で希釈し、そして5%〜7%のCO2で37℃にて6〜24時間、インキュベートする。この細胞を再度、遠心分離し、PBS中で洗浄し、そして10mlの成長培地へ48時間の間、戻す。
【0048】
LPCはまた、リン酸カルシウムトランスフェクション技術を用いて遺伝子的に改変される。標準的なリン酸カルシウムトランスフェクションでは、細胞を、単一の細胞の懸濁液へと機械的に分離し、そして、50%のコンフルーエンス(50,000〜75,000(細胞/cm2))で組織培養用に処理されたシャーレ上にプレートし、一晩付着させる。1つのプロトコルでは、改変されたリン酸カルシウムトランスフェクション手順を、以下のように行う:TEで440μlまで希釈した滅菌TE緩衝液(10mM Tris、0.25mM EDTA、pH7.5)中のDNA(15〜25μg)、および60μlの2M CaCl2(1M HEPES緩衝液でpH5.8まで)を、DNA/TE緩衝液へと加える。合計500μlの2×HeBS(HEPESで緩衝化した生理食塩水;275mM NaCl、10mM KCl、1.4mM Na2HPO4、12mMブドウ糖、40mM HEPES緩衝粉末、pH6.92)をこの混合物へ一滴ずつ加える。この混合物を室温にて20分間、静置する。細胞を1×HeBSで簡単に洗浄し、そして1mlのリン酸カルシウムで沈殿したDNAの溶液を、各プレートへと加え、そしてその細胞を37℃で20分間インキュベートする。このインキュベーションの後に、10mlの培地を、細胞に加え、そしてプレートを、さらに3〜6時間、インキュベーター(37℃、9.5%CO2)中に配置する。DNAおよび培地を、インキュベーション期間の最後に、吸引によって除去し、細胞を3回洗浄し、そしてインキュベーターに戻す。
【0049】
遺伝子改変が生物学的に活性な物質の生成のためである場合、その物質は、所与の肝臓障害の処置に有用な物質であり得る。LPCは、生物学的に活性な因子(例えば、成長因子、成長因子レセプター)を発現するために遺伝子的に改変される。例えば、細胞が増殖誘導成長因子または分化誘導成長因子を分泌するように、細胞を遺伝子的に改変することが望ましくあり得る。肝臓障害の処置において有用な成長因子産物としては、HGF、VEGF、FGF−1、FGF−2、EGF、TGFα、TGF、PDGF、IGF、およびインターロイキンが挙げられる。
【0050】
遺伝子改変されたLPCは、細胞治療または遺伝子治療のために、遺伝子的に改変された細胞によって生成された生物学的に活性な分子を必要としているレシピエントのCNSへと、移植され得る。移植技術は、以下に記述される。
【0051】
あるいは、遺伝子的に改変されたLPCは、移植の前に、インビトロでの種々の分化プロトコルに供され得る。一旦、細胞が分化されると、細胞は、所望のタンパク質の発現のために再度アッセイされる。所望の表現型を有する細胞は、単離され得、そして、遺伝子的に改変された細胞によって発現された、タンパク質または生物学的に活性な分子を必要としているレシピエントへと移植され得る。
【0052】
(肝臓前駆細胞への薬物の効果をスクリーニングするための方法)
LPC培養物は、潜在的な治療組成物のスクリーニングのために使用され得る。例えば、LPCは、肝臓細胞の増殖、分化、または生存をもたらす化合物を同定するために使用される。加えて、LPCは、抗ウィルス化合物を同定しウィルスの感染性を決定するためか、または、試験化合物の代謝産物を同定するために、使用される。これらの試験組成物は、種々の投与量で培養物中の細胞に適用され得る。そして細胞の応答が、種々の期間、モニタリングされる。細胞の物理的特徴は、顕微鏡を用いて細胞の成長および形態を観察することによって分析され得る。タンパク質(例えば、酵素、レセプター、および他の細胞表面分子)または神経伝達物質、アミノ酸、神経ペプチド、および生体アミンの新たな発現レベルまたは増加した発現レベルの誘導は、このような分子のレベルの変化を同定し得る当該分野で公知の任意の技術を用いて、分析され得る。このような技術としては、このような分子に対する抗体を使用する免疫組織化学、または生化学的分析が挙げられる。このような生化学的分析としては、タンパク質アッセイ、酵素的アッセイ、レセプター結合アッセイ、酵素結合イムノソルベントアッセイ(ELISA)、電気泳動分析、高性能液体クロマトグラフィ(HPLC)による分析、ウェスタンブロット、および放射免疫アッセイ(RIA)が挙げられる。核酸分析(例えば、ノーザンブロット)は、これらの分子またはこれらの分子を合成する酵素をコードするmRNAのレベルを検査するために使用され得る。
【0053】
LPCは、肝細胞への生物学的因子の効果を決定する方法において使用され得る。用語「生物学的因子」とは、任意の因子(例えば、ウィルス、タンパク質、ペプチド、アミノ酸、脂質、炭水化物、核酸、ヌクレオチド、薬物、プロドラッグ、または、神経細胞への効果(このような効果が有害であっても、有益であっても、またはその他でも)を有し得る他の因子)をいう。肝細胞に有益である生物学的因子は、本明細書中で「肝臓因子」と呼ばれる。この用語は、肝細胞の増殖、分化もしくは機能、または肝臓疾患もしくは肝臓障害の処置について潜在的に有用であると証明され得る、生物学的または薬学的に活性な任意の物質を含む用語である。
【0054】
LPCの培養物からの肝細胞への潜在的な生物学的因子の効果の決定は、肝臓組織からかまたは、代わりに、肝臓疾患もしくは肝臓障害に罹患している宿主から得られ得る。培養条件の選択は、試験される特定の因子、そして実験者が達成することを望む効果に依存する。一旦、細胞が所望のドナー組織から入手されると、その細胞は、インビトロで増殖される。
【0055】
移植目的のために大量の細胞を生成するのに有用である、LPCの増殖能力を増加する生物学的因子についてスクリーニングすることが可能である。LPCの増殖を阻害する生物学的因子についてスクリーニングすることもまた可能である。LPCは、対象となる生物学的因子の存在下でプレートされ、そして増殖が起こる程度についてアッセイされる。LPCおよびLPCの子孫の分化および生存への、生物学的因子または生物学的因子の組み合わせの効果が、決定され得る。
【0056】
スクリーニングの前に、分化するよう既に誘導されているLPCをスクリーニングすることが可能である。分化の前に、生物学的因子をLPCに適用することによって、分化プロセスへのその生物学的因子の効果を決定することもまた可能である。一般に、生物学的因子は、可溶化され得、そして種々の濃度にて培養培地に添加されて、各用量におけるこの因子の効果を決定し得る。培養培地は、因子の濃度をある程度一定に保つための量で、数日毎に生物学的因子を補充され得る。
【0057】
増殖における変化は、細胞数における増加または減少によって観察される。「制御因子」は、LPCの増殖への制御効果を有する生物学的因子である。例えば、生物学的因子は、増殖誘導成長因子(例えば、EGF)に応答してインビトロで増殖するLPCの数を増加させるかまたは減少させる場合、「制御因子」とみなされる。あるいは、増殖誘導成長因子に応答するLPCの数は同じままであり得るが、制御因子の追加が、LPCの増殖速度に影響を与える。増殖誘導成長因子は、別の増殖誘導成長因子と組合わせて使用される場合に制御因子として作用し得る。
【0058】
このようなスクリーニング方法を使用すると、当業者は、肝細胞の増殖および分化、または分化した肝細胞の生存または機能への生物学的因子の効果について試験することによって、肝細胞への薬物の潜在的な副作用についてスクリーニングし得る。増殖されたLPCは、代表的には、約5〜10×106細胞/mlの密度でプレートされる。分化した特定の細胞型または所与の細胞の組み合わせへの生物学的因子の効果を試験することが望ましい場合、分化の後に得られた肝細胞と胆管細胞との比は、異なる型の細胞と隔てることによって処理され得る。
【0059】
生物学的因子の効果は、コントロール培養物に対する有意差に基づいて同定される。これは、発現された表現型の比、細胞の生存度、および遺伝子発現における変化のような基準に関しての差である。細胞の物理的な特徴は、顕微鏡で細胞の形態および成長を観察することによって分析され得る。タンパク質(例えば、酵素、レセプター、および他の細胞表面分子)の新たな発現レベルまたは増加した発現レベルの誘導は、このような分子のレベルの変化を同定し得る当該分野で公知の任意の技術を用いて、分析され得る。このような技術としては、このような分子に対する抗体を使用する免疫組織化学、または生化学的分析が挙げられる。このような生化学的分析としては、タンパク質アッセイ、酵素的アッセイ、レセプター結合アッセイ、酵素結合イムノソルベントアッセイ(ELISA)、電気泳動分析、高性能液体クロマトグラフィ(HPLC)による分析、ウェスタンブロット、および放射免疫アッセイ(RIA)が挙げられる。核酸分析(例えば、ノーザンブロットおよびPCR)は、これらの分子またはこれらの分子を合成する酵素をコードするmRNAのレベルを検査するために使用され得る。
【0060】
LPCの増殖、およびLPCの子孫の増殖、分化および生存に関与する因子、ならびに生物学的因子に対するそれらの応答は、当該分野で公知の技術を用いて、LPCの発生の異なる段階におけるLPCまたはLPCの子孫から、cDNAライブラリーを構築することによって単離され得る。ある発生段階における細胞からのライブラリーは、発生の異なる段階における細胞のライブラリーと比較されて、発生の間の遺伝子発現の順序を決定し、そして種々の生物学的因子の効果を明らかにするか、または肝臓細胞における遺伝子発現を変化させる新たな生物学的因子を明らかにする。ライブラリーが機能不全組織から調製される場合、機能不全組織からのライブラリーと正常組織からのライブラリーとを比較することによって、原因として機能不全を引き起こすために役割を果たす遺伝因子が同定され得る。この情報は、障害を処置するために治療設計において使用され得る。加えて、プローブが、種々の遺伝障害の診断における使用のためか、または発生における特定の段階で肝細胞を同定することに使用するために同定され得る。
【0061】
本発明は、以下の実施例によってさらに示されるが、それらに限定はされない。
【実施例】
【0062】
(実施例1:一般的な方法)
(ヒト胎児肝細胞の単離)
本研究の許可は、Huddinge大学病院にある現地の倫理委員会から認可された。ヒトFL組織を、スウェーデンの手引きに従って、妊娠の6〜9.5週間目に中絶された胎児から入手した。研究プロトコルは、現地の倫理委員会によって認可された。改変された吸引掻爬術を行った(33)。在胎齢を、妊娠期間の12週間未満の胎児においては特有の解剖学的マーカー(34)に従って推定し、そしてそれより後の胎児においては、超音波大横径測定によって推定した(35)。妊娠齢は、最終月経から算出した妊娠期間として与えられる。中絶を、明らかな異常がない妊娠において行った(つまり、異常がある胎児を含まなかった)。FLを、解体し、そしてRPMI 1640培地(Gibco,Invitrogen Corp.UK)を含む滅菌チューブ中に配置した。次いで、肝臓を、70μmの金属メッシュを通すことによって、単一の細胞の懸濁液へと分解した。単一の細胞の懸濁液を、200gで10分間遠心して、細胞をペレットにした。胎児組織を提供するすべての女性は、梅毒、トキソプラスマ症、風疹、HIV−1、サイトメガロウイルス、B型肝炎、およびC型肝炎、パロウィルス、ならびに1型単純ヘルペスおよび2型単純ヘルペスについて血清学的にスクリーニングされている。
【0063】
(磁気細胞分類による細胞の単離およびインビトロでの培養)
単一の細胞の懸濁液を、妊娠7〜9週目における胎児肝細胞から調製した。細胞をヒト原始前駆細胞濃縮単離キット(Stem cell technologies,Vancouver,Canada)を用いて、その後に、磁気活性化細胞分離磁気ビーズシステム(Stem cell technologies)によって単離した。この方法は、この集団のネガティブ選択に基づいている。この選択には、12個の系統特異的な細胞表面抗原に対する抗体(抗CD2、抗CD3、抗CD14、抗CD16、抗CD19、抗CD24、抗CD36、抗CD38、抗CD45RA、抗CD56、抗CD66b、抗グリコホリンA)を含む枯渇カクテル(depletion cocktail)を使用する。この手順を、販売元によって説明されているように実行した。すべての場合において、回収した前駆細胞を、直ちにフローサイトメトリーによって分析し、lin+細胞の汚染が存在しないことを確かめ、かつ細胞の前駆体の表現型(CD117+/CD34+/lin−)を確認した。手順の最後に、回収された前駆細胞を生存度について試験した。この後で、この細胞を、I型コラーゲンで被覆されたプラスチックのペトリ皿(Biocoat,Becton and Dickinson,New Jersey,USA)に播種した。そして、10%不活化胎児子ウシ血清、ペニシリンおよびストレプトマイシン、5%L−グルタミン、5%最小必須アミノ酸、50ng/mlのHGF(R&D Systems,Abingdon,England)、20ng/mlのEGF(R&D Systems)、および10ng/mlの塩基性繊維芽細胞成長因子(R&D Systems)を含有するダルベッコ改変イーグル培地(DMEM)(GIBCO,Invitrogen,Stockholm,Sweden)中で培養した。三日ごとに毎回、培地を回収し、遠心分離し、滅菌ろ過処理し、そして条件培地(CM)として使用した。その後の全ての継代培養を、20%CMを使用して行った。いくつかの実験において、前駆細胞を、20ng/mlのHGF、10ng/mlのEGF、および50ng/mlのVEGF(R&D Systems)を含有するDMEM培地中で成長させ、そして培養物中で分裂させた。増殖を検出するために、培養物中の細胞を、チミジンアナログBrdU(30mM)とともに30分間、インキュベートした。細胞を洗浄し、そして、ヒトのアルブミンに対するFITC結合ヤギ抗体(Natutec,Frankfurt,Germany)、非結合抗ヒトサイトケラチン19(Neomarker,USA)、および非結合抗BrdU抗体(Sigma,Stockholm,Sweden)を使用して、アルブミン、サイトケラチン19、およびBrdU(30mM)で染色した。表現型決定のために使用された他の抗体は、抗CD45、抗CD14、抗CD90、抗CD117、抗CD34(Pharmingen,USA)、抗Flk−1(ReliaTech,Germany)、ならびに二次サブクラス特異的抗体であるヤギ抗マウスIgG1(FITC/Texas red)およびヤギ抗マウスIgG2a(FITC/Texas red)であった。フローサイトメトリーおよび免疫細胞化学を、前駆細胞を表現型決定するために使用した。この手順を記載されているように行った(36)。
【0064】
新たに単離された前駆細胞(P0)、および第6継代(P6)および第12継代(P12)においてインビトロで拡張された細胞を、移植研究のために使用した。
【0065】
(マウス)
動物を世話し、Huddinge病院にあるThe animal care and use committeが認可した動物プロトコルを使用する。肝障害を、部分的な肝切除の24時間前に、体重1kgあたり0.7gの腹腔内への、GalN(Sigma Chemicals Co.,Stockholm,Sweden)の投与によって、C57ブラック/ヌードマウス(n=16)に誘導した。GalNを、pH7.4のリン酸緩衝生理食塩水(PBS)中に、100mg/mlにて溶解した。部分的な肝切除(PH)を、以前に説明されたように行った(37)。GalNの投与を、PHの後、10日間続けた。
【0066】
肝臓前駆細胞を、これらの動物の脾臓へと移植した。動物を、エーテル下で麻酔し、そして代表的には、200μlのDMEM培地中に懸濁された1×105の新たに単離された細胞(P0)、および第6と第12継代における1×106の細胞を、およそ10〜15秒にわたって脾臓へと注入した。4匹のマウスを、DMEM培地のみで偽移植した。止血を確実にした後、腹腔切開を閉じ、そして動物を回復に近づくまでモニタリングした。
【0067】
(肝臓の調製および凍結切片における蛍光分析)
マウスを、移植の4週間後に殺し、そして肝臓、脾臓および肺を切除した。約2mm2の肝臓の各々から2つか3つの生検を、液体窒素中でショック凍結し、そしてRT−PCR分析を行うためのRNAの単離のために使用した。残りの肝臓組織を、蛍光分析および免疫組織化学的分析のためにショック凍結した。5μmの厚さの凍結切片を風乾し、そして、メタノール中で10分間、冷30%アセトンで固定し、そして免疫組織化学によってさらに分析した。
【0068】
(免疫組織化学)
本発明者らはまず、マウスの実質中のヒト細胞の検出のために2つの方法を試験した;a)ジゴキシゲニン標識総ヒトDNAプローブ(Cytocell,Oxfordshire UK)を使用するインサイチュハブリダイゼーション技術(38)、およびb)マウス抗ヒト核モノクローナル抗体(Chemicon,CA,USA)の後に、ビオチン化ウマ抗マウス二次抗体で染色する。免疫ペルオキシダーゼ手順を、製造元によって説明されているように、Vectastain Elite ABCキット(ImmunKemi,Stockholm,Sweden)を使用して行った。ジアミノベンジジン三塩酸塩(DAB)−ニッケル基質キットを、色展開剤として使用した。二重染色について、DAB(茶色に染色する)および/またはDAB−Ni(黒)および/またはVector NovaRedキットの組み合わせを使用した。使用した他の一次抗体は、ヒトのアルブミンに対する(マウスとは交差反応しない)FITC結合ヤギ抗体(Natutec,Frankfurt,Germany)、非結合抗ヒトサイトケラチン19(Neomarker,USA)、および(ジペプチジルペプチダーゼIVを検出する)非結合マウス抗ヒトCD26(Pharmingen,USA)であった。GGT、G−6−Pおよびグリコーゲンを、以前に説明されたようにインサイチュで示した(39,40)。切片をヘマトキシリンで対比染色し、マウント培地(ImmunKemi,Stockholm,Sweden)中でマウントした。
【0069】
(形態計測分析)
各マウスの肝臓の60枚の連続切片を、DAB−Ni−ポジティブなヒト細胞についてスクリーニングした。移植された細胞の数を、3つのサイズのクラスター(すなわち、単独で配置する細胞、または2以上〜20の細胞のクラスターまたは20より多い細胞のクラスター)で決定した。本発明者らは、移植された動物の全て由来の組織において最低限100の高倍率視野で分析した。
【0070】
(逆転写ポリメラーゼ連鎖反応(RT−PCR))
全RNAを、4匹のヒト−マウスキメラマウスの肝臓組織から、および1匹の正常マウスの肝臓組織から、Micro−FastTrack RNA単離キット(Invitrogen,Groningen,The Netherlands)を使用して、抽出した。本発明者らは、マウスの肝臓における、ヒトのアルブミン、CK19、α−フェトプロテインおよびα1−抗トリプシンの発現を検出するために、ヒト特異的プライマーを使用した。プライマーを、Primer Expressソフトウェア バージョン2.0(Applied Biosystems)を使用することにより、CK−19、α−フェトプロテイン、アルブミン、抗トリプシン、およびグルコース6リン酸デヒドロゲナーゼ(G6PD)について選択した。プライマーセットの各々を、DNAで汚染されずに、cDNAのみを標的とするために設計した。プライマーセットは、CyberGene(Huddinge,Sweden)によって商業的に合成された。
【0071】
【化1】

PCR反応を、96ウェル光学プレートに総容量25μlで、二つ組で行った。各反応物は、2.5μLのcDNA、12.5μLのSYBR Green Master Mix(Applied Biosystems)、および500nMの各プライマーを含んだ。ポジティブコントロールおよびネガティブコントロールを、全ての実行において含んだ。熱サイクル条件は、製造元によって推奨されているように、最初に50℃で2分間、そして95℃で10分間であった。サイクル条件は、95℃で15秒間、そして60℃で1分間の40サイクルであった。ハウスキーピング遺伝子であるG6PDを、内部標準コントロールとして含んだ。これは、RNAの単離および逆転写の成功、ならびに全てのサンプルにおけるRNAの総量を確認するために使用した。RT−PCRからの結果を可視化するために、本発明者らは、使用できる状態の12.5%の非変性ポリアクリルアミドゲル電気泳動システム上でPCR産物を泳動し、そしてそのバンドを、自動化された銀染色(Pharmacia Biotech,Uppsala,Sweden)によって染色した。
【0072】
(統計方法)
データを、平均値±SDのように示す。差異の有意性を、スチューデントt検定および分散分析(ANOVA)を用いて分析した。0.05未満のp値は、有意であるとみなした。
【0073】
(実施例2 CD117+/CD34+/LIN−肝臓前駆細胞は、インビトロで肝細胞および胆管細胞へと分化し得る)
原始造血前駆体を単離するために設計されたキットを使用して、ヒト胎児肝臓由来の細胞集団(gw6〜9)を、任意の関係付けられた造血マーカーを発現しなかった集団から得た。この集団のさらなる表現型決定は、幹細胞マーカーCD117およびCD34の発現を示したが、肝臓マーカー(例えば、アルブミン(肝臓マーカー)およびCK19(胆管細胞マーカー))の発現は示さなかった。Thy−1(CD90)もCD45もまた発現しなかった(図1a)。この集団は、妊娠6〜9週目における全体の胎児肝臓の約0.5%〜0.7%を表す。CD45およびCD90の発現は、細胞の継代培養の間には観察されなかった。培養の際に、これらの細胞が、付着性の集団(約85%)(図1b)と非付着性集団(約15%)の両方の混合物であることを見出した。非付着性集団は、以下で与えられた培養条件の下で、さらに拡張され得なかった。CD117+/CD34+/Lin−細胞およびそれらの子孫は、コロニー中で成長する傾向がある(図1b)。培養物中で2日後に付着前駆細胞によって発現される最初のマーカーは、c−Met(肝細胞成長因子レセプター)であった(図1b)。肝細胞成長因子(HGF)および上皮成長因子(EGF)を含有する培養培地中で2週間、これらの細胞をインビトロで培養することは、4つタイプの細胞の存在を示した:i)アルブミンおよびCK19を発現する約85%のダブルポジティブ細胞、ii)約4%のダブルネガティブ細胞、iii)アルブミンのみを発現する約6%のシングルポジティブ細胞、iv)約5%のCK19のみを発現するシングルポジティブ細胞(図1c)。この表現型は、培養の間、数継代にわたって維持された(図1c)。しかし、P11以降から、ダブルポジティブ細胞(約60%)およびタブルネガティブ(約3%)の数においてわずかな減少があった。一方で、アルブミンまたはCK19についてのシングルポジティブ細胞は増加した。これらのデータは、発生早期の肝臓由来の非造血性原始前駆細胞がインビトロで肝細胞および胆管細胞へと分化し得ることを実証する。
(実施例3 CD117+/CD34+/LIN−肝臓前駆細胞は、血管内皮成長因子の存在下で肝臓類洞内皮細胞へと分化する)
興味深いことに、付着性CD117+/CD34+/Lin−細胞を、50ng/mlの血管内皮成長因子(VEGF)を含有する培養培地中で分化させた場合、本発明者らは、内皮様の形態を有する細胞の大きな集団を観察した。胎児肝臓内皮前駆体に発現されることが公知であるFlk−1を含む肝細胞マーカーを使用する、この細胞集団のさらなる特徴付けは、4つの集団を明らかにした:a)レセプターFlk−1を発現する内皮細胞(約50%)、b)肝細胞(約13%)、c)胆管細胞(約17%)(図1d)、および、d)これらのマーカーのうちのいずれも発現しなかった細胞集団(約20%)(データは示さず)。Flk−1+細胞の類洞表現型を、抗体の大きなパネルを使用して確認した(表1)。ヒト臍静脈内皮細胞を、血管内皮細胞と類洞内皮細胞との間の表現型の差を実証するために使用した(表1)。電子顕微鏡分析は、類洞内皮細胞に特徴的な窓(fenestrne)の存在、およびの基底膜の欠如を明らかにした(データは示さず)。これらのデータは、発生早期の肝臓由来の非造血性原始前駆細胞が、肝細胞、胆管細胞、および内皮細胞へとインビトロで分化し得ることを示す。
【0074】
【表1】
(実施例4 肝臓前駆細胞は、インビトロで広範に継代され得る)
肝臓前駆体の増殖は、培養の間、20%の条件培地(CM)の恒常的な使用に依存した。20%のCMを使用すると、これらの細胞を、少なくとも12継代まで継代し得る。CMを除去すると、得られる細胞継代の数が有意に減少した(p<0.001;図2a)。このことは、細胞の増殖における減少を反映する。高い増殖能力が、BrdUの取り込みによって検出されたように、ダブルポジティブ細胞(ALB+CK19+)およびアルブミンのシングルポジティブ(ALB+CK19−)において観察されたことが見出された(図2bおよび図2c)。この細胞は、安定な表現型を長期間の間、維持され得る。すべてのFLが、急速に増殖し、そして数継代の間、培養物中で維持され得る細胞を生成したわけでないことを言及することは重要である(表2)。長期間、増殖の成功は、得られたFL組織の質に依存した。しかし、試験されたすべてのFLから単離されたCD117+/CD34+/LIN−細胞が肝細胞へと分化したことを見出した。インビトロ研究からのデータは、CD117+/CD34+/LIN−細胞およびそれらの子孫が、広範な複製能力を有し、そして安定な表現型で長期間の間維持され得ることを実証した。
【0075】
【表2】
(実施例5:拡張されたヒト胎児前駆細胞は、GalN処置マウスの肝臓中で成熟肝細胞、成熟胆管細胞、および成熟類洞へと首尾よく移植され、分化する)
新たに単離されたCD117+/CD34+/Lin−細胞(P0)、および培養物中で拡張された細胞(P6およびP12)が、分化する可能性を有しインビボで機能するかどうかを試験するために、これらの細胞を、最初に部分的に肝切除されたマウスへと移植し、次いでDガラアクトサミン(GalN)で処置して急性肝損傷を誘導した。このプロトコルは、急性肝損傷を誘導し、肝臓再分化を促進させる(21)。コントロールの2匹マウスおよび試験グループの2匹は、処置の24時間後に死亡した。細胞移植の4週間後にマウスが生存していた結果が示される。脾臓内の移植の後、初代P0細胞、ならびにP6およびP12継代培養細胞は、GalN処置マウスの肝臓において生存した。移植された細胞を位置決めするための、ヒトのセントロメアプローブおよび抗ヒト核抗体を使用して得られた結果を比較する実験は、同様の結果を示した。ヒトDNAプローブの種特異性(図3aおよび図3b)および抗ヒト核抗体は、ヒト肝臓によるポジティブな結果(図3c)、およびヌードマウスからの偽移植された肝臓によるネガティブな免疫組織化学(図3d)によって実証される。新たに単離されたCD117+/CD34+/Lin−細胞は、移植した場合、移植の4週間後にて、肝細胞(図3e)、胆管細胞(図3f)、および形成された胆管(図3g)へと分化した。P6およびP12における肝臓前駆細胞は、移植した場合、移植され、そして急性的に損傷した肝臓を再構成した(図3h−j)。移植された細胞を、マウスの肝臓中で見出したが(図3k)、例えば、脾臓(図3l)および肺(図3m)のような他の組織において、そして偽移植された動物(図3n)においては見出されなかった。移植されたヒト細胞は、肝臓マーカー(例えば、グルコース6リン酸(G−6−P)(図4a)およびグリコーゲン(図4b))、ならびに胆管マーカー(例えば、ジペプチジルペプチダーゼIV(DPPIV)(図4c)およびγグルタミルトランスペプチダーゼ(GGT)(図4d))を発現した。3匹のマウスでは、ヒト肝細胞によって90%再増殖した、再分化する組織の切片が見出された(図4e)。ヒト前駆体によって完全に再増殖したヒト胆管の透明な領域もまた、観察された。これらの細胞は、CK19およびそして抗ヒト核抗体に対してポジティブに染色した(図4f)。マウスのアルブミンとは交差反応しないヒトのアルブミンに対するモノクローナル抗体を、移植された細胞におけるヒトのアルブミンの発現を検査するために使用した。ヌードマウスからの偽移植された肝臓によるネガティブな免疫組織化学(図4g)は、抗体の種特異性を示した。移植されたマウス10匹すべてにおいて、いくつかのアルブミンポジティブ構造(図4h−j)が見出されたが、偽移植されたマウス(n=2)においてヒトのアルブミン発現する病巣は、同一の条件を使用しても観察されなかった。ヒトのアルブミンは、マウスの肝臓中のみで検出され、しかし脾臓または肺においては観察されなかった。興味深く注目するものとしては、分析されたマウスのいずれにおいても、腫瘍の形成が観察されなかったことである。
【0076】
肝臓の移植が、P0、p6、およびp12におけるヒト肝臓前駆体の間で匹敵する状態であったかどうかを決定するために、マウス肝臓の各々の60枚の連続切片を、DAB−Ni−ポジティブなヒト細胞についてスクリーニングした。移植された細胞の数を、3つのサイズのクラスター(すなわち、単独で配置する細胞、または5以上〜20の細胞のクラスター、または20より多い細胞のクラスター)で決定した。この分析は、P0、P6およびP12細胞を受容する動物の間で、単独で配置する移植された細胞の数において、または2〜20以上の細胞のクラスターにおいて、有意差が存在しなかったことを示した。20より多い細胞のクラスターは、P0細胞を移植されたマウスにおいては何も観察されなかったが、P6およびP12における細胞を受容したマウスにおいては観察された(表3)(p<0.001;ANOVA)。P0細胞を移植されたマウスにおいて、本発明者らは、ヒトの移植された細胞の数における10倍の増加を検出し、一方で、P6細胞およびP12細胞を注入されたマウスにおいては、6倍の増加を検出した(表3)。これらのデータは、新たに単離されたCD117+/CD34+/LIN−細胞が、首尾よく増殖し、そしてインビボで成熟肝細胞、成熟胆管細胞、および成熟類洞内皮細胞へと分化することを実証する。加えて、インビトロで拡張されたCD117+/CD34+/Lin−細胞およびそれらの子孫は、インビボで増殖し、そして、損傷したマウス肝臓を首尾よく再構築する。
【0077】
【表3】
(実施例6:ヒト肝臓前駆細胞を移植したマウスにおけるヒト肝臓特異的遺伝子の転写)
植え付けられた細胞の移植は、移植されたマウスにおけるヒト遺伝子の発現を決定することによって確認した。屠殺したマウス肝臓を、ヒト肝臓特異的遺伝子(アルブミン、α1−抗トリプシン、CK19、およびAFPを含む)について特異的なプライマーを使用するRT−PCRによって、ヒト胎児肝臓前駆細胞の移植の1ヵ月後に、分析した。偽移植されたヌードマウスの肝臓からのRNAは、RNAの完全性のためのコントロールとして使用されたG6PDについて増幅を結果としてもたらした。CK19、アルブミン、およびAFPプライマーは、プライマーが個々のマウス遺伝子を増幅しなかったので、ヒトに特異的な種であった。しかし、α1−抗トリプシンが種特異的であることは見出されなかった(図4k)。これらの結果は、新たに単離された(P0)細胞、および種々の継代(P6およびP12)における細胞の両方が、損傷したマウス肝臓に移植することを示す。
【0078】
(実施例7:肝臓前駆細胞を移植されたマウスにおけるヒト第VIII因子の保護)
ヒト第VIII因子のレベルは、化学的に肝臓を損傷したマウスおよび肝切除したマウスにおいては極僅かであり、同様に、同じ様式で処置された、そしてさらに成長因子HGFおよび/または間質細胞を受容したマウスにおいても、同様に極僅かである。しかし、ヒト肝臓前駆細胞のみ、またはHGF+間質細胞+前駆細胞の組み合わせを受容する化学的に肝臓を損傷したマウスおよび肝切除したマウスは、それらの血漿においてヒト第VIII因子の高いレベルを実証した(図6)。
【0079】
(他の実施形態)
特定の実施形態が本明細書中で詳細に開示されているが、これは、例示目的のために例として開示されており、前に添付された特許請求の範囲に関して限定することは意図されていない。具体的には、種々の置換、変更、および修正が、請求項に規定されるような本発明の精神および範囲から逸脱することなく、本発明に対して行われ得ることが、本発明者らによって企図される。核酸の出発物質、所定のクローン、またはライブラリーのタイプの選択が、本明細書中に記載された実施形態の知識を有する当業者にとって、日常的なことであると考えられる。他の局面、利益、および修正は、上述の特許請求の範囲内にあるとみなした。
【技術分野】
【0001】
(発明の分野)
本発明は、前駆細胞に関する。
【背景技術】
【0002】
(発明の背景)
急性肝不全は、依然として高い死亡率を有する重大な問題である。肝機能障害および肝不全を生じる疾患の発症率が高いにもかかわらず、内科的治療における主な進歩は、現在、特定の形態のウィルス性肝炎の予防および処置に限られている。急性および慢性の肝疾患は、治療的なアプローチよりも対症的なアプローチで、依然として処置されている。同所性肝移植は、今までのところ、末期の肝疾患を有する患者のための唯一利用可能な治療である。残念なことに、ドナーの臓器のアベイラビリティは制限されているので、多くの患者が、肝臓移植を待ちながら毎年亡くなっている。最近では、正常肝細胞を罹患した肝臓へと移植することが、代替治療として用いられている。しかし、臓器ドナーの不足が、肝細胞移植の臨床適用を制限している。
【0003】
幹細胞およびそれらの子孫細胞を用いた細胞治療は、肝疾患を有する患者のための、大部分が未対処の医学的必要性に対する有望な新しいアプローチである。多くの研究が、実験的に誘導した急性肝不全において、(肝外肝臓組織および肝内肝臓組織から得られた)幹/前駆細胞の移植の可能性を調べている。成人の臓器由来の幹/前駆細胞は、機能性の肝細胞を産出し得るが、このような細胞は、稀である。遺伝性疾患および後天性疾患を回復するための肝臓前駆細胞移植に関する臨床治療プロトコルは、完全に発達した機能を依然として保持したままの細胞を大量に生じる能力に、大いに恩恵を受ける。
【発明の概要】
【発明が解決しようとする課題】
【0004】
従って、機能性の肝細胞へと分化し得る肝臓前駆細胞の供給源に対する必要性が存在する。
【課題を解決するための手段】
【0005】
(発明の要旨)
本発明は、肝臓前駆細胞の発見に基づいている。前駆細胞は、多分化能細胞である。前駆細胞は、(例えば、インビボまたはインビトロで)肝細胞(例えば、肝細胞、胆管細胞、または肝臓内皮細胞(すなわち、類洞細胞))へと分化することが可能である。従って、本発明は、肝臓前駆細胞培養物(例えば、インビトロ培養物)を特色とする。この培養物は、付着培養物である。あるいは、培養物中の細胞は、懸濁液中にある。この細胞は、肝臓組織(例えば、胎児肝臓組織)由来である。上記組織は、哺乳動物(例えば、ヒト、霊長類、マウス、ラット、イヌ、ネコ、ウシ、ウマ、ブタ)由来である。上記細胞は、CD117およびCD34に対して免疫反応性であり、Linには免疫反応しない。上記細胞は、インビトロで増殖する。上記細胞は、2回、3回、4回、5回、6回、7回、8回、9回、10回、11回、12回、15回、20回、25回またはそれ以上倍加することが可能であり、そして肝細胞、胆管細胞、または肝臓内皮細胞へと分化する能力を保持する。
【0006】
さらに提供されるものは、1つ以上の分化因子を含有する培養培地中で肝臓前駆細胞培養物を分化させることにより、肝細胞、胆管細胞、または肝臓内皮細胞を生成する方法である。分化因子としては、例えば、血管内皮成長因子(VEGF)、肝細胞成長因子(HGF)および表皮成長因子(EGF)が挙げられる。
【0007】
本発明はさらに、CD117+、CD34+、およびLin−のヒト肝臓前駆細胞を提供し、そしてその細胞を宿主へ移植することにより、(例えば、ヒト、霊長類、マウス、ラット、イヌ、ネコ、ウシ、ウマ、ブタのような哺乳動物である)宿主中に、前駆細胞または前駆細胞の子孫を移植する方法を、特徴とする。必要に応じて、肝臓の間質細胞または間葉細胞は、宿主中に共移植される。宿主は、前駆細胞の前、後、または同時に肝細胞増殖因子を投与される。宿主は、肝臓障害または肝臓組織損傷に罹患している。例えば、被験体は、肝炎、肝硬変、肝臓癌、脂肪肝疾患、ライ症候群、糖原病、肝嚢腫、またはウィルソン病を罹患している。移植は、臨床上の利益(例えば、特定の肝臓障害の1つ以上の症状を軽減すること)を与える。肝臓障害は、当該分野で公知の方法を使用して、医師によって診断される。
【0008】
肝細胞の増殖、分化、または生存をもたらす化合物は、肝臓前駆細胞培養物を試験化合物に接触させ、そして、その化合物が肝臓前駆細胞の増殖、分化、または生存に対する効果を有するかどうかを決定することによって、同定される。同様に、試験化合物の代謝産物が決定される。代謝産物は、培養物を試験化合物に接触させた後、培養培地をスクリーニングすることにより同定される。代謝産物は、当該分野で公知の方法(例えば、HPLC、質量分析、またはゲル電気泳動)によって同定される。試験化合物の抗ウィルス活性は、試験化合物の存在下または非存在下で、ウィルスを前駆細胞培養物へと導入し、その細胞の生存率を決定することによって決定される。試験化合物の非存在下と比較した、試験化合物の存在下における生存率の増加は、この試験化合物が抗ウィルス活性を有することを示す。ウィルスの感染性は、前駆細胞培養物をウィルスに接触させ、そしてこの細胞における増殖または生存の効果を決定することによって決定される。ウィルスと接触させなかった細胞培養物と比較した、生存または増殖の減少は、このウィルスが肝細胞に感染し得ることを示す。その一方、ウィルスと接触させなかった細胞培養物と比較した、生存または増殖における類似性は、このウィルスが肝細胞に感染しないことを示す。
【0009】
必要に応じて、培養物は、その培養物を試験化合物に接触させる前に、分化する。増殖および/または生存は、当該分野で公知の方法(例えば、BrdUアッセイ)によって決定される。分化は、肝細胞表面マーカーを形態学的かまたは組織学的に決定することによって決定される。ウィルスは、肝臓栄養性ウィルス(例えば、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルス)である。
本発明はまた、以下の項目を提供する。
(項目1)
ヒトの肝臓組織由来の細胞を含むインビトロ細胞培養物であって、該培養物中の該細胞は、
a.CD117+、CD34+、およびLin−であり;
b.培養物中で増殖し得;そして
c.インビトロで肝細胞、胆管細胞、または類洞細胞へ分化し得る、
細胞である、培養物。
(項目2)
項目1に記載の培養物であって、前記培養物は、少なくとも6回、倍加し得る、培養物。
(項目3)
項目1に記載の培養物であって、前記培養物は、少なくとも12回、倍加し得る、培養物。
(項目4)
項目1に記載の培養物であって、前記培養物は、付着培養物である、培養物。
(項目5)
インビトロでヒトの肝類洞細胞を生成する方法であって、該方法は、
a.CD117+、CD34+、およびLin−の細胞を含む細胞懸濁物を提供する工程;
b.該細胞懸濁液を培養する工程;ならびに
c.血管内皮成長因子を含有する培地中で該細胞の子孫を分化させる工程
を包含する、方法。
(項目6)
ヒトの肝臓の肝細胞または胆管細胞をインビトロで生成する方法であって、該方法は、
a.CD117+、CD34+、およびLin−の細胞を含む細胞懸濁物を提供する工程;
b.該細胞懸濁液を培養する工程;ならびに
c.EGFおよびHGFを含有する培地中で該細胞の子孫を分化させる工程
を包含する、方法。
(項目7)
インビボで肝細胞、または胆管細胞または類洞細胞へ分化する、ヒトの肝臓前駆細胞の集団を生成する方法であって、CD117+、CD34+、およびLin−である細胞を、ヒトの肝臓由来細胞の集団から選択する工程を包含する、方法。
(項目8)
多分化能肝臓前駆細胞の子孫を宿主へ移植する方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;ならびに
b.該細胞を該宿主中に移植する工程
を包含する、方法。
(項目9)
肝細胞の増殖、分化、または生存をもたらす化合物についてスクリーニングする方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化ための多分化能力を維持する、工程;
b.該培養物を試験化合物に接触させる工程;ならびに
c.該化合物が、肝細胞の増殖、分化、または生存をもたらすかどうかを決定する工程;
を包含する、方法。
(項目10)
項目9に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目11)
試験化合物の代謝産物を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;
b.該培養物を試験化合物に接触させる工程;ならびに
c.該培養物とインキュベーションした後に、該試験化合物由来の代謝産物を同定する工程;
を包含する、方法。
(項目12)
項目11に記載の方法であって、工程bを実施する前に、前記培養物の分化を誘導する工程を包含する、方法。
(項目13)
試験化合物の抗ウィルス活性を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させるための多分化能力を維持する、工程;
b.該培養物をウィルスおよび試験化合物に接触させる工程;ならびに
c.該培養物の生存率をコントロール細胞培養物と比較する工程;
を包含し、該コントロール培養物と比較された該培養物の生存率の増加が、該試験化合物の抗ウィルス活性を示している、方法。
(項目14)
項目13に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目15)
項目13に記載の方法であって、前記ウィルスは、肝臓栄養性ウィルスである、方法。
(項目16)
項目15に記載の方法であって、前記肝臓栄養性ウィルスは、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルスである、方法。
(項目17)
ウィルスの感染性を決定するインビトロでの方法であって、該方法は、
a.多分化能のCD117+、CD34+、およびLin−のヒト肝臓前駆細胞を含む、インビトロ細胞培養物を提供する工程であって、該細胞は、肝細胞、胆管細胞、または類洞細胞へ分化させる多分化能力を維持する、工程;
b.該培養物をウィルスに接触させる工程;ならびに
c.該ウィルスが、該培養物中の該細胞の増殖または生存に対する効果を有するかどうかを決定する工程;
を包含する、方法。
(項目18)
項目17に記載の方法であって、該方法は、工程bを実施する前に、前記培養物の分化を誘導する工程をさらに包含する、方法。
(項目19)
項目17に記載の方法であって、前記ウィルスは、肝臓栄養性ウィルスである、方法。
(項目20)
項目19に記載の方法であって、前記肝臓栄養性ウィルスは、A型肝炎ウィルス、B型肝炎ウィルス、またはC型肝炎ウィルスである、方法。
(項目21)
項目8に記載の方法であって、胎児肝臓間質細胞または胎児肝臓間葉細胞を前記宿主中に共移植する工程をさらに包含する、方法。
(項目22)
項目8に記載の方法であって、前記宿主に肝細胞成長因子を投与する工程をさらに包含する、方法。
【0010】
他に規定されない限り、本明細書中で使用されるすべての技術用語および科学用語は、本発明が属する分野の当業者に一般に理解されるような同じ意味を有する。本明細書中に記載される方法および材料と類似するかまたはそれらと同等である方法および材料が、本発明の実施または試験において使用され得るが、適切な方法および材料が、以下に記載される。本明細書中で言及された他のすべての刊行物、特許出願、特許、および参考文献は、その全体が参考して援用される。矛盾する場合は、定義を含む本明細書が統制する。さらに、材料、方法、実施例は、例示するのみであり、限定されることを意図しない。
本発明の他の特徴および利点は、以下の詳細な説明および上記特許請求の範囲から明かである。
【図面の簡単な説明】
【0011】
【図1A】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Aは、単離の直後に磁気によって分類したFL細胞を、CD117およびCD34については二重染色したが、CD90、CD45、アルブミン、およびサイトケラチン19(CK19)については染色しなかった。
【図1B】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Bは、CD117+/CD34+/Lin−細胞は、培養される際にコロニー内で成長し、そして発現される第1マーカーは、肝細胞成長因子レセプター(c−Met)であった(拡大率:40倍)。
【図1C】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Cは、種々の継代において新たに単離し膨張させた細胞は、ALB−CK19−(白矢印)、ALB+(緑)、CK19+(赤)、ALB+CK19−(緑;肝細胞)およびALB−CK19+(赤;胆管細胞)を生じた(拡大率:60倍)。
【図1D】図1は、ヒト肝臓前駆細胞の特徴づけを示す、一連のFACS分析および写真である。図1Dは、血管内皮成長因子を含有する培養培地中で成長させた際の、磁気によって分類したCD117+/CD34+/Lin−付着細胞は、Flk−1+内皮細胞(約50%)、CK19+胆管細胞(約13%)およびアルブミン+肝細胞(約17%)へと分化した。およそ20%の細胞は、これらのマーカーのうちのいずれも発現しなかった。
【図2A】図2Aは、肝臓前駆体およびそれらの子孫の増殖を示す棒グラフである。20%に調製された培地(CM)の使用は、肝臓前駆細胞の増殖を有意に増加し、そしてCMを使用しないものと比較して数回継代され得る(p<0.001;スチューデントt検定)。
【図2B】図2Bは、フローサイトメトリー分析の結果を示す棒グラフである。このグラフは、高い増殖度(BrdU+細胞)が、アルブミンCK19+、アルブミン+CK19−、およびアルブミン−CK19+細胞において観察されたが、その一方で、両方ともネガティブな(アルブミン−CK19−)細胞は、より静止状態であったことを示す。
【図2C】図2Cは、種々の継代における肝臓前駆細胞を示す一連の写真であり、アルブミン(緑)およびCK19(赤)については、蛍光染色され、そして、BrdUの取り込みの検出については酵素学的に染色した。これらは、種々の部分集団において増殖能力を示した(拡大率:40倍)。
【図3】図3A−3Nは、マウス肝臓中のヒト肝臓前駆細胞の局在化の一連の写真である。a−b;ヒト肝臓(DAB)において核シグナルを示すが、マウス肝臓においては示さない、ヒト動原体プローブを用いたインサイチュハイブリダイゼーション。c−d:免疫組織化学を使用し、抗ヒト核抗体(DAB−Ni)で同様の結果を得た。e−j;新たに単離した細胞(e−g)である、第6継代および第12継代細胞が、D−ガラクトサミン処置(GalN)マウスへ移植される際に、肝細胞、胆管細胞、および内皮細胞への分化を示した(DAB−Ni;矢印の先端)。k;移植された細胞は、肝臓中で観察されたが、l−m;GalN処置マウスの脾臓、肺では、観察されなかった。n;偽移植されたマウスの肝臓を、コントロールとして用いた(拡大率:60倍)。
【図4A−J】図4A−Jは、ヒト肝臓前駆細胞のインビボでの発生運命を示す一連の写真である。a−d;GalN処置マウスへの移植の1ヶ月後のヒト肝臓前駆細胞は、グルコース−6−リン酸(茶)、グリコーゲン(ピンク)、ジペプチジルペプチダーゼIV(赤茶)、γグルタミルトランスペプチダーゼ(茶)を含んだ。抗ヒト核抗体(DAB−Ni;矢印の先端)による二重染色は、ヒト核(黒)を視覚化した。e;ヒト肝臓前駆体(核;黒矢印の先端)によるマウス組織切片全体の再生。f;サイトケラチン19(赤茶)の発現および、g−j;ヒトアルブミン(緑)は、偽移植したマウスでは観察されなかったが、新たに単離した、第6〜第12継代のヒト肝臓前駆細胞を移植した、GalN処置マウスの肝臓中で観察された(矢印の先端)。a−j;(拡大率:60倍)。f;(拡大率:200倍)。
【図4K】図4Kは、マウス肝臓におけるヒト肝臓特異的遺伝子の転写を示す写真である。ヒトサイトケラチン19、αフェトプロテイン、およびアルブミンは、ヒト肝臓前駆細胞を受容したGalN処置マウスの肝臓では検出されたが、偽移植されたマウスの肝臓では検出されなかった。しかし、α1アンチトリプシンは、コントロール中でもまたわずかに増幅した。グルコース−6−リン酸デヒドロゲナーゼを、ハウスキーピング遺伝子として使用した。
【図5】図5は、胎児肝臓間質参謀および胎児肝臓前駆細胞の共移植の効果を示す一連の写真である。A;抗ヒト核抗体で染色した正常ヒト肝臓切片は、陽性染色を示す(黒/茶)。B;正常マウス肝臓切片では、同じ抗体による陽性染色が見られなかったが、抗体の特異性を示した。C;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして肝細胞成長因子(HGF)を皮下注入して、そして胎児肝臓前駆細胞を移植した後のマウスは、高い細胞移植度をもたらさなかった。D;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして胎児肝臓から単離された間質細胞を注入した後のマウスは、高い細胞移植度をもたらさなかった。E−H;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして胎児肝臓間質細胞と前駆/幹細胞との混合物を移植した後のマウスは、高い肝細胞移植度をもたらした。I−K;レトロルシン(30mg/kg)で処置し、部分的に肝切除し、そして肝細胞成長因子(HGF)を皮下注入して、そして胎児肝臓間質細胞および肝臓前駆細胞を移植した後のマウスは、高い細胞移植度をもたらした。
【図6】図6は、種々の方法で処置した正常ヌードC57ブラックマウスおよびC57ブラックマウスにおける、ヒト第VIII因子の検出レベルを示すチャートである。
【図7】図7は、胎児肝臓および成人肝臓に対する、種々の造血細胞、肝細胞、および脾臓細胞の表面マーカーの発現を示すチャートである。
【図8】図8は、ヒトの胎児肝臓および成人肝臓における肝臓マーカーを示す一連の写真である。
【図9A】図9Aは、ヒト肝臓前駆細胞の特徴づけを示す一連のFACS分析である。
【図9B−F】図9B−Fは、異なるマトリックス上のヒト肝臓前駆細胞の形態を示す一連の写真である。
【図10】図10の上図は、ヒト肝臓前駆細胞における肝臓マーカーの遺伝子発現を示すノーザンブロットの写真である。図10の下図は、ヒト肝臓前駆細胞における肝臓マーカーの遺伝子発現を示すチャートである。
【発明を実施するための形態】
【0012】
(詳細な説明)
本発明は、数世代の間インビトロで拡張しかつ祖先の表現型を維持する、ヒト胎児肝臓内の非造血性前駆細胞の規定された集団における予想外の発見に基づいている。これらの細胞は、急性肝損傷を有する動物へと移植される際に、肝細胞、胆管細胞、および類洞細胞への機能的な分化を示した。
【0013】
肝臓疾患としては、世界的にみて顕著な罹患率および死亡率に関連する、広範囲にわたる急性的な状態および慢性的な状態の両方が挙げられる。肝細胞移植は、肝臓疾患の処置において強大な治療的潜在能力を有するが、肝細胞移植の臨床的使用は、ドナー組織の不足によって阻まれている。成人または胎児の肝細胞前駆体からのインビトロでの肝細胞の生成、または成熟肝細胞をインビボで生成し得る前駆体集団の同定は、この問題を解決する。本明細書中に記載されたデータは、ヒト胎児肝臓からの規定された細胞集団の同定を実証する。このヒト胎児肝臓は、エキソビボで数世代の間、首尾よく拡張され、そして急性肝損傷を有する動物へと移植される際に、肝細胞、胆管細胞、および類洞細胞への機能的な分化を示す。肝臓前駆細胞のインビトロで成功した拡張および分化は、肝細胞移植、候補治療薬の代謝試験および毒性試験、ならびに遺伝子治療のビヒクルに有用である。
【0014】
本発明は、インビトロまたはインビボ(すなわち、インサイチュ)で増殖させるためにヒト胎児肝臓組織からの多分化能肝臓前駆細胞を誘導して、肝細胞、胆管細胞、および類洞細胞へと分化することが可能な多分化能ヒト前駆細胞の子孫を大量に生成するための方法を提供する。ヒト肝臓前駆細胞の子孫を分化するための方法もまた、提供される。
【0015】
(ヒト肝臓前駆細胞)
本発明は、ヒト肝臓前駆細胞(本明細書中でLPCと呼ばれる)を提供する。LPC細胞は、本発明の方法を用いて増殖させるために誘導され得る未分化細胞である。LPCは、各細胞分裂において、少なくとも1つの娘細胞がまたLPC細胞であるように、自己保持することが可能である。LPCは、100倍、250倍、500倍、1000倍、2000倍、3000倍、4000倍、5000倍以上、拡張されることが可能である。
【0016】
LPCの表現型決定は、これらの細胞が、どの関係付けられた(committed)肝細胞マーカーも発現しないが、この細胞が幹細胞マーカーを発現することを明らかにする。例えば、LPCは、CD117およびCD34の両方に対して免疫反応性であり、そして系統表面抗原(Lineage surface antigen)(Lin)に対しては非免疫反応性である。LPCは、多分化能前駆細胞である。多分化能前駆細胞とは、この細胞が1つより多くの細胞型へと分化することが可能であることを意味する。例えば、その細胞は、肝細胞、胆管細胞、または類洞細胞へと分化することが可能である。
【0017】
CD117はまた、c−kit(造血幹細胞因子(steel factor)レセプターまたは幹細胞因子レセプター)としても公知である。CD117は、IIIクラスレセプターチロシンキナーゼファミリーに属する、145kDの細胞表面糖タンパク質である。これは、多分化能造血性幹細胞を含む大多数の造血前駆細胞上で発現され、同様に関係付けられた骨髄性前駆細胞、赤血球系前駆細胞、およびリンパ系前駆細胞上でも発現される。CD117はまた、わずかな成熟造血性細胞(例えば、肥満細胞)上で発現される。CD34は、ヒトのリンパ系造血性前駆細胞および骨髄性造血性前駆細胞上で発現された110kDの単鎖膜貫通糖タンパク質である。系統表面抗原とは、異なる14個の細胞表面タンパク質のうちの13個が成熟血液細胞系統のマーカーである場合の混合物である。
【0018】
LPCは、胎児の肝臓組織から得られる。肝臓組織は、肝臓組織を有する任意の動物(例えば、魚、爬虫類、鳥、両生類、および哺乳動物(例えば、好ましくは、マウスのようなげっ歯類およびヒト))から入手され得る。この組織は、少なくとも4週齢、5週齢、6週齢、7週齢、8週齢、9週齢、10週齢以上の胎児から得られる。LPCは、全体の胎児肝臓のうちのおよそ0.5〜0.7%に相当する。
【0019】
LPCは、インビトロの長期培養において維持され得る。LPCは、培養中で2回、3回、4回、5回、6回、7回、8回、9回、10回、11回、12回以上継代されることが可能である。
【0020】
移植の前には、新たに単離されたLPC(CD117+/CD34+/Lin−細胞)は、アルブミンおよびCK19を発現しなかった。しかし、移植後には、これらの細胞は、成熟肝細胞および胆管細胞へと分化された。このことは、ヒトのアルブミン、CK19、G−6−P、GGT、DPPIV、およびグリコーゲンの発現によって決定され、そして、これらの細胞は高い増殖能力を有していた。肝臓前駆細胞は、2つの造血性関連マーカー(c−kitおよびCD34)を発現したが、非造血性細胞から造血性細胞を区別するマーカーであるCD45は、発現しなかった。従って、本明細書中に記載される大規模な増殖能力を有するLPCは、造血性起源ではない。さらに、インビトロで成人造血性幹細胞を拡張する限定された能力とは異なり、胎児肝臓からの肝臓前駆体は、高い増殖能力を有する。単離されたCD117+/CD34+/lin−細胞のすべてが、インビトロでの培養の間に、培養プレートに付着し、そして肝細胞へと分化するわけではなかったという観察は、これらの細胞の部分集団のみが肝細胞の前駆体であることを示す。CD117+/CD34+/lin−細胞は、HGFおよびEGFを含有する培地中で培養した場合、4タイプの細胞(ALB−CK19−肝細胞、ALB+CK19+肝細胞、ALB+CK19−肝細胞、およびALB−CK19+胆管細胞)を生じた。ダブルネガティブ細胞を除いてすべての他の部分集団は、増殖細胞(BrdU+)を高い比率で有した。ALB−CK19−細胞の部分セットは、インビトロで非常に静止状態であったが、インビボでのP0継代において移植された細胞数の10倍の増加は、これらの細胞が、高い増殖能力を有し得ることを示す。肝臓前駆細胞の高い増殖能力にもかかわらず、腫瘍は、ヒトの細胞移植の4週間後にインビボで観察されなかった。興味深いことに、早期胎児肝臓からのCD117+/CD34+/Lin−細胞は、VEGFを含有する倍地中で培養した場合、肝細胞および胆管細胞へと分化しただけでなくまた、類洞内皮細胞へ、分化した。
【0021】
(培養条件)
LPCは、本明細書中に記載された方法を用いて増殖される。細胞は、ドナー組織の連結している細胞外マトリクスから個々の細胞を分離することによって、その組織から入手される。胎児からの組織は、滅菌手順を用いて除去され、そして細胞は、酵素(例えば、トリプシン、コラゲナーゼなど)を用いる処置を含む当該分野で公知の任意の方法を用いてか、または例えば鈍器もしくはホモジナイザーを用いる物理的な分離方法を用いて、分離される。胎児細胞の分離は、組織培養培地中で行われ得る。
【0022】
例えば、細胞の分離は、0.1%トリプシンおよび0.05%DNaseを含むDMEM中で行われ得る。分離された細胞は、200〜2000rpmの間(通常は400〜800rpmの間)の低速で遠心分離され、そして次いで、培養培地中に再懸濁される。肝細胞は、懸濁液中かまたは固定された基質上で培養され得る。分離された細胞懸濁液は、細胞を支持することが可能な任意の容器中(具体的には、培養フラスコ、培養プレート、またはローラー瓶、そしてより具代的には、25cm2培養フラスコのような小さな培養フラスコ中)に播種される。懸濁液中で培養された細胞は、およそ5×104〜2×105(細胞/ml)(例えば、1×105(細胞/ml))で再懸濁される。固定された基質上にプレートされた細胞は、およそ2〜3×103(細胞/cm3)でプレートされる。必要に応じて、培養プレートは、コラーゲンのようなマトリクスタンパク質で被覆される。分離された肝細胞は、細胞増殖を支持することが可能な任意の公知の培養培地(HEM、DMEM、PRMI、F−12などを含む)へと配置され得る。この培地は、細胞代謝に必要な補充物(例えば、グルタミンおよび他のアミノ酸、ビタミン、ミネラル、およびトランスフェリンのようなタンパク質など)を含有する。培養培地はまた、酵母、細菌、および真菌類による汚染を防ぐための抗生物質(例えば、ペニシリン、ストレプトマイシン、ゲンタマイシンなど)を含み得る。培養培地は、ウシ、ウマ、ニワトリなどに由来する血清を含み得る。
【0023】
培養条件は、生理学的条件に近いべきである。培養培地のpHは、生理学的pH(例えば、pH6〜8の間、pH約7〜7.8の間、またはpH7.4)に近いべきである。生理学的温度の範囲は、約30℃〜40℃の間である。LPCは、約32℃〜約38℃の間(例えば、約35℃〜約37℃の間)の温度にて培養される。
【0024】
必要に応じて、培養培地は、少なくとも1つの増殖誘導(「マイトジェン」)成長因子を補充される。「成長因子」とは、LPCに対する成長効果、増殖誘導効果、分化誘導効果、または栄養効果を有する、タンパク質、ペプチド、または他の分子である。「増殖誘導成長因子」は、LPCを増殖させることを可能にする栄養因子であり、細胞の表面上のレセプターと結合して、細胞に栄養効果または成長誘導効果を与える任意の分子を含む。増殖誘導成長因子としては、EGF、アンフィレグリン(amphiregulin)、酸性線維芽細胞成長因子(aFGFまたはFGF−1)、塩基性線維芽細胞成長因子(bFGFまたはFGF−2)、トランスホーミング増殖因子α(TGFα)、VEGFおよびこれらの組み合わせが挙げられる。成長因子は、通常、約1fg/ml〜1mg/mlの間の範囲の濃度で培養培地へと加えられる。約1〜100ng/mlの間の濃度で、通常は十分である。簡単な滴定アッセイは、特定の成長因子の最適濃度を決定するために容易に実施され得る。
【0025】
成長因子および栄養因子の生物学的効果は、一般に、細胞表面のレセプターとの結合を介して仲介される。これら多くの因子に対するレセプターは、同定されてきており、そして特定のレセプターに対する抗体および分子プローブが、利用可能である。LPCは、分化のすべての段階において成長因子レセプターの存在について分析され得る。多くの場合、特定のレセプターの同定は、外因性成長因子または栄養因子の付加を伴う、特定の発生経路に沿ったさらなる細胞の分化における使用のためのストラテジーについての手引きを提供する。
【0026】
一般に、インビトロで約3〜10日後に、培地を吸引し、そして培養フラスコへ新鮮な培地を加えることによって、LPCを増殖する。必要に応じて、吸引された培地は、収集され、フィルタ処理され、そして次に続くLPCの継代のための条件培地として使用される。例えば、10%、20%、30%、40%以上の条件培地が使用される。
【0027】
LPC細胞培養物は、増殖を再開するために用意に継代され得る。例えば、インビトロで約3〜7日後に、培養フラスコは、十分に振盪され、そして次いで、LPCは、50ml遠心管へと移され、低速で遠心される。この培地は吸引され、LPCは、少量の培養培地中に再懸濁される。次いで細胞は、カウントされ、そして増殖を再開するために所望の密度にて再度プレートされる。この手順は、毎週繰り返され、各継代における生存可能な細胞数の対数的な増加をもたらし得る。この手順は、所望の数のLPCが入手されるまで続けられる。
【0028】
LPCおよびLPCの子孫は、それらが必要とされるまで、当該分野における任意の公知の方法によって凍結保存され得る(例えば、米国特許出願第5,071,741号、PCT国際特許出願第WO93/14191号、同WO95/07611号、同WO96/27287号、同WO96/29862号、および同WO98/14058号、Karlssonら、65 Biophysical J.2524−2536(1993)を参照のこと)。LPCは、等張液中、好ましくは、特定の凍結保存剤(cryopreservant)を含有する細胞培養培地中に懸濁され得る。このような凍結保存剤としては、ジメチルスルホキシド(DMSO)、グリセロールなどが挙げられる。これらの凍結保存剤は、5〜5%(例えば、8〜10%)の濃度で使用される。細胞は、−10℃〜−150℃(例えば、−20℃〜−100℃、または−70℃〜−80℃)の温度まで徐々に凍結される。
(ヒト肝臓前駆細胞の分化)
培養条件に依存して、LPCは、肝細胞、胆管細胞、または類洞細胞へと分化され得る。
【0029】
LPCは、HGFおよびEGFを有する培養培地中の固定された基質上でLPCを培養することによって、肝細胞、または胆管細胞へと分化され得る。あるいは、LPCは、VEGFを有する培養培地中の固定された基質上でLPCを培養することによって、類洞細胞へと分化され得る。
【0030】
LPCの分化はまた、成長を引き起こす生物学的事象のカスケードを活性化する当該分野で公知の任意の方法によって誘導され得る。この生物学的事象としては、イノシトール三リン酸塩および細胞内のCa2+の遊離、ジアシルグリセロールの遊離、ならびにプロテインキナーゼCおよび他の細胞キナーゼの活性化などが挙げられる。ホルボールエステル、分化誘導成長因子、および他の化学シグナルによる処置は、分化を誘導し得る。LPCの増殖のための増殖誘導成長因子(上記参照)の代わりに、分化誘導成長因子は、LPCの分化に影響を与えるために培養培地に加えられ得る。成長因子を誘導する他の分化としては、血小板由来成長因子(PDGF)、チロトロピン放出ホルモン(TRH)、トランスホーミング成長因子β(TGF)、インスリン様成長因子(IGF−1)などが挙げられる。
【0031】
分化した肝細胞、胆管細胞、または類洞細胞は、当該分野で公知の免疫細胞化学的技術を用いて検出される。免疫細胞化学(例えば、二重標識免疫蛍光法および免疫ペルオキシダーゼ法)は、肝細胞の細胞特徴または表現型特性を区別するために、細胞タンパク質を検出する抗体を使用する。肝細胞および胆管細胞に対する細胞マーカーとしては、アルブミンおよびCK10が挙げられ、一方で類洞細胞に対する細胞マーカーとしては、Flkが挙げられる。他の適切なマーカーとしては、グルコース6ホスファターゼ、グリコーゲン、ジペピジルペプチダーゼIV、γグルタリルトランスペプチダーゼが挙げられる。
【0032】
免疫細胞化学はまた、例えば、アルブミン、α1−抗トリプシン、CK−19、α胎児タンパク質、またはヒト第VIII因子のような肝臓機能を担う肝臓遺伝子の発現を検出することによって、肝細胞を同定するために使用され得る。
【0033】
インサイチュハイブリダイゼーション組織化学はまた、肝臓遺伝子のmRNAに特異的なcDNAプローブまたはRNAプローブを用いて行われ得る。これらの技術は、特定の表現型の同定を増強するための免疫細胞化学的方法と組み合わされ得る。必要であれば、上で議論された抗体および分子プローブは、ウェスタンブロット手順およびノーザンブロット手順(それぞれが細胞の同定に役立つ)に適用され得る。
【0034】
(ヒト肝臓前駆細胞の移植)
新しい細胞の損傷した肝臓への移植は、損傷した肝臓組織を修復し、それによって肝臓機能を回復する可能性を有する。必要に応じて胎児間質細胞および/またはHGFは、LPCとともに共移植される。しかし、移植目的に適する細胞の欠如は、この手順が満たすべき最大の可能性を妨げている。「適切な」細胞とは、以下の基準:(1)大量に入手され得ること;(2)必要に応じて、遺伝物質の挿入を可能とするためにインビトロで増殖され得ること;(3)無制限に生存することが可能であり、肝臓における移植に対する肝臓修復を促進することが可能であること;そして(4)非免疫原性であり、好ましくは、患者自身の組織からかまたは適合し得るドナーから入手されることを、満たす細胞である。
【0035】
胎児の肝臓組織から入手可能なLPC(これは、本明細書中に記載された培養条件を用いてインビトロで維持される場合に、長期間にわたって分裂することが可能である)は、肝臓移植目的に適する細胞の望ましい必要条件のすべてを満たし、かつ不死化されていない細胞として特に適切な細胞株であり、かつ腫瘍形成の発生源ではない。肝臓障害の処置におけるLPCの使用は、動物モデルの使用によって実証され得る。
【0036】
LPCは、異常な肝臓または肝不全症状を有する任意の動物に投与される。LPCは、宿主に対して異種であるドナー組織から調製され得る。異種移植を成功させるために、移植された組織に対する免疫応答を低減させるか排除する何らかの方法が、通常、利用される。従って、LPCレシピエントは、免疫抑制薬物(例えば、シクロスポリン)の使用を介してか、または局所的に適用された免疫抑制剤を利用する局所的免疫抑制ストラテジーを介して、免疫抑制され得る。局所的免疫抑制は、Gruber、54 Transplantation 1−11(1992)によって開示される。米国特許第5,026,365号は、局所的免疫抑制に適切なカプセル化方法を開示する。
【0037】
免疫抑制技術の利用の代替として、胎児幹細胞における相同組み換えを用いる遺伝子置換法または遺伝子ノックアウト法(Smithiesら、317 Nature 230−234(1985)によって教示され、細胞株における遺伝子置換または遺伝子ノックアウトまで拡張された(Zhengら、88 Proc.Natl.Acad.Sci.8067−8071(1991))が、主要組織適合遺伝子複合体(MHC)遺伝子の切除のために、LPCに適用され得る。MHC発現を欠くLPCは、同種異系を越えて濃縮された肝細胞集団の移植を可能にし、そしておそらくレシピエントを免疫抑制する必要なく異種間の組織適合性の障壁を越えて濃縮された肝細胞集団の移植でさえも可能にする。レシピエントを免疫抑制する必要なく組織適合性の障壁をなくす。ドナー細胞の抗原性を低減させるための組み換え方法の使用についての一般的な総説および引用はまた、Gruber、54 Transplantation 1−11(1992)によって開示される。表面修飾によって移植片の免疫原性を低減する例示的なアプローチは、PCT国際特許出願第WO92/04033号および同第PCT/US99/24630号によって開示される。あるいは、移植片の免疫原性は、MHC抗原を変化させたかまたは欠失させたトランスジェニック動物からLPCを調製することによって低減され得る。
【0038】
LPCは、マイクロカプセル化(例えば、本明細書中で参考として援用される、米国特許出願第4,352,883号;同第4,353,888号;および同第5,084,350号を参照のこと)、およびマクロカプセル化(例えば、それぞれが本明細書中で参考として援用される、米国特許第5,284,761号;同第5,158,881号、同第4,976,859号、および同第4,968,733号、ならびにPCT国際特許出願第WO92/19195号および同第WO95/05452号を参照のこと)を含む、公知のカプセル化技術に従って、因子を宿主へと送達するためにカプセル化され得、かつ使用され得る。マクロカプセル化は、米国特許第5,284,761号;同第5,158,881号;同第4,976,859号;同第4,968,733号;同第5,800,828号およびPCT国際特許出願第WO95/05452号において開示され、それぞれが本明細書中で参考として援用される。デバイス中の細胞数は、変動され得る;好ましくは、各デバイスは、103〜109の間の細胞を含む(例えば、105〜107の細胞)。複数のマクロカプセル化デバイスが、宿主中に移植され得る。
【0039】
レシピエントの組織と同種異系である組織から調製されたLPCは、レシピエントの組織適合性型とほぼ一致させるように、周知の組織型決定方法によって使用のために試験される。
【0040】
LPCは時折、レシピエント自身の肝臓から調製され得る(例えば、腫瘍除去生検の場合)。このような場合、LPCは、分離された組織から生成され得、そして上述の方法を用いてインビトロで増殖され得る。適切な細胞数が増殖された際に、LPCは、収集され得、必要に応じて遺伝的に改変され得、そしてレシピエントの肝臓へと直接注入するために整えられ得る。LPCは、肝臓移植片を形成し得る肝臓領域に投与され、その結果、その細胞は、隣接する肝細胞との通常の結合を形成し、移植されたかまたは既存の肝細胞との接触を維持する。従って、移植されたLPCは、疾患および加齢が原因で損傷している肝臓組織を再構築する。
【0041】
宿主の肝臓組織への移植片の機能的な組み込みは、種々の機能を回復することに関して移植片の有効性を検査することによって評価され得る。この検査としては、アラニントランスアミナーゼ(ALT)、アスパラギン酸トランスアミナーゼ(AST)、アルカリホスフォターゼ(ALP)、アルブミン、総タンパク質、ならびに総ビリルビンおよび直接ビリルビンについての血液検査が挙げられる。
【0042】
移植における使用のためにインビトロでLPCを拡張する能力はまた、エキソビボの遺伝子治療にも有用である。従って、LPCは、エキソビボでの遺伝子治療試行におけるビヒクルとしての使用のために、肝臓細胞を回収および拡張するさらなる方法を提供する。
【0043】
(肝臓前駆細胞の遺伝子改変)
LPCは、形質転換されてない初代細胞であるが、それらは、連続的な細胞株の特徴を保有する。未分化の状態において、LPCは、連続的に分裂し、そしてそれ故に、遺伝子改変の標的となる。いくつかの実施形態において、遺伝子改変された細胞は、上述の任意の方法によって、肝細胞、胆管細胞、または類洞細胞へと分化されるように誘導される。
【0044】
用語「遺伝子改変」とは、外来DNAの意図的な導入によって、LPCの遺伝子型を、安定にかまたは一時的に変更することをいう。DNAは、合成DNAまたは天然に由来するDNAであり得、そして遺伝子、遺伝子部分、または他の有用なDNA配列を含み得る。本明細書中で使用される場合、用語「遺伝子改変」とは、天然に起こる変化(例えば、天然のウィルス活性、天然の遺伝子組み換えなどを介して起こる変化)を含むことを意図しない。
【0045】
細胞の有用な遺伝子改変のいずれもが、本発明の範囲内にある。例えば、LPCは、生物学的に活性な物質(例えば、成長因子など)を生成するか、または生成を増加させるために改変され得る。一実施形態では、生物学的に活性な物質は、転写因子(例えば、遺伝的分化を調節する転写因子)である。代替的な実施形態では、生物学的に活性な物質は、非分裂促進的な増殖因子(例えば、v−myc、SV−40ラージT、またはテロメアーゼ)である。
【0046】
遺伝的改変は、ウィルスベクター(レトロウィルス、改変されたヘルペスウィルス、ヘルペスウィルス、アデノウィルス、アデノ関連ウィルスなど)を用いる感染によってか、または、当該分野において公知の方法を使用するトランスフェクション(リポフェクション、リン酸カルシウムトランスフェクション、DEAE−デキストラン、エレクトロポレーションなど)のいずれかによって行われる(Maniatisら、Molecular Cloning:A Laboratory Manual(Cold Spring Harbor Laboratory,N.Y.,1982)を参照のこと)。例えば、キメラ遺伝子構築物は、ウィルス(例えば、レトロウィルス)の末端反復配列(LTR)、サルウィルス40(SV40)、サイトメガロウイルス(CMV);または哺乳動物細胞特異的プロモーター(例えば、チロシンヒドロキシラーゼ(TH;ドーパミン細胞についてのマーカー))、DBH、フェニルエタノールアミンN−メチルトランスフェラーゼ(PNMT)、ChAT、GFAP、NSE、所望のタンパク質をコードする構造遺伝子の発現を指示するNFタンパク質(NE−L、NF−M、NF−Hなど)を含み得る。加えて、ベクターは、薬物選択マーカー(例えば、E.coliアミノグリコシドホスフォトランスフェラーゼ遺伝子)を含み得、これは、試験遺伝子とともに共感染(co−infected)される際に、タンパク質合成インヒビターであるジェネテシン(G418)に対する耐性を与える。
【0047】
LPCは、発現ベクターによるトランスフェクションを使用して遺伝子的に改変され得る。1つのプロトコルでは、遺伝子を含むベクターDNAを、0.1×TE(1mM Tris(pH8.0)、0.1mM EDTA)中で40μg/mlの濃度まで希釈する。22μlのDNAを、使い捨ての滅菌5mlプラスチックチューブ中の、250μlの2×HBS(280mM NaCl、10mM KCl、1.5mM Na2HPO4、12mMブドウ糖、50mM HEPES)に加える。31μlの2M CaCl2をゆっくりと加え、そして混合物を、室温にて30分間インキュベートする。この30分のインキュベーションの間に、細胞を、800gで5分間、4℃にて遠心分離する。細胞を、20容量の氷冷されたPBS中に再懸濁し、そして1×107の細胞のアリコートに分ける。これらを再度、遠心分離する。細胞のアリコートの各々を、1mlのDNA−CaCl2懸濁液中に再懸濁し、そして室温にて20分間インキュベートする。次いで、細胞を成長培地中で希釈し、そして5%〜7%のCO2で37℃にて6〜24時間、インキュベートする。この細胞を再度、遠心分離し、PBS中で洗浄し、そして10mlの成長培地へ48時間の間、戻す。
【0048】
LPCはまた、リン酸カルシウムトランスフェクション技術を用いて遺伝子的に改変される。標準的なリン酸カルシウムトランスフェクションでは、細胞を、単一の細胞の懸濁液へと機械的に分離し、そして、50%のコンフルーエンス(50,000〜75,000(細胞/cm2))で組織培養用に処理されたシャーレ上にプレートし、一晩付着させる。1つのプロトコルでは、改変されたリン酸カルシウムトランスフェクション手順を、以下のように行う:TEで440μlまで希釈した滅菌TE緩衝液(10mM Tris、0.25mM EDTA、pH7.5)中のDNA(15〜25μg)、および60μlの2M CaCl2(1M HEPES緩衝液でpH5.8まで)を、DNA/TE緩衝液へと加える。合計500μlの2×HeBS(HEPESで緩衝化した生理食塩水;275mM NaCl、10mM KCl、1.4mM Na2HPO4、12mMブドウ糖、40mM HEPES緩衝粉末、pH6.92)をこの混合物へ一滴ずつ加える。この混合物を室温にて20分間、静置する。細胞を1×HeBSで簡単に洗浄し、そして1mlのリン酸カルシウムで沈殿したDNAの溶液を、各プレートへと加え、そしてその細胞を37℃で20分間インキュベートする。このインキュベーションの後に、10mlの培地を、細胞に加え、そしてプレートを、さらに3〜6時間、インキュベーター(37℃、9.5%CO2)中に配置する。DNAおよび培地を、インキュベーション期間の最後に、吸引によって除去し、細胞を3回洗浄し、そしてインキュベーターに戻す。
【0049】
遺伝子改変が生物学的に活性な物質の生成のためである場合、その物質は、所与の肝臓障害の処置に有用な物質であり得る。LPCは、生物学的に活性な因子(例えば、成長因子、成長因子レセプター)を発現するために遺伝子的に改変される。例えば、細胞が増殖誘導成長因子または分化誘導成長因子を分泌するように、細胞を遺伝子的に改変することが望ましくあり得る。肝臓障害の処置において有用な成長因子産物としては、HGF、VEGF、FGF−1、FGF−2、EGF、TGFα、TGF、PDGF、IGF、およびインターロイキンが挙げられる。
【0050】
遺伝子改変されたLPCは、細胞治療または遺伝子治療のために、遺伝子的に改変された細胞によって生成された生物学的に活性な分子を必要としているレシピエントのCNSへと、移植され得る。移植技術は、以下に記述される。
【0051】
あるいは、遺伝子的に改変されたLPCは、移植の前に、インビトロでの種々の分化プロトコルに供され得る。一旦、細胞が分化されると、細胞は、所望のタンパク質の発現のために再度アッセイされる。所望の表現型を有する細胞は、単離され得、そして、遺伝子的に改変された細胞によって発現された、タンパク質または生物学的に活性な分子を必要としているレシピエントへと移植され得る。
【0052】
(肝臓前駆細胞への薬物の効果をスクリーニングするための方法)
LPC培養物は、潜在的な治療組成物のスクリーニングのために使用され得る。例えば、LPCは、肝臓細胞の増殖、分化、または生存をもたらす化合物を同定するために使用される。加えて、LPCは、抗ウィルス化合物を同定しウィルスの感染性を決定するためか、または、試験化合物の代謝産物を同定するために、使用される。これらの試験組成物は、種々の投与量で培養物中の細胞に適用され得る。そして細胞の応答が、種々の期間、モニタリングされる。細胞の物理的特徴は、顕微鏡を用いて細胞の成長および形態を観察することによって分析され得る。タンパク質(例えば、酵素、レセプター、および他の細胞表面分子)または神経伝達物質、アミノ酸、神経ペプチド、および生体アミンの新たな発現レベルまたは増加した発現レベルの誘導は、このような分子のレベルの変化を同定し得る当該分野で公知の任意の技術を用いて、分析され得る。このような技術としては、このような分子に対する抗体を使用する免疫組織化学、または生化学的分析が挙げられる。このような生化学的分析としては、タンパク質アッセイ、酵素的アッセイ、レセプター結合アッセイ、酵素結合イムノソルベントアッセイ(ELISA)、電気泳動分析、高性能液体クロマトグラフィ(HPLC)による分析、ウェスタンブロット、および放射免疫アッセイ(RIA)が挙げられる。核酸分析(例えば、ノーザンブロット)は、これらの分子またはこれらの分子を合成する酵素をコードするmRNAのレベルを検査するために使用され得る。
【0053】
LPCは、肝細胞への生物学的因子の効果を決定する方法において使用され得る。用語「生物学的因子」とは、任意の因子(例えば、ウィルス、タンパク質、ペプチド、アミノ酸、脂質、炭水化物、核酸、ヌクレオチド、薬物、プロドラッグ、または、神経細胞への効果(このような効果が有害であっても、有益であっても、またはその他でも)を有し得る他の因子)をいう。肝細胞に有益である生物学的因子は、本明細書中で「肝臓因子」と呼ばれる。この用語は、肝細胞の増殖、分化もしくは機能、または肝臓疾患もしくは肝臓障害の処置について潜在的に有用であると証明され得る、生物学的または薬学的に活性な任意の物質を含む用語である。
【0054】
LPCの培養物からの肝細胞への潜在的な生物学的因子の効果の決定は、肝臓組織からかまたは、代わりに、肝臓疾患もしくは肝臓障害に罹患している宿主から得られ得る。培養条件の選択は、試験される特定の因子、そして実験者が達成することを望む効果に依存する。一旦、細胞が所望のドナー組織から入手されると、その細胞は、インビトロで増殖される。
【0055】
移植目的のために大量の細胞を生成するのに有用である、LPCの増殖能力を増加する生物学的因子についてスクリーニングすることが可能である。LPCの増殖を阻害する生物学的因子についてスクリーニングすることもまた可能である。LPCは、対象となる生物学的因子の存在下でプレートされ、そして増殖が起こる程度についてアッセイされる。LPCおよびLPCの子孫の分化および生存への、生物学的因子または生物学的因子の組み合わせの効果が、決定され得る。
【0056】
スクリーニングの前に、分化するよう既に誘導されているLPCをスクリーニングすることが可能である。分化の前に、生物学的因子をLPCに適用することによって、分化プロセスへのその生物学的因子の効果を決定することもまた可能である。一般に、生物学的因子は、可溶化され得、そして種々の濃度にて培養培地に添加されて、各用量におけるこの因子の効果を決定し得る。培養培地は、因子の濃度をある程度一定に保つための量で、数日毎に生物学的因子を補充され得る。
【0057】
増殖における変化は、細胞数における増加または減少によって観察される。「制御因子」は、LPCの増殖への制御効果を有する生物学的因子である。例えば、生物学的因子は、増殖誘導成長因子(例えば、EGF)に応答してインビトロで増殖するLPCの数を増加させるかまたは減少させる場合、「制御因子」とみなされる。あるいは、増殖誘導成長因子に応答するLPCの数は同じままであり得るが、制御因子の追加が、LPCの増殖速度に影響を与える。増殖誘導成長因子は、別の増殖誘導成長因子と組合わせて使用される場合に制御因子として作用し得る。
【0058】
このようなスクリーニング方法を使用すると、当業者は、肝細胞の増殖および分化、または分化した肝細胞の生存または機能への生物学的因子の効果について試験することによって、肝細胞への薬物の潜在的な副作用についてスクリーニングし得る。増殖されたLPCは、代表的には、約5〜10×106細胞/mlの密度でプレートされる。分化した特定の細胞型または所与の細胞の組み合わせへの生物学的因子の効果を試験することが望ましい場合、分化の後に得られた肝細胞と胆管細胞との比は、異なる型の細胞と隔てることによって処理され得る。
【0059】
生物学的因子の効果は、コントロール培養物に対する有意差に基づいて同定される。これは、発現された表現型の比、細胞の生存度、および遺伝子発現における変化のような基準に関しての差である。細胞の物理的な特徴は、顕微鏡で細胞の形態および成長を観察することによって分析され得る。タンパク質(例えば、酵素、レセプター、および他の細胞表面分子)の新たな発現レベルまたは増加した発現レベルの誘導は、このような分子のレベルの変化を同定し得る当該分野で公知の任意の技術を用いて、分析され得る。このような技術としては、このような分子に対する抗体を使用する免疫組織化学、または生化学的分析が挙げられる。このような生化学的分析としては、タンパク質アッセイ、酵素的アッセイ、レセプター結合アッセイ、酵素結合イムノソルベントアッセイ(ELISA)、電気泳動分析、高性能液体クロマトグラフィ(HPLC)による分析、ウェスタンブロット、および放射免疫アッセイ(RIA)が挙げられる。核酸分析(例えば、ノーザンブロットおよびPCR)は、これらの分子またはこれらの分子を合成する酵素をコードするmRNAのレベルを検査するために使用され得る。
【0060】
LPCの増殖、およびLPCの子孫の増殖、分化および生存に関与する因子、ならびに生物学的因子に対するそれらの応答は、当該分野で公知の技術を用いて、LPCの発生の異なる段階におけるLPCまたはLPCの子孫から、cDNAライブラリーを構築することによって単離され得る。ある発生段階における細胞からのライブラリーは、発生の異なる段階における細胞のライブラリーと比較されて、発生の間の遺伝子発現の順序を決定し、そして種々の生物学的因子の効果を明らかにするか、または肝臓細胞における遺伝子発現を変化させる新たな生物学的因子を明らかにする。ライブラリーが機能不全組織から調製される場合、機能不全組織からのライブラリーと正常組織からのライブラリーとを比較することによって、原因として機能不全を引き起こすために役割を果たす遺伝因子が同定され得る。この情報は、障害を処置するために治療設計において使用され得る。加えて、プローブが、種々の遺伝障害の診断における使用のためか、または発生における特定の段階で肝細胞を同定することに使用するために同定され得る。
【0061】
本発明は、以下の実施例によってさらに示されるが、それらに限定はされない。
【実施例】
【0062】
(実施例1:一般的な方法)
(ヒト胎児肝細胞の単離)
本研究の許可は、Huddinge大学病院にある現地の倫理委員会から認可された。ヒトFL組織を、スウェーデンの手引きに従って、妊娠の6〜9.5週間目に中絶された胎児から入手した。研究プロトコルは、現地の倫理委員会によって認可された。改変された吸引掻爬術を行った(33)。在胎齢を、妊娠期間の12週間未満の胎児においては特有の解剖学的マーカー(34)に従って推定し、そしてそれより後の胎児においては、超音波大横径測定によって推定した(35)。妊娠齢は、最終月経から算出した妊娠期間として与えられる。中絶を、明らかな異常がない妊娠において行った(つまり、異常がある胎児を含まなかった)。FLを、解体し、そしてRPMI 1640培地(Gibco,Invitrogen Corp.UK)を含む滅菌チューブ中に配置した。次いで、肝臓を、70μmの金属メッシュを通すことによって、単一の細胞の懸濁液へと分解した。単一の細胞の懸濁液を、200gで10分間遠心して、細胞をペレットにした。胎児組織を提供するすべての女性は、梅毒、トキソプラスマ症、風疹、HIV−1、サイトメガロウイルス、B型肝炎、およびC型肝炎、パロウィルス、ならびに1型単純ヘルペスおよび2型単純ヘルペスについて血清学的にスクリーニングされている。
【0063】
(磁気細胞分類による細胞の単離およびインビトロでの培養)
単一の細胞の懸濁液を、妊娠7〜9週目における胎児肝細胞から調製した。細胞をヒト原始前駆細胞濃縮単離キット(Stem cell technologies,Vancouver,Canada)を用いて、その後に、磁気活性化細胞分離磁気ビーズシステム(Stem cell technologies)によって単離した。この方法は、この集団のネガティブ選択に基づいている。この選択には、12個の系統特異的な細胞表面抗原に対する抗体(抗CD2、抗CD3、抗CD14、抗CD16、抗CD19、抗CD24、抗CD36、抗CD38、抗CD45RA、抗CD56、抗CD66b、抗グリコホリンA)を含む枯渇カクテル(depletion cocktail)を使用する。この手順を、販売元によって説明されているように実行した。すべての場合において、回収した前駆細胞を、直ちにフローサイトメトリーによって分析し、lin+細胞の汚染が存在しないことを確かめ、かつ細胞の前駆体の表現型(CD117+/CD34+/lin−)を確認した。手順の最後に、回収された前駆細胞を生存度について試験した。この後で、この細胞を、I型コラーゲンで被覆されたプラスチックのペトリ皿(Biocoat,Becton and Dickinson,New Jersey,USA)に播種した。そして、10%不活化胎児子ウシ血清、ペニシリンおよびストレプトマイシン、5%L−グルタミン、5%最小必須アミノ酸、50ng/mlのHGF(R&D Systems,Abingdon,England)、20ng/mlのEGF(R&D Systems)、および10ng/mlの塩基性繊維芽細胞成長因子(R&D Systems)を含有するダルベッコ改変イーグル培地(DMEM)(GIBCO,Invitrogen,Stockholm,Sweden)中で培養した。三日ごとに毎回、培地を回収し、遠心分離し、滅菌ろ過処理し、そして条件培地(CM)として使用した。その後の全ての継代培養を、20%CMを使用して行った。いくつかの実験において、前駆細胞を、20ng/mlのHGF、10ng/mlのEGF、および50ng/mlのVEGF(R&D Systems)を含有するDMEM培地中で成長させ、そして培養物中で分裂させた。増殖を検出するために、培養物中の細胞を、チミジンアナログBrdU(30mM)とともに30分間、インキュベートした。細胞を洗浄し、そして、ヒトのアルブミンに対するFITC結合ヤギ抗体(Natutec,Frankfurt,Germany)、非結合抗ヒトサイトケラチン19(Neomarker,USA)、および非結合抗BrdU抗体(Sigma,Stockholm,Sweden)を使用して、アルブミン、サイトケラチン19、およびBrdU(30mM)で染色した。表現型決定のために使用された他の抗体は、抗CD45、抗CD14、抗CD90、抗CD117、抗CD34(Pharmingen,USA)、抗Flk−1(ReliaTech,Germany)、ならびに二次サブクラス特異的抗体であるヤギ抗マウスIgG1(FITC/Texas red)およびヤギ抗マウスIgG2a(FITC/Texas red)であった。フローサイトメトリーおよび免疫細胞化学を、前駆細胞を表現型決定するために使用した。この手順を記載されているように行った(36)。
【0064】
新たに単離された前駆細胞(P0)、および第6継代(P6)および第12継代(P12)においてインビトロで拡張された細胞を、移植研究のために使用した。
【0065】
(マウス)
動物を世話し、Huddinge病院にあるThe animal care and use committeが認可した動物プロトコルを使用する。肝障害を、部分的な肝切除の24時間前に、体重1kgあたり0.7gの腹腔内への、GalN(Sigma Chemicals Co.,Stockholm,Sweden)の投与によって、C57ブラック/ヌードマウス(n=16)に誘導した。GalNを、pH7.4のリン酸緩衝生理食塩水(PBS)中に、100mg/mlにて溶解した。部分的な肝切除(PH)を、以前に説明されたように行った(37)。GalNの投与を、PHの後、10日間続けた。
【0066】
肝臓前駆細胞を、これらの動物の脾臓へと移植した。動物を、エーテル下で麻酔し、そして代表的には、200μlのDMEM培地中に懸濁された1×105の新たに単離された細胞(P0)、および第6と第12継代における1×106の細胞を、およそ10〜15秒にわたって脾臓へと注入した。4匹のマウスを、DMEM培地のみで偽移植した。止血を確実にした後、腹腔切開を閉じ、そして動物を回復に近づくまでモニタリングした。
【0067】
(肝臓の調製および凍結切片における蛍光分析)
マウスを、移植の4週間後に殺し、そして肝臓、脾臓および肺を切除した。約2mm2の肝臓の各々から2つか3つの生検を、液体窒素中でショック凍結し、そしてRT−PCR分析を行うためのRNAの単離のために使用した。残りの肝臓組織を、蛍光分析および免疫組織化学的分析のためにショック凍結した。5μmの厚さの凍結切片を風乾し、そして、メタノール中で10分間、冷30%アセトンで固定し、そして免疫組織化学によってさらに分析した。
【0068】
(免疫組織化学)
本発明者らはまず、マウスの実質中のヒト細胞の検出のために2つの方法を試験した;a)ジゴキシゲニン標識総ヒトDNAプローブ(Cytocell,Oxfordshire UK)を使用するインサイチュハブリダイゼーション技術(38)、およびb)マウス抗ヒト核モノクローナル抗体(Chemicon,CA,USA)の後に、ビオチン化ウマ抗マウス二次抗体で染色する。免疫ペルオキシダーゼ手順を、製造元によって説明されているように、Vectastain Elite ABCキット(ImmunKemi,Stockholm,Sweden)を使用して行った。ジアミノベンジジン三塩酸塩(DAB)−ニッケル基質キットを、色展開剤として使用した。二重染色について、DAB(茶色に染色する)および/またはDAB−Ni(黒)および/またはVector NovaRedキットの組み合わせを使用した。使用した他の一次抗体は、ヒトのアルブミンに対する(マウスとは交差反応しない)FITC結合ヤギ抗体(Natutec,Frankfurt,Germany)、非結合抗ヒトサイトケラチン19(Neomarker,USA)、および(ジペプチジルペプチダーゼIVを検出する)非結合マウス抗ヒトCD26(Pharmingen,USA)であった。GGT、G−6−Pおよびグリコーゲンを、以前に説明されたようにインサイチュで示した(39,40)。切片をヘマトキシリンで対比染色し、マウント培地(ImmunKemi,Stockholm,Sweden)中でマウントした。
【0069】
(形態計測分析)
各マウスの肝臓の60枚の連続切片を、DAB−Ni−ポジティブなヒト細胞についてスクリーニングした。移植された細胞の数を、3つのサイズのクラスター(すなわち、単独で配置する細胞、または2以上〜20の細胞のクラスターまたは20より多い細胞のクラスター)で決定した。本発明者らは、移植された動物の全て由来の組織において最低限100の高倍率視野で分析した。
【0070】
(逆転写ポリメラーゼ連鎖反応(RT−PCR))
全RNAを、4匹のヒト−マウスキメラマウスの肝臓組織から、および1匹の正常マウスの肝臓組織から、Micro−FastTrack RNA単離キット(Invitrogen,Groningen,The Netherlands)を使用して、抽出した。本発明者らは、マウスの肝臓における、ヒトのアルブミン、CK19、α−フェトプロテインおよびα1−抗トリプシンの発現を検出するために、ヒト特異的プライマーを使用した。プライマーを、Primer Expressソフトウェア バージョン2.0(Applied Biosystems)を使用することにより、CK−19、α−フェトプロテイン、アルブミン、抗トリプシン、およびグルコース6リン酸デヒドロゲナーゼ(G6PD)について選択した。プライマーセットの各々を、DNAで汚染されずに、cDNAのみを標的とするために設計した。プライマーセットは、CyberGene(Huddinge,Sweden)によって商業的に合成された。
【0071】
【化1】
PCR反応を、96ウェル光学プレートに総容量25μlで、二つ組で行った。各反応物は、2.5μLのcDNA、12.5μLのSYBR Green Master Mix(Applied Biosystems)、および500nMの各プライマーを含んだ。ポジティブコントロールおよびネガティブコントロールを、全ての実行において含んだ。熱サイクル条件は、製造元によって推奨されているように、最初に50℃で2分間、そして95℃で10分間であった。サイクル条件は、95℃で15秒間、そして60℃で1分間の40サイクルであった。ハウスキーピング遺伝子であるG6PDを、内部標準コントロールとして含んだ。これは、RNAの単離および逆転写の成功、ならびに全てのサンプルにおけるRNAの総量を確認するために使用した。RT−PCRからの結果を可視化するために、本発明者らは、使用できる状態の12.5%の非変性ポリアクリルアミドゲル電気泳動システム上でPCR産物を泳動し、そしてそのバンドを、自動化された銀染色(Pharmacia Biotech,Uppsala,Sweden)によって染色した。
【0072】
(統計方法)
データを、平均値±SDのように示す。差異の有意性を、スチューデントt検定および分散分析(ANOVA)を用いて分析した。0.05未満のp値は、有意であるとみなした。
【0073】
(実施例2 CD117+/CD34+/LIN−肝臓前駆細胞は、インビトロで肝細胞および胆管細胞へと分化し得る)
原始造血前駆体を単離するために設計されたキットを使用して、ヒト胎児肝臓由来の細胞集団(gw6〜9)を、任意の関係付けられた造血マーカーを発現しなかった集団から得た。この集団のさらなる表現型決定は、幹細胞マーカーCD117およびCD34の発現を示したが、肝臓マーカー(例えば、アルブミン(肝臓マーカー)およびCK19(胆管細胞マーカー))の発現は示さなかった。Thy−1(CD90)もCD45もまた発現しなかった(図1a)。この集団は、妊娠6〜9週目における全体の胎児肝臓の約0.5%〜0.7%を表す。CD45およびCD90の発現は、細胞の継代培養の間には観察されなかった。培養の際に、これらの細胞が、付着性の集団(約85%)(図1b)と非付着性集団(約15%)の両方の混合物であることを見出した。非付着性集団は、以下で与えられた培養条件の下で、さらに拡張され得なかった。CD117+/CD34+/Lin−細胞およびそれらの子孫は、コロニー中で成長する傾向がある(図1b)。培養物中で2日後に付着前駆細胞によって発現される最初のマーカーは、c−Met(肝細胞成長因子レセプター)であった(図1b)。肝細胞成長因子(HGF)および上皮成長因子(EGF)を含有する培養培地中で2週間、これらの細胞をインビトロで培養することは、4つタイプの細胞の存在を示した:i)アルブミンおよびCK19を発現する約85%のダブルポジティブ細胞、ii)約4%のダブルネガティブ細胞、iii)アルブミンのみを発現する約6%のシングルポジティブ細胞、iv)約5%のCK19のみを発現するシングルポジティブ細胞(図1c)。この表現型は、培養の間、数継代にわたって維持された(図1c)。しかし、P11以降から、ダブルポジティブ細胞(約60%)およびタブルネガティブ(約3%)の数においてわずかな減少があった。一方で、アルブミンまたはCK19についてのシングルポジティブ細胞は増加した。これらのデータは、発生早期の肝臓由来の非造血性原始前駆細胞がインビトロで肝細胞および胆管細胞へと分化し得ることを実証する。
(実施例3 CD117+/CD34+/LIN−肝臓前駆細胞は、血管内皮成長因子の存在下で肝臓類洞内皮細胞へと分化する)
興味深いことに、付着性CD117+/CD34+/Lin−細胞を、50ng/mlの血管内皮成長因子(VEGF)を含有する培養培地中で分化させた場合、本発明者らは、内皮様の形態を有する細胞の大きな集団を観察した。胎児肝臓内皮前駆体に発現されることが公知であるFlk−1を含む肝細胞マーカーを使用する、この細胞集団のさらなる特徴付けは、4つの集団を明らかにした:a)レセプターFlk−1を発現する内皮細胞(約50%)、b)肝細胞(約13%)、c)胆管細胞(約17%)(図1d)、および、d)これらのマーカーのうちのいずれも発現しなかった細胞集団(約20%)(データは示さず)。Flk−1+細胞の類洞表現型を、抗体の大きなパネルを使用して確認した(表1)。ヒト臍静脈内皮細胞を、血管内皮細胞と類洞内皮細胞との間の表現型の差を実証するために使用した(表1)。電子顕微鏡分析は、類洞内皮細胞に特徴的な窓(fenestrne)の存在、およびの基底膜の欠如を明らかにした(データは示さず)。これらのデータは、発生早期の肝臓由来の非造血性原始前駆細胞が、肝細胞、胆管細胞、および内皮細胞へとインビトロで分化し得ることを示す。
【0074】
【表1】
(実施例4 肝臓前駆細胞は、インビトロで広範に継代され得る)
肝臓前駆体の増殖は、培養の間、20%の条件培地(CM)の恒常的な使用に依存した。20%のCMを使用すると、これらの細胞を、少なくとも12継代まで継代し得る。CMを除去すると、得られる細胞継代の数が有意に減少した(p<0.001;図2a)。このことは、細胞の増殖における減少を反映する。高い増殖能力が、BrdUの取り込みによって検出されたように、ダブルポジティブ細胞(ALB+CK19+)およびアルブミンのシングルポジティブ(ALB+CK19−)において観察されたことが見出された(図2bおよび図2c)。この細胞は、安定な表現型を長期間の間、維持され得る。すべてのFLが、急速に増殖し、そして数継代の間、培養物中で維持され得る細胞を生成したわけでないことを言及することは重要である(表2)。長期間、増殖の成功は、得られたFL組織の質に依存した。しかし、試験されたすべてのFLから単離されたCD117+/CD34+/LIN−細胞が肝細胞へと分化したことを見出した。インビトロ研究からのデータは、CD117+/CD34+/LIN−細胞およびそれらの子孫が、広範な複製能力を有し、そして安定な表現型で長期間の間維持され得ることを実証した。
【0075】
【表2】
(実施例5:拡張されたヒト胎児前駆細胞は、GalN処置マウスの肝臓中で成熟肝細胞、成熟胆管細胞、および成熟類洞へと首尾よく移植され、分化する)
新たに単離されたCD117+/CD34+/Lin−細胞(P0)、および培養物中で拡張された細胞(P6およびP12)が、分化する可能性を有しインビボで機能するかどうかを試験するために、これらの細胞を、最初に部分的に肝切除されたマウスへと移植し、次いでDガラアクトサミン(GalN)で処置して急性肝損傷を誘導した。このプロトコルは、急性肝損傷を誘導し、肝臓再分化を促進させる(21)。コントロールの2匹マウスおよび試験グループの2匹は、処置の24時間後に死亡した。細胞移植の4週間後にマウスが生存していた結果が示される。脾臓内の移植の後、初代P0細胞、ならびにP6およびP12継代培養細胞は、GalN処置マウスの肝臓において生存した。移植された細胞を位置決めするための、ヒトのセントロメアプローブおよび抗ヒト核抗体を使用して得られた結果を比較する実験は、同様の結果を示した。ヒトDNAプローブの種特異性(図3aおよび図3b)および抗ヒト核抗体は、ヒト肝臓によるポジティブな結果(図3c)、およびヌードマウスからの偽移植された肝臓によるネガティブな免疫組織化学(図3d)によって実証される。新たに単離されたCD117+/CD34+/Lin−細胞は、移植した場合、移植の4週間後にて、肝細胞(図3e)、胆管細胞(図3f)、および形成された胆管(図3g)へと分化した。P6およびP12における肝臓前駆細胞は、移植した場合、移植され、そして急性的に損傷した肝臓を再構成した(図3h−j)。移植された細胞を、マウスの肝臓中で見出したが(図3k)、例えば、脾臓(図3l)および肺(図3m)のような他の組織において、そして偽移植された動物(図3n)においては見出されなかった。移植されたヒト細胞は、肝臓マーカー(例えば、グルコース6リン酸(G−6−P)(図4a)およびグリコーゲン(図4b))、ならびに胆管マーカー(例えば、ジペプチジルペプチダーゼIV(DPPIV)(図4c)およびγグルタミルトランスペプチダーゼ(GGT)(図4d))を発現した。3匹のマウスでは、ヒト肝細胞によって90%再増殖した、再分化する組織の切片が見出された(図4e)。ヒト前駆体によって完全に再増殖したヒト胆管の透明な領域もまた、観察された。これらの細胞は、CK19およびそして抗ヒト核抗体に対してポジティブに染色した(図4f)。マウスのアルブミンとは交差反応しないヒトのアルブミンに対するモノクローナル抗体を、移植された細胞におけるヒトのアルブミンの発現を検査するために使用した。ヌードマウスからの偽移植された肝臓によるネガティブな免疫組織化学(図4g)は、抗体の種特異性を示した。移植されたマウス10匹すべてにおいて、いくつかのアルブミンポジティブ構造(図4h−j)が見出されたが、偽移植されたマウス(n=2)においてヒトのアルブミン発現する病巣は、同一の条件を使用しても観察されなかった。ヒトのアルブミンは、マウスの肝臓中のみで検出され、しかし脾臓または肺においては観察されなかった。興味深く注目するものとしては、分析されたマウスのいずれにおいても、腫瘍の形成が観察されなかったことである。
【0076】
肝臓の移植が、P0、p6、およびp12におけるヒト肝臓前駆体の間で匹敵する状態であったかどうかを決定するために、マウス肝臓の各々の60枚の連続切片を、DAB−Ni−ポジティブなヒト細胞についてスクリーニングした。移植された細胞の数を、3つのサイズのクラスター(すなわち、単独で配置する細胞、または5以上〜20の細胞のクラスター、または20より多い細胞のクラスター)で決定した。この分析は、P0、P6およびP12細胞を受容する動物の間で、単独で配置する移植された細胞の数において、または2〜20以上の細胞のクラスターにおいて、有意差が存在しなかったことを示した。20より多い細胞のクラスターは、P0細胞を移植されたマウスにおいては何も観察されなかったが、P6およびP12における細胞を受容したマウスにおいては観察された(表3)(p<0.001;ANOVA)。P0細胞を移植されたマウスにおいて、本発明者らは、ヒトの移植された細胞の数における10倍の増加を検出し、一方で、P6細胞およびP12細胞を注入されたマウスにおいては、6倍の増加を検出した(表3)。これらのデータは、新たに単離されたCD117+/CD34+/LIN−細胞が、首尾よく増殖し、そしてインビボで成熟肝細胞、成熟胆管細胞、および成熟類洞内皮細胞へと分化することを実証する。加えて、インビトロで拡張されたCD117+/CD34+/Lin−細胞およびそれらの子孫は、インビボで増殖し、そして、損傷したマウス肝臓を首尾よく再構築する。
【0077】
【表3】
(実施例6:ヒト肝臓前駆細胞を移植したマウスにおけるヒト肝臓特異的遺伝子の転写)
植え付けられた細胞の移植は、移植されたマウスにおけるヒト遺伝子の発現を決定することによって確認した。屠殺したマウス肝臓を、ヒト肝臓特異的遺伝子(アルブミン、α1−抗トリプシン、CK19、およびAFPを含む)について特異的なプライマーを使用するRT−PCRによって、ヒト胎児肝臓前駆細胞の移植の1ヵ月後に、分析した。偽移植されたヌードマウスの肝臓からのRNAは、RNAの完全性のためのコントロールとして使用されたG6PDについて増幅を結果としてもたらした。CK19、アルブミン、およびAFPプライマーは、プライマーが個々のマウス遺伝子を増幅しなかったので、ヒトに特異的な種であった。しかし、α1−抗トリプシンが種特異的であることは見出されなかった(図4k)。これらの結果は、新たに単離された(P0)細胞、および種々の継代(P6およびP12)における細胞の両方が、損傷したマウス肝臓に移植することを示す。
【0078】
(実施例7:肝臓前駆細胞を移植されたマウスにおけるヒト第VIII因子の保護)
ヒト第VIII因子のレベルは、化学的に肝臓を損傷したマウスおよび肝切除したマウスにおいては極僅かであり、同様に、同じ様式で処置された、そしてさらに成長因子HGFおよび/または間質細胞を受容したマウスにおいても、同様に極僅かである。しかし、ヒト肝臓前駆細胞のみ、またはHGF+間質細胞+前駆細胞の組み合わせを受容する化学的に肝臓を損傷したマウスおよび肝切除したマウスは、それらの血漿においてヒト第VIII因子の高いレベルを実証した(図6)。
【0079】
(他の実施形態)
特定の実施形態が本明細書中で詳細に開示されているが、これは、例示目的のために例として開示されており、前に添付された特許請求の範囲に関して限定することは意図されていない。具体的には、種々の置換、変更、および修正が、請求項に規定されるような本発明の精神および範囲から逸脱することなく、本発明に対して行われ得ることが、本発明者らによって企図される。核酸の出発物質、所定のクローン、またはライブラリーのタイプの選択が、本明細書中に記載された実施形態の知識を有する当業者にとって、日常的なことであると考えられる。他の局面、利益、および修正は、上述の特許請求の範囲内にあるとみなした。
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1A】

【図1B】

【図1C】
【図1D】
【図2A】
【図2B】

【図2C】
【図3】

【図4A−J】

【図4K】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B−F】
【図10】
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図3】
【図4A−J】
【図4K】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B−F】
【図10】
【公開番号】特開2011−103884(P2011−103884A)
【公開日】平成23年6月2日(2011.6.2)
【国際特許分類】
【出願番号】特願2010−261849(P2010−261849)
【出願日】平成22年11月24日(2010.11.24)
【分割の表示】特願2006−548478(P2006−548478)の分割
【原出願日】平成17年1月14日(2005.1.14)
【出願人】(506241112)ノーバヘップ アーベー (2)
【Fターム(参考)】
【公開日】平成23年6月2日(2011.6.2)
【国際特許分類】
【出願日】平成22年11月24日(2010.11.24)
【分割の表示】特願2006−548478(P2006−548478)の分割
【原出願日】平成17年1月14日(2005.1.14)
【出願人】(506241112)ノーバヘップ アーベー (2)
【Fターム(参考)】
[ Back to top ]