
ヒトインスリン受容体を介する医薬品の送達
【課題】ヒトインスリン受容体(HIR)に結合するヒト化マウス抗体が提供される。
【解決手段】該ヒト化マウス抗体は、HIRを発現するヒト器官および組織へ医薬品を送達するためのトロイの木馬として使用するために適切である。該ヒト化抗体は、神経医薬品を血流から血液脳関門(BBB)を越えて脳へ輸送するために特に良好に適している。該ヒト化マウス抗体は、遺伝子組換えにより該医薬品に融合させることができ、またはアビジン−ビオチン結合系を使用して該医薬品に結合させることができる。
【解決手段】該ヒト化マウス抗体は、HIRを発現するヒト器官および組織へ医薬品を送達するためのトロイの木馬として使用するために適切である。該ヒト化抗体は、神経医薬品を血流から血液脳関門(BBB)を越えて脳へ輸送するために特に良好に適している。該ヒト化マウス抗体は、遺伝子組換えにより該医薬品に融合させることができ、またはアビジン−ビオチン結合系を使用して該医薬品に結合させることができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般に血流からヒト脳およびその他のヒトインスリン受容体を発現する器官もしくは組織への医薬品(pharmaceutical agents)の送達に関する。より詳細には、本発明は、ヒトインスリン受容体(HIR)に容易に結合できる化合物を形成するために医薬品に付着させることのできる「ヒト化」モノクローナル抗体(MAb)の開発に関する。この化合物は、脳毛細血管内皮上に位置するインスリン受容体によってヒト血液脳関門(BBB)を越えることができる。BBBを越えると、ヒト化モノクローナル抗体/医薬品化合物は、さらにまた脳細胞上に位置するインスリン受容体を介して脳細胞内への受容体媒介性エンドサイトーシスを受けることができる。
【背景技術】
【0002】
本明細書で言及した出版物およびその他の参考文献は、本発明の背景およびその実践に関する追加される詳細を提供するために、参考文献として組み込まれる。便宜上、参考文書は添付の文献目録に著者名および発行日によって分類されている。
【0003】
BBBは、循環中の薬物、タンパク質治療薬、アンチセンス薬物、および遺伝子薬品の脳内取り込みを防止する組織全体に及ぶ障壁膜である。重篤な脳疾患を治療するための薬物もしくは遺伝子は、(a)その薬物もしくは遺伝子を脳内に直接注射する、したがってBBBを迂回する工程によって、または(b)その薬物もしくは遺伝子がBBBを越える経血管的経路を介して脳内に入るようにその薬物もしくは遺伝子を血流内に注射する工程のどちらかによってヒト脳へ送達することができる。薬物の脳内投与を用いる場合は、頭部に穴を穿孔して開頭術を実施する必要がある。この開頭術に基づく脳アプローチによる薬物送達は、高額な費用がかかり高度に侵襲性であることに加えて、その薬物もしくは遺伝子が注射針の先端で脳のほんのわずかな容積にしか送達されないために有効ではない。その薬物もしくは遺伝子を脳内に広く分布させることのできる唯一の方法は、血流への注入後の経血管的経路である。しかしこの後者のアプローチは、BBBを越えて輸送される能力を必要とする。BBBは、安全に越えるのが極めて困難であり、かつ頑強であることが証明されている。
【0004】
以前の研究は、BBB受容体/輸送系に結合する分子版トロイの木馬を使用することでBBBを越えて薬物もしくは遺伝子を運び込めることを証明している。これらのトロイの木馬は、修飾されたタンパク質、内因性ペプチド、またはペプチド模倣(peptidomimetic)モノクローナル抗体(MAb)であってよい。例えば、HIRMAb 83−14はヒトインスリン受容体(HIR)に結合するマウスMAbである。この結合はMAb 83−14(Pardridgeら, 1995)の、およびMAbに付着したいずれかの薬物もしくは遺伝子ペイロード(Wuら, 1997)のBBBを越える輸送を誘発する。
【0005】
BBBを越えて薬物もしくは遺伝子を運び込むための分子版トロイの木馬の使用は米国特許第4,801,575号および第6,372,250号に記載されている。MAb輸送ベクターへの薬物の結合は、アビジン−ビオチンテクノロジーの使用により容易になる。このアプローチでは、薬物もしくはタンパク質治療薬がモノビオチン化されて抗体ベクターとアビジンもしくはストレプトアビジンとの共役結合体に結合される。抗体主体(based)輸送ベクターへの薬物の結合を促進するためのアビジン−ビオチンテクノロジーは、米国特許第6,287,792号に記載されている。薬物が遺伝子組換えによりMAb輸送ベクターに融合されている融合タンパク質もまた使用されている。
【0006】
HIRMAb 83−14は、生きているアカゲザルのBBBを越えて迅速に輸送されることが、ヒトBBBの解剖学的基質である単離ヒト脳毛細血管に強度に結合することが証明されている(Pardridgeら, 1995を参照)。どちらの場合も、霊長類もしくはヒトBBBでの結合および輸送に関するHIRMAb 83−14の活性は、トランスフェリン受容体のような他のBBB受容体をターゲットとしうる他のペプチド模倣MAbの結合または輸送より10倍高い(Pardridge, 1997)。現在のところ、HIRMAb 83−14は既知の最も活性なBBB輸送ベクターである(Pardridge, 1997)。これに基づくと、HIRMAb 83−14は薬物をインビボで霊長類の脳へ送達するために極めて有用な物質であることが証明されており、さらにヒト脳への脳薬物もしくは遺伝子送達にとって高度に活性であろう。
【0007】
HIRMAb 83−14は、このマウスタンパク質が免疫原性であるためにヒトにおいて使用することができない。HIRMAb 83−14の遺伝子組換え形は、キメラ抗体または遺伝子組換え「ヒト化」HIRMAbのどちらかの形態でヒトにおいて使用できよう。しかしキメラ抗体またはヒト化抗体いずれかの遺伝子組換えおよび産生を実施するためには、最初に抗体重鎖の可変領域(VH)および抗体軽鎖の可変領域(VL)をクローニングする必要がある。VHおよびVL遺伝子のクローニングに続いて、遺伝子をシーケンシングし、ヌクレオチド配列からアミノ酸配列を推定しなければならない。このアミノ酸配列と共に、当業者に知られている工学技術(Footeら, 1992)を使用すると、マウスHIRMAb 83−14のヒト化を実施することが可能になる。しかしHIRMAb 83−14は、ヒト化した後には生物活性を損失する可能性がある(Pichlaら, 1997)。このため、生物活性を維持しながらマウスHIRMAbをヒト化できるかどうかは不確実である。
【0008】
HIRMAb 83−14のキメラ形は遺伝子組換えされており、このキメラ抗体はHIRに結合し、霊長類の脳内へ輸送される(Colomaら, 2000)。しかしキメラ抗体はVHおよびVLのどちらについても全マウスFRを保持しており、このため、キメラ抗体は依然としてヒトにおいては免疫原性である(Bruggemannら, 1989)。キメラ抗体とは対照的に、ヒト化抗体はVHおよびVLのどちらについてもヒトFRアミノ酸配列を使用し、HVの3つの相補性決定領域(CDR)およびVLの3つのCDRしか保持していないであろう。全部のマウスMAbをヒト化することはできないが、これはマウスFRがヒトFR配列によって置換されると生物活性の損失が発生するためである(Pichlaら, 1997)。抗体の生物活性は一定のマウスFRアミノ酸を逆置換することによって復元できる(米国特許第5,585,089号を参照)。それでも、FRアミノ酸逆置換を用いた場合でさえ、一定の抗体は生物活性を保持しながらヒト化することができない(Pichlaら, 1997)。このため、主要なマウスCDRおよびFRアミノ酸配列が分かっている場合でさえ、マウスHIRMAb 83−14をヒト化できるかどうかは不確実である。
【発明の概要】
【課題を解決するための手段】
【0009】
本発明によれば、マウスHIRMAb 83−14抗体をヒト化して、ヒトをインビボで治療するための薬物および診断薬と組み合わせて使用できる生物活性のヒト化インスリン受容体(HIR)抗体を提供できることが見いだされた。HIR抗体は、アビジン−ビオチン共役法を使用して薬物または診断薬に共役させることができ、または遺伝子組換え技術を使用して融合タンパク質としてHIR抗体/薬物の組み合わせを調製することができる。HIR抗体は、BBBを越えてヒト脳へ神経薬理学的物質を送達するために特に良好に適している。HIR抗体のヒト化特性は、ヒトにおける免疫学的反応を顕著に低下させる。
【0010】
本発明のヒト化マウス抗体はHIRに結合することができ、どちらも可変領域および定常領域を含むアミノ酸の重鎖(HC)およびアミノ酸の軽鎖(LC)を含有している。HCおよびLCの可変領域には、フレームワーク領域(FR)間に挿入されている相補性決定領域(CDR)が含まれる。
【0011】
HCは可変領域のアミノ末端に位置する第1CDR、HC可変領域のカルボキシル末端に位置する第3CDRならびに前記第1および第3CDRの間に位置する第2CDRを含有している。第1CDR、第2CDR、および第3CDRのアミノ酸配列は各々配列番号31、33および35、ならびにそれらの組み合わせ同等物である。HCフレームワーク領域には、第1CDRのアミノ末端に隣接する第1FR、前記第1および第2CDRの間に位置する第2FR、前記第2および第3CDRの間に位置する第3FR、ならびに前記第3CDRのカルボキシル末端に隣接する第4FRが含まれる。本発明によると、HCの4つのFRは、全抗体がHIRに関して生物活性を維持していて、ヒトにおいて免疫原性ではないようにヒト化される。
【0012】
LCは可変領域のアミノ末端に位置する第1CDR、可変領域のカルボキシル末端に位置する第3CDR、ならびに前記第1および第3CDRの間に位置する第2CDRを含有している。第1CDR、第2CDR、および第3CDRのアミノ酸配列は各々配列番号38、40および42、ならびにそれらの組み合わせ同等物である。LCフレームワーク領域には、前記第1CDRのアミノ末端に隣接する第1FR、前記第1および第2CDRの間に位置する第2FR、前記第2および第3CDRの間に位置する第3FR、ならびに前記第3CDRのカルボキシル末端に隣接て位置する第4FRが含まれる。本発明によると、LCの4つのFRは、全抗体がHIRに関して生物活性を維持していて、ヒトにおいて最小の免疫原性を有するようにヒト化される。
【0013】
マウス抗体の定常領域は、またヒトにおける免疫原性を最小限に抑えるために修飾される。マウスHC定常領域は、IgG1などのヒト免疫グロブリンからのHC定常領域と置換される。マウスLC定常領域は、カッパ(κ)LC定常領域のようなヒト免疫グロブリンのLCからの定常領域と置換される。マウスHCおよびLC定常領域とヒト定常領域との置換は、HIR結合に関してヒト化抗体の生物活性に有害な影響を及ぼさないことが見いだされた。
【0014】
本発明は、ヒト化マウス抗体自体を含むだけではなく、薬物または診断薬に結合したヒト化抗体から構成される医薬組成物も含んでいる。ヒト化抗体は、HIRを介してBBBを越える輸送および/または細胞内へのエンドサイトーシスを提供するために薬物または診断薬をインビボでHIRへ送達する際に有効である。これらの組成物は、神経医薬品を脳へ送達するためにヒトへ静脈内(iv)注入するために特に良好に適している。
【0015】
本発明の上記で考察した、およびその他の多数の特徴および付随する利点は、添付の図面を結び付けて詳細な説明を参照することにより、明確に理解されるであろう。
【図面の簡単な説明】
【0016】
【図1】図1は、マウスVH(配列番号1)およびマウスVL(配列番号2)のヌクレオチド配列ならびにマウスVH(配列番号3)およびマウスVL(配列番号4)の推定アミノ酸配列を示し、これらは、83−14マウスHIRMAbの重鎖(HC)および軽鎖(LC)双方の3つのフレームワーク(FR)領域および4つの相補性決定領域(CDR)を示している。アスタリスク(*)が付けられたアミノ酸は、無傷マウスLCまたは無傷マウスHCのトリプシン消化ペプチドのいずれかのアミノ酸シーケンシングによって確認された;アミノ酸シーケンシングのために、ハイブリドーマ順化培地から無傷マウスIgGを精製した後に無傷マウスHCまたはLCをゲル精製した。
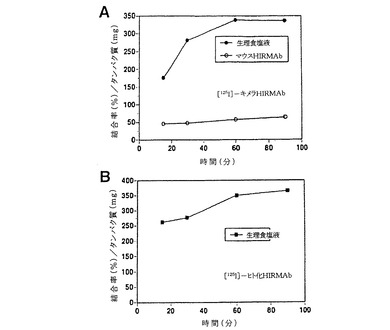
【図2】図2Aおよび2Bは、ヒト剖検脳から機械的均質化法を用いて得られた単離ヒト脳毛細血管について実施した放射性受容体アッセイの結果を示したグラフである。これらの毛細血管は[125I]−標識キメラHIRMAb(Colomaら, 2000)(図2A)または[125I]−バージョン5ヒト化HIRMAb(図2B)と一緒にインキュベートした。これらのデータは、どちらの抗体も同等に、ヒトにおけるBBBの解剖学的基礎を形成するヒト脳毛細血管に良好に結合することを示す。
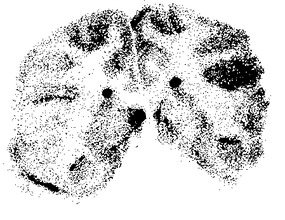
【図3】図3は、本発明によるヒト化モノクローナル抗体を用いて治療されたアカゲザルの脳スキャンを示した図である。[125I]−標識バージョン5 HIRMAbを麻酔したアカゲザルに静脈内注射し、120分後に動物を安楽死させた。迅速に脳を切除し、冠状半球形スラブに切り分け、これらを直ちに冷凍した。クリオスタット切片(20μm)を作製し、X線フィルムに露光させた。このフィルムを走査すると、図3に示した画像が得られた。この画像は霊長類の脳の灰白質と白質との間の明白な境界を示す。灰白質中の方が血管密度が高いために、白質より多くのヒト化HIRMAbの取り込みが見られる。
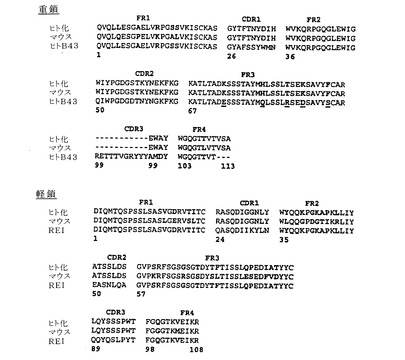
【図4】図4は、以下の重鎖および軽鎖両方の3FRおよび3CDRのアミノ酸配列比較を示す:(a)バージョン5ヒト化HIRMAb、(v)オリジナルのマウス83−14 HIRMAb、および(c)B43ヒトIgGのVHまたはREIヒトIgGのVL。
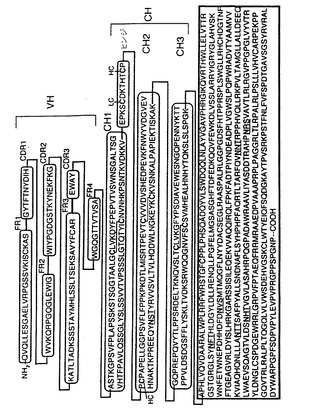
【図5】図5は、ヒトα−L−イヅロニダーゼ(IDUA)(配列番号48)の融合タンパク質のアミノ酸配列であり、これは、ヒトインスリン受容体(HIRMAb)に対するヒト化モノクローナル抗体の重鎖(HC)のカルボキシル末端に融合している。HCは可変領域(VH)および定常領域(CH)から構成されている;CHはさらに3つのサブ領域、CH1(配列番号44)、CH2(配列番号45)、およびCH3(配列番号46)から構成される;CH1およびCH2領域は12アミノ酸ヒンジ領域(配列番号47)によって結合されている。VHは4つのフレームワーク領域(FR1=配列番号30;FR2=配列番号32;FR3=配列番号34;およびFR4=配列番号36)ならびに3つの相補性決定領域(CDR)(CDR1=配列番号31;CDR2=配列番号33;およびCDR3=配列番号35)から構成される。CHについて示されたアミノ酸配列は現行データベース内でよく知られており、ヒトIgG1のCH配列に一致する。CHのCH2領域内のアスパラギン(N)残基上には単一N結合グリコシル化部位が存在し、下線によって示したようにIDUA配列内には6つの潜在的N−結合グリコシル化部位が存在する。
【発明を実施するための形態】
【0017】
本発明は、MAb−83−14として同定されたマウスモノクローナル抗体をヒトにおいてインビボで使用できるようにする前記抗体のヒト化を含んでいる。以前に言及されたように(Pardridge,ら 1995)、MAb 83−14はヒトまたはアカゲザル血液脳関門でヒトインスリン受容体に対する高親和性を有しており、BBBを越えて神経医薬品を輸送するためにトロイの木馬として使用するための候補物質である。本明細書で使用する用語「医薬品」は、ヒトにおける疾患を治療または診断するために使用される薬物、遺伝子または化学物質を含むことが意図されている。用語「神経医薬品」には、脳疾患を治療するために使用される医薬品が含まれる。本発明のヒト化抗体のトロイの木馬は、神経医薬品を血流からBBBを越えて脳へ輸送するために特に良好に適している。
【0018】
マウスMAb−83−14のHCおよびLCの可変領域の完全アミノ酸配列は、実施例1に記載したとおりに決定された。マウスVH(配列番号1)およびマウスVL(配列番号2)を発現する遺伝子のヌクレオチド配列は、図1に表示されている。マウスVH(配列番号3)およびマウスVL(配列番号4)を発現する遺伝子のアミノ酸配列もまた、図1に表示されている。マウスMAb 83−14 VHおよびVLの可変領域のアミノ酸配列(各々、配列番号3および4)もまた、図4に示されている。本発明のヒト化マウス抗体は、HIRに強力に結合する抗体の能力を破壊せずにヒト抗体により密接に類似するようにマウス抗体の可変領域のアミノ酸配列を修飾することによって調製される。さらに、ヒト化抗体には、ヒト抗体にも一致する定常領域が含まれる。
【0019】
ヒト化マウス抗体には、定常領域(CH)および可変領域(VH)から構成されるアミノ酸の重鎖(HC)が含まれる。HCの可変領域はアミノ末端およびカルボキシル末端を有しており、さらに4つのFRの間に挿入された3つのCDRを含んでいる。第1CDR(CDR1)は、VHのアミノ末端の近くに位置しており、第3CDR(CDR3)はHCのカルボキシル末端の近くに位置している。マウスMAb 83−14 HC CDR1、CDR2、およびCDR3のアミノ酸配列は、各々配列番号31、33、および35に表示されている。HC CDRは抗体がHIRへ結合するために不可欠であるので、ヒト化抗体が配列番号31、33および35に一致するアミノ酸配列を備えるHC CDRを有するのが好ましい。しかし、ヒト化抗体には、HC内に配列番号31、33および35に「個別に同等」であるアミノ酸配列を有するCDRを含まれることがある。「個別に同等」なアミノ酸配列は、少なくとも75%の配列同一性を有しており、抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、個別に同等のアミノ酸配列は、配列番号31、33および35との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、少なくとも95%の配列同一性を有する個別に同等のアミノ酸配列である。
【0020】
3つのVH CDRアミノ酸配列は、さらにまたアミノ酸配列(VH CDR1、VH CDR2およびVH CDR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVH CDR配列の組み合わせ群の同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列31、32および35を組み合わせたものと少なくとも75%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号31、33および35において見いだされる組み合わせ配列との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号31、33および35)との少なくとも95%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0021】
VH CDRアミノ酸配列が上記に規定した個別の同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いCDRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のCDRが個別同等性の基準を満たさないことがあるという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。例えば、VH CDR3(配列番号35)はわずか4アミノ酸長である。2つのアミノ酸が変化される場合は、次に個別配列同一性はわずか50%であり、これは上記に規定の個別同等性に対する75%下限の下方にある。しかし、この特定配列は、CDR1、CDR2およびCDR3を組み合わせた配列の配列同一性が群同等性要件を満たすことを前提に、組み合わせ同等VH CDR群の一部として使用するためにさらに適している。
【0022】
ヒト化マウス抗体には、さらにまた定常領域(CL)および可変領域(VL)から構成されるアミノ酸の軽鎖(LC)が含まれる。LCの可変領域はアミノ末端およびカルボキシル末端を有しており、さらに4つのFRの間に挿入された3つのCDRを含んでいる。第1CDR(CDR1)は、VLのアミノ末端の近くに位置しており、第3CDR(CDR3)はVLのカルボキシル末端の近くに位置している。マウスMAb 83−14 LC CDR1、CDR2、およびCDR3のアミノ酸配列は、各々配列番号38、40および42に表示されている。VL CDRは抗体がHIRに結合するためにも重要であるので、ヒト化抗体が配列番号38、40および42と同一であるアミノ酸配列を備えるLC CDRを有するのが好ましい。しかし、ヒト化抗体には、VL内に配列番号38、40および42と「個別に同等」であるアミノ酸配列を有するCDRが含まれることがある。「個別に同等」なアミノ酸配列は、少なくとも75%配列同一性を有しており、抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、個別に同等のアミノ酸配列は、配列番号38、40または42との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、少なくとも95%の配列同一性を有する個別に同等のアミノ酸配列である。
【0023】
3つのVL CDRアミノ酸配列は、さらにまたアミノ酸配列(VL CDR1、VL CDR2およびVL CDR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVL CDR配列の組み合わせ群の同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列38、40および42を組み合わせたものと少なくとも75%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号38、40および42において見いだされる組み合わせ配列との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号38、40および42)との少なくとも95%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0024】
VL CDRアミノ酸配列が上記に規定した個別の同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いCDRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のCDRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。例えば、VH CDR3(配列番号42)は9アミノ酸長である。3つのアミノ酸が変化される場合は、次に個別配列同一性はわずか66%であり、これは上記に規定の個別同等性に対して75%下限の下方にある。しかし、この特定配列は、CDR1、CDR2およびCDR3を組み合わせた配列の配列同一性が群同等性要件を満たすことを前提に、組み合わせ同等VL CDR群の一部として使用するためにさらに適する。
【0025】
VHの第1フレームワーク領域(FR1)はヒト化抗体のアミノ末端に位置する。第4フレームワーク領域(FR4)は、ヒト化抗体のカルボキシル末端の近くに位置する。ヒト化VH FR1、FR2、FR3およびFR4の代表的な好ましいアミノ酸配列は各々配列番号30、32、34および36に表示されており、これらの好ましい配列はバージョン5ヒト化HIRMAbに一致する(表3)。FR2のアミノ酸配列(配列番号32)は、マウスMAb 83−14 VH FR2またはヒトIgG、B43のアミノ酸配列(図4を参照)と同一である。VH FR1およびFR4のアミノ酸配列(配列番号30および36)は、マウスMAb 83−14とは相違するアミノ酸配列を有するB43ヒト抗体フレームワーク領域に一致する(図4)。バージョン5ヒト化HIRMAbのVH
FR3のアミノ酸配列(配列番号34)は、マウス83−14抗体のVH FR3に一致する(表3)。好ましいVH FR配列は抗体の生物活性を破壊せずに修飾することができる。適切なまた別の、または同等のFRには、配列番号30、32、34または36と少なくとも70%の個別配列同一性を有し、結果として生じる抗体がHIRに結合する能力を破壊しないFRが含まれる。好ましくは、また別のFRは、置換される好ましいVH FRとの少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、置換される好ましいVH FRとの少なくとも90%の配列同一性を有するまた別のFRである。
【0026】
4つのVH FRアミノ酸配列は、さらにまたアミノ酸配列(VH FR1、VH FR2およびVH FR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVH FR配列の組み合わせ群の代替物または同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列30、32、34および36を組み合わせたものと少なくとも70%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号30、32、34および36において見いだされる組み合わせ配列との少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号30、32、34および36)との少なくとも90%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0027】
また別のVH FRアミノ酸配列が上記に規定した個別同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いFRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のFRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。
【0028】
LCの第1フレームワーク領域(FR1)はヒト化抗体のVLのアミノ末端に位置する。第4フレームワーク領域(FR4)は、ヒト化抗体のVLのカルボキシル末端の近くに位置する。ヒト化VL FR1、FR2、FR3およびFR4の代表的な好ましいアミノ酸配列は、各々配列番号37、39、41および43に表示されている。VH FR1、FR2、FR3およびFR4のアミノ酸配列(配列番号37、39、41および43)は、マウスMAb 83−14とは相違するアミノ酸配列を有するREIヒト抗体フレームワーク領域に一致する(図4を参照)。好ましいVL FR配列は抗体の生物活性を破壊せずに修飾することができる。適切なまた別の、または同等のFRには、配列番号37、39、41および43と少なくとも70%の個別配列同一性を有し、結果として生じる抗体がHIRに結合する能力を破壊しないFRが含まれる。好ましくは、同等またはまた別のFRは、置換される好ましいVL FRとの少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、置換される好ましいVL FRとの少なくとも90%の配列同一性を有するまた別のFRである。
【0029】
4つのVL FRアミノ酸配列は、さらにまたアミノ酸配列(VL FR1、VL FR2、VL FR3およびVL FR4)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVL FR配列の組み合わせ群の代替物または同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列37、39、41および43を組み合わせたものと少なくとも70%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号37、39、41および43において見いだされる組み合わせ配列との少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号37、39、41および43)との少なくとも90%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0030】
また別のVL FRアミノ酸配列が上記に規定した個別同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いFRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のFRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。
【0031】
バージョン5は、本発明による好ましいヒト化抗体である。バージョン5のVHおよびVLのアミノ酸配列は、各々配列番号5および6に表示されている。バージョン5の調製および同定については、実施例2、表3および図4により詳細に記載されている。バージョン5のVH FRのアミノ酸配列は、上記に記載の好ましいVH FR配列(配列番号30、32、34および36)と一致する。さらに、バージョン5のVL FRのアミノ酸配列は、上記に記載の好ましいVL FR配列(配列番号37、39、41および43)と一致する。バージョン5のVHおよびVL FRは、「ヒト化」されているVHおよびVL LC FRの好ましい例である。「ヒト化」とは、HCまたはLCいずれかにおける4つのフレームワーク領域が、結果として生じる抗体がHIRに結合する能力を破壊せずにヒト抗体(HAb)からのFRとできる限り緊密に適合されていることを意味する。HCのために使用されるヒト抗体モデルはB43抗体であり、LCのために使用されるヒト抗体モデルはREI抗体であり、B43およびREI抗体配列はどちらもよく知られており、公共データベースから入手可能である。HCまたはLC FRがヒト化された場合、1つ以上のFRが選択されたHAbテンプレートと完全には一致せず、マウス抗体との同一性または類似性を保持していることがあり得る。マウスアミノ酸配列がヒト化FR内に残されている程度は、ヒトにおける免疫原性反応の可能性を減少させるためにできる限り低く維持されなければならない。
【0032】
ヒト化されているFRの例は、実施例2および表3で説明されている。マウスMAb 84−13のFRに密接に一致するヒト抗体からのフレームワーク領域が選択される。次にヒトFRはマウスFRに代えてMAb−83−14内に置換される。次に結果として生じる抗体が試験される。1群としてのFRは、修飾された抗体がまだHIR受容体に強力に結合し、ヒトにおける免疫原性を減少させている場合にのみヒト化されていると見なされる。最初の試験が成功しない場合は、ヒトFRがわずかに修飾され、結果として生じる抗体が試験される。MAb 84−13のHC FRをヒト化するために使用できるHC
FRを有する代表的なヒト抗体には、Genbank(受託番号S78322)に預託されているB43ヒトIgG(配列番号12)が含まれており、マウス84−13 VHとのVH相同性を備えるその他のヒトIgG分子は、免疫グロブリン配列のKabatデータベースなどの公共データベースを検索することによって見いだすことができる。MAb 84−13のLC FRをヒト化するために使用できるLC FRを有する代表的なヒト抗体には、Genbank(受託番号1WTLB)に預託されているヒトREI抗体(配列番号13)が含まれており、マウス83−14 VLとのVL相同性を備えるその他のヒトIgG分子は、免疫グロブリン配列のKabatデータベースなどの公共データベースを検索することによって見いだすことができる。
【0033】
ヒト化抗体が適正に機能するためには、HCおよびLCは各々定常領域を含有していなければならない。抗体がHIRに結合する能力を破壊しないことを前提に、あらゆる数の様々なヒト抗体定常領域をヒト化抗体内に組み込むことができる。適切なヒト抗体HC定常領域にはヒトIgG1、IgG2、IgG3、またはIgG4が含まれる。好ましいHC定常領域はヒトIgG1である。適切なヒト抗体LC定常領域にはカッパ(κ)またはλが含まれる。ヒトκLC定常領域が好ましい。
【0034】
ヒト化抗体は、事前にHIRにアクセスすることによって遺伝子、薬物および診断薬を細胞へ送達するために使用されてきた他の抗体ターゲティング剤(トロイの木馬)と同一方法で使用できる。ヒト化抗体は、典型的には薬物または診断用化合物(医薬品)に結合しており、適切な医薬担体と組み合わされて静脈内(iv)投与される。適切な担体を用いると、薬物/ヒト化抗体複合体は、皮下、筋肉内、鼻腔内、クモ膜下、または経口投与することができよう。ヒト化抗体を医薬品へ結合させることのできる多数の方法がある。ヒト化抗体はアビジンもしくはストレプトアビジンに融合させ、抗体トロイの木馬と医薬品とを一緒に結合させるためにアビジン−ビオチン結合を使用する既知の方法によって、モノビオチン化されている医薬品へ結合させることができる。あるいは、ヒト化抗体および医薬品は遺伝子組換え方法を使用して単一融合タンパク質として発現させることができる。
【0035】
それにヒト化抗体を結合させることのできる代表的な医薬品には、遺伝子送達のための小分子、組換えタンパク質、合成ペプチド、アンチセンス物質またはナノ容器が含まれる。代表的な組換えタンパク質には、塩基性線維芽細胞成長因子(bFGF)、ヒトα−L−イヅロニダーゼ(IDUA)、脳由来神経栄養性因子などの他のニューロトロフィン、またはその他のリソソーム酵素が含まれる。本発明のヒト化抗体などの、BBBを越えてbFGFを輸送するためのトロイの木馬の使用は、本特許出願と同一譲受人によって所有されており、本特許出願と同一日に出願された同時係属の米国特許出願(UC Case
2002−094−1、代理人整理番号第0180−0027号)の中に記載されている。
【0036】
ヒト化抗体が医薬品と結合されると、患者に知られている他のコンジュゲート(結合体)または融合タンパク質と同一方法で患者に投与される。特定の用量または治療レジメンは、送達される医薬品および治療される状態に依存して広範囲に変動するであろう。好ましい投与経路は静脈内(iv)である。適切な担体には、Tweenシリーズの界面活性剤由来の1つなどの低濃度の中性洗剤を含む、または含まない酢酸塩、リン酸塩、TRISまたは様々な他のバッファーを用いて緩衝された生理食塩液もしくは水が含まれる。BBBを越えて神経医薬品を送達するためには、好ましくはヒト抗体/医薬品によるトロイの木馬化合物が使用される。しかし、ヒト化されたトロイの木馬は、さらにまた一般にHIRを有するいずれかの器官または組織へ医薬品を送達することもできる。
【0037】
以下の実施例では、本発明によるヒト化モノクローナル抗体が発見された方法およびそれらの作製および使用に関する更なる詳細について記載する。
【実施例1】
【0038】
マウス83−14 VHおよびVL遺伝子のクローニング
ポリA+RNAを83−14ハイブリドーマ細胞系(Soosら, 1986)から単離し、それを使用して逆転写酵素素を用いて相補的DNA(cDNA)を生成した。このcDNAを、マウス抗体遺伝子のVHおよびVLを特異的に増幅させるオリゴデオキシヌクレオチド(ODN)を用いて83−14 VHまたは83−14 VL遺伝子のうちのどちらかのポリメラーゼ連鎖反応(PCR)増幅と一緒に使用したが、類似方法は周知である(Liら, 1999)。これらの遺伝子フラグメントのPCR増幅に適したPCR ODNの配列は周知である(Li., 1999)。PCR産物を1%アガロースゲルで単離し、予想された0.4KbのVHおよびVL遺伝子産物を単離した。一本鎖Fv(ScFv)抗体をコードできるように、VHおよびVL遺伝子フラグメントを連続的に細菌発現プラスミドにサブクローニングした。次にScFv発現プラスミドを使用して大腸菌を形質転換させた。個々のコロニーを寒天プレート上で同定し、LB培地中で液体培養物を生成した。この培地をアカゲザル脳の免疫組織化学検査に使用して、アカゲザル脳微小血管またはBBBへ強力に結合する抗体を産生するクローンを同定した。この免疫組織化学検査は、それらのコロニーが機能的83−14ScFvを分泌することを同定した。83−14 VHおよびVL遺伝子の同定に続いて、自動DNAシーケンシング法を使用してヌクレオチド配列を両方向で決定した。マウス83−14 VH(配列番号1)およびマウスVL(配列番号2)のヌクレオチド配列は、マウスVH(配列番号3)およびマウスVL(配列番号4)の推定アミノ酸配列を生じさせる。アミノ酸配列は、HCおよびLCのマウス83−14 HIRMAb両方の全3つのCDRおよび全4つのFRについて与えられる。図1では、LCの可変領域をVLとし、HCの可変領域をVHとしている。
【実施例2】
【0039】
83−14 HIRMAbの反復ヒト化:バージョン1からバージョン5
83−14 MAbのヒト化は83−14 MAb内のマウスFRはLCおよびHCの両方の可変領域内の適切なヒトFR領域によって置換されるCDR/FRグラフティングによって実施した。KabatデータベースはMatchプログラムを使用してスクリーニングした。マウス83−14 VHまたはVLアミノ酸配列をヒトIgG VHまたはヒトκ軽鎖データベースと比較した。できる限り最小のミスマッチを使用して、マウス83−14 VHおよびVLのアミノ酸配列と高度に相同であるFRアミノ酸配列を含有する可能性のあるいくつかのヒトIgG分子を同定した。最後にマウス83−14 HIRMAbのCDR/FRグラフティングのために、B43ヒトIgG1重鎖およびREIヒトκ軽鎖のフレームワーク領域を選択した。
【0040】
合成ヒト化83−14 VLおよびVH遺伝子を増幅させるために、長さが69−94ヌクレオチドから構成される6種のODNプライマーのセットを設計した(表1および2)。これらのODNプライマーは5’および3’末端のどちらにもおいても24ヌクレオチド重複しており、標準ソフトウエアを用いて二次構造を分析した。46℃を超えるTmを生じる安定な二次構造は、これらの構造の融点を32−46℃へ低下させるために第1、第2、または第3文字のコドンの置換によって修正した。さらに5’および3’末端両方に対応するプライマーもまた設計したが、これらによって人工遺伝子のPCR増幅が可能になった。これらの新規の配列は、アスパラギン残基での如何なるコンセンサスN−グリコシル化部位も欠如している。
【0041】
【表1】

【0042】
【表2】
【0043】
PCRは、各5pモルの6つのオーバーラッピングODN、ヌクレオチド、ならびにTaqおよびTaq伸長DNAポリメラーゼを含有する総量100μL中で実施した。PCR後、ヒト化VHおよびVL遺伝子を細菌発現プラスミド中で個別にライゲートし、大腸菌を形質転換させた。幾つかのクローンを単離し、個別にシーケンシングし、PCR導入シーケンスエラーを含有しないクローンを引き続いて生成させた。
【0044】
制限エンドヌクレアーゼを用いてヒト化VHインサートを細菌発現プラスミドから遊離させ、以前に記載されたように(Colomaら, 1992;米国特許第5,624,659号)真核細胞発現ベクター内にライゲートした。ヒト化VL合成遺伝子に対して類似の方法を実施した。骨髄腫細胞にヒト化軽鎖遺伝子をトランスフェクトし、引き続き、この細胞系にバージョン1のヒト化重鎖遺伝子をトランスフェクトした(表3)。無傷ヒトIgGを分泌するクローンを同定するために、トランスフェクトした骨髄腫細胞を96ウエルELISAでスクリーニングした。複数回試みたが、ヒトIgGを産生する細胞系を同定することはできなかった。これとは反対に、ノーザンブロット分析はトランスフェクトした細胞系が予想されたヒト化83−14 mRNAを産生することを示し、これはこの細胞系のトランスフェクションが成功したことを実証していた。これらの結果は、FRアミノ酸置換を含有していないバージョン1のヒト化HIRMAbがこの細胞から分泌されないことを示しており、このことは、ヒト化HCがヒト化LCと適正には集合しないことを示唆していた。バージョン1は、25Cl’Cl抗体(Bejcekら, 1995)に対応するFRアミノ酸を含有する合成HC遺伝子から誘導された。これにより、新規のHC人工遺伝子が調製され、これは、B43ヒトIgGのアミノ酸配列(Bejcekら, 1995)である異なるヒトIgG配列由来のHC FRアミノ酸を含有しており、これはバージョン2のヒト化HIRMAbを生じた(表3)。しかし、バージョン2ヒト化HIRMAbはトランスフェクトした骨髄腫細胞によって分泌されなかった。バージョン1および2はどちらも、Rechaviら(1983)から誘導された同一のHCシグナルペプチド(表3)を含有している。シグナルペプチドがIgG分泌に及ぼす作用を評価するために、シグナルペプチド配列を、キメラHIRMAbを産生するために使用するシグナルペプチド配列に変更したが(Colomaら, 2000)、このシグナルペプチド配列は表3に示されている。ヒト化HIRMAbのバージョン2および3はそのシグナルペプチドに関してしか相違していなかった(表3)。しかし、バージョン3は骨髄腫細胞からは分泌されず、これはこのシグナルペプチドがヒト化HIRMAbの分泌の欠如の原因ではないことを示している。
【0045】
上記の所見は、ヒトFR領域へのマウス83−14 CDRの単純なグラフティングが適正には集合および分泌できないタンパク質を産生させることを証明した。以前の研究は、HIRMAbのキメラ形がトランスフェクトした骨髄腫細胞中で適切にプロセシングかつ分泌されることを証明していた(Colomaら, 2000)。これは、ヒト化HCまたはLCのFR内の一定のアミノ酸配列がヒト化HIRMAbの適正な集合および分泌を妨害することを示唆した。そこで、キメラ/ヒト化ハイブリッド分子を組換え作製した。バージョン4aはマウスFR1ならびにヒト化FR2、FR3、およびFR4を含有していた;バージョン4bはマウスFR3、およびFR4ならびにヒト化FR1およびFR2を含有していた(表3)。バージョン4aおよび4bのどちらも分泌されたが、バージョン4bはバージョン4aより活性であった。これらの所見は、FR3またはFR4のどちらかの中のアミノ酸がヒト化HIRMAbの分泌の欠如の原因であることを示していた。ヒトおよびマウスFR4は1アミノ酸しか相違しなかった(表3);このため、FR4の配列をヒト配列に対応させるために部位特異的突然変異誘発によって変化させ、このバージョンをバージョン5とした(表3)。バージョン5 HIRMAbは、VHのFR3内のヒト配列とVH内のFR3に対するオリジナルのマウス配列との置換を用いるとオリジナルのCDRグラフティング抗体と一致した。全バージョンのHIRMAbを産生する際には、FR置換を備えていない同一CDRグラフティングLCを使用した。これは、LCにおけるFR変化を必要としない可能性があることを証明した他の研究(Grazianoら, 1995)と一致している。
【0046】
【表3】
【0047】
ヒト25C1C1 IgG重鎖(HC)可変領域(VH)のFRを使用してバージョン1を設計した。
バージョン1は、ノーザンブロット分析によって決定されたHC mRNAが高度に富裕であるにもかかわらず、トランスフェクトした骨髄腫細胞から分泌hIgGを産生しなかった。
【0048】
ヒトB43 IgG HC可変領域のFRを使用してバージョン2を再設計した。バージョン1および2におけるペプチドシグナル#1(MDWTWRVLCLLAVAPGAHS)(配列番号49)をバージョン3におけるシグナルペプチド#2(MGWSWVMLFLLSVTAGKGL)(配列番号50)と取り替えた。バージョン2および3におけるFRおよびCDRは同一であった。バージョン4a、4bおよび5に対してはシグナルペプチド#2を使用した。
バージョン4aはヒトFR2、3および4、ならびにマウスFR1を有している。
バージョン4bはヒトFR1および2、ならびにマウスFR3および4を有している。
バージョン5は、ヒトFR1、2および4、ならびにマウスFR3を使用して生成した。
バージョン4a、4bおよび5は分泌hIgGを産生したが、他方バージョン1、2および3はIgGを分泌しなかった。バージョン4a、4b、および5の中では、含有するマウスフレームワークアミノ酸置換が少ないのでバージョン5が好ましい。
【0049】
このタンパク質のバージョン5形は、トランスフェクトした骨髄腫細胞から無傷で分泌された。分泌されたバージョン5ヒト化HIRMAbはタンパク質Aアフィニティークロマトグラフィーによって精製し、HIRに対するこの抗体の親和性は、以前に記載されたように(Colomaら, 2000)、リガンドとして[125I]−標識マウス83−14 Mabを使用した免疫放射定量アッセイ(IRMA)を用いて試験した。これらの結果は、HIRに対するこの抗体の親和性が維持されていることを証明した。IRMAでは、抗原はトランスフェクトしたCHO細胞から産生し、CHO細胞順化培地のレクチンアフィニティークロマトグラフィーによって精製されたHIRの細胞外ドメインであった。マウスおよびバージョン5ヒト化83−14 HIRMAbの解離定数(KD)は各々2.7±0.4nMおよび3.7±0.4nMであった。これらの結果は、83−14 HIRMAbが、米国特許第5,585,089号に教示されているように、(a)種々のヒト免疫グロブリン分子由来のHCおよびLCのFR領域を入手し、および(b)抗体構造の分子モデリングの使用を必要としない方法を使用することによって良好にヒト化されていることを示している。他の適用(Grazianoら, 1995)と同様に、抗体のLCにおけるFRアミノ酸変化は必要とされなかった。
【実施例3】
【0050】
ヒト化HIRMAbのヒトBBBへの結合
以前の研究は、放射標識マウスHIRMAbが、60〜120分間のインキュベーション時間でタンパク質1mgにつきおよそ400%の結合率でヒト脳毛細血管に強力に結合すると報告している(Pardridgeら, 1995)。この実施例では、放射標識バージョン5ヒト化HIRMAbを用いて類似の所見が記録された。ヒト脳毛細血管を放射受容体アッセイにおいて[125I]バージョン5ヒト化HIRMAbと一緒にインキュベートすると、室温での60分間のインキュベーションで結合率はほぼ400%となり、これは[125I]−キメラHIRMAbのヒト脳毛細血管への結合に似ていた(図2Aおよび2Bを参照)。これとは対照的に、ヒト脳毛細血管への非特異的IgGの結合は、類似のインキュベーション時間ではタンパク質1mgにつき5%未満である(Pardridgeら, 1995)。この実施例は、バージョン5ヒト化HIRMAbがインビボでBBBを形成するヒト脳毛細血管によって強力に結合かつエンドサイトーシスされたことを証明している。
【実施例4】
【0051】
インビボの霊長類BBBを越えるヒト化HIRMAbの輸送
ヒト化バージョン5HIRMAbを125−ヨウ素により放射標識し、成体アカゲザルに静脈内注射した。2時間後に動物を致死させ、脳を切除して冷凍した。クリオスタット切片(20ミクロン)を作製し、x線フィルムに露光させた。フィルムを走査すると、ヒト化HIRMAbの霊長類脳取り込みの画像が生成された(図3)。霊長類脳の白質および灰白質の区域が明瞭に描出され、取り込みは白質より灰白質内の方が多かった。白質と比較して、灰白質内のヒトHIRMAbの高度の取り込みは灰白質内の3倍高い血管密度と一致している。アカゲザルに3倍高い非特異的IgGが注射しても抗体の脳取り込みは生じない(Pardridgeら, 1995)。これらのフィルムオートラジオグラフィー試験は、ヒト化HIRMAbがインビボで霊長類BBBを越えて薬物(ヨウ素)を運べることを証明している。ヒトBBB(図2)へのヒト化HIRMAbの高度の結合に基づくと、ヒトにおいてもインビボでの高い脳取り込みの類似所見が記録されるであろう。
【実施例5】
【0052】
CDRまたはFRアミノ酸置換による抗体の親和性成熟法
バージョン5ヒト化HIRMAb、マウス83−14 HIRMAb、ならびにB43
HCまたはREI LC抗体についてのHCのVHのアミノ酸配列およびLCのVLのアミノ酸配列は図4に示されている。図4におけるCDRアミノ酸を前提にすると、抗体工学術分野の当業者(Schierら, 1996)は、いわゆる「親和性成熟法」または分子進化法と呼ばれる工程において83−14 HCまたはLC CDR配列内で一定のアミノ酸置換を作製することができる。これは無作為に、または抗体のHCもしくはLCのいずれかのFR領域で作製した一本鎖アミノ酸変化に類似する免疫グロブリン構造のX線回折モデルによって誘導して実施することができる(米国特許第5,585,089号)。同様に、図4におけるFRアミノ酸配列が与えられると、当業者はターゲット(標的)HIR抗原に対するHIRMAbの親和性をさらに最適化するために、HCもしくはLC FR領域における一定のアミノ酸置換を作製することができる。これらの置換は、FRおよびCDR領域両方について以前に記載された配列同一性の限界に留意しなければならない。これらの変化は、増加した結合もしくは増加したエンドサイトーシスまたはその両方のいずれかを引き起こすことができる。
【実施例6】
【0053】
ヒト化HIRMAb/α−L−イヅロニダーゼ融合タンパク質
α−L−イヅロニダーゼ(IDUA)は、ハーラー症候群またはI型ムコ多糖体沈着症(MPS)を有する患者において欠如している酵素であり、脳に有害な影響を及ぼす。脳病理は、最終的にはこの遺伝性疾患を有する小児の早期の死亡を生じさせる。I型MPS患者に対するIDUA酵素補充療法(ERT)は脳疾患にとって有効ではないが、それはこの酵素がBBBを越えないからである。これは重篤な問題であり、この疾患を有する小児がERTを受けている場合でさえ早期に死亡することを意味する。この酵素は、この酵素がヒト化HIRMAbなどの分子版トロイの木馬に付着されることを前提に、末梢投与後にはヒトBBBを越えて送達することができよう。IDUAは、アビジン−ビオチン工学技術を用いてヒト化HIRMAbへ付着させることができる。このアプローチでは、IDUA酵素はヒト化HIRMAbおよびアビジンの融合タンパク質産生に平行してモノビオチン化される。さらに、IDUAはアビジン−ビオチン工学技術ではなく、ビオチン化の必要またはアビジンなどの異種タンパク質の使用を回避する遺伝子組換え技術を用いてヒト化HIRMAbへ付着させることもできよう。このアプローチでは、IDUAをコードする遺伝子がHIRMAb重鎖もしくは軽鎖タンパク質のアミノ末端もしくはカルボキシル末端に一致するヒト化HIRMAb重鎖もしくは軽鎖遺伝子の領域に融合される。融合遺伝子の構築および適切な原核細胞もしくは真核細胞発現ベクター内への挿入に続いて、HIRMAb/IDUA融合タンパク質は精製および製造のために大量生産される。典型的MAb/IDUA融合タンパク質のアミノ酸配列および一般構造式は図5に示されている(配列番号48)。この構築物中では、この酵素はヒト化HIRMAbの重鎖(HC)のカルボキシル末端に融合している。図5に示されているIDUAのアミノ酸配列は、成熟した、プロセシングされた酵素のアミノ酸配列である。あるいは、この酵素はHIRMAb HCのアミノ末端またはヒト化HIRMAb軽鎖(LC)のアミノ末端もしくはカルボキシル末端に融合させることができよう。さらに、IDUA配列内の1つ以上のアミノ酸はこの酵素の生物活性を保持しながら修飾することができよう。リソソーム酵素および抗体の融合タンパク質が調製されており、これらの融合タンパク質は生物活性を保持している(Haismaら, 1998)。この融合タンパク質をコードする融合遺伝子はpCEP4などの数種の市販で入手できる永続的発現ベクター内に挿入でき、細胞系を永続的にトランスフェクトしてヒグロマイシンまたはその他の選択剤を用いて選択することができる。順化培地は、組換えヒト化HIRMAb/IDUA融合タンパク質を精製するために濃縮することができる。
【実施例7】
【0054】
HIRMAbのヒトインスリン受容体への結合における軽鎖(LC)の役割
ヒト化HIRMAb軽鎖または、抗ダンシルMAb軽鎖であった「代替軽鎖」(Shin and Morrison, 1990)をコードするプラスミドを用いて、骨髄腫細胞(NSO)をトランスフェクトした。抗ダンシル軽鎖は、抗ダンシルIgGから誘導され、この場合、ダンシルは抗体生成において使用される一般的なハプテンである。ヒト化HIRMAb軽鎖を用いてトランスフェクトした骨髄腫細胞系、および代替軽鎖を用いてトランスフェクトした骨髄腫細胞系のどちらも、引き続いて、キメラHIRMAbの重鎖をコードするプラスミドを用いてトランスフェクトした。1つの細胞系は抗HIRMAbキメラ重鎖および抗HIRMAbヒト化軽鎖から構成されるIgGを分泌し、このIgGをキメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGとする。もう1つの細胞系は、キメラHIRMAb重鎖および抗ダンシル軽鎖から構成されるIgGを分泌し、このIgGをキメラHIRMAb HC/ダンシルLC IgGとする。どちらの細胞系も、骨髄腫細胞上清についてのヒトIgG ELISAを用いて決定されたように、ヒト化HIRMAb軽鎖または抗ダンシル軽鎖のどちらかを用いてプロセシングされたIgGを分泌した。これらのデータは、キメラHIRMAb重鎖が非特異的もしくは代用軽鎖を産生する骨髄腫細胞によってプロセシングかつ分泌され得ることを示した。これらのキメラ抗体とHIRの可溶性細胞外ドメイン(ECD)との反応性はELISAによって決定された。HIR ECDは、以前に記載されたように(Colomaら, 2000)、HIR ECDを用いてトランスフェクトしたCHO細胞の順化培地のレクチンアフィニティークロマトグラフィーによって精製した。HIR ECD ELISAでは、マウス83−14 HIRMAbを陽性コントロールとして使用し、マウスIgG2aを陰性コントロールとして使用した。陰性コントロールはごくわずかなELISAシグナルを生じた;マウス83−14 MAbを用いた検量線は、1μg/mL(マウス83−14 MAb)で飽和状態に到達した吸光度の線形増加を生じさせた。ELISAにおける免疫反応を、分光光度計を用いて定量し、このアッセイにおける405nmでの最大吸光度(A405)は0.9であった。キメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGを分泌するすべての単離骨髄腫細胞クローンは、検量線を最大化した免疫反応性レベルを示してHIR ECD ELISAにおいて陽性であった。さらに、キメラHIRMAb HC/dansyl LC IgGを分泌する骨髄腫細胞クローンはさらにまたHIR ECD ELISAにおいて陽性シグナルを生じ、A405レベルはキメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGを用いて得られたA405レベルの約50%であった。これらの所見は、軽鎖がヒトインスリン受容体の細胞外ドメインであるその標的抗原へのHIRMAbへの結合において小さな役割しか果たさないことを示している。この解釈は、ヒト化HIRMAbのHIR ECDへの活性の結合を可能にするためにヒト化LCにおけるFR置換が必要とされなかったという所見によって支持される(実施例2を参照)。これらの所見は、無傷ヒト化HIRMAbの標的HIR抗原への結合の損失を最小限に抑えながらHIRMAb軽鎖のアミノ酸配列における大きな変化(50%以上)を作製できることを証明している。したがって、広範囲のLCを使用すると、それらがHCと適合性であるならば、本発明によってヒト化抗体を調製することができる。LCは、LCがHCと結合され、結果として生じる抗体がHIRに結合する能力を破壊しない場合は、HCと「適合性」があると見なされる。さらに、LCは、ヒトにおける免疫原性反応が最小限に抑えられるようにヒトまたは十分にヒト化されなければならない。選択されたヒトまたはヒト化LC配列がHCと適合性であるかどうかは、日常的実験を使用して決定できる。
【0055】
本発明の代表的実施形態を記載してきたが、当業者には本開示は典型に過ぎないこと、様々な他の代替、適応および修飾を本発明の範囲内で実施できることに留意されたい。したがって、本発明は上記の実施形態および実施例に限定されず、添付の特許請求項によってのみ限定される。
【0056】
参考文献
Bruggemann M.ら(1989)、「キメラ抗体の免疫原性」、J. Exp. Med., 170:2153−2157.
Coloma M.J., Lee H.J., Kurihara A., Landaw E.M., Boado R.J., Morrison S.L., and Pardridge W.M.(2000)、「遺伝子組換えキメラモノクローナル抗体のヒトインスリン受容体への霊長類血液脳関門を越える輸送」、Pharm. Res., 17:266−274.
Coloma M.J., Hastings A., Wims L.A., and Morrison S.L.(1992)、「ポリメラーゼ連鎖反応によって生成した可変領域を使用して抗体分子を発現させるための新規ベクター」、J. Immunol. Methods, 152:89−104.
Foote J. and Winter G.(1992)、「超可変ループの立体構造に影響を及ぼす抗体フレームワーク残基」、J. Mol. Biol., 224:487−499.
【0057】
Graziano R.F.ら(1995)、「ヒト化抗γ−Ig受容体I型(FcγRI)モノクローナル抗体の構築および特性解析」、J. Immunol., 155:4996−5002.
Haisma J.J.ら(2000)、「抗体指向性酵素プロドラッグ療法のための一本鎖抗CD20抗体およびヒトβ−グルクロニダーゼの融合タンパク質の構築および特性解析」、Blood. 92:184−190.
Li J.Y., Sugimura K., Boado R.J., Lee H.J., Zhang C., Dubel S., and Pardridge W.M.(1999)、「遺伝子組換え脳薬物送達ベクター抗トランスフェリン受容体一本鎖抗体−ストレプトアビジン融合遺伝子およびタンパク質のインビボ適用」、Protein Engineering, 12:787−796.
Miller G.(2002)、「障壁を破壊する」、Science, 297:1116−1118.
Pardridge W.M.(1997)、「脳への薬物送達」、J. Cereb.
Blood Flow Metabol., 17:713−731.
【0058】
Pardridge W.M., Buciak J.L., and Friden P.M.(1991)、「インビボでの血液脳関門を通しての抗トランスフェリン受容体抗体の選択的輸送」、J. Pharmacol. Exp. Ther., 259:66−70.
Pardridge W.M., Kang Y.−S., Bucjak J.L.,and Yang J.(1995)、「ヒトインスリン受容体モノクローナル抗体はインビトロではヒト脳毛細血管への高度の親和性結合および霊長類におけるインビボでは血液脳関門を通る迅速なトランスサイトーシスを受ける」、 Pharm., Res.12:807−816.
Pichla W.L., Murali R., and Burnett R.M.(1997)、「骨髄腫関連性GD2ガングリオシドへのFabフラグメントの結晶構造」、J. Struct. Biol., 119:6−16.
Rechavi G.ら(1983)、「免疫グロブリン重鎖可変領域(VH)遺伝子サブグループの進化の様相」、Proc. Natl. Acad. Sci.(U.S.A.)80:855−859.
Schier R.ら(1996)、「抗体結合部位の中心にある相補性決定領域の分子進化によるピコモル親和性抗c−erbB−2一本鎖Fvの単離」、J. Mol. Biol., 263:551−557.
【0059】
Shin S.U. and Morrison S.L.(1990)、「インスリン様成長因子1へ特異的に結合した抗体結合の発現および特性解析:細胞ターゲティングの潜在的用途」、Proc. Natl. Acad. Sci. U.S.A. 87:5322−5326.
Soos M.A.ら(1986)、「ヒトインスリン受容体上の多数のエピトープと反応するモノクローナル抗体」、Biochem. J. 235:199−208.
米国特許第5,624,659号(発行日:1997年4月29日)、「テネイシンを発現する脳腫瘍を治療する方法」、(発明者:Darell D. BignerおよびMichael R. Zalutsky;譲受人:デューク大学)。
米国特許第6,287,792号(発行日:2001年9月11日)、「アビジン−ビオチン工学技術を使用したインビボ組織および細胞へのアンチセンスオリゴデオキシヌクレオチドおよびペプチドの薬物送達」、(発明者:William M. PardridgeおよびRuben J. Boado;譲受人:カリフォルニア大学)。
【0060】
Vogel C.L.ら(2002)、「HER2−過剰発現性転移性乳癌の第一選択療法における単一薬剤としてのトラスツズマブの有効性および安全性」、J. Clin.
Oncol., 20:719−726.
Wu D., Yang J., and Pardridge W.M.(1997)、「ヒトインスリン受容体に対するモノクローナル抗体を用いてのインビボでの霊長類血液脳関門を越えるペプチド放射性医薬品のドラッグターゲティング」、J. Clin.
Invest., 100:1804−1812.
Zhang Y., Lee H.J., Boado R.J., and Pardridge W.M.(2002)、「ヒト脳癌細胞へのアンチセンス遺伝子の受容体媒介性送達」、J. Gene Med., 4:183−194.
【技術分野】
【0001】
本発明は、一般に血流からヒト脳およびその他のヒトインスリン受容体を発現する器官もしくは組織への医薬品(pharmaceutical agents)の送達に関する。より詳細には、本発明は、ヒトインスリン受容体(HIR)に容易に結合できる化合物を形成するために医薬品に付着させることのできる「ヒト化」モノクローナル抗体(MAb)の開発に関する。この化合物は、脳毛細血管内皮上に位置するインスリン受容体によってヒト血液脳関門(BBB)を越えることができる。BBBを越えると、ヒト化モノクローナル抗体/医薬品化合物は、さらにまた脳細胞上に位置するインスリン受容体を介して脳細胞内への受容体媒介性エンドサイトーシスを受けることができる。
【背景技術】
【0002】
本明細書で言及した出版物およびその他の参考文献は、本発明の背景およびその実践に関する追加される詳細を提供するために、参考文献として組み込まれる。便宜上、参考文書は添付の文献目録に著者名および発行日によって分類されている。
【0003】
BBBは、循環中の薬物、タンパク質治療薬、アンチセンス薬物、および遺伝子薬品の脳内取り込みを防止する組織全体に及ぶ障壁膜である。重篤な脳疾患を治療するための薬物もしくは遺伝子は、(a)その薬物もしくは遺伝子を脳内に直接注射する、したがってBBBを迂回する工程によって、または(b)その薬物もしくは遺伝子がBBBを越える経血管的経路を介して脳内に入るようにその薬物もしくは遺伝子を血流内に注射する工程のどちらかによってヒト脳へ送達することができる。薬物の脳内投与を用いる場合は、頭部に穴を穿孔して開頭術を実施する必要がある。この開頭術に基づく脳アプローチによる薬物送達は、高額な費用がかかり高度に侵襲性であることに加えて、その薬物もしくは遺伝子が注射針の先端で脳のほんのわずかな容積にしか送達されないために有効ではない。その薬物もしくは遺伝子を脳内に広く分布させることのできる唯一の方法は、血流への注入後の経血管的経路である。しかしこの後者のアプローチは、BBBを越えて輸送される能力を必要とする。BBBは、安全に越えるのが極めて困難であり、かつ頑強であることが証明されている。
【0004】
以前の研究は、BBB受容体/輸送系に結合する分子版トロイの木馬を使用することでBBBを越えて薬物もしくは遺伝子を運び込めることを証明している。これらのトロイの木馬は、修飾されたタンパク質、内因性ペプチド、またはペプチド模倣(peptidomimetic)モノクローナル抗体(MAb)であってよい。例えば、HIRMAb 83−14はヒトインスリン受容体(HIR)に結合するマウスMAbである。この結合はMAb 83−14(Pardridgeら, 1995)の、およびMAbに付着したいずれかの薬物もしくは遺伝子ペイロード(Wuら, 1997)のBBBを越える輸送を誘発する。
【0005】
BBBを越えて薬物もしくは遺伝子を運び込むための分子版トロイの木馬の使用は米国特許第4,801,575号および第6,372,250号に記載されている。MAb輸送ベクターへの薬物の結合は、アビジン−ビオチンテクノロジーの使用により容易になる。このアプローチでは、薬物もしくはタンパク質治療薬がモノビオチン化されて抗体ベクターとアビジンもしくはストレプトアビジンとの共役結合体に結合される。抗体主体(based)輸送ベクターへの薬物の結合を促進するためのアビジン−ビオチンテクノロジーは、米国特許第6,287,792号に記載されている。薬物が遺伝子組換えによりMAb輸送ベクターに融合されている融合タンパク質もまた使用されている。
【0006】
HIRMAb 83−14は、生きているアカゲザルのBBBを越えて迅速に輸送されることが、ヒトBBBの解剖学的基質である単離ヒト脳毛細血管に強度に結合することが証明されている(Pardridgeら, 1995を参照)。どちらの場合も、霊長類もしくはヒトBBBでの結合および輸送に関するHIRMAb 83−14の活性は、トランスフェリン受容体のような他のBBB受容体をターゲットとしうる他のペプチド模倣MAbの結合または輸送より10倍高い(Pardridge, 1997)。現在のところ、HIRMAb 83−14は既知の最も活性なBBB輸送ベクターである(Pardridge, 1997)。これに基づくと、HIRMAb 83−14は薬物をインビボで霊長類の脳へ送達するために極めて有用な物質であることが証明されており、さらにヒト脳への脳薬物もしくは遺伝子送達にとって高度に活性であろう。
【0007】
HIRMAb 83−14は、このマウスタンパク質が免疫原性であるためにヒトにおいて使用することができない。HIRMAb 83−14の遺伝子組換え形は、キメラ抗体または遺伝子組換え「ヒト化」HIRMAbのどちらかの形態でヒトにおいて使用できよう。しかしキメラ抗体またはヒト化抗体いずれかの遺伝子組換えおよび産生を実施するためには、最初に抗体重鎖の可変領域(VH)および抗体軽鎖の可変領域(VL)をクローニングする必要がある。VHおよびVL遺伝子のクローニングに続いて、遺伝子をシーケンシングし、ヌクレオチド配列からアミノ酸配列を推定しなければならない。このアミノ酸配列と共に、当業者に知られている工学技術(Footeら, 1992)を使用すると、マウスHIRMAb 83−14のヒト化を実施することが可能になる。しかしHIRMAb 83−14は、ヒト化した後には生物活性を損失する可能性がある(Pichlaら, 1997)。このため、生物活性を維持しながらマウスHIRMAbをヒト化できるかどうかは不確実である。
【0008】
HIRMAb 83−14のキメラ形は遺伝子組換えされており、このキメラ抗体はHIRに結合し、霊長類の脳内へ輸送される(Colomaら, 2000)。しかしキメラ抗体はVHおよびVLのどちらについても全マウスFRを保持しており、このため、キメラ抗体は依然としてヒトにおいては免疫原性である(Bruggemannら, 1989)。キメラ抗体とは対照的に、ヒト化抗体はVHおよびVLのどちらについてもヒトFRアミノ酸配列を使用し、HVの3つの相補性決定領域(CDR)およびVLの3つのCDRしか保持していないであろう。全部のマウスMAbをヒト化することはできないが、これはマウスFRがヒトFR配列によって置換されると生物活性の損失が発生するためである(Pichlaら, 1997)。抗体の生物活性は一定のマウスFRアミノ酸を逆置換することによって復元できる(米国特許第5,585,089号を参照)。それでも、FRアミノ酸逆置換を用いた場合でさえ、一定の抗体は生物活性を保持しながらヒト化することができない(Pichlaら, 1997)。このため、主要なマウスCDRおよびFRアミノ酸配列が分かっている場合でさえ、マウスHIRMAb 83−14をヒト化できるかどうかは不確実である。
【発明の概要】
【課題を解決するための手段】
【0009】
本発明によれば、マウスHIRMAb 83−14抗体をヒト化して、ヒトをインビボで治療するための薬物および診断薬と組み合わせて使用できる生物活性のヒト化インスリン受容体(HIR)抗体を提供できることが見いだされた。HIR抗体は、アビジン−ビオチン共役法を使用して薬物または診断薬に共役させることができ、または遺伝子組換え技術を使用して融合タンパク質としてHIR抗体/薬物の組み合わせを調製することができる。HIR抗体は、BBBを越えてヒト脳へ神経薬理学的物質を送達するために特に良好に適している。HIR抗体のヒト化特性は、ヒトにおける免疫学的反応を顕著に低下させる。
【0010】
本発明のヒト化マウス抗体はHIRに結合することができ、どちらも可変領域および定常領域を含むアミノ酸の重鎖(HC)およびアミノ酸の軽鎖(LC)を含有している。HCおよびLCの可変領域には、フレームワーク領域(FR)間に挿入されている相補性決定領域(CDR)が含まれる。
【0011】
HCは可変領域のアミノ末端に位置する第1CDR、HC可変領域のカルボキシル末端に位置する第3CDRならびに前記第1および第3CDRの間に位置する第2CDRを含有している。第1CDR、第2CDR、および第3CDRのアミノ酸配列は各々配列番号31、33および35、ならびにそれらの組み合わせ同等物である。HCフレームワーク領域には、第1CDRのアミノ末端に隣接する第1FR、前記第1および第2CDRの間に位置する第2FR、前記第2および第3CDRの間に位置する第3FR、ならびに前記第3CDRのカルボキシル末端に隣接する第4FRが含まれる。本発明によると、HCの4つのFRは、全抗体がHIRに関して生物活性を維持していて、ヒトにおいて免疫原性ではないようにヒト化される。
【0012】
LCは可変領域のアミノ末端に位置する第1CDR、可変領域のカルボキシル末端に位置する第3CDR、ならびに前記第1および第3CDRの間に位置する第2CDRを含有している。第1CDR、第2CDR、および第3CDRのアミノ酸配列は各々配列番号38、40および42、ならびにそれらの組み合わせ同等物である。LCフレームワーク領域には、前記第1CDRのアミノ末端に隣接する第1FR、前記第1および第2CDRの間に位置する第2FR、前記第2および第3CDRの間に位置する第3FR、ならびに前記第3CDRのカルボキシル末端に隣接て位置する第4FRが含まれる。本発明によると、LCの4つのFRは、全抗体がHIRに関して生物活性を維持していて、ヒトにおいて最小の免疫原性を有するようにヒト化される。
【0013】
マウス抗体の定常領域は、またヒトにおける免疫原性を最小限に抑えるために修飾される。マウスHC定常領域は、IgG1などのヒト免疫グロブリンからのHC定常領域と置換される。マウスLC定常領域は、カッパ(κ)LC定常領域のようなヒト免疫グロブリンのLCからの定常領域と置換される。マウスHCおよびLC定常領域とヒト定常領域との置換は、HIR結合に関してヒト化抗体の生物活性に有害な影響を及ぼさないことが見いだされた。
【0014】
本発明は、ヒト化マウス抗体自体を含むだけではなく、薬物または診断薬に結合したヒト化抗体から構成される医薬組成物も含んでいる。ヒト化抗体は、HIRを介してBBBを越える輸送および/または細胞内へのエンドサイトーシスを提供するために薬物または診断薬をインビボでHIRへ送達する際に有効である。これらの組成物は、神経医薬品を脳へ送達するためにヒトへ静脈内(iv)注入するために特に良好に適している。
【0015】
本発明の上記で考察した、およびその他の多数の特徴および付随する利点は、添付の図面を結び付けて詳細な説明を参照することにより、明確に理解されるであろう。
【図面の簡単な説明】
【0016】
【図1】図1は、マウスVH(配列番号1)およびマウスVL(配列番号2)のヌクレオチド配列ならびにマウスVH(配列番号3)およびマウスVL(配列番号4)の推定アミノ酸配列を示し、これらは、83−14マウスHIRMAbの重鎖(HC)および軽鎖(LC)双方の3つのフレームワーク(FR)領域および4つの相補性決定領域(CDR)を示している。アスタリスク(*)が付けられたアミノ酸は、無傷マウスLCまたは無傷マウスHCのトリプシン消化ペプチドのいずれかのアミノ酸シーケンシングによって確認された;アミノ酸シーケンシングのために、ハイブリドーマ順化培地から無傷マウスIgGを精製した後に無傷マウスHCまたはLCをゲル精製した。
【図2】図2Aおよび2Bは、ヒト剖検脳から機械的均質化法を用いて得られた単離ヒト脳毛細血管について実施した放射性受容体アッセイの結果を示したグラフである。これらの毛細血管は[125I]−標識キメラHIRMAb(Colomaら, 2000)(図2A)または[125I]−バージョン5ヒト化HIRMAb(図2B)と一緒にインキュベートした。これらのデータは、どちらの抗体も同等に、ヒトにおけるBBBの解剖学的基礎を形成するヒト脳毛細血管に良好に結合することを示す。
【図3】図3は、本発明によるヒト化モノクローナル抗体を用いて治療されたアカゲザルの脳スキャンを示した図である。[125I]−標識バージョン5 HIRMAbを麻酔したアカゲザルに静脈内注射し、120分後に動物を安楽死させた。迅速に脳を切除し、冠状半球形スラブに切り分け、これらを直ちに冷凍した。クリオスタット切片(20μm)を作製し、X線フィルムに露光させた。このフィルムを走査すると、図3に示した画像が得られた。この画像は霊長類の脳の灰白質と白質との間の明白な境界を示す。灰白質中の方が血管密度が高いために、白質より多くのヒト化HIRMAbの取り込みが見られる。
【図4】図4は、以下の重鎖および軽鎖両方の3FRおよび3CDRのアミノ酸配列比較を示す:(a)バージョン5ヒト化HIRMAb、(v)オリジナルのマウス83−14 HIRMAb、および(c)B43ヒトIgGのVHまたはREIヒトIgGのVL。
【図5】図5は、ヒトα−L−イヅロニダーゼ(IDUA)(配列番号48)の融合タンパク質のアミノ酸配列であり、これは、ヒトインスリン受容体(HIRMAb)に対するヒト化モノクローナル抗体の重鎖(HC)のカルボキシル末端に融合している。HCは可変領域(VH)および定常領域(CH)から構成されている;CHはさらに3つのサブ領域、CH1(配列番号44)、CH2(配列番号45)、およびCH3(配列番号46)から構成される;CH1およびCH2領域は12アミノ酸ヒンジ領域(配列番号47)によって結合されている。VHは4つのフレームワーク領域(FR1=配列番号30;FR2=配列番号32;FR3=配列番号34;およびFR4=配列番号36)ならびに3つの相補性決定領域(CDR)(CDR1=配列番号31;CDR2=配列番号33;およびCDR3=配列番号35)から構成される。CHについて示されたアミノ酸配列は現行データベース内でよく知られており、ヒトIgG1のCH配列に一致する。CHのCH2領域内のアスパラギン(N)残基上には単一N結合グリコシル化部位が存在し、下線によって示したようにIDUA配列内には6つの潜在的N−結合グリコシル化部位が存在する。
【発明を実施するための形態】
【0017】
本発明は、MAb−83−14として同定されたマウスモノクローナル抗体をヒトにおいてインビボで使用できるようにする前記抗体のヒト化を含んでいる。以前に言及されたように(Pardridge,ら 1995)、MAb 83−14はヒトまたはアカゲザル血液脳関門でヒトインスリン受容体に対する高親和性を有しており、BBBを越えて神経医薬品を輸送するためにトロイの木馬として使用するための候補物質である。本明細書で使用する用語「医薬品」は、ヒトにおける疾患を治療または診断するために使用される薬物、遺伝子または化学物質を含むことが意図されている。用語「神経医薬品」には、脳疾患を治療するために使用される医薬品が含まれる。本発明のヒト化抗体のトロイの木馬は、神経医薬品を血流からBBBを越えて脳へ輸送するために特に良好に適している。
【0018】
マウスMAb−83−14のHCおよびLCの可変領域の完全アミノ酸配列は、実施例1に記載したとおりに決定された。マウスVH(配列番号1)およびマウスVL(配列番号2)を発現する遺伝子のヌクレオチド配列は、図1に表示されている。マウスVH(配列番号3)およびマウスVL(配列番号4)を発現する遺伝子のアミノ酸配列もまた、図1に表示されている。マウスMAb 83−14 VHおよびVLの可変領域のアミノ酸配列(各々、配列番号3および4)もまた、図4に示されている。本発明のヒト化マウス抗体は、HIRに強力に結合する抗体の能力を破壊せずにヒト抗体により密接に類似するようにマウス抗体の可変領域のアミノ酸配列を修飾することによって調製される。さらに、ヒト化抗体には、ヒト抗体にも一致する定常領域が含まれる。
【0019】
ヒト化マウス抗体には、定常領域(CH)および可変領域(VH)から構成されるアミノ酸の重鎖(HC)が含まれる。HCの可変領域はアミノ末端およびカルボキシル末端を有しており、さらに4つのFRの間に挿入された3つのCDRを含んでいる。第1CDR(CDR1)は、VHのアミノ末端の近くに位置しており、第3CDR(CDR3)はHCのカルボキシル末端の近くに位置している。マウスMAb 83−14 HC CDR1、CDR2、およびCDR3のアミノ酸配列は、各々配列番号31、33、および35に表示されている。HC CDRは抗体がHIRへ結合するために不可欠であるので、ヒト化抗体が配列番号31、33および35に一致するアミノ酸配列を備えるHC CDRを有するのが好ましい。しかし、ヒト化抗体には、HC内に配列番号31、33および35に「個別に同等」であるアミノ酸配列を有するCDRを含まれることがある。「個別に同等」なアミノ酸配列は、少なくとも75%の配列同一性を有しており、抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、個別に同等のアミノ酸配列は、配列番号31、33および35との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、少なくとも95%の配列同一性を有する個別に同等のアミノ酸配列である。
【0020】
3つのVH CDRアミノ酸配列は、さらにまたアミノ酸配列(VH CDR1、VH CDR2およびVH CDR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVH CDR配列の組み合わせ群の同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列31、32および35を組み合わせたものと少なくとも75%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号31、33および35において見いだされる組み合わせ配列との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号31、33および35)との少なくとも95%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0021】
VH CDRアミノ酸配列が上記に規定した個別の同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いCDRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のCDRが個別同等性の基準を満たさないことがあるという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。例えば、VH CDR3(配列番号35)はわずか4アミノ酸長である。2つのアミノ酸が変化される場合は、次に個別配列同一性はわずか50%であり、これは上記に規定の個別同等性に対する75%下限の下方にある。しかし、この特定配列は、CDR1、CDR2およびCDR3を組み合わせた配列の配列同一性が群同等性要件を満たすことを前提に、組み合わせ同等VH CDR群の一部として使用するためにさらに適している。
【0022】
ヒト化マウス抗体には、さらにまた定常領域(CL)および可変領域(VL)から構成されるアミノ酸の軽鎖(LC)が含まれる。LCの可変領域はアミノ末端およびカルボキシル末端を有しており、さらに4つのFRの間に挿入された3つのCDRを含んでいる。第1CDR(CDR1)は、VLのアミノ末端の近くに位置しており、第3CDR(CDR3)はVLのカルボキシル末端の近くに位置している。マウスMAb 83−14 LC CDR1、CDR2、およびCDR3のアミノ酸配列は、各々配列番号38、40および42に表示されている。VL CDRは抗体がHIRに結合するためにも重要であるので、ヒト化抗体が配列番号38、40および42と同一であるアミノ酸配列を備えるLC CDRを有するのが好ましい。しかし、ヒト化抗体には、VL内に配列番号38、40および42と「個別に同等」であるアミノ酸配列を有するCDRが含まれることがある。「個別に同等」なアミノ酸配列は、少なくとも75%配列同一性を有しており、抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、個別に同等のアミノ酸配列は、配列番号38、40または42との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、少なくとも95%の配列同一性を有する個別に同等のアミノ酸配列である。
【0023】
3つのVL CDRアミノ酸配列は、さらにまたアミノ酸配列(VL CDR1、VL CDR2およびVL CDR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVL CDR配列の組み合わせ群の同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列38、40および42を組み合わせたものと少なくとも75%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号38、40および42において見いだされる組み合わせ配列との少なくとも85%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号38、40および42)との少なくとも95%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0024】
VL CDRアミノ酸配列が上記に規定した個別の同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いCDRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のCDRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。例えば、VH CDR3(配列番号42)は9アミノ酸長である。3つのアミノ酸が変化される場合は、次に個別配列同一性はわずか66%であり、これは上記に規定の個別同等性に対して75%下限の下方にある。しかし、この特定配列は、CDR1、CDR2およびCDR3を組み合わせた配列の配列同一性が群同等性要件を満たすことを前提に、組み合わせ同等VL CDR群の一部として使用するためにさらに適する。
【0025】
VHの第1フレームワーク領域(FR1)はヒト化抗体のアミノ末端に位置する。第4フレームワーク領域(FR4)は、ヒト化抗体のカルボキシル末端の近くに位置する。ヒト化VH FR1、FR2、FR3およびFR4の代表的な好ましいアミノ酸配列は各々配列番号30、32、34および36に表示されており、これらの好ましい配列はバージョン5ヒト化HIRMAbに一致する(表3)。FR2のアミノ酸配列(配列番号32)は、マウスMAb 83−14 VH FR2またはヒトIgG、B43のアミノ酸配列(図4を参照)と同一である。VH FR1およびFR4のアミノ酸配列(配列番号30および36)は、マウスMAb 83−14とは相違するアミノ酸配列を有するB43ヒト抗体フレームワーク領域に一致する(図4)。バージョン5ヒト化HIRMAbのVH
FR3のアミノ酸配列(配列番号34)は、マウス83−14抗体のVH FR3に一致する(表3)。好ましいVH FR配列は抗体の生物活性を破壊せずに修飾することができる。適切なまた別の、または同等のFRには、配列番号30、32、34または36と少なくとも70%の個別配列同一性を有し、結果として生じる抗体がHIRに結合する能力を破壊しないFRが含まれる。好ましくは、また別のFRは、置換される好ましいVH FRとの少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、置換される好ましいVH FRとの少なくとも90%の配列同一性を有するまた別のFRである。
【0026】
4つのVH FRアミノ酸配列は、さらにまたアミノ酸配列(VH FR1、VH FR2およびVH FR3)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVH FR配列の組み合わせ群の代替物または同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列30、32、34および36を組み合わせたものと少なくとも70%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号30、32、34および36において見いだされる組み合わせ配列との少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号30、32、34および36)との少なくとも90%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0027】
また別のVH FRアミノ酸配列が上記に規定した個別同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いFRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のFRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。
【0028】
LCの第1フレームワーク領域(FR1)はヒト化抗体のVLのアミノ末端に位置する。第4フレームワーク領域(FR4)は、ヒト化抗体のVLのカルボキシル末端の近くに位置する。ヒト化VL FR1、FR2、FR3およびFR4の代表的な好ましいアミノ酸配列は、各々配列番号37、39、41および43に表示されている。VH FR1、FR2、FR3およびFR4のアミノ酸配列(配列番号37、39、41および43)は、マウスMAb 83−14とは相違するアミノ酸配列を有するREIヒト抗体フレームワーク領域に一致する(図4を参照)。好ましいVL FR配列は抗体の生物活性を破壊せずに修飾することができる。適切なまた別の、または同等のFRには、配列番号37、39、41および43と少なくとも70%の個別配列同一性を有し、結果として生じる抗体がHIRに結合する能力を破壊しないFRが含まれる。好ましくは、同等またはまた別のFRは、置換される好ましいVL FRとの少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、置換される好ましいVL FRとの少なくとも90%の配列同一性を有するまた別のFRである。
【0029】
4つのVL FRアミノ酸配列は、さらにまたアミノ酸配列(VL FR1、VL FR2、VL FR3およびVL FR4)の1つの組み合わせ群と見なすことができる。本発明は、さらにまたVL FR配列の組み合わせ群の代替物または同等物も含んでいる。そのような「組み合わせ同等物」は、アミノ酸配列37、39、41および43を組み合わせたものと少なくとも70%の配列同一性を有しており、その抗体のHIRへの結合に有害な影響を及ぼさないアミノ酸配列である。好ましくは、組み合わせ同等アミノ酸配列は、配列番号37、39、41および43において見いだされる組み合わせ配列との少なくとも80%の配列同一性を有するであろう。よりいっそう好ましいのは、組み合わせアミノ酸配列(配列番号37、39、41および43)との少なくとも90%の配列同一性を有する組み合わせ同等アミノ酸配列である。
【0030】
また別のVL FRアミノ酸配列が上記に規定した個別同等性および組み合わせ同等性の要件をどちらも満たすことが好ましい。しかし、特により短いFRについて、組み合わせ同等性の基準が満たされる場合でさえ1つ以上のFRが個別同等性の基準を満たさないという一定の状況が存在する。そのような状況では、個別同等性の要件は、組み合わせ同等性の要件が満たされることを前提に適用されない。
【0031】
バージョン5は、本発明による好ましいヒト化抗体である。バージョン5のVHおよびVLのアミノ酸配列は、各々配列番号5および6に表示されている。バージョン5の調製および同定については、実施例2、表3および図4により詳細に記載されている。バージョン5のVH FRのアミノ酸配列は、上記に記載の好ましいVH FR配列(配列番号30、32、34および36)と一致する。さらに、バージョン5のVL FRのアミノ酸配列は、上記に記載の好ましいVL FR配列(配列番号37、39、41および43)と一致する。バージョン5のVHおよびVL FRは、「ヒト化」されているVHおよびVL LC FRの好ましい例である。「ヒト化」とは、HCまたはLCいずれかにおける4つのフレームワーク領域が、結果として生じる抗体がHIRに結合する能力を破壊せずにヒト抗体(HAb)からのFRとできる限り緊密に適合されていることを意味する。HCのために使用されるヒト抗体モデルはB43抗体であり、LCのために使用されるヒト抗体モデルはREI抗体であり、B43およびREI抗体配列はどちらもよく知られており、公共データベースから入手可能である。HCまたはLC FRがヒト化された場合、1つ以上のFRが選択されたHAbテンプレートと完全には一致せず、マウス抗体との同一性または類似性を保持していることがあり得る。マウスアミノ酸配列がヒト化FR内に残されている程度は、ヒトにおける免疫原性反応の可能性を減少させるためにできる限り低く維持されなければならない。
【0032】
ヒト化されているFRの例は、実施例2および表3で説明されている。マウスMAb 84−13のFRに密接に一致するヒト抗体からのフレームワーク領域が選択される。次にヒトFRはマウスFRに代えてMAb−83−14内に置換される。次に結果として生じる抗体が試験される。1群としてのFRは、修飾された抗体がまだHIR受容体に強力に結合し、ヒトにおける免疫原性を減少させている場合にのみヒト化されていると見なされる。最初の試験が成功しない場合は、ヒトFRがわずかに修飾され、結果として生じる抗体が試験される。MAb 84−13のHC FRをヒト化するために使用できるHC
FRを有する代表的なヒト抗体には、Genbank(受託番号S78322)に預託されているB43ヒトIgG(配列番号12)が含まれており、マウス84−13 VHとのVH相同性を備えるその他のヒトIgG分子は、免疫グロブリン配列のKabatデータベースなどの公共データベースを検索することによって見いだすことができる。MAb 84−13のLC FRをヒト化するために使用できるLC FRを有する代表的なヒト抗体には、Genbank(受託番号1WTLB)に預託されているヒトREI抗体(配列番号13)が含まれており、マウス83−14 VLとのVL相同性を備えるその他のヒトIgG分子は、免疫グロブリン配列のKabatデータベースなどの公共データベースを検索することによって見いだすことができる。
【0033】
ヒト化抗体が適正に機能するためには、HCおよびLCは各々定常領域を含有していなければならない。抗体がHIRに結合する能力を破壊しないことを前提に、あらゆる数の様々なヒト抗体定常領域をヒト化抗体内に組み込むことができる。適切なヒト抗体HC定常領域にはヒトIgG1、IgG2、IgG3、またはIgG4が含まれる。好ましいHC定常領域はヒトIgG1である。適切なヒト抗体LC定常領域にはカッパ(κ)またはλが含まれる。ヒトκLC定常領域が好ましい。
【0034】
ヒト化抗体は、事前にHIRにアクセスすることによって遺伝子、薬物および診断薬を細胞へ送達するために使用されてきた他の抗体ターゲティング剤(トロイの木馬)と同一方法で使用できる。ヒト化抗体は、典型的には薬物または診断用化合物(医薬品)に結合しており、適切な医薬担体と組み合わされて静脈内(iv)投与される。適切な担体を用いると、薬物/ヒト化抗体複合体は、皮下、筋肉内、鼻腔内、クモ膜下、または経口投与することができよう。ヒト化抗体を医薬品へ結合させることのできる多数の方法がある。ヒト化抗体はアビジンもしくはストレプトアビジンに融合させ、抗体トロイの木馬と医薬品とを一緒に結合させるためにアビジン−ビオチン結合を使用する既知の方法によって、モノビオチン化されている医薬品へ結合させることができる。あるいは、ヒト化抗体および医薬品は遺伝子組換え方法を使用して単一融合タンパク質として発現させることができる。
【0035】
それにヒト化抗体を結合させることのできる代表的な医薬品には、遺伝子送達のための小分子、組換えタンパク質、合成ペプチド、アンチセンス物質またはナノ容器が含まれる。代表的な組換えタンパク質には、塩基性線維芽細胞成長因子(bFGF)、ヒトα−L−イヅロニダーゼ(IDUA)、脳由来神経栄養性因子などの他のニューロトロフィン、またはその他のリソソーム酵素が含まれる。本発明のヒト化抗体などの、BBBを越えてbFGFを輸送するためのトロイの木馬の使用は、本特許出願と同一譲受人によって所有されており、本特許出願と同一日に出願された同時係属の米国特許出願(UC Case
2002−094−1、代理人整理番号第0180−0027号)の中に記載されている。
【0036】
ヒト化抗体が医薬品と結合されると、患者に知られている他のコンジュゲート(結合体)または融合タンパク質と同一方法で患者に投与される。特定の用量または治療レジメンは、送達される医薬品および治療される状態に依存して広範囲に変動するであろう。好ましい投与経路は静脈内(iv)である。適切な担体には、Tweenシリーズの界面活性剤由来の1つなどの低濃度の中性洗剤を含む、または含まない酢酸塩、リン酸塩、TRISまたは様々な他のバッファーを用いて緩衝された生理食塩液もしくは水が含まれる。BBBを越えて神経医薬品を送達するためには、好ましくはヒト抗体/医薬品によるトロイの木馬化合物が使用される。しかし、ヒト化されたトロイの木馬は、さらにまた一般にHIRを有するいずれかの器官または組織へ医薬品を送達することもできる。
【0037】
以下の実施例では、本発明によるヒト化モノクローナル抗体が発見された方法およびそれらの作製および使用に関する更なる詳細について記載する。
【実施例1】
【0038】
マウス83−14 VHおよびVL遺伝子のクローニング
ポリA+RNAを83−14ハイブリドーマ細胞系(Soosら, 1986)から単離し、それを使用して逆転写酵素素を用いて相補的DNA(cDNA)を生成した。このcDNAを、マウス抗体遺伝子のVHおよびVLを特異的に増幅させるオリゴデオキシヌクレオチド(ODN)を用いて83−14 VHまたは83−14 VL遺伝子のうちのどちらかのポリメラーゼ連鎖反応(PCR)増幅と一緒に使用したが、類似方法は周知である(Liら, 1999)。これらの遺伝子フラグメントのPCR増幅に適したPCR ODNの配列は周知である(Li., 1999)。PCR産物を1%アガロースゲルで単離し、予想された0.4KbのVHおよびVL遺伝子産物を単離した。一本鎖Fv(ScFv)抗体をコードできるように、VHおよびVL遺伝子フラグメントを連続的に細菌発現プラスミドにサブクローニングした。次にScFv発現プラスミドを使用して大腸菌を形質転換させた。個々のコロニーを寒天プレート上で同定し、LB培地中で液体培養物を生成した。この培地をアカゲザル脳の免疫組織化学検査に使用して、アカゲザル脳微小血管またはBBBへ強力に結合する抗体を産生するクローンを同定した。この免疫組織化学検査は、それらのコロニーが機能的83−14ScFvを分泌することを同定した。83−14 VHおよびVL遺伝子の同定に続いて、自動DNAシーケンシング法を使用してヌクレオチド配列を両方向で決定した。マウス83−14 VH(配列番号1)およびマウスVL(配列番号2)のヌクレオチド配列は、マウスVH(配列番号3)およびマウスVL(配列番号4)の推定アミノ酸配列を生じさせる。アミノ酸配列は、HCおよびLCのマウス83−14 HIRMAb両方の全3つのCDRおよび全4つのFRについて与えられる。図1では、LCの可変領域をVLとし、HCの可変領域をVHとしている。
【実施例2】
【0039】
83−14 HIRMAbの反復ヒト化:バージョン1からバージョン5
83−14 MAbのヒト化は83−14 MAb内のマウスFRはLCおよびHCの両方の可変領域内の適切なヒトFR領域によって置換されるCDR/FRグラフティングによって実施した。KabatデータベースはMatchプログラムを使用してスクリーニングした。マウス83−14 VHまたはVLアミノ酸配列をヒトIgG VHまたはヒトκ軽鎖データベースと比較した。できる限り最小のミスマッチを使用して、マウス83−14 VHおよびVLのアミノ酸配列と高度に相同であるFRアミノ酸配列を含有する可能性のあるいくつかのヒトIgG分子を同定した。最後にマウス83−14 HIRMAbのCDR/FRグラフティングのために、B43ヒトIgG1重鎖およびREIヒトκ軽鎖のフレームワーク領域を選択した。
【0040】
合成ヒト化83−14 VLおよびVH遺伝子を増幅させるために、長さが69−94ヌクレオチドから構成される6種のODNプライマーのセットを設計した(表1および2)。これらのODNプライマーは5’および3’末端のどちらにもおいても24ヌクレオチド重複しており、標準ソフトウエアを用いて二次構造を分析した。46℃を超えるTmを生じる安定な二次構造は、これらの構造の融点を32−46℃へ低下させるために第1、第2、または第3文字のコドンの置換によって修正した。さらに5’および3’末端両方に対応するプライマーもまた設計したが、これらによって人工遺伝子のPCR増幅が可能になった。これらの新規の配列は、アスパラギン残基での如何なるコンセンサスN−グリコシル化部位も欠如している。
【0041】
【表1】
【0042】
【表2】
【0043】
PCRは、各5pモルの6つのオーバーラッピングODN、ヌクレオチド、ならびにTaqおよびTaq伸長DNAポリメラーゼを含有する総量100μL中で実施した。PCR後、ヒト化VHおよびVL遺伝子を細菌発現プラスミド中で個別にライゲートし、大腸菌を形質転換させた。幾つかのクローンを単離し、個別にシーケンシングし、PCR導入シーケンスエラーを含有しないクローンを引き続いて生成させた。
【0044】
制限エンドヌクレアーゼを用いてヒト化VHインサートを細菌発現プラスミドから遊離させ、以前に記載されたように(Colomaら, 1992;米国特許第5,624,659号)真核細胞発現ベクター内にライゲートした。ヒト化VL合成遺伝子に対して類似の方法を実施した。骨髄腫細胞にヒト化軽鎖遺伝子をトランスフェクトし、引き続き、この細胞系にバージョン1のヒト化重鎖遺伝子をトランスフェクトした(表3)。無傷ヒトIgGを分泌するクローンを同定するために、トランスフェクトした骨髄腫細胞を96ウエルELISAでスクリーニングした。複数回試みたが、ヒトIgGを産生する細胞系を同定することはできなかった。これとは反対に、ノーザンブロット分析はトランスフェクトした細胞系が予想されたヒト化83−14 mRNAを産生することを示し、これはこの細胞系のトランスフェクションが成功したことを実証していた。これらの結果は、FRアミノ酸置換を含有していないバージョン1のヒト化HIRMAbがこの細胞から分泌されないことを示しており、このことは、ヒト化HCがヒト化LCと適正には集合しないことを示唆していた。バージョン1は、25Cl’Cl抗体(Bejcekら, 1995)に対応するFRアミノ酸を含有する合成HC遺伝子から誘導された。これにより、新規のHC人工遺伝子が調製され、これは、B43ヒトIgGのアミノ酸配列(Bejcekら, 1995)である異なるヒトIgG配列由来のHC FRアミノ酸を含有しており、これはバージョン2のヒト化HIRMAbを生じた(表3)。しかし、バージョン2ヒト化HIRMAbはトランスフェクトした骨髄腫細胞によって分泌されなかった。バージョン1および2はどちらも、Rechaviら(1983)から誘導された同一のHCシグナルペプチド(表3)を含有している。シグナルペプチドがIgG分泌に及ぼす作用を評価するために、シグナルペプチド配列を、キメラHIRMAbを産生するために使用するシグナルペプチド配列に変更したが(Colomaら, 2000)、このシグナルペプチド配列は表3に示されている。ヒト化HIRMAbのバージョン2および3はそのシグナルペプチドに関してしか相違していなかった(表3)。しかし、バージョン3は骨髄腫細胞からは分泌されず、これはこのシグナルペプチドがヒト化HIRMAbの分泌の欠如の原因ではないことを示している。
【0045】
上記の所見は、ヒトFR領域へのマウス83−14 CDRの単純なグラフティングが適正には集合および分泌できないタンパク質を産生させることを証明した。以前の研究は、HIRMAbのキメラ形がトランスフェクトした骨髄腫細胞中で適切にプロセシングかつ分泌されることを証明していた(Colomaら, 2000)。これは、ヒト化HCまたはLCのFR内の一定のアミノ酸配列がヒト化HIRMAbの適正な集合および分泌を妨害することを示唆した。そこで、キメラ/ヒト化ハイブリッド分子を組換え作製した。バージョン4aはマウスFR1ならびにヒト化FR2、FR3、およびFR4を含有していた;バージョン4bはマウスFR3、およびFR4ならびにヒト化FR1およびFR2を含有していた(表3)。バージョン4aおよび4bのどちらも分泌されたが、バージョン4bはバージョン4aより活性であった。これらの所見は、FR3またはFR4のどちらかの中のアミノ酸がヒト化HIRMAbの分泌の欠如の原因であることを示していた。ヒトおよびマウスFR4は1アミノ酸しか相違しなかった(表3);このため、FR4の配列をヒト配列に対応させるために部位特異的突然変異誘発によって変化させ、このバージョンをバージョン5とした(表3)。バージョン5 HIRMAbは、VHのFR3内のヒト配列とVH内のFR3に対するオリジナルのマウス配列との置換を用いるとオリジナルのCDRグラフティング抗体と一致した。全バージョンのHIRMAbを産生する際には、FR置換を備えていない同一CDRグラフティングLCを使用した。これは、LCにおけるFR変化を必要としない可能性があることを証明した他の研究(Grazianoら, 1995)と一致している。
【0046】
【表3】
【0047】
ヒト25C1C1 IgG重鎖(HC)可変領域(VH)のFRを使用してバージョン1を設計した。
バージョン1は、ノーザンブロット分析によって決定されたHC mRNAが高度に富裕であるにもかかわらず、トランスフェクトした骨髄腫細胞から分泌hIgGを産生しなかった。
【0048】
ヒトB43 IgG HC可変領域のFRを使用してバージョン2を再設計した。バージョン1および2におけるペプチドシグナル#1(MDWTWRVLCLLAVAPGAHS)(配列番号49)をバージョン3におけるシグナルペプチド#2(MGWSWVMLFLLSVTAGKGL)(配列番号50)と取り替えた。バージョン2および3におけるFRおよびCDRは同一であった。バージョン4a、4bおよび5に対してはシグナルペプチド#2を使用した。
バージョン4aはヒトFR2、3および4、ならびにマウスFR1を有している。
バージョン4bはヒトFR1および2、ならびにマウスFR3および4を有している。
バージョン5は、ヒトFR1、2および4、ならびにマウスFR3を使用して生成した。
バージョン4a、4bおよび5は分泌hIgGを産生したが、他方バージョン1、2および3はIgGを分泌しなかった。バージョン4a、4b、および5の中では、含有するマウスフレームワークアミノ酸置換が少ないのでバージョン5が好ましい。
【0049】
このタンパク質のバージョン5形は、トランスフェクトした骨髄腫細胞から無傷で分泌された。分泌されたバージョン5ヒト化HIRMAbはタンパク質Aアフィニティークロマトグラフィーによって精製し、HIRに対するこの抗体の親和性は、以前に記載されたように(Colomaら, 2000)、リガンドとして[125I]−標識マウス83−14 Mabを使用した免疫放射定量アッセイ(IRMA)を用いて試験した。これらの結果は、HIRに対するこの抗体の親和性が維持されていることを証明した。IRMAでは、抗原はトランスフェクトしたCHO細胞から産生し、CHO細胞順化培地のレクチンアフィニティークロマトグラフィーによって精製されたHIRの細胞外ドメインであった。マウスおよびバージョン5ヒト化83−14 HIRMAbの解離定数(KD)は各々2.7±0.4nMおよび3.7±0.4nMであった。これらの結果は、83−14 HIRMAbが、米国特許第5,585,089号に教示されているように、(a)種々のヒト免疫グロブリン分子由来のHCおよびLCのFR領域を入手し、および(b)抗体構造の分子モデリングの使用を必要としない方法を使用することによって良好にヒト化されていることを示している。他の適用(Grazianoら, 1995)と同様に、抗体のLCにおけるFRアミノ酸変化は必要とされなかった。
【実施例3】
【0050】
ヒト化HIRMAbのヒトBBBへの結合
以前の研究は、放射標識マウスHIRMAbが、60〜120分間のインキュベーション時間でタンパク質1mgにつきおよそ400%の結合率でヒト脳毛細血管に強力に結合すると報告している(Pardridgeら, 1995)。この実施例では、放射標識バージョン5ヒト化HIRMAbを用いて類似の所見が記録された。ヒト脳毛細血管を放射受容体アッセイにおいて[125I]バージョン5ヒト化HIRMAbと一緒にインキュベートすると、室温での60分間のインキュベーションで結合率はほぼ400%となり、これは[125I]−キメラHIRMAbのヒト脳毛細血管への結合に似ていた(図2Aおよび2Bを参照)。これとは対照的に、ヒト脳毛細血管への非特異的IgGの結合は、類似のインキュベーション時間ではタンパク質1mgにつき5%未満である(Pardridgeら, 1995)。この実施例は、バージョン5ヒト化HIRMAbがインビボでBBBを形成するヒト脳毛細血管によって強力に結合かつエンドサイトーシスされたことを証明している。
【実施例4】
【0051】
インビボの霊長類BBBを越えるヒト化HIRMAbの輸送
ヒト化バージョン5HIRMAbを125−ヨウ素により放射標識し、成体アカゲザルに静脈内注射した。2時間後に動物を致死させ、脳を切除して冷凍した。クリオスタット切片(20ミクロン)を作製し、x線フィルムに露光させた。フィルムを走査すると、ヒト化HIRMAbの霊長類脳取り込みの画像が生成された(図3)。霊長類脳の白質および灰白質の区域が明瞭に描出され、取り込みは白質より灰白質内の方が多かった。白質と比較して、灰白質内のヒトHIRMAbの高度の取り込みは灰白質内の3倍高い血管密度と一致している。アカゲザルに3倍高い非特異的IgGが注射しても抗体の脳取り込みは生じない(Pardridgeら, 1995)。これらのフィルムオートラジオグラフィー試験は、ヒト化HIRMAbがインビボで霊長類BBBを越えて薬物(ヨウ素)を運べることを証明している。ヒトBBB(図2)へのヒト化HIRMAbの高度の結合に基づくと、ヒトにおいてもインビボでの高い脳取り込みの類似所見が記録されるであろう。
【実施例5】
【0052】
CDRまたはFRアミノ酸置換による抗体の親和性成熟法
バージョン5ヒト化HIRMAb、マウス83−14 HIRMAb、ならびにB43
HCまたはREI LC抗体についてのHCのVHのアミノ酸配列およびLCのVLのアミノ酸配列は図4に示されている。図4におけるCDRアミノ酸を前提にすると、抗体工学術分野の当業者(Schierら, 1996)は、いわゆる「親和性成熟法」または分子進化法と呼ばれる工程において83−14 HCまたはLC CDR配列内で一定のアミノ酸置換を作製することができる。これは無作為に、または抗体のHCもしくはLCのいずれかのFR領域で作製した一本鎖アミノ酸変化に類似する免疫グロブリン構造のX線回折モデルによって誘導して実施することができる(米国特許第5,585,089号)。同様に、図4におけるFRアミノ酸配列が与えられると、当業者はターゲット(標的)HIR抗原に対するHIRMAbの親和性をさらに最適化するために、HCもしくはLC FR領域における一定のアミノ酸置換を作製することができる。これらの置換は、FRおよびCDR領域両方について以前に記載された配列同一性の限界に留意しなければならない。これらの変化は、増加した結合もしくは増加したエンドサイトーシスまたはその両方のいずれかを引き起こすことができる。
【実施例6】
【0053】
ヒト化HIRMAb/α−L−イヅロニダーゼ融合タンパク質
α−L−イヅロニダーゼ(IDUA)は、ハーラー症候群またはI型ムコ多糖体沈着症(MPS)を有する患者において欠如している酵素であり、脳に有害な影響を及ぼす。脳病理は、最終的にはこの遺伝性疾患を有する小児の早期の死亡を生じさせる。I型MPS患者に対するIDUA酵素補充療法(ERT)は脳疾患にとって有効ではないが、それはこの酵素がBBBを越えないからである。これは重篤な問題であり、この疾患を有する小児がERTを受けている場合でさえ早期に死亡することを意味する。この酵素は、この酵素がヒト化HIRMAbなどの分子版トロイの木馬に付着されることを前提に、末梢投与後にはヒトBBBを越えて送達することができよう。IDUAは、アビジン−ビオチン工学技術を用いてヒト化HIRMAbへ付着させることができる。このアプローチでは、IDUA酵素はヒト化HIRMAbおよびアビジンの融合タンパク質産生に平行してモノビオチン化される。さらに、IDUAはアビジン−ビオチン工学技術ではなく、ビオチン化の必要またはアビジンなどの異種タンパク質の使用を回避する遺伝子組換え技術を用いてヒト化HIRMAbへ付着させることもできよう。このアプローチでは、IDUAをコードする遺伝子がHIRMAb重鎖もしくは軽鎖タンパク質のアミノ末端もしくはカルボキシル末端に一致するヒト化HIRMAb重鎖もしくは軽鎖遺伝子の領域に融合される。融合遺伝子の構築および適切な原核細胞もしくは真核細胞発現ベクター内への挿入に続いて、HIRMAb/IDUA融合タンパク質は精製および製造のために大量生産される。典型的MAb/IDUA融合タンパク質のアミノ酸配列および一般構造式は図5に示されている(配列番号48)。この構築物中では、この酵素はヒト化HIRMAbの重鎖(HC)のカルボキシル末端に融合している。図5に示されているIDUAのアミノ酸配列は、成熟した、プロセシングされた酵素のアミノ酸配列である。あるいは、この酵素はHIRMAb HCのアミノ末端またはヒト化HIRMAb軽鎖(LC)のアミノ末端もしくはカルボキシル末端に融合させることができよう。さらに、IDUA配列内の1つ以上のアミノ酸はこの酵素の生物活性を保持しながら修飾することができよう。リソソーム酵素および抗体の融合タンパク質が調製されており、これらの融合タンパク質は生物活性を保持している(Haismaら, 1998)。この融合タンパク質をコードする融合遺伝子はpCEP4などの数種の市販で入手できる永続的発現ベクター内に挿入でき、細胞系を永続的にトランスフェクトしてヒグロマイシンまたはその他の選択剤を用いて選択することができる。順化培地は、組換えヒト化HIRMAb/IDUA融合タンパク質を精製するために濃縮することができる。
【実施例7】
【0054】
HIRMAbのヒトインスリン受容体への結合における軽鎖(LC)の役割
ヒト化HIRMAb軽鎖または、抗ダンシルMAb軽鎖であった「代替軽鎖」(Shin and Morrison, 1990)をコードするプラスミドを用いて、骨髄腫細胞(NSO)をトランスフェクトした。抗ダンシル軽鎖は、抗ダンシルIgGから誘導され、この場合、ダンシルは抗体生成において使用される一般的なハプテンである。ヒト化HIRMAb軽鎖を用いてトランスフェクトした骨髄腫細胞系、および代替軽鎖を用いてトランスフェクトした骨髄腫細胞系のどちらも、引き続いて、キメラHIRMAbの重鎖をコードするプラスミドを用いてトランスフェクトした。1つの細胞系は抗HIRMAbキメラ重鎖および抗HIRMAbヒト化軽鎖から構成されるIgGを分泌し、このIgGをキメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGとする。もう1つの細胞系は、キメラHIRMAb重鎖および抗ダンシル軽鎖から構成されるIgGを分泌し、このIgGをキメラHIRMAb HC/ダンシルLC IgGとする。どちらの細胞系も、骨髄腫細胞上清についてのヒトIgG ELISAを用いて決定されたように、ヒト化HIRMAb軽鎖または抗ダンシル軽鎖のどちらかを用いてプロセシングされたIgGを分泌した。これらのデータは、キメラHIRMAb重鎖が非特異的もしくは代用軽鎖を産生する骨髄腫細胞によってプロセシングかつ分泌され得ることを示した。これらのキメラ抗体とHIRの可溶性細胞外ドメイン(ECD)との反応性はELISAによって決定された。HIR ECDは、以前に記載されたように(Colomaら, 2000)、HIR ECDを用いてトランスフェクトしたCHO細胞の順化培地のレクチンアフィニティークロマトグラフィーによって精製した。HIR ECD ELISAでは、マウス83−14 HIRMAbを陽性コントロールとして使用し、マウスIgG2aを陰性コントロールとして使用した。陰性コントロールはごくわずかなELISAシグナルを生じた;マウス83−14 MAbを用いた検量線は、1μg/mL(マウス83−14 MAb)で飽和状態に到達した吸光度の線形増加を生じさせた。ELISAにおける免疫反応を、分光光度計を用いて定量し、このアッセイにおける405nmでの最大吸光度(A405)は0.9であった。キメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGを分泌するすべての単離骨髄腫細胞クローンは、検量線を最大化した免疫反応性レベルを示してHIR ECD ELISAにおいて陽性であった。さらに、キメラHIRMAb HC/dansyl LC IgGを分泌する骨髄腫細胞クローンはさらにまたHIR ECD ELISAにおいて陽性シグナルを生じ、A405レベルはキメラHIRMAb重鎖/ヒト化HIRMAb軽鎖IgGを用いて得られたA405レベルの約50%であった。これらの所見は、軽鎖がヒトインスリン受容体の細胞外ドメインであるその標的抗原へのHIRMAbへの結合において小さな役割しか果たさないことを示している。この解釈は、ヒト化HIRMAbのHIR ECDへの活性の結合を可能にするためにヒト化LCにおけるFR置換が必要とされなかったという所見によって支持される(実施例2を参照)。これらの所見は、無傷ヒト化HIRMAbの標的HIR抗原への結合の損失を最小限に抑えながらHIRMAb軽鎖のアミノ酸配列における大きな変化(50%以上)を作製できることを証明している。したがって、広範囲のLCを使用すると、それらがHCと適合性であるならば、本発明によってヒト化抗体を調製することができる。LCは、LCがHCと結合され、結果として生じる抗体がHIRに結合する能力を破壊しない場合は、HCと「適合性」があると見なされる。さらに、LCは、ヒトにおける免疫原性反応が最小限に抑えられるようにヒトまたは十分にヒト化されなければならない。選択されたヒトまたはヒト化LC配列がHCと適合性であるかどうかは、日常的実験を使用して決定できる。
【0055】
本発明の代表的実施形態を記載してきたが、当業者には本開示は典型に過ぎないこと、様々な他の代替、適応および修飾を本発明の範囲内で実施できることに留意されたい。したがって、本発明は上記の実施形態および実施例に限定されず、添付の特許請求項によってのみ限定される。
【0056】
参考文献
Bruggemann M.ら(1989)、「キメラ抗体の免疫原性」、J. Exp. Med., 170:2153−2157.
Coloma M.J., Lee H.J., Kurihara A., Landaw E.M., Boado R.J., Morrison S.L., and Pardridge W.M.(2000)、「遺伝子組換えキメラモノクローナル抗体のヒトインスリン受容体への霊長類血液脳関門を越える輸送」、Pharm. Res., 17:266−274.
Coloma M.J., Hastings A., Wims L.A., and Morrison S.L.(1992)、「ポリメラーゼ連鎖反応によって生成した可変領域を使用して抗体分子を発現させるための新規ベクター」、J. Immunol. Methods, 152:89−104.
Foote J. and Winter G.(1992)、「超可変ループの立体構造に影響を及ぼす抗体フレームワーク残基」、J. Mol. Biol., 224:487−499.
【0057】
Graziano R.F.ら(1995)、「ヒト化抗γ−Ig受容体I型(FcγRI)モノクローナル抗体の構築および特性解析」、J. Immunol., 155:4996−5002.
Haisma J.J.ら(2000)、「抗体指向性酵素プロドラッグ療法のための一本鎖抗CD20抗体およびヒトβ−グルクロニダーゼの融合タンパク質の構築および特性解析」、Blood. 92:184−190.
Li J.Y., Sugimura K., Boado R.J., Lee H.J., Zhang C., Dubel S., and Pardridge W.M.(1999)、「遺伝子組換え脳薬物送達ベクター抗トランスフェリン受容体一本鎖抗体−ストレプトアビジン融合遺伝子およびタンパク質のインビボ適用」、Protein Engineering, 12:787−796.
Miller G.(2002)、「障壁を破壊する」、Science, 297:1116−1118.
Pardridge W.M.(1997)、「脳への薬物送達」、J. Cereb.
Blood Flow Metabol., 17:713−731.
【0058】
Pardridge W.M., Buciak J.L., and Friden P.M.(1991)、「インビボでの血液脳関門を通しての抗トランスフェリン受容体抗体の選択的輸送」、J. Pharmacol. Exp. Ther., 259:66−70.
Pardridge W.M., Kang Y.−S., Bucjak J.L.,and Yang J.(1995)、「ヒトインスリン受容体モノクローナル抗体はインビトロではヒト脳毛細血管への高度の親和性結合および霊長類におけるインビボでは血液脳関門を通る迅速なトランスサイトーシスを受ける」、 Pharm., Res.12:807−816.
Pichla W.L., Murali R., and Burnett R.M.(1997)、「骨髄腫関連性GD2ガングリオシドへのFabフラグメントの結晶構造」、J. Struct. Biol., 119:6−16.
Rechavi G.ら(1983)、「免疫グロブリン重鎖可変領域(VH)遺伝子サブグループの進化の様相」、Proc. Natl. Acad. Sci.(U.S.A.)80:855−859.
Schier R.ら(1996)、「抗体結合部位の中心にある相補性決定領域の分子進化によるピコモル親和性抗c−erbB−2一本鎖Fvの単離」、J. Mol. Biol., 263:551−557.
【0059】
Shin S.U. and Morrison S.L.(1990)、「インスリン様成長因子1へ特異的に結合した抗体結合の発現および特性解析:細胞ターゲティングの潜在的用途」、Proc. Natl. Acad. Sci. U.S.A. 87:5322−5326.
Soos M.A.ら(1986)、「ヒトインスリン受容体上の多数のエピトープと反応するモノクローナル抗体」、Biochem. J. 235:199−208.
米国特許第5,624,659号(発行日:1997年4月29日)、「テネイシンを発現する脳腫瘍を治療する方法」、(発明者:Darell D. BignerおよびMichael R. Zalutsky;譲受人:デューク大学)。
米国特許第6,287,792号(発行日:2001年9月11日)、「アビジン−ビオチン工学技術を使用したインビボ組織および細胞へのアンチセンスオリゴデオキシヌクレオチドおよびペプチドの薬物送達」、(発明者:William M. PardridgeおよびRuben J. Boado;譲受人:カリフォルニア大学)。
【0060】
Vogel C.L.ら(2002)、「HER2−過剰発現性転移性乳癌の第一選択療法における単一薬剤としてのトラスツズマブの有効性および安全性」、J. Clin.
Oncol., 20:719−726.
Wu D., Yang J., and Pardridge W.M.(1997)、「ヒトインスリン受容体に対するモノクローナル抗体を用いてのインビボでの霊長類血液脳関門を越えるペプチド放射性医薬品のドラッグターゲティング」、J. Clin.
Invest., 100:1804−1812.
Zhang Y., Lee H.J., Boado R.J., and Pardridge W.M.(2002)、「ヒト脳癌細胞へのアンチセンス遺伝子の受容体媒介性送達」、J. Gene Med., 4:183−194.
【特許請求の範囲】
【請求項1】
ヒトインスリン受容体と結合するヒト化抗体であって、前記抗体は:
a)定常領域および可変領域を含むアミノ酸の重鎖であって、前記可変領域はアミノ末端およびカルボキシル末端を有し、フレームワーク領域間に挿入された相補性決定領域を含み、前記相補性決定領域は前記可変領域のアミノ末端に位置する第1相補性決定領域、前記可変領域のカルボキシル末端に位置する第3相補性決定領域、ならびに前記第1および第3相補性決定領域の間に位置する第2相補性決定領域を含み、前記第1相補性決定領域、前記第2相補性決定領域および前記第3相補性決定領域のアミノ酸配列は各々配列番号31、配列番号33、配列番号35およびそれらの組み合わせ同等物であり、さらに前記フレームワーク領域は前記第1相補性決定領域のアミノ末端に隣接する第1フレームワーク領域、前記第1および第2相補性決定領域の間に位置する第2フレームワーク領域、前記第2および第3相補性決定領域の間に位置する第3フレームワーク領域ならびに前記第3相補性決定領域のカルボキシル末端に隣接する第4フレームワーク領域を含み、このとき前記フレームワーク領域がヒト化されているアミノ酸の重鎖:および
b)前記重鎖と適合可能なアミノ酸の軽鎖、
を含むヒト化抗体。
【請求項2】
請求項1記載のヒト化抗体であって、前記アミノ酸の軽鎖は定常領域および可変領域を含み、前記可変領域はアミノ末端およびカルボキシル末端を有し且つフレームワーク領域間に挿入された相補性決定領域を含み、前記相補性決定領域は前記可変領域のアミノ末端に位置する第1相補性決定領域、前記可変領域のカルボキシル末端に位置する第3相補性決定領域、ならびに前記第1および第3相補性決定領域の間に位置する第2相補性決定領域を含み、前記第1相補性決定領域、前記第2相補性決定領域および前記第3相補性決定領域のアミノ酸配列は各々配列番号38、40および42ならびにそれらの組み合わせ同等物であり、さらに前記フレームワーク領域は前記第1相補性決定領域のアミノ末端に隣接する第1フレームワーク領域、前記第1および第2相補性決定領域の間に位置する第2フレームワーク領域、前記第2および第3相補性決定領域の間に位置する第3フレームワーク領域ならびに前記第3相補性決定領域のカルボキシル末端に隣接する第4フレームワーク領域を含み、このとき前記フレームワーク領域がヒト化されている、前記ヒトインスリン受容体と結合する請求項1記載のヒト化抗体。
【請求項3】
前記重鎖の前記定常領域はヒトIgG1定常領域である、請求項1記載のヒト化抗体。
【請求項4】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項2記載のヒト化抗体。
【請求項5】
前記重鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号30、32、34および36、ならびにそれらの組み合わせ同等物である、請求項1記載のヒト化抗体。
【請求項6】
前記軽鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号37、39、41および43、ならびにそれらの組み合わせ同等物である、請求項2記載のヒト化抗体。
【請求項7】
前記軽鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号30、32、34および36、ならびにそれらの組み合わせ同等物である、請求項6記載のヒト化抗体。
【請求項8】
前記重鎖の定常領域はヒトIgG1定常領域である、請求項7記載のヒト化抗体。
【請求項9】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項7記載のヒト化抗体。
【請求項10】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項8記載のヒト化抗体。
【請求項11】
ヒトインスリン受容体に結合可能な組成物であって、前記組成物は:
医薬品;および
請求項1記載のヒト化抗体を含むヒトインスリン受容体ターゲティング物質であって、前記ヒトインスリン受容体ターゲティング物質が前記医薬品に結合している当該物質
を含む組成物。
【請求項12】
前記医薬品はビオチン化されており、前記ヒトインスリン受容体ターゲティング物質はアビジンもしくはストレプトアビジンを含み、前記医薬品は少なくとも1つのアビジン−ビオチン結合を介して前記ヒトインスリン受容体ターゲティング物質に結合されている、請求項11記載の組成物。
【請求項13】
前記医薬品はモノビオチン化されている、請求項12記載の組成物。
【請求項14】
前記医薬品は神経医薬品である、請求項11記載の組成物。
【請求項15】
前記ヒトインスリン受容体ターゲティング物質はヒト化マウス抗体から本質的に構成され、また前記ヒト化マウス抗体は前記ヒト化マウス抗体および前記医薬品を含む融合タンパク質を形成するための遺伝子融合法によって前記医薬品へ結合されている、請求項11記載の組成物。
【請求項16】
ヒトへ静脈内投与するための医薬製剤であって、前記医薬製剤は請求項11記載の組成物と、前記医薬製剤を静脈内投与するための前記組成物に容認可能な担体と、を含む医薬製剤。
【請求項17】
前記医薬品は神経医薬品である、請求項16記載の医薬製剤。
【請求項18】
神経医薬品がヒト血液脳関門を越える能力を強化する方法であって、前記神経医薬品を請求項1記載のヒト化マウス抗体へ結合させる工程を含む方法。
【請求項19】
前記神経医薬品はアビジン−ビオチン結合を介して前記ヒト化マウス抗体へ結合されている、請求項18記載の方法。
【請求項20】
前記神経医薬品は遺伝子融合法によって前記ヒト化マウス抗体へ結合されている、請求項18記載の方法。
【請求項21】
インビボで神経医薬品をヒトインスリン受容体にターゲティングさせる方法であって、前記医薬品を請求項1記載のヒト化マウス抗体へ結合させる工程を含む方法。
【請求項22】
前記医薬品はアビジン−ビオチン結合を介して前記ヒト化マウス抗体へ結合される、請求項21記載の方法。
【請求項23】
前記神経医薬品は遺伝子融合法によって前記ヒト化マウス抗体へ結合される、請求項21記載の方法。
【請求項24】
医薬品をヒト患者へ静脈内投与するための方法であって、前記方法は請求項16記載の医薬製剤を前記ヒト患者の血流に注入する工程を含む方法。
【請求項1】
ヒトインスリン受容体と結合するヒト化抗体であって、前記抗体は:
a)定常領域および可変領域を含むアミノ酸の重鎖であって、前記可変領域はアミノ末端およびカルボキシル末端を有し、フレームワーク領域間に挿入された相補性決定領域を含み、前記相補性決定領域は前記可変領域のアミノ末端に位置する第1相補性決定領域、前記可変領域のカルボキシル末端に位置する第3相補性決定領域、ならびに前記第1および第3相補性決定領域の間に位置する第2相補性決定領域を含み、前記第1相補性決定領域、前記第2相補性決定領域および前記第3相補性決定領域のアミノ酸配列は各々配列番号31、配列番号33、配列番号35およびそれらの組み合わせ同等物であり、さらに前記フレームワーク領域は前記第1相補性決定領域のアミノ末端に隣接する第1フレームワーク領域、前記第1および第2相補性決定領域の間に位置する第2フレームワーク領域、前記第2および第3相補性決定領域の間に位置する第3フレームワーク領域ならびに前記第3相補性決定領域のカルボキシル末端に隣接する第4フレームワーク領域を含み、このとき前記フレームワーク領域がヒト化されているアミノ酸の重鎖:および
b)前記重鎖と適合可能なアミノ酸の軽鎖、
を含むヒト化抗体。
【請求項2】
請求項1記載のヒト化抗体であって、前記アミノ酸の軽鎖は定常領域および可変領域を含み、前記可変領域はアミノ末端およびカルボキシル末端を有し且つフレームワーク領域間に挿入された相補性決定領域を含み、前記相補性決定領域は前記可変領域のアミノ末端に位置する第1相補性決定領域、前記可変領域のカルボキシル末端に位置する第3相補性決定領域、ならびに前記第1および第3相補性決定領域の間に位置する第2相補性決定領域を含み、前記第1相補性決定領域、前記第2相補性決定領域および前記第3相補性決定領域のアミノ酸配列は各々配列番号38、40および42ならびにそれらの組み合わせ同等物であり、さらに前記フレームワーク領域は前記第1相補性決定領域のアミノ末端に隣接する第1フレームワーク領域、前記第1および第2相補性決定領域の間に位置する第2フレームワーク領域、前記第2および第3相補性決定領域の間に位置する第3フレームワーク領域ならびに前記第3相補性決定領域のカルボキシル末端に隣接する第4フレームワーク領域を含み、このとき前記フレームワーク領域がヒト化されている、前記ヒトインスリン受容体と結合する請求項1記載のヒト化抗体。
【請求項3】
前記重鎖の前記定常領域はヒトIgG1定常領域である、請求項1記載のヒト化抗体。
【請求項4】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項2記載のヒト化抗体。
【請求項5】
前記重鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号30、32、34および36、ならびにそれらの組み合わせ同等物である、請求項1記載のヒト化抗体。
【請求項6】
前記軽鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号37、39、41および43、ならびにそれらの組み合わせ同等物である、請求項2記載のヒト化抗体。
【請求項7】
前記軽鎖の第1フレームワーク領域、第2フレームワーク領域、第3フレームワーク領域および第4フレームワーク領域のアミノ酸配列は各々配列番号30、32、34および36、ならびにそれらの組み合わせ同等物である、請求項6記載のヒト化抗体。
【請求項8】
前記重鎖の定常領域はヒトIgG1定常領域である、請求項7記載のヒト化抗体。
【請求項9】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項7記載のヒト化抗体。
【請求項10】
前記軽鎖の前記定常領域はヒトκ定常領域である、請求項8記載のヒト化抗体。
【請求項11】
ヒトインスリン受容体に結合可能な組成物であって、前記組成物は:
医薬品;および
請求項1記載のヒト化抗体を含むヒトインスリン受容体ターゲティング物質であって、前記ヒトインスリン受容体ターゲティング物質が前記医薬品に結合している当該物質
を含む組成物。
【請求項12】
前記医薬品はビオチン化されており、前記ヒトインスリン受容体ターゲティング物質はアビジンもしくはストレプトアビジンを含み、前記医薬品は少なくとも1つのアビジン−ビオチン結合を介して前記ヒトインスリン受容体ターゲティング物質に結合されている、請求項11記載の組成物。
【請求項13】
前記医薬品はモノビオチン化されている、請求項12記載の組成物。
【請求項14】
前記医薬品は神経医薬品である、請求項11記載の組成物。
【請求項15】
前記ヒトインスリン受容体ターゲティング物質はヒト化マウス抗体から本質的に構成され、また前記ヒト化マウス抗体は前記ヒト化マウス抗体および前記医薬品を含む融合タンパク質を形成するための遺伝子融合法によって前記医薬品へ結合されている、請求項11記載の組成物。
【請求項16】
ヒトへ静脈内投与するための医薬製剤であって、前記医薬製剤は請求項11記載の組成物と、前記医薬製剤を静脈内投与するための前記組成物に容認可能な担体と、を含む医薬製剤。
【請求項17】
前記医薬品は神経医薬品である、請求項16記載の医薬製剤。
【請求項18】
神経医薬品がヒト血液脳関門を越える能力を強化する方法であって、前記神経医薬品を請求項1記載のヒト化マウス抗体へ結合させる工程を含む方法。
【請求項19】
前記神経医薬品はアビジン−ビオチン結合を介して前記ヒト化マウス抗体へ結合されている、請求項18記載の方法。
【請求項20】
前記神経医薬品は遺伝子融合法によって前記ヒト化マウス抗体へ結合されている、請求項18記載の方法。
【請求項21】
インビボで神経医薬品をヒトインスリン受容体にターゲティングさせる方法であって、前記医薬品を請求項1記載のヒト化マウス抗体へ結合させる工程を含む方法。
【請求項22】
前記医薬品はアビジン−ビオチン結合を介して前記ヒト化マウス抗体へ結合される、請求項21記載の方法。
【請求項23】
前記神経医薬品は遺伝子融合法によって前記ヒト化マウス抗体へ結合される、請求項21記載の方法。
【請求項24】
医薬品をヒト患者へ静脈内投与するための方法であって、前記方法は請求項16記載の医薬製剤を前記ヒト患者の血流に注入する工程を含む方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−144178(P2011−144178A)
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−23913(P2011−23913)
【出願日】平成23年2月7日(2011.2.7)
【分割の表示】特願2004−557217(P2004−557217)の分割
【原出願日】平成15年11月18日(2003.11.18)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【出願番号】特願2011−23913(P2011−23913)
【出願日】平成23年2月7日(2011.2.7)
【分割の表示】特願2004−557217(P2004−557217)の分割
【原出願日】平成15年11月18日(2003.11.18)
【出願人】(592130699)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (364)
【氏名又は名称原語表記】The Regents of The University of California
【Fターム(参考)】
[ Back to top ]