
ヒトエンドカインα
【課題】疾患状態に関与するTNFに類似したサイトカインを提供する。
【解決手段】以下をコードするヌクレオチド配列から選択される配列に少なくとも95%同一な配列を有する単離された核酸分子:(a)特定のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによりコードされる全長エンドカインαポリペプチド;(b)約44〜169残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされる細胞外エンドカインαポリペプチド;(c)約18〜43残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされるエンドカインα膜貫通ドメイン;(d)約1〜17残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされるエンドカインα細胞内ドメイン;および(e)上記のいずれかに相補的なヌクレオチド配列。
【解決手段】以下をコードするヌクレオチド配列から選択される配列に少なくとも95%同一な配列を有する単離された核酸分子:(a)特定のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによりコードされる全長エンドカインαポリペプチド;(b)約44〜169残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされる細胞外エンドカインαポリペプチド;(c)約18〜43残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされるエンドカインα膜貫通ドメイン;(d)約1〜17残基の特定のアミノ酸配列を有するか、または上記cDNAクローンによりコードされるエンドカインα細胞内ドメイン;および(e)上記のいずれかに相補的なヌクレオチド配列。
【発明の詳細な説明】
【技術分野】
【0001】
発明の背景
発明の分野
本発明はエンドカインαタンパク質に関する。詳細には、エンドカインαタンパク質をコードする単離された核酸分子が提供される。エンドカインαポリペプチド、ならびにベクター、宿主細胞およびこれらを産生するための組換え方法が提供される。
【背景技術】
【0002】
関連分野
腫瘍壊死因子-α(TNFα;カケクチンとも呼ばれる)は、内毒素または他の刺激に応答して、17kDのタンパク質サブユニットの可溶性のホモトリマーとして、単球およびマクロファージによって主として分泌される(非特許文献1)。膜に結合したTNFの26kDの前駆体形態もまた記載されている(非特許文献2)。
【0003】
蓄積された証拠により、TNFが多面的な生物学的活性を有する調節サイトカインであることが示されている。これらの活性としては以下が挙げられる:リポタンパク質リパーゼ合成(「カケクチン」活性)(非特許文献3)、多形核白血球の活性化(非特許文献4;非特許文献5)、細胞増殖の阻害または細胞増殖の刺激(非特許文献6;非特許文献7; 非特許文献8)、所定の形質転換された細胞型における細胞傷害作用(非特許文献8; 非特許文献9)、抗ウイルス活性(非特許文献10;非特許文献11)、骨再吸収の刺激(非特許文献12; 非特許文献13)、コラゲナーゼおよびプロスタグランジンE2産生の刺激(非特許文献14);ならびにT細胞(非特許文献15)、B細胞(非特許文献16)、単球(非特許文献17)、胸腺細胞(非特許文献18)の活性化を含む免疫調節作用、ならびに主要組織適合複合体(MHC)クラスIおよびクラスII分子の細胞表面発現の刺激(非特許文献19)。
【0004】
TNFは、組織損傷を生じる炎症誘発性作用(例えば、血管内皮細胞における凝血原活性の誘導(非特許文献20)、好中球およびリンパ球の接着の増大(非特許文献21)、およびマクロファージ、好中球および血管内皮細胞由来の血小板活性化因子の放出の刺激(非特許文献22)について記載されている。
【0005】
最近の証拠は、TNFを、多くの感染の原因(非特許文献23)、免疫不全、腫瘍性疾患、例えば、いくつかの悪性腫瘍に付随する悪液質(非特許文献24)、および自己免疫疾患および移植片対宿主疾患(非特許文献25)に結び付ける。TNFのガンおよび感染性疾患との関連は、宿主の異化状態にしばしば関連する。ガン患者における主要な問題は、体重減少(通常、食欲不振に関連する)である。生じる大規模な体力の消耗は「悪液質」として知られる(非特許文献26)。悪液質としては、進行性の体重減少、食欲不振、および悪性増殖に応答した身体質量の持続性の浸食が挙げられる。従って、悪液質の状態は、有意な罹患率と関連し、そしてガン死亡率の大多数を担う。多数の研究により、TNFがガン、感染性病理、および他の異化状態における悪液質の重要なメディエータであることが示唆された。
【0006】
TNFは、グラム陰性敗血症および内毒素ショックの病態生理学的結果において中心的な役割を果たすと考えられている(非特許文献27; 非特許文献28;非特許文献29)。これには、発熱、倦怠感、食欲不振、および悪液質が挙げられる。内毒素は、強力な単球/マクロファージ活性化因子であり、TNFの産生および分泌(非特許文献30)ならびに他のサイトカインの産生および分泌を刺激する。TNFは、内毒素の多くの生物学的影響を模倣し得るので、内毒素関連疾病の臨床的徴候を担う中心的なメディエータであると結論付けられた。TNFおよび他の単球由来サイトカインは、内毒素に対する代謝性および神経ホルモンの応答を媒介する(非特許文献31)。ヒトボランティアへの内毒素投与は、インフルエンザ様症状症状(発熱、頻脈、増大した代謝速度、およびストレスホルモン放出を含む)を伴う急性疾病を生じる(非特許文献32)。循環TNFのレベルの上昇はまた、グラム陰性敗血症を患う患者において見出されている(非特許文献33;非特許文献34;非特許文献35; 非特許文献36)。
【0007】
TNFを中和することに向けられる受動免疫療法は、グラム陰性敗血症および内毒血症(上記で考察したように、これらの病理状態における増大したTNF産生および上昇したTNFレベルに基づく)において有益な効果を有し得る。
【0008】
カケクチンとして特徴付けられた「モジュラー」物質(後にTNFと同一であることが見出された)に対する抗体は、Ceramiら(特許文献1、1987年3月4日)に開示された。このような抗体は、診断的免疫アッセイおよび細菌感染におけるショックの治療に有用であると考えられた。Rubinら(特許文献2、1987年4月22日)は、ヒトTNFに対するモノクローナル抗体、そのような抗体を分泌するハイブリドーマ、そのような抗体を産生する方法、およびTNFのイムノアッセイにおけるそのような抗体の使用を開示した。Yoneら(特許文献3、1988年10月26日)は、mAbを含む抗TNF抗体、および疾患(特に、川崎病、および細菌感染)のイムノアッセイ診断における有用性を開示した。川崎病(乳児の急性発熱性粘膜皮膚リンパ節症候群;非特許文献37;非特許文献38)を有する患者の体液は、この疾患の進行に関連する上昇したTNFレベルを含むと考えられた(Yoneら、前述)。
【0009】
他の研究者らは、インビトロでの中和活性を有する組換えヒトTNFに特異的なmAbを記載した(非特許文献39;非特許文献40;非特許文献41;非特許文献42;非特許文献43; 非特許文献44)。これらのmAbのいくつかは、ヒトTNFのエピトープをマッピングするため、および酵素イムノアッセイを開発するため(非特許文献41;非特許文献43;非特許文献44)、ならびに組換えTNFの精製を補助するため(非特許文献42)に使用された。しかし、これらの研究は、免疫原性、特異性の欠失および/または医薬の適合性に起因する、ヒトにおけるインビボでの診断的または治療的使用に使用され得るTNF中和抗体を産生するための基礎を提供しない。
【0010】
有害な生理学的変化を抑止し、そして実験的な内毒血症および菌血症における致命的なチャレンジの後の死滅を妨げるための、TNFに対する中和抗血清またはmAbがヒト以外の哺乳動物において示されている。この効果は、例えば、齧歯類の死亡率アッセイおよび霊長類の疾患モデル系において示されている(非特許文献45;非特許文献46; 非特許文献47; 非特許文献48; 非特許文献49; 非特許文献50;非特許文献51)。
【0011】
今日まで、ヒトにおける抗TNF mAb治療を用いる実験は、限定はされているが、(例えば、関節炎および敗血症において)有益な治療結果を示している。例えば、非特許文献52;非特許文献53; 非特許文献54; 非特許文献55;非特許文献56を参照のこと。
【0012】
サイトカインレセプターの配列分析により、膜タンパク質の数種のサブファミリーが規定された:(1)Igスーパーファミリー、(2)ヘマトポイエチン(サイトカインレセプタースーパーファミリー)および(3)腫瘍壊死因子(TNF)/神経増殖因子(NGF)レセプタースーパーファミリー(TNFスーパーファミリーの総説については、非特許文献57および非特許文献58を参照のこと)。TNF/NGFレセプタースーパーファミリーは少なくとも10種の異なるタンパク質を含む。非特許文献57。これらのレセプターに対するリガンドは、同定されており、そして少なくとも2つのサイトカインスーパーファミリーに属する。非特許文献57。
【0013】
従って、疾患状態に関与するTNFに類似したサイトカインを提供する必要がある。このような新規のサイトカインは、TNF様障害の治療のための新規の抗体またはこれらのTNF様サイトカインに結合する他のアンタゴニストを作製するために使用され得る。
【特許文献1】欧州特許第0,212,489号明細書
【特許文献2】欧州特許第0,218,868号明細書
【特許文献3】欧州特許第0,288,088号明細書
【非特許文献1】Smith,R.A.ら、J.Biol.Chem.262:6951-6954 (1987)
【非特許文献2】Kriegler,M.ら、Cell53:45-53(1988)
【非特許文献3】Beutler,B.ら、Nature316:552 (1985)
【非特許文献4】Klebanoff,S.J.ら、J.Immunol. 136:4220(1986)
【非特許文献5】Perussia,B.ら、J.Immunol. 138:765 (1987)
【非特許文献6】Vilcek,J.ら、J.Exp.Med.163:632 (1986)
【非特許文献7】Sugarman,B.J.ら、Science 230:943 (1985)
【非特許文献8】Lachman,L.B.ら、J.Immunol.138:2913 (1987)
【非特許文献9】Darzynkiewicz,Z.ら、Canc.Res.44:83 (1984)
【非特許文献10】Kohase, M.ら、Cell 45:659 (1986)
【非特許文献11】Wong, G.H.W.ら、Nature323:819 (1986)
【非特許文献12】Bertolini, D.R.ら、Nature 319:516 (1986)
【非特許文献13】Saklatvala,J.,Nature 322: 547 (1986)
【非特許文献14】Dayer, J.-M.ら、J.Exp.Med.162:2163 (1985)
【非特許文献15】Yokota, S.ら、J.Immunol. 140:531 (1988)
【非特許文献16】Kehrl, J.H.ら、J.Exp.Med.166:786(1987)
【非特許文献17】Philip, R.ら、Nature 323:86 (1986)
【非特許文献18】Ranges, G.E.ら、J.Exp.Med.167:1472 (1988)
【非特許文献19】Collins,T.ら、Proc.Natl.Acad.Sci.USA 83:446 (1986); Pujol-Borrel,R.ら、Nature326:304(1987)
【非特許文献20】Pober,J.S.ら、J.Immunol. 136:1680 (1986)
【非特許文献21】Pober, J.S.ら、J.Immunol.138:3319 (1987)
【非特許文献22】Camussi, G.ら、J.Exp.Med.166:1390(1987)
【非特許文献23】Cerami,A.ら、Immunol.Today9:28 (1988)
【非特許文献24】Oliff, A.ら、Cell 50:555 (1987)
【非特許文献25】Piguet,P.-F.ら、J.Exp.Med. 166:1280 (1987)
【非特許文献26】Kern,K.A.ら、J.Parent.Enter.Nutr. 12:286-298 (1988)
【非特許文献27】Michie,H.R.ら、Br.J.Surg. 76:670-671 (1989)
【非特許文献28】Debets, J.M.H.ら、Second Vienna Shock Forump.463-466 (1989)
【非特許文献29】Simpson, S.Q.ら、Crit.Care Clin. 5:27-47 (1989)
【非特許文献30】Kornbluth,S.K.ら、J.Immunol. 137:2585-2591 (1986)
【非特許文献31】Michie,H.R.ら、N.Eng.J.Med. 318:1481-1486 (1988)
【非特許文献32】Revhaug,A.ら、Arch.Surg.123:162-170 (1988)
【非特許文献33】Waage, A.ら、Lancet1:355-357 (1987)
【非特許文献34】Hammerle, A.F.ら、Second Vienna Shock Forum p.715-718 (1989)
【非特許文献35】Debets, J.M.H.ら、Crit.Care Med. 17:489-497 (1989)
【非特許文献36】Calandra, T.ら、J.Infec.Dis.161:982-987 (1990)
【非特許文献37】Kawasaki, T., Allergy 16:178 (1967)
【非特許文献38】Kawasaki, T., Shonica (Pediatrics) 26:935(1985)
【非特許文献39】Liang, C-M.ら、Biochem.Biophys.Res.Comm. 137:847-854(1986)
【非特許文献40】Meager, A.ら、Hybridoma 6:305-311 (1987)
【非特許文献41】Fendlyら、Hybridoma 6:359-369(1987)
【非特許文献42】Bringman, T.S.ら、Hybridoma 6:489-507 (1987)
【非特許文献43】Hirai, M.ら、J.Immunol.Meth.96:57-62 (1987)
【非特許文献44】Moller, A.ら(Cytokine 2:162-169 (1990))
【非特許文献45】Mathison,J.C.ら、J.Clin.Invest. 81:1925-1937 (1988)
【非特許文献46】Beutler, B.ら、Science 229:869-871(1985)
【非特許文献47】Tracey, K.J.ら、Nature 330:662-664 (1987)
【非特許文献48】Shimamoto, Y.ら、Immunol.Lett.17:311-318 (1988)
【非特許文献49】Silva, A.T.ら、J.Infect.Dis. 162:421-427 (1990)
【非特許文献50】Opal, S.M.ら、J.Infect.Dis.161:1148-1152 (1990)
【非特許文献51】Hinshaw, L.B.ら、Circ.Shock 30:279-292 (1990)
【非特許文献52】Elliott,M.J.ら、Baillieres Clin. Rheumatol. 9:633-52 (1995)
【非特許文献53】Feldmann M.ら、Ann.N.Y.Acad.Sci.USA766:272-8 (1995)
【非特許文献54】van der Poll,Tら、Shock 3: 1-12 (1995)
【非特許文献55】Wherryら、Crit.Care.Med.21:S436-40 (1993)
【非特許文献56】Tracey K.J.ら、Crit.Care Med. 21:S415-22 (1993)
【非特許文献57】GrussおよびDower,Blood 85(12):3378-3404 (1995)
【非特許文献58】AggarwalおよびNatarajan, Eur.Cytokine Netw.,7(2):93-124 (1996)
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明の目的は、疾患状態に関与するTNFに類似したサイトカインを提供することであ
る。
【課題を解決するための手段】
【0015】
本発明は、以下からなる群から選択される配列に少なくとも95%同一であるヌクレオチド配列を有するポリヌクレオチドを含有する単離された核酸分子を提供する:(a)図1(配列番号2)の完全なアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされる全長エンドカインαポリペプチドをコードするヌクレオチド配列;(b)図1(配列番号2)の約44残基〜約169残基のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされる細胞外エンドカ
インαポリペプチドをコードするヌクレオチド配列;(c)図1(配列番号2)の約18残基〜
約43残基のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクロ
ーンによってコードされるエンドカインα膜貫通ドメインをコードするヌクレオチド配列;(d)図1(配列番号2)の約1残基〜約17残基のアミノ酸配列を有するか、またはATCC受
託番号第97640号に含まれるcDNAクローンによってコードされるエンドカインα細胞内ド
メインをコードするヌクレオチド配列;および(e)(a)、(b)、(c)または(d)のヌクレオチ
ド配列のいずれかに相補的なヌクレオチド配列。
【0016】
1つの実施形態において、ストリンジェントなハイブリダイゼーション条件下で、上記の(a)、(b)、(c)、(d)または(e)のヌクレオチド配列に同一なヌクレオチド配列を有する
ポリヌクレオチドにハイブリダイズするポリヌクレオチドを含む単離された核酸分子であって、このポリヌクレオチドがストリンジェントなハイブリダイゼーション条件下でA残基のみまたはT残基のみからなるヌクレオチド配列を有するポリヌクレオチドにはハイブリダイズしない、単離された核酸分子を提供する。
【0017】
1つの実施形態において、上記の(a)、(b)、(c)、(d)または(e)のアミノ酸配列を有す
るエンドカインαポリペプチドのエピトープ保有部分のアミノ酸配列をコードするポリヌクレオチドを含む、単離された核酸分子を提供する。
【0018】
別の局面において、本発明は、以下からなる群から選択されるエンドカインαポリペプチドのエピトープ保有部分をコードする、上記の単離された核酸分子を提供する:図1(配列番号2)の約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)の
約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約94〜約105のアミノ酸残基を含むポリペプチド;図
1(配列番号2)の約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)の約148〜約158のアミノ酸残基を含むポリペプチド。
【0019】
1つの実施形態において、このポリヌクレオチドがRNAである、上記の単離された核酸
分子を提供する。
【0020】
1つの実施形態において、このポリヌクレオチドがRNAである、上記の単離された核酸
分子を提供する。
【0021】
1つの実施形態において、上記の単離された核酸分子をベクターに挿入する工程を包含する、組換えベクターを作製する方法を提供する。
【0022】
1つの実施形態において、上記の方法によって産生された、組換えベクターを提供する。
【0023】
1つの実施形態において、宿主細胞に上記の組換えベクターを導入する工程を包含する、組換え宿主細胞を作製する方法を提供する。
【0024】
1つの実施形態において、上記の方法によって産生された組換え宿主細胞を提供する。
【0025】
別の局面において、エンドカインαポリペプチドを産生するための組換え方法を提供し、この方法は、上記の組換え宿主細胞を、このポリペプチドが発現されるような条件下で培養し、そしてこのポリペプチドを回収する工程を包含する。
【0026】
別の局面において、以下からなる群から選択される配列に少なくとも95%同一であるアミノ酸配列を有する単離されたポリペプチドを提供する:(a)図1(配列番号2)の全長の
エンドカインαポリペプチド、またはATCC受託番号第97640号に含まれるcDNAクローンに
よってコードされる全長エンドカインαポリペプチドのアミノ酸配列;(b)図1(配列番号2)の約44残基〜約169残基、またはATCC受託番号第97640号に含まれるcDNAクローンによ
ってコードされる細胞外エンドカインαポリペプチドのアミノ酸配列;(c)図1(配列番号2)の約18残基〜約43残基、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされるエンドカインα膜貫通ドメインのアミノ酸配列;(d)図1(配列番号2)の
約1残基〜約17残基、またはATCC受託番号第97640号に含まれるcDNAクローンによってコ
ードされるエンドカインα細胞内ドメインのアミノ酸配列;および(e)(a)、(b)、(c)または(d)のポリペプチド配列のいずれか1つのエピトープ保有部分のアミノ酸配列。
【0027】
別の局面において、エンドカインαのエピトープ保有部分を含む単離されたポリペプチドを提供し、この部分が以下からなる群から選択される、単離されたポリペプチド:図1(配列番号2)の約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)
の約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約94〜約105のアミノ酸残基を含むポリペプチド;
図1(配列番号2)の約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)の約148〜約158のアミノ酸残基を含むポリペプチド。
【0028】
1つの実施形態において、上記のエンドカインαポリペプチドに特異的に結合する、単離された抗体または抗体フラグメントを提供する。
【0029】
別の局面において、エンドカインα活性のレベルの減少を必要とする個体を処置する方法を提供し、この方法は、上記の単離された抗体または抗体フラグメントを含む組成物を該個体に投与する工程を包含する。
【0030】
別の局面において、以下の工程を包含する診断方法を提供する:(a)哺乳動物の細胞ま
たは体液中のエンドカインα遺伝子発現レベルをアッセイする工程;および(b)該エンド
カインα遺伝子発現レベルと標準のエンドカインα遺伝子発現レベルとを比較する工程であって、それによって該標準と比較した該エンドカインα遺伝子発現レベルにおける増大または減少がTNF関連障害の指標となる、工程。
【発明の効果】
【0031】
本発明によって、疾患状態に関与するTNFに類似したサイトカインが提供される。
【発明を実施するための最良の形態】
【0032】
発明の要旨
本発明は、TNFに類似し、そして類似の生物学的効果および活性を有すると考えられる
サイトカインをコードするポリヌクレオチドを含む単離された核酸分子を提供する。このサイトカインは、エンドカインαと呼ばれ、そして図1(配列番号2)のアミノ酸配列、または1996年6月27日にATCC受託番号97640として細菌宿主中で寄託されたcDNAクローン
によりコードされるアミノ酸配列の少なくとも一部を有するエンドカインαポリペプチドを含む。このヌクレオチド配列(寄託されたエンドカインαのcDNAクローンを配列決定することによって決定された)は、約169アミノ酸残基のポリペプチドをコードするオープ
ンリーディングフレームを含む。これは、N末端のメチオニン、約17アミノ酸残基の細胞内ドメイン、約26アミノ酸の膜貫通ドメイン、約126アミノ酸の細胞外ドメイン、および
約19kDaの完全なタンパク質についての推定の分子量を含む。予想される成熟エンドカイ
ンαタンパク質の126アミノ酸配列は、図1(配列番号2)(残基44〜169)に示される。
【0033】
別の局面において、本発明は、1996年6月27日にATCC受託番号97640として寄託された
クローンのcDNAによってコードされるアミノ酸配列を有するエンドカインαポリペプチドをコードする単離された核酸分子を提供する。好ましくは、この核酸分子は、上記の寄託されたcDNAによってコードされる成熟ポリペプチドをコードする。
【0034】
本発明は、さらに、本明細書中に記載の核酸分子の核酸フラグメントに関する。好ましい核酸フラグメントとしては、以下をコードする核酸分子が挙げられる:エンドカインα細胞内ドメイン(図1(配列番号2)の約1〜約17のアミノ酸残基)を含むポリペプチド;エンドカインα膜貫通ドメインを含むポリペプチド(図1(配列番号2)の約18〜約43のアミノ酸残基);およびエンドカインα細胞外ドメインを含むポリペプチド(図1(配列番
号2)の約44〜約169のアミノ酸残基)。
【0035】
本発明のさらなる実施態様は、本明細書中に記載される核酸分子のいずれかのヌクレオチド配列に少なくとも90%同一、そしてより好ましくは、少なくとも95%、96%、97%、98%、または99%同一であるヌクレオチド配列を有する単離された核酸分子を含む。
【0036】
本発明はまた、本発明の単離された核酸分子を含む組換えベクター、その組換えベクターを含有する宿主細胞、および組換え技術によるエンドカインαポリペプチドの産生に関する。
【0037】
本発明のポリペプチドは、寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド(詳細には、成熟ポリペプチド)、ならびに寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド、またはそれらのフラグメントのアミノ酸配列に対して少なくとも90%の類似性、より好ましくは少なくとも95%の類似性を有するアミノ酸配列を有するポリペプチドを含む。本発明のさらなるポリペプチドは、寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド、またはそれらのフラグメントのアミノ酸配列に少なくとも80%同一、より好ましくは少なくとも90%または95%同一であるアミノ酸配列を有するポリペプチドを含む。
【0038】
本発明の好ましいポリペプチドフラグメントは、以下を含むポリペプチドを含む:エンドカインα細胞内ドメイン、エンドカインα膜貫通ドメイン、およびエンドカインα細胞外ドメイン。
【0039】
本発明は、上記のようなアミノ酸を有するエンドカインαポリペプチドに特異的に結合する抗体を単離する方法をさらに提供する。このような抗体は、エンドカインαおよび/
またはTNF関連障害の処置において、アンタゴニストとして診断的または治療的に有用で
あり得る。
【0040】
発明の詳細な説明
本発明は、図1(配列番号2)に示されるアミノ酸配列(クローン化されたcDNAを配列決定することによって決定した)を有するエンドカインαタンパク質をコードするポリヌクレオチドを含む、単離された核酸分子を提供する。エンドカインαは、腫瘍壊死因子(TNF)リガンドファミリーの新規なメンバーであり、そしてヒトTNFαおよび関連するTNFファミリーメンバー(図2)と配列相同性を共有する。図1(配列番号1)に示されるヌクレオチド配列は、cDNAクローンを配列決定することによって得られた。これは、1996年6月27日に、アメリカンタイプカルチャーコレクション,12301 Park Lawn Drive, Rockville, Maryland 20852に寄託され、そして受託番号第97640号を与えられた。寄託されたクローンは、pBluescriptSK(-)プラスミド(Stratagene, LaJolla, CA)中に含まれる。
【0041】
核酸分子
他に示されない限り、本明細書中でDNA分子を配列決定することによって決定されたす
べてのヌクレオチド配列は、自動化DNA配列決定機(例えば、AppliedBiosystems, Inc.からのModel 373)を用いて決定され、そして本明細書中で決定されるDNA分子によってコードされるポリペプチドのすべてのアミノ酸配列は、上記のように決定されるDNA配列の翻
訳によって予想された。従って、この自動化アプローチによって決定された任意のDNA配
列について当該分野において公知のように、本明細書中で決定される任意のヌクレオチド配列はいくつかの誤りを含み得る。自動化によって決定されるヌクレオチド配列は、配列
決定されるDNA分子の実際のヌクレオチド配列に対して、代表的には少なくとも約90%の
同一、より代表的には少なくとも約95%から少なくとも約99.99%の同一である。実際の
配列は、当該分野において周知の手動DNA配列決定方法を含む他のアプローチによってよ
り正確に決定され得る。当該分野においてまた公知のように、実際の配列と比較した、決定されるヌクレオチド配列における単一の挿入または欠失は、ヌクレオチド配列の翻訳におけるフレームシフトを引き起こし、その結果、決定されるヌクレオチド配列によってコードされる予想されるアミノ酸配列は、挿入または欠失のような点にて始まる配列決定されるDNA分子によって実際にコードされるアミノ酸配列とは完全に異なる。
【0042】
他に示されない限り、本明細書中に示される各「ヌクレオチド配列」は、デオキシリボヌクレオチド(A, G, C, およびTと省略される)の配列として示される。しかし、核酸分
子またはポリヌクレオチドの「核酸配列」によって、DNA分子またはポリヌクレオチドに
はデオキシリボヌクレオチドが、そしてRNA分子またはポリヌクレオチドにはリボヌクレ
オチド(A,G, C, およびU)の対応する配列(ここで特定されるデオキシヌクレオチド配列における各チミジンデオキシヌクレオチド(T)は、リボヌクレオチドのウリジン(U)によって置き換えられる)が意図される。例えば、デオキシリボヌクレオチドの略語を用いて示される図1(配列番号1)の配列を有するRNA分子との言及は、配列番号1の各デオキ
シヌクレオチドA,GまたはCが、対応するリボヌクレオチドA, G, またはCによって置き換
えられ、そして各デオキシヌクレオチドTが、リボヌクレオチドUによって置き換えられる配列を有するRNA分子を示すことが意図される。
【0043】
本明細書中で提供される情報(例えば、図1中のヌクレオチド配列)を用いて、エンドカインαポリペプチドをコードする本発明の核酸分子は、標準的なクローニングおよびスクリーニング手順(例えば、開始物質としてmRNAを用いるクローニングcDNAのための手順)を用いて得られ得る。本発明の図のように、図1(配列番号1)において記載される核酸分子は、ヒト脳線条由来のcDNAライブラリー中で発見された。エンドカインαcDNAの一部に対応する発現される配列タグはまた、いくつかの内皮ライブラリーおよび胎児肝臓ライブラリーにおいて見出された。
【0044】
エンドカインα遺伝子は、約169のアミノ酸残基のタンパク質をコードするオープンリ
ーディングフレーム、約17のアミノ酸の細胞内ドメイン(図1(配列番号2)中の約1〜約17のアミノ酸残基)、約26のアミノ酸の膜貫通ドメイン(図1(配列番号2)中の約18〜約43のアミノ酸残基)、約126アミノ酸の細胞外ドメイン(図1(配列番号2)中の約44〜約169のアミノ酸残基);および約19kDaの推定分子量を含む。図1(配列番号2)に
おいて示されるエンドカインαタンパク質は、ヒトTNF-αに対して約30%の類似し、そして約22%同一であり、それは、GenBankに、受託番号第U42764号として利用され得る。
【0045】
当業者が理解するように、上記の配列決定誤差の可能性、ならびに異なる公知のタンパク質におけるリーダーの切断部位の可変性に起因して、寄託されたcDNAによってコードされる実際のエンドカインαポリペプチドは、約169のアミノ酸を含むが、約154〜184の範
囲のアミノ酸で任意の場所であり得る。当業者によって、使用される基準に依存して、上記のエンドカインαタンパク質ドメインの正確な「位置」が異なり得ることもまた理解される。従って、例えば、図1(配列番号2)において示されるエンドカインα細胞内膜貫通ドメインおよび細胞外ドメインの正確な位置は、ドメインを定義するために使用される基準に依存してわずかに変化し得る(例えば、正確な位置は、図1に示されるものと比較して約1〜約5の残基で異なり得る)。
【0046】
示されるように、本発明の核酸分子は、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNA、およびクローニングによって得られるか、または合成的に生成されるゲノムDNAを含む)であり得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAは、コ
ード鎖(センス鎖としても知られる)であり得るか、またはそれは、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0047】
「単離された」核酸分子によって、天然の環境から取り出された核酸分子(DNAまたはRNA)が意図される。例えば、ベクター中に含まれる組換えDNA分子は、本発明の目的のた
めに単離されることが意図される。単離されたDNA分子のさらなる例は、異種の宿主細胞
において維持される組換えDNA分子、または溶液中の精製された(部分的または実質的)DNA分子を含む。単離されたRNA分子は、本発明のDNA分子のインビボまたはインビトロでのRNA転写物を含む。本発明に従って単離された核酸分子は、合成的に生成されるような分
子をさらに含む。
【0048】
本発明の単離された核酸分子は、図1(配列番号1)において示されるオープンリーディングフレーム(ORG)を含むDNA分子を含み、そしてさらに、図1(配列番号1)におい
て示されるORF配列のすべてまたは一部と実質的に異なるが、遺伝コードの縮重に起因し
て、エンドカインαタンパク質またはそのフラグメントをなおコードする核酸分子を含む。当然のことながら、遺伝コードは当該分野において周知である。従って、上記のような縮重した変異体を生成することは、当業者にとって日常的である。
【0049】
別の局面において、本発明は、1996年6月27日にATCC寄託番号97640として寄託された
クローンのcDNAによってコードされるアミノ酸配列を有するエンドカインαポリペプチドをコードする単離された核酸分子を提供する。好ましくは、核酸分子は、上記の寄託されたcDNAクローンによってコードされる成熟ポリペプチドをコードする。本発明は、さらに、図1(配列番号1)に示されるヌクレオチド配列、もしくは上記の寄託されたクローン中に含まれるエンドカインαcDNAのヌクレオチド配列を有する単離された核酸分子、または上記の配列の一つと相補的な配列を有する核酸分子を提供する。そのような単離された分子、特にDNA分子は、染色体とのインサイチュハイブリダイゼーションによって遺伝子
マッピングするため、および例えばノーザンブロット分析によってヒト組織中のエンドカインα遺伝子の発現を検出するためのプローブとして有用である。以下に詳細に記載されるように、特定の組織または体液におけるエンドカインα遺伝子発現の変化を検出することは、特定の障害を示す。
【0050】
本発明は、さらに、本明細書中で記載される単離した核酸分子のフラグメントに関する。寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)に示されるヌクレオチド配列を有する単離された核酸分子のフラグメントによって、本明細書中で考察されるように、診断的プローブおよびプライマーとして有用である、長さが、少なくとも約15nt、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntが意図される。当然のことながら、長さがより大きなフラグメント50〜1500ntもまた、寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)において示されるようなヌクレオチド配列のすべてではなくともほとんどに対応するフラグメントのように、本発明に従って有用である。例えば、長さが少なくとも20ntのフラグメントによって、寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)に示されるようなヌクレオチド配列からの20以上の隣接した塩基を含むフラグメントが意図される。この遺伝子が寄託され、そして図1(配列番号1)において示されるヌクレオチド配列が提供されるので、そのようなDNAフラグメントを生成することは、当
業者にとって日常的である。例えば、制限エンドヌクレアーゼ切断、または超音波処理による剪断は、種々のサイズのフラグメントを生成するために容易に使用され得る。あるいは、そのようなフラグメントは、合成的に生成され得る。
【0051】
本発明の好ましい核酸フラグメントは、以下をコードする核酸分子を含む:エンドカインα細胞内ドメインを含むポリペプチド(図1(配列番号2)中の約1〜約17のアミノ酸
残基);エンドカインα膜貫通ドメインを含むポリペプチド(図1(配列番号2)中の約18〜約43のアミノ酸残基);およびエンドカインα細胞外ドメインを含むポリペプチド(図1(配列番号2)中の約44〜約169のアミノ酸残基)。
【0052】
本発明のさらに好ましい核酸フラグメントは、エンドカインαタンパク質のエピトープ保有部分をコードする核酸分子を含む。特に、本発明のそのような核酸フラグメントは、以下をコードする核酸分子を含む:図1(配列番号2)中の約44〜約158のアミノ酸残基
を含むポリペプチド;図1(配列番号2)中の約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約94〜約105のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約108〜約132
のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約148〜約158のアミノ酸残基を含むポリペプチド。本発明者らは、上記のポリペプチドフラグメントが、エンドカインαタンパク質の抗原性領域であることを決定した。エンドカインαタンパク質の他のそのようなエピトープ保有部分を決定するための方法は、以下に詳細に記載される。
【0053】
別の局面において、本発明は、ストリンジェントなハイブリダイゼーション条件下で、上記の本発明の核酸分子(例えば、1996年6月27日に作成されたATCC寄託第97640号中に
含まれるcDNAクローン)中のポリヌクレオチドの一部にハイブリダイズするポリヌクレオチドを含む単離した核酸分子を提供する。「ストリンジェントなハイブリダイゼーション条件」によって、括弧内を含む溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNA)中42℃での一晩のインキュベ
ーション、続く約65℃にて0.1×SSCにおいてフィルターを洗浄することが意図される。ポリヌクレオチドの「一部」にハイブリダイズするポリヌクレオチドによって、参照のポリヌクレオチドの少なくとも約15ヌクレオチド(nt)、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは約30〜70ntにハイブリダイズするポリヌクレオチド(DNAまたはRNAのいずれか)が意図される。これらは、以上、および以下でより詳細に考察されるような診断用プローブおよびプライマーとして有用である。
【0054】
当然のことながら、参照のポリヌクレオチド(例えば、寄託されたcDNAクローン)のより大きな部分(例えば、長さが50〜500ntの部分)、または参照のポリヌクレオチドの完
全長とまでもハイブリダイズするポリヌクレオチドはまた、寄託されたcDNAのヌクレオチド配列または図1(配列番号1)に示されるようなヌクレオチド配列のすべてではなくともほとんどに対応するポリヌクレオチドのように、本発明に従ったプローブとして有用である。例えば、「長さが少なくとも20nt」のポリヌクレオチドの部分によって、参照のポリヌクレオチドのヌクレオチド配列(例えば、寄託されたcDNA、または図1(配列番号1)に示されるようなヌクレオチド配列)からの20以上の隣接したヌクレオチドが意図される。示されるように、そのような部分は、従来のDNAハイブリダイゼーション技術に従っ
たプローブとして、または例えば、Sambrook,J.ら編、Molecular Cloning, A Laboratory
Manual, 第2版、Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY (1989)(その全開示は、本明細書中において参考として援用される)に記載されるようなポリメラーゼ連鎖反応(PCR)による標的配列の増幅のためのプライマーとしてのいずれかで診
断的に有用である。
【0055】
エンドカインαcDNAクローンは寄託されており、そしてそのヌクレオチド配列が図1(配列番号1)において提供されるので、エンドカインαcDNA分子の一部にハイブリダイズするポリヌクレオチドを生成することは、当業者に日常的である。例えば、エンドカインαcDNAクローンの制限エンドヌクレアーゼ切断または超音波による剪断は、エンドカイン
αcDNA分子の一部にハイブリダイズするポリヌクレオチドである種々のサイズのDNA部分
を生成するために容易に使用され得る。あるいは、本発明のハイブリダイズするポリヌクレオチドは、公知の技術に従って合成的に生成され得る。
【0056】
当然のことながら、ポリA配列(例えば、図1(配列番号1)に示されるエンドカイン
αcDNAの3’末端ポリ(A)付加物)にのみ、またはT(またはU)の相補的な部分(stretch)
にハイブリダイズするポリヌクレオチドは、本発明の核酸の一部にハイブリダイズするために使用される本発明のポリヌクレオチドに含まれない。なぜなら、そのようなポリヌクレオチドは、poly(A)伸長またはその相補物を含む任意の核酸分子(例えば、特に任意の
二本鎖cDNAクローン)にハイブリダイズする。
【0057】
示されるように、エンドカインαタンパク質をコードする本発明の核酸分子は、以下を含むが、それらに限定されない:それ自体によって、成熟ペプチドのアミノ酸配列をコードする核酸分子;成熟したポリペプチドのコード配列およびさらなる配列(例えば、プレタンパク質配列またはプロタンパク質配列またはプレプロタンパク質配列);成熟ポリペプチドのコード配列で、上記のさらなるコード配列を含むかまたは含まず、更なる非コード配列とともに集まっており、この配列はイントロンおよび非コード5’および3’配列(例えば、転写、mRNA(スプライシングおよびポリアデニル化シグナル(例えば、リボソーム結合およびmRNAの安定性)を含む)において役割を担う転写非翻訳配列)を含むがこれらに限定されない;さらなる機能性を提供するようなさらなるアミノ酸をコードするさらなるコード配列。従って、例えば、ポリペプチドをコードする配列は、マーカー配列(例えば、融合されたポリペプチドの精製を容易にするペプチドをコードする配列)に融合され得る。本発明のこの局面の特定の好ましい実施態様において、マーカーアミノ酸配列は、ヘキサ-ヒスチジンペプチド(例えば、pQEベクター(Qiagen,Inc.)において提供されるタグ)、他の中では、それらの多くは公的および/または商業的に入手可能である。例えば、Gentzら、Proc. Natl.Acad. Sci. USA 86: 821-824 (1989)において記載されるよう
に、ヘキサヒスチジンは、融合タンパク質の簡便な精製を提供する。「HA」タグは、インフルエンザ赤血球凝集素(HA)タンパク質由来のエピトープに対応する精製のために有用な別のペプチドであり、それは、Wilsonら、Cell37: 767 (1984)によって記載されている。他のそのような融合タンパク質は、NまたはC末端にてFcに融合されるエンドカインαタンパク質を含む。
【0058】
本発明は、さらに、エンドカインαタンパク質の一部、アナログ、または誘導体をコードする本発明の核酸分子の変異体に関する。変異体は、天然の対立遺伝子変異体のように、天然に生じ得る。「対立遺伝子変異体」によって、生物の染色体上の所定の遺伝子座を占める遺伝子のいくつかの交換可能な形態の1つが意図される。天然に存在しない変異体は、例えば当該分野で公知の変異誘発技術を用いて生成され得る。
【0059】
そのような変異体は、ヌクレオチド置換、欠失、または添加によって生成される変異体を含む。置換、欠失、または添加は、1つ以上のヌクレオチドを含み得る。変異体は、コードもしくは非コード領域、またはその両方において変化され得る。コード領域における変異は、保存性もしくは非保存性アミノ酸置換、欠失、または添加を生成し得る。これらの中で特に好ましいものは、サイレント置換、添加、および欠失であり、これらは、エンドカインαタンパク質またはその一部の特性および活性を変化しない。これらの点において特にまた好ましいものは、保存性置換である。最も高度に好ましいものは、図1(配列番号2)において示されるアミノ酸配列、または寄託されたcDNAクローンによってコードされる成熟エンドカインαアミノ酸配列を有する成熟エンドカインαタンパク質をコードする核酸分子である。
【0060】
本発明のさらなる実施態様は、(a)図1(配列番号2)に示される完全なアミノ酸配列
を有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされる
全長エンドカインαポリペプチドをコードするヌクレオチド配列;(b)図1(配列番号2
)中のほぼ44位からほぼ169位のアミノ酸配列を有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされるエンドカインα細胞外ドメインをコードするヌクレオチド配列;(c)図1(配列番号2)中のほぼ18位からほぼ43位のアミノ酸配列を
有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされるエ
ンドカインα膜貫通ドメインをコードするヌクレオチド配列;(d)図1(配列番号2)中
のほぼ1位からほぼ17位でのアミノ酸配列を有するか、もしくはATCC受託番号97640に含
まれるcDNAクローンによってコードされるエンドカインα細胞内ドメインをコードするヌクレオチド配列;または(e) (a)、(b)、(c)、もしくは(d)のヌクレオチド配列のいずれかと相補的なヌクレオチド配列と、少なくとも90%同一のヌクレオチド配列を有し、そしてより好ましくは少なくとも95%、96%、97%、98%、または99%同一のヌクレオチド配列を有するポリヌクレオチドを含む単離された核酸分子を含む。
【0061】
エンドカインαポリペプチドをコードする参照ヌクレオチド配列と少なくとも、例えば、95%「同一」のヌクレオチド配列を有するポリヌクレオチドによって、ポリヌクレオチド配列が、エンドカインαポリペプチドをコードする参照ヌクレオチド配列の各100ヌク
レオチドあたり5つまでの点変異を含み得ることを除いて、ポリヌクレオチドのヌクレオチド配列が、参照配列と同一であることが意図される。言い換えれば、参照ヌクレオチド配列と少なくとも95%同一であるヌクレオチド配列を有するポリヌクレオチドを得るために、参照配列における5%までのヌクレオチドが欠失され得るかもしくは別のヌクレオチドで置換され得るか、または参照配列において全ヌクレオチドの5%までの多数のヌクレオチドが、参照配列中に挿入され得る。参照配列のこれらの変異は、参照ヌクレオチド配列の5’もしくは3’末端位置でか、または参照配列内のヌクレオチドの中で個々に、もしくは参照配列内の1つ以上の連続した群でのいずれかで散在されて、これらの末端位置の間のどこかで生じ得る。
【0062】
実際には、任意の特定の核酸分子が、図1に示されるヌクレオチド配列または寄託されたcDNAクローンのヌクレオチド配列に少なくとも90%、95%、96%、97%、98%、または99%同一であるかどうかは、例えば、BESTFITプログラム(WisconsinSequence Analysis Package, Version 8 for Unix(登録商標), Genetics Computer Group,University Research Park, 575 Science Drive, Madison, WI 53711)のような公知のコンピュータープログラムを用いて従来通りに決定され得る。BESTFITは、SmithおよびWaterman,Adv. Appl. Math. 2:482-489 (1981)の局所的相同性アルゴリズムを用いて、2つの配列間の相同性の最適のセグメントを見出す。BESTFITまたは任意の他の配列整列プログラムを使用して、
特定の配列が、例えば本発明による参照配列に95%同一であるか否かを決定する場合、パラメーターは、もちろん、同一性の割合が、参照ヌクレオチド配列の全長にわたって計算されるように、そして参照配列の総ヌクレオチド数の5%までの相同性におけるギャップが許容されるように、設定される。
【0063】
本出願は、エンドカインαタンパク質活性を有するポリペプチドをコードするか否かに関係なく、上記の核酸配列と少なくとも90%、95%、96%、97%、98%、または99%同一であるこのような核酸分子に関する。これは、特定の核酸分子がエンドカインα活性を有するポリペプチドをコードしない場合でさえ、当業者は、核酸分子をどのようにして、例えば、ハイブリダイゼーションプローブ、またはポリメラーゼ連鎖反応(PCR)のプライマ
ーとして使用するかをなお知るからである。エンドカインα活性を有するポリペプチドをコードしない本発明の核酸分子の使用としては、とりわけ(1)エンドカインα遺伝子また
はその対立遺伝子変異体をcDNAライブラリーから単離すること;(2)Vermaら,Human Chromosomes: a Manual of Basic Techniques, Pergamon Press, New York(1988)に記載の、エンドカインα遺伝子の正確な染色体位置を提供するための分裂中期染色体展開物に対する
インサイチュハイブリダイゼーション(FISH);および、(3)特定の組織におけるエンドカ
インαmRNA発現を検出するためのノーザンブロット分析、が挙げられる。
【0064】
しかし、実際に、エンドカインαタンパク質活性を有するポリペプチドをコードする上記の核酸配列に少なくとも90%、95%、96%、97%、98%、または99%同一である配列を有するこのような核酸分子が好ましい。「エンドカインα活性を有するポリペプチド」によって、特定の生物学的アッセイにおいて測定される場合、エンドカインαタンパク質と比較して類似する(しかし、必ずしも同一ではない)活性を示すポリペプチドが意図される。エンドカインα活性は、公知の方法に従ってアッセイされ得る。例えば、細胞傷害性アッセイまたは細胞増殖アッセイが使用され得、ここでエンドカインαポリペプチドは培養物中の細胞へ添加され、そして細胞に対するエンドカインの効果が、細胞数の増大または減少を測定することによって決定される。
【0065】
もちろん、遺伝子コードの縮重のために、当業者は、上記の核酸配列に少なくとも90%、95%、96%、97%、98%、または99%同一である配列を有する多数の核酸分子が、「エンドカインαタンパク質活性を有する」ポリペプチドをコードすることを直ちに認識する。実際に、縮重改変体の全ては同じポリペプチドをコードするので、これは上記の比較アッセイを実施することなしでさえ当業者に明らかである。縮重改変体でないそのような核酸分子について、合理的な数がまたエンドカインαタンパク質活性を有するポリペプチドをコードすることが、当該分野でさらに認識される。これは、当業者が、タンパク質の機能により低い有意性で影響しそうであるか、または有意には影響しそうにないかのいずれかのアミノ酸置換(例えば、1つの脂肪族アミノ酸の第二の脂肪族アミノ酸への置換)を完全に知っているからである。
【0066】
例えば、どのように表現型的にサイレントなアミノ酸置換を作製するかに関するガイダンスは、Bowie, J.U.ら, Science 247:1306-1310(1990)に提供される。ここで著者らは、アミノ酸配列の変化に対する寛容を研究するための2つの主要なアプローチが存在することを示す。第一の方法は、進化のプロセスに依存し、ここで変異は、自然淘汰によって受容されるか拒絶されるかのいずれかである。第二のアプローチは、クローン化遺伝子の特定の位置でアミノ酸変化を導入するために遺伝子操作を、そして機能を維持する配列を同定するために選択またはスクリーニングを用いる。著者らが述べるように、これらの研究は、タンパク質が、アミノ酸置換に驚くほど寛容であることを明らかにしている。著者らは、どのアミノ酸の変化が、タンパク質の特定の位置で許容性でありそうであるかをさらに示している。例えば、大部分の埋没したアミノ酸残基は非極性側鎖を必要とし、一方表面の側鎖の特性は一般的にわずかしか保存されていない。他のこのような表現型的にサイレントな置換は、Bowie,J.U.ら,前出およびその中に引用される参考文献に記載される。
【0067】
ベクターおよび宿主細胞
本発明はまた、本発明の単離されたDNA分子を含むベクター、組換えベクターで遺伝子
操作された宿主細胞、および組換え技術によるエンドカインαポリペプチドまたはその部分の産生に関する。
【0068】
組換え構築物は、感染、形質導入、トランスフェクション、トランスベクション(transvection)、エレクトロポレーション、および形質転換のような周知の技術を用いて宿主細胞に導入され得る。ベクターは、例えば、ファージベクター、プラスミドベクター、ウイルスベクター、またはレトロウイルスベクターであり得る。レトロウイルスベクターは、複製可能かまたは複製欠損であり得る。後者の場合、ウイルスの増殖は、一般的に、相補宿主細胞においてのみ生じる。
【0069】
ポリヌクレオチドは、宿主細胞における増殖のための選択マーカーを含むベクターに結
合され得る。一般的に、プラスミドベクターは、リン酸カルシウム沈殿物のような沈殿物中か、または荷電された脂質との複合体中で導入される。ベクターがウイルスである場合、ベクターは、適切なパッケージング細胞株を用いてインビトロでパッケージングされ得、次いで宿主細胞に形質導入され得る。
【0070】
目的のポリヌクレオチドに対するシス作用性制御領域を含むベクターが好ましい。適切なトランス作用性因子は、宿主によって供給される得るか、相補ベクターによって供給され得るか、または宿主への導入の際にベクター自体によって供給され得る。
【0071】
この事に関する特定の好ましい実施態様において、ベクターは、誘導性および/または細胞型特異的であり得る特異的な発現を提供する。このようなベクターの中で特に好ましいベクターは、温度および栄養添加物のような操作することが容易である環境因子によって誘導性のベクターである。
【0072】
本発明において有用な発現ベクターとしては、染色体ベクター、エピソームベクター、およびウイルス由来ベクター(例えば、細菌プラスミド、バクテリオファージ、酵母エピソーム、酵母染色体エレメント、ウイルス(例えば、バキュロウイルス、パポバウイルス、ワクシニアウイルス、アデノウイルス、トリポックスウイルス、仮性狂犬病ウイルス、およびレトロウイルス)、ならびにそれらの組合せに由来するベクター(例えば、コスミドおよびファージミド)が挙げられる。例えば、Ausubel,下記、Sambrook,下記を参照の
こと。
【0073】
DNAインサートは、適切なプロモーター(例えば、少し名を挙げると、ファージλPLプ
ロモーター、E.coli:lacプロモーター、trpプロモーター、およびtacプロモーター、SV40初期プロモーターおよび後期プロモーター、ならびにレトロウイルスLTRのプロモーター
)に作動可能に連結されるべきである。他の適切なプロモーターは、当業者に公知である。発現構築物は、さらに、転写開始、転写終結のための部位、および、転写領域中に翻訳のためのリボゾーム結合部位を含む。構築物によって発現される成熟転写物のコード部分は、翻訳されるべきポリペプチドの始めに転写開始AUGを含み、そして終わりに適切に位
置される終止コドンを含む。
【0074】
示されるように、発現ベクターは、好ましくは少なくとも1つの選択マーカーを含む。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼ耐性またはネオマイシン耐性、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。適切な宿主の代表的な例としては、細菌細胞(例えば、E.coli細胞、Streptomyces細胞、およびSalmonellatyphimurium細胞);真菌細胞(例えば酵母細胞);昆虫細胞(例えば、Drosophila S2細胞およびSpodoptera Sf9細胞);動物細胞(例えば、CHO細胞、COS細胞、およびBowes黒色
腫細胞);ならびに植物細胞が挙げられる。上記の宿主細胞のための適切な培養培地および条件は当該分野で公知である。
【0075】
細菌における使用に好ましいベクターの中には、pQE70、pQE60、およびpQE-9(Qiagen
から入手可能);pBSベクター、Phagescriptベクター、Bluescriptベクター、pNH8A、pNH16a、pNH18A、pNH46A(Stratageneから入手可能);ならびにptrc99a、pKK223-3、pKK233-3、pDR540、pRlT5(Pharmaciaから入手可能)が含まれる。好ましい真核生物ベクターの中には、pWLNEO、pSV2CAT、pOG44、pXT1、およびpSG(Stratageneから入手可能);ならびにpSVK3、pBPV、pMSG、およびpSVL(Pharmacicaから入手可能)ががある。他の適切なベク
ターは、当業者に容易に明らかである。
【0076】
本発明における使用に適した公知の細菌プロモーターの中には、E.coli lacIおよびlac
Zプロモーター、T3プロモーターおよびT7プロモーター、gptプロモーター、λPRプロモーターおよびλPLプロモーター、ならびにtrpプロモーターが含まれる。適切な真核生物プ
ロモーターとしては、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、初期SV40プロモーターおよび後期SV40プロモーター、レトロウイルスLTRのプロモーター(例
えば、ラウス肉腫ウイルス(RSV)のプロモーター)、ならびにメタロチオネインプロモー
ター(例えば、マウスメタロチオネインIプロモーター)が挙げられる。
【0077】
宿主細胞への構築物の導入は、リン酸カルシウムトランスフェクション、DEAEデキストラン媒介トランスフェクション、カチオン性脂質媒介トランスフェクション、エレクトロポレーション、形質導入、感染または他の方法によってもたらされ得る。このような方法は、Davisら,Basic Methods in Molecular Biology(1986)のような多くの標準的研究室マニュアルに記載されている。
【0078】
高等真核生物による本発明のポリペプチドをコードするDNAの転写は、ベクター中にエ
ンハンサー配列を挿入することによって増大させ得る。エンハンサーは、所定の宿主細胞型におけるプロモーターの転写活性を増大するように働く、通常約10〜300bpのDNAのシス作用性エレメントである。エンハンサーの例としては、SV40エンハンサー(これは、複製起点の後期側上の100〜270bpに位置される)、サイトメガロウイルスの初期プロモーターエンハンサー、複製起点の後期側上のポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。
【0079】
翻訳されたタンパク質の小胞体の管腔内へか、周辺質空間内へか、または細胞外環境内への分泌のために、適切な分泌シグナルが、発現されるポリペプチド中に組み込まれ得る。シグナルは、ポリペプチドに対して内因性であり得るか、またはそれらは異種シグナルであり得る。
【0080】
ポリペプチドは、融合タンパク質のような改変された形態で発現され得、そして分泌シグナルだけでなく、付加的な異種の機能的領域も含み得る。さらなる例において、付加的なアミノ酸、特に荷電性アミノ酸の領域が、宿主細胞内での、精製の間の、または続く操作および保存の間の、安定性および持続性を改善するために、ポリペプチドのN末端に付加され得る。また示されるように、領域(単数または複数)はまた、精製を容易にするためにポリペプチドへ付加され得る。そのような領域は、ポリペプチドの最終調製の前に除去され得る。とりわけ、分泌および排出を生じるため、安定性を改善するため、および精製を容易にするためのペプチド部分のポリペプチドへの付加は、当該分野でよく知られており、そして日常的な技術である。好ましい融合タンパク質は、レセプターの可溶化に有用な免疫グロブリン由来の異種領域を含む。例えば、EP A 0,464,533(また、カナダ対応出願2,045,869)は、別のヒトタンパク質またはその一部とともに免疫グロブリン分子の
定常領域の種々の部分を含む融合タンパク質を開示する。多くの場合、融合タンパク質中のFc部分は、治療および診断における使用に十分に有利であり、従って、例えば改善された薬物動態学的特性を生じる(EPA 0,232,262)。一方、いくつかの使用について、融合タ
ンパク質が、記載される有利な様式で、発現され、検出され、および精製された後にFc部分が欠失され得ることが望ましい。これは、Fc部分が、治療および診断における使用の妨害であると判明する場合(例えば、融合タンパク質が免疫のための抗原として使用されるべき場合)である。薬物探索において、例えばhIL-5のようなヒトタンパク質は、アゴニ
スト(例えば、hIL-5)を同定するための高処理能力スクリーニングアッセイの目的でFc
部分と融合されている。D.Bennettら,Journal of Molecular Recognition 8:52-58(1995)、およびK.Johansonら, The Journal ofBiological Chemistry 270(16):9459-9471(1995)を参照のこと。
【0081】
エンドカインαタンパク質は、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまた
は陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む周知の方法によって組換え細胞培養物から回収され、そして精製され得る。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0082】
本発明のポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明のポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
エンドカインαポリペプチドおよびペプチド
本発明はさらに、寄託されたcDNAによってコードされるアミノ酸配列、もしくは図1(配列番号2)のアミノ酸配列を有する単離されたエンドカインαポリペプチド、または上記ポリペプチドの一部を含むペプチドもしくはポリペプチドを提供する。用語「ペプチド」および「オリゴペプチド」は、(通常認識されるように)同義語と考えられ、そして各用語は、文脈がペプチド結合によって結合された少なくとも2つのアミノ酸の鎖を示すことを必要とする場合、交換可能に使用され得る。用語「ポリペプチド」は、10より多いアミノ酸残基を含む鎖に対して本明細書中で使用される。本明細書中の全てのオリゴペプチドおよびポリペプチドの式または配列は、左から右へ、そしてアミノ末端からカルボキシ末端の方向に書かれる。
【0083】
「単離された」ポリペプチドまたはタンパク質によって、その天然の環境から取り出されたポリペプチドまたはタンパク質が意図される。例えば、組換え宿主細胞中で発現された組換え産生されたポリペプチドおよびタンパク質は、例えば、SmithおよびJohnson, Gene 67:31-40 (1988)に記載の一段階法のような任意の適切な技術によって実質的に精製されている天然または組換えのポリペプチドおよびタンパク質がそうであるように、本発明の目的で単離されていると考えられる。
【0084】
エンドカインαポリペプチドのいくつかのアミノ酸配列が、このタンパク質の構造または機能に有意に影響することなく改変され得ることが、当該分野で認識される。配列内のこのような差異が意図される場合、活性を決定するタンパク質の重要な領域が存在することを覚えておくべきである。一般的に、類似の機能を実行する残基が使用される場合、三次構造を形成する残基の置換が可能である。他の例において、残基の型は、変化がタンパク質の重要でない領域で生じる場合、全く重要でないかもしれない。
【0085】
従って、本発明は、さらに、実質的なエンドカインαポリペプチド活性を示すか、または下記に考察されるタンパク質フラグメントのようなエンドカインαタンパク質の領域を含むエンドカインαポリペプチドの改変体を含む。このような変異体は、欠失、挿入、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)を含む。小さな変化、またはこのような「中性」アミノ酸置換は、一般的にほとんど活性に影響しない。
【0086】
代表的に保存的置換と見られるのは、脂肪族アミノ酸Ala、Val、Leu、およびIleの中での1つのアミノ酸の別のアミノ酸への置換;ヒドロキシル残基SerおよびThrの交換、酸性残基AspおよびGluの交換、アミド残基AsnおよびGlnの間の置換、塩基性残基LysおよびArgの交換、ならびに芳香族残基Phe、Tyrの間の置換である。
【0087】
上記に詳細に示されるように、どのアミノ酸の変化が表現型的にサイレントでありそう
か(すなわち、機能に対して有意に有害な効果を有しそうにないか)に関するさらなるガイダンスは、Bowie, J.U.ら「Deciphering the Message in Protein Sequences: Toleranceto Amino Acid Substitutions」,Science 247:1306-1310 (1990)に見出され得る。
【0088】
本発明のポリペプチドは、以下のような、図1(配列番号2)に示されるか、または寄託されたcDNAによってコードされるようなアミノ酸配列を有するポリペプチドを含む:完全長タンパク質、細胞内ドメイン、膜貫通ドメイン、および細胞外ドメイン、ならびに本明細書中に記載のポリペプチドと少なくとも90%の類似性、より好ましくは少なくとも95%の類似性、およびさらにより好ましくは少なくとも96%、97%、98%、または99%の類似性を有するポリペプチド。さらに本発明のポリペプチドは、本明細書中に記載のポリペプチドと、少なくとも80%同一、より好ましくは少なくとも90%または95%同一、さらにより好ましくは少なくとも96%、97%、98%、または99%同一であるポリペプチドを含み、そしてまた、少なくとも30アミノ酸、およびより好ましくは少なくとも50アミノ酸を有するこのようなポリペプチドの部分を含む。
【0089】
2つのポリペプチドについての「%類似性」によって、BESTFITプログラム(Wisconsin
Sequence Analysis Package, Version 8 for UNIX(登録商標), Genetics Computer Group, University Resarch Park, 575 Science Drive,Madison, WI 53711)および類似性を決定するための省略時設定(default setting)を用いて2つのポリペプチドのアミノ酸配
列を比較することによって生じる類似性のスコアが意図される。BESTFITは、SmithおよびWaterman,Adv. Appl. Math. 2:482-489 (1981)の局所的相同性アルゴリズムを、2つの配列間の類似性の最適のセグメントを見出すために使用する。
【0090】
エンドカインαポリペプチドの参照アミノ酸配列に少なくとも、例えば、95%「同一な」アミノ酸配列を有するポリペプチドにより、ポリペプチド配列がエンドカインαポリペプチドの参照アミノ酸配列の各100アミノ酸毎に5つまでのアミノ酸変化を含み得ること
を除いて、ポリペプチドのアミノ酸配列は参照配列と同一であることが意図される。言い換えれば、参照アミノ酸配列に少なくとも95%同一のアミノ酸配列を有するポリペプチドを得るために、参照配列において5%までのアミノ酸残基が、欠失されるかもしくは他のアミノ酸と置換され得、または参照配列における5%までの総アミノ酸残基の多数のアミノ酸が参照配列に挿入され得る。これらの参照配列の改変は、参照アミノ酸配列のアミノ末端位置もしくはカルボキシ末端位置で、または参照配列内または参照配列内の1つ以上の近接した基内の個々の残基間のいずれかで散在したこれらの末端位置の間のどこかで、起こり得る。
【0091】
実際には、任意の特定のポリペプチドが、例えば、図1(配列番号2)に示されるアミノ酸配列、または寄託されたcDNAクローンによりコードされるアミノ酸配列に対して、少なくとも90%、95%、96%、97%、98%、または99%同一であるかどうかは、BESTFITプ
ログラム(WisconsinSequence Analysis Package, Version8 Unix(登録商標)版、Genetics Computer Group, UniversityResearch Park, 575 Science Drive, Madison, WI 53711)のような公知のコンピュータープログラムを使用して慣習的に決定され得る。特定の
配列が、例えば、本発明の参照配列に95%同一であるかどうかを決定するために、BESTFITまたは任意の他の配列アラインメントプログラムを使用する場合、当然、同一性の百分
率が参照アミノ酸配列の全長にわたって計算され、そして参照配列内のアミノ酸残基の総数の5%までの相同性におけるギャップが許容されるようなパラメーターが設定される。
【0092】
下記に詳細に示すように、本発明のポリペプチドは、下記のようにエンドカインαタンパク質発現を検出するための診断アッセイにおいて有用である、またはエンドカインαタンパク質機能を阻害し得るアゴニストおよびアンタゴニストとして、ポリクローナル抗体およびモノクローナル抗体を惹起するために使用され得る。さらに、このようなポリペプ
チドは、本発明の候補アゴニストおよびアンタゴニストでもあるエンドカインαタンパク質結合タンパク質を「捕獲する」ための酵母ツーハイブリッドシステムにおいて使用され得る。酵母ツーハイブリッドシステムは、FieldsおよびSong,Nature 340:245-246(1989
)に記載されている。
【0093】
別の局面において、本発明は、本発明のポリペプチドのエピトープ-保有部分を含む、
ペプチドまたはポリペプチドを提供する。このポリペプチドのエピトープ部分は、本発明のポリペプチドの免疫原性または抗原性エピロープである。「免疫原性エピトープ」は、タンパク質全体が免疫原性である場合、抗体応答を誘発するタンパク質の一部として定義される。これらの免疫原性エピトープは、分子上の2、3の焦点に制限されると考えられている。一方では、抗体が結合し得るタンパク質分子の領域は、「抗原性エピトープ」と定義され得る。タンパク質の免疫原性エピトープの数は、一般には、抗原性エピトープの数よりも少ない。例えば、Geysen,H.M.ら、Proc. Natl. Acad. Sci. USA 81:3998-4002(1984)を参照のこと。
【0094】
抗原性エピトープを保有するペプチドまたはポリペプチド(すなわち、抗体が結合し得るタンパク質分子の領域を含む)の選択に関して、タンパク質配列の一部を模倣する比較的短い合成ペプチドが、部分的に模倣されたタンパク質と反応する抗血清を日常的に誘発し得ることが当該分野で周知である。例えば、Sutcliffe,J.G.ら、Science 219:660-666(1983)を参照のこと。タンパク質-反応性血清を誘発し得るペプチドは、しばしばタンパク質の一次配列に頻繁に存在し、そして単純な化学的法則のセットにより特徴付けられ得、そしてインタクトなタンパク質(すなわち、免疫原性エピトープ)の免疫ドミナント領域にも、アミノ末端またはカルボキシル末端にも、制限されない。極度に疎水性であるペプチドおよび6以下の残基のペプチドは、一般には、模倣タンパク質に結合する抗体の誘導に効果がなく;より長い、可溶性ペプチド、特にプロリン残基を含むペプチドは、通常は有効である。Sutcliffeら、前出、661。例えば、これらのガイドラインに従って設計された30のペプチドのうち18(インフルエンザウイルス赤血球凝集素HA1ポリペプチド鎖の配
列の75%を覆う8-39残基を含む)は、HA1タンパク質またはインタクトなウイルスと反応
する抗体を誘導した;そしてMuLVポリメラーゼからの12/12ペプチドおよび狂犬病糖タン
パク質からの18/18はそれぞれのタンパク質を沈澱する抗体を誘導した。
【0095】
本発明の抗原性エピトープ-保有ペプチドおよびポリペプチドは、それゆえ、本発明の
ポリペプチドに特異的に結合するモノクローナル抗体を含む抗体を惹起するのに有用である。従って、抗原エピトープ-保有ペプチドで免疫化されたドナーからの脾臓細胞の融合
により得られるハイブリドーマの大部分は、一般に天然のタンパク質と反応性がある抗体を分泌する。Sutcliffeら、前出、663。抗原性エピトープ-保有ペプチドまたはポリペプ
チドにより惹起された抗体は、模倣タンパク質を検出するのに有用であり、そして異なるペプチドに対する抗体が、翻訳語プロセシングを受けるタンパク質前駆体の種々の領域の末路を追跡するために使用され得る。免疫沈降アッセイにおいて、短いペプチド(例えば、約9アミノ酸)でさえ、より長いペプチドに結合しそして置換し得ることが示されているので、ペプチドおよび抗ペプチド抗体は、模倣タンパク質についての種々の定性的または定量的アッセイ、例えば、競合的アッセイにおいて使用され得る。例えば、Wilson,I.A.ら、Cell 37:767-778(1984)777を参照のこと。本発明の抗ペプチド抗体もまた、模倣
タンパク質の精製(例えば、当該分野で周知の方法を使用して、吸着クロマトグラフィーにより)に有用である。
【0096】
上記のガンダンスに従って設計された本発明の抗原性エピトープ-保有ペプチドまたは
ポリペプチドは、好ましくは本発明のポリペプチドのアミノ酸配列内に含まれる少なくとも7、より好ましくは少なくとも9、そして最も好ましくは約15〜約30アミノ酸の間の配列を含む。しかし、本発明のポリペプチドのアミノ酸配列の約30〜約50アミノ酸または全
体までの任意の長さおよび全体を含む、本発明のポリペプチドのアミノ酸配列のより大部分を含むペプチドまたはポリペプチドもまた、本発明のエピトープ-保有ペプチドまたは
ポリペプチドであると考えられ、そしてまた模倣タンパク質と反応する抗体を誘導するのに有用である。好ましくは、エピトープ-保有ペプチドのアミノ酸配列は、水溶液中で実
質的な溶解性を提供するように選択され(すなわち、その配列は、比較的親水性残基を含み、そして高度な疎水性配列は好ましくは回避される);そしてプロリン残基を含む配列が特に好ましい。
【0097】
エンドカイン特異的ポリクローナルおよびモノクローナル抗体を産生するために使用され得る抗原性ポリペプチドの非限定的な例には、図1(配列番号2)における約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約94〜約105のアミノ酸残基を含むポリペプチド;図1
(配列番号2)における約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)における約148〜約158のアミノ酸残基を含むポリペプチドが挙げられる。上記のように、本発明者らは、上記のポリペプチドフラグメントがエンドカインαタンパク質の抗原性領域であることを決定した。
【0098】
本発明のエピトープ-保有ペプチドおよびポリペプチドは、本発明の核酸分子を使用す
る組換え手段を含むペプチドまたはポリペプチドを作製するための任意の従来の手段により産生され得る。例えば、短いエピトープ-保有アミノ酸配列は、組換え体産生および精
製の間、ならびに抗ペプチド抗体を産生するための免疫化の間、キャリアとして作用するより大きなポリペプチドに融合され得る。エピトープ-保有ペプチドはまた、化学合成の
公知の方法を使用して合成され得る。例えば、Houghtenは、4週間未満で、調製されそして特徴付けられた(ELISA-タイプ結合研究により)HA1ポリペプチドのセグメントの単一
のアミノ酸改変体を示す10〜20mgの248の異なる13残基ペプチドのような多数のペプチド
の合成のための簡単な方法を記載している。Houghten,R.A., Proc. Natl. Acad. Sci. USA 82:5131-5135 (1985)を参照のこと。この「SimultaneousMultiple Peptide Synthesis(SMPS)」プロセスは、さらにHoughtenら(1986)の米国特許第4,631,211号に記載される。この手順において、種々のペプチドの固相合成のための個々の樹脂は、分離溶媒透過性パケットに含まれ、固相法に関連する多くの同一の反復工程の最適な使用を可能にする。完全なマニュアル手順は、同時に行われる500〜1000以上の合成を可能にする。Houghtenら、前出、5134。
【0099】
本発明のエピトープ保有ペプチドおよびポリペプチドは、当該分野に周知の方法によって抗体を誘導するために使用される。例えば、Sutcliffeら、前出;Wilsonら、前出;Chow, Mら、Proc. Natl. Acad. Sci. USA82:910-914;およびBittle, F. J.ら、J. Gen. Virol. 66:2347-2354(1985)を参照のこと。一般には、動物は遊離ペプチドで免疫化され得る;しかし、抗ペプチド抗体力価はペプチドを高分子キャリア(例えば、キーホールリンペットヘモシアニン(KLH)または破傷風毒素)にカップリングすることにより追加免疫
され得る。例えば、システインを含有するペプチドは、m-マレイミドベンゾイル-N-ヒド
ロキシスクシンイミドエステル(MBS)のようなリンカーを使用してキャリアにカップリ
ングされ得、一方、他のペプチドは、グルタルアルデヒドのようなより一般的な連結剤を使用してキャリアーにカップリングされ得る。
【0100】
ウサギ、ラット、およびマウスのような動物は、遊離またはキャリア-カップリングペ
プチドのいずれかで、例えば、約100μgのペプチドまたはキャリアタンパク質およびFreundのアジュバントを含むエマルジョンの腹腔内および/または皮内注射により免疫化され
る。いくつかの追加免疫注射が、例えば、固体表面に吸着された遊離ペプチドを使用して
ELISAアッセイにより検出され得る有用な力価の抗ペプチド抗体を提供するために、例え
ば、約2週間の間隔で必要とされ得る。免疫化動物からの血清における抗ペプチド抗体の力価は、抗ペプチド抗体の選択により、例えば、当該分野で周知の方法による固体支持体上のペプチドへの吸収および選択された抗体の溶出により増加され得る。
【0101】
本発明の免疫原性エピトープ-保有ペプチド、すなわち、抗体応答を惹起するタンパク
質の部分は、全体のタンパク質が免疫原性である場合、当該分野で公知の方法により同定される。例えば、Geysenら(1984)前出は、酵素-結合免疫吸収アッセイにおける反応に
十分に純粋な何百というペプチドの固体支持体上の迅速な併用的合成の手順を開示する。合成ペプチドの抗体との相互作用は、次いで、それらを支持体から除去することなく容易に検出される。この様式において、所望のタンパク質の免疫原性エピトープを保有するペプチドは、当業者により日常的に同定され得る。
【0102】
例えば、口蹄疫ウイルスの被膜タンパク質における免疫学的に重要なエピトープは、タンパク質の213のアミノ酸配列全体を覆う全ての208の可能なヘキサペプチドの重複セットの合成による7アミノ酸の解明によりGeysenらによって位置付けされた。次いで、全ての20アミノ酸が順にエピトープ内の各位置で置換されたペプチドの完全な置換セットが合成され、そして抗体との反応のための特異性を与える特定のアミノ酸が決定された。従って、本発明のエピトープ-保有ペプチドのペプチドアナログは、この方法により日常的に作
成され得る。Geysen(1987)の米国特許第4,708,781号は、所望のタンパク質の免疫原性
エピトープを保有するペプチドを同定するこの方法をさらに記載している。
【0103】
さらになお、Geysen(1990)の米国特許第5,194,392号は、目的の抗体の特定のパラト
ープ(抗原結合部位)に相補的であるエピトープの位相幾何学的等価物(例えば、「ミモトープ」)であるモノマー(アミノ酸または他の化合物)の配列を検出または決定する一般的な方法を記載する。より一般的には、Geysen(1989)の米国特許第4,433,092号は、
目的の特定のレセプターのリガンド結合部位に相補的であるリガンドの位相等価であるモノマーの配列を検出または決定する方法を記載する。同様に、PeralkylatedOligopeptide
MixturesにおけるHoughten, R. A.ら(1996)の米国特許第5,480,971号は、線状C1-C7-
アルキル過アルキル化(peralkylated)オリゴペプチドおよびセットおよびこのようなペプチドのライブラリー、ならびに目的のアクセプター分子に、優先的に結合する過剰アルキル化オリゴペプチドの配列を決定するためにこのようなオリゴペプチドセットおよびライブラリーを使用する方法を開示する。従って、本発明のエピトープ-保有ペプチドの非
ペプチドアナログはまた、これらの方法により日常的に作成され得る。
【0104】
本発明者らは、エンドカインαタンパク質が、3つの主要な構造ドメインを示す169残
基タンパク質であることを発見した。細胞内ドメインは、図1(配列番号2)における約1〜約17の残基内で同定された。膜貫通ドメインは、図1(配列番号2)における約18〜約43の残基内で同定された。細胞外ドメインは、図1(配列番号2)における約44〜約169の残基内で同定された。従って、本発明は、エンドカインα細胞内ドメイン、膜貫通ド
メイン、およびエンドカインα細胞外ドメインから選択されるポリペプチドを含む好ましいエンドカインαタンパク質フラグメントをさらに提供する。
【0105】
エンドカインαタンパク質の細胞外ドメインは、免疫グロブリン(IgG)の定ドメイン
の一部と結合し得、キメラペプチドを生じる。これらの融合タンパク質は、インビボで増加した半減期を示す。これは、例えば、ヒトCD4-ポリペプチドの最初の2つのドメインおよび哺乳動物免疫グロブリンの重鎖または軽鎖の定常領域の種々のドメインからなるキメラタンパク質について示されている(EPA394,827;Trauneckerら、Nature 331:84-86(1988))。IgG部分によるジスルフィド結合ダイマー構造を有する融合タンパク質はまた、
リガンドの結合および中和において、モノマー細胞外ドメイン単独よりも有効であり得る
(Fountoulakisら、J.Biochem. 270:3958-3964(1995))。
【0106】
「ポリペプチドおよびペプチド」のこの節に引用される各文書の全体の開示は、ここで参考として本明細書中に援用される。
【0107】
エンドカインα関連障害診断
エンドカインαは、サイトカインのTNFファミリーの新規のメンバーである。エンドカ
インα関連障害について、「標準」エンドカインα遺伝子発現レベル(すなわち、障害を有さない個体からの組織または体液におけるエンドカインα発現レベル)と比較して、実質的に変化した(増加または減少した)レベルのエンドカインα遺伝子発現が、このような障害を有する個体から採取された組織または他の細胞または体液(例えば、血清、血漿、尿、滑液、また脊髄液)において検出され得ると考えられる。従って、本発明は、エンドカインα関連障害の診断の間に有用な診断方法を提供する。これは個体からの組織または他の細胞または体液におけるエンドカインαタンパク質をコードする遺伝子の発現レベルを測定すること、および測定した遺伝子発現レベルと標準エンドカインα遺伝子発現レベルとを比較することを含む。それにより、標準と比較された遺伝子発現レベルにおける増加または減少は、エンドカインα関連障害の指標である。
【0108】
個体により、哺乳動物個体、好ましくはヒトが意図される。「エンドカインαタンパク質をコードする遺伝子の発現レベルを測定すること」により、第一の生物学的サンプルにおいて直接的(例えば、絶対的なタンパク質レベルまたはmRNAレベルを測定または評価することにより)または間接的(例えば、第二の生物学的サンプルにおけるエンドカインαタンパク質レベルまたはmRNAレベルと比較することにより)のいずれかで、エンドカインαタンパク質のレベルまたはエンドカインαタンパク質をコードするmRNAレベルの定性的測定もしくは定量的測定または評価が意図される。好ましくは、第一の生物学的サンプルにおけるエンドカインαタンパク質レベルまたはmRNAレベルが測定または評価され、そして標準エンドカインαタンパク質レベルまたはmRNAレベルと比較される。標準は、障害を有さない個体から得られた第二の生物学的サンプルから得られるか、またはエンドカインαを含む障害を有さない個体の集団からの平均レベルにより決定され得る。当該分野において評価されるように、一旦標準エンドカインαタンパク質レベルまたはmRNAレベルが公知になると、それは比較のための標準として繰り返して使用され得る。
【0109】
「生物学的サンプル」によって、個体、体液、細胞株、組織培養物、またはエンドカインαタンパク質もしくはmRNAを含む他の供給源から得られる任意の生物学的サンプルが意図される。示されるように、生物学的サンプルは、分泌された成熟エンドカインαタンパク質を含む体液(例えば、血清、血漿、尿、滑液、および随液)あるいはエンドカインαを発現することが見出された組織供給源を含む。哺乳動物から組織生検および体液を得るための方法は当該分野で周知である。生物学的サンプルがmRNAを含む場合、組織生検が好ましい供給源である。
【0110】
本発明は、当該分野で公知であるかまたは本明細書中で示されるようなTNF様障害と同
様に、哺乳動物(好ましくは、ヒト)の種々のエンドカインα関連障害の診断に有用である。これらは、免疫調節および炎症、細胞増殖、血管新生、腫瘍転移、アポトーシス、敗血症、および内毒素血症に関連する障害を含む。
【0111】
全細胞RNAが、任意の適切な技術(例えば、ChomczynskiおよびSacchi,Anal. Biochem. 162:156-159 (1987)に記載されている一工程のグアニジン-チオシアネート-フェノール-
クロロホルム法)を使用して生物学的サンプルから単離され得る。次いで、エンドカインαポリペプチドをコードするmRNAのレベルが、任意の適切な方法を用いてアッセイされる
。これらは、ノーザンブロット分析、S1ヌクレアーゼマッピング、ポリメラーゼ連鎖反応(PCR)、ポリメラーゼ連鎖反応と組み合わせた逆転写(RT-PCR)、およびリガーゼ連鎖
反応と組み合わせた逆転写(RT-LCR)を含む。
【0112】
ノーザンブロット分析は、Haradaら、Cell 63:303-312 (1990)に記載されているように行われ得る。簡潔には、全RNAが上記のように生物学的サンプルから調製される。ノーザ
ンブロットのためには、RNAは適切な緩衝液(例えば、グリオキサール/ジメチルスルホ
キシド/リン酸ナトリウム緩衝液)中で変性され、アガロースゲル電気泳動に供され、そしてニトロセルロースフィルターに転写される。RNAがUVリンカーによってフィルターに
結合された後、フィルターは、ホルムアミド、SSC、デンハルト溶液、変性させたサケ精
子、SDS、およびリン酸ナトリウム緩衝液を含有する溶液中で予備ハイブリダイゼーショ
ンされる。任意の適切な方法(例えば、32PマルチプライムDNA標識システム(Amersham)
)に従って標識されたエンドカインαタンパク質cDNAが、プローブとして使用される。一晩のハイブリダイゼーション後、フィルターは洗浄され、そしてX線フィルムに曝される。本発明に従うプローブとしての使用のためのcDNAは、上記の節に記載されており、そして少なくとも15bpの長さが好ましい。
【0113】
S1マッピングは、Fujitaら、Cell49:357-367 (1987)に記載されているように行われ得る。S1マッピングでの使用のためのプローブDNAを調製するために、上記のcDNAのセンス
鎖がテンプレートとして使用され、標識されたアンチセンスDNAが合成される。次いで、
アンチセンスDNAが、適切な制限エンドヌクレアーゼを使用して消化され、所望の長さの
さらなるDNAプローブが生成され得る。このようなアンチセンスプローブは、標的mRNA(
すなわち、エンドカインαタンパク質をコードするmRNA)に対応する保護されたバンドを可視化するために有用である。ノーザンブロット分析が、上記のように行われ得る。
【0114】
好ましくは、エンドカインαタンパク質をコードするmRNAのレベルは、Makinoら、Technique 2:295-301 (1990) に記載されるようなRT-PCR法を使用してアッセイされる。この
方法によれば、ポリアクリルアミドゲルバンド中の「アンプリコン」の放射活性は、標的mRNAの初期濃度に直線的に相関する。簡潔には、この方法は、RTプライマーおよび適切な緩衝液を含む反応混合物中の生物学的サンプルから単離された全RNAを添加する工程を包
含する。プライマーのアニーリングのためのインキュベーション後、混合物はRT緩衝液、dNTP、DTT、RNaseインヒビター、および逆転写酵素を補充され得る。RNAの逆転写を達成
するためのインキュベーション後、次いで、RT産物は標識されたプライマーを使用するPCRに供される。あるいは、プライマーを標識するよりもむしろ、標識されたdNTPがPCR反応混合物中に含まれ得る。PCR増幅は、従来技術に従ってDNAサーマルサイクラー中で行われ得る。増幅を達成するための適切な数のラウンド後、PCR反応混合物はポリアクリルアミ
ドゲル上で電気泳動される。ゲルの乾燥後、適切なバンド(エンドカインαタンパク質をコードするmRNAに対応する)の放射活性は、画像解析機を使用して定量される。RTおよびPCR反応の成分および条件、試薬およびゲル濃度、ならびに標識方法は、当該分野で周知
である。RT-PCR法の変形は当業者に明らかである。
【0115】
逆転写された標的のmRNAを増幅する任意のオリゴヌクレオチドプライマーのセットが使用され得、そして上記の節で記載されるように設計され得る。
【0116】
生物学的サンプル中のエンドカインαタンパク質のレベルのアッセイは、任意の当該分野で公知の方法を使用して行われ得る。抗体に基づく技術が、生物学的サンプル中のエンドカインαタンパク質レベルをアッセイするために好ましい。例えば、組織中でのエンドカインαタンパク質の発現は、伝統的な免疫組織学的方法で研究され得る。これらの場合、特異的認識は一次抗体(ポリクローナルまたはモノクローナル)によって提供されるが、二次検出系は、蛍光、酵素、または他の結合された二次抗体を利用し得る。結果として
、病理学試験のための組織切片の免疫組織学的染色が得られる。組織はまた、ウェスタンブロットまたはドット/スロットアッセイ(Jalkanen, M.ら、J. Cell. Biol. 101:976-985 (1985);Jalkanen, Mら、J.Cell. Biol. 105:3087-3096 (1987))のためのエンドカイ
ンαタンパク質の遊離のために、例えば、尿素および中性の界面活性剤を用いて抽出され得る。陽イオン性固相の使用に基づくこの技術において、エンドカインαタンパク質の定量は、単離されたエンドカインαタンパク質を標準として使用して達成され得る。この技術はまた、体液にも適用され得る。このサンプルに関して、エンドカインαタンパク質のモル濃度は、血清、血漿、尿、滑液、随液などのような異なる体液について、エンドカインαタンパク質含量の標準値の設定を補助する。次いで、エンドカインαタンパク質の量の正常値は、健常な個体に由来する値を使用して設定され得、これは、試験被検体から得られる値と比較され得る。
【0117】
エンドカインαタンパク質レベルを検出するために有用な他の抗体に基づく方法は、イムノアッセイ(例えば、酵素結合免疫吸着アッセイ(ELISA)およびラジオイムノアッセ
イ(RIA))を含む。例えば、エンドカインαタンパク質特異的モノクローナル抗体は、
エンドカインαタンパク質を検出および定量するための、免疫吸着剤としておよび酵素標識プローブとしての両方に使用され得る。サンプル中に存在するエンドカインαタンパク質の量は、直線回帰コンピューターアルゴリズムを使用して、標準的な調製物中に存在する量との比較によって算出され得る。腫瘍抗原を検出するためのこのようなELISAは、Iacobelliら、BreastCancer Research and Treatment 11:19-30 (1988) に記載されている。別のELISAアッセイにおいては、2つの異なる特異的なモノクローナル抗体が、体液中の
エンドカインαタンパク質を検出するために使用され得る。このアッセイにおいて、一方の抗体が免疫吸着剤として使用され、そして他方が酵素標識プローブとして使用される。
【0118】
上記の技術は、本質的に、「一工程」または「二工程」アッセイとして行われ得る。「一工程」アッセイは、固定化抗体とエンドカインαタンパク質とを接触させる工程を包含し、そして洗浄する工程、標識された抗体と混合物とを接触させる工程は含まない。「二工程」アッセイは、洗浄する工程、その後、標識された抗体と混合物とを接触させる工程を包含する。他の従来の方法もまた、適切に使用され得る。通常、支持体上にアッセイ系の1つの成分を固定することが所望され、それによって系の他の成分は、その成分との接触およびサンプルからの容易な除去を生じることが可能となる。
【0119】
適切な酵素標識は、例えば、基質との反応による過酸化水素の生成を触媒するオキシダーゼ群由来のものを含む。グルコースオキシダーゼは、それが良好な安定性を有し、そしてその基質(グルコース)が容易に入手できるために、特に好ましい。オキシダーゼ標識の活性は、酵素-標識抗体/基質反応によって形成される過酸化水素の濃度を測定するこ
とによってアッセイされ得る。酵素に加えて、他の適切な標識として、放射性同位元素(例えば、ヨウ素(125I、121I)、炭素(14C)、イオウ(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99mTc))、ならびに蛍光標識(例えば、フルオレセインおよびローダミン)ならびにビオチンが挙げられる。
【0120】
個体から得られる生物学的サンプル中のエンドカインαタンパク質レベルをアッセイすることに加えて、エンドカインαタンパク質はまた、画像解析によってインビボで検出され得る。エンドカインαタンパク質のインビボでの画像解析のための抗体標識またはマーカーとして、X線撮影法、NMR、またはESRによって検出可能なものが挙げられる。X線撮影法については、適切な標識として、検出可能な放射線を放射するが、被検体に対して明らかには有害ではない、バリウムまたはセシウムのような放射性同位元素が挙げられる。NMRおよびESRのための適切なマーカーとして、関連のハイブリドーマの栄養分の標識によって抗体中に取り込まれ得る、重水素のような検出可能な特徴的な回転を有するものが挙げられる。
【0121】
放射性同位元素(例えば、131I、111In、99mTc)、放射性不透明体(radio-opaque)基質、または核磁気共鳴によって検出可能な物質のような適切な検出可能な画像解析部分で標識されている、エンドカインαタンパク質-特異的抗体または抗体部分が、障害につい
て試験される哺乳動物中に(例えば、非経口的、皮下、または静脈内)導入される。被検体の大きさおよび使用される画像解析システムによって、診断用の画像を生じるために必要とされる画像解析部分の量が決定されることが、当該分野で理解される。放射性同位元素部分の場合、ヒト被検体については、注射される放射活性の量は、通常約5〜20ミリキュリーの範囲の99mTcである。次いで、標識抗体または抗体部分は、エンドカインαタン
パク質を含む細胞の位置に優先的に蓄積する。インビボでの腫瘍画像解析は、S.W.Burchielら、「Immunopharmacokinetics of Radiolabeled Antibodies and Their Portions」(TumerImaging 第13章:The Radiochemical Detection of Cancer, Burchiel, S.W.およびRhodes,B.A.編、Masson Publishing Inc. (1982))に記載されている。
【0122】
本発明での使用のためのエンドカインαタンパク質特異的抗体は、完全なエンドカインαタンパク質またはその抗原性ポリペプチド部分に対して惹起され得る。これは、アルブミンのようなキャリアタンパク質と共に、またはそれが充分に長い(少なくとも約25アミノ酸)場合はキャリアを伴わずに、動物系(例えば、ウサギまたはマウス)に対して提示され得る。
【0123】
本明細書中で使用される場合、用語「抗体」(Ab)または「モノクローナル抗体」(Mab)は、エンドカインαタンパク質に特異的に結合し得る完全な分子および抗体タンパク
質(例えば、FabおよびF(ab’)2部分のような)を含むことを意味する。FabおよびF(ab’)2部分は完全な抗体のFc部分を欠いており、循環によってさらに迅速に除去され、そして完全な抗体の非特異的組織結合をほとんど有し得ない(Wahlら、J.Nucl. Med. 24:316-325 (1983))。従って、これらの部分が好ましい。
【0124】
本発明の抗体は、任意の種々の方法によって調製され得る。例えば、エンドカインαタンパク質またはその抗原性部分を発現する細胞は、ポリクローナル抗体を含む血清の産生を誘導するために動物に投与され得る。好ましい方法において、エンドカインαタンパク質の調製物は、それが天然の混入物を実質的に含まないように、上記のように調製され、そして精製される。次いで、このような調製物は、より大きな特異的活性のポリクローナル抗血清を産生するために動物に導入される。
【0125】
最も好ましい方法において、本発明の抗体はモノクローナル抗体(またはそのエンドカインαタンパク質結合部分)である。このようなモノクローナル抗体は、ハイブリドーマ技術(例えば、Colligan, Current Protocols in Immunology, Wiley Interscience, NewYork (1990-1996);HarlowおよびLane, Antibodies:A Laboratory Manual, Chs. 6-9, ColdSpring Harbor Press, Cold Spring Harbor, N.Y.(1988);Ausubel、前出、第11章を参照のこと、これらの参考文献は、その全体が本明細書中に参考として援用される)を用いて調製され得る。
【0126】
一般に、このような手順は、エンドカインαポリペプチド抗原またはエンドカインαポリペプチド発現細胞で動物(好ましくは、マウス)を免疫する工程を包含する。適切な細胞は、抗エンドカインαタンパク質抗体に結合するそれらの能力によって認識され得る。このような細胞は、任意の適切な組織培養培地(例えば、10%ウシ胎児血清(約56℃で不活化した)を補充し、約10μg/lの非必須アミノ酸、約1,000U/mlのペニシリン、および約100μg/mlのストレプトマイシンを補充したEarle改変イーグル培地)中で培養され得る。このようなマウスの脾細胞が抽出され、そして適切な骨髄腫細胞株と融合される。任意の適切な骨髄腫細胞株(例えば、アメリカンタイプカルチャーコレクション(ATCC)(Rock
ville,Maryland, USA)から入手可能な親骨髄腫細胞株(SP2O))が、本発明に従って使用
され得る。融合後、得られたハイブリドーマ細胞はHAT培地中で選択的に維持され、次い
でWandsら、Gastroenterology80:225-232 (1981);HarlowおよびLane、前出、第7章に記載されているような限界希釈によってクローニングされる。次いで、このような選択によって得られたハイブリドーマ細胞は、エンドカインα抗原に結合し得る抗体を分泌するクローンを同定するためにアッセイされる。
【0127】
あるいは、エンドカインαタンパク質抗原に結合し得るさらなる抗体が、抗イディオタイプ抗体の使用を通じて二工程手順で産生され得る。このような方法は、抗体はそれ自体が抗原であるという事実を使用し、従って、二次抗体に結合する抗体を得ることが可能である。この方法に従って、エンドカインαタンパク質特異的抗体は、動物(好ましくは、マウス)を免疫するために使用される。次いで、このような動物の脾細胞はハイブリドーマ細胞を産生するために使用され、そしてハイブリドーマ細胞は、エンドカインαタンパク質特異的抗体に結合する能力がエンドカインαタンパク質抗原によってブロックされ得る抗体を産生するクローンを同定するためにスクリーニングされる。このような抗体は、エンドカインαタンパク質特異的抗体に対する抗イディオタイプ抗体を含み、そしてさらなるエンドカインαタンパク質特異的抗体の形成を誘導するために動物を免疫するために使用され得る。
【0128】
本発明の抗体のFab部分およびF(ab’)2部分および他の部分が、本明細書中に開示される方法に従って用いられ得ることが認識される。このような部分は、代表的には、パパイン(Fab部分を生成するため)またはペプシン(F(ab’)2部分を生成するため)のような酵素を用いるタンパク質分解切断により生成される。あるいは、エンドカインα(endokinealpha)タンパク質結合部分は、組換えDNA技術の適用を通して、または合成化学を通して生成され得る。
【0129】
ヒトにおける診断のために、インビボでのイメージングを用いて、増強されたレベルのエンドカインαタンパク質を検出する場合、「ヒト化」キメラモノクローナル抗体を使用することが好ましくあり得る。このような抗体は、上記のモノクローナル抗体を生成するハイブリドーマ細胞由来の遺伝構築物を用いて生成され得る。キメラ抗体を生成するための方法は、当該分野で公知である。総説については、Morrison, Science 229:1202 (1985); Oiら, BioTechniques 4:214 (1986);Cabillyら, 米国特許第4,816,567号; Taniguchi
ら, EP 171496; Morrisonら, EP 173494;Neubergerら, WO 8601533; Robinsonら, WO 8702671; Boulianneら, Nature 312:643(1984); Neubergerら, Nature 314:268 (1985)を参
照のこと。
【0130】
本発明のエンドカインαタンパク質特異的抗体にさらに適切な標識は、以下に提供される。適切な酵素標識の例は、リンゴ酸デヒドロゲナーゼ、スタフィロコッカスヌクレアーゼ、Δ-5-ステロイドイソメラーゼ、酵母アルコールデヒドロゲナーゼ、α-グリセロールリン酸デヒドロゲナーゼ、トリオースリン酸イソメラーゼ、ペルオキシダーゼ、アルカリホスファターゼ、アスパラギナーゼ、グルコースオキシダーゼ、β-ガラクトシダーゼ、
リボヌクレアーゼ、ウレアーゼ、カタラーゼ、グルコース-6-リン酸デヒドロゲナーゼ、
グルコアミラーゼ、およびアセチルコリンエステラーゼを含む。
【0131】
適切な放射性同位体標識の例は、3H、111In、125I、131I、32P、35S、14C、51Cr、57To、58Co、59Fe、75Se、152Eu、90Y、67Cu、217Ci、211At、212Pb、47Sc、109Pdなどを含む。111Inおよび99mTcは、インビボでのイメージングが用いられる場合に好ましい同位体である。なぜなら、これは、125Iまたは131Iで標識したモノクローナル抗体の肝臓による脱ハロゲン化の問題を回避するからである。さらに、この放射性核種(radionucleotide)は、イメージングのためにより好ましいγ放出エネルギーを有する(Perkinsら
,Eur. J. Nucl. Med. 10:296-301 (1985); Carasquilloら, J. Nucl. Med. 28:281-287(1987))。例えば、1-(p-イソチオシアネートベンジル)-DPTAを用いてモノクローナル抗体
にカップリングした111Inは、非腫瘍性組織(特に肝臓)における取り込みをほとんど示
さなかった。それゆえ、腫瘍局在化の特異性を増強する(Estebanら,J. Nucl. Med. 28:861-870 (1987))。
【0132】
適切な非放射性同位体標識の例は、157Gd、55Mn、162Dy、52Tr、および56Feを含む。
【0133】
適切な蛍光標識の例は、152Eu標識、フルオレセイン標識、イソチオシアネート標識、
ローダミン標識、フィコエリトリン標識、フィコシアニン標識、アロフィコシアニン標識、o-フタルアルデヒド(o-phthaldehide)標識、およびフルオレサミン標識を含む。
【0134】
適切な毒素標識の例は、ジフテリア毒素、リシン、およびコレラ毒素を含む。
【0135】
化学発光標識の例は、ルミナール標識、イソルミナール標識、芳香族アクリジニウムエステル標識、イミダゾール標識、アクリジニウム塩標識、シュウ酸エステル標識、ルシフェリン標識、ルシフェラーゼ標識、およびエクオリン標識を含む。
【0136】
核磁気共鳴コントラスト剤の例は、Gd、Mn、およびFeのような重金属原子核を含む。
【0137】
上記の標識を抗体に結合させるための代表的な技術は、Kennedyら(Clin. Chim. Acta 70:1-31 (1976))およびSchursら(Clin. Chim. Acta81:1-40 (1977))により提供される。後者において言及されるカップリング技術は、グルタルアルデヒド法、過ヨウ素酸法、ジマレイミド法、m-マレイミドベンジル-N-ヒドロキシ-スクシンイミドエステル法であり、これらの方法は全て本明細書中に参考として援用される。
【0138】
染色体アッセイ
本発明の核酸分子はまた、染色体の同定に有用である。配列は、個々のヒト染色体上の特定の位置に特異的に標的化され、そしてその位置にハイブリダイズし得る。さらに、現在は染色体上の特定の部位を同定する必要性がある。現在、実際の配列データ(反復多型)に基づいた染色体標識化試薬は、染色体位置の標識にほとんど利用可能でない。本発明によるDNAの染色体へのマッピングは、これらの配列と疾患に関連する遺伝子との相関付
けにおいて重要な第1工程である。
【0139】
この点における特定の好ましい実施態様において、本明細書中に開示されるcDNAは、エンドカインαタンパク質遺伝子のゲノムDNAをクローニングするために使用される。これ
は、種々の周知の技術および一般に市販されているライブラリーを使用して達成され得る。次いで、ゲノムDNAは、この目的のための周知の技術を使用してインサイチュ染色体マ
ッピングのために使用される。代表的には、染色体マッピングの日常的な手順に従う、いくつかの試行錯誤が、良好なインサイチュハイブリダイゼーションシグナルを与えるゲノムプローブを同定するために必要であり得る。
【0140】
いくつかの場合において、さらに、配列は、cDNAからPCRプライマー(好ましくは15〜25bp)を調製することにより染色体にマップされ得る。遺伝子の3’非翻訳領域のコンピューター解析が、ゲノムDNA内で1より多いエキソンにまたがらず、従って増幅プロセスを
複雑化しないプライマーを迅速に選択するために使用される。次いで、これらのプライマーは、個々のヒト染色体を含む体細胞ハイブリッドのPCRスクリーニングに使用される。
プライマーに対応するヒト遺伝子を含むそれらのハイブリッドのみが増幅部分を生じる。
【0141】
体細胞ハイブリッドのPCRマッピングは、特定の染色体に特定のDNAを割り当てるための迅速な手順である。同じオリゴヌクレオチドプライマーを本発明と共に使用して、特定の染色体由来の部分のパネルまたは類似の様式での大きなゲノムクローンのプールを用いて、準位置決定(sublocalization)が達成され得る。染色体にマップするために同様に使用
され得る他のマッピングストラテジーは、インサイチュハイブリダイゼーション、標識してフロー選別した(flow-sorted)染色体を用いるプレスクリーニング、および染色体特異
的cDNAライブラリーを構築するためのハイブリダイゼーションによる予備選択を含む。
【0142】
cDNAクローンの中期染色体スプレッド(spread)への蛍光インサイチュハイブリダイゼ
ーション(「FISH」)は、1工程で正確な染色体位置を提供するために使用され得る。この技術は、50bpまたは60bpほど短いcDNA由来のプローブを用いて使用され得る。この技術の総説については、Vermaら,Human Chromosomes: A Manual of Basic Techniques, Pergamon Press, New York(1988)を参照のこと。
【0143】
一旦配列が正確な染色体位置にマップされると、配列の染色体上での物理的な位置を遺伝地図のデータと相関させ得る。このようなデータは、例えば、V. McKusick, Mendelian
Inheritance in Man(Johns Hopkins University,Welch Medical Libraryからオンラインで入手可能である)において見出される。次いで、同じ染色体領域にマップされた遺伝子
と疾患との間の関係が、連鎖解析(物理的に隣接した遺伝子の同時遺伝)により同定される。
【0144】
次に、罹患個体と非罹患個体との間のcDNA配列またはゲノム配列における相違を決定する必要がある。変異がいくつかまたは全ての罹患個体に観察されるが、いずれの正常な個体にも観察されない場合、この変異は疾患の原因因子であるようである。
【0145】
物理的マッピング技術および遺伝的マッピング技術の現在の解像度では、疾患に関連する染色体領域に正確に位置決めされたcDNAは、50と500との間の潜在的原因遺伝子の1つ
であり得る。(これは、1メガベースのマッピング解像度、および20kbあたり1遺伝子と仮定する)。
【0146】
エンドカインαタンパク質および抗体治療
上記のように、TNFは、組織損傷をもたらす、その炎症誘発性(pro-inflammatory)作
用(例えば、血管内皮細胞への凝血原活性の誘導(Pober,J.S.ら, J.Immunol. 136:1680 (1986))、好中球およびリンパ球の増加した接着(Pober J.S.ら, J.Immunol.138:3319 (1987))、ならびにマクロファージ、好中球、および血管内皮細胞からの血小板活性化因
子の放出の刺激(Camussi, G.ら, J.Exp. Med. 166:1390 (1987)))について留意される。最近の証拠は、TNFを多くの感染(Cerami A.ら, Immunol.Today 9:28 (1988))、免疫
疾患、腫瘍性疾患(例えば、悪液質を伴ういくつかの悪性疾患(Oliff A.ら, Cell 50:555 (1987))ならびに自己免疫疾患および対宿主性移植片疾患(Piguet,P.-F.ら, J. Exp. Med. 166:1280 (1987)))における病因に関係付けている。多数の研究は、TNFが、ガン
、感染性の疾患、および他の異化状態における悪液質の重要なメディエータであることを示唆した。
【0147】
従って、本発明のエンドカインαタンパク質は、好ましくは、放射性同位体または細胞増殖抑制性薬物との結合後に、腫瘍標的化のために用いられ得る(GrussおよびDower, Blood 85(12):3378-3404 (1995))。エンドカインαは、黒色腫および肉腫を有する患者に
おいて、腫瘍退縮および患者の寿命の延長のために、局所注射を通して、または別々の四肢への灌流において用いられ得る(AggarwalおよびNatarajan,Eur. Cytokine Netw. 7(2):92-124(1996))。
【0148】
本発明のエンドカインαはまた、特定の状況において治療的役割(例えば、ウイルス感染、細菌感染、酵母感染、真菌感染、および他の感染(toxoplasma gondii、schistosoma
mansoni、listeria monocytogens、およびBCGを含む)に対する活性)を有し得る。エンドカインαのこれらの効果は、間接的であり得、従って好ましくは、マクロファージ、好酸球、線維芽細胞、または好中球の活性化を通して媒介される。
【0149】
TNFはまた、発熱、倦怠感、食欲不振、および悪液質を含む、グラム陰性敗血症および
内毒素性ショックの病態生理学的結果において中心的な役割を果たすと考えられる(Michie,H.R.ら, Br. J. Surg. 76:670-671 (1989); Debets, J. M. H.ら, Second Vienna ShockForum, 463-466頁(1989); Simpson, S.Q.ら, Crit. Care Clin. 5:27-47 (1989))。内毒素は、TNF(Kornbluth,S.K.ら, J. Immunol. 137:2585-2591 (1986))および他のサ
イトカインの生成および分泌を刺激する、強力な単球/マクロファージアクチベーターである。上昇したレベルの循環TNFはまた、グラム陰性敗血症を罹患する患者において見出
されている(Waage,A.ら, Lancet 1:355-357 (1987); Hammerle, A.F.ら, Second Vienna
Shock Forum715-718頁 (1989); Debets, J.M.H.ら, Crit. Care Med. 17:489-497 (1989); Calandra,T.ら, J. Infec. Dis. 161:982-987 (1990))。
【0150】
TNFに対する中和抗血清またはmAbは、ヒト以外の哺乳動物において、実験的内毒血症および菌血症における有害な生理学的変化を排除し、そして致死的なチャレンジの後の死を防止することが示されている。この効果は、例えば、齧歯類の致死率アッセイおよび霊長類の病態モデル系において実証されている(Mathison,J.C.ら, J. Clin. Invest. 81:1925-1937 (1988); Beutler, B.ら, Science 229:869-871(1985); Tracey, K.J.ら, Nature 330:662-664 (1987); Shimamoto, Y.ら, Immunol.Lett. 17:311-318 (1988); Silva, A.T.ら, J. Infet. Dis. 162:421-427 (1990); Opal,S.M.ら, J. Infect. Dis. 161:1148-1152 (1990); Hinshaw, L.B.ら, Circ. Shock30:279-292 (1990))。今日まで、ヒトにおける抗TNF mAb治療での経験は、限られていたが、例えば、関節炎および敗血症において有
益な治療結果を示す。例えば、Elliott,M.J.ら, Baillieres Clin. Rheumatol. 9:633-52
(1995); Feldmann, M.ら, Ann. N.Y.Acad. Sci. USA 766:272-8 (1995); van der Poll,
T.ら, Shock 3:1-12 (1995); Wherryら,Crit. Care. Med. 21:S436-40 (1993); Tracey K.J.ら, Crit. Care Med. 21:S415-22(1993)を参照のこと。
【0151】
エンドカインαは、TNFの多くの生物学的効果を示すと考えられるので、本発明はさら
に、上記の障害の1つ以上を処置するために哺乳動物(好ましくはヒト)患者に抗エンドカインα抗体を投与する工程を含む、抗体に基づく治療に関する。抗エンドカインαポリクローナル抗体およびモノクローナル抗体を生成するための方法は、上記に詳細に記載される。このような抗体は、当該分野で公知のように、または本明細書中に記載されるように、薬学的に受容可能な組成物中に提供され得る。
【0152】
本発明の抗体が治療的に用られ得る方法の概要は、エンドカインαを身体内において局所的または全身的に結合させること、または、例えば、補体(CDC)もしくはエフェクタ
ー細胞(ADCC)により媒介されるような抗体の直接細胞傷害性によることを含む。これらのアプローチのいくつかは、下記にさらに詳細に記載される。本明細書中に提供される教示で武装すれば、当業者は、本発明の抗体を診断目的、モニタリング目的、または治療目的のために過度の実験を伴うことなくどのように使用するかがわかる。
【0153】
本発明の薬学的組成物は、それらの意図される目的を達成する任意の手段により投与され得る。抗体、それらのフラグメント、または誘導体の投与のための量および措置は、TNF関連疾患の処置の臨床分野の当業者によって容易に決定され得る。
【0154】
例えば、投与は、非経口、皮下、静脈内、筋肉内、腹腔内、経皮、または頬の経路によってであり得る。あるいは、または同時に、投与は、経口経路によってであり得る。投与される投薬量は、レシピエントの年齢、健康状態、および体重、同時処置の種類(ある場合)、処置の頻度、および所望される効果の性質に依存する。
【0155】
本発明の範囲内の組成物は、抗体、フラグメント、または誘導体が、その意図される目的を達成するために有効な量で含まれる全ての組成物を含む。個々の必要性は異なるが、各成分の有効量の最適な範囲の決定は、当該分野の技量の範囲内である。有効用量は、個々のキメラ抗体またはモノクローナル抗体、結合した治療剤(下記を参照のこと)の存在および性質、患者およびその臨床状態の関数であり、そして約10μg/kg体重〜約5000mg/kg体重で変化し得る。好ましい投薬量は、0.1〜500mg/kg体重を含む。
【0156】
薬理学的に活性な化合物に加えて、新規な薬学的組成物は、適切な薬学的に受容可能なキャリアを含み得る。このキャリアは、薬学的に用いられ得る調製物中での活性な化合物の処理を容易にする賦形剤および助剤を含む。好ましくは、調製物は、約0.01〜99%、好ましくは約20〜75%の活性化合物を、賦形剤とともに含む。
【0157】
同様に、イメージングのための検出可能に標識された形態、または治療のための遊離もしくは結合した形態のような、非経口投与のための本発明のエンドカインα抗体またはフラグメントの調製物は、無菌の水性または非水性の溶液、懸濁液、および乳濁液を含む。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油(例えばオリーブ油)、および注入可能な有機エステル(例えば、オレイン酸エチル)である。水性キャリアは、生理食塩水および緩衝化媒体を含む、水、アルコール/水溶液、乳濁液、または懸濁液、塩化ナトリウム溶液を含む非経口ビヒクル、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル、または固定油を含む。静脈内ビヒクルは、流体補充物および栄養補充物(replenisher)(例えば、リンゲルデキストロース
に基づく補充物)などを含む。保存剤および他の添加剤はまた、例えば、抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなどのように存在し得る。一般的に、Remington’sPharmaceutical Science, 第16版, Mack Publishing Co., Easton, PA, 1980を参照のこ
と。
【0158】
特に、本発明の抗体、フラグメント、および誘導体は、本明細書中に記載されるようなエンドカインα関連障害を有するかまたは発症している被験体を処置するために有用である。このような処置は、抗体、フラグメント、もしくは誘導体、またはその結合体の単回用量または複数回用量を非経口投与する工程を含む。
【0159】
本発明の抗体は、他のモノクローナル抗体もしくはキメラ抗体と、または抗体と相互作用するエフェクター細胞の数もしくは活性を増加させるのに役立つ、リンホカインまたは造血性増殖因子などと組み合わせて有利に利用され得る。
【0160】
エンドカインαの循環濃度は、(TNFと同様に)非敗血症個体において約10pg/mlの範囲、そして敗血症患者において約50pg/mlに到達し、そしてTNFの敗血症症候群において100pg/mlより上で、非常に低い傾向がある(Hammerle,A.F.ら, 1989, 上記)か、またはエン
ドカインα関連障害の部位で検出可能でのみあり得るので、高親和性および/または強力なインビボのエンドカインα阻害抗体および/またはエンドカインα中和抗体、フラグメント、またはその領域を、エンドカインα免疫アッセイおよびエンドカイン関連障害の治療の両方に用いることが好ましい。このような抗体、フラグメント、または領域は、好ましくは、ヒトエンドカインαに、少なくとも108M-1、より好ましくは少なくとも109
M-1、例えば、5×108 M-1、8×108M-1、2×109 M-1、4×109 M-1、6×10
9M-1、8×109 M-1のKaとして表される親和性を有する。
【0161】
ヒトでの治療的使用のために好ましいのは、本発明による、強力なインビボでのエンドカイン阻害活性および/または中和活性を有する、高親和性マウスおよびマウス/ヒト、またはヒト/ヒトのキメラ抗体、ならびにフラグメント、領域、および誘導体である。これらの抗体、ならびにフラグメント、領域、および誘導体は、例えば、エンドカイン誘導性IL-1、IL-6、またはTNF分泌、凝血原活性、細胞接着分子(例えば、ELAM-1およびICAM-1)の発現、および分裂促進活性を、インビボ、インサイチュ、およびインビトロでブロ
ックする。
【0162】
本発明を一般的に記載してきたが、本発明は、例示のために提供され、そして限定することを意図しない以下の実施例を参照することにより、さらに容易に理解される。
【実施例】
【0163】
実施例1:E.coliにおける成熟エンドカインαの発現および精製
寄託されたcDNAにおいて成熟エンドカインαをコードするDNA配列を、エンドカインα
タンパク質のアミノ酸末端配列に特異的なPCRオリゴヌクレオチドプライマーを用いて増
幅する。クローニングを容易にするための制限部位を含むさらなるヌクレオチドを、それぞれ5’および3’の配列に添加する。
【0164】
5’オリゴヌクレオチドプライマーは、配列 GCG CCA TGG CTA AGT TTG GAC CAT(配列番号5)を有し、これは下線を付したNcoI制限部位を含む。
【0165】
3’プライマーは、配列GCG AAG CTT TCA AGT CTC TAG GAG ATG(配列番号6)を有し
、これは下線を付したHindIII制限部位を含む。
【0166】
制限部位は、細菌発現ベクターpQE60の制限酵素部位に対して都合良く、これらの実施
例においてM15/rep4宿主細胞における細菌発現のために使用する(Qiagen,Inc. Chatsworth, CA, 91311)。pQE60は、アンピシリン抗生物質耐性(Ampr)をコードし、細菌の複製起点(ori)、IPTG誘導プロモーター、リボソーム結合部位(「RBS」)、6-Hisタグ、お
よび制限酵素部位を含む。
【0167】
増幅されたエンドカインαタンパク質DNAおよびベクターpQE60の両方をNcoIおよびHindIIIで消化し、次いで消化されたDNAを一緒に連結する。エンドカインαタンパク質DNAの
制限されたpQE60ベクターへの挿入は、エンドカインαタンパク質コード領域を、ベクタ
ーのIPTG誘導プロモーターの下流に、かつこれに作動可能に連結するように、そしてエンドカインαタンパク質の翻訳のために適切に位置された開始ATGにインフレームに配置す
る。
【0168】
連結混合物を、標準的な手順を用いて、コンピテントなE.coli細胞に形質転換する。このような手順は、Sambrookら、Molecular Cloning: A LaboratoryManual、第2版、Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)に記載されている
。多数のコピーのプラスミドpREP4を含有するE.coli株M15/rep4(これは、lacリプレッサーを発現し、そしてまたカナマイシン耐性(Kanr)を付与する)を本明細書に記載の例示的な実施例を行うにあたって使用する。この株(これは、エンドカインαタンパク質を発現するために適切である多くの内の唯一である)は、Qiagenから市販されている。
【0169】
形質転換体を、アンピシリンおよびカナマイシンの存在下でLBプレート上で増殖するそれらの能力により同定する。プラスミドDNAを耐性コロニーから単離し、そしてクローン
化DNAの同定を制限分析により確認する。
【0170】
所望の構築物を含むクローンを、アンピシリン(100μg/ml)とカナマイシン(25μg/ml)との両方を補充したLB培地における液体培養で一晩(「O/N」)増殖する。
【0171】
O/N培養物を用いて約1:100〜1:250の希釈で大規模培養に接種する。細胞を、0.4と0.6
との間の600NMでの吸光度(OD600)にまで増殖させた。次いで、イソプロピル-B-D-チオ
ガラクトピラノシド(「IPTG」)を加えて1mMの最終濃度にし、lacIリプレッサーを不活性化することにより、lacリプレッサー感受性プロモーターからの転写を誘導する。細胞
をさらに3〜4時間引き続きインキュベートする。次いで、標準的方法によって細胞を遠心分離により収集し、そして破壊する。日常的な回収技術を用いて破壊した細胞から封入体を精製し、そしてタンパク質を封入体から8M尿素中へ可溶化する。可溶化したタンパク質を含む8M尿素溶液を、2×リン酸緩衝化生理食塩水(「PBS」)中でPD-10カラムを通し、それによって尿素を除去し、緩衝液を交換し、そしてタンパク質を再び折り畳む。タンパク質をクロマトグラフィーのさらなる工程によって精製してエンドトキシンを除去する。次いで、濾過滅菌する。濾過滅菌したタンパク質調製物を、2×PBS中で保存する
。
【0172】
実施例2:バキュロウイルス発現系における成熟エンドカインαのクローニングおよび発現
寄託されたクローンにおける完全エンドカインαタンパク質をコードするcDNA配列を、この遺伝子の5’配列および3’配列に対応するPCRオリゴヌクレオチドプライマーを用
いて増幅する:
5’プライマーは、配列 GC GGA TCC CGA GAC TGC TAA GGA GCC(配列番号7)を有し
、下線を付したBamHI制限酵素部位を含む、そして図1におけるエンドカインαタンパク
質の部分をコードする配列に相補的な配列を含む。
【0173】
3’プライマーは、配列 GC GGA TCC CTA GGA GAT GAA TTG GGG ATT TG(配列番号8)を有し、下線を付したBamHI制限部位を含み、そして図1におけるエンドカインαタンパ
ク質の部分に相補的な配列を含む。
【0174】
増幅されたフラグメントを、市販のキット(「Geneclean」 BIO 101 Inc., La Jolla, Ca.)を用いて、1%アガロースゲルより単離する。次いで、このフラグメントをBamHIで消化し、そして1%アガロースゲルで再び精製する。このフラグメントを、本明細書中でF2と命名する。
【0175】
ベクターpA2GPを用いて、Summersら、AManual of Methods for Baculovirus Vectors and Insect Cell Culture Procedures,Texas Agricultural Experimental Station Bulletin No. 1555(1987)に記載の標準的な方法を用いて、エンドカインαタンパク質をバキ
ュロウイルス発現系において発現する。この発現ベクターは、Autographacalifornica核
多角体病ウイルス(AcMNPV)の強いポリヘドリンプロモーター、それに続く便利な制限部位を含む。AcMNPV gp67のシグナルペプチド(N末端メチオニンを含む)は、BamHI部位のちょうど上流に位置する。シミアンウイルス40(「SV40」)のポリアデニル化部位を、効率的なポリアデニル化のために用いる。組換えウイルスの容易な選択のために、E.coli由来のβガラクトシダーゼ遺伝子をポリヘドリンプロモーターと同方向に挿入し、その後ポリヘドリン遺伝子のポリアデニル化シグナルが続く。ポリヘドリン配列を、野生型ウイルスDNAとの細胞媒介性相同組換えのためにウイルス配列で両端で隣接させ、クローン化ポ
リヌクレオチドを発現する生存可能なウイルスを生成する。
【0176】
当業者が容易に理解するように、構築が転写、翻訳、輸送などのために適切に位置したシグナル(例えば、必要ならばインフレームでのAUGおよびシグナルペプチド)を提供す
るならば、多くの他のバキュロウイルスベクターが、pA2-GPの代わりに用いられ得る。例えば、pAc373、pVL941、およびpAcIM1である。このようなベクターは、とりわけ、Luckowら、Virology,170:31-39に記載される。
【0177】
当該分野で公知の日常的な手順を用いて、プラスミドを制限酵素XbaIで消化し、次いでウシ腸ホスファターゼを用いて脱リン酸化する。次いで、市販のキット(「Geneclean」BIO 101 Inc.,La Jolla, Ca)を用いてDNAを1%アガロースゲルから単離する。このベク
ターDNAを、本明細書中で「V2」と命名する。
【0178】
フラグメントF2および脱リン酸化プラスミドV2を、T4DNAリガーゼでともに連結する。E.coli HB101細胞を連結混合物で形質転換し、そして培養プレート上に播く。XbaIを用い
て個々のコロニーからのDNAを消化し、次いでゲル電気泳動によって消化産物を分析する
ことによって、ヒトエンドカインα遺伝子を有するプラスミドを含む細菌を同定する。クローン化フラグメントの配列を、DNA配列決定により確認する。
【0179】
5μgのプラスミドを、Felgnerら Proc. Natl. Acad. Sci.USA, 84:7413-7417 (1987)によって記載されるリポフェクション法を用いて、1.0μgの市販の線状化バキュロウイルスDNA(「BaculoGoldTMbaculovirus DNA」, Pharmingen, San Diego, CA.)とともに同時トランスフェクトする。1μgのBaculoGoldTMウイルスDNAおよび5μgのプラスミドを、50μlの無血清グレース培地(Life Technologies Inc.,Gaithersburg, MD)を含むマイクロタイタープレートの無菌ウェル中で混合する。その後、10μlのリポフェクチンおよび90
μlのグレース培地を添加し、混合し、そして室温にて15分間インキュベートする。次い
で、そのトランスフェクション混合物を、無血清グレース培地1mlを有する35mm組織培養プレート内に播種されたSf9昆虫細胞(ATCCCRL 1711)に滴下する。プレートを、新たに
添加した溶液を混合するために、前後に振盪する。次いでプレートを、27℃で5時間インキュベートする。5時間後、トランスフェクション溶液をプレートから除去し、そして10%ウシ胎児血清を補充した1mlのグレース昆虫培地を添加する。プレートをインキュベーターに戻し、そして27℃で4日間培養を続ける。
【0180】
4日後、上清を回収し、そしてSummersおよびSmith(前出)に記載されるようにプラークアッセイを行う。青く染色されたプラークを産生するgal発現クローンの容易な同定お
よび単離を可能にするために、「BlueGal」(Life Technologies Inc., Gaithersburg)
を有するアガロースゲルを用いる。(このタイプの「プラークアッセイ」の詳細な説明はまた、LifeTechnologies Inc.、Gaithersburg、で配布される昆虫細胞培養およびバキュ
ロウイルス学の使用者ガイド(9〜10頁)においても見い出され得る)。
【0181】
連続希釈の4日後、ウイルスを細胞に添加する。適切なインキュベーションの後、青く染色されたプラークをエッペンドルフピペットのチップで拾う。次いで、組換えウイルスを含む寒天を、200μlのグレース培地を含むエッペンドルフチューブ中に再懸濁する。寒天を、短時間の遠心分離により除去し、そして組換えバキュロウイルスを含む上清を、35mmディッシュに播種されたSf9細胞に感染するために用いる。4日後、これらの培養ディ
ッシュの上清を回収し、次いでそれらを4℃で保存する。適切に挿入されたhESSBI、II、およびIIIを含むクローンを、このプラスミド制限マッピングおよび配列決定を含むDNA分析によって同定する。
【0182】
Sf9細胞を、10%熱不活化FBSを補充したグレース培地中で増殖する。細胞を、約2(約1〜約3)の感染多重度(「MOI」)で組換えバキュロウイルスで感染させる。6時間後
、その培地を除去し、そしてメチオニンおよびシステインを除いたSF900II培地(Life Technologies Inc., Gaithersburgから入手可能)に置き換える。42時間後、5μCiの35S-メチオニンおよび5μCiの35S-システイン(Amershamから入手可能)を添加する。細胞
をさらに16時間インキュベートし、次いで細胞を遠心分離により収集し、溶解し、そして標識されたタンパク質をSDS-PAGEおよびオートラジオグラフィーにより可視化する。
【0183】
実施例3:CHO細胞におけるクローニングおよび発現
ベクターpC1を、エンドカインαタンパク質の発現のために使用する。プラスミドpC1は、プラスミドpSV2-dhfr(ATCC受託番号37146)の誘導体である。両方のプラスミドは、SV40初期プロモーターの制御下で、マウスDHFR遺伝子を含む。これらのプラスミドでトランスフェクトされるジヒドロ葉酸活性を欠如するチャイニーズハムスター卵巣細胞または他の細胞は、化学治療剤メトトレキサートを補充した選択培地(αマイナスMEM、LifeTechnologies)中で細胞を増殖させることによって選択され得る。メトトレキサート(MTX)に耐性である細胞におけるDHFR遺伝子の増幅は、よく考証されている(例えば、Alt,F.W.ら、J. Biol. Chem. 253:1357-1370(1978)、Hamlin, J.L.およびMa, C., Biochem. etBiophys. Acta, 1097:107-143(1990)、Page, M.J.およびSydenham, M.A., Biotechnology9:64-68(1991)を参照のこと)。漸増濃度のMTXにおける細胞増殖は、DHFR遺伝子の増幅の結果
として、標的酵素DHFRを過剰産生することによって薬物への耐性を生じる。第二遺伝子がDHFR遺伝子に連結される場合、通常、同時増幅され、そして過剰発現される。細胞株が遺伝子の1,000以上のコピーを有することは、当該水準である。続いて、メトトレキサート
が取り除かれる場合、細胞株は、染色体(単数または複数)に取り込まれる増幅遺伝子を含む。
【0184】
プラスミドpC1は、ラウス肉腫ウイルス(Cullenら、Molecularand Celluar Biology, 438-4470(1985年3月))の長末端反復(LTR)の目的の強力なプロモーターの遺伝子、お
よびヒトサイトメガロウイルス(CMV)(Boshartら、Cell41:521-530(1985))の最初期
遺伝子のエンハンサーから単離されたフラグメントの遺伝子の発現を含む。プロモーターの下流は、遺伝子の取り込みを可能にする以下の1つの制限酵素切断部位である:BamHI
、PvuII、およびNruI。これらのクローニング部位の後ろに、プラスミドは、全ての3つ
のリーディングフレームにおける翻訳停止コドン、続いてラットプレプロインシュリン遺伝子の3’イントロンおよびポリアデニル化部位を含む。他の高効率プロモーターもまた
、発現(例えば、ヒトβアクチンプロモーター、SV40初期もしくは後期プロモーター、または他のレトロウイルス(例えば、HIVおよびHTLVI)からの長末端反復のために使用し得る。mRNAのポリアデニル化のために、他のシグナル(例えば、ヒト成長ホルモンまたはグロビン遺伝子由来)も同様に使用し得る。
【0185】
染色体に挿入した目的の遺伝子を有する安定な細胞株もまた、選択マーカー(例えば、gpt、G418、またはハイグロマイシン)での同時トランスフェクトに基づいて選択し得る
。開始における1つ以上の選択マーカー(例えば、G418およびメトトレキサート)を使用することが有用である。
【0186】
プラスミドpC1を制限酵素BamHIで消化し、ついでウシ小腸ホスファターゼ(phosphate)
を用いて、当該分野で公知の手順によって脱リン酸化する。次いで、ベクターを、1%アガロースゲルから単離する。
【0187】
エンドカインαをコードするDNA配列(ATCC番号97640)を、遺伝子の5’および3’配列に対応するPCRオリゴヌクレオチドプライマーを用いて増幅する:
5’プライマーは、配列5’GCG GGA TCC GCC ATC ATG CCT TTA AGC CAT TC 3’(配列番号9)を有し、これは、下線を付したBamHI制限酵素部位、続いて図1(配列番号1)の
エンドカインαの配列の塩基を含む。発現ベクター(以下に記載)に挿入した、ヒトエンドカインαをコードする増幅したフラグメントの5’末端は、有効なシグナルペプチドを
提供する。真核生物細胞における翻訳の開始のために十分なシグナル(Kozak,M., J. Mol
. Biol. 196:947-950(1987)によって記載される)を、構築物のベクター部分に適切に配
置する。
【0188】
3’プライマーは、配列5’GC GGA TCC CTA GGA GAT GAA TTG GGG ATT TG 3’(配列番
号10)を有し、これは、Asp718制限、続いて図1(配列番号1)に示されるエンドカインαコード配列に相補的なヌクレオチド(停止コドンを含む)を含む。
【0189】
増幅したフラグメントを、上記の1%アガロースゲルから単離し、次いでエンドヌクレアーゼBamHIおよびAsp718で消化し、次いで1%アガロースゲルで再び精製する。
【0190】
次いで、単離したフラグメントおよび脱リン酸化ベクターをT4 DNAリガーゼで連結する。次いで、E.coli HB101細胞を形質転換し、そして制限酵素BamHIを用いて正しい配向に
挿入されたプラスミドpC1を含む細菌を同定する。挿入された遺伝子の配列を、DNA配列決定によって確認する。
【0191】
CHO-DHFR細胞のトランスフェクション
活性なDHFR酵素を欠如するチャイニーズハムスター卵巣細胞を、トランスフェクションのために使用する。5μgの発現プラスミドC1を、リポフェクション法(Felgnerら、前出)を用いて、0.5μgのプラスミドpSVneoとともに同時トランスフェクトする。プラスミドpSV2-neoは優性選択マーカー(G418を含む一群の抗生物質への耐性を与える酵素をコードするTn5由来の遺伝子neo)を含む。細胞を、1mg/mlのG418を補充したαマイナスMEMに播種する。2日後、細胞をトリプシン処理し、そしてハイブリドーマクローニングプレート(Greiner,Germany)に播種し、そして10〜14日間培養する。この期間の後、シングルコ
ロニーをトリプシン処理し、次いで異なる濃度のメトトレキサート(25nM、50nM、100nM
、200nM、400nM)を用いて、6ウェルペトリ皿に播種する。次いで、最も高濃度のメトトレキサートで増殖するクローンを、一様に高濃度のメトトレキート(500nM、1μM、2μM、5μM)を含む新たな6ウェルプレートに移す。同じ手順を、クローンが100μMの濃度で増殖するまで反復する。
【0192】
所望の遺伝子産物の発現を、ウエスタンブロット分析およびSDS-PAGEによって分析する。
【0193】
実施例4:エンドカインα発現の組織分布
ノーザンブロット分析を行って、とりわけSambrookら(前出)によって記載される方法を用いて、ヒト組織におけるエンドカインαタンパク質をコードする遺伝子の発現レベルの試験を行った。本発明のエンドカインαタンパク質の完全ヌクレオチド配列を含むcDMAプローブ(配列番号1)を、rediprimeTMDMA標識系(AmershamLife Science)を用いて、製造業者の説明書に従って、32Pで標識した。標識後、プローブを、CHROMA SPIN-100TMカラム(ClontechLaboratories, Inc.)を用いて、製造業者のプロトコル番号PT1200-1に従って、精製した。次いで、精製した標識化プローブを使用して、エンドカインαタンパク質をコードする遺伝子の発現について、種々のヒト組織を実験した。
【0194】
種々のヒト組織(H)またはヒト免疫系組織(IM)を含む多組織ノーザン(MTN)ブロットをClontechから入手し、そしてExpressHybTMHybridization Solution(Clontech)を用いて、製造業者のプロトコル番号PT1190-1に従って、標識化プローブで試験した。ハイブリダイゼーションおよび洗浄に続いて、ブロットを取り付け、そして-70℃にて一晩フィ
ルムに暴露し、そして標準的な手順に従ってフィルムを現像した。
【0195】
本発明のエンドカインαタンパク質をコードする遺伝子の発現を、ヒト脳線条体(striatrium)および膵臓組織において検出した。
【0196】
本発明が、前述の説明および実施例に詳細に記載される以外に実施され得ることは明白である。
【0197】
本発明の多数の改変および変異が、上記の教示に照らして可能であり、それゆえ添付の請求の範囲内である。
【0198】
本明細書中に参考として援用される全ての特許、特許明細書、および刊行物の開示は、本明細書に参考として援用される。
【産業上の利用可能性】
【0199】
本発明の新規のサイトカインは、TNF様障害の治療のための新規の抗体またはこれらのTNF様サイトカインに結合する他のアンタゴニストを作製するために使用され得る。
【図面の簡単な説明】
【0200】
【図1A】図1Aは、エンドカインαタンパク質のヌクレオチド(配列番号1)および推定アミノ酸(配列番号2)配列を示す。アミノ酸1〜17は、細胞内ドメインを示し、アミノ酸18〜43は、膜貫通ドメイン(下線を付した配列)を示し、そしてアミノ酸44〜169は、細胞外ドメイン(残りの配列)を示す。
【図1B】図1Aの続き。
【図2】図2は、エンドカインαタンパク質(配列番号2)、組織壊死因子α(TNF-α)(配列番号3)、およびTNF-β(配列番号4)のアミノ酸配列間の類似性が存在する領域を示す。J.Hein法を、PAM250残基重量表とともに使用した。四角の囲いは、コンセンサスに正確に一致する残基を示す。
【図3】図3は、エンドカインαアミノ酸配列の分析を提供する。α、β、ターンおよびコイル領域;親水性および疎水性;両親媒性領域;可動性領域;抗原性指数および表面確率が示される。「抗原性指数-Jameson-Wolf」グラフにおいて、図1のアミノ酸残基44〜54、57〜68、69〜78、94〜105、108〜132および148〜158が、示されたエンドカインαタンパク質の抗原性が高い領域に対応する。
【0201】
(配列表)
【0202】
【表1】

【技術分野】
【0001】
発明の背景
発明の分野
本発明はエンドカインαタンパク質に関する。詳細には、エンドカインαタンパク質をコードする単離された核酸分子が提供される。エンドカインαポリペプチド、ならびにベクター、宿主細胞およびこれらを産生するための組換え方法が提供される。
【背景技術】
【0002】
関連分野
腫瘍壊死因子-α(TNFα;カケクチンとも呼ばれる)は、内毒素または他の刺激に応答して、17kDのタンパク質サブユニットの可溶性のホモトリマーとして、単球およびマクロファージによって主として分泌される(非特許文献1)。膜に結合したTNFの26kDの前駆体形態もまた記載されている(非特許文献2)。
【0003】
蓄積された証拠により、TNFが多面的な生物学的活性を有する調節サイトカインであることが示されている。これらの活性としては以下が挙げられる:リポタンパク質リパーゼ合成(「カケクチン」活性)(非特許文献3)、多形核白血球の活性化(非特許文献4;非特許文献5)、細胞増殖の阻害または細胞増殖の刺激(非特許文献6;非特許文献7; 非特許文献8)、所定の形質転換された細胞型における細胞傷害作用(非特許文献8; 非特許文献9)、抗ウイルス活性(非特許文献10;非特許文献11)、骨再吸収の刺激(非特許文献12; 非特許文献13)、コラゲナーゼおよびプロスタグランジンE2産生の刺激(非特許文献14);ならびにT細胞(非特許文献15)、B細胞(非特許文献16)、単球(非特許文献17)、胸腺細胞(非特許文献18)の活性化を含む免疫調節作用、ならびに主要組織適合複合体(MHC)クラスIおよびクラスII分子の細胞表面発現の刺激(非特許文献19)。
【0004】
TNFは、組織損傷を生じる炎症誘発性作用(例えば、血管内皮細胞における凝血原活性の誘導(非特許文献20)、好中球およびリンパ球の接着の増大(非特許文献21)、およびマクロファージ、好中球および血管内皮細胞由来の血小板活性化因子の放出の刺激(非特許文献22)について記載されている。
【0005】
最近の証拠は、TNFを、多くの感染の原因(非特許文献23)、免疫不全、腫瘍性疾患、例えば、いくつかの悪性腫瘍に付随する悪液質(非特許文献24)、および自己免疫疾患および移植片対宿主疾患(非特許文献25)に結び付ける。TNFのガンおよび感染性疾患との関連は、宿主の異化状態にしばしば関連する。ガン患者における主要な問題は、体重減少(通常、食欲不振に関連する)である。生じる大規模な体力の消耗は「悪液質」として知られる(非特許文献26)。悪液質としては、進行性の体重減少、食欲不振、および悪性増殖に応答した身体質量の持続性の浸食が挙げられる。従って、悪液質の状態は、有意な罹患率と関連し、そしてガン死亡率の大多数を担う。多数の研究により、TNFがガン、感染性病理、および他の異化状態における悪液質の重要なメディエータであることが示唆された。
【0006】
TNFは、グラム陰性敗血症および内毒素ショックの病態生理学的結果において中心的な役割を果たすと考えられている(非特許文献27; 非特許文献28;非特許文献29)。これには、発熱、倦怠感、食欲不振、および悪液質が挙げられる。内毒素は、強力な単球/マクロファージ活性化因子であり、TNFの産生および分泌(非特許文献30)ならびに他のサイトカインの産生および分泌を刺激する。TNFは、内毒素の多くの生物学的影響を模倣し得るので、内毒素関連疾病の臨床的徴候を担う中心的なメディエータであると結論付けられた。TNFおよび他の単球由来サイトカインは、内毒素に対する代謝性および神経ホルモンの応答を媒介する(非特許文献31)。ヒトボランティアへの内毒素投与は、インフルエンザ様症状症状(発熱、頻脈、増大した代謝速度、およびストレスホルモン放出を含む)を伴う急性疾病を生じる(非特許文献32)。循環TNFのレベルの上昇はまた、グラム陰性敗血症を患う患者において見出されている(非特許文献33;非特許文献34;非特許文献35; 非特許文献36)。
【0007】
TNFを中和することに向けられる受動免疫療法は、グラム陰性敗血症および内毒血症(上記で考察したように、これらの病理状態における増大したTNF産生および上昇したTNFレベルに基づく)において有益な効果を有し得る。
【0008】
カケクチンとして特徴付けられた「モジュラー」物質(後にTNFと同一であることが見出された)に対する抗体は、Ceramiら(特許文献1、1987年3月4日)に開示された。このような抗体は、診断的免疫アッセイおよび細菌感染におけるショックの治療に有用であると考えられた。Rubinら(特許文献2、1987年4月22日)は、ヒトTNFに対するモノクローナル抗体、そのような抗体を分泌するハイブリドーマ、そのような抗体を産生する方法、およびTNFのイムノアッセイにおけるそのような抗体の使用を開示した。Yoneら(特許文献3、1988年10月26日)は、mAbを含む抗TNF抗体、および疾患(特に、川崎病、および細菌感染)のイムノアッセイ診断における有用性を開示した。川崎病(乳児の急性発熱性粘膜皮膚リンパ節症候群;非特許文献37;非特許文献38)を有する患者の体液は、この疾患の進行に関連する上昇したTNFレベルを含むと考えられた(Yoneら、前述)。
【0009】
他の研究者らは、インビトロでの中和活性を有する組換えヒトTNFに特異的なmAbを記載した(非特許文献39;非特許文献40;非特許文献41;非特許文献42;非特許文献43; 非特許文献44)。これらのmAbのいくつかは、ヒトTNFのエピトープをマッピングするため、および酵素イムノアッセイを開発するため(非特許文献41;非特許文献43;非特許文献44)、ならびに組換えTNFの精製を補助するため(非特許文献42)に使用された。しかし、これらの研究は、免疫原性、特異性の欠失および/または医薬の適合性に起因する、ヒトにおけるインビボでの診断的または治療的使用に使用され得るTNF中和抗体を産生するための基礎を提供しない。
【0010】
有害な生理学的変化を抑止し、そして実験的な内毒血症および菌血症における致命的なチャレンジの後の死滅を妨げるための、TNFに対する中和抗血清またはmAbがヒト以外の哺乳動物において示されている。この効果は、例えば、齧歯類の死亡率アッセイおよび霊長類の疾患モデル系において示されている(非特許文献45;非特許文献46; 非特許文献47; 非特許文献48; 非特許文献49; 非特許文献50;非特許文献51)。
【0011】
今日まで、ヒトにおける抗TNF mAb治療を用いる実験は、限定はされているが、(例えば、関節炎および敗血症において)有益な治療結果を示している。例えば、非特許文献52;非特許文献53; 非特許文献54; 非特許文献55;非特許文献56を参照のこと。
【0012】
サイトカインレセプターの配列分析により、膜タンパク質の数種のサブファミリーが規定された:(1)Igスーパーファミリー、(2)ヘマトポイエチン(サイトカインレセプタースーパーファミリー)および(3)腫瘍壊死因子(TNF)/神経増殖因子(NGF)レセプタースーパーファミリー(TNFスーパーファミリーの総説については、非特許文献57および非特許文献58を参照のこと)。TNF/NGFレセプタースーパーファミリーは少なくとも10種の異なるタンパク質を含む。非特許文献57。これらのレセプターに対するリガンドは、同定されており、そして少なくとも2つのサイトカインスーパーファミリーに属する。非特許文献57。
【0013】
従って、疾患状態に関与するTNFに類似したサイトカインを提供する必要がある。このような新規のサイトカインは、TNF様障害の治療のための新規の抗体またはこれらのTNF様サイトカインに結合する他のアンタゴニストを作製するために使用され得る。
【特許文献1】欧州特許第0,212,489号明細書
【特許文献2】欧州特許第0,218,868号明細書
【特許文献3】欧州特許第0,288,088号明細書
【非特許文献1】Smith,R.A.ら、J.Biol.Chem.262:6951-6954 (1987)
【非特許文献2】Kriegler,M.ら、Cell53:45-53(1988)
【非特許文献3】Beutler,B.ら、Nature316:552 (1985)
【非特許文献4】Klebanoff,S.J.ら、J.Immunol. 136:4220(1986)
【非特許文献5】Perussia,B.ら、J.Immunol. 138:765 (1987)
【非特許文献6】Vilcek,J.ら、J.Exp.Med.163:632 (1986)
【非特許文献7】Sugarman,B.J.ら、Science 230:943 (1985)
【非特許文献8】Lachman,L.B.ら、J.Immunol.138:2913 (1987)
【非特許文献9】Darzynkiewicz,Z.ら、Canc.Res.44:83 (1984)
【非特許文献10】Kohase, M.ら、Cell 45:659 (1986)
【非特許文献11】Wong, G.H.W.ら、Nature323:819 (1986)
【非特許文献12】Bertolini, D.R.ら、Nature 319:516 (1986)
【非特許文献13】Saklatvala,J.,Nature 322: 547 (1986)
【非特許文献14】Dayer, J.-M.ら、J.Exp.Med.162:2163 (1985)
【非特許文献15】Yokota, S.ら、J.Immunol. 140:531 (1988)
【非特許文献16】Kehrl, J.H.ら、J.Exp.Med.166:786(1987)
【非特許文献17】Philip, R.ら、Nature 323:86 (1986)
【非特許文献18】Ranges, G.E.ら、J.Exp.Med.167:1472 (1988)
【非特許文献19】Collins,T.ら、Proc.Natl.Acad.Sci.USA 83:446 (1986); Pujol-Borrel,R.ら、Nature326:304(1987)
【非特許文献20】Pober,J.S.ら、J.Immunol. 136:1680 (1986)
【非特許文献21】Pober, J.S.ら、J.Immunol.138:3319 (1987)
【非特許文献22】Camussi, G.ら、J.Exp.Med.166:1390(1987)
【非特許文献23】Cerami,A.ら、Immunol.Today9:28 (1988)
【非特許文献24】Oliff, A.ら、Cell 50:555 (1987)
【非特許文献25】Piguet,P.-F.ら、J.Exp.Med. 166:1280 (1987)
【非特許文献26】Kern,K.A.ら、J.Parent.Enter.Nutr. 12:286-298 (1988)
【非特許文献27】Michie,H.R.ら、Br.J.Surg. 76:670-671 (1989)
【非特許文献28】Debets, J.M.H.ら、Second Vienna Shock Forump.463-466 (1989)
【非特許文献29】Simpson, S.Q.ら、Crit.Care Clin. 5:27-47 (1989)
【非特許文献30】Kornbluth,S.K.ら、J.Immunol. 137:2585-2591 (1986)
【非特許文献31】Michie,H.R.ら、N.Eng.J.Med. 318:1481-1486 (1988)
【非特許文献32】Revhaug,A.ら、Arch.Surg.123:162-170 (1988)
【非特許文献33】Waage, A.ら、Lancet1:355-357 (1987)
【非特許文献34】Hammerle, A.F.ら、Second Vienna Shock Forum p.715-718 (1989)
【非特許文献35】Debets, J.M.H.ら、Crit.Care Med. 17:489-497 (1989)
【非特許文献36】Calandra, T.ら、J.Infec.Dis.161:982-987 (1990)
【非特許文献37】Kawasaki, T., Allergy 16:178 (1967)
【非特許文献38】Kawasaki, T., Shonica (Pediatrics) 26:935(1985)
【非特許文献39】Liang, C-M.ら、Biochem.Biophys.Res.Comm. 137:847-854(1986)
【非特許文献40】Meager, A.ら、Hybridoma 6:305-311 (1987)
【非特許文献41】Fendlyら、Hybridoma 6:359-369(1987)
【非特許文献42】Bringman, T.S.ら、Hybridoma 6:489-507 (1987)
【非特許文献43】Hirai, M.ら、J.Immunol.Meth.96:57-62 (1987)
【非特許文献44】Moller, A.ら(Cytokine 2:162-169 (1990))
【非特許文献45】Mathison,J.C.ら、J.Clin.Invest. 81:1925-1937 (1988)
【非特許文献46】Beutler, B.ら、Science 229:869-871(1985)
【非特許文献47】Tracey, K.J.ら、Nature 330:662-664 (1987)
【非特許文献48】Shimamoto, Y.ら、Immunol.Lett.17:311-318 (1988)
【非特許文献49】Silva, A.T.ら、J.Infect.Dis. 162:421-427 (1990)
【非特許文献50】Opal, S.M.ら、J.Infect.Dis.161:1148-1152 (1990)
【非特許文献51】Hinshaw, L.B.ら、Circ.Shock 30:279-292 (1990)
【非特許文献52】Elliott,M.J.ら、Baillieres Clin. Rheumatol. 9:633-52 (1995)
【非特許文献53】Feldmann M.ら、Ann.N.Y.Acad.Sci.USA766:272-8 (1995)
【非特許文献54】van der Poll,Tら、Shock 3: 1-12 (1995)
【非特許文献55】Wherryら、Crit.Care.Med.21:S436-40 (1993)
【非特許文献56】Tracey K.J.ら、Crit.Care Med. 21:S415-22 (1993)
【非特許文献57】GrussおよびDower,Blood 85(12):3378-3404 (1995)
【非特許文献58】AggarwalおよびNatarajan, Eur.Cytokine Netw.,7(2):93-124 (1996)
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明の目的は、疾患状態に関与するTNFに類似したサイトカインを提供することであ
る。
【課題を解決するための手段】
【0015】
本発明は、以下からなる群から選択される配列に少なくとも95%同一であるヌクレオチド配列を有するポリヌクレオチドを含有する単離された核酸分子を提供する:(a)図1(配列番号2)の完全なアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされる全長エンドカインαポリペプチドをコードするヌクレオチド配列;(b)図1(配列番号2)の約44残基〜約169残基のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされる細胞外エンドカ
インαポリペプチドをコードするヌクレオチド配列;(c)図1(配列番号2)の約18残基〜
約43残基のアミノ酸配列を有するか、またはATCC受託番号第97640号に含まれるcDNAクロ
ーンによってコードされるエンドカインα膜貫通ドメインをコードするヌクレオチド配列;(d)図1(配列番号2)の約1残基〜約17残基のアミノ酸配列を有するか、またはATCC受
託番号第97640号に含まれるcDNAクローンによってコードされるエンドカインα細胞内ド
メインをコードするヌクレオチド配列;および(e)(a)、(b)、(c)または(d)のヌクレオチ
ド配列のいずれかに相補的なヌクレオチド配列。
【0016】
1つの実施形態において、ストリンジェントなハイブリダイゼーション条件下で、上記の(a)、(b)、(c)、(d)または(e)のヌクレオチド配列に同一なヌクレオチド配列を有する
ポリヌクレオチドにハイブリダイズするポリヌクレオチドを含む単離された核酸分子であって、このポリヌクレオチドがストリンジェントなハイブリダイゼーション条件下でA残基のみまたはT残基のみからなるヌクレオチド配列を有するポリヌクレオチドにはハイブリダイズしない、単離された核酸分子を提供する。
【0017】
1つの実施形態において、上記の(a)、(b)、(c)、(d)または(e)のアミノ酸配列を有す
るエンドカインαポリペプチドのエピトープ保有部分のアミノ酸配列をコードするポリヌクレオチドを含む、単離された核酸分子を提供する。
【0018】
別の局面において、本発明は、以下からなる群から選択されるエンドカインαポリペプチドのエピトープ保有部分をコードする、上記の単離された核酸分子を提供する:図1(配列番号2)の約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)の
約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約94〜約105のアミノ酸残基を含むポリペプチド;図
1(配列番号2)の約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)の約148〜約158のアミノ酸残基を含むポリペプチド。
【0019】
1つの実施形態において、このポリヌクレオチドがRNAである、上記の単離された核酸
分子を提供する。
【0020】
1つの実施形態において、このポリヌクレオチドがRNAである、上記の単離された核酸
分子を提供する。
【0021】
1つの実施形態において、上記の単離された核酸分子をベクターに挿入する工程を包含する、組換えベクターを作製する方法を提供する。
【0022】
1つの実施形態において、上記の方法によって産生された、組換えベクターを提供する。
【0023】
1つの実施形態において、宿主細胞に上記の組換えベクターを導入する工程を包含する、組換え宿主細胞を作製する方法を提供する。
【0024】
1つの実施形態において、上記の方法によって産生された組換え宿主細胞を提供する。
【0025】
別の局面において、エンドカインαポリペプチドを産生するための組換え方法を提供し、この方法は、上記の組換え宿主細胞を、このポリペプチドが発現されるような条件下で培養し、そしてこのポリペプチドを回収する工程を包含する。
【0026】
別の局面において、以下からなる群から選択される配列に少なくとも95%同一であるアミノ酸配列を有する単離されたポリペプチドを提供する:(a)図1(配列番号2)の全長の
エンドカインαポリペプチド、またはATCC受託番号第97640号に含まれるcDNAクローンに
よってコードされる全長エンドカインαポリペプチドのアミノ酸配列;(b)図1(配列番号2)の約44残基〜約169残基、またはATCC受託番号第97640号に含まれるcDNAクローンによ
ってコードされる細胞外エンドカインαポリペプチドのアミノ酸配列;(c)図1(配列番号2)の約18残基〜約43残基、またはATCC受託番号第97640号に含まれるcDNAクローンによってコードされるエンドカインα膜貫通ドメインのアミノ酸配列;(d)図1(配列番号2)の
約1残基〜約17残基、またはATCC受託番号第97640号に含まれるcDNAクローンによってコ
ードされるエンドカインα細胞内ドメインのアミノ酸配列;および(e)(a)、(b)、(c)または(d)のポリペプチド配列のいずれか1つのエピトープ保有部分のアミノ酸配列。
【0027】
別の局面において、エンドカインαのエピトープ保有部分を含む単離されたポリペプチドを提供し、この部分が以下からなる群から選択される、単離されたポリペプチド:図1(配列番号2)の約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)
の約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)の約94〜約105のアミノ酸残基を含むポリペプチド;
図1(配列番号2)の約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)の約148〜約158のアミノ酸残基を含むポリペプチド。
【0028】
1つの実施形態において、上記のエンドカインαポリペプチドに特異的に結合する、単離された抗体または抗体フラグメントを提供する。
【0029】
別の局面において、エンドカインα活性のレベルの減少を必要とする個体を処置する方法を提供し、この方法は、上記の単離された抗体または抗体フラグメントを含む組成物を該個体に投与する工程を包含する。
【0030】
別の局面において、以下の工程を包含する診断方法を提供する:(a)哺乳動物の細胞ま
たは体液中のエンドカインα遺伝子発現レベルをアッセイする工程;および(b)該エンド
カインα遺伝子発現レベルと標準のエンドカインα遺伝子発現レベルとを比較する工程であって、それによって該標準と比較した該エンドカインα遺伝子発現レベルにおける増大または減少がTNF関連障害の指標となる、工程。
【発明の効果】
【0031】
本発明によって、疾患状態に関与するTNFに類似したサイトカインが提供される。
【発明を実施するための最良の形態】
【0032】
発明の要旨
本発明は、TNFに類似し、そして類似の生物学的効果および活性を有すると考えられる
サイトカインをコードするポリヌクレオチドを含む単離された核酸分子を提供する。このサイトカインは、エンドカインαと呼ばれ、そして図1(配列番号2)のアミノ酸配列、または1996年6月27日にATCC受託番号97640として細菌宿主中で寄託されたcDNAクローン
によりコードされるアミノ酸配列の少なくとも一部を有するエンドカインαポリペプチドを含む。このヌクレオチド配列(寄託されたエンドカインαのcDNAクローンを配列決定することによって決定された)は、約169アミノ酸残基のポリペプチドをコードするオープ
ンリーディングフレームを含む。これは、N末端のメチオニン、約17アミノ酸残基の細胞内ドメイン、約26アミノ酸の膜貫通ドメイン、約126アミノ酸の細胞外ドメイン、および
約19kDaの完全なタンパク質についての推定の分子量を含む。予想される成熟エンドカイ
ンαタンパク質の126アミノ酸配列は、図1(配列番号2)(残基44〜169)に示される。
【0033】
別の局面において、本発明は、1996年6月27日にATCC受託番号97640として寄託された
クローンのcDNAによってコードされるアミノ酸配列を有するエンドカインαポリペプチドをコードする単離された核酸分子を提供する。好ましくは、この核酸分子は、上記の寄託されたcDNAによってコードされる成熟ポリペプチドをコードする。
【0034】
本発明は、さらに、本明細書中に記載の核酸分子の核酸フラグメントに関する。好ましい核酸フラグメントとしては、以下をコードする核酸分子が挙げられる:エンドカインα細胞内ドメイン(図1(配列番号2)の約1〜約17のアミノ酸残基)を含むポリペプチド;エンドカインα膜貫通ドメインを含むポリペプチド(図1(配列番号2)の約18〜約43のアミノ酸残基);およびエンドカインα細胞外ドメインを含むポリペプチド(図1(配列番
号2)の約44〜約169のアミノ酸残基)。
【0035】
本発明のさらなる実施態様は、本明細書中に記載される核酸分子のいずれかのヌクレオチド配列に少なくとも90%同一、そしてより好ましくは、少なくとも95%、96%、97%、98%、または99%同一であるヌクレオチド配列を有する単離された核酸分子を含む。
【0036】
本発明はまた、本発明の単離された核酸分子を含む組換えベクター、その組換えベクターを含有する宿主細胞、および組換え技術によるエンドカインαポリペプチドの産生に関する。
【0037】
本発明のポリペプチドは、寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド(詳細には、成熟ポリペプチド)、ならびに寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド、またはそれらのフラグメントのアミノ酸配列に対して少なくとも90%の類似性、より好ましくは少なくとも95%の類似性を有するアミノ酸配列を有するポリペプチドを含む。本発明のさらなるポリペプチドは、寄託されたcDNAによってコードされるポリペプチド、図1(配列番号2)のポリペプチド、またはそれらのフラグメントのアミノ酸配列に少なくとも80%同一、より好ましくは少なくとも90%または95%同一であるアミノ酸配列を有するポリペプチドを含む。
【0038】
本発明の好ましいポリペプチドフラグメントは、以下を含むポリペプチドを含む:エンドカインα細胞内ドメイン、エンドカインα膜貫通ドメイン、およびエンドカインα細胞外ドメイン。
【0039】
本発明は、上記のようなアミノ酸を有するエンドカインαポリペプチドに特異的に結合する抗体を単離する方法をさらに提供する。このような抗体は、エンドカインαおよび/
またはTNF関連障害の処置において、アンタゴニストとして診断的または治療的に有用で
あり得る。
【0040】
発明の詳細な説明
本発明は、図1(配列番号2)に示されるアミノ酸配列(クローン化されたcDNAを配列決定することによって決定した)を有するエンドカインαタンパク質をコードするポリヌクレオチドを含む、単離された核酸分子を提供する。エンドカインαは、腫瘍壊死因子(TNF)リガンドファミリーの新規なメンバーであり、そしてヒトTNFαおよび関連するTNFファミリーメンバー(図2)と配列相同性を共有する。図1(配列番号1)に示されるヌクレオチド配列は、cDNAクローンを配列決定することによって得られた。これは、1996年6月27日に、アメリカンタイプカルチャーコレクション,12301 Park Lawn Drive, Rockville, Maryland 20852に寄託され、そして受託番号第97640号を与えられた。寄託されたクローンは、pBluescriptSK(-)プラスミド(Stratagene, LaJolla, CA)中に含まれる。
【0041】
核酸分子
他に示されない限り、本明細書中でDNA分子を配列決定することによって決定されたす
べてのヌクレオチド配列は、自動化DNA配列決定機(例えば、AppliedBiosystems, Inc.からのModel 373)を用いて決定され、そして本明細書中で決定されるDNA分子によってコードされるポリペプチドのすべてのアミノ酸配列は、上記のように決定されるDNA配列の翻
訳によって予想された。従って、この自動化アプローチによって決定された任意のDNA配
列について当該分野において公知のように、本明細書中で決定される任意のヌクレオチド配列はいくつかの誤りを含み得る。自動化によって決定されるヌクレオチド配列は、配列
決定されるDNA分子の実際のヌクレオチド配列に対して、代表的には少なくとも約90%の
同一、より代表的には少なくとも約95%から少なくとも約99.99%の同一である。実際の
配列は、当該分野において周知の手動DNA配列決定方法を含む他のアプローチによってよ
り正確に決定され得る。当該分野においてまた公知のように、実際の配列と比較した、決定されるヌクレオチド配列における単一の挿入または欠失は、ヌクレオチド配列の翻訳におけるフレームシフトを引き起こし、その結果、決定されるヌクレオチド配列によってコードされる予想されるアミノ酸配列は、挿入または欠失のような点にて始まる配列決定されるDNA分子によって実際にコードされるアミノ酸配列とは完全に異なる。
【0042】
他に示されない限り、本明細書中に示される各「ヌクレオチド配列」は、デオキシリボヌクレオチド(A, G, C, およびTと省略される)の配列として示される。しかし、核酸分
子またはポリヌクレオチドの「核酸配列」によって、DNA分子またはポリヌクレオチドに
はデオキシリボヌクレオチドが、そしてRNA分子またはポリヌクレオチドにはリボヌクレ
オチド(A,G, C, およびU)の対応する配列(ここで特定されるデオキシヌクレオチド配列における各チミジンデオキシヌクレオチド(T)は、リボヌクレオチドのウリジン(U)によって置き換えられる)が意図される。例えば、デオキシリボヌクレオチドの略語を用いて示される図1(配列番号1)の配列を有するRNA分子との言及は、配列番号1の各デオキ
シヌクレオチドA,GまたはCが、対応するリボヌクレオチドA, G, またはCによって置き換
えられ、そして各デオキシヌクレオチドTが、リボヌクレオチドUによって置き換えられる配列を有するRNA分子を示すことが意図される。
【0043】
本明細書中で提供される情報(例えば、図1中のヌクレオチド配列)を用いて、エンドカインαポリペプチドをコードする本発明の核酸分子は、標準的なクローニングおよびスクリーニング手順(例えば、開始物質としてmRNAを用いるクローニングcDNAのための手順)を用いて得られ得る。本発明の図のように、図1(配列番号1)において記載される核酸分子は、ヒト脳線条由来のcDNAライブラリー中で発見された。エンドカインαcDNAの一部に対応する発現される配列タグはまた、いくつかの内皮ライブラリーおよび胎児肝臓ライブラリーにおいて見出された。
【0044】
エンドカインα遺伝子は、約169のアミノ酸残基のタンパク質をコードするオープンリ
ーディングフレーム、約17のアミノ酸の細胞内ドメイン(図1(配列番号2)中の約1〜約17のアミノ酸残基)、約26のアミノ酸の膜貫通ドメイン(図1(配列番号2)中の約18〜約43のアミノ酸残基)、約126アミノ酸の細胞外ドメイン(図1(配列番号2)中の約44〜約169のアミノ酸残基);および約19kDaの推定分子量を含む。図1(配列番号2)に
おいて示されるエンドカインαタンパク質は、ヒトTNF-αに対して約30%の類似し、そして約22%同一であり、それは、GenBankに、受託番号第U42764号として利用され得る。
【0045】
当業者が理解するように、上記の配列決定誤差の可能性、ならびに異なる公知のタンパク質におけるリーダーの切断部位の可変性に起因して、寄託されたcDNAによってコードされる実際のエンドカインαポリペプチドは、約169のアミノ酸を含むが、約154〜184の範
囲のアミノ酸で任意の場所であり得る。当業者によって、使用される基準に依存して、上記のエンドカインαタンパク質ドメインの正確な「位置」が異なり得ることもまた理解される。従って、例えば、図1(配列番号2)において示されるエンドカインα細胞内膜貫通ドメインおよび細胞外ドメインの正確な位置は、ドメインを定義するために使用される基準に依存してわずかに変化し得る(例えば、正確な位置は、図1に示されるものと比較して約1〜約5の残基で異なり得る)。
【0046】
示されるように、本発明の核酸分子は、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNA、およびクローニングによって得られるか、または合成的に生成されるゲノムDNAを含む)であり得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAは、コ
ード鎖(センス鎖としても知られる)であり得るか、またはそれは、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0047】
「単離された」核酸分子によって、天然の環境から取り出された核酸分子(DNAまたはRNA)が意図される。例えば、ベクター中に含まれる組換えDNA分子は、本発明の目的のた
めに単離されることが意図される。単離されたDNA分子のさらなる例は、異種の宿主細胞
において維持される組換えDNA分子、または溶液中の精製された(部分的または実質的)DNA分子を含む。単離されたRNA分子は、本発明のDNA分子のインビボまたはインビトロでのRNA転写物を含む。本発明に従って単離された核酸分子は、合成的に生成されるような分
子をさらに含む。
【0048】
本発明の単離された核酸分子は、図1(配列番号1)において示されるオープンリーディングフレーム(ORG)を含むDNA分子を含み、そしてさらに、図1(配列番号1)におい
て示されるORF配列のすべてまたは一部と実質的に異なるが、遺伝コードの縮重に起因し
て、エンドカインαタンパク質またはそのフラグメントをなおコードする核酸分子を含む。当然のことながら、遺伝コードは当該分野において周知である。従って、上記のような縮重した変異体を生成することは、当業者にとって日常的である。
【0049】
別の局面において、本発明は、1996年6月27日にATCC寄託番号97640として寄託された
クローンのcDNAによってコードされるアミノ酸配列を有するエンドカインαポリペプチドをコードする単離された核酸分子を提供する。好ましくは、核酸分子は、上記の寄託されたcDNAクローンによってコードされる成熟ポリペプチドをコードする。本発明は、さらに、図1(配列番号1)に示されるヌクレオチド配列、もしくは上記の寄託されたクローン中に含まれるエンドカインαcDNAのヌクレオチド配列を有する単離された核酸分子、または上記の配列の一つと相補的な配列を有する核酸分子を提供する。そのような単離された分子、特にDNA分子は、染色体とのインサイチュハイブリダイゼーションによって遺伝子
マッピングするため、および例えばノーザンブロット分析によってヒト組織中のエンドカインα遺伝子の発現を検出するためのプローブとして有用である。以下に詳細に記載されるように、特定の組織または体液におけるエンドカインα遺伝子発現の変化を検出することは、特定の障害を示す。
【0050】
本発明は、さらに、本明細書中で記載される単離した核酸分子のフラグメントに関する。寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)に示されるヌクレオチド配列を有する単離された核酸分子のフラグメントによって、本明細書中で考察されるように、診断的プローブおよびプライマーとして有用である、長さが、少なくとも約15nt、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntが意図される。当然のことながら、長さがより大きなフラグメント50〜1500ntもまた、寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)において示されるようなヌクレオチド配列のすべてではなくともほとんどに対応するフラグメントのように、本発明に従って有用である。例えば、長さが少なくとも20ntのフラグメントによって、寄託されたcDNAのヌクレオチド配列、または図1(配列番号1)に示されるようなヌクレオチド配列からの20以上の隣接した塩基を含むフラグメントが意図される。この遺伝子が寄託され、そして図1(配列番号1)において示されるヌクレオチド配列が提供されるので、そのようなDNAフラグメントを生成することは、当
業者にとって日常的である。例えば、制限エンドヌクレアーゼ切断、または超音波処理による剪断は、種々のサイズのフラグメントを生成するために容易に使用され得る。あるいは、そのようなフラグメントは、合成的に生成され得る。
【0051】
本発明の好ましい核酸フラグメントは、以下をコードする核酸分子を含む:エンドカインα細胞内ドメインを含むポリペプチド(図1(配列番号2)中の約1〜約17のアミノ酸
残基);エンドカインα膜貫通ドメインを含むポリペプチド(図1(配列番号2)中の約18〜約43のアミノ酸残基);およびエンドカインα細胞外ドメインを含むポリペプチド(図1(配列番号2)中の約44〜約169のアミノ酸残基)。
【0052】
本発明のさらに好ましい核酸フラグメントは、エンドカインαタンパク質のエピトープ保有部分をコードする核酸分子を含む。特に、本発明のそのような核酸フラグメントは、以下をコードする核酸分子を含む:図1(配列番号2)中の約44〜約158のアミノ酸残基
を含むポリペプチド;図1(配列番号2)中の約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約94〜約105のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約108〜約132
のアミノ酸残基を含むポリペプチド;図1(配列番号2)中の約148〜約158のアミノ酸残基を含むポリペプチド。本発明者らは、上記のポリペプチドフラグメントが、エンドカインαタンパク質の抗原性領域であることを決定した。エンドカインαタンパク質の他のそのようなエピトープ保有部分を決定するための方法は、以下に詳細に記載される。
【0053】
別の局面において、本発明は、ストリンジェントなハイブリダイゼーション条件下で、上記の本発明の核酸分子(例えば、1996年6月27日に作成されたATCC寄託第97640号中に
含まれるcDNAクローン)中のポリヌクレオチドの一部にハイブリダイズするポリヌクレオチドを含む単離した核酸分子を提供する。「ストリンジェントなハイブリダイゼーション条件」によって、括弧内を含む溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNA)中42℃での一晩のインキュベ
ーション、続く約65℃にて0.1×SSCにおいてフィルターを洗浄することが意図される。ポリヌクレオチドの「一部」にハイブリダイズするポリヌクレオチドによって、参照のポリヌクレオチドの少なくとも約15ヌクレオチド(nt)、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは約30〜70ntにハイブリダイズするポリヌクレオチド(DNAまたはRNAのいずれか)が意図される。これらは、以上、および以下でより詳細に考察されるような診断用プローブおよびプライマーとして有用である。
【0054】
当然のことながら、参照のポリヌクレオチド(例えば、寄託されたcDNAクローン)のより大きな部分(例えば、長さが50〜500ntの部分)、または参照のポリヌクレオチドの完
全長とまでもハイブリダイズするポリヌクレオチドはまた、寄託されたcDNAのヌクレオチド配列または図1(配列番号1)に示されるようなヌクレオチド配列のすべてではなくともほとんどに対応するポリヌクレオチドのように、本発明に従ったプローブとして有用である。例えば、「長さが少なくとも20nt」のポリヌクレオチドの部分によって、参照のポリヌクレオチドのヌクレオチド配列(例えば、寄託されたcDNA、または図1(配列番号1)に示されるようなヌクレオチド配列)からの20以上の隣接したヌクレオチドが意図される。示されるように、そのような部分は、従来のDNAハイブリダイゼーション技術に従っ
たプローブとして、または例えば、Sambrook,J.ら編、Molecular Cloning, A Laboratory
Manual, 第2版、Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY (1989)(その全開示は、本明細書中において参考として援用される)に記載されるようなポリメラーゼ連鎖反応(PCR)による標的配列の増幅のためのプライマーとしてのいずれかで診
断的に有用である。
【0055】
エンドカインαcDNAクローンは寄託されており、そしてそのヌクレオチド配列が図1(配列番号1)において提供されるので、エンドカインαcDNA分子の一部にハイブリダイズするポリヌクレオチドを生成することは、当業者に日常的である。例えば、エンドカインαcDNAクローンの制限エンドヌクレアーゼ切断または超音波による剪断は、エンドカイン
αcDNA分子の一部にハイブリダイズするポリヌクレオチドである種々のサイズのDNA部分
を生成するために容易に使用され得る。あるいは、本発明のハイブリダイズするポリヌクレオチドは、公知の技術に従って合成的に生成され得る。
【0056】
当然のことながら、ポリA配列(例えば、図1(配列番号1)に示されるエンドカイン
αcDNAの3’末端ポリ(A)付加物)にのみ、またはT(またはU)の相補的な部分(stretch)
にハイブリダイズするポリヌクレオチドは、本発明の核酸の一部にハイブリダイズするために使用される本発明のポリヌクレオチドに含まれない。なぜなら、そのようなポリヌクレオチドは、poly(A)伸長またはその相補物を含む任意の核酸分子(例えば、特に任意の
二本鎖cDNAクローン)にハイブリダイズする。
【0057】
示されるように、エンドカインαタンパク質をコードする本発明の核酸分子は、以下を含むが、それらに限定されない:それ自体によって、成熟ペプチドのアミノ酸配列をコードする核酸分子;成熟したポリペプチドのコード配列およびさらなる配列(例えば、プレタンパク質配列またはプロタンパク質配列またはプレプロタンパク質配列);成熟ポリペプチドのコード配列で、上記のさらなるコード配列を含むかまたは含まず、更なる非コード配列とともに集まっており、この配列はイントロンおよび非コード5’および3’配列(例えば、転写、mRNA(スプライシングおよびポリアデニル化シグナル(例えば、リボソーム結合およびmRNAの安定性)を含む)において役割を担う転写非翻訳配列)を含むがこれらに限定されない;さらなる機能性を提供するようなさらなるアミノ酸をコードするさらなるコード配列。従って、例えば、ポリペプチドをコードする配列は、マーカー配列(例えば、融合されたポリペプチドの精製を容易にするペプチドをコードする配列)に融合され得る。本発明のこの局面の特定の好ましい実施態様において、マーカーアミノ酸配列は、ヘキサ-ヒスチジンペプチド(例えば、pQEベクター(Qiagen,Inc.)において提供されるタグ)、他の中では、それらの多くは公的および/または商業的に入手可能である。例えば、Gentzら、Proc. Natl.Acad. Sci. USA 86: 821-824 (1989)において記載されるよう
に、ヘキサヒスチジンは、融合タンパク質の簡便な精製を提供する。「HA」タグは、インフルエンザ赤血球凝集素(HA)タンパク質由来のエピトープに対応する精製のために有用な別のペプチドであり、それは、Wilsonら、Cell37: 767 (1984)によって記載されている。他のそのような融合タンパク質は、NまたはC末端にてFcに融合されるエンドカインαタンパク質を含む。
【0058】
本発明は、さらに、エンドカインαタンパク質の一部、アナログ、または誘導体をコードする本発明の核酸分子の変異体に関する。変異体は、天然の対立遺伝子変異体のように、天然に生じ得る。「対立遺伝子変異体」によって、生物の染色体上の所定の遺伝子座を占める遺伝子のいくつかの交換可能な形態の1つが意図される。天然に存在しない変異体は、例えば当該分野で公知の変異誘発技術を用いて生成され得る。
【0059】
そのような変異体は、ヌクレオチド置換、欠失、または添加によって生成される変異体を含む。置換、欠失、または添加は、1つ以上のヌクレオチドを含み得る。変異体は、コードもしくは非コード領域、またはその両方において変化され得る。コード領域における変異は、保存性もしくは非保存性アミノ酸置換、欠失、または添加を生成し得る。これらの中で特に好ましいものは、サイレント置換、添加、および欠失であり、これらは、エンドカインαタンパク質またはその一部の特性および活性を変化しない。これらの点において特にまた好ましいものは、保存性置換である。最も高度に好ましいものは、図1(配列番号2)において示されるアミノ酸配列、または寄託されたcDNAクローンによってコードされる成熟エンドカインαアミノ酸配列を有する成熟エンドカインαタンパク質をコードする核酸分子である。
【0060】
本発明のさらなる実施態様は、(a)図1(配列番号2)に示される完全なアミノ酸配列
を有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされる
全長エンドカインαポリペプチドをコードするヌクレオチド配列;(b)図1(配列番号2
)中のほぼ44位からほぼ169位のアミノ酸配列を有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされるエンドカインα細胞外ドメインをコードするヌクレオチド配列;(c)図1(配列番号2)中のほぼ18位からほぼ43位のアミノ酸配列を
有するか、もしくはATCC受託番号97640に含まれるcDNAクローンによってコードされるエ
ンドカインα膜貫通ドメインをコードするヌクレオチド配列;(d)図1(配列番号2)中
のほぼ1位からほぼ17位でのアミノ酸配列を有するか、もしくはATCC受託番号97640に含
まれるcDNAクローンによってコードされるエンドカインα細胞内ドメインをコードするヌクレオチド配列;または(e) (a)、(b)、(c)、もしくは(d)のヌクレオチド配列のいずれかと相補的なヌクレオチド配列と、少なくとも90%同一のヌクレオチド配列を有し、そしてより好ましくは少なくとも95%、96%、97%、98%、または99%同一のヌクレオチド配列を有するポリヌクレオチドを含む単離された核酸分子を含む。
【0061】
エンドカインαポリペプチドをコードする参照ヌクレオチド配列と少なくとも、例えば、95%「同一」のヌクレオチド配列を有するポリヌクレオチドによって、ポリヌクレオチド配列が、エンドカインαポリペプチドをコードする参照ヌクレオチド配列の各100ヌク
レオチドあたり5つまでの点変異を含み得ることを除いて、ポリヌクレオチドのヌクレオチド配列が、参照配列と同一であることが意図される。言い換えれば、参照ヌクレオチド配列と少なくとも95%同一であるヌクレオチド配列を有するポリヌクレオチドを得るために、参照配列における5%までのヌクレオチドが欠失され得るかもしくは別のヌクレオチドで置換され得るか、または参照配列において全ヌクレオチドの5%までの多数のヌクレオチドが、参照配列中に挿入され得る。参照配列のこれらの変異は、参照ヌクレオチド配列の5’もしくは3’末端位置でか、または参照配列内のヌクレオチドの中で個々に、もしくは参照配列内の1つ以上の連続した群でのいずれかで散在されて、これらの末端位置の間のどこかで生じ得る。
【0062】
実際には、任意の特定の核酸分子が、図1に示されるヌクレオチド配列または寄託されたcDNAクローンのヌクレオチド配列に少なくとも90%、95%、96%、97%、98%、または99%同一であるかどうかは、例えば、BESTFITプログラム(WisconsinSequence Analysis Package, Version 8 for Unix(登録商標), Genetics Computer Group,University Research Park, 575 Science Drive, Madison, WI 53711)のような公知のコンピュータープログラムを用いて従来通りに決定され得る。BESTFITは、SmithおよびWaterman,Adv. Appl. Math. 2:482-489 (1981)の局所的相同性アルゴリズムを用いて、2つの配列間の相同性の最適のセグメントを見出す。BESTFITまたは任意の他の配列整列プログラムを使用して、
特定の配列が、例えば本発明による参照配列に95%同一であるか否かを決定する場合、パラメーターは、もちろん、同一性の割合が、参照ヌクレオチド配列の全長にわたって計算されるように、そして参照配列の総ヌクレオチド数の5%までの相同性におけるギャップが許容されるように、設定される。
【0063】
本出願は、エンドカインαタンパク質活性を有するポリペプチドをコードするか否かに関係なく、上記の核酸配列と少なくとも90%、95%、96%、97%、98%、または99%同一であるこのような核酸分子に関する。これは、特定の核酸分子がエンドカインα活性を有するポリペプチドをコードしない場合でさえ、当業者は、核酸分子をどのようにして、例えば、ハイブリダイゼーションプローブ、またはポリメラーゼ連鎖反応(PCR)のプライマ
ーとして使用するかをなお知るからである。エンドカインα活性を有するポリペプチドをコードしない本発明の核酸分子の使用としては、とりわけ(1)エンドカインα遺伝子また
はその対立遺伝子変異体をcDNAライブラリーから単離すること;(2)Vermaら,Human Chromosomes: a Manual of Basic Techniques, Pergamon Press, New York(1988)に記載の、エンドカインα遺伝子の正確な染色体位置を提供するための分裂中期染色体展開物に対する
インサイチュハイブリダイゼーション(FISH);および、(3)特定の組織におけるエンドカ
インαmRNA発現を検出するためのノーザンブロット分析、が挙げられる。
【0064】
しかし、実際に、エンドカインαタンパク質活性を有するポリペプチドをコードする上記の核酸配列に少なくとも90%、95%、96%、97%、98%、または99%同一である配列を有するこのような核酸分子が好ましい。「エンドカインα活性を有するポリペプチド」によって、特定の生物学的アッセイにおいて測定される場合、エンドカインαタンパク質と比較して類似する(しかし、必ずしも同一ではない)活性を示すポリペプチドが意図される。エンドカインα活性は、公知の方法に従ってアッセイされ得る。例えば、細胞傷害性アッセイまたは細胞増殖アッセイが使用され得、ここでエンドカインαポリペプチドは培養物中の細胞へ添加され、そして細胞に対するエンドカインの効果が、細胞数の増大または減少を測定することによって決定される。
【0065】
もちろん、遺伝子コードの縮重のために、当業者は、上記の核酸配列に少なくとも90%、95%、96%、97%、98%、または99%同一である配列を有する多数の核酸分子が、「エンドカインαタンパク質活性を有する」ポリペプチドをコードすることを直ちに認識する。実際に、縮重改変体の全ては同じポリペプチドをコードするので、これは上記の比較アッセイを実施することなしでさえ当業者に明らかである。縮重改変体でないそのような核酸分子について、合理的な数がまたエンドカインαタンパク質活性を有するポリペプチドをコードすることが、当該分野でさらに認識される。これは、当業者が、タンパク質の機能により低い有意性で影響しそうであるか、または有意には影響しそうにないかのいずれかのアミノ酸置換(例えば、1つの脂肪族アミノ酸の第二の脂肪族アミノ酸への置換)を完全に知っているからである。
【0066】
例えば、どのように表現型的にサイレントなアミノ酸置換を作製するかに関するガイダンスは、Bowie, J.U.ら, Science 247:1306-1310(1990)に提供される。ここで著者らは、アミノ酸配列の変化に対する寛容を研究するための2つの主要なアプローチが存在することを示す。第一の方法は、進化のプロセスに依存し、ここで変異は、自然淘汰によって受容されるか拒絶されるかのいずれかである。第二のアプローチは、クローン化遺伝子の特定の位置でアミノ酸変化を導入するために遺伝子操作を、そして機能を維持する配列を同定するために選択またはスクリーニングを用いる。著者らが述べるように、これらの研究は、タンパク質が、アミノ酸置換に驚くほど寛容であることを明らかにしている。著者らは、どのアミノ酸の変化が、タンパク質の特定の位置で許容性でありそうであるかをさらに示している。例えば、大部分の埋没したアミノ酸残基は非極性側鎖を必要とし、一方表面の側鎖の特性は一般的にわずかしか保存されていない。他のこのような表現型的にサイレントな置換は、Bowie,J.U.ら,前出およびその中に引用される参考文献に記載される。
【0067】
ベクターおよび宿主細胞
本発明はまた、本発明の単離されたDNA分子を含むベクター、組換えベクターで遺伝子
操作された宿主細胞、および組換え技術によるエンドカインαポリペプチドまたはその部分の産生に関する。
【0068】
組換え構築物は、感染、形質導入、トランスフェクション、トランスベクション(transvection)、エレクトロポレーション、および形質転換のような周知の技術を用いて宿主細胞に導入され得る。ベクターは、例えば、ファージベクター、プラスミドベクター、ウイルスベクター、またはレトロウイルスベクターであり得る。レトロウイルスベクターは、複製可能かまたは複製欠損であり得る。後者の場合、ウイルスの増殖は、一般的に、相補宿主細胞においてのみ生じる。
【0069】
ポリヌクレオチドは、宿主細胞における増殖のための選択マーカーを含むベクターに結
合され得る。一般的に、プラスミドベクターは、リン酸カルシウム沈殿物のような沈殿物中か、または荷電された脂質との複合体中で導入される。ベクターがウイルスである場合、ベクターは、適切なパッケージング細胞株を用いてインビトロでパッケージングされ得、次いで宿主細胞に形質導入され得る。
【0070】
目的のポリヌクレオチドに対するシス作用性制御領域を含むベクターが好ましい。適切なトランス作用性因子は、宿主によって供給される得るか、相補ベクターによって供給され得るか、または宿主への導入の際にベクター自体によって供給され得る。
【0071】
この事に関する特定の好ましい実施態様において、ベクターは、誘導性および/または細胞型特異的であり得る特異的な発現を提供する。このようなベクターの中で特に好ましいベクターは、温度および栄養添加物のような操作することが容易である環境因子によって誘導性のベクターである。
【0072】
本発明において有用な発現ベクターとしては、染色体ベクター、エピソームベクター、およびウイルス由来ベクター(例えば、細菌プラスミド、バクテリオファージ、酵母エピソーム、酵母染色体エレメント、ウイルス(例えば、バキュロウイルス、パポバウイルス、ワクシニアウイルス、アデノウイルス、トリポックスウイルス、仮性狂犬病ウイルス、およびレトロウイルス)、ならびにそれらの組合せに由来するベクター(例えば、コスミドおよびファージミド)が挙げられる。例えば、Ausubel,下記、Sambrook,下記を参照の
こと。
【0073】
DNAインサートは、適切なプロモーター(例えば、少し名を挙げると、ファージλPLプ
ロモーター、E.coli:lacプロモーター、trpプロモーター、およびtacプロモーター、SV40初期プロモーターおよび後期プロモーター、ならびにレトロウイルスLTRのプロモーター
)に作動可能に連結されるべきである。他の適切なプロモーターは、当業者に公知である。発現構築物は、さらに、転写開始、転写終結のための部位、および、転写領域中に翻訳のためのリボゾーム結合部位を含む。構築物によって発現される成熟転写物のコード部分は、翻訳されるべきポリペプチドの始めに転写開始AUGを含み、そして終わりに適切に位
置される終止コドンを含む。
【0074】
示されるように、発現ベクターは、好ましくは少なくとも1つの選択マーカーを含む。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼ耐性またはネオマイシン耐性、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。適切な宿主の代表的な例としては、細菌細胞(例えば、E.coli細胞、Streptomyces細胞、およびSalmonellatyphimurium細胞);真菌細胞(例えば酵母細胞);昆虫細胞(例えば、Drosophila S2細胞およびSpodoptera Sf9細胞);動物細胞(例えば、CHO細胞、COS細胞、およびBowes黒色
腫細胞);ならびに植物細胞が挙げられる。上記の宿主細胞のための適切な培養培地および条件は当該分野で公知である。
【0075】
細菌における使用に好ましいベクターの中には、pQE70、pQE60、およびpQE-9(Qiagen
から入手可能);pBSベクター、Phagescriptベクター、Bluescriptベクター、pNH8A、pNH16a、pNH18A、pNH46A(Stratageneから入手可能);ならびにptrc99a、pKK223-3、pKK233-3、pDR540、pRlT5(Pharmaciaから入手可能)が含まれる。好ましい真核生物ベクターの中には、pWLNEO、pSV2CAT、pOG44、pXT1、およびpSG(Stratageneから入手可能);ならびにpSVK3、pBPV、pMSG、およびpSVL(Pharmacicaから入手可能)ががある。他の適切なベク
ターは、当業者に容易に明らかである。
【0076】
本発明における使用に適した公知の細菌プロモーターの中には、E.coli lacIおよびlac
Zプロモーター、T3プロモーターおよびT7プロモーター、gptプロモーター、λPRプロモーターおよびλPLプロモーター、ならびにtrpプロモーターが含まれる。適切な真核生物プ
ロモーターとしては、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、初期SV40プロモーターおよび後期SV40プロモーター、レトロウイルスLTRのプロモーター(例
えば、ラウス肉腫ウイルス(RSV)のプロモーター)、ならびにメタロチオネインプロモー
ター(例えば、マウスメタロチオネインIプロモーター)が挙げられる。
【0077】
宿主細胞への構築物の導入は、リン酸カルシウムトランスフェクション、DEAEデキストラン媒介トランスフェクション、カチオン性脂質媒介トランスフェクション、エレクトロポレーション、形質導入、感染または他の方法によってもたらされ得る。このような方法は、Davisら,Basic Methods in Molecular Biology(1986)のような多くの標準的研究室マニュアルに記載されている。
【0078】
高等真核生物による本発明のポリペプチドをコードするDNAの転写は、ベクター中にエ
ンハンサー配列を挿入することによって増大させ得る。エンハンサーは、所定の宿主細胞型におけるプロモーターの転写活性を増大するように働く、通常約10〜300bpのDNAのシス作用性エレメントである。エンハンサーの例としては、SV40エンハンサー(これは、複製起点の後期側上の100〜270bpに位置される)、サイトメガロウイルスの初期プロモーターエンハンサー、複製起点の後期側上のポリオーマエンハンサー、およびアデノウイルスエンハンサーが挙げられる。
【0079】
翻訳されたタンパク質の小胞体の管腔内へか、周辺質空間内へか、または細胞外環境内への分泌のために、適切な分泌シグナルが、発現されるポリペプチド中に組み込まれ得る。シグナルは、ポリペプチドに対して内因性であり得るか、またはそれらは異種シグナルであり得る。
【0080】
ポリペプチドは、融合タンパク質のような改変された形態で発現され得、そして分泌シグナルだけでなく、付加的な異種の機能的領域も含み得る。さらなる例において、付加的なアミノ酸、特に荷電性アミノ酸の領域が、宿主細胞内での、精製の間の、または続く操作および保存の間の、安定性および持続性を改善するために、ポリペプチドのN末端に付加され得る。また示されるように、領域(単数または複数)はまた、精製を容易にするためにポリペプチドへ付加され得る。そのような領域は、ポリペプチドの最終調製の前に除去され得る。とりわけ、分泌および排出を生じるため、安定性を改善するため、および精製を容易にするためのペプチド部分のポリペプチドへの付加は、当該分野でよく知られており、そして日常的な技術である。好ましい融合タンパク質は、レセプターの可溶化に有用な免疫グロブリン由来の異種領域を含む。例えば、EP A 0,464,533(また、カナダ対応出願2,045,869)は、別のヒトタンパク質またはその一部とともに免疫グロブリン分子の
定常領域の種々の部分を含む融合タンパク質を開示する。多くの場合、融合タンパク質中のFc部分は、治療および診断における使用に十分に有利であり、従って、例えば改善された薬物動態学的特性を生じる(EPA 0,232,262)。一方、いくつかの使用について、融合タ
ンパク質が、記載される有利な様式で、発現され、検出され、および精製された後にFc部分が欠失され得ることが望ましい。これは、Fc部分が、治療および診断における使用の妨害であると判明する場合(例えば、融合タンパク質が免疫のための抗原として使用されるべき場合)である。薬物探索において、例えばhIL-5のようなヒトタンパク質は、アゴニ
スト(例えば、hIL-5)を同定するための高処理能力スクリーニングアッセイの目的でFc
部分と融合されている。D.Bennettら,Journal of Molecular Recognition 8:52-58(1995)、およびK.Johansonら, The Journal ofBiological Chemistry 270(16):9459-9471(1995)を参照のこと。
【0081】
エンドカインαタンパク質は、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまた
は陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む周知の方法によって組換え細胞培養物から回収され、そして精製され得る。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0082】
本発明のポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明のポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
エンドカインαポリペプチドおよびペプチド
本発明はさらに、寄託されたcDNAによってコードされるアミノ酸配列、もしくは図1(配列番号2)のアミノ酸配列を有する単離されたエンドカインαポリペプチド、または上記ポリペプチドの一部を含むペプチドもしくはポリペプチドを提供する。用語「ペプチド」および「オリゴペプチド」は、(通常認識されるように)同義語と考えられ、そして各用語は、文脈がペプチド結合によって結合された少なくとも2つのアミノ酸の鎖を示すことを必要とする場合、交換可能に使用され得る。用語「ポリペプチド」は、10より多いアミノ酸残基を含む鎖に対して本明細書中で使用される。本明細書中の全てのオリゴペプチドおよびポリペプチドの式または配列は、左から右へ、そしてアミノ末端からカルボキシ末端の方向に書かれる。
【0083】
「単離された」ポリペプチドまたはタンパク質によって、その天然の環境から取り出されたポリペプチドまたはタンパク質が意図される。例えば、組換え宿主細胞中で発現された組換え産生されたポリペプチドおよびタンパク質は、例えば、SmithおよびJohnson, Gene 67:31-40 (1988)に記載の一段階法のような任意の適切な技術によって実質的に精製されている天然または組換えのポリペプチドおよびタンパク質がそうであるように、本発明の目的で単離されていると考えられる。
【0084】
エンドカインαポリペプチドのいくつかのアミノ酸配列が、このタンパク質の構造または機能に有意に影響することなく改変され得ることが、当該分野で認識される。配列内のこのような差異が意図される場合、活性を決定するタンパク質の重要な領域が存在することを覚えておくべきである。一般的に、類似の機能を実行する残基が使用される場合、三次構造を形成する残基の置換が可能である。他の例において、残基の型は、変化がタンパク質の重要でない領域で生じる場合、全く重要でないかもしれない。
【0085】
従って、本発明は、さらに、実質的なエンドカインαポリペプチド活性を示すか、または下記に考察されるタンパク質フラグメントのようなエンドカインαタンパク質の領域を含むエンドカインαポリペプチドの改変体を含む。このような変異体は、欠失、挿入、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)を含む。小さな変化、またはこのような「中性」アミノ酸置換は、一般的にほとんど活性に影響しない。
【0086】
代表的に保存的置換と見られるのは、脂肪族アミノ酸Ala、Val、Leu、およびIleの中での1つのアミノ酸の別のアミノ酸への置換;ヒドロキシル残基SerおよびThrの交換、酸性残基AspおよびGluの交換、アミド残基AsnおよびGlnの間の置換、塩基性残基LysおよびArgの交換、ならびに芳香族残基Phe、Tyrの間の置換である。
【0087】
上記に詳細に示されるように、どのアミノ酸の変化が表現型的にサイレントでありそう
か(すなわち、機能に対して有意に有害な効果を有しそうにないか)に関するさらなるガイダンスは、Bowie, J.U.ら「Deciphering the Message in Protein Sequences: Toleranceto Amino Acid Substitutions」,Science 247:1306-1310 (1990)に見出され得る。
【0088】
本発明のポリペプチドは、以下のような、図1(配列番号2)に示されるか、または寄託されたcDNAによってコードされるようなアミノ酸配列を有するポリペプチドを含む:完全長タンパク質、細胞内ドメイン、膜貫通ドメイン、および細胞外ドメイン、ならびに本明細書中に記載のポリペプチドと少なくとも90%の類似性、より好ましくは少なくとも95%の類似性、およびさらにより好ましくは少なくとも96%、97%、98%、または99%の類似性を有するポリペプチド。さらに本発明のポリペプチドは、本明細書中に記載のポリペプチドと、少なくとも80%同一、より好ましくは少なくとも90%または95%同一、さらにより好ましくは少なくとも96%、97%、98%、または99%同一であるポリペプチドを含み、そしてまた、少なくとも30アミノ酸、およびより好ましくは少なくとも50アミノ酸を有するこのようなポリペプチドの部分を含む。
【0089】
2つのポリペプチドについての「%類似性」によって、BESTFITプログラム(Wisconsin
Sequence Analysis Package, Version 8 for UNIX(登録商標), Genetics Computer Group, University Resarch Park, 575 Science Drive,Madison, WI 53711)および類似性を決定するための省略時設定(default setting)を用いて2つのポリペプチドのアミノ酸配
列を比較することによって生じる類似性のスコアが意図される。BESTFITは、SmithおよびWaterman,Adv. Appl. Math. 2:482-489 (1981)の局所的相同性アルゴリズムを、2つの配列間の類似性の最適のセグメントを見出すために使用する。
【0090】
エンドカインαポリペプチドの参照アミノ酸配列に少なくとも、例えば、95%「同一な」アミノ酸配列を有するポリペプチドにより、ポリペプチド配列がエンドカインαポリペプチドの参照アミノ酸配列の各100アミノ酸毎に5つまでのアミノ酸変化を含み得ること
を除いて、ポリペプチドのアミノ酸配列は参照配列と同一であることが意図される。言い換えれば、参照アミノ酸配列に少なくとも95%同一のアミノ酸配列を有するポリペプチドを得るために、参照配列において5%までのアミノ酸残基が、欠失されるかもしくは他のアミノ酸と置換され得、または参照配列における5%までの総アミノ酸残基の多数のアミノ酸が参照配列に挿入され得る。これらの参照配列の改変は、参照アミノ酸配列のアミノ末端位置もしくはカルボキシ末端位置で、または参照配列内または参照配列内の1つ以上の近接した基内の個々の残基間のいずれかで散在したこれらの末端位置の間のどこかで、起こり得る。
【0091】
実際には、任意の特定のポリペプチドが、例えば、図1(配列番号2)に示されるアミノ酸配列、または寄託されたcDNAクローンによりコードされるアミノ酸配列に対して、少なくとも90%、95%、96%、97%、98%、または99%同一であるかどうかは、BESTFITプ
ログラム(WisconsinSequence Analysis Package, Version8 Unix(登録商標)版、Genetics Computer Group, UniversityResearch Park, 575 Science Drive, Madison, WI 53711)のような公知のコンピュータープログラムを使用して慣習的に決定され得る。特定の
配列が、例えば、本発明の参照配列に95%同一であるかどうかを決定するために、BESTFITまたは任意の他の配列アラインメントプログラムを使用する場合、当然、同一性の百分
率が参照アミノ酸配列の全長にわたって計算され、そして参照配列内のアミノ酸残基の総数の5%までの相同性におけるギャップが許容されるようなパラメーターが設定される。
【0092】
下記に詳細に示すように、本発明のポリペプチドは、下記のようにエンドカインαタンパク質発現を検出するための診断アッセイにおいて有用である、またはエンドカインαタンパク質機能を阻害し得るアゴニストおよびアンタゴニストとして、ポリクローナル抗体およびモノクローナル抗体を惹起するために使用され得る。さらに、このようなポリペプ
チドは、本発明の候補アゴニストおよびアンタゴニストでもあるエンドカインαタンパク質結合タンパク質を「捕獲する」ための酵母ツーハイブリッドシステムにおいて使用され得る。酵母ツーハイブリッドシステムは、FieldsおよびSong,Nature 340:245-246(1989
)に記載されている。
【0093】
別の局面において、本発明は、本発明のポリペプチドのエピトープ-保有部分を含む、
ペプチドまたはポリペプチドを提供する。このポリペプチドのエピトープ部分は、本発明のポリペプチドの免疫原性または抗原性エピロープである。「免疫原性エピトープ」は、タンパク質全体が免疫原性である場合、抗体応答を誘発するタンパク質の一部として定義される。これらの免疫原性エピトープは、分子上の2、3の焦点に制限されると考えられている。一方では、抗体が結合し得るタンパク質分子の領域は、「抗原性エピトープ」と定義され得る。タンパク質の免疫原性エピトープの数は、一般には、抗原性エピトープの数よりも少ない。例えば、Geysen,H.M.ら、Proc. Natl. Acad. Sci. USA 81:3998-4002(1984)を参照のこと。
【0094】
抗原性エピトープを保有するペプチドまたはポリペプチド(すなわち、抗体が結合し得るタンパク質分子の領域を含む)の選択に関して、タンパク質配列の一部を模倣する比較的短い合成ペプチドが、部分的に模倣されたタンパク質と反応する抗血清を日常的に誘発し得ることが当該分野で周知である。例えば、Sutcliffe,J.G.ら、Science 219:660-666(1983)を参照のこと。タンパク質-反応性血清を誘発し得るペプチドは、しばしばタンパク質の一次配列に頻繁に存在し、そして単純な化学的法則のセットにより特徴付けられ得、そしてインタクトなタンパク質(すなわち、免疫原性エピトープ)の免疫ドミナント領域にも、アミノ末端またはカルボキシル末端にも、制限されない。極度に疎水性であるペプチドおよび6以下の残基のペプチドは、一般には、模倣タンパク質に結合する抗体の誘導に効果がなく;より長い、可溶性ペプチド、特にプロリン残基を含むペプチドは、通常は有効である。Sutcliffeら、前出、661。例えば、これらのガイドラインに従って設計された30のペプチドのうち18(インフルエンザウイルス赤血球凝集素HA1ポリペプチド鎖の配
列の75%を覆う8-39残基を含む)は、HA1タンパク質またはインタクトなウイルスと反応
する抗体を誘導した;そしてMuLVポリメラーゼからの12/12ペプチドおよび狂犬病糖タン
パク質からの18/18はそれぞれのタンパク質を沈澱する抗体を誘導した。
【0095】
本発明の抗原性エピトープ-保有ペプチドおよびポリペプチドは、それゆえ、本発明の
ポリペプチドに特異的に結合するモノクローナル抗体を含む抗体を惹起するのに有用である。従って、抗原エピトープ-保有ペプチドで免疫化されたドナーからの脾臓細胞の融合
により得られるハイブリドーマの大部分は、一般に天然のタンパク質と反応性がある抗体を分泌する。Sutcliffeら、前出、663。抗原性エピトープ-保有ペプチドまたはポリペプ
チドにより惹起された抗体は、模倣タンパク質を検出するのに有用であり、そして異なるペプチドに対する抗体が、翻訳語プロセシングを受けるタンパク質前駆体の種々の領域の末路を追跡するために使用され得る。免疫沈降アッセイにおいて、短いペプチド(例えば、約9アミノ酸)でさえ、より長いペプチドに結合しそして置換し得ることが示されているので、ペプチドおよび抗ペプチド抗体は、模倣タンパク質についての種々の定性的または定量的アッセイ、例えば、競合的アッセイにおいて使用され得る。例えば、Wilson,I.A.ら、Cell 37:767-778(1984)777を参照のこと。本発明の抗ペプチド抗体もまた、模倣
タンパク質の精製(例えば、当該分野で周知の方法を使用して、吸着クロマトグラフィーにより)に有用である。
【0096】
上記のガンダンスに従って設計された本発明の抗原性エピトープ-保有ペプチドまたは
ポリペプチドは、好ましくは本発明のポリペプチドのアミノ酸配列内に含まれる少なくとも7、より好ましくは少なくとも9、そして最も好ましくは約15〜約30アミノ酸の間の配列を含む。しかし、本発明のポリペプチドのアミノ酸配列の約30〜約50アミノ酸または全
体までの任意の長さおよび全体を含む、本発明のポリペプチドのアミノ酸配列のより大部分を含むペプチドまたはポリペプチドもまた、本発明のエピトープ-保有ペプチドまたは
ポリペプチドであると考えられ、そしてまた模倣タンパク質と反応する抗体を誘導するのに有用である。好ましくは、エピトープ-保有ペプチドのアミノ酸配列は、水溶液中で実
質的な溶解性を提供するように選択され(すなわち、その配列は、比較的親水性残基を含み、そして高度な疎水性配列は好ましくは回避される);そしてプロリン残基を含む配列が特に好ましい。
【0097】
エンドカイン特異的ポリクローナルおよびモノクローナル抗体を産生するために使用され得る抗原性ポリペプチドの非限定的な例には、図1(配列番号2)における約44〜約158のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約44〜約54のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約57〜約68のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約69〜約78のアミノ酸残基を含むポリペプチド;図1(配列番号2)における約94〜約105のアミノ酸残基を含むポリペプチド;図1
(配列番号2)における約108〜約132のアミノ酸残基を含むポリペプチド;および図1(配列番号2)における約148〜約158のアミノ酸残基を含むポリペプチドが挙げられる。上記のように、本発明者らは、上記のポリペプチドフラグメントがエンドカインαタンパク質の抗原性領域であることを決定した。
【0098】
本発明のエピトープ-保有ペプチドおよびポリペプチドは、本発明の核酸分子を使用す
る組換え手段を含むペプチドまたはポリペプチドを作製するための任意の従来の手段により産生され得る。例えば、短いエピトープ-保有アミノ酸配列は、組換え体産生および精
製の間、ならびに抗ペプチド抗体を産生するための免疫化の間、キャリアとして作用するより大きなポリペプチドに融合され得る。エピトープ-保有ペプチドはまた、化学合成の
公知の方法を使用して合成され得る。例えば、Houghtenは、4週間未満で、調製されそして特徴付けられた(ELISA-タイプ結合研究により)HA1ポリペプチドのセグメントの単一
のアミノ酸改変体を示す10〜20mgの248の異なる13残基ペプチドのような多数のペプチド
の合成のための簡単な方法を記載している。Houghten,R.A., Proc. Natl. Acad. Sci. USA 82:5131-5135 (1985)を参照のこと。この「SimultaneousMultiple Peptide Synthesis(SMPS)」プロセスは、さらにHoughtenら(1986)の米国特許第4,631,211号に記載される。この手順において、種々のペプチドの固相合成のための個々の樹脂は、分離溶媒透過性パケットに含まれ、固相法に関連する多くの同一の反復工程の最適な使用を可能にする。完全なマニュアル手順は、同時に行われる500〜1000以上の合成を可能にする。Houghtenら、前出、5134。
【0099】
本発明のエピトープ保有ペプチドおよびポリペプチドは、当該分野に周知の方法によって抗体を誘導するために使用される。例えば、Sutcliffeら、前出;Wilsonら、前出;Chow, Mら、Proc. Natl. Acad. Sci. USA82:910-914;およびBittle, F. J.ら、J. Gen. Virol. 66:2347-2354(1985)を参照のこと。一般には、動物は遊離ペプチドで免疫化され得る;しかし、抗ペプチド抗体力価はペプチドを高分子キャリア(例えば、キーホールリンペットヘモシアニン(KLH)または破傷風毒素)にカップリングすることにより追加免疫
され得る。例えば、システインを含有するペプチドは、m-マレイミドベンゾイル-N-ヒド
ロキシスクシンイミドエステル(MBS)のようなリンカーを使用してキャリアにカップリ
ングされ得、一方、他のペプチドは、グルタルアルデヒドのようなより一般的な連結剤を使用してキャリアーにカップリングされ得る。
【0100】
ウサギ、ラット、およびマウスのような動物は、遊離またはキャリア-カップリングペ
プチドのいずれかで、例えば、約100μgのペプチドまたはキャリアタンパク質およびFreundのアジュバントを含むエマルジョンの腹腔内および/または皮内注射により免疫化され
る。いくつかの追加免疫注射が、例えば、固体表面に吸着された遊離ペプチドを使用して
ELISAアッセイにより検出され得る有用な力価の抗ペプチド抗体を提供するために、例え
ば、約2週間の間隔で必要とされ得る。免疫化動物からの血清における抗ペプチド抗体の力価は、抗ペプチド抗体の選択により、例えば、当該分野で周知の方法による固体支持体上のペプチドへの吸収および選択された抗体の溶出により増加され得る。
【0101】
本発明の免疫原性エピトープ-保有ペプチド、すなわち、抗体応答を惹起するタンパク
質の部分は、全体のタンパク質が免疫原性である場合、当該分野で公知の方法により同定される。例えば、Geysenら(1984)前出は、酵素-結合免疫吸収アッセイにおける反応に
十分に純粋な何百というペプチドの固体支持体上の迅速な併用的合成の手順を開示する。合成ペプチドの抗体との相互作用は、次いで、それらを支持体から除去することなく容易に検出される。この様式において、所望のタンパク質の免疫原性エピトープを保有するペプチドは、当業者により日常的に同定され得る。
【0102】
例えば、口蹄疫ウイルスの被膜タンパク質における免疫学的に重要なエピトープは、タンパク質の213のアミノ酸配列全体を覆う全ての208の可能なヘキサペプチドの重複セットの合成による7アミノ酸の解明によりGeysenらによって位置付けされた。次いで、全ての20アミノ酸が順にエピトープ内の各位置で置換されたペプチドの完全な置換セットが合成され、そして抗体との反応のための特異性を与える特定のアミノ酸が決定された。従って、本発明のエピトープ-保有ペプチドのペプチドアナログは、この方法により日常的に作
成され得る。Geysen(1987)の米国特許第4,708,781号は、所望のタンパク質の免疫原性
エピトープを保有するペプチドを同定するこの方法をさらに記載している。
【0103】
さらになお、Geysen(1990)の米国特許第5,194,392号は、目的の抗体の特定のパラト
ープ(抗原結合部位)に相補的であるエピトープの位相幾何学的等価物(例えば、「ミモトープ」)であるモノマー(アミノ酸または他の化合物)の配列を検出または決定する一般的な方法を記載する。より一般的には、Geysen(1989)の米国特許第4,433,092号は、
目的の特定のレセプターのリガンド結合部位に相補的であるリガンドの位相等価であるモノマーの配列を検出または決定する方法を記載する。同様に、PeralkylatedOligopeptide
MixturesにおけるHoughten, R. A.ら(1996)の米国特許第5,480,971号は、線状C1-C7-
アルキル過アルキル化(peralkylated)オリゴペプチドおよびセットおよびこのようなペプチドのライブラリー、ならびに目的のアクセプター分子に、優先的に結合する過剰アルキル化オリゴペプチドの配列を決定するためにこのようなオリゴペプチドセットおよびライブラリーを使用する方法を開示する。従って、本発明のエピトープ-保有ペプチドの非
ペプチドアナログはまた、これらの方法により日常的に作成され得る。
【0104】
本発明者らは、エンドカインαタンパク質が、3つの主要な構造ドメインを示す169残
基タンパク質であることを発見した。細胞内ドメインは、図1(配列番号2)における約1〜約17の残基内で同定された。膜貫通ドメインは、図1(配列番号2)における約18〜約43の残基内で同定された。細胞外ドメインは、図1(配列番号2)における約44〜約169の残基内で同定された。従って、本発明は、エンドカインα細胞内ドメイン、膜貫通ド
メイン、およびエンドカインα細胞外ドメインから選択されるポリペプチドを含む好ましいエンドカインαタンパク質フラグメントをさらに提供する。
【0105】
エンドカインαタンパク質の細胞外ドメインは、免疫グロブリン(IgG)の定ドメイン
の一部と結合し得、キメラペプチドを生じる。これらの融合タンパク質は、インビボで増加した半減期を示す。これは、例えば、ヒトCD4-ポリペプチドの最初の2つのドメインおよび哺乳動物免疫グロブリンの重鎖または軽鎖の定常領域の種々のドメインからなるキメラタンパク質について示されている(EPA394,827;Trauneckerら、Nature 331:84-86(1988))。IgG部分によるジスルフィド結合ダイマー構造を有する融合タンパク質はまた、
リガンドの結合および中和において、モノマー細胞外ドメイン単独よりも有効であり得る
(Fountoulakisら、J.Biochem. 270:3958-3964(1995))。
【0106】
「ポリペプチドおよびペプチド」のこの節に引用される各文書の全体の開示は、ここで参考として本明細書中に援用される。
【0107】
エンドカインα関連障害診断
エンドカインαは、サイトカインのTNFファミリーの新規のメンバーである。エンドカ
インα関連障害について、「標準」エンドカインα遺伝子発現レベル(すなわち、障害を有さない個体からの組織または体液におけるエンドカインα発現レベル)と比較して、実質的に変化した(増加または減少した)レベルのエンドカインα遺伝子発現が、このような障害を有する個体から採取された組織または他の細胞または体液(例えば、血清、血漿、尿、滑液、また脊髄液)において検出され得ると考えられる。従って、本発明は、エンドカインα関連障害の診断の間に有用な診断方法を提供する。これは個体からの組織または他の細胞または体液におけるエンドカインαタンパク質をコードする遺伝子の発現レベルを測定すること、および測定した遺伝子発現レベルと標準エンドカインα遺伝子発現レベルとを比較することを含む。それにより、標準と比較された遺伝子発現レベルにおける増加または減少は、エンドカインα関連障害の指標である。
【0108】
個体により、哺乳動物個体、好ましくはヒトが意図される。「エンドカインαタンパク質をコードする遺伝子の発現レベルを測定すること」により、第一の生物学的サンプルにおいて直接的(例えば、絶対的なタンパク質レベルまたはmRNAレベルを測定または評価することにより)または間接的(例えば、第二の生物学的サンプルにおけるエンドカインαタンパク質レベルまたはmRNAレベルと比較することにより)のいずれかで、エンドカインαタンパク質のレベルまたはエンドカインαタンパク質をコードするmRNAレベルの定性的測定もしくは定量的測定または評価が意図される。好ましくは、第一の生物学的サンプルにおけるエンドカインαタンパク質レベルまたはmRNAレベルが測定または評価され、そして標準エンドカインαタンパク質レベルまたはmRNAレベルと比較される。標準は、障害を有さない個体から得られた第二の生物学的サンプルから得られるか、またはエンドカインαを含む障害を有さない個体の集団からの平均レベルにより決定され得る。当該分野において評価されるように、一旦標準エンドカインαタンパク質レベルまたはmRNAレベルが公知になると、それは比較のための標準として繰り返して使用され得る。
【0109】
「生物学的サンプル」によって、個体、体液、細胞株、組織培養物、またはエンドカインαタンパク質もしくはmRNAを含む他の供給源から得られる任意の生物学的サンプルが意図される。示されるように、生物学的サンプルは、分泌された成熟エンドカインαタンパク質を含む体液(例えば、血清、血漿、尿、滑液、および随液)あるいはエンドカインαを発現することが見出された組織供給源を含む。哺乳動物から組織生検および体液を得るための方法は当該分野で周知である。生物学的サンプルがmRNAを含む場合、組織生検が好ましい供給源である。
【0110】
本発明は、当該分野で公知であるかまたは本明細書中で示されるようなTNF様障害と同
様に、哺乳動物(好ましくは、ヒト)の種々のエンドカインα関連障害の診断に有用である。これらは、免疫調節および炎症、細胞増殖、血管新生、腫瘍転移、アポトーシス、敗血症、および内毒素血症に関連する障害を含む。
【0111】
全細胞RNAが、任意の適切な技術(例えば、ChomczynskiおよびSacchi,Anal. Biochem. 162:156-159 (1987)に記載されている一工程のグアニジン-チオシアネート-フェノール-
クロロホルム法)を使用して生物学的サンプルから単離され得る。次いで、エンドカインαポリペプチドをコードするmRNAのレベルが、任意の適切な方法を用いてアッセイされる
。これらは、ノーザンブロット分析、S1ヌクレアーゼマッピング、ポリメラーゼ連鎖反応(PCR)、ポリメラーゼ連鎖反応と組み合わせた逆転写(RT-PCR)、およびリガーゼ連鎖
反応と組み合わせた逆転写(RT-LCR)を含む。
【0112】
ノーザンブロット分析は、Haradaら、Cell 63:303-312 (1990)に記載されているように行われ得る。簡潔には、全RNAが上記のように生物学的サンプルから調製される。ノーザ
ンブロットのためには、RNAは適切な緩衝液(例えば、グリオキサール/ジメチルスルホ
キシド/リン酸ナトリウム緩衝液)中で変性され、アガロースゲル電気泳動に供され、そしてニトロセルロースフィルターに転写される。RNAがUVリンカーによってフィルターに
結合された後、フィルターは、ホルムアミド、SSC、デンハルト溶液、変性させたサケ精
子、SDS、およびリン酸ナトリウム緩衝液を含有する溶液中で予備ハイブリダイゼーショ
ンされる。任意の適切な方法(例えば、32PマルチプライムDNA標識システム(Amersham)
)に従って標識されたエンドカインαタンパク質cDNAが、プローブとして使用される。一晩のハイブリダイゼーション後、フィルターは洗浄され、そしてX線フィルムに曝される。本発明に従うプローブとしての使用のためのcDNAは、上記の節に記載されており、そして少なくとも15bpの長さが好ましい。
【0113】
S1マッピングは、Fujitaら、Cell49:357-367 (1987)に記載されているように行われ得る。S1マッピングでの使用のためのプローブDNAを調製するために、上記のcDNAのセンス
鎖がテンプレートとして使用され、標識されたアンチセンスDNAが合成される。次いで、
アンチセンスDNAが、適切な制限エンドヌクレアーゼを使用して消化され、所望の長さの
さらなるDNAプローブが生成され得る。このようなアンチセンスプローブは、標的mRNA(
すなわち、エンドカインαタンパク質をコードするmRNA)に対応する保護されたバンドを可視化するために有用である。ノーザンブロット分析が、上記のように行われ得る。
【0114】
好ましくは、エンドカインαタンパク質をコードするmRNAのレベルは、Makinoら、Technique 2:295-301 (1990) に記載されるようなRT-PCR法を使用してアッセイされる。この
方法によれば、ポリアクリルアミドゲルバンド中の「アンプリコン」の放射活性は、標的mRNAの初期濃度に直線的に相関する。簡潔には、この方法は、RTプライマーおよび適切な緩衝液を含む反応混合物中の生物学的サンプルから単離された全RNAを添加する工程を包
含する。プライマーのアニーリングのためのインキュベーション後、混合物はRT緩衝液、dNTP、DTT、RNaseインヒビター、および逆転写酵素を補充され得る。RNAの逆転写を達成
するためのインキュベーション後、次いで、RT産物は標識されたプライマーを使用するPCRに供される。あるいは、プライマーを標識するよりもむしろ、標識されたdNTPがPCR反応混合物中に含まれ得る。PCR増幅は、従来技術に従ってDNAサーマルサイクラー中で行われ得る。増幅を達成するための適切な数のラウンド後、PCR反応混合物はポリアクリルアミ
ドゲル上で電気泳動される。ゲルの乾燥後、適切なバンド(エンドカインαタンパク質をコードするmRNAに対応する)の放射活性は、画像解析機を使用して定量される。RTおよびPCR反応の成分および条件、試薬およびゲル濃度、ならびに標識方法は、当該分野で周知
である。RT-PCR法の変形は当業者に明らかである。
【0115】
逆転写された標的のmRNAを増幅する任意のオリゴヌクレオチドプライマーのセットが使用され得、そして上記の節で記載されるように設計され得る。
【0116】
生物学的サンプル中のエンドカインαタンパク質のレベルのアッセイは、任意の当該分野で公知の方法を使用して行われ得る。抗体に基づく技術が、生物学的サンプル中のエンドカインαタンパク質レベルをアッセイするために好ましい。例えば、組織中でのエンドカインαタンパク質の発現は、伝統的な免疫組織学的方法で研究され得る。これらの場合、特異的認識は一次抗体(ポリクローナルまたはモノクローナル)によって提供されるが、二次検出系は、蛍光、酵素、または他の結合された二次抗体を利用し得る。結果として
、病理学試験のための組織切片の免疫組織学的染色が得られる。組織はまた、ウェスタンブロットまたはドット/スロットアッセイ(Jalkanen, M.ら、J. Cell. Biol. 101:976-985 (1985);Jalkanen, Mら、J.Cell. Biol. 105:3087-3096 (1987))のためのエンドカイ
ンαタンパク質の遊離のために、例えば、尿素および中性の界面活性剤を用いて抽出され得る。陽イオン性固相の使用に基づくこの技術において、エンドカインαタンパク質の定量は、単離されたエンドカインαタンパク質を標準として使用して達成され得る。この技術はまた、体液にも適用され得る。このサンプルに関して、エンドカインαタンパク質のモル濃度は、血清、血漿、尿、滑液、随液などのような異なる体液について、エンドカインαタンパク質含量の標準値の設定を補助する。次いで、エンドカインαタンパク質の量の正常値は、健常な個体に由来する値を使用して設定され得、これは、試験被検体から得られる値と比較され得る。
【0117】
エンドカインαタンパク質レベルを検出するために有用な他の抗体に基づく方法は、イムノアッセイ(例えば、酵素結合免疫吸着アッセイ(ELISA)およびラジオイムノアッセ
イ(RIA))を含む。例えば、エンドカインαタンパク質特異的モノクローナル抗体は、
エンドカインαタンパク質を検出および定量するための、免疫吸着剤としておよび酵素標識プローブとしての両方に使用され得る。サンプル中に存在するエンドカインαタンパク質の量は、直線回帰コンピューターアルゴリズムを使用して、標準的な調製物中に存在する量との比較によって算出され得る。腫瘍抗原を検出するためのこのようなELISAは、Iacobelliら、BreastCancer Research and Treatment 11:19-30 (1988) に記載されている。別のELISAアッセイにおいては、2つの異なる特異的なモノクローナル抗体が、体液中の
エンドカインαタンパク質を検出するために使用され得る。このアッセイにおいて、一方の抗体が免疫吸着剤として使用され、そして他方が酵素標識プローブとして使用される。
【0118】
上記の技術は、本質的に、「一工程」または「二工程」アッセイとして行われ得る。「一工程」アッセイは、固定化抗体とエンドカインαタンパク質とを接触させる工程を包含し、そして洗浄する工程、標識された抗体と混合物とを接触させる工程は含まない。「二工程」アッセイは、洗浄する工程、その後、標識された抗体と混合物とを接触させる工程を包含する。他の従来の方法もまた、適切に使用され得る。通常、支持体上にアッセイ系の1つの成分を固定することが所望され、それによって系の他の成分は、その成分との接触およびサンプルからの容易な除去を生じることが可能となる。
【0119】
適切な酵素標識は、例えば、基質との反応による過酸化水素の生成を触媒するオキシダーゼ群由来のものを含む。グルコースオキシダーゼは、それが良好な安定性を有し、そしてその基質(グルコース)が容易に入手できるために、特に好ましい。オキシダーゼ標識の活性は、酵素-標識抗体/基質反応によって形成される過酸化水素の濃度を測定するこ
とによってアッセイされ得る。酵素に加えて、他の適切な標識として、放射性同位元素(例えば、ヨウ素(125I、121I)、炭素(14C)、イオウ(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99mTc))、ならびに蛍光標識(例えば、フルオレセインおよびローダミン)ならびにビオチンが挙げられる。
【0120】
個体から得られる生物学的サンプル中のエンドカインαタンパク質レベルをアッセイすることに加えて、エンドカインαタンパク質はまた、画像解析によってインビボで検出され得る。エンドカインαタンパク質のインビボでの画像解析のための抗体標識またはマーカーとして、X線撮影法、NMR、またはESRによって検出可能なものが挙げられる。X線撮影法については、適切な標識として、検出可能な放射線を放射するが、被検体に対して明らかには有害ではない、バリウムまたはセシウムのような放射性同位元素が挙げられる。NMRおよびESRのための適切なマーカーとして、関連のハイブリドーマの栄養分の標識によって抗体中に取り込まれ得る、重水素のような検出可能な特徴的な回転を有するものが挙げられる。
【0121】
放射性同位元素(例えば、131I、111In、99mTc)、放射性不透明体(radio-opaque)基質、または核磁気共鳴によって検出可能な物質のような適切な検出可能な画像解析部分で標識されている、エンドカインαタンパク質-特異的抗体または抗体部分が、障害につい
て試験される哺乳動物中に(例えば、非経口的、皮下、または静脈内)導入される。被検体の大きさおよび使用される画像解析システムによって、診断用の画像を生じるために必要とされる画像解析部分の量が決定されることが、当該分野で理解される。放射性同位元素部分の場合、ヒト被検体については、注射される放射活性の量は、通常約5〜20ミリキュリーの範囲の99mTcである。次いで、標識抗体または抗体部分は、エンドカインαタン
パク質を含む細胞の位置に優先的に蓄積する。インビボでの腫瘍画像解析は、S.W.Burchielら、「Immunopharmacokinetics of Radiolabeled Antibodies and Their Portions」(TumerImaging 第13章:The Radiochemical Detection of Cancer, Burchiel, S.W.およびRhodes,B.A.編、Masson Publishing Inc. (1982))に記載されている。
【0122】
本発明での使用のためのエンドカインαタンパク質特異的抗体は、完全なエンドカインαタンパク質またはその抗原性ポリペプチド部分に対して惹起され得る。これは、アルブミンのようなキャリアタンパク質と共に、またはそれが充分に長い(少なくとも約25アミノ酸)場合はキャリアを伴わずに、動物系(例えば、ウサギまたはマウス)に対して提示され得る。
【0123】
本明細書中で使用される場合、用語「抗体」(Ab)または「モノクローナル抗体」(Mab)は、エンドカインαタンパク質に特異的に結合し得る完全な分子および抗体タンパク
質(例えば、FabおよびF(ab’)2部分のような)を含むことを意味する。FabおよびF(ab’)2部分は完全な抗体のFc部分を欠いており、循環によってさらに迅速に除去され、そして完全な抗体の非特異的組織結合をほとんど有し得ない(Wahlら、J.Nucl. Med. 24:316-325 (1983))。従って、これらの部分が好ましい。
【0124】
本発明の抗体は、任意の種々の方法によって調製され得る。例えば、エンドカインαタンパク質またはその抗原性部分を発現する細胞は、ポリクローナル抗体を含む血清の産生を誘導するために動物に投与され得る。好ましい方法において、エンドカインαタンパク質の調製物は、それが天然の混入物を実質的に含まないように、上記のように調製され、そして精製される。次いで、このような調製物は、より大きな特異的活性のポリクローナル抗血清を産生するために動物に導入される。
【0125】
最も好ましい方法において、本発明の抗体はモノクローナル抗体(またはそのエンドカインαタンパク質結合部分)である。このようなモノクローナル抗体は、ハイブリドーマ技術(例えば、Colligan, Current Protocols in Immunology, Wiley Interscience, NewYork (1990-1996);HarlowおよびLane, Antibodies:A Laboratory Manual, Chs. 6-9, ColdSpring Harbor Press, Cold Spring Harbor, N.Y.(1988);Ausubel、前出、第11章を参照のこと、これらの参考文献は、その全体が本明細書中に参考として援用される)を用いて調製され得る。
【0126】
一般に、このような手順は、エンドカインαポリペプチド抗原またはエンドカインαポリペプチド発現細胞で動物(好ましくは、マウス)を免疫する工程を包含する。適切な細胞は、抗エンドカインαタンパク質抗体に結合するそれらの能力によって認識され得る。このような細胞は、任意の適切な組織培養培地(例えば、10%ウシ胎児血清(約56℃で不活化した)を補充し、約10μg/lの非必須アミノ酸、約1,000U/mlのペニシリン、および約100μg/mlのストレプトマイシンを補充したEarle改変イーグル培地)中で培養され得る。このようなマウスの脾細胞が抽出され、そして適切な骨髄腫細胞株と融合される。任意の適切な骨髄腫細胞株(例えば、アメリカンタイプカルチャーコレクション(ATCC)(Rock
ville,Maryland, USA)から入手可能な親骨髄腫細胞株(SP2O))が、本発明に従って使用
され得る。融合後、得られたハイブリドーマ細胞はHAT培地中で選択的に維持され、次い
でWandsら、Gastroenterology80:225-232 (1981);HarlowおよびLane、前出、第7章に記載されているような限界希釈によってクローニングされる。次いで、このような選択によって得られたハイブリドーマ細胞は、エンドカインα抗原に結合し得る抗体を分泌するクローンを同定するためにアッセイされる。
【0127】
あるいは、エンドカインαタンパク質抗原に結合し得るさらなる抗体が、抗イディオタイプ抗体の使用を通じて二工程手順で産生され得る。このような方法は、抗体はそれ自体が抗原であるという事実を使用し、従って、二次抗体に結合する抗体を得ることが可能である。この方法に従って、エンドカインαタンパク質特異的抗体は、動物(好ましくは、マウス)を免疫するために使用される。次いで、このような動物の脾細胞はハイブリドーマ細胞を産生するために使用され、そしてハイブリドーマ細胞は、エンドカインαタンパク質特異的抗体に結合する能力がエンドカインαタンパク質抗原によってブロックされ得る抗体を産生するクローンを同定するためにスクリーニングされる。このような抗体は、エンドカインαタンパク質特異的抗体に対する抗イディオタイプ抗体を含み、そしてさらなるエンドカインαタンパク質特異的抗体の形成を誘導するために動物を免疫するために使用され得る。
【0128】
本発明の抗体のFab部分およびF(ab’)2部分および他の部分が、本明細書中に開示される方法に従って用いられ得ることが認識される。このような部分は、代表的には、パパイン(Fab部分を生成するため)またはペプシン(F(ab’)2部分を生成するため)のような酵素を用いるタンパク質分解切断により生成される。あるいは、エンドカインα(endokinealpha)タンパク質結合部分は、組換えDNA技術の適用を通して、または合成化学を通して生成され得る。
【0129】
ヒトにおける診断のために、インビボでのイメージングを用いて、増強されたレベルのエンドカインαタンパク質を検出する場合、「ヒト化」キメラモノクローナル抗体を使用することが好ましくあり得る。このような抗体は、上記のモノクローナル抗体を生成するハイブリドーマ細胞由来の遺伝構築物を用いて生成され得る。キメラ抗体を生成するための方法は、当該分野で公知である。総説については、Morrison, Science 229:1202 (1985); Oiら, BioTechniques 4:214 (1986);Cabillyら, 米国特許第4,816,567号; Taniguchi
ら, EP 171496; Morrisonら, EP 173494;Neubergerら, WO 8601533; Robinsonら, WO 8702671; Boulianneら, Nature 312:643(1984); Neubergerら, Nature 314:268 (1985)を参
照のこと。
【0130】
本発明のエンドカインαタンパク質特異的抗体にさらに適切な標識は、以下に提供される。適切な酵素標識の例は、リンゴ酸デヒドロゲナーゼ、スタフィロコッカスヌクレアーゼ、Δ-5-ステロイドイソメラーゼ、酵母アルコールデヒドロゲナーゼ、α-グリセロールリン酸デヒドロゲナーゼ、トリオースリン酸イソメラーゼ、ペルオキシダーゼ、アルカリホスファターゼ、アスパラギナーゼ、グルコースオキシダーゼ、β-ガラクトシダーゼ、
リボヌクレアーゼ、ウレアーゼ、カタラーゼ、グルコース-6-リン酸デヒドロゲナーゼ、
グルコアミラーゼ、およびアセチルコリンエステラーゼを含む。
【0131】
適切な放射性同位体標識の例は、3H、111In、125I、131I、32P、35S、14C、51Cr、57To、58Co、59Fe、75Se、152Eu、90Y、67Cu、217Ci、211At、212Pb、47Sc、109Pdなどを含む。111Inおよび99mTcは、インビボでのイメージングが用いられる場合に好ましい同位体である。なぜなら、これは、125Iまたは131Iで標識したモノクローナル抗体の肝臓による脱ハロゲン化の問題を回避するからである。さらに、この放射性核種(radionucleotide)は、イメージングのためにより好ましいγ放出エネルギーを有する(Perkinsら
,Eur. J. Nucl. Med. 10:296-301 (1985); Carasquilloら, J. Nucl. Med. 28:281-287(1987))。例えば、1-(p-イソチオシアネートベンジル)-DPTAを用いてモノクローナル抗体
にカップリングした111Inは、非腫瘍性組織(特に肝臓)における取り込みをほとんど示
さなかった。それゆえ、腫瘍局在化の特異性を増強する(Estebanら,J. Nucl. Med. 28:861-870 (1987))。
【0132】
適切な非放射性同位体標識の例は、157Gd、55Mn、162Dy、52Tr、および56Feを含む。
【0133】
適切な蛍光標識の例は、152Eu標識、フルオレセイン標識、イソチオシアネート標識、
ローダミン標識、フィコエリトリン標識、フィコシアニン標識、アロフィコシアニン標識、o-フタルアルデヒド(o-phthaldehide)標識、およびフルオレサミン標識を含む。
【0134】
適切な毒素標識の例は、ジフテリア毒素、リシン、およびコレラ毒素を含む。
【0135】
化学発光標識の例は、ルミナール標識、イソルミナール標識、芳香族アクリジニウムエステル標識、イミダゾール標識、アクリジニウム塩標識、シュウ酸エステル標識、ルシフェリン標識、ルシフェラーゼ標識、およびエクオリン標識を含む。
【0136】
核磁気共鳴コントラスト剤の例は、Gd、Mn、およびFeのような重金属原子核を含む。
【0137】
上記の標識を抗体に結合させるための代表的な技術は、Kennedyら(Clin. Chim. Acta 70:1-31 (1976))およびSchursら(Clin. Chim. Acta81:1-40 (1977))により提供される。後者において言及されるカップリング技術は、グルタルアルデヒド法、過ヨウ素酸法、ジマレイミド法、m-マレイミドベンジル-N-ヒドロキシ-スクシンイミドエステル法であり、これらの方法は全て本明細書中に参考として援用される。
【0138】
染色体アッセイ
本発明の核酸分子はまた、染色体の同定に有用である。配列は、個々のヒト染色体上の特定の位置に特異的に標的化され、そしてその位置にハイブリダイズし得る。さらに、現在は染色体上の特定の部位を同定する必要性がある。現在、実際の配列データ(反復多型)に基づいた染色体標識化試薬は、染色体位置の標識にほとんど利用可能でない。本発明によるDNAの染色体へのマッピングは、これらの配列と疾患に関連する遺伝子との相関付
けにおいて重要な第1工程である。
【0139】
この点における特定の好ましい実施態様において、本明細書中に開示されるcDNAは、エンドカインαタンパク質遺伝子のゲノムDNAをクローニングするために使用される。これ
は、種々の周知の技術および一般に市販されているライブラリーを使用して達成され得る。次いで、ゲノムDNAは、この目的のための周知の技術を使用してインサイチュ染色体マ
ッピングのために使用される。代表的には、染色体マッピングの日常的な手順に従う、いくつかの試行錯誤が、良好なインサイチュハイブリダイゼーションシグナルを与えるゲノムプローブを同定するために必要であり得る。
【0140】
いくつかの場合において、さらに、配列は、cDNAからPCRプライマー(好ましくは15〜25bp)を調製することにより染色体にマップされ得る。遺伝子の3’非翻訳領域のコンピューター解析が、ゲノムDNA内で1より多いエキソンにまたがらず、従って増幅プロセスを
複雑化しないプライマーを迅速に選択するために使用される。次いで、これらのプライマーは、個々のヒト染色体を含む体細胞ハイブリッドのPCRスクリーニングに使用される。
プライマーに対応するヒト遺伝子を含むそれらのハイブリッドのみが増幅部分を生じる。
【0141】
体細胞ハイブリッドのPCRマッピングは、特定の染色体に特定のDNAを割り当てるための迅速な手順である。同じオリゴヌクレオチドプライマーを本発明と共に使用して、特定の染色体由来の部分のパネルまたは類似の様式での大きなゲノムクローンのプールを用いて、準位置決定(sublocalization)が達成され得る。染色体にマップするために同様に使用
され得る他のマッピングストラテジーは、インサイチュハイブリダイゼーション、標識してフロー選別した(flow-sorted)染色体を用いるプレスクリーニング、および染色体特異
的cDNAライブラリーを構築するためのハイブリダイゼーションによる予備選択を含む。
【0142】
cDNAクローンの中期染色体スプレッド(spread)への蛍光インサイチュハイブリダイゼ
ーション(「FISH」)は、1工程で正確な染色体位置を提供するために使用され得る。この技術は、50bpまたは60bpほど短いcDNA由来のプローブを用いて使用され得る。この技術の総説については、Vermaら,Human Chromosomes: A Manual of Basic Techniques, Pergamon Press, New York(1988)を参照のこと。
【0143】
一旦配列が正確な染色体位置にマップされると、配列の染色体上での物理的な位置を遺伝地図のデータと相関させ得る。このようなデータは、例えば、V. McKusick, Mendelian
Inheritance in Man(Johns Hopkins University,Welch Medical Libraryからオンラインで入手可能である)において見出される。次いで、同じ染色体領域にマップされた遺伝子
と疾患との間の関係が、連鎖解析(物理的に隣接した遺伝子の同時遺伝)により同定される。
【0144】
次に、罹患個体と非罹患個体との間のcDNA配列またはゲノム配列における相違を決定する必要がある。変異がいくつかまたは全ての罹患個体に観察されるが、いずれの正常な個体にも観察されない場合、この変異は疾患の原因因子であるようである。
【0145】
物理的マッピング技術および遺伝的マッピング技術の現在の解像度では、疾患に関連する染色体領域に正確に位置決めされたcDNAは、50と500との間の潜在的原因遺伝子の1つ
であり得る。(これは、1メガベースのマッピング解像度、および20kbあたり1遺伝子と仮定する)。
【0146】
エンドカインαタンパク質および抗体治療
上記のように、TNFは、組織損傷をもたらす、その炎症誘発性(pro-inflammatory)作
用(例えば、血管内皮細胞への凝血原活性の誘導(Pober,J.S.ら, J.Immunol. 136:1680 (1986))、好中球およびリンパ球の増加した接着(Pober J.S.ら, J.Immunol.138:3319 (1987))、ならびにマクロファージ、好中球、および血管内皮細胞からの血小板活性化因
子の放出の刺激(Camussi, G.ら, J.Exp. Med. 166:1390 (1987)))について留意される。最近の証拠は、TNFを多くの感染(Cerami A.ら, Immunol.Today 9:28 (1988))、免疫
疾患、腫瘍性疾患(例えば、悪液質を伴ういくつかの悪性疾患(Oliff A.ら, Cell 50:555 (1987))ならびに自己免疫疾患および対宿主性移植片疾患(Piguet,P.-F.ら, J. Exp. Med. 166:1280 (1987)))における病因に関係付けている。多数の研究は、TNFが、ガン
、感染性の疾患、および他の異化状態における悪液質の重要なメディエータであることを示唆した。
【0147】
従って、本発明のエンドカインαタンパク質は、好ましくは、放射性同位体または細胞増殖抑制性薬物との結合後に、腫瘍標的化のために用いられ得る(GrussおよびDower, Blood 85(12):3378-3404 (1995))。エンドカインαは、黒色腫および肉腫を有する患者に
おいて、腫瘍退縮および患者の寿命の延長のために、局所注射を通して、または別々の四肢への灌流において用いられ得る(AggarwalおよびNatarajan,Eur. Cytokine Netw. 7(2):92-124(1996))。
【0148】
本発明のエンドカインαはまた、特定の状況において治療的役割(例えば、ウイルス感染、細菌感染、酵母感染、真菌感染、および他の感染(toxoplasma gondii、schistosoma
mansoni、listeria monocytogens、およびBCGを含む)に対する活性)を有し得る。エンドカインαのこれらの効果は、間接的であり得、従って好ましくは、マクロファージ、好酸球、線維芽細胞、または好中球の活性化を通して媒介される。
【0149】
TNFはまた、発熱、倦怠感、食欲不振、および悪液質を含む、グラム陰性敗血症および
内毒素性ショックの病態生理学的結果において中心的な役割を果たすと考えられる(Michie,H.R.ら, Br. J. Surg. 76:670-671 (1989); Debets, J. M. H.ら, Second Vienna ShockForum, 463-466頁(1989); Simpson, S.Q.ら, Crit. Care Clin. 5:27-47 (1989))。内毒素は、TNF(Kornbluth,S.K.ら, J. Immunol. 137:2585-2591 (1986))および他のサ
イトカインの生成および分泌を刺激する、強力な単球/マクロファージアクチベーターである。上昇したレベルの循環TNFはまた、グラム陰性敗血症を罹患する患者において見出
されている(Waage,A.ら, Lancet 1:355-357 (1987); Hammerle, A.F.ら, Second Vienna
Shock Forum715-718頁 (1989); Debets, J.M.H.ら, Crit. Care Med. 17:489-497 (1989); Calandra,T.ら, J. Infec. Dis. 161:982-987 (1990))。
【0150】
TNFに対する中和抗血清またはmAbは、ヒト以外の哺乳動物において、実験的内毒血症および菌血症における有害な生理学的変化を排除し、そして致死的なチャレンジの後の死を防止することが示されている。この効果は、例えば、齧歯類の致死率アッセイおよび霊長類の病態モデル系において実証されている(Mathison,J.C.ら, J. Clin. Invest. 81:1925-1937 (1988); Beutler, B.ら, Science 229:869-871(1985); Tracey, K.J.ら, Nature 330:662-664 (1987); Shimamoto, Y.ら, Immunol.Lett. 17:311-318 (1988); Silva, A.T.ら, J. Infet. Dis. 162:421-427 (1990); Opal,S.M.ら, J. Infect. Dis. 161:1148-1152 (1990); Hinshaw, L.B.ら, Circ. Shock30:279-292 (1990))。今日まで、ヒトにおける抗TNF mAb治療での経験は、限られていたが、例えば、関節炎および敗血症において有
益な治療結果を示す。例えば、Elliott,M.J.ら, Baillieres Clin. Rheumatol. 9:633-52
(1995); Feldmann, M.ら, Ann. N.Y.Acad. Sci. USA 766:272-8 (1995); van der Poll,
T.ら, Shock 3:1-12 (1995); Wherryら,Crit. Care. Med. 21:S436-40 (1993); Tracey K.J.ら, Crit. Care Med. 21:S415-22(1993)を参照のこと。
【0151】
エンドカインαは、TNFの多くの生物学的効果を示すと考えられるので、本発明はさら
に、上記の障害の1つ以上を処置するために哺乳動物(好ましくはヒト)患者に抗エンドカインα抗体を投与する工程を含む、抗体に基づく治療に関する。抗エンドカインαポリクローナル抗体およびモノクローナル抗体を生成するための方法は、上記に詳細に記載される。このような抗体は、当該分野で公知のように、または本明細書中に記載されるように、薬学的に受容可能な組成物中に提供され得る。
【0152】
本発明の抗体が治療的に用られ得る方法の概要は、エンドカインαを身体内において局所的または全身的に結合させること、または、例えば、補体(CDC)もしくはエフェクタ
ー細胞(ADCC)により媒介されるような抗体の直接細胞傷害性によることを含む。これらのアプローチのいくつかは、下記にさらに詳細に記載される。本明細書中に提供される教示で武装すれば、当業者は、本発明の抗体を診断目的、モニタリング目的、または治療目的のために過度の実験を伴うことなくどのように使用するかがわかる。
【0153】
本発明の薬学的組成物は、それらの意図される目的を達成する任意の手段により投与され得る。抗体、それらのフラグメント、または誘導体の投与のための量および措置は、TNF関連疾患の処置の臨床分野の当業者によって容易に決定され得る。
【0154】
例えば、投与は、非経口、皮下、静脈内、筋肉内、腹腔内、経皮、または頬の経路によってであり得る。あるいは、または同時に、投与は、経口経路によってであり得る。投与される投薬量は、レシピエントの年齢、健康状態、および体重、同時処置の種類(ある場合)、処置の頻度、および所望される効果の性質に依存する。
【0155】
本発明の範囲内の組成物は、抗体、フラグメント、または誘導体が、その意図される目的を達成するために有効な量で含まれる全ての組成物を含む。個々の必要性は異なるが、各成分の有効量の最適な範囲の決定は、当該分野の技量の範囲内である。有効用量は、個々のキメラ抗体またはモノクローナル抗体、結合した治療剤(下記を参照のこと)の存在および性質、患者およびその臨床状態の関数であり、そして約10μg/kg体重〜約5000mg/kg体重で変化し得る。好ましい投薬量は、0.1〜500mg/kg体重を含む。
【0156】
薬理学的に活性な化合物に加えて、新規な薬学的組成物は、適切な薬学的に受容可能なキャリアを含み得る。このキャリアは、薬学的に用いられ得る調製物中での活性な化合物の処理を容易にする賦形剤および助剤を含む。好ましくは、調製物は、約0.01〜99%、好ましくは約20〜75%の活性化合物を、賦形剤とともに含む。
【0157】
同様に、イメージングのための検出可能に標識された形態、または治療のための遊離もしくは結合した形態のような、非経口投与のための本発明のエンドカインα抗体またはフラグメントの調製物は、無菌の水性または非水性の溶液、懸濁液、および乳濁液を含む。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油(例えばオリーブ油)、および注入可能な有機エステル(例えば、オレイン酸エチル)である。水性キャリアは、生理食塩水および緩衝化媒体を含む、水、アルコール/水溶液、乳濁液、または懸濁液、塩化ナトリウム溶液を含む非経口ビヒクル、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル、または固定油を含む。静脈内ビヒクルは、流体補充物および栄養補充物(replenisher)(例えば、リンゲルデキストロース
に基づく補充物)などを含む。保存剤および他の添加剤はまた、例えば、抗菌剤、抗酸化剤、キレート化剤、および不活性ガスなどのように存在し得る。一般的に、Remington’sPharmaceutical Science, 第16版, Mack Publishing Co., Easton, PA, 1980を参照のこ
と。
【0158】
特に、本発明の抗体、フラグメント、および誘導体は、本明細書中に記載されるようなエンドカインα関連障害を有するかまたは発症している被験体を処置するために有用である。このような処置は、抗体、フラグメント、もしくは誘導体、またはその結合体の単回用量または複数回用量を非経口投与する工程を含む。
【0159】
本発明の抗体は、他のモノクローナル抗体もしくはキメラ抗体と、または抗体と相互作用するエフェクター細胞の数もしくは活性を増加させるのに役立つ、リンホカインまたは造血性増殖因子などと組み合わせて有利に利用され得る。
【0160】
エンドカインαの循環濃度は、(TNFと同様に)非敗血症個体において約10pg/mlの範囲、そして敗血症患者において約50pg/mlに到達し、そしてTNFの敗血症症候群において100pg/mlより上で、非常に低い傾向がある(Hammerle,A.F.ら, 1989, 上記)か、またはエン
ドカインα関連障害の部位で検出可能でのみあり得るので、高親和性および/または強力なインビボのエンドカインα阻害抗体および/またはエンドカインα中和抗体、フラグメント、またはその領域を、エンドカインα免疫アッセイおよびエンドカイン関連障害の治療の両方に用いることが好ましい。このような抗体、フラグメント、または領域は、好ましくは、ヒトエンドカインαに、少なくとも108M-1、より好ましくは少なくとも109
M-1、例えば、5×108 M-1、8×108M-1、2×109 M-1、4×109 M-1、6×10
9M-1、8×109 M-1のKaとして表される親和性を有する。
【0161】
ヒトでの治療的使用のために好ましいのは、本発明による、強力なインビボでのエンドカイン阻害活性および/または中和活性を有する、高親和性マウスおよびマウス/ヒト、またはヒト/ヒトのキメラ抗体、ならびにフラグメント、領域、および誘導体である。これらの抗体、ならびにフラグメント、領域、および誘導体は、例えば、エンドカイン誘導性IL-1、IL-6、またはTNF分泌、凝血原活性、細胞接着分子(例えば、ELAM-1およびICAM-1)の発現、および分裂促進活性を、インビボ、インサイチュ、およびインビトロでブロ
ックする。
【0162】
本発明を一般的に記載してきたが、本発明は、例示のために提供され、そして限定することを意図しない以下の実施例を参照することにより、さらに容易に理解される。
【実施例】
【0163】
実施例1:E.coliにおける成熟エンドカインαの発現および精製
寄託されたcDNAにおいて成熟エンドカインαをコードするDNA配列を、エンドカインα
タンパク質のアミノ酸末端配列に特異的なPCRオリゴヌクレオチドプライマーを用いて増
幅する。クローニングを容易にするための制限部位を含むさらなるヌクレオチドを、それぞれ5’および3’の配列に添加する。
【0164】
5’オリゴヌクレオチドプライマーは、配列 GCG CCA TGG CTA AGT TTG GAC CAT(配列番号5)を有し、これは下線を付したNcoI制限部位を含む。
【0165】
3’プライマーは、配列GCG AAG CTT TCA AGT CTC TAG GAG ATG(配列番号6)を有し
、これは下線を付したHindIII制限部位を含む。
【0166】
制限部位は、細菌発現ベクターpQE60の制限酵素部位に対して都合良く、これらの実施
例においてM15/rep4宿主細胞における細菌発現のために使用する(Qiagen,Inc. Chatsworth, CA, 91311)。pQE60は、アンピシリン抗生物質耐性(Ampr)をコードし、細菌の複製起点(ori)、IPTG誘導プロモーター、リボソーム結合部位(「RBS」)、6-Hisタグ、お
よび制限酵素部位を含む。
【0167】
増幅されたエンドカインαタンパク質DNAおよびベクターpQE60の両方をNcoIおよびHindIIIで消化し、次いで消化されたDNAを一緒に連結する。エンドカインαタンパク質DNAの
制限されたpQE60ベクターへの挿入は、エンドカインαタンパク質コード領域を、ベクタ
ーのIPTG誘導プロモーターの下流に、かつこれに作動可能に連結するように、そしてエンドカインαタンパク質の翻訳のために適切に位置された開始ATGにインフレームに配置す
る。
【0168】
連結混合物を、標準的な手順を用いて、コンピテントなE.coli細胞に形質転換する。このような手順は、Sambrookら、Molecular Cloning: A LaboratoryManual、第2版、Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)に記載されている
。多数のコピーのプラスミドpREP4を含有するE.coli株M15/rep4(これは、lacリプレッサーを発現し、そしてまたカナマイシン耐性(Kanr)を付与する)を本明細書に記載の例示的な実施例を行うにあたって使用する。この株(これは、エンドカインαタンパク質を発現するために適切である多くの内の唯一である)は、Qiagenから市販されている。
【0169】
形質転換体を、アンピシリンおよびカナマイシンの存在下でLBプレート上で増殖するそれらの能力により同定する。プラスミドDNAを耐性コロニーから単離し、そしてクローン
化DNAの同定を制限分析により確認する。
【0170】
所望の構築物を含むクローンを、アンピシリン(100μg/ml)とカナマイシン(25μg/ml)との両方を補充したLB培地における液体培養で一晩(「O/N」)増殖する。
【0171】
O/N培養物を用いて約1:100〜1:250の希釈で大規模培養に接種する。細胞を、0.4と0.6
との間の600NMでの吸光度(OD600)にまで増殖させた。次いで、イソプロピル-B-D-チオ
ガラクトピラノシド(「IPTG」)を加えて1mMの最終濃度にし、lacIリプレッサーを不活性化することにより、lacリプレッサー感受性プロモーターからの転写を誘導する。細胞
をさらに3〜4時間引き続きインキュベートする。次いで、標準的方法によって細胞を遠心分離により収集し、そして破壊する。日常的な回収技術を用いて破壊した細胞から封入体を精製し、そしてタンパク質を封入体から8M尿素中へ可溶化する。可溶化したタンパク質を含む8M尿素溶液を、2×リン酸緩衝化生理食塩水(「PBS」)中でPD-10カラムを通し、それによって尿素を除去し、緩衝液を交換し、そしてタンパク質を再び折り畳む。タンパク質をクロマトグラフィーのさらなる工程によって精製してエンドトキシンを除去する。次いで、濾過滅菌する。濾過滅菌したタンパク質調製物を、2×PBS中で保存する
。
【0172】
実施例2:バキュロウイルス発現系における成熟エンドカインαのクローニングおよび発現
寄託されたクローンにおける完全エンドカインαタンパク質をコードするcDNA配列を、この遺伝子の5’配列および3’配列に対応するPCRオリゴヌクレオチドプライマーを用
いて増幅する:
5’プライマーは、配列 GC GGA TCC CGA GAC TGC TAA GGA GCC(配列番号7)を有し
、下線を付したBamHI制限酵素部位を含む、そして図1におけるエンドカインαタンパク
質の部分をコードする配列に相補的な配列を含む。
【0173】
3’プライマーは、配列 GC GGA TCC CTA GGA GAT GAA TTG GGG ATT TG(配列番号8)を有し、下線を付したBamHI制限部位を含み、そして図1におけるエンドカインαタンパ
ク質の部分に相補的な配列を含む。
【0174】
増幅されたフラグメントを、市販のキット(「Geneclean」 BIO 101 Inc., La Jolla, Ca.)を用いて、1%アガロースゲルより単離する。次いで、このフラグメントをBamHIで消化し、そして1%アガロースゲルで再び精製する。このフラグメントを、本明細書中でF2と命名する。
【0175】
ベクターpA2GPを用いて、Summersら、AManual of Methods for Baculovirus Vectors and Insect Cell Culture Procedures,Texas Agricultural Experimental Station Bulletin No. 1555(1987)に記載の標準的な方法を用いて、エンドカインαタンパク質をバキ
ュロウイルス発現系において発現する。この発現ベクターは、Autographacalifornica核
多角体病ウイルス(AcMNPV)の強いポリヘドリンプロモーター、それに続く便利な制限部位を含む。AcMNPV gp67のシグナルペプチド(N末端メチオニンを含む)は、BamHI部位のちょうど上流に位置する。シミアンウイルス40(「SV40」)のポリアデニル化部位を、効率的なポリアデニル化のために用いる。組換えウイルスの容易な選択のために、E.coli由来のβガラクトシダーゼ遺伝子をポリヘドリンプロモーターと同方向に挿入し、その後ポリヘドリン遺伝子のポリアデニル化シグナルが続く。ポリヘドリン配列を、野生型ウイルスDNAとの細胞媒介性相同組換えのためにウイルス配列で両端で隣接させ、クローン化ポ
リヌクレオチドを発現する生存可能なウイルスを生成する。
【0176】
当業者が容易に理解するように、構築が転写、翻訳、輸送などのために適切に位置したシグナル(例えば、必要ならばインフレームでのAUGおよびシグナルペプチド)を提供す
るならば、多くの他のバキュロウイルスベクターが、pA2-GPの代わりに用いられ得る。例えば、pAc373、pVL941、およびpAcIM1である。このようなベクターは、とりわけ、Luckowら、Virology,170:31-39に記載される。
【0177】
当該分野で公知の日常的な手順を用いて、プラスミドを制限酵素XbaIで消化し、次いでウシ腸ホスファターゼを用いて脱リン酸化する。次いで、市販のキット(「Geneclean」BIO 101 Inc.,La Jolla, Ca)を用いてDNAを1%アガロースゲルから単離する。このベク
ターDNAを、本明細書中で「V2」と命名する。
【0178】
フラグメントF2および脱リン酸化プラスミドV2を、T4DNAリガーゼでともに連結する。E.coli HB101細胞を連結混合物で形質転換し、そして培養プレート上に播く。XbaIを用い
て個々のコロニーからのDNAを消化し、次いでゲル電気泳動によって消化産物を分析する
ことによって、ヒトエンドカインα遺伝子を有するプラスミドを含む細菌を同定する。クローン化フラグメントの配列を、DNA配列決定により確認する。
【0179】
5μgのプラスミドを、Felgnerら Proc. Natl. Acad. Sci.USA, 84:7413-7417 (1987)によって記載されるリポフェクション法を用いて、1.0μgの市販の線状化バキュロウイルスDNA(「BaculoGoldTMbaculovirus DNA」, Pharmingen, San Diego, CA.)とともに同時トランスフェクトする。1μgのBaculoGoldTMウイルスDNAおよび5μgのプラスミドを、50μlの無血清グレース培地(Life Technologies Inc.,Gaithersburg, MD)を含むマイクロタイタープレートの無菌ウェル中で混合する。その後、10μlのリポフェクチンおよび90
μlのグレース培地を添加し、混合し、そして室温にて15分間インキュベートする。次い
で、そのトランスフェクション混合物を、無血清グレース培地1mlを有する35mm組織培養プレート内に播種されたSf9昆虫細胞(ATCCCRL 1711)に滴下する。プレートを、新たに
添加した溶液を混合するために、前後に振盪する。次いでプレートを、27℃で5時間インキュベートする。5時間後、トランスフェクション溶液をプレートから除去し、そして10%ウシ胎児血清を補充した1mlのグレース昆虫培地を添加する。プレートをインキュベーターに戻し、そして27℃で4日間培養を続ける。
【0180】
4日後、上清を回収し、そしてSummersおよびSmith(前出)に記載されるようにプラークアッセイを行う。青く染色されたプラークを産生するgal発現クローンの容易な同定お
よび単離を可能にするために、「BlueGal」(Life Technologies Inc., Gaithersburg)
を有するアガロースゲルを用いる。(このタイプの「プラークアッセイ」の詳細な説明はまた、LifeTechnologies Inc.、Gaithersburg、で配布される昆虫細胞培養およびバキュ
ロウイルス学の使用者ガイド(9〜10頁)においても見い出され得る)。
【0181】
連続希釈の4日後、ウイルスを細胞に添加する。適切なインキュベーションの後、青く染色されたプラークをエッペンドルフピペットのチップで拾う。次いで、組換えウイルスを含む寒天を、200μlのグレース培地を含むエッペンドルフチューブ中に再懸濁する。寒天を、短時間の遠心分離により除去し、そして組換えバキュロウイルスを含む上清を、35mmディッシュに播種されたSf9細胞に感染するために用いる。4日後、これらの培養ディ
ッシュの上清を回収し、次いでそれらを4℃で保存する。適切に挿入されたhESSBI、II、およびIIIを含むクローンを、このプラスミド制限マッピングおよび配列決定を含むDNA分析によって同定する。
【0182】
Sf9細胞を、10%熱不活化FBSを補充したグレース培地中で増殖する。細胞を、約2(約1〜約3)の感染多重度(「MOI」)で組換えバキュロウイルスで感染させる。6時間後
、その培地を除去し、そしてメチオニンおよびシステインを除いたSF900II培地(Life Technologies Inc., Gaithersburgから入手可能)に置き換える。42時間後、5μCiの35S-メチオニンおよび5μCiの35S-システイン(Amershamから入手可能)を添加する。細胞
をさらに16時間インキュベートし、次いで細胞を遠心分離により収集し、溶解し、そして標識されたタンパク質をSDS-PAGEおよびオートラジオグラフィーにより可視化する。
【0183】
実施例3:CHO細胞におけるクローニングおよび発現
ベクターpC1を、エンドカインαタンパク質の発現のために使用する。プラスミドpC1は、プラスミドpSV2-dhfr(ATCC受託番号37146)の誘導体である。両方のプラスミドは、SV40初期プロモーターの制御下で、マウスDHFR遺伝子を含む。これらのプラスミドでトランスフェクトされるジヒドロ葉酸活性を欠如するチャイニーズハムスター卵巣細胞または他の細胞は、化学治療剤メトトレキサートを補充した選択培地(αマイナスMEM、LifeTechnologies)中で細胞を増殖させることによって選択され得る。メトトレキサート(MTX)に耐性である細胞におけるDHFR遺伝子の増幅は、よく考証されている(例えば、Alt,F.W.ら、J. Biol. Chem. 253:1357-1370(1978)、Hamlin, J.L.およびMa, C., Biochem. etBiophys. Acta, 1097:107-143(1990)、Page, M.J.およびSydenham, M.A., Biotechnology9:64-68(1991)を参照のこと)。漸増濃度のMTXにおける細胞増殖は、DHFR遺伝子の増幅の結果
として、標的酵素DHFRを過剰産生することによって薬物への耐性を生じる。第二遺伝子がDHFR遺伝子に連結される場合、通常、同時増幅され、そして過剰発現される。細胞株が遺伝子の1,000以上のコピーを有することは、当該水準である。続いて、メトトレキサート
が取り除かれる場合、細胞株は、染色体(単数または複数)に取り込まれる増幅遺伝子を含む。
【0184】
プラスミドpC1は、ラウス肉腫ウイルス(Cullenら、Molecularand Celluar Biology, 438-4470(1985年3月))の長末端反復(LTR)の目的の強力なプロモーターの遺伝子、お
よびヒトサイトメガロウイルス(CMV)(Boshartら、Cell41:521-530(1985))の最初期
遺伝子のエンハンサーから単離されたフラグメントの遺伝子の発現を含む。プロモーターの下流は、遺伝子の取り込みを可能にする以下の1つの制限酵素切断部位である:BamHI
、PvuII、およびNruI。これらのクローニング部位の後ろに、プラスミドは、全ての3つ
のリーディングフレームにおける翻訳停止コドン、続いてラットプレプロインシュリン遺伝子の3’イントロンおよびポリアデニル化部位を含む。他の高効率プロモーターもまた
、発現(例えば、ヒトβアクチンプロモーター、SV40初期もしくは後期プロモーター、または他のレトロウイルス(例えば、HIVおよびHTLVI)からの長末端反復のために使用し得る。mRNAのポリアデニル化のために、他のシグナル(例えば、ヒト成長ホルモンまたはグロビン遺伝子由来)も同様に使用し得る。
【0185】
染色体に挿入した目的の遺伝子を有する安定な細胞株もまた、選択マーカー(例えば、gpt、G418、またはハイグロマイシン)での同時トランスフェクトに基づいて選択し得る
。開始における1つ以上の選択マーカー(例えば、G418およびメトトレキサート)を使用することが有用である。
【0186】
プラスミドpC1を制限酵素BamHIで消化し、ついでウシ小腸ホスファターゼ(phosphate)
を用いて、当該分野で公知の手順によって脱リン酸化する。次いで、ベクターを、1%アガロースゲルから単離する。
【0187】
エンドカインαをコードするDNA配列(ATCC番号97640)を、遺伝子の5’および3’配列に対応するPCRオリゴヌクレオチドプライマーを用いて増幅する:
5’プライマーは、配列5’GCG GGA TCC GCC ATC ATG CCT TTA AGC CAT TC 3’(配列番号9)を有し、これは、下線を付したBamHI制限酵素部位、続いて図1(配列番号1)の
エンドカインαの配列の塩基を含む。発現ベクター(以下に記載)に挿入した、ヒトエンドカインαをコードする増幅したフラグメントの5’末端は、有効なシグナルペプチドを
提供する。真核生物細胞における翻訳の開始のために十分なシグナル(Kozak,M., J. Mol
. Biol. 196:947-950(1987)によって記載される)を、構築物のベクター部分に適切に配
置する。
【0188】
3’プライマーは、配列5’GC GGA TCC CTA GGA GAT GAA TTG GGG ATT TG 3’(配列番
号10)を有し、これは、Asp718制限、続いて図1(配列番号1)に示されるエンドカインαコード配列に相補的なヌクレオチド(停止コドンを含む)を含む。
【0189】
増幅したフラグメントを、上記の1%アガロースゲルから単離し、次いでエンドヌクレアーゼBamHIおよびAsp718で消化し、次いで1%アガロースゲルで再び精製する。
【0190】
次いで、単離したフラグメントおよび脱リン酸化ベクターをT4 DNAリガーゼで連結する。次いで、E.coli HB101細胞を形質転換し、そして制限酵素BamHIを用いて正しい配向に
挿入されたプラスミドpC1を含む細菌を同定する。挿入された遺伝子の配列を、DNA配列決定によって確認する。
【0191】
CHO-DHFR細胞のトランスフェクション
活性なDHFR酵素を欠如するチャイニーズハムスター卵巣細胞を、トランスフェクションのために使用する。5μgの発現プラスミドC1を、リポフェクション法(Felgnerら、前出)を用いて、0.5μgのプラスミドpSVneoとともに同時トランスフェクトする。プラスミドpSV2-neoは優性選択マーカー(G418を含む一群の抗生物質への耐性を与える酵素をコードするTn5由来の遺伝子neo)を含む。細胞を、1mg/mlのG418を補充したαマイナスMEMに播種する。2日後、細胞をトリプシン処理し、そしてハイブリドーマクローニングプレート(Greiner,Germany)に播種し、そして10〜14日間培養する。この期間の後、シングルコ
ロニーをトリプシン処理し、次いで異なる濃度のメトトレキサート(25nM、50nM、100nM
、200nM、400nM)を用いて、6ウェルペトリ皿に播種する。次いで、最も高濃度のメトトレキサートで増殖するクローンを、一様に高濃度のメトトレキート(500nM、1μM、2μM、5μM)を含む新たな6ウェルプレートに移す。同じ手順を、クローンが100μMの濃度で増殖するまで反復する。
【0192】
所望の遺伝子産物の発現を、ウエスタンブロット分析およびSDS-PAGEによって分析する。
【0193】
実施例4:エンドカインα発現の組織分布
ノーザンブロット分析を行って、とりわけSambrookら(前出)によって記載される方法を用いて、ヒト組織におけるエンドカインαタンパク質をコードする遺伝子の発現レベルの試験を行った。本発明のエンドカインαタンパク質の完全ヌクレオチド配列を含むcDMAプローブ(配列番号1)を、rediprimeTMDMA標識系(AmershamLife Science)を用いて、製造業者の説明書に従って、32Pで標識した。標識後、プローブを、CHROMA SPIN-100TMカラム(ClontechLaboratories, Inc.)を用いて、製造業者のプロトコル番号PT1200-1に従って、精製した。次いで、精製した標識化プローブを使用して、エンドカインαタンパク質をコードする遺伝子の発現について、種々のヒト組織を実験した。
【0194】
種々のヒト組織(H)またはヒト免疫系組織(IM)を含む多組織ノーザン(MTN)ブロットをClontechから入手し、そしてExpressHybTMHybridization Solution(Clontech)を用いて、製造業者のプロトコル番号PT1190-1に従って、標識化プローブで試験した。ハイブリダイゼーションおよび洗浄に続いて、ブロットを取り付け、そして-70℃にて一晩フィ
ルムに暴露し、そして標準的な手順に従ってフィルムを現像した。
【0195】
本発明のエンドカインαタンパク質をコードする遺伝子の発現を、ヒト脳線条体(striatrium)および膵臓組織において検出した。
【0196】
本発明が、前述の説明および実施例に詳細に記載される以外に実施され得ることは明白である。
【0197】
本発明の多数の改変および変異が、上記の教示に照らして可能であり、それゆえ添付の請求の範囲内である。
【0198】
本明細書中に参考として援用される全ての特許、特許明細書、および刊行物の開示は、本明細書に参考として援用される。
【産業上の利用可能性】
【0199】
本発明の新規のサイトカインは、TNF様障害の治療のための新規の抗体またはこれらのTNF様サイトカインに結合する他のアンタゴニストを作製するために使用され得る。
【図面の簡単な説明】
【0200】
【図1A】図1Aは、エンドカインαタンパク質のヌクレオチド(配列番号1)および推定アミノ酸(配列番号2)配列を示す。アミノ酸1〜17は、細胞内ドメインを示し、アミノ酸18〜43は、膜貫通ドメイン(下線を付した配列)を示し、そしてアミノ酸44〜169は、細胞外ドメイン(残りの配列)を示す。
【図1B】図1Aの続き。
【図2】図2は、エンドカインαタンパク質(配列番号2)、組織壊死因子α(TNF-α)(配列番号3)、およびTNF-β(配列番号4)のアミノ酸配列間の類似性が存在する領域を示す。J.Hein法を、PAM250残基重量表とともに使用した。四角の囲いは、コンセンサスに正確に一致する残基を示す。
【図3】図3は、エンドカインαアミノ酸配列の分析を提供する。α、β、ターンおよびコイル領域;親水性および疎水性;両親媒性領域;可動性領域;抗原性指数および表面確率が示される。「抗原性指数-Jameson-Wolf」グラフにおいて、図1のアミノ酸残基44〜54、57〜68、69〜78、94〜105、108〜132および148〜158が、示されたエンドカインαタンパク質の抗原性が高い領域に対応する。
【0201】
(配列表)
【0202】
【表1】
【特許請求の範囲】
【請求項1】
本明細書に記載される発明。
【請求項1】
本明細書に記載される発明。
【図1A】


【図1B】

【図2】
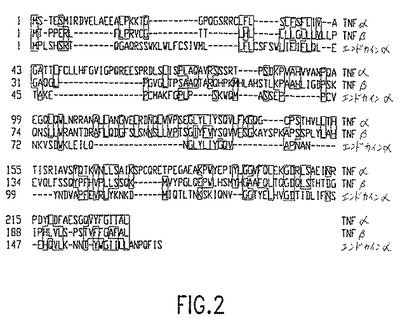
【図3】
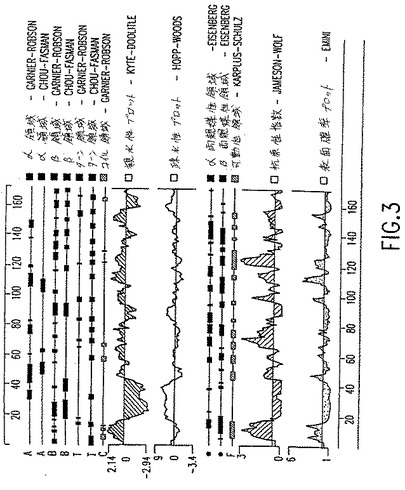
【図1B】
【図2】
【図3】
【公開番号】特開2009−77723(P2009−77723A)
【公開日】平成21年4月16日(2009.4.16)
【国際特許分類】
【出願番号】特願2008−269226(P2008−269226)
【出願日】平成20年10月17日(2008.10.17)
【分割の表示】特願平10−510675の分割
【原出願日】平成8年8月16日(1996.8.16)
【出願人】(597018381)ヒューマン ジノーム サイエンシーズ, インコーポレイテッド (44)
【氏名又は名称原語表記】Human Genome Sciences, Inc.
【Fターム(参考)】
【公開日】平成21年4月16日(2009.4.16)
【国際特許分類】
【出願日】平成20年10月17日(2008.10.17)
【分割の表示】特願平10−510675の分割
【原出願日】平成8年8月16日(1996.8.16)
【出願人】(597018381)ヒューマン ジノーム サイエンシーズ, インコーポレイテッド (44)
【氏名又は名称原語表記】Human Genome Sciences, Inc.
【Fターム(参考)】
[ Back to top ]