
ヒトトール相同体
【課題】出願人は、本出願で(DNA40021によってコードされる)PRO285、(DNA42663によってコードされる)PRO286および(DNA47361によってコードされる)PRO358と命名された新規なヒトトールポリペプチドをコードする3種類の新規なcDNAクローンを同定することを目的とする。
【解決手段】本発明は、ヒトトールたんぱく質PRO285、PRO286またはPRO358をコードする新規DNAの同定および単離、およびこれらのたんぱく質の組換え生産方法と手段に関する。本発明は、またPRO285またはPRO286またはPRO358トールたんぱく質に特異的に結合する抗体に関する。
【解決手段】本発明は、ヒトトールたんぱく質PRO285、PRO286またはPRO358をコードする新規DNAの同定および単離、およびこれらのたんぱく質の組換え生産方法と手段に関する。本発明は、またPRO285またはPRO286またはPRO358トールたんぱく質に特異的に結合する抗体に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的には、ここでDNA40021、DNA42663およびDNA47361と称する新規なDNAの同定と単離、および該DNAによりコードされる新規なヒトトール(Toll)相同体(それぞれPRO285、PRO286およびPRO358と称する)の組換え生産に関する。
【背景技術】
【0002】
膜結合タンパク質およびレセプタは多細胞生物の形成、分化および維持に重要な役割を果たすことができる。多くの各細胞の運命、例えば、増殖、移動、分化または他の細胞との相互作用は、典型的には他の細胞および/または直接の環境から受け取られた情報により支配される。この情報はしばしば分泌ポリペプチド(例えば有糸分裂因子、生存因子、細胞毒因子、分化因子、神経ペプチドおよびホルモン)により伝達された後に多様な細胞レセプタまたは膜結合タンパク質により受け取られ解釈される。そのような膜結合タンパク質および細胞レセプタとしてはサイトカインレセプタ、レセプタキナーゼ、レセプタホスファターゼ、細胞間相互作用に関与するレセプタおよびセレクチン類やインテグリン類のような細胞付着分子が挙げられるが、これらに限定されない。例えば、細胞の増殖と分化を調節するシグナル変換は、一部は多様な細胞タンパク質のリン酸化により調節される。このプロセスを触媒する酵素であるタンパク質チロシンキナーゼは成長因子レセプタとしても働くことができる。具体的な例として繊維芽細胞成長因子レセプタや神経成長因子レセプタが挙げられる。
【0003】
膜結合タンパク質とレセプタ分子は、医薬および診断剤等の多様な工業的応用性を有する。例えば、レセプタ免疫アドヘシンはレセプタリガンド相互作用をブロックする治療剤として用いることができる。膜結合タンパク質は関連するレセプタ/リガンド相互作用の潜在的なペプチドまたは小分子阻害物質のスクリーニングのために用いることもできる。
【0004】
新しい天然レセプタタンパク質を同定するための努力が工業界と大学の両者によりなされている。新規なレセプタタンパク質をコードする配列を同定するために哺乳類の組換えDNAライブラリーのスクリーニングに多くの努力が集められている。
胚背側−腹側パターンの確立に中心的な役割を演じる母性遺伝効果遺伝子であるショウジョウバエのトール遺伝子のクローニングは、Hashimotoら,Cell 52,269−279(1988)により報告されている。ショウジョウバエのトール遺伝子は803個のアミノ酸の細胞質外ドメインと269個のアミノ酸の細胞質ドメインを有する統合膜タンパク質をコードする。この細胞質外ドメインは膜にわたる潜在的なセグメントを有し、多くのトランスメンブレンタンパク質に見られる構造的モチーフであるロイシンに富むセグメントの複数のコピーを有する。トールタンパク質は、ショウジョウバエ胚の背側−腹側パターンを調節し、そのリガンドのスペッツルに結合する際に転写因子ドーサルを活性化するMorisato and Anderson,Cell 76,677−688(1994)。成熟したショウジョウバエにおいて、トール/ドーサルシグナル経路は抗微生物免疫応答に関与するLenaitreら,Cell 86,973−983(1996)。
【0005】
ショウジョウバエのトールタンパク質のヒト相同体はMedzhitovらNature 388、394−397(1997)により記載されている。このヒトのトールは、ショウジョウバエのトールと同じように、21個のタンデムに繰り返されたロイシンに富むモチーフ(ロイシンに富む領域−LRR)からなり、非LRR領域により分離された細胞外ドメインとヒトインターロイキン−1(IL−1)の細胞質ドメインに相同性のある細胞質ドメインとを有するタイプIのトランスメンブレンタンパク質である。ヒト細胞系にトランスフェクトされたヒトトールの構成的に活性のある変異体は、NF−κΒの活性化および炎症サイトカインIL−1、IL−6およびIL−8のNF−κΒ調節遺伝子の発現、ならびに天然のT細胞の活性化に必要とされる構成促進分子B7.1の発現を誘導することが可能なことが示された。トールは免疫系の非クローン性レセプタとしての脊椎動物中で機能し、脊椎動物で固有かつ適応性の免疫応答を活性化するシグナルを誘導できることが示唆されている。上記のMedzhitovらにより報告されたヒトトール遺伝子は脾臓および末梢血液白血球(PBL)において最も強く発現され、筆者らは他の組織中でのその発現は、それが感染の初期警告システムとして働きうるマクロファージおよび樹状突起細胞の存在によるものであることを示唆した。公共のGenBankデータベースは下記のトール配列を有する:トール1(ランダム配列の全長cDNA#HUMRSC786−1と同一であるDNAX# HSU88540−1);トール2(DNAX# HSU88878−1);トール3(DNAX# HSU88879−1);トール4(上記のMedzhitovらにより報告されたDNA配列と同一であるDNAX# HSU88880−1)。部分的なトール配列(トール5)はGenBankからDNA# HSU88881−1として利用できる。
【0006】
トール様レセプタと命名されたショウジョウバエトールタンパク質のさらなるヒト相同体(huTLRs1−5)が最近クローニングされ、ショウジョウバエ対応物の局在構造によく似ることが示された(Rockら,Proc.Natl.Acad.Sci.USA 95,588−593[1998])。あるヒトTLR(トールタンパク質相同体 − 上記のMedzhitovら;TLR4−上記のRockら)の構成的活性のある変異体の過剰発現はNFκΒの活性化および炎症サイトカインと構成促進分子の誘導をもたらす。上記のMedzhitovら。
【特許文献1】米国特許第5,364,934号
【特許文献2】米国特許第5,428,130号
【特許文献3】米国特許第3,959,642号
【非特許文献1】MedzhitovらNature 388、394−397(1997)
【非特許文献2】Rockら,Proc.Natl.Acad.Sci.USA 95,588−593(1998)
【発明の開示】
【発明が解決しようとする課題】
【0007】
出願人は、本出願で(DNA40021によってコードされる)PRO285、(DNA42663によってコードされる)PRO286および(DNA47361によってコードされる)PRO358と命名された新規なヒトトールポリペプチドをコードする3種類の新規なcDNAクローンを同定した。
【課題を解決するための手段】
【0008】
一実施態様において、本発明は、(a)図1(配列番号1)のアミノ酸残基27〜839を有するPRO286ポリペプチドをコードするDNA分子;または(b)図4(配列番号3)のアミノ酸残基27〜825を有するPRO286ポリペプチドをコードするDNA分子、または図18〜19(配列番号13)のアミノ酸20〜575を有するPRO358ポリペプチドをコードするDNA分子に対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも90%の配列同一性、最も好ましくは約95%の配列同一性を有するポリペプチドをコードするDNAからなる単離された核酸分子、または(d)前記(a)(b)または(c)のDNA分子の相補体を提供する。該相補DNA分子はそのようなコード核酸配列に対して少なくとも適度にストリンジェントな条件下で、および任意に高いストリンジェントな条件下で安定的に結合しているのが好ましい。
【0009】
さらなる実施態様において、単離された核酸分子は、図1(配列番号1)のアミノ酸1〜839の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有し;または図4(配列番号3)のアミノ酸1〜1041の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有し;または図18〜19(配列番号13)のアミノ酸1〜811の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有するポリヌクレオチドからなる。
【0010】
特定の実施態様において、本発明は、N末端シグナル配列を有するか有さない、および各全長配列のトランスメンブレン領域を有するか有さない天然または変異体のPRO285、PRO286およびPRO358ポリペプチドをコードするDNAからなる単離された核酸分子を提供する。一つの態様において、単離された核酸は、図1(配列番号1)のアミノ酸残基1〜1049、図4(配列番号3)の1〜1041、および図18〜19(配列番号13)の1〜811を有する成熟した全長の天然PRO285、PRO286またはPRO358ポリペプチドをコードするDNAからなるか、またはそうしたコード核酸配列に相補性のあるものである。別の態様において、本発明はN末端シグナル配列を有さない天然のPRO285、PRO286またはPRO358ポリペプチドをコードするDNAからなる単離された核酸分子またはそうしたコード核酸配列に相補性のある単離された核酸分子に関する。さらに別の実施態様において、本発明は、全長の天然PRO285、PRO286およびPRO358タンパク質のトランスメンブレン−ドメイン欠失または不活性化形をコードする核酸に関する。
【0011】
別の態様において、本発明は、図2と3(配列番号2)のおよその残基85〜およそ3283(当該残基を含む)の核酸の相補体に対して、または図5から7(配列番号4)のおよその残基57〜およそ4199(当該残基を含む)の核酸の相補体に対して、または図21〜22配列番号14)のおよその残基111〜およそ2544(当該残基を含む)の核酸の相補体に対してハイブリダイズするDNAからなる、PRO285、PRO286またはPRO358ポリペプチドをコードする単離された核酸分子に関する。好ましくは、ハイブリダイゼーションはストリンジェントなハイブリダイゼーションと洗浄条件下で起る。
【0012】
別の態様において、本発明は、(a)図1(配列番号1)の残基1〜1049(当該残基を含む)、または図4(配列番号3)のアミノ酸残基1〜1041(当該残基を含む)、または図18〜19(配列番号13)のアミノ酸残基1〜811(当該残基を含む)のアミノ酸配列に比較して少なくとも約80%の正量、好ましくは少なくとも約85%の正量、より好ましくは少なくとも90%の正量、最も好ましくは少なくとも約95%の正量を示すポリペプチドをコードするDNA、または(b)前記(a)のDNAの相補体に関する。
【0013】
別の実施態様において、本発明は、1997年10月17日にATCC No.209389にて寄託されたクローン(DNA40021−1154)、1997年10月17日にATCC No.209386にて寄託されたクローン(DNA42663−1154)、または1997年11月7日にATCC No.209431にて寄託されたクローン(DNA47361−1249)からなる単離された核酸分子を提供する。
【0014】
さらに別の実施態様において、本発明はPRO285、PRO286およびPRO358ポリペプチドまたはそれらの変異体をコードするDNAからなるベクターを提供する。よって、ベクターは上記のいずれの単離核酸分子からなってもよい。
【0015】
特定の実施態様において、本発明は、図18〜19(配列番号13)のアミノ酸20〜811の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも90%の配列同一性、最も好ましくは少なくとも約95%の配列同一性を有するポリヌクレオチドからなるベクター、またはそのようなポリヌクレオチドの相補体からなるベクターを提供する。特定の実施態様において、該ベクターは、N末端シグナル配列(およそアミノ酸1〜19)を有するか有さない新規なトール相同体(PRO358)、またはトランスメンブレン−ドメイン(およそアミノ酸576〜595)が欠損または不活性化したその変異体、または成熟タンパク質の細胞外ドメイン(およそアミノ酸20〜596)、またはこれらの配列のいずれか一つからなるタンパク質をコードするDNAからなる。そのようなベクターを有する宿主細胞も提供される。類似する実施態様が、各シグナル配列を有するか有さないPRO285およびPRO286トール相同体および/またはそれらのトランスメンブレン−ドメイン欠損または不活性化変異体からなるベクターについて明らかであり、具体的にはベクターは成熟PRO85およびPRO286トール相同体の細胞外ドメインをそれぞれ有する。
そのようなベクターを有する宿主も提供される。例えば、宿主細胞はCHO細胞、大腸菌または酵母であってよい。
【0016】
PRO285、PRO286およびPRO358ポリペプチドの製造方法がさらに提供され、該製法はPRO285、PRO286およびPRO358のそれぞれの発現に適する条件下で宿主細胞を培養し、PRO285、PRO286またはPRO358を細胞培養物から回収することからなる。
【0017】
別の実施態様において、本発明は単離されたPRO285、PRO286およびPRO358ポリペプチドを提供する。特に、本発明は、一実施態様において図1と3(配列番号1と3)のそれぞれのアミノ酸残基1〜1049および1〜1041からなるアミノ酸配列を有する単離された天然配列PRO285およびPRO286ポリペプチドを提供する。また、本発明は上記いずれかの単離核酸分子によりコードされるPRO285とPRO286ポリペプチドの変異体も提供する。特定の変異体として、各N末端シグナル配列を欠き、および/または各トランスメンブレンおよび/または細胞質ドメインを欠失または不活性化させた全長の天然配列PRO285とPRO286ポリペプチドの欠失(端を切り取った)変異体が挙げられるが、これらに限定されない。さらに、本発明は単離された天然配列PRO358ポリペプチドまたはその変異体を提供する。特に、本発明は、ある種の実施態様において、図18〜19(配列番号13)の残基20〜575、または20〜811、または1〜811からなるアミノ酸配列を有する単離された天然配列PRO358ポリペプチドを提供する。
【0018】
さらに別の態様において、本発明は、図1(配列番号1)のアミノ酸残基1〜1049;または図4(配列番号3)のアミノ酸残基1〜1041、または図18〜19(配列番号13)のアミノ酸残基1〜811のアミノ酸配列と比較して、少なくとも約80%の正量(positives)、好ましくは少なくとも約85%の正量、より好ましくは少なくとも90%の正量、最も好ましくは少なくとも約95%の正量を示すアミノ酸配列からなる単離されたPRO285、PRO286またはPRO358ポリペプチドに関する。
【0019】
さらに別の態様において、本発明は、(I)(a)図1(配列番号1)のおよそのアミノ酸残基1〜およそ1049(当該残基を含む)のアミノ酸残基、または図4(配列番号3)のおよそのアミノ酸残基1〜およそ1041(当該残基を含む)のアミノ酸残基、または図18〜19(配列番号13)のおよそのアミノ酸残基1〜およそ811(当該残基を含む)のアミノ酸残基の配列を有するPRO285、PRO286またはPRO358ポリペプチドをコードするDNA分子、または(b)前記の(a)のDNA分子の相補体に対して試験DNA分子をストリンジェントな条件下でハイブリダイズさせ、そして試験DNA分子が(a)または(b)に対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも約90%配列同一性、最も好ましくは少なくとも約95%の配列同一性を有する場合、(ii)該試験DNA分子を有する宿主細胞を該ポリペプチドの発現に適する条件下で培養し、(iii)該ポリペプチドを細胞培養物から回収することにより生産されるポリペプチドを提供する。
【0020】
別の実施態様において、本発明は、PRO285、PRO286またはPRO358を異種ポリペプチドまたはアミノ酸配列に融合させてなるキメラ分子を提供する。そのようなキメラ分子の具体例は、PRO285、PRO286またはPRO358ポリペプチドをエピトープタグ配列または免疫グロブリンのFc領域に融合させてなる。そのようなキメラ分子の具体例は、PRO358(シグナルペプチドおよび/またはトランスメンブレン−ドメインおよび任意に細胞内ドメインが欠損したその変異体を含む)をエピトープタグ配列または免疫グロブリンのFc領域に融合させてなる。好ましい実施態様において、該融合物は、少なくともCH2とCH3ドメインを有する免疫グロブリン定常領域に融合させたPRO358の細胞外ドメインを有する。類似の特定の実施態様が存在し、PRO258またはPRO286ポリペプチドからなるキメラ分子についてここで開示される。
【0021】
別の実施態様において、本発明はPRO285、PRO286またはPRO358ポリペプチドに特異的に結合する抗体を提供する。任意に、抗体はモノクローナル抗体である。具体的には、本発明は二重特異性を有する抗体、例えば2つ以上のトールポリペプチドを結合させる二特異的抗体を包含する。
さらに別の実施態様において、本発明は天然のPRO285、PRO286およびPRO358ポリペプチドのアゴニストおよびアンタゴニストに関する。特定の実施態様において、該アゴニストまたはアンタゴニストは抗PRO285、抗PRO286または抗PRO358抗体である。
【0022】
さらに別の実施態様において、本発明は天然PRO285、PRO286およびPRO358ポリペプチドのアゴニストまたはアンタゴニストを同定するスクリーニングアッセイに関する。
【0023】
さらに別の実施態様において、本発明は、薬学的に許容される担体と組み合わせたPRO285、PRO286またはPRO358ポリペプチド、または上記のアゴニストまたはアンタゴニストからなる組成物に関する。
さらに、本発明は、薬学的に許容される担体と組み合わせた、PRO285、PRO286またはPRO358ポリペプチドに特異的に結合する抗体からなる組成物に関する。
【0024】
また、本発明は、効果的な量のPRO285、PRO286またはPRO358ポリペプチドのアンタゴニストを患者に投与することからなる敗血症性ショックの治療方法に関する。具体的な実施態様において、該アンタゴニストは天然のPRO285、PRO286またはPRO358ポリペプチドに特異的に結合する阻止抗体である。
【発明を実施するための最良の形態】
【0025】
I.定義
ここで用いられる「PROポリペプチド」、「PRO286ポリペプチド」、「PRO285」および「PRO286」との用語は、(ここでさらに定義される)天然配列PRO285およびPRO286トールタンパク質および変異体を包含する。該PRO285およびPRO286ポリペプチドは、ヒト組織タイプ等の多様なソースまたは別のソースから単離してもよいし、組換えまたは合成方法により、またはこれらの技術や類似の技術の組合せにより調製してもよい。
【0026】
「天然配列RPO285」または「天然配列PRO286」は天然から得られたPRO285またはPRO286と同一のアミノ酸配列を有するポリペプチドからなる。そのような天然配列トールポリペプチドは天然から単離することができ、または組換えまたは合成手段により作ることができる。具体的には、「天然配列PRO285」および「天然配列PRO286」とは、ここに開示されるPRO285とPRO286ポリペプチドの天然の端切断または分泌された形(例えば、細胞外ドメイン配列)、PRO285とPRO286ポリペプチドの天然の変異体形(例えば、交互にスプライスされた形)および天然の対立遺伝子変異体を包含する。本発明の一実施態様において、天然配列PRO285は、図1(配列番号1)のアミノ酸1〜1049からなる成熟または全長の天然配列PRO285ポリペプチドであり、天然配列PRO286は図4(配列番号3)のアミノ酸1〜1041からなる成熟または全長の天然配列PRO286ポリペプチドである。さらなる実施態様において、天然配列PRO285は、図1(配列番号1)のアミノ酸27〜1049または27〜836、または図4(配列番号3)のアミノ酸27〜1041または27〜825からなる。
【0027】
「PRO285変異体」および「PRO286変異体」との用語は、全長天然配列PRO285に関して図1(配列番号1)に示された類推アミノ酸配列を有するPRO285に対して少なくとも約80%のアミノ酸配列同一性、または全長天然配列PRO286に関しては図4(配列番号3)に示された類推アミノ酸配列を有するPRO286に対して少なくとも約80%のアミノ酸配列同一性を有する以下に定義する活性PRO285またはPRO286ポリペプチドを意味する。そのような変異体としては、例えば、一つ以上のアミノ酸残基が図1と3(配列番号1と3)の配列のN末端またはC末端でそれぞれ付加または欠失したPRO285およびPRO286を包含する。通常、PRO285またはPRO286変異体は、図1または図4(配列番号1と3)のアミノ酸配列に対して少なくとも約80%のアミノ酸同一性、より好ましくは少なくとも約90%のアミノ酸配列同一性、およびさらに好ましくは少なくとも95%のアミノ酸同一性を有するであろう。好ましい変異体は、天然配列PRO285またはPRO286ポリペプチドの細胞外ドメインに対して高度な配列同一性を示すものである。特定の実施態様において、本発明のPRO285およびPRO286変異体は、対応する天然のタンパク質の細胞内ドメインの少なくともC末端部分を保持し、最も好ましくは細胞内および細胞外ドメインのほとんどを保持する。しかし、それらの意図する使用に基づいて、そのような変異体は多様なアミノ酸の変更、例えばこれらの領域内に置換、欠失および/または挿入を有してよい。
【0028】
ここで用いられる「PRO358ポリペプチド」、「PRO358」、「PRO358トール相同体」およびその文字通りの変異体は、(さらにここで定義される)PRO358トールタンパク質と変異体を包含する。PRO358ポリペプチドは例えばヒト組織またはそれ以外のソース等の多様なソースから単離してもよいし、または組換え法または合成方法、またはこれらの技術や類似した技術の組合せにより調製してよい。
【0029】
「天然配列PRO358」は天然から得られたPRO358の同一のアミノ酸配列を有するポリペプチドからなる。そのような天然配列トールポリペプチドは天然から単離できるか、または組換えまたは合成手段により製造することができる。「天然配列PRO358」との用語は、具体的には、ここに開示のPRO368ポリペプチドの天然の端切断または分泌形(例えば、細胞外ドメイン配列)、天然の変異体形(例えば、交互にスプライスされた形)および天然の対立遺伝子変異体を包含する。本発明の一実施態様において、天然配列PRO358は、N末端シグナル配列(アミノ酸1〜19)を有するか有さず、およびN末端メチオニンを有するか有さない図18〜19(配列番号13)のアミノ酸20〜811からなる成熟または全長の天然配列PRO358ポリペプチドである。別の実施態様において、天然配列PRO358は、N末端シグナル配列を有するか有さず、およびN末端メチオニンを有するか有さない全長タンパク質(アミノ酸29〜575)の細胞外ドメインを保持する全長PRO358の可溶形である。
【0030】
「PRO358変異体」との用語は、図18〜19(配列番号13)に示される類推アミノ酸配列を有するPRO358に対して少なくとも約80%、好ましくは少なくとも約85%、より好ましくは少なくとも90%、最も好ましくは少なくとも約95%のアミノ配列同一性を有する下記の活性PRO358ポリペプチドを意味する。そのような変異体としては、例えば、一つ以上のアミノ酸が図18〜19(配列番号13)の配列のN末端またはC末端で付加または欠損したPRO358ポリペプチドを挙げられる。具体的には、変異体は、細胞内ドメインの一部またはすべてが欠損してよい、天然配列PRO358のトランスメンブレン−ドメイン欠失または不活性化変異体が挙げられる。好ましい変異体は天然配列PRO358ポリペプチドの細胞外ドメインに対して高い配列同一性を有するものである。特定の実施態様において、本発明のPRO358変異体は対応する天然タンパク質の細胞内ドメインの少なくともC末端を保持し、最も好ましくは細胞内および細胞外ドメインのほとんどを保持する。しかし、それらの意図する使用に基づいて、そのような変異体は多様なアミノ酸の変更、例えば、これらの領域内での置換、欠失および/または挿入を有してよい。
【0031】
ここで同定されたPRO285、PRO286およびPRO358配列に関する「アミノ酸配列同一度(%)」とは、配列を整列させ、必要であれば空所を導入して配列の最大同一性を得て、配列同一性の一部としての保存置換物は考慮しない後の、PRO285、PRO286またはPRO358配列中のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントと定義される。アミノ酸配列同一度を決める目的のためのアライメントは、例えばBLAST、ALIGNまたはメガライン(DNASTAR)ソフトウエア等の一般に利用可能なコンピュータソフトウエアを用いて当業者の技量内にある多様な方法で行うことができる。当業者は比較される配列の全長にわたる最大アライメントを得るために必要なアルゴリズム等のアライメント測定用の適当なパラメータを決めることができる。ALIGNソフトウエアがアミノ酸配列同一性を決定するために好ましい。
【0032】
具体的な特徴において、ここで同定されたPRO285、PRO286およびPRO358配列に関する「アミノ酸同一度(%)」とは、配列を整列させ、必要であれば空所を導入して配列の最大同一度を得て、配列同一性の一部としての保存置換物は考慮しない後の、PRO285、PRO286またはPRO358配列中のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントと定義される。ここで用いられる同一性値(%)は、[Altschulら,Methods in Enzymology,266:460−480(1996);http://blast.wustl/edu/blast/README.html]から得られたWU−BLAST−2により作られる。WU−BLAST−2は幾つかのサーチパラメータを使用し、そのほとんどがデフォルト値に設定される。調節可能なパラメータは、オーバーラップスパン=1、オーバーラップフラクション=0.125、ワード閾(T)=11の値内で設定される。HSP SとHSP S2パラメータは動的な値で、特定の配列の構成と調べられる興味のある配列に対する特定のデータベースの構成に基づいてプログラム自体により確立される;しかし、その値は感度を高めるために調節してもよい。アミノ酸配列同一性値%は、整列させた領域中の「長い」配列の残基の合計数で割られた、マッチする同一残基の数により決定される。「長い」配列は整列させた領域中の最も実際の残基を有する配列である(アライメントスコアを最大化するためにWU−Blast−2により導入された空所は無視する)。
【0033】
上記のように実施される配列比較の文脈において、「正量(positives)」との用語は、同一ではないが、(例えば保存的置換の結果として)類似の性質を有する比較配列中の残基を包含する。正量の%値は、上記のように、長い配列中の残基の合計数により割られたBLOSUM 62マトリックス中の正量値を示す残基部分により決定される。
【0034】
ここで同定されたDNA40021、DNA42663およびDNA47361配列に関する「核酸配列同一度(%)」は、配列を整列させ、必要であれば空所を導入して配列の最大同一度を得て、DNA40021、DNA42663およびDNA47361配列中のヌクレオチドと同一である候補配列中のヌクレオチドのパーセントと定義される。核酸配列同一度を決める目的のためのアライメントは、例えばBLAST、ALIGNまたはメガライン(DNASTAR)ソフトウエア等の一般に利用可能なコンピュータソフトウエアを用いて当業者の技量内にある多様な方法で行うことができる。当業者は比較される配列の全長にわたる最大アライメントを得るために必要なアルゴリズム等のアライメント測定用の適当なパラメータを決めることができる。ALIGNソフトウエアが核酸配列同一性を決定するために好ましい。
【0035】
具体的には、ここで同定されたPRO285、PRO286およびPRO358ポリペプチドのコード配列に関する「核酸配列同一度(%)」とはPRO285、PRO286またはPRO358のコード配列中のヌクレオチド残基と同一である候補配列中のヌクレオチド残基のパーセントと定義される。ここで用いられる同一性値は、デフォルトパラメータに設定されたWU−BLAST−2のBLASTNモジュールにより作られ、オーバーラップスパンとオーバーラップフラクションをそれぞれ1および0.125に設定した。
【0036】
ここに開示の多様なポリペプチドを記載するために用いられる場合の「単離された」とはその天然の環境の成分から同定され、分離され、および/または回収されたポリペプチドを意味する。その天然環境のきょう雑成分は典型的には該ポリペプチドの診断または治療使用に干渉する材料であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性の溶質が挙げられよう。好ましい実施態様において、該ポリペプチドは、(1)スピンニングカップシークエネータを用いて、N末端または中間アミノ酸配列の少なくとも15残基を得るのに十分な程度まで、または(2)非還元または還元条件下に、クマシーブルーまたは好ましくは銀染色を用いてSDS−PAGEにより均一となるまで精製する。PRO285、PRO286またはPRO358の天然環境の少なくとも一つの構成要素は存在しないために、単離されたポリペプチドは組換え細胞中のin situでのポリペプチドを包含する。しかし、通常は単離されたポリペプチドは少なくとも一つの精製工程で調製されよう。
【0037】
「単離された」DNA40021、DNA42663またはDNA47361核酸分子は、DNA40021、DNA42663またはDNA47361核酸の天然のソースに通常会合している少なくとも一つのきょう雑核酸分子から分離され同定される核酸分子である。単離されたDNA40021、DNA42663またはDNA47361核酸分子は天然に見られるものとは形態や設定において異なる。したがって、単離されたDNA40021、DNA42663またはDNA47361核酸分子は、天然の細胞中で存在するDNA40021、DNA42663またはDNA47361核酸とは区別される。しかし、単離されたDNA40021、DNA42663またはDNA47361核酸分子は、例えば核酸分子が天然の細胞とは異なる遺伝子配置にあるDNA40021、DNA42663またはDNA47361を通常発現する細胞中に含まれるDNA40021、DNA42663およびDNA47361核酸分子を包含する。
【0038】
「トールレセプタ2」、「TLR2」および「huTLR2」は相互に交換可能に用いられ、Rockら,Proc.Natl.Acad.Sci.USA 95,588−593(1998)により「HuTLR2」と命名されたヒトトールレセプタを示している。huTLR2の塩基配列とアミノ酸配列をそれぞれ図16(配列番号11)と図17(配列番号12)に示す。
【0039】
「発現ベクター」との用語は、ここではトール相同タンパク質をコードする核酸が、適当な宿主細胞中でその発現に影響を与えることのできる制御配列に操作可能に結合されたベクターを定義するために用いる。通常、ベクターは複製部位を有する(ただし、染色体統合が起る場合これは必要ではない)。発現ベクターは形質転換細胞において表現型の選択を提供できるマーカー配列も有する。例えば、大腸菌は典型的には大腸菌種に由来するプラスミドであるpBR322を用いて形質転換される(Bolivar,ら,Gene 2:95[1977])。pBR322はアンピシリンおよびテトラサイクリン耐性遺伝子を有しているために、目的がクローニングであろうと、発現であろうと形質転換細胞の同定の容易な手段を提供する。また、発現ベクターは、最も望ましくは、転写と翻訳の制御に有用な配列、例えばプロモータおよびShine−Dalgarno配列(原核生物に関して)またはプロモータとエンハンサ(哺乳類細胞に関して)を有しよう。プロモータとしては誘導性のあるものでよいが、誘導性である必要はなく;哺乳類宿主用のCMVプロモータ等の強力な構成プロモータが宿主細胞毒なしにLHRを産生することがわかっている。発現ベクターは発現の制御、複製配列または選択遺伝子を有する必要はないと思われるが、それらが存在しないと、ハイブリッド形質転換体の同定や高いレベルのハイブリッド免疫グロブリン発現の達成が困難になろう。
【0040】
「制御(control)配列」とは、特定の宿主生物中で、操作可能に結合させたコード配列の発現に必要なDNA配列を指している。原核細胞に適する制御配列とは、例えば、プロモータ、任意にオペレーター配列、およびリボソーム結合部位が挙げられる。真核細胞は、プロモータ、ポリアデ二ル化シグナルおよびエンハンサを利用することが公知である。
【0041】
核酸が別の核酸配列と機能的な関係に置かれているときに、それは「操作可能に(operably)結合」されている。例えば、プレ配列用または分泌リーダー用のDNAがポリペプチド用のDNAに操作可能に結合しているとは、それが該ポリペプチドの分泌に関与する前タンパク質として発現している場合であり;プロモータまたはエンハンサがコード配列に操作可能に結合されているとは、それが該配列の転写に影響を与える場合であり;またはリボソーム結合部位がコード配列に操作可能に結合されているとは、それが翻訳を容易にするように配置されている場合である。一般的に、「操作可能に結合されている」とは、結合されているDNA配列が隣接しており、分泌リーダーの場合、隣接しており、リーディングフェーズにあることを意味する。しかし、エンハンサは隣接している必要はない。結合は便利な制限部位での連結により行われる。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを慣用の実施にしたがって用いる。
【0042】
「抗体」との用語は最も広い意味で用いられ、具体的には単一の抗PRO285、抗PRO286および抗PRO358モノクローナル抗体(アゴニスト、アンタゴニストおよび中和抗体を含む)、および多エピトープ特異性を有する抗PRO285、抗PRO286および抗PRO358抗体組成物を包含する。ここで用いられる「モノクローナル抗体」との用語は実質的に均一な抗体の集団(すなわち、該集団を構成する個々の抗体は少量存在する可能性のある起り得る天然の変異を除けば同一である)から得られた抗体を意味する。
【0043】
「アンタゴニスト」との用語は最も広い意味で用いられ、ここに開示される天然のトールレセプタの生物学的活性を部分的または完全にブロック、阻止、抑制または中和する分子を包含する。同様に、「アゴニスト」との用語は最も広い意味で用いられ、ここに開示される天然のトールレセプタの生物学的活性に類似するか、増強する分子を包含する。適当なアゴニストまたはアンタゴニスト分子は具体的にはアゴニストまたはアンタゴニスト抗体または抗体断片、断片または天然のトールレセプタポリペプチドのアミノ酸配列の変異体、ペプチド、小有機分子等を包含する。
【0044】
ここでの目的のための「活性のある」または「活性」とは、天然または天然に存在するPRO285、PRO286およびPRO358のそれぞれの生物学的および/または免疫学的活性を保持するPRO285、PRO286およびPRO358の形態を意味する。好ましい「活性」は、炎症サイトカインであるIL−1、IL−6およびIL−8のNFκΒおよび/またはNF−κΒ制御遺伝子の活性化を誘導する能力である。別の好ましい「活性」は脊椎動物で固有および/または適合性の免疫応答を活性化する能力である。さらに好ましい「活性」は微生物上に存在する保存された分子構造物の存在を感知する能力であり、具体的にはリポ多糖(LPS)シグナルを仲介する能力である。同じ「活性」の定義がPRO285、PRO286およびPRO358ポリペプチドのアゴニスト(例えば、アゴニスト抗体)に適用される。上記のように、PRO285、PRO286またはPRO358ポリペプチドのアンタゴニスト(アゴニスト抗体を含む)の活性はPRO285、PRO286またはPRO358ポリペプチドの上記同定の活性のいずれに対しても拮抗作用する能力、例えばそれら活性を部分的または完全にブロック、阻止、抑制または中和する能力と定義される。
【0045】
ハイブリダイゼーション反応の「ストリンジェンシー」とは当業者により容易に決定でき、通常、プローブの長さ、洗浄温度および塩濃度に依存する経験的な算定である。一般に、長いプローブは適当なアニーリングのために高温を必要とし、逆に短いプローブは低温を必要とする。相補鎖がそれらの融解温度以下の環境にある場合、ハイブリダイゼーションは通常、変性DNAが再アニールする能力に依存する。プローブと、ハイブリダイゼーションが可能な配列との所望の相同性の程度が高ければ高いほど使用できる相対的温度が高くなる。その結果、さらに高い相対的温度は反応条件をさらにストリンジェントにする傾向があり、他方で低温は厳格性を低くする傾向があるということになる。さらなる詳細とハイブリダイゼーション反応のストリンジェンシーの説明については、Ausubelら,Current Protocols in Molecular Biology(1995)を参照されたい。
【0046】
ここに定義される「ストリンジェントな条件」または「高ストリンジェントな条件」は、(1)洗浄のための低いイオン強度と高い温度、例えば0.015Mの塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で用いる;(2)ハイブリダイゼーション中にホルムアミド等の変性剤、例えば、0.1%ウシ血清アルブミン/0.1% Ficoll/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム(pH6.5)で、750mM塩化ナトリウム、75mMクエン酸を有する42℃の50%(v/v)ホルムアミドを用いる、(3)50%ホルムアミド、5×SSC(0.75M NaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×Denhardtの溶液、超音波処理サケ精子DNA(50μl/ml)、0.1%SDSおよび10%硫酸デキストランを42℃で用い、42℃での0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)および55Cの50%ホルムアミドでの洗浄およびその後の55℃でのEDTA含有0.1×SSCからなる高ストリンジェントな洗浄によって同定されよう。
【0047】
「適度にストリンジェントな条件」とは、Sambrookら,Molecular Cloning:A Laboratory Manual,New York:Cold Spring Harbor Press,1989)により記載されたように同定できるもので、上記の記載のものよりストリンジェンシーの低い洗浄溶液とハイブリダイゼーション条件(例えば、温度、イオン強度およびSDSの%)の使用を伴う。適度にストリンジェントな条件の具体例としては、20%ホルムアルデヒド、5×SSC(150mM NaCl、15mMクエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×Denhartの溶液、10%硫酸デキストラン、および20mg/mlの変性専断サケ精子DNAからなる溶液中で37℃の一晩のインキュベーション後に、ろ紙を約37〜50℃の1×SSCで洗浄することが挙げられる。当業者は、プローブの長さ等の因子を適合させるように、いかに温度、イオン強度等を必要にあわせて調節するかを分かるであろう。
ここで用いられる場合の「エピトープタグ」との用語は、FIZZポリペプチドを「タグポリペプチド」に融合させてなるキメラポリペプチドを意味する。タグポリペプチドは、抗体が作られうるエピトープを提供するのに十分な残基を有するが、それが融合するポリペプチドの活性に干渉しないように短い残基を有する。タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応しないようにかなり独自なものでなければならない。適するタグポリペプチドは、一般的に、少なくとも6アミノ酸残基、通常は約8〜50アミノ酸残基(好ましくは、約10〜20アミノ酸残基)を有する。
【0048】
ここで用いられる「免疫アドヘシン(immunoadhesin)」との用語は、異種タンパク質(「アドヘシン」)の結合特異性を免疫グロブリンの定常ドメインのエフェクター機能に組み合わせている抗体様分子を示す。構造的には、免疫アドヘシンは、抗体(すなわち「異種」のもの)の抗原認識結合部位ではない所望の結合特異性を有するアミノ酸配列と免系グロブリンの定常部ドメイン配列との融合からなる。免疫アドヘシン分子のアドヘシン部分は、典型的には、レセプタまたはリガンドの少なくとも結合部位からなる隣接アミノ酸配列である。免疫アドヘシンにおける免疫グロブリン定常ドメイン配列は、IgG−1、IgG−2、IgG−3またはIgG−4サブタイプ、IgA(IgA−1およびIgA−2等)、IgE、IgDまたはIgM等の免疫グロブリンから得てよい。
【0049】
「治療」とは、その目的が標的とする病理学的状態または疾患を予防または遅らせる(軽減)することにある場合、治療的処置と予防的または防止的対策の両方を意味する。治療を必要とする対象は該疾患を既に有する対象、ならびに該疾患を持ちそうな対象または該疾患を予防しようとする対象である。
「長期(chronic)」投与とは、初期の治療効果(活性)を長期間にわたって保持するように、急性の態様に対する連続的な形態での薬剤の投与を言う。
【0050】
治療目的の「哺乳類」とは、ヒト、家畜および農場動物、および動物園、スポーツまたはペット動物、例えばイヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ等の哺乳類として分類される動物を言う。好ましくは、哺乳類はヒトである。
一つ以上のさらなる治療薬「と組み合わせた」投与とは、同時投与および順番は問わない連続的な投与を含む。
【0051】
「リポ多糖」または「LPS」との用語は「内毒素(endotoxin)」と同義語として用いられている。リポ多糖(LPS)はグラム陰性細菌、例えば大腸菌の外膜の特徴的な成分である。それらは多糖部分とリピドAと呼ばれる脂肪からなる。多糖は細菌の種類により異なっており、(3〜8糖の繰返し単位から作られる)O特異鎖と二部分の核から作られる。リピドAは、事実上、リン酸により修飾された二つのグルコサミン糖と異なる数の脂肪酸を常に含む。さらなる情報に関しては、Rietschel and Brade,Scientific American August 1992,54−61を参照されたい。
【0052】
「敗血症性ショック」との用語はここでは最も広い意味で用いられ、Bone,Ann.Intern Med.114,332−333(1991)に開示されたすべての定義を含む。具体的には、敗血症性ショックは、敗血症と呼ばれる症候群である感染に対する全身応答で始まる。この症候群が低血圧と器官不全をもたらす場合、それは敗血症性ショックと呼ばれる。敗血症性ショックはグラム陽性菌および菌類、ならびに内毒素を有するグラム陰性菌により開始されるだろう。よって、ここでの定義は「内毒素ショック」に限定されない。
【0053】
II.発明の組成物と方法
A.全長PRO285、PRO286およびPRO358
本発明は、本出願でPRO285および8PRO286と称されるポリペプチドをコードする、新しく同定、単離された塩基配列を提供する。特に、出願人は下記の実施例でさらに詳細に開示されるPRO285およびPRO286ポリペプチドをコードするcDNAを同定し、単離した。BLASTおよびFastA配列アライメントコンピュータプログラムを用いて、出願人は、PRO285およびPRO286のコード配列が、GenBankデータベース中のDNA配列HSU88540_1、HSU88878_1、HSU88879_1、HSU88880_1およびHSU88881_1に対して高度に相同性のあることを発見した。
【0054】
さらに、本発明は、本出願でPRO358と称されるポリペプチドをコードする、新しく同定、単離された塩基配列を提供する。特に、出願人は下記の実施例でさらに詳細に開示されるヒトトールポリペプチド(PRO358)をコードするcDNAを同定し、単離した。BLASTおよびFastA配列アライメントコンピュータプログラムを用いて、出願人は、PRO358のコード配列が、GenBankデータベース中のDNA配列HSU88540_1、HSU88878_1、HSU88879_1、HSU88880_1、HS88881_1およびHSU79260_1に対して顕著な相同性を示すことを発見した。HSU79260_1を除けば、示されたタンパク質はヒトトール様レセプタと同定された。
【0055】
したがって、本出願で開示されたPRO285、PRO286およびPRO358タンパク質が、ショウジョウバエタンパク質トールの新しく同定されたヒト相同体であり、適合免疫において重要な役割を果たしていると現在考えられている。さらに具体的にはPRO285、PRO286およびPRO358は、炎症、敗血症性ショックおよび病原体に対する応答にかかわっていると思われ、例えば、糖尿病、ALS、ガン、リューマチ性関節炎および潰瘍等の免疫応答により悪化する多様な医学的な状態に適切な役割を果たしている。微生物上の保存された分子構築物の存在を感知する病原体パターン認識レセプタとしてのPRO285、PRO286およびPRO358の役割は、公知のヒトトール様レセプタであるTLR2がLPSシグナルの直接の仲介物質であることを示している本出願で開示されたデータによってさらに支持される。
【0056】
B.PRO285、PRO286およびPRO358変異体
ここに記載された全長の天然配列PRO285、PRO286およびPRO358に加えて、これらの配列の変異体を調製できることが意図される。PRO285、PRO286およびPRO358変異体は、適当なヌクレオチド変化をPRO285、PRO286またはPRO358DNAに導入することにより、または所望の変異体ポリペプチドの合成により調製することができる。当業者は、PRO285、PRO286またはPRO358ポリペプチドの翻訳後プロセスを、例えばグリコシル化部位の数や位置を変えることにより、または膜固定特性を変更する等により変更できることを理解しよう。
【0057】
天然の全長配列PRO285、PRO286およびPRO358、またはここに記載のPRO285、PRO286またはPRO358の多様なドメインにおける変異は、例えば、米国特許第5,364,934号に示された保存または非保存変異に関する技術とガイドラインを用いて行うことができる。変異は、対応する天然配列ポリペプチドと比較してアミノ酸配列の変化をもたらす、PRO285、PRO286またはPRO358ポリペプチドをコードする一つ以上のコドンの置換、欠失または挿入であってよい。任意に、変異は、少なくとも一つのアミノ酸を、PRO285、PRO286またはPRO358の一つ以上のドメインにおいて他のアミノ酸に置換することによる。どのアミノ酸が所望の活性に悪影響を与えることなく挿入、置換または欠失させてよいかを決定するガイダンスは、PRO285、PRO286またはPRO358の配列を相同性のある公知のタンパク質分子と比較し、高い相同性のある領域でなされたアミノ酸配列の変化の数を最小化することにより発見されよう。アミノ酸置換は、ロイシンのセリンによる置換等、あるアミノ酸の、類似の構造および/または化学的性質を有する別のアミノ酸による置換(すなわち保存アミノ酸の置換)の結果でありうる。挿入または欠失は任意に1〜5アミノ酸の範囲であってよい。認められる変異は、配列中のアミノ酸の挿入、欠失または置換を系統的に行い、得られた変異体の活性を下記の実施例に記載のインビトロアッセイにより調べることにより決定してよい。
【0058】
多様性はオリゴヌクレオチド仲介(部位特異的)変異、アラニンスキャンニングおよびPCR変異等の公知の方法を用いて行うことができる。部位特異的変異[Carterら,Nucl.Acids Res.,13:4331(1986);Zollerら,Nucl.Acids Res.,10:6487(1987)]、カセット変異[Wellsら,Gene,34:315(1985)]、制限選択変異[Wellsら,Philos,Trans.R.Soc.London SerA,317:415(1986)]または他の公知の技術をクローニングされたDNAに行ってPRO285またはPRO286変異体DNAを作ることができる。
【0059】
スキャンニングアミノ酸分析は隣接した配列に沿った一つ以上のアミノ酸を同定するために用いることもできる。比較的小さい中性アミノ酸が好ましいスキャンニングアミノ酸中にある。そのようなアミノ酸としてはアラニン、グリシン、セリンおよびシステインが挙げられる。アラニンはベータ炭素を超えて側鎖を除去し、変異体の主鎖立体配座を変えることが少ないのでこのグループの中でも好ましいスキャンニングアミノ酸である。アラニンは最も共通するアミノ酸であるために、これが典型的には望ましい。さらに、これは、埋め込まれた位置や露出した位置にしばしば見られる[Creighton,The Proteins(W.H.Freeman & Co.,N.Y.);Chothia,J.Mol.Biol.,150:1(1976)]。アラニン置換が十分な量の変異体を作らない場合、イソテリックなアミノ酸を用いることができる。
【0060】
ここに開示されるPRO285、PRO286およびPRO358トールタンパク質の変異体としては、トランスメンブレンドメインが欠失または不活性化されたタンパク質が挙げられる。トランスメンブレン領域は、細胞膜の脂質二層にわたる適当な大きさの高度に疎水性または親油性ドメインである。それらは天然の成熟PRO285、PRO286およびPRO358ポリペプチドを細胞膜に固定すると考えられている。PRO285では、トランスメンブレンドメインはおよそのアミノ酸の位置840からおよそのアミノ酸位置864まで伸びている。PRO286においては、トランスメンブレンドメインはおよそのアミノ酸位置826とおよそのアミノ酸位置848との間にある。PRO358では、トランスメンブレンドメインはおよそのアミノ酸位置576とアミノ酸位置595との間にある。
【0061】
トランスメンブレンドメインの欠失または置換は、PRO285、PRO286およびPRO358ポリペプチドの可溶形の回収を、その細胞または膜の脂質親和性を減少させ、その水溶性を向上させることにより容易にして提供する。トランスメンブレンおよび細胞質ドメインが欠失する場合、体により外来と認識される細胞内ペプチドをさらすか、または潜在的に免疫原性のある異種ポリペプチドの挿入により、潜在的に免疫原性のあるエピトープの導入を避ける。トランスメンブレンドメイン欠失PRO285、PRO286またはPRO358の主要な利点は、それが組換え宿主の培養培地に分泌されることである。この変異体は血液等の体液に可溶性であり、細胞膜脂質に対して容易に感知できる親和性を有さないために、組換え細胞培養物からのその回収を非常に容易にする。
【0062】
前記の考察から、置換、欠失、挿入またはそれらの組合せを導入して最終構築物に到達することは十分に明らかであろう。一般的な問題として、可溶性の変異体は機能的なトランスメンブレンドメインを有さず、好ましくは機能的な細胞質配列を有さないだろう。これは関連するドメインの欠失により通常行われるが、十分な挿入または置換による変異体もこの目的のために効果的である。例えば、トランスメンブレンドメインは、アミノ酸配列、例えば、ともに親水性の水治療法的な特徴を示す約5〜50個のセリン、スレオニン、リジン、アルギニン、グルタミン、アスパラギン酸等の親水性残基のランダムまたは一定の配列によって置換される。欠失(末端切除)PRO285、PRO286およびPRO358変異体のように、これらの変異体は組換え宿主の細胞培地中に分泌される。
【0063】
全長の成熟PRO285、PRO286およびPRO358ポリペプチドのさらなる欠失変異体(またはそれらのトランスメンブレンドメイン欠失〜不活性化形)としては、N末端シグナルペプチド(PRO285およびPRO286についてはアミノ酸1〜19、およびPRO358についてはアミノ酸1〜26として推定上同定されたもの)および/または開始メチオニンが欠失した変異体が挙げられる。天然のシグナル配列は、別のトール様タンパク質、または他のヒトまたは非ヒト(例えば、細菌、酵母または非ヒト哺乳類の)シグナル配列であってよい別の(異種)シグナルペプチドによって置換されてもよい。
【0064】
細胞内領域、および特にそのC末端部分がこれらのポリペプチドの生物学的機能に重要であると思われる。したがって、その目的が対応する天然のトール様タンパク質の生物学的活性を保持する変異体を作ることにあるのなら、これらの領域の少なくとも実質的な部分は保持されるか、またはその変更がある場合、該変更は保存アミノ酸の置換および/または挿入を伴うか、またはアミノ酸が挿入される領域に存在するアミノ酸に性質の類似したアミノ酸を伴うものとする。しかし、天然トールレセプタの生物学的機能の実質的な修飾が必要とされる場合(例えば、それぞれの天然トールポリペプチドのアンタゴニストを作ることが目的の場合)、その変更は、対応する天然のトールポリペプチドにおいて標的とされる位置のアミノ酸とは性質の異なるアミノ酸の置換および/または挿入を伴う。
【0065】
天然のアミノ酸は共通する側鎖の性質に基づいて下記グループに分類される:(1)疎水性:ノルロイシン、メチオニン、アラニン、バリン、ロイシン、イソロイシン;
(2)中性疎水性:システイン、セリン、スレオニン;
(3)酸性:アスパラギン酸、グルタミン酸;
(4)塩基性:アスパラギン、グルタミン、ヒスチジン、リジン、アルギニン;
(5)鎖の配向に影響を与える残基:グリシン、プロリン;および(6)芳香性:トリプトファン、チロシン、フェニルアラニン。
【0066】
保存的置換は一グループ内の一メンバーを同一グループ内の別のメンバーに交換することによるが、非保存的置換はこれらのクラスのうちの一クラスのメンバーを別のものに交換することを必要としよう。非保存的置換により得られる変異体は得られた変異体の生物学的性質/機能にさらに顕著な変化をもたらすことが期待される。
【0067】
アミノ酸挿入は、長さが1残基から百以上の残基を有するポリペプチドまでの範囲のアミノ末端および/またはカルボキシル末端の融合、ならびに単一または複数のアミノ酸残基の配列内挿入を包含する。配列内挿入(すなわち、PRO285、PRO286またはPRO358タンパク質アミノ酸配列内の挿入)は、通常約1〜10残基、より好ましくは1〜5残基、さらに好ましくは1〜3残基の範囲であってよい。末端挿入の具体例としては、N末端のメチオニル残基を有するPRO285、PRO286およびPRO358ポリペプチド、細菌組換え細胞培養物におけるその直接の発現の人工的産物および組換え宿主細胞に由来する成熟I−TRAFタンパク質の分泌を容易にするためにPRO285、PRO286またはPRO358分子のN末端への異種N末端シグナル配列の融合体が挙げられる。そのようなシグナル配列は通常は意図される宿主細胞種から得られるので、該宿主細胞種と同種であろう。適当な配列としては、大腸菌用のSTIIまたはIpp、酵母用のアルファ因子および哺乳類細胞用のヘルペスgD等のウイルスシグナルが挙げられる。
【0068】
ここに開示される天然トール様分子の他の挿入的変異体としては、免疫原性ポリペプチド、例えばベータ−ラクタマーゼ等の細菌ポリペプチドまたは大腸菌trp遺伝子座によりコードされる酵素、または酵母タンパク質に対する天然配列分子のN末端またはC末端の融合体、および1989年4月6日公開のWO89/02922に記載のように免疫グロブリン領域(好ましくは、免疫アドヘシンを生じる免疫グロブリンの定常部)、アルブミンまたはフェリチン等の長い半減期を有するタンパク質とのC末端融合体が挙げられる。免疫グロブリン融合体の生産に関しては、1995年6月27日発行の米国特許第5,428,130号を参照されたい。
【0069】
変異体トール様タンパク質の特性を前もって予想することはしばしば困難なので、スクリーニングは最適な変異体を選択する必要があることが理解されよう。この目的のために、以下に説明するような生化学的または他のスクリーニングアッセイが容易に利用可能となるだろう。
【0070】
C.PRO285、PRO286およびPRO385トールタンパク質の修飾
PRO285、PRO286およびPRO358ヒトトール相同体の共有結合による修飾が本発明の範囲に含まれる。一つの種類の共有修飾は、PRO285、PRO286およびPRO358タンパク質の標的アミノ酸残基を、選択された側鎖またはNまたはC末端残基と反応可能な有機誘導体化剤と反応させることによる。二官能剤による誘導体化が、例えば、PRO285、PRO286またはPRO358を抗PRO285、抗PRO286または抗PRO358抗体の精製法に用いられる非水溶性の支持体マトリックスまたは表面に対して架橋させるか、またはその逆にマトリックスまたは表面を該タンパク質に架橋させるために有用である。
【0071】
通常用いられる架橋剤としては、例えば1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステル、例えば、4−アジドサリチル酸とのエステル、ホモ二官能性イミドエステル、例えば3,3’−ジチオビス−(スクシンイミジルプロピオネート)等のジスクシンイミジルエステル、ビス−N−マレイミド−1,8−オクタン等の二官能性マレイミドおよびメチル−3−[(p−アジドフェニル)ジチオ]プロピオイミデート等の薬剤が挙げられる。
【0072】
他の修飾としては、グルタミニルおよびアルパラギニル残基の、それらの対応するグルタミルおよびアスパルチル残基へのそれぞれの脱アミノ化、プロリンおよびリジンのヒドロキシル化、セリルまたはスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニンおよびヒスチジン側鎖のα−アミノ基のメチル化[T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,San Francisco,pp.79−86(1983)]、N末端アミンのアセチル化、およびC末端カルボキシル基のアミド化が挙げられる。
【0073】
二官能性剤による誘導体化は、ここにおけるトール様レセプタとポリペプチドとの細胞内会合体の調製、ならびにアッセイまたはアフィニティー精製に使用される非水溶性支持体マトリックスまたは表面に対するこれらポリペプチドの架橋のために有用である。さらに、鎖間の架橋の研究は配座構造に関する直接の情報を提供しよう。通常用いられる架橋剤としては、1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステル、ホモ二官能性イミドエステルおよび二官能性マレイミドが挙げられる。メチル−3−[(p−アジドフェニル)ジチオ]プロピオイミデートは、光の存在下に架橋を形成できる光活性化可能な中間体を作る。もしくは、臭化シアン活性化炭水化物等の反応性非水溶性マトリックスおよび米国特許第3,959,642号、同第3,969,287号、同第3,691,016号、同第4,195,128号、同第4,247,642号、同第4,229,537号、同第4,055,635号および同第4,330,440号に記載されたシステム反応性の非水溶性基質がタンパク質固定化と架橋に用いられる。
【0074】
本発明の範囲に含まれるPRO285、PRO286およびPRO358ポリペプチドの別の種類の共有修飾はポリペプチドの天然グリコシル化パターンを変えることからなる。「天然グリコシル化パターンを変える」とは、ここでの目的のために、(内在するグリコシル化部位を除去するか、または化学的および/または酵素的手段によりグリコシル化を欠失させることにより)天然配列中に見られる一つ以上の炭水化物部分を欠失させ、および/または天然配列中には存在しない一つ以上のグリコシル化部位を付加させることを意味する。さらに、この語句は、存在する炭水化物の性質と比率の変化を伴う天然タンパク質のグリコシル化の定性的変化を包含する。
【0075】
(DNA40021によりコードされる)天然の全長PRO285は、アミノ酸位置66、69、167、202、215、361、413、488、523、534、590、679、720、799および942で潜在的なN結合グリコシル化部位を有する。(DNA42663によりコードされる)天然の全長PRO286は、アミノ酸位置29、42、80、88、115、160、247、285、293、358、362、395、416、443、511、546、582、590、640、680、752,937および1026で潜在的なN結合グリコシル化部位を有する。
【0076】
PRO285、PRO286およびPRO358ポリペプチドに対するグリコシル化部位の付加はアミノ酸配列を変更することによって行ってよい。その変更は例えば天然配列に対して一つ以上のセリンまたはスレオニン残基の付加または置換により行ってよい(O結合グリコシル化部位の場合)。アミノ酸配列は、特にPRO285、PRO286およびPRO358ポリペプチドをコードするDNAを、所望のアミノ酸に翻訳するコドンが作られるように前選択された塩基において変異させることによるDNAレベルでの変化により任意に変更してよい。
【0077】
PRO285、PRO286およびPRO358ポリペプチド上の炭水化物の数を増加させる別の手段はグルコシドのポリペプチドへの化学的または酵素的な結合による。そのような方法は、例えば1987年9月11日公告のWO87/05330およびAplinとWriston,CRC Crit.Rev.Biochem.,pp.259−306(1981)に記載されている。
【0078】
PRO285、PRO286およびPRO358ポリペプチド上に存在する炭水化物部分の除去は化学的または酵素的に、またはグリコシル化の標的として働くアミノ酸残基をコードするコドンの変異置換により行ってよい。化学的脱グリコシル技術は公知であり、例えばHakimuddinら,Arch.Biochem.Biophys.,259:52(1987)およびEdgeら,Anal.Biochem.,118:131(1981)により記載されている。ポリペプチド上の炭水化物部分の酵素的切断はThotakuraら,Meth.Enzymol.,138:350(1987)に記載されたように多様なエンド−およびエキソ−グリコシダーゼを用いて行うことができる。
【0079】
別の共有修飾は、PRO285、PRO286およびPRO358ポリペプチドを多様な非タンパク質性ポリマー、例えばポリエチレングリコール(PEG)、ポリプロピレングリコールまたはポリオキシアルキレンに米国特許第4,640,835号、同第4,406,689号、同第4,801,144号、同第4,670,417号、同第4,791,192号または同第4,179,337号に記載された方法で連結することからなる。
【0080】
本発明のPRO285、PRO286およびPRO358ポリペプチドは、PRO285、PRO286、PRO358またはその断片を別の異種ポリペプチドまたはアミノ酸配列に融合させてなるキメラ分子を形成する方法で修飾してもよい。一実施態様において、そのようなキメラ分子は、抗タグ抗体が選択的に結合することのできるエピトープを提供するタグポリペプチドに対するPRO285、PRO286またはPRO358ポリペプチドの融合体からなる。エピトープタグは通常は天然または変異体PRO285、PRO286またはPRO358分子のN末端またはC末端に置かれる。そのようなエピトープのタグ付加形の存在はタグポリペプチドに対する抗体を用いることにより検出することができる。また、エピトープタグの提供により、PRO285、PRO286またはPRO358ポリペプチドが、エピトープタグに結合する抗タグ抗体または別の種類のアフィニティーマトリックスを用いるアフィニティー精製により容易に精製できる。
【0081】
多様なタグポリペプチドおよびそれらの抗体は当業界で公知である。具体例としては、ポリ−ヒスチジン(poly−his)またはポリ−ヒスチジン−グリシン(poly−his−gly)タグ;カゼHAタグポリペプチドおよびその抗体12CA5[Fieldら,Mol.Cell.Biol.,8:2159−2165(1988)];c−mycタグおよびそれに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evanら,Molecular and Cellular Biology,5:3610−3616(1985)];および単純ヘルペスウイルス糖タンパク質D(gD)タグおよびその抗体[Paborskyら,Protein Engineering,3(6):547−553(1990)]が挙げられる。他のタグポリペプチドとしてはFlagペプチド[Hoppら,BioTechnology,6:1204−1210(1988)];KT3エピトープペプチド[Martinら,Science,255:192−194(1992)];α−チュブリンエピトープペプチド[Skinnerら,J.Biol.Chem.,266:15163−15166(1991)];およびT7遺伝子10タンパク質ペプチドタグ[Lutz−freyermuthら,Proc.Natl.Acad.Sci.USA,87:6393−6397(1990)]が挙げられる。
【0082】
さらなる実施態様において、キメラ分子は、PRO285、PRO286またはPRO358ポリペプチドまたはその断片と免疫グロブリンまたは免疫グロブリンの特定の領域との融合体からなってよい。二価形のキメラ分子のために、そのような融合体はIgG分子等のIgのFc領域に対する融合であってよい。Ig融合体は、好ましくはIg分子内の少なくとも一つの可変部の代わりとしてPRO285、PRO286またはPRO358ポリペプチドの可溶の(トランスメンブレンドメイン欠失または不活性化)ポリペプチドを置換させた物を包含する。免疫グロブリン融合体の製造については、1995年6月27日発行の米国特許第5,428,130号も参照されたい。
【0083】
D.PRO285、PRO286およびPRO358ポリペプチドの調製
以下の説明は、主に、これらのタンパク質をコードする核酸(例えば、DNA40021、DNA42663およびDNA47361のそれぞれ)を有するベクターを用いて形質転換またはトランスフェクトした細胞を培養することによるPRO285、PRO286およびPRO358トール相同体の生産に関する。勿論、当分野で当業界で公知である別法を用いてPRO285、PRO286、PRO358またはそれらの変異体を調製することも意図される。例えば、PRO285、PRO286またはPRO358配列、またはそれらの一部を、固相技術を用いる直接のペプチド合成により作ってもよい[例えば、Stewartら,Solid−Phase Peptide Synthesis,W.H.Freeman Co.,San Fransisco,CA(1969);Merrifield,J.Am.Chem.Soc.,85:2149−2154(1963)を参照されたい]。インビトロのタンパク質合成は手動による技術を用いて、または自動化により行ってもよい。自動化された合成は例えばApplied Biosystemsペプチド合成機(米国、カリフォルニア州、フォスター市)を製造業者の指示にしたがって用いて行ってもよい。PRO285、PRO286またはPRO358の多様な部分は個々に化学合成し、それらを化学的または酵素的合成法を用いて組み合わせて全長PRO285、PRO286またはPRO358を作ってもよい。
【0084】
1.PRO285、PRO286またはPRO358をコードするDNAの単離
PRO285、PRO286またはPRO358をコードするDNAはPRO285、PRO286またはPRO358mRNAを有し、それを検出可能なレベルで発現すると思われる組織から調製されたcDNAライブラリーから得てもよい。したがって、PRO285、PRO286またはPRO358DNAは、実施例に記載されるようにヒト組織から調製されたcDNAライブラリーから便利に得ることができる。内在する遺伝子は染色体ライブラリーから、またはオリゴヌクレオチド合成によって得てもよい。実施例に記載のライブラリーに加えて、本発明のヒトトールタンパク質をコードするDNAは例えば脾臓細胞または抹消血液白血球(PBL)から単離することができる。
【0085】
ライブラリーは、対象とする遺伝子またはそれによってコードされるタンパク質を同定するように設計されたプローブ(例えば、PRO285、PRO286またはPRO358のタンパク質または少なくとも約20〜80塩基のオリゴヌクレオチドに対する抗体等)を用いてスクリーニングすることができる。選択されたプローブによるcDNAまたは染色体ライブラリーのスクリーニングは、例えばSambrookら,Molecular Cloning:A Laboratory Manual(New York:Cold Spring Harbor Laboratory Press,1989)によって記載されたような標準的な手法を用いて行うことができる。PRO285、PRO286またはPRO358をコードする遺伝子を単離する別の手段はPCR法を用いることである[上記のSambrookら;Dieffenbachら,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press,1995)]。
【0086】
下記の実施例はcDNAライブラリーのスクリーニング技術を説明する。プローブとして選択されたオリゴヌクレオチド配列は、偽陽性を最小化するように十分な長さを有し、十分に明確なものでなければならない。オリゴヌクレオチドは、スクリーニングするライブラリー中のDNAとのハイブリダイゼーションの際に検出できるように好ましくは標識する。標識方法はよく知られており、32P標識ATP、ビオチニル化または酵素標識等の放射能標識の利用を包含する。適度なストリンジェンシーや高いストリンジェンシー等のハイブリダイゼーション条件は上記のSambrookらにより提供されている。
【0087】
そのようなライブラリースクリーニング法で同定される配列は、GenBank等の公共のデータベースまたは他の民間の配列データベースに寄託され利用可能な配列と比較し、アライメントさせることができる。分子の定められた領域内または全長配列に沿う(アミノ酸またはヌクレオチドレベルでの)配列の同一性は、相同性/配列同一性を測定するための多様なアルゴリズムを使用するALIGN、DNAstarおよびINHERIT等のコンピュータソフトウエアプログラムを用いる配列アラインメントにより決定することができる。
【0088】
タンパク質をコードする配列を有する核酸は、cDNAに逆転写されていないmRNAの前駆体またはプロセス中間体を検出するために、ここで最初に開示された類推アミノ酸配列を用いて、必要であれば上記のSambrookらに記載された慣用のプライマー伸長法を用いて、選択されたcDNAまたは染色体ライブラリーをスクリーニングすることによって得てもよい。
【0089】
2.宿主細胞の選択と形質転換
ヒトトールタンパク質の生産のためにここに記載された発現ベクターまたはクローニングベクターを用いて宿主細胞をトランスフェクトまたは形質転換し、プロモータを誘導し、形質転換体を選択し、かつ所望の配列をコードする遺伝子を増幅するために適切に改良された慣用の栄養培地で培養する。培地、温度、pH等の培養条件は当業者らにより過度な実験なしに選択することができる。通常、細胞培養物の生産性を最大化する原理、方法および実用的な技術は、Mammalian Cell Biotechnology:a Practical Approach,M.Butler,編(IRLプレス、1991)および上記のSambrookらに見ることができる。
【0090】
トランスフェクション法は当業者に知られており、例えばCaPO4およびエレクトロポレーションがある。使用される宿主細胞に基づいて、形質転換はそのような細胞に適する標準的な技術を用いて行う。上記のSambrookらにより記載された塩化カルシウムを用いるカルシウム処理、またはエレクトロポレーションが、実質的な細胞壁障壁を有する原核細胞または他の細胞に一般的に用いられる。Agrobacterium tumefaciensによる感染は、Shawら,Gene,23:315(1983)および1989年6月29日公開のWO89/05859に記載されているように、ある種の植物細胞の形質転換に用いられる。そのような細胞壁のない哺乳類の細胞に関しては、Grahamとvander編のリン酸カルシウム法,Virology,52:456−457(1978)を用いることができる。哺乳類細胞宿主システム形質転換の一般的な特徴は米国特許第4,399,216号に記載されている。酵母の形質転換は典型的にはVan Solingenら,J.Bact.,130:946(1977)およびHsiaoら,Proc.Natl.Acad.Sci.(USA),76:3829(1979)の方法にしたがって実施する。しかし、核マイクロインジェクション、エレクトロポレーション、そのままの細胞との細菌プロトプラスト融合、またはポリブレン、ポリオルニチン等のポリカチオン等によりDNAを細胞に導入する他の方法も用いてよい。哺乳類細胞の形質転換の多様な方法に関しては、Keownら,Methods in Enzymology,185:527−537(1990)およびMansourら,Nature,336:348−352(1988)を参照されたい。
【0091】
ここでのベクター中のDNAのクローニングまたは発現に適する宿主細胞として、原核細胞、酵母または高等真核細胞が挙げられる。適当な原核細胞としては、真正細菌、例えばグラム陰性、またはグラム陽性細菌、例えば大腸菌等のEnterobacteriaceaeが挙げられるが、これらに限定されない。大腸菌K12株MM294(ATCC 31,446);大腸菌X1776(ATCC 31,537);大腸菌株W3110(ATCC 27,325)およびK5 772(ATCC 53,635)等の多様な大腸菌株が一般に利用可能である。
【0092】
原核細胞に加えて、糸状菌または酵母等の真核微生物が、ヒトトールをコードするベクターの適当なクローニング宿主または発現宿主である。Sacccharomyces cerevisiaeが通常用いられる下等真核宿主微生物である。
【0093】
グリコシル化ヒトトールタンパク質の発現に適する宿主細胞は多細胞生物から得られる。無脊椎動物細胞の具体例としては、ショウジョウバエS2およびSpodotera Sf9等の昆虫細胞、ならびに植物細胞が挙げられる。有用な宿主細胞系としてはチャイニーズハムスター卵巣(CHO)およびCOS細胞が挙げられる。さらに具体的な例としては、SV40で形質転換されたサル腎臓CV1細胞系(COS−7、ATCC CRL 1651);ヒト胎児腎臓細胞系(懸濁培養物中での増殖のためにサブクローニングされた293または293細胞、Grahamら,J.Gen Virol.,36:59(1977);チャイニーズハムスター卵巣細胞/−DHFR(CHO,Urlaub and Chasin,Proc.Natl.Acad.Sci.USA,77:4216(1980);マウスセルトリー細胞(TM4,Mather,Biol.Reprod.,23:243−251(1980);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝臓細胞(HEPG2,HB 8065);およびマウスの乳せん腫瘍(MMT 060562,ATCC CCL51)が挙げられる。適当な宿主細胞の選択は当業者の技量内にあるであろう。
【0094】
3.複製可能なベクターの選択と使用
PRO285、PRO286またはPRO358をコードする核酸(例えばcDNAまたは染色体DNA)はクローニング(DNAの増幅)用または発現用の複製可能なベクターに挿入してよい。多様なベクターが一般に利用できる。例えば、ベクターはプラスミド、コスミド、ウイルス粒子またはファージの形であってよい。適当な核酸配列はベクターに多様な方法により挿入してよい。一般的に、DNAは公知の方法を用いて適当な制限エンドヌクレアーゼ部位に挿入させる。通常、ベクター成分としてはシグナル配列、複製開始部位、一種類以上のマーカー遺伝子、エンハンサエレメント、プロモータおよび転写終止配列が挙げられるが、これらに限定されない。これらの成分を一つ以上を有する適当なベクターの構築は当業者に公知の標準的な連結技術を用いる。
【0095】
PRO285、PRO286およびPRO358タンパク質は、組換えにより直接に作ってもよいのみならず、成熟タンパク質またはポリペプチドのN末端に特定の切断部位を有するシグナル配列または他のポリペプチドであってよい異種ポリペプチドとの融合ポリペプチドとして作ってもよい。通常、シグナル配列はベクターの一成分であってよく、またはベクターに挿入されたPRO285、PRO286またはPRO358DNAの一部であってよい。シグナル配列は、例えば、アルカリホスファターゼ、ペニシニリナーゼ、lpp、または熱安定エンテロトキシンIIリーダーの群から選択される原核生物シグナル配列であってよい。酵母での分泌のために、シグナル配列は、例えば酵母インベルターゼリーダー、アルファ因子リーダー(SaccharomycesとKluyveromycesのα因子リーダー等であり、後者は米国特許第5,010,182号に記載されている)、または酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー(1990年4月4日公開のEP362,179)、または1990年11月15日公開のWO90/13646に記載のシグナルであってよい。哺乳類細胞での発現において、タンパク質分泌を指令するため、同一または関連種の分泌ポリペプチドのシグナル配列等の哺乳類のシグナル配列ならびにウイルス分泌リーダー用いてよい。
【0096】
発現ベクターおよびクローニングベクターは両者ともに、一種類以上の選択された宿主細胞中でベクターの複製を可能とする核酸配列を有する。そのような配列は多様な細菌、酵母およびウイルスに関して当業界で公知である。プラスミドpBR322の複製開始部位はほとんどのグラム陰性細菌に適し、2μプラスミド開始部位は酵母に適し、多様なウイルス開始部位(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)は哺乳動物細胞中のクローニングベクターに有用である。
【0097】
発現ベクターとクローニングベクターは典型的には、選択マーカーとも呼ばれる選択遺伝子を有しよう。典型的な選択遺伝子は、(a)抗生物質または他の毒素、例えば、アンピシリン、ネオマイシン、メトトレキセートまたはテトラサイクリンに対する耐性を付与するか、(b)栄養要求性欠損を相補するか、または(c)複合培地からは利用できない重要な栄養素を供給するタンパク質をコードするもので、例えばBacilliのDアラニンラセマーゼをコードする遺伝子がある。
【0098】
哺乳類細胞の適当な選択マーカーの具体例は、PRO285、PRO286またはPRO358核酸を取り込む能力のある細胞の同定を可能とするもので、例えばDHFRまたはチミジンキナーゼである。野生型DHFRが用いられる場合の適当な宿主細胞は、Urlaubら,Proc.Natl.Acad.Sci.USA,77:4216(1980)により記載されているように調製され、増殖させたDHFR活性を欠損したCHO細胞系である。酵母に用いられる適当な選択遺伝子は酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら,Nature,282:39(1979);Kingsmanら,Gene,7:141(1979);Tschemerら,Gene,10:157(1980)]。該trp1遺伝子は、トリプトファンでの生育能を欠く酵母変異株、例えばATCC No.44076またはPEP4−1用の選択マーカーを提供する[Jones,Genetics,85:12(1977)]。
【0099】
発現ベクターおよびクローニングベクターは、通常は、mRNA合成を指令するために、PRO285、PRO286またはPRO358タンパク質をコードする核酸配列に操作可能に連結させたプロモータを有する。多様な潜在的宿主細胞に認識されるプロモータは当業界で公知である。原核生物宿主での使用に適するプロモータとしては、β−ラクタマーゼおよびラクトースプロモータシステム[Changら,Nature,275:615(1978);Goeddelら,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモータシステム[Goeddel,Nucleic Acids Res.,8:4057(1980);EP36,776]およびtacプロモータ等のハイブリッドプロモータ[deBoerら,Proc.Natl.Acad.Sci.USA,80:21−25(1983)]が挙げられる。細菌システムに使用されるプロモータもPRO285、PRO286またはPRO358をコードするDNAに操作可能に連結させたShine−Dalgarno(S.D.)配列を有しよう。
【0100】
酵母宿主に使用される適当なプロモータ配列の具体例としては、3−ホスホグリセレートキナーゼのプロモータ[Hitzemanら,J.Biol.Chem.,255:2073(1980)]または他の解糖系酵素[Hessら,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)]、例えばエノラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ヘキソキナーゼ、ピルベートデカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−ホスフェートイソメラーゼ、3−ホスホグリセレートムターゼ、ピルベートキナーゼ、トリオースホスフェートイソメラーゼ、ホスホグルコースイソメラーゼおよびグルコキナーゼが挙げられる。
【0101】
他の酵母プロモータは成長条件により制御を受ける転写の付加的な利点を有する誘導性プロモータであって、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸性ホスファターゼ、窒素代謝に関連した分解酵素、メタロチオネイン、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼおよびマルトースやガラクトースの利用に関わる酵素のプロモータ領域である。酵母発現に使用される適当なベクターとプロモータはEP73,657にさらに記載されている。
【0102】
哺乳類の宿主細胞中のベクターからのPRO285、PRO286またはPRO358転写は、例えばポリオーマウイルス、鶏痘ウイルス(1989年7月5日発行のUK2,211,504)、アデノウイルス(アデノウイルス2等)、ウシ乳頭腫ウイルス(SV40)、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびサルウイルス40(SV40)等のウイルスのゲノムから得られるプロモータ、異種哺乳類プロモータ、例えばアクチンプロモータまたは免疫グロブリンプロモータおよび熱ショックプロモータにより制御を受ける(但し、そのようなプロモータが宿主細胞系に適合性がある場合である)。
【0103】
PRO285、PRO286またはPRO358ポリペプチドをコードするDNAの高等真核生物による転写はヘンハンサー配列をベクターに挿入することにより高めてもよい。エンハンサは、通常約10〜300bpのDNAのシス作用エレメントであり、プロモータに作用してその転写を高める。哺乳類の(グロビン、エラスターゼ、アルブミン、α−フェトタンパク質およびインシュリン)遺伝子に由来する多くのエンハンサ配列が現在公知である。しかし、典型的には、真核細胞ウイルスのエンハンサが用いられよう。具体例としては、複製起源の後期側に位置するSV40(bp100〜270)、サイトメガロウイルス初期プロモータエンハンサ、複製開始部位の後期側に位置するポリオーマエンハンサおよびアデノウイルスエンハンサが挙げられる。エンハンサは、PRO285、PRO286またはPRO358をコードする配列に対して5’または3’の位置でベクター中にスプライスさせてよいが、好ましくはプロモータから5’の側に配置する。
【0104】
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、または他の多核生物に由来する有核細胞)に用いられる発現ベクターは転写の終止およびmRNAの安定化に必要な配列も有しよう。そのような配列は、通常は真核生物またはウイルスDNAまたはcDNAの5’および場合によっては3’の非翻訳領域から利用可能である。これらの領域はPRO285、PRO286またはPRO358をコードするmRNAの非翻訳部分のポリアデニル化断片として転写されるヌクレオチドセグメントを有する。
【0105】
組換え脊椎動物細胞培養物中でのPRO285、PRO286またはPRO358の合成に適合に適するさらに別の方法、ベクターおよび宿主細胞はGethingら,Nature,293:620−625(1981);Manteiら,Nature,281:40−46(1979);EP117,060;およびEP117,058に記載されている。
【0106】
4.遺伝子増幅の検出/発現
試料中の遺伝子増幅および/または発現は、ここに提供される配列に基づく適当に標識されたプローブを用いて、例えば、mRNAの転写を定量する慣用のサザンブロッティング、ノーザンブロッティング[Thomas,Proc.Natl.Acad Sci.USA,77:5201−5205(1980)]、ドットブロッティング(DNA合成)、またはin situハイブリダイゼーションにより直接測定してもよい。もしくは、特定の二本鎖、例えばDNA二本鎖、RNA二本鎖およびDNA−RNAハイブリッド二本鎖またはDNA−タンパク質二本鎖を認識できる抗体を用いてもよい。さらに、該抗体を標識してもよく、アッセイを行って二本鎖を表面に結合させ、該表面上での二本鎖の形成によって、二本鎖に結合した抗体の存在が検出できる。
【0107】
もしくは、遺伝子発現は、細胞または組織切片の免疫組織化学染色、細胞培養物または体液のアッセイ等の免疫学的方法により測定して、遺伝子産物の発現を直接に定量してもよい。試料液の免疫組織化学染色および/またはアッセイに有用な抗体はモノクローナルまたはポリクローナルのいずれでもよく、哺乳類中に作ってもよい。便利には、天然配列PRO285、PRO286またはPRO358ポリペプチドに対して、またはここに提供されたDNA配列に基づく合成ペプチドに対して、またはPRO285、PRO286またはPRO358DNAに融合させ、かつ特定の抗体エピトープをコードする外因性配列に対して調製してもよい。
【0108】
5.ポリペプチドの精製
PRO285、PRO286またはPRO358の形態は培養培地または宿主細胞溶解物から回収してよい。膜に結合している場合、適当な変性溶液(例えば、Triton−X100)または酵素による切断を用いて膜から放出させることができる。PRO285、PRO286またはPRO358の発現に用いられる細胞は、凍結/解凍の繰返し、超音波処理、機械的破壊または細胞溶解剤等の多様な物理的または化学的手段により破壊することができる。
【0109】
組換え細胞タンパク質またはポリペプチドからPRO285、PRO286またはPRO358を精製することも望ましいであろう。下記の方法は適当な精製方法の例示である:イオン交換カラムによる分画;エタノール沈殿;逆相HPLC;シリカまたはカチオン交換樹脂、例えばDEAEによるクロマトグラフィー;クロマトフォーカシング;SDS−PAGE;硫安沈殿;例えばSephadex G−75を用いるゲルろ過;IgG等の不純物を除去するためのプロテインAセファロースカラム;およびトールタンパク質のエピトープタグ付加の形態に結合する金属キレートカラム。タンパク質精製の多様な方法を用いてよく、そのような方法は公知であり、例えばDeutscher,Methods in Enzymology,182(1990)およびScopes,Protein Purification:Principles and Practics,Springer−Verlag,New York(1982)に記載されている。選択される精製方法は、例えば使用される精製法および生産される特定のトールタンパク質の性質に依存しよう。
【0110】
E.トールタンパク質に関する使用とコードする核酸
本発明のトールタンパク質をコードする塩基配列(またはそれらの相補物)は、ハイブリダイゼーションプローブとして、染色体および遺伝子地図作成において、およびアンチセンスRNAおよびDNAの生成における使用等、分子生物学の技術において多様な利用性を有する。トール核酸は、ここに記載の組換え技術によるPRO285、PRO286およびPRO358ポリペプチドの調製にも有用であろう。
【0111】
PRO285、PRO286およびPRO358をそれぞれコードする全長の天然配列DNA40021、DNA42663およびDNA47361遺伝子、またはそれらの部分は、cDNAライブラリーのハイブリダイゼーションプローブとして用いて、該全長遺伝子を単離するか、または図1、3および12A〜Bにそれぞれに開示されたPRO285、PRO286またはPRO358配列に対して所望の配列同一性を有するさらに別の遺伝子(例えば、PRO285、PRO286またはPRO358の天然変異体、またはそれらのさらなるヒト相同体、または他の種からの相同体をコードする遺伝子)を単離することができる。プローブの長さは任意に約20〜約50塩基である。ハイブリダイゼーションプローブは図2と3(配列番号2)、または図5から7(配列番号4)、または図21〜22(配列番号14)の塩基配列または天然配列のプロモータ、エンハンサエレメントおよびイントロン等のゲノム配列から得てもよい。例えば、スクリーニング法は、公知のDNA配列を用いるPRO285、またはPRO286、またはPRO358遺伝子のコード領域を単離して約40塩基の選択プローブを合成することからなるだろう。ハイブリダイゼーションプローブは、32Pまたは35S等のラジオヌクレオチド、またはアビジン/ビオチン結合システムによりプローブに結合させたアルカリホスファターゼ等の酵素標識等の多様な標識により標識してよい。本発明のPRO285、PRO286またはPRO358遺伝子(DNA40021、42663および47361)に相補的な配列を有する標識プローブを用いて、ヒトcDNA、染色体DNAまたはmRNAのライブラリーをスクリーニングして、該ライブラリーのどのメンバーに該プローブがハイブリダイズするかを決定する。ハイブリダイゼーション技術は下記の実施例でさらに詳細に説明する。
【0112】
該プローブをPCR技術に用いて密接に関連するトール配列の同定用の配列プールを作ってよい。
ここでトールタンパク質をコードするヌクレオチド配列を用いて、該トールタンパク質をコードする遺伝子の地図作成および遺伝病を有する個人の遺伝的分析のためにハイブリダイゼーションプローブを構築することもできる。ここで提供されるヌクレオチド配列は、公知の技術、例えばin situハイブリダイゼーション、公知の染色体マーカーに対する連結分析およびライブラリーに対するハイブリダイゼーションスクリーニングを用いて染色体および染色体の特別な領域に対して地図を作成してよい。
【0113】
本発明のヒトトールタンパク質はトール仲介シグナル変換に関与する他のタンパク質または分子を同定するアッセイにも用いることができる。例えば、PRO285、PRO286およびPRO358は、ヒトトールの現時点では未知の天然リガンド、またはキナーゼ関連の潜在的トールレセプタ等のヒトトールレセプタによる活性化またはシグナルに(直接または間接的に)関与する他の因子を同定するのに有用である。さらに、レセプタ/リガンド結合相互作用の阻害物質を同定できる。そのような結合相互作用に関与するタンパク質は、結合相互作用のペプチドまたは小分子の阻害物質またはアゴニストのスクリーニングに用いることもできる。スクリーニングアッセイを設計して、天然トールポリペプチドまたは天然のトールポリペプチドのリガンドの生物学的活性に似た主要な化合物を発見することができる。そのようなスクリーニングアッセイは、化学的ライブラリーの高処理量のスクリーニングが可能なアッセイを包含するために、アッセイは小分子医薬候補の同定に特に適する。意図される小分子は合成または無機化合物を包含する。これらのアッセイは、当分野でよく特徴付けられているタンパク質−タンパク質結合アッセイ、生化学スクリーニングアッセイ、免疫アッセイおよび細胞に基づくアッセイ等の多様な形態で実施することができる。
【0114】
インビトロアッセイはトールレセプタポリペプチド等の成分の混合物を用い、該ポリペプチドは別のペプチドまたはポリペプチド、例えば検出用または固定用タグ等との融合生成物の一部であってよい。アッセイ混合物は(結合アッセイのために)天然の細胞内または細胞外のトールが結合する標的(すなわち、トールリガンド、またはトールレセプタにより活性化し、および/またはシグナルを発することの公知の別の分子)をさらに有してもよい。天然の結合標的を用いてよいが、そうした天然の結合標的(例えばペプチド)部分がアッセイで便利に測定可能な対象トールタンパク質に対して結合親和性と親和力を提供する場合、そうした部分を用いることがしばしば望ましい。アッセイ混合物は候補薬理剤も含有する。候補試薬は多様な薬品の種類を包含するが、それらは典型的には有機化合物であり、好ましくは小有機化合物であり、合成または天然化合物のライブラリー等の多様なソースから得られる。塩、緩衝液、中性タンパク質、例えばアルブミン、変性剤、プロテアーゼインヒビター、ヌクレアーゼインヒビター、殺菌剤等の他の多様な試薬も該混合物に含有させてもよい。
【0115】
インビトロ結合アッセイにおいて、得られた混合物を、候補分子の存在を除けば、トールタンパク質が参照結合親和性により細胞結合標的、部分または相同体に特異的に結合する条件下にインキュベートする。該混合物成分は、必要な結合を提供するいかなる順番でも加えることができ、インキュベーションは最適な結合を容易にするいかなる温度で実施してもよい。インキュベーション時間は、最適な結合のために選択するが、迅速な高処理量のスクリーニングを容易にするように最小の時間とする。
【0116】
インキュベーション後、トールタンパク質と一つ以上の結合標的との試薬バイアス結合を慣用の技術により検出する。無細胞結合タイプのアッセイのために、結合物を未結合物から分離するために分離工程がよく用いられる。分離は沈殿(例えば、TCA沈殿、免疫沈殿等)、固定化(例えば固体基質への固定化)等の後に例えば膜によるろ過(例えばワットマンP−18イオン交換紙、Polyfitronicの疎水GFC膜等)、ゲルクロマトグラフィー(例えばゲルろ過、アフィニティー等)による洗浄によって行ってもよい。トール依存転写アッセイについては、結合をトール依存レポーターの発現の変化により検出する。
【0117】
検出は慣用の方法によって行えばよい。無細胞結合アッセイのために、通常、成分の一つは標識からなるか、また標識に結合させる。該標識は放射性、発光、光学的または電子密度等としての直接の検出、またはエピトープタグ、酵素等の間接的な検出を提供しよう。多様な方法が、標識の性質および他のアッセイ成分に基いて、例えば、光学的または電子密度、放射性放出、非放射性エネルギー転移等を用いて標識を検出してよく、または抗体複合物によって間接的に検出してもよい。
【0118】
PRO285、PRO286またはPRO358をコードする核酸、またはそれらの修飾形を用いてトランスジェニック動物または「ノックアウト」動物を作ることもでき、これらの動物は治療に有用な試薬の開発やスクリーニングに有用である。トランスジェニック動物(例えば、マウスまたはラット)はトランスジーンを含有する細胞を有する動物であり、該トランスジーンは該動物に導入されたか、または該動物の親等に胎児期で、例えば胚の段階で導入されたものである。トランスジーンはトランスジェニック動物を作った細胞のゲノムに組み込まれるDNAである。一実施態様において、確立された技術にしたがって、PRO285またはPRO286をコードするcDNAを用いてPRO285、PRO286またはPRO358をコードするゲノムDNAをクローニングでき、ゲノム配列を用いて、PRO285、PRO286またはPRO358をコードするDNAを発現する細胞を有するトランスジェニック動物を作ることができる。トランスジェニック動物、特にマウスまたはラット等の動物を作る方法は当分野では慣用となっており、例えば米国特許第4,736,866号と同第4,870,009号に記載されている。典型的には、特定の細胞が組織特異的エンハンサによるトランスジーン導入の標的となる。胚の段階に動物の生殖細胞系列に導入されたPRO285、PRO286またはPRO358をコードするトランスジーンのコピーを有するトランスジェニック動物を用いて、PRO285、PRO286またはPRO358をコードするDNAの増強した発現の効果を調べることができる。そのような動物は、例えばその過剰発現に関連した病理状態からの保護を付与すると考えられる試薬に関する試験動物として用いることができる。本発明のこの特徴にしたがって、動物を試薬で処置し、トランスジーンを有する未処置の動物と比較した場合の病理状態の低下率が該病理状態についての潜在的な治療的介在を示すものであろう。
【0119】
もしくは、PRO285またはPRO286またはPRO358の非ヒト脊椎動物(例えば哺乳類)の相同体を用いて、PRO285、PRO286またはPRO358タンパク質をコードする内在遺伝子と、該動物の胚細胞に導入されたPRO285、PRO286またはPRO358をコードする変更染色体遺伝子との均一組換えの結果として、PRO285またはPRO286またはPRO358をコードする、欠陥または変更遺伝子を有する「ノックアウト」動物を作ることができる。例えば、PRO285、PRO286またはPRO358をコードするcDNAを用いて、PRO285、PRO286またはPRO358をコードする染色体DNAを確立された技術にしたがってクローニングすることができる。PRO285、PRO286またはPRO358をコードする染色体DNAの一部を欠失させるか、または別の遺伝子、例えば統合をモニターするために用いることのできる選択マーカーをコードする遺伝子に置換させることができる。典型的には、(5’と3’の両末端において)数キロベースの未変更の隣接DNAをベクターに含有させる[例えば、均一組換えベクターの記載に関しては、例えばThomas and Chepecchi,Cell,51:503(1987)を参照されたい]。ベクターを(例えばエレクトロポレーションにより)胚幹細胞系に導入し、この導入されたDNAが内在DNAと均一組換えした細胞を選択する[例えば、Liら,Cell,69:915(1992)を参照されたい]。次に、選択された細胞を動物(例えばマウスまたはラット)の胚盤胞に注入して集合体キメラを形成する[例えば、Bradley,Teratocarcinomas and Embryonic Stem Cells:A Practical Approach,E.J.Robertson,編(IRL,Oxford,1987),pp.113−152を参照されたい]。キメラ胚を次に適当な疑妊娠メス親動物に移植し、胚を生育させて「ノックアウト」動物を創出する。均一組換えDNAを生殖細胞に有する子孫は標準的な技術をにより同定することができ、これを用いて、動物のすべての細胞が均一組換えDNAを有する動物を繁殖させる。ノックアウト動物は、例えば、ある種の病理状態に対する防御能やPRO285、PRO286またはPRO358ポリペプチドがないことによる病理状態の進展のために特徴付けることができる。
【0120】
ここに開示されたトールポリペプチドをコードする核酸は遺伝子治療に用いることもできる。遺伝子治療への利用において、遺伝子を細胞に導入して、例えば欠陥遺伝子の置換のために治療効果のある遺伝子産物のインビボ合成を行う。「遺伝子治療」とは、持続する効果が一回の治療により得られる通常の遺伝子治療と、治療効果のあるDNAまたはmRNAの一回または繰り返しの投与をともなう遺伝治療剤の投与との両方を含む。アンチセンスRNAおよびDNAはある種の遺伝子の発現をインビボでブロックする治療薬として用いることができる。短いアンチセンスオリゴヌクレオチドを細胞に移入させて、細胞内で細胞膜の限定された取込みにより引き起こされたそれらの低い細胞内濃度にもかかわらず阻害物質として作用させることができることが既に示されている(Zamecnikら,Proc.Natl.Acad.Sci.USA 83,4143−4146[1986])。これらのオリゴヌクレオチドは、例えばそれらの負電荷ホスホジエステルグループを未電荷のグループに置換することによりそれらの取込みを高めるように修飾することができる。
【0121】
核酸を生きた細胞に導入するのに利用することのできる多様な技術が存在する。これらの技術は、核酸を培養細胞にインビボで移すのか、または意図される宿主の細胞にインビボで移すのかにしたがって変えられる。核酸を哺乳類細胞にインビトロで移すのに適する技術としては、リポソームの利用、エレクトロポレーション、マイクロインジェクション、細胞融合、DEAE−デキストラン、リン酸カルシウム沈殿法等が挙げられる。現在好ましいインビボ遺伝子転移技術としては、ウイルス(典型的にはレトロウイルス)ベクターによるトランスフェクションとウイルスコートタンパク質−リポソーム仲介トランスフェクションとが挙げられる(Dzauら,Trends in Biotechnology 11,205−210[1993])。ある場合では、細胞表面膜タンパク質または標的細胞に対して特異性のある抗体、標的細胞上のレセプタに対するリガンド等、標的細胞を標的とする試薬を核酸ソースに提供することが望ましい。リポソームを使用する場合、エンドサイトーシスに関連する細胞表面膜タンパク質に結合するタンパク質は、標的化および/または例えば特定の細胞タイプに引き付けられるキャプシドタンパク質またはその断片、サイクリングで内在化を受けるタンパク質に対する抗体、細胞内局在化を標的とし、細胞内半減期を向上させるタンパク質の取り込みを容易するために用いてよい。レセプタ仲介エンドサイトーシスの技術は、例えば、Wuら,J.Biol.Chem.262,4429−4432(1987)およびWagnerら,Proc.Natl.Acad.Sci.USA 87,3410−3414(1990)によって記載されている。最近公知の遺伝子製造と遺伝子治療方法の概説については、Andersonら,Science 256,808−813(1992)を参照されたい。
トールタンパク質に関連してここで挙げた多様な利用は、天然トールレセプタの少なくとも一つの生物学的機能に似た、天然トールレセプタのアゴニストにも利用できる。
【0122】
F.抗トールタンパク質抗体
さらに、本発明は抗トールタンパク質抗体を提供する。例示的な抗体として、ポリクローナル、モノクローナル、ヒト化、二特異性およびヘテロ複合抗体が挙げられる。
【0123】
1.ポリクローナル抗体
抗トールタンパク質抗体はポリクローナル抗体からなってもよい。ポリクローナル抗体の調製方法は当業者で公知である。ポリクローナル抗体は、例えば免疫剤および所望であればアジュバントの一回以上の注射により哺乳類中に生じさせることができる。典型的には、免疫剤および/またはアジュバントを複数回の皮下または腹腔内注射により哺乳類に注射する。免疫剤はPRO285およびPRO286ポリペプチドまたはその融合タンパク質を含有してよい。免疫剤を、免疫される哺乳動物で免疫原性のあることが公知のタンパク質に複合させることが有用であろう。そのような免疫原性タンパク質の具体例としては、キーホールリンペットヘモシアニン、血清アルブミン、ウシチログロビンおよび大豆トリプシンインヒビターが挙げられるが、これらに限定されない。使用できるアジュバントの具体例としてはフロイントの完全アジュバントおよびMPL−TDMアジュバント(モノホスホリルリピドA、合成トレハロースジコリノミコレート)が挙げられる。免疫法は当業者によって過度な実験なしに選択できよう。
【0124】
2.モノクローナル抗体
もしくは、抗トールタンパク質抗体はモノクローナル抗体であってよい。モノクローナル抗体は、KohlerとMilstein,Nature,256:495(1975)によって記載されたようなハイブリドーマ法を用いて調製してよい。ハイブリドーマ法において、マウス、ハムスターまたは他の適当な宿主動物を典型的には免疫剤で免疫して、該免疫剤に特異的に結合する抗体を産生するか、もしくは産生することのできるリンパ球を誘引する。あるいは、リンパ球をインビトロで免疫しうる。
【0125】
免疫剤としては、典型的には、PRO285、PRO286またはPRO358ポリペプチドまたはそれらの融合タンパク質が挙げられる。一般的には、ヒト起源の細胞を望む場合、抹消血液リンパ球(「PBL」)を用い、または非ヒト哺乳類ソースを望む場合、脾臓細胞またはリンパ節細胞を用いる。次に、該リンパ球を、ポリエチレングリコール等の適当な融合試薬を用いて不死化細胞系に融合させてハイブリドーマ細胞を形成させる[Goding,Monoclonal Antibodies:Principles and Practice,Academic Press,(1986)pp.59−103]。不死化細胞系は通常は悪性化した哺乳類細胞、特にネズミ、ウシおよびヒト起源の骨髄腫細胞である。通常は、ラットもしくはネズミ骨髄腫細胞系を用いる。該ハイブリドーマ細胞を、好ましくは、未融合の不死化細胞の増殖や生存を妨げるために一種類以上の物質を含む適当な培養培地で培養してよい。例えば、親細胞がヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマの培養培地は典型的にはHGPRT欠損細胞の成長を妨げる物質であるヒポキサンチン、アミノプテリンおよびチミジン(「HAT培地」)を含有するものとなろう。
【0126】
好ましい不死化細胞系は、選択された抗体産生細胞による安定で高レベルの抗体発現を効率的に支持し、HAT培地等の培地に感受性のあるものである。さらに好ましい不死化細胞系は、例えば米国カリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerまたはメリーランド州ロックビルのAmerican Type Culture Collectionから入手することのできるネズミ骨髄腫細胞系である。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞系もヒトモノクローナル抗体の生産に関して記載されている[Kozbor,J.Immunol.,133:3001(1984);Brodeurら,Monoclonal Antibody Production Techniques and Applications,Marcel Dekker,Ink.,New York(1987)pp.51−63]。
【0127】
ハイブリドーマ細胞が培養される細胞培地は、次にPRO285、PRO286またはPRO385に対するモノクローナル抗体の存在についてアッセイすることができる。好ましくは、ハイブリドーマ細胞によって作られるモノクローナル抗体の結合特異性は、免疫沈殿またはラジオイムノアッセイ(RIA)または酵素結合免疫吸着法(ELISA)等のインビトロアッセイにより測定される。そうした技術やアッセイは公知である。モノクローナル抗体の結合親和性は、例えば、MunsonおよびPollardのSatchard分析、Anal.Biochem.,107:220(1980)によって測定することができる。
【0128】
所望のハイブリドーマ細胞が同定されたら、クローンを限界希釈法によりサブクローニングし、標準的な方法により増殖させる[上記のGoding]。この目的の培養培地としては、例えば、ダルベッコの改良イーグル培地およびRPMI1640培地が挙げられる。もしくは、ハイブリドーマ細胞を哺乳類中の腹水としてインビボで増殖させてもよい。
【0129】
サブクローンにより分泌されたモノクローナル抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィー等の慣用の免疫グロブリン精製方法により培養培地または腹水から単離し、精製してよい。
【0130】
モノクローナル抗体は、組換えDNA法、例えば米国特許第4,816,567号に記載された方法により作ってもよい。本発明のモノクローナル抗体をコードするDNAは慣用の方法を用いて(例えば、げっ歯類の抗体の重鎖と軽鎖をコードする遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを用いることにより)容易に単離して配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましいソースとして働く。いったん単離されたら、DNAを発現ベクターに入れてよく、次にそれ自体では免疫グロブリンタンパク質を作らないサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞または骨髄腫細胞に該ベクターをトランスフェクトして、組換え宿主細胞中でモノクローナル抗体の合成を得る。該DNAは例えばげっ歯類の相同配列の代わりにヒトの重鎖または軽鎖定常ドメインに置き換えることにより[米国特許弟4,816,567号;上記のMorrisonら]、または該免疫グロブリンをコードする配列に非免疫グロブリンポリペプチドのコード配列の全部または一部を共有結合させることにより修飾してもよい。そのような非免疫グロブリンポリペプチドは、本発明抗体の定常ドメインの代わりとするか、または本発明抗体の抗原結合部位の可変ドメインの代わりとしてキメラ二価抗体を作ることができる。
【0131】
抗体は一価抗体であってよい。一価抗体の調製方法は当業界で公知である。例えば、ある方法では免疫グロブリンの軽鎖と修飾重鎖の組換え発現をともなう。重鎖は、通常、重鎖の架橋を妨げるようにFc領域のいずれかの位置で端が切り取られている。もしくは、関連するシステイン残基を別のアミノ酸残基によって置換するか、または架橋を妨げるように欠失させる。
インビトロ方法も一価抗体の調製に適する。抗体を消化し、その断片、特にFab断片を作るのは日常的な公知の方法を用いて行うことができる。
【0132】
3.ヒト化およびヒト抗体
本発明の抗トール抗体はさらにヒト化抗体またはヒト抗体からなってよい。非ヒト(例えば、ネズミ)抗体のヒト化形態は、非ヒト免疫グロブリンに由来する最小の配列を有するキメラ免系グロブリン、その免疫グロブリン鎖またはその断片(例えば、抗体のFv、Fab、Fab’、F(ab’)2または他の抗原結合副配列)である。ヒト化抗体としては、受容者の相補性決定部位(CDR)の残基が、所望の特異性、親和性および能力を有するマウス、ラットまたはウサギ等の非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(受容者抗体)が挙げられる。ある場合では、ヒト免疫グロブリンのFv枠残基が対応する非ヒト残基で置換される。ヒト化抗体は、受容者抗体にも移入CDRにも枠配列にも見られない残基からなってもよい。一般的に、ヒト化抗体は、すべてまたは実質的にすべてのCDR領域が非ヒト免疫グロブリンのものと対応し、すべてまたは実質的にすべてのFR領域がヒト免疫グロブリン共通配列のものである少なくとも一つまたは二つの実質的にすべての可変ドメインからなるだろう。ヒト化抗体は最適には免疫グロブリン定常部(Fc)の少なくとも一部、典型的にはヒト免疫グロブリンの該領域からなってもよいであろう[Jonesら,Nature,321:522−525(1986);Reichmannら,Nature,332:323−329(1988);およびPresta,Curr.Op.Struct.Biol.,2:593−596(1992)]。
【0133】
非ヒト抗体をヒト化する方法は当業界で公知である。通常、ヒト化抗体は非ヒトであるソースからの一つ以上のアミノ酸残基を導入している。これらの非ヒトアミノ酸残基は、典型的には「輸入」可変ドメインから取られた「輸入」残基としばしば称される。ヒト化は、基本的には、Winterと共同研究者の方法にしたがって[Jonesら,Nature,321:522−525(1986);Riechmannら,Nature,332:323−327(1988);Verhoeyenら,Science,239:1534−1536(1988)]、げっ歯類のCDRまたはCDR配列をヒト抗体の対応する配列の代わりとすることにより行うことができる。したがって、そのような「ヒト化」抗体は、実質的にそのままのヒト可変ドメインが非ヒト種に由来する対応配列によって置換されたキメラ抗体である(米国特許第4,816,567号)。実際には、ヒト化抗体は典型的には、一部のCDR残基およびおそらく一部のFR残基がネズミ抗体の相同部位の残基によって置換されているヒト抗体である。
【0134】
ヒト抗体は、ファージディスプレイライブラリー等の公知の多様な技術を用いて作ることもできる[Hoogenboom and Winter,J.Mol.Biol.,227:381(1991);Marksら,J.Mol.Biol.,222:581(1991)]。ColeらおよびBoernerらの技術もヒトモノクローナル抗体の調製に利用できる[Coleら,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77(1985)and Boernerら,J.Immunol.,147(1):86−95(1991)]。同様に、ヒト抗体は、ヒト免疫グロブリン遺伝子座をトランスジェニック動物、例えば内在性免疫グロブリン遺伝子が部分的または完全に不活性化されたマウスに導入することにより作ることができる。誘発により、ヒト抗体の産生が観察され、これは遺伝子再配置、構築および抗体レパートリー等のすべての点においてヒトで見られるものと非常に似ている。このアプローチは、例えば米国特許第5,545,807号、同第5,545,806号、同第5,569,825号、同第5,625,126号、同第5,633,425号、同第5,661,016号;および以下の科学出版物:Marksら,Bio/Technology 10,779−783(1992);Lonbergら,Nature 868 856−859(1994);Morrison,Nature 368,812−13(1994);Fishwildら Nature Biotechnology 14,845−51(1996);Neuberger,Nature Biotechnology 14,826(1996);Lonberg and Huszar,Intern.Rev,Immunol.,13 65−93(1995)に記載されている。
【0135】
4.二特異性抗体
二特異性抗体は、少なくとも二つの異なる抗原に対する結合特異性を有するモノクローナルの、好ましくはヒトまたはヒト化抗体である。この場合、結合特異性の一つはPRO285、PRO286またはPRO358タンパク質に対するものであり、別のものは他の抗原に対するもので、好ましくは細胞表面タンパク質またはレセプタまたはレセプタサブユニットに対するものである。二つの異なるトール様タンパク質、例えば本出願で開示されたトール相同体のいずれか二つ、またはここに開示されたトールタンパク質、および公知のトールタンパク質、例えばTLR2に対して特異性を有する二特異性抗体を調製することも可能である。そのような二特異性抗体はトールレセプタによる異なる病原体パターンの認識をブロックすることができるために、敗血症または敗血症性ショックの治療に顕著な利点を有することが期待される。
【0136】
二特異性抗体の製造方法は公知である。伝統的には、二種類の重鎖が異なる特異性を有する二種類の免疫グロブリンの重鎖/軽鎖対の共発現に基づく[Milstein and Cuello,Nature,305:537−539(1983)]。免疫グロブリンの重鎖と軽鎖のランダムな集合のために、これらのハイブリドーマ(クアドローマ)は10種類の異なる抗体分子の潜在的な混合物を産生し、このうちわずかに一種類のみが正しい二特異性構造を有する。正しい分子の精製は、通常、アフィニティークロマトグラフィー工程により行われる。同様な方法が、1993年5月13日公開のWO93/08829およびTrauneckerら,EMBO J.,10:3655−3659(1991)に開示されている。
【0137】
所望の結合特異性(抗体−抗原結合部位)を有する抗体可変ドメインは免疫グロブリンの定常ドメイン配列に融合させることができる。この融合は、少なくともヒンジ部分のCH2とCH3領域からなる免疫グロブリンの重鎖定常ドメインに対する融合であるのが好ましい。該融合の少なくとも一つに存在する軽鎖結合に必要な部位を有する第一の重鎖定常部(CH1)を有することが好ましい。重鎖形態の免疫グロブリンをコードするDNAおよび所望であれば免疫グロブリン軽鎖を別々の発現ベクターに挿入し、適当な宿主生物にともにトランスフェクトさせる。二特異性抗体の作成のさらなる詳細については、例えばSureshら,Methods in Enzymology,121:210(1986)を参照されたい。
【0138】
5.ヘテロ複合抗体
ヘテロ複合抗体も本発明の範囲内にある。ヘテロ複合抗体は二つの共有結合した抗体からなる。例えば、そのような抗体は免疫系細胞の標的を不要な細胞に向けさせるために[米国特許第4,676,980号]、およびHIV感染の治療のために[WO91/00360;WO92/200373;EP03089]報告されている。該抗体は、合成タンパク質化学の公知の方法、例えば架橋剤をともなう方法を用いてインビトロで調製してよい。例えば、免疫毒素はジスルフィド交換反応を用いるか、またはチオエーテル結合を形成することにより構築してよい。この目的に適する試薬の具体例としてはイミノチオレートおよびメチル−4−メルカプトブチリミデートおよび米国特許第4,676,980号に開示のものが挙げられる。
【0139】
G.抗トールタンパク質抗体に関する使用
本発明の抗トール抗体は多様な有用性を有する。例えば、抗PRO285、抗PRO286、抗PRO358および抗TLR2抗体は、PRO285、PRO286、PRO358またはTLR2に関して、例えば特定細胞、組織または血清中でのその発現を検出する診断アッセイに用いることができる。不均一相または均一相のいずれかで行われる競合結合アッセイ、直接または間接サンドイッチアッセイおよび免疫沈殿アッセイ等の公知の多様な診断アッセイ技術を用いてよい[Zola,Monoclonal Antibodies:A Manual of Techniques,CRC Press,Inc.(1987)pp.147−158]。診断アッセイで用いられる抗体は検出可能な部分で標識してよい。検出可能な部分とは直接的または間接的に検出可能なシグナルを作ることのできるものでなければならない。例えば、検出可能な部分とは、放射性同位元素、例えば3H、14C、32P、35Sまたは125I、蛍光または化学発光化合物、例えばフルオレセインイソチオシアネート、ローダミンまたはルシフェリン、または酵素、例えばアルカリホスファターゼ、ベータ−ガラクトシダーゼまたは西洋ワサビパーオキシダーゼであってよい。Hunterら,Nature,144:945(1962)、Davidら,Biotechnology,13:1014(1974)、Painら,J.Immunol.Meth.40:219(1981)およびNygren,J.Histochem.and Cytochem.,30:407(1982)により記載された公知のいずれかの方法により抗体を該検出可能な部分に複合化させてよい。
【0140】
抗PRO285、抗PRO286、抗PRO358または抗TLR2抗体も組換え細胞培養物または天然のソースからのこれらタンパク質のアフィニティー精製に有用である。このプロセスにおいて、これらのTrollタンパク質に対する抗体を、Sephadex樹脂またはろ紙等の支持体に公知の方法を用いて固定化する。次に、この固定化抗体を、精製すべき該タンパク質を有する試料に接触させ、その後、固定化抗体に結合したPRO285、PRO286、PRO358またはTLR2タンパク質以外の試料中の実質的にすべての物質を除去する適当な溶媒で支持体を洗浄する。最後に、該タンパク質を抗体から解離させる別の適当な溶媒で支持体を洗浄する。
【0141】
抗トールレセプタ(すなわち、抗PRO285、抗PRO286、抗PRO358または抗TLR2抗体)は各トールレセプタの生物活性をブロックするのに有用でもあろう。トールレセプタファミリーの主要な機能は、微生物上に存在する保存された分子パターンの存在を察知する病原体パターン認識レセプタとして作用することであると考えられる。多様な細菌により作られる潜在的な致死をもたらす分子であるリポ多糖(LPS、内毒素としても知られる)は血液中でリポ多糖結合タンパク質(LBP)に結合する。この形成された複合体は次にCD14として知られるレセプタを活性化する。次に何が起るかについては未だにコンセンサスが得られていない。ある仮定によれば、CD14は、低分子量の前炎症仲介体の産生に関与するサイトカイン、細胞付着タンパク質および酵素を作るようにマクロファージを直接に指令するのではなく、むしろLPSに第二レセプタ活性化ができるようにする。もしくは、LPSはLBPまたはCD14からの助けなくしてある種のレセプタを直接に活性化することも示唆されている。本出願で開示されるデータは、ヒトトール様レセプタはLBPとCD14が応答するようにLPSによって活性化されるシグナルレセプタであることを示す。この機能は、病態生理学的条件下で敗血症性ショックと呼ばれるしばしば致命的な症候群を導きうるので、(他のトールレセプタンタゴニストとちょうど同じように)抗トールレセプタ抗体は敗血症性ショックの治療に有用であろう。異なるトールレセプタは異なる病原体、例えばグラム陰性またはグラム陽性細菌の多様な株を認識できると予想される。したがって、異なる状況では、異なるトールレセプタに特異的に結合する抗体混合物または二特異性抗トール抗体との併用治療が望ましいであろう。
【0142】
抗huTLR2抗体がこのレセプタのLPSによる誘導をブロックするのに特に有用であると考えられることがここで具体的に示されている。LPSに対する露出が敗血症性ショックを導き得ることが示されているように(Parrillo,N.Engl.J.Med.328,1471−1477[1993])、抗huTLR2抗体は敗血症性ショックの治療に潜在的に有用である。
抗トールレセプタ抗体と関連して挙げた上記の治療および診断利用も他のトールアンタゴニスト、すなわち、トールレセプタ活性化および/またはトールレセプタにより仲介されるシグナル変換をブロックする他の分子(タンパク質、ペプチド、小有機分子等)に対しても適用することができる。
【0143】
それらの治療潜在能力を鑑みて、トールタンパク質(天然のトール相同体の変異体を含む)およびそれらのアゴニストおよびアンタゴニスト(抗トール抗体を含むがこれに限定されない)を治療利用に適する組成物に配合する。治療組成物は、所望の純度を有する有効成分を任意の薬理学的に許容される担体、賦形剤または安定剤に混合し、凍結乾燥配合物または水性溶液の形で保存用として調製する(Remington’s Pharmaceutical Science 第16版,Osol,A.編 1980)。許容される担体、賦形剤または安定剤は、使用される用量と濃度では受容者(受容動物)に対して毒性がないものとし、リン酸塩、クエン酸塩および他の有機酸等の緩衝液;アスコルビン酸等の抗酸化剤;低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチンまたは免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性高分子、グリシン、グルタミン、アスパラギン、アルギニンまたはリジン等のアミノ酸;グルコース、マンノース、またはデキストリン等の単糖類、二糖類および他の炭水化物;EDTA等のキレート剤;マンニトールやソルビトール等の糖アルコール;ナトリウム等の塩形成対イオン;および/またはTween、PluronicsまたはPEG等の非イオン性界面活性剤が挙げられる。
【0144】
有効成分は、例えば、コアセルベーション技術または界面合成により調製された、例えばヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリレート)マイクロカプセル等のそれぞれのマイクロカプセル、またはコロイド医薬輸送システム(例えば、リポソーム、アルブミンミクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)に、またはマイクロエマルジョンに封じ込めてもよい。そのような技術は上記のRemingtons’s Pharmaceutical Sciencesに開示されている。
【0145】
インビボ投与に用いられる配合物は無菌でなければならない。これは、凍結乾燥および再構成の前後に滅菌ろ過膜を用いたろ過により容易になされる。
ここでの治療組成物は、滅菌アクセス孔を有する容器、例えば、皮下注射針で穴が開けられるストッパーを有する静脈内溶液バックまたはバイアルに入れる。
投与経路は、例えば注射または静脈内、腹腔内、脳内、筋肉内、眼内、動脈内または病変内の経路、局所投与、または徐放システム等の公知の方法にしたがう。
【0146】
徐放性調製物の適当な実例としては、形付けられた製品、例えばフィルムまたはマイクロカプセルの形にした半透性のポリマーマトリックスが挙げられる。徐放性マトリックスとしては、ポリエステル、ポリラクチド(米国特許3,773,919、EP58,481)、L−グルタミン酸とガンマLグルタミン酸エチルとの徐放性の共重合体(U.Sidmanら,Biopolymers 22(1):547−556[1993])、ポリ(2−ヒドロキシエチル−メタクリレート)(R.Langer,ら,J.Bimed.Mater.Res.15:167−277[1881]およびR.Langer,Chem.Tech.12:98−105[1982])、エチレンビニルアセテート(R.Langerら,Id.)またはポリ−D−(−)−3−ヒドロキシ酪酸(EP133,988)が挙げられる。徐放性の組成物はリポソームも包含する。本発明の範囲内の分子を含有するリポソームはそれ自体公知の方法で調製される[DE 3,218,121;Epsteinら,Proc.Natl.Acad.Sci.USA 82:3688−3692(1985);Hwangら,Proc.Natl.Acad.Sci.USA 77:4030−4034(1980);EP52322;EP36676A;EP88046;EP143949;EP142641;日本国特許出願83−118008;米国特許4,485,045および4,544,545;およびEP102,324]。通常、リポソームは、脂質含量が約30mol%コレステロールを超える小さい(約200〜800オングストローム)単層タイプであり、選択される比率は最適なNT−4治療に適合させる。
【0147】
有効成分の効果的な量は、例えば治療の目的、投与経路および患者の状態に依存しよう。したがって、治療者は、最適な治療効果を得るために必要とされる投与量を評価し、投与経路を修正する必要がある。典型的な一日あたりの投与量は、上記のファクターに依存しながら約1μg/kg〜100mg/kgまたはそれ以上である。典型的には、臨床医は必要とされる生物学的効果を提供する投与量に達するまで本発明の分子を投与しよう。この治療の進展は慣用のアッセイにより容易にモニターされる。
【0148】
以下の実施例は、例示の目的のみで提示されるものであり、本発明の範囲を制限する意図は全くない。
本発明の明細書で引用されるすべての特許および文献の参照は、その全てが参照としてここに取り込まれる。
【実施例】
【0149】
別記した以外は、実施例で参照される市販の試薬は、製造者の指示に従って用いた。以下の実施例中、および明細書を通しての、ATCC受託番号により示される起源は、アメリカンタイプカルチャーコレクション、Rockville、Marylandである。
【0150】
[実施例1]
ヒトPRO285をコードするcDNAクローンの単離
私有の発現配列タグ(EST)DNAデータベース(LIFESEQ(商標)、Incyte Pharmaceuticals,Palo Alto,CA)を検索し、ショウジョウバエ トールたんぱく質に相同性を示すEST(#2243209)を同定した。 ESTに基づき、PCRプライマーのペア(フォワードおよびリバース)
TAAAGACCCAGCTGTGACCG(配列番号5)
ATCCATGAGCCTCTGATGGG(配列番号6)
と、プローブ
ATTTATGTCTCGAGGAAAGGGACTGGTTACCAGGGCAGCCAGTTC(配列番号7)
を合成した。
【0151】
cDNAライブラリーの構築のためのmRNAは、ヒト胎盤組織より単離した。cDNAクローンを単離するために用いられたcDNAライブラリーは、Invitrogen、サンディエゴ、CA(Fast Track 2)からの試薬などの市販の試薬を用いた標準的な方法により構築された。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそサイズを合わせ、定められた方向でクローニングベクターpCR2.1(Invtrigen,Inc.)内に、Life Technologies,Gaithersburg,MD(Super Script Plasmid System)からの試薬とプロトコールを用いてクローン化した。二重鎖cDNAは、1000bpを超えるサイズに合わせ、そのcDNAをBamHI/NotI開裂ベクターにクローン化した。pCR2.1は市販されているプラスミドであり、PCRフラグメントのクローン化を容易にするようにデザインされており、選択のためにAmpRとKanR遺伝子、青白選択のためにLacZを有している。
【0152】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO285遺伝子をコードするクローンの単離のために用いた。
【0153】
cDNAクローンはその全てを配列決定した。DNA40021(PRO285をコードしている)の全ヌクレオチド配列を、図2と3(配列番号2)に示す。クローンDNA40021は、ヌクレオチド位置61-63に明らかな翻訳開始部位を有するシングルオペロンリーディングフレームを含む(図2と3)。予想されるポリペプチド前駆体は、1049アミノ酸の長さであり、推定シグナルペプチドをアミノ酸位置1-29に、推定膜間ドメインをアミノ酸位置837-860の間に、およびロイシンジッパーパターンをアミノ酸位置132-153と704-725に、各々含む。なお、示された境界はおおよそのものであり、示された領域の実際の限界は、数アミノ酸の差で異なっているかもしれない。クローンDNA40021は、ATCCに寄託され(表示DNA40021-1154)、ATCC寄託番号ATCC209389が付与されている。
【0154】
全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0155】
[実施例2]
ヒトPRO286をコードするcDNAクローンの単離
所有されている発現配列タグ(EST)DNAデータベース(LIFESEQ(商標)、Incyte Pharmaceutical,Palo Alto,CA)を検索し、ショウジョウバエ トールたんぱく質に相同性を示すEST(#694401)を同定した。 ESTに基づき、PCRプライマーのペア(フォワードおよびリバース)
GCCGAGACAAAAACGTTCTCC(配列番号8)
CATCCATGTTCTCATCCATTAGCC(配列番号9)
と、プローブ
TCGACAACCTCATGCAGAGCATCAACCAAAGCAAGAAAACAGTATT(配列番号10)
を合成した。
【0156】
cDNAライブラリーの構築のためのmRNAは、ヒト胎盤組織より単離した。
このRNAは、Life Technologies,Gaithersburg,MD(Super Script Plasmid System)からの試薬とプロトコールを用いて、ベクターpRK5D中にオリゴdTプライム化cDNAを生成させるために用いた。pRK5Dはsp6転写開始部位、それに続くSfiI制源酵素部位、さらにそれに続くXhoI/NotI cDNAクローニング部位を有するクローニングベクターである。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそ1000bpを超えるサイズを合わせ、定められた方向でXhoI/NotI開裂pRK5Dにクローン化した。
【0157】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO286遺伝子をコードするクローンの単離のために用いた。
【0158】
cDNAクローンはその全てを配列決定した。DNA42663(PRO286をコードしている)の全ヌクレオチド配列を、図5から7(配列番号4)に示す。クローンDNA42663は、ヌクレオチド位置57-59に明らかな翻訳開始部位を有するシングルオペロンリーディングフレームを含む(図5から7)。予想されるポリペプチド前駆体は、1041アミノ酸の長さであり、推定シグナルペプチドをアミノ酸位置1-26に、推定膜間ドメインをアミノ酸位置826-848に、およびロイシンジッパーパターンをアミノ酸位置130-151、206-227、662-684、669-690、および693-614に、各々含む。なお、示された境界はおおよそのものであり、示された領域の実際の限界は、数アミノ酸の差で異なっているかもしれない。
クローンDNA42663は、ATCCに寄託され(表示DNA42663-1154)、ATCC寄託番号ATCC209386が付与されている。
【0159】
PRO286の全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0160】
[実施例3]
ヒトトール受容体ファミリーの既知のメンバーからの細胞外ドメイン(ECD)配列(存在する場合、分泌シグナル配列を含む)をESTデータベースの検索に用いた。ESTデータベースはパブリックデータべース(例えば、GenBank)、および所有されているデータベース(例えばLIFESEQ(商標)、Incyte Pharmaceuticals、Palo Alto、CA)を含んでいた。検索は、コンピュータープログラムBLASTまたはBLAST2(Altschul、およびGish、Methods in Enzymology 266:460-80(1996))を用いて、EST配列の6フレーム翻訳に対するECDたんぱく質配列の比較として行なわれた。既知のたんぱく質をコードしていない70(あるいは、あるケースでは90)以上のBlastスコアとのそれらの比較を、プログラム”pharap”を用いて、コンセンサスDNA配列に集めて組み立てた。(Phil Green、ワシントン大学、シアトル、ワシントン)。
【0161】
ESTは、Incyteデータベースで同定した(INC3115949)。
そのEST配列に基づき、PCRにより対象の配列を含むcDNAライブラリーを同定し、PRO358の全長コード配列のクローンを単離するためのプローブとして使用するために、オリゴヌクレオチドを合成した。
【0162】
PCRプライマーのペア(フォワードおよびリバース)を合成した:TCCCACCAGGTATCATAAACTGAA(配列番号15) TTATAGACAATCTGTTCTCATCAGAGA(配列番号16)
プローブも合成した:
AAAAAGCATACTTGGAATGGCCCAAGGATAGGTGTAAATG(配列番号17)
【0163】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO358遺伝子をコードするクローンの単離のために用いた。
【0164】
cDNAライブラリーの構築のためのRNAは、ヒト骨髄組織より単離した(LIB256)。cDNAクローンを単離するために用いられたcDNAライブラリーは、Invitrogen、サンディエゴ、CAからの試薬などの市販の試薬を用いた標準的な方法により構築された。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそサイズを合わせ、定められた方向で適当なクローニングベクター(例えばpRKBまたはpRKD;pRK5Bは、SiI部位を含まないpRK5Dの前駆体である;Holmesら、Science,253:1278-1280(1991))内にユニークなXhoIとNotI部位にクローン化した。
【0165】
上述の単離されたクローンのDNA配列決定は、PRO358の全長DNA配列(図21および22、配列番号14)とPRO358の誘導たんぱく質配列(図18および19、配列番号13)を与えた。
同定されたクローン(DNA47361)の全ヌクレオチド配列を、図21〜22(配列番号14)に示す。クローンDNA47361は、図21と22の下線のヌクレオチド位置に明らかな翻訳開始部位(ATG開始シグナル)を有するシングルオペロンリーディングフレームを含む(図2と3)。予想されるポリペプチド前駆体は、811アミノ酸の長さであり、推定シグナルペプチド(アミノ酸1-19)、細胞外ドメイン(アミノ酸20-575、位置55から位置575までの領域のロイシンリッチ反復を含む)、推定膜間ドメイン(アミノ酸576-595)を含む。クローンDNA47361は、ATCCに寄託され(表示DNA47361-1249)、ATCC寄託番号ATCC209431が付与されている。
【0166】
PRO286の全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0167】
[実施例4]
PRO285、PRO286およびPRO358DNAの、ハイブリダイゼーションプローブとしての使用。
以下の方法は、PRO285、PRO286またはPRO358ポリペプチドをコードするヌクレオチド配列の、ハイブリダイゼーションプローブとしての使用を記載している。
トール相同体のコード配列を含むDNAは、ヒト組織cDNAライブラリー又はヒト組織ゲノムライブラリーにおける相同性DNA(例えばこれらの特定のトールたんぱく質の天然由来の変異体をコードするもの)をスクリーニングするためのプローブとして用いることができる。
【0168】
どちらかのライブラリーDNAを含むフィルターのハイブリダイゼーションと洗浄は、以下の高ストリンジェントな条件下で行われる。放射性標識したトール相同体由来プローブの、フィルターへのハイブリダイゼーションは、50%ホルムアミド、5xSSC、0.1%SDS、0.1%ピロリン酸ナトリウム、50mMリン酸ナトリウム、pH 6.8、2xDenhardt溶液、及び10%硫酸デキストランの溶液内で、42°C、20時間行う。このフィルターの洗浄は、0.1xSSC及び0.1%SDSの水溶液内で42°Cで行う。
【0169】
全長の未変性配列トール相同体をコードするDNAについて所望の配列相同性を有するDNAは、次いで、当該技術分野で知られる標準的な技術により同定することができる。
【0170】
[実施例5]
大腸菌(E.coli)内でのPRO285、PRO286およびPRO358の発現
この実施例は、大腸菌内での組換発現による、PRO285、PRO286またはPRO358(”トール相同体”)の非グリコシル化形態での調製を例示するものである。
トール相同体をコードするDNA配列をまず、選択したPCRプリマーを使用して増幅させる。このプライマーは、選択した発現ベクター上の制限酵素部位に対応する制限酵素部位を含む必要がある。種々の発現ベクターを使用することができる。適切なベクターの例としては、pBR322(大腸菌由来;Bolivarら、Gene, 2:95(1977)を参照)であって、アンピシリン及びテトラサイクリンの耐性遺伝子を含むものである。ベクターは、制限酵素で消化して脱リン酸化する。PCRで増幅した配列を次いでベクターへ連結する。ベクターは好ましくは抗生物質耐性遺伝子、trpプロモーター、ポリhisリーダー(最初の6つのSTIIコドン、ポリhis配列、及びエンテロキナーゼ開裂部位を含む)、PRO285コード領域、ラムダ転写終了符号、及びargU遺伝子をコードする配列を含むであろう。
【0171】
次いで連結混合物を使用して、選択した大腸菌株へ形質転換するが、これにはSambrookら(上記)に記載の方法を使用する。形質転換体は、LBプレート上での増殖能力により同定し、抗生物質耐性のコロニーを選択する。プラスミドDNAは、制限分析及びDNA配列決定により同定及び確認することができる。
【0172】
選択したクローンを、抗生物質を補ったLBブロス等の液体培地中で一晩増殖させることもできる。この一晩培養したものを次いで、より大スケールの培養用の接種用に使用することができる。細胞を次いで所望の吸光度まで増殖させ、その間に、発現プロモーターをオンにする。
【0173】
細胞を更に数時間培養した後、細胞を遠心により回収する。遠心して得られた細胞の沈殿物を、当該技術分野で知られる種々の薬剤を使用して可溶化し、可溶化したトール相同体を次いで、金属キレート性カラムを蛋白質との強固な結合を許容する条件下で使用して精製する。
【0174】
[実施例6]
ほ乳類細胞におけるPRO285、PRO286およびPRO358の発現
本実施例は、ほ乳類細胞内での組換え発現による、PRO285、PRO286およびPRO358(“トール相同体”)のグリコシル化された形態での調製を例示するものである。
【0175】
pRK5ベクター(1989年3月15日公開のEP 307,247を参照)を発現ベクターとして使用する。適宜、Sambrookら(上記)に記載されるものなどの連結法を利用して、選択した制限酵素を使用してトール相同体をコードするDNAを挿入して、トール相同体をコードするDNAをpRK5に連結する。得られるベクターは、場合により、pRK5−PRO285、PRO286、またはPRO358と呼ぶ。
【0176】
1つの態様においては、293細胞を宿主細胞として選択することができる。ヒトの293細胞(ATCC CCL1573)は、ウシ胎児血清及び適宜栄養成分及び/又は抗生物質で補ったDMEM等の培養液で、組織培養プレート内で周密的になるまで増殖させた。約10μgのpRK5−PRO285、PRO286、またはPRO358DNAを、VA RNA遺伝子(Thimmappayaら, Cell, 31:543 (1982))をコードする約1μgのDNAと混合し、500μlの1mMトリス−HCl、0.1mM EDTA、0.227M CaCl2に溶解した。この混合物に対して、500μlの50mM HEPES(pH7.35)、280mM NaCl、1.5mM NaPO4、を滴加し、25°Cで10分間、沈殿を形成させる。この沈殿物を再懸濁し、293細胞へ添加し、約4時間、37°Cで静置する。培養液を吸引除去し、2mlのPBS中20%グリセロールを30秒間添加する。この293細胞を次いで無血清培地で洗浄し、新鮮な培地を添加して細胞を約5日間インキュベーションする。
【0177】
形質移入の約24時間後、培養培地を除去して培養培地(のみ)で置換するか、又は200μCi/mlの35S−システイン及び200μCi/mlの35Sメチオニンを含有する培養培地で置換する。12時間のインキュベーション後、馴化培地を回収し、スピンフィルターで濃縮し、15%SDS−ゲルにかける。処理したゲルは、乾燥して設定した時間、フィルムを露出してPRO285ポリペプチドの存在を明らかにすることができる。形質移入細胞を含む培養物はさらに、(無血清培地で)インキュベーションして、この培地を選択したバイオアッセイで試験することもできる。
【0178】
代替技術において、トール相同体DNAは、Somparyracら(Proc. Natl. Acad. Sci., 12:7575 (1981))の記載する硫酸デキストラン法を使用して、293細胞内に一過性導入することができる。293細胞を、スピナーフラスコ内で最大濃度まで増殖させ、700μgのpRK5−PRO(285)/(286)/(358)DNAを添加する。細胞をまず、スピナーフラスコから遠心により濃縮して、PBSで洗浄する。DNA−デキストラン沈殿物を細胞のペレット上で4時間インキュベーションする。細胞を20%のグリセロールで90秒間処理し、組織培養培地で洗浄し、細胞培地、5μg/mlウシインシュリン、及び0.1μg/mlのウシトランスフェリンを含有するスピナーフラスコに再導入する。約4日後、馴化培地を遠心して細胞及びその破砕片を除去する。対応する発現トール相同体を含む試料は、次いで、例えば透析及び/又はカラムクロマトグラフィー等の選択した方法により濃縮・精製することができる。
【0179】
別の態様において、トール相同体は、CHO細胞内で発現することもできる。pRK5ベクターを、例えばCaPO4またはDEAEデキストラン等の既知の試薬を使用してCHO細胞へ形質移入することができる。上記したように、細胞培養物をインキュベーションして、培地を培養培地(のみ)、又は35S−メチオニン等の放射性標識を含む培地で置換する。PRO285、PRO286、またはPRO358ポリペプチドの存在を確認した後、培地を無血清培地で置換する。好ましくは、培養物は約6時間インキュベーションし、次いで馴化培地を回収する。発現したトール相同体を含む培地を次いで、何らかの選択した方法を選択して濃縮・精製することができる。
【0180】
エピトープタグ化トール相同体もまた、CHOの宿主細胞内で発現することができる。このトール相同体DNAは、pRK5ベクターからサブクローニングすることもできる。このサブクローンインサートは、PCRにより選択したエピトープ、例えばポリ−hisのタグとフレームを合わせ、バキュロウイルス発現ベクターへ融合させることができる。このポリ−hisタグ化挿入物は次いで、DHFR等の、安定クローンの選択用の選択標識を含む、SV40駆動性ベクターへサブクローニングすることができる。最後に、このCHO細胞をSV40駆動性のベクターで(上記したようにして)形質移入することができる。上記したように、発現を変化させるために標識を行なっても良い。発現したポリ−Hisタグ化トール相同体を含む培養培地は次いで、例えばNi2+-キレートアフィニティクロマトグラフィー等の何らかの選択した方法により濃縮・精製することができる。
【0181】
[実施例7]
酵母中でのPRO285、PRO286およびPRO358の発現
以下の方法は、酵母内での、PRO285、PRO286またはPRO358(”トール相同体”)の組換発現を記載したものである。
まず、酵母発現ベクターを、ADH2/GAPDHプロモーターから、トール相同体を細胞内生産又は分泌させるために構築する。所望のトール相同体をコードするDNA、選択したシグナルペプチド、及びプロモーターを選択したプラスミド中の適切な制限酵素部位に挿入して、細胞内発現をさせる。分泌用には、選択したトール相同体をコードするDNAを選択したプラスミドへ、ADH2/GAPDHプロモーター、酵母アルファ因子分泌シグナル/リーダー配列、及び(必要ならば)発現用のリンカー配列とともにクローニングすることができる。
【0182】
例えば酵母株AB110等の酵母細胞は、上記したような発現プラスミドで形質転換し、選択した培養培地で培養することができる。形質転換した酵母の上清は、10%トリクロロ酢酸で沈殿してSDS−PAGEによる分離、それに引き続くゲルのクマシーブルー染料での染色により分析することができる。
【0183】
組換体トール相同体は引き続いて、酵母細胞を培養液より遠心で除去し、選択したカートリッジフィルターで濃縮することにより単離・精製することができる。トール相同体を含有する濃縮物は、選択したカラムクロマトグラフィーの樹脂を用いて更に精製することができる。
【0184】
[実施例8]
バキュロウイルス感染昆虫細胞内におけるPRO285、PRO286およびPRO358の発現
以下の方法は、バキュロウイルス感染昆虫細胞でのPRO285、PRO286またはPRO358(”トール相同体”)の組換発現を記載している。
トール相同体コード配列を、バキュロウイルス発現ベクターで含まれるエピトープタグの上流に融合させる。このようなエイトープタグには、ポリ−hisタグや(IgGのFc領域のような)免疫グロブリンタグがある。種々のプラスミドを使用することができるが、これには商業的に入手可能なプラスミド、例えばpVL1393(Novagen)などに由来するものがある。簡潔にいうと、トール相同体コード配列またはコード配列の所望の部分(例えば細胞外ドメインをコードする配列等)を5’及び3’領域に相補的なプライマーでPCRにより増幅させる。5’プライマーは、フランキング(選択した)制限酵素部位を含むことができる。その産物を次いで、それらの選択した制限酵素で消化し、発現ベクター中へサブクローニングする。
【0185】
組換バキュロウイルスは、上記のプラスミドとBaculoGold(商標)ウイルスDNA(Pharmingen)とをSpodoptera frugipedra(“Sf9”)細胞(ATCC CRL1711)へ、リポフェクチン(GIBCO−BRLより商業的に入手可能)を使用して共形質転換して作り出される。28°Cで4乃至5日間インキュベーションした後、放出されたウイルスを回収して更なる増殖に使用する。ウイルス感染及び蛋白質発現は、O'Reilleyら(Baculovirus expression vectors: A laboratory Manual, Oxford: Oxford University Press (1994))に記載されるようにして行う。
【0186】
発現したポリ−hisタグ化トール相同体は次いで、例えばNi2+−キレートアフィニティクロマトグラフィーで、以下のようにして精製することができる。抽出物は組換ウイルスに感染したSf9細胞より、Rupertら、Nature, 362:175-179 (1993)により記載されたようにして調製する。簡潔にいうと、Sf9細胞を洗浄し、超音波処理緩衝液(25mL Hepes pH7.9、12.5mM MgCl2、0.1mM EDTA、10%グリセロール、0.1%NP−40、0.4M KCl)中に再懸濁し、氷上で20秒間、2回、超音波処理する。超音波処理物を遠心して、その上清を充填用緩衝液(50mMリン酸塩、300mM NaCl、10%グリセロール、pH7.8)で50倍に希釈し、0.45μmのフィルターに通す。Ni2+−NTAアガロースカラム(Qiagenより商業的に入手可能)を5mLのベッド容量で調製し、25mLの水で洗浄し、25mLの充填用緩衝液で平衡化する。濾過済の細胞抽出物を毎分0.5mLでカラムにのせる。カラムは、 A280がベースラインになるまで充填用緩衝液で洗浄し、その点から画分の回収を開始する。次に、カラムを二次洗浄緩衝液(50mMリン酸塩、300mM NaCl、10%グリセロール、pH6.0)で洗浄するが、これにより非特異的に結合した蛋白質が溶出する。再びA280のベースラインになった後、カラムを二次洗浄緩衝液中の0乃至500mMのイミダゾール勾配で展開する。1mLの画分を回収してSDS−PAGE及び銀染色又はアルカリホスファターゼにコンジュゲートしたNi2+−NTA(Qiagen)を利用したウェスタンブロッティングで分析する。溶出したHis10のタグ化PRO285をプールし、充填用緩衝液に対して透析する。
【0187】
或いは、IgGタグ化(又はFcタグ化)トール相同体の精製は、例えばプロテインAやプロテインGカラムクロマトグラフィーを含む、既知のクロマトグラフィー技術を利用して行うこともできる。
【0188】
[実施例9]
NF-κBアッセイ NF-κB経路を介したトールたんぱく質シグナルとして、それらの生理活性は、NF-κBアッセイでテストすることができる。このアッセイにおいて、Jurkat細胞は、製造者の指示に従って、Lipofectamine試薬(Gibco BRL)を用いて一過性形質導入される。NF-κB駆動ルシフェラーゼ遺伝子を含む、1μgのpB2XLucプラスミドを、PRO285またはPRO286をコードする挿入を有するあるいは有していない、1μgのpSRαN発現ベクターを用いて構築する。陽性対照として、細胞はPMA(酢酸フォルボールミリスチル;20ng/ml)とPHA(フィトヘマグルチニン(phytohaemaglutinin))を用いて、3から4時間処理される。細胞は、Promegaからの試薬を用いてルシフェラーゼ活性の測定のために2から3日後に溶解させる。
【0189】
[実施例10]
PRO285、PRO286またはPRO358に結合する抗体の調製
この実施例は、PRO285、PRO286またはPRO358(”トール相同体”)に特異的に結合することができるモノクローナル抗体の調製を例証する。
モノクローナル抗体を生産する技術は、当該技術分野において周知であり、例えば、Goding(上掲)に記載されている。用いられる免疫原は、精製されたトール相同体、トール相同体を含む融合タンパク、および細胞表面に組み換えトール相同体を発現するトール相同体を含む。免疫原の選択は、過度の実験を要することなく当業者によって実施されることができる。
【0190】
Balb/c等のマウスを、完全Freund'sアジュバント中に乳化されたトール相同体免疫原で免疫して、1−100マイクログラムの量で皮下または腹膜内に注射する。あるいは、この免疫原はMPL-TDMアジュバント(Ribi Immunochemical Research, Hamilton, MT)中に乳化され、動物の後ろ足に接種する。免疫されたマウスは、選択されたアジュバントに乳化されたさらなる免疫原を用いて10ないし12日後にブースト(boost)される。その後、数週間、マウスは、さらなる免疫注射でブーストされてもよい。血清サンプルは、PRO285抗体を検出するためにELISAアッセイで試験するための後眼窩採血(retro-orbital bleeding)によりマウスから定期的に得ることができる。
【0191】
適切な抗体価が検出された後、抗体に“陽性”の動物は、トール相同体の最終的な静脈注射により注射されてもよい。3−4日後、このマウスがと殺され、脾臓細胞が回収される。脾臓細胞は、ATCC、No.CRL1597から入手できるP3X63AgU.1のような選択されたマウスミエローマセルラインに融合(35%ポリエチレングリコールを用いて)される。融合体はハイブリドーマ細胞を生じ、非融合細胞、ミエローマハイブリッド、および脾臓細胞ハイブリッドの増殖を阻害するためにHAT(ヒポキサンチン、アミノプテリン、およびチミジン)培地を含む96ウェル組織培養プレートにプレートされ得る。
【0192】
ハイブリドーマ細胞は、対応するトール相同体に対する反応性についてELISAでスクリーニングされる。トール相同体に対する所望のモノクローナル抗体を分泌する“陽性”ハイブリドーマ細胞の決定は、当業者の技術範囲である。
陽性ハイブリドーマ細胞は、抗トール相同体モノクローナル抗体を含む腹水を産生するために同系のBalb/cマウスに腹膜内接種されてもよい。あるいは、このハイブリドーマ細胞は、組織培養フラスコまたはローラーボトルで生育することもできる。腹水中に産生されたモノクローナル抗体の精製は、硫安沈殿、ゲル排除クロマトグラフィーを用いて実施することができる。あるいは、プロテインAまたはプロテインGに対する抗体の結合に基づくアフィニティークロマトグラフィーを用いることができる。
【0193】
[実施例11]
HuTLR2は、リポ多糖(LPS)誘導細胞シグナルの媒体となる。
方法試薬 [3H]標識化、非標識化、LCD25およびS.minnesotaR595LPSは、List Biochemicals(キャンベル、CA)から、および他のすべてのLPSはSigma Chemical Co.(セントルイス、MO)から得た。LPは、ヒトLBP発現ベクターでトランスフェクトされた293細胞からの条件付培地として供給された。TLR2-Fc融合たんぱく質は、バキュロウィルス系で生産し、上述したように精製した。Markら,J.Biol.Chem 269,10720-10728(1994)。
【0194】
発現プラスミドの構築 ヒトTLR2をコードするcDNAは、ヒト胎児肺ライブラリーよりクローン化した。予測されるアミノ酸配列は、アミノ酸726がgluからaspに置換されているのを除いて、以前発表された配列(Rockら、上述)のそれと一致した。TLR2(dG.TLR2)のアミノ酸末端エピト−プタグバージョンを、位置17のロイシン(TLR2の予想される成熟型の最初のアミノ酸)のすぐ上流にXhoI制現部位を加え、これを上述したように単純ヘルペスウィルスタイプ1の糖たんぱく質Dのアミノ酸1-53に結合することにより構築した。Markら、上述。PCR産物を配列決定し、プロマイシン耐性遺伝子を含む哺乳類の発現ベクターにサブクローン化した。gD.TLR2のC末端トランケーション(truncation)変異体は、細胞内ドメインのコード配列に存在するBlpI(変異体△1)またはNsiI(変異体△2)部位での、および発現ベクターの3’ポリリンカーに存在するNotI部位でのcDNAの消化と、その後のオリゴヌクレオチドリンカーの結さつにより構築した。
【0195】
Δ1:5'-TCA GCG GTA AGC-3'(配列番号18)および 5'-GGC CGC TTA CCG C-3'(配列番号19)
Δ2:5'-TAA GCT TAA CG-3'(配列番号20)および 5'-GGC CGC TTA AGC TTA TGC A-3'(配列番号21)
【0196】
CD4/TLR2キメラはPCRにより構築し、CD4とTLR2配列の連結部にリンカーでコードされたバリンを有して、ヒトTLR2のアミノ酸588-784(膜間および細胞内ドメイン)に融合した、ヒトCD4のアミノ酸1-205(シグナルペプチドと2つの免疫グロブリン様ドメイン)を含んでいた。pGL3.ELAM.tkレポータープラスミドは、ルシフェラーゼレポータープラスミドpGL3(Promega)のSacIとHindIII部位の間に挿入された、以下の配列5'-GGT ACC TCC TGA CAT CAT TGT AAT TTT AAG CAT CGT GGA TAT TCC CGG GAA AGT TTT TGG ATG CCA TTG GGG ATT TCC TCT TTA GAT CTG GCG CGG TCC CAG GTCCAC TTC GCA TAT TAA GGT GAC GCG TGT GGC CTC GAA CAC CGA GCG ACC CTG CAGCGA CCC GCA AGC TT-3'(配列番号22)
を含んでいた。
【0197】
LBPのC末端エピト-プタグバージョン(LBP-FLAG)をPCRにより、ネイティブなストップコドンに変えてAsc1部位の付加とこのフラグメントのpRK5-FLAGへのサブクローニングによって構築し、アミノ酸GRADYKDDDDK(配列番号23)のC末端付加を得た。
【0198】
安定なセルライン/プール 293のヒト胚芽腎細胞を、10%FBS、2mMグルタミンおよびペニシリン/ストレプトマイシンが添加されたLGDMEM/HEM's F12(50:50)培地中で増殖させた。gD.TLR2の安定な発現のために、細胞をgD.TLR2発現ベクターで形質導入し、最終濃度1μg/mlのプロマイシン耐性で選択した。細胞の安定なプール(293-TLR2 クローン1)は、上述のMarkらに記載されたように、FACSとウェスタンブロット分析により特徴付けられた。
【0199】
ルシフェラーゼレポーターアッセイおよび電気泳動移動度シフトアッセイ(EMSA)
29332の親または安定な細胞(ウェル当たり2×105細胞)を6ウェルプレート中にシードし、後日、0.5μgのルシフェラーゼレポータープラスミドpGL3-ELAM.tkと内部コントロールとして0.05μgのRenillaルシフェラーゼレポート化ベクターと共に発現プラスミドで形質導入した。24時間後、細胞はLPS、LBP、またはLPSとLBPの両方で処理し、レポーター遺伝子活性を測定した。データを、蛍光ルシフェラーゼ活性をRenilla Luciferaseのそれで除した相対ルシフェラーゼ活性として示す。EMSAとしては、核抽出物を調製し、コンセンサスNF-κB結合部位を含む5’-[32P]-放射標識化オリゴヌクレオチドとのDNA-結合反応に用いた(Santa Cruz Biotechnology,sc-2511)。複合体中のNF-κBの同定は、NF-κBに対する抗体によるスーパーシフトにより確かめた(データは示さず)。
【0200】
RNA発現
組織ノザンブロットは、Clontechより購入し、TLR2の細胞外ドメインを囲むプローブにハイブリダイズさせた。ポリアデニル化mRNAを293細胞または293−TLR2細胞より単離し、ノザンブロットをヒトIL-8 cDNAフラグメントでプローブさせた。TLR2発現を、定量的PCRで、実質的に に記載されているリアルタイム”taqman(商標)”技術を用いて測定し、モデル770配列検出器(Applied Biosystems, Foster City, CA, USA)で、実質的に(Luohら、J.Mol.Endocrinol.18,77―85[1997])に記載されているように分析した。
【0201】
フォワードとリバースプライマー5'-GCG GGA AGG ATT TTG GGT AA-3'(配列番号24)
5'-GAT CCC AAC TAG ACA AAG ACT GGT C-3'(配列番号25)
を、レポーター染色FAMで5’ヌクレオチドを、消光(quenching)染色TAMRAで3’ヌクレオチドを標識したハイブリダイゼーションプローブ5'-TGA GAG CTG CGA TAA AGT CCT AGG TTC CCA TAT-3'(配列番号26)
と共に用いた。
マクロファージ/単球を1μg/mlのLPSで16時間処理した。
【0202】
レセプター結合アッセイ 直接の結合を測定するために、20ngの[3H]-LPSを、15μlのプロテインAセファロースを含む100μlの結合バッファー(150mM NaCl,20mM Hepes,0.03% BSA)中の600ngのTLR2―Fcと混合した。室温での3時間のインキュベーションの後、プロテインAセファロースサンプルは、冷PBS/0.1% NP-40で2回洗浄し、1%SDSと25mM EDTAを含む結合バッファー中に再懸濁し、カウントした。
【0203】
結果
ショウジョウバエで、トール受容体は、胚芽の背腹方向の(dorso-vental)パターン形成に必要であり、成ハエ(adult fly)での抗真菌免疫反応にも関与する。BelvinおよびAnderson,Ann.Rev.Cell.Biol.12,393-416(1996);Lemaitreら、Cell 86,973-983(1996)。トールは、多重ロイシンリッチ反復(LRRs)を有する細胞外ドメインと、インターロイキン1受容体(IL-1R)に配列相同性を有するサイトプラスマドメインを含むタイプIの膜間たんぱく質であり、いくつかの植物病耐性たんぱく質である。トールの活性化は、NF-κB経路の活性化を通して遺伝子の誘導を引き起こす。前述の通り、クローン化されたいくつかのヒト相同体があり、それらのいくつかは、本出願中に新規たんぱく質として開示されている。これらのヒトたんぱく質は、それらのショウジョウバエ複製(counterpart)の局所的構造を反映している。1つのヒトTLR(TLR4)の構成的活性変異体の過剰発現が、NF-κBの活性化と、炎症性サイトカインとコンスティミュラトリー(constimulatory)分子の誘導をもたらすことが示されている(Medzhitovら、およびRockら、上述)。
【0204】
ヒトTLRがLPS-誘導細胞活性化に関与しているかもしれないことを調べるために、我々は最初に、TLRsの発現を種々の免疫組織で調べた。TLRsの1つであるTLR2は、調べた全てのリンパ様の組織で発現されることが見出され、最高の発現は末梢の血液白血球であった(図8a)。TLR2の発現は、単球/マクロファージ、主なCD14−発現およびLPS反応性細胞で、富化される。興味深いことに、tLR2は、LPS(図8b)での単離された単球/マクロファージの刺激に関して、CD14で報告されたように(Matsuuraら、Eur.J.Immunol.22,1663-1665[1992];CrostonJ.Biol.Chem.270,16514-16517[1995])、上方制御(up-regulated)される。
【0205】
この結果は、TLR2がLPS−仲介細胞シグナルに関与しているかどうかを測定することを我々に促した。我々は、アミノ末端エピトープタグを含むTLR2(gD−TLR2)のバージョンを発現するヒト胎芽腎293細胞を作製した。個々のクローンと同様に、クローンの安定したプールを単離し、約105kDaの新規たんぱく質が発現することを示し(図10)、TLR2(〜89kDa)の予想サイズとN−結合グリコシルかの4つの可能性のある部位の存在に一致した。我々は、E−セレクチン遺伝子のNF−κB応答性エンハンサにより駆動される報告された遺伝子の発現を測定することにより、293または293−TLR2細胞とLBPの応答を、調べた(Clostonら、上述)。LPSでもLBPでも単独処理では、大きな遺伝子活性化がおこらないが、LPSとLBPの両方の添加は、TLR2を発現する細胞内でレポーター遺伝子活性の実質的な誘導を起こすが、対照の293細胞では起こらない(図9)。さらに、電気泳動移動度シフトアッセイ(EMSA)を用いて、我々はLPSが、LBPと組み合わさって、TLR2発現細胞でNK-κB活性を誘導することを見出した(図11)。293細胞でのLPS-誘導NF-κB活性のキネテッィクスは、ミエロイドと非ミエロイド細胞のそれと類似しており(Deludeら、J.Biol.Chem.269,22253-22260[1994];Leeら,Proc.Natl.Acad.Sci.USA,90,9930-9934[1993])、NF-κBの核の位置は、LPSに曝した後、30分間以内で最大である。シクロヘキシミド(図11)またはアクチノマイシンD(示さず)による前処理はNF-κB活性化を阻害しないために、293TLR2細胞中のLPS/LBPによるNF-κBの活性化は、de novoたんぱく質合成を必要としない。
【0206】
ミエロイド細胞に存在するCD14の膜結合型(mCD14)と、プラスマに存在する可溶性CD14(sCD14)(Bazilら、Eur.J.Immunol.16,1583-1589[1986])は、LPSに対する細胞の応答性を増進させることが示された。我々は293細胞がそれらの表面でほとんどあるは全くCD14を発現しないことを観察した(データは示さず)。しかしながら、293細胞の一過性の形質導入は、mCD14がTLR2-媒介LPS応答性の感受性と規模を増加した(図12)。
【0207】
上に示されたデータは、TLR2が、LPSのためのシグナルトランスデューサーとして機能するかもしれないことを示唆した。LPS反応の媒介におけるTLR2の細胞内ドメインの役割を調べるために、我々は、13(TLR-△1)または141アミノ酸(TLR2-△2)のC-末端トランケーション(truncation)を有するTLR2変異体が、一過性の形質導入293細胞でELAMレポーターを制御できるかどうか測定した。我々は、FACS分析により(示さず)およびウェスタンブロットにより(図13c)、細胞表面でこれらのレポーターの発現を検出できたが、両方のC末端のトランケーション変異体が、レポーター遺伝子の活性化を欠損したことを観察した。TLR2‐△1で欠失した細胞内ドメインの領域は、IL-1R-結合キナーゼIRAK(Crostonら、上述)との会合に要求されるIL-1R細胞内ドメインの領域と著しい類似性を有している(図13b)。我々はまた、TLR2の細胞外ドメイン(ECD)が、TLR2のECDがCD4のECDの一部に置換されたTLR2変異体もまた応答できないLPS-応答性に要求されることを示した(図13aおよび13b)。
【0208】
LPSは、近位の疎水性リピドA部分と、遠位の親水性O抗原多糖領域と、リピドAとO抗原構造を連結するコアオリゴ糖からなる複雑な糖脂質である。リピドA部分とは異なり、異なるグラム陰性細菌からO抗原構造には非常に多様性が有る。リピドAはLPS応答に必要であり、リピドAの脂肪酸側鎖を除く処理はLPSを不活性化する。我々は、アルカリ加水分解により”無毒化された”LPSと同様に、種々のグラム陰性細菌から調製されるLPSの効果を比較した。我々は、Escherichia coliセロタイプLCD25から単離されたLPSが、TLR2を活性化するために血清的に異なるEscherichia coli 055:B5 LPSよりほとんど2つのオーダーの規模で効果が高いことを観察した(図14a)。S.minnesota R595 LPSから調製されたLPSは、TLR2活性の可能性の有るインデューサーであるが、TLR2は”無毒化LPS”に応答できない。
【0209】
我々は、可溶型のTLR2細胞外ドメイン(TLR2‐Fc)が3H標識化LPSがin vitroアッセイで結合するかどうかを測定することにより、TLR2がLPSに結合するかどうかを調べた。我々は、3H−LCD25 LPSがTLR2−Fc融合たんぱく質に結合するが、Fc単独あるいは、いくつかのほかのレセプターのECDを含む融合たんぱく質には結合しないことを観察した(図14b)。この結合は特異的に、冷LCD25LPSにコンピートされたが、無毒化LPSではされなかった。LPSのTLR2−Fcへの結合の予備的分析は、Kdが比較的低い(500-700nM)ことと結合のキネティックスがを非常に遅いこと(データ示さず)を示唆する。我々は、他のたんぱく質、例えばLBPは、LBPがLPSをフリー、CD14との結合(ミセル型)から移す作用をするのと非常に類似して、LPSのTLR2へのin vivo結合の増進に作用するかもしれないと推測する。このことは、LBPがLPSに対するTLR2の感受性応答を得るために必要であることを示す我々のin vivoの結果と一致する(図9)。
【0210】
マクロファージのILS処理は、多くの炎症性サイトカインの発現をもたらす。同様にTLR2の293細胞での発現は、LPS/LBPと応答してIL-8 mRNAの>100倍の誘導をもたらすが、無毒化LPSはこのアッセイで不活性である(図15)。
【0211】
これらのデータはTLR2が、先天性免疫応答においてセンチネル(sentinel)役割、細菌の病原性に対する防御の最初のラインを果たすことを示唆する。TLR2とCD14は共にメロイド細胞で発現され、それらの誘導は、協働してLPS処理で誘導される。非ミエロイド細胞中のTLR2の発現は、LBPに依存し、mCD14の発現により増進されるメカニズムにより、通常の非応答性細胞にLPS応答性を与える。TLR2発現細胞のLPS処理は、NF-κBの活性化とそれにつづくIL-8などの適応反応を開始する遺伝子の誘導をもたらす(図15)。我々のデータは、インタクトな細胞外および細胞内ドメインがLPS応答に必要であるために、TLR2がLPSの存在を感知することとこの情報をプラスマ膜を通して伝えることの両方に関与していることを示唆する。さらに、IRAKとの会合に必要なIL-1Rの一部に相同性を有するTLR2のC末端尾領域は、NF-κB活性化に必要である。
【0212】
ショウジョウバエのトールおよびトール関連レセプター18Wheelerは、細菌と真菌に対する応答において、各々抗菌性ペプチドの誘導において重要な役割をはたす。Medzhitovら、上述。遺伝データは、トールに対するリガンドとしてのSpatzleが、背腹方向の(dorsovental)パターンに関与していることを示唆しており、Spatzleの相同体が免疫応答においてヒトTLRsの制御に重要かもしれないとの推測をもたらす。TLR2のLPSによる活性化がシクロヘキシミドにより阻害されないこと、TLR2の細胞外ドメインがLPSに対するin vitroでの低い親和性のレセプターであることの我々の観察は、TLR2がLPS認識に関与しているというモデルと一致する。我々のデータは、他のたんぱく質(例えばSpatzle相同体)がLPSに対するTLR2の応答を修正するかもしれないという可能性を排除しない。我々は、TLR2とショウジョウバエ トールの細胞外ドメインが共にLPRsを含むにもかかわらず、アミノ酸同一性が20%未満であることに注目する。第2に、LPRたんぱく質は、たんぱく質、ペプチドおよび炭水化物などの種々のタイプの分子に対するパターン認識レセプター(PRRs)である。Danglら、Cell 91,17-24(1997)。第3に、ショウジョウバエ免疫応答におけるSpatzleに対する要求は、トールのそれに比べて明らかではない。トールの欠損とは異なり、Spatzle変異体は、抗菌ペプチドのDefensinおよびAttacinの通常のレベルを誘導し、真菌の攻撃の後のCecropinA発現の部分的欠損だけであり、外傷の応答におけるDorsalの活性化で欠損していない。Lemaitreら,Cell 86,973-983(1996);Lemaitreら、EMBO J.14,536-545(1995)。
【0213】
前に記したように、TLRは、特に本出願で開示されている、3つの新規レセプター(DNA40021、DNA42663、およびDNA47361に各々コードされた)を含むヒトトール関連レセプターの大きなファミリーのメンバーである。この実施例で提示されたデータは,TLR4のNF-κB応答遺伝子の活性化における関与の証拠と同様に、レセプターのこのファミリーの主要な機能は、Janewayとその共同研究者によって最初に示唆されたように、微生物に存在する保存された分子構造の存在を感知する病原体パターン認識レセプターとして作用することであることを示唆している(Medzhitovら、上述)。ヒトTLRファミリーは、敗血症性ショックの治療のための治療方法におけるターゲットかもしれない。
【0214】
[実施例12]
in situハイブリダイゼーション in situハイブリダイゼーションは、細胞または組織調製物内における核酸配列の検出および位置決定のための強力かつ用途の広い技術である。例えば、遺伝子発現部位を同定するため、転写の組織分布を分析するため、ウイルス感染を同定および位置決定するため、特異的なmRNA合成の変化を追求するため、並びに染色体地図作製を支援するために使用できる。
【0215】
in situハイブリダイゼーションは、PCR産生33P-ラベル化リボプローブを用いて、LuとGillett, Cell Vision 1:169-176 (1994)によるプロトコールの最適化されたバージョンに従って行われた。簡単に説明すると、ホルマリン固定、パラフィン包埋されたヒト組織が切断され、パラフィンが除かれ、プロテイナーゼK(20g/ml)で15分間37℃でタンパク除去され、上記LuとGillettに記載されているようにin situハイブリダイゼーションのために処理される。[33−P]UTP-ラベルされたアンチセンスリボプローブがPCR産物から産生され、55℃で一晩ハイブリダイズされた。このスライドを、Kodak NTB2核トラックエマルジョンに浸して、4週間感光させた。
【0216】
33P-リボプローブ合成 6.0μl(125mCi)の33P-UTP(Amersham BF 1002, SA<2000 Ci/mmol)を乾燥させた。乾燥した33P-UTPを含む各チューブに、以下の成分を添加した: 2.0μl 5x 転写バッファー 1.0μl DTT(100mM)
2.0μl NTP混合物(2.5mM:10μ;それぞれ10mMのGTP、CTP&ATP+10μl H2O)
1.0μl UTP(50μM)
1.0μl Rnasin 1.0μl DNA鋳型(1μg)
1.0μl H2O 1.0μl RNAポリメラーゼ(通常、PCR産物T3=AS、T7=S)
【0217】
このチューブを、37℃で1時間インキュベートした。1.0μlのRQ1 DNアーゼを添加し、その後に37℃で15分間インキュベートした。90μlのTE(10mM Tris pH7.6/1mM EDTA pH8.0)を添加し、この混合物をDE81ペーパーにピペットで移した。残りの溶液を、Microcon-50限外濾過ユニットに充填し、プログラム10を用いて遠心した(6分)。濾過ユニットを第二チューブにあけ、プログラム2を用いて遠心した(3分)。最終的な回収遠心の後に、100μlのTEを添加した。1μlの最終産物をDE81ペーパーにピペットで移し、6mlのBiofluor IIにおいてカウントした。
【0218】
このプローブをTBE/尿素ゲルにかけた。1−3μlのプローブまたは5μlのRNA Mrk IIIを3μlの充填バッファーに添加した。95℃のヒートブロックで3分間熱した後、このゲルを直ちに氷に移した。ゲルのウェルを洗い、サンプルを充填し、45分間180−250ボルトで流した。このゲルをサランラップで覆い、−70℃のフリーザーの中で1時間から一晩の間、増感紙を用いてXARフィルムに感光させた。
【0219】
33P-ハイブリダイゼーション冷凍切片の前処理
このスライドをフリーザーから取り出し、アルミニウムのトレイに移し、室温で5分間解凍した。このトレイを、凝縮を下げるために、5分間55℃のインキュベーターに置いた。このスライドをフュームフードにおいて氷上で4%パラホルムアルデヒド中で10分間固定し、0.5xSSCで5分間、室温で洗浄した(25ml 20xSSC+975ml SQH2O)。10分間37℃で0.5μg/mlのプロテイナーゼK中で脱タンパクした後(250mlの予め温められたRNアーゼを含まないRNAバッファー中の10mg/mlストックの12.5μl)、この切片を0.5xSSCで10分間室温で洗浄した。この切片を70%、95%、100%のエタノール、2分間でそれぞれ脱水した。
【0220】
パラフィン-包埋された切片の前処理
このスライドを脱パラフィン化し、SQ H2Oに移し、2xSSC、室温、5分間でそれぞれ二回洗浄した。この切片を、20μg/mlのプロテイナーゼK(250mlのRNアーゼを含まないRNアーゼバッファー中の10mg/mlの500μl;37℃、15分)−ヒト胎芽、あるいは、8xのプロテイナーゼK(250mlのRNaseバッファー中の100μl;37℃、30分)−ホルマリン組織において脱タンパク化した。次いで、上述したように0.5xSSCですすぎ、脱水を行った。
【0221】
プレハイブリダイゼーション
このスライドを、Boxバッファー(4xSSC、50%ホルムアミド)−飽和フィルターペーパーを備えたプラスチックボックスに配置した。この組織を50μlのハイブリダイゼーションバッファー(3.75g デキストランスルファート+6mlSQ H2O)で被覆し、ボルテックスし、キャップを緩めて2分間マイクロ波で加熱した。氷上で冷却した後、18.75mlのホルムアミド、3.75mlの20xSSCおよび9mlのSQ H2Oを添加し、組織をよくボルテックスし、42℃で1−4時間インキュベートした。
【0222】
ハイブリダイゼーション
スライド当たり、1.0x106cpmプローブと1.0μltRNA(50mg/mlストック)を3分間95℃で加熱した。このスライドを氷冷し、スライド当たり48μlのハイブリダイゼーションバッファーを添加した。ボルテックスした後、50μlの33P混合物を、スライドの50μlのプレハイブリダイゼーションに添加した。このスライドを55℃で一晩インキュベートした。
【0223】
洗浄
洗浄を2x10分間、2xSSC、EDTAを用いて室温で行い(400ml 20xSSC+16ml 0.25M EDTA、Vf=4L)、その後に37℃で30分間RNアーゼA処理した(250mlのRNアーゼバッファー中の10mg/mlの500μl=20μg/ml)。このスライドを2x10分間、2xSSC、EDTAを用いて室温で洗浄した。ストリンジェントな洗浄条件は、55℃で2時間、0.1xSSC、EDTA(20ml 20xSSC+16ml EDTA、Vf=4L)であった。
【0224】
結果PRO285(DNA40021)
ヒト成人および胎児組織でのPRO285(DNA40021)の発現パターンを調べた。全長DNA40021配列に基づき合成した以下のプローブを用いた:オリゴ1:GGA TTC TAA TAC GAC TCA CTA TAG GGC AAA CTC TGC CCT GTG ATG TCA(配列番号27)オリゴ2:CTA TGA AAT TAA CCC TCA CTA AAG GGA ACG AGG GCA ATT TCC ACT TAG (配列番号28)
【0225】
この実験で、胎盤において、およびマウス胎児肝臓の造血細胞全般において低レベルの発現が観察された。ヒト胎児、成人またはチンパンジーリンパヌードではいずれも発現は検出されず、ヒト胎児またはヒト成人脾臓でも発現は検出されなかった。これらのデータは、おそらくin situハイブリダーゼーションアッセイでは感度が足りないため、ノザンブロットまたはPCRデータと完全には一致していない。より感度の高い条件下で、さらなる組織がいくらかの発現を示す可能性がある。
【0226】
PRO358(DNA47361)
ヒト成人および胎児組織でのPRO358(DNA47361)の発現パターンを調べた。全長DNA47361配列に基づき合成した以下のプローブを用いた:オリゴ1:GGA TTC TAA TAC GAC TCA CTA TAG GGC TGG CAA TAA ACT GGA GAC ACT (配列番号29)オリゴ2:CTA TGA AAT TAA CCC TCA CTA AAG GGA TTG AGT TGT TCT TGG GTT GTT(配列番号30)
【0227】
この実験で、消化管関連リンパ球組織および胎児における発達中の脾白髄質において、発現が観察された。正常成人脾臓のpALS領域で低いレベルの発現が観察された。他の全ての組織では陰性であったが、より感度の高い条件下で他の組織型において低いレベルの発現が観察できる可能性がある。
【0228】
材料の寄託
以下の材料は、American Type Culture Collection, 2301 Parklawn Drive, Rockville, MD,USA (ATCC)に寄託されている。
材料 ATCC寄託番号 寄託日DNA40021-1154 209389 1997年10月17日(PRO285をコードしている)
DNA42663-1154 209386 1997年10月17日(PRO286をコードしている)
DNA47361-1249 209431 1997年11月7日
【0229】
これらの寄託物は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定およびそれに基づく規則の下で作製された。これは、寄託日から30年間、寄託物の生存可能な培養物の維持を保証する。寄託物はブダペスト条約の下でATCCにより入手可能であり、Genentech, Inc.とATCCとの間の合意を得ることを条件として、直接関係する米国特許の発行、もしくは米国またはその他の国の特許出願の公開に基づいて、公衆に寄託物の培養物の子孫の永久的かつ無制限の利用可能性を保証し、そして、その子孫の入手を、35 USC 第122条およびそれに準ずる長官規則(特に886OG638に関して37 CFR 第1.14条を含む)に従って付与される米国特許商標庁長官によって定められた者に保証する。
【0230】
本願の譲受人は、寄託物の培養物が適切な条件下で培養された場合に死滅または損なわれた場合には、この材料を速やかに同一物で置き換えることに同意した。寄託材料の利用可能性は、その国の特許法に従って政府の権力の下に付与された権利に違反して本発明を実施するライセンスとして解釈されない。
【0231】
上記明細書は、当業者が本発明を十分に実施できるものと解する。寄託された実施態様は、本発明の一部の態様の単なる例示に過ぎないこと、並びに、機能的に等価なあらゆる構成物が本発明の範囲に含まれることから、本発明は、寄託された構成物により、範囲が制限されるものではない。ここに言う材料の寄託は、上記記述が、その最良の形態を含めた本発明のあらゆる態様の実施を可能にするのに不適切であることの承認、あるいは、それが示す特定の例示に請求の範囲を限定すると解釈されるとの承認を構成するものではない。実際に、ここに記載または示されたものに加えて、本発明の種々の変更が、上記記載から当業者にとって明白になり、添付された請求の範囲内に含まれる。
【図面の簡単な説明】
【0232】
【図1】PRO285と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号1)を示す。
【図2】PRO285をコードする、天然配列ヒトトールタンパク質のDNA40021と命名されたcDNA(配列番号2)の塩基配列を示す。
【図3】図2の配列の続きである。
【図4】PRO286と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号3)を示す。
【図5】PRO286をコードする、天然配列ヒトトールタンパク質のDNA42663と命名されたcDNA(配列番号4)の塩基配列を示す。
【図6】図5の配列の続きである。
【図7】図6の配列の続きである。
【図8】ヒトトールレセプタ2(huTLR2)の発現パターンを示す(上記のRockら)。a.TLR2プローブで検出されたヒトの複数の免疫組織のノーザン分析。PRL(抹消血液白血球)。b.マクロファージにおけるTLR2の豊富な発現、およびLPSに対するTLR2の転写の上流制御。定量的RT−PCRを用いて、PBL、T細胞、マクロファージ(MΦ)およびLPS促進マクロファージ(MΦ+LPS)におけるTLR2の相対的発現レベルを測定した。
【図9】TLR2はLPS誘導シグナルを仲介する。a.TLR2を安定して発現する293細胞はLPS応答性を得る。gD.TLR2を発現する安定クローンの集団(293−TLR2pop1)またはgD.TLR2を発現する単一クローン(293−TLR2クローン1)または発現ベクターのみで安定にトランスフェクトされたコントロール細胞(293−MSCV)は、pGL3.ELAM.tkで過渡的にトランスフェクトし、無血清培地中でLBPを存在または存在させずに6時間、1μg/mlの055:B5エンハンサで促進した。ELAMエンハンサの活性化は実施例に記載のように測定した。二つの独立した実験から結果を得た。ELAMエンハンサを欠いたコントロールレポータープラスミドを用いた場合、促進は観察されなかった(データは示さず)。レポータープラスミドの発現は未処理細胞またはLBPのみで処理された細胞において同一であった(データは示さず)。
【図10】293細胞においてエピトープ付加されたTLR2の発現を示すウェスタンブロット。
【図11】NF−κΒのTLR2依存性LPS促進活性化とトランスロケーションのタイムコース。核抽出物は055:B5 LPS(10μg/ml)とLBPで示された時間で処理された細胞(上部)、または1μMシクロヘキシミド(CHX)で1時間前処理され、無血清培地中で1μg/mlのLPSの存在下で1時間促進された細胞(低部)から調製した。
【図12】TLR2によるNF−κΒ活性化に対するmCD14の効果。ベクターコントロール(193−MSCV)または293−TLR2pop1細胞にレポータープラスミド、およびCD14発現ベクター(+mCD14)またはベクターコントロール(−mCD14)をそれぞれをトランスフェクトした。24時間後、トランスフェクトされた細胞を、無血清培地中でLBSの存在下に055:B5 LPSで6時間促進した。示されたデータは3つの独立した実験からのものを表わす。
【図13】シグナルにおけるTLR2のドメイン機能。a.多様なTLR2構築物の模式図。TLR2−WT、全長エピトープタグ付加形のTLR2、TLR2−Δ1およびΔ2はカルボキシル末端での13個または141個のアミノ酸の末端切断をそれぞれ表わす。CD4−TLR2、TLR2の細胞外ドメインをヒトCD4のアミノ酸1〜205により置き換えたヒトCD4−TLR2キメラ。ECD、細胞外ドメイン;TM、トランスメンブレン領域;ICD、細胞内ドメイン。b.IL−1RおよびTLR2シグナル変換に重要なC末端残基。残基番号は各タンパク質の右に示されている。矢印はTLR2−Δ1末端切断の位置を示した。*、IL−1Rシグナルに重要な残基(Heguyら,J.Biol.Chem.267,2605−2609[1992];Crostonら,J.Biol.Chem.270,16514−16517[1995])1 I、同一アミノ酸;:、保存的変化。c.TLR−R2変異体はLPSとLBPに対してNF−κΒを誘導することはない。293個の細胞は、示されたように全長TLR2またはTLR2変異体をコードする発現ベクターとpGL3.FLAM.tkで過渡的にトランスフェクトされた。また、細胞はCD14発現プラスミド(+mCD14)またはコントロールプラスミド(−mCD14)でトランスフェクトした。各タンパク質の等しい発現は、抗gDまたはCD4抗体を用いるウェスタンブロッティングにより確認される(下)。ルシフェラーゼアッセイは実施例に記載のように行った。データは二回の実験から得られた。
【図14】大腸菌K12 LPS(LCD25)の高い潜在力とそのTLR2への結合を示す。a.多様なLPS調製物の投与量応答曲線。b.TLR2の細胞外ドメインに対する[3H]−LPS(LCD25)の特異的相互作用。特異的結合はTLR2−Fcに対して観察されたが、Fcのみ、またはRse、Axl、Her2またはHer4の細胞外ドメインを有する融合タンパク質に対しては観察されなかった。TLR2−Fcに対する結合はLCD25 LPSと特異的に競合したが、脱毒素化されたLPSでは競合しなかった。
【図15】TLR2はLPS誘導IL−8発現に必要とされる。mCD14を過渡的に発現する293−MSCVベクターコントロールと293−TLR2細胞をLBPのみ、または示された種類のLPSとともに、無血清培地中1μg/mlの濃度で6時間促進した。等しい量のpoly−(A)RNAをノーザン分析のために用いた。
【図16】huTLR2をコードする塩基配列(配列番号11)。
【図17】huTLR2のアミノ酸配列(配列番号12)。
【図18】PRO358と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号13)を示す。この図において、アミノ酸1〜19は推定シグナル配列を形成し、アミノ酸20〜575は、アミノ酸20〜54がロイシンに富む繰返しの特徴を有する推定細胞外ドメインであり、アミノ酸576〜595は推定トランスメンブレンドメインであるが、アミノ酸596〜811は細胞内ドメインを形成する。
【図19】図18の配列の続きである。
【図20】図19の配列の続きである。
【図21】(配列番号14)成熟した全長トールタンパク質PRO358をコードするDNA47361と命名された天然配列ヒトトールタンパク質cDNAの塩基配列を示す。示された配列は外来性配列を有するので、ATG開始コドンに下線を付し、TAA停止コドンは四角で囲んだ。
【図22】図21の配列の続きである。
【技術分野】
【0001】
本発明は、一般的には、ここでDNA40021、DNA42663およびDNA47361と称する新規なDNAの同定と単離、および該DNAによりコードされる新規なヒトトール(Toll)相同体(それぞれPRO285、PRO286およびPRO358と称する)の組換え生産に関する。
【背景技術】
【0002】
膜結合タンパク質およびレセプタは多細胞生物の形成、分化および維持に重要な役割を果たすことができる。多くの各細胞の運命、例えば、増殖、移動、分化または他の細胞との相互作用は、典型的には他の細胞および/または直接の環境から受け取られた情報により支配される。この情報はしばしば分泌ポリペプチド(例えば有糸分裂因子、生存因子、細胞毒因子、分化因子、神経ペプチドおよびホルモン)により伝達された後に多様な細胞レセプタまたは膜結合タンパク質により受け取られ解釈される。そのような膜結合タンパク質および細胞レセプタとしてはサイトカインレセプタ、レセプタキナーゼ、レセプタホスファターゼ、細胞間相互作用に関与するレセプタおよびセレクチン類やインテグリン類のような細胞付着分子が挙げられるが、これらに限定されない。例えば、細胞の増殖と分化を調節するシグナル変換は、一部は多様な細胞タンパク質のリン酸化により調節される。このプロセスを触媒する酵素であるタンパク質チロシンキナーゼは成長因子レセプタとしても働くことができる。具体的な例として繊維芽細胞成長因子レセプタや神経成長因子レセプタが挙げられる。
【0003】
膜結合タンパク質とレセプタ分子は、医薬および診断剤等の多様な工業的応用性を有する。例えば、レセプタ免疫アドヘシンはレセプタリガンド相互作用をブロックする治療剤として用いることができる。膜結合タンパク質は関連するレセプタ/リガンド相互作用の潜在的なペプチドまたは小分子阻害物質のスクリーニングのために用いることもできる。
【0004】
新しい天然レセプタタンパク質を同定するための努力が工業界と大学の両者によりなされている。新規なレセプタタンパク質をコードする配列を同定するために哺乳類の組換えDNAライブラリーのスクリーニングに多くの努力が集められている。
胚背側−腹側パターンの確立に中心的な役割を演じる母性遺伝効果遺伝子であるショウジョウバエのトール遺伝子のクローニングは、Hashimotoら,Cell 52,269−279(1988)により報告されている。ショウジョウバエのトール遺伝子は803個のアミノ酸の細胞質外ドメインと269個のアミノ酸の細胞質ドメインを有する統合膜タンパク質をコードする。この細胞質外ドメインは膜にわたる潜在的なセグメントを有し、多くのトランスメンブレンタンパク質に見られる構造的モチーフであるロイシンに富むセグメントの複数のコピーを有する。トールタンパク質は、ショウジョウバエ胚の背側−腹側パターンを調節し、そのリガンドのスペッツルに結合する際に転写因子ドーサルを活性化するMorisato and Anderson,Cell 76,677−688(1994)。成熟したショウジョウバエにおいて、トール/ドーサルシグナル経路は抗微生物免疫応答に関与するLenaitreら,Cell 86,973−983(1996)。
【0005】
ショウジョウバエのトールタンパク質のヒト相同体はMedzhitovらNature 388、394−397(1997)により記載されている。このヒトのトールは、ショウジョウバエのトールと同じように、21個のタンデムに繰り返されたロイシンに富むモチーフ(ロイシンに富む領域−LRR)からなり、非LRR領域により分離された細胞外ドメインとヒトインターロイキン−1(IL−1)の細胞質ドメインに相同性のある細胞質ドメインとを有するタイプIのトランスメンブレンタンパク質である。ヒト細胞系にトランスフェクトされたヒトトールの構成的に活性のある変異体は、NF−κΒの活性化および炎症サイトカインIL−1、IL−6およびIL−8のNF−κΒ調節遺伝子の発現、ならびに天然のT細胞の活性化に必要とされる構成促進分子B7.1の発現を誘導することが可能なことが示された。トールは免疫系の非クローン性レセプタとしての脊椎動物中で機能し、脊椎動物で固有かつ適応性の免疫応答を活性化するシグナルを誘導できることが示唆されている。上記のMedzhitovらにより報告されたヒトトール遺伝子は脾臓および末梢血液白血球(PBL)において最も強く発現され、筆者らは他の組織中でのその発現は、それが感染の初期警告システムとして働きうるマクロファージおよび樹状突起細胞の存在によるものであることを示唆した。公共のGenBankデータベースは下記のトール配列を有する:トール1(ランダム配列の全長cDNA#HUMRSC786−1と同一であるDNAX# HSU88540−1);トール2(DNAX# HSU88878−1);トール3(DNAX# HSU88879−1);トール4(上記のMedzhitovらにより報告されたDNA配列と同一であるDNAX# HSU88880−1)。部分的なトール配列(トール5)はGenBankからDNA# HSU88881−1として利用できる。
【0006】
トール様レセプタと命名されたショウジョウバエトールタンパク質のさらなるヒト相同体(huTLRs1−5)が最近クローニングされ、ショウジョウバエ対応物の局在構造によく似ることが示された(Rockら,Proc.Natl.Acad.Sci.USA 95,588−593[1998])。あるヒトTLR(トールタンパク質相同体 − 上記のMedzhitovら;TLR4−上記のRockら)の構成的活性のある変異体の過剰発現はNFκΒの活性化および炎症サイトカインと構成促進分子の誘導をもたらす。上記のMedzhitovら。
【特許文献1】米国特許第5,364,934号
【特許文献2】米国特許第5,428,130号
【特許文献3】米国特許第3,959,642号
【非特許文献1】MedzhitovらNature 388、394−397(1997)
【非特許文献2】Rockら,Proc.Natl.Acad.Sci.USA 95,588−593(1998)
【発明の開示】
【発明が解決しようとする課題】
【0007】
出願人は、本出願で(DNA40021によってコードされる)PRO285、(DNA42663によってコードされる)PRO286および(DNA47361によってコードされる)PRO358と命名された新規なヒトトールポリペプチドをコードする3種類の新規なcDNAクローンを同定した。
【課題を解決するための手段】
【0008】
一実施態様において、本発明は、(a)図1(配列番号1)のアミノ酸残基27〜839を有するPRO286ポリペプチドをコードするDNA分子;または(b)図4(配列番号3)のアミノ酸残基27〜825を有するPRO286ポリペプチドをコードするDNA分子、または図18〜19(配列番号13)のアミノ酸20〜575を有するPRO358ポリペプチドをコードするDNA分子に対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも90%の配列同一性、最も好ましくは約95%の配列同一性を有するポリペプチドをコードするDNAからなる単離された核酸分子、または(d)前記(a)(b)または(c)のDNA分子の相補体を提供する。該相補DNA分子はそのようなコード核酸配列に対して少なくとも適度にストリンジェントな条件下で、および任意に高いストリンジェントな条件下で安定的に結合しているのが好ましい。
【0009】
さらなる実施態様において、単離された核酸分子は、図1(配列番号1)のアミノ酸1〜839の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有し;または図4(配列番号3)のアミノ酸1〜1041の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有し;または図18〜19(配列番号13)のアミノ酸1〜811の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約90%の配列同一性、好ましくは少なくとも約95%の配列同一性を有するポリヌクレオチドからなる。
【0010】
特定の実施態様において、本発明は、N末端シグナル配列を有するか有さない、および各全長配列のトランスメンブレン領域を有するか有さない天然または変異体のPRO285、PRO286およびPRO358ポリペプチドをコードするDNAからなる単離された核酸分子を提供する。一つの態様において、単離された核酸は、図1(配列番号1)のアミノ酸残基1〜1049、図4(配列番号3)の1〜1041、および図18〜19(配列番号13)の1〜811を有する成熟した全長の天然PRO285、PRO286またはPRO358ポリペプチドをコードするDNAからなるか、またはそうしたコード核酸配列に相補性のあるものである。別の態様において、本発明はN末端シグナル配列を有さない天然のPRO285、PRO286またはPRO358ポリペプチドをコードするDNAからなる単離された核酸分子またはそうしたコード核酸配列に相補性のある単離された核酸分子に関する。さらに別の実施態様において、本発明は、全長の天然PRO285、PRO286およびPRO358タンパク質のトランスメンブレン−ドメイン欠失または不活性化形をコードする核酸に関する。
【0011】
別の態様において、本発明は、図2と3(配列番号2)のおよその残基85〜およそ3283(当該残基を含む)の核酸の相補体に対して、または図5から7(配列番号4)のおよその残基57〜およそ4199(当該残基を含む)の核酸の相補体に対して、または図21〜22配列番号14)のおよその残基111〜およそ2544(当該残基を含む)の核酸の相補体に対してハイブリダイズするDNAからなる、PRO285、PRO286またはPRO358ポリペプチドをコードする単離された核酸分子に関する。好ましくは、ハイブリダイゼーションはストリンジェントなハイブリダイゼーションと洗浄条件下で起る。
【0012】
別の態様において、本発明は、(a)図1(配列番号1)の残基1〜1049(当該残基を含む)、または図4(配列番号3)のアミノ酸残基1〜1041(当該残基を含む)、または図18〜19(配列番号13)のアミノ酸残基1〜811(当該残基を含む)のアミノ酸配列に比較して少なくとも約80%の正量、好ましくは少なくとも約85%の正量、より好ましくは少なくとも90%の正量、最も好ましくは少なくとも約95%の正量を示すポリペプチドをコードするDNA、または(b)前記(a)のDNAの相補体に関する。
【0013】
別の実施態様において、本発明は、1997年10月17日にATCC No.209389にて寄託されたクローン(DNA40021−1154)、1997年10月17日にATCC No.209386にて寄託されたクローン(DNA42663−1154)、または1997年11月7日にATCC No.209431にて寄託されたクローン(DNA47361−1249)からなる単離された核酸分子を提供する。
【0014】
さらに別の実施態様において、本発明はPRO285、PRO286およびPRO358ポリペプチドまたはそれらの変異体をコードするDNAからなるベクターを提供する。よって、ベクターは上記のいずれの単離核酸分子からなってもよい。
【0015】
特定の実施態様において、本発明は、図18〜19(配列番号13)のアミノ酸20〜811の配列からなるポリペプチドをコードするポリヌクレオチドに対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも90%の配列同一性、最も好ましくは少なくとも約95%の配列同一性を有するポリヌクレオチドからなるベクター、またはそのようなポリヌクレオチドの相補体からなるベクターを提供する。特定の実施態様において、該ベクターは、N末端シグナル配列(およそアミノ酸1〜19)を有するか有さない新規なトール相同体(PRO358)、またはトランスメンブレン−ドメイン(およそアミノ酸576〜595)が欠損または不活性化したその変異体、または成熟タンパク質の細胞外ドメイン(およそアミノ酸20〜596)、またはこれらの配列のいずれか一つからなるタンパク質をコードするDNAからなる。そのようなベクターを有する宿主細胞も提供される。類似する実施態様が、各シグナル配列を有するか有さないPRO285およびPRO286トール相同体および/またはそれらのトランスメンブレン−ドメイン欠損または不活性化変異体からなるベクターについて明らかであり、具体的にはベクターは成熟PRO85およびPRO286トール相同体の細胞外ドメインをそれぞれ有する。
そのようなベクターを有する宿主も提供される。例えば、宿主細胞はCHO細胞、大腸菌または酵母であってよい。
【0016】
PRO285、PRO286およびPRO358ポリペプチドの製造方法がさらに提供され、該製法はPRO285、PRO286およびPRO358のそれぞれの発現に適する条件下で宿主細胞を培養し、PRO285、PRO286またはPRO358を細胞培養物から回収することからなる。
【0017】
別の実施態様において、本発明は単離されたPRO285、PRO286およびPRO358ポリペプチドを提供する。特に、本発明は、一実施態様において図1と3(配列番号1と3)のそれぞれのアミノ酸残基1〜1049および1〜1041からなるアミノ酸配列を有する単離された天然配列PRO285およびPRO286ポリペプチドを提供する。また、本発明は上記いずれかの単離核酸分子によりコードされるPRO285とPRO286ポリペプチドの変異体も提供する。特定の変異体として、各N末端シグナル配列を欠き、および/または各トランスメンブレンおよび/または細胞質ドメインを欠失または不活性化させた全長の天然配列PRO285とPRO286ポリペプチドの欠失(端を切り取った)変異体が挙げられるが、これらに限定されない。さらに、本発明は単離された天然配列PRO358ポリペプチドまたはその変異体を提供する。特に、本発明は、ある種の実施態様において、図18〜19(配列番号13)の残基20〜575、または20〜811、または1〜811からなるアミノ酸配列を有する単離された天然配列PRO358ポリペプチドを提供する。
【0018】
さらに別の態様において、本発明は、図1(配列番号1)のアミノ酸残基1〜1049;または図4(配列番号3)のアミノ酸残基1〜1041、または図18〜19(配列番号13)のアミノ酸残基1〜811のアミノ酸配列と比較して、少なくとも約80%の正量(positives)、好ましくは少なくとも約85%の正量、より好ましくは少なくとも90%の正量、最も好ましくは少なくとも約95%の正量を示すアミノ酸配列からなる単離されたPRO285、PRO286またはPRO358ポリペプチドに関する。
【0019】
さらに別の態様において、本発明は、(I)(a)図1(配列番号1)のおよそのアミノ酸残基1〜およそ1049(当該残基を含む)のアミノ酸残基、または図4(配列番号3)のおよそのアミノ酸残基1〜およそ1041(当該残基を含む)のアミノ酸残基、または図18〜19(配列番号13)のおよそのアミノ酸残基1〜およそ811(当該残基を含む)のアミノ酸残基の配列を有するPRO285、PRO286またはPRO358ポリペプチドをコードするDNA分子、または(b)前記の(a)のDNA分子の相補体に対して試験DNA分子をストリンジェントな条件下でハイブリダイズさせ、そして試験DNA分子が(a)または(b)に対して少なくとも約80%の配列同一性、好ましくは少なくとも約85%の配列同一性、より好ましくは少なくとも約90%配列同一性、最も好ましくは少なくとも約95%の配列同一性を有する場合、(ii)該試験DNA分子を有する宿主細胞を該ポリペプチドの発現に適する条件下で培養し、(iii)該ポリペプチドを細胞培養物から回収することにより生産されるポリペプチドを提供する。
【0020】
別の実施態様において、本発明は、PRO285、PRO286またはPRO358を異種ポリペプチドまたはアミノ酸配列に融合させてなるキメラ分子を提供する。そのようなキメラ分子の具体例は、PRO285、PRO286またはPRO358ポリペプチドをエピトープタグ配列または免疫グロブリンのFc領域に融合させてなる。そのようなキメラ分子の具体例は、PRO358(シグナルペプチドおよび/またはトランスメンブレン−ドメインおよび任意に細胞内ドメインが欠損したその変異体を含む)をエピトープタグ配列または免疫グロブリンのFc領域に融合させてなる。好ましい実施態様において、該融合物は、少なくともCH2とCH3ドメインを有する免疫グロブリン定常領域に融合させたPRO358の細胞外ドメインを有する。類似の特定の実施態様が存在し、PRO258またはPRO286ポリペプチドからなるキメラ分子についてここで開示される。
【0021】
別の実施態様において、本発明はPRO285、PRO286またはPRO358ポリペプチドに特異的に結合する抗体を提供する。任意に、抗体はモノクローナル抗体である。具体的には、本発明は二重特異性を有する抗体、例えば2つ以上のトールポリペプチドを結合させる二特異的抗体を包含する。
さらに別の実施態様において、本発明は天然のPRO285、PRO286およびPRO358ポリペプチドのアゴニストおよびアンタゴニストに関する。特定の実施態様において、該アゴニストまたはアンタゴニストは抗PRO285、抗PRO286または抗PRO358抗体である。
【0022】
さらに別の実施態様において、本発明は天然PRO285、PRO286およびPRO358ポリペプチドのアゴニストまたはアンタゴニストを同定するスクリーニングアッセイに関する。
【0023】
さらに別の実施態様において、本発明は、薬学的に許容される担体と組み合わせたPRO285、PRO286またはPRO358ポリペプチド、または上記のアゴニストまたはアンタゴニストからなる組成物に関する。
さらに、本発明は、薬学的に許容される担体と組み合わせた、PRO285、PRO286またはPRO358ポリペプチドに特異的に結合する抗体からなる組成物に関する。
【0024】
また、本発明は、効果的な量のPRO285、PRO286またはPRO358ポリペプチドのアンタゴニストを患者に投与することからなる敗血症性ショックの治療方法に関する。具体的な実施態様において、該アンタゴニストは天然のPRO285、PRO286またはPRO358ポリペプチドに特異的に結合する阻止抗体である。
【発明を実施するための最良の形態】
【0025】
I.定義
ここで用いられる「PROポリペプチド」、「PRO286ポリペプチド」、「PRO285」および「PRO286」との用語は、(ここでさらに定義される)天然配列PRO285およびPRO286トールタンパク質および変異体を包含する。該PRO285およびPRO286ポリペプチドは、ヒト組織タイプ等の多様なソースまたは別のソースから単離してもよいし、組換えまたは合成方法により、またはこれらの技術や類似の技術の組合せにより調製してもよい。
【0026】
「天然配列RPO285」または「天然配列PRO286」は天然から得られたPRO285またはPRO286と同一のアミノ酸配列を有するポリペプチドからなる。そのような天然配列トールポリペプチドは天然から単離することができ、または組換えまたは合成手段により作ることができる。具体的には、「天然配列PRO285」および「天然配列PRO286」とは、ここに開示されるPRO285とPRO286ポリペプチドの天然の端切断または分泌された形(例えば、細胞外ドメイン配列)、PRO285とPRO286ポリペプチドの天然の変異体形(例えば、交互にスプライスされた形)および天然の対立遺伝子変異体を包含する。本発明の一実施態様において、天然配列PRO285は、図1(配列番号1)のアミノ酸1〜1049からなる成熟または全長の天然配列PRO285ポリペプチドであり、天然配列PRO286は図4(配列番号3)のアミノ酸1〜1041からなる成熟または全長の天然配列PRO286ポリペプチドである。さらなる実施態様において、天然配列PRO285は、図1(配列番号1)のアミノ酸27〜1049または27〜836、または図4(配列番号3)のアミノ酸27〜1041または27〜825からなる。
【0027】
「PRO285変異体」および「PRO286変異体」との用語は、全長天然配列PRO285に関して図1(配列番号1)に示された類推アミノ酸配列を有するPRO285に対して少なくとも約80%のアミノ酸配列同一性、または全長天然配列PRO286に関しては図4(配列番号3)に示された類推アミノ酸配列を有するPRO286に対して少なくとも約80%のアミノ酸配列同一性を有する以下に定義する活性PRO285またはPRO286ポリペプチドを意味する。そのような変異体としては、例えば、一つ以上のアミノ酸残基が図1と3(配列番号1と3)の配列のN末端またはC末端でそれぞれ付加または欠失したPRO285およびPRO286を包含する。通常、PRO285またはPRO286変異体は、図1または図4(配列番号1と3)のアミノ酸配列に対して少なくとも約80%のアミノ酸同一性、より好ましくは少なくとも約90%のアミノ酸配列同一性、およびさらに好ましくは少なくとも95%のアミノ酸同一性を有するであろう。好ましい変異体は、天然配列PRO285またはPRO286ポリペプチドの細胞外ドメインに対して高度な配列同一性を示すものである。特定の実施態様において、本発明のPRO285およびPRO286変異体は、対応する天然のタンパク質の細胞内ドメインの少なくともC末端部分を保持し、最も好ましくは細胞内および細胞外ドメインのほとんどを保持する。しかし、それらの意図する使用に基づいて、そのような変異体は多様なアミノ酸の変更、例えばこれらの領域内に置換、欠失および/または挿入を有してよい。
【0028】
ここで用いられる「PRO358ポリペプチド」、「PRO358」、「PRO358トール相同体」およびその文字通りの変異体は、(さらにここで定義される)PRO358トールタンパク質と変異体を包含する。PRO358ポリペプチドは例えばヒト組織またはそれ以外のソース等の多様なソースから単離してもよいし、または組換え法または合成方法、またはこれらの技術や類似した技術の組合せにより調製してよい。
【0029】
「天然配列PRO358」は天然から得られたPRO358の同一のアミノ酸配列を有するポリペプチドからなる。そのような天然配列トールポリペプチドは天然から単離できるか、または組換えまたは合成手段により製造することができる。「天然配列PRO358」との用語は、具体的には、ここに開示のPRO368ポリペプチドの天然の端切断または分泌形(例えば、細胞外ドメイン配列)、天然の変異体形(例えば、交互にスプライスされた形)および天然の対立遺伝子変異体を包含する。本発明の一実施態様において、天然配列PRO358は、N末端シグナル配列(アミノ酸1〜19)を有するか有さず、およびN末端メチオニンを有するか有さない図18〜19(配列番号13)のアミノ酸20〜811からなる成熟または全長の天然配列PRO358ポリペプチドである。別の実施態様において、天然配列PRO358は、N末端シグナル配列を有するか有さず、およびN末端メチオニンを有するか有さない全長タンパク質(アミノ酸29〜575)の細胞外ドメインを保持する全長PRO358の可溶形である。
【0030】
「PRO358変異体」との用語は、図18〜19(配列番号13)に示される類推アミノ酸配列を有するPRO358に対して少なくとも約80%、好ましくは少なくとも約85%、より好ましくは少なくとも90%、最も好ましくは少なくとも約95%のアミノ配列同一性を有する下記の活性PRO358ポリペプチドを意味する。そのような変異体としては、例えば、一つ以上のアミノ酸が図18〜19(配列番号13)の配列のN末端またはC末端で付加または欠損したPRO358ポリペプチドを挙げられる。具体的には、変異体は、細胞内ドメインの一部またはすべてが欠損してよい、天然配列PRO358のトランスメンブレン−ドメイン欠失または不活性化変異体が挙げられる。好ましい変異体は天然配列PRO358ポリペプチドの細胞外ドメインに対して高い配列同一性を有するものである。特定の実施態様において、本発明のPRO358変異体は対応する天然タンパク質の細胞内ドメインの少なくともC末端を保持し、最も好ましくは細胞内および細胞外ドメインのほとんどを保持する。しかし、それらの意図する使用に基づいて、そのような変異体は多様なアミノ酸の変更、例えば、これらの領域内での置換、欠失および/または挿入を有してよい。
【0031】
ここで同定されたPRO285、PRO286およびPRO358配列に関する「アミノ酸配列同一度(%)」とは、配列を整列させ、必要であれば空所を導入して配列の最大同一性を得て、配列同一性の一部としての保存置換物は考慮しない後の、PRO285、PRO286またはPRO358配列中のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントと定義される。アミノ酸配列同一度を決める目的のためのアライメントは、例えばBLAST、ALIGNまたはメガライン(DNASTAR)ソフトウエア等の一般に利用可能なコンピュータソフトウエアを用いて当業者の技量内にある多様な方法で行うことができる。当業者は比較される配列の全長にわたる最大アライメントを得るために必要なアルゴリズム等のアライメント測定用の適当なパラメータを決めることができる。ALIGNソフトウエアがアミノ酸配列同一性を決定するために好ましい。
【0032】
具体的な特徴において、ここで同定されたPRO285、PRO286およびPRO358配列に関する「アミノ酸同一度(%)」とは、配列を整列させ、必要であれば空所を導入して配列の最大同一度を得て、配列同一性の一部としての保存置換物は考慮しない後の、PRO285、PRO286またはPRO358配列中のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントと定義される。ここで用いられる同一性値(%)は、[Altschulら,Methods in Enzymology,266:460−480(1996);http://blast.wustl/edu/blast/README.html]から得られたWU−BLAST−2により作られる。WU−BLAST−2は幾つかのサーチパラメータを使用し、そのほとんどがデフォルト値に設定される。調節可能なパラメータは、オーバーラップスパン=1、オーバーラップフラクション=0.125、ワード閾(T)=11の値内で設定される。HSP SとHSP S2パラメータは動的な値で、特定の配列の構成と調べられる興味のある配列に対する特定のデータベースの構成に基づいてプログラム自体により確立される;しかし、その値は感度を高めるために調節してもよい。アミノ酸配列同一性値%は、整列させた領域中の「長い」配列の残基の合計数で割られた、マッチする同一残基の数により決定される。「長い」配列は整列させた領域中の最も実際の残基を有する配列である(アライメントスコアを最大化するためにWU−Blast−2により導入された空所は無視する)。
【0033】
上記のように実施される配列比較の文脈において、「正量(positives)」との用語は、同一ではないが、(例えば保存的置換の結果として)類似の性質を有する比較配列中の残基を包含する。正量の%値は、上記のように、長い配列中の残基の合計数により割られたBLOSUM 62マトリックス中の正量値を示す残基部分により決定される。
【0034】
ここで同定されたDNA40021、DNA42663およびDNA47361配列に関する「核酸配列同一度(%)」は、配列を整列させ、必要であれば空所を導入して配列の最大同一度を得て、DNA40021、DNA42663およびDNA47361配列中のヌクレオチドと同一である候補配列中のヌクレオチドのパーセントと定義される。核酸配列同一度を決める目的のためのアライメントは、例えばBLAST、ALIGNまたはメガライン(DNASTAR)ソフトウエア等の一般に利用可能なコンピュータソフトウエアを用いて当業者の技量内にある多様な方法で行うことができる。当業者は比較される配列の全長にわたる最大アライメントを得るために必要なアルゴリズム等のアライメント測定用の適当なパラメータを決めることができる。ALIGNソフトウエアが核酸配列同一性を決定するために好ましい。
【0035】
具体的には、ここで同定されたPRO285、PRO286およびPRO358ポリペプチドのコード配列に関する「核酸配列同一度(%)」とはPRO285、PRO286またはPRO358のコード配列中のヌクレオチド残基と同一である候補配列中のヌクレオチド残基のパーセントと定義される。ここで用いられる同一性値は、デフォルトパラメータに設定されたWU−BLAST−2のBLASTNモジュールにより作られ、オーバーラップスパンとオーバーラップフラクションをそれぞれ1および0.125に設定した。
【0036】
ここに開示の多様なポリペプチドを記載するために用いられる場合の「単離された」とはその天然の環境の成分から同定され、分離され、および/または回収されたポリペプチドを意味する。その天然環境のきょう雑成分は典型的には該ポリペプチドの診断または治療使用に干渉する材料であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性の溶質が挙げられよう。好ましい実施態様において、該ポリペプチドは、(1)スピンニングカップシークエネータを用いて、N末端または中間アミノ酸配列の少なくとも15残基を得るのに十分な程度まで、または(2)非還元または還元条件下に、クマシーブルーまたは好ましくは銀染色を用いてSDS−PAGEにより均一となるまで精製する。PRO285、PRO286またはPRO358の天然環境の少なくとも一つの構成要素は存在しないために、単離されたポリペプチドは組換え細胞中のin situでのポリペプチドを包含する。しかし、通常は単離されたポリペプチドは少なくとも一つの精製工程で調製されよう。
【0037】
「単離された」DNA40021、DNA42663またはDNA47361核酸分子は、DNA40021、DNA42663またはDNA47361核酸の天然のソースに通常会合している少なくとも一つのきょう雑核酸分子から分離され同定される核酸分子である。単離されたDNA40021、DNA42663またはDNA47361核酸分子は天然に見られるものとは形態や設定において異なる。したがって、単離されたDNA40021、DNA42663またはDNA47361核酸分子は、天然の細胞中で存在するDNA40021、DNA42663またはDNA47361核酸とは区別される。しかし、単離されたDNA40021、DNA42663またはDNA47361核酸分子は、例えば核酸分子が天然の細胞とは異なる遺伝子配置にあるDNA40021、DNA42663またはDNA47361を通常発現する細胞中に含まれるDNA40021、DNA42663およびDNA47361核酸分子を包含する。
【0038】
「トールレセプタ2」、「TLR2」および「huTLR2」は相互に交換可能に用いられ、Rockら,Proc.Natl.Acad.Sci.USA 95,588−593(1998)により「HuTLR2」と命名されたヒトトールレセプタを示している。huTLR2の塩基配列とアミノ酸配列をそれぞれ図16(配列番号11)と図17(配列番号12)に示す。
【0039】
「発現ベクター」との用語は、ここではトール相同タンパク質をコードする核酸が、適当な宿主細胞中でその発現に影響を与えることのできる制御配列に操作可能に結合されたベクターを定義するために用いる。通常、ベクターは複製部位を有する(ただし、染色体統合が起る場合これは必要ではない)。発現ベクターは形質転換細胞において表現型の選択を提供できるマーカー配列も有する。例えば、大腸菌は典型的には大腸菌種に由来するプラスミドであるpBR322を用いて形質転換される(Bolivar,ら,Gene 2:95[1977])。pBR322はアンピシリンおよびテトラサイクリン耐性遺伝子を有しているために、目的がクローニングであろうと、発現であろうと形質転換細胞の同定の容易な手段を提供する。また、発現ベクターは、最も望ましくは、転写と翻訳の制御に有用な配列、例えばプロモータおよびShine−Dalgarno配列(原核生物に関して)またはプロモータとエンハンサ(哺乳類細胞に関して)を有しよう。プロモータとしては誘導性のあるものでよいが、誘導性である必要はなく;哺乳類宿主用のCMVプロモータ等の強力な構成プロモータが宿主細胞毒なしにLHRを産生することがわかっている。発現ベクターは発現の制御、複製配列または選択遺伝子を有する必要はないと思われるが、それらが存在しないと、ハイブリッド形質転換体の同定や高いレベルのハイブリッド免疫グロブリン発現の達成が困難になろう。
【0040】
「制御(control)配列」とは、特定の宿主生物中で、操作可能に結合させたコード配列の発現に必要なDNA配列を指している。原核細胞に適する制御配列とは、例えば、プロモータ、任意にオペレーター配列、およびリボソーム結合部位が挙げられる。真核細胞は、プロモータ、ポリアデ二ル化シグナルおよびエンハンサを利用することが公知である。
【0041】
核酸が別の核酸配列と機能的な関係に置かれているときに、それは「操作可能に(operably)結合」されている。例えば、プレ配列用または分泌リーダー用のDNAがポリペプチド用のDNAに操作可能に結合しているとは、それが該ポリペプチドの分泌に関与する前タンパク質として発現している場合であり;プロモータまたはエンハンサがコード配列に操作可能に結合されているとは、それが該配列の転写に影響を与える場合であり;またはリボソーム結合部位がコード配列に操作可能に結合されているとは、それが翻訳を容易にするように配置されている場合である。一般的に、「操作可能に結合されている」とは、結合されているDNA配列が隣接しており、分泌リーダーの場合、隣接しており、リーディングフェーズにあることを意味する。しかし、エンハンサは隣接している必要はない。結合は便利な制限部位での連結により行われる。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーを慣用の実施にしたがって用いる。
【0042】
「抗体」との用語は最も広い意味で用いられ、具体的には単一の抗PRO285、抗PRO286および抗PRO358モノクローナル抗体(アゴニスト、アンタゴニストおよび中和抗体を含む)、および多エピトープ特異性を有する抗PRO285、抗PRO286および抗PRO358抗体組成物を包含する。ここで用いられる「モノクローナル抗体」との用語は実質的に均一な抗体の集団(すなわち、該集団を構成する個々の抗体は少量存在する可能性のある起り得る天然の変異を除けば同一である)から得られた抗体を意味する。
【0043】
「アンタゴニスト」との用語は最も広い意味で用いられ、ここに開示される天然のトールレセプタの生物学的活性を部分的または完全にブロック、阻止、抑制または中和する分子を包含する。同様に、「アゴニスト」との用語は最も広い意味で用いられ、ここに開示される天然のトールレセプタの生物学的活性に類似するか、増強する分子を包含する。適当なアゴニストまたはアンタゴニスト分子は具体的にはアゴニストまたはアンタゴニスト抗体または抗体断片、断片または天然のトールレセプタポリペプチドのアミノ酸配列の変異体、ペプチド、小有機分子等を包含する。
【0044】
ここでの目的のための「活性のある」または「活性」とは、天然または天然に存在するPRO285、PRO286およびPRO358のそれぞれの生物学的および/または免疫学的活性を保持するPRO285、PRO286およびPRO358の形態を意味する。好ましい「活性」は、炎症サイトカインであるIL−1、IL−6およびIL−8のNFκΒおよび/またはNF−κΒ制御遺伝子の活性化を誘導する能力である。別の好ましい「活性」は脊椎動物で固有および/または適合性の免疫応答を活性化する能力である。さらに好ましい「活性」は微生物上に存在する保存された分子構造物の存在を感知する能力であり、具体的にはリポ多糖(LPS)シグナルを仲介する能力である。同じ「活性」の定義がPRO285、PRO286およびPRO358ポリペプチドのアゴニスト(例えば、アゴニスト抗体)に適用される。上記のように、PRO285、PRO286またはPRO358ポリペプチドのアンタゴニスト(アゴニスト抗体を含む)の活性はPRO285、PRO286またはPRO358ポリペプチドの上記同定の活性のいずれに対しても拮抗作用する能力、例えばそれら活性を部分的または完全にブロック、阻止、抑制または中和する能力と定義される。
【0045】
ハイブリダイゼーション反応の「ストリンジェンシー」とは当業者により容易に決定でき、通常、プローブの長さ、洗浄温度および塩濃度に依存する経験的な算定である。一般に、長いプローブは適当なアニーリングのために高温を必要とし、逆に短いプローブは低温を必要とする。相補鎖がそれらの融解温度以下の環境にある場合、ハイブリダイゼーションは通常、変性DNAが再アニールする能力に依存する。プローブと、ハイブリダイゼーションが可能な配列との所望の相同性の程度が高ければ高いほど使用できる相対的温度が高くなる。その結果、さらに高い相対的温度は反応条件をさらにストリンジェントにする傾向があり、他方で低温は厳格性を低くする傾向があるということになる。さらなる詳細とハイブリダイゼーション反応のストリンジェンシーの説明については、Ausubelら,Current Protocols in Molecular Biology(1995)を参照されたい。
【0046】
ここに定義される「ストリンジェントな条件」または「高ストリンジェントな条件」は、(1)洗浄のための低いイオン強度と高い温度、例えば0.015Mの塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で用いる;(2)ハイブリダイゼーション中にホルムアミド等の変性剤、例えば、0.1%ウシ血清アルブミン/0.1% Ficoll/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム(pH6.5)で、750mM塩化ナトリウム、75mMクエン酸を有する42℃の50%(v/v)ホルムアミドを用いる、(3)50%ホルムアミド、5×SSC(0.75M NaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×Denhardtの溶液、超音波処理サケ精子DNA(50μl/ml)、0.1%SDSおよび10%硫酸デキストランを42℃で用い、42℃での0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)および55Cの50%ホルムアミドでの洗浄およびその後の55℃でのEDTA含有0.1×SSCからなる高ストリンジェントな洗浄によって同定されよう。
【0047】
「適度にストリンジェントな条件」とは、Sambrookら,Molecular Cloning:A Laboratory Manual,New York:Cold Spring Harbor Press,1989)により記載されたように同定できるもので、上記の記載のものよりストリンジェンシーの低い洗浄溶液とハイブリダイゼーション条件(例えば、温度、イオン強度およびSDSの%)の使用を伴う。適度にストリンジェントな条件の具体例としては、20%ホルムアルデヒド、5×SSC(150mM NaCl、15mMクエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×Denhartの溶液、10%硫酸デキストラン、および20mg/mlの変性専断サケ精子DNAからなる溶液中で37℃の一晩のインキュベーション後に、ろ紙を約37〜50℃の1×SSCで洗浄することが挙げられる。当業者は、プローブの長さ等の因子を適合させるように、いかに温度、イオン強度等を必要にあわせて調節するかを分かるであろう。
ここで用いられる場合の「エピトープタグ」との用語は、FIZZポリペプチドを「タグポリペプチド」に融合させてなるキメラポリペプチドを意味する。タグポリペプチドは、抗体が作られうるエピトープを提供するのに十分な残基を有するが、それが融合するポリペプチドの活性に干渉しないように短い残基を有する。タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応しないようにかなり独自なものでなければならない。適するタグポリペプチドは、一般的に、少なくとも6アミノ酸残基、通常は約8〜50アミノ酸残基(好ましくは、約10〜20アミノ酸残基)を有する。
【0048】
ここで用いられる「免疫アドヘシン(immunoadhesin)」との用語は、異種タンパク質(「アドヘシン」)の結合特異性を免疫グロブリンの定常ドメインのエフェクター機能に組み合わせている抗体様分子を示す。構造的には、免疫アドヘシンは、抗体(すなわち「異種」のもの)の抗原認識結合部位ではない所望の結合特異性を有するアミノ酸配列と免系グロブリンの定常部ドメイン配列との融合からなる。免疫アドヘシン分子のアドヘシン部分は、典型的には、レセプタまたはリガンドの少なくとも結合部位からなる隣接アミノ酸配列である。免疫アドヘシンにおける免疫グロブリン定常ドメイン配列は、IgG−1、IgG−2、IgG−3またはIgG−4サブタイプ、IgA(IgA−1およびIgA−2等)、IgE、IgDまたはIgM等の免疫グロブリンから得てよい。
【0049】
「治療」とは、その目的が標的とする病理学的状態または疾患を予防または遅らせる(軽減)することにある場合、治療的処置と予防的または防止的対策の両方を意味する。治療を必要とする対象は該疾患を既に有する対象、ならびに該疾患を持ちそうな対象または該疾患を予防しようとする対象である。
「長期(chronic)」投与とは、初期の治療効果(活性)を長期間にわたって保持するように、急性の態様に対する連続的な形態での薬剤の投与を言う。
【0050】
治療目的の「哺乳類」とは、ヒト、家畜および農場動物、および動物園、スポーツまたはペット動物、例えばイヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ等の哺乳類として分類される動物を言う。好ましくは、哺乳類はヒトである。
一つ以上のさらなる治療薬「と組み合わせた」投与とは、同時投与および順番は問わない連続的な投与を含む。
【0051】
「リポ多糖」または「LPS」との用語は「内毒素(endotoxin)」と同義語として用いられている。リポ多糖(LPS)はグラム陰性細菌、例えば大腸菌の外膜の特徴的な成分である。それらは多糖部分とリピドAと呼ばれる脂肪からなる。多糖は細菌の種類により異なっており、(3〜8糖の繰返し単位から作られる)O特異鎖と二部分の核から作られる。リピドAは、事実上、リン酸により修飾された二つのグルコサミン糖と異なる数の脂肪酸を常に含む。さらなる情報に関しては、Rietschel and Brade,Scientific American August 1992,54−61を参照されたい。
【0052】
「敗血症性ショック」との用語はここでは最も広い意味で用いられ、Bone,Ann.Intern Med.114,332−333(1991)に開示されたすべての定義を含む。具体的には、敗血症性ショックは、敗血症と呼ばれる症候群である感染に対する全身応答で始まる。この症候群が低血圧と器官不全をもたらす場合、それは敗血症性ショックと呼ばれる。敗血症性ショックはグラム陽性菌および菌類、ならびに内毒素を有するグラム陰性菌により開始されるだろう。よって、ここでの定義は「内毒素ショック」に限定されない。
【0053】
II.発明の組成物と方法
A.全長PRO285、PRO286およびPRO358
本発明は、本出願でPRO285および8PRO286と称されるポリペプチドをコードする、新しく同定、単離された塩基配列を提供する。特に、出願人は下記の実施例でさらに詳細に開示されるPRO285およびPRO286ポリペプチドをコードするcDNAを同定し、単離した。BLASTおよびFastA配列アライメントコンピュータプログラムを用いて、出願人は、PRO285およびPRO286のコード配列が、GenBankデータベース中のDNA配列HSU88540_1、HSU88878_1、HSU88879_1、HSU88880_1およびHSU88881_1に対して高度に相同性のあることを発見した。
【0054】
さらに、本発明は、本出願でPRO358と称されるポリペプチドをコードする、新しく同定、単離された塩基配列を提供する。特に、出願人は下記の実施例でさらに詳細に開示されるヒトトールポリペプチド(PRO358)をコードするcDNAを同定し、単離した。BLASTおよびFastA配列アライメントコンピュータプログラムを用いて、出願人は、PRO358のコード配列が、GenBankデータベース中のDNA配列HSU88540_1、HSU88878_1、HSU88879_1、HSU88880_1、HS88881_1およびHSU79260_1に対して顕著な相同性を示すことを発見した。HSU79260_1を除けば、示されたタンパク質はヒトトール様レセプタと同定された。
【0055】
したがって、本出願で開示されたPRO285、PRO286およびPRO358タンパク質が、ショウジョウバエタンパク質トールの新しく同定されたヒト相同体であり、適合免疫において重要な役割を果たしていると現在考えられている。さらに具体的にはPRO285、PRO286およびPRO358は、炎症、敗血症性ショックおよび病原体に対する応答にかかわっていると思われ、例えば、糖尿病、ALS、ガン、リューマチ性関節炎および潰瘍等の免疫応答により悪化する多様な医学的な状態に適切な役割を果たしている。微生物上の保存された分子構築物の存在を感知する病原体パターン認識レセプタとしてのPRO285、PRO286およびPRO358の役割は、公知のヒトトール様レセプタであるTLR2がLPSシグナルの直接の仲介物質であることを示している本出願で開示されたデータによってさらに支持される。
【0056】
B.PRO285、PRO286およびPRO358変異体
ここに記載された全長の天然配列PRO285、PRO286およびPRO358に加えて、これらの配列の変異体を調製できることが意図される。PRO285、PRO286およびPRO358変異体は、適当なヌクレオチド変化をPRO285、PRO286またはPRO358DNAに導入することにより、または所望の変異体ポリペプチドの合成により調製することができる。当業者は、PRO285、PRO286またはPRO358ポリペプチドの翻訳後プロセスを、例えばグリコシル化部位の数や位置を変えることにより、または膜固定特性を変更する等により変更できることを理解しよう。
【0057】
天然の全長配列PRO285、PRO286およびPRO358、またはここに記載のPRO285、PRO286またはPRO358の多様なドメインにおける変異は、例えば、米国特許第5,364,934号に示された保存または非保存変異に関する技術とガイドラインを用いて行うことができる。変異は、対応する天然配列ポリペプチドと比較してアミノ酸配列の変化をもたらす、PRO285、PRO286またはPRO358ポリペプチドをコードする一つ以上のコドンの置換、欠失または挿入であってよい。任意に、変異は、少なくとも一つのアミノ酸を、PRO285、PRO286またはPRO358の一つ以上のドメインにおいて他のアミノ酸に置換することによる。どのアミノ酸が所望の活性に悪影響を与えることなく挿入、置換または欠失させてよいかを決定するガイダンスは、PRO285、PRO286またはPRO358の配列を相同性のある公知のタンパク質分子と比較し、高い相同性のある領域でなされたアミノ酸配列の変化の数を最小化することにより発見されよう。アミノ酸置換は、ロイシンのセリンによる置換等、あるアミノ酸の、類似の構造および/または化学的性質を有する別のアミノ酸による置換(すなわち保存アミノ酸の置換)の結果でありうる。挿入または欠失は任意に1〜5アミノ酸の範囲であってよい。認められる変異は、配列中のアミノ酸の挿入、欠失または置換を系統的に行い、得られた変異体の活性を下記の実施例に記載のインビトロアッセイにより調べることにより決定してよい。
【0058】
多様性はオリゴヌクレオチド仲介(部位特異的)変異、アラニンスキャンニングおよびPCR変異等の公知の方法を用いて行うことができる。部位特異的変異[Carterら,Nucl.Acids Res.,13:4331(1986);Zollerら,Nucl.Acids Res.,10:6487(1987)]、カセット変異[Wellsら,Gene,34:315(1985)]、制限選択変異[Wellsら,Philos,Trans.R.Soc.London SerA,317:415(1986)]または他の公知の技術をクローニングされたDNAに行ってPRO285またはPRO286変異体DNAを作ることができる。
【0059】
スキャンニングアミノ酸分析は隣接した配列に沿った一つ以上のアミノ酸を同定するために用いることもできる。比較的小さい中性アミノ酸が好ましいスキャンニングアミノ酸中にある。そのようなアミノ酸としてはアラニン、グリシン、セリンおよびシステインが挙げられる。アラニンはベータ炭素を超えて側鎖を除去し、変異体の主鎖立体配座を変えることが少ないのでこのグループの中でも好ましいスキャンニングアミノ酸である。アラニンは最も共通するアミノ酸であるために、これが典型的には望ましい。さらに、これは、埋め込まれた位置や露出した位置にしばしば見られる[Creighton,The Proteins(W.H.Freeman & Co.,N.Y.);Chothia,J.Mol.Biol.,150:1(1976)]。アラニン置換が十分な量の変異体を作らない場合、イソテリックなアミノ酸を用いることができる。
【0060】
ここに開示されるPRO285、PRO286およびPRO358トールタンパク質の変異体としては、トランスメンブレンドメインが欠失または不活性化されたタンパク質が挙げられる。トランスメンブレン領域は、細胞膜の脂質二層にわたる適当な大きさの高度に疎水性または親油性ドメインである。それらは天然の成熟PRO285、PRO286およびPRO358ポリペプチドを細胞膜に固定すると考えられている。PRO285では、トランスメンブレンドメインはおよそのアミノ酸の位置840からおよそのアミノ酸位置864まで伸びている。PRO286においては、トランスメンブレンドメインはおよそのアミノ酸位置826とおよそのアミノ酸位置848との間にある。PRO358では、トランスメンブレンドメインはおよそのアミノ酸位置576とアミノ酸位置595との間にある。
【0061】
トランスメンブレンドメインの欠失または置換は、PRO285、PRO286およびPRO358ポリペプチドの可溶形の回収を、その細胞または膜の脂質親和性を減少させ、その水溶性を向上させることにより容易にして提供する。トランスメンブレンおよび細胞質ドメインが欠失する場合、体により外来と認識される細胞内ペプチドをさらすか、または潜在的に免疫原性のある異種ポリペプチドの挿入により、潜在的に免疫原性のあるエピトープの導入を避ける。トランスメンブレンドメイン欠失PRO285、PRO286またはPRO358の主要な利点は、それが組換え宿主の培養培地に分泌されることである。この変異体は血液等の体液に可溶性であり、細胞膜脂質に対して容易に感知できる親和性を有さないために、組換え細胞培養物からのその回収を非常に容易にする。
【0062】
前記の考察から、置換、欠失、挿入またはそれらの組合せを導入して最終構築物に到達することは十分に明らかであろう。一般的な問題として、可溶性の変異体は機能的なトランスメンブレンドメインを有さず、好ましくは機能的な細胞質配列を有さないだろう。これは関連するドメインの欠失により通常行われるが、十分な挿入または置換による変異体もこの目的のために効果的である。例えば、トランスメンブレンドメインは、アミノ酸配列、例えば、ともに親水性の水治療法的な特徴を示す約5〜50個のセリン、スレオニン、リジン、アルギニン、グルタミン、アスパラギン酸等の親水性残基のランダムまたは一定の配列によって置換される。欠失(末端切除)PRO285、PRO286およびPRO358変異体のように、これらの変異体は組換え宿主の細胞培地中に分泌される。
【0063】
全長の成熟PRO285、PRO286およびPRO358ポリペプチドのさらなる欠失変異体(またはそれらのトランスメンブレンドメイン欠失〜不活性化形)としては、N末端シグナルペプチド(PRO285およびPRO286についてはアミノ酸1〜19、およびPRO358についてはアミノ酸1〜26として推定上同定されたもの)および/または開始メチオニンが欠失した変異体が挙げられる。天然のシグナル配列は、別のトール様タンパク質、または他のヒトまたは非ヒト(例えば、細菌、酵母または非ヒト哺乳類の)シグナル配列であってよい別の(異種)シグナルペプチドによって置換されてもよい。
【0064】
細胞内領域、および特にそのC末端部分がこれらのポリペプチドの生物学的機能に重要であると思われる。したがって、その目的が対応する天然のトール様タンパク質の生物学的活性を保持する変異体を作ることにあるのなら、これらの領域の少なくとも実質的な部分は保持されるか、またはその変更がある場合、該変更は保存アミノ酸の置換および/または挿入を伴うか、またはアミノ酸が挿入される領域に存在するアミノ酸に性質の類似したアミノ酸を伴うものとする。しかし、天然トールレセプタの生物学的機能の実質的な修飾が必要とされる場合(例えば、それぞれの天然トールポリペプチドのアンタゴニストを作ることが目的の場合)、その変更は、対応する天然のトールポリペプチドにおいて標的とされる位置のアミノ酸とは性質の異なるアミノ酸の置換および/または挿入を伴う。
【0065】
天然のアミノ酸は共通する側鎖の性質に基づいて下記グループに分類される:(1)疎水性:ノルロイシン、メチオニン、アラニン、バリン、ロイシン、イソロイシン;
(2)中性疎水性:システイン、セリン、スレオニン;
(3)酸性:アスパラギン酸、グルタミン酸;
(4)塩基性:アスパラギン、グルタミン、ヒスチジン、リジン、アルギニン;
(5)鎖の配向に影響を与える残基:グリシン、プロリン;および(6)芳香性:トリプトファン、チロシン、フェニルアラニン。
【0066】
保存的置換は一グループ内の一メンバーを同一グループ内の別のメンバーに交換することによるが、非保存的置換はこれらのクラスのうちの一クラスのメンバーを別のものに交換することを必要としよう。非保存的置換により得られる変異体は得られた変異体の生物学的性質/機能にさらに顕著な変化をもたらすことが期待される。
【0067】
アミノ酸挿入は、長さが1残基から百以上の残基を有するポリペプチドまでの範囲のアミノ末端および/またはカルボキシル末端の融合、ならびに単一または複数のアミノ酸残基の配列内挿入を包含する。配列内挿入(すなわち、PRO285、PRO286またはPRO358タンパク質アミノ酸配列内の挿入)は、通常約1〜10残基、より好ましくは1〜5残基、さらに好ましくは1〜3残基の範囲であってよい。末端挿入の具体例としては、N末端のメチオニル残基を有するPRO285、PRO286およびPRO358ポリペプチド、細菌組換え細胞培養物におけるその直接の発現の人工的産物および組換え宿主細胞に由来する成熟I−TRAFタンパク質の分泌を容易にするためにPRO285、PRO286またはPRO358分子のN末端への異種N末端シグナル配列の融合体が挙げられる。そのようなシグナル配列は通常は意図される宿主細胞種から得られるので、該宿主細胞種と同種であろう。適当な配列としては、大腸菌用のSTIIまたはIpp、酵母用のアルファ因子および哺乳類細胞用のヘルペスgD等のウイルスシグナルが挙げられる。
【0068】
ここに開示される天然トール様分子の他の挿入的変異体としては、免疫原性ポリペプチド、例えばベータ−ラクタマーゼ等の細菌ポリペプチドまたは大腸菌trp遺伝子座によりコードされる酵素、または酵母タンパク質に対する天然配列分子のN末端またはC末端の融合体、および1989年4月6日公開のWO89/02922に記載のように免疫グロブリン領域(好ましくは、免疫アドヘシンを生じる免疫グロブリンの定常部)、アルブミンまたはフェリチン等の長い半減期を有するタンパク質とのC末端融合体が挙げられる。免疫グロブリン融合体の生産に関しては、1995年6月27日発行の米国特許第5,428,130号を参照されたい。
【0069】
変異体トール様タンパク質の特性を前もって予想することはしばしば困難なので、スクリーニングは最適な変異体を選択する必要があることが理解されよう。この目的のために、以下に説明するような生化学的または他のスクリーニングアッセイが容易に利用可能となるだろう。
【0070】
C.PRO285、PRO286およびPRO385トールタンパク質の修飾
PRO285、PRO286およびPRO358ヒトトール相同体の共有結合による修飾が本発明の範囲に含まれる。一つの種類の共有修飾は、PRO285、PRO286およびPRO358タンパク質の標的アミノ酸残基を、選択された側鎖またはNまたはC末端残基と反応可能な有機誘導体化剤と反応させることによる。二官能剤による誘導体化が、例えば、PRO285、PRO286またはPRO358を抗PRO285、抗PRO286または抗PRO358抗体の精製法に用いられる非水溶性の支持体マトリックスまたは表面に対して架橋させるか、またはその逆にマトリックスまたは表面を該タンパク質に架橋させるために有用である。
【0071】
通常用いられる架橋剤としては、例えば1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステル、例えば、4−アジドサリチル酸とのエステル、ホモ二官能性イミドエステル、例えば3,3’−ジチオビス−(スクシンイミジルプロピオネート)等のジスクシンイミジルエステル、ビス−N−マレイミド−1,8−オクタン等の二官能性マレイミドおよびメチル−3−[(p−アジドフェニル)ジチオ]プロピオイミデート等の薬剤が挙げられる。
【0072】
他の修飾としては、グルタミニルおよびアルパラギニル残基の、それらの対応するグルタミルおよびアスパルチル残基へのそれぞれの脱アミノ化、プロリンおよびリジンのヒドロキシル化、セリルまたはスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニンおよびヒスチジン側鎖のα−アミノ基のメチル化[T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,San Francisco,pp.79−86(1983)]、N末端アミンのアセチル化、およびC末端カルボキシル基のアミド化が挙げられる。
【0073】
二官能性剤による誘導体化は、ここにおけるトール様レセプタとポリペプチドとの細胞内会合体の調製、ならびにアッセイまたはアフィニティー精製に使用される非水溶性支持体マトリックスまたは表面に対するこれらポリペプチドの架橋のために有用である。さらに、鎖間の架橋の研究は配座構造に関する直接の情報を提供しよう。通常用いられる架橋剤としては、1,1−ビス(ジアゾアセチル)−2−フェニルエタン、グルタルアルデヒド、N−ヒドロキシスクシンイミドエステル、ホモ二官能性イミドエステルおよび二官能性マレイミドが挙げられる。メチル−3−[(p−アジドフェニル)ジチオ]プロピオイミデートは、光の存在下に架橋を形成できる光活性化可能な中間体を作る。もしくは、臭化シアン活性化炭水化物等の反応性非水溶性マトリックスおよび米国特許第3,959,642号、同第3,969,287号、同第3,691,016号、同第4,195,128号、同第4,247,642号、同第4,229,537号、同第4,055,635号および同第4,330,440号に記載されたシステム反応性の非水溶性基質がタンパク質固定化と架橋に用いられる。
【0074】
本発明の範囲に含まれるPRO285、PRO286およびPRO358ポリペプチドの別の種類の共有修飾はポリペプチドの天然グリコシル化パターンを変えることからなる。「天然グリコシル化パターンを変える」とは、ここでの目的のために、(内在するグリコシル化部位を除去するか、または化学的および/または酵素的手段によりグリコシル化を欠失させることにより)天然配列中に見られる一つ以上の炭水化物部分を欠失させ、および/または天然配列中には存在しない一つ以上のグリコシル化部位を付加させることを意味する。さらに、この語句は、存在する炭水化物の性質と比率の変化を伴う天然タンパク質のグリコシル化の定性的変化を包含する。
【0075】
(DNA40021によりコードされる)天然の全長PRO285は、アミノ酸位置66、69、167、202、215、361、413、488、523、534、590、679、720、799および942で潜在的なN結合グリコシル化部位を有する。(DNA42663によりコードされる)天然の全長PRO286は、アミノ酸位置29、42、80、88、115、160、247、285、293、358、362、395、416、443、511、546、582、590、640、680、752,937および1026で潜在的なN結合グリコシル化部位を有する。
【0076】
PRO285、PRO286およびPRO358ポリペプチドに対するグリコシル化部位の付加はアミノ酸配列を変更することによって行ってよい。その変更は例えば天然配列に対して一つ以上のセリンまたはスレオニン残基の付加または置換により行ってよい(O結合グリコシル化部位の場合)。アミノ酸配列は、特にPRO285、PRO286およびPRO358ポリペプチドをコードするDNAを、所望のアミノ酸に翻訳するコドンが作られるように前選択された塩基において変異させることによるDNAレベルでの変化により任意に変更してよい。
【0077】
PRO285、PRO286およびPRO358ポリペプチド上の炭水化物の数を増加させる別の手段はグルコシドのポリペプチドへの化学的または酵素的な結合による。そのような方法は、例えば1987年9月11日公告のWO87/05330およびAplinとWriston,CRC Crit.Rev.Biochem.,pp.259−306(1981)に記載されている。
【0078】
PRO285、PRO286およびPRO358ポリペプチド上に存在する炭水化物部分の除去は化学的または酵素的に、またはグリコシル化の標的として働くアミノ酸残基をコードするコドンの変異置換により行ってよい。化学的脱グリコシル技術は公知であり、例えばHakimuddinら,Arch.Biochem.Biophys.,259:52(1987)およびEdgeら,Anal.Biochem.,118:131(1981)により記載されている。ポリペプチド上の炭水化物部分の酵素的切断はThotakuraら,Meth.Enzymol.,138:350(1987)に記載されたように多様なエンド−およびエキソ−グリコシダーゼを用いて行うことができる。
【0079】
別の共有修飾は、PRO285、PRO286およびPRO358ポリペプチドを多様な非タンパク質性ポリマー、例えばポリエチレングリコール(PEG)、ポリプロピレングリコールまたはポリオキシアルキレンに米国特許第4,640,835号、同第4,406,689号、同第4,801,144号、同第4,670,417号、同第4,791,192号または同第4,179,337号に記載された方法で連結することからなる。
【0080】
本発明のPRO285、PRO286およびPRO358ポリペプチドは、PRO285、PRO286、PRO358またはその断片を別の異種ポリペプチドまたはアミノ酸配列に融合させてなるキメラ分子を形成する方法で修飾してもよい。一実施態様において、そのようなキメラ分子は、抗タグ抗体が選択的に結合することのできるエピトープを提供するタグポリペプチドに対するPRO285、PRO286またはPRO358ポリペプチドの融合体からなる。エピトープタグは通常は天然または変異体PRO285、PRO286またはPRO358分子のN末端またはC末端に置かれる。そのようなエピトープのタグ付加形の存在はタグポリペプチドに対する抗体を用いることにより検出することができる。また、エピトープタグの提供により、PRO285、PRO286またはPRO358ポリペプチドが、エピトープタグに結合する抗タグ抗体または別の種類のアフィニティーマトリックスを用いるアフィニティー精製により容易に精製できる。
【0081】
多様なタグポリペプチドおよびそれらの抗体は当業界で公知である。具体例としては、ポリ−ヒスチジン(poly−his)またはポリ−ヒスチジン−グリシン(poly−his−gly)タグ;カゼHAタグポリペプチドおよびその抗体12CA5[Fieldら,Mol.Cell.Biol.,8:2159−2165(1988)];c−mycタグおよびそれに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evanら,Molecular and Cellular Biology,5:3610−3616(1985)];および単純ヘルペスウイルス糖タンパク質D(gD)タグおよびその抗体[Paborskyら,Protein Engineering,3(6):547−553(1990)]が挙げられる。他のタグポリペプチドとしてはFlagペプチド[Hoppら,BioTechnology,6:1204−1210(1988)];KT3エピトープペプチド[Martinら,Science,255:192−194(1992)];α−チュブリンエピトープペプチド[Skinnerら,J.Biol.Chem.,266:15163−15166(1991)];およびT7遺伝子10タンパク質ペプチドタグ[Lutz−freyermuthら,Proc.Natl.Acad.Sci.USA,87:6393−6397(1990)]が挙げられる。
【0082】
さらなる実施態様において、キメラ分子は、PRO285、PRO286またはPRO358ポリペプチドまたはその断片と免疫グロブリンまたは免疫グロブリンの特定の領域との融合体からなってよい。二価形のキメラ分子のために、そのような融合体はIgG分子等のIgのFc領域に対する融合であってよい。Ig融合体は、好ましくはIg分子内の少なくとも一つの可変部の代わりとしてPRO285、PRO286またはPRO358ポリペプチドの可溶の(トランスメンブレンドメイン欠失または不活性化)ポリペプチドを置換させた物を包含する。免疫グロブリン融合体の製造については、1995年6月27日発行の米国特許第5,428,130号も参照されたい。
【0083】
D.PRO285、PRO286およびPRO358ポリペプチドの調製
以下の説明は、主に、これらのタンパク質をコードする核酸(例えば、DNA40021、DNA42663およびDNA47361のそれぞれ)を有するベクターを用いて形質転換またはトランスフェクトした細胞を培養することによるPRO285、PRO286およびPRO358トール相同体の生産に関する。勿論、当分野で当業界で公知である別法を用いてPRO285、PRO286、PRO358またはそれらの変異体を調製することも意図される。例えば、PRO285、PRO286またはPRO358配列、またはそれらの一部を、固相技術を用いる直接のペプチド合成により作ってもよい[例えば、Stewartら,Solid−Phase Peptide Synthesis,W.H.Freeman Co.,San Fransisco,CA(1969);Merrifield,J.Am.Chem.Soc.,85:2149−2154(1963)を参照されたい]。インビトロのタンパク質合成は手動による技術を用いて、または自動化により行ってもよい。自動化された合成は例えばApplied Biosystemsペプチド合成機(米国、カリフォルニア州、フォスター市)を製造業者の指示にしたがって用いて行ってもよい。PRO285、PRO286またはPRO358の多様な部分は個々に化学合成し、それらを化学的または酵素的合成法を用いて組み合わせて全長PRO285、PRO286またはPRO358を作ってもよい。
【0084】
1.PRO285、PRO286またはPRO358をコードするDNAの単離
PRO285、PRO286またはPRO358をコードするDNAはPRO285、PRO286またはPRO358mRNAを有し、それを検出可能なレベルで発現すると思われる組織から調製されたcDNAライブラリーから得てもよい。したがって、PRO285、PRO286またはPRO358DNAは、実施例に記載されるようにヒト組織から調製されたcDNAライブラリーから便利に得ることができる。内在する遺伝子は染色体ライブラリーから、またはオリゴヌクレオチド合成によって得てもよい。実施例に記載のライブラリーに加えて、本発明のヒトトールタンパク質をコードするDNAは例えば脾臓細胞または抹消血液白血球(PBL)から単離することができる。
【0085】
ライブラリーは、対象とする遺伝子またはそれによってコードされるタンパク質を同定するように設計されたプローブ(例えば、PRO285、PRO286またはPRO358のタンパク質または少なくとも約20〜80塩基のオリゴヌクレオチドに対する抗体等)を用いてスクリーニングすることができる。選択されたプローブによるcDNAまたは染色体ライブラリーのスクリーニングは、例えばSambrookら,Molecular Cloning:A Laboratory Manual(New York:Cold Spring Harbor Laboratory Press,1989)によって記載されたような標準的な手法を用いて行うことができる。PRO285、PRO286またはPRO358をコードする遺伝子を単離する別の手段はPCR法を用いることである[上記のSambrookら;Dieffenbachら,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press,1995)]。
【0086】
下記の実施例はcDNAライブラリーのスクリーニング技術を説明する。プローブとして選択されたオリゴヌクレオチド配列は、偽陽性を最小化するように十分な長さを有し、十分に明確なものでなければならない。オリゴヌクレオチドは、スクリーニングするライブラリー中のDNAとのハイブリダイゼーションの際に検出できるように好ましくは標識する。標識方法はよく知られており、32P標識ATP、ビオチニル化または酵素標識等の放射能標識の利用を包含する。適度なストリンジェンシーや高いストリンジェンシー等のハイブリダイゼーション条件は上記のSambrookらにより提供されている。
【0087】
そのようなライブラリースクリーニング法で同定される配列は、GenBank等の公共のデータベースまたは他の民間の配列データベースに寄託され利用可能な配列と比較し、アライメントさせることができる。分子の定められた領域内または全長配列に沿う(アミノ酸またはヌクレオチドレベルでの)配列の同一性は、相同性/配列同一性を測定するための多様なアルゴリズムを使用するALIGN、DNAstarおよびINHERIT等のコンピュータソフトウエアプログラムを用いる配列アラインメントにより決定することができる。
【0088】
タンパク質をコードする配列を有する核酸は、cDNAに逆転写されていないmRNAの前駆体またはプロセス中間体を検出するために、ここで最初に開示された類推アミノ酸配列を用いて、必要であれば上記のSambrookらに記載された慣用のプライマー伸長法を用いて、選択されたcDNAまたは染色体ライブラリーをスクリーニングすることによって得てもよい。
【0089】
2.宿主細胞の選択と形質転換
ヒトトールタンパク質の生産のためにここに記載された発現ベクターまたはクローニングベクターを用いて宿主細胞をトランスフェクトまたは形質転換し、プロモータを誘導し、形質転換体を選択し、かつ所望の配列をコードする遺伝子を増幅するために適切に改良された慣用の栄養培地で培養する。培地、温度、pH等の培養条件は当業者らにより過度な実験なしに選択することができる。通常、細胞培養物の生産性を最大化する原理、方法および実用的な技術は、Mammalian Cell Biotechnology:a Practical Approach,M.Butler,編(IRLプレス、1991)および上記のSambrookらに見ることができる。
【0090】
トランスフェクション法は当業者に知られており、例えばCaPO4およびエレクトロポレーションがある。使用される宿主細胞に基づいて、形質転換はそのような細胞に適する標準的な技術を用いて行う。上記のSambrookらにより記載された塩化カルシウムを用いるカルシウム処理、またはエレクトロポレーションが、実質的な細胞壁障壁を有する原核細胞または他の細胞に一般的に用いられる。Agrobacterium tumefaciensによる感染は、Shawら,Gene,23:315(1983)および1989年6月29日公開のWO89/05859に記載されているように、ある種の植物細胞の形質転換に用いられる。そのような細胞壁のない哺乳類の細胞に関しては、Grahamとvander編のリン酸カルシウム法,Virology,52:456−457(1978)を用いることができる。哺乳類細胞宿主システム形質転換の一般的な特徴は米国特許第4,399,216号に記載されている。酵母の形質転換は典型的にはVan Solingenら,J.Bact.,130:946(1977)およびHsiaoら,Proc.Natl.Acad.Sci.(USA),76:3829(1979)の方法にしたがって実施する。しかし、核マイクロインジェクション、エレクトロポレーション、そのままの細胞との細菌プロトプラスト融合、またはポリブレン、ポリオルニチン等のポリカチオン等によりDNAを細胞に導入する他の方法も用いてよい。哺乳類細胞の形質転換の多様な方法に関しては、Keownら,Methods in Enzymology,185:527−537(1990)およびMansourら,Nature,336:348−352(1988)を参照されたい。
【0091】
ここでのベクター中のDNAのクローニングまたは発現に適する宿主細胞として、原核細胞、酵母または高等真核細胞が挙げられる。適当な原核細胞としては、真正細菌、例えばグラム陰性、またはグラム陽性細菌、例えば大腸菌等のEnterobacteriaceaeが挙げられるが、これらに限定されない。大腸菌K12株MM294(ATCC 31,446);大腸菌X1776(ATCC 31,537);大腸菌株W3110(ATCC 27,325)およびK5 772(ATCC 53,635)等の多様な大腸菌株が一般に利用可能である。
【0092】
原核細胞に加えて、糸状菌または酵母等の真核微生物が、ヒトトールをコードするベクターの適当なクローニング宿主または発現宿主である。Sacccharomyces cerevisiaeが通常用いられる下等真核宿主微生物である。
【0093】
グリコシル化ヒトトールタンパク質の発現に適する宿主細胞は多細胞生物から得られる。無脊椎動物細胞の具体例としては、ショウジョウバエS2およびSpodotera Sf9等の昆虫細胞、ならびに植物細胞が挙げられる。有用な宿主細胞系としてはチャイニーズハムスター卵巣(CHO)およびCOS細胞が挙げられる。さらに具体的な例としては、SV40で形質転換されたサル腎臓CV1細胞系(COS−7、ATCC CRL 1651);ヒト胎児腎臓細胞系(懸濁培養物中での増殖のためにサブクローニングされた293または293細胞、Grahamら,J.Gen Virol.,36:59(1977);チャイニーズハムスター卵巣細胞/−DHFR(CHO,Urlaub and Chasin,Proc.Natl.Acad.Sci.USA,77:4216(1980);マウスセルトリー細胞(TM4,Mather,Biol.Reprod.,23:243−251(1980);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝臓細胞(HEPG2,HB 8065);およびマウスの乳せん腫瘍(MMT 060562,ATCC CCL51)が挙げられる。適当な宿主細胞の選択は当業者の技量内にあるであろう。
【0094】
3.複製可能なベクターの選択と使用
PRO285、PRO286またはPRO358をコードする核酸(例えばcDNAまたは染色体DNA)はクローニング(DNAの増幅)用または発現用の複製可能なベクターに挿入してよい。多様なベクターが一般に利用できる。例えば、ベクターはプラスミド、コスミド、ウイルス粒子またはファージの形であってよい。適当な核酸配列はベクターに多様な方法により挿入してよい。一般的に、DNAは公知の方法を用いて適当な制限エンドヌクレアーゼ部位に挿入させる。通常、ベクター成分としてはシグナル配列、複製開始部位、一種類以上のマーカー遺伝子、エンハンサエレメント、プロモータおよび転写終止配列が挙げられるが、これらに限定されない。これらの成分を一つ以上を有する適当なベクターの構築は当業者に公知の標準的な連結技術を用いる。
【0095】
PRO285、PRO286およびPRO358タンパク質は、組換えにより直接に作ってもよいのみならず、成熟タンパク質またはポリペプチドのN末端に特定の切断部位を有するシグナル配列または他のポリペプチドであってよい異種ポリペプチドとの融合ポリペプチドとして作ってもよい。通常、シグナル配列はベクターの一成分であってよく、またはベクターに挿入されたPRO285、PRO286またはPRO358DNAの一部であってよい。シグナル配列は、例えば、アルカリホスファターゼ、ペニシニリナーゼ、lpp、または熱安定エンテロトキシンIIリーダーの群から選択される原核生物シグナル配列であってよい。酵母での分泌のために、シグナル配列は、例えば酵母インベルターゼリーダー、アルファ因子リーダー(SaccharomycesとKluyveromycesのα因子リーダー等であり、後者は米国特許第5,010,182号に記載されている)、または酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー(1990年4月4日公開のEP362,179)、または1990年11月15日公開のWO90/13646に記載のシグナルであってよい。哺乳類細胞での発現において、タンパク質分泌を指令するため、同一または関連種の分泌ポリペプチドのシグナル配列等の哺乳類のシグナル配列ならびにウイルス分泌リーダー用いてよい。
【0096】
発現ベクターおよびクローニングベクターは両者ともに、一種類以上の選択された宿主細胞中でベクターの複製を可能とする核酸配列を有する。そのような配列は多様な細菌、酵母およびウイルスに関して当業界で公知である。プラスミドpBR322の複製開始部位はほとんどのグラム陰性細菌に適し、2μプラスミド開始部位は酵母に適し、多様なウイルス開始部位(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)は哺乳動物細胞中のクローニングベクターに有用である。
【0097】
発現ベクターとクローニングベクターは典型的には、選択マーカーとも呼ばれる選択遺伝子を有しよう。典型的な選択遺伝子は、(a)抗生物質または他の毒素、例えば、アンピシリン、ネオマイシン、メトトレキセートまたはテトラサイクリンに対する耐性を付与するか、(b)栄養要求性欠損を相補するか、または(c)複合培地からは利用できない重要な栄養素を供給するタンパク質をコードするもので、例えばBacilliのDアラニンラセマーゼをコードする遺伝子がある。
【0098】
哺乳類細胞の適当な選択マーカーの具体例は、PRO285、PRO286またはPRO358核酸を取り込む能力のある細胞の同定を可能とするもので、例えばDHFRまたはチミジンキナーゼである。野生型DHFRが用いられる場合の適当な宿主細胞は、Urlaubら,Proc.Natl.Acad.Sci.USA,77:4216(1980)により記載されているように調製され、増殖させたDHFR活性を欠損したCHO細胞系である。酵母に用いられる適当な選択遺伝子は酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら,Nature,282:39(1979);Kingsmanら,Gene,7:141(1979);Tschemerら,Gene,10:157(1980)]。該trp1遺伝子は、トリプトファンでの生育能を欠く酵母変異株、例えばATCC No.44076またはPEP4−1用の選択マーカーを提供する[Jones,Genetics,85:12(1977)]。
【0099】
発現ベクターおよびクローニングベクターは、通常は、mRNA合成を指令するために、PRO285、PRO286またはPRO358タンパク質をコードする核酸配列に操作可能に連結させたプロモータを有する。多様な潜在的宿主細胞に認識されるプロモータは当業界で公知である。原核生物宿主での使用に適するプロモータとしては、β−ラクタマーゼおよびラクトースプロモータシステム[Changら,Nature,275:615(1978);Goeddelら,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモータシステム[Goeddel,Nucleic Acids Res.,8:4057(1980);EP36,776]およびtacプロモータ等のハイブリッドプロモータ[deBoerら,Proc.Natl.Acad.Sci.USA,80:21−25(1983)]が挙げられる。細菌システムに使用されるプロモータもPRO285、PRO286またはPRO358をコードするDNAに操作可能に連結させたShine−Dalgarno(S.D.)配列を有しよう。
【0100】
酵母宿主に使用される適当なプロモータ配列の具体例としては、3−ホスホグリセレートキナーゼのプロモータ[Hitzemanら,J.Biol.Chem.,255:2073(1980)]または他の解糖系酵素[Hessら,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)]、例えばエノラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ、ヘキソキナーゼ、ピルベートデカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−ホスフェートイソメラーゼ、3−ホスホグリセレートムターゼ、ピルベートキナーゼ、トリオースホスフェートイソメラーゼ、ホスホグルコースイソメラーゼおよびグルコキナーゼが挙げられる。
【0101】
他の酵母プロモータは成長条件により制御を受ける転写の付加的な利点を有する誘導性プロモータであって、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸性ホスファターゼ、窒素代謝に関連した分解酵素、メタロチオネイン、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼおよびマルトースやガラクトースの利用に関わる酵素のプロモータ領域である。酵母発現に使用される適当なベクターとプロモータはEP73,657にさらに記載されている。
【0102】
哺乳類の宿主細胞中のベクターからのPRO285、PRO286またはPRO358転写は、例えばポリオーマウイルス、鶏痘ウイルス(1989年7月5日発行のUK2,211,504)、アデノウイルス(アデノウイルス2等)、ウシ乳頭腫ウイルス(SV40)、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびサルウイルス40(SV40)等のウイルスのゲノムから得られるプロモータ、異種哺乳類プロモータ、例えばアクチンプロモータまたは免疫グロブリンプロモータおよび熱ショックプロモータにより制御を受ける(但し、そのようなプロモータが宿主細胞系に適合性がある場合である)。
【0103】
PRO285、PRO286またはPRO358ポリペプチドをコードするDNAの高等真核生物による転写はヘンハンサー配列をベクターに挿入することにより高めてもよい。エンハンサは、通常約10〜300bpのDNAのシス作用エレメントであり、プロモータに作用してその転写を高める。哺乳類の(グロビン、エラスターゼ、アルブミン、α−フェトタンパク質およびインシュリン)遺伝子に由来する多くのエンハンサ配列が現在公知である。しかし、典型的には、真核細胞ウイルスのエンハンサが用いられよう。具体例としては、複製起源の後期側に位置するSV40(bp100〜270)、サイトメガロウイルス初期プロモータエンハンサ、複製開始部位の後期側に位置するポリオーマエンハンサおよびアデノウイルスエンハンサが挙げられる。エンハンサは、PRO285、PRO286またはPRO358をコードする配列に対して5’または3’の位置でベクター中にスプライスさせてよいが、好ましくはプロモータから5’の側に配置する。
【0104】
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、または他の多核生物に由来する有核細胞)に用いられる発現ベクターは転写の終止およびmRNAの安定化に必要な配列も有しよう。そのような配列は、通常は真核生物またはウイルスDNAまたはcDNAの5’および場合によっては3’の非翻訳領域から利用可能である。これらの領域はPRO285、PRO286またはPRO358をコードするmRNAの非翻訳部分のポリアデニル化断片として転写されるヌクレオチドセグメントを有する。
【0105】
組換え脊椎動物細胞培養物中でのPRO285、PRO286またはPRO358の合成に適合に適するさらに別の方法、ベクターおよび宿主細胞はGethingら,Nature,293:620−625(1981);Manteiら,Nature,281:40−46(1979);EP117,060;およびEP117,058に記載されている。
【0106】
4.遺伝子増幅の検出/発現
試料中の遺伝子増幅および/または発現は、ここに提供される配列に基づく適当に標識されたプローブを用いて、例えば、mRNAの転写を定量する慣用のサザンブロッティング、ノーザンブロッティング[Thomas,Proc.Natl.Acad Sci.USA,77:5201−5205(1980)]、ドットブロッティング(DNA合成)、またはin situハイブリダイゼーションにより直接測定してもよい。もしくは、特定の二本鎖、例えばDNA二本鎖、RNA二本鎖およびDNA−RNAハイブリッド二本鎖またはDNA−タンパク質二本鎖を認識できる抗体を用いてもよい。さらに、該抗体を標識してもよく、アッセイを行って二本鎖を表面に結合させ、該表面上での二本鎖の形成によって、二本鎖に結合した抗体の存在が検出できる。
【0107】
もしくは、遺伝子発現は、細胞または組織切片の免疫組織化学染色、細胞培養物または体液のアッセイ等の免疫学的方法により測定して、遺伝子産物の発現を直接に定量してもよい。試料液の免疫組織化学染色および/またはアッセイに有用な抗体はモノクローナルまたはポリクローナルのいずれでもよく、哺乳類中に作ってもよい。便利には、天然配列PRO285、PRO286またはPRO358ポリペプチドに対して、またはここに提供されたDNA配列に基づく合成ペプチドに対して、またはPRO285、PRO286またはPRO358DNAに融合させ、かつ特定の抗体エピトープをコードする外因性配列に対して調製してもよい。
【0108】
5.ポリペプチドの精製
PRO285、PRO286またはPRO358の形態は培養培地または宿主細胞溶解物から回収してよい。膜に結合している場合、適当な変性溶液(例えば、Triton−X100)または酵素による切断を用いて膜から放出させることができる。PRO285、PRO286またはPRO358の発現に用いられる細胞は、凍結/解凍の繰返し、超音波処理、機械的破壊または細胞溶解剤等の多様な物理的または化学的手段により破壊することができる。
【0109】
組換え細胞タンパク質またはポリペプチドからPRO285、PRO286またはPRO358を精製することも望ましいであろう。下記の方法は適当な精製方法の例示である:イオン交換カラムによる分画;エタノール沈殿;逆相HPLC;シリカまたはカチオン交換樹脂、例えばDEAEによるクロマトグラフィー;クロマトフォーカシング;SDS−PAGE;硫安沈殿;例えばSephadex G−75を用いるゲルろ過;IgG等の不純物を除去するためのプロテインAセファロースカラム;およびトールタンパク質のエピトープタグ付加の形態に結合する金属キレートカラム。タンパク質精製の多様な方法を用いてよく、そのような方法は公知であり、例えばDeutscher,Methods in Enzymology,182(1990)およびScopes,Protein Purification:Principles and Practics,Springer−Verlag,New York(1982)に記載されている。選択される精製方法は、例えば使用される精製法および生産される特定のトールタンパク質の性質に依存しよう。
【0110】
E.トールタンパク質に関する使用とコードする核酸
本発明のトールタンパク質をコードする塩基配列(またはそれらの相補物)は、ハイブリダイゼーションプローブとして、染色体および遺伝子地図作成において、およびアンチセンスRNAおよびDNAの生成における使用等、分子生物学の技術において多様な利用性を有する。トール核酸は、ここに記載の組換え技術によるPRO285、PRO286およびPRO358ポリペプチドの調製にも有用であろう。
【0111】
PRO285、PRO286およびPRO358をそれぞれコードする全長の天然配列DNA40021、DNA42663およびDNA47361遺伝子、またはそれらの部分は、cDNAライブラリーのハイブリダイゼーションプローブとして用いて、該全長遺伝子を単離するか、または図1、3および12A〜Bにそれぞれに開示されたPRO285、PRO286またはPRO358配列に対して所望の配列同一性を有するさらに別の遺伝子(例えば、PRO285、PRO286またはPRO358の天然変異体、またはそれらのさらなるヒト相同体、または他の種からの相同体をコードする遺伝子)を単離することができる。プローブの長さは任意に約20〜約50塩基である。ハイブリダイゼーションプローブは図2と3(配列番号2)、または図5から7(配列番号4)、または図21〜22(配列番号14)の塩基配列または天然配列のプロモータ、エンハンサエレメントおよびイントロン等のゲノム配列から得てもよい。例えば、スクリーニング法は、公知のDNA配列を用いるPRO285、またはPRO286、またはPRO358遺伝子のコード領域を単離して約40塩基の選択プローブを合成することからなるだろう。ハイブリダイゼーションプローブは、32Pまたは35S等のラジオヌクレオチド、またはアビジン/ビオチン結合システムによりプローブに結合させたアルカリホスファターゼ等の酵素標識等の多様な標識により標識してよい。本発明のPRO285、PRO286またはPRO358遺伝子(DNA40021、42663および47361)に相補的な配列を有する標識プローブを用いて、ヒトcDNA、染色体DNAまたはmRNAのライブラリーをスクリーニングして、該ライブラリーのどのメンバーに該プローブがハイブリダイズするかを決定する。ハイブリダイゼーション技術は下記の実施例でさらに詳細に説明する。
【0112】
該プローブをPCR技術に用いて密接に関連するトール配列の同定用の配列プールを作ってよい。
ここでトールタンパク質をコードするヌクレオチド配列を用いて、該トールタンパク質をコードする遺伝子の地図作成および遺伝病を有する個人の遺伝的分析のためにハイブリダイゼーションプローブを構築することもできる。ここで提供されるヌクレオチド配列は、公知の技術、例えばin situハイブリダイゼーション、公知の染色体マーカーに対する連結分析およびライブラリーに対するハイブリダイゼーションスクリーニングを用いて染色体および染色体の特別な領域に対して地図を作成してよい。
【0113】
本発明のヒトトールタンパク質はトール仲介シグナル変換に関与する他のタンパク質または分子を同定するアッセイにも用いることができる。例えば、PRO285、PRO286およびPRO358は、ヒトトールの現時点では未知の天然リガンド、またはキナーゼ関連の潜在的トールレセプタ等のヒトトールレセプタによる活性化またはシグナルに(直接または間接的に)関与する他の因子を同定するのに有用である。さらに、レセプタ/リガンド結合相互作用の阻害物質を同定できる。そのような結合相互作用に関与するタンパク質は、結合相互作用のペプチドまたは小分子の阻害物質またはアゴニストのスクリーニングに用いることもできる。スクリーニングアッセイを設計して、天然トールポリペプチドまたは天然のトールポリペプチドのリガンドの生物学的活性に似た主要な化合物を発見することができる。そのようなスクリーニングアッセイは、化学的ライブラリーの高処理量のスクリーニングが可能なアッセイを包含するために、アッセイは小分子医薬候補の同定に特に適する。意図される小分子は合成または無機化合物を包含する。これらのアッセイは、当分野でよく特徴付けられているタンパク質−タンパク質結合アッセイ、生化学スクリーニングアッセイ、免疫アッセイおよび細胞に基づくアッセイ等の多様な形態で実施することができる。
【0114】
インビトロアッセイはトールレセプタポリペプチド等の成分の混合物を用い、該ポリペプチドは別のペプチドまたはポリペプチド、例えば検出用または固定用タグ等との融合生成物の一部であってよい。アッセイ混合物は(結合アッセイのために)天然の細胞内または細胞外のトールが結合する標的(すなわち、トールリガンド、またはトールレセプタにより活性化し、および/またはシグナルを発することの公知の別の分子)をさらに有してもよい。天然の結合標的を用いてよいが、そうした天然の結合標的(例えばペプチド)部分がアッセイで便利に測定可能な対象トールタンパク質に対して結合親和性と親和力を提供する場合、そうした部分を用いることがしばしば望ましい。アッセイ混合物は候補薬理剤も含有する。候補試薬は多様な薬品の種類を包含するが、それらは典型的には有機化合物であり、好ましくは小有機化合物であり、合成または天然化合物のライブラリー等の多様なソースから得られる。塩、緩衝液、中性タンパク質、例えばアルブミン、変性剤、プロテアーゼインヒビター、ヌクレアーゼインヒビター、殺菌剤等の他の多様な試薬も該混合物に含有させてもよい。
【0115】
インビトロ結合アッセイにおいて、得られた混合物を、候補分子の存在を除けば、トールタンパク質が参照結合親和性により細胞結合標的、部分または相同体に特異的に結合する条件下にインキュベートする。該混合物成分は、必要な結合を提供するいかなる順番でも加えることができ、インキュベーションは最適な結合を容易にするいかなる温度で実施してもよい。インキュベーション時間は、最適な結合のために選択するが、迅速な高処理量のスクリーニングを容易にするように最小の時間とする。
【0116】
インキュベーション後、トールタンパク質と一つ以上の結合標的との試薬バイアス結合を慣用の技術により検出する。無細胞結合タイプのアッセイのために、結合物を未結合物から分離するために分離工程がよく用いられる。分離は沈殿(例えば、TCA沈殿、免疫沈殿等)、固定化(例えば固体基質への固定化)等の後に例えば膜によるろ過(例えばワットマンP−18イオン交換紙、Polyfitronicの疎水GFC膜等)、ゲルクロマトグラフィー(例えばゲルろ過、アフィニティー等)による洗浄によって行ってもよい。トール依存転写アッセイについては、結合をトール依存レポーターの発現の変化により検出する。
【0117】
検出は慣用の方法によって行えばよい。無細胞結合アッセイのために、通常、成分の一つは標識からなるか、また標識に結合させる。該標識は放射性、発光、光学的または電子密度等としての直接の検出、またはエピトープタグ、酵素等の間接的な検出を提供しよう。多様な方法が、標識の性質および他のアッセイ成分に基いて、例えば、光学的または電子密度、放射性放出、非放射性エネルギー転移等を用いて標識を検出してよく、または抗体複合物によって間接的に検出してもよい。
【0118】
PRO285、PRO286またはPRO358をコードする核酸、またはそれらの修飾形を用いてトランスジェニック動物または「ノックアウト」動物を作ることもでき、これらの動物は治療に有用な試薬の開発やスクリーニングに有用である。トランスジェニック動物(例えば、マウスまたはラット)はトランスジーンを含有する細胞を有する動物であり、該トランスジーンは該動物に導入されたか、または該動物の親等に胎児期で、例えば胚の段階で導入されたものである。トランスジーンはトランスジェニック動物を作った細胞のゲノムに組み込まれるDNAである。一実施態様において、確立された技術にしたがって、PRO285またはPRO286をコードするcDNAを用いてPRO285、PRO286またはPRO358をコードするゲノムDNAをクローニングでき、ゲノム配列を用いて、PRO285、PRO286またはPRO358をコードするDNAを発現する細胞を有するトランスジェニック動物を作ることができる。トランスジェニック動物、特にマウスまたはラット等の動物を作る方法は当分野では慣用となっており、例えば米国特許第4,736,866号と同第4,870,009号に記載されている。典型的には、特定の細胞が組織特異的エンハンサによるトランスジーン導入の標的となる。胚の段階に動物の生殖細胞系列に導入されたPRO285、PRO286またはPRO358をコードするトランスジーンのコピーを有するトランスジェニック動物を用いて、PRO285、PRO286またはPRO358をコードするDNAの増強した発現の効果を調べることができる。そのような動物は、例えばその過剰発現に関連した病理状態からの保護を付与すると考えられる試薬に関する試験動物として用いることができる。本発明のこの特徴にしたがって、動物を試薬で処置し、トランスジーンを有する未処置の動物と比較した場合の病理状態の低下率が該病理状態についての潜在的な治療的介在を示すものであろう。
【0119】
もしくは、PRO285またはPRO286またはPRO358の非ヒト脊椎動物(例えば哺乳類)の相同体を用いて、PRO285、PRO286またはPRO358タンパク質をコードする内在遺伝子と、該動物の胚細胞に導入されたPRO285、PRO286またはPRO358をコードする変更染色体遺伝子との均一組換えの結果として、PRO285またはPRO286またはPRO358をコードする、欠陥または変更遺伝子を有する「ノックアウト」動物を作ることができる。例えば、PRO285、PRO286またはPRO358をコードするcDNAを用いて、PRO285、PRO286またはPRO358をコードする染色体DNAを確立された技術にしたがってクローニングすることができる。PRO285、PRO286またはPRO358をコードする染色体DNAの一部を欠失させるか、または別の遺伝子、例えば統合をモニターするために用いることのできる選択マーカーをコードする遺伝子に置換させることができる。典型的には、(5’と3’の両末端において)数キロベースの未変更の隣接DNAをベクターに含有させる[例えば、均一組換えベクターの記載に関しては、例えばThomas and Chepecchi,Cell,51:503(1987)を参照されたい]。ベクターを(例えばエレクトロポレーションにより)胚幹細胞系に導入し、この導入されたDNAが内在DNAと均一組換えした細胞を選択する[例えば、Liら,Cell,69:915(1992)を参照されたい]。次に、選択された細胞を動物(例えばマウスまたはラット)の胚盤胞に注入して集合体キメラを形成する[例えば、Bradley,Teratocarcinomas and Embryonic Stem Cells:A Practical Approach,E.J.Robertson,編(IRL,Oxford,1987),pp.113−152を参照されたい]。キメラ胚を次に適当な疑妊娠メス親動物に移植し、胚を生育させて「ノックアウト」動物を創出する。均一組換えDNAを生殖細胞に有する子孫は標準的な技術をにより同定することができ、これを用いて、動物のすべての細胞が均一組換えDNAを有する動物を繁殖させる。ノックアウト動物は、例えば、ある種の病理状態に対する防御能やPRO285、PRO286またはPRO358ポリペプチドがないことによる病理状態の進展のために特徴付けることができる。
【0120】
ここに開示されたトールポリペプチドをコードする核酸は遺伝子治療に用いることもできる。遺伝子治療への利用において、遺伝子を細胞に導入して、例えば欠陥遺伝子の置換のために治療効果のある遺伝子産物のインビボ合成を行う。「遺伝子治療」とは、持続する効果が一回の治療により得られる通常の遺伝子治療と、治療効果のあるDNAまたはmRNAの一回または繰り返しの投与をともなう遺伝治療剤の投与との両方を含む。アンチセンスRNAおよびDNAはある種の遺伝子の発現をインビボでブロックする治療薬として用いることができる。短いアンチセンスオリゴヌクレオチドを細胞に移入させて、細胞内で細胞膜の限定された取込みにより引き起こされたそれらの低い細胞内濃度にもかかわらず阻害物質として作用させることができることが既に示されている(Zamecnikら,Proc.Natl.Acad.Sci.USA 83,4143−4146[1986])。これらのオリゴヌクレオチドは、例えばそれらの負電荷ホスホジエステルグループを未電荷のグループに置換することによりそれらの取込みを高めるように修飾することができる。
【0121】
核酸を生きた細胞に導入するのに利用することのできる多様な技術が存在する。これらの技術は、核酸を培養細胞にインビボで移すのか、または意図される宿主の細胞にインビボで移すのかにしたがって変えられる。核酸を哺乳類細胞にインビトロで移すのに適する技術としては、リポソームの利用、エレクトロポレーション、マイクロインジェクション、細胞融合、DEAE−デキストラン、リン酸カルシウム沈殿法等が挙げられる。現在好ましいインビボ遺伝子転移技術としては、ウイルス(典型的にはレトロウイルス)ベクターによるトランスフェクションとウイルスコートタンパク質−リポソーム仲介トランスフェクションとが挙げられる(Dzauら,Trends in Biotechnology 11,205−210[1993])。ある場合では、細胞表面膜タンパク質または標的細胞に対して特異性のある抗体、標的細胞上のレセプタに対するリガンド等、標的細胞を標的とする試薬を核酸ソースに提供することが望ましい。リポソームを使用する場合、エンドサイトーシスに関連する細胞表面膜タンパク質に結合するタンパク質は、標的化および/または例えば特定の細胞タイプに引き付けられるキャプシドタンパク質またはその断片、サイクリングで内在化を受けるタンパク質に対する抗体、細胞内局在化を標的とし、細胞内半減期を向上させるタンパク質の取り込みを容易するために用いてよい。レセプタ仲介エンドサイトーシスの技術は、例えば、Wuら,J.Biol.Chem.262,4429−4432(1987)およびWagnerら,Proc.Natl.Acad.Sci.USA 87,3410−3414(1990)によって記載されている。最近公知の遺伝子製造と遺伝子治療方法の概説については、Andersonら,Science 256,808−813(1992)を参照されたい。
トールタンパク質に関連してここで挙げた多様な利用は、天然トールレセプタの少なくとも一つの生物学的機能に似た、天然トールレセプタのアゴニストにも利用できる。
【0122】
F.抗トールタンパク質抗体
さらに、本発明は抗トールタンパク質抗体を提供する。例示的な抗体として、ポリクローナル、モノクローナル、ヒト化、二特異性およびヘテロ複合抗体が挙げられる。
【0123】
1.ポリクローナル抗体
抗トールタンパク質抗体はポリクローナル抗体からなってもよい。ポリクローナル抗体の調製方法は当業者で公知である。ポリクローナル抗体は、例えば免疫剤および所望であればアジュバントの一回以上の注射により哺乳類中に生じさせることができる。典型的には、免疫剤および/またはアジュバントを複数回の皮下または腹腔内注射により哺乳類に注射する。免疫剤はPRO285およびPRO286ポリペプチドまたはその融合タンパク質を含有してよい。免疫剤を、免疫される哺乳動物で免疫原性のあることが公知のタンパク質に複合させることが有用であろう。そのような免疫原性タンパク質の具体例としては、キーホールリンペットヘモシアニン、血清アルブミン、ウシチログロビンおよび大豆トリプシンインヒビターが挙げられるが、これらに限定されない。使用できるアジュバントの具体例としてはフロイントの完全アジュバントおよびMPL−TDMアジュバント(モノホスホリルリピドA、合成トレハロースジコリノミコレート)が挙げられる。免疫法は当業者によって過度な実験なしに選択できよう。
【0124】
2.モノクローナル抗体
もしくは、抗トールタンパク質抗体はモノクローナル抗体であってよい。モノクローナル抗体は、KohlerとMilstein,Nature,256:495(1975)によって記載されたようなハイブリドーマ法を用いて調製してよい。ハイブリドーマ法において、マウス、ハムスターまたは他の適当な宿主動物を典型的には免疫剤で免疫して、該免疫剤に特異的に結合する抗体を産生するか、もしくは産生することのできるリンパ球を誘引する。あるいは、リンパ球をインビトロで免疫しうる。
【0125】
免疫剤としては、典型的には、PRO285、PRO286またはPRO358ポリペプチドまたはそれらの融合タンパク質が挙げられる。一般的には、ヒト起源の細胞を望む場合、抹消血液リンパ球(「PBL」)を用い、または非ヒト哺乳類ソースを望む場合、脾臓細胞またはリンパ節細胞を用いる。次に、該リンパ球を、ポリエチレングリコール等の適当な融合試薬を用いて不死化細胞系に融合させてハイブリドーマ細胞を形成させる[Goding,Monoclonal Antibodies:Principles and Practice,Academic Press,(1986)pp.59−103]。不死化細胞系は通常は悪性化した哺乳類細胞、特にネズミ、ウシおよびヒト起源の骨髄腫細胞である。通常は、ラットもしくはネズミ骨髄腫細胞系を用いる。該ハイブリドーマ細胞を、好ましくは、未融合の不死化細胞の増殖や生存を妨げるために一種類以上の物質を含む適当な培養培地で培養してよい。例えば、親細胞がヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマの培養培地は典型的にはHGPRT欠損細胞の成長を妨げる物質であるヒポキサンチン、アミノプテリンおよびチミジン(「HAT培地」)を含有するものとなろう。
【0126】
好ましい不死化細胞系は、選択された抗体産生細胞による安定で高レベルの抗体発現を効率的に支持し、HAT培地等の培地に感受性のあるものである。さらに好ましい不死化細胞系は、例えば米国カリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerまたはメリーランド州ロックビルのAmerican Type Culture Collectionから入手することのできるネズミ骨髄腫細胞系である。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞系もヒトモノクローナル抗体の生産に関して記載されている[Kozbor,J.Immunol.,133:3001(1984);Brodeurら,Monoclonal Antibody Production Techniques and Applications,Marcel Dekker,Ink.,New York(1987)pp.51−63]。
【0127】
ハイブリドーマ細胞が培養される細胞培地は、次にPRO285、PRO286またはPRO385に対するモノクローナル抗体の存在についてアッセイすることができる。好ましくは、ハイブリドーマ細胞によって作られるモノクローナル抗体の結合特異性は、免疫沈殿またはラジオイムノアッセイ(RIA)または酵素結合免疫吸着法(ELISA)等のインビトロアッセイにより測定される。そうした技術やアッセイは公知である。モノクローナル抗体の結合親和性は、例えば、MunsonおよびPollardのSatchard分析、Anal.Biochem.,107:220(1980)によって測定することができる。
【0128】
所望のハイブリドーマ細胞が同定されたら、クローンを限界希釈法によりサブクローニングし、標準的な方法により増殖させる[上記のGoding]。この目的の培養培地としては、例えば、ダルベッコの改良イーグル培地およびRPMI1640培地が挙げられる。もしくは、ハイブリドーマ細胞を哺乳類中の腹水としてインビボで増殖させてもよい。
【0129】
サブクローンにより分泌されたモノクローナル抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィー等の慣用の免疫グロブリン精製方法により培養培地または腹水から単離し、精製してよい。
【0130】
モノクローナル抗体は、組換えDNA法、例えば米国特許第4,816,567号に記載された方法により作ってもよい。本発明のモノクローナル抗体をコードするDNAは慣用の方法を用いて(例えば、げっ歯類の抗体の重鎖と軽鎖をコードする遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを用いることにより)容易に単離して配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましいソースとして働く。いったん単離されたら、DNAを発現ベクターに入れてよく、次にそれ自体では免疫グロブリンタンパク質を作らないサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞または骨髄腫細胞に該ベクターをトランスフェクトして、組換え宿主細胞中でモノクローナル抗体の合成を得る。該DNAは例えばげっ歯類の相同配列の代わりにヒトの重鎖または軽鎖定常ドメインに置き換えることにより[米国特許弟4,816,567号;上記のMorrisonら]、または該免疫グロブリンをコードする配列に非免疫グロブリンポリペプチドのコード配列の全部または一部を共有結合させることにより修飾してもよい。そのような非免疫グロブリンポリペプチドは、本発明抗体の定常ドメインの代わりとするか、または本発明抗体の抗原結合部位の可変ドメインの代わりとしてキメラ二価抗体を作ることができる。
【0131】
抗体は一価抗体であってよい。一価抗体の調製方法は当業界で公知である。例えば、ある方法では免疫グロブリンの軽鎖と修飾重鎖の組換え発現をともなう。重鎖は、通常、重鎖の架橋を妨げるようにFc領域のいずれかの位置で端が切り取られている。もしくは、関連するシステイン残基を別のアミノ酸残基によって置換するか、または架橋を妨げるように欠失させる。
インビトロ方法も一価抗体の調製に適する。抗体を消化し、その断片、特にFab断片を作るのは日常的な公知の方法を用いて行うことができる。
【0132】
3.ヒト化およびヒト抗体
本発明の抗トール抗体はさらにヒト化抗体またはヒト抗体からなってよい。非ヒト(例えば、ネズミ)抗体のヒト化形態は、非ヒト免疫グロブリンに由来する最小の配列を有するキメラ免系グロブリン、その免疫グロブリン鎖またはその断片(例えば、抗体のFv、Fab、Fab’、F(ab’)2または他の抗原結合副配列)である。ヒト化抗体としては、受容者の相補性決定部位(CDR)の残基が、所望の特異性、親和性および能力を有するマウス、ラットまたはウサギ等の非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(受容者抗体)が挙げられる。ある場合では、ヒト免疫グロブリンのFv枠残基が対応する非ヒト残基で置換される。ヒト化抗体は、受容者抗体にも移入CDRにも枠配列にも見られない残基からなってもよい。一般的に、ヒト化抗体は、すべてまたは実質的にすべてのCDR領域が非ヒト免疫グロブリンのものと対応し、すべてまたは実質的にすべてのFR領域がヒト免疫グロブリン共通配列のものである少なくとも一つまたは二つの実質的にすべての可変ドメインからなるだろう。ヒト化抗体は最適には免疫グロブリン定常部(Fc)の少なくとも一部、典型的にはヒト免疫グロブリンの該領域からなってもよいであろう[Jonesら,Nature,321:522−525(1986);Reichmannら,Nature,332:323−329(1988);およびPresta,Curr.Op.Struct.Biol.,2:593−596(1992)]。
【0133】
非ヒト抗体をヒト化する方法は当業界で公知である。通常、ヒト化抗体は非ヒトであるソースからの一つ以上のアミノ酸残基を導入している。これらの非ヒトアミノ酸残基は、典型的には「輸入」可変ドメインから取られた「輸入」残基としばしば称される。ヒト化は、基本的には、Winterと共同研究者の方法にしたがって[Jonesら,Nature,321:522−525(1986);Riechmannら,Nature,332:323−327(1988);Verhoeyenら,Science,239:1534−1536(1988)]、げっ歯類のCDRまたはCDR配列をヒト抗体の対応する配列の代わりとすることにより行うことができる。したがって、そのような「ヒト化」抗体は、実質的にそのままのヒト可変ドメインが非ヒト種に由来する対応配列によって置換されたキメラ抗体である(米国特許第4,816,567号)。実際には、ヒト化抗体は典型的には、一部のCDR残基およびおそらく一部のFR残基がネズミ抗体の相同部位の残基によって置換されているヒト抗体である。
【0134】
ヒト抗体は、ファージディスプレイライブラリー等の公知の多様な技術を用いて作ることもできる[Hoogenboom and Winter,J.Mol.Biol.,227:381(1991);Marksら,J.Mol.Biol.,222:581(1991)]。ColeらおよびBoernerらの技術もヒトモノクローナル抗体の調製に利用できる[Coleら,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77(1985)and Boernerら,J.Immunol.,147(1):86−95(1991)]。同様に、ヒト抗体は、ヒト免疫グロブリン遺伝子座をトランスジェニック動物、例えば内在性免疫グロブリン遺伝子が部分的または完全に不活性化されたマウスに導入することにより作ることができる。誘発により、ヒト抗体の産生が観察され、これは遺伝子再配置、構築および抗体レパートリー等のすべての点においてヒトで見られるものと非常に似ている。このアプローチは、例えば米国特許第5,545,807号、同第5,545,806号、同第5,569,825号、同第5,625,126号、同第5,633,425号、同第5,661,016号;および以下の科学出版物:Marksら,Bio/Technology 10,779−783(1992);Lonbergら,Nature 868 856−859(1994);Morrison,Nature 368,812−13(1994);Fishwildら Nature Biotechnology 14,845−51(1996);Neuberger,Nature Biotechnology 14,826(1996);Lonberg and Huszar,Intern.Rev,Immunol.,13 65−93(1995)に記載されている。
【0135】
4.二特異性抗体
二特異性抗体は、少なくとも二つの異なる抗原に対する結合特異性を有するモノクローナルの、好ましくはヒトまたはヒト化抗体である。この場合、結合特異性の一つはPRO285、PRO286またはPRO358タンパク質に対するものであり、別のものは他の抗原に対するもので、好ましくは細胞表面タンパク質またはレセプタまたはレセプタサブユニットに対するものである。二つの異なるトール様タンパク質、例えば本出願で開示されたトール相同体のいずれか二つ、またはここに開示されたトールタンパク質、および公知のトールタンパク質、例えばTLR2に対して特異性を有する二特異性抗体を調製することも可能である。そのような二特異性抗体はトールレセプタによる異なる病原体パターンの認識をブロックすることができるために、敗血症または敗血症性ショックの治療に顕著な利点を有することが期待される。
【0136】
二特異性抗体の製造方法は公知である。伝統的には、二種類の重鎖が異なる特異性を有する二種類の免疫グロブリンの重鎖/軽鎖対の共発現に基づく[Milstein and Cuello,Nature,305:537−539(1983)]。免疫グロブリンの重鎖と軽鎖のランダムな集合のために、これらのハイブリドーマ(クアドローマ)は10種類の異なる抗体分子の潜在的な混合物を産生し、このうちわずかに一種類のみが正しい二特異性構造を有する。正しい分子の精製は、通常、アフィニティークロマトグラフィー工程により行われる。同様な方法が、1993年5月13日公開のWO93/08829およびTrauneckerら,EMBO J.,10:3655−3659(1991)に開示されている。
【0137】
所望の結合特異性(抗体−抗原結合部位)を有する抗体可変ドメインは免疫グロブリンの定常ドメイン配列に融合させることができる。この融合は、少なくともヒンジ部分のCH2とCH3領域からなる免疫グロブリンの重鎖定常ドメインに対する融合であるのが好ましい。該融合の少なくとも一つに存在する軽鎖結合に必要な部位を有する第一の重鎖定常部(CH1)を有することが好ましい。重鎖形態の免疫グロブリンをコードするDNAおよび所望であれば免疫グロブリン軽鎖を別々の発現ベクターに挿入し、適当な宿主生物にともにトランスフェクトさせる。二特異性抗体の作成のさらなる詳細については、例えばSureshら,Methods in Enzymology,121:210(1986)を参照されたい。
【0138】
5.ヘテロ複合抗体
ヘテロ複合抗体も本発明の範囲内にある。ヘテロ複合抗体は二つの共有結合した抗体からなる。例えば、そのような抗体は免疫系細胞の標的を不要な細胞に向けさせるために[米国特許第4,676,980号]、およびHIV感染の治療のために[WO91/00360;WO92/200373;EP03089]報告されている。該抗体は、合成タンパク質化学の公知の方法、例えば架橋剤をともなう方法を用いてインビトロで調製してよい。例えば、免疫毒素はジスルフィド交換反応を用いるか、またはチオエーテル結合を形成することにより構築してよい。この目的に適する試薬の具体例としてはイミノチオレートおよびメチル−4−メルカプトブチリミデートおよび米国特許第4,676,980号に開示のものが挙げられる。
【0139】
G.抗トールタンパク質抗体に関する使用
本発明の抗トール抗体は多様な有用性を有する。例えば、抗PRO285、抗PRO286、抗PRO358および抗TLR2抗体は、PRO285、PRO286、PRO358またはTLR2に関して、例えば特定細胞、組織または血清中でのその発現を検出する診断アッセイに用いることができる。不均一相または均一相のいずれかで行われる競合結合アッセイ、直接または間接サンドイッチアッセイおよび免疫沈殿アッセイ等の公知の多様な診断アッセイ技術を用いてよい[Zola,Monoclonal Antibodies:A Manual of Techniques,CRC Press,Inc.(1987)pp.147−158]。診断アッセイで用いられる抗体は検出可能な部分で標識してよい。検出可能な部分とは直接的または間接的に検出可能なシグナルを作ることのできるものでなければならない。例えば、検出可能な部分とは、放射性同位元素、例えば3H、14C、32P、35Sまたは125I、蛍光または化学発光化合物、例えばフルオレセインイソチオシアネート、ローダミンまたはルシフェリン、または酵素、例えばアルカリホスファターゼ、ベータ−ガラクトシダーゼまたは西洋ワサビパーオキシダーゼであってよい。Hunterら,Nature,144:945(1962)、Davidら,Biotechnology,13:1014(1974)、Painら,J.Immunol.Meth.40:219(1981)およびNygren,J.Histochem.and Cytochem.,30:407(1982)により記載された公知のいずれかの方法により抗体を該検出可能な部分に複合化させてよい。
【0140】
抗PRO285、抗PRO286、抗PRO358または抗TLR2抗体も組換え細胞培養物または天然のソースからのこれらタンパク質のアフィニティー精製に有用である。このプロセスにおいて、これらのTrollタンパク質に対する抗体を、Sephadex樹脂またはろ紙等の支持体に公知の方法を用いて固定化する。次に、この固定化抗体を、精製すべき該タンパク質を有する試料に接触させ、その後、固定化抗体に結合したPRO285、PRO286、PRO358またはTLR2タンパク質以外の試料中の実質的にすべての物質を除去する適当な溶媒で支持体を洗浄する。最後に、該タンパク質を抗体から解離させる別の適当な溶媒で支持体を洗浄する。
【0141】
抗トールレセプタ(すなわち、抗PRO285、抗PRO286、抗PRO358または抗TLR2抗体)は各トールレセプタの生物活性をブロックするのに有用でもあろう。トールレセプタファミリーの主要な機能は、微生物上に存在する保存された分子パターンの存在を察知する病原体パターン認識レセプタとして作用することであると考えられる。多様な細菌により作られる潜在的な致死をもたらす分子であるリポ多糖(LPS、内毒素としても知られる)は血液中でリポ多糖結合タンパク質(LBP)に結合する。この形成された複合体は次にCD14として知られるレセプタを活性化する。次に何が起るかについては未だにコンセンサスが得られていない。ある仮定によれば、CD14は、低分子量の前炎症仲介体の産生に関与するサイトカイン、細胞付着タンパク質および酵素を作るようにマクロファージを直接に指令するのではなく、むしろLPSに第二レセプタ活性化ができるようにする。もしくは、LPSはLBPまたはCD14からの助けなくしてある種のレセプタを直接に活性化することも示唆されている。本出願で開示されるデータは、ヒトトール様レセプタはLBPとCD14が応答するようにLPSによって活性化されるシグナルレセプタであることを示す。この機能は、病態生理学的条件下で敗血症性ショックと呼ばれるしばしば致命的な症候群を導きうるので、(他のトールレセプタンタゴニストとちょうど同じように)抗トールレセプタ抗体は敗血症性ショックの治療に有用であろう。異なるトールレセプタは異なる病原体、例えばグラム陰性またはグラム陽性細菌の多様な株を認識できると予想される。したがって、異なる状況では、異なるトールレセプタに特異的に結合する抗体混合物または二特異性抗トール抗体との併用治療が望ましいであろう。
【0142】
抗huTLR2抗体がこのレセプタのLPSによる誘導をブロックするのに特に有用であると考えられることがここで具体的に示されている。LPSに対する露出が敗血症性ショックを導き得ることが示されているように(Parrillo,N.Engl.J.Med.328,1471−1477[1993])、抗huTLR2抗体は敗血症性ショックの治療に潜在的に有用である。
抗トールレセプタ抗体と関連して挙げた上記の治療および診断利用も他のトールアンタゴニスト、すなわち、トールレセプタ活性化および/またはトールレセプタにより仲介されるシグナル変換をブロックする他の分子(タンパク質、ペプチド、小有機分子等)に対しても適用することができる。
【0143】
それらの治療潜在能力を鑑みて、トールタンパク質(天然のトール相同体の変異体を含む)およびそれらのアゴニストおよびアンタゴニスト(抗トール抗体を含むがこれに限定されない)を治療利用に適する組成物に配合する。治療組成物は、所望の純度を有する有効成分を任意の薬理学的に許容される担体、賦形剤または安定剤に混合し、凍結乾燥配合物または水性溶液の形で保存用として調製する(Remington’s Pharmaceutical Science 第16版,Osol,A.編 1980)。許容される担体、賦形剤または安定剤は、使用される用量と濃度では受容者(受容動物)に対して毒性がないものとし、リン酸塩、クエン酸塩および他の有機酸等の緩衝液;アスコルビン酸等の抗酸化剤;低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチンまたは免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性高分子、グリシン、グルタミン、アスパラギン、アルギニンまたはリジン等のアミノ酸;グルコース、マンノース、またはデキストリン等の単糖類、二糖類および他の炭水化物;EDTA等のキレート剤;マンニトールやソルビトール等の糖アルコール;ナトリウム等の塩形成対イオン;および/またはTween、PluronicsまたはPEG等の非イオン性界面活性剤が挙げられる。
【0144】
有効成分は、例えば、コアセルベーション技術または界面合成により調製された、例えばヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリレート)マイクロカプセル等のそれぞれのマイクロカプセル、またはコロイド医薬輸送システム(例えば、リポソーム、アルブミンミクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)に、またはマイクロエマルジョンに封じ込めてもよい。そのような技術は上記のRemingtons’s Pharmaceutical Sciencesに開示されている。
【0145】
インビボ投与に用いられる配合物は無菌でなければならない。これは、凍結乾燥および再構成の前後に滅菌ろ過膜を用いたろ過により容易になされる。
ここでの治療組成物は、滅菌アクセス孔を有する容器、例えば、皮下注射針で穴が開けられるストッパーを有する静脈内溶液バックまたはバイアルに入れる。
投与経路は、例えば注射または静脈内、腹腔内、脳内、筋肉内、眼内、動脈内または病変内の経路、局所投与、または徐放システム等の公知の方法にしたがう。
【0146】
徐放性調製物の適当な実例としては、形付けられた製品、例えばフィルムまたはマイクロカプセルの形にした半透性のポリマーマトリックスが挙げられる。徐放性マトリックスとしては、ポリエステル、ポリラクチド(米国特許3,773,919、EP58,481)、L−グルタミン酸とガンマLグルタミン酸エチルとの徐放性の共重合体(U.Sidmanら,Biopolymers 22(1):547−556[1993])、ポリ(2−ヒドロキシエチル−メタクリレート)(R.Langer,ら,J.Bimed.Mater.Res.15:167−277[1881]およびR.Langer,Chem.Tech.12:98−105[1982])、エチレンビニルアセテート(R.Langerら,Id.)またはポリ−D−(−)−3−ヒドロキシ酪酸(EP133,988)が挙げられる。徐放性の組成物はリポソームも包含する。本発明の範囲内の分子を含有するリポソームはそれ自体公知の方法で調製される[DE 3,218,121;Epsteinら,Proc.Natl.Acad.Sci.USA 82:3688−3692(1985);Hwangら,Proc.Natl.Acad.Sci.USA 77:4030−4034(1980);EP52322;EP36676A;EP88046;EP143949;EP142641;日本国特許出願83−118008;米国特許4,485,045および4,544,545;およびEP102,324]。通常、リポソームは、脂質含量が約30mol%コレステロールを超える小さい(約200〜800オングストローム)単層タイプであり、選択される比率は最適なNT−4治療に適合させる。
【0147】
有効成分の効果的な量は、例えば治療の目的、投与経路および患者の状態に依存しよう。したがって、治療者は、最適な治療効果を得るために必要とされる投与量を評価し、投与経路を修正する必要がある。典型的な一日あたりの投与量は、上記のファクターに依存しながら約1μg/kg〜100mg/kgまたはそれ以上である。典型的には、臨床医は必要とされる生物学的効果を提供する投与量に達するまで本発明の分子を投与しよう。この治療の進展は慣用のアッセイにより容易にモニターされる。
【0148】
以下の実施例は、例示の目的のみで提示されるものであり、本発明の範囲を制限する意図は全くない。
本発明の明細書で引用されるすべての特許および文献の参照は、その全てが参照としてここに取り込まれる。
【実施例】
【0149】
別記した以外は、実施例で参照される市販の試薬は、製造者の指示に従って用いた。以下の実施例中、および明細書を通しての、ATCC受託番号により示される起源は、アメリカンタイプカルチャーコレクション、Rockville、Marylandである。
【0150】
[実施例1]
ヒトPRO285をコードするcDNAクローンの単離
私有の発現配列タグ(EST)DNAデータベース(LIFESEQ(商標)、Incyte Pharmaceuticals,Palo Alto,CA)を検索し、ショウジョウバエ トールたんぱく質に相同性を示すEST(#2243209)を同定した。 ESTに基づき、PCRプライマーのペア(フォワードおよびリバース)
TAAAGACCCAGCTGTGACCG(配列番号5)
ATCCATGAGCCTCTGATGGG(配列番号6)
と、プローブ
ATTTATGTCTCGAGGAAAGGGACTGGTTACCAGGGCAGCCAGTTC(配列番号7)
を合成した。
【0151】
cDNAライブラリーの構築のためのmRNAは、ヒト胎盤組織より単離した。cDNAクローンを単離するために用いられたcDNAライブラリーは、Invitrogen、サンディエゴ、CA(Fast Track 2)からの試薬などの市販の試薬を用いた標準的な方法により構築された。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそサイズを合わせ、定められた方向でクローニングベクターpCR2.1(Invtrigen,Inc.)内に、Life Technologies,Gaithersburg,MD(Super Script Plasmid System)からの試薬とプロトコールを用いてクローン化した。二重鎖cDNAは、1000bpを超えるサイズに合わせ、そのcDNAをBamHI/NotI開裂ベクターにクローン化した。pCR2.1は市販されているプラスミドであり、PCRフラグメントのクローン化を容易にするようにデザインされており、選択のためにAmpRとKanR遺伝子、青白選択のためにLacZを有している。
【0152】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO285遺伝子をコードするクローンの単離のために用いた。
【0153】
cDNAクローンはその全てを配列決定した。DNA40021(PRO285をコードしている)の全ヌクレオチド配列を、図2と3(配列番号2)に示す。クローンDNA40021は、ヌクレオチド位置61-63に明らかな翻訳開始部位を有するシングルオペロンリーディングフレームを含む(図2と3)。予想されるポリペプチド前駆体は、1049アミノ酸の長さであり、推定シグナルペプチドをアミノ酸位置1-29に、推定膜間ドメインをアミノ酸位置837-860の間に、およびロイシンジッパーパターンをアミノ酸位置132-153と704-725に、各々含む。なお、示された境界はおおよそのものであり、示された領域の実際の限界は、数アミノ酸の差で異なっているかもしれない。クローンDNA40021は、ATCCに寄託され(表示DNA40021-1154)、ATCC寄託番号ATCC209389が付与されている。
【0154】
全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0155】
[実施例2]
ヒトPRO286をコードするcDNAクローンの単離
所有されている発現配列タグ(EST)DNAデータベース(LIFESEQ(商標)、Incyte Pharmaceutical,Palo Alto,CA)を検索し、ショウジョウバエ トールたんぱく質に相同性を示すEST(#694401)を同定した。 ESTに基づき、PCRプライマーのペア(フォワードおよびリバース)
GCCGAGACAAAAACGTTCTCC(配列番号8)
CATCCATGTTCTCATCCATTAGCC(配列番号9)
と、プローブ
TCGACAACCTCATGCAGAGCATCAACCAAAGCAAGAAAACAGTATT(配列番号10)
を合成した。
【0156】
cDNAライブラリーの構築のためのmRNAは、ヒト胎盤組織より単離した。
このRNAは、Life Technologies,Gaithersburg,MD(Super Script Plasmid System)からの試薬とプロトコールを用いて、ベクターpRK5D中にオリゴdTプライム化cDNAを生成させるために用いた。pRK5Dはsp6転写開始部位、それに続くSfiI制源酵素部位、さらにそれに続くXhoI/NotI cDNAクローニング部位を有するクローニングベクターである。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそ1000bpを超えるサイズを合わせ、定められた方向でXhoI/NotI開裂pRK5Dにクローン化した。
【0157】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO286遺伝子をコードするクローンの単離のために用いた。
【0158】
cDNAクローンはその全てを配列決定した。DNA42663(PRO286をコードしている)の全ヌクレオチド配列を、図5から7(配列番号4)に示す。クローンDNA42663は、ヌクレオチド位置57-59に明らかな翻訳開始部位を有するシングルオペロンリーディングフレームを含む(図5から7)。予想されるポリペプチド前駆体は、1041アミノ酸の長さであり、推定シグナルペプチドをアミノ酸位置1-26に、推定膜間ドメインをアミノ酸位置826-848に、およびロイシンジッパーパターンをアミノ酸位置130-151、206-227、662-684、669-690、および693-614に、各々含む。なお、示された境界はおおよそのものであり、示された領域の実際の限界は、数アミノ酸の差で異なっているかもしれない。
クローンDNA42663は、ATCCに寄託され(表示DNA42663-1154)、ATCC寄託番号ATCC209386が付与されている。
【0159】
PRO286の全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0160】
[実施例3]
ヒトトール受容体ファミリーの既知のメンバーからの細胞外ドメイン(ECD)配列(存在する場合、分泌シグナル配列を含む)をESTデータベースの検索に用いた。ESTデータベースはパブリックデータべース(例えば、GenBank)、および所有されているデータベース(例えばLIFESEQ(商標)、Incyte Pharmaceuticals、Palo Alto、CA)を含んでいた。検索は、コンピュータープログラムBLASTまたはBLAST2(Altschul、およびGish、Methods in Enzymology 266:460-80(1996))を用いて、EST配列の6フレーム翻訳に対するECDたんぱく質配列の比較として行なわれた。既知のたんぱく質をコードしていない70(あるいは、あるケースでは90)以上のBlastスコアとのそれらの比較を、プログラム”pharap”を用いて、コンセンサスDNA配列に集めて組み立てた。(Phil Green、ワシントン大学、シアトル、ワシントン)。
【0161】
ESTは、Incyteデータベースで同定した(INC3115949)。
そのEST配列に基づき、PCRにより対象の配列を含むcDNAライブラリーを同定し、PRO358の全長コード配列のクローンを単離するためのプローブとして使用するために、オリゴヌクレオチドを合成した。
【0162】
PCRプライマーのペア(フォワードおよびリバース)を合成した:TCCCACCAGGTATCATAAACTGAA(配列番号15) TTATAGACAATCTGTTCTCATCAGAGA(配列番号16)
プローブも合成した:
AAAAAGCATACTTGGAATGGCCCAAGGATAGGTGTAAATG(配列番号17)
【0163】
全長クローン源としていくつかのライブラリーをスクリーニングするために、ライブライリーからのDNAを、上記PCRプライマーペアを用いて、PCR増幅によりスクリーニングした。ついでポジティブライブラリーを、プローブオリゴヌクレオチドとPCRプライマーの一方を用いて、PRO358遺伝子をコードするクローンの単離のために用いた。
【0164】
cDNAライブラリーの構築のためのRNAは、ヒト骨髄組織より単離した(LIB256)。cDNAクローンを単離するために用いられたcDNAライブラリーは、Invitrogen、サンディエゴ、CAからの試薬などの市販の試薬を用いた標準的な方法により構築された。cDNAは、NotI部位を含むオリゴdTにより開始し(prime)、ブラントでSalIヘミキナーゼ化(hemikinased)アダプターに連結し、NotIで切断し、ゲル電気泳動でおおよそサイズを合わせ、定められた方向で適当なクローニングベクター(例えばpRKBまたはpRKD;pRK5Bは、SiI部位を含まないpRK5Dの前駆体である;Holmesら、Science,253:1278-1280(1991))内にユニークなXhoIとNotI部位にクローン化した。
【0165】
上述の単離されたクローンのDNA配列決定は、PRO358の全長DNA配列(図21および22、配列番号14)とPRO358の誘導たんぱく質配列(図18および19、配列番号13)を与えた。
同定されたクローン(DNA47361)の全ヌクレオチド配列を、図21〜22(配列番号14)に示す。クローンDNA47361は、図21と22の下線のヌクレオチド位置に明らかな翻訳開始部位(ATG開始シグナル)を有するシングルオペロンリーディングフレームを含む(図2と3)。予想されるポリペプチド前駆体は、811アミノ酸の長さであり、推定シグナルペプチド(アミノ酸1-19)、細胞外ドメイン(アミノ酸20-575、位置55から位置575までの領域のロイシンリッチ反復を含む)、推定膜間ドメイン(アミノ酸576-595)を含む。クローンDNA47361は、ATCCに寄託され(表示DNA47361-1249)、ATCC寄託番号ATCC209431が付与されている。
【0166】
PRO286の全長配列のBLASTおよびFastA配列アラインメント分析(ALIGNコンピュータープログラムを用いて)によれば、それはショウジョウバエ トールたんぱく質のヒトアナログであり、以下のヒトトールプロモーターに相同性を有している:Toll1(DNAX# HSU88540-1、これはランダム配列化全長cDNA #HUMRSC786-1と同一);Toll2(DNAX# HSU88878-1);Toll3(DNAX# HSU88879-1);およびToll4(DNAX# HSU88880-1)。
【0167】
[実施例4]
PRO285、PRO286およびPRO358DNAの、ハイブリダイゼーションプローブとしての使用。
以下の方法は、PRO285、PRO286またはPRO358ポリペプチドをコードするヌクレオチド配列の、ハイブリダイゼーションプローブとしての使用を記載している。
トール相同体のコード配列を含むDNAは、ヒト組織cDNAライブラリー又はヒト組織ゲノムライブラリーにおける相同性DNA(例えばこれらの特定のトールたんぱく質の天然由来の変異体をコードするもの)をスクリーニングするためのプローブとして用いることができる。
【0168】
どちらかのライブラリーDNAを含むフィルターのハイブリダイゼーションと洗浄は、以下の高ストリンジェントな条件下で行われる。放射性標識したトール相同体由来プローブの、フィルターへのハイブリダイゼーションは、50%ホルムアミド、5xSSC、0.1%SDS、0.1%ピロリン酸ナトリウム、50mMリン酸ナトリウム、pH 6.8、2xDenhardt溶液、及び10%硫酸デキストランの溶液内で、42°C、20時間行う。このフィルターの洗浄は、0.1xSSC及び0.1%SDSの水溶液内で42°Cで行う。
【0169】
全長の未変性配列トール相同体をコードするDNAについて所望の配列相同性を有するDNAは、次いで、当該技術分野で知られる標準的な技術により同定することができる。
【0170】
[実施例5]
大腸菌(E.coli)内でのPRO285、PRO286およびPRO358の発現
この実施例は、大腸菌内での組換発現による、PRO285、PRO286またはPRO358(”トール相同体”)の非グリコシル化形態での調製を例示するものである。
トール相同体をコードするDNA配列をまず、選択したPCRプリマーを使用して増幅させる。このプライマーは、選択した発現ベクター上の制限酵素部位に対応する制限酵素部位を含む必要がある。種々の発現ベクターを使用することができる。適切なベクターの例としては、pBR322(大腸菌由来;Bolivarら、Gene, 2:95(1977)を参照)であって、アンピシリン及びテトラサイクリンの耐性遺伝子を含むものである。ベクターは、制限酵素で消化して脱リン酸化する。PCRで増幅した配列を次いでベクターへ連結する。ベクターは好ましくは抗生物質耐性遺伝子、trpプロモーター、ポリhisリーダー(最初の6つのSTIIコドン、ポリhis配列、及びエンテロキナーゼ開裂部位を含む)、PRO285コード領域、ラムダ転写終了符号、及びargU遺伝子をコードする配列を含むであろう。
【0171】
次いで連結混合物を使用して、選択した大腸菌株へ形質転換するが、これにはSambrookら(上記)に記載の方法を使用する。形質転換体は、LBプレート上での増殖能力により同定し、抗生物質耐性のコロニーを選択する。プラスミドDNAは、制限分析及びDNA配列決定により同定及び確認することができる。
【0172】
選択したクローンを、抗生物質を補ったLBブロス等の液体培地中で一晩増殖させることもできる。この一晩培養したものを次いで、より大スケールの培養用の接種用に使用することができる。細胞を次いで所望の吸光度まで増殖させ、その間に、発現プロモーターをオンにする。
【0173】
細胞を更に数時間培養した後、細胞を遠心により回収する。遠心して得られた細胞の沈殿物を、当該技術分野で知られる種々の薬剤を使用して可溶化し、可溶化したトール相同体を次いで、金属キレート性カラムを蛋白質との強固な結合を許容する条件下で使用して精製する。
【0174】
[実施例6]
ほ乳類細胞におけるPRO285、PRO286およびPRO358の発現
本実施例は、ほ乳類細胞内での組換え発現による、PRO285、PRO286およびPRO358(“トール相同体”)のグリコシル化された形態での調製を例示するものである。
【0175】
pRK5ベクター(1989年3月15日公開のEP 307,247を参照)を発現ベクターとして使用する。適宜、Sambrookら(上記)に記載されるものなどの連結法を利用して、選択した制限酵素を使用してトール相同体をコードするDNAを挿入して、トール相同体をコードするDNAをpRK5に連結する。得られるベクターは、場合により、pRK5−PRO285、PRO286、またはPRO358と呼ぶ。
【0176】
1つの態様においては、293細胞を宿主細胞として選択することができる。ヒトの293細胞(ATCC CCL1573)は、ウシ胎児血清及び適宜栄養成分及び/又は抗生物質で補ったDMEM等の培養液で、組織培養プレート内で周密的になるまで増殖させた。約10μgのpRK5−PRO285、PRO286、またはPRO358DNAを、VA RNA遺伝子(Thimmappayaら, Cell, 31:543 (1982))をコードする約1μgのDNAと混合し、500μlの1mMトリス−HCl、0.1mM EDTA、0.227M CaCl2に溶解した。この混合物に対して、500μlの50mM HEPES(pH7.35)、280mM NaCl、1.5mM NaPO4、を滴加し、25°Cで10分間、沈殿を形成させる。この沈殿物を再懸濁し、293細胞へ添加し、約4時間、37°Cで静置する。培養液を吸引除去し、2mlのPBS中20%グリセロールを30秒間添加する。この293細胞を次いで無血清培地で洗浄し、新鮮な培地を添加して細胞を約5日間インキュベーションする。
【0177】
形質移入の約24時間後、培養培地を除去して培養培地(のみ)で置換するか、又は200μCi/mlの35S−システイン及び200μCi/mlの35Sメチオニンを含有する培養培地で置換する。12時間のインキュベーション後、馴化培地を回収し、スピンフィルターで濃縮し、15%SDS−ゲルにかける。処理したゲルは、乾燥して設定した時間、フィルムを露出してPRO285ポリペプチドの存在を明らかにすることができる。形質移入細胞を含む培養物はさらに、(無血清培地で)インキュベーションして、この培地を選択したバイオアッセイで試験することもできる。
【0178】
代替技術において、トール相同体DNAは、Somparyracら(Proc. Natl. Acad. Sci., 12:7575 (1981))の記載する硫酸デキストラン法を使用して、293細胞内に一過性導入することができる。293細胞を、スピナーフラスコ内で最大濃度まで増殖させ、700μgのpRK5−PRO(285)/(286)/(358)DNAを添加する。細胞をまず、スピナーフラスコから遠心により濃縮して、PBSで洗浄する。DNA−デキストラン沈殿物を細胞のペレット上で4時間インキュベーションする。細胞を20%のグリセロールで90秒間処理し、組織培養培地で洗浄し、細胞培地、5μg/mlウシインシュリン、及び0.1μg/mlのウシトランスフェリンを含有するスピナーフラスコに再導入する。約4日後、馴化培地を遠心して細胞及びその破砕片を除去する。対応する発現トール相同体を含む試料は、次いで、例えば透析及び/又はカラムクロマトグラフィー等の選択した方法により濃縮・精製することができる。
【0179】
別の態様において、トール相同体は、CHO細胞内で発現することもできる。pRK5ベクターを、例えばCaPO4またはDEAEデキストラン等の既知の試薬を使用してCHO細胞へ形質移入することができる。上記したように、細胞培養物をインキュベーションして、培地を培養培地(のみ)、又は35S−メチオニン等の放射性標識を含む培地で置換する。PRO285、PRO286、またはPRO358ポリペプチドの存在を確認した後、培地を無血清培地で置換する。好ましくは、培養物は約6時間インキュベーションし、次いで馴化培地を回収する。発現したトール相同体を含む培地を次いで、何らかの選択した方法を選択して濃縮・精製することができる。
【0180】
エピトープタグ化トール相同体もまた、CHOの宿主細胞内で発現することができる。このトール相同体DNAは、pRK5ベクターからサブクローニングすることもできる。このサブクローンインサートは、PCRにより選択したエピトープ、例えばポリ−hisのタグとフレームを合わせ、バキュロウイルス発現ベクターへ融合させることができる。このポリ−hisタグ化挿入物は次いで、DHFR等の、安定クローンの選択用の選択標識を含む、SV40駆動性ベクターへサブクローニングすることができる。最後に、このCHO細胞をSV40駆動性のベクターで(上記したようにして)形質移入することができる。上記したように、発現を変化させるために標識を行なっても良い。発現したポリ−Hisタグ化トール相同体を含む培養培地は次いで、例えばNi2+-キレートアフィニティクロマトグラフィー等の何らかの選択した方法により濃縮・精製することができる。
【0181】
[実施例7]
酵母中でのPRO285、PRO286およびPRO358の発現
以下の方法は、酵母内での、PRO285、PRO286またはPRO358(”トール相同体”)の組換発現を記載したものである。
まず、酵母発現ベクターを、ADH2/GAPDHプロモーターから、トール相同体を細胞内生産又は分泌させるために構築する。所望のトール相同体をコードするDNA、選択したシグナルペプチド、及びプロモーターを選択したプラスミド中の適切な制限酵素部位に挿入して、細胞内発現をさせる。分泌用には、選択したトール相同体をコードするDNAを選択したプラスミドへ、ADH2/GAPDHプロモーター、酵母アルファ因子分泌シグナル/リーダー配列、及び(必要ならば)発現用のリンカー配列とともにクローニングすることができる。
【0182】
例えば酵母株AB110等の酵母細胞は、上記したような発現プラスミドで形質転換し、選択した培養培地で培養することができる。形質転換した酵母の上清は、10%トリクロロ酢酸で沈殿してSDS−PAGEによる分離、それに引き続くゲルのクマシーブルー染料での染色により分析することができる。
【0183】
組換体トール相同体は引き続いて、酵母細胞を培養液より遠心で除去し、選択したカートリッジフィルターで濃縮することにより単離・精製することができる。トール相同体を含有する濃縮物は、選択したカラムクロマトグラフィーの樹脂を用いて更に精製することができる。
【0184】
[実施例8]
バキュロウイルス感染昆虫細胞内におけるPRO285、PRO286およびPRO358の発現
以下の方法は、バキュロウイルス感染昆虫細胞でのPRO285、PRO286またはPRO358(”トール相同体”)の組換発現を記載している。
トール相同体コード配列を、バキュロウイルス発現ベクターで含まれるエピトープタグの上流に融合させる。このようなエイトープタグには、ポリ−hisタグや(IgGのFc領域のような)免疫グロブリンタグがある。種々のプラスミドを使用することができるが、これには商業的に入手可能なプラスミド、例えばpVL1393(Novagen)などに由来するものがある。簡潔にいうと、トール相同体コード配列またはコード配列の所望の部分(例えば細胞外ドメインをコードする配列等)を5’及び3’領域に相補的なプライマーでPCRにより増幅させる。5’プライマーは、フランキング(選択した)制限酵素部位を含むことができる。その産物を次いで、それらの選択した制限酵素で消化し、発現ベクター中へサブクローニングする。
【0185】
組換バキュロウイルスは、上記のプラスミドとBaculoGold(商標)ウイルスDNA(Pharmingen)とをSpodoptera frugipedra(“Sf9”)細胞(ATCC CRL1711)へ、リポフェクチン(GIBCO−BRLより商業的に入手可能)を使用して共形質転換して作り出される。28°Cで4乃至5日間インキュベーションした後、放出されたウイルスを回収して更なる増殖に使用する。ウイルス感染及び蛋白質発現は、O'Reilleyら(Baculovirus expression vectors: A laboratory Manual, Oxford: Oxford University Press (1994))に記載されるようにして行う。
【0186】
発現したポリ−hisタグ化トール相同体は次いで、例えばNi2+−キレートアフィニティクロマトグラフィーで、以下のようにして精製することができる。抽出物は組換ウイルスに感染したSf9細胞より、Rupertら、Nature, 362:175-179 (1993)により記載されたようにして調製する。簡潔にいうと、Sf9細胞を洗浄し、超音波処理緩衝液(25mL Hepes pH7.9、12.5mM MgCl2、0.1mM EDTA、10%グリセロール、0.1%NP−40、0.4M KCl)中に再懸濁し、氷上で20秒間、2回、超音波処理する。超音波処理物を遠心して、その上清を充填用緩衝液(50mMリン酸塩、300mM NaCl、10%グリセロール、pH7.8)で50倍に希釈し、0.45μmのフィルターに通す。Ni2+−NTAアガロースカラム(Qiagenより商業的に入手可能)を5mLのベッド容量で調製し、25mLの水で洗浄し、25mLの充填用緩衝液で平衡化する。濾過済の細胞抽出物を毎分0.5mLでカラムにのせる。カラムは、 A280がベースラインになるまで充填用緩衝液で洗浄し、その点から画分の回収を開始する。次に、カラムを二次洗浄緩衝液(50mMリン酸塩、300mM NaCl、10%グリセロール、pH6.0)で洗浄するが、これにより非特異的に結合した蛋白質が溶出する。再びA280のベースラインになった後、カラムを二次洗浄緩衝液中の0乃至500mMのイミダゾール勾配で展開する。1mLの画分を回収してSDS−PAGE及び銀染色又はアルカリホスファターゼにコンジュゲートしたNi2+−NTA(Qiagen)を利用したウェスタンブロッティングで分析する。溶出したHis10のタグ化PRO285をプールし、充填用緩衝液に対して透析する。
【0187】
或いは、IgGタグ化(又はFcタグ化)トール相同体の精製は、例えばプロテインAやプロテインGカラムクロマトグラフィーを含む、既知のクロマトグラフィー技術を利用して行うこともできる。
【0188】
[実施例9]
NF-κBアッセイ NF-κB経路を介したトールたんぱく質シグナルとして、それらの生理活性は、NF-κBアッセイでテストすることができる。このアッセイにおいて、Jurkat細胞は、製造者の指示に従って、Lipofectamine試薬(Gibco BRL)を用いて一過性形質導入される。NF-κB駆動ルシフェラーゼ遺伝子を含む、1μgのpB2XLucプラスミドを、PRO285またはPRO286をコードする挿入を有するあるいは有していない、1μgのpSRαN発現ベクターを用いて構築する。陽性対照として、細胞はPMA(酢酸フォルボールミリスチル;20ng/ml)とPHA(フィトヘマグルチニン(phytohaemaglutinin))を用いて、3から4時間処理される。細胞は、Promegaからの試薬を用いてルシフェラーゼ活性の測定のために2から3日後に溶解させる。
【0189】
[実施例10]
PRO285、PRO286またはPRO358に結合する抗体の調製
この実施例は、PRO285、PRO286またはPRO358(”トール相同体”)に特異的に結合することができるモノクローナル抗体の調製を例証する。
モノクローナル抗体を生産する技術は、当該技術分野において周知であり、例えば、Goding(上掲)に記載されている。用いられる免疫原は、精製されたトール相同体、トール相同体を含む融合タンパク、および細胞表面に組み換えトール相同体を発現するトール相同体を含む。免疫原の選択は、過度の実験を要することなく当業者によって実施されることができる。
【0190】
Balb/c等のマウスを、完全Freund'sアジュバント中に乳化されたトール相同体免疫原で免疫して、1−100マイクログラムの量で皮下または腹膜内に注射する。あるいは、この免疫原はMPL-TDMアジュバント(Ribi Immunochemical Research, Hamilton, MT)中に乳化され、動物の後ろ足に接種する。免疫されたマウスは、選択されたアジュバントに乳化されたさらなる免疫原を用いて10ないし12日後にブースト(boost)される。その後、数週間、マウスは、さらなる免疫注射でブーストされてもよい。血清サンプルは、PRO285抗体を検出するためにELISAアッセイで試験するための後眼窩採血(retro-orbital bleeding)によりマウスから定期的に得ることができる。
【0191】
適切な抗体価が検出された後、抗体に“陽性”の動物は、トール相同体の最終的な静脈注射により注射されてもよい。3−4日後、このマウスがと殺され、脾臓細胞が回収される。脾臓細胞は、ATCC、No.CRL1597から入手できるP3X63AgU.1のような選択されたマウスミエローマセルラインに融合(35%ポリエチレングリコールを用いて)される。融合体はハイブリドーマ細胞を生じ、非融合細胞、ミエローマハイブリッド、および脾臓細胞ハイブリッドの増殖を阻害するためにHAT(ヒポキサンチン、アミノプテリン、およびチミジン)培地を含む96ウェル組織培養プレートにプレートされ得る。
【0192】
ハイブリドーマ細胞は、対応するトール相同体に対する反応性についてELISAでスクリーニングされる。トール相同体に対する所望のモノクローナル抗体を分泌する“陽性”ハイブリドーマ細胞の決定は、当業者の技術範囲である。
陽性ハイブリドーマ細胞は、抗トール相同体モノクローナル抗体を含む腹水を産生するために同系のBalb/cマウスに腹膜内接種されてもよい。あるいは、このハイブリドーマ細胞は、組織培養フラスコまたはローラーボトルで生育することもできる。腹水中に産生されたモノクローナル抗体の精製は、硫安沈殿、ゲル排除クロマトグラフィーを用いて実施することができる。あるいは、プロテインAまたはプロテインGに対する抗体の結合に基づくアフィニティークロマトグラフィーを用いることができる。
【0193】
[実施例11]
HuTLR2は、リポ多糖(LPS)誘導細胞シグナルの媒体となる。
方法試薬 [3H]標識化、非標識化、LCD25およびS.minnesotaR595LPSは、List Biochemicals(キャンベル、CA)から、および他のすべてのLPSはSigma Chemical Co.(セントルイス、MO)から得た。LPは、ヒトLBP発現ベクターでトランスフェクトされた293細胞からの条件付培地として供給された。TLR2-Fc融合たんぱく質は、バキュロウィルス系で生産し、上述したように精製した。Markら,J.Biol.Chem 269,10720-10728(1994)。
【0194】
発現プラスミドの構築 ヒトTLR2をコードするcDNAは、ヒト胎児肺ライブラリーよりクローン化した。予測されるアミノ酸配列は、アミノ酸726がgluからaspに置換されているのを除いて、以前発表された配列(Rockら、上述)のそれと一致した。TLR2(dG.TLR2)のアミノ酸末端エピト−プタグバージョンを、位置17のロイシン(TLR2の予想される成熟型の最初のアミノ酸)のすぐ上流にXhoI制現部位を加え、これを上述したように単純ヘルペスウィルスタイプ1の糖たんぱく質Dのアミノ酸1-53に結合することにより構築した。Markら、上述。PCR産物を配列決定し、プロマイシン耐性遺伝子を含む哺乳類の発現ベクターにサブクローン化した。gD.TLR2のC末端トランケーション(truncation)変異体は、細胞内ドメインのコード配列に存在するBlpI(変異体△1)またはNsiI(変異体△2)部位での、および発現ベクターの3’ポリリンカーに存在するNotI部位でのcDNAの消化と、その後のオリゴヌクレオチドリンカーの結さつにより構築した。
【0195】
Δ1:5'-TCA GCG GTA AGC-3'(配列番号18)および 5'-GGC CGC TTA CCG C-3'(配列番号19)
Δ2:5'-TAA GCT TAA CG-3'(配列番号20)および 5'-GGC CGC TTA AGC TTA TGC A-3'(配列番号21)
【0196】
CD4/TLR2キメラはPCRにより構築し、CD4とTLR2配列の連結部にリンカーでコードされたバリンを有して、ヒトTLR2のアミノ酸588-784(膜間および細胞内ドメイン)に融合した、ヒトCD4のアミノ酸1-205(シグナルペプチドと2つの免疫グロブリン様ドメイン)を含んでいた。pGL3.ELAM.tkレポータープラスミドは、ルシフェラーゼレポータープラスミドpGL3(Promega)のSacIとHindIII部位の間に挿入された、以下の配列5'-GGT ACC TCC TGA CAT CAT TGT AAT TTT AAG CAT CGT GGA TAT TCC CGG GAA AGT TTT TGG ATG CCA TTG GGG ATT TCC TCT TTA GAT CTG GCG CGG TCC CAG GTCCAC TTC GCA TAT TAA GGT GAC GCG TGT GGC CTC GAA CAC CGA GCG ACC CTG CAGCGA CCC GCA AGC TT-3'(配列番号22)
を含んでいた。
【0197】
LBPのC末端エピト-プタグバージョン(LBP-FLAG)をPCRにより、ネイティブなストップコドンに変えてAsc1部位の付加とこのフラグメントのpRK5-FLAGへのサブクローニングによって構築し、アミノ酸GRADYKDDDDK(配列番号23)のC末端付加を得た。
【0198】
安定なセルライン/プール 293のヒト胚芽腎細胞を、10%FBS、2mMグルタミンおよびペニシリン/ストレプトマイシンが添加されたLGDMEM/HEM's F12(50:50)培地中で増殖させた。gD.TLR2の安定な発現のために、細胞をgD.TLR2発現ベクターで形質導入し、最終濃度1μg/mlのプロマイシン耐性で選択した。細胞の安定なプール(293-TLR2 クローン1)は、上述のMarkらに記載されたように、FACSとウェスタンブロット分析により特徴付けられた。
【0199】
ルシフェラーゼレポーターアッセイおよび電気泳動移動度シフトアッセイ(EMSA)
29332の親または安定な細胞(ウェル当たり2×105細胞)を6ウェルプレート中にシードし、後日、0.5μgのルシフェラーゼレポータープラスミドpGL3-ELAM.tkと内部コントロールとして0.05μgのRenillaルシフェラーゼレポート化ベクターと共に発現プラスミドで形質導入した。24時間後、細胞はLPS、LBP、またはLPSとLBPの両方で処理し、レポーター遺伝子活性を測定した。データを、蛍光ルシフェラーゼ活性をRenilla Luciferaseのそれで除した相対ルシフェラーゼ活性として示す。EMSAとしては、核抽出物を調製し、コンセンサスNF-κB結合部位を含む5’-[32P]-放射標識化オリゴヌクレオチドとのDNA-結合反応に用いた(Santa Cruz Biotechnology,sc-2511)。複合体中のNF-κBの同定は、NF-κBに対する抗体によるスーパーシフトにより確かめた(データは示さず)。
【0200】
RNA発現
組織ノザンブロットは、Clontechより購入し、TLR2の細胞外ドメインを囲むプローブにハイブリダイズさせた。ポリアデニル化mRNAを293細胞または293−TLR2細胞より単離し、ノザンブロットをヒトIL-8 cDNAフラグメントでプローブさせた。TLR2発現を、定量的PCRで、実質的に に記載されているリアルタイム”taqman(商標)”技術を用いて測定し、モデル770配列検出器(Applied Biosystems, Foster City, CA, USA)で、実質的に(Luohら、J.Mol.Endocrinol.18,77―85[1997])に記載されているように分析した。
【0201】
フォワードとリバースプライマー5'-GCG GGA AGG ATT TTG GGT AA-3'(配列番号24)
5'-GAT CCC AAC TAG ACA AAG ACT GGT C-3'(配列番号25)
を、レポーター染色FAMで5’ヌクレオチドを、消光(quenching)染色TAMRAで3’ヌクレオチドを標識したハイブリダイゼーションプローブ5'-TGA GAG CTG CGA TAA AGT CCT AGG TTC CCA TAT-3'(配列番号26)
と共に用いた。
マクロファージ/単球を1μg/mlのLPSで16時間処理した。
【0202】
レセプター結合アッセイ 直接の結合を測定するために、20ngの[3H]-LPSを、15μlのプロテインAセファロースを含む100μlの結合バッファー(150mM NaCl,20mM Hepes,0.03% BSA)中の600ngのTLR2―Fcと混合した。室温での3時間のインキュベーションの後、プロテインAセファロースサンプルは、冷PBS/0.1% NP-40で2回洗浄し、1%SDSと25mM EDTAを含む結合バッファー中に再懸濁し、カウントした。
【0203】
結果
ショウジョウバエで、トール受容体は、胚芽の背腹方向の(dorso-vental)パターン形成に必要であり、成ハエ(adult fly)での抗真菌免疫反応にも関与する。BelvinおよびAnderson,Ann.Rev.Cell.Biol.12,393-416(1996);Lemaitreら、Cell 86,973-983(1996)。トールは、多重ロイシンリッチ反復(LRRs)を有する細胞外ドメインと、インターロイキン1受容体(IL-1R)に配列相同性を有するサイトプラスマドメインを含むタイプIの膜間たんぱく質であり、いくつかの植物病耐性たんぱく質である。トールの活性化は、NF-κB経路の活性化を通して遺伝子の誘導を引き起こす。前述の通り、クローン化されたいくつかのヒト相同体があり、それらのいくつかは、本出願中に新規たんぱく質として開示されている。これらのヒトたんぱく質は、それらのショウジョウバエ複製(counterpart)の局所的構造を反映している。1つのヒトTLR(TLR4)の構成的活性変異体の過剰発現が、NF-κBの活性化と、炎症性サイトカインとコンスティミュラトリー(constimulatory)分子の誘導をもたらすことが示されている(Medzhitovら、およびRockら、上述)。
【0204】
ヒトTLRがLPS-誘導細胞活性化に関与しているかもしれないことを調べるために、我々は最初に、TLRsの発現を種々の免疫組織で調べた。TLRsの1つであるTLR2は、調べた全てのリンパ様の組織で発現されることが見出され、最高の発現は末梢の血液白血球であった(図8a)。TLR2の発現は、単球/マクロファージ、主なCD14−発現およびLPS反応性細胞で、富化される。興味深いことに、tLR2は、LPS(図8b)での単離された単球/マクロファージの刺激に関して、CD14で報告されたように(Matsuuraら、Eur.J.Immunol.22,1663-1665[1992];CrostonJ.Biol.Chem.270,16514-16517[1995])、上方制御(up-regulated)される。
【0205】
この結果は、TLR2がLPS−仲介細胞シグナルに関与しているかどうかを測定することを我々に促した。我々は、アミノ末端エピトープタグを含むTLR2(gD−TLR2)のバージョンを発現するヒト胎芽腎293細胞を作製した。個々のクローンと同様に、クローンの安定したプールを単離し、約105kDaの新規たんぱく質が発現することを示し(図10)、TLR2(〜89kDa)の予想サイズとN−結合グリコシルかの4つの可能性のある部位の存在に一致した。我々は、E−セレクチン遺伝子のNF−κB応答性エンハンサにより駆動される報告された遺伝子の発現を測定することにより、293または293−TLR2細胞とLBPの応答を、調べた(Clostonら、上述)。LPSでもLBPでも単独処理では、大きな遺伝子活性化がおこらないが、LPSとLBPの両方の添加は、TLR2を発現する細胞内でレポーター遺伝子活性の実質的な誘導を起こすが、対照の293細胞では起こらない(図9)。さらに、電気泳動移動度シフトアッセイ(EMSA)を用いて、我々はLPSが、LBPと組み合わさって、TLR2発現細胞でNK-κB活性を誘導することを見出した(図11)。293細胞でのLPS-誘導NF-κB活性のキネテッィクスは、ミエロイドと非ミエロイド細胞のそれと類似しており(Deludeら、J.Biol.Chem.269,22253-22260[1994];Leeら,Proc.Natl.Acad.Sci.USA,90,9930-9934[1993])、NF-κBの核の位置は、LPSに曝した後、30分間以内で最大である。シクロヘキシミド(図11)またはアクチノマイシンD(示さず)による前処理はNF-κB活性化を阻害しないために、293TLR2細胞中のLPS/LBPによるNF-κBの活性化は、de novoたんぱく質合成を必要としない。
【0206】
ミエロイド細胞に存在するCD14の膜結合型(mCD14)と、プラスマに存在する可溶性CD14(sCD14)(Bazilら、Eur.J.Immunol.16,1583-1589[1986])は、LPSに対する細胞の応答性を増進させることが示された。我々は293細胞がそれらの表面でほとんどあるは全くCD14を発現しないことを観察した(データは示さず)。しかしながら、293細胞の一過性の形質導入は、mCD14がTLR2-媒介LPS応答性の感受性と規模を増加した(図12)。
【0207】
上に示されたデータは、TLR2が、LPSのためのシグナルトランスデューサーとして機能するかもしれないことを示唆した。LPS反応の媒介におけるTLR2の細胞内ドメインの役割を調べるために、我々は、13(TLR-△1)または141アミノ酸(TLR2-△2)のC-末端トランケーション(truncation)を有するTLR2変異体が、一過性の形質導入293細胞でELAMレポーターを制御できるかどうか測定した。我々は、FACS分析により(示さず)およびウェスタンブロットにより(図13c)、細胞表面でこれらのレポーターの発現を検出できたが、両方のC末端のトランケーション変異体が、レポーター遺伝子の活性化を欠損したことを観察した。TLR2‐△1で欠失した細胞内ドメインの領域は、IL-1R-結合キナーゼIRAK(Crostonら、上述)との会合に要求されるIL-1R細胞内ドメインの領域と著しい類似性を有している(図13b)。我々はまた、TLR2の細胞外ドメイン(ECD)が、TLR2のECDがCD4のECDの一部に置換されたTLR2変異体もまた応答できないLPS-応答性に要求されることを示した(図13aおよび13b)。
【0208】
LPSは、近位の疎水性リピドA部分と、遠位の親水性O抗原多糖領域と、リピドAとO抗原構造を連結するコアオリゴ糖からなる複雑な糖脂質である。リピドA部分とは異なり、異なるグラム陰性細菌からO抗原構造には非常に多様性が有る。リピドAはLPS応答に必要であり、リピドAの脂肪酸側鎖を除く処理はLPSを不活性化する。我々は、アルカリ加水分解により”無毒化された”LPSと同様に、種々のグラム陰性細菌から調製されるLPSの効果を比較した。我々は、Escherichia coliセロタイプLCD25から単離されたLPSが、TLR2を活性化するために血清的に異なるEscherichia coli 055:B5 LPSよりほとんど2つのオーダーの規模で効果が高いことを観察した(図14a)。S.minnesota R595 LPSから調製されたLPSは、TLR2活性の可能性の有るインデューサーであるが、TLR2は”無毒化LPS”に応答できない。
【0209】
我々は、可溶型のTLR2細胞外ドメイン(TLR2‐Fc)が3H標識化LPSがin vitroアッセイで結合するかどうかを測定することにより、TLR2がLPSに結合するかどうかを調べた。我々は、3H−LCD25 LPSがTLR2−Fc融合たんぱく質に結合するが、Fc単独あるいは、いくつかのほかのレセプターのECDを含む融合たんぱく質には結合しないことを観察した(図14b)。この結合は特異的に、冷LCD25LPSにコンピートされたが、無毒化LPSではされなかった。LPSのTLR2−Fcへの結合の予備的分析は、Kdが比較的低い(500-700nM)ことと結合のキネティックスがを非常に遅いこと(データ示さず)を示唆する。我々は、他のたんぱく質、例えばLBPは、LBPがLPSをフリー、CD14との結合(ミセル型)から移す作用をするのと非常に類似して、LPSのTLR2へのin vivo結合の増進に作用するかもしれないと推測する。このことは、LBPがLPSに対するTLR2の感受性応答を得るために必要であることを示す我々のin vivoの結果と一致する(図9)。
【0210】
マクロファージのILS処理は、多くの炎症性サイトカインの発現をもたらす。同様にTLR2の293細胞での発現は、LPS/LBPと応答してIL-8 mRNAの>100倍の誘導をもたらすが、無毒化LPSはこのアッセイで不活性である(図15)。
【0211】
これらのデータはTLR2が、先天性免疫応答においてセンチネル(sentinel)役割、細菌の病原性に対する防御の最初のラインを果たすことを示唆する。TLR2とCD14は共にメロイド細胞で発現され、それらの誘導は、協働してLPS処理で誘導される。非ミエロイド細胞中のTLR2の発現は、LBPに依存し、mCD14の発現により増進されるメカニズムにより、通常の非応答性細胞にLPS応答性を与える。TLR2発現細胞のLPS処理は、NF-κBの活性化とそれにつづくIL-8などの適応反応を開始する遺伝子の誘導をもたらす(図15)。我々のデータは、インタクトな細胞外および細胞内ドメインがLPS応答に必要であるために、TLR2がLPSの存在を感知することとこの情報をプラスマ膜を通して伝えることの両方に関与していることを示唆する。さらに、IRAKとの会合に必要なIL-1Rの一部に相同性を有するTLR2のC末端尾領域は、NF-κB活性化に必要である。
【0212】
ショウジョウバエのトールおよびトール関連レセプター18Wheelerは、細菌と真菌に対する応答において、各々抗菌性ペプチドの誘導において重要な役割をはたす。Medzhitovら、上述。遺伝データは、トールに対するリガンドとしてのSpatzleが、背腹方向の(dorsovental)パターンに関与していることを示唆しており、Spatzleの相同体が免疫応答においてヒトTLRsの制御に重要かもしれないとの推測をもたらす。TLR2のLPSによる活性化がシクロヘキシミドにより阻害されないこと、TLR2の細胞外ドメインがLPSに対するin vitroでの低い親和性のレセプターであることの我々の観察は、TLR2がLPS認識に関与しているというモデルと一致する。我々のデータは、他のたんぱく質(例えばSpatzle相同体)がLPSに対するTLR2の応答を修正するかもしれないという可能性を排除しない。我々は、TLR2とショウジョウバエ トールの細胞外ドメインが共にLPRsを含むにもかかわらず、アミノ酸同一性が20%未満であることに注目する。第2に、LPRたんぱく質は、たんぱく質、ペプチドおよび炭水化物などの種々のタイプの分子に対するパターン認識レセプター(PRRs)である。Danglら、Cell 91,17-24(1997)。第3に、ショウジョウバエ免疫応答におけるSpatzleに対する要求は、トールのそれに比べて明らかではない。トールの欠損とは異なり、Spatzle変異体は、抗菌ペプチドのDefensinおよびAttacinの通常のレベルを誘導し、真菌の攻撃の後のCecropinA発現の部分的欠損だけであり、外傷の応答におけるDorsalの活性化で欠損していない。Lemaitreら,Cell 86,973-983(1996);Lemaitreら、EMBO J.14,536-545(1995)。
【0213】
前に記したように、TLRは、特に本出願で開示されている、3つの新規レセプター(DNA40021、DNA42663、およびDNA47361に各々コードされた)を含むヒトトール関連レセプターの大きなファミリーのメンバーである。この実施例で提示されたデータは,TLR4のNF-κB応答遺伝子の活性化における関与の証拠と同様に、レセプターのこのファミリーの主要な機能は、Janewayとその共同研究者によって最初に示唆されたように、微生物に存在する保存された分子構造の存在を感知する病原体パターン認識レセプターとして作用することであることを示唆している(Medzhitovら、上述)。ヒトTLRファミリーは、敗血症性ショックの治療のための治療方法におけるターゲットかもしれない。
【0214】
[実施例12]
in situハイブリダイゼーション in situハイブリダイゼーションは、細胞または組織調製物内における核酸配列の検出および位置決定のための強力かつ用途の広い技術である。例えば、遺伝子発現部位を同定するため、転写の組織分布を分析するため、ウイルス感染を同定および位置決定するため、特異的なmRNA合成の変化を追求するため、並びに染色体地図作製を支援するために使用できる。
【0215】
in situハイブリダイゼーションは、PCR産生33P-ラベル化リボプローブを用いて、LuとGillett, Cell Vision 1:169-176 (1994)によるプロトコールの最適化されたバージョンに従って行われた。簡単に説明すると、ホルマリン固定、パラフィン包埋されたヒト組織が切断され、パラフィンが除かれ、プロテイナーゼK(20g/ml)で15分間37℃でタンパク除去され、上記LuとGillettに記載されているようにin situハイブリダイゼーションのために処理される。[33−P]UTP-ラベルされたアンチセンスリボプローブがPCR産物から産生され、55℃で一晩ハイブリダイズされた。このスライドを、Kodak NTB2核トラックエマルジョンに浸して、4週間感光させた。
【0216】
33P-リボプローブ合成 6.0μl(125mCi)の33P-UTP(Amersham BF 1002, SA<2000 Ci/mmol)を乾燥させた。乾燥した33P-UTPを含む各チューブに、以下の成分を添加した: 2.0μl 5x 転写バッファー 1.0μl DTT(100mM)
2.0μl NTP混合物(2.5mM:10μ;それぞれ10mMのGTP、CTP&ATP+10μl H2O)
1.0μl UTP(50μM)
1.0μl Rnasin 1.0μl DNA鋳型(1μg)
1.0μl H2O 1.0μl RNAポリメラーゼ(通常、PCR産物T3=AS、T7=S)
【0217】
このチューブを、37℃で1時間インキュベートした。1.0μlのRQ1 DNアーゼを添加し、その後に37℃で15分間インキュベートした。90μlのTE(10mM Tris pH7.6/1mM EDTA pH8.0)を添加し、この混合物をDE81ペーパーにピペットで移した。残りの溶液を、Microcon-50限外濾過ユニットに充填し、プログラム10を用いて遠心した(6分)。濾過ユニットを第二チューブにあけ、プログラム2を用いて遠心した(3分)。最終的な回収遠心の後に、100μlのTEを添加した。1μlの最終産物をDE81ペーパーにピペットで移し、6mlのBiofluor IIにおいてカウントした。
【0218】
このプローブをTBE/尿素ゲルにかけた。1−3μlのプローブまたは5μlのRNA Mrk IIIを3μlの充填バッファーに添加した。95℃のヒートブロックで3分間熱した後、このゲルを直ちに氷に移した。ゲルのウェルを洗い、サンプルを充填し、45分間180−250ボルトで流した。このゲルをサランラップで覆い、−70℃のフリーザーの中で1時間から一晩の間、増感紙を用いてXARフィルムに感光させた。
【0219】
33P-ハイブリダイゼーション冷凍切片の前処理
このスライドをフリーザーから取り出し、アルミニウムのトレイに移し、室温で5分間解凍した。このトレイを、凝縮を下げるために、5分間55℃のインキュベーターに置いた。このスライドをフュームフードにおいて氷上で4%パラホルムアルデヒド中で10分間固定し、0.5xSSCで5分間、室温で洗浄した(25ml 20xSSC+975ml SQH2O)。10分間37℃で0.5μg/mlのプロテイナーゼK中で脱タンパクした後(250mlの予め温められたRNアーゼを含まないRNAバッファー中の10mg/mlストックの12.5μl)、この切片を0.5xSSCで10分間室温で洗浄した。この切片を70%、95%、100%のエタノール、2分間でそれぞれ脱水した。
【0220】
パラフィン-包埋された切片の前処理
このスライドを脱パラフィン化し、SQ H2Oに移し、2xSSC、室温、5分間でそれぞれ二回洗浄した。この切片を、20μg/mlのプロテイナーゼK(250mlのRNアーゼを含まないRNアーゼバッファー中の10mg/mlの500μl;37℃、15分)−ヒト胎芽、あるいは、8xのプロテイナーゼK(250mlのRNaseバッファー中の100μl;37℃、30分)−ホルマリン組織において脱タンパク化した。次いで、上述したように0.5xSSCですすぎ、脱水を行った。
【0221】
プレハイブリダイゼーション
このスライドを、Boxバッファー(4xSSC、50%ホルムアミド)−飽和フィルターペーパーを備えたプラスチックボックスに配置した。この組織を50μlのハイブリダイゼーションバッファー(3.75g デキストランスルファート+6mlSQ H2O)で被覆し、ボルテックスし、キャップを緩めて2分間マイクロ波で加熱した。氷上で冷却した後、18.75mlのホルムアミド、3.75mlの20xSSCおよび9mlのSQ H2Oを添加し、組織をよくボルテックスし、42℃で1−4時間インキュベートした。
【0222】
ハイブリダイゼーション
スライド当たり、1.0x106cpmプローブと1.0μltRNA(50mg/mlストック)を3分間95℃で加熱した。このスライドを氷冷し、スライド当たり48μlのハイブリダイゼーションバッファーを添加した。ボルテックスした後、50μlの33P混合物を、スライドの50μlのプレハイブリダイゼーションに添加した。このスライドを55℃で一晩インキュベートした。
【0223】
洗浄
洗浄を2x10分間、2xSSC、EDTAを用いて室温で行い(400ml 20xSSC+16ml 0.25M EDTA、Vf=4L)、その後に37℃で30分間RNアーゼA処理した(250mlのRNアーゼバッファー中の10mg/mlの500μl=20μg/ml)。このスライドを2x10分間、2xSSC、EDTAを用いて室温で洗浄した。ストリンジェントな洗浄条件は、55℃で2時間、0.1xSSC、EDTA(20ml 20xSSC+16ml EDTA、Vf=4L)であった。
【0224】
結果PRO285(DNA40021)
ヒト成人および胎児組織でのPRO285(DNA40021)の発現パターンを調べた。全長DNA40021配列に基づき合成した以下のプローブを用いた:オリゴ1:GGA TTC TAA TAC GAC TCA CTA TAG GGC AAA CTC TGC CCT GTG ATG TCA(配列番号27)オリゴ2:CTA TGA AAT TAA CCC TCA CTA AAG GGA ACG AGG GCA ATT TCC ACT TAG (配列番号28)
【0225】
この実験で、胎盤において、およびマウス胎児肝臓の造血細胞全般において低レベルの発現が観察された。ヒト胎児、成人またはチンパンジーリンパヌードではいずれも発現は検出されず、ヒト胎児またはヒト成人脾臓でも発現は検出されなかった。これらのデータは、おそらくin situハイブリダーゼーションアッセイでは感度が足りないため、ノザンブロットまたはPCRデータと完全には一致していない。より感度の高い条件下で、さらなる組織がいくらかの発現を示す可能性がある。
【0226】
PRO358(DNA47361)
ヒト成人および胎児組織でのPRO358(DNA47361)の発現パターンを調べた。全長DNA47361配列に基づき合成した以下のプローブを用いた:オリゴ1:GGA TTC TAA TAC GAC TCA CTA TAG GGC TGG CAA TAA ACT GGA GAC ACT (配列番号29)オリゴ2:CTA TGA AAT TAA CCC TCA CTA AAG GGA TTG AGT TGT TCT TGG GTT GTT(配列番号30)
【0227】
この実験で、消化管関連リンパ球組織および胎児における発達中の脾白髄質において、発現が観察された。正常成人脾臓のpALS領域で低いレベルの発現が観察された。他の全ての組織では陰性であったが、より感度の高い条件下で他の組織型において低いレベルの発現が観察できる可能性がある。
【0228】
材料の寄託
以下の材料は、American Type Culture Collection, 2301 Parklawn Drive, Rockville, MD,USA (ATCC)に寄託されている。
材料 ATCC寄託番号 寄託日DNA40021-1154 209389 1997年10月17日(PRO285をコードしている)
DNA42663-1154 209386 1997年10月17日(PRO286をコードしている)
DNA47361-1249 209431 1997年11月7日
【0229】
これらの寄託物は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の規定およびそれに基づく規則の下で作製された。これは、寄託日から30年間、寄託物の生存可能な培養物の維持を保証する。寄託物はブダペスト条約の下でATCCにより入手可能であり、Genentech, Inc.とATCCとの間の合意を得ることを条件として、直接関係する米国特許の発行、もしくは米国またはその他の国の特許出願の公開に基づいて、公衆に寄託物の培養物の子孫の永久的かつ無制限の利用可能性を保証し、そして、その子孫の入手を、35 USC 第122条およびそれに準ずる長官規則(特に886OG638に関して37 CFR 第1.14条を含む)に従って付与される米国特許商標庁長官によって定められた者に保証する。
【0230】
本願の譲受人は、寄託物の培養物が適切な条件下で培養された場合に死滅または損なわれた場合には、この材料を速やかに同一物で置き換えることに同意した。寄託材料の利用可能性は、その国の特許法に従って政府の権力の下に付与された権利に違反して本発明を実施するライセンスとして解釈されない。
【0231】
上記明細書は、当業者が本発明を十分に実施できるものと解する。寄託された実施態様は、本発明の一部の態様の単なる例示に過ぎないこと、並びに、機能的に等価なあらゆる構成物が本発明の範囲に含まれることから、本発明は、寄託された構成物により、範囲が制限されるものではない。ここに言う材料の寄託は、上記記述が、その最良の形態を含めた本発明のあらゆる態様の実施を可能にするのに不適切であることの承認、あるいは、それが示す特定の例示に請求の範囲を限定すると解釈されるとの承認を構成するものではない。実際に、ここに記載または示されたものに加えて、本発明の種々の変更が、上記記載から当業者にとって明白になり、添付された請求の範囲内に含まれる。
【図面の簡単な説明】
【0232】
【図1】PRO285と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号1)を示す。
【図2】PRO285をコードする、天然配列ヒトトールタンパク質のDNA40021と命名されたcDNA(配列番号2)の塩基配列を示す。
【図3】図2の配列の続きである。
【図4】PRO286と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号3)を示す。
【図5】PRO286をコードする、天然配列ヒトトールタンパク質のDNA42663と命名されたcDNA(配列番号4)の塩基配列を示す。
【図6】図5の配列の続きである。
【図7】図6の配列の続きである。
【図8】ヒトトールレセプタ2(huTLR2)の発現パターンを示す(上記のRockら)。a.TLR2プローブで検出されたヒトの複数の免疫組織のノーザン分析。PRL(抹消血液白血球)。b.マクロファージにおけるTLR2の豊富な発現、およびLPSに対するTLR2の転写の上流制御。定量的RT−PCRを用いて、PBL、T細胞、マクロファージ(MΦ)およびLPS促進マクロファージ(MΦ+LPS)におけるTLR2の相対的発現レベルを測定した。
【図9】TLR2はLPS誘導シグナルを仲介する。a.TLR2を安定して発現する293細胞はLPS応答性を得る。gD.TLR2を発現する安定クローンの集団(293−TLR2pop1)またはgD.TLR2を発現する単一クローン(293−TLR2クローン1)または発現ベクターのみで安定にトランスフェクトされたコントロール細胞(293−MSCV)は、pGL3.ELAM.tkで過渡的にトランスフェクトし、無血清培地中でLBPを存在または存在させずに6時間、1μg/mlの055:B5エンハンサで促進した。ELAMエンハンサの活性化は実施例に記載のように測定した。二つの独立した実験から結果を得た。ELAMエンハンサを欠いたコントロールレポータープラスミドを用いた場合、促進は観察されなかった(データは示さず)。レポータープラスミドの発現は未処理細胞またはLBPのみで処理された細胞において同一であった(データは示さず)。
【図10】293細胞においてエピトープ付加されたTLR2の発現を示すウェスタンブロット。
【図11】NF−κΒのTLR2依存性LPS促進活性化とトランスロケーションのタイムコース。核抽出物は055:B5 LPS(10μg/ml)とLBPで示された時間で処理された細胞(上部)、または1μMシクロヘキシミド(CHX)で1時間前処理され、無血清培地中で1μg/mlのLPSの存在下で1時間促進された細胞(低部)から調製した。
【図12】TLR2によるNF−κΒ活性化に対するmCD14の効果。ベクターコントロール(193−MSCV)または293−TLR2pop1細胞にレポータープラスミド、およびCD14発現ベクター(+mCD14)またはベクターコントロール(−mCD14)をそれぞれをトランスフェクトした。24時間後、トランスフェクトされた細胞を、無血清培地中でLBSの存在下に055:B5 LPSで6時間促進した。示されたデータは3つの独立した実験からのものを表わす。
【図13】シグナルにおけるTLR2のドメイン機能。a.多様なTLR2構築物の模式図。TLR2−WT、全長エピトープタグ付加形のTLR2、TLR2−Δ1およびΔ2はカルボキシル末端での13個または141個のアミノ酸の末端切断をそれぞれ表わす。CD4−TLR2、TLR2の細胞外ドメインをヒトCD4のアミノ酸1〜205により置き換えたヒトCD4−TLR2キメラ。ECD、細胞外ドメイン;TM、トランスメンブレン領域;ICD、細胞内ドメイン。b.IL−1RおよびTLR2シグナル変換に重要なC末端残基。残基番号は各タンパク質の右に示されている。矢印はTLR2−Δ1末端切断の位置を示した。*、IL−1Rシグナルに重要な残基(Heguyら,J.Biol.Chem.267,2605−2609[1992];Crostonら,J.Biol.Chem.270,16514−16517[1995])1 I、同一アミノ酸;:、保存的変化。c.TLR−R2変異体はLPSとLBPに対してNF−κΒを誘導することはない。293個の細胞は、示されたように全長TLR2またはTLR2変異体をコードする発現ベクターとpGL3.FLAM.tkで過渡的にトランスフェクトされた。また、細胞はCD14発現プラスミド(+mCD14)またはコントロールプラスミド(−mCD14)でトランスフェクトした。各タンパク質の等しい発現は、抗gDまたはCD4抗体を用いるウェスタンブロッティングにより確認される(下)。ルシフェラーゼアッセイは実施例に記載のように行った。データは二回の実験から得られた。
【図14】大腸菌K12 LPS(LCD25)の高い潜在力とそのTLR2への結合を示す。a.多様なLPS調製物の投与量応答曲線。b.TLR2の細胞外ドメインに対する[3H]−LPS(LCD25)の特異的相互作用。特異的結合はTLR2−Fcに対して観察されたが、Fcのみ、またはRse、Axl、Her2またはHer4の細胞外ドメインを有する融合タンパク質に対しては観察されなかった。TLR2−Fcに対する結合はLCD25 LPSと特異的に競合したが、脱毒素化されたLPSでは競合しなかった。
【図15】TLR2はLPS誘導IL−8発現に必要とされる。mCD14を過渡的に発現する293−MSCVベクターコントロールと293−TLR2細胞をLBPのみ、または示された種類のLPSとともに、無血清培地中1μg/mlの濃度で6時間促進した。等しい量のpoly−(A)RNAをノーザン分析のために用いた。
【図16】huTLR2をコードする塩基配列(配列番号11)。
【図17】huTLR2のアミノ酸配列(配列番号12)。
【図18】PRO358と命名された天然配列ヒトトールタンパク質の得られたアミノ酸配列(配列番号13)を示す。この図において、アミノ酸1〜19は推定シグナル配列を形成し、アミノ酸20〜575は、アミノ酸20〜54がロイシンに富む繰返しの特徴を有する推定細胞外ドメインであり、アミノ酸576〜595は推定トランスメンブレンドメインであるが、アミノ酸596〜811は細胞内ドメインを形成する。
【図19】図18の配列の続きである。
【図20】図19の配列の続きである。
【図21】(配列番号14)成熟した全長トールタンパク質PRO358をコードするDNA47361と命名された天然配列ヒトトールタンパク質cDNAの塩基配列を示す。示された配列は外来性配列を有するので、ATG開始コドンに下線を付し、TAA停止コドンは四角で囲んだ。
【図22】図21の配列の続きである。
【特許請求の範囲】
【請求項1】
(a)配列番号3のアミノ酸残基1から825を有し、かつNF−κBの活性化を誘導する能力を有するPRO286ポリペプチドをコードするDNA分子、または上記DNA分子の相補体;あるいは、
(b)配列番号4のヒトトール蛋白質cDNAによりコードされ、かつNF−κBの活性化を誘導する能力を有する同じポリペプチドをコードするDNA分子、または上記DNA分子の相補体;
を含む、単離された核酸分子。
【請求項2】
配列番号3のアミノ酸残基1から1041を有するPRO286ポリペプチドをコードするDNAを含む、請求項1記載の単離された核酸分子。
【請求項3】
配列番号3のアミノ酸残基1から825と、849から1041を有するPRO286ポリペプチドをコードするDNAを含む、請求項1記載の単離された核酸分子。
【請求項4】
上記DNAが配列番号4のヌクレオチド位置57から始まり、ヌクレオチド位置4199で終結するヌクレオチド配列、またはその相補体を含む、請求項1記載の核酸分子。
【請求項5】
請求項1から4のいずれか一項記載の核酸分子を含むベクター。
【請求項6】
上記ベクターで形質転換された宿主細胞により認識される制御配列に操作可能に結合された、請求項5記載のベクター。
【請求項7】
請求項5または6記載のベクターを含む宿主細胞。
【請求項8】
上記細胞がCHO細胞である、請求項7記載の宿主細胞。
【請求項9】
上記細胞が大腸菌である、請求項7記載の宿主細胞。
【請求項10】
上記細胞が酵母細胞である、請求項7記載の宿主細胞。
【請求項11】
請求項7から10のいずれか一項記載の宿主細胞を、請求項1から4のいずれか一項記載の核酸分子によりコードされるポリペプチドの発現に適した条件下で培養し、上記ポリペプチドを回収することを含むトールポリペプチドの製造方法。
【請求項12】
配列番号3で示されるアミノ酸配列における一もしくは数個のアミノ酸残基の置換、欠失または挿入によって得られるアミノ酸からなり、かつNF−κBの活性化を誘導する能力を有する、単離されたポリペプチド。
【請求項13】
配列番号3のアミノ酸残基1から825を有する、請求項12記載のポリペプチド。
【請求項14】
配列番号3のアミノ酸残基1から1041を有する、請求項12記載のポリペプチド。
【請求項15】
配列番号3のアミノ酸残基1から825と、849から1041を有する、請求項12記載のポリペプチド。
【請求項16】
配列番号4のヌクレオチド位置57から始まるヌクレオチド配列によりコードされる、請求項12記載のポリペプチド。
【請求項17】
ヘテロなアミノ酸配列に融合している、請求項12から16のいずれか一項記載のPRO286を含むキメラ分子。
【請求項18】
上記ヘテロなアミノ酸配列がエピトープタグ配列である、請求項17記載のキメラ分子。
【請求項19】
上記ヘテロなアミノ酸配列が免疫グロブリンのFc領域である、請求項18記載のキメラ分子。
【請求項20】
配列番号3に示されるアミノ酸配列を有するポリペプチドに特異的に結合する抗体。
【請求項21】
上記抗体がモノクローナル抗体である、請求項20記載の抗体。
【請求項1】
(a)配列番号3のアミノ酸残基1から825を有し、かつNF−κBの活性化を誘導する能力を有するPRO286ポリペプチドをコードするDNA分子、または上記DNA分子の相補体;あるいは、
(b)配列番号4のヒトトール蛋白質cDNAによりコードされ、かつNF−κBの活性化を誘導する能力を有する同じポリペプチドをコードするDNA分子、または上記DNA分子の相補体;
を含む、単離された核酸分子。
【請求項2】
配列番号3のアミノ酸残基1から1041を有するPRO286ポリペプチドをコードするDNAを含む、請求項1記載の単離された核酸分子。
【請求項3】
配列番号3のアミノ酸残基1から825と、849から1041を有するPRO286ポリペプチドをコードするDNAを含む、請求項1記載の単離された核酸分子。
【請求項4】
上記DNAが配列番号4のヌクレオチド位置57から始まり、ヌクレオチド位置4199で終結するヌクレオチド配列、またはその相補体を含む、請求項1記載の核酸分子。
【請求項5】
請求項1から4のいずれか一項記載の核酸分子を含むベクター。
【請求項6】
上記ベクターで形質転換された宿主細胞により認識される制御配列に操作可能に結合された、請求項5記載のベクター。
【請求項7】
請求項5または6記載のベクターを含む宿主細胞。
【請求項8】
上記細胞がCHO細胞である、請求項7記載の宿主細胞。
【請求項9】
上記細胞が大腸菌である、請求項7記載の宿主細胞。
【請求項10】
上記細胞が酵母細胞である、請求項7記載の宿主細胞。
【請求項11】
請求項7から10のいずれか一項記載の宿主細胞を、請求項1から4のいずれか一項記載の核酸分子によりコードされるポリペプチドの発現に適した条件下で培養し、上記ポリペプチドを回収することを含むトールポリペプチドの製造方法。
【請求項12】
配列番号3で示されるアミノ酸配列における一もしくは数個のアミノ酸残基の置換、欠失または挿入によって得られるアミノ酸からなり、かつNF−κBの活性化を誘導する能力を有する、単離されたポリペプチド。
【請求項13】
配列番号3のアミノ酸残基1から825を有する、請求項12記載のポリペプチド。
【請求項14】
配列番号3のアミノ酸残基1から1041を有する、請求項12記載のポリペプチド。
【請求項15】
配列番号3のアミノ酸残基1から825と、849から1041を有する、請求項12記載のポリペプチド。
【請求項16】
配列番号4のヌクレオチド位置57から始まるヌクレオチド配列によりコードされる、請求項12記載のポリペプチド。
【請求項17】
ヘテロなアミノ酸配列に融合している、請求項12から16のいずれか一項記載のPRO286を含むキメラ分子。
【請求項18】
上記ヘテロなアミノ酸配列がエピトープタグ配列である、請求項17記載のキメラ分子。
【請求項19】
上記ヘテロなアミノ酸配列が免疫グロブリンのFc領域である、請求項18記載のキメラ分子。
【請求項20】
配列番号3に示されるアミノ酸配列を有するポリペプチドに特異的に結合する抗体。
【請求項21】
上記抗体がモノクローナル抗体である、請求項20記載の抗体。
【図1】



【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図9】
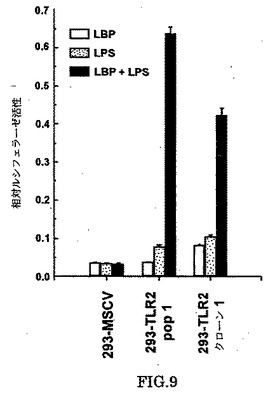
【図12】
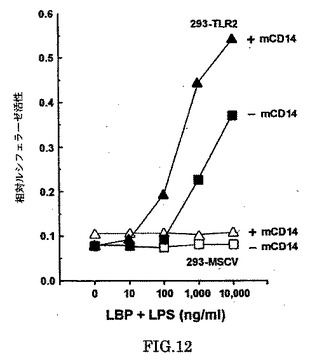
【図14】
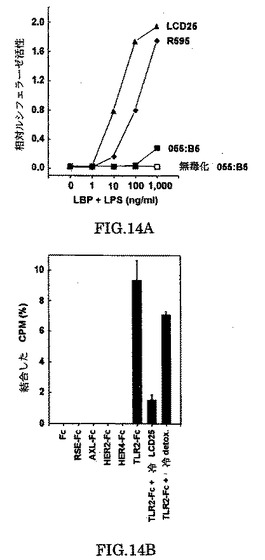
【図16】

【図17】

【図18】

【図19】

【図20】

【図21】

【図22】

【図8】
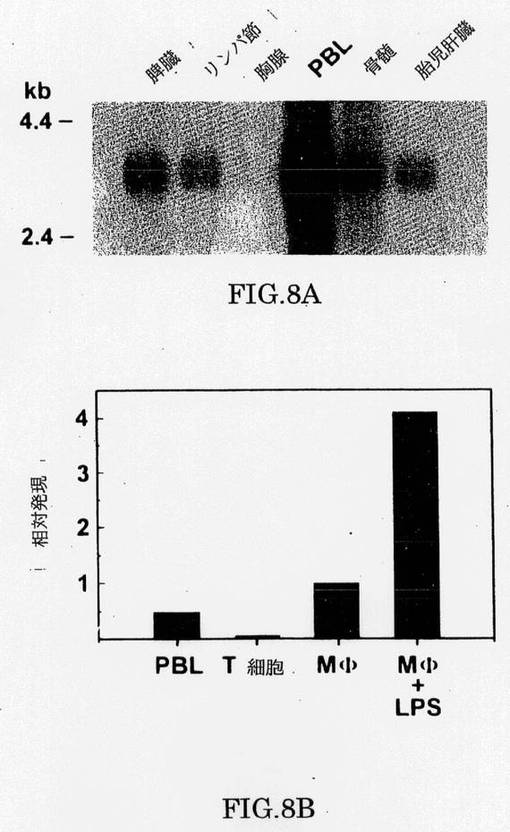
【図10】

【図11】
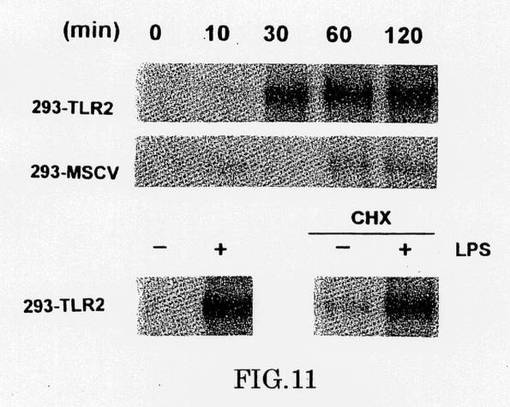
【図13】
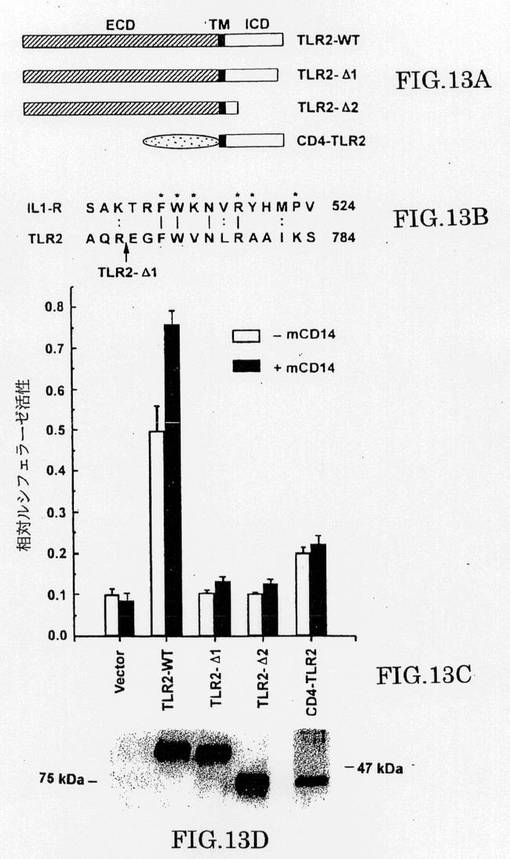
【図15】
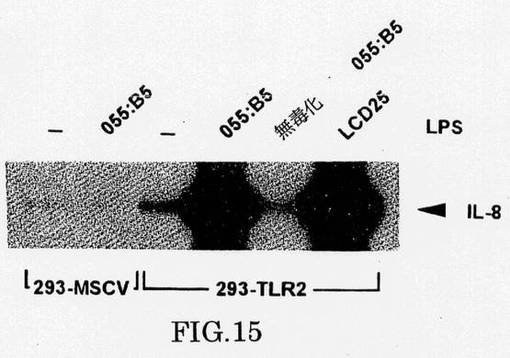
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図9】
【図12】
【図14】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図8】
【図10】
【図11】
【図13】
【図15】
【公開番号】特開2009−131265(P2009−131265A)
【公開日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願番号】特願2008−328570(P2008−328570)
【出願日】平成20年12月24日(2008.12.24)
【分割の表示】特願2000−517077(P2000−517077)の分割
【原出願日】平成10年10月7日(1998.10.7)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願日】平成20年12月24日(2008.12.24)
【分割の表示】特願2000−517077(P2000−517077)の分割
【原出願日】平成10年10月7日(1998.10.7)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]