
ヒトニューロンニコチン性アセチルコリン受容体組成物及び該組成物を使用する方法
【課題】ヒトニューロンニコチン性アセチルコリン受容体のα及びβサブユニットをコードする核酸分子、該核酸分子を含む哺乳動物及び両生動物細胞、並びにα及びβサブユニットの製造方法を提供する。
【解決手段】ヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットをコードする核酸分子及びβ3サブユニットをコードする分子。また、複数のサブユニットの組合わせ、例えばα1、α2、α3、α4、α5、α6及び/又はα7サブユニットのうちの一つ以上とβ3サブユニットの一つ以上との組合わせ、又はβ2、β3及び/又はβ4サブユニットとα6サブユニットとの組合せ。
【解決手段】ヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットをコードする核酸分子及びβ3サブユニットをコードする分子。また、複数のサブユニットの組合わせ、例えばα1、α2、α3、α4、α5、α6及び/又はα7サブユニットのうちの一つ以上とβ3サブユニットの一つ以上との組合わせ、又はβ2、β3及び/又はβ4サブユニットとα6サブユニットとの組合せ。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願明細書は、米国内向けでは、1995年6月7日出願のElliottらによる米国特許出願明細書第08/484,722号“HUMAN NEURONAL NICOTINIC ACETYLCHOLINE RECEPTOR COMPOSITIONS AND METHODS EMPLOYING SAME(ヒトニューロンニコチン性アセチルコリン受容体組成物及び該組成物を使用する方法)”の一部継続出願である。米国特許出願明細書第08/484,722号の論題は全体が本出願明細書に参考として包含される。
【0002】
本出願明細書は、米国特許明細書第5,369,028号及び米国特許出願明細書第08/028,031号、第08/149,503号、第08/496,855号、第07/938,154号、第08/467,574号、第08/466,589号、第08/487,596号にも関連している。これらの米国特許出願明細書及び米国特許明細書の論題は本出願明細書に参考として包含される。
【0003】
発明の分野
本発明は、ヒトニューロンニコチン性アセチルコリン受容体タンパク質サブユニットをコードする核酸分子、及びコードされたタンパク質に関する。特に、ヒトニューロンニコチン性アセチルコリン受容体αサブユニットをコードするDNA及びRNA、αサブユニットタンパク質、βサブユニットをコードするDNA及びRNA、βサブユニットタンパク質、並びにこれらの組合わせを提供する。
【0004】
発明の背景
リガンド依存性イオンチャンネルは中枢神経系の細胞間の情報伝達手段を提供する。これらのチャンネルは、ある細胞から放出された信号(例えば神経伝達物質と称する化学物質)を、標的細胞膜に沿って伝搬する電気信号に変換する。中枢及び末梢神経系には種々の神経伝達物質及び神経伝達物質受容体が存在する。神経筋及びニューロン由来のニコチン性アセチルコリン受容体(nAChR)を含む5種類のリガンド依存性受容体類が同定された(Stroudら,1990 Biochemistry 29:11009−11023)。しかしながら、種々の受容体が神経系の種々の部分で神経伝達物質又は他の調節リガンドに対する種々の応答を発生させるメカニズムは殆ど解明されていない。
【0005】
ニコチン性アセチルコリン受容体(nAChR)は、神経筋及びニューロン由来のマルチサブユニットタンパク質である。これらの受容体は、神経伝達物質アセチルコリン(ACh)との相互作用時に、神経と筋肉との間、及びニューロン間のシナプス伝達を仲介するリガンド依存性イオンチャンネルを形成する。様々なニコチン性アセチルコリン受容体(nAChR)サブユニットが存在するため、様々なnAChR組成物(即ちサブユニットの組合わせ)が存在する。異なるnAChR組成物は種々のリガンドに対して異なる特異性を示すため、薬理学的に識別可能である。脊椎動物神経筋接合部、脊椎動物交感神経節及び脊椎動物中枢神経系で発現されるニコチン性アセチルコリン受容体は、種々のnAChR組成物に結合する種々のリガンドの作用に基づいて識別された。例えば、神経筋接合部のニコチン性アセチルコリン受容体の活性化を阻止するメガネヘビα神経毒は、幾つかの異なるニューロン由来細胞系で発現される一部のニューロンニコチン性アセチルコリン受容体の活性化は阻止しない。
【0006】
筋肉nAChRは、化学量論(α)2β(γ又はε)δで5種類のサブユニットからなる糖タンパク質である。各サブユニットは約50〜60キロドルトン(kd)の質量を有し、異なる遺伝子によってコードされる。(α)2β(γ又はε)δ複合体は、二つのリガンド結合部位と一つのリガンド依存性トランスメンブランチャンネルとを含む機能性受容体を形成する。筋肉ニコチン性nAChRは、コリン作動性アゴニストとの相互作用時にナトリウムイオンを伝導する。ナトリウムイオンの流入は、プラズマ膜の内外で維持されていた正常なイオン勾配を即座に破壊し、その結果、プラズマ膜が脱分極する。膜の内外の電位差を減少させると、神経筋接合部で化学信号が電気信号に変換され、筋肉の収縮を誘発する。
【0007】
機能性筋肉ニコチン性アセチルコリン受容体は、αβδγサブユニット、αβγサブユニット,αβδサブユニット、αδγサブユニットで形成されたが、一つのサブユニットだけでは形成されていない(例えば、Kurosakiら(1987)FEBS Lett.214 253−248;Comachoら(1993)J.Neuroscience 13:605−613参照)。これに対し、機能性ニューロンnAChRは、αサブユニットのみ、又はα及びβサブユニットの組合わせによって形成できる。より大きいαサブユニットは一般的にACh結合サブユニットと考えられており、より小さい分子量のβサブユニットは一般的に構造サブユニットと考えられているが、βサブユニットがAChに結合する能力を有していない、又はACh結合部位の形成に関与しないという決定的な証拠はない。機能性イオンチャンネルの形成に関与するサブユニットはいずれも、結果として得られるチャンネルの構造に寄与するという範囲で、ACh結合能力(又は不能)にかかわりなく「構造」サブユニットである。リガンド依存性イオンチャンネルでもあるニューロンnAChRは、自律神経系及び中枢神経系の神経節(ニューロンnAChRが信号伝達を仲介する場所)、並びにシナプス前及びシナプス外位置(ニューロンnAChRが神経伝達を調節し、別の機能も果たし得る場所)で発現される。Wonnacottら(1990)Progress in Brain Research,A.Nordberら編,Elsevier,Amsterdam,157−163。
【0008】
nAChRをコードするDNAは幾つかの由来源から分離された。この種の研究から得られる情報に基づいて、しばらくの間、筋肉、自律神経節及び中枢神経系で発現されるnAChRがそれぞれ機能的に異なることは明らかだとされていた。この機能的多様性は、少なくとも部分的には、存在する種々のnAChRサブユニットの数が多いことに起因し得る。しかしながら、特にニューロン細胞で、(どの)nAChRサブユニットがどのように組合わせられて独自のnAChRサブタイプを形成するのかは完全には解明されていない。実際、特定のサブタイプのみがアルツハイマー病のような疾患に関与し得ることは証明されている。また、類似の組織又は細胞種類に由来するnAChRが類似の交差種であるかどうかは不明である。
【0009】
従って、各ヒトニューロンnAChRサブユニット、この種のサブユニットを含む組換え細胞、及びこれらから形成された受容体をコードするDNAを分離し、特徴を解明する必要がある。ヒトニューロンnAChRの機能を調べ、疾患特異的な薬理学的活性物質を得るために、分離(好ましくは精製)ヒトニューロンnAChR及び分離(好ましくは精製)ヒトニューロンnAChRサブユニットを得る必要もある。また、このような薬理学的活性物質を同定するためのアッセイを開発する必要もある。
【0010】
前述のような核酸、細胞、受容体サブユニット及び受容体組成物を利用できれば、非ヒトnAChRデータ、又はヒトもしくは非ヒト筋肉もしくは神経節nAChRデータから導き出した予測に基づくヒトニューロンnAChRの構造及び機能に関する推測の不確実さが除去される。
【0011】
従って本発明は、ヒトニューロンニコチン性アセチルコリン受容体のサブユニットをコードするDNAを分離し特徴を解明することを目的とする。本発明は、ヒトニューロンニコチン性アセチルコリン受容体サブユニットを組換えにより製造する方法を提供することも目的とする。本発明は更に、精製受容体サブユニットを提供すること、並びにヒトニューロンnAChRの活性を調節する化合物を同定するために化合物をスクリーニングする方法を提供することも目的とする。
【0012】
これらの目的及び他の目的は、本明細書の本文及び「特許請求の範囲」を更に検討することにより当業者に理解されよう。
【0013】
発明の概要
ヒトニューロンnAChRα及びβサブユニットをコードする分離核酸分子を提供する。特に、ヒトニューロンnAChRα6及びβ3サブユニットをコードする分離DNA及びRNAを提供する。前記DNAによってコードされるメッセンジャーRNA及びポリペプチドも提供する。
【0014】
α6及びβ3サブユニットを含む組換えヒトニューロンニコチン性nAChRサブユニット、並びにその製造方法も提供する。更に、少なくとも一つのヒトニューロンニコチン性nAChRサブユニットを含む組換えヒトニューロンニコチン性アセチルコリン受容体、及びその製造方法も提供する。また、宿主細胞でコードされた一つ以上のnAChRサブユニットと外来DNAもしくはRNA(即ち宿主細胞内に導入した本明細書に記載のDNAもしくはRNA)でコードされた一つ以上のnAChRサブユニットとの混合物を含む組換えニューロンニコチン性nAChR、及びその製造方法も提供する。
【0015】
前述のサブユニットをコードするDNAを含むプラスミドも提供する。本発明は、前述のDNA、mRNA又はプラスミドを含む組換え細胞も提供する。この種の細胞は、例えばDNAの複製、ヒトnAChRサブユニット及び組換え受容体の製造、並びに一つ以上のヒトサブユニットを含む受容体を発現する細胞の製造に有用である。
【0016】
本発明が提供するDNA、RNA、ベクター、受容体サブユニット、受容体サブユニットの組合わせ及び細胞は、選択したニューロンニコチン性nAChR受容体サブタイプ、及びこれらのサブタイプの特定の組合わせ、並びに受容体サブユニットに対する抗体の製造を可能にする。これは、その存在が単一nAChRサブタイプの分析を妨害し得る他の多くの受容体タンパク質が実質的に混入していない合成もしくは組換え受容体及び受容体サブユニットの製造手段を提供するものである。所望の受容体サブタイプを入手できれば、薬剤物質が特定の受容体サブタイプに与える作用を観察することが可能になり、従ってヒト及びヒトニューロンニコチン性nAChRsサブタイプに特異的な試験システムで薬剤物質の初期in vitroスクリーニングを実施することが可能になる。
【0017】
本発明は、ここに開示するDNA分子の一部と、該DNAでコードされたタンパク質に特異的に結合する抗体とを含む一本鎖プローブも提供する。また、配列番号15のヌクレオチド98−211を含む分離核酸分子も提供する。
【0018】
前記DNAでコードされたタンパク質も提供する。これらのタンパク質は、前記DNAを適当な原核又は真核宿主細胞内で発現させ、その結果産生されたタンパク質を分離することにより製造し得る。
【0019】
機能性ニューロンニコチン性アセチルコリン受容体サブユニット及びこれらの組合わせを同定する方法も提供する。
【0020】
ヒトニコチン性アセチルコリン受容体の活性を調節する化合物を同定するアッセイも提供する。特定の受容体組成物に対する薬剤の作用を調べるために薬剤物質をin vitroスクリーニングすることができれば、受容体サブタイプ特異的又は疾患特異的薬剤の開発及びスクリーニングが可能になる。種々の潜在的アゴニスト又はアンタゴニストを用いて単一受容体サブユニット又は特定受容体サブタイプの組合わせを試験すれば、個々のサブユニットの機能及び活性に関して更に情報が得られ、受容体サブユニット又は受容体サブタイプの一つ以上と極めて特異的に相互作用することができる化合物の同定及び設計が可能になる。得られる薬剤は、種々のサブタイプを発現する細胞を用いてスクリーニングすることにより同定した薬剤と比べて、望ましくない副作用が少ない。
【0021】
更に、薬剤の開発及び種々の疾患状態の治療に関連して、ヒトニューロンnAChRサブユニットをコードするDNA及びRNAを利用できれば、特定の疾患状態の発生と相関し得るような遺伝子中の任意の変化(例えば突然変異)を同定する手段が得られる。また、このような突然変異を、後で作用を調べるために実験動物又はin vitroアッセイシステム中に導入し得る合成DNA配列中に特異的に導入することにより、前述のような疾患状態の動物モデルの製造が可能になる。
【0022】
図面の簡単な説明
第1図は、α6コーディングフラグメントをEcoRI挿入物として含むサイトメガロウイルス(CMV)プロモーターをベースとするベクターpcDNA3−KEアルファ6.3の制限地図を示す説明図である。
【0023】
第2図は、β3コーディングフラグメントをEcoRI挿入物として含むCMVプローモーターをベースとするベクターpcDNA3−KBベータ3.2の制限地図を示す説明図である。
【0024】
好ましい実施態様の詳細な説明
定義
特に指示がない限り、本明細書で使用する総ての技術及び科学用語は、本発明が属する分野の当業者によって一般的に理解されているものと同じ意味を表す。本明細書で言及する総ての特許及び出版物は、特に指示がない限り、全体が参考として本明細書に包含される。
【0025】
本明細書で核酸分子、DNA、RNA、ポリペプチド又はタンパク質を形容する語として使用する「分離」(又は実質的に精製した、もしくは純粋な)という用語は、このように形容されるDNA、RNA、ポリペプチド又はタンパク質が人の手を介してin vivo細胞環境から分離されたものであることを意味する。従って、例えば本明細書中の「分離(又は実質的に純粋な)DNA」とは、当業者に使用されている標準的方法(例えばManiatisら(1982)Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY参照)で精製したDNAフラグメントを意味する。
【0026】
同様にして、本明細書でDNA、RNA、ポリペプチド又はタンパク質を形容する語として使用する「組換え」という用語は、このように形容されるDNA、RNA、ポリペプチド又はタンパク質が、例えばクローニング、組換え発現といったような方法で人為的に製造されたものであることを意味する。従って、例えば本明細書中の組換えタンパク質とは、人為的に加えられたDNAを含みこれを発現する組換え宿主により産生されたタンパク質を意味する。
【0027】
本明細書で使用するベクター(又はプラスミド)という用語は、外来DNAをその発現又は複製のために細胞内に導入するのに使用される別個のエレメントを意味する。このようなビヒクルの選択及び使用は、当業者が随意に実施できる範囲のものである。発現ベクターには、調節配列と作用的に結合するDNAを発現することができるベクター、例えば前記のようなDNAフラグメントの発現を生起させることができるプローモー領域が含まれる。従って、発現ベクターとは、組換えDNAもしくはRNA構築物、例えばプラスミド、ファージ、組換えウイルス、又は、宿主細胞内に導入されるとベクター上の適当な部位内にクローニングされたDNAを発現させる他のベクターを意味する。適当な発現ベクターは当業者に良く知られており、真核細胞及び/又は原核細胞内で複製できるもの、並びにエピソーム状態を維持するもの、又は宿主細胞ゲノム内に組込まれるものがある。真核宿主細胞、特に哺乳動物細胞内でnAChRサブニットを発現させるための、現時点で好ましいプラスミドの非限定的具体例としては、サイトメガロウイルス(CMV)、サルウイルス40(SV40)及びマウス乳癌ウイルス(MMTV)プロモーター含有ベクター、例えば、pCMV、pcDNA1,pcDNA3、pZeoSV、pCEP4、pMAMneo及びpMAMhygが挙げられる。
【0028】
本明細書で使用するプロモーター領域という用語は、DNAに作用的に結合してその転写を制御するDNAセグメントを意味する。プロモーター領域は、RNAポリメラーゼの認識、結合及び転写開始に十分な特異的配列を含む。プロモーター領域のこの部分をプロモーターと称する。プロモーター領域は更に、RNAポリメラーゼの認識、結合及び転写開始活性を調節する配列も含む。これらの配列はシス作用性であり得、又はトランス作用因子に応答し得る。プロモーターは、調節の種類に応じて構成性であり得、又は調節される。本発明で使用し得るプロモーターの具体例としては、SV40初期プロモーター、サイトメガロウイルス(CMV)プロモーター、マウス乳癌ウイルス(MMTV)ステロイド誘導プロモーター及びモロネーマウス白血病ウイルス(MMLV)プロモーター、並びに他の適当なプロモーターが挙げられる。
【0029】
本明細書で使用する「作用的に結合」という用語は、DNAと調節及びエフェクターヌクレオチド配列、例えばプロモーター、エンハンサー、転写及び翻訳開始及び終結部位、並びに他のシグナル配列との機能的関係を指す。例えば、DNAとプロモーターとの作用的結合は、DNAとプロモーターとの物理的及び機能的関係であって、DNAの転写が、該DNAを特異的に認識し、これに結合し且つこれを転写するRNAポリメラーゼによりプロモーターから開始されるような関係を意味する。発現及び/又はin vitro転写を最適化するためには、クローンの5’非翻訳部分を除去又は変化させて、余計な潜在的な別の翻訳開始コドン、又は転写もしくは翻訳のレベルで発現を妨害もしくは低下させる別の配列を除去する必要があり得る。あるいは、コンセンサスリボソーム結合部位(例えばKozak(1991)J.Biol.Chem.266:19867−19870参照)を開始コドンの5’に隣接させて挿入して、発現を促進することもできる。このような修飾が望ましい(又は必要である)かどうかは、経験的に決定し得る。
【0030】
本明細書で使用する発現という用語は、ポリ核酸がmRNAに転写され、ペプチド、ポリペプチド又はタンパク質に翻訳されるプロセスを意味する。ポリ核酸がゲノムDNAに由来する場合には、適当な真核宿主細胞又は生物を選択すれば、発現はmRNAのスプライシングを含み得る。
【0031】
哺乳動物細胞のトランスフェクションに特に好ましいベクターはSV40プロモーターをベースとする発現ベクター、例えばpZeoSV(Invitrogen,San Diego,CA)、CMVプロモーターをベースとするベクター、例えばpcDNA1、pcDNA3、pCEP4(Invitrogen,San Diego,CA)及びMMTVプロモーターをベースとするベクター、例えばpMAMneo(Clontech,Inc.)が挙げられる。
【0032】
本明細書中のヒトαサブユニット遺伝子とは、ヒトニューロンニコチン性アセチルコリン受容体のαサブユニットをコードする遺伝子である。ヒトnAChRのαサブユニットは典型的には、デンキナマズ(Torpedo)αサブユニット(Nodaら(1982)Nature 299:793−797参照)のシステイン192及び193と相同である、サブユニットの推定細胞外ドメイン内の隣接システイン残基の保存を示す。
【0033】
本明細書で使用するαサブユニットサブタイプという用語は、ここに開示するニューロンnAChRαサブユニットコーディングDNAクローンのうちの少なくとも一つに高ストリンジェンシー条件下でハイブリダイズするDNAでコードされるヒトニューロンnAChRサブユニットを意味する。αサブユニットは通常、生理学的条件下で、生理学的濃度で、任意的なβサブユニットの存在下で(即ち、あるαサブユニットは単独で機能するが、別のαサブユニットはβサブユニットの存在を必要とする)AChに結合し、通常は、本明細書に記載の方法又は当業者に公知の方法で評価して、機能性のnAChRを形成する。
【0034】
前述のようなαサブユニットをコードするが、遺伝暗号の縮重に起因して、指定のハイブリダイゼーション条件下で開示のDNAに必ずしもハイブリダイズしないDNA分子でコードされるαサブユニットも考えられる。このようなサブユニットも、通常は一つ以上のβサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性の受容体を形成する。典型的には、αサブユニットが別のスプライシングから生じるRNA(即ちスプライス変種(splice variant))でコードされる場合を除いて、αをコードするDNA及び該DNAでコードされたαサブユニットは、本明細書に記載のαサブユニットDNA(及び該DNAでコードされたタンパク質)の少なくとも一つに対して実質的な配列相同を有する。ここで理解されることとして、スプライス変種をコードするDNA又はRNAは全体で本発明が提供するDNA又はRNAに対して90%以下の相同を有し得るが、本明細書に開示のDNAフラグメントに対してほぼ100%相同の領域を含んでおり、開始及び終止コドンを含み機能性αサブユニットをコードする読取り枠をコードする。
【0035】
本明細書中のヒトβサブユニット遺伝子とは、ヒトニューロンニコチン性アセチルコリン受容体のβサブユニットをコードする遺伝子である。推定上のニューロンnAChRサブユニットに「β」という名称を付けた理由は、隣接システイン残基の欠失にある(これらの残基はαサブユニットに特徴的なものである)。βサブユニットはしばしば構成性nAChRサブユニットと呼ばれる(但しβサブユニットがACh結合特性を有する可能性もある)。適当なβサブユニットと適当なαサブユニットとを組合わせれば、機能性受容体が形成される。
【0036】
本明細書中のβサブユニットサブタイプとは、本明細書に開示のニューロンnAChRコーディングDNAの少なくとも一つに高ストリンジェンシー条件下でハイブリダイズするDNAでコードされるニューロンnAChRサブユニットを意味する。βサブユニットは適当なαサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性のnAChRを形成し得る。
【0037】
前述のようなβサブタイプをコードするが、遺伝暗号の縮重に起因して、指定のハイブリダイゼーション条件下で必ずしも開示のDNAとハイブリダイズしないDNAにコードされるβサブユニットも考えられる。この種のサブユニットも適当なαサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性の受容体を形成し得る。典型的には、βサブユニットがスプライス変種として生じるRNAでコードされる場合を除いて、βコーディングDNA及び該DNAにコードされたβサブユニットは、本明細書に記載のβコーディングDNA及びβサブユニットタンパク質に対して実質的な配列相同を共有する。ここで理解されることとして、スプライス変種をコードするDNA又はRNAは全体で本発明が提供するDNA又はRNAに対して90%以下の相同を有し得るが、このようなDNAは本明細書に開示のDNAに対してほぼ100%相同の領域を含む。
【0038】
本明細書で使用するnAChRサブタイプという用語は、α及び/又はβサブユニットサブタイプの特定の組合わせを含むニコチン性アセチルコリン受容体、例えばヒトnAChRα6及びβ3サブユニットを含む受容体を意味する。
【0039】
本明細書中のスプライス変異体とは、1種類以上のmRNAを形成させることになるゲノムDNAの一次転写体の分別プロセシングによって生じる変異体nAChRサブユニットコーディング核酸を意味する。分別的にプロセシングされたゲノムDNAに由来するcDNAは、完全に同じアミノ酸を有する部分と、異なるアミノ酸配列を有する部分とを有するnAChRサブユニットをコードする。従って、同一のゲノム配列が多数の関連mRNA及びタンパク質の産生を生起し得る。その結果得られるmRNA及びタンパク質を「スプライス変異体」と称する。
【0040】
本明細書中の外来(heterologous又はforeign)DNA及びRNAとは、それを含んでいるゲノムの一部としては生来存在しない、又は生来の位置とは異なるゲノム内位置に存在するDNA又はRNAを意味する。典型的には、細胞に対して内生ではなく、人為的に細胞内に導入されたDNA又はRNAである。外来DNAの非限定的具体例としては、ヒトnAChRサブユニットをコードするDNA、及び転写、翻訳又は他の調節可能な生化学的プロセスに作用することにより内生DNAの発現を仲介又は変化させるRNA又はタンパク質をコードするDNAが挙げられる。ヒトnAChRサブユニットをコードするDNAのような外来DNAを発現する細胞は、同じ又は異なるニコチン性アセチルコリン受容体サブユニットをコードするDNAを含み得る。外来DNAは発現される必要はなく、宿主細胞ゲノム内に組込まれるように、又はエピソーム状に維持されるように導入し得る。
【0041】
本明細書では、ハイブリダイゼーションのストリンジェンシーとは、ポリ核酸ハイブリッドを安定にしておく条件を意味する。当業者に公知のように、ハイブリッドの安定性はハイブリッドの融解温度(Tm)に反映される。Tmは下記の式:
【0042】
【化1】

によって概算できる。前記式中、lはヌクレオチドで表されるハイブリッドの長さである。Tmは配列相同が1%減少する毎に約1〜1.5℃低下する。一般的には、ハイブリッドの安定性はナトリウムイオン濃度及び温度の関数である。典型的には、ハイブリダイゼーション反応を低ストリンジェンシー条件下で実施し、次いで異なる、但しより高いストリンジェンシーで洗浄を行う。ハイブリダイゼーションストリンジェンシーという用語は、このような洗浄条件について使用される。
【0043】
本明細書では、(1)高ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関しては、0.018M NaCl中65℃で安定なハイブリッドを形成する核酸配列だけがハイブリダイズできる条件を意味する(即ち、あるハイブリッドが0.018M NaCl中65℃で安定でなければ、そのハイブリッドは本明細書で考慮する高ストリンジェンシー条件下で安定ではない)。高ストリンジェンシー条件は例えば、50%ホルムアミド、5×デンハーツ(Denhardt)溶液、5×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで0.1×SSPE及び0.1%SDS中65℃で洗浄することにより与えることができる。(2)中間ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関して、50%ホルムアミド、5×デンハーツ溶液、5×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで0.2×SSPE及び0.2%SDS中60℃で洗浄することに等しい条件を意味する。(3)低ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関して、10%ホルムアミド、5×デンハーツ溶液、6×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNAでハイブリダイセーションを行い、次いで1×SSPE及び0.2%SDS中50℃で洗浄することに等しい条件を意味する。(4)高ストリンジェンシー条件は、オリゴヌクレオチド(即ち長さ約30ヌクレオチド以下の合成DNA)に関しては、10%ホルムアミド、5×デンハーツ溶液、6×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで1×SSPE及び0.2%SDS中50℃で洗浄することに等しい条件を意味する。
【0044】
尚、これらの条件は種々の緩衝液及び温度を使用して再現し得、必ずしも厳密ではない。
【0045】
デンハーツ溶液及びSSPEは(例えばSambrookら,(1989)Molecular Cloning:A Laboratory Manual,第二版,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY)、他の適当なハイブリダイセーション緩衝液と同様に当業者に良く知られている。例えば、SSPEはpH7.4のリン酸緩衝0.18M NaClである。SSPEは例えば、175.3gのNaClと27.6gのNaH2PO4と7.4gのEDTAとを800mlの水に溶解し、pHを7.4に調整し、次いで水を1lになるまで加えることにより20×原液として調製できる。デンハーツ溶液(Denhardt(1966)Biochem.Biophys.Res.Commun.23:641)は例えば、5gのFicoll(タイプ400,Pharmacia LKB Biotechnology,INC.,Piscataway,NJ)と5gのポリビニルピロリドンと5gのウシ血清アルブミン(フラクションV;Sigma,St.Louis,MO)とを混合し、水を500mlになるまで加え、粒状物を除去すべく濾過することにより、50×原液として調製できる。
【0046】
本明細書で使用する「実質的な配列相同」という表現は、同一度が約90%以上の二つのヌクレオチド配列、及び典型的には95%以上のアミノ酸が同一であるアミノ酸配列に関するものである。しかしながら、スプライス変異体として生じる前記レベルより低い相同を有する、又は保守的(conservative)アミノ酸置換(もしくは縮重コドンの置換)によって修飾されたタンパク質(及びこのようなタンパク質をコードするDNAもしくはmRNA)も期待される。
【0047】
「実質的に同じ」という表現は、本明細書では、ここに開示する実際の配列と比べて僅かな重要ではない配列変化を有するDNAヌクレオチド配列、RNAリボヌクレオチド配列又はアミノ酸配列もしくはタンパク質について使用されている。実質的に同じ種は、開示の配列と機能的に同等であるとみなされる。従って、本明細書中の機能的に同等の核酸分子又はタンパク質とは、実質的に同じ結果を得るために実質的に同じ方法で機能するように十分に類似している核酸分子又はタンパク質である。
【0048】
本明細書で使用する「僅かな重要ではない配列変化」とは、ここに開示するDNA、RNA又はタンパク質と実質的に同じである配列が、ここに開示するヒト由来配列と機能的に同等であることを意味する。機能的に同等の配列は、実質的に同じ方法で機能して、ここに開示するヒト由来核酸及びアミノ酸組成物と実質的に同じ組成物を産生する。特に、機能的に同等のDNA分子は、ここに開示するものと同じ、又は保守的アミノ酸変化、例えばある非極性残基による別の非極性残基の置換、もしくはある帯電残基による類似の帯電残基の置換(例えば表1参照)を有するヒト由来タンパク質をコードする。これらの変化には、当業者によってタンパク質の三次構造を実質的に変化させないと認識されているものが含まれる。
【0049】
適当なアミノ酸保存置換は当業者に公知であり、通常は結果として得られる分子の生物学的活性を変えずに実施し得る。当業者に認識されているように、一般的には、ポリペプチドの非必須領域における単一アミノ酸の置換は生物学的活性を実質的に変化させない(例えばWatsonら、Molecular Biology of the Gene,第四版,1987,The Benjamin/Cummings Pub.Co.,p.244参照)。このような置換は、下記の表1に示すように実施するのが好ましい。
【0050】
【表1】
【0051】
別の置換も許容し得、経験的に決定し得るか、又は既知の保存置換と一致し得る。ポリペプチドのこの種の修飾はいずれも当業者に公知の任意の方法で実施し得る。
【0052】
本明細書中のヒトニューロンnAChRの活性とは、nAChRに特有の任意の活性を意味する。このような活性は典型的には一つ以上のin vitro方法で測定することができ、しばしばヒトニューロンnAChRのin vivo活性に対応する。このような活性は当業者に公知の任意の方法、例えば刺激に応答して組換え体チャンネルに流れる電流の量を測定することにより測定し得る。
【0053】
ヒトニューロンnAChRの存在及び/又は活性を決定する方法の非限定的具体例としては、ニコチン結合、86Rbイオン流、Ca2+流入、細胞の電気生理学的応答、RNAを導入した卵母細胞の電気生理学的応答を測定するアッセイが挙げられる。ここでは特に、DNA又はmRNAを含む細胞を試験化合物と接触させた時のnAChR仲介応答を測定又は検出する方法を提供する。
【0054】
本明細書中の組換え又は外来ヒトニューロンnAChRとは、受容体タンパク質を発現することができる細胞内に導入され発現された外来DNAにコードされるサブユニットを一つ以上含む受容体を意味する。組換えヒトニューロンnAChRは、宿主細胞にとって内生のDNAにより産生されるサブユニットも含み得る。ある実施態様では、組換え又は外来ヒトニューロンnAChRは、外来DNAにコードされるサブユニットのみを含み得る。
【0055】
本明細書中の機能性ニューロンnAChRとは、ここに開示する、又は当業者に公知のin vitro又はin vivoアッセイで評価して、ニューロンニコチン性nAChR活性を示す受容体を意味する。当業者に公知の、及び本発明で提供する任意の方法で評価し得る前述の活性を所有していれば、受容体は機能性とみなすことができる。nAChRタンパク質及び/又は活性の検出方法の非限定的具体例としては、ニコチン結合、86Rbイオン流、Ca2+流入、及び一つ以上の受容体サブユニットサブタイプをコードする外来DNA又はmRNAを含む細胞の電気生理学的応答を測定するアッセイが挙げられる。α及びβサブユニットの総ての組合わせが機能性受容体を形成し得るとは限らないため、このような受容体を産生する特定のサブユニット及び細胞の特徴を十分に解明するために、α及びβサブユニットの組合わせを多数試験し得る。従って、本明細書で組換え又は外来ヒトニューロンnAChRに関して使用する「機能性」という用語は、受容体チャンネルが刺激に応答して例えばNa+、K+、Ca2+又はBa2+のようなヒトニューロンnAChR浸透性イオンを提供し且つその流入を調節することができ、及び/又は受容体に対する親和性を有するリガンドと結合できることを意味する。このようなヒトニューロンnAChR活性は、電気生理学的方法、薬理的方法又は当業者の公知の別の方法により、宿主細胞によって生じ得る内因性nAChR活性から識別されることが好ましい。
【0056】
本明細書中のある種類の「対照」細胞又は「対照」培養物は、対照培養物は試験化合物に暴露しないという点を除いて、試験化合物に暴露した細胞又は培養物と実質的に同様に処理した細胞又は培養物である。別の種類の「対照」細胞又は「対照」培養物は、対照培養に使用した細胞が機能性ニコチン性アセチルコリン受容体を発現しないという点を除いて、トランスフェクションした細胞と同一である細胞又は細胞培養物であり得る。この場合は、細胞又は各種類の細胞の培養物をアッセイすべき化合物の存在下で実質的に同じ反応条件に暴露した時の、試験化合物に対するニコチン性アセチルコリン受容体陰性細胞の応答(又は応答欠失)と、試験化合物に対する試験細胞の応答とを比較する。
【0057】
本明細書中の「ニューロンnAChRの活性を調節する」化合物又はシグナルとは、前記化合物又はシグナルの存在下と不在下とではnAChRの活性が異なるようにnAChRの活性を変化させる化合物又はシグナルを意味する。このような化合物又はシグナルとしては、特にアゴニスト及びアンタゴニストが挙げられる。アゴニストという用語は、受容体の機能を活性化する物質又はシグナル、例えばAChを意味し、アンタゴニストという用語は、受容体の機能を妨害する物質を意味する。典型的には、アンタゴニストの作用は、アゴニストによる活性化の阻止として観察される。アンタゴニストには競合アンタゴニスト及び非競合アンタゴニストがある。競合アンタゴニスト(又は競合遮断剤)は、同じ又は近傍の部位に関してアゴニスト(例えばリガンド又は神経伝達物質)に特異的な部位又はその近傍と相互作用する。非競合アンタゴニスト又は遮断剤は、アゴニストと相互作用する部位以外の部位と相互作用することにより受容体の機能を不活化する。
【0058】
A. 分離DNAクローン
ヒトニューロンnAChRα及びβサブユニットをコードするDNA分子を提供する。特定的には、ヒトニューロンnAChRのα6及びβ3サブユニットをコードする分離DNA分子を開示する。前述のDNAでコードされる組換えメッセンジャーRNA(mRNA)及び組換えポリペプチドも提供する。
【0059】
本明細書で使用する「α6サブユニットをコードする(コーディング)核酸」という用語は、同じ名称のニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNA又はRNAを意味する。このような核酸分子は多くの方法で特徴を調べることができ、例えばDNAのヌクレオチド(又はRNAのリボヌクレオチド)は配列番号10又は配列番号20に示すアミノ酸配列をコードし得る。
【0060】
現時点で好ましいα6コーディング核酸としては、高ストリンジェンシー条件下で、配列番号9に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド143−1624)、又は配列番号19に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド143−1579)にハイブリダイズするDNA又はRNAが挙げられる。
【0061】
特に好ましいα6コーディング核酸分子は、配列番号10又は配列番号20に示すものと実質的に同じアミノ酸配列を有する(即ち保守的アミノ酸置換のみを有する)タンパク質をコードする分子である。最も好ましい分子は、配列番号9に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド143−1624)又は配列番号19に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド143−1579)を有するヌクレオチド(又はTがUで置換されたリボヌクレオチド)配列である。
【0062】
典型的には、α6サブユニットがスプライス変異体として生じる場合を除いて、α6コーディングDNAはここに開示するα6コーディング核酸分子に対して実質的な配列相同(即ち約90%以上)を有する。スプライス変異体をコードするDNA又はRNAはここに開示するDNA又はRNAに対して90%以下の総合的配列相同を有し得るが、このようなスプライス変異体は、ここに開示する核酸分子の一つ以上に対してほぼ100%相同の領域を含む。
【0063】
本発明は、「β3サブユニットコーディング核酸」も提供する。これには、同じ名称のニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNA又はRNA分子が含まれる。このような核酸分子は多くの方法で特徴を調べることができ、例えばDNAのヌクレオチド(又はRNAのリボヌクレオチド)は配列番号16に示すアミノ酸配列をコードし得る。
【0064】
現時点で好ましいβ3コーディング核酸としては、高ストリンジェンシー条件下で、配列番号15に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド98−1471)にハイブリダイズするDNA又はRNAが挙げられる。より好ましいのは、配列番号16に示すアミノ酸配列(又は実質的に保守的アミノ酸置換のみを有する前記アミノ酸配列)を含むタンパク質をコードする核酸である。本発明が提供する特に好ましいβ3コーディング核酸分子は、配列番号15に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド98−1471)を有する。
【0065】
典型的には、β3サブユニットがスプライス変異体として生じる場合以外は、β3コーディング核酸は開示のβ3コーディング核酸分子に対して実質的な配列相同(約90%以上)を有する。スプライス変異体をコードするDNA又はRNAは開示のDNA又はRNAに対して90%以下の総合的配列相同を有し得るが、このような核酸は、前述の核酸分子の一つ以上に対してほぼ100相同の領域を含む。
【0066】
B. プローブ
ヒトニューロンニコチン性nAChRα及びβサブユニットをコードするDNAは、ここに開示するDNA(配列番号9又は15に由来するヌクレオチドを含む)を用いて、適当なハイブリダイゼーション条件下で適当なヒトcDNA又はヒトゲノムライブラリーをスクリーニングすることにより分離し得る。適当なライブラリーは、ニューロン組織試料、脳底神経節、視床及び視床下部組織から製造し得る。ライブラリーは、DNAのサブユニットコーディング配列全体を含むDNA部分を用いてスクリーニングするのが好ましく、又はライブラリーは適当なプローブを用いてスクリーニングし得る。典型的には、プローブを、同定可能なタグ、例えば放射性標識、酵素又は他の当業者に公知のタグで標識する。
【0067】
α6及びβ3コーディング核酸を分離する方法で使用するためのプローブも提供する。例えばヒトα6サブユニットに関しては、プローブは、配列番号9又は配列番号19に示す任意の27個の塩基と同じ(又はその相補体)である少なくとも27個の連続塩基を含むヌクレオチド配列を有する一本鎖DNA又はRNA分子である。
【0068】
ヒトβ3サブユニットに関しては、プローブは、配列番号15に示すシグナル配列/コーディング配列の最初の105個のヌクレオチドに由来する任意の28個の塩基と同じ(又はその相補体)である少なくとも28個の連続塩基を含むヌクレオチド配列を有する一本鎖DNA又はRNAである。
【0069】
プローブを構築するための好ましい領域の非限定的具体例としては、5’及び/又は3’コーディング配列、トランスメンブランドメインをコードすると予測される配列を含む領域、細胞質ループをコードすると予測される配列を含む領域、シグナル配列、並びにアセチルコリン(ACh)及びα−ブンガロトキシン(α−bgtx)結合部位が挙げられる。ACh及びα−bgtx結合には、典型的には、デンキナマズnAChRαサブユニットの残基190−198に対応するアミノ酸(例えばKarlin(1993)Curr.Opin.Neurobiol.3:299−309参照)が関与する。別のプローブに関して、前述のような領域を含む大凡のアミノ酸残基を下記の表2に示す。
【0070】
【表2】
【0071】
あるいは、DNAの一部分を、特定ライブラリー内の選択したフラグメントを増幅するためのプライマーとして使用することもできる。
【0072】
C. ヒトニューロンニコチン性アセチルコリン受容体のα6及びβ3サブユニットをコードするクローンの分離
前述のプローブを使用して適当なライブラリーをスクリーニングする。各サブユニットをコードするDNAを得るのに適したライブラリーの非限定的具体例としては、ヒトα6コーディングDNAを分離する場合の黒質、視床又は視床下部、並びにヒトβ3コーディングDNAを分離する場合の黒質又は視床が挙げられる。
【0073】
ライブラリーのスクリーニング後、ハイブリダイゼーションシグナルの検出によって陽性クローンを同定する。同定されたクローンは、制限酵素地図及び/又はDNA配列分析よって特徴を調べ、次いで開示の配列と比較して、完全なα又はβサブユニットをコードするDNAを含んでいるかどうかを確認する。選択したクローンが不完全であれば、これらのクローンを使用して、重複クローンを得るために同じ又は異なるライブラリーを再スクリーニングし得る。所望であれば、ライブラリーは、完全なα又はβサブユニットをコードする重複クローンが得られるまで陽性クローンを用いて繰り返しスクリーニングし得る。ライブラリーがcDNAライブラリーの場合には、重複クローンは読取り枠を含む。ライブラリーがゲノムライブラリーの場合には、重複クローンはエキソン及びイントロンを含み得る。完全クローンは、本発明が提供するDNA及びコードされたタンパク質との比較によって同定し得る。
【0074】
ヒトニューロンnAChRα及びβサブユニットの種々のサブタイプをコードする相補的DNAクローンが分離された。サブユニットの各サブタイプは、異なる遺伝子でコードされると思われる。ここに開示するDNAクローンは、各サブタイプをコードするゲノムクローンの分離、並びに種々の神経組織から製造したライブラリーのスクリーニングによる任意のスプライス変異体の分離に使用し得る。ヒトニューロンnAChRサブユニットのスプライス変異体は、当業者に良く知られている核酸増幅方法を使用して検出することができる。この操作は、分岐(divergent)配列を包囲するDNA配列をベースとするオリゴヌクレオチドをヒトRNA又はゲノムDNA増幅用プライマーとして使用して行う。増幅産物の大きさ及び配列決定により、スプライス変異体の存在を明らかにすることができる。また、ハイブリダイゼーションによってヒトゲノムDNA配列を分離すると、ヒトニューロンnAChRサブユニットをコードする転写体の種々のスプライス変異体に対応する、イントロンで分離された複数のエキソンを含むDNAが得られる。
【0075】
総てのサブユニットサブタイプが総ての神経組織又は総ての脳部分で発現されるとは限らないことが判明した。従って、特定のサブユニットサブタイプ又はこのようなサブタイプのスプライス変異体をコードするcDNAを分離するためには、種々のニューロン又は神経組織から製造したライブラリーをスクリーニングすることが好ましい。
【0076】
D. α6及びβ3コーディング核酸を含む細胞及びベクター
ヒトnAChRサブユニットをコードする前述の核酸分子は、更に操作を行うためにベクター内に導入できる。適当な発現ベクター内へのクローン化DNAの導入、一つ以上の別個の遺伝子をコードする単一の発現構築物もしくは複数の発現構築物の組合わせ又は直鎖状DNAの真核細胞内へのトランスフェクション、並びにトランスフェクションした細胞の選択は当業者に良く知られている(例えばSambrookら(1989)Molecular Cloning:A Laboratory Manual,第二版,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY参照)。外来DNAは、当業者に公知の任意の方法、例えばCaPO4沈降により外来DNAをコードする発現構築物をトランスフェクションする方法で宿主細胞内に導入し得る(例えば、Wiglerら(1979)Proc.Natl.Acad.Sci.U.S.A.76:1373−1376参照)。次いで組換え細胞を、DNAでコードされたサブユニットを発現させる条件下で培養し得る。好ましい細胞の非限定的具体例としては、哺乳動物細胞(例えばHEK293、CHO及びLtk細胞)、酵母細胞(例えばPichia pastorisのようなメチロトロフィック(methylotrophic)酵母細胞)及び細菌細胞(例えば大腸菌)が挙げられる。
【0077】
α6及びβ3サブユニットをコードする核酸は、更に操作を行うために、個々に、又は別のニコチン性アセチルコリン受容体をコードする核酸と組合わせてベクター内に導入できる。ヒトニューロンnAChRサブユニットをコードする完全長DNAクローンが、ベクターpcDNA3に、即ちCMVプロモーター/エンハンサーと、CMVプロモーター/エンハンサーの下流のポリリンカーと、これに次ぐウシ成長ホルモン(BGH)ポリアデニル化シグナルとを含むpUC19ベースの哺乳動物細胞発現ベクター内に、挿入された。nAChRサブユニットをコードするDNAをCMVプロモーターとBGHポリアデニル化シグナルとの間に配置すると、構築物をトランスフェクションした哺乳動物細胞内でDNAの構成的発現が得られる。哺乳動物細胞内でのヒトnAChRサブユニットコーディングDNAの誘導発現のためには、DNAをpMAMneoのようなプラスミド内に挿入し得る。このプラスミドは、作用的に結合した外来DNAのステロイド誘導発現のためのマウス乳癌ウイルス(MMTV)プロモーターを含む。宿主細胞が、細胞内へのグルココルチコイド(即ちMMTVプロモーターの誘発物質)の取込みに必要な内生グルココルチコイド受容体を発現しない場合には、更にグルココルチコイド受容体をコードするDNA(ATCC受託番号67200)を細胞にトランスフェクションする必要がある。
【0078】
別の実施態様では、前述のポリ核酸(即ちDNA又はmRNA)を含む細胞を提供する。DNAの複製及びnAChRサブユニットの製造には、細菌、酵母及び哺乳動物細胞のような宿主細胞を使用し得る。発現ベクターを構築し、in vitro転写体を形成し、DNAを哺乳動物細胞内にトランスフェクションし、卵母細胞に注入し、ここに記載のような受容体発現及び機能を評価するための電気生理学的分析及び他の分析を実施する方法は、PCT出願公開明細書PCT/US91/02311号、PCT/US94/02447号、PCT/US91/05625号及びPCT/US92/11090号、米国特許明細書第5,369,028号並びに同時係属米国特許出願明細書第07/563,751号及び第07/812,254号にも記述されている。これらの出願明細書の論題はその全体が参考として本明細書に包含される。
【0079】
本発明が提供するDNAは、酵母細胞(例えばPichia、特にPichia pastoris(米国特許明細書第4,882,279号、第4,837,148号、第4,929,555号及び第4,855,231号参照)、Saccharomyces cerevisiae,Candida tropicalis,Hansenula polymorpha及び他の酵母細胞)を含む任意の真核細胞内で発現し得、現時点で好ましいのは、本発明が提供するヒトニューロンニコチン性nAChRサブユニットをコードするDNAを発現するための、市販の系及び他の当業者に公知の系を含む哺乳動物発現系である。DNAのRNA転写体の発現にはアフリカツメガエル卵母細胞が好ましい。
【0080】
任意のヒトニューロンニコチン性nAChRサブユニットをコードするクローン化完全長DNAは、真核細胞内での発現のためにプラスミドベクター内に導入し得る。このようなDNAはゲノムDNA又はcDNAであってよい。各々が少なくとも一つのヒトニューロンnAChRサブユニットをコードする単一のプラスミド又は複数のプラスミドの組合わせを宿主細胞にトランスフェクションし得る。外来DNAは細胞内でエピソームエレメントとして維持され得、又は細胞の染色体DNA中に組込まれ得る。
【0081】
DNA又はRNAを導入し得る真核細胞としては、このようなDNAもしくはRNAをトランスフェクション又は注入し得る任意の細胞が挙げられる。好ましい細胞は、一時的又は安定的にトランスフェクションすることができ、DNA及びRNAを発現することもできる細胞である。現時点で最も好ましい細胞は、外来DNAでコードされたサブユニットを一つ以上含む組換え又は外来ヒトニューロンニコチン性nAChRを形成することができる細胞である。このような細胞は経験的に同定し得、又はトランスフェクションもしくは注入が容易なことが知られているものの中から選択し得る。
【0082】
DNAを導入するための細胞の非限定的具体例としては、哺乳動物由来の細胞(例えばCOS細胞、マウスL細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト胎児腎臓(HEK)細胞、GH3細胞及び他の当業者に公知の細胞)、両生動物細胞(例えばXenopus laevis卵母細胞)及び酵母細胞(例えばSaccharomyces cerevisiae,Pichia pastoris)が挙げられる。注入RNA転写体を発現するための代表的な細胞はXenopus laevis卵母細胞である。DNAのトランスフェクションに好ましい細胞は当業者の公知であり、又は経験的に同定し得、具体例としてはHEK293(受託番号#CRL 1573でATCCから入手可能)、Ltk−細胞(受託番号#CCL1.3でATCCから入手可能)、COS−7細胞(受託番号#CRL 1651でATCCから入手可能)及びGH3細胞(受託番号#CCL82.1でATCCから入手可能)が挙げられる。現時点で好ましい細胞としては、GH3細胞及びHEK293細胞、特に懸濁液増殖用に調整され、液体窒素中で冷凍し、その後解凍して再増殖することができるHEK293細胞が挙げられる。HEK293細胞は、例えばGormanの米国特許明細書第5,024,939号に記載されている(Stillmanら(1985)Mol.Cell Biol.5:2051−2060も参照)。
【0083】
DNAは当業者に公知の方法で細胞内に安定に導入することができ、又は一時的に導入し得る。安定にトランスフェクションした哺乳動物細胞は、nAChRサブユニットをコードするDNAを含む一つ以上の発現構築物及び選択可能マーカー遺伝子(非限定的具体例としてネオマイシン耐性、ゼオシン耐性もしくはハイグロマイシン耐性用の遺伝子)を有する別個の発現ベクターを細胞にトランスフェクションするか、又はnAChRサブユニットをコードするDNAと選択可能マーカーとを含む一つ以上の発現構築物を細胞にトランスフェクションし、トランスフェクションした細胞を、マーカー遺伝子発現細胞に関して選択的な条件下で増殖させることにより製造し得る。このような細胞を製造するためには、外来DNAでコードされたヒトサブユニットを含むヒトニューロンnAChRを形成するのに十分な濃度のサブユニットコーディング核酸を細胞にトランスフェクションしなければならない。サブユニットをコードするDNAの正確な量及び比率は経験的に決定し得、サブユニット、細胞及びアッセイ条件の特定の組合わせに関して最適化し得る。特に好ましいのは、外来DNA又はRNAのみによってコードされたサブユニットを含むニューロンnAChRを発現する組換え細胞である。
【0084】
E. 組換えnAChR及びnAChRサブユニットタンパク質
本発明は、実質的に純粋なヒトnAChRサブユニットタンパク質、特にヒトα6及びβ3サブユニットタンパク質を提供する。前記ヒトnAChRサブユニットタンパク質の少なくとも一つを含む組換えnAChRも提供する。従って、本発明の別の実施態様は、組換えヒトnAChRサブユニット及び該サブユニットを含む受容体の製造方法を含む。
【0085】
好ましい実施態様では、ヒトnAChRサブユニット、特にヒトnAChRα6及びβ3サブユニットをコードするDNAをベクター内に連結し、得られた構築物を適当な宿主細胞内に導入して、特定のヒトニューロンnAChR受容体サブタイプ又はサブタイプの特定の組合わせを発現する形質転換細胞系を形成する。得られた細胞系は次いで、受容体機能に対する薬剤の作用の再現可能な定量分析を行うために量的に製造し得る。別の実施態様では、各サブユニットをコードするDNAのin vitro転写によりmRNAを製造し得る。単一サブユニットクローン又は複数のクローンの組合わせに由来する前記mRNAは、次いでアフリカツメガエル卵母細胞内に注入し得、そこでmRNAがヒト受容体サブユニットの合成を指示し、これらのサブユニットが機能性受容体を形成する。あるいは、機能性受容体を発現させるために、サブユニットをコードするDNAを卵母細胞に直接注入することもできる。トランスフェクションを行った哺乳動物細胞、又は注入を行った卵母細胞は、次いで本明細書に記載の薬剤スクリーニング方法で使用し得る。
【0086】
得られた組換え細胞は培養し得、又はこのような培養物又はその継代培養物から継代培養(subculture、又は哺乳動物の場合はpassage)し得る。トランスフェクション、注入及び組換え細胞培養方法は当業者に公知である。また、ヒトニューロンニコチン性nAChRサブユニットは、当業者に公知のタンパク質精製方法を用いて精製し得る。例えば、サブユニットの一つ以上に特異的に結合する抗体又は他のリガンドを、サブユニット又はサブユニット含有ヒトニューロンnAChRのアフィニティ精製に使用し得る。
【0087】
ある実施態様では、ヒトニューロンnAChRサブユニット及び機能性受容体を発現する細胞の製造方法も提供する。これらの方法の一つでは、ニューロンnAChRの少なくとも一つのαサブユニット及び少なくとも一つのβサブユニットをコードするDNAを宿主細胞にトランスフェクションする。α及び/又はβサブユニットをコードするDNA又はRNAを含むトランスフェクション細胞は、ノーザンブロット又はスロットブロット分析のような方法を用いて選択できる。トランスフェクション細胞は、nAChRタンパク質を発現する細胞を同定するための分析にもかける。分析は例えば、細胞がアセチルコリン、ニコチン又はnAChRアゴニストに結合する能力を測定し、非トランスフェクション宿主細胞又は他の適当な対照細胞のニコチン結合能力と比較するか、又はnAChRアゴニストに応答して細胞膜に流れる電流を電気生理学的に観察することにより実施できる。
【0088】
特に好ましいこととして、外来DNAを含み、このDNAを発現し、組換え機能性ニューロンnAChRを形成する真核細胞が得られる。より好ましいこととして、組換えニューロンnAChR活性を容易に検出することができる。なぜなら、これは非トランスフェクション宿主細胞に存在しないか、又は非トランスフェクション細胞内で示されない量を有する種類のものだからである。組換え受容体を含むような細胞は例えば、ヒトニューロンニコチン性nAChRα6及びβ3をサブユニットをコードするDNAで形質転換した細胞に、一つ以上のα及び/又はβnAChRサブユニットの存在下又は不在下で、対応するタンパク質を発現させることにより製造し得る。得られる合成又は組換え受容体はα6及びβ3nAChRサブユニットを含む。このような受容体は、種々の用途、例えば非ヒト受容体又はヒト組織調製物を使用する先行技術のアッセイシステムでしばしば見られる妨害がないアッセイシステムの一部として有用である。また、単一受容体サブユニットを種々の潜在的アゴニスト又はアンタゴニストを用いて試験すれば、個々のサブユニットの機能及び活性に関して更に情報が得られる。このような情報は、受容体サブユニットの一つ以上と極めて特異的に相互作用することができる化合物の同定につながり得る。このような特異性は、医療用途で極めて有用であり得る。
【0089】
一つ以上のヒトニューロンnAChRサブユニットをコードするDNAは、個々のサブユニット及び機能性nAChRを発現するために、適当な宿主細胞(例えば真核又は原核細胞)内に導入し得る。好ましくは、αサブユニット及びβサブユニットの組合わせを細胞内に導入し得る。このような組合わせは、α2、α3、α4、α5、α6及びα7とβ2、β3及び/又はβ4との任意の一つ以上の組合わせを含む。これらのサブユニットの各々に関する配列情報は添付の配列表に示す。α5の配列情報はProc.Natl.Acad.Sci.USA(1992)89:1572−1576にも記載されており、α2、α3、α4、α7、β2及びβ4の配列情報は、本明細書に参考として包含されるPCT出願公開明細書第94/20617号にも記載されている。現時点で好ましいサブユニットの組合わせとしては、α6及び/又はβ3とα2、α3、α4、α5、β2又はβ4のうちのいずれか一つ以上との組合わせが挙げられる。一部のサブユニットは別のサブユニットの不在下でイオン輸送機能を有し得、他のサブユニットはイオン輸送機能を示すために二つ以上のサブユニットの組合わせを必要とすることが認識されている。例えば、α7サブユニットは任意のβサブユニットを付加しない状態で機能する。また、一部のサブユニットは単独又は組合わせ状態で機能性nAChRを形成し得ないが、他のnAChRサブユニットの組合わせの特性を調節し得る。
【0090】
ある実施態様では、細胞内で翻訳される少なくとも一つのRNA転写体を含む第一の組成物をヒトニューロンnAChRのサブユニット内に導入することによって、外来ヒトニューロンnAChRを含む真核細胞を製造する。好ましい実施態様では、翻訳されるサブユニットがヒトニューロンnAChRのαサブユニットを含む。より好ましくは、導入する組成物は、αサブユニットをコードするRNA転写体を含み、ヒトニューロンnAChRのβサブユニットをコードするRNA転写体も含む。RNA転写体は、ヒトニューロンnAChRをコードするDNAをトランスフェクションした細胞から得ることができ、又はサブユニットをコードするDNAのin vitro転写によって得ることができる。クローン化DNAをin vitro転写させ、得られたmRNAを真核細胞内に注入する方法は当業者に良く知られている。ヒトニューロンnAChRDNAクローンのin vitro転写体の発現には両生動物卵母細胞が特に好ましい。イオンチャンネルを研究するためのアフリカツメガエル卵母細胞の使用については、例えばDascal(1989)CRC Crit.Rev.Biochem.22:317−387を参照し得る。
【0091】
一つ以上のαサブタイプ及び一つ以上のβサブタイプをコードするDNA又はRNAを段階的に細胞内に導入することが可能である。得られた細胞は、機能性nAChR活性を検出するために、本明細書に記載の方法又は当業者に公知の方法で試験し得る。このような試験は、機能性nAChRを形成するα及びβサブユニットサブタイプの組合わせ、並びに機能性nAChRを形成する個々のサブユニットの同定を可能にする。
【0092】
組換え真核細胞の表面に存在する組換え受容体は、ヒトニューロンnAChRサブユニットをコードするDNA又はmRNAでコードされた一つ以上のサブユニットを含み得、又は宿主細胞にコードされたサブユニットと外来DNAもしくはmRNAにコードされたサブユニットとの混合物を含み得る。組換え受容体は均質であり得、又はサブタイプ混合物であり得る。種々の由来源、例えばマウス及びヒトに由来する受容体をコードするDNA又はmRNAの混合物も細胞内に導入し得る。このようにして、α6及びβ3サブユニットのみを含む、又はここに開示する任意の別のα及びβサブユニットとの組合わせを含む組換え受容体を発現する細胞を製造し得る。例えば、ここに記載のα6及びβ3サブユニットのいずれか又は両方を、α2、α3、α4、α5、α7、β2及び/又はβ4受容体サブユニットと一緒に発現できる。前述のように、一部のニューロンnAChRサブユニットは、別のサブユニットの不在下で機能性受容体を形成する能力を有し得、従って機能性受容体を形成するために同時発現が必ず必要であるとは限らない。また、一部のnAChRサブユニットは、機能性受容体の形成に参加するために、二つ以上のnAChRサブユニットとの同時発現を必要とし得る。
【0093】
F. アッセイ
本発明の実施態様の一つでは、組換えヒトニューロンnAChR発現哺乳動物細胞又は卵母細胞を試験化合物と接触させ、次いで試験化合物の存在下及び不在下でのnAChR仲介応答を比較するか、又は試験化合物の存在に対する試験細胞もしくは対照細胞のnAChR仲介応答を比較することにより、試験化合物の調節作用を評価することができる。
【0094】
当業者に理解されているように、ヒトニューロンnAChR活性を調節する化合物(例えばアゴニスト及びアンタゴニスト)を同定するためのアッセイ方法は通常、対照との比較を必要とする。前述のように、ある種類の「対照」細胞又は「対照」培養物は、対照培養物は試験化合物に暴露しないという点を除いて、試験化合物に暴露した細胞又は培養物と実質的に同じ方法で処理した細胞又は培養物である。例えば、電圧固定電気生理学的操作を使用する方法では、細胞を浸漬する外部溶液を変えるだけで、同じ細胞を試験化合物の存在下及び不在下で試験することができる。別の種類の「対照」細胞又は「対照」培養物は、対照培養物に使用した細胞が機能性ヒトニューロンnAChRを発現しないという点を除いて、トランスフェクション細胞と同じである細胞又は細胞培養物であり得る。この場合は、細胞又は各種類の細胞の培養物を、アッセイすべき化合物の存在下で実質的に同じ反応条件に暴露した時の、試験化合物に対する受容体陰性(対照)細胞の応答(又は応答欠如)と、試験化合物に対する試験細胞の応答とを比較する。
【0095】
機能性組換えヒトニューロンnAChRはヒトニューロンnAChRの少なくとも一つのαサブユニット、又は少なくとも一つのαサブユニット及びβサブユニットを含む。これらのサブユニットを発現する真核細胞は、RNA転写体の注入及びDNAのトランスフェクションによって製造された。これらの細胞は、外来ヒトニューロンnAChRサブユニットの一つ以上を含むヒトニューロンnAChRに起因し得るnAChR活性を示した。
【0096】
機能性外来ヒトニューロンnAChRの活性の測定については、内生nAChR活性、そして所望であれば、内生宿主細胞サブユニットと外来サブユニットとの混合物を含むnAChRの活性を、可能であれば、化学的、薬理学的及び電気生理学的方法で大幅に阻害すべきである。
【0097】
G. 抗体
本発明は、前述のnAChRサブユニット又はその一部分に対して産生される抗体も提供する。この種の抗体は、受容体組織の検出、サブタイプ組成、機能ドメインの構造、受容体の精製の評価、並びに診断及び治療用途で使用し得る。好ましくは治療用途では、使用する抗体はモノクローナル抗体である。
【0098】
前述の抗体は、本明細書に記載のnAChRサブユニットタンパク質又はその一部分を抗体産生用の抗原として使用して、当業者に良く知られている標準的方法で製造できる。抗ペプチド抗体及び抗融合タンパク質抗体の両方を使用し得る[例えば、Bahouthら(1991)Trends Pharmacol.Sci.12:338−343;Current Protocols in Molecular Biology(Ausubelら編),John Wiley and Sons,New York(1989)参照]。免疫原として(合成ペプチド又は組換えにより産生した細菌融合タンパク質として)使用するためのnAChRサブユニットの部分の選択で考慮すべき要素としては、抗原性、アクセスの容易さ(即ち細胞外及び細胞質ドメイン)、特定サブタイプに対する特有性、及び他の当業者に公知の要素が挙げられる。
【0099】
サブタイプ特異的抗体を入手できれば、免疫化学の手法を種々のサブタイプの分布及び発現密度(例えば正常及び疾患脳組織内)の観察に適用することが可能になる。ヒトnAChRサブユニットを免疫原として製造した抗体は、種々の特性の中でも特に、生物学的試料又はこの種の試料に由来する溶液中に存在し得るヒトnAChR又はそのサブユニットに特異的且つ優先的に結合する能力、及び/又は前記ヒトnAChRもしくはそのサブユニットを免疫沈降させる能力を有する。このような抗体は、これらの抗体が特異的に結合するサブユニットを含むヒトnAChRを発現する細胞を選択的に分離するためにも使用し得る。これらの抗体は診断及び治療用途でも使用し得る。更に別の実施態様では、受容体を有効量の前記抗体と接触させることにより、nAChRのイオンチャンネル活性を調節する方法も提供する。
【0100】
本明細書に記載の抗体は、標準的方法、例えば腹腔内、筋内、静脈内もしくは皮下注射、移植又は経皮による投与方法を用いて対象に投与し得る。当業者は、使用する投与方法に基づいて、投薬形態、治療計画等を容易に決定し得る。
【0101】
以下の実施例は本発明を例示するためのものであって、本発明の範囲を限定するものではない。
【実施例1】
【0102】
ヒトnAChRα6サブユニットをコードするDNAの分離
ヒト黒質cDNAライブラリー(Clontech Laboratories,Inc.)を、マウスnAChRα6サブユニットcDNAのフラグメントへのハイブリダイゼーションについてスクリーニングした。分離したプラークをニトロセルロースフィルターに移し、ハイブリダイゼーションを5×デンハーツ、5×SSPE、50%ホルムアミド、200μg/ml変性サケ精子DNA及び0.2%SDS中で42℃で実施した。洗浄を0.2×SSPE、0.2%SDS中60℃で実施した。
【0103】
五つのハイブリッド形成クローンをプラーク精製し、制限エンドヌクレアーゼ地図及びDNA配列決定分析により特徴を調べた。市販のλgt10前進及びリバースオリゴヌクレオチドプライマーを使用して、cDNA挿入物の5’及び3’末端のDNA配列を決定した。五つのcDNA挿入物のDNA配列の分析の結果、三つのクローンが、翻訳開始コドンと完全長α読取り枠(配列番号9のヌクレオチド143−1624)と翻訳終止コドンと5’−非翻訳配列の更に142個のヌクレオチドと3’−非翻訳配列の116個のヌクレオチドとを含んでいた。完全長クローンのヌクレオチド配列から推定されるアミノ酸配列は、マウスnAChRα6サブユニットDNAから推定されるアミノ酸配列に対して〜82%の同一性を有する。推定マウス及びヒトα6アミノ酸配列の幾つかの領域は著しく相違している:アミノ酸1−30(ヒトシグナル配列はマウス配列に対して〜56%の同一性しか有していない)、アミノ酸31−50(ヒト配列はマウス配列に対して〜70%の同一性しか有していない)、アミノ酸344−391(ヒト配列はマウス配列に対して〜40%の同一性しか有していない)、アミノ酸401−428(ヒト配列はマウス配列に対して〜64%の同一性しか有していない)。
【0104】
また、単一クローンKEα6.5の挿入DNAは、α6コーディング配列の45個のヌクレオチドを欠失しており、その結果、推定アミノ酸配列の15個のアミノ酸残基(配列番号10の残基74−88)が枠内欠失(in−frame deletion)していると決定された。この配列を欠失している
α6サブユニット変異体のヌクレオチド配列は配列番号19に示し、該ヌクレオチド配列から推定されるアミノ酸配列は配列番号20に示す。興味深いことに、欠失部位のすぐ下流の推定アミノ酸配列は、推定マウスα6アミノ酸配列に対して〜58%のアミノ酸同一性しか有していない(配列番号10のアミノ酸89−100)。
【実施例2】
【0105】
ヒトnAChRβ3サブユニットをコードするDNAの分離
ヒト黒質cDNAライブラリー(Clontech Laboratories,Inc.)を、ヒトニコチン性nAChRβ3サブユニットcDNAに相補的な合成オリゴヌクレオチドへのハイブリダイゼーションについてスクリーニングした。分離したプラークをニトロセルロースフィルターに移し、オリゴヌクレオチドに対して高ストリンジェンシーの条件下で(洗浄条件1×SSPE、0.2%SDS、50℃)、配列番号15のヌクレオチド212−230及び1442−1469を含むヒトβ3nAChRサブユニットcDNAの配列に対して相補的な合成オリゴヌクレオチドとハイブリダイズさせた。
【0106】
二つのハイブリッド形成クローンをプラーク精製し、制限エンドヌクレアーゼ地図で特徴を調べた。市販のT7及びSP6オリゴヌクレオチドプライマーを使用して、cDNA挿入物の5’及び3’末端のDNA配列を決定した。クローンKBβ3.2の完全配列が決定された。クローンKBβ3.2は、完全長β3nAChRサブユニットをコードする1,377ヌクレオチド読取り枠(配列番号15のヌクレオチド98−1471)と、5’非翻訳配列の97個のヌクレオチド及び3’非翻訳配列の454個のヌクレオチドとを含む1927bpのcDNA挿入物を含んでいる。完全長クローンのヌクレオチド配列から推定されるアミノ酸配列は、マウスニコチン性nAChRβ3サブユニットDNAから推定されるアミノ酸配列に対して〜81%の同一性を有する。推定マウス及びヒトβ3アミノ酸配列の幾つかの領域は著しく相違している:アミノ酸1−28(ヒトシグナル配列はマウス配列に対して〜25%の同一性しか有していない)、アミノ酸347−393(ヒト配列はマウス配列に対して〜55%の同一性しか有していない)、アミノ酸440−464(ヒト配列はマウス配列に対して〜68%の同一性しか有していない)。
【実施例3】
【0107】
組換えヒトニューロンnAChRサブユニットを発現するための構築物の製造
ヒトニューロンnAChRサブユニットをコードする分離cDNAを、哺乳動物宿主細胞内でのサブユニットの発現に使用され、アフリカツメガエル卵母細胞内で発現すべきDNAからのin vitro転写体の形成に使用されるベクターに導入した。構築物の製造には下記のベクターを使用した。
【0108】
A. ヒトnAChRα6サブユニットを発現するための構築物
完全長nAChRα6サブユニットをコードする1,743bp EcoRIフラグメントを標準的方法でKEα6.3から分離し、ベクターpcDNA3のEcoRIポリリンカー部位に連結して、pcDNA3−KEα6.3を形成した(第1図参照)。プラスミドpcDNA3(第1図参照)は、CMVプロモーター/エンハンサーと、CMVプロモーター/エンハンサーの下流に位置するT7バクテリオファージRNAポリメラーゼプロモーターと、T7プロモーターの下流のウシ成長ホルモン(BGH)ポリアデニル化シグナルと、T7プロモーター及びBGHポリアデニル化シグナル間のポリリンカーとを含むpUC19ベースのベクターである。従ってこのベクターは、ポリリンカー位にベクター内に導入された外来DNAを哺乳動物細胞内で発現させるのに必要な調節エレメントを総て含む。更に、T7プロモーターがポリリンカーのすぐ上流に位置するため、このプラスミドはポリリンカーレベル位にベクター内にサブクローニングされた外来DNAのin vitro転写体の合成に使用できる。このプラスミドは更に、トランスフェクション時に選択可能マーカーとして使用されるネオマイシン耐性をコードする遺伝子を含む。
【0109】
第1図はpcDNA3−KEα6.3の部分的制限地図も示している。
【0110】
完全長ヒトnAChRα6サブユニットの発現は、翻訳開始コドンの前にコンセンサスリボソーム結合部位[RBS;例えばKozak(1991)J.Biol Chem.266:19867−19870参照]を導入することによって最適化した。既存の5’非翻訳領域を、DNA鋳型としてプラスミドpcDNA3−KEα6.3と、適当なヌクレオチドRBS置換体及び非翻訳5’HindIII及びEcoRI部位を含む相補上流オリゴヌクレオチドと、翻訳開始コドン下流のα6コーディング〜450ヌクレオチド配列に相補的なオリゴヌクレオチドとを使用して、PCR突然変異誘発により修飾した。その結果得られた増幅産物は、HindIII及びEcoRI部位と、これに続くコンセンサスRBS及びヒトnAChRα6コーディング配列ヌクレオチド1−459(配列番号9のヌクレオチド143−602)とを含んでいた。増幅DNAをHindIII及びBamHIで消化し、308bpのHindIII−BamHIフラグメントを分離し、pcDNA3−KEα6.3の5.3kb BamHI−PvuIフラグメント及びpcDNA3由来の1.4kb PvuI−HindIIIフラグメントに連結して、〜7.0kbプラスミドpcDNA3−KEα6RBSを形成した。
【0111】
B. ヒトニューロンnAChRβ3サブユニットを発現させるための構築物
完全長ニコチン性AChRβ3サブユニットをコードする〜2.0kb EcoRIフラグメントを標準的方法でKEβ3.2から分離し、ベクターpcDNA3のEcoRIポリリンカー部位に連結して、pcDNA3−KEβ3.2を形成した(第2図参照)。第2図はpcDNA3.KEβ3.2の部分的制限地図も示している。
【0112】
完全長ヒトニコチン性nAChRβ3サブユニットの発現は、翻訳開始コドンの前にコンセンサスリボソーム結合部位(RBS)を導入することによって最適化する。α6nAChRサブユニットについて上述したものと類似の方法を用いてPCR突然変異誘発により既存の5’非翻訳領域を修飾し、pcDNA3−KEβ3RBSを形成した。
【実施例4】
【0113】
アフリカツメガエル内での組換えヒトニューロンnAChRの発現
アフリカツメガエル卵母細胞に、α6及びβ3サブユニットをコードするDNAを含む構築物から製造したin vitro転写体を注入する。二電極電圧固定法を使用して卵母細胞トランスメンブラン電流の電気生理学的測定を行う(例えばStuhmer(1992)Meth.Enzymol.207:310−339参照)。
【0114】
1. in vitro転写体の調製
mMessage及びmMachine in vitro転写キットを製造業者のキャップ化転写体プロトコル通りに使用して(AMBION,Inc.,Austin,TXのカタログ1344)、直線化プラスミドからpcDNA3−KEαRBS及びpcDNA3−KEβ3RBSの組換えキャップ化転写体を合成する。合成した転写体の質量をUV吸光率によって測定し、転写体の完全性をアガロースゲル電気泳動で調べる。
【0115】
2. 電気生理学
アフリカツメガエル卵母細胞に、卵母細胞当たり12.5、50又は125ngの1種類以上のヒトニコチン性nAChRα及びβサブユニット転写体を注入する。卵母細胞の調製及び注入操作は、Dascal(1987),Crit.Rev.Biochem.22:317:387に記載のように実施する。mRNAの注入から2〜6日後、二電極電圧固定法を用いて卵母細胞を調べる。1μMのアトロピンを含み、100μMのd−チュボキュラリンを含む又は含まないリンガー液(115mM NaCl、2.5mM KCl、1.8mM CaCl2、10mM HEPES、pH7.3)に細胞を浸漬する。細胞を−60〜−80mVで電圧固定する。Axotapeソフトウエアで2〜5Hzでデータを得る。アゴニストアセチルコリン(ACh)、ニコチン及びシチシンを0.1μM〜100μMの濃度で加える。
【実施例5】
【0116】
哺乳動物細胞内でのヒトnAChRサブユニットの組換え発現
ヒト胎児腎臓(HEK)293細胞に、ヒトニューロンニコチン性nAChRα6及びβ3サブユニットをコードするDNAを一時的及び安定的にトランスフェクションする。種々のアッセイ、例えば電気生理学的方法、Ca2++感受性蛍光指示薬をベースとするアッセイを用いて、一時的トランスフェクション細胞をニコチン性nAChRの発現について分析する。
【0117】
1. HEK細胞の一時的トランスフェクション
HEK細胞に、一つ以上のαサブユニット及び/又は一つ以上のβサブユニットをコードするDNAを一時的に同時トランスフェクションする。約2×106のHEK細胞に18μgの指定プラスミドを、標準的CaPO4トランスフェクション方法で(Wiglerら(1979)Proc.Natl.Acad.Sci.U.S.A.76:1373−1376)、又はリポフェクタミン(lipofectamine)を製造業者(Bethesda Research Laboratory(BRL),Gaitherburg,MD)の指示通りに使用して、一時的にトランスフェクションする。更に、トランスフェクション効率を観察するために、CMVプロモーターに融合した大腸菌βガラクトシダーゼ遺伝子を含むプラスミドpCMVβgal(Clontech Laboratories,Palo Alto,CA)2μgをリポーター遺伝子として同時トランスフェクションする。βガラクトシダーゼ活性の測定により、トランスフェクション細胞をβガラクトシダーゼ発現について分析する[Miller(1972)Experiments in Molecular Genetics,pp.352−355,Cold Spring Harbor Press]。トランスフェクション細胞は、βガラクトシダーゼ及びX−gal基質が関与する反応の生成物を直接染色することにより、βガラクトシダーゼ発現についても分析し得る[Jones(1986)EMBO 5:133−3142]。
【0118】
2. HEK細胞の安定なトランスフェクション
リン酸カルシウムトランスフェクション法[Current Protocols in Molecular Biology,Vol.1,Wiley Inter−Science,Supplement 14,Unit 9.1.1−9.1.9(1990)]を使用してHEK細胞のトランスフェクションを行う。所望のα及びβサブユニットをコードするDNAとpSV2neo(選択可能マーカー)とを含むDNA/リン酸カルシウム沈降物1mlをHEK細胞にトランスフェクションする。1μg/mlのG418を含む培地で14日間増殖させた後、形成されたコロニーをクローニングシリンダーを用いて個々に分離する。分離物を限界希釈にかけ、後述のように最大レベルのnAChRを発現するものを同定するためにスクリーニングする。
【実施例6】
【0119】
ヒトニューロンnAChRを発現する細胞系の特徴分析
ヒトニューロンnAChRサブユニットをコードするDNAをトランスフェクションすることにより形成した実施例5に記載のような組換え細胞系は、下記の方法の一つ以上を用いて更に特徴を分析し得る。
【0120】
A. α及び/又はβサブユニットコーディングメッセージの発現に関するノーザン又はスロットブロット分析
〜1×107個の細胞から全RNAを分離し、各細胞種類のRNAを10〜15μg使用してノーザンブロット又はスロットブロットハイブリダイゼーション分析を行う。ヒトニューロンnAChRコーディングプラスミド由来の挿入物は、ニックトランスレーションにかけてプローブとして使用し得る。また、各ブロット上のRNAの存在又は不在を確認し、細胞系間のα又はβ特異的mRNAレベルの差の定量に使用する大体の標準を与えるために、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ(GAPD)遺伝子配列(Tsoら(1985)Nucleic Acids Res.13:2485)をニックトランスレーションにかけ、二重フィルター上で対照プローブとして使用し得る。典型的なノーザンブロット及びスロットブロットハイブリダイゼーション並びに洗浄条件は下記の通りである:5×SSPE、5×デンハーツ溶液、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、50%ホルムアミド中42℃でハイブリダイズし、次いで0.1×SSPE、0.1%SDS中65℃で洗浄する。
【0121】
B. 結合アッセイ
ヒトニューロンnAChRのα又はα及びβサブユニットをコードするDNAのトランスフェクションによって製造した細胞系は、例えばニコチン又は他のアゴニストに結合する能力について、対照細胞系、例えばニューロン由来細胞系PC12(Boulterら(1986)Nature 319:368−374;ATCC#CRL1721)及びIMR32(Clementiら(1986)Int.J.Neurochem.47:291−297;ATCC#CCL127)、並びに筋肉由来細胞系BC3H1(Patrickら(1977)J.Biol.Chem.252:2143−2153)と比較して分析し得る。陰性対照細胞(即ち、トランスフェクション細胞の形成に使用した宿主細胞)もアッセイに含まれる。アッセイは次のように実施する:アッセイにかける直前に、トランスフェクション細胞をスクラッピングによってプレートから除去する。使用する陽性対照細胞は、PC12、BC3H1及びIMR32である(これらの細胞には数日間新しい培地を与えなかった)。37℃のアッセイ緩衝液(50mMトリス/HCl、1mM MgCl2、2mM CaCl2、120mM NaCl、3mM EDTA、2mg/ml BSA及び0.1%アプロチニン、pH7.4)中で濯ぐことにより対照細胞系を除去する。細胞を洗浄し、1×106/250μlの濃度に再懸濁する。各プラスチックアッセイ管に、250μlの細胞溶液と、15nM 3H−ニコチンと、1mM非標識ニコチン有または無と、最終容量を500μlにする量のアッセイ緩衝液とを加える。トランスフェクション細胞系のアッセイを室温で30分間インキュベートする。陽性対照細胞のアッセイは1℃で2分間インキュベートする。適当な時間にわたってインキュベートした後、アッセイ容量の450μlアリコートを、0.05%ポリエチレンイミン中4℃で24時間インキュベートすることにより予備処理したWhatman GF/Cグラスファイバーフィルターで濾過する。次いで、フィルターを各回4mlの氷冷アッセイ緩衝液を使用して2回洗浄する。洗浄後、フィルターを乾燥し、5mlのシンチレーション液を入れたバイアルに加え、放射能を測定する。
【0122】
C. 86Rbイオン流アッセイ
ニコチン又はnAChRアゴニスト及びアンタゴニストがトランスフェクション細胞及び対照細胞内への86Rbの流入を仲介する能力は、細胞表面上の機能性nAChRの存在を指示するものであることが分かっている。86Rbイオン流アッセイは下記のように実施する:
1. 実験の前夜、ウェル数6のポリリシンコーティングしたプレートにウェル当たり2×106個の細胞(即ちウェル当たり2ml)を播く。
2. 培養培地をデカントし、プレートを2mlのアッセイ緩衝液(50mM HEPES、260mMスクロース、5.4mM KCl、1.8mM CaCl2、0.8mM MgSO4、5.5mMグルコース)で室温で洗浄する。
3. アッセイ緩衝液をデカントし、3μCi/mlの86Rbを5mMウワバイン及び最大応答を与える濃度のアゴニスト又はアンタゴニストと共に含むアッセイ緩衝液を1ml加える。
4. プレートを氷上1℃で4分間インキュベートする。
5. 緩衝液を廃棄物容器にデカントし、各ウェルを3mlのアッセイ緩衝液で洗浄し、次いで各回2mlで2回洗浄する。
6. 細胞をウェル当たり2×0.5mlの0.2%SDSで溶解し、5mlのシンチレーション液を入れたシンチレーションバイアルに移す。
7. 各バイアル5に含まれている放射能を測定し、データを計算する。このアッセイでは陽性対照細胞から下記のデータが得られた:
【0123】
【表3】
【0124】
D. ヒトニューロンnAChRサブユニットをコードするDNAをトランスフェクションした哺乳動物細胞の電気生理学的分析
組換え受容体の活性の評価、又は試験化合物がリガンド依存性組換えnAChRを介してカチオン流の規模及び持続時間を助長、拮抗又は調節する能力の評価には、電気生理学的測定を使用し得る。発現したニューロンnAChRの機能は、二電極電圧固定法及びパッチ固定法を含む種々の電気生理学的方法で評価できる。nAChRに固有のカチオン伝導チャンネルは、アセチルコリン(ACh)又は他のニコチン性コリン作動性アゴニストに応答して開き、その結果、生理学的条件下で主にナトリウム及びカリウムイオンにより担われているトランスメンブラン電流が流れる。この電流は電圧固定法で直接観察することができる。好ましい実施態様では、トランスフェクションを行った哺乳動物細胞又は注入を行った卵母細胞を、nAChRアゴニスト依存電流の存在に関して電気生理学的に分析する。
【0125】
E. 蛍光指示薬をベースとするアッセイ
アゴニストによりリガンド依存性nAChRが活性化されると、Ca++を含むカチオンが受容体チャンネルを介して流入する。チャンネルを介するCa++の細胞内流入は、細胞内ストアに含まれていたカルシウムの放出を誘発し得る。一価のカチオンがチャンネルを介して細胞内に流入すると、膜の脱分極及びその結果として生じる電圧依存カルシウムチャンネルの活性化を通して、細胞質Ca++レベルが増加し得る。従って、細胞内カルシウム濃度の一時的増加を検出する方法は、機能性ニコチン性nAChR発現の分析に適用できる。細胞内カルシウム濃度を測定する方法の一つは、カルシウム感受性蛍光指示薬の使用をベースとする。
【0126】
カルシウム感受性指示薬、例えばフルオ(fluo)−3(カタログ番号FO1241、Molecular Probes,Inc.,Eugene,OR)は、膜透過性であるアセトキシメチルエステルとして入手できる。アセトキシメチルエステル形態の指示薬が細胞内に入ると、エステル基が細胞質ゾルエステラーゼによって除去され、その結果細胞質ゾル内に遊離指示薬が捕捉される。遊離指示薬とカルシウムとが相互作用すると、指示薬の蛍光が増加する。従って、指示薬を含んでいる細胞の細胞内Ca2+濃度の増加は、蛍光の増加として直接示すことができる。ニコチン性nAChRをアッセイするための自動化蛍光検出システムは文献に記述されている(米国特許出願明細書第08/229,150号、第08/244,985号、第08/434,511号及び第08/434,968号並びに対応する国際特許出願公開明細書US92/11090号参照。国際特許出願公開明細書第96/05488号も参照)。
【0127】
自動化蛍光指示薬ベースアッセイを用いて、適当なα及び/又はβサブユニット並びにα6及びβ3サブユニットをコードするDNAを一時的又は安定的に同時トランスフェクションしたHEK細胞を、機能性組換えnAChRの発現について分析する。アッセイの手順は次の通りである。非トランスフェクションHEK細胞と適当なα及び/又はβサブユニットをコードするDNAを同時トランスフェクションしたHEK細胞とを、ウェル数96のミクロタイター皿のウェル内に播き、HBS(125mM NaCl、5mM KCl、1.8mM CaCl2、0.62mMm MgSO4、6mMグルコース、20mM HEPES、pH7.4)中20μMフルオ−3、0.2%Pluronic F−127を含む培地中で20℃で2時間インキュベートすることによりフルオ−3を充填する。次いで、細胞をアッセイ緩衝液(即ちHBS)で洗浄する。アンタゴニストd−ツボクラリンを一部のウェルに最終濃度10μMで加える。次いで、ミクロタイター皿を蛍光プレート読取り器内に配置し、各ウェルの基本蛍光を測定し、記録した後、アゴニスト、例えば200μMニコチンをウェルに加える。ニコチンの添加後、約60秒間にわたり、ウェルの蛍光を繰り返し観察する。
【0128】
非トランスフェクションHEK細胞の蛍光は、ニコチン添加後にも変化しない。これに対し、同時トランスフェクションを行った細胞の蛍光は、d−ツボクラリンの不在下では、ウェルへのニコチンの添加後に急増する。このニコチン刺激による蛍光の増加は、アンタゴニストd−ツボクラリンに暴露した同時トランスフェクション細胞では観察されない。このような結果は、同時トランスフェクション細胞が、ニコチンによって活性化され且つd−ツボクラリンによって阻害される機能性組換えnAChRを発現することを示すものである。
【0129】
好ましい実施例を挙げて本発明を詳細に説明してきたが、種々の変形が本発明の思想及び範囲を逸脱せずに可能である。
【0130】
変形例は当業者に明らかであるため、本発明は後述の「請求の範囲」によってのみ限定される。
【0131】
【表4】
【0132】
配列の要約
配列番号1はヒトニューロンニコチン性アセチルコリン受容体のα2サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0133】
配列番号2は配列番号1に示すヒトニューロンニコチン性アセチルコリン受容体のα2pサブユニットのアミノ酸配列である。
【0134】
配列番号3はヒトニューロンニコチン性アセチルコリン受容体のα3サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0135】
配列番号4は配列番号3に示すヒトニューロンニコチン性アセチルコリン受容体のα3サブユニットのアミノ酸配列である。
【0136】
配列番号5はヒトニューロンニコチン性アセチルコリン受容体のα4サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0137】
配列番号6は配列番号5に示すヒトニューロンニコチン性アセチルコリン受容体のα4サブユニットのアミノ酸配列である。 配列番号7はヒトニューロンニコチン性アセチルコリン受容体のα5サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0138】
配列番号8は配列番号7に示すヒトニューロンニコチン性アセチルコリン受容体のα5サブユニットのアミノ酸配列である。
【0139】
配列番号9はヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0140】
配列番号10は配列番号9に示すヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットのアミノ酸配列である。
【0141】
配列番号11はヒトニューロンニコチン性アセチルコリン受容体のα7サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0142】
配列番号12は配列番号11に示すヒトニューロンニコチン性アセチルコリン受容体のα7サブユニットのアミノ酸配列である。
【0143】
配列番号13はヒトニューロンニコチン性アセチルコリン受容体のβ2サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0144】
配列番号14は配列番号13に示すヒトニューロンニコチン性アセチルコリン受容体のβ2サブユニットのアミノ酸配列である。
【0145】
配列番号15はヒトニューロンニコチン性アセチルコリン受容体のβ3サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0146】
配列番号16は配列番号15に示すヒトニューロンニコチン性アセチルコリン受容体のβ3サブユニットのアミノ酸配列である。
【0147】
配列番号17はヒトニューロンニコチン性アセチルコリン受容体のβ4サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0148】
配列番号18は配列番号17に示すヒトニューロンニコチン性アセチルコリン受容体のβ4サブユニットのアミノ酸配列である。
【0149】
配列番号19はヒトニューロンニコチン性アセチルコリン受容体の変異体α6サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0150】
配列番号20は配列番号19に示すヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットのアミノ酸配列である。
【図面の簡単な説明】
【0151】
【図1】第1図は、α6コーディングフラグメントをEcoRI挿入物として含むサイトメガロウイルス(CMV)プロモーターをベースとするベクターpcDNA3−KEアルファ6.3の制限地図を示す説明図である。
【図2】第2図は、β3コーディングフラグメントをEcoRI挿入物として含むCMVプローモーターをベースとするベクターpcDNA3−KBベータ3.2の制限地図を示す説明図である。
【技術分野】
【0001】
関連出願
本出願明細書は、米国内向けでは、1995年6月7日出願のElliottらによる米国特許出願明細書第08/484,722号“HUMAN NEURONAL NICOTINIC ACETYLCHOLINE RECEPTOR COMPOSITIONS AND METHODS EMPLOYING SAME(ヒトニューロンニコチン性アセチルコリン受容体組成物及び該組成物を使用する方法)”の一部継続出願である。米国特許出願明細書第08/484,722号の論題は全体が本出願明細書に参考として包含される。
【0002】
本出願明細書は、米国特許明細書第5,369,028号及び米国特許出願明細書第08/028,031号、第08/149,503号、第08/496,855号、第07/938,154号、第08/467,574号、第08/466,589号、第08/487,596号にも関連している。これらの米国特許出願明細書及び米国特許明細書の論題は本出願明細書に参考として包含される。
【0003】
発明の分野
本発明は、ヒトニューロンニコチン性アセチルコリン受容体タンパク質サブユニットをコードする核酸分子、及びコードされたタンパク質に関する。特に、ヒトニューロンニコチン性アセチルコリン受容体αサブユニットをコードするDNA及びRNA、αサブユニットタンパク質、βサブユニットをコードするDNA及びRNA、βサブユニットタンパク質、並びにこれらの組合わせを提供する。
【0004】
発明の背景
リガンド依存性イオンチャンネルは中枢神経系の細胞間の情報伝達手段を提供する。これらのチャンネルは、ある細胞から放出された信号(例えば神経伝達物質と称する化学物質)を、標的細胞膜に沿って伝搬する電気信号に変換する。中枢及び末梢神経系には種々の神経伝達物質及び神経伝達物質受容体が存在する。神経筋及びニューロン由来のニコチン性アセチルコリン受容体(nAChR)を含む5種類のリガンド依存性受容体類が同定された(Stroudら,1990 Biochemistry 29:11009−11023)。しかしながら、種々の受容体が神経系の種々の部分で神経伝達物質又は他の調節リガンドに対する種々の応答を発生させるメカニズムは殆ど解明されていない。
【0005】
ニコチン性アセチルコリン受容体(nAChR)は、神経筋及びニューロン由来のマルチサブユニットタンパク質である。これらの受容体は、神経伝達物質アセチルコリン(ACh)との相互作用時に、神経と筋肉との間、及びニューロン間のシナプス伝達を仲介するリガンド依存性イオンチャンネルを形成する。様々なニコチン性アセチルコリン受容体(nAChR)サブユニットが存在するため、様々なnAChR組成物(即ちサブユニットの組合わせ)が存在する。異なるnAChR組成物は種々のリガンドに対して異なる特異性を示すため、薬理学的に識別可能である。脊椎動物神経筋接合部、脊椎動物交感神経節及び脊椎動物中枢神経系で発現されるニコチン性アセチルコリン受容体は、種々のnAChR組成物に結合する種々のリガンドの作用に基づいて識別された。例えば、神経筋接合部のニコチン性アセチルコリン受容体の活性化を阻止するメガネヘビα神経毒は、幾つかの異なるニューロン由来細胞系で発現される一部のニューロンニコチン性アセチルコリン受容体の活性化は阻止しない。
【0006】
筋肉nAChRは、化学量論(α)2β(γ又はε)δで5種類のサブユニットからなる糖タンパク質である。各サブユニットは約50〜60キロドルトン(kd)の質量を有し、異なる遺伝子によってコードされる。(α)2β(γ又はε)δ複合体は、二つのリガンド結合部位と一つのリガンド依存性トランスメンブランチャンネルとを含む機能性受容体を形成する。筋肉ニコチン性nAChRは、コリン作動性アゴニストとの相互作用時にナトリウムイオンを伝導する。ナトリウムイオンの流入は、プラズマ膜の内外で維持されていた正常なイオン勾配を即座に破壊し、その結果、プラズマ膜が脱分極する。膜の内外の電位差を減少させると、神経筋接合部で化学信号が電気信号に変換され、筋肉の収縮を誘発する。
【0007】
機能性筋肉ニコチン性アセチルコリン受容体は、αβδγサブユニット、αβγサブユニット,αβδサブユニット、αδγサブユニットで形成されたが、一つのサブユニットだけでは形成されていない(例えば、Kurosakiら(1987)FEBS Lett.214 253−248;Comachoら(1993)J.Neuroscience 13:605−613参照)。これに対し、機能性ニューロンnAChRは、αサブユニットのみ、又はα及びβサブユニットの組合わせによって形成できる。より大きいαサブユニットは一般的にACh結合サブユニットと考えられており、より小さい分子量のβサブユニットは一般的に構造サブユニットと考えられているが、βサブユニットがAChに結合する能力を有していない、又はACh結合部位の形成に関与しないという決定的な証拠はない。機能性イオンチャンネルの形成に関与するサブユニットはいずれも、結果として得られるチャンネルの構造に寄与するという範囲で、ACh結合能力(又は不能)にかかわりなく「構造」サブユニットである。リガンド依存性イオンチャンネルでもあるニューロンnAChRは、自律神経系及び中枢神経系の神経節(ニューロンnAChRが信号伝達を仲介する場所)、並びにシナプス前及びシナプス外位置(ニューロンnAChRが神経伝達を調節し、別の機能も果たし得る場所)で発現される。Wonnacottら(1990)Progress in Brain Research,A.Nordberら編,Elsevier,Amsterdam,157−163。
【0008】
nAChRをコードするDNAは幾つかの由来源から分離された。この種の研究から得られる情報に基づいて、しばらくの間、筋肉、自律神経節及び中枢神経系で発現されるnAChRがそれぞれ機能的に異なることは明らかだとされていた。この機能的多様性は、少なくとも部分的には、存在する種々のnAChRサブユニットの数が多いことに起因し得る。しかしながら、特にニューロン細胞で、(どの)nAChRサブユニットがどのように組合わせられて独自のnAChRサブタイプを形成するのかは完全には解明されていない。実際、特定のサブタイプのみがアルツハイマー病のような疾患に関与し得ることは証明されている。また、類似の組織又は細胞種類に由来するnAChRが類似の交差種であるかどうかは不明である。
【0009】
従って、各ヒトニューロンnAChRサブユニット、この種のサブユニットを含む組換え細胞、及びこれらから形成された受容体をコードするDNAを分離し、特徴を解明する必要がある。ヒトニューロンnAChRの機能を調べ、疾患特異的な薬理学的活性物質を得るために、分離(好ましくは精製)ヒトニューロンnAChR及び分離(好ましくは精製)ヒトニューロンnAChRサブユニットを得る必要もある。また、このような薬理学的活性物質を同定するためのアッセイを開発する必要もある。
【0010】
前述のような核酸、細胞、受容体サブユニット及び受容体組成物を利用できれば、非ヒトnAChRデータ、又はヒトもしくは非ヒト筋肉もしくは神経節nAChRデータから導き出した予測に基づくヒトニューロンnAChRの構造及び機能に関する推測の不確実さが除去される。
【0011】
従って本発明は、ヒトニューロンニコチン性アセチルコリン受容体のサブユニットをコードするDNAを分離し特徴を解明することを目的とする。本発明は、ヒトニューロンニコチン性アセチルコリン受容体サブユニットを組換えにより製造する方法を提供することも目的とする。本発明は更に、精製受容体サブユニットを提供すること、並びにヒトニューロンnAChRの活性を調節する化合物を同定するために化合物をスクリーニングする方法を提供することも目的とする。
【0012】
これらの目的及び他の目的は、本明細書の本文及び「特許請求の範囲」を更に検討することにより当業者に理解されよう。
【0013】
発明の概要
ヒトニューロンnAChRα及びβサブユニットをコードする分離核酸分子を提供する。特に、ヒトニューロンnAChRα6及びβ3サブユニットをコードする分離DNA及びRNAを提供する。前記DNAによってコードされるメッセンジャーRNA及びポリペプチドも提供する。
【0014】
α6及びβ3サブユニットを含む組換えヒトニューロンニコチン性nAChRサブユニット、並びにその製造方法も提供する。更に、少なくとも一つのヒトニューロンニコチン性nAChRサブユニットを含む組換えヒトニューロンニコチン性アセチルコリン受容体、及びその製造方法も提供する。また、宿主細胞でコードされた一つ以上のnAChRサブユニットと外来DNAもしくはRNA(即ち宿主細胞内に導入した本明細書に記載のDNAもしくはRNA)でコードされた一つ以上のnAChRサブユニットとの混合物を含む組換えニューロンニコチン性nAChR、及びその製造方法も提供する。
【0015】
前述のサブユニットをコードするDNAを含むプラスミドも提供する。本発明は、前述のDNA、mRNA又はプラスミドを含む組換え細胞も提供する。この種の細胞は、例えばDNAの複製、ヒトnAChRサブユニット及び組換え受容体の製造、並びに一つ以上のヒトサブユニットを含む受容体を発現する細胞の製造に有用である。
【0016】
本発明が提供するDNA、RNA、ベクター、受容体サブユニット、受容体サブユニットの組合わせ及び細胞は、選択したニューロンニコチン性nAChR受容体サブタイプ、及びこれらのサブタイプの特定の組合わせ、並びに受容体サブユニットに対する抗体の製造を可能にする。これは、その存在が単一nAChRサブタイプの分析を妨害し得る他の多くの受容体タンパク質が実質的に混入していない合成もしくは組換え受容体及び受容体サブユニットの製造手段を提供するものである。所望の受容体サブタイプを入手できれば、薬剤物質が特定の受容体サブタイプに与える作用を観察することが可能になり、従ってヒト及びヒトニューロンニコチン性nAChRsサブタイプに特異的な試験システムで薬剤物質の初期in vitroスクリーニングを実施することが可能になる。
【0017】
本発明は、ここに開示するDNA分子の一部と、該DNAでコードされたタンパク質に特異的に結合する抗体とを含む一本鎖プローブも提供する。また、配列番号15のヌクレオチド98−211を含む分離核酸分子も提供する。
【0018】
前記DNAでコードされたタンパク質も提供する。これらのタンパク質は、前記DNAを適当な原核又は真核宿主細胞内で発現させ、その結果産生されたタンパク質を分離することにより製造し得る。
【0019】
機能性ニューロンニコチン性アセチルコリン受容体サブユニット及びこれらの組合わせを同定する方法も提供する。
【0020】
ヒトニコチン性アセチルコリン受容体の活性を調節する化合物を同定するアッセイも提供する。特定の受容体組成物に対する薬剤の作用を調べるために薬剤物質をin vitroスクリーニングすることができれば、受容体サブタイプ特異的又は疾患特異的薬剤の開発及びスクリーニングが可能になる。種々の潜在的アゴニスト又はアンタゴニストを用いて単一受容体サブユニット又は特定受容体サブタイプの組合わせを試験すれば、個々のサブユニットの機能及び活性に関して更に情報が得られ、受容体サブユニット又は受容体サブタイプの一つ以上と極めて特異的に相互作用することができる化合物の同定及び設計が可能になる。得られる薬剤は、種々のサブタイプを発現する細胞を用いてスクリーニングすることにより同定した薬剤と比べて、望ましくない副作用が少ない。
【0021】
更に、薬剤の開発及び種々の疾患状態の治療に関連して、ヒトニューロンnAChRサブユニットをコードするDNA及びRNAを利用できれば、特定の疾患状態の発生と相関し得るような遺伝子中の任意の変化(例えば突然変異)を同定する手段が得られる。また、このような突然変異を、後で作用を調べるために実験動物又はin vitroアッセイシステム中に導入し得る合成DNA配列中に特異的に導入することにより、前述のような疾患状態の動物モデルの製造が可能になる。
【0022】
図面の簡単な説明
第1図は、α6コーディングフラグメントをEcoRI挿入物として含むサイトメガロウイルス(CMV)プロモーターをベースとするベクターpcDNA3−KEアルファ6.3の制限地図を示す説明図である。
【0023】
第2図は、β3コーディングフラグメントをEcoRI挿入物として含むCMVプローモーターをベースとするベクターpcDNA3−KBベータ3.2の制限地図を示す説明図である。
【0024】
好ましい実施態様の詳細な説明
定義
特に指示がない限り、本明細書で使用する総ての技術及び科学用語は、本発明が属する分野の当業者によって一般的に理解されているものと同じ意味を表す。本明細書で言及する総ての特許及び出版物は、特に指示がない限り、全体が参考として本明細書に包含される。
【0025】
本明細書で核酸分子、DNA、RNA、ポリペプチド又はタンパク質を形容する語として使用する「分離」(又は実質的に精製した、もしくは純粋な)という用語は、このように形容されるDNA、RNA、ポリペプチド又はタンパク質が人の手を介してin vivo細胞環境から分離されたものであることを意味する。従って、例えば本明細書中の「分離(又は実質的に純粋な)DNA」とは、当業者に使用されている標準的方法(例えばManiatisら(1982)Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY参照)で精製したDNAフラグメントを意味する。
【0026】
同様にして、本明細書でDNA、RNA、ポリペプチド又はタンパク質を形容する語として使用する「組換え」という用語は、このように形容されるDNA、RNA、ポリペプチド又はタンパク質が、例えばクローニング、組換え発現といったような方法で人為的に製造されたものであることを意味する。従って、例えば本明細書中の組換えタンパク質とは、人為的に加えられたDNAを含みこれを発現する組換え宿主により産生されたタンパク質を意味する。
【0027】
本明細書で使用するベクター(又はプラスミド)という用語は、外来DNAをその発現又は複製のために細胞内に導入するのに使用される別個のエレメントを意味する。このようなビヒクルの選択及び使用は、当業者が随意に実施できる範囲のものである。発現ベクターには、調節配列と作用的に結合するDNAを発現することができるベクター、例えば前記のようなDNAフラグメントの発現を生起させることができるプローモー領域が含まれる。従って、発現ベクターとは、組換えDNAもしくはRNA構築物、例えばプラスミド、ファージ、組換えウイルス、又は、宿主細胞内に導入されるとベクター上の適当な部位内にクローニングされたDNAを発現させる他のベクターを意味する。適当な発現ベクターは当業者に良く知られており、真核細胞及び/又は原核細胞内で複製できるもの、並びにエピソーム状態を維持するもの、又は宿主細胞ゲノム内に組込まれるものがある。真核宿主細胞、特に哺乳動物細胞内でnAChRサブニットを発現させるための、現時点で好ましいプラスミドの非限定的具体例としては、サイトメガロウイルス(CMV)、サルウイルス40(SV40)及びマウス乳癌ウイルス(MMTV)プロモーター含有ベクター、例えば、pCMV、pcDNA1,pcDNA3、pZeoSV、pCEP4、pMAMneo及びpMAMhygが挙げられる。
【0028】
本明細書で使用するプロモーター領域という用語は、DNAに作用的に結合してその転写を制御するDNAセグメントを意味する。プロモーター領域は、RNAポリメラーゼの認識、結合及び転写開始に十分な特異的配列を含む。プロモーター領域のこの部分をプロモーターと称する。プロモーター領域は更に、RNAポリメラーゼの認識、結合及び転写開始活性を調節する配列も含む。これらの配列はシス作用性であり得、又はトランス作用因子に応答し得る。プロモーターは、調節の種類に応じて構成性であり得、又は調節される。本発明で使用し得るプロモーターの具体例としては、SV40初期プロモーター、サイトメガロウイルス(CMV)プロモーター、マウス乳癌ウイルス(MMTV)ステロイド誘導プロモーター及びモロネーマウス白血病ウイルス(MMLV)プロモーター、並びに他の適当なプロモーターが挙げられる。
【0029】
本明細書で使用する「作用的に結合」という用語は、DNAと調節及びエフェクターヌクレオチド配列、例えばプロモーター、エンハンサー、転写及び翻訳開始及び終結部位、並びに他のシグナル配列との機能的関係を指す。例えば、DNAとプロモーターとの作用的結合は、DNAとプロモーターとの物理的及び機能的関係であって、DNAの転写が、該DNAを特異的に認識し、これに結合し且つこれを転写するRNAポリメラーゼによりプロモーターから開始されるような関係を意味する。発現及び/又はin vitro転写を最適化するためには、クローンの5’非翻訳部分を除去又は変化させて、余計な潜在的な別の翻訳開始コドン、又は転写もしくは翻訳のレベルで発現を妨害もしくは低下させる別の配列を除去する必要があり得る。あるいは、コンセンサスリボソーム結合部位(例えばKozak(1991)J.Biol.Chem.266:19867−19870参照)を開始コドンの5’に隣接させて挿入して、発現を促進することもできる。このような修飾が望ましい(又は必要である)かどうかは、経験的に決定し得る。
【0030】
本明細書で使用する発現という用語は、ポリ核酸がmRNAに転写され、ペプチド、ポリペプチド又はタンパク質に翻訳されるプロセスを意味する。ポリ核酸がゲノムDNAに由来する場合には、適当な真核宿主細胞又は生物を選択すれば、発現はmRNAのスプライシングを含み得る。
【0031】
哺乳動物細胞のトランスフェクションに特に好ましいベクターはSV40プロモーターをベースとする発現ベクター、例えばpZeoSV(Invitrogen,San Diego,CA)、CMVプロモーターをベースとするベクター、例えばpcDNA1、pcDNA3、pCEP4(Invitrogen,San Diego,CA)及びMMTVプロモーターをベースとするベクター、例えばpMAMneo(Clontech,Inc.)が挙げられる。
【0032】
本明細書中のヒトαサブユニット遺伝子とは、ヒトニューロンニコチン性アセチルコリン受容体のαサブユニットをコードする遺伝子である。ヒトnAChRのαサブユニットは典型的には、デンキナマズ(Torpedo)αサブユニット(Nodaら(1982)Nature 299:793−797参照)のシステイン192及び193と相同である、サブユニットの推定細胞外ドメイン内の隣接システイン残基の保存を示す。
【0033】
本明細書で使用するαサブユニットサブタイプという用語は、ここに開示するニューロンnAChRαサブユニットコーディングDNAクローンのうちの少なくとも一つに高ストリンジェンシー条件下でハイブリダイズするDNAでコードされるヒトニューロンnAChRサブユニットを意味する。αサブユニットは通常、生理学的条件下で、生理学的濃度で、任意的なβサブユニットの存在下で(即ち、あるαサブユニットは単独で機能するが、別のαサブユニットはβサブユニットの存在を必要とする)AChに結合し、通常は、本明細書に記載の方法又は当業者に公知の方法で評価して、機能性のnAChRを形成する。
【0034】
前述のようなαサブユニットをコードするが、遺伝暗号の縮重に起因して、指定のハイブリダイゼーション条件下で開示のDNAに必ずしもハイブリダイズしないDNA分子でコードされるαサブユニットも考えられる。このようなサブユニットも、通常は一つ以上のβサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性の受容体を形成する。典型的には、αサブユニットが別のスプライシングから生じるRNA(即ちスプライス変種(splice variant))でコードされる場合を除いて、αをコードするDNA及び該DNAでコードされたαサブユニットは、本明細書に記載のαサブユニットDNA(及び該DNAでコードされたタンパク質)の少なくとも一つに対して実質的な配列相同を有する。ここで理解されることとして、スプライス変種をコードするDNA又はRNAは全体で本発明が提供するDNA又はRNAに対して90%以下の相同を有し得るが、本明細書に開示のDNAフラグメントに対してほぼ100%相同の領域を含んでおり、開始及び終止コドンを含み機能性αサブユニットをコードする読取り枠をコードする。
【0035】
本明細書中のヒトβサブユニット遺伝子とは、ヒトニューロンニコチン性アセチルコリン受容体のβサブユニットをコードする遺伝子である。推定上のニューロンnAChRサブユニットに「β」という名称を付けた理由は、隣接システイン残基の欠失にある(これらの残基はαサブユニットに特徴的なものである)。βサブユニットはしばしば構成性nAChRサブユニットと呼ばれる(但しβサブユニットがACh結合特性を有する可能性もある)。適当なβサブユニットと適当なαサブユニットとを組合わせれば、機能性受容体が形成される。
【0036】
本明細書中のβサブユニットサブタイプとは、本明細書に開示のニューロンnAChRコーディングDNAの少なくとも一つに高ストリンジェンシー条件下でハイブリダイズするDNAでコードされるニューロンnAChRサブユニットを意味する。βサブユニットは適当なαサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性のnAChRを形成し得る。
【0037】
前述のようなβサブタイプをコードするが、遺伝暗号の縮重に起因して、指定のハイブリダイゼーション条件下で必ずしも開示のDNAとハイブリダイズしないDNAにコードされるβサブユニットも考えられる。この種のサブユニットも適当なαサブユニットサブタイプと一緒に、本明細書に記載の方法又は当業者に公知の方法で評価して機能性の受容体を形成し得る。典型的には、βサブユニットがスプライス変種として生じるRNAでコードされる場合を除いて、βコーディングDNA及び該DNAにコードされたβサブユニットは、本明細書に記載のβコーディングDNA及びβサブユニットタンパク質に対して実質的な配列相同を共有する。ここで理解されることとして、スプライス変種をコードするDNA又はRNAは全体で本発明が提供するDNA又はRNAに対して90%以下の相同を有し得るが、このようなDNAは本明細書に開示のDNAに対してほぼ100%相同の領域を含む。
【0038】
本明細書で使用するnAChRサブタイプという用語は、α及び/又はβサブユニットサブタイプの特定の組合わせを含むニコチン性アセチルコリン受容体、例えばヒトnAChRα6及びβ3サブユニットを含む受容体を意味する。
【0039】
本明細書中のスプライス変異体とは、1種類以上のmRNAを形成させることになるゲノムDNAの一次転写体の分別プロセシングによって生じる変異体nAChRサブユニットコーディング核酸を意味する。分別的にプロセシングされたゲノムDNAに由来するcDNAは、完全に同じアミノ酸を有する部分と、異なるアミノ酸配列を有する部分とを有するnAChRサブユニットをコードする。従って、同一のゲノム配列が多数の関連mRNA及びタンパク質の産生を生起し得る。その結果得られるmRNA及びタンパク質を「スプライス変異体」と称する。
【0040】
本明細書中の外来(heterologous又はforeign)DNA及びRNAとは、それを含んでいるゲノムの一部としては生来存在しない、又は生来の位置とは異なるゲノム内位置に存在するDNA又はRNAを意味する。典型的には、細胞に対して内生ではなく、人為的に細胞内に導入されたDNA又はRNAである。外来DNAの非限定的具体例としては、ヒトnAChRサブユニットをコードするDNA、及び転写、翻訳又は他の調節可能な生化学的プロセスに作用することにより内生DNAの発現を仲介又は変化させるRNA又はタンパク質をコードするDNAが挙げられる。ヒトnAChRサブユニットをコードするDNAのような外来DNAを発現する細胞は、同じ又は異なるニコチン性アセチルコリン受容体サブユニットをコードするDNAを含み得る。外来DNAは発現される必要はなく、宿主細胞ゲノム内に組込まれるように、又はエピソーム状に維持されるように導入し得る。
【0041】
本明細書では、ハイブリダイゼーションのストリンジェンシーとは、ポリ核酸ハイブリッドを安定にしておく条件を意味する。当業者に公知のように、ハイブリッドの安定性はハイブリッドの融解温度(Tm)に反映される。Tmは下記の式:
【0042】
【化1】
によって概算できる。前記式中、lはヌクレオチドで表されるハイブリッドの長さである。Tmは配列相同が1%減少する毎に約1〜1.5℃低下する。一般的には、ハイブリッドの安定性はナトリウムイオン濃度及び温度の関数である。典型的には、ハイブリダイゼーション反応を低ストリンジェンシー条件下で実施し、次いで異なる、但しより高いストリンジェンシーで洗浄を行う。ハイブリダイゼーションストリンジェンシーという用語は、このような洗浄条件について使用される。
【0043】
本明細書では、(1)高ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関しては、0.018M NaCl中65℃で安定なハイブリッドを形成する核酸配列だけがハイブリダイズできる条件を意味する(即ち、あるハイブリッドが0.018M NaCl中65℃で安定でなければ、そのハイブリッドは本明細書で考慮する高ストリンジェンシー条件下で安定ではない)。高ストリンジェンシー条件は例えば、50%ホルムアミド、5×デンハーツ(Denhardt)溶液、5×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで0.1×SSPE及び0.1%SDS中65℃で洗浄することにより与えることができる。(2)中間ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関して、50%ホルムアミド、5×デンハーツ溶液、5×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで0.2×SSPE及び0.2%SDS中60℃で洗浄することに等しい条件を意味する。(3)低ストリンジェンシー条件とは、フラグメントのハイブリダイセーションに関して、10%ホルムアミド、5×デンハーツ溶液、6×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNAでハイブリダイセーションを行い、次いで1×SSPE及び0.2%SDS中50℃で洗浄することに等しい条件を意味する。(4)高ストリンジェンシー条件は、オリゴヌクレオチド(即ち長さ約30ヌクレオチド以下の合成DNA)に関しては、10%ホルムアミド、5×デンハーツ溶液、6×SSPE、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、42℃でハイブリダイセーションを行い、次いで1×SSPE及び0.2%SDS中50℃で洗浄することに等しい条件を意味する。
【0044】
尚、これらの条件は種々の緩衝液及び温度を使用して再現し得、必ずしも厳密ではない。
【0045】
デンハーツ溶液及びSSPEは(例えばSambrookら,(1989)Molecular Cloning:A Laboratory Manual,第二版,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY)、他の適当なハイブリダイセーション緩衝液と同様に当業者に良く知られている。例えば、SSPEはpH7.4のリン酸緩衝0.18M NaClである。SSPEは例えば、175.3gのNaClと27.6gのNaH2PO4と7.4gのEDTAとを800mlの水に溶解し、pHを7.4に調整し、次いで水を1lになるまで加えることにより20×原液として調製できる。デンハーツ溶液(Denhardt(1966)Biochem.Biophys.Res.Commun.23:641)は例えば、5gのFicoll(タイプ400,Pharmacia LKB Biotechnology,INC.,Piscataway,NJ)と5gのポリビニルピロリドンと5gのウシ血清アルブミン(フラクションV;Sigma,St.Louis,MO)とを混合し、水を500mlになるまで加え、粒状物を除去すべく濾過することにより、50×原液として調製できる。
【0046】
本明細書で使用する「実質的な配列相同」という表現は、同一度が約90%以上の二つのヌクレオチド配列、及び典型的には95%以上のアミノ酸が同一であるアミノ酸配列に関するものである。しかしながら、スプライス変異体として生じる前記レベルより低い相同を有する、又は保守的(conservative)アミノ酸置換(もしくは縮重コドンの置換)によって修飾されたタンパク質(及びこのようなタンパク質をコードするDNAもしくはmRNA)も期待される。
【0047】
「実質的に同じ」という表現は、本明細書では、ここに開示する実際の配列と比べて僅かな重要ではない配列変化を有するDNAヌクレオチド配列、RNAリボヌクレオチド配列又はアミノ酸配列もしくはタンパク質について使用されている。実質的に同じ種は、開示の配列と機能的に同等であるとみなされる。従って、本明細書中の機能的に同等の核酸分子又はタンパク質とは、実質的に同じ結果を得るために実質的に同じ方法で機能するように十分に類似している核酸分子又はタンパク質である。
【0048】
本明細書で使用する「僅かな重要ではない配列変化」とは、ここに開示するDNA、RNA又はタンパク質と実質的に同じである配列が、ここに開示するヒト由来配列と機能的に同等であることを意味する。機能的に同等の配列は、実質的に同じ方法で機能して、ここに開示するヒト由来核酸及びアミノ酸組成物と実質的に同じ組成物を産生する。特に、機能的に同等のDNA分子は、ここに開示するものと同じ、又は保守的アミノ酸変化、例えばある非極性残基による別の非極性残基の置換、もしくはある帯電残基による類似の帯電残基の置換(例えば表1参照)を有するヒト由来タンパク質をコードする。これらの変化には、当業者によってタンパク質の三次構造を実質的に変化させないと認識されているものが含まれる。
【0049】
適当なアミノ酸保存置換は当業者に公知であり、通常は結果として得られる分子の生物学的活性を変えずに実施し得る。当業者に認識されているように、一般的には、ポリペプチドの非必須領域における単一アミノ酸の置換は生物学的活性を実質的に変化させない(例えばWatsonら、Molecular Biology of the Gene,第四版,1987,The Benjamin/Cummings Pub.Co.,p.244参照)。このような置換は、下記の表1に示すように実施するのが好ましい。
【0050】
【表1】
【0051】
別の置換も許容し得、経験的に決定し得るか、又は既知の保存置換と一致し得る。ポリペプチドのこの種の修飾はいずれも当業者に公知の任意の方法で実施し得る。
【0052】
本明細書中のヒトニューロンnAChRの活性とは、nAChRに特有の任意の活性を意味する。このような活性は典型的には一つ以上のin vitro方法で測定することができ、しばしばヒトニューロンnAChRのin vivo活性に対応する。このような活性は当業者に公知の任意の方法、例えば刺激に応答して組換え体チャンネルに流れる電流の量を測定することにより測定し得る。
【0053】
ヒトニューロンnAChRの存在及び/又は活性を決定する方法の非限定的具体例としては、ニコチン結合、86Rbイオン流、Ca2+流入、細胞の電気生理学的応答、RNAを導入した卵母細胞の電気生理学的応答を測定するアッセイが挙げられる。ここでは特に、DNA又はmRNAを含む細胞を試験化合物と接触させた時のnAChR仲介応答を測定又は検出する方法を提供する。
【0054】
本明細書中の組換え又は外来ヒトニューロンnAChRとは、受容体タンパク質を発現することができる細胞内に導入され発現された外来DNAにコードされるサブユニットを一つ以上含む受容体を意味する。組換えヒトニューロンnAChRは、宿主細胞にとって内生のDNAにより産生されるサブユニットも含み得る。ある実施態様では、組換え又は外来ヒトニューロンnAChRは、外来DNAにコードされるサブユニットのみを含み得る。
【0055】
本明細書中の機能性ニューロンnAChRとは、ここに開示する、又は当業者に公知のin vitro又はin vivoアッセイで評価して、ニューロンニコチン性nAChR活性を示す受容体を意味する。当業者に公知の、及び本発明で提供する任意の方法で評価し得る前述の活性を所有していれば、受容体は機能性とみなすことができる。nAChRタンパク質及び/又は活性の検出方法の非限定的具体例としては、ニコチン結合、86Rbイオン流、Ca2+流入、及び一つ以上の受容体サブユニットサブタイプをコードする外来DNA又はmRNAを含む細胞の電気生理学的応答を測定するアッセイが挙げられる。α及びβサブユニットの総ての組合わせが機能性受容体を形成し得るとは限らないため、このような受容体を産生する特定のサブユニット及び細胞の特徴を十分に解明するために、α及びβサブユニットの組合わせを多数試験し得る。従って、本明細書で組換え又は外来ヒトニューロンnAChRに関して使用する「機能性」という用語は、受容体チャンネルが刺激に応答して例えばNa+、K+、Ca2+又はBa2+のようなヒトニューロンnAChR浸透性イオンを提供し且つその流入を調節することができ、及び/又は受容体に対する親和性を有するリガンドと結合できることを意味する。このようなヒトニューロンnAChR活性は、電気生理学的方法、薬理的方法又は当業者の公知の別の方法により、宿主細胞によって生じ得る内因性nAChR活性から識別されることが好ましい。
【0056】
本明細書中のある種類の「対照」細胞又は「対照」培養物は、対照培養物は試験化合物に暴露しないという点を除いて、試験化合物に暴露した細胞又は培養物と実質的に同様に処理した細胞又は培養物である。別の種類の「対照」細胞又は「対照」培養物は、対照培養に使用した細胞が機能性ニコチン性アセチルコリン受容体を発現しないという点を除いて、トランスフェクションした細胞と同一である細胞又は細胞培養物であり得る。この場合は、細胞又は各種類の細胞の培養物をアッセイすべき化合物の存在下で実質的に同じ反応条件に暴露した時の、試験化合物に対するニコチン性アセチルコリン受容体陰性細胞の応答(又は応答欠失)と、試験化合物に対する試験細胞の応答とを比較する。
【0057】
本明細書中の「ニューロンnAChRの活性を調節する」化合物又はシグナルとは、前記化合物又はシグナルの存在下と不在下とではnAChRの活性が異なるようにnAChRの活性を変化させる化合物又はシグナルを意味する。このような化合物又はシグナルとしては、特にアゴニスト及びアンタゴニストが挙げられる。アゴニストという用語は、受容体の機能を活性化する物質又はシグナル、例えばAChを意味し、アンタゴニストという用語は、受容体の機能を妨害する物質を意味する。典型的には、アンタゴニストの作用は、アゴニストによる活性化の阻止として観察される。アンタゴニストには競合アンタゴニスト及び非競合アンタゴニストがある。競合アンタゴニスト(又は競合遮断剤)は、同じ又は近傍の部位に関してアゴニスト(例えばリガンド又は神経伝達物質)に特異的な部位又はその近傍と相互作用する。非競合アンタゴニスト又は遮断剤は、アゴニストと相互作用する部位以外の部位と相互作用することにより受容体の機能を不活化する。
【0058】
A. 分離DNAクローン
ヒトニューロンnAChRα及びβサブユニットをコードするDNA分子を提供する。特定的には、ヒトニューロンnAChRのα6及びβ3サブユニットをコードする分離DNA分子を開示する。前述のDNAでコードされる組換えメッセンジャーRNA(mRNA)及び組換えポリペプチドも提供する。
【0059】
本明細書で使用する「α6サブユニットをコードする(コーディング)核酸」という用語は、同じ名称のニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNA又はRNAを意味する。このような核酸分子は多くの方法で特徴を調べることができ、例えばDNAのヌクレオチド(又はRNAのリボヌクレオチド)は配列番号10又は配列番号20に示すアミノ酸配列をコードし得る。
【0060】
現時点で好ましいα6コーディング核酸としては、高ストリンジェンシー条件下で、配列番号9に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド143−1624)、又は配列番号19に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド143−1579)にハイブリダイズするDNA又はRNAが挙げられる。
【0061】
特に好ましいα6コーディング核酸分子は、配列番号10又は配列番号20に示すものと実質的に同じアミノ酸配列を有する(即ち保守的アミノ酸置換のみを有する)タンパク質をコードする分子である。最も好ましい分子は、配列番号9に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド143−1624)又は配列番号19に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド143−1579)を有するヌクレオチド(又はTがUで置換されたリボヌクレオチド)配列である。
【0062】
典型的には、α6サブユニットがスプライス変異体として生じる場合を除いて、α6コーディングDNAはここに開示するα6コーディング核酸分子に対して実質的な配列相同(即ち約90%以上)を有する。スプライス変異体をコードするDNA又はRNAはここに開示するDNA又はRNAに対して90%以下の総合的配列相同を有し得るが、このようなスプライス変異体は、ここに開示する核酸分子の一つ以上に対してほぼ100%相同の領域を含む。
【0063】
本発明は、「β3サブユニットコーディング核酸」も提供する。これには、同じ名称のニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNA又はRNA分子が含まれる。このような核酸分子は多くの方法で特徴を調べることができ、例えばDNAのヌクレオチド(又はRNAのリボヌクレオチド)は配列番号16に示すアミノ酸配列をコードし得る。
【0064】
現時点で好ましいβ3コーディング核酸としては、高ストリンジェンシー条件下で、配列番号15に示すコーディング配列(好ましくは該コーディング配列の実質的に全部、即ちヌクレオチド98−1471)にハイブリダイズするDNA又はRNAが挙げられる。より好ましいのは、配列番号16に示すアミノ酸配列(又は実質的に保守的アミノ酸置換のみを有する前記アミノ酸配列)を含むタンパク質をコードする核酸である。本発明が提供する特に好ましいβ3コーディング核酸分子は、配列番号15に示すものと実質的に同じヌクレオチド配列(即ち、特に該配列のヌクレオチド98−1471)を有する。
【0065】
典型的には、β3サブユニットがスプライス変異体として生じる場合以外は、β3コーディング核酸は開示のβ3コーディング核酸分子に対して実質的な配列相同(約90%以上)を有する。スプライス変異体をコードするDNA又はRNAは開示のDNA又はRNAに対して90%以下の総合的配列相同を有し得るが、このような核酸は、前述の核酸分子の一つ以上に対してほぼ100相同の領域を含む。
【0066】
B. プローブ
ヒトニューロンニコチン性nAChRα及びβサブユニットをコードするDNAは、ここに開示するDNA(配列番号9又は15に由来するヌクレオチドを含む)を用いて、適当なハイブリダイゼーション条件下で適当なヒトcDNA又はヒトゲノムライブラリーをスクリーニングすることにより分離し得る。適当なライブラリーは、ニューロン組織試料、脳底神経節、視床及び視床下部組織から製造し得る。ライブラリーは、DNAのサブユニットコーディング配列全体を含むDNA部分を用いてスクリーニングするのが好ましく、又はライブラリーは適当なプローブを用いてスクリーニングし得る。典型的には、プローブを、同定可能なタグ、例えば放射性標識、酵素又は他の当業者に公知のタグで標識する。
【0067】
α6及びβ3コーディング核酸を分離する方法で使用するためのプローブも提供する。例えばヒトα6サブユニットに関しては、プローブは、配列番号9又は配列番号19に示す任意の27個の塩基と同じ(又はその相補体)である少なくとも27個の連続塩基を含むヌクレオチド配列を有する一本鎖DNA又はRNA分子である。
【0068】
ヒトβ3サブユニットに関しては、プローブは、配列番号15に示すシグナル配列/コーディング配列の最初の105個のヌクレオチドに由来する任意の28個の塩基と同じ(又はその相補体)である少なくとも28個の連続塩基を含むヌクレオチド配列を有する一本鎖DNA又はRNAである。
【0069】
プローブを構築するための好ましい領域の非限定的具体例としては、5’及び/又は3’コーディング配列、トランスメンブランドメインをコードすると予測される配列を含む領域、細胞質ループをコードすると予測される配列を含む領域、シグナル配列、並びにアセチルコリン(ACh)及びα−ブンガロトキシン(α−bgtx)結合部位が挙げられる。ACh及びα−bgtx結合には、典型的には、デンキナマズnAChRαサブユニットの残基190−198に対応するアミノ酸(例えばKarlin(1993)Curr.Opin.Neurobiol.3:299−309参照)が関与する。別のプローブに関して、前述のような領域を含む大凡のアミノ酸残基を下記の表2に示す。
【0070】
【表2】
【0071】
あるいは、DNAの一部分を、特定ライブラリー内の選択したフラグメントを増幅するためのプライマーとして使用することもできる。
【0072】
C. ヒトニューロンニコチン性アセチルコリン受容体のα6及びβ3サブユニットをコードするクローンの分離
前述のプローブを使用して適当なライブラリーをスクリーニングする。各サブユニットをコードするDNAを得るのに適したライブラリーの非限定的具体例としては、ヒトα6コーディングDNAを分離する場合の黒質、視床又は視床下部、並びにヒトβ3コーディングDNAを分離する場合の黒質又は視床が挙げられる。
【0073】
ライブラリーのスクリーニング後、ハイブリダイゼーションシグナルの検出によって陽性クローンを同定する。同定されたクローンは、制限酵素地図及び/又はDNA配列分析よって特徴を調べ、次いで開示の配列と比較して、完全なα又はβサブユニットをコードするDNAを含んでいるかどうかを確認する。選択したクローンが不完全であれば、これらのクローンを使用して、重複クローンを得るために同じ又は異なるライブラリーを再スクリーニングし得る。所望であれば、ライブラリーは、完全なα又はβサブユニットをコードする重複クローンが得られるまで陽性クローンを用いて繰り返しスクリーニングし得る。ライブラリーがcDNAライブラリーの場合には、重複クローンは読取り枠を含む。ライブラリーがゲノムライブラリーの場合には、重複クローンはエキソン及びイントロンを含み得る。完全クローンは、本発明が提供するDNA及びコードされたタンパク質との比較によって同定し得る。
【0074】
ヒトニューロンnAChRα及びβサブユニットの種々のサブタイプをコードする相補的DNAクローンが分離された。サブユニットの各サブタイプは、異なる遺伝子でコードされると思われる。ここに開示するDNAクローンは、各サブタイプをコードするゲノムクローンの分離、並びに種々の神経組織から製造したライブラリーのスクリーニングによる任意のスプライス変異体の分離に使用し得る。ヒトニューロンnAChRサブユニットのスプライス変異体は、当業者に良く知られている核酸増幅方法を使用して検出することができる。この操作は、分岐(divergent)配列を包囲するDNA配列をベースとするオリゴヌクレオチドをヒトRNA又はゲノムDNA増幅用プライマーとして使用して行う。増幅産物の大きさ及び配列決定により、スプライス変異体の存在を明らかにすることができる。また、ハイブリダイゼーションによってヒトゲノムDNA配列を分離すると、ヒトニューロンnAChRサブユニットをコードする転写体の種々のスプライス変異体に対応する、イントロンで分離された複数のエキソンを含むDNAが得られる。
【0075】
総てのサブユニットサブタイプが総ての神経組織又は総ての脳部分で発現されるとは限らないことが判明した。従って、特定のサブユニットサブタイプ又はこのようなサブタイプのスプライス変異体をコードするcDNAを分離するためには、種々のニューロン又は神経組織から製造したライブラリーをスクリーニングすることが好ましい。
【0076】
D. α6及びβ3コーディング核酸を含む細胞及びベクター
ヒトnAChRサブユニットをコードする前述の核酸分子は、更に操作を行うためにベクター内に導入できる。適当な発現ベクター内へのクローン化DNAの導入、一つ以上の別個の遺伝子をコードする単一の発現構築物もしくは複数の発現構築物の組合わせ又は直鎖状DNAの真核細胞内へのトランスフェクション、並びにトランスフェクションした細胞の選択は当業者に良く知られている(例えばSambrookら(1989)Molecular Cloning:A Laboratory Manual,第二版,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY参照)。外来DNAは、当業者に公知の任意の方法、例えばCaPO4沈降により外来DNAをコードする発現構築物をトランスフェクションする方法で宿主細胞内に導入し得る(例えば、Wiglerら(1979)Proc.Natl.Acad.Sci.U.S.A.76:1373−1376参照)。次いで組換え細胞を、DNAでコードされたサブユニットを発現させる条件下で培養し得る。好ましい細胞の非限定的具体例としては、哺乳動物細胞(例えばHEK293、CHO及びLtk細胞)、酵母細胞(例えばPichia pastorisのようなメチロトロフィック(methylotrophic)酵母細胞)及び細菌細胞(例えば大腸菌)が挙げられる。
【0077】
α6及びβ3サブユニットをコードする核酸は、更に操作を行うために、個々に、又は別のニコチン性アセチルコリン受容体をコードする核酸と組合わせてベクター内に導入できる。ヒトニューロンnAChRサブユニットをコードする完全長DNAクローンが、ベクターpcDNA3に、即ちCMVプロモーター/エンハンサーと、CMVプロモーター/エンハンサーの下流のポリリンカーと、これに次ぐウシ成長ホルモン(BGH)ポリアデニル化シグナルとを含むpUC19ベースの哺乳動物細胞発現ベクター内に、挿入された。nAChRサブユニットをコードするDNAをCMVプロモーターとBGHポリアデニル化シグナルとの間に配置すると、構築物をトランスフェクションした哺乳動物細胞内でDNAの構成的発現が得られる。哺乳動物細胞内でのヒトnAChRサブユニットコーディングDNAの誘導発現のためには、DNAをpMAMneoのようなプラスミド内に挿入し得る。このプラスミドは、作用的に結合した外来DNAのステロイド誘導発現のためのマウス乳癌ウイルス(MMTV)プロモーターを含む。宿主細胞が、細胞内へのグルココルチコイド(即ちMMTVプロモーターの誘発物質)の取込みに必要な内生グルココルチコイド受容体を発現しない場合には、更にグルココルチコイド受容体をコードするDNA(ATCC受託番号67200)を細胞にトランスフェクションする必要がある。
【0078】
別の実施態様では、前述のポリ核酸(即ちDNA又はmRNA)を含む細胞を提供する。DNAの複製及びnAChRサブユニットの製造には、細菌、酵母及び哺乳動物細胞のような宿主細胞を使用し得る。発現ベクターを構築し、in vitro転写体を形成し、DNAを哺乳動物細胞内にトランスフェクションし、卵母細胞に注入し、ここに記載のような受容体発現及び機能を評価するための電気生理学的分析及び他の分析を実施する方法は、PCT出願公開明細書PCT/US91/02311号、PCT/US94/02447号、PCT/US91/05625号及びPCT/US92/11090号、米国特許明細書第5,369,028号並びに同時係属米国特許出願明細書第07/563,751号及び第07/812,254号にも記述されている。これらの出願明細書の論題はその全体が参考として本明細書に包含される。
【0079】
本発明が提供するDNAは、酵母細胞(例えばPichia、特にPichia pastoris(米国特許明細書第4,882,279号、第4,837,148号、第4,929,555号及び第4,855,231号参照)、Saccharomyces cerevisiae,Candida tropicalis,Hansenula polymorpha及び他の酵母細胞)を含む任意の真核細胞内で発現し得、現時点で好ましいのは、本発明が提供するヒトニューロンニコチン性nAChRサブユニットをコードするDNAを発現するための、市販の系及び他の当業者に公知の系を含む哺乳動物発現系である。DNAのRNA転写体の発現にはアフリカツメガエル卵母細胞が好ましい。
【0080】
任意のヒトニューロンニコチン性nAChRサブユニットをコードするクローン化完全長DNAは、真核細胞内での発現のためにプラスミドベクター内に導入し得る。このようなDNAはゲノムDNA又はcDNAであってよい。各々が少なくとも一つのヒトニューロンnAChRサブユニットをコードする単一のプラスミド又は複数のプラスミドの組合わせを宿主細胞にトランスフェクションし得る。外来DNAは細胞内でエピソームエレメントとして維持され得、又は細胞の染色体DNA中に組込まれ得る。
【0081】
DNA又はRNAを導入し得る真核細胞としては、このようなDNAもしくはRNAをトランスフェクション又は注入し得る任意の細胞が挙げられる。好ましい細胞は、一時的又は安定的にトランスフェクションすることができ、DNA及びRNAを発現することもできる細胞である。現時点で最も好ましい細胞は、外来DNAでコードされたサブユニットを一つ以上含む組換え又は外来ヒトニューロンニコチン性nAChRを形成することができる細胞である。このような細胞は経験的に同定し得、又はトランスフェクションもしくは注入が容易なことが知られているものの中から選択し得る。
【0082】
DNAを導入するための細胞の非限定的具体例としては、哺乳動物由来の細胞(例えばCOS細胞、マウスL細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト胎児腎臓(HEK)細胞、GH3細胞及び他の当業者に公知の細胞)、両生動物細胞(例えばXenopus laevis卵母細胞)及び酵母細胞(例えばSaccharomyces cerevisiae,Pichia pastoris)が挙げられる。注入RNA転写体を発現するための代表的な細胞はXenopus laevis卵母細胞である。DNAのトランスフェクションに好ましい細胞は当業者の公知であり、又は経験的に同定し得、具体例としてはHEK293(受託番号#CRL 1573でATCCから入手可能)、Ltk−細胞(受託番号#CCL1.3でATCCから入手可能)、COS−7細胞(受託番号#CRL 1651でATCCから入手可能)及びGH3細胞(受託番号#CCL82.1でATCCから入手可能)が挙げられる。現時点で好ましい細胞としては、GH3細胞及びHEK293細胞、特に懸濁液増殖用に調整され、液体窒素中で冷凍し、その後解凍して再増殖することができるHEK293細胞が挙げられる。HEK293細胞は、例えばGormanの米国特許明細書第5,024,939号に記載されている(Stillmanら(1985)Mol.Cell Biol.5:2051−2060も参照)。
【0083】
DNAは当業者に公知の方法で細胞内に安定に導入することができ、又は一時的に導入し得る。安定にトランスフェクションした哺乳動物細胞は、nAChRサブユニットをコードするDNAを含む一つ以上の発現構築物及び選択可能マーカー遺伝子(非限定的具体例としてネオマイシン耐性、ゼオシン耐性もしくはハイグロマイシン耐性用の遺伝子)を有する別個の発現ベクターを細胞にトランスフェクションするか、又はnAChRサブユニットをコードするDNAと選択可能マーカーとを含む一つ以上の発現構築物を細胞にトランスフェクションし、トランスフェクションした細胞を、マーカー遺伝子発現細胞に関して選択的な条件下で増殖させることにより製造し得る。このような細胞を製造するためには、外来DNAでコードされたヒトサブユニットを含むヒトニューロンnAChRを形成するのに十分な濃度のサブユニットコーディング核酸を細胞にトランスフェクションしなければならない。サブユニットをコードするDNAの正確な量及び比率は経験的に決定し得、サブユニット、細胞及びアッセイ条件の特定の組合わせに関して最適化し得る。特に好ましいのは、外来DNA又はRNAのみによってコードされたサブユニットを含むニューロンnAChRを発現する組換え細胞である。
【0084】
E. 組換えnAChR及びnAChRサブユニットタンパク質
本発明は、実質的に純粋なヒトnAChRサブユニットタンパク質、特にヒトα6及びβ3サブユニットタンパク質を提供する。前記ヒトnAChRサブユニットタンパク質の少なくとも一つを含む組換えnAChRも提供する。従って、本発明の別の実施態様は、組換えヒトnAChRサブユニット及び該サブユニットを含む受容体の製造方法を含む。
【0085】
好ましい実施態様では、ヒトnAChRサブユニット、特にヒトnAChRα6及びβ3サブユニットをコードするDNAをベクター内に連結し、得られた構築物を適当な宿主細胞内に導入して、特定のヒトニューロンnAChR受容体サブタイプ又はサブタイプの特定の組合わせを発現する形質転換細胞系を形成する。得られた細胞系は次いで、受容体機能に対する薬剤の作用の再現可能な定量分析を行うために量的に製造し得る。別の実施態様では、各サブユニットをコードするDNAのin vitro転写によりmRNAを製造し得る。単一サブユニットクローン又は複数のクローンの組合わせに由来する前記mRNAは、次いでアフリカツメガエル卵母細胞内に注入し得、そこでmRNAがヒト受容体サブユニットの合成を指示し、これらのサブユニットが機能性受容体を形成する。あるいは、機能性受容体を発現させるために、サブユニットをコードするDNAを卵母細胞に直接注入することもできる。トランスフェクションを行った哺乳動物細胞、又は注入を行った卵母細胞は、次いで本明細書に記載の薬剤スクリーニング方法で使用し得る。
【0086】
得られた組換え細胞は培養し得、又はこのような培養物又はその継代培養物から継代培養(subculture、又は哺乳動物の場合はpassage)し得る。トランスフェクション、注入及び組換え細胞培養方法は当業者に公知である。また、ヒトニューロンニコチン性nAChRサブユニットは、当業者に公知のタンパク質精製方法を用いて精製し得る。例えば、サブユニットの一つ以上に特異的に結合する抗体又は他のリガンドを、サブユニット又はサブユニット含有ヒトニューロンnAChRのアフィニティ精製に使用し得る。
【0087】
ある実施態様では、ヒトニューロンnAChRサブユニット及び機能性受容体を発現する細胞の製造方法も提供する。これらの方法の一つでは、ニューロンnAChRの少なくとも一つのαサブユニット及び少なくとも一つのβサブユニットをコードするDNAを宿主細胞にトランスフェクションする。α及び/又はβサブユニットをコードするDNA又はRNAを含むトランスフェクション細胞は、ノーザンブロット又はスロットブロット分析のような方法を用いて選択できる。トランスフェクション細胞は、nAChRタンパク質を発現する細胞を同定するための分析にもかける。分析は例えば、細胞がアセチルコリン、ニコチン又はnAChRアゴニストに結合する能力を測定し、非トランスフェクション宿主細胞又は他の適当な対照細胞のニコチン結合能力と比較するか、又はnAChRアゴニストに応答して細胞膜に流れる電流を電気生理学的に観察することにより実施できる。
【0088】
特に好ましいこととして、外来DNAを含み、このDNAを発現し、組換え機能性ニューロンnAChRを形成する真核細胞が得られる。より好ましいこととして、組換えニューロンnAChR活性を容易に検出することができる。なぜなら、これは非トランスフェクション宿主細胞に存在しないか、又は非トランスフェクション細胞内で示されない量を有する種類のものだからである。組換え受容体を含むような細胞は例えば、ヒトニューロンニコチン性nAChRα6及びβ3をサブユニットをコードするDNAで形質転換した細胞に、一つ以上のα及び/又はβnAChRサブユニットの存在下又は不在下で、対応するタンパク質を発現させることにより製造し得る。得られる合成又は組換え受容体はα6及びβ3nAChRサブユニットを含む。このような受容体は、種々の用途、例えば非ヒト受容体又はヒト組織調製物を使用する先行技術のアッセイシステムでしばしば見られる妨害がないアッセイシステムの一部として有用である。また、単一受容体サブユニットを種々の潜在的アゴニスト又はアンタゴニストを用いて試験すれば、個々のサブユニットの機能及び活性に関して更に情報が得られる。このような情報は、受容体サブユニットの一つ以上と極めて特異的に相互作用することができる化合物の同定につながり得る。このような特異性は、医療用途で極めて有用であり得る。
【0089】
一つ以上のヒトニューロンnAChRサブユニットをコードするDNAは、個々のサブユニット及び機能性nAChRを発現するために、適当な宿主細胞(例えば真核又は原核細胞)内に導入し得る。好ましくは、αサブユニット及びβサブユニットの組合わせを細胞内に導入し得る。このような組合わせは、α2、α3、α4、α5、α6及びα7とβ2、β3及び/又はβ4との任意の一つ以上の組合わせを含む。これらのサブユニットの各々に関する配列情報は添付の配列表に示す。α5の配列情報はProc.Natl.Acad.Sci.USA(1992)89:1572−1576にも記載されており、α2、α3、α4、α7、β2及びβ4の配列情報は、本明細書に参考として包含されるPCT出願公開明細書第94/20617号にも記載されている。現時点で好ましいサブユニットの組合わせとしては、α6及び/又はβ3とα2、α3、α4、α5、β2又はβ4のうちのいずれか一つ以上との組合わせが挙げられる。一部のサブユニットは別のサブユニットの不在下でイオン輸送機能を有し得、他のサブユニットはイオン輸送機能を示すために二つ以上のサブユニットの組合わせを必要とすることが認識されている。例えば、α7サブユニットは任意のβサブユニットを付加しない状態で機能する。また、一部のサブユニットは単独又は組合わせ状態で機能性nAChRを形成し得ないが、他のnAChRサブユニットの組合わせの特性を調節し得る。
【0090】
ある実施態様では、細胞内で翻訳される少なくとも一つのRNA転写体を含む第一の組成物をヒトニューロンnAChRのサブユニット内に導入することによって、外来ヒトニューロンnAChRを含む真核細胞を製造する。好ましい実施態様では、翻訳されるサブユニットがヒトニューロンnAChRのαサブユニットを含む。より好ましくは、導入する組成物は、αサブユニットをコードするRNA転写体を含み、ヒトニューロンnAChRのβサブユニットをコードするRNA転写体も含む。RNA転写体は、ヒトニューロンnAChRをコードするDNAをトランスフェクションした細胞から得ることができ、又はサブユニットをコードするDNAのin vitro転写によって得ることができる。クローン化DNAをin vitro転写させ、得られたmRNAを真核細胞内に注入する方法は当業者に良く知られている。ヒトニューロンnAChRDNAクローンのin vitro転写体の発現には両生動物卵母細胞が特に好ましい。イオンチャンネルを研究するためのアフリカツメガエル卵母細胞の使用については、例えばDascal(1989)CRC Crit.Rev.Biochem.22:317−387を参照し得る。
【0091】
一つ以上のαサブタイプ及び一つ以上のβサブタイプをコードするDNA又はRNAを段階的に細胞内に導入することが可能である。得られた細胞は、機能性nAChR活性を検出するために、本明細書に記載の方法又は当業者に公知の方法で試験し得る。このような試験は、機能性nAChRを形成するα及びβサブユニットサブタイプの組合わせ、並びに機能性nAChRを形成する個々のサブユニットの同定を可能にする。
【0092】
組換え真核細胞の表面に存在する組換え受容体は、ヒトニューロンnAChRサブユニットをコードするDNA又はmRNAでコードされた一つ以上のサブユニットを含み得、又は宿主細胞にコードされたサブユニットと外来DNAもしくはmRNAにコードされたサブユニットとの混合物を含み得る。組換え受容体は均質であり得、又はサブタイプ混合物であり得る。種々の由来源、例えばマウス及びヒトに由来する受容体をコードするDNA又はmRNAの混合物も細胞内に導入し得る。このようにして、α6及びβ3サブユニットのみを含む、又はここに開示する任意の別のα及びβサブユニットとの組合わせを含む組換え受容体を発現する細胞を製造し得る。例えば、ここに記載のα6及びβ3サブユニットのいずれか又は両方を、α2、α3、α4、α5、α7、β2及び/又はβ4受容体サブユニットと一緒に発現できる。前述のように、一部のニューロンnAChRサブユニットは、別のサブユニットの不在下で機能性受容体を形成する能力を有し得、従って機能性受容体を形成するために同時発現が必ず必要であるとは限らない。また、一部のnAChRサブユニットは、機能性受容体の形成に参加するために、二つ以上のnAChRサブユニットとの同時発現を必要とし得る。
【0093】
F. アッセイ
本発明の実施態様の一つでは、組換えヒトニューロンnAChR発現哺乳動物細胞又は卵母細胞を試験化合物と接触させ、次いで試験化合物の存在下及び不在下でのnAChR仲介応答を比較するか、又は試験化合物の存在に対する試験細胞もしくは対照細胞のnAChR仲介応答を比較することにより、試験化合物の調節作用を評価することができる。
【0094】
当業者に理解されているように、ヒトニューロンnAChR活性を調節する化合物(例えばアゴニスト及びアンタゴニスト)を同定するためのアッセイ方法は通常、対照との比較を必要とする。前述のように、ある種類の「対照」細胞又は「対照」培養物は、対照培養物は試験化合物に暴露しないという点を除いて、試験化合物に暴露した細胞又は培養物と実質的に同じ方法で処理した細胞又は培養物である。例えば、電圧固定電気生理学的操作を使用する方法では、細胞を浸漬する外部溶液を変えるだけで、同じ細胞を試験化合物の存在下及び不在下で試験することができる。別の種類の「対照」細胞又は「対照」培養物は、対照培養物に使用した細胞が機能性ヒトニューロンnAChRを発現しないという点を除いて、トランスフェクション細胞と同じである細胞又は細胞培養物であり得る。この場合は、細胞又は各種類の細胞の培養物を、アッセイすべき化合物の存在下で実質的に同じ反応条件に暴露した時の、試験化合物に対する受容体陰性(対照)細胞の応答(又は応答欠如)と、試験化合物に対する試験細胞の応答とを比較する。
【0095】
機能性組換えヒトニューロンnAChRはヒトニューロンnAChRの少なくとも一つのαサブユニット、又は少なくとも一つのαサブユニット及びβサブユニットを含む。これらのサブユニットを発現する真核細胞は、RNA転写体の注入及びDNAのトランスフェクションによって製造された。これらの細胞は、外来ヒトニューロンnAChRサブユニットの一つ以上を含むヒトニューロンnAChRに起因し得るnAChR活性を示した。
【0096】
機能性外来ヒトニューロンnAChRの活性の測定については、内生nAChR活性、そして所望であれば、内生宿主細胞サブユニットと外来サブユニットとの混合物を含むnAChRの活性を、可能であれば、化学的、薬理学的及び電気生理学的方法で大幅に阻害すべきである。
【0097】
G. 抗体
本発明は、前述のnAChRサブユニット又はその一部分に対して産生される抗体も提供する。この種の抗体は、受容体組織の検出、サブタイプ組成、機能ドメインの構造、受容体の精製の評価、並びに診断及び治療用途で使用し得る。好ましくは治療用途では、使用する抗体はモノクローナル抗体である。
【0098】
前述の抗体は、本明細書に記載のnAChRサブユニットタンパク質又はその一部分を抗体産生用の抗原として使用して、当業者に良く知られている標準的方法で製造できる。抗ペプチド抗体及び抗融合タンパク質抗体の両方を使用し得る[例えば、Bahouthら(1991)Trends Pharmacol.Sci.12:338−343;Current Protocols in Molecular Biology(Ausubelら編),John Wiley and Sons,New York(1989)参照]。免疫原として(合成ペプチド又は組換えにより産生した細菌融合タンパク質として)使用するためのnAChRサブユニットの部分の選択で考慮すべき要素としては、抗原性、アクセスの容易さ(即ち細胞外及び細胞質ドメイン)、特定サブタイプに対する特有性、及び他の当業者に公知の要素が挙げられる。
【0099】
サブタイプ特異的抗体を入手できれば、免疫化学の手法を種々のサブタイプの分布及び発現密度(例えば正常及び疾患脳組織内)の観察に適用することが可能になる。ヒトnAChRサブユニットを免疫原として製造した抗体は、種々の特性の中でも特に、生物学的試料又はこの種の試料に由来する溶液中に存在し得るヒトnAChR又はそのサブユニットに特異的且つ優先的に結合する能力、及び/又は前記ヒトnAChRもしくはそのサブユニットを免疫沈降させる能力を有する。このような抗体は、これらの抗体が特異的に結合するサブユニットを含むヒトnAChRを発現する細胞を選択的に分離するためにも使用し得る。これらの抗体は診断及び治療用途でも使用し得る。更に別の実施態様では、受容体を有効量の前記抗体と接触させることにより、nAChRのイオンチャンネル活性を調節する方法も提供する。
【0100】
本明細書に記載の抗体は、標準的方法、例えば腹腔内、筋内、静脈内もしくは皮下注射、移植又は経皮による投与方法を用いて対象に投与し得る。当業者は、使用する投与方法に基づいて、投薬形態、治療計画等を容易に決定し得る。
【0101】
以下の実施例は本発明を例示するためのものであって、本発明の範囲を限定するものではない。
【実施例1】
【0102】
ヒトnAChRα6サブユニットをコードするDNAの分離
ヒト黒質cDNAライブラリー(Clontech Laboratories,Inc.)を、マウスnAChRα6サブユニットcDNAのフラグメントへのハイブリダイゼーションについてスクリーニングした。分離したプラークをニトロセルロースフィルターに移し、ハイブリダイゼーションを5×デンハーツ、5×SSPE、50%ホルムアミド、200μg/ml変性サケ精子DNA及び0.2%SDS中で42℃で実施した。洗浄を0.2×SSPE、0.2%SDS中60℃で実施した。
【0103】
五つのハイブリッド形成クローンをプラーク精製し、制限エンドヌクレアーゼ地図及びDNA配列決定分析により特徴を調べた。市販のλgt10前進及びリバースオリゴヌクレオチドプライマーを使用して、cDNA挿入物の5’及び3’末端のDNA配列を決定した。五つのcDNA挿入物のDNA配列の分析の結果、三つのクローンが、翻訳開始コドンと完全長α読取り枠(配列番号9のヌクレオチド143−1624)と翻訳終止コドンと5’−非翻訳配列の更に142個のヌクレオチドと3’−非翻訳配列の116個のヌクレオチドとを含んでいた。完全長クローンのヌクレオチド配列から推定されるアミノ酸配列は、マウスnAChRα6サブユニットDNAから推定されるアミノ酸配列に対して〜82%の同一性を有する。推定マウス及びヒトα6アミノ酸配列の幾つかの領域は著しく相違している:アミノ酸1−30(ヒトシグナル配列はマウス配列に対して〜56%の同一性しか有していない)、アミノ酸31−50(ヒト配列はマウス配列に対して〜70%の同一性しか有していない)、アミノ酸344−391(ヒト配列はマウス配列に対して〜40%の同一性しか有していない)、アミノ酸401−428(ヒト配列はマウス配列に対して〜64%の同一性しか有していない)。
【0104】
また、単一クローンKEα6.5の挿入DNAは、α6コーディング配列の45個のヌクレオチドを欠失しており、その結果、推定アミノ酸配列の15個のアミノ酸残基(配列番号10の残基74−88)が枠内欠失(in−frame deletion)していると決定された。この配列を欠失している
α6サブユニット変異体のヌクレオチド配列は配列番号19に示し、該ヌクレオチド配列から推定されるアミノ酸配列は配列番号20に示す。興味深いことに、欠失部位のすぐ下流の推定アミノ酸配列は、推定マウスα6アミノ酸配列に対して〜58%のアミノ酸同一性しか有していない(配列番号10のアミノ酸89−100)。
【実施例2】
【0105】
ヒトnAChRβ3サブユニットをコードするDNAの分離
ヒト黒質cDNAライブラリー(Clontech Laboratories,Inc.)を、ヒトニコチン性nAChRβ3サブユニットcDNAに相補的な合成オリゴヌクレオチドへのハイブリダイゼーションについてスクリーニングした。分離したプラークをニトロセルロースフィルターに移し、オリゴヌクレオチドに対して高ストリンジェンシーの条件下で(洗浄条件1×SSPE、0.2%SDS、50℃)、配列番号15のヌクレオチド212−230及び1442−1469を含むヒトβ3nAChRサブユニットcDNAの配列に対して相補的な合成オリゴヌクレオチドとハイブリダイズさせた。
【0106】
二つのハイブリッド形成クローンをプラーク精製し、制限エンドヌクレアーゼ地図で特徴を調べた。市販のT7及びSP6オリゴヌクレオチドプライマーを使用して、cDNA挿入物の5’及び3’末端のDNA配列を決定した。クローンKBβ3.2の完全配列が決定された。クローンKBβ3.2は、完全長β3nAChRサブユニットをコードする1,377ヌクレオチド読取り枠(配列番号15のヌクレオチド98−1471)と、5’非翻訳配列の97個のヌクレオチド及び3’非翻訳配列の454個のヌクレオチドとを含む1927bpのcDNA挿入物を含んでいる。完全長クローンのヌクレオチド配列から推定されるアミノ酸配列は、マウスニコチン性nAChRβ3サブユニットDNAから推定されるアミノ酸配列に対して〜81%の同一性を有する。推定マウス及びヒトβ3アミノ酸配列の幾つかの領域は著しく相違している:アミノ酸1−28(ヒトシグナル配列はマウス配列に対して〜25%の同一性しか有していない)、アミノ酸347−393(ヒト配列はマウス配列に対して〜55%の同一性しか有していない)、アミノ酸440−464(ヒト配列はマウス配列に対して〜68%の同一性しか有していない)。
【実施例3】
【0107】
組換えヒトニューロンnAChRサブユニットを発現するための構築物の製造
ヒトニューロンnAChRサブユニットをコードする分離cDNAを、哺乳動物宿主細胞内でのサブユニットの発現に使用され、アフリカツメガエル卵母細胞内で発現すべきDNAからのin vitro転写体の形成に使用されるベクターに導入した。構築物の製造には下記のベクターを使用した。
【0108】
A. ヒトnAChRα6サブユニットを発現するための構築物
完全長nAChRα6サブユニットをコードする1,743bp EcoRIフラグメントを標準的方法でKEα6.3から分離し、ベクターpcDNA3のEcoRIポリリンカー部位に連結して、pcDNA3−KEα6.3を形成した(第1図参照)。プラスミドpcDNA3(第1図参照)は、CMVプロモーター/エンハンサーと、CMVプロモーター/エンハンサーの下流に位置するT7バクテリオファージRNAポリメラーゼプロモーターと、T7プロモーターの下流のウシ成長ホルモン(BGH)ポリアデニル化シグナルと、T7プロモーター及びBGHポリアデニル化シグナル間のポリリンカーとを含むpUC19ベースのベクターである。従ってこのベクターは、ポリリンカー位にベクター内に導入された外来DNAを哺乳動物細胞内で発現させるのに必要な調節エレメントを総て含む。更に、T7プロモーターがポリリンカーのすぐ上流に位置するため、このプラスミドはポリリンカーレベル位にベクター内にサブクローニングされた外来DNAのin vitro転写体の合成に使用できる。このプラスミドは更に、トランスフェクション時に選択可能マーカーとして使用されるネオマイシン耐性をコードする遺伝子を含む。
【0109】
第1図はpcDNA3−KEα6.3の部分的制限地図も示している。
【0110】
完全長ヒトnAChRα6サブユニットの発現は、翻訳開始コドンの前にコンセンサスリボソーム結合部位[RBS;例えばKozak(1991)J.Biol Chem.266:19867−19870参照]を導入することによって最適化した。既存の5’非翻訳領域を、DNA鋳型としてプラスミドpcDNA3−KEα6.3と、適当なヌクレオチドRBS置換体及び非翻訳5’HindIII及びEcoRI部位を含む相補上流オリゴヌクレオチドと、翻訳開始コドン下流のα6コーディング〜450ヌクレオチド配列に相補的なオリゴヌクレオチドとを使用して、PCR突然変異誘発により修飾した。その結果得られた増幅産物は、HindIII及びEcoRI部位と、これに続くコンセンサスRBS及びヒトnAChRα6コーディング配列ヌクレオチド1−459(配列番号9のヌクレオチド143−602)とを含んでいた。増幅DNAをHindIII及びBamHIで消化し、308bpのHindIII−BamHIフラグメントを分離し、pcDNA3−KEα6.3の5.3kb BamHI−PvuIフラグメント及びpcDNA3由来の1.4kb PvuI−HindIIIフラグメントに連結して、〜7.0kbプラスミドpcDNA3−KEα6RBSを形成した。
【0111】
B. ヒトニューロンnAChRβ3サブユニットを発現させるための構築物
完全長ニコチン性AChRβ3サブユニットをコードする〜2.0kb EcoRIフラグメントを標準的方法でKEβ3.2から分離し、ベクターpcDNA3のEcoRIポリリンカー部位に連結して、pcDNA3−KEβ3.2を形成した(第2図参照)。第2図はpcDNA3.KEβ3.2の部分的制限地図も示している。
【0112】
完全長ヒトニコチン性nAChRβ3サブユニットの発現は、翻訳開始コドンの前にコンセンサスリボソーム結合部位(RBS)を導入することによって最適化する。α6nAChRサブユニットについて上述したものと類似の方法を用いてPCR突然変異誘発により既存の5’非翻訳領域を修飾し、pcDNA3−KEβ3RBSを形成した。
【実施例4】
【0113】
アフリカツメガエル内での組換えヒトニューロンnAChRの発現
アフリカツメガエル卵母細胞に、α6及びβ3サブユニットをコードするDNAを含む構築物から製造したin vitro転写体を注入する。二電極電圧固定法を使用して卵母細胞トランスメンブラン電流の電気生理学的測定を行う(例えばStuhmer(1992)Meth.Enzymol.207:310−339参照)。
【0114】
1. in vitro転写体の調製
mMessage及びmMachine in vitro転写キットを製造業者のキャップ化転写体プロトコル通りに使用して(AMBION,Inc.,Austin,TXのカタログ1344)、直線化プラスミドからpcDNA3−KEαRBS及びpcDNA3−KEβ3RBSの組換えキャップ化転写体を合成する。合成した転写体の質量をUV吸光率によって測定し、転写体の完全性をアガロースゲル電気泳動で調べる。
【0115】
2. 電気生理学
アフリカツメガエル卵母細胞に、卵母細胞当たり12.5、50又は125ngの1種類以上のヒトニコチン性nAChRα及びβサブユニット転写体を注入する。卵母細胞の調製及び注入操作は、Dascal(1987),Crit.Rev.Biochem.22:317:387に記載のように実施する。mRNAの注入から2〜6日後、二電極電圧固定法を用いて卵母細胞を調べる。1μMのアトロピンを含み、100μMのd−チュボキュラリンを含む又は含まないリンガー液(115mM NaCl、2.5mM KCl、1.8mM CaCl2、10mM HEPES、pH7.3)に細胞を浸漬する。細胞を−60〜−80mVで電圧固定する。Axotapeソフトウエアで2〜5Hzでデータを得る。アゴニストアセチルコリン(ACh)、ニコチン及びシチシンを0.1μM〜100μMの濃度で加える。
【実施例5】
【0116】
哺乳動物細胞内でのヒトnAChRサブユニットの組換え発現
ヒト胎児腎臓(HEK)293細胞に、ヒトニューロンニコチン性nAChRα6及びβ3サブユニットをコードするDNAを一時的及び安定的にトランスフェクションする。種々のアッセイ、例えば電気生理学的方法、Ca2++感受性蛍光指示薬をベースとするアッセイを用いて、一時的トランスフェクション細胞をニコチン性nAChRの発現について分析する。
【0117】
1. HEK細胞の一時的トランスフェクション
HEK細胞に、一つ以上のαサブユニット及び/又は一つ以上のβサブユニットをコードするDNAを一時的に同時トランスフェクションする。約2×106のHEK細胞に18μgの指定プラスミドを、標準的CaPO4トランスフェクション方法で(Wiglerら(1979)Proc.Natl.Acad.Sci.U.S.A.76:1373−1376)、又はリポフェクタミン(lipofectamine)を製造業者(Bethesda Research Laboratory(BRL),Gaitherburg,MD)の指示通りに使用して、一時的にトランスフェクションする。更に、トランスフェクション効率を観察するために、CMVプロモーターに融合した大腸菌βガラクトシダーゼ遺伝子を含むプラスミドpCMVβgal(Clontech Laboratories,Palo Alto,CA)2μgをリポーター遺伝子として同時トランスフェクションする。βガラクトシダーゼ活性の測定により、トランスフェクション細胞をβガラクトシダーゼ発現について分析する[Miller(1972)Experiments in Molecular Genetics,pp.352−355,Cold Spring Harbor Press]。トランスフェクション細胞は、βガラクトシダーゼ及びX−gal基質が関与する反応の生成物を直接染色することにより、βガラクトシダーゼ発現についても分析し得る[Jones(1986)EMBO 5:133−3142]。
【0118】
2. HEK細胞の安定なトランスフェクション
リン酸カルシウムトランスフェクション法[Current Protocols in Molecular Biology,Vol.1,Wiley Inter−Science,Supplement 14,Unit 9.1.1−9.1.9(1990)]を使用してHEK細胞のトランスフェクションを行う。所望のα及びβサブユニットをコードするDNAとpSV2neo(選択可能マーカー)とを含むDNA/リン酸カルシウム沈降物1mlをHEK細胞にトランスフェクションする。1μg/mlのG418を含む培地で14日間増殖させた後、形成されたコロニーをクローニングシリンダーを用いて個々に分離する。分離物を限界希釈にかけ、後述のように最大レベルのnAChRを発現するものを同定するためにスクリーニングする。
【実施例6】
【0119】
ヒトニューロンnAChRを発現する細胞系の特徴分析
ヒトニューロンnAChRサブユニットをコードするDNAをトランスフェクションすることにより形成した実施例5に記載のような組換え細胞系は、下記の方法の一つ以上を用いて更に特徴を分析し得る。
【0120】
A. α及び/又はβサブユニットコーディングメッセージの発現に関するノーザン又はスロットブロット分析
〜1×107個の細胞から全RNAを分離し、各細胞種類のRNAを10〜15μg使用してノーザンブロット又はスロットブロットハイブリダイゼーション分析を行う。ヒトニューロンnAChRコーディングプラスミド由来の挿入物は、ニックトランスレーションにかけてプローブとして使用し得る。また、各ブロット上のRNAの存在又は不在を確認し、細胞系間のα又はβ特異的mRNAレベルの差の定量に使用する大体の標準を与えるために、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ(GAPD)遺伝子配列(Tsoら(1985)Nucleic Acids Res.13:2485)をニックトランスレーションにかけ、二重フィルター上で対照プローブとして使用し得る。典型的なノーザンブロット及びスロットブロットハイブリダイゼーション並びに洗浄条件は下記の通りである:5×SSPE、5×デンハーツ溶液、0.2%SDS、200μg/ml変性音波処理ニシン精子DNA、50%ホルムアミド中42℃でハイブリダイズし、次いで0.1×SSPE、0.1%SDS中65℃で洗浄する。
【0121】
B. 結合アッセイ
ヒトニューロンnAChRのα又はα及びβサブユニットをコードするDNAのトランスフェクションによって製造した細胞系は、例えばニコチン又は他のアゴニストに結合する能力について、対照細胞系、例えばニューロン由来細胞系PC12(Boulterら(1986)Nature 319:368−374;ATCC#CRL1721)及びIMR32(Clementiら(1986)Int.J.Neurochem.47:291−297;ATCC#CCL127)、並びに筋肉由来細胞系BC3H1(Patrickら(1977)J.Biol.Chem.252:2143−2153)と比較して分析し得る。陰性対照細胞(即ち、トランスフェクション細胞の形成に使用した宿主細胞)もアッセイに含まれる。アッセイは次のように実施する:アッセイにかける直前に、トランスフェクション細胞をスクラッピングによってプレートから除去する。使用する陽性対照細胞は、PC12、BC3H1及びIMR32である(これらの細胞には数日間新しい培地を与えなかった)。37℃のアッセイ緩衝液(50mMトリス/HCl、1mM MgCl2、2mM CaCl2、120mM NaCl、3mM EDTA、2mg/ml BSA及び0.1%アプロチニン、pH7.4)中で濯ぐことにより対照細胞系を除去する。細胞を洗浄し、1×106/250μlの濃度に再懸濁する。各プラスチックアッセイ管に、250μlの細胞溶液と、15nM 3H−ニコチンと、1mM非標識ニコチン有または無と、最終容量を500μlにする量のアッセイ緩衝液とを加える。トランスフェクション細胞系のアッセイを室温で30分間インキュベートする。陽性対照細胞のアッセイは1℃で2分間インキュベートする。適当な時間にわたってインキュベートした後、アッセイ容量の450μlアリコートを、0.05%ポリエチレンイミン中4℃で24時間インキュベートすることにより予備処理したWhatman GF/Cグラスファイバーフィルターで濾過する。次いで、フィルターを各回4mlの氷冷アッセイ緩衝液を使用して2回洗浄する。洗浄後、フィルターを乾燥し、5mlのシンチレーション液を入れたバイアルに加え、放射能を測定する。
【0122】
C. 86Rbイオン流アッセイ
ニコチン又はnAChRアゴニスト及びアンタゴニストがトランスフェクション細胞及び対照細胞内への86Rbの流入を仲介する能力は、細胞表面上の機能性nAChRの存在を指示するものであることが分かっている。86Rbイオン流アッセイは下記のように実施する:
1. 実験の前夜、ウェル数6のポリリシンコーティングしたプレートにウェル当たり2×106個の細胞(即ちウェル当たり2ml)を播く。
2. 培養培地をデカントし、プレートを2mlのアッセイ緩衝液(50mM HEPES、260mMスクロース、5.4mM KCl、1.8mM CaCl2、0.8mM MgSO4、5.5mMグルコース)で室温で洗浄する。
3. アッセイ緩衝液をデカントし、3μCi/mlの86Rbを5mMウワバイン及び最大応答を与える濃度のアゴニスト又はアンタゴニストと共に含むアッセイ緩衝液を1ml加える。
4. プレートを氷上1℃で4分間インキュベートする。
5. 緩衝液を廃棄物容器にデカントし、各ウェルを3mlのアッセイ緩衝液で洗浄し、次いで各回2mlで2回洗浄する。
6. 細胞をウェル当たり2×0.5mlの0.2%SDSで溶解し、5mlのシンチレーション液を入れたシンチレーションバイアルに移す。
7. 各バイアル5に含まれている放射能を測定し、データを計算する。このアッセイでは陽性対照細胞から下記のデータが得られた:
【0123】
【表3】
【0124】
D. ヒトニューロンnAChRサブユニットをコードするDNAをトランスフェクションした哺乳動物細胞の電気生理学的分析
組換え受容体の活性の評価、又は試験化合物がリガンド依存性組換えnAChRを介してカチオン流の規模及び持続時間を助長、拮抗又は調節する能力の評価には、電気生理学的測定を使用し得る。発現したニューロンnAChRの機能は、二電極電圧固定法及びパッチ固定法を含む種々の電気生理学的方法で評価できる。nAChRに固有のカチオン伝導チャンネルは、アセチルコリン(ACh)又は他のニコチン性コリン作動性アゴニストに応答して開き、その結果、生理学的条件下で主にナトリウム及びカリウムイオンにより担われているトランスメンブラン電流が流れる。この電流は電圧固定法で直接観察することができる。好ましい実施態様では、トランスフェクションを行った哺乳動物細胞又は注入を行った卵母細胞を、nAChRアゴニスト依存電流の存在に関して電気生理学的に分析する。
【0125】
E. 蛍光指示薬をベースとするアッセイ
アゴニストによりリガンド依存性nAChRが活性化されると、Ca++を含むカチオンが受容体チャンネルを介して流入する。チャンネルを介するCa++の細胞内流入は、細胞内ストアに含まれていたカルシウムの放出を誘発し得る。一価のカチオンがチャンネルを介して細胞内に流入すると、膜の脱分極及びその結果として生じる電圧依存カルシウムチャンネルの活性化を通して、細胞質Ca++レベルが増加し得る。従って、細胞内カルシウム濃度の一時的増加を検出する方法は、機能性ニコチン性nAChR発現の分析に適用できる。細胞内カルシウム濃度を測定する方法の一つは、カルシウム感受性蛍光指示薬の使用をベースとする。
【0126】
カルシウム感受性指示薬、例えばフルオ(fluo)−3(カタログ番号FO1241、Molecular Probes,Inc.,Eugene,OR)は、膜透過性であるアセトキシメチルエステルとして入手できる。アセトキシメチルエステル形態の指示薬が細胞内に入ると、エステル基が細胞質ゾルエステラーゼによって除去され、その結果細胞質ゾル内に遊離指示薬が捕捉される。遊離指示薬とカルシウムとが相互作用すると、指示薬の蛍光が増加する。従って、指示薬を含んでいる細胞の細胞内Ca2+濃度の増加は、蛍光の増加として直接示すことができる。ニコチン性nAChRをアッセイするための自動化蛍光検出システムは文献に記述されている(米国特許出願明細書第08/229,150号、第08/244,985号、第08/434,511号及び第08/434,968号並びに対応する国際特許出願公開明細書US92/11090号参照。国際特許出願公開明細書第96/05488号も参照)。
【0127】
自動化蛍光指示薬ベースアッセイを用いて、適当なα及び/又はβサブユニット並びにα6及びβ3サブユニットをコードするDNAを一時的又は安定的に同時トランスフェクションしたHEK細胞を、機能性組換えnAChRの発現について分析する。アッセイの手順は次の通りである。非トランスフェクションHEK細胞と適当なα及び/又はβサブユニットをコードするDNAを同時トランスフェクションしたHEK細胞とを、ウェル数96のミクロタイター皿のウェル内に播き、HBS(125mM NaCl、5mM KCl、1.8mM CaCl2、0.62mMm MgSO4、6mMグルコース、20mM HEPES、pH7.4)中20μMフルオ−3、0.2%Pluronic F−127を含む培地中で20℃で2時間インキュベートすることによりフルオ−3を充填する。次いで、細胞をアッセイ緩衝液(即ちHBS)で洗浄する。アンタゴニストd−ツボクラリンを一部のウェルに最終濃度10μMで加える。次いで、ミクロタイター皿を蛍光プレート読取り器内に配置し、各ウェルの基本蛍光を測定し、記録した後、アゴニスト、例えば200μMニコチンをウェルに加える。ニコチンの添加後、約60秒間にわたり、ウェルの蛍光を繰り返し観察する。
【0128】
非トランスフェクションHEK細胞の蛍光は、ニコチン添加後にも変化しない。これに対し、同時トランスフェクションを行った細胞の蛍光は、d−ツボクラリンの不在下では、ウェルへのニコチンの添加後に急増する。このニコチン刺激による蛍光の増加は、アンタゴニストd−ツボクラリンに暴露した同時トランスフェクション細胞では観察されない。このような結果は、同時トランスフェクション細胞が、ニコチンによって活性化され且つd−ツボクラリンによって阻害される機能性組換えnAChRを発現することを示すものである。
【0129】
好ましい実施例を挙げて本発明を詳細に説明してきたが、種々の変形が本発明の思想及び範囲を逸脱せずに可能である。
【0130】
変形例は当業者に明らかであるため、本発明は後述の「請求の範囲」によってのみ限定される。
【0131】
【表4】
【0132】
配列の要約
配列番号1はヒトニューロンニコチン性アセチルコリン受容体のα2サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0133】
配列番号2は配列番号1に示すヒトニューロンニコチン性アセチルコリン受容体のα2pサブユニットのアミノ酸配列である。
【0134】
配列番号3はヒトニューロンニコチン性アセチルコリン受容体のα3サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0135】
配列番号4は配列番号3に示すヒトニューロンニコチン性アセチルコリン受容体のα3サブユニットのアミノ酸配列である。
【0136】
配列番号5はヒトニューロンニコチン性アセチルコリン受容体のα4サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0137】
配列番号6は配列番号5に示すヒトニューロンニコチン性アセチルコリン受容体のα4サブユニットのアミノ酸配列である。 配列番号7はヒトニューロンニコチン性アセチルコリン受容体のα5サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0138】
配列番号8は配列番号7に示すヒトニューロンニコチン性アセチルコリン受容体のα5サブユニットのアミノ酸配列である。
【0139】
配列番号9はヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0140】
配列番号10は配列番号9に示すヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットのアミノ酸配列である。
【0141】
配列番号11はヒトニューロンニコチン性アセチルコリン受容体のα7サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0142】
配列番号12は配列番号11に示すヒトニューロンニコチン性アセチルコリン受容体のα7サブユニットのアミノ酸配列である。
【0143】
配列番号13はヒトニューロンニコチン性アセチルコリン受容体のβ2サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0144】
配列番号14は配列番号13に示すヒトニューロンニコチン性アセチルコリン受容体のβ2サブユニットのアミノ酸配列である。
【0145】
配列番号15はヒトニューロンニコチン性アセチルコリン受容体のβ3サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0146】
配列番号16は配列番号15に示すヒトニューロンニコチン性アセチルコリン受容体のβ3サブユニットのアミノ酸配列である。
【0147】
配列番号17はヒトニューロンニコチン性アセチルコリン受容体のβ4サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0148】
配列番号18は配列番号17に示すヒトニューロンニコチン性アセチルコリン受容体のβ4サブユニットのアミノ酸配列である。
【0149】
配列番号19はヒトニューロンニコチン性アセチルコリン受容体の変異体α6サブユニットをコードするヌクレオチド配列及びその推定アミノ酸配列である。
【0150】
配列番号20は配列番号19に示すヒトニューロンニコチン性アセチルコリン受容体のα6サブユニットのアミノ酸配列である。
【図面の簡単な説明】
【0151】
【図1】第1図は、α6コーディングフラグメントをEcoRI挿入物として含むサイトメガロウイルス(CMV)プロモーターをベースとするベクターpcDNA3−KEアルファ6.3の制限地図を示す説明図である。
【図2】第2図は、β3コーディングフラグメントをEcoRI挿入物として含むCMVプローモーターをベースとするベクターpcDNA3−KBベータ3.2の制限地図を示す説明図である。
【特許請求の範囲】
【請求項1】
ヒトニューロンニコチン性アセチルコリン受容体の全長β3サブユニットをコードする分離核酸分子であって、
β3サブユニットをコードするヌクレオチド配列は、以下のaからcのヌクレオチド配列から選択される前記核酸分子:
a.配列番号15に示すヌクレオチド98−1471を含むヌクレオチド配列;
b.配列番号16を含むアミノ酸配列をコードするヌクレオチド配列;
c.aまたはbに示された配列に縮重するヌクレオチド配列。
【請求項2】
配列番号15に示すヌクレオチド98−1471を含む請求項1の核酸分子。
【請求項3】
配列番号15に示す最初の105個のヌクレオチドからなる翻訳配列に示されている任意の28個の連続塩基又はその相補体を含む少なくとも28塩基の長さを有する一本鎖核酸分子。
【請求項4】
標識されている請求項3に記載の核酸分子。
【請求項5】
32Pで標識されている請求項4に記載の核酸分子。
【請求項6】
ヒトニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNAを分離するための方法であって、請求項4に記載の核酸分子を用いてライブラリーをスクリーニングし、少なくとも低ストリンジェンシー条件下で請求項4に記載の核酸にハイブリダイズするクローンを分離する操作を含む前記方法。
【請求項7】
分離クローンが高ストリンジェンシー条件下でハイブリダイズする請求項6に記載の方法。
【請求項8】
細胞が原核細胞又は真核細胞であり、核酸分子が細胞に対して外来性である請求項1に記載の核酸分子を含む細胞。
【請求項9】
哺乳動物細胞又は両生動物卵母細胞である請求項8に記載の細胞。
【請求項10】
DNAである請求項1に記載の分子。
【請求項11】
RNAである請求項1に記載の分子。
【請求項12】
ヒトニューロンニコチン性アセチルコリン受容体の活性を調節する化合物を同定するために化合物をスクリーニングする方法であって、該方法は、請求項8に記載の細胞内のニューロンニコチン性アセチルコリン受容体活性に対する試験化合物の作用を決定し、これを対照細胞に対する作用又は化合物の不在下における細胞のニューロンニコチン性アセチルコリン受容体活性と比較することからなる前記方法。
【請求項13】
配列番号16に示されたアミノ酸配列と90%の同一性を有する、実質的に純粋なヒトニューロンニコチン性アセチルコリン受容体β3サブユニット。
【請求項14】
配列番号16に示されたアミノ酸配列と90%の同一性を有するヒトニューロンニコチン性アセチルコリン受容体β3サブユニットを含む実質的に純粋な組換えヒトニューロンニコチン性アセチルコリン受容体。
【請求項15】
機能性ニューロンニコチン性アセチルコリン受容体サブユニット及びこれらの組合わせを同定するための方法であって、(a)請求項1に記載の核酸分子を真核細胞に導入し、(b)ステップ(a)の細胞内のニューロンニコチン性アセチルコリン受容体活性を検出する操作を含み、前記活性が、導入分子でコードされたサブユニットを含む受容体によって仲介される前記方法。
【請求項16】
mRNAである請求項1に記載の核酸。
【請求項17】
請求項16に記載のmRNAを含む分離細胞。
【請求項18】
配列番号15のヌクレオチド98−211を含む分離核酸分子。
【請求項19】
配列番号15に示すヌクレオチド配列を含む請求項1に記載の核酸分子。
【請求項20】
ニューロンニコチン性アセチルコリン受容体を発現する請求項8に記載の細胞。
【請求項21】
配列番号16のアミノ酸1−28をコードするヌクレオチド配列を含む分離核酸分子。
【請求項22】
配列番号16のアミノ酸1−20をコードするヌクレオチド配列を含む分離核酸分子。
【請求項23】
ヒトcDNAライブラリーから得られる請求項1に記載の核酸分子。
【請求項24】
配列番号15に示すヌクレオチド98−211と高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列を含む請求項1に記載の核酸分子。
【請求項1】
ヒトニューロンニコチン性アセチルコリン受容体の全長β3サブユニットをコードする分離核酸分子であって、
β3サブユニットをコードするヌクレオチド配列は、以下のaからcのヌクレオチド配列から選択される前記核酸分子:
a.配列番号15に示すヌクレオチド98−1471を含むヌクレオチド配列;
b.配列番号16を含むアミノ酸配列をコードするヌクレオチド配列;
c.aまたはbに示された配列に縮重するヌクレオチド配列。
【請求項2】
配列番号15に示すヌクレオチド98−1471を含む請求項1の核酸分子。
【請求項3】
配列番号15に示す最初の105個のヌクレオチドからなる翻訳配列に示されている任意の28個の連続塩基又はその相補体を含む少なくとも28塩基の長さを有する一本鎖核酸分子。
【請求項4】
標識されている請求項3に記載の核酸分子。
【請求項5】
32Pで標識されている請求項4に記載の核酸分子。
【請求項6】
ヒトニューロンニコチン性アセチルコリン受容体サブユニットをコードするDNAを分離するための方法であって、請求項4に記載の核酸分子を用いてライブラリーをスクリーニングし、少なくとも低ストリンジェンシー条件下で請求項4に記載の核酸にハイブリダイズするクローンを分離する操作を含む前記方法。
【請求項7】
分離クローンが高ストリンジェンシー条件下でハイブリダイズする請求項6に記載の方法。
【請求項8】
細胞が原核細胞又は真核細胞であり、核酸分子が細胞に対して外来性である請求項1に記載の核酸分子を含む細胞。
【請求項9】
哺乳動物細胞又は両生動物卵母細胞である請求項8に記載の細胞。
【請求項10】
DNAである請求項1に記載の分子。
【請求項11】
RNAである請求項1に記載の分子。
【請求項12】
ヒトニューロンニコチン性アセチルコリン受容体の活性を調節する化合物を同定するために化合物をスクリーニングする方法であって、該方法は、請求項8に記載の細胞内のニューロンニコチン性アセチルコリン受容体活性に対する試験化合物の作用を決定し、これを対照細胞に対する作用又は化合物の不在下における細胞のニューロンニコチン性アセチルコリン受容体活性と比較することからなる前記方法。
【請求項13】
配列番号16に示されたアミノ酸配列と90%の同一性を有する、実質的に純粋なヒトニューロンニコチン性アセチルコリン受容体β3サブユニット。
【請求項14】
配列番号16に示されたアミノ酸配列と90%の同一性を有するヒトニューロンニコチン性アセチルコリン受容体β3サブユニットを含む実質的に純粋な組換えヒトニューロンニコチン性アセチルコリン受容体。
【請求項15】
機能性ニューロンニコチン性アセチルコリン受容体サブユニット及びこれらの組合わせを同定するための方法であって、(a)請求項1に記載の核酸分子を真核細胞に導入し、(b)ステップ(a)の細胞内のニューロンニコチン性アセチルコリン受容体活性を検出する操作を含み、前記活性が、導入分子でコードされたサブユニットを含む受容体によって仲介される前記方法。
【請求項16】
mRNAである請求項1に記載の核酸。
【請求項17】
請求項16に記載のmRNAを含む分離細胞。
【請求項18】
配列番号15のヌクレオチド98−211を含む分離核酸分子。
【請求項19】
配列番号15に示すヌクレオチド配列を含む請求項1に記載の核酸分子。
【請求項20】
ニューロンニコチン性アセチルコリン受容体を発現する請求項8に記載の細胞。
【請求項21】
配列番号16のアミノ酸1−28をコードするヌクレオチド配列を含む分離核酸分子。
【請求項22】
配列番号16のアミノ酸1−20をコードするヌクレオチド配列を含む分離核酸分子。
【請求項23】
ヒトcDNAライブラリーから得られる請求項1に記載の核酸分子。
【請求項24】
配列番号15に示すヌクレオチド98−211と高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列を含む請求項1に記載の核酸分子。
【図1】

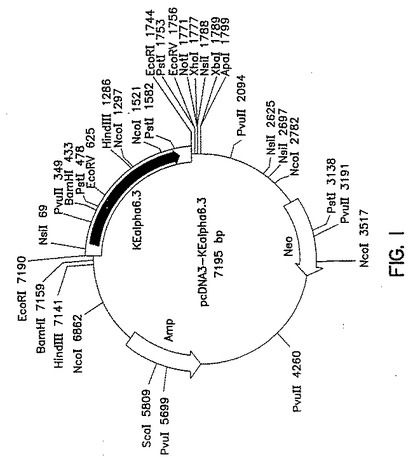
【図2】
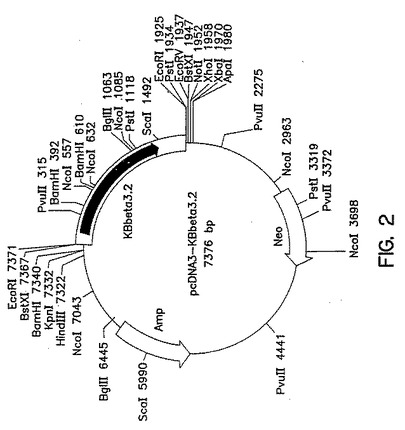
【図2】
【公開番号】特開2009−50270(P2009−50270A)
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願番号】特願2008−258987(P2008−258987)
【出願日】平成20年10月3日(2008.10.3)
【分割の表示】特願平9−503220の分割
【原出願日】平成8年6月7日(1996.6.7)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願日】平成20年10月3日(2008.10.3)
【分割の表示】特願平9−503220の分割
【原出願日】平成8年6月7日(1996.6.7)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
[ Back to top ]