
ヒト・インターフェロン−ガンマ中和活性を持つ、完全ヒト抗体Fab断片
本発明は、インターフェロン−ガンマ(IFNγ)の選択的結合剤を提供する。より具体的には、本発明は、IFNγに選択的に結合し、そして関節リウマチ、全身性エリテマトーデスおよび多発性硬化症などの自己免疫および炎症性疾患に関連する状態を防止するかまたは治療するのに使用可能な、抗体および抗原結合ドメインを提供する。前記の抗体および抗原結合ドメインをコードする核酸分子、並びに該核酸分子を産生するための発現ベクターおよび宿主細胞もまた、提供する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
発明の分野
本発明は、ヒト・インターフェロン・ガンマ(hIFNγ)に結合し、そして該分子と同族(cognate)受容体、IFNγ−Rとの相互作用を阻害し、そして/またはIFNγに誘発される生物学的作用を修飾する、新規完全ヒト抗体Fab断片に関する。より具体的には、本発明は、ファージ・ディスプレー・ライブラリーであって、ユニークなFab断片を含有し、該断片が次いで、全長ヒトIgG抗体に変換される、前記ファージ・ディスプレー・ライブラリーのhIFNγ親和性選択を通じて単離される中和Fab断片(Fab)に関する。hIFNγ中和活性、高親和性、およびin vivoでの長い半減期という望ましい特質を有する、hIFNγに対するこれらの新規完全ヒト抗体を用いて、多様な自己免疫および炎症性疾患を防止するかまたは治療することも可能である。本発明の完全ヒトFabを産生するための核酸分子、ベクターおよび宿主細胞もまた提供する。
【0002】
発明の背景
抗体は、生物薬剤学研究および薬剤発見の試みにおいて、長年、非常に重要な役割を果たしてきた。ヒト疾患の治療のための療法剤としての抗体の有用性は、抗体が:(a)in vivoでの半減期が長く;(b)高い親和性および特異性で標的(単数または複数)に結合可能であり;そして(c)免疫エフェクター機能(補体結合および抗体依存性細胞傷害作用など)を仲介する能力を持ちうるため、長年、理想とされてきた。
【0003】
しかし、今までは、非ヒト種から得た抗体に、不都合な免疫原性があり、抗体の長期臨床的有用性が制限されることから、療法抗体の概念を実行することは、大幅に制限されていた。最近の技術的進歩によって、免疫原性がより低く、そして長期の療法潜在能力を持つ、完全ヒト抗体を得る手段を提供することによって、これらの制限を克服する新規の方法が提供されてきている。さらに、コンビナトリアル・ライブラリー法および抗体操作における発展によって、抗体親和性、半減期、および/またはエフェクター機能を修飾する機会が得られるようになってきた。
【0004】
高親和性、高特異性であり、そしてin vivoでアゴニスト活性またはアンタゴニスト活性を持つ抗体を発見するのに、ファージ・コートタンパク質に融合された抗体断片の繊維状ファージ・ディスプレー・コンビナトリアル・ライブラリー(いわゆる「ファージ・ディスプレー・ライブラリー」)を使用する、こうした技術の1つが有効に用いられてきている。
【0005】
ヒト・インターフェロン・ガンマ(hIFNγ)は、活性化Tリンパ球およびナチュラルキラー細胞に産生されるリンホカインである。該分子は、抗増殖活性、抗ウイルス活性および免疫調節活性を示し、そして免疫系の大部分の初代細胞上にあるヘテロ二量体受容体、hIFNγ−Rに結合し;Langerら, Immunology Today, 9:393(1988)、そして炎症につながる事象カスケードを誘発する。IFNγの抗ウイルス活性および免疫調節活性は、いくつかの臨床状態に有益な影響を有することが知られる。しかし、IFNγ活性が有害な影響を有することが知られる、多くの臨床設定がある。例えば、自己免疫疾患は、高レベルの血中hIFNγと関連し、そして現在、IFNγの隔絶が、関節リウマチ(RA)、全身性エリテマトーデス(SLE)および多発性硬化症(MS)などの自己免疫疾患の症状軽減と関連することを示唆する証拠がある:例えばSkurkovichら, Intern. Journal of Immunotherapy, 14:23−32(1998);Gerezら, Clin. Exp. Immunol., 109:296−303(1997)を参照されたい。IFNγ活性はまた、悪液質、敗血症ショックおよびクローン病などの疾患状態にも関連付けられてきている。
【0006】
hIFNγのその受容体への相互作用を遮断することは、これに関連する最も上流の段階に介入することに相当するため、hIFNγ中和活性を持つ完全ヒト抗体は、魅力的な療法製品候補に相当する。ヒト・インターフェロン−ガンマ(hIFNγ)を療法標的として用いる抗体を同定するのに、ファージ・ディスプレー・ライブラリー技術を使用することが、本発明の目的である。
【0007】
発明の概要
本発明は、ヒト・インターフェロン−ガンマ(hIFNγ)に結合する、新規完全ヒト抗体Fab断片を提供する。1つの態様において、完全ヒト抗体Fab断片は、hIFNγとその同族受容体、hIFNγ−Rとの相互作用を、部分的にまたは完全に阻害し、そしてそれによって、hIFNγ活性を、部分的にまたは完全に阻害する方式で、hIFNγに結合する;すなわち抗体は、hIFNγのアンタゴニストである。好ましくは、hIFNγは哺乳動物hIFNγである。より好ましくは、hIFNγは、可溶性型または細胞表面結合型であることも可能なヒトhIFNγ、あるいはその断片、誘導体および変異体(variant)である。
【0008】
本発明の抗体は、ネズミまたはヒトhIFNγなどのhIFNγ、好ましくはヒトhIFNγ、あるいはその免疫原性断片、誘導体または変異体で、動物を免疫することによって、調製可能である。さらに、hIFNγがトランスフェクション細胞で発現されて、そしてその表面に結合するように、hIFNγをコードする核酸分子を含有するベクターでトランスフェクションされた細胞を用いて、動物を免疫することも可能である。あるいは、hIFNγへの結合に関して、抗体または抗原結合ドメイン配列を含むライブラリーをスクリーニングすることによって、抗体を得ることも可能である。こうしたライブラリーは、組み立てられたファージ粒子表面上に発現されるバクテリオファージ・コートタンパク質へのタンパク質融合体またはペプチド融合体、およびファージ粒子内に含有されるコードDNA配列として、バクテリオファージ中で、好適に調製される(いわゆる「ファージ・ディスプレー・ライブラリー」)。1つの例において、ファージ・ディスプレー・ライブラリーは、可変軽鎖および重鎖などのヒト抗体をコードするDNA配列を含有する。
【0009】
抗体または抗原結合ドメインは、hIFNγに結合可能であり、そして部分的にまたは完全にhIFNγ活性を中和する、天然抗体に類似の四量体糖タンパク質であることも可能であるし、あるいは一本鎖抗体;Fv、Fab、Fab’またはF(ab)’断片、二重特異性抗体、異種抗体、あるいはそれらの他の断片、変異体、または誘導体であることも可能である。抗体または抗原結合ドメインを、ハイブリドーマ細胞株(例えばマウス骨髄腫細胞に融合させた脾臓細胞などの、抗体産生細胞)で産生することも可能であるし、あるいは前記抗体または抗原結合ドメインをコードする核酸分子でトランスフェクションした異種細胞株で産生することも可能である。
【0010】
本発明の抗体または抗原結合ドメインは:
(a)図3〜13(配列番号65〜配列番号86)に示すようなFab重鎖アミノ酸配列;
(b)(a)の配列の保存的アミノ酸置換を含む、重鎖アミノ酸配列;
(c)(a)の配列に少なくとも約80%同一である、重鎖アミノ酸配列;あるいは
(d)(a)、(b)または(c)の断片または誘導体
を含み、hIFNγに選択的に結合する。
【0011】
別の態様において、本発明の抗体または抗原結合ドメインは、上記と同一の図3〜13に示すようなFab重鎖アミノ酸配列(配列番号65〜配列番号86)および図14〜24に示すようなFab軽鎖アミノ酸配列(配列番号87〜配列番号108)を含む抗体または抗原結合ドメインに認識される、ヒトhIFNγ上のエピトープを認識する。
【0012】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:
各Vl鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vl)、CDR2(Vl)およびCDR3(Vl)と称されるCDRアミノ酸配列を含み、CDR1(Vl)が:
【0013】
【化1】
【0014】
からなる群より選択され、
CDR2(Vl)が:
【0015】
【化2】
【0016】
からなる群より選択され、
そしてCDR3(Vl)が:
【0017】
【化3】
【0018】
からなる群より選択され、
CDR1(Vl)、CDR2(Vl)およびCDR3(Vl)が互いに独立に選択され;そして
各Vh鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vh)、CDR2(Vh)およびCDR3(Vh)と称されるCDRアミノ酸配列を含み、CDR1(Vh)が:
【0019】
【化4】
【0020】
からなる群より選択され、
CDR2(Vh)が:
【0021】
【化5】
【0022】
からなる群より選択され、
CDR3(Vh)が:
【0023】
【化6】
【0024】
からなる群より選択され;
CDR1(Vh)、CDR2(Vh)およびCDR3(Vh)が互いに独立に選択される。
【0025】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0026】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIASNYVQ(配列番号02)を有するCDR1、配列EDNQRPS(配列番号13)を有するCDR2、および配列QSYDGSAWV(配列番号24)を有するCDR3を含み;そして
Vh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列TSWNAGGPIDY(配列番号55)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0027】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TRSSGSIASYYVQ(配列番号03)を有するCDR1、配列EDDQRPS(配列番号14)を有するCDR2、および配列QSYDRNSLV(配列番号25)を有するCDR3を含み;そして
Vh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列DRVGYSSSLLDY(配列番号56)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0028】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RATQSLLHGNGHNYLD(配列番号04)を有するCDR1、配列MGSNRAS(配列番号15)を有するCDR2、および配列MQALQLPPT(配列番号26)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号36)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号46)を有するCDR2、および配列DKGSRITIFGVVGSAGFDY(配列番号57)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0029】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLVHSDGNTYLS(配列番号05)を有するCDR1、配列KISNRFS(配列番号16)を有するCDR2、および配列MQATQLPYT(配列番号27)を有するCDR3を含み;そして
Vh鎖が、配列NARMGVS(配列番号37)を有するCDR1、配列HIFSNDEESYSTSLKS(配列番号47)を有するCDR2、および配列LLLYEGFDP(配列番号58)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0030】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列SGDVLARKYAR(配列番号06)を有するCDR1、配列KDRERPS(配列番号17)を有するCDR2、および配列YSAADNRGV(配列番号28)を有するCDR3を含み;そして
Vh鎖が、配列SYAMH(配列番号38)を有するCDR1、配列VISYDGSNKYYADSVKG(配列番号48)を有するCDR2、および配列DLVLTMTSRRAAFDI(配列番号59)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0031】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列GGDNLGGKSLH(配列番号07)を有するCDR1、配列DDSDRPS(配列番号18)を有するCDR2、および配列QVWDGSSDQRV(配列番号29)を有するCDR3を含み;そして
Vh鎖が、配列SYSMN(配列番号39)を有するCDR1、配列SISSGSSYRYDADSVKG(配列番号49)を有するCDR2、および配列DQWGTISGNDY(配列番号60)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0032】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLLHTNEYNYLD(配列番号08)を有するCDR1、配列LGSNRAP(配列番号19)を有するCDR2、および配列MQALQTPRT(配列番号30)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号40)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号50)を有するCDR2、および配列GWPTYVWGSYRPKGYFDY(配列番号61)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0033】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIANNYVH(配列番号09)を有するCDR1、配列EDDQRPS(配列番号20)を有するCDR2、および配列QSYDNSNSFVV(配列番号31)を有するCDR3を含み;そして
Vh鎖が、配列SGGYSWS(配列番号41)を有するCDR1、配列YIYHSGSTYYNPSLKS(配列番号51)を有するCDR2、および配列GDWGYFDY(配列番号62)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0034】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RASQYVSSNSLA(配列番号10)を有するCDR1、配列GASNRAT(配列番号21)を有するCDR2、および配列QQYGSSPIT(配列番号32)を有するCDR3を含み;そして
Vh鎖が、配列SNYMS(配列番号42)を有するCDR1、配列VIYSGGSTYYADSVKG(配列番号52)を有するCDR2、および配列DADGGDYGY(配列番号63)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0035】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLLRSNGYNYLA(配列番号11)を有するCDR1、配列LASNRAS(配列番号22)を有するCDR2、および配列VHGVHIPYT(配列番号33)を有するCDR3を含み;そして
Vh鎖が、配列SNEAGVG(配列番号43)を有するCDR1、配列LLYWDDDKRYSPSLRS(配列番号53)を有するCDR2、および配列RLVRYGGYSTGGFDV(配列番号64)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0036】
本発明の抗体および抗原結合ドメインは、軽鎖および重鎖アミノ酸配列をコードするゲノムDNAに存在する生殖系列核酸由来である。抗体は、生殖系列配列再編成突然変異および体細胞突然変異の産物である核酸配列にコードされる。
【0037】
1つの態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含み;そして抗体は、IFNγポリペプチドに選択的に結合する。
【0038】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図34におけるようなVH1生殖系列遺伝子(配列番号123)の再編成変異体または体細胞変異体を含む。
【0039】
別の態様において、Vl鎖が、図42におけるようなVκ2生殖系列遺伝子(配列番号131)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図35におけるようなVH2生殖系列遺伝子(配列番号124)の再編成変異体または体細胞変異体を含む。
【0040】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含む。
【0041】
別の態様において、Vl鎖が、図44におけるようなVλ3生殖系列遺伝子(配列番号133)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図36におけるようなVH3生殖系列遺伝子(配列番号125)の再編成変異体または体細胞変異体を含む。
【0042】
別の態様において、Vl鎖が、図45におけるようなVλ3生殖系列遺伝子(配列番号134)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図37におけるようなVH3生殖系列遺伝子(配列番号126)の再編成変異体または体細胞変異体を含む。
【0043】
別の態様において、Vl鎖が、図46におけるようなVκ3生殖系列遺伝子(配列番号135)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図38におけるようなVH3生殖系列遺伝子(配列番号127)の再編成変異体または体細胞変異体を含む。
【0044】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図39におけるようなVH4生殖系列遺伝子(配列番号128)の再編成変異体または体細胞変異体を含む。
【0045】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含む。
【0046】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図40におけるようなVH2生殖系列遺伝子(配列番号129)の再編成変異体または体細胞変異体を含む。
【0047】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図34におけるようなVH1生殖系列遺伝子(配列番号123)の再編成変異体または体細胞変異体を含む。
【0048】
本発明の選択的結合剤(抗体または抗原結合ドメイン)は、IFNγの少なくとも1つの活性、例えばIFNγのIFNγ−Rへの結合を、部分的にまたは完全に阻害する。
1つの態様において、IFNγアンタゴニスト、例えば抗体または抗原ドメインは、狼瘡様疾患、関節炎、または多発性硬化症様症候群を経験しているか、またはこれらを発展させるリスクがある動物に投与される。IFNγアンタゴニストを用いて、ループス腎炎、関節リウマチ、および/または多発性硬化症を防止し、そして/または治療することも可能である。
【0049】
本発明の抗体または抗原結合ドメインおよび薬学的に許容しうるキャリアーを含む組成物もまた、提供する。
【0050】
発明の詳細な説明
本発明は、ヒト・インターフェロン−ガンマ・タンパク質(hIFNγ)に選択的に結合する剤(「選択的結合剤」)を提供する。好ましくは、該剤は、IFNγアンタゴニストまたは阻害剤であり、IFNγの少なくとも1つの活性、例えばIFNγのその同族受容体への結合を、部分的にまたは完全に阻害する。1つの態様において、完全ヒト抗体断片は、IFNγのその同族受容体への結合を、部分的にまたは完全に遮断し、そしてIFNγ活性を、部分的にまたは完全に阻害するように、IFNγに選択的に結合する。
【0051】
用語「選択的結合剤」は、IFNγに優先的に結合する分子を指す。選択的結合剤には、タンパク質、ペプチド、核酸、炭水化物、脂質、または小分子量化合物が含まれることも可能である。好ましい態様において、選択的結合剤は抗体であり、例えば可溶性型または結合型で、標識されていることも可能である、ポリクローナル抗体、モノクローナル抗体(mAb)、キメラ抗体、CDR移植抗体、抗体に対する抗イディオタイプ(抗Id)抗体とともに、限定されるわけではないが、酵素切断、ペプチド合成または組換え技術を含む既知の技術によって提供される、それらの断片、領域または誘導体である。本発明の抗IFNγ選択的結合剤は、IFNγの部分に結合することが可能であり、IFNγのIFNγ−R受容体への結合を阻害する。
【0052】
本発明の抗体および抗原結合ドメインは、IFNγに選択的に結合し、すなわち、これらは他の抗原より高い結合親和性で、優先的にIFNγに結合する。抗体は、ヒトIFNγに選択的に結合するが、ネズミIFNγなどの非ヒトIFNγにもまた検出可能であるように結合することも可能である。あるいは、抗体は、非ヒトIFNγに選択的に結合するが、ヒトIFNγにもまた検出可能であるように結合することも可能である。あるいは、抗体は、もっぱらヒトIFNγに結合し、非ヒトIFNγには検出可能に結合しないことも可能である。
【0053】
用語「モノクローナル抗体」は、実質的に均質な抗体の集団から得られる抗体を指し、ここで、各モノクローナル抗体は、典型的には、抗原上の単一のエピトープを認識するであろう。用語「モノクローナル」は、抗体を作成するいかなる特定の方法にも限定されない。例えば、本発明のモノクローナル抗体は、Kohlerら;Nature, 256:495(1975)に記載されるようなハイブリドーマ法によって作成することも可能であるし、また例えば本明細書記載の技術を用いて、ファージ・ライブラリーから単離することも可能である。
【0054】
用語「抗原結合ドメイン」または「抗原結合領域」は、選択的結合剤(例えば抗体分子)の部分であって、抗原と相互作用し、そして該結合剤に、抗原に対する特異性および親和性を与えるアミノ酸残基を含有する、前記部分を指す。好ましくは、抗原結合領域は、ヒト起源のものであろう。他の態様において、抗原結合領域は、他の動物種、特にウサギ、ラットまたはハムスターなどのげっ歯類に由来することも可能である。
【0055】
用語「エピトープ」は、1以上の結合剤の抗原結合領域で、選択的結合剤(例えば抗体)によって認識され、そして結合されることが可能な分子いずれかの部分を指す。エピトープは通常、アミノ酸または糖側鎖などの、分子の化学的に活性である表面集団からなり、そして特定の三次元構造特性とともに特定の電荷特性を有する。「阻害性エピトープおよび/または中和エピトープ」は、選択的結合剤に結合された際、そのエピトープを含有する分子または生物のin vivo、in vitro、またはin situでの、より好ましくはin vivoでの、IFNγのその受容体への結合を含めた、生物学的活性の損失を生じるエピトープを意図する。用語「軽鎖」は、抗体に関して用いた場合、定常ドメインのアミノ酸配列に基づいて、カッパ(κ)またはラムダ(λ)と呼ばれる2つの別個の種類を指す。
【0056】
用語「重鎖」は、抗体に関して用いた場合、重鎖定常ドメインのアミノ酸配列に基づいて、アルファ、デルタ、イプシロン、ガンマおよびミューと呼ばれる5つの別個の種類を指す。重鎖のこれらの別個の種類は、抗体の5つのクラス、それぞれ、IgA、IgD、IgE、IgGおよびIgMを生じさせ、これには、IgGの4つのサブクラス、すなわちIgG1、IgG2、IgG3およびIgG4が含まれる。
【0057】
用語「可変領域」または「可変ドメイン」は、典型的には、重鎖ではアミノ末端の約120〜130アミノ酸、そして軽鎖では約100〜110アミノ酸であり、抗体間で配列が大規模に異なり、そして特定の抗原に対する特定の各抗体の結合および特異性において用いられる、軽鎖の部分および重鎖の部分を指す。配列の可変性は、相補性決定領域(CDR)と呼ばれる領域に集中しており、一方、可変ドメイン中のよりよく保存される領域は、フレームワーク領域(FR)と呼ばれる。軽鎖のCDRおよび重鎖のCDRは、抗原と抗体の相互作用に関与する。
【0058】
用語「定常領域」または「定常ドメイン」は、抗体の抗原への結合に直接関与していないが、Fc受容体との相互作用など、多様なエフェクター機能を示す、軽鎖のカルボキシ末端部分および重鎖のカルボキシ末端部分を指す。
【0059】
用語「ヒト・インターフェロン−ガンマ」または「ヒト・インターフェロン−ガンマ・ポリペプチド」は、その開示が本明細書に援用されるPCT公報WO 83/04053に記載されるアミノ酸配列を含むポリペプチド、および関連ポリペプチドを指す。関連ポリペプチドには、対立遺伝子変異体;スプライス変異体;断片;誘導体;置換変異体、欠失変異体、および挿入変異体;融合ポリペプチド;並びに種間相同体(homolog)が含まれる。免疫反応を生じるのに十分なIFNγの可溶性型もまた含まれる。IFNγは、本明細書に定義するような成熟ポリペプチドであることも可能であり、そして調製した方法に応じて、アミノ末端メチオニン残基を有することもまた持たないことも可能である。
【0060】
用語「断片」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、全長アミノ酸配列に満たない配列を含むペプチドまたはポリペプチドを指す。こうした断片は、例えばアミノ末端の一部切除(truncation)、カルボキシ末端の一部切除、および/またはアミノ酸配列からの残基(単数または複数)の内部欠失から生じることも可能である。断片は、選択的RNAスプライシング、またはin vivoプロテアーゼ活性から生じることも可能である。
【0061】
用語「変異体」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、天然配列または非修飾配列に比較した際に、1以上のアミノ酸配列置換、欠失、および/または付加を含むペプチドまたはポリペプチドを指す。例えば、IFNγ変異体は、天然IFNγのアミノ酸配列に対する1以上の変化から生じることも可能である。やはり例として、IFNγの選択的結合剤の変異体は、天然のまたはあらかじめ修飾されていない選択的結合剤のアミノ酸配列に対する1以上の変化から生じることも可能である。変異体は、対立遺伝子変異体またはスプライス変異体などの天然存在変異体であることも可能であるし、あるいは人工的に構築することも可能である。ポリペプチド変異体は、前記変異体をコードする、対応する核酸分子から調製することも可能である。
【0062】
用語「誘導体」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、化学的に修飾されているポリペプチドまたはペプチド、あるいはその変異体、断片または誘導体を指す。例には、1以上のポリマー、例えば水溶性ポリマー、N連結、またはO連結炭水化物、糖、リン酸、および/または他のこうした分子の共有結合が含まれる。誘導体は、付着する分子の種類または位置いずれかが、天然存在または出発ペプチドまたはポリペプチドと異なる方式で修飾されている。誘導体には、ペプチドまたはポリペプチド上に天然に存在する1以上の化学基の欠失がさらに含まれる。
【0063】
用語「融合」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、ペプチドまたはポリペプチド、あるいはその断片、変異体および/または誘導体と、異種ペプチドまたはポリペプチドとの連結を指す。
【0064】
用語「生物学的に活性」は、IFNγに関連して、またはタンパク質性選択的結合剤に関連して用いた場合、IFNγまたは選択的結合剤の少なくとも1つの活性特性を有するペプチドまたはポリペプチドを指す。IFNγの選択的結合剤は、IFNγの少なくとも1つの生物学的活性に関して、アゴニスト活性、アンタゴニスト活性、あるいは中和活性または遮断活性を有することも可能である。
【0065】
用語「天然存在」は、核酸分子、ポリペプチド、宿主細胞等の生物学的材料に関連して用いた場合、天然に見出され、そして人間によって操作されていないものを指す。
用語「単離」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、天然環境で見出される混入ポリペプチドを少なくとも1つ含まないペプチドまたはポリペプチドを指し、そして好ましくは、療法使用および診断使用に干渉するであろう、いかなる他の混入哺乳動物ポリペプチドも実質的に含まないペプチドまたはポリペプチドを指す。
【0066】
用語「成熟」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、リーダー配列を欠くペプチドまたはポリペプチドを指す。該用語はまた、アミノ末端(リーダー配列を含むまたは含まない)および/またはカルボキシ末端のタンパク質分解的プロセシング、より大きい前駆体からのより小さいポリペプチドの切断、N連結および/またはO連結グリコシル化等のペプチドまたはポリペプチドの他の修飾も含むことも可能である。
【0067】
用語「有効量」および「療法的有効量」は、IFNγの選択的結合剤に関連して用いた場合、IFNγの1以上の生物学的活性レベルの観察可能な変化を支持するのに有用であるかまたは必要である、選択的結合剤の量を指す。前記変化は、IFNγ活性レベルの増加または減少いずれであることも可能である。
【0068】
用語「保存的アミノ酸置換」は、その位でのアミノ酸残基の極性または電荷にほとんどまたはまったく影響しないような、天然アミノ酸残基の非天然残基での置換を指す。例えば、保存的置換は、ポリペプチド中の非極性残基を他の非極性残基いずれかで置換することから生じる。さらに、アラニン・スキャニング突然変異誘発に関して、先に記載されているように、ポリペプチド中の天然残基いずれかをアラニンで置換することもまた可能である;Cunninghamら, Science, 244:1081−1085(1989)。保存的アミノ酸置換の典型的な規則を表Iに示す。
【0069】
表I
保存的アミノ酸置換
【0070】
【表1】
【0071】
保存的アミノ酸置換はまた、典型的には、生物学的系における合成よりも化学的ペプチド合成によって取り込まれる、非天然存在アミノ酸残基も含む。これらには、ペプチド模倣体、およびアミノ酸部分の他の逆転型または反転型が含まれる。
【0072】
アミノ酸配列に対する保存的修飾(およびコードするヌクレオチドに対する対応する修飾)は、天然存在IFNγまたは選択的結合剤と類似の機能特性および化学特性を有するIFNγポリペプチド(およびそのタンパク質性選択的結合剤)を産生可能である。対照的に、(a)置換領域における分子主鎖の構造、例えばシートまたはらせんコンホメーション、(b)標的部位での、分子の電荷または疎水性、あるいは(c)側鎖バルクの維持に対する影響が有意に異なる置換を選択することによって、IFNγ(およびそのタンパク質性選択的結合剤)の機能特性および/または化学特性の実質的な修飾を達成することも可能である。天然存在残基は、共通の側鎖特性に基づいて、グループに分けることも可能である:
1)疎水性:ノルロイシン、Met、Ala、Val、Leu、Ile;
2)中性親水性:Cys、Ser、Thr;
3)酸性:Asp、Glu;
4)塩基性:Asn、Gln、His、Lys、Arg;
5)鎖配向に影響を及ぼす残基:Gly、Pro;および
6)芳香族:Trp、Tyr、Phe。
【0073】
非保存的置換は、これらの種類の1つのメンバーと別の種類のメンバーとの交換を伴うことも可能である。
2以上の核酸分子および/またはポリペプチドの「同一性または類似性」は、2以上の別個の配列の関連性の測定値を提供する。用語「同一性」は、2つの別個のアミノ酸配列の対応する位で同一であるアミノ酸を指す。用語「類似性」は、2つの別個のアミノ酸配列の対応する位で、同一であるか、または上に定義するような保存的置換であるアミノ酸を指す。
【0074】
同一性または類似性の度合いは、限定されるわけではないが、Computational Molecular Biology, Lesk, A.M.監修, Oxford University Press, ニューヨーク, 1988;Biocomputing:Informatics and Genome Projects, Smith, D.W.監修, Academic Press, ニューヨーク, 1993;Computer Analysis of Sequence Data, Part 1, Griffin, A.M.およびGriffin, H.G.監修, Humana Press, ニュージャージー, 1994;Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987;Sequence Analysis Primer, Gribskov, M.およびDevereux, J.監修, M. Stockton Press, ニューヨーク, 1991;およびCarilloら, SIAM J. Applied Math., 48:1073(1988)に記載されるものを含む、既知の方法によって、容易に計算可能である。
【0075】
同一性および/または類似性を決定する好ましい方法を設計して、試験する配列間の最大のマッチを得る。同一性および類似性を決定する方法は、公的に入手可能なコンピュータプログラムに体系化されている。2つの配列間の同一性および類似性を決定する典型的なコンピュータプログラム法には、限定されるわけではないが、GAP;Devereuxら, Nucleic Acids Research, 12:387(1984);遺伝学コンピュータグループ、ウィスコンシン大学、ウィスコンシン州マディソン;BLASTP、BLASTNおよびFASTA、Altschulら,J. Mol. Biol., 215:403−410(1990)を含むGCGプログラムパッケージが含まれる。BLAST Xプログラムは、米国バイオテクノロジー情報センター(NCBI)および他の供給源(BLAST Manual, Altschulら NCB NLM NIH メリーランド州ベセスダ)から公的に入手可能である。周知のSmith Watermanアルゴリズムを用いて、同一性を決定することもまた可能である。
【0076】
IFNγポリペプチド
本発明の選択的結合剤をスクリーニングし、そして同定するため、IFNγポリペプチド、並びにその断片、変異体および誘導体を標的分子として用いる。選択的結合剤として抗体を調製することが望ましい場合、IFNγポリペプチドは、好ましくは免疫原性であり、すなわち、これらは動物に投与された際に、免疫応答を誘発する。あるいは、in vitro技術によって抗体を調製する場合、標的分子として用いられるIFNγポリペプチドは、抗体または抗原結合ドメインと検出可能に結合することが可能である。
【0077】
生物学的方法または化学的方法によって、IFNγポリペプチドを調製する。組換えIFNγをコードするDNA配列の発現などの生物学的な方法が当該技術分野に知られる;例えば、Sambrookら、上記を参照されたい。Merrifieldら, J. Am. Chem. Soc., 85:2149(1963)、Houghtenら, Proc Natl Acad. Sci. USA, 82:5132(1985)、並びにStewartおよびYoung, Solid phase peptide synthesis, Pierce Chemical Co., イリノイ州ロックフォード(1984)に示されるような化学合成法を用いて、本発明のIFNγポリペプチドを調製することもまた可能である。こうしたポリペプチドを、アミノ末端メチオニンを伴い、または伴わずに合成することも可能である。化学的に合成したIFNγポリペプチド、あるいはその断片または変異体を、これらの参考文献に示される方法を用いて酸化して、ジスルフィド架橋を形成することも可能である。化学合成によって調製された本発明のIFNγポリペプチドは、組換えによって産生されたか、または天然供給源から精製された、対応するIFNγポリペプチドに匹敵する生物学的活性を、少なくとも1つ有するであろう。
【0078】
IFNγポリペプチドを、IFNγポリペプチドが天然に見出される供給源組織および/または体液などの生物学的試料から単離することによって、得ることも可能である。IFNγポリペプチドの供給源は、起源が、ヒトまたは非ヒトであることも可能である。電気泳動による分離、その後の電気的溶出、多様な種類のクロマトグラフィー(アフィニティー、免疫親和性、分子ふるい、および/またはイオン交換)、および/または高圧液体クロマトグラフィーなどの、当該技術分野に知られる方法を用いて、天然存在IFNγポリペプチドの単離を達成することも可能である。例えば、組換えで産生したIFNγポリペプチドまたはそのペプチド断片に対して調製した抗体を用いて、精製中のIFNγポリペプチドの存在を監視することも可能である。
【0079】
本発明のポリペプチドには、単離IFNγポリペプチド、並びに上に定義するような、断片、変異体、融合ポリペプチド、および誘導体を含む、IFNγポリペプチドに関連するポリペプチドが含まれる。本発明のIFNγ断片は、アミノ末端(リーダー配列を含むまたは含まない)の一部切除、カルボキシ末端の一部切除、および/またはポリペプチド内部の欠失から生じることも可能である。こうしたIFNγポリペプチド断片は、所望によって、アミノ末端メチオニン残基を含むことも可能である。本発明のポリペプチドは、抗体応答を誘発可能である点で、免疫原性であろう。
【0080】
本発明のIFNγポリペプチド変異体には、天然IFNγアミノ酸配列に比較した際、1以上のアミノ酸置換、付加および/または欠失が含まれる。アミノ酸置換は、上に定義するように保存的であることも可能であるし、または非保存的であることも、あるいはその組み合わせいずれかであることも可能である。変異体は、カルボキシ末端またはアミノ末端いずれかに、アミノ酸残基の付加を有することも可能である(この場合、アミノ末端は、リーダー配列を含んでもよいし、また含まなくてもよい)。
【0081】
本発明の態様には、IFNγグリコシル化変異体およびシステイン変異体が含まれる。IFNγグリコシル化変異体には、天然IFNγポリペプチドに比較して、グリコシル化部位の数および/または種類が改変されている変異体が含まれる。1つの態様において、IFNγグリコシル化変異体は、天然IFNγに比較して、より多くのまたはより少ない数のN連結グリコシル化部位を含む。
【0082】
N連結炭水化物鎖の再編成を含むIFNγグリコシル化変異体であって、1以上のN連結グリコシル化部位(典型的には天然存在であるもの)が除去され、そして1以上の新規N連結部位が生成されている、前記変異体もまた提供する。IFNγシステイン変異体は、天然IFNγに比較して、より多い数の、あるいはより少ない数のシステイン残基を含む。1つの態様において、1以上のシステイン残基が欠失されるか、または別のアミノ酸(例えばセリン)で置換される。IFNγのシステイン変異体は、変性状態から単離された後、IFNγが生物学的に活性であるコンホメーションに再フォールディングするのを補助することによって、生物学的に活性であるIFNγの回収を改善することも可能である。
【0083】
IFNγポリペプチド変異体の調製は、当該技術分野の技術レベル内である。1つのアプローチにおいて、天然IFNγに1以上のアミノ酸置換、欠失および/または付加を導入することも可能であり、ここでIFNγ変異体は、IFNγの天然構造および/または少なくとも1つの生物学的活性を保持する。1つのアプローチは、比較的低いそして高い同一性および/または類似性の領域を同定するため、多様な異なる種由来のIFNγポリペプチドの配列を比較することである。比較的低い同一性および/または類似性を有するIFNγポリペプチドの領域は、構造および活性に必須である可能性はより低く、そしてしたがって、アミノ酸改変、特に非保存性である改変に、より耐性でありうると認識される。比較的保存される領域であっても、活性を保持しつつ、保存的アミノ酸置換を導入可能であることもまた認識される。
【0084】
別のアプローチにおいて、構造−機能関連性を用いて、類似のポリペプチドにおいて、活性または構造に重要な残基を同定することも可能である。例えば、IFNγ、および構造−機能解析が利用可能な腫瘍壊死因子ファミリーの他のメンバーの間で保存されるアミノ酸残基を比較し、そしてこうした比較に基づいて、IFNγ中のどのアミノ酸残基が活性または構造に重要であるかを予測することも可能である。当業者は、こうして予測されたIFNγの重要なアミノ酸残基に関して、化学的に類似のアミノ酸置換を選択することも可能である。
【0085】
さらに別のアプローチにおいて、IFNγの二次構造または三次構造(IFNγ結晶のX線回折によって、または構造予測法によって決定されるものいずれか)の解析を行って、IFNγポリペプチド内の実際の構造または予測される構造に関して、特定のアミノ酸残基の位置を決定することも可能である。この情報を用いて、IFNγポリペプチドの二次構造および/または三次構造をできるだけ多く保持することを模索する方式で、アミノ酸変化を導入することも可能である。さらに別のアプローチにおいて、アミノ酸置換を導入し、そして本明細書に記載するアッセイを用いて、生物学的活性に関して、改変されたIFNγポリペプチドを試験することによって、特定の位でアミノ酸を改変する影響を実験的に試験することも可能である。
【0086】
アラニン・スキャニング突然変異誘発などの技術(Cunninghamら、上記)が、このアプローチに特によく適している。IFNγ中の多様なアミノ酸位で、多くの置換を導入し、そしてファージ・ディスプレー・ライブラリーの一部として、改変されたポリペプチド集団をスクリーニングすることによって、多くの改変された配列を好適に試験することも可能である。このアプローチを用いて、活性に必須のIFNγポリペプチドの領域を容易に決定することも可能である。
【0087】
上記方法は、天然構造を保持するIFNγ変異体を生成するのに有用である。したがって、各変異体に対して作成された抗体は、IFNγの天然構造決定基またはエピトープを認識する可能性があり、そしてまた、天然IFNγに結合する可能性もある。しかし、いくつかの場合、天然IFNγ構造を保持しないか、あるいは部分的にまたは完全に空白である(unfilled)IFNγ変異体を産生することが望ましい可能性もある。こうしたタンパク質に対して作成された抗体は、IFNγ上の覆い隠されたエピトープを認識するであろう。
【0088】
本発明はまた、異種ペプチドまたはタンパク質に融合された、IFNγポリペプチド、並びにその断片、変異体、および誘導体を含む、IFNγ融合ポリペプチドも提供する。異種ペプチドおよびタンパク質には、限定されるわけではないが:IFNγ融合ポリペプチドの検出および/または単離を可能にするエピトープ;膜貫通受容体タンパク質またはその部分、例えば細胞外ドメイン、または膜貫通ドメインおよび細胞内ドメイン;膜貫通受容体タンパク質に結合するリガンドまたはその部分;触媒として活性である酵素またはその部分;オリゴマー化を促進するタンパク質またはペプチド、例えばロイシンジッパードメイン;および安定性を増加させるタンパク質またはペプチド、例えば免疫グロブリン定常領域が含まれる。
【0089】
IFNγポリペプチドをそれ自体、あるいはその断片、変異体、または誘導体と融合させることも可能である。融合は、IFNγポリペプチドのアミノ末端またはカルボキシ末端いずれで行うことも可能であるし、そしてリンカー分子またはアダプター分子を伴わずに直接であることも、あるいはリンカー分子またはアダプター分子を介したものであることも可能である。また、リンカー分子またはアダプター分子を、DNA制限エンドヌクレアーゼまたはプロテアーゼの切断部位を含んで設計して、融合部分の分離を可能にすることも可能である。本発明のさらなる態様において、IFNγポリペプチド、断片、変異体および/または誘導体をヒトIgGのFc領域に融合させる。1つの例において、当業者に知られる方法を用いて、IFNγポリペプチドのN末端またはC末端いずれかに、ヒトIgGヒンジ、ch2およびch3領域を融合させることも可能である。別の例において、ヒンジ領域、並びにch2およびch3領域の部分を融合させることも可能である。タンパク質アフィニティーカラムを使用することによって、こうして産生したIFNγ・Fc融合ポリペプチドを精製することも可能である。さらに、Fc領域に融合させたペプチドおよびタンパク質は、融合させていない対応物よりも、実質的により長いin vivo半減期を示すことも見出されてきている。また、Fc領域への融合は、融合ポリペプチドの二量体化/多量体化も可能にする。Fc領域は、天然存在Fc領域であることも可能であるし、また療法特質や循環時間などの特定の特質を改善し、凝集を減少させるように改変することなども可能である。
【0090】
IFNγポリペプチド誘導体は、本発明の範囲内に含まれる。こうした誘導体は、IFNγポリペプチドがポリマーに連結された、化学的に修飾されたIFNγポリペプチド組成物である。選択されるポリマーは、付着するタンパク質が、生理学的環境などの水性環境において沈殿しないように、典型的には、水溶性である。ポリマーは、いかなる分子量のものであることも可能であるし、そして分枝していてもまたは分枝していなくてもよい。IFNγポリペプチドポリマーの範囲内に含まれるのは、ポリマー混合物である。好ましくは、最終産物調製物を療法的に使用するため、ポリマーは薬学的に許容しうるものであろう。
【0091】
水溶性ポリマーまたはその混合物は、例えば、ポリエチレングリコール(PEG)、モノメトキシ−ポリエチレングリコール、デキストラン(例えば約6kDの低分子量デキストランなど)、セルロース、または他の炭水化物に基づくポリマー、ポリ−(N−ビニルピロリドン)ポリエチレングリコール、プロピレングリコール・ホモポリマー、ポリプロピレンオキシド/エチレンオキシド・コポリマー、ポリオキシエチル化ポリオール類(例えばグリセロール)およびポリビニルアルコールであることも可能である。
【0092】
好ましい水溶性ポリマーはポリエチレングリコールである。本明細書において、ポリエチレングリコールは、他のタンパク質を誘導体化するのに用いられてきたPEGのいかなる型も、例えばモノ−(C1−C10)アルコキシ−、またはアリールオキシ−ポリエチレングリコールも含むよう意図される。本発明にやはり含まれるのは、共有結合IFNγ多量体を調製するのに使用可能な二官能性PEG架橋分子である。
【0093】
化学的に誘導体化されたIFNγポリペプチドを調製する方法が当該技術分野に知られる。例えば、Francisら, Focus on Growth Factors, 3:4−10(1992);EP 0 154 316;およびEP 0 401 384に記載される方法を用いて、PEGでのIFNγポリペプチドの誘導体化を行うことも可能である。好ましい態様において、IFNγポリペプチド誘導体は、アミノ末端に単一のPEG部分を有するであろう;本明細書に援用される米国特許第5,985,265号を参照されたい。
【0094】
本明細書に開示するIFNγポリペプチド誘導体は、非修飾ポリペプチドに比較して、IFNγの少なくとも1つの生物学的活性の増進または減少を示すことも可能であるし、あるいは増加したまたは減少した半減期または安定性を示すことも可能である。
【0095】
IFNγ選択的結合剤
IFNγポリペプチド、およびその断片、変異体および誘導体を用いて、IFNγの選択的結合剤を同定することも可能である。上に定義するように、IFNγの選択的結合剤は、タンパク質性結合剤および非タンパク質性結合剤を両方含み、そして本発明の1つの好ましい態様において、選択的結合剤はタンパク質性である。さらに別の好ましい態様において、選択的結合剤は、IFNγ、好ましくはヒトIFNγに結合する抗体またはその断片である。本発明の抗体は、IFNγの少なくとも1つの生物学的活性のレベルを増進するアゴニスト抗体;またはIFNγの少なくとも1つの生物学的活性のレベルを減少させるアンタゴニスト抗体であることも可能である。IFNγのアンタゴニスト抗体を、IFNγの阻害性抗体または中和抗体と呼ぶこともまた可能である。こうした抗体は本発明の好ましい態様であるが、IFNγ活性のアゴニストまたはアンタゴニストである他のタンパク質性選択的結合剤もまた、本発明に含まれることが理解される。
【0096】
以下の実施例に記載するように、IFNγの少なくとも1つの活性を阻害する抗IFNγ抗体および抗原結合ドメインが同定されてきている。本発明の態様には、図3〜13のいずれかに示すような重鎖Fab配列を含み、そしてカッパ軽鎖配列またはラムダ軽鎖配列をさらに含む抗体が含まれる。軽鎖Fab配列は、図14〜24に示すとおりであってもよい。例えば、「BS−A」抗体は、それぞれ、図14および図3の軽鎖配列および重鎖配列を有し;「BS−B」抗体は、それぞれ、図15および図4の軽鎖配列および重鎖配列を有し;「RD−B1」抗体は、それぞれ、図16および図5の軽鎖配列および重鎖配列を有し;「RD−A2」抗体は、それぞれ、図17および図6の軽鎖配列および重鎖配列を有し;「58C」抗体は、それぞれ、図18および図7の軽鎖配列および重鎖配列を有し;「GP−A」抗体は、それぞれ、図19および図8の軽鎖配列および重鎖配列を有し;「57D」抗体は、それぞれ、図20および図9の軽鎖配列および重鎖配列を有し;「57E」抗体は、それぞれ、図21および図10の軽鎖配列および重鎖配列を有し;「IFN−A」抗体は、それぞれ、図22および図11の軽鎖配列および重鎖配列を有し;「67C」抗体は、それぞれ、図23および図12の軽鎖配列および重鎖配列を有し;そして「59−A2」抗体は、それぞれ、図24および図13の軽鎖配列および重鎖配列を有する。本発明の抗体は、アイソタイプ、IgG、IgM、IgA、IgE、またはIgDいずれか由来のヒトFc領域をさらに含む。好ましくは、Fc領域は、ヒトIgG、例えばIgG1、IgG2、IgG3、またはIgG4由来である。
【0097】
本発明はまた、本明細書に開示するFab配列の断片、変異体、または誘導体を含む抗体または抗原結合ドメインも提供する。断片には、軽鎖または重鎖のFab配列いずれかの可変ドメインが含まれ、これらは、典型的には軽鎖または重鎖の定常ドメインに連結されている。変異体には、図14〜24のいずれか1つのFab配列に、少なくとも約80%、85%、90%、95%、98%または99%同一または類似である軽鎖Fab配列、または対応する可変ドメインを含む抗体、あるいは図3〜13のいずれか1つのFab配列に、少なくとも約80%、85%、90%、95%、98%または99%同一または類似である重鎖Fab配列、または対応する可変ドメインを含む抗体が含まれる。抗体は、典型的には、重鎖および軽鎖の定常領域と会合して、全長抗体を形成することも可能である。
【0098】
本発明の抗体および抗原結合ドメイン、並びにその断片、変異体および誘導体は、IFNγポリペプチド、好ましくはヒトIFNγポリペプチドに選択的に結合する能力を保持するであろう。1つの態様において、抗体は、約1nM以下、あるいは0.1nM以下、あるいは10pM以下、あるいは10pM未満の解離定数(KD)でIFNγポリペプチドに結合するであろう。
【0099】
本発明の抗体には、ポリクローナル抗体、単一特異性ポリクローナル抗体、モノクローナル抗体、組換え抗体、キメラ抗体、ヒト化抗体、完全ヒト抗体、一本鎖抗体および/または二重特異性抗体が含まれる。抗体断片には、IFNγポリペプチド上のエピトープに結合する抗IFNγ抗体の部分が含まれる。こうした断片の例には、Fab、F(ab’)、F(ab)’、Fv、およびsFv断片が含まれる。抗体は、全長抗体の酵素切断によって、または組換えDNA技術によって、例えば抗体可変領域をコードする核酸配列を含有する組換えプラスミドの発現などによって、生成可能である。
【0100】
ポリクローナル抗体は、抗原で免疫した動物の血清由来の抗体分子の異種集団である。抗原は、抗体に結合されることが可能であり、さらに、動物が、その抗原のエピトープに結合可能な抗体を産生するよう誘導可能である、分子または分子の部分である。抗原は、1以上のエピトープを有することも可能である。上述の特異的反応は、抗原が、非常に選択的な方式で、対応する抗体と反応し、そして他の抗原に誘発されうる多数の他の抗体とは反応しないであろうことを示すよう意味する。
【0101】
IFNγポリペプチドに対して向けられるポリクローナル抗体は、一般的に、IFNγおよびアジュバントの多数回の皮下注射または腹腔内注射によって、動物(例えばウサギまたはマウス)において作成される。本発明にしたがって、IFNγポリペプチド、あるいはその変異体、断片、または誘導体を、免疫しようとする種において免疫原性であるキャリアータンパク質、例えばキーホールリンペット(keyhole limpet)・ヘモシアニン、血清、アルブミン、ウシ・チログロブリン、またはダイズ(soybean)・トリプシン阻害剤にコンジュゲート化することが有用である可能性もある。また、ミョウバンなどの凝集剤を用いて、免疫応答を増進する。免疫後、動物から出血させ、そして抗IFNγ抗体力価に関して、血清をアッセイする。
【0102】
モノクローナル抗体(mAb)は、抗原に特異的な抗体の実質的に均質な集団を含有し、この集団は、実質的に類似のエピトープ結合部位を含有する。こうした抗体は、IgG、IgM、IgE、IgA、IgDおよびそのサブクラスいずれかを含む、免疫グロブリンクラスいずれのものであることも可能である。本発明のモノクローナル抗体を産生するハイブリドーマを、in vitro、in situまたはin vivoで培養することも可能である。in vivoまたはin situでの高力価の産生が好ましい産生法である。IFNγに対して向けられるモノクローナル抗体は、培養中の連続細胞株による抗体分子の産生を提供する方法いずれかを用いて産生される。モノクローナル抗体を調製するのに適した方法の例には、Kohlerら, Nature, 256:495−497(1975)のハイブリドーマ法、並びにヒトB細胞ハイブリドーマ法、Kozbor, J. Immunol,. 133:3001(1984);Brodeurら, Monoclonal Antibody Production Techniques and Applications, pp.51−63(Marcel Dekker, Inc. , ニューヨーク, 1987);並びにHarlowおよびLane, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory(1988)が含まれ;これら参考文献の内容は、本明細書に完全に援用される。
【0103】
好ましい抗IFNγ選択的結合剤には、ヒトIFNγのその同族受容体、hIFNγ−Rへの結合を、部分的にまたは完全に阻害するであろうモノクローナル抗体、あるいは実質的に同一の特異的結合特性を有する抗体とともに、その断片および領域が含まれる。競合的阻害によって、モノクローナル抗体特異性および親和性を決定するのに好ましい方法は、Harlowら, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー, 1988);Colliganら監修, Current Protocols in Immunology, Greene Publishing Assoc. and Wiley Interscience, ニューヨーク, (1992, 1993);およびMuller, Meth. Enzymol., 92:589−601(1983)に見出されることも可能である。これらの参考文献は、本明細書に援用される。本発明にやはり提供するのは、IFNγポリペプチドと反応するモノクローナル抗体を産生するハイブリドーマ細胞株である。
【0104】
キメラ抗体は、異なる部分が異なる動物種に由来する分子であり、例えばネズミ・モノクローナル抗体由来の可変領域およびヒト免疫グロブリン定常領域を有するものがある。キメラ抗体は、主に、適用中の免疫原性を減少させ、そして産生中の収率を増加させるように用いられ、例えば、ネズミ・モノクローナル抗体が、ハイブリドーマからはより高い収率で得られるが、ヒトにおいてより免疫原性である場合、ヒト/ネズミ・キメラモノクローナル抗体を用いる。
【0105】
キメラ抗体およびその産生法が当該技術分野に知られる。Cabillyら, Proc. Natl. Acad.Sci. USA, 81:3273−3277(1984);Morrisonら, Proc. Natl. Acad. Sci. USA, 81:6851−6855(1984);Boulianneら, Nature, 312:643−646(1984);Neubergerら, Nature, 314:268−270(1985);Liuら, Proc. Natl. Acad. Sci. USA, 84:3439−3443(1987);並びにHarlowおよびLane Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory(1988)。これらの参考文献は、本明細書に援用される。
【0106】
例えば、本発明のキメラモノクローナル抗体を療法剤として用いることも可能である。こうしたキメラ抗体では、望ましい生物学的活性を示す限り、重鎖および/または軽鎖の部分が、特定の種に由来するか、または1つの特定の抗体クラスもしくはサブクラスに属する抗体の対応する配列と同一であるかまたは相同であり、一方、鎖(単数または複数)の残りが、別の種に由来するか、または別の抗体クラスもしくはサブクラスに属する抗体とともにこうした抗体の断片の対応する配列と同一であるかまたは相同である;例えば、米国特許第4,816,567号およびMorrisonら, Proc. Natl. Acad. Sci., 81:6851−6855(1985)を参照されたい。
【0107】
本明細書において、用語「キメラ抗体」には、一価、二価または多価免疫グロブリンが含まれる。一価キメラ抗体は、キメラL鎖とのジスルフィド架橋を通じて会合するキメラH鎖によって形成される二量体(HL)である。二価キメラ抗体は、少なくとも1つのジスルフィド架橋を通じて会合する2つのHL二量体によって形成される四量体(H2L2)である。多価キメラ抗体もまた、例えば凝集するCH領域(例えばIgM H鎖、またはμ鎖由来のもの)を使用することによって産生可能である。
【0108】
本発明のネズミ抗体およびキメラ抗体、断片および領域は、個々の重鎖(H)および/または軽鎖(L)免疫グロブリン鎖を含むことも可能である。キメラH鎖は、IFNγに特異的な非ヒト抗体のH鎖由来の抗原結合領域を含み、これがヒトH鎖C領域(CH)の少なくとも部分、例えばCH1またはCH2に連結される。
【0109】
本発明記載のキメラL鎖は、ヒトL鎖C領域(CL)の少なくとも部分に連結された、IFNγに特異的な非ヒト抗体のL鎖由来の抗原結合領域を含む。
既知の方法工程にしたがって、個々のポリペプチド鎖を適切に会合させることによって、選択的結合剤、例えば同一のまたは異なる可変領域結合特異性を持つキメラH鎖およびL鎖を有する抗体、断片、または誘導体もまた調製可能である;例えば、Ausubelら監修 Current Protocols in Molecular Biology, Wiley Interscience, ニューヨーク(1993)およびHarlowら, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1988)を参照されたい。これらの参考文献の内容は、本明細書に完全に援用される。このアプローチを用いて、キメラH鎖(またはその誘導体)を発現する宿主を、キメラL鎖(またはその誘導体)を発現する宿主と別個に培養し、そして免疫グロブリン鎖を別個に回収して、そして次いで会合させる。あるいは、宿主を同時培養して、そして鎖を培地中で自発的に会合可能にし、その後、組み立てられた免疫グロブリン、断片または誘導体を回収することも可能である。
【0110】
例として、本発明の選択的結合剤(例えばキメラ抗体)の抗原結合領域は、好ましくは、ヒトIFNγに特異的な非ヒト抗体由来である。こうした非ヒト抗体をコードするDNAの好ましい供給源には、抗体を産生する細胞株、例えば一般的にハイブリドーマとして知られるハイブリッド細胞株が含まれる。
【0111】
本発明はまた、抗IFNγ抗体の断片、変異体および誘導体も提供し、ここで用語「断片」、「変異体」、「誘導体」および「融合体」は、本明細書に定義される。本発明は、非修飾抗IFNγ抗体に機能的に類似である、すなわち非修飾抗体の活性を少なくとも1つ保持する、抗IFNγ抗体の断片、変異体、誘導体、および融合体を含む。上述の修飾に加えて、植物毒素および細菌毒素などの細胞毒性タンパク質をコードする遺伝子配列の付加も含まれる。抗IFNγ抗体の断片、変異体、誘導体および融合体を、本発明の宿主のいずれから産生することも可能である。
【0112】
適切な断片には、例えば、Fab、Fab’、F(ab’)2、FvおよびscFvが含まれる。これらの断片は、損なわれていない(intact)抗体のFc断片を欠き、循環からより迅速に一掃され、そして損なわれていない抗体よりも非特異性組織結合がより少なくなりうる;Wahlら, J. Nucl. Med., 24:316−325(1983)。これらの断片は、当該技術分野に周知の方法を用いて、例えばパパイン(Fab断片を産生する)またはペプシン(F(ab’)2断片を産生する)などの酵素でのタンパク質分解切断によって、損なわれていない抗体から産生される。これらの抗原結合領域および/または本発明のモノクローナル抗体に認識されるエピトープを同定すると、本出願の態様と同等である、類似の結合特性および療法的有用性または診断有用性を持つさらなるモノクローナル抗体を生成するのに必要な情報が提供される。
【0113】
選択的結合剤の変異体もまた提供する。1つの態様において、抗体および抗原結合ドメインの変異体は、軽鎖および/または重鎖アミノ酸配列中に変化を含み、これらの変化は天然存在であるか、または組換えDNA技術を用いた天然配列のin vitro操作によって導入されている。天然存在変異体には、外来(foreign)抗原に対する抗体応答の生成中、対応する生殖系列ヌクレオチド配列において、in vivoで生成される「体細胞」変異体が含まれる。配列において、本発明の典型的なFabを生成する、生殖系列可変軽鎖および重鎖配列中の体細胞突然変異にコードされる変異体は、Fab「BS−A」に関しては図33および図41に、Fab「BS−B」に関しては図34および図41に、Fab「RD−B1」に関しては図35および図42に、Fab「RD−B1」に関しては図35および図42に、Fab「RD−A2」に関しては図33および図43に、Fab「58C」に関しては図36および図44に、Fab「GP−A」に関しては図37および図45に、Fab「57D」に関しては図38および図46に、Fab「57E」に関しては図39および図41に、Fab「IFN−A」に関しては図33および図43に、Fab「67C」に関しては図40および図43に、そしてFab「59−A2」に関しては図34および図41に示される。
【0114】
抗IFNγ抗体および抗原結合ドメインの変異体はまた、当該技術分野に知られる突然変異誘発技術によっても調製可能である。1つの例において、抗体コード領域全体で、無作為にアミノ酸変化を導入することも可能であり、そして望ましい活性、例えばIFNγに対する結合親和性に関して、生じた変異体をスクリーニングすることも可能である。あるいは、IFNγ抗体の選択される領域に、例えば軽鎖および/または重鎖CDR、およびフレームワーク領域に、アミノ酸変化を導入することも可能であり、そしてIFNγへの結合または他の何らかの活性に関して、生じた抗体をスクリーニングすることも可能である。アミノ酸変化は、単一アミノ酸相違から、既定のCDR、例えばCDR3内のアミノ酸のありうるすべての順列(permutation)の範囲で、CDR中に1以上のアミノ酸置換を含む。別の方法において、CDR内の少なくとも1つの残基をアラニンで置換することによって、CDR内の各残基のIFNγ結合への寄与を評価することも可能である;Lewisら, Mol. Immunol., 32:1065−1072(1995)。次いで、より最適な配列を決定するため、IFNγへの結合に最適でない残基を変化させることも可能である。アミノ酸を挿入して、CDR、例えばCDR3のサイズを増加させることによって生成される変異体もまた、含まれる。例えば、大部分の軽鎖CDR3配列は、長さ9アミノ酸である。9残基より短い、抗体中の軽鎖CDR3配列は、適切なアミノ酸を挿入して、CDRの長さを増加させることによって、IFNγへの結合に関して最適化されることも可能である。
【0115】
1つの態様において、抗体または抗原結合ドメイン変異体は、1以上の重鎖または軽鎖CDR1、CDR2またはCDR3に、そして所望によって1以上の重鎖または軽鎖フレームワーク領域FR1、FR2またはFR3に、1以上のアミノ酸変化を含む。アミノ酸変化は、アミノ酸残基の置換、欠失および/または挿入を含む。典型的な変異体には、配列GYYWS(配列番号34);EINHSGSTNYNPSLKS(配列番号44);またはGRARNWRSRFDY(配列番号54)に1以上のアミノ酸変化を持つ「BS−A」重鎖可変領域変異体、あるいは配列TGSSGSIASHYVQ(配列番号01);EDKERPS(配列番号12);またはQSYDSSNQWV(配列番号23)に1以上のアミノ酸変化を持つ「BS−A」軽鎖可変領域変異体が含まれる。前述の「BS−A」重鎖および軽鎖可変領域変異体は、フレームワーク領域に1以上のアミノ酸変化をさらに含むことも可能である。
【0116】
1つの例において、1以上のアミノ酸変化を導入して、体細胞突然変異フレームワーク残基を、その位の生殖系列残基で置換することも可能である。前述のアミノ酸変化が置換である場合、変化は保存的置換でもまた非保存的置換でもよい。変異体を、軽鎖または重鎖いずれかの「鎖シャフリング」によって調製することもまた可能である;Marksら Biotechnology, 10:779−783(1992)。典型的には、単一の軽鎖(または重鎖)を、重鎖(または軽鎖)のレパートリーを有するライブラリーと組み合わせて、そして生じた集団を、IFNγへの結合などの望ましい活性に関してスクリーニングする。この技術は、重鎖および軽鎖両方のレパートリーを含むライブラリーで可能であるよりも、単一の軽鎖(または重鎖)と組み合わされた異なる重鎖(または軽鎖)のより多数の試料をスクリーニングすることを可能にする。
【0117】
本発明の選択的結合剤は、二重特異性であることも可能である。本発明の二重特異性選択的結合剤は、いくつかの立体配置のものであることも可能である。例えば、二重特異性抗体は、一本鎖抗体(または抗体断片)に似ているが、2つの異なる抗原結合部位(可変領域)を有する。二重特異性抗体は、化学的技術によって;例えば、Kranzら, Proc. Natl. Acad. Sci. USA, 78:5807(1981)を参照されたい;「ポリドーマ(polydoma)」技術によって;米国特許第4,474,893号;または組換えDNA技術によって、産生可能である。
【0118】
本発明の選択的結合剤はまた、異種抗体であることも可能である。異種抗体は、ともに連結された、各々異なる特異性を有する、2以上の抗体、または抗体結合断片(Fab)である。
【0119】
本発明はまた、「ヒト化」抗体にも関する。非ヒト抗体をヒト化する方法は、当該技術分野に周知である。一般的に、ヒト化抗体は、非ヒトである供給源からヒト抗体に導入された1以上のアミノ酸残基を有する。一般的に、非ヒト残基は、CDRに存在するであろう。ヒト化は、当該技術分野に知られる方法にしたがって、げっ歯類相補性決定領域(CDR)をヒト抗体の対応する領域に対して置換することによって、実行可能である;Jonesら, Nature 321:522−525(1986);Riechmannら, Nature, 332:323−327(1988);Verhoeyenら, Science, 239:1534−1536(1988)。
【0120】
キメラ抗体、CDR移植抗体、およびヒト化抗体を含む、本発明の選択的結合剤は、当該技術分野に知られる組換え法によっても産生可能である。抗体をコードする核酸を宿主細胞に導入し、そして本明細書に記載し、そして当該技術分野に知られる、材料および方法を用いて発現させる。好ましい態様において、CHO細胞などの哺乳動物宿主細胞において、抗体を産生する。宿主細胞にトランスフェクションした組換えDNAの発現によって、または上述のようなハイブリドーマ細胞における発現によって、完全ヒト抗体を産生することも可能である。
【0121】
モノクローナル抗体の生成を回避して、抗体分子の抗原結合領域の組換えDNA型を生成する技術が本発明の実施内に含まれる。これを行うため、免疫動物から採取した免疫系細胞から、抗体特異的メッセンジャーRNA分子を抽出し、そして相補DNA(cDNA)に転写する。次いで、cDNAを細菌発現系にクローニングする。本発明を実施するのに適した技術の一例は、発現されたFabタンパク質を細胞膜周辺腔(細菌細胞膜および細胞壁の間)に移動させるか、または分泌させる、リーダー配列を有する、繊維状バクテリオファージM13由来ファージミド・ベクター系を用いる。抗原に結合するものに関して、非常に多数の機能するFab断片を迅速に生成し、そしてスクリーニングすることも可能である。こうしたIFNγ選択的結合剤(IFNγポリペプチドに対して特異性を持つFab断片)は、本明細書に定義され、議論され、そして請求される際、用語「抗体」に明確に含まれる。
【0122】
やはり本発明の範囲内にあるのは、適切な抗原特異性を持つマウス抗体分子由来の遺伝子を、適切な生物学的活性、例えばヒト補体を活性化し、そしてADCCを仲介する能力を持つヒト抗体分子由来の遺伝子とともにスプライシングすることによって、キメラ抗体を産生するために開発された技術である;Morrisonら, Proc. Natl. Acad. Sci., 81:6851(1984);Neubergerら, Nature, 312:604(1984)。1つの例は、Fc領域の、異なるアイソタイプのFc領域との交換である。この技術によって産生される抗体などの選択的結合剤は、本発明の範囲内である。
【0123】
本発明の好ましい態様において、抗IFNγ抗体は、完全ヒト抗体である。したがって、IFNγポリペプチドに結合し、そしてヒト生殖系列免疫グロブリン核酸配列、並びにその断片、合成変異体、誘導体および融合体の天然存在体細胞変異体である核酸配列にコードされる抗体が本発明に含まれる。こうした抗体は、当該技術分野に知られる方法いずれによって産生してもよい。典型的な方法には、内因性免疫グロブリン産生を伴わずに、ヒト抗体レパートリーを産生することが可能なトランスジェニック動物(例えばマウス)を、IFNγ抗原(免疫応答を誘発することが可能であり、そして所望によってキャリアーにコンジュゲート化されているIFNγポリペプチドいずれか)で免疫することが含まれる;例えばJakobovitsら, Proc. Natl. Acad. Sci., 90:2551−2555(1993);Jakobovitsら, Nature, 362:255−258(1993);Bruggermannら, Year in Immunol., 7:33(1993)を参照されたい。
【0124】
あるいは、ファージ・ディスプレー抗体ライブラリーのin vitroスクリーニングを通じて、ヒト抗体を生成することも可能である;例えば、本明細書に援用される、Hoogenboomら, J. Mol. Biol., 227:381(1991);Marksら, J. Mol. Biol., 222:581(1991)を参照されたい。多様な抗体含有ファージ・ディスプレー・ライブラリーが記載されており、そして当業者によって容易に調製可能である。ライブラリーは、適切な標的に対してスクリーニング可能な、多様なヒト抗体配列、例えばヒトFab、Fv、およびscFv断片を含有することも可能である。実施例2は、IFNγに選択的に結合する分子を同定するための、IFNγに対するFabファージ・ライブラリーのスクリーニングを記載する。ファージ・ディスプレー・ライブラリーが、IFNγの選択的結合剤を同定するためにスクリーニングすることが可能な抗体以外のペプチドまたはタンパク質を含んでもよいことが認識されるであろう。
【0125】
抗イディオタイプ(抗Id)抗体は、一般的に抗体の抗原結合部位と会合するユニークな決定基を認識する抗体である。Id抗体は、抗Idを調製しようとするモノクローナル抗体で、該モノクローナル抗体の供給源と同一種および同一遺伝子型(例えばマウス系統)の動物を免疫することによって、調製可能である。免疫された動物は、これらのイディオタイプ決定基に対する抗体(抗Id抗体)を産生することによって、免疫抗体のイディオタイプ決定基を認識し、そして応答するであろう;例えば本明細書に完全に援用される米国特許第4,699,880号を参照されたい。抗Id抗体はまた、さらに別の動物において免疫応答を誘導し、いわゆる抗−抗Id抗体を産生する、「免疫原」としても使用可能である。抗−抗Idは、抗Idを誘導した元来のモノクローナル抗体と、エピトープが同一であることも可能である。したがって、mAbのイディオタイプ決定基に対する抗体を用いることによって、同一の特異性の抗体を発現する他のクローンを同定することが可能である。
【0126】
IFNγの選択的結合剤の産生
調製しようとするIFNγの選択的結合剤が、タンパク質性選択的結合剤、例えば抗体または抗原結合ドメインである場合、前記剤を産生するため、多様な生物学的方法または化学的方法が利用可能である。
【0127】
療法使用のために十分な量の選択的結合剤を産生するには、生物学的方法が好ましい。本発明の抗体および抗原結合ドメインの産生には、標準的組換えDNA技術が特に有用である。発現産物の回収のための典型的な発現ベクター、宿主細胞および方法を以下に記載する。
【0128】
標準的連結技術を用いて、IFNγ抗体または抗原結合ドメインをコードする核酸分子を適切な発現ベクターに挿入する。ベクターは、典型的には、使用する特定の宿主細胞で機能するように選択される(すなわち、ベクターは、遺伝子の増幅および/または遺伝子の発現が起こりうるように、宿主細胞機構と適合する)。抗IFNγ抗体をコードする核酸分子を、原核宿主細胞、酵母宿主細胞、昆虫宿主細胞(バキュロウイルス系)および/または真核宿主細胞で増幅/発現することも可能である。宿主細胞の選択は、部分的に、抗IFNγ抗体を翻訳後に(post−transitionally)修飾(例えばグリコシル化および/またはリン酸化)しようとしているかどうかに応じるであろう。修飾するのであれば、酵母宿主細胞、昆虫宿主細胞、または哺乳動物宿主細胞が好ましい。発現ベクターの概説に関しては、Meth. Enz. V. 185, D.V. Goeddel監修 Academic Press Inc., カリフォルニア州サンディエゴ(1990)を参照されたい。
【0129】
典型的には、いかなる宿主細胞で用いられる発現ベクターも、以下の構成要素の1以上を含有するであろう:プロモーター、1以上のエンハンサー配列、複製起点、転写終結配列、ドナーおよびアクセプターのスプライス部位を含有する完全イントロン配列、分泌のためのリーダー配列、リボソーム結合部位、ポリアデニル化配列、発現しようとするポリペプチドをコードする核酸を挿入するためのポリリンカー領域、および選択可能マーカー要素。これらの配列は各々、以下により詳細に論じられる。
【0130】
ベクター構成要素は、相同配列(すなわち宿主細胞と同一種および/または同一系統由来)、異種配列(すなわち宿主細胞種以外の種または系統由来)、ハイブリッド(すなわち1より多い供給源由来の異なる配列の組み合わせ)、合成配列、または通常、免疫グロブリン発現を制御するように機能する天然配列であることも可能である。こうしたものとして、ベクター構成要素の供給源は、宿主細胞機構において機能し、そして該機構によって活性化可能である限り、いかなる原核生物または真核生物、いかなる脊椎動物または無脊椎動物、またはいかなる植物であることも可能である。
【0131】
複製起点は、発現に用いられる宿主細胞の種類に基づいて選択される。例えば、プラスミドpBR322(製品番号303−3s、New England Biolabs、マサチューセッツ州ビバリー)由来の複製起点は、大部分のグラム陰性細菌に適しており、一方、SV40、ポリオーマ、アデノウイルス、水疱性口内炎ウイルス(VSV)またはパピローマウイルス(HPVまたはBPVなど)由来の多様な起点は、哺乳動物細胞におけるクローニングベクターに有用である。一般的に、複製起点構成要素は哺乳動物発現ベクターには必要ない(例えば、SV40起点がしばしば用いられるが、これはただ初期プロモーターを含有するからである)。
【0132】
転写終結配列は、典型的にはポリペプチドコード領域の3’端に位置し、そして転写を終結させるように働く。通常、原核細胞中の転写終結配列には、G−Cリッチ断片に続いてポリT配列がある。配列を、ライブラリーから容易にクローニングするか、またはベクターの一部として商業的に購入してもよいが、上述のものなどの核酸合成法を用いて、容易に合成することも可能である。
【0133】
選択可能マーカー遺伝子要素は、選択培地中で増殖する宿主細胞の生存および増殖に必要なタンパク質をコードする。典型的な選択マーカー遺伝子は(a)抗生物質または他の毒素、例えば原核宿主細胞ではアンピシリン、テトラサイクリン、またはカナマイシンに対する耐性を与えるタンパク質、(b)細胞の栄養要求不全を補足するタンパク質;または(c)複合培地から入手可能でない非常に重要な栄養素を供給するタンパク質をコードする。好ましい選択可能マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、およびテトラサイクリン耐性遺伝子である。原核宿主細胞および真核宿主細胞において、ネオマイシン耐性遺伝子もまた使用可能である。
【0134】
他の選択遺伝子を用いて、発現しようとする遺伝子を増幅することも可能である。増幅は、増殖に決定的に重要なタンパク質の産生に対する要求がより高い遺伝子が、組換え細胞の代々の世代の染色体内で、タンデムに反復されるプロセスである。哺乳動物細胞に適した選択可能マーカーの例には、ジヒドロ葉酸レダクターゼ(DHFR)およびチミジンキナーゼが含まれる。哺乳動物細胞形質転換体を、ベクターに存在するマーカーのため、形質転換体のみがユニークに適応して生存する選択圧下に置く。培地中の選択剤の濃度を連続的に変化させて、それによって選択遺伝子および抗IFNγ抗体をコードするDNA両方の増幅が導かれる条件下で、形質転換細胞を培養することによって、選択圧を課す。その結果、増幅されたDNAから、増加した量の抗体が合成される。
【0135】
リボソーム結合部位は、通常、mRNAの翻訳開始に必要であり、そしてシャイン・ダルガノ配列(原核生物)またはコザック配列(真核生物)によって特徴付けられる。これら要素は、典型的には、発現しようとするポリペプチドのプロモーターの3’で、そしてコード配列の5’に位置する。シャイン・ダルガノ配列は多様であるが、典型的にはポリプリンである(すなわち高いA−G含量を有する)。多くのシャイン・ダルガノ配列が同定されてきており、各々、上述の方法を用いて、容易に合成可能であり、そして原核ベクターで使用可能である。
【0136】
リーダー配列またはシグナル配列は、ポリペプチドの分泌を指示するのに用いられる。シグナル配列はポリペプチドコード領域内にまたはその5’端に直接配置されることも可能である。多くのシグナル配列が同定されてきており、そして発現に用いる宿主細胞に基づいて選択可能である。本発明において、シグナル配列は、抗IFNγ抗体または抗原結合ドメインをコードする核酸配列に相同である(天然存在である)か、またはこれら配列に対して異種であることも可能である。選択される異種シグナル配列は、シグナルペプチダーゼにより、宿主細胞によって、認識され、そしてプロセシングされる、すなわち切断されるものであるべきである。天然免疫グロブリンシグナル配列を認識せず、そしてプロセシングしない原核宿主細胞では、シグナル配列を、例えばアルカリホスファターゼ、ペニシリナーゼ、または熱安定性内毒素IIのリーダーの群から選択される原核シグナル配列によって置換する。酵母分泌のため、天然免疫グロブリンシグナル配列を、酵母インベルターゼ、アルファ因子、または酸ホスファターゼのリーダーによって置換することも可能である。哺乳動物細胞発現では、天然シグナル配列で十分だが、他の哺乳動物シグナル配列が適切である可能性もある。
【0137】
大部分の場合、宿主細胞から抗IFNγ抗体または抗原結合ドメインが分泌される結果、抗体からシグナルペプチドが除去されるであろう。したがって、成熟抗体は、いかなるリーダー配列またはシグナル配列も欠くであろう。
【0138】
真核宿主細胞発現系におけるグリコシル化が望ましいなどのいくつかの場合、グリコシル化または収率を改善するため、多様なプレ配列を操作することも可能である。例えば、特定のシグナルペプチドのペプチダーゼ切断部位を改変することも可能であるし、またはグリコシル化に影響を及ぼすことも可能なプロ配列を付加することも可能である。最終タンパク質産物は、−1位(成熟タンパク質の最初のアミノ酸に比較して)に、完全には除去されなかった可能性もある、発現によって生じる1以上のさらなるアミノ酸を有することも可能である。例えば、最終タンパク質産物は、N末端に付着して、ペプチダーゼ切断部位に見られる1つまたは2つのアミノ酸を有することも可能である。あるいは、いくつかの酵素切断部位を用いることによって、望ましいIFNγポリペプチドのわずかに一部切除された型を生じることも可能であり、これは酵素が成熟ポリペプチド内のこうした領域を切断する場合である。
【0139】
本発明の発現ベクターは、典型的には、宿主生物に認識され、そして抗IFNγ抗体または抗原結合ドメインをコードする核酸分子に機能可能であるように連結されているプロモーターを含有するであろう。発現に用いる宿主細胞および望ましいタンパク質収率に応じて、天然プロモーターまたは異種プロモーターを使用することも可能である。
【0140】
原核宿主で用いるのに適したプロモーターには、ベータ−ラクタマーゼおよびラクトースプロモーター系;アルカリホスファターゼ、トリプトファン(trp)プロモーター系;およびtacプロモーターなどのハイブリッドプロモーターが含まれる。他の既知の細菌プロモーターもまた、適している。これらの配列は公表されており、それによって、当業者が、必要に応じて、必要な制限部位いずれかを供給するリンカーまたはアダプターを用いて、これらプロモーターを望ましいDNA配列(単数または複数)に連結することが可能になる。
【0141】
酵母宿主で用いるのに適したプロモーターもまた、当該技術分野に周知である。酵母エンハンサーは、酵母プロモーターとともに、好適に用いられる。哺乳動物宿主細胞で用いるのに適したプロモーターが周知であり、そしてポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシ・パピローマウイルス、鳥・肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および最も好ましくはサル・ウイルス40(SV40)などのウイルスのゲノムから得られるものが含まれる。他の適切な哺乳動物プロモーターには、異種哺乳動物プロモーター、例えば熱ショックプロモーターおよびアクチン・プロモーターが含まれる。
【0142】
本発明の選択的結合剤を発現するのに使用可能なさらなるプロモーターには、限定されるわけではないが:SV40初期プロモーター領域;BernoistおよびChambon, Nature, 290:304−310(1981)、CMVプロモーター、ラウス肉腫ウイルスの3’末端反復配列(long terminal repeat)に含有されるプロモーター;Yamamotoら, Cell, 22:787−797(1980)、ヘルペス・チミジンキナーゼ・プロモーター;Wagnerら, Proc. Natl. Acad. Sci. U.S.A., 78:144−1445(1981)、メタロチオネイン遺伝子の制御配列;Brinsterら, Nature, 296:39−42(1982)、ベータ−ラクタマーゼ・プロモーターなどの原核生物発現ベクター;Villa−Kamaroffら, Proc. Natl. Acad. Sci. U.S.A., 75:3727−3731(1978)、またはtacプロモーター;DeBoerら, Proc. Natl. Acad. Sci. U.S.A., 80:21−25(1983)が含まれる。
【0143】
やはり重要なのは、組織特異性を有し、そしてトランスジェニック動物において利用されてきている、以下の動物転写調節領域である:膵臓腺房細胞で活性であるエラスターゼI遺伝子調節領域;Swiftら, Cell, 38:639−646(1984);Ornitzら, Cold Spring Harbor Symp. Quant. Biol. 50:399−409(1986);MacDonald, Hepatology, 7:425−515(1987)、膵臓ベータ細胞で活性であるインスリン遺伝子調節領域;Hanahan, Nature, 315:115−122(1985)、リンパ球で活性である免疫グロブリン遺伝子調節領域;Grosschedlら, Cell, 38:647−658(1984);Adamesら, Nature, 318:533−538(1985);Alexanderら, Mol. Cell. Biol., 7:1436−1444(1987)、精巣細胞、乳房細胞、リンパ球およびマスト細胞で活性であるマウス乳腺腫瘍ウイルス調節領域;Lederら, Cell, 45:485−495(1986)、肝臓で活性であるアルブミン遺伝子調節領域;Pinkertら, Genes and Devel., 1:268−276(1987)、肝臓で活性であるアルファ・フェトプロテイン遺伝子調節領域;Krumlaufら, Mol. Cell. Biol., 5:1639−1648(1985);Hammerら, Science, 235:53−58(1987)、肝臓で活性であるアルファ1−アンチトリプシン遺伝子調節領域;Kelseyら, Genes and Devel., 1:161−171(1987);骨髄細胞で活性であるベータ−グロビン遺伝子調節領域;Mogramら, Nature, 315:338−340(1985);Kolliasら, Cell, 46:89−94(1986)、脳のオリゴデンドロサイトで活性であるミエリン塩基性タンパク質遺伝子調節領域;Readheadら, Cell, 48:703−712(1987)、骨格筋で活性であるミオシン軽鎖−2遺伝子調節領域;Sani, Nature, 314:283−286(1985)、および視床下部で活性である性腺刺激ホルモン放出ホルモン遺伝子調節領域;Masonら, Science, 234:1372−1378(1986)。
【0144】
エンハンサー配列をベクターに挿入して、真核宿主細胞において転写を増加させることも可能である。哺乳動物遺伝子で利用可能ないくつかのエンハンサー配列が知られる(例えばグロビン、エラスターゼ、アルブミン、アルファ−フェト−プロテインおよびインスリン)。しかし、典型的には、ウイルス由来のエンハンサーが用いられるであろう。SV40エンハンサー、サイトメガロウイルス初期プロモーター・エンハンサー、ポリオーマ・エンハンサー、およびアデノウイルス・エンハンサーは、真核プロモーター活性化のための典型的な増進要素である。エンハンサーは、ポリペプチドコード領域の5’または3’の位で、ベクターにスプライシングすることも可能であるが、典型的には、プロモーターの5’の部位に位置する。
【0145】
本発明を実施するための好ましいベクターは、細菌細胞、昆虫細胞、および哺乳動物宿主細胞と適合するものである。こうしたベクターには、とりわけ、pCRII、pCR3、およびpcDNA3.1(Invitrogen社、カリフォルニア州サンディエゴ)、pBSII(Stratagene社、カリフォルニア州ラホヤ)、pET15(Novagen、ウィスコンシン州マディソン)、pGEX(Pharmacia Biotech、ニュージャージー州ピスカタウェイ)、pEGFP−N2(Clontech、カリフォルニア州パロアルト)、pETL(BlueBacII;Invitrogen、カリフォルニア州カールスバッド)、pDSR−アルファ(PCR公報第WO90/14363)およびpFastBacDual(Gibco/BRL、ニューヨーク州グランドアイランド)が含まれる。
【0146】
さらなる可能なベクターには、限定されるわけではないが、コスミド、プラスミドまたは修飾ウイルスが含まれるが、ベクター系は、選択した宿主細胞に適合しなければならない。こうしたベクターには、限定されるわけではないが、Bluescript(登録商標)プラスミド誘導体(ColE1に基づく高コピー数ファージミド、Stratagene Cloning Systems Inc.、カリフォルニア州ラホヤ)、Taq増幅PCR産物をクローニングするために設計されたPCRクローニングプラスミド(例えばTOPOTM TAクローニング(登録商標)キット、PCR2.1(登録商標)プラスミド誘導体、Invitrogen、カリフォルニア州カールスバッド)、および哺乳動物ベクター、酵母ベクターまたはウイルスベクター、例えばバキュロウイルス発現系(pBacPAKプラスミド誘導体、Clontech、カリフォルニア州パロアルト)が含まれる。組換え分子は、形質転換、トランスフェクション、感染、エレクトロポレーション、または他の既知の技術を介して、宿主細胞に導入可能である。
【0147】
本発明の宿主細胞は、原核宿主細胞(例えば大腸菌(E. coli))または真核宿主細胞(例えば酵母細胞、昆虫細胞、または脊椎動物細胞)であることも可能である。宿主細胞は、適切な条件下で培養すると、本発明の抗体または抗原結合ドメインを発現し、これを続いて培地から収集する(宿主細胞がこれらを培地に分泌する場合)か、またはこれらを産生する宿主細胞から直接収集する(これらが分泌されない場合)ことも可能である。適切な宿主細胞の選択は、望ましい発現レベル、活性に望ましいかまたは必要なポリペプチド修飾、例えばグリコシル化またはリン酸化、および生物学的活性分子にフォールディングするのが容易であることなどの多様な要因に応じるであろう。
【0148】
いくつかの適切な宿主細胞が当該技術分野に知られ、そして多くがアメリカン・タイプ・カルチャー・コレクション(ATCC)、バージニア州マナサスから入手可能である。例には、チャイニーズハムスター卵巣細胞(CHO)(ATCC番号CCL61)、CHO DHFR細胞;Urlaubら, Proc. Natl. Acad. Sci. USA, 97:4216−4220(1980)、ヒト胎児由来腎臓(HEK)293細胞または293T細胞(ATCC番号CRL1573)、あるいは3T3細胞(ATCC番号CCL92)などの哺乳動物細胞が含まれる。形質転換、培養、増幅、スクリーニング、並びに産物産生および産物精製に適した哺乳動物宿主細胞および方法の選択は当該技術分野に知られる。他の適切な哺乳動物細胞株は、サルCOS−1(ATCC番号CRL1650)およびCOS−7細胞株(ATCC第CRL1651)、およびCV−1細胞株(ATCC番号CCL70)である。さらなる典型的な哺乳動物宿主細胞には、霊長類細胞株およびげっ歯類細胞株が含まれ、形質転換細胞株が含まれる。正常な二倍体細胞、初代組織のin vitro培養から得られる細胞株とともに、初代外植片もまた適切である。候補細胞は、選択遺伝子の遺伝子型が不完全であることも可能であるし、また優性に作用する選択遺伝子を含有することも可能である。他の適切な哺乳動物細胞株には、限定されるわけではないが、マウス神経芽細胞腫N2A細胞、HeLa細胞、マウスL−929細胞、Swissマウス、Balb−cマウスまたはNIHマウス由来の3T3細胞株、BHKまたはHaKハムスター細胞株が含まれ、これらはアメリカン・タイプ・カルチャー・コレクション(バージニア州マナサス)から入手可能である。これら細胞株は各々、タンパク質発現の技術分野の当業者に知られ、そして入手可能である。
【0149】
本発明に適した宿主細胞として、同様に有用であるのは、細菌細胞である。例えば、大腸菌の多様な株(例えばHB101(ATCC番号33694)、DH5α、DH10、およびMC1061(ATCC番号53338))は、バイオテクノロジーの分野で、宿主細胞として周知である。枯草菌(B. subtilis)、シュードモナス属(Pseudomonas)種、他のバチルス属(Bacillus)種、ストレプトミセス属(Streptomyces)種等の多様な株もまた、この方法で使用可能である。
【0150】
当業者に知られる酵母細胞の多くの株もまた、本発明のポリペプチドを発現するための宿主細胞として、入手可能である。好ましい酵母細胞には、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisae)が含まれる。
【0151】
さらに、望ましい場合、本発明の方法には、昆虫細胞系が利用可能である。こうした系は、例えば、Kittsら, Biotechniques, 14:810−817(1993)、Lucklow, Curr. Opin. Biotechnol., 4:564−572(1993)およびLucklowら, J. Virol., 67:4566−4579(1993)に記載される。好ましい昆虫細胞は、Sf−9およびHi5(Invitrogen、カリフォルニア州カールスバッド)である。
【0152】
抗IFNγ抗体または抗原結合ドメインをコードする核酸分子の、選択した宿主細胞への形質転換またはトランスフェクションは、塩化カルシウム法、エレクトロポレーション法、マイクロインジェクション法、リポフェクション法またはDEAEデキストラン法などの方法を含む、周知の方法によって達成可能である。選択される方法は、部分的に、用いる宿主細胞の種類と相関関係にあるであろう。これらの方法および他の適切な方法が当業者に周知であり、そして例えばSambrookら、上記に示される。
【0153】
トランスジェニック動物を使用して、グリコシル化選択的結合剤、例えば抗体および抗原結合ドメインを発現することも可能である。例えば、ミルクを産生するトランスジェニック動物(例えばウシまたはヤギ)を用いて、そして動物のミルク中にグリコシル化された結合剤を得ることも可能である。あるいは、植物を用いて、グリコシル化された選択的結合剤を産生することも可能である。
【0154】
当業者に周知の標準的培地を用いて、IFNγの選択的結合剤をコードする発現ベクターを含む(すなわち形質転換されたかまたはトランスフェクションされた)宿主細胞を培養してもよい。培地は、通常、細胞の増殖および生存に必要なすべての栄養素を含有するであろう。大腸菌細胞を培養するのに適した培地は、例えば、ルリア・ブロス(LB)および/またはテリフィック・ブロス(TB)である。真核細胞を培養するのに適した培地は、RPMI1640、MEM、DMEMであり、これらすべてには、培養中の特定の細胞株に必要とされるような血清および/または増殖因子を補うことも可能である。昆虫培養に適した培地は、必要に応じて、イーストレート、ラクトアルブミン加水分解物、および/またはウシ胎児血清を補ったグレース培地である。
【0155】
典型的には、トランスフェクション細胞および形質転換細胞の選択的増殖に有用な抗生物質または他の化合物が、培地の補充剤として添加される。使用すべき化合物は、宿主細胞を形質転換するプラスミド上に存在する選択可能マーカー要素によって、決定されるであろう。例えば、選択可能マーカー要素が、カナマイシン耐性である場合、培地に添加される化合物はカナマイシンであろう。選択的増殖のための他の化合物には、アンピシリン、テトラサイクリンおよびネオマイシンが含まれる。
【0156】
当該技術分野に知られる標準法を用いて、宿主細胞に産生される抗IFNγ抗体または抗原結合ドメインの量を評価することも可能である。こうした方法には、限定なしに、ウェスタンブロット解析、SDS−ポリアクリルアミドゲル電気泳動、非変性ゲル電気泳動、HPLC分離、免疫沈降、および/または活性アッセイが含まれる。
【0157】
細胞培地に分泌されている抗IFNγ抗体または抗原結合ドメインの精製は、アフィニティークロマトグラフィー、免疫親和性クロマトグラフィーまたはイオン交換クロマトグラフィー、分子ふるいクロマトグラフィー、分取用ゲル電気泳動または等電点電気泳動、クロマトフォーカシング、および高圧液体クロマトグラフィーを含む、多様な技術を用いて、達成可能である。例えば、Fc領域に選択的に結合するプロテインAを用いたアフィニティークロマトグラフィーによって、Fc領域を含む抗体を好適に精製することも可能である。カルボキシル末端またはアミノ末端いずれかの、ヘキサヒスチジン、あるいはFLAG(Eastman Kodak Co.、コネチカット州ニューヘブン)またはmyc(Invitrogen)などの他の小ペプチドなどのアフィニティータグを用いて、抗体または抗原結合ドメインの修飾型を調製し、そして一工程アフィニティーカラムによって精製することも可能である。例えば、ポリヒスチジンは、高い親和性および特異性でニッケルに結合し、したがってニッケルのアフィニティーカラム(例えばQiagen(登録商標)ニッケルカラム)を、ポリヒスチジンでタグ化した選択的結合剤の精製に用いることも可能である;例えばAusubelら監修, Current Protocols in Molecular Biology, セクション10.11.8, John Wiley & Sons, ニューヨーク(1993)を参照されたい。いくつかの例では、1より多い精製工程が必要である可能性もある。
【0158】
原核宿主細胞で発現される本発明の選択的結合剤は、細胞膜周辺腔または細胞質いずれかで、可溶性型で、あるいは細胞内封入体の一部として、不溶性型で、存在することも可能である。選択的結合剤は、当業者に知られる標準的技術いずれかを用いて、宿主細胞から抽出することも可能である。例えば、フレンチプレス、ホモジナイズ、および/または超音波処理の後、遠心分離することによって、宿主細胞を溶解して、周辺質/細胞質の内容物を放出させることも可能である。
【0159】
当該技術分野に知られる方法を用いて、細胞質に存在するか、または細胞膜周辺腔から放出されるか、いずれかの抗IFNγ抗体または抗原結合ドメインの可溶性型をさらに精製することも可能であり、例えば浸透圧ショック技術によって、細菌細胞膜周辺腔からFab断片を放出させる。抗体または抗原結合ドメインが、封入体を形成した場合、これらはしばしば、内部細胞膜および/または外部細胞膜に結合することも可能であり、そしてしたがって、遠心分離後、主にペレット成分中に見られるであろう。次いで、ペレット成分を、アルカリpHではジチオスレイトールなどの還元剤の存在下で、または酸pHではTrisカルボキシエチルホスフィンなどの還元剤の存在下で、極端なpHで処理するか、あるいは界面活性剤、グアニジン、グアニジン誘導体、尿素、または尿素誘導体などのカオトロピック剤で処理して、封入体を放出させ、粉々にし、そして可溶化することも可能である。次いで、ゲル電気泳動、免疫沈降等を用いて、可溶性選択的結合剤を解析することも可能である。可溶化抗体または抗原結合ドメインを単離することが望ましい場合、以下に示す方法およびMarstonら, Meth. Enz., 182:264−275(1990)に示される方法などの標準法を用いて、単離を達成することも可能である。
【0160】
いくつかの場合、抗体または抗原結合ドメインは、単離の際には、生物学的に活性でなくてもよい。ポリペプチドを三次構造に「再フォールディング」するか、または変換して、そしてジスルフィド連結を生成するための、多様な方法を用いて、生物学的活性を回復することも可能である。こうした方法には、可溶化ポリペプチドを、通常7を超えるpHに、そして特定の濃度のカオトロープの存在下で、曝露することが含まれる。カオトロープの選択は、封入体可溶化に用いる選択と非常に類似であるが、通常、カオトロープは、より低い濃度で用いられ、そして可溶化に用いるカオトロープと必ずしも同一ではない。大部分の場合、再フォールディング/酸化溶液はまた、還元剤を含有するか、または還元剤に加えて特定の比で酸化型を含有して、特定の酸化還元電位を生じ、ジスルフィド・シャフリングを可能にして、タンパク質のシステイン架橋(単数または複数)の形成を生じる。通常用いられるレドックス対には、システイン/シスタミン、グルタチオン(GSH)/ジチオビスGSH、塩化第二銅、ジチオスレイトール(DTT)/ジチアンDTT、および2−メルカプトエタノール(bME)/ジチオ−b(ME)が含まれる。多くの場合、共溶媒(cosolvent)を用いることも可能であるし、また再フォールディングの効率を増加させるのに共溶媒が必要とされる可能性もあり、そしてこの目的に用いられる、より一般的な試薬には、グリセロール、多様な分子量のポリエチレングリコール、アルギニン等が含まれる。
【0161】
本発明の抗体および抗原結合ドメインはまた、Merrifieldら, J. Am. Chem. Soc., 85:2149(1963);Houghtenら, Proc Natl Acad. Sci. USA, 82:5132(1985)、並びにStewartおよびYoung, Solid Phase Peptide Synthesis, Pierce Chemical Co., イリノイ州ロックフォード(1984)に示されるものなどの当該技術分野に知られる技術を用いた化学合成法(例えば固相ペプチド合成)によっても調製可能である。こうしたポリペプチドを、アミノ末端にメチオニンを含み、または含まずに合成することも可能である。これらの参考文献に示される方法を用いて、化学的に合成された抗体および抗原結合ドメインを酸化して、ジスルフィド架橋を形成することも可能である。こうして調製された抗体は、天然のまたは組換えによって産生された抗opgbp抗体または抗原結合ドメインと関連する生物学的活性を少なくとも1つ保持するであろう。
【0162】
IFNγの選択的結合剤に関するアッセイ
IFNγの少なくとも1つの生物学的活性を部分的にまたは完全に阻害する選択的結合剤を同定するためのスクリーニング法を本発明に提供する。IFNγの生物学的活性の阻害には、限定されるわけではないが、IFNγのその同族受容体、IFNγ−Rへの結合の阻害、in vitroでのA549に対するIFNγの抗増殖活性の阻害、並びにin vitroおよびin vivoでのIFNγによる単球の活性化の阻害が含まれる。本発明の選択的結合剤には、抗IFNγ抗体、並びにその断片、変異体、誘導体および融合体、ペプチド、ペプチド模倣体化合物または有機模倣体(organo−mimetic)化合物が含まれる。IFNγの生物学的活性を部分的にまたは完全に阻害可能な選択的結合剤を同定するためのスクリーニング法には、in vitroアッセイまたはin vivoアッセイが含まれることも可能である。in vitroアッセイには、IFNγのIFNγ−Rへの結合を検出するものが含まれ、そしてこうしたアッセイを用いて、IFNγのIFNγ−Rへの結合の速度または度合いを増加させるかまたは減少させる能力に関して、IFNγの選択的結合剤をスクリーニングすることも可能である。1つの種類のアッセイでは、IFNγポリペプチド、好ましくは細胞外ドメインなどのIFNγの可溶性型を固体支持体(例えばアガロースまたはアクリルビーズ)に固定し、そしてIFNγ−Rポリペプチドを、IFNγの選択的結合剤の存在下または非存在下いずれかで添加する。選択的結合剤の存在下または非存在下でIFNγおよびIFNγ−Rの結合の度合いを測定する。例えば放射標識、蛍光標識または酵素反応によって、結合を検出することも可能である。
【0163】
あるいは、BIAcoreアッセイ系(Pharmacia、ニュージャージー州ピスカタウェイ)などの表面プラズモン共鳴検出装置系を用いて、結合反応を行うことも可能である。結合反応は、製造者のプロトコルにしたがって、実行可能である。
【0164】
上述のものなどのin vitroアッセイを好適に用いて、IFNγのIFNγ−Rへの結合に対する影響に関して、迅速に多数の選択的結合剤をスクリーニングすることも可能である。アッセイを自動化して、ファージ・ディスプレー・ライブラリー、合成ペプチドライブラリーおよび化学合成ライブラリーで生成される化合物をスクリーニングすることも可能である。
【0165】
IFNγのIFNγ−Rへの結合を増加させるかまたは減少させる選択的結合剤を、どちらかのポリペプチドを発現する細胞および細胞株を用いて、細胞培養中でスクリーニングすることもまた可能である。細胞および細胞株は、いかなる哺乳動物から得ることも可能であるが、好ましくは、ヒトまたは他の霊長類、イヌまたはげっ歯類供給源由来であろう。例えば、選択的結合剤の存在下または非存在下で、表面にIFNγ−Rを発現している細胞に対するIFNγの結合を評価して、そして例えばIFNγに対するビオチン化抗体を用いたフローサイトメトリーによって、結合の度合いを測定することも可能である。
【0166】
in vitro活性アッセイを用いて、IFNγ活性を阻害する選択的結合剤を同定することもまた可能である。アッセイの例には、A549細胞増殖アッセイおよびTHP−1 HLA−DR発現アッセイが含まれる。
【0167】
選択的結合剤が、NZBxNZW F1マウスモデルにおいて、タンパク尿の発生を遅延させ、そして生存時間を増加させることが可能であるかどうかを決定するため、in vivoアッセイもまた利用可能である。
【0168】
診断適用のため、特定の態様において、IFNγの選択的結合剤、例えば抗体およびその抗原結合ドメインを、典型的には、検出可能部分で標識する。検出可能部分は、直接または間接的に、検出可能シグナルを産生可能なものいずれであってもよい。例えば、検出可能部分は、放射性同位体、例えば3H、14C、32P、35S、または125I、蛍光化合物または化学発光化合物、例えばフルオレセイン・イソチオシアネート、ローダミン、またはルシフェリン;あるいは酵素、例えばアルカリホスファターゼ、β−ガラクトシダーゼまたは西洋ワサビ(horseradish)ペルオキシダーゼであってもよい;Bayerら, Meth. Enz., 184:138−163(1990)。
【0169】
本発明の選択的結合剤は、いかなる既知のアッセイ法で使用することも可能であり、こうした方法には、例えばIFNγポリペプチドを検出し、そして定量化するための、ラジオイムノアッセイ、競合的結合アッセイ、直接および間接的サンドイッチアッセイ(ELISA)、および免疫沈降アッセイがある(Sola, Monoclonal Antibodies:A Manual of Techniques, pp.147−158(CRC Press, 1987))。抗体は、使用中のアッセイ法に適した親和性で、IFNγポリペプチドに結合するであろう。
【0170】
本発明の選択的結合剤はまた、in vivo画像化にも有用であり、ここで、例えば検出可能部分で標識した選択的結合剤を、動物の好ましくは血流中に投与し、そして宿主中の標識抗体の存在および位置をアッセイする。剤を、核磁気共鳴によってであれ、放射線によってであれ、または当該技術分野に知られる他の手段によってであれ、動物中で検出可能ないずれの部分で標識することも可能である。
【0171】
本発明はまた、抗体または抗原結合ドメインなどのIFNγの選択的結合剤、および生物学的試料中のIFNγレベルを検出するのに有用な他の試薬を含むキットにも関する。こうした試薬には、二次活性、検出可能標識、ブロッキング血清、陽性対照試料および陰性対照試料、および検出試薬が含まれることも可能である。
【0172】
IFNγ選択的結合剤の療法使用
本発明の選択的結合剤は、療法剤として使用可能である。療法選択的結合剤は、IFNγアゴニストまたはアンタゴニストであることも可能であり、そして1つの態様において、in vitroまたはin vivoでIFNγポリペプチドの生物学的活性の少なくとも1つを阻害する抗IFNγアンタゴニスト抗体である。例えば、IFNγのアンタゴニストは、IFNγのIFNγ−Rへの結合を阻害するであろう。あるいは、IFNγアンタゴニストは、実施例1に記載するものなどのA549細胞増殖アッセイにおいて、測定可能なND50(50%増殖を生じる濃度)によって示されるようなin vitroでのヒト肺癌の増殖を刺激するであろう。
【0173】
IFNγアンタゴニスト、例えば抗IFNγアンタゴニスト抗体および抗原結合ドメインを用いて、限定されるわけではないが、以下の:急性膵炎;ALS;アルツハイマー病;AIDS誘導悪疫質を含む悪疫質/摂食障害;喘息および他の肺疾患;アテローム性動脈硬化症;慢性疲労症候群;クロストリジウム属(Clostridium)関連下痢を含むクロストリジウム属関連疾病;うっ血性心不全、冠状動脈再狭窄、心筋梗塞、心筋機能不全(例えば敗血症に関連するもの)、および冠状動脈バイパス移植を含む冠状動脈状態および徴候;癌、例えば多発性骨髄腫、並びに骨髄性白血病(例えばAMLおよびCML)および他の白血病とともに腫瘍転位;発熱;糸球体腎炎;移植片対宿主病/移植拒絶;出血性ショック;例えば角膜移植に関与することもありうるような、炎症性眼疾患;脳虚血(例えば、各々神経変性につながりうる、外傷、癲癇、出血または脳卒中などの結果の脳傷害)を含む虚血;学習障害;多発性硬化症;筋障害(例えば筋タンパク質代謝、特に敗血症におけるもの);神経毒性(例えばHIVに誘導されるようなもの);骨粗鬆症;癌関連疼痛を含む疼痛;パーキンソン病;歯周病;神経毒性;早期陣痛;乾癬;再灌流傷害;敗血症ショック;放射療法の副作用;顎関節疾患;睡眠障害;ブドウ膜炎;または挫傷(strain)、捻挫、軟骨損傷、外傷、整形手術、感染もしくは他の疾患過程から生じる炎症状態;若年発症1型糖尿病およびインスリン耐性糖尿病(例えば肥満に関連するようなもの)を含む糖尿病;子宮内膜症、子宮内膜炎、および関連状態;線維筋痛症または痛覚脱失症;痛覚過敏;クローン病を含む炎症性腸疾患;肺疾患(例えば成人呼吸困難症候群、および肺線維症);神経炎症性疾患;眼変性およびブドウ膜炎を含む、眼疾患および状態;毛孔性紅色粃糠疹(PRP);前立腺炎(細菌性または非細菌性)および関連状態;乾癬および関連状態;肺線維症;再灌流傷害;骨関節炎、関節リウマチ、若年性(リウマチ様)関節炎、血清陰性多発性関節炎、強直性脊椎炎、ライター症候群および反応性関節炎、スティル病、乾癬性関節炎、腸疾患に基づく関節炎、多発性筋炎、皮膚筋炎、強皮症、全身性硬化症、脈管炎(例えば川崎病)、脳脈管炎、ライム病、ブドウ球菌(staphylococcal)誘導(「敗血症」)関節炎、シェーグレン症候群、リウマチ熱、多発性軟骨炎およびリウマチ性多発性筋痛および巨細胞性動脈炎を含む、関節の炎症状態およびリウマチ疾患;敗血症ショック;全身性エリテマトーデス(SLE)腎炎;放射療法による副作用;顎関節疾患;甲状腺炎;組織移植、または挫傷、捻挫、軟骨損傷、外傷、および整形手術から生じる炎症状態を含む、自己免疫疾患および炎症状態を防止するかまたは治療することも可能である。
【0174】
より具体的には、IFNγアンタゴニスト、例えば抗IFNγアンタゴニスト抗体および抗原結合ドメインを用いて、関節炎(特に関節リウマチ)、全身性エリテマトーデス(SLE)、移植片対宿主病(GvHD)、多発性硬化症および糖尿病を防止するかまたは治療することも可能である。
【0175】
アンタゴニスト抗体および抗原結合ドメインを含む、本発明のIFNγアンタゴニストを、単独で、または抗IFNγアンタゴニスト抗体および抗原結合ドメインなどの他の療法剤IFNγアンタゴニストと組み合わせて投与して、これらを用いて、多様な炎症状態、自己免疫状態、および骨損失につながる他の状態を防止するかまたは治療することも可能である。状態および望ましい治療レベルに応じて、2、3、またはそれより多い剤を投与することも可能である。同一処方に含むか、または治療キットに含むことによって、これらの剤をともに提供することも可能であり、あるいは別個に提供することも可能である。遺伝子治療によって投与する際、タンパク質剤をコードする遺伝子を、所望によって同一プロモーター領域の制御下で同一ベクターに含めてもよいし、または別個のベクターに含めてもよい。前述の種類の特に好ましい分子は以下のとおりである。
・IL−1阻害剤:IL−1raタンパク質および可溶性IL−1受容体。最も好ましいIL−1阻害剤はアナキンラである。
・TNF−α阻害剤:可溶性腫瘍壊死因子I型受容体(sTNF−RI;−RIはまた、p55受容体とも呼ばれる);可溶性腫瘍壊死因子II型受容体(p75受容体とも呼ばれる);およびTNF受容体に結合するモノクローナル抗体。最も好ましいのは、WO 98/24463に記載されるようなsTNF−RI、エタネルセプト(エンブレル(登録商標))、およびアバカイン(登録商標)である。典型的なTNF−α阻害剤は、EP 422 339、EP 308 378、EP 393 438、EP 398 327、およびEP 418 014に記載される。
・セリンプロテアーゼ阻害剤:SLPI、ALP、MPI、HUSI−I、BMI、およびCUSI。SLPIがLPS反応を阻害することが示されたように、これらの阻害剤を、典型的なLPS調節因子として見ることもまた可能である。Jinら(1997), Cell 88(3):417−26(本明細書に援用される)。
【0176】
薬剤組成物
IFNγ選択的結合剤の薬剤組成物は、本発明の範囲内にある。こうした組成物は、療法的または予防的に有効な量のIFNγ選択的結合剤、例えば抗体、あるいはその断片、変異体、誘導体または融合体を、薬学的に許容しうる剤と混合して含む。好ましい態様において、薬剤組成物は、IFNγの少なくとも1つの生物学的活性を部分的にまたは完全に阻害する抗IFNγアンタゴニスト抗体を、薬学的に許容しうる剤と混合して含む。典型的には、抗体は、動物への投与のため、十分に精製されているであろう。
【0177】
本発明の組成物に使用するための薬学的に許容しうる剤には、当該技術分野に周知であるような、キャリアー、賦形剤、希釈剤、酸化防止剤、保存剤、着色剤、フレーバー剤および希釈剤、乳化剤、懸濁剤、溶媒、増量剤(filler)、充填剤(bulking agent)、緩衝剤、搬送ビヒクル、等張化剤、共溶媒、湿潤剤、錯化剤、緩衝化剤、抗菌剤および界面活性剤が含まれる。
【0178】
中性緩衝生理食塩水または血清アルブミンと混合した生理食塩水が、典型的な適切なキャリアーである。組成物にやはり含まれるのは、アスコルビン酸などの酸化防止剤;低分子量ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニンまたはリジンなどのアミノ酸;単糖、二糖、およびグルコース、マンノースまたはデキストリン類を含む他の炭水化物;EDTAなどのキレート剤;マンニトールまたはソルビトールなどの糖アルコール;ナトリウムなどの塩形成対イオン;および/またはTween、プルロニック類またはポリエチレングリコールなどの非イオン性界面活性剤である。やはり例として、適切な等張化増進剤には、ハロゲン化アルカリ金属(好ましくは塩化ナトリウムまたは塩化カリウム)、マンニトール、ソルビトール等が含まれる。適切な保存剤には、限定されるわけではないが、塩化ベンザルコニウム、チメロサール、フェネチル・アルコール、メチルパラベン、プロピルパラベン、クロルヘキシジン、ソルビン酸等が含まれる。過酸化水素もまた、保存剤として使用可能である。
【0179】
適切な共溶媒は、例えばグリセリン、プロピレングリコール、およびポリエチレングリコールである。適切な錯化剤は、例えばカフェイン、ポリビニルピロリドン、ベータ−シクロデキストリンまたはヒドロキシ−プロピル−ベータ−シクロデキストリンである。適切な界面活性剤または湿潤剤には、ソルビタンエステル類、ポリソルベート80などのポリソルベート類、トロメタミン、レシチン、コレステロール、チロキサポール(tyloxapal)等が含まれる。緩衝剤は、酢酸、ホウ酸、クエン酸、リン酸、炭酸水素、またはTris−HClなどの慣用的緩衝剤であることも可能である。酢酸緩衝液はpH4.0〜5.5程度であることも可能であり、そしてTris緩衝液はpH7.0〜8.5程度であることも可能である。さらなる薬学的剤が、Remington’s Pharmaceutical Sciences, 第18版, A.R. Gennaro監修, Mack Publishing社、1990に示されており、該文献の相当する部分は、本明細書に援用される。
【0180】
組成物は、液体型であっても、あるいは凍結乾燥型またはフリーズドライ型であってもよい。凍結乾燥型には、スクロースなどの賦形剤が含まれうる。本発明の組成物は、非経口投与に適している。好ましい態様において、組成物は、当業者に利用可能ないずれかの経路によって、例えば皮下経路、静脈内経路、筋内経路、腹腔内経路、大脳内(実質内)経路、脳室内経路、筋内経路、眼内経路、動脈内経路、または病変内経路によって、動物に注射するかまたは注入するのに適している。非経口処方は、典型的には、無菌、病原体不含の等張化水性溶液であり、所望によって薬学的に許容しうる保存剤を含有するであろう。
【0181】
最適な薬剤処方は、意図される投与経路、搬送形式および望ましい投薬量に応じて、当業者が容易に決定可能である。
他の処方もまた本発明に意図される。薬剤組成物はまた、ポリ乳酸、ポリグリコール酸などのポリマー化合物の粒子性調製物、またはリポソームへのIFNγ選択的結合剤(抗体など)の導入も含むことも可能である。ヒアルロン酸もまた使用可能であり、そしてこの酸は、循環中にある期間の持続を促進する効果も有しうる。薬剤組成物は、IFNγ選択的結合剤(例えば抗体)と、注射可能微小球体、生体侵食性粒子またはビーズ、あるいはリポソームとの処方もまた含み、こうした処方は、選択的結合剤の制御放出または徐放を提供し、こうした処方を、次いで、蓄積注射として搬送してもよい。他の適切な搬送手段には、移植可能搬送装置が含まれる。
【0182】
IFNγ選択的結合剤(抗体など)を含む薬剤組成物は、吸入用の乾燥粉末としても処方可能である。こうした吸入溶液はまた、エアロゾル搬送用の液化噴霧剤中にも処方可能である。さらに別の処方において、溶液を噴霧することも可能である。IFNγ選択的結合剤を含有する特定の処方を経口投与してもよいこともまた意図される。この方式で投与される処方を、錠剤およびカプセルなどの固形投薬型の配合に習慣的に用いられるキャリアーを伴い、または伴わずに、処方することも可能である。例えば、生物学的利用能が最大となりそして前全身分解が最小となる胃腸管で、処方の活性部分が放出されるように、カプセルを設計することも可能である。選択的結合剤の吸収を促進する、さらなる剤を含むことも可能である。希釈剤、フレーバー剤、低融点ワックス、植物油、滑沢剤、懸濁剤、錠剤崩壊剤、および結合剤もまた、使用可能である。
【0183】
別の調製は、有効量のIFNγ選択的結合剤を、錠剤の製造に適した非毒性賦形剤と混合して含むことも可能である。錠剤を滅菌水、または別の適切なビヒクルに溶解することによって、単位用量型の溶液を調製することも可能である。適切な賦形剤には、限定されるわけではないが、不活性希釈剤、例えば炭酸カルシウム、炭酸ナトリウムもしくは炭酸水素ナトリウム、ラクトース、またはリン酸カルシウム;あるいは結合剤、例えばデンプン、ゼラチン、またはアラビアゴム(acacia);あるいは滑沢剤、例えばステアリン酸マグネシウム、ステアリン酸、またはタルクが含まれる。
【0184】
さらなる処方が当業者には明らかであり、こうした処方には、IFNγ選択的結合剤を、1以上の他の療法剤と組み合わせて含む処方が含まれる。多様な他の徐放または制御搬送手段、例えばリポソームキャリアー、生体侵食性微小粒子または多孔ビーズおよび蓄積注射を処方するための技術もまた、当業者に知られる。例えば、その開示が本明細書に援用される、Supersaxoら、薬剤組成物搬送用の制御放出多孔ポリマー微小粒子の説明(WO 93/15722(PCT/US93/00829)を参照されたい)を参照されたい。
【0185】
投与方式に関わらず、体重、体表面積または臓器サイズにしたがって、特定の用量を計算することも可能である。上述の処方各々を伴う治療の適切な投薬量を決定するのに必要な計算のさらなる微調整は、当業者によって日常的に行われ、そして当業者が日常的に行う仕事の範囲内である。適切な用量−反応データの使用を通じて、適切な投薬量を確認することも可能である。
【0186】
さらに、肺投与によって、本発明の薬剤組成物を投与することも可能であり、例えば化学的に修飾されたタンパク質の肺搬送を開示し、本明細書に援用される、PCT WO94/20069を参照されたい。肺搬送のため、粒子サイズは、遠位肺への搬送に適したものでなければならない。例えば、粒子サイズは1μm〜5μmであることも可能であるが、例えば各粒子がかなり多孔性であれば、より大きい粒子も使用可能である。
【0187】
あるいはまたはさらに、IFNγ選択的結合剤が吸収されているか、または被包されている、膜、スポンジ、または他の適切な材料を、罹患領域に移植することを介して、組成物を局所投与することも可能である。移植装置を用いる場合、適切な組織または臓器いずれかに装置を移植し、そしてIFNγ選択的結合剤の搬送は、多量(bolus)投与を介して、または連続投与を介して、または連続注入を用い、カテーテルを介して、装置から直接なされることも可能である。
【0188】
本発明の薬剤組成物は、徐放処方または徐放調製物中でもまた投与可能である。徐放調製物の適切な例には、成形物品の形の半透性ポリマーマトリックス、例えばフィルム、または微小カプセルが含まれる。徐放マトリックスには、ポリエステル、ヒドロゲル、ポリ乳酸(例えば米国特許第3,773,919号、EP 58,481)、L−グルタミン酸およびガンマ・エチル−L−グルタメートのコポリマー(Sidmanら, Biopolymers, 22:547−556(1983))、ポリ(2−ヒドロキシエチル−メタクリレート)(Langerら, J. Biomed. Mater. Res., 15:167−277(1981)およびLanger, Chem. Tech., 12:98−105(1982))、エチレン酢酸ビニル、またはポリ−D(−)−3−ヒドロキシ酪酸が含まれる。徐放組成物にはまた、当該技術分野に知られるいくつかの方法のいずれかによって調製可能なリポソームが含まれることも可能である。例えばEppsteinら, Proc. Natl. Acad. Sci. USA, 82:3688−3692(1985);EP 36,676;EP 88,046;およびEP 143,949を参照されたい。
【0189】
いくつかの例では、IFNγ選択的結合剤組成物を含む薬剤組成物を、ex vivo方式で使用することが望ましい可能性もある。この場合、患者から取り除いた細胞、組織または臓器を、IFNγ選択的結合剤を含む薬剤組成物に曝露し、その後、続いて、細胞、組織および/または臓器を患者に移植しなおす。
【0190】
他の場合では、本明細書に記載するものなどの方法を用いて、遺伝子操作した特定の細胞を患者に移植して、ポリペプチド、選択的結合剤、断片、変異体、または誘導体を発現させ分泌させることによって、IFNγ選択的結合剤を含む組成物を搬送することも可能である。こうした細胞は、動物細胞またはヒト細胞であってもよく、そして患者自身の組織あるいはヒトまたは非ヒトいずれかの別の供給源由来であってもよい。所望によって、細胞を不死化してもよい。しかし、免疫応答の可能性を減少させるため、細胞を被包して、周囲組織への浸潤を回避することが好ましい。被包材料は、典型的には、タンパク質産物(単数または複数)の放出を許すが、患者の免疫系による、または周囲組織由来の他の有害因子による分解を防止する、生体適合性の半透性ポリマー被包体(enclosure)または膜である。
【0191】
細胞の膜被包に用いられる方法は当業者に周知であり、そして被包細胞の調製および該細胞の患者への移植は、過度の実験を伴わずに達成可能である。例えば米国特許第4,892,538号、第5,011,472号、および第5,106,627号を参照されたい。生存細胞を被包する系がPCT WO 91/10425(Aebischerら)に記載される。他の多様な徐放または制御搬送手段、例えばリポソームキャリアー、生体侵食性粒子またはビーズを処方するための技術もまた、当該技術分野に知られ、そして記載されている。細胞を被包し、また被包せずに、患者の適切な体組織または臓器に移植してもよい。
【0192】
IFNγ選択的結合剤(例えば抗IFNγ抗体、あるいはその断片、変異体、誘導体、および融合体)を含む薬剤組成物の療法的または予防的に有効な量は、例えば該組成物を用いる徴候などの療法目的、投与経路、および被験者の状態に応じるであろう。本発明のIFNγアンタゴニスト抗体または抗原結合ドメインは、自己免疫および/または炎症状態を防止し、そして/または治療するため、療法的または予防的に有効な量で投与される。
【0193】
以下の実施例は、本発明をさらに十分に例示するために提供され、そして本発明の範囲を限定するとは解釈されない。
【0194】
(実施例1)
試薬およびアッセイ
これらの研究に用いるスクリーニング標的を:1)EP 0423845またはPCT公報WO 83/04053に記載されるような、hIFNγをコードするcDNAの大腸菌における発現;または2)以下のような、hIFNγをコードするcDNAのCHO宿主細胞における発現から調製した:PCR(標準的条件)を用いて、テンプレートとしてヒト脾臓Marathon Ready cDNA(Clonetech)を用い、ヒトINFγをコードする全長配列を増幅した。発現プラスミドに配列をサブクローニングし、そしてDH10B細胞(Gibco Life Sciences)をDNAで形質転換し、DNAを調製し、そしてリン酸カルシウム法(Speciality Media, Inc.)によって、CHO細胞にトランスフェクションした。高発現細胞株クローンを用いて、血清不含馴化培地を生成した。
【0195】
hINFγを含有するCHO細胞馴化培地を濃縮し、透析し、そして次いで、いくつかのクロマトグラフィー工程を通じて精製した。最初の工程は、非グリコシル化hIFNγ型に対して高グリコシル化型を分離する、標準的NaCl勾配を用いたQ−HP(Pharmacia)クロマトグラフィーであった。コムギ(wheat)胚芽凝集素クロマトグラフィー(EY Laboratories)を通して、Q−HPプールをさらに精製した。精製物質は、クーマシー−ブルーおよび銀染色SDS−PAGE両方によって判断すると、95%より純粋であった。物質は、ゲル凝固法(カブトガニ(Limulus)アメーバ様細胞溶解物)によってアッセイした際、内毒素が低レベルであった。R&D Systemsのヤギ(goat)抗hIFNγ中和抗体(カタログ番号AF−285−NA、ロット番号ZW019011)を用いて、ウェスタンブロットによって、hINFγの同一性を確認した。吸光係数法(0.66)を用いて、最終タンパク質濃度を決定した。物質2ロットをそれぞれ生成した。収率は40mg/lであった。最終物質をPBS中に配合した。
【0196】
CHO細胞におけるヒトIFNγR1−Fcタンパク質の発現
これらの研究において、標的からファージ抗体を溶出するのに用いたヒトIFNγR1−Fcタンパク質を以下のように調製した:PCR(標準的条件)を用いて、テンプレートとしてヒトリンパ球Marathon Ready cDNA(Clonetechより購入)を用い、ヒトIFNγR1をコードする全長配列を増幅した。PCR(標準的条件)を用いて、ヒトIgG1のFc部分をコードする配列を増幅した。重複PCRを用いて、IFNγR1−Fc融合構築物をコードする配列(IFNγR1のアミノ酸1〜Ser246)を生成して、そして発現プラスミドに該配列をサブクローニングした。DH10B細胞(Gibco Life Sciences)をDNAで形質転換し、DNAを調製し、そしてリン酸カルシウム法(Speciality Media, Inc.)によって、CHO細胞にトランスフェクションした。高発現細胞株クローンを用いて、血清不含馴化培地を生成した。
【0197】
標準的プロテインG Fast−Flowカラム(Pharmacia)を通して、hINFγR1−Fcを含有するCHO細胞馴化培地を濃縮し、そして精製した。吸光係数として1.44を用いて、A280によって最終濃度を決定した。N末端配列決定解析を通じて、精製試料の同一性を確認した。この物質をPBS中に配合した。
【0198】
抗体
モノクローナル抗hIFNγ抗体、クローン2578.111をR&D Systemsから購入した(カタログ番号MAB285、ロット番号KW07)。モノクローナル抗hIFNγ抗体、クローンMMHG−1をBiosourceから購入した(カタログ番号AHC4834、ロット番号10803−015)。組換えヒトIFNγ受容体1(rhIFNγ R1)をR&D Systemsから購入した(カタログ番号673−IR)。rhIFNγ R1の計算上の分子量は25,000ダルトンである。グリコシル化の結果、組換えタンパク質は、SDS−PAGE上、40〜50kDaタンパク質として移動する。
【0199】
A549細胞増殖アッセイ
IFNγの抗体中和を評価するために用いたA549細胞増殖アッセイは、96ウェルアッセイであり、そして一般的に以下のように記載される:第1日、1)アッセイ培地(F12K、5%FBS、1xPen/Strep L−グルタミン)中の最高濃度からAbを1:2で連続希釈する。アッセイにおいて望ましい濃度の4倍で、全部で10回の希釈を行う。2つ組のため、各希釈物は少なくとも最終200μlが必要である;2)用量反応曲線中の有効用量の90%に基づいて、スパイク用にIFNγを適切な濃度に希釈する。アッセイにおいて望ましい濃度の4倍で、IFNγスパイクを作成する;3)Titertek試験管中で、各4xAb希釈物150μlを、4xINFγスパイク150μlと合わせる。ピペッティングによって混合する。ふたをして、室温で1時間インキュベーションする。(注:AbおよびIFNγの濃度は、ここで、アッセイ濃度の2倍である);4)(所望による)AbおよびIFNγをインキュベーションしている間、力価決定(titration)曲線用にIFNγを希釈する。4000ng/mlから始めて、1:3希釈を12回行う。アッセイにおいて、3つ組には300μlが必要であるため、希釈終了時に必要な体積は少なくとも400μlであるはずである。必要となるまで、4℃で保存する;5)インキュベーションが完了する前に、5mlトリプシン中で、A549細胞をトリプシン処理する。フラスコに20mlのアッセイ培地を添加し、そして50mlコニカルに移して、そしてIEC中、室温で1/2〜3/4の速度で遠心分離する;6)細胞を吸引する。7.5mlのアッセイ培地に再懸濁する。トリパンブルー中、1:1で計数する;7)アッセイ培地中、細胞を2.5x104細胞/mlに希釈する。各試料について、2.5x103細胞/ウェルを、96ウェルFalconに0.1mlで植え付ける;8)1時間後、2つ組の2つのウェル各々に、100μl Ab/IFNγ混合物を添加する;そして9)37℃、5%CO2および高湿度で5日間インキュベーションする。第5日:1)ウェルあたり20μlのAlamar Blueを添加する。37℃、5%CO2および高湿度で3〜4時間インキュベーションする;2)FL500蛍光プレート読取装置のスイッチを入れる。プレートからふたを取り除き、そしてふたなしで10分間振盪する;そして3)FL500を読み取る。設定:中程度に3秒間振盪、励起530/25、発光590/35、感度34。
【0200】
(実施例2)
ヒトFabライブラリーのスクリーニング
スクリーニング法
ヒトFabライブラリーを構築し、そしてスクリーニングする、一般的な方法は、de Haardら(Advanced Drug Delivery Reviews, 31:5−31(1998);J. Biol. Chem., 274:18218−18230(1999))に記載された。以下の方法によって、hIFNγに結合するFab断片をスクリーニングした。
【0201】
Nuncイムノチューブを、0.1M炭酸Na、pH9.6中、0.39μg/mlのhIFNγ 4mlで、Nutator上、室温で2時間コーティングした。Target Quest、NV(オランダ・アムステルダム)凍結ファージライブラリーストック(試験管あたり750μl中、4x1012pfu)のアリコットを融解後、1/5体積(150μl)のPEG溶液(20%ポリエチレングリコール8000、2.5M NaCl、オートクレーブ)を添加し、そして試験管を氷上に1時間放置してファージを沈殿させることによって、グリセロール(15%)を取り除いた。沈殿したファージ粒子を4000rpm、4℃で15分間遠心してペレットとし、次いで500μl PBS、pH7.4に再懸濁した。IFNγでコーティングしたイムノチューブを4ml PBSで3回洗浄し、そしてNutator上、4mlの2%MPBSを用いて、室温で1時間ブロッキングした。同時に、500μlの4%MPBSをファージ懸濁物に添加し、そして室温で30分間〜1時間インキュベーションして、ファージ粒子のプレ・ブロッキングを可能にした。ブロッキングしたイムノチューブをPBST(PBS中の0.1%Tween20)で2回洗浄し、そしてPBSで2回洗浄した。プレ・ブロッキングしたファージ混合物を、3mlの2%MPBSを含有する、洗浄したイムノチューブに添加した。回転装置上で30分間インキュベーションし、その後、室温で1.5時間、静置インキュベーションした後、ファージ混合物を廃棄した。試験管をまずPBSTで20回、次いでPBSで20回洗浄した。回転装置上、0.4%MPBS、pH7.4中、1μMの特異的溶出試薬(それぞれhIFNγ、GPNA、RDMA、BSMA、またはrhIFNγ R1)1mlとインキュベーションすることによって、結合したファージ粒子を溶出した。溶出したファージ粒子を無菌50mlコニカル・ポリプロピレン試験管に移し、そして氷上で保存した。各ファージ溶出物、約20μlを、力価決定のため、取り置いた。増幅のため、残った溶出ファージ粒子を、5mlのTG1培養物(OD590約0.5)および4mlの2xYTを含有する50mlコニカル試験管に添加した。IFN感染(IFNection)混合物を振盪せずに37℃で30分間インキュベーションし、次いで、3500rpmで20分間回転させた。細胞ペレットを1500μlの2xYT−AGブロスに懸濁し、そして5つのSOBCGプレート上、300μl/プレートで蒔いた。プレートを30℃で一晩インキュベーションした。20時間インキュベーションした後、プレートから細胞スクレーパーで細胞を回収し、これにプレートあたり4mlの2xYT−AGを添加した。この工程を3回反復した。回収細胞のごく一部をファージ救出に用いた(以下を参照されたい)。残った細胞懸濁物を3500rpmで20分間回転させた。細胞ペレットをペレットサイズの1/2体積の50%グリセロールに再懸濁して、グリセロールストックを作成し、そして−80℃で保存した。
【0202】
増幅細胞懸濁物からのファージ救出を以下のように行った。蒔いて増幅させた約0.5mlの回収細胞懸濁物を用いて、約0.3のOD590になるように50mlの2xYT−AGに接種した。振盪装置上、OD590が0.5になるまで培養物を37℃でインキュベーションした。M.O.I.20のM13KO7ヘルパーファージ(GIBCO BRL、カタログ番号18311−019、1.1x1011pfu/ml)1mlで、10mlの培養物をIFN感染させて、そして37℃で30分間、インキュベーター中でインキュベーションした。IFN感染した細胞を4000rpmで20分間回転させて落とした。細胞ペレットを50mlの2xYT−AKに再懸濁し、250mlフラスコに移して、そして270rpmで20時間、振盪しながら30℃でインキュベーションした。一晩培養物を4000rpmで20分間回転させて、細胞破片を取り除いた。上清を再び遠心分離して、細胞破片が除去されることを確実にした。PEG溶液(20%PEG8000、2.5M NaCl)約1/5体積を上清に添加して、ファージ粒子を沈殿させた。混合物を氷上で少なくとも1時間インキュベーションし、次いで4000rpmで20分間遠心分離して、沈殿したファージ粒子を収集した。ファージペレットを1mlのPBSに再懸濁し、そして微量遠心分離試験管に移した。ファージ懸濁物を氷上で1時間放置して、ファージ粒子の完全な懸濁を可能にし、次いで、14,000rpmで2分間回転させて、残った細胞破片を取り除いた。ファージ沈殿工程を反復した。最終ファージペレットを1.1mlのPBSに懸濁し、そして延長した期間、氷上に放置して、ファージ粒子の完全な懸濁を確実にした。ファージ懸濁物を14,000rpmで2分間遠心分離して、残った細胞破片を取り除いた。500μlの救出したファージ懸濁物を用いて、250μlの50%グリセロールを添加することによって、グリセロールストックを作成した。100μlの救出ファージ懸濁物をファージプールELISA(以下を参照されたい)用に取り置いた。残った500μlの救出ファージを次の周期のパニングに用いた。
【0203】
ファージプールELISA
ファージプールELISAを以下のように行った:大腸菌で発現したhIFNγを、Nunc MaxiSorb Immunoプレートにおいて、0.1M炭酸Na、pH9.6中、0.39μg/ml、100μl/ウェルで、穏やかに振動させながら、室温で2時間プレーティングした。コーティングしたプレートをPBSで3回洗浄し、次いで、振動装置上、室温で1時間、2%MPBS 300μl/ウェルを用いてブロッキングした。陰性対照として、抗原でコーティングされていない別のNunc Immunoプレートもまた、2%MPBSでブロッキングした。その間に、120μlの救出された各ファージプールを、96ウェルCostar 3790プレート中、120μlの0.8%MTBSでプレ・ブロッキングし、そして使用準備が整うまで、室温で放置した。ブロッキングしたプレートをどちらも、0.1%TBST(TBS:10mM Tris−HCl、pH7.5、1mM EDTA、150mM NaCl;Tween−20 0.1%)で5回洗浄した。プレ・ブロッキングしたファージ希釈物を、抗原でコーティングしたプレートおよび陰性対照プレート両方に分配し(100μl/ウェル)、そして振動装置上、室温で1時間インキュベーションした。記載するようにプレートを洗浄した後、0.4%MTBS中、1:1000倍希釈したHRP/抗M13モノクローナル・コンジュゲート(Amersham Pharmacia Biotech、カタログ番号27−9421−01)100μl/ウェルを分配し、そして振動装置上、室温で1時間インキュベーションした。記載するようにプレートを洗浄した。基質、1−StepTM ABTS(Pierce、カタログ番号37615)100μl/ウェルを添加した後、プレートを1時間インキュベーションした。シグナル検出のため、OD405を測定した。さらなる解析のため、個々のクローンの供給源として、ELISA陽性ファージプールを用いた。
【0204】
(実施例3)
IFNγに結合するFabファージクローンの同定
DNAフィンガープリンティング
ELISA陽性ファージプール由来の重鎖および軽鎖両方を含有する全長クローンを同定するため、96ウェルThermowellプレート中でポリメラーゼ連鎖反応(PCR)を行った。典型的には、各ウェルは、25μlのPCR反応混合物(2.5μl 10xPCR緩衝液、21.625μlの水、25mMの0.25μl dNTP類、10pmol/μlの0.25μlプライマー870−02(以下を参照されたい)、10pmol/μlの0.25μlプライマー2182−83(以下を参照されたい)、5単位/μlの0.125μl Taqポリメラーゼ)を含有する。
【0205】
【化7】
【0206】
個々のコロニーを摘み取り、そしてまずPCRプレート中のウェルに再懸濁し、次いで300μl/ウェルの2xYT−AGブロス(2xYTブロス:100μg/mlアンピシリンおよび2%グルコースを含有する、水1lあたり、10g酵母エキス、16gバクト−トリプトン、5g NaCl)を満たした96ウェル深底ウェルブロックの対応するウェルに再懸濁する。PCR反応条件は、94℃5分間の変性周期1回、40周期の94℃45秒間、55℃45秒間、72℃1.5分間、その後、72℃10分間の伸長周期1回であった。PCR反応が完了した後、3μl/ウェルのPCR反応混合物を、0.5μl/mlのエチジウムブロミドを含有する、余分に長い4x(24+2)の1%TAEゲル(Embi Tec、カタログ番号GE−3820)上で、120ボルトで1時間泳動した。1kb plus DNAラダー(Gibco BRL、カタログ番号10787−018)に比較することによって、1.6kbより大きい挿入物を持つクローンを全長クローンと同定した。
【0207】
ユニークな全長クローンの同定を以下のように行った:同定された全長クローンのPCR増幅した挿入物に対して、BstNI消化を行った。96ウェルThermowellプレート中の試料あたり16μlのPCR反応混合物に、3μl 10x緩衝液2(NEBL)、10mg/mlの0.3μl BSA、10μlの水および0.7μlのBstNI(NEBL)を含有する14μlのBstNI消化マスター溶液を添加した。プレートを60℃で3時間インキュベーションした。消化した試料の各々13μlを、0.5μl/mlのエチジウムブロミドを含有する、余分に長い2x(24+2)の4%TAEゲル(Embi Tec、カタログ番号GE−3817)上で、100ボルトで3時間泳動した。BstNI断片パターンの相違に基づいて、ユニークなクローンを同定した。
【0208】
クローンファージELISA
同定されたユニークな全長クローンのFabファージを96ウェル形式で救出した。96ウェルの2ml深底ウェルブロックにおいて、480μl/ウェル 2xYTAGブロスに、選択したユニークな全長クローンの一晩培養物20μlを接種し、次いで、300rpm、37℃で3時間インキュベーションした。細胞をIFN感染させるため、各ウェルに、100μlの1:10希釈したM13KO7ヘルパーファージ希釈物を添加した。ブロックを振盪せずに37℃で30分間インキュベーションし、次いで、150rpmでさらに30分間、穏やかに振盪した。ブロックを3600rpmで20分間遠心分離して、IFN感染させた細胞をペレットにした。各ウェルの細胞ペレットを480μlの2xYTAK(100μg/mlアンピシリンおよび40μg/mlカナマイシンを含有する2xYTブロス)に懸濁し、次いで約20時間に渡って、30℃で一晩インキュベーションした。3600rpmで20分間遠心分離することによって、細胞破片を分離した。救出したファージ上清を注意深く別の無菌96ウェルブロックに移した。救出したファージを用いて、実施例2のファージプールELISAに記載したのと正確に同じように、クローンファージELISAを行った。0.2以上の純OD405を生じるクローンをIFNγ結合候補と見なした。
【0209】
大規模ファージ救出ELISA
ELISAにおいて、FabファージがIFNγに濃度依存的に結合することを立証することによって、同定したユニークなFabファージクローンが特異的にIFNγに結合することを確認した。大規模救出によってFabファージを得た。
【0210】
個々のクローンの大規模救出を、以下のように行った:同定したユニークなIFNγ結合クローンのFabファージを大規模に救出した。250mlの無菌フラスコ中で、50mlの2xYT−AGブロスに、選択したIFNγ結合クローンの一晩培養物200μlを接種し、そして培養物のOD590が0.5に達するまで、37℃、2700rpmでインキュベーションした。5mlのM13KO7ヘルパーファージ(GIBCO BRL、カタログ番号18311−019、1.1x1011pfu/ml)をM.O.I.20で添加して、細胞を感染させた。細胞/ヘルパーファージ混合物を振盪せずに37℃で30分間インキュベーションし、次いで4000rpmで20分間遠心分離して、感染細胞をペレットにした。細胞ペレットを50mlの2xYTAKブロス(100μg/mlアンピシリンおよび40μg/mlカナマイシンを含有する2xYTブロス)に懸濁し、次いで270rpmで振盪しながら、約20時間に渡って、30℃で一晩インキュベーションした。一晩培養物を4000rpmで20分間遠心分離することによって、細胞破片を取り除いた。上清を再び遠心分離して、細胞破片が除去されていることを確実にした。上清に、10ml(1/5体積)のPEG溶液(20%PEG8000、2.5M NaCl)を添加して、ファージ粒子を沈殿させた。混合物を氷上で少なくとも1時間インキュベーションし、そして4000rpmで20分間遠心分離して、沈殿したファージ粒子を収集した。ファージペレットを1mlのPBSに再懸濁して、そして微量遠心分離試験管に移した。ファージ懸濁物を氷上で1時間放置して、ファージ粒子の完全な懸濁を可能にし、次いで、14,000rpmで2分間回転させて、残った細胞破片を取り除いた。ファージ沈殿工程を反復した。最終ファージペレットを1mlのPBSに懸濁し、そして延長した期間、氷上に放置して、ファージ粒子の完全な懸濁を確実にした。ファージ懸濁物を14,000rpmで2分間遠心分離して、残った細胞破片を取り除いた。最終ファージ懸濁物を4℃で保存した。実施例2のファージプールELISAに記載するように、ファージELISAを行った。典型的には1x109pfu/ウェル〜1x1011pfu/ウェルの、少なくとも6つの異なる濃度の大規模救出ファージを、対応するウェルに添加した。
【0211】
総数11のFabクローンを同定した。大腸菌hIFN−γ溶出を用いて、ファージプールからFabクローン「IFN−A」、「57E」、および「57D」を同定した。GPNA溶出を用いて、ファージプールからFabクローン「GP−A」および「58C」を同定した。RDMA溶出を用いて、ファージプールからFabクローン「RD−A2」、「RD−B」および「59−A2」を同定した。BSMA溶出を用いて、ファージプールからFabクローン「BS−A」および「BS−B」を同定した。hIFN−γ R1溶出を用いて、ファージプールからFabクローン「67C」を同定した。大規模救出ファージ調製物に対して、9つのユニークなクローンの濃度依存的クローンファージELISAを行って、そしてこれを図1および図2に例示した。これらのFabファージは、ELISAプロフィールに基づいて、3つのグループに分けることも可能である。グループAには、Fabクローン「GP−A」および「BS−B」が含まれる。これらの2つのFabファージは強い結合剤であり、ELISAシグナルは5E9 pfu/ウェルで飽和に到達する。グループBには、Fabクローン「BS−A」、「RD−A2」、「INF−A」および「57E」が含まれる。これらは強い結合剤から穏やかな結合剤であり、優れた濃度依存性結合曲線を示す。グループCには、Fabクローン「57D」、「58C」、「RD−B」および「67C」が含まれる。これらは弱いが特異的であり、そして濃度依存性のIFNγ結合剤である。
【0212】
Fabクローンの配列解析
ユニークなIFNγ結合Fabファージクローンの確認を以下のように行った:QIAfilterTMプラスミド・ミディ・キット(Qiagen、カタログ番号12245)を用いて、ユニークなFab BstNI消化パターン各々の代表のプラスミドDNAを調製し、そして配列決定に送った。Fab BstNIパターンすべての配列がユニークであることが確認され、そして個々の重鎖配列および軽鎖配列が明らかになった(図3〜24を参照されたい)。
【0213】
Fab「BS−A」、「BS−B」、「RD−B1」、「RD−A2」、「58C」、「GP−A」、「57D」、「57E」、「IFN−A」、「67C」および「59−A2」の重鎖のDNAおよび予測されるアミノ酸配列(それぞれ配列番号65〜86)を、それぞれ図3〜13に示した。Fab「BS−A」、「BS−B」、「RD−B1」、「RD−A2」、「58C」、「GP−A」、「57D」、「57E」、「IFN−A」、「67C」および「59−A2」の軽鎖のDNAおよび予測されるアミノ酸配列(それぞれ配列番号87〜108)を、それぞれ図14〜24に示した。図31に示すように、11のFabすべての重鎖および軽鎖のアミノ酸配列を比較した。GCGの「BestFit」プログラムを用いて、Fab各対間の同一性および類似性のパーセンテージを得た。重鎖中の最も密接なマッチは、「BS−A」、「RD−A2」および「IFN−A」中である。「BS−A」および「RD−A2」の重鎖配列は、同一のフレームワークおよびCDR1およびCDR2を有する。これらはCDR3のみが異なり、そして92.6%の同一性および93.4%の類似性を有する。最初のアミノ酸を例外として、「BS−A」および「IFN−A」の重鎖配列は、同一のフレームワークおよびCDR1およびCDR2を有し、そして異なるCDR3を有する。これらは93.5%の同一性および95.1%の類似性を共有する。同じことが「IFN−A」および「RD−A2」の重鎖配列にも当てはまり、これらは90.5%の同一性および92.1%の類似性を持つ。Fab「57E」の重鎖のアミノ酸配列は、「BS−A」の重鎖配列に88.1%の同一性および89.0%の類似性を示し、「RD−A2」の重鎖配列に88.8%の同一性および90.0%の類似性を示し、そしてIFN−Aの重鎖配列に89.0%の同一性および90.7%の類似性を示す。Fab「BS−B」の重鎖のアミノ酸配列は、「57D」の重鎖配列に88.6%の同一性および90.4%の類似性を示し、「GP−A」の重鎖配列に81.7%の同一性および83.3%の類似性を示し、そして58Cの重鎖配列に83.9%の同一性および84.7%の類似性を示す。軽鎖中の最も密接なマッチは、「59−A2」および「BS−A」間の90.8%の同一性および91.7%の類似性、「BS−A」および「BS−B」間の89.0%の同一性および90.9%の類似性、「57E」および「BS−A」間の88.2%の同一性および90.9%の類似性、「57E」および「BS−B」間の89.7%の同一性および91.5%の類似性、そして「59−A2」および「BS−B」間の88.2%の同一性および88.2%の類似性である。Fabの3対、「59−A2」/「BS−B」、「57E」/「BS−A」および「IFN−A」/「RD−A2」のみが、重鎖および軽鎖両方で密接にマッチしている。
【0214】
相補性決定領域(CDR)のアミノ酸配列の比較を図25に示す。11の抗IFNγ Fabの重鎖CDR3は、類似性をほとんど共有しない。図32に示すように、重鎖CDRまたは軽鎖CDRのいずれの類似性にしたがって、Fabをグループ分けすることも可能である。クローン「BS−A」、「IFN−A」および「RD−A2」は、同一の重鎖CDR1およびCDR2を有する。しかし、最後の3つの残基が同一である(FDY)ことを除いて、これらの重鎖CDR3は、非常に異なる。興味深いことに、IFN−AおよびRD−A2はまた、緊密にマッチする軽鎖CDR1(11/16の同一残基)、CDR2(5/7の同一残基)、およびCDR3(8/9の同一残基)も共有する。クローン「BS−B」、「59−A2」、「GP−A」、および「57D」は、類似の重鎖CDR1およびCDR2を有する。クローン「BS−B」および「59−A2」は、同一の重鎖CDR1およびCDR2を有するが、非常に異なる重鎖CDR3を有する。クローン「59−A2」、「BS−A」、「BS−B」および「57E」の3つの軽鎖CDRはすべて、非常に類似である。
【0215】
(実施例4)
可溶性Fabの発現および精製
大腸菌株HB2151(Pharmacia)を、ユニークな結合剤のプラスミドDNAで形質転換した。形質転換HB2151の一晩培養物を2xYT−AGブロス中、30℃で増殖させた。100μg/mlアンピシリンおよび0.1%グルコースを含有する750mlの2xYTに、一晩培養物7.5mlを接種し、そして37℃で約2時間、振盪しながら(270rpm)インキュベーションした。OD590が0.8〜1.0に到達したら、誘導のため、IPTGを1mM添加した。培養物を振盪しながら30℃で4時間増殖させ続けた。培養物を4000rpmで20分間遠心分離し、そして上清を廃棄した。浸透圧ショックアプローチを用いて、Fabの周辺腔放出を達成した。8mlの氷冷TES(0.2M Tris、0.5mM EDTA、17.1%スクロース、pH8.0)に細胞を懸濁し、そしてときどき穏やかに振盪しながら氷上で5〜10分間インキュベーションした。空の試験管を8.8mlのTES/H2O(1:3)でリンスし、これを細胞懸濁物にプールした。
【0216】
細胞懸濁物を氷上でさらに20分間インキュベーションし、そして4,000rpmで15分間遠心分離した。上清を注意深く別の試験管に移し、そして8000rpmで20分間、再度遠心分離した。生じた上清は、TES放出周辺腔分画であった。細胞ペレットを10ml TES/15mM MgSO4に再懸濁し、氷上で15分間インキュベーションし、次いで上述のように2回遠心分離した。最後の上清は、Mg放出周辺腔分画であり、そしてTES放出周辺腔分画にともにプールした。
【0217】
周辺腔分画に、最終濃度1mg/mlとなるようにキャリアーおよび安定化剤としてBSAを添加した。プロテアーゼ阻害剤を加えた、2lの超音波処理緩衝液(20mM Tris−HCl/0.1M NaCl、pH8.5)に対して、1回交換しつつ、周辺腔分画を透析した。1/10体積のあらかじめ平衡化したTALON樹脂(Clontech)に周辺腔分画を添加し、そして穏やかに振動させながら4℃で1時間インキュベーションした。樹脂混合物を1300rpmで3分間遠心分離し、そして上清をできるだけ多く取り除いた。樹脂を10体積の超音波処理緩衝液で洗浄し、次いで1300rpmで3分間遠心分離した。上清を廃棄した。洗浄した樹脂を1ベッド体積の超音波処理緩衝液に懸濁し、そしてカラムに充填し、これを3ベッド体積の超音波処理緩衝液で洗浄した。2ベッド体積の200mMイミダゾールを用いて、Fabを溶出させた。精製したFabをPBS、pH7.4に透析した。
【0218】
(実施例5)
全長ヒトIFNγ抗体のクローニングおよび発現
以下の方法によって、FAbクローンを全長抗体に変換した。
【0219】
pDSRα19:hIgG1 CHの構築
プラスミドpDSRα19:抗ヒトOPGL IgG1をHindIIIおよびBsmBIで消化して、抗ヒトOPGL可変領域のコード領域を除去した。1.0kbpヒトIgG1定常領域ドメイン(CH1、ヒンジ、CH2およびCH3ドメイン)を含有する直鎖プラスミドpDSRα19:hIgG1 CHをゲル単離し、そしてFAb由来抗IFN−ガンマ可変領域を受け入れさせるのに用いた。
【0220】
pDSRα19:抗IFNガンマBS−A重鎖の構築
抗IFN−ガンマFAb重鎖cDNAをpDSRα19:hIgG1 CHにクローニングして、FAbを全長IgGに変換した。「BS−A」重鎖をコードするプラスミドの構築を本明細書に記載する。類似の方法を用いて、他のFAb重鎖をクローニングした。シグナル配列を持つFAbを生成するため、3工程PCRを行った。まず、FAb cDNAテンプレートとともにプライマー2485−51(以下に示す)および2465−68(以下に示す)を用いた。条件は:Pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。次いで、プライマー2148−98(以下に示す)および2465−68(以下に示す)を用いてPCR産物を増幅し、その後、プライマー2489−36(以下に示す)および2465−68(以下に示す)を用いて増幅した。最終PCR産物をQiagenで精製し、HindIIIおよびBsmBIで切断し、そしてQiagenで精製した。5’コザック(翻訳開始)部位および哺乳動物発現のための以下のシグナル配列:
【0221】
【化8】
【0222】
とともにFAbを含有するこの断片を、pDSRα19:hIgG1 CHに連結した。
【0223】
【化9】
【0224】
pDSRα19:抗IFNガンマBS−A重鎖の構築
FAb軽鎖cDNAをpDSRα19にクローニングして、FAbを全長抗体に変換した。「BS−A」軽鎖をコードするプラスミドの構築を本明細書に記載する。類似の方法を用いて、他のFAbをクローニングした。シグナル配列を持つFAb「BS−A」を生成するため、3工程PCRを行った。まず、FAb cDNAテンプレートとともにプライマー2525−43(以下に示す)および2578−27(以下に示す)を用いた。PCR条件は:pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。次いで、PCR産物をゲル精製し、そして次いで、プライマー2148−98(以下に示す)および2578−27(以下に示す)で増幅した。次に、再び、FAb cDNAテンプレートとともに、プライマー2578−26(以下に示す)および2469−67(以下に示す)を用いた。PCR条件は:Pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。PCR産物をゲル単離し、そして同じ条件を用いて再増幅した。最後にゲル単離したPCR産物をプライマー2489−36(以下に示す)および2469−67(以下に示す)と混合し、そして増幅した。最終PCR産物をQiagenで精製し、XbaIおよびSalIで切断し、そしてQiagenで精製した。5’コザック(翻訳開始)部位および哺乳動物発現のための以下のシグナル配列:
【0225】
【化10】
【0226】
とともにFAbを含有するこの断片を、pDSRα19に連結した。
【0227】
【化11】
【0228】
抗体調製
重鎖および軽鎖全長抗体をコードするcDNAを含有する発現ベクターをCHO細胞にトランスフェクションし、そして重鎖および軽鎖の発現、並びに細胞培地への分泌を可能にする条件下で培養した。0.45μmの酢酸セルロース・フィルター(Corning、マサチューセッツ州アクトン)を通して馴化培地をろ過し、そして塩化カルシウム不含で、そして塩化マグネシウム不含である、PBS−ダルベッコのリン酸緩衝生理食塩水(Gibco BRL Products、ニューヨーク州グランドアイランド)で平衡化しておいたプロテインGセファロース(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)カラムに適用した。試料を適用した後、280nmの吸光度がベースラインに達するまで、カラムをPBSで洗浄した。100mMグリシン、pH2.5を用いて、タンパク質の溶出を達成した。分画を収集し、そして1M Tris−HCl、pH9.2を添加することによって、直ちに中和した。クーマシー染色によってSDS−ポリアクリルアミドゲルを視覚化することによって抗体を検出した。
【0229】
抗体を含有する分画をプールし、Centricon 10(Amicon)または体積がより多い場合は、Centriprep 10(Amicon)いずれかを用いて、濃縮し、そしてPBS中に透析ろ過(diafilter)した。
【0230】
単離抗体を、Superose 6(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)上のゲルろ過によって性質決定し、そして該抗体は、単量体IgGとして移動することが示された。
【0231】
(実施例6)
FabおよびIgGの親和性測定
表面プラズモン共鳴技術(BIAcore、Pharmacia、ニュージャージー州ピスカタウェイ)によって、結合定数(Kd)、結合速度定数(ka)および解離速度定数(kd)を決定した。Fabおよび抗体のBIAcore解析を以下のように行った:BIACORE2000(BIACORE Inc.)を室温で用いて、実験を行った。CHOが発現したhIFNγをCM5チップに固定した。多様な濃度のFabまたはFab IgGをhu−IFNγ表面に注入した。BIAEVALUATION 3.1ソフトウェア(BIACORE, Inc.)を用いてデータを解析した。結果を図30に示す。
【0232】
(実施例7)
FabおよびIgGの活性測定
BIAcore中和アッセイ
IgGに変換されたFabの中和活性をBIAcore上で試験した(実施例6を参照されたい)。結果を図29に示す。BS−B IgGに関して、9nMのIC50で、IFNγ−R1へのhu IFNγ結合の濃度依存性阻害が観察された。
【0233】
A549細胞増殖アッセイ
A549増殖アッセイ(実施例1に記載)において、FabおよびIgGの中和活性測定を以下のように行った:A549細胞を、標的化されたFabまたはIgG(多様な濃度)およびCHOに発現されたhIFNγ(2ng/mlまたは5ng/ml)の混合物で処理した。Fab濃度は、0.3〜150μg/mlの範囲であった。IgG濃度は0.1〜100μg/mlの範囲であった。陽性対照Ab(Pharmingen B27)濃度は、0.01〜5μg/mlの範囲であった。処理5日後、Alamar Blueで細胞を染色し、そして染色4時間後、FL500プレート読取装置上で解析した。BS−A Fab、BS−B FabおよびGP−A Fabに関しては図26に、そしてBS−A IgGおよびBS−B IgGに関しては図27に結果を示す。図26のBS−A FabおよびBS−B Fab、並びに図27のBS−A IgGおよびBS−B IgGは、増殖活性として測定した際、高濃度で、陽性対照よりも約2桁高い中和活性を有することが示された。
【0234】
本発明は、好ましい態様に関して記載されてきているが、当業者には多様な変型および修飾が思い浮かぶであろうことが理解された。したがって、付随する請求項が、請求された際に本発明の範囲内に属するであろう、こうした同等の変型をすべて含むと意図された。
【図面の簡単な説明】
【0235】
【図1】図1は、主なファージFabクローンのhIFNγに対する反応性に関するELISAの結果を示すグラフである。2%MPBSであらかじめブロッキングしたファージ懸濁物を、ウェルあたり最大100μl用いて、ファージ希釈を行い、ELISAにおいて典型的な109〜1011ファージ/ウェルの範囲とした。実施例3に記載するように、ELISA用のファージストックを調製した。値は、単一点測定によるものであり、そしてシグナル検出のため、OD405を測定した。
【図2】図2は、主なFab「GP−A」および「BS−B」クローンのhIFNγに対する反応性に関する、用量依存性クローンファージELISAの結果を示すグラフである。2%MPBSであらかじめブロッキングしたファージ懸濁物を、ウェルあたり最大100μl用いて、ファージ希釈を行い、ELISAにおいて典型的な109〜1011ファージ/ウェルの範囲とした。実施例3に記載するように、ELISA用のファージストックを調製した。値は、単一点測定によるものであり、そしてシグナル検出のため、OD405を測定した。
【図3】図3は、Fab「BS−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図4】図4は、Fab「BS−B」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図5】図5は、Fab「RD−B1」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図6】図6は、Fab「RD−A2」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図7】図7は、Fab「58C」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図8】図8は、Fab「GP−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図9】図9は、Fab「57D」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図10】図10は、Fab「57E」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図11】図11は、Fab「IFN−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図12】図12は、Fab「67C」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図13】図13は、Fab「59−A2」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図14】図14は、Fab「BS−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図15】図15は、Fab「BS−B」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図16】図16は、Fab「RD−B1」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図17】図17は、Fab「RD−A2」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図18】図18は、Fab「58C」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図19】図19は、Fab「GP−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図20】図20は、Fab「57D」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図21】図21は、Fab「57E」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図22】図22は、Fab「IFN−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図23】図23は、Fab「67C」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図24】図24は、Fab「59−A2」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図25】図25は、Fab「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」の重鎖および軽鎖相補性決定領域(CDR)のアミノ酸配列の比較を示す。
【図26】図26は、A549細胞アッセイで測定した際のFab「BS−A」および「BS−B」の中和活性を示すグラフである。実施例4に記載するようにFabを精製し、そして0.3〜150μg/mlのFab濃度範囲で添加した。Pharmingen B27 Ab(0.01〜5μg/mlの濃度範囲)を陽性対照として用いた。処理5日後、細胞をAlamar Blueで染色し、そしてFL500プレート読取装置上で、染色4時間後に解析した。
【図27】図27は、A549細胞アッセイで測定した際の「BS−A」IgGおよび「BS−B」IgGの中和活性を示すグラフである。実施例4に記載するようにIgGを精製し、そして0.1〜100μg/mlのIgG濃度範囲で添加した。Pharmingen B27 Ab(0.01〜5μg/mlの濃度範囲)を陽性対照として用いた。hIFNγと反応しない無関係なAb、AT−IgG(0.01〜5μg/mlの濃度範囲)を陰性対照として用いた。処理5日後、細胞をAlamar Blueで染色し、そしてFL500プレート読取装置上で、染色4時間後に解析した。
【図28】図28は、BiaCoreによって、そしてA549細胞アッセイにおいて測定した際の、「BS−A」、「BS−B」、「RD−A2」、「RD−B」、「IFN−A」、「57E」、「57D」、「GP−A」、「58C」および「67C」IgGの親和性および中和活性の比較を提供する表である。BIAEVALUATIONを用いて、BIACOREデータを解析した。
【図29】図29は、BIACoreで測定した際の「BS−A」IgGおよび「BS−B」IgGの中和活性を示すグラフである。相対結合反応(%)を、試料濃度(nM)に対してプロットしている。
【図30】図30は、BIACoreによって測定した際の、抗IFNγ Fab「BS−A」、「BS−B」、「IFN−A」および「GP−A」、並びに対応するIgGの親和性の比較を提供する表である。
【図31】図31は、図3〜24に示すFabアミノ酸配列の比較を示す。重鎖および軽鎖Fab「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」の予測されるアミノ酸配列を、同一性および類似性に関して比較した。GCGの「BestFit」プログラムを用いて、Fabの各対間の同一性および類似性パーセントを得た。
【図32】図32は、重鎖および軽鎖「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」Fabの相補性決定領域(CDR)の並列を示す。
【図33】図33は、予測されるFab「BS−A」、「RD−A2」および「IFN−A」重鎖アミノ酸配列(それぞれ、図3、6および11の両端を含む残基1〜120、1〜127および1〜126)と、VH4ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列4−34、D領域配列1−1、3−3または3−16、およびJ領域配列JH4を含む。FR1、FR2およびFR3は、3つのフレームワーク領域を指定し、CDR1、CDR2およびCDR3は、3つの相補性決定領域を指定し、そしてH1、H2およびH3は、フレームワーク領域およびCDRの間の対応する接合部配列を指定する。「BS−A」、「RD−A2」、「IFN−A」および生殖系列V、D、またはJ配列の間の相違を太字で示す。図33〜46の生殖系列アミノ酸残基の番号付けは、Kabatら, Sequences of Proteins of Immunological Interest, 米国保健社会福祉省, 第4版(1991)に記載されるとおりである。
【図34】図34は、予測されるFab「BS−B」、および「59−A2」重鎖アミノ酸配列(それぞれ、図4および13の両端を含む残基1〜121および1〜120)と、VH1ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列1−18、D領域配列6−13、1−1または1−7、およびJ領域配列JH4を含む。
【図35】図35は、予測されるFab「RD−B1」重鎖アミノ酸配列(図5の両端を含む残基1〜119)と、VH2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−26、D領域配列3−22、およびJ領域配列JH5を含む。
【図36】図36は、予測されるFab「58C」重鎖アミノ酸配列(図7の両端を含む残基1〜119)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−21、未知のD領域配列、およびJ領域配列JH4を含む。
【図37】図37は、予測されるFab「GP−A」重鎖アミノ酸配列(図8の両端を含む残基1〜124)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−30.3、D領域配列3−10、およびJ領域配列JH3を含む。
【図38】図38は、予測されるFab「57D」重鎖アミノ酸配列(図9の両端を含む残基1〜117)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−53、D領域配列3−16、および未知のJ領域配列を含む。
【図39】図39は、予測されるFab「57E」重鎖アミノ酸配列(図10の両端を含む残基1〜118)と、VH4ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列4−61、D領域配列7−27、およびJ領域配列JH4を含む。
【図40】図40は、予測されるFab「67C」重鎖アミノ酸配列(図12の両端を含む残基1〜119)と、VH2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−05、D領域配列5−18、およびJ領域配列JH6を含む。
【図41】図41は、予測されるFab「BS−A」、「BS−B」、「57E」および「59−A2」軽鎖アミノ酸配列(それぞれ、図14、15、21および24の両端を含む残基1〜111、1〜110、1〜112および1〜110)と、Vλ6ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列6aおよび未知のJ領域配列あるいはJL2またはJL3を含む。
【図42】図42は、予測されるFab「RD−B1」軽鎖アミノ酸配列(図16の両端を含む残基1〜112)と、Vκ2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A23およびJ領域配列JK2を含む。
【図43】図43は、予測されるFab「RD−A2」、「IFN−A」および「67C」軽鎖アミノ酸配列(それぞれ、図17、22および23の両端を含む残基1〜112、1〜110、1〜112および1〜110)と、Vκ2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A19、並びにそれぞれJ領域配列JK3、JK2およびJK2を含む。
【図44】図44は、予測されるFab「58C」軽鎖アミノ酸配列(図18の両端を含む残基1〜108)と、Vλ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3hおよび未知のJ領域配列を含む。
【図45】図45は、予測されるFab「GP−A」軽鎖アミノ酸配列(図19の両端を含む残基1〜106)と、Vλ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−19およびJ領域配列JL2またはJL3を含む。
【図46】図46は、予測されるFab「57D」軽鎖アミノ酸配列(図20の両端を含む残基1〜108)と、Vκ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A27およびJ領域配列JK5を含む。
【図47】図47は、Fabクラスの比較を示す。多配列比較解析用のGCG(遺伝学コンピュータグループ, 575 Science Drive, Madison, WI 53711)PileUpプログラムを用いて、Fabクラス比較を行った。記号(**)は、既知の生殖系列配列には関連していたが、最も密接なマッチング多様性(D)領域または連結領域(J)が決定不能であったことを示す。記号(*)は、同定された連結領域に比較して、1、2または3残基の変異が起きたことを示す。
【配列表】
【発明の詳細な説明】
【0001】
発明の分野
本発明は、ヒト・インターフェロン・ガンマ(hIFNγ)に結合し、そして該分子と同族(cognate)受容体、IFNγ−Rとの相互作用を阻害し、そして/またはIFNγに誘発される生物学的作用を修飾する、新規完全ヒト抗体Fab断片に関する。より具体的には、本発明は、ファージ・ディスプレー・ライブラリーであって、ユニークなFab断片を含有し、該断片が次いで、全長ヒトIgG抗体に変換される、前記ファージ・ディスプレー・ライブラリーのhIFNγ親和性選択を通じて単離される中和Fab断片(Fab)に関する。hIFNγ中和活性、高親和性、およびin vivoでの長い半減期という望ましい特質を有する、hIFNγに対するこれらの新規完全ヒト抗体を用いて、多様な自己免疫および炎症性疾患を防止するかまたは治療することも可能である。本発明の完全ヒトFabを産生するための核酸分子、ベクターおよび宿主細胞もまた提供する。
【0002】
発明の背景
抗体は、生物薬剤学研究および薬剤発見の試みにおいて、長年、非常に重要な役割を果たしてきた。ヒト疾患の治療のための療法剤としての抗体の有用性は、抗体が:(a)in vivoでの半減期が長く;(b)高い親和性および特異性で標的(単数または複数)に結合可能であり;そして(c)免疫エフェクター機能(補体結合および抗体依存性細胞傷害作用など)を仲介する能力を持ちうるため、長年、理想とされてきた。
【0003】
しかし、今までは、非ヒト種から得た抗体に、不都合な免疫原性があり、抗体の長期臨床的有用性が制限されることから、療法抗体の概念を実行することは、大幅に制限されていた。最近の技術的進歩によって、免疫原性がより低く、そして長期の療法潜在能力を持つ、完全ヒト抗体を得る手段を提供することによって、これらの制限を克服する新規の方法が提供されてきている。さらに、コンビナトリアル・ライブラリー法および抗体操作における発展によって、抗体親和性、半減期、および/またはエフェクター機能を修飾する機会が得られるようになってきた。
【0004】
高親和性、高特異性であり、そしてin vivoでアゴニスト活性またはアンタゴニスト活性を持つ抗体を発見するのに、ファージ・コートタンパク質に融合された抗体断片の繊維状ファージ・ディスプレー・コンビナトリアル・ライブラリー(いわゆる「ファージ・ディスプレー・ライブラリー」)を使用する、こうした技術の1つが有効に用いられてきている。
【0005】
ヒト・インターフェロン・ガンマ(hIFNγ)は、活性化Tリンパ球およびナチュラルキラー細胞に産生されるリンホカインである。該分子は、抗増殖活性、抗ウイルス活性および免疫調節活性を示し、そして免疫系の大部分の初代細胞上にあるヘテロ二量体受容体、hIFNγ−Rに結合し;Langerら, Immunology Today, 9:393(1988)、そして炎症につながる事象カスケードを誘発する。IFNγの抗ウイルス活性および免疫調節活性は、いくつかの臨床状態に有益な影響を有することが知られる。しかし、IFNγ活性が有害な影響を有することが知られる、多くの臨床設定がある。例えば、自己免疫疾患は、高レベルの血中hIFNγと関連し、そして現在、IFNγの隔絶が、関節リウマチ(RA)、全身性エリテマトーデス(SLE)および多発性硬化症(MS)などの自己免疫疾患の症状軽減と関連することを示唆する証拠がある:例えばSkurkovichら, Intern. Journal of Immunotherapy, 14:23−32(1998);Gerezら, Clin. Exp. Immunol., 109:296−303(1997)を参照されたい。IFNγ活性はまた、悪液質、敗血症ショックおよびクローン病などの疾患状態にも関連付けられてきている。
【0006】
hIFNγのその受容体への相互作用を遮断することは、これに関連する最も上流の段階に介入することに相当するため、hIFNγ中和活性を持つ完全ヒト抗体は、魅力的な療法製品候補に相当する。ヒト・インターフェロン−ガンマ(hIFNγ)を療法標的として用いる抗体を同定するのに、ファージ・ディスプレー・ライブラリー技術を使用することが、本発明の目的である。
【0007】
発明の概要
本発明は、ヒト・インターフェロン−ガンマ(hIFNγ)に結合する、新規完全ヒト抗体Fab断片を提供する。1つの態様において、完全ヒト抗体Fab断片は、hIFNγとその同族受容体、hIFNγ−Rとの相互作用を、部分的にまたは完全に阻害し、そしてそれによって、hIFNγ活性を、部分的にまたは完全に阻害する方式で、hIFNγに結合する;すなわち抗体は、hIFNγのアンタゴニストである。好ましくは、hIFNγは哺乳動物hIFNγである。より好ましくは、hIFNγは、可溶性型または細胞表面結合型であることも可能なヒトhIFNγ、あるいはその断片、誘導体および変異体(variant)である。
【0008】
本発明の抗体は、ネズミまたはヒトhIFNγなどのhIFNγ、好ましくはヒトhIFNγ、あるいはその免疫原性断片、誘導体または変異体で、動物を免疫することによって、調製可能である。さらに、hIFNγがトランスフェクション細胞で発現されて、そしてその表面に結合するように、hIFNγをコードする核酸分子を含有するベクターでトランスフェクションされた細胞を用いて、動物を免疫することも可能である。あるいは、hIFNγへの結合に関して、抗体または抗原結合ドメイン配列を含むライブラリーをスクリーニングすることによって、抗体を得ることも可能である。こうしたライブラリーは、組み立てられたファージ粒子表面上に発現されるバクテリオファージ・コートタンパク質へのタンパク質融合体またはペプチド融合体、およびファージ粒子内に含有されるコードDNA配列として、バクテリオファージ中で、好適に調製される(いわゆる「ファージ・ディスプレー・ライブラリー」)。1つの例において、ファージ・ディスプレー・ライブラリーは、可変軽鎖および重鎖などのヒト抗体をコードするDNA配列を含有する。
【0009】
抗体または抗原結合ドメインは、hIFNγに結合可能であり、そして部分的にまたは完全にhIFNγ活性を中和する、天然抗体に類似の四量体糖タンパク質であることも可能であるし、あるいは一本鎖抗体;Fv、Fab、Fab’またはF(ab)’断片、二重特異性抗体、異種抗体、あるいはそれらの他の断片、変異体、または誘導体であることも可能である。抗体または抗原結合ドメインを、ハイブリドーマ細胞株(例えばマウス骨髄腫細胞に融合させた脾臓細胞などの、抗体産生細胞)で産生することも可能であるし、あるいは前記抗体または抗原結合ドメインをコードする核酸分子でトランスフェクションした異種細胞株で産生することも可能である。
【0010】
本発明の抗体または抗原結合ドメインは:
(a)図3〜13(配列番号65〜配列番号86)に示すようなFab重鎖アミノ酸配列;
(b)(a)の配列の保存的アミノ酸置換を含む、重鎖アミノ酸配列;
(c)(a)の配列に少なくとも約80%同一である、重鎖アミノ酸配列;あるいは
(d)(a)、(b)または(c)の断片または誘導体
を含み、hIFNγに選択的に結合する。
【0011】
別の態様において、本発明の抗体または抗原結合ドメインは、上記と同一の図3〜13に示すようなFab重鎖アミノ酸配列(配列番号65〜配列番号86)および図14〜24に示すようなFab軽鎖アミノ酸配列(配列番号87〜配列番号108)を含む抗体または抗原結合ドメインに認識される、ヒトhIFNγ上のエピトープを認識する。
【0012】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:
各Vl鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vl)、CDR2(Vl)およびCDR3(Vl)と称されるCDRアミノ酸配列を含み、CDR1(Vl)が:
【0013】
【化1】
【0014】
からなる群より選択され、
CDR2(Vl)が:
【0015】
【化2】
【0016】
からなる群より選択され、
そしてCDR3(Vl)が:
【0017】
【化3】
【0018】
からなる群より選択され、
CDR1(Vl)、CDR2(Vl)およびCDR3(Vl)が互いに独立に選択され;そして
各Vh鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vh)、CDR2(Vh)およびCDR3(Vh)と称されるCDRアミノ酸配列を含み、CDR1(Vh)が:
【0019】
【化4】
【0020】
からなる群より選択され、
CDR2(Vh)が:
【0021】
【化5】
【0022】
からなる群より選択され、
CDR3(Vh)が:
【0023】
【化6】
【0024】
からなる群より選択され;
CDR1(Vh)、CDR2(Vh)およびCDR3(Vh)が互いに独立に選択される。
【0025】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0026】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIASNYVQ(配列番号02)を有するCDR1、配列EDNQRPS(配列番号13)を有するCDR2、および配列QSYDGSAWV(配列番号24)を有するCDR3を含み;そして
Vh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列TSWNAGGPIDY(配列番号55)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0027】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TRSSGSIASYYVQ(配列番号03)を有するCDR1、配列EDDQRPS(配列番号14)を有するCDR2、および配列QSYDRNSLV(配列番号25)を有するCDR3を含み;そして
Vh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列DRVGYSSSLLDY(配列番号56)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0028】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RATQSLLHGNGHNYLD(配列番号04)を有するCDR1、配列MGSNRAS(配列番号15)を有するCDR2、および配列MQALQLPPT(配列番号26)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号36)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号46)を有するCDR2、および配列DKGSRITIFGVVGSAGFDY(配列番号57)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0029】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLVHSDGNTYLS(配列番号05)を有するCDR1、配列KISNRFS(配列番号16)を有するCDR2、および配列MQATQLPYT(配列番号27)を有するCDR3を含み;そして
Vh鎖が、配列NARMGVS(配列番号37)を有するCDR1、配列HIFSNDEESYSTSLKS(配列番号47)を有するCDR2、および配列LLLYEGFDP(配列番号58)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0030】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列SGDVLARKYAR(配列番号06)を有するCDR1、配列KDRERPS(配列番号17)を有するCDR2、および配列YSAADNRGV(配列番号28)を有するCDR3を含み;そして
Vh鎖が、配列SYAMH(配列番号38)を有するCDR1、配列VISYDGSNKYYADSVKG(配列番号48)を有するCDR2、および配列DLVLTMTSRRAAFDI(配列番号59)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0031】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列GGDNLGGKSLH(配列番号07)を有するCDR1、配列DDSDRPS(配列番号18)を有するCDR2、および配列QVWDGSSDQRV(配列番号29)を有するCDR3を含み;そして
Vh鎖が、配列SYSMN(配列番号39)を有するCDR1、配列SISSGSSYRYDADSVKG(配列番号49)を有するCDR2、および配列DQWGTISGNDY(配列番号60)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0032】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLLHTNEYNYLD(配列番号08)を有するCDR1、配列LGSNRAP(配列番号19)を有するCDR2、および配列MQALQTPRT(配列番号30)を有するCDR3を含み;そして
Vh鎖が、配列GYYWS(配列番号40)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号50)を有するCDR2、および配列GWPTYVWGSYRPKGYFDY(配列番号61)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0033】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列TGSSGSIANNYVH(配列番号09)を有するCDR1、配列EDDQRPS(配列番号20)を有するCDR2、および配列QSYDNSNSFVV(配列番号31)を有するCDR3を含み;そして
Vh鎖が、配列SGGYSWS(配列番号41)を有するCDR1、配列YIYHSGSTYYNPSLKS(配列番号51)を有するCDR2、および配列GDWGYFDY(配列番号62)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0034】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RASQYVSSNSLA(配列番号10)を有するCDR1、配列GASNRAT(配列番号21)を有するCDR2、および配列QQYGSSPIT(配列番号32)を有するCDR3を含み;そして
Vh鎖が、配列SNYMS(配列番号42)を有するCDR1、配列VIYSGGSTYYADSVKG(配列番号52)を有するCDR2、および配列DADGGDYGY(配列番号63)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0035】
別の態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み:Vl鎖が、配列RSSQSLLRSNGYNYLA(配列番号11)を有するCDR1、配列LASNRAS(配列番号22)を有するCDR2、および配列VHGVHIPYT(配列番号33)を有するCDR3を含み;そして
Vh鎖が、配列SNEAGVG(配列番号43)を有するCDR1、配列LLYWDDDKRYSPSLRS(配列番号53)を有するCDR2、および配列RLVRYGGYSTGGFDV(配列番号64)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている。
【0036】
本発明の抗体および抗原結合ドメインは、軽鎖および重鎖アミノ酸配列をコードするゲノムDNAに存在する生殖系列核酸由来である。抗体は、生殖系列配列再編成突然変異および体細胞突然変異の産物である核酸配列にコードされる。
【0037】
1つの態様において、本発明の抗体または抗原結合ドメインは、Vl鎖およびVh鎖を含み、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含み;そして抗体は、IFNγポリペプチドに選択的に結合する。
【0038】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図34におけるようなVH1生殖系列遺伝子(配列番号123)の再編成変異体または体細胞変異体を含む。
【0039】
別の態様において、Vl鎖が、図42におけるようなVκ2生殖系列遺伝子(配列番号131)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図35におけるようなVH2生殖系列遺伝子(配列番号124)の再編成変異体または体細胞変異体を含む。
【0040】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含む。
【0041】
別の態様において、Vl鎖が、図44におけるようなVλ3生殖系列遺伝子(配列番号133)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図36におけるようなVH3生殖系列遺伝子(配列番号125)の再編成変異体または体細胞変異体を含む。
【0042】
別の態様において、Vl鎖が、図45におけるようなVλ3生殖系列遺伝子(配列番号134)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図37におけるようなVH3生殖系列遺伝子(配列番号126)の再編成変異体または体細胞変異体を含む。
【0043】
別の態様において、Vl鎖が、図46におけるようなVκ3生殖系列遺伝子(配列番号135)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図38におけるようなVH3生殖系列遺伝子(配列番号127)の再編成変異体または体細胞変異体を含む。
【0044】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図39におけるようなVH4生殖系列遺伝子(配列番号128)の再編成変異体または体細胞変異体を含む。
【0045】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図33におけるようなVH4生殖系列遺伝子(配列番号122)の再編成変異体または体細胞変異体を含む。
【0046】
別の態様において、Vl鎖が、図43におけるようなVκ2生殖系列遺伝子(配列番号132)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図40におけるようなVH2生殖系列遺伝子(配列番号129)の再編成変異体または体細胞変異体を含む。
【0047】
別の態様において、Vl鎖が、図41におけるようなVλ6生殖系列遺伝子(配列番号130)の再編成変異体または体細胞変異体を含み;そしてVh鎖が、図34におけるようなVH1生殖系列遺伝子(配列番号123)の再編成変異体または体細胞変異体を含む。
【0048】
本発明の選択的結合剤(抗体または抗原結合ドメイン)は、IFNγの少なくとも1つの活性、例えばIFNγのIFNγ−Rへの結合を、部分的にまたは完全に阻害する。
1つの態様において、IFNγアンタゴニスト、例えば抗体または抗原ドメインは、狼瘡様疾患、関節炎、または多発性硬化症様症候群を経験しているか、またはこれらを発展させるリスクがある動物に投与される。IFNγアンタゴニストを用いて、ループス腎炎、関節リウマチ、および/または多発性硬化症を防止し、そして/または治療することも可能である。
【0049】
本発明の抗体または抗原結合ドメインおよび薬学的に許容しうるキャリアーを含む組成物もまた、提供する。
【0050】
発明の詳細な説明
本発明は、ヒト・インターフェロン−ガンマ・タンパク質(hIFNγ)に選択的に結合する剤(「選択的結合剤」)を提供する。好ましくは、該剤は、IFNγアンタゴニストまたは阻害剤であり、IFNγの少なくとも1つの活性、例えばIFNγのその同族受容体への結合を、部分的にまたは完全に阻害する。1つの態様において、完全ヒト抗体断片は、IFNγのその同族受容体への結合を、部分的にまたは完全に遮断し、そしてIFNγ活性を、部分的にまたは完全に阻害するように、IFNγに選択的に結合する。
【0051】
用語「選択的結合剤」は、IFNγに優先的に結合する分子を指す。選択的結合剤には、タンパク質、ペプチド、核酸、炭水化物、脂質、または小分子量化合物が含まれることも可能である。好ましい態様において、選択的結合剤は抗体であり、例えば可溶性型または結合型で、標識されていることも可能である、ポリクローナル抗体、モノクローナル抗体(mAb)、キメラ抗体、CDR移植抗体、抗体に対する抗イディオタイプ(抗Id)抗体とともに、限定されるわけではないが、酵素切断、ペプチド合成または組換え技術を含む既知の技術によって提供される、それらの断片、領域または誘導体である。本発明の抗IFNγ選択的結合剤は、IFNγの部分に結合することが可能であり、IFNγのIFNγ−R受容体への結合を阻害する。
【0052】
本発明の抗体および抗原結合ドメインは、IFNγに選択的に結合し、すなわち、これらは他の抗原より高い結合親和性で、優先的にIFNγに結合する。抗体は、ヒトIFNγに選択的に結合するが、ネズミIFNγなどの非ヒトIFNγにもまた検出可能であるように結合することも可能である。あるいは、抗体は、非ヒトIFNγに選択的に結合するが、ヒトIFNγにもまた検出可能であるように結合することも可能である。あるいは、抗体は、もっぱらヒトIFNγに結合し、非ヒトIFNγには検出可能に結合しないことも可能である。
【0053】
用語「モノクローナル抗体」は、実質的に均質な抗体の集団から得られる抗体を指し、ここで、各モノクローナル抗体は、典型的には、抗原上の単一のエピトープを認識するであろう。用語「モノクローナル」は、抗体を作成するいかなる特定の方法にも限定されない。例えば、本発明のモノクローナル抗体は、Kohlerら;Nature, 256:495(1975)に記載されるようなハイブリドーマ法によって作成することも可能であるし、また例えば本明細書記載の技術を用いて、ファージ・ライブラリーから単離することも可能である。
【0054】
用語「抗原結合ドメイン」または「抗原結合領域」は、選択的結合剤(例えば抗体分子)の部分であって、抗原と相互作用し、そして該結合剤に、抗原に対する特異性および親和性を与えるアミノ酸残基を含有する、前記部分を指す。好ましくは、抗原結合領域は、ヒト起源のものであろう。他の態様において、抗原結合領域は、他の動物種、特にウサギ、ラットまたはハムスターなどのげっ歯類に由来することも可能である。
【0055】
用語「エピトープ」は、1以上の結合剤の抗原結合領域で、選択的結合剤(例えば抗体)によって認識され、そして結合されることが可能な分子いずれかの部分を指す。エピトープは通常、アミノ酸または糖側鎖などの、分子の化学的に活性である表面集団からなり、そして特定の三次元構造特性とともに特定の電荷特性を有する。「阻害性エピトープおよび/または中和エピトープ」は、選択的結合剤に結合された際、そのエピトープを含有する分子または生物のin vivo、in vitro、またはin situでの、より好ましくはin vivoでの、IFNγのその受容体への結合を含めた、生物学的活性の損失を生じるエピトープを意図する。用語「軽鎖」は、抗体に関して用いた場合、定常ドメインのアミノ酸配列に基づいて、カッパ(κ)またはラムダ(λ)と呼ばれる2つの別個の種類を指す。
【0056】
用語「重鎖」は、抗体に関して用いた場合、重鎖定常ドメインのアミノ酸配列に基づいて、アルファ、デルタ、イプシロン、ガンマおよびミューと呼ばれる5つの別個の種類を指す。重鎖のこれらの別個の種類は、抗体の5つのクラス、それぞれ、IgA、IgD、IgE、IgGおよびIgMを生じさせ、これには、IgGの4つのサブクラス、すなわちIgG1、IgG2、IgG3およびIgG4が含まれる。
【0057】
用語「可変領域」または「可変ドメイン」は、典型的には、重鎖ではアミノ末端の約120〜130アミノ酸、そして軽鎖では約100〜110アミノ酸であり、抗体間で配列が大規模に異なり、そして特定の抗原に対する特定の各抗体の結合および特異性において用いられる、軽鎖の部分および重鎖の部分を指す。配列の可変性は、相補性決定領域(CDR)と呼ばれる領域に集中しており、一方、可変ドメイン中のよりよく保存される領域は、フレームワーク領域(FR)と呼ばれる。軽鎖のCDRおよび重鎖のCDRは、抗原と抗体の相互作用に関与する。
【0058】
用語「定常領域」または「定常ドメイン」は、抗体の抗原への結合に直接関与していないが、Fc受容体との相互作用など、多様なエフェクター機能を示す、軽鎖のカルボキシ末端部分および重鎖のカルボキシ末端部分を指す。
【0059】
用語「ヒト・インターフェロン−ガンマ」または「ヒト・インターフェロン−ガンマ・ポリペプチド」は、その開示が本明細書に援用されるPCT公報WO 83/04053に記載されるアミノ酸配列を含むポリペプチド、および関連ポリペプチドを指す。関連ポリペプチドには、対立遺伝子変異体;スプライス変異体;断片;誘導体;置換変異体、欠失変異体、および挿入変異体;融合ポリペプチド;並びに種間相同体(homolog)が含まれる。免疫反応を生じるのに十分なIFNγの可溶性型もまた含まれる。IFNγは、本明細書に定義するような成熟ポリペプチドであることも可能であり、そして調製した方法に応じて、アミノ末端メチオニン残基を有することもまた持たないことも可能である。
【0060】
用語「断片」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、全長アミノ酸配列に満たない配列を含むペプチドまたはポリペプチドを指す。こうした断片は、例えばアミノ末端の一部切除(truncation)、カルボキシ末端の一部切除、および/またはアミノ酸配列からの残基(単数または複数)の内部欠失から生じることも可能である。断片は、選択的RNAスプライシング、またはin vivoプロテアーゼ活性から生じることも可能である。
【0061】
用語「変異体」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、天然配列または非修飾配列に比較した際に、1以上のアミノ酸配列置換、欠失、および/または付加を含むペプチドまたはポリペプチドを指す。例えば、IFNγ変異体は、天然IFNγのアミノ酸配列に対する1以上の変化から生じることも可能である。やはり例として、IFNγの選択的結合剤の変異体は、天然のまたはあらかじめ修飾されていない選択的結合剤のアミノ酸配列に対する1以上の変化から生じることも可能である。変異体は、対立遺伝子変異体またはスプライス変異体などの天然存在変異体であることも可能であるし、あるいは人工的に構築することも可能である。ポリペプチド変異体は、前記変異体をコードする、対応する核酸分子から調製することも可能である。
【0062】
用語「誘導体」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、化学的に修飾されているポリペプチドまたはペプチド、あるいはその変異体、断片または誘導体を指す。例には、1以上のポリマー、例えば水溶性ポリマー、N連結、またはO連結炭水化物、糖、リン酸、および/または他のこうした分子の共有結合が含まれる。誘導体は、付着する分子の種類または位置いずれかが、天然存在または出発ペプチドまたはポリペプチドと異なる方式で修飾されている。誘導体には、ペプチドまたはポリペプチド上に天然に存在する1以上の化学基の欠失がさらに含まれる。
【0063】
用語「融合」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、ペプチドまたはポリペプチド、あるいはその断片、変異体および/または誘導体と、異種ペプチドまたはポリペプチドとの連結を指す。
【0064】
用語「生物学的に活性」は、IFNγに関連して、またはタンパク質性選択的結合剤に関連して用いた場合、IFNγまたは選択的結合剤の少なくとも1つの活性特性を有するペプチドまたはポリペプチドを指す。IFNγの選択的結合剤は、IFNγの少なくとも1つの生物学的活性に関して、アゴニスト活性、アンタゴニスト活性、あるいは中和活性または遮断活性を有することも可能である。
【0065】
用語「天然存在」は、核酸分子、ポリペプチド、宿主細胞等の生物学的材料に関連して用いた場合、天然に見出され、そして人間によって操作されていないものを指す。
用語「単離」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、天然環境で見出される混入ポリペプチドを少なくとも1つ含まないペプチドまたはポリペプチドを指し、そして好ましくは、療法使用および診断使用に干渉するであろう、いかなる他の混入哺乳動物ポリペプチドも実質的に含まないペプチドまたはポリペプチドを指す。
【0066】
用語「成熟」は、IFNγに関連して、またはIFNγのタンパク質性選択的結合剤に関連して用いた場合、リーダー配列を欠くペプチドまたはポリペプチドを指す。該用語はまた、アミノ末端(リーダー配列を含むまたは含まない)および/またはカルボキシ末端のタンパク質分解的プロセシング、より大きい前駆体からのより小さいポリペプチドの切断、N連結および/またはO連結グリコシル化等のペプチドまたはポリペプチドの他の修飾も含むことも可能である。
【0067】
用語「有効量」および「療法的有効量」は、IFNγの選択的結合剤に関連して用いた場合、IFNγの1以上の生物学的活性レベルの観察可能な変化を支持するのに有用であるかまたは必要である、選択的結合剤の量を指す。前記変化は、IFNγ活性レベルの増加または減少いずれであることも可能である。
【0068】
用語「保存的アミノ酸置換」は、その位でのアミノ酸残基の極性または電荷にほとんどまたはまったく影響しないような、天然アミノ酸残基の非天然残基での置換を指す。例えば、保存的置換は、ポリペプチド中の非極性残基を他の非極性残基いずれかで置換することから生じる。さらに、アラニン・スキャニング突然変異誘発に関して、先に記載されているように、ポリペプチド中の天然残基いずれかをアラニンで置換することもまた可能である;Cunninghamら, Science, 244:1081−1085(1989)。保存的アミノ酸置換の典型的な規則を表Iに示す。
【0069】
表I
保存的アミノ酸置換
【0070】
【表1】
【0071】
保存的アミノ酸置換はまた、典型的には、生物学的系における合成よりも化学的ペプチド合成によって取り込まれる、非天然存在アミノ酸残基も含む。これらには、ペプチド模倣体、およびアミノ酸部分の他の逆転型または反転型が含まれる。
【0072】
アミノ酸配列に対する保存的修飾(およびコードするヌクレオチドに対する対応する修飾)は、天然存在IFNγまたは選択的結合剤と類似の機能特性および化学特性を有するIFNγポリペプチド(およびそのタンパク質性選択的結合剤)を産生可能である。対照的に、(a)置換領域における分子主鎖の構造、例えばシートまたはらせんコンホメーション、(b)標的部位での、分子の電荷または疎水性、あるいは(c)側鎖バルクの維持に対する影響が有意に異なる置換を選択することによって、IFNγ(およびそのタンパク質性選択的結合剤)の機能特性および/または化学特性の実質的な修飾を達成することも可能である。天然存在残基は、共通の側鎖特性に基づいて、グループに分けることも可能である:
1)疎水性:ノルロイシン、Met、Ala、Val、Leu、Ile;
2)中性親水性:Cys、Ser、Thr;
3)酸性:Asp、Glu;
4)塩基性:Asn、Gln、His、Lys、Arg;
5)鎖配向に影響を及ぼす残基:Gly、Pro;および
6)芳香族:Trp、Tyr、Phe。
【0073】
非保存的置換は、これらの種類の1つのメンバーと別の種類のメンバーとの交換を伴うことも可能である。
2以上の核酸分子および/またはポリペプチドの「同一性または類似性」は、2以上の別個の配列の関連性の測定値を提供する。用語「同一性」は、2つの別個のアミノ酸配列の対応する位で同一であるアミノ酸を指す。用語「類似性」は、2つの別個のアミノ酸配列の対応する位で、同一であるか、または上に定義するような保存的置換であるアミノ酸を指す。
【0074】
同一性または類似性の度合いは、限定されるわけではないが、Computational Molecular Biology, Lesk, A.M.監修, Oxford University Press, ニューヨーク, 1988;Biocomputing:Informatics and Genome Projects, Smith, D.W.監修, Academic Press, ニューヨーク, 1993;Computer Analysis of Sequence Data, Part 1, Griffin, A.M.およびGriffin, H.G.監修, Humana Press, ニュージャージー, 1994;Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987;Sequence Analysis Primer, Gribskov, M.およびDevereux, J.監修, M. Stockton Press, ニューヨーク, 1991;およびCarilloら, SIAM J. Applied Math., 48:1073(1988)に記載されるものを含む、既知の方法によって、容易に計算可能である。
【0075】
同一性および/または類似性を決定する好ましい方法を設計して、試験する配列間の最大のマッチを得る。同一性および類似性を決定する方法は、公的に入手可能なコンピュータプログラムに体系化されている。2つの配列間の同一性および類似性を決定する典型的なコンピュータプログラム法には、限定されるわけではないが、GAP;Devereuxら, Nucleic Acids Research, 12:387(1984);遺伝学コンピュータグループ、ウィスコンシン大学、ウィスコンシン州マディソン;BLASTP、BLASTNおよびFASTA、Altschulら,J. Mol. Biol., 215:403−410(1990)を含むGCGプログラムパッケージが含まれる。BLAST Xプログラムは、米国バイオテクノロジー情報センター(NCBI)および他の供給源(BLAST Manual, Altschulら NCB NLM NIH メリーランド州ベセスダ)から公的に入手可能である。周知のSmith Watermanアルゴリズムを用いて、同一性を決定することもまた可能である。
【0076】
IFNγポリペプチド
本発明の選択的結合剤をスクリーニングし、そして同定するため、IFNγポリペプチド、並びにその断片、変異体および誘導体を標的分子として用いる。選択的結合剤として抗体を調製することが望ましい場合、IFNγポリペプチドは、好ましくは免疫原性であり、すなわち、これらは動物に投与された際に、免疫応答を誘発する。あるいは、in vitro技術によって抗体を調製する場合、標的分子として用いられるIFNγポリペプチドは、抗体または抗原結合ドメインと検出可能に結合することが可能である。
【0077】
生物学的方法または化学的方法によって、IFNγポリペプチドを調製する。組換えIFNγをコードするDNA配列の発現などの生物学的な方法が当該技術分野に知られる;例えば、Sambrookら、上記を参照されたい。Merrifieldら, J. Am. Chem. Soc., 85:2149(1963)、Houghtenら, Proc Natl Acad. Sci. USA, 82:5132(1985)、並びにStewartおよびYoung, Solid phase peptide synthesis, Pierce Chemical Co., イリノイ州ロックフォード(1984)に示されるような化学合成法を用いて、本発明のIFNγポリペプチドを調製することもまた可能である。こうしたポリペプチドを、アミノ末端メチオニンを伴い、または伴わずに合成することも可能である。化学的に合成したIFNγポリペプチド、あるいはその断片または変異体を、これらの参考文献に示される方法を用いて酸化して、ジスルフィド架橋を形成することも可能である。化学合成によって調製された本発明のIFNγポリペプチドは、組換えによって産生されたか、または天然供給源から精製された、対応するIFNγポリペプチドに匹敵する生物学的活性を、少なくとも1つ有するであろう。
【0078】
IFNγポリペプチドを、IFNγポリペプチドが天然に見出される供給源組織および/または体液などの生物学的試料から単離することによって、得ることも可能である。IFNγポリペプチドの供給源は、起源が、ヒトまたは非ヒトであることも可能である。電気泳動による分離、その後の電気的溶出、多様な種類のクロマトグラフィー(アフィニティー、免疫親和性、分子ふるい、および/またはイオン交換)、および/または高圧液体クロマトグラフィーなどの、当該技術分野に知られる方法を用いて、天然存在IFNγポリペプチドの単離を達成することも可能である。例えば、組換えで産生したIFNγポリペプチドまたはそのペプチド断片に対して調製した抗体を用いて、精製中のIFNγポリペプチドの存在を監視することも可能である。
【0079】
本発明のポリペプチドには、単離IFNγポリペプチド、並びに上に定義するような、断片、変異体、融合ポリペプチド、および誘導体を含む、IFNγポリペプチドに関連するポリペプチドが含まれる。本発明のIFNγ断片は、アミノ末端(リーダー配列を含むまたは含まない)の一部切除、カルボキシ末端の一部切除、および/またはポリペプチド内部の欠失から生じることも可能である。こうしたIFNγポリペプチド断片は、所望によって、アミノ末端メチオニン残基を含むことも可能である。本発明のポリペプチドは、抗体応答を誘発可能である点で、免疫原性であろう。
【0080】
本発明のIFNγポリペプチド変異体には、天然IFNγアミノ酸配列に比較した際、1以上のアミノ酸置換、付加および/または欠失が含まれる。アミノ酸置換は、上に定義するように保存的であることも可能であるし、または非保存的であることも、あるいはその組み合わせいずれかであることも可能である。変異体は、カルボキシ末端またはアミノ末端いずれかに、アミノ酸残基の付加を有することも可能である(この場合、アミノ末端は、リーダー配列を含んでもよいし、また含まなくてもよい)。
【0081】
本発明の態様には、IFNγグリコシル化変異体およびシステイン変異体が含まれる。IFNγグリコシル化変異体には、天然IFNγポリペプチドに比較して、グリコシル化部位の数および/または種類が改変されている変異体が含まれる。1つの態様において、IFNγグリコシル化変異体は、天然IFNγに比較して、より多くのまたはより少ない数のN連結グリコシル化部位を含む。
【0082】
N連結炭水化物鎖の再編成を含むIFNγグリコシル化変異体であって、1以上のN連結グリコシル化部位(典型的には天然存在であるもの)が除去され、そして1以上の新規N連結部位が生成されている、前記変異体もまた提供する。IFNγシステイン変異体は、天然IFNγに比較して、より多い数の、あるいはより少ない数のシステイン残基を含む。1つの態様において、1以上のシステイン残基が欠失されるか、または別のアミノ酸(例えばセリン)で置換される。IFNγのシステイン変異体は、変性状態から単離された後、IFNγが生物学的に活性であるコンホメーションに再フォールディングするのを補助することによって、生物学的に活性であるIFNγの回収を改善することも可能である。
【0083】
IFNγポリペプチド変異体の調製は、当該技術分野の技術レベル内である。1つのアプローチにおいて、天然IFNγに1以上のアミノ酸置換、欠失および/または付加を導入することも可能であり、ここでIFNγ変異体は、IFNγの天然構造および/または少なくとも1つの生物学的活性を保持する。1つのアプローチは、比較的低いそして高い同一性および/または類似性の領域を同定するため、多様な異なる種由来のIFNγポリペプチドの配列を比較することである。比較的低い同一性および/または類似性を有するIFNγポリペプチドの領域は、構造および活性に必須である可能性はより低く、そしてしたがって、アミノ酸改変、特に非保存性である改変に、より耐性でありうると認識される。比較的保存される領域であっても、活性を保持しつつ、保存的アミノ酸置換を導入可能であることもまた認識される。
【0084】
別のアプローチにおいて、構造−機能関連性を用いて、類似のポリペプチドにおいて、活性または構造に重要な残基を同定することも可能である。例えば、IFNγ、および構造−機能解析が利用可能な腫瘍壊死因子ファミリーの他のメンバーの間で保存されるアミノ酸残基を比較し、そしてこうした比較に基づいて、IFNγ中のどのアミノ酸残基が活性または構造に重要であるかを予測することも可能である。当業者は、こうして予測されたIFNγの重要なアミノ酸残基に関して、化学的に類似のアミノ酸置換を選択することも可能である。
【0085】
さらに別のアプローチにおいて、IFNγの二次構造または三次構造(IFNγ結晶のX線回折によって、または構造予測法によって決定されるものいずれか)の解析を行って、IFNγポリペプチド内の実際の構造または予測される構造に関して、特定のアミノ酸残基の位置を決定することも可能である。この情報を用いて、IFNγポリペプチドの二次構造および/または三次構造をできるだけ多く保持することを模索する方式で、アミノ酸変化を導入することも可能である。さらに別のアプローチにおいて、アミノ酸置換を導入し、そして本明細書に記載するアッセイを用いて、生物学的活性に関して、改変されたIFNγポリペプチドを試験することによって、特定の位でアミノ酸を改変する影響を実験的に試験することも可能である。
【0086】
アラニン・スキャニング突然変異誘発などの技術(Cunninghamら、上記)が、このアプローチに特によく適している。IFNγ中の多様なアミノ酸位で、多くの置換を導入し、そしてファージ・ディスプレー・ライブラリーの一部として、改変されたポリペプチド集団をスクリーニングすることによって、多くの改変された配列を好適に試験することも可能である。このアプローチを用いて、活性に必須のIFNγポリペプチドの領域を容易に決定することも可能である。
【0087】
上記方法は、天然構造を保持するIFNγ変異体を生成するのに有用である。したがって、各変異体に対して作成された抗体は、IFNγの天然構造決定基またはエピトープを認識する可能性があり、そしてまた、天然IFNγに結合する可能性もある。しかし、いくつかの場合、天然IFNγ構造を保持しないか、あるいは部分的にまたは完全に空白である(unfilled)IFNγ変異体を産生することが望ましい可能性もある。こうしたタンパク質に対して作成された抗体は、IFNγ上の覆い隠されたエピトープを認識するであろう。
【0088】
本発明はまた、異種ペプチドまたはタンパク質に融合された、IFNγポリペプチド、並びにその断片、変異体、および誘導体を含む、IFNγ融合ポリペプチドも提供する。異種ペプチドおよびタンパク質には、限定されるわけではないが:IFNγ融合ポリペプチドの検出および/または単離を可能にするエピトープ;膜貫通受容体タンパク質またはその部分、例えば細胞外ドメイン、または膜貫通ドメインおよび細胞内ドメイン;膜貫通受容体タンパク質に結合するリガンドまたはその部分;触媒として活性である酵素またはその部分;オリゴマー化を促進するタンパク質またはペプチド、例えばロイシンジッパードメイン;および安定性を増加させるタンパク質またはペプチド、例えば免疫グロブリン定常領域が含まれる。
【0089】
IFNγポリペプチドをそれ自体、あるいはその断片、変異体、または誘導体と融合させることも可能である。融合は、IFNγポリペプチドのアミノ末端またはカルボキシ末端いずれで行うことも可能であるし、そしてリンカー分子またはアダプター分子を伴わずに直接であることも、あるいはリンカー分子またはアダプター分子を介したものであることも可能である。また、リンカー分子またはアダプター分子を、DNA制限エンドヌクレアーゼまたはプロテアーゼの切断部位を含んで設計して、融合部分の分離を可能にすることも可能である。本発明のさらなる態様において、IFNγポリペプチド、断片、変異体および/または誘導体をヒトIgGのFc領域に融合させる。1つの例において、当業者に知られる方法を用いて、IFNγポリペプチドのN末端またはC末端いずれかに、ヒトIgGヒンジ、ch2およびch3領域を融合させることも可能である。別の例において、ヒンジ領域、並びにch2およびch3領域の部分を融合させることも可能である。タンパク質アフィニティーカラムを使用することによって、こうして産生したIFNγ・Fc融合ポリペプチドを精製することも可能である。さらに、Fc領域に融合させたペプチドおよびタンパク質は、融合させていない対応物よりも、実質的により長いin vivo半減期を示すことも見出されてきている。また、Fc領域への融合は、融合ポリペプチドの二量体化/多量体化も可能にする。Fc領域は、天然存在Fc領域であることも可能であるし、また療法特質や循環時間などの特定の特質を改善し、凝集を減少させるように改変することなども可能である。
【0090】
IFNγポリペプチド誘導体は、本発明の範囲内に含まれる。こうした誘導体は、IFNγポリペプチドがポリマーに連結された、化学的に修飾されたIFNγポリペプチド組成物である。選択されるポリマーは、付着するタンパク質が、生理学的環境などの水性環境において沈殿しないように、典型的には、水溶性である。ポリマーは、いかなる分子量のものであることも可能であるし、そして分枝していてもまたは分枝していなくてもよい。IFNγポリペプチドポリマーの範囲内に含まれるのは、ポリマー混合物である。好ましくは、最終産物調製物を療法的に使用するため、ポリマーは薬学的に許容しうるものであろう。
【0091】
水溶性ポリマーまたはその混合物は、例えば、ポリエチレングリコール(PEG)、モノメトキシ−ポリエチレングリコール、デキストラン(例えば約6kDの低分子量デキストランなど)、セルロース、または他の炭水化物に基づくポリマー、ポリ−(N−ビニルピロリドン)ポリエチレングリコール、プロピレングリコール・ホモポリマー、ポリプロピレンオキシド/エチレンオキシド・コポリマー、ポリオキシエチル化ポリオール類(例えばグリセロール)およびポリビニルアルコールであることも可能である。
【0092】
好ましい水溶性ポリマーはポリエチレングリコールである。本明細書において、ポリエチレングリコールは、他のタンパク質を誘導体化するのに用いられてきたPEGのいかなる型も、例えばモノ−(C1−C10)アルコキシ−、またはアリールオキシ−ポリエチレングリコールも含むよう意図される。本発明にやはり含まれるのは、共有結合IFNγ多量体を調製するのに使用可能な二官能性PEG架橋分子である。
【0093】
化学的に誘導体化されたIFNγポリペプチドを調製する方法が当該技術分野に知られる。例えば、Francisら, Focus on Growth Factors, 3:4−10(1992);EP 0 154 316;およびEP 0 401 384に記載される方法を用いて、PEGでのIFNγポリペプチドの誘導体化を行うことも可能である。好ましい態様において、IFNγポリペプチド誘導体は、アミノ末端に単一のPEG部分を有するであろう;本明細書に援用される米国特許第5,985,265号を参照されたい。
【0094】
本明細書に開示するIFNγポリペプチド誘導体は、非修飾ポリペプチドに比較して、IFNγの少なくとも1つの生物学的活性の増進または減少を示すことも可能であるし、あるいは増加したまたは減少した半減期または安定性を示すことも可能である。
【0095】
IFNγ選択的結合剤
IFNγポリペプチド、およびその断片、変異体および誘導体を用いて、IFNγの選択的結合剤を同定することも可能である。上に定義するように、IFNγの選択的結合剤は、タンパク質性結合剤および非タンパク質性結合剤を両方含み、そして本発明の1つの好ましい態様において、選択的結合剤はタンパク質性である。さらに別の好ましい態様において、選択的結合剤は、IFNγ、好ましくはヒトIFNγに結合する抗体またはその断片である。本発明の抗体は、IFNγの少なくとも1つの生物学的活性のレベルを増進するアゴニスト抗体;またはIFNγの少なくとも1つの生物学的活性のレベルを減少させるアンタゴニスト抗体であることも可能である。IFNγのアンタゴニスト抗体を、IFNγの阻害性抗体または中和抗体と呼ぶこともまた可能である。こうした抗体は本発明の好ましい態様であるが、IFNγ活性のアゴニストまたはアンタゴニストである他のタンパク質性選択的結合剤もまた、本発明に含まれることが理解される。
【0096】
以下の実施例に記載するように、IFNγの少なくとも1つの活性を阻害する抗IFNγ抗体および抗原結合ドメインが同定されてきている。本発明の態様には、図3〜13のいずれかに示すような重鎖Fab配列を含み、そしてカッパ軽鎖配列またはラムダ軽鎖配列をさらに含む抗体が含まれる。軽鎖Fab配列は、図14〜24に示すとおりであってもよい。例えば、「BS−A」抗体は、それぞれ、図14および図3の軽鎖配列および重鎖配列を有し;「BS−B」抗体は、それぞれ、図15および図4の軽鎖配列および重鎖配列を有し;「RD−B1」抗体は、それぞれ、図16および図5の軽鎖配列および重鎖配列を有し;「RD−A2」抗体は、それぞれ、図17および図6の軽鎖配列および重鎖配列を有し;「58C」抗体は、それぞれ、図18および図7の軽鎖配列および重鎖配列を有し;「GP−A」抗体は、それぞれ、図19および図8の軽鎖配列および重鎖配列を有し;「57D」抗体は、それぞれ、図20および図9の軽鎖配列および重鎖配列を有し;「57E」抗体は、それぞれ、図21および図10の軽鎖配列および重鎖配列を有し;「IFN−A」抗体は、それぞれ、図22および図11の軽鎖配列および重鎖配列を有し;「67C」抗体は、それぞれ、図23および図12の軽鎖配列および重鎖配列を有し;そして「59−A2」抗体は、それぞれ、図24および図13の軽鎖配列および重鎖配列を有する。本発明の抗体は、アイソタイプ、IgG、IgM、IgA、IgE、またはIgDいずれか由来のヒトFc領域をさらに含む。好ましくは、Fc領域は、ヒトIgG、例えばIgG1、IgG2、IgG3、またはIgG4由来である。
【0097】
本発明はまた、本明細書に開示するFab配列の断片、変異体、または誘導体を含む抗体または抗原結合ドメインも提供する。断片には、軽鎖または重鎖のFab配列いずれかの可変ドメインが含まれ、これらは、典型的には軽鎖または重鎖の定常ドメインに連結されている。変異体には、図14〜24のいずれか1つのFab配列に、少なくとも約80%、85%、90%、95%、98%または99%同一または類似である軽鎖Fab配列、または対応する可変ドメインを含む抗体、あるいは図3〜13のいずれか1つのFab配列に、少なくとも約80%、85%、90%、95%、98%または99%同一または類似である重鎖Fab配列、または対応する可変ドメインを含む抗体が含まれる。抗体は、典型的には、重鎖および軽鎖の定常領域と会合して、全長抗体を形成することも可能である。
【0098】
本発明の抗体および抗原結合ドメイン、並びにその断片、変異体および誘導体は、IFNγポリペプチド、好ましくはヒトIFNγポリペプチドに選択的に結合する能力を保持するであろう。1つの態様において、抗体は、約1nM以下、あるいは0.1nM以下、あるいは10pM以下、あるいは10pM未満の解離定数(KD)でIFNγポリペプチドに結合するであろう。
【0099】
本発明の抗体には、ポリクローナル抗体、単一特異性ポリクローナル抗体、モノクローナル抗体、組換え抗体、キメラ抗体、ヒト化抗体、完全ヒト抗体、一本鎖抗体および/または二重特異性抗体が含まれる。抗体断片には、IFNγポリペプチド上のエピトープに結合する抗IFNγ抗体の部分が含まれる。こうした断片の例には、Fab、F(ab’)、F(ab)’、Fv、およびsFv断片が含まれる。抗体は、全長抗体の酵素切断によって、または組換えDNA技術によって、例えば抗体可変領域をコードする核酸配列を含有する組換えプラスミドの発現などによって、生成可能である。
【0100】
ポリクローナル抗体は、抗原で免疫した動物の血清由来の抗体分子の異種集団である。抗原は、抗体に結合されることが可能であり、さらに、動物が、その抗原のエピトープに結合可能な抗体を産生するよう誘導可能である、分子または分子の部分である。抗原は、1以上のエピトープを有することも可能である。上述の特異的反応は、抗原が、非常に選択的な方式で、対応する抗体と反応し、そして他の抗原に誘発されうる多数の他の抗体とは反応しないであろうことを示すよう意味する。
【0101】
IFNγポリペプチドに対して向けられるポリクローナル抗体は、一般的に、IFNγおよびアジュバントの多数回の皮下注射または腹腔内注射によって、動物(例えばウサギまたはマウス)において作成される。本発明にしたがって、IFNγポリペプチド、あるいはその変異体、断片、または誘導体を、免疫しようとする種において免疫原性であるキャリアータンパク質、例えばキーホールリンペット(keyhole limpet)・ヘモシアニン、血清、アルブミン、ウシ・チログロブリン、またはダイズ(soybean)・トリプシン阻害剤にコンジュゲート化することが有用である可能性もある。また、ミョウバンなどの凝集剤を用いて、免疫応答を増進する。免疫後、動物から出血させ、そして抗IFNγ抗体力価に関して、血清をアッセイする。
【0102】
モノクローナル抗体(mAb)は、抗原に特異的な抗体の実質的に均質な集団を含有し、この集団は、実質的に類似のエピトープ結合部位を含有する。こうした抗体は、IgG、IgM、IgE、IgA、IgDおよびそのサブクラスいずれかを含む、免疫グロブリンクラスいずれのものであることも可能である。本発明のモノクローナル抗体を産生するハイブリドーマを、in vitro、in situまたはin vivoで培養することも可能である。in vivoまたはin situでの高力価の産生が好ましい産生法である。IFNγに対して向けられるモノクローナル抗体は、培養中の連続細胞株による抗体分子の産生を提供する方法いずれかを用いて産生される。モノクローナル抗体を調製するのに適した方法の例には、Kohlerら, Nature, 256:495−497(1975)のハイブリドーマ法、並びにヒトB細胞ハイブリドーマ法、Kozbor, J. Immunol,. 133:3001(1984);Brodeurら, Monoclonal Antibody Production Techniques and Applications, pp.51−63(Marcel Dekker, Inc. , ニューヨーク, 1987);並びにHarlowおよびLane, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory(1988)が含まれ;これら参考文献の内容は、本明細書に完全に援用される。
【0103】
好ましい抗IFNγ選択的結合剤には、ヒトIFNγのその同族受容体、hIFNγ−Rへの結合を、部分的にまたは完全に阻害するであろうモノクローナル抗体、あるいは実質的に同一の特異的結合特性を有する抗体とともに、その断片および領域が含まれる。競合的阻害によって、モノクローナル抗体特異性および親和性を決定するのに好ましい方法は、Harlowら, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー, 1988);Colliganら監修, Current Protocols in Immunology, Greene Publishing Assoc. and Wiley Interscience, ニューヨーク, (1992, 1993);およびMuller, Meth. Enzymol., 92:589−601(1983)に見出されることも可能である。これらの参考文献は、本明細書に援用される。本発明にやはり提供するのは、IFNγポリペプチドと反応するモノクローナル抗体を産生するハイブリドーマ細胞株である。
【0104】
キメラ抗体は、異なる部分が異なる動物種に由来する分子であり、例えばネズミ・モノクローナル抗体由来の可変領域およびヒト免疫グロブリン定常領域を有するものがある。キメラ抗体は、主に、適用中の免疫原性を減少させ、そして産生中の収率を増加させるように用いられ、例えば、ネズミ・モノクローナル抗体が、ハイブリドーマからはより高い収率で得られるが、ヒトにおいてより免疫原性である場合、ヒト/ネズミ・キメラモノクローナル抗体を用いる。
【0105】
キメラ抗体およびその産生法が当該技術分野に知られる。Cabillyら, Proc. Natl. Acad.Sci. USA, 81:3273−3277(1984);Morrisonら, Proc. Natl. Acad. Sci. USA, 81:6851−6855(1984);Boulianneら, Nature, 312:643−646(1984);Neubergerら, Nature, 314:268−270(1985);Liuら, Proc. Natl. Acad. Sci. USA, 84:3439−3443(1987);並びにHarlowおよびLane Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory(1988)。これらの参考文献は、本明細書に援用される。
【0106】
例えば、本発明のキメラモノクローナル抗体を療法剤として用いることも可能である。こうしたキメラ抗体では、望ましい生物学的活性を示す限り、重鎖および/または軽鎖の部分が、特定の種に由来するか、または1つの特定の抗体クラスもしくはサブクラスに属する抗体の対応する配列と同一であるかまたは相同であり、一方、鎖(単数または複数)の残りが、別の種に由来するか、または別の抗体クラスもしくはサブクラスに属する抗体とともにこうした抗体の断片の対応する配列と同一であるかまたは相同である;例えば、米国特許第4,816,567号およびMorrisonら, Proc. Natl. Acad. Sci., 81:6851−6855(1985)を参照されたい。
【0107】
本明細書において、用語「キメラ抗体」には、一価、二価または多価免疫グロブリンが含まれる。一価キメラ抗体は、キメラL鎖とのジスルフィド架橋を通じて会合するキメラH鎖によって形成される二量体(HL)である。二価キメラ抗体は、少なくとも1つのジスルフィド架橋を通じて会合する2つのHL二量体によって形成される四量体(H2L2)である。多価キメラ抗体もまた、例えば凝集するCH領域(例えばIgM H鎖、またはμ鎖由来のもの)を使用することによって産生可能である。
【0108】
本発明のネズミ抗体およびキメラ抗体、断片および領域は、個々の重鎖(H)および/または軽鎖(L)免疫グロブリン鎖を含むことも可能である。キメラH鎖は、IFNγに特異的な非ヒト抗体のH鎖由来の抗原結合領域を含み、これがヒトH鎖C領域(CH)の少なくとも部分、例えばCH1またはCH2に連結される。
【0109】
本発明記載のキメラL鎖は、ヒトL鎖C領域(CL)の少なくとも部分に連結された、IFNγに特異的な非ヒト抗体のL鎖由来の抗原結合領域を含む。
既知の方法工程にしたがって、個々のポリペプチド鎖を適切に会合させることによって、選択的結合剤、例えば同一のまたは異なる可変領域結合特異性を持つキメラH鎖およびL鎖を有する抗体、断片、または誘導体もまた調製可能である;例えば、Ausubelら監修 Current Protocols in Molecular Biology, Wiley Interscience, ニューヨーク(1993)およびHarlowら, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1988)を参照されたい。これらの参考文献の内容は、本明細書に完全に援用される。このアプローチを用いて、キメラH鎖(またはその誘導体)を発現する宿主を、キメラL鎖(またはその誘導体)を発現する宿主と別個に培養し、そして免疫グロブリン鎖を別個に回収して、そして次いで会合させる。あるいは、宿主を同時培養して、そして鎖を培地中で自発的に会合可能にし、その後、組み立てられた免疫グロブリン、断片または誘導体を回収することも可能である。
【0110】
例として、本発明の選択的結合剤(例えばキメラ抗体)の抗原結合領域は、好ましくは、ヒトIFNγに特異的な非ヒト抗体由来である。こうした非ヒト抗体をコードするDNAの好ましい供給源には、抗体を産生する細胞株、例えば一般的にハイブリドーマとして知られるハイブリッド細胞株が含まれる。
【0111】
本発明はまた、抗IFNγ抗体の断片、変異体および誘導体も提供し、ここで用語「断片」、「変異体」、「誘導体」および「融合体」は、本明細書に定義される。本発明は、非修飾抗IFNγ抗体に機能的に類似である、すなわち非修飾抗体の活性を少なくとも1つ保持する、抗IFNγ抗体の断片、変異体、誘導体、および融合体を含む。上述の修飾に加えて、植物毒素および細菌毒素などの細胞毒性タンパク質をコードする遺伝子配列の付加も含まれる。抗IFNγ抗体の断片、変異体、誘導体および融合体を、本発明の宿主のいずれから産生することも可能である。
【0112】
適切な断片には、例えば、Fab、Fab’、F(ab’)2、FvおよびscFvが含まれる。これらの断片は、損なわれていない(intact)抗体のFc断片を欠き、循環からより迅速に一掃され、そして損なわれていない抗体よりも非特異性組織結合がより少なくなりうる;Wahlら, J. Nucl. Med., 24:316−325(1983)。これらの断片は、当該技術分野に周知の方法を用いて、例えばパパイン(Fab断片を産生する)またはペプシン(F(ab’)2断片を産生する)などの酵素でのタンパク質分解切断によって、損なわれていない抗体から産生される。これらの抗原結合領域および/または本発明のモノクローナル抗体に認識されるエピトープを同定すると、本出願の態様と同等である、類似の結合特性および療法的有用性または診断有用性を持つさらなるモノクローナル抗体を生成するのに必要な情報が提供される。
【0113】
選択的結合剤の変異体もまた提供する。1つの態様において、抗体および抗原結合ドメインの変異体は、軽鎖および/または重鎖アミノ酸配列中に変化を含み、これらの変化は天然存在であるか、または組換えDNA技術を用いた天然配列のin vitro操作によって導入されている。天然存在変異体には、外来(foreign)抗原に対する抗体応答の生成中、対応する生殖系列ヌクレオチド配列において、in vivoで生成される「体細胞」変異体が含まれる。配列において、本発明の典型的なFabを生成する、生殖系列可変軽鎖および重鎖配列中の体細胞突然変異にコードされる変異体は、Fab「BS−A」に関しては図33および図41に、Fab「BS−B」に関しては図34および図41に、Fab「RD−B1」に関しては図35および図42に、Fab「RD−B1」に関しては図35および図42に、Fab「RD−A2」に関しては図33および図43に、Fab「58C」に関しては図36および図44に、Fab「GP−A」に関しては図37および図45に、Fab「57D」に関しては図38および図46に、Fab「57E」に関しては図39および図41に、Fab「IFN−A」に関しては図33および図43に、Fab「67C」に関しては図40および図43に、そしてFab「59−A2」に関しては図34および図41に示される。
【0114】
抗IFNγ抗体および抗原結合ドメインの変異体はまた、当該技術分野に知られる突然変異誘発技術によっても調製可能である。1つの例において、抗体コード領域全体で、無作為にアミノ酸変化を導入することも可能であり、そして望ましい活性、例えばIFNγに対する結合親和性に関して、生じた変異体をスクリーニングすることも可能である。あるいは、IFNγ抗体の選択される領域に、例えば軽鎖および/または重鎖CDR、およびフレームワーク領域に、アミノ酸変化を導入することも可能であり、そしてIFNγへの結合または他の何らかの活性に関して、生じた抗体をスクリーニングすることも可能である。アミノ酸変化は、単一アミノ酸相違から、既定のCDR、例えばCDR3内のアミノ酸のありうるすべての順列(permutation)の範囲で、CDR中に1以上のアミノ酸置換を含む。別の方法において、CDR内の少なくとも1つの残基をアラニンで置換することによって、CDR内の各残基のIFNγ結合への寄与を評価することも可能である;Lewisら, Mol. Immunol., 32:1065−1072(1995)。次いで、より最適な配列を決定するため、IFNγへの結合に最適でない残基を変化させることも可能である。アミノ酸を挿入して、CDR、例えばCDR3のサイズを増加させることによって生成される変異体もまた、含まれる。例えば、大部分の軽鎖CDR3配列は、長さ9アミノ酸である。9残基より短い、抗体中の軽鎖CDR3配列は、適切なアミノ酸を挿入して、CDRの長さを増加させることによって、IFNγへの結合に関して最適化されることも可能である。
【0115】
1つの態様において、抗体または抗原結合ドメイン変異体は、1以上の重鎖または軽鎖CDR1、CDR2またはCDR3に、そして所望によって1以上の重鎖または軽鎖フレームワーク領域FR1、FR2またはFR3に、1以上のアミノ酸変化を含む。アミノ酸変化は、アミノ酸残基の置換、欠失および/または挿入を含む。典型的な変異体には、配列GYYWS(配列番号34);EINHSGSTNYNPSLKS(配列番号44);またはGRARNWRSRFDY(配列番号54)に1以上のアミノ酸変化を持つ「BS−A」重鎖可変領域変異体、あるいは配列TGSSGSIASHYVQ(配列番号01);EDKERPS(配列番号12);またはQSYDSSNQWV(配列番号23)に1以上のアミノ酸変化を持つ「BS−A」軽鎖可変領域変異体が含まれる。前述の「BS−A」重鎖および軽鎖可変領域変異体は、フレームワーク領域に1以上のアミノ酸変化をさらに含むことも可能である。
【0116】
1つの例において、1以上のアミノ酸変化を導入して、体細胞突然変異フレームワーク残基を、その位の生殖系列残基で置換することも可能である。前述のアミノ酸変化が置換である場合、変化は保存的置換でもまた非保存的置換でもよい。変異体を、軽鎖または重鎖いずれかの「鎖シャフリング」によって調製することもまた可能である;Marksら Biotechnology, 10:779−783(1992)。典型的には、単一の軽鎖(または重鎖)を、重鎖(または軽鎖)のレパートリーを有するライブラリーと組み合わせて、そして生じた集団を、IFNγへの結合などの望ましい活性に関してスクリーニングする。この技術は、重鎖および軽鎖両方のレパートリーを含むライブラリーで可能であるよりも、単一の軽鎖(または重鎖)と組み合わされた異なる重鎖(または軽鎖)のより多数の試料をスクリーニングすることを可能にする。
【0117】
本発明の選択的結合剤は、二重特異性であることも可能である。本発明の二重特異性選択的結合剤は、いくつかの立体配置のものであることも可能である。例えば、二重特異性抗体は、一本鎖抗体(または抗体断片)に似ているが、2つの異なる抗原結合部位(可変領域)を有する。二重特異性抗体は、化学的技術によって;例えば、Kranzら, Proc. Natl. Acad. Sci. USA, 78:5807(1981)を参照されたい;「ポリドーマ(polydoma)」技術によって;米国特許第4,474,893号;または組換えDNA技術によって、産生可能である。
【0118】
本発明の選択的結合剤はまた、異種抗体であることも可能である。異種抗体は、ともに連結された、各々異なる特異性を有する、2以上の抗体、または抗体結合断片(Fab)である。
【0119】
本発明はまた、「ヒト化」抗体にも関する。非ヒト抗体をヒト化する方法は、当該技術分野に周知である。一般的に、ヒト化抗体は、非ヒトである供給源からヒト抗体に導入された1以上のアミノ酸残基を有する。一般的に、非ヒト残基は、CDRに存在するであろう。ヒト化は、当該技術分野に知られる方法にしたがって、げっ歯類相補性決定領域(CDR)をヒト抗体の対応する領域に対して置換することによって、実行可能である;Jonesら, Nature 321:522−525(1986);Riechmannら, Nature, 332:323−327(1988);Verhoeyenら, Science, 239:1534−1536(1988)。
【0120】
キメラ抗体、CDR移植抗体、およびヒト化抗体を含む、本発明の選択的結合剤は、当該技術分野に知られる組換え法によっても産生可能である。抗体をコードする核酸を宿主細胞に導入し、そして本明細書に記載し、そして当該技術分野に知られる、材料および方法を用いて発現させる。好ましい態様において、CHO細胞などの哺乳動物宿主細胞において、抗体を産生する。宿主細胞にトランスフェクションした組換えDNAの発現によって、または上述のようなハイブリドーマ細胞における発現によって、完全ヒト抗体を産生することも可能である。
【0121】
モノクローナル抗体の生成を回避して、抗体分子の抗原結合領域の組換えDNA型を生成する技術が本発明の実施内に含まれる。これを行うため、免疫動物から採取した免疫系細胞から、抗体特異的メッセンジャーRNA分子を抽出し、そして相補DNA(cDNA)に転写する。次いで、cDNAを細菌発現系にクローニングする。本発明を実施するのに適した技術の一例は、発現されたFabタンパク質を細胞膜周辺腔(細菌細胞膜および細胞壁の間)に移動させるか、または分泌させる、リーダー配列を有する、繊維状バクテリオファージM13由来ファージミド・ベクター系を用いる。抗原に結合するものに関して、非常に多数の機能するFab断片を迅速に生成し、そしてスクリーニングすることも可能である。こうしたIFNγ選択的結合剤(IFNγポリペプチドに対して特異性を持つFab断片)は、本明細書に定義され、議論され、そして請求される際、用語「抗体」に明確に含まれる。
【0122】
やはり本発明の範囲内にあるのは、適切な抗原特異性を持つマウス抗体分子由来の遺伝子を、適切な生物学的活性、例えばヒト補体を活性化し、そしてADCCを仲介する能力を持つヒト抗体分子由来の遺伝子とともにスプライシングすることによって、キメラ抗体を産生するために開発された技術である;Morrisonら, Proc. Natl. Acad. Sci., 81:6851(1984);Neubergerら, Nature, 312:604(1984)。1つの例は、Fc領域の、異なるアイソタイプのFc領域との交換である。この技術によって産生される抗体などの選択的結合剤は、本発明の範囲内である。
【0123】
本発明の好ましい態様において、抗IFNγ抗体は、完全ヒト抗体である。したがって、IFNγポリペプチドに結合し、そしてヒト生殖系列免疫グロブリン核酸配列、並びにその断片、合成変異体、誘導体および融合体の天然存在体細胞変異体である核酸配列にコードされる抗体が本発明に含まれる。こうした抗体は、当該技術分野に知られる方法いずれによって産生してもよい。典型的な方法には、内因性免疫グロブリン産生を伴わずに、ヒト抗体レパートリーを産生することが可能なトランスジェニック動物(例えばマウス)を、IFNγ抗原(免疫応答を誘発することが可能であり、そして所望によってキャリアーにコンジュゲート化されているIFNγポリペプチドいずれか)で免疫することが含まれる;例えばJakobovitsら, Proc. Natl. Acad. Sci., 90:2551−2555(1993);Jakobovitsら, Nature, 362:255−258(1993);Bruggermannら, Year in Immunol., 7:33(1993)を参照されたい。
【0124】
あるいは、ファージ・ディスプレー抗体ライブラリーのin vitroスクリーニングを通じて、ヒト抗体を生成することも可能である;例えば、本明細書に援用される、Hoogenboomら, J. Mol. Biol., 227:381(1991);Marksら, J. Mol. Biol., 222:581(1991)を参照されたい。多様な抗体含有ファージ・ディスプレー・ライブラリーが記載されており、そして当業者によって容易に調製可能である。ライブラリーは、適切な標的に対してスクリーニング可能な、多様なヒト抗体配列、例えばヒトFab、Fv、およびscFv断片を含有することも可能である。実施例2は、IFNγに選択的に結合する分子を同定するための、IFNγに対するFabファージ・ライブラリーのスクリーニングを記載する。ファージ・ディスプレー・ライブラリーが、IFNγの選択的結合剤を同定するためにスクリーニングすることが可能な抗体以外のペプチドまたはタンパク質を含んでもよいことが認識されるであろう。
【0125】
抗イディオタイプ(抗Id)抗体は、一般的に抗体の抗原結合部位と会合するユニークな決定基を認識する抗体である。Id抗体は、抗Idを調製しようとするモノクローナル抗体で、該モノクローナル抗体の供給源と同一種および同一遺伝子型(例えばマウス系統)の動物を免疫することによって、調製可能である。免疫された動物は、これらのイディオタイプ決定基に対する抗体(抗Id抗体)を産生することによって、免疫抗体のイディオタイプ決定基を認識し、そして応答するであろう;例えば本明細書に完全に援用される米国特許第4,699,880号を参照されたい。抗Id抗体はまた、さらに別の動物において免疫応答を誘導し、いわゆる抗−抗Id抗体を産生する、「免疫原」としても使用可能である。抗−抗Idは、抗Idを誘導した元来のモノクローナル抗体と、エピトープが同一であることも可能である。したがって、mAbのイディオタイプ決定基に対する抗体を用いることによって、同一の特異性の抗体を発現する他のクローンを同定することが可能である。
【0126】
IFNγの選択的結合剤の産生
調製しようとするIFNγの選択的結合剤が、タンパク質性選択的結合剤、例えば抗体または抗原結合ドメインである場合、前記剤を産生するため、多様な生物学的方法または化学的方法が利用可能である。
【0127】
療法使用のために十分な量の選択的結合剤を産生するには、生物学的方法が好ましい。本発明の抗体および抗原結合ドメインの産生には、標準的組換えDNA技術が特に有用である。発現産物の回収のための典型的な発現ベクター、宿主細胞および方法を以下に記載する。
【0128】
標準的連結技術を用いて、IFNγ抗体または抗原結合ドメインをコードする核酸分子を適切な発現ベクターに挿入する。ベクターは、典型的には、使用する特定の宿主細胞で機能するように選択される(すなわち、ベクターは、遺伝子の増幅および/または遺伝子の発現が起こりうるように、宿主細胞機構と適合する)。抗IFNγ抗体をコードする核酸分子を、原核宿主細胞、酵母宿主細胞、昆虫宿主細胞(バキュロウイルス系)および/または真核宿主細胞で増幅/発現することも可能である。宿主細胞の選択は、部分的に、抗IFNγ抗体を翻訳後に(post−transitionally)修飾(例えばグリコシル化および/またはリン酸化)しようとしているかどうかに応じるであろう。修飾するのであれば、酵母宿主細胞、昆虫宿主細胞、または哺乳動物宿主細胞が好ましい。発現ベクターの概説に関しては、Meth. Enz. V. 185, D.V. Goeddel監修 Academic Press Inc., カリフォルニア州サンディエゴ(1990)を参照されたい。
【0129】
典型的には、いかなる宿主細胞で用いられる発現ベクターも、以下の構成要素の1以上を含有するであろう:プロモーター、1以上のエンハンサー配列、複製起点、転写終結配列、ドナーおよびアクセプターのスプライス部位を含有する完全イントロン配列、分泌のためのリーダー配列、リボソーム結合部位、ポリアデニル化配列、発現しようとするポリペプチドをコードする核酸を挿入するためのポリリンカー領域、および選択可能マーカー要素。これらの配列は各々、以下により詳細に論じられる。
【0130】
ベクター構成要素は、相同配列(すなわち宿主細胞と同一種および/または同一系統由来)、異種配列(すなわち宿主細胞種以外の種または系統由来)、ハイブリッド(すなわち1より多い供給源由来の異なる配列の組み合わせ)、合成配列、または通常、免疫グロブリン発現を制御するように機能する天然配列であることも可能である。こうしたものとして、ベクター構成要素の供給源は、宿主細胞機構において機能し、そして該機構によって活性化可能である限り、いかなる原核生物または真核生物、いかなる脊椎動物または無脊椎動物、またはいかなる植物であることも可能である。
【0131】
複製起点は、発現に用いられる宿主細胞の種類に基づいて選択される。例えば、プラスミドpBR322(製品番号303−3s、New England Biolabs、マサチューセッツ州ビバリー)由来の複製起点は、大部分のグラム陰性細菌に適しており、一方、SV40、ポリオーマ、アデノウイルス、水疱性口内炎ウイルス(VSV)またはパピローマウイルス(HPVまたはBPVなど)由来の多様な起点は、哺乳動物細胞におけるクローニングベクターに有用である。一般的に、複製起点構成要素は哺乳動物発現ベクターには必要ない(例えば、SV40起点がしばしば用いられるが、これはただ初期プロモーターを含有するからである)。
【0132】
転写終結配列は、典型的にはポリペプチドコード領域の3’端に位置し、そして転写を終結させるように働く。通常、原核細胞中の転写終結配列には、G−Cリッチ断片に続いてポリT配列がある。配列を、ライブラリーから容易にクローニングするか、またはベクターの一部として商業的に購入してもよいが、上述のものなどの核酸合成法を用いて、容易に合成することも可能である。
【0133】
選択可能マーカー遺伝子要素は、選択培地中で増殖する宿主細胞の生存および増殖に必要なタンパク質をコードする。典型的な選択マーカー遺伝子は(a)抗生物質または他の毒素、例えば原核宿主細胞ではアンピシリン、テトラサイクリン、またはカナマイシンに対する耐性を与えるタンパク質、(b)細胞の栄養要求不全を補足するタンパク質;または(c)複合培地から入手可能でない非常に重要な栄養素を供給するタンパク質をコードする。好ましい選択可能マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、およびテトラサイクリン耐性遺伝子である。原核宿主細胞および真核宿主細胞において、ネオマイシン耐性遺伝子もまた使用可能である。
【0134】
他の選択遺伝子を用いて、発現しようとする遺伝子を増幅することも可能である。増幅は、増殖に決定的に重要なタンパク質の産生に対する要求がより高い遺伝子が、組換え細胞の代々の世代の染色体内で、タンデムに反復されるプロセスである。哺乳動物細胞に適した選択可能マーカーの例には、ジヒドロ葉酸レダクターゼ(DHFR)およびチミジンキナーゼが含まれる。哺乳動物細胞形質転換体を、ベクターに存在するマーカーのため、形質転換体のみがユニークに適応して生存する選択圧下に置く。培地中の選択剤の濃度を連続的に変化させて、それによって選択遺伝子および抗IFNγ抗体をコードするDNA両方の増幅が導かれる条件下で、形質転換細胞を培養することによって、選択圧を課す。その結果、増幅されたDNAから、増加した量の抗体が合成される。
【0135】
リボソーム結合部位は、通常、mRNAの翻訳開始に必要であり、そしてシャイン・ダルガノ配列(原核生物)またはコザック配列(真核生物)によって特徴付けられる。これら要素は、典型的には、発現しようとするポリペプチドのプロモーターの3’で、そしてコード配列の5’に位置する。シャイン・ダルガノ配列は多様であるが、典型的にはポリプリンである(すなわち高いA−G含量を有する)。多くのシャイン・ダルガノ配列が同定されてきており、各々、上述の方法を用いて、容易に合成可能であり、そして原核ベクターで使用可能である。
【0136】
リーダー配列またはシグナル配列は、ポリペプチドの分泌を指示するのに用いられる。シグナル配列はポリペプチドコード領域内にまたはその5’端に直接配置されることも可能である。多くのシグナル配列が同定されてきており、そして発現に用いる宿主細胞に基づいて選択可能である。本発明において、シグナル配列は、抗IFNγ抗体または抗原結合ドメインをコードする核酸配列に相同である(天然存在である)か、またはこれら配列に対して異種であることも可能である。選択される異種シグナル配列は、シグナルペプチダーゼにより、宿主細胞によって、認識され、そしてプロセシングされる、すなわち切断されるものであるべきである。天然免疫グロブリンシグナル配列を認識せず、そしてプロセシングしない原核宿主細胞では、シグナル配列を、例えばアルカリホスファターゼ、ペニシリナーゼ、または熱安定性内毒素IIのリーダーの群から選択される原核シグナル配列によって置換する。酵母分泌のため、天然免疫グロブリンシグナル配列を、酵母インベルターゼ、アルファ因子、または酸ホスファターゼのリーダーによって置換することも可能である。哺乳動物細胞発現では、天然シグナル配列で十分だが、他の哺乳動物シグナル配列が適切である可能性もある。
【0137】
大部分の場合、宿主細胞から抗IFNγ抗体または抗原結合ドメインが分泌される結果、抗体からシグナルペプチドが除去されるであろう。したがって、成熟抗体は、いかなるリーダー配列またはシグナル配列も欠くであろう。
【0138】
真核宿主細胞発現系におけるグリコシル化が望ましいなどのいくつかの場合、グリコシル化または収率を改善するため、多様なプレ配列を操作することも可能である。例えば、特定のシグナルペプチドのペプチダーゼ切断部位を改変することも可能であるし、またはグリコシル化に影響を及ぼすことも可能なプロ配列を付加することも可能である。最終タンパク質産物は、−1位(成熟タンパク質の最初のアミノ酸に比較して)に、完全には除去されなかった可能性もある、発現によって生じる1以上のさらなるアミノ酸を有することも可能である。例えば、最終タンパク質産物は、N末端に付着して、ペプチダーゼ切断部位に見られる1つまたは2つのアミノ酸を有することも可能である。あるいは、いくつかの酵素切断部位を用いることによって、望ましいIFNγポリペプチドのわずかに一部切除された型を生じることも可能であり、これは酵素が成熟ポリペプチド内のこうした領域を切断する場合である。
【0139】
本発明の発現ベクターは、典型的には、宿主生物に認識され、そして抗IFNγ抗体または抗原結合ドメインをコードする核酸分子に機能可能であるように連結されているプロモーターを含有するであろう。発現に用いる宿主細胞および望ましいタンパク質収率に応じて、天然プロモーターまたは異種プロモーターを使用することも可能である。
【0140】
原核宿主で用いるのに適したプロモーターには、ベータ−ラクタマーゼおよびラクトースプロモーター系;アルカリホスファターゼ、トリプトファン(trp)プロモーター系;およびtacプロモーターなどのハイブリッドプロモーターが含まれる。他の既知の細菌プロモーターもまた、適している。これらの配列は公表されており、それによって、当業者が、必要に応じて、必要な制限部位いずれかを供給するリンカーまたはアダプターを用いて、これらプロモーターを望ましいDNA配列(単数または複数)に連結することが可能になる。
【0141】
酵母宿主で用いるのに適したプロモーターもまた、当該技術分野に周知である。酵母エンハンサーは、酵母プロモーターとともに、好適に用いられる。哺乳動物宿主細胞で用いるのに適したプロモーターが周知であり、そしてポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシ・パピローマウイルス、鳥・肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および最も好ましくはサル・ウイルス40(SV40)などのウイルスのゲノムから得られるものが含まれる。他の適切な哺乳動物プロモーターには、異種哺乳動物プロモーター、例えば熱ショックプロモーターおよびアクチン・プロモーターが含まれる。
【0142】
本発明の選択的結合剤を発現するのに使用可能なさらなるプロモーターには、限定されるわけではないが:SV40初期プロモーター領域;BernoistおよびChambon, Nature, 290:304−310(1981)、CMVプロモーター、ラウス肉腫ウイルスの3’末端反復配列(long terminal repeat)に含有されるプロモーター;Yamamotoら, Cell, 22:787−797(1980)、ヘルペス・チミジンキナーゼ・プロモーター;Wagnerら, Proc. Natl. Acad. Sci. U.S.A., 78:144−1445(1981)、メタロチオネイン遺伝子の制御配列;Brinsterら, Nature, 296:39−42(1982)、ベータ−ラクタマーゼ・プロモーターなどの原核生物発現ベクター;Villa−Kamaroffら, Proc. Natl. Acad. Sci. U.S.A., 75:3727−3731(1978)、またはtacプロモーター;DeBoerら, Proc. Natl. Acad. Sci. U.S.A., 80:21−25(1983)が含まれる。
【0143】
やはり重要なのは、組織特異性を有し、そしてトランスジェニック動物において利用されてきている、以下の動物転写調節領域である:膵臓腺房細胞で活性であるエラスターゼI遺伝子調節領域;Swiftら, Cell, 38:639−646(1984);Ornitzら, Cold Spring Harbor Symp. Quant. Biol. 50:399−409(1986);MacDonald, Hepatology, 7:425−515(1987)、膵臓ベータ細胞で活性であるインスリン遺伝子調節領域;Hanahan, Nature, 315:115−122(1985)、リンパ球で活性である免疫グロブリン遺伝子調節領域;Grosschedlら, Cell, 38:647−658(1984);Adamesら, Nature, 318:533−538(1985);Alexanderら, Mol. Cell. Biol., 7:1436−1444(1987)、精巣細胞、乳房細胞、リンパ球およびマスト細胞で活性であるマウス乳腺腫瘍ウイルス調節領域;Lederら, Cell, 45:485−495(1986)、肝臓で活性であるアルブミン遺伝子調節領域;Pinkertら, Genes and Devel., 1:268−276(1987)、肝臓で活性であるアルファ・フェトプロテイン遺伝子調節領域;Krumlaufら, Mol. Cell. Biol., 5:1639−1648(1985);Hammerら, Science, 235:53−58(1987)、肝臓で活性であるアルファ1−アンチトリプシン遺伝子調節領域;Kelseyら, Genes and Devel., 1:161−171(1987);骨髄細胞で活性であるベータ−グロビン遺伝子調節領域;Mogramら, Nature, 315:338−340(1985);Kolliasら, Cell, 46:89−94(1986)、脳のオリゴデンドロサイトで活性であるミエリン塩基性タンパク質遺伝子調節領域;Readheadら, Cell, 48:703−712(1987)、骨格筋で活性であるミオシン軽鎖−2遺伝子調節領域;Sani, Nature, 314:283−286(1985)、および視床下部で活性である性腺刺激ホルモン放出ホルモン遺伝子調節領域;Masonら, Science, 234:1372−1378(1986)。
【0144】
エンハンサー配列をベクターに挿入して、真核宿主細胞において転写を増加させることも可能である。哺乳動物遺伝子で利用可能ないくつかのエンハンサー配列が知られる(例えばグロビン、エラスターゼ、アルブミン、アルファ−フェト−プロテインおよびインスリン)。しかし、典型的には、ウイルス由来のエンハンサーが用いられるであろう。SV40エンハンサー、サイトメガロウイルス初期プロモーター・エンハンサー、ポリオーマ・エンハンサー、およびアデノウイルス・エンハンサーは、真核プロモーター活性化のための典型的な増進要素である。エンハンサーは、ポリペプチドコード領域の5’または3’の位で、ベクターにスプライシングすることも可能であるが、典型的には、プロモーターの5’の部位に位置する。
【0145】
本発明を実施するための好ましいベクターは、細菌細胞、昆虫細胞、および哺乳動物宿主細胞と適合するものである。こうしたベクターには、とりわけ、pCRII、pCR3、およびpcDNA3.1(Invitrogen社、カリフォルニア州サンディエゴ)、pBSII(Stratagene社、カリフォルニア州ラホヤ)、pET15(Novagen、ウィスコンシン州マディソン)、pGEX(Pharmacia Biotech、ニュージャージー州ピスカタウェイ)、pEGFP−N2(Clontech、カリフォルニア州パロアルト)、pETL(BlueBacII;Invitrogen、カリフォルニア州カールスバッド)、pDSR−アルファ(PCR公報第WO90/14363)およびpFastBacDual(Gibco/BRL、ニューヨーク州グランドアイランド)が含まれる。
【0146】
さらなる可能なベクターには、限定されるわけではないが、コスミド、プラスミドまたは修飾ウイルスが含まれるが、ベクター系は、選択した宿主細胞に適合しなければならない。こうしたベクターには、限定されるわけではないが、Bluescript(登録商標)プラスミド誘導体(ColE1に基づく高コピー数ファージミド、Stratagene Cloning Systems Inc.、カリフォルニア州ラホヤ)、Taq増幅PCR産物をクローニングするために設計されたPCRクローニングプラスミド(例えばTOPOTM TAクローニング(登録商標)キット、PCR2.1(登録商標)プラスミド誘導体、Invitrogen、カリフォルニア州カールスバッド)、および哺乳動物ベクター、酵母ベクターまたはウイルスベクター、例えばバキュロウイルス発現系(pBacPAKプラスミド誘導体、Clontech、カリフォルニア州パロアルト)が含まれる。組換え分子は、形質転換、トランスフェクション、感染、エレクトロポレーション、または他の既知の技術を介して、宿主細胞に導入可能である。
【0147】
本発明の宿主細胞は、原核宿主細胞(例えば大腸菌(E. coli))または真核宿主細胞(例えば酵母細胞、昆虫細胞、または脊椎動物細胞)であることも可能である。宿主細胞は、適切な条件下で培養すると、本発明の抗体または抗原結合ドメインを発現し、これを続いて培地から収集する(宿主細胞がこれらを培地に分泌する場合)か、またはこれらを産生する宿主細胞から直接収集する(これらが分泌されない場合)ことも可能である。適切な宿主細胞の選択は、望ましい発現レベル、活性に望ましいかまたは必要なポリペプチド修飾、例えばグリコシル化またはリン酸化、および生物学的活性分子にフォールディングするのが容易であることなどの多様な要因に応じるであろう。
【0148】
いくつかの適切な宿主細胞が当該技術分野に知られ、そして多くがアメリカン・タイプ・カルチャー・コレクション(ATCC)、バージニア州マナサスから入手可能である。例には、チャイニーズハムスター卵巣細胞(CHO)(ATCC番号CCL61)、CHO DHFR細胞;Urlaubら, Proc. Natl. Acad. Sci. USA, 97:4216−4220(1980)、ヒト胎児由来腎臓(HEK)293細胞または293T細胞(ATCC番号CRL1573)、あるいは3T3細胞(ATCC番号CCL92)などの哺乳動物細胞が含まれる。形質転換、培養、増幅、スクリーニング、並びに産物産生および産物精製に適した哺乳動物宿主細胞および方法の選択は当該技術分野に知られる。他の適切な哺乳動物細胞株は、サルCOS−1(ATCC番号CRL1650)およびCOS−7細胞株(ATCC第CRL1651)、およびCV−1細胞株(ATCC番号CCL70)である。さらなる典型的な哺乳動物宿主細胞には、霊長類細胞株およびげっ歯類細胞株が含まれ、形質転換細胞株が含まれる。正常な二倍体細胞、初代組織のin vitro培養から得られる細胞株とともに、初代外植片もまた適切である。候補細胞は、選択遺伝子の遺伝子型が不完全であることも可能であるし、また優性に作用する選択遺伝子を含有することも可能である。他の適切な哺乳動物細胞株には、限定されるわけではないが、マウス神経芽細胞腫N2A細胞、HeLa細胞、マウスL−929細胞、Swissマウス、Balb−cマウスまたはNIHマウス由来の3T3細胞株、BHKまたはHaKハムスター細胞株が含まれ、これらはアメリカン・タイプ・カルチャー・コレクション(バージニア州マナサス)から入手可能である。これら細胞株は各々、タンパク質発現の技術分野の当業者に知られ、そして入手可能である。
【0149】
本発明に適した宿主細胞として、同様に有用であるのは、細菌細胞である。例えば、大腸菌の多様な株(例えばHB101(ATCC番号33694)、DH5α、DH10、およびMC1061(ATCC番号53338))は、バイオテクノロジーの分野で、宿主細胞として周知である。枯草菌(B. subtilis)、シュードモナス属(Pseudomonas)種、他のバチルス属(Bacillus)種、ストレプトミセス属(Streptomyces)種等の多様な株もまた、この方法で使用可能である。
【0150】
当業者に知られる酵母細胞の多くの株もまた、本発明のポリペプチドを発現するための宿主細胞として、入手可能である。好ましい酵母細胞には、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisae)が含まれる。
【0151】
さらに、望ましい場合、本発明の方法には、昆虫細胞系が利用可能である。こうした系は、例えば、Kittsら, Biotechniques, 14:810−817(1993)、Lucklow, Curr. Opin. Biotechnol., 4:564−572(1993)およびLucklowら, J. Virol., 67:4566−4579(1993)に記載される。好ましい昆虫細胞は、Sf−9およびHi5(Invitrogen、カリフォルニア州カールスバッド)である。
【0152】
抗IFNγ抗体または抗原結合ドメインをコードする核酸分子の、選択した宿主細胞への形質転換またはトランスフェクションは、塩化カルシウム法、エレクトロポレーション法、マイクロインジェクション法、リポフェクション法またはDEAEデキストラン法などの方法を含む、周知の方法によって達成可能である。選択される方法は、部分的に、用いる宿主細胞の種類と相関関係にあるであろう。これらの方法および他の適切な方法が当業者に周知であり、そして例えばSambrookら、上記に示される。
【0153】
トランスジェニック動物を使用して、グリコシル化選択的結合剤、例えば抗体および抗原結合ドメインを発現することも可能である。例えば、ミルクを産生するトランスジェニック動物(例えばウシまたはヤギ)を用いて、そして動物のミルク中にグリコシル化された結合剤を得ることも可能である。あるいは、植物を用いて、グリコシル化された選択的結合剤を産生することも可能である。
【0154】
当業者に周知の標準的培地を用いて、IFNγの選択的結合剤をコードする発現ベクターを含む(すなわち形質転換されたかまたはトランスフェクションされた)宿主細胞を培養してもよい。培地は、通常、細胞の増殖および生存に必要なすべての栄養素を含有するであろう。大腸菌細胞を培養するのに適した培地は、例えば、ルリア・ブロス(LB)および/またはテリフィック・ブロス(TB)である。真核細胞を培養するのに適した培地は、RPMI1640、MEM、DMEMであり、これらすべてには、培養中の特定の細胞株に必要とされるような血清および/または増殖因子を補うことも可能である。昆虫培養に適した培地は、必要に応じて、イーストレート、ラクトアルブミン加水分解物、および/またはウシ胎児血清を補ったグレース培地である。
【0155】
典型的には、トランスフェクション細胞および形質転換細胞の選択的増殖に有用な抗生物質または他の化合物が、培地の補充剤として添加される。使用すべき化合物は、宿主細胞を形質転換するプラスミド上に存在する選択可能マーカー要素によって、決定されるであろう。例えば、選択可能マーカー要素が、カナマイシン耐性である場合、培地に添加される化合物はカナマイシンであろう。選択的増殖のための他の化合物には、アンピシリン、テトラサイクリンおよびネオマイシンが含まれる。
【0156】
当該技術分野に知られる標準法を用いて、宿主細胞に産生される抗IFNγ抗体または抗原結合ドメインの量を評価することも可能である。こうした方法には、限定なしに、ウェスタンブロット解析、SDS−ポリアクリルアミドゲル電気泳動、非変性ゲル電気泳動、HPLC分離、免疫沈降、および/または活性アッセイが含まれる。
【0157】
細胞培地に分泌されている抗IFNγ抗体または抗原結合ドメインの精製は、アフィニティークロマトグラフィー、免疫親和性クロマトグラフィーまたはイオン交換クロマトグラフィー、分子ふるいクロマトグラフィー、分取用ゲル電気泳動または等電点電気泳動、クロマトフォーカシング、および高圧液体クロマトグラフィーを含む、多様な技術を用いて、達成可能である。例えば、Fc領域に選択的に結合するプロテインAを用いたアフィニティークロマトグラフィーによって、Fc領域を含む抗体を好適に精製することも可能である。カルボキシル末端またはアミノ末端いずれかの、ヘキサヒスチジン、あるいはFLAG(Eastman Kodak Co.、コネチカット州ニューヘブン)またはmyc(Invitrogen)などの他の小ペプチドなどのアフィニティータグを用いて、抗体または抗原結合ドメインの修飾型を調製し、そして一工程アフィニティーカラムによって精製することも可能である。例えば、ポリヒスチジンは、高い親和性および特異性でニッケルに結合し、したがってニッケルのアフィニティーカラム(例えばQiagen(登録商標)ニッケルカラム)を、ポリヒスチジンでタグ化した選択的結合剤の精製に用いることも可能である;例えばAusubelら監修, Current Protocols in Molecular Biology, セクション10.11.8, John Wiley & Sons, ニューヨーク(1993)を参照されたい。いくつかの例では、1より多い精製工程が必要である可能性もある。
【0158】
原核宿主細胞で発現される本発明の選択的結合剤は、細胞膜周辺腔または細胞質いずれかで、可溶性型で、あるいは細胞内封入体の一部として、不溶性型で、存在することも可能である。選択的結合剤は、当業者に知られる標準的技術いずれかを用いて、宿主細胞から抽出することも可能である。例えば、フレンチプレス、ホモジナイズ、および/または超音波処理の後、遠心分離することによって、宿主細胞を溶解して、周辺質/細胞質の内容物を放出させることも可能である。
【0159】
当該技術分野に知られる方法を用いて、細胞質に存在するか、または細胞膜周辺腔から放出されるか、いずれかの抗IFNγ抗体または抗原結合ドメインの可溶性型をさらに精製することも可能であり、例えば浸透圧ショック技術によって、細菌細胞膜周辺腔からFab断片を放出させる。抗体または抗原結合ドメインが、封入体を形成した場合、これらはしばしば、内部細胞膜および/または外部細胞膜に結合することも可能であり、そしてしたがって、遠心分離後、主にペレット成分中に見られるであろう。次いで、ペレット成分を、アルカリpHではジチオスレイトールなどの還元剤の存在下で、または酸pHではTrisカルボキシエチルホスフィンなどの還元剤の存在下で、極端なpHで処理するか、あるいは界面活性剤、グアニジン、グアニジン誘導体、尿素、または尿素誘導体などのカオトロピック剤で処理して、封入体を放出させ、粉々にし、そして可溶化することも可能である。次いで、ゲル電気泳動、免疫沈降等を用いて、可溶性選択的結合剤を解析することも可能である。可溶化抗体または抗原結合ドメインを単離することが望ましい場合、以下に示す方法およびMarstonら, Meth. Enz., 182:264−275(1990)に示される方法などの標準法を用いて、単離を達成することも可能である。
【0160】
いくつかの場合、抗体または抗原結合ドメインは、単離の際には、生物学的に活性でなくてもよい。ポリペプチドを三次構造に「再フォールディング」するか、または変換して、そしてジスルフィド連結を生成するための、多様な方法を用いて、生物学的活性を回復することも可能である。こうした方法には、可溶化ポリペプチドを、通常7を超えるpHに、そして特定の濃度のカオトロープの存在下で、曝露することが含まれる。カオトロープの選択は、封入体可溶化に用いる選択と非常に類似であるが、通常、カオトロープは、より低い濃度で用いられ、そして可溶化に用いるカオトロープと必ずしも同一ではない。大部分の場合、再フォールディング/酸化溶液はまた、還元剤を含有するか、または還元剤に加えて特定の比で酸化型を含有して、特定の酸化還元電位を生じ、ジスルフィド・シャフリングを可能にして、タンパク質のシステイン架橋(単数または複数)の形成を生じる。通常用いられるレドックス対には、システイン/シスタミン、グルタチオン(GSH)/ジチオビスGSH、塩化第二銅、ジチオスレイトール(DTT)/ジチアンDTT、および2−メルカプトエタノール(bME)/ジチオ−b(ME)が含まれる。多くの場合、共溶媒(cosolvent)を用いることも可能であるし、また再フォールディングの効率を増加させるのに共溶媒が必要とされる可能性もあり、そしてこの目的に用いられる、より一般的な試薬には、グリセロール、多様な分子量のポリエチレングリコール、アルギニン等が含まれる。
【0161】
本発明の抗体および抗原結合ドメインはまた、Merrifieldら, J. Am. Chem. Soc., 85:2149(1963);Houghtenら, Proc Natl Acad. Sci. USA, 82:5132(1985)、並びにStewartおよびYoung, Solid Phase Peptide Synthesis, Pierce Chemical Co., イリノイ州ロックフォード(1984)に示されるものなどの当該技術分野に知られる技術を用いた化学合成法(例えば固相ペプチド合成)によっても調製可能である。こうしたポリペプチドを、アミノ末端にメチオニンを含み、または含まずに合成することも可能である。これらの参考文献に示される方法を用いて、化学的に合成された抗体および抗原結合ドメインを酸化して、ジスルフィド架橋を形成することも可能である。こうして調製された抗体は、天然のまたは組換えによって産生された抗opgbp抗体または抗原結合ドメインと関連する生物学的活性を少なくとも1つ保持するであろう。
【0162】
IFNγの選択的結合剤に関するアッセイ
IFNγの少なくとも1つの生物学的活性を部分的にまたは完全に阻害する選択的結合剤を同定するためのスクリーニング法を本発明に提供する。IFNγの生物学的活性の阻害には、限定されるわけではないが、IFNγのその同族受容体、IFNγ−Rへの結合の阻害、in vitroでのA549に対するIFNγの抗増殖活性の阻害、並びにin vitroおよびin vivoでのIFNγによる単球の活性化の阻害が含まれる。本発明の選択的結合剤には、抗IFNγ抗体、並びにその断片、変異体、誘導体および融合体、ペプチド、ペプチド模倣体化合物または有機模倣体(organo−mimetic)化合物が含まれる。IFNγの生物学的活性を部分的にまたは完全に阻害可能な選択的結合剤を同定するためのスクリーニング法には、in vitroアッセイまたはin vivoアッセイが含まれることも可能である。in vitroアッセイには、IFNγのIFNγ−Rへの結合を検出するものが含まれ、そしてこうしたアッセイを用いて、IFNγのIFNγ−Rへの結合の速度または度合いを増加させるかまたは減少させる能力に関して、IFNγの選択的結合剤をスクリーニングすることも可能である。1つの種類のアッセイでは、IFNγポリペプチド、好ましくは細胞外ドメインなどのIFNγの可溶性型を固体支持体(例えばアガロースまたはアクリルビーズ)に固定し、そしてIFNγ−Rポリペプチドを、IFNγの選択的結合剤の存在下または非存在下いずれかで添加する。選択的結合剤の存在下または非存在下でIFNγおよびIFNγ−Rの結合の度合いを測定する。例えば放射標識、蛍光標識または酵素反応によって、結合を検出することも可能である。
【0163】
あるいは、BIAcoreアッセイ系(Pharmacia、ニュージャージー州ピスカタウェイ)などの表面プラズモン共鳴検出装置系を用いて、結合反応を行うことも可能である。結合反応は、製造者のプロトコルにしたがって、実行可能である。
【0164】
上述のものなどのin vitroアッセイを好適に用いて、IFNγのIFNγ−Rへの結合に対する影響に関して、迅速に多数の選択的結合剤をスクリーニングすることも可能である。アッセイを自動化して、ファージ・ディスプレー・ライブラリー、合成ペプチドライブラリーおよび化学合成ライブラリーで生成される化合物をスクリーニングすることも可能である。
【0165】
IFNγのIFNγ−Rへの結合を増加させるかまたは減少させる選択的結合剤を、どちらかのポリペプチドを発現する細胞および細胞株を用いて、細胞培養中でスクリーニングすることもまた可能である。細胞および細胞株は、いかなる哺乳動物から得ることも可能であるが、好ましくは、ヒトまたは他の霊長類、イヌまたはげっ歯類供給源由来であろう。例えば、選択的結合剤の存在下または非存在下で、表面にIFNγ−Rを発現している細胞に対するIFNγの結合を評価して、そして例えばIFNγに対するビオチン化抗体を用いたフローサイトメトリーによって、結合の度合いを測定することも可能である。
【0166】
in vitro活性アッセイを用いて、IFNγ活性を阻害する選択的結合剤を同定することもまた可能である。アッセイの例には、A549細胞増殖アッセイおよびTHP−1 HLA−DR発現アッセイが含まれる。
【0167】
選択的結合剤が、NZBxNZW F1マウスモデルにおいて、タンパク尿の発生を遅延させ、そして生存時間を増加させることが可能であるかどうかを決定するため、in vivoアッセイもまた利用可能である。
【0168】
診断適用のため、特定の態様において、IFNγの選択的結合剤、例えば抗体およびその抗原結合ドメインを、典型的には、検出可能部分で標識する。検出可能部分は、直接または間接的に、検出可能シグナルを産生可能なものいずれであってもよい。例えば、検出可能部分は、放射性同位体、例えば3H、14C、32P、35S、または125I、蛍光化合物または化学発光化合物、例えばフルオレセイン・イソチオシアネート、ローダミン、またはルシフェリン;あるいは酵素、例えばアルカリホスファターゼ、β−ガラクトシダーゼまたは西洋ワサビ(horseradish)ペルオキシダーゼであってもよい;Bayerら, Meth. Enz., 184:138−163(1990)。
【0169】
本発明の選択的結合剤は、いかなる既知のアッセイ法で使用することも可能であり、こうした方法には、例えばIFNγポリペプチドを検出し、そして定量化するための、ラジオイムノアッセイ、競合的結合アッセイ、直接および間接的サンドイッチアッセイ(ELISA)、および免疫沈降アッセイがある(Sola, Monoclonal Antibodies:A Manual of Techniques, pp.147−158(CRC Press, 1987))。抗体は、使用中のアッセイ法に適した親和性で、IFNγポリペプチドに結合するであろう。
【0170】
本発明の選択的結合剤はまた、in vivo画像化にも有用であり、ここで、例えば検出可能部分で標識した選択的結合剤を、動物の好ましくは血流中に投与し、そして宿主中の標識抗体の存在および位置をアッセイする。剤を、核磁気共鳴によってであれ、放射線によってであれ、または当該技術分野に知られる他の手段によってであれ、動物中で検出可能ないずれの部分で標識することも可能である。
【0171】
本発明はまた、抗体または抗原結合ドメインなどのIFNγの選択的結合剤、および生物学的試料中のIFNγレベルを検出するのに有用な他の試薬を含むキットにも関する。こうした試薬には、二次活性、検出可能標識、ブロッキング血清、陽性対照試料および陰性対照試料、および検出試薬が含まれることも可能である。
【0172】
IFNγ選択的結合剤の療法使用
本発明の選択的結合剤は、療法剤として使用可能である。療法選択的結合剤は、IFNγアゴニストまたはアンタゴニストであることも可能であり、そして1つの態様において、in vitroまたはin vivoでIFNγポリペプチドの生物学的活性の少なくとも1つを阻害する抗IFNγアンタゴニスト抗体である。例えば、IFNγのアンタゴニストは、IFNγのIFNγ−Rへの結合を阻害するであろう。あるいは、IFNγアンタゴニストは、実施例1に記載するものなどのA549細胞増殖アッセイにおいて、測定可能なND50(50%増殖を生じる濃度)によって示されるようなin vitroでのヒト肺癌の増殖を刺激するであろう。
【0173】
IFNγアンタゴニスト、例えば抗IFNγアンタゴニスト抗体および抗原結合ドメインを用いて、限定されるわけではないが、以下の:急性膵炎;ALS;アルツハイマー病;AIDS誘導悪疫質を含む悪疫質/摂食障害;喘息および他の肺疾患;アテローム性動脈硬化症;慢性疲労症候群;クロストリジウム属(Clostridium)関連下痢を含むクロストリジウム属関連疾病;うっ血性心不全、冠状動脈再狭窄、心筋梗塞、心筋機能不全(例えば敗血症に関連するもの)、および冠状動脈バイパス移植を含む冠状動脈状態および徴候;癌、例えば多発性骨髄腫、並びに骨髄性白血病(例えばAMLおよびCML)および他の白血病とともに腫瘍転位;発熱;糸球体腎炎;移植片対宿主病/移植拒絶;出血性ショック;例えば角膜移植に関与することもありうるような、炎症性眼疾患;脳虚血(例えば、各々神経変性につながりうる、外傷、癲癇、出血または脳卒中などの結果の脳傷害)を含む虚血;学習障害;多発性硬化症;筋障害(例えば筋タンパク質代謝、特に敗血症におけるもの);神経毒性(例えばHIVに誘導されるようなもの);骨粗鬆症;癌関連疼痛を含む疼痛;パーキンソン病;歯周病;神経毒性;早期陣痛;乾癬;再灌流傷害;敗血症ショック;放射療法の副作用;顎関節疾患;睡眠障害;ブドウ膜炎;または挫傷(strain)、捻挫、軟骨損傷、外傷、整形手術、感染もしくは他の疾患過程から生じる炎症状態;若年発症1型糖尿病およびインスリン耐性糖尿病(例えば肥満に関連するようなもの)を含む糖尿病;子宮内膜症、子宮内膜炎、および関連状態;線維筋痛症または痛覚脱失症;痛覚過敏;クローン病を含む炎症性腸疾患;肺疾患(例えば成人呼吸困難症候群、および肺線維症);神経炎症性疾患;眼変性およびブドウ膜炎を含む、眼疾患および状態;毛孔性紅色粃糠疹(PRP);前立腺炎(細菌性または非細菌性)および関連状態;乾癬および関連状態;肺線維症;再灌流傷害;骨関節炎、関節リウマチ、若年性(リウマチ様)関節炎、血清陰性多発性関節炎、強直性脊椎炎、ライター症候群および反応性関節炎、スティル病、乾癬性関節炎、腸疾患に基づく関節炎、多発性筋炎、皮膚筋炎、強皮症、全身性硬化症、脈管炎(例えば川崎病)、脳脈管炎、ライム病、ブドウ球菌(staphylococcal)誘導(「敗血症」)関節炎、シェーグレン症候群、リウマチ熱、多発性軟骨炎およびリウマチ性多発性筋痛および巨細胞性動脈炎を含む、関節の炎症状態およびリウマチ疾患;敗血症ショック;全身性エリテマトーデス(SLE)腎炎;放射療法による副作用;顎関節疾患;甲状腺炎;組織移植、または挫傷、捻挫、軟骨損傷、外傷、および整形手術から生じる炎症状態を含む、自己免疫疾患および炎症状態を防止するかまたは治療することも可能である。
【0174】
より具体的には、IFNγアンタゴニスト、例えば抗IFNγアンタゴニスト抗体および抗原結合ドメインを用いて、関節炎(特に関節リウマチ)、全身性エリテマトーデス(SLE)、移植片対宿主病(GvHD)、多発性硬化症および糖尿病を防止するかまたは治療することも可能である。
【0175】
アンタゴニスト抗体および抗原結合ドメインを含む、本発明のIFNγアンタゴニストを、単独で、または抗IFNγアンタゴニスト抗体および抗原結合ドメインなどの他の療法剤IFNγアンタゴニストと組み合わせて投与して、これらを用いて、多様な炎症状態、自己免疫状態、および骨損失につながる他の状態を防止するかまたは治療することも可能である。状態および望ましい治療レベルに応じて、2、3、またはそれより多い剤を投与することも可能である。同一処方に含むか、または治療キットに含むことによって、これらの剤をともに提供することも可能であり、あるいは別個に提供することも可能である。遺伝子治療によって投与する際、タンパク質剤をコードする遺伝子を、所望によって同一プロモーター領域の制御下で同一ベクターに含めてもよいし、または別個のベクターに含めてもよい。前述の種類の特に好ましい分子は以下のとおりである。
・IL−1阻害剤:IL−1raタンパク質および可溶性IL−1受容体。最も好ましいIL−1阻害剤はアナキンラである。
・TNF−α阻害剤:可溶性腫瘍壊死因子I型受容体(sTNF−RI;−RIはまた、p55受容体とも呼ばれる);可溶性腫瘍壊死因子II型受容体(p75受容体とも呼ばれる);およびTNF受容体に結合するモノクローナル抗体。最も好ましいのは、WO 98/24463に記載されるようなsTNF−RI、エタネルセプト(エンブレル(登録商標))、およびアバカイン(登録商標)である。典型的なTNF−α阻害剤は、EP 422 339、EP 308 378、EP 393 438、EP 398 327、およびEP 418 014に記載される。
・セリンプロテアーゼ阻害剤:SLPI、ALP、MPI、HUSI−I、BMI、およびCUSI。SLPIがLPS反応を阻害することが示されたように、これらの阻害剤を、典型的なLPS調節因子として見ることもまた可能である。Jinら(1997), Cell 88(3):417−26(本明細書に援用される)。
【0176】
薬剤組成物
IFNγ選択的結合剤の薬剤組成物は、本発明の範囲内にある。こうした組成物は、療法的または予防的に有効な量のIFNγ選択的結合剤、例えば抗体、あるいはその断片、変異体、誘導体または融合体を、薬学的に許容しうる剤と混合して含む。好ましい態様において、薬剤組成物は、IFNγの少なくとも1つの生物学的活性を部分的にまたは完全に阻害する抗IFNγアンタゴニスト抗体を、薬学的に許容しうる剤と混合して含む。典型的には、抗体は、動物への投与のため、十分に精製されているであろう。
【0177】
本発明の組成物に使用するための薬学的に許容しうる剤には、当該技術分野に周知であるような、キャリアー、賦形剤、希釈剤、酸化防止剤、保存剤、着色剤、フレーバー剤および希釈剤、乳化剤、懸濁剤、溶媒、増量剤(filler)、充填剤(bulking agent)、緩衝剤、搬送ビヒクル、等張化剤、共溶媒、湿潤剤、錯化剤、緩衝化剤、抗菌剤および界面活性剤が含まれる。
【0178】
中性緩衝生理食塩水または血清アルブミンと混合した生理食塩水が、典型的な適切なキャリアーである。組成物にやはり含まれるのは、アスコルビン酸などの酸化防止剤;低分子量ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニンまたはリジンなどのアミノ酸;単糖、二糖、およびグルコース、マンノースまたはデキストリン類を含む他の炭水化物;EDTAなどのキレート剤;マンニトールまたはソルビトールなどの糖アルコール;ナトリウムなどの塩形成対イオン;および/またはTween、プルロニック類またはポリエチレングリコールなどの非イオン性界面活性剤である。やはり例として、適切な等張化増進剤には、ハロゲン化アルカリ金属(好ましくは塩化ナトリウムまたは塩化カリウム)、マンニトール、ソルビトール等が含まれる。適切な保存剤には、限定されるわけではないが、塩化ベンザルコニウム、チメロサール、フェネチル・アルコール、メチルパラベン、プロピルパラベン、クロルヘキシジン、ソルビン酸等が含まれる。過酸化水素もまた、保存剤として使用可能である。
【0179】
適切な共溶媒は、例えばグリセリン、プロピレングリコール、およびポリエチレングリコールである。適切な錯化剤は、例えばカフェイン、ポリビニルピロリドン、ベータ−シクロデキストリンまたはヒドロキシ−プロピル−ベータ−シクロデキストリンである。適切な界面活性剤または湿潤剤には、ソルビタンエステル類、ポリソルベート80などのポリソルベート類、トロメタミン、レシチン、コレステロール、チロキサポール(tyloxapal)等が含まれる。緩衝剤は、酢酸、ホウ酸、クエン酸、リン酸、炭酸水素、またはTris−HClなどの慣用的緩衝剤であることも可能である。酢酸緩衝液はpH4.0〜5.5程度であることも可能であり、そしてTris緩衝液はpH7.0〜8.5程度であることも可能である。さらなる薬学的剤が、Remington’s Pharmaceutical Sciences, 第18版, A.R. Gennaro監修, Mack Publishing社、1990に示されており、該文献の相当する部分は、本明細書に援用される。
【0180】
組成物は、液体型であっても、あるいは凍結乾燥型またはフリーズドライ型であってもよい。凍結乾燥型には、スクロースなどの賦形剤が含まれうる。本発明の組成物は、非経口投与に適している。好ましい態様において、組成物は、当業者に利用可能ないずれかの経路によって、例えば皮下経路、静脈内経路、筋内経路、腹腔内経路、大脳内(実質内)経路、脳室内経路、筋内経路、眼内経路、動脈内経路、または病変内経路によって、動物に注射するかまたは注入するのに適している。非経口処方は、典型的には、無菌、病原体不含の等張化水性溶液であり、所望によって薬学的に許容しうる保存剤を含有するであろう。
【0181】
最適な薬剤処方は、意図される投与経路、搬送形式および望ましい投薬量に応じて、当業者が容易に決定可能である。
他の処方もまた本発明に意図される。薬剤組成物はまた、ポリ乳酸、ポリグリコール酸などのポリマー化合物の粒子性調製物、またはリポソームへのIFNγ選択的結合剤(抗体など)の導入も含むことも可能である。ヒアルロン酸もまた使用可能であり、そしてこの酸は、循環中にある期間の持続を促進する効果も有しうる。薬剤組成物は、IFNγ選択的結合剤(例えば抗体)と、注射可能微小球体、生体侵食性粒子またはビーズ、あるいはリポソームとの処方もまた含み、こうした処方は、選択的結合剤の制御放出または徐放を提供し、こうした処方を、次いで、蓄積注射として搬送してもよい。他の適切な搬送手段には、移植可能搬送装置が含まれる。
【0182】
IFNγ選択的結合剤(抗体など)を含む薬剤組成物は、吸入用の乾燥粉末としても処方可能である。こうした吸入溶液はまた、エアロゾル搬送用の液化噴霧剤中にも処方可能である。さらに別の処方において、溶液を噴霧することも可能である。IFNγ選択的結合剤を含有する特定の処方を経口投与してもよいこともまた意図される。この方式で投与される処方を、錠剤およびカプセルなどの固形投薬型の配合に習慣的に用いられるキャリアーを伴い、または伴わずに、処方することも可能である。例えば、生物学的利用能が最大となりそして前全身分解が最小となる胃腸管で、処方の活性部分が放出されるように、カプセルを設計することも可能である。選択的結合剤の吸収を促進する、さらなる剤を含むことも可能である。希釈剤、フレーバー剤、低融点ワックス、植物油、滑沢剤、懸濁剤、錠剤崩壊剤、および結合剤もまた、使用可能である。
【0183】
別の調製は、有効量のIFNγ選択的結合剤を、錠剤の製造に適した非毒性賦形剤と混合して含むことも可能である。錠剤を滅菌水、または別の適切なビヒクルに溶解することによって、単位用量型の溶液を調製することも可能である。適切な賦形剤には、限定されるわけではないが、不活性希釈剤、例えば炭酸カルシウム、炭酸ナトリウムもしくは炭酸水素ナトリウム、ラクトース、またはリン酸カルシウム;あるいは結合剤、例えばデンプン、ゼラチン、またはアラビアゴム(acacia);あるいは滑沢剤、例えばステアリン酸マグネシウム、ステアリン酸、またはタルクが含まれる。
【0184】
さらなる処方が当業者には明らかであり、こうした処方には、IFNγ選択的結合剤を、1以上の他の療法剤と組み合わせて含む処方が含まれる。多様な他の徐放または制御搬送手段、例えばリポソームキャリアー、生体侵食性微小粒子または多孔ビーズおよび蓄積注射を処方するための技術もまた、当業者に知られる。例えば、その開示が本明細書に援用される、Supersaxoら、薬剤組成物搬送用の制御放出多孔ポリマー微小粒子の説明(WO 93/15722(PCT/US93/00829)を参照されたい)を参照されたい。
【0185】
投与方式に関わらず、体重、体表面積または臓器サイズにしたがって、特定の用量を計算することも可能である。上述の処方各々を伴う治療の適切な投薬量を決定するのに必要な計算のさらなる微調整は、当業者によって日常的に行われ、そして当業者が日常的に行う仕事の範囲内である。適切な用量−反応データの使用を通じて、適切な投薬量を確認することも可能である。
【0186】
さらに、肺投与によって、本発明の薬剤組成物を投与することも可能であり、例えば化学的に修飾されたタンパク質の肺搬送を開示し、本明細書に援用される、PCT WO94/20069を参照されたい。肺搬送のため、粒子サイズは、遠位肺への搬送に適したものでなければならない。例えば、粒子サイズは1μm〜5μmであることも可能であるが、例えば各粒子がかなり多孔性であれば、より大きい粒子も使用可能である。
【0187】
あるいはまたはさらに、IFNγ選択的結合剤が吸収されているか、または被包されている、膜、スポンジ、または他の適切な材料を、罹患領域に移植することを介して、組成物を局所投与することも可能である。移植装置を用いる場合、適切な組織または臓器いずれかに装置を移植し、そしてIFNγ選択的結合剤の搬送は、多量(bolus)投与を介して、または連続投与を介して、または連続注入を用い、カテーテルを介して、装置から直接なされることも可能である。
【0188】
本発明の薬剤組成物は、徐放処方または徐放調製物中でもまた投与可能である。徐放調製物の適切な例には、成形物品の形の半透性ポリマーマトリックス、例えばフィルム、または微小カプセルが含まれる。徐放マトリックスには、ポリエステル、ヒドロゲル、ポリ乳酸(例えば米国特許第3,773,919号、EP 58,481)、L−グルタミン酸およびガンマ・エチル−L−グルタメートのコポリマー(Sidmanら, Biopolymers, 22:547−556(1983))、ポリ(2−ヒドロキシエチル−メタクリレート)(Langerら, J. Biomed. Mater. Res., 15:167−277(1981)およびLanger, Chem. Tech., 12:98−105(1982))、エチレン酢酸ビニル、またはポリ−D(−)−3−ヒドロキシ酪酸が含まれる。徐放組成物にはまた、当該技術分野に知られるいくつかの方法のいずれかによって調製可能なリポソームが含まれることも可能である。例えばEppsteinら, Proc. Natl. Acad. Sci. USA, 82:3688−3692(1985);EP 36,676;EP 88,046;およびEP 143,949を参照されたい。
【0189】
いくつかの例では、IFNγ選択的結合剤組成物を含む薬剤組成物を、ex vivo方式で使用することが望ましい可能性もある。この場合、患者から取り除いた細胞、組織または臓器を、IFNγ選択的結合剤を含む薬剤組成物に曝露し、その後、続いて、細胞、組織および/または臓器を患者に移植しなおす。
【0190】
他の場合では、本明細書に記載するものなどの方法を用いて、遺伝子操作した特定の細胞を患者に移植して、ポリペプチド、選択的結合剤、断片、変異体、または誘導体を発現させ分泌させることによって、IFNγ選択的結合剤を含む組成物を搬送することも可能である。こうした細胞は、動物細胞またはヒト細胞であってもよく、そして患者自身の組織あるいはヒトまたは非ヒトいずれかの別の供給源由来であってもよい。所望によって、細胞を不死化してもよい。しかし、免疫応答の可能性を減少させるため、細胞を被包して、周囲組織への浸潤を回避することが好ましい。被包材料は、典型的には、タンパク質産物(単数または複数)の放出を許すが、患者の免疫系による、または周囲組織由来の他の有害因子による分解を防止する、生体適合性の半透性ポリマー被包体(enclosure)または膜である。
【0191】
細胞の膜被包に用いられる方法は当業者に周知であり、そして被包細胞の調製および該細胞の患者への移植は、過度の実験を伴わずに達成可能である。例えば米国特許第4,892,538号、第5,011,472号、および第5,106,627号を参照されたい。生存細胞を被包する系がPCT WO 91/10425(Aebischerら)に記載される。他の多様な徐放または制御搬送手段、例えばリポソームキャリアー、生体侵食性粒子またはビーズを処方するための技術もまた、当該技術分野に知られ、そして記載されている。細胞を被包し、また被包せずに、患者の適切な体組織または臓器に移植してもよい。
【0192】
IFNγ選択的結合剤(例えば抗IFNγ抗体、あるいはその断片、変異体、誘導体、および融合体)を含む薬剤組成物の療法的または予防的に有効な量は、例えば該組成物を用いる徴候などの療法目的、投与経路、および被験者の状態に応じるであろう。本発明のIFNγアンタゴニスト抗体または抗原結合ドメインは、自己免疫および/または炎症状態を防止し、そして/または治療するため、療法的または予防的に有効な量で投与される。
【0193】
以下の実施例は、本発明をさらに十分に例示するために提供され、そして本発明の範囲を限定するとは解釈されない。
【0194】
(実施例1)
試薬およびアッセイ
これらの研究に用いるスクリーニング標的を:1)EP 0423845またはPCT公報WO 83/04053に記載されるような、hIFNγをコードするcDNAの大腸菌における発現;または2)以下のような、hIFNγをコードするcDNAのCHO宿主細胞における発現から調製した:PCR(標準的条件)を用いて、テンプレートとしてヒト脾臓Marathon Ready cDNA(Clonetech)を用い、ヒトINFγをコードする全長配列を増幅した。発現プラスミドに配列をサブクローニングし、そしてDH10B細胞(Gibco Life Sciences)をDNAで形質転換し、DNAを調製し、そしてリン酸カルシウム法(Speciality Media, Inc.)によって、CHO細胞にトランスフェクションした。高発現細胞株クローンを用いて、血清不含馴化培地を生成した。
【0195】
hINFγを含有するCHO細胞馴化培地を濃縮し、透析し、そして次いで、いくつかのクロマトグラフィー工程を通じて精製した。最初の工程は、非グリコシル化hIFNγ型に対して高グリコシル化型を分離する、標準的NaCl勾配を用いたQ−HP(Pharmacia)クロマトグラフィーであった。コムギ(wheat)胚芽凝集素クロマトグラフィー(EY Laboratories)を通して、Q−HPプールをさらに精製した。精製物質は、クーマシー−ブルーおよび銀染色SDS−PAGE両方によって判断すると、95%より純粋であった。物質は、ゲル凝固法(カブトガニ(Limulus)アメーバ様細胞溶解物)によってアッセイした際、内毒素が低レベルであった。R&D Systemsのヤギ(goat)抗hIFNγ中和抗体(カタログ番号AF−285−NA、ロット番号ZW019011)を用いて、ウェスタンブロットによって、hINFγの同一性を確認した。吸光係数法(0.66)を用いて、最終タンパク質濃度を決定した。物質2ロットをそれぞれ生成した。収率は40mg/lであった。最終物質をPBS中に配合した。
【0196】
CHO細胞におけるヒトIFNγR1−Fcタンパク質の発現
これらの研究において、標的からファージ抗体を溶出するのに用いたヒトIFNγR1−Fcタンパク質を以下のように調製した:PCR(標準的条件)を用いて、テンプレートとしてヒトリンパ球Marathon Ready cDNA(Clonetechより購入)を用い、ヒトIFNγR1をコードする全長配列を増幅した。PCR(標準的条件)を用いて、ヒトIgG1のFc部分をコードする配列を増幅した。重複PCRを用いて、IFNγR1−Fc融合構築物をコードする配列(IFNγR1のアミノ酸1〜Ser246)を生成して、そして発現プラスミドに該配列をサブクローニングした。DH10B細胞(Gibco Life Sciences)をDNAで形質転換し、DNAを調製し、そしてリン酸カルシウム法(Speciality Media, Inc.)によって、CHO細胞にトランスフェクションした。高発現細胞株クローンを用いて、血清不含馴化培地を生成した。
【0197】
標準的プロテインG Fast−Flowカラム(Pharmacia)を通して、hINFγR1−Fcを含有するCHO細胞馴化培地を濃縮し、そして精製した。吸光係数として1.44を用いて、A280によって最終濃度を決定した。N末端配列決定解析を通じて、精製試料の同一性を確認した。この物質をPBS中に配合した。
【0198】
抗体
モノクローナル抗hIFNγ抗体、クローン2578.111をR&D Systemsから購入した(カタログ番号MAB285、ロット番号KW07)。モノクローナル抗hIFNγ抗体、クローンMMHG−1をBiosourceから購入した(カタログ番号AHC4834、ロット番号10803−015)。組換えヒトIFNγ受容体1(rhIFNγ R1)をR&D Systemsから購入した(カタログ番号673−IR)。rhIFNγ R1の計算上の分子量は25,000ダルトンである。グリコシル化の結果、組換えタンパク質は、SDS−PAGE上、40〜50kDaタンパク質として移動する。
【0199】
A549細胞増殖アッセイ
IFNγの抗体中和を評価するために用いたA549細胞増殖アッセイは、96ウェルアッセイであり、そして一般的に以下のように記載される:第1日、1)アッセイ培地(F12K、5%FBS、1xPen/Strep L−グルタミン)中の最高濃度からAbを1:2で連続希釈する。アッセイにおいて望ましい濃度の4倍で、全部で10回の希釈を行う。2つ組のため、各希釈物は少なくとも最終200μlが必要である;2)用量反応曲線中の有効用量の90%に基づいて、スパイク用にIFNγを適切な濃度に希釈する。アッセイにおいて望ましい濃度の4倍で、IFNγスパイクを作成する;3)Titertek試験管中で、各4xAb希釈物150μlを、4xINFγスパイク150μlと合わせる。ピペッティングによって混合する。ふたをして、室温で1時間インキュベーションする。(注:AbおよびIFNγの濃度は、ここで、アッセイ濃度の2倍である);4)(所望による)AbおよびIFNγをインキュベーションしている間、力価決定(titration)曲線用にIFNγを希釈する。4000ng/mlから始めて、1:3希釈を12回行う。アッセイにおいて、3つ組には300μlが必要であるため、希釈終了時に必要な体積は少なくとも400μlであるはずである。必要となるまで、4℃で保存する;5)インキュベーションが完了する前に、5mlトリプシン中で、A549細胞をトリプシン処理する。フラスコに20mlのアッセイ培地を添加し、そして50mlコニカルに移して、そしてIEC中、室温で1/2〜3/4の速度で遠心分離する;6)細胞を吸引する。7.5mlのアッセイ培地に再懸濁する。トリパンブルー中、1:1で計数する;7)アッセイ培地中、細胞を2.5x104細胞/mlに希釈する。各試料について、2.5x103細胞/ウェルを、96ウェルFalconに0.1mlで植え付ける;8)1時間後、2つ組の2つのウェル各々に、100μl Ab/IFNγ混合物を添加する;そして9)37℃、5%CO2および高湿度で5日間インキュベーションする。第5日:1)ウェルあたり20μlのAlamar Blueを添加する。37℃、5%CO2および高湿度で3〜4時間インキュベーションする;2)FL500蛍光プレート読取装置のスイッチを入れる。プレートからふたを取り除き、そしてふたなしで10分間振盪する;そして3)FL500を読み取る。設定:中程度に3秒間振盪、励起530/25、発光590/35、感度34。
【0200】
(実施例2)
ヒトFabライブラリーのスクリーニング
スクリーニング法
ヒトFabライブラリーを構築し、そしてスクリーニングする、一般的な方法は、de Haardら(Advanced Drug Delivery Reviews, 31:5−31(1998);J. Biol. Chem., 274:18218−18230(1999))に記載された。以下の方法によって、hIFNγに結合するFab断片をスクリーニングした。
【0201】
Nuncイムノチューブを、0.1M炭酸Na、pH9.6中、0.39μg/mlのhIFNγ 4mlで、Nutator上、室温で2時間コーティングした。Target Quest、NV(オランダ・アムステルダム)凍結ファージライブラリーストック(試験管あたり750μl中、4x1012pfu)のアリコットを融解後、1/5体積(150μl)のPEG溶液(20%ポリエチレングリコール8000、2.5M NaCl、オートクレーブ)を添加し、そして試験管を氷上に1時間放置してファージを沈殿させることによって、グリセロール(15%)を取り除いた。沈殿したファージ粒子を4000rpm、4℃で15分間遠心してペレットとし、次いで500μl PBS、pH7.4に再懸濁した。IFNγでコーティングしたイムノチューブを4ml PBSで3回洗浄し、そしてNutator上、4mlの2%MPBSを用いて、室温で1時間ブロッキングした。同時に、500μlの4%MPBSをファージ懸濁物に添加し、そして室温で30分間〜1時間インキュベーションして、ファージ粒子のプレ・ブロッキングを可能にした。ブロッキングしたイムノチューブをPBST(PBS中の0.1%Tween20)で2回洗浄し、そしてPBSで2回洗浄した。プレ・ブロッキングしたファージ混合物を、3mlの2%MPBSを含有する、洗浄したイムノチューブに添加した。回転装置上で30分間インキュベーションし、その後、室温で1.5時間、静置インキュベーションした後、ファージ混合物を廃棄した。試験管をまずPBSTで20回、次いでPBSで20回洗浄した。回転装置上、0.4%MPBS、pH7.4中、1μMの特異的溶出試薬(それぞれhIFNγ、GPNA、RDMA、BSMA、またはrhIFNγ R1)1mlとインキュベーションすることによって、結合したファージ粒子を溶出した。溶出したファージ粒子を無菌50mlコニカル・ポリプロピレン試験管に移し、そして氷上で保存した。各ファージ溶出物、約20μlを、力価決定のため、取り置いた。増幅のため、残った溶出ファージ粒子を、5mlのTG1培養物(OD590約0.5)および4mlの2xYTを含有する50mlコニカル試験管に添加した。IFN感染(IFNection)混合物を振盪せずに37℃で30分間インキュベーションし、次いで、3500rpmで20分間回転させた。細胞ペレットを1500μlの2xYT−AGブロスに懸濁し、そして5つのSOBCGプレート上、300μl/プレートで蒔いた。プレートを30℃で一晩インキュベーションした。20時間インキュベーションした後、プレートから細胞スクレーパーで細胞を回収し、これにプレートあたり4mlの2xYT−AGを添加した。この工程を3回反復した。回収細胞のごく一部をファージ救出に用いた(以下を参照されたい)。残った細胞懸濁物を3500rpmで20分間回転させた。細胞ペレットをペレットサイズの1/2体積の50%グリセロールに再懸濁して、グリセロールストックを作成し、そして−80℃で保存した。
【0202】
増幅細胞懸濁物からのファージ救出を以下のように行った。蒔いて増幅させた約0.5mlの回収細胞懸濁物を用いて、約0.3のOD590になるように50mlの2xYT−AGに接種した。振盪装置上、OD590が0.5になるまで培養物を37℃でインキュベーションした。M.O.I.20のM13KO7ヘルパーファージ(GIBCO BRL、カタログ番号18311−019、1.1x1011pfu/ml)1mlで、10mlの培養物をIFN感染させて、そして37℃で30分間、インキュベーター中でインキュベーションした。IFN感染した細胞を4000rpmで20分間回転させて落とした。細胞ペレットを50mlの2xYT−AKに再懸濁し、250mlフラスコに移して、そして270rpmで20時間、振盪しながら30℃でインキュベーションした。一晩培養物を4000rpmで20分間回転させて、細胞破片を取り除いた。上清を再び遠心分離して、細胞破片が除去されることを確実にした。PEG溶液(20%PEG8000、2.5M NaCl)約1/5体積を上清に添加して、ファージ粒子を沈殿させた。混合物を氷上で少なくとも1時間インキュベーションし、次いで4000rpmで20分間遠心分離して、沈殿したファージ粒子を収集した。ファージペレットを1mlのPBSに再懸濁し、そして微量遠心分離試験管に移した。ファージ懸濁物を氷上で1時間放置して、ファージ粒子の完全な懸濁を可能にし、次いで、14,000rpmで2分間回転させて、残った細胞破片を取り除いた。ファージ沈殿工程を反復した。最終ファージペレットを1.1mlのPBSに懸濁し、そして延長した期間、氷上に放置して、ファージ粒子の完全な懸濁を確実にした。ファージ懸濁物を14,000rpmで2分間遠心分離して、残った細胞破片を取り除いた。500μlの救出したファージ懸濁物を用いて、250μlの50%グリセロールを添加することによって、グリセロールストックを作成した。100μlの救出ファージ懸濁物をファージプールELISA(以下を参照されたい)用に取り置いた。残った500μlの救出ファージを次の周期のパニングに用いた。
【0203】
ファージプールELISA
ファージプールELISAを以下のように行った:大腸菌で発現したhIFNγを、Nunc MaxiSorb Immunoプレートにおいて、0.1M炭酸Na、pH9.6中、0.39μg/ml、100μl/ウェルで、穏やかに振動させながら、室温で2時間プレーティングした。コーティングしたプレートをPBSで3回洗浄し、次いで、振動装置上、室温で1時間、2%MPBS 300μl/ウェルを用いてブロッキングした。陰性対照として、抗原でコーティングされていない別のNunc Immunoプレートもまた、2%MPBSでブロッキングした。その間に、120μlの救出された各ファージプールを、96ウェルCostar 3790プレート中、120μlの0.8%MTBSでプレ・ブロッキングし、そして使用準備が整うまで、室温で放置した。ブロッキングしたプレートをどちらも、0.1%TBST(TBS:10mM Tris−HCl、pH7.5、1mM EDTA、150mM NaCl;Tween−20 0.1%)で5回洗浄した。プレ・ブロッキングしたファージ希釈物を、抗原でコーティングしたプレートおよび陰性対照プレート両方に分配し(100μl/ウェル)、そして振動装置上、室温で1時間インキュベーションした。記載するようにプレートを洗浄した後、0.4%MTBS中、1:1000倍希釈したHRP/抗M13モノクローナル・コンジュゲート(Amersham Pharmacia Biotech、カタログ番号27−9421−01)100μl/ウェルを分配し、そして振動装置上、室温で1時間インキュベーションした。記載するようにプレートを洗浄した。基質、1−StepTM ABTS(Pierce、カタログ番号37615)100μl/ウェルを添加した後、プレートを1時間インキュベーションした。シグナル検出のため、OD405を測定した。さらなる解析のため、個々のクローンの供給源として、ELISA陽性ファージプールを用いた。
【0204】
(実施例3)
IFNγに結合するFabファージクローンの同定
DNAフィンガープリンティング
ELISA陽性ファージプール由来の重鎖および軽鎖両方を含有する全長クローンを同定するため、96ウェルThermowellプレート中でポリメラーゼ連鎖反応(PCR)を行った。典型的には、各ウェルは、25μlのPCR反応混合物(2.5μl 10xPCR緩衝液、21.625μlの水、25mMの0.25μl dNTP類、10pmol/μlの0.25μlプライマー870−02(以下を参照されたい)、10pmol/μlの0.25μlプライマー2182−83(以下を参照されたい)、5単位/μlの0.125μl Taqポリメラーゼ)を含有する。
【0205】
【化7】
【0206】
個々のコロニーを摘み取り、そしてまずPCRプレート中のウェルに再懸濁し、次いで300μl/ウェルの2xYT−AGブロス(2xYTブロス:100μg/mlアンピシリンおよび2%グルコースを含有する、水1lあたり、10g酵母エキス、16gバクト−トリプトン、5g NaCl)を満たした96ウェル深底ウェルブロックの対応するウェルに再懸濁する。PCR反応条件は、94℃5分間の変性周期1回、40周期の94℃45秒間、55℃45秒間、72℃1.5分間、その後、72℃10分間の伸長周期1回であった。PCR反応が完了した後、3μl/ウェルのPCR反応混合物を、0.5μl/mlのエチジウムブロミドを含有する、余分に長い4x(24+2)の1%TAEゲル(Embi Tec、カタログ番号GE−3820)上で、120ボルトで1時間泳動した。1kb plus DNAラダー(Gibco BRL、カタログ番号10787−018)に比較することによって、1.6kbより大きい挿入物を持つクローンを全長クローンと同定した。
【0207】
ユニークな全長クローンの同定を以下のように行った:同定された全長クローンのPCR増幅した挿入物に対して、BstNI消化を行った。96ウェルThermowellプレート中の試料あたり16μlのPCR反応混合物に、3μl 10x緩衝液2(NEBL)、10mg/mlの0.3μl BSA、10μlの水および0.7μlのBstNI(NEBL)を含有する14μlのBstNI消化マスター溶液を添加した。プレートを60℃で3時間インキュベーションした。消化した試料の各々13μlを、0.5μl/mlのエチジウムブロミドを含有する、余分に長い2x(24+2)の4%TAEゲル(Embi Tec、カタログ番号GE−3817)上で、100ボルトで3時間泳動した。BstNI断片パターンの相違に基づいて、ユニークなクローンを同定した。
【0208】
クローンファージELISA
同定されたユニークな全長クローンのFabファージを96ウェル形式で救出した。96ウェルの2ml深底ウェルブロックにおいて、480μl/ウェル 2xYTAGブロスに、選択したユニークな全長クローンの一晩培養物20μlを接種し、次いで、300rpm、37℃で3時間インキュベーションした。細胞をIFN感染させるため、各ウェルに、100μlの1:10希釈したM13KO7ヘルパーファージ希釈物を添加した。ブロックを振盪せずに37℃で30分間インキュベーションし、次いで、150rpmでさらに30分間、穏やかに振盪した。ブロックを3600rpmで20分間遠心分離して、IFN感染させた細胞をペレットにした。各ウェルの細胞ペレットを480μlの2xYTAK(100μg/mlアンピシリンおよび40μg/mlカナマイシンを含有する2xYTブロス)に懸濁し、次いで約20時間に渡って、30℃で一晩インキュベーションした。3600rpmで20分間遠心分離することによって、細胞破片を分離した。救出したファージ上清を注意深く別の無菌96ウェルブロックに移した。救出したファージを用いて、実施例2のファージプールELISAに記載したのと正確に同じように、クローンファージELISAを行った。0.2以上の純OD405を生じるクローンをIFNγ結合候補と見なした。
【0209】
大規模ファージ救出ELISA
ELISAにおいて、FabファージがIFNγに濃度依存的に結合することを立証することによって、同定したユニークなFabファージクローンが特異的にIFNγに結合することを確認した。大規模救出によってFabファージを得た。
【0210】
個々のクローンの大規模救出を、以下のように行った:同定したユニークなIFNγ結合クローンのFabファージを大規模に救出した。250mlの無菌フラスコ中で、50mlの2xYT−AGブロスに、選択したIFNγ結合クローンの一晩培養物200μlを接種し、そして培養物のOD590が0.5に達するまで、37℃、2700rpmでインキュベーションした。5mlのM13KO7ヘルパーファージ(GIBCO BRL、カタログ番号18311−019、1.1x1011pfu/ml)をM.O.I.20で添加して、細胞を感染させた。細胞/ヘルパーファージ混合物を振盪せずに37℃で30分間インキュベーションし、次いで4000rpmで20分間遠心分離して、感染細胞をペレットにした。細胞ペレットを50mlの2xYTAKブロス(100μg/mlアンピシリンおよび40μg/mlカナマイシンを含有する2xYTブロス)に懸濁し、次いで270rpmで振盪しながら、約20時間に渡って、30℃で一晩インキュベーションした。一晩培養物を4000rpmで20分間遠心分離することによって、細胞破片を取り除いた。上清を再び遠心分離して、細胞破片が除去されていることを確実にした。上清に、10ml(1/5体積)のPEG溶液(20%PEG8000、2.5M NaCl)を添加して、ファージ粒子を沈殿させた。混合物を氷上で少なくとも1時間インキュベーションし、そして4000rpmで20分間遠心分離して、沈殿したファージ粒子を収集した。ファージペレットを1mlのPBSに再懸濁して、そして微量遠心分離試験管に移した。ファージ懸濁物を氷上で1時間放置して、ファージ粒子の完全な懸濁を可能にし、次いで、14,000rpmで2分間回転させて、残った細胞破片を取り除いた。ファージ沈殿工程を反復した。最終ファージペレットを1mlのPBSに懸濁し、そして延長した期間、氷上に放置して、ファージ粒子の完全な懸濁を確実にした。ファージ懸濁物を14,000rpmで2分間遠心分離して、残った細胞破片を取り除いた。最終ファージ懸濁物を4℃で保存した。実施例2のファージプールELISAに記載するように、ファージELISAを行った。典型的には1x109pfu/ウェル〜1x1011pfu/ウェルの、少なくとも6つの異なる濃度の大規模救出ファージを、対応するウェルに添加した。
【0211】
総数11のFabクローンを同定した。大腸菌hIFN−γ溶出を用いて、ファージプールからFabクローン「IFN−A」、「57E」、および「57D」を同定した。GPNA溶出を用いて、ファージプールからFabクローン「GP−A」および「58C」を同定した。RDMA溶出を用いて、ファージプールからFabクローン「RD−A2」、「RD−B」および「59−A2」を同定した。BSMA溶出を用いて、ファージプールからFabクローン「BS−A」および「BS−B」を同定した。hIFN−γ R1溶出を用いて、ファージプールからFabクローン「67C」を同定した。大規模救出ファージ調製物に対して、9つのユニークなクローンの濃度依存的クローンファージELISAを行って、そしてこれを図1および図2に例示した。これらのFabファージは、ELISAプロフィールに基づいて、3つのグループに分けることも可能である。グループAには、Fabクローン「GP−A」および「BS−B」が含まれる。これらの2つのFabファージは強い結合剤であり、ELISAシグナルは5E9 pfu/ウェルで飽和に到達する。グループBには、Fabクローン「BS−A」、「RD−A2」、「INF−A」および「57E」が含まれる。これらは強い結合剤から穏やかな結合剤であり、優れた濃度依存性結合曲線を示す。グループCには、Fabクローン「57D」、「58C」、「RD−B」および「67C」が含まれる。これらは弱いが特異的であり、そして濃度依存性のIFNγ結合剤である。
【0212】
Fabクローンの配列解析
ユニークなIFNγ結合Fabファージクローンの確認を以下のように行った:QIAfilterTMプラスミド・ミディ・キット(Qiagen、カタログ番号12245)を用いて、ユニークなFab BstNI消化パターン各々の代表のプラスミドDNAを調製し、そして配列決定に送った。Fab BstNIパターンすべての配列がユニークであることが確認され、そして個々の重鎖配列および軽鎖配列が明らかになった(図3〜24を参照されたい)。
【0213】
Fab「BS−A」、「BS−B」、「RD−B1」、「RD−A2」、「58C」、「GP−A」、「57D」、「57E」、「IFN−A」、「67C」および「59−A2」の重鎖のDNAおよび予測されるアミノ酸配列(それぞれ配列番号65〜86)を、それぞれ図3〜13に示した。Fab「BS−A」、「BS−B」、「RD−B1」、「RD−A2」、「58C」、「GP−A」、「57D」、「57E」、「IFN−A」、「67C」および「59−A2」の軽鎖のDNAおよび予測されるアミノ酸配列(それぞれ配列番号87〜108)を、それぞれ図14〜24に示した。図31に示すように、11のFabすべての重鎖および軽鎖のアミノ酸配列を比較した。GCGの「BestFit」プログラムを用いて、Fab各対間の同一性および類似性のパーセンテージを得た。重鎖中の最も密接なマッチは、「BS−A」、「RD−A2」および「IFN−A」中である。「BS−A」および「RD−A2」の重鎖配列は、同一のフレームワークおよびCDR1およびCDR2を有する。これらはCDR3のみが異なり、そして92.6%の同一性および93.4%の類似性を有する。最初のアミノ酸を例外として、「BS−A」および「IFN−A」の重鎖配列は、同一のフレームワークおよびCDR1およびCDR2を有し、そして異なるCDR3を有する。これらは93.5%の同一性および95.1%の類似性を共有する。同じことが「IFN−A」および「RD−A2」の重鎖配列にも当てはまり、これらは90.5%の同一性および92.1%の類似性を持つ。Fab「57E」の重鎖のアミノ酸配列は、「BS−A」の重鎖配列に88.1%の同一性および89.0%の類似性を示し、「RD−A2」の重鎖配列に88.8%の同一性および90.0%の類似性を示し、そしてIFN−Aの重鎖配列に89.0%の同一性および90.7%の類似性を示す。Fab「BS−B」の重鎖のアミノ酸配列は、「57D」の重鎖配列に88.6%の同一性および90.4%の類似性を示し、「GP−A」の重鎖配列に81.7%の同一性および83.3%の類似性を示し、そして58Cの重鎖配列に83.9%の同一性および84.7%の類似性を示す。軽鎖中の最も密接なマッチは、「59−A2」および「BS−A」間の90.8%の同一性および91.7%の類似性、「BS−A」および「BS−B」間の89.0%の同一性および90.9%の類似性、「57E」および「BS−A」間の88.2%の同一性および90.9%の類似性、「57E」および「BS−B」間の89.7%の同一性および91.5%の類似性、そして「59−A2」および「BS−B」間の88.2%の同一性および88.2%の類似性である。Fabの3対、「59−A2」/「BS−B」、「57E」/「BS−A」および「IFN−A」/「RD−A2」のみが、重鎖および軽鎖両方で密接にマッチしている。
【0214】
相補性決定領域(CDR)のアミノ酸配列の比較を図25に示す。11の抗IFNγ Fabの重鎖CDR3は、類似性をほとんど共有しない。図32に示すように、重鎖CDRまたは軽鎖CDRのいずれの類似性にしたがって、Fabをグループ分けすることも可能である。クローン「BS−A」、「IFN−A」および「RD−A2」は、同一の重鎖CDR1およびCDR2を有する。しかし、最後の3つの残基が同一である(FDY)ことを除いて、これらの重鎖CDR3は、非常に異なる。興味深いことに、IFN−AおよびRD−A2はまた、緊密にマッチする軽鎖CDR1(11/16の同一残基)、CDR2(5/7の同一残基)、およびCDR3(8/9の同一残基)も共有する。クローン「BS−B」、「59−A2」、「GP−A」、および「57D」は、類似の重鎖CDR1およびCDR2を有する。クローン「BS−B」および「59−A2」は、同一の重鎖CDR1およびCDR2を有するが、非常に異なる重鎖CDR3を有する。クローン「59−A2」、「BS−A」、「BS−B」および「57E」の3つの軽鎖CDRはすべて、非常に類似である。
【0215】
(実施例4)
可溶性Fabの発現および精製
大腸菌株HB2151(Pharmacia)を、ユニークな結合剤のプラスミドDNAで形質転換した。形質転換HB2151の一晩培養物を2xYT−AGブロス中、30℃で増殖させた。100μg/mlアンピシリンおよび0.1%グルコースを含有する750mlの2xYTに、一晩培養物7.5mlを接種し、そして37℃で約2時間、振盪しながら(270rpm)インキュベーションした。OD590が0.8〜1.0に到達したら、誘導のため、IPTGを1mM添加した。培養物を振盪しながら30℃で4時間増殖させ続けた。培養物を4000rpmで20分間遠心分離し、そして上清を廃棄した。浸透圧ショックアプローチを用いて、Fabの周辺腔放出を達成した。8mlの氷冷TES(0.2M Tris、0.5mM EDTA、17.1%スクロース、pH8.0)に細胞を懸濁し、そしてときどき穏やかに振盪しながら氷上で5〜10分間インキュベーションした。空の試験管を8.8mlのTES/H2O(1:3)でリンスし、これを細胞懸濁物にプールした。
【0216】
細胞懸濁物を氷上でさらに20分間インキュベーションし、そして4,000rpmで15分間遠心分離した。上清を注意深く別の試験管に移し、そして8000rpmで20分間、再度遠心分離した。生じた上清は、TES放出周辺腔分画であった。細胞ペレットを10ml TES/15mM MgSO4に再懸濁し、氷上で15分間インキュベーションし、次いで上述のように2回遠心分離した。最後の上清は、Mg放出周辺腔分画であり、そしてTES放出周辺腔分画にともにプールした。
【0217】
周辺腔分画に、最終濃度1mg/mlとなるようにキャリアーおよび安定化剤としてBSAを添加した。プロテアーゼ阻害剤を加えた、2lの超音波処理緩衝液(20mM Tris−HCl/0.1M NaCl、pH8.5)に対して、1回交換しつつ、周辺腔分画を透析した。1/10体積のあらかじめ平衡化したTALON樹脂(Clontech)に周辺腔分画を添加し、そして穏やかに振動させながら4℃で1時間インキュベーションした。樹脂混合物を1300rpmで3分間遠心分離し、そして上清をできるだけ多く取り除いた。樹脂を10体積の超音波処理緩衝液で洗浄し、次いで1300rpmで3分間遠心分離した。上清を廃棄した。洗浄した樹脂を1ベッド体積の超音波処理緩衝液に懸濁し、そしてカラムに充填し、これを3ベッド体積の超音波処理緩衝液で洗浄した。2ベッド体積の200mMイミダゾールを用いて、Fabを溶出させた。精製したFabをPBS、pH7.4に透析した。
【0218】
(実施例5)
全長ヒトIFNγ抗体のクローニングおよび発現
以下の方法によって、FAbクローンを全長抗体に変換した。
【0219】
pDSRα19:hIgG1 CHの構築
プラスミドpDSRα19:抗ヒトOPGL IgG1をHindIIIおよびBsmBIで消化して、抗ヒトOPGL可変領域のコード領域を除去した。1.0kbpヒトIgG1定常領域ドメイン(CH1、ヒンジ、CH2およびCH3ドメイン)を含有する直鎖プラスミドpDSRα19:hIgG1 CHをゲル単離し、そしてFAb由来抗IFN−ガンマ可変領域を受け入れさせるのに用いた。
【0220】
pDSRα19:抗IFNガンマBS−A重鎖の構築
抗IFN−ガンマFAb重鎖cDNAをpDSRα19:hIgG1 CHにクローニングして、FAbを全長IgGに変換した。「BS−A」重鎖をコードするプラスミドの構築を本明細書に記載する。類似の方法を用いて、他のFAb重鎖をクローニングした。シグナル配列を持つFAbを生成するため、3工程PCRを行った。まず、FAb cDNAテンプレートとともにプライマー2485−51(以下に示す)および2465−68(以下に示す)を用いた。条件は:Pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。次いで、プライマー2148−98(以下に示す)および2465−68(以下に示す)を用いてPCR産物を増幅し、その後、プライマー2489−36(以下に示す)および2465−68(以下に示す)を用いて増幅した。最終PCR産物をQiagenで精製し、HindIIIおよびBsmBIで切断し、そしてQiagenで精製した。5’コザック(翻訳開始)部位および哺乳動物発現のための以下のシグナル配列:
【0221】
【化8】
【0222】
とともにFAbを含有するこの断片を、pDSRα19:hIgG1 CHに連結した。
【0223】
【化9】
【0224】
pDSRα19:抗IFNガンマBS−A重鎖の構築
FAb軽鎖cDNAをpDSRα19にクローニングして、FAbを全長抗体に変換した。「BS−A」軽鎖をコードするプラスミドの構築を本明細書に記載する。類似の方法を用いて、他のFAbをクローニングした。シグナル配列を持つFAb「BS−A」を生成するため、3工程PCRを行った。まず、FAb cDNAテンプレートとともにプライマー2525−43(以下に示す)および2578−27(以下に示す)を用いた。PCR条件は:pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。次いで、PCR産物をゲル精製し、そして次いで、プライマー2148−98(以下に示す)および2578−27(以下に示す)で増幅した。次に、再び、FAb cDNAテンプレートとともに、プライマー2578−26(以下に示す)および2469−67(以下に示す)を用いた。PCR条件は:Pfuポリメラーゼ、並びに適切な緩衝液およびヌクレオチドを用いて、94℃1分間、(94℃20秒間、48℃30秒間、74℃30秒間)を4周期、(94℃20秒間、66℃30秒間、74℃30秒間)を25周期、および74℃5分間であった。PCR産物をゲル単離し、そして同じ条件を用いて再増幅した。最後にゲル単離したPCR産物をプライマー2489−36(以下に示す)および2469−67(以下に示す)と混合し、そして増幅した。最終PCR産物をQiagenで精製し、XbaIおよびSalIで切断し、そしてQiagenで精製した。5’コザック(翻訳開始)部位および哺乳動物発現のための以下のシグナル配列:
【0225】
【化10】
【0226】
とともにFAbを含有するこの断片を、pDSRα19に連結した。
【0227】
【化11】
【0228】
抗体調製
重鎖および軽鎖全長抗体をコードするcDNAを含有する発現ベクターをCHO細胞にトランスフェクションし、そして重鎖および軽鎖の発現、並びに細胞培地への分泌を可能にする条件下で培養した。0.45μmの酢酸セルロース・フィルター(Corning、マサチューセッツ州アクトン)を通して馴化培地をろ過し、そして塩化カルシウム不含で、そして塩化マグネシウム不含である、PBS−ダルベッコのリン酸緩衝生理食塩水(Gibco BRL Products、ニューヨーク州グランドアイランド)で平衡化しておいたプロテインGセファロース(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)カラムに適用した。試料を適用した後、280nmの吸光度がベースラインに達するまで、カラムをPBSで洗浄した。100mMグリシン、pH2.5を用いて、タンパク質の溶出を達成した。分画を収集し、そして1M Tris−HCl、pH9.2を添加することによって、直ちに中和した。クーマシー染色によってSDS−ポリアクリルアミドゲルを視覚化することによって抗体を検出した。
【0229】
抗体を含有する分画をプールし、Centricon 10(Amicon)または体積がより多い場合は、Centriprep 10(Amicon)いずれかを用いて、濃縮し、そしてPBS中に透析ろ過(diafilter)した。
【0230】
単離抗体を、Superose 6(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)上のゲルろ過によって性質決定し、そして該抗体は、単量体IgGとして移動することが示された。
【0231】
(実施例6)
FabおよびIgGの親和性測定
表面プラズモン共鳴技術(BIAcore、Pharmacia、ニュージャージー州ピスカタウェイ)によって、結合定数(Kd)、結合速度定数(ka)および解離速度定数(kd)を決定した。Fabおよび抗体のBIAcore解析を以下のように行った:BIACORE2000(BIACORE Inc.)を室温で用いて、実験を行った。CHOが発現したhIFNγをCM5チップに固定した。多様な濃度のFabまたはFab IgGをhu−IFNγ表面に注入した。BIAEVALUATION 3.1ソフトウェア(BIACORE, Inc.)を用いてデータを解析した。結果を図30に示す。
【0232】
(実施例7)
FabおよびIgGの活性測定
BIAcore中和アッセイ
IgGに変換されたFabの中和活性をBIAcore上で試験した(実施例6を参照されたい)。結果を図29に示す。BS−B IgGに関して、9nMのIC50で、IFNγ−R1へのhu IFNγ結合の濃度依存性阻害が観察された。
【0233】
A549細胞増殖アッセイ
A549増殖アッセイ(実施例1に記載)において、FabおよびIgGの中和活性測定を以下のように行った:A549細胞を、標的化されたFabまたはIgG(多様な濃度)およびCHOに発現されたhIFNγ(2ng/mlまたは5ng/ml)の混合物で処理した。Fab濃度は、0.3〜150μg/mlの範囲であった。IgG濃度は0.1〜100μg/mlの範囲であった。陽性対照Ab(Pharmingen B27)濃度は、0.01〜5μg/mlの範囲であった。処理5日後、Alamar Blueで細胞を染色し、そして染色4時間後、FL500プレート読取装置上で解析した。BS−A Fab、BS−B FabおよびGP−A Fabに関しては図26に、そしてBS−A IgGおよびBS−B IgGに関しては図27に結果を示す。図26のBS−A FabおよびBS−B Fab、並びに図27のBS−A IgGおよびBS−B IgGは、増殖活性として測定した際、高濃度で、陽性対照よりも約2桁高い中和活性を有することが示された。
【0234】
本発明は、好ましい態様に関して記載されてきているが、当業者には多様な変型および修飾が思い浮かぶであろうことが理解された。したがって、付随する請求項が、請求された際に本発明の範囲内に属するであろう、こうした同等の変型をすべて含むと意図された。
【図面の簡単な説明】
【0235】
【図1】図1は、主なファージFabクローンのhIFNγに対する反応性に関するELISAの結果を示すグラフである。2%MPBSであらかじめブロッキングしたファージ懸濁物を、ウェルあたり最大100μl用いて、ファージ希釈を行い、ELISAにおいて典型的な109〜1011ファージ/ウェルの範囲とした。実施例3に記載するように、ELISA用のファージストックを調製した。値は、単一点測定によるものであり、そしてシグナル検出のため、OD405を測定した。
【図2】図2は、主なFab「GP−A」および「BS−B」クローンのhIFNγに対する反応性に関する、用量依存性クローンファージELISAの結果を示すグラフである。2%MPBSであらかじめブロッキングしたファージ懸濁物を、ウェルあたり最大100μl用いて、ファージ希釈を行い、ELISAにおいて典型的な109〜1011ファージ/ウェルの範囲とした。実施例3に記載するように、ELISA用のファージストックを調製した。値は、単一点測定によるものであり、そしてシグナル検出のため、OD405を測定した。
【図3】図3は、Fab「BS−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図4】図4は、Fab「BS−B」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図5】図5は、Fab「RD−B1」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図6】図6は、Fab「RD−A2」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図7】図7は、Fab「58C」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図8】図8は、Fab「GP−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図9】図9は、Fab「57D」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図10】図10は、Fab「57E」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図11】図11は、Fab「IFN−A」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図12】図12は、Fab「67C」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図13】図13は、Fab「59−A2」重鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図14】図14は、Fab「BS−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図15】図15は、Fab「BS−B」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図16】図16は、Fab「RD−B1」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図17】図17は、Fab「RD−A2」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図18】図18は、Fab「58C」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図19】図19は、Fab「GP−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図20】図20は、Fab「57D」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図21】図21は、Fab「57E」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図22】図22は、Fab「IFN−A」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図23】図23は、Fab「67C」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図24】図24は、Fab「59−A2」軽鎖のヌクレオチド配列およびアミノ酸配列を示す。
【図25】図25は、Fab「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」の重鎖および軽鎖相補性決定領域(CDR)のアミノ酸配列の比較を示す。
【図26】図26は、A549細胞アッセイで測定した際のFab「BS−A」および「BS−B」の中和活性を示すグラフである。実施例4に記載するようにFabを精製し、そして0.3〜150μg/mlのFab濃度範囲で添加した。Pharmingen B27 Ab(0.01〜5μg/mlの濃度範囲)を陽性対照として用いた。処理5日後、細胞をAlamar Blueで染色し、そしてFL500プレート読取装置上で、染色4時間後に解析した。
【図27】図27は、A549細胞アッセイで測定した際の「BS−A」IgGおよび「BS−B」IgGの中和活性を示すグラフである。実施例4に記載するようにIgGを精製し、そして0.1〜100μg/mlのIgG濃度範囲で添加した。Pharmingen B27 Ab(0.01〜5μg/mlの濃度範囲)を陽性対照として用いた。hIFNγと反応しない無関係なAb、AT−IgG(0.01〜5μg/mlの濃度範囲)を陰性対照として用いた。処理5日後、細胞をAlamar Blueで染色し、そしてFL500プレート読取装置上で、染色4時間後に解析した。
【図28】図28は、BiaCoreによって、そしてA549細胞アッセイにおいて測定した際の、「BS−A」、「BS−B」、「RD−A2」、「RD−B」、「IFN−A」、「57E」、「57D」、「GP−A」、「58C」および「67C」IgGの親和性および中和活性の比較を提供する表である。BIAEVALUATIONを用いて、BIACOREデータを解析した。
【図29】図29は、BIACoreで測定した際の「BS−A」IgGおよび「BS−B」IgGの中和活性を示すグラフである。相対結合反応(%)を、試料濃度(nM)に対してプロットしている。
【図30】図30は、BIACoreによって測定した際の、抗IFNγ Fab「BS−A」、「BS−B」、「IFN−A」および「GP−A」、並びに対応するIgGの親和性の比較を提供する表である。
【図31】図31は、図3〜24に示すFabアミノ酸配列の比較を示す。重鎖および軽鎖Fab「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」の予測されるアミノ酸配列を、同一性および類似性に関して比較した。GCGの「BestFit」プログラムを用いて、Fabの各対間の同一性および類似性パーセントを得た。
【図32】図32は、重鎖および軽鎖「BS−A」、「BS−B」、「RD−A2」、「RD−B1」、「IFN−A」、「57E」、「57D」、「GP−A」、「58−C」、「67C」および「59−A2」Fabの相補性決定領域(CDR)の並列を示す。
【図33】図33は、予測されるFab「BS−A」、「RD−A2」および「IFN−A」重鎖アミノ酸配列(それぞれ、図3、6および11の両端を含む残基1〜120、1〜127および1〜126)と、VH4ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列4−34、D領域配列1−1、3−3または3−16、およびJ領域配列JH4を含む。FR1、FR2およびFR3は、3つのフレームワーク領域を指定し、CDR1、CDR2およびCDR3は、3つの相補性決定領域を指定し、そしてH1、H2およびH3は、フレームワーク領域およびCDRの間の対応する接合部配列を指定する。「BS−A」、「RD−A2」、「IFN−A」および生殖系列V、D、またはJ配列の間の相違を太字で示す。図33〜46の生殖系列アミノ酸残基の番号付けは、Kabatら, Sequences of Proteins of Immunological Interest, 米国保健社会福祉省, 第4版(1991)に記載されるとおりである。
【図34】図34は、予測されるFab「BS−B」、および「59−A2」重鎖アミノ酸配列(それぞれ、図4および13の両端を含む残基1〜121および1〜120)と、VH1ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列1−18、D領域配列6−13、1−1または1−7、およびJ領域配列JH4を含む。
【図35】図35は、予測されるFab「RD−B1」重鎖アミノ酸配列(図5の両端を含む残基1〜119)と、VH2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−26、D領域配列3−22、およびJ領域配列JH5を含む。
【図36】図36は、予測されるFab「58C」重鎖アミノ酸配列(図7の両端を含む残基1〜119)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−21、未知のD領域配列、およびJ領域配列JH4を含む。
【図37】図37は、予測されるFab「GP−A」重鎖アミノ酸配列(図8の両端を含む残基1〜124)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−30.3、D領域配列3−10、およびJ領域配列JH3を含む。
【図38】図38は、予測されるFab「57D」重鎖アミノ酸配列(図9の両端を含む残基1〜117)と、VH3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3−53、D領域配列3−16、および未知のJ領域配列を含む。
【図39】図39は、予測されるFab「57E」重鎖アミノ酸配列(図10の両端を含む残基1〜118)と、VH4ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列4−61、D領域配列7−27、およびJ領域配列JH4を含む。
【図40】図40は、予測されるFab「67C」重鎖アミノ酸配列(図12の両端を含む残基1〜119)と、VH2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−05、D領域配列5−18、およびJ領域配列JH6を含む。
【図41】図41は、予測されるFab「BS−A」、「BS−B」、「57E」および「59−A2」軽鎖アミノ酸配列(それぞれ、図14、15、21および24の両端を含む残基1〜111、1〜110、1〜112および1〜110)と、Vλ6ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列6aおよび未知のJ領域配列あるいはJL2またはJL3を含む。
【図42】図42は、予測されるFab「RD−B1」軽鎖アミノ酸配列(図16の両端を含む残基1〜112)と、Vκ2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A23およびJ領域配列JK2を含む。
【図43】図43は、予測されるFab「RD−A2」、「IFN−A」および「67C」軽鎖アミノ酸配列(それぞれ、図17、22および23の両端を含む残基1〜112、1〜110、1〜112および1〜110)と、Vκ2ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A19、並びにそれぞれJ領域配列JK3、JK2およびJK2を含む。
【図44】図44は、予測されるFab「58C」軽鎖アミノ酸配列(図18の両端を含む残基1〜108)と、Vλ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列3hおよび未知のJ領域配列を含む。
【図45】図45は、予測されるFab「GP−A」軽鎖アミノ酸配列(図19の両端を含む残基1〜106)と、Vλ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列2−19およびJ領域配列JL2またはJL3を含む。
【図46】図46は、予測されるFab「57D」軽鎖アミノ酸配列(図20の両端を含む残基1〜108)と、Vκ3ファミリー由来の生殖系列配列の比較を示す。生殖系列配列は、V領域配列A27およびJ領域配列JK5を含む。
【図47】図47は、Fabクラスの比較を示す。多配列比較解析用のGCG(遺伝学コンピュータグループ, 575 Science Drive, Madison, WI 53711)PileUpプログラムを用いて、Fabクラス比較を行った。記号(**)は、既知の生殖系列配列には関連していたが、最も密接なマッチング多様性(D)領域または連結領域(J)が決定不能であったことを示す。記号(*)は、同定された連結領域に比較して、1、2または3残基の変異が起きたことを示す。
【配列表】
【特許請求の範囲】
【請求項1】
インターフェロン−ガンマ・タンパク質に結合し、そしてアンタゴニスト抗体である、抗体または抗原結合ドメイン、あるいはその断片、変異体(variant)または誘導体。
【請求項2】
インターフェロン−ガンマ・タンパク質が哺乳動物インターフェロン−ガンマ・タンパク質である、請求項1の抗体。
【請求項3】
インターフェロン−ガンマ・タンパク質がヒト・インターフェロン−ガンマ・タンパク質またはその免疫原性断片である、請求項2の抗体。
【請求項4】
免疫原性断片が、ヒト・インターフェロン−ガンマ・タンパク質の細胞外ドメインの少なくとも一部を含む、請求項3の抗体。
【請求項5】
インターフェロン−ガンマ・タンパク質のインターフェロン−ガンマ受容体への結合を阻害する、請求項1の抗体。
【請求項6】
炎症を阻害する、請求項1の抗体。
【請求項7】
阻害する、請求項1の抗体。
【請求項8】
Fv、scFv、Fab、Fab’およびF(ab’)2からなる群より選択される、請求項1の抗体。
【請求項9】
ヒト抗体である、請求項1の抗体。
【請求項10】
抗体または抗原結合ドメインであって:
(a)図3(配列番号66)、図4(配列番号68)、図5(配列番号70)、図6(配列番号72)、図7(配列番号74)、図8(配列番号76)、図9(配列番号78)、図10(配列番号80)、図11(配列番号82)、図12(配列番号84)または図13(配列番号86)に示すようなFab重鎖アミノ酸配列;
(b)(a)の配列の保存的アミノ酸置換を含む、重鎖アミノ酸配列;
(c)(a)の配列に少なくとも約80%同一である、重鎖アミノ酸配列;あるいは
(d)(a)、(b)または(c)の断片または誘導体
を含み、IFNγに選択的に結合する、前記抗体または抗原結合ドメイン。
【請求項11】
カッパ軽鎖またはラムダ軽鎖をさらに含む、請求項10の抗体。
【請求項12】
ヒトFc領域をさらに含む、請求項10の抗体。
【請求項13】
図3(配列番号66)、図4(配列番号68)、図5(配列番号70)、図6(配列番号72)、図7(配列番号74)、図8(配列番号76)、図9(配列番号78)、図10(配列番号80)、図11(配列番号82)、図12(配列番号84)または図13(配列番号86)に示すようなFab重鎖アミノ酸配列、および図14(配列番号88)、図15(配列番号90)、図16(配列番号92)、図17(配列番号94)、図18(配列番号96)、図19(配列番号98)、図20(配列番号100)、図21(配列番号102)、図22(配列番号104)、図23(配列番号106)または図24(配列番号108)に示すようなFab軽鎖アミノ酸配列を含む抗体または抗原結合ドメインに認識される、ヒトIFNγ上のエピトープを認識する、抗体または抗原結合ドメイン。
【請求項14】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体または抗原結合ドメインであって:
各Vl鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vl)、CDR2(Vl)およびCDR3(Vl)と称されるCDRアミノ酸配列を含み、CDR1(Vl)が:
【化1】
からなる群より選択され、
CDR2(Vl)が:
【化2】
からなる群より選択され、
そしてCDR3(Vl)が:
【化3】
からなる群より選択され、
CDR1(Vl)、CDR2(Vl)およびCDR3(Vl)が互いに独立に選択され;そして
各Vh鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vh)、CDR2(Vh)およびCDR3(Vh)と称されるCDRアミノ酸配列を含み、CDR1(Vh)が:
【化4】
からなる群より選択され、
CDR2(Vh)が:
【化5】
からなる群より選択され、
CDR3(Vh)が:
【化6】
からなる群より選択され;
CDR1(Vh)、CDR2(Vh)およびCDR3(Vh)が互いに独立に選択される、
前記抗体または抗原結合ドメイン。
【請求項15】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む、請求項14の抗体であって:
Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIASNYVQ(配列番号02)を有するCDR1、配列EDNQRPS(配列番号13)を有するCDR2、および配列QSYDGSAWV(配列番号24)を有するCDR3を含み;そしてVh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列TSWNAGGPIDY(配列番号55)を有するCDR3を含むか;または
Vl鎖が、配列TRSSGSIASYYVQ(配列番号03)を有するCDR1、配列EDDQRPS(配列番号14)を有するCDR2、および配列QSYDRNSLV(配列番号25)を有するCDR3を含み;そしてVh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列DRVGYSSSLLDY(配列番号56)を有するCDR3を含むか;または
Vl鎖が、配列RATQSLLHGNGHNYLD(配列番号04)を有するCDR1、配列MGSNRAS(配列番号15)を有するCDR2、および配列MQALQLPPT(配列番号26)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号36)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号46)を有するCDR2、および配列DKGSRITIFGVVGSAGFDY(配列番号57)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLVHSDGNTYLS(配列番号05)を有するCDR1、配列KISNRFS(配列番号16)を有するCDR2、および配列MQATQLPYT(配列番号27)を有するCDR3を含み;そしてVh鎖が、配列NARMGVS(配列番号37)を有するCDR1、配列HIFSNDEESYSTSLKS(配列番号47)を有するCDR2、および配列LLLYEGFDP(配列番号58)を有するCDR3を含むか;または
Vl鎖が、配列SGDVLARKYAR(配列番号06)を有するCDR1、配列KDRERPS(配列番号17)を有するCDR2、および配列YSAADNRGV(配列番号28)を有するCDR3を含み;そしてVh鎖が、配列SYAMH(配列番号38)を有するCDR1、配列VISYDGSNKYYADSVKG(配列番号48)を有するCDR2、および配列DLVLTMTSRRAAFDI(配列番号59)を有するCDR3を含むか;または
Vl鎖が、配列GGDNLGGKSLH(配列番号07)を有するCDR1、配列DDSDRPS(配列番号18)を有するCDR2、および配列QVWDGSSDQRV(配列番号29)を有するCDR3を含み;そしてVh鎖が、配列SYSMN(配列番号39)を有するCDR1、配列SISSGSSYRYDADSVKG(配列番号49)を有するCDR2、および配列DQWGTISGNDY(配列番号60)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLLHTNEYNYLD(配列番号08)を有するCDR1、配列LGSNRAP(配列番号19)を有するCDR2、および配列MQALQTPRT(配列番号30)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号40)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号50)を有するCDR2、および配列GWPTYVWGSYRPKGYFDY(配列番号61)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIANNYVH(配列番号09)を有するCDR1、配列EDDQRPS(配列番号20)を有するCDR2、および配列QSYDNSNSFVV(配列番号31)を有するCDR3を含み;そしてVh鎖が、配列SGGYSWS(配列番号41)を有するCDR1、配列YIYHSGSTYYNPSLKS(配列番号51)を有するCDR2、および配列GDWGYFDY(配列番号62)を有するCDR3を含むか;または
Vl鎖が、配列RASQYVSSNSLA(配列番号10)を有するCDR1、配列GASNRAT(配列番号21)を有するCDR2、および配列QQYGSSPIT(配列番号32)を有するCDR3を含み;そしてVh鎖が、配列SNYMS(配列番号42)を有するCDR1、配列VIYSGGSTYYADSVKG(配列番号52)を有するCDR2、および配列DADGGDYGY(配列番号63)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLLRSNGYNYLA(配列番号11)を有するCDR1、配列LASNRAS(配列番号22)を有するCDR2、および配列VHGVHIPYT(配列番号33)を有するCDR3を含み;そしてVh鎖が、配列SNEAGVG(配列番号43)を有するCDR1、配列LLYWDDDKRYSPSLRS(配列番号53)を有するCDR2、および配列RLVRYGGYSTGGFDV(配列番号64)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている、前記抗体。
【請求項16】
ヒトFc領域をさらに含む、請求項14または15の抗体。
【請求項17】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図33の生殖系列配列(配列番号122)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項18】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図34の生殖系列配列(配列番号123)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項19】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図42の生殖系列配列(配列番号131)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図35の生殖系列配列(配列番号124)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項20】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図43の生殖系列配列(配列番号132)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図33の生殖系列配列(配列番号122)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項21】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図44の生殖系列配列(配列番号133)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図36の生殖系列配列(配列番号125)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項22】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図45の生殖系列配列(配列番号134)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図37の生殖系列配列(配列番号126)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項23】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図46の生殖系列配列(配列番号135)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図38の生殖系列配列(配列番号127)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項24】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図39の生殖系列配列(配列番号128)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項25】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図43の生殖系列配列(配列番号132)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図40の生殖系列配列(配列番号129)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項26】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図34の生殖系列配列(配列番号123)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項27】
モノクローナル抗体、ヒト化抗体、二重特異性抗体、一本鎖抗体、または異種抗体である、請求項1の抗体。
【請求項28】
請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27のいずれかの抗体をコードする、単離核酸分子。
【請求項29】
請求項28の核酸分子を含む発現ベクター。
【請求項30】
請求項29の発現ベクターを含む宿主細胞。
【請求項31】
CHO細胞である、請求項30の宿主細胞。
【請求項32】
核酸分子の発現を可能にする条件下で、請求項31の宿主細胞を培養することを含む、抗体産生法。
【請求項33】
IgGアイソタイプがIgG、IgM、IgA、IgEおよびIgDから選択される、請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27の抗体。
【請求項34】
アイソタイプがIgG1、IgG2、IgG3またはIgG4である、請求項33の抗体。
【請求項35】
請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27のいずれかの抗体または抗原結合ドメイン、あるいはその断片、変異体または誘導体、および薬学的に許容しうるキャリアーを含む組成物。
【請求項36】
請求項35の組成物の有効量を哺乳動物に投与することを含む、自己免疫疾患を防止するかまたは治療する方法。
【請求項37】
請求項35の組成物の有効量を哺乳動物に投与することを含む、炎症状態を防止するかまたは治療する方法。
【請求項1】
インターフェロン−ガンマ・タンパク質に結合し、そしてアンタゴニスト抗体である、抗体または抗原結合ドメイン、あるいはその断片、変異体(variant)または誘導体。
【請求項2】
インターフェロン−ガンマ・タンパク質が哺乳動物インターフェロン−ガンマ・タンパク質である、請求項1の抗体。
【請求項3】
インターフェロン−ガンマ・タンパク質がヒト・インターフェロン−ガンマ・タンパク質またはその免疫原性断片である、請求項2の抗体。
【請求項4】
免疫原性断片が、ヒト・インターフェロン−ガンマ・タンパク質の細胞外ドメインの少なくとも一部を含む、請求項3の抗体。
【請求項5】
インターフェロン−ガンマ・タンパク質のインターフェロン−ガンマ受容体への結合を阻害する、請求項1の抗体。
【請求項6】
炎症を阻害する、請求項1の抗体。
【請求項7】
阻害する、請求項1の抗体。
【請求項8】
Fv、scFv、Fab、Fab’およびF(ab’)2からなる群より選択される、請求項1の抗体。
【請求項9】
ヒト抗体である、請求項1の抗体。
【請求項10】
抗体または抗原結合ドメインであって:
(a)図3(配列番号66)、図4(配列番号68)、図5(配列番号70)、図6(配列番号72)、図7(配列番号74)、図8(配列番号76)、図9(配列番号78)、図10(配列番号80)、図11(配列番号82)、図12(配列番号84)または図13(配列番号86)に示すようなFab重鎖アミノ酸配列;
(b)(a)の配列の保存的アミノ酸置換を含む、重鎖アミノ酸配列;
(c)(a)の配列に少なくとも約80%同一である、重鎖アミノ酸配列;あるいは
(d)(a)、(b)または(c)の断片または誘導体
を含み、IFNγに選択的に結合する、前記抗体または抗原結合ドメイン。
【請求項11】
カッパ軽鎖またはラムダ軽鎖をさらに含む、請求項10の抗体。
【請求項12】
ヒトFc領域をさらに含む、請求項10の抗体。
【請求項13】
図3(配列番号66)、図4(配列番号68)、図5(配列番号70)、図6(配列番号72)、図7(配列番号74)、図8(配列番号76)、図9(配列番号78)、図10(配列番号80)、図11(配列番号82)、図12(配列番号84)または図13(配列番号86)に示すようなFab重鎖アミノ酸配列、および図14(配列番号88)、図15(配列番号90)、図16(配列番号92)、図17(配列番号94)、図18(配列番号96)、図19(配列番号98)、図20(配列番号100)、図21(配列番号102)、図22(配列番号104)、図23(配列番号106)または図24(配列番号108)に示すようなFab軽鎖アミノ酸配列を含む抗体または抗原結合ドメインに認識される、ヒトIFNγ上のエピトープを認識する、抗体または抗原結合ドメイン。
【請求項14】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体または抗原結合ドメインであって:
各Vl鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vl)、CDR2(Vl)およびCDR3(Vl)と称されるCDRアミノ酸配列を含み、CDR1(Vl)が:
【化1】
からなる群より選択され、
CDR2(Vl)が:
【化2】
からなる群より選択され、
そしてCDR3(Vl)が:
【化3】
からなる群より選択され、
CDR1(Vl)、CDR2(Vl)およびCDR3(Vl)が互いに独立に選択され;そして
各Vh鎖が、フレームワーク・アミノ酸配列によって分離されるCDR1(Vh)、CDR2(Vh)およびCDR3(Vh)と称されるCDRアミノ酸配列を含み、CDR1(Vh)が:
【化4】
からなる群より選択され、
CDR2(Vh)が:
【化5】
からなる群より選択され、
CDR3(Vh)が:
【化6】
からなる群より選択され;
CDR1(Vh)、CDR2(Vh)およびCDR3(Vh)が互いに独立に選択される、
前記抗体または抗原結合ドメイン。
【請求項15】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む、請求項14の抗体であって:
Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIASHYVQ(配列番号01)を有するCDR1、配列EDKERPS(配列番号12)を有するCDR2、および配列QSYDSSNQWV(配列番号23)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号34)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号44)を有するCDR2、および配列GRARNWRSRFDY(配列番号54)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIASNYVQ(配列番号02)を有するCDR1、配列EDNQRPS(配列番号13)を有するCDR2、および配列QSYDGSAWV(配列番号24)を有するCDR3を含み;そしてVh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列TSWNAGGPIDY(配列番号55)を有するCDR3を含むか;または
Vl鎖が、配列TRSSGSIASYYVQ(配列番号03)を有するCDR1、配列EDDQRPS(配列番号14)を有するCDR2、および配列QSYDRNSLV(配列番号25)を有するCDR3を含み;そしてVh鎖が、配列SYAMS(配列番号35)を有するCDR1、配列AISGSGGSTYYADSVKG(配列番号45)を有するCDR2、および配列DRVGYSSSLLDY(配列番号56)を有するCDR3を含むか;または
Vl鎖が、配列RATQSLLHGNGHNYLD(配列番号04)を有するCDR1、配列MGSNRAS(配列番号15)を有するCDR2、および配列MQALQLPPT(配列番号26)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号36)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号46)を有するCDR2、および配列DKGSRITIFGVVGSAGFDY(配列番号57)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLVHSDGNTYLS(配列番号05)を有するCDR1、配列KISNRFS(配列番号16)を有するCDR2、および配列MQATQLPYT(配列番号27)を有するCDR3を含み;そしてVh鎖が、配列NARMGVS(配列番号37)を有するCDR1、配列HIFSNDEESYSTSLKS(配列番号47)を有するCDR2、および配列LLLYEGFDP(配列番号58)を有するCDR3を含むか;または
Vl鎖が、配列SGDVLARKYAR(配列番号06)を有するCDR1、配列KDRERPS(配列番号17)を有するCDR2、および配列YSAADNRGV(配列番号28)を有するCDR3を含み;そしてVh鎖が、配列SYAMH(配列番号38)を有するCDR1、配列VISYDGSNKYYADSVKG(配列番号48)を有するCDR2、および配列DLVLTMTSRRAAFDI(配列番号59)を有するCDR3を含むか;または
Vl鎖が、配列GGDNLGGKSLH(配列番号07)を有するCDR1、配列DDSDRPS(配列番号18)を有するCDR2、および配列QVWDGSSDQRV(配列番号29)を有するCDR3を含み;そしてVh鎖が、配列SYSMN(配列番号39)を有するCDR1、配列SISSGSSYRYDADSVKG(配列番号49)を有するCDR2、および配列DQWGTISGNDY(配列番号60)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLLHTNEYNYLD(配列番号08)を有するCDR1、配列LGSNRAP(配列番号19)を有するCDR2、および配列MQALQTPRT(配列番号30)を有するCDR3を含み;そしてVh鎖が、配列GYYWS(配列番号40)を有するCDR1、配列EINHSGSTNYNPSLKS(配列番号50)を有するCDR2、および配列GWPTYVWGSYRPKGYFDY(配列番号61)を有するCDR3を含むか;または
Vl鎖が、配列TGSSGSIANNYVH(配列番号09)を有するCDR1、配列EDDQRPS(配列番号20)を有するCDR2、および配列QSYDNSNSFVV(配列番号31)を有するCDR3を含み;そしてVh鎖が、配列SGGYSWS(配列番号41)を有するCDR1、配列YIYHSGSTYYNPSLKS(配列番号51)を有するCDR2、および配列GDWGYFDY(配列番号62)を有するCDR3を含むか;または
Vl鎖が、配列RASQYVSSNSLA(配列番号10)を有するCDR1、配列GASNRAT(配列番号21)を有するCDR2、および配列QQYGSSPIT(配列番号32)を有するCDR3を含み;そしてVh鎖が、配列SNYMS(配列番号42)を有するCDR1、配列VIYSGGSTYYADSVKG(配列番号52)を有するCDR2、および配列DADGGDYGY(配列番号63)を有するCDR3を含むか;または
Vl鎖が、配列RSSQSLLRSNGYNYLA(配列番号11)を有するCDR1、配列LASNRAS(配列番号22)を有するCDR2、および配列VHGVHIPYT(配列番号33)を有するCDR3を含み;そしてVh鎖が、配列SNEAGVG(配列番号43)を有するCDR1、配列LLYWDDDKRYSPSLRS(配列番号53)を有するCDR2、および配列RLVRYGGYSTGGFDV(配列番号64)を有するCDR3を含み;
各Vl鎖およびVh鎖上のCDR1、CDR2およびCDR3が、フレームワーク・アミノ酸配列によって分離されている、前記抗体。
【請求項16】
ヒトFc領域をさらに含む、請求項14または15の抗体。
【請求項17】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図33の生殖系列配列(配列番号122)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項18】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図34の生殖系列配列(配列番号123)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項19】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図42の生殖系列配列(配列番号131)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図35の生殖系列配列(配列番号124)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項20】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図43の生殖系列配列(配列番号132)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図33の生殖系列配列(配列番号122)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項21】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図44の生殖系列配列(配列番号133)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図36の生殖系列配列(配列番号125)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項22】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図45の生殖系列配列(配列番号134)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図37の生殖系列配列(配列番号126)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項23】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図46の生殖系列配列(配列番号135)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図38の生殖系列配列(配列番号127)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項24】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図39の生殖系列配列(配列番号128)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項25】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図43の生殖系列配列(配列番号132)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図40の生殖系列配列(配列番号129)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項26】
可変軽鎖(Vl鎖)および可変重鎖(Vh鎖)を含む抗体であって:
Vl鎖が、図41の生殖系列配列(配列番号130)の再編成変異体または体細胞変異体を含み;そして
Vh鎖が、図34の生殖系列配列(配列番号123)の再編成変異体または体細胞変異体を含み;そしてインターフェロン−ガンマ・タンパク質に選択的に結合する、前記抗体。
【請求項27】
モノクローナル抗体、ヒト化抗体、二重特異性抗体、一本鎖抗体、または異種抗体である、請求項1の抗体。
【請求項28】
請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27のいずれかの抗体をコードする、単離核酸分子。
【請求項29】
請求項28の核酸分子を含む発現ベクター。
【請求項30】
請求項29の発現ベクターを含む宿主細胞。
【請求項31】
CHO細胞である、請求項30の宿主細胞。
【請求項32】
核酸分子の発現を可能にする条件下で、請求項31の宿主細胞を培養することを含む、抗体産生法。
【請求項33】
IgGアイソタイプがIgG、IgM、IgA、IgEおよびIgDから選択される、請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27の抗体。
【請求項34】
アイソタイプがIgG1、IgG2、IgG3またはIgG4である、請求項33の抗体。
【請求項35】
請求項1、10、13、14、15、17、18、19、20、21、22、23、24、25、26または27のいずれかの抗体または抗原結合ドメイン、あるいはその断片、変異体または誘導体、および薬学的に許容しうるキャリアーを含む組成物。
【請求項36】
請求項35の組成物の有効量を哺乳動物に投与することを含む、自己免疫疾患を防止するかまたは治療する方法。
【請求項37】
請求項35の組成物の有効量を哺乳動物に投与することを含む、炎症状態を防止するかまたは治療する方法。
【図1】

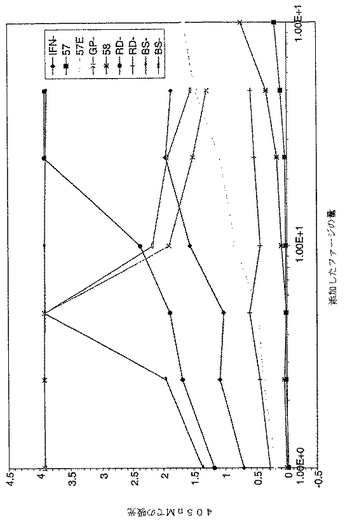
【図2】
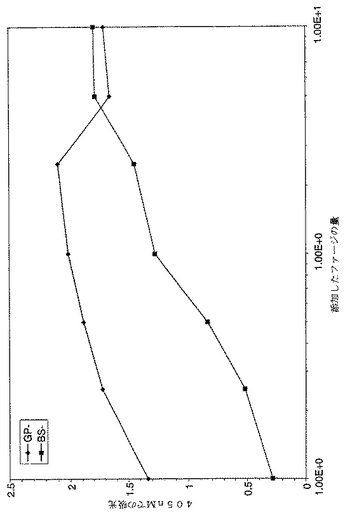
【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図14】

【図15】

【図16】

【図17】

【図18】

【図19】

【図20】

【図21】

【図22】

【図23】

【図24】

【図25】

【図26】
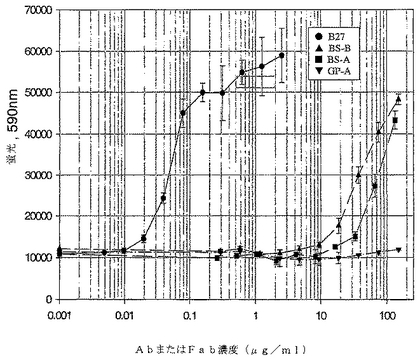
【図27】
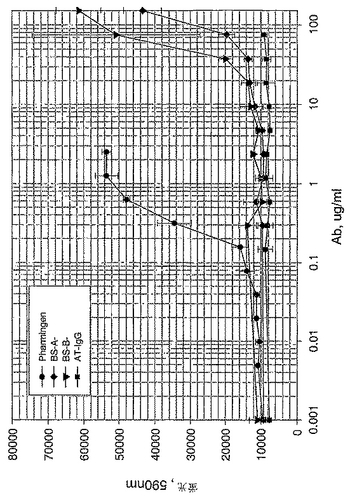
【図28】
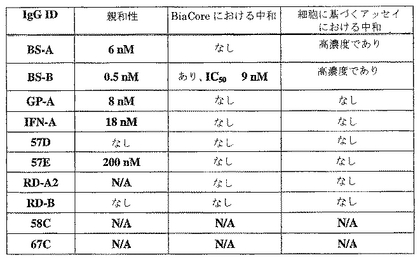
【図29】
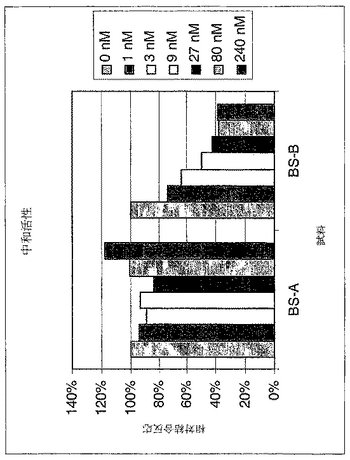
【図30】

【図31】
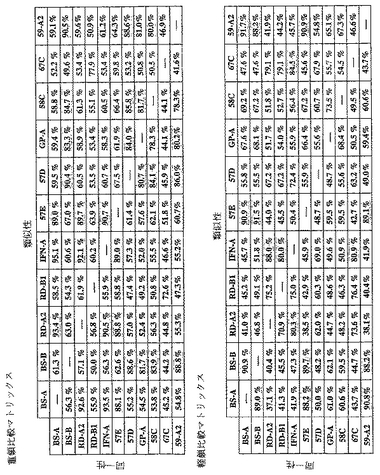
【図32】

【図33】
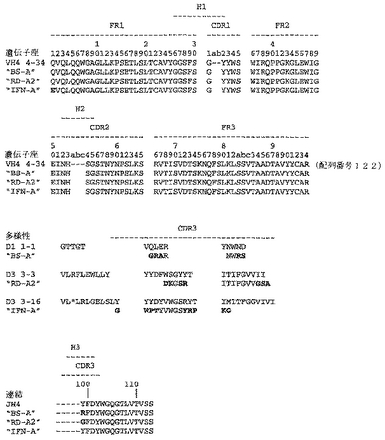
【図34】
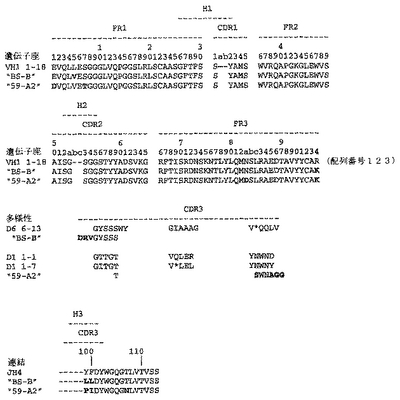
【図35】
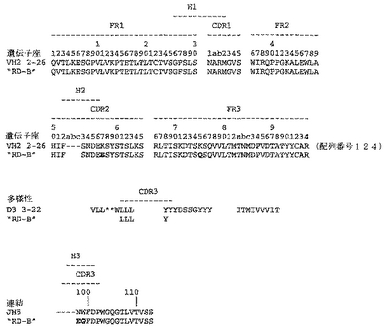
【図36】
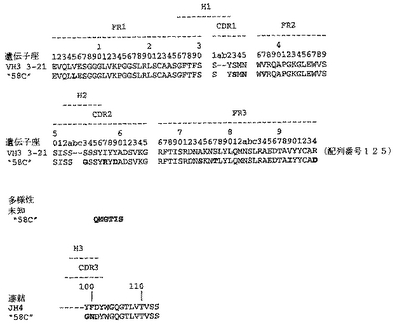
【図37】
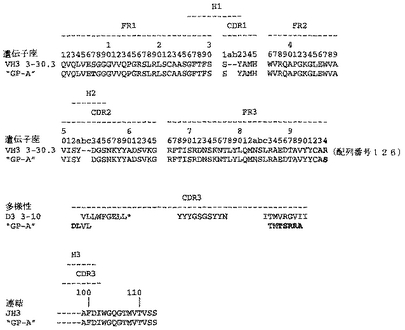
【図38】
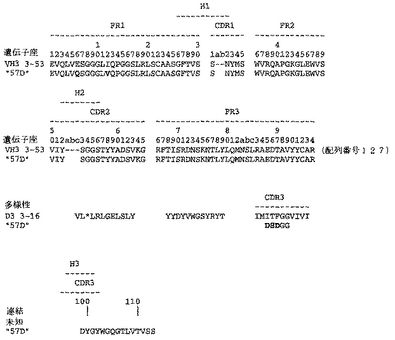
【図39】
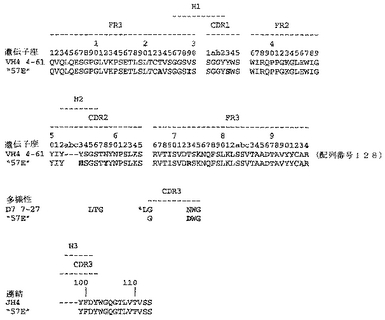
【図40】
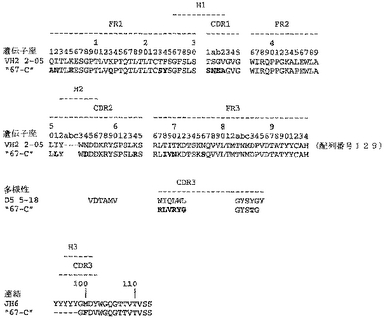
【図41】
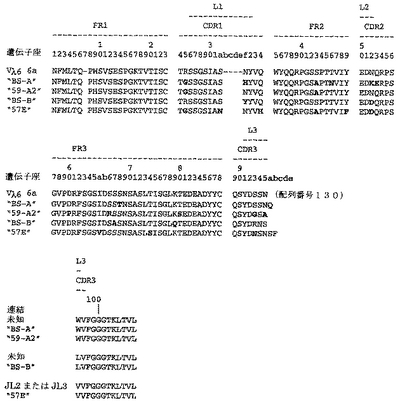
【図42】
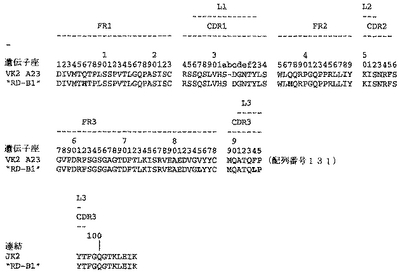
【図43】
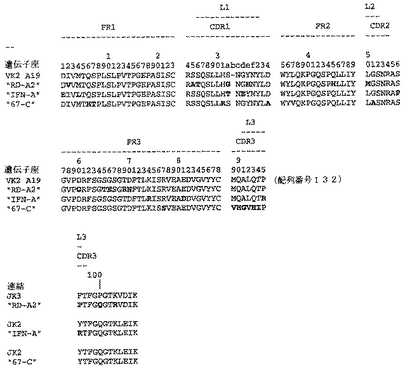
【図44】
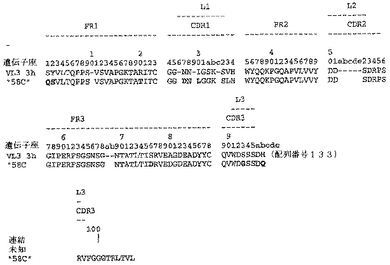
【図45】
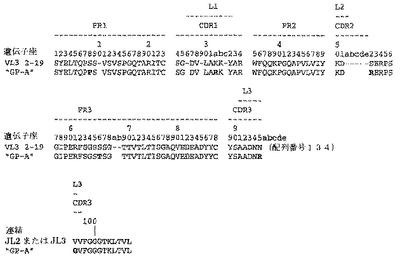
【図46】
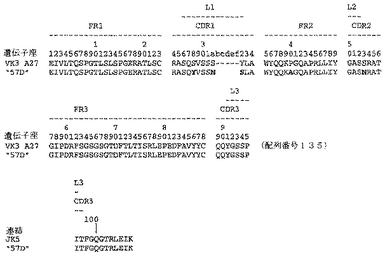
【図47】
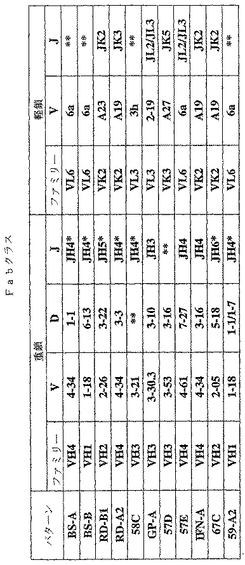
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【公表番号】特表2006−502739(P2006−502739A)
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願番号】特願2004−553382(P2004−553382)
【出願日】平成14年10月17日(2002.10.17)
【国際出願番号】PCT/US2002/033556
【国際公開番号】WO2004/046306
【国際公開日】平成16年6月3日(2004.6.3)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願日】平成14年10月17日(2002.10.17)
【国際出願番号】PCT/US2002/033556
【国際公開番号】WO2004/046306
【国際公開日】平成16年6月3日(2004.6.3)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
[ Back to top ]