
ヒト以外のトランスジェニック動物における内因性免疫グロブリン発現の抑制
本発明は1つまたはいくつかのヒトまたはヒト化免疫グロブリン導入遺伝子座を発現する遺伝子操作されたヒト以外のトランスジェニック動物における内因性抗体発現の抑制に関する新規研究法を提供する。内因性免疫グロブリン発現するB細胞においてのみ毒素のような自殺遺伝子を選択発現するが、ヒト(化)免疫グロブリンを発現するB細胞では発現しないことにより、ヒト以外のトランスジェニック動物における内因性免疫グロブリン発現を抑制する。この方法によりトランスジェニック動物の血液、乳または卵中のヒト化またはヒト抗体をコードする導入遺伝子座の優勢な発現が可能になる。

【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は内因性免疫グロブリンを発現するB細胞では自殺遺伝子を選択的に発現するが、ヒトまたはヒト化免疫グロブリンまたは免疫グロブリン鎖のような外因性免疫グロブリンまたは免疫グロブリン鎖を発現するB細胞では発現しないことにより、ヒト以外のトランスジェニック動物における内因性免疫グロブリン発現を抑制する方法に関する。この方法により例えばヒト以外のトランスジェニック動物の血液、乳および卵中のヒトまたはヒト化抗体の優勢な発現が可能になる。
【背景技術】
【0002】
発明の背景
ヒトマウスキメラ抗体を発現するマウスの作製は非特許文献1に記載されている。ヒト免疫グロブリンポリペプチドを発現するマウスの作製は非特許文献2;非特許文献3;および非特許文献4により記載されており、ならびにBACクローンを用いるトランスジェニックマウスの作製は非特許文献5に記載されている。ヒト抗体を発現するウシの作製は非特許文献6に記載されている。
【0003】
動物における遺伝子導入は非特許文献7に記載されている。トランスジェニックウサギの作製は非特許文献8;および非特許文献9に記載されている。トランスジェニックニワトリの作製は非特許文献10;および非特許文献11;および非特許文献12に記載されている。
【0004】
免疫グロブリン発現の損傷を有するウサギは非特許文献13;ならびに非特許文献14に記載されている。ガンマグロブリン血症のニワトリは非特許文献15;および非特許文献16に記載されている。
【0005】
細胞からの動物のクローニングは非特許文献17;非特許文献18;非特許文献19;非特許文献20;および非特許文献21に記載されている。ウサギの核移植クローニングは非特許文献22、非特許文献23に記載されている。
【0006】
ヒト(化)免疫グロブリン導入遺伝子座を発現するヒト以外のトランスジェニック動物の生成およびかかるトランスジェニック動物からの抗体の生成は特許文献1、特許文献2ならびに特許文献3、特許文献4;および特許文献5に詳細に記載されている。キメラ哺乳動物宿主の相同組換えは特許文献6に例示されている。DNAを胚に導入するための方法は特許文献7に記載されている。胚性幹細胞の維持および増殖は特許文献8に記載されている。
【0007】
毒素基盤の研究法を用いる自殺遺伝子は非特許文献24;非特許文献25;非特許文献26;非特許文献27;非特許文献28;非特許文献29;非特許文献30;非特許文献31;非特許文献32;非特許文献33に記載されている。無毒性プロドラッグ酵素研究法を用いる自殺遺伝子は非特許文献34に記載されている。
【0008】
2Aペプチド配列を含有するウイルス性タンパク質の切断活性は非特許文献35、非特許文献36、非特許文献37、非特許文献38、非特許文献39に記載されている。
【0009】
リコンビナーゼおよびその特性は非特許文献40に記載されている。2つの核酸の非常に特異的な配列間の相同組換えを認識および触媒する部位特異的リコンビナーゼが公知である。例えば部位特異的リコンビナーゼのインテグラーゼファミリーに属するφC31およびR4が公知である(非特許文献41;非特許文献42)。ヒトおよびマウスゲノムを含むいくつかのゲノムでは偽attP部位が天然にある(非特許文献43)。8.9kbの大型のVIIコラーゲンcDNAの一次前駆患者皮膚細胞へのインビトロでのφC31インテグラーゼ媒介遺伝子移入は非特許文献44に報告されている。トランスジェニック哺乳動物の操作におけるφC31、TP901−1およびR4ファージインテグラーゼの使用は非特許文献45で実証されている。
【0010】
B細胞を含む細胞のアブレーションは非特許文献46、非特許文献47および非特許文献48に記載されている。
【0011】
【特許文献1】国際公開第92/03918号パンフレット
【特許文献2】国際公開第02/12437号パンフレット
【特許文献3】米国特許第5545807号明細書
【特許文献4】米国特許第5814318号明細書
【特許文献5】米国特許第5570429号明細書
【特許文献6】米国特許第5416260号明細書
【特許文献7】米国特許第5567607号明細書
【特許文献8】米国特許第5453357号明細書
【非特許文献1】Pluschkeら、Journal of Immunological Methods 1998年、215:27−37
【非特許文献2】Neubergerら、Nature 1989年、338:350−2
【非特許文献3】Lonbergら、Int.Rev.Immunol.1995年、13(1):65−93
【非特許文献4】Bruggemannら、Curr.Opin.Biotechnol.1997年、8(4):455−8
【非特許文献5】Yangら、Nat.Biotechnol.1997年、15:859−65
【非特許文献6】Kuroiwaら、Nature Biotech 2002年、20(9):889−894
【非特許文献7】Wall RJ、Theriogenology 2002年、57(1):189−201
【非特許文献8】Fan,J.ら、Pathol Int.1999年、49:583−94
【非特許文献9】Bremら、Mol.Reprod.Dev.1996年、44:56−62
【非特許文献10】Etchesら、Methods in Molecular Biology 1997年、62:433−450
【非特許文献11】Painら、Cells Tissues Organs 1999年、165(3−4):212−9
【非特許文献12】Shermanら、Nature Biotech 1998年、16:1050−1053
【非特許文献13】Chenら、J.Immunol.1993年、150:2783−2793
【非特許文献14】Lamoyi EおよびMage RG.、J.Exp.Med.1985年、162:1149−1160
【非特許文献15】Frommelら、J.Immunol.1970年、105(l):1−6
【非特許文献16】Benedictら、Adv.Exp.Med.Biol.1977年、88(2):197−205
【非特許文献17】T.Wakayamaら、Nature 1998年、394:369−374
【非特許文献18】J.B.Cibelliら、Science 1998年、280:1256−1258
【非特許文献19】J.B.Cibelliら、Nature Biotechnology 1998年、16:642−646
【非特許文献20】A.E.Schniekeら、Science 1997年、278:2130−2133
【非特許文献21】K.H.Campbellら、Nature 1996年、380:64−66
【非特許文献22】Sticeら、Biology of Reproduction 1988年、39:657−664
【非特許文献23】Challah−Jacquesら、Cloning and Stem Cells 2003年、8(4):295−299
【非特許文献24】Leongら、Science 1983年、220:515−7
【非特許文献25】Maxwellら、Cancer Research 1986年、46:4660−4664
【非特許文献26】Palmiterら、Cell 1987年、50:435−443
【非特許文献27】Maxwellら、Cell 1991年、51:4299−4304
【非特許文献28】Maxwellら、Leukemia and Lymphoma 1992年、7:457−462
【非特許文献29】Aguilaら、Proc.Natl.Acad.Sci.1995年、92:10192−10196
【非特許文献30】Grieshammerら、Developmental Biology 1998年、197:234−247
【非特許文献31】Bartellら、Biology of Reproduction 2000年、63:409−416
【非特許文献32】Erlandssonら、J.Exp.Med.2001年、194:557−570
【非特許文献33】Leeら、Human Gene Therapy 2002年、13:533−542
【非特許文献34】Methods in Molecular Medicine:Suicide Gene Therapy, Methods and Reviews、Caroline J Springer編、Human Press、2004年
【非特許文献35】Palmenbergら、Virology 1992年、190:754−762
【非特許文献36】Ryanら、J Gen Virol 72:2727−2732(1991)
【非特許文献37】Donnellyら、J Gen Virol 2001年、82:1027−1041
【非特許文献38】Donnellyら、J Gen Virol 2001年、82:1013−1025
【非特許文献39】Szymaczakら、Nature Biotech 2004年、22(5):589−594
【非特許文献40】Kolb A.F.、Cloning Stem Cells 2002年、41:65−80
【非特許文献41】Grothら、Proc.Natl.Acad.Sci.2000年、97:5995−6000
【非特許文献42】Olivaresら、Nature Biotechnol.2002年、20(11):1124−8
【非特許文献43】Thyagarajanら、Mol.and Cell.Biol.2001年、21:3926−3934
【非特許文献44】Ortiz−Urdaら、Nature Medicine 8:1166−1170(2002)
【非特許文献45】Hollisら、Repro.Biol.and Endocrinol.2003年、1:79
【非特許文献46】Erlandssonら、J Exp Med 194(5):557−570(2001)
【非特許文献47】Maxwellら、Cancer Research 1991年、51:4299−4304
【非特許文献48】Palmiterら、Cell 1987年、50:435−443
【発明の開示】
【課題を解決するための手段】
【0012】
発明の要旨
本発明はトランスジェニック動物における内因性免疫グロブリン生成を抑制するための方法に関する。方法は内因性免疫グロブリンを発現するB細胞において自殺遺伝子を選択的に発現するが、ヒトまたはヒト化免疫グロブリンを発現するB細胞では自殺遺伝子を発現しないことを伴う。とりわけ本発明は1つまたはいくつかのヒトまたはヒト(化)免疫グロブリン導入遺伝子座を含有するヒト以外のトランスジェニック動物において内因性免疫グロブリン遺伝子座の発現を抑制するための方法に関係する。結果的にヒト(化)導入遺伝子座はヒト以外のトランスジェニック動物において遺伝子再構成および変異過程を被り、実質的に内因性免疫グロブリン生成の不在下で多様化されたヒト(化)抗体レパートリーを生成することができる。
【0013】
とりわけ本発明はヒト以外のトランスジェニック動物の内因性免疫グロブリンを生成するB細胞では少なくとも1つの自殺遺伝子を選択的に発現するが、外因性免疫グロブリンを生成するB細胞では発現しないことを含み、それにより内因性免疫グロブリンを生成するB細胞は枯渇し、そして内因性免疫グロブリンの生成は抑制されるが、外因性免疫グロブリンの生成は抑制されない、外因性免疫グロブリン導入遺伝子座を担持するヒト以外のトランスジェニック動物のB細胞における内因性免疫グロブリン(Ig)生成の選択的抑制のための方法に関係する。好ましい実施態様では、外因性免疫グロブリンはヒト化免疫グロブリン重および/または軽鎖配列である。
【0014】
さらなる態様では、ヒト以外のトランスジェニック動物のB細胞に導入された自殺遺伝子はB細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼ配列によりフランキングされている。
【0015】
なお別のさらなる態様では、さらに該組換え配列を認識するリコンビナーゼをコードする発現構築物の一部としてヒト以外のトランスジェニック動物のB細胞にヒト(化)免疫グロブリン鎖導入遺伝子座を導入し、ここで自殺遺伝子の発現はヒト化免疫グロブリン導入遺伝子座を発現するB細胞におけるリコンビナーゼの発現により活性化される。
【0016】
特定の態様では、自殺遺伝子は細菌性、真菌性、昆虫性および植物毒素からなる群から選択される。好ましい実施態様では、自殺遺伝子はジフテリア毒素A鎖である。
【0017】
別の実施態様では、自殺遺伝子はプロドラッグ変換酵素である。この実施態様の1つの態様では、プロドラッグ変換酵素は非哺乳動物起源である。さらなる態様では非哺乳動物プロドラッグ変換酵素はウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、β−ラクタマーゼ、多剤活性化酵素(MDAE)、β−ガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)からなる群から選択される。
【0018】
さらに別の実施態様では、プロドラッグ変換酵素はヒト起源である。さらなる態様では、ヒトプロドラッグ変換酵素はデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)からなる群から選択される。
【0019】
別の態様では、リコンビナーゼCre、Cre様、F1p、φC31、λインテグラーゼ、ファージR4リコンビナーゼ、TP901−1リコンビナーゼ、原核細胞性トランスポーゼス、真核細胞性トランスポーゼス、ウイルス性レトロトランスポーゼス、ショウジョウバエコピア様レトロトランスポーゼスおよび非ウイルス性レトロトランスポーゼスからなる群から選択される。さらなる実施態様では、トランスポーゼスまたはレトロトランスポーゼスはTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される。
【0020】
1つの実施態様では、ヒト以外のトランスジェニック動物は若年期に遺伝子再構成により抗体多様化を実質的に停止する。さらなる実施態様では、ヒト以外のトランスジェニック動物は生まれて1か月以内に抗体多様化を実質的に停止する。
【0021】
別の実施態様では、ヒト以外のトランスジェニック動物はげっ歯類、ウサギ、鳥類、ウシ、ブタ、ヒツジ、ヤギおよびウマからなる群から選択される。好ましい実施態様では、げっ歯類はマウスまたはラットである。
【0022】
特定の態様では、本発明は、ヒトまたはヒト化免疫グロブリン重および/または軽鎖導入遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0023】
別の態様では、本発明は、B細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0024】
1つの実施態様では、本発明はヒトまたはヒト化免疫グロブリン重および/または軽鎖遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子、ならびにB細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0025】
1つの実施態様では、前記した遺伝子導入発現構築物はCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4およびTP901−1リコンビナーゼからなる群から選択される部位特異的リコンビナーゼを含む。
【0026】
別の実施態様では、前記した遺伝子導入発現構築物は原核細胞性または真核細胞性のいずれかのトランスポーゼスであるリコンビナーゼを含む。
【0027】
さらに別の実施態様では、前記した遺伝子導入発現構築物はウイルス性、ショウジョウバエコピア様または非ウイルス性レトロトランスポーゼスのいずれかであるリコンビナーゼを含む。
【0028】
さらなる実施態様では、前記した遺伝子導入発現構築物はTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択されるリコンビナーゼを含む。
【0029】
特定の実施態様では、前記した遺伝子導入発現構築物はlox P部位、FRT部位、細菌性ゲノム組換え部位およびファージ組換え部位からなる群から選択される組換え部位を含む。
【0030】
さらなる実施態様では、細菌性ゲノム組換え部位はattBであり、そしてファージ組換え部位はattPまたは偽attPまたは偽attB部位である。
【0031】
特定の実施態様では、前記した遺伝子導入発現構築物はウイルス性2A/2Bまたは2A様/2B配列から得られる自己切断ペプチドを含む。
【0032】
さらなる実施態様では、ウイルスはピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーからなる群、またはC型ロタウイルスファミリーから選択される。さらに別の実施態様では、ウイルスは口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルスまたはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)からなる群から選択される。
【0033】
特定の実施態様では、前記した遺伝子導入発現構築物はCD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b、mb−1、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖および免疫グロブリンJ−鎖またはその修飾体からなる群から選択されるプロモーター/エンハンサーを用いてB細胞において特異的に発現される自殺遺伝子を含む。好ましい実施態様では、B細胞特異的プロモーター/エンハンサーはカッパ軽鎖遺伝子プロモーターまたはその修飾体である。
【0034】
1つの態様では本発明は、内因性免疫グロブリン生成が内因性免疫グロブリンを生成するB細胞における自殺遺伝子の選択的発現により抑制されるが、ヒト(化)免疫グロブリンを発現するB細胞は自殺遺伝子を発現せず、そしてしたがって増殖するヒト以外の動物を生成する方法に関係する。
【0035】
とりわけ本発明は前記した遺伝子導入発現構築物を発現するヒト以外のトランスジェニック動物に関係する。
【0036】
1つの実施態様では、ヒト以外のトランスジェニック動物は実質的に遺伝子変換により抗体多様性を生じる。
【0037】
全ての態様において、好ましいヒト以外の動物には、限定するものではないがげっ歯類(例えばマウス、ラット)、ウサギ、鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)、ウシ、ブタ、ヒツジ、ヤギ、ウマ、ロバおよびその他の家畜が含まれる。好ましい実施態様では、ヒト以外のトランスジェニック動物はマウスまたはラットのいずれかである。
【発明を実施するための最良の形態】
【0038】
発明の詳細な説明
定義
特記しない場合、本明細書で用いる技術的および科学的用語は本発明が属する分野の当業者により一般に理解されるのと同一の意味を有する。Singletonら、Dictionary of Microbiology and Molecular Biology、第2版、J.Wiley & Sons(ニューヨーク、ニューヨーク州、1994年)およびMarch、Advanced Organic Chemistry Reactions, Mechanisms and Structure、第4版、John Wiley & Sons(ニューヨーク、ニューヨーク州、1992年)は本出願で用いる多くの用語に対する一般的な手引きを当業者に提供する。
【0039】
当業者は本明細書に記載したものに類似するかまたは均等である多くの方法および材料を認識し、それを本発明の実施にあたり用いることができる。実際に本発明は記載した方法および材料に何ら限定されるものではない。本発明の目的のために以下の用語を後記で定義する。
【0040】
「B細胞」は免疫グロブリン遺伝子セグメントの再構成を被り、そしてそのライフサイクルのいくつかの段階で免疫グロブリン遺伝子を発現することができるB系統細胞として定義される。これらの細胞には、限定するものではないが初期プロB細胞、後期プロB細胞、大型プレB細胞、小型プレB細胞、未成熟B細胞、成熟B細胞、記憶B細胞、形質細胞等が含まれる。
【0041】
「抗体」(Ab)および「免疫グロブリン」(Ig)は同一の構造特性を有する糖タンパク質である。抗体は特異的抗原に対して結合特異性を呈するが、免疫グロブリンは抗体および抗原特異性を欠如するその他の抗体様分子の双方を含む。「抗体」なる用語は本明細書では最も広義で用いられ、そして具体的には、限定するものではないがモノクローナル抗体(モノクローナル抗体全長を含む)、ポリクローナル抗体、多重特異性抗体(例えば二重特異性抗体)およびそれが望ましい特異性を呈する限り抗体フラグメントに及ぶ。
【0042】
本明細書で用いる「Ig遺伝子セグメント」なる用語はIg分子の種々の部分をコードするDNAのセグメントを意味し、それは動物およびヒトの生殖細胞系列に存在し、そしてB細胞で一緒になって再構成されたIg遺伝子を形成する。したがって、本明細書で用いるIg遺伝子セグメントはV遺伝子セグメント、D遺伝子セグメント、J遺伝子セグメントおよびC領域遺伝子セグメントを含む。VDJまたはVJセグメントの機能的再構成の結果、免疫グロブリン重または軽鎖の発現に至る。
【0043】
本明細書で用いる「ヒトIg遺伝子導入遺伝子座または遺伝子座またはセグメント」なる用語はヒトIg遺伝子座またはそのセグメントの天然発生配列、ヒトIg遺伝子座またはそのセグメントの天然発生配列の変性形態、およびヒトIg遺伝子座またはそのセグメントの天然発生配列によりコードされるポリペプチドに実質的に同一のポリペプチド配列をコードする合成配列の双方を含む。この局面で、「実質的に」とはアミノ酸配列同一性の程度が少なくとも約85%〜95%、または少なくとも約90%〜95%、または少なくとも約95%、または少なくとも約98%であることを意味する。特定の好ましい実施態様では、ヒトIg遺伝子セグメントはヒトにおいて免疫グロブリン分子に非免疫原性にする。ここでは「ヒトまたはヒト化免疫グロブリン(Ig)重および/または軽鎖遺伝子座」または「ヒトまたはヒト化Ig遺伝子座」なる用語は互換的に用いられる。
【0044】
「ヒト抗体」および「ヒト免疫グロブリン」なる用語は完全なヒト配列を含む抗体および免疫グロブリン分子を意味するために用いられる。
【0045】
本明細書で用いる「ヒト化抗体」および「ヒト化免疫グロブリン」なる用語はヒト免疫グロブリンポリペプチド配列(またはヒト免疫グロブリン遺伝子セグメントによりコードされるポリペプチド配列)の少なくとも一部を含む免疫グロブリン分子を意味する。本発明のヒト化免疫グロブリン分子を、ヒト化免疫グロブリン分子を生成するように操作されたヒト以外のトランスジェニック動物から単離することができる。かかるヒト化免疫グロブリン分子は、動物から調製された、または動物に由来する細胞から調製された非ヒト化免疫グロブリン分子に相対して、霊長類、特にヒトに対して免疫原性が低い。ヒト化免疫グロブリンまたは抗体は遺伝子変換および遺伝子変換動物における体細胞超変異によりさらに多様化された免疫グロブリン(Ig)および抗体を含む。かかるヒト化Igまたは抗体はヒトにより天然に作成されたものではないので(ヒトは遺伝子変換によりその抗体レパートリーを多様化しないので)、それらは「ヒト」ではなく、そしてしかもヒト化Igまたは抗体はその構造でヒトIg配列を有するので、それらはヒトに対して免疫原性ではない。
【0046】
「導入遺伝子または導入遺伝子構築物」は、天然または動物または動物の細胞において通常見出されない合成タンパク質をコードする配列を有するDNAフラグメントである。本明細書で用いる「導入遺伝子構築物」なる用語は、構造的な「目的の遺伝子」および遺伝子移入を促進するその他の配列を含有するポリヌクレオチド分子を意味する。本発明は2つの導入遺伝子構築物:(1)ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼおよび(2)免疫細胞特異的自殺導入遺伝子構築物に言及する。
【0047】
「遺伝子導入発現構築物」は、ヒト以外のトランスジェニック動物の特異的な細胞内で一時的または細胞特異的または目的の導入遺伝子の発現強化のいずれかのために必要なその他の調節DNA配列と一緒に、本発明の1つまたはいくつかの導入遺伝子構築物をコードする配列を伴うDNAフラグメントを意味する。
【0048】
「ヒト(化)Ig遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子または導入遺伝子構築物」は、単一のmRNAに転写される導入遺伝子構築物を意味し、それは後記で論じる自己切断メカニズムのために2つのポリペプチド、主にヒト(化)免疫グロブリン鎖およびリコンビナーゼに翻訳される。
【0049】
本明細書で用いる「自己切断ペプチド」なる用語は、ペプチド配列自体の中の2つのアミノ酸残基の間に生じる切断活性を随伴するペプチド配列を意味する。例えば2A/2Bペプチドまたは2A/2B様ペプチドでは、切断は2Aペプチド上のグリシン残基と2Bペプチド上のプロリン残基との間で生じる。これは2A/2Bペプチドの2Aグリシン残基と2Bプロリン残基との間の正常なペプチド形成が損なわれている翻訳の間の「リボソームスキップ機構」によって生じ、2Bペプチドの残りの翻訳には影響することはない。かかるリボソームスキップ機構は当分野において周知であり、そして単一のメッセンジャーRNAによりコードされるいくつかのタンパク質の発現のためのいくつかのウイルスにより用いられることが解っている。
【0050】
本明細書で用いる「リコンビナーゼ」なる用語は、「組換え部位」と称される定義された部位間の部位特異的組換えを促進することができる酵素の群を意味し、ここで2つの組換え部位は単一の核酸分子内で物理的に分かれているか、または別個の核酸分子上にある。2つの定義された組換え部位の配列は同一である必要はない。リコンビナーゼの群の中には「インテグラーゼ」(例えばCre、Cre様、FLPおよびλインテグラーゼのような部位特異的リコンビナーゼ)および「リゾルベース/インベルターゼ」(例えばφC31インテグラーゼ、R4インテグラーゼおよびTP−901インテグラーゼ)を含むいくつかのサブファミリーがある。「リコンビナーゼ」なる用語にはまた、限定するものではないが原核細胞性または真核細胞性トランスポーゼス、ウイルス性またはショウジョウバエコピア様または哺乳動物レトロトランスポーゼスを含む非ウイルス性レトロトランスポーゼスも含まれる。原核細胞性トランスポーゼスの実例には、Tn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。真核細胞性トランスポーゼスにはショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAcおよびDsエレメント等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。レトロトランスポーゼスにはL1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノス等でコードされるものが含まれる。トランスポーゼスをMp、Spm、En、ドット、Mu、I転位性エレメントからなる群から選択することもできる。
【0051】
本明細書で用いる「野生型組換え部位」なる用語は通常インテグラーゼのようなリコンビナーゼにより用いられる組換え部位を意味する。
【0052】
「偽組換え部位」とは、部位がその野生型組換え部位の配列と同一の配列を有していなくても、リコンビナーゼが組換えを促進することができる部位を意味する。
【0053】
本明細書で用いる「自殺遺伝子または自殺導入遺伝子」なる用語は、発現が遺伝子を発現する細胞の死亡に至るタンパク質をコードする遺伝子を意味する。例えばタンパク質は毒素(例えばジフテリア毒素A鎖)または無毒性のプロドラッグを毒性生成物に変換する酵素(例えばチミジンキナーゼ、カルボキシルエステラーゼ、カルボキシペプチダーゼ、チトクロームP450アイソザイム、デオキシリボヌクレオチドキナーゼ、ニトロリダクターゼ等)でよい。自殺遺伝子がプロドラッグ変換酵素をコードする場合、それを発現する細胞はプロドラッグに暴露された時点で死亡する。本明細書で用いる「自殺遺伝子生成物」なる用語は「自殺遺伝子」によりコードされるタンパク質を意味する。自殺遺伝子発現は免疫細胞特異的プロモーターにより、好ましくはB細胞プロモーターにより駆動される。
【0054】
「自殺遺伝子発現の不活性化を可能にする配列」なる用語はリコンビナーゼにより認識される自殺遺伝子をフランキングする組換え部位を意味する。
【0055】
「プロドラッグ」なる用語は代謝によりインビボで毒性生成物、代謝物または薬物に変換可能である化合物を意味する。
【0056】
「内因性Ig(免疫グロブリン)発現B細胞」および「内因性B細胞」なる用語は互換的に用いられ、そして動物の内因性免疫グロブリン遺伝子座を発現するこれらのB細胞を意味する。本発明のB細胞はそのゲノムに自殺遺伝子を含有する。内因性B細胞は自殺遺伝子を発現し、そしてその結果、最終的に枯渇し、そしてしたがって動物の内因性Ig発現が抑制される。
【0057】
「外因性Ig(免疫グロブリン)発現B細胞」および「外因性B細胞」なる用語は、かかるB細胞に導入された外因性ヒト(化)Ig導入遺伝子座の産生性再構成を被るヒト以外の動物のこれらのB細胞を意味する。ヒト(化)Ig遺伝子座は、これもまた部位特異的リコンビナーゼをコードする発現構築物の一部としてかかるB細胞に導入される。ヒト(化)Ig遺伝子座の産生性再構成の結果、ヒト(化)Igおよび導入遺伝子コード化リコンビナーゼの発現に至る。結果として、B細胞のゲノムから自殺遺伝子が切除され、そして細胞は細胞死を免れる。したがって好ましくは、トランスジェニック動物により発現されるIg生成物はヒト(化)免疫グロブリンである。
【0058】
「自殺遺伝子の選択的発現」とは好ましくは免疫細胞内での、さらに好ましくはB細胞内での、そして最も好ましくは内因性B細胞内での自殺遺伝子生成物の発現を意味する。免疫細胞またはB細胞内での自殺遺伝子の選択的発現は各々免疫特異的またはB細胞特異的プロモーターを用いることにより達成されて自殺遺伝子発現を駆動する。
【0059】
「選択的不活性化」なる用語は、外因性B細胞のゲノムからの自殺遺伝子またはその部分の選択的切除を意味する。自殺遺伝子は外因性B細胞において発現される導入遺伝子コード化リコンビナーゼにより認識される組換え部位によりフランキングされるので、自殺遺伝子は切除されるかまたは不活性化される。
【0060】
Ig生成細胞の「枯渇」は、ヒト以外のまたは非ヒト化Igを発現する内因性B細胞集団の部分的または完全な死滅、瀕死および/または除去として定義される。さらに後記で説明するように、選択されるトランスジェニック動物が、抗体再構成が若年期に停止するものである場合、内因性B細胞の枯渇はさらに有効であり得る。
【0061】
「内因性免疫グロブリン生成の選択的抑制」は、自殺遺伝子を発現する内因性B細胞の枯渇による、ヒト以外のトランスジェニック動物の内因性免疫グロブリンの生成の選択的抑制を意味する。したがって、トランスジェニック動物により優勢に発現される免疫グロブリン生成物はヒト(化)免疫グロブリンである。
【0062】
「抗体多様性」および「抗体レパートリー」なる用語は互換的に用いられ、そして生物が発現できる全抗体特異性全体を意味する。
【0063】
遺伝子再構成および遺伝子変換を行う能力を有するIg遺伝子座はまた本明細書では「機能的」Ig遺伝子座をも意味し、そして機能的Ig遺伝子座により作成された多様性を有する抗体はまた本明細書では「機能的」抗体または抗体の「機能的」レパートリーをも意味する。
【0064】
「モノクローナル抗体」なる用語はB細胞の単一のクローンにより合成された抗体分子を意味する。
【0065】
「ポリクローナル抗体」なる用語はB細胞の集団により合成された抗体分子の集団を意味するために用いられる。
【0066】
「ポリヌクレオチド」および「核酸」なる用語は互換的に用いられ、そして単数または複数で用いられる場合、一般に任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドを意味し、これは未修飾RNAもしくDNAまたは修飾RNAもしくはDNAでよい。したがって、例えば本明細書で定義するようなポリヌクレオチドには、限定するものではないが一本鎖および二本鎖DNA、一本鎖および二本鎖領域を含むDNA、一本鎖および二本鎖RNA、ならびに一本鎖および二本鎖領域を含むRNA、一本鎖もしくはさらに典型的には二本鎖でよいか、または一本鎖および二本鎖領域を含んでよいDNAおよびRNAを含むハイブリッド分子が含まれる。加えて、本明細書で用いる「ポリヌクレオチド」なる用語はRNAもしくはDNAまたはRNAおよびDNAの双方を含む三本鎖領域を意味する。かかる領域の鎖は同一の分子から、または異なる分子からでよい。領域は1つまたはそれより多い分子全てを含んでよいが、さらに典型的にはいくつかの分子のある領域のみを伴う。三本鎖らせん領域の分子の1つはオリゴヌクレオチドである。「ポリヌクレオチド」なる用語は具体的にはcDNAを含む。その用語は1つまたはそれより多い修飾塩基を含有するDNA(cDNAを含む)およびRNAを含む。したがって、安定性またはその他の理由のために修飾された骨格を有するDNAまたはRNAは、その用語が本明細書で意図されるような「ポリヌクレオチド」である。さらに、イノシンのような通常でない塩基またはトリチウム化塩基のような修飾塩基を含むDNAまたはRNAは本明細書で定義する「ポリヌクレオチド」の用語に含まれる。一般に「ポリヌクレオチド」なる用語は未修飾ポリヌクレオチドの全ての化学的、酵素的および/または代謝的修飾形態、ならびにウイルスおよび単純細胞および複雑型細胞を含む細胞のDNAおよびRNA特性の化学的形態を包含する。
【0067】
本明細書で用いる「ヒト以外の(トランスジェニック)動物」には、限定するものではないが例えばヒト以外の霊長類、げっ歯類(例えばマウスおよびラット)、例えばウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマおよびロバのような非げっ歯類哺乳動物、ならびに鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)のような哺乳動物が含まれる。本明細書で用いる「非霊長類動物」なる用語には、限定するものではないが前記で具体的に列挙した哺乳動物を含む霊長類以外の哺乳動物が含まれる。
【0068】
「一次抗体レパートリーを創成するために実質的に遺伝子変換および/または体細胞超変異により抗体多様性を創成する動物」または「遺伝子変換動物」なる語句および文法的な均等は抗体多様化の主要な機構が、遺伝子再構成と対照的に遺伝子変換および/または超変異であるかかる動物を意味するために用いられる。かかる動物には、限定するものではないがウサギ、鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)ウシおよびブタが含まれる。特に好ましいヒト以外の動物はウサギおよびニワトリである。
【0069】
動物による「若年期の抗体遺伝子再構成の停止」とは、典型的には免疫グロブリン遺伝子の再構成が生まれて1か月以内に停止するこれらの動物を意味する。かかる動物の実例には、限定するものではないがウサギ、鳥類(例えばニワトリ)、ヒツジ、ヤギ、ウシ、ブタおよびウマである。
【0070】
詳細な説明
本発明は、例えば動物を(複数の)ヒト(化)免疫グロブリンの発現にさらに適当なものにする目的で、ヒト以外の動物における内因性免疫グロブリン生成を抑制するための方法を提供する。
【0071】
本発明によれば、内因性免疫グロブリンを発現するB細胞における自殺遺伝子の選択的発現により、内因性免疫グロブリン生成は(複数の)ヒト(化)免疫グロブリンのような外因性免疫グロブリン配列を発現するヒト以外のトランスジェニック動物において選択的に抑制される。自殺遺伝子は導入遺伝子として動物のゲノムに組み込まれ、そして例えばヒト(化)Ig導入遺伝子座をも導入する遺伝子導入発現構築物の一部として、または別個に例えば別個の遺伝子導入発現構築物を用いて導入され得る。後者の場合、2つの発現構築物を同時にまたは異なる時間でトランスジェニック動物に導入することができる。
【0072】
自殺遺伝子は免疫特異的プロモーター、好ましくはB細胞特異的プロモーターを用いて動物のB細胞において発現され、そしてリコンビナーゼにより認識される組換え配列によりフランキングされる。したがって、組換え配列によりフランキングされた自殺遺伝子は初期には動物の全てのB細胞に存在する。「外因性B細胞」では、ヒト(化)免疫グロブリン−自己切断ペプチド−リコンビナーゼ分子をコードする外因性免疫グロブリン導入遺伝子座の産生性再構成はかかるB細胞におけるリコンビナーゼの選択的発現に至る。リコンビナーゼはかかる細胞において自殺遺伝子をフランキングする組換え部位を認識する。結果的に外因性B細胞では自殺遺伝子は選択的に切除され、それは結果的に細胞死を免れる。対照的に内因性B細胞の内因性免疫グロブリン遺伝子座の産生性再構成はリコンビナーゼの発現に至らない。結果的に内因性B細胞における自殺遺伝子発現はこの細胞集団の死亡に至り、そしてその結果ヒト以外のトランスジェニック動物によるヒト(化)免疫グロブリンの発現を抑制することなく内因性免疫グロブリン生成を抑制する。
【0073】
導入遺伝子は1つまたはいくつかの天然または、通常動物もしくは動物の細胞において見出されない合成タンパク質をコードする配列を有するDNAフラグメントである。前核のマイクロインジェクション、トランスフェクション、核移植クローニング、精子媒介の遺伝子移入、精巣媒介の遺伝子移入等を含む種々の技術により、(複数の)DNAフラグメントを動物のゲノムに導入することができる。本発明は2つの導入遺伝子または導入遺伝子構築物(1)ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子および(2)免疫細胞特異的自殺導入遺伝子に言及する。各々の導入遺伝子はその独自の調節配列に作動可能なように連結されている。例えば自殺遺伝子の発現はB細胞特異的プロモーターにより駆動され得る。2つの導入遺伝子構築物は2つの別個のベクターまたは同一のベクター(プラスミド)に存在し得る。1つの実施態様では、2つの導入遺伝子構築物を別個の時点で導入することができる。これに代えて、双方の導入遺伝子構築物を同時に動物に導入することができる。好ましい実施態様では、自殺導入遺伝子の発現は、重鎖再構成を生じた後に起こるように調節される。後記で論じる機構から明白であるように、これは外因性B細胞におけるリコンビナーゼの発現のための時間、そしてしたがって外因性B細胞のゲノムからの自殺遺伝子のリコンビナーゼ媒介の切除、したがってかかる細胞における自殺遺伝子発現の停止のための時間を見越している。加えて、本発明の方法で用いるベクターは選択を可能にするためにゲンタマイシン、ネオマイシン、カナマイシン等のような抗生物質選択マーカーをコードするDNA配列を含有できる。
【0074】
本発明の1つの態様では、導入遺伝子は自己切断ペプチド(例えば2Aペプチドまたは2A様ペプチド)をコードするDNA配列を含む。導入遺伝子の免疫グロブリンコード化配列とリコンビナーゼコード化配列の間の自己切断ペプチドコード化配列の挿入により1つのメッセンジャーRNAの生成に至る。しかしながらこのmRNAの翻訳の結果、ペプチドの自己切断機構のために2つの別個のタンパク質、(複数の)免疫グロブリンおよびリコンビナーゼに至る。したがってリコンビナーゼの発現をVDJまたはVJセグメントの機能的再構成と結びつけることができる。
【0075】
本発明の1つのかかる実施態様では、自己切断はピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーを含む、またはC型ロタウイルスファミリーからのウイルスの2A/2Bペプチドまたは2A様/2B配列により媒介される。ピコルナウイルス科ウイルスファミリーにはエンテロ、ライノ、カルディオおよびアフトならびに口蹄疫(FMDV)ウイルスが含まれる。ピコルナウイルス様昆虫ウイルスファミリーには感染性フラシェリー病ウイルス(IFV)、ショウジョウバエCウイルス(DCV)、急性ミツバチ麻痺病ウイルス(ABPV)およびコオロギ麻痺病ウイルス(CrPV)ならびに昆虫ウイルスゾセア・アシグナ(Thosea asigna)ウイルス(TaV)のようなウイルスが含まれる。C型ロタウイルスファミリーにはウシ、ブタおよびヒトC型ロタウイルが含まれる。さらなる実施態様では、切断配列はポリオウイルス、ライノウイルス、コクサッキーウイルス、脳心筋炎ウイルス(EMCV)、メンゴウイルス、ブタテシオウイルス−1、またはタイラーネズミ脳炎ウイルス(TMEV)等のいずれかからの2A様/2B配列を含み得る。好ましい実施態様では、自己切断タンパク質配列は口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルスまたはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)のいずれかの2A/2Bペプチドである。Palmenbergら、Virology 190:754−762(1992)、Ryanら、J Gen Virol 72:2727−2732(1991)、Donnellyら、J Gen Virol 82:1027−1041(2001)、Donnellyら、J Gen Virol 82:1013−1025(2001)、Szymaczakら、Nature Biotech 22(5):589−594(2004)。
【0076】
本発明の方法で用いるその他の導入遺伝子は部位特異的リコンビナーゼをコードする。部位特異的リコンビナーゼは2つの核酸、例えばDNAセグメント間の相同組換えを触媒する。これらのリコンビナーゼは組換えの双方のパートナーの非常に特異的な配列を認識する。触媒の機構は様々な型の部位特異的リコンビナーゼで異なってよいが、それらは全て、基礎を成す機構に関わらず本明細書に含まれ、そして本発明の実施に適当である。
【0077】
特定の実施態様では、リコンビナーゼは例えばCre、Flpリコンビナーゼ等でよい。CreおよびFlpは2つの最も一般的に用いられる酵素であり、それは非常に特異的なDNA配列に作用するのみである。Creは2つの34塩基対長のloxP部位間でDNAの組換えを触媒するが、Flpはfrt部位を標的とする。真核細胞のDNAの部位特異的組換えのためのCreリコンビナーゼの使用は米国特許第4959317号に記載されている。真核細胞のトランスフェクションのための部位特異的リコンビナーゼの使用は米国特許第6632672号に記載されている。部位特異的組換えは一般に米国特許第4673640号に記載されている。Cre/loxP基盤のクローニング系は例えばBD Biosciences−Clontech(パロアルト、カリフォルニア州)(Creator(商標))またはInvitrogen(カールスバッド、カリフォルニア州)(Echo(商標))から市販により入手可能である。
【0078】
別の実施態様では、リコンビナーゼはλインテグラーゼ、φC31、TP901−1およびR4からなる群から選択されるファージによりコードされる部位特異的リコンビナーゼでよい。φC31およびR4は部位特異的リコンビナーゼのインテグラーゼファミリーに属するが、TP901−1は拡張されたリゾルベースファミリーに属する。R4インテグラーゼはストレプトミセス・パルブルスのファージR4のゲノムから誘導された部位特異的一方向性リコンビナーゼである。部位特異的インテグラーゼTP901−1はラクトコッカス・ラクティス亜種クレモリスのファージTP901−1によりコードされる。λは大腸菌に感染する鋳型バクテリオファージである。ファージは組換えのための1つの結合部位(attP)を有し、そして大腸菌細菌ゲノムは組換えのための1つの結合部位(attB)を有する。本発明の局面では、野生型組換え部位を例えば相同系から誘導し、そして異種性配列に随伴させることができる。したがってattB部位をその他の系に置いてインテグラーゼの基質として作用させることができる。さらに別の実施態様では、リコンビナーゼは細菌ゲノム組換え部位(attB)およびファージゲノム組換え部位(attP)の間の組換えを触媒することができるか、または第1の部位は偽attB部位を含むことができ、そして/もしくは第2の部位は偽attP部位を含むことができるかまたはその逆である(Grothら、Proc.Nat.Acad.Sci.97:5995−6000(2000);Olivaresら、Nature Biotechnol.20(11):1124−8(2002));(Thyagarajanら、Mol. and Cell.Biol.21:3926−3934(2001));Hollisら、Repro.Biol.and Endocrinol.1:79(2003))。「偽組換え部位」とは、たとえ部位がその野生型組換え部位の配列と同一の配列を有せずとも、リコンビナーゼが組換えを促進できる部位を意味する。
【0079】
さらに別の実施態様では、リコンビナーゼはトランスポーゼスまたはレトロトランスポーゼスでよい。トランスポゾンまたはレトロトランスポーゼスはカットアンドペースト機構によりその転位を触媒する酵素であり、そして任意の導入遺伝子の移入または挿入に用いることができる。これらはヒト、鳥類、げっ歯類等のような脊椎動物を含む広範な種のゲノムへの任意のDNA配列の挿入または移入のための非ウイルス性および非相同性の方法を提供する。例えばショウジョウバエエレメントマリナーはそれ自体をニワトリ生殖細胞系に転位させることが示された(Shermanら、Nature Biotechnol.16:1050−1053(1998))。sleeping beautyトランスポーゼス系を用いるマウスおよびヒトゲノムのような哺乳動物系への長期間の導入遺伝子発現またはトランスポゾンDNAの効率的な挿入はYantら、Nature Genetics 25:35−41(2000);Dupuyら、Proc.Nat.Acad.Sci.99:4495−4499(2002)およびGeurtsら、Mol.Therapy 8:108−117(2003)により実証されている。L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)、ミノスのようなその他のトランスポーゼスは脊椎動物種において活性であることが示されており、そしてしたがって遺伝子移入に、または挿入変異ベクターとして有用である(Largaespada,David A.、Repro.Biol.and Endocrinol.1:80(2003))。トランスポーゼスの実例としては、限定するものではないが原核細胞性または真核細胞性トランスポーゼス、ウイルス性、ショウジョウバエコピア様または哺乳動物レトロトランスポーゼス等を含む非ウイルス性レトロトランスポーゼスが含まれる。原核細胞性トランスポーゼスには、Tn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。真核細胞性トランスポーゼスにはショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAcおよびDsエレメント等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。レトロトランスポーゼスにはL1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)、ミノス等でコードされるものが含まれる。トランスポーゼスをMp、Spm、En、ドット、Mu、およびI転位性エレメントからなる群から選択することもできる。
【0080】
本発明の1つの態様では、前記した部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子を用いて内因性B細胞を選択的に死滅させる。自殺遺伝子を組換え部位でフランキングすることにより、外因性B細胞においてのみリコンビナーゼまたはトランスポゾンの発現時の自殺遺伝子の不活性化に至る。自殺遺伝子発現の不活性化は自殺遺伝子またはその部分の切除により達成することができる。これに代えて、自殺遺伝子の発現に必要な配列を切除の標的とすることができる。別の研究法によれば、DNAフラグメントの反転または挿入によりトランスポゾンジャンピング、トランスポゾン挿入または反転のために自殺遺伝子発現を不活性化することができる。
【0081】
本発明の実施において用いる自殺遺伝子には毒素遺伝子および毒性生成物を生じる無毒性プロドラッグ変換酵素が含まれる。本発明の方法において用いることができる活性毒素およびそのフラグメントには、例えばジフテリア毒素A鎖(DTA)のような細菌性毒素もしくはそのフラグメント、シグマ毒素、エキソトキシンA鎖(緑膿菌由来)等またはリシンA鎖、アブリンA鎖、モデクシンA鎖、アルファサルシン、シナアブラギリタンパク質、ジアンチンタンパク質、ヨウシュヤマゴボウタンパク質(PAPI、PAPAIIおよびPAP−S)、ツルレイシ阻害剤、クルシン、クロチン、サボンソウ阻害剤、ゲロニン、ミトゲリン、レストリクトシン、フェノマイシン、エノマイシンおよびトリコテセン、殺虫性毒素、は虫類毒液等のような植物もしくは真菌毒素およびその非結合活性フラグメントが含まれる。好ましい実施態様では、用いる毒素はジフテリア毒素A鎖(DTA)である。
【0082】
標的B細胞に導入された自殺遺伝子がプロドラッグ変換酵素をコードする場合、酵素は特異的無毒性プロドラッグを活性化して最終的に標的B細胞を死滅させる毒性代謝物を創成する。この研究法では、内因性B細胞生成を抑制するために2工程処理方法を設計することができる。第1工程では、当分野において公知の種々の方式で外来酵素のための遺伝子を発現させるかまたはB細胞に分配することができる。第2工程では、動物に投与されたプロドラッグは最終的にそれを死滅させる酵素を発現するB細胞により活性化されて毒性代謝物になる。
【0083】
自殺遺伝子として有用なプロドラッグ変換酵素は一般に2つの主要なクラスで見出されている。第1のクラスはヒト対応物を伴うかまたは伴わない非哺乳動物起源の酵素である。実例にはウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、βラクタマーゼ、多剤活性化酵素(MDAE)、βガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)が含まれる。第2のクラスはヒト起源の酵素からなる。これにはデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)アイソザイムが含まれる。酵素プロドラッグ系のさらなる実例はMethods in Molecular Medicine:Suicide Gene Therapy,Methods and Reviews(Caroline J Springer編、Human Press、2004年)の表1に列挙されており、それを出典明示により本明細書の一部とする。したがって本発明で自殺遺伝子として用いるプロドラッグ変換酵素には、限定するものではないが前記したように非哺乳動物、非ヒト起源およびヒト起源の酵素が含まれる。
【0084】
本発明で有用である適当なプロドラッグには、限定するものではないがガンシクロビル、アシクロビル、5−(アジリジン−1−イル)−2,3ジニトロベナミド、カペシタビン、イリノテカン、カルバマート基盤の20(S)−カンプトテシン、ジニトロベンズアミドアジリジンCB1954およびそのナイトロジェンマスタード類似体SN23862、2−アミノアントラセン(2−AA)および4−イポメアノール(4−IM)等が含まれる。本発明で有用であるプロドラッグのさらなる実例は対応するその活性化酵素と共にMethods in Molecular Medicine:Suicide Gene Therapy, Methods and Reviews(Caroline J Springer編、Human Press、2004年)の表1に列挙されており、それを出典明示により本明細書の一部とする。酵素活性化プロドラッグはしばしば、例えばエタニダゾール、フルオソール、ミソニダゾール、ニモラゾール、テモポルフィン、トリアパザミンのような放射線増感薬、またはカスパーゼのようなその他のアポトーシス剤の発現を用いるその他の細胞死滅方法と組み合わせて用いられることがあり、標的内因性Ig発現B細胞を死滅させる相乗効果を導く。
【0085】
したがって自殺遺伝子の発現は種々の機構により内因性免疫グロブリンを発現するB細胞を排除することができる。例えばDTAの発現によりタンパク質合成の阻止および続く細胞死に至る。チミジンキナーゼはガンシクロビルおよびアシクロビルをその対応する一リン酸形態にリン酸化し、それは続いて細胞性キナーゼにより毒性の三リン酸誘導体に変換される。毒性三リン酸の分裂細胞DNAへの取り込みにより細胞死に至る。ニトロリダクターゼは5−(アジリジン−1−イル)−2,3ジニトロベナミドを2−および4−ヒドロキシルアミノ誘導体に変換し、そのときに細胞性チオエステルを有する4−ヒドロキシルアミノ誘導体の非酵素反応によりDNAを架橋することができる強力な細胞毒性二価性アルキル化剤が作成される。当分野において公知の自殺遺伝子は本発明において有用であるが、これらの自殺遺伝子が内因性B細胞を排除するように作用する(複数の)機構に束縛または限定されることはない。
【0086】
本発明の好ましい実施態様では、自殺遺伝子は野生型または偽組換え部位(例えばCreにより認識されるloxP部位またはFlpにより認識されるFRT部位)によりフランキングされたジフテリア毒素A鎖(DTA)をコードする。
【0087】
本発明の別の実施態様では、自殺遺伝子は 野生型または偽組換え部位によりフランキングされたチミジンキナーゼをコードする。
【0088】
1つの実施態様では、部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子を、ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子の導入の前、同時または後のいずれかに、別個の遺伝子導入ベクターで動物に導入する。別の実施態様では、部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子をヒトIg遺伝子座−リコンビナーゼ導入遺伝子のためのものとして同一の遺伝子導入ベクターで動物に導入する。全ての態様で、自殺遺伝子は動物ゲノムに組み込まれ、そして免疫細胞においてのみ特異的に発現されることを確実にする免疫細胞特異的プロモーターにより、および好ましくはB細胞においてのみ特異的に発現されることを確実にし、その他の細胞型では発現されないB細胞特異的プロモーターにより、その発現が駆動される。
【0089】
自殺遺伝子の発現はB細胞特異的プロモーターにより制御されて、その発現は非B細胞またはヒト以外のトランスジェニック動物の組織では「スイッチオフ」になる。B細胞特異的遺伝子の発現を制御するプロモーター(およびエンハンサー)、またはその変種もしくは操作された部分は自殺遺伝子のかかるB細胞特異的発現に有用である。B細胞特異的遺伝子のプロモーター/エンハンサーの実例には、限定するものではないがCD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b(B29またはIgベータとしても公知である)、mb−1(Igアルファとしても公知)、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖、免疫グロブリンJ−鎖等のプロモーター/エンハンサーが含まれる。好ましい実施態様では、カッパ軽鎖遺伝子プロモーター/エンハンサーが自殺遺伝子のB細胞特異的発現を駆動する。
【0090】
したがって、内因性免疫グロブリン生成の抑制の結果、ヒト(化)Ig導入遺伝子座の優勢な発現に至る。換言すれば、内因性B細胞の枯渇によりヒト(化)抗体の富化に至る。好ましくは、内因性B細胞の富化は100%に近くなる。
【0091】
本発明のさらに別の態様では、導入遺伝子は免疫グロブリン重鎖および/もしくは免疫グロブリン軽鎖またはその部分をコードする。遺伝子座は生殖細胞系の立体配置または再構成形態でよい。コード化配列またはその部分はヒト免疫グロブリンをコードすることができ、その結果ヒト(化)抗体の発現に至る。
【0092】
ヒト(化)抗体をコードする(複数の)導入遺伝子は1つまたはいくつかのヒトIgセグメント(例えばヒトIgV、D、JまたはC遺伝子セグメント)を含有するIg遺伝子座またはIg遺伝子座の大部分を含有する。これに代えて、導入遺伝子はヒト免疫グロブリン遺伝子座またはその大部分である。かかるヒトIg遺伝子座またはかかる修飾されたIg遺伝子座またはIg遺伝子座の修飾された部分を含有する導入遺伝子もまた本明細書では「ヒト(化)Ig導入遺伝子座」と称され、ヒト以外のトランスジェニック動物において再構成を被ることができ、それによりヒト免疫グロブリンポリペプチド配列の少なくとも一部を有する抗体の多様化されたレパートリーが生成される。
【0093】
免疫グロブリン重および軽鎖遺伝子は個々の遺伝子によりコードされ、そしてイントロン配列により分けられたいくつかのセグメントを含む。したがってヒト免疫グロブリン重鎖のための遺伝子は第14染色体で見出される。重鎖(VH)の可変領域は3つの遺伝子セグメント(V、DおよびJセグメント)、続いてC領域をコード化する同義遺伝子を含む。V領域は大型のスペーサーによってC領域から分けられ、そしてV、DおよびJセグメントをコードする個々の遺伝子もまたスペーサーによって分けられる。
【0094】
2つの型の免疫グロブリン軽鎖:κおよびλがある。ヒトκ軽鎖の遺伝子は第2染色体に見出され、そしてヒトλ軽鎖の遺伝子は第22染色体に見出される。抗体軽鎖の可変領域は別個の遺伝子セグメントによりコードされるVセグメントおよびJセグメントを含む。κ軽鎖遺伝子の生殖細胞系立体配置では、およそ100〜200V領域遺伝子が直線的な配置で存在し、各遺伝子はそれ独自のリーダー配列、続いておよそ5J遺伝子セグメントおよびC領域遺伝子セグメントを有する。全てのV領域はイントロンにより分けられ、そして同様にV、JおよびC領域遺伝子セグメントを分けるイントロンが存在する。
【0095】
感染に対して保護する免疫系の能力は、抗体の多様なレパートリーを創成するために特殊化された遺伝的な仕組みにある。B細胞の抗体コード化遺伝子は可変(V)領域の結合部位の無数の組み合わせを可能にする様式で組み立てられる。1012より多い可能な結合構造がかかる機構から生じると推測される。ヒトを含む全ての動物において抗体産生過程は免疫グロブリン(Ig)遺伝子座の可変(V)、多様性(D)および連結(J)セグメントを組換えることにより始まる。この工程の後に、動物種に依存して2つの一般的な機構を用いて抗体の多様な結合構造を生成する。
【0096】
ヒトおよびマウスのようないくつかの動物では、免疫グロブリン重鎖遺伝子座にV、DおよびJセグメントの複数のコピー、ならびに免疫グロブリン軽鎖伝子座にVおよびJセグメントの複数のコピーが存在する。これらの動物における抗体多様性は一次的には遺伝子再構成、すなわち再構成された重鎖可変領域および軽鎖可変領域を形成するための遺伝子セグメントの異なる組み合わせにより作成される。しかしながら別の動物(例えばウサギ、鳥類、例えばニワトリ、ガチョウおよびアヒル、ヒツジ、ヤギならびにウシ)では、遺伝子再構成は抗体多様性の作成においてあまり大きな役割を果たさない。例えば、ウサギでは、非常に限定された数のV遺伝子セグメントのみ、もっともよくあるのはV領域の3’末端のV遺伝子セグメントが遺伝子再構成に用いられて近接するVDJセグメントを形成する。ニワトリでは、1つのV遺伝子セグメント(D領域に隣接するもの、すなわち「3’近位V遺伝子セグメント」)のみ、1つのDセグメントおよび1つのJセグメントのみを重鎖再構成で用い、そして1つのV遺伝子セグメント(3’近位V遺伝子セグメント)および1つのJセグメントのみを軽鎖再構成で用いる。したがってこれらの動物では初期に再構成された可変領域配列の中では多様性が少ししかなく、それは連結部多様化の結果である。再構成されたIg遺伝子のさらなる多様化は、上流V遺伝子セグメントから誘導される短い配列が、再構成されたIg遺伝子のV遺伝子セグメント内の短い配列を置き換える過程である遺伝子変換により達成される。抗体配列のさらなる多様化を超変異により作成することができる。
【0097】
免疫グロブリン(抗体)は5つのクラス(IgG、IgM、IgA、IgEおよびIgD)に属し、各々は免疫防御において異なる生物学的役割を有している。血液中に最も豊富であり、そして感染に対する応答において強力であるのはIgGクラスである。ヒトIgGクラス内にはFcドメインを含む重鎖定常領域の構造により決定される4つのサブクラス(IgG1、IgG2、IgG3およびIgG4アイソタイプ)がある。抗体のF(ab)ドメインは抗原の特異的配列(エピトープ)に結合するが、抗体のFcドメインは抗原を排除するために免疫系のその他の構成成分を補充および活性化する。
【0098】
元来の抗体および免疫グロブリンは通常約150000ダルトンのヘテロ四量体糖タンパク質であり、2つの同一の軽(L)鎖および2つの同一の重(H)鎖からなる。各軽鎖は(複数の)共有結合性ジスルフィド結合により重鎖に連結されているが、ジスルフィド結合の数は様々な免疫グロブリンアイソタイプの重鎖間で異なる。各重および軽鎖はまた規則正しい間隔の鎖内ジスルフィド架橋を有する。各重鎖は一方の末端で可変ドメイン(VH)を有し、多くの定常ドメインがそれに続く。各軽鎖は一方の末端で可変ドメイン(VL)、およびその他方の末端で定常ドメインを有し、軽鎖の定常ドメインは重鎖の第1の定常ドメインとアラインされ、そして軽鎖可変ドメインは重鎖の可変ドメインとアラインされている。特定のアミノ酸残基は軽鎖および重鎖可変ドメイン間のインターフェースを形成すると考えられている(Chothiaら、J.Mol.Biol.186:651(1985);NovotnyおよびHaber、Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0099】
「可変」なる用語は可変ドメインの特定の部分が抗体間の配列で大きく異なるという事実を意味し、そしてその特定の抗原に関する特定の各抗体の結合および特異性において用いられる。しかしながら可変性は抗体の可変ドメインにわたって均一に分布されるわけではない。それは双方共に軽鎖および重鎖可変ドメインの相補性決定領域(CDR)または超可変領域と称される3つのセグメントに集中している。可変ドメインのさらに高度に保存された部分はフレームワーク(FR)と称される。元来の重および軽鎖の可変ドメインは各々3つのCDRによりつながれた4つのFR領域を含む。各鎖のCDRはFR領域により近接近してまとまり、そしてその他の鎖のCDRと一緒に抗体の抗原結合部位の形成に寄与する(Kabatら、Sequences of Proteins of Immunological Interest、第5版、National Institute of Health,、ベセスダ、メリーランド州(1991年)参照)。定常ドメインは抗体の抗原に対する結合に直接関与しないが、抗体依存性細胞毒性における抗体の関与のような種々のエフェクター機能を呈する。
【0100】
ヒト動物導入遺伝子座の創成により高収率で多様化された高親和性ヒト(化)(ポリクローナル)抗体を発現するトランスジェニック動物の創成が可能になる。一般にヒト以外の動物における免疫グロブリン(Ig)遺伝子座のヒト化は1つまたはそれより多いヒトIg遺伝子セグメントの動物のゲノムへの組み込みを伴い、ヒト(化)免疫グロブリン遺伝子座を創成する。したがってヒト(化)Ig重鎖遺伝子座の創成は1つまたはそれより多いVおよび/もしくはDおよび/もしくはJセグメント、ならびに/またはC領域セグメントの動物ゲノムへの組み込みを伴う。同様に、ヒト化Ig軽鎖遺伝子座の創成は1つまたはそれより多いVおよび/もしくはJセグメント、ならびに/またはC領域セグメントの動物ゲノムへの組み込みを伴う。
【0101】
染色体位置に関わらず、本発明のヒト(化)Ig遺伝子座はヒト以外の動物において遺伝子再構成および遺伝子変換および超変異を被る能力を有し、それによりヒト(化)Ig分子の多様化されたレパートリーを生成する。遺伝子再構成および遺伝子変換を被る能力を有するIg遺伝子座は「機能的」Ig遺伝子座とも称され、そして機能的Ig遺伝子座により作成された多様性を有する抗体は「機能的」抗体または抗体分子の「機能的」レパートリーとも称される。
【0102】
1つの態様では、抗体レパートリーの多様化が若年期に停止する動物が本発明において有用である。B細胞は造血幹細胞から発達する。抗原暴露の前にB細胞は一連の成熟工程を被り、その最終生成物は成熟B細胞であり、それはその他の細胞表面シグナリング分子と一緒にその細胞表面に独特な膜結合性IgMおよびしばしばIgDを発現する。ヒトでは遺伝子再構成による抗体多様化は一生にわたって生じるが、その他の動物では、抗体レパートリーの多様化は若年期に、典型的には生まれて1か月以内に停止する。
【0103】
免疫グロブリン遺伝子の再構成が若年期に停止する動物では、この限定された期間に生成された全てまたはほとんどのB細胞を死滅させることにより効果的に内因性免疫グロブリン生成の持続的または永続的な抑止に至る。1つまたはいくつかのヒトまたはヒト化免疫グロブリン導入遺伝子座を含有するトランスジェニック動物では、これにより動物の内因性免疫グロブリン生成の不在下でのヒトまたはヒト化免疫グロブリンの生成が可能になる。このように遺伝子再構成が若年期に停止する動物では(複数の)内因性免疫グロブリンの発現が効果的に抑制され得る。かかる動物の実例は、限定するものではないがウサギ、鳥類(例えばニワトリ)、ヒツジ、ヤギ、ウシ、ブタおよびウマである。
【0104】
本発明によれば、ヒト(化)免疫グロブリンを作ることができるトランスジェニック動物は、動物の(1つまたは複数の)レシピエント細胞に本明細書に前記した1つまたはそれより多い遺伝子導入ベクターを導入することにより作られ、その1つはヒト(化)Ig遺伝子座を担持し、そして遺伝的に修飾された(1つまたは複数の)レシピエント細胞から動物を誘導する。
【0105】
レシピエント細胞は例えば遺伝子変換および/または超変異により抗体多様性を作成するヒト以外の動物、例えば鳥類(例えばニワトリ)、ウサギ、ウシ等由来でよい。かかる動物では3’近位V遺伝子セグメントは免疫グロブリンの生成に優先的に用いられる。動物の3’近位V遺伝子セグメントを置換することか、または3’近位V遺伝子セグメントの近接近で配置されることのいずれかにより、導入遺伝子ベクターのIg遺伝子座へのヒトV遺伝子セグメントの組み込むことで、大部分の免疫グロブリンでのヒトV領域ポリペプチド配列の発現に至る。これに代えて、再構成されたヒトV(D)Jセグメントを導入遺伝子ベクターの免疫グロブリン遺伝子座のJ遺伝子座に挿入することができる。
【0106】
無作為組み込みによるかまたは標的組み込みにより、ヒト(化)Ig遺伝子座および自殺遺伝子を含有する目的の遺伝子を含有する遺伝子導入ベクターを(1つまたは複数の)レシピエント細胞に導入し、そして次に(1つまたは複数の)レシピエント細胞のゲノムに組み込むことができる。
【0107】
無作為組み込みのために、標準的な遺伝子導入技術によりヒト(化)Ig遺伝子座を含有する遺伝子導入ベクターを動物のレシピエント細胞に導入することができる。例えば遺伝子導入ベクターを受精卵母細胞の前核に直接注射することができる。また卵母細胞の受精の前に精子の遺伝子導入ベクターとのコインキュベーションにより遺伝子導入ベクターを導入することもできる。受精卵母細胞からトランスジェニック動物を発達させることができる。遺伝子導入ベクターを導入する別の方式は、胚性幹細胞をトランスフェクトし、そして続いて遺伝的に修飾された胚性幹細胞を発達中の胚に注射することによる。これに代えて遺伝子導入ベクター(裸でまたは促進剤と組み合わせて)を発達中の胚に直接注射することができる。最終的に、トランスジェニック動物の少なくともいくつかの体細胞のゲノムに組み込まれたヒト(化)Ig導入遺伝子を含有する胚からキメラトランスジェニック動物を生成する。
【0108】
特定の実施態様では、ヒト(化)Ig遺伝子座を含有する導入遺伝子は内因性免疫グロブリン遺伝子の発現が損なわれた動物株から誘導されたレシピエント細胞(例えば受精卵母細胞または発達中の胚)のゲノムにランダムに組み込まれる。かかる動物株の使用によりヒト(化)遺伝子導入Ig遺伝子座からの免疫グロブリン分子の優先的な発現が可能になる。かかる動物の実例には、アリシアおよびバジレアウサギ株、ならびに無ガンマグロブリン血症ニワトリ株、ならびに免疫グロブリンノックアウトマウスが含まれる。これに代えて、ヒト(化)免疫グロブリン導入遺伝子または遺伝子座を有するトランスジェニック動物を内因性免疫グロブリンの発現が損なわれた動物株と交配させることができる。損なわれた内因性Ig遺伝子座およびヒト(化)遺伝子導入Ig遺伝子座に関してホモ接合性の子孫を得ることができる。
【0109】
標的組み込みのために、遺伝子導入ベクターを胚性幹細胞または既に分化した体細胞のような適切な動物レシピエント細胞に導入することができる。その後、導入遺伝子が動物ゲノムに組み込まれ、そして相同組換えにより対応する内因性Ig遺伝子座を置換した細胞を標準的な方法により選択することができる。例えばKuroiwaら、Nature Genetics、2004年6月6日を参照のこと。次いで選択した細胞を除核した核移植単位細胞、例えば卵母細胞または胚性幹細胞と融合することができ、その細胞は全能性であり、そして機能的新生仔を形成することができる。十分に確立されている従来技術に従って融合を実施する。また卵母細胞の除核および核移植を注入ピペットを用いる顕微手術により実施することもできる。(例えばWakayamaら、Nature 394:369(1998)参照。)得られた卵細胞を次いで適切な培地中で培養し、そして同期化したレシピエントに移入してトランスジェニック動物を作成する。これに代えて選択された遺伝的に修飾された細胞を発達中の胚に注射することができ、それを続いてキメラ動物に発達させる。
【0110】
さらに本発明によれば、その1つが内因性Ig遺伝子セグメントのフランキング配列に相同である5’および3’フランキング配列に連結されたヒトIg遺伝子セグメントを担持する本明細書にて前記した1つまたはそれより多い組換えベクターを(1つまたは複数の)レシピエント細胞に導入し、次いで相同組換えにより内因性Ig遺伝子セグメントがヒトIg遺伝子セグメントにより置き換えられている細胞を選択し、そして選択された遺伝的に修飾された(1つまたは複数の)レシピエント細胞から動物を誘導することにより、ヒト(化)免疫グロブリンを生成できるトランスジェニック動物を作ることもできる。
【0111】
遺伝子導入ベクターの標的挿入に類似して、この研究法においてレシピエント細胞として使用するのに適切な細胞には胚性幹細胞または既に分化した体細胞が含まれる。任意の実行可能な手段、例えばトランスフェクションによりヒトIg遺伝子セグメントを担持する組換えベクターをかかるレシピエント細胞に導入することができる。その後相同組換えによりヒトIg遺伝子セグメントが対応する内因性Ig遺伝子セグメントと置き換わっている細胞を標準的な方法により選択することができる。これらの遺伝的に修飾された細胞をトランスジェニック動物のクローニングのための核移植手順における核ドナー細胞として提供することができる。これに代えて選択された遺伝的に修飾された胚性幹細胞を発達中の胚に注射することができ、それを続いてキメラ動物に発達させることができる。
【0112】
具体的な実施態様では、胚期の間に導入遺伝子を胚に直接注射することにより、またはそれらを妊娠母体もしくは産卵するメンドリに間接的に注射することにより本発明の導入遺伝子構築物をトランスジェニック動物に導入することができる。結果的に動物の免疫グロブリン分子を発現する内因性B細胞は枯渇し得て、そして故にトランスジェニック子孫が抗原での免疫に応答してヒト(化)抗体を優勢に生成する。
【0113】
前記の方法のいずれかにより生成されたトランスジェニック動物は本発明の別の実施態様を形成する。トランスジェニック動物は少なくとも1つの、すなわち1つまたはそれより多い、そこからヒト(化)抗体の機能的レパートリーが生成されるヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有する。
【0114】
具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有するトランスジェニックウサギを提供する。本発明のトランスジェニックウサギはヒト(化)Ig遺伝子座を再構成および遺伝子変換し、そしてヒト(化)抗体の機能的レパートリーを発現することができる。
【0115】
別の具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有するトランスジェニックニワトリを提供する。本発明のトランスジェニックニワトリはヒト(化)Ig遺伝子座を再構成および遺伝子変換し、そしてヒト(化)抗体の機能的レパートリーを発現することができる。別の具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)V領域および自殺遺伝子をゲノムに有するトランスジェニックマウスを提供する。ヒト(化)V領域はヒト以外のスペーサー配列によりフランキングされた少なくとも2つのヒトV遺伝子セグメントを含む。トランスジェニックマウスはヒトVエレメントを再構成し、そして抗体の機能的レパートリーを発現することができる。
【0116】
抗原での免疫が該トランスジェニック動物における同一抗原に対するヒト(化)抗体の生成を招く。
【0117】
本発明の好ましい実施態様は、内因性B細胞を枯渇させるためにヒト(化)Ig遺伝子座および少なくとも1つの自殺遺伝子を有し、そしてヒト(化)ポリクローナル抗血清を生成するトランスジェニック動物に向けられているが、霊長類化Ig遺伝子座および霊長類化ポリクローナル抗血清を有するトランスジェニック動物もまた本発明の精神内にあることは理解されよう。ヒト(化)ポリクローナル抗血清組成物に類似して霊長類化ポリクローナル抗血清組成物は、ヒト個体における免疫原性が低下している可能性がある。
【0118】
一度多様化されたヒト(化)免疫グロブリン分子を生成することができるヒト以外のトランスジェニック動物が作られると(さらに後記で説明するように)、抗原で動物を免疫することによりヒト(化)免疫グロブリンおよび抗原に対するヒト(化)抗体調製物を容易に得ることができる。種々の抗原を用いてトランスジェニック宿主動物を免疫することができる。かかる抗原には、微生物、例えばウイルスおよび単細胞生物(例えば細菌および菌類)、生存、弱毒化もしくは死亡した、微生物のフラグメント、または微生物から単離された抗原分子が含まれる。
【0119】
動物を免疫するのに用いるための好ましい細菌性抗原には、莢膜多糖類5および8型のような黄色ブドウ球菌からの精製抗原、アルファ毒素のような毒性因子の組換え体、アドヘシン結合タンパク質、コラーゲン結合タンパク質、およびフィブロネクチン結合タンパク質が含まれる。好ましい細菌性抗原にはまた黄色ブドウ球菌、緑膿菌、腸球菌、エンテロバクターおよび肺炎桿菌の弱毒体、またはこれらの細菌細胞からの培養上澄みも含まれる。免疫に用いることができるその他の細菌性抗原には精製リポ多糖類(LPS)、莢膜抗原、莢膜多糖類および/または外膜タンパク質の組換え体、フィブロネクチン結合タンパク質、内毒素、ならびに緑膿菌、腸球菌、エンテロバクターおよび肺炎桿菌の外毒素が含まれる。
【0120】
真菌に対する抗体の作成のための好ましい抗原には、真菌の弱毒体またはその外膜タンパク質が含まれ、その真菌には、限定するものではないがカンジダ・アルビカンス、カンジダ・パラプシローシス、カンジダ・トロピカリスおよびクリプトコッカス・ネオホルマンスが含まれる。
【0121】
ウイルスに対する抗体を作成するために免疫で用いるための好ましい抗原には、エンベロープタンパク質およびウイルスの弱毒体が含まれ、そのウイルスには、限定するものではないが呼吸器多核体ウイルス(RSV)(特にFプロテイン)、C型肝炎ウイルス(HCV)、B型肝炎ウイルス(HBV)、サイトメガロウイルス(CMV)、EBVおよびHSVが含まれる。
【0122】
単離された腫瘍細胞または腫瘍細胞系を有するトランスジェニック動物を免疫することにより癌の処置のための治療用抗体を作成することができ、腫瘍関連抗原には、限定するものではないがHer−2−neu抗原(それに対する抗体は乳癌の処置に有用である);CD19、CD20、CD22およびCD53抗原(それに対する抗体はB細胞リンパ腫の処置に有用である)、(3)前立腺特異的膜抗原(PMSA)(それに対する抗体は前立腺癌の処置に有用である)、および17−1A分子(それに対する抗体は結腸癌の処置に有用である)が含まれる。
【0123】
任意の都合のよい様式で、補助剤を伴うかまたは伴わずに抗原をトランスジェニック宿主動物に投与することができ、そして予め決定されたスケジュールに従って投与することができる。
【0124】
免疫後、抗原に特異的な医薬品等級のポリクローナル抗体の精製のために免疫したトランスジェニック動物からの血清または乳を分画することができる。トランスジェニック鳥類の場合、卵黄を分画することにより抗体を作ることもできる。クロマトグラフィー(親和性、イオン交換、ゲルろ過等)、硫酸アンモニウムのような塩、エタノールのような有機溶媒またはポリエチレングリコールのようなポリマーを用いる選択的沈殿により、濃縮され、精製された免疫グロブリン分画を得ることができる。
【0125】
分画されたヒト(化)抗体をヒトでの静脈内投与に適当な無毒性のパイロジェン不含培地、例えば滅菌緩衝食塩水に溶解または希釈することができる。
【0126】
投与に用いられる抗体調製物は一般に、免疫グロブリン濃度が0.1から100mg/ml、さらに通常的には1から10mg/mlであることを特徴とする。抗体調製物は種々のアイソタイプの免疫グロブリンを含有できる。これに代えて、抗体調製物はただ1つのアイソタイプ、または多くの選択されたアイソタイプの抗体を含有することができる。
【0127】
ヒト(化)モノクローナル抗体を作るために、その動物の内因性免疫グロブリンを発現するB細胞が枯渇している、免疫トランスジェニック動物から脾臓細胞を単離する。単離した脾臓細胞を、ハイブリドーマの生成のために形質転換された細胞系との細胞融合に用いるか、または抗体コード化cDNAを標準的な分子生物学技術によりクローン化し、そしてトランスフェクトされた細胞中で発現させる。モノクローナル抗体を作るための手順は当分野において十分に確立されている。例えば欧州特許出願第0583980Al号(「Method For Generating Monoclonal Antibodies From Rabbits」)、米国特許第4977081号(「Stable Rabbit−Mouse Hybridomas And Secretion Products Thereof」)、国際公開公報第97/16537号(「Stable Chicken B−cell Line And Method of Use Thereof」)、および欧州特許第0491057B1号(「Hybridoma Which Produces Avian Specific Immunoglobulin G」)(それらの開示は出典明示により本明細書の一部とする)参照。クローン化されたcDNAからのモノクローナル抗体のインビトロ生成はAndris−Widhopfら、「Methods for the generation of chicken monoclonal antibody fragments by phage display」、J lmmunol Methods 242:159(2000)およびBurton, D. R.、「Phage display」、Immunotechnology 1:87(1995)(それらの開示は出典明示により本明細書の一部とする)により記載されている。
【0128】
たいていの場合、抗体調製物は未修飾免疫グロブリン、すなわち例えば化学物質または酵素によりさらなる修飾を加えていない動物から調製したヒト(化)抗体からなる。これに代えて免疫グロブリン分画を酵素消化(例えばペプシン、パパイン、プラスミン、グリコシダーゼ、ヌクレアーゼ等で)、加熱等のような処理に供する、および/またはさらに分画することができる。
【0129】
本発明の好ましい実施態様はヒト化抗体を生成するヒト以外のトランスジェニック動物における内因性免疫グロブリン生成の抑制のための方法に向けられており、それにより望ましいヒト(化)免疫グロブリンの富化が可能になる。1つの実施態様では、当分野において公知の方法を用いてヒト(化)免疫グロブリン遺伝子を含む導入遺伝子、自己切断ペプチドおよびリコンビナーゼをトランスジェニック動物に導入し、外因性B細胞と称されるB細胞における付随するヒト(化)免疫グロブリンおよびリコンビナーゼ遺伝子の発現を確実にする。本明細書に記載した、または当分野において周知の任意の種々のリコンビナーゼ、自己切断ペプチドまたは免疫グロブリン遺伝子を導入遺伝子で用いることができる。さらにこの実施態様では、内因性免疫グロブリンを発現するB細胞における自殺遺伝子の選択的発現により内因性免疫グロブリン生成の抑制が達成され、そしてしたがって細胞死により枯渇する。対応して、導入遺伝子の発現によりリコンビナーゼ媒介機構を介して自殺遺伝子は外因性B細胞のゲノムから切除される。したがって外因性B細胞は生存し、そしてヒト(化)免疫グロブリンをコードする導入遺伝子を産生的に生成する。以前に記載した、および当分野において公知の様々な自殺遺伝子は本発明の実施態様である。この実施態様の1つの態様では、導入遺伝子を介して自殺遺伝子をトランスジェニック動物のゲノムに導入し、そしてその発現を免疫細胞特異的プロモーターにより、好ましくはB細胞特異的プロモーターにより駆動してB細胞において自殺遺伝子を選択的に発現させ、したがって非B細胞集団の不必要な細胞死を防御する。本明細書にて記載した、または当分野において周知の種々の免疫細胞およびB細胞特異的プロモーターを用いてB細胞において自殺遺伝子を選択的に発現させることができる。別の実施態様では、本発明で用いるトランスジェニック動物は遺伝子変換動物であるか、または若年期に停止する遺伝子再構成により抗体多様化を被ることができる。さらに前記した方法を用いて作成した遺伝子導入ベクターおよびトランスジェニック動物もまた本発明の実施態様である。
【0130】
本発明をさらに説明するが、決して以下の実施例により限定されるものではない。
【実施例】
【0131】
実施例1
loxPが導入されたDT−A発現ベクターの構築
DT−A自殺遺伝子のB細胞特異的発現を達成するために、ウサギカッパ1をコードするBACクローン179L1(ジェンバンク受け入れ番号AY495827)を修飾する。V1の下流のスペーサーからJ遺伝子座を通るカッパ1遺伝子座、イントロンエンハンサー、定常領域をコードするエクソン、3’エンハンサーの下流の配列に対する3’エンハンサーを含むBACクローンの46kbフラグメントをETクローニングによりサブクローニングする。さらなるゲンタマイシン選択カセットを有するpBELOBACベクター骨格をBAC179L1に相同な50bpを有するプライマーでPCR増幅する。加えて順方向プライマーはAttBインテグラーゼ認識部位およびPvuI制限酵素認識部位を有する。加えて逆方向プライマーはPvuI制限酵素認識部位を有する。
【0132】
【化1】
【0133】
【化2】
ETクローニングのために、PCR生成物をBAC179L1ならびに誘導ラムダファージ組換え酵素Redα、Redβおよびγを含有するストレプトマイシン抵抗性大腸菌株に形質転換する。これらの組換えタンパク質を同時トランスフェクトしたプラスミド(プラスミドpSC101−γβαを有するDH10B大腸菌)またはゲノムに組み込まれたラムダプロファージ(DY380大腸菌株)のいずれかから発現させる。ゲンタマイシンを用いて陽性クローン179L1(46kb)を選択し、そして制限酵素消化により確認する。
【0134】
5’から3’の2つのSV40ポリA部位−loxP部位−ウサギカッパ1プロモーター−DT−A−FRT部位−loxP部位−2つのSV40ポリA部位からなるDNAフラグメントを化学的に合成する。FLP媒介の組み込みによりカナマイシン選択カセットをFRT部位に導入する。サブクローニングされた179L1の46kbフラグメント(46kb)に相同な50bpを有するプライマーで合成フラグメントをPCR増幅する。ETクローニングによりJ遺伝子座を合成DT−Aフラグメントに対して変化させる。カナマイシンを用いて陽性クローンを選択し、そして制限酵素分析により確認する。
【0135】
再度FLP媒介組換えによりカナマイシン選択カセットを除去する。カナマイシン抵抗性の喪失により陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより確認する。
【0136】
最終構築物はV1の下流のスペーサー、合成DT−Aフラグメント、イントロンエンハンサー、定常領域をコードするエクソン、3’エンハンサーの下流の配列に対する3’エンハンサーを含む。
【0137】
構築物をトランスジェニック動物の作成に用いる。
【0138】
実施例2
IgMの膜形態、2A自己切断ペプチド、Creリコンビナーゼからなる融合タンパク質をコードするヒト(化)重鎖遺伝子座の構築
定常、可変、および連結遺伝子セグメントまたは3’エンハンサー領域に特異的なプローブを用いてゲノムDNAライブラリーからウサギ免疫グロブリン重鎖遺伝子座配列を含有するBACおよびフォスミドクローンを単離した。単離したBAC27N5(ジェンバンク受け入れ番号AY386696)、219D23(ジェンバンク受け入れ番号AY386695)、225P18(ジェンバンク受け入れ番号AY386697)、38A2(ジェンバンク受け入れ番号AY386694)およびフォスミドFos15B(ジェンバンク受け入れ番号AY3866968)をシークエンシングした(Rosら、Gene 330:49−59)。
【0139】
ETクローニングによる大腸菌における相同組換えにより選択した免疫グロブリンコード化配列を対応するヒト対応物と交換した(E−Chiang Leeら、Genomics 73:56−65;Daiguan Yuら、PNAS 97:5978−5983;Muyrersら、Nucleic Acids Research 27:1555−1557;Zhangら、Nature Biotechnology 18:1314−1317)。
【0140】
これに代えて、インビトロでライゲーションおよび続く大腸菌の形質転換によりDNAフラグメントを組換えした。インビトロライゲーションおよび形質転換、ETクローニングにより、またはCreリコンビナーゼ媒介組み込みによりBACおよび/またはFosl5Bまたはその部分を組み合わせた。
【0141】
ETクローニングのために標的配列を含有するベクターを、誘導ラムダファージ組換え酵素Redα、Redβおよびγを含有するストレプトマイシン抵抗性大腸菌株に形質転換した。同時トランスフェクトしたプラスミド(プラスミドpSC101−γβαを有するDH10B大腸菌)またはゲノムに組み込まれたラムダプロファージ(DY380大腸菌株)のいずれかからこれらの組換えタンパク質を発現させた。ETクローニング手順は2つの相同組換え工程を包含した。
【0142】
第1の工程では、標的遺伝子座を選択−対抗選択カセット(例えばネオマイシンに対する抵抗性(neo)およびストレプトマイシンに対する感受性(rpsL)を付与するneo−rpsL)により置き換えた。neo−抵抗性コロニーを単離した後、相同組換えによる選択カセットの挿入を制限酵素分析および部分的シークエンシングにより確認した。
【0143】
第2の工程では、rpsL−neo選択カセットを新しい配列と交換した。制限分析およびシークエンシングによりストレプトマイシン抵抗性クローンを分析した。ETクローニング手順に用いたフラグメントは20から50bp長のフランキング配列を有し、それは標的配列と同一であった。ライゲーションに用いた配列はその3’および5’末端で適切な制限酵素部位を有した。これらの部位は天然発生部位であるか、または適切な部位を含有するプライマーを用いるPCRにより導入されたかのいずれかであった。
【0144】
これに代えて配列を合成により作成した。
【0145】
ETクローニングによりウサギJH、BAC219D23のCμおよびBAC27N5のCγの対応するそのヒト対応物との置き換えによりヒト化重鎖を構築した。ETクローニング手順に用いたヒト配列をヒトゲノムDNAからPCRにより増幅した。
【0146】
ヒトCμ、CγおよびJH遺伝子セグメントをウサギ標的配列に相同な50bpを有するプライマーを用いて増幅した。
【0147】
【化3】
BACクローン225P18のクローン219D23との、およびBAC27N5のフォスミド15Bとのライゲーションの後、ライゲートした構築物を大腸菌に形質転換し、そしてCreリコンビナーゼ媒介挿入によりつないだ。これにより18ウサギ可変遺伝子、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCε、ウサギCα4および3’エンハンサーエレメントからなる機能的遺伝子座に至った。
【0148】
自己切断F2Aペプチドの配列およびCREリコンビナーゼのコドン最適化コード化配列(iCRE)を含むDNAフラグメントを化学的に合成する。
【0149】
【化4】
ETクローニングによりIgMのM2膜エクソンを合成M2−F2A−iCREフラグメントに対して変化させる。
【0150】
ETクローニング手順は2つの相同組換え工程を包含した。第1の工程では、標的遺伝子座を選択−対抗選択カセット(例えばネオマイシンに対する抵抗性(neo)およびストレプトマイシンに対する感受性(rpsL)を付与するneo−rpsL)により置き換える。このためにrpsL−neo選択カセットを標的配列に相同な50bpを有するプライマーを用いてPCR増幅する。neo−抵抗性コロニーを単離した後、相同組換えによる選択カセットの挿入を制限酵素分析および部分的シークエンシングにより確認する。
【0151】
第2の工程では、rpsL−neo選択カセットを合成M2−F2A−iCREフラグメントと交換する。ストレプトマイシン抵抗性により陽性クローンを同定し、そして制限分析およびシークエンシングにより分析する。
【0152】
得られたBACをトランスジェニック動物作成に使用する。
【0153】
実施例3
ヒト化軽鎖遺伝子座の構築
ウサギゲノムBACライブラリーのスクリーニングによりウサギ軽鎖K1遺伝子セグメント(ジェンバンク受け入れ番号AY495827、AY495826)を含有する2つのBAC(179L1および215M22)の同定に至った。
【0154】
前記したようにETクローニングによりウサギCκ1をヒトCκアロタイプKm3と交換した。
【0155】
ヒトCκ(アロタイプKm3)を標的配列に相同な50bpフランキング配列を有するプライマーを用いてPCRにより増幅した。
【0156】
【化5】
ホモロジーアームはウサギ生殖細胞系カッパ(b5;ジェンバンク受け入れ番号K01363)の公開されている配列に基づいて設計され、そしてCκのイントロン−エクソン境界に適合した。
【0157】
BAC179L1におけるヒトCκに対するウサギCκの交換をシークエンシングにより確認した。
【0158】
BAC179L1−huCkを2つのETクローニングにより修飾した。BAC179L1に相同な50bpを有するプライマーを用いてネオマイシン選択カセットを増幅した。加えて順方向プライマーはi−CeuIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。ネオマイシンで陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。BAC179L1に相同な50bp配列を含有するプライマーを用いてゼオシン選択カセットを増幅した。加えて順方向プライマーはi−SceIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。ゼオシンで陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0159】
BAC215M22をETクローニングにより修飾した。BAC215M22に相同な50bpを有するプライマーを用いてゲンタマイシン抵抗性遺伝子を増幅した。加えて順方向プライマーはi−CeuIメガヌクレアーゼ部位を有し、そして逆方向プライマーはi−SceIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。得られたクローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0160】
【化6】
【0161】
【化7】
修飾されたBAC179L1および225M22をi−CeuIおよびi−SceIで切断した。98kbおよび132kbのフラグメントを精製し、そしてライゲートした。得られたクローンをカナマイシンおよびクロラムフェニコールで選択し、そして制限酵素消化、i−SceIおよびi−CeuI制限部位を含有する領域のPCR、ならびにシークエンシングにより正確性を確かめた。得られたBACを179−215−huCkと称した。
【0162】
ETクローニングによりBAC179−215−huCkのウサギJk1およびJk2を合成ヒト再構成カッパ1VJ遺伝子により置き換えた。ウサギプロモーター、ウサギリーダー、ウサギイントロンおよびヒトVJ遺伝子を有するDNAフラグメントを化学的に合成した。合成ヒトVJのコドン使用を最適化してウサギVカッパ遺伝子に対して最高のDNA配列相同性を達成した。
【0163】
BAC179L1に相同な50bpを有する順方向プライマーならびにゲンタマイシン抵抗性遺伝子およびFRT部位に相同な逆方向プライマーを用いて合成ヒトVJをPCR増幅した。FRT部位を有する順方向プライマーならびにBAC179L1およびFRT部位に相同な50bpを有する逆方向プライマーを用いてゲンタマイシン抵抗性遺伝子を増幅した。合成ヒトVJ遺伝子のための順方向プライマーおよびゲンタマイシン抵抗性遺伝子のための逆方向プライマーを用いてオーバーラップ伸長PCRによりヒト合成ヒトVJおよびゲンタマイシン抵抗性遺伝子を組み合わせた。得られたフラグメントをETクローニングに使用した。陽性クローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0164】
【化8】
部位特異的組換えによりFlpリコンビナーゼの発現を介してゲンタマイシン抵抗性遺伝子を除去した。組換えの後、1つのFRTが残った。ETクローニングによりFRT部位を削除した。合成ヒトVJからの232bpフラグメントをPCRにより増幅し、そしてETクローニングに使用した。得られたコロニーをFRT部位の喪失に関してPCRによりスクリーニングし、そしてシークエンシングにより確認した。
【0165】
ETクローニングによりBAC179−215−huCkのネオマイシン抵抗性遺伝子を置き換えた。BAC179−215−huCkに相同な50bpを有するプライマーを用いてゲンタマイシン抵抗性(pRep−Genta;Genebridges)遺伝子をPCRにより増幅した。加えて順方向プライマーはloxP部位、attB部位およびPvuI制限部位を有した。得られたクローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0166】
得られたBACをトランスジェニック動物の作成に使用した。
【0167】
【化9】
実施例4
ヒト化重鎖免疫グロブリンを発現するトランスジェニックマウスおよびウサギの作成
カッパ軽鎖プロモーター/エンハンサー配列の制御下でヒト化重および軽鎖免疫グロブリン遺伝子座およびloxPが導入されたジフテリア毒素A遺伝子を含有するトランスジェニックウサギおよびマウスを、受精卵母細胞の前核へのDNAの注射および、続く里親への胚の移入により作成する。トランスジェニック創始動物をPCRにより同定する。ヒト(化)免疫グロブリンMおよびGの発現をELISAにより測定する。ヒト化IgGの発現は1〜5mg/mlである。マウスおよびウサギIgGの発現は各々1〜5μg/mlである。
【0168】
実施例5
ヒト化重鎖免疫グロブリンを発現するトランスジェニックニワトリの作成
精巣媒介の遺伝子移入によりトランスジェニックニワトリを作成する。DNA構築物を
(50μg)を0.9%NaCl 500μl中リポフェクション試薬(スーパーフェクト)250μlと混合し、そしてオンドリの精巣に注射する。3〜4週後、トランスジェニック精子を有するオンドリをPCR分析により同定し、そしてメンドリと交配させる。トランスジェニック子孫をPCRにより同定する。ヒト化IgGの発現は1〜5mg/mlである。ニワトリIgYの発現は各々1〜5μg/mlである。
【0169】
本開示全体にわたって引用した全ての参照文献はそこに引用された参照文献と一緒に出典明示により本明細書の一部とする。
【0170】
本発明を特定の実施態様を参照することにより説明したが、そのように限定されるものではない。種々の修飾を容易に利用でき、そして本発明が取り組む方式を実質的に変化させることなく実施できることは当業者には理解されよう。かかる修飾の全てが、具体的には本明細書にて特許請求される本発明の範囲内であると意図される。
【図面の簡単な説明】
【0171】
【図1】外因性免疫グロブリン(Ig)発現B細胞および内因性免疫グロブリン(Ig)発現B細胞で生じる事象の説明図である。2つの導入遺伝子(Ig重鎖遺伝子座およびDTA)を有する細胞では、遺伝子導入免疫グロブリン重鎖遺伝子座の産生性再構成の結果、重鎖(HC)およびCreリコンビナーゼの発現に至る。続いて、Cre媒介の組換えの結果、DTA発現カセットのループアウトに至る。したがって、DTAは発現されず、そして遺伝子導入免疫グロブリンHC遺伝子座を発現するかかる外因性B細胞は生存する。内因性HC遺伝子座の産生性再構成の結果、Creリコンビナーゼの発現には至らない。したがって、DTA発現カセットは活性化され、そしてかかる内因性B細胞は死亡する。
【技術分野】
【0001】
発明の分野
本発明は内因性免疫グロブリンを発現するB細胞では自殺遺伝子を選択的に発現するが、ヒトまたはヒト化免疫グロブリンまたは免疫グロブリン鎖のような外因性免疫グロブリンまたは免疫グロブリン鎖を発現するB細胞では発現しないことにより、ヒト以外のトランスジェニック動物における内因性免疫グロブリン発現を抑制する方法に関する。この方法により例えばヒト以外のトランスジェニック動物の血液、乳および卵中のヒトまたはヒト化抗体の優勢な発現が可能になる。
【背景技術】
【0002】
発明の背景
ヒトマウスキメラ抗体を発現するマウスの作製は非特許文献1に記載されている。ヒト免疫グロブリンポリペプチドを発現するマウスの作製は非特許文献2;非特許文献3;および非特許文献4により記載されており、ならびにBACクローンを用いるトランスジェニックマウスの作製は非特許文献5に記載されている。ヒト抗体を発現するウシの作製は非特許文献6に記載されている。
【0003】
動物における遺伝子導入は非特許文献7に記載されている。トランスジェニックウサギの作製は非特許文献8;および非特許文献9に記載されている。トランスジェニックニワトリの作製は非特許文献10;および非特許文献11;および非特許文献12に記載されている。
【0004】
免疫グロブリン発現の損傷を有するウサギは非特許文献13;ならびに非特許文献14に記載されている。ガンマグロブリン血症のニワトリは非特許文献15;および非特許文献16に記載されている。
【0005】
細胞からの動物のクローニングは非特許文献17;非特許文献18;非特許文献19;非特許文献20;および非特許文献21に記載されている。ウサギの核移植クローニングは非特許文献22、非特許文献23に記載されている。
【0006】
ヒト(化)免疫グロブリン導入遺伝子座を発現するヒト以外のトランスジェニック動物の生成およびかかるトランスジェニック動物からの抗体の生成は特許文献1、特許文献2ならびに特許文献3、特許文献4;および特許文献5に詳細に記載されている。キメラ哺乳動物宿主の相同組換えは特許文献6に例示されている。DNAを胚に導入するための方法は特許文献7に記載されている。胚性幹細胞の維持および増殖は特許文献8に記載されている。
【0007】
毒素基盤の研究法を用いる自殺遺伝子は非特許文献24;非特許文献25;非特許文献26;非特許文献27;非特許文献28;非特許文献29;非特許文献30;非特許文献31;非特許文献32;非特許文献33に記載されている。無毒性プロドラッグ酵素研究法を用いる自殺遺伝子は非特許文献34に記載されている。
【0008】
2Aペプチド配列を含有するウイルス性タンパク質の切断活性は非特許文献35、非特許文献36、非特許文献37、非特許文献38、非特許文献39に記載されている。
【0009】
リコンビナーゼおよびその特性は非特許文献40に記載されている。2つの核酸の非常に特異的な配列間の相同組換えを認識および触媒する部位特異的リコンビナーゼが公知である。例えば部位特異的リコンビナーゼのインテグラーゼファミリーに属するφC31およびR4が公知である(非特許文献41;非特許文献42)。ヒトおよびマウスゲノムを含むいくつかのゲノムでは偽attP部位が天然にある(非特許文献43)。8.9kbの大型のVIIコラーゲンcDNAの一次前駆患者皮膚細胞へのインビトロでのφC31インテグラーゼ媒介遺伝子移入は非特許文献44に報告されている。トランスジェニック哺乳動物の操作におけるφC31、TP901−1およびR4ファージインテグラーゼの使用は非特許文献45で実証されている。
【0010】
B細胞を含む細胞のアブレーションは非特許文献46、非特許文献47および非特許文献48に記載されている。
【0011】
【特許文献1】国際公開第92/03918号パンフレット
【特許文献2】国際公開第02/12437号パンフレット
【特許文献3】米国特許第5545807号明細書
【特許文献4】米国特許第5814318号明細書
【特許文献5】米国特許第5570429号明細書
【特許文献6】米国特許第5416260号明細書
【特許文献7】米国特許第5567607号明細書
【特許文献8】米国特許第5453357号明細書
【非特許文献1】Pluschkeら、Journal of Immunological Methods 1998年、215:27−37
【非特許文献2】Neubergerら、Nature 1989年、338:350−2
【非特許文献3】Lonbergら、Int.Rev.Immunol.1995年、13(1):65−93
【非特許文献4】Bruggemannら、Curr.Opin.Biotechnol.1997年、8(4):455−8
【非特許文献5】Yangら、Nat.Biotechnol.1997年、15:859−65
【非特許文献6】Kuroiwaら、Nature Biotech 2002年、20(9):889−894
【非特許文献7】Wall RJ、Theriogenology 2002年、57(1):189−201
【非特許文献8】Fan,J.ら、Pathol Int.1999年、49:583−94
【非特許文献9】Bremら、Mol.Reprod.Dev.1996年、44:56−62
【非特許文献10】Etchesら、Methods in Molecular Biology 1997年、62:433−450
【非特許文献11】Painら、Cells Tissues Organs 1999年、165(3−4):212−9
【非特許文献12】Shermanら、Nature Biotech 1998年、16:1050−1053
【非特許文献13】Chenら、J.Immunol.1993年、150:2783−2793
【非特許文献14】Lamoyi EおよびMage RG.、J.Exp.Med.1985年、162:1149−1160
【非特許文献15】Frommelら、J.Immunol.1970年、105(l):1−6
【非特許文献16】Benedictら、Adv.Exp.Med.Biol.1977年、88(2):197−205
【非特許文献17】T.Wakayamaら、Nature 1998年、394:369−374
【非特許文献18】J.B.Cibelliら、Science 1998年、280:1256−1258
【非特許文献19】J.B.Cibelliら、Nature Biotechnology 1998年、16:642−646
【非特許文献20】A.E.Schniekeら、Science 1997年、278:2130−2133
【非特許文献21】K.H.Campbellら、Nature 1996年、380:64−66
【非特許文献22】Sticeら、Biology of Reproduction 1988年、39:657−664
【非特許文献23】Challah−Jacquesら、Cloning and Stem Cells 2003年、8(4):295−299
【非特許文献24】Leongら、Science 1983年、220:515−7
【非特許文献25】Maxwellら、Cancer Research 1986年、46:4660−4664
【非特許文献26】Palmiterら、Cell 1987年、50:435−443
【非特許文献27】Maxwellら、Cell 1991年、51:4299−4304
【非特許文献28】Maxwellら、Leukemia and Lymphoma 1992年、7:457−462
【非特許文献29】Aguilaら、Proc.Natl.Acad.Sci.1995年、92:10192−10196
【非特許文献30】Grieshammerら、Developmental Biology 1998年、197:234−247
【非特許文献31】Bartellら、Biology of Reproduction 2000年、63:409−416
【非特許文献32】Erlandssonら、J.Exp.Med.2001年、194:557−570
【非特許文献33】Leeら、Human Gene Therapy 2002年、13:533−542
【非特許文献34】Methods in Molecular Medicine:Suicide Gene Therapy, Methods and Reviews、Caroline J Springer編、Human Press、2004年
【非特許文献35】Palmenbergら、Virology 1992年、190:754−762
【非特許文献36】Ryanら、J Gen Virol 72:2727−2732(1991)
【非特許文献37】Donnellyら、J Gen Virol 2001年、82:1027−1041
【非特許文献38】Donnellyら、J Gen Virol 2001年、82:1013−1025
【非特許文献39】Szymaczakら、Nature Biotech 2004年、22(5):589−594
【非特許文献40】Kolb A.F.、Cloning Stem Cells 2002年、41:65−80
【非特許文献41】Grothら、Proc.Natl.Acad.Sci.2000年、97:5995−6000
【非特許文献42】Olivaresら、Nature Biotechnol.2002年、20(11):1124−8
【非特許文献43】Thyagarajanら、Mol.and Cell.Biol.2001年、21:3926−3934
【非特許文献44】Ortiz−Urdaら、Nature Medicine 8:1166−1170(2002)
【非特許文献45】Hollisら、Repro.Biol.and Endocrinol.2003年、1:79
【非特許文献46】Erlandssonら、J Exp Med 194(5):557−570(2001)
【非特許文献47】Maxwellら、Cancer Research 1991年、51:4299−4304
【非特許文献48】Palmiterら、Cell 1987年、50:435−443
【発明の開示】
【課題を解決するための手段】
【0012】
発明の要旨
本発明はトランスジェニック動物における内因性免疫グロブリン生成を抑制するための方法に関する。方法は内因性免疫グロブリンを発現するB細胞において自殺遺伝子を選択的に発現するが、ヒトまたはヒト化免疫グロブリンを発現するB細胞では自殺遺伝子を発現しないことを伴う。とりわけ本発明は1つまたはいくつかのヒトまたはヒト(化)免疫グロブリン導入遺伝子座を含有するヒト以外のトランスジェニック動物において内因性免疫グロブリン遺伝子座の発現を抑制するための方法に関係する。結果的にヒト(化)導入遺伝子座はヒト以外のトランスジェニック動物において遺伝子再構成および変異過程を被り、実質的に内因性免疫グロブリン生成の不在下で多様化されたヒト(化)抗体レパートリーを生成することができる。
【0013】
とりわけ本発明はヒト以外のトランスジェニック動物の内因性免疫グロブリンを生成するB細胞では少なくとも1つの自殺遺伝子を選択的に発現するが、外因性免疫グロブリンを生成するB細胞では発現しないことを含み、それにより内因性免疫グロブリンを生成するB細胞は枯渇し、そして内因性免疫グロブリンの生成は抑制されるが、外因性免疫グロブリンの生成は抑制されない、外因性免疫グロブリン導入遺伝子座を担持するヒト以外のトランスジェニック動物のB細胞における内因性免疫グロブリン(Ig)生成の選択的抑制のための方法に関係する。好ましい実施態様では、外因性免疫グロブリンはヒト化免疫グロブリン重および/または軽鎖配列である。
【0014】
さらなる態様では、ヒト以外のトランスジェニック動物のB細胞に導入された自殺遺伝子はB細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼ配列によりフランキングされている。
【0015】
なお別のさらなる態様では、さらに該組換え配列を認識するリコンビナーゼをコードする発現構築物の一部としてヒト以外のトランスジェニック動物のB細胞にヒト(化)免疫グロブリン鎖導入遺伝子座を導入し、ここで自殺遺伝子の発現はヒト化免疫グロブリン導入遺伝子座を発現するB細胞におけるリコンビナーゼの発現により活性化される。
【0016】
特定の態様では、自殺遺伝子は細菌性、真菌性、昆虫性および植物毒素からなる群から選択される。好ましい実施態様では、自殺遺伝子はジフテリア毒素A鎖である。
【0017】
別の実施態様では、自殺遺伝子はプロドラッグ変換酵素である。この実施態様の1つの態様では、プロドラッグ変換酵素は非哺乳動物起源である。さらなる態様では非哺乳動物プロドラッグ変換酵素はウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、β−ラクタマーゼ、多剤活性化酵素(MDAE)、β−ガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)からなる群から選択される。
【0018】
さらに別の実施態様では、プロドラッグ変換酵素はヒト起源である。さらなる態様では、ヒトプロドラッグ変換酵素はデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)からなる群から選択される。
【0019】
別の態様では、リコンビナーゼCre、Cre様、F1p、φC31、λインテグラーゼ、ファージR4リコンビナーゼ、TP901−1リコンビナーゼ、原核細胞性トランスポーゼス、真核細胞性トランスポーゼス、ウイルス性レトロトランスポーゼス、ショウジョウバエコピア様レトロトランスポーゼスおよび非ウイルス性レトロトランスポーゼスからなる群から選択される。さらなる実施態様では、トランスポーゼスまたはレトロトランスポーゼスはTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される。
【0020】
1つの実施態様では、ヒト以外のトランスジェニック動物は若年期に遺伝子再構成により抗体多様化を実質的に停止する。さらなる実施態様では、ヒト以外のトランスジェニック動物は生まれて1か月以内に抗体多様化を実質的に停止する。
【0021】
別の実施態様では、ヒト以外のトランスジェニック動物はげっ歯類、ウサギ、鳥類、ウシ、ブタ、ヒツジ、ヤギおよびウマからなる群から選択される。好ましい実施態様では、げっ歯類はマウスまたはラットである。
【0022】
特定の態様では、本発明は、ヒトまたはヒト化免疫グロブリン重および/または軽鎖導入遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0023】
別の態様では、本発明は、B細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0024】
1つの実施態様では、本発明はヒトまたはヒト化免疫グロブリン重および/または軽鎖遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子、ならびにB細胞特異的プロモーターの制御下にあり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物に関係する。
【0025】
1つの実施態様では、前記した遺伝子導入発現構築物はCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4およびTP901−1リコンビナーゼからなる群から選択される部位特異的リコンビナーゼを含む。
【0026】
別の実施態様では、前記した遺伝子導入発現構築物は原核細胞性または真核細胞性のいずれかのトランスポーゼスであるリコンビナーゼを含む。
【0027】
さらに別の実施態様では、前記した遺伝子導入発現構築物はウイルス性、ショウジョウバエコピア様または非ウイルス性レトロトランスポーゼスのいずれかであるリコンビナーゼを含む。
【0028】
さらなる実施態様では、前記した遺伝子導入発現構築物はTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択されるリコンビナーゼを含む。
【0029】
特定の実施態様では、前記した遺伝子導入発現構築物はlox P部位、FRT部位、細菌性ゲノム組換え部位およびファージ組換え部位からなる群から選択される組換え部位を含む。
【0030】
さらなる実施態様では、細菌性ゲノム組換え部位はattBであり、そしてファージ組換え部位はattPまたは偽attPまたは偽attB部位である。
【0031】
特定の実施態様では、前記した遺伝子導入発現構築物はウイルス性2A/2Bまたは2A様/2B配列から得られる自己切断ペプチドを含む。
【0032】
さらなる実施態様では、ウイルスはピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーからなる群、またはC型ロタウイルスファミリーから選択される。さらに別の実施態様では、ウイルスは口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルスまたはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)からなる群から選択される。
【0033】
特定の実施態様では、前記した遺伝子導入発現構築物はCD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b、mb−1、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖および免疫グロブリンJ−鎖またはその修飾体からなる群から選択されるプロモーター/エンハンサーを用いてB細胞において特異的に発現される自殺遺伝子を含む。好ましい実施態様では、B細胞特異的プロモーター/エンハンサーはカッパ軽鎖遺伝子プロモーターまたはその修飾体である。
【0034】
1つの態様では本発明は、内因性免疫グロブリン生成が内因性免疫グロブリンを生成するB細胞における自殺遺伝子の選択的発現により抑制されるが、ヒト(化)免疫グロブリンを発現するB細胞は自殺遺伝子を発現せず、そしてしたがって増殖するヒト以外の動物を生成する方法に関係する。
【0035】
とりわけ本発明は前記した遺伝子導入発現構築物を発現するヒト以外のトランスジェニック動物に関係する。
【0036】
1つの実施態様では、ヒト以外のトランスジェニック動物は実質的に遺伝子変換により抗体多様性を生じる。
【0037】
全ての態様において、好ましいヒト以外の動物には、限定するものではないがげっ歯類(例えばマウス、ラット)、ウサギ、鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)、ウシ、ブタ、ヒツジ、ヤギ、ウマ、ロバおよびその他の家畜が含まれる。好ましい実施態様では、ヒト以外のトランスジェニック動物はマウスまたはラットのいずれかである。
【発明を実施するための最良の形態】
【0038】
発明の詳細な説明
定義
特記しない場合、本明細書で用いる技術的および科学的用語は本発明が属する分野の当業者により一般に理解されるのと同一の意味を有する。Singletonら、Dictionary of Microbiology and Molecular Biology、第2版、J.Wiley & Sons(ニューヨーク、ニューヨーク州、1994年)およびMarch、Advanced Organic Chemistry Reactions, Mechanisms and Structure、第4版、John Wiley & Sons(ニューヨーク、ニューヨーク州、1992年)は本出願で用いる多くの用語に対する一般的な手引きを当業者に提供する。
【0039】
当業者は本明細書に記載したものに類似するかまたは均等である多くの方法および材料を認識し、それを本発明の実施にあたり用いることができる。実際に本発明は記載した方法および材料に何ら限定されるものではない。本発明の目的のために以下の用語を後記で定義する。
【0040】
「B細胞」は免疫グロブリン遺伝子セグメントの再構成を被り、そしてそのライフサイクルのいくつかの段階で免疫グロブリン遺伝子を発現することができるB系統細胞として定義される。これらの細胞には、限定するものではないが初期プロB細胞、後期プロB細胞、大型プレB細胞、小型プレB細胞、未成熟B細胞、成熟B細胞、記憶B細胞、形質細胞等が含まれる。
【0041】
「抗体」(Ab)および「免疫グロブリン」(Ig)は同一の構造特性を有する糖タンパク質である。抗体は特異的抗原に対して結合特異性を呈するが、免疫グロブリンは抗体および抗原特異性を欠如するその他の抗体様分子の双方を含む。「抗体」なる用語は本明細書では最も広義で用いられ、そして具体的には、限定するものではないがモノクローナル抗体(モノクローナル抗体全長を含む)、ポリクローナル抗体、多重特異性抗体(例えば二重特異性抗体)およびそれが望ましい特異性を呈する限り抗体フラグメントに及ぶ。
【0042】
本明細書で用いる「Ig遺伝子セグメント」なる用語はIg分子の種々の部分をコードするDNAのセグメントを意味し、それは動物およびヒトの生殖細胞系列に存在し、そしてB細胞で一緒になって再構成されたIg遺伝子を形成する。したがって、本明細書で用いるIg遺伝子セグメントはV遺伝子セグメント、D遺伝子セグメント、J遺伝子セグメントおよびC領域遺伝子セグメントを含む。VDJまたはVJセグメントの機能的再構成の結果、免疫グロブリン重または軽鎖の発現に至る。
【0043】
本明細書で用いる「ヒトIg遺伝子導入遺伝子座または遺伝子座またはセグメント」なる用語はヒトIg遺伝子座またはそのセグメントの天然発生配列、ヒトIg遺伝子座またはそのセグメントの天然発生配列の変性形態、およびヒトIg遺伝子座またはそのセグメントの天然発生配列によりコードされるポリペプチドに実質的に同一のポリペプチド配列をコードする合成配列の双方を含む。この局面で、「実質的に」とはアミノ酸配列同一性の程度が少なくとも約85%〜95%、または少なくとも約90%〜95%、または少なくとも約95%、または少なくとも約98%であることを意味する。特定の好ましい実施態様では、ヒトIg遺伝子セグメントはヒトにおいて免疫グロブリン分子に非免疫原性にする。ここでは「ヒトまたはヒト化免疫グロブリン(Ig)重および/または軽鎖遺伝子座」または「ヒトまたはヒト化Ig遺伝子座」なる用語は互換的に用いられる。
【0044】
「ヒト抗体」および「ヒト免疫グロブリン」なる用語は完全なヒト配列を含む抗体および免疫グロブリン分子を意味するために用いられる。
【0045】
本明細書で用いる「ヒト化抗体」および「ヒト化免疫グロブリン」なる用語はヒト免疫グロブリンポリペプチド配列(またはヒト免疫グロブリン遺伝子セグメントによりコードされるポリペプチド配列)の少なくとも一部を含む免疫グロブリン分子を意味する。本発明のヒト化免疫グロブリン分子を、ヒト化免疫グロブリン分子を生成するように操作されたヒト以外のトランスジェニック動物から単離することができる。かかるヒト化免疫グロブリン分子は、動物から調製された、または動物に由来する細胞から調製された非ヒト化免疫グロブリン分子に相対して、霊長類、特にヒトに対して免疫原性が低い。ヒト化免疫グロブリンまたは抗体は遺伝子変換および遺伝子変換動物における体細胞超変異によりさらに多様化された免疫グロブリン(Ig)および抗体を含む。かかるヒト化Igまたは抗体はヒトにより天然に作成されたものではないので(ヒトは遺伝子変換によりその抗体レパートリーを多様化しないので)、それらは「ヒト」ではなく、そしてしかもヒト化Igまたは抗体はその構造でヒトIg配列を有するので、それらはヒトに対して免疫原性ではない。
【0046】
「導入遺伝子または導入遺伝子構築物」は、天然または動物または動物の細胞において通常見出されない合成タンパク質をコードする配列を有するDNAフラグメントである。本明細書で用いる「導入遺伝子構築物」なる用語は、構造的な「目的の遺伝子」および遺伝子移入を促進するその他の配列を含有するポリヌクレオチド分子を意味する。本発明は2つの導入遺伝子構築物:(1)ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼおよび(2)免疫細胞特異的自殺導入遺伝子構築物に言及する。
【0047】
「遺伝子導入発現構築物」は、ヒト以外のトランスジェニック動物の特異的な細胞内で一時的または細胞特異的または目的の導入遺伝子の発現強化のいずれかのために必要なその他の調節DNA配列と一緒に、本発明の1つまたはいくつかの導入遺伝子構築物をコードする配列を伴うDNAフラグメントを意味する。
【0048】
「ヒト(化)Ig遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子または導入遺伝子構築物」は、単一のmRNAに転写される導入遺伝子構築物を意味し、それは後記で論じる自己切断メカニズムのために2つのポリペプチド、主にヒト(化)免疫グロブリン鎖およびリコンビナーゼに翻訳される。
【0049】
本明細書で用いる「自己切断ペプチド」なる用語は、ペプチド配列自体の中の2つのアミノ酸残基の間に生じる切断活性を随伴するペプチド配列を意味する。例えば2A/2Bペプチドまたは2A/2B様ペプチドでは、切断は2Aペプチド上のグリシン残基と2Bペプチド上のプロリン残基との間で生じる。これは2A/2Bペプチドの2Aグリシン残基と2Bプロリン残基との間の正常なペプチド形成が損なわれている翻訳の間の「リボソームスキップ機構」によって生じ、2Bペプチドの残りの翻訳には影響することはない。かかるリボソームスキップ機構は当分野において周知であり、そして単一のメッセンジャーRNAによりコードされるいくつかのタンパク質の発現のためのいくつかのウイルスにより用いられることが解っている。
【0050】
本明細書で用いる「リコンビナーゼ」なる用語は、「組換え部位」と称される定義された部位間の部位特異的組換えを促進することができる酵素の群を意味し、ここで2つの組換え部位は単一の核酸分子内で物理的に分かれているか、または別個の核酸分子上にある。2つの定義された組換え部位の配列は同一である必要はない。リコンビナーゼの群の中には「インテグラーゼ」(例えばCre、Cre様、FLPおよびλインテグラーゼのような部位特異的リコンビナーゼ)および「リゾルベース/インベルターゼ」(例えばφC31インテグラーゼ、R4インテグラーゼおよびTP−901インテグラーゼ)を含むいくつかのサブファミリーがある。「リコンビナーゼ」なる用語にはまた、限定するものではないが原核細胞性または真核細胞性トランスポーゼス、ウイルス性またはショウジョウバエコピア様または哺乳動物レトロトランスポーゼスを含む非ウイルス性レトロトランスポーゼスも含まれる。原核細胞性トランスポーゼスの実例には、Tn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。真核細胞性トランスポーゼスにはショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAcおよびDsエレメント等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。レトロトランスポーゼスにはL1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノス等でコードされるものが含まれる。トランスポーゼスをMp、Spm、En、ドット、Mu、I転位性エレメントからなる群から選択することもできる。
【0051】
本明細書で用いる「野生型組換え部位」なる用語は通常インテグラーゼのようなリコンビナーゼにより用いられる組換え部位を意味する。
【0052】
「偽組換え部位」とは、部位がその野生型組換え部位の配列と同一の配列を有していなくても、リコンビナーゼが組換えを促進することができる部位を意味する。
【0053】
本明細書で用いる「自殺遺伝子または自殺導入遺伝子」なる用語は、発現が遺伝子を発現する細胞の死亡に至るタンパク質をコードする遺伝子を意味する。例えばタンパク質は毒素(例えばジフテリア毒素A鎖)または無毒性のプロドラッグを毒性生成物に変換する酵素(例えばチミジンキナーゼ、カルボキシルエステラーゼ、カルボキシペプチダーゼ、チトクロームP450アイソザイム、デオキシリボヌクレオチドキナーゼ、ニトロリダクターゼ等)でよい。自殺遺伝子がプロドラッグ変換酵素をコードする場合、それを発現する細胞はプロドラッグに暴露された時点で死亡する。本明細書で用いる「自殺遺伝子生成物」なる用語は「自殺遺伝子」によりコードされるタンパク質を意味する。自殺遺伝子発現は免疫細胞特異的プロモーターにより、好ましくはB細胞プロモーターにより駆動される。
【0054】
「自殺遺伝子発現の不活性化を可能にする配列」なる用語はリコンビナーゼにより認識される自殺遺伝子をフランキングする組換え部位を意味する。
【0055】
「プロドラッグ」なる用語は代謝によりインビボで毒性生成物、代謝物または薬物に変換可能である化合物を意味する。
【0056】
「内因性Ig(免疫グロブリン)発現B細胞」および「内因性B細胞」なる用語は互換的に用いられ、そして動物の内因性免疫グロブリン遺伝子座を発現するこれらのB細胞を意味する。本発明のB細胞はそのゲノムに自殺遺伝子を含有する。内因性B細胞は自殺遺伝子を発現し、そしてその結果、最終的に枯渇し、そしてしたがって動物の内因性Ig発現が抑制される。
【0057】
「外因性Ig(免疫グロブリン)発現B細胞」および「外因性B細胞」なる用語は、かかるB細胞に導入された外因性ヒト(化)Ig導入遺伝子座の産生性再構成を被るヒト以外の動物のこれらのB細胞を意味する。ヒト(化)Ig遺伝子座は、これもまた部位特異的リコンビナーゼをコードする発現構築物の一部としてかかるB細胞に導入される。ヒト(化)Ig遺伝子座の産生性再構成の結果、ヒト(化)Igおよび導入遺伝子コード化リコンビナーゼの発現に至る。結果として、B細胞のゲノムから自殺遺伝子が切除され、そして細胞は細胞死を免れる。したがって好ましくは、トランスジェニック動物により発現されるIg生成物はヒト(化)免疫グロブリンである。
【0058】
「自殺遺伝子の選択的発現」とは好ましくは免疫細胞内での、さらに好ましくはB細胞内での、そして最も好ましくは内因性B細胞内での自殺遺伝子生成物の発現を意味する。免疫細胞またはB細胞内での自殺遺伝子の選択的発現は各々免疫特異的またはB細胞特異的プロモーターを用いることにより達成されて自殺遺伝子発現を駆動する。
【0059】
「選択的不活性化」なる用語は、外因性B細胞のゲノムからの自殺遺伝子またはその部分の選択的切除を意味する。自殺遺伝子は外因性B細胞において発現される導入遺伝子コード化リコンビナーゼにより認識される組換え部位によりフランキングされるので、自殺遺伝子は切除されるかまたは不活性化される。
【0060】
Ig生成細胞の「枯渇」は、ヒト以外のまたは非ヒト化Igを発現する内因性B細胞集団の部分的または完全な死滅、瀕死および/または除去として定義される。さらに後記で説明するように、選択されるトランスジェニック動物が、抗体再構成が若年期に停止するものである場合、内因性B細胞の枯渇はさらに有効であり得る。
【0061】
「内因性免疫グロブリン生成の選択的抑制」は、自殺遺伝子を発現する内因性B細胞の枯渇による、ヒト以外のトランスジェニック動物の内因性免疫グロブリンの生成の選択的抑制を意味する。したがって、トランスジェニック動物により優勢に発現される免疫グロブリン生成物はヒト(化)免疫グロブリンである。
【0062】
「抗体多様性」および「抗体レパートリー」なる用語は互換的に用いられ、そして生物が発現できる全抗体特異性全体を意味する。
【0063】
遺伝子再構成および遺伝子変換を行う能力を有するIg遺伝子座はまた本明細書では「機能的」Ig遺伝子座をも意味し、そして機能的Ig遺伝子座により作成された多様性を有する抗体はまた本明細書では「機能的」抗体または抗体の「機能的」レパートリーをも意味する。
【0064】
「モノクローナル抗体」なる用語はB細胞の単一のクローンにより合成された抗体分子を意味する。
【0065】
「ポリクローナル抗体」なる用語はB細胞の集団により合成された抗体分子の集団を意味するために用いられる。
【0066】
「ポリヌクレオチド」および「核酸」なる用語は互換的に用いられ、そして単数または複数で用いられる場合、一般に任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドを意味し、これは未修飾RNAもしくDNAまたは修飾RNAもしくはDNAでよい。したがって、例えば本明細書で定義するようなポリヌクレオチドには、限定するものではないが一本鎖および二本鎖DNA、一本鎖および二本鎖領域を含むDNA、一本鎖および二本鎖RNA、ならびに一本鎖および二本鎖領域を含むRNA、一本鎖もしくはさらに典型的には二本鎖でよいか、または一本鎖および二本鎖領域を含んでよいDNAおよびRNAを含むハイブリッド分子が含まれる。加えて、本明細書で用いる「ポリヌクレオチド」なる用語はRNAもしくはDNAまたはRNAおよびDNAの双方を含む三本鎖領域を意味する。かかる領域の鎖は同一の分子から、または異なる分子からでよい。領域は1つまたはそれより多い分子全てを含んでよいが、さらに典型的にはいくつかの分子のある領域のみを伴う。三本鎖らせん領域の分子の1つはオリゴヌクレオチドである。「ポリヌクレオチド」なる用語は具体的にはcDNAを含む。その用語は1つまたはそれより多い修飾塩基を含有するDNA(cDNAを含む)およびRNAを含む。したがって、安定性またはその他の理由のために修飾された骨格を有するDNAまたはRNAは、その用語が本明細書で意図されるような「ポリヌクレオチド」である。さらに、イノシンのような通常でない塩基またはトリチウム化塩基のような修飾塩基を含むDNAまたはRNAは本明細書で定義する「ポリヌクレオチド」の用語に含まれる。一般に「ポリヌクレオチド」なる用語は未修飾ポリヌクレオチドの全ての化学的、酵素的および/または代謝的修飾形態、ならびにウイルスおよび単純細胞および複雑型細胞を含む細胞のDNAおよびRNA特性の化学的形態を包含する。
【0067】
本明細書で用いる「ヒト以外の(トランスジェニック)動物」には、限定するものではないが例えばヒト以外の霊長類、げっ歯類(例えばマウスおよびラット)、例えばウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマおよびロバのような非げっ歯類哺乳動物、ならびに鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)のような哺乳動物が含まれる。本明細書で用いる「非霊長類動物」なる用語には、限定するものではないが前記で具体的に列挙した哺乳動物を含む霊長類以外の哺乳動物が含まれる。
【0068】
「一次抗体レパートリーを創成するために実質的に遺伝子変換および/または体細胞超変異により抗体多様性を創成する動物」または「遺伝子変換動物」なる語句および文法的な均等は抗体多様化の主要な機構が、遺伝子再構成と対照的に遺伝子変換および/または超変異であるかかる動物を意味するために用いられる。かかる動物には、限定するものではないがウサギ、鳥類(例えばニワトリ、シチメンチョウ、アヒル、ガチョウ等)ウシおよびブタが含まれる。特に好ましいヒト以外の動物はウサギおよびニワトリである。
【0069】
動物による「若年期の抗体遺伝子再構成の停止」とは、典型的には免疫グロブリン遺伝子の再構成が生まれて1か月以内に停止するこれらの動物を意味する。かかる動物の実例には、限定するものではないがウサギ、鳥類(例えばニワトリ)、ヒツジ、ヤギ、ウシ、ブタおよびウマである。
【0070】
詳細な説明
本発明は、例えば動物を(複数の)ヒト(化)免疫グロブリンの発現にさらに適当なものにする目的で、ヒト以外の動物における内因性免疫グロブリン生成を抑制するための方法を提供する。
【0071】
本発明によれば、内因性免疫グロブリンを発現するB細胞における自殺遺伝子の選択的発現により、内因性免疫グロブリン生成は(複数の)ヒト(化)免疫グロブリンのような外因性免疫グロブリン配列を発現するヒト以外のトランスジェニック動物において選択的に抑制される。自殺遺伝子は導入遺伝子として動物のゲノムに組み込まれ、そして例えばヒト(化)Ig導入遺伝子座をも導入する遺伝子導入発現構築物の一部として、または別個に例えば別個の遺伝子導入発現構築物を用いて導入され得る。後者の場合、2つの発現構築物を同時にまたは異なる時間でトランスジェニック動物に導入することができる。
【0072】
自殺遺伝子は免疫特異的プロモーター、好ましくはB細胞特異的プロモーターを用いて動物のB細胞において発現され、そしてリコンビナーゼにより認識される組換え配列によりフランキングされる。したがって、組換え配列によりフランキングされた自殺遺伝子は初期には動物の全てのB細胞に存在する。「外因性B細胞」では、ヒト(化)免疫グロブリン−自己切断ペプチド−リコンビナーゼ分子をコードする外因性免疫グロブリン導入遺伝子座の産生性再構成はかかるB細胞におけるリコンビナーゼの選択的発現に至る。リコンビナーゼはかかる細胞において自殺遺伝子をフランキングする組換え部位を認識する。結果的に外因性B細胞では自殺遺伝子は選択的に切除され、それは結果的に細胞死を免れる。対照的に内因性B細胞の内因性免疫グロブリン遺伝子座の産生性再構成はリコンビナーゼの発現に至らない。結果的に内因性B細胞における自殺遺伝子発現はこの細胞集団の死亡に至り、そしてその結果ヒト以外のトランスジェニック動物によるヒト(化)免疫グロブリンの発現を抑制することなく内因性免疫グロブリン生成を抑制する。
【0073】
導入遺伝子は1つまたはいくつかの天然または、通常動物もしくは動物の細胞において見出されない合成タンパク質をコードする配列を有するDNAフラグメントである。前核のマイクロインジェクション、トランスフェクション、核移植クローニング、精子媒介の遺伝子移入、精巣媒介の遺伝子移入等を含む種々の技術により、(複数の)DNAフラグメントを動物のゲノムに導入することができる。本発明は2つの導入遺伝子または導入遺伝子構築物(1)ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子および(2)免疫細胞特異的自殺導入遺伝子に言及する。各々の導入遺伝子はその独自の調節配列に作動可能なように連結されている。例えば自殺遺伝子の発現はB細胞特異的プロモーターにより駆動され得る。2つの導入遺伝子構築物は2つの別個のベクターまたは同一のベクター(プラスミド)に存在し得る。1つの実施態様では、2つの導入遺伝子構築物を別個の時点で導入することができる。これに代えて、双方の導入遺伝子構築物を同時に動物に導入することができる。好ましい実施態様では、自殺導入遺伝子の発現は、重鎖再構成を生じた後に起こるように調節される。後記で論じる機構から明白であるように、これは外因性B細胞におけるリコンビナーゼの発現のための時間、そしてしたがって外因性B細胞のゲノムからの自殺遺伝子のリコンビナーゼ媒介の切除、したがってかかる細胞における自殺遺伝子発現の停止のための時間を見越している。加えて、本発明の方法で用いるベクターは選択を可能にするためにゲンタマイシン、ネオマイシン、カナマイシン等のような抗生物質選択マーカーをコードするDNA配列を含有できる。
【0074】
本発明の1つの態様では、導入遺伝子は自己切断ペプチド(例えば2Aペプチドまたは2A様ペプチド)をコードするDNA配列を含む。導入遺伝子の免疫グロブリンコード化配列とリコンビナーゼコード化配列の間の自己切断ペプチドコード化配列の挿入により1つのメッセンジャーRNAの生成に至る。しかしながらこのmRNAの翻訳の結果、ペプチドの自己切断機構のために2つの別個のタンパク質、(複数の)免疫グロブリンおよびリコンビナーゼに至る。したがってリコンビナーゼの発現をVDJまたはVJセグメントの機能的再構成と結びつけることができる。
【0075】
本発明の1つのかかる実施態様では、自己切断はピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーを含む、またはC型ロタウイルスファミリーからのウイルスの2A/2Bペプチドまたは2A様/2B配列により媒介される。ピコルナウイルス科ウイルスファミリーにはエンテロ、ライノ、カルディオおよびアフトならびに口蹄疫(FMDV)ウイルスが含まれる。ピコルナウイルス様昆虫ウイルスファミリーには感染性フラシェリー病ウイルス(IFV)、ショウジョウバエCウイルス(DCV)、急性ミツバチ麻痺病ウイルス(ABPV)およびコオロギ麻痺病ウイルス(CrPV)ならびに昆虫ウイルスゾセア・アシグナ(Thosea asigna)ウイルス(TaV)のようなウイルスが含まれる。C型ロタウイルスファミリーにはウシ、ブタおよびヒトC型ロタウイルが含まれる。さらなる実施態様では、切断配列はポリオウイルス、ライノウイルス、コクサッキーウイルス、脳心筋炎ウイルス(EMCV)、メンゴウイルス、ブタテシオウイルス−1、またはタイラーネズミ脳炎ウイルス(TMEV)等のいずれかからの2A様/2B配列を含み得る。好ましい実施態様では、自己切断タンパク質配列は口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルスまたはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)のいずれかの2A/2Bペプチドである。Palmenbergら、Virology 190:754−762(1992)、Ryanら、J Gen Virol 72:2727−2732(1991)、Donnellyら、J Gen Virol 82:1027−1041(2001)、Donnellyら、J Gen Virol 82:1013−1025(2001)、Szymaczakら、Nature Biotech 22(5):589−594(2004)。
【0076】
本発明の方法で用いるその他の導入遺伝子は部位特異的リコンビナーゼをコードする。部位特異的リコンビナーゼは2つの核酸、例えばDNAセグメント間の相同組換えを触媒する。これらのリコンビナーゼは組換えの双方のパートナーの非常に特異的な配列を認識する。触媒の機構は様々な型の部位特異的リコンビナーゼで異なってよいが、それらは全て、基礎を成す機構に関わらず本明細書に含まれ、そして本発明の実施に適当である。
【0077】
特定の実施態様では、リコンビナーゼは例えばCre、Flpリコンビナーゼ等でよい。CreおよびFlpは2つの最も一般的に用いられる酵素であり、それは非常に特異的なDNA配列に作用するのみである。Creは2つの34塩基対長のloxP部位間でDNAの組換えを触媒するが、Flpはfrt部位を標的とする。真核細胞のDNAの部位特異的組換えのためのCreリコンビナーゼの使用は米国特許第4959317号に記載されている。真核細胞のトランスフェクションのための部位特異的リコンビナーゼの使用は米国特許第6632672号に記載されている。部位特異的組換えは一般に米国特許第4673640号に記載されている。Cre/loxP基盤のクローニング系は例えばBD Biosciences−Clontech(パロアルト、カリフォルニア州)(Creator(商標))またはInvitrogen(カールスバッド、カリフォルニア州)(Echo(商標))から市販により入手可能である。
【0078】
別の実施態様では、リコンビナーゼはλインテグラーゼ、φC31、TP901−1およびR4からなる群から選択されるファージによりコードされる部位特異的リコンビナーゼでよい。φC31およびR4は部位特異的リコンビナーゼのインテグラーゼファミリーに属するが、TP901−1は拡張されたリゾルベースファミリーに属する。R4インテグラーゼはストレプトミセス・パルブルスのファージR4のゲノムから誘導された部位特異的一方向性リコンビナーゼである。部位特異的インテグラーゼTP901−1はラクトコッカス・ラクティス亜種クレモリスのファージTP901−1によりコードされる。λは大腸菌に感染する鋳型バクテリオファージである。ファージは組換えのための1つの結合部位(attP)を有し、そして大腸菌細菌ゲノムは組換えのための1つの結合部位(attB)を有する。本発明の局面では、野生型組換え部位を例えば相同系から誘導し、そして異種性配列に随伴させることができる。したがってattB部位をその他の系に置いてインテグラーゼの基質として作用させることができる。さらに別の実施態様では、リコンビナーゼは細菌ゲノム組換え部位(attB)およびファージゲノム組換え部位(attP)の間の組換えを触媒することができるか、または第1の部位は偽attB部位を含むことができ、そして/もしくは第2の部位は偽attP部位を含むことができるかまたはその逆である(Grothら、Proc.Nat.Acad.Sci.97:5995−6000(2000);Olivaresら、Nature Biotechnol.20(11):1124−8(2002));(Thyagarajanら、Mol. and Cell.Biol.21:3926−3934(2001));Hollisら、Repro.Biol.and Endocrinol.1:79(2003))。「偽組換え部位」とは、たとえ部位がその野生型組換え部位の配列と同一の配列を有せずとも、リコンビナーゼが組換えを促進できる部位を意味する。
【0079】
さらに別の実施態様では、リコンビナーゼはトランスポーゼスまたはレトロトランスポーゼスでよい。トランスポゾンまたはレトロトランスポーゼスはカットアンドペースト機構によりその転位を触媒する酵素であり、そして任意の導入遺伝子の移入または挿入に用いることができる。これらはヒト、鳥類、げっ歯類等のような脊椎動物を含む広範な種のゲノムへの任意のDNA配列の挿入または移入のための非ウイルス性および非相同性の方法を提供する。例えばショウジョウバエエレメントマリナーはそれ自体をニワトリ生殖細胞系に転位させることが示された(Shermanら、Nature Biotechnol.16:1050−1053(1998))。sleeping beautyトランスポーゼス系を用いるマウスおよびヒトゲノムのような哺乳動物系への長期間の導入遺伝子発現またはトランスポゾンDNAの効率的な挿入はYantら、Nature Genetics 25:35−41(2000);Dupuyら、Proc.Nat.Acad.Sci.99:4495−4499(2002)およびGeurtsら、Mol.Therapy 8:108−117(2003)により実証されている。L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)、ミノスのようなその他のトランスポーゼスは脊椎動物種において活性であることが示されており、そしてしたがって遺伝子移入に、または挿入変異ベクターとして有用である(Largaespada,David A.、Repro.Biol.and Endocrinol.1:80(2003))。トランスポーゼスの実例としては、限定するものではないが原核細胞性または真核細胞性トランスポーゼス、ウイルス性、ショウジョウバエコピア様または哺乳動物レトロトランスポーゼス等を含む非ウイルス性レトロトランスポーゼスが含まれる。原核細胞性トランスポーゼスには、Tn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。真核細胞性トランスポーゼスにはショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAcおよびDsエレメント等の転位可能なエレメントでコードされるトランスポーゼスが含まれる。レトロトランスポーゼスにはL1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)、ミノス等でコードされるものが含まれる。トランスポーゼスをMp、Spm、En、ドット、Mu、およびI転位性エレメントからなる群から選択することもできる。
【0080】
本発明の1つの態様では、前記した部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子を用いて内因性B細胞を選択的に死滅させる。自殺遺伝子を組換え部位でフランキングすることにより、外因性B細胞においてのみリコンビナーゼまたはトランスポゾンの発現時の自殺遺伝子の不活性化に至る。自殺遺伝子発現の不活性化は自殺遺伝子またはその部分の切除により達成することができる。これに代えて、自殺遺伝子の発現に必要な配列を切除の標的とすることができる。別の研究法によれば、DNAフラグメントの反転または挿入によりトランスポゾンジャンピング、トランスポゾン挿入または反転のために自殺遺伝子発現を不活性化することができる。
【0081】
本発明の実施において用いる自殺遺伝子には毒素遺伝子および毒性生成物を生じる無毒性プロドラッグ変換酵素が含まれる。本発明の方法において用いることができる活性毒素およびそのフラグメントには、例えばジフテリア毒素A鎖(DTA)のような細菌性毒素もしくはそのフラグメント、シグマ毒素、エキソトキシンA鎖(緑膿菌由来)等またはリシンA鎖、アブリンA鎖、モデクシンA鎖、アルファサルシン、シナアブラギリタンパク質、ジアンチンタンパク質、ヨウシュヤマゴボウタンパク質(PAPI、PAPAIIおよびPAP−S)、ツルレイシ阻害剤、クルシン、クロチン、サボンソウ阻害剤、ゲロニン、ミトゲリン、レストリクトシン、フェノマイシン、エノマイシンおよびトリコテセン、殺虫性毒素、は虫類毒液等のような植物もしくは真菌毒素およびその非結合活性フラグメントが含まれる。好ましい実施態様では、用いる毒素はジフテリア毒素A鎖(DTA)である。
【0082】
標的B細胞に導入された自殺遺伝子がプロドラッグ変換酵素をコードする場合、酵素は特異的無毒性プロドラッグを活性化して最終的に標的B細胞を死滅させる毒性代謝物を創成する。この研究法では、内因性B細胞生成を抑制するために2工程処理方法を設計することができる。第1工程では、当分野において公知の種々の方式で外来酵素のための遺伝子を発現させるかまたはB細胞に分配することができる。第2工程では、動物に投与されたプロドラッグは最終的にそれを死滅させる酵素を発現するB細胞により活性化されて毒性代謝物になる。
【0083】
自殺遺伝子として有用なプロドラッグ変換酵素は一般に2つの主要なクラスで見出されている。第1のクラスはヒト対応物を伴うかまたは伴わない非哺乳動物起源の酵素である。実例にはウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、βラクタマーゼ、多剤活性化酵素(MDAE)、βガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)が含まれる。第2のクラスはヒト起源の酵素からなる。これにはデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)アイソザイムが含まれる。酵素プロドラッグ系のさらなる実例はMethods in Molecular Medicine:Suicide Gene Therapy,Methods and Reviews(Caroline J Springer編、Human Press、2004年)の表1に列挙されており、それを出典明示により本明細書の一部とする。したがって本発明で自殺遺伝子として用いるプロドラッグ変換酵素には、限定するものではないが前記したように非哺乳動物、非ヒト起源およびヒト起源の酵素が含まれる。
【0084】
本発明で有用である適当なプロドラッグには、限定するものではないがガンシクロビル、アシクロビル、5−(アジリジン−1−イル)−2,3ジニトロベナミド、カペシタビン、イリノテカン、カルバマート基盤の20(S)−カンプトテシン、ジニトロベンズアミドアジリジンCB1954およびそのナイトロジェンマスタード類似体SN23862、2−アミノアントラセン(2−AA)および4−イポメアノール(4−IM)等が含まれる。本発明で有用であるプロドラッグのさらなる実例は対応するその活性化酵素と共にMethods in Molecular Medicine:Suicide Gene Therapy, Methods and Reviews(Caroline J Springer編、Human Press、2004年)の表1に列挙されており、それを出典明示により本明細書の一部とする。酵素活性化プロドラッグはしばしば、例えばエタニダゾール、フルオソール、ミソニダゾール、ニモラゾール、テモポルフィン、トリアパザミンのような放射線増感薬、またはカスパーゼのようなその他のアポトーシス剤の発現を用いるその他の細胞死滅方法と組み合わせて用いられることがあり、標的内因性Ig発現B細胞を死滅させる相乗効果を導く。
【0085】
したがって自殺遺伝子の発現は種々の機構により内因性免疫グロブリンを発現するB細胞を排除することができる。例えばDTAの発現によりタンパク質合成の阻止および続く細胞死に至る。チミジンキナーゼはガンシクロビルおよびアシクロビルをその対応する一リン酸形態にリン酸化し、それは続いて細胞性キナーゼにより毒性の三リン酸誘導体に変換される。毒性三リン酸の分裂細胞DNAへの取り込みにより細胞死に至る。ニトロリダクターゼは5−(アジリジン−1−イル)−2,3ジニトロベナミドを2−および4−ヒドロキシルアミノ誘導体に変換し、そのときに細胞性チオエステルを有する4−ヒドロキシルアミノ誘導体の非酵素反応によりDNAを架橋することができる強力な細胞毒性二価性アルキル化剤が作成される。当分野において公知の自殺遺伝子は本発明において有用であるが、これらの自殺遺伝子が内因性B細胞を排除するように作用する(複数の)機構に束縛または限定されることはない。
【0086】
本発明の好ましい実施態様では、自殺遺伝子は野生型または偽組換え部位(例えばCreにより認識されるloxP部位またはFlpにより認識されるFRT部位)によりフランキングされたジフテリア毒素A鎖(DTA)をコードする。
【0087】
本発明の別の実施態様では、自殺遺伝子は 野生型または偽組換え部位によりフランキングされたチミジンキナーゼをコードする。
【0088】
1つの実施態様では、部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子を、ヒトIg遺伝子座−自己切断ペプチド−リコンビナーゼ導入遺伝子の導入の前、同時または後のいずれかに、別個の遺伝子導入ベクターで動物に導入する。別の実施態様では、部位特異的リコンビナーゼ認識部位によりフランキングされた自殺遺伝子をヒトIg遺伝子座−リコンビナーゼ導入遺伝子のためのものとして同一の遺伝子導入ベクターで動物に導入する。全ての態様で、自殺遺伝子は動物ゲノムに組み込まれ、そして免疫細胞においてのみ特異的に発現されることを確実にする免疫細胞特異的プロモーターにより、および好ましくはB細胞においてのみ特異的に発現されることを確実にし、その他の細胞型では発現されないB細胞特異的プロモーターにより、その発現が駆動される。
【0089】
自殺遺伝子の発現はB細胞特異的プロモーターにより制御されて、その発現は非B細胞またはヒト以外のトランスジェニック動物の組織では「スイッチオフ」になる。B細胞特異的遺伝子の発現を制御するプロモーター(およびエンハンサー)、またはその変種もしくは操作された部分は自殺遺伝子のかかるB細胞特異的発現に有用である。B細胞特異的遺伝子のプロモーター/エンハンサーの実例には、限定するものではないがCD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b(B29またはIgベータとしても公知である)、mb−1(Igアルファとしても公知)、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖、免疫グロブリンJ−鎖等のプロモーター/エンハンサーが含まれる。好ましい実施態様では、カッパ軽鎖遺伝子プロモーター/エンハンサーが自殺遺伝子のB細胞特異的発現を駆動する。
【0090】
したがって、内因性免疫グロブリン生成の抑制の結果、ヒト(化)Ig導入遺伝子座の優勢な発現に至る。換言すれば、内因性B細胞の枯渇によりヒト(化)抗体の富化に至る。好ましくは、内因性B細胞の富化は100%に近くなる。
【0091】
本発明のさらに別の態様では、導入遺伝子は免疫グロブリン重鎖および/もしくは免疫グロブリン軽鎖またはその部分をコードする。遺伝子座は生殖細胞系の立体配置または再構成形態でよい。コード化配列またはその部分はヒト免疫グロブリンをコードすることができ、その結果ヒト(化)抗体の発現に至る。
【0092】
ヒト(化)抗体をコードする(複数の)導入遺伝子は1つまたはいくつかのヒトIgセグメント(例えばヒトIgV、D、JまたはC遺伝子セグメント)を含有するIg遺伝子座またはIg遺伝子座の大部分を含有する。これに代えて、導入遺伝子はヒト免疫グロブリン遺伝子座またはその大部分である。かかるヒトIg遺伝子座またはかかる修飾されたIg遺伝子座またはIg遺伝子座の修飾された部分を含有する導入遺伝子もまた本明細書では「ヒト(化)Ig導入遺伝子座」と称され、ヒト以外のトランスジェニック動物において再構成を被ることができ、それによりヒト免疫グロブリンポリペプチド配列の少なくとも一部を有する抗体の多様化されたレパートリーが生成される。
【0093】
免疫グロブリン重および軽鎖遺伝子は個々の遺伝子によりコードされ、そしてイントロン配列により分けられたいくつかのセグメントを含む。したがってヒト免疫グロブリン重鎖のための遺伝子は第14染色体で見出される。重鎖(VH)の可変領域は3つの遺伝子セグメント(V、DおよびJセグメント)、続いてC領域をコード化する同義遺伝子を含む。V領域は大型のスペーサーによってC領域から分けられ、そしてV、DおよびJセグメントをコードする個々の遺伝子もまたスペーサーによって分けられる。
【0094】
2つの型の免疫グロブリン軽鎖:κおよびλがある。ヒトκ軽鎖の遺伝子は第2染色体に見出され、そしてヒトλ軽鎖の遺伝子は第22染色体に見出される。抗体軽鎖の可変領域は別個の遺伝子セグメントによりコードされるVセグメントおよびJセグメントを含む。κ軽鎖遺伝子の生殖細胞系立体配置では、およそ100〜200V領域遺伝子が直線的な配置で存在し、各遺伝子はそれ独自のリーダー配列、続いておよそ5J遺伝子セグメントおよびC領域遺伝子セグメントを有する。全てのV領域はイントロンにより分けられ、そして同様にV、JおよびC領域遺伝子セグメントを分けるイントロンが存在する。
【0095】
感染に対して保護する免疫系の能力は、抗体の多様なレパートリーを創成するために特殊化された遺伝的な仕組みにある。B細胞の抗体コード化遺伝子は可変(V)領域の結合部位の無数の組み合わせを可能にする様式で組み立てられる。1012より多い可能な結合構造がかかる機構から生じると推測される。ヒトを含む全ての動物において抗体産生過程は免疫グロブリン(Ig)遺伝子座の可変(V)、多様性(D)および連結(J)セグメントを組換えることにより始まる。この工程の後に、動物種に依存して2つの一般的な機構を用いて抗体の多様な結合構造を生成する。
【0096】
ヒトおよびマウスのようないくつかの動物では、免疫グロブリン重鎖遺伝子座にV、DおよびJセグメントの複数のコピー、ならびに免疫グロブリン軽鎖伝子座にVおよびJセグメントの複数のコピーが存在する。これらの動物における抗体多様性は一次的には遺伝子再構成、すなわち再構成された重鎖可変領域および軽鎖可変領域を形成するための遺伝子セグメントの異なる組み合わせにより作成される。しかしながら別の動物(例えばウサギ、鳥類、例えばニワトリ、ガチョウおよびアヒル、ヒツジ、ヤギならびにウシ)では、遺伝子再構成は抗体多様性の作成においてあまり大きな役割を果たさない。例えば、ウサギでは、非常に限定された数のV遺伝子セグメントのみ、もっともよくあるのはV領域の3’末端のV遺伝子セグメントが遺伝子再構成に用いられて近接するVDJセグメントを形成する。ニワトリでは、1つのV遺伝子セグメント(D領域に隣接するもの、すなわち「3’近位V遺伝子セグメント」)のみ、1つのDセグメントおよび1つのJセグメントのみを重鎖再構成で用い、そして1つのV遺伝子セグメント(3’近位V遺伝子セグメント)および1つのJセグメントのみを軽鎖再構成で用いる。したがってこれらの動物では初期に再構成された可変領域配列の中では多様性が少ししかなく、それは連結部多様化の結果である。再構成されたIg遺伝子のさらなる多様化は、上流V遺伝子セグメントから誘導される短い配列が、再構成されたIg遺伝子のV遺伝子セグメント内の短い配列を置き換える過程である遺伝子変換により達成される。抗体配列のさらなる多様化を超変異により作成することができる。
【0097】
免疫グロブリン(抗体)は5つのクラス(IgG、IgM、IgA、IgEおよびIgD)に属し、各々は免疫防御において異なる生物学的役割を有している。血液中に最も豊富であり、そして感染に対する応答において強力であるのはIgGクラスである。ヒトIgGクラス内にはFcドメインを含む重鎖定常領域の構造により決定される4つのサブクラス(IgG1、IgG2、IgG3およびIgG4アイソタイプ)がある。抗体のF(ab)ドメインは抗原の特異的配列(エピトープ)に結合するが、抗体のFcドメインは抗原を排除するために免疫系のその他の構成成分を補充および活性化する。
【0098】
元来の抗体および免疫グロブリンは通常約150000ダルトンのヘテロ四量体糖タンパク質であり、2つの同一の軽(L)鎖および2つの同一の重(H)鎖からなる。各軽鎖は(複数の)共有結合性ジスルフィド結合により重鎖に連結されているが、ジスルフィド結合の数は様々な免疫グロブリンアイソタイプの重鎖間で異なる。各重および軽鎖はまた規則正しい間隔の鎖内ジスルフィド架橋を有する。各重鎖は一方の末端で可変ドメイン(VH)を有し、多くの定常ドメインがそれに続く。各軽鎖は一方の末端で可変ドメイン(VL)、およびその他方の末端で定常ドメインを有し、軽鎖の定常ドメインは重鎖の第1の定常ドメインとアラインされ、そして軽鎖可変ドメインは重鎖の可変ドメインとアラインされている。特定のアミノ酸残基は軽鎖および重鎖可変ドメイン間のインターフェースを形成すると考えられている(Chothiaら、J.Mol.Biol.186:651(1985);NovotnyおよびHaber、Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0099】
「可変」なる用語は可変ドメインの特定の部分が抗体間の配列で大きく異なるという事実を意味し、そしてその特定の抗原に関する特定の各抗体の結合および特異性において用いられる。しかしながら可変性は抗体の可変ドメインにわたって均一に分布されるわけではない。それは双方共に軽鎖および重鎖可変ドメインの相補性決定領域(CDR)または超可変領域と称される3つのセグメントに集中している。可変ドメインのさらに高度に保存された部分はフレームワーク(FR)と称される。元来の重および軽鎖の可変ドメインは各々3つのCDRによりつながれた4つのFR領域を含む。各鎖のCDRはFR領域により近接近してまとまり、そしてその他の鎖のCDRと一緒に抗体の抗原結合部位の形成に寄与する(Kabatら、Sequences of Proteins of Immunological Interest、第5版、National Institute of Health,、ベセスダ、メリーランド州(1991年)参照)。定常ドメインは抗体の抗原に対する結合に直接関与しないが、抗体依存性細胞毒性における抗体の関与のような種々のエフェクター機能を呈する。
【0100】
ヒト動物導入遺伝子座の創成により高収率で多様化された高親和性ヒト(化)(ポリクローナル)抗体を発現するトランスジェニック動物の創成が可能になる。一般にヒト以外の動物における免疫グロブリン(Ig)遺伝子座のヒト化は1つまたはそれより多いヒトIg遺伝子セグメントの動物のゲノムへの組み込みを伴い、ヒト(化)免疫グロブリン遺伝子座を創成する。したがってヒト(化)Ig重鎖遺伝子座の創成は1つまたはそれより多いVおよび/もしくはDおよび/もしくはJセグメント、ならびに/またはC領域セグメントの動物ゲノムへの組み込みを伴う。同様に、ヒト化Ig軽鎖遺伝子座の創成は1つまたはそれより多いVおよび/もしくはJセグメント、ならびに/またはC領域セグメントの動物ゲノムへの組み込みを伴う。
【0101】
染色体位置に関わらず、本発明のヒト(化)Ig遺伝子座はヒト以外の動物において遺伝子再構成および遺伝子変換および超変異を被る能力を有し、それによりヒト(化)Ig分子の多様化されたレパートリーを生成する。遺伝子再構成および遺伝子変換を被る能力を有するIg遺伝子座は「機能的」Ig遺伝子座とも称され、そして機能的Ig遺伝子座により作成された多様性を有する抗体は「機能的」抗体または抗体分子の「機能的」レパートリーとも称される。
【0102】
1つの態様では、抗体レパートリーの多様化が若年期に停止する動物が本発明において有用である。B細胞は造血幹細胞から発達する。抗原暴露の前にB細胞は一連の成熟工程を被り、その最終生成物は成熟B細胞であり、それはその他の細胞表面シグナリング分子と一緒にその細胞表面に独特な膜結合性IgMおよびしばしばIgDを発現する。ヒトでは遺伝子再構成による抗体多様化は一生にわたって生じるが、その他の動物では、抗体レパートリーの多様化は若年期に、典型的には生まれて1か月以内に停止する。
【0103】
免疫グロブリン遺伝子の再構成が若年期に停止する動物では、この限定された期間に生成された全てまたはほとんどのB細胞を死滅させることにより効果的に内因性免疫グロブリン生成の持続的または永続的な抑止に至る。1つまたはいくつかのヒトまたはヒト化免疫グロブリン導入遺伝子座を含有するトランスジェニック動物では、これにより動物の内因性免疫グロブリン生成の不在下でのヒトまたはヒト化免疫グロブリンの生成が可能になる。このように遺伝子再構成が若年期に停止する動物では(複数の)内因性免疫グロブリンの発現が効果的に抑制され得る。かかる動物の実例は、限定するものではないがウサギ、鳥類(例えばニワトリ)、ヒツジ、ヤギ、ウシ、ブタおよびウマである。
【0104】
本発明によれば、ヒト(化)免疫グロブリンを作ることができるトランスジェニック動物は、動物の(1つまたは複数の)レシピエント細胞に本明細書に前記した1つまたはそれより多い遺伝子導入ベクターを導入することにより作られ、その1つはヒト(化)Ig遺伝子座を担持し、そして遺伝的に修飾された(1つまたは複数の)レシピエント細胞から動物を誘導する。
【0105】
レシピエント細胞は例えば遺伝子変換および/または超変異により抗体多様性を作成するヒト以外の動物、例えば鳥類(例えばニワトリ)、ウサギ、ウシ等由来でよい。かかる動物では3’近位V遺伝子セグメントは免疫グロブリンの生成に優先的に用いられる。動物の3’近位V遺伝子セグメントを置換することか、または3’近位V遺伝子セグメントの近接近で配置されることのいずれかにより、導入遺伝子ベクターのIg遺伝子座へのヒトV遺伝子セグメントの組み込むことで、大部分の免疫グロブリンでのヒトV領域ポリペプチド配列の発現に至る。これに代えて、再構成されたヒトV(D)Jセグメントを導入遺伝子ベクターの免疫グロブリン遺伝子座のJ遺伝子座に挿入することができる。
【0106】
無作為組み込みによるかまたは標的組み込みにより、ヒト(化)Ig遺伝子座および自殺遺伝子を含有する目的の遺伝子を含有する遺伝子導入ベクターを(1つまたは複数の)レシピエント細胞に導入し、そして次に(1つまたは複数の)レシピエント細胞のゲノムに組み込むことができる。
【0107】
無作為組み込みのために、標準的な遺伝子導入技術によりヒト(化)Ig遺伝子座を含有する遺伝子導入ベクターを動物のレシピエント細胞に導入することができる。例えば遺伝子導入ベクターを受精卵母細胞の前核に直接注射することができる。また卵母細胞の受精の前に精子の遺伝子導入ベクターとのコインキュベーションにより遺伝子導入ベクターを導入することもできる。受精卵母細胞からトランスジェニック動物を発達させることができる。遺伝子導入ベクターを導入する別の方式は、胚性幹細胞をトランスフェクトし、そして続いて遺伝的に修飾された胚性幹細胞を発達中の胚に注射することによる。これに代えて遺伝子導入ベクター(裸でまたは促進剤と組み合わせて)を発達中の胚に直接注射することができる。最終的に、トランスジェニック動物の少なくともいくつかの体細胞のゲノムに組み込まれたヒト(化)Ig導入遺伝子を含有する胚からキメラトランスジェニック動物を生成する。
【0108】
特定の実施態様では、ヒト(化)Ig遺伝子座を含有する導入遺伝子は内因性免疫グロブリン遺伝子の発現が損なわれた動物株から誘導されたレシピエント細胞(例えば受精卵母細胞または発達中の胚)のゲノムにランダムに組み込まれる。かかる動物株の使用によりヒト(化)遺伝子導入Ig遺伝子座からの免疫グロブリン分子の優先的な発現が可能になる。かかる動物の実例には、アリシアおよびバジレアウサギ株、ならびに無ガンマグロブリン血症ニワトリ株、ならびに免疫グロブリンノックアウトマウスが含まれる。これに代えて、ヒト(化)免疫グロブリン導入遺伝子または遺伝子座を有するトランスジェニック動物を内因性免疫グロブリンの発現が損なわれた動物株と交配させることができる。損なわれた内因性Ig遺伝子座およびヒト(化)遺伝子導入Ig遺伝子座に関してホモ接合性の子孫を得ることができる。
【0109】
標的組み込みのために、遺伝子導入ベクターを胚性幹細胞または既に分化した体細胞のような適切な動物レシピエント細胞に導入することができる。その後、導入遺伝子が動物ゲノムに組み込まれ、そして相同組換えにより対応する内因性Ig遺伝子座を置換した細胞を標準的な方法により選択することができる。例えばKuroiwaら、Nature Genetics、2004年6月6日を参照のこと。次いで選択した細胞を除核した核移植単位細胞、例えば卵母細胞または胚性幹細胞と融合することができ、その細胞は全能性であり、そして機能的新生仔を形成することができる。十分に確立されている従来技術に従って融合を実施する。また卵母細胞の除核および核移植を注入ピペットを用いる顕微手術により実施することもできる。(例えばWakayamaら、Nature 394:369(1998)参照。)得られた卵細胞を次いで適切な培地中で培養し、そして同期化したレシピエントに移入してトランスジェニック動物を作成する。これに代えて選択された遺伝的に修飾された細胞を発達中の胚に注射することができ、それを続いてキメラ動物に発達させる。
【0110】
さらに本発明によれば、その1つが内因性Ig遺伝子セグメントのフランキング配列に相同である5’および3’フランキング配列に連結されたヒトIg遺伝子セグメントを担持する本明細書にて前記した1つまたはそれより多い組換えベクターを(1つまたは複数の)レシピエント細胞に導入し、次いで相同組換えにより内因性Ig遺伝子セグメントがヒトIg遺伝子セグメントにより置き換えられている細胞を選択し、そして選択された遺伝的に修飾された(1つまたは複数の)レシピエント細胞から動物を誘導することにより、ヒト(化)免疫グロブリンを生成できるトランスジェニック動物を作ることもできる。
【0111】
遺伝子導入ベクターの標的挿入に類似して、この研究法においてレシピエント細胞として使用するのに適切な細胞には胚性幹細胞または既に分化した体細胞が含まれる。任意の実行可能な手段、例えばトランスフェクションによりヒトIg遺伝子セグメントを担持する組換えベクターをかかるレシピエント細胞に導入することができる。その後相同組換えによりヒトIg遺伝子セグメントが対応する内因性Ig遺伝子セグメントと置き換わっている細胞を標準的な方法により選択することができる。これらの遺伝的に修飾された細胞をトランスジェニック動物のクローニングのための核移植手順における核ドナー細胞として提供することができる。これに代えて選択された遺伝的に修飾された胚性幹細胞を発達中の胚に注射することができ、それを続いてキメラ動物に発達させることができる。
【0112】
具体的な実施態様では、胚期の間に導入遺伝子を胚に直接注射することにより、またはそれらを妊娠母体もしくは産卵するメンドリに間接的に注射することにより本発明の導入遺伝子構築物をトランスジェニック動物に導入することができる。結果的に動物の免疫グロブリン分子を発現する内因性B細胞は枯渇し得て、そして故にトランスジェニック子孫が抗原での免疫に応答してヒト(化)抗体を優勢に生成する。
【0113】
前記の方法のいずれかにより生成されたトランスジェニック動物は本発明の別の実施態様を形成する。トランスジェニック動物は少なくとも1つの、すなわち1つまたはそれより多い、そこからヒト(化)抗体の機能的レパートリーが生成されるヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有する。
【0114】
具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有するトランスジェニックウサギを提供する。本発明のトランスジェニックウサギはヒト(化)Ig遺伝子座を再構成および遺伝子変換し、そしてヒト(化)抗体の機能的レパートリーを発現することができる。
【0115】
別の具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)Ig遺伝子座および自殺遺伝子をゲノムに有するトランスジェニックニワトリを提供する。本発明のトランスジェニックニワトリはヒト(化)Ig遺伝子座を再構成および遺伝子変換し、そしてヒト(化)抗体の機能的レパートリーを発現することができる。別の具体的な実施態様では、本発明は1つまたはそれより多いヒト(化)V領域および自殺遺伝子をゲノムに有するトランスジェニックマウスを提供する。ヒト(化)V領域はヒト以外のスペーサー配列によりフランキングされた少なくとも2つのヒトV遺伝子セグメントを含む。トランスジェニックマウスはヒトVエレメントを再構成し、そして抗体の機能的レパートリーを発現することができる。
【0116】
抗原での免疫が該トランスジェニック動物における同一抗原に対するヒト(化)抗体の生成を招く。
【0117】
本発明の好ましい実施態様は、内因性B細胞を枯渇させるためにヒト(化)Ig遺伝子座および少なくとも1つの自殺遺伝子を有し、そしてヒト(化)ポリクローナル抗血清を生成するトランスジェニック動物に向けられているが、霊長類化Ig遺伝子座および霊長類化ポリクローナル抗血清を有するトランスジェニック動物もまた本発明の精神内にあることは理解されよう。ヒト(化)ポリクローナル抗血清組成物に類似して霊長類化ポリクローナル抗血清組成物は、ヒト個体における免疫原性が低下している可能性がある。
【0118】
一度多様化されたヒト(化)免疫グロブリン分子を生成することができるヒト以外のトランスジェニック動物が作られると(さらに後記で説明するように)、抗原で動物を免疫することによりヒト(化)免疫グロブリンおよび抗原に対するヒト(化)抗体調製物を容易に得ることができる。種々の抗原を用いてトランスジェニック宿主動物を免疫することができる。かかる抗原には、微生物、例えばウイルスおよび単細胞生物(例えば細菌および菌類)、生存、弱毒化もしくは死亡した、微生物のフラグメント、または微生物から単離された抗原分子が含まれる。
【0119】
動物を免疫するのに用いるための好ましい細菌性抗原には、莢膜多糖類5および8型のような黄色ブドウ球菌からの精製抗原、アルファ毒素のような毒性因子の組換え体、アドヘシン結合タンパク質、コラーゲン結合タンパク質、およびフィブロネクチン結合タンパク質が含まれる。好ましい細菌性抗原にはまた黄色ブドウ球菌、緑膿菌、腸球菌、エンテロバクターおよび肺炎桿菌の弱毒体、またはこれらの細菌細胞からの培養上澄みも含まれる。免疫に用いることができるその他の細菌性抗原には精製リポ多糖類(LPS)、莢膜抗原、莢膜多糖類および/または外膜タンパク質の組換え体、フィブロネクチン結合タンパク質、内毒素、ならびに緑膿菌、腸球菌、エンテロバクターおよび肺炎桿菌の外毒素が含まれる。
【0120】
真菌に対する抗体の作成のための好ましい抗原には、真菌の弱毒体またはその外膜タンパク質が含まれ、その真菌には、限定するものではないがカンジダ・アルビカンス、カンジダ・パラプシローシス、カンジダ・トロピカリスおよびクリプトコッカス・ネオホルマンスが含まれる。
【0121】
ウイルスに対する抗体を作成するために免疫で用いるための好ましい抗原には、エンベロープタンパク質およびウイルスの弱毒体が含まれ、そのウイルスには、限定するものではないが呼吸器多核体ウイルス(RSV)(特にFプロテイン)、C型肝炎ウイルス(HCV)、B型肝炎ウイルス(HBV)、サイトメガロウイルス(CMV)、EBVおよびHSVが含まれる。
【0122】
単離された腫瘍細胞または腫瘍細胞系を有するトランスジェニック動物を免疫することにより癌の処置のための治療用抗体を作成することができ、腫瘍関連抗原には、限定するものではないがHer−2−neu抗原(それに対する抗体は乳癌の処置に有用である);CD19、CD20、CD22およびCD53抗原(それに対する抗体はB細胞リンパ腫の処置に有用である)、(3)前立腺特異的膜抗原(PMSA)(それに対する抗体は前立腺癌の処置に有用である)、および17−1A分子(それに対する抗体は結腸癌の処置に有用である)が含まれる。
【0123】
任意の都合のよい様式で、補助剤を伴うかまたは伴わずに抗原をトランスジェニック宿主動物に投与することができ、そして予め決定されたスケジュールに従って投与することができる。
【0124】
免疫後、抗原に特異的な医薬品等級のポリクローナル抗体の精製のために免疫したトランスジェニック動物からの血清または乳を分画することができる。トランスジェニック鳥類の場合、卵黄を分画することにより抗体を作ることもできる。クロマトグラフィー(親和性、イオン交換、ゲルろ過等)、硫酸アンモニウムのような塩、エタノールのような有機溶媒またはポリエチレングリコールのようなポリマーを用いる選択的沈殿により、濃縮され、精製された免疫グロブリン分画を得ることができる。
【0125】
分画されたヒト(化)抗体をヒトでの静脈内投与に適当な無毒性のパイロジェン不含培地、例えば滅菌緩衝食塩水に溶解または希釈することができる。
【0126】
投与に用いられる抗体調製物は一般に、免疫グロブリン濃度が0.1から100mg/ml、さらに通常的には1から10mg/mlであることを特徴とする。抗体調製物は種々のアイソタイプの免疫グロブリンを含有できる。これに代えて、抗体調製物はただ1つのアイソタイプ、または多くの選択されたアイソタイプの抗体を含有することができる。
【0127】
ヒト(化)モノクローナル抗体を作るために、その動物の内因性免疫グロブリンを発現するB細胞が枯渇している、免疫トランスジェニック動物から脾臓細胞を単離する。単離した脾臓細胞を、ハイブリドーマの生成のために形質転換された細胞系との細胞融合に用いるか、または抗体コード化cDNAを標準的な分子生物学技術によりクローン化し、そしてトランスフェクトされた細胞中で発現させる。モノクローナル抗体を作るための手順は当分野において十分に確立されている。例えば欧州特許出願第0583980Al号(「Method For Generating Monoclonal Antibodies From Rabbits」)、米国特許第4977081号(「Stable Rabbit−Mouse Hybridomas And Secretion Products Thereof」)、国際公開公報第97/16537号(「Stable Chicken B−cell Line And Method of Use Thereof」)、および欧州特許第0491057B1号(「Hybridoma Which Produces Avian Specific Immunoglobulin G」)(それらの開示は出典明示により本明細書の一部とする)参照。クローン化されたcDNAからのモノクローナル抗体のインビトロ生成はAndris−Widhopfら、「Methods for the generation of chicken monoclonal antibody fragments by phage display」、J lmmunol Methods 242:159(2000)およびBurton, D. R.、「Phage display」、Immunotechnology 1:87(1995)(それらの開示は出典明示により本明細書の一部とする)により記載されている。
【0128】
たいていの場合、抗体調製物は未修飾免疫グロブリン、すなわち例えば化学物質または酵素によりさらなる修飾を加えていない動物から調製したヒト(化)抗体からなる。これに代えて免疫グロブリン分画を酵素消化(例えばペプシン、パパイン、プラスミン、グリコシダーゼ、ヌクレアーゼ等で)、加熱等のような処理に供する、および/またはさらに分画することができる。
【0129】
本発明の好ましい実施態様はヒト化抗体を生成するヒト以外のトランスジェニック動物における内因性免疫グロブリン生成の抑制のための方法に向けられており、それにより望ましいヒト(化)免疫グロブリンの富化が可能になる。1つの実施態様では、当分野において公知の方法を用いてヒト(化)免疫グロブリン遺伝子を含む導入遺伝子、自己切断ペプチドおよびリコンビナーゼをトランスジェニック動物に導入し、外因性B細胞と称されるB細胞における付随するヒト(化)免疫グロブリンおよびリコンビナーゼ遺伝子の発現を確実にする。本明細書に記載した、または当分野において周知の任意の種々のリコンビナーゼ、自己切断ペプチドまたは免疫グロブリン遺伝子を導入遺伝子で用いることができる。さらにこの実施態様では、内因性免疫グロブリンを発現するB細胞における自殺遺伝子の選択的発現により内因性免疫グロブリン生成の抑制が達成され、そしてしたがって細胞死により枯渇する。対応して、導入遺伝子の発現によりリコンビナーゼ媒介機構を介して自殺遺伝子は外因性B細胞のゲノムから切除される。したがって外因性B細胞は生存し、そしてヒト(化)免疫グロブリンをコードする導入遺伝子を産生的に生成する。以前に記載した、および当分野において公知の様々な自殺遺伝子は本発明の実施態様である。この実施態様の1つの態様では、導入遺伝子を介して自殺遺伝子をトランスジェニック動物のゲノムに導入し、そしてその発現を免疫細胞特異的プロモーターにより、好ましくはB細胞特異的プロモーターにより駆動してB細胞において自殺遺伝子を選択的に発現させ、したがって非B細胞集団の不必要な細胞死を防御する。本明細書にて記載した、または当分野において周知の種々の免疫細胞およびB細胞特異的プロモーターを用いてB細胞において自殺遺伝子を選択的に発現させることができる。別の実施態様では、本発明で用いるトランスジェニック動物は遺伝子変換動物であるか、または若年期に停止する遺伝子再構成により抗体多様化を被ることができる。さらに前記した方法を用いて作成した遺伝子導入ベクターおよびトランスジェニック動物もまた本発明の実施態様である。
【0130】
本発明をさらに説明するが、決して以下の実施例により限定されるものではない。
【実施例】
【0131】
実施例1
loxPが導入されたDT−A発現ベクターの構築
DT−A自殺遺伝子のB細胞特異的発現を達成するために、ウサギカッパ1をコードするBACクローン179L1(ジェンバンク受け入れ番号AY495827)を修飾する。V1の下流のスペーサーからJ遺伝子座を通るカッパ1遺伝子座、イントロンエンハンサー、定常領域をコードするエクソン、3’エンハンサーの下流の配列に対する3’エンハンサーを含むBACクローンの46kbフラグメントをETクローニングによりサブクローニングする。さらなるゲンタマイシン選択カセットを有するpBELOBACベクター骨格をBAC179L1に相同な50bpを有するプライマーでPCR増幅する。加えて順方向プライマーはAttBインテグラーゼ認識部位およびPvuI制限酵素認識部位を有する。加えて逆方向プライマーはPvuI制限酵素認識部位を有する。
【0132】
【化1】
【0133】
【化2】
ETクローニングのために、PCR生成物をBAC179L1ならびに誘導ラムダファージ組換え酵素Redα、Redβおよびγを含有するストレプトマイシン抵抗性大腸菌株に形質転換する。これらの組換えタンパク質を同時トランスフェクトしたプラスミド(プラスミドpSC101−γβαを有するDH10B大腸菌)またはゲノムに組み込まれたラムダプロファージ(DY380大腸菌株)のいずれかから発現させる。ゲンタマイシンを用いて陽性クローン179L1(46kb)を選択し、そして制限酵素消化により確認する。
【0134】
5’から3’の2つのSV40ポリA部位−loxP部位−ウサギカッパ1プロモーター−DT−A−FRT部位−loxP部位−2つのSV40ポリA部位からなるDNAフラグメントを化学的に合成する。FLP媒介の組み込みによりカナマイシン選択カセットをFRT部位に導入する。サブクローニングされた179L1の46kbフラグメント(46kb)に相同な50bpを有するプライマーで合成フラグメントをPCR増幅する。ETクローニングによりJ遺伝子座を合成DT−Aフラグメントに対して変化させる。カナマイシンを用いて陽性クローンを選択し、そして制限酵素分析により確認する。
【0135】
再度FLP媒介組換えによりカナマイシン選択カセットを除去する。カナマイシン抵抗性の喪失により陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより確認する。
【0136】
最終構築物はV1の下流のスペーサー、合成DT−Aフラグメント、イントロンエンハンサー、定常領域をコードするエクソン、3’エンハンサーの下流の配列に対する3’エンハンサーを含む。
【0137】
構築物をトランスジェニック動物の作成に用いる。
【0138】
実施例2
IgMの膜形態、2A自己切断ペプチド、Creリコンビナーゼからなる融合タンパク質をコードするヒト(化)重鎖遺伝子座の構築
定常、可変、および連結遺伝子セグメントまたは3’エンハンサー領域に特異的なプローブを用いてゲノムDNAライブラリーからウサギ免疫グロブリン重鎖遺伝子座配列を含有するBACおよびフォスミドクローンを単離した。単離したBAC27N5(ジェンバンク受け入れ番号AY386696)、219D23(ジェンバンク受け入れ番号AY386695)、225P18(ジェンバンク受け入れ番号AY386697)、38A2(ジェンバンク受け入れ番号AY386694)およびフォスミドFos15B(ジェンバンク受け入れ番号AY3866968)をシークエンシングした(Rosら、Gene 330:49−59)。
【0139】
ETクローニングによる大腸菌における相同組換えにより選択した免疫グロブリンコード化配列を対応するヒト対応物と交換した(E−Chiang Leeら、Genomics 73:56−65;Daiguan Yuら、PNAS 97:5978−5983;Muyrersら、Nucleic Acids Research 27:1555−1557;Zhangら、Nature Biotechnology 18:1314−1317)。
【0140】
これに代えて、インビトロでライゲーションおよび続く大腸菌の形質転換によりDNAフラグメントを組換えした。インビトロライゲーションおよび形質転換、ETクローニングにより、またはCreリコンビナーゼ媒介組み込みによりBACおよび/またはFosl5Bまたはその部分を組み合わせた。
【0141】
ETクローニングのために標的配列を含有するベクターを、誘導ラムダファージ組換え酵素Redα、Redβおよびγを含有するストレプトマイシン抵抗性大腸菌株に形質転換した。同時トランスフェクトしたプラスミド(プラスミドpSC101−γβαを有するDH10B大腸菌)またはゲノムに組み込まれたラムダプロファージ(DY380大腸菌株)のいずれかからこれらの組換えタンパク質を発現させた。ETクローニング手順は2つの相同組換え工程を包含した。
【0142】
第1の工程では、標的遺伝子座を選択−対抗選択カセット(例えばネオマイシンに対する抵抗性(neo)およびストレプトマイシンに対する感受性(rpsL)を付与するneo−rpsL)により置き換えた。neo−抵抗性コロニーを単離した後、相同組換えによる選択カセットの挿入を制限酵素分析および部分的シークエンシングにより確認した。
【0143】
第2の工程では、rpsL−neo選択カセットを新しい配列と交換した。制限分析およびシークエンシングによりストレプトマイシン抵抗性クローンを分析した。ETクローニング手順に用いたフラグメントは20から50bp長のフランキング配列を有し、それは標的配列と同一であった。ライゲーションに用いた配列はその3’および5’末端で適切な制限酵素部位を有した。これらの部位は天然発生部位であるか、または適切な部位を含有するプライマーを用いるPCRにより導入されたかのいずれかであった。
【0144】
これに代えて配列を合成により作成した。
【0145】
ETクローニングによりウサギJH、BAC219D23のCμおよびBAC27N5のCγの対応するそのヒト対応物との置き換えによりヒト化重鎖を構築した。ETクローニング手順に用いたヒト配列をヒトゲノムDNAからPCRにより増幅した。
【0146】
ヒトCμ、CγおよびJH遺伝子セグメントをウサギ標的配列に相同な50bpを有するプライマーを用いて増幅した。
【0147】
【化3】
BACクローン225P18のクローン219D23との、およびBAC27N5のフォスミド15Bとのライゲーションの後、ライゲートした構築物を大腸菌に形質転換し、そしてCreリコンビナーゼ媒介挿入によりつないだ。これにより18ウサギ可変遺伝子、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCε、ウサギCα4および3’エンハンサーエレメントからなる機能的遺伝子座に至った。
【0148】
自己切断F2Aペプチドの配列およびCREリコンビナーゼのコドン最適化コード化配列(iCRE)を含むDNAフラグメントを化学的に合成する。
【0149】
【化4】
ETクローニングによりIgMのM2膜エクソンを合成M2−F2A−iCREフラグメントに対して変化させる。
【0150】
ETクローニング手順は2つの相同組換え工程を包含した。第1の工程では、標的遺伝子座を選択−対抗選択カセット(例えばネオマイシンに対する抵抗性(neo)およびストレプトマイシンに対する感受性(rpsL)を付与するneo−rpsL)により置き換える。このためにrpsL−neo選択カセットを標的配列に相同な50bpを有するプライマーを用いてPCR増幅する。neo−抵抗性コロニーを単離した後、相同組換えによる選択カセットの挿入を制限酵素分析および部分的シークエンシングにより確認する。
【0151】
第2の工程では、rpsL−neo選択カセットを合成M2−F2A−iCREフラグメントと交換する。ストレプトマイシン抵抗性により陽性クローンを同定し、そして制限分析およびシークエンシングにより分析する。
【0152】
得られたBACをトランスジェニック動物作成に使用する。
【0153】
実施例3
ヒト化軽鎖遺伝子座の構築
ウサギゲノムBACライブラリーのスクリーニングによりウサギ軽鎖K1遺伝子セグメント(ジェンバンク受け入れ番号AY495827、AY495826)を含有する2つのBAC(179L1および215M22)の同定に至った。
【0154】
前記したようにETクローニングによりウサギCκ1をヒトCκアロタイプKm3と交換した。
【0155】
ヒトCκ(アロタイプKm3)を標的配列に相同な50bpフランキング配列を有するプライマーを用いてPCRにより増幅した。
【0156】
【化5】
ホモロジーアームはウサギ生殖細胞系カッパ(b5;ジェンバンク受け入れ番号K01363)の公開されている配列に基づいて設計され、そしてCκのイントロン−エクソン境界に適合した。
【0157】
BAC179L1におけるヒトCκに対するウサギCκの交換をシークエンシングにより確認した。
【0158】
BAC179L1−huCkを2つのETクローニングにより修飾した。BAC179L1に相同な50bpを有するプライマーを用いてネオマイシン選択カセットを増幅した。加えて順方向プライマーはi−CeuIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。ネオマイシンで陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。BAC179L1に相同な50bp配列を含有するプライマーを用いてゼオシン選択カセットを増幅した。加えて順方向プライマーはi−SceIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。ゼオシンで陽性クローンを選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0159】
BAC215M22をETクローニングにより修飾した。BAC215M22に相同な50bpを有するプライマーを用いてゲンタマイシン抵抗性遺伝子を増幅した。加えて順方向プライマーはi−CeuIメガヌクレアーゼ部位を有し、そして逆方向プライマーはi−SceIメガヌクレアーゼ部位を有した。PCR生成物をETクローニングに使用した。得られたクローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0160】
【化6】
【0161】
【化7】
修飾されたBAC179L1および225M22をi−CeuIおよびi−SceIで切断した。98kbおよび132kbのフラグメントを精製し、そしてライゲートした。得られたクローンをカナマイシンおよびクロラムフェニコールで選択し、そして制限酵素消化、i−SceIおよびi−CeuI制限部位を含有する領域のPCR、ならびにシークエンシングにより正確性を確かめた。得られたBACを179−215−huCkと称した。
【0162】
ETクローニングによりBAC179−215−huCkのウサギJk1およびJk2を合成ヒト再構成カッパ1VJ遺伝子により置き換えた。ウサギプロモーター、ウサギリーダー、ウサギイントロンおよびヒトVJ遺伝子を有するDNAフラグメントを化学的に合成した。合成ヒトVJのコドン使用を最適化してウサギVカッパ遺伝子に対して最高のDNA配列相同性を達成した。
【0163】
BAC179L1に相同な50bpを有する順方向プライマーならびにゲンタマイシン抵抗性遺伝子およびFRT部位に相同な逆方向プライマーを用いて合成ヒトVJをPCR増幅した。FRT部位を有する順方向プライマーならびにBAC179L1およびFRT部位に相同な50bpを有する逆方向プライマーを用いてゲンタマイシン抵抗性遺伝子を増幅した。合成ヒトVJ遺伝子のための順方向プライマーおよびゲンタマイシン抵抗性遺伝子のための逆方向プライマーを用いてオーバーラップ伸長PCRによりヒト合成ヒトVJおよびゲンタマイシン抵抗性遺伝子を組み合わせた。得られたフラグメントをETクローニングに使用した。陽性クローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0164】
【化8】
部位特異的組換えによりFlpリコンビナーゼの発現を介してゲンタマイシン抵抗性遺伝子を除去した。組換えの後、1つのFRTが残った。ETクローニングによりFRT部位を削除した。合成ヒトVJからの232bpフラグメントをPCRにより増幅し、そしてETクローニングに使用した。得られたコロニーをFRT部位の喪失に関してPCRによりスクリーニングし、そしてシークエンシングにより確認した。
【0165】
ETクローニングによりBAC179−215−huCkのネオマイシン抵抗性遺伝子を置き換えた。BAC179−215−huCkに相同な50bpを有するプライマーを用いてゲンタマイシン抵抗性(pRep−Genta;Genebridges)遺伝子をPCRにより増幅した。加えて順方向プライマーはloxP部位、attB部位およびPvuI制限部位を有した。得られたクローンをゲンタマイシンで選択し、そして制限酵素消化およびシークエンシングにより正確性を確かめた。
【0166】
得られたBACをトランスジェニック動物の作成に使用した。
【0167】
【化9】
実施例4
ヒト化重鎖免疫グロブリンを発現するトランスジェニックマウスおよびウサギの作成
カッパ軽鎖プロモーター/エンハンサー配列の制御下でヒト化重および軽鎖免疫グロブリン遺伝子座およびloxPが導入されたジフテリア毒素A遺伝子を含有するトランスジェニックウサギおよびマウスを、受精卵母細胞の前核へのDNAの注射および、続く里親への胚の移入により作成する。トランスジェニック創始動物をPCRにより同定する。ヒト(化)免疫グロブリンMおよびGの発現をELISAにより測定する。ヒト化IgGの発現は1〜5mg/mlである。マウスおよびウサギIgGの発現は各々1〜5μg/mlである。
【0168】
実施例5
ヒト化重鎖免疫グロブリンを発現するトランスジェニックニワトリの作成
精巣媒介の遺伝子移入によりトランスジェニックニワトリを作成する。DNA構築物を
(50μg)を0.9%NaCl 500μl中リポフェクション試薬(スーパーフェクト)250μlと混合し、そしてオンドリの精巣に注射する。3〜4週後、トランスジェニック精子を有するオンドリをPCR分析により同定し、そしてメンドリと交配させる。トランスジェニック子孫をPCRにより同定する。ヒト化IgGの発現は1〜5mg/mlである。ニワトリIgYの発現は各々1〜5μg/mlである。
【0169】
本開示全体にわたって引用した全ての参照文献はそこに引用された参照文献と一緒に出典明示により本明細書の一部とする。
【0170】
本発明を特定の実施態様を参照することにより説明したが、そのように限定されるものではない。種々の修飾を容易に利用でき、そして本発明が取り組む方式を実質的に変化させることなく実施できることは当業者には理解されよう。かかる修飾の全てが、具体的には本明細書にて特許請求される本発明の範囲内であると意図される。
【図面の簡単な説明】
【0171】
【図1】外因性免疫グロブリン(Ig)発現B細胞および内因性免疫グロブリン(Ig)発現B細胞で生じる事象の説明図である。2つの導入遺伝子(Ig重鎖遺伝子座およびDTA)を有する細胞では、遺伝子導入免疫グロブリン重鎖遺伝子座の産生性再構成の結果、重鎖(HC)およびCreリコンビナーゼの発現に至る。続いて、Cre媒介の組換えの結果、DTA発現カセットのループアウトに至る。したがって、DTAは発現されず、そして遺伝子導入免疫グロブリンHC遺伝子座を発現するかかる外因性B細胞は生存する。内因性HC遺伝子座の産生性再構成の結果、Creリコンビナーゼの発現には至らない。したがって、DTA発現カセットは活性化され、そしてかかる内因性B細胞は死亡する。
【特許請求の範囲】
【請求項1】
ヒト以外のトランスジェニック動物の内因性免疫グロブリンを生成するB細胞において少なくとも1つの自殺遺伝子を選択的に発現するが、外因性免疫グロブリンを生成するB細胞では発現しないこと含み、それにより内因性免疫グロブリンを生成するB細胞は枯渇し、そして内因性免疫グロブリンの生成は抑制されるが該外因性免疫グロブリンの生成は抑制されない外因性免疫グロブリン導入遺伝子座を担持する該ヒト以外のトランスジェニック動物のB細胞における内因性免疫グロブリン生成の選択的抑制のための方法。
【請求項2】
該外因性免疫グロブリンがヒト(化)免疫グロブリン重および/または軽鎖配列である請求項1に記載の方法。
【請求項3】
該ヒト以外のトランスジェニック動物のB細胞に導入された該自殺遺伝子がB細胞特異的プロモーターの制御下にあり、そして組換え配列によりフランキングされる請求項1または請求項2に記載の方法。
【請求項4】
該ヒト(化)免疫グロブリン鎖導入遺伝子座をさらに該組換え配列を認識するリコンビナーゼをコードする発現構築物の一部として該ヒト以外のトランスジェニック動物のB細胞に導入し、ここで該自殺遺伝子の発現が該ヒト(化)免疫グロブリン導入遺伝子座を発現するB細胞における該リコンビナーゼの発現により不活性化される請求項3に記載の方法。
【請求項5】
該自殺遺伝子が細菌性、真菌性、殺虫性および植物の毒素からなる群から選択される請求項1または請求項2に記載の方法。
【請求項6】
該自殺遺伝子がジフテリア毒素A鎖である請求項1または請求項2に記載の方法。
【請求項7】
該自殺遺伝子がプロドラッグ変換酵素である請求項1または請求項2に記載の方法。
【請求項8】
プロドラッグ変換酵素が非哺乳動物起源である請求項7に記載の方法。
【請求項9】
非哺乳動物起源の該プロドラッグ変換酵素がウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、β−ラクタマーゼ、多剤活性化酵素(MDAE)、β−ガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)からなる群から選択される請求項8に記載の方法。
【請求項10】
プロドラッグ変換酵素がヒト起源である請求項7に記載の方法。
【請求項11】
ヒト起源のプロドラッグ変換酵素がデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)からなる群から選択される請求項10に記載の方法。
【請求項12】
該リコンビナーゼがCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4リコンビナーゼ、TP901−1リコンビナーゼ、原核細胞性トランスポーゼス、真核細胞性トランスポーゼス、ウイルス性レトロトランスポーゼス、ショウジョウバエコピア様レトロトランスポーゼスおよび非ウイルス性レトロトランスポーゼスからなる群から選択される請求項4に記載の方法。
【請求項13】
該トランスポーゼスまたはレトロトランスポーゼスがTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される請求項12に記載の方法。
【請求項14】
該ヒト以外のトランスジェニック動物が若年期の遺伝子再構成により抗体多様化を実質的に停止する請求項1または請求項2に記載の方法。
【請求項15】
該ヒト以外のトランスジェニック動物が生まれて1か月以内に抗体多様化を実質的に停止する請求項14に記載の方法。
【請求項16】
該ヒト以外のトランスジェニック動物がげっ歯類、ウサギ、鳥類、ウシ、ブタ、ヒツジ、ヤギおよびウマからなる群から選択される請求項1または請求項2に記載の方法。
【請求項17】
該げっ歯類がマウスまたはラットである請求項16に記載の方法。
【請求項18】
(1)導入遺伝子、(2)ヒトまたはヒト化免疫グロブリン重および/または軽鎖導入遺伝子座(3)自己切断ペプチドならびに(4)リコンビナーゼを含む遺伝子導入発現構築物。
【請求項19】
(1)導入遺伝子および(2)B細胞特異的プロモーターの制御下であり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子を含む遺伝子導入発現構築物。
【請求項20】
ヒトまたはヒト化免疫グロブリン重および/または軽鎖遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子ならびにB細胞特異的プロモーターの制御下であり、そして該リコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物。
【請求項21】
該リコンビナーゼがCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4リコンビナーゼおよびTP901−1リコンビナーゼからなる群から選択される部位特異的リコンビナーゼである請求項18、19または20に記載の遺伝子導入発現構築物。
【請求項22】
該リコンビナーゼが原核細胞性または真核細胞性トランスポーゼスのいずれかである請求項18、19または20に記載の遺伝子導入発現構築物。
【請求項23】
該リコンビナーゼがウイルス性、ショウジョウバエコピア様または非ウイルス性レトロトランスポーゼスのいずれかである請求項22に記載の遺伝子導入発現構築物。
【請求項24】
該レトロトランスポーゼスがTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される請求項23に記載の遺伝子導入発現構築物。
【請求項25】
該組換え部位がloxP部位、FRT部位、細菌性ゲノム組換え部位およびファージ組換え部位からなる群から選択される請求項19または20に記載の遺伝子導入発現構築物。
【請求項26】
該細菌性ゲノム組換え部位がattBであり、そして該ファージ組換え部位がattPまたは偽attPまたは偽attB部位である請求項25に記載の遺伝子導入発現構築物。
【請求項27】
該自己切断ペプチドがウイルス性2A/2Bまたは2A様/2B配列から得られる請求項18または20に記載の遺伝子導入発現構築物。
【請求項28】
該ウイルスがピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーからなる群からまたはC型ロタウイルスファミリーから選択される請求項27に記載の遺伝子導入発現構築物。
【請求項29】
該ウイルスが口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルス、またはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)からなる群から選択される請求項27に記載の遺伝子導入発現構築物。
【請求項30】
CD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b、mb−1、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖および免疫グロブリンJ鎖またはその修飾体からなる群から選択されるプロモーター/エンハンサーを用いてB細胞において該自殺遺伝子が特異的に発現される請求項19または20に記載の遺伝子導入発現構築物。
【請求項31】
該B細胞特異的プロモーター/エンハンサーがカッパ軽鎖遺伝子プロモーターまたはその修飾体である請求項30に記載の遺伝子導入発現構築物。
【請求項32】
請求項18および19または20に記載の遺伝子導入発現構築物を発現するヒト以外のトランスジェニック動物。
【請求項33】
実質的に遺伝子変換により抗体多様性を作成する請求項32に記載のヒト以外のトランスジェニック動物。
【請求項34】
げっ歯類、ウサギ、ニワトリ、シチメンチョウ、アヒルおよびガチョウを含む鳥類からなる群から選択される請求項32に記載のヒト以外のトランスジェニック動物。
【請求項35】
マウスまたはラットのいずれかである請求項34に記載のヒト以外のトランスジェニック動物。
【請求項1】
ヒト以外のトランスジェニック動物の内因性免疫グロブリンを生成するB細胞において少なくとも1つの自殺遺伝子を選択的に発現するが、外因性免疫グロブリンを生成するB細胞では発現しないこと含み、それにより内因性免疫グロブリンを生成するB細胞は枯渇し、そして内因性免疫グロブリンの生成は抑制されるが該外因性免疫グロブリンの生成は抑制されない外因性免疫グロブリン導入遺伝子座を担持する該ヒト以外のトランスジェニック動物のB細胞における内因性免疫グロブリン生成の選択的抑制のための方法。
【請求項2】
該外因性免疫グロブリンがヒト(化)免疫グロブリン重および/または軽鎖配列である請求項1に記載の方法。
【請求項3】
該ヒト以外のトランスジェニック動物のB細胞に導入された該自殺遺伝子がB細胞特異的プロモーターの制御下にあり、そして組換え配列によりフランキングされる請求項1または請求項2に記載の方法。
【請求項4】
該ヒト(化)免疫グロブリン鎖導入遺伝子座をさらに該組換え配列を認識するリコンビナーゼをコードする発現構築物の一部として該ヒト以外のトランスジェニック動物のB細胞に導入し、ここで該自殺遺伝子の発現が該ヒト(化)免疫グロブリン導入遺伝子座を発現するB細胞における該リコンビナーゼの発現により不活性化される請求項3に記載の方法。
【請求項5】
該自殺遺伝子が細菌性、真菌性、殺虫性および植物の毒素からなる群から選択される請求項1または請求項2に記載の方法。
【請求項6】
該自殺遺伝子がジフテリア毒素A鎖である請求項1または請求項2に記載の方法。
【請求項7】
該自殺遺伝子がプロドラッグ変換酵素である請求項1または請求項2に記載の方法。
【請求項8】
プロドラッグ変換酵素が非哺乳動物起源である請求項7に記載の方法。
【請求項9】
非哺乳動物起源の該プロドラッグ変換酵素がウイルス性チミジンキナーゼ(TK)、細菌性サイトシンデアミナーゼ(CD)、細菌性カルボキシペプチダーゼG2(CPG2)、プリンヌクレオチドホスホリラーゼ(PNP)、チミジンホスホリラーゼ(TP)、ニトロリダクターゼ(NR)、D−アミノ酸オキシダーゼ(DAAO)、キサンチン−グアニンホスホリボシルトランスフェラーゼ(XGPRT)、ペニシリン−Gアミダーゼ(PGA)、β−ラクタマーゼ、多剤活性化酵素(MDAE)、β−ガラクトシダーゼ(β−Gal)、西洋ワサビベルオキシダーゼ(HRP)およびデオキシリボヌクレオチドキナーゼ(DRNK)からなる群から選択される請求項8に記載の方法。
【請求項10】
プロドラッグ変換酵素がヒト起源である請求項7に記載の方法。
【請求項11】
ヒト起源のプロドラッグ変換酵素がデオキシシチジンキナーゼ(dCK)、カルボキシルエステラーゼ(CE)、カルボキシペプチダーゼA(CPA)、β−グルクロニダーゼ(−Glu)およびチトクロームP450(CYP)からなる群から選択される請求項10に記載の方法。
【請求項12】
該リコンビナーゼがCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4リコンビナーゼ、TP901−1リコンビナーゼ、原核細胞性トランスポーゼス、真核細胞性トランスポーゼス、ウイルス性レトロトランスポーゼス、ショウジョウバエコピア様レトロトランスポーゼスおよび非ウイルス性レトロトランスポーゼスからなる群から選択される請求項4に記載の方法。
【請求項13】
該トランスポーゼスまたはレトロトランスポーゼスがTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される請求項12に記載の方法。
【請求項14】
該ヒト以外のトランスジェニック動物が若年期の遺伝子再構成により抗体多様化を実質的に停止する請求項1または請求項2に記載の方法。
【請求項15】
該ヒト以外のトランスジェニック動物が生まれて1か月以内に抗体多様化を実質的に停止する請求項14に記載の方法。
【請求項16】
該ヒト以外のトランスジェニック動物がげっ歯類、ウサギ、鳥類、ウシ、ブタ、ヒツジ、ヤギおよびウマからなる群から選択される請求項1または請求項2に記載の方法。
【請求項17】
該げっ歯類がマウスまたはラットである請求項16に記載の方法。
【請求項18】
(1)導入遺伝子、(2)ヒトまたはヒト化免疫グロブリン重および/または軽鎖導入遺伝子座(3)自己切断ペプチドならびに(4)リコンビナーゼを含む遺伝子導入発現構築物。
【請求項19】
(1)導入遺伝子および(2)B細胞特異的プロモーターの制御下であり、そしてリコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子を含む遺伝子導入発現構築物。
【請求項20】
ヒトまたはヒト化免疫グロブリン重および/または軽鎖遺伝子座、自己切断ペプチドおよびリコンビナーゼをさらに含む第1の導入遺伝子ならびにB細胞特異的プロモーターの制御下であり、そして該リコンビナーゼにより認識される組換え部位によりフランキングされた自殺遺伝子をさらに含む第2の導入遺伝子を含む遺伝子導入発現構築物。
【請求項21】
該リコンビナーゼがCre、Cre様、Flp、φC31、λインテグラーゼ、ファージR4リコンビナーゼおよびTP901−1リコンビナーゼからなる群から選択される部位特異的リコンビナーゼである請求項18、19または20に記載の遺伝子導入発現構築物。
【請求項22】
該リコンビナーゼが原核細胞性または真核細胞性トランスポーゼスのいずれかである請求項18、19または20に記載の遺伝子導入発現構築物。
【請求項23】
該リコンビナーゼがウイルス性、ショウジョウバエコピア様または非ウイルス性レトロトランスポーゼスのいずれかである請求項22に記載の遺伝子導入発現構築物。
【請求項24】
該レトロトランスポーゼスがTn1、Tn2、Tn3、Tn4、Tn5、Tn6、Tn9、Tn10、Tn30、Tn101、Tn501、Tn903、Tnl000、Tn1681、Tn2901、ショウジョウバエマリナー、sleeping beautyトランスポーゼス、ショウジョウバエPエレメント、トウモロコシAc、Ds、Mp、Spm、En、ドット、Mu、I、L1、Tol2 Tc1、Tc3、マリナー(Himar1)、マリナー(mos1)およびミノスからなる群から選択される請求項23に記載の遺伝子導入発現構築物。
【請求項25】
該組換え部位がloxP部位、FRT部位、細菌性ゲノム組換え部位およびファージ組換え部位からなる群から選択される請求項19または20に記載の遺伝子導入発現構築物。
【請求項26】
該細菌性ゲノム組換え部位がattBであり、そして該ファージ組換え部位がattPまたは偽attPまたは偽attB部位である請求項25に記載の遺伝子導入発現構築物。
【請求項27】
該自己切断ペプチドがウイルス性2A/2Bまたは2A様/2B配列から得られる請求項18または20に記載の遺伝子導入発現構築物。
【請求項28】
該ウイルスがピコルナウイルス科ウイルスファミリー、ウマ鼻炎A(ERAV)ウイルスファミリー、ピコルナウイルス様昆虫ウイルスファミリーからなる群からまたはC型ロタウイルスファミリーから選択される請求項27に記載の遺伝子導入発現構築物。
【請求項29】
該ウイルスが口蹄疫ウイルス(FMDV)、ウマ鼻炎A(ERAV)ウイルス、またはゾセア・アシグナ(Thosea asigna)ウイルス(TaV)からなる群から選択される請求項27に記載の遺伝子導入発現構築物。
【請求項30】
CD19、CD20、CD21、CD22、CD23、CD24、CD40、CD72、Blimp−1、CD79b、mb−1、チロシンキナーゼblk、VpreB、免疫グロブリンカッパ軽鎖、免疫グロブリンラムダ軽鎖および免疫グロブリンJ鎖またはその修飾体からなる群から選択されるプロモーター/エンハンサーを用いてB細胞において該自殺遺伝子が特異的に発現される請求項19または20に記載の遺伝子導入発現構築物。
【請求項31】
該B細胞特異的プロモーター/エンハンサーがカッパ軽鎖遺伝子プロモーターまたはその修飾体である請求項30に記載の遺伝子導入発現構築物。
【請求項32】
請求項18および19または20に記載の遺伝子導入発現構築物を発現するヒト以外のトランスジェニック動物。
【請求項33】
実質的に遺伝子変換により抗体多様性を作成する請求項32に記載のヒト以外のトランスジェニック動物。
【請求項34】
げっ歯類、ウサギ、ニワトリ、シチメンチョウ、アヒルおよびガチョウを含む鳥類からなる群から選択される請求項32に記載のヒト以外のトランスジェニック動物。
【請求項35】
マウスまたはラットのいずれかである請求項34に記載のヒト以外のトランスジェニック動物。
【図1】

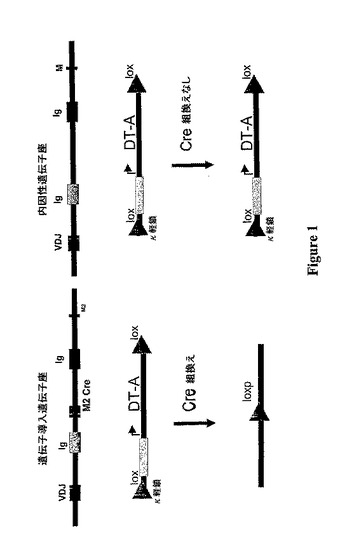
【公表番号】特表2008−517600(P2008−517600A)
【公表日】平成20年5月29日(2008.5.29)
【国際特許分類】
【出願番号】特願2007−538110(P2007−538110)
【出願日】平成17年10月21日(2005.10.21)
【国際出願番号】PCT/US2005/038095
【国際公開番号】WO2006/047367
【国際公開日】平成18年5月4日(2006.5.4)
【出願人】(506011504)セラピューティック ヒューマン ポリクローナルズ, インコーポレイテッド (6)
【Fターム(参考)】
【公表日】平成20年5月29日(2008.5.29)
【国際特許分類】
【出願日】平成17年10月21日(2005.10.21)
【国際出願番号】PCT/US2005/038095
【国際公開番号】WO2006/047367
【国際公開日】平成18年5月4日(2006.5.4)
【出願人】(506011504)セラピューティック ヒューマン ポリクローナルズ, インコーポレイテッド (6)
【Fターム(参考)】
[ Back to top ]