
ヒト化モノクローナル抗体のアミノ酸配列およびヌクレオチド配列、並びにヒト化モノクローナル抗体
【課題】ヒト化モノクローナル抗体のアミノ酸配列およびヌクレオチド配列、並びにその用途を提供する。
【解決手段】腫瘍壊死因子αに結合するヒト化モノクローナル抗体。特定のアミノ酸配列を含む軽鎖の可変領域、及び特定のアミノ酸配列を含む重鎖の可変領域を含み、前記アミノ酸配列は、少なくとも一つのアミノ酸置換を有し、アミノ酸置換は、軽鎖可変領域の第10アミノ酸のイソロイシンのトレオニンへの置換、第18アミノ酸のリジンのアルギニンへの置換、また、重鎖可変領域配列の第2アミノ酸のリジンのグルタミンへの置換、第10アミノ酸のトリプトファンのロイシンへの置換、第18アミノ酸のリジンのアルギニンへの置換、および第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つである、前記ヒト化モノクローナル抗体。
【解決手段】腫瘍壊死因子αに結合するヒト化モノクローナル抗体。特定のアミノ酸配列を含む軽鎖の可変領域、及び特定のアミノ酸配列を含む重鎖の可変領域を含み、前記アミノ酸配列は、少なくとも一つのアミノ酸置換を有し、アミノ酸置換は、軽鎖可変領域の第10アミノ酸のイソロイシンのトレオニンへの置換、第18アミノ酸のリジンのアルギニンへの置換、また、重鎖可変領域配列の第2アミノ酸のリジンのグルタミンへの置換、第10アミノ酸のトリプトファンのロイシンへの置換、第18アミノ酸のリジンのアルギニンへの置換、および第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つである、前記ヒト化モノクローナル抗体。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗腫瘍壊死因子αの抗体に関し、特に、腫瘍壊死因子αを高度に中和することが可能なヒト化モノクローナル抗体およびそのアミノ酸配列に関するものである。
【背景技術】
【0002】
腫瘍壊死因子α(tumor necrosis factor−alpha、TNF−α)は、炎症誘発性サイトカインであり、マクロファージ細胞と単球を含む免疫システム細胞により最初に生成される。腫瘍壊死因子αは、ホモ三量体蛋白質として存在し、各サブユニット中で、最初、一個の26kDaの膜貫通前駆体蛋白質として翻訳される。腫瘍壊死因子α変換酵素(TACE)により、TNF−αの膜貫通領域に近い位置で切割を行なった後、腫瘍壊死因子αの可溶三量体を放出し、エフェクター細胞上の構造区分タイプIとタイプIIの腫瘍壊死因子レセプター(TNFRIとTNFRII)を結合することにより、その活性を発揮する。
【0003】
膜貫通形式の腫瘍壊死因子αも、独特の生物機能が知られており、例えば、細胞間の接触方式における、細胞毒性活性とポリクローナルB細胞の活性化(Mitomaら、2008)が挙げられる。腫瘍壊死因子αが自己免疫過程に対し、特定の影響を有することが既に証明されており、多くの自己免疫疾病の主要な治療目標となっている(Feldmann、2001)。今までのところ、ある抗腫瘍壊死因子αの試薬、例えば、エタネルセプト(etanercept)、アダリムマブ(adalimumab)、及びインフリキシマブ(infliximab)は、既に、米国食品医薬局(FDA)により承認されており、いずれも、主要な薬理学的メカニズムの作用として、可溶形の腫瘍壊死因子αを効果的に中和する能力を有する。しかし、これらの拮抗薬の膜貫通形式の腫瘍壊死因子αに対する結合作用は異なり、臨床疾病上で、異なる結果をもたらす(Taylor、2010)。例えば、エタネルセプトは、膜貫通形式の腫瘍壊死因子αが重要な役割を果たす肉芽腫性疾患の発病に対して臨床的に有効ではない(2008、Mitoma)。よって、抗腫瘍壊死因子αの試薬が、膜貫通形式の腫瘍壊死因子αと結合して、抗体依存性細胞媒介性障害(ADCC)、補体依存性細胞障害(CDC)、アポトーシス、及び外から内へのシグナル伝達メカニズムをトリガーすることができるかどうかが必要条件である。
【0004】
臨床実験におけるマウスモノクローナル抗体の使用による主な障害は、患者にヒト抗マウス抗体反応(HAMA)が生じる可能性があるということである(Owens and Young、1994;Sandhu、1992;Schroffら、1985)。よって、臨床使用の効果を改善するため、遺伝子工程技術を使用して、マウスに属するアミノ酸残基をヒトに対応するアミノ酸残基に置換して、患者の免疫原生の誘発の可能性を減少させることが行なわれている。
【0005】
理想的な抗体のヒト型化は、抗原に対する特異性と親和性を維持すると共に、できる限り、免疫原性を低下させることができる。これまでのところ、抗体のヒト型化は、既に、多くの方式により行なわれており、例えば、マウス抗原と可変領域を結合して、ヒト抗体の定常領域に遺伝子的に融合してなるキメラ抗体が最も早期に試され、免疫原性を減少させている(Morrisonら、1984)。しかし、キメラ抗体には、所望でない抗可変領域反応が生じる(Bruggemannら、1989)。もう一つの方式は、相補性決定領域移植法(CDR−gtafting)であり、齧歯目抗体の相補性決定領域を、ヒト抗体のFvフレームワーク(FR)に転移することを含む(Verhoeyenら、1988)。残念ながら、相補性決定領域とFvフレームワークとの接触面の変化により、抗原との結合が激しく妨害される。最初の相補性決定領域移植抗体は、親の結合親和性を失う傾向にあるので、相補性決定領域のループ構造にとって極めて重要であると見なされるマウスフレームワークアミノ酸の復帰突然変異のための別の作業が必要である(Queenら、1989)。可変領域表面再構成(variable domain resurfacing)によるヒト化は、親抗体の特異性と結合親和性を維持することができる別のアプローチであり、マウスFvフレームワーク中の表面に露出する残基を、一般のヒト抗体中に見出される表面露出残基に代替させることにより、抗体の免疫原性を減少させることができる(Fontayneら、2006;Padlan、1991;Roguskaら、1994;Staelensら、2006;Zhangら、2005)。現在の分子生物技術は、このアミノ酸を変化させる方法を更に容易にしているが、特に、信頼できる抗体のコンピュータモデルが必要であるとき、溶剤中の表面に露出する残基を確認することは依然として困難である(Fontayneら、2006)。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、膜貫通型腫瘍壊死因子αと結合する能力を有するヒト化モノクローナル抗体のアミノ酸配列およびヌクレオチド配列、並びにヒト化モノクローナル抗体を提供することにある。更に本発明の他の目的は、抗体依存性細胞媒介性細胞障害を生じさせ、膜貫通型腫瘍壊死因子αを中和し得、膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物の調製に用いられるヒト化モノクローナル抗体を提供することにある。
【課題を解決するための手段】
【0007】
本発明に係るヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の配列を含む軽鎖の可変領域のアミノ酸配列、及び配列番号2の配列を含む重鎖の可変領域のアミノ酸配列を含み、前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0008】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0009】
また、本発明に係る他のヒト化モノクローナル抗体のアミノ酸配列は、配列番号5の配列を含む軽鎖の可変領域のアミノ酸配列、及び配列番号6の配列を含む重鎖の可変領域のアミノ酸配列を含み、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0010】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0011】
本発明に係るヒト化モノクローナル抗体のヌクレオチド配列は、配列番号7の配列を含む軽鎖の可変領域のヌクレオチド配列、及び配列番号8の配列を含む重鎖の可変領域のヌクレオチド配列を含み、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0012】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0013】
本発明のヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号1の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号2の配列を含む重鎖を含み、前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0014】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0015】
上記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含むことが好ましい。
【0016】
上記ヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMであることが好ましい。
本発明の他のヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含み、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0017】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0018】
上記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含むことが好ましい。
【0019】
上記ヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMであることが好ましい。
【0020】
また、本発明に係る膜貫通型腫瘍壊死因子αを中和する方法は、ヒト化モノクローナル抗体を膜貫通型腫瘍壊死因子αに結合させる工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含むところに要旨を有するものである。
【0021】
また、本発明に係る抗体依存性細胞媒介性細胞障害を生じさせる方法は、ヒト化モノクローナル抗体を対象の膜貫通型腫瘍壊死因子αに結合させる工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含むところに要旨を有するものである。
【0022】
また、本発明に係る膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法は、ヒト化モノクローナル抗体を提供する工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含み、且つ、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【発明の効果】
【0023】
本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αと結合する能力を有し、抗体依存性細胞媒介性細胞障害を生じさせることができる。従って、本発明のヒト化モノクローナル抗体は、抗体依存性細胞媒介性細胞障害を生じさせる用途や、膜貫通型腫瘍壊死因子αを中和する用途や、膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物の用途などに応用することができる。
【図面の簡単な説明】
【0024】
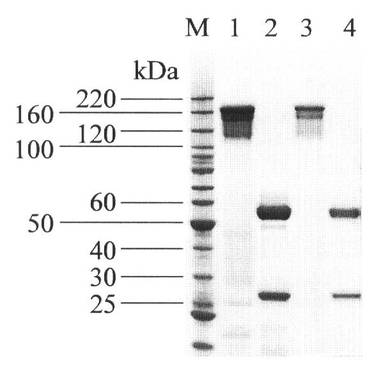
【図1】図1は、精製されたm357IgGとh357IgGのSDS−PAGE分析結果を示す図である。m357IgG(レーン1と2)とh357IgG(レーン3およびレーン4)は、マウス骨髄腫NS0細胞で発現され、蛋白質Aカラムにより培養基から精製された。サンプルは、非還元状態(レーン1と3)と還元状態(レーン2と4)下で、MOPSバッファ溶液により、4〜12%SDS/Bis−Trisポリアクリルアミドゲルで電気泳動された。ゲルはクアシンブルーで染色された。Mは分子量標準品である。
【図2】図2は、m357IgGの軽鎖cDNAとm357IgGの重鎖cDNAの電気泳動分析の結果をそれぞれ示す図である。
【図3A】図3Aは、m357IgGの重鎖可変領域および軽鎖可変領域について、最終の正確な構造を示す図である。
【図3B】図3Bは、m357IgGの分子表面モデルを示す図である。
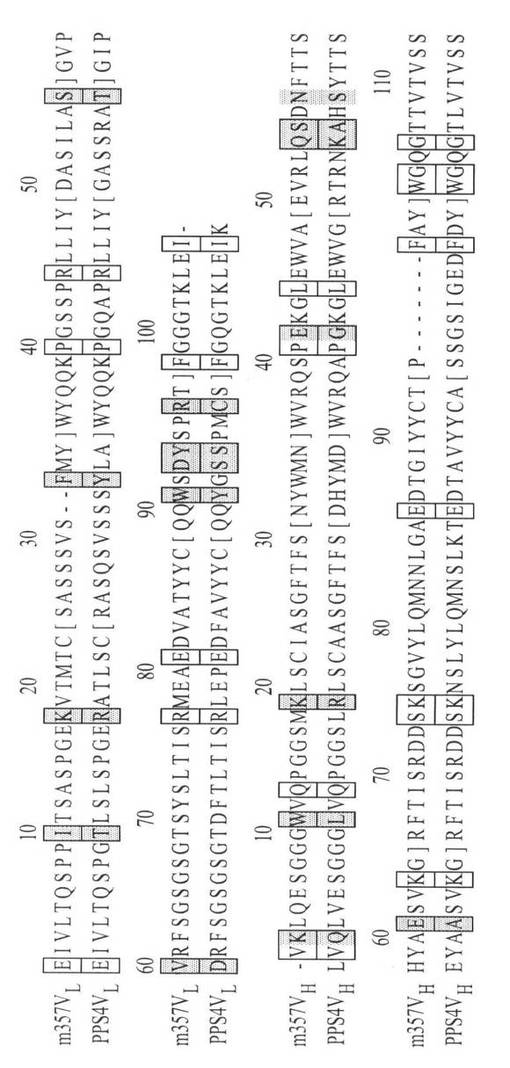
【図4】図4は、m357IgGとPPS4の重鎖可変領域VH(A)、及び軽鎖可変領域VL(B)のアミノ酸配列を示す図である。m357IgGのヒト化のためのヒト表面殘基のアクセプタとして用いたm357IgG Fvと最も相同性が高かったPPS4 Fv配列を、比較のため、PPS4VLおよびPPS4VHとして示す。CDR殘基を括弧([ ])中に示し、保存表面殘基を白色フレームで表示し、非保存表面殘基を灰色フレームで表示した。Kabatの慣例(Kabat、1991)に基づいて、アミノ酸配列をナンバリングした。
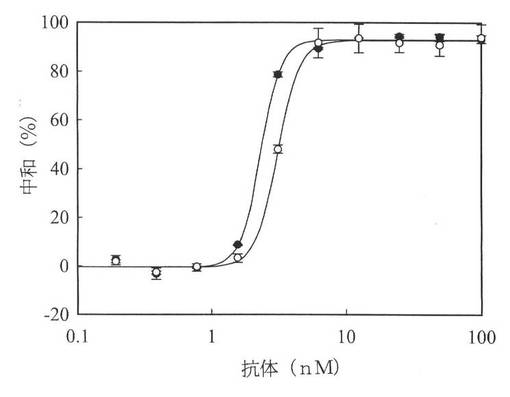
【図5】図5は、m357−IgG抗体とh357−IgG抗体による、L929細胞中の腫瘍壊死因子α媒介細胞毒性の中和を示す図である。種々の濃度のm357−IgG(●)抗体とh357−IgG(○)抗体をL929細胞に加え、100ng/mlのヒト腫瘍壊死因子αと共に培養した。37℃で16時間培養した後、比色MTT分析により、細胞の生存を分析した。
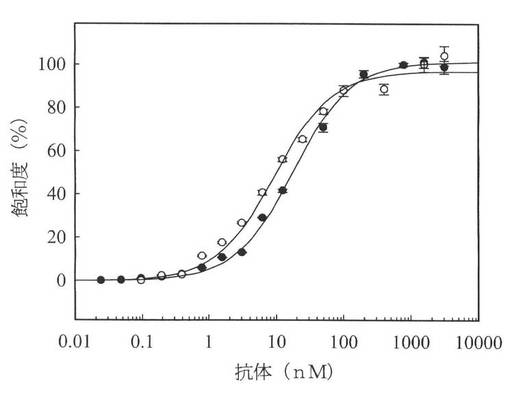
【図6】図6は、m357−IgG抗体とh357−IgG抗体の細胞表面上の膜貫通型腫瘍壊死因子αに対する飽和結合分析を示す図である。膜貫通型腫瘤壞死因子αトランスフェクトNS0細胞を、m357−IgG(○)またはh357−IgG(●)抗体の連続希釈物(対数希釈)と共に4℃で1時間培養した。これらの細胞を3回洗浄し、それぞれ、m357−IgGに反応するAlexa Fluor 488ヤギ抗マウスIgG(H+L)と、h357−IgGに反応するAlexa Fluor 647ヤギ抗ヒトIgG(H+L)と共に4℃で1時間培養した。その後、細胞を洗浄し、FACSCaliburフローサイトメータで分析した。
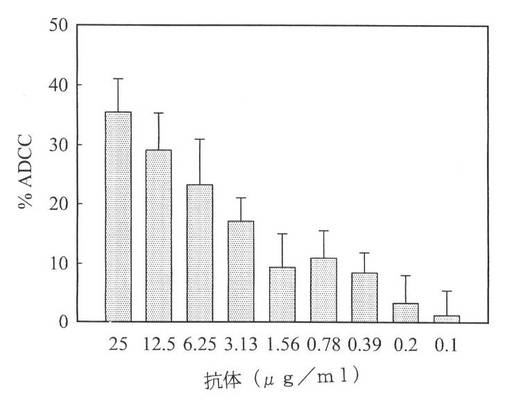
【図7】図7は、膜貫通型腫瘍壊死因子α発現細胞に対するh357IgGの抗体依存性細胞媒介性細胞傷害を示す図である。膜貫通型腫瘍壊死因子α発現細胞を異なる濃度のh357IgG抗体の存在下で1時間培養した。その後、ヒト末梢血単核球PBMCをエフェクター細胞として用い、膜貫通型腫瘍壊死因子α発現細胞を目標細胞として用いた。細胞質ゾルから上澄み液に遊離するLDH量を測定することにより、細胞毒性を計算した。この結果を、溶解バッファ処理細胞群に対して100%溶解した細胞溶解率として示す。
【発明を実施するための形態】
【0025】
本発明の一態様において、本発明は、ヒト化モノクローナル抗体のアミノ酸配列を提供する。上記ヒト化モノクローナル抗体は腫瘍壊死因子α(TNF−α)と結合し、腫瘍壊死因子αはヒト腫瘍壊死因子αであっても良い。上述のヒト化モノクローナル抗体のアミノ酸配列は、軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列を含む。一実施形態において、本発明によるヒト化モノクローナル抗体のアミノ酸配列は、免疫グロブリンG(IgG)抗体のアミノ酸配列であり、軽鎖可変領域のアミノ酸配列、重鎖可変領域のアミノ酸配列、およびヒト免疫グロブリンG抗体の定常部のアミノ酸配列を含む。
【0026】
一実施形態において、本発明によるヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列は、腫瘍壊死因子αと結合することができる非ヒトモノクローナル抗体に対し、可変領域表面再構成を行なうことにより得られる。
【0027】
まず、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列を得る。一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体は、マウスモノクローナル抗体を含んでいても良い。このマウスモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、重鎖可変領域アミノ酸配列は、配列番号2の配列を含む。
【0028】
続いて、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列に基づいて、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列の分子モデルを構築すると共に、非定常表面残基をラベルする。一実施形態において、分子モデリングプロセスは、コンピュータアシストホモロジーモデリングにより行なうことができる。
【0029】
その後、非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)と同一性が最も高いヒト配列を探すと共に、両者を比較して、非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)アミノ酸配列の置換可能な残基を決定する。最後に、この置換可能な残基は、非ヒトモノクローナル抗体の可変領域の置換可能な残基の位置に対応する位置に存在する、ヒト配列に対応するアミノ酸残基に置換されて、腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列を獲得する。
【0030】
一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、且つ、重鎖可変領域アミノ酸配列は、配列番号2の配列を含んでいる。非ヒトモノクローナル抗体の可変領域と最も相同性が高い配列を有するヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。この実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のアミノ酸配列に対して行われる置換は、少なくとも以下の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。よって、得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の軽鎖可変領域アミノ酸配列、及び配列番号2の重鎖可変領域アミノ酸配列を含み、且つ、配列番号1の配列と配列番号2の配列は少なくとも以下の置換の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。
【0031】
別の実施形態において、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、非ヒトモノクローナル抗体の重鎖可変領域アミノ酸配列は、配列番号2の配列を含んでいる。非ヒトモノクローナル抗体の可変領域と最も同一性が高いヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。上述した配列比較実行後に得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列は、配列番号5を有し、重鎖可変領域のアミノ酸配列は、配列番号6を含む。
【0032】
よって、本発明の別の態様によれば、本発明はまた、上述したヒト化モノクローナル抗体のヌクレオチド配列も提供する。本発明のヒト化モノクローナル抗体のヌクレオチド配列は、軽鎖可変領域のヌクレオチド配列と重鎖可変領域のヌクレオチド配列を含む。一実施形態において、本発明によるヒト化モノクローナル抗体のヌクレオチド配列は、免疫グロブリンG(IgG)抗体のヌクレオチド配列であり、軽鎖可変領域のヌクレオチド配列、重鎖可変領域のヌクレオチド配列、およびヒト免疫グロブリンG抗体の定常領域のヌクレオチド配列を含む。
【0033】
一実施形態において、ヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド配列は、配列番号5の配列をエンコードするヌクレオチド配列を含み、重鎖可変領域ヌクレオチド配列は、配列番号6の配列をエンコードするヌクレオチド配列を含む。配列番号5の配列をエンコードするヌクレオチド配列は配列番号7の配列であり、配列番号6の配列をエンコードするヌクレオチド配列は配列番号8の配列である。
【0034】
本発明の別の態様において、本発明は更に、ヒト化モノクローナル抗体を提供する。このヒト化モノクローナル抗体は腫瘍壊死因子αと結合し、腫瘍壊死因子αは、ヒトの腫瘍壊死因子αであっても良い。ヒト化モノクローナル抗体は、軽鎖と重鎖を含む。一実施形態において、本発明によるヒト化モノクローナル抗体は免疫グロブリンG(IgG)抗体であり、一本鎖は、軽鎖可変領域、重鎖可変領域、およびヒト免疫グロブリンG定常領域を含む。
【0035】
一実施形態において、本発明によるヒト化モノクローナル抗体は、以下のステップにより得られる。
【0036】
まず、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域ヌクレオチド断片およびその配列と、重鎖可変領域ヌクレオチド断片およびその配列を得る。一実施形態において、上述の非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド断片(cDNA)は、非ヒトモノクローナル抗体を生成することができるハイブリドーマの総RNAを抽出した後、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のそれぞれに対応する二個のプライマー対により、総RNAに対して逆転写ポリメラーゼ連鎖反応(RT−PCR)を行なうことによって得られる。次いで、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド断片の配列をそれぞれ決定した後、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド配列を得る。一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体はマウス抗体を含む。このマウス抗体の軽鎖可変領域ヌクレオチド配列は、配列番号9の配列を有し、重鎖可変領域ヌクレオチド配列は、配列番号10の配列を有する。
【0037】
続いて、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド配列に基づいて、それぞれ、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列を獲得する。一実施形態において、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を有し、非ヒトモノクローナル抗体の重鎖可変領域アミノ酸配列は、配列番号2の配列を有する。
【0038】
その後、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列により、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域の分子モデルを構築すると共に、非定常表面残基をラベルする。一実施形態において、分子モデリングプロセスは、コンピュータアシストホモロジーモデリングにより行なう。
【0039】
続いて、本発明による腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域アミノ酸配列と最も同一性が高いヒト配列を探すと共に、両者の比較を実行して、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)アミノ酸配列の置換可能な残基を決定する。最後に、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域アミノ酸配列のヌクレオチド配列を、置換可能な残基の対応位置でエンコードして、点変異により、この残基位置に対応するヒト配列の残基をエンコードするヌクレオチドに置換して、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド断片と重鎖可変領域のヌクレオチド断片を獲得する。
【0040】
その後、本技術領域で熟知される方法により、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド断片と重鎖可変領域のヌクレオチド断片と、既知のヒト抗体定常領域ヌクレオチド断片を、それぞれ、適切な発現ベクターにクローン化し、この発現ベクターを適切な宿主細胞にトランスフェクトすると、宿主細胞は、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体を発現する。
【0041】
一実施形態において、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を有し、且つ、重鎖可変領域アミノ酸配列は、配列番号2の配列を有する。前述の非ヒトモノクローナル抗体の可変領域(軽鎖可変領域アミノ酸配列および重鎖可変領域)アミノ酸配列と相同性が最も高いヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。この実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のアミノ酸配列に対して行なわれる置換は、少なくとも以下の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンがアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。よって、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の軽鎖可変領域アミノ酸配列、および配列番号2の重鎖可変領域アミノ酸配列を含み、且つ、配列番号1の配列と配列番号2の配列は、少なくとも一つの以下のアミノ酸置換を含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸がグリシンに置換される。
【0042】
別の実施形態において、本発明による腫瘍壊死因子αと結合するモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、重鎖可変領域アミノ酸配列は、配列番号2の配列を含む。本発明による腫瘍壊死因子αと結合するモノクローナル抗体の可変領域(軽鎖可変領域および重鎖可変領域)アミノ酸配列と相同性が最も高いヒト配列は、PPS4の軽鎖可変領域配列(配列番号3)と重鎖可変領域配列(配列番号4)である。上記のようにして配列を比較した後、得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号5の軽鎖可変領域を含み、配列番号6の重鎖可変領域のアミノ酸配列を含む。よって、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体は、配列番号5の軽鎖可変領域アミノ酸配列、及び配列番号6の重鎖可変領域アミノ酸配列を含む。
【0043】
本発明のヒト化モノクローナル抗体と結合する腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む。本発明のヒト化モノクローナル抗体の放出型腫瘍壊死因子αに対する結合親和性は、約20−40nMであり、好ましくは、約10−20nMである。
【0044】
更に本発明のヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は約20−40nMであり、好ましくは、約10−20nMである。一実施形態において、本発明のヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は約16.8nMである。
【0045】
更に一実施形態において、本発明のヒト化モノクローナル抗体は、細胞上で発現される膜貫通型腫瘍壊死因子αと結合することにより、抗体依存性細胞媒介性細胞障害(ADCC)を生じる。
【0046】
現在分かっていることは、膜貫通型腫瘍壊死因子αと多くの疾病が関係しているということである。例えば、細胞表面が発現する膜貫通型腫瘍壊死因子αのマクロファージ細胞と単球細胞は、例えば、クローン病やウェゲナー肉芽腫症などの肉芽腫性疾患で重要な役割を果たしており、抗体依存性細胞媒介性細胞障害により、細胞を直接滅亡させる(Beenhouwertsら、2004)。
【0047】
このように本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αと結合する能力を有し、且つ、抗体依存性細胞媒介性細胞障害を生じさせる能力があるので、本発明の更なる態様において、本発明は、膜貫通型腫瘍壊死因子αを中和するヒト化モノクローナル抗体の用途、および抗体依存性細胞媒介性細胞障害を生じさせるヒト化モノクローナル抗体の用途も提供する。
【0048】
更に本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αに相関する疾病を治療する薬物の用途にも応用できる。
【実施例】
【0049】
材料と方法
A.ヒト腫瘍壊死因子α(TNF−α)と結合するマウスモノクローナル抗体
1.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体の獲得と精製
ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体を産生し得るハイブリドーマ細胞株357−104−4(ECACC No.92030603)(ヨーロピアン・コレクション・オブ・セル・カルチャーから購入)をマウス腹腔内に注射し、マウスに腫瘍と腹水を生じさせた。生成したマウス腹水は、大量のヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)を含む。上述のステップは、GlycoNEX Inc.(http://www.glyconex.com.tw/)に委託して行なった。
【0050】
続いて、この腹水をマウスから取り出し、腹水をプロテインAカラム(GEヘルスケア)に通して精製し、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)を獲得した。
【0051】
2.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のcDNA配列の獲得
【0052】
(1)ハイブリドーマ細胞株357−104−4(ECACC No.92030603)の総RNAを、QIAGEN RNeasy Mini Kitにより抽出した。
【0053】
(2)アマシャム(Amersham)社製のLight Primer MixとHeavy Primersの二個のプライマー対により、総RNAに対して逆転写ポリメラーゼ連鎖反応(RT−PCR)を行い、m357IgGの軽鎖と重鎖のcDNAをそれぞれ、獲得した。
【0054】
(3)この軽鎖と重鎖のcDNAを、それぞれ、TOPOベクター上にクローン化して、二個のクローンを得た。
【0055】
(4)DNA配列決定後、制限酵素部位を有する二個のプライマー対を設計した。
【0056】
(5)制限酵素部位を有する二個のプライマー対を合成した。
ここで、m357IgGの軽鎖のcDNAに用いるプライマー対は以下のとおりである:
357VL’Ascプライマー:
5’-CAGGCGCGCCGAAATTGTGCTGACCCAGTC-3’(配列番号11)
357VL3’Glinkプライマー:
5’-CCAGAGCCACCTCCGCCTGAACCGCCTCCACCCAATTTCCAGCTTGC-3’(配列番号12)。
【0057】
m357IgGの重鎖のcDNAに用いるプライマー対は以下のとおりである:
357VH5’Glinkプライマー:
5’-TCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGGTGAAACTGCAGGA-3’(配列番号13)
357VH3’Not:
5’-CAGCGGCCGCTGAGGAGACGGTGACCGTGGT-3’(配列番号14)。
【0058】
その後、ステップ(3)で形成された2つのクローン中でポリメラーゼ連鎖反応を行なって、m357IgGの軽鎖と重鎖のcDNAを増幅させた。
【0059】
B.再構成のための、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のコンピューターモデリング
Discovery Studioモデリング2.1(Accelrys社、サンディエゴ、カナダ)を使用して、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のホモロジーモデリングプロセスを行なった。m357IgGの軽鎖可変領域(VL)と重鎖可変領域(VH)に対し、二個の別々のBLASTP検索を行なった。蛋白質データバンク(http://www.rcsb.org/pdb/home/home.do)中で検索することによって配列を分析し、m357蛋白質配列のホモロジーを同定した。マウス抗乳癌抗体Fab断片の重鎖可変領域SM−3[PDB entry:1SM3](Dokurnoら、1998)と抗サーマス・アクタチクス(thermus aquaticus)DNA ポリメラーゼIモノクローナル抗体Fab断片の軽鎖可変領域構造[PDB entry:1AY1](Muraliら、1998)のホモロジーモデリングに基づいて、m357IgG Fv断片の三次元(3D)構造を構築した。最終的な三次元モデルは、MODELLERモジュール(Saliら、1995)により生成され、空間的拘束を満足させる(satisfaction of spatial restraints)ことにより、比較蛋白質構造モデリングの自動方法を行なった。m357IgGに対し、蛋白質ホモロジーモデリングおよびループモデリングを自動的に行なった。モデル抗体ループ(Model antibody Loops)モジュールを使用することにより、最高の配列鑑別度で、PDBデータベースからテンプレート構造を選択してCDRループのモデル構築を行い、、且つ、ループ精製モジュールを使用して、CDRループのモデル構築を完全にし、立体衝突(steric clash)を最小化すると共に、正確な結合距離と角度を確認した。その後、Discovery Studioモデリング2.1中のCHARMm(B.R.Brooks、1983)プログラムと、Accelrys CHARMm forcefieldにより、構造のエネルギーを最小化して、このモデル全体を更に純化した。構造のエネルギー最小化は、二つのステップで行なった。まず、抑制最急降下最小化(restrained steepest descent minimization)を5000ステップいった後、続いて、フレームワーク(framework)のαカーボンが、適切な位置で維持固定されるまで、共役勾配最小化(conjugated gradient minimization)を5000ステップ行なった。AREAIMOLプログラム(CCP4、1994)により、三次元モデル上で、m357IgG残基の溶媒接近表面積(solvent−accessible surface areas)を計算した。相対接近が30%以上の残基は、接近可能であると定義した。
【0060】
C.IgGの構築、発現および精製
オーバーラッピングPCRにより、m357IgGのヒト化可変領域(軽鎖可変領域および重鎖可変領域)をエンコードするcDNA断片を獲得した。続いて、ヒトIgGからの定常領域配列とオーバーラッピングPCRにより合成される可変領域配列を、哺乳動物発現ベクターpSecTag2/Hygro(重鎖)(Invitrogen)とpcDNA3.3−TOPO TA(軽鎖)(Invitrogen)中で、サブクローニングした。その後、EcoRV制限酵素認識部位を利用して、二個の構築体(construct)を融合させ、pSec−pcDNA−h357−IgGを生成した。メーカーの使用説明書により、エフェクテン(Effectene、Qiagen)を使用し、重鎖と軽鎖遺伝子を含むプラスミドを、マウス骨髄腫NS0細胞(ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャー、Salisbury、Wiltshire、英国)中でトランスフェクトさせた。ハイグロマイシン(Hygromycin)(400μg/ml)で4週間抽出した後、安定したクローンを、無血清の既知組成培地HyQNS0(Hyclone)中で、5×105cells/mlの初期播種密度にてシェーカー中で培養した。37℃で5日間後の培養基を取り出し、蛋白質A(GE Health−care)クロマトグラフィにより、抗体を上澄みから精製した。
【0061】
D.抗腫瘍壊死因子αの中和効力分析
既知の方法(Matthews N、1987)に従って、アクチノマイシンDで処理されたマウス線維芽細胞L929細胞(ATCC Cat.No.CCL−1)により、ヒト腫瘍壊死因子αに対するm357IgGおよびh357IgGの中和活性を測定した。要約すると、L929細胞を、96ウェルプレート中に3×105cells/wellで三回播種すると共に、10%(v/v)ウシ胎仔血清を添加したRPMI 1640培地中で16時間培養した。その後、幾つかの抗体希釈液を調製し、アクチノマイシンD(2μg/ml)と腫瘍壊死因子α(TNF−α)(100ng/ml)を含む培地中に入れ、37℃で16時間培養した。上清を除去した後、3−(4,5−ジメチル−2−チアゾリル)−2,5−ジフェニルテトラゾリウムブロミド(MTT)(5mg/ml)(Sigma−Aldrich)を加え、37℃で4時間培養した。その後、SDS溶液(10%)をウェル中に加えた。室温で24時間培養した後、色度計により、各ウェル中の色が紫色に変化するのを記録した。570nmで測定される溶出の光学密度(OD)を測定した結果、生存細胞の数量と正の相関が見られた。ブランクの対照群(培養物のみ)、腫瘍壊死因子αの対照群(腫瘍壊死因子αのみ)、および抗体の対照群(抗体のみ)もまた、上記実験中で行なわれた。シグマプロットソフトウェア(Systat software,Inc、Richmond、カナダ)を用いて複雜なS状非線形回帰(complex sigmoid non−linear regression)分析によりED50値を計算した。
【0062】
E.NS0細胞上の膜貫通型腫瘍壊死因子αの安定した発現
既知の技術(Perezら、1990)で示されるように、抵抗腫瘍壊死因子α変換酵素(TNF−α converting enzyme、TACE)媒介開裂に対して耐性がある欠失変異体膜貫通腫瘍壊死因子αを、部位特異的変異導入法によって生成した。非開裂形式の膜貫通腫瘍壊死因子αでは、天然の膜貫通腫瘍壊死因子αのアミノ酸+1〜+12が削除された。非開裂形式の膜貫通腫瘍壊死因子α遺伝子は、pSecTag2/Hygro哺乳動物発現ベクター(Invitrogen)にクローン化されると共に、エフェクテンにより、マウス骨髄腫NS0細胞にトランスフェクトされ、細胞表面上で膜貫通腫瘍壊死因子αを発現させた。
【0063】
F.m357−IgGとh357−IgGの膜貫通腫瘍壊死因子αに対する飽和結合分析
膜貫通腫瘍壊死因子αトランスフェクトNS0細胞を、m357−IgGとh357−IgGの連続希釈物(対数希釈)と共に、2%のウシ胎仔血清を含むリン酸緩衝生理食塩水(蛍光活性化細胞分類[FACS]バッファ)中にて4℃で1時間培養した。この細胞を、蛍光活性化細胞分類バッファで3回洗浄した後、それぞれ、m357−IgGのAlexa Fluor 488ヤギ抗マウスIgG(H+L)とh357−IgGのAlexa Fluor 647ヤギ抗ヒトIgG(H+L)により、4℃で1時間染色した。FACSCaliburフロー血球計算器(Becton Dickinson、San Jose、カナダ)により蛍光強度を測定した。
【0064】
G.抗体依存性細胞媒介性細胞障害(ADCC)の分析
目標物質であるh357−IgGの抗体依存性細胞媒介性細胞障害(ADCC)の活性を、LDH細胞毒性検出キット(Clontech)により測定し、製造メーカーの取扱説明書に基づいて、損傷した細胞のサイトゾルから遊離するLDH活性を測定した。要約すると、高度に表現された膜貫通腫瘍壊死因子αの細胞を、異なる濃度のh357抗体の存在下で、5%CO2インキュベーターのアッセイ培地(1%FBS含有DEME)中で1時間培養した後、エフェクター細胞として梢血単核細胞(PBMC)を加えた(エフェクター細胞:目標細胞=20:1)。更に37℃で16時間培養した後、ウェル毎に100μlの上澄みを採取し、新しい96−ウェルの平底プレート中に移した。LDH基質(100μl)を各ウェルに加え、避光下にて室温で30分間培養した。サンプルの吸光度を、ELISAリーダーにより490nmで測定した。溶解バッファにより最大放出(release)を決定した。以下の式により、特定の溶解百分率を計算した:%細胞毒性=[実験例の放出−自然放出]/[最大放出−自然放出]×100。
【0065】
結果
A.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)
【0066】
1.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体の分析
精製したm357IgGに対してSDS−PAGE分析を行なった。この結果を図1のレーン1とレーン2に示す。レーン1は非還元状態における精製したm357IgGのSDS−PAGE分析結果であり、レーン2は還元状態における精製したm357IgGのSDS−PAGE分析結果である。
【0067】
2.m357IgGの軽鎖と重鎖のcDNA
ポリメラーゼ連鎖反応により得られたm357IgGの軽鎖と重鎖のcDNAについて電気泳動分析を行なった。この結果を図2に示す。レーン1はm357IgGの軽鎖のcDNA(配列番号9)であり、レーン2はm357IgGの重鎖のcDNA(配列番号10)である。
【0068】
B.m357可変断片の分子モデリング
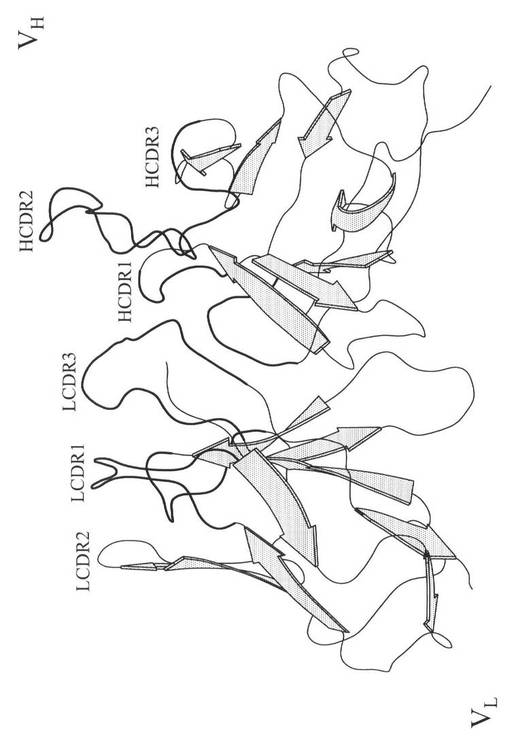
RT−PCRにより、ハイブリドーマ細胞株357−101−4(ECACC No. 92030603)からの抗腫瘍壊死因子αモノクローナル抗体m357IgGの重鎖可変領域(VH)および軽鎖可変領域(VL)をエンコードしたcDNA(データは示さず)を得た。「材料と方法」中に記載したホモロジーモデリング(program MODELLER)により、それぞれ、m357IgGの重鎖可変領域VH、および軽鎖可変領域VLの推定アミノ酸配列の三次元構造を構築した。その後、PDBからの1SM3(配列同一性および相似性が、それぞれ、87%と93%)テンプレート構造が1.95Å、PDBからの1AY1(配列同一性および相似性が、それぞれ、85%と91%)テンプレート構造が2.20Åの解析度で、m357IgGの重鎖可変領域および軽鎖可変領域をそれぞれ、モデリングした。Discovery Studioモデリング2.1プログラムにより、m357IgGの重鎖可変領域VHおよび軽鎖可変領域VLの最終の正確な構造を獲得した。この結果を図3Aに示す。
【0069】
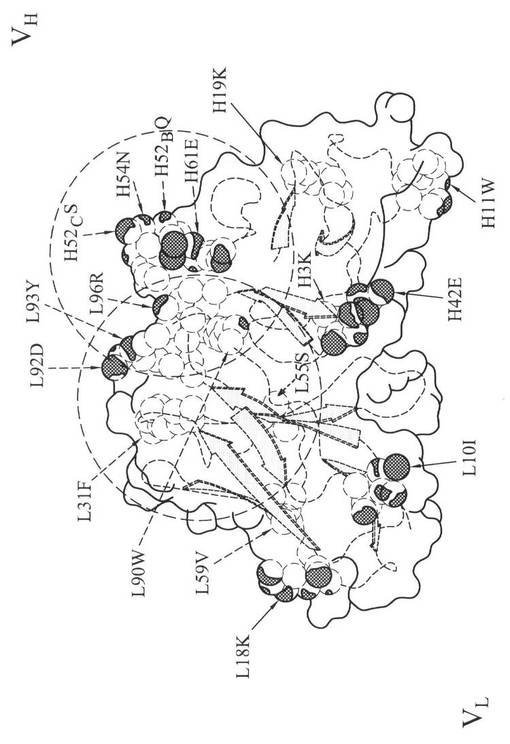
図3Aは、m357IgGの可変領域を示す図である。m357IgGの三次元構造は、それぞれ、重鎖可変領域に対応する1SM3と軽鎖可変領域に対応する1AY1の結晶構造を比較するホモロジーモデリングにより生成した。CDRループを太線で示す。図3Bは、m357IgGの分子表面モデルを示す図であり、非ヒト様(non−human like)表面残基とCDRループの関係を示す。相対溶媒接近が30%以上の17個の残基は、非ヒト様の表面残基であり、太い矢印で示す。CDRループを太線で示す。5Å内のCDRループに近い残基(点線の円)を点線矢印で示す。好ましくは「ヒト」アミノ酸に突然変異する6個の残基を実線矢印で示す。重鎖可変領域フレームワーク中の4個の残基をヒト化のために決定し、軽鎖可変領域フレームワーク中の2個の残基のみヒト化のために決定した。
【0070】
C.m357IgG可変領域断片のヒト化
可変領域再構成によるヒト化は、最初にPadlanが1991年に提案した。この方法では、抗体のフレームワーク領域中の潜在抗原部位は排除され、抗体の抗原親和性に影響しない(Padlan、1991)。この方法は、ヒト抗マウス抗体(HAMA)が、表面残基から派生する可変領域だけに反応することを前提としており、他の研究者(Fontayneら、2006;O’Connorら、1998;Staelensら、2006)により採用され改変されてきた。本実施例では、以下の三つのステップにより、m357IgGの可変領域再構成を行なった。まず、m357IgGの重鎖可変領域と軽鎖可変領域をそれぞれ構築した。次に、AREAIMOLプログラムを使用して溶媒接近可能残基を計算し、非ヒト様のフレームワーク表面残基を同定した。最後に、ヒトフレームワークの配列アライメント結果に基づいて、これらの表面残基を、ヒトの対応表面残基に突然変異させた。可変領域上で、マウスm357IgGの非ヒト様フレームワーク表面残基を置換するため、ヒト目標配列から、一組の高度に相同性を有する表面残基を選択した。IMGTデータベース(http://imgt.cines.fr/)中で検索を行い、検索結果から、ファージ提示法、またはヒト化抗体の配列を排除して、m357IgGと対応する可変領域に対し、相同性が最も高いヒト重鎖可変領域と軽鎖可変領域の配列ペアを同定した。ヒト配列で見出される最も同一性の高い表面残基は、PPS4の可変領域である(フレームワーク領域中の配列同一性は、それぞれ、重鎖可変領域;76%、軽鎖可変領域:73%)(図4)。AREAIMOLプログラムの計算結果に基づいて、重鎖可変領域中の20個の表面残基のうち8個は、ヒトとマウスの配列間で非保存的であり、軽鎖可変領域中の16個の表面残基のうち9個は非保存的であった。これら17個(8個+9個)の表面残基は、置換の候補者であった。しかし、CDR領域中の重鎖H52BQ、重鎖H52CS、重鎖H54N、重鎖H61E、軽鎖L31F、軽鎖L55S、軽鎖L90W、軽鎖L92D、軽鎖L93Y、および軽鎖L96Rの置換は、CDRのコンフォメーションを潜在的に変化させ、軽鎖中のCDR2近くの5Å内の余分な非定常表面残基であるL59Vも、結合親和性に潜在的に影響する(図3B)。その結果、これらの11個のマウス残基は維持され、抗原親和性を保持する。最後に、残りの6個の残基を選択して、ヒト保存残基に置換した。PPS4と比較すると、軽鎖可変領域の第10の比較位置におけるイソロイシンはトレオニンに置換され(m357IgGの軽鎖可変領域の第10アミノ酸のイソロイシンがトレオニンに置換される)、第18の比較位置のリジンはアルギニンに置換される(m357軽鎖可変領域の第18のアミノ酸のリジンがアルギニンに置換される)。PPS4と比較すると、重鎖可変領域の第3の比較位置のリジンはグルタミンに置換され(m357重鎖可変領域の第2アミノ酸のリジンがグルタミンに置換される)、第11の比較位置のトリプトファンがロイシンに置換され(m357重鎖可変領域の第10アミノ酸のトリプトファンがロイシンに置換される)、第19の比較位置のリジンがアルギニンに置換され(重鎖可変領域の第18アミノ酸のリジンがアルギニンに置換される)、第42の比較位置のグルタミン酸がグリシンに置換される(重鎖可変領域の第41アミノ酸のグルタミン酸がグリシンに置換される)(図4)。
【0071】
D.ヒト化357(h357)IgG1の構築と発現。
m357IgGのヒト化重鎖可変領域と軽鎖可変領域のアミノ酸配列を、インフレームで、ヒトIgG1の重鎖とカッパ軽鎖定常領域に融合させた。完全なヒト化357(h357)IgG1分子を発現するため、二種の哺乳動物発現ベクターであるpSecTag2/HygroとpcDNA3.3−TOPO TAを使用し、h357IgGのヒト化重鎖と軽鎖をそれぞれ、導入した。その後、軽鎖発現カセットを重鎖発現pSecTag2/Hygroベクターに連接させ、単一の発現ベクターを得た。組み換えh357IgGの発現レベルは約14mg/Lであった。蛋白質Aクロマトグラフィにより、m357IgGとh357IgGを含む培養基をそれぞれ、精製すると共に、SDS−PAGEにより蛋白質精製物を決定した(図1)。図1に示すように、非還元状態下で、二つの抗体は、155kDaの分子量を有する単一バンド(レーン1と3)を示した。還元状態下では、二つの抗体は、それぞれ55kDa(重鎖)と26kDa(軽鎖)の分子量を有する二つの蛋白質バンド(レーン2と4)を生成した。
【0072】
E.m357IgGとh357IgGによる腫瘍壊死因子α媒介細胞毒性の中和
抗腫瘍壊死因子α抗体の機能活性を調べるために、可溶性腫瘍壊死因子α活性を抑制する抗体の能力をテストした。腫瘍壊死因子αは、マウスL929細胞に対して細胞毒性を生じさせる。組み換えヒト腫瘍壊死因子α抗体および上記細胞の共培養により、L929アッセイでm357IgGとh357IgGを両方評価した。図5に示すように、100ng/mlのヒト腫瘍壊死因子αで処理されたL929細胞中の腫瘍壊死因子α媒介細胞毒性は、m357IgGとh357IgGの両方により、用量依存的に効果的に中和され、ED50値はそれぞれ、3.07nMと2.30nMであった。この結果は、ヒト化357IgGは、マウス357IgGと同程度の濃度で、腫瘍壊死因子αの中和活性を維持することを示している。
【0073】
F.h357IgG抗体と膜貫通腫瘍壊死因子αとの結合活性
腫瘍壊死因子αは、膜関連の前駆体(膜貫通腫瘍壊死因子α)として存在し、腫瘍壊死因子α変換酵素(TNF変換酵素、TACE)により媒介される蛋白分解切断により成熟した可溶体が放出される。多くの研究により、腫瘍壊死因子α拮抗薬は、アポトーシス、抗体依存性細胞媒介性細胞障害(ADCC)、補体依存性細胞障害活性(CDC)、外側から内側へのシグナリングメカニズムにより、細胞溶解を生じることが調査されてきた(Aroraら、2009;Caronら、1999;Mitomaら、2008;Scallonら、1995)。h357IgGと膜貫通腫瘍壊死因子αとの結合能力を分析するため、膜貫通腫瘍壊死因子αの非開裂形式のcDNAを、NS0細胞にトランスフェクトして細胞膜上で発現させ、フロー血球計算器を用いてm357IgGとh357IgGの両方の結合活性を評価した。図6のデータは、m357IgGとh357IgGは共に、濃度依存的に膜貫通腫瘍壊死因子αと結合することを示しており、KD値はそれぞれ、12.0nMと16.8nMである。同程度のKD値は、ヒト化過程において、膜貫通TNF−αの結合親和性を変化させないことを示す。h357IgGがナノモル範囲の結合親和性を有することは、h357IgGが、膜貫通腫瘍壊死因子αを介して、更に多くのエフェクター機能、またはアポトーシスメカニズムを潜在的にトリガーすることを示唆している。
【0074】
G.h357IgGの抗体依存性細胞媒介性細胞障害に対する能力
既知の文献で示されるように、インフリキシマブおよびアダリムマブの膜貫通腫瘍壊死因子αに対する結合親和性は、エタネルセプトよりも高く、抗体依存性細胞媒介性細胞障害(ADCC)、補体依存性細胞障害活性(CDC)、またはアポトーシスにより、細胞表面発現膜貫通腫瘍壊死因子αの細胞死滅作用に影響するが、これは、臨床疾病で異なる効果が生じる原因のひとつである(Taylor、2010)。細胞表面発現腫瘍壊死因子α様マクロファージと単球は、例えばクローン病やウェゲナー肉芽腫症などの肉芽腫性疾患において非常に重要な役割を果たしており、抗体依存性細胞媒介性細胞障害により、細胞を直接、死滅させる(Beenhouwerら、2004)。腫瘍壊死因子α拮抗薬が膜貫通型腫瘍壊死因子α発現細胞に結合すると、これらの細胞は、ナチュラルキラー細胞のターゲットとされる。ヒトIgG1由来のFc領域からなるh357IgGは、腫瘍壊死因子α産生細胞中で、潜在的に細胞溶解を生じる。よって、h357IgGの、膜貫通腫瘍壊死因子α発現目標細胞に対するADCC仲介能力を評価するため、分離した末梢血単核球PMBCを抗体依存性細胞媒介性細胞障害の分析に用いた。エフェクター細胞:目標細胞(E:T)は20:1である。h357IgGの濃度が6.25ug/mlのときに、腫瘍壊死因子α目標細胞の20%以上が溶解した(図7)。これらのデータは、h357IgGが、細胞表面で発現された膜貫通腫瘍壊死因子αとの結合によって抗体依存性細胞媒介性細胞障害を仲介することを示唆している。従って、h357IgGは、ADCC能を有する治療抗体と同様、更に効果的な腫瘍壊死因子α中和抗体に発展する可能性を有している。
【0075】
本発明では好ましい実施例を上記の通り開示したが、本発明は上記実施例に限定されず、当業者であれば、本発明の精神と領域を逸脱しない範囲内で各種の改変や調整を行なうことができ、このようなものも本発明の範囲内に包含される。
【0076】
本願明細書中に記載した文献の詳細は、以下のとおりである。
【0077】
【表1】

【0078】
【表2】
【0079】
【表3】
【0080】
【表4】
【技術分野】
【0001】
本発明は、抗腫瘍壊死因子αの抗体に関し、特に、腫瘍壊死因子αを高度に中和することが可能なヒト化モノクローナル抗体およびそのアミノ酸配列に関するものである。
【背景技術】
【0002】
腫瘍壊死因子α(tumor necrosis factor−alpha、TNF−α)は、炎症誘発性サイトカインであり、マクロファージ細胞と単球を含む免疫システム細胞により最初に生成される。腫瘍壊死因子αは、ホモ三量体蛋白質として存在し、各サブユニット中で、最初、一個の26kDaの膜貫通前駆体蛋白質として翻訳される。腫瘍壊死因子α変換酵素(TACE)により、TNF−αの膜貫通領域に近い位置で切割を行なった後、腫瘍壊死因子αの可溶三量体を放出し、エフェクター細胞上の構造区分タイプIとタイプIIの腫瘍壊死因子レセプター(TNFRIとTNFRII)を結合することにより、その活性を発揮する。
【0003】
膜貫通形式の腫瘍壊死因子αも、独特の生物機能が知られており、例えば、細胞間の接触方式における、細胞毒性活性とポリクローナルB細胞の活性化(Mitomaら、2008)が挙げられる。腫瘍壊死因子αが自己免疫過程に対し、特定の影響を有することが既に証明されており、多くの自己免疫疾病の主要な治療目標となっている(Feldmann、2001)。今までのところ、ある抗腫瘍壊死因子αの試薬、例えば、エタネルセプト(etanercept)、アダリムマブ(adalimumab)、及びインフリキシマブ(infliximab)は、既に、米国食品医薬局(FDA)により承認されており、いずれも、主要な薬理学的メカニズムの作用として、可溶形の腫瘍壊死因子αを効果的に中和する能力を有する。しかし、これらの拮抗薬の膜貫通形式の腫瘍壊死因子αに対する結合作用は異なり、臨床疾病上で、異なる結果をもたらす(Taylor、2010)。例えば、エタネルセプトは、膜貫通形式の腫瘍壊死因子αが重要な役割を果たす肉芽腫性疾患の発病に対して臨床的に有効ではない(2008、Mitoma)。よって、抗腫瘍壊死因子αの試薬が、膜貫通形式の腫瘍壊死因子αと結合して、抗体依存性細胞媒介性障害(ADCC)、補体依存性細胞障害(CDC)、アポトーシス、及び外から内へのシグナル伝達メカニズムをトリガーすることができるかどうかが必要条件である。
【0004】
臨床実験におけるマウスモノクローナル抗体の使用による主な障害は、患者にヒト抗マウス抗体反応(HAMA)が生じる可能性があるということである(Owens and Young、1994;Sandhu、1992;Schroffら、1985)。よって、臨床使用の効果を改善するため、遺伝子工程技術を使用して、マウスに属するアミノ酸残基をヒトに対応するアミノ酸残基に置換して、患者の免疫原生の誘発の可能性を減少させることが行なわれている。
【0005】
理想的な抗体のヒト型化は、抗原に対する特異性と親和性を維持すると共に、できる限り、免疫原性を低下させることができる。これまでのところ、抗体のヒト型化は、既に、多くの方式により行なわれており、例えば、マウス抗原と可変領域を結合して、ヒト抗体の定常領域に遺伝子的に融合してなるキメラ抗体が最も早期に試され、免疫原性を減少させている(Morrisonら、1984)。しかし、キメラ抗体には、所望でない抗可変領域反応が生じる(Bruggemannら、1989)。もう一つの方式は、相補性決定領域移植法(CDR−gtafting)であり、齧歯目抗体の相補性決定領域を、ヒト抗体のFvフレームワーク(FR)に転移することを含む(Verhoeyenら、1988)。残念ながら、相補性決定領域とFvフレームワークとの接触面の変化により、抗原との結合が激しく妨害される。最初の相補性決定領域移植抗体は、親の結合親和性を失う傾向にあるので、相補性決定領域のループ構造にとって極めて重要であると見なされるマウスフレームワークアミノ酸の復帰突然変異のための別の作業が必要である(Queenら、1989)。可変領域表面再構成(variable domain resurfacing)によるヒト化は、親抗体の特異性と結合親和性を維持することができる別のアプローチであり、マウスFvフレームワーク中の表面に露出する残基を、一般のヒト抗体中に見出される表面露出残基に代替させることにより、抗体の免疫原性を減少させることができる(Fontayneら、2006;Padlan、1991;Roguskaら、1994;Staelensら、2006;Zhangら、2005)。現在の分子生物技術は、このアミノ酸を変化させる方法を更に容易にしているが、特に、信頼できる抗体のコンピュータモデルが必要であるとき、溶剤中の表面に露出する残基を確認することは依然として困難である(Fontayneら、2006)。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の目的は、膜貫通型腫瘍壊死因子αと結合する能力を有するヒト化モノクローナル抗体のアミノ酸配列およびヌクレオチド配列、並びにヒト化モノクローナル抗体を提供することにある。更に本発明の他の目的は、抗体依存性細胞媒介性細胞障害を生じさせ、膜貫通型腫瘍壊死因子αを中和し得、膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物の調製に用いられるヒト化モノクローナル抗体を提供することにある。
【課題を解決するための手段】
【0007】
本発明に係るヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の配列を含む軽鎖の可変領域のアミノ酸配列、及び配列番号2の配列を含む重鎖の可変領域のアミノ酸配列を含み、前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0008】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0009】
また、本発明に係る他のヒト化モノクローナル抗体のアミノ酸配列は、配列番号5の配列を含む軽鎖の可変領域のアミノ酸配列、及び配列番号6の配列を含む重鎖の可変領域のアミノ酸配列を含み、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0010】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0011】
本発明に係るヒト化モノクローナル抗体のヌクレオチド配列は、配列番号7の配列を含む軽鎖の可変領域のヌクレオチド配列、及び配列番号8の配列を含む重鎖の可変領域のヌクレオチド配列を含み、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0012】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0013】
本発明のヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号1の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号2の配列を含む重鎖を含み、前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、前記モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0014】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0015】
上記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含むことが好ましい。
【0016】
上記ヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMであることが好ましい。
本発明の他のヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含み、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【0017】
上記腫瘍壊死因子αは、ヒト腫瘍壊死因子αであることが好ましい。
【0018】
上記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含むことが好ましい。
【0019】
上記ヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMであることが好ましい。
【0020】
また、本発明に係る膜貫通型腫瘍壊死因子αを中和する方法は、ヒト化モノクローナル抗体を膜貫通型腫瘍壊死因子αに結合させる工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含むところに要旨を有するものである。
【0021】
また、本発明に係る抗体依存性細胞媒介性細胞障害を生じさせる方法は、ヒト化モノクローナル抗体を対象の膜貫通型腫瘍壊死因子αに結合させる工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含むところに要旨を有するものである。
【0022】
また、本発明に係る膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法は、ヒト化モノクローナル抗体を提供する工程を含み、前記ヒト化モノクローナル抗体は、軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖を含み、且つ、前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合するところに要旨を有するものである。
【発明の効果】
【0023】
本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αと結合する能力を有し、抗体依存性細胞媒介性細胞障害を生じさせることができる。従って、本発明のヒト化モノクローナル抗体は、抗体依存性細胞媒介性細胞障害を生じさせる用途や、膜貫通型腫瘍壊死因子αを中和する用途や、膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物の用途などに応用することができる。
【図面の簡単な説明】
【0024】
【図1】図1は、精製されたm357IgGとh357IgGのSDS−PAGE分析結果を示す図である。m357IgG(レーン1と2)とh357IgG(レーン3およびレーン4)は、マウス骨髄腫NS0細胞で発現され、蛋白質Aカラムにより培養基から精製された。サンプルは、非還元状態(レーン1と3)と還元状態(レーン2と4)下で、MOPSバッファ溶液により、4〜12%SDS/Bis−Trisポリアクリルアミドゲルで電気泳動された。ゲルはクアシンブルーで染色された。Mは分子量標準品である。
【図2】図2は、m357IgGの軽鎖cDNAとm357IgGの重鎖cDNAの電気泳動分析の結果をそれぞれ示す図である。
【図3A】図3Aは、m357IgGの重鎖可変領域および軽鎖可変領域について、最終の正確な構造を示す図である。
【図3B】図3Bは、m357IgGの分子表面モデルを示す図である。
【図4】図4は、m357IgGとPPS4の重鎖可変領域VH(A)、及び軽鎖可変領域VL(B)のアミノ酸配列を示す図である。m357IgGのヒト化のためのヒト表面殘基のアクセプタとして用いたm357IgG Fvと最も相同性が高かったPPS4 Fv配列を、比較のため、PPS4VLおよびPPS4VHとして示す。CDR殘基を括弧([ ])中に示し、保存表面殘基を白色フレームで表示し、非保存表面殘基を灰色フレームで表示した。Kabatの慣例(Kabat、1991)に基づいて、アミノ酸配列をナンバリングした。
【図5】図5は、m357−IgG抗体とh357−IgG抗体による、L929細胞中の腫瘍壊死因子α媒介細胞毒性の中和を示す図である。種々の濃度のm357−IgG(●)抗体とh357−IgG(○)抗体をL929細胞に加え、100ng/mlのヒト腫瘍壊死因子αと共に培養した。37℃で16時間培養した後、比色MTT分析により、細胞の生存を分析した。
【図6】図6は、m357−IgG抗体とh357−IgG抗体の細胞表面上の膜貫通型腫瘍壊死因子αに対する飽和結合分析を示す図である。膜貫通型腫瘤壞死因子αトランスフェクトNS0細胞を、m357−IgG(○)またはh357−IgG(●)抗体の連続希釈物(対数希釈)と共に4℃で1時間培養した。これらの細胞を3回洗浄し、それぞれ、m357−IgGに反応するAlexa Fluor 488ヤギ抗マウスIgG(H+L)と、h357−IgGに反応するAlexa Fluor 647ヤギ抗ヒトIgG(H+L)と共に4℃で1時間培養した。その後、細胞を洗浄し、FACSCaliburフローサイトメータで分析した。
【図7】図7は、膜貫通型腫瘍壊死因子α発現細胞に対するh357IgGの抗体依存性細胞媒介性細胞傷害を示す図である。膜貫通型腫瘍壊死因子α発現細胞を異なる濃度のh357IgG抗体の存在下で1時間培養した。その後、ヒト末梢血単核球PBMCをエフェクター細胞として用い、膜貫通型腫瘍壊死因子α発現細胞を目標細胞として用いた。細胞質ゾルから上澄み液に遊離するLDH量を測定することにより、細胞毒性を計算した。この結果を、溶解バッファ処理細胞群に対して100%溶解した細胞溶解率として示す。
【発明を実施するための形態】
【0025】
本発明の一態様において、本発明は、ヒト化モノクローナル抗体のアミノ酸配列を提供する。上記ヒト化モノクローナル抗体は腫瘍壊死因子α(TNF−α)と結合し、腫瘍壊死因子αはヒト腫瘍壊死因子αであっても良い。上述のヒト化モノクローナル抗体のアミノ酸配列は、軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列を含む。一実施形態において、本発明によるヒト化モノクローナル抗体のアミノ酸配列は、免疫グロブリンG(IgG)抗体のアミノ酸配列であり、軽鎖可変領域のアミノ酸配列、重鎖可変領域のアミノ酸配列、およびヒト免疫グロブリンG抗体の定常部のアミノ酸配列を含む。
【0026】
一実施形態において、本発明によるヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列は、腫瘍壊死因子αと結合することができる非ヒトモノクローナル抗体に対し、可変領域表面再構成を行なうことにより得られる。
【0027】
まず、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列を得る。一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体は、マウスモノクローナル抗体を含んでいても良い。このマウスモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、重鎖可変領域アミノ酸配列は、配列番号2の配列を含む。
【0028】
続いて、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列に基づいて、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列の分子モデルを構築すると共に、非定常表面残基をラベルする。一実施形態において、分子モデリングプロセスは、コンピュータアシストホモロジーモデリングにより行なうことができる。
【0029】
その後、非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)と同一性が最も高いヒト配列を探すと共に、両者を比較して、非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)アミノ酸配列の置換可能な残基を決定する。最後に、この置換可能な残基は、非ヒトモノクローナル抗体の可変領域の置換可能な残基の位置に対応する位置に存在する、ヒト配列に対応するアミノ酸残基に置換されて、腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列と重鎖可変領域のアミノ酸配列を獲得する。
【0030】
一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、且つ、重鎖可変領域アミノ酸配列は、配列番号2の配列を含んでいる。非ヒトモノクローナル抗体の可変領域と最も相同性が高い配列を有するヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。この実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のアミノ酸配列に対して行われる置換は、少なくとも以下の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。よって、得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の軽鎖可変領域アミノ酸配列、及び配列番号2の重鎖可変領域アミノ酸配列を含み、且つ、配列番号1の配列と配列番号2の配列は少なくとも以下の置換の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。
【0031】
別の実施形態において、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、非ヒトモノクローナル抗体の重鎖可変領域アミノ酸配列は、配列番号2の配列を含んでいる。非ヒトモノクローナル抗体の可変領域と最も同一性が高いヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。上述した配列比較実行後に得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域のアミノ酸配列は、配列番号5を有し、重鎖可変領域のアミノ酸配列は、配列番号6を含む。
【0032】
よって、本発明の別の態様によれば、本発明はまた、上述したヒト化モノクローナル抗体のヌクレオチド配列も提供する。本発明のヒト化モノクローナル抗体のヌクレオチド配列は、軽鎖可変領域のヌクレオチド配列と重鎖可変領域のヌクレオチド配列を含む。一実施形態において、本発明によるヒト化モノクローナル抗体のヌクレオチド配列は、免疫グロブリンG(IgG)抗体のヌクレオチド配列であり、軽鎖可変領域のヌクレオチド配列、重鎖可変領域のヌクレオチド配列、およびヒト免疫グロブリンG抗体の定常領域のヌクレオチド配列を含む。
【0033】
一実施形態において、ヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド配列は、配列番号5の配列をエンコードするヌクレオチド配列を含み、重鎖可変領域ヌクレオチド配列は、配列番号6の配列をエンコードするヌクレオチド配列を含む。配列番号5の配列をエンコードするヌクレオチド配列は配列番号7の配列であり、配列番号6の配列をエンコードするヌクレオチド配列は配列番号8の配列である。
【0034】
本発明の別の態様において、本発明は更に、ヒト化モノクローナル抗体を提供する。このヒト化モノクローナル抗体は腫瘍壊死因子αと結合し、腫瘍壊死因子αは、ヒトの腫瘍壊死因子αであっても良い。ヒト化モノクローナル抗体は、軽鎖と重鎖を含む。一実施形態において、本発明によるヒト化モノクローナル抗体は免疫グロブリンG(IgG)抗体であり、一本鎖は、軽鎖可変領域、重鎖可変領域、およびヒト免疫グロブリンG定常領域を含む。
【0035】
一実施形態において、本発明によるヒト化モノクローナル抗体は、以下のステップにより得られる。
【0036】
まず、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域ヌクレオチド断片およびその配列と、重鎖可変領域ヌクレオチド断片およびその配列を得る。一実施形態において、上述の非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド断片(cDNA)は、非ヒトモノクローナル抗体を生成することができるハイブリドーマの総RNAを抽出した後、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のそれぞれに対応する二個のプライマー対により、総RNAに対して逆転写ポリメラーゼ連鎖反応(RT−PCR)を行なうことによって得られる。次いで、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド断片の配列をそれぞれ決定した後、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド配列を得る。一実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体はマウス抗体を含む。このマウス抗体の軽鎖可変領域ヌクレオチド配列は、配列番号9の配列を有し、重鎖可変領域ヌクレオチド配列は、配列番号10の配列を有する。
【0037】
続いて、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のヌクレオチド配列に基づいて、それぞれ、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列を獲得する。一実施形態において、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を有し、非ヒトモノクローナル抗体の重鎖可変領域アミノ酸配列は、配列番号2の配列を有する。
【0038】
その後、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列と重鎖可変領域アミノ酸配列により、非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域の分子モデルを構築すると共に、非定常表面残基をラベルする。一実施形態において、分子モデリングプロセスは、コンピュータアシストホモロジーモデリングにより行なう。
【0039】
続いて、本発明による腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域アミノ酸配列と最も同一性が高いヒト配列を探すと共に、両者の比較を実行して、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域(軽鎖可変領域と重鎖可変領域)アミノ酸配列の置換可能な残基を決定する。最後に、前述の腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の可変領域アミノ酸配列のヌクレオチド配列を、置換可能な残基の対応位置でエンコードして、点変異により、この残基位置に対応するヒト配列の残基をエンコードするヌクレオチドに置換して、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド断片と重鎖可変領域のヌクレオチド断片を獲得する。
【0040】
その後、本技術領域で熟知される方法により、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体の軽鎖可変領域ヌクレオチド断片と重鎖可変領域のヌクレオチド断片と、既知のヒト抗体定常領域ヌクレオチド断片を、それぞれ、適切な発現ベクターにクローン化し、この発現ベクターを適切な宿主細胞にトランスフェクトすると、宿主細胞は、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体を発現する。
【0041】
一実施形態において、非ヒトモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を有し、且つ、重鎖可変領域アミノ酸配列は、配列番号2の配列を有する。前述の非ヒトモノクローナル抗体の可変領域(軽鎖可変領域アミノ酸配列および重鎖可変領域)アミノ酸配列と相同性が最も高いヒト配列は、PPS4の軽鎖可変領域アミノ酸配列(配列番号3)と重鎖可変領域アミノ酸配列(配列番号4)である。この実施形態において、腫瘍壊死因子αと結合する非ヒトモノクローナル抗体の軽鎖可変領域と重鎖可変領域のアミノ酸配列に対して行なわれる置換は、少なくとも以下の一つを含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンがアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸はグリシンに置換される。よって、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号1の軽鎖可変領域アミノ酸配列、および配列番号2の重鎖可変領域アミノ酸配列を含み、且つ、配列番号1の配列と配列番号2の配列は、少なくとも一つの以下のアミノ酸置換を含む:配列番号1の第10アミノ酸のイソロイシンはトレオニンに置換され、配列番号1の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第2アミノ酸のリジンはグルタミンに置換され、配列番号2の第10アミノ酸のトリプトファンはロイシンに置換され、配列番号2の第18アミノ酸のリジンはアルギニンに置換され、配列番号2の第41アミノ酸のグルタミン酸がグリシンに置換される。
【0042】
別の実施形態において、本発明による腫瘍壊死因子αと結合するモノクローナル抗体の軽鎖可変領域アミノ酸配列は、配列番号1の配列を含み、重鎖可変領域アミノ酸配列は、配列番号2の配列を含む。本発明による腫瘍壊死因子αと結合するモノクローナル抗体の可変領域(軽鎖可変領域および重鎖可変領域)アミノ酸配列と相同性が最も高いヒト配列は、PPS4の軽鎖可変領域配列(配列番号3)と重鎖可変領域配列(配列番号4)である。上記のようにして配列を比較した後、得られた腫瘍壊死因子αと結合するヒト化モノクローナル抗体のアミノ酸配列は、配列番号5の軽鎖可変領域を含み、配列番号6の重鎖可変領域のアミノ酸配列を含む。よって、本発明による腫瘍壊死因子αと結合するヒト化モノクローナル抗体は、配列番号5の軽鎖可変領域アミノ酸配列、及び配列番号6の重鎖可変領域アミノ酸配列を含む。
【0043】
本発明のヒト化モノクローナル抗体と結合する腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む。本発明のヒト化モノクローナル抗体の放出型腫瘍壊死因子αに対する結合親和性は、約20−40nMであり、好ましくは、約10−20nMである。
【0044】
更に本発明のヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は約20−40nMであり、好ましくは、約10−20nMである。一実施形態において、本発明のヒト化モノクローナル抗体の膜貫通型腫瘍壊死因子αに対する結合親和性は約16.8nMである。
【0045】
更に一実施形態において、本発明のヒト化モノクローナル抗体は、細胞上で発現される膜貫通型腫瘍壊死因子αと結合することにより、抗体依存性細胞媒介性細胞障害(ADCC)を生じる。
【0046】
現在分かっていることは、膜貫通型腫瘍壊死因子αと多くの疾病が関係しているということである。例えば、細胞表面が発現する膜貫通型腫瘍壊死因子αのマクロファージ細胞と単球細胞は、例えば、クローン病やウェゲナー肉芽腫症などの肉芽腫性疾患で重要な役割を果たしており、抗体依存性細胞媒介性細胞障害により、細胞を直接滅亡させる(Beenhouwertsら、2004)。
【0047】
このように本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αと結合する能力を有し、且つ、抗体依存性細胞媒介性細胞障害を生じさせる能力があるので、本発明の更なる態様において、本発明は、膜貫通型腫瘍壊死因子αを中和するヒト化モノクローナル抗体の用途、および抗体依存性細胞媒介性細胞障害を生じさせるヒト化モノクローナル抗体の用途も提供する。
【0048】
更に本発明のヒト化モノクローナル抗体は、膜貫通型腫瘍壊死因子αに相関する疾病を治療する薬物の用途にも応用できる。
【実施例】
【0049】
材料と方法
A.ヒト腫瘍壊死因子α(TNF−α)と結合するマウスモノクローナル抗体
1.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体の獲得と精製
ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体を産生し得るハイブリドーマ細胞株357−104−4(ECACC No.92030603)(ヨーロピアン・コレクション・オブ・セル・カルチャーから購入)をマウス腹腔内に注射し、マウスに腫瘍と腹水を生じさせた。生成したマウス腹水は、大量のヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)を含む。上述のステップは、GlycoNEX Inc.(http://www.glyconex.com.tw/)に委託して行なった。
【0050】
続いて、この腹水をマウスから取り出し、腹水をプロテインAカラム(GEヘルスケア)に通して精製し、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)を獲得した。
【0051】
2.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のcDNA配列の獲得
【0052】
(1)ハイブリドーマ細胞株357−104−4(ECACC No.92030603)の総RNAを、QIAGEN RNeasy Mini Kitにより抽出した。
【0053】
(2)アマシャム(Amersham)社製のLight Primer MixとHeavy Primersの二個のプライマー対により、総RNAに対して逆転写ポリメラーゼ連鎖反応(RT−PCR)を行い、m357IgGの軽鎖と重鎖のcDNAをそれぞれ、獲得した。
【0054】
(3)この軽鎖と重鎖のcDNAを、それぞれ、TOPOベクター上にクローン化して、二個のクローンを得た。
【0055】
(4)DNA配列決定後、制限酵素部位を有する二個のプライマー対を設計した。
【0056】
(5)制限酵素部位を有する二個のプライマー対を合成した。
ここで、m357IgGの軽鎖のcDNAに用いるプライマー対は以下のとおりである:
357VL’Ascプライマー:
5’-CAGGCGCGCCGAAATTGTGCTGACCCAGTC-3’(配列番号11)
357VL3’Glinkプライマー:
5’-CCAGAGCCACCTCCGCCTGAACCGCCTCCACCCAATTTCCAGCTTGC-3’(配列番号12)。
【0057】
m357IgGの重鎖のcDNAに用いるプライマー対は以下のとおりである:
357VH5’Glinkプライマー:
5’-TCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGGTGAAACTGCAGGA-3’(配列番号13)
357VH3’Not:
5’-CAGCGGCCGCTGAGGAGACGGTGACCGTGGT-3’(配列番号14)。
【0058】
その後、ステップ(3)で形成された2つのクローン中でポリメラーゼ連鎖反応を行なって、m357IgGの軽鎖と重鎖のcDNAを増幅させた。
【0059】
B.再構成のための、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のコンピューターモデリング
Discovery Studioモデリング2.1(Accelrys社、サンディエゴ、カナダ)を使用して、ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)のホモロジーモデリングプロセスを行なった。m357IgGの軽鎖可変領域(VL)と重鎖可変領域(VH)に対し、二個の別々のBLASTP検索を行なった。蛋白質データバンク(http://www.rcsb.org/pdb/home/home.do)中で検索することによって配列を分析し、m357蛋白質配列のホモロジーを同定した。マウス抗乳癌抗体Fab断片の重鎖可変領域SM−3[PDB entry:1SM3](Dokurnoら、1998)と抗サーマス・アクタチクス(thermus aquaticus)DNA ポリメラーゼIモノクローナル抗体Fab断片の軽鎖可変領域構造[PDB entry:1AY1](Muraliら、1998)のホモロジーモデリングに基づいて、m357IgG Fv断片の三次元(3D)構造を構築した。最終的な三次元モデルは、MODELLERモジュール(Saliら、1995)により生成され、空間的拘束を満足させる(satisfaction of spatial restraints)ことにより、比較蛋白質構造モデリングの自動方法を行なった。m357IgGに対し、蛋白質ホモロジーモデリングおよびループモデリングを自動的に行なった。モデル抗体ループ(Model antibody Loops)モジュールを使用することにより、最高の配列鑑別度で、PDBデータベースからテンプレート構造を選択してCDRループのモデル構築を行い、、且つ、ループ精製モジュールを使用して、CDRループのモデル構築を完全にし、立体衝突(steric clash)を最小化すると共に、正確な結合距離と角度を確認した。その後、Discovery Studioモデリング2.1中のCHARMm(B.R.Brooks、1983)プログラムと、Accelrys CHARMm forcefieldにより、構造のエネルギーを最小化して、このモデル全体を更に純化した。構造のエネルギー最小化は、二つのステップで行なった。まず、抑制最急降下最小化(restrained steepest descent minimization)を5000ステップいった後、続いて、フレームワーク(framework)のαカーボンが、適切な位置で維持固定されるまで、共役勾配最小化(conjugated gradient minimization)を5000ステップ行なった。AREAIMOLプログラム(CCP4、1994)により、三次元モデル上で、m357IgG残基の溶媒接近表面積(solvent−accessible surface areas)を計算した。相対接近が30%以上の残基は、接近可能であると定義した。
【0060】
C.IgGの構築、発現および精製
オーバーラッピングPCRにより、m357IgGのヒト化可変領域(軽鎖可変領域および重鎖可変領域)をエンコードするcDNA断片を獲得した。続いて、ヒトIgGからの定常領域配列とオーバーラッピングPCRにより合成される可変領域配列を、哺乳動物発現ベクターpSecTag2/Hygro(重鎖)(Invitrogen)とpcDNA3.3−TOPO TA(軽鎖)(Invitrogen)中で、サブクローニングした。その後、EcoRV制限酵素認識部位を利用して、二個の構築体(construct)を融合させ、pSec−pcDNA−h357−IgGを生成した。メーカーの使用説明書により、エフェクテン(Effectene、Qiagen)を使用し、重鎖と軽鎖遺伝子を含むプラスミドを、マウス骨髄腫NS0細胞(ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャー、Salisbury、Wiltshire、英国)中でトランスフェクトさせた。ハイグロマイシン(Hygromycin)(400μg/ml)で4週間抽出した後、安定したクローンを、無血清の既知組成培地HyQNS0(Hyclone)中で、5×105cells/mlの初期播種密度にてシェーカー中で培養した。37℃で5日間後の培養基を取り出し、蛋白質A(GE Health−care)クロマトグラフィにより、抗体を上澄みから精製した。
【0061】
D.抗腫瘍壊死因子αの中和効力分析
既知の方法(Matthews N、1987)に従って、アクチノマイシンDで処理されたマウス線維芽細胞L929細胞(ATCC Cat.No.CCL−1)により、ヒト腫瘍壊死因子αに対するm357IgGおよびh357IgGの中和活性を測定した。要約すると、L929細胞を、96ウェルプレート中に3×105cells/wellで三回播種すると共に、10%(v/v)ウシ胎仔血清を添加したRPMI 1640培地中で16時間培養した。その後、幾つかの抗体希釈液を調製し、アクチノマイシンD(2μg/ml)と腫瘍壊死因子α(TNF−α)(100ng/ml)を含む培地中に入れ、37℃で16時間培養した。上清を除去した後、3−(4,5−ジメチル−2−チアゾリル)−2,5−ジフェニルテトラゾリウムブロミド(MTT)(5mg/ml)(Sigma−Aldrich)を加え、37℃で4時間培養した。その後、SDS溶液(10%)をウェル中に加えた。室温で24時間培養した後、色度計により、各ウェル中の色が紫色に変化するのを記録した。570nmで測定される溶出の光学密度(OD)を測定した結果、生存細胞の数量と正の相関が見られた。ブランクの対照群(培養物のみ)、腫瘍壊死因子αの対照群(腫瘍壊死因子αのみ)、および抗体の対照群(抗体のみ)もまた、上記実験中で行なわれた。シグマプロットソフトウェア(Systat software,Inc、Richmond、カナダ)を用いて複雜なS状非線形回帰(complex sigmoid non−linear regression)分析によりED50値を計算した。
【0062】
E.NS0細胞上の膜貫通型腫瘍壊死因子αの安定した発現
既知の技術(Perezら、1990)で示されるように、抵抗腫瘍壊死因子α変換酵素(TNF−α converting enzyme、TACE)媒介開裂に対して耐性がある欠失変異体膜貫通腫瘍壊死因子αを、部位特異的変異導入法によって生成した。非開裂形式の膜貫通腫瘍壊死因子αでは、天然の膜貫通腫瘍壊死因子αのアミノ酸+1〜+12が削除された。非開裂形式の膜貫通腫瘍壊死因子α遺伝子は、pSecTag2/Hygro哺乳動物発現ベクター(Invitrogen)にクローン化されると共に、エフェクテンにより、マウス骨髄腫NS0細胞にトランスフェクトされ、細胞表面上で膜貫通腫瘍壊死因子αを発現させた。
【0063】
F.m357−IgGとh357−IgGの膜貫通腫瘍壊死因子αに対する飽和結合分析
膜貫通腫瘍壊死因子αトランスフェクトNS0細胞を、m357−IgGとh357−IgGの連続希釈物(対数希釈)と共に、2%のウシ胎仔血清を含むリン酸緩衝生理食塩水(蛍光活性化細胞分類[FACS]バッファ)中にて4℃で1時間培養した。この細胞を、蛍光活性化細胞分類バッファで3回洗浄した後、それぞれ、m357−IgGのAlexa Fluor 488ヤギ抗マウスIgG(H+L)とh357−IgGのAlexa Fluor 647ヤギ抗ヒトIgG(H+L)により、4℃で1時間染色した。FACSCaliburフロー血球計算器(Becton Dickinson、San Jose、カナダ)により蛍光強度を測定した。
【0064】
G.抗体依存性細胞媒介性細胞障害(ADCC)の分析
目標物質であるh357−IgGの抗体依存性細胞媒介性細胞障害(ADCC)の活性を、LDH細胞毒性検出キット(Clontech)により測定し、製造メーカーの取扱説明書に基づいて、損傷した細胞のサイトゾルから遊離するLDH活性を測定した。要約すると、高度に表現された膜貫通腫瘍壊死因子αの細胞を、異なる濃度のh357抗体の存在下で、5%CO2インキュベーターのアッセイ培地(1%FBS含有DEME)中で1時間培養した後、エフェクター細胞として梢血単核細胞(PBMC)を加えた(エフェクター細胞:目標細胞=20:1)。更に37℃で16時間培養した後、ウェル毎に100μlの上澄みを採取し、新しい96−ウェルの平底プレート中に移した。LDH基質(100μl)を各ウェルに加え、避光下にて室温で30分間培養した。サンプルの吸光度を、ELISAリーダーにより490nmで測定した。溶解バッファにより最大放出(release)を決定した。以下の式により、特定の溶解百分率を計算した:%細胞毒性=[実験例の放出−自然放出]/[最大放出−自然放出]×100。
【0065】
結果
A.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体(m357IgG)
【0066】
1.ヒト腫瘍壊死因子αと結合するマウスモノクローナル抗体の分析
精製したm357IgGに対してSDS−PAGE分析を行なった。この結果を図1のレーン1とレーン2に示す。レーン1は非還元状態における精製したm357IgGのSDS−PAGE分析結果であり、レーン2は還元状態における精製したm357IgGのSDS−PAGE分析結果である。
【0067】
2.m357IgGの軽鎖と重鎖のcDNA
ポリメラーゼ連鎖反応により得られたm357IgGの軽鎖と重鎖のcDNAについて電気泳動分析を行なった。この結果を図2に示す。レーン1はm357IgGの軽鎖のcDNA(配列番号9)であり、レーン2はm357IgGの重鎖のcDNA(配列番号10)である。
【0068】
B.m357可変断片の分子モデリング
RT−PCRにより、ハイブリドーマ細胞株357−101−4(ECACC No. 92030603)からの抗腫瘍壊死因子αモノクローナル抗体m357IgGの重鎖可変領域(VH)および軽鎖可変領域(VL)をエンコードしたcDNA(データは示さず)を得た。「材料と方法」中に記載したホモロジーモデリング(program MODELLER)により、それぞれ、m357IgGの重鎖可変領域VH、および軽鎖可変領域VLの推定アミノ酸配列の三次元構造を構築した。その後、PDBからの1SM3(配列同一性および相似性が、それぞれ、87%と93%)テンプレート構造が1.95Å、PDBからの1AY1(配列同一性および相似性が、それぞれ、85%と91%)テンプレート構造が2.20Åの解析度で、m357IgGの重鎖可変領域および軽鎖可変領域をそれぞれ、モデリングした。Discovery Studioモデリング2.1プログラムにより、m357IgGの重鎖可変領域VHおよび軽鎖可変領域VLの最終の正確な構造を獲得した。この結果を図3Aに示す。
【0069】
図3Aは、m357IgGの可変領域を示す図である。m357IgGの三次元構造は、それぞれ、重鎖可変領域に対応する1SM3と軽鎖可変領域に対応する1AY1の結晶構造を比較するホモロジーモデリングにより生成した。CDRループを太線で示す。図3Bは、m357IgGの分子表面モデルを示す図であり、非ヒト様(non−human like)表面残基とCDRループの関係を示す。相対溶媒接近が30%以上の17個の残基は、非ヒト様の表面残基であり、太い矢印で示す。CDRループを太線で示す。5Å内のCDRループに近い残基(点線の円)を点線矢印で示す。好ましくは「ヒト」アミノ酸に突然変異する6個の残基を実線矢印で示す。重鎖可変領域フレームワーク中の4個の残基をヒト化のために決定し、軽鎖可変領域フレームワーク中の2個の残基のみヒト化のために決定した。
【0070】
C.m357IgG可変領域断片のヒト化
可変領域再構成によるヒト化は、最初にPadlanが1991年に提案した。この方法では、抗体のフレームワーク領域中の潜在抗原部位は排除され、抗体の抗原親和性に影響しない(Padlan、1991)。この方法は、ヒト抗マウス抗体(HAMA)が、表面残基から派生する可変領域だけに反応することを前提としており、他の研究者(Fontayneら、2006;O’Connorら、1998;Staelensら、2006)により採用され改変されてきた。本実施例では、以下の三つのステップにより、m357IgGの可変領域再構成を行なった。まず、m357IgGの重鎖可変領域と軽鎖可変領域をそれぞれ構築した。次に、AREAIMOLプログラムを使用して溶媒接近可能残基を計算し、非ヒト様のフレームワーク表面残基を同定した。最後に、ヒトフレームワークの配列アライメント結果に基づいて、これらの表面残基を、ヒトの対応表面残基に突然変異させた。可変領域上で、マウスm357IgGの非ヒト様フレームワーク表面残基を置換するため、ヒト目標配列から、一組の高度に相同性を有する表面残基を選択した。IMGTデータベース(http://imgt.cines.fr/)中で検索を行い、検索結果から、ファージ提示法、またはヒト化抗体の配列を排除して、m357IgGと対応する可変領域に対し、相同性が最も高いヒト重鎖可変領域と軽鎖可変領域の配列ペアを同定した。ヒト配列で見出される最も同一性の高い表面残基は、PPS4の可変領域である(フレームワーク領域中の配列同一性は、それぞれ、重鎖可変領域;76%、軽鎖可変領域:73%)(図4)。AREAIMOLプログラムの計算結果に基づいて、重鎖可変領域中の20個の表面残基のうち8個は、ヒトとマウスの配列間で非保存的であり、軽鎖可変領域中の16個の表面残基のうち9個は非保存的であった。これら17個(8個+9個)の表面残基は、置換の候補者であった。しかし、CDR領域中の重鎖H52BQ、重鎖H52CS、重鎖H54N、重鎖H61E、軽鎖L31F、軽鎖L55S、軽鎖L90W、軽鎖L92D、軽鎖L93Y、および軽鎖L96Rの置換は、CDRのコンフォメーションを潜在的に変化させ、軽鎖中のCDR2近くの5Å内の余分な非定常表面残基であるL59Vも、結合親和性に潜在的に影響する(図3B)。その結果、これらの11個のマウス残基は維持され、抗原親和性を保持する。最後に、残りの6個の残基を選択して、ヒト保存残基に置換した。PPS4と比較すると、軽鎖可変領域の第10の比較位置におけるイソロイシンはトレオニンに置換され(m357IgGの軽鎖可変領域の第10アミノ酸のイソロイシンがトレオニンに置換される)、第18の比較位置のリジンはアルギニンに置換される(m357軽鎖可変領域の第18のアミノ酸のリジンがアルギニンに置換される)。PPS4と比較すると、重鎖可変領域の第3の比較位置のリジンはグルタミンに置換され(m357重鎖可変領域の第2アミノ酸のリジンがグルタミンに置換される)、第11の比較位置のトリプトファンがロイシンに置換され(m357重鎖可変領域の第10アミノ酸のトリプトファンがロイシンに置換される)、第19の比較位置のリジンがアルギニンに置換され(重鎖可変領域の第18アミノ酸のリジンがアルギニンに置換される)、第42の比較位置のグルタミン酸がグリシンに置換される(重鎖可変領域の第41アミノ酸のグルタミン酸がグリシンに置換される)(図4)。
【0071】
D.ヒト化357(h357)IgG1の構築と発現。
m357IgGのヒト化重鎖可変領域と軽鎖可変領域のアミノ酸配列を、インフレームで、ヒトIgG1の重鎖とカッパ軽鎖定常領域に融合させた。完全なヒト化357(h357)IgG1分子を発現するため、二種の哺乳動物発現ベクターであるpSecTag2/HygroとpcDNA3.3−TOPO TAを使用し、h357IgGのヒト化重鎖と軽鎖をそれぞれ、導入した。その後、軽鎖発現カセットを重鎖発現pSecTag2/Hygroベクターに連接させ、単一の発現ベクターを得た。組み換えh357IgGの発現レベルは約14mg/Lであった。蛋白質Aクロマトグラフィにより、m357IgGとh357IgGを含む培養基をそれぞれ、精製すると共に、SDS−PAGEにより蛋白質精製物を決定した(図1)。図1に示すように、非還元状態下で、二つの抗体は、155kDaの分子量を有する単一バンド(レーン1と3)を示した。還元状態下では、二つの抗体は、それぞれ55kDa(重鎖)と26kDa(軽鎖)の分子量を有する二つの蛋白質バンド(レーン2と4)を生成した。
【0072】
E.m357IgGとh357IgGによる腫瘍壊死因子α媒介細胞毒性の中和
抗腫瘍壊死因子α抗体の機能活性を調べるために、可溶性腫瘍壊死因子α活性を抑制する抗体の能力をテストした。腫瘍壊死因子αは、マウスL929細胞に対して細胞毒性を生じさせる。組み換えヒト腫瘍壊死因子α抗体および上記細胞の共培養により、L929アッセイでm357IgGとh357IgGを両方評価した。図5に示すように、100ng/mlのヒト腫瘍壊死因子αで処理されたL929細胞中の腫瘍壊死因子α媒介細胞毒性は、m357IgGとh357IgGの両方により、用量依存的に効果的に中和され、ED50値はそれぞれ、3.07nMと2.30nMであった。この結果は、ヒト化357IgGは、マウス357IgGと同程度の濃度で、腫瘍壊死因子αの中和活性を維持することを示している。
【0073】
F.h357IgG抗体と膜貫通腫瘍壊死因子αとの結合活性
腫瘍壊死因子αは、膜関連の前駆体(膜貫通腫瘍壊死因子α)として存在し、腫瘍壊死因子α変換酵素(TNF変換酵素、TACE)により媒介される蛋白分解切断により成熟した可溶体が放出される。多くの研究により、腫瘍壊死因子α拮抗薬は、アポトーシス、抗体依存性細胞媒介性細胞障害(ADCC)、補体依存性細胞障害活性(CDC)、外側から内側へのシグナリングメカニズムにより、細胞溶解を生じることが調査されてきた(Aroraら、2009;Caronら、1999;Mitomaら、2008;Scallonら、1995)。h357IgGと膜貫通腫瘍壊死因子αとの結合能力を分析するため、膜貫通腫瘍壊死因子αの非開裂形式のcDNAを、NS0細胞にトランスフェクトして細胞膜上で発現させ、フロー血球計算器を用いてm357IgGとh357IgGの両方の結合活性を評価した。図6のデータは、m357IgGとh357IgGは共に、濃度依存的に膜貫通腫瘍壊死因子αと結合することを示しており、KD値はそれぞれ、12.0nMと16.8nMである。同程度のKD値は、ヒト化過程において、膜貫通TNF−αの結合親和性を変化させないことを示す。h357IgGがナノモル範囲の結合親和性を有することは、h357IgGが、膜貫通腫瘍壊死因子αを介して、更に多くのエフェクター機能、またはアポトーシスメカニズムを潜在的にトリガーすることを示唆している。
【0074】
G.h357IgGの抗体依存性細胞媒介性細胞障害に対する能力
既知の文献で示されるように、インフリキシマブおよびアダリムマブの膜貫通腫瘍壊死因子αに対する結合親和性は、エタネルセプトよりも高く、抗体依存性細胞媒介性細胞障害(ADCC)、補体依存性細胞障害活性(CDC)、またはアポトーシスにより、細胞表面発現膜貫通腫瘍壊死因子αの細胞死滅作用に影響するが、これは、臨床疾病で異なる効果が生じる原因のひとつである(Taylor、2010)。細胞表面発現腫瘍壊死因子α様マクロファージと単球は、例えばクローン病やウェゲナー肉芽腫症などの肉芽腫性疾患において非常に重要な役割を果たしており、抗体依存性細胞媒介性細胞障害により、細胞を直接、死滅させる(Beenhouwerら、2004)。腫瘍壊死因子α拮抗薬が膜貫通型腫瘍壊死因子α発現細胞に結合すると、これらの細胞は、ナチュラルキラー細胞のターゲットとされる。ヒトIgG1由来のFc領域からなるh357IgGは、腫瘍壊死因子α産生細胞中で、潜在的に細胞溶解を生じる。よって、h357IgGの、膜貫通腫瘍壊死因子α発現目標細胞に対するADCC仲介能力を評価するため、分離した末梢血単核球PMBCを抗体依存性細胞媒介性細胞障害の分析に用いた。エフェクター細胞:目標細胞(E:T)は20:1である。h357IgGの濃度が6.25ug/mlのときに、腫瘍壊死因子α目標細胞の20%以上が溶解した(図7)。これらのデータは、h357IgGが、細胞表面で発現された膜貫通腫瘍壊死因子αとの結合によって抗体依存性細胞媒介性細胞障害を仲介することを示唆している。従って、h357IgGは、ADCC能を有する治療抗体と同様、更に効果的な腫瘍壊死因子α中和抗体に発展する可能性を有している。
【0075】
本発明では好ましい実施例を上記の通り開示したが、本発明は上記実施例に限定されず、当業者であれば、本発明の精神と領域を逸脱しない範囲内で各種の改変や調整を行なうことができ、このようなものも本発明の範囲内に包含される。
【0076】
本願明細書中に記載した文献の詳細は、以下のとおりである。
【0077】
【表1】
【0078】
【表2】
【0079】
【表3】
【0080】
【表4】
【特許請求の範囲】
【請求項1】
ヒト化モノクローナル抗体のアミノ酸配列であって、
配列番号1の配列を含む軽鎖の可変領域のアミノ酸配列、及び
配列番号2の配列を含む重鎖の可変領域のアミノ酸配列
を含み、
前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、
前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のアミノ酸配列。
【請求項2】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項1に記載のヒト化モノクローナル抗体のアミノ酸配列。
【請求項3】
ヒト化モノクローナル抗体のアミノ酸配列であって、
配列番号5の配列を含む軽鎖の可変領域のアミノ酸配列、及び
配列番号6の配列を含む重鎖の可変領域のアミノ酸配列
を含み、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のアミノ酸配列。
【請求項4】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項3に記載のヒト化モノクローナル抗体のアミノ酸配列。
【請求項5】
ヒト化モノクローナル抗体のヌクレオチド配列であって、
配列番号7の配列を含む軽鎖の可変領域のヌクレオチド配列、及び
配列番号8の配列を含む重鎖の可変領域のヌクレオチド配列
を含み、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のヌクレオチド配列。
【請求項6】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項5に記載のヒト化モノクローナル抗体のヌクレオチド配列。
【請求項7】
ヒト化モノクローナル抗体であって、
軽鎖の可変領域のアミノ酸配列が配列番号1の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号2の配列を含む重鎖
を含み、
前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、
前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体。
【請求項8】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項7に記載のヒト化モノクローナル抗体。
【請求項9】
前記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む請求項7に記載のヒト化モノクローナル抗体。
【請求項10】
前記ヒト化モノクローナル抗体の前記膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMである請求項9に記載のヒト化モノクローナル抗体。
【請求項11】
ヒト化モノクローナル抗体であって、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含み、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体。
【請求項12】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項11に記載のヒト化モノクローナル抗体。
【請求項13】
前記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む請求項11に記載のヒト化モノクローナル抗体。
【請求項14】
前記ヒト化モノクローナル抗体の前記膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMである請求項13に記載のヒト化モノクローナル抗体。
【請求項15】
膜貫通型腫瘍壊死因子αを中和する方法であって、
ヒト化モノクローナル抗体を膜貫通型腫瘍壊死因子αに結合させる工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含むことを特徴とする膜貫通型腫瘍壊死因子αを中和する方法。
【請求項16】
抗体依存性細胞媒介性細胞障害を生じさせる方法であって、
ヒト化モノクローナル抗体を対象の膜貫通型腫瘍壊死因子αに結合させる工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含むことを特徴とする抗体依存性細胞媒介性細胞障害を生じさせる方法。
【請求項17】
膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法であって、
ヒト化モノクローナル抗体を提供する工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含み、且つ、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とする膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法。
【請求項1】
ヒト化モノクローナル抗体のアミノ酸配列であって、
配列番号1の配列を含む軽鎖の可変領域のアミノ酸配列、及び
配列番号2の配列を含む重鎖の可変領域のアミノ酸配列
を含み、
前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、
前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のアミノ酸配列。
【請求項2】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項1に記載のヒト化モノクローナル抗体のアミノ酸配列。
【請求項3】
ヒト化モノクローナル抗体のアミノ酸配列であって、
配列番号5の配列を含む軽鎖の可変領域のアミノ酸配列、及び
配列番号6の配列を含む重鎖の可変領域のアミノ酸配列
を含み、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のアミノ酸配列。
【請求項4】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項3に記載のヒト化モノクローナル抗体のアミノ酸配列。
【請求項5】
ヒト化モノクローナル抗体のヌクレオチド配列であって、
配列番号7の配列を含む軽鎖の可変領域のヌクレオチド配列、及び
配列番号8の配列を含む重鎖の可変領域のヌクレオチド配列
を含み、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体のヌクレオチド配列。
【請求項6】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項5に記載のヒト化モノクローナル抗体のヌクレオチド配列。
【請求項7】
ヒト化モノクローナル抗体であって、
軽鎖の可変領域のアミノ酸配列が配列番号1の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号2の配列を含む重鎖
を含み、
前記配列番号1の配列と前記配列番号2の配列は、少なくとも一つのアミノ酸置換を有し、
前記アミノ酸置換は、配列番号1の第10アミノ酸のイソロイシンのトレオニンへの置換、配列番号1の第18アミノ酸のリジンのアルギニンへの置換、配列番号2の第2アミノ酸のリジンのグルタミンへの置換、配列番号2の第10アミノ酸のトリプトファンのロイシンへの置換、配列番号2の第18アミノ酸のリジンのアルギニンへの置換、および配列番号2の第41アミノ酸のグルタミン酸のグリシンへの置換よりなる群から選択される少なくとも一つであり、且つ、
前記モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体。
【請求項8】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項7に記載のヒト化モノクローナル抗体。
【請求項9】
前記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む請求項7に記載のヒト化モノクローナル抗体。
【請求項10】
前記ヒト化モノクローナル抗体の前記膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMである請求項9に記載のヒト化モノクローナル抗体。
【請求項11】
ヒト化モノクローナル抗体であって、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含み、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とするヒト化モノクローナル抗体。
【請求項12】
前記腫瘍壊死因子αは、ヒト腫瘍壊死因子αである請求項11に記載のヒト化モノクローナル抗体。
【請求項13】
前記腫瘍壊死因子αは、放出型腫瘍壊死因子α、または膜貫通型腫瘍壊死因子αを含む請求項11に記載のヒト化モノクローナル抗体。
【請求項14】
前記ヒト化モノクローナル抗体の前記膜貫通型腫瘍壊死因子αに対する結合親和性は、約20−40nMである請求項13に記載のヒト化モノクローナル抗体。
【請求項15】
膜貫通型腫瘍壊死因子αを中和する方法であって、
ヒト化モノクローナル抗体を膜貫通型腫瘍壊死因子αに結合させる工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含むことを特徴とする膜貫通型腫瘍壊死因子αを中和する方法。
【請求項16】
抗体依存性細胞媒介性細胞障害を生じさせる方法であって、
ヒト化モノクローナル抗体を対象の膜貫通型腫瘍壊死因子αに結合させる工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含むことを特徴とする抗体依存性細胞媒介性細胞障害を生じさせる方法。
【請求項17】
膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法であって、
ヒト化モノクローナル抗体を提供する工程を含み、
前記ヒト化モノクローナル抗体は、
軽鎖の可変領域のアミノ酸配列が配列番号5の配列を含む軽鎖、及び
重鎖の可変領域のアミノ酸配列が配列番号6の配列を含む重鎖
を含み、且つ、
前記ヒト化モノクローナル抗体は、腫瘍壊死因子αに結合することを特徴とする膜貫通型腫瘍壊死因子αに関する疾病を治療する薬物を調製する方法。
【図1】

【図2】

【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2012−85632(P2012−85632A)
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願番号】特願2010−291151(P2010−291151)
【出願日】平成22年12月27日(2010.12.27)
【出願人】(390023582)財団法人工業技術研究院 (524)
【氏名又は名称原語表記】INDUSTRIAL TECHNOLOGY RESEARCH INSTITUTE
【住所又は居所原語表記】195 Chung Hsing Rd.,Sec.4,Chutung,Hsin−Chu,Taiwan R.O.C
【Fターム(参考)】
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願日】平成22年12月27日(2010.12.27)
【出願人】(390023582)財団法人工業技術研究院 (524)
【氏名又は名称原語表記】INDUSTRIAL TECHNOLOGY RESEARCH INSTITUTE
【住所又は居所原語表記】195 Chung Hsing Rd.,Sec.4,Chutung,Hsin−Chu,Taiwan R.O.C
【Fターム(参考)】
[ Back to top ]