
ヒト化非ヒト哺乳動物を製造する方法
ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で、機能的ヒト血液細胞系統を再構成する方法を、本明細書で提供する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト血液細胞系統に分化する条件下で維持され、それによって非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成する。また、非ヒト哺乳動物で免疫原に対するヒト抗体を産生する方法、モノクローナル抗体を分泌するハイブリドーマならびにB細胞により産生される抗体(たとえば、ポリクローナル抗体類;モノクローナル抗体類)および本方法で作られる非ヒト哺乳動物も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、2009年6月29日提出の米国仮特許出願第61/221,438号の利益を主張する。上記出願の教示全体を参照により本明細書に援用する。
【背景技術】
【0002】
発明の背景
病原体感染に対するヒト免疫応答を、小動物モデルで、体系的かつ制御された方法で研究することが強く求められている。過去20年にわたり、Tリンパ球およびBリンパ球が欠如し、ヒト血液系細胞を有する、重症複合免疫不全(scid)マウスの再構成に大変な努力が向けられてきた(Shultz LD,Ishikawa F,Greiner DL(2007)Nat RevImmunol 7:118−130)。しかし、レシピエントマウスで、生着が不十分であり、ヒトT細胞およびB細胞が急速に消失し、あるいは造血器悪性腫瘍が急速に発症したため、初期の試みは不成功に終わった。scid変異かまたは組換え活性化遺伝子(Rag)の変異のいずれかのためにT細胞およびB細胞が欠損しているばかりでなく、共通γ鎖(γcまたはIl2rg)欠失のためにナチュラルキラー(NK)細胞も欠損しているレシピエントマウスを使用することにより、大きな進歩が達成された(Hiramatsu H,et al.(2003)Blood 102:873−880;Traggiai E,et al.(2004)Science 304:104−107)。NOD−scidIl2rg−/−(NSG)レシピエントまたはBALB/c−Rag2−/−Il2rg−/−レシピエントのいずれかへのヒト造血幹細胞(HSC)の養子移入は、レシピエント骨髄(BM)における安定したHSCの長期生着および末梢における全てのヒト血液系細胞の生成(ヒト化マウスまたはヒューマウス(humice))をもたらす(Hiramatsu H,et al.(2003)Blood 102:873−880;Traggiai E,et al.(2004)Science 304:104−107)。
【0003】
既存のヒト化マウスモデルは、ヒト病原体による感染(Davis PH,Stanley SL,Jr.(2003)Cell Microbiol 5:849-860;Bente DA,et al.(2005)J Virol 79:13797-13799;Islas−Ohlmayer M,et al.(2004)J Virol 78:13891-13900;Guirado E,et al.(2006)Microbes Infect 8:1252-1259;Kneteman NM,et al.(2006)Hepatology 43:1346-1353;Jiang Q,et al.(2008)Blood 112:2858-2868)、特にヒト血液系細胞を感染させるものを研究するための重要なツールを提供する。これらのモデルはまた、小動物モデルでの病原体に対するヒト免疫応答の調査研究を可能にする。しかし、現在利用可能なモデルは、最適なものから程遠い。たとえば、ヒト細胞再構成のレベルは、異なる細胞系統間で著しく異なる。細胞の再構成は骨が折れ、T細胞の再構成は合理的であるが、B細胞およびT細胞は機能的ではない。加えて、NK細胞および骨髄系細胞の再構成は一般に不十分であり、検出不能である。
【0004】
したがって、ヒト免疫系の改善された非ヒトモデルおよびそれらを製造する方法が強く求められている。
【発明の概要】
【0005】
ヒト造血幹細胞(HSC)によるヒト血液細胞系統の再構成不良は主として、非ヒト哺乳動物でこれらの細胞系統を発生および維持するのに必要な、適切なヒトサイトカイン類の欠乏の結果であることを、本明細書に示す。ヒトIL−15およびFlt−3/Flk−2リガンドをコード化するプラスミドDNAがヒト化マウスに送達されたとき(たとえば、流体力学的尾静脈注入による)、ヒトサイトカイン類の発現は2〜3週間持続し、またNK細胞レベル上昇は1カ月を超える期間誘導された。サイトカイン誘導性NK細胞は、活性化受容体と抑制性受容体の両者を発現し、インビトロで標的細胞を死滅させ、またインビボでウイルス感染に強く応答した。同様に、ヒトGM−CSFおよびIL−4、マクロファージコロニー刺激因子、またはエリスロポエチンおよびIL−3の発現は、それぞれ樹枝状細胞、単球類/マクロファージ類、または赤血球の再構成を有意に強化した(参照により本明細書に援用されるChen,Q.,et al.,Proc.Natl.Acad.Sci.,USA,106:21783−21788(2009)を参照されたい)。また、GM−CSFおよびIL−4はヒトT細胞およびヒトB細胞の再構成も強化した。このように、(たとえば、流体力学的送達により)ヒトサイトカイン遺伝子発現をヒトHSCと一緒に使用することは、ヒト化マウスにおける特定のヒト血液細胞系統の再構成を改善するための簡単で効率のよい方法であり、ヒト疾病およびそれらの進行をモデル化し、また小動物モデルにおけるヒト免疫応答を研究するための重要なツールを提供する。
【0006】
したがって、本明細書では、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成し、それによってヒト化非ヒト哺乳動物を製造する方法を提供する。特定の実施態様で、ヒト化マウス(ヒューマウス)が作られる。
【0007】
一態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する(1つまたは複数の)核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成する方法を対象とする。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト血液細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成する。
【0008】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトNK細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトNK細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物におけるヒトNK細胞の再構成を強化する。
【0009】
また別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト樹枝状細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト樹枝状細胞の再構成を強化する。
【0010】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する。
【0011】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト赤血球への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト赤血球に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト赤血球を再構成する。
【0012】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する。
【0013】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコードする核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で、免疫原に対するヒト抗体を産生する方法を対象とし、ここで、ヒトサイトカイン類は、ヒトHSCの機能的ヒトT細胞および機能的ヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞および機能的ヒトB細胞に分化する条件下で維持される。非ヒト哺乳動物は、免疫原で免疫され、また、ヒトB細胞が非ヒト哺乳動物で免疫原に対するヒト抗体を産生する条件下で維持され、それによって、非ヒト哺乳動物で免疫原に対するヒト抗体を産生する。免疫原に対する抗体を産生するB細胞は、さらに単離して、免疫原に対するモノクローナル抗体を分泌するハイブリドーマの産生に使用することができる。
【0014】
モノクローナル抗体を分泌するハイブリドーマならびにB細胞により産生される抗体類(たとえば、ポリクローナル抗体類;モノクローナル抗体類)もまた、本発明に包含される。本明細書で提供される方法により作られる非ヒト哺乳動物もまた、本発明に包含される。
【0015】
本明細書に記載の方法は、非ヒト哺乳動物(たとえば、ヒト化マウス)で機能的ヒト血液細胞系統(たとえば、骨髄性細胞;リンパ球系細胞)を再構成するため、および非ヒト哺乳動物でヒト抗体を産生するための、簡単で効率のよい方法を提供する。
【図面の簡単な説明】
【0016】
【図1】ヒューマウスの骨髄(BM)由来のヒトCD34+細胞は、インビトロで刺激してNK細胞に分化させることができる。(図1A)ヒューマウスのBM由来(左)およびMACSにより抗CD34ビーズで精製した後(右)の単核細胞の、CD34とCD133の染色プロフィールの比較。表示されるイベントは、ヒトCD45+細胞にプレゲート設定する。数値は、ゲートリージョン内の細胞のパーセンテージを示す。マウス4匹のうち1匹からの代表的なデーターを示す。(図1B)IL−15およびFLの非存在下(Ctrl)または存在下で7日間培養したヒトCD34+細胞のNKp46の染色特性対CD56の染色特性。数値は、CD56+NKp46+細胞のパーセンテージを示す。
【図2】流体力学的注入介在性遺伝子送達は、マウスで、全身的ヒトサイトカイン環境を創る。ヒトIL−15およびFLを発現するpcDNAベクターを混合し(各50μg)、ヒューマウスに流体力学的に注入した。表示の時点に、マウス血清中のヒトIL−15およびFLのレベルをELISAで分析した(各時点ごとにn=3)。
【図3】インビボで、IL−15およびFLの発現は、ヒトNK細胞発生を刺激する。空のpcDNAベクター(Ctrl)またはIL−15および/またはFLを発現するpcDNAベクターを、ヒト化マウスに流体力学的に注入した。9日後、表示の組織から細胞を調製し、ヒトCD45、CD3、およびCD56の染色をした。(図3A)ドットプロットは、CD45+細胞にゲート設定したCD3の染色特性対CD56の染色特性を示す。数値は、ゲートリージョンのCD56+CD3−細胞のパーセンテージを示す。群当たりマウス5匹のうち1匹からの代表的なデーターを示す。異なる3つのドナーHSCで、ヒューマウスを構築した。(図3B)サイトカイン遺伝子送達の9日後の、様々な器官におけるCD45+ヒト白血球内のCD56+CD3−NK細胞の発生頻度(平均値±SEM)の比較(n=5)。(図3C)経時的な、血中のCD45+ヒト白血球のCD56+CD3−NK細胞の発生頻度(平均値±SEM)(n=3)。
【図4A−D】ヒトNK細胞は機能的である。(図4A)ヒューマウスにおける、アデノウイルス感染後の、NK細胞介在性肝損傷。サイトカイン遺伝子送達の9日後、PBSまたは複製欠損−アデノウイルス(Ad)を、ヒューマウスに流体力学的に注入した。感染の3日後、H&E染色用にヒューマウスの肝臓を採取した(各群ごとにn=3)。矢印は、壊死および白血球浸潤の領域を示す。Ctrl,サイトカイン遺伝子を送達しないヒューマウス;IL−15/FL,サイトカイン遺伝子を送達したヒューマウス。拡大図を示す。(図4B)血清中のALTレベルの比較。注入の5日後にアデノウイルスまたはPBS処理ヒューマウスから血清を採取し、ALT活性の分析をした(平均値±SEM,群当たりn=3)。IL−15/FL/Adと他群との間でP<0.05。(図4C)アデノウイルス感染ヒューマウスにおける血清IFN−γレベル。アデノウイルス感染ヒューマウスから血清を採取し、ELISAでIFN−γを測定した。平均値±SEMを示す(群当たりn=3)。P<0.05。(図4D)様々なヒューマウスの肝臓におけるMNCの数。アデノウイルス感染の5日後に肝臓を採取した。肝MNC総数を計数し、フローサイトメトリー分析によりヒトCD45+細胞数を決定した(平均値±SEM、群当たりn=3)。IL−15/FL/Adと他群との間でP<0.05。
【図4E】(図4E)CD56+ヒトNK細胞の、肝臓における病変への局在化。肝臓組織をパラフィンに包埋し、薄切し、CD56の染色をし、顕微鏡法で分析した。矢印は病変の領域を示す。マウス3匹のうち1匹からの代表的な画像を示す。
【図5】対応するヒトサイトカイン遺伝子送達による、特定のヒト血液系細胞の誘導。(図5A)樹枝状細胞の再構成改善。空のpcDNAベクターまたは指定されたヒトサイトカイン遺伝子を発現するpcDNAベクターを、ヒューマウスに流体力学的に注入した。注入の9日後、様々な器官から単一細胞懸濁液を調製し、ヒトCD45、CD11c、およびCD209の染色をした。CD45+ヒト細胞にゲート設定したCD209の染色特性対CD11cの染色特性を示す。マウス5匹のうち1匹からの代表的なデーターを示す。(図5B)単球/マクロファージの再構成改善。実験は、pcDNA−コード化M−CSFを注入したこと、および細胞をヒトCD45およびCD14について染色したこと以外、(図5A)の場合と同様に実施した。CD45+ヒト細胞にゲート設定したCD14の染色特性対CD45の染色特性を示す。マウス3匹のうち1匹からの代表的なデーターを示す。(図5C)赤血球の再構成改善。実験は、pcDNA−コード化EPOおよびIL−3を注入したこと、および注入の7日後および30日後に、血液をヒトCD235abについて染色したこと以外、(図5A)の場合と同様に実施した。全血液細胞のCD235abの染色特性対DAPIの染色特性を示す。マウス3匹のうち1匹からの代表的なデーターを示す。数値は、ゲートリージョン内の細胞のパーセンテージを示す。
【図6】ヒト化マウスにおけるヒトNK細胞の再構成。(図6A)HSC生着の12週後、末梢血の単核細胞におけるヒト細胞系統の再構成を、フローサイトメトリーで分析した。ドットプロットは、生存有核細胞にゲート設定したヒトCD45の染色特性対マウスCD45染色特性、あるいはCD45+ヒト細胞にゲート設定したCD3の染色特性対CD19の染色特性またはCD14の染色特性対CD56の染色特性を表す。(図6B)ヒト化マウスの血中、骨髄、脾臓、肺、および肝臓におけるヒトCD45+細胞の、CD14の染色特性対CD56の染色特性。数値は、ゲートリージョン内の細胞のパーセンテージを示す。マウス6匹のうち1匹からの代表的なデーターを示す。
【図7】IL−2シグナルペプチド配列を使用して発現されたときの、循環におけるIL−15レベル上昇。(図7A)IL−15発現ベクターの模式図。内在性シグナル配列(SP)またはIL−2SPのいずれかが付いたIL−15遺伝子を、CMVプロモーターを含むpcDNAベクターにクローニングした。(図7B)血清IL−15レベルの比較。空のpcDNAベクター(Ctrl)、IL−15をコード化するpcDNAベクター、およびIL−2シグナル配列が付いたIL−15をコード化するpcDNAベクターを、NSGマウスに流体力学的に注入した。注入の7日後に、血清を採取し、IL−15レベルをELISAで検定した。
【図8】IL−15およびFL発現後の、ヒト細胞数増加。(図8A〜8G)空のpcDNAベクター(Ctrl)、またはIL−15とFLの両者を発現するpcDNAベクターを、ヒト化マウスに流体力学的に注入した。9日後、表示の組織から細胞を調製し、ヒトCD45に加えてCD3、CD56、CD14、CD11c、CD1c、ILT7、CD303、およびCD19の染色をした。様々な器官における、ヒトCD45+白血球、CD56+NK細胞、CD11c+CD1c+樹枝状細胞、ILT7+CD303+形質細胞様樹状細胞、CD14+単球/マクロファージ、CD3+T細胞、およびCD19+B細胞の絶対数は、総細胞数に特定の細胞型の発生頻度を乗じることによって算出した。平均値±SEMを示す(n=3)。骨髄(BM)における細胞数は大腿骨2本からであった。
【図9】IL−15およびFL処理ヒューマウスにおけるヒトNK細胞の細胞表面表現型。IL−15遺伝子およびFL遺伝子の送達後9日に、表示の器官から細胞を調製し、ヒトCD45、CD56に加えてNKG2D、NKG2A、CD7、CD69、CD94、NKp46、KIR、またはCD16の染色をした。CD45+ヒト細胞にゲート設定した、CD56の染色特性対NKG2D、NKG2A、CD7、CD69、CD94、NKp46、KIR、またはCD16の染色特性を示す。数値は、ゲートリージョン内の細胞のパーセンテージを示す。
【図10】IL−15およびFL処理ヒューマウス由来のヒトNK細胞の細胞傷害性および刺激。(図10A)NK細胞は、細胞溶解性である。サイトカイン遺伝子送達の9日後、ヒトNK細胞をBMおよび脾臓から精製し、異なるエフェクター対標的(E:T)比でK562細胞と混合し、4時間培養した。NK細胞の細胞溶解活性は、上澄の乳酸デヒドロゲナーゼ酵素活性を測定することによって決定した。(図10B)NK細胞は、インビトロで、poly(I:C)刺激後、IFN−γを産生する。精製NK細胞(5×105)を単独で、またはpoly(I:C)(50μg/ml)の存在下で、またはpoly(I:C)およびインビトロで分化したヒトDC(5×105)の存在下で、培養した(材料と方法参照)。24時間後に、ELISAで、ヒトIFN−γについて上澄を分析した。(図10C)NK細胞は、インビボでpoly(I:C)刺激後、IFN−γを産生する。ヒューマウスに、poly(I:C)(マウス当たり200pg)を静脈内注入した。注入の24時間後に血清を採取し、ELISAでヒトIFN−γの検定をした(n=4)。P<0.05。
【図11】インビトロでの、ヒトCD34+細胞の分化。CD34+ヒト細胞を、ヒューマウスのBMから精製し(左)、GM−CSFに加えてIL−4、またはM−CSFの存在下で7日間、あるいはEPOに加えてIL−3の存在下で20日間、培養した。次いで細胞を、CD45に加えてCD209およびCD11c、またはCD14およびCD33、またはCD235abについて検定した。CD45+細胞について、CD209の染色特性対CD11cの染色特性およびCD14の染色特性対CD33の染色特性を示す。CD235ab発現をヒストグラム(太線)で示す。EPOおよびIL−3無しで培養した精製CD34+細胞をコントロールとして使用した(細線)。
【図12】ヒト化マウスにおける、破傷風トキソイドワクチン免疫後の、ヒト細胞増殖。(図12A)免疫の実験的流れ:第0日に、類似したヒト白血球再構成(50〜80%)を有する12週齢のヒト化マウスに、ヒトGM−CSFおよびIL−4をコード化するプラスミドまたは空のpcDNAベクター(ベクター)を、流体力学的に注入した。7日後、これらのマウスを、破傷風トキソイド(TT)で3回、免疫した。第1回の免疫は、2l.f.の破傷風トキソイドワクチンの腹腔内注射で実施し、続いて2〜3週間隔でさらに2回、ブースターを実施した。第2回ブースターの2週間後、マウスを分析した。(図12B)GM−CSFおよびIL−4処理マウス由来の脾臓は、有意に腫大していた。(図12C)様々なヒューマウスの脾臓における単核細胞(MNC)数。脾臓を、免疫後に採取した。脾臓のMNC総数を数え、フローサイトメトリー分析でヒトCD45+細胞数を決定した(平均値±SEM,群当たりn=3)。
【図13】GM−CSFおよびIL−4処理した、TT免疫ヒューマウスにおける、ヒトB細胞の細胞表面表現型。細胞を脾臓から調製し、ヒトCD45、マウスCD45、CD19に加えてIgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色をした。CD45+ヒト細胞にゲート設定した、CD19の染色特性対IgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色特性を示す。
【図14】GM−CSFおよびIL−4処理した、TT免疫ヒューマウスにおける、ヒトT細胞の細胞表面表現型。細胞を脾臓から調製し、ヒトCD45、マウスCD45、CD19に加えてIgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色をした。CD45+ヒト細胞にゲート設定した、CD3の染色特性対T細胞活性化マーカー:HLA−DRおよびCD40Lの染色特性を示す。
【図15】TT免疫ヒューマウスにおける、ヒトIgG、IgMおよびTT特異的ヒトIgGの血清レベル。TT免疫ヒューマウスから血清を採取し、ヒトIgG、IgMおよびTT特異的ヒトIgGをELISAで測定した。(図15A)血清中のヒト総IgG。GM−CSF+IL−4処理マウスは、ベクター処理マウスより有意に高いヒト総IgGレベルを生じさせた。(図15B)血清中のヒト総IgM。GM−CSF+IL−4処理マウスはまた、より高い総ヒトIgM血清レベルを有する。(図15C)血清中のヒトTT特異的IgG。
【図16】サイトカイン処理マウスにおけるTT特異的ヒトT細胞反応。第3回免疫の2週間後に、脾臓を採取し、ヒトT細胞のパーセンテージをフローサイトメトリーで測定した。ELISPOT検定の場合、同数(5×105)の、異なる試料からのヒトT細胞を、抗ヒトIFN−γまたは抗ヒトIL−4抗体で被覆したウェルに播種し、3種類の条件下で24時間培養した:培地のみ(ctrl)、PMAの存在下またはTT−特異的ペプチドの存在下。ELISPOTを現像した。(図16A)免疫マウス由来の脾細胞を含む代表的なヒトIFN−γELISPOTウェル。(図16B)免疫マウス由来の脾細胞を含む代表的なヒトIL−4ELISPOTウェル。2つの独立した実験の1つからのデータを示す。
【図17】ヒトIL−15、FL、GM−CSF、IL−4およびM−CSFをコード化するDNAプラスミドの混合物(各50μg)を、PBSに溶解し、12週齢のヒト化マウスに注入した(n=2)。7日後、血清を採取し、ELISAで、これらのヒトサイトカイン類の分析をした。
【発明を実施するための形態】
【0017】
近年、ヒト胎仔胸腺、肝臓、および自己ヒトCD34+細胞を移植されたNOD−scidマウス(BLTマウス)で、ヒト樹枝状細胞(DC)および単球/マクロファージの意義深い再構成が報告された(Wege AK,et al.(2008)Curr Top Microbiol Immunol 324:149−165)。しかしまだ、BLTマウスにヒトNK細胞は存在しなかった。NK細胞および骨髄性細胞は、自然免疫反応において重要な役割を果たすため、これらの細胞型の十分な再構成レベルを有するヒト非ヒト哺乳動物、たとえばヒト化マウス等の発生は、感染病調査研究および血液系細胞を含む他の調査研究(たとえば、貧血症、免疫不全症、癌等の、血液病調査)におけるヒト化マウスモデルの可能性を十分に実現するために重要である。
【0018】
全ての血液細胞系統は、共通のヒト造血幹細胞(HSC)に由来する。サイトカイン類は、それらの分化および維持の間、重要な役割を果たす。たとえば、IL−15は、NK細胞の発生および生存に必要であり(Mrozek E,et al.(1996)Blood 87:2632−2640)、GM−CSFおよびIL−4は樹枝状細胞(DC)発生に(Rosenzwajg M,et al.(1996)Blood 87:535−544)、マクロファージコロニー刺激因子(M−CSF)は単球/マクロファージの発生および維持に(Stec M,et al.(2007)J Leukoc Biol 82:594−602)、またエリスロポエチン(EPO)およびIL−3は赤血球発生に必要である(Giarratana MC,et al.(2005)Nat Biotechnol 23:69−74)。しかし、ヒトとマウスとの間の進化的分岐のため、これらのサイトカイン類は種特異的である(すなわち、マウスサイトカイン類はヒト細胞に対して作用しない)。たとえば、マウスIL−15は、ヒトNK細胞および前駆体に対して何の作用も持たず(Eisenman J,et al.(2002)Cytokine 20:121-129)、ヒューマウスにおけるヒトNK細胞の再構成不良という結果に終わる(Huntington ND,et al.(2009)J Exp Med 206:25−34;Kalberer CP,et al.(2003)Blood 102:127−135)。同様に、マウスGM−CSF、IL−4(Metcalf D(1986)Blood 67:257−267;Mosmann TR,et al.(1987)J Immunol 138:1813−1816)、M−CSF(Fixe P,Praloran V(1997)Eur Cytokine Netw 8:125−136)、およびIL−3(Stevenson LM,Jones DG(1994)J CompPathol111:99−106)は全て、ヒト細胞に対して何の作用もないと報告されている。
【0019】
ヒューマウスにおけるNK細胞および骨髄性細胞の不十分な再構成および機能は、特定のヒトサイトカイン類の欠如に起因するのかどうかを調査研究した。再構成されたマウスにおけるヒトサイトカイン類の発現が、特定のヒト血液系細胞の分化、生存、および機能を刺激するかどうかを究明するための実験を、本明細書に記述する。
【0020】
この調査研究が、ヒト化マウスを使用して実証されるような、ヒト化非ヒト哺乳動物における特定のヒト血液系細胞の再構成を改善するための簡単で効率のよい方法の開発につながった。ヒトIL−15およびFlt−3/Flk−2リガンド(FL)をコード化する核酸を送達した際に、特定のヒトサイトカイン類が、ヒューマウスの循環中に2〜3週間検出された。結果として、有意に増加した数のヒトNK細胞が、様々な器官で1カ月以上確認された。サイトカイン誘導性NK細胞は、インビトロおよびインビボの両方で、完全に機能した。同じ戦略を使用して、ヒューマウスにおけるヒト樹枝状細胞、単球/マクロファージ、および赤血球の再構成レベルも大幅に強化された。従前のヒト化マウスモデルにおけるNK細胞および骨髄性細胞の再構成不良は、それらの分化および維持に必要な適切なヒトサイトカイン類の欠如の結果であること、およびヒトサイトカイン遺伝子の送達(たとえば、流体力学的送達)は、これらの細胞系統の再構成不良を克服するための簡単で効率のよい方法であることを、本明細書に記載の研究は示す。
【0021】
したがって、一態様で、本発明は、機能的ヒト血液系細胞(たとえば、単一のヒト血液系細胞(たとえば、NK細胞);多様なヒト血液系細胞(たとえば、NK細胞、樹枝状細胞、T細胞、B細胞等々)を、また幾つかの実施態様で、全てのヒト血液系細胞)を、非ヒト哺乳動物で再構成する方法を対象とする。この実施態様で、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入される。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCで非ヒト哺乳動物が再構成される条件下で維持され、それによって、非ヒト哺乳動物でヒト造血幹細胞(HSC)を再構成する。
【0022】
本明細書で使用されるとき、HSC(たとえば、ヒトHSC)は、レシピエントに移植されるとき、「移植レシピエント(たとえば、非ヒト哺乳動物;免疫不全非ヒト哺乳動物)の造血システムを「再生息させる」または「再構成する」ことができ、またレシピエントで(たとえば、長期)造血を持続できる、自己再生幹細胞である。HSCは、骨髄性(たとえば、単球およびマクロファージ、好中球、好塩基球、好酸球、赤血球、巨核球/血小板、樹枝状細胞)およびリンパ球系(たとえば、T−細胞、B−細胞、NK−細胞)を含む、血液細胞型を生じさせる(に分化する)多能性幹細胞である。本明細書に記載の方法に示す通り、再構成されたヒトHSCは、非ヒト哺乳動物で、ヒトNK細胞、ヒト単球、ヒトマクロファージ、ヒト樹枝状細胞、ヒト赤血球、ヒトB細胞、ヒトT細胞またはそれらの組合せに分化することができる。
【0023】
HSCは細胞マーカーCD34を発現するので、通例「CD34+」と呼ばれる。当業者には明らかなように、HSCはまた他の細胞マーカー、たとえばCD133および/またはCD90等も発現できる(「CD133+」、「CD90+」)。場合によっては、HSCは、発現されないマーカー、たとえば、CD38を特徴とする。したがって、本発明の一実施態様で、本明細書に記載の方法で使用されるヒトHSCは、CD34+、CD90+、CD133+、CD34+CD38−、CD34+CD90+、CD34+CD133+CD38−、CD133+CD38−、CD133+CD90+CD38−、CD34+CD133+CD90+CD38−、またはそれらの任意の組合せである。ある特定の実施態様で、HSCは、本明細書で「二重陽性」細胞または「DP」細胞あるいは「DPC」とも呼ばれる、CD34(「CD34+」)とCD133+(「CD133+」)の両方である。別の実施態様で、HSCはCD34+CD133+であり、そしてCD38−および/またはCD90+をさらに含んでもよい。
【0024】
HSCは、ドナー(たとえば、人類、霊長類、ブタ、マウス、等々を含む、哺乳動物等の脊椎動物)の大腿骨、股関節、肋骨、胸骨、および他の骨等の骨髄に見られる。臨床使用および科学的使用のための、他のHSC源としては、臍帯血、胎盤、胎仔肝臓、動員末梢血、非動員(non−mobilized)(または不動員(unmobilized))末梢血、胎仔肝臓、胎仔脾臓、胚性幹細胞、および大動脈−性腺−中腎(AGM)、またはそれらの組合せなどがある。
【0025】
当業者に理解されるであろうが、動員末梢血は、HSC(たとえば、CD34+細胞)を強化した末梢血を指す。化学療法剤および/またはG−CSF等の化学物質の投与は、幹細胞を骨髄から末梢循環に動員する。たとえば、少なくとも5日間の、または約5日間の、顆粒球コロニー刺激因子(G−CSF)の投与は、CD34+細胞を末梢血に動員する。循環するCD34+細胞の30倍富化が確認され、そのピーク値は、G−CSF投与開始後第5日に現れる。末梢血の動員がなければ、循環するCD34+細胞の数は非常に少なく、総単核血球の0.01〜0.05%と推定される。
【0026】
本方法で使用するためのヒトHSCは、単一ドナーから得ても多数のドナーから得てもよい。加えて、本明細書に記載の方法で使用されるHSCは、用時単離されたHSC、凍結保存HSCS、またはそれらの組合せであってもよい。
【0027】
当該技術分野で周知の通り、HSCは、当該技術分野で周知の種々の方法を使用して、これらの供給源から得ることができる。たとえば、HSCは、針と注射器を使用して、たとえば、股関節部、大腿骨、等々の骨髄から除去することにより直接得ることもでき、あるいは骨髄コンパートメントから細胞を放出させる、顆粒球コロニー刺激因子(G−CSF)等のサイトカイン類でドナーを前処置した後、血液から得ることもできる。
【0028】
本発明の方法で使用するためのHSCは、得られたまま(たとえば、未増殖で)またはHSCを非ヒト哺乳動物に導入する前に操作して(たとえば、増殖して)、非ヒト哺乳動物に直接導入することができる。一実施態様で、HSCを非ヒト哺乳動物に導入する前に、HSCを増殖する。当業者に理解されるであろうが、HSCを増殖させるために使用できる方法は種々ある(たとえば、Zhang,Y.,et al.,Tissue Engineering,12(8):2161−2170(2006);Zhang CC,et al.,Blood,111(7):3415−3423(2008)参照)。ある特定の実施態様で、成長因子類(たとえば、アンギオポエチン様5(Angplt5)成長因子、IGF−結合蛋白質2(IGFBP2)、幹細胞因子(SCF)、線維芽細胞成長因子(FGF)、トロンボポエチン(TPO)、またはそれらの組合せ)の存在下で、HSCを間葉幹細胞(MSC)と共培養して細胞培養をすることにより、HSCの個体群を増殖することができる。細胞培養は、増殖したHSC個体群が産生される条件下で維持される(たとえば、Maroun,K.,et al.,ISSCR,7th Annual Meeting,Abstract No.1401(July8−11,2009)、_____________として公開された、2010年5月28日に提出された、PCT出願第PCT/US2010/036664号(代理人整理番号No.4471.1000−001)(参照により本明細書に援用する)を参照)。
【0029】
本明細書に記載の方法では、ヒトHSCの機能的ヒト細胞への分化を誘導するために1つまたは複数のヒトサイトカイン類をコード化する(1つまたは複数の)核酸(たとえば、DNA、RNA)も非ヒト哺乳動物に導入される。当該技術分野で周知の通り、サイトカイン類は、免疫細胞の分化、増殖または機能を刺激または抑制する蛋白質である。多数のヒトサイトカイン類の核酸配列も、当該技術分野で周知である(たとえば、www.ncbi.nlm.nih.gov参照)。1つまたは複数のサイトカイン類をコード化する核酸を入手するための方法は、当該技術分野でルーチンであり、種々の供給源(たとえば、血清)から核酸を単離すること(たとえば、クローニング)、組換えで核酸を産生することまたは商業的供給源から核酸を入手することを含む。
【0030】
本発明の方法で使用することができる種々のヒトサイトカイン類がある。このようなヒトサイトカイン類の例としては、インターロイキン−12(IL−12)、インターロイキン−15(IL−15)、Fms関連チロシンキナーゼ3リガンド(Flt3L)、Flt3L/Flk2リガンド(FL)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン−4(IL−4)、インターロイキン−3(IL−3)、マクロファージコロニー刺激因子(M−CSF)、エリスロポエチン(EPO)およびそれらの組合せなどがある。本明細書に記載の方法で使用するのに好適な他のサイトカイン類の例を一覧表にした。非ヒト哺乳動物に導入されるサイトカインのタイプおよびサイトカイン類の数は、ヒトHSCの分化が非ヒト哺乳動物で起こるとき、どのヒト血液細胞系統が再構成されるべきかによって異なる。たとえば、実施例1に示す通り、ヒトIL−15およびFlt−3/Flk−2リガンドをコード化する核酸が(たとえば、流体力学的尾静脈注入により)ヒト化マウスに導入されたとき、ヒトサイトカイン類の発現は2〜3週間持続し、上昇したヒトNK細胞レベルは1カ月以上にわたって誘導された。サイトカイン誘導性NK細胞は、活性化受容体および抑制性受容体の両者を発現し、インビトロで標的細胞を死滅させ、インビボでウイルス感染に強く応答した。同様に、ヒトGM−CSFおよびIL−4の発現は、ヒト樹枝状細胞の有意に強化された再構成を招き;マクロファージコロニー刺激因子の発現は、ヒト単球/マクロファージの有意に強化された再構成を招き;またエリスロポエチンおよびIL−3の発現は、ヒト赤血球の有意に強化された再構成を招いた(参照により本明細書に援用される、Chen,Q.,et al.,Proc.Natl.Acad.Sci.,USA,106:21783−21788(2009)参照)。実施例2に示す通り、GM−CSFおよびIL−4の発現は、機能的ヒトT細胞およびヒトB細胞の再構を強化した。
【0031】
幾つかの態様で、少なくとも1種のサイトカイン、少なくとも2種のサイトカイン類、少なくとも3種のサイトカイン類、少なくとも4種のサイトカイン類、少なくとも5種のサイトカイン類、少なくとも6種のサイトカイン類、少なくとも7種のサイトカイン類、少なくとも8種のサイトカイン、少なくとも9種のサイトカイン類、少なくとも10種のサイトカイン類、少なくとも11種のサイトカイン類、少なくとも12種のサイトカイン類、少なくとも13種のサイトカイン類、少なくとも14種のサイトカイン類、少なくとも15種のサイトカイン類、少なくとも16種のサイトカイン類、少なくとも17種のサイトカイン類、少なくとも18種のサイトカイン類、少なくとも19種のサイトカイン類、または少なくとも20種のサイトカイン類(を含む)が、非ヒト哺乳動物に導入される。他の態様で、1種のサイトカイン、2種のサイトカイン類、3種のサイトカイン類、4種のサイトカイン類、5種のサイトカイン類、6種のサイトカイン類、7種のサイトカイン類、8種のサイトカイン、9種のサイトカイン類、10種のサイトカイン類、11種のサイトカイン類、12種のサイトカイン類、13種のサイトカイン類、14種のサイトカイン類、15種のサイトカイン類、16種のサイトカイン類、17種のサイトカイン類、18種のサイトカイン類、19種のサイトカイン類、または20種のサイトカイン類のみ(から成る、から本質的に成る)が、非ヒト哺乳動物に導入される。各ヒトサイトカインをコード化する核酸を同時にまたは順次導入してもよい(たとえば、複数のサイトカインを非ヒト哺乳動物で発現させるべき場合、各サイトカインをコード化する各核酸を、独自の1つのプラスミドまたはベクターで導入してもよく、または多数のプラスミド類またはベクター類で導入してもよい;あるいは、導入されるべきサイトカイン類をコード化する全ての核酸を、1種のプラスミドまたはベクターで導入してもよい)。
【0032】
本発明の方法では、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入される。本明細書で使用されるとき、用語「哺乳動物」および「哺乳類」は、それらの子に授乳する、生存する子を出産する(真獣類または有胎盤哺乳動物)かまたは産卵する(後獣類または無胎盤哺乳動物)かいずれかの、単孔類、有袋類および有胎盤動物を含む、任意の脊椎動物を指す。本明細書に記載の方法で使用することができる哺乳類種の例としては、非ヒト霊長類(たとえば、サル、チンパンジー)、齧歯類(たとえば、ラット、マウス、モルモット)、イヌ、ネコ、および反芻動物(たとえば、ウシ、ブタ、ウマ)などがある。一実施態様において、非ヒト哺乳動物はマウスである。本明細書に記載の方法で使用される非ヒト哺乳動物は、成体、新生仔(たとえば、<48時間齢;子)または子宮内であってもよい。
【0033】
特定の実施態様で、非ヒト哺乳動物は免疫不全非ヒト哺乳動物である、すなわち、その免疫系に1つまたは複数の欠損があり(たとえば、NSGまたはNOD scid γ(NOD.Cg−Prkdcscid Il2rgtm1Wjl/SzJ)マウス)、結果として、ヒトHSCが導入されるとき、ヒト血液細胞系統の再構成を許容する、非ヒト哺乳動物である。たとえば、その非ヒト哺乳動物は、それ自身のT細胞、B細胞、NK細胞またはそれらの組合せを欠く。特定の実施態様で、非ヒト哺乳動物は、たとえば重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、またサイトカイン−受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scid IL2Rγ−/−マウス);およびBalb/crag−/−γc−/−マウス等の、免疫不全マウスである。
【0034】
免疫不全マウスの他の具体例としては、重症複合免疫不全(scid)マウス、非肥満糖尿病(NOD)−scidマウス、IL2rg−/−マウス(たとえば、NOD/LySz−scidIL2rg−/−マウス、NOD/Shi−scidIL2rg−/−マウス(NOGマウス)、BALB/c−Rag−/−IL2rg−/−マウス、H2d−Rag−/−IL2rg−/−マウス)、NOD/Rag−/−IL2rg−/−マウスなどがあるが、その限りではない。
【0035】
幾つかの実施態様で、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸の導入前に(たとえば、ヒトHSCの再構成をさらに強化するために)、非ヒト哺乳動物を処置または操作する。たとえば、非ヒト哺乳動物を操作して、ヒトHSCの生着および/または再構成をさらに強化することができる。一実施態様で、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸の導入前に、非ヒト哺乳動物を照射する。別の実施態様で、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸の導入前に、1つまたは複数の化学療法剤が非ヒト哺乳動物に投与される。
【0036】
やはり当業者に理解されるであろうが、HSCおよびサイトカイン類をコード化する核酸を非ヒト哺乳動物に導入する種々の方法がある。そのような方法の例としては、皮内、筋肉内、免疫、眼内、大腿内、脳室内、頭蓋内、髄腔内、静脈内、心臓内、肝臓内、骨髄内、皮下、局所、経口および鼻腔内投与経路などがあるが、その限りではない。他の好適な導入方法としては、子宮内注入、流体力学的遺伝子送達、遺伝子療法、充電式または生分解性装置、粒子加速器(「遺伝子銃」)および徐放性ポリマー装置も挙げることができる。
【0037】
任意のそのような投与経路等々を使用して、HSCを非ヒトに導入することができる。ある特定の実施態様で、HSCを非ヒト哺乳動物に心臓内注入する。
【0038】
1つまたは複数のサイトカイン類をコード化する核酸はまた、その核酸(類)が非ヒト哺乳動物で発現される限り、任意のそのような投与経路を使用して導入することもできる。たとえば、1つまたは複数のサイトカイン類をコード化する核酸を、プラスミド(たとえば、pcDNA3.1(+))またはウイルスベクター(たとえば、アデノウイルス、アデノ随伴ウイルス、レンチウイルス、レトロウイルス等々)で、裸の核酸(裸のDNA)として導入することができる。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、(たとえば、非ヒト哺乳動物の尾静脈への)流体力学的注入を使用して、プラスミドで導入される。
【0039】
HSCおよび1つまたは複数のサイトカイン類をコード化する核酸は、同時にまたは順次導入することができ、また当業者に理解されるであろうが、使用しようとしている非ヒト哺乳動物のタイプ、発現させようとしているサイトカイン類およびヒトHSCの分化が非ヒト哺乳動物で起こるとき、どのヒト血液系細胞が発現されるかおよび/または強化されるか等の因子によって異なる。ある特定の実施態様で、HSCは新生仔(たとえば、約48時間齢)に導入され、またサイトカイン類をコード化する核酸は約1か月、約2カ月、約3カ月、約4カ月、約5カ月、約6カ月、約7カ月、約8カ月、約9カ月、約10カ月、約11カ月、約12カ月後に導入される。
【0040】
いったんHSCおよび1つまたは複数のサイトカイン類をコード化する核酸が導入されると、非ヒト哺乳動物は、核酸が発現されて、HSCで非ヒトが再構成される条件下で維持される。本発明の非ヒト動物が維持されるそのような条件は、当業者に周知の通り、哺乳動物の基本的欲求を満たすこと(たとえば、食糧、水、光)である。
【0041】
本明細書に記載の方法は、核酸が発現されるかどうか、ヒトHSCが存在するかどうかおよび/またはヒトHSCが、1つまたは複数のヒト血液系細胞に分化しているかどうかを究明することをさらに含む。核酸が発現されるかどうかおよび/またはHSCで非ヒトが再構成されるかどうかを究明する方法は、本明細書に提供され、また当業者に周知である。たとえば、ヒトHSCの表面細胞マーカーに特異的な抗体を使用するフローサイトメトリー分析法を使用して、ヒトHSCが非ヒト哺乳動物に存在することを検出できる。加えて、血清を非ヒト哺乳動物から採取して、ヒトサイトカイン類の存在を検定することができる。分化したHSC(たとえば、NK細胞、樹枝状細胞、T細胞、B細胞、単球/マクロファージ、赤血球)の機能を評価するための検定法も使用することができる。そのような検定法は本明細書にも記述されており、また当業者に周知である。たとえば、本明細書に記載の通り、サイトカイン誘導性ヒトNK細胞は、インビトロ検定法(乳酸デヒドロゲナーゼ検定法)で標的細胞を死滅させ、またインビボでウイルス感染に強く応答した。
【0042】
1つまたは複数のヒトサイトカイン遺伝子をコード化する核酸の送達によって、1つまたは複数のヒト血液細胞系統を非ヒト哺乳動物(たとえば、ヒト化マウス)で再構成できることは、種々の方法で使用できる。
【0043】
たとえば、一態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトNK細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトNK細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトNK細を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトIL−15およびヒトFlt−3/Flk−2リガンドをコード化する。
【0044】
別の実施態様で、非ヒト哺乳動物の末梢血の白血球の約3%〜約25%が、ヒトNK細胞(たとえば、機能的NK細胞)である。他の実施態様で、非ヒト哺乳動物の末梢血の白血球の約3%、約4%、約5%、約6%、約7%、約8%、約9%、約10%、約11%、約12%、約13%、約14%、約15%、約16%、約17%、約18%、約19%、約20%、約21%、約22%、約23%、約24%、または約25%が、ヒトNK細胞である。
【0045】
さらに他の実施態様で、ヒトNK細胞(たとえば、機能的NK細胞)の発現は約1〜約30日間、維持され(また、場合によっては、たとえば好適なコントロールと比較して強化され)、そして特定の実施態様で、ヒトNK細胞の発現は約1日間、約2日間、約3日間、約4日間、約5日間、約6日間、約7日間、約8日間、約9日間、約10日間、約11日間、約12日間、約13日間、約14日間、約15日間、約16日間、約17日間、約18日間、約19日間、約20日間、約21日間、約22日間、約23日間、約24日間、約25日間、約26日間、約27日間、約28日間、約29日間、約30日間、または約31日間維持される。
【0046】
幾つかの実施態様で、非ヒト哺乳動物におけるヒトNK細胞は、ヒトで見られる通常の(野生型)NK細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。そのような発現は、ヒトNK細胞が非ヒト哺乳動物で確かに機能することを示す。たとえば一実施態様で、ヒトNK細胞はCD56+NK細胞細胞である。他の実施態様で、ヒトNK細胞は、NKG2D、NKG2A、CD94、KIR、NKp46、CD7、CD69、Cd16、またはそれらの組合せを発現する。
【0047】
さらに他の実施態様で、非ヒト哺乳動物におけるヒトNK細胞は、適切な刺激(たとえば、Toll様受容体3アゴニスト・ポリ(I:C);ヒト樹枝状細胞;アデノウイルス)を受けて、標的細胞を死滅させたりIFN−γを発現したりすることができる。
【0048】
別の態様で、本発明は、免疫不全非ヒト哺乳動物に、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を導入することを含む、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法を対象とし、ここでヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト樹枝状細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトGM−CSFおよびヒトIL−4をコード化する。別の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトGM−CSF、ヒトIL−4およびヒトFlt−3/Flk−2リガンドをコード化する。
【0049】
他の実施態様で、非ヒト哺乳動物におけるヒト樹枝状細胞は、ヒトで見られる通常の(野生型)樹枝状細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、非ヒト哺乳動物におけるヒト樹枝状細胞は、CD11c+CD209骨髄性樹枝状細胞(たとえば、血液、脾臓、骨髄、肺、肝臓で発現される)、ILT7+CD303+形質細胞様樹状細胞またはそれらの組合せである。
【0050】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現され、ヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトマクロファージコロニー刺激因子をコード化する。
【0051】
他の実施態様で、非ヒト哺乳動物におけるヒト単球/マクロファージは、ヒトで見られる通常の(野生型)単球/マクロファージの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、ヒト単球/マクロファージはCD14+を発現する。
【0052】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト赤血球への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト赤血球に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト赤血球を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトエリスロポエチンおよびIL−3をコード化する。
【0053】
別の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、ヒトで見られる通常の(野生型)赤血球の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、ヒト赤血球はCD235ab+を発現する。
【0054】
さらに他の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、非ヒト哺乳動物における全赤血球の約1%〜約10%、または約3%〜約5%を構成する。特定の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、非ヒト哺乳動物における全赤血球の約1%、約2%、約3%、約4%、約5%、約6%、約7%、約8%、約9%または約10%を構成する。
【0055】
ある特定の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカインをコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法を対象とし、ここでヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する。一実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、GM−CSFおよびIL−4をコード化する。本方法は、非ヒト哺乳動物を免疫原で免疫すること、および非ヒト哺乳動物が免疫原に対するヒト抗体を産生する条件下で非ヒト動物を維持することを、さらに含んでもよい。
【0056】
また別の特定の態様で、本発明は、非ヒト哺乳動物で、免疫原に対するヒト抗体を生成する方法を対象とする。本方法で、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入されるが、ここでヒトサイトカイン類は、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞および機能的ヒトB細胞に分化する条件下で維持される。非ヒト哺乳動物は免疫原で免疫され、またヒトB細胞が非ヒト哺乳動物で免疫原に対するヒト抗体を産生する条件下で維持され、それによって、非ヒト哺乳動物で免疫原に対するヒト抗体を生成する。一実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、GM−CSFおよびIL−4をコード化する。
【0057】
当該技術分野で周知の通り、「免疫原」は、免疫応答を誘導し、抗体産生を増進することができる物質である。本法で使用するための種々の免疫原は、当該技術分野で周知である。たとえば、免疫原は、下記の全部または免疫原性部分であってもよい:ヒトまたは他種由来の蛋白質、(たとえば、正常細胞または腫瘍細胞等の疾病細胞の)細胞表面蛋白質、生物(たとえば、生物の被膜、莢膜、細胞壁、鞭毛、線毛および毒素を含む、生物の免疫原性部分)、ウイルス蛋白質、細菌蛋白質、毒素、多糖、リポ蛋白質、(たとえば、アセチル化、メチル化、グリコシル化された)修飾蛋白質、核酸(たとえば、DNA、ペプチド、蛋白質または多糖と結合されるときのRNA)、化学的エピトープ、等々。
【0058】
これらの方法は、ヒト抗体を産生するヒトB細胞を、非ヒト哺乳動物から単離することをさらに含むことができる。B細胞を非ヒト哺乳動物から単離する方法は、当該技術分野で周知である。たとえば、フローサイトメトリーまたはB細胞特異的蛋白質に特異的な抗体に基づく磁気精製を使用する細胞選別(たとえば、John Wiley and Sons,Inc.ed.John E.Coligan et al.によるCurrent Protocols in Immunology,Copyright(著作権)2010参照)。
【0059】
当該技術分野で周知の通り、「抗体」または「免疫グロブリン」は、血中を循環するB細胞により産生される免疫系の蛋白質成分であり、細菌やウイルスのような免疫原を認識して、それらを無力化する。免疫原への暴露後、抗体は血中を循環し続け、将来その抗原に暴露されることを防ぐ。本明細書に記載の方法を使用して、ヒトB細胞により産生されるあらゆるタイプの抗体を得ることができる。モノクローナル抗体はポリクローナルであってもよく、モノクローナル抗体であってもよい。そのような抗体の例は当該技術分野で周知であり、IgG(たとえば、IgG1、IgG2、IgG3、IgG4)、IgM、IgDおよびIgAなどがある。
【0060】
本方法は、単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;およびヒトB細胞と不死化細胞が融合して、免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、その組合せを維持することを、さらに含むことができる。
【0061】
当該技術分野で周知の通り、免疫後適切なときに、たとえば、抗体価が最高のときに、抗体産生細胞を対象から得て、標準的な技術、たとえば最初にKohler and Milstein,Nature 256:495−497(1975)によって記述されたハイブリドーマ技術、ヒトB細胞ハイブリドーマ技術(Kozbor et al.,Immunol.Today 4:72(1983)),EBV−ハイブリドーマ技術(Cole et al.,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,Inc.,pp.77−96(1985))またはトリオーマ技術等により、モノクローナル抗体の調製に使用することができる。ハイブリドーマを作るためのテクノロジーは周知である(一般にCurrent Protocols in Immunology,Coliganet al.,(eds.)John Wiley & Sons,Inc.,New York,NY(1994)参照)。簡単に記述すると、上述の通り、不死化細胞株(一般に骨髄腫)を、免疫原で免疫された哺乳動物由来のリンパ球(一般に脾細胞)に融合させ、本発明のポリペプチドを結合するモノクローナル抗体を産生するハイブリドーマを同定するために、結果として得られたハイブリドーマ細胞の培養上澄をスクリーニングする。
【0062】
リンパ球と不死化細胞株を融合させるために使用される多くの周知のプロトコールのいずれも、免疫原に対するモノクローナル抗体を生成するために利用することができる(たとえば、Current Protocols in Immunology,supra;Galfre et al.,Nature,266:55052(1977);R.H.Kenneth,in Monoclonal Antibodies:A New Dimension In Biological Analyses,Plenum Publishing Corp.,New York,New York(1980);およびLerner,Yale J.Biol.Med.54:387−402(1981)参照)。
【0063】
さらに、一般の当業者は、そのような方法の多くの変形ならびにヒトB細胞により産生される抗体を得るために使用することができる他の方法があることを理解するであろう。たとえば、ヒトB細胞により発現されるヒト抗体の全部または機能的部分をコード化する配列を、既知の技術を使用してクローニングすることができる。一般に、本手法は、抗原特異的B細胞の単離(たとえば、蛍光色素標識抗原で染色される、フローサイトメトリーによる選別)および変性プライマーおよび単一細胞ポリメラーゼ連鎖反応(PCR)を使用する抗体遺伝子のVDJ部分の増幅を含む。クローニングされ、配列決定された抗体遺伝子のVDJ部分を定常領域遺伝子セグメントと結合し、CHO細胞株等の細胞株で抗体を作る(たとえば、Hahn,S.,et al.,Cell Mol.Life Sci.(2000),57(1):96−105参照)。
【0064】
非ヒト哺乳動物により産生されるヒト抗体を産生および/または単離する方法のもう1つの例は、ヒトB細胞をウイルスにより不死化することを含む。本方法では、たとえば、エプスタイン−バールウイルス(Epstein−Barr Virus)(EBV)または修飾EBVを使用してB細胞を不死化することができる(たとえば、Lanzavecchia,A.,Curr.opin.Biotechnol.(2007) 18(6):523−528参照)。
【0065】
当業者に理解されるであろうが、サイトカイン類に加えて、成長因子等の蛋白質(たとえば、ヒト蛋白質;ヒト分泌蛋白質)、ステロイド類、および/または小分子の発現を本方法で使用して、血液系細胞以上に、ヒト細胞の再構成および/または機能を改善することができる。たとえば、ヒトサイトカイン類の1つまたは複数のアゴニストを非ヒト哺乳動物に導入して、HSCの再構成を強化することができる。
【0066】
当業者に理解されるであろうが、「機能的(または「生物学的に活性な」または「成熟した」)ヒトNK細胞」、「機能的ヒト樹枝状細胞」、「機能的ヒト単球/マクロファージ」、「機能的ヒト赤血球、「機能的ヒトT細胞」および「機能的ヒトB細胞」は全て、分化した細胞(ヒトNK細胞、ヒト樹枝状細胞、ヒト単球/マクロファージ、ヒト赤血球、ヒトT細胞、またはヒトB細胞であっても)が、対応する、ヒトで見られる通常の(野生型)細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全部を発現し、そして結果として、それらがヒトで機能するのと同様に非ヒト哺乳動物で機能するということを指す。
【0067】
非ヒト哺乳動物におけるヒト血液系細胞の機能を測定するための検定法は、当業者に周知であり、また本明細書に記述されている。たとえば、NK細胞傷害性検定法を使用して、非ヒト哺乳動物におけるNK細胞の機能を測定することができる。
【0068】
本発明のある態様で、非ヒト哺乳動物におけるヒト血液細胞系統および/またはある特定のヒト細胞系統(たとえば、NK細胞、樹枝状細胞、単球/マクロファージ、赤血球、T細胞、B細胞)の再構成が強化される。再構成強化は、たとえば、適当なコントロールと比較して強化された細胞型の発現(たとえば、1つまたは複数のヒト血液系細胞数の増加;細胞型が発現される期間の増加(たとえば、>30日))を指す。そのようなコントロールは当業者に明白である。適当なコントロールの例は、1つまたは複数のサイトカイン類をコード化する核酸ではなく、ヒトHSCが導入された非ヒト哺乳動物である。
【0069】
本発明の他の態様は、組成物を含む。一態様で、本発明は、本明細書に記載の方法により作られる非ヒト動物を含む。
【0070】
他の態様で、本発明は、本明細書に記載の方法により産生されるハイブリドーマ(単離ハイブリドーマ)およびそのハイブリドーマにより産生されるモノクローナル抗体(単離モノクローナル抗体)を含む。
【0071】
本明細書で使用されるとき、「単離された」(たとえば、「単離されたB細胞」;「単離されたハイブリドーマ」、「単離されたモノクローナル抗体」)は、それが自然に存在する複雑な(たとえば、細胞の)環境、または器官、身体、組織、血液または培地に関して実質的に単離されていることを指す。場合によっては、単離された材料は、組成物の一部(たとえば、他の物質を含む粗抽出物)、バッファー系、培養系または試薬混合物を形成する。状況が違えば、材料は、本質的に均質になるまで精製することができる。単離されたB細胞個体群は、(全細胞数を基準として)存在する全細胞の少なくとも約50%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約99%を構成する。一実施態様で、本発明は、本明細書に記載の方法で産生される、単離された、または実質的に単離された(または精製された、本質的に精製された)B細胞、ハイブリドーマおよび/またはモノクローナル抗体を対象とする。
【0072】
したがって、本明細書に示す通り、ヒト化マウスで確認されるヒト生来の応答および適応応答が不十分な主な理由は、適切なヒトサイトカイン発現が再構成マウスで欠如しているために、特定のヒト血液系細胞の再構成が低レベル、および/または特定のヒト血液細胞系統が機能的に成熟不良であることに起因する。再構成非ヒト哺乳動物(たとえば、マウス)で様々なヒトサイトカイン類を発現し、また結果として得られる非ヒト哺乳動物におけるヒト血液系細胞の再構成を著しく改善するための、効率よくかつ汎用的な方法を、本明細書に記述する。
【0073】
非ヒト哺乳動物でヒトサイトカイン類を発現するために、流体力学的注入(たとえば、7秒で体重の10%)を使用して、ヒトサイトカイン類をコード化するDNAベクターを哺乳動物に導入した。この実施態様で、注入されたDNAの一部が肝細胞によって取り込まれ、マウスでヒトサイトカインが発現する結果となった。ヒトサイトカイン類をコード化する核酸をこのようにして導入することにより、特定のヒト細胞サブセットの発生が向上し、ヒト細胞数および免疫応答の増加がもたらされた。
【0074】
具体的には、ヒトサイトカイン類を、ベクターにクローニングした。操作したヒトインターロイキン15(IL−15)遺伝子およびFms−関連チロシンキナーゼ3リガンド(Flt3L)遺伝子を、個々にpcDNA3.1(+)ベクターにクローニングした。
【0075】
pcDNA3.1(+)プラスミドをインビボ遺伝子送達用ベクターとして使用した。インビボでNK細胞を誘導するために、pcDNA−IL2/IL15およびpcDNA−Flt3Lプラスミドを構築した。簡単に記述すると、IL−15のシグナルペプチドは通常長く(48aa)、またIL−15の分泌を制限するため、ヒトIL−15のシグナルペプチド配列を、ヒトIL−2のそれで置き換えた。IL−15に応答するNK細胞前駆体の発生頻度を高めるために、速効性サイトカインflt3リガンド(FL)を使用した。これらの2つの組換え遺伝子配列をpcDNA3.1(+)ベクターに挿入した。
【0076】
流体力学に基づく遺伝子導入法で、尾静脈を介して大量の溶液を急速注入することにより、プラスミドをマウスに投与した。簡単に記述すると、8〜12週齢のヒト化マウスに、各プラスミド50μgを含む生理食塩水1.8mlを、7秒以内で静脈注入した。
【0077】
注入後第7日に、これらのマウスの尾静脈から採血して、主として血中のヒト免疫細胞の再構成を分析した。次いで、数匹のマウスを第9日および第16日に屠殺して、肝臓、肺、脾臓、骨髄、およびリンパ節におけるヒト細胞再構成の動力学を分析した。血清を異なる時点に採取し、循環血液中のヒトサイトカインレベルをELISAで分析した。
【0078】
流体力学的注入後、血清中に有意なレベルのIL−15およびFlt3Lが2〜3週間検出された。相応して、血中、脾臓、骨髄、肺および肝臓におけるCD56+CD3−ヒトナチュラルキラー(NK)細胞の再構成は、ヒト器官で確認されるものに匹敵するレベルまで有意に上昇した。CD45+ヒト細胞の絶対数も、全ての器官で増加した。
【0079】
この方法によってヒト化マウスで生成したNK細胞は、通常のNK細胞表現型を有し、またインビトロでもインビボでも、polyI:CおよびLPS刺激に応答したインターフェロン−γ(IFN−γ)産生に関して機能的であった。ヒト化マウスから精製されたNK細胞は、インビトロで、K562標的細胞に対して細胞傷害活性を示した。さらに、NK細胞は、インビボで、ウイルス感染に応答した。同様の手法を使用して、GM−CSFおよびIL−4を発現することにより樹枝状細胞再構成を強化し、MCSFを発現することによりマクロファージおよび単球再構成を強化し、EPOおよびIL3を発現することによりヒト赤血球再構成を強化した。
【0080】
裸のDNAの流体力学的注入は、血清中で2〜3週間、ヒトサイトカイン類が検出される結果となった。当業者に明白な通り、ヒト遺伝子はまた、ウイルスベクター(たとえば、アデノウイルス介在性遺伝子発現;レンチウイルス介在性遺伝子発現)によって発現させることもでき、これはマウスで高レベルかつ長期にわたるヒト遺伝子発現をもたらすことができる。
【0081】
一部の種類のヒト免疫細胞の再構成は、ヒト造血幹細胞移植NOD/SCID IL2Rγ−/−マウスで非常に低く、これはマウスサイトカイン環境に対する反応性が不十分なためである。免疫細胞発生および成熟に不可欠な多くのサイトカイン類は、NK細胞に対するIL−15、DCに対するGM−CSF/IL−4、およびマクロファージに対するM−CSF等のような、種特異的活性を示す。有意に高レベルの全身性外因性ヒトサイトカインの発現をもたらし、結果としてヒト化マウスでのヒト免疫細胞発生を促進することになる、裸のサイトカイン発現プラスミドの投与を使用する流体力学に基づくインビボトランスフェクション手順を、本明細書に記述する。
【0082】
本方法および組成物は、ヒトHSC由来の血液細胞系統を有する非ヒト哺乳動物を再構成するために使用される現行の方法に優る多数の利点を提供する。たとえば、サイトカインをコード化する核酸は、所望の結果を達成するために、一度導入しさえすればよい;サイトカイン類をコード化する核酸を使用すると、大規模調製がはるかにし易く、はるかに安定して長期保存し易く、またはるかに遺伝子操作し易い;3週間にわたる全身性ヒトサイトカイン発現で、流体力学的に基づく注入を実施することができ、またサイトカインを導入するためにアデノウイルスベクターおよび/またはレンチウイルスベクターを使用して、発現をより長く持続できると思われる;多数の遺伝子構築物を組み合わせることができる(図17参照);本方法は、ヒト化マウスでHSC(たとえば、骨髄性細胞)を再構成するための簡単で効率のよい方法を提供する。
【実施例】
【0083】
例示
実施例1
ヒトサイトカイン類の発現は、ヒト化マウス(ヒューマウス)における、特定のヒト血液系細胞の再構成を劇的に改善する。
【0084】
材料と方法
HSC単離、ヒト化マウスの構築、および流体力学的遺伝子送達。ヒト臍帯血は、Singapore Cord Blood Bankから入手した。臍帯血単核細胞(MNC)は、Ficoll−Hypaque密度勾配により分離した。CD34+細胞は、RosetteSep(登録商標)システムで、製造業者のプロトコール((Stem Cell Technologies)に従って精製した。CD34+細胞の純度は>95%であった。HSCを増殖させるために、精製CD34+細胞を、規定の因子の存在下、無血清培地で11〜14日間培養した(Zhang CC,Kaba M,Iizuka S,Huynh H,Lodish HF(2008)Blood 111:3415−3423)。非増殖HSCと増殖HSCの両方を使用して、ヒト化マウスを作った。
【0085】
NSGマウスは、Jackson Laboratoriesから購入し、Nanyang Technological UniversityおよびNational University of Singaporeの動物施設で、特定病原体不在条件下で維持した。マウスを再構成するために、γ線源を使用して100cGyを新生仔(48時間齢未満)に照射し、CD34+CD133+細胞(1×105細胞/レシピエント)を心内注入した。ヒトサイトカイン遺伝子は、pcDNA3.1(+)ベクター(Invitrogen)に別々にクローニングした。プラスミドDNAは、Maxi−prep Kit(Qiagen)で精製した。流体力学的遺伝子送達のために、12週齢のヒューマウスに、各プラスミド50μgを含む総量1.8mlの生理食塩水を、27ゲージの針を使用して7秒以内で注入した。試料およびマウスを用いた調査研究は全て、National University of SingaporeおよびNanyang Technological Universityの施設ガイドラインを順守して実施した。
【0086】
単一細胞調製、抗体、およびフローサイトメトリー。単一細胞懸濁液は、標準手順により、脾臓および骨髄(BM)から調製した。MNCをヒューマウス肝臓から単離するために、肝臓を、200ゲージのステンレススチールメッシュを押し通し、50xgで5分間の遠心分離により残屑を除去した。MNCを含む上澄を回収し、PBSで洗浄し、40%Percoll(Sigma)を含むRPMI培地1640中に再懸濁した。細胞懸濁液を、70%Percollの上に静かに重ね、750xgで20分間、遠心分離した。MNCを相間から回収し、PBSで2回洗浄した。MNCを肺から単離するために、肺を細かく刻み、0.05%コラゲナーゼ(Sigma)および0.01%DNase I(Sigma)を含む培地中に懸濁し、37℃で20分間インキュベートした。肺試料を、200ゲージのステンレススチールメッシュを押し通し、上述の通り、Percoll遠心分離でMNCを単離した。
【0087】
下記の抗体を使用した:Becton−DicksonからのCD3(SK7)、CD34(581)、CD19(HIB19)、NKG2D(1D11)、NKp46(9E2)、CD94(DX22)、CD16(3G8)、CD56(B159)、HLA−DR(L243)、CD14(M5E2)、CD11c(B−ly6)、CD209(DCN46)、CD7(M−T701)、CD45(2D1)、CD69(L78)、CD33(WM53);BioLegendからのKIR2DL2/L3(DX27)、ILT7(17G10.2)およびCD235ab(HIR2);Miltenyi BiotecからのCD303(AC144);Beckman CoulterからのCD159a(NKG2A;Z199);およびeBioscienceからのCD133(EMK08)およびマウスCD45.1(A20)。細胞を、氷上で、0.2%BSAおよび0.05%アジ化ナトリウムを含有するPBS 100μl中の適切な抗体で30分間染色した。フローサイトメトリーは、FACSDivaソフトウェアを使用したLSRIIフローサイトメーター(Becton,Dickinson and Co.)で実施した。試料当たり10,000〜1,000,000の事象を収集し、Flowjoソフトウェアを使用して解析した。
【0088】
インビトロでのヒトCD34+細胞の分化。BM MNCを12週齢のヒューマウスから単離した。CD34+細胞を、MACS(登録商標)マイクロビーズ(Miltenyi Biotec)で濃縮した。精製細胞を、RPMI 1640、10%FCS中、37℃および5%CO2で培養した。NK細胞、DC、単球/マクロファージ、および赤血球の分化用に、それぞれ、50ng/ml SCF、50ng/ml FLおよび50ng/ml IL−15;50ng/ml SCF、20ng/ml GM−CSFおよび50ng/ml IL−4;50ng/ml SCFおよび30ng/ml M−CSF;および100ng/ml SCF、5ng/ml IL−3および3U/ml EPOを使用した。サイトカイン類は全て、R&D Systemsから購入した。
【0089】
NK細胞細胞傷害性検定および刺激。遺伝子送達の9日後、幹細胞PE選択キット(Stem Cell Technologies)を使用して正の選択により、CD56+NK細胞を脾臓およびBMから精製した。細胞を洗浄し、2%FCSを含むIMDM中に再懸濁し、NK感受性標的K562(ATCC)に対する細胞傷害性を、4時間乳酸デヒドロゲナーゼ放出検定法(CytoTox96;Promega)で測定した。
【0090】
インビトロ刺激用に、精製NK細胞を、ヒトDCを含む、または含まない、RPMI 1640、10%FCS、L−グルタミン2mM、ピルビン酸ナトリウム1mM、ペニシリン、およびストレプトマイシン中、37℃および5%CO2で24時間、培養した。ヒトDCは、記述の通り、臍帯血CD34+細胞から分化させた(Rosenzwajg M,Canque B,Gluckman JC(1996)Blood 87:535−544)。次に、インビトロでNK細胞を刺激するために、50pg/ml poly(I:C)(Sigma)を培養に添加した。インビボ刺激用に、ヒューマウスにpoly(I:C)200μgを静脈内注入した。血清中または培養上澄中のIFN−γレベルを、ELISA Kits(R&D Systems)で測定した。
【0091】
アデノウイルス感染、ALT、および組織学。複製欠損、E1およびE3欠失、タイプ5緑色蛍光蛋白質発現アデノ−Xウイルス(AdGFP)は、Clontechから購入した。AdGFPを、HEK293細胞中で増殖させ、CsCl不連続密度勾配遠心分離により精製した。4×109pfu AdGFPウイルスを、流体力学的注入により尾静脈を介してヒューマウスに投与した。アデノウイルス感染の5日後、血清を採取し、コバス(cobas) c 111アナライザー(Roche Diagnostics Ltd.)を使用して、ALT活性について分析した。
【0092】
組織学的分析用に、肝臓を摘除し、パラフィン中に包埋して、厚さ5μmの切片を作製した。パラフィン切片をH&Eで染色し、光学顕微鏡で分析した。2色蛍光免疫染色用には、非特異的染色のブロッキング後、脱パラフィン処理切片を、PE−コンジュゲート抗ヒトCD56抗体(MEM−188;Biolegend)の最適希釈物で染色した。切片を、MIRAX MIDI蛍光顕微鏡(Zeiss)で分析した。
【0093】
統計解析。データを、平均値および平均値の標準誤差として示す。群間差をStudent t検定で解析した。<0.05のP値を、統計学的有意と考えた。計算は全て、Origin8.0ソフトウェアパッケージを使用して実施した。
【0094】
結果
インビトロでの、ヒトサイトカイン類によるNK細胞分化の刺激。ヒト化マウスを構築するために、ヒト臍帯血から単離したCD34+HSCを、亜致死照射したNSG仔に養子移入した。再構成の12週間後、末梢血由来の単核細胞(MNC)を、ヒトCD45およびマウスCD45に特異的な抗体で染色した(図6A〜6B)。平均再構成率は、血液で約50%であった[再構成率=%CD45+ヒト細胞/(%CD45+ヒト細胞+%CD45+マウス細胞)]。CD45+ヒト白血球の中で、CD19+B細胞のレベルは40〜85%の範囲であり、CD3+T細胞のレベルは10〜50%の範囲であった。NK細胞は、血中、BM、脾臓、肺、および肝臓で検出されたが、それらの発生頻度は、対応するヒト組織またはマウス組織における発生頻度より有意に低かった(図6A〜6B参照)。
【0095】
ヒューマウスにおけるNK細胞再構成不良の根底となる原因を究明するために、我々は、ヒューマウスのBMに由来するヒトCD34+細胞が、インビトロで、ヒトIL−15およびFLにより刺激されて、NK細胞に分化するかどうかを試験した。FLは、IL−15に応答できるCD34+NK前駆細胞を含む、多様な造血細胞系統の分化を刺激する(Yu H,et al.(1998)Blood 92:3647−3657)。FLとIL−15の組合せは、CD34+前駆体の、NK細胞方向への分化を促進すると予期される。したがって、ヒューマウスBM由来の精製ヒトCD34+細胞(>80%)(図1A)を、FLおよびIL−15の存在下で7日間培養し、NK細胞マーカーCD56およびNKp46の発現について分析した。サイトカイン類の存在下では、細胞の約11%がCD56およびNKp46の両者について陽性であったが、サイトカイン類の非存在下では非常に少ない細胞が陽性であった(図1B)。これらの結果から、ヒューマウスのBM中のCD34+ヒト細胞は、適切なサイトカイン環境が提供されれば、NK細胞に分化することができることが示唆される。
【0096】
プラスミドDNAの流体力学的注入による、マウスにおけるヒトサイトカイン類の発現。IL−15およびFLの存在下では、インビトロでヒトNK細胞が発生するという知見から、これらのヒトサイトカイン類は、ヒューマウスにおけるNK細胞発生も刺激し得ることが示唆される。ヒトサイトカイン類をマウスに導入する一つの方法は、組換え蛋白質を毎日注入することによる。この方法は厄介で高価なため、我々はサイトカインコード化DNAプラスミドの流体力学的送達により、マウスで、ヒトサイトカイン類を発現させた。ヒトIL−15は、異常に長いシグナルペプチド配列を有し(45aa残基)、これは、IL−15の分泌不良につながることが知られている(Meazza R,et al.(1997)Eur J Immunol 27:1049−1054)。IL−15分泌のレベルを高めるために、我々は、IL−15シグナルペプチドがヒトIL−2のシグナルペプチドに置き換えられた、IL−15発現ベクターを構築した(図7A〜7B)。この置換は、血清IL−15レベルを約100倍上昇させた(図7B)。IL−15コード化プラスミド50μgの単回流体力学的注入で、高レベルのIL−15が、注入の1日後に血清中で検出され、有意なレベルが14日間維持された(図2)。同様に、FLコード化プラスミドの単回注入は、FLが血清で21日間発現するという結果をもたらした。したがって、サイトカイン遺伝子の流体力学的送達は、マウスでヒトサイトカイン類を導入するための簡単で効率のよい方法である。
【0097】
IL−15およびFL遺伝子送達後のヒトNK細胞の再構成強化。IL−15およびFL発現がNK細胞発生に及ぼす影響を測定するために、遺伝子送達の9日後に、ヒューマウスの様々な器官におけるNK細胞再構成についてフローサイトメトリーで分析した。空のpcDNAベクターまたはFLコード化ベクターの注入は、CD56+NK細胞の発生頻度に有意な影響を及ぼさなかった(図3A)。しかし、IL−15の発現は、血中、脾臓、BM、肺、および肝臓におけるCD56+NK細胞の発生頻度を有意に増加させた(図3Aおよび3B)。NK細胞の発生頻度の増加は、IL−15とFLの両者がヒューマウスで発現されたとき、さらに劇的であり、通常のヒト末梢血(白血球の5%〜21%)(Maurice RG,O’Gorman ADD(2008)Handbook of Human Immunology(CRC Press,Boca Raton))および通常のマウス組織(Zhang J,et al.(2005)Cell Mol Immunol 2:271−280)で確認されるレベルに達した。NK細胞の発生頻度上昇に対応して、NK細胞の絶対数が脾臓およびBMで著しく増加した(図S3B)。さらに、血中でのCD56+NK細胞の高い発生頻度は、遺伝子送達後少なくとも30日間維持された(図3C)。加えて、サイトカイン誘導性NK細胞は、活性化受容体NKG2D、抑制性受容体NKG2A、CD94、およびKIR、細胞傷害誘発受容体NKp46、NK細胞マーカーCD7、早期活性化マーカーCD69、およびFC受容体CD16を含め、NK細胞機能に重要であることが知られている細胞表面受容体の多くを発現した(図9)。これらの結果から、サイトカイン誘導性NK細胞は通常のNK細胞の特徴的な表面表現型を示すことが分かる。
【0098】
NK細胞発生の刺激に加えて、FLとIL−15の両者は、他の造血細胞系統にも効果を発揮することが知られている(Diener KR,Moldenhauer LM,Lyons AB,Brown MP,Hayball JD(2008)Exp Hematol 36:51−60;Dong J,McPherson CM,Stambrook PJ(2002)Cancer Biol Ther 1:486−489;Blom B,Ho S,Antonenko S,Liu YJ(2000)J Exp Med 192:1785−1796;Armitage RJ,Macduff BM,Eisenman J,Paxton R,Grabstein KH(1995)J Immunol 154:483−490)。したがって、ヒューマウスの脾臓、BM、肺、および肝臓由来の細胞を列挙し、フローサイトメトリーで分析した。IL−15およびFLの発現はまた、脾臓およびBMにおけるCD14+単球/マクロファージ、CD11c+CD1c+骨髄性樹枝状細胞、ILT7+CD303+形質細胞様樹状細胞、およびCD19+B細胞の有意な増加も誘導した(図8A〜8G参照)。これらの結果は、ヒトIL−15およびFLの発現が、ヒト化マウスにおけるNK細胞ならびに他の骨髄性細胞およびリンパ球系細胞の再構成を劇的に改善することを示す。
【0099】
サイトカイン誘導性NK細胞は機能的である。我々は、サイトカイン誘導性ヒトNK細胞が機能的である(すなわち、標的細胞を死滅させることができ、また適切な刺激後にIFN−γを発現できる)かどうかを調査研究した。CD56+NK細胞は、IL−15およびFL処理マウスのBMおよび脾臓から精製した。MHCクラスI欠損標的細胞K562と混合したとき、添加したNK細胞が増加するにつれて、高レベルの標的細胞溶解が確認された(図10A)。NK細胞を刺激して炎症性サイトカイン類を産生させることが知られている、Toll様受容体3アゴニストpoly(I:C)(Schmidt KN,et al.(2004)J Immunol 172:138−143)で精製NK細胞を刺激したとき、IFN−γが培養上澄で検出された(図10B)。ヒトDCの存在下で、IFN−γ分泌レベルはさらに上昇した。poly(I:C)をヒト化マウスに注入したとき、サイトカインコード化DNAを注入したヒューマウスの血清で、非注入ヒューマウスと比較して有意に高いレベルのIFN−γが検出された(図10C)。
【0100】
我々はまた、NK細胞依存性肝損傷を引き起こすことが知られているアデノウイルス(Chen Q,Wei H,Sun R,Zhang J,Tian Z(2008)Hepatology 47:648−658)を、ヒューマウスに投与した。サイトカイン遺伝子送達の9日後、複製欠損アデノウイルスをヒューマウスに静脈内注入した。3日後、肝臓を採取し、H&Eで染色した。大量の白血球浸潤および広域の壊死が、IL−15およびFL処理したアデノウイルス感染ヒューマウスの肝臓で確認された。しかし、未処理のアデノウイルス感染ヒューマウスは、軽度の細胞浸潤および損傷を示しただけであった(図4A)。相応して、IL−15およびFL処理したアデノウイルス感染ヒューマウスで、血清アラニンアミノトランスフェラーゼ(ALT)レベルが有意に上昇していた(図4B)。この上昇は、肝臓における、血清IFN−γレベルの約4倍の増加(図4C)および、浸潤性ヒト白血球の約5倍の増加(図4D)と相関性があった。肝薄片の免疫組織化学分析は、CD56+NK細胞の病変内局在化を裏付けた(図4E)。これらの結果から、サイトカイン誘導性ヒトNK細胞は機能的であることが強く示唆される。
【0101】
他のヒト血液細胞系統の再構成の改善。我々は、サイトカイン遺伝子送達が、ヒューマウスにおける特定のヒト血液細胞系統の再構成を改善するための一般的方法として使用できるかどうかを試験した。培養で、ヒューマウスのBMから精製したヒトCD34+細胞を、GM−CSFおよびIL−4で刺激してCD11c+CD209+DCに、M−CSFで刺激してCD14+単球/マクロファージに、またEPOおよびIL−3で刺激してCD235ab+赤血球に、分化させた(図11)。インビボで、GM−CSF、IL−4、およびFLを発現するDNAベクターをヒューマウスに流体力学的に送達することにより、血中、脾臓、BM、肺、および肝臓でD11c+CD209+DCの発生頻度が著しく増加する結果となった(図5A)。同様に、M−CSFの発現は、リンパ器官と非リンパ器官の両者におけるCD14+単球/マクロファージの再構成の改善へと導いた(図5B)。EPOおよびIL−3の発現は、血中でのCD235ab+ヒト赤血球の出現をもたらし(図5C)、全赤血球の3〜5%に達した。したがって、DNAプラスミドの流体力学的注入によるサイトカイン遺伝子発現は、ヒューマウスにおける特定のヒト血液細胞系統の再構成を改善するための一般的で効率のよい方法である。
【0102】
考察
NK細胞および骨髄性細胞の再構成は、NSGまたはBALB/c−Rag2−/−Il2rg−/−マウスをレシピエントとして使用するヒト化マウスモデルでは、一般に不十分である。BLTマウスで、DCおよび単球/マクロファージがかなり再構成されるにも拘わらず、ヒトNK細胞およびRBCは存在しない。NK細胞または様々な骨髄性細胞の発生および維持に必要な、IL−15、GM−CSF、IL−4、M−CSF、およびIL−3を含む多くのサイトカイン類は、ヒトとマウスとの間で有意な配列分散を示すことが分かった。以前の研究は、これらのネズミサイトカイン類が、適切なヒト細胞型に殆ど影響しないことを証明していた。これらのサイトカイン類は、主として非造血細胞によって産生されるため、これらのヒトサイトカイン類の欠如によって、ヒューマウスにおけるNK細胞および骨髄性細胞の再構成不良を説明し得るであろう。
【0103】
この解釈を裏付けるために、ヒューマウスのBMから単離されたヒトCD34+前駆細胞を刺激して、インビトロで、NK細胞、DC、単球/マクロファージ、および赤血球に分化できることを示した。サイトカインコード化プラスミドDNAの流体力学的送達により、ヒト化マウスで適切なヒトサイトカイン類が導入されるとき、有意に高いレベルのNK細胞、DC、単球/マクロファージ、および赤血球が誘導される。サイトカイン類の血清レベルが低下するにつれて、再構成レベルも低下する。したがって、ヒューマウスにおけるNK細胞および骨髄性細胞の再構成不良は、それらの分化および維持に通常必要な適切なヒトサイトカイン類が欠如した結果である。適切なサイトカイン類の導入は、ヒューマウスにおけるこれらのヒト血液細胞系統の再構成レベルの劇的な上昇につながる。
【0104】
流体力学的遺伝子送達は、マウスで、高レベルの、一過性の肝および全身性導入遺伝子発現を生じさせるために広く使用される(Suda T,LiuD(2007)Mol Ther 15:2063−2069)。本方法は、短時間(6〜8秒)で、大量(体重の10%)のDNAの尾静脈注入を含む。流体力学的圧力は、肝臓損傷を引き起こし、肝細胞によるDNAの取り込みにつながる(Suda T,Liu D(2007)Mol Ther 15:2063−2069)。転写および翻訳の後、サイトカイン類が循環内に分泌されて、BMまたは他の器官の標的細胞に到達することができる。したがって、サイトカインコード化DNAの単回注入で、IL−15は血清中で2週間にわたって検出され、FLは3週間にわたって検出された。IL−15とFLとの血清持続期間の差は、恐らく蛋白質の半減期の差のためであるか、またはIL−15が通常はIL−15Ra鎖を介して細胞表面上に結合されているためである(Mortier E,Woo T,Advincula R,Gozalo S,Ma A(2008)J Exp Med 205:1213−1225)。単回のDNA注入によって産生されるIL−15およびFLの量は、少なくとも30日間、NK細胞レベルの著しい上昇を誘導するのに明らかに十分である。サイトカイン類が循環中でもはや検出されなかったとき、NK細胞が存続することは、サイトカイン類の重要な役割が、分化の初期段階に発揮されることを示す。NK細胞は、いったん生成すると、サイトカイン類が循環で検出できなくなった後も、長時間にわたって生存することができる。NK細胞は、多様な血液細胞系統に影響を及ぼすため、FLおよびIL−15の発現はまた、脾臓およびBMでの、単球/マクロファージ、DC、およびB細胞のレベル上昇をもたらすが、T細胞のレベル上昇はもたらさない。さらに、流体力学的遺伝子送達による適切なサイトカイン類の発現はまた、DC、単球/マクロファージ、および赤血球を含む、特定の骨髄系細胞の再構成を著しく強化し、本手法の汎用性を示す。ヒトサイトカイン遺伝子送達の7日後に明白になる、NK細胞、DCおよび単球/マクロファージの改善した再構成と比較して、赤血球の改善した再構成は、サイトカイン遺伝子送達の30日後までピークレベルに到達しなかった。これは、一方ではヒトWBCとマウスWBCの比率の差によって説明でき、また他方ではヒトRBCとマウスRBCの比率の差によって説明できる。マウスRBCは大量なため、類似したパーセンテージに到達するために十分な数のヒトRBCを産生するには長時間を要する。以前に、2つのグループが、組換えヒトIL−15をNOD−scidマウスまたはBALB/c−Rag2−/−Il2rg−/−マウスに注入することによって強化されたNK細胞発生について報告した(Huntington ND,et al.(2009)J Exp Med 206:25−34;Kalberer CP,Siegler U,Wodnar−Filipowicz A(2003)Blood 102:127−135)。厄介で高価な、毎日のサイトカイン注入と比較して、流体力学的遺伝子送達によるサイトカイン遺伝子の発現は、単回注入で、30日間以上にわたって特定の血液細胞系統の再構成上昇をもたらすため、手頃で、簡単、かつ効率のよい方法である。
【0105】
サイトカイン誘導性NK細胞は、通常の表面表現型および機能を示す。NOD−scidマウスで、毎日の組換えIL−15の注入後にヒトNK細胞が生成された、以前の観察結果と対照的に、この細胞はNKp46を発現したが、NKG2DおよびNKG2Aは発現しなかった(Kalberer CP,Siegler U,Wodnar−Filipowicz A(2003)Blood 102:127−135)。本研究で、サイトカイン誘導性ヒトNK細胞は、活性化受容体NKG2D、抑制性受容体NKG2AおよびKIR、ならびに細胞傷害誘発受容体NKp46を含む、NK受容体の3つの主要なファミリー全てを発現した。一貫して、サイトカイン誘導性NK細胞は、インビトロでもインビボでも、poly(I:C)刺激のすぐ後で、MHCクラスI−欠損標的細胞を溶解することができ、またIFN−γを分泌することができる。さらに、IL−15およびFL処理ヒューマウスにおける、広範囲の肝臓壊死および高レベルの血清ALTで示されるように、サイトカイン誘導性NK細胞は、アデノウイルス感染に対する強い反応を開始することができる。IFN−γ分泌により肝炎を仲介する、マウスにおける野生型NK細胞(Chen Q,Wei H,Sun R,Zhan gJ,Tian Z(2008)Hepatology 47:648−658;Rosenberger CM,Clark AE,Treuting PM,Johnson CD,Aderem A(2008)Proc Natl Acad Sci USA 105:2544−2549)と同様、IL−15およびFL処理アデノウイルス感染ヒューマウスの血清IFN−γレベルは有意に上昇していた。これらの結果から、サイトカイン誘導性NK細胞は、表面表現型でも機能でも、正常であることが示唆される。
【0106】
実施例2
ヒトサイトカイン類の発現は、ヒト化マウスにおけるヒトT細胞およびB細胞の再構成および機能を改善する。
ヒトT細胞およびB細胞の再構成は、ヒト化マウスで妥当であるが、最適な機能を示さない。たとえば、ヒトCD8+T細胞反応は、ウイルス投与後に検出されてきたが、CD4+T細胞の機能は異常である;ヒト化マウスで、ヒトB細胞介在性抗体反応は存在しない。本明細書に示す通り、ヒトT細胞およびB細胞応答の異常はまた、マウスにおける、マウスサイトカイン類とヒト細胞との間の不十分な交差反応にも起因する。ヒトGM−CSFおよびIL−4コード化プラスミドのヒューマウスへの注入が、T細胞に対する主要抗原提示細胞であると考えられる、ヒトCD209+樹枝状細胞の再構成改善に導いたことを、本明細書に示す。さらに、IL−4は、ヒトB細胞における細胞増殖、生存、ならびにIgGおよびIgEへの免疫グロブリンクラススイッチ、ならびにナイーブCD4+T細胞によるTh2表現型の獲得を促進することも証明された。トキソイド(TT)ワクチンを使用して、GM−CSFおよびIL−4処理ヒューマウスを免疫し、これらのマウスがTT特異的抗体応答を引き起こすかどうかを決定した。
【0107】
本明細書に記載の通り、類似したヒト白血球再構成(50〜80%)を有する12週齢のヒト化マウスに、ヒトGM−CSFおよびIL−4をコード化するプラスミドまたは空のpcDNAベクター(ベクター)を、流体力学的に注入した。7日後、これらのマウスに、破傷風トキソイド(TT)を、3週間の投与間隔で3回免疫した(図12A)。第3回の免疫の2週間後に、脾臓および血清を採取した。図12Bは、GM−CSFおよびIL−4処理マウスからの脾臓は、ベクター処理マウスと比較して、有意に拡張したことを示す。相応して、GM−CSFおよびIL−4処理マウスの脾臓で、ヒト単核細胞(MNC)の劇的な増殖(約20倍)が認められた(図12C)。脾臓におけるヒトB細胞およびT細胞の細胞表面表現型別の結果から、ヒトB細胞は、サイトカイン処理および免疫の全過程を通して通常のヒト特性と同一である成熟した抗体産生期(CD19lowCD20−)まで成長した(図13);が、ヒトT細胞は、HLA−DRおよびCD40Lの発現を上方制御することにより活性化されることも分かった(図14)。
【0108】
免疫した、GM−CSFおよびIL−4処理マウス由来の血清中のヒトIgGおよびIgM総レベルは、それぞれ、1.3mgおよび140μgという高値に達し(図15A,15B)、ヒトでのレベル(それぞれ4mgおよび1mg)に類似していた。最も重要なことは、抗原特異的ヒト抗体応答が、ヒューマウスで初めて首尾よく実証されたことである。ヒトTT特異的IgGはベクター処理マウスで検出できなかったが、サイトカイン類処理マウスでは、0.16IU/mlという平均値に達した(図15C)。免疫化後、ヒトで、0.1IU/mlという抗破傷風トキソイド抗体は、個体を感染から守るのに十分である。さらに、GM−CSF+IL−4処理マウスにおけるヒトT細胞応答も、TT抗原に対して特異性を示した(図16A〜16B)。破傷風毒素ペプチド(830−843)を使用して、脾臓T細胞を刺激した。GM−CSF+IL−4処理マウス由来のT細胞は、ベクター処理マウス由来の細胞と比較して、TT特異的刺激後に、有意レベルのヒトIFN−γおよびIL−4を産生することができた。
【0109】
本明細書に記載の方法を使用して、有意レベルのヒト抗原特異的抗体応答を、マウスで実証することができた。したがって、本明細書に記載の方法は、ワクチン類を試験するため、および治療目的のヒト抗体を産生するための、有用な基盤を提供する。
【0110】
【表1】

【0111】
【表2】
【0112】
【表3】
【0113】
全ての特許、公開された出願および本明細書に引用された参考文献の教示は、参照によりその全体を援用する。
【0114】
本発明は具体的に示され、またその例示的な実施態様を参照しながら記述されているが、添付の請求の範囲に含まれる本発明の範囲から逸脱せずに、形状および詳細の様々な変更を加えられることを、当業者は理解するであろう。
【技術分野】
【0001】
関連出願
本出願は、2009年6月29日提出の米国仮特許出願第61/221,438号の利益を主張する。上記出願の教示全体を参照により本明細書に援用する。
【背景技術】
【0002】
発明の背景
病原体感染に対するヒト免疫応答を、小動物モデルで、体系的かつ制御された方法で研究することが強く求められている。過去20年にわたり、Tリンパ球およびBリンパ球が欠如し、ヒト血液系細胞を有する、重症複合免疫不全(scid)マウスの再構成に大変な努力が向けられてきた(Shultz LD,Ishikawa F,Greiner DL(2007)Nat RevImmunol 7:118−130)。しかし、レシピエントマウスで、生着が不十分であり、ヒトT細胞およびB細胞が急速に消失し、あるいは造血器悪性腫瘍が急速に発症したため、初期の試みは不成功に終わった。scid変異かまたは組換え活性化遺伝子(Rag)の変異のいずれかのためにT細胞およびB細胞が欠損しているばかりでなく、共通γ鎖(γcまたはIl2rg)欠失のためにナチュラルキラー(NK)細胞も欠損しているレシピエントマウスを使用することにより、大きな進歩が達成された(Hiramatsu H,et al.(2003)Blood 102:873−880;Traggiai E,et al.(2004)Science 304:104−107)。NOD−scidIl2rg−/−(NSG)レシピエントまたはBALB/c−Rag2−/−Il2rg−/−レシピエントのいずれかへのヒト造血幹細胞(HSC)の養子移入は、レシピエント骨髄(BM)における安定したHSCの長期生着および末梢における全てのヒト血液系細胞の生成(ヒト化マウスまたはヒューマウス(humice))をもたらす(Hiramatsu H,et al.(2003)Blood 102:873−880;Traggiai E,et al.(2004)Science 304:104−107)。
【0003】
既存のヒト化マウスモデルは、ヒト病原体による感染(Davis PH,Stanley SL,Jr.(2003)Cell Microbiol 5:849-860;Bente DA,et al.(2005)J Virol 79:13797-13799;Islas−Ohlmayer M,et al.(2004)J Virol 78:13891-13900;Guirado E,et al.(2006)Microbes Infect 8:1252-1259;Kneteman NM,et al.(2006)Hepatology 43:1346-1353;Jiang Q,et al.(2008)Blood 112:2858-2868)、特にヒト血液系細胞を感染させるものを研究するための重要なツールを提供する。これらのモデルはまた、小動物モデルでの病原体に対するヒト免疫応答の調査研究を可能にする。しかし、現在利用可能なモデルは、最適なものから程遠い。たとえば、ヒト細胞再構成のレベルは、異なる細胞系統間で著しく異なる。細胞の再構成は骨が折れ、T細胞の再構成は合理的であるが、B細胞およびT細胞は機能的ではない。加えて、NK細胞および骨髄系細胞の再構成は一般に不十分であり、検出不能である。
【0004】
したがって、ヒト免疫系の改善された非ヒトモデルおよびそれらを製造する方法が強く求められている。
【発明の概要】
【0005】
ヒト造血幹細胞(HSC)によるヒト血液細胞系統の再構成不良は主として、非ヒト哺乳動物でこれらの細胞系統を発生および維持するのに必要な、適切なヒトサイトカイン類の欠乏の結果であることを、本明細書に示す。ヒトIL−15およびFlt−3/Flk−2リガンドをコード化するプラスミドDNAがヒト化マウスに送達されたとき(たとえば、流体力学的尾静脈注入による)、ヒトサイトカイン類の発現は2〜3週間持続し、またNK細胞レベル上昇は1カ月を超える期間誘導された。サイトカイン誘導性NK細胞は、活性化受容体と抑制性受容体の両者を発現し、インビトロで標的細胞を死滅させ、またインビボでウイルス感染に強く応答した。同様に、ヒトGM−CSFおよびIL−4、マクロファージコロニー刺激因子、またはエリスロポエチンおよびIL−3の発現は、それぞれ樹枝状細胞、単球類/マクロファージ類、または赤血球の再構成を有意に強化した(参照により本明細書に援用されるChen,Q.,et al.,Proc.Natl.Acad.Sci.,USA,106:21783−21788(2009)を参照されたい)。また、GM−CSFおよびIL−4はヒトT細胞およびヒトB細胞の再構成も強化した。このように、(たとえば、流体力学的送達により)ヒトサイトカイン遺伝子発現をヒトHSCと一緒に使用することは、ヒト化マウスにおける特定のヒト血液細胞系統の再構成を改善するための簡単で効率のよい方法であり、ヒト疾病およびそれらの進行をモデル化し、また小動物モデルにおけるヒト免疫応答を研究するための重要なツールを提供する。
【0006】
したがって、本明細書では、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成し、それによってヒト化非ヒト哺乳動物を製造する方法を提供する。特定の実施態様で、ヒト化マウス(ヒューマウス)が作られる。
【0007】
一態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する(1つまたは複数の)核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成する方法を対象とする。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト血液細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト血液細胞系統を再構成する。
【0008】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトNK細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトNK細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物におけるヒトNK細胞の再構成を強化する。
【0009】
また別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト樹枝状細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト樹枝状細胞の再構成を強化する。
【0010】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する。
【0011】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト赤血球への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト赤血球に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト赤血球を再構成する。
【0012】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する。
【0013】
別の態様で、本発明は、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコードする核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で、免疫原に対するヒト抗体を産生する方法を対象とし、ここで、ヒトサイトカイン類は、ヒトHSCの機能的ヒトT細胞および機能的ヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞および機能的ヒトB細胞に分化する条件下で維持される。非ヒト哺乳動物は、免疫原で免疫され、また、ヒトB細胞が非ヒト哺乳動物で免疫原に対するヒト抗体を産生する条件下で維持され、それによって、非ヒト哺乳動物で免疫原に対するヒト抗体を産生する。免疫原に対する抗体を産生するB細胞は、さらに単離して、免疫原に対するモノクローナル抗体を分泌するハイブリドーマの産生に使用することができる。
【0014】
モノクローナル抗体を分泌するハイブリドーマならびにB細胞により産生される抗体類(たとえば、ポリクローナル抗体類;モノクローナル抗体類)もまた、本発明に包含される。本明細書で提供される方法により作られる非ヒト哺乳動物もまた、本発明に包含される。
【0015】
本明細書に記載の方法は、非ヒト哺乳動物(たとえば、ヒト化マウス)で機能的ヒト血液細胞系統(たとえば、骨髄性細胞;リンパ球系細胞)を再構成するため、および非ヒト哺乳動物でヒト抗体を産生するための、簡単で効率のよい方法を提供する。
【図面の簡単な説明】
【0016】
【図1】ヒューマウスの骨髄(BM)由来のヒトCD34+細胞は、インビトロで刺激してNK細胞に分化させることができる。(図1A)ヒューマウスのBM由来(左)およびMACSにより抗CD34ビーズで精製した後(右)の単核細胞の、CD34とCD133の染色プロフィールの比較。表示されるイベントは、ヒトCD45+細胞にプレゲート設定する。数値は、ゲートリージョン内の細胞のパーセンテージを示す。マウス4匹のうち1匹からの代表的なデーターを示す。(図1B)IL−15およびFLの非存在下(Ctrl)または存在下で7日間培養したヒトCD34+細胞のNKp46の染色特性対CD56の染色特性。数値は、CD56+NKp46+細胞のパーセンテージを示す。
【図2】流体力学的注入介在性遺伝子送達は、マウスで、全身的ヒトサイトカイン環境を創る。ヒトIL−15およびFLを発現するpcDNAベクターを混合し(各50μg)、ヒューマウスに流体力学的に注入した。表示の時点に、マウス血清中のヒトIL−15およびFLのレベルをELISAで分析した(各時点ごとにn=3)。
【図3】インビボで、IL−15およびFLの発現は、ヒトNK細胞発生を刺激する。空のpcDNAベクター(Ctrl)またはIL−15および/またはFLを発現するpcDNAベクターを、ヒト化マウスに流体力学的に注入した。9日後、表示の組織から細胞を調製し、ヒトCD45、CD3、およびCD56の染色をした。(図3A)ドットプロットは、CD45+細胞にゲート設定したCD3の染色特性対CD56の染色特性を示す。数値は、ゲートリージョンのCD56+CD3−細胞のパーセンテージを示す。群当たりマウス5匹のうち1匹からの代表的なデーターを示す。異なる3つのドナーHSCで、ヒューマウスを構築した。(図3B)サイトカイン遺伝子送達の9日後の、様々な器官におけるCD45+ヒト白血球内のCD56+CD3−NK細胞の発生頻度(平均値±SEM)の比較(n=5)。(図3C)経時的な、血中のCD45+ヒト白血球のCD56+CD3−NK細胞の発生頻度(平均値±SEM)(n=3)。
【図4A−D】ヒトNK細胞は機能的である。(図4A)ヒューマウスにおける、アデノウイルス感染後の、NK細胞介在性肝損傷。サイトカイン遺伝子送達の9日後、PBSまたは複製欠損−アデノウイルス(Ad)を、ヒューマウスに流体力学的に注入した。感染の3日後、H&E染色用にヒューマウスの肝臓を採取した(各群ごとにn=3)。矢印は、壊死および白血球浸潤の領域を示す。Ctrl,サイトカイン遺伝子を送達しないヒューマウス;IL−15/FL,サイトカイン遺伝子を送達したヒューマウス。拡大図を示す。(図4B)血清中のALTレベルの比較。注入の5日後にアデノウイルスまたはPBS処理ヒューマウスから血清を採取し、ALT活性の分析をした(平均値±SEM,群当たりn=3)。IL−15/FL/Adと他群との間でP<0.05。(図4C)アデノウイルス感染ヒューマウスにおける血清IFN−γレベル。アデノウイルス感染ヒューマウスから血清を採取し、ELISAでIFN−γを測定した。平均値±SEMを示す(群当たりn=3)。P<0.05。(図4D)様々なヒューマウスの肝臓におけるMNCの数。アデノウイルス感染の5日後に肝臓を採取した。肝MNC総数を計数し、フローサイトメトリー分析によりヒトCD45+細胞数を決定した(平均値±SEM、群当たりn=3)。IL−15/FL/Adと他群との間でP<0.05。
【図4E】(図4E)CD56+ヒトNK細胞の、肝臓における病変への局在化。肝臓組織をパラフィンに包埋し、薄切し、CD56の染色をし、顕微鏡法で分析した。矢印は病変の領域を示す。マウス3匹のうち1匹からの代表的な画像を示す。
【図5】対応するヒトサイトカイン遺伝子送達による、特定のヒト血液系細胞の誘導。(図5A)樹枝状細胞の再構成改善。空のpcDNAベクターまたは指定されたヒトサイトカイン遺伝子を発現するpcDNAベクターを、ヒューマウスに流体力学的に注入した。注入の9日後、様々な器官から単一細胞懸濁液を調製し、ヒトCD45、CD11c、およびCD209の染色をした。CD45+ヒト細胞にゲート設定したCD209の染色特性対CD11cの染色特性を示す。マウス5匹のうち1匹からの代表的なデーターを示す。(図5B)単球/マクロファージの再構成改善。実験は、pcDNA−コード化M−CSFを注入したこと、および細胞をヒトCD45およびCD14について染色したこと以外、(図5A)の場合と同様に実施した。CD45+ヒト細胞にゲート設定したCD14の染色特性対CD45の染色特性を示す。マウス3匹のうち1匹からの代表的なデーターを示す。(図5C)赤血球の再構成改善。実験は、pcDNA−コード化EPOおよびIL−3を注入したこと、および注入の7日後および30日後に、血液をヒトCD235abについて染色したこと以外、(図5A)の場合と同様に実施した。全血液細胞のCD235abの染色特性対DAPIの染色特性を示す。マウス3匹のうち1匹からの代表的なデーターを示す。数値は、ゲートリージョン内の細胞のパーセンテージを示す。
【図6】ヒト化マウスにおけるヒトNK細胞の再構成。(図6A)HSC生着の12週後、末梢血の単核細胞におけるヒト細胞系統の再構成を、フローサイトメトリーで分析した。ドットプロットは、生存有核細胞にゲート設定したヒトCD45の染色特性対マウスCD45染色特性、あるいはCD45+ヒト細胞にゲート設定したCD3の染色特性対CD19の染色特性またはCD14の染色特性対CD56の染色特性を表す。(図6B)ヒト化マウスの血中、骨髄、脾臓、肺、および肝臓におけるヒトCD45+細胞の、CD14の染色特性対CD56の染色特性。数値は、ゲートリージョン内の細胞のパーセンテージを示す。マウス6匹のうち1匹からの代表的なデーターを示す。
【図7】IL−2シグナルペプチド配列を使用して発現されたときの、循環におけるIL−15レベル上昇。(図7A)IL−15発現ベクターの模式図。内在性シグナル配列(SP)またはIL−2SPのいずれかが付いたIL−15遺伝子を、CMVプロモーターを含むpcDNAベクターにクローニングした。(図7B)血清IL−15レベルの比較。空のpcDNAベクター(Ctrl)、IL−15をコード化するpcDNAベクター、およびIL−2シグナル配列が付いたIL−15をコード化するpcDNAベクターを、NSGマウスに流体力学的に注入した。注入の7日後に、血清を採取し、IL−15レベルをELISAで検定した。
【図8】IL−15およびFL発現後の、ヒト細胞数増加。(図8A〜8G)空のpcDNAベクター(Ctrl)、またはIL−15とFLの両者を発現するpcDNAベクターを、ヒト化マウスに流体力学的に注入した。9日後、表示の組織から細胞を調製し、ヒトCD45に加えてCD3、CD56、CD14、CD11c、CD1c、ILT7、CD303、およびCD19の染色をした。様々な器官における、ヒトCD45+白血球、CD56+NK細胞、CD11c+CD1c+樹枝状細胞、ILT7+CD303+形質細胞様樹状細胞、CD14+単球/マクロファージ、CD3+T細胞、およびCD19+B細胞の絶対数は、総細胞数に特定の細胞型の発生頻度を乗じることによって算出した。平均値±SEMを示す(n=3)。骨髄(BM)における細胞数は大腿骨2本からであった。
【図9】IL−15およびFL処理ヒューマウスにおけるヒトNK細胞の細胞表面表現型。IL−15遺伝子およびFL遺伝子の送達後9日に、表示の器官から細胞を調製し、ヒトCD45、CD56に加えてNKG2D、NKG2A、CD7、CD69、CD94、NKp46、KIR、またはCD16の染色をした。CD45+ヒト細胞にゲート設定した、CD56の染色特性対NKG2D、NKG2A、CD7、CD69、CD94、NKp46、KIR、またはCD16の染色特性を示す。数値は、ゲートリージョン内の細胞のパーセンテージを示す。
【図10】IL−15およびFL処理ヒューマウス由来のヒトNK細胞の細胞傷害性および刺激。(図10A)NK細胞は、細胞溶解性である。サイトカイン遺伝子送達の9日後、ヒトNK細胞をBMおよび脾臓から精製し、異なるエフェクター対標的(E:T)比でK562細胞と混合し、4時間培養した。NK細胞の細胞溶解活性は、上澄の乳酸デヒドロゲナーゼ酵素活性を測定することによって決定した。(図10B)NK細胞は、インビトロで、poly(I:C)刺激後、IFN−γを産生する。精製NK細胞(5×105)を単独で、またはpoly(I:C)(50μg/ml)の存在下で、またはpoly(I:C)およびインビトロで分化したヒトDC(5×105)の存在下で、培養した(材料と方法参照)。24時間後に、ELISAで、ヒトIFN−γについて上澄を分析した。(図10C)NK細胞は、インビボでpoly(I:C)刺激後、IFN−γを産生する。ヒューマウスに、poly(I:C)(マウス当たり200pg)を静脈内注入した。注入の24時間後に血清を採取し、ELISAでヒトIFN−γの検定をした(n=4)。P<0.05。
【図11】インビトロでの、ヒトCD34+細胞の分化。CD34+ヒト細胞を、ヒューマウスのBMから精製し(左)、GM−CSFに加えてIL−4、またはM−CSFの存在下で7日間、あるいはEPOに加えてIL−3の存在下で20日間、培養した。次いで細胞を、CD45に加えてCD209およびCD11c、またはCD14およびCD33、またはCD235abについて検定した。CD45+細胞について、CD209の染色特性対CD11cの染色特性およびCD14の染色特性対CD33の染色特性を示す。CD235ab発現をヒストグラム(太線)で示す。EPOおよびIL−3無しで培養した精製CD34+細胞をコントロールとして使用した(細線)。
【図12】ヒト化マウスにおける、破傷風トキソイドワクチン免疫後の、ヒト細胞増殖。(図12A)免疫の実験的流れ:第0日に、類似したヒト白血球再構成(50〜80%)を有する12週齢のヒト化マウスに、ヒトGM−CSFおよびIL−4をコード化するプラスミドまたは空のpcDNAベクター(ベクター)を、流体力学的に注入した。7日後、これらのマウスを、破傷風トキソイド(TT)で3回、免疫した。第1回の免疫は、2l.f.の破傷風トキソイドワクチンの腹腔内注射で実施し、続いて2〜3週間隔でさらに2回、ブースターを実施した。第2回ブースターの2週間後、マウスを分析した。(図12B)GM−CSFおよびIL−4処理マウス由来の脾臓は、有意に腫大していた。(図12C)様々なヒューマウスの脾臓における単核細胞(MNC)数。脾臓を、免疫後に採取した。脾臓のMNC総数を数え、フローサイトメトリー分析でヒトCD45+細胞数を決定した(平均値±SEM,群当たりn=3)。
【図13】GM−CSFおよびIL−4処理した、TT免疫ヒューマウスにおける、ヒトB細胞の細胞表面表現型。細胞を脾臓から調製し、ヒトCD45、マウスCD45、CD19に加えてIgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色をした。CD45+ヒト細胞にゲート設定した、CD19の染色特性対IgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色特性を示す。
【図14】GM−CSFおよびIL−4処理した、TT免疫ヒューマウスにおける、ヒトT細胞の細胞表面表現型。細胞を脾臓から調製し、ヒトCD45、マウスCD45、CD19に加えてIgM、IgD、CD10、CD268、CD5、CD21、CD27、IgG、またはCD20の染色をした。CD45+ヒト細胞にゲート設定した、CD3の染色特性対T細胞活性化マーカー:HLA−DRおよびCD40Lの染色特性を示す。
【図15】TT免疫ヒューマウスにおける、ヒトIgG、IgMおよびTT特異的ヒトIgGの血清レベル。TT免疫ヒューマウスから血清を採取し、ヒトIgG、IgMおよびTT特異的ヒトIgGをELISAで測定した。(図15A)血清中のヒト総IgG。GM−CSF+IL−4処理マウスは、ベクター処理マウスより有意に高いヒト総IgGレベルを生じさせた。(図15B)血清中のヒト総IgM。GM−CSF+IL−4処理マウスはまた、より高い総ヒトIgM血清レベルを有する。(図15C)血清中のヒトTT特異的IgG。
【図16】サイトカイン処理マウスにおけるTT特異的ヒトT細胞反応。第3回免疫の2週間後に、脾臓を採取し、ヒトT細胞のパーセンテージをフローサイトメトリーで測定した。ELISPOT検定の場合、同数(5×105)の、異なる試料からのヒトT細胞を、抗ヒトIFN−γまたは抗ヒトIL−4抗体で被覆したウェルに播種し、3種類の条件下で24時間培養した:培地のみ(ctrl)、PMAの存在下またはTT−特異的ペプチドの存在下。ELISPOTを現像した。(図16A)免疫マウス由来の脾細胞を含む代表的なヒトIFN−γELISPOTウェル。(図16B)免疫マウス由来の脾細胞を含む代表的なヒトIL−4ELISPOTウェル。2つの独立した実験の1つからのデータを示す。
【図17】ヒトIL−15、FL、GM−CSF、IL−4およびM−CSFをコード化するDNAプラスミドの混合物(各50μg)を、PBSに溶解し、12週齢のヒト化マウスに注入した(n=2)。7日後、血清を採取し、ELISAで、これらのヒトサイトカイン類の分析をした。
【発明を実施するための形態】
【0017】
近年、ヒト胎仔胸腺、肝臓、および自己ヒトCD34+細胞を移植されたNOD−scidマウス(BLTマウス)で、ヒト樹枝状細胞(DC)および単球/マクロファージの意義深い再構成が報告された(Wege AK,et al.(2008)Curr Top Microbiol Immunol 324:149−165)。しかしまだ、BLTマウスにヒトNK細胞は存在しなかった。NK細胞および骨髄性細胞は、自然免疫反応において重要な役割を果たすため、これらの細胞型の十分な再構成レベルを有するヒト非ヒト哺乳動物、たとえばヒト化マウス等の発生は、感染病調査研究および血液系細胞を含む他の調査研究(たとえば、貧血症、免疫不全症、癌等の、血液病調査)におけるヒト化マウスモデルの可能性を十分に実現するために重要である。
【0018】
全ての血液細胞系統は、共通のヒト造血幹細胞(HSC)に由来する。サイトカイン類は、それらの分化および維持の間、重要な役割を果たす。たとえば、IL−15は、NK細胞の発生および生存に必要であり(Mrozek E,et al.(1996)Blood 87:2632−2640)、GM−CSFおよびIL−4は樹枝状細胞(DC)発生に(Rosenzwajg M,et al.(1996)Blood 87:535−544)、マクロファージコロニー刺激因子(M−CSF)は単球/マクロファージの発生および維持に(Stec M,et al.(2007)J Leukoc Biol 82:594−602)、またエリスロポエチン(EPO)およびIL−3は赤血球発生に必要である(Giarratana MC,et al.(2005)Nat Biotechnol 23:69−74)。しかし、ヒトとマウスとの間の進化的分岐のため、これらのサイトカイン類は種特異的である(すなわち、マウスサイトカイン類はヒト細胞に対して作用しない)。たとえば、マウスIL−15は、ヒトNK細胞および前駆体に対して何の作用も持たず(Eisenman J,et al.(2002)Cytokine 20:121-129)、ヒューマウスにおけるヒトNK細胞の再構成不良という結果に終わる(Huntington ND,et al.(2009)J Exp Med 206:25−34;Kalberer CP,et al.(2003)Blood 102:127−135)。同様に、マウスGM−CSF、IL−4(Metcalf D(1986)Blood 67:257−267;Mosmann TR,et al.(1987)J Immunol 138:1813−1816)、M−CSF(Fixe P,Praloran V(1997)Eur Cytokine Netw 8:125−136)、およびIL−3(Stevenson LM,Jones DG(1994)J CompPathol111:99−106)は全て、ヒト細胞に対して何の作用もないと報告されている。
【0019】
ヒューマウスにおけるNK細胞および骨髄性細胞の不十分な再構成および機能は、特定のヒトサイトカイン類の欠如に起因するのかどうかを調査研究した。再構成されたマウスにおけるヒトサイトカイン類の発現が、特定のヒト血液系細胞の分化、生存、および機能を刺激するかどうかを究明するための実験を、本明細書に記述する。
【0020】
この調査研究が、ヒト化マウスを使用して実証されるような、ヒト化非ヒト哺乳動物における特定のヒト血液系細胞の再構成を改善するための簡単で効率のよい方法の開発につながった。ヒトIL−15およびFlt−3/Flk−2リガンド(FL)をコード化する核酸を送達した際に、特定のヒトサイトカイン類が、ヒューマウスの循環中に2〜3週間検出された。結果として、有意に増加した数のヒトNK細胞が、様々な器官で1カ月以上確認された。サイトカイン誘導性NK細胞は、インビトロおよびインビボの両方で、完全に機能した。同じ戦略を使用して、ヒューマウスにおけるヒト樹枝状細胞、単球/マクロファージ、および赤血球の再構成レベルも大幅に強化された。従前のヒト化マウスモデルにおけるNK細胞および骨髄性細胞の再構成不良は、それらの分化および維持に必要な適切なヒトサイトカイン類の欠如の結果であること、およびヒトサイトカイン遺伝子の送達(たとえば、流体力学的送達)は、これらの細胞系統の再構成不良を克服するための簡単で効率のよい方法であることを、本明細書に記載の研究は示す。
【0021】
したがって、一態様で、本発明は、機能的ヒト血液系細胞(たとえば、単一のヒト血液系細胞(たとえば、NK細胞);多様なヒト血液系細胞(たとえば、NK細胞、樹枝状細胞、T細胞、B細胞等々)を、また幾つかの実施態様で、全てのヒト血液系細胞)を、非ヒト哺乳動物で再構成する方法を対象とする。この実施態様で、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入される。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCで非ヒト哺乳動物が再構成される条件下で維持され、それによって、非ヒト哺乳動物でヒト造血幹細胞(HSC)を再構成する。
【0022】
本明細書で使用されるとき、HSC(たとえば、ヒトHSC)は、レシピエントに移植されるとき、「移植レシピエント(たとえば、非ヒト哺乳動物;免疫不全非ヒト哺乳動物)の造血システムを「再生息させる」または「再構成する」ことができ、またレシピエントで(たとえば、長期)造血を持続できる、自己再生幹細胞である。HSCは、骨髄性(たとえば、単球およびマクロファージ、好中球、好塩基球、好酸球、赤血球、巨核球/血小板、樹枝状細胞)およびリンパ球系(たとえば、T−細胞、B−細胞、NK−細胞)を含む、血液細胞型を生じさせる(に分化する)多能性幹細胞である。本明細書に記載の方法に示す通り、再構成されたヒトHSCは、非ヒト哺乳動物で、ヒトNK細胞、ヒト単球、ヒトマクロファージ、ヒト樹枝状細胞、ヒト赤血球、ヒトB細胞、ヒトT細胞またはそれらの組合せに分化することができる。
【0023】
HSCは細胞マーカーCD34を発現するので、通例「CD34+」と呼ばれる。当業者には明らかなように、HSCはまた他の細胞マーカー、たとえばCD133および/またはCD90等も発現できる(「CD133+」、「CD90+」)。場合によっては、HSCは、発現されないマーカー、たとえば、CD38を特徴とする。したがって、本発明の一実施態様で、本明細書に記載の方法で使用されるヒトHSCは、CD34+、CD90+、CD133+、CD34+CD38−、CD34+CD90+、CD34+CD133+CD38−、CD133+CD38−、CD133+CD90+CD38−、CD34+CD133+CD90+CD38−、またはそれらの任意の組合せである。ある特定の実施態様で、HSCは、本明細書で「二重陽性」細胞または「DP」細胞あるいは「DPC」とも呼ばれる、CD34(「CD34+」)とCD133+(「CD133+」)の両方である。別の実施態様で、HSCはCD34+CD133+であり、そしてCD38−および/またはCD90+をさらに含んでもよい。
【0024】
HSCは、ドナー(たとえば、人類、霊長類、ブタ、マウス、等々を含む、哺乳動物等の脊椎動物)の大腿骨、股関節、肋骨、胸骨、および他の骨等の骨髄に見られる。臨床使用および科学的使用のための、他のHSC源としては、臍帯血、胎盤、胎仔肝臓、動員末梢血、非動員(non−mobilized)(または不動員(unmobilized))末梢血、胎仔肝臓、胎仔脾臓、胚性幹細胞、および大動脈−性腺−中腎(AGM)、またはそれらの組合せなどがある。
【0025】
当業者に理解されるであろうが、動員末梢血は、HSC(たとえば、CD34+細胞)を強化した末梢血を指す。化学療法剤および/またはG−CSF等の化学物質の投与は、幹細胞を骨髄から末梢循環に動員する。たとえば、少なくとも5日間の、または約5日間の、顆粒球コロニー刺激因子(G−CSF)の投与は、CD34+細胞を末梢血に動員する。循環するCD34+細胞の30倍富化が確認され、そのピーク値は、G−CSF投与開始後第5日に現れる。末梢血の動員がなければ、循環するCD34+細胞の数は非常に少なく、総単核血球の0.01〜0.05%と推定される。
【0026】
本方法で使用するためのヒトHSCは、単一ドナーから得ても多数のドナーから得てもよい。加えて、本明細書に記載の方法で使用されるHSCは、用時単離されたHSC、凍結保存HSCS、またはそれらの組合せであってもよい。
【0027】
当該技術分野で周知の通り、HSCは、当該技術分野で周知の種々の方法を使用して、これらの供給源から得ることができる。たとえば、HSCは、針と注射器を使用して、たとえば、股関節部、大腿骨、等々の骨髄から除去することにより直接得ることもでき、あるいは骨髄コンパートメントから細胞を放出させる、顆粒球コロニー刺激因子(G−CSF)等のサイトカイン類でドナーを前処置した後、血液から得ることもできる。
【0028】
本発明の方法で使用するためのHSCは、得られたまま(たとえば、未増殖で)またはHSCを非ヒト哺乳動物に導入する前に操作して(たとえば、増殖して)、非ヒト哺乳動物に直接導入することができる。一実施態様で、HSCを非ヒト哺乳動物に導入する前に、HSCを増殖する。当業者に理解されるであろうが、HSCを増殖させるために使用できる方法は種々ある(たとえば、Zhang,Y.,et al.,Tissue Engineering,12(8):2161−2170(2006);Zhang CC,et al.,Blood,111(7):3415−3423(2008)参照)。ある特定の実施態様で、成長因子類(たとえば、アンギオポエチン様5(Angplt5)成長因子、IGF−結合蛋白質2(IGFBP2)、幹細胞因子(SCF)、線維芽細胞成長因子(FGF)、トロンボポエチン(TPO)、またはそれらの組合せ)の存在下で、HSCを間葉幹細胞(MSC)と共培養して細胞培養をすることにより、HSCの個体群を増殖することができる。細胞培養は、増殖したHSC個体群が産生される条件下で維持される(たとえば、Maroun,K.,et al.,ISSCR,7th Annual Meeting,Abstract No.1401(July8−11,2009)、_____________として公開された、2010年5月28日に提出された、PCT出願第PCT/US2010/036664号(代理人整理番号No.4471.1000−001)(参照により本明細書に援用する)を参照)。
【0029】
本明細書に記載の方法では、ヒトHSCの機能的ヒト細胞への分化を誘導するために1つまたは複数のヒトサイトカイン類をコード化する(1つまたは複数の)核酸(たとえば、DNA、RNA)も非ヒト哺乳動物に導入される。当該技術分野で周知の通り、サイトカイン類は、免疫細胞の分化、増殖または機能を刺激または抑制する蛋白質である。多数のヒトサイトカイン類の核酸配列も、当該技術分野で周知である(たとえば、www.ncbi.nlm.nih.gov参照)。1つまたは複数のサイトカイン類をコード化する核酸を入手するための方法は、当該技術分野でルーチンであり、種々の供給源(たとえば、血清)から核酸を単離すること(たとえば、クローニング)、組換えで核酸を産生することまたは商業的供給源から核酸を入手することを含む。
【0030】
本発明の方法で使用することができる種々のヒトサイトカイン類がある。このようなヒトサイトカイン類の例としては、インターロイキン−12(IL−12)、インターロイキン−15(IL−15)、Fms関連チロシンキナーゼ3リガンド(Flt3L)、Flt3L/Flk2リガンド(FL)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン−4(IL−4)、インターロイキン−3(IL−3)、マクロファージコロニー刺激因子(M−CSF)、エリスロポエチン(EPO)およびそれらの組合せなどがある。本明細書に記載の方法で使用するのに好適な他のサイトカイン類の例を一覧表にした。非ヒト哺乳動物に導入されるサイトカインのタイプおよびサイトカイン類の数は、ヒトHSCの分化が非ヒト哺乳動物で起こるとき、どのヒト血液細胞系統が再構成されるべきかによって異なる。たとえば、実施例1に示す通り、ヒトIL−15およびFlt−3/Flk−2リガンドをコード化する核酸が(たとえば、流体力学的尾静脈注入により)ヒト化マウスに導入されたとき、ヒトサイトカイン類の発現は2〜3週間持続し、上昇したヒトNK細胞レベルは1カ月以上にわたって誘導された。サイトカイン誘導性NK細胞は、活性化受容体および抑制性受容体の両者を発現し、インビトロで標的細胞を死滅させ、インビボでウイルス感染に強く応答した。同様に、ヒトGM−CSFおよびIL−4の発現は、ヒト樹枝状細胞の有意に強化された再構成を招き;マクロファージコロニー刺激因子の発現は、ヒト単球/マクロファージの有意に強化された再構成を招き;またエリスロポエチンおよびIL−3の発現は、ヒト赤血球の有意に強化された再構成を招いた(参照により本明細書に援用される、Chen,Q.,et al.,Proc.Natl.Acad.Sci.,USA,106:21783−21788(2009)参照)。実施例2に示す通り、GM−CSFおよびIL−4の発現は、機能的ヒトT細胞およびヒトB細胞の再構を強化した。
【0031】
幾つかの態様で、少なくとも1種のサイトカイン、少なくとも2種のサイトカイン類、少なくとも3種のサイトカイン類、少なくとも4種のサイトカイン類、少なくとも5種のサイトカイン類、少なくとも6種のサイトカイン類、少なくとも7種のサイトカイン類、少なくとも8種のサイトカイン、少なくとも9種のサイトカイン類、少なくとも10種のサイトカイン類、少なくとも11種のサイトカイン類、少なくとも12種のサイトカイン類、少なくとも13種のサイトカイン類、少なくとも14種のサイトカイン類、少なくとも15種のサイトカイン類、少なくとも16種のサイトカイン類、少なくとも17種のサイトカイン類、少なくとも18種のサイトカイン類、少なくとも19種のサイトカイン類、または少なくとも20種のサイトカイン類(を含む)が、非ヒト哺乳動物に導入される。他の態様で、1種のサイトカイン、2種のサイトカイン類、3種のサイトカイン類、4種のサイトカイン類、5種のサイトカイン類、6種のサイトカイン類、7種のサイトカイン類、8種のサイトカイン、9種のサイトカイン類、10種のサイトカイン類、11種のサイトカイン類、12種のサイトカイン類、13種のサイトカイン類、14種のサイトカイン類、15種のサイトカイン類、16種のサイトカイン類、17種のサイトカイン類、18種のサイトカイン類、19種のサイトカイン類、または20種のサイトカイン類のみ(から成る、から本質的に成る)が、非ヒト哺乳動物に導入される。各ヒトサイトカインをコード化する核酸を同時にまたは順次導入してもよい(たとえば、複数のサイトカインを非ヒト哺乳動物で発現させるべき場合、各サイトカインをコード化する各核酸を、独自の1つのプラスミドまたはベクターで導入してもよく、または多数のプラスミド類またはベクター類で導入してもよい;あるいは、導入されるべきサイトカイン類をコード化する全ての核酸を、1種のプラスミドまたはベクターで導入してもよい)。
【0032】
本発明の方法では、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入される。本明細書で使用されるとき、用語「哺乳動物」および「哺乳類」は、それらの子に授乳する、生存する子を出産する(真獣類または有胎盤哺乳動物)かまたは産卵する(後獣類または無胎盤哺乳動物)かいずれかの、単孔類、有袋類および有胎盤動物を含む、任意の脊椎動物を指す。本明細書に記載の方法で使用することができる哺乳類種の例としては、非ヒト霊長類(たとえば、サル、チンパンジー)、齧歯類(たとえば、ラット、マウス、モルモット)、イヌ、ネコ、および反芻動物(たとえば、ウシ、ブタ、ウマ)などがある。一実施態様において、非ヒト哺乳動物はマウスである。本明細書に記載の方法で使用される非ヒト哺乳動物は、成体、新生仔(たとえば、<48時間齢;子)または子宮内であってもよい。
【0033】
特定の実施態様で、非ヒト哺乳動物は免疫不全非ヒト哺乳動物である、すなわち、その免疫系に1つまたは複数の欠損があり(たとえば、NSGまたはNOD scid γ(NOD.Cg−Prkdcscid Il2rgtm1Wjl/SzJ)マウス)、結果として、ヒトHSCが導入されるとき、ヒト血液細胞系統の再構成を許容する、非ヒト哺乳動物である。たとえば、その非ヒト哺乳動物は、それ自身のT細胞、B細胞、NK細胞またはそれらの組合せを欠く。特定の実施態様で、非ヒト哺乳動物は、たとえば重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、またサイトカイン−受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scid IL2Rγ−/−マウス);およびBalb/crag−/−γc−/−マウス等の、免疫不全マウスである。
【0034】
免疫不全マウスの他の具体例としては、重症複合免疫不全(scid)マウス、非肥満糖尿病(NOD)−scidマウス、IL2rg−/−マウス(たとえば、NOD/LySz−scidIL2rg−/−マウス、NOD/Shi−scidIL2rg−/−マウス(NOGマウス)、BALB/c−Rag−/−IL2rg−/−マウス、H2d−Rag−/−IL2rg−/−マウス)、NOD/Rag−/−IL2rg−/−マウスなどがあるが、その限りではない。
【0035】
幾つかの実施態様で、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸の導入前に(たとえば、ヒトHSCの再構成をさらに強化するために)、非ヒト哺乳動物を処置または操作する。たとえば、非ヒト哺乳動物を操作して、ヒトHSCの生着および/または再構成をさらに強化することができる。一実施態様で、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸の導入前に、非ヒト哺乳動物を照射する。別の実施態様で、HSCおよび1つまたは複数のサイトカイン類をコード化する核酸の導入前に、1つまたは複数の化学療法剤が非ヒト哺乳動物に投与される。
【0036】
やはり当業者に理解されるであろうが、HSCおよびサイトカイン類をコード化する核酸を非ヒト哺乳動物に導入する種々の方法がある。そのような方法の例としては、皮内、筋肉内、免疫、眼内、大腿内、脳室内、頭蓋内、髄腔内、静脈内、心臓内、肝臓内、骨髄内、皮下、局所、経口および鼻腔内投与経路などがあるが、その限りではない。他の好適な導入方法としては、子宮内注入、流体力学的遺伝子送達、遺伝子療法、充電式または生分解性装置、粒子加速器(「遺伝子銃」)および徐放性ポリマー装置も挙げることができる。
【0037】
任意のそのような投与経路等々を使用して、HSCを非ヒトに導入することができる。ある特定の実施態様で、HSCを非ヒト哺乳動物に心臓内注入する。
【0038】
1つまたは複数のサイトカイン類をコード化する核酸はまた、その核酸(類)が非ヒト哺乳動物で発現される限り、任意のそのような投与経路を使用して導入することもできる。たとえば、1つまたは複数のサイトカイン類をコード化する核酸を、プラスミド(たとえば、pcDNA3.1(+))またはウイルスベクター(たとえば、アデノウイルス、アデノ随伴ウイルス、レンチウイルス、レトロウイルス等々)で、裸の核酸(裸のDNA)として導入することができる。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、(たとえば、非ヒト哺乳動物の尾静脈への)流体力学的注入を使用して、プラスミドで導入される。
【0039】
HSCおよび1つまたは複数のサイトカイン類をコード化する核酸は、同時にまたは順次導入することができ、また当業者に理解されるであろうが、使用しようとしている非ヒト哺乳動物のタイプ、発現させようとしているサイトカイン類およびヒトHSCの分化が非ヒト哺乳動物で起こるとき、どのヒト血液系細胞が発現されるかおよび/または強化されるか等の因子によって異なる。ある特定の実施態様で、HSCは新生仔(たとえば、約48時間齢)に導入され、またサイトカイン類をコード化する核酸は約1か月、約2カ月、約3カ月、約4カ月、約5カ月、約6カ月、約7カ月、約8カ月、約9カ月、約10カ月、約11カ月、約12カ月後に導入される。
【0040】
いったんHSCおよび1つまたは複数のサイトカイン類をコード化する核酸が導入されると、非ヒト哺乳動物は、核酸が発現されて、HSCで非ヒトが再構成される条件下で維持される。本発明の非ヒト動物が維持されるそのような条件は、当業者に周知の通り、哺乳動物の基本的欲求を満たすこと(たとえば、食糧、水、光)である。
【0041】
本明細書に記載の方法は、核酸が発現されるかどうか、ヒトHSCが存在するかどうかおよび/またはヒトHSCが、1つまたは複数のヒト血液系細胞に分化しているかどうかを究明することをさらに含む。核酸が発現されるかどうかおよび/またはHSCで非ヒトが再構成されるかどうかを究明する方法は、本明細書に提供され、また当業者に周知である。たとえば、ヒトHSCの表面細胞マーカーに特異的な抗体を使用するフローサイトメトリー分析法を使用して、ヒトHSCが非ヒト哺乳動物に存在することを検出できる。加えて、血清を非ヒト哺乳動物から採取して、ヒトサイトカイン類の存在を検定することができる。分化したHSC(たとえば、NK細胞、樹枝状細胞、T細胞、B細胞、単球/マクロファージ、赤血球)の機能を評価するための検定法も使用することができる。そのような検定法は本明細書にも記述されており、また当業者に周知である。たとえば、本明細書に記載の通り、サイトカイン誘導性ヒトNK細胞は、インビトロ検定法(乳酸デヒドロゲナーゼ検定法)で標的細胞を死滅させ、またインビボでウイルス感染に強く応答した。
【0042】
1つまたは複数のヒトサイトカイン遺伝子をコード化する核酸の送達によって、1つまたは複数のヒト血液細胞系統を非ヒト哺乳動物(たとえば、ヒト化マウス)で再構成できることは、種々の方法で使用できる。
【0043】
たとえば、一態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトNK細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトNK細胞系統に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトNK細を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトIL−15およびヒトFlt−3/Flk−2リガンドをコード化する。
【0044】
別の実施態様で、非ヒト哺乳動物の末梢血の白血球の約3%〜約25%が、ヒトNK細胞(たとえば、機能的NK細胞)である。他の実施態様で、非ヒト哺乳動物の末梢血の白血球の約3%、約4%、約5%、約6%、約7%、約8%、約9%、約10%、約11%、約12%、約13%、約14%、約15%、約16%、約17%、約18%、約19%、約20%、約21%、約22%、約23%、約24%、または約25%が、ヒトNK細胞である。
【0045】
さらに他の実施態様で、ヒトNK細胞(たとえば、機能的NK細胞)の発現は約1〜約30日間、維持され(また、場合によっては、たとえば好適なコントロールと比較して強化され)、そして特定の実施態様で、ヒトNK細胞の発現は約1日間、約2日間、約3日間、約4日間、約5日間、約6日間、約7日間、約8日間、約9日間、約10日間、約11日間、約12日間、約13日間、約14日間、約15日間、約16日間、約17日間、約18日間、約19日間、約20日間、約21日間、約22日間、約23日間、約24日間、約25日間、約26日間、約27日間、約28日間、約29日間、約30日間、または約31日間維持される。
【0046】
幾つかの実施態様で、非ヒト哺乳動物におけるヒトNK細胞は、ヒトで見られる通常の(野生型)NK細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。そのような発現は、ヒトNK細胞が非ヒト哺乳動物で確かに機能することを示す。たとえば一実施態様で、ヒトNK細胞はCD56+NK細胞細胞である。他の実施態様で、ヒトNK細胞は、NKG2D、NKG2A、CD94、KIR、NKp46、CD7、CD69、Cd16、またはそれらの組合せを発現する。
【0047】
さらに他の実施態様で、非ヒト哺乳動物におけるヒトNK細胞は、適切な刺激(たとえば、Toll様受容体3アゴニスト・ポリ(I:C);ヒト樹枝状細胞;アデノウイルス)を受けて、標的細胞を死滅させたりIFN−γを発現したりすることができる。
【0048】
別の態様で、本発明は、免疫不全非ヒト哺乳動物に、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を導入することを含む、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法を対象とし、ここでヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト樹枝状細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトGM−CSFおよびヒトIL−4をコード化する。別の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトGM−CSF、ヒトIL−4およびヒトFlt−3/Flk−2リガンドをコード化する。
【0049】
他の実施態様で、非ヒト哺乳動物におけるヒト樹枝状細胞は、ヒトで見られる通常の(野生型)樹枝状細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、非ヒト哺乳動物におけるヒト樹枝状細胞は、CD11c+CD209骨髄性樹枝状細胞(たとえば、血液、脾臓、骨髄、肺、肝臓で発現される)、ILT7+CD303+形質細胞様樹状細胞またはそれらの組合せである。
【0050】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現され、ヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトマクロファージコロニー刺激因子をコード化する。
【0051】
他の実施態様で、非ヒト哺乳動物におけるヒト単球/マクロファージは、ヒトで見られる通常の(野生型)単球/マクロファージの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、ヒト単球/マクロファージはCD14+を発現する。
【0052】
別の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカイン類をコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法を対象とし、ここで、ヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒト赤血球への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒト赤血球に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒト赤血球を再構成する。ある特定の実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、ヒトエリスロポエチンおよびIL−3をコード化する。
【0053】
別の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、ヒトで見られる通常の(野生型)赤血球の細胞表面マーカーの、1つまたは複数、および場合によっては全てを発現する。一実施態様で、ヒト赤血球はCD235ab+を発現する。
【0054】
さらに他の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、非ヒト哺乳動物における全赤血球の約1%〜約10%、または約3%〜約5%を構成する。特定の実施態様で、非ヒト哺乳動物におけるヒト赤血球は、非ヒト哺乳動物における全赤血球の約1%、約2%、約3%、約4%、約5%、約6%、約7%、約8%、約9%または約10%を構成する。
【0055】
ある特定の態様で、本発明は、ヒトHSCおよび1つまたは複数のヒトサイトカインをコード化する核酸を免疫不全非ヒト哺乳動物に導入することを含む、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法を対象とし、ここでヒトサイトカイン類は、非ヒト哺乳動物で発現されるとき、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持され、それによって、非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する。一実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、GM−CSFおよびIL−4をコード化する。本方法は、非ヒト哺乳動物を免疫原で免疫すること、および非ヒト哺乳動物が免疫原に対するヒト抗体を産生する条件下で非ヒト動物を維持することを、さらに含んでもよい。
【0056】
また別の特定の態様で、本発明は、非ヒト哺乳動物で、免疫原に対するヒト抗体を生成する方法を対象とする。本方法で、ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸が非ヒト哺乳動物に導入されるが、ここでヒトサイトカイン類は、ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する。非ヒト哺乳動物は、非ヒト哺乳動物で核酸が発現されてヒトHSCが機能的ヒトT細胞および機能的ヒトB細胞に分化する条件下で維持される。非ヒト哺乳動物は免疫原で免疫され、またヒトB細胞が非ヒト哺乳動物で免疫原に対するヒト抗体を産生する条件下で維持され、それによって、非ヒト哺乳動物で免疫原に対するヒト抗体を生成する。一実施態様で、1つまたは複数のサイトカイン類をコード化する核酸は、GM−CSFおよびIL−4をコード化する。
【0057】
当該技術分野で周知の通り、「免疫原」は、免疫応答を誘導し、抗体産生を増進することができる物質である。本法で使用するための種々の免疫原は、当該技術分野で周知である。たとえば、免疫原は、下記の全部または免疫原性部分であってもよい:ヒトまたは他種由来の蛋白質、(たとえば、正常細胞または腫瘍細胞等の疾病細胞の)細胞表面蛋白質、生物(たとえば、生物の被膜、莢膜、細胞壁、鞭毛、線毛および毒素を含む、生物の免疫原性部分)、ウイルス蛋白質、細菌蛋白質、毒素、多糖、リポ蛋白質、(たとえば、アセチル化、メチル化、グリコシル化された)修飾蛋白質、核酸(たとえば、DNA、ペプチド、蛋白質または多糖と結合されるときのRNA)、化学的エピトープ、等々。
【0058】
これらの方法は、ヒト抗体を産生するヒトB細胞を、非ヒト哺乳動物から単離することをさらに含むことができる。B細胞を非ヒト哺乳動物から単離する方法は、当該技術分野で周知である。たとえば、フローサイトメトリーまたはB細胞特異的蛋白質に特異的な抗体に基づく磁気精製を使用する細胞選別(たとえば、John Wiley and Sons,Inc.ed.John E.Coligan et al.によるCurrent Protocols in Immunology,Copyright(著作権)2010参照)。
【0059】
当該技術分野で周知の通り、「抗体」または「免疫グロブリン」は、血中を循環するB細胞により産生される免疫系の蛋白質成分であり、細菌やウイルスのような免疫原を認識して、それらを無力化する。免疫原への暴露後、抗体は血中を循環し続け、将来その抗原に暴露されることを防ぐ。本明細書に記載の方法を使用して、ヒトB細胞により産生されるあらゆるタイプの抗体を得ることができる。モノクローナル抗体はポリクローナルであってもよく、モノクローナル抗体であってもよい。そのような抗体の例は当該技術分野で周知であり、IgG(たとえば、IgG1、IgG2、IgG3、IgG4)、IgM、IgDおよびIgAなどがある。
【0060】
本方法は、単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;およびヒトB細胞と不死化細胞が融合して、免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、その組合せを維持することを、さらに含むことができる。
【0061】
当該技術分野で周知の通り、免疫後適切なときに、たとえば、抗体価が最高のときに、抗体産生細胞を対象から得て、標準的な技術、たとえば最初にKohler and Milstein,Nature 256:495−497(1975)によって記述されたハイブリドーマ技術、ヒトB細胞ハイブリドーマ技術(Kozbor et al.,Immunol.Today 4:72(1983)),EBV−ハイブリドーマ技術(Cole et al.,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,Inc.,pp.77−96(1985))またはトリオーマ技術等により、モノクローナル抗体の調製に使用することができる。ハイブリドーマを作るためのテクノロジーは周知である(一般にCurrent Protocols in Immunology,Coliganet al.,(eds.)John Wiley & Sons,Inc.,New York,NY(1994)参照)。簡単に記述すると、上述の通り、不死化細胞株(一般に骨髄腫)を、免疫原で免疫された哺乳動物由来のリンパ球(一般に脾細胞)に融合させ、本発明のポリペプチドを結合するモノクローナル抗体を産生するハイブリドーマを同定するために、結果として得られたハイブリドーマ細胞の培養上澄をスクリーニングする。
【0062】
リンパ球と不死化細胞株を融合させるために使用される多くの周知のプロトコールのいずれも、免疫原に対するモノクローナル抗体を生成するために利用することができる(たとえば、Current Protocols in Immunology,supra;Galfre et al.,Nature,266:55052(1977);R.H.Kenneth,in Monoclonal Antibodies:A New Dimension In Biological Analyses,Plenum Publishing Corp.,New York,New York(1980);およびLerner,Yale J.Biol.Med.54:387−402(1981)参照)。
【0063】
さらに、一般の当業者は、そのような方法の多くの変形ならびにヒトB細胞により産生される抗体を得るために使用することができる他の方法があることを理解するであろう。たとえば、ヒトB細胞により発現されるヒト抗体の全部または機能的部分をコード化する配列を、既知の技術を使用してクローニングすることができる。一般に、本手法は、抗原特異的B細胞の単離(たとえば、蛍光色素標識抗原で染色される、フローサイトメトリーによる選別)および変性プライマーおよび単一細胞ポリメラーゼ連鎖反応(PCR)を使用する抗体遺伝子のVDJ部分の増幅を含む。クローニングされ、配列決定された抗体遺伝子のVDJ部分を定常領域遺伝子セグメントと結合し、CHO細胞株等の細胞株で抗体を作る(たとえば、Hahn,S.,et al.,Cell Mol.Life Sci.(2000),57(1):96−105参照)。
【0064】
非ヒト哺乳動物により産生されるヒト抗体を産生および/または単離する方法のもう1つの例は、ヒトB細胞をウイルスにより不死化することを含む。本方法では、たとえば、エプスタイン−バールウイルス(Epstein−Barr Virus)(EBV)または修飾EBVを使用してB細胞を不死化することができる(たとえば、Lanzavecchia,A.,Curr.opin.Biotechnol.(2007) 18(6):523−528参照)。
【0065】
当業者に理解されるであろうが、サイトカイン類に加えて、成長因子等の蛋白質(たとえば、ヒト蛋白質;ヒト分泌蛋白質)、ステロイド類、および/または小分子の発現を本方法で使用して、血液系細胞以上に、ヒト細胞の再構成および/または機能を改善することができる。たとえば、ヒトサイトカイン類の1つまたは複数のアゴニストを非ヒト哺乳動物に導入して、HSCの再構成を強化することができる。
【0066】
当業者に理解されるであろうが、「機能的(または「生物学的に活性な」または「成熟した」)ヒトNK細胞」、「機能的ヒト樹枝状細胞」、「機能的ヒト単球/マクロファージ」、「機能的ヒト赤血球、「機能的ヒトT細胞」および「機能的ヒトB細胞」は全て、分化した細胞(ヒトNK細胞、ヒト樹枝状細胞、ヒト単球/マクロファージ、ヒト赤血球、ヒトT細胞、またはヒトB細胞であっても)が、対応する、ヒトで見られる通常の(野生型)細胞の細胞表面マーカーの、1つまたは複数、および場合によっては全部を発現し、そして結果として、それらがヒトで機能するのと同様に非ヒト哺乳動物で機能するということを指す。
【0067】
非ヒト哺乳動物におけるヒト血液系細胞の機能を測定するための検定法は、当業者に周知であり、また本明細書に記述されている。たとえば、NK細胞傷害性検定法を使用して、非ヒト哺乳動物におけるNK細胞の機能を測定することができる。
【0068】
本発明のある態様で、非ヒト哺乳動物におけるヒト血液細胞系統および/またはある特定のヒト細胞系統(たとえば、NK細胞、樹枝状細胞、単球/マクロファージ、赤血球、T細胞、B細胞)の再構成が強化される。再構成強化は、たとえば、適当なコントロールと比較して強化された細胞型の発現(たとえば、1つまたは複数のヒト血液系細胞数の増加;細胞型が発現される期間の増加(たとえば、>30日))を指す。そのようなコントロールは当業者に明白である。適当なコントロールの例は、1つまたは複数のサイトカイン類をコード化する核酸ではなく、ヒトHSCが導入された非ヒト哺乳動物である。
【0069】
本発明の他の態様は、組成物を含む。一態様で、本発明は、本明細書に記載の方法により作られる非ヒト動物を含む。
【0070】
他の態様で、本発明は、本明細書に記載の方法により産生されるハイブリドーマ(単離ハイブリドーマ)およびそのハイブリドーマにより産生されるモノクローナル抗体(単離モノクローナル抗体)を含む。
【0071】
本明細書で使用されるとき、「単離された」(たとえば、「単離されたB細胞」;「単離されたハイブリドーマ」、「単離されたモノクローナル抗体」)は、それが自然に存在する複雑な(たとえば、細胞の)環境、または器官、身体、組織、血液または培地に関して実質的に単離されていることを指す。場合によっては、単離された材料は、組成物の一部(たとえば、他の物質を含む粗抽出物)、バッファー系、培養系または試薬混合物を形成する。状況が違えば、材料は、本質的に均質になるまで精製することができる。単離されたB細胞個体群は、(全細胞数を基準として)存在する全細胞の少なくとも約50%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約99%を構成する。一実施態様で、本発明は、本明細書に記載の方法で産生される、単離された、または実質的に単離された(または精製された、本質的に精製された)B細胞、ハイブリドーマおよび/またはモノクローナル抗体を対象とする。
【0072】
したがって、本明細書に示す通り、ヒト化マウスで確認されるヒト生来の応答および適応応答が不十分な主な理由は、適切なヒトサイトカイン発現が再構成マウスで欠如しているために、特定のヒト血液系細胞の再構成が低レベル、および/または特定のヒト血液細胞系統が機能的に成熟不良であることに起因する。再構成非ヒト哺乳動物(たとえば、マウス)で様々なヒトサイトカイン類を発現し、また結果として得られる非ヒト哺乳動物におけるヒト血液系細胞の再構成を著しく改善するための、効率よくかつ汎用的な方法を、本明細書に記述する。
【0073】
非ヒト哺乳動物でヒトサイトカイン類を発現するために、流体力学的注入(たとえば、7秒で体重の10%)を使用して、ヒトサイトカイン類をコード化するDNAベクターを哺乳動物に導入した。この実施態様で、注入されたDNAの一部が肝細胞によって取り込まれ、マウスでヒトサイトカインが発現する結果となった。ヒトサイトカイン類をコード化する核酸をこのようにして導入することにより、特定のヒト細胞サブセットの発生が向上し、ヒト細胞数および免疫応答の増加がもたらされた。
【0074】
具体的には、ヒトサイトカイン類を、ベクターにクローニングした。操作したヒトインターロイキン15(IL−15)遺伝子およびFms−関連チロシンキナーゼ3リガンド(Flt3L)遺伝子を、個々にpcDNA3.1(+)ベクターにクローニングした。
【0075】
pcDNA3.1(+)プラスミドをインビボ遺伝子送達用ベクターとして使用した。インビボでNK細胞を誘導するために、pcDNA−IL2/IL15およびpcDNA−Flt3Lプラスミドを構築した。簡単に記述すると、IL−15のシグナルペプチドは通常長く(48aa)、またIL−15の分泌を制限するため、ヒトIL−15のシグナルペプチド配列を、ヒトIL−2のそれで置き換えた。IL−15に応答するNK細胞前駆体の発生頻度を高めるために、速効性サイトカインflt3リガンド(FL)を使用した。これらの2つの組換え遺伝子配列をpcDNA3.1(+)ベクターに挿入した。
【0076】
流体力学に基づく遺伝子導入法で、尾静脈を介して大量の溶液を急速注入することにより、プラスミドをマウスに投与した。簡単に記述すると、8〜12週齢のヒト化マウスに、各プラスミド50μgを含む生理食塩水1.8mlを、7秒以内で静脈注入した。
【0077】
注入後第7日に、これらのマウスの尾静脈から採血して、主として血中のヒト免疫細胞の再構成を分析した。次いで、数匹のマウスを第9日および第16日に屠殺して、肝臓、肺、脾臓、骨髄、およびリンパ節におけるヒト細胞再構成の動力学を分析した。血清を異なる時点に採取し、循環血液中のヒトサイトカインレベルをELISAで分析した。
【0078】
流体力学的注入後、血清中に有意なレベルのIL−15およびFlt3Lが2〜3週間検出された。相応して、血中、脾臓、骨髄、肺および肝臓におけるCD56+CD3−ヒトナチュラルキラー(NK)細胞の再構成は、ヒト器官で確認されるものに匹敵するレベルまで有意に上昇した。CD45+ヒト細胞の絶対数も、全ての器官で増加した。
【0079】
この方法によってヒト化マウスで生成したNK細胞は、通常のNK細胞表現型を有し、またインビトロでもインビボでも、polyI:CおよびLPS刺激に応答したインターフェロン−γ(IFN−γ)産生に関して機能的であった。ヒト化マウスから精製されたNK細胞は、インビトロで、K562標的細胞に対して細胞傷害活性を示した。さらに、NK細胞は、インビボで、ウイルス感染に応答した。同様の手法を使用して、GM−CSFおよびIL−4を発現することにより樹枝状細胞再構成を強化し、MCSFを発現することによりマクロファージおよび単球再構成を強化し、EPOおよびIL3を発現することによりヒト赤血球再構成を強化した。
【0080】
裸のDNAの流体力学的注入は、血清中で2〜3週間、ヒトサイトカイン類が検出される結果となった。当業者に明白な通り、ヒト遺伝子はまた、ウイルスベクター(たとえば、アデノウイルス介在性遺伝子発現;レンチウイルス介在性遺伝子発現)によって発現させることもでき、これはマウスで高レベルかつ長期にわたるヒト遺伝子発現をもたらすことができる。
【0081】
一部の種類のヒト免疫細胞の再構成は、ヒト造血幹細胞移植NOD/SCID IL2Rγ−/−マウスで非常に低く、これはマウスサイトカイン環境に対する反応性が不十分なためである。免疫細胞発生および成熟に不可欠な多くのサイトカイン類は、NK細胞に対するIL−15、DCに対するGM−CSF/IL−4、およびマクロファージに対するM−CSF等のような、種特異的活性を示す。有意に高レベルの全身性外因性ヒトサイトカインの発現をもたらし、結果としてヒト化マウスでのヒト免疫細胞発生を促進することになる、裸のサイトカイン発現プラスミドの投与を使用する流体力学に基づくインビボトランスフェクション手順を、本明細書に記述する。
【0082】
本方法および組成物は、ヒトHSC由来の血液細胞系統を有する非ヒト哺乳動物を再構成するために使用される現行の方法に優る多数の利点を提供する。たとえば、サイトカインをコード化する核酸は、所望の結果を達成するために、一度導入しさえすればよい;サイトカイン類をコード化する核酸を使用すると、大規模調製がはるかにし易く、はるかに安定して長期保存し易く、またはるかに遺伝子操作し易い;3週間にわたる全身性ヒトサイトカイン発現で、流体力学的に基づく注入を実施することができ、またサイトカインを導入するためにアデノウイルスベクターおよび/またはレンチウイルスベクターを使用して、発現をより長く持続できると思われる;多数の遺伝子構築物を組み合わせることができる(図17参照);本方法は、ヒト化マウスでHSC(たとえば、骨髄性細胞)を再構成するための簡単で効率のよい方法を提供する。
【実施例】
【0083】
例示
実施例1
ヒトサイトカイン類の発現は、ヒト化マウス(ヒューマウス)における、特定のヒト血液系細胞の再構成を劇的に改善する。
【0084】
材料と方法
HSC単離、ヒト化マウスの構築、および流体力学的遺伝子送達。ヒト臍帯血は、Singapore Cord Blood Bankから入手した。臍帯血単核細胞(MNC)は、Ficoll−Hypaque密度勾配により分離した。CD34+細胞は、RosetteSep(登録商標)システムで、製造業者のプロトコール((Stem Cell Technologies)に従って精製した。CD34+細胞の純度は>95%であった。HSCを増殖させるために、精製CD34+細胞を、規定の因子の存在下、無血清培地で11〜14日間培養した(Zhang CC,Kaba M,Iizuka S,Huynh H,Lodish HF(2008)Blood 111:3415−3423)。非増殖HSCと増殖HSCの両方を使用して、ヒト化マウスを作った。
【0085】
NSGマウスは、Jackson Laboratoriesから購入し、Nanyang Technological UniversityおよびNational University of Singaporeの動物施設で、特定病原体不在条件下で維持した。マウスを再構成するために、γ線源を使用して100cGyを新生仔(48時間齢未満)に照射し、CD34+CD133+細胞(1×105細胞/レシピエント)を心内注入した。ヒトサイトカイン遺伝子は、pcDNA3.1(+)ベクター(Invitrogen)に別々にクローニングした。プラスミドDNAは、Maxi−prep Kit(Qiagen)で精製した。流体力学的遺伝子送達のために、12週齢のヒューマウスに、各プラスミド50μgを含む総量1.8mlの生理食塩水を、27ゲージの針を使用して7秒以内で注入した。試料およびマウスを用いた調査研究は全て、National University of SingaporeおよびNanyang Technological Universityの施設ガイドラインを順守して実施した。
【0086】
単一細胞調製、抗体、およびフローサイトメトリー。単一細胞懸濁液は、標準手順により、脾臓および骨髄(BM)から調製した。MNCをヒューマウス肝臓から単離するために、肝臓を、200ゲージのステンレススチールメッシュを押し通し、50xgで5分間の遠心分離により残屑を除去した。MNCを含む上澄を回収し、PBSで洗浄し、40%Percoll(Sigma)を含むRPMI培地1640中に再懸濁した。細胞懸濁液を、70%Percollの上に静かに重ね、750xgで20分間、遠心分離した。MNCを相間から回収し、PBSで2回洗浄した。MNCを肺から単離するために、肺を細かく刻み、0.05%コラゲナーゼ(Sigma)および0.01%DNase I(Sigma)を含む培地中に懸濁し、37℃で20分間インキュベートした。肺試料を、200ゲージのステンレススチールメッシュを押し通し、上述の通り、Percoll遠心分離でMNCを単離した。
【0087】
下記の抗体を使用した:Becton−DicksonからのCD3(SK7)、CD34(581)、CD19(HIB19)、NKG2D(1D11)、NKp46(9E2)、CD94(DX22)、CD16(3G8)、CD56(B159)、HLA−DR(L243)、CD14(M5E2)、CD11c(B−ly6)、CD209(DCN46)、CD7(M−T701)、CD45(2D1)、CD69(L78)、CD33(WM53);BioLegendからのKIR2DL2/L3(DX27)、ILT7(17G10.2)およびCD235ab(HIR2);Miltenyi BiotecからのCD303(AC144);Beckman CoulterからのCD159a(NKG2A;Z199);およびeBioscienceからのCD133(EMK08)およびマウスCD45.1(A20)。細胞を、氷上で、0.2%BSAおよび0.05%アジ化ナトリウムを含有するPBS 100μl中の適切な抗体で30分間染色した。フローサイトメトリーは、FACSDivaソフトウェアを使用したLSRIIフローサイトメーター(Becton,Dickinson and Co.)で実施した。試料当たり10,000〜1,000,000の事象を収集し、Flowjoソフトウェアを使用して解析した。
【0088】
インビトロでのヒトCD34+細胞の分化。BM MNCを12週齢のヒューマウスから単離した。CD34+細胞を、MACS(登録商標)マイクロビーズ(Miltenyi Biotec)で濃縮した。精製細胞を、RPMI 1640、10%FCS中、37℃および5%CO2で培養した。NK細胞、DC、単球/マクロファージ、および赤血球の分化用に、それぞれ、50ng/ml SCF、50ng/ml FLおよび50ng/ml IL−15;50ng/ml SCF、20ng/ml GM−CSFおよび50ng/ml IL−4;50ng/ml SCFおよび30ng/ml M−CSF;および100ng/ml SCF、5ng/ml IL−3および3U/ml EPOを使用した。サイトカイン類は全て、R&D Systemsから購入した。
【0089】
NK細胞細胞傷害性検定および刺激。遺伝子送達の9日後、幹細胞PE選択キット(Stem Cell Technologies)を使用して正の選択により、CD56+NK細胞を脾臓およびBMから精製した。細胞を洗浄し、2%FCSを含むIMDM中に再懸濁し、NK感受性標的K562(ATCC)に対する細胞傷害性を、4時間乳酸デヒドロゲナーゼ放出検定法(CytoTox96;Promega)で測定した。
【0090】
インビトロ刺激用に、精製NK細胞を、ヒトDCを含む、または含まない、RPMI 1640、10%FCS、L−グルタミン2mM、ピルビン酸ナトリウム1mM、ペニシリン、およびストレプトマイシン中、37℃および5%CO2で24時間、培養した。ヒトDCは、記述の通り、臍帯血CD34+細胞から分化させた(Rosenzwajg M,Canque B,Gluckman JC(1996)Blood 87:535−544)。次に、インビトロでNK細胞を刺激するために、50pg/ml poly(I:C)(Sigma)を培養に添加した。インビボ刺激用に、ヒューマウスにpoly(I:C)200μgを静脈内注入した。血清中または培養上澄中のIFN−γレベルを、ELISA Kits(R&D Systems)で測定した。
【0091】
アデノウイルス感染、ALT、および組織学。複製欠損、E1およびE3欠失、タイプ5緑色蛍光蛋白質発現アデノ−Xウイルス(AdGFP)は、Clontechから購入した。AdGFPを、HEK293細胞中で増殖させ、CsCl不連続密度勾配遠心分離により精製した。4×109pfu AdGFPウイルスを、流体力学的注入により尾静脈を介してヒューマウスに投与した。アデノウイルス感染の5日後、血清を採取し、コバス(cobas) c 111アナライザー(Roche Diagnostics Ltd.)を使用して、ALT活性について分析した。
【0092】
組織学的分析用に、肝臓を摘除し、パラフィン中に包埋して、厚さ5μmの切片を作製した。パラフィン切片をH&Eで染色し、光学顕微鏡で分析した。2色蛍光免疫染色用には、非特異的染色のブロッキング後、脱パラフィン処理切片を、PE−コンジュゲート抗ヒトCD56抗体(MEM−188;Biolegend)の最適希釈物で染色した。切片を、MIRAX MIDI蛍光顕微鏡(Zeiss)で分析した。
【0093】
統計解析。データを、平均値および平均値の標準誤差として示す。群間差をStudent t検定で解析した。<0.05のP値を、統計学的有意と考えた。計算は全て、Origin8.0ソフトウェアパッケージを使用して実施した。
【0094】
結果
インビトロでの、ヒトサイトカイン類によるNK細胞分化の刺激。ヒト化マウスを構築するために、ヒト臍帯血から単離したCD34+HSCを、亜致死照射したNSG仔に養子移入した。再構成の12週間後、末梢血由来の単核細胞(MNC)を、ヒトCD45およびマウスCD45に特異的な抗体で染色した(図6A〜6B)。平均再構成率は、血液で約50%であった[再構成率=%CD45+ヒト細胞/(%CD45+ヒト細胞+%CD45+マウス細胞)]。CD45+ヒト白血球の中で、CD19+B細胞のレベルは40〜85%の範囲であり、CD3+T細胞のレベルは10〜50%の範囲であった。NK細胞は、血中、BM、脾臓、肺、および肝臓で検出されたが、それらの発生頻度は、対応するヒト組織またはマウス組織における発生頻度より有意に低かった(図6A〜6B参照)。
【0095】
ヒューマウスにおけるNK細胞再構成不良の根底となる原因を究明するために、我々は、ヒューマウスのBMに由来するヒトCD34+細胞が、インビトロで、ヒトIL−15およびFLにより刺激されて、NK細胞に分化するかどうかを試験した。FLは、IL−15に応答できるCD34+NK前駆細胞を含む、多様な造血細胞系統の分化を刺激する(Yu H,et al.(1998)Blood 92:3647−3657)。FLとIL−15の組合せは、CD34+前駆体の、NK細胞方向への分化を促進すると予期される。したがって、ヒューマウスBM由来の精製ヒトCD34+細胞(>80%)(図1A)を、FLおよびIL−15の存在下で7日間培養し、NK細胞マーカーCD56およびNKp46の発現について分析した。サイトカイン類の存在下では、細胞の約11%がCD56およびNKp46の両者について陽性であったが、サイトカイン類の非存在下では非常に少ない細胞が陽性であった(図1B)。これらの結果から、ヒューマウスのBM中のCD34+ヒト細胞は、適切なサイトカイン環境が提供されれば、NK細胞に分化することができることが示唆される。
【0096】
プラスミドDNAの流体力学的注入による、マウスにおけるヒトサイトカイン類の発現。IL−15およびFLの存在下では、インビトロでヒトNK細胞が発生するという知見から、これらのヒトサイトカイン類は、ヒューマウスにおけるNK細胞発生も刺激し得ることが示唆される。ヒトサイトカイン類をマウスに導入する一つの方法は、組換え蛋白質を毎日注入することによる。この方法は厄介で高価なため、我々はサイトカインコード化DNAプラスミドの流体力学的送達により、マウスで、ヒトサイトカイン類を発現させた。ヒトIL−15は、異常に長いシグナルペプチド配列を有し(45aa残基)、これは、IL−15の分泌不良につながることが知られている(Meazza R,et al.(1997)Eur J Immunol 27:1049−1054)。IL−15分泌のレベルを高めるために、我々は、IL−15シグナルペプチドがヒトIL−2のシグナルペプチドに置き換えられた、IL−15発現ベクターを構築した(図7A〜7B)。この置換は、血清IL−15レベルを約100倍上昇させた(図7B)。IL−15コード化プラスミド50μgの単回流体力学的注入で、高レベルのIL−15が、注入の1日後に血清中で検出され、有意なレベルが14日間維持された(図2)。同様に、FLコード化プラスミドの単回注入は、FLが血清で21日間発現するという結果をもたらした。したがって、サイトカイン遺伝子の流体力学的送達は、マウスでヒトサイトカイン類を導入するための簡単で効率のよい方法である。
【0097】
IL−15およびFL遺伝子送達後のヒトNK細胞の再構成強化。IL−15およびFL発現がNK細胞発生に及ぼす影響を測定するために、遺伝子送達の9日後に、ヒューマウスの様々な器官におけるNK細胞再構成についてフローサイトメトリーで分析した。空のpcDNAベクターまたはFLコード化ベクターの注入は、CD56+NK細胞の発生頻度に有意な影響を及ぼさなかった(図3A)。しかし、IL−15の発現は、血中、脾臓、BM、肺、および肝臓におけるCD56+NK細胞の発生頻度を有意に増加させた(図3Aおよび3B)。NK細胞の発生頻度の増加は、IL−15とFLの両者がヒューマウスで発現されたとき、さらに劇的であり、通常のヒト末梢血(白血球の5%〜21%)(Maurice RG,O’Gorman ADD(2008)Handbook of Human Immunology(CRC Press,Boca Raton))および通常のマウス組織(Zhang J,et al.(2005)Cell Mol Immunol 2:271−280)で確認されるレベルに達した。NK細胞の発生頻度上昇に対応して、NK細胞の絶対数が脾臓およびBMで著しく増加した(図S3B)。さらに、血中でのCD56+NK細胞の高い発生頻度は、遺伝子送達後少なくとも30日間維持された(図3C)。加えて、サイトカイン誘導性NK細胞は、活性化受容体NKG2D、抑制性受容体NKG2A、CD94、およびKIR、細胞傷害誘発受容体NKp46、NK細胞マーカーCD7、早期活性化マーカーCD69、およびFC受容体CD16を含め、NK細胞機能に重要であることが知られている細胞表面受容体の多くを発現した(図9)。これらの結果から、サイトカイン誘導性NK細胞は通常のNK細胞の特徴的な表面表現型を示すことが分かる。
【0098】
NK細胞発生の刺激に加えて、FLとIL−15の両者は、他の造血細胞系統にも効果を発揮することが知られている(Diener KR,Moldenhauer LM,Lyons AB,Brown MP,Hayball JD(2008)Exp Hematol 36:51−60;Dong J,McPherson CM,Stambrook PJ(2002)Cancer Biol Ther 1:486−489;Blom B,Ho S,Antonenko S,Liu YJ(2000)J Exp Med 192:1785−1796;Armitage RJ,Macduff BM,Eisenman J,Paxton R,Grabstein KH(1995)J Immunol 154:483−490)。したがって、ヒューマウスの脾臓、BM、肺、および肝臓由来の細胞を列挙し、フローサイトメトリーで分析した。IL−15およびFLの発現はまた、脾臓およびBMにおけるCD14+単球/マクロファージ、CD11c+CD1c+骨髄性樹枝状細胞、ILT7+CD303+形質細胞様樹状細胞、およびCD19+B細胞の有意な増加も誘導した(図8A〜8G参照)。これらの結果は、ヒトIL−15およびFLの発現が、ヒト化マウスにおけるNK細胞ならびに他の骨髄性細胞およびリンパ球系細胞の再構成を劇的に改善することを示す。
【0099】
サイトカイン誘導性NK細胞は機能的である。我々は、サイトカイン誘導性ヒトNK細胞が機能的である(すなわち、標的細胞を死滅させることができ、また適切な刺激後にIFN−γを発現できる)かどうかを調査研究した。CD56+NK細胞は、IL−15およびFL処理マウスのBMおよび脾臓から精製した。MHCクラスI欠損標的細胞K562と混合したとき、添加したNK細胞が増加するにつれて、高レベルの標的細胞溶解が確認された(図10A)。NK細胞を刺激して炎症性サイトカイン類を産生させることが知られている、Toll様受容体3アゴニストpoly(I:C)(Schmidt KN,et al.(2004)J Immunol 172:138−143)で精製NK細胞を刺激したとき、IFN−γが培養上澄で検出された(図10B)。ヒトDCの存在下で、IFN−γ分泌レベルはさらに上昇した。poly(I:C)をヒト化マウスに注入したとき、サイトカインコード化DNAを注入したヒューマウスの血清で、非注入ヒューマウスと比較して有意に高いレベルのIFN−γが検出された(図10C)。
【0100】
我々はまた、NK細胞依存性肝損傷を引き起こすことが知られているアデノウイルス(Chen Q,Wei H,Sun R,Zhang J,Tian Z(2008)Hepatology 47:648−658)を、ヒューマウスに投与した。サイトカイン遺伝子送達の9日後、複製欠損アデノウイルスをヒューマウスに静脈内注入した。3日後、肝臓を採取し、H&Eで染色した。大量の白血球浸潤および広域の壊死が、IL−15およびFL処理したアデノウイルス感染ヒューマウスの肝臓で確認された。しかし、未処理のアデノウイルス感染ヒューマウスは、軽度の細胞浸潤および損傷を示しただけであった(図4A)。相応して、IL−15およびFL処理したアデノウイルス感染ヒューマウスで、血清アラニンアミノトランスフェラーゼ(ALT)レベルが有意に上昇していた(図4B)。この上昇は、肝臓における、血清IFN−γレベルの約4倍の増加(図4C)および、浸潤性ヒト白血球の約5倍の増加(図4D)と相関性があった。肝薄片の免疫組織化学分析は、CD56+NK細胞の病変内局在化を裏付けた(図4E)。これらの結果から、サイトカイン誘導性ヒトNK細胞は機能的であることが強く示唆される。
【0101】
他のヒト血液細胞系統の再構成の改善。我々は、サイトカイン遺伝子送達が、ヒューマウスにおける特定のヒト血液細胞系統の再構成を改善するための一般的方法として使用できるかどうかを試験した。培養で、ヒューマウスのBMから精製したヒトCD34+細胞を、GM−CSFおよびIL−4で刺激してCD11c+CD209+DCに、M−CSFで刺激してCD14+単球/マクロファージに、またEPOおよびIL−3で刺激してCD235ab+赤血球に、分化させた(図11)。インビボで、GM−CSF、IL−4、およびFLを発現するDNAベクターをヒューマウスに流体力学的に送達することにより、血中、脾臓、BM、肺、および肝臓でD11c+CD209+DCの発生頻度が著しく増加する結果となった(図5A)。同様に、M−CSFの発現は、リンパ器官と非リンパ器官の両者におけるCD14+単球/マクロファージの再構成の改善へと導いた(図5B)。EPOおよびIL−3の発現は、血中でのCD235ab+ヒト赤血球の出現をもたらし(図5C)、全赤血球の3〜5%に達した。したがって、DNAプラスミドの流体力学的注入によるサイトカイン遺伝子発現は、ヒューマウスにおける特定のヒト血液細胞系統の再構成を改善するための一般的で効率のよい方法である。
【0102】
考察
NK細胞および骨髄性細胞の再構成は、NSGまたはBALB/c−Rag2−/−Il2rg−/−マウスをレシピエントとして使用するヒト化マウスモデルでは、一般に不十分である。BLTマウスで、DCおよび単球/マクロファージがかなり再構成されるにも拘わらず、ヒトNK細胞およびRBCは存在しない。NK細胞または様々な骨髄性細胞の発生および維持に必要な、IL−15、GM−CSF、IL−4、M−CSF、およびIL−3を含む多くのサイトカイン類は、ヒトとマウスとの間で有意な配列分散を示すことが分かった。以前の研究は、これらのネズミサイトカイン類が、適切なヒト細胞型に殆ど影響しないことを証明していた。これらのサイトカイン類は、主として非造血細胞によって産生されるため、これらのヒトサイトカイン類の欠如によって、ヒューマウスにおけるNK細胞および骨髄性細胞の再構成不良を説明し得るであろう。
【0103】
この解釈を裏付けるために、ヒューマウスのBMから単離されたヒトCD34+前駆細胞を刺激して、インビトロで、NK細胞、DC、単球/マクロファージ、および赤血球に分化できることを示した。サイトカインコード化プラスミドDNAの流体力学的送達により、ヒト化マウスで適切なヒトサイトカイン類が導入されるとき、有意に高いレベルのNK細胞、DC、単球/マクロファージ、および赤血球が誘導される。サイトカイン類の血清レベルが低下するにつれて、再構成レベルも低下する。したがって、ヒューマウスにおけるNK細胞および骨髄性細胞の再構成不良は、それらの分化および維持に通常必要な適切なヒトサイトカイン類が欠如した結果である。適切なサイトカイン類の導入は、ヒューマウスにおけるこれらのヒト血液細胞系統の再構成レベルの劇的な上昇につながる。
【0104】
流体力学的遺伝子送達は、マウスで、高レベルの、一過性の肝および全身性導入遺伝子発現を生じさせるために広く使用される(Suda T,LiuD(2007)Mol Ther 15:2063−2069)。本方法は、短時間(6〜8秒)で、大量(体重の10%)のDNAの尾静脈注入を含む。流体力学的圧力は、肝臓損傷を引き起こし、肝細胞によるDNAの取り込みにつながる(Suda T,Liu D(2007)Mol Ther 15:2063−2069)。転写および翻訳の後、サイトカイン類が循環内に分泌されて、BMまたは他の器官の標的細胞に到達することができる。したがって、サイトカインコード化DNAの単回注入で、IL−15は血清中で2週間にわたって検出され、FLは3週間にわたって検出された。IL−15とFLとの血清持続期間の差は、恐らく蛋白質の半減期の差のためであるか、またはIL−15が通常はIL−15Ra鎖を介して細胞表面上に結合されているためである(Mortier E,Woo T,Advincula R,Gozalo S,Ma A(2008)J Exp Med 205:1213−1225)。単回のDNA注入によって産生されるIL−15およびFLの量は、少なくとも30日間、NK細胞レベルの著しい上昇を誘導するのに明らかに十分である。サイトカイン類が循環中でもはや検出されなかったとき、NK細胞が存続することは、サイトカイン類の重要な役割が、分化の初期段階に発揮されることを示す。NK細胞は、いったん生成すると、サイトカイン類が循環で検出できなくなった後も、長時間にわたって生存することができる。NK細胞は、多様な血液細胞系統に影響を及ぼすため、FLおよびIL−15の発現はまた、脾臓およびBMでの、単球/マクロファージ、DC、およびB細胞のレベル上昇をもたらすが、T細胞のレベル上昇はもたらさない。さらに、流体力学的遺伝子送達による適切なサイトカイン類の発現はまた、DC、単球/マクロファージ、および赤血球を含む、特定の骨髄系細胞の再構成を著しく強化し、本手法の汎用性を示す。ヒトサイトカイン遺伝子送達の7日後に明白になる、NK細胞、DCおよび単球/マクロファージの改善した再構成と比較して、赤血球の改善した再構成は、サイトカイン遺伝子送達の30日後までピークレベルに到達しなかった。これは、一方ではヒトWBCとマウスWBCの比率の差によって説明でき、また他方ではヒトRBCとマウスRBCの比率の差によって説明できる。マウスRBCは大量なため、類似したパーセンテージに到達するために十分な数のヒトRBCを産生するには長時間を要する。以前に、2つのグループが、組換えヒトIL−15をNOD−scidマウスまたはBALB/c−Rag2−/−Il2rg−/−マウスに注入することによって強化されたNK細胞発生について報告した(Huntington ND,et al.(2009)J Exp Med 206:25−34;Kalberer CP,Siegler U,Wodnar−Filipowicz A(2003)Blood 102:127−135)。厄介で高価な、毎日のサイトカイン注入と比較して、流体力学的遺伝子送達によるサイトカイン遺伝子の発現は、単回注入で、30日間以上にわたって特定の血液細胞系統の再構成上昇をもたらすため、手頃で、簡単、かつ効率のよい方法である。
【0105】
サイトカイン誘導性NK細胞は、通常の表面表現型および機能を示す。NOD−scidマウスで、毎日の組換えIL−15の注入後にヒトNK細胞が生成された、以前の観察結果と対照的に、この細胞はNKp46を発現したが、NKG2DおよびNKG2Aは発現しなかった(Kalberer CP,Siegler U,Wodnar−Filipowicz A(2003)Blood 102:127−135)。本研究で、サイトカイン誘導性ヒトNK細胞は、活性化受容体NKG2D、抑制性受容体NKG2AおよびKIR、ならびに細胞傷害誘発受容体NKp46を含む、NK受容体の3つの主要なファミリー全てを発現した。一貫して、サイトカイン誘導性NK細胞は、インビトロでもインビボでも、poly(I:C)刺激のすぐ後で、MHCクラスI−欠損標的細胞を溶解することができ、またIFN−γを分泌することができる。さらに、IL−15およびFL処理ヒューマウスにおける、広範囲の肝臓壊死および高レベルの血清ALTで示されるように、サイトカイン誘導性NK細胞は、アデノウイルス感染に対する強い反応を開始することができる。IFN−γ分泌により肝炎を仲介する、マウスにおける野生型NK細胞(Chen Q,Wei H,Sun R,Zhan gJ,Tian Z(2008)Hepatology 47:648−658;Rosenberger CM,Clark AE,Treuting PM,Johnson CD,Aderem A(2008)Proc Natl Acad Sci USA 105:2544−2549)と同様、IL−15およびFL処理アデノウイルス感染ヒューマウスの血清IFN−γレベルは有意に上昇していた。これらの結果から、サイトカイン誘導性NK細胞は、表面表現型でも機能でも、正常であることが示唆される。
【0106】
実施例2
ヒトサイトカイン類の発現は、ヒト化マウスにおけるヒトT細胞およびB細胞の再構成および機能を改善する。
ヒトT細胞およびB細胞の再構成は、ヒト化マウスで妥当であるが、最適な機能を示さない。たとえば、ヒトCD8+T細胞反応は、ウイルス投与後に検出されてきたが、CD4+T細胞の機能は異常である;ヒト化マウスで、ヒトB細胞介在性抗体反応は存在しない。本明細書に示す通り、ヒトT細胞およびB細胞応答の異常はまた、マウスにおける、マウスサイトカイン類とヒト細胞との間の不十分な交差反応にも起因する。ヒトGM−CSFおよびIL−4コード化プラスミドのヒューマウスへの注入が、T細胞に対する主要抗原提示細胞であると考えられる、ヒトCD209+樹枝状細胞の再構成改善に導いたことを、本明細書に示す。さらに、IL−4は、ヒトB細胞における細胞増殖、生存、ならびにIgGおよびIgEへの免疫グロブリンクラススイッチ、ならびにナイーブCD4+T細胞によるTh2表現型の獲得を促進することも証明された。トキソイド(TT)ワクチンを使用して、GM−CSFおよびIL−4処理ヒューマウスを免疫し、これらのマウスがTT特異的抗体応答を引き起こすかどうかを決定した。
【0107】
本明細書に記載の通り、類似したヒト白血球再構成(50〜80%)を有する12週齢のヒト化マウスに、ヒトGM−CSFおよびIL−4をコード化するプラスミドまたは空のpcDNAベクター(ベクター)を、流体力学的に注入した。7日後、これらのマウスに、破傷風トキソイド(TT)を、3週間の投与間隔で3回免疫した(図12A)。第3回の免疫の2週間後に、脾臓および血清を採取した。図12Bは、GM−CSFおよびIL−4処理マウスからの脾臓は、ベクター処理マウスと比較して、有意に拡張したことを示す。相応して、GM−CSFおよびIL−4処理マウスの脾臓で、ヒト単核細胞(MNC)の劇的な増殖(約20倍)が認められた(図12C)。脾臓におけるヒトB細胞およびT細胞の細胞表面表現型別の結果から、ヒトB細胞は、サイトカイン処理および免疫の全過程を通して通常のヒト特性と同一である成熟した抗体産生期(CD19lowCD20−)まで成長した(図13);が、ヒトT細胞は、HLA−DRおよびCD40Lの発現を上方制御することにより活性化されることも分かった(図14)。
【0108】
免疫した、GM−CSFおよびIL−4処理マウス由来の血清中のヒトIgGおよびIgM総レベルは、それぞれ、1.3mgおよび140μgという高値に達し(図15A,15B)、ヒトでのレベル(それぞれ4mgおよび1mg)に類似していた。最も重要なことは、抗原特異的ヒト抗体応答が、ヒューマウスで初めて首尾よく実証されたことである。ヒトTT特異的IgGはベクター処理マウスで検出できなかったが、サイトカイン類処理マウスでは、0.16IU/mlという平均値に達した(図15C)。免疫化後、ヒトで、0.1IU/mlという抗破傷風トキソイド抗体は、個体を感染から守るのに十分である。さらに、GM−CSF+IL−4処理マウスにおけるヒトT細胞応答も、TT抗原に対して特異性を示した(図16A〜16B)。破傷風毒素ペプチド(830−843)を使用して、脾臓T細胞を刺激した。GM−CSF+IL−4処理マウス由来のT細胞は、ベクター処理マウス由来の細胞と比較して、TT特異的刺激後に、有意レベルのヒトIFN−γおよびIL−4を産生することができた。
【0109】
本明細書に記載の方法を使用して、有意レベルのヒト抗原特異的抗体応答を、マウスで実証することができた。したがって、本明細書に記載の方法は、ワクチン類を試験するため、および治療目的のヒト抗体を産生するための、有用な基盤を提供する。
【0110】
【表1】
【0111】
【表2】
【0112】
【表3】
【0113】
全ての特許、公開された出願および本明細書に引用された参考文献の教示は、参照によりその全体を援用する。
【0114】
本発明は具体的に示され、またその例示的な実施態様を参照しながら記述されているが、添付の請求の範囲に含まれる本発明の範囲から逸脱せずに、形状および詳細の様々な変更を加えられることを、当業者は理解するであろう。
【特許請求の範囲】
【請求項1】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入すること;および
b)非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが機能的ヒト血液細胞系統に分化する条件下で、維持すること;
を含み、
それによって、前記非ヒト哺乳動物で、機能的ヒト血液細胞系統を再構成する、
非ヒト哺乳動物で、機能的ヒト血液細胞系統を再構成する方法。
【請求項2】
前期HSCが前記非ヒト哺乳動物導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項1に記載の方法。
【請求項3】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで導入される、請求項1に記載の方法。
【請求項4】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項3に記載の方法。
【請求項5】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項4に記載の方法。
【請求項6】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用してプラスミドDNAとして導入される、請求項1に記載の方法。
【請求項7】
前記1つまたは複数のヒトサイトカイン類が、インターロイキン−12(IL−12)、インターロイキン−15(IL−15)、Flt3L(Fms関連チロシンキナーゼ3リガンド)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン−4(IL−4)、インターロイキン−3(IL−3)、マクロファージコロニー刺激因子(M−CSF)、エリスロポエチン(EPO)およびそれらの組合せからなる群から選択される、請求項1に記載の方法。
【請求項8】
前記非ヒト哺乳動物がマウスである、請求項1に記載の方法。
【請求項9】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、かつサイトカイン受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scid IL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項8に記載の方法。
【請求項10】
再構成される前記機能的ヒト血液細胞系統が、機能的ヒト骨髄性細胞、機能ヒトリンパ球系細胞またはそれらの組合せである、請求項1に記載の方法。
【請求項11】
前記ヒト骨髄性細胞が、ヒト単球、ヒトマクロファージ、ヒト樹枝状細胞、ヒト赤血球、およびそれらの組合せであり;また前記ヒトリンパ球系細胞が、ヒトNK細胞、ヒトB細胞、ヒトT細胞、およびそれらの組合せである、請求項10に記載の方法。
【請求項12】
前記1つまたは複数のヒトサイトカイン類が、インターロイキン−15(IL−15)およびFms関連チロシンキナーゼ3リガンド(Flt3L)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統がヒトNK細胞である、請求項1に記載の方法。
【請求項13】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン4(IL−4)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト樹枝状細胞である、請求項1に記載の方法。
【請求項14】
前記1つまたは複数のサイトカイン類が、マクロファージコロニー刺激因子(MCSF)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト単球/マクロファージである、請求項1に記載の方法。
【請求項15】
前記1つまたは複数のサイトカイン類が、エリスロポエチン(EPO)およびインターロイキン3(IL−3)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト赤血球である、請求項1に記載の方法。
【請求項16】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒトNK細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒトNK細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物におけるヒトNK細胞の再構成を強化する、
非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法。
【請求項17】
前記核酸が、ヒトIL−15およびヒトFlt−3/Flk−2リガンドをコード化する、請求項16に記載の方法。
【請求項18】
前記非ヒト哺乳動物の末梢血中の白血球の約5%〜約21%がヒトNK細胞である、請求項17に記載の方法。
【請求項19】
前記ヒトNK細胞の発現が、前記非ヒト哺乳動物で約30日間維持される、請求項18に記載の方法。
【請求項20】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒト樹枝状細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト樹枝状細胞の再構成を強化する、
非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法。
【請求項21】
前記核酸が、ヒトGM−CSFおよびヒトIL−4をコード化する、請求項20に記載の方法。
【請求項22】
ヒトFlt−3/Flk−2リガンドをコード化する核酸を導入することをさらに含む、請求項21に記載の方法。
【請求項23】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持すること:
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト単球/マクロファージの再構成を強化する、
非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法。
【請求項24】
前記核酸が、ヒトマクロファージコロニー刺激因子をコード化する、請求項23に記載の方法。
【請求項25】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト赤血球への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒト赤血球に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト赤血球の再構成を強化する、
非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法。
【請求項26】
前記核酸が、ヒトエリスロポエチンおよびヒトIL−3をコード化する、請求項25に記載の方法。
【請求項27】
前記赤血球が、前記非ヒト哺乳動物における全赤血球の約3%〜約5%を構成する、請求項26に記載の方法。
【請求項28】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物で、機能的ヒトT細胞およびヒトB細胞を再構成する、
非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法。
【請求項29】
前記HSCが前記非ヒト哺乳動物に導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項28に記載の方法。
【請求項30】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで、導入される、請求項28に記載の方法。
【請求項31】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項30に記載の方法。
【請求項32】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項31に記載の方法。
【請求項33】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用して、プラスミドDNAとして導入される、請求項28に記載の方法。
【請求項34】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン−4(IL−4)である、請求項28に記載の方法。
【請求項35】
前記非ヒト哺乳動物がマウスである、請求項28に記載の方法。
【請求項36】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、サイトカイン受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scidIL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項35に記載の方法。
【請求項37】
前記ヒト抗体が、ヒトIgG、ヒトIgMまたはそれらの組合せである、請求項28に記載の方法。
【請求項38】
c)前記非ヒト哺乳動物を免疫原で免疫すること;および
d)前記非ヒト動物を、前記非ヒト哺乳動物が前記免疫原に対するヒト抗体を産生する条件下で維持すること;
をさらに含む、請求項28に記載の方法。
【請求項39】
前記ヒト抗体を産生するヒトB細胞を、前記非ヒト哺乳動物から単離することをさらに含み、それによって単離されたヒトB細胞を作る、請求項38に記載の方法。
【請求項40】
前記単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;および前記ヒトB細胞と前記不死化細胞が融合して、前記免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、前記組合せを維持すること;をさらに含む、請求項39に記載の方法。
【請求項41】
前記ヒトB細胞により発現されるヒト抗体の全部または機能的部分をコード化する配列をクローニングすることをさらに含む、請求項39に記載の方法。
【請求項42】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する、核酸を導入すること;
b)前記非ヒト哺乳動物で前記核酸が発現されて前記HSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で、前記非ヒト哺乳動物を維持すること;
c)前記非ヒト哺乳動物を、前記免疫原で免疫すること;および
d)前記非ヒト動物を、前記非ヒト哺乳動物で前記ヒトB細胞が前記免疫原に対するヒト抗体を産生する条件下で維持すること;を含み、
それによって、前記非ヒト哺乳動物で、前記免疫原に対するヒト抗体を生成する、
非ヒト哺乳動物で免疫原に対するヒト抗体を生成する方法。
【請求項43】
前記HSCが前記非ヒト哺乳動物に導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項42に記載の方法。
【請求項44】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで、導入される、請求項42に記載の方法。
【請求項45】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項44に記載の方法。
【請求項46】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項45に記載の方法。
【請求項47】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用して、プラスミドDNAとして導入される、請求項42に記載の方法。
【請求項48】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン−4(IL−4)である、請求項42に記載の方法。
【請求項49】
前記非ヒト哺乳動物がマウスである、請求項42に記載の方法。
【請求項50】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、前記サイトカイン−受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scidIL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項49に記載の方法。
【請求項51】
前記ヒト抗体が、ヒトIgG、ヒトIgMまたはそれらの組合せである、請求項42に記載の方法。
【請求項52】
前記ヒト抗体を産生するヒトB細胞を前記非ヒト哺乳動物から単離することをさらに含み、それによって単離されたヒトB細胞を作る、請求項42に記載の方法。
【請求項53】
前記単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;および前記ヒトB細胞および前記不死化細胞が融合して、前記免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、前記組合せを維持すること;をさらに含む、請求項52に記載の方法。
【請求項54】
前記免疫原が破傷風トキソイドである、請求項42に記載の方法。
【請求項55】
前記ヒトB細胞により発現される前記ヒト抗体の全部または機能的部分をコード化する配列をクローニングすることをさらに含む、請求項52に記載の方法。
【請求項56】
請求項1に記載の方法で作られる非ヒト哺乳動物。
【請求項57】
請求項16に記載の方法で作られる非ヒト哺乳動物。
【請求項58】
請求項20に記載の方法で作られる非ヒト哺乳動物。
【請求項59】
請求項23に記載の方法で作られる非ヒト哺乳動物。
【請求項60】
請求項25に記載の方法で作られる非ヒト哺乳動物。
【請求項61】
請求項28に記載の方法で作られる非ヒト哺乳動物。
【請求項62】
請求項40に記載の方法で作られるハイブリドーマ。
【請求項63】
請求項62に記載のハイブリドーマにより分泌されるモノクローナル抗体。
【請求項64】
請求項53に記載の方法で作られるハイブリドーマ。
【請求項65】
請求項64に記載のハイブリドーマにより分泌されるモノクローナル抗体。
【請求項1】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入すること;および
b)非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが機能的ヒト血液細胞系統に分化する条件下で、維持すること;
を含み、
それによって、前記非ヒト哺乳動物で、機能的ヒト血液細胞系統を再構成する、
非ヒト哺乳動物で、機能的ヒト血液細胞系統を再構成する方法。
【請求項2】
前期HSCが前記非ヒト哺乳動物導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項1に記載の方法。
【請求項3】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで導入される、請求項1に記載の方法。
【請求項4】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項3に記載の方法。
【請求項5】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項4に記載の方法。
【請求項6】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用してプラスミドDNAとして導入される、請求項1に記載の方法。
【請求項7】
前記1つまたは複数のヒトサイトカイン類が、インターロイキン−12(IL−12)、インターロイキン−15(IL−15)、Flt3L(Fms関連チロシンキナーゼ3リガンド)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン−4(IL−4)、インターロイキン−3(IL−3)、マクロファージコロニー刺激因子(M−CSF)、エリスロポエチン(EPO)およびそれらの組合せからなる群から選択される、請求項1に記載の方法。
【請求項8】
前記非ヒト哺乳動物がマウスである、請求項1に記載の方法。
【請求項9】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、かつサイトカイン受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scid IL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項8に記載の方法。
【請求項10】
再構成される前記機能的ヒト血液細胞系統が、機能的ヒト骨髄性細胞、機能ヒトリンパ球系細胞またはそれらの組合せである、請求項1に記載の方法。
【請求項11】
前記ヒト骨髄性細胞が、ヒト単球、ヒトマクロファージ、ヒト樹枝状細胞、ヒト赤血球、およびそれらの組合せであり;また前記ヒトリンパ球系細胞が、ヒトNK細胞、ヒトB細胞、ヒトT細胞、およびそれらの組合せである、請求項10に記載の方法。
【請求項12】
前記1つまたは複数のヒトサイトカイン類が、インターロイキン−15(IL−15)およびFms関連チロシンキナーゼ3リガンド(Flt3L)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統がヒトNK細胞である、請求項1に記載の方法。
【請求項13】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン4(IL−4)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト樹枝状細胞である、請求項1に記載の方法。
【請求項14】
前記1つまたは複数のサイトカイン類が、マクロファージコロニー刺激因子(MCSF)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト単球/マクロファージである、請求項1に記載の方法。
【請求項15】
前記1つまたは複数のサイトカイン類が、エリスロポエチン(EPO)およびインターロイキン3(IL−3)であり、前記非ヒト哺乳動物で再構成される前記機能的血液細胞系統が、ヒト赤血球である、請求項1に記載の方法。
【請求項16】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒトNK細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒトNK細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物におけるヒトNK細胞の再構成を強化する、
非ヒト哺乳動物で機能的ヒトNK細胞を再構成する方法。
【請求項17】
前記核酸が、ヒトIL−15およびヒトFlt−3/Flk−2リガンドをコード化する、請求項16に記載の方法。
【請求項18】
前記非ヒト哺乳動物の末梢血中の白血球の約5%〜約21%がヒトNK細胞である、請求項17に記載の方法。
【請求項19】
前記ヒトNK細胞の発現が、前記非ヒト哺乳動物で約30日間維持される、請求項18に記載の方法。
【請求項20】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト樹枝状細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒト樹枝状細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト樹枝状細胞の再構成を強化する、
非ヒト哺乳動物で機能的ヒト樹枝状細胞を再構成する方法。
【請求項21】
前記核酸が、ヒトGM−CSFおよびヒトIL−4をコード化する、請求項20に記載の方法。
【請求項22】
ヒトFlt−3/Flk−2リガンドをコード化する核酸を導入することをさらに含む、請求項21に記載の方法。
【請求項23】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト単球/マクロファージへの分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが機能的ヒト単球/マクロファージに分化する条件下で維持すること:
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト単球/マクロファージの再構成を強化する、
非ヒト哺乳動物で機能的ヒト単球/マクロファージを再構成する方法。
【請求項24】
前記核酸が、ヒトマクロファージコロニー刺激因子をコード化する、請求項23に記載の方法。
【請求項25】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒト赤血球への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒト赤血球に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物における機能的ヒト赤血球の再構成を強化する、
非ヒト哺乳動物で機能的ヒト赤血球を再構成する方法。
【請求項26】
前記核酸が、ヒトエリスロポエチンおよびヒトIL−3をコード化する、請求項25に記載の方法。
【請求項27】
前記赤血球が、前記非ヒト哺乳動物における全赤血球の約3%〜約5%を構成する、請求項26に記載の方法。
【請求項28】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカインをコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記非ヒト哺乳動物で発現されるとき、前記ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する、核酸を導入すること;および
b)前記非ヒト哺乳動物を、前記非ヒト哺乳動物で前記核酸が発現されて前記ヒトHSCが前記非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞に分化する条件下で維持すること;
を含み、
それによって、前記非ヒト哺乳動物で、機能的ヒトT細胞およびヒトB細胞を再構成する、
非ヒト哺乳動物で機能的ヒトT細胞およびヒトB細胞を再構成する方法。
【請求項29】
前記HSCが前記非ヒト哺乳動物に導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項28に記載の方法。
【請求項30】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで、導入される、請求項28に記載の方法。
【請求項31】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項30に記載の方法。
【請求項32】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項31に記載の方法。
【請求項33】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用して、プラスミドDNAとして導入される、請求項28に記載の方法。
【請求項34】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン−4(IL−4)である、請求項28に記載の方法。
【請求項35】
前記非ヒト哺乳動物がマウスである、請求項28に記載の方法。
【請求項36】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、サイトカイン受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scidIL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項35に記載の方法。
【請求項37】
前記ヒト抗体が、ヒトIgG、ヒトIgMまたはそれらの組合せである、請求項28に記載の方法。
【請求項38】
c)前記非ヒト哺乳動物を免疫原で免疫すること;および
d)前記非ヒト動物を、前記非ヒト哺乳動物が前記免疫原に対するヒト抗体を産生する条件下で維持すること;
をさらに含む、請求項28に記載の方法。
【請求項39】
前記ヒト抗体を産生するヒトB細胞を、前記非ヒト哺乳動物から単離することをさらに含み、それによって単離されたヒトB細胞を作る、請求項38に記載の方法。
【請求項40】
前記単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;および前記ヒトB細胞と前記不死化細胞が融合して、前記免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、前記組合せを維持すること;をさらに含む、請求項39に記載の方法。
【請求項41】
前記ヒトB細胞により発現されるヒト抗体の全部または機能的部分をコード化する配列をクローニングすることをさらに含む、請求項39に記載の方法。
【請求項42】
a)ヒト造血幹細胞(HSC)および1つまたは複数のヒトサイトカイン類をコード化する核酸を、免疫不全非ヒト哺乳動物に導入することであって、前記ヒトサイトカイン類は、前記ヒトHSCの機能的ヒトT細胞およびヒトB細胞への分化を促進する、核酸を導入すること;
b)前記非ヒト哺乳動物で前記核酸が発現されて前記HSCが機能的ヒトT細胞およびヒトB細胞に分化する条件下で、前記非ヒト哺乳動物を維持すること;
c)前記非ヒト哺乳動物を、前記免疫原で免疫すること;および
d)前記非ヒト動物を、前記非ヒト哺乳動物で前記ヒトB細胞が前記免疫原に対するヒト抗体を産生する条件下で維持すること;を含み、
それによって、前記非ヒト哺乳動物で、前記免疫原に対するヒト抗体を生成する、
非ヒト哺乳動物で免疫原に対するヒト抗体を生成する方法。
【請求項43】
前記HSCが前記非ヒト哺乳動物に導入された後に、前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が導入される、請求項42に記載の方法。
【請求項44】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、裸のDNAとして、またはベクターで、導入される、請求項42に記載の方法。
【請求項45】
前記ベクターが、プラスミド、ウイルスベクター、またはそれらの組合せである、請求項44に記載の方法。
【請求項46】
前記ウイルスベクターが、レンチウイルスウイルスまたはアデノウイルスベクターである、請求項45に記載の方法。
【請求項47】
前記1つまたは複数のヒトサイトカイン類をコード化する前記核酸が、流体力学的注入を使用して、プラスミドDNAとして導入される、請求項42に記載の方法。
【請求項48】
前記1つまたは複数のヒトサイトカイン類が、顆粒球マクロファージコロニー刺激因子(GM−CSF)およびインターロイキン−4(IL−4)である、請求項42に記載の方法。
【請求項49】
前記非ヒト哺乳動物がマウスである、請求項42に記載の方法。
【請求項50】
前記マウスが、重症複合免疫不全変異を担持する非肥満糖尿病マウス(NOD/scidマウス);重症複合免疫不全変異を担持し、前記サイトカイン−受容体γ鎖の遺伝子を欠く非肥満糖尿病マウス(NOD/scidIL2Rγ−/−マウス)およびBalb/crag−/−γc−/−マウスからなる群から選択される、請求項49に記載の方法。
【請求項51】
前記ヒト抗体が、ヒトIgG、ヒトIgMまたはそれらの組合せである、請求項42に記載の方法。
【請求項52】
前記ヒト抗体を産生するヒトB細胞を前記非ヒト哺乳動物から単離することをさらに含み、それによって単離されたヒトB細胞を作る、請求項42に記載の方法。
【請求項53】
前記単離されたヒトB細胞を不死化細胞と接触させ、それによって組合せを作ること;および前記ヒトB細胞および前記不死化細胞が融合して、前記免疫原に対するモノクローナル抗体を産生するハイブリドーマを形成する条件下で、前記組合せを維持すること;をさらに含む、請求項52に記載の方法。
【請求項54】
前記免疫原が破傷風トキソイドである、請求項42に記載の方法。
【請求項55】
前記ヒトB細胞により発現される前記ヒト抗体の全部または機能的部分をコード化する配列をクローニングすることをさらに含む、請求項52に記載の方法。
【請求項56】
請求項1に記載の方法で作られる非ヒト哺乳動物。
【請求項57】
請求項16に記載の方法で作られる非ヒト哺乳動物。
【請求項58】
請求項20に記載の方法で作られる非ヒト哺乳動物。
【請求項59】
請求項23に記載の方法で作られる非ヒト哺乳動物。
【請求項60】
請求項25に記載の方法で作られる非ヒト哺乳動物。
【請求項61】
請求項28に記載の方法で作られる非ヒト哺乳動物。
【請求項62】
請求項40に記載の方法で作られるハイブリドーマ。
【請求項63】
請求項62に記載のハイブリドーマにより分泌されるモノクローナル抗体。
【請求項64】
請求項53に記載の方法で作られるハイブリドーマ。
【請求項65】
請求項64に記載のハイブリドーマにより分泌されるモノクローナル抗体。
【図1】

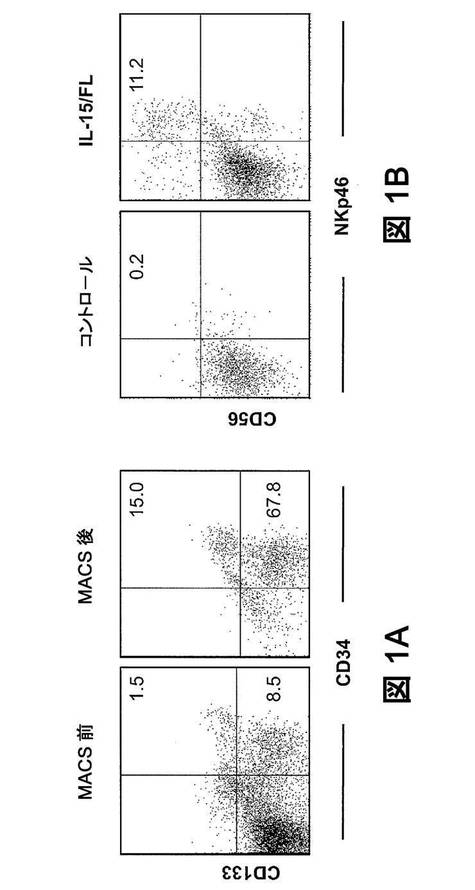
【図2】
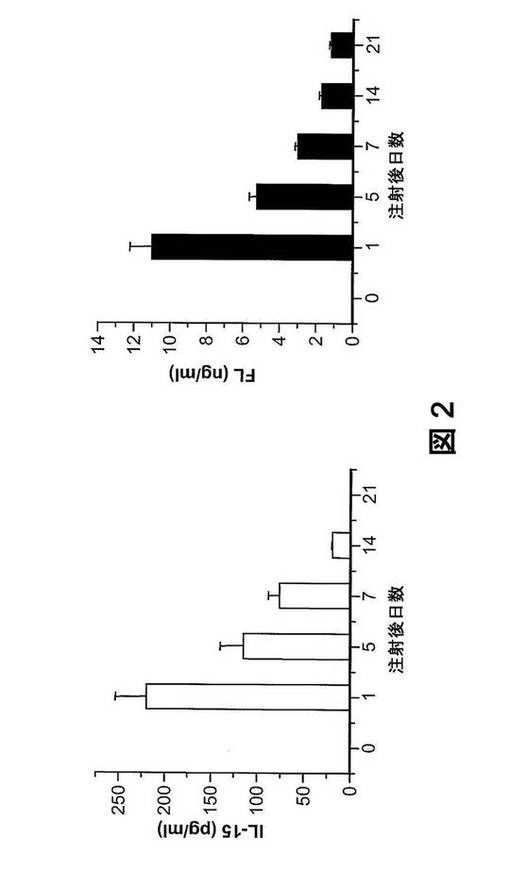
【図3】
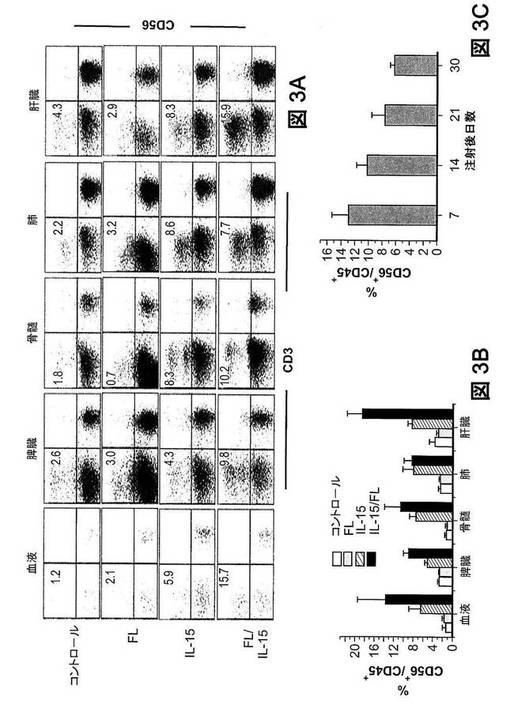
【図4A−D】
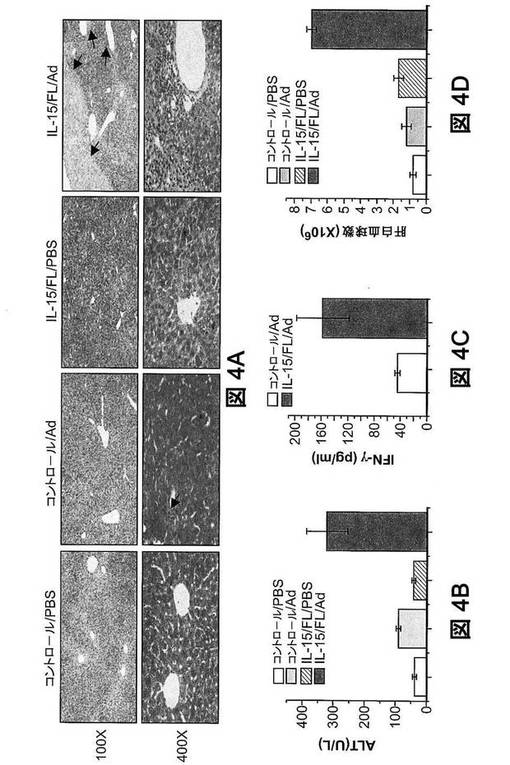
【図4E】

【図5】

【図6】
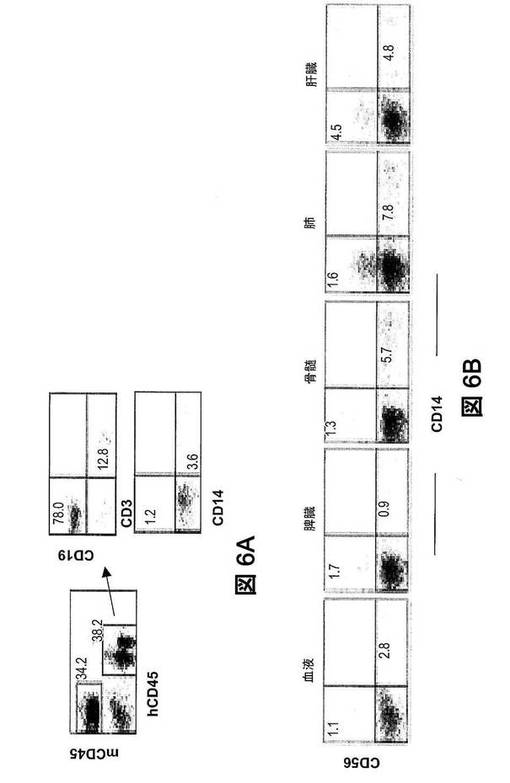
【図7】

【図8】
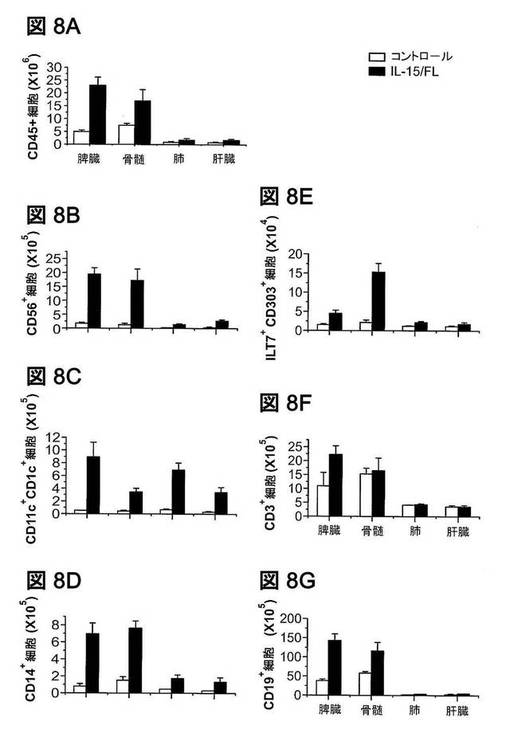
【図9】

【図10】
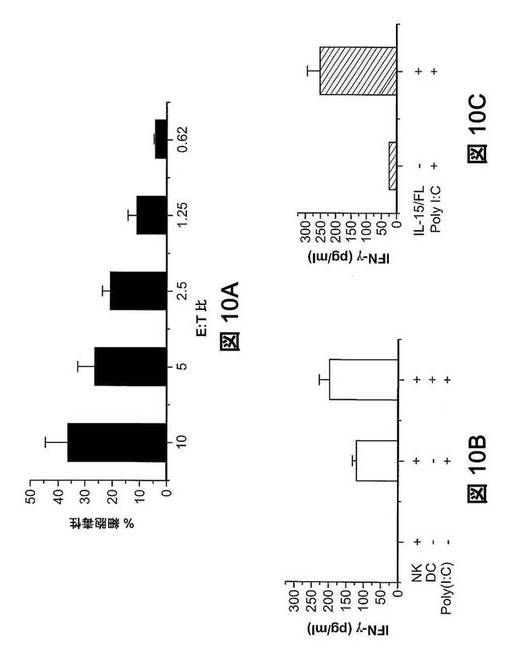
【図11】
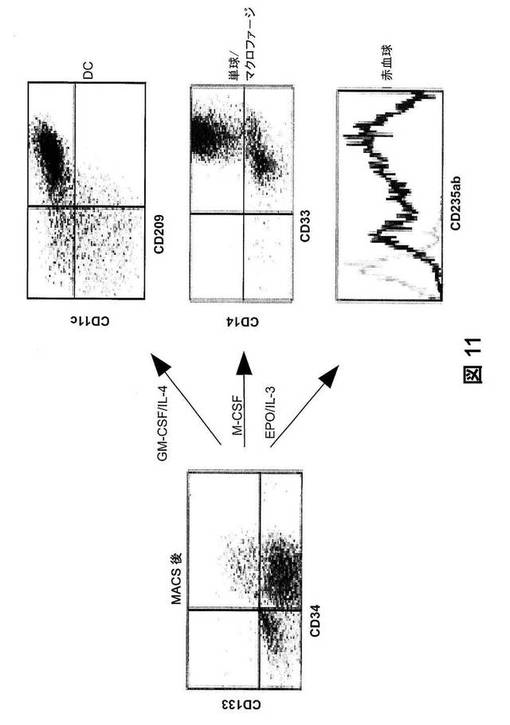
【図12】
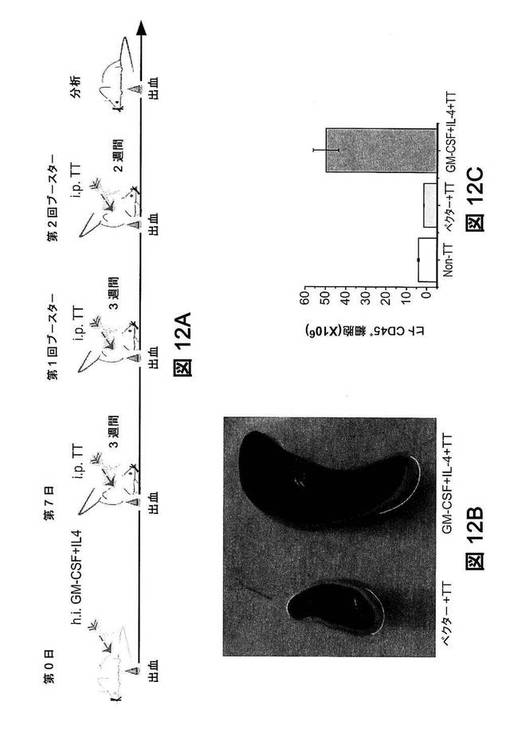
【図13】
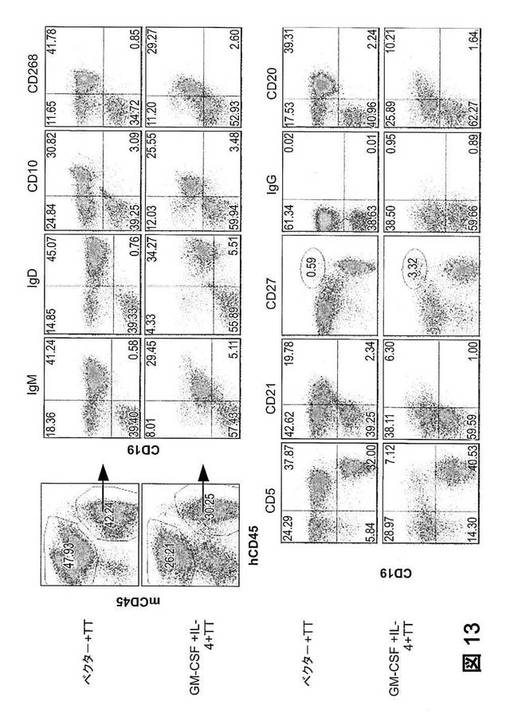
【図14】
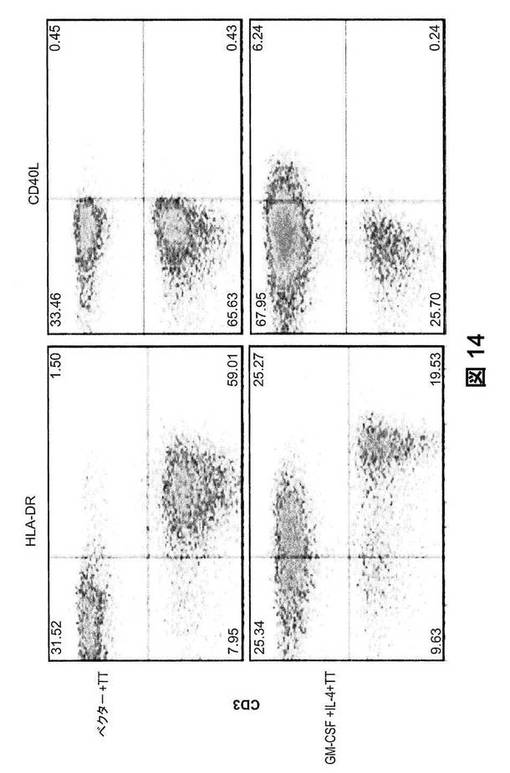
【図15】
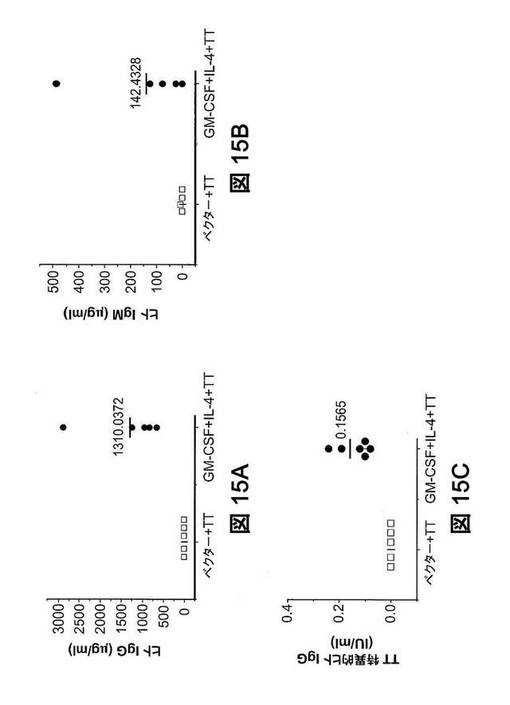
【図16】
【図17】

【図2】
【図3】
【図4A−D】
【図4E】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2012−531896(P2012−531896A)
【公表日】平成24年12月13日(2012.12.13)
【国際特許分類】
【出願番号】特願2012−517837(P2012−517837)
【出願日】平成22年6月28日(2010.6.28)
【国際出願番号】PCT/US2010/040260
【国際公開番号】WO2011/002727
【国際公開日】平成23年1月6日(2011.1.6)
【出願人】(512000499)
【出願人】(512000466)
【Fターム(参考)】
【公表日】平成24年12月13日(2012.12.13)
【国際特許分類】
【出願日】平成22年6月28日(2010.6.28)
【国際出願番号】PCT/US2010/040260
【国際公開番号】WO2011/002727
【国際公開日】平成23年1月6日(2011.1.6)
【出願人】(512000499)
【出願人】(512000466)
【Fターム(参考)】
[ Back to top ]