
ヒト心血管前駆細胞
本発明は、ヒト心血管前駆細胞の集団、前記細胞を作製する方法、ならびに心血管コロニーの集団及び心筋細胞の集団を産生するために前記細胞を使用する方法を提供する。心筋細胞置換療法の方法をも提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2008年3月27日に出願された米国特許仮出願第61/040,049号の利益を主張する。
【0002】
(連邦政府による資金提供を受けた試験開発の記載)
本発明は、米国立衛生試験所によって授与された助成金番号第R01HL71800−01号及びP20−GM07501号の下で政府による後援を受けて実施された。米国政府は、本発明に関して所定の権利を有することができる。
【0003】
(技術分野)
胚発生中、身体の組織は、3つの主要な細胞集団である外胚葉、中胚葉及び胚性内胚葉から形成される。これらの細胞集団は、原発胚細胞層としても公知であり、原腸形成として公知のプロセスを通して形成される。原腸形成後に、各原発胚細胞層は特定セットの細胞集団及び組織を生成する。中胚葉は血球、内皮細胞、心筋及び骨格筋、ならびに脂肪細胞を生じさせる。胚性内胚葉は、肝臓、膵臓及び肺を生成する。外胚葉は、神経系、皮膚及び副腎組織を生じさせる。これらの胚細胞層からの組織発達のプロセスは、複雑な分子変化を反映する複数の分化工程を含んでいる。中胚葉及びその誘導体に関して、3つの別個のステージが規定されてきた。その第1は、胚盤葉上層として公知の構造内の細胞からの中胚葉の誘導である。新生中胚葉としても公知である新規に形成された中胚葉は、初期胚内の将来の組織発達の部位となる様々な位置へ移動する。このプロセスは、パターニングとして公知であり、特定組織に向かう初期分化ステージを反映する可能性が高い一部の分子変化を必要とする。最終ステージは、特異化として公知であり、パターン化された中胚葉亜集団からの別個の組織の発生を含んでいる。
【背景技術】
【0004】
これまでに得られた証拠は、中胚葉が別個の発生能を備える亜集団を表す連続する波のように誘導されることを示唆している。最初に形成された中胚葉は胚外領域へ移動して造血細胞及び内皮細胞を生じさせるが、次の集団は発生中の胚内で前方へ移動して心臓及び頭部の間葉となる。これらの分化系列関係は、最初は組織学的分析を通して規定され、その後は主として細胞追跡試験によって確証されてきた。造血への関与に関しては、現在ではES細胞分化モデルからの研究、及びマウス胚上での研究から、最初に同定できる前駆細胞は血管芽細胞として公知の細胞であり、血管能力もまた示す細胞であるという強力な証拠が得られている(非特許文献1、非特許文献2)。この前駆細胞についての分析は、この前駆細胞が中胚葉遺伝子ブラキュリ及び受容体チロシンキナーゼFlk−1を共発現することを明らかにしたが、これは中胚葉の亜集団が造血及び血管分化系列へ関与することを表すことを示している(非特許文献3)。分化系列追跡試験は、心臓がFlk−1+集団から発達することを証明したが、これは心血管系のための同等の多能性細胞が存在する可能性があることを示唆している(非特許文献4)。ES細胞分化培養の分析は、心臓及び内皮潜在力を備えるFlk−1+前駆細胞の存在についての証拠を提供した(非特許文献5)。近年の試験もまた、複数の心血管分化系列を生じさせることのできるマウス心血管前駆細胞の存在を支持している(非特許文献6、非特許文献7、非特許文献8)。ヒト心血管前駆細胞集団は、これまで同定されていない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Choi et al.(1998);Development 125:725−732
【非特許文献2】Huber et al.(2004)Nature 432:625−30
【非特許文献3】Fehling et al.(2003)Development 130:4217−4227
【非特許文献4】Ema et al.(2006)Blood 107:111−117
【非特許文献5】Yamashita et al.(2005)FASEB 19:1534−1536
【非特許文献6】Kattman et al.(2006)Dev. Cell 11:723−732
【非特許文献7】Moretti et al.(2006)Cell 127:1151−1165
【非特許文献8】Wu et al.,(2006)Cell 127:1137−1150
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、ヒト心血管前駆細胞の濃縮集団を提供する。ヒト心血管前駆細胞は、細胞表面マーカーであるKDRの存在及び細胞表面マーカーであるC−KITの非存在を特徴とする。ヒト心血管前駆細胞は、in vitro及びin vivoで心筋細胞、内皮細胞、ならびに血管平滑筋細胞に分化することができる。
【課題を解決するための手段】
【0007】
また別の実施形態では、本発明は、ヒト胚様体(EB)からヒト心血管前駆細胞を産生する方法であって、前記EBをアクチビン及び骨形成因子(BMP)及び任意で塩基性線維芽細胞成長因子(bFGF)の存在下で培養する工程と、引き続いてヒト心血管前駆細胞を提供するためにWnt成長因子の阻害剤及び任意で血管内皮細胞成長因子(VEGF)の存在下で培養する工程と、ならびに前記細胞培養から前記細胞を収集する工程とを含む方法を提供する。
【0008】
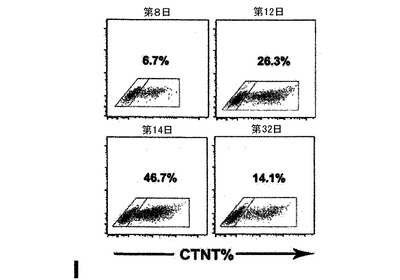
また別の実施形態では、本発明は、少なくとも約30%及び好ましくは少なくとも約40%、及びより好ましくは少なくとも約50%のヒト心筋細胞を含有する細胞集団を発生させる方法であって、前記ヒト心血管前駆細胞をWntの阻害剤及び任意でVEGFの存在下で培養する工程を含む方法を提供する。一実施形態では、細胞は、単層として培養される。また別の実施形態では、細胞は、凝集体として培養される。心筋細胞の存在は、細胞の収縮活性を評価する工程によって、または例えば心トロポニンT(CTNT)などの心筋細胞の指標となる遺伝子の発現を測定する工程によって決定できる。
【0009】
本発明は、心筋細胞、内皮細胞及び血管平滑筋細胞を含有するヒト心血管コロニーを発生させる方法であって、前記ヒト心血管前駆細胞をVEGF、bFGF、及びWntの阻害剤の存在下で培養する工程を含む方法をさらに提供する。好ましい実施形態では、細胞は、メチルセルロース中で培養される。
【0010】
ヒト心血管前駆細胞及びそれに由来する分化細胞は、心血管前駆細胞、心筋細胞、内皮細胞、血管平滑筋細胞、及び心血管組織に影響を及ぼす薬剤についてスクリーニングする方法において有用である。
【0011】
ヒト心血管前駆細胞は、また増殖及び貯蔵のため、ならびに心筋細胞、内皮細胞及び血管平滑筋細胞の起源としても有用である。
【0012】
ヒト心血管前駆細胞及びそれに由来する分化細胞は、また哺乳動物における心臓機能を改善もしくは再構成するため、及び移植のためにin vitroで心血管組織を操作するための方法においても有用である。
【図面の簡単な説明】
【0013】
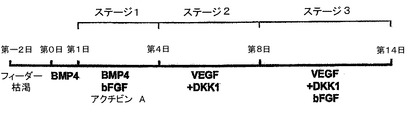
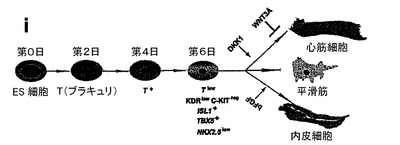
【図1】本発明の一実施形態によってヒト胚幹細胞(hESC)から心臓分化系列への分化を方向付けるために使用されるプロトコルの略図である。
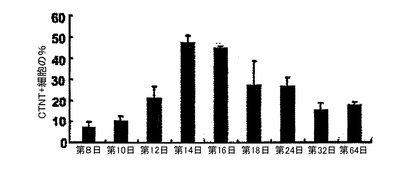
【図2】BMP4、bFGF及びアクチビンAの組み合わせを用いて誘導されたEB内でのCTNT+細胞発達の動態を示す。EBを、表示時点に収集し、CTNTを発現する細胞の頻度について細胞内フローサイトメトリーによって分析した。
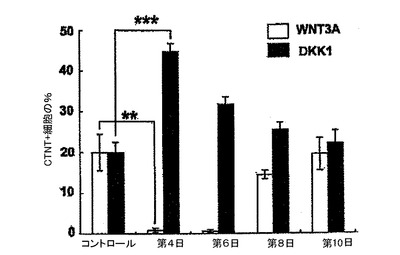
【図3】異なるステージでのWntシグナル伝達経路の操作後の14日齢EB内でのCTNT+細胞の頻度を示す。
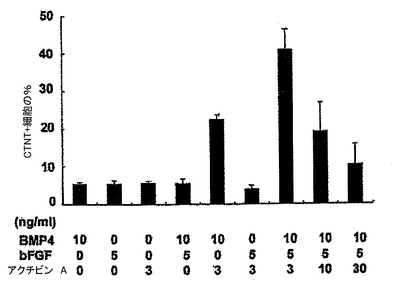
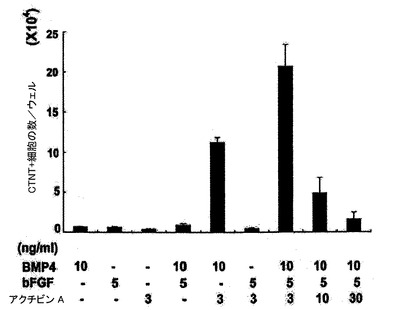
【図4】誘導ステージ中のBMP4、アクチビンA及びbFGFの組み合わせにおける培養後の、14日齢EB内でのCTNT+細胞の頻度を示す。
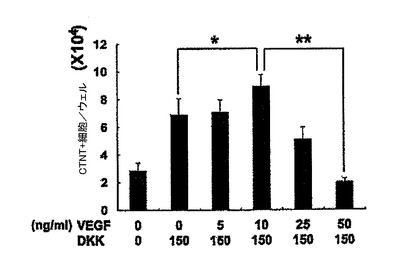
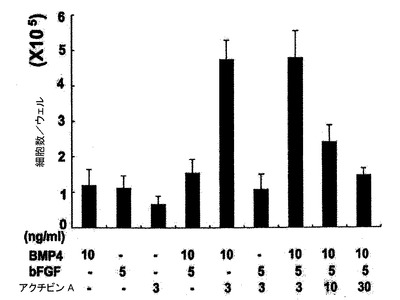
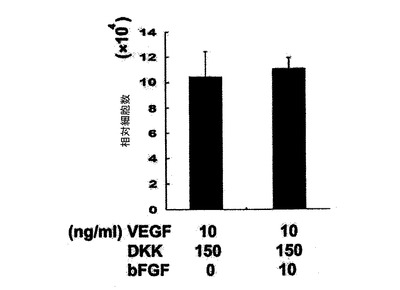
【図5】VEGF及びDKK1の表示した組み合わせを用いて処置した14日齢EBについての24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。図1〜5については、バーは、3回の独立実験の標準誤差または平均値を表す。統計的分析は、対応のないt検定を用いて実施した。*P=0.07、**P<0.01、***P<0.001。
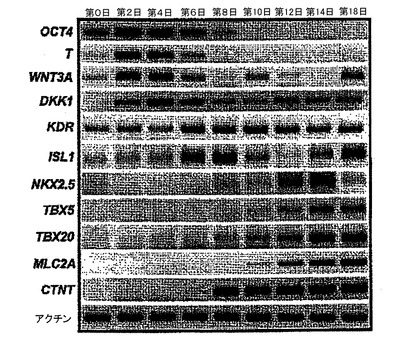
【図6】異なる発達ステージでのEBの遺伝子発現分析を示す。
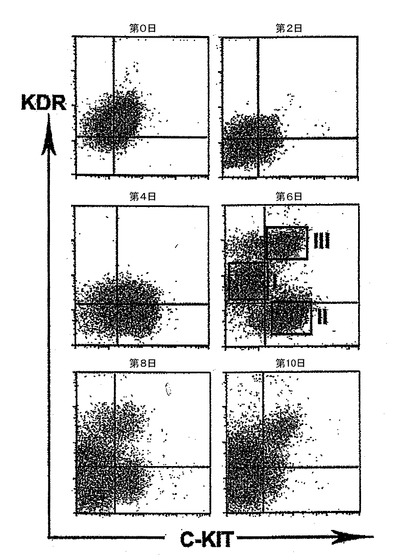
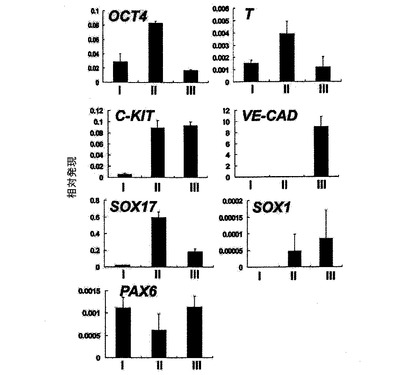
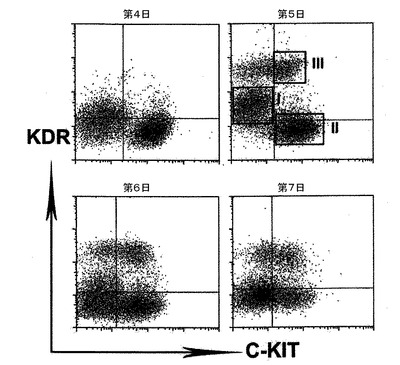
【図7】異なる日齢でのEBのフローサイトメトリー分析を示す。
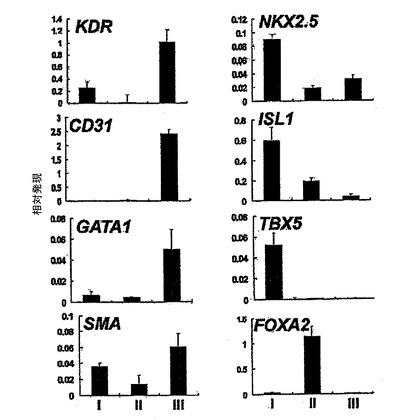
【図8】6日齢EBから単離した3つの集団の定量的RT−PCR遺伝子分析を示す。
【図9】3つの6日齢集団の心臓電位を示す。
【図10】6日齢KDRlow/C−KITneg集団の心臓電位を示す。
【図11】6日齢EB由来KDRlow/C−KITneg分画から発生した付着性集団の定量的RT−PCR発現分析を示す。
【図12】KDRlow/C−KITneg集団の培養の10〜12日後のCTNT+、CD31+及びSMA+細胞の存在についての分析を示す。
【図13】混合RFP/GFP培養から単離された4日齢心血管コロニーの発現分析を示す。
【図14】プレーティングしたKDRlow/C−KITneg由来細胞の数と発達する心血管コロニーの数との関係を示す細胞用量反応を示す。
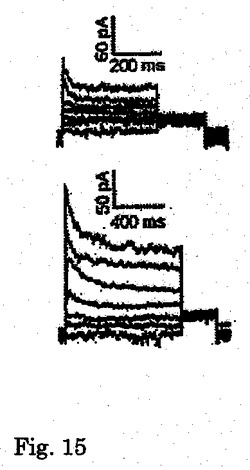
【図15】KDRlow/C−KITneg由来心筋細胞中で自然に発現した一過性外向きK+電流(Ito)の全細胞電位固定記録の代表的追跡を示す。
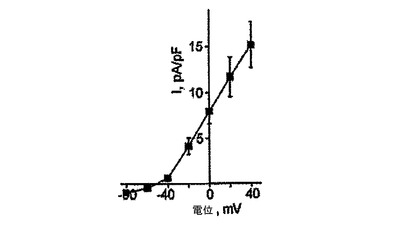
【図16】図15に示したように、細胞についての平均電流密度−電位関係を示す。1バッチの細胞10個中8個はIto電流を示し、平均値±SEM電流密度はこれらの8個の細胞からの追跡を用いてプロットした。
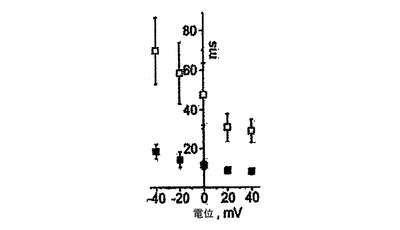
【図17】図16におけるような細胞についてのピーク電流までの平均時間(黒い四角)及び不活性化τ(白い四角)を示す(n=8)。
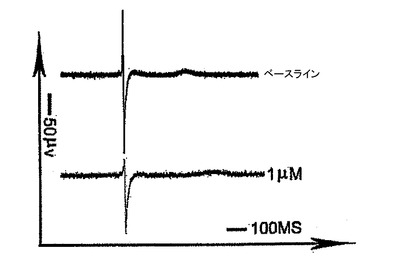
【図18】2〜3週間にわたってMEA(Multi Channel Systems)培養皿内で培養した6日齢EBから単離したKDRlow/C−KITneg細胞についての電気活動を示す。細胞外電気活動は、1μMキニジン(Sigma)を用いて、または用いずに記録した。
【図19】hESC培養内での心血管前駆細胞の発達を示すモデルを示す。
【図20】未分化hES2細胞、4日齢EB及び収縮性心筋細胞を備える14日齢EBの画像を示す(倍率200倍)。
【図21】EB内でのCTNT+細胞発達の動態を示す。
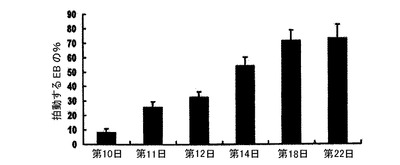
【図22】図1に示したプロトコルを用いて誘導された収縮性EB発達の動態を示す。150〜200個のEBが、収縮性EBの存在について各時点でスコア付けされた。バーは、3回の独立実験の平均値の標準誤差を表す。
【図23】グラフの下方に示した因子の組み合わせを用いて誘導した後の14日齢EB内での24ウェルプレートの1ウェル当たりの細胞総数を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
【図24】グラフの下方に示した因子の組み合わせを用いて誘導した後の14日齢EB内での24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
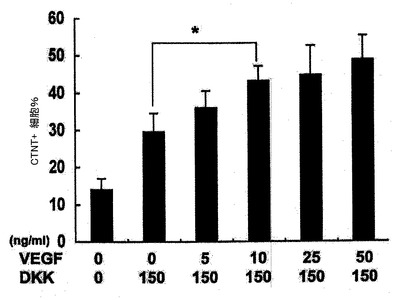
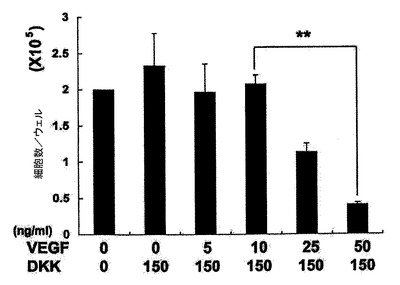
【図25】VEGF及びDKKの表示した組み合わせを用いて処置した14日齢EB由来のCTNT+細胞の頻度を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
【図26】VEGF及びDKKの表示した組み合わせを用いて処置した14日齢EBについての24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。
【図27】6日齢のEBから単離した3つの集団の定量的RT−PCR分析を示す。シクロフィニン(cyclophinin)に対して標準化した平均発現を示した。バーは、3回の独立実験の平均値の標準誤差を表す。
【図28】6日齢EBから単離されたKDRlow/C−KITneg集団の単層培養中での10〜14日後の相対細胞数のグラフである。96ウェルプレートの1ウェル当たり40,000個の細胞は、グラフの下方に示した因子の組み合わせを用いて誘導した。バーは、3回の独立実験の平均値の標準誤差を表す。
【図29】H1 hESCから発生したEB内での3つの異なるKDR/C−KIT集団の発生を証明しているフローサイトメトリー分析を示す。
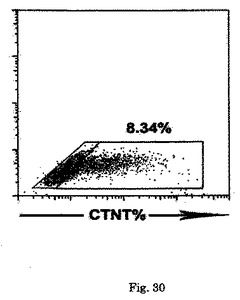
【図30】7日間の培養後のH1 hESC由来KDRlow/C−KITneg分画からのCTNT+細胞の発達を示す。
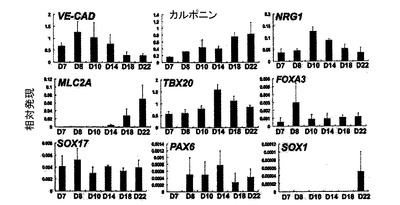
【図31】VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)の存在下で培養されたKDRlow/C−KITneg由来付着性集団の定量的RT−PCR発現分析を示す。培養は規定時点に収集され、表示した遺伝子の発現について分析された。シクロフィニンに対して標準化した平均発現を示した。
【図32】hES2由来KDRlow/C−KITneg集団から発生した個々の4日齢心血管コロニーの発現分析を示す。
【発明を実施するための形態】
【0014】
本発明は、ヒト心血管前駆細胞の集団を提供する。本発明のヒト心血管前駆細胞は、適切な条件下では、心筋細胞、内皮細胞、及び血管平滑筋細胞へ分化することができる。ヒト心血管前駆細胞は、チロシンキナーゼ受容体KDRの発現及びチロシンキナーゼ受容体C−KITの発現の欠如を特徴とする。
【0015】
本発明の一実施形態では、ヒト心血管前駆細胞は、胚様体(EB)から入手される。EBは、広範囲の分化系列から発達する集団を含有する三次元コロニーである。胚幹細胞(ESC)からEBを形成するための条件は、当技術分野において公知である。例えば、胚フィーダー細胞上に維持されたヒトESCは、例えばMatrigel(商標)(BD Biosciences、マサチューセッツ州ベッドフォード)などの基底膜マトリックスの層上で培養し、小クラスターを形成するために解離させ、そしてBMPの存在下で無血清培地中においてプレーティングすることによってフィーダーを枯渇させることができる。本発明の好ましい一実施形態では、フィーダーを枯渇させたヒトESCがEBを形成するために約1日間にわたりBMP4の存在下で無血清培地中にて培養される。また別の好ましい実施形態では、BMP4の濃度は、約0.1ng/ml〜約1.0ng/ml、及びより好ましくは約0.5ng/mlである。
【0016】
ヒトESCは、市販で、または当技術分野において公知の方法によって入手できる。例えば、ヒトESCは、当技術分野において公知の方法、及び例えばThomson et al.の米国特許第5,843,780号明細書に開示された方法、及びReubinoff et al.(2000)Nature Biotech 18:399による方法によって胚盤胞から調製できる。
【0017】
EBは、当技術分野において公知の方法、及び例えばTakahashi et al.(2007)Cell 131:861及びMali et al.(2008)Stem Cells 26:1998によって開示された方法によって誘導多能性幹細胞(iPS)から形成することもできる。
【0018】
本発明のヒト心血管前駆細胞は、ヒトEBをアクチビン及びBMPの存在下で無血清培地中において培養する工程と、次に心血管前駆細胞の集団を提供するためにWntの阻害剤の存在下で培養する工程と、及び任意で前記培養から細胞を収集する工程と、さらに任意で心血管前駆細胞のための収集された細胞集団を濃縮する工程とによって入手することができる。好ましい実施形態では、アクチビンはアクチビンAであり、BMPはBMP4であり、Wntの阻害剤はDickkopf−1(DKK−1)である。
【0019】
本発明のまた別の実施形態は、ヒト心血管前駆細胞を産出するための方法であって、ヒトEBを約1〜約4日間にわたりアクチビンA及びBMP4及び任意のbFGFの存在下で無血清培地中において培養する工程と、及び引き続き前記細胞を約1〜2日間にわたりDKK−1及び任意のVEGFの存在下で無血清培地中において培養する工程と、細胞集団を収集する工程と、ならびに任意でヒト心血管前駆細胞について前記集団を濃縮する工程とを含む方法を提供する。
【0020】
好ましい実施形態では、EBは、アクチビンA及びBMP4及び任意のbFGFの存在下で約4日間にわたり培養される。アクチビンA、BMP4及び任意のbFGFの濃度は、T及びWNT3Aの発現のアップレギュレーションを特徴とする細胞の原始線条様集団を誘導するために十分である。その後の工程では、DKK−1及び任意のVEGFの濃度は、心血管前駆細胞の集団を提供するために十分である。
【0021】
好ましい実施形態では、アクチビンAの濃度は、約0.5ng/ml〜約30ng/ml、及びより好ましくは約1.0ng/ml〜約5.0ng/mlである。最も好ましい実施形態では、アクチビンAの濃度は、約3.0ng/mlである。
【0022】
好ましい実施形態では、BMP4の濃度は、約1.0ng/ml〜約20.0ng/ml、及びより好ましくは約5.0ng/ml〜約15ng/mlである。最も好ましい実施形態では、BMP4の濃度は、約10ng/mlである。
【0023】
好ましい実施形態では、bFGFの濃度は、約1.0ng/ml〜約10.0ng/ml、及びより好ましくは約3.0ng/ml〜約6.0ng/mlである。最も好ましい実施形態では、bFGFの濃度は、約5.0ng/mlである。
【0024】
好ましい実施形態では、DKK1の濃度は、約100ng/ml〜約200ng/ml、及びより好ましくは約125ng/ml〜約175ng/mlである。最も好ましい実施形態では、DKK1の濃度は、約150ng/mlである。
【0025】
別の好ましい実施形態では、VEGFの濃度は、約1.0ng/ml〜約50.0ng/ml、及びより好ましくは約1.0ng/ml〜約25.0ng/mlである。最も好ましい実施形態では、VEGFの濃度は、約10.0ng/mlである。
【0026】
本発明のまた別の好ましい実施形態では、ヒトEBは、約4日間にわたり無血清培地中、約3.0ng/mlのアクチビンA、約10.0ng/mlのBMP4及び約5.0ng/mlのbFGFの存在下で培養されるが、その時点に約150ng/mlのDKK1及び約10.0ng/mlのVEGFが加えられ、細胞は心血管前駆細胞を産生するために約1〜2日間にわたり培養される。また別の好ましい実施形態では、前記集団は、例えばKDRを発現するがC−KITを発現しない細胞を選別することによって、心血管前駆細胞に濃縮される。好ましい実施形態では、この選別は、免疫選択によって、例えばフローサイトメトリーによって行われる。
【0027】
また別の実施形態では、本発明は、ヒト心筋細胞の集団を発生させる方法であって、前記ヒト心血管前駆細胞を約7〜約10日間にわたりWntの阻害剤及び任意にVEGFの存在下で培養する工程と、及び培養からヒト心筋細胞の集団を収集する工程とを含む方法を提供する。好ましい実施形態では、Wntの阻害剤はDKK1であり、DKK1及びVEGFの好ましい濃度は、上記に記載したとおりである。1つの好ましい実施形態では、細胞は、ゼラチンコーティングウェル内の単層としてプレーティングされる。また別の実施形態では、細胞は、凝集体として低クラスター培養皿内で培養される。ヒト心筋細胞の集団は、少なくとも約30%、好ましくは少なくとも約40%、及びより好ましくは少なくとも約50%の心筋細胞を含有している。他の実施形態では、ヒト心筋細胞の集団は、少なくとも約70%、好ましくは少なくとも約80%、より好ましくは少なくとも約90%、及び100%までの心筋細胞を含有している。心筋細胞は、細胞の収縮活性を評価する工程によって、または例えばCTNTなどの心筋細胞の指標となる遺伝子の発現を測定する工程によって同定できる。
【0028】
本発明のヒト心血管前駆細胞はさらにまた、当業者には公知の分化条件を用いて、例えば心房、心室、及びペースメーカー細胞を含む心筋細胞の亜集団を発生させるために有用である。
【0029】
また別の実施形態では、本発明は、心筋細胞、内皮細胞及び血管平滑筋細胞を含有する心血管コロニーを発生させる方法であって、前記ヒト心血管前駆細胞を約3〜約7日間にわたりVEGF、bFGF、及びWntの阻害剤、好ましくはDKK1の存在下で培養する工程を含む方法を提供する。好ましい実施形態では、細胞は、メチルセルロース中にプレーティングされる。VEGF、bFGF及びDKKの好ましい濃度は、上記に記載したとおりである。心筋細胞、内皮細胞、及び血管平滑筋細胞の存在は、例えばCTNTなどの心筋細胞の指標となる遺伝子、及び例えばCD31、VE−カドヘリンなどの内皮細胞の指標となる遺伝子ならびに例えばSMA及びカルポニンなどの血管平滑筋細胞の指標となる遺伝子の発現を測定する工程によって決定できる。
【0030】
本発明は、ヒト心血管前駆細胞、心血管コロニー、心筋細胞、内皮細胞及び血管平滑筋細胞に作用を及ぼす薬剤についてスクリーニングするための方法をさらに提供する。本方法は、上記に記載した細胞集団の1つに由来する細胞を候補薬と接触させる工程と、及び前記薬剤が前記細胞集団に作用を及ぼすかどうかを決定する工程とを含んでいる。試験対象の薬剤は、天然もしくは合成、1つの化合物もしくは混合物、低分子またはポリペプチド、多糖類、ポリヌクレオチドなどを含むポリマー、抗体もしくはそのフラグメント、1ライブラリーの天然もしくは合成化合物からの化合物、理論的薬物設計から得られた化合物、例えば細胞培養条件などの条件、または細胞集団に及ぼす作用を当技術分野において公知のアッセイを用いて評価できる任意の薬剤であってよい。細胞集団に及ぼす作用は、例えば、マーカー発現、受容体結合、収縮能力、電気生理学、細胞生存能力、生存率、形態、またはDNA合成もしくは修復についてのアッセイを含む、表現型もしくは活性についての任意の標準アッセイによって決定することができる。標準増殖及び分化アッセイは、米国特許第6,110,739号明細書に記載されている。そのような薬剤は、in vivo及びin vitroでの細胞成長、分化及び生存率の制御、ならびに組織の維持、再生及び修復のために有用である。
【0031】
本発明は、ヒト心血管前駆細胞の集団を含む組成物及びヒト心血管コロニーの集団を含む組成物をさらに提供する。本組成物は、薬学的に許容される担体及び希釈剤を含むことができる。本組成物は、移植を促進する成分をさらに含むことができる。これらの集団を含む組成物は、細胞及び組織の置換及び修復のため、ならびに心筋細胞、内皮細胞、及び血管平滑筋細胞の集団をin vitro及びin vivoで発生させるために有用である。ヒト心血管前駆細胞を含む組成物は、前駆細胞集団を拡張させるために有用である。本組成物は、心臓の状態を治療するための医薬品または送達装置として調製できる。
【0032】
また別の実施形態では、本発明は、例えば、先天性心疾患、冠動脈性心疾患、心筋症、心内膜炎及びうっ血性心不全を含む不十分な心臓機能を特徴とする障害を治療するために有用な細胞置換方法及び組織置換方法を提供する。分化細胞及び心血管前駆細胞はどちらも置換療法のために有用であるが、それは前駆細胞集団が心筋細胞、内皮細胞及び血管平滑筋細胞分化系列へin vivoで分化できるためである。これらの細胞はさらにin vitroで心血管細胞を発生させるためにも有用である。心臓組織を操作するための方法は当技術分野において公知であり、例えば、Birlaによって“Stem Cell Therapy and Tissue Engineering for Cardiovascular Repair” Springer, 2006で概説されている。
【0033】
したがって、一実施形態では、本発明は、心筋細胞置換療法のための方法であって、当該治療を必要とする被験者に本発明によって得られたヒト心血管前駆細胞が濃縮された細胞集団から単離された心筋細胞を含む組成物を投与する工程を含む方法を提供する。また別の実施形態では、本発明は、不十分な心臓機能を特徴とする障害を治療する方法であって、当該治療を必要とする被験者にヒト心血管前駆細胞を含む組成物を投与する工程を含む方法を提供する。好ましい実施形態では、被験者は、ヒトである。本組成物は、心臓組織への送達もしくは遊走を生じさせる、例えば注射もしくは移植を含む経路によって、そして少なくとも1つの有害作用もしくは症状または障害の減少を生じさせる条件下で投与することができる。
【0034】
本明細書に引用した全参考文献は、開示内容全体が参考として本明細書で援用される。
【実施例】
【0035】
以下の実施例は、本発明をさらに具体的に説明するために役立つ。
【0036】
(実施例1)
材料及び方法
ヒトES細胞の維持
WiCell Research Institute(ウィスコンシン州マディソン)からのH1(NIHコード番号WA01)及びESI International(シンガポール国)からのhES2(NIHコード番号ES02)を、6ウェル組織培養プレート内で20%ノックアウト血清代替物(SR)、100μM非必須アミノ酸、2mMグルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン(Invitrogen、ニューヨーク州グランドアイランド)、10−4M β−メルカプトエタノール(Sigma、ミズーリ州セントルイス)及び20ng/mlのhbFGF(R&D Systems、ミネソタ州ミネアポリス)が補充されたダルベッコ変法イーグル培地(DMEM)/F12(50:50;Mediatech、バージニア州ハーンドン)からなるhESC培地中にて、照射マウス胚フィーダー細胞上で維持した。AAVS1標的化hES2細胞系は、親hES2細胞のAAV2−TRUF11(CAG−GFP−TK−neo)及び野生型AAV2両方の106ウイルス粒子との共感染によって発生させた。G418選択後、GFP陽性細胞を選別し、サブクローンを単離した。トランス遺伝子の標的化組込みは、ライゲーション媒介ポリメラーゼ連鎖反応(LM PCR)によって確認された。野生型AAV配列は、GFP陽性クローン中では検出されなかった。RFP発現hES細胞は、Irion et al.(2007)Nat. Biotechnol. 12:1477−1482によって記載されたようなhES2細胞のRosa26遺伝子座へRFP cDNAを標的化する工程によって生成した。
【0037】
ヒトES細胞の分化
EBを発生させる前に、hESCは、フィーダーを枯渇させるためにMatrigel(商標)(BD Biosciences、マサチューセッツ州ベッドフォード)の層上で48時間にわたり培養した。Matrigel(商標)培養からの細胞はコラゲナーゼB(1mg/ml、Roche、インディアナ州インディアナポリス)を用いた20分間の処理、その後のトリプシン−EDTA(0.05%)を用いた2分間の処理によって小クラスターに解離させた。第0〜1日から、EBは2ml塩基性培地(StemPro−34(Invitrogen)、2mMのグルタミン、4×10−4Mのモノチオグリセロール(MTG)、及び50μg/mlのアスコルビン酸(Sigma)+0.5ng/mlのBMP4(R&D Systems)中にプレーティングした。以下の濃度の因子をEB形成、中胚葉誘導及び心臓特異化のために使用した。BMP4−10ng/ml、hbFGF−5ng/ml、アクチビンA−3ng/ml、hDKK1−150ng/ml、hVEGF−10ng/ml。これらの因子は、以下の順序で加えた。第1〜4日には、BMP4、hbFGF及びアクチビンA、第4〜8日には、VEGF及びDKK1、第8日以降には、VEGF、DKK1、及びbFGF。培地は、第8日以降には4日毎に取り替えた。全ヒト因子及び阻害剤は、R&D Systems(ミネソタ州ミネアポリス)から購入した。培養は、培養の初期10〜12日間は5%CO2、5%O2、及び90%N2環境内で維持し、その後は5%CO2/気体環境内で維持した。
【0038】
フローサイトメトリーによる選別及びMatrigel(商標)上での管形成
EBを収集し、トリプシン(0.25%トリプシン−EDTA)を用いて単一の細胞に解離させた。トリプシン処理後、Facscaliburフローサイトメーター(Becton Dickinson)を用いて分析を実施した。細胞は、MoFlo(Dako Cytomation、コロラド州フォートコリンズ)セルソーターを用いて選別した。フローサイトメトリーデータは、FlowJo(Treestar、カリフォルニア州サンカルロス)ソフトウエアプログラムを用いて分析した。抗KDR−PE、抗−C−KIT−APCは、R&D Systemsから購入した。選別したKDRlow/C−KITneg集団は、VEGF(25ng/ml)及びbFGF(25ng/ml)を補充したStempro34培地中で分化させた。管状構造は、Matrigel(商標)コーティングガラス製カバースリップへ移してから24時間以内に形成された。
【0039】
免疫蛍光
解離した細胞は、2日間にわたりガラス製カバースリップ上で培養した。この時点で、細胞を4%PFAで固定し、次に染色した。以下の抗体を免疫染色のために使用した。R&D Systemsからの抗ヒトCD31、抗ヒトVE−カドヘリン、Lab Vision(カリフォルニア州フリーモント)からの抗マウストロポニンT及び抗ヒト平滑筋アクチン、Chemicon(カリフォルニア州テメキュラ)からの抗ヒトANP、コネキシン43及び抗ヒトα/β MHC抗体、Sigma(ミズーリ州セントルイス)からの抗ヒトα−アクチニン。抗ヒト平滑筋ミオシン重鎖(SMHC)、カルデスモン、フォン・ヴィレブランド因子抗体は、DakoCytomation(カリフォルニア州カーピンテリア)から購入した。Cy2、Cy3及びCy5共役二次抗体は、Jackson ImmunoResearch(ペンシルベニア州ウェストグローブ)から入手した。蛍光は、Leica DMRA2蛍光顕微鏡(Leica、ドイツ国ウェッツラー)を用いて視認し、画像はデジタル式Hamamatsu CCDカメラ(日本国浜松市)を使用して記録した。
【0040】
コロニーアッセイ
心血管コロニーを発生させるために、6日齢EBから単離したKDRlow/C−KITneg細胞は、VEGF(25ng/ml)、bFGF(10ng/ml)及びDKK1(150ng/ml)の存在下で2〜3日間にわたり凝集させた。このステージに、凝集体を解離させ、細胞は5%CO2、5%O2、及び90%N2環境内においてVEGF(25ng/ml)、bFGF(25ng/ml)及びDKK1(150ng/ml)を含有するメチルセルロース中で培養した。コロニーは、コロニー頻度分析のために4〜6日間の培養後にスコア付けした。
【0041】
RT−PCR
発現試験のためには、個々のコロニーをメチルセルロース培養から単離し、Brady et al.(1993)Methods Enzymol. 225:611−623によって以前に記載されたプロトコルの修正版を用いて分析した。増幅cDNAを次に標準PCRにかけた。リアルタイム定量的PCRは、MasterCycler EP RealPlex(Eppendort)上で実施した。実験は白金SYBR GreenER qPCR SuperMix(Invitrogen)を用いて3回ずつ実施した。全プライマーは、表1に記載した。全アニーリング反応は、60℃で実施した。
【0042】
【表1−1】
【0043】
【表1−2】
【0044】
【表1−3】
【0045】
KDRlow/C−KITneg由来集団のin vivo分析
hESCを発現するGFP由来のKDRlow/C−KITneg細胞を移植前5〜10日間にわたって、VEGF(10ng/ml)、bFGF(10ng/ml)及びDKK1(150ng/ml)の存在下で培養した。NOD/SCIDγマウスに麻酔をかけ、気管挿管し、100,000個のドナー細胞を開胸手術において左心室壁内へ直接注射した。手術の2〜11週後に心臓を収集し、カコジル酸バッファー中の1%パラホルムアルデヒド中で固定し、300μmでビブラトーム(Vibrotome)にて切片化を行った。移植領域を同定し、10μmで凍結切片化を行った。免疫組織化学検査は、GFP抗体(Chemicon、AB3080、1:100;Vector ABC及びDABキット)、α−アクチニン抗体(Sigma、A7811、1:500;Chemicon AQ300R二次抗体、1:20)、CD31抗体(Dako、M0823、1:50;Vector ABC及びDABキット)、ならびに平滑筋MHC抗体(Biomedical Technologies、BT−562、1:300;Alexa 555二次抗体、1:200)について実施した。共焦点像は、ImageJ and Pierre Bourdoncleのプラグインをデフォルト設定で使用して共局在化について分析した。マウス梗塞モデルにおける評価のためには、心筋梗塞はRubert et al.(2007)Nature Biotechnol. 25:993−994によって記載された技術を使用して、直接的な冠状動脈結紮によってSCIDベージュマウスにおいて誘導した。10〜20分後に、マウスに500,000個のKDRlow/C−KITneg由来細胞(n=9)または同等量の無血清培地(n=12)を注射した。全注射は、梗塞の境界帯で試みた。2週間後、心室機能の評価は、9.4テスラの磁気共鳴イメージングを用いて実施した。
【0046】
パッチクランプ
全細胞パッチクランプ記録は、IX50倒立顕微鏡(Olympus)、Multiclamp 700A増幅器、Digidata 1300アナログ/デジタル変換器及びpClamp9.1ソフトウエア(Axon Instruments、カリフォルニア州フォスターシティ)を備えるPCを用いて室温で実施した。浴溶液(単位:mM)は、NaCl 136、KCl 4、CaCl2 1、MgCl2 2、CoCl2 5、HEPES 10、グルコース10、及びテトロドトキシン(TTX)0.02(pH7.4)であった。ピペット(単位:mM)は、KCl 135、EGTA 10、HEPES 10、及びグルコース5(pH7.2)を含有する細胞内溶液を充填すると3〜5MΩ抵抗のピペットであった。細胞を、−30mVのテールパルス(図4におけるような持続期間)前には、−80mVから+40mVへの電位を20mV増分で試験するために−80mVの保持電位から段階付けた。データは、pClamp9.1ソフトウエア(Axon Instruments)を用いて分析した。電流振幅は、細胞サイズに標準化した(全細胞膜容量)。不活性化τ値は、電流減衰の指数関数的適合を用いて計算した。
【0047】
電場電位の記録
6日齢EBから単離したKDRlow/C−KITneg細胞は、2〜4週間にわたりStemPro−34+10ng/mlのVEGF及び150ng/mlのDKK1を備えるMEA(Multi Channel Systems)培養皿内で培養した。記録の2日前に、細胞は15%FBSを備えるDMEM(細胞増殖用)へ切り換えた。細胞外電気活動は、60チャンネルから同時に記録し、ソフトウエアMC Rack(Multi Channel Systems)を用いて分析した。
【0048】
(実施例2)
hESC分化培養中での心臓発達の調節
hESCから心臓分化系列への分化を方向付けるために、原始線条様集団の誘導、心臓中胚葉のパターニング及び心血管分化系列の特異化を含む段階的プロトコルは、様々なモデル系において中胚葉誘導及び心臓特異化で役割を果たすことが公知である因子のBMP4、アクチビンA、VEGF、bFGF及びDKK1の組み合わせを用いて設計した。プロトコルの概略は、図1に示した。本明細書に例示したプロトコルを用いると、hESCは、EBの形成を可能にするために24時間にわたりBMP4の存在下で最初に分化された。第1ステージでのアクチビンとBMP4との組み合わせは、マウスにおけるこれらの集団内で発現することが公知の遺伝子であるT(ブラキュリ)及びWNT3aのアップレギュレーション及び一過性発現によって証明されるように、原始線条様集団及び中胚葉を誘導した(図6)。第2ステージでは、VEGF及びWnt阻害剤であるDKK1が中胚葉を更にパターン化し、KDR+前駆細胞集団の拡張及び成熟を促進した。分化の第8日には、発達中の心血管分化系列の持続的拡張を支援するためにbFGFが再び加えられた。このプロトコルは、収縮細胞ならびに心臓トロポニンT(CTNT)(図2)、α−アクチニン、α/βミオシン重鎖、ANP及びコネキシン43(図20、21)を発現した細胞の出現によって証明されたように、EB内の心臓発達を支援する。CTNT+細胞の最高頻度は培養の第14〜16日に定期的に観察されたが(図2、21)、この集団はEB内で2カ月間の期間にわたって維持することができた。収縮性EBの発達の動態は、CTNT+集団の出現と平行していた(図22)。
【0049】
(実施例3)
ヒトES細胞由来の心臓分化系列の発生にWntシグナル伝達が果たす役割
Wntシグナル伝達経路がヒトES細胞由来の心臓分化系列の出現に果たす役割を、原始線条誘導を越える段階(第2ステージ)に特に焦点を当てて調査した。EBは、プロトコルに略説したように、BMP4、bFGF及びアクチビンAの存在下で誘導した。Wnt3AまたはDKK1のいずれかを図3に示した日に培養へ加えた。これらの因子は、14日間を通して培養中で維持された。VEGF及びbFGFは、指示した時点に加えた。第14日に、EBを収集し、上述したように分析した。対照は、WNTまたはDKK1を受容しなかった培養であった。分化第4日におけるEBへのDKK1の添加は、培養第14日のCTNT+細胞の頻度の2倍を超える(40%までの)増加をもたらした(図3)。DKK1の作用は、第6または8日に加えられた場合はこれほど顕著ではなく、添加が分化第10日に添加された場合はCTNT+細胞の頻度の増強は見られなかった。Wnt3Aは反対の作用を有しており、第4または6日に培養に加えられた場合はCTNT+細胞の発達を完全に抑制した。これらの所見は、古典的Wnt経路のステージ特異的阻害がBMP4/アクチビン誘導性PS集団の心臓特異化を促進するために不可欠であることを示している。
【0050】
(実施例4)
心臓分化系列の誘導においてBMP4、アクチビンA及びbFGFが果たす役割
hESC分化培養中での心臓分化系列の誘導においてBMP4、アクチビンA及びbFGFが果たす役割を評価するために、誘導ステージ(第1〜4日)中に単一因子ならびに様々な組み合わせを試験した。第4日には、図1に略説したように、因子を除去し、EBを処理した。第4〜14日に、EBは図5、25、及び26に示したようにVEGF及びDKK1の組み合わせを用いて処理し、第8日以降にはbFGFを添加した。EBは第14日に収集し、各組み合わせについてのCTNT+細胞の頻度及び24ウェルプレートの1ウェル当たりのCTNT+細胞の総数について分析した。BMP4、bFGFもしくはアクチビンA単独またはBMP4及びFGFまたはアクチビンA及びbFGFの組み合わせは、低頻度(図4)及び生成されたCTNT+細胞の総数(図23、24)によって証明されたように心臓発達の貧弱な誘導因子であった。BMP4及びアクチビンAは有意な数のCTNT+細胞を誘導するが、これら3つの因子の組み合わせは最も強力であり、心臓細胞の最高の頻度及び総数を発生し、40%〜50%のCTNT+細胞からなる培養を常に生産した。VEGFは心臓発達のために必須ではなかったが、10ng/mlのこの因子の添加は発達したCTNT+細胞の総数を増加させた。この濃度を超えてVEGFの濃度が増加すると培養中の細胞数を減少させ、これは発生したCTNT+細胞の総数を有意に減少させた(図5、25及び26)。
【0051】
発達中のEBの分子分析は、原始線条様集団の確立後に発現パターンにおける動的変化を明らかにした。T及びWNT3Aと一緒に、DKK1の発現は早期にアップレギュレートされ、18日間の経過を通して持続した(図6)。分化の第2〜4日の内因性DKK1の発現は、心臓中胚葉を特異化するように機能することができ、外因性阻害剤の添加の不在下で心臓分化系列を確立させる(図3)。KDRは、未分化ES細胞内で発現した。この発現レベルは、第4〜6日に増加すると思われ、その後12日間にわたって持続された。初期胚中の2次心臓形成領域の前駆細胞をマークする遺伝子であるISL1は、分化第8日に最初に検出された心臓転写因子Nkx2.5の発現に先行して第6〜8日にEB中で発現した。適正な心臓発達に必要とされる2つのTBX転写因子であるTBX5及びTBX20、ならびに収縮性タンパク質MLC2A及びCTNTの発現は、分化の第8〜10日にアップレギュレートされ、これは心臓発達の開始を反映していた。
【0052】
(実施例5)
心血管電位を用いたKDRlow/C−KITneg EB集団の同定
発達中のEBをKDR及びC−KITの発現について分析した。C−KITを別個の集団を規定するための電位マーカーとして使用したが、それはマウスEB内でのC−KITの発現が最初期血球血管芽細胞由来造血前駆細胞及び血管前駆細胞ならびに前原始線条及び発達中の内胚葉を同定するからである。図7は、異なる日齢のEBのフローサイトメトリー分析を示す。図7に示すように、3つの別個の集団であるKDRhigh/C−KIT+、KDRlow/C−KITneg及びKDRneg/C−KIT+は、6日齢EB内で分解される可能性がある。3つの集団の発達は、BMP4及びアクチビンAの両方を用いた誘導に依存した(図示しない)。3つの集団を細胞選別によって単離し、遺伝子発現パターン及び心臓電位について分析した。図8は、6日齢のEBから単離した3つの集団の定量的RT−PCR遺伝子発現分析を示す。平均発現をシクロフィニンへ標準化した。KDRhigh/C−KIT+集団は、血管発達に関連する遺伝子であるCD31、VE−カドヘリン及びSMA、ならびに造血に関与することの指標となる遺伝子であるGATA−1を発現した(図8、27)。心臓発達に関係する、NKX2.5、ISL1及びTBX5を含む遺伝子は、KDRlow/C−KITneg分画内では最高レベルで発現した。この分画はSMA、極めて低レベルのGATA−1もまた発現したが、VE−カドヘリンもしくはCD31は検出しなかった。SMAは早期に心臓分化系列ならびにVSM分化系列内で発現するので、この発現は心臓発達の開始を反映できた。KDRneg/C−KIT+細胞は最高レベルの3分画のOCT4及びTを発現したが、これは残留未分化ES細胞ならびに原始線条様細胞の存在を示す。この分画内の比較的高レベルのFOXA2及びSOX17は、それが内胚葉分化系列に関与する細胞もまた含有していることを示唆している。SOX1及びPAX6は極めて低レベルでしか検出されず、これはこの発達ステージではEB内で神経外胚葉分化系列への分化があったとしてもほんのわずかに過ぎないことを示唆している(図27)。これらをまとめると、これらの発現パターンは、KDRhigh/C−KIT+集団が造血及び血管前駆細胞を含有する、KDRlow/C−KITneg集団が心臓前駆細胞を含有する、そしてKDRneg/C−KIT+集団が未分化ES細胞、原始線条細胞及び内胚葉細胞の混合物からなることを示している。
【0053】
心臓電位をモニターするために、細胞はゼラチンコーティングウェル上の単層としてプレーティングするか、または低クラスター培養皿内で再凝集させ、凝集体として培養した。詳細には、細胞は6日齢EBから単離し、VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)と共に単層としてゼラチンコーティングウェル上で培養した。CTNT+細胞は、10日間の培養後に分析した。結果を図9に示す。6日齢EBからのKDRlow/C−KITneg細胞を選別し、単層として、またはVEGF(10ng/ml)及びDKK1(150ng/ml)とともに低クラスターウェル内の凝集体として培養した。培養を収集し、7〜10日間にわたる分化後にCTNT+細胞の存在について分析した。結果を図10に示す。数(%)は、3回の独立実験の平均値±標準誤差を表す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.01、***P<0.001。KDRlow/C−KITneg集団は最大の心筋細胞電位(図9)を提示しており、懸濁培養中の凝集体内ならびに付着性単層内の両方において容易にCTNT+細胞及び収縮性細胞の集団を発生した(図10)。凝集体内のおよそ40%の細胞がCTNT+であったが、単層培養の50%超は一貫してこのマーカーを発現した。単層培養内の心筋細胞の高頻度は、同調塊(synchronous mass)として収縮する細胞層の発達を常にもたらした。単離されたKDRlow/C−KITneg細胞は、凝集体としてはおよそ1.5倍(データは示していない)、及び単層培養内では3倍に拡張した(図28)。略説した誘導プロトコル及び選別ストラテジーを用いると、4インプットのhESCに付き1の心筋細胞のアウトプットが予測された。典型的な実験では、2×106個のhESCは、1×106個の6日齢EB細胞を発生したが、そのうちの30%(3.0×9 05細胞)はKDRlow/C−KITneg分画を提示した。この集団は、単層培養フォーマット内で3倍(1×106細胞)に拡張し、およそ50%の心筋細胞(5×105)からなる集団を産出した。2次hESC系であるH1から発生したEBの動態分析は、分化第6日ではなく第5日に3KDR/C−KIT集団の発達を証明した。5日齢KDRlow/C−KITneg集団の分析は、それが心臓電位もまた示すことを示した(図29、30)。
【0054】
(実施例6)
KDRlow/C−KITneg由来分化系列の特性解析
VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)とともにプレーティングした6日齢EB由来KDRlow/C−KITneg分画から発生した付着性集団についての定量的RT−PCR発現分析を実施した。培養は図11に示した日に収集し、細胞を発現について分析した(D7はプレーティングの1日後の集団を表す)。シクロフィニンに対して標準化した平均発現を示した。バーは、3回の独立実験の平均値の標準誤差を表す。
【0055】
プレーティングから異なる日数後でのKDRlow/C−KITneg由来付着性集団についての発現分析(qPCR)は、内皮(CD31、VE−カドヘリン)、VSM(カルポニン、SMA)、心臓発達(NKX2.5、ISL1、TBX5、TBX20)及び心臓成熟(CTNT、MLC2A)に関連する遺伝子のアップレギュレーションを証明した(図11、31)。NFATC及びニューレグリン 1(NRGl)の発現は、培養中の心内膜の存在を示唆している。低レベルのNEUD、PAX6、SOX1、FOXA2、FOXA3、SOX17及びMEOX1の発現は、これらの培養が神経外胚葉、内胚葉または体中胚葉で汚染されていないことを示している(図11、31)。
【0056】
KDRlow/C−KITneg細胞を6日齢EBから単離し、図12のグラフの下方に指した因子の様々な組み合わせを用いて単層として培養した。細胞を収集し、10〜12日間にわたる培養後にCTNT+、CD31+及びSMA+細胞の存在について分析した。SMA抗体は、心筋細胞及び平滑筋細胞の両方を染色することができる。数(%)は、3回の独立実験の平均値の標準誤差を表す。
【0057】
VEGF及びDKK1中で10〜12日間培養したKDRlow/C−KITneg由来付着性細胞のフローサイトメトリー分析は、ほぼ90%がSMAを発現し、50%がCTNTを発現し、4%がCD31を発現することを明らかにした(図12)。培養へのbFGFの添加は、CTNT+細胞の比率を30%へ減少させ、CD31+集団を30%へ増加させた。SMA+細胞の比率は、bFGFの存在下で平均80%へ穏やかに減少した。bFGFの添加は、単層培養中で細胞数に有意には影響を及ぼさなかった(図28)。これらの所見は、KDRlow/C−KITneg由来集団内の細胞の大多数が心血管分化系列の細胞であることを明らかに示している。それらはさらに、bFGFがこの集団内で発達する心筋細胞及び内皮細胞の比率に影響を及ぼすことができることもまた示している。
【0058】
VEGF、DKK1及びbFGF中で培養したKDRlow/C−KITneg由来集団の免疫染色分析は、CD31+、VE−カドヘリン+及びフォン・ヴィレブランド因子+内皮細胞の存在、CTNT+心筋細胞の存在ならびにVSM発達の指標となるSMA+、SMHC+及びカルデスモン+細胞の存在を証明した。集団内の未成熟心筋細胞はCTNT及びSMAの両方を発現したが、他方VSM細胞はSMAしか発現しなかった。KDRlow/C−KITneg由来内皮細胞についてさらに特性解析するために、この集団をVEGF及びbFGFの存在下で拡張させ、次にMatrigel(商標)コーティングカバースリップ上で培養した。これらの条件下では、細胞は管状構造への内皮細胞の組織化の指標である格子を形成した。これらの構造内の細胞はCD31を発現し、Dil−AC−LDLを取り込む能力を示して、それらの内皮細胞表現型を確証した。免疫染色試験からの所見は、フローサイトメトリー分析所見と一致しており、KDRlow/C−KITneg由来集団が心臓、内皮及び血管平滑筋分化系列の細胞からなることを証明している。
【0059】
KDRlow/C−KITneg由来細胞のin vivoでの発達可能性を証明するために、それらをさらにNOD/SCIDマウスの心臓内へ移植した。これらの試験のためには、hES2細胞系のGFP発現バージョンを使用した。組織学的分析は、移植後に心臓内のGFP+細胞の存在を明らかにした。抗体を用いたGFPの検出は、落射蛍光によって視認された集団と同一集団を同定したが、これはGFPシグナルがドナー細胞の存在を表し、バックグラウンド蛍光の結果としては生じないことを示している。α−アクチン、CD31もしくはSMHCのいずれかを共発現するGFP+集団が移植片内で検出されたが、これは心臓、内皮及び血管平滑筋分化系列へのin vivoでの分化を示している。移植された動物のいずれにおいても奇形腫は検出されなかった(N>10)。この集団のin vivoでの潜在性をさらに評価するために、KDRlow/C−KITneg由来細胞をSCIDベージュマウスの梗塞心臓内へ直接的に移植した。2週間後に分析すると、KDRlow/C−KITneg由来心血管集団が移植された動物は、培地単独が注射された集団より31%高い駆出率を有していた(56%±3.6%対39%±4.8%、p=0.008)。これらの所見は、以前の報告書と一致しており、hESC由来心筋細胞の移植が心筋梗塞の齧歯類モデルにおける心臓機能の改善をもたらすことを証明している。
【0060】
(実施例7)
心血管コロニー形成細胞の同定及び特性解析
VEGF、bFGF及びDKK1の存在下においてメチルセルロース中でプレーティングすると、KDRlow/C−KITneg由来細胞は培養4日間以内に小さな小型コロニーを生成した。個々の4日齢コロニーのPCR分析は、心臓(CTNT)、血管(CD31及び/またはVE−カドヘリン)及びVSM(SMA及び/またはカルポニン)の発達の指標となるマーカーの共発現を示した(図13、32)。さらに6日間にわたり培養中で維持すると、これらのコロニーの一部分が収縮細胞を発生し、それらが心筋細胞を含有することを確証した。ISL1及びTBX5は、典型的には同一コロニーでは発現しなかったので、これはそれらの発現が異なる心臓形成領域から心臓細胞を発達する別個の亜集団を含有するコロニーを規定できることを示唆している。カバースリップ上で増殖した個々のコロニー由来の付着性集団の免疫染色は、SMA+、CTNT+及びVE−CAD+細胞の存在を明らかにし、これらのコロニーが心臓、内皮及びVSM潜在性を有することを確証した。
【0061】
2つの異なるアプローチを使用して、心血管コロニーがクローン性であるかどうかを決定した。最初に、hESCを発現するGFP由来のKDRlow/C−KITneg由来細胞を、メチセルロースコロニーアッセイにおいて赤色蛍光タンパク質(RFP)を発現するように組換えられたhESC由来のKDRlow/C−KITneg由来細胞とメチセルロースアッセイにおいて混合した。4日後に分析されたコロニーは、GFPもしくはRFPのいずれかを発現するが両方は発現しないことが見出されたが(図13)、この所見はそれらが単細胞から発生するのであり、細胞凝集の結果として形成されるものではないという解釈と一致する所見である。第2のアプローチとして、様々な数のKDRlow/C−KITneg由来細胞を心臓メチルセルロース培養中でプレーティングする細胞用量応答実験を実施した。発達するコロニーの数とプレーティングされた細胞数との関係は、1に近い勾配を持つ線形であり、これはこれらのコロニーが単細胞に由来するという意見をさらに支持している(図14)。これらをまとめると、これらの所見は、これらのコロニーが心血管コロニー形成細胞(hCV−CFC)に由来する心血管細胞のクローンを表すことを示している。
【0062】
KDRlow/C−KITneg由来心筋細胞の機能的電位は、全細胞電流及び電場電位測定値を用いて評価した。全細胞電圧固定分析では、試験した細胞の80%が優勢な電位依存性の一過性の外向きカリウム電流を発現した(図15、16)。この電流の電位依存性、密度及びゲーティング動態(図16、17)は、ヒト心房及び心室心筋において見出されるItoカリウム電流のものと似ていた。微小電極を使用して培養細胞から記録した電場電位は、ヒト心血管前駆細胞由来心臓細胞が電気的に相互に共役していることを明らかにした。さらに、予想されたように、Vaugn Williamsクラス1a薬剤であるキニジンは、測定T波振幅を減少させ、QT間隔を増加させた(図19)。
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2008年3月27日に出願された米国特許仮出願第61/040,049号の利益を主張する。
【0002】
(連邦政府による資金提供を受けた試験開発の記載)
本発明は、米国立衛生試験所によって授与された助成金番号第R01HL71800−01号及びP20−GM07501号の下で政府による後援を受けて実施された。米国政府は、本発明に関して所定の権利を有することができる。
【0003】
(技術分野)
胚発生中、身体の組織は、3つの主要な細胞集団である外胚葉、中胚葉及び胚性内胚葉から形成される。これらの細胞集団は、原発胚細胞層としても公知であり、原腸形成として公知のプロセスを通して形成される。原腸形成後に、各原発胚細胞層は特定セットの細胞集団及び組織を生成する。中胚葉は血球、内皮細胞、心筋及び骨格筋、ならびに脂肪細胞を生じさせる。胚性内胚葉は、肝臓、膵臓及び肺を生成する。外胚葉は、神経系、皮膚及び副腎組織を生じさせる。これらの胚細胞層からの組織発達のプロセスは、複雑な分子変化を反映する複数の分化工程を含んでいる。中胚葉及びその誘導体に関して、3つの別個のステージが規定されてきた。その第1は、胚盤葉上層として公知の構造内の細胞からの中胚葉の誘導である。新生中胚葉としても公知である新規に形成された中胚葉は、初期胚内の将来の組織発達の部位となる様々な位置へ移動する。このプロセスは、パターニングとして公知であり、特定組織に向かう初期分化ステージを反映する可能性が高い一部の分子変化を必要とする。最終ステージは、特異化として公知であり、パターン化された中胚葉亜集団からの別個の組織の発生を含んでいる。
【背景技術】
【0004】
これまでに得られた証拠は、中胚葉が別個の発生能を備える亜集団を表す連続する波のように誘導されることを示唆している。最初に形成された中胚葉は胚外領域へ移動して造血細胞及び内皮細胞を生じさせるが、次の集団は発生中の胚内で前方へ移動して心臓及び頭部の間葉となる。これらの分化系列関係は、最初は組織学的分析を通して規定され、その後は主として細胞追跡試験によって確証されてきた。造血への関与に関しては、現在ではES細胞分化モデルからの研究、及びマウス胚上での研究から、最初に同定できる前駆細胞は血管芽細胞として公知の細胞であり、血管能力もまた示す細胞であるという強力な証拠が得られている(非特許文献1、非特許文献2)。この前駆細胞についての分析は、この前駆細胞が中胚葉遺伝子ブラキュリ及び受容体チロシンキナーゼFlk−1を共発現することを明らかにしたが、これは中胚葉の亜集団が造血及び血管分化系列へ関与することを表すことを示している(非特許文献3)。分化系列追跡試験は、心臓がFlk−1+集団から発達することを証明したが、これは心血管系のための同等の多能性細胞が存在する可能性があることを示唆している(非特許文献4)。ES細胞分化培養の分析は、心臓及び内皮潜在力を備えるFlk−1+前駆細胞の存在についての証拠を提供した(非特許文献5)。近年の試験もまた、複数の心血管分化系列を生じさせることのできるマウス心血管前駆細胞の存在を支持している(非特許文献6、非特許文献7、非特許文献8)。ヒト心血管前駆細胞集団は、これまで同定されていない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Choi et al.(1998);Development 125:725−732
【非特許文献2】Huber et al.(2004)Nature 432:625−30
【非特許文献3】Fehling et al.(2003)Development 130:4217−4227
【非特許文献4】Ema et al.(2006)Blood 107:111−117
【非特許文献5】Yamashita et al.(2005)FASEB 19:1534−1536
【非特許文献6】Kattman et al.(2006)Dev. Cell 11:723−732
【非特許文献7】Moretti et al.(2006)Cell 127:1151−1165
【非特許文献8】Wu et al.,(2006)Cell 127:1137−1150
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、ヒト心血管前駆細胞の濃縮集団を提供する。ヒト心血管前駆細胞は、細胞表面マーカーであるKDRの存在及び細胞表面マーカーであるC−KITの非存在を特徴とする。ヒト心血管前駆細胞は、in vitro及びin vivoで心筋細胞、内皮細胞、ならびに血管平滑筋細胞に分化することができる。
【課題を解決するための手段】
【0007】
また別の実施形態では、本発明は、ヒト胚様体(EB)からヒト心血管前駆細胞を産生する方法であって、前記EBをアクチビン及び骨形成因子(BMP)及び任意で塩基性線維芽細胞成長因子(bFGF)の存在下で培養する工程と、引き続いてヒト心血管前駆細胞を提供するためにWnt成長因子の阻害剤及び任意で血管内皮細胞成長因子(VEGF)の存在下で培養する工程と、ならびに前記細胞培養から前記細胞を収集する工程とを含む方法を提供する。
【0008】
また別の実施形態では、本発明は、少なくとも約30%及び好ましくは少なくとも約40%、及びより好ましくは少なくとも約50%のヒト心筋細胞を含有する細胞集団を発生させる方法であって、前記ヒト心血管前駆細胞をWntの阻害剤及び任意でVEGFの存在下で培養する工程を含む方法を提供する。一実施形態では、細胞は、単層として培養される。また別の実施形態では、細胞は、凝集体として培養される。心筋細胞の存在は、細胞の収縮活性を評価する工程によって、または例えば心トロポニンT(CTNT)などの心筋細胞の指標となる遺伝子の発現を測定する工程によって決定できる。
【0009】
本発明は、心筋細胞、内皮細胞及び血管平滑筋細胞を含有するヒト心血管コロニーを発生させる方法であって、前記ヒト心血管前駆細胞をVEGF、bFGF、及びWntの阻害剤の存在下で培養する工程を含む方法をさらに提供する。好ましい実施形態では、細胞は、メチルセルロース中で培養される。
【0010】
ヒト心血管前駆細胞及びそれに由来する分化細胞は、心血管前駆細胞、心筋細胞、内皮細胞、血管平滑筋細胞、及び心血管組織に影響を及ぼす薬剤についてスクリーニングする方法において有用である。
【0011】
ヒト心血管前駆細胞は、また増殖及び貯蔵のため、ならびに心筋細胞、内皮細胞及び血管平滑筋細胞の起源としても有用である。
【0012】
ヒト心血管前駆細胞及びそれに由来する分化細胞は、また哺乳動物における心臓機能を改善もしくは再構成するため、及び移植のためにin vitroで心血管組織を操作するための方法においても有用である。
【図面の簡単な説明】
【0013】
【図1】本発明の一実施形態によってヒト胚幹細胞(hESC)から心臓分化系列への分化を方向付けるために使用されるプロトコルの略図である。
【図2】BMP4、bFGF及びアクチビンAの組み合わせを用いて誘導されたEB内でのCTNT+細胞発達の動態を示す。EBを、表示時点に収集し、CTNTを発現する細胞の頻度について細胞内フローサイトメトリーによって分析した。
【図3】異なるステージでのWntシグナル伝達経路の操作後の14日齢EB内でのCTNT+細胞の頻度を示す。
【図4】誘導ステージ中のBMP4、アクチビンA及びbFGFの組み合わせにおける培養後の、14日齢EB内でのCTNT+細胞の頻度を示す。
【図5】VEGF及びDKK1の表示した組み合わせを用いて処置した14日齢EBについての24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。図1〜5については、バーは、3回の独立実験の標準誤差または平均値を表す。統計的分析は、対応のないt検定を用いて実施した。*P=0.07、**P<0.01、***P<0.001。
【図6】異なる発達ステージでのEBの遺伝子発現分析を示す。
【図7】異なる日齢でのEBのフローサイトメトリー分析を示す。
【図8】6日齢EBから単離した3つの集団の定量的RT−PCR遺伝子分析を示す。
【図9】3つの6日齢集団の心臓電位を示す。
【図10】6日齢KDRlow/C−KITneg集団の心臓電位を示す。
【図11】6日齢EB由来KDRlow/C−KITneg分画から発生した付着性集団の定量的RT−PCR発現分析を示す。
【図12】KDRlow/C−KITneg集団の培養の10〜12日後のCTNT+、CD31+及びSMA+細胞の存在についての分析を示す。
【図13】混合RFP/GFP培養から単離された4日齢心血管コロニーの発現分析を示す。
【図14】プレーティングしたKDRlow/C−KITneg由来細胞の数と発達する心血管コロニーの数との関係を示す細胞用量反応を示す。
【図15】KDRlow/C−KITneg由来心筋細胞中で自然に発現した一過性外向きK+電流(Ito)の全細胞電位固定記録の代表的追跡を示す。
【図16】図15に示したように、細胞についての平均電流密度−電位関係を示す。1バッチの細胞10個中8個はIto電流を示し、平均値±SEM電流密度はこれらの8個の細胞からの追跡を用いてプロットした。
【図17】図16におけるような細胞についてのピーク電流までの平均時間(黒い四角)及び不活性化τ(白い四角)を示す(n=8)。
【図18】2〜3週間にわたってMEA(Multi Channel Systems)培養皿内で培養した6日齢EBから単離したKDRlow/C−KITneg細胞についての電気活動を示す。細胞外電気活動は、1μMキニジン(Sigma)を用いて、または用いずに記録した。
【図19】hESC培養内での心血管前駆細胞の発達を示すモデルを示す。
【図20】未分化hES2細胞、4日齢EB及び収縮性心筋細胞を備える14日齢EBの画像を示す(倍率200倍)。
【図21】EB内でのCTNT+細胞発達の動態を示す。
【図22】図1に示したプロトコルを用いて誘導された収縮性EB発達の動態を示す。150〜200個のEBが、収縮性EBの存在について各時点でスコア付けされた。バーは、3回の独立実験の平均値の標準誤差を表す。
【図23】グラフの下方に示した因子の組み合わせを用いて誘導した後の14日齢EB内での24ウェルプレートの1ウェル当たりの細胞総数を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
【図24】グラフの下方に示した因子の組み合わせを用いて誘導した後の14日齢EB内での24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
【図25】VEGF及びDKKの表示した組み合わせを用いて処置した14日齢EB由来のCTNT+細胞の頻度を示す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.05、**P<0.01。
【図26】VEGF及びDKKの表示した組み合わせを用いて処置した14日齢EBについての24ウェルプレートの1ウェル当たりのCTNT+細胞の総数を示す。
【図27】6日齢のEBから単離した3つの集団の定量的RT−PCR分析を示す。シクロフィニン(cyclophinin)に対して標準化した平均発現を示した。バーは、3回の独立実験の平均値の標準誤差を表す。
【図28】6日齢EBから単離されたKDRlow/C−KITneg集団の単層培養中での10〜14日後の相対細胞数のグラフである。96ウェルプレートの1ウェル当たり40,000個の細胞は、グラフの下方に示した因子の組み合わせを用いて誘導した。バーは、3回の独立実験の平均値の標準誤差を表す。
【図29】H1 hESCから発生したEB内での3つの異なるKDR/C−KIT集団の発生を証明しているフローサイトメトリー分析を示す。
【図30】7日間の培養後のH1 hESC由来KDRlow/C−KITneg分画からのCTNT+細胞の発達を示す。
【図31】VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)の存在下で培養されたKDRlow/C−KITneg由来付着性集団の定量的RT−PCR発現分析を示す。培養は規定時点に収集され、表示した遺伝子の発現について分析された。シクロフィニンに対して標準化した平均発現を示した。
【図32】hES2由来KDRlow/C−KITneg集団から発生した個々の4日齢心血管コロニーの発現分析を示す。
【発明を実施するための形態】
【0014】
本発明は、ヒト心血管前駆細胞の集団を提供する。本発明のヒト心血管前駆細胞は、適切な条件下では、心筋細胞、内皮細胞、及び血管平滑筋細胞へ分化することができる。ヒト心血管前駆細胞は、チロシンキナーゼ受容体KDRの発現及びチロシンキナーゼ受容体C−KITの発現の欠如を特徴とする。
【0015】
本発明の一実施形態では、ヒト心血管前駆細胞は、胚様体(EB)から入手される。EBは、広範囲の分化系列から発達する集団を含有する三次元コロニーである。胚幹細胞(ESC)からEBを形成するための条件は、当技術分野において公知である。例えば、胚フィーダー細胞上に維持されたヒトESCは、例えばMatrigel(商標)(BD Biosciences、マサチューセッツ州ベッドフォード)などの基底膜マトリックスの層上で培養し、小クラスターを形成するために解離させ、そしてBMPの存在下で無血清培地中においてプレーティングすることによってフィーダーを枯渇させることができる。本発明の好ましい一実施形態では、フィーダーを枯渇させたヒトESCがEBを形成するために約1日間にわたりBMP4の存在下で無血清培地中にて培養される。また別の好ましい実施形態では、BMP4の濃度は、約0.1ng/ml〜約1.0ng/ml、及びより好ましくは約0.5ng/mlである。
【0016】
ヒトESCは、市販で、または当技術分野において公知の方法によって入手できる。例えば、ヒトESCは、当技術分野において公知の方法、及び例えばThomson et al.の米国特許第5,843,780号明細書に開示された方法、及びReubinoff et al.(2000)Nature Biotech 18:399による方法によって胚盤胞から調製できる。
【0017】
EBは、当技術分野において公知の方法、及び例えばTakahashi et al.(2007)Cell 131:861及びMali et al.(2008)Stem Cells 26:1998によって開示された方法によって誘導多能性幹細胞(iPS)から形成することもできる。
【0018】
本発明のヒト心血管前駆細胞は、ヒトEBをアクチビン及びBMPの存在下で無血清培地中において培養する工程と、次に心血管前駆細胞の集団を提供するためにWntの阻害剤の存在下で培養する工程と、及び任意で前記培養から細胞を収集する工程と、さらに任意で心血管前駆細胞のための収集された細胞集団を濃縮する工程とによって入手することができる。好ましい実施形態では、アクチビンはアクチビンAであり、BMPはBMP4であり、Wntの阻害剤はDickkopf−1(DKK−1)である。
【0019】
本発明のまた別の実施形態は、ヒト心血管前駆細胞を産出するための方法であって、ヒトEBを約1〜約4日間にわたりアクチビンA及びBMP4及び任意のbFGFの存在下で無血清培地中において培養する工程と、及び引き続き前記細胞を約1〜2日間にわたりDKK−1及び任意のVEGFの存在下で無血清培地中において培養する工程と、細胞集団を収集する工程と、ならびに任意でヒト心血管前駆細胞について前記集団を濃縮する工程とを含む方法を提供する。
【0020】
好ましい実施形態では、EBは、アクチビンA及びBMP4及び任意のbFGFの存在下で約4日間にわたり培養される。アクチビンA、BMP4及び任意のbFGFの濃度は、T及びWNT3Aの発現のアップレギュレーションを特徴とする細胞の原始線条様集団を誘導するために十分である。その後の工程では、DKK−1及び任意のVEGFの濃度は、心血管前駆細胞の集団を提供するために十分である。
【0021】
好ましい実施形態では、アクチビンAの濃度は、約0.5ng/ml〜約30ng/ml、及びより好ましくは約1.0ng/ml〜約5.0ng/mlである。最も好ましい実施形態では、アクチビンAの濃度は、約3.0ng/mlである。
【0022】
好ましい実施形態では、BMP4の濃度は、約1.0ng/ml〜約20.0ng/ml、及びより好ましくは約5.0ng/ml〜約15ng/mlである。最も好ましい実施形態では、BMP4の濃度は、約10ng/mlである。
【0023】
好ましい実施形態では、bFGFの濃度は、約1.0ng/ml〜約10.0ng/ml、及びより好ましくは約3.0ng/ml〜約6.0ng/mlである。最も好ましい実施形態では、bFGFの濃度は、約5.0ng/mlである。
【0024】
好ましい実施形態では、DKK1の濃度は、約100ng/ml〜約200ng/ml、及びより好ましくは約125ng/ml〜約175ng/mlである。最も好ましい実施形態では、DKK1の濃度は、約150ng/mlである。
【0025】
別の好ましい実施形態では、VEGFの濃度は、約1.0ng/ml〜約50.0ng/ml、及びより好ましくは約1.0ng/ml〜約25.0ng/mlである。最も好ましい実施形態では、VEGFの濃度は、約10.0ng/mlである。
【0026】
本発明のまた別の好ましい実施形態では、ヒトEBは、約4日間にわたり無血清培地中、約3.0ng/mlのアクチビンA、約10.0ng/mlのBMP4及び約5.0ng/mlのbFGFの存在下で培養されるが、その時点に約150ng/mlのDKK1及び約10.0ng/mlのVEGFが加えられ、細胞は心血管前駆細胞を産生するために約1〜2日間にわたり培養される。また別の好ましい実施形態では、前記集団は、例えばKDRを発現するがC−KITを発現しない細胞を選別することによって、心血管前駆細胞に濃縮される。好ましい実施形態では、この選別は、免疫選択によって、例えばフローサイトメトリーによって行われる。
【0027】
また別の実施形態では、本発明は、ヒト心筋細胞の集団を発生させる方法であって、前記ヒト心血管前駆細胞を約7〜約10日間にわたりWntの阻害剤及び任意にVEGFの存在下で培養する工程と、及び培養からヒト心筋細胞の集団を収集する工程とを含む方法を提供する。好ましい実施形態では、Wntの阻害剤はDKK1であり、DKK1及びVEGFの好ましい濃度は、上記に記載したとおりである。1つの好ましい実施形態では、細胞は、ゼラチンコーティングウェル内の単層としてプレーティングされる。また別の実施形態では、細胞は、凝集体として低クラスター培養皿内で培養される。ヒト心筋細胞の集団は、少なくとも約30%、好ましくは少なくとも約40%、及びより好ましくは少なくとも約50%の心筋細胞を含有している。他の実施形態では、ヒト心筋細胞の集団は、少なくとも約70%、好ましくは少なくとも約80%、より好ましくは少なくとも約90%、及び100%までの心筋細胞を含有している。心筋細胞は、細胞の収縮活性を評価する工程によって、または例えばCTNTなどの心筋細胞の指標となる遺伝子の発現を測定する工程によって同定できる。
【0028】
本発明のヒト心血管前駆細胞はさらにまた、当業者には公知の分化条件を用いて、例えば心房、心室、及びペースメーカー細胞を含む心筋細胞の亜集団を発生させるために有用である。
【0029】
また別の実施形態では、本発明は、心筋細胞、内皮細胞及び血管平滑筋細胞を含有する心血管コロニーを発生させる方法であって、前記ヒト心血管前駆細胞を約3〜約7日間にわたりVEGF、bFGF、及びWntの阻害剤、好ましくはDKK1の存在下で培養する工程を含む方法を提供する。好ましい実施形態では、細胞は、メチルセルロース中にプレーティングされる。VEGF、bFGF及びDKKの好ましい濃度は、上記に記載したとおりである。心筋細胞、内皮細胞、及び血管平滑筋細胞の存在は、例えばCTNTなどの心筋細胞の指標となる遺伝子、及び例えばCD31、VE−カドヘリンなどの内皮細胞の指標となる遺伝子ならびに例えばSMA及びカルポニンなどの血管平滑筋細胞の指標となる遺伝子の発現を測定する工程によって決定できる。
【0030】
本発明は、ヒト心血管前駆細胞、心血管コロニー、心筋細胞、内皮細胞及び血管平滑筋細胞に作用を及ぼす薬剤についてスクリーニングするための方法をさらに提供する。本方法は、上記に記載した細胞集団の1つに由来する細胞を候補薬と接触させる工程と、及び前記薬剤が前記細胞集団に作用を及ぼすかどうかを決定する工程とを含んでいる。試験対象の薬剤は、天然もしくは合成、1つの化合物もしくは混合物、低分子またはポリペプチド、多糖類、ポリヌクレオチドなどを含むポリマー、抗体もしくはそのフラグメント、1ライブラリーの天然もしくは合成化合物からの化合物、理論的薬物設計から得られた化合物、例えば細胞培養条件などの条件、または細胞集団に及ぼす作用を当技術分野において公知のアッセイを用いて評価できる任意の薬剤であってよい。細胞集団に及ぼす作用は、例えば、マーカー発現、受容体結合、収縮能力、電気生理学、細胞生存能力、生存率、形態、またはDNA合成もしくは修復についてのアッセイを含む、表現型もしくは活性についての任意の標準アッセイによって決定することができる。標準増殖及び分化アッセイは、米国特許第6,110,739号明細書に記載されている。そのような薬剤は、in vivo及びin vitroでの細胞成長、分化及び生存率の制御、ならびに組織の維持、再生及び修復のために有用である。
【0031】
本発明は、ヒト心血管前駆細胞の集団を含む組成物及びヒト心血管コロニーの集団を含む組成物をさらに提供する。本組成物は、薬学的に許容される担体及び希釈剤を含むことができる。本組成物は、移植を促進する成分をさらに含むことができる。これらの集団を含む組成物は、細胞及び組織の置換及び修復のため、ならびに心筋細胞、内皮細胞、及び血管平滑筋細胞の集団をin vitro及びin vivoで発生させるために有用である。ヒト心血管前駆細胞を含む組成物は、前駆細胞集団を拡張させるために有用である。本組成物は、心臓の状態を治療するための医薬品または送達装置として調製できる。
【0032】
また別の実施形態では、本発明は、例えば、先天性心疾患、冠動脈性心疾患、心筋症、心内膜炎及びうっ血性心不全を含む不十分な心臓機能を特徴とする障害を治療するために有用な細胞置換方法及び組織置換方法を提供する。分化細胞及び心血管前駆細胞はどちらも置換療法のために有用であるが、それは前駆細胞集団が心筋細胞、内皮細胞及び血管平滑筋細胞分化系列へin vivoで分化できるためである。これらの細胞はさらにin vitroで心血管細胞を発生させるためにも有用である。心臓組織を操作するための方法は当技術分野において公知であり、例えば、Birlaによって“Stem Cell Therapy and Tissue Engineering for Cardiovascular Repair” Springer, 2006で概説されている。
【0033】
したがって、一実施形態では、本発明は、心筋細胞置換療法のための方法であって、当該治療を必要とする被験者に本発明によって得られたヒト心血管前駆細胞が濃縮された細胞集団から単離された心筋細胞を含む組成物を投与する工程を含む方法を提供する。また別の実施形態では、本発明は、不十分な心臓機能を特徴とする障害を治療する方法であって、当該治療を必要とする被験者にヒト心血管前駆細胞を含む組成物を投与する工程を含む方法を提供する。好ましい実施形態では、被験者は、ヒトである。本組成物は、心臓組織への送達もしくは遊走を生じさせる、例えば注射もしくは移植を含む経路によって、そして少なくとも1つの有害作用もしくは症状または障害の減少を生じさせる条件下で投与することができる。
【0034】
本明細書に引用した全参考文献は、開示内容全体が参考として本明細書で援用される。
【実施例】
【0035】
以下の実施例は、本発明をさらに具体的に説明するために役立つ。
【0036】
(実施例1)
材料及び方法
ヒトES細胞の維持
WiCell Research Institute(ウィスコンシン州マディソン)からのH1(NIHコード番号WA01)及びESI International(シンガポール国)からのhES2(NIHコード番号ES02)を、6ウェル組織培養プレート内で20%ノックアウト血清代替物(SR)、100μM非必須アミノ酸、2mMグルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン(Invitrogen、ニューヨーク州グランドアイランド)、10−4M β−メルカプトエタノール(Sigma、ミズーリ州セントルイス)及び20ng/mlのhbFGF(R&D Systems、ミネソタ州ミネアポリス)が補充されたダルベッコ変法イーグル培地(DMEM)/F12(50:50;Mediatech、バージニア州ハーンドン)からなるhESC培地中にて、照射マウス胚フィーダー細胞上で維持した。AAVS1標的化hES2細胞系は、親hES2細胞のAAV2−TRUF11(CAG−GFP−TK−neo)及び野生型AAV2両方の106ウイルス粒子との共感染によって発生させた。G418選択後、GFP陽性細胞を選別し、サブクローンを単離した。トランス遺伝子の標的化組込みは、ライゲーション媒介ポリメラーゼ連鎖反応(LM PCR)によって確認された。野生型AAV配列は、GFP陽性クローン中では検出されなかった。RFP発現hES細胞は、Irion et al.(2007)Nat. Biotechnol. 12:1477−1482によって記載されたようなhES2細胞のRosa26遺伝子座へRFP cDNAを標的化する工程によって生成した。
【0037】
ヒトES細胞の分化
EBを発生させる前に、hESCは、フィーダーを枯渇させるためにMatrigel(商標)(BD Biosciences、マサチューセッツ州ベッドフォード)の層上で48時間にわたり培養した。Matrigel(商標)培養からの細胞はコラゲナーゼB(1mg/ml、Roche、インディアナ州インディアナポリス)を用いた20分間の処理、その後のトリプシン−EDTA(0.05%)を用いた2分間の処理によって小クラスターに解離させた。第0〜1日から、EBは2ml塩基性培地(StemPro−34(Invitrogen)、2mMのグルタミン、4×10−4Mのモノチオグリセロール(MTG)、及び50μg/mlのアスコルビン酸(Sigma)+0.5ng/mlのBMP4(R&D Systems)中にプレーティングした。以下の濃度の因子をEB形成、中胚葉誘導及び心臓特異化のために使用した。BMP4−10ng/ml、hbFGF−5ng/ml、アクチビンA−3ng/ml、hDKK1−150ng/ml、hVEGF−10ng/ml。これらの因子は、以下の順序で加えた。第1〜4日には、BMP4、hbFGF及びアクチビンA、第4〜8日には、VEGF及びDKK1、第8日以降には、VEGF、DKK1、及びbFGF。培地は、第8日以降には4日毎に取り替えた。全ヒト因子及び阻害剤は、R&D Systems(ミネソタ州ミネアポリス)から購入した。培養は、培養の初期10〜12日間は5%CO2、5%O2、及び90%N2環境内で維持し、その後は5%CO2/気体環境内で維持した。
【0038】
フローサイトメトリーによる選別及びMatrigel(商標)上での管形成
EBを収集し、トリプシン(0.25%トリプシン−EDTA)を用いて単一の細胞に解離させた。トリプシン処理後、Facscaliburフローサイトメーター(Becton Dickinson)を用いて分析を実施した。細胞は、MoFlo(Dako Cytomation、コロラド州フォートコリンズ)セルソーターを用いて選別した。フローサイトメトリーデータは、FlowJo(Treestar、カリフォルニア州サンカルロス)ソフトウエアプログラムを用いて分析した。抗KDR−PE、抗−C−KIT−APCは、R&D Systemsから購入した。選別したKDRlow/C−KITneg集団は、VEGF(25ng/ml)及びbFGF(25ng/ml)を補充したStempro34培地中で分化させた。管状構造は、Matrigel(商標)コーティングガラス製カバースリップへ移してから24時間以内に形成された。
【0039】
免疫蛍光
解離した細胞は、2日間にわたりガラス製カバースリップ上で培養した。この時点で、細胞を4%PFAで固定し、次に染色した。以下の抗体を免疫染色のために使用した。R&D Systemsからの抗ヒトCD31、抗ヒトVE−カドヘリン、Lab Vision(カリフォルニア州フリーモント)からの抗マウストロポニンT及び抗ヒト平滑筋アクチン、Chemicon(カリフォルニア州テメキュラ)からの抗ヒトANP、コネキシン43及び抗ヒトα/β MHC抗体、Sigma(ミズーリ州セントルイス)からの抗ヒトα−アクチニン。抗ヒト平滑筋ミオシン重鎖(SMHC)、カルデスモン、フォン・ヴィレブランド因子抗体は、DakoCytomation(カリフォルニア州カーピンテリア)から購入した。Cy2、Cy3及びCy5共役二次抗体は、Jackson ImmunoResearch(ペンシルベニア州ウェストグローブ)から入手した。蛍光は、Leica DMRA2蛍光顕微鏡(Leica、ドイツ国ウェッツラー)を用いて視認し、画像はデジタル式Hamamatsu CCDカメラ(日本国浜松市)を使用して記録した。
【0040】
コロニーアッセイ
心血管コロニーを発生させるために、6日齢EBから単離したKDRlow/C−KITneg細胞は、VEGF(25ng/ml)、bFGF(10ng/ml)及びDKK1(150ng/ml)の存在下で2〜3日間にわたり凝集させた。このステージに、凝集体を解離させ、細胞は5%CO2、5%O2、及び90%N2環境内においてVEGF(25ng/ml)、bFGF(25ng/ml)及びDKK1(150ng/ml)を含有するメチルセルロース中で培養した。コロニーは、コロニー頻度分析のために4〜6日間の培養後にスコア付けした。
【0041】
RT−PCR
発現試験のためには、個々のコロニーをメチルセルロース培養から単離し、Brady et al.(1993)Methods Enzymol. 225:611−623によって以前に記載されたプロトコルの修正版を用いて分析した。増幅cDNAを次に標準PCRにかけた。リアルタイム定量的PCRは、MasterCycler EP RealPlex(Eppendort)上で実施した。実験は白金SYBR GreenER qPCR SuperMix(Invitrogen)を用いて3回ずつ実施した。全プライマーは、表1に記載した。全アニーリング反応は、60℃で実施した。
【0042】
【表1−1】
【0043】
【表1−2】
【0044】
【表1−3】
【0045】
KDRlow/C−KITneg由来集団のin vivo分析
hESCを発現するGFP由来のKDRlow/C−KITneg細胞を移植前5〜10日間にわたって、VEGF(10ng/ml)、bFGF(10ng/ml)及びDKK1(150ng/ml)の存在下で培養した。NOD/SCIDγマウスに麻酔をかけ、気管挿管し、100,000個のドナー細胞を開胸手術において左心室壁内へ直接注射した。手術の2〜11週後に心臓を収集し、カコジル酸バッファー中の1%パラホルムアルデヒド中で固定し、300μmでビブラトーム(Vibrotome)にて切片化を行った。移植領域を同定し、10μmで凍結切片化を行った。免疫組織化学検査は、GFP抗体(Chemicon、AB3080、1:100;Vector ABC及びDABキット)、α−アクチニン抗体(Sigma、A7811、1:500;Chemicon AQ300R二次抗体、1:20)、CD31抗体(Dako、M0823、1:50;Vector ABC及びDABキット)、ならびに平滑筋MHC抗体(Biomedical Technologies、BT−562、1:300;Alexa 555二次抗体、1:200)について実施した。共焦点像は、ImageJ and Pierre Bourdoncleのプラグインをデフォルト設定で使用して共局在化について分析した。マウス梗塞モデルにおける評価のためには、心筋梗塞はRubert et al.(2007)Nature Biotechnol. 25:993−994によって記載された技術を使用して、直接的な冠状動脈結紮によってSCIDベージュマウスにおいて誘導した。10〜20分後に、マウスに500,000個のKDRlow/C−KITneg由来細胞(n=9)または同等量の無血清培地(n=12)を注射した。全注射は、梗塞の境界帯で試みた。2週間後、心室機能の評価は、9.4テスラの磁気共鳴イメージングを用いて実施した。
【0046】
パッチクランプ
全細胞パッチクランプ記録は、IX50倒立顕微鏡(Olympus)、Multiclamp 700A増幅器、Digidata 1300アナログ/デジタル変換器及びpClamp9.1ソフトウエア(Axon Instruments、カリフォルニア州フォスターシティ)を備えるPCを用いて室温で実施した。浴溶液(単位:mM)は、NaCl 136、KCl 4、CaCl2 1、MgCl2 2、CoCl2 5、HEPES 10、グルコース10、及びテトロドトキシン(TTX)0.02(pH7.4)であった。ピペット(単位:mM)は、KCl 135、EGTA 10、HEPES 10、及びグルコース5(pH7.2)を含有する細胞内溶液を充填すると3〜5MΩ抵抗のピペットであった。細胞を、−30mVのテールパルス(図4におけるような持続期間)前には、−80mVから+40mVへの電位を20mV増分で試験するために−80mVの保持電位から段階付けた。データは、pClamp9.1ソフトウエア(Axon Instruments)を用いて分析した。電流振幅は、細胞サイズに標準化した(全細胞膜容量)。不活性化τ値は、電流減衰の指数関数的適合を用いて計算した。
【0047】
電場電位の記録
6日齢EBから単離したKDRlow/C−KITneg細胞は、2〜4週間にわたりStemPro−34+10ng/mlのVEGF及び150ng/mlのDKK1を備えるMEA(Multi Channel Systems)培養皿内で培養した。記録の2日前に、細胞は15%FBSを備えるDMEM(細胞増殖用)へ切り換えた。細胞外電気活動は、60チャンネルから同時に記録し、ソフトウエアMC Rack(Multi Channel Systems)を用いて分析した。
【0048】
(実施例2)
hESC分化培養中での心臓発達の調節
hESCから心臓分化系列への分化を方向付けるために、原始線条様集団の誘導、心臓中胚葉のパターニング及び心血管分化系列の特異化を含む段階的プロトコルは、様々なモデル系において中胚葉誘導及び心臓特異化で役割を果たすことが公知である因子のBMP4、アクチビンA、VEGF、bFGF及びDKK1の組み合わせを用いて設計した。プロトコルの概略は、図1に示した。本明細書に例示したプロトコルを用いると、hESCは、EBの形成を可能にするために24時間にわたりBMP4の存在下で最初に分化された。第1ステージでのアクチビンとBMP4との組み合わせは、マウスにおけるこれらの集団内で発現することが公知の遺伝子であるT(ブラキュリ)及びWNT3aのアップレギュレーション及び一過性発現によって証明されるように、原始線条様集団及び中胚葉を誘導した(図6)。第2ステージでは、VEGF及びWnt阻害剤であるDKK1が中胚葉を更にパターン化し、KDR+前駆細胞集団の拡張及び成熟を促進した。分化の第8日には、発達中の心血管分化系列の持続的拡張を支援するためにbFGFが再び加えられた。このプロトコルは、収縮細胞ならびに心臓トロポニンT(CTNT)(図2)、α−アクチニン、α/βミオシン重鎖、ANP及びコネキシン43(図20、21)を発現した細胞の出現によって証明されたように、EB内の心臓発達を支援する。CTNT+細胞の最高頻度は培養の第14〜16日に定期的に観察されたが(図2、21)、この集団はEB内で2カ月間の期間にわたって維持することができた。収縮性EBの発達の動態は、CTNT+集団の出現と平行していた(図22)。
【0049】
(実施例3)
ヒトES細胞由来の心臓分化系列の発生にWntシグナル伝達が果たす役割
Wntシグナル伝達経路がヒトES細胞由来の心臓分化系列の出現に果たす役割を、原始線条誘導を越える段階(第2ステージ)に特に焦点を当てて調査した。EBは、プロトコルに略説したように、BMP4、bFGF及びアクチビンAの存在下で誘導した。Wnt3AまたはDKK1のいずれかを図3に示した日に培養へ加えた。これらの因子は、14日間を通して培養中で維持された。VEGF及びbFGFは、指示した時点に加えた。第14日に、EBを収集し、上述したように分析した。対照は、WNTまたはDKK1を受容しなかった培養であった。分化第4日におけるEBへのDKK1の添加は、培養第14日のCTNT+細胞の頻度の2倍を超える(40%までの)増加をもたらした(図3)。DKK1の作用は、第6または8日に加えられた場合はこれほど顕著ではなく、添加が分化第10日に添加された場合はCTNT+細胞の頻度の増強は見られなかった。Wnt3Aは反対の作用を有しており、第4または6日に培養に加えられた場合はCTNT+細胞の発達を完全に抑制した。これらの所見は、古典的Wnt経路のステージ特異的阻害がBMP4/アクチビン誘導性PS集団の心臓特異化を促進するために不可欠であることを示している。
【0050】
(実施例4)
心臓分化系列の誘導においてBMP4、アクチビンA及びbFGFが果たす役割
hESC分化培養中での心臓分化系列の誘導においてBMP4、アクチビンA及びbFGFが果たす役割を評価するために、誘導ステージ(第1〜4日)中に単一因子ならびに様々な組み合わせを試験した。第4日には、図1に略説したように、因子を除去し、EBを処理した。第4〜14日に、EBは図5、25、及び26に示したようにVEGF及びDKK1の組み合わせを用いて処理し、第8日以降にはbFGFを添加した。EBは第14日に収集し、各組み合わせについてのCTNT+細胞の頻度及び24ウェルプレートの1ウェル当たりのCTNT+細胞の総数について分析した。BMP4、bFGFもしくはアクチビンA単独またはBMP4及びFGFまたはアクチビンA及びbFGFの組み合わせは、低頻度(図4)及び生成されたCTNT+細胞の総数(図23、24)によって証明されたように心臓発達の貧弱な誘導因子であった。BMP4及びアクチビンAは有意な数のCTNT+細胞を誘導するが、これら3つの因子の組み合わせは最も強力であり、心臓細胞の最高の頻度及び総数を発生し、40%〜50%のCTNT+細胞からなる培養を常に生産した。VEGFは心臓発達のために必須ではなかったが、10ng/mlのこの因子の添加は発達したCTNT+細胞の総数を増加させた。この濃度を超えてVEGFの濃度が増加すると培養中の細胞数を減少させ、これは発生したCTNT+細胞の総数を有意に減少させた(図5、25及び26)。
【0051】
発達中のEBの分子分析は、原始線条様集団の確立後に発現パターンにおける動的変化を明らかにした。T及びWNT3Aと一緒に、DKK1の発現は早期にアップレギュレートされ、18日間の経過を通して持続した(図6)。分化の第2〜4日の内因性DKK1の発現は、心臓中胚葉を特異化するように機能することができ、外因性阻害剤の添加の不在下で心臓分化系列を確立させる(図3)。KDRは、未分化ES細胞内で発現した。この発現レベルは、第4〜6日に増加すると思われ、その後12日間にわたって持続された。初期胚中の2次心臓形成領域の前駆細胞をマークする遺伝子であるISL1は、分化第8日に最初に検出された心臓転写因子Nkx2.5の発現に先行して第6〜8日にEB中で発現した。適正な心臓発達に必要とされる2つのTBX転写因子であるTBX5及びTBX20、ならびに収縮性タンパク質MLC2A及びCTNTの発現は、分化の第8〜10日にアップレギュレートされ、これは心臓発達の開始を反映していた。
【0052】
(実施例5)
心血管電位を用いたKDRlow/C−KITneg EB集団の同定
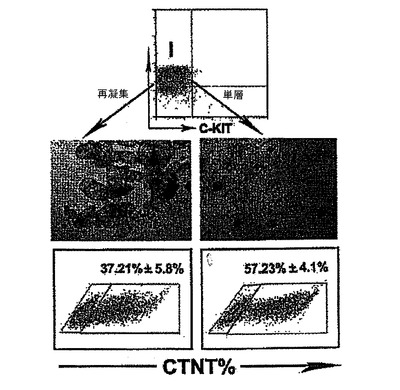
発達中のEBをKDR及びC−KITの発現について分析した。C−KITを別個の集団を規定するための電位マーカーとして使用したが、それはマウスEB内でのC−KITの発現が最初期血球血管芽細胞由来造血前駆細胞及び血管前駆細胞ならびに前原始線条及び発達中の内胚葉を同定するからである。図7は、異なる日齢のEBのフローサイトメトリー分析を示す。図7に示すように、3つの別個の集団であるKDRhigh/C−KIT+、KDRlow/C−KITneg及びKDRneg/C−KIT+は、6日齢EB内で分解される可能性がある。3つの集団の発達は、BMP4及びアクチビンAの両方を用いた誘導に依存した(図示しない)。3つの集団を細胞選別によって単離し、遺伝子発現パターン及び心臓電位について分析した。図8は、6日齢のEBから単離した3つの集団の定量的RT−PCR遺伝子発現分析を示す。平均発現をシクロフィニンへ標準化した。KDRhigh/C−KIT+集団は、血管発達に関連する遺伝子であるCD31、VE−カドヘリン及びSMA、ならびに造血に関与することの指標となる遺伝子であるGATA−1を発現した(図8、27)。心臓発達に関係する、NKX2.5、ISL1及びTBX5を含む遺伝子は、KDRlow/C−KITneg分画内では最高レベルで発現した。この分画はSMA、極めて低レベルのGATA−1もまた発現したが、VE−カドヘリンもしくはCD31は検出しなかった。SMAは早期に心臓分化系列ならびにVSM分化系列内で発現するので、この発現は心臓発達の開始を反映できた。KDRneg/C−KIT+細胞は最高レベルの3分画のOCT4及びTを発現したが、これは残留未分化ES細胞ならびに原始線条様細胞の存在を示す。この分画内の比較的高レベルのFOXA2及びSOX17は、それが内胚葉分化系列に関与する細胞もまた含有していることを示唆している。SOX1及びPAX6は極めて低レベルでしか検出されず、これはこの発達ステージではEB内で神経外胚葉分化系列への分化があったとしてもほんのわずかに過ぎないことを示唆している(図27)。これらをまとめると、これらの発現パターンは、KDRhigh/C−KIT+集団が造血及び血管前駆細胞を含有する、KDRlow/C−KITneg集団が心臓前駆細胞を含有する、そしてKDRneg/C−KIT+集団が未分化ES細胞、原始線条細胞及び内胚葉細胞の混合物からなることを示している。
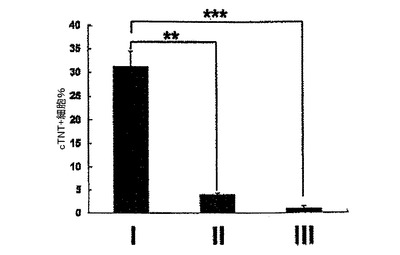
【0053】
心臓電位をモニターするために、細胞はゼラチンコーティングウェル上の単層としてプレーティングするか、または低クラスター培養皿内で再凝集させ、凝集体として培養した。詳細には、細胞は6日齢EBから単離し、VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)と共に単層としてゼラチンコーティングウェル上で培養した。CTNT+細胞は、10日間の培養後に分析した。結果を図9に示す。6日齢EBからのKDRlow/C−KITneg細胞を選別し、単層として、またはVEGF(10ng/ml)及びDKK1(150ng/ml)とともに低クラスターウェル内の凝集体として培養した。培養を収集し、7〜10日間にわたる分化後にCTNT+細胞の存在について分析した。結果を図10に示す。数(%)は、3回の独立実験の平均値±標準誤差を表す。バーは、3回の独立実験の平均値の標準誤差を表す。統計的分析は、対応のないt検定を用いて実施した。*P<0.01、***P<0.001。KDRlow/C−KITneg集団は最大の心筋細胞電位(図9)を提示しており、懸濁培養中の凝集体内ならびに付着性単層内の両方において容易にCTNT+細胞及び収縮性細胞の集団を発生した(図10)。凝集体内のおよそ40%の細胞がCTNT+であったが、単層培養の50%超は一貫してこのマーカーを発現した。単層培養内の心筋細胞の高頻度は、同調塊(synchronous mass)として収縮する細胞層の発達を常にもたらした。単離されたKDRlow/C−KITneg細胞は、凝集体としてはおよそ1.5倍(データは示していない)、及び単層培養内では3倍に拡張した(図28)。略説した誘導プロトコル及び選別ストラテジーを用いると、4インプットのhESCに付き1の心筋細胞のアウトプットが予測された。典型的な実験では、2×106個のhESCは、1×106個の6日齢EB細胞を発生したが、そのうちの30%(3.0×9 05細胞)はKDRlow/C−KITneg分画を提示した。この集団は、単層培養フォーマット内で3倍(1×106細胞)に拡張し、およそ50%の心筋細胞(5×105)からなる集団を産出した。2次hESC系であるH1から発生したEBの動態分析は、分化第6日ではなく第5日に3KDR/C−KIT集団の発達を証明した。5日齢KDRlow/C−KITneg集団の分析は、それが心臓電位もまた示すことを示した(図29、30)。
【0054】
(実施例6)
KDRlow/C−KITneg由来分化系列の特性解析
VEGF(10ng/ml)、DKK1(150ng/ml)及びbFGF(10ng/ml)とともにプレーティングした6日齢EB由来KDRlow/C−KITneg分画から発生した付着性集団についての定量的RT−PCR発現分析を実施した。培養は図11に示した日に収集し、細胞を発現について分析した(D7はプレーティングの1日後の集団を表す)。シクロフィニンに対して標準化した平均発現を示した。バーは、3回の独立実験の平均値の標準誤差を表す。
【0055】
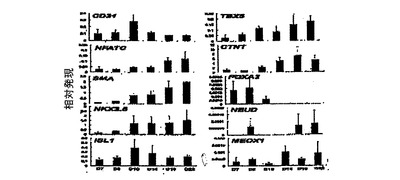
プレーティングから異なる日数後でのKDRlow/C−KITneg由来付着性集団についての発現分析(qPCR)は、内皮(CD31、VE−カドヘリン)、VSM(カルポニン、SMA)、心臓発達(NKX2.5、ISL1、TBX5、TBX20)及び心臓成熟(CTNT、MLC2A)に関連する遺伝子のアップレギュレーションを証明した(図11、31)。NFATC及びニューレグリン 1(NRGl)の発現は、培養中の心内膜の存在を示唆している。低レベルのNEUD、PAX6、SOX1、FOXA2、FOXA3、SOX17及びMEOX1の発現は、これらの培養が神経外胚葉、内胚葉または体中胚葉で汚染されていないことを示している(図11、31)。
【0056】
KDRlow/C−KITneg細胞を6日齢EBから単離し、図12のグラフの下方に指した因子の様々な組み合わせを用いて単層として培養した。細胞を収集し、10〜12日間にわたる培養後にCTNT+、CD31+及びSMA+細胞の存在について分析した。SMA抗体は、心筋細胞及び平滑筋細胞の両方を染色することができる。数(%)は、3回の独立実験の平均値の標準誤差を表す。
【0057】
VEGF及びDKK1中で10〜12日間培養したKDRlow/C−KITneg由来付着性細胞のフローサイトメトリー分析は、ほぼ90%がSMAを発現し、50%がCTNTを発現し、4%がCD31を発現することを明らかにした(図12)。培養へのbFGFの添加は、CTNT+細胞の比率を30%へ減少させ、CD31+集団を30%へ増加させた。SMA+細胞の比率は、bFGFの存在下で平均80%へ穏やかに減少した。bFGFの添加は、単層培養中で細胞数に有意には影響を及ぼさなかった(図28)。これらの所見は、KDRlow/C−KITneg由来集団内の細胞の大多数が心血管分化系列の細胞であることを明らかに示している。それらはさらに、bFGFがこの集団内で発達する心筋細胞及び内皮細胞の比率に影響を及ぼすことができることもまた示している。
【0058】
VEGF、DKK1及びbFGF中で培養したKDRlow/C−KITneg由来集団の免疫染色分析は、CD31+、VE−カドヘリン+及びフォン・ヴィレブランド因子+内皮細胞の存在、CTNT+心筋細胞の存在ならびにVSM発達の指標となるSMA+、SMHC+及びカルデスモン+細胞の存在を証明した。集団内の未成熟心筋細胞はCTNT及びSMAの両方を発現したが、他方VSM細胞はSMAしか発現しなかった。KDRlow/C−KITneg由来内皮細胞についてさらに特性解析するために、この集団をVEGF及びbFGFの存在下で拡張させ、次にMatrigel(商標)コーティングカバースリップ上で培養した。これらの条件下では、細胞は管状構造への内皮細胞の組織化の指標である格子を形成した。これらの構造内の細胞はCD31を発現し、Dil−AC−LDLを取り込む能力を示して、それらの内皮細胞表現型を確証した。免疫染色試験からの所見は、フローサイトメトリー分析所見と一致しており、KDRlow/C−KITneg由来集団が心臓、内皮及び血管平滑筋分化系列の細胞からなることを証明している。
【0059】
KDRlow/C−KITneg由来細胞のin vivoでの発達可能性を証明するために、それらをさらにNOD/SCIDマウスの心臓内へ移植した。これらの試験のためには、hES2細胞系のGFP発現バージョンを使用した。組織学的分析は、移植後に心臓内のGFP+細胞の存在を明らかにした。抗体を用いたGFPの検出は、落射蛍光によって視認された集団と同一集団を同定したが、これはGFPシグナルがドナー細胞の存在を表し、バックグラウンド蛍光の結果としては生じないことを示している。α−アクチン、CD31もしくはSMHCのいずれかを共発現するGFP+集団が移植片内で検出されたが、これは心臓、内皮及び血管平滑筋分化系列へのin vivoでの分化を示している。移植された動物のいずれにおいても奇形腫は検出されなかった(N>10)。この集団のin vivoでの潜在性をさらに評価するために、KDRlow/C−KITneg由来細胞をSCIDベージュマウスの梗塞心臓内へ直接的に移植した。2週間後に分析すると、KDRlow/C−KITneg由来心血管集団が移植された動物は、培地単独が注射された集団より31%高い駆出率を有していた(56%±3.6%対39%±4.8%、p=0.008)。これらの所見は、以前の報告書と一致しており、hESC由来心筋細胞の移植が心筋梗塞の齧歯類モデルにおける心臓機能の改善をもたらすことを証明している。
【0060】
(実施例7)
心血管コロニー形成細胞の同定及び特性解析
VEGF、bFGF及びDKK1の存在下においてメチルセルロース中でプレーティングすると、KDRlow/C−KITneg由来細胞は培養4日間以内に小さな小型コロニーを生成した。個々の4日齢コロニーのPCR分析は、心臓(CTNT)、血管(CD31及び/またはVE−カドヘリン)及びVSM(SMA及び/またはカルポニン)の発達の指標となるマーカーの共発現を示した(図13、32)。さらに6日間にわたり培養中で維持すると、これらのコロニーの一部分が収縮細胞を発生し、それらが心筋細胞を含有することを確証した。ISL1及びTBX5は、典型的には同一コロニーでは発現しなかったので、これはそれらの発現が異なる心臓形成領域から心臓細胞を発達する別個の亜集団を含有するコロニーを規定できることを示唆している。カバースリップ上で増殖した個々のコロニー由来の付着性集団の免疫染色は、SMA+、CTNT+及びVE−CAD+細胞の存在を明らかにし、これらのコロニーが心臓、内皮及びVSM潜在性を有することを確証した。
【0061】
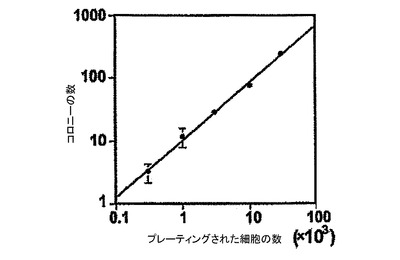
2つの異なるアプローチを使用して、心血管コロニーがクローン性であるかどうかを決定した。最初に、hESCを発現するGFP由来のKDRlow/C−KITneg由来細胞を、メチセルロースコロニーアッセイにおいて赤色蛍光タンパク質(RFP)を発現するように組換えられたhESC由来のKDRlow/C−KITneg由来細胞とメチセルロースアッセイにおいて混合した。4日後に分析されたコロニーは、GFPもしくはRFPのいずれかを発現するが両方は発現しないことが見出されたが(図13)、この所見はそれらが単細胞から発生するのであり、細胞凝集の結果として形成されるものではないという解釈と一致する所見である。第2のアプローチとして、様々な数のKDRlow/C−KITneg由来細胞を心臓メチルセルロース培養中でプレーティングする細胞用量応答実験を実施した。発達するコロニーの数とプレーティングされた細胞数との関係は、1に近い勾配を持つ線形であり、これはこれらのコロニーが単細胞に由来するという意見をさらに支持している(図14)。これらをまとめると、これらの所見は、これらのコロニーが心血管コロニー形成細胞(hCV−CFC)に由来する心血管細胞のクローンを表すことを示している。
【0062】
KDRlow/C−KITneg由来心筋細胞の機能的電位は、全細胞電流及び電場電位測定値を用いて評価した。全細胞電圧固定分析では、試験した細胞の80%が優勢な電位依存性の一過性の外向きカリウム電流を発現した(図15、16)。この電流の電位依存性、密度及びゲーティング動態(図16、17)は、ヒト心房及び心室心筋において見出されるItoカリウム電流のものと似ていた。微小電極を使用して培養細胞から記録した電場電位は、ヒト心血管前駆細胞由来心臓細胞が電気的に相互に共役していることを明らかにした。さらに、予想されたように、Vaugn Williamsクラス1a薬剤であるキニジンは、測定T波振幅を減少させ、QT間隔を増加させた(図19)。
【特許請求の範囲】
【請求項1】
ヒト心血管前駆細胞の集団であって、前記前駆細胞はKDRを発現するがC−KITを発現しないヒト心血管前駆細胞の集団。
【請求項2】
前記前駆細胞が、心筋細胞、内皮細胞、及び血管平滑筋細胞の前駆細胞である、請求項1に記載のヒト心血管前駆細胞の集団。
【請求項3】
ヒト胚様体をアクチビン及びBMPの存在下で無血清培地中において培養する工程と、その後に前記培養にWntの阻害剤を加える工程とによって得られる、請求項1に記載のヒト心血管前駆細胞の集団。
【請求項4】
前記アクチビンが、アクチビンAである請求項3に記載のヒト心血管前駆細胞の集団。
【請求項5】
前記BMPが、BMP4である請求項3に記載のヒト心血管前駆細胞の集団。
【請求項6】
前記Wntの阻害剤が、DKK−1である請求項3に記載のヒト心血管前駆細胞の集団。
【請求項7】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体をアクチビン及びBMPの存在下で無血清培地中において培養する工程、及び
(b)前記培地へWntの阻害剤を加え、ヒト心血管前駆細胞の集団を提供するために培養する工程を含む方法。
【請求項8】
(c)収集した細胞集団を提供するために前記ヒト心血管前駆細胞を培養から収集する工程をさらに含む、請求項7に記載の方法。
【請求項9】
(d)前記収集した細胞集団を心血管前駆細胞に濃縮する工程をさらに含む、請求項8に記載の方法。
【請求項10】
前記アクチビンが、アクチビンAである請求項7に記載の方法。
【請求項11】
前記BMPが、BMP4である請求項7に記載の方法。
【請求項12】
前記Wntの阻害剤が、DKK1である請求項7に記載の方法。
【請求項13】
前記工程(a)が、bFGFの存在をさらに含む請求項7に記載の方法。
【請求項14】
前記工程(b)が、VEGFを加える工程をさらに含む請求項7に記載の方法。
【請求項15】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体を約1〜4日間にわたりアクチビンA及びBMP4の存在下で無血清培地中において培養する工程、
(b)DKK1を前記無血清培地へ加え、約1〜2日間にわたり培養する工程、及び、
(c)ヒト心血管前駆細胞の集団を収集する工程を含む方法。
【請求項16】
前記工程(a)が、bFGFの存在をさらに含む請求項15に記載の方法。
【請求項17】
前記工程(b)が、VEGFを加える工程をさらに含む請求項15に記載の方法。
【請求項18】
前記アクチビンAの濃度が、約0.5ng/ml〜約30ng/mlである請求項15に記載の方法。
【請求項19】
前記BMP4の濃度が、約1.0ng/ml〜約20ng/mlである請求項15に記載の方法。
【請求項20】
bFGFの濃度が、約1.0ng/ml〜約10ng/mlである請求項15に記載の方法。
【請求項21】
前記DKK1の濃度が、約100ng/ml〜約200ng/mlである請求項15に記載の方法。
【請求項22】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項17に記載の方法。
【請求項23】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体を約4日間にわたり約3.0ng/mlのアクチビンA、約10.0ng/mlのBMP4及び約5.0ng/mlのbFGFの存在下にて無血清培地中で培養する工程、及び
(b)約150ng/mlのDKK1及び約10.0ng/mlのVEGFを無血清培地へ加え、心血管前駆細胞を生成するために約1〜2日間にわたり培養する工程を含む方法。
【請求項24】
(c)KDRを発現するがC−KITを発現しない細胞を選別する工程をさらに含む請求項23に記載の方法。
【請求項25】
前記選別が、免疫選択による請求項24に記載の方法。
【請求項26】
前記選別が、フローサイトメトリーによる請求項24に記載の方法。
【請求項27】
ヒト心筋細胞の集団を発生する方法であって、KDRを発現するがC−KITを発現しないヒト心血管前駆細胞の集団を約7〜約10日間にわたりWntの阻害剤の存在下で培養する工程と、ヒト心筋細胞の集団を収集する工程とを含む方法。
【請求項28】
前記ヒト心血管前駆細胞をVEGF及びWntの阻害剤の存在下で培養する工程を含む請求項27に記載の方法。
【請求項29】
前記Wntの阻害剤が、DKK1である請求項27に記載の方法。
【請求項30】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項28に記載の方法。
【請求項31】
DKK2の濃度が、約100ng/ml〜約200ng/mlである請求項27に記載の方法。
【請求項32】
前記ヒト心血管前駆細胞が、単層として培養される請求項27に記載の方法。
【請求項33】
前記ヒト心血管前駆細胞が、凝集体として培養される請求項27に記載の方法。
【請求項34】
心筋細胞、内皮細胞及び血管平滑筋細胞を含有する心血管コロニーを発生する方法であって、ヒト心血管前駆細胞を約3〜約7日間にわたりVEGF、bFGF、及びWntの阻害剤の存在下で培養する工程を含む方法。
【請求項35】
前記Wntの阻害剤が、DKK1である請求項34に記載の方法。
【請求項36】
前記DKK1の濃度が、約100ng/ml〜約200ng/mlである請求項35に記載の方法。
【請求項37】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項34に記載の方法。
【請求項38】
前記bFGFの濃度が、約1.0ng/ml〜約10.0ng/mlである請求項34に記載の方法。
【請求項39】
ヒト心血管前駆細胞の集団を含む組成物。
【請求項40】
請求項34の方法によって産出されたヒト心血管コロニーの集団を含む組成物。
【請求項41】
心筋細胞を置換する方法であって、当該置換を必要とする被験者に請求項27に記載の方法によって産生されたヒト心筋細胞を含む組成物を投与する工程を含む方法。
【請求項42】
心機能不全を特徴とする障害を治療する方法であって、前記治療を必要とする被験者にヒト心血管前駆細胞を含む組成物を投与する工程を含む方法。
【請求項1】
ヒト心血管前駆細胞の集団であって、前記前駆細胞はKDRを発現するがC−KITを発現しないヒト心血管前駆細胞の集団。
【請求項2】
前記前駆細胞が、心筋細胞、内皮細胞、及び血管平滑筋細胞の前駆細胞である、請求項1に記載のヒト心血管前駆細胞の集団。
【請求項3】
ヒト胚様体をアクチビン及びBMPの存在下で無血清培地中において培養する工程と、その後に前記培養にWntの阻害剤を加える工程とによって得られる、請求項1に記載のヒト心血管前駆細胞の集団。
【請求項4】
前記アクチビンが、アクチビンAである請求項3に記載のヒト心血管前駆細胞の集団。
【請求項5】
前記BMPが、BMP4である請求項3に記載のヒト心血管前駆細胞の集団。
【請求項6】
前記Wntの阻害剤が、DKK−1である請求項3に記載のヒト心血管前駆細胞の集団。
【請求項7】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体をアクチビン及びBMPの存在下で無血清培地中において培養する工程、及び
(b)前記培地へWntの阻害剤を加え、ヒト心血管前駆細胞の集団を提供するために培養する工程を含む方法。
【請求項8】
(c)収集した細胞集団を提供するために前記ヒト心血管前駆細胞を培養から収集する工程をさらに含む、請求項7に記載の方法。
【請求項9】
(d)前記収集した細胞集団を心血管前駆細胞に濃縮する工程をさらに含む、請求項8に記載の方法。
【請求項10】
前記アクチビンが、アクチビンAである請求項7に記載の方法。
【請求項11】
前記BMPが、BMP4である請求項7に記載の方法。
【請求項12】
前記Wntの阻害剤が、DKK1である請求項7に記載の方法。
【請求項13】
前記工程(a)が、bFGFの存在をさらに含む請求項7に記載の方法。
【請求項14】
前記工程(b)が、VEGFを加える工程をさらに含む請求項7に記載の方法。
【請求項15】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体を約1〜4日間にわたりアクチビンA及びBMP4の存在下で無血清培地中において培養する工程、
(b)DKK1を前記無血清培地へ加え、約1〜2日間にわたり培養する工程、及び、
(c)ヒト心血管前駆細胞の集団を収集する工程を含む方法。
【請求項16】
前記工程(a)が、bFGFの存在をさらに含む請求項15に記載の方法。
【請求項17】
前記工程(b)が、VEGFを加える工程をさらに含む請求項15に記載の方法。
【請求項18】
前記アクチビンAの濃度が、約0.5ng/ml〜約30ng/mlである請求項15に記載の方法。
【請求項19】
前記BMP4の濃度が、約1.0ng/ml〜約20ng/mlである請求項15に記載の方法。
【請求項20】
bFGFの濃度が、約1.0ng/ml〜約10ng/mlである請求項15に記載の方法。
【請求項21】
前記DKK1の濃度が、約100ng/ml〜約200ng/mlである請求項15に記載の方法。
【請求項22】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項17に記載の方法。
【請求項23】
ヒト心血管前駆細胞を産生するための方法であって、
(a)ヒト胚様体を約4日間にわたり約3.0ng/mlのアクチビンA、約10.0ng/mlのBMP4及び約5.0ng/mlのbFGFの存在下にて無血清培地中で培養する工程、及び
(b)約150ng/mlのDKK1及び約10.0ng/mlのVEGFを無血清培地へ加え、心血管前駆細胞を生成するために約1〜2日間にわたり培養する工程を含む方法。
【請求項24】
(c)KDRを発現するがC−KITを発現しない細胞を選別する工程をさらに含む請求項23に記載の方法。
【請求項25】
前記選別が、免疫選択による請求項24に記載の方法。
【請求項26】
前記選別が、フローサイトメトリーによる請求項24に記載の方法。
【請求項27】
ヒト心筋細胞の集団を発生する方法であって、KDRを発現するがC−KITを発現しないヒト心血管前駆細胞の集団を約7〜約10日間にわたりWntの阻害剤の存在下で培養する工程と、ヒト心筋細胞の集団を収集する工程とを含む方法。
【請求項28】
前記ヒト心血管前駆細胞をVEGF及びWntの阻害剤の存在下で培養する工程を含む請求項27に記載の方法。
【請求項29】
前記Wntの阻害剤が、DKK1である請求項27に記載の方法。
【請求項30】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項28に記載の方法。
【請求項31】
DKK2の濃度が、約100ng/ml〜約200ng/mlである請求項27に記載の方法。
【請求項32】
前記ヒト心血管前駆細胞が、単層として培養される請求項27に記載の方法。
【請求項33】
前記ヒト心血管前駆細胞が、凝集体として培養される請求項27に記載の方法。
【請求項34】
心筋細胞、内皮細胞及び血管平滑筋細胞を含有する心血管コロニーを発生する方法であって、ヒト心血管前駆細胞を約3〜約7日間にわたりVEGF、bFGF、及びWntの阻害剤の存在下で培養する工程を含む方法。
【請求項35】
前記Wntの阻害剤が、DKK1である請求項34に記載の方法。
【請求項36】
前記DKK1の濃度が、約100ng/ml〜約200ng/mlである請求項35に記載の方法。
【請求項37】
前記VEGFの濃度が、約1.0ng/ml〜約50.0ng/mlである請求項34に記載の方法。
【請求項38】
前記bFGFの濃度が、約1.0ng/ml〜約10.0ng/mlである請求項34に記載の方法。
【請求項39】
ヒト心血管前駆細胞の集団を含む組成物。
【請求項40】
請求項34の方法によって産出されたヒト心血管コロニーの集団を含む組成物。
【請求項41】
心筋細胞を置換する方法であって、当該置換を必要とする被験者に請求項27に記載の方法によって産生されたヒト心筋細胞を含む組成物を投与する工程を含む方法。
【請求項42】
心機能不全を特徴とする障害を治療する方法であって、前記治療を必要とする被験者にヒト心血管前駆細胞を含む組成物を投与する工程を含む方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】

【図13】

【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】

【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【公表番号】特表2011−517563(P2011−517563A)
【公表日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願番号】特願2011−502014(P2011−502014)
【出願日】平成21年3月25日(2009.3.25)
【国際出願番号】PCT/US2009/038220
【国際公開番号】WO2009/120762
【国際公開日】平成21年10月1日(2009.10.1)
【出願人】(502375437)マウント シナイ スクール オブ メディスン オブ ニューヨーク ユニバーシティー (11)
【Fターム(参考)】
【公表日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願日】平成21年3月25日(2009.3.25)
【国際出願番号】PCT/US2009/038220
【国際公開番号】WO2009/120762
【国際公開日】平成21年10月1日(2009.10.1)
【出願人】(502375437)マウント シナイ スクール オブ メディスン オブ ニューヨーク ユニバーシティー (11)
【Fターム(参考)】
[ Back to top ]