
ヒト癌化細胞の作製方法
【課題】安全性に問題がない、ヒト細胞を癌化させる方法、前記方法により得られる細胞、前記方法に用いるためのキット、および前記方法により得られた細胞を抗癌剤の薬効評価に用いる方法を提供すること。
【解決手段】ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、およびヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法。
【解決手段】ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、およびヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト癌化細胞の作製方法に関する。さらに詳しくは、ヒト28S rRNA由来アンチセンスRNAを用いたヒト細胞の癌化細胞の作製方法、該方法により得られる細胞、該方法に用いるためのキット、及び該方法により得られた細胞を抗癌剤の薬効評価に用いる方法に関する。
【背景技術】
【0002】
シミアンウイルス40(SV40)ラージT抗原、アデノウイルスE1A、E1B、ヒトパピローマウイルスE6、E7等のDNA腫瘍ウイルスタンパク質をコードする腫瘍遺伝子を用いたヒト細胞の研究から、ヒト培養細胞の悪性転換は、不死化過程と腫瘍形成過程の2つのフェーズからなるものであるという発癌モデルが仮定される。
【0003】
不死化過程では、テロメラーゼ活性によって、染色体末端部にテロメア配列が付加されることにより、無限の寿命を取得することが可能となる。従って、不死化が起こると、細胞はクローン性増殖を起こすことにより、遺伝子に何らかの変化がもたらされる時間が増えて、発癌形質に変化する可能性が高くなる。また、腫瘍形成過程では、不死化された細胞が、特定の成長因子の非存在下で自立増殖が可能となる。
【0004】
一般的に、癌細胞は、正常細胞とは異なる不特定な表現型、例えば、ゲノム不安定型、化学信号非応答型、非分化型等を発現する。腫瘍形成過程では、特定のタンパク質に関係する遺伝子変異が累積したり、又は、あまり知られていないが、腫瘍形成初期に活性化された因子によって遺伝子発現が連鎖したりするが、前記変異型が生じる原因がこのような過程に関係するかどうかは依然不明である。
【0005】
近年、ヒト正常細胞が、ヒトテロメラーゼ触媒サブユニット(hTERT)、SV40初期領域(ER)、及び活性型H-ras遺伝子を発現させることによって形質転換され得ることが報告されている(非特許文献1参照)。それによると、hTERT存在下では、SV40ラージT抗原(LT)がレチノブラストーマタンパク質(pRB)腫瘍抑制回路とp53腫瘍抑制回路を不活化し、SV40スモールT抗原(ST)がフォスファターゼ2Aを機能不全にする。また、活性型ras遺伝子は、形質転換細胞の腫瘍形成に関係することが知られている。従って、ヒト細胞の悪性転換には、このような遺伝子が最低でも必要であることが示唆される。
【0006】
また、特許文献1では、hTERTとSV40ラージT抗原(LT)が存在する状態で、Srcを発現させる、ヒト由来細胞の癌化誘導方法が開示されている。
【0007】
一方、本発明者らは、核内低分子RNA U5の二次構造のファーストステム(first stem)の3'末端側の特定の塩基配列に、ポリ(A)鎖を付加したもの〔トランスフォーミングRNA(TR)ともいう〕を、ノンコーディングmRNAとして発現させることにより、ラット線維芽細胞3Y1を腫瘍細胞に誘導することができることを報告している(非特許文献2参照)。3Y1細胞は、元来、自発的に不死化して永久に増殖することができる特性を有する。従って、TRが不死化ではなく腫瘍形成に関係することが推察されたので、さらに検討したところ、前記形質転換が、TRの特定のポリプリン配列、GGAGAGGAA(配列番号3)に大きく依存するものであることを見出した。
【0008】
またさらに、ウサギ網赤血球抽出液を用いて検討した結果、in vitroで合成されたTR配列が、ペプチド合成の鎖伸長初期に、リボソームの28S rRNAに直接結合して分泌シグナルペプチドに関連する翻訳に影響を及ぼすことを報告している(非特許文献3参照)。そして、前記ポリプリン配列を含むオリゴデオキシヌクレオチド(ODN)は、28S rRNA上のCUCC配列(配列番号4)と結合し、一方、前記ポリプリン配列のアンチセンス配列を含むオリゴデオキシヌクレオチド(ODN)は、シグナル認識顆粒(SRP)の7SL RNA上のGAGG(塩基48-51)(配列番号5)に結合することも見出されている。このことから、鎖伸長阻害は、28S rRNAとSRP RNAの間の生理的作用に影響を及ぼしていると考えられる。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特開2008−109889号公報
【非特許文献】
【0010】
【非特許文献1】W.C.Hahn et al, “Creation of human tumour cells withdefined genetic elements.”letter, Nature, 29 July 1999
【非特許文献2】K.Hamada, “Morphological transformation caused by a partial sequence of U5 small nuclear RNA.” Mol. Carcinog., 1997, 20, p175-188
【非特許文献3】K.Hamada et al, “Effect of Transforming RNA on the Synthesis of a Protein with a Secretory Signal Sequence in Vitro.” J. Biol. Chem., 1999, 274(22), p15786-15796
【発明の概要】
【発明が解決しようとする課題】
【0011】
しかしながら、前記TRは、hTERTの存在下では、c-myc、活性型H-ras、又はSV40 Large T等をコードする遺伝子と組み合わせることによっては、ヒト細胞を形質転換して癌化細胞に誘導できないことが判明した。
【0012】
本発明の課題は、安全性に問題がない、ヒト細胞を癌化させる方法、該方法により得られる細胞、該方法に用いるためのキット、及び該方法により得られた細胞を抗癌剤の薬効評価に用いる方法を提供することにある。
【課題を解決するための手段】
【0013】
本発明者らは、上記課題を解決する為に検討を重ねた結果、ヒト特異的なトランスフォーミングRNA(hTR)として、ヒト28S rRNAに相補的な特定のアンチセンス短鎖RNAが、リボソーム表面で結合して、ヒト細胞を癌化させることが出来ることを見出し、本発明を完成するに至った。
【0014】
即ち、本発明は、
〔1〕 ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法、
〔2〕 ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、同一又は異なるベクターに組み込み、ヒト由来細胞に導入する、ヒト細胞を癌化する遺伝子の導入方法、
〔3〕 前記〔1〕記載の方法により得られる、ヒト細胞の癌化細胞、
〔4〕 ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有してなる、前記〔1〕記載の方法に用いるためのキット、ならびに
〔5〕 前記〔3〕記載の癌化細胞に候補化合物を接触させる工程、及び、前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に、候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程を含む、抗癌剤のスクリーニング方法
に関する。
【発明の効果】
【0015】
本発明のヒト細胞の癌化細胞の作製方法によると、ヒト細胞を、より安全に、かつ、強力な癌化形質を有する癌化細胞に誘導できるものである。
【図面の簡単な説明】
【0016】
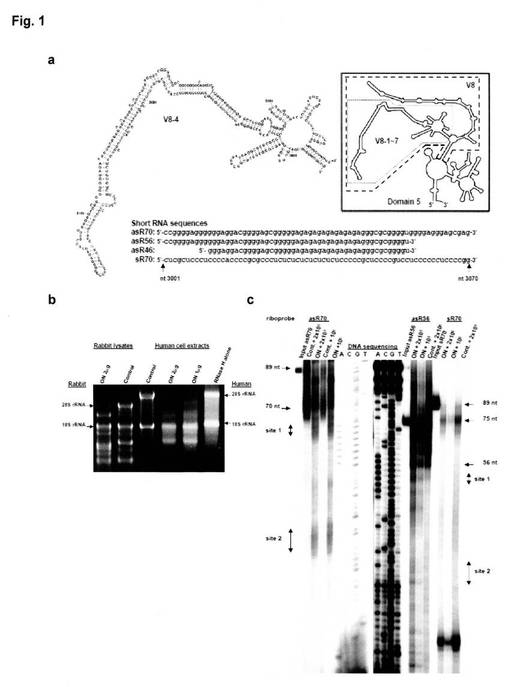
【図1】図1は、ヒト28S rRNAとantisense short RNAsの相互作用の結果を示す図である。図1−aは、ヒト28S rRNAのドメイン5の推定二次構造を示すものであり、左がV8-4領域、右がドメイン5の全体を示し、下段にはasR70(配列番号1)、asR56(配列番号6)、asR46(配列番号2)、sR70(配列番号7)の塩基配列を示す。図1−bは、ONと共培養した際のRNase Hによる切断評価の結果を示すものであり、左がウサギ溶解液について、右がヒド細胞抽出液について結果を示す。図1−cは、asR70、asR56、及びsR70の結合特性を示し、asR70については、左から、asR70プローブのみ、未処理RNA(2.0×105cpm asR70プローブ添加)、ODN処理RNA(2.0×105cpm asR70プローブ添加)、未処理RNA(1.0×105cpm asR70プローブ添加)、ODN処理RNA(1.0×105cpm asR70プローブ添加)の結果を、asR56については、左から、asR56プローブのみ、ODN処理RNA(2.0×105cpm asR56プローブ添加)、ODN処理RNA(1.0×105cpm asR56プローブ添加)、未処理RNA(2.0×105cpm asR56プローブ添加)の結果を、sR70については、左から、sR70プローブのみ、ODN処理RNA(2.0×105cpm sR70プローブ添加)、ODN処理RNA(1.0×105cpm sR70プローブ添加)、未処理RNA(2.0×105cpm sR70プローブ添加)を示す。なお、total RNAを20unitsのRNaseHで処理したものも併せて示す。
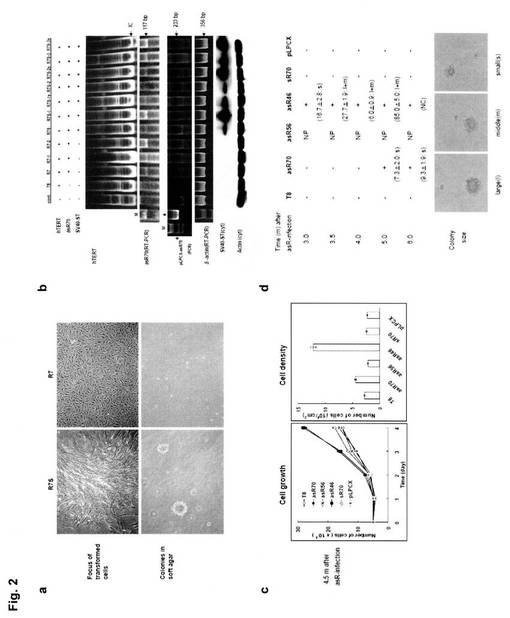
【図2】図2−aは、形質転換細胞の形態、及び軟寒天培地でのコロニー形成を示す図である。左がhTERT、SV40ST、及びasR70によって形質転換された細胞、右がhTERT、及びasR70によって形質転換された細胞であり、上段が細胞形態図、下段がコロニー形成を示す図である。図2−bは、上段より、hTERTについての結果(TRAPアッセイ)、asR70についての結果(RT-PCR)を、SV40STについての結果(イムノブロッティング)を示す図である。図2−cは、リボソーム由来オリゴヌクレオチドの形質転換細胞の成長に及ぼす影響を示す図である。導入後4.5ヵ月後の細胞状態を示し、左図が細胞成長を、右図が細胞密度を示す。図2−dは、形質転換細胞のコロニー形成試験の結果を示す。写真は、左から、大コロニー、中コロニー、小コロニーについて、それぞれ一般的な例を示す。
【図3】図3は、形質転換細胞の核型分析の結果を示す図である。
【図4】図4は、Bipタンパク合成の抑制及び小胞体ストレス反応誘導の結果を示す図である。図4−aは、35Sメチオニンを用いたパルスラベル・免疫沈降法によるBip、フィブロネクチン、インテグリンβ1の合成速度を調べた結果を示す図である。図4−b、cは、Bip、eIF2α、リン酸化eIF2α、ATF4のイムノブロッティングの結果を示す図である。
【発明を実施するための形態】
【0017】
本発明のヒト細胞の癌化細胞の作製方法は、ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、及びSV40スモールT抗原(SV40ST)遺伝子を導入する工程を含むものであるが、その際に、ヒト28S rRNA由来アンチセンスオリゴヌクレオチドを併用することを大きな特徴とする。
【0018】
ヒト28S rRNAの5'末端から約3kb領域には、ドメイン5のV8-4領域が存在する(図1−a参照)。V8領域は、例えば、シャペロン、修飾酵素、SRP又はトランスロコン(小胞体膜上にあるタンパク質輸送装置)等の非リボソーム性因子を、リボソーム中のL1サイドからトンネル出口の方に配置する作用を有すると報告されているイーストES27に相当する。また、V8-4領域は、5'末端側の後半部分に、CUCC配列(配列番号4)を6個有する。従って、本発明者らは、これらの事象を総合的に判断した結果、ヒト28S rRNAのES27に相補的な短鎖RNAが、ヒトトランスファーミングRNA(hTR)として用いることができると考え、hTRをhTERT及びSV40STと併用したところ、驚くべきことにも、ヒト細胞を形質転換させることができることを見出した。得られた形質転換細胞は、足場非依存性増殖能を有し、染色体も異常であることから、hTRを用いることで、hTERT及びSV40STの導入が可能となり、ヒト細胞を癌化細胞に形質転換できることが示唆される。
【0019】
本発明のヒト細胞の癌化細胞の作製方法は、ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む。
【0020】
本明細書において、「遺伝子」とは、生物の遺伝情報を担う因子であり、DNA及びRNAを含む意味で用いられる。また、「ヌクレオチド」とはDNA及びRNAを含む意味で用いられる。また、「アンチセンスオリゴヌクレオチド」とは、特定の塩基配列(以下、センス配列という)に対し相補的な塩基配列を有し、かつセンス配列にハイブリダイズし得るオリゴヌクレオチドをいう。
【0021】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子としては、特に限定はなく、当該分野で公知のものが挙げられる。テロメラーゼは、細胞分裂によるテロメア長の短縮に拮抗して、テロメア長を維持する酵素である。テロメラーゼは、テロメア配列の鋳型となるRNA、及び、逆転写酵素を構成要素に含み、この逆転写酵素部分がhTERTである。
【0022】
SV40スモールT抗原(SV40ST)遺伝子としては、特に限定はなく、当該分野で公知のものが挙げられる。SV40(simian virus 40)はポリオーマウイルス科のウイルスで、アカゲザル腎細胞から分離されたものである。このウイルスは、細胞への感染初期(DNA合成が起きる前)に、初期タンパク質として、ラージT抗原及びスモールT抗原を産生するが、本発明においては、スモールT抗原を抗原ウイルスとして用いる。
【0023】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドとしては、ヒト28S rRNAの拡張セグメント27(ES27)領域中にあるセンス配列のアンチセンスオリゴヌクレオチドが挙げられる。なかでも、配列番号1で示される塩基配列を含むオリゴヌクレオチド、配列番号2で示される塩基配列を含むオリゴヌクレオチド、及び配列番号6で示される塩基配列を含むオリゴヌクレオチドが好ましく、実質的に配列番号1で示される塩基配列からなるオリゴヌクレオチド、実質的に配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び実質的に配列番号6で示される塩基配列からなるオリゴヌクレオチドがより好ましく、配列番号1で示される塩基配列からなるオリゴヌクレオチド、及び配列番号2で示される塩基配列からなるオリゴヌクレオチドがさらに好ましい。
【0024】
また、本発明においては、前記オリゴヌクレオチドとして、前記塩基配列において1もしくは数個のヌクレオチドの欠失、付加、挿入又は置換を有するオリゴヌクレオチドも、好適に用いられる。かかるオリゴヌクレオチドとしては、具体的には、前記塩基配列で示されるオリゴヌクレオチドに対して少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有するオリゴヌクレオチド、又はそれを含むオリゴヌクレオチドを挙げることができる。また、塩基長としては、特に限定されないが、好ましくは40〜200、より好ましくは40〜150、さらに好ましくは40〜100、さらに好ましくは40〜90、さらに好ましくは40〜80、さらに好ましくは46〜70である。このようなオリゴヌクレオチドは、配列番号1で示される塩基配列からなるオリゴヌクレオチド、配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び配列番号6で示される塩基配列からなるオリゴヌクレオチドと同様の効果を奏するものである。なお、本明細書において、相同性は、例えば、Altschulら(The Journal of Molecular Biology,215,403−410(1990))の開発したアルゴリズムを使用した検索プログラムBLASTを用いて求めることができる。
【0025】
配列番号1で示される塩基配列からなるオリゴヌクレオチド、配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び配列番号6で示される塩基配列からなるオリゴヌクレオチドの模式図を図1に示す。これらは、ヒト28S rRNAのドメイン5に位置する、3001〜3070塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR70ともいう)、3015〜3060塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR46ともいう)、及び3015〜3070塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR56ともいう)である。いずれのオリゴヌクレオチドも、拡張セグメント27(エクスパンジョンセグメント27、ES27、2889〜3551塩基)領域に存在するセンス配列に相補的であり、ES27の機能、即ち、増殖に関係することが示唆される。なお、本明細書において、asRとは、アンチセンス短鎖RNA(antisense short RNA)を意味し、sRとは、センス短鎖RNA(sense short RNA)を意味する。
【0026】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドの合成方法としては、特に限定されず、公知のオリゴヌクレオチド合成機を用いたホスホロアミダイト法、ホスホロチオエート法、ホスホトリエステル法等を用いることができる。
【0027】
前記遺伝子及びアンチセンスオリゴヌクレオチドは、安定性や細胞に対する親和性を高めるために、その活性を著しく低下させない範囲で、公知の方法に従って、修飾体、置換体としてもよく、例えば、リン酸エステル基又はリボース部分の水酸基を他の安定な基に置換した誘導体として用いることも可能である。
【0028】
このような遺伝子及びアンチセンスオリゴヌクレオチドが導入されるヒト由来細胞としては、特に限定はなく、ヒト線維芽細胞、ヒト内皮細胞、ヒト上皮細胞等が挙げられる。なお、前記細胞の由来、組織は特に限定されない。
【0029】
ヒト由来細胞へ、前記遺伝子及びアンチセンスオリゴヌクレオチドを導入する方法としては、特に限定はなく、例えば、前記遺伝子及びアンチセンスオリゴヌクレオチドを任意のベクターに組み込んだものを用いることができる。
【0030】
ベクターは、宿主細胞において自立複製可能であると同時に、前記遺伝子及びアンチセンスオリゴヌクレオチド以外に、プロモーター、及び転写終結配列を含有することが好ましい。また、プロモーターを制御する遺伝子が含まれていてもよい。なお、プロモーターとしては、前記遺伝子及びアンチセンスオリゴヌクレオチドを、宿主細胞中に発現させることが可能であれば特に限定はない。
【0031】
本発明に用いられる好適なベクターとしては、例えば、アデノウイルスベクター、ワクシニアウイルスベクター、レトロウイルスベクター等が挙げられる。
【0032】
前記遺伝子及びアンチセンスオリゴヌクレオチドは、同時に一括又は個々に導入してもよく、異なるタイミングにそれぞれを導入してもよく、前記遺伝子及びアンチセンスオリゴヌクレオチドを別々のベクターに組み込んだものを用いてもよく、同一のベクターに組み込んだものを用いてもよい。別々のベクターに組み込んだものを用いる場合、例えば、hTERT遺伝子はpBABEベクターに、SV40ST遺伝子はpLHCXベクターに、アンチセンスオリゴヌクレオチドはpLPCXベクターに組み込んだものを用いることができる。また、同一のベクターに組み込んだものを用いる場合、好適なベクターとしてはレトロウイルスベクターが挙げられ、前記遺伝子及びアンチセンスオリゴヌクレオチドの該ベクター内での配置は、特に限定はなく、例えば、プロモーターと転写終結配列の間に、プロモーター側から、hTERT遺伝子、SV40ST遺伝子、アンチセンスオリゴヌクレオチドの順で配置される。
【0033】
ベクターの導入方法としては、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0034】
また、前記ベクターに導入細胞への感染用ウイルスDNAをさらに組み込んだベクターを調製して、ベクターが導入されたウイルスをヒト細胞に感染させることで、ベクター導入を行ってもよい。感染用ウイルスとしては、アデノウイルス、アデノ随伴ウイルス、レトロウイルス等が挙げられる。
【0035】
なお、かかるベクターの構築方法、具体的な用法等については、例えば、Sambrook, J., et. al., Molecular cloning: A Laboratory Mannual; 2nd Ed., Cold Spring Harbor Laboratory. Cold Spring Harbor. NY., 1989.等の成書を参照すればよい。
【0036】
かくして、ヒト由来細胞に、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入することができる。得られた細胞は、hTERT及びSV40STによってコードされるタンパク質を発現すると共に、ヒト28S rRNA由来アンチセンスオリゴヌクレオチドを発現する、形質転換細胞となる。従って、本発明は、また、前記hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入することにより得られる細胞を提供する。該細胞は、細胞に適した条件下で培養し、集団倍加を繰り返して、公知の薬剤による選択を行ってもよい。なお、本発明においては、hTERT遺伝子が既に組み込まれたhTERT不死化細胞に、SV40ST遺伝子及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入したものも、本発明の範囲内に含まれる。hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドによって癌化が誘導されたヒト細胞の状態については、後述の実施例に記載の方法に従って、確認することができる。
【0037】
また、本発明では、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを組み合わせて用いることで、ヒト細胞を癌化できることから、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有するキットを、ヒト細胞の癌化用に用いることができる。本発明は、また、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有するキットを提供する。
【0038】
またさらに、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを組み合わせて用いることで、ヒト細胞の癌化が容易であり、また、癌化程度も一定であることから、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを用いて癌化されたヒト細胞を、抗癌剤のスクリーニングに用いることができる。従って、本発明は、また、抗癌剤のスクリーニング方法を提供する。前記方法は、ヒトの系でのin vitro評価であるため、in vivoへの想定が容易となり、より簡便、かつ、迅速に有効物質を見出すことが可能となる。
【0039】
具体的な方法としては、
hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、ヒト細胞に導入して得られた癌化細胞に候補化合物を接触させる工程、ならびに、
前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程
を含む方法が挙げられる。
【0040】
hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドが導入されたヒト細胞に抗癌剤を接触させる方法としては、特に限定はなく、当該分野で公知の方法を用いることができる。
【0041】
また、抗癌剤の接触前後のヒト細胞の癌化程度の判断としては、例えば、同じ実験系において、接触後の一部の細胞が他の細胞に比べて、トランスフォーム形態消失、アポトーシスの誘導、及び足場非依存性の消失等の変化が認められる場合には、該細胞に接触させた候補化合物が、抗癌剤としての作用を有し、癌化を縮小する効果があると判定することができる。
【実施例】
【0042】
以下、本発明を実施例に基づいて説明するが、本発明はこれらの実施例等によりなんら限定されるものではない。なお、hTERT遺伝子は、Dr. F.Ishikaw又はaddgene1774より入手したものを、SV40ST遺伝子は、JCRB gene bank(pMTI0D)より入手したものを、ヒト28S rRNA由来オリゴヌクレオチドは、合成したもの又はTakara Bio社より入手したものを用いた。また、ヒト線維芽細胞(MJ90細胞)はDr. O.M.Pereira-Smithより、ヒト線維芽細胞(TIG3細胞)はRB cell bankより入手したものを用いた。
【0043】
参考例1<GGAGAGGAA配列(配列番号3)の結合特性>
10μLのウサギ溶解液又は20μLのヒト細胞抽出液に、トランスファーミングRNA(TR)の一部であるポリプリン配列、GGAGAGGAA(配列番号3)を添加し、RNase Hの存在下で反応させた。その後、total RNAを回収して、ポリアクリルアミドゲル電気泳動にかけた結果を図1−bに示す。結果、GGAGAGGAA配列(配列番号3)は、28S rRNAに結合していることが分かる。
【0044】
参考例2<asR70、asR56、及びsR70の結合特性>
asR70、asR56、及びsR70の結合特性を以下の方法に従って調べた。結果を図1−cに示す。具体的には、先ず、HindIII-Xbalで切断されたプラスミドpGEM3(Promega社製)に挿入して作製したプラスミドpG-asR70、pG-asR56、及びpG-sR70から、SP6 RNA polymerase(Takara Bio社製)と[α-32P]-GTP(Perkin-Elmer社製)を用いて、asR70、asR56、及びsR70の標識RNAプローブ(riboprobe)を作製した。得られたプローブをポリアクリルアミドゲル電気泳動にかけたところ、asR70プローブが89塩基(左から1番目のレーン)、asR56プローブが75塩基(右から8番目のレーン)、及びsR70プローブが89塩基(右から4番目のレーン)の長さを有するプローブであった。
【0045】
次に、20μLのヒト細胞溶解液と4μgのオリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)を混合してRNase Hで切断後、回収したtotal RNAをasR70、asR56、及びsR70のリボプローブと混合し、RNase A 及び RNase T1で処理した。なお、比較として、オリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)を用いない例についても、同様の処理を行った。
【0046】
オリゴデオキシヌクレオチド処理total RNAはasR70プローブとの結合において二箇所の切断部位を示した。即ち、オリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)は、28S rRNAの3001〜3070塩基内に結合部位を有することが判明した(左から3番目と5番目のレーン)。具体的には、28S rRNAの3063〜3066塩基であるCUCC配列(配列番号4、サイト1)と、3035〜3040塩基であるCUCUCU配列(配列番号8、サイト2)と結合し得ることが分かった。また、14ヌクレオチド短いasR56についても、同じサイトにおいて結合することが判明した(右から6番目と7番目のレーン)。このサイト1とサイト2は、約23〜24ヌクレオチド離れて位置し、サイト1はV8-4領域のヘアピンループに位置し、一方、サイト2は膨らみ部分に位置するものであり、asR70及びasR56は、前記2箇所のサイトで28S rRNAと結合することが示唆される。一方、sR70プローブと28S rRNAとの結合は確認されなかった(右から2番目と3番目のレーン)。また、非特許文献3において、配列番号3で示されるオリゴデオキシヌクレオチドの修飾配列であるGAGAGAGAG配列(配列番号9)は、分泌タンパクの合成を抑制することが出来ないことが示されていることから、V8-4領域では、サイト2よりもサイト1においてSRPのRNAと相互作用することでタンパク質合成を制御していることが示唆される。なお、asR56には、サイト2に結合するasR70中の配列が含まれているが、異所性結合を伴って結合が弱いと推定される。
【0047】
実施例1<形質転換細胞の形態、癌化形質>
hTERT、asR70、及びSV40STによって形質転換されたMJ90細胞(hTERT+asR70+SV40ST、R7S)、ならびに、hTERT及びasR70によって形質転換されたMJ90細胞(hTERT+asR70、R7)について、位相差顕微鏡を用いて細胞形態を観察した。また、軟寒天培地でのコロニー形成試験も行った。結果を図2−aに示す。
【0048】
なお、形質転換細胞は次のようにして調製したものを用いた。具体的には、先ず、asR70を挿入したpGEM3(Promega社製)をHindIII-EcoRIで切断した断片をpLPCX(Clontech社製)に挿入したベクターを調製した(pLPCX-asR70)。PCRで増幅したSV40ST断片をpLHCX(Clontech社製)に挿入したベクターを調製した(pLHCX-ST)。hTERT発現用プラスミドは、pcDNAhTERTn2(Dr. F.Ishikawより提供)及びpBABE-neo-hTERT(Addgene plasmid 1774)を用いた。次に、pLPCX-asR70及びpLHCX-STは、レトロウイルスパッケイジングプラスミドpCL-10A1(Imgenex社製)と共に、293T 細胞(Riken Cell Bankから入手)にトランスフェクトした後、ウイルス回収の上、MJ90細胞に感染させた。pcDNAhTERTn2を用いる場合は、lipofectamine 2000を用いてMJ90細胞にトランスフェクトした。pBABE-neo-hTERTを用いる場合は、ウイルス作製後、TIG3細胞に感染させた。得られた細胞は、薬剤選択を行って親株よりもライフスパンの長いものを選択して実験に供した。
【0049】
図2−aより、hTERT、SV40ST、及びasR70によって形質転換された細胞(R7S)は、細胞と細胞の境界が不明瞭となり、軟寒天培地でも移植後14〜18日にはコロニーを形成していたため、足場非依存性増殖能を有し癌化していることがわかる。
【0050】
実施例2<形質転換細胞におけるhTERT、SV40ST、asR70の発現>
正常ヒト線維芽細胞(intact、MJ90)、hTERT不死化細胞(T8)、ならびに、hTERT及びasR70を共トランスフェクションした細胞(R7)、そのクローン(R7-1、R7-2)を準備した。また、細胞(R7)にSV40STを導入した細胞(R7S)、そのクローン(R7S-1、R7S-2、R7S-3)、及びサブクローン(R7S-1s、R7S-2s、R7S-3s)も準備した。なお、準備した細胞は全て実施例1と同様にして選択された株を用いた。また、全細胞株について、実施例1と同様にしてコロニー形成試験を行った結果、細胞(R7Sシリーズ)は多くのコロニー形成が確認されたが、細胞(R7、R7-1、R7-2)、細胞(MJ90)、及び細胞(T8)では全くコロニーが形成されなかった。
【0051】
各細胞の抽出液(1μg)を用いて、PCRを用いたTRAP(telomeric repeat amplification protocol)アッセイを行って、テロメラーゼ活性(hTERT活性)を調べた。結果を図2-b(PCR結果の一番上)に示す。細胞(MJ90)以外は、テロメラーゼ活性があることが示された。なお、図中の「IC」は、PCR用の内部標準物質である。
【0052】
asR70については、asR70特異的な下記に示すプライマーを用いたRT-PCRを行って調べた(図2-bのPCR結果の上から2番目)。その結果、細胞(MJ90)及び細胞(T8)以外の細胞において、増幅産物(117bp)が認められた。
pLPCX-retroF:5'-CGCTAGCGCTACCGGACTC-3'(配列番号10)
asR70-R:5'-ATAAAGCTTCTCGCTCCCTCCCCACCC-3'(配列番号11)
【0053】
また、増幅産物(117bp)がasR70 cDNAに特異的なものであることを確かめるために、前記pLPCX-retroFプライマーと、asR70のDNAを含むプラスミドの非転写領域に特異的なプライマーとして、
pLPCX-seqR:5'-TGGGGTCTTTCATTCCC-3'(配列番号12)
を用いたPCR増幅を行ったが、増幅産物(233bp)が確認されず、asR70がプラスミドに由来するものではなく組み込まれたものであることが確認できた(図2-bのPCR結果の上から3番目)。さらに、増幅量が等量であることを示すために、下記プライマーを用いて、β-アクチン(356bp)の増幅量を調べた(図2-bのPCR結果の一番下)。なお、図2-b中の「M」は、分子量マーカー(HaeIII digest marker)を示し、「*」は、プラスミドpLPCX-asR70を増幅した結果を示す。
forward:5'-TGAAGTGTGACGTGGACATCCGC-3'(配列番号13)
reverse:5'-GCCAATCTCATCTTGTTTTCTGCGC-3'(配列番号14)
【0054】
以上より、細胞(MJ90)及び細胞(T8)以外は、asR70の発現が確認され、それらはプラスミド由来ではなく、形質転換により組み込まれたものに由来することが明らかになった。
【0055】
また、SV40STについては、細胞質のイムノブロッティングにより調べた。結果を図2-bの下段(イムノブロッティング結果)に示す。その結果、細胞(R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)において、SV40ST(15〜20kD)の発現が確認された。なお、全ての細胞において、コントロールとして、Actinの発現も確認した(図2-bのイムノブロッティング結果下)。
【0056】
実施例3<ヒト28S rRNA由来オリゴヌクレオチドの形質転換細胞の成長に及ぼす影響>
実施例2で用いたhTERT不死化細胞(T8)を13ヶ月培養した後、液体窒素中で保存した。その後、解凍し、数回複製してから、以下の実験に用いた。
【0057】
前記細胞(T8)に、intactのpLPCX(pLPCX)、asR70を組み込んだpLPCX(asR70)、asR56を組み込んだpLPCX(asR56)、asR46を組み込んだpLPCX(asR46)、又はsR70を組み込んだpLPCX(sR70)をそれぞれ導入し、ピューロマイシンによる薬剤選択を行って、該導入細胞(T8、pLPCX、asR70、asR56、asR46、sR70)を選択した。導入4.5ヵ月後の細胞株について、細胞成長と細胞密度を調べた(n=3)。結果を図2-cに示す。なお、細胞成長は、一定数に調整された細胞の経時的な細胞数増加を測定することにより調べ、細胞密度は、一定面積に占める細胞数を測定することにより調べることができる。
【0058】
図2-cより、導入4.5ヵ月後の細胞では、asR70を導入した細胞及びasR46を導入した細胞が、細胞成長速度が大きく、また細胞密度も高いものであった。このことから、asR70及びasR46は、細胞増殖メカニズムに影響を及ぼすものであることが示唆される。
【0059】
実施例4<形質転換細胞のコロニー形成>
実施例2で用いたhTERT不死化細胞(T8)に、intactのpLPCX(pLPCX)、asR70を組み込んだpLPCX(asR70)、asR56を組み込んだpLPCX(asR56)、asR46を組み込んだpLPCX(asR46)、又はsR70を組み込んだpLPCX(sR70)をそれぞれ導入後、intactのpLPCXを導入した細胞以外には、さらにSV40STを組み込んだpLPCXを導入後、ピューロマイシンによる薬剤選択を行って、該導入細胞(T8、pLPCX、asR70、asR56、asR46、sR70)を選択した。得られた細胞を培養し、経時的に、コロニー形成試験を行った(n=3)。また、コロニー形成が認められた場合は、コロニーを大きさ「大(large、細胞の10倍以上の大きさ)」、「中(middle、細胞の5〜9倍程度の大きさ)」、「小(small、細胞の3〜4倍程度の大きさ)」によって分類し、大コロニーと中コロニーの合計コロニー数(l+m)、及び小コロニー数(s)を計数し、平均値を求めた。結果を図2-dに示す。なお、図2-dにおいて「NP」とは、コロニーが認められるものの増殖が確認されない場合を示し、「NC」とはコロニー計数を行わなかった場合を意味する。
【0060】
図2-dより、細胞(asR70)及び細胞(asR46)では、軟寒天中でのコロニー形成が確認され、いずれの細胞も癌化していることが分かる。また、細胞(asR46)は、細胞(asR70)より多くのコロニーを形成していることから、癌化効率が高いことが示唆される。これらより、asR70及びasR46は、より高い形質転換効果を奏することが示唆される。また、asR46、hTERT、及びSV40STを用いることで、ヒト胎児肺由来二倍体線維芽細胞TIG3を形質転換できることも確認できた(結果示さず)。なお、前記細胞(asR46)(以降R4Sともいう)を単離して、そのクローン(R4S-1、R4S-2、R4S-3)を調製し、以降の実験に用いた。
【0061】
実施例5<形質転換細胞の核型分析>
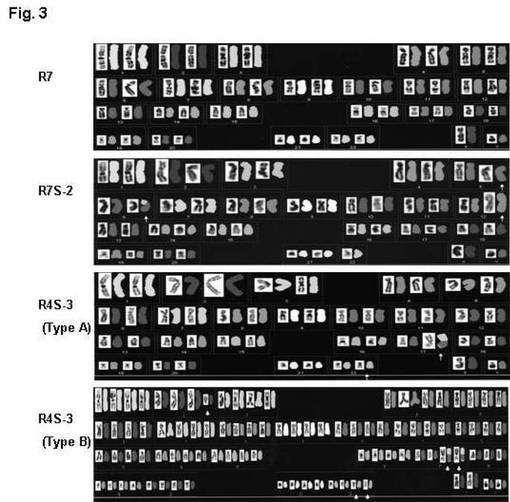
実施例2で用いた、hTERT不死化細胞(T8)、hTERT及びasR70を共トランスフェクションした細胞(R7)、細胞(T8)にSV40STを導入した細胞(T8+ST)、細胞(R7)にSV40STを導入した細胞のクローン(R7S-1、R7S-2、R7S-3)、ならびに実施例4で用いた、hTERT不死化細胞(T8)にasR46とSV40STを導入した細胞のクローン(R4S-1、R4S-2、R4S-3)のそれぞれについて、5細胞ずつ、核型分析(reverse DAPI banding - spectral karyotyping analysis、SKY)を行った。結果を表1に、また代表的なSKY染色図を図3に示す。
【0062】
【表1】

【0063】
表1及び図3より、細胞(T8)、細胞(R7)、及び細胞(T8+ST)は、46本の染色体が確認され、正常の二倍体細胞であることが分かる。一方、細胞(R7S-1)と細胞(R7S-3)は、46本の染色体が認められたものの、1箇所でゲノム再編成が行われていることが明らかとなった。また、細胞(R7S-2)は、染色体数が45本と正常より1本少なく、4箇所でゲノム再編成が確認された。また、細胞(R4S-1、R4S-2、R4S-3)はいずれも、同じ株であったとしても、染色体数やゲノム再編成が行われている箇所などが様々であり、種々のタイプが確認された。なかでも、細胞(R4S-1、R4S-2、R4S-3)は細胞(R7S-1、R7S-2、R7S-3)より、変異がより大きいものであるが、細胞(R4S-1、R4S-2)には正常な細胞も存在することから、これらの変異は、形質転換後に生じていることが示唆される。これより、hTERT、SV40ST及びasR70形質転換細胞は、細胞内の遺伝情報が、正常細胞やhTERT及びasR70形質転換細胞とは異なることが示唆される。
【0064】
実施例6<形質転換細胞における小胞体ストレス反応誘導>
実施例4と同様にして得られた細胞(T8、pLPCX、asR70、asR56、asR46、sR70)について、小胞体ストレス応答を示す物質の発現を確認した。また、細胞(asR70)については、実施例2と同様にして得られた細胞、即ち、細胞(T8、R7、R7-1、R7-2、R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)について同様の実験を行った。
【0065】
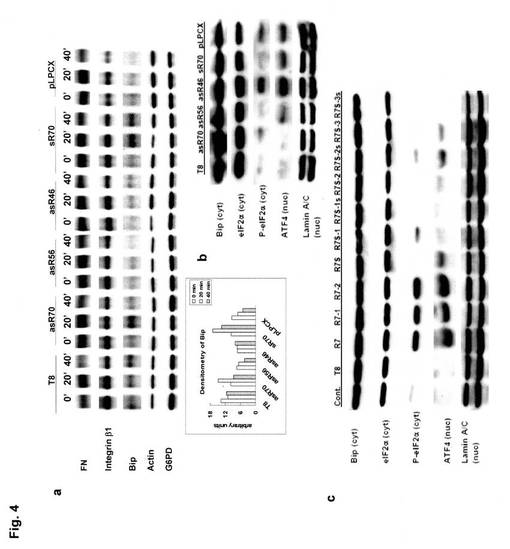
具体的には、35Sメチオニンを用いたパルスラベル・免疫沈降法に従って、細胞外マトリックスのフィブロネクチン(FN)、膜透過型レセプターのインテグリン ベータ1(integrin β1)、及び小胞体局在シャペロンのBipの発現量を各細胞について測定した。細胞(T8、pLPCX、asR70、asR56、asR46、sR70)に関する結果を図4−aに示す。
【0066】
図4−aより、Bipの発現は、細胞(asR70)、細胞(asR56)、細胞(asR46)及び細胞(pLPCX)で経時的に減少していることが分かった(図4−aの枠内グラフ参照)。一方、FN及びintegrin β1の発現は、シグナル非依存性のアクチンやグルコース6リン酸デヒドロゲナーゼと同様の傾向を示し、殆ど変化が認められなかった。FN及びintegrin β1は、細胞内で生産性が高いため変化が認められなかったが、Bipの発現量に変化が認められたことから、細胞(asR70)、細胞(asR56)、細胞(asR46)のリボゾームにおいてBipの合成が抑制されていることが示唆される。
【0067】
また、Bipの影響について更に詳しく検討するために、細胞質タンパク質について同様の検討を行った。具体的には、30μgの細胞質タンパク質を10% SDS/PAGEで分離し、Bip、eIF2a(eukaryotic initiation factor 2a)、リン酸化eIF2a(P-eIF2a)、及びATF4(activating transcription factor 4)のイムノブロッティングを行った。細胞(T8、pLPCX、asR70、asR56、asR46、sR70)に関する結果を図4−b、細胞(T8、R7、R7-1、R7-2、R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)に関する結果を図4−cに示す。なお、Bipの合成が抑制されると、eIF2aがリン酸化されてタンパク合成が抑制され、リン酸化されたeIF2aがATF4の翻訳を開始するという、小胞体のストレス反応が誘導される。
【0068】
図4−b、cより、Bip、eIF2aの発現量は、いずれの細胞においても同程度であったが、細胞(asR46)、細胞(R7)、細胞(R7-1)、細胞(R7-2)において、リン酸化eIF2aの発現がより顕著に確認され、また、付随するATF4の誘導も確認された。このことから、asR46とasR70は、Bip合成を抑制することで、小胞体ストレス応答を誘導していることが示唆される。
【0069】
実施例7<抗癌剤のスクリーニング>
hTERT、SV40ST、及びヒト28S rRNA由来オリゴヌクレオチドが導入された細胞を細胞培養器具にて培養し、そこに、候補化合物を溶媒に溶解したものを添加して、さらに培養する。一方、対照として、候補化合物を添加しない群も設定する。候補化合物を添加しない群をコントロールとして、添加しない群よりも細胞のトランスフォーム形態の消失、アポトーシスの誘導、及び足場非依存性増殖の消失等の変化が認められる場合に、該細胞に添加した候補化合物は、抗癌剤としての作用を有する可能性が高いと判断することができる。
【産業上の利用可能性】
【0070】
本発明のヒト細胞の癌化細胞の作製方法は、癌発症メカニズムの解明のため、様々なヒト正常細胞に癌化を誘導する際に利用することができ、新たな創薬ターゲット分子の探索に有効に利用することができる。
【配列表フリーテキスト】
【0071】
配列表の配列番号1は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR70)である。
配列表の配列番号2は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR46)である。
配列表の配列番号3は、トランスファーミングRNAの部分塩基配列である。
配列表の配列番号4は、ヒト28S rRNAの部分塩基配列である。
配列表の配列番号5は、ヒトSRP RNAの部分塩基配列である。
配列表の配列番号6は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR56)である。
配列表の配列番号7は、ヒト28S rRNA由来オリゴヌクレオチド(sR70)である。
配列表の配列番号8は、ヒト28S rRNAの部分塩基配列である。
配列表の配列番号9は、トランスファーミングRNAの部分塩基配列を修飾した配列である。
配列表の配列番号10は、pLPCX-retroF用プライマーである。
配列表の配列番号11は、asR70 cDNA用プライマーである。
配列表の配列番号12は、pLPCX-seqR用プライマーである。
配列表の配列番号13は、β-アクチン用プライマーである。
配列表の配列番号14は、β-アクチン用プライマーである。
【技術分野】
【0001】
本発明は、ヒト癌化細胞の作製方法に関する。さらに詳しくは、ヒト28S rRNA由来アンチセンスRNAを用いたヒト細胞の癌化細胞の作製方法、該方法により得られる細胞、該方法に用いるためのキット、及び該方法により得られた細胞を抗癌剤の薬効評価に用いる方法に関する。
【背景技術】
【0002】
シミアンウイルス40(SV40)ラージT抗原、アデノウイルスE1A、E1B、ヒトパピローマウイルスE6、E7等のDNA腫瘍ウイルスタンパク質をコードする腫瘍遺伝子を用いたヒト細胞の研究から、ヒト培養細胞の悪性転換は、不死化過程と腫瘍形成過程の2つのフェーズからなるものであるという発癌モデルが仮定される。
【0003】
不死化過程では、テロメラーゼ活性によって、染色体末端部にテロメア配列が付加されることにより、無限の寿命を取得することが可能となる。従って、不死化が起こると、細胞はクローン性増殖を起こすことにより、遺伝子に何らかの変化がもたらされる時間が増えて、発癌形質に変化する可能性が高くなる。また、腫瘍形成過程では、不死化された細胞が、特定の成長因子の非存在下で自立増殖が可能となる。
【0004】
一般的に、癌細胞は、正常細胞とは異なる不特定な表現型、例えば、ゲノム不安定型、化学信号非応答型、非分化型等を発現する。腫瘍形成過程では、特定のタンパク質に関係する遺伝子変異が累積したり、又は、あまり知られていないが、腫瘍形成初期に活性化された因子によって遺伝子発現が連鎖したりするが、前記変異型が生じる原因がこのような過程に関係するかどうかは依然不明である。
【0005】
近年、ヒト正常細胞が、ヒトテロメラーゼ触媒サブユニット(hTERT)、SV40初期領域(ER)、及び活性型H-ras遺伝子を発現させることによって形質転換され得ることが報告されている(非特許文献1参照)。それによると、hTERT存在下では、SV40ラージT抗原(LT)がレチノブラストーマタンパク質(pRB)腫瘍抑制回路とp53腫瘍抑制回路を不活化し、SV40スモールT抗原(ST)がフォスファターゼ2Aを機能不全にする。また、活性型ras遺伝子は、形質転換細胞の腫瘍形成に関係することが知られている。従って、ヒト細胞の悪性転換には、このような遺伝子が最低でも必要であることが示唆される。
【0006】
また、特許文献1では、hTERTとSV40ラージT抗原(LT)が存在する状態で、Srcを発現させる、ヒト由来細胞の癌化誘導方法が開示されている。
【0007】
一方、本発明者らは、核内低分子RNA U5の二次構造のファーストステム(first stem)の3'末端側の特定の塩基配列に、ポリ(A)鎖を付加したもの〔トランスフォーミングRNA(TR)ともいう〕を、ノンコーディングmRNAとして発現させることにより、ラット線維芽細胞3Y1を腫瘍細胞に誘導することができることを報告している(非特許文献2参照)。3Y1細胞は、元来、自発的に不死化して永久に増殖することができる特性を有する。従って、TRが不死化ではなく腫瘍形成に関係することが推察されたので、さらに検討したところ、前記形質転換が、TRの特定のポリプリン配列、GGAGAGGAA(配列番号3)に大きく依存するものであることを見出した。
【0008】
またさらに、ウサギ網赤血球抽出液を用いて検討した結果、in vitroで合成されたTR配列が、ペプチド合成の鎖伸長初期に、リボソームの28S rRNAに直接結合して分泌シグナルペプチドに関連する翻訳に影響を及ぼすことを報告している(非特許文献3参照)。そして、前記ポリプリン配列を含むオリゴデオキシヌクレオチド(ODN)は、28S rRNA上のCUCC配列(配列番号4)と結合し、一方、前記ポリプリン配列のアンチセンス配列を含むオリゴデオキシヌクレオチド(ODN)は、シグナル認識顆粒(SRP)の7SL RNA上のGAGG(塩基48-51)(配列番号5)に結合することも見出されている。このことから、鎖伸長阻害は、28S rRNAとSRP RNAの間の生理的作用に影響を及ぼしていると考えられる。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特開2008−109889号公報
【非特許文献】
【0010】
【非特許文献1】W.C.Hahn et al, “Creation of human tumour cells withdefined genetic elements.”letter, Nature, 29 July 1999
【非特許文献2】K.Hamada, “Morphological transformation caused by a partial sequence of U5 small nuclear RNA.” Mol. Carcinog., 1997, 20, p175-188
【非特許文献3】K.Hamada et al, “Effect of Transforming RNA on the Synthesis of a Protein with a Secretory Signal Sequence in Vitro.” J. Biol. Chem., 1999, 274(22), p15786-15796
【発明の概要】
【発明が解決しようとする課題】
【0011】
しかしながら、前記TRは、hTERTの存在下では、c-myc、活性型H-ras、又はSV40 Large T等をコードする遺伝子と組み合わせることによっては、ヒト細胞を形質転換して癌化細胞に誘導できないことが判明した。
【0012】
本発明の課題は、安全性に問題がない、ヒト細胞を癌化させる方法、該方法により得られる細胞、該方法に用いるためのキット、及び該方法により得られた細胞を抗癌剤の薬効評価に用いる方法を提供することにある。
【課題を解決するための手段】
【0013】
本発明者らは、上記課題を解決する為に検討を重ねた結果、ヒト特異的なトランスフォーミングRNA(hTR)として、ヒト28S rRNAに相補的な特定のアンチセンス短鎖RNAが、リボソーム表面で結合して、ヒト細胞を癌化させることが出来ることを見出し、本発明を完成するに至った。
【0014】
即ち、本発明は、
〔1〕 ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法、
〔2〕 ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、同一又は異なるベクターに組み込み、ヒト由来細胞に導入する、ヒト細胞を癌化する遺伝子の導入方法、
〔3〕 前記〔1〕記載の方法により得られる、ヒト細胞の癌化細胞、
〔4〕 ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有してなる、前記〔1〕記載の方法に用いるためのキット、ならびに
〔5〕 前記〔3〕記載の癌化細胞に候補化合物を接触させる工程、及び、前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に、候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程を含む、抗癌剤のスクリーニング方法
に関する。
【発明の効果】
【0015】
本発明のヒト細胞の癌化細胞の作製方法によると、ヒト細胞を、より安全に、かつ、強力な癌化形質を有する癌化細胞に誘導できるものである。
【図面の簡単な説明】
【0016】
【図1】図1は、ヒト28S rRNAとantisense short RNAsの相互作用の結果を示す図である。図1−aは、ヒト28S rRNAのドメイン5の推定二次構造を示すものであり、左がV8-4領域、右がドメイン5の全体を示し、下段にはasR70(配列番号1)、asR56(配列番号6)、asR46(配列番号2)、sR70(配列番号7)の塩基配列を示す。図1−bは、ONと共培養した際のRNase Hによる切断評価の結果を示すものであり、左がウサギ溶解液について、右がヒド細胞抽出液について結果を示す。図1−cは、asR70、asR56、及びsR70の結合特性を示し、asR70については、左から、asR70プローブのみ、未処理RNA(2.0×105cpm asR70プローブ添加)、ODN処理RNA(2.0×105cpm asR70プローブ添加)、未処理RNA(1.0×105cpm asR70プローブ添加)、ODN処理RNA(1.0×105cpm asR70プローブ添加)の結果を、asR56については、左から、asR56プローブのみ、ODN処理RNA(2.0×105cpm asR56プローブ添加)、ODN処理RNA(1.0×105cpm asR56プローブ添加)、未処理RNA(2.0×105cpm asR56プローブ添加)の結果を、sR70については、左から、sR70プローブのみ、ODN処理RNA(2.0×105cpm sR70プローブ添加)、ODN処理RNA(1.0×105cpm sR70プローブ添加)、未処理RNA(2.0×105cpm sR70プローブ添加)を示す。なお、total RNAを20unitsのRNaseHで処理したものも併せて示す。
【図2】図2−aは、形質転換細胞の形態、及び軟寒天培地でのコロニー形成を示す図である。左がhTERT、SV40ST、及びasR70によって形質転換された細胞、右がhTERT、及びasR70によって形質転換された細胞であり、上段が細胞形態図、下段がコロニー形成を示す図である。図2−bは、上段より、hTERTについての結果(TRAPアッセイ)、asR70についての結果(RT-PCR)を、SV40STについての結果(イムノブロッティング)を示す図である。図2−cは、リボソーム由来オリゴヌクレオチドの形質転換細胞の成長に及ぼす影響を示す図である。導入後4.5ヵ月後の細胞状態を示し、左図が細胞成長を、右図が細胞密度を示す。図2−dは、形質転換細胞のコロニー形成試験の結果を示す。写真は、左から、大コロニー、中コロニー、小コロニーについて、それぞれ一般的な例を示す。
【図3】図3は、形質転換細胞の核型分析の結果を示す図である。
【図4】図4は、Bipタンパク合成の抑制及び小胞体ストレス反応誘導の結果を示す図である。図4−aは、35Sメチオニンを用いたパルスラベル・免疫沈降法によるBip、フィブロネクチン、インテグリンβ1の合成速度を調べた結果を示す図である。図4−b、cは、Bip、eIF2α、リン酸化eIF2α、ATF4のイムノブロッティングの結果を示す図である。
【発明を実施するための形態】
【0017】
本発明のヒト細胞の癌化細胞の作製方法は、ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、及びSV40スモールT抗原(SV40ST)遺伝子を導入する工程を含むものであるが、その際に、ヒト28S rRNA由来アンチセンスオリゴヌクレオチドを併用することを大きな特徴とする。
【0018】
ヒト28S rRNAの5'末端から約3kb領域には、ドメイン5のV8-4領域が存在する(図1−a参照)。V8領域は、例えば、シャペロン、修飾酵素、SRP又はトランスロコン(小胞体膜上にあるタンパク質輸送装置)等の非リボソーム性因子を、リボソーム中のL1サイドからトンネル出口の方に配置する作用を有すると報告されているイーストES27に相当する。また、V8-4領域は、5'末端側の後半部分に、CUCC配列(配列番号4)を6個有する。従って、本発明者らは、これらの事象を総合的に判断した結果、ヒト28S rRNAのES27に相補的な短鎖RNAが、ヒトトランスファーミングRNA(hTR)として用いることができると考え、hTRをhTERT及びSV40STと併用したところ、驚くべきことにも、ヒト細胞を形質転換させることができることを見出した。得られた形質転換細胞は、足場非依存性増殖能を有し、染色体も異常であることから、hTRを用いることで、hTERT及びSV40STの導入が可能となり、ヒト細胞を癌化細胞に形質転換できることが示唆される。
【0019】
本発明のヒト細胞の癌化細胞の作製方法は、ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む。
【0020】
本明細書において、「遺伝子」とは、生物の遺伝情報を担う因子であり、DNA及びRNAを含む意味で用いられる。また、「ヌクレオチド」とはDNA及びRNAを含む意味で用いられる。また、「アンチセンスオリゴヌクレオチド」とは、特定の塩基配列(以下、センス配列という)に対し相補的な塩基配列を有し、かつセンス配列にハイブリダイズし得るオリゴヌクレオチドをいう。
【0021】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子としては、特に限定はなく、当該分野で公知のものが挙げられる。テロメラーゼは、細胞分裂によるテロメア長の短縮に拮抗して、テロメア長を維持する酵素である。テロメラーゼは、テロメア配列の鋳型となるRNA、及び、逆転写酵素を構成要素に含み、この逆転写酵素部分がhTERTである。
【0022】
SV40スモールT抗原(SV40ST)遺伝子としては、特に限定はなく、当該分野で公知のものが挙げられる。SV40(simian virus 40)はポリオーマウイルス科のウイルスで、アカゲザル腎細胞から分離されたものである。このウイルスは、細胞への感染初期(DNA合成が起きる前)に、初期タンパク質として、ラージT抗原及びスモールT抗原を産生するが、本発明においては、スモールT抗原を抗原ウイルスとして用いる。
【0023】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドとしては、ヒト28S rRNAの拡張セグメント27(ES27)領域中にあるセンス配列のアンチセンスオリゴヌクレオチドが挙げられる。なかでも、配列番号1で示される塩基配列を含むオリゴヌクレオチド、配列番号2で示される塩基配列を含むオリゴヌクレオチド、及び配列番号6で示される塩基配列を含むオリゴヌクレオチドが好ましく、実質的に配列番号1で示される塩基配列からなるオリゴヌクレオチド、実質的に配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び実質的に配列番号6で示される塩基配列からなるオリゴヌクレオチドがより好ましく、配列番号1で示される塩基配列からなるオリゴヌクレオチド、及び配列番号2で示される塩基配列からなるオリゴヌクレオチドがさらに好ましい。
【0024】
また、本発明においては、前記オリゴヌクレオチドとして、前記塩基配列において1もしくは数個のヌクレオチドの欠失、付加、挿入又は置換を有するオリゴヌクレオチドも、好適に用いられる。かかるオリゴヌクレオチドとしては、具体的には、前記塩基配列で示されるオリゴヌクレオチドに対して少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有するオリゴヌクレオチド、又はそれを含むオリゴヌクレオチドを挙げることができる。また、塩基長としては、特に限定されないが、好ましくは40〜200、より好ましくは40〜150、さらに好ましくは40〜100、さらに好ましくは40〜90、さらに好ましくは40〜80、さらに好ましくは46〜70である。このようなオリゴヌクレオチドは、配列番号1で示される塩基配列からなるオリゴヌクレオチド、配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び配列番号6で示される塩基配列からなるオリゴヌクレオチドと同様の効果を奏するものである。なお、本明細書において、相同性は、例えば、Altschulら(The Journal of Molecular Biology,215,403−410(1990))の開発したアルゴリズムを使用した検索プログラムBLASTを用いて求めることができる。
【0025】
配列番号1で示される塩基配列からなるオリゴヌクレオチド、配列番号2で示される塩基配列からなるオリゴヌクレオチド、及び配列番号6で示される塩基配列からなるオリゴヌクレオチドの模式図を図1に示す。これらは、ヒト28S rRNAのドメイン5に位置する、3001〜3070塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR70ともいう)、3015〜3060塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR46ともいう)、及び3015〜3070塩基で示されるオリゴヌクレオチドに対するアンチセンスオリゴヌクレオチド(以降、asR56ともいう)である。いずれのオリゴヌクレオチドも、拡張セグメント27(エクスパンジョンセグメント27、ES27、2889〜3551塩基)領域に存在するセンス配列に相補的であり、ES27の機能、即ち、増殖に関係することが示唆される。なお、本明細書において、asRとは、アンチセンス短鎖RNA(antisense short RNA)を意味し、sRとは、センス短鎖RNA(sense short RNA)を意味する。
【0026】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドの合成方法としては、特に限定されず、公知のオリゴヌクレオチド合成機を用いたホスホロアミダイト法、ホスホロチオエート法、ホスホトリエステル法等を用いることができる。
【0027】
前記遺伝子及びアンチセンスオリゴヌクレオチドは、安定性や細胞に対する親和性を高めるために、その活性を著しく低下させない範囲で、公知の方法に従って、修飾体、置換体としてもよく、例えば、リン酸エステル基又はリボース部分の水酸基を他の安定な基に置換した誘導体として用いることも可能である。
【0028】
このような遺伝子及びアンチセンスオリゴヌクレオチドが導入されるヒト由来細胞としては、特に限定はなく、ヒト線維芽細胞、ヒト内皮細胞、ヒト上皮細胞等が挙げられる。なお、前記細胞の由来、組織は特に限定されない。
【0029】
ヒト由来細胞へ、前記遺伝子及びアンチセンスオリゴヌクレオチドを導入する方法としては、特に限定はなく、例えば、前記遺伝子及びアンチセンスオリゴヌクレオチドを任意のベクターに組み込んだものを用いることができる。
【0030】
ベクターは、宿主細胞において自立複製可能であると同時に、前記遺伝子及びアンチセンスオリゴヌクレオチド以外に、プロモーター、及び転写終結配列を含有することが好ましい。また、プロモーターを制御する遺伝子が含まれていてもよい。なお、プロモーターとしては、前記遺伝子及びアンチセンスオリゴヌクレオチドを、宿主細胞中に発現させることが可能であれば特に限定はない。
【0031】
本発明に用いられる好適なベクターとしては、例えば、アデノウイルスベクター、ワクシニアウイルスベクター、レトロウイルスベクター等が挙げられる。
【0032】
前記遺伝子及びアンチセンスオリゴヌクレオチドは、同時に一括又は個々に導入してもよく、異なるタイミングにそれぞれを導入してもよく、前記遺伝子及びアンチセンスオリゴヌクレオチドを別々のベクターに組み込んだものを用いてもよく、同一のベクターに組み込んだものを用いてもよい。別々のベクターに組み込んだものを用いる場合、例えば、hTERT遺伝子はpBABEベクターに、SV40ST遺伝子はpLHCXベクターに、アンチセンスオリゴヌクレオチドはpLPCXベクターに組み込んだものを用いることができる。また、同一のベクターに組み込んだものを用いる場合、好適なベクターとしてはレトロウイルスベクターが挙げられ、前記遺伝子及びアンチセンスオリゴヌクレオチドの該ベクター内での配置は、特に限定はなく、例えば、プロモーターと転写終結配列の間に、プロモーター側から、hTERT遺伝子、SV40ST遺伝子、アンチセンスオリゴヌクレオチドの順で配置される。
【0033】
ベクターの導入方法としては、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0034】
また、前記ベクターに導入細胞への感染用ウイルスDNAをさらに組み込んだベクターを調製して、ベクターが導入されたウイルスをヒト細胞に感染させることで、ベクター導入を行ってもよい。感染用ウイルスとしては、アデノウイルス、アデノ随伴ウイルス、レトロウイルス等が挙げられる。
【0035】
なお、かかるベクターの構築方法、具体的な用法等については、例えば、Sambrook, J., et. al., Molecular cloning: A Laboratory Mannual; 2nd Ed., Cold Spring Harbor Laboratory. Cold Spring Harbor. NY., 1989.等の成書を参照すればよい。
【0036】
かくして、ヒト由来細胞に、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入することができる。得られた細胞は、hTERT及びSV40STによってコードされるタンパク質を発現すると共に、ヒト28S rRNA由来アンチセンスオリゴヌクレオチドを発現する、形質転換細胞となる。従って、本発明は、また、前記hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入することにより得られる細胞を提供する。該細胞は、細胞に適した条件下で培養し、集団倍加を繰り返して、公知の薬剤による選択を行ってもよい。なお、本発明においては、hTERT遺伝子が既に組み込まれたhTERT不死化細胞に、SV40ST遺伝子及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入したものも、本発明の範囲内に含まれる。hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドによって癌化が誘導されたヒト細胞の状態については、後述の実施例に記載の方法に従って、確認することができる。
【0037】
また、本発明では、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを組み合わせて用いることで、ヒト細胞を癌化できることから、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有するキットを、ヒト細胞の癌化用に用いることができる。本発明は、また、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有するキットを提供する。
【0038】
またさらに、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを組み合わせて用いることで、ヒト細胞の癌化が容易であり、また、癌化程度も一定であることから、hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを用いて癌化されたヒト細胞を、抗癌剤のスクリーニングに用いることができる。従って、本発明は、また、抗癌剤のスクリーニング方法を提供する。前記方法は、ヒトの系でのin vitro評価であるため、in vivoへの想定が容易となり、より簡便、かつ、迅速に有効物質を見出すことが可能となる。
【0039】
具体的な方法としては、
hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、ヒト細胞に導入して得られた癌化細胞に候補化合物を接触させる工程、ならびに、
前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程
を含む方法が挙げられる。
【0040】
hTERT遺伝子、SV40ST遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドが導入されたヒト細胞に抗癌剤を接触させる方法としては、特に限定はなく、当該分野で公知の方法を用いることができる。
【0041】
また、抗癌剤の接触前後のヒト細胞の癌化程度の判断としては、例えば、同じ実験系において、接触後の一部の細胞が他の細胞に比べて、トランスフォーム形態消失、アポトーシスの誘導、及び足場非依存性の消失等の変化が認められる場合には、該細胞に接触させた候補化合物が、抗癌剤としての作用を有し、癌化を縮小する効果があると判定することができる。
【実施例】
【0042】
以下、本発明を実施例に基づいて説明するが、本発明はこれらの実施例等によりなんら限定されるものではない。なお、hTERT遺伝子は、Dr. F.Ishikaw又はaddgene1774より入手したものを、SV40ST遺伝子は、JCRB gene bank(pMTI0D)より入手したものを、ヒト28S rRNA由来オリゴヌクレオチドは、合成したもの又はTakara Bio社より入手したものを用いた。また、ヒト線維芽細胞(MJ90細胞)はDr. O.M.Pereira-Smithより、ヒト線維芽細胞(TIG3細胞)はRB cell bankより入手したものを用いた。
【0043】
参考例1<GGAGAGGAA配列(配列番号3)の結合特性>
10μLのウサギ溶解液又は20μLのヒト細胞抽出液に、トランスファーミングRNA(TR)の一部であるポリプリン配列、GGAGAGGAA(配列番号3)を添加し、RNase Hの存在下で反応させた。その後、total RNAを回収して、ポリアクリルアミドゲル電気泳動にかけた結果を図1−bに示す。結果、GGAGAGGAA配列(配列番号3)は、28S rRNAに結合していることが分かる。
【0044】
参考例2<asR70、asR56、及びsR70の結合特性>
asR70、asR56、及びsR70の結合特性を以下の方法に従って調べた。結果を図1−cに示す。具体的には、先ず、HindIII-Xbalで切断されたプラスミドpGEM3(Promega社製)に挿入して作製したプラスミドpG-asR70、pG-asR56、及びpG-sR70から、SP6 RNA polymerase(Takara Bio社製)と[α-32P]-GTP(Perkin-Elmer社製)を用いて、asR70、asR56、及びsR70の標識RNAプローブ(riboprobe)を作製した。得られたプローブをポリアクリルアミドゲル電気泳動にかけたところ、asR70プローブが89塩基(左から1番目のレーン)、asR56プローブが75塩基(右から8番目のレーン)、及びsR70プローブが89塩基(右から4番目のレーン)の長さを有するプローブであった。
【0045】
次に、20μLのヒト細胞溶解液と4μgのオリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)を混合してRNase Hで切断後、回収したtotal RNAをasR70、asR56、及びsR70のリボプローブと混合し、RNase A 及び RNase T1で処理した。なお、比較として、オリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)を用いない例についても、同様の処理を行った。
【0046】
オリゴデオキシヌクレオチド処理total RNAはasR70プローブとの結合において二箇所の切断部位を示した。即ち、オリゴデオキシヌクレオチドGGAGAGGAA(配列番号3)は、28S rRNAの3001〜3070塩基内に結合部位を有することが判明した(左から3番目と5番目のレーン)。具体的には、28S rRNAの3063〜3066塩基であるCUCC配列(配列番号4、サイト1)と、3035〜3040塩基であるCUCUCU配列(配列番号8、サイト2)と結合し得ることが分かった。また、14ヌクレオチド短いasR56についても、同じサイトにおいて結合することが判明した(右から6番目と7番目のレーン)。このサイト1とサイト2は、約23〜24ヌクレオチド離れて位置し、サイト1はV8-4領域のヘアピンループに位置し、一方、サイト2は膨らみ部分に位置するものであり、asR70及びasR56は、前記2箇所のサイトで28S rRNAと結合することが示唆される。一方、sR70プローブと28S rRNAとの結合は確認されなかった(右から2番目と3番目のレーン)。また、非特許文献3において、配列番号3で示されるオリゴデオキシヌクレオチドの修飾配列であるGAGAGAGAG配列(配列番号9)は、分泌タンパクの合成を抑制することが出来ないことが示されていることから、V8-4領域では、サイト2よりもサイト1においてSRPのRNAと相互作用することでタンパク質合成を制御していることが示唆される。なお、asR56には、サイト2に結合するasR70中の配列が含まれているが、異所性結合を伴って結合が弱いと推定される。
【0047】
実施例1<形質転換細胞の形態、癌化形質>
hTERT、asR70、及びSV40STによって形質転換されたMJ90細胞(hTERT+asR70+SV40ST、R7S)、ならびに、hTERT及びasR70によって形質転換されたMJ90細胞(hTERT+asR70、R7)について、位相差顕微鏡を用いて細胞形態を観察した。また、軟寒天培地でのコロニー形成試験も行った。結果を図2−aに示す。
【0048】
なお、形質転換細胞は次のようにして調製したものを用いた。具体的には、先ず、asR70を挿入したpGEM3(Promega社製)をHindIII-EcoRIで切断した断片をpLPCX(Clontech社製)に挿入したベクターを調製した(pLPCX-asR70)。PCRで増幅したSV40ST断片をpLHCX(Clontech社製)に挿入したベクターを調製した(pLHCX-ST)。hTERT発現用プラスミドは、pcDNAhTERTn2(Dr. F.Ishikawより提供)及びpBABE-neo-hTERT(Addgene plasmid 1774)を用いた。次に、pLPCX-asR70及びpLHCX-STは、レトロウイルスパッケイジングプラスミドpCL-10A1(Imgenex社製)と共に、293T 細胞(Riken Cell Bankから入手)にトランスフェクトした後、ウイルス回収の上、MJ90細胞に感染させた。pcDNAhTERTn2を用いる場合は、lipofectamine 2000を用いてMJ90細胞にトランスフェクトした。pBABE-neo-hTERTを用いる場合は、ウイルス作製後、TIG3細胞に感染させた。得られた細胞は、薬剤選択を行って親株よりもライフスパンの長いものを選択して実験に供した。
【0049】
図2−aより、hTERT、SV40ST、及びasR70によって形質転換された細胞(R7S)は、細胞と細胞の境界が不明瞭となり、軟寒天培地でも移植後14〜18日にはコロニーを形成していたため、足場非依存性増殖能を有し癌化していることがわかる。
【0050】
実施例2<形質転換細胞におけるhTERT、SV40ST、asR70の発現>
正常ヒト線維芽細胞(intact、MJ90)、hTERT不死化細胞(T8)、ならびに、hTERT及びasR70を共トランスフェクションした細胞(R7)、そのクローン(R7-1、R7-2)を準備した。また、細胞(R7)にSV40STを導入した細胞(R7S)、そのクローン(R7S-1、R7S-2、R7S-3)、及びサブクローン(R7S-1s、R7S-2s、R7S-3s)も準備した。なお、準備した細胞は全て実施例1と同様にして選択された株を用いた。また、全細胞株について、実施例1と同様にしてコロニー形成試験を行った結果、細胞(R7Sシリーズ)は多くのコロニー形成が確認されたが、細胞(R7、R7-1、R7-2)、細胞(MJ90)、及び細胞(T8)では全くコロニーが形成されなかった。
【0051】
各細胞の抽出液(1μg)を用いて、PCRを用いたTRAP(telomeric repeat amplification protocol)アッセイを行って、テロメラーゼ活性(hTERT活性)を調べた。結果を図2-b(PCR結果の一番上)に示す。細胞(MJ90)以外は、テロメラーゼ活性があることが示された。なお、図中の「IC」は、PCR用の内部標準物質である。
【0052】
asR70については、asR70特異的な下記に示すプライマーを用いたRT-PCRを行って調べた(図2-bのPCR結果の上から2番目)。その結果、細胞(MJ90)及び細胞(T8)以外の細胞において、増幅産物(117bp)が認められた。
pLPCX-retroF:5'-CGCTAGCGCTACCGGACTC-3'(配列番号10)
asR70-R:5'-ATAAAGCTTCTCGCTCCCTCCCCACCC-3'(配列番号11)
【0053】
また、増幅産物(117bp)がasR70 cDNAに特異的なものであることを確かめるために、前記pLPCX-retroFプライマーと、asR70のDNAを含むプラスミドの非転写領域に特異的なプライマーとして、
pLPCX-seqR:5'-TGGGGTCTTTCATTCCC-3'(配列番号12)
を用いたPCR増幅を行ったが、増幅産物(233bp)が確認されず、asR70がプラスミドに由来するものではなく組み込まれたものであることが確認できた(図2-bのPCR結果の上から3番目)。さらに、増幅量が等量であることを示すために、下記プライマーを用いて、β-アクチン(356bp)の増幅量を調べた(図2-bのPCR結果の一番下)。なお、図2-b中の「M」は、分子量マーカー(HaeIII digest marker)を示し、「*」は、プラスミドpLPCX-asR70を増幅した結果を示す。
forward:5'-TGAAGTGTGACGTGGACATCCGC-3'(配列番号13)
reverse:5'-GCCAATCTCATCTTGTTTTCTGCGC-3'(配列番号14)
【0054】
以上より、細胞(MJ90)及び細胞(T8)以外は、asR70の発現が確認され、それらはプラスミド由来ではなく、形質転換により組み込まれたものに由来することが明らかになった。
【0055】
また、SV40STについては、細胞質のイムノブロッティングにより調べた。結果を図2-bの下段(イムノブロッティング結果)に示す。その結果、細胞(R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)において、SV40ST(15〜20kD)の発現が確認された。なお、全ての細胞において、コントロールとして、Actinの発現も確認した(図2-bのイムノブロッティング結果下)。
【0056】
実施例3<ヒト28S rRNA由来オリゴヌクレオチドの形質転換細胞の成長に及ぼす影響>
実施例2で用いたhTERT不死化細胞(T8)を13ヶ月培養した後、液体窒素中で保存した。その後、解凍し、数回複製してから、以下の実験に用いた。
【0057】
前記細胞(T8)に、intactのpLPCX(pLPCX)、asR70を組み込んだpLPCX(asR70)、asR56を組み込んだpLPCX(asR56)、asR46を組み込んだpLPCX(asR46)、又はsR70を組み込んだpLPCX(sR70)をそれぞれ導入し、ピューロマイシンによる薬剤選択を行って、該導入細胞(T8、pLPCX、asR70、asR56、asR46、sR70)を選択した。導入4.5ヵ月後の細胞株について、細胞成長と細胞密度を調べた(n=3)。結果を図2-cに示す。なお、細胞成長は、一定数に調整された細胞の経時的な細胞数増加を測定することにより調べ、細胞密度は、一定面積に占める細胞数を測定することにより調べることができる。
【0058】
図2-cより、導入4.5ヵ月後の細胞では、asR70を導入した細胞及びasR46を導入した細胞が、細胞成長速度が大きく、また細胞密度も高いものであった。このことから、asR70及びasR46は、細胞増殖メカニズムに影響を及ぼすものであることが示唆される。
【0059】
実施例4<形質転換細胞のコロニー形成>
実施例2で用いたhTERT不死化細胞(T8)に、intactのpLPCX(pLPCX)、asR70を組み込んだpLPCX(asR70)、asR56を組み込んだpLPCX(asR56)、asR46を組み込んだpLPCX(asR46)、又はsR70を組み込んだpLPCX(sR70)をそれぞれ導入後、intactのpLPCXを導入した細胞以外には、さらにSV40STを組み込んだpLPCXを導入後、ピューロマイシンによる薬剤選択を行って、該導入細胞(T8、pLPCX、asR70、asR56、asR46、sR70)を選択した。得られた細胞を培養し、経時的に、コロニー形成試験を行った(n=3)。また、コロニー形成が認められた場合は、コロニーを大きさ「大(large、細胞の10倍以上の大きさ)」、「中(middle、細胞の5〜9倍程度の大きさ)」、「小(small、細胞の3〜4倍程度の大きさ)」によって分類し、大コロニーと中コロニーの合計コロニー数(l+m)、及び小コロニー数(s)を計数し、平均値を求めた。結果を図2-dに示す。なお、図2-dにおいて「NP」とは、コロニーが認められるものの増殖が確認されない場合を示し、「NC」とはコロニー計数を行わなかった場合を意味する。
【0060】
図2-dより、細胞(asR70)及び細胞(asR46)では、軟寒天中でのコロニー形成が確認され、いずれの細胞も癌化していることが分かる。また、細胞(asR46)は、細胞(asR70)より多くのコロニーを形成していることから、癌化効率が高いことが示唆される。これらより、asR70及びasR46は、より高い形質転換効果を奏することが示唆される。また、asR46、hTERT、及びSV40STを用いることで、ヒト胎児肺由来二倍体線維芽細胞TIG3を形質転換できることも確認できた(結果示さず)。なお、前記細胞(asR46)(以降R4Sともいう)を単離して、そのクローン(R4S-1、R4S-2、R4S-3)を調製し、以降の実験に用いた。
【0061】
実施例5<形質転換細胞の核型分析>
実施例2で用いた、hTERT不死化細胞(T8)、hTERT及びasR70を共トランスフェクションした細胞(R7)、細胞(T8)にSV40STを導入した細胞(T8+ST)、細胞(R7)にSV40STを導入した細胞のクローン(R7S-1、R7S-2、R7S-3)、ならびに実施例4で用いた、hTERT不死化細胞(T8)にasR46とSV40STを導入した細胞のクローン(R4S-1、R4S-2、R4S-3)のそれぞれについて、5細胞ずつ、核型分析(reverse DAPI banding - spectral karyotyping analysis、SKY)を行った。結果を表1に、また代表的なSKY染色図を図3に示す。
【0062】
【表1】
【0063】
表1及び図3より、細胞(T8)、細胞(R7)、及び細胞(T8+ST)は、46本の染色体が確認され、正常の二倍体細胞であることが分かる。一方、細胞(R7S-1)と細胞(R7S-3)は、46本の染色体が認められたものの、1箇所でゲノム再編成が行われていることが明らかとなった。また、細胞(R7S-2)は、染色体数が45本と正常より1本少なく、4箇所でゲノム再編成が確認された。また、細胞(R4S-1、R4S-2、R4S-3)はいずれも、同じ株であったとしても、染色体数やゲノム再編成が行われている箇所などが様々であり、種々のタイプが確認された。なかでも、細胞(R4S-1、R4S-2、R4S-3)は細胞(R7S-1、R7S-2、R7S-3)より、変異がより大きいものであるが、細胞(R4S-1、R4S-2)には正常な細胞も存在することから、これらの変異は、形質転換後に生じていることが示唆される。これより、hTERT、SV40ST及びasR70形質転換細胞は、細胞内の遺伝情報が、正常細胞やhTERT及びasR70形質転換細胞とは異なることが示唆される。
【0064】
実施例6<形質転換細胞における小胞体ストレス反応誘導>
実施例4と同様にして得られた細胞(T8、pLPCX、asR70、asR56、asR46、sR70)について、小胞体ストレス応答を示す物質の発現を確認した。また、細胞(asR70)については、実施例2と同様にして得られた細胞、即ち、細胞(T8、R7、R7-1、R7-2、R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)について同様の実験を行った。
【0065】
具体的には、35Sメチオニンを用いたパルスラベル・免疫沈降法に従って、細胞外マトリックスのフィブロネクチン(FN)、膜透過型レセプターのインテグリン ベータ1(integrin β1)、及び小胞体局在シャペロンのBipの発現量を各細胞について測定した。細胞(T8、pLPCX、asR70、asR56、asR46、sR70)に関する結果を図4−aに示す。
【0066】
図4−aより、Bipの発現は、細胞(asR70)、細胞(asR56)、細胞(asR46)及び細胞(pLPCX)で経時的に減少していることが分かった(図4−aの枠内グラフ参照)。一方、FN及びintegrin β1の発現は、シグナル非依存性のアクチンやグルコース6リン酸デヒドロゲナーゼと同様の傾向を示し、殆ど変化が認められなかった。FN及びintegrin β1は、細胞内で生産性が高いため変化が認められなかったが、Bipの発現量に変化が認められたことから、細胞(asR70)、細胞(asR56)、細胞(asR46)のリボゾームにおいてBipの合成が抑制されていることが示唆される。
【0067】
また、Bipの影響について更に詳しく検討するために、細胞質タンパク質について同様の検討を行った。具体的には、30μgの細胞質タンパク質を10% SDS/PAGEで分離し、Bip、eIF2a(eukaryotic initiation factor 2a)、リン酸化eIF2a(P-eIF2a)、及びATF4(activating transcription factor 4)のイムノブロッティングを行った。細胞(T8、pLPCX、asR70、asR56、asR46、sR70)に関する結果を図4−b、細胞(T8、R7、R7-1、R7-2、R7S、R7S-1、R7S-2、R7S-3、R7S-1s、R7S-2s、R7S-3s)に関する結果を図4−cに示す。なお、Bipの合成が抑制されると、eIF2aがリン酸化されてタンパク合成が抑制され、リン酸化されたeIF2aがATF4の翻訳を開始するという、小胞体のストレス反応が誘導される。
【0068】
図4−b、cより、Bip、eIF2aの発現量は、いずれの細胞においても同程度であったが、細胞(asR46)、細胞(R7)、細胞(R7-1)、細胞(R7-2)において、リン酸化eIF2aの発現がより顕著に確認され、また、付随するATF4の誘導も確認された。このことから、asR46とasR70は、Bip合成を抑制することで、小胞体ストレス応答を誘導していることが示唆される。
【0069】
実施例7<抗癌剤のスクリーニング>
hTERT、SV40ST、及びヒト28S rRNA由来オリゴヌクレオチドが導入された細胞を細胞培養器具にて培養し、そこに、候補化合物を溶媒に溶解したものを添加して、さらに培養する。一方、対照として、候補化合物を添加しない群も設定する。候補化合物を添加しない群をコントロールとして、添加しない群よりも細胞のトランスフォーム形態の消失、アポトーシスの誘導、及び足場非依存性増殖の消失等の変化が認められる場合に、該細胞に添加した候補化合物は、抗癌剤としての作用を有する可能性が高いと判断することができる。
【産業上の利用可能性】
【0070】
本発明のヒト細胞の癌化細胞の作製方法は、癌発症メカニズムの解明のため、様々なヒト正常細胞に癌化を誘導する際に利用することができ、新たな創薬ターゲット分子の探索に有効に利用することができる。
【配列表フリーテキスト】
【0071】
配列表の配列番号1は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR70)である。
配列表の配列番号2は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR46)である。
配列表の配列番号3は、トランスファーミングRNAの部分塩基配列である。
配列表の配列番号4は、ヒト28S rRNAの部分塩基配列である。
配列表の配列番号5は、ヒトSRP RNAの部分塩基配列である。
配列表の配列番号6は、ヒト28S rRNA由来アンチセンスオリゴヌクレオチド(asR56)である。
配列表の配列番号7は、ヒト28S rRNA由来オリゴヌクレオチド(sR70)である。
配列表の配列番号8は、ヒト28S rRNAの部分塩基配列である。
配列表の配列番号9は、トランスファーミングRNAの部分塩基配列を修飾した配列である。
配列表の配列番号10は、pLPCX-retroF用プライマーである。
配列表の配列番号11は、asR70 cDNA用プライマーである。
配列表の配列番号12は、pLPCX-seqR用プライマーである。
配列表の配列番号13は、β-アクチン用プライマーである。
配列表の配列番号14は、β-アクチン用プライマーである。
【特許請求の範囲】
【請求項1】
ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法。
【請求項2】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドが、配列番号1で示される塩基配列を含むオリゴヌクレオチド、又は配列番号1で示される塩基配列に対して70%以上の相同性を有するオリゴヌクレオチドを含むオリゴヌクレオチドである、請求項1記載の方法。
【請求項3】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドが、配列番号2で示される塩基配列を含むオリゴヌクレオチド、又は配列番号2で示される塩基配列に対して70%以上の相同性を有するオリゴヌクレオチドを含むオリゴヌクレオチドである、請求項1記載の方法。
【請求項4】
ヒト由来細胞がヒト線維芽細胞である、請求項1〜3いずれか記載の方法。
【請求項5】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、同一又は異なるベクターに組み込み、ヒト由来細胞に導入する、ヒト細胞を癌化する遺伝子の導入方法。
【請求項6】
請求項1〜4いずれか記載の方法により得られる、ヒト細胞の癌化細胞。
【請求項7】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有してなる、請求項1〜4いずれか記載の方法に用いるためのキット。
【請求項8】
請求項6記載の癌化細胞に候補化合物を接触させる工程、及び、前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に、候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程を含む、抗癌剤のスクリーニング方法。
【請求項1】
ヒト由来細胞に、ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを導入する工程を含む、ヒト細胞の癌化細胞の作製方法。
【請求項2】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドが、配列番号1で示される塩基配列を含むオリゴヌクレオチド、又は配列番号1で示される塩基配列に対して70%以上の相同性を有するオリゴヌクレオチドを含むオリゴヌクレオチドである、請求項1記載の方法。
【請求項3】
ヒト28S rRNA由来アンチセンスオリゴヌクレオチドが、配列番号2で示される塩基配列を含むオリゴヌクレオチド、又は配列番号2で示される塩基配列に対して70%以上の相同性を有するオリゴヌクレオチドを含むオリゴヌクレオチドである、請求項1記載の方法。
【請求項4】
ヒト由来細胞がヒト線維芽細胞である、請求項1〜3いずれか記載の方法。
【請求項5】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを、同一又は異なるベクターに組み込み、ヒト由来細胞に導入する、ヒト細胞を癌化する遺伝子の導入方法。
【請求項6】
請求項1〜4いずれか記載の方法により得られる、ヒト細胞の癌化細胞。
【請求項7】
ヒトテロメラーゼ触媒サブユニット(hTERT)遺伝子、SV40スモールT抗原(SV40ST)遺伝子、及びヒト28S rRNA由来アンチセンスオリゴヌクレオチドを含有してなる、請求項1〜4いずれか記載の方法に用いるためのキット。
【請求項8】
請求項6記載の癌化細胞に候補化合物を接触させる工程、及び、前記候補化合物の接触前に比べて接触後の細胞の方が癌化程度が小さいと認められる場合に、候補化合物が抗癌剤としての作用を有する可能性が高いと判定する工程を含む、抗癌剤のスクリーニング方法。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−239726(P2011−239726A)
【公開日】平成23年12月1日(2011.12.1)
【国際特許分類】
【出願番号】特願2010−114535(P2010−114535)
【出願日】平成22年5月18日(2010.5.18)
【出願人】(510137467)株式会社TRDIGM (1)
【Fターム(参考)】
【公開日】平成23年12月1日(2011.12.1)
【国際特許分類】
【出願日】平成22年5月18日(2010.5.18)
【出願人】(510137467)株式会社TRDIGM (1)
【Fターム(参考)】
[ Back to top ]