
ヒト胚性および人工多能性幹細胞の分化
本発明は、多能性未分化hESCまたはiPSCを軟骨形成細胞系譜の細胞に段階的かつ均一に分化させることができる培養系、培養方法および培養条件に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般的に、ヒト胚性および人工多能性幹細胞の分化を軟骨系譜に誘導する方法に関する。
【背景技術】
【0002】
米国で最も多く見られる消耗性の慢性健康障害の1つに軟骨の変性疾患、たとえば、変形性関節症がある。軟骨変性疾患の処置は、組織の自己修復能が乏しいため特に臨床上の課題になっている。ヒト胚性幹細胞(hESC)は、複数の細胞型に分化する能力および無限の自己複製能を有するので、軟骨欠損を修復する強力なツールになる可能性がある。hESCが病変および損傷軟骨を修復する可能性を実現するには、hESCの軟骨形成細胞系譜(chondrogenic lineage)への分化を誘導する方法を開発する必要がある。
【0003】
ESC由来の細胞が様々な程度で軟骨細胞に分化するいくつかの培養系が開発されているが、これらの系では軟骨細胞分化(chondrogenic differentiation)が均一ではなく、軟骨細胞は、分化する細胞の亜集団のみであり、軟骨修復のために細胞集団を利用することが難しくなっている。以下に記載する本発明の以前には、軟骨細胞分化のプロトコルは、ESC由来の胚様体(EB)の細胞を利用していた。こうした従来の方法の欠点は、EB由来の細胞の細胞の不均一性により、軟骨修復に使用し得る軟骨形成細胞の均質な集団が得にくくなることである。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、こうした従来の方法に伴う問題の多くを解決するものであり、1つには、前もってEB形成を形成せずに、幹細胞、たとえば未分化多能性ヒト胚性幹細胞(hESC)または人工多能性幹細胞(iPSCまたはiPS細胞)が軟骨形成細胞系譜へと段階的に実質的に均一に分化するのを迅速かつ直接誘導する培養系および培養条件に基づくものである。また、この方法では臍帯血または羊水から採取した幹細胞も使用する。この方法を用いて作製される、規定の分化段階の実質的に純粋な軟骨形成細胞は、従来の方法に伴う細胞不均一性(組織型および分化の段階に関する)の問題および欠点に対する解決策となる。本明細書に記載の細胞集団は、非軟骨形成細胞を実質的に含まない。たとえば、この集団は、軟骨形成細胞を少なくとも85%、95〜98%、および最大99%または100%含む。本発明はまた、ESCの軟骨形成細胞系譜への分化の進行を促進し、かつ軟骨細胞分化の均一性の達成に寄与する高密度培養条件、および追加手順も提供する。
【0005】
胚様体は、ESCまたはiPSC由来の細胞の凝集塊である。こうした凝集塊、すなわちEBは、培養物の目視検査で確認される。凝集(EBの形成)すると、分化が開始される。EBは、成人生体に存在する細胞型の全部を生じさせる胚の3胚葉すべてに由来する複数の細胞型に分化する。本発明の以前、hESC由来の軟骨形成前駆細胞の集団を産生する従来の方法の欠点は、非軟骨形成細胞の混入、すなわち、EBが所望の標的系譜(軟骨形成細胞系譜)以外の系譜に分化して産生された非軟骨形成細胞の混入であった。本明細書に記載の方法はEB形成を回避するため、得られる軟骨形成前駆細胞には、実質的に他の系譜の細胞が混入しない。この方法は、培養中のEBが非軟骨形成組織/細胞に分化するという問題を解決する。従来の方法は、混入している望ましくない細胞を除去する必要があったため、本明細書に記載のプロセスは、従来の方法に比べより効率的かつ高速であるうえ、費用効果に優れている。
【課題を解決するための手段】
【0006】
ヒト胚性幹細胞(hESC)を軟骨形成細胞に直接分化させる方法は、EBを実質的に含まないhESCの集団を支持体上に用意し、hESCを前記支持体から剥離して前記hESCを分離し、hESCの高密度マイクロマス培養を樹立し、集団を骨形成タンパク質と接触させることにより、hESCが軟骨形成細胞の実質的に均一な集団に直接分化することにより行われる。同様に、人工多能性幹細胞(iPSC)を軟骨形成細胞に直接分化させる方法は、EBを実質的に含まないiPSCの集団を支持体上に用意し、iPSCを支持体から剥離してiPSCを分離し、iPSCの高密度培養を樹立し、iPSCを骨形成タンパク質と接触させてiPSCの軟骨形成細胞系譜への分化を誘導することにより行われる。iPSCは軟骨形成細胞の実質的に均一な集団に直接分化する。
【0007】
一態様では、本明細書に記載の培養系および培養条件は、実質的に均一な軟骨分化を約1週間または2週間未満の内に、たとえば、約2日〜約3日未満、約3日〜約4日未満、約5日〜約6日未満、約6日〜約7日未満、約7日〜約8日未満、約8日〜約9日未満、約9日〜約10日未満、約10日〜約11日未満、約11日〜約12日未満、約12日〜約13日未満、または約13日〜約14日未満の内に促進する。別の態様では、本発明の培養系および培養条件は、未分化多能性ヒト胚性幹細胞の少なくとも約50〜100%が軟骨形成細胞系譜に分化するように、たとえば、未分化多能性ヒト胚性幹細胞の少なくとも約50〜60%、少なくとも約60〜70%、少なくとも約70〜80%、少なくとも約80〜90%、または少なくとも約90〜100%が軟骨形成細胞系譜に分化するように非常に均一な分化を促進する。好ましくは、この集団は、軟骨形成細胞を95%含み、一層好ましくは、集団は軟骨形成細胞を98%含む。
【0008】
分化の段階は、時間的に、あるいは、一連の遺伝子マーカーの発現により説明される。時間(日数)は、hESCまたはiPSCの高密度培養の樹立後の期間を示すものである。ゼロ時は、細胞濃度または密度を10μl当たり少なくとも1×105細胞に調節する時点である。「2日」の細胞は、高密度コロニーの樹立後2日間培養した細胞である。高密度培養は、10μl当たり少なくとも1×105の濃度で細胞を含む。たとえば、2〜3日の細胞は、軟骨系譜(すなわち、軟骨前駆体)に入ったばかりであることを特徴とし、4日目の細胞は、軟骨細胞分化の初期相(すなわち、初期の軟骨細胞)にあり、7〜14日の細胞は、軟骨細胞への明らかな分化を均一に示すものの、肥大成熟化を示していない(すなわち、細胞は、十分に分化した軟骨細胞であることを特徴とする)(図14)。
【0009】
本発明は、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に及ぶ様々な相の軟骨形成細胞系譜におけるhESC由来の前駆体細胞の特徴付けについて記載する。様々な相の軟骨形成細胞系譜におけるhESC由来の前駆体細胞について、細胞ベースの組織工学治療により軟骨を修復する能力を解析する。
【0010】
hESCの軟骨細胞分化を促進する条件は、人工多能性幹細胞(iPSC)の均一な軟骨分化も促進する。本発明はまた、患者由来のiPSCの軟骨細胞分化を誘導する方法も提供する。たとえば、患者個人を考慮した治療アプローチにiPSC由来の軟骨形成前駆細胞(自家細胞)を使用する。別の例では、本方法を用いて細胞および細胞株を作製して、疾患のメカニズムを研究するため、軟骨異形成症などの軟骨の遺伝子性障害の基本的なメカニズムを解明する研究ツールとして使用するほか、細胞を使用して治療薬のスクリーニングも行う。
【0011】
上記で論じたように、hESCの軟骨細胞分化を促進する条件は、人工多能性幹細胞(iPSC)、すなわち、再プログラム化されて多能性状態にある体細胞の均一な軟骨分化も促進する。iPSCが軟骨形成細胞系譜に信頼性高く直接かつ均一に分化すれば、患者特異的な自家細胞治療が可能になる。したがって、軟骨欠損の修復には、hESC由来の
前駆体(precursor)だけでなく、iPSC由来の軟骨形成前駆体細胞も使用される。
【0012】
本発明の培養系および培養条件は、胚様体(EB)由来の細胞を細胞ベースの軟骨修復治療に使用しにくくしている細胞の不均一性を回避する。この方法は直接かつ均一な分化を促進し、EB段階を回避するため、規定の分化段階、すなわち、初期と後期の細胞集団を確実に生じさせる。遺伝子発現プロファイリングを行えば、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に及ぶ様々な段階の軟骨形成細胞系譜におけるhESC由来の前駆体細胞を同定することができる。初期相の軟骨形成細胞系譜の細胞(たとえば、Sox9などの初期軟骨マーカーを含む)は、分化を続けるだけでなく、移植患者に移植された後にin situで局所微小環境のシグナルに応答するため、系譜の後期の細胞と比べ、軟骨修復に関与するのに望ましい。軟骨形成細胞系譜の後期の細胞(たとえば、細胞がより明らかに分化したことを示すマーカー、アグリカン(aggrecan)などの軟骨マーカーを含む)は、軟骨修復/再建手順に好適である。このため、本方法は、様々な分化の段階および異なる系譜の不均一な細胞集団を生じる従来の方法と異なり、段階的に生じた細胞の供給源を提供し、処置される病状に最も適した分化の段階に基づき治療を個々に合わせることを可能にする。
【0013】
本発明はまた、hESCおよびiPSCの軟骨形成細胞系譜への分化の進行を促進し、かつ軟骨細胞分化の均一性の達成に寄与する条件および追加手順も提供する。こうした改変には、(1)高密度培養から分離したhESCの維持(EBは存在しない)、(2)増殖因子、特にBMP−2およびTGFβ−1を含む培養物の補充、(3)マウス胚性線維芽細胞のフィーダー層上ではなく、高分子もしくはゼラチン支持体またはスキャフォールド上、あるいは高分子もしくはゼラチン支持体またはスキャフォールド内でのhESCおよびiPSCの培養、(4)マイクロマス培養物を調製する前にトリプシンではなく、Accutase、TrypLE Selectまたは他の酵素を使用してhESCを分離すること、ならびに、(5)マイクロマス培養の樹立において分離に伴うアポトーシスを減弱するRho結合キナーゼ(ROCK)阻害剤の適用が含まれる。好適な高分子もしくはゼラチン支持体またはスキャフォールド材料として、基底膜支持体(たとえばマトリゲル、Chondrogide)、コラーゲンもしくはゼラチン、ヒアルロナン(たとえばHyaffl 1またはHyStemC)もしくはキトサンを含むヒドロゲルまたはスポンジ、またはPEG(ポリエチレングリコール)もしくはPLGA(ポリラクチカシドグリオリト酸)スキャフォールド、あるいは細胞外マトリックス成分を含む任意の支持体が挙げられる。
【0014】
具体的には、本発明は、無血清培地を用いてhESCを支持体上で培養し、hESCを支持体から剥離してhESCを分離し、無血清培地を用いてhESCを高密度培養(マイクロマスまたはペレット)として培養し、骨形成タンパク質−2(BMP2;NG_023233(GI:300068920))、参照によって本明細書に援用する)をマイクロマス培養基に投与することにより、ヒト胚性幹細胞(hESC)の軟骨形成細胞系譜への分化を誘導する方法を提供する。hESCは、EBを実質的に含まない。好ましくは、hESCは、EBをまったく含まない。BMP2は、hESCの高密度マイクロマス培養から12時間後、24時間後、48時間後、72時間後、または96時間後に投与する。好ましくは、増殖因子(単数または複数)は、高密度細胞培養の樹立から48時間後に加える。あるいは、BMP−2およびトランスフォーミング増殖因子β(TGFβ;NM_000660(GI:260655621))、参照によって本明細書に援用する)を一緒に加える。BMP2の濃度は、25〜200ng/mlの範囲であり、TFGβ1の濃度は、2.5〜20ng/mlの範囲である。BMPとTGFとの比率は約5:1〜20:1であり、好ましくは約10:1である。
【0015】
hESCは、高密度マイクロマス培養としてhESCを培養する前にトリプシン、TrypLE SelectまたはAccutaseなどの分離剤と接触させる。任意に、hESC培養の支持体は、マウス胚性線維芽細胞(MEF)を含むフィーダー層である。あるいは、hESC培養の支持体は、人工基底膜材料(たとえば、マトリゲル)などのゼラチン組成物、または他の好適な支持体またはスキャフォールドである。一態様では、この方法は、トランスフォーミング増殖因子β−1(TGFβ1)をマイクロマス培養基に投与することをさらに含む。任意に、この方法は、Rho結合キナーゼ(ROCK)阻害剤の投与をさらに含む。
【0016】
一態様では、hESCは、軟骨形成細胞系譜の細胞に約14日以内に、たとえば、約10日以内、約7日以内、約4日以内または約3日以内に分化する。本明細書に記載の方法は、hESCの少なくとも約85%、たとえば、少なくとも約90%、少なくとも約95%、少なくとも約98%、少なくとも約99%または約100%が軟骨形成細胞系譜の細胞に分化するように細胞の分化を誘導する。分化は、好ましくはウシ関節軟骨細胞、たとえば、分化したウシ関節軟骨細胞の非存在下で行われる。また、このプロセスは、肝細胞の癌細胞株などの細胞または細胞株の条件培地の非存在下でも行われる。
【0017】
本発明はまた、無血清培地を用いてiPSCを支持体上で培養し、iPSCを支持体からしてiPSCを分離し、無血清培地を用いてiPSCを高密度マイクロマス培養として培養し、骨形成タンパク質−2(BMP2)をマイクロマス培養に投与することにより、人工多能性幹細胞(iPSC)の軟骨形成細胞系譜への分化を誘導する方法も提供する。iPSCは、EBを実質的に含まない。好ましくは、iPSCはEBをまったく含まない。一態様では、iPSCは、軟骨障害の患者由来である。一態様では、軟骨障害は軟骨異形成症である。
【0018】
上述の方法は、規定の分化の段階にあり、かつEB由来の細胞、たとえば、非軟骨形成細胞系譜の細胞を含まないことを特徴とする軟骨形成細胞を生じさせる。たとえば、軟骨形成細胞の集団は非軟骨形成細胞を実質的に含まず、集団は、上記のようにESCまたはiPSCから分化している。別の例では、細胞集団は、同調的に生じるか、または段階的に生じる。均一に分化した軟骨形成細胞の集団は、細胞の少なくとも85%(もしくは98%、99%、または100%)が単一の規定の分化の段階にあることを特徴とする。一部の臨床応用では、分化の段階は比較的初期であり、たとえば、集団は十分に分化した軟骨細胞を実質的に含まない。たとえば、この段階は、3日目の軟骨形成細胞または4日目の軟骨形成細胞からなる群から選択される。系譜の初期相の細胞を利用すると、移植後に関節軟骨の修復を促進する局所環境シグナルに応答しやすくなる。以上のように、軟骨を修復または再建する方法は、損傷または病変軟骨を上述の細胞集団のいずれかと接触させることにより行われる。同様に、関節炎を予防または処置する方法は、個体(ヒトまたは他の動物、たとえば、イヌ、ネコ、ウマ)の関節接合部(articulating joint)または関節腔に、記載した軟骨形成細胞の集団のいずれかを投与することにより行われる。
【0019】
本発明は、傷害または変性に起因するような軟骨障害または欠損を処置する方法を含む。処置される障害として、関節接合部の外傷性障害(たとえば、前十字靭帯(ACL)または他の靭帯を含む膝の損傷または断裂、半月板の損傷または骨折)、肘の外傷性障害、肩の外傷性障害(たとえば、回旋筋腱板損傷)、顎の外傷性障害(顎関節、すなわちTMJ疾患)、または手指の外傷性障害、および他の軟骨または骨格損傷のほか、変形性関節症(OA)など加齢に伴い発症する関節炎、または関節リウマチ(RA)など炎症を伴う関節炎、または外傷性障害による関節炎などの慢性状態、および関節接合部のいずれかが侵される慢性状態が挙げられる。この細胞は、傷害または慢性疾患による関節軟骨の損傷の予防に有用である。また、この細胞は、線維軟骨、靭帯(ligment)もしくは半
月板の障害または変性の修復にも有用である。治療方法は、本明細書の方法により産生された細胞を、軟骨障害、たとえば、軟骨傷害または変形性関節症などの関節炎の患者に投与するステップを含む。また、治療には、本方法により産生された細胞を導入して、傷害または疾患による慢性関節軟骨変性の発症を遅延させるかまたは予防することも含まれる。本方法により産生された軟骨形成細胞で軟骨を修復する方法は、移植、注射または注入により関節接合部または他の標的部位に細胞を投与することを含んでいた。たとえば、細胞は、関節鏡視下で投与する。場合によっては、関連する欠損または状態を修復する外科的または関節鏡視下手順の前、その後、またはその最中に細胞を投与または移植する。たとえば、ACL修復手順、半月板修復、回旋筋腱板修復、または他の手順と併用して関節腔に細胞を投与する。また、細胞は、事故もしくは軍事衝突が原因の外傷性障害、または先天性欠損症により失われた手指または四肢を再生する方法にも有用である。
【0020】
本方法の大きな利点は、規定の分化の段階で、たとえば、細胞が分化し始めたばかり(たとえば、2日目の細胞、3日目の細胞もしくは4日目の細胞)、または十分に分化した軟骨細胞(たとえば、7日目〜14日目の細胞)の段階で細胞の実質的に純粋な集団を単離することを可能にすることである。十分に分化した軟骨細胞は、多くの軟骨修復および再建治療に有用である。分化の初期段階の細胞は、同じ目的で使用されるか、または他の目的で使用され、その場合、コロニー形成し、さらに分化するだけでなく、in situで局所環境シグナルに応答するという別の利点が存在する。
【0021】
また、上述の方法を用いて産生された細胞は、疾患の遺伝学的基盤の判定、および処置に使用される治療薬のスクリーニング、または軟骨障害の重症度の軽減にも有用である。たとえば、軟骨障害の発生に関係する遺伝子を同定する方法は、既知の分化の段階(たとえば、十分に分化した細胞の2日目〜3日目の細胞、4日目の細胞)の実質的に均一な細胞集団を用意し、正常対照細胞と比較した遺伝子発現の増加または減少を検出することにより行う。遺伝子発現の増加または減少から、差次的に発現する遺伝子は、軟骨障害の発生に関係することが示される。たとえば、この細胞は、軟骨異形成症または軟骨無形成症と診断された患者から採取されたiPSCである。
【0022】
軟骨障害の重症度を処置または軽減する治療薬(たとえば、軟骨促進剤)を同定する例示的なスクリーニング方法は、既知の分化の段階(たとえば、十分に分化した細胞の2日目〜3日目の細胞、4日目の細胞)の実質的に均一な細胞集団を用意し、集団を候補化合物と接触させ、軟骨形成を検出することにより行う。前記化合物の存在下での軟骨形成が化合物の非存在下と比較して増加すれば、化合物が軟骨の形成を促進することが示される。軟骨形成が減少すれば、化合物が軟骨の形成を阻害することが示される。
【0023】
本発明の他の特徴と利点については、以下のその好ましい実施形態の説明、および特許請求の範囲から明らかになるであろう。他に定義しない限り、本明細書で使用する技術用語および科学用語はすべて、本発明が属する技術分野の当業者が一般に理解しているのと同じ意味を持つ。本発明の実施または試験には、本明細書に記載したのと類似または同等の方法および材料を使用してもよいが、以下に好適な方法および材料について記載する。本明細書に引用する公開された外国特許および特許出願はすべて、参照によって本明細書に援用する。本明細書に引用する、受託番号で表されるGenBankおよびNCBIの寄託物(submissions)は、参照によって本明細書に援用する。本明細書に引用する他の公表された参考文献、文書、原稿および科学文献はすべて、参照によって本明細書に援用する。矛盾がある場合、定義を含め本明細書が優先する。さらに、材料、方法、および例は、単に例示のためのものであり、限定的であることを意図するものではない。
【0024】
本発明の他の特徴と利点は、以下の詳細な説明および特許請求の範囲から明らかになる
であろう。
【図面の簡単な説明】
【0025】
【図1】A:放射線照射マウス胚性線維芽細胞のフィーダー層上で培養された未分化多能性H9ヒト胚性幹細胞(hESC)のコロニーを示す顕微鏡写真、B:形成された翌日のhESCコロニーに由来する胚様体(EB)を示す顕微鏡写真。
【図2】A:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、未処理の対照培養物を示す顕微鏡写真、B:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、骨形成タンパク質−2(BMP2)処理した培養物を示す顕微鏡写真、C:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、トランスフォーミング増殖因子β1(TGFβ1)処理した培養物を示す顕微鏡写真、D:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物を示す顕微鏡写真について、BMP2とTGFβ1との両方で処理された培養物を示す顕微鏡写真。
【図3】A:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、B:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、C:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、D:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真。
【図4】A:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、アルシアンブルー染色された14日目の未処理の対照培養物を示す顕微鏡写真、B:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、アルシアンブルー染色された14日目のBMP2処理培養物を示す顕微鏡写真、C:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、II型コラーゲン抗体で免疫染色した14日目のBMP2処理培養物の矢状断面。
【図5】A:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、B:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、C:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、D:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、E:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、F:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真。
【図6】A:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、未処理の対照培養物を示す顕微鏡写真、B:図未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、BMP2処理培養物を示す顕微鏡写真、C:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、TGFβ1処理培養物を示す顕微鏡写真、D:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、BMP2とTGFβ1との両方で処理された培養物を示す顕微鏡写真。
【図7】A:培養から48時間でBMP2を供給し、リアルタイム定量RT−PCRにより測定した3日目(D3)、4日目(D4)、7日目(D7)、および14日目(D14)のhESCマイクロマス培養物におけるSox9転写物の発現、アグリカン転写物の発現、Col2a1転写物の発現、Col10a1転写物の発現およびオステオポンチン(OPN)転写物の発現(A)を示す棒グラフ。発現レベルは、内部対照としてGAPDHを用いてΔΔCt法により判定した。相対量は、−2ΔΔCtとして計算し、2日目の発現レベルに対して補正して、これを値1に設定した。各値は、3つの測定値の平均値(±SEM)(4日目、7日目、および14日目)、または2つの測定値の平均(±range)(3日目)である、B:培養から48時間でBMP2を供給し、リアルタイム定量RT−PCRにより測定した3日目(D3)、4日目(D4)、7日目(D7)、および14日目(D14)のhESCマイクロマス培養物におけるBrachyury転写物の発現(B)を示す棒グラフ。発現レベルは、内部対照としてGAPDHを用いてΔΔCt法により判定した。相対量は、−2ΔΔCtとして計算し、2日目の発現レベルに対して補正して、これを値1に設定した。各値は、3つの測定値の平均値(±SEM)(4日目、7日目、および14日目)、または2つの測定値の平均(±range)(3日目)である、C:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるBrachyuryの発現レベルD:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるSox9の発現レベル、E:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるアグリカンの発現レベル。
【図8】アグリカンの球間ドメインに対する抗体で免疫染色した14日目のBMP2処理培養物の断面の顕微鏡写真。細胞内アグリカン染色を示す細胞が培養物の全体に存在する。これは、II型コラーゲン細胞外マトリックスで囲まれた細胞の実質的にすべてが(図4のA〜Cに示す)、細胞自律性(cell autonomous)マーカーを用いた免疫染色によるアッセイされる軟骨マーカー、アグリカンを発現していることを示す。
【図9】A:BMP2で処理された14日目のhESCマイクロマス培養物のアルシアンブルー染色およびNuclear Fast Red染色矢状断面を示す顕微鏡写真。アルシアンブルーで染色可能な細胞外マトリックスで囲まれた細胞は、BMP2処理培養物の断面の実質的に全範囲にわたり存在し、さらに、少数の管状構造物も存在する(矢印)、B:BMP2およびTGF−β1の両方で処理されたhESCマイクロマス培養物の断面を示す顕微鏡写真。細胞の実質的にすべてがアルシアンブルー軟骨基質で囲まれており、非軟骨形成組織は、ほとんどあるいはまったく検出できない、C:BMP2とTGF−β1とで処理されたhESC培養物の断面を示す顕微鏡写真。広範囲の軟骨組織に加えて小管(矢印)も存在する。こうした管は、BMP2とTGF−β1とで処理された培養物の非常に小さな割合を占めるだけであり、培養物の大部分は軟骨分化を示す。図9Cの下のテーブル1は、図9Aに示した14日目のBMP2処理培養物の断面中に存在する細胞の88%がアルシアンブルー陽性基質で囲まれていることを示す。図9Bおよび図9Cに示したBMP2およびTGFβ1処理培養物の断面では、細胞の97.3%がアルシアンブルー陽性基質で囲まれている。これらの結果から、本方法のBMP2処理、およびBMP2/TGFβ1処理hESC培養による軟骨細胞分化の実質的な均一性が明らかにされる。
【図10】胚様体(EB)細胞から樹立され、BMP2とTGF−β1とで処理されたマイクロマス培養物のアルシアンブルー染色およびNuclear Fast Red染色矢状断面を示す顕微鏡写真。アルシアンブルー染色された軟骨形成組織だけでなく、大量の脂肪組織(Ad)、さらには管(矢印)も存在する。
【図11】本明細書に記載の方法に供されたヒト包皮線維芽細胞に由来するiPS細胞株の軟骨基質をアルシアンブルー染色することによりアッセイされる軟骨細胞分化を示す顕微鏡写真。図11の(A)は、BMP2を補充せずに維持したiPSCの14日目の対照マイクロマス培養物を示す。アルシアンブルー陽性基質はほとんど存在しない。図11の(B)は、14日目のBMP2処理iPSCマイクロマス培養物を示す。強く広範囲のアルシアンブルー染色が培養物の全体に存在する。
【図12】A:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。トリプシン(A)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、B:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。TrypLE Select(B)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、C:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真である。Accutase(C)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、D:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない、E:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない、F:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない。
【図13】A:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2の存在下で維持されたiPSCの14日目のマイクロマス培養物を示す。BMP2を補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積する、B:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2とTGFβ1との組み合わせの存在下で維持された14日目のiPSCマイクロマス培養物を示す。BMP2とTGFβ1との組み合わせを補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積する、C:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2またはTGFβ1の非存在下で維持された14日目のiPSCマイクロマス培養物を示す。BMP2、またはBMP2とTGFβ1との組み合わせを補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積するのに対し、BMP2補充を受けなかったiPSCマイクロマス培養物は、アルシアンブルー陽性基質がほとんど蓄積されない。
【図14】A:マウス関節軟骨損傷モデルを示す顕微鏡写真。対照無手術マウス膝の前額断を示す。内側半月(mm)が確認される。靭帯の外科的切除および部分的な半月板切除(meniscetomy)による膝の不安定化から8週後のマウス膝の前額断を示す、B:マウス関節軟骨損傷モデルを示す顕微鏡写真。靭帯の外科的切除および部分的な半月板切除(meniscetomy)による膝の不安定化から8週後のマウス膝の前額断を示す。内側半月が存在しないことをアスタリスクで示す。関節軟骨損傷の局在化した領域が存在する(矢印)、C:Aの拡大図、D:Bの拡大図。
【図15】A:マウス指先再生モデルを示す顕微鏡写真。骨がアリザリンレッドで染色されている対照無手術マウスの指先を示す、B:マウス指先再生モデルを示す顕微鏡写真。6週間前に切断されたマウス指先を示す。すべての構造が完全かつ自然に再生されている、C:マウス指先再生モデルを示す顕微鏡写真。軟組織および硬組織の存在を示す対照指先の断面を示す、D:マウス指先再生モデルを示す顕微鏡写真。6週間前に切断されたマウス指先を示す。すべての構造が完全かつ自然に再生されている。
【図16】A:遺伝性軟骨障害(軟骨異形成症)の患者のヒト線維芽細胞からのiPSCの誘導を示す顕微鏡写真。未分化多能性幹細胞に特徴的な外観を持つコロニーを含む生きている培養物を示す(10×)、B:遺伝性軟骨障害(軟骨異形成症)の患者のヒト線維芽細胞からのiPSCの誘導を示す顕微鏡写真。iPSCの多能性幹細胞のマーカー(SSEA−4およびNanog)に対する免疫細胞化学的染色を示す。
【図17】未分化ヒト胚性幹細胞または人工ヒト多能性幹細胞から軟骨形成細胞を作製するためのステップを示すフロー図。
【発明を実施するための形態】
【0026】
軟骨の変性疾患、たとえば、変形性関節症は、多く見られる消耗性の慢性健康障害であり、成人の生活の質を低下させる主な原因の1つである(Magne D et al.,2005 Trends in Molecular Medicine,11:519−526)。変形性関節症(OA)は、関節軟骨の分解および変性を特徴とする非炎症性の変性関節疾患である。OAは、65歳を超える大部分の人が罹患し、40歳を超える集団の90%が、関節に何らかの形で痛みおよび不動を引き起こす軟骨変性を呈すると推定される(Song L et al.,2004 Cytotherapy,6:596−601)。軟骨変性疾患は、組織の自己修復能および再生能が乏しいため、その処置は特別な課題であり、軟骨病変の処置は、主要な臨床上の問題になっている。胚盤胞の内細胞塊に由来するヒト胚性幹細胞(hESC)は、無限の自己複製能を有しつつも、複数の細胞型に分化する能力を維持しているため、細胞ベースの組織工学治療により軟骨欠損を修復する強力なツールになる。実際、下記のように、hESCは、軟骨修復のための前駆体細胞を無限に供給する。
【0027】
軟骨修復にhESCを使用するには、病変軟骨の修復に関与し得る前駆体細胞へのhESCの分化を十分に誘導する必要がある。本発明の以前に、ESC由来の細胞が様々な程度で軟骨細胞に分化するいくつかの培養系が開発されてきた(;Elisseeff J
et al.,2005 Orthod Craniofac Res,8:150−161;Harkness L et al.,2009 Stem Cell Rev
5;353−368;Heng BC et al.,2004 Stem Cells,22:1152−1167;Hoben GM et al 2009 Stem Cells Dev 18:283−292;Hwang NS et al.,2006 Stem Cells,24:284−291;Hwang NS et al.,2006 Tissue Eng,12:2695−2706;Jukes et al.,2008 Tissue Engineering Part A,14:135−147;Kawaguchi J et al.,2005 Bone,36:758−769;Koay EJ et al.,2007 Stem Cells 25:2183−2190;Koay EJ and Athanasiou KA 2009 tissue Eng Part A 15:2249−2257;Kramer J et al.,2000 Mechanisms of Development,92:193−205;Kramer J et al.,2005 Cell Biology International,29:139−146;Kramer J et a
l.,2005 Anat Embryol,210:175−185;Ofek G et al.,2009 J Biomech Eng 131:061011;Sui
YP et al.,2003 Differentiation,71:578−585;Toh WS et al.,2007 Stem Cells,25:950−960;Toh WS et al.,2009 J Cell Mol Med 13B:3570−3590;Vats A et al.,2006 Tissue Eng 12:1687−1697;zur Nieden NI et al.,2005
BMC Dev Biol,5:5−15)。しかしながら、これらの系では軟骨細胞は、分化する細胞の亜集団のみであり、軟骨修復のために細胞集団を利用することが難しくなっている。
【0028】
さらに、軟骨細胞分化プロトコルの大部分は、ESC由来の胚様体(EB)の細胞を利用する(Elisseeff J et al.,2005 Orthod Craniofac Res,8:150−161;Harkness L et al.,2009 Stem Cell Rev 5:353−368;Heng BC et al.,2004 Stem Cells,22:1152−1167;Hoben GM et al 2009 Stem Cells Dev 18:283−292;Hwang NS et al.,2006 Stem Cells,24:284−291;Hwang NS et al.,2006 Tissue Eng,12:2695−2706;Jukes et al.,2008 Tissue Engineering
Part A,14:135−147;Kawaguchi J et al.,2005 Bone,36:758−769;Koay EJ et al.,2007 Stem Cells 25:2183−2190;Koay EJ and Athanasiou KA 2009 tissue Eng Part A 15:2249−2257;Kramer J et al.,2000 Mechanisms of Development,92:193−205;Kramer J et al.,2005 Cell Biology International,29:139−146;Kramer J et al.,2005 Anat Embryol,210:175−185;Ofek G et al.,2009 J Biomech Eng
131:061011;Sui YP et al.,2003 Differentiation,71:578−585;Toh WS et al.,2007 Stem Cells,25:950−960;Toh WS et al.,2009 J Cell Mol Med 13B:3570−3590;Vats A et al.,2006 Tissue Eng 12:1687−1697;zur Nieden
NI et al.,2005 BMC Dev Biol,5:5−15;Waese and Stanford 2010 Stem Cell Res Sept 6)。EBは、未分化なESCまたはコロニーの懸濁液を非接着性支持体上でインキュベートすることにより形成される細胞の三次元凝集塊である。EBの細胞は、胚の3胚葉に由来するすべての細胞型で構成される半ば系統化された組織(semi−organized tissues)に分化することができる。EBで生じる細胞環境および相互作用は、正確に制御することが難しく、細胞の不均一性を助長している。このため、EB由来の細胞を利用すると、組織修復に使用することができる軟骨形成細胞の均質な集団を得にくくなる。
【0029】
前もってEB形成せずにhESCの軟骨細胞分化が試みられた、いくつかのプロトコルが報告されている。1つのプロトコルは、分化したウシ関節軟骨細胞とhESCを長期間(数週間)共培養するものであった(Hwang NS et al.,2008 PLoS ONE,3:1−10)。別のプロトコルは、明確に定義されていない肝細胞の癌細胞株の条件培地でESCを前培養するものであった(Hwang YS et al.,2008 Stem Cells Dev,17:971−978)が、この研究では
、軟骨細胞分化が限定的であり、非均一であった(Hwang NS et al.,2008 PLoS ONE,3:1−10)。
【0030】
別の研究では、前もってEB形成せずにマウスES細胞のマイクロマス培養物を利用して軟骨分化を獲得しようとした(Yamashita A et al.,2009,Cell Death and Differentiation 16:278−286)。Yamashitaのプロトコルは、培養開始時に増殖因子を加え、その結果かなりの細胞死が引き起され、その後培養支持体からの剥離が行われるものであった。さらに、この培養物は、斑状の非均一なアルシアンブルー染色のみを示し、RT−PCRで検出可能な信頼できる軟骨マーカー、アグリカンをほとんど発現しないか、まったく発現しない。培養物は、肥大軟骨マーカー10型コラーゲンを発現し、石灰化が起こることが報告されており、培養中の細胞が培養期間の初期に肥大軟骨細胞に成熟したことが示された。肥大軟骨細胞の成熟は変形性関節症と関連するため、これらの特徴は、この方法の大きな欠点である。また、この培養物は、血管マーカーflk1を発現することも報告されたことから、培養の不均一性が示された。形成される比較的少量の軟骨形成組織が肥大成熟化し、軟骨形成組織を軟骨修復に不適切なものにする。実際、同じ著者らによる第2の研究(Yamashita A et al.,2010 PLoS One 5:el0998)では、培養物がインビトロでさらに肥大し、最終的に骨様組織を形成することが報告された。
【0031】
これに対し、本明細書に記載の方法は、組織学的解析および分子解析によりアッセイされる実質的に均一な軟骨細胞分化を生じさせる。この方法は、アグリカンおよび他の軟骨マーカーの高レベルの発現を示す細胞を生じさせる。これらの培養物が示す段階的な分化により、軟骨修復に使用してもよい、個々の分化段階の比較的純粋な細胞の集団を得ることができる。さらに、この細胞は肥大成熟化を起こさないことから、軟骨修復に特に好ましい。
【0032】
こうした相違は、1つには増殖因子を加えるタイミングに関係している。Yamashitaは増殖因子を高密度培養の開始時(ゼロ時)に加えたのに対し、本明細書に記載の方法では、増殖因子を培養物に加えるのは高密度培養の樹立から24時間後または48時間後である。マイクロマス培養の軟骨形成前駆体を培養の開始時(0時)にBMPで処理すると、細胞死が引き起こされ、軟骨細胞分化が阻害されるため、増殖因子を補充するタイミングは、軟骨細胞分化にとって重要である(Fisher et al 2007)。他の研究者は、インビボでBMPにより軟骨形成前駆体を処理すると、大量の細胞死が引き起こされることを明らかにしている(Macias et al 1997)。また、hESCの生存を促進するには、培養物にROCK阻害剤を加える(Watanabe
K et al.,2007;Li X 2009)。好適な阻害剤として、Y27632およびH1152(どちらもCalbiochemから入手可能)、AR−12286(Aerie Pharmaceuticals)のほか、HA−1100.HCl([ヒドロキシファスジル;1−ヒドロキシ−5−イソキノリンスルホニル)ホモ−ピペラジン]、3−(4−ピリジル)−1H−インドール、H−1152.2HCl([H−1152P;(S)−(+)−2−メチル−1−[(4−メチル−5−イソキノリニル)スルホニル]ホモピペラジン、およびN−(4−ピリジル)−N’−(2,4,6−トリクロロフェニル)尿素(これらのうち最後の4つは、Alexis Biotchemicalsから入手可能)が挙げられる。
【0033】
また、間葉系幹細胞様中間体(mesenchymal stem cell−like intermediate)を利用してhESCを軟骨細胞に間接的に分化するプロトコルも報告されている。しかしながら、これらのプロトコルの一部は、EBを利用して間葉系幹細胞様中間体を産生する(Lee EJ et al.,2010 Tissu
e Eng A 16:705−717;Brown SE et al.,2008,Cells Tiss Org 189:256−260;Mahmood A et al.,2010 J Bone Min Res 25:1216−1233;Hwang et al 2008;Cohen S et al.,2010 Tissue
Eng A 16:3119−3139;Harkness L et al.,2009 Stem Cell Rev 5:353−368)ため、上記で論じたように、細胞の不均一性が生じる。同様に、ヒト生殖隆起から単離したヒト幹細胞由来の間葉系幹細胞中間体からの軟骨細胞分化を報告したプロトコルでも、EBのステップを使用している(hEG細胞、Varghese S et al.,2010 Stem Cells 28:765−774)。他のプロトコルでは、間葉系中間体を産生するため、マウス骨髄由来間質細胞株であるOP9細胞との共培養(Barberi T et al.,2005 PLoS Med 2 e161)、あるいはヒト関節軟骨細胞との共培養(Bigdeli N et al.,2009 Stem Cells 27:1812−1821)を必要とした。いくつかのプロトコルでは、紡錘形の線維芽細胞/間葉系様細胞を産生させ、その後様々な軟骨細胞分化プロトコルに付すため、間葉系幹細胞マーカーを発現しているhESCの亜集団を同定すべくFACSソーティングを使用する(Lian Q et al.,2007 Stem Cells 25:425−436;Kopher RA et al.,2010 Bone 47:718−728;Stavropoulos ME et al.,2009 Curr Prot Stem
Cell Biol 9:1F.8.1−1F.8.10;Barberi T et
al.,2005 PLoS Med 2 el61;Barberi T et al.,2007 Nature Med 13:642−651)か、あるいは、継代の繰り返し、および低密度でのhESCの大規模かつ長期的(数週間)培養を行う(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692;Arpornmeaklong P et al.,2009 Stem Cells Dev 18:955−968;Boyd NL et al.,2009 Tissue Eng A 15:1897−1908)ことにより、間葉系幹細胞様中間体の細胞集団を得た。しかしながら、これらのプロトコルで得られた軟骨細胞分化は、II型コラーゲンの斑状の免疫染色に示されるように不均一なものであり(Arpornmeaklong P et al.,2009 Stem Cells Dev 18:955−968)、組織に均一性がなく(Kopher et al 2010)、不均一にII型コラーゲンおよびSox9が分布し(Barberi T et al.,2007 Nature Med 13:642−651;Stavropoulos ME et al.,2009 Curr Prot Stem Cell Biol 9:1F.8.1−1F.8.10)、アルシアンブルー染色をほとんど示さない不均一な組織が形成され(Lian Q et al.,2007 Stem Cells 25:425−436)、培養プレートから剥離した、特徴付けが十分になされていない集団が産生された(Boyd NL et al.,2009 Tissue Eng A 15:1897−1908)。また、BMP7の存在下でhESC由来の間葉系線維芽細胞様細胞をペレット培養に供したプロトコル(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692)でも不均一な軟骨形成の分化が起こり、培養物は広範囲に及び未分化な中核を示し、軟骨細胞分化は、ペレットの周辺の細胞に限定されていた。また、BMP7とTGFβ1とで処理したペレットの軟骨細胞分化(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692)も均一ではなく、未分化な中核があり、周辺のII型コラーゲン免疫染色が全体として弱かった。これらの培養物での、信頼できる軟骨マーカー、アグリカンの発現は、未処理対照ペレットと比較して大きく増加しなかった。
【0034】
これらの従来の方法の結果は、実質的に均一な細胞を段階的に直接生じさせる、本明細
書に記載の方法を用いたhESCによる軟骨細胞分化とは異なる。もう1つの利点として、本発明の方法は、分化の個々の段階で同調的に細胞を生じさせる。たとえば、軟骨前駆体(2〜3日の細胞、軟骨系譜に入ったばかりであることを特徴とする)、初期の軟骨細胞(4日目の細胞、軟骨細胞分化の初期相にあることを特徴とする)、または十分に分化した軟骨細胞(7〜14日の細胞、軟骨細胞への明らかな分化を均一に示すものの、肥大成熟化を示していない)を採取するため、別々の時点で細胞が回収される。
【0035】
【表1】
上述の各細胞集団は、理想的には特定の臨床状態の処置に適している。たとえば、軟骨前駆体(chondroprogentor)(2日目〜3日目の細胞)は、四肢または手指の再生、半月板(mensical)または靭帯の修復、および骨折修復に優先的に使用される。初期の軟骨細胞(4日目の細胞)は、傷害による関節軟骨欠損の修復、慢性変性疾患(OAまたはRA)による関節軟骨の修復または予防、外傷または先天性欠損により失われた軟骨組織の再生および置換、半月板または骨折の修復、ならびに顎関節の線維軟骨および軟骨の修復に使用される。十分に分化した軟骨細胞は、急性もしくは外傷性障害後の関節軟骨の修復、または慢性OAもしくはRAによる関節軟骨損傷の修復もしくは予防のほか、内軟骨性骨化による骨折修復に使用される。各段階は、軟骨発生および分化のモデリングに有用であり、iPSC由来の細胞は、患者個人を考慮した修復および再生のほか、ヒト疾患のモデリング、標的治療の設計および試験、ならびに遺伝的に傷害された軟骨組織の疾患特異的修復に特に有用である。
【0036】
iPSCの軟骨細胞分化については、既に報告されている(Medvedev SP et al.,2010 Stem Cells Dev Oct 17;Lian Q
et al.,2010 Circulation 121:1113−1123)。しかしながら、一方のプロトコルは、iPSCからのEBの形成を必要とし(Medvedev SP et al.,2010 Stem Cells Dev Oct 17)、もう一方のプロトコルは、間葉系幹細胞様中間体の細胞集団を採取するため、軟骨細胞分化を誘導する前に蛍光活性化セルソーター(FACS)によりiPSCを分取して間葉系幹細胞マーカーを発現している亜集団を同定する必要がある(Lian Q et al.,2010 Circulation 121:1113−1123)。このプロトコルを用いた軟骨細胞分化の結果、II型コラーゲンに免疫反応を示す細胞は、約60%に過ぎなかった(Lian Q et al.,2010 Circulation 121:1113−1123)。さらに本方法のもう1つの利点は、FACS(時間がかかるうえ、混入のリスクが生じ得る)の使用が回避されることである。
【0037】
本明細書に記載の方法は、EBを利用しない。本方法は、hESCまたはiPSCと、分化したウシ関節軟骨細胞、ヒト関節軟骨細胞、またはマウスOP9細胞との共培養を必要としない。本方法は、肝細胞の癌細胞由来の培地との前培養もしくは共培養も必要としない。さらに、本方法は、単層(monoloayer)での長期の継代および培養、またはFACSによる細胞分取も必要としない。本方法は、より高速である(3週間未満、
2週間未満または1週間未満、たとえば、細胞は、高密度細胞培養の樹立から2〜3日後という短期間で軟骨形成細胞系譜に入る)。本方法により、実質的に均一な軟骨細胞分化が段階的に起こり、軟骨修復および他の使用に合わせて別々の分化の段階で細胞の集団を産生することが可能になる。
【0038】
前もって胚様体(EB)を形成せずに、未分化多能性hESCの軟骨形成細胞系譜への段階的かつ均一な分化を迅速に直接促進する本発明の培養系および培養条件について、以下に記載する。未分化多能性hESC、およびhESC由来の胚様体(EB)の細胞を高密度マイクロマス培養条件に付し、胚の肢芽間葉系細胞の軟骨細胞への分化が誘導された(Gay SW and Kosher RA,1984 J Exp Zool,232:317−326;Kosher RA et al.,1986 Dev Biol,118:112−117;Kosher RA et al.,1986 J Cell Biol,102:1151−1156;Kulyk WM et al.,1991 Matrix,11:282−288)。この高密度マイクロマス培養系は、胚の四肢の発生において間葉系前駆体細胞の軟骨細胞分化の開始を特徴付ける細胞の密な並列、および細胞間相互作用を刺激する。BMP2単独で処理(またはBMP2とTGFβとを組み合わせて一緒に投与)した未分化多能性hESCのマイクロマス培養物は、前もってEB形成をせずに軟骨形成細胞系譜への段階的かつ実質的に均一な(たとえば、少なくとも88%)分化を直接示す。
【0039】
BMP2およびTGFβ1を補充する別のアプローチとして、骨形成タンパク質−4(BMP4、NG_009215(GI:219521814)、参照によって本明細書に援用する)、骨形成タンパク質−7(BMP7、NM_001719(GI:187608319)、参照によって本明細書に援用する)、増殖分化因子−5(GDF5、NG_008076(GI:193083169)、参照によって本明細書に援用する)、トランスフォーミング増殖因子β−3(TGFβ3、NG_011715(GI:225735563)、参照によって本明細書に援用する)、またはインスリン様成長因子−1(IGF−I、NG_011713(GI:225735562)、参照によって本明細書に援用する)などの他の増殖因子を培養物に補充する。これらの各因子は、軟骨形成細胞系譜への分化を促進する(Waese EY and Stanford WL 2010
Stem Cell Res Sept 6;Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692;An C et al.,2010 Ann Biomed Eng 38:1647−1654;Moore YR et al.,2010 J Clin Periodontol 37:288−298)。
【0040】
マイクロマス培養の別のアプローチとしては、hESCをペレット培養に付す。この方法では、分離したhESCまたはiPSC(約1×105〜1×107細胞)をマイクロフュージチューブでペレット状にする。次いでマイクロマス培養のため、このペレットを上記と同じ軟骨細胞分化プロトコルに付す。ペレットに増殖因子(単数または複数)を加え、ペレットに拡散させて細胞の分化を誘導する。その後治療のため、ペレットそのものを、関節もしくは関節腔または他の修復組織への移植、注入、または他の投与手段に使用する。このプロトコルによっても、EBを形成せずに、hESCの段階的かつ実質的に均一な軟骨細胞分化が直接達成される。また、このペレット培養プロトコルは、iPSCの軟骨形成細胞系譜への分化の誘導にも使用される。
【0041】
細胞培養系:系Iおよび系II
細胞は、2つの系の1つを用いて調製される。系Iでは、高密度培養を樹立し、24時間培養し、この時点で細胞にBMP2またはBMP2とTGFβとの組み合わせを含む軟骨形成培地を加える。系IIでは、高密度培養を樹立し、24時間培養し、この時点で増
殖因子(単数または複数)を用いずに細胞に軟骨形成培地を加える。3日目の初めに、細胞にBMP2またはBMP2とTGFβとの組み合わせを加える。1日後(3日目の終わりに)、遺伝子発現プロファイルおよび物理的特性/組織学的特徴から明らかなように、細胞は、実質的に均一に軟骨形成細胞系譜に入る。培養の時期、増殖因子の添加、および分化の段階を図示するフローチャートを図17に示す。
【0042】
下記のように、分化の進行過程の様々な時期に回収されたhESC由来の細胞の遺伝子発現プロファイルを用いることで、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に至る様々な相の軟骨形成細胞系譜の細胞を同定することができる。本明細書に記載の方法を用いれば、様々な相の軟骨形成細胞系譜のhESC由来の前駆体細胞が、細胞ベースの組織工学治療により軟骨を修復する能力を解析することができる。また、均一かつ段階的な軟骨細胞分化が起これば、軟骨形成細胞系譜に分化することが決定した前駆体細胞への未分化多能性hESCの段階的な変化に関係する遺伝子、シグナル、および制御ネットワークを同定しやすくなる。
【0043】
治療方法
細胞を投与するアプローチとして、自家軟骨細胞移植の場合(ACI,Zazlav K et al.,2009 Am J Sports Med,37:42−55)、またはスキャフォールドフリーアプローチにより関節鏡視下の場合(Jubel A et al.,2008 Am J Sports Med 36:1555−1564;Erggelet C et al 2003 Arthroscopy 19:108−110)には、膝を開く手術などの外科的手段により、マイクロマス培養物またはペレット培養物を損傷部位に直接移植する。一部の手順では、培養物をコラゲナーゼ消化または他の手段によりさらに分離し、細胞をスキャフォールドまたは他の支持体に播種する。こうした支持体として、コラーゲン膜(たとえばChondrogide)、ヒアルロナン含有マトリックス(たとえばHyaff−11)、ポリグラクチンフリース、または他のマトリックスがある。次いで細胞を播種したマトリックスを、マトリックス誘導自家軟骨細胞修復の場合(MACI:matrix−induced autologous chondrocyte repair)(Brittberg,2010 Am J Sports Med,38:1259−1271)、またはHyalgraftCの場合(Gobbi A et al.,2006:Am J Sports Med 34:1763−1773)には、外科的に移植するか(Kon et al.,2009 Am J Sports Med 37:156S−166S)、あるいは、関節鏡視下で移植する(Gianni E et al.,2008 Am J Sports Med 36:873−880;Nixon AJ 2002 Clinical Tech Equine Practice 1:257−269)。また、分離した細胞を、コラーゲン、キトサン、アガロースまたはヒアルロナンを含むゲル中に封入し、その細胞−ゲル混合物を用いて、外科的手順で損傷領域に充填する(Funayama A et
al.,2008 J Orthop Surg 13:225−232;Hoemann C et al.,2005 OsteoArthritis Cart 13:318−329;Emans PJ et al.,2010 PNAS 107:3418−3423)。これらの方法では、使用する細胞の濃度は、(1×106〜2×107細胞/ml)である。
【0044】
あるいは、非侵襲的に関節内注射により、分離した細胞を膝に直接注射または注入する。導入される細胞数は、(1×106〜2×107細胞/ml)である。自家間葉系幹細胞(MSC)を傷害された膝または関節炎の膝に導入する場合(Centeno CJ et al.,2008 Pain Physician 11:343−353)、またはMSCを軟骨が損傷したラット、ブタおよびヤギの関節に注射する場合(Horie
M et al.,2009 Stem Cells 27:878−887;Lee
KB et al.,2007 Stem Cells 25:2964−2971;Murphy JM et al.,2003 Arthritis Rheum 48:3464−3474)には、細胞は食塩水を用いて注射する。別のアプローチでは、細胞は、臨床的関節炎痛および不動(immmobility)を軽減する関節内補充療法に使用されるヒアルロナン調製物(たとえばSynvisc、Supartz)などの他の物質と併用して(Petrella RJ and Petrella M 2006,J Rheumatol 33:951−956;Brander VA and Stadler TS 2009 Phys Sports Med 37:38−48)、あるいは、骨関節炎の進行を遅らせるBMP7などの増殖因子と併用して(Hunter DJ et al.,2010,BMC Musculoskelet Disord 11:232;Hayashi M et al.,2010 J Orthop Res 28:1502−1506)注射する。
【0045】
細胞送達の1つのアプローチは、関節鏡視下での投与である。場合によって、関連する欠損または状態を修復する外科的手順または関節鏡視下手順の前、その後、またはその最中に細胞を投与または移植する。たとえば、ACL修復手順、半月板修復、回旋筋腱板修復、または他の手順と併用して関節腔に細胞を投与する。このアプローチは、動物における間葉系幹細胞の半月板損傷の修復を評価するために使用されており(Horie M et al.,2009 Stem Cells 27:878−887)、ヒトに好適である(Centano CJ et al.,2008 Med Hypotheses 71:900−908.)。
【0046】
本細胞および方法は、スポーツ関連、戦闘関連、および他の骨、軟骨および四肢の外傷性障害の修復を支援するのに有用である(Goldstein 2006 J Am Acad Orthop Surg 14:S152−156;Sundelacruz S and Kaplan DL 2009 Sem Cell Dev Biol 20:646−655)。本細胞は、長骨骨折の修復に有用であり、内軟骨性骨化、すなわち軟骨を鋳型に骨化形成が起こり正常な一連の発生現象(normal developmental sequence)を再現させるプロセスにより修復を行う(Kronenberg HM 2003 Nature 423:332−336)。間葉系幹細胞を利用する方法に記載したように、細胞は、注射または移植により骨折の領域に導入されると、リモデリングが起こり修復のための骨を形成する(Kallai I et al.,2010 J Biomech 43:2315−2320;Scotti C et al.,2009 PNAS 107:7251−7256)。
【0047】
また、細胞は、事故もしくは軍事衝突が原因の外傷性障害または先天性欠損症により失われた手指または四肢を再生する方法にも有用である。イモリおよびカエルなどの下等動物は、四肢を再生する能力を有するものの、成体哺乳動物では、再生反応を行うのに必要な細胞およびシグナルが不十分であるため(Gurtner GC et al.,2007 Ann Rev Med 58:299−312;Muneoka K et al.,2008 Sci Am 298:56−63)、この能力の大部分が失われている(Muller TL et al.,1999 Sem Cell Dev Biol 10:405−413)。
【0048】
マウスの四肢再生モデルが樹立された。このモデルでは、切断された指尖部は自然な再生反応を示すのに対し、切断された指基部は再生することができない(Han M et
al.,2008 Dev Biol 315:125−135)。このモデルは、hESCおよびiPSC由来の軟骨前駆体細胞、特に骨格および軟骨形成細胞系譜(たとえば、2日目、3日目または4日目の細胞)に入ったばかりの細胞が哺乳動物の四肢再生に関与または促進する能力を試験する系となる。四肢(腕もしくは脚、手もしくは足)また
は指(手指もしくは足指)の再生治療のための送達方法は、本方法により産生された細胞を指または四肢に注射または移植により導入することを含む。細胞は、増殖因子を使用してあるいは使用せずにスキャフォールドまたはゲルを用いて導入する。この指再生モデルを図15のA〜Dに示す。
【0049】
疾患モデル
ヒト疾患のインビトロでのモデリングに際して、さらに標的治療法の設計および試験のための系として、遺伝性疾患の患者から採取したiPSCが使用される(Amabile
G and Meissner A 2009 Trends Molec Med 15:59−68;Laustriat D et al.,2010 Biochem
Soc Trans 38:1051−1057)。本発明も、遺伝性軟骨疾患のインビトロでのモデリング、および治療法の設計および試験に有用である。軟骨異形成症および軟骨無形成症(achondoplasia)を含むこれらの障害は、外観が変形して障害を生み出す低身長、さらには死を引き起こす(Krakow D and Rimoin D 2010 Genet Med 12:327−341)。このため、軟骨に影響を与える遺伝子性障害(たとえば、軟骨異形成症、軟骨無形成症または他の症状)のある個体から採取した線維芽細胞または他の細胞型からiPSCを得て、本明細書に記載の方法を適用して軟骨細胞分化を誘導する。このiPSC由来の軟骨形成細胞は、インビトロで疾患プロセスを再現し、疾患の病理学的変化の機構研究、ならびに標的治療法(therapueutics)の設計および試験を可能にする。このアプローチは、たとえば細胞外マトリックスタンパク質の合成または硫酸化;代謝酵素、イオンチャネルもしくはトランスポーター;分子のフォールディング、プロセシングもしくは分解;ホルモン、増殖因子、受容体もしくはシグナル伝達物質;転写因子;RNAプロセシング;または細胞骨格タンパク質の欠損などにより引き起こされるような、メカニズムの大部分が不明である軟骨異形成症に適用される(Krakow D and Rimoin D 2010 Genet Med 12:327−341)。このアプローチは、正常もしくは異常な軟骨機能に重要な新規な遺伝子または治療の新規な標的を同定するための遺伝子プロファイリングにも適用される(Pogue R et al.,2004 Matrix
Biol 23:299−307)。このアプローチを遺伝子ターゲティングと組み合わせて単一遺伝子(mongenic)軟骨異形成症の患者特異的細胞による治療を試みてもよい(Wong GK and Chiu AT 2010 Biotechnol
Adv July 24)。
【0050】
軟骨修復の評価には、大型および小型動物の実験モデルを使用する(Chu CR et al.,2010 Tissue Eng Part B 16:105−115)。あるモデルでは、外科的な膝靭帯の切断および/または部分的な半月板切除(meniscetomy)により膝関節を不安定にして、荷重を不適切に集中させ、限局性の関節軟骨の破壊を引き越す。このモデルで起こる軟骨欠損は非常に再現性がよく、その軟骨破壊の発生および進行において、慢性ヒト変形性関節症で見られる状態が忠実に再現される。このモデルは、様々な動物の関節に間葉系幹細胞を注射することにより、損傷した関節軟骨の細胞による修復を評価するのに使用されている(たとえばMurphy JM et al.,2003 Arthritis Rheum 48:3464−3474;Kubo S et al.,2009 Arthritis Rheum 60:155−165)。このモデルは、齧歯動物にも応用されている(Kamekura S et al.,2005 Osteoarthritis Cart 13:632−641;Glasson SS et al.,2007 Osteoarthritis Cart 15:1061−1069;Welch ID et al.,2009 Arthritis Res Ther 11:R14)。このモデルは現在、免疫能が低下したマウスで樹立され、hESCおよびiPSC由来の軟骨形成細胞による修復のインビボ試験に使用されている(図14のA〜D)。
【0051】
軟骨修復の評価に使用される別のモデルは、外科的に作製した関節軟骨欠損を利用する(Ahern BJ et al.,2009 Osteoarthritis Cart 17:705−713)。このモデルでは、大腿骨膝蓋溝の非荷重部を露出させ、大腿骨を切除するか、または軟骨表面から軟骨下骨に及ぶ穴を開けて全層欠損を機械的に作製する。このアプローチは大型動物に使用され、ラット(Dausse Y et al.,2003 Osteoarthrtis Cart 11:16−28)、およびマウス(Eltawil NM et al.,2009 Osteoarthritis
Cart 17:695−704;Osteoarthritis Cart 6:695−704)にも適合されている。このモデルは、急性傷害後の関節軟骨損傷を刺激し、ヒトの場合と同様に、欠損を治癒できないと、損傷部位の外側で軟骨変性がさらに起こり、明らかなOAに至る。
【0052】
実施例1:hESCの培養および胚様体形成
WiCell Research Instituteで作製されたH9 hESC株(Thomson JA et al.,1998 Science,282:1145−1147)。University of Connecticut Stem Cell Coreにより標準的なプロトコルを用いて調製された放射線照射CF1マウス胚性線維芽細胞(MEF)をフィーダー層とし、以前に記載された無血清hESC培地(Toh WS et al.,2007 Stem Cells,25:950−960)でH9細胞を培養した(図1のAを参照)。1mg/mlのコラゲナーゼIV(Invitrogen)で処理してから4〜6日ごとにコロニーを継代した。本研究では、継代数36〜45のhESCを使用した。
【0053】
EBを作製するため、0.1%ディスパーゼ(Invitrogen)で37℃にて20〜30分間処理してhESCのコロニーをMEFフィーダー層から剥離し、T75 Ultra−Low Attachment Flask(Corning,Lowell,MA)中のEB形成培地に懸濁し(Toh WS et al.,2007 Stem
Cells,25:950−960)、5日間インキュベートした。培地は、48時間ごとに交換した。図1のBに示すように、懸濁されたhESCコロニーは、細胞の三次元凝集塊からなるEBを形成する。
【0054】
EB細胞および未分化hESC由来のマイクロマス培養物の調製
5日目のEB(図1のB)および未分化hESCコロニー(図1のA)から分離した両方の細胞からマイクロマス培養物を調製した。分離の前に、hESCコロニーを上記のようにMEFフィーダー層から剥離した。剥離したhESCコロニーおよびEBを0.05%トリプシン/EDTA(Invitrogen)で処理することにより単細胞に分離し、続いて細胞懸濁液を40μmのセルストレーナー(BD Biosciences,Franklin lakes,NJ)に通した。分離した細胞を、10%FBS(FBS,Hyclone)および10%>KSRを補充した高グルコースDMEMに2×107細胞/mlで懸濁した。マイクロマス培養物は、10μlの細胞懸濁液(2×105細胞)を24ウェル組織培養ディッシュ(Nunc;Fisher)の各ウェルにスポットして調製した。細胞接着を促進するため加湿5%CO2インキュベーター中で37℃にて2時間インキュベートした後、0.5mlの同じ培地を各ウェルに加えた。24時間後(培養の2日目の開始)、培地を除去し、培養物に無血清軟骨形成培地を供給した(Toh WS et al.,2007 Stem Cells,25:950−960)。軟骨形成培地は、ITS(6.25μg/mlのインスリン、6.25μg/mlのトランスフェリン、6.25μg/mlのセレン)、1.25mg/mlのウシ血清アルブミン、5.35μg/mlのリノール酸(BD Biosciences)、1%KSR、40μg/mlのL−プロリン(Sigma−Aldrich)、50μg/mlのアスコル
ビン酸2−ホスフェート(Sigma−Aldrich)、1%非必須アミノ酸(Invitrogen)、10−7M デキサメタゾン(Sigma−Aldrich)、および100単位/100μgのペニシリン/ストレプトマイシン(Invitrogen)を補充した高グルコースDMEMを含む。次いで細胞を、100ng/mlの組換えヒトBMP2(R&D Systems,Minneapolis)、10ng/mlの組換えヒトTGF−β1(R&D Systems,Minneapolis)、あるいは、100ng/mlの組換えヒトBMP2と10ng/mlの組換えヒトTGF−β1との組み合わせのいずれかの存在下または非存在下で培養する。一部の実験では、hESCのマイクロマス培養物を、増殖因子を含まない軟骨形成培地で24時間(2日目を通して)培養し、その後(培養の3日目の始めに)、培養物に、上記のような増殖因子を補充した新鮮な軟骨形成培地を供給した。増殖因子を含む培地は、培養期間を通して48時間ごとに交換した。
【0055】
アルシアンブルー染色、II型コラーゲンおよびアグリカンの免疫染色
マイクロマス培養物をアルシアンブルー、pH1.0で以前に記載されたように染色して、軟骨基質の蓄積を組織化学的にモニターした(Gay SW and Kosher
RA,1984 J Exp Zool,232:317−326)。II型コラーゲンの免疫染色では、4%パラホルムアルデヒドを含むリン酸塩緩衝生理食塩水(PBS)で30分間マイクロマス培養物を固定し、PBSで洗浄し、組織培養プレートから掻き取り、脱水し、パラフィンに包埋し、7μmの矢状断切片を作製した。免疫組織化学的検査については、Vectastain Elite ABCキット(Vector,Burlingame,CA)と、II型コラーゲンに対するモノクローナル抗体(抗II型コラーゲン、クローン6B3;Chemicon)、および軟骨マーカー、アグリカンの球間ドメインに対するモノクローナル抗体(6−B−4;Abeam)とを用いて行った。切片を脱パラフィンし、0.5%H2O2メタノール溶液で15分間クエンチし、ペプシン(Labvision)とインキュベートすることにより抗原賦活化を行った。切片を、5%正常ウマ血清を含むTBSでブロッキングしてから、1:200希釈II型コラーゲン抗体と1時間インキュベートした。次いでこの切片を、1:200希釈ビオチン化ウマ抗マウスIgGと室温にて1時間インキュベートし、アビジン−ビオチン西洋わさびペルオキシダーゼ複合体(Vectastain ABCキット)で処理し、ジアミノベンジジン/H2O2(Vector)で発色させた。陰性対照は、一次抗体とインキュベートしないものであり、ほとんど、あるいはまったく染色を示さなかった。
【0056】
リアルタイム逆転写ポリメラーゼ連鎖反応(リアルタイムRT−PCR)
Qiagen RNeasy Miniキット(Qiagen,Chatsworth,CA)を用いてマイクロマス培養物から全RNAを抽出し、RNase−free DNase(Ambion,Austin,TX)で処理して混入する恐れがあるゲノムDNAを除去した。High Capacity cDNA Reverse Transcription Kit(Applied Biosystems,Foster City,CA)を用いて、20μlの反応容量に対して2μgのRNAをcDNAに逆転写した。リアルタイムRT−PCRは、Sox9(アッセイID Hs00165814_ml)、アグリカン(アッセイID Hs00153936_ml)、II型コラーゲン(Col2a1)(アッセイID Hs00264051_ml)、X型コラーゲン(Col10a1)(アッセイID Hs00166657_ml)、オステオポンチン(opn)(アッセイID Hs00960942_ml)、Indian hedgehog(Ihh)(アッセイID Hs01081801_ml)、Brachyury(アッセイID Hs00610080_ml)、およびグリエルアルデヒド−3−リン酸デヒドロゲナーゼ)(GAPDH)(Hs99999905_ml)について、ABI Prism 7900 Sequence Detection System,TaqMan Gene Expression Master Mix、およびTaqMan
Gene Expression Assays (Applied Biosystems)を用いて行った。遺伝子発現レベルは、内部標準遺伝子としてGAPDHを用いてΔΔCt法により判定した。各遺伝子の相対量は、解析対象の各処理および時点ごとに2〜3つの独立したマイクロマス培養物それぞれから調製したサンプル2つずつの−2ΔΔCtとして計算した。サーマルサイクリング条件は、50℃で2分、95℃で10分、その後94℃で15秒の変性を40サイクル、60℃で1分の伸長反応であった。
【0057】
実施例2:未分化多能性hESCの直接かつ段階的な軟骨細胞分化
EBの特徴を示す細胞環境および固有の細胞多様性が明確に定義されていないため、未分化多能性H9 hESC(図1のA)が高密度培養に供されたときに軟骨形成細胞系譜に直接分化する能力を調べた。高密度培養物は、濃度が培地の10μl当たり1×105細胞を超える。たとえば、細胞の濃度は、10μl当たり1〜4×105細胞、たとえば、10μl当たり2×105細胞である。より大規模な培養が望ましい場合、体積を増やすが、細胞密度/比率は、上述の範囲内にとどめるものとする。
【0058】
分離したH9細胞のマイクロマス培養の樹立の翌日、100ng/mlのBMP2の存在下および非存在下で無血清「軟骨形成培地」を補充した。図4のAに示すように、分離した未分化なH9 hESC細胞から直接樹立した14日目の未処理のマイクロマス培養では、ごくわずかな小さなアルシアンブルー染色領域のみが存在した。外来性のBMP2の存在下では、14日目のBMP2処理hESCマイクロマス培養物中に実質的に均一で強く染色されたアルシアンブルー陽性基質が蓄積する(図4のB)。均一なアルシアンブルー染色だけでなく、コラーゲン抗体を用いた免疫染色により検出可能で軟骨に特徴的なII型コラーゲンII型が、BMP2処理培養物の全体に存在した(図4のC)。
【0059】
上記のように培養した未分化なH9細胞マイクロマス培養物でアルシアンブルー基質の蓄積が進行するのを図5のA〜Cに示す。培養から3日後では、検出可能なアルシアンブルー染色はほとんど認められなかったが(図5のA)、7日目には、比較的均一なアルシアンブルー染色が培養物全体で検出可能であった(図5のB)。14日目では、より強く染色されたアルシアンブルー陽性基質が培養物全体に存在した(図5のC)。これらの結果から、未分化多能性hESCは、適切な細胞環境(高密度マイクロマス培養)、およびBMP2などの外来性のシグナル伝達分子が供給されると、EB段階を経ずに直接かつ極めて均一な軟骨形成を示すことが示唆される。
【0060】
上述の軟骨細胞分化プロトコルを改変すると、hESCの軟骨形成細胞系譜への分化の進行がさらに促進される。この改変プロトコルでは、hESCマイクロマス培養の樹立の翌日、BMP2を含まない無血清「軟骨形成培地」を補充し、次いで24時間後に100ng/mlのBMP2を補充する。図5のD〜Fに示すように、3日目の培養物では検出可能なアルシアンブルー陽性基質がほとんど認められないが、7日目には、培養物の全体に均一でかなり強いアルシアンブルー基質が存在した。14日目には、培養物の全体に非常に強く染色されたアルシアンブルー基質が存在した(図5のF)。外来性のBMP2を3日目に供給したこの改変プロトコルを利用すると(図5のD〜F)、BMP2を2日目にhESC培養物に補充したプロトコルと比較して(図5のA〜C)、アルシアンブルー染色の範囲および強さが大きくなった。
【0061】
実施例3:hESCマイクロマス培養物における様々な相の軟骨形成細胞系譜の細胞に関する遺伝子発現プロファイリングによる特徴付け
多能性hESCの軟骨形成細胞系譜への分化の進行を確認および定量し、さらに様々な相の系譜の前駆体細胞の特徴付けを行うため、軟骨形成細胞系譜の様々な段階に特徴的なマーカー遺伝子の発現をリアルタイム定量RT−PCRにより調べた。上記の図5のD〜Fに示すようなアルシアンブルー基質染色によりアッセイされる段階的な軟骨細胞分化を
誘導するように改変した軟骨細胞分化プロトコルを行った多能性hESCのBMP2処理マイクロマス培養物における遺伝子発現パターンについて、何回も解析した。
【0062】
図7のAに示すように、外来性のBMP2の添加から24時間後の培養の3日目の終了時に、軟骨形成の転写因子Sox9の発現が7倍と著しくアップレギュレートされる。Sox9は、軟骨分化の「マスター制御遺伝子」として特徴付けられており、軟骨形成の実質的にすべての初期相の決定的なレギュレーターである(Akiyama H et al.,2002 Genes and Development 16:2813−2828;Lefebvre V and Smits P,2005 Birth Defects Res C Embryo Today,75:200−212)。したがって、3日目の培養物におけるSox9発現の著しいアップレギュレーションは、細胞が軟骨形成細胞系譜に入ったことと整合する。また、3日目の培養物で転写因子Brachyuryの発現が24倍のアップレギュレーションを示すことも注目に値する(図7のB)。Brachyuryは、中胚葉系譜のマーカーであり、発生中の四肢に軟骨を形成する前軟骨形成中胚葉細胞にも発現する(Herrmann BG,1995 Seminars in Developmental Biology,6:385−394;Hoffmann A et al.,2002 J Cell Sci,115:769−781;Liu C et al.,2003 Development,130:1327−1337)。したがって、Sox9と共にBrachyuryの発現がアップレギュレートされたことは、3日目の培養が軟骨形成細胞系譜に入ったことをさらに示唆する。3日目の培養物はSox9およびBrachyuryの発現のアップレギュレーションを示すものの、この培養物は、軟骨マーカー、アグリカンの発現のアップレギュレーションを示さない(図7のA)。3日目の培養物では、アグリカンは2日目と同じ低い基準レベルで発現する。アグリカンは、軟骨基質の主要な硫酸化プロテオグリカンであり、明らかに分化した軟骨細胞の信頼できる高度に特異的な分子マーカーである(Han Y and Lefebvre V,2008 Molecular and Cellular Biology,28:4999−5013)。これらの結果から、3日目の培養物は軟骨形成細胞系譜に入っているものの、まだ軟骨細胞への明らかな分化を示していないことが示唆される。また、3日目の培養物はアグリカン発現のアップレギュレーションを示していないが、軟骨に特徴的なII型コラーゲンをコードするCol2a1の転写物の発現が約4倍のアップレギュレーションを示すことにも留意されたい(図7のA)。しかしながら、Col2a1は、アグリカンと異なり、発生中の四肢の軟骨形成細胞系譜にある初期の前軟骨形成間葉系細胞だけでなく、分化した軟骨細胞にも発現する(Han Y
and Lefebvre V,2008 Molecular and Cellular Biology,28:4999−5013;Nah HD and Upholt WB,1991 J Biol Chem,266:23446−23452;Sakai K et al.,2001 Matrix Biol,19:761−767)。この遺伝子発現パターンからは、3日目の培養物が軟骨形成細胞系譜には入っているものの、まだ軟骨細胞への明らかな分化を示していないことが示唆される。
【0063】
図7のAおよびBに示すように、4日目の培養物は、転写制御因子Sox9およびBrachyuryの発現のさらなるアップレギュレーションを示す。4日目の培養物では、3日目の培養物よりSox9発現が2倍を超えて高く、Brachyuryの発現は5倍高くなっている(図7のAおよびB)。さらに、4日目の培養物は、信頼できる軟骨マーカー、アグリカンの発現の4倍を超えるアップレギュレーションを示す(図7のA)。アグリカンの発現のアップレギュレーションから、4日目の培養物が軟骨細胞への明らかな分化の初期相にあることが示唆される。
【0064】
培養の7日目には、Sox9およびBrachyuryの発現が著しくダウンレギュレートされる(図7のAおよびB)。7日目では4日目と比較して、Sox9の発現が約1
0倍(図7のA)低下し、Brachyuryの発現が約24倍低下する(図7のB)。14日目には、Sox9の発現は低下し続けて非常に低レベルになり、14日目のBrachyuryの発現は無視できる程度である(図7のAおよびB)。7日目および14日目のSox9およびBrachyuryの発現のダウンレギュレーションは、軟骨マーカー、アグリカンおよびCol2a1の発現の約8倍のアップレギュレーションと同時に起こる。上記のような、7日目および14日目のアグリカンおよびCol2a1発現のアップレギュレーションは、免疫染色により検出可能なアルシアンブルー陽性基質およびII型コラーゲンの均一な蓄積と相関する。これらの遺伝子発現および染色のパターンから、7日目および14日目の培養物は、軟骨細胞への非常に明らかな分化を均一に示していることが示唆される。
【0065】
注目されるのは、明らかな軟骨分化を示した7日目および14日目の培養物が肥大軟骨細胞成熟のマーカーを発現していないことである。7日目または14日目の培養物では、肥大軟骨細胞の信頼できる分子マーカーCol10a1の発現が検出されず、最終肥大成熟化のマーカー、オステオポンチン(OPN)の発現が無視できる程度である(図7のA)。また、7日目および14日目の培養物は、成熟を開始させる前肥大軟骨細胞のマーカーIndian hedgehog(Ihh)の発現も無視できる程度である。これらの結果から、7日目および14日目の培養物を構成する軟骨細胞は肥大軟骨細胞への成熟を示していないことが示唆される。
【0066】
不適切な肥大成熟化は変形性関節症の特徴であるため、ヒト(または他の哺乳動物)の治療に使用される細胞集団にこの表現型が存在することは、望ましくない。一部の系では、BMPにより誘導される軟骨形成培養物が、X型コラーゲン(COL)およびIndian hedgehog(IHH)など軟骨細胞肥大に関連する遺伝子の高い発現を示す。こうした肥大関連の変化は、変形性関節症などの病状に見られる。こうした肥大関連マーカーが非存在であるか、またはその発現が無視できる程度であることは、本明細書に記載の細胞が他の調製物に対して利点を有しており、理想的には病変または損傷した関節軟骨組織への治療投与に適していることが示唆される。
【0067】
図7のC〜Eに示すように、BMP2単独で処理したhESCマイクロマス培養物の軟骨系譜マーカーの発現を、TGFβ1単独処理した培養物、またはTGFβ1とBMP2との組み合わせで処理した培養物の発現と比較した。TGFβ1は、BMP2と異なり単独では軟骨マーカーの発現を刺激しない。実際、TGFβ1処理培養物および未処理の対照培養物は、アグリカン、Sox9、およびBrachyuryの発現が同程度の低レベルを示す(図7のC〜E)。
【0068】
実施例4:様々な相の軟骨形成細胞系譜における未分化多能性hESCの分化/未分化多能性hESCの前駆体細胞への直接かつ段階的な分化
変形性関節症などの軟骨の変性疾患を処置するためのhESCを実現するには、hESCの軟骨形成細胞系譜への分化を誘導する方法を開発する必要がある。hESCの軟骨形成細胞系譜への段階的かつ均一な分化を可能にする培養系および培養条件については上述した。上記のように、BMP2(またはBMP2とTGFβとの両方)の存在下で未分化多能性hESCをマイクロマス培養に供すと、前もってEB形成をせずに軟骨形成細胞系譜への段階的かつ非常に均一な分化が直接起こる。
【0069】
多能性未分化hESCが示す直接かつ段階的な軟骨細胞分化により、軟骨形成細胞系譜の様々な相にある前駆体細胞の特徴付けが可能になり、細胞ベースの組織工学治療を用いて損傷または病変したヒト軟骨を修復する能力を試験および比較することができるようになった。培養の様々な時期に得られた、BMP2処理hESCマイクロマス培養物の遺伝子発現プロファイリングにより、軟骨形成細胞系譜に入ったばかりの細胞(3日目の培養
物、すなわち、軟骨前駆体)、明らかな軟骨細胞分化の初期相にある細胞(4日目の培養物、すなわち、初期の軟骨細胞)、および軟骨細胞への明らかな分化を均一に示すが、肥大成熟化を示していない細胞(7日目および14日目の培養物、すなわち、十分に分化した軟骨細胞)を同定した。
【0070】
特に、3日目のhESCマイクロマス培養物は転写制御因子Sox9およびBrachyuryの発現のアップレギュレーションを特徴とするが、明らかに分化した軟骨細胞の信頼できる高度に特異的な分子マーカー、アグリカンの発現のアップレギュレーションを示さず、アルシアンブルー陽性基質もほとんど、またはまったく示さない。このため、3日目の培養物は、軟骨形成細胞系譜に入ってはいるものの、まだ軟骨細胞への明らかな分化を示してない。こうした軟骨形成細胞系譜の初期でのこの前駆体細胞の特徴は、軟骨欠損の修復に有用である。非常に初期相の軟骨形成細胞系譜の細胞は、系譜の後期の細胞と比較して修復への関与を促進するシグナルにより応答しやすい。
【0071】
発生の特定の段階にある細胞集団は、ある種の臨床徴候に好ましい。たとえば、2日目〜4日目の細胞(軟骨形成前駆体または初期の軟骨細胞)は、損傷した組織および軟骨に融合しやすく、より優れた長期にわたる修復を誘発する。
【0072】
軟骨前駆体(2日目〜3日目の細胞)は、四肢または手指の再生、半月板(mensical)または靭帯の修復、および骨折修復に優先的に使用される。初期の軟骨細胞(4日目の細胞)は、傷害による関節軟骨欠損の修復、慢性疾患(OAまたはRA)による関節軟骨損傷の修復または予防、外傷または先天性欠損により失われた軟骨組織の再生および置換、半月板の修復、ならびに顎関節の線維軟骨の修復に使用される。より十分に分化した軟骨細胞は、急性もしくは外傷性障害後の関節軟骨の修復、または疾患(OAまたはRA)による関節軟骨の修復もしくは予防のほか、内軟骨性骨化による骨折修復などへの適用に好適である。各段階は、軟骨発生および分化のモデリングに有用であり、iPSC由来の細胞は、患者個人を考慮した修復および再生のほか、ヒト疾患のモデリング、標的治療の設計および試験、ならびに遺伝的に傷害された軟骨組織の疾患特異的修復に特に有用である。
【0073】
4日目のBMP2処理hESCマイクロマス培養物は、Sox9およびBrachyuryの発現のさらなるアップレギュレーションを示し、信頼できる軟骨マーカー、アグリカンの発現の4倍を超えるアップレギュレーションを示すことから、4日目の培養物が軟骨細胞への明らかな分化の初期相にあることが示唆される。7日目および14日目の培養物は、Sox9およびBrachyuryの発現のダウンレギュレーションが、軟骨マーカー、アグリカンおよびCol2a1の発現の約8倍のアップレギュレーションと同時に起こることを特徴とする。また、均一なアルシアンブルー陽性基質も7日目および14日目の培養物の全体に存在し、免疫染色により検出可能なII型コラーゲンも培養物の全体に存在する。7日目および14日目の培養物は、Col10a1、Ihhもしくはオステオポンチンなどの肥大軟骨細胞成熟のマーカーを発現しない。このため、7日目および14日目の培養物は、軟骨細胞への明らかな分化を示しているが、肥大成熟化を示していない。このことは、関節軟骨を構成する軟骨細胞が肥大成熟化を示さないのが通常であり、実際に関節軟骨細胞の不適切な肥大成熟化は変形性関節症に特徴的なものであるため、関節軟骨の修復に細胞を利用することを考慮する際に重要である。7日目および/または14日目の培養物の軟骨細胞が肥大成熟化を示していないということにより、これらの細胞は軟骨欠損の修復の好ましい候補となる。
【0074】
前駆体細胞を軟骨形成細胞系譜に誘導する決定プロセスに関係する因子を同定するためのhESCのマイクロマス培養物
軟骨分化の開始を制御するSox9など一部の制御遺伝子が同定されているとはいえ、
胚性前駆体細胞の軟骨形成細胞系譜への段階的な決定またはコミットメントに関係する遺伝子および他の因子についてはほとんど知られていない。BMP2の存在下でのマイクロマス培養におけるhESCの段階的分化は、こうした初期の決定現象に関係する因子を明らかにする好適な系となる。未分化多能性hESCから軟骨細胞への系譜進行の様々な段階における細胞のトランスクリプトームを解析すれば、初期の軟骨形成細胞系譜のコミットメント現象を制御する遺伝子、シグナル伝達分子、および制御ネットワークを同定し、決定プロセスに関係する未知の遺伝子を明らかにしやすくなる。
【0075】
上述の遺伝子発現プロファイリングからは、細胞を軟骨形成細胞系譜に誘導する際の転写因子Brachyuryの役割が示される。これまでの研究から、Tボックス転写因子であるBrachyuryは、原腸形成において高度に発現し、初期の(primary)中胚葉形成で不可欠な役割を果たしていることが明らかになっている(Herrmann BG,1995 Seminars in Developmental Biology,6:385−394)。発生の後期には、Brachyuryは、肢芽形成の開始時に側部中胚葉に発現し、その後発生中の四肢に軟骨を形成する前軟骨形成間葉系細胞に発現する(Liu C et al.,2003 Development,130:1327−1337)。上記のように、Brachyuryの発現は、BMP2で処理したhESCマイクロマス培養物が、軟骨形成の転写因子Sox9の発現のアップレギュレーションによりアッセイされる軟骨形成細胞系譜に入ったばかりのときにアップレギュレートされる(24倍)。BrachyuryおよびSox9の発現のアップレギュレーションは、信頼できる軟骨マーカー、アグリカンの発現のアップレギュレーションの前に同時に起こる。次いでBMP2処理hESCマイクロマス培養物でアグリカンの発現のアップレギュレーションが開始されると、BrachyuryおよびSox9の発現が同時にさらに増加する。軟骨形成の開始後、明らかな軟骨分化が起こると、BrachyuryおよびSox9の発現は同時に無視できるレベルまで低下する。BMP2処理未分化多能性hESCの軟骨細胞への段階的分化におけるこの発現パターンから、Brachyuryが中胚葉性前駆体細胞の軟骨形成細胞系譜への初期コミットメントの調節に関係していることが示唆される。
【0076】
Brachyuryの発現は、BMP2シグナル伝達に応答して間葉系細胞株C3H10T1/2の軟骨細胞分化の開始時にアップレギュレートされ(Hoffmann A et al.,2002 J Cell Sci,115:769−781)、Brachyuryの強制発現によりSox9の発現およびC3H10T1/2細胞の軟骨細胞分化が促進される(Hoffmann A et al.,2002 J Cell Sci,115:769−781)ことが明らかになっている。さらに、ドミナントネガティブ型のBrachyuryは、C3H10T1/2細胞のBMP2刺激による軟骨細胞分化を障害する(Hoffmann A et al.,2002 J Cell Sci,115:769−781)。Brachyuryは、C3H101/2細胞などの前軟骨形成前駆体細胞にBMP2に応答して軟骨細胞分化を示す能力を与える可能性があることが示唆されている(Hoffmann A et al.,2002 J Cell Sci,115:769−781)。上述の結果は、Brachyuryの発現が、BMP2に応答してhESCが軟骨形成細胞系譜に入るとアップレギュレートされることを示唆し、以前の研究と整合している。これらの研究は、Brachyuryが、前駆体細胞を軟骨形成細胞系譜に誘導する決定プロセスに関係していることを示す。
【0077】
実施例5:ヒト胚性幹細胞の軟骨形成細胞系譜への分化
図4のCに示すように、BMP2処理hESCマイクロマス培養物全体に細胞外軟骨に特徴的なII型コラーゲンが存在することが、培養物の矢状断面をII型コラーゲン抗体で免疫染色することにより検出される。さらに、軟骨マーカー、アグリカンの球状ドメインに対するモノクローナル抗体で細胞質が染色される細胞も、BMP−2処理hESCマ
イクロマス培養物全体に存在する(図9)ことから、II型コラーゲン細胞外マトリックスで囲まれた細胞の実質的にすべてが、細胞自律性(cell autonomous)マーカーで免疫染色することによりアッセイされる軟骨マーカー、アグリカンを発現していることが示される。
【0078】
培養物における軟骨基質の蓄積は通常、基質の硫酸化プロテオグリカンを染色するアルシアンブルー、pH1.0を用いたインタクトな培養物のホールマウント染色によりモニターする。BMP2単独、またはBMP2とTGF−β1とにより処理したhESCマイクロマス培養物ではホールマウント染色により、アルシアンブルーで染色可能な基質の非常に均一な蓄積が検出可能である(図4〜図6)。軟骨分化の程度を組織学的に評価するため、hESCマイクロマス培養物の矢状断面をアルシアンブルーで染色し、核染色のNuclear Fast Redで対比染色した。アルシアンブルーにて染色可能な細胞外マトリックスで囲まれた細胞は、BMP2の単独処理(図10A)、あるいはBMP2とTGF−β1とで処理(図10Bおよび図10C)したhESCマイクロマス培養物の組織学的切片の実質的に全範囲にわたり存在する。BMP2処理hESC培養物は、広範囲に及ぶ軟骨組織に加えて少数の管状構造物を含んでいた(図10A;矢印)。培養物にTGF−β1およびBMP2を供給すると、検出可能な管の数はかなり減少し、管は培養物の非常に小さな割合を占めるだけである。したがって、BMP2およびTGF−β1の両方で処理したhESCマイクロマス培養物では、軟骨細胞分化が、完全には均質ではないが非常に均一に起こり、以前に報告された他の培養系の場合よりかなり安定して広範囲に及ぶようになる。実際、胚様体(EB)細胞から樹立された、BMP2+TGF−β処理マイクロマス培養物は、非常に不均一であり、軟骨形成組織だけでなく、大量の脂肪組織、さらには管からなる(図11)。
【0079】
実施例6:再生医療およびヒト疾患モデリングのためのiPSC由来軟骨細胞
iPSCは患者自身の細胞から採取されるため、患者特異的細胞を用いた治療および再生医療の手段となる(Amabile G and Meissner A 2009 Trends Mol Med 15:59−68)。図11、図12、および図13に示すように、iPSCは、本明細書に記載の方法により誘導され軟骨形成細胞系譜への段階的な分化を示す。
【0080】
図11(A)は、BMP2を補充せずに14日間維持されたiPSCの対照マイクロマス培養物を示し、アルシアンブルー陽性基質の蓄積をほとんど示していない。図11(B)は、14日目のBMP2処理iPSCマイクロマス培養物を示し、培養物全体に強くて広範囲のアルシアンブルー染色が存在することを示す。
【0081】
BMP2補充iPSC由来マイクロマス培養物による軟骨細胞分化の進行を確認するため、Brachyury、Sox9、Col2a1およびアグリカンなど軟骨形成細胞系譜に特徴的なマーカー遺伝子のmRNA発現のリアルタイム定量RT−PCRを使用した。これらのマーカー遺伝子のレベルは、培養期間中、BMP2を補充したhESCマイクロマス培養物の場合と同様のアップレギュレーションを示した。さらに、BMP2補充iPSCマイクロマス培養物では、肥大軟骨細胞のマーカー、オステオポンチンmRNAの発現が無視できる程度であった。したがって、本方法を用いたiPSCのマイクロマス培養物は、インビトロで軟骨形成細胞系譜への段階的分化を示し、さらに肥大を示さない。
【0082】
3種の酵素的アプローチ、トリプシン−EDTA、AccutaseまたはTrypLE Selectによって単細胞に分離したiPSCから樹立したマイクロマス培養物別に軟骨細胞分化を比較する研究を行った。図12に示すように、7日後、トリプシン−EDTA(図12のA)、TrypLE Select(図12のB)、あるいはAccutase(図12のC)のいずれかで予め分離したiPSCから樹立したBMP2補充マ
イクロマス培養物はそれぞれ、強く染色された広範囲のアルシアンブルー陽性基質の蓄積を同程度に示すことから、各分離方法は、BMP2に応答してこのプロトコルで軟骨細胞分化をうまく示すことができる単一のiPSCを作製するのに同等に有効である。BMP2を補充せずに維持した培養物には、アルシアンブルー陽性基質がほとんど存在しない(図12のD〜F)。
【0083】
図13に示すように、iPSCマイクロマス培養物は、BMP2(図13のA)、およびBMP2とTGFβ1との組み合わせ(図13のB)に応答して培養から14日後に広範囲の強いアルシアンブルー染色によりアッセイされる軟骨細胞分化を同程度に示す。これに対し、BMP2補充を受けなかったiPSCマイクロマス培養物は、アルシアンブルー陽性基質がほとんど蓄積されない(図13のC)。
【0084】
したがって、本方法は、上述のアプローチを用いて、傷害またはOAもしくはRAなどの慢性疾患による関節の関節軟骨欠損修復、および四肢再生、半月板もしくは靭帯欠損の修復、および骨折修復などの患者個人を考慮した軟骨修復および再建治療のためのiPSCの作製に有用である。
【0085】
遺伝子性障害の個体から採取されたiPSCは、疾患モデリングと、標的治療の設計および試験のスクリーニングとの手段となる(Laustriat D et al.,2010 Biochem Soc Trans 38:1051−1057;Lengner CJ 2010 Ann NY Acad Sci 1192:38−44)。iPSCを遺伝子ターゲティングと組み合わせると、単一遺伝子疾患を処置するアプローチとなる(Wong GKY and Chiu AT 2010 Biotechnol
Adv 28:715−724)。
【0086】
図16に示すようなiPSCを、軟骨異形成症のヒト患者から作製した。遺伝性軟骨障害のヒト患者由来のiPSCは、本明細書に記載の方法を用いて軟骨形成細胞系譜に分化するように誘導される。hESCおよびiPSCの軟骨細胞分化の誘導に使用した高密度マイクロマスアプローチなどの高密度培養法は、毒性試験として(Piersma AH
2004 Toxicol Lett 149:147−153;Ponce RA 2001 Curr Protoc Toxicol 13:13.3)、軟骨発生の新規な遺伝子制御因子を同定するためのゲノムスクリーニングとして(James CG et al.,2005 Mol Biol Cell 16:5316−5333)、軟骨形成に影響を与えるデベロプメンタルトキシン(developmental toxin)の検出のため(Hanse et al 2001 Free Radic Biol Med 31:1582−1592)、および有望な骨格形成異常治療法の機構評価のため(Woods A et al.,2007 Endocrinol 148:5030−5041)に使用するのに好適である。軟骨の遺伝子性障害(たとえば、軟骨異形成症または軟骨無形成症)の患者に由来し、本明細書に記載の方法により軟骨細胞分化を示すように誘導されたiPSCは、疾患プロセスのモデリングと、薬物療法の設計および試験とに有用である。軟骨異形成を示すiPSC由来の軟骨形成細胞の遺伝子プロファイリングを用いれば、疾患プロセスに関わり、治療介入のための疾患特異的な標的となるキー遺伝子および因子が同定される。疾患の標的治療剤については、インビトロでのその薬に対する、本方法により産生された病変したiPSC由来の軟骨形成細胞の応答を判定することにより、インビトロで試験される。また、遺伝的に病変した患者に正常な軟骨構造または機能を再建するアプローチとして、本方法により産生されたiPSC由来の疾患特異的な軟骨形成細胞を遺伝子治療と併用する。
【0087】
本明細書に言及した特許および科学文献は、当業者に入手可能な知識を明らかにするものである。本明細書に引用する米国特許、および公開もしくは未公開米国特許出願はすべ
て、参照によって援用する。本明細書に引用する公開された外国特許および特許出願はすべて、参照によって本明細書に援用する。本明細書に引用する他の公表された参考文献、文書、原稿および科学文献についても、参照によって本明細書に援用する。
【0088】
本発明についてその好ましい実施形態を参照しながら詳細に図示して記載してきたが、本発明添付の特許請求の範囲により包含される範囲から逸脱しない範囲で、本発明の形式および細部に様々な変更が可能であることが当業者には理解されよう。
【技術分野】
【0001】
本発明は一般的に、ヒト胚性および人工多能性幹細胞の分化を軟骨系譜に誘導する方法に関する。
【背景技術】
【0002】
米国で最も多く見られる消耗性の慢性健康障害の1つに軟骨の変性疾患、たとえば、変形性関節症がある。軟骨変性疾患の処置は、組織の自己修復能が乏しいため特に臨床上の課題になっている。ヒト胚性幹細胞(hESC)は、複数の細胞型に分化する能力および無限の自己複製能を有するので、軟骨欠損を修復する強力なツールになる可能性がある。hESCが病変および損傷軟骨を修復する可能性を実現するには、hESCの軟骨形成細胞系譜(chondrogenic lineage)への分化を誘導する方法を開発する必要がある。
【0003】
ESC由来の細胞が様々な程度で軟骨細胞に分化するいくつかの培養系が開発されているが、これらの系では軟骨細胞分化(chondrogenic differentiation)が均一ではなく、軟骨細胞は、分化する細胞の亜集団のみであり、軟骨修復のために細胞集団を利用することが難しくなっている。以下に記載する本発明の以前には、軟骨細胞分化のプロトコルは、ESC由来の胚様体(EB)の細胞を利用していた。こうした従来の方法の欠点は、EB由来の細胞の細胞の不均一性により、軟骨修復に使用し得る軟骨形成細胞の均質な集団が得にくくなることである。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、こうした従来の方法に伴う問題の多くを解決するものであり、1つには、前もってEB形成を形成せずに、幹細胞、たとえば未分化多能性ヒト胚性幹細胞(hESC)または人工多能性幹細胞(iPSCまたはiPS細胞)が軟骨形成細胞系譜へと段階的に実質的に均一に分化するのを迅速かつ直接誘導する培養系および培養条件に基づくものである。また、この方法では臍帯血または羊水から採取した幹細胞も使用する。この方法を用いて作製される、規定の分化段階の実質的に純粋な軟骨形成細胞は、従来の方法に伴う細胞不均一性(組織型および分化の段階に関する)の問題および欠点に対する解決策となる。本明細書に記載の細胞集団は、非軟骨形成細胞を実質的に含まない。たとえば、この集団は、軟骨形成細胞を少なくとも85%、95〜98%、および最大99%または100%含む。本発明はまた、ESCの軟骨形成細胞系譜への分化の進行を促進し、かつ軟骨細胞分化の均一性の達成に寄与する高密度培養条件、および追加手順も提供する。
【0005】
胚様体は、ESCまたはiPSC由来の細胞の凝集塊である。こうした凝集塊、すなわちEBは、培養物の目視検査で確認される。凝集(EBの形成)すると、分化が開始される。EBは、成人生体に存在する細胞型の全部を生じさせる胚の3胚葉すべてに由来する複数の細胞型に分化する。本発明の以前、hESC由来の軟骨形成前駆細胞の集団を産生する従来の方法の欠点は、非軟骨形成細胞の混入、すなわち、EBが所望の標的系譜(軟骨形成細胞系譜)以外の系譜に分化して産生された非軟骨形成細胞の混入であった。本明細書に記載の方法はEB形成を回避するため、得られる軟骨形成前駆細胞には、実質的に他の系譜の細胞が混入しない。この方法は、培養中のEBが非軟骨形成組織/細胞に分化するという問題を解決する。従来の方法は、混入している望ましくない細胞を除去する必要があったため、本明細書に記載のプロセスは、従来の方法に比べより効率的かつ高速であるうえ、費用効果に優れている。
【課題を解決するための手段】
【0006】
ヒト胚性幹細胞(hESC)を軟骨形成細胞に直接分化させる方法は、EBを実質的に含まないhESCの集団を支持体上に用意し、hESCを前記支持体から剥離して前記hESCを分離し、hESCの高密度マイクロマス培養を樹立し、集団を骨形成タンパク質と接触させることにより、hESCが軟骨形成細胞の実質的に均一な集団に直接分化することにより行われる。同様に、人工多能性幹細胞(iPSC)を軟骨形成細胞に直接分化させる方法は、EBを実質的に含まないiPSCの集団を支持体上に用意し、iPSCを支持体から剥離してiPSCを分離し、iPSCの高密度培養を樹立し、iPSCを骨形成タンパク質と接触させてiPSCの軟骨形成細胞系譜への分化を誘導することにより行われる。iPSCは軟骨形成細胞の実質的に均一な集団に直接分化する。
【0007】
一態様では、本明細書に記載の培養系および培養条件は、実質的に均一な軟骨分化を約1週間または2週間未満の内に、たとえば、約2日〜約3日未満、約3日〜約4日未満、約5日〜約6日未満、約6日〜約7日未満、約7日〜約8日未満、約8日〜約9日未満、約9日〜約10日未満、約10日〜約11日未満、約11日〜約12日未満、約12日〜約13日未満、または約13日〜約14日未満の内に促進する。別の態様では、本発明の培養系および培養条件は、未分化多能性ヒト胚性幹細胞の少なくとも約50〜100%が軟骨形成細胞系譜に分化するように、たとえば、未分化多能性ヒト胚性幹細胞の少なくとも約50〜60%、少なくとも約60〜70%、少なくとも約70〜80%、少なくとも約80〜90%、または少なくとも約90〜100%が軟骨形成細胞系譜に分化するように非常に均一な分化を促進する。好ましくは、この集団は、軟骨形成細胞を95%含み、一層好ましくは、集団は軟骨形成細胞を98%含む。
【0008】
分化の段階は、時間的に、あるいは、一連の遺伝子マーカーの発現により説明される。時間(日数)は、hESCまたはiPSCの高密度培養の樹立後の期間を示すものである。ゼロ時は、細胞濃度または密度を10μl当たり少なくとも1×105細胞に調節する時点である。「2日」の細胞は、高密度コロニーの樹立後2日間培養した細胞である。高密度培養は、10μl当たり少なくとも1×105の濃度で細胞を含む。たとえば、2〜3日の細胞は、軟骨系譜(すなわち、軟骨前駆体)に入ったばかりであることを特徴とし、4日目の細胞は、軟骨細胞分化の初期相(すなわち、初期の軟骨細胞)にあり、7〜14日の細胞は、軟骨細胞への明らかな分化を均一に示すものの、肥大成熟化を示していない(すなわち、細胞は、十分に分化した軟骨細胞であることを特徴とする)(図14)。
【0009】
本発明は、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に及ぶ様々な相の軟骨形成細胞系譜におけるhESC由来の前駆体細胞の特徴付けについて記載する。様々な相の軟骨形成細胞系譜におけるhESC由来の前駆体細胞について、細胞ベースの組織工学治療により軟骨を修復する能力を解析する。
【0010】
hESCの軟骨細胞分化を促進する条件は、人工多能性幹細胞(iPSC)の均一な軟骨分化も促進する。本発明はまた、患者由来のiPSCの軟骨細胞分化を誘導する方法も提供する。たとえば、患者個人を考慮した治療アプローチにiPSC由来の軟骨形成前駆細胞(自家細胞)を使用する。別の例では、本方法を用いて細胞および細胞株を作製して、疾患のメカニズムを研究するため、軟骨異形成症などの軟骨の遺伝子性障害の基本的なメカニズムを解明する研究ツールとして使用するほか、細胞を使用して治療薬のスクリーニングも行う。
【0011】
上記で論じたように、hESCの軟骨細胞分化を促進する条件は、人工多能性幹細胞(iPSC)、すなわち、再プログラム化されて多能性状態にある体細胞の均一な軟骨分化も促進する。iPSCが軟骨形成細胞系譜に信頼性高く直接かつ均一に分化すれば、患者特異的な自家細胞治療が可能になる。したがって、軟骨欠損の修復には、hESC由来の
前駆体(precursor)だけでなく、iPSC由来の軟骨形成前駆体細胞も使用される。
【0012】
本発明の培養系および培養条件は、胚様体(EB)由来の細胞を細胞ベースの軟骨修復治療に使用しにくくしている細胞の不均一性を回避する。この方法は直接かつ均一な分化を促進し、EB段階を回避するため、規定の分化段階、すなわち、初期と後期の細胞集団を確実に生じさせる。遺伝子発現プロファイリングを行えば、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に及ぶ様々な段階の軟骨形成細胞系譜におけるhESC由来の前駆体細胞を同定することができる。初期相の軟骨形成細胞系譜の細胞(たとえば、Sox9などの初期軟骨マーカーを含む)は、分化を続けるだけでなく、移植患者に移植された後にin situで局所微小環境のシグナルに応答するため、系譜の後期の細胞と比べ、軟骨修復に関与するのに望ましい。軟骨形成細胞系譜の後期の細胞(たとえば、細胞がより明らかに分化したことを示すマーカー、アグリカン(aggrecan)などの軟骨マーカーを含む)は、軟骨修復/再建手順に好適である。このため、本方法は、様々な分化の段階および異なる系譜の不均一な細胞集団を生じる従来の方法と異なり、段階的に生じた細胞の供給源を提供し、処置される病状に最も適した分化の段階に基づき治療を個々に合わせることを可能にする。
【0013】
本発明はまた、hESCおよびiPSCの軟骨形成細胞系譜への分化の進行を促進し、かつ軟骨細胞分化の均一性の達成に寄与する条件および追加手順も提供する。こうした改変には、(1)高密度培養から分離したhESCの維持(EBは存在しない)、(2)増殖因子、特にBMP−2およびTGFβ−1を含む培養物の補充、(3)マウス胚性線維芽細胞のフィーダー層上ではなく、高分子もしくはゼラチン支持体またはスキャフォールド上、あるいは高分子もしくはゼラチン支持体またはスキャフォールド内でのhESCおよびiPSCの培養、(4)マイクロマス培養物を調製する前にトリプシンではなく、Accutase、TrypLE Selectまたは他の酵素を使用してhESCを分離すること、ならびに、(5)マイクロマス培養の樹立において分離に伴うアポトーシスを減弱するRho結合キナーゼ(ROCK)阻害剤の適用が含まれる。好適な高分子もしくはゼラチン支持体またはスキャフォールド材料として、基底膜支持体(たとえばマトリゲル、Chondrogide)、コラーゲンもしくはゼラチン、ヒアルロナン(たとえばHyaffl 1またはHyStemC)もしくはキトサンを含むヒドロゲルまたはスポンジ、またはPEG(ポリエチレングリコール)もしくはPLGA(ポリラクチカシドグリオリト酸)スキャフォールド、あるいは細胞外マトリックス成分を含む任意の支持体が挙げられる。
【0014】
具体的には、本発明は、無血清培地を用いてhESCを支持体上で培養し、hESCを支持体から剥離してhESCを分離し、無血清培地を用いてhESCを高密度培養(マイクロマスまたはペレット)として培養し、骨形成タンパク質−2(BMP2;NG_023233(GI:300068920))、参照によって本明細書に援用する)をマイクロマス培養基に投与することにより、ヒト胚性幹細胞(hESC)の軟骨形成細胞系譜への分化を誘導する方法を提供する。hESCは、EBを実質的に含まない。好ましくは、hESCは、EBをまったく含まない。BMP2は、hESCの高密度マイクロマス培養から12時間後、24時間後、48時間後、72時間後、または96時間後に投与する。好ましくは、増殖因子(単数または複数)は、高密度細胞培養の樹立から48時間後に加える。あるいは、BMP−2およびトランスフォーミング増殖因子β(TGFβ;NM_000660(GI:260655621))、参照によって本明細書に援用する)を一緒に加える。BMP2の濃度は、25〜200ng/mlの範囲であり、TFGβ1の濃度は、2.5〜20ng/mlの範囲である。BMPとTGFとの比率は約5:1〜20:1であり、好ましくは約10:1である。
【0015】
hESCは、高密度マイクロマス培養としてhESCを培養する前にトリプシン、TrypLE SelectまたはAccutaseなどの分離剤と接触させる。任意に、hESC培養の支持体は、マウス胚性線維芽細胞(MEF)を含むフィーダー層である。あるいは、hESC培養の支持体は、人工基底膜材料(たとえば、マトリゲル)などのゼラチン組成物、または他の好適な支持体またはスキャフォールドである。一態様では、この方法は、トランスフォーミング増殖因子β−1(TGFβ1)をマイクロマス培養基に投与することをさらに含む。任意に、この方法は、Rho結合キナーゼ(ROCK)阻害剤の投与をさらに含む。
【0016】
一態様では、hESCは、軟骨形成細胞系譜の細胞に約14日以内に、たとえば、約10日以内、約7日以内、約4日以内または約3日以内に分化する。本明細書に記載の方法は、hESCの少なくとも約85%、たとえば、少なくとも約90%、少なくとも約95%、少なくとも約98%、少なくとも約99%または約100%が軟骨形成細胞系譜の細胞に分化するように細胞の分化を誘導する。分化は、好ましくはウシ関節軟骨細胞、たとえば、分化したウシ関節軟骨細胞の非存在下で行われる。また、このプロセスは、肝細胞の癌細胞株などの細胞または細胞株の条件培地の非存在下でも行われる。
【0017】
本発明はまた、無血清培地を用いてiPSCを支持体上で培養し、iPSCを支持体からしてiPSCを分離し、無血清培地を用いてiPSCを高密度マイクロマス培養として培養し、骨形成タンパク質−2(BMP2)をマイクロマス培養に投与することにより、人工多能性幹細胞(iPSC)の軟骨形成細胞系譜への分化を誘導する方法も提供する。iPSCは、EBを実質的に含まない。好ましくは、iPSCはEBをまったく含まない。一態様では、iPSCは、軟骨障害の患者由来である。一態様では、軟骨障害は軟骨異形成症である。
【0018】
上述の方法は、規定の分化の段階にあり、かつEB由来の細胞、たとえば、非軟骨形成細胞系譜の細胞を含まないことを特徴とする軟骨形成細胞を生じさせる。たとえば、軟骨形成細胞の集団は非軟骨形成細胞を実質的に含まず、集団は、上記のようにESCまたはiPSCから分化している。別の例では、細胞集団は、同調的に生じるか、または段階的に生じる。均一に分化した軟骨形成細胞の集団は、細胞の少なくとも85%(もしくは98%、99%、または100%)が単一の規定の分化の段階にあることを特徴とする。一部の臨床応用では、分化の段階は比較的初期であり、たとえば、集団は十分に分化した軟骨細胞を実質的に含まない。たとえば、この段階は、3日目の軟骨形成細胞または4日目の軟骨形成細胞からなる群から選択される。系譜の初期相の細胞を利用すると、移植後に関節軟骨の修復を促進する局所環境シグナルに応答しやすくなる。以上のように、軟骨を修復または再建する方法は、損傷または病変軟骨を上述の細胞集団のいずれかと接触させることにより行われる。同様に、関節炎を予防または処置する方法は、個体(ヒトまたは他の動物、たとえば、イヌ、ネコ、ウマ)の関節接合部(articulating joint)または関節腔に、記載した軟骨形成細胞の集団のいずれかを投与することにより行われる。
【0019】
本発明は、傷害または変性に起因するような軟骨障害または欠損を処置する方法を含む。処置される障害として、関節接合部の外傷性障害(たとえば、前十字靭帯(ACL)または他の靭帯を含む膝の損傷または断裂、半月板の損傷または骨折)、肘の外傷性障害、肩の外傷性障害(たとえば、回旋筋腱板損傷)、顎の外傷性障害(顎関節、すなわちTMJ疾患)、または手指の外傷性障害、および他の軟骨または骨格損傷のほか、変形性関節症(OA)など加齢に伴い発症する関節炎、または関節リウマチ(RA)など炎症を伴う関節炎、または外傷性障害による関節炎などの慢性状態、および関節接合部のいずれかが侵される慢性状態が挙げられる。この細胞は、傷害または慢性疾患による関節軟骨の損傷の予防に有用である。また、この細胞は、線維軟骨、靭帯(ligment)もしくは半
月板の障害または変性の修復にも有用である。治療方法は、本明細書の方法により産生された細胞を、軟骨障害、たとえば、軟骨傷害または変形性関節症などの関節炎の患者に投与するステップを含む。また、治療には、本方法により産生された細胞を導入して、傷害または疾患による慢性関節軟骨変性の発症を遅延させるかまたは予防することも含まれる。本方法により産生された軟骨形成細胞で軟骨を修復する方法は、移植、注射または注入により関節接合部または他の標的部位に細胞を投与することを含んでいた。たとえば、細胞は、関節鏡視下で投与する。場合によっては、関連する欠損または状態を修復する外科的または関節鏡視下手順の前、その後、またはその最中に細胞を投与または移植する。たとえば、ACL修復手順、半月板修復、回旋筋腱板修復、または他の手順と併用して関節腔に細胞を投与する。また、細胞は、事故もしくは軍事衝突が原因の外傷性障害、または先天性欠損症により失われた手指または四肢を再生する方法にも有用である。
【0020】
本方法の大きな利点は、規定の分化の段階で、たとえば、細胞が分化し始めたばかり(たとえば、2日目の細胞、3日目の細胞もしくは4日目の細胞)、または十分に分化した軟骨細胞(たとえば、7日目〜14日目の細胞)の段階で細胞の実質的に純粋な集団を単離することを可能にすることである。十分に分化した軟骨細胞は、多くの軟骨修復および再建治療に有用である。分化の初期段階の細胞は、同じ目的で使用されるか、または他の目的で使用され、その場合、コロニー形成し、さらに分化するだけでなく、in situで局所環境シグナルに応答するという別の利点が存在する。
【0021】
また、上述の方法を用いて産生された細胞は、疾患の遺伝学的基盤の判定、および処置に使用される治療薬のスクリーニング、または軟骨障害の重症度の軽減にも有用である。たとえば、軟骨障害の発生に関係する遺伝子を同定する方法は、既知の分化の段階(たとえば、十分に分化した細胞の2日目〜3日目の細胞、4日目の細胞)の実質的に均一な細胞集団を用意し、正常対照細胞と比較した遺伝子発現の増加または減少を検出することにより行う。遺伝子発現の増加または減少から、差次的に発現する遺伝子は、軟骨障害の発生に関係することが示される。たとえば、この細胞は、軟骨異形成症または軟骨無形成症と診断された患者から採取されたiPSCである。
【0022】
軟骨障害の重症度を処置または軽減する治療薬(たとえば、軟骨促進剤)を同定する例示的なスクリーニング方法は、既知の分化の段階(たとえば、十分に分化した細胞の2日目〜3日目の細胞、4日目の細胞)の実質的に均一な細胞集団を用意し、集団を候補化合物と接触させ、軟骨形成を検出することにより行う。前記化合物の存在下での軟骨形成が化合物の非存在下と比較して増加すれば、化合物が軟骨の形成を促進することが示される。軟骨形成が減少すれば、化合物が軟骨の形成を阻害することが示される。
【0023】
本発明の他の特徴と利点については、以下のその好ましい実施形態の説明、および特許請求の範囲から明らかになるであろう。他に定義しない限り、本明細書で使用する技術用語および科学用語はすべて、本発明が属する技術分野の当業者が一般に理解しているのと同じ意味を持つ。本発明の実施または試験には、本明細書に記載したのと類似または同等の方法および材料を使用してもよいが、以下に好適な方法および材料について記載する。本明細書に引用する公開された外国特許および特許出願はすべて、参照によって本明細書に援用する。本明細書に引用する、受託番号で表されるGenBankおよびNCBIの寄託物(submissions)は、参照によって本明細書に援用する。本明細書に引用する他の公表された参考文献、文書、原稿および科学文献はすべて、参照によって本明細書に援用する。矛盾がある場合、定義を含め本明細書が優先する。さらに、材料、方法、および例は、単に例示のためのものであり、限定的であることを意図するものではない。
【0024】
本発明の他の特徴と利点は、以下の詳細な説明および特許請求の範囲から明らかになる
であろう。
【図面の簡単な説明】
【0025】
【図1】A:放射線照射マウス胚性線維芽細胞のフィーダー層上で培養された未分化多能性H9ヒト胚性幹細胞(hESC)のコロニーを示す顕微鏡写真、B:形成された翌日のhESCコロニーに由来する胚様体(EB)を示す顕微鏡写真。
【図2】A:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、未処理の対照培養物を示す顕微鏡写真、B:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、骨形成タンパク質−2(BMP2)処理した培養物を示す顕微鏡写真、C:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物について、トランスフォーミング増殖因子β1(TGFβ1)処理した培養物を示す顕微鏡写真、D:EBの細胞から樹立されて21日目のアルシアンブルー染色されたマイクロマス培養物を示す顕微鏡写真について、BMP2とTGFβ1との両方で処理された培養物を示す顕微鏡写真。
【図3】A:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、B:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、C:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、D:BMP2とTGFβ1との両方で処理したEB細胞マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真。
【図4】A:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、アルシアンブルー染色された14日目の未処理の対照培養物を示す顕微鏡写真、B:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、アルシアンブルー染色された14日目のBMP2処理培養物を示す顕微鏡写真、C:未分化多能性hESCから直接樹立されたマイクロマス培養物におけるアルシアンブルー軟骨基質およびII型コラーゲンの蓄積に対するBMP2の作用について、II型コラーゲン抗体で免疫染色した14日目のBMP2処理培養物の矢状断面。
【図5】A:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、B:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、C:2日目にBMP2を含む「軟骨形成培地」を培養物に補充した、未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、D:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、E:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真、F:2日目にBMP2を含まない「軟骨形成培地」を培養物に供給し、その後3日目にBMP2を補充した未分化多能性hESCから樹立されたBMP2処理マイクロマス培養物のアルシアンブルー基質の蓄積の経時的変化を示す顕微鏡写真。
【図6】A:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、未処理の対照培養物を示す顕微鏡写真、B:図未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、BMP2処理培養物を示す顕微鏡写真、C:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、TGFβ1処理培養物を示す顕微鏡写真、D:未分化多能性hESCから樹立され、アルシアンブルー染色された14日目のマイクロマス培養物について、BMP2とTGFβ1との両方で処理された培養物を示す顕微鏡写真。
【図7】A:培養から48時間でBMP2を供給し、リアルタイム定量RT−PCRにより測定した3日目(D3)、4日目(D4)、7日目(D7)、および14日目(D14)のhESCマイクロマス培養物におけるSox9転写物の発現、アグリカン転写物の発現、Col2a1転写物の発現、Col10a1転写物の発現およびオステオポンチン(OPN)転写物の発現(A)を示す棒グラフ。発現レベルは、内部対照としてGAPDHを用いてΔΔCt法により判定した。相対量は、−2ΔΔCtとして計算し、2日目の発現レベルに対して補正して、これを値1に設定した。各値は、3つの測定値の平均値(±SEM)(4日目、7日目、および14日目)、または2つの測定値の平均(±range)(3日目)である、B:培養から48時間でBMP2を供給し、リアルタイム定量RT−PCRにより測定した3日目(D3)、4日目(D4)、7日目(D7)、および14日目(D14)のhESCマイクロマス培養物におけるBrachyury転写物の発現(B)を示す棒グラフ。発現レベルは、内部対照としてGAPDHを用いてΔΔCt法により判定した。相対量は、−2ΔΔCtとして計算し、2日目の発現レベルに対して補正して、これを値1に設定した。各値は、3つの測定値の平均値(±SEM)(4日目、7日目、および14日目)、または2つの測定値の平均(±range)(3日目)である、C:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるBrachyuryの発現レベルD:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるSox9の発現レベル、E:3日目(D3)、4日目(D4)、および7日目(D7)の未処理の対照(BT−、青)、BMP2処理(B+、赤)、BMP2とTGFβ1とにより処理(BT+、緑)、およびTGFβ1処理(T+、紫)hESCマイクロマス培養物におけるアグリカンの発現レベル。
【図8】アグリカンの球間ドメインに対する抗体で免疫染色した14日目のBMP2処理培養物の断面の顕微鏡写真。細胞内アグリカン染色を示す細胞が培養物の全体に存在する。これは、II型コラーゲン細胞外マトリックスで囲まれた細胞の実質的にすべてが(図4のA〜Cに示す)、細胞自律性(cell autonomous)マーカーを用いた免疫染色によるアッセイされる軟骨マーカー、アグリカンを発現していることを示す。
【図9】A:BMP2で処理された14日目のhESCマイクロマス培養物のアルシアンブルー染色およびNuclear Fast Red染色矢状断面を示す顕微鏡写真。アルシアンブルーで染色可能な細胞外マトリックスで囲まれた細胞は、BMP2処理培養物の断面の実質的に全範囲にわたり存在し、さらに、少数の管状構造物も存在する(矢印)、B:BMP2およびTGF−β1の両方で処理されたhESCマイクロマス培養物の断面を示す顕微鏡写真。細胞の実質的にすべてがアルシアンブルー軟骨基質で囲まれており、非軟骨形成組織は、ほとんどあるいはまったく検出できない、C:BMP2とTGF−β1とで処理されたhESC培養物の断面を示す顕微鏡写真。広範囲の軟骨組織に加えて小管(矢印)も存在する。こうした管は、BMP2とTGF−β1とで処理された培養物の非常に小さな割合を占めるだけであり、培養物の大部分は軟骨分化を示す。図9Cの下のテーブル1は、図9Aに示した14日目のBMP2処理培養物の断面中に存在する細胞の88%がアルシアンブルー陽性基質で囲まれていることを示す。図9Bおよび図9Cに示したBMP2およびTGFβ1処理培養物の断面では、細胞の97.3%がアルシアンブルー陽性基質で囲まれている。これらの結果から、本方法のBMP2処理、およびBMP2/TGFβ1処理hESC培養による軟骨細胞分化の実質的な均一性が明らかにされる。
【図10】胚様体(EB)細胞から樹立され、BMP2とTGF−β1とで処理されたマイクロマス培養物のアルシアンブルー染色およびNuclear Fast Red染色矢状断面を示す顕微鏡写真。アルシアンブルー染色された軟骨形成組織だけでなく、大量の脂肪組織(Ad)、さらには管(矢印)も存在する。
【図11】本明細書に記載の方法に供されたヒト包皮線維芽細胞に由来するiPS細胞株の軟骨基質をアルシアンブルー染色することによりアッセイされる軟骨細胞分化を示す顕微鏡写真。図11の(A)は、BMP2を補充せずに維持したiPSCの14日目の対照マイクロマス培養物を示す。アルシアンブルー陽性基質はほとんど存在しない。図11の(B)は、14日目のBMP2処理iPSCマイクロマス培養物を示す。強く広範囲のアルシアンブルー染色が培養物の全体に存在する。
【図12】A:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。トリプシン(A)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、B:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。TrypLE Select(B)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、C:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真である。Accutase(C)で予め分離し、培養から7日後にアルシアンブルーでホールマウント染色したBMP2補充iPSCのマイクロマス培養物を示す。各培養物では、アルシアンブルー蓄積が同程度に顕著である、D:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない、E:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない、F:アルシアンブルー染色によりアッセイされるマイクロマス培養物の軟骨細胞分化に対するiPSC分離法の作用を示す顕微鏡写真。BMP2の非存在下で維持されたiPSC培養物を示し、使用した分離法を問わず、アルシアンブルー陽性基質がほとんど存在しない。
【図13】A:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2の存在下で維持されたiPSCの14日目のマイクロマス培養物を示す。BMP2を補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積する、B:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2とTGFβ1との組み合わせの存在下で維持された14日目のiPSCマイクロマス培養物を示す。BMP2とTGFβ1との組み合わせを補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積する、C:BMP2およびTGFB1の組み合わせに応答したiPSCマイクロマス培養物の軟骨細胞分化を示す顕微鏡写真。BMP2またはTGFβ1の非存在下で維持された14日目のiPSCマイクロマス培養物を示す。BMP2、またはBMP2とTGFβ1との組み合わせを補充した培養物は、強く広範囲のアルシアンブルー陽性基質が同程度に蓄積するのに対し、BMP2補充を受けなかったiPSCマイクロマス培養物は、アルシアンブルー陽性基質がほとんど蓄積されない。
【図14】A:マウス関節軟骨損傷モデルを示す顕微鏡写真。対照無手術マウス膝の前額断を示す。内側半月(mm)が確認される。靭帯の外科的切除および部分的な半月板切除(meniscetomy)による膝の不安定化から8週後のマウス膝の前額断を示す、B:マウス関節軟骨損傷モデルを示す顕微鏡写真。靭帯の外科的切除および部分的な半月板切除(meniscetomy)による膝の不安定化から8週後のマウス膝の前額断を示す。内側半月が存在しないことをアスタリスクで示す。関節軟骨損傷の局在化した領域が存在する(矢印)、C:Aの拡大図、D:Bの拡大図。
【図15】A:マウス指先再生モデルを示す顕微鏡写真。骨がアリザリンレッドで染色されている対照無手術マウスの指先を示す、B:マウス指先再生モデルを示す顕微鏡写真。6週間前に切断されたマウス指先を示す。すべての構造が完全かつ自然に再生されている、C:マウス指先再生モデルを示す顕微鏡写真。軟組織および硬組織の存在を示す対照指先の断面を示す、D:マウス指先再生モデルを示す顕微鏡写真。6週間前に切断されたマウス指先を示す。すべての構造が完全かつ自然に再生されている。
【図16】A:遺伝性軟骨障害(軟骨異形成症)の患者のヒト線維芽細胞からのiPSCの誘導を示す顕微鏡写真。未分化多能性幹細胞に特徴的な外観を持つコロニーを含む生きている培養物を示す(10×)、B:遺伝性軟骨障害(軟骨異形成症)の患者のヒト線維芽細胞からのiPSCの誘導を示す顕微鏡写真。iPSCの多能性幹細胞のマーカー(SSEA−4およびNanog)に対する免疫細胞化学的染色を示す。
【図17】未分化ヒト胚性幹細胞または人工ヒト多能性幹細胞から軟骨形成細胞を作製するためのステップを示すフロー図。
【発明を実施するための形態】
【0026】
軟骨の変性疾患、たとえば、変形性関節症は、多く見られる消耗性の慢性健康障害であり、成人の生活の質を低下させる主な原因の1つである(Magne D et al.,2005 Trends in Molecular Medicine,11:519−526)。変形性関節症(OA)は、関節軟骨の分解および変性を特徴とする非炎症性の変性関節疾患である。OAは、65歳を超える大部分の人が罹患し、40歳を超える集団の90%が、関節に何らかの形で痛みおよび不動を引き起こす軟骨変性を呈すると推定される(Song L et al.,2004 Cytotherapy,6:596−601)。軟骨変性疾患は、組織の自己修復能および再生能が乏しいため、その処置は特別な課題であり、軟骨病変の処置は、主要な臨床上の問題になっている。胚盤胞の内細胞塊に由来するヒト胚性幹細胞(hESC)は、無限の自己複製能を有しつつも、複数の細胞型に分化する能力を維持しているため、細胞ベースの組織工学治療により軟骨欠損を修復する強力なツールになる。実際、下記のように、hESCは、軟骨修復のための前駆体細胞を無限に供給する。
【0027】
軟骨修復にhESCを使用するには、病変軟骨の修復に関与し得る前駆体細胞へのhESCの分化を十分に誘導する必要がある。本発明の以前に、ESC由来の細胞が様々な程度で軟骨細胞に分化するいくつかの培養系が開発されてきた(;Elisseeff J
et al.,2005 Orthod Craniofac Res,8:150−161;Harkness L et al.,2009 Stem Cell Rev
5;353−368;Heng BC et al.,2004 Stem Cells,22:1152−1167;Hoben GM et al 2009 Stem Cells Dev 18:283−292;Hwang NS et al.,2006 Stem Cells,24:284−291;Hwang NS et al.,2006 Tissue Eng,12:2695−2706;Jukes et al.,2008 Tissue Engineering Part A,14:135−147;Kawaguchi J et al.,2005 Bone,36:758−769;Koay EJ et al.,2007 Stem Cells 25:2183−2190;Koay EJ and Athanasiou KA 2009 tissue Eng Part A 15:2249−2257;Kramer J et al.,2000 Mechanisms of Development,92:193−205;Kramer J et al.,2005 Cell Biology International,29:139−146;Kramer J et a
l.,2005 Anat Embryol,210:175−185;Ofek G et al.,2009 J Biomech Eng 131:061011;Sui
YP et al.,2003 Differentiation,71:578−585;Toh WS et al.,2007 Stem Cells,25:950−960;Toh WS et al.,2009 J Cell Mol Med 13B:3570−3590;Vats A et al.,2006 Tissue Eng 12:1687−1697;zur Nieden NI et al.,2005
BMC Dev Biol,5:5−15)。しかしながら、これらの系では軟骨細胞は、分化する細胞の亜集団のみであり、軟骨修復のために細胞集団を利用することが難しくなっている。
【0028】
さらに、軟骨細胞分化プロトコルの大部分は、ESC由来の胚様体(EB)の細胞を利用する(Elisseeff J et al.,2005 Orthod Craniofac Res,8:150−161;Harkness L et al.,2009 Stem Cell Rev 5:353−368;Heng BC et al.,2004 Stem Cells,22:1152−1167;Hoben GM et al 2009 Stem Cells Dev 18:283−292;Hwang NS et al.,2006 Stem Cells,24:284−291;Hwang NS et al.,2006 Tissue Eng,12:2695−2706;Jukes et al.,2008 Tissue Engineering
Part A,14:135−147;Kawaguchi J et al.,2005 Bone,36:758−769;Koay EJ et al.,2007 Stem Cells 25:2183−2190;Koay EJ and Athanasiou KA 2009 tissue Eng Part A 15:2249−2257;Kramer J et al.,2000 Mechanisms of Development,92:193−205;Kramer J et al.,2005 Cell Biology International,29:139−146;Kramer J et al.,2005 Anat Embryol,210:175−185;Ofek G et al.,2009 J Biomech Eng
131:061011;Sui YP et al.,2003 Differentiation,71:578−585;Toh WS et al.,2007 Stem Cells,25:950−960;Toh WS et al.,2009 J Cell Mol Med 13B:3570−3590;Vats A et al.,2006 Tissue Eng 12:1687−1697;zur Nieden
NI et al.,2005 BMC Dev Biol,5:5−15;Waese and Stanford 2010 Stem Cell Res Sept 6)。EBは、未分化なESCまたはコロニーの懸濁液を非接着性支持体上でインキュベートすることにより形成される細胞の三次元凝集塊である。EBの細胞は、胚の3胚葉に由来するすべての細胞型で構成される半ば系統化された組織(semi−organized tissues)に分化することができる。EBで生じる細胞環境および相互作用は、正確に制御することが難しく、細胞の不均一性を助長している。このため、EB由来の細胞を利用すると、組織修復に使用することができる軟骨形成細胞の均質な集団を得にくくなる。
【0029】
前もってEB形成せずにhESCの軟骨細胞分化が試みられた、いくつかのプロトコルが報告されている。1つのプロトコルは、分化したウシ関節軟骨細胞とhESCを長期間(数週間)共培養するものであった(Hwang NS et al.,2008 PLoS ONE,3:1−10)。別のプロトコルは、明確に定義されていない肝細胞の癌細胞株の条件培地でESCを前培養するものであった(Hwang YS et al.,2008 Stem Cells Dev,17:971−978)が、この研究では
、軟骨細胞分化が限定的であり、非均一であった(Hwang NS et al.,2008 PLoS ONE,3:1−10)。
【0030】
別の研究では、前もってEB形成せずにマウスES細胞のマイクロマス培養物を利用して軟骨分化を獲得しようとした(Yamashita A et al.,2009,Cell Death and Differentiation 16:278−286)。Yamashitaのプロトコルは、培養開始時に増殖因子を加え、その結果かなりの細胞死が引き起され、その後培養支持体からの剥離が行われるものであった。さらに、この培養物は、斑状の非均一なアルシアンブルー染色のみを示し、RT−PCRで検出可能な信頼できる軟骨マーカー、アグリカンをほとんど発現しないか、まったく発現しない。培養物は、肥大軟骨マーカー10型コラーゲンを発現し、石灰化が起こることが報告されており、培養中の細胞が培養期間の初期に肥大軟骨細胞に成熟したことが示された。肥大軟骨細胞の成熟は変形性関節症と関連するため、これらの特徴は、この方法の大きな欠点である。また、この培養物は、血管マーカーflk1を発現することも報告されたことから、培養の不均一性が示された。形成される比較的少量の軟骨形成組織が肥大成熟化し、軟骨形成組織を軟骨修復に不適切なものにする。実際、同じ著者らによる第2の研究(Yamashita A et al.,2010 PLoS One 5:el0998)では、培養物がインビトロでさらに肥大し、最終的に骨様組織を形成することが報告された。
【0031】
これに対し、本明細書に記載の方法は、組織学的解析および分子解析によりアッセイされる実質的に均一な軟骨細胞分化を生じさせる。この方法は、アグリカンおよび他の軟骨マーカーの高レベルの発現を示す細胞を生じさせる。これらの培養物が示す段階的な分化により、軟骨修復に使用してもよい、個々の分化段階の比較的純粋な細胞の集団を得ることができる。さらに、この細胞は肥大成熟化を起こさないことから、軟骨修復に特に好ましい。
【0032】
こうした相違は、1つには増殖因子を加えるタイミングに関係している。Yamashitaは増殖因子を高密度培養の開始時(ゼロ時)に加えたのに対し、本明細書に記載の方法では、増殖因子を培養物に加えるのは高密度培養の樹立から24時間後または48時間後である。マイクロマス培養の軟骨形成前駆体を培養の開始時(0時)にBMPで処理すると、細胞死が引き起こされ、軟骨細胞分化が阻害されるため、増殖因子を補充するタイミングは、軟骨細胞分化にとって重要である(Fisher et al 2007)。他の研究者は、インビボでBMPにより軟骨形成前駆体を処理すると、大量の細胞死が引き起こされることを明らかにしている(Macias et al 1997)。また、hESCの生存を促進するには、培養物にROCK阻害剤を加える(Watanabe
K et al.,2007;Li X 2009)。好適な阻害剤として、Y27632およびH1152(どちらもCalbiochemから入手可能)、AR−12286(Aerie Pharmaceuticals)のほか、HA−1100.HCl([ヒドロキシファスジル;1−ヒドロキシ−5−イソキノリンスルホニル)ホモ−ピペラジン]、3−(4−ピリジル)−1H−インドール、H−1152.2HCl([H−1152P;(S)−(+)−2−メチル−1−[(4−メチル−5−イソキノリニル)スルホニル]ホモピペラジン、およびN−(4−ピリジル)−N’−(2,4,6−トリクロロフェニル)尿素(これらのうち最後の4つは、Alexis Biotchemicalsから入手可能)が挙げられる。
【0033】
また、間葉系幹細胞様中間体(mesenchymal stem cell−like intermediate)を利用してhESCを軟骨細胞に間接的に分化するプロトコルも報告されている。しかしながら、これらのプロトコルの一部は、EBを利用して間葉系幹細胞様中間体を産生する(Lee EJ et al.,2010 Tissu
e Eng A 16:705−717;Brown SE et al.,2008,Cells Tiss Org 189:256−260;Mahmood A et al.,2010 J Bone Min Res 25:1216−1233;Hwang et al 2008;Cohen S et al.,2010 Tissue
Eng A 16:3119−3139;Harkness L et al.,2009 Stem Cell Rev 5:353−368)ため、上記で論じたように、細胞の不均一性が生じる。同様に、ヒト生殖隆起から単離したヒト幹細胞由来の間葉系幹細胞中間体からの軟骨細胞分化を報告したプロトコルでも、EBのステップを使用している(hEG細胞、Varghese S et al.,2010 Stem Cells 28:765−774)。他のプロトコルでは、間葉系中間体を産生するため、マウス骨髄由来間質細胞株であるOP9細胞との共培養(Barberi T et al.,2005 PLoS Med 2 e161)、あるいはヒト関節軟骨細胞との共培養(Bigdeli N et al.,2009 Stem Cells 27:1812−1821)を必要とした。いくつかのプロトコルでは、紡錘形の線維芽細胞/間葉系様細胞を産生させ、その後様々な軟骨細胞分化プロトコルに付すため、間葉系幹細胞マーカーを発現しているhESCの亜集団を同定すべくFACSソーティングを使用する(Lian Q et al.,2007 Stem Cells 25:425−436;Kopher RA et al.,2010 Bone 47:718−728;Stavropoulos ME et al.,2009 Curr Prot Stem
Cell Biol 9:1F.8.1−1F.8.10;Barberi T et
al.,2005 PLoS Med 2 el61;Barberi T et al.,2007 Nature Med 13:642−651)か、あるいは、継代の繰り返し、および低密度でのhESCの大規模かつ長期的(数週間)培養を行う(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692;Arpornmeaklong P et al.,2009 Stem Cells Dev 18:955−968;Boyd NL et al.,2009 Tissue Eng A 15:1897−1908)ことにより、間葉系幹細胞様中間体の細胞集団を得た。しかしながら、これらのプロトコルで得られた軟骨細胞分化は、II型コラーゲンの斑状の免疫染色に示されるように不均一なものであり(Arpornmeaklong P et al.,2009 Stem Cells Dev 18:955−968)、組織に均一性がなく(Kopher et al 2010)、不均一にII型コラーゲンおよびSox9が分布し(Barberi T et al.,2007 Nature Med 13:642−651;Stavropoulos ME et al.,2009 Curr Prot Stem Cell Biol 9:1F.8.1−1F.8.10)、アルシアンブルー染色をほとんど示さない不均一な組織が形成され(Lian Q et al.,2007 Stem Cells 25:425−436)、培養プレートから剥離した、特徴付けが十分になされていない集団が産生された(Boyd NL et al.,2009 Tissue Eng A 15:1897−1908)。また、BMP7の存在下でhESC由来の間葉系線維芽細胞様細胞をペレット培養に供したプロトコル(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692)でも不均一な軟骨形成の分化が起こり、培養物は広範囲に及び未分化な中核を示し、軟骨細胞分化は、ペレットの周辺の細胞に限定されていた。また、BMP7とTGFβ1とで処理したペレットの軟骨細胞分化(Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692)も均一ではなく、未分化な中核があり、周辺のII型コラーゲン免疫染色が全体として弱かった。これらの培養物での、信頼できる軟骨マーカー、アグリカンの発現は、未処理対照ペレットと比較して大きく増加しなかった。
【0034】
これらの従来の方法の結果は、実質的に均一な細胞を段階的に直接生じさせる、本明細
書に記載の方法を用いたhESCによる軟骨細胞分化とは異なる。もう1つの利点として、本発明の方法は、分化の個々の段階で同調的に細胞を生じさせる。たとえば、軟骨前駆体(2〜3日の細胞、軟骨系譜に入ったばかりであることを特徴とする)、初期の軟骨細胞(4日目の細胞、軟骨細胞分化の初期相にあることを特徴とする)、または十分に分化した軟骨細胞(7〜14日の細胞、軟骨細胞への明らかな分化を均一に示すものの、肥大成熟化を示していない)を採取するため、別々の時点で細胞が回収される。
【0035】
【表1】
上述の各細胞集団は、理想的には特定の臨床状態の処置に適している。たとえば、軟骨前駆体(chondroprogentor)(2日目〜3日目の細胞)は、四肢または手指の再生、半月板(mensical)または靭帯の修復、および骨折修復に優先的に使用される。初期の軟骨細胞(4日目の細胞)は、傷害による関節軟骨欠損の修復、慢性変性疾患(OAまたはRA)による関節軟骨の修復または予防、外傷または先天性欠損により失われた軟骨組織の再生および置換、半月板または骨折の修復、ならびに顎関節の線維軟骨および軟骨の修復に使用される。十分に分化した軟骨細胞は、急性もしくは外傷性障害後の関節軟骨の修復、または慢性OAもしくはRAによる関節軟骨損傷の修復もしくは予防のほか、内軟骨性骨化による骨折修復に使用される。各段階は、軟骨発生および分化のモデリングに有用であり、iPSC由来の細胞は、患者個人を考慮した修復および再生のほか、ヒト疾患のモデリング、標的治療の設計および試験、ならびに遺伝的に傷害された軟骨組織の疾患特異的修復に特に有用である。
【0036】
iPSCの軟骨細胞分化については、既に報告されている(Medvedev SP et al.,2010 Stem Cells Dev Oct 17;Lian Q
et al.,2010 Circulation 121:1113−1123)。しかしながら、一方のプロトコルは、iPSCからのEBの形成を必要とし(Medvedev SP et al.,2010 Stem Cells Dev Oct 17)、もう一方のプロトコルは、間葉系幹細胞様中間体の細胞集団を採取するため、軟骨細胞分化を誘導する前に蛍光活性化セルソーター(FACS)によりiPSCを分取して間葉系幹細胞マーカーを発現している亜集団を同定する必要がある(Lian Q et al.,2010 Circulation 121:1113−1123)。このプロトコルを用いた軟骨細胞分化の結果、II型コラーゲンに免疫反応を示す細胞は、約60%に過ぎなかった(Lian Q et al.,2010 Circulation 121:1113−1123)。さらに本方法のもう1つの利点は、FACS(時間がかかるうえ、混入のリスクが生じ得る)の使用が回避されることである。
【0037】
本明細書に記載の方法は、EBを利用しない。本方法は、hESCまたはiPSCと、分化したウシ関節軟骨細胞、ヒト関節軟骨細胞、またはマウスOP9細胞との共培養を必要としない。本方法は、肝細胞の癌細胞由来の培地との前培養もしくは共培養も必要としない。さらに、本方法は、単層(monoloayer)での長期の継代および培養、またはFACSによる細胞分取も必要としない。本方法は、より高速である(3週間未満、
2週間未満または1週間未満、たとえば、細胞は、高密度細胞培養の樹立から2〜3日後という短期間で軟骨形成細胞系譜に入る)。本方法により、実質的に均一な軟骨細胞分化が段階的に起こり、軟骨修復および他の使用に合わせて別々の分化の段階で細胞の集団を産生することが可能になる。
【0038】
前もって胚様体(EB)を形成せずに、未分化多能性hESCの軟骨形成細胞系譜への段階的かつ均一な分化を迅速に直接促進する本発明の培養系および培養条件について、以下に記載する。未分化多能性hESC、およびhESC由来の胚様体(EB)の細胞を高密度マイクロマス培養条件に付し、胚の肢芽間葉系細胞の軟骨細胞への分化が誘導された(Gay SW and Kosher RA,1984 J Exp Zool,232:317−326;Kosher RA et al.,1986 Dev Biol,118:112−117;Kosher RA et al.,1986 J Cell Biol,102:1151−1156;Kulyk WM et al.,1991 Matrix,11:282−288)。この高密度マイクロマス培養系は、胚の四肢の発生において間葉系前駆体細胞の軟骨細胞分化の開始を特徴付ける細胞の密な並列、および細胞間相互作用を刺激する。BMP2単独で処理(またはBMP2とTGFβとを組み合わせて一緒に投与)した未分化多能性hESCのマイクロマス培養物は、前もってEB形成をせずに軟骨形成細胞系譜への段階的かつ実質的に均一な(たとえば、少なくとも88%)分化を直接示す。
【0039】
BMP2およびTGFβ1を補充する別のアプローチとして、骨形成タンパク質−4(BMP4、NG_009215(GI:219521814)、参照によって本明細書に援用する)、骨形成タンパク質−7(BMP7、NM_001719(GI:187608319)、参照によって本明細書に援用する)、増殖分化因子−5(GDF5、NG_008076(GI:193083169)、参照によって本明細書に援用する)、トランスフォーミング増殖因子β−3(TGFβ3、NG_011715(GI:225735563)、参照によって本明細書に援用する)、またはインスリン様成長因子−1(IGF−I、NG_011713(GI:225735562)、参照によって本明細書に援用する)などの他の増殖因子を培養物に補充する。これらの各因子は、軟骨形成細胞系譜への分化を促進する(Waese EY and Stanford WL 2010
Stem Cell Res Sept 6;Nakagawa T et al.,2009 Arthritis Rheum 60:3686−3692;An C et al.,2010 Ann Biomed Eng 38:1647−1654;Moore YR et al.,2010 J Clin Periodontol 37:288−298)。
【0040】
マイクロマス培養の別のアプローチとしては、hESCをペレット培養に付す。この方法では、分離したhESCまたはiPSC(約1×105〜1×107細胞)をマイクロフュージチューブでペレット状にする。次いでマイクロマス培養のため、このペレットを上記と同じ軟骨細胞分化プロトコルに付す。ペレットに増殖因子(単数または複数)を加え、ペレットに拡散させて細胞の分化を誘導する。その後治療のため、ペレットそのものを、関節もしくは関節腔または他の修復組織への移植、注入、または他の投与手段に使用する。このプロトコルによっても、EBを形成せずに、hESCの段階的かつ実質的に均一な軟骨細胞分化が直接達成される。また、このペレット培養プロトコルは、iPSCの軟骨形成細胞系譜への分化の誘導にも使用される。
【0041】
細胞培養系:系Iおよび系II
細胞は、2つの系の1つを用いて調製される。系Iでは、高密度培養を樹立し、24時間培養し、この時点で細胞にBMP2またはBMP2とTGFβとの組み合わせを含む軟骨形成培地を加える。系IIでは、高密度培養を樹立し、24時間培養し、この時点で増
殖因子(単数または複数)を用いずに細胞に軟骨形成培地を加える。3日目の初めに、細胞にBMP2またはBMP2とTGFβとの組み合わせを加える。1日後(3日目の終わりに)、遺伝子発現プロファイルおよび物理的特性/組織学的特徴から明らかなように、細胞は、実質的に均一に軟骨形成細胞系譜に入る。培養の時期、増殖因子の添加、および分化の段階を図示するフローチャートを図17に示す。
【0042】
下記のように、分化の進行過程の様々な時期に回収されたhESC由来の細胞の遺伝子発現プロファイルを用いることで、軟骨形成細胞系譜に入ったばかりの細胞から、明らかに分化した軟骨細胞に至る様々な相の軟骨形成細胞系譜の細胞を同定することができる。本明細書に記載の方法を用いれば、様々な相の軟骨形成細胞系譜のhESC由来の前駆体細胞が、細胞ベースの組織工学治療により軟骨を修復する能力を解析することができる。また、均一かつ段階的な軟骨細胞分化が起これば、軟骨形成細胞系譜に分化することが決定した前駆体細胞への未分化多能性hESCの段階的な変化に関係する遺伝子、シグナル、および制御ネットワークを同定しやすくなる。
【0043】
治療方法
細胞を投与するアプローチとして、自家軟骨細胞移植の場合(ACI,Zazlav K et al.,2009 Am J Sports Med,37:42−55)、またはスキャフォールドフリーアプローチにより関節鏡視下の場合(Jubel A et al.,2008 Am J Sports Med 36:1555−1564;Erggelet C et al 2003 Arthroscopy 19:108−110)には、膝を開く手術などの外科的手段により、マイクロマス培養物またはペレット培養物を損傷部位に直接移植する。一部の手順では、培養物をコラゲナーゼ消化または他の手段によりさらに分離し、細胞をスキャフォールドまたは他の支持体に播種する。こうした支持体として、コラーゲン膜(たとえばChondrogide)、ヒアルロナン含有マトリックス(たとえばHyaff−11)、ポリグラクチンフリース、または他のマトリックスがある。次いで細胞を播種したマトリックスを、マトリックス誘導自家軟骨細胞修復の場合(MACI:matrix−induced autologous chondrocyte repair)(Brittberg,2010 Am J Sports Med,38:1259−1271)、またはHyalgraftCの場合(Gobbi A et al.,2006:Am J Sports Med 34:1763−1773)には、外科的に移植するか(Kon et al.,2009 Am J Sports Med 37:156S−166S)、あるいは、関節鏡視下で移植する(Gianni E et al.,2008 Am J Sports Med 36:873−880;Nixon AJ 2002 Clinical Tech Equine Practice 1:257−269)。また、分離した細胞を、コラーゲン、キトサン、アガロースまたはヒアルロナンを含むゲル中に封入し、その細胞−ゲル混合物を用いて、外科的手順で損傷領域に充填する(Funayama A et
al.,2008 J Orthop Surg 13:225−232;Hoemann C et al.,2005 OsteoArthritis Cart 13:318−329;Emans PJ et al.,2010 PNAS 107:3418−3423)。これらの方法では、使用する細胞の濃度は、(1×106〜2×107細胞/ml)である。
【0044】
あるいは、非侵襲的に関節内注射により、分離した細胞を膝に直接注射または注入する。導入される細胞数は、(1×106〜2×107細胞/ml)である。自家間葉系幹細胞(MSC)を傷害された膝または関節炎の膝に導入する場合(Centeno CJ et al.,2008 Pain Physician 11:343−353)、またはMSCを軟骨が損傷したラット、ブタおよびヤギの関節に注射する場合(Horie
M et al.,2009 Stem Cells 27:878−887;Lee
KB et al.,2007 Stem Cells 25:2964−2971;Murphy JM et al.,2003 Arthritis Rheum 48:3464−3474)には、細胞は食塩水を用いて注射する。別のアプローチでは、細胞は、臨床的関節炎痛および不動(immmobility)を軽減する関節内補充療法に使用されるヒアルロナン調製物(たとえばSynvisc、Supartz)などの他の物質と併用して(Petrella RJ and Petrella M 2006,J Rheumatol 33:951−956;Brander VA and Stadler TS 2009 Phys Sports Med 37:38−48)、あるいは、骨関節炎の進行を遅らせるBMP7などの増殖因子と併用して(Hunter DJ et al.,2010,BMC Musculoskelet Disord 11:232;Hayashi M et al.,2010 J Orthop Res 28:1502−1506)注射する。
【0045】
細胞送達の1つのアプローチは、関節鏡視下での投与である。場合によって、関連する欠損または状態を修復する外科的手順または関節鏡視下手順の前、その後、またはその最中に細胞を投与または移植する。たとえば、ACL修復手順、半月板修復、回旋筋腱板修復、または他の手順と併用して関節腔に細胞を投与する。このアプローチは、動物における間葉系幹細胞の半月板損傷の修復を評価するために使用されており(Horie M et al.,2009 Stem Cells 27:878−887)、ヒトに好適である(Centano CJ et al.,2008 Med Hypotheses 71:900−908.)。
【0046】
本細胞および方法は、スポーツ関連、戦闘関連、および他の骨、軟骨および四肢の外傷性障害の修復を支援するのに有用である(Goldstein 2006 J Am Acad Orthop Surg 14:S152−156;Sundelacruz S and Kaplan DL 2009 Sem Cell Dev Biol 20:646−655)。本細胞は、長骨骨折の修復に有用であり、内軟骨性骨化、すなわち軟骨を鋳型に骨化形成が起こり正常な一連の発生現象(normal developmental sequence)を再現させるプロセスにより修復を行う(Kronenberg HM 2003 Nature 423:332−336)。間葉系幹細胞を利用する方法に記載したように、細胞は、注射または移植により骨折の領域に導入されると、リモデリングが起こり修復のための骨を形成する(Kallai I et al.,2010 J Biomech 43:2315−2320;Scotti C et al.,2009 PNAS 107:7251−7256)。
【0047】
また、細胞は、事故もしくは軍事衝突が原因の外傷性障害または先天性欠損症により失われた手指または四肢を再生する方法にも有用である。イモリおよびカエルなどの下等動物は、四肢を再生する能力を有するものの、成体哺乳動物では、再生反応を行うのに必要な細胞およびシグナルが不十分であるため(Gurtner GC et al.,2007 Ann Rev Med 58:299−312;Muneoka K et al.,2008 Sci Am 298:56−63)、この能力の大部分が失われている(Muller TL et al.,1999 Sem Cell Dev Biol 10:405−413)。
【0048】
マウスの四肢再生モデルが樹立された。このモデルでは、切断された指尖部は自然な再生反応を示すのに対し、切断された指基部は再生することができない(Han M et
al.,2008 Dev Biol 315:125−135)。このモデルは、hESCおよびiPSC由来の軟骨前駆体細胞、特に骨格および軟骨形成細胞系譜(たとえば、2日目、3日目または4日目の細胞)に入ったばかりの細胞が哺乳動物の四肢再生に関与または促進する能力を試験する系となる。四肢(腕もしくは脚、手もしくは足)また
は指(手指もしくは足指)の再生治療のための送達方法は、本方法により産生された細胞を指または四肢に注射または移植により導入することを含む。細胞は、増殖因子を使用してあるいは使用せずにスキャフォールドまたはゲルを用いて導入する。この指再生モデルを図15のA〜Dに示す。
【0049】
疾患モデル
ヒト疾患のインビトロでのモデリングに際して、さらに標的治療法の設計および試験のための系として、遺伝性疾患の患者から採取したiPSCが使用される(Amabile
G and Meissner A 2009 Trends Molec Med 15:59−68;Laustriat D et al.,2010 Biochem
Soc Trans 38:1051−1057)。本発明も、遺伝性軟骨疾患のインビトロでのモデリング、および治療法の設計および試験に有用である。軟骨異形成症および軟骨無形成症(achondoplasia)を含むこれらの障害は、外観が変形して障害を生み出す低身長、さらには死を引き起こす(Krakow D and Rimoin D 2010 Genet Med 12:327−341)。このため、軟骨に影響を与える遺伝子性障害(たとえば、軟骨異形成症、軟骨無形成症または他の症状)のある個体から採取した線維芽細胞または他の細胞型からiPSCを得て、本明細書に記載の方法を適用して軟骨細胞分化を誘導する。このiPSC由来の軟骨形成細胞は、インビトロで疾患プロセスを再現し、疾患の病理学的変化の機構研究、ならびに標的治療法(therapueutics)の設計および試験を可能にする。このアプローチは、たとえば細胞外マトリックスタンパク質の合成または硫酸化;代謝酵素、イオンチャネルもしくはトランスポーター;分子のフォールディング、プロセシングもしくは分解;ホルモン、増殖因子、受容体もしくはシグナル伝達物質;転写因子;RNAプロセシング;または細胞骨格タンパク質の欠損などにより引き起こされるような、メカニズムの大部分が不明である軟骨異形成症に適用される(Krakow D and Rimoin D 2010 Genet Med 12:327−341)。このアプローチは、正常もしくは異常な軟骨機能に重要な新規な遺伝子または治療の新規な標的を同定するための遺伝子プロファイリングにも適用される(Pogue R et al.,2004 Matrix
Biol 23:299−307)。このアプローチを遺伝子ターゲティングと組み合わせて単一遺伝子(mongenic)軟骨異形成症の患者特異的細胞による治療を試みてもよい(Wong GK and Chiu AT 2010 Biotechnol
Adv July 24)。
【0050】
軟骨修復の評価には、大型および小型動物の実験モデルを使用する(Chu CR et al.,2010 Tissue Eng Part B 16:105−115)。あるモデルでは、外科的な膝靭帯の切断および/または部分的な半月板切除(meniscetomy)により膝関節を不安定にして、荷重を不適切に集中させ、限局性の関節軟骨の破壊を引き越す。このモデルで起こる軟骨欠損は非常に再現性がよく、その軟骨破壊の発生および進行において、慢性ヒト変形性関節症で見られる状態が忠実に再現される。このモデルは、様々な動物の関節に間葉系幹細胞を注射することにより、損傷した関節軟骨の細胞による修復を評価するのに使用されている(たとえばMurphy JM et al.,2003 Arthritis Rheum 48:3464−3474;Kubo S et al.,2009 Arthritis Rheum 60:155−165)。このモデルは、齧歯動物にも応用されている(Kamekura S et al.,2005 Osteoarthritis Cart 13:632−641;Glasson SS et al.,2007 Osteoarthritis Cart 15:1061−1069;Welch ID et al.,2009 Arthritis Res Ther 11:R14)。このモデルは現在、免疫能が低下したマウスで樹立され、hESCおよびiPSC由来の軟骨形成細胞による修復のインビボ試験に使用されている(図14のA〜D)。
【0051】
軟骨修復の評価に使用される別のモデルは、外科的に作製した関節軟骨欠損を利用する(Ahern BJ et al.,2009 Osteoarthritis Cart 17:705−713)。このモデルでは、大腿骨膝蓋溝の非荷重部を露出させ、大腿骨を切除するか、または軟骨表面から軟骨下骨に及ぶ穴を開けて全層欠損を機械的に作製する。このアプローチは大型動物に使用され、ラット(Dausse Y et al.,2003 Osteoarthrtis Cart 11:16−28)、およびマウス(Eltawil NM et al.,2009 Osteoarthritis
Cart 17:695−704;Osteoarthritis Cart 6:695−704)にも適合されている。このモデルは、急性傷害後の関節軟骨損傷を刺激し、ヒトの場合と同様に、欠損を治癒できないと、損傷部位の外側で軟骨変性がさらに起こり、明らかなOAに至る。
【0052】
実施例1:hESCの培養および胚様体形成
WiCell Research Instituteで作製されたH9 hESC株(Thomson JA et al.,1998 Science,282:1145−1147)。University of Connecticut Stem Cell Coreにより標準的なプロトコルを用いて調製された放射線照射CF1マウス胚性線維芽細胞(MEF)をフィーダー層とし、以前に記載された無血清hESC培地(Toh WS et al.,2007 Stem Cells,25:950−960)でH9細胞を培養した(図1のAを参照)。1mg/mlのコラゲナーゼIV(Invitrogen)で処理してから4〜6日ごとにコロニーを継代した。本研究では、継代数36〜45のhESCを使用した。
【0053】
EBを作製するため、0.1%ディスパーゼ(Invitrogen)で37℃にて20〜30分間処理してhESCのコロニーをMEFフィーダー層から剥離し、T75 Ultra−Low Attachment Flask(Corning,Lowell,MA)中のEB形成培地に懸濁し(Toh WS et al.,2007 Stem
Cells,25:950−960)、5日間インキュベートした。培地は、48時間ごとに交換した。図1のBに示すように、懸濁されたhESCコロニーは、細胞の三次元凝集塊からなるEBを形成する。
【0054】
EB細胞および未分化hESC由来のマイクロマス培養物の調製
5日目のEB(図1のB)および未分化hESCコロニー(図1のA)から分離した両方の細胞からマイクロマス培養物を調製した。分離の前に、hESCコロニーを上記のようにMEFフィーダー層から剥離した。剥離したhESCコロニーおよびEBを0.05%トリプシン/EDTA(Invitrogen)で処理することにより単細胞に分離し、続いて細胞懸濁液を40μmのセルストレーナー(BD Biosciences,Franklin lakes,NJ)に通した。分離した細胞を、10%FBS(FBS,Hyclone)および10%>KSRを補充した高グルコースDMEMに2×107細胞/mlで懸濁した。マイクロマス培養物は、10μlの細胞懸濁液(2×105細胞)を24ウェル組織培養ディッシュ(Nunc;Fisher)の各ウェルにスポットして調製した。細胞接着を促進するため加湿5%CO2インキュベーター中で37℃にて2時間インキュベートした後、0.5mlの同じ培地を各ウェルに加えた。24時間後(培養の2日目の開始)、培地を除去し、培養物に無血清軟骨形成培地を供給した(Toh WS et al.,2007 Stem Cells,25:950−960)。軟骨形成培地は、ITS(6.25μg/mlのインスリン、6.25μg/mlのトランスフェリン、6.25μg/mlのセレン)、1.25mg/mlのウシ血清アルブミン、5.35μg/mlのリノール酸(BD Biosciences)、1%KSR、40μg/mlのL−プロリン(Sigma−Aldrich)、50μg/mlのアスコル
ビン酸2−ホスフェート(Sigma−Aldrich)、1%非必須アミノ酸(Invitrogen)、10−7M デキサメタゾン(Sigma−Aldrich)、および100単位/100μgのペニシリン/ストレプトマイシン(Invitrogen)を補充した高グルコースDMEMを含む。次いで細胞を、100ng/mlの組換えヒトBMP2(R&D Systems,Minneapolis)、10ng/mlの組換えヒトTGF−β1(R&D Systems,Minneapolis)、あるいは、100ng/mlの組換えヒトBMP2と10ng/mlの組換えヒトTGF−β1との組み合わせのいずれかの存在下または非存在下で培養する。一部の実験では、hESCのマイクロマス培養物を、増殖因子を含まない軟骨形成培地で24時間(2日目を通して)培養し、その後(培養の3日目の始めに)、培養物に、上記のような増殖因子を補充した新鮮な軟骨形成培地を供給した。増殖因子を含む培地は、培養期間を通して48時間ごとに交換した。
【0055】
アルシアンブルー染色、II型コラーゲンおよびアグリカンの免疫染色
マイクロマス培養物をアルシアンブルー、pH1.0で以前に記載されたように染色して、軟骨基質の蓄積を組織化学的にモニターした(Gay SW and Kosher
RA,1984 J Exp Zool,232:317−326)。II型コラーゲンの免疫染色では、4%パラホルムアルデヒドを含むリン酸塩緩衝生理食塩水(PBS)で30分間マイクロマス培養物を固定し、PBSで洗浄し、組織培養プレートから掻き取り、脱水し、パラフィンに包埋し、7μmの矢状断切片を作製した。免疫組織化学的検査については、Vectastain Elite ABCキット(Vector,Burlingame,CA)と、II型コラーゲンに対するモノクローナル抗体(抗II型コラーゲン、クローン6B3;Chemicon)、および軟骨マーカー、アグリカンの球間ドメインに対するモノクローナル抗体(6−B−4;Abeam)とを用いて行った。切片を脱パラフィンし、0.5%H2O2メタノール溶液で15分間クエンチし、ペプシン(Labvision)とインキュベートすることにより抗原賦活化を行った。切片を、5%正常ウマ血清を含むTBSでブロッキングしてから、1:200希釈II型コラーゲン抗体と1時間インキュベートした。次いでこの切片を、1:200希釈ビオチン化ウマ抗マウスIgGと室温にて1時間インキュベートし、アビジン−ビオチン西洋わさびペルオキシダーゼ複合体(Vectastain ABCキット)で処理し、ジアミノベンジジン/H2O2(Vector)で発色させた。陰性対照は、一次抗体とインキュベートしないものであり、ほとんど、あるいはまったく染色を示さなかった。
【0056】
リアルタイム逆転写ポリメラーゼ連鎖反応(リアルタイムRT−PCR)
Qiagen RNeasy Miniキット(Qiagen,Chatsworth,CA)を用いてマイクロマス培養物から全RNAを抽出し、RNase−free DNase(Ambion,Austin,TX)で処理して混入する恐れがあるゲノムDNAを除去した。High Capacity cDNA Reverse Transcription Kit(Applied Biosystems,Foster City,CA)を用いて、20μlの反応容量に対して2μgのRNAをcDNAに逆転写した。リアルタイムRT−PCRは、Sox9(アッセイID Hs00165814_ml)、アグリカン(アッセイID Hs00153936_ml)、II型コラーゲン(Col2a1)(アッセイID Hs00264051_ml)、X型コラーゲン(Col10a1)(アッセイID Hs00166657_ml)、オステオポンチン(opn)(アッセイID Hs00960942_ml)、Indian hedgehog(Ihh)(アッセイID Hs01081801_ml)、Brachyury(アッセイID Hs00610080_ml)、およびグリエルアルデヒド−3−リン酸デヒドロゲナーゼ)(GAPDH)(Hs99999905_ml)について、ABI Prism 7900 Sequence Detection System,TaqMan Gene Expression Master Mix、およびTaqMan
Gene Expression Assays (Applied Biosystems)を用いて行った。遺伝子発現レベルは、内部標準遺伝子としてGAPDHを用いてΔΔCt法により判定した。各遺伝子の相対量は、解析対象の各処理および時点ごとに2〜3つの独立したマイクロマス培養物それぞれから調製したサンプル2つずつの−2ΔΔCtとして計算した。サーマルサイクリング条件は、50℃で2分、95℃で10分、その後94℃で15秒の変性を40サイクル、60℃で1分の伸長反応であった。
【0057】
実施例2:未分化多能性hESCの直接かつ段階的な軟骨細胞分化
EBの特徴を示す細胞環境および固有の細胞多様性が明確に定義されていないため、未分化多能性H9 hESC(図1のA)が高密度培養に供されたときに軟骨形成細胞系譜に直接分化する能力を調べた。高密度培養物は、濃度が培地の10μl当たり1×105細胞を超える。たとえば、細胞の濃度は、10μl当たり1〜4×105細胞、たとえば、10μl当たり2×105細胞である。より大規模な培養が望ましい場合、体積を増やすが、細胞密度/比率は、上述の範囲内にとどめるものとする。
【0058】
分離したH9細胞のマイクロマス培養の樹立の翌日、100ng/mlのBMP2の存在下および非存在下で無血清「軟骨形成培地」を補充した。図4のAに示すように、分離した未分化なH9 hESC細胞から直接樹立した14日目の未処理のマイクロマス培養では、ごくわずかな小さなアルシアンブルー染色領域のみが存在した。外来性のBMP2の存在下では、14日目のBMP2処理hESCマイクロマス培養物中に実質的に均一で強く染色されたアルシアンブルー陽性基質が蓄積する(図4のB)。均一なアルシアンブルー染色だけでなく、コラーゲン抗体を用いた免疫染色により検出可能で軟骨に特徴的なII型コラーゲンII型が、BMP2処理培養物の全体に存在した(図4のC)。
【0059】
上記のように培養した未分化なH9細胞マイクロマス培養物でアルシアンブルー基質の蓄積が進行するのを図5のA〜Cに示す。培養から3日後では、検出可能なアルシアンブルー染色はほとんど認められなかったが(図5のA)、7日目には、比較的均一なアルシアンブルー染色が培養物全体で検出可能であった(図5のB)。14日目では、より強く染色されたアルシアンブルー陽性基質が培養物全体に存在した(図5のC)。これらの結果から、未分化多能性hESCは、適切な細胞環境(高密度マイクロマス培養)、およびBMP2などの外来性のシグナル伝達分子が供給されると、EB段階を経ずに直接かつ極めて均一な軟骨形成を示すことが示唆される。
【0060】
上述の軟骨細胞分化プロトコルを改変すると、hESCの軟骨形成細胞系譜への分化の進行がさらに促進される。この改変プロトコルでは、hESCマイクロマス培養の樹立の翌日、BMP2を含まない無血清「軟骨形成培地」を補充し、次いで24時間後に100ng/mlのBMP2を補充する。図5のD〜Fに示すように、3日目の培養物では検出可能なアルシアンブルー陽性基質がほとんど認められないが、7日目には、培養物の全体に均一でかなり強いアルシアンブルー基質が存在した。14日目には、培養物の全体に非常に強く染色されたアルシアンブルー基質が存在した(図5のF)。外来性のBMP2を3日目に供給したこの改変プロトコルを利用すると(図5のD〜F)、BMP2を2日目にhESC培養物に補充したプロトコルと比較して(図5のA〜C)、アルシアンブルー染色の範囲および強さが大きくなった。
【0061】
実施例3:hESCマイクロマス培養物における様々な相の軟骨形成細胞系譜の細胞に関する遺伝子発現プロファイリングによる特徴付け
多能性hESCの軟骨形成細胞系譜への分化の進行を確認および定量し、さらに様々な相の系譜の前駆体細胞の特徴付けを行うため、軟骨形成細胞系譜の様々な段階に特徴的なマーカー遺伝子の発現をリアルタイム定量RT−PCRにより調べた。上記の図5のD〜Fに示すようなアルシアンブルー基質染色によりアッセイされる段階的な軟骨細胞分化を
誘導するように改変した軟骨細胞分化プロトコルを行った多能性hESCのBMP2処理マイクロマス培養物における遺伝子発現パターンについて、何回も解析した。
【0062】
図7のAに示すように、外来性のBMP2の添加から24時間後の培養の3日目の終了時に、軟骨形成の転写因子Sox9の発現が7倍と著しくアップレギュレートされる。Sox9は、軟骨分化の「マスター制御遺伝子」として特徴付けられており、軟骨形成の実質的にすべての初期相の決定的なレギュレーターである(Akiyama H et al.,2002 Genes and Development 16:2813−2828;Lefebvre V and Smits P,2005 Birth Defects Res C Embryo Today,75:200−212)。したがって、3日目の培養物におけるSox9発現の著しいアップレギュレーションは、細胞が軟骨形成細胞系譜に入ったことと整合する。また、3日目の培養物で転写因子Brachyuryの発現が24倍のアップレギュレーションを示すことも注目に値する(図7のB)。Brachyuryは、中胚葉系譜のマーカーであり、発生中の四肢に軟骨を形成する前軟骨形成中胚葉細胞にも発現する(Herrmann BG,1995 Seminars in Developmental Biology,6:385−394;Hoffmann A et al.,2002 J Cell Sci,115:769−781;Liu C et al.,2003 Development,130:1327−1337)。したがって、Sox9と共にBrachyuryの発現がアップレギュレートされたことは、3日目の培養が軟骨形成細胞系譜に入ったことをさらに示唆する。3日目の培養物はSox9およびBrachyuryの発現のアップレギュレーションを示すものの、この培養物は、軟骨マーカー、アグリカンの発現のアップレギュレーションを示さない(図7のA)。3日目の培養物では、アグリカンは2日目と同じ低い基準レベルで発現する。アグリカンは、軟骨基質の主要な硫酸化プロテオグリカンであり、明らかに分化した軟骨細胞の信頼できる高度に特異的な分子マーカーである(Han Y and Lefebvre V,2008 Molecular and Cellular Biology,28:4999−5013)。これらの結果から、3日目の培養物は軟骨形成細胞系譜に入っているものの、まだ軟骨細胞への明らかな分化を示していないことが示唆される。また、3日目の培養物はアグリカン発現のアップレギュレーションを示していないが、軟骨に特徴的なII型コラーゲンをコードするCol2a1の転写物の発現が約4倍のアップレギュレーションを示すことにも留意されたい(図7のA)。しかしながら、Col2a1は、アグリカンと異なり、発生中の四肢の軟骨形成細胞系譜にある初期の前軟骨形成間葉系細胞だけでなく、分化した軟骨細胞にも発現する(Han Y
and Lefebvre V,2008 Molecular and Cellular Biology,28:4999−5013;Nah HD and Upholt WB,1991 J Biol Chem,266:23446−23452;Sakai K et al.,2001 Matrix Biol,19:761−767)。この遺伝子発現パターンからは、3日目の培養物が軟骨形成細胞系譜には入っているものの、まだ軟骨細胞への明らかな分化を示していないことが示唆される。
【0063】
図7のAおよびBに示すように、4日目の培養物は、転写制御因子Sox9およびBrachyuryの発現のさらなるアップレギュレーションを示す。4日目の培養物では、3日目の培養物よりSox9発現が2倍を超えて高く、Brachyuryの発現は5倍高くなっている(図7のAおよびB)。さらに、4日目の培養物は、信頼できる軟骨マーカー、アグリカンの発現の4倍を超えるアップレギュレーションを示す(図7のA)。アグリカンの発現のアップレギュレーションから、4日目の培養物が軟骨細胞への明らかな分化の初期相にあることが示唆される。
【0064】
培養の7日目には、Sox9およびBrachyuryの発現が著しくダウンレギュレートされる(図7のAおよびB)。7日目では4日目と比較して、Sox9の発現が約1
0倍(図7のA)低下し、Brachyuryの発現が約24倍低下する(図7のB)。14日目には、Sox9の発現は低下し続けて非常に低レベルになり、14日目のBrachyuryの発現は無視できる程度である(図7のAおよびB)。7日目および14日目のSox9およびBrachyuryの発現のダウンレギュレーションは、軟骨マーカー、アグリカンおよびCol2a1の発現の約8倍のアップレギュレーションと同時に起こる。上記のような、7日目および14日目のアグリカンおよびCol2a1発現のアップレギュレーションは、免疫染色により検出可能なアルシアンブルー陽性基質およびII型コラーゲンの均一な蓄積と相関する。これらの遺伝子発現および染色のパターンから、7日目および14日目の培養物は、軟骨細胞への非常に明らかな分化を均一に示していることが示唆される。
【0065】
注目されるのは、明らかな軟骨分化を示した7日目および14日目の培養物が肥大軟骨細胞成熟のマーカーを発現していないことである。7日目または14日目の培養物では、肥大軟骨細胞の信頼できる分子マーカーCol10a1の発現が検出されず、最終肥大成熟化のマーカー、オステオポンチン(OPN)の発現が無視できる程度である(図7のA)。また、7日目および14日目の培養物は、成熟を開始させる前肥大軟骨細胞のマーカーIndian hedgehog(Ihh)の発現も無視できる程度である。これらの結果から、7日目および14日目の培養物を構成する軟骨細胞は肥大軟骨細胞への成熟を示していないことが示唆される。
【0066】
不適切な肥大成熟化は変形性関節症の特徴であるため、ヒト(または他の哺乳動物)の治療に使用される細胞集団にこの表現型が存在することは、望ましくない。一部の系では、BMPにより誘導される軟骨形成培養物が、X型コラーゲン(COL)およびIndian hedgehog(IHH)など軟骨細胞肥大に関連する遺伝子の高い発現を示す。こうした肥大関連の変化は、変形性関節症などの病状に見られる。こうした肥大関連マーカーが非存在であるか、またはその発現が無視できる程度であることは、本明細書に記載の細胞が他の調製物に対して利点を有しており、理想的には病変または損傷した関節軟骨組織への治療投与に適していることが示唆される。
【0067】
図7のC〜Eに示すように、BMP2単独で処理したhESCマイクロマス培養物の軟骨系譜マーカーの発現を、TGFβ1単独処理した培養物、またはTGFβ1とBMP2との組み合わせで処理した培養物の発現と比較した。TGFβ1は、BMP2と異なり単独では軟骨マーカーの発現を刺激しない。実際、TGFβ1処理培養物および未処理の対照培養物は、アグリカン、Sox9、およびBrachyuryの発現が同程度の低レベルを示す(図7のC〜E)。
【0068】
実施例4:様々な相の軟骨形成細胞系譜における未分化多能性hESCの分化/未分化多能性hESCの前駆体細胞への直接かつ段階的な分化
変形性関節症などの軟骨の変性疾患を処置するためのhESCを実現するには、hESCの軟骨形成細胞系譜への分化を誘導する方法を開発する必要がある。hESCの軟骨形成細胞系譜への段階的かつ均一な分化を可能にする培養系および培養条件については上述した。上記のように、BMP2(またはBMP2とTGFβとの両方)の存在下で未分化多能性hESCをマイクロマス培養に供すと、前もってEB形成をせずに軟骨形成細胞系譜への段階的かつ非常に均一な分化が直接起こる。
【0069】
多能性未分化hESCが示す直接かつ段階的な軟骨細胞分化により、軟骨形成細胞系譜の様々な相にある前駆体細胞の特徴付けが可能になり、細胞ベースの組織工学治療を用いて損傷または病変したヒト軟骨を修復する能力を試験および比較することができるようになった。培養の様々な時期に得られた、BMP2処理hESCマイクロマス培養物の遺伝子発現プロファイリングにより、軟骨形成細胞系譜に入ったばかりの細胞(3日目の培養
物、すなわち、軟骨前駆体)、明らかな軟骨細胞分化の初期相にある細胞(4日目の培養物、すなわち、初期の軟骨細胞)、および軟骨細胞への明らかな分化を均一に示すが、肥大成熟化を示していない細胞(7日目および14日目の培養物、すなわち、十分に分化した軟骨細胞)を同定した。
【0070】
特に、3日目のhESCマイクロマス培養物は転写制御因子Sox9およびBrachyuryの発現のアップレギュレーションを特徴とするが、明らかに分化した軟骨細胞の信頼できる高度に特異的な分子マーカー、アグリカンの発現のアップレギュレーションを示さず、アルシアンブルー陽性基質もほとんど、またはまったく示さない。このため、3日目の培養物は、軟骨形成細胞系譜に入ってはいるものの、まだ軟骨細胞への明らかな分化を示してない。こうした軟骨形成細胞系譜の初期でのこの前駆体細胞の特徴は、軟骨欠損の修復に有用である。非常に初期相の軟骨形成細胞系譜の細胞は、系譜の後期の細胞と比較して修復への関与を促進するシグナルにより応答しやすい。
【0071】
発生の特定の段階にある細胞集団は、ある種の臨床徴候に好ましい。たとえば、2日目〜4日目の細胞(軟骨形成前駆体または初期の軟骨細胞)は、損傷した組織および軟骨に融合しやすく、より優れた長期にわたる修復を誘発する。
【0072】
軟骨前駆体(2日目〜3日目の細胞)は、四肢または手指の再生、半月板(mensical)または靭帯の修復、および骨折修復に優先的に使用される。初期の軟骨細胞(4日目の細胞)は、傷害による関節軟骨欠損の修復、慢性疾患(OAまたはRA)による関節軟骨損傷の修復または予防、外傷または先天性欠損により失われた軟骨組織の再生および置換、半月板の修復、ならびに顎関節の線維軟骨の修復に使用される。より十分に分化した軟骨細胞は、急性もしくは外傷性障害後の関節軟骨の修復、または疾患(OAまたはRA)による関節軟骨の修復もしくは予防のほか、内軟骨性骨化による骨折修復などへの適用に好適である。各段階は、軟骨発生および分化のモデリングに有用であり、iPSC由来の細胞は、患者個人を考慮した修復および再生のほか、ヒト疾患のモデリング、標的治療の設計および試験、ならびに遺伝的に傷害された軟骨組織の疾患特異的修復に特に有用である。
【0073】
4日目のBMP2処理hESCマイクロマス培養物は、Sox9およびBrachyuryの発現のさらなるアップレギュレーションを示し、信頼できる軟骨マーカー、アグリカンの発現の4倍を超えるアップレギュレーションを示すことから、4日目の培養物が軟骨細胞への明らかな分化の初期相にあることが示唆される。7日目および14日目の培養物は、Sox9およびBrachyuryの発現のダウンレギュレーションが、軟骨マーカー、アグリカンおよびCol2a1の発現の約8倍のアップレギュレーションと同時に起こることを特徴とする。また、均一なアルシアンブルー陽性基質も7日目および14日目の培養物の全体に存在し、免疫染色により検出可能なII型コラーゲンも培養物の全体に存在する。7日目および14日目の培養物は、Col10a1、Ihhもしくはオステオポンチンなどの肥大軟骨細胞成熟のマーカーを発現しない。このため、7日目および14日目の培養物は、軟骨細胞への明らかな分化を示しているが、肥大成熟化を示していない。このことは、関節軟骨を構成する軟骨細胞が肥大成熟化を示さないのが通常であり、実際に関節軟骨細胞の不適切な肥大成熟化は変形性関節症に特徴的なものであるため、関節軟骨の修復に細胞を利用することを考慮する際に重要である。7日目および/または14日目の培養物の軟骨細胞が肥大成熟化を示していないということにより、これらの細胞は軟骨欠損の修復の好ましい候補となる。
【0074】
前駆体細胞を軟骨形成細胞系譜に誘導する決定プロセスに関係する因子を同定するためのhESCのマイクロマス培養物
軟骨分化の開始を制御するSox9など一部の制御遺伝子が同定されているとはいえ、
胚性前駆体細胞の軟骨形成細胞系譜への段階的な決定またはコミットメントに関係する遺伝子および他の因子についてはほとんど知られていない。BMP2の存在下でのマイクロマス培養におけるhESCの段階的分化は、こうした初期の決定現象に関係する因子を明らかにする好適な系となる。未分化多能性hESCから軟骨細胞への系譜進行の様々な段階における細胞のトランスクリプトームを解析すれば、初期の軟骨形成細胞系譜のコミットメント現象を制御する遺伝子、シグナル伝達分子、および制御ネットワークを同定し、決定プロセスに関係する未知の遺伝子を明らかにしやすくなる。
【0075】
上述の遺伝子発現プロファイリングからは、細胞を軟骨形成細胞系譜に誘導する際の転写因子Brachyuryの役割が示される。これまでの研究から、Tボックス転写因子であるBrachyuryは、原腸形成において高度に発現し、初期の(primary)中胚葉形成で不可欠な役割を果たしていることが明らかになっている(Herrmann BG,1995 Seminars in Developmental Biology,6:385−394)。発生の後期には、Brachyuryは、肢芽形成の開始時に側部中胚葉に発現し、その後発生中の四肢に軟骨を形成する前軟骨形成間葉系細胞に発現する(Liu C et al.,2003 Development,130:1327−1337)。上記のように、Brachyuryの発現は、BMP2で処理したhESCマイクロマス培養物が、軟骨形成の転写因子Sox9の発現のアップレギュレーションによりアッセイされる軟骨形成細胞系譜に入ったばかりのときにアップレギュレートされる(24倍)。BrachyuryおよびSox9の発現のアップレギュレーションは、信頼できる軟骨マーカー、アグリカンの発現のアップレギュレーションの前に同時に起こる。次いでBMP2処理hESCマイクロマス培養物でアグリカンの発現のアップレギュレーションが開始されると、BrachyuryおよびSox9の発現が同時にさらに増加する。軟骨形成の開始後、明らかな軟骨分化が起こると、BrachyuryおよびSox9の発現は同時に無視できるレベルまで低下する。BMP2処理未分化多能性hESCの軟骨細胞への段階的分化におけるこの発現パターンから、Brachyuryが中胚葉性前駆体細胞の軟骨形成細胞系譜への初期コミットメントの調節に関係していることが示唆される。
【0076】
Brachyuryの発現は、BMP2シグナル伝達に応答して間葉系細胞株C3H10T1/2の軟骨細胞分化の開始時にアップレギュレートされ(Hoffmann A et al.,2002 J Cell Sci,115:769−781)、Brachyuryの強制発現によりSox9の発現およびC3H10T1/2細胞の軟骨細胞分化が促進される(Hoffmann A et al.,2002 J Cell Sci,115:769−781)ことが明らかになっている。さらに、ドミナントネガティブ型のBrachyuryは、C3H10T1/2細胞のBMP2刺激による軟骨細胞分化を障害する(Hoffmann A et al.,2002 J Cell Sci,115:769−781)。Brachyuryは、C3H101/2細胞などの前軟骨形成前駆体細胞にBMP2に応答して軟骨細胞分化を示す能力を与える可能性があることが示唆されている(Hoffmann A et al.,2002 J Cell Sci,115:769−781)。上述の結果は、Brachyuryの発現が、BMP2に応答してhESCが軟骨形成細胞系譜に入るとアップレギュレートされることを示唆し、以前の研究と整合している。これらの研究は、Brachyuryが、前駆体細胞を軟骨形成細胞系譜に誘導する決定プロセスに関係していることを示す。
【0077】
実施例5:ヒト胚性幹細胞の軟骨形成細胞系譜への分化
図4のCに示すように、BMP2処理hESCマイクロマス培養物全体に細胞外軟骨に特徴的なII型コラーゲンが存在することが、培養物の矢状断面をII型コラーゲン抗体で免疫染色することにより検出される。さらに、軟骨マーカー、アグリカンの球状ドメインに対するモノクローナル抗体で細胞質が染色される細胞も、BMP−2処理hESCマ
イクロマス培養物全体に存在する(図9)ことから、II型コラーゲン細胞外マトリックスで囲まれた細胞の実質的にすべてが、細胞自律性(cell autonomous)マーカーで免疫染色することによりアッセイされる軟骨マーカー、アグリカンを発現していることが示される。
【0078】
培養物における軟骨基質の蓄積は通常、基質の硫酸化プロテオグリカンを染色するアルシアンブルー、pH1.0を用いたインタクトな培養物のホールマウント染色によりモニターする。BMP2単独、またはBMP2とTGF−β1とにより処理したhESCマイクロマス培養物ではホールマウント染色により、アルシアンブルーで染色可能な基質の非常に均一な蓄積が検出可能である(図4〜図6)。軟骨分化の程度を組織学的に評価するため、hESCマイクロマス培養物の矢状断面をアルシアンブルーで染色し、核染色のNuclear Fast Redで対比染色した。アルシアンブルーにて染色可能な細胞外マトリックスで囲まれた細胞は、BMP2の単独処理(図10A)、あるいはBMP2とTGF−β1とで処理(図10Bおよび図10C)したhESCマイクロマス培養物の組織学的切片の実質的に全範囲にわたり存在する。BMP2処理hESC培養物は、広範囲に及ぶ軟骨組織に加えて少数の管状構造物を含んでいた(図10A;矢印)。培養物にTGF−β1およびBMP2を供給すると、検出可能な管の数はかなり減少し、管は培養物の非常に小さな割合を占めるだけである。したがって、BMP2およびTGF−β1の両方で処理したhESCマイクロマス培養物では、軟骨細胞分化が、完全には均質ではないが非常に均一に起こり、以前に報告された他の培養系の場合よりかなり安定して広範囲に及ぶようになる。実際、胚様体(EB)細胞から樹立された、BMP2+TGF−β処理マイクロマス培養物は、非常に不均一であり、軟骨形成組織だけでなく、大量の脂肪組織、さらには管からなる(図11)。
【0079】
実施例6:再生医療およびヒト疾患モデリングのためのiPSC由来軟骨細胞
iPSCは患者自身の細胞から採取されるため、患者特異的細胞を用いた治療および再生医療の手段となる(Amabile G and Meissner A 2009 Trends Mol Med 15:59−68)。図11、図12、および図13に示すように、iPSCは、本明細書に記載の方法により誘導され軟骨形成細胞系譜への段階的な分化を示す。
【0080】
図11(A)は、BMP2を補充せずに14日間維持されたiPSCの対照マイクロマス培養物を示し、アルシアンブルー陽性基質の蓄積をほとんど示していない。図11(B)は、14日目のBMP2処理iPSCマイクロマス培養物を示し、培養物全体に強くて広範囲のアルシアンブルー染色が存在することを示す。
【0081】
BMP2補充iPSC由来マイクロマス培養物による軟骨細胞分化の進行を確認するため、Brachyury、Sox9、Col2a1およびアグリカンなど軟骨形成細胞系譜に特徴的なマーカー遺伝子のmRNA発現のリアルタイム定量RT−PCRを使用した。これらのマーカー遺伝子のレベルは、培養期間中、BMP2を補充したhESCマイクロマス培養物の場合と同様のアップレギュレーションを示した。さらに、BMP2補充iPSCマイクロマス培養物では、肥大軟骨細胞のマーカー、オステオポンチンmRNAの発現が無視できる程度であった。したがって、本方法を用いたiPSCのマイクロマス培養物は、インビトロで軟骨形成細胞系譜への段階的分化を示し、さらに肥大を示さない。
【0082】
3種の酵素的アプローチ、トリプシン−EDTA、AccutaseまたはTrypLE Selectによって単細胞に分離したiPSCから樹立したマイクロマス培養物別に軟骨細胞分化を比較する研究を行った。図12に示すように、7日後、トリプシン−EDTA(図12のA)、TrypLE Select(図12のB)、あるいはAccutase(図12のC)のいずれかで予め分離したiPSCから樹立したBMP2補充マ
イクロマス培養物はそれぞれ、強く染色された広範囲のアルシアンブルー陽性基質の蓄積を同程度に示すことから、各分離方法は、BMP2に応答してこのプロトコルで軟骨細胞分化をうまく示すことができる単一のiPSCを作製するのに同等に有効である。BMP2を補充せずに維持した培養物には、アルシアンブルー陽性基質がほとんど存在しない(図12のD〜F)。
【0083】
図13に示すように、iPSCマイクロマス培養物は、BMP2(図13のA)、およびBMP2とTGFβ1との組み合わせ(図13のB)に応答して培養から14日後に広範囲の強いアルシアンブルー染色によりアッセイされる軟骨細胞分化を同程度に示す。これに対し、BMP2補充を受けなかったiPSCマイクロマス培養物は、アルシアンブルー陽性基質がほとんど蓄積されない(図13のC)。
【0084】
したがって、本方法は、上述のアプローチを用いて、傷害またはOAもしくはRAなどの慢性疾患による関節の関節軟骨欠損修復、および四肢再生、半月板もしくは靭帯欠損の修復、および骨折修復などの患者個人を考慮した軟骨修復および再建治療のためのiPSCの作製に有用である。
【0085】
遺伝子性障害の個体から採取されたiPSCは、疾患モデリングと、標的治療の設計および試験のスクリーニングとの手段となる(Laustriat D et al.,2010 Biochem Soc Trans 38:1051−1057;Lengner CJ 2010 Ann NY Acad Sci 1192:38−44)。iPSCを遺伝子ターゲティングと組み合わせると、単一遺伝子疾患を処置するアプローチとなる(Wong GKY and Chiu AT 2010 Biotechnol
Adv 28:715−724)。
【0086】
図16に示すようなiPSCを、軟骨異形成症のヒト患者から作製した。遺伝性軟骨障害のヒト患者由来のiPSCは、本明細書に記載の方法を用いて軟骨形成細胞系譜に分化するように誘導される。hESCおよびiPSCの軟骨細胞分化の誘導に使用した高密度マイクロマスアプローチなどの高密度培養法は、毒性試験として(Piersma AH
2004 Toxicol Lett 149:147−153;Ponce RA 2001 Curr Protoc Toxicol 13:13.3)、軟骨発生の新規な遺伝子制御因子を同定するためのゲノムスクリーニングとして(James CG et al.,2005 Mol Biol Cell 16:5316−5333)、軟骨形成に影響を与えるデベロプメンタルトキシン(developmental toxin)の検出のため(Hanse et al 2001 Free Radic Biol Med 31:1582−1592)、および有望な骨格形成異常治療法の機構評価のため(Woods A et al.,2007 Endocrinol 148:5030−5041)に使用するのに好適である。軟骨の遺伝子性障害(たとえば、軟骨異形成症または軟骨無形成症)の患者に由来し、本明細書に記載の方法により軟骨細胞分化を示すように誘導されたiPSCは、疾患プロセスのモデリングと、薬物療法の設計および試験とに有用である。軟骨異形成を示すiPSC由来の軟骨形成細胞の遺伝子プロファイリングを用いれば、疾患プロセスに関わり、治療介入のための疾患特異的な標的となるキー遺伝子および因子が同定される。疾患の標的治療剤については、インビトロでのその薬に対する、本方法により産生された病変したiPSC由来の軟骨形成細胞の応答を判定することにより、インビトロで試験される。また、遺伝的に病変した患者に正常な軟骨構造または機能を再建するアプローチとして、本方法により産生されたiPSC由来の疾患特異的な軟骨形成細胞を遺伝子治療と併用する。
【0087】
本明細書に言及した特許および科学文献は、当業者に入手可能な知識を明らかにするものである。本明細書に引用する米国特許、および公開もしくは未公開米国特許出願はすべ
て、参照によって援用する。本明細書に引用する公開された外国特許および特許出願はすべて、参照によって本明細書に援用する。本明細書に引用する他の公表された参考文献、文書、原稿および科学文献についても、参照によって本明細書に援用する。
【0088】
本発明についてその好ましい実施形態を参照しながら詳細に図示して記載してきたが、本発明添付の特許請求の範囲により包含される範囲から逸脱しない範囲で、本発明の形式および細部に様々な変更が可能であることが当業者には理解されよう。
【特許請求の範囲】
【請求項1】
ヒト胚性幹細胞(hESC)を軟骨形成細胞に直接分化させる方法であって、
胚様体(EB)を実質的に含まないhESCの集団を支持体上に用意する工程と、
前記hESCを前記支持体から剥離し、前記hESCを分離する工程と、
前記hESCの高密度マイクロマス培養を樹立する工程と、
前記集団を骨形成タンパク質と接触させる工程と
を含み、
前記hESCは軟骨形成細胞の実質的に均一な集団に直接分化する
方法。
【請求項2】
前記支持体はマウス胚性線維芽細胞(MEF)を含むフィーダー層である、請求項1に記載の方法。
【請求項3】
前記支持体はマトリゲル、あるいは、他の高分子もしくはゼラチン支持体またはスキャフォールドである、請求項1に記載の方法。
【請求項4】
前記hESCにトランスフォーミング増殖因子βを投与することをさらに含む、請求項1に記載の方法。
【請求項5】
前記BMPはBMP−2を含み、前記BMP−2は前記高密度マイクロマス培養の樹立から約24時間後に投与される、請求項1に記載の方法。
【請求項6】
前記BMPはBMP−2を含み、前記BMP−2は前記高密度マイクロマス培養の樹立から約48時間後に投与される、請求項1に記載の方法。
【請求項7】
前記hESCは約14日以内に十分に分化した軟骨細胞に分化する、請求項1に記載の方法。
【請求項8】
前記hESCは約7日以内に十分に分化した軟骨細胞に分化する、請求項1に記載の方法。
【請求項9】
前記hESCは約2〜3日以内に軟骨前駆体細胞に分化する、請求項1に記載の方法。
【請求項10】
前記hESCは約4日以内に初期の軟骨細胞に分化する、請求項1に記載の方法。
【請求項11】
前記hESCの少なくとも約85%は軟骨形成細胞系譜の細胞に分化する、請求項1に記載の方法。
【請求項12】
前記hESCの少なくとも約95%は軟骨形成細胞系譜の細胞に分化する、請求項1に記載の方法。
【請求項13】
前記hESCはトリプシン、TrypLE SelectまたはAccutaseによって分離される、請求項1に記載の方法。
【請求項14】
Rho結合キナーゼ(ROCK)阻害剤を投与する工程をさらに含む、請求項1に記載の方法。
【請求項15】
人工多能性幹細胞(iPSC)を軟骨形成細胞に直接分化させる方法であって、
EBを実質的に含まないiPSCの集団を支持体上に用意する工程と、
前記iPSCを前記支持体から剥離し、前記iPSCを分離する工程と、
前記iPSCの高密度培養を樹立する工程と、
前記iPSCを骨形成タンパク質と接触させることにより、iPSCの軟骨形成細胞系譜への分化を誘導する工程と
を含む方法。
【請求項16】
hESCはマイクロマス懸濁液培養またはペレット培養の形態である、請求項1または15に記載の方法。
【請求項17】
非軟骨形成細胞を実質的に含まず、かつ請求項1に記載の方法によりESCから分化した軟骨形成細胞の集団。
【請求項18】
非軟骨形成細胞を実質的に含まず、かつ請求項15に記載の方法によりiPSCから分化した軟骨形成細胞の集団。
【請求項19】
均一に分化した軟骨形成細胞の集団であって、前記細胞の少なくとも95%は単一の規定の分化の段階にあり、前記集団は十分に分化した軟骨細胞を実質的に含まない軟骨形成細胞の集団。
【請求項20】
前記段階は2日目もしくは3日目の軟骨形成細胞(軟骨前駆体)、または4日目の軟骨形成細胞(初期の軟骨細胞)からなる群から選択される、請求項19に記載の集団。
【請求項21】
損傷または病変軟骨を請求項17、18、19、20に記載の集団と接触させる工程を含む、軟骨を修復または再建する方法。
【請求項22】
請求項17、18、19、20に記載の集団を関節接合部に投与する工程を含む、関節炎を予防または処置する方法。
【請求項23】
骨折部位を請求項17、18、19、20に記載の集団と接触させる工程を含む、骨折修復を処置する方法。
【請求項24】
請求項19または20に記載の集団を関節接合部に投与する工程を含む、損傷もしくは断裂した靭帯または半月板を処置または修復する方法。
【請求項25】
請求項19または20に記載の集団によって手指または四肢を再生する方法。
【請求項26】
軟骨障害の発生に関係する遺伝子を同定する方法であって、請求項18、19、20に記載の集団を用意する工程と、正常対照細胞と比較した遺伝子発現の増加または減少を検出する工程とを含み、前記増加または減少から前記遺伝子が前記軟骨障害の発生に関係することが示唆される方法。
【請求項27】
前記軟骨障害は軟骨異形成症または軟骨無形成症を含む、請求項26に記載の方法。
【請求項28】
軟骨促進剤を同定する方法であって、請求項18、19または20に記載の集団を候補化合物と接触させる工程と、軟骨形成を検出する工程とを含み、前記化合物の非存在下と比較した前記前記化合物の存在下での軟骨形成の増加から前記化合物が軟骨の形成を促進することが示唆される方法。
【請求項29】
前記支持体はマウス胚性線維芽細胞(MEF)を含むフィーダー層である、請求項15に記載の方法。
【請求項30】
前記支持体はマトリゲル、あるいは、他の高分子もしくはゼラチン支持体またはスキャフォールドである、請求項15に記載の方法。
【請求項31】
前記hESCにトランスフォーミング増殖因子βを投与する工程をさらに含む、請求項15に記載の方法。
【請求項32】
BMP−2はiPSCの高密度マイクロマス培養の樹立から約24時間後に投与される、請求項15に記載の方法。
【請求項33】
BMP−2は前記iPSCの高密度マイクロマス培養の開始から48時間後に投与される、請求項15に記載の方法。
【請求項34】
前記iPSCは約14日以内に十分に分化した軟骨細胞に分化する、請求項15に記載の方法。
【請求項35】
前記iPSCは7日以内に十分に分化した軟骨細胞に分化する、請求項15に記載の方法。
【請求項36】
前記iPSCは約2〜3日以内に軟骨前駆体細胞に分化する、請求項15に記載の方法。
【請求項37】
前記iPSCは約4日以内に初期の軟骨細胞に分化する、請求項15に記載の方法。
【請求項38】
前記iPSCの少なくとも約85%は軟骨形成細胞系譜の細胞に分化する、請求項15に記載の方法。
【請求項39】
前記iPSCの少なくとも約95%は軟骨形成細胞系譜の細胞に分化する、請求項15に記載の方法。
【請求項40】
前記iPSCはトリプシン、TrypLE SelectまたはAccutaseによって分離される、請求項15に記載の方法。
【請求項41】
Rho結合キナーゼ(ROCK)阻害剤を投与する工程をさらに含む、請求項15に記載の方法。
【請求項1】
ヒト胚性幹細胞(hESC)を軟骨形成細胞に直接分化させる方法であって、
胚様体(EB)を実質的に含まないhESCの集団を支持体上に用意する工程と、
前記hESCを前記支持体から剥離し、前記hESCを分離する工程と、
前記hESCの高密度マイクロマス培養を樹立する工程と、
前記集団を骨形成タンパク質と接触させる工程と
を含み、
前記hESCは軟骨形成細胞の実質的に均一な集団に直接分化する
方法。
【請求項2】
前記支持体はマウス胚性線維芽細胞(MEF)を含むフィーダー層である、請求項1に記載の方法。
【請求項3】
前記支持体はマトリゲル、あるいは、他の高分子もしくはゼラチン支持体またはスキャフォールドである、請求項1に記載の方法。
【請求項4】
前記hESCにトランスフォーミング増殖因子βを投与することをさらに含む、請求項1に記載の方法。
【請求項5】
前記BMPはBMP−2を含み、前記BMP−2は前記高密度マイクロマス培養の樹立から約24時間後に投与される、請求項1に記載の方法。
【請求項6】
前記BMPはBMP−2を含み、前記BMP−2は前記高密度マイクロマス培養の樹立から約48時間後に投与される、請求項1に記載の方法。
【請求項7】
前記hESCは約14日以内に十分に分化した軟骨細胞に分化する、請求項1に記載の方法。
【請求項8】
前記hESCは約7日以内に十分に分化した軟骨細胞に分化する、請求項1に記載の方法。
【請求項9】
前記hESCは約2〜3日以内に軟骨前駆体細胞に分化する、請求項1に記載の方法。
【請求項10】
前記hESCは約4日以内に初期の軟骨細胞に分化する、請求項1に記載の方法。
【請求項11】
前記hESCの少なくとも約85%は軟骨形成細胞系譜の細胞に分化する、請求項1に記載の方法。
【請求項12】
前記hESCの少なくとも約95%は軟骨形成細胞系譜の細胞に分化する、請求項1に記載の方法。
【請求項13】
前記hESCはトリプシン、TrypLE SelectまたはAccutaseによって分離される、請求項1に記載の方法。
【請求項14】
Rho結合キナーゼ(ROCK)阻害剤を投与する工程をさらに含む、請求項1に記載の方法。
【請求項15】
人工多能性幹細胞(iPSC)を軟骨形成細胞に直接分化させる方法であって、
EBを実質的に含まないiPSCの集団を支持体上に用意する工程と、
前記iPSCを前記支持体から剥離し、前記iPSCを分離する工程と、
前記iPSCの高密度培養を樹立する工程と、
前記iPSCを骨形成タンパク質と接触させることにより、iPSCの軟骨形成細胞系譜への分化を誘導する工程と
を含む方法。
【請求項16】
hESCはマイクロマス懸濁液培養またはペレット培養の形態である、請求項1または15に記載の方法。
【請求項17】
非軟骨形成細胞を実質的に含まず、かつ請求項1に記載の方法によりESCから分化した軟骨形成細胞の集団。
【請求項18】
非軟骨形成細胞を実質的に含まず、かつ請求項15に記載の方法によりiPSCから分化した軟骨形成細胞の集団。
【請求項19】
均一に分化した軟骨形成細胞の集団であって、前記細胞の少なくとも95%は単一の規定の分化の段階にあり、前記集団は十分に分化した軟骨細胞を実質的に含まない軟骨形成細胞の集団。
【請求項20】
前記段階は2日目もしくは3日目の軟骨形成細胞(軟骨前駆体)、または4日目の軟骨形成細胞(初期の軟骨細胞)からなる群から選択される、請求項19に記載の集団。
【請求項21】
損傷または病変軟骨を請求項17、18、19、20に記載の集団と接触させる工程を含む、軟骨を修復または再建する方法。
【請求項22】
請求項17、18、19、20に記載の集団を関節接合部に投与する工程を含む、関節炎を予防または処置する方法。
【請求項23】
骨折部位を請求項17、18、19、20に記載の集団と接触させる工程を含む、骨折修復を処置する方法。
【請求項24】
請求項19または20に記載の集団を関節接合部に投与する工程を含む、損傷もしくは断裂した靭帯または半月板を処置または修復する方法。
【請求項25】
請求項19または20に記載の集団によって手指または四肢を再生する方法。
【請求項26】
軟骨障害の発生に関係する遺伝子を同定する方法であって、請求項18、19、20に記載の集団を用意する工程と、正常対照細胞と比較した遺伝子発現の増加または減少を検出する工程とを含み、前記増加または減少から前記遺伝子が前記軟骨障害の発生に関係することが示唆される方法。
【請求項27】
前記軟骨障害は軟骨異形成症または軟骨無形成症を含む、請求項26に記載の方法。
【請求項28】
軟骨促進剤を同定する方法であって、請求項18、19または20に記載の集団を候補化合物と接触させる工程と、軟骨形成を検出する工程とを含み、前記化合物の非存在下と比較した前記前記化合物の存在下での軟骨形成の増加から前記化合物が軟骨の形成を促進することが示唆される方法。
【請求項29】
前記支持体はマウス胚性線維芽細胞(MEF)を含むフィーダー層である、請求項15に記載の方法。
【請求項30】
前記支持体はマトリゲル、あるいは、他の高分子もしくはゼラチン支持体またはスキャフォールドである、請求項15に記載の方法。
【請求項31】
前記hESCにトランスフォーミング増殖因子βを投与する工程をさらに含む、請求項15に記載の方法。
【請求項32】
BMP−2はiPSCの高密度マイクロマス培養の樹立から約24時間後に投与される、請求項15に記載の方法。
【請求項33】
BMP−2は前記iPSCの高密度マイクロマス培養の開始から48時間後に投与される、請求項15に記載の方法。
【請求項34】
前記iPSCは約14日以内に十分に分化した軟骨細胞に分化する、請求項15に記載の方法。
【請求項35】
前記iPSCは7日以内に十分に分化した軟骨細胞に分化する、請求項15に記載の方法。
【請求項36】
前記iPSCは約2〜3日以内に軟骨前駆体細胞に分化する、請求項15に記載の方法。
【請求項37】
前記iPSCは約4日以内に初期の軟骨細胞に分化する、請求項15に記載の方法。
【請求項38】
前記iPSCの少なくとも約85%は軟骨形成細胞系譜の細胞に分化する、請求項15に記載の方法。
【請求項39】
前記iPSCの少なくとも約95%は軟骨形成細胞系譜の細胞に分化する、請求項15に記載の方法。
【請求項40】
前記iPSCはトリプシン、TrypLE SelectまたはAccutaseによって分離される、請求項15に記載の方法。
【請求項41】
Rho結合キナーゼ(ROCK)阻害剤を投与する工程をさらに含む、請求項15に記載の方法。
【図8】


【図10】
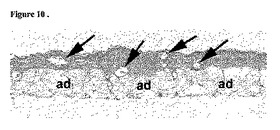
【図13】
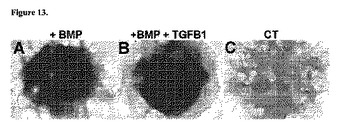
【図17】

【図1】
【図2】
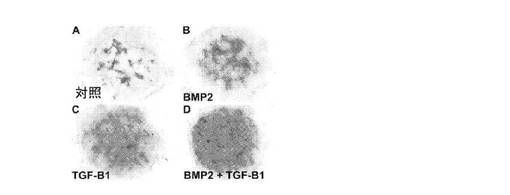
【図3】
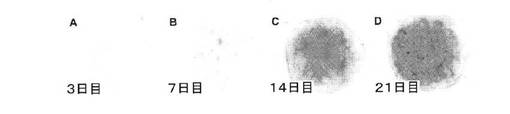
【図4】
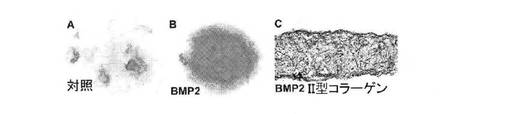
【図5】
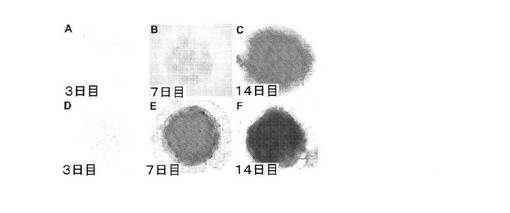
【図6】
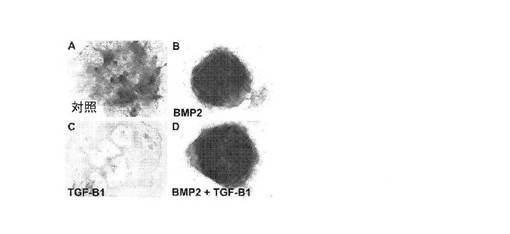
【図7】
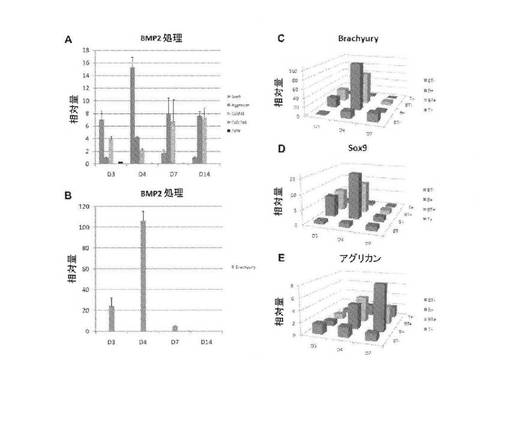
【図9】
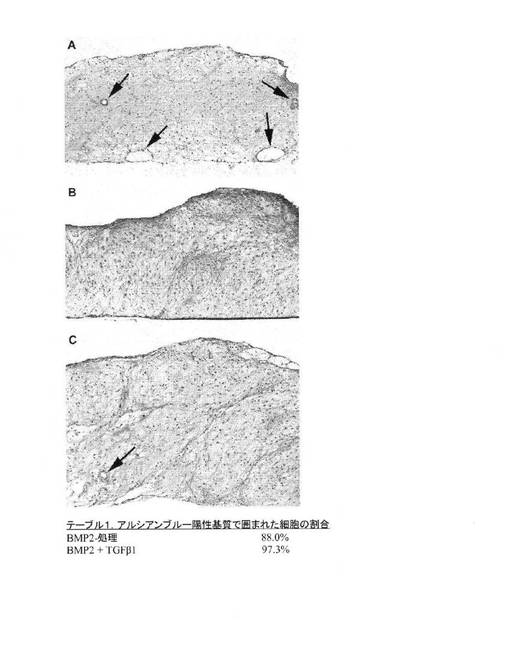
【図11】
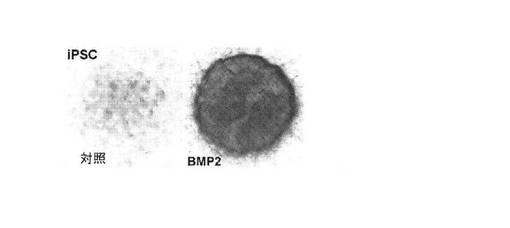
【図12】
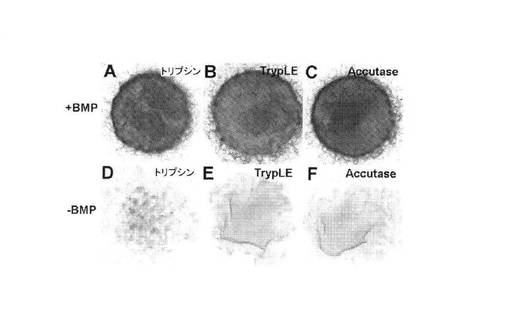
【図14】
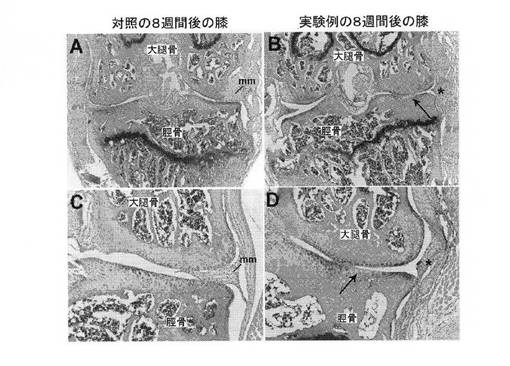
【図15】
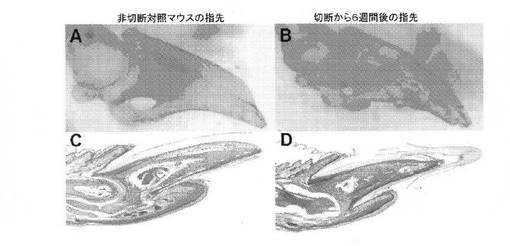
【図16】

【図10】
【図13】
【図17】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図9】
【図11】
【図12】
【図14】
【図15】
【図16】
【公表番号】特表2013−511987(P2013−511987A)
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−541196(P2012−541196)
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/US2010/058024
【国際公開番号】WO2011/066403
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(501315876)ユニバーシティ オブ コネチカット (22)
【Fターム(参考)】
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/US2010/058024
【国際公開番号】WO2011/066403
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(501315876)ユニバーシティ オブ コネチカット (22)
【Fターム(参考)】
[ Back to top ]