
ヒト胚性幹細胞の膵内分泌系への分化
本発明は、膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、米国特許出願シリアル番号第61/110,287号(2008年10月31日出願)に対する優先権を請求する。
【0002】
(発明の分野)
本発明は、多能性幹細胞の分化を促進する方法を提供する。具体的には本発明は、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【背景技術】
【0003】
I型糖尿病の細胞置換療法における進歩及び移植可能なランゲルハンス島の不足のため、生着に適したインスリン産生細胞すなわちβ細胞の供給源の開発に注目が集まっている。1つの手法は、例えば、胚性幹細胞のような多能性幹細胞から機能性のβ細胞を生成することである。
【0004】
脊椎動物の胚発生において、多能性細胞は、原腸形成として公知のプロセスにて、3つの胚葉(外胚葉、中胚葉、及び内胚葉)を含む細胞のグループを生じる。例えば、甲状腺、胸腺、膵臓、腸、及び肝臓等の組織は、内胚葉から中間ステージを経て発達する。このプロセスにおける中間ステージは、胚体内胚葉の形成である。胚体内胚葉細胞はHNF−3β、GATA4、MIXL1、CXCR4及びSOX17などの多数のマーカーを発現する。
【0005】
膵臓の形成は、胚体内胚葉の膵臓内胚葉への分化により起こる。膵臓内胚葉の細胞は膵臓−十二指腸ホメオボックス遺伝子、PDX1を発現する。PDX1が存在しない場合、膵臓は、腹側芽及び背側芽の形成を越えて発達しない。したがって、PDX1の発現は、膵臓器官形成において重要な工程となっている。成熟した膵臓は、他の細胞型間で外分泌組織及び内分泌組織を含有する。外分泌組織及び内分泌組織は、膵臓内胚葉の分化によって生じる。
【0006】
島細胞の特徴を保持する細胞がマウスの胚細胞から誘導されたことが報告されている。例えば、Lumelskyら(Science 292:1389、2001年)は、マウスの胚性幹細胞の、膵島と同様のインスリン分泌構造への分化を報告している。Soriaら(Diabetes 49:157、2000年)は、ストレプトゾトシン糖尿病のマウスにおいて、マウスの胚性幹細胞から誘導されたインスリン分泌細胞が糖血症を正常化することを報告している。
【0007】
一例において、Horiら(PNAS 99:16105,2002)は、ホスホイノシチド3−キナーゼ(LY294002)の阻害剤でマウス胚性幹細胞を処理することにより、β細胞に類似した細胞が生じたことを開示している。
【0008】
他の例では、Blyszczukら(PNAS 100:998、2003年)が、Pax4を構成的に発現しているマウス胚性幹細胞からのインスリン産生細胞の生成を報告している。
【0009】
Micallefらは、レチノイン酸が、胚性幹細胞のPdx1陽性膵臓内胚葉の形成に対する関与を制御できることを報告している。レチノイン酸は、胚における原腸形成の終了時に対応する期間中の、胚性幹細胞分化の4日目に培養液に添加すると、Pdx1発現の誘発に最も効果的である(Diabetes 54:301、2005年)。
【0010】
Miyazakiらは、Pdx1を過剰発現しているマウス胚性幹細胞株を報告している。Miyazakiらの結果は、外因性のPdx1発現が、得られた分化細胞内でインスリン、ソマトスタチン、グルコキナーゼ、ニューロゲニン3、P48、Pax6、及びHNF6遺伝子の発現を明らかに増加させたことを示している(Diabetes 53:1030、2004年)。
【0011】
Skoudyらは、マウス胚性幹細胞内で、アクチビンA(TGF−βスーパーファミリーのメンバー)が、膵臓外分泌遺伝子(p48及びアミラーゼ)、並びに内分泌遺伝子(Pdx1、インスリン及びグルカゴン)の発現を上方制御することを報告している。最大の効果は、1nMアクチビンAを使用した場合に認められた。Skoudyらはまた、インスリン及びPdx1 mRNAの発現レベルはレチノイン酸により影響されなかったが、3nMのFGF7による処理によりPdx1の転写産物のレベルが増大したことも観察している(Biochem.J.379:749,2004)。
【0012】
Shirakiらは、胚性幹細胞のPdx1陽性細胞への分化を特異的に増大させる増殖因子の効果を研究した。彼らは、TGF−β2によってPdx1陽性細胞が高い比率で再現可能に得られたことを観察している(Genes Cells.2005 Jun;10(6):503〜16.)。
【0013】
Gordonらは、血清の非存在下、かつアクチビンとWntシグナル伝達阻害剤の存在下で、マウス胚性幹細胞からbrachyury+/HNF−3β+内胚葉細胞が誘導されることを示している(米国特許出願公開2006/0003446(A1)号)。
【0014】
Gordonら(PNAS,Vol 103,page 16806,2006)は、「Wnt及びTGF−β/nodal/アクチビンの同時シグナル伝達が前原始線条の形成には必要であった」と述べている。
【0015】
しかしながら、胚性幹細胞発達のマウスモデルは、例えば、ヒトなどのより高等な哺乳動物における発達プログラムを正確には模倣しない恐れがある。
【0016】
Thomsonらは、ヒト胚盤胞から胚性幹細胞を単離した(Science 282:114,1998年)。同時に、Gearhart及び共同研究者は、胎児性腺組織から、ヒト胚性生殖(hEG)細胞株を誘導した(Shamblottら、Proc.Natl.Acad.Sci.USA 95:13726,1998)。白血病抑制因子(LIF)と共に培養するのみでも分化を阻止し得るマウス胚性幹細胞とは異なり、ヒト胚性幹細胞は、非常に特殊な条件下で維持する必要がある(米国特許第6,200,806号、国際公開第99/20741号;同第01/51616号)。
【0017】
D’Amourらは、高濃度のアクチビン及び低濃度の血清の存在下で、ヒト胚性幹細胞由来の胚体内胚葉の濃縮化された培養物が調製されたことを述べている(Nature Biotechnology 2005)。これらの細胞を、マウスの腎臓被膜下に移植することにより、一部の内胚葉性器官の特徴を有する、より成熟した細胞への分化が得られた。ヒト胚性幹細胞由来の胚体内胚葉細胞は、FGF−10の添加後、PDx1陽性細胞に更に分化することができる(米国特許出願公開第2005/0266554(A1)号)。
【0018】
D’Amourら(Nature Biotechnology−24,1392〜1401(2006))は、「我々は、ヒト胚性幹細胞(hES)を、インスリン、グルカゴン、ソマトスタチン、膵臓ポリペプチド及びグレリンといった膵臓ホルモンを合成可能な内分泌細胞に転換させる分化プロセスを開発した。このプロセスは、胚体内胚葉、腸管内胚葉、膵臓内胚葉及び内分泌前駆体が、内分泌ホルモンを発現する細胞へと向かう段階に類似した段階を通して細胞を指向させることにより、インビボでの膵臓器官形成を模倣する。」と述べている。
【0019】
別の例において、Fiskらは、ヒト胚性幹細胞から膵島細胞を産生するシステムを報告している(米国特許出願公開第2006/0040387(A1)号)。この場合、分化経路は3つのステージに分割された。先ず、ヒト胚性幹細胞を、酪酸ナトリウムとアクチビンAの組み合わせを用いて内胚葉に分化した。次に細胞をノギンなどのTGF−βアンタゴニストとEGF又はベータセルリンの組み合わせと培養してPDx1陽性細胞を生成する。最終分化は、ニコチンアミドにより誘発された。
【0020】
1つの例において、Benvenistryらは、「我々は、Pdx1の過剰発現が膵臓に多く見られる遺伝子の発現を高めたことを結論付ける。インスリン発現の誘導には、インビボでのみ存在する更なるシグナルを必要とする可能性がある。」と述べている(Benvenistryら、Stem Cells 2006;24:1923〜1930)。
【0021】
サイクリンはβ細胞機能に関係していると見なされてきた。例えば、Liljaらは、「インスリンを分泌する膵臓β細胞にはCdk5が存在する」と報告する(J.Biol.Chem.,Vol.276,Issue 36,34199〜34205,September 7,2001)。Liljaらは、「Cdk5はβ細胞中に存在し、インスリン開口分泌の正の調節因子として作用する。」と記載する。
【0022】
他の例では、Marzoらは「Cdk4ノックインマウスではβ細胞集団が有意に増加し、かつCdk4ノックインマウスは、Cdk4がI型糖尿病において膵臓β細胞集団を再生するのに潜在的な標的であることを示す、生理学的機能を有する」と記載する(Diabetalogia,Vol.47,Number4,686〜694,April 1,2004.)。
【0023】
他の例では、Ubedaらは、サイクリン依存性キナーゼ5活性の阻害が、膵臓β細胞を糖毒性から保護するということを報告する(J.Biol.Chem.,Vol.281,Issue 39,28858〜28864,September 29,2006)。
【0024】
他の例では、Weiらは、グルコース刺激型のインスリン分泌の、Cdk5依存性の制御を報告する(Nature Medicine 11,1104〜1108(1 October 2005.))。
【0025】
他の例では、Vanderfordらは以下のように記載している。「MafAは膵臓のβ細胞内で発現される塩基性ロイシンジッパー型転写制御因子であり、β細胞のバイオロジーの様々な態様に関与していることから正常なグルコース恒常性を維持するために必要とされる。MafAたんぱく質レベルは、未だ十分には特徴づけられていないメカニズムによって、高濃度のブドウ糖に反応して増加することが知られている。我々は、別々の細胞内シグナル伝達イベントがmafA発現を制御するのか否かを調査した。我々は、一般的なキナーゼ阻害剤であるスタウロスポリンが、タンパク質の安定性を変化させることなくmafA発現を誘導することを見出した。MAPキナーゼJNKの阻害は、mafA発現に対するスタウロスポリンの効果を模倣する。カルモジュリンキナーゼとカルシウムシグナル伝達も同様に、高濃度のブドウ糖によりmafA発現を刺激するのに重要である。しかしながら、スタウロスポリン、JNK、及びカルモジュリンキナーゼは、インスリン発現の誘導に対して異なる効果を有する。これらのデータは、MafAレベルが、複数のキナーゼ経路の協調作用により厳密に制御されていることを明らかにした。」(Archives of Biochemistry and Biophysics(2008),doi:10.1016/j.abb.2008.10.001)。
【発明の概要】
【発明が解決しようとする課題】
【0026】
したがって、多能性幹細胞を膵内分泌細胞、膵臓ホルモン発現細胞、又は膵臓ホルモン分泌細胞へと分化させるための方法の開発に対して、未だに有意な必要性が存在している。本発明は、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【課題を解決するための手段】
【0027】
一実施形態では、本発明は、膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【図面の簡単な説明】
【0028】
【図1】パネルaは、膵内分泌系に特徴的なマーカーを発現している細胞でのインスリン発現とグルカゴン発現の比に対する、EMD Calbiochemキナーゼ阻害剤ライブラリ由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。英数字ラベルは表1に示される同一の化合物と対応する。パネルbは、膵内分泌系に特徴的なマーカーを発現している細胞でのMAFA発現とARX4発現の比に対する、EMD Calbiochemキナーゼ阻害剤ライブラリ由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。英数字ラベルは表1に示される同一の化合物と対応する。
【図2A】実施例1に記載の方法に従って処理した、ステージ6処理の4日目の細胞の4x顕微鏡像を示す。
【図2B】0.5μMの化合物PubChemID# 5330812で処理した、処理の4日目の細胞の4x顕微鏡像を示す。
【図2C】1μMの化合物PubChemID# 5330812で処理した、処理の4日目の細胞の4x顕微鏡像を示す。
【図2D】実施例1に記載の方法に従って処理した、ステージ6処理の6日目の細胞の20x顕微鏡像を示す。
【図2E】0.5μMの化合物PubChemID #5330812で処理した、処理の6日目の細胞の20x顕微鏡像を示す。
【図2F】1μMの化合物PubChemID# 5330812で処理した、処理の6日目の細胞の20x顕微鏡像を示す。
【図3】0.5μM(濃色のバー)又は1.0μM(淡色のバー)の化合物PubChem ID#5330812での5日間の処理後の、膵内分泌系に特徴的なマーカーを発現している細胞での、23種の遺伝子の発現を示す。発現レベルは0日目、2日目及び5日目で測定した。
【図4】実施例4に記載の分化プロトコルの、ステージ7で処理した細胞における、膵内分泌系に特徴的なマーカーの発現に対する、CDK阻害剤III処理の効果を示す。
【図5】島様のクラスタのジチゾン(dithazone)染色に対する、CDK阻害剤III処理の効果を示す。
【図6】実施例5に記載の方法に従って生産されたインスリン産生細胞中の、インスリン、シナプトフィジン及びグルカゴンの発現を示す。記載のたんぱく質の発現はFACSにより測定した。
【図7】実施例5に記載の方法に従って生産されたインスリン産生細胞中の、インスリン、シナプトフィジン及びグルカゴンの発現を示す。記載のたんぱく質の発現はFACSにより測定した。
【図8】本発明の方法により生産された、インスリン産生細胞中のMAFA(パネルa)及びインスリン(パネルb)の発現を示す。1、2、3及び4日目の細胞サンプルを、PCR解析に用いた。CDK阻害剤での処理の4日後に、CDK阻害剤を培養物から除去し、DMEM−F12+1%B27+20ng/mLアクチビンA中で更に4日間培養した。4日間の終了時に、PCR解析のためにサンプルを三つ組複製で回収した。
【図9】膵内分泌系に特徴的なマーカーを発現している細胞でのMAFA発現に対する、EMD Calbiochemキナーゼ阻害剤ライブラリI由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。
【図10】膵内分泌系に特徴的なマーカーを発現している細胞での、インスリン、グルカゴン、ソマトスタチン及びMAFAのmRNA発現に対するゲニステイン(genestein)の効果を、リアルタイムPCRにより測定されたものとして示す。
【発明を実施するための形態】
【0029】
開示を明確にするために、本発明の「発明を実施するための形態」を、限定を目的とすることなく、本発明の特定の特徴、実施形態、又は応用を説明若しくは図示した以下の小項目に分ける。
【0030】
定義
幹細胞は、単一の細胞レベルにて自己複製し、分化して後代細胞を生成する、それら両方の能力で定義される未分化細胞であり、後代細胞には、自己複製前駆細胞、非再生前駆細胞、及び最終分化細胞が含まれる。幹細胞はまた、インビトロで複数の胚葉(内胚葉、中胚葉及び外胚葉)から様々な細胞系の機能的細胞に分化する能力によって、また移植後に複数の胚葉の組織を生じ、胚盤胞への注入後、全部ではないとしても殆どの組織を提供する能力によっても、特徴付けられる。
【0031】
幹細胞は、発生上の能力によって、(1)全ての胚性又は胚体外細胞のタイプを生ずる能力を有することを意味する、分化全能性、(2)全ての胚性細胞のタイプを生ずる能力を有することを意味する、分化万能性、(3)細胞系のサブセットを生ずる能力を有するが、それらが全て特定の組織、臓器、又は生理学的システムのものであるような、分化多能性(例えば、造血幹細胞(HSC)は、HSC(自己再生性)、血球限定的寡能性前駆細胞、及び、血液の通常の成分である全ての細胞種及び要素(例えば、血小板)を生じうる)、(4)多能性幹細胞よりも限定された細胞系のサブセットを生ずる能力を有することを意味する、分化寡能性、及び(5)単一の細胞系(例えば、精原幹細胞)を生ずる能力を有することを意味する、分化単一性に分類される。
【0032】
分化は、非特殊化の(「中立の」)又は比較的特殊化されていない細胞が、例えば、神経細胞又は筋細胞などの特殊化した細胞の特徴を獲得するプロセスである。分化した、又は分化を誘発された細胞は、細胞系内でより特殊化した(「傾倒した」)状況を呈している細胞である。分化プロセスに適用した際の用語「傾倒した」は、通常の環境下で特定の細胞型又は細胞型の小集合に分化し続ける分化経路の地点に進行しており、通常の環境下で異なる細胞型に分化し、又はより分化していない細胞型に戻ることができない細胞を指す。脱分化は、細胞が細胞系内で比較的特殊化されて(又は傾倒して)いない状況に戻るプロセスを指す。本明細書で使用するとき、細胞系は、細胞の遺伝、すなわち、その細胞がどの細胞から来たか、またどの細胞を生じ得るかを規定する。細胞系は、細胞を発達及び分化の遺伝的スキーム内に配置する。系特異的なマーカーは、対象とする系の細胞の表現型に特異的に関連した特徴を指し、中立細胞の対象とする系への分化を評価する際に使用することができる。
【0033】
「β−細胞系」は、転写因子PDX−1、及び以下の転写因子、すなわち、NGN3、NKX2.2、NKX6.1、NEUROD、ISL1、HNF3 β、MAFA、PAX4、又はPAX6の少なくとも1つについて遺伝子の発現が陽性である細胞を指す。β細胞系に特徴的なマーカーを発現している細胞としては、β細胞が挙げられる。
【0034】
本明細書で使用するとき、「胚体内胚葉系に特徴的なマーカーを発現している細胞」とは、以下のマーカー、すなわちSOX17、GATA−4、HNF−3 β、GSC、CER1、Nodal、FGF8、Brachyury、Mix様ホメオボックスタンパク質、FGF4 CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA−6、CXCR4、C−Kit、CD99又はOTX2のうちの少なくとも1つを発現する細胞を指す。胚体内胚葉系に特徴的なマーカーを発現している細胞としては、原始線条前駆体細胞、原始線条細胞、中内胚葉細胞及び胚体内胚葉細胞が挙げられる。
【0035】
本明細書で使用するとき、「膵臓内胚葉系に特徴的なマーカーを発現している細胞」とは、以下のマーカー、すなわち、PDX1、HNF1 β、PTF1 α、HNF−、又はHB9のうちの少なくとも1つを発現している細胞を指す。膵臓内胚葉系に特徴的なマーカーを発現している細胞としては、膵臓内胚葉細胞、原腸管細胞、後部前腸細胞が挙げられる。
【0036】
本明細書で使用するとき、「膵内分泌系に特徴的なマーカーを発現している細胞」は、以下のマーカー、すなわち、NGN3、NEUROD、ISL1、PDX1、NKX6.1、PAX4、NGN3、又はPTF1 αのうちの少なくとも1つを発現している細胞を指す。膵臓内分泌系に特徴的なマーカーを発現する細胞としては、膵臓内分泌細胞、膵臓ホルモン発現細胞及び膵臓ホルモン分泌細胞、並びにβ細胞系の細胞が挙げられる。
【0037】
本明細書で使用するとき、「胚体内胚葉」は、原腸形成中、胚盤葉上層から生じ、胃腸管及びその誘導体を形成する細胞の特徴を保持する細胞を指す。胚体内胚葉細胞は、以下のマーカー、すなわち、HNF−3 β、GATA4、SOX17、Cerberus、OTX2、グースコイド、C−Kit、CD99、及びMIXL1を発現する。
【0038】
本明細書で使用するとき、「胚体外内胚葉」は、以下のマーカー、すなわち、SOX7、AFP、又はSPARCのうちの少なくとも1つを発現する細胞の集団を指す。
【0039】
本明細書で使用するとき、「マーカー」とは、対象とする細胞で差異的に発現される核酸又はポリペプチド分子である。本文脈において、差異的な発現は、陽性マーカーのレベルの増大及び陰性マーカーのレベルの減少を意味する。検出可能なレベルのマーカー核酸又はポリペプチドは、他の細胞と比較して対象とする細胞内で十分高く又は低く、そのため当該技術分野において既知の多様な方法のいずれかを使用して、対象とする細胞を他の細胞から識別及び区別することができる。
【0040】
本明細書で使用するとき、「中内胚葉細胞」は、以下のマーカー、すなわちCD48、eomesodermin(EOMES)、SOX17、DKK4、HNF3 β、GSC、FGF17、又はGATA6のうちの少なくとも1つを発現している細胞を指す。
【0041】
本明細書で使用するとき、「膵臓内分泌細胞」又は「膵臓ホルモン発現細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、又は膵臓ポリペプチドのうちの少なくとも1つを発現することができる細胞を指す。
【0042】
本明細書で使用するとき、「膵臓内胚葉細胞」は、以下のマーカー、すなわち、NGN3、NEUROD、ISL1、PDX1、PAX4又はNKX2.2のうちの少なくとも1つを発現することが可能な細胞を指す。
【0043】
本明細書で使用するとき、「膵臓ホルモン産生細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、又は膵臓ポリペプチドのうちの少なくとも1つを産生することが可能な細胞を指す。
【0044】
本明細書で言うところの「膵臓ホルモン分泌細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを分泌することが可能な細胞を指して言う。
【0045】
本明細書で使用するとき、「前腸後部細胞」は、以下のマーカー、すなわちPDX1、HNF1、PTF1 α、HNF6、HB9、又はPROX1のうちの少なくとも1つを分泌することができる細胞を指す。
【0046】
本明細書で使用するとき、「前原始線条細胞」とは、以下のマーカー、すなわち、Nodal、又はFGF8のうちの少なくとも1つを発現する細胞を指す。
【0047】
本明細書で使用するとき、「原腸管細胞」とは、以下のマーカー、すなわち、HNF−1又はHNF4Aのうちの少なくとも1つを分泌することができる細胞を指す。
【0048】
本明細書で使用するとき、「原始線条細胞」とは、以下のマーカー、すなわち、Brachyury、Mix様ホメオボックスタンパク質、又はFGF4のうちの少なくとも1つを発現する細胞を指す。
【0049】
多能性幹細胞の単離、増殖及び培養
多能性幹細胞の特徴付け
多能性幹細胞は、ステージ特異的胚抗原(SSEA)3及び4の1以上、並びにTra−1−60及びTra−1−81と呼ばれる抗体によって検出可能なマーカーを発現し得る(Thomsonら、Science 282:1145,1998)。インビトロで多能性幹細胞を分化させると、SSEA−4、Tra−1−60、及びTra−1−81の発現が消失し(存在する場合)、SSEA−1の発現が増大する。未分化の多能性幹細胞は通常アルカリホスファターゼ活性を有し、これは、細胞を4%パラホルムアルデヒドで固定した後、製造業者(Vector Laboratories(Burlingame Calif.))によって述べられるようにVectorRedを基質として現像することによって検出することができる。未分化多能性幹細胞はまた、RT−PCRで検出されるように、一般にOct−4及びTERTも発現する。
【0050】
増殖させた多能性幹細胞の別の望ましい表現型は、内胚葉、中胚葉、及び外胚葉組織の3胚葉のすべての細胞に分化する能力である。多能性幹細胞の多能性は、例えば、細胞を重症複合免疫不全症(SCID)マウスに注入し、形成される奇形腫を4%パラホルムアルデヒドで固定し、次いでこれを3つの胚細胞層からの細胞種の証拠について組織学的に調べることによって確認することができる。代替的に、多能性は、胚様体を形成させ、この胚様体を3つの胚葉に関連したマーカーの存在に関して評価することにより決定することができる。
【0051】
増殖した多能性幹細胞株は、標準的なGバンド法を使用して核型を決定することができ、確立された対応する霊長類種の核型と比較される。「正常な核型」を有する細胞を獲得することが望ましく、これは細胞が正倍数体であることを意味し、全ヒト染色体が存在し、かつ、著しく変更されてはいない。
【0052】
多能性幹細胞の源
使用が可能な多能性幹細胞の種類としては、妊娠期間中の任意の時期(必ずしもではないが、通常は妊娠約10〜12週よりも前)に採取した前胚性組織(例えば胚盤胞等)、胚性組織、胎児組織などの、妊娠後に形成される組織に由来する多能性細胞の株化細胞系が含まれる。非限定的な例は、例えばヒト胚幹細胞株H1、H7、及びH9(WiCell)などのヒト胚幹細胞又はヒト胚生殖細胞の確立株である。それらの細胞の最初の樹立又は安定化中に本開示の組成物を使用することも想定され、その場合、源となる細胞は、源となる組織から直接採取した一次多能性細胞である。フィーダー細胞の不在下で既に培養された多能性幹細胞集団から採取した細胞も好適である。例えば、BG01v(BresaGen,Athens,GA)などの変異ヒト胚性幹細胞株も好適である。
【0053】
一実施形態では、ヒト胚性幹細胞は、Thomsonら(米国特許第5,843,780号、Science 282:1145,1998、Curr.Top.Dev.Biol.38:133 ff.,1998、Proc.Natl.Acad.Sci.U.S.A.92:7844,1995)に記載されているように調製される。
【0054】
多能性幹細胞の培養
一実施形態では、多能性幹細胞は、一般にフィーダー細胞の層上で培養され、このフィーダー細胞は、多能性幹細胞を様々な方法で支持する。あるいは、多能性幹細胞を、フィーダー細胞を基本的に含まないにも関わらず、細胞を実質的に分化させることなく多能性幹細胞の増殖を支持するような培養システム中で培養する。フィーダー細胞不含培養における多能性幹細胞の分化を伴わない増殖は、あらかじめ他の細胞種を培養することにより条件づけした培地を使用して支持される。あるいはフィーダー細胞不含培養における多能性幹細胞の分化を伴わない増殖は、合成培地を使用して支持される。
【0055】
例えば、Reubinoffら(Nature Biotechnology 18:399〜404(2000))及びThompsonら(Science 6 November 1998:Vol.282.no.5391,pp.1145〜1147)は、マウス胚性繊維芽フィーダー細胞層を用いてヒト胚盤胞からの多能性幹細胞系を培養することについて開示している。
【0056】
Richardsら(Stem Cells 21:546〜556,2003)は、11種類の異なるヒトの成人、胎児、及び新生児フィーダー細胞層についてヒト多能性幹細胞の培養を支持する能力の評価を行っている。Richardsらは、「成人の皮膚線維芽フィーダー細胞上で培養したヒト胚性幹細胞系は、ヒト胚性幹細胞の形態を有し、多能性を維持する」と述べている。
【0057】
米国特許出願公開第20020072117号は、無フィーダー細胞培養中で霊長類の多能性幹細胞の増殖を支持する培地を生成する細胞系を開示している。使用される細胞系は、胚性組織から得られるかあるいは胚性幹細胞から分化した間葉系かつ繊維芽細胞様の細胞系である。米国特許出願公開第20020072117号は、この細胞系の1次フィーダー細胞層としての使用を更に開示している。
【0058】
別の例として、Wangら(Stem Cells 23:1221〜1227,2005)は、ヒト胚性幹細胞由来のフィーダー細胞層上でヒト多能性幹細胞を長期にわたって増殖させるための方法を開示している。
【0059】
別の例として、Stojkovicら(Stem Cells 2005 23:306〜314,2005)は、ヒト胚性幹細胞の自然分化により誘導されたフィーダー細胞システムを開示している。
【0060】
更なる別の例として、Miyamotoら(Stem Cells 22:433〜440,2004)は、ヒトの胎盤から得られたフィーダー細胞の供給源を開示している。
【0061】
Amitら(Biol.Reprod 68:2150〜2156、2003年)は、ヒト包皮に由来するフィーダー細胞層を開示している。
【0062】
別の例として、Inzunzaら(Stem Cells 23:544〜549,2005)は、ヒトの出生直後産児の包皮線維芽細胞から得られたフィーダー細胞層を開示している。
【0063】
米国特許第6642048号は、無フィーダー細胞培養中での霊長類の多能性幹(pPS)細胞の増殖を支持する培地、及びこうした培地の製造に有用な細胞系を開示している。米国特許第6642048号は、「本発明は、胚性組織から得られるかあるいは胚性幹細胞から分化した間葉系かつ繊維芽細胞様の細胞系を含む。本開示では、こうした細胞系を誘導し、培地を調整し、この馴化培地を用いて幹細胞を増殖させるための方法を説明及び図示する」と述べている。
【0064】
別の例として、国際公開第2005014799号は、哺乳動物細胞の維持、増殖及び分化のための条件培地を開示している。国際公開第2005014799号は、「本発明に基づいて製造される培地は、マウス細胞、特にMMH(Metマウス肝細胞)と称される分化及び不死化したトランスジェニック肝細胞の細胞分泌活性によって馴化される」と述べている。
【0065】
別の例として、Xuら(Stem Cells 22:972〜980,2004)は、ヒトテロメラーゼ逆転写酵素を過剰発現するように遺伝子改変されたヒト胚性幹細胞由来細胞から得られた馴化培地を開示している。
【0066】
別の例において、米国特許出願公開第20070010011号は、多能性幹細胞を維持するための合成培地を開示している。
【0067】
代替的な一培養システムでは、胚性幹細胞の増殖を促進することが可能な増殖因子を添加した無血清培地を使用している。例えば、Cheonら(BioReprod DOI:10.1095/biolreprod.105.046870,October 19,2005)は、胚性幹細胞の自己再生を誘発することが可能な異なる増殖因子を添加した非馴化血清補充(SR)培地中に胚性幹細胞が維持された、無フィーダー細胞かつ無血清の培養システムを開示している。
【0068】
別の例において、Levensteinら(Stem Cells 24:568〜574,2006)は、線維芽細胞又は馴化培地の非存在下で、bFGFを添加した培地を使用して、胚幹細胞を長期間培養する方法を開示している。
【0069】
別の例において、米国特許出願公開第20050148070号は、血清及び繊維芽細胞フィーダー細胞方法を含まない合成培地中でのヒト胚幹細胞の培養方法を開示し、同方法は、アルブミン、アミノ酸、ビタミン、無機物、少なくとも1つのトランスフェリン又はトランスフェリン代替物、少なくとも1つのインスリン又はインスリン代替物を含有する培地中で細胞を培養し、この培地は、本質的に哺乳動物胎児血清を含有せず、線維芽細胞増殖因子シグナル伝達受容体を活性化できる少なくとも約100ng/mLの線維芽細胞増殖因子を含有し、ここで増殖因子は、線維芽細胞フィーダー層のみでなく他の源からも供給され、培地はフィーダー細胞又は馴化培地なしで、未分化状態の幹細胞の増殖を支持した。
【0070】
別の例において、米国特許出願公開第20050233446号は、未分化の霊長類始原幹細胞を含む幹細胞の培養に有用な合成培地を開示している。溶液において、培地は、培養されている幹細胞と比較して実質的に等張である。所定の培養おいて、特定の培地は、基本培地と、実質的に未分化の始原幹細胞の増殖の支持に必要な、ある量のbFGF、インスリン、及びアスコルビン酸の各々とを含有する。
【0071】
別の例として、米国特許第6800480号は、「1つの実施形態において、実質的に未分化状態の霊長類由来の始原幹細胞を増殖させるための細胞培地であって、霊長類由来の始原幹細胞の増殖を支持するうえで効果的な低浸透圧、低エンドトキシンの基礎培地を含む細胞培地を提供する。この基本培地は、霊長類由来の始原幹細胞の増殖を支持するうえで効果的な栄養素血清、並びに、フィーダー細胞、及びフィーダー細胞から誘導される細胞外基質成分からなる群から選択される基質と組み合わされる。培地は更に、非必須アミノ酸、抗酸化剤、並びに、ヌクレオシド及びピルビン酸塩からなる群から選択される第1の増殖因子を含む。」と述べている。
【0072】
別の例では、米国特許出願公開第20050244962号は、「1つの態様において本発明は、霊長類の胚性幹細胞を培養する方法を提供する。哺乳動物の胎児血清を基本的に含まない(好ましくはあらゆる動物の血清をも基本的に含まない)培養中で、単に繊維芽フィーダー細胞層以外の供給源から供給される線維芽細胞増殖因子の存在下で幹細胞を培養する。好ましい1つの形態では、充分な量の繊維芽増殖因子を添加することによって、幹細胞の培養を維持するために従来必要とされていた繊維芽フィーダー細胞層の必要性がなくなる。」と述べている。
【0073】
更なる例として、国際特許出願公開第2005065354号は、基本的に無フィーダー細胞かつ無血清の合成等張培地であって、a.基礎培地、b.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のbFGF、c.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のインスリン、及びd.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のアスコルビン酸、を含む培地を開示している。
【0074】
別の例として、国際公開第2005086845号は、幹細胞を、細胞を未分化な状態に維持するのに十分な量の、トランスフォーミング増殖因子β(TGF−β)ファミリータンパク質のメンバー、線維芽細胞増殖因子(FGF)ファミリータンパク質のメンバー、又はニコチンアミド(NIC)に、所望の結果を得るのに充分な時間曝露することを含む、未分化の幹細胞を維持するための方法を開示している。
【0075】
多能性幹細胞は、好適な培養基質上に播くことができる。一実施形態では、好適な培養基質は、例えば基底膜から誘導されたもの、又は接着分子受容体−リガンド結合の一部を形成し得るものなどの細胞外マトリックス成分である。一実施形態において、好適な培養基質は、MATRIGEL(登録商標)(Becton Dickenson)である。MATRIGEL(登録商標)は、Engelbreth−Holm Swarm腫瘍細胞からの可溶性製剤であり、室温でゲル化して再構成基底膜を形成する。
【0076】
他の細胞外マトリックス成分及び成分混合物は代替物として好適である。増殖させる細胞型に応じて、これは、ラミニン、フィブロネクチン、プロテオグリカン、エンタクチン、ヘパラン硫塩、及び同様物を、単独で又は様々な組み合わせで含み得る。
【0077】
多能性幹細胞は、細胞の生存、増殖、及び所望の特徴の維持を促進する培地の存在下、基質上に好適な分布にて播かれてもよい。これら全特徴は、播種分布に細心の注意を払うことから利益を得、当業者は容易に決定することができる。
【0078】
好適な培地は、以下の成分、例えば、ダルベッコ変法イーグル培地(DMEM)、Gibco No.11965−092;ノックアウトダルベッコ変法イーグル培地(KO DMEM)、Gibco No.10829−018;ハムF12/50% DMEM基礎培地、200mM L−グルタミン、Gibco No.15039−027;非不可欠アミノ酸溶液、Gibco No.11140−050;β−メルカプトエタノール、Sigma No.7522;ヒト組み換え塩基性線維芽細胞増殖因子(bFGF)、Gibco No.13256−029などから調製することができる。
【0079】
多能性幹細胞からの膵臓ホルモン産生細胞の形成
一実施形態では、本発明は、多能性幹細胞から膵臓ホルモン産生細胞を作製するための方法を提供し、かかる方法は、
a.多能性幹細胞を培養する工程と、
b.多能性幹細胞を、胚体内胚葉系に特徴的なマーカーを発現している細胞に分化させる工程と、
c.胚体内胚葉系に特徴的なマーカーを発現している細胞を、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化させる工程と、
d.膵臓内胚葉系に特徴的なマーカーを発現している細胞を、膵内分泌系に特徴的なマーカーを発現している細胞に分化させる工程と、を含む。
【0080】
本発明での使用に好適な多能性幹細胞としては、例えばヒト胚性幹細胞株H9(NIH code:WA09)、ヒト胚性幹細胞株H1(NIH code:WA01)、ヒト胚性幹細胞株H7(NIH code:WA07)、及びヒト胚性幹細胞株SA002(Cellartis,Sweden)が挙げられる。多能性細胞に特徴的な以下のマ−カー、すなわち、ABCG2、cripto、CD9、FOXD3、コネキシン43、コネキシン45、OCT4、SOX2、NANOG、hTERT、UTF1、ZFP42、SSEA3、SSEA4、Tra1−60又はTra1−81のうちの少なくとも1つを発現する細胞も本発明での使用に適している。
【0081】
胚体内胚葉系に特徴的なマーカーは、SOX17、GATA4、HNF3 β、GSC、CER1、NODAL、FGF8、Brachyury、Mix−様ホメオボックスタンパク質、FGF4 CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、及びOTX2からなる群より選択される。本発明での使用に好適なものは、胚体内胚葉系に特徴的なマーカーのうちの少なくとも1つを発現している細胞である。本発明の一態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、原始線条前駆体細胞である。別の態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、中内胚葉細胞である。別の態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、胚体内胚葉細胞である。
【0082】
膵臓内胚葉系に特徴的なマーカーは、PDX1、HNF−1 β、PTF1 α、HNF6、HB9及びPROX1からなる群から選択される。本発明での使用に好適なものは、膵臓内胚葉系の特徴を示す少なくとも1つのマーカーを発現している細胞である。本発明の一態様において、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉細胞である。
【0083】
膵内分泌系に特徴的なマーカーは、NGN3、NEUROD、ISL1、PDX1、NKX6.1、PAX4、及びPTF−1 αからなる群から選択される。一実施形態では、膵内分泌細胞は、以下のホルモン:インスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを発現することができる。本発明で使用するのに好適なものは、膵内分泌系の特徴を示すマーカーを少なくとも1つ発現する細胞である。本発明の一態様において、膵内分泌系に特徴的なマーカーを発現している細胞は、膵内分泌細胞である。膵内分泌細胞は、膵臓ホルモン発現細胞であってもよい。代替的に、膵内分泌細胞は、膵臓ホルモン分泌細胞であってもよい。
【0084】
本発明の一態様では、膵臓内分泌細胞は、β細胞系統に特徴的なマーカーを発現する細胞である。β細胞系に特徴的なマーカーを発現している細胞は、PDx1と、以下の転写因子、すなわち、NGN3、NKX2.2、NKX6.1、NEUROD、ISL1、HNF−3 β、MAFA、PAX4、又はPAX6のうちの少なくとも1つを発現する。本発明の一態様では、β細胞系統に特徴的なマーカーを発現する細胞は、β細胞である。
【0085】
胚体内胚葉系に特徴的なマーカーを発現している細胞の形成
多能性幹細胞は、当該技術分野のいかなる方法、又は本発明で提案されるいかなる方法によって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させてもよい。
【0086】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 23,1534〜1541(2005)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0087】
例えば、多能性幹細胞は、Shinozakiら、Development 131,1651〜1662(2004)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0088】
例えば、多能性幹細胞は、McLeanら、Stem Cells 25,29〜38(2007)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0089】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0090】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンA及び血清と培養し、次いで細胞をアクチビンA及び異なる濃度の血清と培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の一例は、Nature Biotechnology 23,1534〜1541(2005)に開示されている。
【0091】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンAと、別の濃度の血清の存在下で培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の例は、D’Amourら、Nature Biotechnology,2005に開示されている。
【0092】
例えば、多能性幹細胞は、アクチビンA及びWntリガンドを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いでWntリガンドを除去し、細胞をアクチビンAと、血清の存在下で培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0093】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0094】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0095】
例えば、多能性幹細胞は、米国特許出願第60/990,529号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0096】
例えば、多能性幹細胞は、米国特許出願第61/076,889号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0097】
例えば、多能性幹細胞は、米国特許出願第61/076,900号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0098】
例えば、多能性幹細胞は、米国特許出願第61/076,908号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0099】
例えば、多能性幹細胞は、米国特許出願第61/076,915号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0100】
胚体内胚葉系に特徴的なマーカーを発現している細胞の分化
胚体内胚葉系に特徴的なマーカーを発現している細胞の形成は、以下の特定のプロトコルの前後に、マーカーの存在に関して試験することにより決定することができる。多能性幹細胞は、一般にかかるマーカーを発現しない。したがって、多能性細胞の分化は、細胞がそれらの発現を開始した際に検出される。
【0101】
分化効率は、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより測定することができる。
【0102】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0103】
多能性幹細胞の特徴は当業者に周知であり、多能性幹細胞の更なる特徴は、継続して同定されている。例えば、多能性幹細胞マーカーとしては、以下のもの:ABCG2、cripto、FOXD3、コネキシン43、コネキシン45、OCT4、SOX2、NANOG、hTERT、UTF1、ZFP42、SSEA3、SSEA4、Tra1−60、又はTra1−81のうちの1つ以上の発現が挙げられる。
【0104】
多能性幹細胞を本発明の方法で処理した後、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現している細胞により発現される、例えばCXCR4などのタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより、分化した細胞を精製することができる。
【0105】
膵臓内胚葉系に特徴的なマーカーを発現している細胞の形成
胚体内胚葉系に特徴的なマーカーを発現している細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化され得る。
【0106】
例えば、胚体内胚葉系に特徴的なマーカーを発現している細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示されている方法に従って、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化され得る。
【0107】
例えば、胚体内胚葉系に特徴的なマーカーを発現している細胞は、胚体内胚葉系に特徴的なマーカーを発現している細胞を、線維芽細胞増殖因子及びヘッジホッグシグナル伝達経路阻害剤KAAD−シクロパミンで処理した後、線維芽細胞増殖因子及びKAAD−シクロパミンを含有する培地を除去し、続いて細胞をレチノイン酸、線維芽細胞増殖因子及びKAAD−シクロパミンを含有する培地中で培養することにより、膵臓内胚葉系に特徴的なマーカーを発現している細胞に更に分化させられる。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0108】
本発明の一態様では、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞をレチノイン酸及び少なくとも1種類の線維芽細胞増殖因子で所定の時間処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞を膵臓内胚葉系に特徴的なマーカーを発現する細胞に更に分化させる。
【0109】
本発明の一態様では、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞をレチノイン酸及び少なくとも1種類の線維芽細胞増殖因子で所定の時間処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞を膵臓内胚葉系に特徴的なマーカーを発現する細胞に更に分化させる。
【0110】
本発明の1つの態様では、胚体内胚葉系に特徴的なマーカーを発現している細胞を米国特許出願第60/990,529号に記載の方法に従って処理することにより、膵臓内胚葉系に特徴的なマーカーを発現している細胞へと胚体内胚葉系に特徴的なマーカーを発現している細胞を更に分化させる。
【0111】
膵臓内胚葉系に特徴的なマーカーを発現する細胞の検出
膵臓内胚葉系に特徴的なマーカーは、当業者に周知であり、膵臓内胚葉系の特徴を示す追加のマーカーが、継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して膵臓内胚葉系の特徴を示す性質を獲得したことを確認するために使用され得る。膵臓内胚葉系に特異的なマーカーとしては、例えば、HLXB9、PTF−1 α、PDX1、HNF6、HNF−1 βなどの転写因子の1つ以上のものの発現が挙げられる。
【0112】
分化効率は、膵臓内胚葉系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、処理した細胞集団を曝露することにより測定することができる。
【0113】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0114】
膵内分泌系に特徴的なマーカーを発現している細胞の形成
膵臓内胚葉系に特徴的なマーカーを発現している細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵内分泌系に特徴的なマーカーを発現している細胞へと分化させることができる。
【0115】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示されている方法に従って、膵内分泌系に特徴的なマーカーを発現している細胞に分化され得る。
【0116】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をDAPT及びエキセンディン4を含有する培地中で培養し、次にDAPT及びエキセンディン4を含有する培地を除去し、続いて細胞をエキセンディン1、IGF−1及びHGFを含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0117】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をエキセンディン4を含有する培地中で培養し、次にエキセンディン4を含有する培地を除去し、続いて細胞をエキセンディン1、IGF−1及びHGFを含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’ Amourら、Nature Biotechnology,2006に開示されている。
【0118】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をDAPT及びエキセンディン4を含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’Amourら、Nature Biotechnology,2006に開示されている。
【0119】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をエキセンディン4を含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’Amourら、Nature Biotechnology,2006に開示されている。
【0120】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0121】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0122】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第60/953,178号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0123】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、米国特許出願第60/990,529号に開示された方法に従って処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0124】
本発明の一態様では、本発明は、膵内分泌系と関連付けられるマーカーの発現量を増加させる方法を提供し、この方法は、膵内分泌系に特徴的なマーカーを発現している細胞を、米国特許出願第61/110,278号に示された方法に従って、十分な量のTGF−β受容体作動剤を含む培地で処理することで、膵内分泌系に関連付けられるマーカーの発現量を増加させることを含む。
【0125】
膵内分泌系に特徴的なマーカーを発現している細胞の検出
膵内分泌系に特徴的なマーカーは当業者に周知であり、膵内分泌系の特徴を示す追加のマーカーが継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して膵内分泌系の特徴を示す性質を獲得したことを確認するために使用され得る。膵内分泌系に特異的なマーカーとしては、例えばNGN3、NEUROD又はISL1などの転写因子の1種以上の発現が挙げられる。
【0126】
β細胞系の細胞に特徴的なマーカーは当業者に周知であり、β細胞系の細胞の特徴を示す追加のマーカーが継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して、β−細胞系の特徴を示す性質を獲得したことを確認するために使用することができる。β細胞系に特異的な特徴としては、例えば、特にPDX1、NKX2.2、NKX6.1、ISL1、PAX6、PAX4、NEUROD、HNF1 β、HNF6、HNF3 β又はMAFAなどの転写因子の1つ以上の発現が挙げられる。これらの転写因子は、内分泌細胞の同定について当該技術分野で十分に確立されている。例えば、Edlund(Nature Reviews genetics 3:524〜632(2002年))を参照されたい。
【0127】
分化効率は、膵内分泌系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、被処理細胞集団を曝露することにより測定することができる。あるいは分化効率は、β細胞系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、被処理細胞集団を曝露することにより測定することができる。
【0128】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0129】
本発明の一態様では、分化効率は、処理後の所定の細胞培養物中のインスリン陽性細胞の百分率を測定することによって求められる。一実施形態では、本発明の方法は、所定の培養物中で約100%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約90%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約80%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約70%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約60%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約50%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約40%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約30%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約20%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約10%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約5%のインスリン陽性細胞を生成する。
【0130】
本発明の一態様では、分化効率は、グルコース刺激によるインスリン分泌を、細胞が放出するC−ペプチドの量を測定することで検出し、測定することにより求められる。一実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約1000ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約900ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約800ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約700ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約600ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約500ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約400ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約500ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約400ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約300ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約200ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約100ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約90ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約80ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約70ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約60ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約50ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約40ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約30ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約20ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約10ngのC−ペプチドを産生する。
【0131】
膵内分泌系に特徴的なマーカーを発現している細胞でのMAFAの発現の増加
一実施形態では、本発明は、膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【0132】
サイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ1を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ2を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ4を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ5を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ9を阻害し得る。あるいは、サイクリン依存性キナーゼ阻害剤は、これらの任意の組み合わせの、サイクリン依存性キナーゼの複数のイソ型を阻害し得る。
【0133】
サイクリン依存性キナーゼ阻害剤はタンパク質であり得る。あるいはサイクリン依存性キナーゼ阻害剤はペプチドであり得る。あるいはサイクリン依存性キナーゼ阻害剤は低分子であり得る。一実施形態では、低分子サイクリン依存性キナーゼ阻害剤は、7−n−ブチル−6−(4−ヒドロキシフェニル)[5H]ピロロ[2,3−b]ピラジン、9−ニトロ−7,12−ジヒドロインドロ[3,2−d][1]ベンゾアゼピン−6(5H)−オン、3−(6−オキソ−9−ニトロ−5,6,7,12−テトラヒドロインドロ[3,2−d][1]ベンゾアゼピン−2−イル)プロピオニトリル、(2R)−2−((6−((3−アミノ−5−クロロフェニル)アミノ)−9−(1−メチルエチル)−9H−プリン−2−イル)アミノ)−3−メチル−1−ブタノール、アルシリアフラビンA、[6−ベンジルアミノ−2−(3−ヒドロキシプロピルアミノ)−9−イソプロピルプリン、ブチロラクトンI、(Z)−1−(3−エチル−5−メトキシ−2,3−ジヒドロベンゾチアゾール−2−イリデン)プロパン−2−オン、2−(3−ヒドロキシプロピルアミノ)−6−(o−ヒドロキシベンジルアミノ)−9−イソプロピルプリン、1−(2,6−ジクロロフェニル)−1,5−ジヒドロ−6−((4−(2−ヒドロキシエトキシ)フェニル)メチル)−3−(1−メチルエチル)−4H−ピラゾロ[3,4−d]ピリミジン−4−オン、Cdk/サイクリン阻害ペプチドIII、3−(2−クロロ−3−インドリルメチレン)−1,3−ジヒドロインドール−2−オン、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレート、RO−3306、N−(cis−2−アミノシクロヘキシル)−N−(3−クロロフェニル)−9−エチル−9H−プリン−2,6−ジアミン、6−シクロヘキシルメトキシ−2−(4’−スルファモイルアニリノ)プリン、5−アミノ−3−((4−(アミノスルホニル)フェニル)アミノ)−N−(2,6−ジフルオロフェニル)−1H−1,2,4−トリアゾール−1−カルボチオアミド、3−アミノ−1H−ピラゾロ[3,4−b]キノキサリン、Cdk2阻害剤I、Cdk2阻害剤II、2(ビス−(ヒドロキシエチル)アミノ)−6−(4−メトキシベンジルアミノ)−9−イソプロピルプリン、4−(6−シクロヘキシルメトキシ−9H−プリン−2−イルアミノ)−N,N−ジエチルベンズアミド、N4−(6−アミノピリミジン−4−イル)−スルファニルアミド、(4−(2−アミノ−4−メチルチアゾール−5−イル)ピリミジン−2−イル)−(3−ニトロフェニル)アミン、2−ブロモ−12,13−ジヒドロ−5H−インドロ[2,3−a]ピロロ[3,4−c]カルバゾール−5,7(6H)−ジオン、1,4−ジメトキシアクリジン−9(10H)−チオン、5−(N−(4−メチルフェニル)アミノ)−2−メチル−4,7−ジオキソベンゾチアゾール、4−(3,5−ジアミノ−1Hピラゾール−4−イルアゾ)−フェノール、2−(2−ヒドロキシエチルアミノ)−6−(3−クロロアニリノ)−9−イソプロピルプリン、ファスカプリシン、インジルビン−3’−モノオキシム、インジルビン−3’−モノオキシム、5−ヨード−,インジルビン−3’−モノオキシム−5−スルホン酸、イソグラヌラチミド、2−(2−ヒドロキシエチルアミノ)−6−ベンジルアミノ−9−メチルプリン、6−(2−ヒドロキシベンジルアミノ)−2−((1R)−(ヒドロキシメチル)プロピル)アミノ)−9−イソプロピルプリン、5−ブロモ−3−(2−(4−フルオロフェニル)−2−オキソエチリジン)−1,3−ジヒドロインドール−2−オン、N6,N6−ジメチルアデニン、2−(1R−イソプロピル−2−ヒドロキシエチルアミノ)−6−(3−クロロアニリノ)−9−イソプロピル−プリン、ラパマイシン、2−(R)−(1−エチル−2−ヒドロキシエチルアミノ)−6−ベンジルアミノ−9−イソプロピルプリン、スキトネミン、3−[1−(3H−イミダゾール−4−イル)−メタ−(Z)−イリデン]−5−メトキシ−1,3−ジヒドロインドール−2−オン、及び4−(3’−ヒドロキシフェニル)アミノ−6,7−ジメトキシキナゾリンからなる群から選択される。
【0134】
一実施形態では、サイクリン依存性キナーゼはエチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートである。一実施形態では、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートを、内分泌腺系に特徴的なマーカーを発現している細胞に約0.1μM〜約10μMの範囲の濃度で、約1日〜7日間にわたって加える。
【0135】
一実施形態では、内分泌腺系に特徴的なマーカーを発現している細胞を、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートで約1日〜約7日間にわたって処理する。
【0136】
以下の実施例により本発明を更に例示するが、本発明はこれらの実施例により限定されるものではない。
【実施例】
【0137】
(実施例1)
ウシ胎児血清の非存在下での、ヒト胚性幹細胞の細胞株H1の膵内分泌細胞への分化
ヒト胚性幹細胞株H1の継代数52の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと分化させるために、以下の分化プロトコルを実施した。
a.2% BSA(カタログ#152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ#1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ#100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R&D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ#565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ#616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理した(ステージ5)。
【0138】
培地は毎日交換した。各ステージにおいて血球計数器を用いて細胞数を計算し、PCR分析用にRNAを採取した。試料はすべて3つ組複製で採取した。
【0139】
(実施例2)
実施例1に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリII由来の化合物の効果のスクリーニング
継代数44のヒト胚性幹細胞株H1の細胞を、MATRIGEL(商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従ってステージ5まで分化させた。これに続き、細胞をEMD Calbiochem化合物ライブラリ(カタログ#539745,Calbiochem,San Diego,Ca)由来の化合物を最終濃度1μMで含有しているDMEM/F12+1% B27で、4日間にわたって処理した。ビヒクルを含有しているウェルを対照として含めた。プロトコルを通して、培地は毎日交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びArx4の発現について解析した。結果は、リアルタイムPCRでの測定結果により、処理サンプル対未処理対照の、インスリン/グルカゴン比(図1、パネルa)、又はMAFAとARX4の比(図1、パネルb)として表わされる。各ウェル#について対応するPubChem化合物ID#は、表1に列挙する。
【0140】
1μMの濃度の化合物A6、B7、B8、又はC2での、膵内分泌系に特徴的なマーカーを発現している細胞の処理は、およそ3.0の、あるいはそれよりも大きいインスリン/グルカゴン発現比をもたらした(図1のパネルaを参照されたい)。
【0141】
次いで、MAFA/ARX4の比に対するこれらの化合物の効果を調べたところ、数個の化合物での膵内分泌系に特徴的なマーカーを発現している細胞の処理は、ライブラリ内の他の試験化合物よりも、MAFAとARX4の比に、より大きな変化をもたらしたことを観察した:化合物C2で処理した細胞は、約1000のMAFA/ARX4比を示した。膵内分泌系に特徴的なマーカーを発現している細胞の化合物C2での処理は、約100のMAFA/ARX4比をもたらした(図1のパネルbを参照されたい)。
【0142】
(実施例3)
実施例1に概説された分化プロトコルに従って処理した細胞でのインスリン及びMAFA発現に対する、サイクリン依存性キナーゼ阻害剤処理の効果
実施例2でインスリン発現とグルカゴン発現の比、又はMAFAとARX4の比を増加させたいくつかの化合物は、サイクリン依存性キナーゼ阻害剤であった。このような化合物の1つはPubChem化合物ID# 5330797(5−アミノ−3−((4−(アミノスルホニル)フェニル)アミノ)−N−(2,6−ジフルオロフェニル)−1H−1,2,4−トリアゾール−1−カルボチオアミド)(カタログ# 217714;Calbiochem,San Diego,Ca)である。これらの観察結果を確証づけるために、MATRIGEL(登録商標)をコートした10cm2のディッシュ上で、継代数42のヒト胚性幹細胞株H1の細胞を培養し、実施例1に記載の方法に従って、ステージ5まで処理した。ステージ5以降は、1% B27と1μMのPubChem化合物ID# 5330797を含有するDMEM/F12で、細胞を6日間にわたって処理した。培地は1日おきに交換した。リアルタイムPCR用の細胞サンプルを化合物での処理前、化合物処理の2日目及び5日目に採取した。
【0143】
化合物処理対未処理対照の4日目又は6日目の細胞の特徴的な顕微鏡写真を図2に示す。未処理細胞は高密度で充填され(図3、パネルa及びd)、個別の細胞を区別することは困難である。しかしながら、0.5μM又は1μMのPubChem化合物ID#5330797での6日間にわたる処理後には、未処理対照(図2、パネルd)と比較してそれぞれの核が目に見えるようになり(図2、パネルe及びf)、細胞集団に分化が生じたことを示している。これはまた、図2のパネルb及びcに示されるような細胞層中の間隙により見出され得る、一部の細胞死を伴う。
【0144】
PubChem化合物ID# 5330797での細胞処理は、程度は異なるもののインスリン、グルカゴン、MAFA、MAFB及びソマトスタチンの発現上昇をもたらした。培養0日目(処理前)と比較した場合の、処理あたりの遺伝子発現の相対的な誘導を図3のパネルa〜vに示す。1μMのPubChem化合物ID# 5330797で処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的に処理の48時間の時点でのグルカゴン発現がおよそ1.5倍に上昇した。この発現量は、処理の5日以降に処理前のレベル未満に下降した。0.5μMのPubChem化合物ID# 5330797での処理では、グルカゴン発現の上昇は観察されなかった(図3、パネルaを参照されたい)。
【0145】
1μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にインスリン発現がおよそ1.5倍に上昇した(図3、パネルbを参照されたい)。
【0146】
1μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にMAFA発現がおよそ200倍に上昇した(図3、パネルdを参照されたい)。
【0147】
0.5μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にMAFB発現がおよそ1.5倍に上昇した(図3、パネルcを参照されたい)。ソマトスタチンの発現では、用量依存的な上昇が観察された(図3、パネルe)。
【0148】
PubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、アミラーゼの発現量は変化しなかった(図3、パネルfを参照されたい)。しかしながら、PAX4(図3、パネルh)、NKX6.1(図3、パネルk)、PDX1(図3、パネルl)、NEUROD(図3、パネルo)、及びBRN4(図3、パネルq)では発現レベルに減少が観察された。
【0149】
(実施例4)
サイクリン依存性キナーゼ阻害剤処理は、島様クラスタにおいてMAFA発現を上昇させた
継代数52のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、実施例1に記載の方法に従って分化させた。膵内分泌系に特徴的なマーカーを発現している細胞を更に成熟させるために、追加のステージ(ステージ6)を加えた。この実施例では、ステージ6はDMEM/F12+1% B27(Invitrogen,CA)での7日間の処理から構成される。培地は毎日交換した。
【0150】
ステージ6後に、細胞は1Xアキュターゼ(Sigma,MO)で、室温で5分にわたって処理した。アキュターゼを除去し、細胞にDMEM/12+1% B27を加えた。接着した細胞をセルスクレーパーを用いて取り外し、穏やかに再懸濁し、40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート(カタログ# 3471,Corning,Ma)上で懸濁液の状態で培養した。次いで細胞を以下のように処理した:細胞を、20ng/mLのアクチビンA(AA)と、1μmのCDK阻害剤III(カタログ#217714,Calbiochem,Ca)を含有しているDMEM/F12+1% B27で10日間にわたって培養した(ステージ7)。ビヒクルで処理した細胞を対照として含めた。PCR解析用に、7日目〜10日目にサンプルを回収し、ジチゾン染色した。本実施例で概説された方法に従って、懸濁状態で培養した細胞は、形態的に膵臓の島クラスタに類似であると推測された。CDK阻害剤IIIでの処理は、島様クラスタの形態に影響を与えるようには見えなかった。
【0151】
図4のパネルa〜iは、細胞クラスタの遺伝子発現プロファイルに対する、CDK阻害剤III処理の効果を示す。CDK阻害剤IIIでの処理は、膵内分泌系に関連付けられるマーカーの発現を上昇させ、特にプロインスリン転写因子のMAFAの発現を上昇させた。
【0152】
図5のパネルa〜bは、クラスタのジチゾン(dithazone)(DTZ)染色に対する、CDK阻害剤IIIの効果を示す。CDK阻害剤で処理し、かつDTZで染色した細胞クラスタは、CDK阻害剤IIIで処理していないクラスタと比較したときに、より赤みの強い染色パターンを示した。
【0153】
(実施例5)
本発明の方法により作製されたインスリン産生細胞の、FACS解析
継代数42のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたプレート上で培養し、以下のプロトコルを使用してインスリン産生細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ# 565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理し(ステージ5)、次いで
f.DMEM/F12+1% B27で7日間にわたって処理し(ステージ6)、次いで
g.アキュターゼで5分間にわたって処理した後に、接着しているあらゆる残りの細胞を掻き取りにより取り外した。次いで細胞懸濁液を40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート上で、DMEM−高グルコース(カタログ# 11995−073,Invitrogen,Ca)+1% B27+20ng/mLのアクチビンA(AA)、1μmのCDK阻害剤III(カタログ# 217714,Calbiochem,Ca)中に懸濁した状態で、5日間にわたって培養した(ステージ7)。
【0154】
TrypLE Express(Invitrogen,Carlsbad,CA)を用いて、島様クラスタを単一細胞へと分散させ、冷PBSで洗浄した。固定のため、細胞を200〜300μLのCytofix/Cytoperm緩衝液(BD 554722,BD,Ca)中に再懸濁し、4℃で30分間にわたってインキュベートした。細胞を1mLのPerm/Wash緩衝液(BD 554723)で2回洗浄し、Perm/Wash緩衝液中に2%正常ヤギ血清を含有する100μLの染色/ブロッキング溶液中に再懸濁した。フローサイトメトリー解析のために、細胞は以下の一次抗体で染色した:抗−インスリン(ウサギmAb,Cell Signaling No.C27C9;1:100希釈);抗−グルカゴン(マウスMab,Sigma No.G2654,1:100);抗−シナプトフィジン(ウサギポリクローナル抗体,DakoCytomation No A0010,1:50)。細胞を4℃で30分にわたってインキュベートし、Perm/Wash緩衝液で2回洗浄し、更に30分間にわたって、以下のような適切な2次抗体中でインキュベートした:ヤギ抗−ウサギAlexa 647(Invitrogen No.A21246)又はヤギ抗−マウス647(Invitrogen No.A21235);ヤギ抗−ウサギR−PE(BioSource No.ALI4407)。二次抗体はすべて1:200の希釈率で使用した。細胞は、少なくとも1回Perm/Wash緩衝液中で洗浄し、BD FACSアレイを使用して分析した。少なくとも10,000事象を分析のために得た。対照には未分化のH1細胞とβ−TC(CRL−11506(商標)ATCC,VA)細胞株を含んだ。
【0155】
図6のパネルa〜cは、ビヒクルを含有している培地での、ステージ7での処理後の細胞に占める、インスリン陽性細胞、シナプトフィジン(synapthophysin)陽性細胞、及びグルカゴン陽性細胞の割合を示す。図7のパネルa〜cは、1μMのCDK阻害剤IIIを含有している培地での、5日間にわたるステージ7での処理後の、インスリン陽性細胞、シナプトフィジン(synapthophysin)陽性細胞、及びグルカゴン陽性細胞の割合を示す。インスリンホルモン単独の陽性細胞の数は、CDK阻害剤での処理後に3%から8%へと上昇した。加えて、多ホルモン(インスリン及びグルカゴン陽性)細胞の割合は、CDK阻害剤での処理後に減少した。
【0156】
(実施例6)
CDK阻害剤誘導型のMAFA発現の動態
継代数42のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたプレート上で培養し、以下のプロトコルを使用してインスリン産生細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ# 565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理し(ステージ5)、次いで
f.DMEM/F12+1% B27で7日間にわたって処理し(ステージ6)、次いで
g.アキュターゼで5分間にわたって処理した後に、接着しているあらゆる残りの細胞を掻き取りにより取り外した。次いで細胞懸濁液を40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート上で、DMEM−高グルコース(カタログ# 11995−073,Invitrogen,Ca)+1% B27+20ng/mLのアクチビンA(AA)、2μmのCDK阻害剤III(カタログ# 217714,Calbiochem,Ca)中に懸濁した状態で、1〜8日間にわたって培養した(ステージ7)。
【0157】
1、2、3及び4日目のサンプルを、PCR解析のために回収した。CDK阻害剤での処理の4日後に、CDK阻害剤を培養物から除去し、DMEM−F12+1% B27+20ng/mLアクチビンA中で更に4日間培養した。4日間の終了時に、PCR解析のためにサンプルを三つ組複製で回収した。
【0158】
図8のパネルa〜bは、ステージ7の様々な時点でのMAFA及びインスリンの発現パターンを示す。CDK阻害剤処理は、MAFA発現とインスリン発現に有意な上昇をもたらし、これらは時間関数として上昇した。しかしながら、CDK阻害剤の除去は、化合物除去の4日後に得たサンプルにおいてMAFA及びインスリン発現の両方で有意な低下をもたらした。
【0159】
(実施例7)
実施例1に概説された分化プロトコルに従って処理された細胞に対する、BIOMOL(商標)キナーゼ阻害剤ライブラリ由来の化合物の効果のスクリーニング
継代数51のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従って、ステージ5まで分化させた。これに続き、細胞をDMEM/F12+1% B27中で1日間生育させ、次いでBIOMOL(商標)化合物ライブラリ(カタログ# 2832,BIOMOL,Plymouth Meeting,Pa)由来の化合物を最終濃度4μMで含有しているDMEM/F12+1% B27中で、6日間にわたって処理した。ビヒクルを含有しているウェルを対照として含めた。処理プロトコルを通し、ビヒクル又は化合物を含有している培地は1日おきに交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びARX4の発現について解析した。結果は、リアルタイムPCRでの測定結果により、処理サンプル対未処理対照の、インスリン/グルカゴン比(表2)、又はMAFAとArx4の比(表2)として表わされる。各英数字ウェル#に対する、対応するカタログ#、CAS#、及び化合物名又はID数を表3に列挙する。
【0160】
4μMの濃度の化合物C8又はF1での、膵内分泌系に特徴的なマーカーを発現している細胞の処理は、およそ10.0の、あるいはそれよりも大きいインスリン/グルカゴン発現比をもたらした。D9で処理した細胞は、およそ1840.0のインスリン/グルカゴン発現比を有する(表2)。
【0161】
次いで、MAFA/ARX4の比に対するこれらの化合物の効果を調べたところ、数個の化合物での膵内分泌系に特徴的なマーカーを発現している細胞の処理は、ライブラリ内の他の試験化合物よりも、MAFAとARX4の比に、より大きな変化をもたらしたことを観察した:化合物B6又はF1で処理した細胞は、およそ10を超えるMAFA/ARX4比を示した。膵内分泌系に特徴的なマーカーを発現している細胞の化合物C8での処理は、約84のMAFA/ARX4比をもたらした一方で、D9で処理した細胞は約212のMAFA/ARX4比を有した(表2)。
【0162】
(実施例8)
実施例1に概説された分化プロトコルに従って処理した細胞でのインスリン及びMAFA発現に対する、サイクリン依存性キナーゼ阻害剤の効果
継代数51のヒト胚性幹細胞株H1の細胞を、MATRIGEL(商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従ってステージ5まで分化させた。これに続き、細胞をDMEM/F12+1 %B27中で8日間にわたって生育させ、次いでサイクリン依存性キナーゼ阻害剤を最終濃度0.6125、1.25、又は5.0μMで含有しているDMEM/F12+1 %B27中で、4日間にわたって処理した。6種の阻害剤を試験した:PubChem ID# 5330812(EMD cat# 217714)、PubChem ID# 4566(EMD cat#217713)、PubChem ID# 5330797(EMD cat# 219476)、PubChem ID# 73292(EMD cat#341251)、PubChem ID #4592(EMD cat#495620)、及びPubChem ID# 160355(EMD cat #557360)。ビヒクルを含有しているウェルを対照として含めた。処理プロトコルを通し、ビヒクル又は化合物を含有している培地は1日おきに交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びARX4の発現について解析した。結果は、リアルタイムPCRにより測定されたものとして、ビヒクル処理対照に対する倍数変化として記載される。
【0163】
化合物PubChem ID# 5330812、PubChem ID# 4566、PubChem ID# 5330797、及びPubChem ID# 73292の全てが、試験濃度でMAFA発現を刺激したことを観察した(表4)。PubChem ID# 4592及びPubChem ID# 160355は試験濃度ではMAFAを刺激しなかった(表4)。化合物PubChem ID# 5330812、PubChem ID# 4566、PubChem ID# 5330797、PubChem ID# 4592、及びPubChem ID# 160355の全てが、インスリン発現を刺激したことは明らかであった(表4)。化合物(PubChem ID# 5330797)はグルカゴン及びArx4発現(表4)の両方を減少させた一方で、MAFA発現を刺激した。
【0164】
(実施例9)
ウシ胎児血清を欠き、25mMグルコースを含有しているDMEM(DMEM−HG)での、ヒト胚性幹細胞の細胞株H1の膵内分泌細胞への分化
ヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、以下のプロトコルを用いて、膵内分泌系に特徴的なマーカーを発現している細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)を加えたRPMI培地で2日間にわたって処理し(ステージ2)、次いで
c.DMEM−HG+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で6日間にわたって処理し(ステージ3)、次いで
d.DMEM−HG+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM−HG+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理した(ステージ5)。
【0165】
培地は毎日交換した。各ステージにおいて血球計数器を用いて細胞数を計算し、PCR解析用にRNAを採取した。試料はすべて3つ組複製で採取した。
【0166】
(実施例10)
実施例9に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリI由来の化合物の効果のスクリーニング
継代数45のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例9に記載の方法に従ってステージ5まで分化させた。これに続き細胞を供給し、ステージ5の1、3及び5日目に、DMEM−HG、1% B27(Invitrogen,CA)と、1μMのALK5阻害剤II(Calbiochem,Ca)と、DMSO(カタログ# 539744,Calbiochem,San Diego,Ca)に溶解し、2.5μMの最終濃度で処理したEMD Calbiochem化合物ライブラリ1由来の化合物とを含む培地で処理した。ビヒクルを含有しているウェルを対照として含めた。培地はステージ5では1日おきに交換したことを除き、プロトコルを通して毎日交換した。全てのサンプルは2つ組複製で処理した。
【0167】
この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、MAFAの発現について解析した。結果はリアルタイムPCRにより測定されたものとして、MAFA発現H1ヒト胚性幹細胞対未処理H1ヒト胚性幹細胞の倍数増加として表わす(表5)。
【0168】
膵内分泌系に特徴的なマーカーを発現している細胞の、2.5μM濃度の化合物A4(Cat#,124001,Akt阻害剤IV)、E8(Cat# 527450,PKR阻害剤)、及びF9(Cat#539648,スタウロスポリン,N−ベンゾイル−)での処理は、MAFA発現の、ビヒクル処理対照を少なくとも4倍の上昇をもたらした(表5)。2.5μM濃度の化合物E6(Cat# 521233,PDGF受容体チロシンナーゼ阻害剤IV)での処理は、ビヒクル処理対照を少なくとも2.5倍超えるMAFA発現の上昇をもたらした(表5)。
【0169】
(実施例11)
実施例9に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリII由来の化合物の効果のスクリーニング
継代数46のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例9に記載の方法に従ってステージ5まで分化させた。これに続き細胞を供給し、ステージ5の1、3及び5日目に、DMEM−HGと、1% B27(Invitrogen,CA)と、1μMのALK5阻害剤II(Calbiochem,Ca)(ステージ5)と、DMSO(表1及び6,Calbiochem,San Diego,Ca)に溶解させ最終濃度2.5μMに処理したEMD Calbiochem化合物ライブラリII由来の化合物と、を含む培地で処理した。ビヒクルを含有しているウェルを対照として含めた。培地はステージ5では1日おきに交換したことを除き、プロトコルを通して毎日交換した。全てのサンプルは2つ組複製で処理した。
【0170】
この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、MAFAの発現について解析した。リアルタイムPCRにより測定されたものとして、MAFA発現を刺激した化合物に関する結果を示し、かつMAFA発現対対照サンプルにおける倍数上昇として表わす(図9)。
【0171】
膵内分泌系に特徴的なマーカーを発現している細胞の、2.5μM濃度の:アルステルパウロン、2−シアノエチル;SU9516;アルステルパウロン;Cdk1/2阻害剤III;カゼインキナーゼI阻害剤、D4476;又はMEK1/2阻害剤のいずれかでの処理は、未処理対照に対してMAFA発現の4.5倍の上昇をもたらした(表7)。
【0172】
(実施例12)
膵内分泌系に特徴的なマーカーを発現している細胞でMAFA発現を促進する低分子阻害剤での、膵内分泌系に特徴的なマーカーを発現している細胞の細胞周期の進行の阻害
細胞周期の進行に由来して生じる細胞増殖は、細胞外増殖因子で細胞を刺激することにより活性化及び維持され得る。増殖因子は増殖因子受容体の細胞外ドメインへと結合し、受容体の細胞内ドメインの立体構造スイッチを誘導する。このシフトは受容体二量体化及び受容体の細胞内ドメイン上に位置するチロシンキナーゼの活性化を開始し、下流の複数のセリン/スレオニンキナーゼのリン酸化及び活性化を導き、最終的には細胞周期の進行と細胞増殖をもたらす。
【0173】
通常の生理学的な条件下では、インスリンと転写因子MAFAの発現とにより特徴づけられる成熟した膵臓β細胞は無活動状態であり、細胞周期のG0に留まる傾向がある。しかしながら、機能性臓器を形成するのに十分な細胞を生成し、成熟した動物に関する要求を満たす、本発明の膵内分泌系に特徴的なマーカーを発現している細胞は、細胞周期を循環している必要がある。その結果、本発明の膵内分泌系に特徴的なマーカーを発現している細胞は胚発生のある時点でβ細胞へと分化し、活動的に細胞周期により増殖している細胞から、休眠細胞へと移行する。
【0174】
我々のデータは、低分子キナーゼ阻害剤によりシグナルカスケードをブロックして細胞周期の進行を阻害することで、膵内分泌系に特徴的なマーカーを発現している細胞に、成熟膵臓β細胞のマーカーであるMAFAの発現を誘導させ得ることを示す。増殖因子受容体(PDGF受容体チロシンキナーゼ阻害剤IV)に標的とされるキナーゼ阻害剤、又はチロシンキナーゼ受容体の下流のキナーゼを中断する阻害剤(MEK1/2阻害剤、PKR阻害剤、又はAkt阻害剤IV)は、増殖性の、増殖因子/キナーゼに基づくシグナル伝達を中断して細胞周期の休止をもたらし、MAFA発現を誘導する。スタウロスポリンのような広域スペクトル阻害剤の使用は、効果的にMAFAを誘導できるが、有効濃度では細胞毒性でもある。サイクリン依存性キナーゼ阻害剤(アルステルパウロン、2−シアノエチル;SU9516;アルステルパウロン;又はCdk1/2阻害剤III)のような、より直接的な化合物は、スタウロスポリンのような広域スペクトル阻害剤と比較して、より少ない細胞毒性でMAFAを誘導する。
【0175】
広域スペクトルのキナーゼ阻害剤が、MAFA発現と、本発明の、膵内分泌系に特徴的なマーカーを発現している、より成熟した表現型の細胞とを誘導し得るか判定するために、H1ヒトES細胞を実施例9に記載の方法に従って分化させ、ステージ5の1、3、及び5日目に、タンパク質チロシンキナーゼ阻害剤のゲニステイン(ヒト及びマウス細胞株のG2期を休止させ、複数のキナーゼを阻害することが示されている)で処理した。内分泌腺ホルモンのインスリン、ソマトスタチン、及び転写因子MAFAの全てが、投与量10及び30ng/mLで未処理対照に対して発現量の増加を示した一方で、内分泌腺ホルモングルカゴンは10ng/mLで発現を増加させた(図10)。我々は投与量100ng/mLでのゲニステインで、インスリン、グルカゴン、及びソマトスタチン発現の損失と関連付けられる有意な毒性を観察した。
【0176】
これらのデータは、増殖因子受容体チロシンキナーゼから細胞内シグナル伝達キナーゼを介して核及びサイクリン依存性キナーゼに向かうシグナル伝達を阻害するために、標的にされる低分子量のキナーゼ阻害剤により、シグナルカスケードをブロックすることで、細胞周期の進行を阻害することにより、本発明の、膵内分泌系に特徴的なマーカーを発現している細胞に、成熟膵臓β細胞のマーカーであるMAFAを発現させることができることを示す。
【0177】
本明細書を通して引用された刊行物は、その全体が参照により本明細書に組み込まれる。以上、本発明の様々な態様を実施例及び好ましい実施形態を参照して説明したが、本発明の範囲は、上記の説明文によってではなく、特許法の原則の下で適切に解釈される以下の特許請求の範囲によって定義されるものである点は認識されるであろう。
【0178】
【表1】

【0179】
【表2−1】
【0180】
【表2−2】
【0181】
【表2−3】
【0182】
【表3−1】
【0183】
【表3−2】
【0184】
【表3−3】
【0185】
【表4】
【0186】
【表5−1】
【0187】
【表5−2】
【0188】
【表5−3】
【0189】
【表6−1】
【0190】
【表6−2】
【0191】
【表6−3】
【0192】
【表7】
【図1−1】
【図1−2】
【技術分野】
【0001】
本発明は、米国特許出願シリアル番号第61/110,287号(2008年10月31日出願)に対する優先権を請求する。
【0002】
(発明の分野)
本発明は、多能性幹細胞の分化を促進する方法を提供する。具体的には本発明は、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【背景技術】
【0003】
I型糖尿病の細胞置換療法における進歩及び移植可能なランゲルハンス島の不足のため、生着に適したインスリン産生細胞すなわちβ細胞の供給源の開発に注目が集まっている。1つの手法は、例えば、胚性幹細胞のような多能性幹細胞から機能性のβ細胞を生成することである。
【0004】
脊椎動物の胚発生において、多能性細胞は、原腸形成として公知のプロセスにて、3つの胚葉(外胚葉、中胚葉、及び内胚葉)を含む細胞のグループを生じる。例えば、甲状腺、胸腺、膵臓、腸、及び肝臓等の組織は、内胚葉から中間ステージを経て発達する。このプロセスにおける中間ステージは、胚体内胚葉の形成である。胚体内胚葉細胞はHNF−3β、GATA4、MIXL1、CXCR4及びSOX17などの多数のマーカーを発現する。
【0005】
膵臓の形成は、胚体内胚葉の膵臓内胚葉への分化により起こる。膵臓内胚葉の細胞は膵臓−十二指腸ホメオボックス遺伝子、PDX1を発現する。PDX1が存在しない場合、膵臓は、腹側芽及び背側芽の形成を越えて発達しない。したがって、PDX1の発現は、膵臓器官形成において重要な工程となっている。成熟した膵臓は、他の細胞型間で外分泌組織及び内分泌組織を含有する。外分泌組織及び内分泌組織は、膵臓内胚葉の分化によって生じる。
【0006】
島細胞の特徴を保持する細胞がマウスの胚細胞から誘導されたことが報告されている。例えば、Lumelskyら(Science 292:1389、2001年)は、マウスの胚性幹細胞の、膵島と同様のインスリン分泌構造への分化を報告している。Soriaら(Diabetes 49:157、2000年)は、ストレプトゾトシン糖尿病のマウスにおいて、マウスの胚性幹細胞から誘導されたインスリン分泌細胞が糖血症を正常化することを報告している。
【0007】
一例において、Horiら(PNAS 99:16105,2002)は、ホスホイノシチド3−キナーゼ(LY294002)の阻害剤でマウス胚性幹細胞を処理することにより、β細胞に類似した細胞が生じたことを開示している。
【0008】
他の例では、Blyszczukら(PNAS 100:998、2003年)が、Pax4を構成的に発現しているマウス胚性幹細胞からのインスリン産生細胞の生成を報告している。
【0009】
Micallefらは、レチノイン酸が、胚性幹細胞のPdx1陽性膵臓内胚葉の形成に対する関与を制御できることを報告している。レチノイン酸は、胚における原腸形成の終了時に対応する期間中の、胚性幹細胞分化の4日目に培養液に添加すると、Pdx1発現の誘発に最も効果的である(Diabetes 54:301、2005年)。
【0010】
Miyazakiらは、Pdx1を過剰発現しているマウス胚性幹細胞株を報告している。Miyazakiらの結果は、外因性のPdx1発現が、得られた分化細胞内でインスリン、ソマトスタチン、グルコキナーゼ、ニューロゲニン3、P48、Pax6、及びHNF6遺伝子の発現を明らかに増加させたことを示している(Diabetes 53:1030、2004年)。
【0011】
Skoudyらは、マウス胚性幹細胞内で、アクチビンA(TGF−βスーパーファミリーのメンバー)が、膵臓外分泌遺伝子(p48及びアミラーゼ)、並びに内分泌遺伝子(Pdx1、インスリン及びグルカゴン)の発現を上方制御することを報告している。最大の効果は、1nMアクチビンAを使用した場合に認められた。Skoudyらはまた、インスリン及びPdx1 mRNAの発現レベルはレチノイン酸により影響されなかったが、3nMのFGF7による処理によりPdx1の転写産物のレベルが増大したことも観察している(Biochem.J.379:749,2004)。
【0012】
Shirakiらは、胚性幹細胞のPdx1陽性細胞への分化を特異的に増大させる増殖因子の効果を研究した。彼らは、TGF−β2によってPdx1陽性細胞が高い比率で再現可能に得られたことを観察している(Genes Cells.2005 Jun;10(6):503〜16.)。
【0013】
Gordonらは、血清の非存在下、かつアクチビンとWntシグナル伝達阻害剤の存在下で、マウス胚性幹細胞からbrachyury+/HNF−3β+内胚葉細胞が誘導されることを示している(米国特許出願公開2006/0003446(A1)号)。
【0014】
Gordonら(PNAS,Vol 103,page 16806,2006)は、「Wnt及びTGF−β/nodal/アクチビンの同時シグナル伝達が前原始線条の形成には必要であった」と述べている。
【0015】
しかしながら、胚性幹細胞発達のマウスモデルは、例えば、ヒトなどのより高等な哺乳動物における発達プログラムを正確には模倣しない恐れがある。
【0016】
Thomsonらは、ヒト胚盤胞から胚性幹細胞を単離した(Science 282:114,1998年)。同時に、Gearhart及び共同研究者は、胎児性腺組織から、ヒト胚性生殖(hEG)細胞株を誘導した(Shamblottら、Proc.Natl.Acad.Sci.USA 95:13726,1998)。白血病抑制因子(LIF)と共に培養するのみでも分化を阻止し得るマウス胚性幹細胞とは異なり、ヒト胚性幹細胞は、非常に特殊な条件下で維持する必要がある(米国特許第6,200,806号、国際公開第99/20741号;同第01/51616号)。
【0017】
D’Amourらは、高濃度のアクチビン及び低濃度の血清の存在下で、ヒト胚性幹細胞由来の胚体内胚葉の濃縮化された培養物が調製されたことを述べている(Nature Biotechnology 2005)。これらの細胞を、マウスの腎臓被膜下に移植することにより、一部の内胚葉性器官の特徴を有する、より成熟した細胞への分化が得られた。ヒト胚性幹細胞由来の胚体内胚葉細胞は、FGF−10の添加後、PDx1陽性細胞に更に分化することができる(米国特許出願公開第2005/0266554(A1)号)。
【0018】
D’Amourら(Nature Biotechnology−24,1392〜1401(2006))は、「我々は、ヒト胚性幹細胞(hES)を、インスリン、グルカゴン、ソマトスタチン、膵臓ポリペプチド及びグレリンといった膵臓ホルモンを合成可能な内分泌細胞に転換させる分化プロセスを開発した。このプロセスは、胚体内胚葉、腸管内胚葉、膵臓内胚葉及び内分泌前駆体が、内分泌ホルモンを発現する細胞へと向かう段階に類似した段階を通して細胞を指向させることにより、インビボでの膵臓器官形成を模倣する。」と述べている。
【0019】
別の例において、Fiskらは、ヒト胚性幹細胞から膵島細胞を産生するシステムを報告している(米国特許出願公開第2006/0040387(A1)号)。この場合、分化経路は3つのステージに分割された。先ず、ヒト胚性幹細胞を、酪酸ナトリウムとアクチビンAの組み合わせを用いて内胚葉に分化した。次に細胞をノギンなどのTGF−βアンタゴニストとEGF又はベータセルリンの組み合わせと培養してPDx1陽性細胞を生成する。最終分化は、ニコチンアミドにより誘発された。
【0020】
1つの例において、Benvenistryらは、「我々は、Pdx1の過剰発現が膵臓に多く見られる遺伝子の発現を高めたことを結論付ける。インスリン発現の誘導には、インビボでのみ存在する更なるシグナルを必要とする可能性がある。」と述べている(Benvenistryら、Stem Cells 2006;24:1923〜1930)。
【0021】
サイクリンはβ細胞機能に関係していると見なされてきた。例えば、Liljaらは、「インスリンを分泌する膵臓β細胞にはCdk5が存在する」と報告する(J.Biol.Chem.,Vol.276,Issue 36,34199〜34205,September 7,2001)。Liljaらは、「Cdk5はβ細胞中に存在し、インスリン開口分泌の正の調節因子として作用する。」と記載する。
【0022】
他の例では、Marzoらは「Cdk4ノックインマウスではβ細胞集団が有意に増加し、かつCdk4ノックインマウスは、Cdk4がI型糖尿病において膵臓β細胞集団を再生するのに潜在的な標的であることを示す、生理学的機能を有する」と記載する(Diabetalogia,Vol.47,Number4,686〜694,April 1,2004.)。
【0023】
他の例では、Ubedaらは、サイクリン依存性キナーゼ5活性の阻害が、膵臓β細胞を糖毒性から保護するということを報告する(J.Biol.Chem.,Vol.281,Issue 39,28858〜28864,September 29,2006)。
【0024】
他の例では、Weiらは、グルコース刺激型のインスリン分泌の、Cdk5依存性の制御を報告する(Nature Medicine 11,1104〜1108(1 October 2005.))。
【0025】
他の例では、Vanderfordらは以下のように記載している。「MafAは膵臓のβ細胞内で発現される塩基性ロイシンジッパー型転写制御因子であり、β細胞のバイオロジーの様々な態様に関与していることから正常なグルコース恒常性を維持するために必要とされる。MafAたんぱく質レベルは、未だ十分には特徴づけられていないメカニズムによって、高濃度のブドウ糖に反応して増加することが知られている。我々は、別々の細胞内シグナル伝達イベントがmafA発現を制御するのか否かを調査した。我々は、一般的なキナーゼ阻害剤であるスタウロスポリンが、タンパク質の安定性を変化させることなくmafA発現を誘導することを見出した。MAPキナーゼJNKの阻害は、mafA発現に対するスタウロスポリンの効果を模倣する。カルモジュリンキナーゼとカルシウムシグナル伝達も同様に、高濃度のブドウ糖によりmafA発現を刺激するのに重要である。しかしながら、スタウロスポリン、JNK、及びカルモジュリンキナーゼは、インスリン発現の誘導に対して異なる効果を有する。これらのデータは、MafAレベルが、複数のキナーゼ経路の協調作用により厳密に制御されていることを明らかにした。」(Archives of Biochemistry and Biophysics(2008),doi:10.1016/j.abb.2008.10.001)。
【発明の概要】
【発明が解決しようとする課題】
【0026】
したがって、多能性幹細胞を膵内分泌細胞、膵臓ホルモン発現細胞、又は膵臓ホルモン分泌細胞へと分化させるための方法の開発に対して、未だに有意な必要性が存在している。本発明は、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【課題を解決するための手段】
【0027】
一実施形態では、本発明は、膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【図面の簡単な説明】
【0028】
【図1】パネルaは、膵内分泌系に特徴的なマーカーを発現している細胞でのインスリン発現とグルカゴン発現の比に対する、EMD Calbiochemキナーゼ阻害剤ライブラリ由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。英数字ラベルは表1に示される同一の化合物と対応する。パネルbは、膵内分泌系に特徴的なマーカーを発現している細胞でのMAFA発現とARX4発現の比に対する、EMD Calbiochemキナーゼ阻害剤ライブラリ由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。英数字ラベルは表1に示される同一の化合物と対応する。
【図2A】実施例1に記載の方法に従って処理した、ステージ6処理の4日目の細胞の4x顕微鏡像を示す。
【図2B】0.5μMの化合物PubChemID# 5330812で処理した、処理の4日目の細胞の4x顕微鏡像を示す。
【図2C】1μMの化合物PubChemID# 5330812で処理した、処理の4日目の細胞の4x顕微鏡像を示す。
【図2D】実施例1に記載の方法に従って処理した、ステージ6処理の6日目の細胞の20x顕微鏡像を示す。
【図2E】0.5μMの化合物PubChemID #5330812で処理した、処理の6日目の細胞の20x顕微鏡像を示す。
【図2F】1μMの化合物PubChemID# 5330812で処理した、処理の6日目の細胞の20x顕微鏡像を示す。
【図3】0.5μM(濃色のバー)又は1.0μM(淡色のバー)の化合物PubChem ID#5330812での5日間の処理後の、膵内分泌系に特徴的なマーカーを発現している細胞での、23種の遺伝子の発現を示す。発現レベルは0日目、2日目及び5日目で測定した。
【図4】実施例4に記載の分化プロトコルの、ステージ7で処理した細胞における、膵内分泌系に特徴的なマーカーの発現に対する、CDK阻害剤III処理の効果を示す。
【図5】島様のクラスタのジチゾン(dithazone)染色に対する、CDK阻害剤III処理の効果を示す。
【図6】実施例5に記載の方法に従って生産されたインスリン産生細胞中の、インスリン、シナプトフィジン及びグルカゴンの発現を示す。記載のたんぱく質の発現はFACSにより測定した。
【図7】実施例5に記載の方法に従って生産されたインスリン産生細胞中の、インスリン、シナプトフィジン及びグルカゴンの発現を示す。記載のたんぱく質の発現はFACSにより測定した。
【図8】本発明の方法により生産された、インスリン産生細胞中のMAFA(パネルa)及びインスリン(パネルb)の発現を示す。1、2、3及び4日目の細胞サンプルを、PCR解析に用いた。CDK阻害剤での処理の4日後に、CDK阻害剤を培養物から除去し、DMEM−F12+1%B27+20ng/mLアクチビンA中で更に4日間培養した。4日間の終了時に、PCR解析のためにサンプルを三つ組複製で回収した。
【図9】膵内分泌系に特徴的なマーカーを発現している細胞でのMAFA発現に対する、EMD Calbiochemキナーゼ阻害剤ライブラリI由来の化合物の効果を、リアルタイムPCRにより測定されたものとして示す。
【図10】膵内分泌系に特徴的なマーカーを発現している細胞での、インスリン、グルカゴン、ソマトスタチン及びMAFAのmRNA発現に対するゲニステイン(genestein)の効果を、リアルタイムPCRにより測定されたものとして示す。
【発明を実施するための形態】
【0029】
開示を明確にするために、本発明の「発明を実施するための形態」を、限定を目的とすることなく、本発明の特定の特徴、実施形態、又は応用を説明若しくは図示した以下の小項目に分ける。
【0030】
定義
幹細胞は、単一の細胞レベルにて自己複製し、分化して後代細胞を生成する、それら両方の能力で定義される未分化細胞であり、後代細胞には、自己複製前駆細胞、非再生前駆細胞、及び最終分化細胞が含まれる。幹細胞はまた、インビトロで複数の胚葉(内胚葉、中胚葉及び外胚葉)から様々な細胞系の機能的細胞に分化する能力によって、また移植後に複数の胚葉の組織を生じ、胚盤胞への注入後、全部ではないとしても殆どの組織を提供する能力によっても、特徴付けられる。
【0031】
幹細胞は、発生上の能力によって、(1)全ての胚性又は胚体外細胞のタイプを生ずる能力を有することを意味する、分化全能性、(2)全ての胚性細胞のタイプを生ずる能力を有することを意味する、分化万能性、(3)細胞系のサブセットを生ずる能力を有するが、それらが全て特定の組織、臓器、又は生理学的システムのものであるような、分化多能性(例えば、造血幹細胞(HSC)は、HSC(自己再生性)、血球限定的寡能性前駆細胞、及び、血液の通常の成分である全ての細胞種及び要素(例えば、血小板)を生じうる)、(4)多能性幹細胞よりも限定された細胞系のサブセットを生ずる能力を有することを意味する、分化寡能性、及び(5)単一の細胞系(例えば、精原幹細胞)を生ずる能力を有することを意味する、分化単一性に分類される。
【0032】
分化は、非特殊化の(「中立の」)又は比較的特殊化されていない細胞が、例えば、神経細胞又は筋細胞などの特殊化した細胞の特徴を獲得するプロセスである。分化した、又は分化を誘発された細胞は、細胞系内でより特殊化した(「傾倒した」)状況を呈している細胞である。分化プロセスに適用した際の用語「傾倒した」は、通常の環境下で特定の細胞型又は細胞型の小集合に分化し続ける分化経路の地点に進行しており、通常の環境下で異なる細胞型に分化し、又はより分化していない細胞型に戻ることができない細胞を指す。脱分化は、細胞が細胞系内で比較的特殊化されて(又は傾倒して)いない状況に戻るプロセスを指す。本明細書で使用するとき、細胞系は、細胞の遺伝、すなわち、その細胞がどの細胞から来たか、またどの細胞を生じ得るかを規定する。細胞系は、細胞を発達及び分化の遺伝的スキーム内に配置する。系特異的なマーカーは、対象とする系の細胞の表現型に特異的に関連した特徴を指し、中立細胞の対象とする系への分化を評価する際に使用することができる。
【0033】
「β−細胞系」は、転写因子PDX−1、及び以下の転写因子、すなわち、NGN3、NKX2.2、NKX6.1、NEUROD、ISL1、HNF3 β、MAFA、PAX4、又はPAX6の少なくとも1つについて遺伝子の発現が陽性である細胞を指す。β細胞系に特徴的なマーカーを発現している細胞としては、β細胞が挙げられる。
【0034】
本明細書で使用するとき、「胚体内胚葉系に特徴的なマーカーを発現している細胞」とは、以下のマーカー、すなわちSOX17、GATA−4、HNF−3 β、GSC、CER1、Nodal、FGF8、Brachyury、Mix様ホメオボックスタンパク質、FGF4 CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA−6、CXCR4、C−Kit、CD99又はOTX2のうちの少なくとも1つを発現する細胞を指す。胚体内胚葉系に特徴的なマーカーを発現している細胞としては、原始線条前駆体細胞、原始線条細胞、中内胚葉細胞及び胚体内胚葉細胞が挙げられる。
【0035】
本明細書で使用するとき、「膵臓内胚葉系に特徴的なマーカーを発現している細胞」とは、以下のマーカー、すなわち、PDX1、HNF1 β、PTF1 α、HNF−、又はHB9のうちの少なくとも1つを発現している細胞を指す。膵臓内胚葉系に特徴的なマーカーを発現している細胞としては、膵臓内胚葉細胞、原腸管細胞、後部前腸細胞が挙げられる。
【0036】
本明細書で使用するとき、「膵内分泌系に特徴的なマーカーを発現している細胞」は、以下のマーカー、すなわち、NGN3、NEUROD、ISL1、PDX1、NKX6.1、PAX4、NGN3、又はPTF1 αのうちの少なくとも1つを発現している細胞を指す。膵臓内分泌系に特徴的なマーカーを発現する細胞としては、膵臓内分泌細胞、膵臓ホルモン発現細胞及び膵臓ホルモン分泌細胞、並びにβ細胞系の細胞が挙げられる。
【0037】
本明細書で使用するとき、「胚体内胚葉」は、原腸形成中、胚盤葉上層から生じ、胃腸管及びその誘導体を形成する細胞の特徴を保持する細胞を指す。胚体内胚葉細胞は、以下のマーカー、すなわち、HNF−3 β、GATA4、SOX17、Cerberus、OTX2、グースコイド、C−Kit、CD99、及びMIXL1を発現する。
【0038】
本明細書で使用するとき、「胚体外内胚葉」は、以下のマーカー、すなわち、SOX7、AFP、又はSPARCのうちの少なくとも1つを発現する細胞の集団を指す。
【0039】
本明細書で使用するとき、「マーカー」とは、対象とする細胞で差異的に発現される核酸又はポリペプチド分子である。本文脈において、差異的な発現は、陽性マーカーのレベルの増大及び陰性マーカーのレベルの減少を意味する。検出可能なレベルのマーカー核酸又はポリペプチドは、他の細胞と比較して対象とする細胞内で十分高く又は低く、そのため当該技術分野において既知の多様な方法のいずれかを使用して、対象とする細胞を他の細胞から識別及び区別することができる。
【0040】
本明細書で使用するとき、「中内胚葉細胞」は、以下のマーカー、すなわちCD48、eomesodermin(EOMES)、SOX17、DKK4、HNF3 β、GSC、FGF17、又はGATA6のうちの少なくとも1つを発現している細胞を指す。
【0041】
本明細書で使用するとき、「膵臓内分泌細胞」又は「膵臓ホルモン発現細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、又は膵臓ポリペプチドのうちの少なくとも1つを発現することができる細胞を指す。
【0042】
本明細書で使用するとき、「膵臓内胚葉細胞」は、以下のマーカー、すなわち、NGN3、NEUROD、ISL1、PDX1、PAX4又はNKX2.2のうちの少なくとも1つを発現することが可能な細胞を指す。
【0043】
本明細書で使用するとき、「膵臓ホルモン産生細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、又は膵臓ポリペプチドのうちの少なくとも1つを産生することが可能な細胞を指す。
【0044】
本明細書で言うところの「膵臓ホルモン分泌細胞」とは、以下のホルモン、すなわちインスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを分泌することが可能な細胞を指して言う。
【0045】
本明細書で使用するとき、「前腸後部細胞」は、以下のマーカー、すなわちPDX1、HNF1、PTF1 α、HNF6、HB9、又はPROX1のうちの少なくとも1つを分泌することができる細胞を指す。
【0046】
本明細書で使用するとき、「前原始線条細胞」とは、以下のマーカー、すなわち、Nodal、又はFGF8のうちの少なくとも1つを発現する細胞を指す。
【0047】
本明細書で使用するとき、「原腸管細胞」とは、以下のマーカー、すなわち、HNF−1又はHNF4Aのうちの少なくとも1つを分泌することができる細胞を指す。
【0048】
本明細書で使用するとき、「原始線条細胞」とは、以下のマーカー、すなわち、Brachyury、Mix様ホメオボックスタンパク質、又はFGF4のうちの少なくとも1つを発現する細胞を指す。
【0049】
多能性幹細胞の単離、増殖及び培養
多能性幹細胞の特徴付け
多能性幹細胞は、ステージ特異的胚抗原(SSEA)3及び4の1以上、並びにTra−1−60及びTra−1−81と呼ばれる抗体によって検出可能なマーカーを発現し得る(Thomsonら、Science 282:1145,1998)。インビトロで多能性幹細胞を分化させると、SSEA−4、Tra−1−60、及びTra−1−81の発現が消失し(存在する場合)、SSEA−1の発現が増大する。未分化の多能性幹細胞は通常アルカリホスファターゼ活性を有し、これは、細胞を4%パラホルムアルデヒドで固定した後、製造業者(Vector Laboratories(Burlingame Calif.))によって述べられるようにVectorRedを基質として現像することによって検出することができる。未分化多能性幹細胞はまた、RT−PCRで検出されるように、一般にOct−4及びTERTも発現する。
【0050】
増殖させた多能性幹細胞の別の望ましい表現型は、内胚葉、中胚葉、及び外胚葉組織の3胚葉のすべての細胞に分化する能力である。多能性幹細胞の多能性は、例えば、細胞を重症複合免疫不全症(SCID)マウスに注入し、形成される奇形腫を4%パラホルムアルデヒドで固定し、次いでこれを3つの胚細胞層からの細胞種の証拠について組織学的に調べることによって確認することができる。代替的に、多能性は、胚様体を形成させ、この胚様体を3つの胚葉に関連したマーカーの存在に関して評価することにより決定することができる。
【0051】
増殖した多能性幹細胞株は、標準的なGバンド法を使用して核型を決定することができ、確立された対応する霊長類種の核型と比較される。「正常な核型」を有する細胞を獲得することが望ましく、これは細胞が正倍数体であることを意味し、全ヒト染色体が存在し、かつ、著しく変更されてはいない。
【0052】
多能性幹細胞の源
使用が可能な多能性幹細胞の種類としては、妊娠期間中の任意の時期(必ずしもではないが、通常は妊娠約10〜12週よりも前)に採取した前胚性組織(例えば胚盤胞等)、胚性組織、胎児組織などの、妊娠後に形成される組織に由来する多能性細胞の株化細胞系が含まれる。非限定的な例は、例えばヒト胚幹細胞株H1、H7、及びH9(WiCell)などのヒト胚幹細胞又はヒト胚生殖細胞の確立株である。それらの細胞の最初の樹立又は安定化中に本開示の組成物を使用することも想定され、その場合、源となる細胞は、源となる組織から直接採取した一次多能性細胞である。フィーダー細胞の不在下で既に培養された多能性幹細胞集団から採取した細胞も好適である。例えば、BG01v(BresaGen,Athens,GA)などの変異ヒト胚性幹細胞株も好適である。
【0053】
一実施形態では、ヒト胚性幹細胞は、Thomsonら(米国特許第5,843,780号、Science 282:1145,1998、Curr.Top.Dev.Biol.38:133 ff.,1998、Proc.Natl.Acad.Sci.U.S.A.92:7844,1995)に記載されているように調製される。
【0054】
多能性幹細胞の培養
一実施形態では、多能性幹細胞は、一般にフィーダー細胞の層上で培養され、このフィーダー細胞は、多能性幹細胞を様々な方法で支持する。あるいは、多能性幹細胞を、フィーダー細胞を基本的に含まないにも関わらず、細胞を実質的に分化させることなく多能性幹細胞の増殖を支持するような培養システム中で培養する。フィーダー細胞不含培養における多能性幹細胞の分化を伴わない増殖は、あらかじめ他の細胞種を培養することにより条件づけした培地を使用して支持される。あるいはフィーダー細胞不含培養における多能性幹細胞の分化を伴わない増殖は、合成培地を使用して支持される。
【0055】
例えば、Reubinoffら(Nature Biotechnology 18:399〜404(2000))及びThompsonら(Science 6 November 1998:Vol.282.no.5391,pp.1145〜1147)は、マウス胚性繊維芽フィーダー細胞層を用いてヒト胚盤胞からの多能性幹細胞系を培養することについて開示している。
【0056】
Richardsら(Stem Cells 21:546〜556,2003)は、11種類の異なるヒトの成人、胎児、及び新生児フィーダー細胞層についてヒト多能性幹細胞の培養を支持する能力の評価を行っている。Richardsらは、「成人の皮膚線維芽フィーダー細胞上で培養したヒト胚性幹細胞系は、ヒト胚性幹細胞の形態を有し、多能性を維持する」と述べている。
【0057】
米国特許出願公開第20020072117号は、無フィーダー細胞培養中で霊長類の多能性幹細胞の増殖を支持する培地を生成する細胞系を開示している。使用される細胞系は、胚性組織から得られるかあるいは胚性幹細胞から分化した間葉系かつ繊維芽細胞様の細胞系である。米国特許出願公開第20020072117号は、この細胞系の1次フィーダー細胞層としての使用を更に開示している。
【0058】
別の例として、Wangら(Stem Cells 23:1221〜1227,2005)は、ヒト胚性幹細胞由来のフィーダー細胞層上でヒト多能性幹細胞を長期にわたって増殖させるための方法を開示している。
【0059】
別の例として、Stojkovicら(Stem Cells 2005 23:306〜314,2005)は、ヒト胚性幹細胞の自然分化により誘導されたフィーダー細胞システムを開示している。
【0060】
更なる別の例として、Miyamotoら(Stem Cells 22:433〜440,2004)は、ヒトの胎盤から得られたフィーダー細胞の供給源を開示している。
【0061】
Amitら(Biol.Reprod 68:2150〜2156、2003年)は、ヒト包皮に由来するフィーダー細胞層を開示している。
【0062】
別の例として、Inzunzaら(Stem Cells 23:544〜549,2005)は、ヒトの出生直後産児の包皮線維芽細胞から得られたフィーダー細胞層を開示している。
【0063】
米国特許第6642048号は、無フィーダー細胞培養中での霊長類の多能性幹(pPS)細胞の増殖を支持する培地、及びこうした培地の製造に有用な細胞系を開示している。米国特許第6642048号は、「本発明は、胚性組織から得られるかあるいは胚性幹細胞から分化した間葉系かつ繊維芽細胞様の細胞系を含む。本開示では、こうした細胞系を誘導し、培地を調整し、この馴化培地を用いて幹細胞を増殖させるための方法を説明及び図示する」と述べている。
【0064】
別の例として、国際公開第2005014799号は、哺乳動物細胞の維持、増殖及び分化のための条件培地を開示している。国際公開第2005014799号は、「本発明に基づいて製造される培地は、マウス細胞、特にMMH(Metマウス肝細胞)と称される分化及び不死化したトランスジェニック肝細胞の細胞分泌活性によって馴化される」と述べている。
【0065】
別の例として、Xuら(Stem Cells 22:972〜980,2004)は、ヒトテロメラーゼ逆転写酵素を過剰発現するように遺伝子改変されたヒト胚性幹細胞由来細胞から得られた馴化培地を開示している。
【0066】
別の例において、米国特許出願公開第20070010011号は、多能性幹細胞を維持するための合成培地を開示している。
【0067】
代替的な一培養システムでは、胚性幹細胞の増殖を促進することが可能な増殖因子を添加した無血清培地を使用している。例えば、Cheonら(BioReprod DOI:10.1095/biolreprod.105.046870,October 19,2005)は、胚性幹細胞の自己再生を誘発することが可能な異なる増殖因子を添加した非馴化血清補充(SR)培地中に胚性幹細胞が維持された、無フィーダー細胞かつ無血清の培養システムを開示している。
【0068】
別の例において、Levensteinら(Stem Cells 24:568〜574,2006)は、線維芽細胞又は馴化培地の非存在下で、bFGFを添加した培地を使用して、胚幹細胞を長期間培養する方法を開示している。
【0069】
別の例において、米国特許出願公開第20050148070号は、血清及び繊維芽細胞フィーダー細胞方法を含まない合成培地中でのヒト胚幹細胞の培養方法を開示し、同方法は、アルブミン、アミノ酸、ビタミン、無機物、少なくとも1つのトランスフェリン又はトランスフェリン代替物、少なくとも1つのインスリン又はインスリン代替物を含有する培地中で細胞を培養し、この培地は、本質的に哺乳動物胎児血清を含有せず、線維芽細胞増殖因子シグナル伝達受容体を活性化できる少なくとも約100ng/mLの線維芽細胞増殖因子を含有し、ここで増殖因子は、線維芽細胞フィーダー層のみでなく他の源からも供給され、培地はフィーダー細胞又は馴化培地なしで、未分化状態の幹細胞の増殖を支持した。
【0070】
別の例において、米国特許出願公開第20050233446号は、未分化の霊長類始原幹細胞を含む幹細胞の培養に有用な合成培地を開示している。溶液において、培地は、培養されている幹細胞と比較して実質的に等張である。所定の培養おいて、特定の培地は、基本培地と、実質的に未分化の始原幹細胞の増殖の支持に必要な、ある量のbFGF、インスリン、及びアスコルビン酸の各々とを含有する。
【0071】
別の例として、米国特許第6800480号は、「1つの実施形態において、実質的に未分化状態の霊長類由来の始原幹細胞を増殖させるための細胞培地であって、霊長類由来の始原幹細胞の増殖を支持するうえで効果的な低浸透圧、低エンドトキシンの基礎培地を含む細胞培地を提供する。この基本培地は、霊長類由来の始原幹細胞の増殖を支持するうえで効果的な栄養素血清、並びに、フィーダー細胞、及びフィーダー細胞から誘導される細胞外基質成分からなる群から選択される基質と組み合わされる。培地は更に、非必須アミノ酸、抗酸化剤、並びに、ヌクレオシド及びピルビン酸塩からなる群から選択される第1の増殖因子を含む。」と述べている。
【0072】
別の例では、米国特許出願公開第20050244962号は、「1つの態様において本発明は、霊長類の胚性幹細胞を培養する方法を提供する。哺乳動物の胎児血清を基本的に含まない(好ましくはあらゆる動物の血清をも基本的に含まない)培養中で、単に繊維芽フィーダー細胞層以外の供給源から供給される線維芽細胞増殖因子の存在下で幹細胞を培養する。好ましい1つの形態では、充分な量の繊維芽増殖因子を添加することによって、幹細胞の培養を維持するために従来必要とされていた繊維芽フィーダー細胞層の必要性がなくなる。」と述べている。
【0073】
更なる例として、国際特許出願公開第2005065354号は、基本的に無フィーダー細胞かつ無血清の合成等張培地であって、a.基礎培地、b.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のbFGF、c.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のインスリン、及びd.実質的に未分化の哺乳動物幹細胞の増殖を支持するうえで充分な量のアスコルビン酸、を含む培地を開示している。
【0074】
別の例として、国際公開第2005086845号は、幹細胞を、細胞を未分化な状態に維持するのに十分な量の、トランスフォーミング増殖因子β(TGF−β)ファミリータンパク質のメンバー、線維芽細胞増殖因子(FGF)ファミリータンパク質のメンバー、又はニコチンアミド(NIC)に、所望の結果を得るのに充分な時間曝露することを含む、未分化の幹細胞を維持するための方法を開示している。
【0075】
多能性幹細胞は、好適な培養基質上に播くことができる。一実施形態では、好適な培養基質は、例えば基底膜から誘導されたもの、又は接着分子受容体−リガンド結合の一部を形成し得るものなどの細胞外マトリックス成分である。一実施形態において、好適な培養基質は、MATRIGEL(登録商標)(Becton Dickenson)である。MATRIGEL(登録商標)は、Engelbreth−Holm Swarm腫瘍細胞からの可溶性製剤であり、室温でゲル化して再構成基底膜を形成する。
【0076】
他の細胞外マトリックス成分及び成分混合物は代替物として好適である。増殖させる細胞型に応じて、これは、ラミニン、フィブロネクチン、プロテオグリカン、エンタクチン、ヘパラン硫塩、及び同様物を、単独で又は様々な組み合わせで含み得る。
【0077】
多能性幹細胞は、細胞の生存、増殖、及び所望の特徴の維持を促進する培地の存在下、基質上に好適な分布にて播かれてもよい。これら全特徴は、播種分布に細心の注意を払うことから利益を得、当業者は容易に決定することができる。
【0078】
好適な培地は、以下の成分、例えば、ダルベッコ変法イーグル培地(DMEM)、Gibco No.11965−092;ノックアウトダルベッコ変法イーグル培地(KO DMEM)、Gibco No.10829−018;ハムF12/50% DMEM基礎培地、200mM L−グルタミン、Gibco No.15039−027;非不可欠アミノ酸溶液、Gibco No.11140−050;β−メルカプトエタノール、Sigma No.7522;ヒト組み換え塩基性線維芽細胞増殖因子(bFGF)、Gibco No.13256−029などから調製することができる。
【0079】
多能性幹細胞からの膵臓ホルモン産生細胞の形成
一実施形態では、本発明は、多能性幹細胞から膵臓ホルモン産生細胞を作製するための方法を提供し、かかる方法は、
a.多能性幹細胞を培養する工程と、
b.多能性幹細胞を、胚体内胚葉系に特徴的なマーカーを発現している細胞に分化させる工程と、
c.胚体内胚葉系に特徴的なマーカーを発現している細胞を、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化させる工程と、
d.膵臓内胚葉系に特徴的なマーカーを発現している細胞を、膵内分泌系に特徴的なマーカーを発現している細胞に分化させる工程と、を含む。
【0080】
本発明での使用に好適な多能性幹細胞としては、例えばヒト胚性幹細胞株H9(NIH code:WA09)、ヒト胚性幹細胞株H1(NIH code:WA01)、ヒト胚性幹細胞株H7(NIH code:WA07)、及びヒト胚性幹細胞株SA002(Cellartis,Sweden)が挙げられる。多能性細胞に特徴的な以下のマ−カー、すなわち、ABCG2、cripto、CD9、FOXD3、コネキシン43、コネキシン45、OCT4、SOX2、NANOG、hTERT、UTF1、ZFP42、SSEA3、SSEA4、Tra1−60又はTra1−81のうちの少なくとも1つを発現する細胞も本発明での使用に適している。
【0081】
胚体内胚葉系に特徴的なマーカーは、SOX17、GATA4、HNF3 β、GSC、CER1、NODAL、FGF8、Brachyury、Mix−様ホメオボックスタンパク質、FGF4 CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、及びOTX2からなる群より選択される。本発明での使用に好適なものは、胚体内胚葉系に特徴的なマーカーのうちの少なくとも1つを発現している細胞である。本発明の一態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、原始線条前駆体細胞である。別の態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、中内胚葉細胞である。別の態様において、胚体内胚葉系に特徴的なマーカーを発現している細胞は、胚体内胚葉細胞である。
【0082】
膵臓内胚葉系に特徴的なマーカーは、PDX1、HNF−1 β、PTF1 α、HNF6、HB9及びPROX1からなる群から選択される。本発明での使用に好適なものは、膵臓内胚葉系の特徴を示す少なくとも1つのマーカーを発現している細胞である。本発明の一態様において、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉細胞である。
【0083】
膵内分泌系に特徴的なマーカーは、NGN3、NEUROD、ISL1、PDX1、NKX6.1、PAX4、及びPTF−1 αからなる群から選択される。一実施形態では、膵内分泌細胞は、以下のホルモン:インスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを発現することができる。本発明で使用するのに好適なものは、膵内分泌系の特徴を示すマーカーを少なくとも1つ発現する細胞である。本発明の一態様において、膵内分泌系に特徴的なマーカーを発現している細胞は、膵内分泌細胞である。膵内分泌細胞は、膵臓ホルモン発現細胞であってもよい。代替的に、膵内分泌細胞は、膵臓ホルモン分泌細胞であってもよい。
【0084】
本発明の一態様では、膵臓内分泌細胞は、β細胞系統に特徴的なマーカーを発現する細胞である。β細胞系に特徴的なマーカーを発現している細胞は、PDx1と、以下の転写因子、すなわち、NGN3、NKX2.2、NKX6.1、NEUROD、ISL1、HNF−3 β、MAFA、PAX4、又はPAX6のうちの少なくとも1つを発現する。本発明の一態様では、β細胞系統に特徴的なマーカーを発現する細胞は、β細胞である。
【0085】
胚体内胚葉系に特徴的なマーカーを発現している細胞の形成
多能性幹細胞は、当該技術分野のいかなる方法、又は本発明で提案されるいかなる方法によって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させてもよい。
【0086】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 23,1534〜1541(2005)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0087】
例えば、多能性幹細胞は、Shinozakiら、Development 131,1651〜1662(2004)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0088】
例えば、多能性幹細胞は、McLeanら、Stem Cells 25,29〜38(2007)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0089】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示される方法に従って胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0090】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンA及び血清と培養し、次いで細胞をアクチビンA及び異なる濃度の血清と培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の一例は、Nature Biotechnology 23,1534〜1541(2005)に開示されている。
【0091】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンAと、別の濃度の血清の存在下で培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の例は、D’Amourら、Nature Biotechnology,2005に開示されている。
【0092】
例えば、多能性幹細胞は、アクチビンA及びWntリガンドを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いでWntリガンドを除去し、細胞をアクチビンAと、血清の存在下で培養することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0093】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0094】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0095】
例えば、多能性幹細胞は、米国特許出願第60/990,529号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0096】
例えば、多能性幹細胞は、米国特許出願第61/076,889号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0097】
例えば、多能性幹細胞は、米国特許出願第61/076,900号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0098】
例えば、多能性幹細胞は、米国特許出願第61/076,908号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0099】
例えば、多能性幹細胞は、米国特許出願第61/076,915号に開示される方法に従って多能性幹細胞を処理することによって胚体内胚葉系に特徴的なマーカーを発現する細胞に分化させることができる。
【0100】
胚体内胚葉系に特徴的なマーカーを発現している細胞の分化
胚体内胚葉系に特徴的なマーカーを発現している細胞の形成は、以下の特定のプロトコルの前後に、マーカーの存在に関して試験することにより決定することができる。多能性幹細胞は、一般にかかるマーカーを発現しない。したがって、多能性細胞の分化は、細胞がそれらの発現を開始した際に検出される。
【0101】
分化効率は、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより測定することができる。
【0102】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0103】
多能性幹細胞の特徴は当業者に周知であり、多能性幹細胞の更なる特徴は、継続して同定されている。例えば、多能性幹細胞マーカーとしては、以下のもの:ABCG2、cripto、FOXD3、コネキシン43、コネキシン45、OCT4、SOX2、NANOG、hTERT、UTF1、ZFP42、SSEA3、SSEA4、Tra1−60、又はTra1−81のうちの1つ以上の発現が挙げられる。
【0104】
多能性幹細胞を本発明の方法で処理した後、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現している細胞により発現される、例えばCXCR4などのタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより、分化した細胞を精製することができる。
【0105】
膵臓内胚葉系に特徴的なマーカーを発現している細胞の形成
胚体内胚葉系に特徴的なマーカーを発現している細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化され得る。
【0106】
例えば、胚体内胚葉系に特徴的なマーカーを発現している細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示されている方法に従って、膵臓内胚葉系に特徴的なマーカーを発現している細胞に分化され得る。
【0107】
例えば、胚体内胚葉系に特徴的なマーカーを発現している細胞は、胚体内胚葉系に特徴的なマーカーを発現している細胞を、線維芽細胞増殖因子及びヘッジホッグシグナル伝達経路阻害剤KAAD−シクロパミンで処理した後、線維芽細胞増殖因子及びKAAD−シクロパミンを含有する培地を除去し、続いて細胞をレチノイン酸、線維芽細胞増殖因子及びKAAD−シクロパミンを含有する培地中で培養することにより、膵臓内胚葉系に特徴的なマーカーを発現している細胞に更に分化させられる。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0108】
本発明の一態様では、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞をレチノイン酸及び少なくとも1種類の線維芽細胞増殖因子で所定の時間処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞を膵臓内胚葉系に特徴的なマーカーを発現する細胞に更に分化させる。
【0109】
本発明の一態様では、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞をレチノイン酸及び少なくとも1種類の線維芽細胞増殖因子で所定の時間処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞を膵臓内胚葉系に特徴的なマーカーを発現する細胞に更に分化させる。
【0110】
本発明の1つの態様では、胚体内胚葉系に特徴的なマーカーを発現している細胞を米国特許出願第60/990,529号に記載の方法に従って処理することにより、膵臓内胚葉系に特徴的なマーカーを発現している細胞へと胚体内胚葉系に特徴的なマーカーを発現している細胞を更に分化させる。
【0111】
膵臓内胚葉系に特徴的なマーカーを発現する細胞の検出
膵臓内胚葉系に特徴的なマーカーは、当業者に周知であり、膵臓内胚葉系の特徴を示す追加のマーカーが、継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して膵臓内胚葉系の特徴を示す性質を獲得したことを確認するために使用され得る。膵臓内胚葉系に特異的なマーカーとしては、例えば、HLXB9、PTF−1 α、PDX1、HNF6、HNF−1 βなどの転写因子の1つ以上のものの発現が挙げられる。
【0112】
分化効率は、膵臓内胚葉系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、処理した細胞集団を曝露することにより測定することができる。
【0113】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0114】
膵内分泌系に特徴的なマーカーを発現している細胞の形成
膵臓内胚葉系に特徴的なマーカーを発現している細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵内分泌系に特徴的なマーカーを発現している細胞へと分化させることができる。
【0115】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示されている方法に従って、膵内分泌系に特徴的なマーカーを発現している細胞に分化され得る。
【0116】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をDAPT及びエキセンディン4を含有する培地中で培養し、次にDAPT及びエキセンディン4を含有する培地を除去し、続いて細胞をエキセンディン1、IGF−1及びHGFを含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、Nature Biotechnology 24、1392〜1401(2006年)に開示されている。
【0117】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をエキセンディン4を含有する培地中で培養し、次にエキセンディン4を含有する培地を除去し、続いて細胞をエキセンディン1、IGF−1及びHGFを含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’ Amourら、Nature Biotechnology,2006に開示されている。
【0118】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をDAPT及びエキセンディン4を含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’Amourら、Nature Biotechnology,2006に開示されている。
【0119】
例えば、膵臓内胚葉系に特徴的なマーカーを発現している細胞は、膵臓内胚葉系に特徴的なマーカーを発現している細胞をエキセンディン4を含有する培地中で培養することにより、膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化する。この方法の例は、D’Amourら、Nature Biotechnology,2006に開示されている。
【0120】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0121】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0122】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、LifeScan,Inc.に譲渡された米国特許出願第60/953,178号に開示された方法に従って、ノッチシグナル伝達経路を阻害する因子で処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0123】
本発明の一態様では、膵臓内胚葉系に特徴的なマーカーを発現している細胞を、米国特許出願第60/990,529号に開示された方法に従って処理することで、膵臓内胚葉系に特徴的なマーカーを発現している細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと更に分化させる。
【0124】
本発明の一態様では、本発明は、膵内分泌系と関連付けられるマーカーの発現量を増加させる方法を提供し、この方法は、膵内分泌系に特徴的なマーカーを発現している細胞を、米国特許出願第61/110,278号に示された方法に従って、十分な量のTGF−β受容体作動剤を含む培地で処理することで、膵内分泌系に関連付けられるマーカーの発現量を増加させることを含む。
【0125】
膵内分泌系に特徴的なマーカーを発現している細胞の検出
膵内分泌系に特徴的なマーカーは当業者に周知であり、膵内分泌系の特徴を示す追加のマーカーが継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して膵内分泌系の特徴を示す性質を獲得したことを確認するために使用され得る。膵内分泌系に特異的なマーカーとしては、例えばNGN3、NEUROD又はISL1などの転写因子の1種以上の発現が挙げられる。
【0126】
β細胞系の細胞に特徴的なマーカーは当業者に周知であり、β細胞系の細胞の特徴を示す追加のマーカーが継続して同定されている。これらのマーカーは、本発明に従って処理された細胞が分化して、β−細胞系の特徴を示す性質を獲得したことを確認するために使用することができる。β細胞系に特異的な特徴としては、例えば、特にPDX1、NKX2.2、NKX6.1、ISL1、PAX6、PAX4、NEUROD、HNF1 β、HNF6、HNF3 β又はMAFAなどの転写因子の1つ以上の発現が挙げられる。これらの転写因子は、内分泌細胞の同定について当該技術分野で十分に確立されている。例えば、Edlund(Nature Reviews genetics 3:524〜632(2002年))を参照されたい。
【0127】
分化効率は、膵内分泌系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、被処理細胞集団を曝露することにより測定することができる。あるいは分化効率は、β細胞系に特徴的なマーカーを発現している細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に、被処理細胞集団を曝露することにより測定することができる。
【0128】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット法、インサイツハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら、eds.2001 supplement)参照)、及び切片材料の免疫組織学的解析などの免疫測定法、ウェスタンブロット、及び無傷細胞内に到達出来るマーカーに関して、フローサイトメトリー解析(FACS)(例えば、Harlow及びLane、Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0129】
本発明の一態様では、分化効率は、処理後の所定の細胞培養物中のインスリン陽性細胞の百分率を測定することによって求められる。一実施形態では、本発明の方法は、所定の培養物中で約100%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約90%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約80%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約70%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約60%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約50%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約40%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約30%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約20%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約10%のインスリン陽性細胞を生成する。別の実施形態では、本発明の方法は、所定の培養物中で約5%のインスリン陽性細胞を生成する。
【0130】
本発明の一態様では、分化効率は、グルコース刺激によるインスリン分泌を、細胞が放出するC−ペプチドの量を測定することで検出し、測定することにより求められる。一実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約1000ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約900ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約800ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約700ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約600ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約500ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約400ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約500ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約400ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約300ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約200ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約100ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約90ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約80ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約70ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約60ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約50ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約40ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約30ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約20ngのC−ペプチドを産生する。別の実施形態では、本発明の方法により生成される細胞は、DNA 1pg当たり約10ngのC−ペプチドを産生する。
【0131】
膵内分泌系に特徴的なマーカーを発現している細胞でのMAFAの発現の増加
一実施形態では、本発明は、膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法を提供する。
【0132】
サイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ1を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ2を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ4を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ5を阻害し得る。あるいはサイクリン依存性キナーゼ阻害剤はサイクリン依存性キナーゼ9を阻害し得る。あるいは、サイクリン依存性キナーゼ阻害剤は、これらの任意の組み合わせの、サイクリン依存性キナーゼの複数のイソ型を阻害し得る。
【0133】
サイクリン依存性キナーゼ阻害剤はタンパク質であり得る。あるいはサイクリン依存性キナーゼ阻害剤はペプチドであり得る。あるいはサイクリン依存性キナーゼ阻害剤は低分子であり得る。一実施形態では、低分子サイクリン依存性キナーゼ阻害剤は、7−n−ブチル−6−(4−ヒドロキシフェニル)[5H]ピロロ[2,3−b]ピラジン、9−ニトロ−7,12−ジヒドロインドロ[3,2−d][1]ベンゾアゼピン−6(5H)−オン、3−(6−オキソ−9−ニトロ−5,6,7,12−テトラヒドロインドロ[3,2−d][1]ベンゾアゼピン−2−イル)プロピオニトリル、(2R)−2−((6−((3−アミノ−5−クロロフェニル)アミノ)−9−(1−メチルエチル)−9H−プリン−2−イル)アミノ)−3−メチル−1−ブタノール、アルシリアフラビンA、[6−ベンジルアミノ−2−(3−ヒドロキシプロピルアミノ)−9−イソプロピルプリン、ブチロラクトンI、(Z)−1−(3−エチル−5−メトキシ−2,3−ジヒドロベンゾチアゾール−2−イリデン)プロパン−2−オン、2−(3−ヒドロキシプロピルアミノ)−6−(o−ヒドロキシベンジルアミノ)−9−イソプロピルプリン、1−(2,6−ジクロロフェニル)−1,5−ジヒドロ−6−((4−(2−ヒドロキシエトキシ)フェニル)メチル)−3−(1−メチルエチル)−4H−ピラゾロ[3,4−d]ピリミジン−4−オン、Cdk/サイクリン阻害ペプチドIII、3−(2−クロロ−3−インドリルメチレン)−1,3−ジヒドロインドール−2−オン、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレート、RO−3306、N−(cis−2−アミノシクロヘキシル)−N−(3−クロロフェニル)−9−エチル−9H−プリン−2,6−ジアミン、6−シクロヘキシルメトキシ−2−(4’−スルファモイルアニリノ)プリン、5−アミノ−3−((4−(アミノスルホニル)フェニル)アミノ)−N−(2,6−ジフルオロフェニル)−1H−1,2,4−トリアゾール−1−カルボチオアミド、3−アミノ−1H−ピラゾロ[3,4−b]キノキサリン、Cdk2阻害剤I、Cdk2阻害剤II、2(ビス−(ヒドロキシエチル)アミノ)−6−(4−メトキシベンジルアミノ)−9−イソプロピルプリン、4−(6−シクロヘキシルメトキシ−9H−プリン−2−イルアミノ)−N,N−ジエチルベンズアミド、N4−(6−アミノピリミジン−4−イル)−スルファニルアミド、(4−(2−アミノ−4−メチルチアゾール−5−イル)ピリミジン−2−イル)−(3−ニトロフェニル)アミン、2−ブロモ−12,13−ジヒドロ−5H−インドロ[2,3−a]ピロロ[3,4−c]カルバゾール−5,7(6H)−ジオン、1,4−ジメトキシアクリジン−9(10H)−チオン、5−(N−(4−メチルフェニル)アミノ)−2−メチル−4,7−ジオキソベンゾチアゾール、4−(3,5−ジアミノ−1Hピラゾール−4−イルアゾ)−フェノール、2−(2−ヒドロキシエチルアミノ)−6−(3−クロロアニリノ)−9−イソプロピルプリン、ファスカプリシン、インジルビン−3’−モノオキシム、インジルビン−3’−モノオキシム、5−ヨード−,インジルビン−3’−モノオキシム−5−スルホン酸、イソグラヌラチミド、2−(2−ヒドロキシエチルアミノ)−6−ベンジルアミノ−9−メチルプリン、6−(2−ヒドロキシベンジルアミノ)−2−((1R)−(ヒドロキシメチル)プロピル)アミノ)−9−イソプロピルプリン、5−ブロモ−3−(2−(4−フルオロフェニル)−2−オキソエチリジン)−1,3−ジヒドロインドール−2−オン、N6,N6−ジメチルアデニン、2−(1R−イソプロピル−2−ヒドロキシエチルアミノ)−6−(3−クロロアニリノ)−9−イソプロピル−プリン、ラパマイシン、2−(R)−(1−エチル−2−ヒドロキシエチルアミノ)−6−ベンジルアミノ−9−イソプロピルプリン、スキトネミン、3−[1−(3H−イミダゾール−4−イル)−メタ−(Z)−イリデン]−5−メトキシ−1,3−ジヒドロインドール−2−オン、及び4−(3’−ヒドロキシフェニル)アミノ−6,7−ジメトキシキナゾリンからなる群から選択される。
【0134】
一実施形態では、サイクリン依存性キナーゼはエチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートである。一実施形態では、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートを、内分泌腺系に特徴的なマーカーを発現している細胞に約0.1μM〜約10μMの範囲の濃度で、約1日〜7日間にわたって加える。
【0135】
一実施形態では、内分泌腺系に特徴的なマーカーを発現している細胞を、エチル−(6−ヒドロキシ−4−フェニルベンゾ[4,5]フロ[2,3−b])ピリジン−3−カルボキシレートで約1日〜約7日間にわたって処理する。
【0136】
以下の実施例により本発明を更に例示するが、本発明はこれらの実施例により限定されるものではない。
【実施例】
【0137】
(実施例1)
ウシ胎児血清の非存在下での、ヒト胚性幹細胞の細胞株H1の膵内分泌細胞への分化
ヒト胚性幹細胞株H1の継代数52の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、細胞を膵内分泌系に特徴的なマーカーを発現している細胞へと分化させるために、以下の分化プロトコルを実施した。
a.2% BSA(カタログ#152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ#1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ#100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R&D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ#565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ#616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理した(ステージ5)。
【0138】
培地は毎日交換した。各ステージにおいて血球計数器を用いて細胞数を計算し、PCR分析用にRNAを採取した。試料はすべて3つ組複製で採取した。
【0139】
(実施例2)
実施例1に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリII由来の化合物の効果のスクリーニング
継代数44のヒト胚性幹細胞株H1の細胞を、MATRIGEL(商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従ってステージ5まで分化させた。これに続き、細胞をEMD Calbiochem化合物ライブラリ(カタログ#539745,Calbiochem,San Diego,Ca)由来の化合物を最終濃度1μMで含有しているDMEM/F12+1% B27で、4日間にわたって処理した。ビヒクルを含有しているウェルを対照として含めた。プロトコルを通して、培地は毎日交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びArx4の発現について解析した。結果は、リアルタイムPCRでの測定結果により、処理サンプル対未処理対照の、インスリン/グルカゴン比(図1、パネルa)、又はMAFAとARX4の比(図1、パネルb)として表わされる。各ウェル#について対応するPubChem化合物ID#は、表1に列挙する。
【0140】
1μMの濃度の化合物A6、B7、B8、又はC2での、膵内分泌系に特徴的なマーカーを発現している細胞の処理は、およそ3.0の、あるいはそれよりも大きいインスリン/グルカゴン発現比をもたらした(図1のパネルaを参照されたい)。
【0141】
次いで、MAFA/ARX4の比に対するこれらの化合物の効果を調べたところ、数個の化合物での膵内分泌系に特徴的なマーカーを発現している細胞の処理は、ライブラリ内の他の試験化合物よりも、MAFAとARX4の比に、より大きな変化をもたらしたことを観察した:化合物C2で処理した細胞は、約1000のMAFA/ARX4比を示した。膵内分泌系に特徴的なマーカーを発現している細胞の化合物C2での処理は、約100のMAFA/ARX4比をもたらした(図1のパネルbを参照されたい)。
【0142】
(実施例3)
実施例1に概説された分化プロトコルに従って処理した細胞でのインスリン及びMAFA発現に対する、サイクリン依存性キナーゼ阻害剤処理の効果
実施例2でインスリン発現とグルカゴン発現の比、又はMAFAとARX4の比を増加させたいくつかの化合物は、サイクリン依存性キナーゼ阻害剤であった。このような化合物の1つはPubChem化合物ID# 5330797(5−アミノ−3−((4−(アミノスルホニル)フェニル)アミノ)−N−(2,6−ジフルオロフェニル)−1H−1,2,4−トリアゾール−1−カルボチオアミド)(カタログ# 217714;Calbiochem,San Diego,Ca)である。これらの観察結果を確証づけるために、MATRIGEL(登録商標)をコートした10cm2のディッシュ上で、継代数42のヒト胚性幹細胞株H1の細胞を培養し、実施例1に記載の方法に従って、ステージ5まで処理した。ステージ5以降は、1% B27と1μMのPubChem化合物ID# 5330797を含有するDMEM/F12で、細胞を6日間にわたって処理した。培地は1日おきに交換した。リアルタイムPCR用の細胞サンプルを化合物での処理前、化合物処理の2日目及び5日目に採取した。
【0143】
化合物処理対未処理対照の4日目又は6日目の細胞の特徴的な顕微鏡写真を図2に示す。未処理細胞は高密度で充填され(図3、パネルa及びd)、個別の細胞を区別することは困難である。しかしながら、0.5μM又は1μMのPubChem化合物ID#5330797での6日間にわたる処理後には、未処理対照(図2、パネルd)と比較してそれぞれの核が目に見えるようになり(図2、パネルe及びf)、細胞集団に分化が生じたことを示している。これはまた、図2のパネルb及びcに示されるような細胞層中の間隙により見出され得る、一部の細胞死を伴う。
【0144】
PubChem化合物ID# 5330797での細胞処理は、程度は異なるもののインスリン、グルカゴン、MAFA、MAFB及びソマトスタチンの発現上昇をもたらした。培養0日目(処理前)と比較した場合の、処理あたりの遺伝子発現の相対的な誘導を図3のパネルa〜vに示す。1μMのPubChem化合物ID# 5330797で処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的に処理の48時間の時点でのグルカゴン発現がおよそ1.5倍に上昇した。この発現量は、処理の5日以降に処理前のレベル未満に下降した。0.5μMのPubChem化合物ID# 5330797での処理では、グルカゴン発現の上昇は観察されなかった(図3、パネルaを参照されたい)。
【0145】
1μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にインスリン発現がおよそ1.5倍に上昇した(図3、パネルbを参照されたい)。
【0146】
1μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にMAFA発現がおよそ200倍に上昇した(図3、パネルdを参照されたい)。
【0147】
0.5μMのPubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、結果的にMAFB発現がおよそ1.5倍に上昇した(図3、パネルcを参照されたい)。ソマトスタチンの発現では、用量依存的な上昇が観察された(図3、パネルe)。
【0148】
PubChem化合物ID# 5330797で5日間にわたって処理した、膵内分泌系に特徴的なマーカーを発現している細胞では、アミラーゼの発現量は変化しなかった(図3、パネルfを参照されたい)。しかしながら、PAX4(図3、パネルh)、NKX6.1(図3、パネルk)、PDX1(図3、パネルl)、NEUROD(図3、パネルo)、及びBRN4(図3、パネルq)では発現レベルに減少が観察された。
【0149】
(実施例4)
サイクリン依存性キナーゼ阻害剤処理は、島様クラスタにおいてMAFA発現を上昇させた
継代数52のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、実施例1に記載の方法に従って分化させた。膵内分泌系に特徴的なマーカーを発現している細胞を更に成熟させるために、追加のステージ(ステージ6)を加えた。この実施例では、ステージ6はDMEM/F12+1% B27(Invitrogen,CA)での7日間の処理から構成される。培地は毎日交換した。
【0150】
ステージ6後に、細胞は1Xアキュターゼ(Sigma,MO)で、室温で5分にわたって処理した。アキュターゼを除去し、細胞にDMEM/12+1% B27を加えた。接着した細胞をセルスクレーパーを用いて取り外し、穏やかに再懸濁し、40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート(カタログ# 3471,Corning,Ma)上で懸濁液の状態で培養した。次いで細胞を以下のように処理した:細胞を、20ng/mLのアクチビンA(AA)と、1μmのCDK阻害剤III(カタログ#217714,Calbiochem,Ca)を含有しているDMEM/F12+1% B27で10日間にわたって培養した(ステージ7)。ビヒクルで処理した細胞を対照として含めた。PCR解析用に、7日目〜10日目にサンプルを回収し、ジチゾン染色した。本実施例で概説された方法に従って、懸濁状態で培養した細胞は、形態的に膵臓の島クラスタに類似であると推測された。CDK阻害剤IIIでの処理は、島様クラスタの形態に影響を与えるようには見えなかった。
【0151】
図4のパネルa〜iは、細胞クラスタの遺伝子発現プロファイルに対する、CDK阻害剤III処理の効果を示す。CDK阻害剤IIIでの処理は、膵内分泌系に関連付けられるマーカーの発現を上昇させ、特にプロインスリン転写因子のMAFAの発現を上昇させた。
【0152】
図5のパネルa〜bは、クラスタのジチゾン(dithazone)(DTZ)染色に対する、CDK阻害剤IIIの効果を示す。CDK阻害剤で処理し、かつDTZで染色した細胞クラスタは、CDK阻害剤IIIで処理していないクラスタと比較したときに、より赤みの強い染色パターンを示した。
【0153】
(実施例5)
本発明の方法により作製されたインスリン産生細胞の、FACS解析
継代数42のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたプレート上で培養し、以下のプロトコルを使用してインスリン産生細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ# 565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理し(ステージ5)、次いで
f.DMEM/F12+1% B27で7日間にわたって処理し(ステージ6)、次いで
g.アキュターゼで5分間にわたって処理した後に、接着しているあらゆる残りの細胞を掻き取りにより取り外した。次いで細胞懸濁液を40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート上で、DMEM−高グルコース(カタログ# 11995−073,Invitrogen,Ca)+1% B27+20ng/mLのアクチビンA(AA)、1μmのCDK阻害剤III(カタログ# 217714,Calbiochem,Ca)中に懸濁した状態で、5日間にわたって培養した(ステージ7)。
【0154】
TrypLE Express(Invitrogen,Carlsbad,CA)を用いて、島様クラスタを単一細胞へと分散させ、冷PBSで洗浄した。固定のため、細胞を200〜300μLのCytofix/Cytoperm緩衝液(BD 554722,BD,Ca)中に再懸濁し、4℃で30分間にわたってインキュベートした。細胞を1mLのPerm/Wash緩衝液(BD 554723)で2回洗浄し、Perm/Wash緩衝液中に2%正常ヤギ血清を含有する100μLの染色/ブロッキング溶液中に再懸濁した。フローサイトメトリー解析のために、細胞は以下の一次抗体で染色した:抗−インスリン(ウサギmAb,Cell Signaling No.C27C9;1:100希釈);抗−グルカゴン(マウスMab,Sigma No.G2654,1:100);抗−シナプトフィジン(ウサギポリクローナル抗体,DakoCytomation No A0010,1:50)。細胞を4℃で30分にわたってインキュベートし、Perm/Wash緩衝液で2回洗浄し、更に30分間にわたって、以下のような適切な2次抗体中でインキュベートした:ヤギ抗−ウサギAlexa 647(Invitrogen No.A21246)又はヤギ抗−マウス647(Invitrogen No.A21235);ヤギ抗−ウサギR−PE(BioSource No.ALI4407)。二次抗体はすべて1:200の希釈率で使用した。細胞は、少なくとも1回Perm/Wash緩衝液中で洗浄し、BD FACSアレイを使用して分析した。少なくとも10,000事象を分析のために得た。対照には未分化のH1細胞とβ−TC(CRL−11506(商標)ATCC,VA)細胞株を含んだ。
【0155】
図6のパネルa〜cは、ビヒクルを含有している培地での、ステージ7での処理後の細胞に占める、インスリン陽性細胞、シナプトフィジン(synapthophysin)陽性細胞、及びグルカゴン陽性細胞の割合を示す。図7のパネルa〜cは、1μMのCDK阻害剤IIIを含有している培地での、5日間にわたるステージ7での処理後の、インスリン陽性細胞、シナプトフィジン(synapthophysin)陽性細胞、及びグルカゴン陽性細胞の割合を示す。インスリンホルモン単独の陽性細胞の数は、CDK阻害剤での処理後に3%から8%へと上昇した。加えて、多ホルモン(インスリン及びグルカゴン陽性)細胞の割合は、CDK阻害剤での処理後に減少した。
【0156】
(実施例6)
CDK阻害剤誘導型のMAFA発現の動態
継代数42のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたプレート上で培養し、以下のプロトコルを使用してインスリン産生細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.DMEM/F12+2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)で2日間にわたって処理し(ステージ2)、次いで
c.DMEM/F12+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で4日間にわたって処理し(ステージ3)、次いで
d.DMEM/F12+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのDAPT(γセクレターゼ阻害剤)(カタログ# 565784,Calbiochem,CA)+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)+100ng/mLのネトリン−4(R&D Systems,MN)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM/F12+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理し(ステージ5)、次いで
f.DMEM/F12+1% B27で7日間にわたって処理し(ステージ6)、次いで
g.アキュターゼで5分間にわたって処理した後に、接着しているあらゆる残りの細胞を掻き取りにより取り外した。次いで細胞懸濁液を40μmのセルストレイナーで濾過した。ストレイナー上に残った細胞を基本培地ですすぐことで取り外し、超低接着性の培養プレート上で、DMEM−高グルコース(カタログ# 11995−073,Invitrogen,Ca)+1% B27+20ng/mLのアクチビンA(AA)、2μmのCDK阻害剤III(カタログ# 217714,Calbiochem,Ca)中に懸濁した状態で、1〜8日間にわたって培養した(ステージ7)。
【0157】
1、2、3及び4日目のサンプルを、PCR解析のために回収した。CDK阻害剤での処理の4日後に、CDK阻害剤を培養物から除去し、DMEM−F12+1% B27+20ng/mLアクチビンA中で更に4日間培養した。4日間の終了時に、PCR解析のためにサンプルを三つ組複製で回収した。
【0158】
図8のパネルa〜bは、ステージ7の様々な時点でのMAFA及びインスリンの発現パターンを示す。CDK阻害剤処理は、MAFA発現とインスリン発現に有意な上昇をもたらし、これらは時間関数として上昇した。しかしながら、CDK阻害剤の除去は、化合物除去の4日後に得たサンプルにおいてMAFA及びインスリン発現の両方で有意な低下をもたらした。
【0159】
(実施例7)
実施例1に概説された分化プロトコルに従って処理された細胞に対する、BIOMOL(商標)キナーゼ阻害剤ライブラリ由来の化合物の効果のスクリーニング
継代数51のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従って、ステージ5まで分化させた。これに続き、細胞をDMEM/F12+1% B27中で1日間生育させ、次いでBIOMOL(商標)化合物ライブラリ(カタログ# 2832,BIOMOL,Plymouth Meeting,Pa)由来の化合物を最終濃度4μMで含有しているDMEM/F12+1% B27中で、6日間にわたって処理した。ビヒクルを含有しているウェルを対照として含めた。処理プロトコルを通し、ビヒクル又は化合物を含有している培地は1日おきに交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びARX4の発現について解析した。結果は、リアルタイムPCRでの測定結果により、処理サンプル対未処理対照の、インスリン/グルカゴン比(表2)、又はMAFAとArx4の比(表2)として表わされる。各英数字ウェル#に対する、対応するカタログ#、CAS#、及び化合物名又はID数を表3に列挙する。
【0160】
4μMの濃度の化合物C8又はF1での、膵内分泌系に特徴的なマーカーを発現している細胞の処理は、およそ10.0の、あるいはそれよりも大きいインスリン/グルカゴン発現比をもたらした。D9で処理した細胞は、およそ1840.0のインスリン/グルカゴン発現比を有する(表2)。
【0161】
次いで、MAFA/ARX4の比に対するこれらの化合物の効果を調べたところ、数個の化合物での膵内分泌系に特徴的なマーカーを発現している細胞の処理は、ライブラリ内の他の試験化合物よりも、MAFAとARX4の比に、より大きな変化をもたらしたことを観察した:化合物B6又はF1で処理した細胞は、およそ10を超えるMAFA/ARX4比を示した。膵内分泌系に特徴的なマーカーを発現している細胞の化合物C8での処理は、約84のMAFA/ARX4比をもたらした一方で、D9で処理した細胞は約212のMAFA/ARX4比を有した(表2)。
【0162】
(実施例8)
実施例1に概説された分化プロトコルに従って処理した細胞でのインスリン及びMAFA発現に対する、サイクリン依存性キナーゼ阻害剤の効果
継代数51のヒト胚性幹細胞株H1の細胞を、MATRIGEL(商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例1に記載の方法に従ってステージ5まで分化させた。これに続き、細胞をDMEM/F12+1 %B27中で8日間にわたって生育させ、次いでサイクリン依存性キナーゼ阻害剤を最終濃度0.6125、1.25、又は5.0μMで含有しているDMEM/F12+1 %B27中で、4日間にわたって処理した。6種の阻害剤を試験した:PubChem ID# 5330812(EMD cat# 217714)、PubChem ID# 4566(EMD cat#217713)、PubChem ID# 5330797(EMD cat# 219476)、PubChem ID# 73292(EMD cat#341251)、PubChem ID #4592(EMD cat#495620)、及びPubChem ID# 160355(EMD cat #557360)。ビヒクルを含有しているウェルを対照として含めた。処理プロトコルを通し、ビヒクル又は化合物を含有している培地は1日おきに交換した。全てのサンプルは2つ組複製で処理した。この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、インスリン、グルカゴン、MAFA、及びARX4の発現について解析した。結果は、リアルタイムPCRにより測定されたものとして、ビヒクル処理対照に対する倍数変化として記載される。
【0163】
化合物PubChem ID# 5330812、PubChem ID# 4566、PubChem ID# 5330797、及びPubChem ID# 73292の全てが、試験濃度でMAFA発現を刺激したことを観察した(表4)。PubChem ID# 4592及びPubChem ID# 160355は試験濃度ではMAFAを刺激しなかった(表4)。化合物PubChem ID# 5330812、PubChem ID# 4566、PubChem ID# 5330797、PubChem ID# 4592、及びPubChem ID# 160355の全てが、インスリン発現を刺激したことは明らかであった(表4)。化合物(PubChem ID# 5330797)はグルカゴン及びArx4発現(表4)の両方を減少させた一方で、MAFA発現を刺激した。
【0164】
(実施例9)
ウシ胎児血清を欠き、25mMグルコースを含有しているDMEM(DMEM−HG)での、ヒト胚性幹細胞の細胞株H1の膵内分泌細胞への分化
ヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートしたディッシュ(1:30希釈)上で培養し、以下のプロトコルを用いて、膵内分泌系に特徴的なマーカーを発現している細胞へと分化させた:
a.2% BSA(カタログ# 152401,MP Biomedical,Ohio)、100ng/mLのアクチビンA(R&D Systems,MN)、20ng/mLのWNT−3a(カタログ# 1324−WN−002,R&D Systems,MN)、8ng/mLのbFGF(カタログ# 100−18B,PeproTech,NJ)を加えたRPMI培地で1日処理した後に、2% BSA、100ng/mLのアクチビンA、8ng/mLのbFGFを加えたRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
b.2% BSA+50ng/mLのFGF7+0.25μMシクロパミン−KAAD(#239804,Calbiochem,CA)を加えたRPMI培地で2日間にわたって処理し(ステージ2)、次いで
c.DMEM−HG+1% B27(Invitrogen,CA)+50ng/mLのFGF7+0.25μMシクロパミン−KAAD+2μMレチノイン酸(RA)(Sigma,MO)+100ng/mLのノギン(R & D Systems,MN)で6日間にわたって処理し(ステージ3)、次いで
d.DMEM−HG+1% B27(Invitrogen,CA)+100ng/mLノギン+1μMのALK5阻害剤II(カタログ# 616452,Calbiochem,Ca)で3日間にわたって処理し(ステージ4)、次いで
e.DMEM−HG+1% B27(Invitrogen,CA)+1μMのALK5阻害剤II(Calbiochem,Ca)で7日間にわたって処理した(ステージ5)。
【0165】
培地は毎日交換した。各ステージにおいて血球計数器を用いて細胞数を計算し、PCR解析用にRNAを採取した。試料はすべて3つ組複製で採取した。
【0166】
(実施例10)
実施例9に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリI由来の化合物の効果のスクリーニング
継代数45のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例9に記載の方法に従ってステージ5まで分化させた。これに続き細胞を供給し、ステージ5の1、3及び5日目に、DMEM−HG、1% B27(Invitrogen,CA)と、1μMのALK5阻害剤II(Calbiochem,Ca)と、DMSO(カタログ# 539744,Calbiochem,San Diego,Ca)に溶解し、2.5μMの最終濃度で処理したEMD Calbiochem化合物ライブラリ1由来の化合物とを含む培地で処理した。ビヒクルを含有しているウェルを対照として含めた。培地はステージ5では1日おきに交換したことを除き、プロトコルを通して毎日交換した。全てのサンプルは2つ組複製で処理した。
【0167】
この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、MAFAの発現について解析した。結果はリアルタイムPCRにより測定されたものとして、MAFA発現H1ヒト胚性幹細胞対未処理H1ヒト胚性幹細胞の倍数増加として表わす(表5)。
【0168】
膵内分泌系に特徴的なマーカーを発現している細胞の、2.5μM濃度の化合物A4(Cat#,124001,Akt阻害剤IV)、E8(Cat# 527450,PKR阻害剤)、及びF9(Cat#539648,スタウロスポリン,N−ベンゾイル−)での処理は、MAFA発現の、ビヒクル処理対照を少なくとも4倍の上昇をもたらした(表5)。2.5μM濃度の化合物E6(Cat# 521233,PDGF受容体チロシンナーゼ阻害剤IV)での処理は、ビヒクル処理対照を少なくとも2.5倍超えるMAFA発現の上昇をもたらした(表5)。
【0169】
(実施例11)
実施例9に概説された分化プロトコルに従って処理された細胞に対する、EMDキナーゼ阻害剤ライブラリII由来の化合物の効果のスクリーニング
継代数46のヒト胚性幹細胞株H1の細胞を、MATRIGEL(登録商標)をコートした24ウェルディッシュ(1:30希釈)上に播種し、実施例9に記載の方法に従ってステージ5まで分化させた。これに続き細胞を供給し、ステージ5の1、3及び5日目に、DMEM−HGと、1% B27(Invitrogen,CA)と、1μMのALK5阻害剤II(Calbiochem,Ca)(ステージ5)と、DMSO(表1及び6,Calbiochem,San Diego,Ca)に溶解させ最終濃度2.5μMに処理したEMD Calbiochem化合物ライブラリII由来の化合物と、を含む培地で処理した。ビヒクルを含有しているウェルを対照として含めた。培地はステージ5では1日おきに交換したことを除き、プロトコルを通して毎日交換した。全てのサンプルは2つ組複製で処理した。
【0170】
この処理の最後に、PCR解析用にRNAを回収した。サンプルはリアルタイムPCRにより、MAFAの発現について解析した。リアルタイムPCRにより測定されたものとして、MAFA発現を刺激した化合物に関する結果を示し、かつMAFA発現対対照サンプルにおける倍数上昇として表わす(図9)。
【0171】
膵内分泌系に特徴的なマーカーを発現している細胞の、2.5μM濃度の:アルステルパウロン、2−シアノエチル;SU9516;アルステルパウロン;Cdk1/2阻害剤III;カゼインキナーゼI阻害剤、D4476;又はMEK1/2阻害剤のいずれかでの処理は、未処理対照に対してMAFA発現の4.5倍の上昇をもたらした(表7)。
【0172】
(実施例12)
膵内分泌系に特徴的なマーカーを発現している細胞でMAFA発現を促進する低分子阻害剤での、膵内分泌系に特徴的なマーカーを発現している細胞の細胞周期の進行の阻害
細胞周期の進行に由来して生じる細胞増殖は、細胞外増殖因子で細胞を刺激することにより活性化及び維持され得る。増殖因子は増殖因子受容体の細胞外ドメインへと結合し、受容体の細胞内ドメインの立体構造スイッチを誘導する。このシフトは受容体二量体化及び受容体の細胞内ドメイン上に位置するチロシンキナーゼの活性化を開始し、下流の複数のセリン/スレオニンキナーゼのリン酸化及び活性化を導き、最終的には細胞周期の進行と細胞増殖をもたらす。
【0173】
通常の生理学的な条件下では、インスリンと転写因子MAFAの発現とにより特徴づけられる成熟した膵臓β細胞は無活動状態であり、細胞周期のG0に留まる傾向がある。しかしながら、機能性臓器を形成するのに十分な細胞を生成し、成熟した動物に関する要求を満たす、本発明の膵内分泌系に特徴的なマーカーを発現している細胞は、細胞周期を循環している必要がある。その結果、本発明の膵内分泌系に特徴的なマーカーを発現している細胞は胚発生のある時点でβ細胞へと分化し、活動的に細胞周期により増殖している細胞から、休眠細胞へと移行する。
【0174】
我々のデータは、低分子キナーゼ阻害剤によりシグナルカスケードをブロックして細胞周期の進行を阻害することで、膵内分泌系に特徴的なマーカーを発現している細胞に、成熟膵臓β細胞のマーカーであるMAFAの発現を誘導させ得ることを示す。増殖因子受容体(PDGF受容体チロシンキナーゼ阻害剤IV)に標的とされるキナーゼ阻害剤、又はチロシンキナーゼ受容体の下流のキナーゼを中断する阻害剤(MEK1/2阻害剤、PKR阻害剤、又はAkt阻害剤IV)は、増殖性の、増殖因子/キナーゼに基づくシグナル伝達を中断して細胞周期の休止をもたらし、MAFA発現を誘導する。スタウロスポリンのような広域スペクトル阻害剤の使用は、効果的にMAFAを誘導できるが、有効濃度では細胞毒性でもある。サイクリン依存性キナーゼ阻害剤(アルステルパウロン、2−シアノエチル;SU9516;アルステルパウロン;又はCdk1/2阻害剤III)のような、より直接的な化合物は、スタウロスポリンのような広域スペクトル阻害剤と比較して、より少ない細胞毒性でMAFAを誘導する。
【0175】
広域スペクトルのキナーゼ阻害剤が、MAFA発現と、本発明の、膵内分泌系に特徴的なマーカーを発現している、より成熟した表現型の細胞とを誘導し得るか判定するために、H1ヒトES細胞を実施例9に記載の方法に従って分化させ、ステージ5の1、3、及び5日目に、タンパク質チロシンキナーゼ阻害剤のゲニステイン(ヒト及びマウス細胞株のG2期を休止させ、複数のキナーゼを阻害することが示されている)で処理した。内分泌腺ホルモンのインスリン、ソマトスタチン、及び転写因子MAFAの全てが、投与量10及び30ng/mLで未処理対照に対して発現量の増加を示した一方で、内分泌腺ホルモングルカゴンは10ng/mLで発現を増加させた(図10)。我々は投与量100ng/mLでのゲニステインで、インスリン、グルカゴン、及びソマトスタチン発現の損失と関連付けられる有意な毒性を観察した。
【0176】
これらのデータは、増殖因子受容体チロシンキナーゼから細胞内シグナル伝達キナーゼを介して核及びサイクリン依存性キナーゼに向かうシグナル伝達を阻害するために、標的にされる低分子量のキナーゼ阻害剤により、シグナルカスケードをブロックすることで、細胞周期の進行を阻害することにより、本発明の、膵内分泌系に特徴的なマーカーを発現している細胞に、成熟膵臓β細胞のマーカーであるMAFAを発現させることができることを示す。
【0177】
本明細書を通して引用された刊行物は、その全体が参照により本明細書に組み込まれる。以上、本発明の様々な態様を実施例及び好ましい実施形態を参照して説明したが、本発明の範囲は、上記の説明文によってではなく、特許法の原則の下で適切に解釈される以下の特許請求の範囲によって定義されるものである点は認識されるであろう。
【0178】
【表1】
【0179】
【表2−1】
【0180】
【表2−2】
【0181】
【表2−3】
【0182】
【表3−1】
【0183】
【表3−2】
【0184】
【表3−3】
【0185】
【表4】
【0186】
【表5−1】
【0187】
【表5−2】
【0188】
【表5−3】
【0189】
【表6−1】
【0190】
【表6−2】
【0191】
【表6−3】
【0192】
【表7】
【図1−1】
【図1−2】
【特許請求の範囲】
【請求項1】
膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法。
【請求項1】
膵内分泌系に特徴的なマーカーを発現している細胞を、十分な量のサイクリン依存性キナーゼ阻害剤を含む培地中で培養することで、MAFAの発現の増加を生じさせる工程を含む、膵内分泌系に特徴的なマーカーを発現している細胞でMAFAの発現を増加させるための方法。
【図2A】

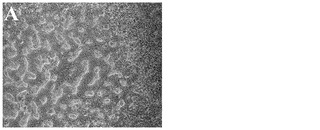
【図2B】
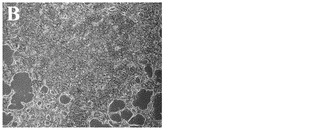
【図2C】
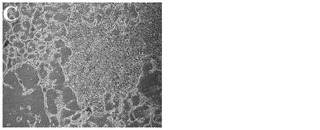
【図2D】
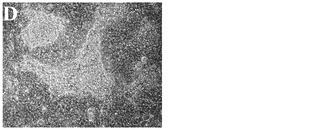
【図2E】
【図2F】

【図3−1】
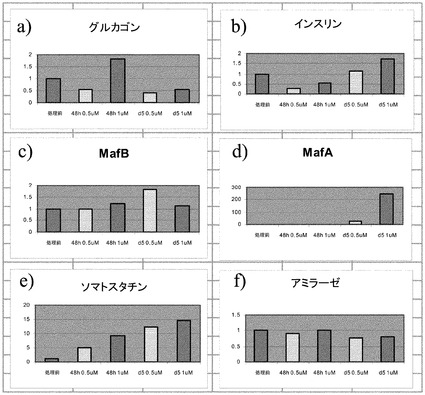
【図3−2】
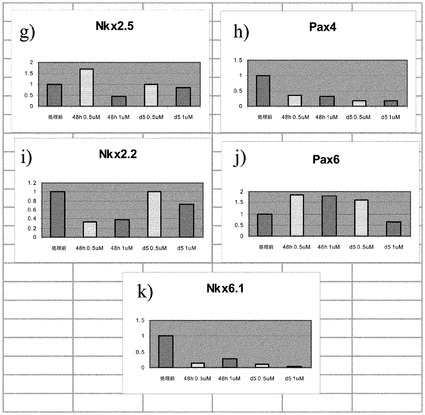
【図3−3】
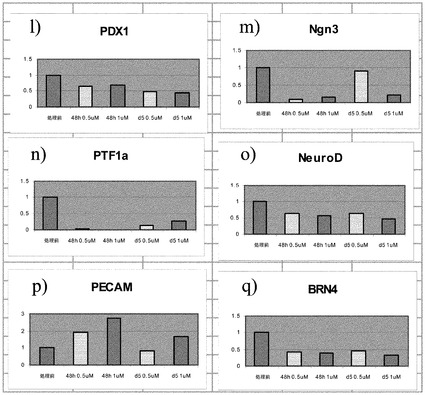
【図3−4】
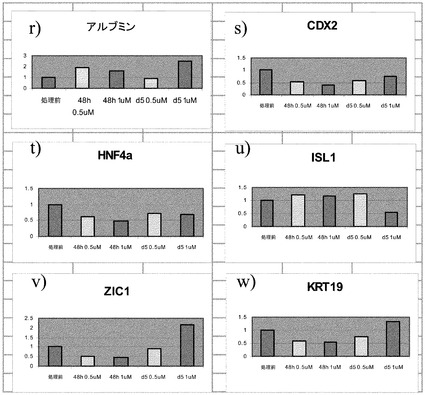
【図4−1】
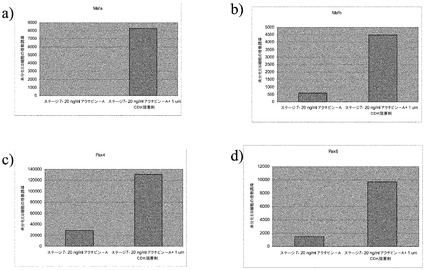
【図4−2】
【図4−3】
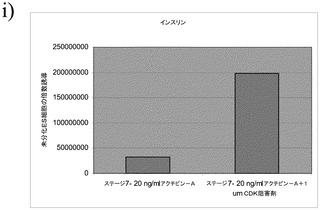
【図5】
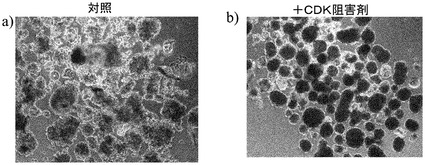
【図6】
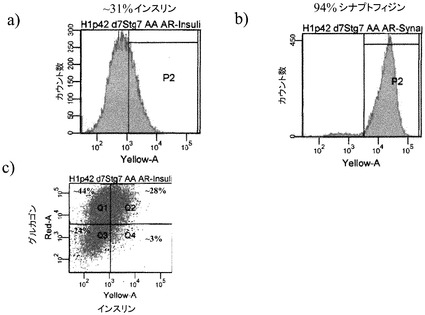
【図7】
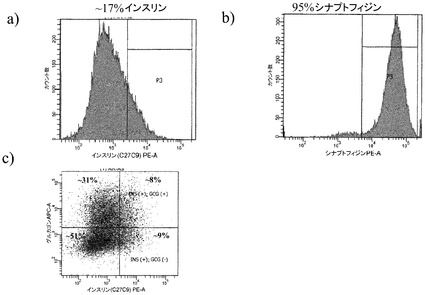
【図8】
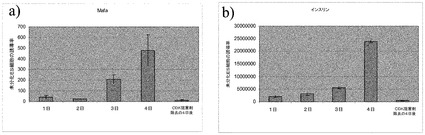
【図9】
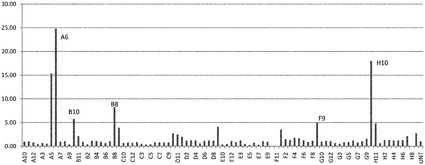
【図10】
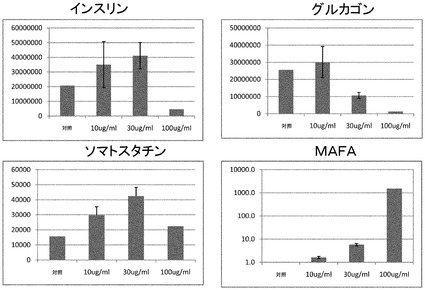
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図4−1】
【図4−2】
【図4−3】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2012−507292(P2012−507292A)
【公表日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願番号】特願2011−534638(P2011−534638)
【出願日】平成21年10月23日(2009.10.23)
【国際出願番号】PCT/US2009/061774
【国際公開番号】WO2010/051223
【国際公開日】平成22年5月6日(2010.5.6)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成24年3月29日(2012.3.29)
【国際特許分類】
【出願日】平成21年10月23日(2009.10.23)
【国際出願番号】PCT/US2009/061774
【国際公開番号】WO2010/051223
【国際公開日】平成22年5月6日(2010.5.6)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]