
ヒト腫瘍抗原からの免疫原性Tヘルパーエピトープおよび前記エピトープを使用する免疫療法の方法
【課題】癌、とりわけ腎癌の免疫療法、ならびに免疫療法で使用するための分子および細胞の提供。
【解決手段】ペプチドがHLA−DRに結合する場合に、結合した該ペプチドが、特定な配列からなるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識する細胞傷害性Tリンパ球(CTL)の産生を誘導可能である、ヒト主要組織適合抗原複合体(MHC)クラスIIの分子のHLA−DR4に結合する能力を有する腫瘍関連ペプチド。
【解決手段】ペプチドがHLA−DRに結合する場合に、結合した該ペプチドが、特定な配列からなるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識する細胞傷害性Tリンパ球(CTL)の産生を誘導可能である、ヒト主要組織適合抗原複合体(MHC)クラスIIの分子のHLA−DR4に結合する能力を有する腫瘍関連ペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、免疫療法の方法、ならびに免疫療法の方法での使用のための分子および細胞に関する。とりわけ、本発明は癌、とりわけ腎癌の免疫療法に関する。本発明はさらに、抗腫瘍免疫応答を刺激するワクチン組成物の薬学的有効成分としてはたらく、単独の若しくは他の腫瘍関連ペプチドと組合せの腫瘍関連Tヘルパー細胞ペプチドエピトープに関する。とりわけ、本発明は、抗腫瘍免疫応答を導き出すためのワクチン組成物で使用し得る、ヒト腫瘍細胞株のHLAクラスII分子由来の338種の新規ペプチド配列に関する。
本発明の目的上、本明細書に引用されるところの全部の参考文献は、そっくりそのまま引用することにより組み込まれる。
【背景技術】
【0002】
免疫応答の刺激は、宿主免疫系により外来として認識される抗原の存在に依存する。腫瘍関連抗原の存在の発見は、今や、腫瘍増殖に介入するため宿主の免疫系を使用する可能性を生じた。免疫系の体液性および細胞性双方の部門の利用の多様な機構が、現在、癌免疫療法のため探求されている。
【0003】
細胞性免疫応答の特定の要素が、腫瘍細胞を特異的に認識かつ破壊することが可能である。腫瘍浸潤細胞集団若しくは末梢血からの細胞傷害性T細胞(CTL)の単離は、こうした細胞が癌に対する自然免疫防御において重要な役割を演じていることを示唆する(非特許文献1)。サイトゾル中に位置するタンパク質由来の通常8ないし10残基の主要組織適合遺伝子複合体(MHC)のクラスI分子をもつペプチドを認識する、CD8+ T細胞(TCD8+)はとりわけ、この応答で重要な役割を演じている。ヒトのMHC分子はヒト白血球抗原(HLA)とも命名されている。
【0004】
MHC分子には2つのクラスがある:MHC−I分子は、内因性タンパク質およびより大きなペプチドのタンパク質分解性切断から生じるペプチドを提示し、核を有するほとんどの細胞上で見出し得る。MHC−II分子は、専門的抗原提示細胞(APC)上でのみ見出し得、そしてエンドサイトーシスの経過中にAPCにより取り込まれ、そして消化される外因性タンパク質のペプチドを提示する。ペプチドとMHC−Iの複合体はCD8陽性の細胞傷害性Tリンパ球により認識され、ペプチドとMHC−II分子の複合体はCD4+−ヘルパーT細胞により認識される。
【0005】
ペプチドが細胞性免疫応答を誘発する(導き出す)ためには、MHC分子に結合しなければならない。この過程はMHC分子のアリルおよびペプチドのアミノ酸配列の特定の多型に依存する。MHCクラスIに結合するペプチドは、長さが通常8〜10残基であり、そして、MHC分子の対応する結合溝と相互作用する、それらの配列中の2個の保存された残基(「アンカー」)を含有する。
【0006】
現在、腫瘍細胞を特異的に認識しかつ養子移入後にいくつかの場合には完全寛解を誘導する治療活性を有する、マウスおよびヒト双方のTCD8+の多数の例が存在する。しかしながら、腫瘍を根絶するT細胞の潜在能力にもかかわらず、多くの腫瘍がin vivoでTCD8+による認識を逃れることが、大部分の癌の進行性増殖からみても明らかである。多様な腫瘍が免疫原性であることが見出されているとは言え、効果的な抗腫瘍免疫応答の刺激は立証することが困難であった。
【0007】
腫瘍特異的細胞傷害性Tリンパ球により認識される抗原、すなわちそれらのエピトープは、酵素、受容体、転写因子などのような全部のタンパク質分類由来の分子であり得る。さらに、例えば、腫瘍関連抗原はまた、例えば変異した遺伝子の産物として腫瘍細胞中のみに存在し得る。腫瘍関連抗原の別の重要な分類は、多様な種類の腫瘍および健康な精巣組織で発現されるCT(「癌精巣」)抗原のような組織特異的構造である。
【0008】
多様な腫瘍関連抗原が同定されている。さらに、多くの研究の労力が付加的な腫瘍関連抗原を同定するために費やされている。腫瘍特異的抗原ともまた当該技術分野で称される、いくつかの群の腫瘍関連抗原は組織特異的である。例として、これに限定されるものでないが、黒色腫のチロシナーゼ、前立腺癌のPSAおよびPSMA、ならびにリンパ腫におけるbcr/ablのような染色体交差が含まれる。しかしながら、同定された多くの腫瘍関連抗原は複数の腫瘍型に存在し、そして、形質転換を実際に引き起こす癌タンパク質および/若しくは腫瘍抑制遺伝子(腫瘍抑制遺伝子は、例えば腎癌について非特許文献2に総説されている)のような数種は、ほぼ全腫瘍型に存在する。例えば、p53(腫瘍抑制遺伝子の一例である)、ras、c−met、myc、pRB、VHLおよびHER−2/neuのような、細胞の増殖および分化を制御する正常細胞タンパク質は、これらの遺伝子産物の発現の上方制御をもたらす突然変異を蓄積してそれによりそれらを発癌性にし得る(非特許文献3;非特許文献4)。これらの変異体タンパク質は、複数の型の癌での腫瘍特異的免疫応答の標的となり得る。
【0009】
腫瘍抑制遺伝子は、多細胞生物体の細胞が腫瘍細胞に変わることができる確率を低下させる遺伝子である。こうした遺伝子の突然変異若しくは欠失は腫瘍の確率を増大させることになる。その点で腫瘍抑制遺伝子は癌遺伝子に類似している。腫瘍抑制遺伝子、若しくはより正確にはそれらがコードするタンパク質は、細胞周期の調節を弱くする又は抑制する効果を有する。これは、基本的に、腫瘍抑制遺伝子/タンパク質により3つの方法により行われる:1.細胞周期の継続に不可欠である遺伝子の抑制。これらの遺伝子が発現されない場合、細胞周期は継続することができず、細胞分裂を効果的に阻害する。2.細胞周期のDNA損傷への結合。細胞中に損傷を受けたDNAが存在する限り、分裂しないはずである。該損傷が修復され得る場合に細胞周期が継続し得る。3.損傷が修復され得ない場合、該細胞はアポトーシスすなわちプログラムされた細胞死を開始して、該生物体のより大きな利益に対してそれが与える脅威を除去するはずである。発見された最初の腫瘍抑制タンパク質はヒト網膜芽種におけるpRbタンパク質であった。重要な腫瘍抑制因子はp53遺伝子(上を参照されたい)である。
【0010】
HPV由来のE6およびE7、若しくはエプスタイン−バーウイルス(EBV)由来のEBNA1のような腫瘍ウイルス由来のトランスフォーミングタンパク質もまた、多くの腫瘍型に存在し、かつ、複数の型の癌で腫瘍特異的免疫応答の標的となり得る(非特許文献5;非特許文献6;非特許文献7;非特許文献8)。MAGEおよびMUCファミリーのような非腫瘍性宿主タンパク質もまた広範に分布している。とりわけ、MAGEファミリーの抗原は、乳癌、肺癌、食道癌、肝癌、甲状腺癌、神経芽細胞腫、胃癌、多発性骨髄腫および黒色腫を包含する多くの多様な癌で見出されている(非特許文献9)。MUCファミリーの抗原は、卵巣および子宮内膜癌、乳癌、多発性骨髄腫、膵癌ならびに結腸および直腸癌と関連づけられている(非特許文献10)。
【0011】
さらに、大部分の癌は1種以上の抗原が関連する。1種以上の腫瘍抗原を発現する腫瘍の例は、これに限定されるものでないが、MUC−1、HER−2/neu、MAGE、p53、T/TnおよびCEAに関連することが示されている乳癌、MUC−2およびMUC−4、CEA、p53ならびにMAGEファミリーに関連することが示されている結腸癌、MAGEファミリーのメンバー、MART−1およびgp100に関連することが示されている黒色腫、ならびにGM2、Tn、sTn、Thompson−Friedenreich抗原(TF)、MUC1、MUC2、ヒト絨毛性ゴナドトロピンのβ鎖(hCGβ)、HER2/neu、PSMAおよびPSAと関連づけられている前立腺癌を含む。事実、腫瘍の抗原欠失バリアントが免疫系の機能下で増殖し得るという事実を代償するために、一連の抗原が、癌に対する免疫療法での使用に提案された(非特許文献11;非特許文献12)。
【0012】
タンパク質が腫瘍特異的抗原として細胞傷害性Tリンパ球により認識されるために、また、治療で使用されるために、特定の前提条件が満たされなければならない。抗原は主として腫瘍細胞により発現され、そして正常の健康な組織により発現されないか若しくは少量で発現するはずである。それぞれの抗原が1つの型の腫瘍にのみならず、高濃度(例えば、細胞あたりのコピー数)でも存在することがさらに望ましい。腫瘍関連抗原由来であるこうしたペプチド(「免疫原性ペプチド」)はin vitro若しくはin vivoのT細胞応答につながるはずであるため、抗原のアミノ酸配列中のエピトープの存在が必須である。
【0013】
現在まで、クラスIIのプロセシング経路中の抗原を標的とするための多数の戦略が記述された。抗原提示細胞(APC)を、取り込まれかつプロセシングされるために目的の抗原とインキュベートすることが可能である(非特許文献13)。他の戦略はリソソーム標的配列を含有する融合タンパク質を使用する。APC中で発現され、こうした融合タンパク質は、抗原をクラスIIのプロセシング区画に向かうことになる(非特許文献14;非特許文献15)。
【0014】
ヘルパーT細胞は、抗腫瘍免疫におけるCTLのエフェクター機能の統合において重要な役割を演じている。Th1型のヘルパーT細胞応答を誘発するヘルパーT細胞エピトープは、それらの細胞表面上に腫瘍関連ペプチド/MHC複合体を表示する腫瘍細胞に向けられた細胞傷害機能を包含するCD8+キラーT細胞のエフェクター機能を支援する。こうして、腫瘍関連ヘルパーT細胞ペプチドエピトープは、単独で若しくは他の腫瘍関連ペプチドとともに、抗腫瘍免疫応答を刺激するワクチン組成物の薬学的有効成分としてはたらき得る。
【0015】
腫瘍ワクチンの開発における主要な責務は、従って、CD4++ CTLにより認識され得る新規腫瘍関連抗原およびそれ由来の免疫原性Tヘルパーエピトープの同定および特徴付けである。従って、ヒト主要組織適合遺伝子複合体(MHC)クラスIIの分子に結合する能力を有するこうしたペプチドの新規アミノ酸配列を提供することが、本発明の一目的である。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第4,440,859号
【特許文献2】米国特許第4,530,901号
【特許文献3】米国特許第4,582,800号
【特許文献4】米国特許第4,677,063号
【特許文献5】米国特許第4,678,751号
【特許文献6】米国特許第4,704,362号
【特許文献7】米国特許第4,710,463号
【特許文献8】米国特許第4,757,006号
【特許文献9】米国特許第4,766,075号
【特許文献10】米国特許第4,810,648号
【特許文献11】WO 95/18145号
【特許文献12】WO 96/03144号
【特許文献13】WO 97/26328号
【特許文献14】WO 93/17095号
【特許文献15】米国特許第4,844,893号
【特許文献16】米国特許第4,690,915号
【非特許文献】
【0017】
【非特許文献1】Cheeverら、Annals N.Y.Acad.Sci.1993 690:101−112
【非特許文献2】Linehan WM、Walther MM、Zbar B.The genetic basis of cancer of the kidney.J Urol.2003 Dec;170(6 Pt 1):2163−72
【非特許文献3】McCarteyら Cancer Research 1998 15:58 2601−5
【非特許文献4】Disisら Ciba Found Symp.1994 187:198−211
【非特許文献5】McKaigら Head Neck 1998 20(3):250−65
【非特許文献6】Punwaneyら Head Neck 1999 21(1):21−9
【非特許文献7】Serthら Cancer Res.1999 15:59(4):823−5
【非特許文献8】Pagano,J.S.Proc.Assoc.Am.Physicians 1999 111(6):573−80
【非特許文献9】Gillespie,A.M.とColeman,R.E.Cancer Treat.Rev.1999 25(4):219−27
【非特許文献10】Segal−Eiras,A.とCroce,M.V.Allergol.Immunopathol.1997 25(4):176−81
【非特許文献11】Zhangら Clin.Cancer Res.1998 4:2669
【非特許文献12】Kawashimaら Hum.Immunol.1998 59:1
【非特許文献13】Chaux,P.、Vantomme,V.、Stroobant,V.、Thielemans,K.、Corthals,J.、Luiten,R.、Eggermont,A.M.、Boon,T.とvan der,B.P.(1999)J.Exp.Med.189、767−778
【非特許文献14】Marks,M.S.、Roche,P.A.、van Donselaar,E.、Woodruff,L.、Peters,P.J.とBonifacino,J.S.(1995)J.Cell Biol.131、351−369
【非特許文献15】Rodriguez,F.、Harkins,S.、Redwine,J.M.、de Pereda,J.M.とWhitton,J.L.(2001)J.Virol.75、10421−10430
【非特許文献16】Strubin,M.、Mach,BとLong,E.O.The comletesequence of the mRNA for the HLA−DR−associated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J.3(4)、869−872(1984)
【非特許文献17】Rammensee H、Bachmann J、Emmerich NP、Bachor OA、Stevanovic S.SYFPEITHI:detabase for MHC ligands and peptide motifs.Immunogenetics.1999 Nov;50(3−4):213−9.総説
【非特許文献18】Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21
【非特許文献19】Endlら J Clin Invest.1997 May 15;99(10):2405−15
【非特許文献20】ten Boschら Blood.1996 Nov;88(9):3522−7
【非特許文献21】Diepolderら J Virol.1997 Aug;71(8):6011−9
【非特許文献22】van der Burgら J Immunol.1999 Jan 1;162(1):152−60
【非特許文献23】Friedeら、Biochim Biophys Acta.1996 Jun 7;1316(2):85−101
【非特許文献24】Seeteら J Immunol.1993 Sep 15;151(6):3163−70.
【非特許文献25】Hammerら Cell.1993 Jul 16;74(1):197−203.
【非特許文献26】Hammerら J Exp Med.1995 May 1;181(5):1847−55
【非特許文献27】Meziereら(1997)J.Immunol.159、3230−3237
【非特許文献28】Luら(1981)J.Org.Chem.46、3433
【非特許文献29】Saikiら(1988)Science 239、487−491
【非特許文献30】Cohenら(1972)Proc.Natl.Acad.Sci.USA 69、2110
【非特許文献31】Sambrookら(1989)Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバー
【非特許文献32】Shermanら(1986)Methods In Yeast Genetics,A Laboratory Manual、ニューヨーク州コールドスプリングハーバー
【非特許文献33】Beggs(1978)Nature 275、104−109
【非特許文献34】Southern(1975)J.Mol.Biol.98、503
【非特許文献35】Berentら(1985)Biotech.3、208
【非特許文献36】Longeneckerら(1993)Ann.NY Acad.Sci.690、276−291
【非特許文献37】Zhouら(1995)Blood 86、3295−3301
【非特許文献38】Rothら(1996)Scand.J.Immunology 43、646−651
【非特許文献39】Conryら(1996)Seminars in Oncology 23、135−147
【非特許文献40】Condonら(1996)Nature Medicine 2、1122−1127
【非特許文献41】Gongら(1997)Nature Medicine 3、558−561
【非特許文献42】Zhaiら(1996)J.Immunol.156、700−710
【非特許文献43】Grahamら(1996)Int J.Cancer 65、664−670
【非特許文献44】Breast Cancer,Advances in biology and therapeutics、Calvoら(編)、John Libbey Eurotext中、Burchellら(1996)pp 309−313
【非特許文献45】KarreとLjunggren(1985)J.Exp.Med.162、1745
【非特許文献46】Thomsonら(1996)J.Immunol.157、822−826
【非特許文献47】Peoplesら(1995)Proc.Natl.Acad.Sci.USA 92、432−436
【非特許文献48】Kawakamiら(1992)J.Immunol.148、638643
【非特許文献49】Plebanskiら(1995)Eur.J.Immunol.25、1783−1787
【非特許文献50】Jochmusら(1997)J.Gen.Virol.78、1689−1695
【非特許文献51】Hillら(1995)J.Exp.Med.181、2221−2228
【非特許文献52】Jeromeら(1993)J.Immunol.151、1654−1662
【非特許文献53】Portaら(1994)Virology 202、449−955
【非特許文献54】Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658
【非特許文献55】Finerら(1994)Blood 83、43
【非特許文献56】Robertsら(1994)Blood 84、2878−2889
【非特許文献57】Moritzら(1994)Proc.Natl.Acad.Sci.USA 91、4318−4322
【非特許文献58】Eshharら(1993)Proc.Natl.Acad.Sci.USA 90、720−724
【非特許文献59】Hwuら(1993)J.Exp.Med.178、361−366
【非特許文献60】Murphyら(1996)The Prostate 29、371−380
【非特許文献61】Tjuaら(1997)The Prostate 32、272−278
【非特許文献62】Gongら(1997)Gene Ther.4、1023−1038
【非特許文献63】Wanら(1997)Hum.Gene Ther.8、1355−1363
【非特許文献64】Spechtら(1997)J.Exp.Med.186、1213−1221
【非特許文献65】Szabolcsら(1997)Blood
【非特許文献66】Tutingら(1997)Eur.J.Immunol.27、2702−2707
【非特許文献67】Ashleyら(1997)J.Exp.Med.186、1177 1182
【非特許文献68】Methods Enzymol.(1986)、121、Hybridoma technology and monoclonal antibodies
【非特許文献69】A.Kibbe、Handbook of Pharmaceutical Excipients、第3版、2000、American Pharmaceutical Association and pharmaceutical press
【非特許文献70】Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221
【非特許文献71】Wang、R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354
【非特許文献72】Hedberg,Y.、Davoodi,E.、Roos,G.、Ljungberg,B.とLandberg,G.(1999)Int.J.Cancer 84、268−272
【非特許文献73】Vasef,M.A.、Brynes,R.K.、Sturm、M.、Bromley,C.とRobinson,R.A.(1999)Mod.Pathol.12、412−416
【非特許文献74】Troussard,X.、Avet−Loiseau,J.、Macro,M.、Mellerin,M.P.、Malet,M.、Roussel,M.とSola,B.(2000)Hematol.J.1、181−185
【非特許文献75】Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827
【非特許文献76】Anderton,S.M.、Viner,N.J.、Matharu,P.、Lowrey,P.A.とWraith,D.C.(2002)Nat.Immunol.3、175−181
【非特許文献77】Lampson,L.AとLevy,R.(1980)J.Immunol.125、293−299
【非特許文献78】Brodsky,F.M.とParham,P.(1982)J.Immunol.128、129−135
【非特許文献79】Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533
【非特許文献80】Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625
【非特許文献81】Shevchenko,A.、Wilm,M.、Vorm,O.とMann,M.(1996)Anal.Chem.68、850−858
【非特許文献82】Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225
【非特許文献83】Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999
【非特許文献84】Bender,A.、Sapp,M.、Schuler,G.、Steinman,R.M.とBhardwaj,N.(1996)J.Immunol.Methods.196、121−135
【非特許文献85】Warmerdam,P.A.、Long,E.O.とRoche,P.A.(1996)J.Cell.Biol.133、281−291
【非特許文献86】Kaplan,J.とKeogh,E.A.(1981)Cell 24、925−932
【非特許文献87】Tietze,C.、Schlesinger,P.とStahl,P.(1982)J.Cell.Biol.92、417−424
【非特許文献88】Seglen,P.O.(1983)Methods Enzymol.96:737−64.、737−764
【発明の概要】
【発明が解決しようとする課題】
【0018】
本発明により、この目的は、添付された配列表の配列番号1ないし配列番号338のいずれかの少なくとも1の配列を含んでなるペプチドの群から選択される腫瘍関連ペプチドを提供することにより解決されるものであって、該ペプチドはヒト主要組織適合遺伝子複合体(MHC)クラスIIの分子に結合する能力を有する。
【課題を解決するための手段】
【0019】
本発明はさらに、抗腫瘍免疫応答を導き出すためのワクチン組成物で使用し得る、ヒト腫瘍細胞株、とりわけ腎癌細胞株のHLAクラスII分子由来の338種の新規ペプチド配列に関する。該新規ペプチド配列は、定義された(例えば腫瘍関連)抗原の未知の天然にプロセシングされたHLAクラスIIリガンドの同定のための新たなかつ一般に応用可能な組合せアプローチにより同定された。従って、ペプチドに基づく免疫療法の新たなかつ有望な候補が、著しく興味深い腫瘍抗原の選択およびそれに由来するペプチド配列の慎重な決定を包含する様式で同定された。
【0020】
本発明の第一の局面は、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかのアミノ酸配列を含んでなるペプチドを提供するが、但し、該ペプチドは、該アミノ酸配列が由来する無傷のヒトポリペプチド(すなわち遺伝子座連関ID(受入番号、下の付属の表を参照されたい)に列挙されるところの完全長配列の1つ)でない。
【0021】
下に本明細書に下述されるとおり、本発明の基礎を形成するペプチドは、MHCクラスIIをもつ細胞(Awells細胞)により提示されるとして全部が同定された。従って、これらの特定のペプチド、ならびに該配列を含有する他のペプチド(すなわち派生ペプチド)は、こうした応答が誘導されるであろう程度が個々のペプチドごとに変動するかもしれないが、おそらく全部が特異的T細胞応答を導き出すであろう。差異は、例えば前記ペプチド中の突然変異により引き起こされ得る(下を参照されたい)。当業者は、とりわけ本明細書の実施例およびそれぞれの文献を参照して、個々のペプチドにより応答が誘導される程度を決定するために適用し得る方法を知っている。
好ましくは、本発明のペプチドは、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかのアミノ酸配列より本質的になる。
【0022】
「より本質的になること」は、本発明のペプチドが、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかの配列に加えて、結合モチーフを含んでなるペプチドのコア配列および免疫原性Tヘルパーエピトープとして機能する、ペプチドの形成部分で必須ではないアミノ酸の付加的なNおよび/若しくはC末端に位置するひと配列(stretch)を含有することを意味している。
【0023】
にもかかわらず、これらのひと配列は、本発明のペプチドの細胞中への効率的な導入を提供するために重要であり得る。本発明の一態様において、本発明のペプチドは、NCBI、GenBank受入番号X00497(Strubin,M.、Mach,BとLong,E.O.The comletesequence of the mRNA for the HLA−DR−associated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J.3(4)、869−872(1984))に由来するところのHLA−DR抗原関連インバリアント鎖(p33、以下で「Ii」)の80のN末端アミノ酸を含んでなる。
【0024】
所定のアミノ酸配列の「バリアント」により、われわれは、ペプチドが、該所定のアミノ酸配列よりなるペプチドと実質的に同じ方法でHLA分子に結合することがなお可能であるような、アミノ酸残基の例えば1若しくは2個の側鎖が(例えばそれらを別の天然に存在するアミノ酸残基の側鎖若しくは何らかの他の側鎖で置換することにより)変えられることを意味している。例えば、ペプチドは、それが、HLA−DRのような適するMHC分子と相互作用しかつそれを結合する能力を改良しない場合は少なくとも維持するように、また、それが、本発明の局面で定義されるところのアミノ酸配列を含有するポリペプチドを異常に発現する細胞を認識かつ死滅させ得る活性化型CTLを生成させる能力を改良しない場合は少なくとも維持するように、改変しうる。以下で記述されるところのデータベースに由来し得るとおり、HLA−DR結合ペプチドのある位置は、典型的に、HLAの結合溝の結合モチーフに嵌るコア配列を形成するアンカー残基である。これらおよびHLA−DRの結合に関与する他の残基の改変は、CTL認識を変えることなく結合を高めうる。
【0025】
T細胞受容体と相互作用するのに不可欠でないアミノ酸残基は、その組み込みがT細胞反応性を実質的に遂げずかつ関連するMHCへの結合を除外しない別のアミノ酸での置換により改変し得る。従って、与えられる条件から離れて、本発明のペプチドは、与えられるところのアミノ酸配列またはそれらの一部分若しくはバリアントを包含するいかなるペプチド(その用語によりオリゴペプチド若しくはポリペプチドを包含する)でもありうる。
【0026】
これらのペプチドが、ある種のHLA特異的アミノ酸モチーフ、ならびに場合によってはコア配列の機能を妨害しない(すなわちペプチドおよびT細胞の相互作用にとって不適切と思われない)Nおよび/若しくはC末端の延長を有する「コア配列」から構成されることが、MHCクラスIIで提示されるペプチドについてさらに既知である。該Nおよび/若しくはC末端の延長は、それぞれ長さが1ないし10アミノ酸の間であり得る。従って、本発明の好ましいペプチドは、9と30アミノ酸の間の全体長さを表す。これらのペプチドを、MHCクラスII分子を添加するために直接使用し得るか、若しくは、該配列を下の記述によるベクターに該配列をクローン化し得るかのいずれかである。これらのペプチドは、細胞内でより長いペプチドのプロセシングの最終産物を形成するため、より長いペプチドを同様に使用し得る。本発明のペプチドはいかなる大きさのものであってもよいが、しかし、典型的にはそれらは分子量が100000未満、好ましくは50000未満、より好ましくは10000未満、および典型的には約5000でありうる。アミノ酸残基の数に関して、本発明のペプチドは、1000残基未満、好ましくは500残基未満、より好ましくは100残基未満を有しうる。
【0027】
約12アミノ酸残基より大きいペプチドを使用してMHC分子に直接結合する場合は、コアHLA結合領域に隣接する残基が、MHC分子の結合溝に特異的に結合するか若しくはCTLにペプチドを提示する該ペプチドの能力に実質的に影響を及ぼさないものであることが好ましい。しかしながら、これらのより大きいペプチドは抗原提示細胞により断片化されうるため、上で既に示したとおり、とりわけポリヌクレオチドによりコードされる場合は、より大きいペプチドを使用しうることが認識されるであろう。
【0028】
MHCリガンド、モチーフ、バリアントのペプチドの例、ならびにNおよび/若しくはC末端の延長のある種の例は、例えば、http://syfpeithi.bmi−heidelberg.com/のデータベースSYFPEITHI(Rammensee H、Bachmann J、Emmerich NP、Bachor OA、Stevanovic S.SYFPEITHI:detabase for MHC ligands and peptide motifs.Immunogenetics.1999 Nov;50(3−4):213−9.総説)、および本明細書に引用されるところの参考文献由来であり得る。
【0029】
制限しない例として、該データベース中のHLA−DRのある種のペプチドは、Ig κ鎖の188−203由来のKHKVYACEVTHQGLSS(Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21);Ig κ鎖の145−159由来のKVQWKVDNALQSGNS(Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21)、GAD65の270−283由来のLPRLIAFTSEHSHF(Endlら J Clin Invest.1997 May 15;99(10):2405−15)、若しくはGAD65の556−575由来のFFRMVISNPAATHQDIDFLI(Endlら J Clin Invest.1997 May 15;99(10):2405−15)である。加えて、ペプチドは、bcr−ablの210kDの融合タンパク質由来のATGFKQSSKALQRPVAS(ten Boschら Blood.1996 Nov;88(9):3522−7)、HCV−1 N33の28−41 Diepolderら J Virol.1997 Aug;71(8):6011−9由来のGYKVLVLNPSVAAT、若しくはHIV−1(HXB2)RTの326−345由来のFRKQNPDIVIQYMDDLYVG(van der Burgら J Immunol.1999 Jan 1;162(1):152−60)の場合でのような、抗原の変異された配列にもまた由来し得る。全部の「アンカー」アミノ酸(Friedeら、Biochim Biophys Acta.1996 Jun 7;1316(2):85−101;Seeteら J Immunol.1993 Sep 15;151(6):3163−70.;Hammerら Cell.1993 Jul 16;74(1):197−203.、およびHammerら J Exp Med.1995 May 1;181(5):1847−55を参照されたい。HLA−DR4の例として)を太字で示し、推定のコア配列に下線を付けている。
【0030】
全部の上述されたペプチドは、所定のアミノ酸配列の「バリアント」という用語により包含される。
【0031】
「ペプチド」により、われわれは、アミノ酸がペプチド(−CO−NH−)結合により結合されている分子のみならず、しかしまたペプチド結合が反転されている分子も包含する。こうしたretro−inversoペプチド模倣物は、例えばMeziereら(1997)J.Immunol.159、3230−3237(引用することにより本明細書に組み込まれる)に記述されるもののような当該技術分野で既知の方法を使用して作成しうる。このアプローチは、バックボーンを伴いかつ側鎖の指向を伴わない変更を含有する擬似ペプチドを作成することを必要とする。Meziereら(1997)は、少なくともMHCクラスIIおよびTヘルパー細胞応答にこれらの擬似ペプチドが有用であることを示す。CO−NHペプチド結合の代わりにNH−CO結合を含有するretro−invesoペプチドは、タンパク質分解に対しはるかにより抵抗性である。
【0032】
典型的には、本発明のペプチドは、抗原提示細胞中で発現される場合に、適切なMHC分子に結合することが可能でありそして適する細胞により提示されかつ適するT細胞応答を導き出しうるフラグメントが生じられるように、プロセシングされうるものである。該ペプチドから生じられるフラグメントもまた本発明のペプチドでありうることが認識されるであろう。便宜的に、本発明のペプチドは、所定のアミノ酸配列またはそれらの一部分若しくはバリアントを包含する一部分、および何らかの所望の特性を賦与するさらなる一部分を含有する。例えば、さらなる部分は、さらなるT細胞エピトープ(第一のT細胞エピトープ含有部分と同一のポリペプチド由来であろうとそうでなかろうと)を包含しうるか、または、それは担体タンパク質若しくはペプチドを包含しうる。従って、一態様において、本発明のペプチドは、切断型のヒトタンパク質、または、タンパク質フラグメントおよび別のポリペプチド部分の融合タンパク質であるが、但し、ヒト部分は1種もしくはそれ以上の発明のアミノ酸配列を包含する。
【0033】
とりわけ好ましい一態様において、本発明のペプチドは、本発明のアミノ酸配列、および最低1個のさらなるT細胞エピトープを包含し、該さらなるT細胞エピトープは、腫瘍関連抗原を異常に発現する腫瘍の型に向けられたT細胞応答の産生を助長することが可能である。従って、本発明のペプチドは、ワクチンとしてもまた使用し得るいわゆる「数珠状構造(beads on a string)」ポリペプチドを包含する。
【0034】
いくつかの応用において、本発明のペプチドを直接使用しうる(すなわち、それらは患者の細胞若しくは患者に与えられる細胞中でのポリヌクレオチドの発現により産生されない)ことが以下から認識されるであろう。こうした応用において、ペプチドが100残基未満を有することが好ましい。
【0035】
本発明のペプチドがHLA−DRに結合することが可能である場合が好ましい。ペプチドがHLA−DR4に選択的に結合する場合がとりわけ好ましい。
【0036】
本発明のペプチドは、本発明の本ペプチドの基礎を形成するポリペプチドを異常に発現する細胞を標的としかつ死滅させるための免疫療法の方法でとりわけ有用である。所定のアミノ酸配列よりなるこれらの特定のペプチドはHLA−DRに結合するため、本発明のペプチドが、HLA−DRに結合し、かつ、そのように結合された場合に、HLA−DR−ペプチド複合体が、適する抗原提示細胞の表面上に存在する場合に所定のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するCTLの産生を導き出すことが可能であることが好ましい。
【0037】
本発明の一態様において、本発明のペプチドは、NCBI、GenBank受入番号X00497に由来するところのHLA−DR抗原関連インバリアント鎖(p33、以下で「Ii」)の80のN末端アミノ酸を含んでなる。
【0038】
「異常に発現される」により、われわれは、該ポリペプチドが正常な発現レベルに比較して過剰発現されている、若しくは、腫瘍が由来する組織中で該遺伝子がサイレントであるがしかし腫瘍中でそれが発現されるという意味を包含する。「過剰発現される」により、われわれは、該ポリペプチドが正常組織中に存在するレベルの最低1.2倍;好ましくは正常組織中に存在するレベルの最低2倍、およびより好ましくは最低5若しくは10倍のレベルで存在することを意味している。
【0039】
ペプチド(少なくともアミノ酸残基間のペプチド結合を含有するもの)は、Luら(1981)J.Org.Chem.46、3433およびその中の参考文献により開示されたところの固相ペプチド合成のFmoc−ポリアミドモードにより合成しうる。一時的なNアミノ基保護は9−フルオレニルメチルオキシカルボニル(Fmoc)基により提供される。この高度に塩基不安定性の保護基の反復切断は、N,N−ジメチルホルムアミド中20%ピペリジンを使用して遂げられる。側鎖の官能性は、それらのブチルエーテル(セリン、トレオニンおよびチロシンの場合)、ブチルエステル(グルタミン酸およびアスパラギン酸の場合)、ブチルオキシカルボニル誘導体(リシンおよびのヒスチジンの場合)、トリチル誘導体(システインの場合)ならびに4−メトキシ−2,3,6−トリメチルベンゼンスルホニル誘導体(アルギニンの場合)として保護しうる。グルタミン若しくはアスパラギンがC末端残基である場合、側鎖のアミド官能性の保護に4,4'−ジメトキシベンズヒドリル基が利用される。固相支持体は、3種の単量体、ジメチルアクリルアミド(バックボーン単量体)、ビスアクリロイルエチレンジアミン(架橋剤)およびアクリロイルサルコシンメチルエステル(官能性化剤)から構成されるポリジメチル−アクリルアミドポリマーに基づく。使用されるペプチドから樹脂の切断可能な結合剤は、酸不安定性の4−ヒドロキシメチル−フェノキシ酢酸誘導体である。全部のアミノ酸誘導体は、アスパラギンおよびグルタミン(反転N,N−ジシクロヘキシル−カルボジイミド/1ヒドロキシベンゾトリアゾールに媒介されるカップリング手順を使用して付加される)を除き、それらの予め形成された対称性の無水物誘導体として付加される。全部のカップリングおよび脱保護反応は、ニンヒドリン、トリニトロベンゼンスルホン酸若しくはイソチン試験手順を使用してモニターする。合成の完了に際し、50%スカベンジャー混合物を含有する95%トリフルオロ酢酸での処理による側鎖保護基の同時の除去で、樹脂支持体からペプチドを切断する。一般に使用されるスカベンジャーは、エタンジチオール、フェノール、アニソールおよび水であり、正確な選択は合成されているペプチドの構成アミノ酸に依存する。
【0040】
トリフルオロ酢酸は、ジエチルエーテルとのその後の摩砕を用いる真空中での蒸発により除去されて、粗ペプチドを提供する。存在するいかなるスカベンジャーも、水相の凍結乾燥に際してスカベンジャーを含まない粗ペプチドを提供する単純な抽出手順により除去される。ペプチド合成のための試薬は、一般に、Calbiochem−Novabiochem(UK)Ltd.、Nottingham NG7 2QJ、UKから入手可能である。
【0041】
精製は、サイズ排除クロマトグラフィー、イオン交換クロマトグラフィー、および(通常は)逆相高速液体クロマトグラフィーのようないずれか1つの技術若しくはそれらの組合せにより遂げることができる。
【0042】
ペプチドの分析は、酸加水分解後の薄層クロマトグラフィー、逆相高速液体クロマトグラフィー、アミノ酸分析を使用して、ならびに、高速原子衝撃(FAB)質量分析、ならびにMALDIおよびESI−Q−TOF質量分析により実施しうる。
【0043】
本発明のさらなる一局面は、本発明のペプチドをコードする核酸(例えばポリヌクレオチド)を提供する。ポリヌクレオチドは、DNA、cDNA、PNA、CNA、RNA若しくはそれらの組合せであることができ、また、それは、それが該ペプチドをコードする限りはイントロンを含有しても若しくはしなくてもよい。もちろん、それは、ポリヌクレオチドによりコード可能である、天然に存在するペプチド結合により結合された天然に存在するアミノ酸を含有するペプチドのみである。本発明のなおさらなる一局面は、本発明のポリペプチドを発現することが可能な発現ベクターを提供する。
【0044】
例えば相補的付着端を介してベクターにポリヌクレオチド、とりわけDNAを作動可能に連結するための多様な方法が開発された。例えば、相補的ホモポリマー領域を、ベクターDNAに挿入されるべきDNAセグメントに付加し得る。ベクターおよびDNAセグメントをその後、相補的ホモポリマー尾部間の水素結合により結合して組換えDNA分子を形成する。
【0045】
1個若しくはそれ以上の制限部位を含有する合成リンカーは、ベクターへのDNAセグメントの一代替結合方法を提供する。前に記述されたところのエンドヌクレアーゼ制限消化により生成されるDNAセグメントを、バクテリオファージT4 DNAポリメラーゼ若しくは大腸菌(E.coli)DNAポリメラーゼIで処理し、これらの酵素は突出している3'一本鎖末端をそれらの3'−5'−エキソヌクレアーゼ活性で除去し、そしてそれらのポリメラーゼ活性でくぼんだ3'端を埋める。
【0046】
これらの活性の組合せは従って平滑端のDNAセグメントを生成する。該平滑端セグメントをその後、バクテリオファージT4 DNAリガーゼのような平滑端DNA分子の核酸連結を触媒することが可能である酵素の存在下で、モル濃度大過剰のリンカー分子とともにインキュベートする。従って、該反応の生成物はそれらの端にポリマーリンカー配列を運搬するDNAセグメントである。これらのDNAセグメントをその後、適切な制限酵素で切断し、そして、該DNAセグメントの末端と適合する末端を生じる酵素で切断した発現ベクターに連結する。
【0047】
多様な制限酵素部位を含有する合成リンカーは、International Biotechnologies Inc、米国コネチカット州ニューヘイブンを包含する多数の供給源から商業的に入手可能である。
【0048】
本発明のポリペプチドをコードするDNAの望ましい一改変方法は、Saikiら(1988)Science 239、487−491により開示されたところのポリメラーゼ連鎖反応を使用することである。この方法は、例えば適する制限部位で設計することにより、適するベクターにDNAを導入するのに使用しうるか、若しくは、それを使用して、当該技術分野で既知であるところの他の有用な方法でDNAを改変しうる。この方法において、酵素で増幅されるべきDNAは、該増幅されたDNAにそれら自身が組み込まれる2種の特異的プライマーにより隣接される。前記特異的プライマーは、当該技術分野で既知の方法を使用して発現ベクターにクローン化するのに使用し得る、制限酵素認識部位を含有しうる。
【0049】
DNA(若しくはレトロウイルスベクターの場合はRNA)がその後、適する宿主中で発現されて、本発明の化合物を含んでなるポリペプチドを産生する。従って、本発明の化合物を構成するポリペプチドをコードするDNAを、本明細書に含有される教示を鑑みて適切に改変された既知技術に従って使用して発現ベクターを構築することができ、この発現ベクターをその後、本発明のポリペプチドの発現および産生のため適切な宿主細胞を形質転換するのに使用する。こうした技術は、Rutterらに1984年4月3日に発行された米国特許第4,440,859号、Weissmanに1985年7月23日に発行された同第4,530,901号、Crowlに1986年4月15日に発行された同第4,582,800号、Markらに1987年6月30日に発行された同第4,677,063号、Goeddelに1987年7月7日に発行された同第4,678,751号、Itakuraらに1987年11月3日に発行された同第4,704,362号、Murrayに1987年12月1日に発行された同第4,710,463号、Toole,Jr.らに1988年7月12日に発行された同第4,757,006号、Goeddelらに1988年8月23日に発行された同第4,766,075号、およびStalkerに1989年3月7日に発行された同第4,810,648号明細書(それらの全部は引用することにより本明細書に組み込まれる)に開示されるものを包含する。
【0050】
本発明の化合物を構成するポリペプチドをコードするDNA(若しくはレトロウイルスベクターの場合はRNA)は、適切な宿主への導入のために多様な他のDNA配列に結合しうる。伴(companion)DNAは、宿主の性質、宿主細胞への該DNAの導入の様式、およびエピソーム維持が望ましいか若しくは組込みが望ましいかに依存することができる。
【0051】
一般に、DNAは、プラスミドのような発現ベクターに、発現のための適正な向きおよび正しい読み枠で挿入する。必要な場合は、DNAは所望の宿主により認識される適切な転写および翻訳調節性の制御ヌクレオチド配列に連結しうるが、こうした制御配列は発現ベクターで一般に利用可能である。ベクターをその後、標準的技術により宿主に導入する。一般に、宿主の全部がベクターにより形質転換されるわけではない。従って、形質転換された宿主細胞を選択することが必要となる。一選択技術は、抗生物質耐性のような、形質転換された細胞中の選択可能な特質をコードするいずれかの必要な調節領域とともに、DNA配列を発現ベクターに組み込むことを伴う。
あるいは、こうした選択可能な特質の遺伝子は、所望の宿主細胞を共形質転換するのに使用する別のベクター上にあってよい。
【0052】
本発明の組換えDNAにより形質転換された宿主細胞をその後、ポリペプチドの発現を可能にするための本明細書に開示される教示を鑑み、十分な時間、および当業者に既知の適切な条件下で培養し、ポリペプチドをその後回収してよい。
【0053】
細菌(例えば大腸菌(E.coli)および枯草菌(Bacillus subtilis)、)酵母(例えば出芽酵母(Saccharomyces cerevisiae))、糸状菌(例えばアスペルギルス属(Aspergillus))、植物細胞、動物細胞および昆虫細胞を包含する多くの発現系が既知である。好ましくは、該系はAwells細胞であってよい。
【0054】
プロモーターは、RNAポリメラーゼの結合および転写を起こさせる、DNA配列により形成される発現調節領域である。模範的な細菌宿主と適合性のあるプロモーター配列は、典型的に、本発明のDNAセグメントの挿入に都合のよい制限部位を含有するプラスミドベクター中に提供されている。典型的な原核生物ベクタープラスミドは、Biorad Laboratoris、(米国カリフォルニア州リッチモンド)から入手可能なpUC18、pUC19、pBR322およびpBR329、ならびにPharmacia、米国ニュージャージー州ピスカタウェイから入手可能なpTrc99AおよびpKK223−3である。
【0055】
典型的な哺乳動物細胞ベクタープラスミドは、Pharmacia、米国ニュージャージー州ピスカタウェイから入手可能なpSVLである。このベクターは、クローン化した遺伝子の発現を駆動するのにSV40後期プロモーターを使用し、最高レベルの発現はCOS−1細胞のようなT抗原産生細胞で見出される。誘導可能な哺乳動物発現ベクターの一例は、またPharmaciaから入手可能なpMSGである。このベクターは、クローン化した遺伝子の発現を駆動するために、マウス乳癌ウイルス末端反復配列のグルココルチコイドで誘導可能なプロモーターを使用する。有用な酵母プラスミドベクターはpRS403−406およびpRS413−416であり、そして一般にStratagene Cloning Systems、La Jolla、CA 92037、USAから入手可能である。プラスミドpRS403、pRS404、pRS405およびpRS406は酵母組込み型プラスミド(YIp)であり、そして酵母の選択可能なマーカー、HIS3、TRP1、LEU2およびURA3を組み込む。プラスミドpRS413−416は酵母セントロメアプラスミド(Ycp)である。多様な宿主細胞との使用のための他のベクターおよび発現系が当該技術分野で公知である。
【0056】
本発明はまた、本発明のポリヌクレオチドベクター構築物で形質転換された宿主細胞にも関する。該宿主細胞は原核生物若しくは真核生物のいずれかであり得る。細菌細胞はいくつかの環境で好ましい原核生物宿主細胞であることができ、そして、典型的には、例えば、Bethesda Research Laboratories Inc.、米国メリーランド州ベセスダから入手可能な大腸菌(E.coli)株DH5、および米国メリーランド州ロックビルのAmerican Type Culture Collection(ATCC)から入手可能なRR1(No ATCC 31343)のような大腸菌(E.coli)の株である。好ましい真核生物宿主細胞は、酵母、昆虫および哺乳動物細胞、好ましくはマウス、ラット、サル若しくはヒト線維芽および腎細胞株からのもののような脊椎動物細胞を包含する。酵母宿主細胞は、Stratagene Cloning Systems、La Jolla、CA 92037、米国から一般に入手可能であるYPH499、YPH500およびYPH501を包含する。好ましい哺乳動物宿主細胞は、CCL61としてATCCから入手可能なチャイニーズハムスター卵巣(CHO)細胞、CRL 1658としてATCCから入手可能なNIH Swissマウス胚細胞NIH/3T3、CRL 1650としてATCCから入手可能なサル腎由来COS−1細胞、およびヒト胎児由来腎臓細胞である293細胞を包含する。好ましい昆虫細胞はバキュロウイルス発現ベクターでトランスフェクトし得るSf9細胞である。
【0057】
本発明のDNA構築物での適切な細胞宿主の形質転換は、使用されるベクターの型に典型的に依存する公知の方法により達成される。原核生物宿主細胞の形質転換に関しては、例えば、Cohenら(1972)Proc.Natl.Acad.Sci.USA 69、2110およびSambrookら(1989)Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバーを参照されたい。酵母細胞の形質転換は、Shermanら(1986)Methods In Yeast Genetics,A Laboratory Manual、ニューヨーク州コールドスプリングハーバーに記述されている。Beggs(1978)Nature 275、104−109の方法もまた有用である。脊椎動物細胞に関しては、こうした細胞のトランスフェクションにおいて有用な試薬、例えばリン酸カルシウムおよびDEAE−デキストラン若しくはリポソーム製剤が、Stratagene Cloning Systems、若しくはLife Technologies Inc.、Gaithersburg、MD 20877、米国から入手可能である。電気穿孔法もまた細胞を形質転換および/若しくはトランスフェクトするのに有用であり、また、酵母細胞、細菌細胞、昆虫細胞および脊椎動物細胞を形質転換するために当該技術分野で公知である。
【0058】
成功裏に形質転換された細胞、すなわち本発明のDNA構築物を含有する細胞は、公知の技術により同定し得る。例えば、本発明の発現構築物の導入から生じる細胞を、本発明のポリペプチドを産生させるために増殖させ得る。細胞を収集かつ溶解し得、そしてそれらのDNA内容物を、Southern(1975)J.Mol.Biol.98、503若しくはBerentら(1985)Biotech.3、208により記述されたもののような方法を使用して、該DNAの存在について検査し得る。あるいは、上清中の外タンパク質の存在を、下述されるところの抗体を使用して検出し得る。
【0059】
組換えDNAの存在について直接アッセイすることに加え、成功裏の形質転換は、組換えDNAがタンパク質の発現を指図することが可能である場合に、公知の免疫学的方法により確認し得る。例えば、発現ベクターで成功裏に形質転換された細胞は、適切な抗原性を表すタンパク質を産生する。形質転換されていることが疑われる細胞のサンプルを収集し、そして、適する抗体を使用してタンパク質についてアッセイする。従って、形質転換された宿主細胞それら自身に加え、本発明はまた、栄養培地中のそれらの細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、若しくはモノクローナル培養物由来の培養物も企図している。
【0060】
本発明のある種の宿主細胞、例えば細菌、酵母および昆虫細胞が、本発明のペプチドの製造で有用であることが認識されるであろう。しかしながら、他の宿主細胞がある種の治療方法で有用でありうる。例えば、樹状細胞のような抗原提示細胞は、それらが適切なMHC分子に負荷されうるような本発明のペプチドを発現させるために有用に使用しうる。
【0061】
本発明のさらなる一局面は、静脈内(i.v.)注入、皮下(s.c.)注入、皮内(i,d.)注入、腹腔内(i.p.)注入、筋肉内(i.m.)注入のためのペプチドの製造方法を提供する。好ましいペプチド注入方法はs.c.、i.p.およびi.v.である。好ましいDNA注入方法は、i.d.、i.m.、s.c.、i.p.およびi.v.である。1から500mgのペプチド若しくはDNAの用量を与えうる。
【0062】
本発明のさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、有効量の本発明のペプチド、または有効量の前記ペプチドをコードするポリヌクレオチド若しくは発現ベクターを患者に投与することを含んでなり、前記ペプチドの該量または前記ポリヌクレオチド若しくは発現ベクターの量は、前記患者で抗標的細胞免疫応答を惹起するのに有効である。標的細胞は、典型的に腫瘍若しくは癌細胞である。
【0063】
ペプチド若しくはペプチドをコードする核酸は腫瘍若しくは癌ワクチンを構成する。それは患者に、冒された器官に直接投与しうるか、またはその後患者に投与される該患者由来の細胞若しくはヒト細胞株にex vivoで適用しうるか、または、その後患者に再投与される該患者由来の免疫細胞からの亜集団を選択するためにin vitroで使用しうる。核酸をin vitroで細胞に投与する場合、インターロイキン−2のような免疫刺激サイトカインを共発現するようにトランスフェクトすることが細胞に有用でありうる。該ペプチドは、実質的に純粋であっても、若しくはDetoxのような免疫刺激アジュバントと組合せても、若しくは免疫刺激サイトカインとともに使用しても、若しくは適する送達系、例えばリポソームを用いて投与してもよい。該ペプチドはまた、キーホールリンペットヘモシアニン(KLH)若しくはマンナンのような適する担体にも複合しうる(WO 95/18145号、およびLongeneckerら(1993)Ann.NY Acad.Sci.690、276−291を参照されたい)。該ペプチドはまた、標識しても、若しくは融合タンパク質であっても、若しくはハイブリッド分子であってもよい。その配列が本発明で示されるペプチドが発現されてCD4 CTLを刺激する。しかしながら、刺激は、CD4+ T細胞により提供される援助の存在下でより効率的である。従って、ハイブリッド分子の融合パートナー若しくは区分は、CD4+ T細胞を刺激するエピトープを適切に提供する。CD4+を刺激するエピトープは当該技術分野で公知であり、そして破傷風トキソイド中で同定されたものを包含する。該ポリヌクレオチドは、実質的に純粋であっても、または適するベクター若しくは送達系に含有されてもよい。
【0064】
適するベクターおよび送達系は、アデノウイルス、ワクシニアウイルス、レトロウイルス、ヘルペスウイルス、アデノ随伴ウイルス、若しくは1種以上のウイルスの要素を含有するハイブリッドに基づく系のようなウイルスを包含する。ウイルス以外の送達系は、DNA送達の技術分野で公知であるところの陽イオン性脂質および陽イオン性ポリマーを包含する。「遺伝子銃」を介するような物理的送達もまた使用しうる。該ペプチド、若しくは該核酸によりコードされるペプチドは、例えば、CD4+ T細胞を刺激する破傷風トキソイドからのエピトープをもつ融合タンパク質でありうる。
【0065】
癌ワクチンでの使用のためのペプチドはいかなる適するペプチドでもありうる。とりわけ、それは適する9merペプチド、あるいは適する7mer若しくは8mer若しくは10mer若しくは11merペプチドまたは12merでありうる。より長いペプチドもまた適しうるが、しかし付属の表1に記述されるところの9mer若しくは10merペプチドが好ましい。
【0066】
適切には、患者に投与されるいかなる核酸も無菌でありかつ発熱性物質を含まない。裸のDNAは筋肉内若しくは皮内若しくは皮下に与えうる。ペプチドは筋肉内、皮内若しくは皮下に与えうる。
【0067】
ワクチン接種は専門の抗原提示細胞により刺激されるCTL応答をもたらし、CTLが一旦刺激されれば、腫瘍細胞中でのMHC発現の増強において一利点が存在しうる。
【0068】
注入の部位、ターゲッティングベクターおよび送達系の使用、または患者からのこうした細胞集団の選択的精製およびペプチド若しくは核酸のex vivo投与のいずれかにより、ワクチンの標的を特定の細胞集団、例えば抗原提示細胞に向けることもまた有用でありうる(例えば、樹状細胞を、Zhouら(1995)Blood 86、3295−3301;Rothら(1996)Scand.J.Immunology 43、646−651に記述されるとおり選別しうる)。例えば、ターゲッティングベクターは、適する場所での抗原の発現を指図する組織若しくは腫瘍特異的プロモーターを含みうる。
【0069】
本発明のさらなる一局面は、従って、有効量の本発明のペプチドを含んでなるか若しくはこうしたペプチドをコードする核酸を含んでなる、癌、または癌若しくは腫瘍細胞に対し有効なワクチンを提供する。ワクチンが核酸ワクチンである場合がとりわけ好ましい。ポリペプチドをコードするDNAワクチンのような核酸ワクチンでの接種がT細胞応答につながることが既知である。
【0070】
便宜上、核酸ワクチンは、いかなる適する核酸送達手段も含みうる。核酸、好ましくはDNAは裸でありうる(すなわち投与されるべき他の成分を実質的に含まない)か、または、それはリポソーム中で若しくはウイルスベクター送達系の一部として送達されうる。
【0071】
樹状細胞による核酸の取り込みおよびコードされるポリペプチドの発現が、免疫応答のプライミングの機構でありうると考えられるが、しかしながら、樹状細胞はトランスフェクトされなくてもよいがしかしなお重要である。それらは、トランスフェクトされた細胞からの発現されたペプチドを組織中に取り込みうるからである。
【0072】
DNAワクチンのようなワクチンが筋中に投与される場合が好ましい。ワクチンが皮膚に投与される場合もまた好ましい。核酸ワクチンはアジュバントを伴わずに投与してもよい。核酸ワクチンはまた、BCG若しくは明礬のようなアジュバントとともにも投与しうる。他の適するアジュバントは、サポニン由来であるAquilaのQS21スティミュロン(stimulon)(Aquila Biotech、米国マサチューセッツ州ウースター)、ミコバクテリウム抽出物および合成細菌細胞壁模倣物、ならびにRibiのDetoxのような特許のアジュバントを包含する。別のサポニン由来アジュバント、Quil Aもまた使用しうる(Superfos、デンマーク)。核酸ワクチンがアジュバントを伴わずに投与される場合が好ましい。フロイントのアジュバントのような他のアジュバントもまた有用でありうる。キーボールリンペットヘモシアニンに、好ましくはまたアジュバントと複合させたペプチドを与えることもまた有用でありうる。
【0073】
癌のペプチドヌクレオチド媒介性の免疫療法は、Conryら(1996)Seminars in Oncology 23、135−147;Condonら(1996)Nature Medicine 2、1122−1127;Gongら(1997)Nature Medicine 3、558−561;Zhaiら(1996)J.Immunol.156、700−710;Grahamら(1996)Int J.Cancer 65、664−670;およびBreast Cancer,Advances in biology and therapeutics、Calvoら(編)、John Libbey Eurotext中、Burchellら(1996)pp 309−313(それらの全部は引用することにより本明細書に組み込まれる)に記述されている。
【0074】
本発明のなおさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞を死滅させるための医薬品の製造における、本発明のペプチド、またはこうしたペプチドをコードするポリヌクレオチド若しくは発現ベクターの使用を提供する。
【0075】
本発明のさらなる一局面は、in vitroでの活性化型細胞傷害性Tリンパ球(CTL)の製造方法を提供し、該方法は、CTLを、適する抗原提示細胞の表面上に発現された、抗原負荷ヒトクラスIIMHC分子と、前記CTLを抗原(該抗原は本発明のペプチドである)特異的様式で活性化するのに十分な時間、in vitroで接触させることを含んでなる。
【0076】
適しては、CTLは、好ましくはTH1型のCD4+ヘルパー細胞である。MHCクラスII分子は、いかなる適する細胞の表面上に発現されてもよく、また、細胞がMHCクラスII分子を天然に発現しない(その場合、該細胞はこうした分子を発現するようトランスフェクトされる)か、またはそれが発現する場合はそれが抗原プロセシング若しくは抗原提示経路に欠陥があるものである場合が好ましい。こうして、MHCクラスII分子を発現する細胞が、CTLを活性化する前に選ばれたペプチド抗原で実質的に完全に予備刺激されることが可能である。
【0077】
抗原提示細胞(若しくは刺激体(stimulator)細胞)は、典型的にその表面上に1個のMHCクラスII分子を有し、そして、好ましくは、前記MHCクラスII分子に選択された抗原をそれ自身負荷することが実質的に不可能である。より詳細に下述されるとおり、MHCクラスII分子は、選択された抗原をin vitroで容易に負荷されうる。
【0078】
好ましくは、該哺乳動物細胞は、TAPペプチド輸送体を欠くか、若しくはその低下されたレベルを有するか、若しくはその低下された機能を有する。TAPペプチド輸送体を欠く適する細胞は、T2、RMA−Sおよびショウジョウバエ(Drosophila)細胞を包含する。TAPは、抗原プロセシングに関与する輸送体(Transporter Associated with antigen Processing)である。
【0079】
ヒトペプチド負荷欠損細胞株(human peptide loading deficient cell line)T2は、カタログ番号CRL 1992で、American Type Culture Collection、12301 Parklawn Drive、Rockville、Maryland 20852、USAから入手可能であり;ショウジョウバエ(Drosophila)細胞株Schneider株2は、カタログ番号CRL 19863でATCCから入手可能であり;マウスRMA−S細胞株は、KarreとLjunggren(1985)J.Exp.Med.162、1745(引用することにより本明細書に組み込まれる)に記述されている。
【0080】
便宜上、前記宿主細胞は、トランスフェクション前にMHCクラスI分子を実質的に発現しない。刺激体細胞が、B7.1、B7.2、ICAM−1およびLFA3のいずれかのようなT細胞共刺激に重要な分子を発現する場合もまた好ましい。
【0081】
多数のMHCクラスII分子および共刺激体分子の核酸配列は、GenBankおよびEMBLデータベースから公的に入手可能である。
【0082】
さらなる一態様において、HLA分子の組合せもまた使用しうる。
【0083】
複数のCD8+ CTLエピトープの送達のための組換え多エピトープワクチンの使用は、Thomsonら(1996)J.Immunol.157、822−826およびWO 96/03144号(それらの双方は引用することにより本明細書に組み込まれる)に記述されている。本発明に関して、単一のワクチン中に1種のペプチド(若しくは1種のペプチドをコードする核酸)を包含することが望ましいことができ、該ペプチドは、本発明のアミノ酸配列およびCD4+ T細胞を刺激するエピトープ(破傷風トキソイドのような)をいずれかの順序で包含する。こうしたワクチンは、癌を処置するのにとりわけ有用であるとみられる。こうした「数珠状構造」ワクチンは、典型的にはDNAワクチンである。
【0084】
in vitroでCTLを生成するのに多数の他の方法を使用しうる。例えば、Peoplesら(1995)Proc.Natl.Acad.Sci.USA 92、432−436およびKawakamiら(1992)J.Immunol.148、638643に記述される方法は、CTLの生成で自己の腫瘍浸潤リンパ球を使用する。Plebanskiら(1995)Eur.J.Immunol.25、1783−1787は、CTLの製造で自己の末梢血リンパ球(PLB)を利用する。Jochmusら(1997)J.Gen.Virol.78、1689−1695は、ペプチド若しくはポリペプチドで樹状細胞をパルスすることを使用することによるか、または組換えウイルスへの感染を介する、自己CTLの製造を記述する。Hillら(1995)J.Exp.Med.181、2221−2228およびJeromeら(1993)J.Immunol.151、1654−1662は、自己CTLの製造でB細胞を利用する。加えて、ペプチド若しくはポリペプチドでパルスしたか、または組換えウイルスに感染させたマクロファージを、自己CTLの製造で使用しうる。
【0085】
同種異系細胞もまたCTLの製造に使用することができ、そして、この方法はWO 97/26328号(引用することにより本明細書に組み込まれる)に詳細に記述されている。例えば、ショウジョウバエ(Drosophila)細胞およびT2細胞に加え、CHO細胞、バキュロウイルスに感染させた昆虫細胞、細菌、酵母、ワクシニアに感染させた標的細胞のような他の細胞を使用して抗原を提示しうる。加えて、植物ウイルスを使用しうる(例えば、外来ペプチドの提示のための多収性の系としてのササゲモザイクウイルスの開発を記述するPortaら(1994)Virology 202、449−955を参照されたい)。
【0086】
本発明のペプチドに向けられる活性化型CTLは治療で有用である。従って、本発明のさらなる一局面は、本発明の前述の方法により得ることができる活性化型CTLを提供する。
【0087】
本発明のなおさらなる一態様は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を選択的に認識する活性化型CTLを提供する。好ましくは、該CTLはHLA/ペプチド複合体と相互作用(例えば結合)することにより前記細胞を認識する。該CTLは、有効な数の活性化型CTLを患者に投与する、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法で有用である。患者に投与されるCTLは、患者由来でありかつ上述されたとおり活性化しうる(すなわちそれらは自己CTLである)。あるいは、該CTLは患者からでないが、しかし別の個体からである。もちろん、該個体が健康な個体である場合が好ましい。「健康な個体」により、われわれは、個体が全般として良好な健康状態にある、好ましくは適確な免疫系を有する、およびより好ましくは、容易に試験かつ検出され得るいかなる疾患にも罹っていないことを意味している。
【0088】
活性化型CTLは、異常なポリペプチドを発現する細胞の認識に関与するT細胞受容体(TCR)を発現する。TCRをコードするcDNAが活性化型CTLからクローン化されかつ発現のためさらなるCTLに移入される場合が有用である。
【0089】
in vivoで、本発明のCD4+ CTLの標的細胞は、腫瘍の細胞(ときにMHCクラスIIを発現する)および/若しくは腫瘍(腫瘍細胞)を取り囲む間質細胞(ときにまたMHCクラスIIを発現する)であり得る。
【0090】
本発明のペプチドに特異的な本発明のCTLクローンのTCRをクローン化する。CTLクローンでのTCRの使用は、(i)TCR可変領域特異的モノクローナル抗体、ならびに(ii)VaおよびVp遺伝子ファミリーに特異的なプライマーを用いるRT PCRを使用して決定する。CTLクローンから抽出したポリA mRNAからcDNAライブラリーを調製する。TCR aおよびP鎖のC末端部分、ならびに同定されたVaおよびPセグメントのN末端部分に特異的なプライマーを使用する。TCR aおよび鎖の完全なcDNAを高忠実度DNAポリメラーゼで増幅し、そして、増幅された産物を適するクローニングベクターにクローン化する。クローン化されたaおよびP鎖遺伝子を、Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658により記述されるところの方法により、一本鎖TCRに集成しうる。この一本鎖構築物中で、VaJセグメントにV DJセグメントが続き、次いでCpセグメント、次いでCD3鎖の膜貫通および細胞質セグメントが続く。この一本鎖TCRをその後レトロウイルス発現ベクターに挿入する(一団のベクターを、成熟ヒトCD8+ Tリンパ球を感染させかつ遺伝子発現を媒介するそれらの能力に基づいて使用しうる。すなわち、レトロウイルスベクター系Katが1つの好ましい可能性である(Finerら(1994)Blood 83、43を参照されたい)。高力価の両指向性レトロウイルスを使用して、腫瘍患者の末梢血から単離した、精製したCD8+若しくはCD4+ Tリンパ球を(Robertsら(1994)Blood 84、2878−2889(引用することにより本明細書に組み込まれる)により公表されたプロトコルに従って)感染させる。抗CD3抗体を使用して、精製したCD8+ T細胞の増殖を誘発し、それは一本鎖TCRのレトロウイルス組込みおよび安定な発現を助長する。レトロウイルス形質導入の効率は、一本鎖TCRに特異的な抗体での感染させたCD8+ T細胞の染色により決定する。形質導入されたCD8+ T細胞のin vitro分析は、該TCR鎖が最初にクローン化された同種拘束性(allo−restricted)CTLクローンで見られると同一の腫瘍特異的死滅をそれらが表すことを確立する。期待された特異性をもつ形質導入されたCD8+ T細胞の集団を、腫瘍患者の養子免疫療法に使用しうる。患者は、108ないし1011の間の自己の形質導入されたCTLで処置しうる。CD8+と同様に、関連する構築物を運搬する形質導入されたCD4+ Tヘルパー細胞を生成させ得る。
【0091】
遺伝子をCTLに導入するための他の適する系は、Moritzら(1994)Proc.Natl.Acad.Sci.USA 91、4318−4322(引用することにより本明細書に組み込まれる)に記述されている。Eshharら(1993)Proc.Natl.Acad.Sci.USA 90、720−724、およびHwuら(1993)J.Exp.Med.178、361−366もまた、CTLのトランスフェクションを記述している。従って、本発明のさらなる局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するTCRを提供し、該TCRは活性化型CTLから得ることができる。
【0092】
TCRだけでなく、TCRに機能上同等な分子が本発明に包含される。これらは、TCRと同一の機能を演じ得る、TCRに機能上同等であるいかなる分子も包含する。とりわけ、こうした分子は、Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658(引用することにより本明細書に組み込まれかつ上に言及された)により記述されかつ上に引用される方法により作成されるところの、遺伝子的に操作された3ドメインの一本鎖TCRを包含する。本発明はまた、TCR若しくは機能上同等な分子をコードするポリヌクレオチド、および該TCR若しくはその機能上同等な分子をコードする発現ベクターも包含する。本発明のTCRを発現するのに適する発現ベクターは、本発明のペプチドの発現に関して上述されたものを包含する。
【0093】
しかしながら、発現ベクターがトランスフェクション後にCTL中でTCRを発現することが可能であるものであることが好ましい。
【0094】
本発明のなおさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、(1)患者からCTLを得る段階;(2)TCR若しくは上で定義されたところの機能上同等な分子をコードするポリヌクレオチドを前記細胞に導入する段階;および(3)段階(2)で製造した細胞を患者に導入する段階を含んでなる。
【0095】
本発明のなおさらなる一局面は、本発明の第一若しくは第二若しくは第三の局面で定義されるところのアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、(1)前記患者から樹状細胞のような抗原提示細胞を得る段階;(2)前記抗原提示細胞を、本発明の第一若しくは第二若しくは第三の局面で定義されるところのペプチド、またはこうしたペプチドをコードするポリヌクレオチドとex vivoで接触させる段階;および(3)そのように処理した抗原提示細胞を患者に再導入する段階を含んでなる。
【0096】
好ましくは、抗原提示細胞は樹状細胞である。適切には、樹状細胞は、抗原ペプチドでパルスされている自己の樹状細胞である。抗原ペプチドは、適切なT細胞応答を生じさせるいかなる適する抗原ペプチドであってもよい。腫瘍関連抗原からのペプチドでパルスした自己の樹状細胞を使用するT細胞療法は、Murphyら(1996)The Prostate 29、371−380およびTjuaら(1997)The Prostate 32、272−278に開示されている。
【0097】
さらなる一態様において、樹状細胞のような抗原提示細胞を、本発明のペプチドをコードするポリヌクレオチドと接触させる。該ポリヌクレオチドはいかなる適するポリヌクレオチドであってもよく、そして、それが樹状細胞を形質導入し、従ってペプチドの提示および免疫の誘導をもたらすことが可能であることが好ましい。
【0098】
便宜上、ポリヌクレオチドはウイルスポリヌクレオチド若しくはウイルスに含まれうる。例えば、アデノウイルスで形質導入した樹状細胞は、MUC1に関して抗原特異的抗腫瘍免疫を誘導することが示された(Gongら(1997)Gene Ther.4、1023−1038を参照されたい)。同様に、アデノウイルスに基づく系を使用することができ(例えば、Wanら(1997)Hum.Gene Ther.8、1355−1363を参照されたい);レトロウイスル系を使用することができ(Spechtら(1997)J.Exp.Med.186、1213−1221、およびSzabolcsら(1997)Blood)、樹状細胞への粒子媒介性移入もまた使用することができ(Tutingら(1997)Eur.J.Immunol.27、2702−2707);そしてRNAもまた使用しうる(Ashleyら(1997)J.Exp.Med.186、1177 1182)。
【0099】
患者における標的細胞の死滅方法に関して、標的細胞が癌細胞、より好ましくは腎癌細胞であることがとりわけ好ましいことが認識されるであろう。
【0100】
本発明の方法により処置される患者がHLA−DRハプロタイプを有する場合がとりわけ好ましい。従って、好ましい一態様において、患者のHLAハプロタイプを処置の前に決定する。HLAハプロタイピングは、いずれの適する方法を使用して実施してもよく、こうした方法は当該技術分野で公知である。
【0101】
本発明は、とりわけ、活性のin vivoワクチン接種のため;in vitroでの自己の樹状細胞の操作、次いでCTL応答を活性化するためのそのように操作した樹状細胞のin vivoでの導入のため;in vitroで自己CTLを活性化し、次いで養子療法のため(すなわちそのように操作したCTLを患者に導入する);および健康なドナー(MHCマッチ若しくはミスマッチの)からのCTLをin vitroで活性化し、次いで養子療法のための、本発明のペプチド(若しくはそれらをコードするポリヌクレオチド)の使用を包含する。
【0102】
好ましい一態様において、本発明のワクチンは、腫瘍の形成を阻害若しくは抑制するために、単独で、若しくは別の癌治療との組合せのいずれかで宿主に投与する。
【0103】
ペプチドワクチンはアジュバントを伴わずに投与しうる。ペプチドワクチンはまた、BCG若しくは明礬のようなアジュバントとともに投与してもよい。他の適するアジュバントは、サポニン由来であるAquilaのQS21スティミュロン(stimulon)(Aquila Biotech、米国マサチューセッツ州ウースター)、ミコバクテリウム抽出物、および合成の細菌細胞壁模倣物、ならびにRibiのDetoxのような特許のアジュバントを包含する。別のサポニン由来アジュバント、Quil Aもまた使用しうる(Superfos、デンマーク)。フロイントのアジュバント若しくはGMCSFのような他のアジュバントもまた有用でありうる。好ましくはまたアジュバントとともにのキーホールリンペットヘモシアニンに複合させたペプチドを与えることもまた有用でありうる。
【0104】
本発明のペプチドはまた診断試薬としても使用し得る。該ペプチドを使用して、CTL集団中に、ペプチドに特異的に向けられているCTLが存在するかどうか、若しくはCTLが治療により誘導されるかどうかを分析し得る。さらに、前駆T細胞の増大を、規定されたペプチドに対する反応性を有するペプチドを用いて試験し得る。さらに、該ペプチドを、該ペプチドが由来する前記抗原を発現する腫瘍の疾患の進行をモニターするためのマーカーとして使用し得る。
【0105】
付属の表1に、同定されたところのペプチドを列挙する。加えて、該表に、ペプチドが由来するタンパク質、およびそれぞれのタンパク質中のペプチドのそれぞれの位置を指定する。さらに、国立衛生研究所の「国立バイオテクノロジー情報センター(National Centre for Biotechnology Information)」のGenbankに関するそれぞれの受入番号を示す(http://www.ncbi.nlm.nih.govを参照されたい)。
【0106】
別の好ましい態様において、該ペプチドを白血球、とりわけTリンパ球の染色に使用する。この使用は、CTL集団中に、ペプチドに向けられている特異的CTLが存在するかどうかが証明されるはずである場合にとりわけ有利である。さらに、該ペプチドを、腫瘍性疾患若しくは障害における治療の進行を決定するためのマーカーとして使用し得る。
【0107】
別の好ましい態様において、該ペプチドを抗体の製造に使用する。ポリクローナル抗体は、該ペプチドの注入を介する動物の免疫化、および免疫グロブリンのその後の精製により、標準的様式で得ることができる。モノクローナル抗体は、例えば、Methods Enzymol.(1986)、121、Hybridoma technology and monoclonal antibodiesに記述されるような標準的プロトコルに従って製造し得る。
【0108】
本発明は、さらなる一局面において、本発明の前記ペプチドの1種若しくはそれ以上を含有する医薬組成物に関する。本組成物は、皮下、皮内、筋肉内のような非経口投与若しくは経口投与に使用する。このため、ペプチドを、薬学的に許容される、好ましくは水性の担体に溶解若しくは懸濁する。加えて、組成物は、緩衝剤、結合剤、膨潤剤、希釈剤、着香料、滑沢剤などのような賦形剤を含有し得る。ペプチドはまた、サイトカインのような免疫刺激物質と一緒になっても投与し得る。こうした組成物中で使用し得る賦形剤の広範囲の一覧は、例えば、A.Kibbe、Handbook of Pharmaceutical Excipients、第3版、2000、American Pharmaceutical Association and pharmaceutical pressから採用し得る。組成物は、腫瘍性疾患の予防(prevention)、予防(prophylaxis)および/若しくは治療に使用し得る。
【0109】
配列番号1ないし配列番号388のいずれかを含んでなる本発明のペプチドの最低1種を含有する医薬組成物を、それぞれのペプチド若しくは抗原と関連づけられる腫瘍性疾患に罹っている患者に投与する。これにより、CTL特異的免疫応答を誘発し得る。
【0110】
本発明の別の局面において、2種若しくはそれ以上の本発明のペプチドの組合せを、直接の組合せで若しくは同一処置レジメン内でのいずれかでワクチンとして使用し得る。さらに、他のペプチド、例えばMHCクラスI特異的ペプチドとの組合せを使用し得る。当業者は、例えばin vitroでのT細胞の生成、ならびに、それらの効率および全体的な存在、ある種のペプチドに対するある種のT細胞の増殖(proliferation)、親和性および増殖(expansion)、ならびに例えばIFN−γ産生を分析することによるT細胞の機能性を試験することにより、免疫原性ペプチドの好ましい組合せを選択することが可能であろう(下の実施例もまた参照されたい)。通常、最も効率的なペプチドをその後、上述されたところの目的上、ワクチンとして組み合わせる。
【0111】
適するワクチンは、2、3、4、5、6、7、8若しくは10種の異なるペプチド、好ましくは4、5、6若しくは7種の異なるペプチド、および最も好ましくは6種の異なるペプチドを含有することができる。
【0112】
最後に、ワクチンは、処置されるべき患者が罹っている癌の特定の型、ならびに、疾患の状態、以前の処置レジメン、患者の免疫状態、およびもちろん該患者のHLAハプロタイプに依存し得る。
【0113】
Iiの80のN末端アミノ酸が、タンパク質をクラスIIプロセシング経路に向けるのに十分であることが示された(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221、Wang、R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354)。本発明の概念の例示的一証明を提供するため、発明者は、Iiの80のN末端アミノ酸および多様な悪性度と関連する例示的1抗原、サイクリンD1(CCND1)よりなる融合タンパク質を生成した。
【0114】
サイクリンD1は、サイクリン依存性キナーゼとの相互作用によりG1−S移行に関与する細胞周期調節物質である。さらに、サイクリンD1は癌原遺伝子であり、かつ、数種の腫瘍型で過剰発現されることが示された(Hedberg,Y.、Davoodi,E.、Roos,G.、Ljungberg,B.とLandberg,G.(1999)Int.J.Cancer 84、268−272、Vasef,M.A.、Brynes,R.K.、Sturm、M.、Bromley,C.とRobinson,R.A.(1999)Mod.Pathol.12、412−416、−Troussard,X.、Avet−Loiseau,J.、Macro,M.、Mellerin,M.P.、Malet,M.、Roussel,M.とSola,B.(2000)Hematol.J.1、181−185)一方、それは、肝および高位腹部大動脈平滑筋を除くいかなる特定の分布も伴わず、大きな一団の健康な器官および組織中で低レベルで発現される(Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827)。示差的質量分析のアプローチにおいて、発明者は、トランスフェクトしたおよびトランスフェクトしていない細胞からの精製したHLAペプチドの質量スペクトルを比較し、そして、これらのHLAクラスIIリガンドの免疫原性の特徴を証明するために、目的の生じるペプチドをin vitro CD4+ Tヘルパー細胞プライミング実験で使用した。
【0115】
腫瘍関連抗原のTヘルパー細胞エピトープの同定は、抗腫瘍免疫療法における重要な責務のままである。ここに、われわれは、腫瘍関連抗原の天然にプロセシングかつ提示されるMHCクラスIIリガンドを同定するための、MSによる示差的ペプチド分析から派生した、新しくかつ一般に応用可能な方法およびペプチドを報告する。このアプローチは、Ii鎖と目的のAgの間の融合タンパク質をコードするベクターでのAPCのトランスフェクション段階、HLAに結合したペプチドの溶出、および、トランスフェクトされない細胞との比較による、トランスフェクタントにより提示されるAg由来ペプチドのMS同定を初めて組合せる。さらに、われわれは、同定されたペプチドに対し誘導されたT細胞が、同族のAgを過剰発現するトランスフェクタントを特異的に認識することを示すことにより、該方法を検証し得た。同定されたペプチドはなお、in vivoでそれらの免疫原性について試験されなければならないが、われわれのアプローチは、天然にプロセシングされるMHCクラスIIリガンドの正確な特徴付けに至る。従って、発明者は、腫瘍関連抗原の合成の重なるペプチド、若しくはクラスIエプトープ予測に比較して精度の低いエピトープ予測により選択された広範なペプチドのいずれかを試験することを回避する。in vivoでのT細胞活性化を誘導することが不可能な潜在性T細胞エピトープの同定につながりうる困難なT細胞アッセイ(Anderton,S.M.、Viner,N.J.、Matharu,P.、Lowrey,P.A.とWraith,D.C.(2002)Nat.Immunol.3、175−181)と対照的に、該研究は、提示されることが見出されている数種のペプチドに集中し得る。さらに、この方法を使用すれば、該ペプチドが天然にプロセシングされることを証明するために組換えAgを産生するか若しくはAgを発現する腫瘍細胞株を有することが必要でない。
【0116】
発明者は、EBVで形質転換したB細胞のクラスIIプロセシング区画中に腫瘍関連抗原を向けるためにIiのN末端を使用した。これを達成するため、われわれは、いかなる抗原もIiとの融合タンパク質として発現し得かつウエスタンブロット分析によりトランスフェクトされた細胞中のタンパク質の発現レベルを測定するためにわれわれを補助する、融通のきくベクターを構築した。IiのN末端が、タンパク質の標的をクラスIIプロセシング区画中に定めるのに十分であることが既に示されている。しかし、今日まで、これは、融合タンパク質をコードするcDNAライブラリーを使用して未知のAgを同定するため(Wang,R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354)、若しくは既知のT細胞クローンの特異性を確認するため(Chaux,P.、Vantomme,V.、Stroobant,V.、Thielemans,K.、Corthals,J.、Luiten,R.、Eggermont,A.M.、Boon,T.とvan der,B.P.(1999)J.Exp.Med.189、767−778)に、卵アルブミンを使用したモデルでのみ記述された(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221)。われわれの知る限り、この方法は、既知の腫瘍関連抗原の天然に提示されたMHCクラスIIに結合したペプチドを同定するために以前に使用されていなかった。MALDI−MASによるトランスフェクトされたおよびトランスフェクトされていない細胞のクラスIIリガンドの示差的分析、ならびにESI−MSによる差次的に発現されたペプチドのさらなる特徴付けは、目的の抗原のクラスIIリガンドの簡単な同定方法をもたらす。ケラチン18融合タンパク質での細胞のトランスフェクションは、われわれの方法が目的の抗原に一般に応用可能であることを証明し、再度、われわれはまた、モデル導入遺伝子ケラチン18からのHLA−DRに提示されるペプチドを記述することも可能であった。
【0117】
発明者は、配列NPPSMVAAGSVVAAV(配列番号1)を有する免疫原性のHLA−DR4に提示されるサイクリンD1ペプチド抗原、ならびにトランスフェクトしたヒト腫瘍細胞株から溶出された337種の他のHLAクラスII関連ペプチドを同定した。サイクリンD1ペプチド、およびヒト腫瘍細胞株から同定した他のペプチドは、ヒト癌の処置のための治療的ワクチンの臨床開発のための可能な候補である(Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827)。
【0118】
本明細書に開示かつ記述されるところの本発明の特徴が、示されるところのそれぞれの組合せでのみならず、しかしまた、本発明の意図される範囲から離れることなく単独の様式でも使用し得ることが、理解されるべきである。
本発明は今や、以下の図面、配列表および実施例を参照してより詳細に記述されることができる。以下の実施例は、具体的説明の目的上のみ提供され、そして本発明を制限することを意図していない。
配列番号1ないし配列番号338は、本発明の、MHCクラスIIにより提示されるペプチドを含有するT細胞エピトープのペプチド配列を示す。
配列番号339ないし配列番号350は、実施例で使用されるところのプライマーの配列を示す。
【図面の簡単な説明】
【0119】
【図1】実験の経過中に作製した融合タンパク質の第一のウエスタンブロット分析を示す。
【図2】実験の経過中に作製した融合タンパク質の第二のウエスタンブロット分析を示す。
【図3】トランスフェクトされない細胞株およびAwells−Ii−サイクリンD1クローンのHPLCクロマトグラムを示す。
【図4】際立つ個々のシグナルを有する好ましい画分の質量分析を示す。
【図5】際立つ個々のシグナルを有する好ましい画分の質量分析を示す。
【図6】ペプチドNPPSMVAAGSVVAAV(配列番号1)(サイクリンD1198−212)の同定を示す。
【図7】サイクリンD1ペプチドによるT細胞の特異的刺激(上図)、および該ペプチドに応答してのT細胞のサイトカイン産生(下図)を示す。
【図8】HLA−DR4/サイクリンD1198−212に特異的なT細胞受容体でのCD4+ Tヘルパー細胞の結果として起こりうる非特異的活性化の陰性対照を示す。
【発明を実施するための形態】
【0120】
本開示および本実施例に際して当業者に明らかであろうように、本発明はまた、免疫原性のHLA−DR4に提示された、配列NPPSMVAAGSVVAAV(配列番号1)を有するサイクリンD1ペプチド抗原およびケラチン18に関する以下の実施例で特異的に使用されるところの腫瘍関連抗原の他のエピトープにも関する。該実施例は、本発明を完全に可能にするために本発明の腫瘍関連抗原の全エピトープ(すなわち配列番号1ないし配列番号338の配列のいずれかを含んでなる)に当業者により容易に適用され得る、一般的例および概要として見られるべきである。
【0121】
本出願を通じて使用される略語:
Ab:抗体
Ag:抗原
APC:抗原提示細胞
CD:表面抗原分類
cpm:1分あたりカウント
DC:樹状細胞
EBV:エプスタイン−バーウイルス
ESI:エレクトロスプレーイオン化
HLA:ヒト白血球抗原
HPLC:高速液体クロマトグラフィー
IFN:インターフェロン
Ii:インバリアント鎖(CD74)
IL:インターロイキン
MALDI:マトリックス支援レーザー脱離/イオン化
MHC:主要組織適合遺伝子複合体
MS:質量分析法
OD450:450nmの波長での光学密度
PBMC:末梢血単核細胞
PCR:ポリメラーゼ連鎖反応
PHA:フィトヘマグルチニン
SDS−PAGE:ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動
S.I.:刺激指数
TOF:飛行時間
【実施例】
【0122】
細胞および抗体
ヒトBリンパ芽球様細胞株Awells(IHW−No.9090;HLA−DRB1*0401、HLA−DRB4*0101)を、10%FCS(Pan、独国アイデンバッハ)を含有しかつ2mM L−グルタミン(BioWhittaker、ベルギー・ヴェルヴィエ)、100U/mlペニシリンおよび100μg/mlストレプトマイシン(BioWhittaker)を補充したRPMI 1640(C.C.Pro、独国ノイシュタット)培地中で維持した。トランスフェクトした細胞クローンに関しては0.8mg/mlのG418(PAA Laboratories、オーストリア・リンツ)を添加した。安定形質転換体をAwells細胞の電気穿孔(280V、975μF;Gene Pulser II、Biorad、独国ミュンヘン)により作製し、次いで制限希釈法を使用してクローン化した。抗体L243(抗HLA−DR)(Lampson,L.AとLevy,R.(1980)J.Immunol.125、293−299)およびW6/32(抗HLAクラスI)(Brodsky,F.M.とParham,P.(1982)J.Immunol.128、129−135)を、プロテインA−セファロースビーズ(Pharmacia、スウェーデン・ウプサラ)を使用してハイブリドーマ培養上清から精製した。Th細胞株は、10%ヒトAB血清(Pel−Freez Clinical Systems、LLC、米国ウィスコンシン州ミルウォーキー)を含有しかつ100U/mlペニシリンおよび100μg/mlストレプトマイシン、2mM L−グルタミンならびに50μM β−メルカプトエタノールを補充したIMDM(BioWhittaker)中で誘導かつ培養した。フローサイトメトリー分析で使用した抗体はPharMingen(米国カリフォルニア州サンディエゴ)からであった。
【0123】
プラスミドDNA構築物
Iiの80のN末端アミノ酸を含有するcDNA(NCBI、GenBank受入番号X00497)を、A.Melms(Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533)から得たベクターpBluescript II KS(+)41−1(Stratagene、独国ハイデルベルク)からPCR反応で増幅し、そして、5'プライマーATCGAAGCTTCCAAGATGCACAGGAGGAGAAGC(配列番号339)および3'プライマーATCGGGATCCTTTGTCCAGCCGGCCCTGCTG(配列番号340)を使用して、pcDNA3のHind IIIおよびBamH I部位にサブクローニングした(pcDNA3−Ii;Invitrogen、独国カールスルーエ)。目的の遺伝子は、それぞれ、ケラチン18(NCBI、GenBank受入番号X12881)について5'プライマーATCGGAATTCTGAGCTTCACCACTCGCTCC(配列番号341)および3'プライマーATCGGCGGCCGCTTAATGCCTCAGAACTTTGGT(配列番号342)、ならびにサイクリンD1(NCBI、GenBankX59798)について5'プライマーATCGGAATTCTGGAACACCAGCTCCTGTGC(配列番号343)および3'プライマーATCGGCGGCCGCTCAGATGTCCACGTCCCGCAC(配列番号344)を使用して、悪性腎組織からのcDNAからPCR反応で増幅した。得られたcDNAを、TOPO TAクローニング(Invitrogen、独国カールスルーエ)を使用してサブクローニングし、そして最後に、Ii配列とインフレームでpcDNA3−IiのEcoR 1およびNot I部位に挿入した。
【0124】
リアルタイム定量的PCR
細胞からのRNAを、製造元の推奨に従ってTRIzol試薬(Invitrogen、独国カールスルーエ)を使用して単離した。cDNAを1μgの全RNAから合成した。リアルタイム定量的PCR(qPCR)を、ABI PRISM 7000配列検出装置(Applied Biosystems、独国ダルムシュタット)を使用して実施した。SYBR Green PCRマスターミックス(Applied Biosystems)を、PCR増幅およびPCR産物のリアルタイム検出に使用した。プライマー配列は後に続くとおりである:18S rRNA、5'プライマーCGGCTACCACATCCAAGGAA(配列番号345)および3'プライマーGCTGGAATTACCGCGGCT(配列番号346);ケラチン18、5'プライマーGAGCCTGGAGACCGAGAAC(配列番号347)および3'プライマーTTGCGAAGATCTGAGCCC(配列番号348);サイクリンD1、5'プライマーCACGATTTCATTGAACACTTCC(配列番号349)および3'プライマーTGAACTTCACCATCTGTGGCAC(配列番号350)。PCR反応は300nMの各プライマーを含む20μl中で実施した(18Sリバースプライマー:50nMのみ)。全部のサンプルを二検体ずつ増幅した。多様な遺伝子についてのトランスフェクトした細胞と野性型細胞の間の発現の差違を、比較閾値周期(comparative threshold cycle)(CT)法(http://docs.appliedbiosystems.com/pebiodocs/04303859.pdfで読み出し可能)を使用する相対定量により、PCR増幅曲線から計算した。18SリボソームRNAを標準化のための参照遺伝子として選んだ。
【0125】
融合タンパク質の検出
融合タンパク質は、Iiのアミノ酸残基12−28に結合するmAb PIN.1(Stressgen,Biomol、独国ハンブルク)を使用するウエスタンブロット分析により検出した。簡潔には、細胞を記述された(Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625)とおり溶解し、ライセートをLaemmli添加液中で沸騰させ、12%SDS−PAGEで分離し、そしてニトロセルロースメンブレンに転写した。BSAでの飽和段階後に、メンブレンをmAb PIN.1(1μg/ml)とともに室温で1時間インキュベートした。タンパク質を、ペルオキシダーゼ結合ヒツジ抗マウスIgG(Amersham Pharmacia、独国フライブルク)を使用して可視化した。いくつかの場合には、トランスフェクトした細胞を、融合タンパク質のエンドソーム/リソソームターゲッティングを検討するために、10〜100μMのクロロキン(Sigma、独国シュタインハイム)の存在下で培養した。細胞をその後溶解し、そしてタンパク質を上述されたとおりウエスタンブロットにより検出した。
【0126】
Ii−ケラチン18もまた、免疫沈降、およびトリプシンでのその後のin situ消化、次いで質量分析により検出した。簡潔には、細胞を記述された(Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625)とおり溶解し、そして、mAb PIN.1およびプロテインA−セファロースビーズ(Pharmacia、スウェーデン・ウプサラ)とともにインキュベートした。セファロースペレットを洗浄し、Laemmli添加液中で沸騰させ、そして12%SDS−PAGE上で泳動した。タンパク質のスポットをゲルから切り出し、そして、本質的には記述された(Shevchenko,A.、Wilm,M.、Vorm,O.とMann,M.(1996)Anal.Chem.68、850−858)とおり、トリプシンでin situで消化した。その後分析は、MALDI−TOF質量分析計(Reflex III、Bruker Daltonik、独国ブレーメン)で実施した。タンパク質の同一性を、ハイブリッド四重極直交加速型飛行時間タンデム質量分析計(QStar Pulsar i Qqoa Tof;Applied Biosystems−MDS Sciex、独国ヴァイターシュタット)に記録したタンデムMSスペクトルにより確認した。
【0127】
MHCクラスIIに結合したペプチドの溶出
凍結細胞ペレット(3.5ないし5×1010細胞)を、以前に記述された(Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225)とおり処理し、そして、ペプチドを、標準的プロトコル(Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999)に従い、HLA−DR特異的mAb L243(Lampson,L.A.とLevy,R.(1980)J.Immunol.125、293−299)を使用して単離した。
【0128】
DR溶出したペプチドの分子分析
ペプチドを逆相高速液体クロマトグラフィー(HPLC、SMART装置、μRPC C2/C18 SC 2.1/10;Amersham Pharmacia Biotech、独国フライブルク)により分離し、そして、Bruker Reflex III質量分析計(Bruker Daltonik)を使用するMALDI−TOF質量分析(MS)により画分を分析した。差次的に提示されたペプチドを、記述された(Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225)ところのハイブリッド四重極直交加速型飛行時間タンデム質量分析計(Q−TOF;Micromass、英国マンチェスター)でのナノESI(エレクトロスプレーイオン化)タンデムMSによりさらに分析した。
【0129】
ペプチドの合成および分析
ペプチドは、Fmoc/tBuストラテジーに従って、自動ペプチド合成機EPS221(Abimed、独国ランゲンフェルト)で合成した。TFA/フェノール/エタンジチオール/チオアニソール/水(容量で90/3.75/1.25/2.5/2.5)での1時間若しくは3時間(アルギニン含有ペプチド)の処理による樹脂の除去後に、ペプチドをメチル−ターシャリブチルエーテル(methyl−tert. butyl ether)から沈殿し、メチル−ターシャリブチルエーテルで1回およびジエチルエーテルで2回洗浄し、そして凍結乾燥前に水に再懸濁した。合成産物をHPLC(Varian star、Zinsser analytics、独国ミュンヘン)およびMALDI−TOF質量分析(future、GSG、独国ブルフザール)により分析した。80%未満の純度のペプチドを分取用HPLC(preparative HPLC)により精製した。
【0130】
単球由来樹状細胞
末梢血単核細胞(PBMC)を、古典的手順に従い、HLA−DRB1*0408、HLA−DRB1*1101、HLA−DRB3*0202、HLA−DRB4*01陽性ドナーから調製した。樹状細胞(DC)は、使用した培地が血清を含まないX−VIVO 15(BioWhittaker)であったことを除き、以前に記述された(Bender,A.、Sapp,M.、Schuler,G.、Steinman,R.M.とBhardwaj,N.(1996)J.Immunol.Methods.196、121−135)とおり、GM−CSFおよびIL−4の存在下で6日間培養したプラスチック付着性PBMCから得た。第6日に、未熟DCを、CD1a、CD11c、CD14、CD40、CD83、CD86、ならびにHLA−DRの細胞表面発現について、CELLQuestソフトウェアを伴うFACScalibur装置(Becton Dickinson、カリフォルニア州マウンテンビュー)でのフローサイトメトリーにより分析した。DCをその後、50μg/mlポリイノシン酸−ポリシチジル酸(ポリI/C、Amersham Pharmacia、スウェーデン・ウプサラ)および10ng/ml TNF−α(PharMingen)の存在下で更に2日間成熟させ、そして、CD14、CD80、CD83およびCD86の細胞表面発現についてフローサイトメトリーにより再度分析した。成熟DCは、CD80、CD83およびCD86分子の明瞭な上方制御を示した。
【0131】
ペプチド特異的Tヘルパー細胞の生成
3×105の成熟DCに、10μMのペプチドNPPSMVAAGSVVAAVを24ウェルプレート中で2時間添加し、そして十分に洗浄した。その後、4×106の新鮮自己PBMCを、Th−1発生に有利であるために10ng/mlのIL−12p70の存在下でDCに添加した。PBMCを、10U/mlのIL−2および5ng/mlのIL−7の存在下で、ペプチドを負荷した照射した自己PBMCで毎週再刺激した。3および5回の再刺激後にT細胞を合わせ、そしてペプチドの存在下で自己PBMCに対し試験した。Tヘルパー細胞株をその後、1μg/mlのPHA、25〜50U/mlのIL−2および5ng/mlのIL−7の存在下に、照射した同種PBMCで1〜2週ごとに増幅し、そしてその後、トランスフェクトした細胞株の認識について試験した。3ないし4週ごとに、Tヘルパー細胞株を、10μMのペプチド、10U/mlのIL−2および5ng/mlのIL−7の存在下に、照射した自己PBMCで再刺激した。
【0132】
Tヘルパー細胞株の機能アッセイおよび特徴付け
Tヘルパー細胞活性化は、チミジン取り込みならびにサイトカイン分泌により推定されるところの細胞増殖により試験した。簡潔には、2×105細胞を、10μMのペプチド若しくは3μg/mlのPHAの存在若しくは非存在下に3検体ずつ96ウェルプレート中で2×105の照射した自己PBMCとともにインキュベートした。24時間後に2部分の50μlの上清を収集かつ凍結し、そして50μlの新鮮培地を細胞に添加した。54時間後に、50μlのトリチウム化チミジン含有培地(0.074MBq/ウェル、Hartmann Analytic、独国ブラウンシュヴァイク)を添加し、そして、チミジンの取り込みを、シンチレーションカウンター(Microbeta、Wallac、独国フライブルク)を使用して72時間に測定した。細胞増殖は、比:(刺激したT細胞のcpmの平均)/(未刺激のT細胞のcpmの平均)に対応する刺激指数(S.I.)として表す。IL−2分泌は、IL−2依存性CTLL−2細胞株を使用して測定した。簡潔には、104細胞を上清の存在下に20〜24時間インキュベートした。その後、チミジン含有培地(0.055MBq/ウェル)を追加の7〜8時間添加し、そしてチミジンの取り込みを上述したとおり測定した。結果はまたS.I.として表す。
【0133】
IFN−γ、IL−4およびIL−6分泌は、PharMingenからの抗体対およびペルオキシダーゼ結合ストレプトアビジンを使用しかつ製造元の推奨に従ったサンドイッチELISAにより測定した。われわれは基質としてSupersensitive TMB(Sigma、独国ドライゼンホーフェン)を使用し、また、反応は2M H2SO4溶液を使用して停止した。OD450をその後測定し、そして結果は標準に従いpg/mlで表した。
【0134】
ペプチド特異的Tヘルパー細胞株によるサイクリンD1でトランスフェクトした細胞の認識
トランスフェクタントに対するT細胞の検出可能な非自己反応の非存在を、多様な細胞数の照射したトランスフェクタントの存在下で該Tヘルパー株を生成させるのに使用したドナーからの新鮮PBMCを共培養することにより立証した。細胞増殖ならびにIL−2、IL−4、IL−6およびIFN−γ分泌を上述したとおり測定し、そして、さらなる実験で使用されるべきエフェクター/標的比をかように決定した。
【0135】
サイクリンD1由来の天然にプロセシングされたペプチドの認識は、サイクリンD1若しくは陰性対照としてのケラチン18のいずれかをコードするプラスミドでトランスフェクトした照射したAwells(4×104細胞)の存在下で、かつ、上で決定された細胞比に従って、ペプチド特異的Tヘルパー細胞株(2×105細胞)を共培養することにより試験した。10μMのペプチド若しくは3μg/mlのPHAの存在下の照射した自己PBMCが陽性対照としてはたらいた。細胞増殖ならびにIL−2およびIFN−γ分泌を上述したとおり測定した。いくつかの実験では、細胞を20μg/mlの精製したL243抗体の存在下で培養した。
【0136】
結果
融合タンパク質を発現する細胞クローンの生成
われわれは、挿入物の3'端の後に一般的クローニング部位(GCS)が続いたような方法で、ベクターpcDNA3中にIiの80のN末端アミノ酸をコードするcDNAをクローン化した。これは、われわれに、Iiおよび目的の遺伝子の融合タンパク質を発現するための融通の利くベクターを与えた。Iiとインフレームで、われわれは、サイクリンD1のcDNA、ならびに対照としてのケラチン18をクローン化した。
【0137】
Awells細胞株を、電気穿孔法を使用して該2種の融合タンパク質をコードするベクターで安定にトランスフェクトした。その後、単細胞クローンを生成し、そして、それらの抗原発現についてmRNAおよびタンパク質レベルで試験した。野性型(トランスフェクトされていない細胞株)に比較して、最良のIi−ケラチン18クローンは、リアルタイム定量的PCR分析により測定されたとおり、5,700倍より多いケラチン18を発現し、また、最良のIi−サイクリンD1クローンは1,200倍より多いサイクリンD1を発現した。該データは18SリボソームRNAで標準化した。
【0138】
ウエスタンブロット分析において、PIN.1抗体を使用してIi融合タンパク質を検出した。Iiの2種のアイソフォームに対応する2本のバンドのみが検出された野性型(Warmerdam,P.A.、Long,E.O.とRoche,P.A.(1996)J.Cell.Biol.133、281−291)と比較して、期待された分子量を伴うIi−ケラチン18若しくはIi−サイクリンD1融合タンパク質を表す2個の付加的なバンドを各クローンで観察し得た(図1)。クローンAwells−Ii−ケラチン18の場合には、融合タンパク質が、免疫沈降、次いでトリプシンでのin situ消化およびその後のMS/MS分析を介してもまた同定された。
【0139】
該クローンはまた、トランスフェクションおよびクローニング手順がそれを妨害したかどうかを決定するため、フローサイトメトリーによりそれらのHLAクラスIおよびクラスII細胞表面発現レベルについても試験した。双方のクローンは、トランスフェクトされていない細胞株と比較して、HLAクラスIおよびクラスII分子の正常な発現レベルを示した。
【0140】
融合タンパク質のエンドソーム/リソソームターゲティング
IiのN末端が、タンパク質の標的をMHCクラスII経路に向けるのに十分であることが既に記述されている(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221、Wang,R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354、Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533)。われわれの構築物がAgプロセシングのその経路に実際に従ったかどうかを試験するため、われわれは、トランスフェクタントを、増大する量のクロロキン(リソソームのpHを上げることによりタンパク質のリソソーム分解を阻害する細胞傷害性の薬物)(Kaplan,J.とKeogh,E.A.(1981)Cell 24、925−932、Tietze,C.、Schlesinger,P.とStahl,P.(1982)J.Cell.Biol.92、417−424、Seglen,P.O.(1983)Methods Enzymol.96:737−64.、737−764)とともに4時間インキュベートした。細胞をその後溶解し、そして融合タンパク質をウエスタンブロットにより検出した。図2は、タンパク質バンドが、クロロキンの量を増大させるにつれてますます強くなることを示し、融合タンパク質の量がクロロキン濃度とともに増大することを示し、そして従って融合タンパク質がMHCクラスIIのタンパク質分解経路に従うことを提供する。
【0141】
HLA−DRに結合したペプチドの示差的質量分析
各クローンおよびトランスフェクトされていない細胞株からの3.5ないし5×1010細胞を増殖させ、そして、HLA−DRに結合したペプチドを、以前に記述された(Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999)とおりにHPLCを介して単離かつ分離した。トランスフェクトされていない細胞株およびAwells−Ii−サイクリンD1クローンのHPLCクロマトグラムを比較した(図3)。HLA−DRで提示されるペプチドレパートリーの小さなほとんど定量的な差違は、図3に示されるところのわずかに異なるUV痕跡になった。われわれの実験から期待されたとおり、明瞭なUVシグナルは、トランスフェクタントにより独占的に提示されるペプチドに割り当て得なかった。Awellsとトランスフェクトした株の間のHLA−DR拘束性ペプチド提示のただわずかな差違は、HPLC画分の大部分が同一のパターンを含有したMALDI−TOF分析によってもまた可視的になった。図4は、ケラチン18でトランスフェクトした株から溶出されたペプチド混合物中にのみ存在しかつケラチン18由来ペプチドを表した顕著な個別のシグナル(m/z:1732.96)を含む唯一の画分の質量分析を示す。図5では、1370.1のm/zシグナルは、サイクリンD1トランスフェクタントからの独占的に提示される1ペプチドを示す。双方のペプチドをナノフローESI MS/MS(タンデムMS)によりより詳細に分析した。発明者は、それぞれ2種のトランスフェクトされた融合タンパク質由来のペプチドNPPSMVAAGSVVAAV(配列番号1)(サイクリンD1198−212)(図6)およびSHYFKIIEDLRAQI(配列番号2)(ケラチン18126−139)を同定することが可能であった。該配列を、対応する合成ペプチドの質量スペクトルにより確認した。
【0142】
ペプチド特異的Tヘルパー細胞株の生成および特徴付け
新規のHLAクラスII由来の腫瘍関連および推定で免疫原性のペプチドの同定のための概念の証明を確立するために、同定されたサイクリンD1ペプチドに特異的なT細胞を、HLA−DR4+樹状細胞に負荷した対応する合成ペプチドでのin vitro刺激により誘導した。それぞれ第3および5回のin vitro(再)刺激後に、Tヘルパー細胞株の特異性を試験した。T細胞を、図7(上図)に示されるとおりサイクリンD1ペプチドにより特異的に刺激した。T細胞は、サイクリンD1ペプチドを負荷した自己PBMCに応答して増殖した(S.I.=5.4)。陽性対照として、PHAは強いT細胞増殖を誘導した(S.I.=330)。発明者は次に、どの型のTヘルパー細胞(Th1対Th2)がペプチドに応答して刺激されたかを、サイトカインプロファイルを検査することにより分析した。図7(下図)に示されるとおり、T細胞はペプチドに応答して低い程度までとは言えIL−2を産生した一方、それらはPHA刺激に対しなお感受性であった。対照的に、ペプチドで誘導したT細胞刺激は、強いIFN−γ分泌(3250pg/ml、図7C)をもたらしたが、しかしIL−4若しくはIL−6分泌をもたらさなかった。とは言え、T細胞はなお、PHAで誘導されるサイトカイン分泌に対しそれぞれ高度におよび中程度に感受性であった。HLA−DR4/サイクリンD1198−212に特異的なT細胞受容体でのCD4+ Tヘルパー細胞の最終的な非特異的活性化についての陰性対照としての無関係のペプチドでの刺激を表8に提示する。結論として、確立されたCD4+ Tヘルパー細胞株は、サイクリンD1198−212ペプチドに特異的でありかつTh1型のものである。この型は、腫瘍細胞の排除における特異的CD8+キラーT細胞の援助においてとりわけ重要である。
【0143】
ペプチド特異的Tヘルパー細胞株は、サイクリンD1でトランスフェクトした細胞もまた認識する
Awells細胞およびT細胞株は完全にHLAが適合しているわけではないため、われわれは最初に、双方を共培養することによりいずれかの非自己反応が生じ得るかどうかを試験した。簡潔には、多様な細胞数の照射したAwells−Ii−ケラチン18若しくはAwells−Ii−サイクリンD1トランスフェクタントを、T細胞ドナーからの固定した数のPBMCの存在下で共培養し、そして、細胞増殖ならびにIL−2、IL−4、IL−6およびIFN−γ分泌を測定した。中程度のT細胞増殖が高細胞数の双方のトランスフェクタントにより誘導されたが、しかし、サイトカイン分泌は観察されなかった(データは示されない)。従って、われわれは、サイトカイン分泌の非存在下でわずかなT細胞増殖のみが観察された5/1という(エフェクターT細胞)/(標的細胞)比を使用することを決定した。結果として、サイトカイン分泌をもたらすT細胞活性化は、トランスフェクタントにより提示される同族の抗原によってのみ特異的に誘導され得た。
【0144】
サイクリンD1ペプチドに特異的なTヘルパー細胞株は、サイクリンD1タンパク質を過剰発現しそして該サイクリンD1ペプチドを天然にプロセシングしかつHLA−DR分子と共同して提示する、トランスフェクトされた細胞を認識することが可能であった。図8に示されるとおり、照射したAwells−Ii−サイクリンD1トランスフェクタントは、IL−2分泌により観察されたとおり、Tヘルパー細胞株を特異的に活性化することが可能であった(S.I.=4.0)。対照的に、Awells−Ii−ケラチン18トランスフェクタント(T細胞刺激の陰性対照として使用され、かつ、HLA−DRと共同して無関係のケラチン18ペプチド126−139を提示することが知られている)はT細胞活性化を誘導せず、ペプチド特異的Tヘルパー細胞株が同族の抗原を特異的に認識することを示した。さらに、これらの結果は、本発明で使用されるサイクリンD1ペプチドが、T細胞エピトープを含有する天然にプロセシングされたペプチドであることを証明する。この活性化は、HLA−DR特異的なブロッキングL243Abの存在により阻害され得た(71.2%)。
【0145】
クラスII分子およびペプチド抗原サイクリンD1198−212を使用する活性化型細胞傷害性リンパ球(CTL)の製造ならびにそれらの投与
活性化型細胞傷害性Tリンパ球(CTL)は、HLA−DRクラスII分子、およびサイクリンD1198−212(配列番号1)からのペプチドを使用して製造する。
一般に、PCT特許出願WO 93/17095号に記述される方法を使用してCTLを製造する。Awells細胞を使用してペプチド抗原をCTLに提示する。HLA−DR分子はAwells細胞により発現される。試験されるべきペプチドを、Applied Biosystemsの合成機、ABI 431A(米国カリフォルニア州フォスターシティ)で合成し、そしてその後HPLCにより精製する。WO 93/17095号に詳細に記述されるとおり、特異的細胞傷害性T細胞の生成のためin vitro条件を最適化するために、刺激体細胞の培養物を適切な培地中で維持する。刺激体細胞は、好ましくは無血清培地(例えばExcell 400)中で維持されるAwells細胞である。
【0146】
活性化されるべき細胞、例えば前駆体CD4+細胞との刺激体細胞のインキュベーション前に、ある量の抗原ペプチドを十分な量の刺激体細胞培養物に添加し、刺激体細胞の表面上で発現されるべきヒトクラスII分子に負荷されたようにする。十分な量のペプチドは、約200、および好ましくは200若しくはそれ以上のペプチドを負荷したヒトクラスIIMHC分子が、各刺激体細胞の表面上で発現されることを可能にする量である。刺激体細胞は、典型的に、20pg/ml超のペプチドとともにインキュベートする。
【0147】
休止期若しくは前駆体CD4+細胞をその後、適切な刺激体細胞を含む培養物中で、CD4+細胞を活性化するのに十分な時間インキュベートする。CD4+細胞はかように抗原特異的様式で活性化される。刺激体細胞に対する休止期若しくは前駆体CD4+(エフェクター)細胞の比は個体ごとに異なることがあり、また、個体のリンパ球の培養条件への従順さのような変数にさらに依存しうる。リンパ球:刺激体細胞(Awells細胞)の比は、典型的に約30:1ないし300:1の範囲にある。例えば、3×101のヒトPBLおよび1×106の生存Awells細胞を混合し、そして20mlのRPMI 1640培地中で維持する。
【0148】
エフェクター/刺激体の培養物は、治療上使用可能な若しくは有効な数のCD4+細胞を刺激するのに必要であるだけの時間維持する。最適な時間は典型的に約1と5日の間であり、「プラトー」すなわち「最大の」特異的CD4+活性化レベルは一般に5日の培養後に観察される。CD4+細胞のin vitro活性化は、典型的に、細胞株のトランスフェクション後短時間内に検出される。CD4+細胞を活性化することが可能な、トランスフェクトされた細胞株での一過性発現は、トランスフェクション48時間以内に検出可能である。これは、ヒトクラスIIMHC分子を発現する形質転換細胞の安定若しくは一過性培養物のいずれも、CD4+細胞の活性化において有効であることを明確に示す。
【0149】
活性化型CD4+細胞は、それらの適切な相補リガンドに結合させるため、刺激体(Awells)細胞、刺激体細胞に負荷したペプチド、またはCD4+細胞(若しくはそのセグメント)に特異的なモノクローナル抗体を使用して、刺激体細胞から効果的に単離しうる。抗体標識した分子をその後、免疫沈降若しくはイムノアッセイ法を介して、刺激体とエフェクター細胞の混合状態から抽出する。
【0150】
活性化型CD4+細胞の有効な細胞傷害性の量は、in vitroとin vivoの使用の間で、ならびにこれらのキラー細胞の最終的標的である細胞の量および型で変動し得、約1×106と1×1012の間の活性化型CTLを成人のヒトに使用する。
活性化型CD4+細胞は、処置されている個体への該CD4+細胞の投与前に、Awells細胞培養物から収集する。
Honsikへの米国特許第4,844,893号明細書およびRosenbergへの米国特許第4,690,915号明細書に例示されるもののような細胞成分の再導入方法を使用する。例えば、静脈内注入を介する活性化型CD8細胞の投与が適切である。
【0151】
表1:本発明で同定されたところの腫瘍関連Tヘルパー細胞ペプチドエピトープ
【表1−1】

【表1−2】
【表1−3】
【表1−4】
【表1−5】
【表1−6】
【表1−7】
【表1−8】
【表1−9】
【表1−10】
【表1−11】
【表1−12】
【表1−13】
【表1−14】
【表1−15】
【技術分野】
【0001】
本発明は、免疫療法の方法、ならびに免疫療法の方法での使用のための分子および細胞に関する。とりわけ、本発明は癌、とりわけ腎癌の免疫療法に関する。本発明はさらに、抗腫瘍免疫応答を刺激するワクチン組成物の薬学的有効成分としてはたらく、単独の若しくは他の腫瘍関連ペプチドと組合せの腫瘍関連Tヘルパー細胞ペプチドエピトープに関する。とりわけ、本発明は、抗腫瘍免疫応答を導き出すためのワクチン組成物で使用し得る、ヒト腫瘍細胞株のHLAクラスII分子由来の338種の新規ペプチド配列に関する。
本発明の目的上、本明細書に引用されるところの全部の参考文献は、そっくりそのまま引用することにより組み込まれる。
【背景技術】
【0002】
免疫応答の刺激は、宿主免疫系により外来として認識される抗原の存在に依存する。腫瘍関連抗原の存在の発見は、今や、腫瘍増殖に介入するため宿主の免疫系を使用する可能性を生じた。免疫系の体液性および細胞性双方の部門の利用の多様な機構が、現在、癌免疫療法のため探求されている。
【0003】
細胞性免疫応答の特定の要素が、腫瘍細胞を特異的に認識かつ破壊することが可能である。腫瘍浸潤細胞集団若しくは末梢血からの細胞傷害性T細胞(CTL)の単離は、こうした細胞が癌に対する自然免疫防御において重要な役割を演じていることを示唆する(非特許文献1)。サイトゾル中に位置するタンパク質由来の通常8ないし10残基の主要組織適合遺伝子複合体(MHC)のクラスI分子をもつペプチドを認識する、CD8+ T細胞(TCD8+)はとりわけ、この応答で重要な役割を演じている。ヒトのMHC分子はヒト白血球抗原(HLA)とも命名されている。
【0004】
MHC分子には2つのクラスがある:MHC−I分子は、内因性タンパク質およびより大きなペプチドのタンパク質分解性切断から生じるペプチドを提示し、核を有するほとんどの細胞上で見出し得る。MHC−II分子は、専門的抗原提示細胞(APC)上でのみ見出し得、そしてエンドサイトーシスの経過中にAPCにより取り込まれ、そして消化される外因性タンパク質のペプチドを提示する。ペプチドとMHC−Iの複合体はCD8陽性の細胞傷害性Tリンパ球により認識され、ペプチドとMHC−II分子の複合体はCD4+−ヘルパーT細胞により認識される。
【0005】
ペプチドが細胞性免疫応答を誘発する(導き出す)ためには、MHC分子に結合しなければならない。この過程はMHC分子のアリルおよびペプチドのアミノ酸配列の特定の多型に依存する。MHCクラスIに結合するペプチドは、長さが通常8〜10残基であり、そして、MHC分子の対応する結合溝と相互作用する、それらの配列中の2個の保存された残基(「アンカー」)を含有する。
【0006】
現在、腫瘍細胞を特異的に認識しかつ養子移入後にいくつかの場合には完全寛解を誘導する治療活性を有する、マウスおよびヒト双方のTCD8+の多数の例が存在する。しかしながら、腫瘍を根絶するT細胞の潜在能力にもかかわらず、多くの腫瘍がin vivoでTCD8+による認識を逃れることが、大部分の癌の進行性増殖からみても明らかである。多様な腫瘍が免疫原性であることが見出されているとは言え、効果的な抗腫瘍免疫応答の刺激は立証することが困難であった。
【0007】
腫瘍特異的細胞傷害性Tリンパ球により認識される抗原、すなわちそれらのエピトープは、酵素、受容体、転写因子などのような全部のタンパク質分類由来の分子であり得る。さらに、例えば、腫瘍関連抗原はまた、例えば変異した遺伝子の産物として腫瘍細胞中のみに存在し得る。腫瘍関連抗原の別の重要な分類は、多様な種類の腫瘍および健康な精巣組織で発現されるCT(「癌精巣」)抗原のような組織特異的構造である。
【0008】
多様な腫瘍関連抗原が同定されている。さらに、多くの研究の労力が付加的な腫瘍関連抗原を同定するために費やされている。腫瘍特異的抗原ともまた当該技術分野で称される、いくつかの群の腫瘍関連抗原は組織特異的である。例として、これに限定されるものでないが、黒色腫のチロシナーゼ、前立腺癌のPSAおよびPSMA、ならびにリンパ腫におけるbcr/ablのような染色体交差が含まれる。しかしながら、同定された多くの腫瘍関連抗原は複数の腫瘍型に存在し、そして、形質転換を実際に引き起こす癌タンパク質および/若しくは腫瘍抑制遺伝子(腫瘍抑制遺伝子は、例えば腎癌について非特許文献2に総説されている)のような数種は、ほぼ全腫瘍型に存在する。例えば、p53(腫瘍抑制遺伝子の一例である)、ras、c−met、myc、pRB、VHLおよびHER−2/neuのような、細胞の増殖および分化を制御する正常細胞タンパク質は、これらの遺伝子産物の発現の上方制御をもたらす突然変異を蓄積してそれによりそれらを発癌性にし得る(非特許文献3;非特許文献4)。これらの変異体タンパク質は、複数の型の癌での腫瘍特異的免疫応答の標的となり得る。
【0009】
腫瘍抑制遺伝子は、多細胞生物体の細胞が腫瘍細胞に変わることができる確率を低下させる遺伝子である。こうした遺伝子の突然変異若しくは欠失は腫瘍の確率を増大させることになる。その点で腫瘍抑制遺伝子は癌遺伝子に類似している。腫瘍抑制遺伝子、若しくはより正確にはそれらがコードするタンパク質は、細胞周期の調節を弱くする又は抑制する効果を有する。これは、基本的に、腫瘍抑制遺伝子/タンパク質により3つの方法により行われる:1.細胞周期の継続に不可欠である遺伝子の抑制。これらの遺伝子が発現されない場合、細胞周期は継続することができず、細胞分裂を効果的に阻害する。2.細胞周期のDNA損傷への結合。細胞中に損傷を受けたDNAが存在する限り、分裂しないはずである。該損傷が修復され得る場合に細胞周期が継続し得る。3.損傷が修復され得ない場合、該細胞はアポトーシスすなわちプログラムされた細胞死を開始して、該生物体のより大きな利益に対してそれが与える脅威を除去するはずである。発見された最初の腫瘍抑制タンパク質はヒト網膜芽種におけるpRbタンパク質であった。重要な腫瘍抑制因子はp53遺伝子(上を参照されたい)である。
【0010】
HPV由来のE6およびE7、若しくはエプスタイン−バーウイルス(EBV)由来のEBNA1のような腫瘍ウイルス由来のトランスフォーミングタンパク質もまた、多くの腫瘍型に存在し、かつ、複数の型の癌で腫瘍特異的免疫応答の標的となり得る(非特許文献5;非特許文献6;非特許文献7;非特許文献8)。MAGEおよびMUCファミリーのような非腫瘍性宿主タンパク質もまた広範に分布している。とりわけ、MAGEファミリーの抗原は、乳癌、肺癌、食道癌、肝癌、甲状腺癌、神経芽細胞腫、胃癌、多発性骨髄腫および黒色腫を包含する多くの多様な癌で見出されている(非特許文献9)。MUCファミリーの抗原は、卵巣および子宮内膜癌、乳癌、多発性骨髄腫、膵癌ならびに結腸および直腸癌と関連づけられている(非特許文献10)。
【0011】
さらに、大部分の癌は1種以上の抗原が関連する。1種以上の腫瘍抗原を発現する腫瘍の例は、これに限定されるものでないが、MUC−1、HER−2/neu、MAGE、p53、T/TnおよびCEAに関連することが示されている乳癌、MUC−2およびMUC−4、CEA、p53ならびにMAGEファミリーに関連することが示されている結腸癌、MAGEファミリーのメンバー、MART−1およびgp100に関連することが示されている黒色腫、ならびにGM2、Tn、sTn、Thompson−Friedenreich抗原(TF)、MUC1、MUC2、ヒト絨毛性ゴナドトロピンのβ鎖(hCGβ)、HER2/neu、PSMAおよびPSAと関連づけられている前立腺癌を含む。事実、腫瘍の抗原欠失バリアントが免疫系の機能下で増殖し得るという事実を代償するために、一連の抗原が、癌に対する免疫療法での使用に提案された(非特許文献11;非特許文献12)。
【0012】
タンパク質が腫瘍特異的抗原として細胞傷害性Tリンパ球により認識されるために、また、治療で使用されるために、特定の前提条件が満たされなければならない。抗原は主として腫瘍細胞により発現され、そして正常の健康な組織により発現されないか若しくは少量で発現するはずである。それぞれの抗原が1つの型の腫瘍にのみならず、高濃度(例えば、細胞あたりのコピー数)でも存在することがさらに望ましい。腫瘍関連抗原由来であるこうしたペプチド(「免疫原性ペプチド」)はin vitro若しくはin vivoのT細胞応答につながるはずであるため、抗原のアミノ酸配列中のエピトープの存在が必須である。
【0013】
現在まで、クラスIIのプロセシング経路中の抗原を標的とするための多数の戦略が記述された。抗原提示細胞(APC)を、取り込まれかつプロセシングされるために目的の抗原とインキュベートすることが可能である(非特許文献13)。他の戦略はリソソーム標的配列を含有する融合タンパク質を使用する。APC中で発現され、こうした融合タンパク質は、抗原をクラスIIのプロセシング区画に向かうことになる(非特許文献14;非特許文献15)。
【0014】
ヘルパーT細胞は、抗腫瘍免疫におけるCTLのエフェクター機能の統合において重要な役割を演じている。Th1型のヘルパーT細胞応答を誘発するヘルパーT細胞エピトープは、それらの細胞表面上に腫瘍関連ペプチド/MHC複合体を表示する腫瘍細胞に向けられた細胞傷害機能を包含するCD8+キラーT細胞のエフェクター機能を支援する。こうして、腫瘍関連ヘルパーT細胞ペプチドエピトープは、単独で若しくは他の腫瘍関連ペプチドとともに、抗腫瘍免疫応答を刺激するワクチン組成物の薬学的有効成分としてはたらき得る。
【0015】
腫瘍ワクチンの開発における主要な責務は、従って、CD4++ CTLにより認識され得る新規腫瘍関連抗原およびそれ由来の免疫原性Tヘルパーエピトープの同定および特徴付けである。従って、ヒト主要組織適合遺伝子複合体(MHC)クラスIIの分子に結合する能力を有するこうしたペプチドの新規アミノ酸配列を提供することが、本発明の一目的である。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第4,440,859号
【特許文献2】米国特許第4,530,901号
【特許文献3】米国特許第4,582,800号
【特許文献4】米国特許第4,677,063号
【特許文献5】米国特許第4,678,751号
【特許文献6】米国特許第4,704,362号
【特許文献7】米国特許第4,710,463号
【特許文献8】米国特許第4,757,006号
【特許文献9】米国特許第4,766,075号
【特許文献10】米国特許第4,810,648号
【特許文献11】WO 95/18145号
【特許文献12】WO 96/03144号
【特許文献13】WO 97/26328号
【特許文献14】WO 93/17095号
【特許文献15】米国特許第4,844,893号
【特許文献16】米国特許第4,690,915号
【非特許文献】
【0017】
【非特許文献1】Cheeverら、Annals N.Y.Acad.Sci.1993 690:101−112
【非特許文献2】Linehan WM、Walther MM、Zbar B.The genetic basis of cancer of the kidney.J Urol.2003 Dec;170(6 Pt 1):2163−72
【非特許文献3】McCarteyら Cancer Research 1998 15:58 2601−5
【非特許文献4】Disisら Ciba Found Symp.1994 187:198−211
【非特許文献5】McKaigら Head Neck 1998 20(3):250−65
【非特許文献6】Punwaneyら Head Neck 1999 21(1):21−9
【非特許文献7】Serthら Cancer Res.1999 15:59(4):823−5
【非特許文献8】Pagano,J.S.Proc.Assoc.Am.Physicians 1999 111(6):573−80
【非特許文献9】Gillespie,A.M.とColeman,R.E.Cancer Treat.Rev.1999 25(4):219−27
【非特許文献10】Segal−Eiras,A.とCroce,M.V.Allergol.Immunopathol.1997 25(4):176−81
【非特許文献11】Zhangら Clin.Cancer Res.1998 4:2669
【非特許文献12】Kawashimaら Hum.Immunol.1998 59:1
【非特許文献13】Chaux,P.、Vantomme,V.、Stroobant,V.、Thielemans,K.、Corthals,J.、Luiten,R.、Eggermont,A.M.、Boon,T.とvan der,B.P.(1999)J.Exp.Med.189、767−778
【非特許文献14】Marks,M.S.、Roche,P.A.、van Donselaar,E.、Woodruff,L.、Peters,P.J.とBonifacino,J.S.(1995)J.Cell Biol.131、351−369
【非特許文献15】Rodriguez,F.、Harkins,S.、Redwine,J.M.、de Pereda,J.M.とWhitton,J.L.(2001)J.Virol.75、10421−10430
【非特許文献16】Strubin,M.、Mach,BとLong,E.O.The comletesequence of the mRNA for the HLA−DR−associated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J.3(4)、869−872(1984)
【非特許文献17】Rammensee H、Bachmann J、Emmerich NP、Bachor OA、Stevanovic S.SYFPEITHI:detabase for MHC ligands and peptide motifs.Immunogenetics.1999 Nov;50(3−4):213−9.総説
【非特許文献18】Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21
【非特許文献19】Endlら J Clin Invest.1997 May 15;99(10):2405−15
【非特許文献20】ten Boschら Blood.1996 Nov;88(9):3522−7
【非特許文献21】Diepolderら J Virol.1997 Aug;71(8):6011−9
【非特許文献22】van der Burgら J Immunol.1999 Jan 1;162(1):152−60
【非特許文献23】Friedeら、Biochim Biophys Acta.1996 Jun 7;1316(2):85−101
【非特許文献24】Seeteら J Immunol.1993 Sep 15;151(6):3163−70.
【非特許文献25】Hammerら Cell.1993 Jul 16;74(1):197−203.
【非特許文献26】Hammerら J Exp Med.1995 May 1;181(5):1847−55
【非特許文献27】Meziereら(1997)J.Immunol.159、3230−3237
【非特許文献28】Luら(1981)J.Org.Chem.46、3433
【非特許文献29】Saikiら(1988)Science 239、487−491
【非特許文献30】Cohenら(1972)Proc.Natl.Acad.Sci.USA 69、2110
【非特許文献31】Sambrookら(1989)Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバー
【非特許文献32】Shermanら(1986)Methods In Yeast Genetics,A Laboratory Manual、ニューヨーク州コールドスプリングハーバー
【非特許文献33】Beggs(1978)Nature 275、104−109
【非特許文献34】Southern(1975)J.Mol.Biol.98、503
【非特許文献35】Berentら(1985)Biotech.3、208
【非特許文献36】Longeneckerら(1993)Ann.NY Acad.Sci.690、276−291
【非特許文献37】Zhouら(1995)Blood 86、3295−3301
【非特許文献38】Rothら(1996)Scand.J.Immunology 43、646−651
【非特許文献39】Conryら(1996)Seminars in Oncology 23、135−147
【非特許文献40】Condonら(1996)Nature Medicine 2、1122−1127
【非特許文献41】Gongら(1997)Nature Medicine 3、558−561
【非特許文献42】Zhaiら(1996)J.Immunol.156、700−710
【非特許文献43】Grahamら(1996)Int J.Cancer 65、664−670
【非特許文献44】Breast Cancer,Advances in biology and therapeutics、Calvoら(編)、John Libbey Eurotext中、Burchellら(1996)pp 309−313
【非特許文献45】KarreとLjunggren(1985)J.Exp.Med.162、1745
【非特許文献46】Thomsonら(1996)J.Immunol.157、822−826
【非特許文献47】Peoplesら(1995)Proc.Natl.Acad.Sci.USA 92、432−436
【非特許文献48】Kawakamiら(1992)J.Immunol.148、638643
【非特許文献49】Plebanskiら(1995)Eur.J.Immunol.25、1783−1787
【非特許文献50】Jochmusら(1997)J.Gen.Virol.78、1689−1695
【非特許文献51】Hillら(1995)J.Exp.Med.181、2221−2228
【非特許文献52】Jeromeら(1993)J.Immunol.151、1654−1662
【非特許文献53】Portaら(1994)Virology 202、449−955
【非特許文献54】Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658
【非特許文献55】Finerら(1994)Blood 83、43
【非特許文献56】Robertsら(1994)Blood 84、2878−2889
【非特許文献57】Moritzら(1994)Proc.Natl.Acad.Sci.USA 91、4318−4322
【非特許文献58】Eshharら(1993)Proc.Natl.Acad.Sci.USA 90、720−724
【非特許文献59】Hwuら(1993)J.Exp.Med.178、361−366
【非特許文献60】Murphyら(1996)The Prostate 29、371−380
【非特許文献61】Tjuaら(1997)The Prostate 32、272−278
【非特許文献62】Gongら(1997)Gene Ther.4、1023−1038
【非特許文献63】Wanら(1997)Hum.Gene Ther.8、1355−1363
【非特許文献64】Spechtら(1997)J.Exp.Med.186、1213−1221
【非特許文献65】Szabolcsら(1997)Blood
【非特許文献66】Tutingら(1997)Eur.J.Immunol.27、2702−2707
【非特許文献67】Ashleyら(1997)J.Exp.Med.186、1177 1182
【非特許文献68】Methods Enzymol.(1986)、121、Hybridoma technology and monoclonal antibodies
【非特許文献69】A.Kibbe、Handbook of Pharmaceutical Excipients、第3版、2000、American Pharmaceutical Association and pharmaceutical press
【非特許文献70】Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221
【非特許文献71】Wang、R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354
【非特許文献72】Hedberg,Y.、Davoodi,E.、Roos,G.、Ljungberg,B.とLandberg,G.(1999)Int.J.Cancer 84、268−272
【非特許文献73】Vasef,M.A.、Brynes,R.K.、Sturm、M.、Bromley,C.とRobinson,R.A.(1999)Mod.Pathol.12、412−416
【非特許文献74】Troussard,X.、Avet−Loiseau,J.、Macro,M.、Mellerin,M.P.、Malet,M.、Roussel,M.とSola,B.(2000)Hematol.J.1、181−185
【非特許文献75】Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827
【非特許文献76】Anderton,S.M.、Viner,N.J.、Matharu,P.、Lowrey,P.A.とWraith,D.C.(2002)Nat.Immunol.3、175−181
【非特許文献77】Lampson,L.AとLevy,R.(1980)J.Immunol.125、293−299
【非特許文献78】Brodsky,F.M.とParham,P.(1982)J.Immunol.128、129−135
【非特許文献79】Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533
【非特許文献80】Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625
【非特許文献81】Shevchenko,A.、Wilm,M.、Vorm,O.とMann,M.(1996)Anal.Chem.68、850−858
【非特許文献82】Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225
【非特許文献83】Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999
【非特許文献84】Bender,A.、Sapp,M.、Schuler,G.、Steinman,R.M.とBhardwaj,N.(1996)J.Immunol.Methods.196、121−135
【非特許文献85】Warmerdam,P.A.、Long,E.O.とRoche,P.A.(1996)J.Cell.Biol.133、281−291
【非特許文献86】Kaplan,J.とKeogh,E.A.(1981)Cell 24、925−932
【非特許文献87】Tietze,C.、Schlesinger,P.とStahl,P.(1982)J.Cell.Biol.92、417−424
【非特許文献88】Seglen,P.O.(1983)Methods Enzymol.96:737−64.、737−764
【発明の概要】
【発明が解決しようとする課題】
【0018】
本発明により、この目的は、添付された配列表の配列番号1ないし配列番号338のいずれかの少なくとも1の配列を含んでなるペプチドの群から選択される腫瘍関連ペプチドを提供することにより解決されるものであって、該ペプチドはヒト主要組織適合遺伝子複合体(MHC)クラスIIの分子に結合する能力を有する。
【課題を解決するための手段】
【0019】
本発明はさらに、抗腫瘍免疫応答を導き出すためのワクチン組成物で使用し得る、ヒト腫瘍細胞株、とりわけ腎癌細胞株のHLAクラスII分子由来の338種の新規ペプチド配列に関する。該新規ペプチド配列は、定義された(例えば腫瘍関連)抗原の未知の天然にプロセシングされたHLAクラスIIリガンドの同定のための新たなかつ一般に応用可能な組合せアプローチにより同定された。従って、ペプチドに基づく免疫療法の新たなかつ有望な候補が、著しく興味深い腫瘍抗原の選択およびそれに由来するペプチド配列の慎重な決定を包含する様式で同定された。
【0020】
本発明の第一の局面は、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかのアミノ酸配列を含んでなるペプチドを提供するが、但し、該ペプチドは、該アミノ酸配列が由来する無傷のヒトポリペプチド(すなわち遺伝子座連関ID(受入番号、下の付属の表を参照されたい)に列挙されるところの完全長配列の1つ)でない。
【0021】
下に本明細書に下述されるとおり、本発明の基礎を形成するペプチドは、MHCクラスIIをもつ細胞(Awells細胞)により提示されるとして全部が同定された。従って、これらの特定のペプチド、ならびに該配列を含有する他のペプチド(すなわち派生ペプチド)は、こうした応答が誘導されるであろう程度が個々のペプチドごとに変動するかもしれないが、おそらく全部が特異的T細胞応答を導き出すであろう。差異は、例えば前記ペプチド中の突然変異により引き起こされ得る(下を参照されたい)。当業者は、とりわけ本明細書の実施例およびそれぞれの文献を参照して、個々のペプチドにより応答が誘導される程度を決定するために適用し得る方法を知っている。
好ましくは、本発明のペプチドは、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかのアミノ酸配列より本質的になる。
【0022】
「より本質的になること」は、本発明のペプチドが、配列番号1ないし配列番号338若しくはそれらのバリアントのいずれかの配列に加えて、結合モチーフを含んでなるペプチドのコア配列および免疫原性Tヘルパーエピトープとして機能する、ペプチドの形成部分で必須ではないアミノ酸の付加的なNおよび/若しくはC末端に位置するひと配列(stretch)を含有することを意味している。
【0023】
にもかかわらず、これらのひと配列は、本発明のペプチドの細胞中への効率的な導入を提供するために重要であり得る。本発明の一態様において、本発明のペプチドは、NCBI、GenBank受入番号X00497(Strubin,M.、Mach,BとLong,E.O.The comletesequence of the mRNA for the HLA−DR−associated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J.3(4)、869−872(1984))に由来するところのHLA−DR抗原関連インバリアント鎖(p33、以下で「Ii」)の80のN末端アミノ酸を含んでなる。
【0024】
所定のアミノ酸配列の「バリアント」により、われわれは、ペプチドが、該所定のアミノ酸配列よりなるペプチドと実質的に同じ方法でHLA分子に結合することがなお可能であるような、アミノ酸残基の例えば1若しくは2個の側鎖が(例えばそれらを別の天然に存在するアミノ酸残基の側鎖若しくは何らかの他の側鎖で置換することにより)変えられることを意味している。例えば、ペプチドは、それが、HLA−DRのような適するMHC分子と相互作用しかつそれを結合する能力を改良しない場合は少なくとも維持するように、また、それが、本発明の局面で定義されるところのアミノ酸配列を含有するポリペプチドを異常に発現する細胞を認識かつ死滅させ得る活性化型CTLを生成させる能力を改良しない場合は少なくとも維持するように、改変しうる。以下で記述されるところのデータベースに由来し得るとおり、HLA−DR結合ペプチドのある位置は、典型的に、HLAの結合溝の結合モチーフに嵌るコア配列を形成するアンカー残基である。これらおよびHLA−DRの結合に関与する他の残基の改変は、CTL認識を変えることなく結合を高めうる。
【0025】
T細胞受容体と相互作用するのに不可欠でないアミノ酸残基は、その組み込みがT細胞反応性を実質的に遂げずかつ関連するMHCへの結合を除外しない別のアミノ酸での置換により改変し得る。従って、与えられる条件から離れて、本発明のペプチドは、与えられるところのアミノ酸配列またはそれらの一部分若しくはバリアントを包含するいかなるペプチド(その用語によりオリゴペプチド若しくはポリペプチドを包含する)でもありうる。
【0026】
これらのペプチドが、ある種のHLA特異的アミノ酸モチーフ、ならびに場合によってはコア配列の機能を妨害しない(すなわちペプチドおよびT細胞の相互作用にとって不適切と思われない)Nおよび/若しくはC末端の延長を有する「コア配列」から構成されることが、MHCクラスIIで提示されるペプチドについてさらに既知である。該Nおよび/若しくはC末端の延長は、それぞれ長さが1ないし10アミノ酸の間であり得る。従って、本発明の好ましいペプチドは、9と30アミノ酸の間の全体長さを表す。これらのペプチドを、MHCクラスII分子を添加するために直接使用し得るか、若しくは、該配列を下の記述によるベクターに該配列をクローン化し得るかのいずれかである。これらのペプチドは、細胞内でより長いペプチドのプロセシングの最終産物を形成するため、より長いペプチドを同様に使用し得る。本発明のペプチドはいかなる大きさのものであってもよいが、しかし、典型的にはそれらは分子量が100000未満、好ましくは50000未満、より好ましくは10000未満、および典型的には約5000でありうる。アミノ酸残基の数に関して、本発明のペプチドは、1000残基未満、好ましくは500残基未満、より好ましくは100残基未満を有しうる。
【0027】
約12アミノ酸残基より大きいペプチドを使用してMHC分子に直接結合する場合は、コアHLA結合領域に隣接する残基が、MHC分子の結合溝に特異的に結合するか若しくはCTLにペプチドを提示する該ペプチドの能力に実質的に影響を及ぼさないものであることが好ましい。しかしながら、これらのより大きいペプチドは抗原提示細胞により断片化されうるため、上で既に示したとおり、とりわけポリヌクレオチドによりコードされる場合は、より大きいペプチドを使用しうることが認識されるであろう。
【0028】
MHCリガンド、モチーフ、バリアントのペプチドの例、ならびにNおよび/若しくはC末端の延長のある種の例は、例えば、http://syfpeithi.bmi−heidelberg.com/のデータベースSYFPEITHI(Rammensee H、Bachmann J、Emmerich NP、Bachor OA、Stevanovic S.SYFPEITHI:detabase for MHC ligands and peptide motifs.Immunogenetics.1999 Nov;50(3−4):213−9.総説)、および本明細書に引用されるところの参考文献由来であり得る。
【0029】
制限しない例として、該データベース中のHLA−DRのある種のペプチドは、Ig κ鎖の188−203由来のKHKVYACEVTHQGLSS(Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21);Ig κ鎖の145−159由来のKVQWKVDNALQSGNS(Kovatsら Eur J Immunol.1997 Apr;27(4):1014−21)、GAD65の270−283由来のLPRLIAFTSEHSHF(Endlら J Clin Invest.1997 May 15;99(10):2405−15)、若しくはGAD65の556−575由来のFFRMVISNPAATHQDIDFLI(Endlら J Clin Invest.1997 May 15;99(10):2405−15)である。加えて、ペプチドは、bcr−ablの210kDの融合タンパク質由来のATGFKQSSKALQRPVAS(ten Boschら Blood.1996 Nov;88(9):3522−7)、HCV−1 N33の28−41 Diepolderら J Virol.1997 Aug;71(8):6011−9由来のGYKVLVLNPSVAAT、若しくはHIV−1(HXB2)RTの326−345由来のFRKQNPDIVIQYMDDLYVG(van der Burgら J Immunol.1999 Jan 1;162(1):152−60)の場合でのような、抗原の変異された配列にもまた由来し得る。全部の「アンカー」アミノ酸(Friedeら、Biochim Biophys Acta.1996 Jun 7;1316(2):85−101;Seeteら J Immunol.1993 Sep 15;151(6):3163−70.;Hammerら Cell.1993 Jul 16;74(1):197−203.、およびHammerら J Exp Med.1995 May 1;181(5):1847−55を参照されたい。HLA−DR4の例として)を太字で示し、推定のコア配列に下線を付けている。
【0030】
全部の上述されたペプチドは、所定のアミノ酸配列の「バリアント」という用語により包含される。
【0031】
「ペプチド」により、われわれは、アミノ酸がペプチド(−CO−NH−)結合により結合されている分子のみならず、しかしまたペプチド結合が反転されている分子も包含する。こうしたretro−inversoペプチド模倣物は、例えばMeziereら(1997)J.Immunol.159、3230−3237(引用することにより本明細書に組み込まれる)に記述されるもののような当該技術分野で既知の方法を使用して作成しうる。このアプローチは、バックボーンを伴いかつ側鎖の指向を伴わない変更を含有する擬似ペプチドを作成することを必要とする。Meziereら(1997)は、少なくともMHCクラスIIおよびTヘルパー細胞応答にこれらの擬似ペプチドが有用であることを示す。CO−NHペプチド結合の代わりにNH−CO結合を含有するretro−invesoペプチドは、タンパク質分解に対しはるかにより抵抗性である。
【0032】
典型的には、本発明のペプチドは、抗原提示細胞中で発現される場合に、適切なMHC分子に結合することが可能でありそして適する細胞により提示されかつ適するT細胞応答を導き出しうるフラグメントが生じられるように、プロセシングされうるものである。該ペプチドから生じられるフラグメントもまた本発明のペプチドでありうることが認識されるであろう。便宜的に、本発明のペプチドは、所定のアミノ酸配列またはそれらの一部分若しくはバリアントを包含する一部分、および何らかの所望の特性を賦与するさらなる一部分を含有する。例えば、さらなる部分は、さらなるT細胞エピトープ(第一のT細胞エピトープ含有部分と同一のポリペプチド由来であろうとそうでなかろうと)を包含しうるか、または、それは担体タンパク質若しくはペプチドを包含しうる。従って、一態様において、本発明のペプチドは、切断型のヒトタンパク質、または、タンパク質フラグメントおよび別のポリペプチド部分の融合タンパク質であるが、但し、ヒト部分は1種もしくはそれ以上の発明のアミノ酸配列を包含する。
【0033】
とりわけ好ましい一態様において、本発明のペプチドは、本発明のアミノ酸配列、および最低1個のさらなるT細胞エピトープを包含し、該さらなるT細胞エピトープは、腫瘍関連抗原を異常に発現する腫瘍の型に向けられたT細胞応答の産生を助長することが可能である。従って、本発明のペプチドは、ワクチンとしてもまた使用し得るいわゆる「数珠状構造(beads on a string)」ポリペプチドを包含する。
【0034】
いくつかの応用において、本発明のペプチドを直接使用しうる(すなわち、それらは患者の細胞若しくは患者に与えられる細胞中でのポリヌクレオチドの発現により産生されない)ことが以下から認識されるであろう。こうした応用において、ペプチドが100残基未満を有することが好ましい。
【0035】
本発明のペプチドがHLA−DRに結合することが可能である場合が好ましい。ペプチドがHLA−DR4に選択的に結合する場合がとりわけ好ましい。
【0036】
本発明のペプチドは、本発明の本ペプチドの基礎を形成するポリペプチドを異常に発現する細胞を標的としかつ死滅させるための免疫療法の方法でとりわけ有用である。所定のアミノ酸配列よりなるこれらの特定のペプチドはHLA−DRに結合するため、本発明のペプチドが、HLA−DRに結合し、かつ、そのように結合された場合に、HLA−DR−ペプチド複合体が、適する抗原提示細胞の表面上に存在する場合に所定のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するCTLの産生を導き出すことが可能であることが好ましい。
【0037】
本発明の一態様において、本発明のペプチドは、NCBI、GenBank受入番号X00497に由来するところのHLA−DR抗原関連インバリアント鎖(p33、以下で「Ii」)の80のN末端アミノ酸を含んでなる。
【0038】
「異常に発現される」により、われわれは、該ポリペプチドが正常な発現レベルに比較して過剰発現されている、若しくは、腫瘍が由来する組織中で該遺伝子がサイレントであるがしかし腫瘍中でそれが発現されるという意味を包含する。「過剰発現される」により、われわれは、該ポリペプチドが正常組織中に存在するレベルの最低1.2倍;好ましくは正常組織中に存在するレベルの最低2倍、およびより好ましくは最低5若しくは10倍のレベルで存在することを意味している。
【0039】
ペプチド(少なくともアミノ酸残基間のペプチド結合を含有するもの)は、Luら(1981)J.Org.Chem.46、3433およびその中の参考文献により開示されたところの固相ペプチド合成のFmoc−ポリアミドモードにより合成しうる。一時的なNアミノ基保護は9−フルオレニルメチルオキシカルボニル(Fmoc)基により提供される。この高度に塩基不安定性の保護基の反復切断は、N,N−ジメチルホルムアミド中20%ピペリジンを使用して遂げられる。側鎖の官能性は、それらのブチルエーテル(セリン、トレオニンおよびチロシンの場合)、ブチルエステル(グルタミン酸およびアスパラギン酸の場合)、ブチルオキシカルボニル誘導体(リシンおよびのヒスチジンの場合)、トリチル誘導体(システインの場合)ならびに4−メトキシ−2,3,6−トリメチルベンゼンスルホニル誘導体(アルギニンの場合)として保護しうる。グルタミン若しくはアスパラギンがC末端残基である場合、側鎖のアミド官能性の保護に4,4'−ジメトキシベンズヒドリル基が利用される。固相支持体は、3種の単量体、ジメチルアクリルアミド(バックボーン単量体)、ビスアクリロイルエチレンジアミン(架橋剤)およびアクリロイルサルコシンメチルエステル(官能性化剤)から構成されるポリジメチル−アクリルアミドポリマーに基づく。使用されるペプチドから樹脂の切断可能な結合剤は、酸不安定性の4−ヒドロキシメチル−フェノキシ酢酸誘導体である。全部のアミノ酸誘導体は、アスパラギンおよびグルタミン(反転N,N−ジシクロヘキシル−カルボジイミド/1ヒドロキシベンゾトリアゾールに媒介されるカップリング手順を使用して付加される)を除き、それらの予め形成された対称性の無水物誘導体として付加される。全部のカップリングおよび脱保護反応は、ニンヒドリン、トリニトロベンゼンスルホン酸若しくはイソチン試験手順を使用してモニターする。合成の完了に際し、50%スカベンジャー混合物を含有する95%トリフルオロ酢酸での処理による側鎖保護基の同時の除去で、樹脂支持体からペプチドを切断する。一般に使用されるスカベンジャーは、エタンジチオール、フェノール、アニソールおよび水であり、正確な選択は合成されているペプチドの構成アミノ酸に依存する。
【0040】
トリフルオロ酢酸は、ジエチルエーテルとのその後の摩砕を用いる真空中での蒸発により除去されて、粗ペプチドを提供する。存在するいかなるスカベンジャーも、水相の凍結乾燥に際してスカベンジャーを含まない粗ペプチドを提供する単純な抽出手順により除去される。ペプチド合成のための試薬は、一般に、Calbiochem−Novabiochem(UK)Ltd.、Nottingham NG7 2QJ、UKから入手可能である。
【0041】
精製は、サイズ排除クロマトグラフィー、イオン交換クロマトグラフィー、および(通常は)逆相高速液体クロマトグラフィーのようないずれか1つの技術若しくはそれらの組合せにより遂げることができる。
【0042】
ペプチドの分析は、酸加水分解後の薄層クロマトグラフィー、逆相高速液体クロマトグラフィー、アミノ酸分析を使用して、ならびに、高速原子衝撃(FAB)質量分析、ならびにMALDIおよびESI−Q−TOF質量分析により実施しうる。
【0043】
本発明のさらなる一局面は、本発明のペプチドをコードする核酸(例えばポリヌクレオチド)を提供する。ポリヌクレオチドは、DNA、cDNA、PNA、CNA、RNA若しくはそれらの組合せであることができ、また、それは、それが該ペプチドをコードする限りはイントロンを含有しても若しくはしなくてもよい。もちろん、それは、ポリヌクレオチドによりコード可能である、天然に存在するペプチド結合により結合された天然に存在するアミノ酸を含有するペプチドのみである。本発明のなおさらなる一局面は、本発明のポリペプチドを発現することが可能な発現ベクターを提供する。
【0044】
例えば相補的付着端を介してベクターにポリヌクレオチド、とりわけDNAを作動可能に連結するための多様な方法が開発された。例えば、相補的ホモポリマー領域を、ベクターDNAに挿入されるべきDNAセグメントに付加し得る。ベクターおよびDNAセグメントをその後、相補的ホモポリマー尾部間の水素結合により結合して組換えDNA分子を形成する。
【0045】
1個若しくはそれ以上の制限部位を含有する合成リンカーは、ベクターへのDNAセグメントの一代替結合方法を提供する。前に記述されたところのエンドヌクレアーゼ制限消化により生成されるDNAセグメントを、バクテリオファージT4 DNAポリメラーゼ若しくは大腸菌(E.coli)DNAポリメラーゼIで処理し、これらの酵素は突出している3'一本鎖末端をそれらの3'−5'−エキソヌクレアーゼ活性で除去し、そしてそれらのポリメラーゼ活性でくぼんだ3'端を埋める。
【0046】
これらの活性の組合せは従って平滑端のDNAセグメントを生成する。該平滑端セグメントをその後、バクテリオファージT4 DNAリガーゼのような平滑端DNA分子の核酸連結を触媒することが可能である酵素の存在下で、モル濃度大過剰のリンカー分子とともにインキュベートする。従って、該反応の生成物はそれらの端にポリマーリンカー配列を運搬するDNAセグメントである。これらのDNAセグメントをその後、適切な制限酵素で切断し、そして、該DNAセグメントの末端と適合する末端を生じる酵素で切断した発現ベクターに連結する。
【0047】
多様な制限酵素部位を含有する合成リンカーは、International Biotechnologies Inc、米国コネチカット州ニューヘイブンを包含する多数の供給源から商業的に入手可能である。
【0048】
本発明のポリペプチドをコードするDNAの望ましい一改変方法は、Saikiら(1988)Science 239、487−491により開示されたところのポリメラーゼ連鎖反応を使用することである。この方法は、例えば適する制限部位で設計することにより、適するベクターにDNAを導入するのに使用しうるか、若しくは、それを使用して、当該技術分野で既知であるところの他の有用な方法でDNAを改変しうる。この方法において、酵素で増幅されるべきDNAは、該増幅されたDNAにそれら自身が組み込まれる2種の特異的プライマーにより隣接される。前記特異的プライマーは、当該技術分野で既知の方法を使用して発現ベクターにクローン化するのに使用し得る、制限酵素認識部位を含有しうる。
【0049】
DNA(若しくはレトロウイルスベクターの場合はRNA)がその後、適する宿主中で発現されて、本発明の化合物を含んでなるポリペプチドを産生する。従って、本発明の化合物を構成するポリペプチドをコードするDNAを、本明細書に含有される教示を鑑みて適切に改変された既知技術に従って使用して発現ベクターを構築することができ、この発現ベクターをその後、本発明のポリペプチドの発現および産生のため適切な宿主細胞を形質転換するのに使用する。こうした技術は、Rutterらに1984年4月3日に発行された米国特許第4,440,859号、Weissmanに1985年7月23日に発行された同第4,530,901号、Crowlに1986年4月15日に発行された同第4,582,800号、Markらに1987年6月30日に発行された同第4,677,063号、Goeddelに1987年7月7日に発行された同第4,678,751号、Itakuraらに1987年11月3日に発行された同第4,704,362号、Murrayに1987年12月1日に発行された同第4,710,463号、Toole,Jr.らに1988年7月12日に発行された同第4,757,006号、Goeddelらに1988年8月23日に発行された同第4,766,075号、およびStalkerに1989年3月7日に発行された同第4,810,648号明細書(それらの全部は引用することにより本明細書に組み込まれる)に開示されるものを包含する。
【0050】
本発明の化合物を構成するポリペプチドをコードするDNA(若しくはレトロウイルスベクターの場合はRNA)は、適切な宿主への導入のために多様な他のDNA配列に結合しうる。伴(companion)DNAは、宿主の性質、宿主細胞への該DNAの導入の様式、およびエピソーム維持が望ましいか若しくは組込みが望ましいかに依存することができる。
【0051】
一般に、DNAは、プラスミドのような発現ベクターに、発現のための適正な向きおよび正しい読み枠で挿入する。必要な場合は、DNAは所望の宿主により認識される適切な転写および翻訳調節性の制御ヌクレオチド配列に連結しうるが、こうした制御配列は発現ベクターで一般に利用可能である。ベクターをその後、標準的技術により宿主に導入する。一般に、宿主の全部がベクターにより形質転換されるわけではない。従って、形質転換された宿主細胞を選択することが必要となる。一選択技術は、抗生物質耐性のような、形質転換された細胞中の選択可能な特質をコードするいずれかの必要な調節領域とともに、DNA配列を発現ベクターに組み込むことを伴う。
あるいは、こうした選択可能な特質の遺伝子は、所望の宿主細胞を共形質転換するのに使用する別のベクター上にあってよい。
【0052】
本発明の組換えDNAにより形質転換された宿主細胞をその後、ポリペプチドの発現を可能にするための本明細書に開示される教示を鑑み、十分な時間、および当業者に既知の適切な条件下で培養し、ポリペプチドをその後回収してよい。
【0053】
細菌(例えば大腸菌(E.coli)および枯草菌(Bacillus subtilis)、)酵母(例えば出芽酵母(Saccharomyces cerevisiae))、糸状菌(例えばアスペルギルス属(Aspergillus))、植物細胞、動物細胞および昆虫細胞を包含する多くの発現系が既知である。好ましくは、該系はAwells細胞であってよい。
【0054】
プロモーターは、RNAポリメラーゼの結合および転写を起こさせる、DNA配列により形成される発現調節領域である。模範的な細菌宿主と適合性のあるプロモーター配列は、典型的に、本発明のDNAセグメントの挿入に都合のよい制限部位を含有するプラスミドベクター中に提供されている。典型的な原核生物ベクタープラスミドは、Biorad Laboratoris、(米国カリフォルニア州リッチモンド)から入手可能なpUC18、pUC19、pBR322およびpBR329、ならびにPharmacia、米国ニュージャージー州ピスカタウェイから入手可能なpTrc99AおよびpKK223−3である。
【0055】
典型的な哺乳動物細胞ベクタープラスミドは、Pharmacia、米国ニュージャージー州ピスカタウェイから入手可能なpSVLである。このベクターは、クローン化した遺伝子の発現を駆動するのにSV40後期プロモーターを使用し、最高レベルの発現はCOS−1細胞のようなT抗原産生細胞で見出される。誘導可能な哺乳動物発現ベクターの一例は、またPharmaciaから入手可能なpMSGである。このベクターは、クローン化した遺伝子の発現を駆動するために、マウス乳癌ウイルス末端反復配列のグルココルチコイドで誘導可能なプロモーターを使用する。有用な酵母プラスミドベクターはpRS403−406およびpRS413−416であり、そして一般にStratagene Cloning Systems、La Jolla、CA 92037、USAから入手可能である。プラスミドpRS403、pRS404、pRS405およびpRS406は酵母組込み型プラスミド(YIp)であり、そして酵母の選択可能なマーカー、HIS3、TRP1、LEU2およびURA3を組み込む。プラスミドpRS413−416は酵母セントロメアプラスミド(Ycp)である。多様な宿主細胞との使用のための他のベクターおよび発現系が当該技術分野で公知である。
【0056】
本発明はまた、本発明のポリヌクレオチドベクター構築物で形質転換された宿主細胞にも関する。該宿主細胞は原核生物若しくは真核生物のいずれかであり得る。細菌細胞はいくつかの環境で好ましい原核生物宿主細胞であることができ、そして、典型的には、例えば、Bethesda Research Laboratories Inc.、米国メリーランド州ベセスダから入手可能な大腸菌(E.coli)株DH5、および米国メリーランド州ロックビルのAmerican Type Culture Collection(ATCC)から入手可能なRR1(No ATCC 31343)のような大腸菌(E.coli)の株である。好ましい真核生物宿主細胞は、酵母、昆虫および哺乳動物細胞、好ましくはマウス、ラット、サル若しくはヒト線維芽および腎細胞株からのもののような脊椎動物細胞を包含する。酵母宿主細胞は、Stratagene Cloning Systems、La Jolla、CA 92037、米国から一般に入手可能であるYPH499、YPH500およびYPH501を包含する。好ましい哺乳動物宿主細胞は、CCL61としてATCCから入手可能なチャイニーズハムスター卵巣(CHO)細胞、CRL 1658としてATCCから入手可能なNIH Swissマウス胚細胞NIH/3T3、CRL 1650としてATCCから入手可能なサル腎由来COS−1細胞、およびヒト胎児由来腎臓細胞である293細胞を包含する。好ましい昆虫細胞はバキュロウイルス発現ベクターでトランスフェクトし得るSf9細胞である。
【0057】
本発明のDNA構築物での適切な細胞宿主の形質転換は、使用されるベクターの型に典型的に依存する公知の方法により達成される。原核生物宿主細胞の形質転換に関しては、例えば、Cohenら(1972)Proc.Natl.Acad.Sci.USA 69、2110およびSambrookら(1989)Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバーを参照されたい。酵母細胞の形質転換は、Shermanら(1986)Methods In Yeast Genetics,A Laboratory Manual、ニューヨーク州コールドスプリングハーバーに記述されている。Beggs(1978)Nature 275、104−109の方法もまた有用である。脊椎動物細胞に関しては、こうした細胞のトランスフェクションにおいて有用な試薬、例えばリン酸カルシウムおよびDEAE−デキストラン若しくはリポソーム製剤が、Stratagene Cloning Systems、若しくはLife Technologies Inc.、Gaithersburg、MD 20877、米国から入手可能である。電気穿孔法もまた細胞を形質転換および/若しくはトランスフェクトするのに有用であり、また、酵母細胞、細菌細胞、昆虫細胞および脊椎動物細胞を形質転換するために当該技術分野で公知である。
【0058】
成功裏に形質転換された細胞、すなわち本発明のDNA構築物を含有する細胞は、公知の技術により同定し得る。例えば、本発明の発現構築物の導入から生じる細胞を、本発明のポリペプチドを産生させるために増殖させ得る。細胞を収集かつ溶解し得、そしてそれらのDNA内容物を、Southern(1975)J.Mol.Biol.98、503若しくはBerentら(1985)Biotech.3、208により記述されたもののような方法を使用して、該DNAの存在について検査し得る。あるいは、上清中の外タンパク質の存在を、下述されるところの抗体を使用して検出し得る。
【0059】
組換えDNAの存在について直接アッセイすることに加え、成功裏の形質転換は、組換えDNAがタンパク質の発現を指図することが可能である場合に、公知の免疫学的方法により確認し得る。例えば、発現ベクターで成功裏に形質転換された細胞は、適切な抗原性を表すタンパク質を産生する。形質転換されていることが疑われる細胞のサンプルを収集し、そして、適する抗体を使用してタンパク質についてアッセイする。従って、形質転換された宿主細胞それら自身に加え、本発明はまた、栄養培地中のそれらの細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、若しくはモノクローナル培養物由来の培養物も企図している。
【0060】
本発明のある種の宿主細胞、例えば細菌、酵母および昆虫細胞が、本発明のペプチドの製造で有用であることが認識されるであろう。しかしながら、他の宿主細胞がある種の治療方法で有用でありうる。例えば、樹状細胞のような抗原提示細胞は、それらが適切なMHC分子に負荷されうるような本発明のペプチドを発現させるために有用に使用しうる。
【0061】
本発明のさらなる一局面は、静脈内(i.v.)注入、皮下(s.c.)注入、皮内(i,d.)注入、腹腔内(i.p.)注入、筋肉内(i.m.)注入のためのペプチドの製造方法を提供する。好ましいペプチド注入方法はs.c.、i.p.およびi.v.である。好ましいDNA注入方法は、i.d.、i.m.、s.c.、i.p.およびi.v.である。1から500mgのペプチド若しくはDNAの用量を与えうる。
【0062】
本発明のさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、有効量の本発明のペプチド、または有効量の前記ペプチドをコードするポリヌクレオチド若しくは発現ベクターを患者に投与することを含んでなり、前記ペプチドの該量または前記ポリヌクレオチド若しくは発現ベクターの量は、前記患者で抗標的細胞免疫応答を惹起するのに有効である。標的細胞は、典型的に腫瘍若しくは癌細胞である。
【0063】
ペプチド若しくはペプチドをコードする核酸は腫瘍若しくは癌ワクチンを構成する。それは患者に、冒された器官に直接投与しうるか、またはその後患者に投与される該患者由来の細胞若しくはヒト細胞株にex vivoで適用しうるか、または、その後患者に再投与される該患者由来の免疫細胞からの亜集団を選択するためにin vitroで使用しうる。核酸をin vitroで細胞に投与する場合、インターロイキン−2のような免疫刺激サイトカインを共発現するようにトランスフェクトすることが細胞に有用でありうる。該ペプチドは、実質的に純粋であっても、若しくはDetoxのような免疫刺激アジュバントと組合せても、若しくは免疫刺激サイトカインとともに使用しても、若しくは適する送達系、例えばリポソームを用いて投与してもよい。該ペプチドはまた、キーホールリンペットヘモシアニン(KLH)若しくはマンナンのような適する担体にも複合しうる(WO 95/18145号、およびLongeneckerら(1993)Ann.NY Acad.Sci.690、276−291を参照されたい)。該ペプチドはまた、標識しても、若しくは融合タンパク質であっても、若しくはハイブリッド分子であってもよい。その配列が本発明で示されるペプチドが発現されてCD4 CTLを刺激する。しかしながら、刺激は、CD4+ T細胞により提供される援助の存在下でより効率的である。従って、ハイブリッド分子の融合パートナー若しくは区分は、CD4+ T細胞を刺激するエピトープを適切に提供する。CD4+を刺激するエピトープは当該技術分野で公知であり、そして破傷風トキソイド中で同定されたものを包含する。該ポリヌクレオチドは、実質的に純粋であっても、または適するベクター若しくは送達系に含有されてもよい。
【0064】
適するベクターおよび送達系は、アデノウイルス、ワクシニアウイルス、レトロウイルス、ヘルペスウイルス、アデノ随伴ウイルス、若しくは1種以上のウイルスの要素を含有するハイブリッドに基づく系のようなウイルスを包含する。ウイルス以外の送達系は、DNA送達の技術分野で公知であるところの陽イオン性脂質および陽イオン性ポリマーを包含する。「遺伝子銃」を介するような物理的送達もまた使用しうる。該ペプチド、若しくは該核酸によりコードされるペプチドは、例えば、CD4+ T細胞を刺激する破傷風トキソイドからのエピトープをもつ融合タンパク質でありうる。
【0065】
癌ワクチンでの使用のためのペプチドはいかなる適するペプチドでもありうる。とりわけ、それは適する9merペプチド、あるいは適する7mer若しくは8mer若しくは10mer若しくは11merペプチドまたは12merでありうる。より長いペプチドもまた適しうるが、しかし付属の表1に記述されるところの9mer若しくは10merペプチドが好ましい。
【0066】
適切には、患者に投与されるいかなる核酸も無菌でありかつ発熱性物質を含まない。裸のDNAは筋肉内若しくは皮内若しくは皮下に与えうる。ペプチドは筋肉内、皮内若しくは皮下に与えうる。
【0067】
ワクチン接種は専門の抗原提示細胞により刺激されるCTL応答をもたらし、CTLが一旦刺激されれば、腫瘍細胞中でのMHC発現の増強において一利点が存在しうる。
【0068】
注入の部位、ターゲッティングベクターおよび送達系の使用、または患者からのこうした細胞集団の選択的精製およびペプチド若しくは核酸のex vivo投与のいずれかにより、ワクチンの標的を特定の細胞集団、例えば抗原提示細胞に向けることもまた有用でありうる(例えば、樹状細胞を、Zhouら(1995)Blood 86、3295−3301;Rothら(1996)Scand.J.Immunology 43、646−651に記述されるとおり選別しうる)。例えば、ターゲッティングベクターは、適する場所での抗原の発現を指図する組織若しくは腫瘍特異的プロモーターを含みうる。
【0069】
本発明のさらなる一局面は、従って、有効量の本発明のペプチドを含んでなるか若しくはこうしたペプチドをコードする核酸を含んでなる、癌、または癌若しくは腫瘍細胞に対し有効なワクチンを提供する。ワクチンが核酸ワクチンである場合がとりわけ好ましい。ポリペプチドをコードするDNAワクチンのような核酸ワクチンでの接種がT細胞応答につながることが既知である。
【0070】
便宜上、核酸ワクチンは、いかなる適する核酸送達手段も含みうる。核酸、好ましくはDNAは裸でありうる(すなわち投与されるべき他の成分を実質的に含まない)か、または、それはリポソーム中で若しくはウイルスベクター送達系の一部として送達されうる。
【0071】
樹状細胞による核酸の取り込みおよびコードされるポリペプチドの発現が、免疫応答のプライミングの機構でありうると考えられるが、しかしながら、樹状細胞はトランスフェクトされなくてもよいがしかしなお重要である。それらは、トランスフェクトされた細胞からの発現されたペプチドを組織中に取り込みうるからである。
【0072】
DNAワクチンのようなワクチンが筋中に投与される場合が好ましい。ワクチンが皮膚に投与される場合もまた好ましい。核酸ワクチンはアジュバントを伴わずに投与してもよい。核酸ワクチンはまた、BCG若しくは明礬のようなアジュバントとともにも投与しうる。他の適するアジュバントは、サポニン由来であるAquilaのQS21スティミュロン(stimulon)(Aquila Biotech、米国マサチューセッツ州ウースター)、ミコバクテリウム抽出物および合成細菌細胞壁模倣物、ならびにRibiのDetoxのような特許のアジュバントを包含する。別のサポニン由来アジュバント、Quil Aもまた使用しうる(Superfos、デンマーク)。核酸ワクチンがアジュバントを伴わずに投与される場合が好ましい。フロイントのアジュバントのような他のアジュバントもまた有用でありうる。キーボールリンペットヘモシアニンに、好ましくはまたアジュバントと複合させたペプチドを与えることもまた有用でありうる。
【0073】
癌のペプチドヌクレオチド媒介性の免疫療法は、Conryら(1996)Seminars in Oncology 23、135−147;Condonら(1996)Nature Medicine 2、1122−1127;Gongら(1997)Nature Medicine 3、558−561;Zhaiら(1996)J.Immunol.156、700−710;Grahamら(1996)Int J.Cancer 65、664−670;およびBreast Cancer,Advances in biology and therapeutics、Calvoら(編)、John Libbey Eurotext中、Burchellら(1996)pp 309−313(それらの全部は引用することにより本明細書に組み込まれる)に記述されている。
【0074】
本発明のなおさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞を死滅させるための医薬品の製造における、本発明のペプチド、またはこうしたペプチドをコードするポリヌクレオチド若しくは発現ベクターの使用を提供する。
【0075】
本発明のさらなる一局面は、in vitroでの活性化型細胞傷害性Tリンパ球(CTL)の製造方法を提供し、該方法は、CTLを、適する抗原提示細胞の表面上に発現された、抗原負荷ヒトクラスIIMHC分子と、前記CTLを抗原(該抗原は本発明のペプチドである)特異的様式で活性化するのに十分な時間、in vitroで接触させることを含んでなる。
【0076】
適しては、CTLは、好ましくはTH1型のCD4+ヘルパー細胞である。MHCクラスII分子は、いかなる適する細胞の表面上に発現されてもよく、また、細胞がMHCクラスII分子を天然に発現しない(その場合、該細胞はこうした分子を発現するようトランスフェクトされる)か、またはそれが発現する場合はそれが抗原プロセシング若しくは抗原提示経路に欠陥があるものである場合が好ましい。こうして、MHCクラスII分子を発現する細胞が、CTLを活性化する前に選ばれたペプチド抗原で実質的に完全に予備刺激されることが可能である。
【0077】
抗原提示細胞(若しくは刺激体(stimulator)細胞)は、典型的にその表面上に1個のMHCクラスII分子を有し、そして、好ましくは、前記MHCクラスII分子に選択された抗原をそれ自身負荷することが実質的に不可能である。より詳細に下述されるとおり、MHCクラスII分子は、選択された抗原をin vitroで容易に負荷されうる。
【0078】
好ましくは、該哺乳動物細胞は、TAPペプチド輸送体を欠くか、若しくはその低下されたレベルを有するか、若しくはその低下された機能を有する。TAPペプチド輸送体を欠く適する細胞は、T2、RMA−Sおよびショウジョウバエ(Drosophila)細胞を包含する。TAPは、抗原プロセシングに関与する輸送体(Transporter Associated with antigen Processing)である。
【0079】
ヒトペプチド負荷欠損細胞株(human peptide loading deficient cell line)T2は、カタログ番号CRL 1992で、American Type Culture Collection、12301 Parklawn Drive、Rockville、Maryland 20852、USAから入手可能であり;ショウジョウバエ(Drosophila)細胞株Schneider株2は、カタログ番号CRL 19863でATCCから入手可能であり;マウスRMA−S細胞株は、KarreとLjunggren(1985)J.Exp.Med.162、1745(引用することにより本明細書に組み込まれる)に記述されている。
【0080】
便宜上、前記宿主細胞は、トランスフェクション前にMHCクラスI分子を実質的に発現しない。刺激体細胞が、B7.1、B7.2、ICAM−1およびLFA3のいずれかのようなT細胞共刺激に重要な分子を発現する場合もまた好ましい。
【0081】
多数のMHCクラスII分子および共刺激体分子の核酸配列は、GenBankおよびEMBLデータベースから公的に入手可能である。
【0082】
さらなる一態様において、HLA分子の組合せもまた使用しうる。
【0083】
複数のCD8+ CTLエピトープの送達のための組換え多エピトープワクチンの使用は、Thomsonら(1996)J.Immunol.157、822−826およびWO 96/03144号(それらの双方は引用することにより本明細書に組み込まれる)に記述されている。本発明に関して、単一のワクチン中に1種のペプチド(若しくは1種のペプチドをコードする核酸)を包含することが望ましいことができ、該ペプチドは、本発明のアミノ酸配列およびCD4+ T細胞を刺激するエピトープ(破傷風トキソイドのような)をいずれかの順序で包含する。こうしたワクチンは、癌を処置するのにとりわけ有用であるとみられる。こうした「数珠状構造」ワクチンは、典型的にはDNAワクチンである。
【0084】
in vitroでCTLを生成するのに多数の他の方法を使用しうる。例えば、Peoplesら(1995)Proc.Natl.Acad.Sci.USA 92、432−436およびKawakamiら(1992)J.Immunol.148、638643に記述される方法は、CTLの生成で自己の腫瘍浸潤リンパ球を使用する。Plebanskiら(1995)Eur.J.Immunol.25、1783−1787は、CTLの製造で自己の末梢血リンパ球(PLB)を利用する。Jochmusら(1997)J.Gen.Virol.78、1689−1695は、ペプチド若しくはポリペプチドで樹状細胞をパルスすることを使用することによるか、または組換えウイルスへの感染を介する、自己CTLの製造を記述する。Hillら(1995)J.Exp.Med.181、2221−2228およびJeromeら(1993)J.Immunol.151、1654−1662は、自己CTLの製造でB細胞を利用する。加えて、ペプチド若しくはポリペプチドでパルスしたか、または組換えウイルスに感染させたマクロファージを、自己CTLの製造で使用しうる。
【0085】
同種異系細胞もまたCTLの製造に使用することができ、そして、この方法はWO 97/26328号(引用することにより本明細書に組み込まれる)に詳細に記述されている。例えば、ショウジョウバエ(Drosophila)細胞およびT2細胞に加え、CHO細胞、バキュロウイルスに感染させた昆虫細胞、細菌、酵母、ワクシニアに感染させた標的細胞のような他の細胞を使用して抗原を提示しうる。加えて、植物ウイルスを使用しうる(例えば、外来ペプチドの提示のための多収性の系としてのササゲモザイクウイルスの開発を記述するPortaら(1994)Virology 202、449−955を参照されたい)。
【0086】
本発明のペプチドに向けられる活性化型CTLは治療で有用である。従って、本発明のさらなる一局面は、本発明の前述の方法により得ることができる活性化型CTLを提供する。
【0087】
本発明のなおさらなる一態様は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を選択的に認識する活性化型CTLを提供する。好ましくは、該CTLはHLA/ペプチド複合体と相互作用(例えば結合)することにより前記細胞を認識する。該CTLは、有効な数の活性化型CTLを患者に投与する、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法で有用である。患者に投与されるCTLは、患者由来でありかつ上述されたとおり活性化しうる(すなわちそれらは自己CTLである)。あるいは、該CTLは患者からでないが、しかし別の個体からである。もちろん、該個体が健康な個体である場合が好ましい。「健康な個体」により、われわれは、個体が全般として良好な健康状態にある、好ましくは適確な免疫系を有する、およびより好ましくは、容易に試験かつ検出され得るいかなる疾患にも罹っていないことを意味している。
【0088】
活性化型CTLは、異常なポリペプチドを発現する細胞の認識に関与するT細胞受容体(TCR)を発現する。TCRをコードするcDNAが活性化型CTLからクローン化されかつ発現のためさらなるCTLに移入される場合が有用である。
【0089】
in vivoで、本発明のCD4+ CTLの標的細胞は、腫瘍の細胞(ときにMHCクラスIIを発現する)および/若しくは腫瘍(腫瘍細胞)を取り囲む間質細胞(ときにまたMHCクラスIIを発現する)であり得る。
【0090】
本発明のペプチドに特異的な本発明のCTLクローンのTCRをクローン化する。CTLクローンでのTCRの使用は、(i)TCR可変領域特異的モノクローナル抗体、ならびに(ii)VaおよびVp遺伝子ファミリーに特異的なプライマーを用いるRT PCRを使用して決定する。CTLクローンから抽出したポリA mRNAからcDNAライブラリーを調製する。TCR aおよびP鎖のC末端部分、ならびに同定されたVaおよびPセグメントのN末端部分に特異的なプライマーを使用する。TCR aおよび鎖の完全なcDNAを高忠実度DNAポリメラーゼで増幅し、そして、増幅された産物を適するクローニングベクターにクローン化する。クローン化されたaおよびP鎖遺伝子を、Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658により記述されるところの方法により、一本鎖TCRに集成しうる。この一本鎖構築物中で、VaJセグメントにV DJセグメントが続き、次いでCpセグメント、次いでCD3鎖の膜貫通および細胞質セグメントが続く。この一本鎖TCRをその後レトロウイルス発現ベクターに挿入する(一団のベクターを、成熟ヒトCD8+ Tリンパ球を感染させかつ遺伝子発現を媒介するそれらの能力に基づいて使用しうる。すなわち、レトロウイルスベクター系Katが1つの好ましい可能性である(Finerら(1994)Blood 83、43を参照されたい)。高力価の両指向性レトロウイルスを使用して、腫瘍患者の末梢血から単離した、精製したCD8+若しくはCD4+ Tリンパ球を(Robertsら(1994)Blood 84、2878−2889(引用することにより本明細書に組み込まれる)により公表されたプロトコルに従って)感染させる。抗CD3抗体を使用して、精製したCD8+ T細胞の増殖を誘発し、それは一本鎖TCRのレトロウイルス組込みおよび安定な発現を助長する。レトロウイルス形質導入の効率は、一本鎖TCRに特異的な抗体での感染させたCD8+ T細胞の染色により決定する。形質導入されたCD8+ T細胞のin vitro分析は、該TCR鎖が最初にクローン化された同種拘束性(allo−restricted)CTLクローンで見られると同一の腫瘍特異的死滅をそれらが表すことを確立する。期待された特異性をもつ形質導入されたCD8+ T細胞の集団を、腫瘍患者の養子免疫療法に使用しうる。患者は、108ないし1011の間の自己の形質導入されたCTLで処置しうる。CD8+と同様に、関連する構築物を運搬する形質導入されたCD4+ Tヘルパー細胞を生成させ得る。
【0091】
遺伝子をCTLに導入するための他の適する系は、Moritzら(1994)Proc.Natl.Acad.Sci.USA 91、4318−4322(引用することにより本明細書に組み込まれる)に記述されている。Eshharら(1993)Proc.Natl.Acad.Sci.USA 90、720−724、およびHwuら(1993)J.Exp.Med.178、361−366もまた、CTLのトランスフェクションを記述している。従って、本発明のさらなる局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するTCRを提供し、該TCRは活性化型CTLから得ることができる。
【0092】
TCRだけでなく、TCRに機能上同等な分子が本発明に包含される。これらは、TCRと同一の機能を演じ得る、TCRに機能上同等であるいかなる分子も包含する。とりわけ、こうした分子は、Chungら(1994)Proc.Natl.Acad.Sci.USA 91、12654−12658(引用することにより本明細書に組み込まれかつ上に言及された)により記述されかつ上に引用される方法により作成されるところの、遺伝子的に操作された3ドメインの一本鎖TCRを包含する。本発明はまた、TCR若しくは機能上同等な分子をコードするポリヌクレオチド、および該TCR若しくはその機能上同等な分子をコードする発現ベクターも包含する。本発明のTCRを発現するのに適する発現ベクターは、本発明のペプチドの発現に関して上述されたものを包含する。
【0093】
しかしながら、発現ベクターがトランスフェクション後にCTL中でTCRを発現することが可能であるものであることが好ましい。
【0094】
本発明のなおさらなる一局面は、本発明のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、(1)患者からCTLを得る段階;(2)TCR若しくは上で定義されたところの機能上同等な分子をコードするポリヌクレオチドを前記細胞に導入する段階;および(3)段階(2)で製造した細胞を患者に導入する段階を含んでなる。
【0095】
本発明のなおさらなる一局面は、本発明の第一若しくは第二若しくは第三の局面で定義されるところのアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を標的とする、患者における標的細胞の死滅方法を提供し、該方法は、(1)前記患者から樹状細胞のような抗原提示細胞を得る段階;(2)前記抗原提示細胞を、本発明の第一若しくは第二若しくは第三の局面で定義されるところのペプチド、またはこうしたペプチドをコードするポリヌクレオチドとex vivoで接触させる段階;および(3)そのように処理した抗原提示細胞を患者に再導入する段階を含んでなる。
【0096】
好ましくは、抗原提示細胞は樹状細胞である。適切には、樹状細胞は、抗原ペプチドでパルスされている自己の樹状細胞である。抗原ペプチドは、適切なT細胞応答を生じさせるいかなる適する抗原ペプチドであってもよい。腫瘍関連抗原からのペプチドでパルスした自己の樹状細胞を使用するT細胞療法は、Murphyら(1996)The Prostate 29、371−380およびTjuaら(1997)The Prostate 32、272−278に開示されている。
【0097】
さらなる一態様において、樹状細胞のような抗原提示細胞を、本発明のペプチドをコードするポリヌクレオチドと接触させる。該ポリヌクレオチドはいかなる適するポリヌクレオチドであってもよく、そして、それが樹状細胞を形質導入し、従ってペプチドの提示および免疫の誘導をもたらすことが可能であることが好ましい。
【0098】
便宜上、ポリヌクレオチドはウイルスポリヌクレオチド若しくはウイルスに含まれうる。例えば、アデノウイルスで形質導入した樹状細胞は、MUC1に関して抗原特異的抗腫瘍免疫を誘導することが示された(Gongら(1997)Gene Ther.4、1023−1038を参照されたい)。同様に、アデノウイルスに基づく系を使用することができ(例えば、Wanら(1997)Hum.Gene Ther.8、1355−1363を参照されたい);レトロウイスル系を使用することができ(Spechtら(1997)J.Exp.Med.186、1213−1221、およびSzabolcsら(1997)Blood)、樹状細胞への粒子媒介性移入もまた使用することができ(Tutingら(1997)Eur.J.Immunol.27、2702−2707);そしてRNAもまた使用しうる(Ashleyら(1997)J.Exp.Med.186、1177 1182)。
【0099】
患者における標的細胞の死滅方法に関して、標的細胞が癌細胞、より好ましくは腎癌細胞であることがとりわけ好ましいことが認識されるであろう。
【0100】
本発明の方法により処置される患者がHLA−DRハプロタイプを有する場合がとりわけ好ましい。従って、好ましい一態様において、患者のHLAハプロタイプを処置の前に決定する。HLAハプロタイピングは、いずれの適する方法を使用して実施してもよく、こうした方法は当該技術分野で公知である。
【0101】
本発明は、とりわけ、活性のin vivoワクチン接種のため;in vitroでの自己の樹状細胞の操作、次いでCTL応答を活性化するためのそのように操作した樹状細胞のin vivoでの導入のため;in vitroで自己CTLを活性化し、次いで養子療法のため(すなわちそのように操作したCTLを患者に導入する);および健康なドナー(MHCマッチ若しくはミスマッチの)からのCTLをin vitroで活性化し、次いで養子療法のための、本発明のペプチド(若しくはそれらをコードするポリヌクレオチド)の使用を包含する。
【0102】
好ましい一態様において、本発明のワクチンは、腫瘍の形成を阻害若しくは抑制するために、単独で、若しくは別の癌治療との組合せのいずれかで宿主に投与する。
【0103】
ペプチドワクチンはアジュバントを伴わずに投与しうる。ペプチドワクチンはまた、BCG若しくは明礬のようなアジュバントとともに投与してもよい。他の適するアジュバントは、サポニン由来であるAquilaのQS21スティミュロン(stimulon)(Aquila Biotech、米国マサチューセッツ州ウースター)、ミコバクテリウム抽出物、および合成の細菌細胞壁模倣物、ならびにRibiのDetoxのような特許のアジュバントを包含する。別のサポニン由来アジュバント、Quil Aもまた使用しうる(Superfos、デンマーク)。フロイントのアジュバント若しくはGMCSFのような他のアジュバントもまた有用でありうる。好ましくはまたアジュバントとともにのキーホールリンペットヘモシアニンに複合させたペプチドを与えることもまた有用でありうる。
【0104】
本発明のペプチドはまた診断試薬としても使用し得る。該ペプチドを使用して、CTL集団中に、ペプチドに特異的に向けられているCTLが存在するかどうか、若しくはCTLが治療により誘導されるかどうかを分析し得る。さらに、前駆T細胞の増大を、規定されたペプチドに対する反応性を有するペプチドを用いて試験し得る。さらに、該ペプチドを、該ペプチドが由来する前記抗原を発現する腫瘍の疾患の進行をモニターするためのマーカーとして使用し得る。
【0105】
付属の表1に、同定されたところのペプチドを列挙する。加えて、該表に、ペプチドが由来するタンパク質、およびそれぞれのタンパク質中のペプチドのそれぞれの位置を指定する。さらに、国立衛生研究所の「国立バイオテクノロジー情報センター(National Centre for Biotechnology Information)」のGenbankに関するそれぞれの受入番号を示す(http://www.ncbi.nlm.nih.govを参照されたい)。
【0106】
別の好ましい態様において、該ペプチドを白血球、とりわけTリンパ球の染色に使用する。この使用は、CTL集団中に、ペプチドに向けられている特異的CTLが存在するかどうかが証明されるはずである場合にとりわけ有利である。さらに、該ペプチドを、腫瘍性疾患若しくは障害における治療の進行を決定するためのマーカーとして使用し得る。
【0107】
別の好ましい態様において、該ペプチドを抗体の製造に使用する。ポリクローナル抗体は、該ペプチドの注入を介する動物の免疫化、および免疫グロブリンのその後の精製により、標準的様式で得ることができる。モノクローナル抗体は、例えば、Methods Enzymol.(1986)、121、Hybridoma technology and monoclonal antibodiesに記述されるような標準的プロトコルに従って製造し得る。
【0108】
本発明は、さらなる一局面において、本発明の前記ペプチドの1種若しくはそれ以上を含有する医薬組成物に関する。本組成物は、皮下、皮内、筋肉内のような非経口投与若しくは経口投与に使用する。このため、ペプチドを、薬学的に許容される、好ましくは水性の担体に溶解若しくは懸濁する。加えて、組成物は、緩衝剤、結合剤、膨潤剤、希釈剤、着香料、滑沢剤などのような賦形剤を含有し得る。ペプチドはまた、サイトカインのような免疫刺激物質と一緒になっても投与し得る。こうした組成物中で使用し得る賦形剤の広範囲の一覧は、例えば、A.Kibbe、Handbook of Pharmaceutical Excipients、第3版、2000、American Pharmaceutical Association and pharmaceutical pressから採用し得る。組成物は、腫瘍性疾患の予防(prevention)、予防(prophylaxis)および/若しくは治療に使用し得る。
【0109】
配列番号1ないし配列番号388のいずれかを含んでなる本発明のペプチドの最低1種を含有する医薬組成物を、それぞれのペプチド若しくは抗原と関連づけられる腫瘍性疾患に罹っている患者に投与する。これにより、CTL特異的免疫応答を誘発し得る。
【0110】
本発明の別の局面において、2種若しくはそれ以上の本発明のペプチドの組合せを、直接の組合せで若しくは同一処置レジメン内でのいずれかでワクチンとして使用し得る。さらに、他のペプチド、例えばMHCクラスI特異的ペプチドとの組合せを使用し得る。当業者は、例えばin vitroでのT細胞の生成、ならびに、それらの効率および全体的な存在、ある種のペプチドに対するある種のT細胞の増殖(proliferation)、親和性および増殖(expansion)、ならびに例えばIFN−γ産生を分析することによるT細胞の機能性を試験することにより、免疫原性ペプチドの好ましい組合せを選択することが可能であろう(下の実施例もまた参照されたい)。通常、最も効率的なペプチドをその後、上述されたところの目的上、ワクチンとして組み合わせる。
【0111】
適するワクチンは、2、3、4、5、6、7、8若しくは10種の異なるペプチド、好ましくは4、5、6若しくは7種の異なるペプチド、および最も好ましくは6種の異なるペプチドを含有することができる。
【0112】
最後に、ワクチンは、処置されるべき患者が罹っている癌の特定の型、ならびに、疾患の状態、以前の処置レジメン、患者の免疫状態、およびもちろん該患者のHLAハプロタイプに依存し得る。
【0113】
Iiの80のN末端アミノ酸が、タンパク質をクラスIIプロセシング経路に向けるのに十分であることが示された(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221、Wang、R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354)。本発明の概念の例示的一証明を提供するため、発明者は、Iiの80のN末端アミノ酸および多様な悪性度と関連する例示的1抗原、サイクリンD1(CCND1)よりなる融合タンパク質を生成した。
【0114】
サイクリンD1は、サイクリン依存性キナーゼとの相互作用によりG1−S移行に関与する細胞周期調節物質である。さらに、サイクリンD1は癌原遺伝子であり、かつ、数種の腫瘍型で過剰発現されることが示された(Hedberg,Y.、Davoodi,E.、Roos,G.、Ljungberg,B.とLandberg,G.(1999)Int.J.Cancer 84、268−272、Vasef,M.A.、Brynes,R.K.、Sturm、M.、Bromley,C.とRobinson,R.A.(1999)Mod.Pathol.12、412−416、−Troussard,X.、Avet−Loiseau,J.、Macro,M.、Mellerin,M.P.、Malet,M.、Roussel,M.とSola,B.(2000)Hematol.J.1、181−185)一方、それは、肝および高位腹部大動脈平滑筋を除くいかなる特定の分布も伴わず、大きな一団の健康な器官および組織中で低レベルで発現される(Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827)。示差的質量分析のアプローチにおいて、発明者は、トランスフェクトしたおよびトランスフェクトしていない細胞からの精製したHLAペプチドの質量スペクトルを比較し、そして、これらのHLAクラスIIリガンドの免疫原性の特徴を証明するために、目的の生じるペプチドをin vitro CD4+ Tヘルパー細胞プライミング実験で使用した。
【0115】
腫瘍関連抗原のTヘルパー細胞エピトープの同定は、抗腫瘍免疫療法における重要な責務のままである。ここに、われわれは、腫瘍関連抗原の天然にプロセシングかつ提示されるMHCクラスIIリガンドを同定するための、MSによる示差的ペプチド分析から派生した、新しくかつ一般に応用可能な方法およびペプチドを報告する。このアプローチは、Ii鎖と目的のAgの間の融合タンパク質をコードするベクターでのAPCのトランスフェクション段階、HLAに結合したペプチドの溶出、および、トランスフェクトされない細胞との比較による、トランスフェクタントにより提示されるAg由来ペプチドのMS同定を初めて組合せる。さらに、われわれは、同定されたペプチドに対し誘導されたT細胞が、同族のAgを過剰発現するトランスフェクタントを特異的に認識することを示すことにより、該方法を検証し得た。同定されたペプチドはなお、in vivoでそれらの免疫原性について試験されなければならないが、われわれのアプローチは、天然にプロセシングされるMHCクラスIIリガンドの正確な特徴付けに至る。従って、発明者は、腫瘍関連抗原の合成の重なるペプチド、若しくはクラスIエプトープ予測に比較して精度の低いエピトープ予測により選択された広範なペプチドのいずれかを試験することを回避する。in vivoでのT細胞活性化を誘導することが不可能な潜在性T細胞エピトープの同定につながりうる困難なT細胞アッセイ(Anderton,S.M.、Viner,N.J.、Matharu,P.、Lowrey,P.A.とWraith,D.C.(2002)Nat.Immunol.3、175−181)と対照的に、該研究は、提示されることが見出されている数種のペプチドに集中し得る。さらに、この方法を使用すれば、該ペプチドが天然にプロセシングされることを証明するために組換えAgを産生するか若しくはAgを発現する腫瘍細胞株を有することが必要でない。
【0116】
発明者は、EBVで形質転換したB細胞のクラスIIプロセシング区画中に腫瘍関連抗原を向けるためにIiのN末端を使用した。これを達成するため、われわれは、いかなる抗原もIiとの融合タンパク質として発現し得かつウエスタンブロット分析によりトランスフェクトされた細胞中のタンパク質の発現レベルを測定するためにわれわれを補助する、融通のきくベクターを構築した。IiのN末端が、タンパク質の標的をクラスIIプロセシング区画中に定めるのに十分であることが既に示されている。しかし、今日まで、これは、融合タンパク質をコードするcDNAライブラリーを使用して未知のAgを同定するため(Wang,R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354)、若しくは既知のT細胞クローンの特異性を確認するため(Chaux,P.、Vantomme,V.、Stroobant,V.、Thielemans,K.、Corthals,J.、Luiten,R.、Eggermont,A.M.、Boon,T.とvan der,B.P.(1999)J.Exp.Med.189、767−778)に、卵アルブミンを使用したモデルでのみ記述された(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221)。われわれの知る限り、この方法は、既知の腫瘍関連抗原の天然に提示されたMHCクラスIIに結合したペプチドを同定するために以前に使用されていなかった。MALDI−MASによるトランスフェクトされたおよびトランスフェクトされていない細胞のクラスIIリガンドの示差的分析、ならびにESI−MSによる差次的に発現されたペプチドのさらなる特徴付けは、目的の抗原のクラスIIリガンドの簡単な同定方法をもたらす。ケラチン18融合タンパク質での細胞のトランスフェクションは、われわれの方法が目的の抗原に一般に応用可能であることを証明し、再度、われわれはまた、モデル導入遺伝子ケラチン18からのHLA−DRに提示されるペプチドを記述することも可能であった。
【0117】
発明者は、配列NPPSMVAAGSVVAAV(配列番号1)を有する免疫原性のHLA−DR4に提示されるサイクリンD1ペプチド抗原、ならびにトランスフェクトしたヒト腫瘍細胞株から溶出された337種の他のHLAクラスII関連ペプチドを同定した。サイクリンD1ペプチド、およびヒト腫瘍細胞株から同定した他のペプチドは、ヒト癌の処置のための治療的ワクチンの臨床開発のための可能な候補である(Weinschenk,T.、Gouttefangeas,C.、Schirle,M.、Obermayr,F.、Walter,S.、Schoor,O.、Kurek,R.、Loeser,W.、Bichler,K.H.、Wernet,D.ら(2002)Cancer Res.62、5818−5827)。
【0118】
本明細書に開示かつ記述されるところの本発明の特徴が、示されるところのそれぞれの組合せでのみならず、しかしまた、本発明の意図される範囲から離れることなく単独の様式でも使用し得ることが、理解されるべきである。
本発明は今や、以下の図面、配列表および実施例を参照してより詳細に記述されることができる。以下の実施例は、具体的説明の目的上のみ提供され、そして本発明を制限することを意図していない。
配列番号1ないし配列番号338は、本発明の、MHCクラスIIにより提示されるペプチドを含有するT細胞エピトープのペプチド配列を示す。
配列番号339ないし配列番号350は、実施例で使用されるところのプライマーの配列を示す。
【図面の簡単な説明】
【0119】
【図1】実験の経過中に作製した融合タンパク質の第一のウエスタンブロット分析を示す。
【図2】実験の経過中に作製した融合タンパク質の第二のウエスタンブロット分析を示す。
【図3】トランスフェクトされない細胞株およびAwells−Ii−サイクリンD1クローンのHPLCクロマトグラムを示す。
【図4】際立つ個々のシグナルを有する好ましい画分の質量分析を示す。
【図5】際立つ個々のシグナルを有する好ましい画分の質量分析を示す。
【図6】ペプチドNPPSMVAAGSVVAAV(配列番号1)(サイクリンD1198−212)の同定を示す。
【図7】サイクリンD1ペプチドによるT細胞の特異的刺激(上図)、および該ペプチドに応答してのT細胞のサイトカイン産生(下図)を示す。
【図8】HLA−DR4/サイクリンD1198−212に特異的なT細胞受容体でのCD4+ Tヘルパー細胞の結果として起こりうる非特異的活性化の陰性対照を示す。
【発明を実施するための形態】
【0120】
本開示および本実施例に際して当業者に明らかであろうように、本発明はまた、免疫原性のHLA−DR4に提示された、配列NPPSMVAAGSVVAAV(配列番号1)を有するサイクリンD1ペプチド抗原およびケラチン18に関する以下の実施例で特異的に使用されるところの腫瘍関連抗原の他のエピトープにも関する。該実施例は、本発明を完全に可能にするために本発明の腫瘍関連抗原の全エピトープ(すなわち配列番号1ないし配列番号338の配列のいずれかを含んでなる)に当業者により容易に適用され得る、一般的例および概要として見られるべきである。
【0121】
本出願を通じて使用される略語:
Ab:抗体
Ag:抗原
APC:抗原提示細胞
CD:表面抗原分類
cpm:1分あたりカウント
DC:樹状細胞
EBV:エプスタイン−バーウイルス
ESI:エレクトロスプレーイオン化
HLA:ヒト白血球抗原
HPLC:高速液体クロマトグラフィー
IFN:インターフェロン
Ii:インバリアント鎖(CD74)
IL:インターロイキン
MALDI:マトリックス支援レーザー脱離/イオン化
MHC:主要組織適合遺伝子複合体
MS:質量分析法
OD450:450nmの波長での光学密度
PBMC:末梢血単核細胞
PCR:ポリメラーゼ連鎖反応
PHA:フィトヘマグルチニン
SDS−PAGE:ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動
S.I.:刺激指数
TOF:飛行時間
【実施例】
【0122】
細胞および抗体
ヒトBリンパ芽球様細胞株Awells(IHW−No.9090;HLA−DRB1*0401、HLA−DRB4*0101)を、10%FCS(Pan、独国アイデンバッハ)を含有しかつ2mM L−グルタミン(BioWhittaker、ベルギー・ヴェルヴィエ)、100U/mlペニシリンおよび100μg/mlストレプトマイシン(BioWhittaker)を補充したRPMI 1640(C.C.Pro、独国ノイシュタット)培地中で維持した。トランスフェクトした細胞クローンに関しては0.8mg/mlのG418(PAA Laboratories、オーストリア・リンツ)を添加した。安定形質転換体をAwells細胞の電気穿孔(280V、975μF;Gene Pulser II、Biorad、独国ミュンヘン)により作製し、次いで制限希釈法を使用してクローン化した。抗体L243(抗HLA−DR)(Lampson,L.AとLevy,R.(1980)J.Immunol.125、293−299)およびW6/32(抗HLAクラスI)(Brodsky,F.M.とParham,P.(1982)J.Immunol.128、129−135)を、プロテインA−セファロースビーズ(Pharmacia、スウェーデン・ウプサラ)を使用してハイブリドーマ培養上清から精製した。Th細胞株は、10%ヒトAB血清(Pel−Freez Clinical Systems、LLC、米国ウィスコンシン州ミルウォーキー)を含有しかつ100U/mlペニシリンおよび100μg/mlストレプトマイシン、2mM L−グルタミンならびに50μM β−メルカプトエタノールを補充したIMDM(BioWhittaker)中で誘導かつ培養した。フローサイトメトリー分析で使用した抗体はPharMingen(米国カリフォルニア州サンディエゴ)からであった。
【0123】
プラスミドDNA構築物
Iiの80のN末端アミノ酸を含有するcDNA(NCBI、GenBank受入番号X00497)を、A.Melms(Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533)から得たベクターpBluescript II KS(+)41−1(Stratagene、独国ハイデルベルク)からPCR反応で増幅し、そして、5'プライマーATCGAAGCTTCCAAGATGCACAGGAGGAGAAGC(配列番号339)および3'プライマーATCGGGATCCTTTGTCCAGCCGGCCCTGCTG(配列番号340)を使用して、pcDNA3のHind IIIおよびBamH I部位にサブクローニングした(pcDNA3−Ii;Invitrogen、独国カールスルーエ)。目的の遺伝子は、それぞれ、ケラチン18(NCBI、GenBank受入番号X12881)について5'プライマーATCGGAATTCTGAGCTTCACCACTCGCTCC(配列番号341)および3'プライマーATCGGCGGCCGCTTAATGCCTCAGAACTTTGGT(配列番号342)、ならびにサイクリンD1(NCBI、GenBankX59798)について5'プライマーATCGGAATTCTGGAACACCAGCTCCTGTGC(配列番号343)および3'プライマーATCGGCGGCCGCTCAGATGTCCACGTCCCGCAC(配列番号344)を使用して、悪性腎組織からのcDNAからPCR反応で増幅した。得られたcDNAを、TOPO TAクローニング(Invitrogen、独国カールスルーエ)を使用してサブクローニングし、そして最後に、Ii配列とインフレームでpcDNA3−IiのEcoR 1およびNot I部位に挿入した。
【0124】
リアルタイム定量的PCR
細胞からのRNAを、製造元の推奨に従ってTRIzol試薬(Invitrogen、独国カールスルーエ)を使用して単離した。cDNAを1μgの全RNAから合成した。リアルタイム定量的PCR(qPCR)を、ABI PRISM 7000配列検出装置(Applied Biosystems、独国ダルムシュタット)を使用して実施した。SYBR Green PCRマスターミックス(Applied Biosystems)を、PCR増幅およびPCR産物のリアルタイム検出に使用した。プライマー配列は後に続くとおりである:18S rRNA、5'プライマーCGGCTACCACATCCAAGGAA(配列番号345)および3'プライマーGCTGGAATTACCGCGGCT(配列番号346);ケラチン18、5'プライマーGAGCCTGGAGACCGAGAAC(配列番号347)および3'プライマーTTGCGAAGATCTGAGCCC(配列番号348);サイクリンD1、5'プライマーCACGATTTCATTGAACACTTCC(配列番号349)および3'プライマーTGAACTTCACCATCTGTGGCAC(配列番号350)。PCR反応は300nMの各プライマーを含む20μl中で実施した(18Sリバースプライマー:50nMのみ)。全部のサンプルを二検体ずつ増幅した。多様な遺伝子についてのトランスフェクトした細胞と野性型細胞の間の発現の差違を、比較閾値周期(comparative threshold cycle)(CT)法(http://docs.appliedbiosystems.com/pebiodocs/04303859.pdfで読み出し可能)を使用する相対定量により、PCR増幅曲線から計算した。18SリボソームRNAを標準化のための参照遺伝子として選んだ。
【0125】
融合タンパク質の検出
融合タンパク質は、Iiのアミノ酸残基12−28に結合するmAb PIN.1(Stressgen,Biomol、独国ハンブルク)を使用するウエスタンブロット分析により検出した。簡潔には、細胞を記述された(Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625)とおり溶解し、ライセートをLaemmli添加液中で沸騰させ、12%SDS−PAGEで分離し、そしてニトロセルロースメンブレンに転写した。BSAでの飽和段階後に、メンブレンをmAb PIN.1(1μg/ml)とともに室温で1時間インキュベートした。タンパク質を、ペルオキシダーゼ結合ヒツジ抗マウスIgG(Amersham Pharmacia、独国フライブルク)を使用して可視化した。いくつかの場合には、トランスフェクトした細胞を、融合タンパク質のエンドソーム/リソソームターゲッティングを検討するために、10〜100μMのクロロキン(Sigma、独国シュタインハイム)の存在下で培養した。細胞をその後溶解し、そしてタンパク質を上述されたとおりウエスタンブロットにより検出した。
【0126】
Ii−ケラチン18もまた、免疫沈降、およびトリプシンでのその後のin situ消化、次いで質量分析により検出した。簡潔には、細胞を記述された(Hsieh,C.S.、deRoos,P.、Honey,K.、Beers,C.とRudensky,A.Y.(2002)J.Immunol.168、2618−2625)とおり溶解し、そして、mAb PIN.1およびプロテインA−セファロースビーズ(Pharmacia、スウェーデン・ウプサラ)とともにインキュベートした。セファロースペレットを洗浄し、Laemmli添加液中で沸騰させ、そして12%SDS−PAGE上で泳動した。タンパク質のスポットをゲルから切り出し、そして、本質的には記述された(Shevchenko,A.、Wilm,M.、Vorm,O.とMann,M.(1996)Anal.Chem.68、850−858)とおり、トリプシンでin situで消化した。その後分析は、MALDI−TOF質量分析計(Reflex III、Bruker Daltonik、独国ブレーメン)で実施した。タンパク質の同一性を、ハイブリッド四重極直交加速型飛行時間タンデム質量分析計(QStar Pulsar i Qqoa Tof;Applied Biosystems−MDS Sciex、独国ヴァイターシュタット)に記録したタンデムMSスペクトルにより確認した。
【0127】
MHCクラスIIに結合したペプチドの溶出
凍結細胞ペレット(3.5ないし5×1010細胞)を、以前に記述された(Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225)とおり処理し、そして、ペプチドを、標準的プロトコル(Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999)に従い、HLA−DR特異的mAb L243(Lampson,L.A.とLevy,R.(1980)J.Immunol.125、293−299)を使用して単離した。
【0128】
DR溶出したペプチドの分子分析
ペプチドを逆相高速液体クロマトグラフィー(HPLC、SMART装置、μRPC C2/C18 SC 2.1/10;Amersham Pharmacia Biotech、独国フライブルク)により分離し、そして、Bruker Reflex III質量分析計(Bruker Daltonik)を使用するMALDI−TOF質量分析(MS)により画分を分析した。差次的に提示されたペプチドを、記述された(Schirle,M.、Keilholz,W.、Weber,B.、Gouttefangeas,C.、Dumrese,T.、Becker,H.D.、Stevanovic,S.とRammensee,H.G.(2000)Eur.J.Immunol.30、2216−2225)ところのハイブリッド四重極直交加速型飛行時間タンデム質量分析計(Q−TOF;Micromass、英国マンチェスター)でのナノESI(エレクトロスプレーイオン化)タンデムMSによりさらに分析した。
【0129】
ペプチドの合成および分析
ペプチドは、Fmoc/tBuストラテジーに従って、自動ペプチド合成機EPS221(Abimed、独国ランゲンフェルト)で合成した。TFA/フェノール/エタンジチオール/チオアニソール/水(容量で90/3.75/1.25/2.5/2.5)での1時間若しくは3時間(アルギニン含有ペプチド)の処理による樹脂の除去後に、ペプチドをメチル−ターシャリブチルエーテル(methyl−tert. butyl ether)から沈殿し、メチル−ターシャリブチルエーテルで1回およびジエチルエーテルで2回洗浄し、そして凍結乾燥前に水に再懸濁した。合成産物をHPLC(Varian star、Zinsser analytics、独国ミュンヘン)およびMALDI−TOF質量分析(future、GSG、独国ブルフザール)により分析した。80%未満の純度のペプチドを分取用HPLC(preparative HPLC)により精製した。
【0130】
単球由来樹状細胞
末梢血単核細胞(PBMC)を、古典的手順に従い、HLA−DRB1*0408、HLA−DRB1*1101、HLA−DRB3*0202、HLA−DRB4*01陽性ドナーから調製した。樹状細胞(DC)は、使用した培地が血清を含まないX−VIVO 15(BioWhittaker)であったことを除き、以前に記述された(Bender,A.、Sapp,M.、Schuler,G.、Steinman,R.M.とBhardwaj,N.(1996)J.Immunol.Methods.196、121−135)とおり、GM−CSFおよびIL−4の存在下で6日間培養したプラスチック付着性PBMCから得た。第6日に、未熟DCを、CD1a、CD11c、CD14、CD40、CD83、CD86、ならびにHLA−DRの細胞表面発現について、CELLQuestソフトウェアを伴うFACScalibur装置(Becton Dickinson、カリフォルニア州マウンテンビュー)でのフローサイトメトリーにより分析した。DCをその後、50μg/mlポリイノシン酸−ポリシチジル酸(ポリI/C、Amersham Pharmacia、スウェーデン・ウプサラ)および10ng/ml TNF−α(PharMingen)の存在下で更に2日間成熟させ、そして、CD14、CD80、CD83およびCD86の細胞表面発現についてフローサイトメトリーにより再度分析した。成熟DCは、CD80、CD83およびCD86分子の明瞭な上方制御を示した。
【0131】
ペプチド特異的Tヘルパー細胞の生成
3×105の成熟DCに、10μMのペプチドNPPSMVAAGSVVAAVを24ウェルプレート中で2時間添加し、そして十分に洗浄した。その後、4×106の新鮮自己PBMCを、Th−1発生に有利であるために10ng/mlのIL−12p70の存在下でDCに添加した。PBMCを、10U/mlのIL−2および5ng/mlのIL−7の存在下で、ペプチドを負荷した照射した自己PBMCで毎週再刺激した。3および5回の再刺激後にT細胞を合わせ、そしてペプチドの存在下で自己PBMCに対し試験した。Tヘルパー細胞株をその後、1μg/mlのPHA、25〜50U/mlのIL−2および5ng/mlのIL−7の存在下に、照射した同種PBMCで1〜2週ごとに増幅し、そしてその後、トランスフェクトした細胞株の認識について試験した。3ないし4週ごとに、Tヘルパー細胞株を、10μMのペプチド、10U/mlのIL−2および5ng/mlのIL−7の存在下に、照射した自己PBMCで再刺激した。
【0132】
Tヘルパー細胞株の機能アッセイおよび特徴付け
Tヘルパー細胞活性化は、チミジン取り込みならびにサイトカイン分泌により推定されるところの細胞増殖により試験した。簡潔には、2×105細胞を、10μMのペプチド若しくは3μg/mlのPHAの存在若しくは非存在下に3検体ずつ96ウェルプレート中で2×105の照射した自己PBMCとともにインキュベートした。24時間後に2部分の50μlの上清を収集かつ凍結し、そして50μlの新鮮培地を細胞に添加した。54時間後に、50μlのトリチウム化チミジン含有培地(0.074MBq/ウェル、Hartmann Analytic、独国ブラウンシュヴァイク)を添加し、そして、チミジンの取り込みを、シンチレーションカウンター(Microbeta、Wallac、独国フライブルク)を使用して72時間に測定した。細胞増殖は、比:(刺激したT細胞のcpmの平均)/(未刺激のT細胞のcpmの平均)に対応する刺激指数(S.I.)として表す。IL−2分泌は、IL−2依存性CTLL−2細胞株を使用して測定した。簡潔には、104細胞を上清の存在下に20〜24時間インキュベートした。その後、チミジン含有培地(0.055MBq/ウェル)を追加の7〜8時間添加し、そしてチミジンの取り込みを上述したとおり測定した。結果はまたS.I.として表す。
【0133】
IFN−γ、IL−4およびIL−6分泌は、PharMingenからの抗体対およびペルオキシダーゼ結合ストレプトアビジンを使用しかつ製造元の推奨に従ったサンドイッチELISAにより測定した。われわれは基質としてSupersensitive TMB(Sigma、独国ドライゼンホーフェン)を使用し、また、反応は2M H2SO4溶液を使用して停止した。OD450をその後測定し、そして結果は標準に従いpg/mlで表した。
【0134】
ペプチド特異的Tヘルパー細胞株によるサイクリンD1でトランスフェクトした細胞の認識
トランスフェクタントに対するT細胞の検出可能な非自己反応の非存在を、多様な細胞数の照射したトランスフェクタントの存在下で該Tヘルパー株を生成させるのに使用したドナーからの新鮮PBMCを共培養することにより立証した。細胞増殖ならびにIL−2、IL−4、IL−6およびIFN−γ分泌を上述したとおり測定し、そして、さらなる実験で使用されるべきエフェクター/標的比をかように決定した。
【0135】
サイクリンD1由来の天然にプロセシングされたペプチドの認識は、サイクリンD1若しくは陰性対照としてのケラチン18のいずれかをコードするプラスミドでトランスフェクトした照射したAwells(4×104細胞)の存在下で、かつ、上で決定された細胞比に従って、ペプチド特異的Tヘルパー細胞株(2×105細胞)を共培養することにより試験した。10μMのペプチド若しくは3μg/mlのPHAの存在下の照射した自己PBMCが陽性対照としてはたらいた。細胞増殖ならびにIL−2およびIFN−γ分泌を上述したとおり測定した。いくつかの実験では、細胞を20μg/mlの精製したL243抗体の存在下で培養した。
【0136】
結果
融合タンパク質を発現する細胞クローンの生成
われわれは、挿入物の3'端の後に一般的クローニング部位(GCS)が続いたような方法で、ベクターpcDNA3中にIiの80のN末端アミノ酸をコードするcDNAをクローン化した。これは、われわれに、Iiおよび目的の遺伝子の融合タンパク質を発現するための融通の利くベクターを与えた。Iiとインフレームで、われわれは、サイクリンD1のcDNA、ならびに対照としてのケラチン18をクローン化した。
【0137】
Awells細胞株を、電気穿孔法を使用して該2種の融合タンパク質をコードするベクターで安定にトランスフェクトした。その後、単細胞クローンを生成し、そして、それらの抗原発現についてmRNAおよびタンパク質レベルで試験した。野性型(トランスフェクトされていない細胞株)に比較して、最良のIi−ケラチン18クローンは、リアルタイム定量的PCR分析により測定されたとおり、5,700倍より多いケラチン18を発現し、また、最良のIi−サイクリンD1クローンは1,200倍より多いサイクリンD1を発現した。該データは18SリボソームRNAで標準化した。
【0138】
ウエスタンブロット分析において、PIN.1抗体を使用してIi融合タンパク質を検出した。Iiの2種のアイソフォームに対応する2本のバンドのみが検出された野性型(Warmerdam,P.A.、Long,E.O.とRoche,P.A.(1996)J.Cell.Biol.133、281−291)と比較して、期待された分子量を伴うIi−ケラチン18若しくはIi−サイクリンD1融合タンパク質を表す2個の付加的なバンドを各クローンで観察し得た(図1)。クローンAwells−Ii−ケラチン18の場合には、融合タンパク質が、免疫沈降、次いでトリプシンでのin situ消化およびその後のMS/MS分析を介してもまた同定された。
【0139】
該クローンはまた、トランスフェクションおよびクローニング手順がそれを妨害したかどうかを決定するため、フローサイトメトリーによりそれらのHLAクラスIおよびクラスII細胞表面発現レベルについても試験した。双方のクローンは、トランスフェクトされていない細胞株と比較して、HLAクラスIおよびクラスII分子の正常な発現レベルを示した。
【0140】
融合タンパク質のエンドソーム/リソソームターゲティング
IiのN末端が、タンパク質の標的をMHCクラスII経路に向けるのに十分であることが既に記述されている(Sanderson,S.、Frauwirth,K.とShastri,N.(1995)Proc.Natl.Acad.Sci.U.S.A 92、7217−7221、Wang,R.F.、Wang,X.、Atwood,A.C.、Topalian,S.L.とRosenberg,S.A.(1999)Science 284、1351−1354、Malcherek,G.、Wirblich,C.、Willcox,N.、Rammensee,H.G.、Trowsdale,J.とMelms,A.(1998)Eur.J.Immunol.28、1524−1533)。われわれの構築物がAgプロセシングのその経路に実際に従ったかどうかを試験するため、われわれは、トランスフェクタントを、増大する量のクロロキン(リソソームのpHを上げることによりタンパク質のリソソーム分解を阻害する細胞傷害性の薬物)(Kaplan,J.とKeogh,E.A.(1981)Cell 24、925−932、Tietze,C.、Schlesinger,P.とStahl,P.(1982)J.Cell.Biol.92、417−424、Seglen,P.O.(1983)Methods Enzymol.96:737−64.、737−764)とともに4時間インキュベートした。細胞をその後溶解し、そして融合タンパク質をウエスタンブロットにより検出した。図2は、タンパク質バンドが、クロロキンの量を増大させるにつれてますます強くなることを示し、融合タンパク質の量がクロロキン濃度とともに増大することを示し、そして従って融合タンパク質がMHCクラスIIのタンパク質分解経路に従うことを提供する。
【0141】
HLA−DRに結合したペプチドの示差的質量分析
各クローンおよびトランスフェクトされていない細胞株からの3.5ないし5×1010細胞を増殖させ、そして、HLA−DRに結合したペプチドを、以前に記述された(Seeger,F.H.、Schirle,M.、Keilholz,W.、Rammensee,H.G.とStevanovic,S.(1999)Immunogenetics 49、996−999)とおりにHPLCを介して単離かつ分離した。トランスフェクトされていない細胞株およびAwells−Ii−サイクリンD1クローンのHPLCクロマトグラムを比較した(図3)。HLA−DRで提示されるペプチドレパートリーの小さなほとんど定量的な差違は、図3に示されるところのわずかに異なるUV痕跡になった。われわれの実験から期待されたとおり、明瞭なUVシグナルは、トランスフェクタントにより独占的に提示されるペプチドに割り当て得なかった。Awellsとトランスフェクトした株の間のHLA−DR拘束性ペプチド提示のただわずかな差違は、HPLC画分の大部分が同一のパターンを含有したMALDI−TOF分析によってもまた可視的になった。図4は、ケラチン18でトランスフェクトした株から溶出されたペプチド混合物中にのみ存在しかつケラチン18由来ペプチドを表した顕著な個別のシグナル(m/z:1732.96)を含む唯一の画分の質量分析を示す。図5では、1370.1のm/zシグナルは、サイクリンD1トランスフェクタントからの独占的に提示される1ペプチドを示す。双方のペプチドをナノフローESI MS/MS(タンデムMS)によりより詳細に分析した。発明者は、それぞれ2種のトランスフェクトされた融合タンパク質由来のペプチドNPPSMVAAGSVVAAV(配列番号1)(サイクリンD1198−212)(図6)およびSHYFKIIEDLRAQI(配列番号2)(ケラチン18126−139)を同定することが可能であった。該配列を、対応する合成ペプチドの質量スペクトルにより確認した。
【0142】
ペプチド特異的Tヘルパー細胞株の生成および特徴付け
新規のHLAクラスII由来の腫瘍関連および推定で免疫原性のペプチドの同定のための概念の証明を確立するために、同定されたサイクリンD1ペプチドに特異的なT細胞を、HLA−DR4+樹状細胞に負荷した対応する合成ペプチドでのin vitro刺激により誘導した。それぞれ第3および5回のin vitro(再)刺激後に、Tヘルパー細胞株の特異性を試験した。T細胞を、図7(上図)に示されるとおりサイクリンD1ペプチドにより特異的に刺激した。T細胞は、サイクリンD1ペプチドを負荷した自己PBMCに応答して増殖した(S.I.=5.4)。陽性対照として、PHAは強いT細胞増殖を誘導した(S.I.=330)。発明者は次に、どの型のTヘルパー細胞(Th1対Th2)がペプチドに応答して刺激されたかを、サイトカインプロファイルを検査することにより分析した。図7(下図)に示されるとおり、T細胞はペプチドに応答して低い程度までとは言えIL−2を産生した一方、それらはPHA刺激に対しなお感受性であった。対照的に、ペプチドで誘導したT細胞刺激は、強いIFN−γ分泌(3250pg/ml、図7C)をもたらしたが、しかしIL−4若しくはIL−6分泌をもたらさなかった。とは言え、T細胞はなお、PHAで誘導されるサイトカイン分泌に対しそれぞれ高度におよび中程度に感受性であった。HLA−DR4/サイクリンD1198−212に特異的なT細胞受容体でのCD4+ Tヘルパー細胞の最終的な非特異的活性化についての陰性対照としての無関係のペプチドでの刺激を表8に提示する。結論として、確立されたCD4+ Tヘルパー細胞株は、サイクリンD1198−212ペプチドに特異的でありかつTh1型のものである。この型は、腫瘍細胞の排除における特異的CD8+キラーT細胞の援助においてとりわけ重要である。
【0143】
ペプチド特異的Tヘルパー細胞株は、サイクリンD1でトランスフェクトした細胞もまた認識する
Awells細胞およびT細胞株は完全にHLAが適合しているわけではないため、われわれは最初に、双方を共培養することによりいずれかの非自己反応が生じ得るかどうかを試験した。簡潔には、多様な細胞数の照射したAwells−Ii−ケラチン18若しくはAwells−Ii−サイクリンD1トランスフェクタントを、T細胞ドナーからの固定した数のPBMCの存在下で共培養し、そして、細胞増殖ならびにIL−2、IL−4、IL−6およびIFN−γ分泌を測定した。中程度のT細胞増殖が高細胞数の双方のトランスフェクタントにより誘導されたが、しかし、サイトカイン分泌は観察されなかった(データは示されない)。従って、われわれは、サイトカイン分泌の非存在下でわずかなT細胞増殖のみが観察された5/1という(エフェクターT細胞)/(標的細胞)比を使用することを決定した。結果として、サイトカイン分泌をもたらすT細胞活性化は、トランスフェクタントにより提示される同族の抗原によってのみ特異的に誘導され得た。
【0144】
サイクリンD1ペプチドに特異的なTヘルパー細胞株は、サイクリンD1タンパク質を過剰発現しそして該サイクリンD1ペプチドを天然にプロセシングしかつHLA−DR分子と共同して提示する、トランスフェクトされた細胞を認識することが可能であった。図8に示されるとおり、照射したAwells−Ii−サイクリンD1トランスフェクタントは、IL−2分泌により観察されたとおり、Tヘルパー細胞株を特異的に活性化することが可能であった(S.I.=4.0)。対照的に、Awells−Ii−ケラチン18トランスフェクタント(T細胞刺激の陰性対照として使用され、かつ、HLA−DRと共同して無関係のケラチン18ペプチド126−139を提示することが知られている)はT細胞活性化を誘導せず、ペプチド特異的Tヘルパー細胞株が同族の抗原を特異的に認識することを示した。さらに、これらの結果は、本発明で使用されるサイクリンD1ペプチドが、T細胞エピトープを含有する天然にプロセシングされたペプチドであることを証明する。この活性化は、HLA−DR特異的なブロッキングL243Abの存在により阻害され得た(71.2%)。
【0145】
クラスII分子およびペプチド抗原サイクリンD1198−212を使用する活性化型細胞傷害性リンパ球(CTL)の製造ならびにそれらの投与
活性化型細胞傷害性Tリンパ球(CTL)は、HLA−DRクラスII分子、およびサイクリンD1198−212(配列番号1)からのペプチドを使用して製造する。
一般に、PCT特許出願WO 93/17095号に記述される方法を使用してCTLを製造する。Awells細胞を使用してペプチド抗原をCTLに提示する。HLA−DR分子はAwells細胞により発現される。試験されるべきペプチドを、Applied Biosystemsの合成機、ABI 431A(米国カリフォルニア州フォスターシティ)で合成し、そしてその後HPLCにより精製する。WO 93/17095号に詳細に記述されるとおり、特異的細胞傷害性T細胞の生成のためin vitro条件を最適化するために、刺激体細胞の培養物を適切な培地中で維持する。刺激体細胞は、好ましくは無血清培地(例えばExcell 400)中で維持されるAwells細胞である。
【0146】
活性化されるべき細胞、例えば前駆体CD4+細胞との刺激体細胞のインキュベーション前に、ある量の抗原ペプチドを十分な量の刺激体細胞培養物に添加し、刺激体細胞の表面上で発現されるべきヒトクラスII分子に負荷されたようにする。十分な量のペプチドは、約200、および好ましくは200若しくはそれ以上のペプチドを負荷したヒトクラスIIMHC分子が、各刺激体細胞の表面上で発現されることを可能にする量である。刺激体細胞は、典型的に、20pg/ml超のペプチドとともにインキュベートする。
【0147】
休止期若しくは前駆体CD4+細胞をその後、適切な刺激体細胞を含む培養物中で、CD4+細胞を活性化するのに十分な時間インキュベートする。CD4+細胞はかように抗原特異的様式で活性化される。刺激体細胞に対する休止期若しくは前駆体CD4+(エフェクター)細胞の比は個体ごとに異なることがあり、また、個体のリンパ球の培養条件への従順さのような変数にさらに依存しうる。リンパ球:刺激体細胞(Awells細胞)の比は、典型的に約30:1ないし300:1の範囲にある。例えば、3×101のヒトPBLおよび1×106の生存Awells細胞を混合し、そして20mlのRPMI 1640培地中で維持する。
【0148】
エフェクター/刺激体の培養物は、治療上使用可能な若しくは有効な数のCD4+細胞を刺激するのに必要であるだけの時間維持する。最適な時間は典型的に約1と5日の間であり、「プラトー」すなわち「最大の」特異的CD4+活性化レベルは一般に5日の培養後に観察される。CD4+細胞のin vitro活性化は、典型的に、細胞株のトランスフェクション後短時間内に検出される。CD4+細胞を活性化することが可能な、トランスフェクトされた細胞株での一過性発現は、トランスフェクション48時間以内に検出可能である。これは、ヒトクラスIIMHC分子を発現する形質転換細胞の安定若しくは一過性培養物のいずれも、CD4+細胞の活性化において有効であることを明確に示す。
【0149】
活性化型CD4+細胞は、それらの適切な相補リガンドに結合させるため、刺激体(Awells)細胞、刺激体細胞に負荷したペプチド、またはCD4+細胞(若しくはそのセグメント)に特異的なモノクローナル抗体を使用して、刺激体細胞から効果的に単離しうる。抗体標識した分子をその後、免疫沈降若しくはイムノアッセイ法を介して、刺激体とエフェクター細胞の混合状態から抽出する。
【0150】
活性化型CD4+細胞の有効な細胞傷害性の量は、in vitroとin vivoの使用の間で、ならびにこれらのキラー細胞の最終的標的である細胞の量および型で変動し得、約1×106と1×1012の間の活性化型CTLを成人のヒトに使用する。
活性化型CD4+細胞は、処置されている個体への該CD4+細胞の投与前に、Awells細胞培養物から収集する。
Honsikへの米国特許第4,844,893号明細書およびRosenbergへの米国特許第4,690,915号明細書に例示されるもののような細胞成分の再導入方法を使用する。例えば、静脈内注入を介する活性化型CD8細胞の投与が適切である。
【0151】
表1:本発明で同定されたところの腫瘍関連Tヘルパー細胞ペプチドエピトープ
【表1−1】
【表1−2】
【表1−3】
【表1−4】
【表1−5】
【表1−6】
【表1−7】
【表1−8】
【表1−9】
【表1−10】
【表1−11】
【表1−12】
【表1−13】
【表1−14】
【表1−15】
【特許請求の範囲】
【請求項1】
配列番号273に記載の連続したアミノ酸配列を含む、15〜30アミノ酸長の腫瘍関連ペプチド。
【請求項2】
ペプチドが、配列番号273のアミノ酸配列からなる、請求項1に記載の腫瘍関連ペプチド。
【請求項3】
ヒト主要組織適合抗原複合体(MHC)クラスIIの分子のHLA−DR4に結合する能力を有する、請求項1または2に記載の腫瘍関連ペプチド。
【請求項4】
ペプチドがHLA−DRに結合する場合に、結合した該ペプチドが、配列番号273に記載のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識する細胞傷害性Tリンパ球(CTL)の産生を誘導可能である、請求項3に記載の腫瘍関連ペプチド。
【請求項5】
該ペプチドが非ペプチド結合を包含する、請求項1〜4のいずれかに記載の腫瘍関連ペプチド。
【請求項6】
該ペプチドが、HLA−DR抗原関連インバリアント鎖(Ii)のN末端アミノ酸を含んでなる融合タンパク質である、請求項1〜5のいずれかに記載の腫瘍関連ペプチド。
【請求項7】
請求項1〜6のいずれか1つに記載ペプチドをコードする核酸。
【請求項8】
DNA、cDNA、PNA、CNA、RNA若しくはそれらの組合せである、請求項7に記載の核酸。
【請求項9】
請求項7または8に記載の核酸を発現することが可能な発現ベクター。
【請求項10】
請求項7または8に記載の核酸または請求項9に記載の発現ベクターを含んでなる宿主細胞。
【請求項11】
組換えAwells細胞である、請求項10に記載の宿主細胞。
【請求項12】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドの製造方法であって、請求項11に記載の宿主細胞を培養すること、および該ペプチドを宿主細胞若しくはその培地から単離することを含んでなる、製造方法。
【請求項13】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチド、および薬学的に許容される担体を含んでなる医薬組成物。
【請求項14】
医薬品での使用のための、請求項1〜6のいずれか1つに記載の腫瘍関連ペプチド。
【請求項15】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドを含んでなる癌ワクチン。
【請求項16】
請求項1〜4のいずれかに示されるところのアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、有効量患者に投与されることにより抗標的細胞免疫応答を誘導する請求項1〜6のいずれか1つに記載のペプチドを含む、標的細胞の死滅剤。
【請求項17】
請求項1〜4のいずれかに示されるところのアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤の製造における、請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドの使用。
【請求項18】
活性化型細胞傷害性Tリンパ球(CTL)のin vitroの製造方法であって、前記CTLを抗原特異的様式で活性化するのに十分な時間、適する抗原提示細胞の表面上に発現された抗原を負荷したヒトクラスII MHC分子とCTLをin vitroで接触させることを含んでなり、該抗原が請求項1〜6のいずれか1つに記載のペプチドである、製造方法。
【請求項19】
該抗原が、十分な量の該抗原を抗原提示細胞と接触させることにより、適する抗原提示細胞の表面上で発現されたクラスII MHC分子に負荷される、請求項18に記載の方法。
【請求項20】
抗原提示細胞が請求項9に記載の発現ベクターを含んでなる、請求項18に記載の方法。
【請求項21】
クラスII MHC分子がHLA−DR4である、請求項18〜20のいずれか1つに記載の方法。
【請求項22】
請求項1〜4のいずれか1つに示されるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を選択的に認識する、請求項18〜21のいずれか1つに記載の方法により製造された活性化型細胞傷害性Tリンパ球(CTL)。
【請求項23】
請求項1〜4のいずれか1つに示されるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するT細胞受容体(TCR)であって、該TCRが、請求項21若しくは22の細胞傷害性Tリンパ球(CTL)から得られるか、または該TCRと機能上同等な分子である、TCR。
【請求項24】
請求項22または23に記載のT細胞受容体(TCR)をコードする核酸。
【請求項25】
請求項22または23に記載のT細胞受容体(TCR)を発現することが可能な発現ベクター。
【請求項26】
請求項1〜4のいずれか1つに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、有効量患者に投与される請求項22若しくは23に定義されるところの細胞傷害性Tリンパ球(CTL)を含む、標的細胞の死滅剤。
【請求項27】
請求項1〜4のいずれかに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、請求項22若しくは23に定義されるところのT細胞受容体(TCR)または当該TCRと機能上同等な分子をコードする核酸が導入された、患者から得られた細胞傷害性Tリンパ球(CTL)を含む、標的細胞の死滅剤。
【請求項28】
標的細胞が癌細胞である、請求項16、26および27のいずれか1つに記載の患者における標的細胞の死滅剤。
【請求項29】
癌が、請求項1〜4のいずれかに記載のアミノ酸配列を含んでなるポリペプチドを異常に発現する腎癌である、請求項28に記載の患者における標的細胞の死滅剤。
【請求項30】
請求項1〜4のいずれか1つに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤の製造における、請求項22または23に定義されるところの細胞傷害性Tリンパ球(CTL)の使用。
【請求項1】
配列番号273に記載の連続したアミノ酸配列を含む、15〜30アミノ酸長の腫瘍関連ペプチド。
【請求項2】
ペプチドが、配列番号273のアミノ酸配列からなる、請求項1に記載の腫瘍関連ペプチド。
【請求項3】
ヒト主要組織適合抗原複合体(MHC)クラスIIの分子のHLA−DR4に結合する能力を有する、請求項1または2に記載の腫瘍関連ペプチド。
【請求項4】
ペプチドがHLA−DRに結合する場合に、結合した該ペプチドが、配列番号273に記載のアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識する細胞傷害性Tリンパ球(CTL)の産生を誘導可能である、請求項3に記載の腫瘍関連ペプチド。
【請求項5】
該ペプチドが非ペプチド結合を包含する、請求項1〜4のいずれかに記載の腫瘍関連ペプチド。
【請求項6】
該ペプチドが、HLA−DR抗原関連インバリアント鎖(Ii)のN末端アミノ酸を含んでなる融合タンパク質である、請求項1〜5のいずれかに記載の腫瘍関連ペプチド。
【請求項7】
請求項1〜6のいずれか1つに記載ペプチドをコードする核酸。
【請求項8】
DNA、cDNA、PNA、CNA、RNA若しくはそれらの組合せである、請求項7に記載の核酸。
【請求項9】
請求項7または8に記載の核酸を発現することが可能な発現ベクター。
【請求項10】
請求項7または8に記載の核酸または請求項9に記載の発現ベクターを含んでなる宿主細胞。
【請求項11】
組換えAwells細胞である、請求項10に記載の宿主細胞。
【請求項12】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドの製造方法であって、請求項11に記載の宿主細胞を培養すること、および該ペプチドを宿主細胞若しくはその培地から単離することを含んでなる、製造方法。
【請求項13】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチド、および薬学的に許容される担体を含んでなる医薬組成物。
【請求項14】
医薬品での使用のための、請求項1〜6のいずれか1つに記載の腫瘍関連ペプチド。
【請求項15】
請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドを含んでなる癌ワクチン。
【請求項16】
請求項1〜4のいずれかに示されるところのアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、有効量患者に投与されることにより抗標的細胞免疫応答を誘導する請求項1〜6のいずれか1つに記載のペプチドを含む、標的細胞の死滅剤。
【請求項17】
請求項1〜4のいずれかに示されるところのアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤の製造における、請求項1〜6のいずれか1つに記載の腫瘍関連ペプチドの使用。
【請求項18】
活性化型細胞傷害性Tリンパ球(CTL)のin vitroの製造方法であって、前記CTLを抗原特異的様式で活性化するのに十分な時間、適する抗原提示細胞の表面上に発現された抗原を負荷したヒトクラスII MHC分子とCTLをin vitroで接触させることを含んでなり、該抗原が請求項1〜6のいずれか1つに記載のペプチドである、製造方法。
【請求項19】
該抗原が、十分な量の該抗原を抗原提示細胞と接触させることにより、適する抗原提示細胞の表面上で発現されたクラスII MHC分子に負荷される、請求項18に記載の方法。
【請求項20】
抗原提示細胞が請求項9に記載の発現ベクターを含んでなる、請求項18に記載の方法。
【請求項21】
クラスII MHC分子がHLA−DR4である、請求項18〜20のいずれか1つに記載の方法。
【請求項22】
請求項1〜4のいずれか1つに示されるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を選択的に認識する、請求項18〜21のいずれか1つに記載の方法により製造された活性化型細胞傷害性Tリンパ球(CTL)。
【請求項23】
請求項1〜4のいずれか1つに示されるアミノ酸配列を含んでなるポリペプチドを異常に発現する細胞を認識するT細胞受容体(TCR)であって、該TCRが、請求項21若しくは22の細胞傷害性Tリンパ球(CTL)から得られるか、または該TCRと機能上同等な分子である、TCR。
【請求項24】
請求項22または23に記載のT細胞受容体(TCR)をコードする核酸。
【請求項25】
請求項22または23に記載のT細胞受容体(TCR)を発現することが可能な発現ベクター。
【請求項26】
請求項1〜4のいずれか1つに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、有効量患者に投与される請求項22若しくは23に定義されるところの細胞傷害性Tリンパ球(CTL)を含む、標的細胞の死滅剤。
【請求項27】
請求項1〜4のいずれかに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤であって、請求項22若しくは23に定義されるところのT細胞受容体(TCR)または当該TCRと機能上同等な分子をコードする核酸が導入された、患者から得られた細胞傷害性Tリンパ球(CTL)を含む、標的細胞の死滅剤。
【請求項28】
標的細胞が癌細胞である、請求項16、26および27のいずれか1つに記載の患者における標的細胞の死滅剤。
【請求項29】
癌が、請求項1〜4のいずれかに記載のアミノ酸配列を含んでなるポリペプチドを異常に発現する腎癌である、請求項28に記載の患者における標的細胞の死滅剤。
【請求項30】
請求項1〜4のいずれか1つに記載のアミノ酸配列を含んでなるポリペプチドを患者中で異常に発現している標的細胞の死滅剤の製造における、請求項22または23に定義されるところの細胞傷害性Tリンパ球(CTL)の使用。
【図3】

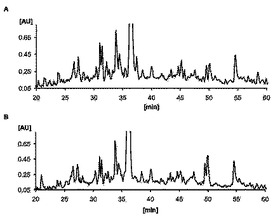
【図4】
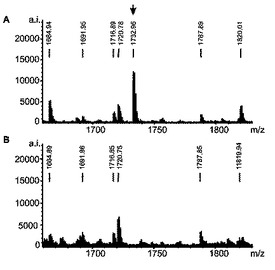
【図5】
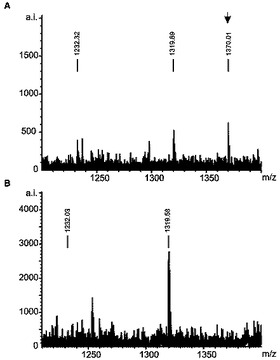
【図6】
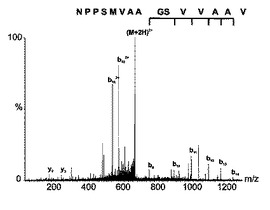
【図7】
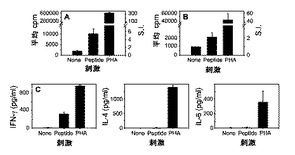
【図8】
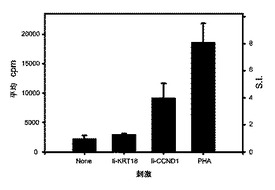
【図1】
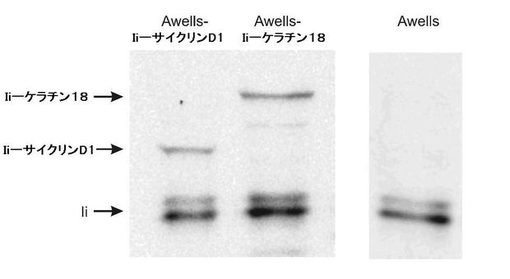
【図2】
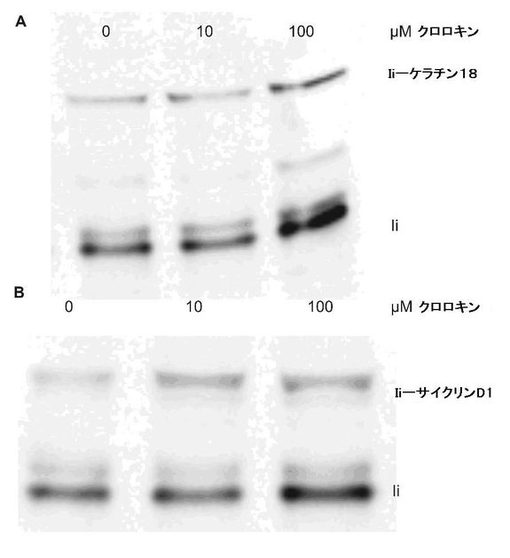
【図4】
【図5】
【図6】
【図7】
【図8】
【図1】
【図2】
【公開番号】特開2011−200231(P2011−200231A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−94758(P2011−94758)
【出願日】平成23年4月21日(2011.4.21)
【分割の表示】特願2007−533893(P2007−533893)の分割
【原出願日】平成17年9月9日(2005.9.9)
【出願人】(506258073)イマティクス バイオテクノロジーズ ゲーエムベーハー (24)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成23年4月21日(2011.4.21)
【分割の表示】特願2007−533893(P2007−533893)の分割
【原出願日】平成17年9月9日(2005.9.9)
【出願人】(506258073)イマティクス バイオテクノロジーズ ゲーエムベーハー (24)
【Fターム(参考)】
[ Back to top ]