
ヒト酸性繊維芽細胞増殖因子の改変ペプチド
【課題】ヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドを提供する。
【解決手段】ヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドであって、該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含み、相対的に高い安定性を持つ。更に、該単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【解決手段】ヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドであって、該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含み、相対的に高い安定性を持つ。更に、該単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、良好な安定性を有する酸性繊維芽細胞増殖因子の改変ペプチドに関する。
【背景技術】
【0002】
(本発明の背景)
イン・ビトロで様々な細胞型の増殖および分化に影響を及ぼす酸性繊維芽細胞増殖因子(aFGF)は、全脳および視床下部などの神経組織由来の一本鎖タンパク質として最初に単離された。aFGFは、ヘパリン依存性の有糸分裂促進物質であり、4つ全ての型の既知FGFレセプターおよびそのスプライス形態に強固に結合することができる。それは、運動機能および感覚機能と関連のあるニューロンの特定サブセット内に局在し、成人の脳から精製され得る。精製aFGFは、神経芽細胞に対する有糸分裂促進物質であり、脊髄ニューロンから伸張する軸索を促進する。
【発明の概要】
【発明が解決しようとする課題】
【0003】
ヒトaFGFのネイティブペプチドはヒトの脳から単離され、154個のアミノ酸から構成される。しかし、ネイティブヒトaFGFのN末端の19個のアミノ酸は、ヒトインターロイキン-1(IL-1)と相同であると同定された。ヒトaFGFとIL-1と類似したポリペプチドのドメインは、例えば、マクロファージの活性化および調節される細胞増殖の停止などと同一の内生免疫応答を引き起こし得る(G. Venkataraman et al., P.N.A.S., 96:3658-63, 1999)。さらに、炎症誘導性サイトカインIL-1およびFGF-1(aFGF)/FGF-2(bFGF)は、同じ構造スキャッホルドを共有し、チロシンキナーゼドメインの同じ受容体結合部位に競合する(A. J. Minter et al., J. Cell Physil., 167:229-37, 1996)。
【課題を解決するための手段】
【0004】
本発明の要旨
本発明は、aFGF135と称するヒト酸性繊維芽細胞増殖因子(aFGF)改変ペプチドを提供するものであって、ここで該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含んでいる。特に、ペプチドaFGF135は、比較的高い安定性を持っており、また既知のaFGFペプチドよりも優れている配列番号:1のアミノ酸配列を含む。
【発明の効果】
【0005】
本発明は、本発明のペプチドaFGF135および医薬上許容し得る担体を含む医薬組成物をさらに提供する。
【0006】
図面の簡単な説明
前記した発明の要旨、ならびに本発明の下記詳細な説明は、添付の図面と併せて読む場合に十分に理解されるであろう。しかし、本発明は、示した正確な配置および手段に限定する意図はないと理解されるべきであろう。
【発明の詳細な説明】
【0007】
(本発明の詳細な説明)
この発明は、aFGF135と称するヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドを提供するものであって、該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含んでいる。特に、ペプチドaFGF135は、配列番号:1のアミノ酸配列を含む。予想外に、ペプチドaFGF135の改変ペプチドは、温度の生理機能の下で比較的高い安定性を有し、その他の既知aFGFとは異なる構造を有する。
【0008】
本発明に従って、ペプチドaFGF135は、比較的高い安定性をもつ配列番号:1のアミノ酸配列を有する。ネイティブなヒトaFGFと比較すると、ペプチドaFGF135は、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失し(「短化aFGF」と呼ぶ)、かつこの20個のa.a.が欠失したaFGFの前にアラニン(Ala)が付加されている。本発明に従って、20個のアミノ酸をネイティブなヒトaFGFから欠失させて、一般経路を介するIL-1様効果を回避させた。これは、ヒトaFGFの最初の19個のアミノ酸がヒトインターロイキン-1(IL-1)と相同であると同定されており、そして該アミノ酸を欠失したためである。予想外の発見は、該ペプチドaFGFは、生理的温度下での優れた安定性を有しており、ヒトaFGFなどの既知aFGFよりもさらに優れている点であった。本発明の実施例に示したとおり、ペプチドaFGF135は、少なくとも48時間のインキュベーション中に体温(例えば、約37℃)で変性しないか、または加水分解しない正しいホールディング状態にあった(図2Aに示したとおりである)。本発明の別の実施態様において、ペプチドaFGF135は、1時間のインキュベーション中に、自己促進分解温度(例えば、約54℃)でその完全な構造を維持した(図2Bに示したとおり)。従って、ペプチドaFGF135は、140個のアミノ酸および127個のアミノ酸を有する各既知aFGF、例えば既知のヒトaFGFと比較して、相対的に高い安定性を有する。
【0009】
ペプチドaFGF135および既知aFGFとの間の差違を示すために、NMR分光器を使用して、ペプチドaFGF135の実際の三次元構造を計算して、既知aFGFと比較した。
【0010】
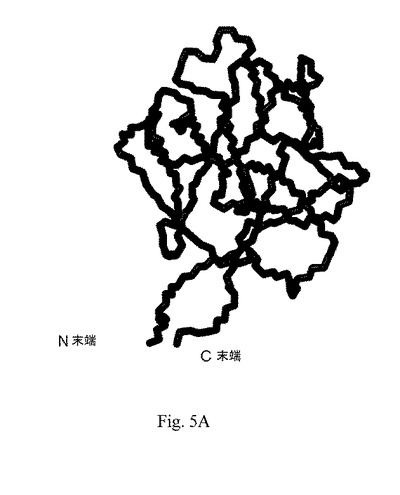
Bernett MJら(Protein, 57(3):626-34, 2004)により刊行されたタンパク質データバンク(PDB)のコード1RG8として140個のアミノ酸を有するヒトaFGFと、Lozano RM.ら(Biochemistry, 2;39(17):4982-93, 2000)により刊行されたPDBのコード1DZDとして127個のアミノ酸を有するヒトaFGFとの比較した場合、ペプチドaFGF135は、図5Aに示したように、例えばC末端およびN末端の露呈および下方左部の結合ループの差違などを示す異なる構造特徴を有する。改変配列を有するペプチドaFGF(20個のアミノ酸の欠失およびAlaの付加などを含む)は、他の既知の組換または天然aFGFよりも高い安定な構造をもたらすことを示唆している。本発明の一実施態様において、分解に関する安定性の比較を行った。そして、市販組換えaFGF(Promega Corporaton)は、1時間54℃でのインキュベーション後のウェスタンブロッティングでは分解バンドを示したが、図3に示したようにペプチドaFGF135は安定であることが判った。
【0011】
本発明は、本発明のペプチドFGF135および医薬上許容し得る担体を含む医薬組成物をさらに提供する。
【0012】
本発明の医薬組成物は、1以上の医薬上許容し得る担体と共に従来の既知方法により製造され得る。本明細書に使用した用語「医薬上許容し得る担体」には、標準的な医薬上許容し得るあらゆる担体を含む。かかる担体は、生理食塩水、生理緩衝食塩水、デキストロース、水、グリセロール、エタノールおよびその組合せ物が挙げられ得るが、これらに限定するものではない。
【0013】
本発明の医薬組成物は、選択された投与様式に好適あらゆる形態に構成され得る。好ましくは、組成物は、外科的領域に直接適用される。
【0014】
本発明は、さらに下記の実施例により説明されるが、これらは限定というよりもむしろ実証を目的として提供される。
【図面の簡単な説明】
【0015】
【図1】図1は、LC-MSMSアッセイにより15281Daと決定されたペプチドaFGF135の分子量を示す略図である。
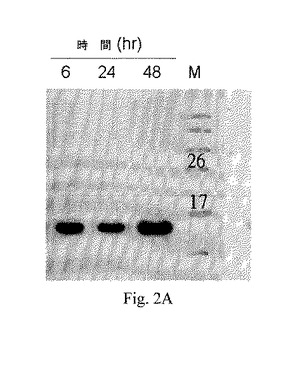
【図2A】図2Aは、48時間、37℃でのインキュベーション後のペプチドaFGF135の分解を示すウェスタンブロッティングの画像である;ここで、レーン1-3は、各々6時間、24時間および48時間のインキュベーションの後の結果であり、レーン4は分子量マーカーであり、各バンド上部の数字は分子量を示す。
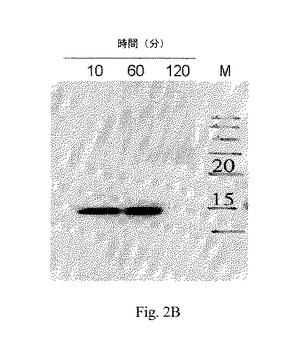
【図2B】図2Bは、120分間(2時間)、54℃でのインキュベーション後のペプチドaFGF135の分解を示すウェスタンブロッティングの画像である;ここで、レーン1から3は、各々10分間、60分間および120分間のインキュベーション後の結果であり、レーン4は分子量マーカーであって、各バンド上部の数字は分子量を示す。
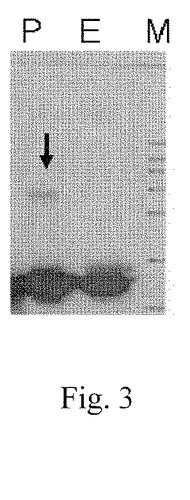
【図3】図3は、1時間、54℃でのインキュベーション後の、ペプチドaFGF135(レーン「E」として示す)および140個のアミノ酸を有する市販ヒトaFGF(レーン「P」として示す)の分解物に関する比較を示すウェスタンブロッティングの画像である;ここで、レーン「M」は分子量マーカーであり、市販ヒトaFGFの分解は黒色矢印により示したレーンPに提示される。
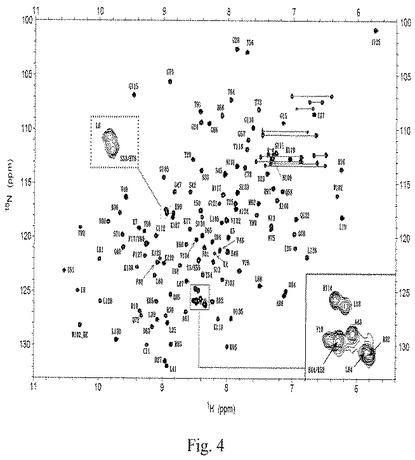
【図4】図4は、液体−状態のタンパク質NMR分光法により検出されたペプチドaFGF135の1H-15N-HSQCスペクトルを示す略図である。
【図5A】図5Aは、図4に示した1H-15N-HSQCスペクトルに従って予測したようなペプチドaFGF135の3D構造略図である。
【図5B】図5Bは、PDB Code 1RG8として140個のアミノ酸を有するヒトaFGFの3D構造の略図である。
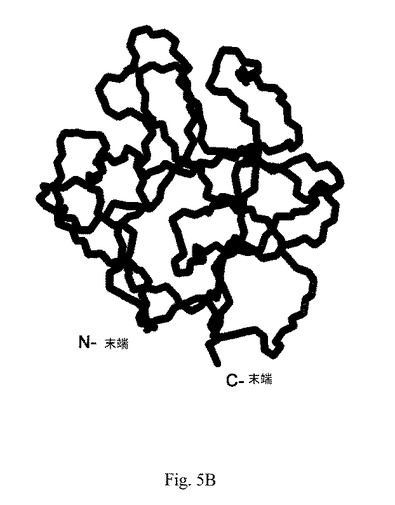
【図5C】図5Cは、PDB Code 1DZDとして127個のアミノ酸を有するヒトaFGFの3D構造の略図である。
【0016】
実施例1:ペプチドaFGF135のクローニング
完全長のヒトaFGFは、Clontech Laboratories, Incから購入したQuick Clone cDNAの生成物であった。構築前に、2つの特異的プライマー配列を下記のように作成した:
配列番号:2: 5’-ACTG▼AATTCATGGCTGAAGGGGAAATCA-3’
配列番号:3: 5’-AAGA▼AGCTTCAATCAGAAGAGACTGGCAGG-3’
【0017】
配列番号:2にはEcoR1制限部位が存在しているが(配列番号:2内に▼として表示した)、配列番号:3にはHindIII制限部位が存在する(配列番号:3内に▼として表示した)。完全長産物を、上記したプライマーと共にPCR増幅に使用し、485塩基対のPCR産物を得た。組換えcDNAがEcoR1およびHindIIIの制限酵素と反応した後に、該切断フラグメントを、同じ制限部位を有するpUC18ベクターに挿入した。正しい配列を含有する組換えベクターpUC18-haFGFを、DNA配列決定のための分析の後に得た。
【0018】
pUC-haFGFのテンプレートに従って、2つの特異的なプライマー配列を下記のとおりに作成した:
配列番号:4:5’-GGCA▼TATGGCTAATTACAAGAAGCCC-3’
配列番号:5:5’-AAGA▼GATCTCTTTAATCAGAAGAGACTGGCAGG-3’
【0019】
配列番号:4にはNde I制限部位が存在しているが(配列番号:4内に▼として表示した)、配列番号:5にはBgl II制限部位がある(配列番号:5内に▼として表示した)。配列番号:4および配列番号:5により増幅したcDNAの長さは、完全長から57個の塩基対分短かった。ペプチドaFGF135は、135個のアミノ酸のみを有し、aFGFの主な機能ドメインを保存した。さらに、第二にアミノ酸-グリシン(G)をN末端中のアラニン(A)に変更した。その部分を次のように示した:配列番号:1中のANYKKPKLLY。pUC-haFGFにより増幅したcDNAフラグメントを、NdeIおよびBglIIの制限酵素と反応させ、同じ制限部位をもつpET3c(Novagen)ベクターに、該切断フラグメントを挿入した。その結果として、pET3c-haFGFを構築した。
【0020】
実施例2:ペプチドaFGF135の発現および単離
pET3c-haFGFの増幅後に、ベクターを、BL21(DE3)(Novagen, Germany)コンピテント細胞を用いてDNAに形質転換した。アンピシリン耐性のE.coliコロニーを、1mM IPTG(イソプロピル β-D-1-チオガラクトピラノシド)の終濃度を用いて誘導する前に、LB培地で培養してOD600=0.3まで増殖させた。16時間(±2時間)のインキュベーション後に、細菌を回収し、27000 xgで遠心分離して、上清を除去した。集菌した細菌をPBSで2回洗浄し、次いで、高圧ホモジナイザーにより溶菌した(Niro Soavi Model NS2006L, Daken Stainless Products Ltd., UK)。溶菌サンプルを、0.22μmの孔サイズを有するシーブに透過させて、タンパク質単離のために準備した。
【0021】
ヒトaFGF135のペプチドを、次のクロマトグラフィーにより単離した:(1)カチオン交換クロマトグラフィー(CMFF column, RM197, GE Healthcare Bio-Sciences USA Corp.);(2)ヘパリンに特異的なアフィニティークロマトグラフィー(ヘパリン FF column, RM 244, GE Healthcare Bio-Sciences USA Corp.);および(3)サイズ排除クロマトグラフィー(Superdex 75 pre-grade column, RM245, GE Healthcare Bio-Sciences USA Corp.)。上記カラムに使用した緩衝液は、リン酸塩溶液であった(0.1% EDTA-Naを有するNa2PO4:NaHPO3=51:49、pH 6.8-7.2)。得られたように最終生成物は、本発明の標的ペプチドであった。ペプチドaFGF135の分子量は、LC-MSMS アッセイにより決定したとおり15281Daであった。
【0022】
実施例3:ペプチドaFGF135の安定性試験
(I)ウェスタンブロッティング分析
粗製ペプチドを、4-20または10-20%の勾配ゲルにおけるSDS-PAGEに供し、次いで電気的転移(Polyblot Transfer System, Model SBD-1000; American Bionetics, Emeryville, CA)により、ニトロセルロース膜(0.05 Am; Schleicher & Schuell, Inc., Keene, NH)に移し取った。aFGFペプチドの安定性を調べるために、8M 尿素を添加するか、または添加しない(潜在的凝集物を分解するまで)Laemmli緩衝溶液[1 M Tris(2.4 ml)pH 6.8、0.8 g SDS-ストック、100%グリセロール(4 ml)、0.01%のブロモフェノールブルー、0.02% β-メルカプトエタノール(電気泳動グレード)(1 ml)、および水(2.8 ml)]を、0.0625M トリス塩、SDSストック1%、および15 mMジチオスレイトルを含むスタッキングゲル緩衝溶液として調製した。サンプルを、室温で1時間保持し、その後4℃で終夜保持した。それを煮沸した後に、SDS-PAGE上で泳動させた。
【0023】
移し取り、そして非特異的タンパク質-結合部位を、3% 乾燥ミルクのTBS溶液によりブロッキングした後に、該ニトロセルロース膜を、洗浄緩衝液(10 mM トリス-HCI、pH 8.0、0.15 M NaCl、0.05% Tween-20)で好適に希釈した(aFGF 抗体の1:500希釈, R&D Systems, Inc.)種々の抗体と共に終夜4℃でインキュベートした。抗原-抗体複合体を、該膜と好適な二次抗体とインキュベートし、それをProtoBlot Western Blot AP System(Promega, Madison, WI)により発色させて可視化した。
【0024】
(II)分解試験
インタクトなaFGFペプチドを、37℃および54℃で各々インキュベートした。この54℃は自己促進性分解温度である。その後、サンプルをウェスタンブロッティングに供した。37℃でのインキュベーション後のサンプルに対する結果を図2Aに示した。該結果は、レーン1-3は各々インキュベーションが6時間、24時間および48時間後の結果であり、レーン4は分子量マーカーであって、また各バンド上部の数字は分子量を表す。54℃で120分間(2時間)のインキュベーション後のペプチドaFGF135に対する結果を図2Bに示した;ここで、レーン1−3は各々10分間、60分間および120分間のインキュベーション後の結果であり、レーン4は分子量マーカーであり、また各バンド上部の数字は分子量を表す。
【0025】
図2Aに示したとおり、該ペプチドaFGF135は、体温(約37℃)にて少なくとも48時間、そのインタクトな構造を維持した。ペプチドaFGF135は、より長期的な神経保護効果かつより良好な安定性を備えていることを示した。
【0026】
図2Bに示したとおり、ペプチドaFGF135は、54℃で少なくとも1時間そのインタクトな構造を維持する。ペプチドaFGF135を、54℃で2時間インキュベートした場合、他の構造に形態を変えたというよりもむしろ完全に分解しており、これは予測不可能な副作用のリスクをもたらし得る。次に、ペプチドaFGFは安全な貯蔵性または移動性を提供した。
【0027】
同じ実験条件下で、安定性試験を、140個のアミノ酸を有する市販組換えヒトaFGF(Promega Corporationからの「Promega aFGF」)について行った。図3に示したとおり、黒色の矢印により示した第二のバンドがPromega aFGFのバンドに表れた;これに対して、ペプチドaFGF135は分解せずにその構造を維持した。一方、マイクロプレートにサンプルを添加すると、Promega aFGFは明らかに白色沈殿物を呈したが、ペプチドaFGF135は透明を呈した。該ペプチドaFGF135は、市販のaFGFよりも良好な安定性を有することを示した。
【0028】
実施例4:1H-15N-NMR構造の特徴分析
現在、NMR分光法は、原子分解にて生物学的マクロ分子様タンパク質および核酸の構造を決定することが出来るよく知られている技術である。ペプチドaFGF135についての構造の特徴分析を、High Field Nuclear Magnetic Resonance Center (Academia Sinica, Taiwan)により行った。
【0029】
要約すると、スペクトルをBruker AVANCE 600で298Kにて記録した。CおよびNの化学シフトを、バックボーンのねじれ角を得るためにプログラムTALOSに使用した。バックボーンについての配列-特異的な共鳴帰属(resonance assignment)を、HNCO、HN(CA)CO、HACNCB、CBCA(CO)NH、HNCAおよびHN(CO)CAの実験を用いて得て、15N-NOESY-HSQCおよび13C-NOESY-HSQCを組み合わせて、15N-TOCSY-HSQCおよび13C-NOESY-HSQCを用いる側鎖配置を得た。バックボーンスペクトルを図4に示した。
【0030】
構造計算のために、二面角をプログラムTALOSにより得て、距離限界を、D2O交換実験の併用にて15N-NOESY-HSQCおよび13C-NOESY-HSQCにより得て、26個の起こり得る水素結合を見出した。ペプチドaFGF135の3D構造特徴を、図5Aに示したとおりプログラムCYANAにより得た。Afgf135についてのNMR拘束および構造計算の統計値を表1に記載した。
【0031】
表1.Afgf135aについてのNMR拘束および構造計算の統計値
【表1】

a. 所与の値は、CYANAによる溶液構造を表す20以上の配座異性体の平均に対応している。
b. 二次構造領域はMOLMO選択を基にしている:7-11、16-21、25-29、39-43、48-53、59-62、68-71、80-85、89-94、127-131。
c. Lozano RMら(Biochemistry, 2;39(17):4982-93, 2000)にて選択された領域に対応する選択領域。
【0032】
ペプチドaFGF135および2つの既知ヒトaFGFの3D構造を、比較のために図5Bおよび図5Cに示した。PDB Code 1RG8として配列番号:6の140個のアミノ酸を有するaFGFを図5Bに示した;ならびに1DZDとして配列番号:7の127個のアミノ酸を有するaFGFを図5Cに示した。両方は、図5Aに示したとおりペプチドaFGF135とは構造的に異なり、ペプチド内のN末端およびC末端は内側に埋没しており、下方左部の結合ループが欠落している。改変アミノ酸配列および独特な構造を有するペプチドaFGF135は、構造的に新規であり、既知aFGFペプチドとは異なることを示した。
【0033】
本発明の広範囲の概念から逸脱せずに上記実施態様に変更を為し得ることは、当分野の技術者には理解されよう。それ故に、本発明は、特定の実施態様に制限されるものではなく、添付の請求の範囲により規定したような本発明の精神および範囲内にある修飾をカバーすることを意図すると解される。
【技術分野】
【0001】
本発明は、良好な安定性を有する酸性繊維芽細胞増殖因子の改変ペプチドに関する。
【背景技術】
【0002】
(本発明の背景)
イン・ビトロで様々な細胞型の増殖および分化に影響を及ぼす酸性繊維芽細胞増殖因子(aFGF)は、全脳および視床下部などの神経組織由来の一本鎖タンパク質として最初に単離された。aFGFは、ヘパリン依存性の有糸分裂促進物質であり、4つ全ての型の既知FGFレセプターおよびそのスプライス形態に強固に結合することができる。それは、運動機能および感覚機能と関連のあるニューロンの特定サブセット内に局在し、成人の脳から精製され得る。精製aFGFは、神経芽細胞に対する有糸分裂促進物質であり、脊髄ニューロンから伸張する軸索を促進する。
【発明の概要】
【発明が解決しようとする課題】
【0003】
ヒトaFGFのネイティブペプチドはヒトの脳から単離され、154個のアミノ酸から構成される。しかし、ネイティブヒトaFGFのN末端の19個のアミノ酸は、ヒトインターロイキン-1(IL-1)と相同であると同定された。ヒトaFGFとIL-1と類似したポリペプチドのドメインは、例えば、マクロファージの活性化および調節される細胞増殖の停止などと同一の内生免疫応答を引き起こし得る(G. Venkataraman et al., P.N.A.S., 96:3658-63, 1999)。さらに、炎症誘導性サイトカインIL-1およびFGF-1(aFGF)/FGF-2(bFGF)は、同じ構造スキャッホルドを共有し、チロシンキナーゼドメインの同じ受容体結合部位に競合する(A. J. Minter et al., J. Cell Physil., 167:229-37, 1996)。
【課題を解決するための手段】
【0004】
本発明の要旨
本発明は、aFGF135と称するヒト酸性繊維芽細胞増殖因子(aFGF)改変ペプチドを提供するものであって、ここで該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含んでいる。特に、ペプチドaFGF135は、比較的高い安定性を持っており、また既知のaFGFペプチドよりも優れている配列番号:1のアミノ酸配列を含む。
【発明の効果】
【0005】
本発明は、本発明のペプチドaFGF135および医薬上許容し得る担体を含む医薬組成物をさらに提供する。
【0006】
図面の簡単な説明
前記した発明の要旨、ならびに本発明の下記詳細な説明は、添付の図面と併せて読む場合に十分に理解されるであろう。しかし、本発明は、示した正確な配置および手段に限定する意図はないと理解されるべきであろう。
【発明の詳細な説明】
【0007】
(本発明の詳細な説明)
この発明は、aFGF135と称するヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドを提供するものであって、該改変ペプチドは、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失することにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含んでいる。特に、ペプチドaFGF135は、配列番号:1のアミノ酸配列を含む。予想外に、ペプチドaFGF135の改変ペプチドは、温度の生理機能の下で比較的高い安定性を有し、その他の既知aFGFとは異なる構造を有する。
【0008】
本発明に従って、ペプチドaFGF135は、比較的高い安定性をもつ配列番号:1のアミノ酸配列を有する。ネイティブなヒトaFGFと比較すると、ペプチドaFGF135は、ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失し(「短化aFGF」と呼ぶ)、かつこの20個のa.a.が欠失したaFGFの前にアラニン(Ala)が付加されている。本発明に従って、20個のアミノ酸をネイティブなヒトaFGFから欠失させて、一般経路を介するIL-1様効果を回避させた。これは、ヒトaFGFの最初の19個のアミノ酸がヒトインターロイキン-1(IL-1)と相同であると同定されており、そして該アミノ酸を欠失したためである。予想外の発見は、該ペプチドaFGFは、生理的温度下での優れた安定性を有しており、ヒトaFGFなどの既知aFGFよりもさらに優れている点であった。本発明の実施例に示したとおり、ペプチドaFGF135は、少なくとも48時間のインキュベーション中に体温(例えば、約37℃)で変性しないか、または加水分解しない正しいホールディング状態にあった(図2Aに示したとおりである)。本発明の別の実施態様において、ペプチドaFGF135は、1時間のインキュベーション中に、自己促進分解温度(例えば、約54℃)でその完全な構造を維持した(図2Bに示したとおり)。従って、ペプチドaFGF135は、140個のアミノ酸および127個のアミノ酸を有する各既知aFGF、例えば既知のヒトaFGFと比較して、相対的に高い安定性を有する。
【0009】
ペプチドaFGF135および既知aFGFとの間の差違を示すために、NMR分光器を使用して、ペプチドaFGF135の実際の三次元構造を計算して、既知aFGFと比較した。
【0010】
Bernett MJら(Protein, 57(3):626-34, 2004)により刊行されたタンパク質データバンク(PDB)のコード1RG8として140個のアミノ酸を有するヒトaFGFと、Lozano RM.ら(Biochemistry, 2;39(17):4982-93, 2000)により刊行されたPDBのコード1DZDとして127個のアミノ酸を有するヒトaFGFとの比較した場合、ペプチドaFGF135は、図5Aに示したように、例えばC末端およびN末端の露呈および下方左部の結合ループの差違などを示す異なる構造特徴を有する。改変配列を有するペプチドaFGF(20個のアミノ酸の欠失およびAlaの付加などを含む)は、他の既知の組換または天然aFGFよりも高い安定な構造をもたらすことを示唆している。本発明の一実施態様において、分解に関する安定性の比較を行った。そして、市販組換えaFGF(Promega Corporaton)は、1時間54℃でのインキュベーション後のウェスタンブロッティングでは分解バンドを示したが、図3に示したようにペプチドaFGF135は安定であることが判った。
【0011】
本発明は、本発明のペプチドFGF135および医薬上許容し得る担体を含む医薬組成物をさらに提供する。
【0012】
本発明の医薬組成物は、1以上の医薬上許容し得る担体と共に従来の既知方法により製造され得る。本明細書に使用した用語「医薬上許容し得る担体」には、標準的な医薬上許容し得るあらゆる担体を含む。かかる担体は、生理食塩水、生理緩衝食塩水、デキストロース、水、グリセロール、エタノールおよびその組合せ物が挙げられ得るが、これらに限定するものではない。
【0013】
本発明の医薬組成物は、選択された投与様式に好適あらゆる形態に構成され得る。好ましくは、組成物は、外科的領域に直接適用される。
【0014】
本発明は、さらに下記の実施例により説明されるが、これらは限定というよりもむしろ実証を目的として提供される。
【図面の簡単な説明】
【0015】
【図1】図1は、LC-MSMSアッセイにより15281Daと決定されたペプチドaFGF135の分子量を示す略図である。
【図2A】図2Aは、48時間、37℃でのインキュベーション後のペプチドaFGF135の分解を示すウェスタンブロッティングの画像である;ここで、レーン1-3は、各々6時間、24時間および48時間のインキュベーションの後の結果であり、レーン4は分子量マーカーであり、各バンド上部の数字は分子量を示す。
【図2B】図2Bは、120分間(2時間)、54℃でのインキュベーション後のペプチドaFGF135の分解を示すウェスタンブロッティングの画像である;ここで、レーン1から3は、各々10分間、60分間および120分間のインキュベーション後の結果であり、レーン4は分子量マーカーであって、各バンド上部の数字は分子量を示す。
【図3】図3は、1時間、54℃でのインキュベーション後の、ペプチドaFGF135(レーン「E」として示す)および140個のアミノ酸を有する市販ヒトaFGF(レーン「P」として示す)の分解物に関する比較を示すウェスタンブロッティングの画像である;ここで、レーン「M」は分子量マーカーであり、市販ヒトaFGFの分解は黒色矢印により示したレーンPに提示される。
【図4】図4は、液体−状態のタンパク質NMR分光法により検出されたペプチドaFGF135の1H-15N-HSQCスペクトルを示す略図である。
【図5A】図5Aは、図4に示した1H-15N-HSQCスペクトルに従って予測したようなペプチドaFGF135の3D構造略図である。
【図5B】図5Bは、PDB Code 1RG8として140個のアミノ酸を有するヒトaFGFの3D構造の略図である。
【図5C】図5Cは、PDB Code 1DZDとして127個のアミノ酸を有するヒトaFGFの3D構造の略図である。
【0016】
実施例1:ペプチドaFGF135のクローニング
完全長のヒトaFGFは、Clontech Laboratories, Incから購入したQuick Clone cDNAの生成物であった。構築前に、2つの特異的プライマー配列を下記のように作成した:
配列番号:2: 5’-ACTG▼AATTCATGGCTGAAGGGGAAATCA-3’
配列番号:3: 5’-AAGA▼AGCTTCAATCAGAAGAGACTGGCAGG-3’
【0017】
配列番号:2にはEcoR1制限部位が存在しているが(配列番号:2内に▼として表示した)、配列番号:3にはHindIII制限部位が存在する(配列番号:3内に▼として表示した)。完全長産物を、上記したプライマーと共にPCR増幅に使用し、485塩基対のPCR産物を得た。組換えcDNAがEcoR1およびHindIIIの制限酵素と反応した後に、該切断フラグメントを、同じ制限部位を有するpUC18ベクターに挿入した。正しい配列を含有する組換えベクターpUC18-haFGFを、DNA配列決定のための分析の後に得た。
【0018】
pUC-haFGFのテンプレートに従って、2つの特異的なプライマー配列を下記のとおりに作成した:
配列番号:4:5’-GGCA▼TATGGCTAATTACAAGAAGCCC-3’
配列番号:5:5’-AAGA▼GATCTCTTTAATCAGAAGAGACTGGCAGG-3’
【0019】
配列番号:4にはNde I制限部位が存在しているが(配列番号:4内に▼として表示した)、配列番号:5にはBgl II制限部位がある(配列番号:5内に▼として表示した)。配列番号:4および配列番号:5により増幅したcDNAの長さは、完全長から57個の塩基対分短かった。ペプチドaFGF135は、135個のアミノ酸のみを有し、aFGFの主な機能ドメインを保存した。さらに、第二にアミノ酸-グリシン(G)をN末端中のアラニン(A)に変更した。その部分を次のように示した:配列番号:1中のANYKKPKLLY。pUC-haFGFにより増幅したcDNAフラグメントを、NdeIおよびBglIIの制限酵素と反応させ、同じ制限部位をもつpET3c(Novagen)ベクターに、該切断フラグメントを挿入した。その結果として、pET3c-haFGFを構築した。
【0020】
実施例2:ペプチドaFGF135の発現および単離
pET3c-haFGFの増幅後に、ベクターを、BL21(DE3)(Novagen, Germany)コンピテント細胞を用いてDNAに形質転換した。アンピシリン耐性のE.coliコロニーを、1mM IPTG(イソプロピル β-D-1-チオガラクトピラノシド)の終濃度を用いて誘導する前に、LB培地で培養してOD600=0.3まで増殖させた。16時間(±2時間)のインキュベーション後に、細菌を回収し、27000 xgで遠心分離して、上清を除去した。集菌した細菌をPBSで2回洗浄し、次いで、高圧ホモジナイザーにより溶菌した(Niro Soavi Model NS2006L, Daken Stainless Products Ltd., UK)。溶菌サンプルを、0.22μmの孔サイズを有するシーブに透過させて、タンパク質単離のために準備した。
【0021】
ヒトaFGF135のペプチドを、次のクロマトグラフィーにより単離した:(1)カチオン交換クロマトグラフィー(CMFF column, RM197, GE Healthcare Bio-Sciences USA Corp.);(2)ヘパリンに特異的なアフィニティークロマトグラフィー(ヘパリン FF column, RM 244, GE Healthcare Bio-Sciences USA Corp.);および(3)サイズ排除クロマトグラフィー(Superdex 75 pre-grade column, RM245, GE Healthcare Bio-Sciences USA Corp.)。上記カラムに使用した緩衝液は、リン酸塩溶液であった(0.1% EDTA-Naを有するNa2PO4:NaHPO3=51:49、pH 6.8-7.2)。得られたように最終生成物は、本発明の標的ペプチドであった。ペプチドaFGF135の分子量は、LC-MSMS アッセイにより決定したとおり15281Daであった。
【0022】
実施例3:ペプチドaFGF135の安定性試験
(I)ウェスタンブロッティング分析
粗製ペプチドを、4-20または10-20%の勾配ゲルにおけるSDS-PAGEに供し、次いで電気的転移(Polyblot Transfer System, Model SBD-1000; American Bionetics, Emeryville, CA)により、ニトロセルロース膜(0.05 Am; Schleicher & Schuell, Inc., Keene, NH)に移し取った。aFGFペプチドの安定性を調べるために、8M 尿素を添加するか、または添加しない(潜在的凝集物を分解するまで)Laemmli緩衝溶液[1 M Tris(2.4 ml)pH 6.8、0.8 g SDS-ストック、100%グリセロール(4 ml)、0.01%のブロモフェノールブルー、0.02% β-メルカプトエタノール(電気泳動グレード)(1 ml)、および水(2.8 ml)]を、0.0625M トリス塩、SDSストック1%、および15 mMジチオスレイトルを含むスタッキングゲル緩衝溶液として調製した。サンプルを、室温で1時間保持し、その後4℃で終夜保持した。それを煮沸した後に、SDS-PAGE上で泳動させた。
【0023】
移し取り、そして非特異的タンパク質-結合部位を、3% 乾燥ミルクのTBS溶液によりブロッキングした後に、該ニトロセルロース膜を、洗浄緩衝液(10 mM トリス-HCI、pH 8.0、0.15 M NaCl、0.05% Tween-20)で好適に希釈した(aFGF 抗体の1:500希釈, R&D Systems, Inc.)種々の抗体と共に終夜4℃でインキュベートした。抗原-抗体複合体を、該膜と好適な二次抗体とインキュベートし、それをProtoBlot Western Blot AP System(Promega, Madison, WI)により発色させて可視化した。
【0024】
(II)分解試験
インタクトなaFGFペプチドを、37℃および54℃で各々インキュベートした。この54℃は自己促進性分解温度である。その後、サンプルをウェスタンブロッティングに供した。37℃でのインキュベーション後のサンプルに対する結果を図2Aに示した。該結果は、レーン1-3は各々インキュベーションが6時間、24時間および48時間後の結果であり、レーン4は分子量マーカーであって、また各バンド上部の数字は分子量を表す。54℃で120分間(2時間)のインキュベーション後のペプチドaFGF135に対する結果を図2Bに示した;ここで、レーン1−3は各々10分間、60分間および120分間のインキュベーション後の結果であり、レーン4は分子量マーカーであり、また各バンド上部の数字は分子量を表す。
【0025】
図2Aに示したとおり、該ペプチドaFGF135は、体温(約37℃)にて少なくとも48時間、そのインタクトな構造を維持した。ペプチドaFGF135は、より長期的な神経保護効果かつより良好な安定性を備えていることを示した。
【0026】
図2Bに示したとおり、ペプチドaFGF135は、54℃で少なくとも1時間そのインタクトな構造を維持する。ペプチドaFGF135を、54℃で2時間インキュベートした場合、他の構造に形態を変えたというよりもむしろ完全に分解しており、これは予測不可能な副作用のリスクをもたらし得る。次に、ペプチドaFGFは安全な貯蔵性または移動性を提供した。
【0027】
同じ実験条件下で、安定性試験を、140個のアミノ酸を有する市販組換えヒトaFGF(Promega Corporationからの「Promega aFGF」)について行った。図3に示したとおり、黒色の矢印により示した第二のバンドがPromega aFGFのバンドに表れた;これに対して、ペプチドaFGF135は分解せずにその構造を維持した。一方、マイクロプレートにサンプルを添加すると、Promega aFGFは明らかに白色沈殿物を呈したが、ペプチドaFGF135は透明を呈した。該ペプチドaFGF135は、市販のaFGFよりも良好な安定性を有することを示した。
【0028】
実施例4:1H-15N-NMR構造の特徴分析
現在、NMR分光法は、原子分解にて生物学的マクロ分子様タンパク質および核酸の構造を決定することが出来るよく知られている技術である。ペプチドaFGF135についての構造の特徴分析を、High Field Nuclear Magnetic Resonance Center (Academia Sinica, Taiwan)により行った。
【0029】
要約すると、スペクトルをBruker AVANCE 600で298Kにて記録した。CおよびNの化学シフトを、バックボーンのねじれ角を得るためにプログラムTALOSに使用した。バックボーンについての配列-特異的な共鳴帰属(resonance assignment)を、HNCO、HN(CA)CO、HACNCB、CBCA(CO)NH、HNCAおよびHN(CO)CAの実験を用いて得て、15N-NOESY-HSQCおよび13C-NOESY-HSQCを組み合わせて、15N-TOCSY-HSQCおよび13C-NOESY-HSQCを用いる側鎖配置を得た。バックボーンスペクトルを図4に示した。
【0030】
構造計算のために、二面角をプログラムTALOSにより得て、距離限界を、D2O交換実験の併用にて15N-NOESY-HSQCおよび13C-NOESY-HSQCにより得て、26個の起こり得る水素結合を見出した。ペプチドaFGF135の3D構造特徴を、図5Aに示したとおりプログラムCYANAにより得た。Afgf135についてのNMR拘束および構造計算の統計値を表1に記載した。
【0031】
表1.Afgf135aについてのNMR拘束および構造計算の統計値
【表1】
a. 所与の値は、CYANAによる溶液構造を表す20以上の配座異性体の平均に対応している。
b. 二次構造領域はMOLMO選択を基にしている:7-11、16-21、25-29、39-43、48-53、59-62、68-71、80-85、89-94、127-131。
c. Lozano RMら(Biochemistry, 2;39(17):4982-93, 2000)にて選択された領域に対応する選択領域。
【0032】
ペプチドaFGF135および2つの既知ヒトaFGFの3D構造を、比較のために図5Bおよび図5Cに示した。PDB Code 1RG8として配列番号:6の140個のアミノ酸を有するaFGFを図5Bに示した;ならびに1DZDとして配列番号:7の127個のアミノ酸を有するaFGFを図5Cに示した。両方は、図5Aに示したとおりペプチドaFGF135とは構造的に異なり、ペプチド内のN末端およびC末端は内側に埋没しており、下方左部の結合ループが欠落している。改変アミノ酸配列および独特な構造を有するペプチドaFGF135は、構造的に新規であり、既知aFGFペプチドとは異なることを示した。
【0033】
本発明の広範囲の概念から逸脱せずに上記実施態様に変更を為し得ることは、当分野の技術者には理解されよう。それ故に、本発明は、特定の実施態様に制限されるものではなく、添付の請求の範囲により規定したような本発明の精神および範囲内にある修飾をカバーすることを意図すると解される。
【特許請求の範囲】
【請求項1】
ヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドであって、
ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失させることにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含む、ペプチド。
【請求項2】
配列番号:1のアミノ酸配列を含む、請求項1記載のペプチド。
【請求項3】
図5Aに示した3D構造を特徴とする、請求項1記載のペプチド。
【請求項4】
相対的に高い安定性を持っている、請求項1記載のペプチド。
【請求項5】
請求項1の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項6】
請求項2の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項7】
請求項3の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項8】
請求項4の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項1】
ヒト酸性繊維芽細胞増殖因子(aFGF)の改変ペプチドであって、
ネイティブなヒトaFGFのN末端から20個のアミノ酸を欠失させることにより短化したネイティブヒトaFGF、および該短化ネイティブaFGFの前に付加されたアラニン(Ala)を含む、ペプチド。
【請求項2】
配列番号:1のアミノ酸配列を含む、請求項1記載のペプチド。
【請求項3】
図5Aに示した3D構造を特徴とする、請求項1記載のペプチド。
【請求項4】
相対的に高い安定性を持っている、請求項1記載のペプチド。
【請求項5】
請求項1の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項6】
請求項2の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項7】
請求項3の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【請求項8】
請求項4の単離ペプチドおよび医薬上許容し得る担体を含む、医薬組成物。
【図1】


【図2A】
【図2B】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図2A】
【図2B】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【公開番号】特開2011−121882(P2011−121882A)
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【外国語出願】
【出願番号】特願2009−279515(P2009−279515)
【出願日】平成21年12月9日(2009.12.9)
【出願人】(509339005)イーユー・ソル・バイオテック・カンパニー・リミテッド (2)
【氏名又は名称原語表記】EU Sol Biotech Co., Ltd.
【Fターム(参考)】
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願番号】特願2009−279515(P2009−279515)
【出願日】平成21年12月9日(2009.12.9)
【出願人】(509339005)イーユー・ソル・バイオテック・カンパニー・リミテッド (2)
【氏名又は名称原語表記】EU Sol Biotech Co., Ltd.
【Fターム(参考)】
[ Back to top ]