
ヒト骨髄由来内皮前駆細胞の同定、及び虚血性傷害後の心筋細胞の機能を改善するためのヒト骨髄由来内皮前駆細胞の使用
【課題】患者の心筋梗塞によって損傷を受けた組織における血管形成を刺激する方法の提供
【解決手段】(a)前記患者のある部位から幹細胞を採取することと、(b)前記幹細胞から内皮前駆細胞を回収することと、(c)前記前駆細胞が前記組織に遊走し、前記組織の新血管新生を刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。あるいは、虚血性傷害によって損傷を受けた組織の部位へのヒト骨髄由来内皮細胞前駆体の輸送を選択的に増加させる方法であって、(a)内皮前駆細胞を患者に投与することと、(b)前記患者にケモカインを投与することによって、前記虚血組織に内皮細胞前駆体を誘引することとを備えた方法。さらに、患者の心筋梗塞による損傷を受けた組織の血管形成又は血管新生を刺激する方法であって、患者に同種異系の幹細胞を注入することを備えた方法。
【解決手段】(a)前記患者のある部位から幹細胞を採取することと、(b)前記幹細胞から内皮前駆細胞を回収することと、(c)前記前駆細胞が前記組織に遊走し、前記組織の新血管新生を刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。あるいは、虚血性傷害によって損傷を受けた組織の部位へのヒト骨髄由来内皮細胞前駆体の輸送を選択的に増加させる方法であって、(a)内皮前駆細胞を患者に投与することと、(b)前記患者にケモカインを投与することによって、前記虚血組織に内皮細胞前駆体を誘引することとを備えた方法。さらに、患者の心筋梗塞による損傷を受けた組織の血管形成又は血管新生を刺激する方法であって、患者に同種異系の幹細胞を注入することを備えた方法。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
本出願は、2000年6月5日に出願された米国特許出願第09/587,441号の一部継続出願であり、その優先権を主張するものである。この親出願を、本明細書の一部として本願に援用する。
【0002】
本出願を通じて、様々な文献が括弧内に引用されている。本発明が属する分野の技術水準をより完全に説明するために、本出願への参照によって、これらの文献全体の開示内容を援用する。これらの参照文献の完全な書誌的引用は、本明細書の末尾に記載されている。
【発明の背景】
【0003】
心筋梗塞後の左心室の組織修復は、その後の心不全及び死亡の主原因である。毛細血管網は、肥大化しているが生存能力を有する心筋による要求の増加に追いつくことができず、心筋の死滅と繊維性置換が起こる。以下に記載されている本発明の第一群の実験は、成人の骨髄が胚血管芽細胞の表現型及び機能的な特徴を備えた内皮細胞前駆細胞を含有しており、これらの前駆細胞は動員、増殖させることが可能であって、実験的な心筋梗塞の後に、梗塞床(infarct bed)への血管形成(vasculogenesis)を誘導するために使用できることを示している。新血管新生は、生存可能な心筋組織の有意且つ持続的な増加、コラーゲンの沈着の減少、及び心筋機能の改善をもたらす。単独で、又は現在用いられている療法と組合せて、サイトカインによって動員されたヒト骨髄由来の自家血管芽細胞を心筋梗塞組織の血行再建術のために使用すると、心筋梗塞後の左心室の組織修復に伴う罹患率及び死亡率が有意に減少する可能性がある。
【0004】
狭い時間枠のうちに迅速な再潅流を行うと、急性心筋梗塞の初期死亡率が著しく減少するが、梗塞後の心不全が増加、多発する(1)。初期の梗塞部位の拡大、梗塞を取り囲む壁の厚さの漸減、及び左心室内腔の膨張を特徴とする心筋梗塞後の左心室の組織修復が、その後の心不全に対する主要な予後因子であることが明らかとされている(2、3)。このプロセスには、本来胎児の段階でしか発現されない遺伝子の転写、心臓の繊維芽細胞によるコラーゲン分泌の急速且つ進行性の増加、心室壁内への繊維組織の沈着、壁の硬さの増大、並びに弛緩期及び収縮期の両期における機能不全が伴う(4,5)。低酸素状態は心臓の繊維芽細胞によるコラーゲンの分泌を直接刺激するが、DNA合成と細胞の増殖を抑制する(6)。動物モデルでは、実験的な心筋梗塞の後、心筋の撤去が終わった時点に遅延して再潅流を行うと、組織修復に極めて有益である(7)。さらに、梗塞に付随する開存性動脈の存在は、常に、梗塞後の期間におけるヒトの生存率の改善を伴う(8)。これは、心室の組織修復過程を改善し、壁の運動の異常な変化を抑制する梗塞血管床が十分に再潅流されることに起因するものと思われる(9)。
【0005】
循環系の細胞因子又は骨髄由来の細胞因子のうち何れかを用いることによって、実験動物モデル中の虚血状態にした非心臓組織の再潅流を首尾よく行い得ることが最近実証された(10−13)。これらの研究では、これらの細胞の正確な性状は明らかにはされていないが、成人の循環系と骨髄の両者に、機能的な内皮細胞に分化する能力(血管形成と称されるプロセス(14−16))を有する前駆細胞が存在することが示されている。出生前期においては、ヒト及びヒトより下等な種の動脈の腹側内皮に由来する前駆細胞は、血管形成と血球新生の両プロセスに関与する細胞因子を生じることが示されている(17、18)。これらの細胞は、胚血管芽細胞(embryonic hemangioblast)と名付けられ、CD34、CD117(幹細胞因子受容体)、Flk−1(血管内皮細胞増殖因子受容体−2、VEGFR−2)、及びTie−2(アンジオポエチン受容体)の発現を特徴とし、VEGFに反応して、芽細胞コロニーを形成する高い増殖能を有することが示された(19−22)。GATA−2をノックアウトした胚幹細胞は、胎児の肝臓及び骨髄による終局的な造血(hematopoiesis)及び造血の開始(seeding)を完全に遮断するので(23)、この後に生じる胚血管芽細胞の成人型多能性幹細胞への増殖及び分化には、GATA−2転写因子の同時発現が関連しているものと思われる。さらに、胚腹側内皮(embryonic ventral endothelium)から分岐した造血及び内皮両細胞系列の最も初期の前駆体は、GATA−2とα4−インテグリンに加え、VEGF受容体を発現していることが示された(24)。本発明の第一群の実験は、成人の骨髄にもGATA−2陽性幹細胞前駆体が存在し、血管芽細胞の特性を表しており、血管形成を誘導するために使用することによって、実験的な心筋梗塞における組織修復と心不全を抑えることが可能であることを示している。
【0006】
既存の成熟内皮から新しい血管が増殖することは血管新生(angiogenesis)と称されており、ある種のCXCケモカインを含む数多くの因子によって制御され得る(47−50)。これに対して、血管形成(vasculogenesis)は、胚血管芽細胞の表現型的特長と血管内皮増殖因子(VEGF)等の受容体チロシンキナーゼによって制御される増殖/分化特性とを有する(54−57)骨髄由来の内皮前駆体(51−53)によって媒介される。治療的血管形成(58−61)は虚血性組織の潅流を改善させ得る能力を有するが、組織虚血の部位への内皮前駆体の選択的輸送に関与する受容体/リガンド相互作用は不明である。以下に記載されている本発明の第二群の実験は、ヒト骨髄由来の血管芽細胞上のCXC受容体とIL−8及びGro−αを含む虚血によって誘導されるELR陽性CXCケモカインとの相互作用の結果、梗塞を起こした心筋中に血管形成が生じ得ることを示している。さらに、CXCR4/SDF−1相互作用を遮断することによって、血管芽細胞を骨髄から虚血性心筋へと再誘導して輸送させることが可能となり、血管形成の増加、心筋細胞の死滅と繊維性置換(fibrous replacement)の減少、及び心機能の改善がもたらされる。前記実験の結果は、IL−8、Gro−α、及び間質由来因子−1(SDF−1、stromal−derived factor−1)を含むCXCケモカインが成人での血管形成を制御する上で中心的な役割を果たしていることを示しており、CXCケモカインと骨髄由来の血管芽細胞上に存在するそれらの受容体との相互作用を操作すると、最適な治療的血管形成及び虚血性組織の撤去(salvage)をもたらし得ることを示唆している。以下に記載されている第三群の実験は、血管芽細胞の虚血性心筋への化学走性(chemotaxis)を媒介する上でもCCケモカインが役割を果たしていることを示している。
【0007】
創傷治癒又は炎症中の血管新生反応は、既存の脈間構造中の内皮細胞と局所的に産生された因子によって調節される細胞外マトリックスとの接着性相互作用の変化に起因し、これによって、内皮細胞の移動、増殖、再構築、及び微小血管の形成が生じる(70)ものと考えられている。ヒトCXCケモカインファミリーは、内皮細胞に結合し、該細胞に対する強力な化学走性活性を有する小さな(<10kD)ヘパリン結合ポリペプチドからなる。N末端の3つのアミノ酸残基(Glu−Leu−Arg、ELRモチーフ)は、内皮細胞上に存在するCXC受容体1及び2へのIL−8及びGro−αなどのCXCケモカインの結合を決定して(49、71)、内皮の化学走性と血管新生を促進する(47−48)。これに対して、ELRモチーフを欠如するCXCケモカインは異なるCXC受容体に結合し、増殖因子によって媒介される血管新生を阻害する(49−72)。SDF−1(ELR陰性のCXCケモカイン)は、CXCR4との相互作用を介した内皮の化学走性の強力な誘導物質であるが(73)、SDF−1の血管新生効果は、発育中の消化管の血管系に限定されるようである(50)。
【0008】
血管形成は、まず出生前期の間に起こり、ヒトの腹側大動脈由来の血管芽細胞が内皮性及び造血性の両細胞因子を生じる(74、75)。近年、成人の骨髄中にも、同様の内皮前駆細胞が同定されており(51−53)、虚血性組織中での血管形成を誘導する能力を有することが示されている(59−61)。しかしながら、このような骨髄由来の前駆体の化学遊走(chemoattraction)に必要とされる虚血性部位からのシグナル、及びこれらの部位への選択的輸送(trafficking)のためにこれらの細胞によって使用される受容体は不明である。心筋梗塞の後には、新血管新生(neoangiogenesis)プロセスが起こる(62、63)が、代償性の肥大が生じている生存可能な組織を維持するには不十分であって、さらなる細胞死、最初の梗塞領域の拡大、及びコラーゲン置換をもたらす(64−66)。組織修復(remodeling)と称されるこのプロセスは、進行性の心不全をもたらす(67−69)。以下に記載されている実験では、ELRモチーフを含有するCXCケモカインがヒト骨髄由来血管芽細胞の組織虚血部位への遊走(migration)を制御するかどうかを調べるために、心筋梗塞のヌードラットモデルを使用した。さらに、選択的な骨髄のホーミングと造血性前駆体の移植は、骨髄の中に恒常的に発現されているSDF−1へのCXCR4の結合に依存するので(76−78)、CXCR4/SDF−1相互作用の阻害が、組織虚血部位へのヒト骨髄由来血管芽細胞の輸送を再誘導することによって、治療的血管形成を増強できるかどうかを調べた。該実験の結果は、IL−8、Gro−α、及びSDF−1を含むCXCケモカインが成人の骨髄依存性血管形成を制御する上で中心的な役割を果たしていることを示している。さらに、以下に記載されている第四群の実験は、幹細胞が梗塞周囲の組織に血管新生を誘導することができることを示している。
【発明の概要】
【0009】
本発明は、虚血によって損傷を受けた患者の組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法を提供する。
【0010】
本発明は、工程(b)の後であり且つ工程(c)の前に、前記内皮前駆細胞を増殖因子(growth factor)と接触させることによって、前記内皮前駆細胞を増殖させる前出の方法も提供する。
【0011】
本発明は、前記増殖因子がサイトカインである前出の方法も提供する。
【0012】
本発明は、前記サイトカインがVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである前出の方法も提供する。
【0013】
本発明は、前記増殖因子がケモカインである前出の方法も提供する。
【0014】
本発明は、前記ケモカインがインターロイキン−8である前出の方法も提供する。
【0015】
本発明は、前記内皮前駆細胞が、増殖(expansion)の前に、他の幹細胞から分離される前出の方法も提供する。
【0016】
本発明は、前記虚血によって損傷を受けた組織が心筋である前出の方法も提供する。
【0017】
本発明は、前記虚血によって損傷を受けた組織が神経系組織である前出の方法も提供する。
【0018】
本発明は、前記幹細胞が前記患者の骨髄から採取される前出の方法も提供する。
【0019】
本発明は、前記骨髄からの前記幹細胞の採取が、前記患者の骨髄からの吸引によって為される前出の方法も提供する。
【0020】
本発明は、前記患者からの前記幹細胞の採取が、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員する(mobilize)ことと、
(b)前記幹細胞を含有する血液の試料を前記患者から採取することと
を備えた方法によって為される前出の方法も提供する。
【0021】
本発明は、前記増殖因子が、皮下、経口、静脈内、又は筋肉内から患者に導入される前出の方法も提供する。
【0022】
本発明は、前記増殖因子が動員を誘導するケモカインである前出の方法も提供する。
【0023】
本発明は、前記ケモカインがインターロイキン−8である前出の方法も提供する。
【0024】
本発明は、前記増殖因子がサイトカインである前出の方法も提供する。
【0025】
本発明は、前記サイトカインがG−CSF、M−CSF、又はGM−CSFである前出の方法も提供する。
【0026】
本発明は、前記内皮前駆細胞が、CD117の発現に基づいて回収される前出の方法も提供する。
【0027】
本発明は、前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される前出の方法も提供する。
【0028】
本発明は、前記内皮前駆細胞が、CD34、VEGF−R、Tie−2,GATA−3、又はAC133のうちの1以上の発現に基づいて回収される前出の方法も提供する。
【0029】
本発明は、前記患者が、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢(limb ischemia)、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している前出の方法も提供する。
【0030】
本発明は、前記患者が虚血によって損傷を受けた組織を患う前に工程(a)が行われ、前記患者が虚血によって損傷を受けた組織を患った後に工程(c)が行われる前出の方法も提供する。
【0031】
本発明は、工程(b)と(c)の間の一期間に、前記内皮前駆細胞が凍結される前出の方法も提供する。
【0032】
本発明は、増殖された後であり且つ工程(c)が行われる前の一期間に、前記内皮前駆細胞が凍結される前出の方法も提供する。
【0033】
本発明は、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記内皮前駆細胞が前記患者に導入される前出の方法も提供する。
【0034】
本発明は、以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を前記患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う前出の方法も提供する。
【0035】
本発明は、患者の梗塞周囲組織における血管新生を刺激する方法であって、 (a)患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記患者の梗塞周囲組織における血管新生を前記内皮前駆細胞が刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた前出の方法も提供する。
【0036】
本発明は、虚血によって損傷を受けた患者の組織への内皮前駆細胞の輸送を選択的に増加させる方法であって、
(a)内皮前駆細胞を患者に投与することと、
(b)ケモカインを前記患者に投与することによって、前記虚血によって損傷を受けた組織に前記内皮前駆細胞を誘引することを備えた前出の方法も提供する。
【0037】
本発明は、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0038】
本発明は、前記内皮前駆細胞と同時に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0039】
本発明は、前記内皮前駆細胞を投与した後に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0040】
本発明は、前記ケモカインがCXCケモカインである前出の方法も提供する。
【0041】
本発明は、前記CXCケモカインがインターロイキン−8、Gro−α、又は間質由来因子−1からなる群から選択される前出の方法も提供する。
【0042】
本発明は、前記ケモカインがCCケモカインである前出の方法も提供する。
【0043】
前記CCケモカインがRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4からなる群から選択される請求項34の方法。
【0044】
本発明は、前記患者の末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織への注入(injection)によって、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0045】
本発明は、患者の中の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法も提供する。
【0046】
本発明は、間質由来因子−1(SDF−1)とCXCR4との相互作用が、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される前出の方法も提供する。
【0047】
本発明は、アンギオテンシン変換酵素阻害剤、AT1−受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備えた前出の方法も提供する。
【0048】
本発明は、患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法も提供する。
【0049】
本発明は、SDF−1の産生が、患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される前出の方法も提供する。
【0050】
本発明は、患者の癌を治療する方法であって、前記癌に関連して増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させ、それにより前記患者の前記癌を治療することを備えた方法も提供する。
【0051】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生されたケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を患者に投与して、このような増殖細胞への内皮前駆細胞の輸送を減少させることにより前記患者の前記癌を治療することを備えた方法も提供する。
【0052】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0053】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0054】
本発明は、前記受容体がCD117受容体である前出の方法も提供する。
【0055】
本発明は、内皮前駆細胞又はマスト前駆細胞中の目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法も提供する。
【0056】
本発明は、前記ベクターがトランスフェクションによって前記細胞中に挿入される前出の方法も提供する。
【0057】
本発明は、前記プロモーターがプレプロエンドセリン−1プロモーターである前出の方法も提供する。
【0058】
本発明は、前記プロモーターが哺乳動物起源である前出の方法も提供する。
【0059】
本発明は、前記プロモーターがヒト起源である前出の方法も提供する。
【0060】
本発明は、癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させるのに有効な一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物を提供する。
【0061】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で回収した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)GATA−2を発現しているとして工程(c)で回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0062】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で回収したマスト前駆細胞の中からGATA−2を発現しているマスト前駆細胞を回収することと、
(d)GATA−2を発現しているとして工程(c)で回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0063】
本発明は、前記異常が虚血によって損傷を受けた組織である前記方法を提供する。
【0064】
本発明は、前記遺伝子産物がプロエンドセリンである前記方法を提供する。
【0065】
本発明は、前記遺伝子産物がエンドセリンである前記方法を提供する。
【0066】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善させる方法であって、 (a)前記患者のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法を提供する。
【0067】
本発明は、前記患者が哺乳動物起源である前記方法を提供する。
【0068】
本発明は、前記哺乳動物がヒト起源である前記方法を提供する。
【0069】
本発明は、患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系(allogeneic)の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が虚血によって損傷を受けた患者の前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法も提供する。
【0070】
本発明は、前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として取得される前記方法を提供する。
【0071】
本発明は、患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞中の内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が虚血によって損傷を受けた患者の前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法を提供する。
【0072】
本発明は、前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として取得される前記方法を提供する。
【0073】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法も提供する。
【0074】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法も提供する。
【0075】
本発明は、内皮前駆細胞を前記患者に導入することをさらに備えた前出の方法も提供する。
【0076】
本発明は、前記患者にG−CSFを導入して内皮前駆細胞を動員することをさらに備えた前出の方法も提供する。
【図面の簡単な説明】
【0077】
【図1A】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0078】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0079】
A.単核細胞の免疫選択後(25)、95%を超える生細胞がCD34を発現する。
【図1B】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0080】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0081】
B.CD34+CD117dimのサブセットは、成熟した血管内皮の表現型特性を備える集団を含んでいる。
【図1C】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0082】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0083】
C.CD34+CD117brightのサブセットは、マウス及びヒトの胚形成が急激に進行する間に生ずる初期の血管芽細胞に特徴的なマーカーを発現する集団を含んでいる。
【図1D】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0084】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0085】
D.GATA−2とGATA−3を同時発現するCD34+CD117bright細胞は、血管芽細胞となり得る造血細胞を規定する別のマーカーであるAC133も発現する。
【図2A】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0086】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0087】
A.正常血清における培養と比較して、VEGF又は虚血の血清に対する増殖応答は、CD117brightGATA−2posBAが同一ドナーのCD117dimGATA−2negBMECに比べて著しく高かった(共にP<0.01)。
【図2B】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0088】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0089】
B.VEGF又は虚血の血清の何れかを用いた培養によって増殖し、多パラメーターのフローサイトメトリー分析によってCD117brightGATA−2posとして特徴付けられる集団は、高い前方散乱(fsc)によって示されるように大きな芽細胞からなっていた。
【図2C】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0090】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0091】
C.CD117brightGATA−2pos細胞の増殖した集団では、正常な培地を用いた培養に比べ、VEGFを用いた培養後での成熟内皮細胞のマーカーの表面発現が増加せず、分化のない芽細胞増殖が示唆された。
【図3A】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0092】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
A.細胞の増殖及び成長の過増殖による玉石模様、
【図3B】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0093】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
B.DiI標識されたアセチル化LDLの均一な取込み、
【図3C】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0094】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
C.フルオレセインと複合されたmAbを用いた免疫蛍光法によって測定したCD34の発現、
【図3D】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0095】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
D.ビオチンと複合されたmAbを用いた免疫ペルオキシダーゼ法によって測定した第VIII因子の発現、及び
【図3E】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0096】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
E.特異的プローブを用いたインサイチューハイブリダイゼーションによって決定されるeNOSの発現、を示す。
【図4−1】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性 A〜C.DiI標識されたヒトCD34を豊富に含む2×106の細胞(CD34純度>95%)、2×106のCD34陰性細胞(CD34純度<5%)、又は2×106の伏在静脈内皮細胞(SVEC)を、冠動脈結紮及び梗塞後のヌードラットに静脈内注入。各ヒト細胞集団は、48時間で、梗塞されたラットの心筋内にほぼ同程度の浸潤を起こした。
【0097】
D.非梗塞ラットの心臓内にヒト細胞が見られない擬似処置
【図4−2】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性 E.CD34陽性細胞(CD34純度>95%)又はCD34陰性細胞(CD34純度<5%)の何れかを与えた梗塞ラットの骨髄及び心臓におけるヒトGATA−2のmRNA発現の測定(GAPDHの発現によって測定した全ヒトRNAに対して規格化した)。虚血組織内のGATA−2mRNAを、虚血のない同一実験条件下で存在するGATA−2mRNAに対する増加倍数として表す。CD34+又はCD34−の何れかの細胞を注入した虚血ラットの骨髄は、ほぼ同じレベルのヒトGATA−2mRNAを含有し、虚血後のGATA−2mRNA発現においてほぼ同じ倍率の誘導を示した。これに対して、CD34+細胞を注入したラットの虚血した心臓は、CD34−細胞を注入したものよりも極めて高いレベルのヒトGATA−2mRNAを含有していた。さらに、梗塞後のGATA−2mRNAの発現の増加率は、CD34−細胞と比較して、CD34+細胞が浸潤した心臓の方が2.6倍高く、CD34+フラクション内のGATA2+細胞が虚血心筋内に選択的に輸送されることが示唆された。
【図4−3】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性F.ヒトCD34+細胞を注入してから2週後のヌードラットの梗塞床内にある血管の連続断面。この血管は、DiIの同時発現によって規定されるヒト内皮細胞、フルオレセイン複合mAbを用いた免疫蛍光法によって測定されるHLA class I、及びビオチン複合mAbを用いた免疫ペルオキシダーゼ法によって測定される第VIII因子を取り込む。
【図5A】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0098】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 A.左心室駆出率(LVEF)は、レシピエントの各グループにおいてLAD結紮後に極度に低下したが、G−CSFによって動員された成人のCD34+細胞の注入のみが、LVEFの有意かつ持続的な回復を伴った(p<0.001)。LVEFの回復は、LAD結紮後のLVEFと梗塞前のLVEFとの平均改善%として計算した。
【図5B】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0099】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 B.同様に、左心室収縮末期の面積(LVAs)は、レシピエントの各グループにおいてLAD結紮後に著しく増加したが、G−CSFによって動員された成人のCD34+細胞の注入のみが、LVAsの有意かつ持続的な減少を伴った(p<0.001)。LVAsの減少は、LAD結紮後のLVAsと梗塞前のLVAsとの平均改善%として計算した。
【図5C】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0100】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 C.各グループの代表的な心臓エコー検査の例を示す。LAD結紮後48時間で弛緩期の機能が各ラットにおいて極度に損なわれる。注入後2週では、弛緩期の機能はCD34+細胞を注入したラットにおいてのみ改善する。この効果は15週の時点で持続している。
【図5D】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0101】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 D.梗塞後15週では、CD34+細胞を注入したラットは他の各グループよりも、正常ラットに対する平均心係数の低下が有意に抑えられた(p<0.001)。
【図6A】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0102】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0103】
A.ヒトCD34+細胞を注入したラットの梗塞域は、肉芽組織の微小血管分布及び細胞性の顕著な増加、多数の毛細血管(矢印の頭)、栄養供給血管(矢印)、並びにマトリックス沈着及び繊維症の減少を示す(×200)。
【図6B】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0104】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0105】
B.これに対して、生理食塩水を注入したコントロールラットの梗塞域は、乏細胞性の密な繊維組識(矢印)からなる心筋瘢痕を示す(×200)。
【図6C】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0106】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0107】
C.ヒトCD34+細胞を注入したラットの虚血心筋は、多数の第VIII因子陽性な間質内の血管芽細胞(矢印)を示し、さらに第VIII因子陽性な毛細血管(矢印の頭)の広汎な増加を示す(×400)。
【図6D】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0108】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0109】
D.生理食塩水を注入したラットの虚血心筋は、第VIII因子陽性血管芽細胞(矢印)を含有せず、第VIII因子陽性血管分布を有する肉芽組織の局所領域(矢印の頭)のみを示す(×400)。
【図6E】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0110】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0111】
E.生理食塩水を注入したラットにおける梗塞後15週のラット心筋のトリクローム染色(×25)。左心室前壁(ant.)内のコラーゲンが豊富な心筋瘢痕は、青く染色され、生きた心筋は赤く染色される。コラーゲン沈着の限局的な島(青)は、左心室後壁(post)にも存在する。前壁の心筋の塊が広範に喪失しており、コラーゲンの沈着及び瘢痕の生成が左心室壁の厚みのほぼ全体を通じて広がり、その結果、動脈瘤の膨張及び典型的なEKG異常(持続性ST部分の上昇)を起こしている。
【図6F】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0112】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0113】
F.これに対して、高度に精製したCD34+細胞を注入したラット中に存在する梗塞後15週のラット心筋のトリクローム染色(×25)では、前壁(ant.)内の生きた心筋の塊が増加しており、EKGが正常であるとともに、梗塞域の大きさが顕著に減少したことが示されている。梗塞域と生きた心筋の接合部では、多数の血管が明瞭である。左心室後壁(post)には、限局性コラーゲンの沈着はない。
【図6G】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0114】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0115】
G.CD34+細胞を与えたラットは、他の各グループに比べ、正常な左心室心筋に対する瘢痕組織の平均サイズが著しく減少した(p<0.01)。心外膜及び心内膜の両領域を含む梗塞の大きさを、デジタルプラニメーター画像解析装置を用いて測定し、所与の切片での心室総外周に対する百分率で示した。各動物に対し、最終的な梗塞サイズを10〜15個の切片の平均値として計算した。
【図7A】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0116】
A.代表的なドナー成人から白血球フェレーシスによって採取した、G−CSFによって動員された骨髄由来細胞の4パラメーターフローサイトメトリーによる表現型キャラクタリゼーション。7−AAD染色によって規定される生細胞のみを分析した。使用した各マーカーに対して、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。CD34+CD117brightのサブセットは、マウス及びヒトの胚形成が急激に進行する間に生ずる初期の血管芽細胞に特徴的なマーカーを発現する集団を含むが、成熟内皮のマーカーを含まない。これらの細胞は、CXCケモカイン受容体も発現する。
【図7B】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0117】
B.ヌードラットへ静脈内注入したDiI標識ヒトCD34を豊富に含む細胞(CD34純度>98%)は、冠動脈を結紮及び梗塞した後にラットの心筋に浸潤するが、擬似手術後48時間では浸潤しない。
【図7C】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0118】
C.純度が2%、40%、又は98%のCD34+の、G−CSFによって動員された2.0×106のヒト骨髄由来細胞を与え、マッソンのトリクローム又は免疫ペルオキシダーゼ法で染色した、代表的なラットから得られたLAD結紮後2週の心筋梗塞床。2%又は40%の何れかの純度のCD34+細胞を与えたラットの梗塞域は、青く染色された乏細胞性の密な繊維組識からなる心筋瘢痕を示す(×400)。これに対し、純度98%のヒトCD34+細胞を注入したラットの梗塞域は、肉芽組織の微小血管分布及び細胞性の顕著な増加、多数の毛細血管、及び最小限のマトリックス沈着と最小限の繊維症を示す(×400)。さらに、抗第VIII因子mAbの結合後の免疫ペルオキシダーゼ染色によって、純度98%のCD34+細胞を注入したラットの梗塞床では第VIII因子陽性な毛細血管の数が著しく増加することが明らかになったが、これは他のラットのいずれにも見られない(×400)。
【図8A】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0119】
A、B. LAD結紮を行っているラットのGAPDHに対する、ラット心筋のIL−8及びGro−αmRNA発現の時間依存的な増加。
【図8B】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0120】
A、B. LAD結紮を行っているラットのGAPDHに対する、ラット心筋のIL−8及びGro−αmRNA発現の時間依存的な増加。
【図8C】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0121】
C.代表的な動物から得られた、基準時間、LAD結紮後12時間、48時間でのIL−8、Gro−α、及びGAPDHのmRNA発現。
【図8D】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0122】
D. LAD結紮を行っているラットの血清中のラットIL−8/Gro−αタンパク質の時間依存的測定。ヒト骨髄由来CD34+細胞の虚血ラット心筋への遊走は、ラットIL−8又はIL−8/Groケモカインファミリー受容体CXCR1及びCXCR2の何れかに対するmAbによって阻害されるが(全てp<0.01)、VEGF又はその受容体のFlk−1に対するmAbによっては阻害されない(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図8E】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0123】
E. IL−8又はGro−αの何れかに対する、又はこれらの血管新生誘発性ケモカインの表面受容体(CXCR1又はCXCR2)に対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋での輸送を対照抗体に比較して40〜60%低下させた(p<0.01)。
【図9A】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0124】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0125】
A.IL−8は、用量に依存して化学走性を誘導する(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図9B】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0126】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0127】
B.化学走性は、IL−8及びSDF−1α/βに応答して増大するが、VEGF又はSCFでは増大しない。
【図9C】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0128】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0129】
C.代表的な蛍光顕微鏡法によって、静脈内に注入したDiI標識されたヒトCD34+細胞(純度98%)のラット心臓への浸潤が、生理食塩水を注入した場合に比べIL−8を心臓内に注入した後に増加することが示された。
【図9D】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0130】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0131】
D.1mg/mlのIL−8を心臓内に注入すると、生理食塩水、VEGF,又は幹細胞因子(SCF)の注入に比べ、DiI標識されたヒトCD34+細胞(純度98%)のラット心臓へのインビボでの化学走性が著しく増大する(p<0.01、結果を異なる3実験の平均値±平均値の標準誤差として表す)。
【図10A】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0132】
A.図示したのは、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地で96時間培養した単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両方で定量化した。虚血の血清は、他の各条件と比較して、CD117brightGATA−2pos細胞のより大きな増殖性応答を誘導した(いずれもp<0.01)。
【図10B】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0133】
B.静脈内注入後2〜14日したラットの骨髄中のヒトCD34+細胞の比率は、LAD結紮によって誘導される虚血後に著しく増加する(結果を、3つの動物における骨髄調査の各時点での平均値±平均値の標準誤差として表す)。
【図10C】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0134】
C. CXCR4、SDF−1,又は抗CD34に対するmAbの、LAD結紮後のラット骨髄及び心筋へのヒトCD34+細胞輸送に対する効果。抗CXCR4又は抗SDF−1を同時投与すると、純度98%のCD34+細胞のラット骨髄への輸送が48時間で著しく低下し、虚血心筋への輸送が増加した(結果を、3つのLAD結紮した動物において注入後48時間に行なった骨髄及び心臓の研究の平均値±平均値の標準誤差として表す)。
【図10D】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0135】
D. CXCR4、SDF−1,又は抗CD34に対するmAbの、LAD結紮後のラット骨髄及び心筋へのヒトCD34+細胞輸送に対する効果。抗CXCR4又は抗SDF−1を同時投与すると、純度98%のCD34+細胞のラット骨髄への輸送が48時間で著しく低下し、虚血心筋への輸送が増加した(結果を、3つのLAD結紮した動物において注入後48時間に行なった骨髄及び心臓の研究の平均値±平均値の標準誤差として表す)。
【図11A】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0136】
A、B.心筋梗塞後の、ヒトCD34+細胞のLVAsの低下に対する効果(A)とLVEFの改善に対する効果(B)。生理食塩水を注入した動物と比較して、純度98%のCD34+を含む2.0×106のヒト細胞を注入すると、LVEFは有意に改善しLVAsを有意に減少させたものの(いずれもp<0.01)、純度2%及び40%のCD34+を含む2.0×106のヒト細胞を注入しても、これらのパラメーターにいかなる効果も与えなかった。しかし、抗CXCR4を純度40%のCD34+細胞と共に同時投与すると、LVEFを有意に改善しLVAsを減少させて(いずれもp<0.01)、純度98%のCD34+細胞を用いたレベルに近づいた。
【図11B】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0137】
A、B.心筋梗塞後の、ヒトCD34+細胞のLVAsの低下に対する効果(A)とLVEFの改善に対する効果(B)。生理食塩水を注入した動物と比較して、純度98%のCD34+を含む2.0×106のヒト細胞を注入すると、LVEFは有意に改善しLVAsを有意に減少させたものの(いずれもp<0.01)、純度2%及び40%のCD34+を含む2.0×106のヒト細胞を注入しても、これらのパラメーターにいかなる効果も与えなかった。しかし、抗CXCR4を純度40%のCD34+細胞と共に同時投与すると、LVEFを有意に改善しLVAsを減少させて(いずれもp<0.01)、純度98%のCD34+細胞を用いたレベルに近づいた。
【図11C】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0138】
C.LAD結紮し、純度2%、40%又は98%のCD34+を含む2.0×106のヒト細胞を注入してから15週後のマッソントリクロームで染色したラット心臓の切片。純度2%及び40%のCD34+細胞を注入したラットの心臓は、純度98%のCD34+細胞を注入したラットの心臓に比べ、前壁の塊が大量に失われ、コラーゲンが沈着(青)し、中隔が肥大した。抗CXCR4mAbを純度40%のCD34+細胞と共に同時投与すると、左心室壁の塊が増加し、コラーゲン沈着が減少した。
【図11D】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0139】
D.98%を超える純度のCD34+細胞を与えたラット、又は抗CXCR4mAbと共に純度40%のCD34+細胞を与えたラットにおける左心室心筋の平均瘢痕/正常比率は、純度2%及び40%のCD34+細胞を与えたラットと比較して著しく低下する(p<0.01)ことを示している。(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図12】CD34+CD117bright血管芽細胞をLAD結紮したラットの血清と共に培養すると、CCR1及びCCR2の表面発現が増加するのに対し、CCR3及びCCR5の表面発現は不変である。
【図13】梗塞した心筋では、CCRに結合した幾つかのケモカインのmRNA発現が時間に依存して増加することを示している。
【図14】MCP−1、MCP−3、及びRANTESに対する、あるいはエオタキシンに対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋への輸送を、対照抗体に比べて40〜60%だけ減少させた(p<0.01)。
【図15】エオタキシンを非梗塞の心臓内に注入すると、CD34+血管芽細胞の輸送を1.5〜1.7倍に増加させたが、他方、増殖因子VEGF及び幹細胞因子の注入は、血管芽細胞の増殖を増加させたものの化学走性には効果がなかった(図示せず)。
【発明を実施するための形態】
【0140】
本明細書において使用する次の各用語は、別段の記述がない限り、以下に述べる定義に従うものとする。
【0141】
本明細書において使用する「BMEC」は、骨髄由来の内皮細胞と定義される。
【0142】
本明細書において使用する血管形成は、骨髄由来の内皮細胞前駆細胞などの「プレ血液(pre−blood)」細胞からの新しい血管の生成と定義される。
【0143】
本明細書において使用する動員は、骨髄由来の内皮細胞前駆細胞が骨髄を離れて末梢循環に入るように誘導することと定義される。当業者であれば、動員された幹細胞を白血球フェレーシスによって身体から採取し得ることが理解できるであろう。
【0144】
本明細書において使用する虚血は、局所的な部位への血管が閉塞したために、その部位への血液供給(循環)が不十分であることと定義される。
【0145】
本明細書において使用するサイトカインは、細胞を増殖又は活性化させる因子と定義される。
【0146】
本明細書において使用するケモカインは、細胞を体内の異なる部位に移動させる因子と定義される。
【0147】
本明細書において使用する虚血性心疾患は、心臓への血液供給量が減少するあらゆる病状と定義される。
【0148】
本明細書において使用する「血管新生」は、既存の血管細胞からの血管の生成と定義される。
【0149】
本明細書において使用する虚血性心疾患は、心臓への血液供給量が減少するあらゆる病状と定義される。
【0150】
本明細書において使用する「VEGF」は、血管内皮増殖因子と定義される。「VEGF−R」は、血管内皮増殖因子受容体と定義される。「FGF」は、繊維芽細胞増殖因子と定義される。「IGF」は、インシュリン様増殖因子と定義される。「SCF」は、幹細胞因子と定義される。「G−CSF」は、顆粒球コロニー刺激因子と定義される。「M−CSF」は、マクロファージコロニー刺激因子と定義される。「GM−CSF」は、顆粒球マクロファージコロニー刺激因子と定義される。「MCP」は、単球走化性タンパク質と定義される。
【0151】
本明細書において使用する「CXC」ケモカインは、ケモカインの構造と定義される。各「C」はシステインを表し、「X」は任意のアミノ酸を表す。
【0152】
本明細書において使用する「CC」ケモカインは、ケモカインの構造と定義される。各「C」はシステインを表す。
【0153】
本明細書において使用する「回収された」は、CD117、GATA−2、GATA−3、CD34を含む(これらには限定されない)細胞上の特異的マーカーに対して作られる、検出可能に標識された抗体を結合する回収可能な細胞に基づいて細胞を検出し、取得することを意味する。
【0154】
本明細書に記載されている、患者に投与するケモカインは、タンパク質の形態又は核酸の形態とすることができる。
【0155】
本発明は、虚血によって損傷を受けた患者の組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法を提供する。
【0156】
さらなる態様において、前記内皮前駆細胞は、工程(b)と工程(c)の間の一期間に凍結される。ある態様において、前記虚血によって損傷を受けた組織は心筋である。別の態様において、前記虚血によって損傷を受けた組織は神経系組織である。
【0157】
ある態様において、工程(b)の後であり且つ工程(c)の前に、増殖因子と接触させることによって、前記内皮前駆細胞を増殖させる。さらなる態様において、前記増殖因子はサイトカインである。さらなる態様において、前記サイトカインはVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである。別の態様において、前記増殖因子はケモカインである。さらなる態様において、前記ケモカインはインターロイキン−8である。ある態様において、前記内皮前駆細胞は、増殖の前に、他の幹細胞から分離される。さらなる態様において、前記内皮前駆細胞は、増殖後であり且つ工程(c)の前に、一定期間凍結される。
【0158】
ある態様において、工程(a)は、前記患者が虚血によって損傷を受けた組織を患う前に行われ、工程(c)は、前記患者が虚血によって損傷を受けた組織を患った後に行われる。
【0159】
ある態様において、前記幹細胞は、前記患者の骨髄から直接採取される。さらなる態様において、前記幹細胞は、前記患者の骨髄からの吸引によって採取される。ある態様において、前記患者からの前記幹細胞は、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員することと、
(b)続いて、幹細胞を含有する血液の試料を前記患者から採取することとを備えた方法によって採取される。
【0160】
さらなる態様において、前記増殖因子は、皮下、経口、静脈内、又は筋肉内から導入される。ある態様において、前記増殖因子は動員を誘導するケモカインである。さらなる態様において、前記ケモカインはインターロイキン−8である。ある態様において、前記増殖因子はサイトカインである。さらなる態様において、前記サイトカインは、G−CSF、M−CSF、又はGM−CSFである。
【0161】
本発明は、前記内皮前駆細胞が、CD117の発現に基づいて回収される前出の方法も提供する。
【0162】
本発明は、前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される前出の方法も提供する。ある態様において、前記遺伝子産物は、以下の群:プレプロエンドセリン−1、ビッグエンドセリン、エンドセリン−1から選択される。
【0163】
ある態様において、前記内皮前駆細胞はGATA−2を発現し、前記内皮前駆細胞は、細胞内GATA−2の発現又はそれらの細胞中のGATA−2活性を検出することによって回収される。
【0164】
ある態様において、前記患者は、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している。
【0165】
ある態様において、前記内皮前駆細胞は、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記患者に導入される。
【0166】
ある態様において、前記方法は、以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を前記患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う。
【0167】
本発明は、患者の梗塞周囲組織における血管新生を刺激する方法であって、 (a)患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記内皮前駆細胞が前記患者の梗塞周囲組織における血管新生を刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法も提供する。
【0168】
本発明は、患者の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を選択的に増加させる方法であって、
(a)内皮前駆細胞を前記患者に投与することと、
(b)ケモカインを前記患者に投与することによって、前記虚血によって損傷を受けた組織に前記内皮前駆細胞を誘引することを備えた方法も提供する。
【0169】
ある態様では、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0170】
別の態様では、前記ケモカインは、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される。別の態様では、前記ケモカインは、前記内皮前駆細胞と同時に前記患者に投与される。別の態様では、前記ケモカインは、前記内皮前駆細胞を投与した後に、前記患者に投与される。ある態様において、前記ケモカインはCXCケモカインである。さらなる態様において、前記CXCケモカインは、インターロイキン−8、Gro−α、又は間質由来因子−1からなる群から選択される。ある態様において、前記ケモカインはCCケモカインである。さらなる態様において、前記CCケモカインはRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4からなる群から選択される。
【0171】
ある態様において、前記ケモカインは、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織への注入によって、前記患者に投与される。
【0172】
本発明は、患者の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法も提供する。
【0173】
ある態様において、前記間質由来因子−1とCXCR4との前記相互作用は、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される。ある態様において、前記方法は、ACE阻害剤、AT受容体遮断薬、又はβ遮断薬、ng酵素阻害剤(ng enzyme inhibitor)、AT1受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備える。
【0174】
本発明は、患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法も提供する。ある態様において、前記SDF−1の産生は、抗SDF−1又は抗CXCR−4モノクローナル抗体を前記患者に投与することによって阻害される。
【0175】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることによって前記患者の前記癌を治療することを備えた方法も提供する。
【0176】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生されたケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への前記内皮前駆細胞の輸送を減少させ、それにより前記患者の前記癌を治療することを備えた方法も提供する。
【0177】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0178】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0179】
本発明は、内皮前駆細胞又はマスト前駆細胞中に存在する目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法も提供する。
【0180】
本発明は、前記ベクターがトランスフェクションによって前記細胞中に挿入される前出の方法も提供する。
【0181】
本発明は、前記プロモーターがプレプロエンドセリン−1プロモーターである前出の方法も提供する。
【0182】
本発明は、前記プロモーターが哺乳動物起源である前出の方法も提供する。
【0183】
本発明は、前記プロモーターがヒト起源である前出の方法も提供する。
【0184】
本発明は、癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させる効果を有する一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物を提供する。
【0185】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で採取した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0186】
ある態様において、前記異常は、虚血によって損傷を受けた組織である。ある態様において、前記遺伝子産物はプロエンドセリンである。ある態様において、前記遺伝子産物はエンドセリンである。ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0187】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で回収したマスト前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0188】
ある態様において、前記異常は、虚血によって損傷を受けた組織である。ある態様において、前記遺伝子産物はプロエンドセリンである。ある態様において、前記遺伝子産物はエンドセリンである。ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0189】
本発明は、心筋梗塞を患った患者の心筋の機能を改善する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法を提供する。
【0190】
ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0191】
本発明は、患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法も提供する。
【0192】
別の態様において、前記同種異系の幹細胞は、胚、胎児、又は臍帯血を起源として採取される。
【0193】
本発明は、患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法を提供する。
【0194】
別の態様において、前記同種異系幹細胞は、胚、胎児、又は臍帯血を起源として採取される。
【0195】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法も提供する。
【0196】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法も提供する。ある態様において、前記方法は、前記患者に内皮前駆細胞を導入することをさらに備える。ある態様において、前記方法は、前記患者にG−CSFを導入して、内皮前駆細胞を動員することをさらに備える。
【0197】
以下に記載されている実験の詳細を参照することによって、本発明をよりよく理解することができるであろう。しかしながら、詳細に記載されている具体的な実験は、その後に記載されている特許請求の範囲においてより完全に記載されている本発明を説明するためのものにすぎないことが、当業者には容易に理解できるであろう。
【実験の詳細】
【0198】
第一群の実験
実験方法及び結果
1.骨髄由来細胞の動員と同定
G−CSF動員後、高純度ヒトCD34細胞(>90%陽性)の60〜80%が幹細胞因子受容体CD117を同時発現し(図1a)、そのうち15−25%はCD117の発現が明瞭で、75〜85%はCD117の発現が不明瞭であった。4種類のパラメーターに関する解析により、VEGFR−2(Flk−1)を発現する2種類のCD34細胞集団が確認された。1つ目はCDdim細胞の20〜30%を占め、VEGFR−2を高濃度に発現し、2つ目はCD117brightの10〜15%を占め、VEGFR−2発現は低濃度であった(図1b)。CD34+117dim細胞集団中のVEGFR−2陽性細胞であって、CD34+117bright分画中には含まれない細胞は、Tie−2、ecNOS、vWF、E−セレクチン(CD62E)及びICAM(CD54)の高発現を含む成熟血管内皮の表現型特性を示した。一方で、図1cで示すように、CD34+CD117bright細胞分画中に属するものであってCD34+CD117dim細胞分画に属さないVEGFR−2陽性細胞は、マウス及びヒト胚形成中に出現するGATA−2、GATA−3及び低レベルのTie−2を含む原始血管芽細胞に特徴的なマーカーを発現した。さらに、GATA−2及びGATA−3を同時発現したCD117bright細胞はまた、強いAC133陽性であった(AC133は、血管芽細胞になり得る造血細胞集団であることを確定するもう一つのマーカーであることが近年示唆されている(2)(図1d)。しかしながら、AC133発現は、GATA−2及びGATA−3が陰性であるCD117dim細胞の集団においても検出されたので、胚性骨髄由来血管芽細胞(BA)表現型の同定には、AC133に加えてGATA−2、GATA−3及びCD117brightが付随して発現することが必要であると本発明者らは結論する。このように、G−CSF処理によって、成熟骨髄由来内皮細胞(BMEC)の大きな集団、及び胚性血管芽細胞(BA)の表現型特性を有するこれより小さな骨髄由来の集団が末梢循環に動員される。
【0199】
2.骨髄由来細胞の増殖
VEGF(27)又は局所的虚血(10−13)の何れかにより動物モデルで循環する内皮細胞前駆体の出現率が増加することが示されたので、次に、VEGF及び虚血性血清中の因子(28)に対するBA及びBMECの増殖応答を比較した。図2aで示すように、VEGF又は虚血性血清の何れかを添加して96時間培養を行った後、CD117brightGATA−2posBAは、同じドナー由来のCD117dimGATA−2negBMECと比較して有意に高い増殖応答を示した。VEGFを添加した場合、BMECはベースラインを超えて1.2倍の増殖増加を示したのに対し、BAの増殖は2.9倍に増加を示し(p<0.01)、その一方で、心筋梗塞のLewラット由来の虚血性血清を添加した場合は、BMECの増殖は正常血清を超えて1.7倍の増加を示したのに対し、BAの増殖は4.3倍の増加を示した(p<0.01)。VEGF又は虚血性血清の何れかを添加して培養すると、GATA−2、GATA−3、及びCD117brightを含む未成熟マーカーは継続的に発現するが、成熟内皮細胞のマーカーは発現しない大型芽細胞のBA集団が著しく増殖したので(図2b)、芽細胞が分化せずに増殖したことが示唆される。内皮増殖培地を用いてフィブロネクチン上でCD34陽性単層培養を7日間行ったところ(29)、増殖の著しい敷石状パターンが観察され(図3a)、ほとんどの接着単層(>95%)において、アセチル化LDLの一定の取り込み、及びCD34、第VIII因子およびeNOSの同時発現を含む内皮細胞に特徴的な特性が認められた(図3b−e)。BMEC集団はVEGF又は虚血性血清中のサイトカインに対する増殖応答が低いため、培養中に著しく過剰増殖した内皮細胞は、おそらくGATA−2、GATA−3及びCD117brightの細胞表面への発現によって特徴付けられるBA集団が起源であろう。
【0200】
3.インビボにおける、骨髄由来CD34+細胞の局所的虚血部位への遊走
次に、局所的虚血誘導後、インビボにおける遊走及び増殖特性について、骨髄由来ヒト細胞と末梢血管由来ヒト細胞との比較を行った。図4a〜cで示すように、冠動脈結紮を行い、梗塞を起こさせたヌードラットに対して、2X106個のDiI−標識ヒトCD34陽性細胞(>95% CD34純度)、CD34陰性細胞(<5% CD34純度)、又は伏在静脈内皮細胞(SVEC)を静脈注射すると、48時間後にラット心筋において同程度の浸潤が観察された(30)。局所的梗塞を伴う心臓の非疾患部位においてはほとんどDiI標識細胞は検出されず(データを示さない)、G−CSF動員CD34+細胞、成熟ヒト内皮細胞いずれも正常心筋部位には浸潤しなかったので(図4d)、細胞輸送は特に梗塞領域へと向けられた。CD34+及びCD34−細胞の虚血性心筋への遊走は同数であったが、正常心筋に対して虚血性心筋におけるヒトGATA−2のmRNA発現の比例増加率(31)は、CD34−細胞の場合と比較してCD34が非常に富む細胞を注射した場合は2.6倍に増加した(p<0.001)(図4e)。さらに、ヒトCD34+細胞を注射して2週間後に、DiI、HLAクラスI、及び第VII因子の同時発現によって定義されるヒト内皮細胞を取り込む血管を検出し得たが、CD34−細胞又はSVEcの注射後には検出されなかった(図4f)。これらの結果を考え合わせると、成人骨髄由来ヒトCD34+細胞には、局所的虚血部位からのインビボのシグナルに選択的に応答して遊走性、局在性、及び内皮分化の増大を示す集団が含有されることが示される。
【0201】
4.梗塞ラット心筋へのG−CSF動員ヒトCD34+細胞注射の影響
次に、G−CSF動員ヒトCD34+(>95%)細胞、CD34−(<5%)細胞、末梢伏在静脈細胞、又は生理的食塩水注射による梗塞ラット心筋への機能的影響を比較した。LAD結紮後、それぞれの群の受容動物において、左心室駆出率(LVEF)が25〜43%低下し、左心室収縮終期領域は44〜90%増大し、左心室機能が著しく抑制された(図5a及びb)。驚いたことには、G−CSF動員成人ヒトCD34+細胞を注射して2週間以内に、LVEFが22±6%(p<0.001)回復した(図5a)。この影響は長期間続き、フォローアップ終了時である15週間後までに、34±4%まで増加した。その一方、G−CSF動員ヒトCD34−細胞、伏在静脈内皮細胞、又は生理的食塩水を注射しても、LVEFに対する影響は皆無であった。同様にして、G−CSF動員ヒトCD34+細胞を注射することにより、左心室収縮終期領域が、2週間後までに26±8%、15週間後までに37±6%減少したが、他の受容動物群ではこのような影響は見られなかった(p<0.001%)(図5b)。各群の代表的な心エコー例を図5Cに示す。さらに、15週間後のCD34+細胞を注射したラットの梗塞後の平均心係数は、正常ラットに対して26±8%減少したにすぎなかったが、他の各群の平均心係数は48〜59%減少した(p<0.001)(図5d)。
【0202】
梗塞して2週間後の組織学的検査(33)によると、CD34+細胞注射によって、対照群と比較すると梗塞部位内において肉芽組織の微小血管形成及び細胞密度の有意な増加とマトリックス沈着及び繊維形成の減少が伴うことが明らかとなった(図6a及びb)。さらに、ヒトCD34+細胞を注射したラットの虚血性心筋には、対照ラットの虚血性心筋と比較して、有意に多数の第VIII因子陽性間質血管芽細胞及び毛細血管が含まれていた(図6c及びd)。毛細血管数の定量によって、CD34+細胞を投与されたラットの梗塞部位内では新血管新生の有意な増加が示された(高倍率視野あたりの第VIII因子陽性毛細血管平均数は、92±5、対して生理食塩水対照では51±4、p<0.01)が、正常心筋においては示されなかった(高倍率視野当たりの毛細血管数、36±2対37±3)。CD34−細胞又はSVECが浸潤した虚血性ラット心筋では、毛細血管数の増加は全く見られなかった。梗塞して15週間後に、高純度CD34+細胞を投与したラットにおいては、他の各群と比較して、前自由壁(anterior free wall)内の生存能力を有する心筋量が増加すると共に梗塞部位の有意な減少が示された(図6e及びf)。CD34+細胞が浸潤した組織の梗塞部位と生存能力を有する心筋との連結部位において、多数の血管がはっきり示された。対照群では、コラーゲン沈着及び瘢痕形成が左心室壁厚のほとんど全体に広がり、動脈瘤拡大及び典型的なEKG異常が認められる一方、CD34+細胞を投与したラットでは、梗塞瘢痕は左心室壁厚のわずか20〜50%にのみ広がっていた。さらに、CD34+細胞を投与したラットでは、非−梗塞部位における病的なコラーゲン沈着が顕著に減少していた。全体的に、瘢痕/正常左心室心筋の平均比率は、CD34+細胞を投与されたラットでは13%であり、それに対して他の各群では36〜45%であった(P<0.01)(図6g)。
【0203】
考察
前記の実験により、ヒト骨髄由来内皮細胞前駆体による梗塞床の新血管新生によって、心筋虚血の齧歯類モデルにおいて、瘢痕発生を予防し、生存能力を有する心筋を維持し、心室機能を向上させることが示される。梗塞後、梗塞部位に接する生存能力を有する心筋組織が有意な程度に肥大を起こす(5、34〜35)。梗塞組織内の新血管新生は組織修復過程において不可欠な要素であると考えられているにも関わらず(36、37)、正常な状態下では、毛細血管ネットワークは組織成長に対応できず、肥大化しているが生存能力を有する心筋によるより大きい要求を支持することが不可能で、その後酸素供給及び栄養供給が不十分となりアポトーシスが引き起こされる。心筋梗塞瘢痕内の新血管新生の進展には、浸潤白血球上で発現しているウロキナーゼ型プラスミノーゲン活性化因子(u−PA)によってプラスミノーゲンを活性化した後、潜在性のコラゲナーゼ及び他のプロテイナーゼを活性化することが必要であると考えられる(38)。u−PA−/−マウスにおいて、欠損している梗塞後心筋血行再建がコンジェニック(cogenic)野生型系統マウスからの骨髄移植により回復したことから、この過程において骨髄由来内皮前駆体が重要であることが示された(38)。ヒト単核細胞及び腫瘍細胞系列において、u−PA mRNA転写及び蛋白質分解活性は、コロニー刺激因子、G−CSF、M−CSF、及びGM−CSFによって有意に促進されるので(39−41)、このことは、治療によって梗塞部位の再血管形成を行うために、これらのサイトカインをインビボ又はエクスビボで使用して多数のヒト成人骨髄由来血管芽細胞を動員し、分化させ得ることの論拠となる。転写因子GATA−2の細胞表面及びRNA発現によって、虚血性部位からのシグナルに応じて増殖して梗塞部位へと移動し、その後新血管新生プロセスに関与するヒト成人骨髄由来血管芽細胞が選択的に同定されると考えられる。特に興味深いことに、GATA−2は、ET−1(強力な血管収縮物質であり、肥大作用を有する自己分泌ペプチドである)の前駆体分子であるプレプロエンドセリン−1(ppET−1)の内皮細胞転写のコファクターである(42)。ppET−1転写は、心筋梗塞後にレニン−アンジオテンシン神経ホルモン系の活性化の結果産生されるアンジオテンシンIIによっても増加するため(43)、アンジオテンシンII細胞表面受容体シグナリングとGATA−2転写活性化との相乗的作用により、梗塞床に浸潤している血管芽細胞は高レベルのET−1を分泌している可能性がある。梗塞瘢痕内に新たに形成された血管が、正常心筋内の血管よりも厚い壁を有し、より強い血管作動物質に対しても低い血管拡張反応を示すという所見(44)は、自己分泌ET−1活性促進による影響と一致し、新血管新生脈管構造が浸潤したGATA−2陽性血管芽細胞由来である可能性を支持する。
【0204】
前記実験の結果を総合すると、胚性血管芽細胞の表現型特性及び機能的特性を持つG−CSF動員成人ヒトCD34+細胞を注射することによって、梗塞血管床において新血管新生を刺激することが可能であり、したがって心筋梗塞においてコラーゲン沈着及び瘢痕形成が減少することが示される。新血管新生によりもたらされる心筋組織修復の減少程度は注目すべきものであるが、ヒト血管芽細胞の注入とACE阻害又はAT1−受容体阻害とを組み合わせることによってアンジオテンシンII依存性心筋繊維芽細胞増殖、コラーゲン分泌、及びプラスミノーゲン活性因子阻害因子(PAI)産生を低下させ、心筋機能をさらに向上させることが可能である(45、46)。心筋梗塞組織の血行再建のために、サイトカインによって動員されたヒト骨髄由来の自家血管芽細胞を現在用いられている治療と併用することによって(47−49)、心筋梗塞後の左心室の組織修復に伴う罹患率及び死亡率を有意に減少させる可能性がもたらされる。
【0205】
第二群の実験
方法
1.サイトカインによって動員されたヒトCD34+細胞の精製
10mg/kgの組換えG−CSF(Amgen、CA)を4日間毎日皮下注射(SC)処置したヒトから、単一ドナーの白血球フェレーシス生成物を採取した。単核細胞をフィコール−ハイパック法によって分離し、高度に精製されたCD34+細胞(>98%陽性)を抗CD34モノクロナール抗体(mAb)(Miltenyi Biotech Ltd、CA)で被覆された磁気ビーズを用いて採取した。精製されたCD34細胞を、CD34、CD117、VEGFR−2、Tie−2、GATA−2、GATA−3、AC133、vWF、eNOS、CD54、CD62E、CXCR1、CXCR2、CXCR4に対するフルオレセイン複合mAbで染色し、FACScan(Becton Dickinson, CA)を用いた4パラメーター蛍光によって分析した。
【0206】
2.ヒト内皮前駆細胞の増殖の研究
20%の正常ラットの血清、虚血ラットの血清、又は20ng/mlのVEGFの何れかを含むRPMI培地内で、単一ドナーのCD34陽性細胞を96時間培養し、次いで、[3H]チミジン(Amersham Life Science Inc、IL、米国)(1mlCi/ウェル)を適用し、LK Betaplate液体シンチレーションカウンター(Wallace、Inc.、Gaithersburg、MD)で取込みを測定した。各条件で96時間培養した後、CD117brightGATA−2pos細胞 の比率もフローサイトメトリー法によって定量化した。
【0207】
3.ヒト骨髄由来の内皮前駆細胞の化学走性
メンブレン(8mm孔)(Neuro Probe、MD)を装着した48穴の走化性チャンバー内に、高度に精製したCD34+細胞(>98%陽性)を播種した。37℃にて2時間インキュベートした後、チャンバーを反転させ、0.2、1.0、及び5.0mg/mlのIL−8、1.0mg/mlのSDF−1α/β、VEGF、及びSCFを含有する培地中で3時間にわたり細胞を培養した。前記メンブレンをメタノールを用いて固定し、Leukostat(Fischer Scientific、Ill)で染色した。10高倍率視野内で遊走する細胞を数えて化学走性を計算した。
【0208】
4.動物、外科的処置、ヒト細胞の注入、及び組織への細胞遊走の定量化
Rowett(rnu/rnu)胸腺欠損ヌードラット(Harlan Sprague Dawley、Indianapolis、Indiana)を、「Columbia University Institute for Animal Care and Use Committee」によって承認された研究に使用した。麻酔後、左開胸を施行し、心膜を開き、左前下行(LAD)冠動脈を結紮した。擬似手術したラットには、冠動脈の周囲を縫合しないで同様の外科的処置を施行した。LAD結紮から48時間後に、CXCR1、CXCR2、CXCR4、CD34、ラットIL−8(ImmunoLaboratories、日本)及びラットSDF−1(R)&D Systems、MN)、又はアイソタイプ対照抗体に対して既知の阻害活性を有するmAbの存在下又は不在下で、G−CSF動員後に単一ドナーから取り出したDiI標識された2.0×106のヒトCD34+細胞(純度>95%、40%、<2%)を尾静脈に注入した。対照動物にはLAD結紮後に生理食塩水を与えた。各グループは6〜10匹のラットからなる。注入後2日して屠殺したラットの心臓内のDiI蛍光を評価することによって、ヒト細胞を注入した後の心筋の浸潤の定量化を実施した(1高倍率視野当たりのDiI陽性な細胞の数として表され、1サンプル当たり最低5視野を調べた)。12匹のラットにおいて、基準日、2、7、及び14日目に、ラット骨髄の総集団に対するHLAクラスI陽性細胞の比率を、フローサイトメトリー法及びRT−PCR法を用いて分析することによってラット骨髄のヒト細胞による浸潤の定量化を実施した。
【0209】
5.心筋機能の分析
細胞又は生理食塩水の注入後2、6、及び15週で、高周波線形配列の変換器(SONOS 5500、Hewlett Packard、Andover、MA)を用いて、基準時間(LAD結紮から48時間後)に心臓エコー検査を実施した。乳頭の中間と頂端のレベルで2D画像を撮影した。2方向の面積長さ法(bi−plane area−length method)によって、弛緩期末期(EDV)及び収縮期末期(ESV)の左心室容積を測定し、左心室駆出率%(LVEF)を[(EDV−ESV)/EDV]×100として計算した。収縮期末期における左心室面積(LVAs)を、僧帽弁のレベルで心臓エコー検査法によって測定した。LVEFの回復及びLVAsの減少は、LAD結紮後の異なる時刻でのLVEF又はLVAsのそれぞれの数値の梗塞前数値に対する平均改善度として計算した。
【0210】
6.組織学及び免疫組織化学
ヒト細胞又は生理食塩水を注入後2週及び15週に、外植されたラットの心臓に対して組織学の研究を実施した。切除後、各実験動物からの左心室を頂端から底部まで10〜15の横断切片にスライスした。代表的な切片を組織検査のためにホルマリンに入れ、免疫ペルオキシダーゼ法を用いて抗第VIII因子mAbで新たに染色して、毛細血管の密度を定量し、又はマッソントリクロームで染色して検鏡板にのせた。デジタルプラニメーター画像解析装置を用いて、心外膜及び心内膜の両領域を含めた梗塞された表面の長さを測定し、心室総外周に対する百分率で表した。各心臓からの全切片の平均値として、最終的な梗塞サイズを計算した。
【0211】
7.ラットのCXCケモカインmRNA及びタンパク質発現の測定
3匹の正常なラット及び12匹のLAD結紮したラットの心臓から、標準的な方法でPoly(A)+mRNAを抽出した。RT−PCR法を使用して、基準時間、LAD結紮してから6、12、24、及び48時間のラットIL−8及びGro−αのmRNAの心筋での発現を、GAPDHの発現によって測定されるラットの全RNAに対して規格化した後に数量化した。オリゴ(dT)15マー及びランダムヘキサマーでプライミングし、Monoleyマウスのリンパ増殖性ウイルス逆転写酵素(Invitrogen、Carlsbad、CA、米国)で逆転写した後、Taqポリメラーゼ(Invitrogen、Carlsbad、CA、米国)、放射標識ジデオキシヌクレオチド([α32p]−ddATP:3、000Ci/mmol、Amersham、Arlington Heights、IL)、並びにラットIL−8、Gro−α及びGAPDHに対するプライマー(Fisher Genosys、CA)を用いて、ポリメラーゼ連鎖反応(PCR)法でcDNAを増幅した。ラットのIL−8、Gro−α及びGAPDHに対するプライマーペア(センス/アンチセンス)は、それぞれ、gaagatagattgcaccgatg(配列番号1)/catagcctctcacatttc(配列番号2)、gcgcccgtccgccaatgagctgcgc(配列番号3)/cttggggacacccttcagcatcttttgg(配列番号4)、及びctctacccacggcaagttcaa(配列番号5)/gggatgaccttgcccacagc(配列番号6)であった。2%アガロースゲルに標識された試料を加え、電気泳動法によって分離し、X線撮影のために−70℃で6時間にわたり露光した。ラットIL−8/GroホモログCINC(ImmunoLaboratories、日本)に対するポリクロナール抗体を用いた市販のELISA法によって、基準時間、LAD結紮後6、12、24、及び48時間目に、4匹のラットにおいて、ラットIL−8/Gro−αの血清量を測定した。ラットIL−8/Gro−αタンパク質の既知の量に対して作成された光学濃度(OD)値の標準曲線に従って、各血清試料中のタンパク質の量を計算した。
【0212】
実験手順及び結果
1.内皮前駆細胞の選択的輸送
98%を超える純度になるように、G−CSFによって動員されたヒトCD34細胞に免疫的選択を施した後、60〜80%が幹細胞因子受容体CD117を同時発現した。四重パラメーター解析によって(図7a)、CD117bright細胞の10〜15%が胚血管芽細胞の表現型特性を示し、転写因子GATA−2とGATA−3、及び内皮前駆細胞の指標となることが最近示された(79)AC133と同様に、VEGFR−2及びTie−2の低レベルの表面発現を伴うことが見出された。これらの細胞は、vWF、eNOS及びE−セレクチンなどの成熟内皮細胞のマーカーを発現していなかったが、CXCケモカイン受容体1、2、及び4に対しては陽性であった。DiI標識された2.0×106のヒトCD34+細胞(純度>98%、40%、及び2%)をLAD結紮したRowettヌードラットに静脈内注入すると、48時間でラット心筋の稠密な浸潤を伴った(図7b)。心臓の局所梗塞に影響されない部位にはDiI標識細胞はほとんど検出されず(図示せず)、DiI標識細胞は擬似手術したラットからの心筋には浸潤しなかった(図7b)ことから、これらのDiI標識細胞は特に梗塞部位に向けて輸送されていた。98%を超える純度のヒトCD34+細胞を与えたラットは、注入後2週間までに、梗塞床の微小血管分布が増加し、マトリックス沈着及び繊維症が減少することを示した(図7c)。98%を超える純度のCD34+を含む2.0×106の細胞を与えたラットの梗塞床においては、2%又は40%の何れかの純度のCD34+を含む2.0×106の細胞を与えたラットの類似の領域においてよりも、1高倍率視野当たりの第VIII因子陽性な毛細血管の数が3倍以上多くなった(p<0.01、図7c)。さらに、これらの毛細血管の大多数は、HLAクラスI分子を発現したことからヒト由来であった(図示せず)。このように、ヒト骨髄由来の様々な細胞集団が梗塞床に遊走するが、血管形成には臨界数の内皮前駆細胞が選択的に輸送されることが必要であると考えられる。
【0213】
2.梗塞された心筋によるCXCケモカインの産生に対する虚血の影響
ヒト白血球の化学走性及び組織の浸潤は、特異的なケモカインとCXC細胞表面受容体との相互作用によって制御されるため、本発明者らは次に、梗塞されたラット心筋によって産生されるCXCケモカインへの虚血の影響を検討した。図8a〜cに見られるように、梗塞された心筋では、CXCR1/2に結合したELR陽性のケモカインであるIL−8及びGro−αのmRNA発現が時間に依存した増加を示し、LAD結紮後6〜12時間で発現が最大となった。非梗塞の心筋と比較して、LAD結紮後の組織は、mRNAの総含量に対して規格化した後で、これらのELR陽性の血管新生誘発性ケモカインの7.2〜7.5倍高いmRNA量を発現した(p<0.001)。さらに、血清のIL−8のレベルは、LAD結紮後6〜12時間以内に8〜10倍増加し(p<0.001)、48時間でもなお増加したままであった(図8d)。IL−8又はGro−αの何れかに対する、又はこれらの血管新生誘発性ケモカインの表面受容体(CXCR1又はCXCR2)に対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋での輸送を対照抗体に比較して40〜60%低下させた(p<0.01、図8E)。
【0214】
3.ケモカインに対するヒト骨髄由来CD34+血管芽細胞の化学走性応答
その後の実験で、本発明者らはヒト骨髄由来CD34+血管芽細胞のIL−8に対するインビトロ及びインビボでの化学走性応答を直接測定した。図9aに示すように、ヒトCD34+細胞のインビトロでの化学走性は、濃度0.2〜5μ/mlのIL−8によって、用量に依存して誘導された。骨髄間質細胞によって恒常的に産生されるELR−ケモカインであるSDF−1は、IL−8と類似の濃度にて、ほぼ同程度のCD34+細胞の化学走性を誘導した(図9b)。これに対して、増殖因子VEGF又は幹細胞因子(SCF)によっては、化学走性は誘導されなかった。さらに、IL−8を1μg/mlだけ非梗塞の心臓内に注入すると、CD34+細胞のインビボでの化学走性を誘導したが(図9c)、対照として用いたVEGF及びSCFは何れもインビボでの化学走性に何ら効果がなかった(図9d)。これらの結果を総合すると、ELR陽性なケモカインの組織発現が亢進されると、骨髄由来の内皮前駆細胞の組織虚血部位への化学走性を誘導することによって、インビボでの血管形成が増強されることが示唆される。
【0215】
4.骨髄から心筋へのヒトCD34陽性細胞の輸送を再誘導するためのCXCR4/SDF−1相互作用の遮断
LAD結紮したラットからの虚血性の血清は、静脈内に注入したヒトCD34+血管芽細胞が損傷した心筋へ輸送されるのを増強するうえに、CD34+CD117bright血管芽細胞集団の循環を急速に増大させ、同時にこれらの細胞の骨髄への輸送を増加させた。図10aに示すように、VEGF又は虚血の血清の何れかを2日間培養すると、CD34+CD117bright血管芽細胞の増殖を、それぞれ2.8倍及び4.3倍に増加させた(p<0.01)。さらに、図10bに示すように、2×106のヒトCD34陽性細胞(純度>95%)を静脈内に注入後2〜14日で、LAD結紮後の虚血ラットから得た骨髄は、正常なラットの骨髄と比較して、5〜8倍高いレベルのヒトCD34+CD117bright血管芽細胞を含有していた(p<0.001)。SDF−1は骨髄間質細胞によって恒常的に発現され、活発に循環している循環CD34+細胞の骨髄での遊走を優先的に促進するため(80)、本発明者らは、ヒトCD34+CD117bright血管芽細胞の虚血ラット骨髄へのホーミングの増加が、SDF−1/CXCR4相互作用の増大のためかどうかを検討した。図10cに示すように、ヒトCXCR4又はラットSDF−1に対するmAbの同時投与は、抗CD34対照抗体と比較して、静脈内投与されたCD34+ヒト血管芽細胞の虚血ラット骨髄への遊走を著しく阻害した(いずれもp<0.001)。さらに、ヒトCXCR4又はラットSDF−1に対するmAbの同時投与は、CD34+ヒト血管芽細胞の虚血ラット心筋への輸送を、それぞれ平均24%及び17%増加させた(いずれもp<0.001、図10d)。2週までには、ヒトCD34+細胞を抗CXCR4mAbと組合せて与えたラットの心筋梗塞床は、CD34+細胞をアイソタイプの対照抗体と組合せて与えたラットの心筋梗塞床と比較して、微小血管分布が3倍超の増加を示した。これらの結果から、静脈内注入したCD34+血管芽細胞は梗塞心筋へ輸送され、ELR+のケモカインの産生が増大するのに応答して血管形成を誘導するが、このプロセスの効率は、血管芽細胞の骨髄へのSDF−1応答性の遊走が伴うことによって著しく低下することが示唆される。CXCR4/SDF−1相互作用を遮断すると、梗塞後に、拡大され循環するヒトCD34陽性細胞集団の骨髄から心筋への輸送を再誘導し、その結果、梗塞床の新血管新生を増強させる。
【0216】
5.心筋機能の改善
LAD結紮後、左心室機能は極度に低下するが、98%を超える純度のCD34+細胞を注入すると2週間以内に左心室の大きさと機能に顕著な回復を伴い、これらの効果は経過観察した15週の全期間にわたり持続した(図11a及びb)。98%を超える純度のCD34+細胞を与えたラットでは、左心室の収縮期末期の面積が梗塞直後に比べて15週までに平均37±6%だけ減少し(図11a)、左心室駆出率(LVEF)は、15週までに平均34±4%だけ回復した(図11b、p<0.001)。純度2%又は40%のCD34+を含む、G−CSFによって動員された2×106のヒト細胞の静脈内注入が、虚血心筋へのほぼ同程度の輸送にもかかわらず心筋機能を顕著には改善しなかったことから、これらのパラメーターの改善は、注入されたCD34+細胞の数に依存する(図11a及びb)。しかし、純度40%のCD34+を含む、G−CSFによって動員されたヒト骨髄由来の細胞と共に、抗CXCR4mAbを同時投与すると、98%を超える純度のCD34+で観察されるレベルまで、LVEFの回復が顕著に改善しLVAsが顕著に減少した。トリクローム染色によって、左心室の塊及びコラーゲン沈着における顕著な差異がこれらのグループ間に観察された(図11c)。純度2%のCD34を含む2×106のヒト細胞を与えたラットでは、左心室の前壁が繊維組織によって完全に入れ替わり、中隔の著しい代償性肥大が存在した。純度40%のCD34を含む2×106のヒト細胞を与えたラットの心臓でも同様の変化が見られた。これに対して、98%を超える純度のCD34+を含む2×106のヒト細胞を与えたラットの心臓では、これより著しく大きな前壁の塊が維持され、中隔の大きさは正常であり、コラーゲンの沈着は最小限であった。特に興味深いことには、純度40%のCD34を含む2×106のヒト細胞を抗CXCR4mAbと共に与えたラットの心臓も、同様に、心筋前壁の塊の増加、中隔肥大の減少、及びコラーゲン沈着の減少を示した。総合すると、繊維性瘢痕/正常左心室心筋の平均比率は、純度2%及び40%のCD34+細胞を与えたラットが36〜45%であるのに対して、98%を超える純度のCD34+細胞を与えたラット、又は純度40%のCD34+細胞を抗CXCR4mAbと共に注入したラットでは、それぞれ、13%及び21%であった(p<0.01、図11d)。したがって、臨界数の内皮前駆細胞の選択的な輸送を増大することによって梗塞床の血管形成が増強されると、組織修復のプロセスがさらに抑制され、生存可能な心筋を救出し、心臓機能を改善することになる。
【0217】
考察
本研究は、虚血組織によって産生されるELR+ケモカインが、骨髄由来の内皮細胞前駆細胞に対して化学遊走物質の濃度勾配を生じることによって、虚血部位での代償的な血管形成の進展を制御することを実証するものである。ELR+CXCケモカインであるIL−8とELR−CXCケモカインであるSDF−1は何れも、CD34+内皮前駆細胞の化学走性、並びに成熟内皮に対して同様の効果を示すが(73)、異なる血管外の部位で発現すると、内皮前駆細胞の方向性の出現に対してそれらは生物学的に相反する影響を及ぼし、その結果、組織の新血管新生に対してもそれらは生物学的に相反する影響を及ぼす。これらの相互作用を理解することで、本発明者らは、心筋での輸送を増加させ、梗塞床の血管形成を誘導し、虚血後の心室の組織修復を抑制し、心筋機能を改善するために、ヒト骨髄由来のCD34+細胞の特異的なサブセットの化学走性特性を操作し増強することができた。
【0218】
基底膜を通る骨髄由来の前駆細胞の遊走は、メタロプロテイナーゼ−9(MMP−9、ゼラチナーゼB)などのタンパク質分解酵素の分泌に依存するため(81)、心臓内のメタロプロテイナーゼの活性が、血管芽細胞の循環からの血管外遊出及び梗塞域内への経内皮遊走を決定する重要な要因かもしれない。IL−8は、好中球中の細胞内貯蔵顆粒から潜在型MMP−9を迅速に(20分以内)放出するように誘導し(82〜83)、非ヒト霊長類におけるインビボでの静脈内投与後、血清のMMP−9のレベルを最大1000倍増加させる(84)。IL−8は骨髄前駆細胞におけるMMP−9の発現を顕著に増加させ(81)、MCP−9に対する中和抗体は骨髄前駆細胞の遊走を抑制するため(85)、本発明者らの研究の結果は、血管芽細胞の浸潤とその後の梗塞床の血管形成が、IL−8に依存したMMP−9分泌の増加に起因するかもしれないことを示唆している。
【0219】
潜在型MMP−9の活性化及びこれと同時に起こるマウスの心筋梗塞瘢痕組織内の新血管新生は、梗塞床に浸潤する骨髄前駆細胞によって同時発現されるウロキナーゼタイプのプラスミノゲン活性化因子(u−PA)に依存することが示された(81)。ヒト細胞内のu−PAの転写及びタンパク質分解活性は、G−CSF及び他のコロニー刺激因子によって顕著に増加する(86〜88)。IL−8によって誘導される化学走性及び前駆細胞の遊走には、機能性G−CSF受容体を介して送達されるシグナルの存在がさらに必要とされるため(89)、IL−8によって媒介される虚血部位への血管芽細胞の輸送にはu−PA活性の増加が必要である可能性がある。このことは、IL−8が高レベルで産生されたにもかかわらず、心筋梗塞後に通常観察される梗塞床における新血管新生の程度が限られていたこと(62、63)を説明すると思われるし、さらに虚血組織の血行再建術治療に使用するためにコロニー刺激因子をインビボ投与又はエクソビボ投与してヒト骨髄由来の血管芽細胞を動員し分化させることに理論的根拠を与える。
【0220】
CXCケモカインであるSDF−1の骨髄間質細胞による恒常的な産生は、造血前駆細胞の骨髄へのホーミング及び移植にとって必須であると考えられる(76〜78)。さらに、SDF−1 −/−マウスは胃腸管の血管新生(50)及び心室中隔の形成(90)の両者に欠陥があるため、非造血組織におけるSDF−1の発現は発達中の血管系において役割を果たす。骨髄由来の内皮前駆細胞はCXCR4を発現し(80)、本明細書で示すようにSDF−1に対して化学走性の応答を示すため、胚形成中又は組織損傷後に、非造血部位でのSDF−1の発現が誘導されることは、組織の新血管新生のプロセスにおいて重要な要素であるかもしれない(91)。CXCR4/SDF−1相互作用の遮断によって、本発明者らがヒト骨髄由来の血管芽細胞の組織虚血部位への輸送を再誘導することができたということは、SDF−1がヒト内皮前駆細胞に対する生物学的に活性な化学走性因子であること、並びにそれが非造血部位で発現されると血管新生誘発活性を有するかもしれないことを強く主張するものである。虚血組織部位におけるSDF−1及び他のケモカインの発現の増大及び局在化が、血管形成を増強する上でELR+CXCケモカインと相乗的であり得るか否かは、将来の研究課題であろう。総合すると、本研究の結果は、IL−8、Gro−α、及びSDF−1を含むCXCケモカインがヒト骨髄に依存する血管形成を制御する際に中心的な役割を果たすこと、さらに、これらのケモカインとヒト骨髄由来の自家血管芽細胞上に存在するそれらの受容体との相互作用を操作することは、組織虚血後の治療的血管形成の潜在的な効力を増強できることを示唆するものである。
【0221】
第3群の実験
実験方法と結果
1.心筋虚血は、CCケモカイン産生を誘導してヒトCD34+血管芽細胞のCCケモカイン受容体の発現を増加させる
ヒト単核細胞の化学走性及び組織浸潤は細胞表面受容体と特定のケモカインリガンドとの相互作用によって制御されるので、血管芽細胞のCCケモカイン受容体発現及び梗塞ラット心筋によるCCケモカイン産生の速度論に対する虚血の影響を研究した。図12に示したように、LAD結紮ラットの血清と共にCD34+CD117bright血管芽細胞を培養すると、CCR1及びCCR2の表面発現は増加したが、CCR3及びCCR5の表面発現に変化はなかった。
【0222】
図13に示したように、梗塞した心筋では、時間に応じていくつかのCCR−結合ケモカインのmRNA発現が増加することが示された。梗塞した心筋では、総mRNA含量で正規化したところ(全てp<0.001)、CCR−2結合CCケモカインMCP−1が8倍を上回る濃度で発現し、MCP−3およびRANTESのmRNAが3〜3.5倍高い濃度で発現し、CCR−3結合ケモカインエオタキシンも発現することが発見された。研究した梗塞組織全てにおいて、これらのCCケモカイン類の発現増加が示されたが、CCR5−結合CCケモカインMIP−1アルファ又はMIP−1ベータの発現誘導は示されなかったので、この遺伝子発現パターンは比較的特異的なものと考えられる。
【0223】
2.虚血性心筋へのヒトCD34+血管芽細胞の輸送は、CC及びCXCケモカイン類の誘導発現によって調節される
次に、虚血性心筋へのヒトの血管芽細胞の輸送が、前記で確認したCCケモカインの誘導発現と関連するかどうかについて研究した。MCP−1、MCP−3、及びRANTESに対する遮断mAb、又はエオタキシンに対する遮断mAbを同時投与すると、ヒトの血管芽細胞の心筋への輸送が対照抗体と比較して40−60%減少した(p<0.01)、(図14)。CCケモカインが血管芽細胞の虚血性心筋への化学走性を媒介することを証明するために、エオタキシンに応じた血管芽細胞の化学走性をインビボで測定した。図15に示したように、心臓の非梗塞部にエオタキシンを心臓内注射すると、CD34+血管芽細胞輸送の1.5−1.7倍の増加が誘導されたが、成長因子VEGF及び幹細胞因子を注射しても、血管芽細胞増殖は増加するものの化学走性に何ら影響はなかった(図示せず)。
【0224】
第4群の実験
筋細胞の大きさの決定。正常なラットの心臓及び梗塞組織切片の梗塞部位並びに梗塞周囲縁、遠位部の筋細胞をトリクローム染色して、大きさを測定した。10−15倍の高倍率視野の各々について100〜200個の筋細胞の横径及び縦径(mm)をイメージプロプラスソフトウェアを用いて400xで測定した。
【0225】
パラフィン組織切片のDNA末端標識による筋細胞アポトーシスの測定
単一細胞レベルにおけるアポトーシスをin situで検出するために、デオキシヌクレオチジルトランスフェラーゼ(TdT)によるDNA末端標識、TUNEL法を用いた(ベーリンガーマンハイム、ドイツ・マンハイム)。生理食塩液又はCD34+ヒト細胞のいずれかを注射して2週間後のLAD結紮ラット及び陰性対照として健康なラットから、ラットの心筋組織切片を切除した。簡単に説明すると、組織を脱パラフィン化してプロティナーゼKで消化し、37℃で60分間、浸潤雰囲気中でTdT及びフルオレセイン標識dUTPとインキュベートした。フルオレセイン特異的抗体複合アルカリホスファターゼで30分間インキュベーションした後、DNAが断片化された核を青色に染色して、TUNEL染色を視覚化した。
【0226】
1.新血管新生は肥大した心筋をアポトーシスから保護する
新血管新生を誘導して心臓機能の改善をもたらす機構を研究した。LAD結紮後2週間経つと、生理食塩水対照群の梗塞周囲縁の筋細胞の外観はゆがんで形が不規則となり、筋細胞の直径は梗塞のないラットと同様となることが(0.020mm+/−0.002 対 0.019mm+/−0.001)結果から示された。一方、CD34+細胞を投与したラットの梗塞周囲縁の筋細胞の形は規則正しい楕円形をしており、対照群ラットの筋細胞より有意に大きかった(直径0.036mm+/−0.004 対 0.019mm+/−0.001、p<0.01)。筋細胞特異的マーカーであるデスミンの染色及びDNA末端標識を同時に行うと、CD34+細胞を注射したラットの梗塞した左心室では、生理食塩液対照群と比較すると、検出されたアポトーシスした筋細胞の数は6分の1であった(アポトーシス指数1.2+0.6対7.1+0.7、p<0.01)。これらの差は梗塞周囲縁内において特に顕著で、生理食塩水処理対照群の形状の不規則な小筋細胞は、アポトーシス核指数が最高であった。また、生理食塩水対照群では左心室壁の75〜80%に筋細胞のアポトーシスが広がっていたが、CD34+細胞を注射したラットでは、筋細胞のアポトーシスは梗塞部から遠位の左心室の最大20〜25%に検出されたにすぎなかった。これらの結果を総合すると、CD34+細胞注射によって誘導された梗塞領域の血管形成及び梗塞周囲の血管新生は、生理食塩水対照群において明らかな遠心性に広がるアポトーシス誘発プロセスを阻止して、梗塞周囲領域内の肥大した心筋の生存を可能にし、心筋機能を改善する。
【0227】
2.早期の新血管新生は後期の心筋組織修復を予防する
最後の実験群によって、対照群および実験群(生理食塩液対CD34+細胞)における2週間後の梗塞周囲縁の筋細胞のアポトーシスの程度と、その後4ヶ月にわたる心筋の組織修復の進行との比較を明らかにした。LVEFが初期に減少し、LVASが増加することは同じであるが、2週間後のコラーゲン沈着又は瘢痕組織/正常な左心室心筋の平均比率は、マッソントリクローム染色によって確認したところ、生理食塩水を投与したラットでは12%であったのに対しCD34+細胞を投与されたラットでは3%であった。梗塞して15週間後では、瘢痕/正常な左心室心筋の平均比率は、CD34+細胞を投与したラットでは13%であったのに対して、それ以外の実験群(生理食塩水、CD34−、SVEC)ではいずれも36−45%であった(p<0.01)。CD34+細胞を投与したラットでは、ラット由来の筋細胞のみを含む前自由壁内に、生存可能な心筋量の有意な増加が示され、ヒトではなくラットのMHC分子を発現していることから、筋細胞はヒト幹細胞前駆体から再分化するのではなく本来の筋細胞を再利用することが確かめられた。対照群では、左心室壁厚のほとんど全体に渡ってコラーゲン沈着および瘢痕形成が広がっており、動脈瘤性拡張および典型的なEKG異常をともなっているが、CD34+細胞を投与したラットでは、左心室壁の壁厚の20〜50%のみに広がっていた。さらに、非梗塞領域における病的なコラーゲン沈着は、CD34+細胞を投与したラットでは著しく減少した。これらの結果を考え合わせると、2週間後に梗塞周囲の筋細胞においてアポトーシスの減少が観察されたことによって、肥大しているが生存可能な筋細胞の生存が延長され、15週間後までに心筋とコラーゲン及び線維組織との置換が阻止されることが示された。
【0228】
考察
梗塞周囲縁および筋細胞間で増殖した毛細血管がラット由来であることが観察され、血管形成に加えて、ヒトの血管芽細胞又は同時投与したその他の骨髄由来因子は、血管新生誘発因子の豊富な発生源となることが可能で、既存の脈管系からさらに血管新生を誘導することを可能にすることを示している。
【参照文献】
【0229】

【発明の詳細な説明】
【0001】
本出願は、2000年6月5日に出願された米国特許出願第09/587,441号の一部継続出願であり、その優先権を主張するものである。この親出願を、本明細書の一部として本願に援用する。
【0002】
本出願を通じて、様々な文献が括弧内に引用されている。本発明が属する分野の技術水準をより完全に説明するために、本出願への参照によって、これらの文献全体の開示内容を援用する。これらの参照文献の完全な書誌的引用は、本明細書の末尾に記載されている。
【発明の背景】
【0003】
心筋梗塞後の左心室の組織修復は、その後の心不全及び死亡の主原因である。毛細血管網は、肥大化しているが生存能力を有する心筋による要求の増加に追いつくことができず、心筋の死滅と繊維性置換が起こる。以下に記載されている本発明の第一群の実験は、成人の骨髄が胚血管芽細胞の表現型及び機能的な特徴を備えた内皮細胞前駆細胞を含有しており、これらの前駆細胞は動員、増殖させることが可能であって、実験的な心筋梗塞の後に、梗塞床(infarct bed)への血管形成(vasculogenesis)を誘導するために使用できることを示している。新血管新生は、生存可能な心筋組織の有意且つ持続的な増加、コラーゲンの沈着の減少、及び心筋機能の改善をもたらす。単独で、又は現在用いられている療法と組合せて、サイトカインによって動員されたヒト骨髄由来の自家血管芽細胞を心筋梗塞組織の血行再建術のために使用すると、心筋梗塞後の左心室の組織修復に伴う罹患率及び死亡率が有意に減少する可能性がある。
【0004】
狭い時間枠のうちに迅速な再潅流を行うと、急性心筋梗塞の初期死亡率が著しく減少するが、梗塞後の心不全が増加、多発する(1)。初期の梗塞部位の拡大、梗塞を取り囲む壁の厚さの漸減、及び左心室内腔の膨張を特徴とする心筋梗塞後の左心室の組織修復が、その後の心不全に対する主要な予後因子であることが明らかとされている(2、3)。このプロセスには、本来胎児の段階でしか発現されない遺伝子の転写、心臓の繊維芽細胞によるコラーゲン分泌の急速且つ進行性の増加、心室壁内への繊維組織の沈着、壁の硬さの増大、並びに弛緩期及び収縮期の両期における機能不全が伴う(4,5)。低酸素状態は心臓の繊維芽細胞によるコラーゲンの分泌を直接刺激するが、DNA合成と細胞の増殖を抑制する(6)。動物モデルでは、実験的な心筋梗塞の後、心筋の撤去が終わった時点に遅延して再潅流を行うと、組織修復に極めて有益である(7)。さらに、梗塞に付随する開存性動脈の存在は、常に、梗塞後の期間におけるヒトの生存率の改善を伴う(8)。これは、心室の組織修復過程を改善し、壁の運動の異常な変化を抑制する梗塞血管床が十分に再潅流されることに起因するものと思われる(9)。
【0005】
循環系の細胞因子又は骨髄由来の細胞因子のうち何れかを用いることによって、実験動物モデル中の虚血状態にした非心臓組織の再潅流を首尾よく行い得ることが最近実証された(10−13)。これらの研究では、これらの細胞の正確な性状は明らかにはされていないが、成人の循環系と骨髄の両者に、機能的な内皮細胞に分化する能力(血管形成と称されるプロセス(14−16))を有する前駆細胞が存在することが示されている。出生前期においては、ヒト及びヒトより下等な種の動脈の腹側内皮に由来する前駆細胞は、血管形成と血球新生の両プロセスに関与する細胞因子を生じることが示されている(17、18)。これらの細胞は、胚血管芽細胞(embryonic hemangioblast)と名付けられ、CD34、CD117(幹細胞因子受容体)、Flk−1(血管内皮細胞増殖因子受容体−2、VEGFR−2)、及びTie−2(アンジオポエチン受容体)の発現を特徴とし、VEGFに反応して、芽細胞コロニーを形成する高い増殖能を有することが示された(19−22)。GATA−2をノックアウトした胚幹細胞は、胎児の肝臓及び骨髄による終局的な造血(hematopoiesis)及び造血の開始(seeding)を完全に遮断するので(23)、この後に生じる胚血管芽細胞の成人型多能性幹細胞への増殖及び分化には、GATA−2転写因子の同時発現が関連しているものと思われる。さらに、胚腹側内皮(embryonic ventral endothelium)から分岐した造血及び内皮両細胞系列の最も初期の前駆体は、GATA−2とα4−インテグリンに加え、VEGF受容体を発現していることが示された(24)。本発明の第一群の実験は、成人の骨髄にもGATA−2陽性幹細胞前駆体が存在し、血管芽細胞の特性を表しており、血管形成を誘導するために使用することによって、実験的な心筋梗塞における組織修復と心不全を抑えることが可能であることを示している。
【0006】
既存の成熟内皮から新しい血管が増殖することは血管新生(angiogenesis)と称されており、ある種のCXCケモカインを含む数多くの因子によって制御され得る(47−50)。これに対して、血管形成(vasculogenesis)は、胚血管芽細胞の表現型的特長と血管内皮増殖因子(VEGF)等の受容体チロシンキナーゼによって制御される増殖/分化特性とを有する(54−57)骨髄由来の内皮前駆体(51−53)によって媒介される。治療的血管形成(58−61)は虚血性組織の潅流を改善させ得る能力を有するが、組織虚血の部位への内皮前駆体の選択的輸送に関与する受容体/リガンド相互作用は不明である。以下に記載されている本発明の第二群の実験は、ヒト骨髄由来の血管芽細胞上のCXC受容体とIL−8及びGro−αを含む虚血によって誘導されるELR陽性CXCケモカインとの相互作用の結果、梗塞を起こした心筋中に血管形成が生じ得ることを示している。さらに、CXCR4/SDF−1相互作用を遮断することによって、血管芽細胞を骨髄から虚血性心筋へと再誘導して輸送させることが可能となり、血管形成の増加、心筋細胞の死滅と繊維性置換(fibrous replacement)の減少、及び心機能の改善がもたらされる。前記実験の結果は、IL−8、Gro−α、及び間質由来因子−1(SDF−1、stromal−derived factor−1)を含むCXCケモカインが成人での血管形成を制御する上で中心的な役割を果たしていることを示しており、CXCケモカインと骨髄由来の血管芽細胞上に存在するそれらの受容体との相互作用を操作すると、最適な治療的血管形成及び虚血性組織の撤去(salvage)をもたらし得ることを示唆している。以下に記載されている第三群の実験は、血管芽細胞の虚血性心筋への化学走性(chemotaxis)を媒介する上でもCCケモカインが役割を果たしていることを示している。
【0007】
創傷治癒又は炎症中の血管新生反応は、既存の脈間構造中の内皮細胞と局所的に産生された因子によって調節される細胞外マトリックスとの接着性相互作用の変化に起因し、これによって、内皮細胞の移動、増殖、再構築、及び微小血管の形成が生じる(70)ものと考えられている。ヒトCXCケモカインファミリーは、内皮細胞に結合し、該細胞に対する強力な化学走性活性を有する小さな(<10kD)ヘパリン結合ポリペプチドからなる。N末端の3つのアミノ酸残基(Glu−Leu−Arg、ELRモチーフ)は、内皮細胞上に存在するCXC受容体1及び2へのIL−8及びGro−αなどのCXCケモカインの結合を決定して(49、71)、内皮の化学走性と血管新生を促進する(47−48)。これに対して、ELRモチーフを欠如するCXCケモカインは異なるCXC受容体に結合し、増殖因子によって媒介される血管新生を阻害する(49−72)。SDF−1(ELR陰性のCXCケモカイン)は、CXCR4との相互作用を介した内皮の化学走性の強力な誘導物質であるが(73)、SDF−1の血管新生効果は、発育中の消化管の血管系に限定されるようである(50)。
【0008】
血管形成は、まず出生前期の間に起こり、ヒトの腹側大動脈由来の血管芽細胞が内皮性及び造血性の両細胞因子を生じる(74、75)。近年、成人の骨髄中にも、同様の内皮前駆細胞が同定されており(51−53)、虚血性組織中での血管形成を誘導する能力を有することが示されている(59−61)。しかしながら、このような骨髄由来の前駆体の化学遊走(chemoattraction)に必要とされる虚血性部位からのシグナル、及びこれらの部位への選択的輸送(trafficking)のためにこれらの細胞によって使用される受容体は不明である。心筋梗塞の後には、新血管新生(neoangiogenesis)プロセスが起こる(62、63)が、代償性の肥大が生じている生存可能な組織を維持するには不十分であって、さらなる細胞死、最初の梗塞領域の拡大、及びコラーゲン置換をもたらす(64−66)。組織修復(remodeling)と称されるこのプロセスは、進行性の心不全をもたらす(67−69)。以下に記載されている実験では、ELRモチーフを含有するCXCケモカインがヒト骨髄由来血管芽細胞の組織虚血部位への遊走(migration)を制御するかどうかを調べるために、心筋梗塞のヌードラットモデルを使用した。さらに、選択的な骨髄のホーミングと造血性前駆体の移植は、骨髄の中に恒常的に発現されているSDF−1へのCXCR4の結合に依存するので(76−78)、CXCR4/SDF−1相互作用の阻害が、組織虚血部位へのヒト骨髄由来血管芽細胞の輸送を再誘導することによって、治療的血管形成を増強できるかどうかを調べた。該実験の結果は、IL−8、Gro−α、及びSDF−1を含むCXCケモカインが成人の骨髄依存性血管形成を制御する上で中心的な役割を果たしていることを示している。さらに、以下に記載されている第四群の実験は、幹細胞が梗塞周囲の組織に血管新生を誘導することができることを示している。
【発明の概要】
【0009】
本発明は、虚血によって損傷を受けた患者の組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法を提供する。
【0010】
本発明は、工程(b)の後であり且つ工程(c)の前に、前記内皮前駆細胞を増殖因子(growth factor)と接触させることによって、前記内皮前駆細胞を増殖させる前出の方法も提供する。
【0011】
本発明は、前記増殖因子がサイトカインである前出の方法も提供する。
【0012】
本発明は、前記サイトカインがVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである前出の方法も提供する。
【0013】
本発明は、前記増殖因子がケモカインである前出の方法も提供する。
【0014】
本発明は、前記ケモカインがインターロイキン−8である前出の方法も提供する。
【0015】
本発明は、前記内皮前駆細胞が、増殖(expansion)の前に、他の幹細胞から分離される前出の方法も提供する。
【0016】
本発明は、前記虚血によって損傷を受けた組織が心筋である前出の方法も提供する。
【0017】
本発明は、前記虚血によって損傷を受けた組織が神経系組織である前出の方法も提供する。
【0018】
本発明は、前記幹細胞が前記患者の骨髄から採取される前出の方法も提供する。
【0019】
本発明は、前記骨髄からの前記幹細胞の採取が、前記患者の骨髄からの吸引によって為される前出の方法も提供する。
【0020】
本発明は、前記患者からの前記幹細胞の採取が、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員する(mobilize)ことと、
(b)前記幹細胞を含有する血液の試料を前記患者から採取することと
を備えた方法によって為される前出の方法も提供する。
【0021】
本発明は、前記増殖因子が、皮下、経口、静脈内、又は筋肉内から患者に導入される前出の方法も提供する。
【0022】
本発明は、前記増殖因子が動員を誘導するケモカインである前出の方法も提供する。
【0023】
本発明は、前記ケモカインがインターロイキン−8である前出の方法も提供する。
【0024】
本発明は、前記増殖因子がサイトカインである前出の方法も提供する。
【0025】
本発明は、前記サイトカインがG−CSF、M−CSF、又はGM−CSFである前出の方法も提供する。
【0026】
本発明は、前記内皮前駆細胞が、CD117の発現に基づいて回収される前出の方法も提供する。
【0027】
本発明は、前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される前出の方法も提供する。
【0028】
本発明は、前記内皮前駆細胞が、CD34、VEGF−R、Tie−2,GATA−3、又はAC133のうちの1以上の発現に基づいて回収される前出の方法も提供する。
【0029】
本発明は、前記患者が、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢(limb ischemia)、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している前出の方法も提供する。
【0030】
本発明は、前記患者が虚血によって損傷を受けた組織を患う前に工程(a)が行われ、前記患者が虚血によって損傷を受けた組織を患った後に工程(c)が行われる前出の方法も提供する。
【0031】
本発明は、工程(b)と(c)の間の一期間に、前記内皮前駆細胞が凍結される前出の方法も提供する。
【0032】
本発明は、増殖された後であり且つ工程(c)が行われる前の一期間に、前記内皮前駆細胞が凍結される前出の方法も提供する。
【0033】
本発明は、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記内皮前駆細胞が前記患者に導入される前出の方法も提供する。
【0034】
本発明は、以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を前記患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う前出の方法も提供する。
【0035】
本発明は、患者の梗塞周囲組織における血管新生を刺激する方法であって、 (a)患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記患者の梗塞周囲組織における血管新生を前記内皮前駆細胞が刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた前出の方法も提供する。
【0036】
本発明は、虚血によって損傷を受けた患者の組織への内皮前駆細胞の輸送を選択的に増加させる方法であって、
(a)内皮前駆細胞を患者に投与することと、
(b)ケモカインを前記患者に投与することによって、前記虚血によって損傷を受けた組織に前記内皮前駆細胞を誘引することを備えた前出の方法も提供する。
【0037】
本発明は、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0038】
本発明は、前記内皮前駆細胞と同時に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0039】
本発明は、前記内皮前駆細胞を投与した後に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0040】
本発明は、前記ケモカインがCXCケモカインである前出の方法も提供する。
【0041】
本発明は、前記CXCケモカインがインターロイキン−8、Gro−α、又は間質由来因子−1からなる群から選択される前出の方法も提供する。
【0042】
本発明は、前記ケモカインがCCケモカインである前出の方法も提供する。
【0043】
前記CCケモカインがRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4からなる群から選択される請求項34の方法。
【0044】
本発明は、前記患者の末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織への注入(injection)によって、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0045】
本発明は、患者の中の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法も提供する。
【0046】
本発明は、間質由来因子−1(SDF−1)とCXCR4との相互作用が、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される前出の方法も提供する。
【0047】
本発明は、アンギオテンシン変換酵素阻害剤、AT1−受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備えた前出の方法も提供する。
【0048】
本発明は、患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法も提供する。
【0049】
本発明は、SDF−1の産生が、患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される前出の方法も提供する。
【0050】
本発明は、患者の癌を治療する方法であって、前記癌に関連して増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させ、それにより前記患者の前記癌を治療することを備えた方法も提供する。
【0051】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生されたケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を患者に投与して、このような増殖細胞への内皮前駆細胞の輸送を減少させることにより前記患者の前記癌を治療することを備えた方法も提供する。
【0052】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0053】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0054】
本発明は、前記受容体がCD117受容体である前出の方法も提供する。
【0055】
本発明は、内皮前駆細胞又はマスト前駆細胞中の目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法も提供する。
【0056】
本発明は、前記ベクターがトランスフェクションによって前記細胞中に挿入される前出の方法も提供する。
【0057】
本発明は、前記プロモーターがプレプロエンドセリン−1プロモーターである前出の方法も提供する。
【0058】
本発明は、前記プロモーターが哺乳動物起源である前出の方法も提供する。
【0059】
本発明は、前記プロモーターがヒト起源である前出の方法も提供する。
【0060】
本発明は、癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させるのに有効な一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物を提供する。
【0061】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で回収した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)GATA−2を発現しているとして工程(c)で回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0062】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で回収したマスト前駆細胞の中からGATA−2を発現しているマスト前駆細胞を回収することと、
(d)GATA−2を発現しているとして工程(c)で回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0063】
本発明は、前記異常が虚血によって損傷を受けた組織である前記方法を提供する。
【0064】
本発明は、前記遺伝子産物がプロエンドセリンである前記方法を提供する。
【0065】
本発明は、前記遺伝子産物がエンドセリンである前記方法を提供する。
【0066】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善させる方法であって、 (a)前記患者のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法を提供する。
【0067】
本発明は、前記患者が哺乳動物起源である前記方法を提供する。
【0068】
本発明は、前記哺乳動物がヒト起源である前記方法を提供する。
【0069】
本発明は、患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系(allogeneic)の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が虚血によって損傷を受けた患者の前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法も提供する。
【0070】
本発明は、前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として取得される前記方法を提供する。
【0071】
本発明は、患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞中の内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が虚血によって損傷を受けた患者の前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法を提供する。
【0072】
本発明は、前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として取得される前記方法を提供する。
【0073】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法も提供する。
【0074】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法も提供する。
【0075】
本発明は、内皮前駆細胞を前記患者に導入することをさらに備えた前出の方法も提供する。
【0076】
本発明は、前記患者にG−CSFを導入して内皮前駆細胞を動員することをさらに備えた前出の方法も提供する。
【図面の簡単な説明】
【0077】
【図1A】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0078】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0079】
A.単核細胞の免疫選択後(25)、95%を超える生細胞がCD34を発現する。
【図1B】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0080】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0081】
B.CD34+CD117dimのサブセットは、成熟した血管内皮の表現型特性を備える集団を含んでいる。
【図1C】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0082】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0083】
C.CD34+CD117brightのサブセットは、マウス及びヒトの胚形成が急激に進行する間に生ずる初期の血管芽細胞に特徴的なマーカーを発現する集団を含んでいる。
【図1D】G−CSFはVEGF受容体を発現する2つのヒト骨髄由来の集団を動員する1つは成熟した内皮細胞の特性を備えたものであり、もう1つは胚血管芽細胞の特性を備えたものである。
【0084】
A〜Dは、代表的なドナー成人から白血球フェレーシス(leucopharesis)によって採取され、G−CSFによって動員された骨髄由来の細胞の4パラメーターフローサイトメトリーによる表現型のキャラクタリゼーションを示す(25)。生細胞(7−AAD染色によって判定する)のみを分析した。使用した各マーカーにおいて、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。
【0085】
D.GATA−2とGATA−3を同時発現するCD34+CD117bright細胞は、血管芽細胞となり得る造血細胞を規定する別のマーカーであるAC133も発現する。
【図2A】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0086】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0087】
A.正常血清における培養と比較して、VEGF又は虚血の血清に対する増殖応答は、CD117brightGATA−2posBAが同一ドナーのCD117dimGATA−2negBMECに比べて著しく高かった(共にP<0.01)。
【図2B】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0088】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0089】
B.VEGF又は虚血の血清の何れかを用いた培養によって増殖し、多パラメーターのフローサイトメトリー分析によってCD117brightGATA−2posとして特徴付けられる集団は、高い前方散乱(fsc)によって示されるように大きな芽細胞からなっていた。
【図2C】骨髄由来の血管芽細胞(BA)は、骨髄由来の内皮細胞(BMEC)よりも、VEGFと虚血の血清の両者に応答してより大きな増殖活性を有する。
【0090】
図示されているのは、蛍光GATA−2mAbによって選別され、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地内で96時間培養された単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞及びCD117dimGATA−2neg細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両者で定量化した。
【0091】
C.CD117brightGATA−2pos細胞の増殖した集団では、正常な培地を用いた培養に比べ、VEGFを用いた培養後での成熟内皮細胞のマーカーの表面発現が増加せず、分化のない芽細胞増殖が示唆された。
【図3A】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0092】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
A.細胞の増殖及び成長の過増殖による玉石模様、
【図3B】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0093】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
B.DiI標識されたアセチル化LDLの均一な取込み、
【図3C】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0094】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
C.フルオレセインと複合されたmAbを用いた免疫蛍光法によって測定したCD34の発現、
【図3D】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0095】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
D.ビオチンと複合されたmAbを用いた免疫ペルオキシダーゼ法によって測定した第VIII因子の発現、及び
【図3E】高度に精製されたヒト骨髄由来CD34細胞は、インビトロでの培養後に内皮細胞に分化する。
【0096】
高度に精製されたCD34+ヒト細胞を7日間内皮増殖培地中で培養すると、成熟した内皮細胞単層の形態的かつ独特な特徴を有する細胞が成長することになる。単層の大部分(>90%)が、
E.特異的プローブを用いたインサイチューハイブリダイゼーションによって決定されるeNOSの発現、を示す。
【図4−1】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性 A〜C.DiI標識されたヒトCD34を豊富に含む2×106の細胞(CD34純度>95%)、2×106のCD34陰性細胞(CD34純度<5%)、又は2×106の伏在静脈内皮細胞(SVEC)を、冠動脈結紮及び梗塞後のヌードラットに静脈内注入。各ヒト細胞集団は、48時間で、梗塞されたラットの心筋内にほぼ同程度の浸潤を起こした。
【0097】
D.非梗塞ラットの心臓内にヒト細胞が見られない擬似処置
【図4−2】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性 E.CD34陽性細胞(CD34純度>95%)又はCD34陰性細胞(CD34純度<5%)の何れかを与えた梗塞ラットの骨髄及び心臓におけるヒトGATA−2のmRNA発現の測定(GAPDHの発現によって測定した全ヒトRNAに対して規格化した)。虚血組織内のGATA−2mRNAを、虚血のない同一実験条件下で存在するGATA−2mRNAに対する増加倍数として表す。CD34+又はCD34−の何れかの細胞を注入した虚血ラットの骨髄は、ほぼ同じレベルのヒトGATA−2mRNAを含有し、虚血後のGATA−2mRNA発現においてほぼ同じ倍率の誘導を示した。これに対して、CD34+細胞を注入したラットの虚血した心臓は、CD34−細胞を注入したものよりも極めて高いレベルのヒトGATA−2mRNAを含有していた。さらに、梗塞後のGATA−2mRNAの発現の増加率は、CD34−細胞と比較して、CD34+細胞が浸潤した心臓の方が2.6倍高く、CD34+フラクション内のGATA2+細胞が虚血心筋内に選択的に輸送されることが示唆された。
【図4−3】心筋虚血誘導後の骨髄及び末梢血管由来のヒト細胞のインビボでの遊走特性及び増殖特性F.ヒトCD34+細胞を注入してから2週後のヌードラットの梗塞床内にある血管の連続断面。この血管は、DiIの同時発現によって規定されるヒト内皮細胞、フルオレセイン複合mAbを用いた免疫蛍光法によって測定されるHLA class I、及びビオチン複合mAbを用いた免疫ペルオキシダーゼ法によって測定される第VIII因子を取り込む。
【図5A】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0098】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 A.左心室駆出率(LVEF)は、レシピエントの各グループにおいてLAD結紮後に極度に低下したが、G−CSFによって動員された成人のCD34+細胞の注入のみが、LVEFの有意かつ持続的な回復を伴った(p<0.001)。LVEFの回復は、LAD結紮後のLVEFと梗塞前のLVEFとの平均改善%として計算した。
【図5B】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0099】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 B.同様に、左心室収縮末期の面積(LVAs)は、レシピエントの各グループにおいてLAD結紮後に著しく増加したが、G−CSFによって動員された成人のCD34+細胞の注入のみが、LVAsの有意かつ持続的な減少を伴った(p<0.001)。LVAsの減少は、LAD結紮後のLVAsと梗塞前のLVAsとの平均改善%として計算した。
【図5C】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0100】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 C.各グループの代表的な心臓エコー検査の例を示す。LAD結紮後48時間で弛緩期の機能が各ラットにおいて極度に損なわれる。注入後2週では、弛緩期の機能はCD34+細胞を注入したラットにおいてのみ改善する。この効果は15週の時点で持続している。
【図5D】G−CSFに動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、心筋機能が向上する。
【0101】
A〜Dは、G−CSFに動員された2×106のヒトCD34+(純度>95%)細胞、CD34−(純度<5%)細胞、末梢伏在静脈の細胞、又は生理食塩水を梗塞ラットの心筋へ注入することの機能上の効果を比較したものである。 D.梗塞後15週では、CD34+細胞を注入したラットは他の各グループよりも、正常ラットに対する平均心係数の低下が有意に抑えられた(p<0.001)。
【図6A】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0102】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0103】
A.ヒトCD34+細胞を注入したラットの梗塞域は、肉芽組織の微小血管分布及び細胞性の顕著な増加、多数の毛細血管(矢印の頭)、栄養供給血管(矢印)、並びにマトリックス沈着及び繊維症の減少を示す(×200)。
【図6B】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0104】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0105】
B.これに対して、生理食塩水を注入したコントロールラットの梗塞域は、乏細胞性の密な繊維組識(矢印)からなる心筋瘢痕を示す(×200)。
【図6C】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0106】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0107】
C.ヒトCD34+細胞を注入したラットの虚血心筋は、多数の第VIII因子陽性な間質内の血管芽細胞(矢印)を示し、さらに第VIII因子陽性な毛細血管(矢印の頭)の広汎な増加を示す(×400)。
【図6D】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0108】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0109】
D.生理食塩水を注入したラットの虚血心筋は、第VIII因子陽性血管芽細胞(矢印)を含有せず、第VIII因子陽性血管分布を有する肉芽組織の局所領域(矢印の頭)のみを示す(×400)。
【図6E】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0110】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0111】
E.生理食塩水を注入したラットにおける梗塞後15週のラット心筋のトリクローム染色(×25)。左心室前壁(ant.)内のコラーゲンが豊富な心筋瘢痕は、青く染色され、生きた心筋は赤く染色される。コラーゲン沈着の限局的な島(青)は、左心室後壁(post)にも存在する。前壁の心筋の塊が広範に喪失しており、コラーゲンの沈着及び瘢痕の生成が左心室壁の厚みのほぼ全体を通じて広がり、その結果、動脈瘤の膨張及び典型的なEKG異常(持続性ST部分の上昇)を起こしている。
【図6F】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0112】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0113】
F.これに対して、高度に精製したCD34+細胞を注入したラット中に存在する梗塞後15週のラット心筋のトリクローム染色(×25)では、前壁(ant.)内の生きた心筋の塊が増加しており、EKGが正常であるとともに、梗塞域の大きさが顕著に減少したことが示されている。梗塞域と生きた心筋の接合部では、多数の血管が明瞭である。左心室後壁(post)には、限局性コラーゲンの沈着はない。
【図6G】G−CSFによって動員されたヒトCD34+細胞を急性梗塞のラットに注入すると、新血管新生を誘導し、心筋の組織修復の過程を修正する。
【0114】
A〜Dは、ヘマトキシリン及びエオシン(A、B)又は抗第VIII因子mAb結合後の免疫ペルオキシダーゼ法(C、D)で染色した代表的な実験動物及び対照動物から得た、LAD結紮から2週後の梗塞ラット心筋を示す。E、Fは、LAD結紮から15週後の代表的な対照動物及び実験動物から得た梗塞ラット心筋のマッソン(Mason)トリクローム染色を示す。Gは、15週の左心室組織の瘢痕/正常%のグループ間差を示す。
【0115】
G.CD34+細胞を与えたラットは、他の各グループに比べ、正常な左心室心筋に対する瘢痕組織の平均サイズが著しく減少した(p<0.01)。心外膜及び心内膜の両領域を含む梗塞の大きさを、デジタルプラニメーター画像解析装置を用いて測定し、所与の切片での心室総外周に対する百分率で示した。各動物に対し、最終的な梗塞サイズを10〜15個の切片の平均値として計算した。
【図7A】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0116】
A.代表的なドナー成人から白血球フェレーシスによって採取した、G−CSFによって動員された骨髄由来細胞の4パラメーターフローサイトメトリーによる表現型キャラクタリゼーション。7−AAD染色によって規定される生細胞のみを分析した。使用した各マーカーに対して、陰影付きの部分は、アイソタイプ対照抗体に対するバックグラウンドの蛍光の対数を表す。CD34+CD117brightのサブセットは、マウス及びヒトの胚形成が急激に進行する間に生ずる初期の血管芽細胞に特徴的なマーカーを発現する集団を含むが、成熟内皮のマーカーを含まない。これらの細胞は、CXCケモカイン受容体も発現する。
【図7B】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0117】
B.ヌードラットへ静脈内注入したDiI標識ヒトCD34を豊富に含む細胞(CD34純度>98%)は、冠動脈を結紮及び梗塞した後にラットの心筋に浸潤するが、擬似手術後48時間では浸潤しない。
【図7C】成人骨髄由来の内皮前駆細胞は、虚血心筋に浸潤して、梗塞床の新血管新生を誘導し、コラーゲンの沈着を抑制する。
【0118】
C.純度が2%、40%、又は98%のCD34+の、G−CSFによって動員された2.0×106のヒト骨髄由来細胞を与え、マッソンのトリクローム又は免疫ペルオキシダーゼ法で染色した、代表的なラットから得られたLAD結紮後2週の心筋梗塞床。2%又は40%の何れかの純度のCD34+細胞を与えたラットの梗塞域は、青く染色された乏細胞性の密な繊維組識からなる心筋瘢痕を示す(×400)。これに対し、純度98%のヒトCD34+細胞を注入したラットの梗塞域は、肉芽組織の微小血管分布及び細胞性の顕著な増加、多数の毛細血管、及び最小限のマトリックス沈着と最小限の繊維症を示す(×400)。さらに、抗第VIII因子mAbの結合後の免疫ペルオキシダーゼ染色によって、純度98%のCD34+細胞を注入したラットの梗塞床では第VIII因子陽性な毛細血管の数が著しく増加することが明らかになったが、これは他のラットのいずれにも見られない(×400)。
【図8A】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0119】
A、B. LAD結紮を行っているラットのGAPDHに対する、ラット心筋のIL−8及びGro−αmRNA発現の時間依存的な増加。
【図8B】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0120】
A、B. LAD結紮を行っているラットのGAPDHに対する、ラット心筋のIL−8及びGro−αmRNA発現の時間依存的な増加。
【図8C】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0121】
C.代表的な動物から得られた、基準時間、LAD結紮後12時間、48時間でのIL−8、Gro−α、及びGAPDHのmRNA発現。
【図8D】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0122】
D. LAD結紮を行っているラットの血清中のラットIL−8/Gro−αタンパク質の時間依存的測定。ヒト骨髄由来CD34+細胞の虚血ラット心筋への遊走は、ラットIL−8又はIL−8/Groケモカインファミリー受容体CXCR1及びCXCR2の何れかに対するmAbによって阻害されるが(全てp<0.01)、VEGF又はその受容体のFlk−1に対するmAbによっては阻害されない(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図8E】ヒト骨髄由来内皮前駆細胞の梗塞部位への遊走は、心筋虚血によって誘導されるCXCR1/2とIL−8/Gro−αの相互作用に依存する。
【0123】
E. IL−8又はGro−αの何れかに対する、又はこれらの血管新生誘発性ケモカインの表面受容体(CXCR1又はCXCR2)に対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋での輸送を対照抗体に比較して40〜60%低下させた(p<0.01)。
【図9A】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0124】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0125】
A.IL−8は、用量に依存して化学走性を誘導する(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図9B】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0126】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0127】
B.化学走性は、IL−8及びSDF−1α/βに応答して増大するが、VEGF又はSCFでは増大しない。
【図9C】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0128】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0129】
C.代表的な蛍光顕微鏡法によって、静脈内に注入したDiI標識されたヒトCD34+細胞(純度98%)のラット心臓への浸潤が、生理食塩水を注入した場合に比べIL−8を心臓内に注入した後に増加することが示された。
【図9D】CXCケモカインは、骨髄由来ヒトCD34+細胞のラット心筋への化学走性を直接誘導する。
【0130】
A及びBは、48穴の走化性チャンバー(Neuro Probe、MD)を用いた、純度98%のヒトCD34+細胞の様々な条件でのインビトロ化学走性の結果を示す。化学走性は、試験した1条件当たり10高倍率視野(hpf)を検討したのちの1hpf当たりの遊走細胞数として定義される。
【0131】
D.1mg/mlのIL−8を心臓内に注入すると、生理食塩水、VEGF,又は幹細胞因子(SCF)の注入に比べ、DiI標識されたヒトCD34+細胞(純度98%)のラット心臓へのインビボでの化学走性が著しく増大する(p<0.01、結果を異なる3実験の平均値±平均値の標準誤差として表す)。
【図10A】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0132】
A.図示したのは、20%の正常なラットの血清、虚血ラットの血清、又は20ng/mlのVEGFを含むRPMI培地で96時間培養した単一ドナーのCD34陽性ヒト細胞の応答である。CD117brightGATA−2pos細胞の数は、[3H]チミジン取込み及びフローサイトメトリーの両方で定量化した。虚血の血清は、他の各条件と比較して、CD117brightGATA−2pos細胞のより大きな増殖性応答を誘導した(いずれもp<0.01)。
【図10B】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0133】
B.静脈内注入後2〜14日したラットの骨髄中のヒトCD34+細胞の比率は、LAD結紮によって誘導される虚血後に著しく増加する(結果を、3つの動物における骨髄調査の各時点での平均値±平均値の標準誤差として表す)。
【図10C】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0134】
C. CXCR4、SDF−1,又は抗CD34に対するmAbの、LAD結紮後のラット骨髄及び心筋へのヒトCD34+細胞輸送に対する効果。抗CXCR4又は抗SDF−1を同時投与すると、純度98%のCD34+細胞のラット骨髄への輸送が48時間で著しく低下し、虚血心筋への輸送が増加した(結果を、3つのLAD結紮した動物において注入後48時間に行なった骨髄及び心臓の研究の平均値±平均値の標準誤差として表す)。
【図10D】CXCR4/SDF−1相互作用を遮断すると、静脈内に注入したヒトCD34+血管芽細胞が骨髄から虚血心筋へ再誘導される。
【0135】
D. CXCR4、SDF−1,又は抗CD34に対するmAbの、LAD結紮後のラット骨髄及び心筋へのヒトCD34+細胞輸送に対する効果。抗CXCR4又は抗SDF−1を同時投与すると、純度98%のCD34+細胞のラット骨髄への輸送が48時間で著しく低下し、虚血心筋への輸送が増加した(結果を、3つのLAD結紮した動物において注入後48時間に行なった骨髄及び心臓の研究の平均値±平均値の標準誤差として表す)。
【図11A】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0136】
A、B.心筋梗塞後の、ヒトCD34+細胞のLVAsの低下に対する効果(A)とLVEFの改善に対する効果(B)。生理食塩水を注入した動物と比較して、純度98%のCD34+を含む2.0×106のヒト細胞を注入すると、LVEFは有意に改善しLVAsを有意に減少させたものの(いずれもp<0.01)、純度2%及び40%のCD34+を含む2.0×106のヒト細胞を注入しても、これらのパラメーターにいかなる効果も与えなかった。しかし、抗CXCR4を純度40%のCD34+細胞と共に同時投与すると、LVEFを有意に改善しLVAsを減少させて(いずれもp<0.01)、純度98%のCD34+細胞を用いたレベルに近づいた。
【図11B】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0137】
A、B.心筋梗塞後の、ヒトCD34+細胞のLVAsの低下に対する効果(A)とLVEFの改善に対する効果(B)。生理食塩水を注入した動物と比較して、純度98%のCD34+を含む2.0×106のヒト細胞を注入すると、LVEFは有意に改善しLVAsを有意に減少させたものの(いずれもp<0.01)、純度2%及び40%のCD34+を含む2.0×106のヒト細胞を注入しても、これらのパラメーターにいかなる効果も与えなかった。しかし、抗CXCR4を純度40%のCD34+細胞と共に同時投与すると、LVEFを有意に改善しLVAsを減少させて(いずれもp<0.01)、純度98%のCD34+細胞を用いたレベルに近づいた。
【図11C】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0138】
C.LAD結紮し、純度2%、40%又は98%のCD34+を含む2.0×106のヒト細胞を注入してから15週後のマッソントリクロームで染色したラット心臓の切片。純度2%及び40%のCD34+細胞を注入したラットの心臓は、純度98%のCD34+細胞を注入したラットの心臓に比べ、前壁の塊が大量に失われ、コラーゲンが沈着(青)し、中隔が肥大した。抗CXCR4mAbを純度40%のCD34+細胞と共に同時投与すると、左心室壁の塊が増加し、コラーゲン沈着が減少した。
【図11D】ヒトCD34+血管芽細胞の梗塞部位への輸送が再誘導されると、組織修復を抑制し、心筋機能を改善する。
【0139】
D.98%を超える純度のCD34+細胞を与えたラット、又は抗CXCR4mAbと共に純度40%のCD34+細胞を与えたラットにおける左心室心筋の平均瘢痕/正常比率は、純度2%及び40%のCD34+細胞を与えたラットと比較して著しく低下する(p<0.01)ことを示している。(結果を、異なる3実験の平均値±平均値の標準誤差として表す)。
【図12】CD34+CD117bright血管芽細胞をLAD結紮したラットの血清と共に培養すると、CCR1及びCCR2の表面発現が増加するのに対し、CCR3及びCCR5の表面発現は不変である。
【図13】梗塞した心筋では、CCRに結合した幾つかのケモカインのmRNA発現が時間に依存して増加することを示している。
【図14】MCP−1、MCP−3、及びRANTESに対する、あるいはエオタキシンに対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋への輸送を、対照抗体に比べて40〜60%だけ減少させた(p<0.01)。
【図15】エオタキシンを非梗塞の心臓内に注入すると、CD34+血管芽細胞の輸送を1.5〜1.7倍に増加させたが、他方、増殖因子VEGF及び幹細胞因子の注入は、血管芽細胞の増殖を増加させたものの化学走性には効果がなかった(図示せず)。
【発明を実施するための形態】
【0140】
本明細書において使用する次の各用語は、別段の記述がない限り、以下に述べる定義に従うものとする。
【0141】
本明細書において使用する「BMEC」は、骨髄由来の内皮細胞と定義される。
【0142】
本明細書において使用する血管形成は、骨髄由来の内皮細胞前駆細胞などの「プレ血液(pre−blood)」細胞からの新しい血管の生成と定義される。
【0143】
本明細書において使用する動員は、骨髄由来の内皮細胞前駆細胞が骨髄を離れて末梢循環に入るように誘導することと定義される。当業者であれば、動員された幹細胞を白血球フェレーシスによって身体から採取し得ることが理解できるであろう。
【0144】
本明細書において使用する虚血は、局所的な部位への血管が閉塞したために、その部位への血液供給(循環)が不十分であることと定義される。
【0145】
本明細書において使用するサイトカインは、細胞を増殖又は活性化させる因子と定義される。
【0146】
本明細書において使用するケモカインは、細胞を体内の異なる部位に移動させる因子と定義される。
【0147】
本明細書において使用する虚血性心疾患は、心臓への血液供給量が減少するあらゆる病状と定義される。
【0148】
本明細書において使用する「血管新生」は、既存の血管細胞からの血管の生成と定義される。
【0149】
本明細書において使用する虚血性心疾患は、心臓への血液供給量が減少するあらゆる病状と定義される。
【0150】
本明細書において使用する「VEGF」は、血管内皮増殖因子と定義される。「VEGF−R」は、血管内皮増殖因子受容体と定義される。「FGF」は、繊維芽細胞増殖因子と定義される。「IGF」は、インシュリン様増殖因子と定義される。「SCF」は、幹細胞因子と定義される。「G−CSF」は、顆粒球コロニー刺激因子と定義される。「M−CSF」は、マクロファージコロニー刺激因子と定義される。「GM−CSF」は、顆粒球マクロファージコロニー刺激因子と定義される。「MCP」は、単球走化性タンパク質と定義される。
【0151】
本明細書において使用する「CXC」ケモカインは、ケモカインの構造と定義される。各「C」はシステインを表し、「X」は任意のアミノ酸を表す。
【0152】
本明細書において使用する「CC」ケモカインは、ケモカインの構造と定義される。各「C」はシステインを表す。
【0153】
本明細書において使用する「回収された」は、CD117、GATA−2、GATA−3、CD34を含む(これらには限定されない)細胞上の特異的マーカーに対して作られる、検出可能に標識された抗体を結合する回収可能な細胞に基づいて細胞を検出し、取得することを意味する。
【0154】
本明細書に記載されている、患者に投与するケモカインは、タンパク質の形態又は核酸の形態とすることができる。
【0155】
本発明は、虚血によって損傷を受けた患者の組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法を提供する。
【0156】
さらなる態様において、前記内皮前駆細胞は、工程(b)と工程(c)の間の一期間に凍結される。ある態様において、前記虚血によって損傷を受けた組織は心筋である。別の態様において、前記虚血によって損傷を受けた組織は神経系組織である。
【0157】
ある態様において、工程(b)の後であり且つ工程(c)の前に、増殖因子と接触させることによって、前記内皮前駆細胞を増殖させる。さらなる態様において、前記増殖因子はサイトカインである。さらなる態様において、前記サイトカインはVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである。別の態様において、前記増殖因子はケモカインである。さらなる態様において、前記ケモカインはインターロイキン−8である。ある態様において、前記内皮前駆細胞は、増殖の前に、他の幹細胞から分離される。さらなる態様において、前記内皮前駆細胞は、増殖後であり且つ工程(c)の前に、一定期間凍結される。
【0158】
ある態様において、工程(a)は、前記患者が虚血によって損傷を受けた組織を患う前に行われ、工程(c)は、前記患者が虚血によって損傷を受けた組織を患った後に行われる。
【0159】
ある態様において、前記幹細胞は、前記患者の骨髄から直接採取される。さらなる態様において、前記幹細胞は、前記患者の骨髄からの吸引によって採取される。ある態様において、前記患者からの前記幹細胞は、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員することと、
(b)続いて、幹細胞を含有する血液の試料を前記患者から採取することとを備えた方法によって採取される。
【0160】
さらなる態様において、前記増殖因子は、皮下、経口、静脈内、又は筋肉内から導入される。ある態様において、前記増殖因子は動員を誘導するケモカインである。さらなる態様において、前記ケモカインはインターロイキン−8である。ある態様において、前記増殖因子はサイトカインである。さらなる態様において、前記サイトカインは、G−CSF、M−CSF、又はGM−CSFである。
【0161】
本発明は、前記内皮前駆細胞が、CD117の発現に基づいて回収される前出の方法も提供する。
【0162】
本発明は、前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される前出の方法も提供する。ある態様において、前記遺伝子産物は、以下の群:プレプロエンドセリン−1、ビッグエンドセリン、エンドセリン−1から選択される。
【0163】
ある態様において、前記内皮前駆細胞はGATA−2を発現し、前記内皮前駆細胞は、細胞内GATA−2の発現又はそれらの細胞中のGATA−2活性を検出することによって回収される。
【0164】
ある態様において、前記患者は、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している。
【0165】
ある態様において、前記内皮前駆細胞は、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記患者に導入される。
【0166】
ある態様において、前記方法は、以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を前記患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う。
【0167】
本発明は、患者の梗塞周囲組織における血管新生を刺激する方法であって、 (a)患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記内皮前駆細胞が前記患者の梗塞周囲組織における血管新生を刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法も提供する。
【0168】
本発明は、患者の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を選択的に増加させる方法であって、
(a)内皮前駆細胞を前記患者に投与することと、
(b)ケモカインを前記患者に投与することによって、前記虚血によって損傷を受けた組織に前記内皮前駆細胞を誘引することを備えた方法も提供する。
【0169】
ある態様では、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される前出の方法も提供する。
【0170】
別の態様では、前記ケモカインは、前記内皮前駆細胞を投与する前に、前記ケモカインが前記患者に投与される。別の態様では、前記ケモカインは、前記内皮前駆細胞と同時に前記患者に投与される。別の態様では、前記ケモカインは、前記内皮前駆細胞を投与した後に、前記患者に投与される。ある態様において、前記ケモカインはCXCケモカインである。さらなる態様において、前記CXCケモカインは、インターロイキン−8、Gro−α、又は間質由来因子−1からなる群から選択される。ある態様において、前記ケモカインはCCケモカインである。さらなる態様において、前記CCケモカインはRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4からなる群から選択される。
【0171】
ある態様において、前記ケモカインは、末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織への注入によって、前記患者に投与される。
【0172】
本発明は、患者の虚血によって損傷を受けた組織への内皮前駆細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法も提供する。
【0173】
ある態様において、前記間質由来因子−1とCXCR4との前記相互作用は、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される。ある態様において、前記方法は、ACE阻害剤、AT受容体遮断薬、又はβ遮断薬、ng酵素阻害剤(ng enzyme inhibitor)、AT1受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備える。
【0174】
本発明は、患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法も提供する。ある態様において、前記SDF−1の産生は、抗SDF−1又は抗CXCR−4モノクローナル抗体を前記患者に投与することによって阻害される。
【0175】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることによって前記患者の前記癌を治療することを備えた方法も提供する。
【0176】
本発明は、患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生されたケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への前記内皮前駆細胞の輸送を減少させ、それにより前記患者の前記癌を治療することを備えた方法も提供する。
【0177】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0178】
本発明は、患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法も提供する。
【0179】
本発明は、内皮前駆細胞又はマスト前駆細胞中に存在する目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法も提供する。
【0180】
本発明は、前記ベクターがトランスフェクションによって前記細胞中に挿入される前出の方法も提供する。
【0181】
本発明は、前記プロモーターがプレプロエンドセリン−1プロモーターである前出の方法も提供する。
【0182】
本発明は、前記プロモーターが哺乳動物起源である前出の方法も提供する。
【0183】
本発明は、前記プロモーターがヒト起源である前出の方法も提供する。
【0184】
本発明は、癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させる効果を有する一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物を提供する。
【0185】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で採取した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0186】
ある態様において、前記異常は、虚血によって損傷を受けた組織である。ある態様において、前記遺伝子産物はプロエンドセリンである。ある態様において、前記遺伝子産物はエンドセリンである。ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0187】
本発明は、患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で回収したマスト前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法を提供する。
【0188】
ある態様において、前記異常は、虚血によって損傷を受けた組織である。ある態様において、前記遺伝子産物はプロエンドセリンである。ある態様において、前記遺伝子産物はエンドセリンである。ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0189】
本発明は、心筋梗塞を患った患者の心筋の機能を改善する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法を提供する。
【0190】
ある態様において、前記患者は哺乳動物である。さらなる態様において、前記哺乳動物はヒトである。
【0191】
本発明は、患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法も提供する。
【0192】
別の態様において、前記同種異系の幹細胞は、胚、胎児、又は臍帯血を起源として採取される。
【0193】
本発明は、患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法を提供する。
【0194】
別の態様において、前記同種異系幹細胞は、胚、胎児、又は臍帯血を起源として採取される。
【0195】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法も提供する。
【0196】
本発明は、心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法も提供する。ある態様において、前記方法は、前記患者に内皮前駆細胞を導入することをさらに備える。ある態様において、前記方法は、前記患者にG−CSFを導入して、内皮前駆細胞を動員することをさらに備える。
【0197】
以下に記載されている実験の詳細を参照することによって、本発明をよりよく理解することができるであろう。しかしながら、詳細に記載されている具体的な実験は、その後に記載されている特許請求の範囲においてより完全に記載されている本発明を説明するためのものにすぎないことが、当業者には容易に理解できるであろう。
【実験の詳細】
【0198】
第一群の実験
実験方法及び結果
1.骨髄由来細胞の動員と同定
G−CSF動員後、高純度ヒトCD34細胞(>90%陽性)の60〜80%が幹細胞因子受容体CD117を同時発現し(図1a)、そのうち15−25%はCD117の発現が明瞭で、75〜85%はCD117の発現が不明瞭であった。4種類のパラメーターに関する解析により、VEGFR−2(Flk−1)を発現する2種類のCD34細胞集団が確認された。1つ目はCDdim細胞の20〜30%を占め、VEGFR−2を高濃度に発現し、2つ目はCD117brightの10〜15%を占め、VEGFR−2発現は低濃度であった(図1b)。CD34+117dim細胞集団中のVEGFR−2陽性細胞であって、CD34+117bright分画中には含まれない細胞は、Tie−2、ecNOS、vWF、E−セレクチン(CD62E)及びICAM(CD54)の高発現を含む成熟血管内皮の表現型特性を示した。一方で、図1cで示すように、CD34+CD117bright細胞分画中に属するものであってCD34+CD117dim細胞分画に属さないVEGFR−2陽性細胞は、マウス及びヒト胚形成中に出現するGATA−2、GATA−3及び低レベルのTie−2を含む原始血管芽細胞に特徴的なマーカーを発現した。さらに、GATA−2及びGATA−3を同時発現したCD117bright細胞はまた、強いAC133陽性であった(AC133は、血管芽細胞になり得る造血細胞集団であることを確定するもう一つのマーカーであることが近年示唆されている(2)(図1d)。しかしながら、AC133発現は、GATA−2及びGATA−3が陰性であるCD117dim細胞の集団においても検出されたので、胚性骨髄由来血管芽細胞(BA)表現型の同定には、AC133に加えてGATA−2、GATA−3及びCD117brightが付随して発現することが必要であると本発明者らは結論する。このように、G−CSF処理によって、成熟骨髄由来内皮細胞(BMEC)の大きな集団、及び胚性血管芽細胞(BA)の表現型特性を有するこれより小さな骨髄由来の集団が末梢循環に動員される。
【0199】
2.骨髄由来細胞の増殖
VEGF(27)又は局所的虚血(10−13)の何れかにより動物モデルで循環する内皮細胞前駆体の出現率が増加することが示されたので、次に、VEGF及び虚血性血清中の因子(28)に対するBA及びBMECの増殖応答を比較した。図2aで示すように、VEGF又は虚血性血清の何れかを添加して96時間培養を行った後、CD117brightGATA−2posBAは、同じドナー由来のCD117dimGATA−2negBMECと比較して有意に高い増殖応答を示した。VEGFを添加した場合、BMECはベースラインを超えて1.2倍の増殖増加を示したのに対し、BAの増殖は2.9倍に増加を示し(p<0.01)、その一方で、心筋梗塞のLewラット由来の虚血性血清を添加した場合は、BMECの増殖は正常血清を超えて1.7倍の増加を示したのに対し、BAの増殖は4.3倍の増加を示した(p<0.01)。VEGF又は虚血性血清の何れかを添加して培養すると、GATA−2、GATA−3、及びCD117brightを含む未成熟マーカーは継続的に発現するが、成熟内皮細胞のマーカーは発現しない大型芽細胞のBA集団が著しく増殖したので(図2b)、芽細胞が分化せずに増殖したことが示唆される。内皮増殖培地を用いてフィブロネクチン上でCD34陽性単層培養を7日間行ったところ(29)、増殖の著しい敷石状パターンが観察され(図3a)、ほとんどの接着単層(>95%)において、アセチル化LDLの一定の取り込み、及びCD34、第VIII因子およびeNOSの同時発現を含む内皮細胞に特徴的な特性が認められた(図3b−e)。BMEC集団はVEGF又は虚血性血清中のサイトカインに対する増殖応答が低いため、培養中に著しく過剰増殖した内皮細胞は、おそらくGATA−2、GATA−3及びCD117brightの細胞表面への発現によって特徴付けられるBA集団が起源であろう。
【0200】
3.インビボにおける、骨髄由来CD34+細胞の局所的虚血部位への遊走
次に、局所的虚血誘導後、インビボにおける遊走及び増殖特性について、骨髄由来ヒト細胞と末梢血管由来ヒト細胞との比較を行った。図4a〜cで示すように、冠動脈結紮を行い、梗塞を起こさせたヌードラットに対して、2X106個のDiI−標識ヒトCD34陽性細胞(>95% CD34純度)、CD34陰性細胞(<5% CD34純度)、又は伏在静脈内皮細胞(SVEC)を静脈注射すると、48時間後にラット心筋において同程度の浸潤が観察された(30)。局所的梗塞を伴う心臓の非疾患部位においてはほとんどDiI標識細胞は検出されず(データを示さない)、G−CSF動員CD34+細胞、成熟ヒト内皮細胞いずれも正常心筋部位には浸潤しなかったので(図4d)、細胞輸送は特に梗塞領域へと向けられた。CD34+及びCD34−細胞の虚血性心筋への遊走は同数であったが、正常心筋に対して虚血性心筋におけるヒトGATA−2のmRNA発現の比例増加率(31)は、CD34−細胞の場合と比較してCD34が非常に富む細胞を注射した場合は2.6倍に増加した(p<0.001)(図4e)。さらに、ヒトCD34+細胞を注射して2週間後に、DiI、HLAクラスI、及び第VII因子の同時発現によって定義されるヒト内皮細胞を取り込む血管を検出し得たが、CD34−細胞又はSVEcの注射後には検出されなかった(図4f)。これらの結果を考え合わせると、成人骨髄由来ヒトCD34+細胞には、局所的虚血部位からのインビボのシグナルに選択的に応答して遊走性、局在性、及び内皮分化の増大を示す集団が含有されることが示される。
【0201】
4.梗塞ラット心筋へのG−CSF動員ヒトCD34+細胞注射の影響
次に、G−CSF動員ヒトCD34+(>95%)細胞、CD34−(<5%)細胞、末梢伏在静脈細胞、又は生理的食塩水注射による梗塞ラット心筋への機能的影響を比較した。LAD結紮後、それぞれの群の受容動物において、左心室駆出率(LVEF)が25〜43%低下し、左心室収縮終期領域は44〜90%増大し、左心室機能が著しく抑制された(図5a及びb)。驚いたことには、G−CSF動員成人ヒトCD34+細胞を注射して2週間以内に、LVEFが22±6%(p<0.001)回復した(図5a)。この影響は長期間続き、フォローアップ終了時である15週間後までに、34±4%まで増加した。その一方、G−CSF動員ヒトCD34−細胞、伏在静脈内皮細胞、又は生理的食塩水を注射しても、LVEFに対する影響は皆無であった。同様にして、G−CSF動員ヒトCD34+細胞を注射することにより、左心室収縮終期領域が、2週間後までに26±8%、15週間後までに37±6%減少したが、他の受容動物群ではこのような影響は見られなかった(p<0.001%)(図5b)。各群の代表的な心エコー例を図5Cに示す。さらに、15週間後のCD34+細胞を注射したラットの梗塞後の平均心係数は、正常ラットに対して26±8%減少したにすぎなかったが、他の各群の平均心係数は48〜59%減少した(p<0.001)(図5d)。
【0202】
梗塞して2週間後の組織学的検査(33)によると、CD34+細胞注射によって、対照群と比較すると梗塞部位内において肉芽組織の微小血管形成及び細胞密度の有意な増加とマトリックス沈着及び繊維形成の減少が伴うことが明らかとなった(図6a及びb)。さらに、ヒトCD34+細胞を注射したラットの虚血性心筋には、対照ラットの虚血性心筋と比較して、有意に多数の第VIII因子陽性間質血管芽細胞及び毛細血管が含まれていた(図6c及びd)。毛細血管数の定量によって、CD34+細胞を投与されたラットの梗塞部位内では新血管新生の有意な増加が示された(高倍率視野あたりの第VIII因子陽性毛細血管平均数は、92±5、対して生理食塩水対照では51±4、p<0.01)が、正常心筋においては示されなかった(高倍率視野当たりの毛細血管数、36±2対37±3)。CD34−細胞又はSVECが浸潤した虚血性ラット心筋では、毛細血管数の増加は全く見られなかった。梗塞して15週間後に、高純度CD34+細胞を投与したラットにおいては、他の各群と比較して、前自由壁(anterior free wall)内の生存能力を有する心筋量が増加すると共に梗塞部位の有意な減少が示された(図6e及びf)。CD34+細胞が浸潤した組織の梗塞部位と生存能力を有する心筋との連結部位において、多数の血管がはっきり示された。対照群では、コラーゲン沈着及び瘢痕形成が左心室壁厚のほとんど全体に広がり、動脈瘤拡大及び典型的なEKG異常が認められる一方、CD34+細胞を投与したラットでは、梗塞瘢痕は左心室壁厚のわずか20〜50%にのみ広がっていた。さらに、CD34+細胞を投与したラットでは、非−梗塞部位における病的なコラーゲン沈着が顕著に減少していた。全体的に、瘢痕/正常左心室心筋の平均比率は、CD34+細胞を投与されたラットでは13%であり、それに対して他の各群では36〜45%であった(P<0.01)(図6g)。
【0203】
考察
前記の実験により、ヒト骨髄由来内皮細胞前駆体による梗塞床の新血管新生によって、心筋虚血の齧歯類モデルにおいて、瘢痕発生を予防し、生存能力を有する心筋を維持し、心室機能を向上させることが示される。梗塞後、梗塞部位に接する生存能力を有する心筋組織が有意な程度に肥大を起こす(5、34〜35)。梗塞組織内の新血管新生は組織修復過程において不可欠な要素であると考えられているにも関わらず(36、37)、正常な状態下では、毛細血管ネットワークは組織成長に対応できず、肥大化しているが生存能力を有する心筋によるより大きい要求を支持することが不可能で、その後酸素供給及び栄養供給が不十分となりアポトーシスが引き起こされる。心筋梗塞瘢痕内の新血管新生の進展には、浸潤白血球上で発現しているウロキナーゼ型プラスミノーゲン活性化因子(u−PA)によってプラスミノーゲンを活性化した後、潜在性のコラゲナーゼ及び他のプロテイナーゼを活性化することが必要であると考えられる(38)。u−PA−/−マウスにおいて、欠損している梗塞後心筋血行再建がコンジェニック(cogenic)野生型系統マウスからの骨髄移植により回復したことから、この過程において骨髄由来内皮前駆体が重要であることが示された(38)。ヒト単核細胞及び腫瘍細胞系列において、u−PA mRNA転写及び蛋白質分解活性は、コロニー刺激因子、G−CSF、M−CSF、及びGM−CSFによって有意に促進されるので(39−41)、このことは、治療によって梗塞部位の再血管形成を行うために、これらのサイトカインをインビボ又はエクスビボで使用して多数のヒト成人骨髄由来血管芽細胞を動員し、分化させ得ることの論拠となる。転写因子GATA−2の細胞表面及びRNA発現によって、虚血性部位からのシグナルに応じて増殖して梗塞部位へと移動し、その後新血管新生プロセスに関与するヒト成人骨髄由来血管芽細胞が選択的に同定されると考えられる。特に興味深いことに、GATA−2は、ET−1(強力な血管収縮物質であり、肥大作用を有する自己分泌ペプチドである)の前駆体分子であるプレプロエンドセリン−1(ppET−1)の内皮細胞転写のコファクターである(42)。ppET−1転写は、心筋梗塞後にレニン−アンジオテンシン神経ホルモン系の活性化の結果産生されるアンジオテンシンIIによっても増加するため(43)、アンジオテンシンII細胞表面受容体シグナリングとGATA−2転写活性化との相乗的作用により、梗塞床に浸潤している血管芽細胞は高レベルのET−1を分泌している可能性がある。梗塞瘢痕内に新たに形成された血管が、正常心筋内の血管よりも厚い壁を有し、より強い血管作動物質に対しても低い血管拡張反応を示すという所見(44)は、自己分泌ET−1活性促進による影響と一致し、新血管新生脈管構造が浸潤したGATA−2陽性血管芽細胞由来である可能性を支持する。
【0204】
前記実験の結果を総合すると、胚性血管芽細胞の表現型特性及び機能的特性を持つG−CSF動員成人ヒトCD34+細胞を注射することによって、梗塞血管床において新血管新生を刺激することが可能であり、したがって心筋梗塞においてコラーゲン沈着及び瘢痕形成が減少することが示される。新血管新生によりもたらされる心筋組織修復の減少程度は注目すべきものであるが、ヒト血管芽細胞の注入とACE阻害又はAT1−受容体阻害とを組み合わせることによってアンジオテンシンII依存性心筋繊維芽細胞増殖、コラーゲン分泌、及びプラスミノーゲン活性因子阻害因子(PAI)産生を低下させ、心筋機能をさらに向上させることが可能である(45、46)。心筋梗塞組織の血行再建のために、サイトカインによって動員されたヒト骨髄由来の自家血管芽細胞を現在用いられている治療と併用することによって(47−49)、心筋梗塞後の左心室の組織修復に伴う罹患率及び死亡率を有意に減少させる可能性がもたらされる。
【0205】
第二群の実験
方法
1.サイトカインによって動員されたヒトCD34+細胞の精製
10mg/kgの組換えG−CSF(Amgen、CA)を4日間毎日皮下注射(SC)処置したヒトから、単一ドナーの白血球フェレーシス生成物を採取した。単核細胞をフィコール−ハイパック法によって分離し、高度に精製されたCD34+細胞(>98%陽性)を抗CD34モノクロナール抗体(mAb)(Miltenyi Biotech Ltd、CA)で被覆された磁気ビーズを用いて採取した。精製されたCD34細胞を、CD34、CD117、VEGFR−2、Tie−2、GATA−2、GATA−3、AC133、vWF、eNOS、CD54、CD62E、CXCR1、CXCR2、CXCR4に対するフルオレセイン複合mAbで染色し、FACScan(Becton Dickinson, CA)を用いた4パラメーター蛍光によって分析した。
【0206】
2.ヒト内皮前駆細胞の増殖の研究
20%の正常ラットの血清、虚血ラットの血清、又は20ng/mlのVEGFの何れかを含むRPMI培地内で、単一ドナーのCD34陽性細胞を96時間培養し、次いで、[3H]チミジン(Amersham Life Science Inc、IL、米国)(1mlCi/ウェル)を適用し、LK Betaplate液体シンチレーションカウンター(Wallace、Inc.、Gaithersburg、MD)で取込みを測定した。各条件で96時間培養した後、CD117brightGATA−2pos細胞 の比率もフローサイトメトリー法によって定量化した。
【0207】
3.ヒト骨髄由来の内皮前駆細胞の化学走性
メンブレン(8mm孔)(Neuro Probe、MD)を装着した48穴の走化性チャンバー内に、高度に精製したCD34+細胞(>98%陽性)を播種した。37℃にて2時間インキュベートした後、チャンバーを反転させ、0.2、1.0、及び5.0mg/mlのIL−8、1.0mg/mlのSDF−1α/β、VEGF、及びSCFを含有する培地中で3時間にわたり細胞を培養した。前記メンブレンをメタノールを用いて固定し、Leukostat(Fischer Scientific、Ill)で染色した。10高倍率視野内で遊走する細胞を数えて化学走性を計算した。
【0208】
4.動物、外科的処置、ヒト細胞の注入、及び組織への細胞遊走の定量化
Rowett(rnu/rnu)胸腺欠損ヌードラット(Harlan Sprague Dawley、Indianapolis、Indiana)を、「Columbia University Institute for Animal Care and Use Committee」によって承認された研究に使用した。麻酔後、左開胸を施行し、心膜を開き、左前下行(LAD)冠動脈を結紮した。擬似手術したラットには、冠動脈の周囲を縫合しないで同様の外科的処置を施行した。LAD結紮から48時間後に、CXCR1、CXCR2、CXCR4、CD34、ラットIL−8(ImmunoLaboratories、日本)及びラットSDF−1(R)&D Systems、MN)、又はアイソタイプ対照抗体に対して既知の阻害活性を有するmAbの存在下又は不在下で、G−CSF動員後に単一ドナーから取り出したDiI標識された2.0×106のヒトCD34+細胞(純度>95%、40%、<2%)を尾静脈に注入した。対照動物にはLAD結紮後に生理食塩水を与えた。各グループは6〜10匹のラットからなる。注入後2日して屠殺したラットの心臓内のDiI蛍光を評価することによって、ヒト細胞を注入した後の心筋の浸潤の定量化を実施した(1高倍率視野当たりのDiI陽性な細胞の数として表され、1サンプル当たり最低5視野を調べた)。12匹のラットにおいて、基準日、2、7、及び14日目に、ラット骨髄の総集団に対するHLAクラスI陽性細胞の比率を、フローサイトメトリー法及びRT−PCR法を用いて分析することによってラット骨髄のヒト細胞による浸潤の定量化を実施した。
【0209】
5.心筋機能の分析
細胞又は生理食塩水の注入後2、6、及び15週で、高周波線形配列の変換器(SONOS 5500、Hewlett Packard、Andover、MA)を用いて、基準時間(LAD結紮から48時間後)に心臓エコー検査を実施した。乳頭の中間と頂端のレベルで2D画像を撮影した。2方向の面積長さ法(bi−plane area−length method)によって、弛緩期末期(EDV)及び収縮期末期(ESV)の左心室容積を測定し、左心室駆出率%(LVEF)を[(EDV−ESV)/EDV]×100として計算した。収縮期末期における左心室面積(LVAs)を、僧帽弁のレベルで心臓エコー検査法によって測定した。LVEFの回復及びLVAsの減少は、LAD結紮後の異なる時刻でのLVEF又はLVAsのそれぞれの数値の梗塞前数値に対する平均改善度として計算した。
【0210】
6.組織学及び免疫組織化学
ヒト細胞又は生理食塩水を注入後2週及び15週に、外植されたラットの心臓に対して組織学の研究を実施した。切除後、各実験動物からの左心室を頂端から底部まで10〜15の横断切片にスライスした。代表的な切片を組織検査のためにホルマリンに入れ、免疫ペルオキシダーゼ法を用いて抗第VIII因子mAbで新たに染色して、毛細血管の密度を定量し、又はマッソントリクロームで染色して検鏡板にのせた。デジタルプラニメーター画像解析装置を用いて、心外膜及び心内膜の両領域を含めた梗塞された表面の長さを測定し、心室総外周に対する百分率で表した。各心臓からの全切片の平均値として、最終的な梗塞サイズを計算した。
【0211】
7.ラットのCXCケモカインmRNA及びタンパク質発現の測定
3匹の正常なラット及び12匹のLAD結紮したラットの心臓から、標準的な方法でPoly(A)+mRNAを抽出した。RT−PCR法を使用して、基準時間、LAD結紮してから6、12、24、及び48時間のラットIL−8及びGro−αのmRNAの心筋での発現を、GAPDHの発現によって測定されるラットの全RNAに対して規格化した後に数量化した。オリゴ(dT)15マー及びランダムヘキサマーでプライミングし、Monoleyマウスのリンパ増殖性ウイルス逆転写酵素(Invitrogen、Carlsbad、CA、米国)で逆転写した後、Taqポリメラーゼ(Invitrogen、Carlsbad、CA、米国)、放射標識ジデオキシヌクレオチド([α32p]−ddATP:3、000Ci/mmol、Amersham、Arlington Heights、IL)、並びにラットIL−8、Gro−α及びGAPDHに対するプライマー(Fisher Genosys、CA)を用いて、ポリメラーゼ連鎖反応(PCR)法でcDNAを増幅した。ラットのIL−8、Gro−α及びGAPDHに対するプライマーペア(センス/アンチセンス)は、それぞれ、gaagatagattgcaccgatg(配列番号1)/catagcctctcacatttc(配列番号2)、gcgcccgtccgccaatgagctgcgc(配列番号3)/cttggggacacccttcagcatcttttgg(配列番号4)、及びctctacccacggcaagttcaa(配列番号5)/gggatgaccttgcccacagc(配列番号6)であった。2%アガロースゲルに標識された試料を加え、電気泳動法によって分離し、X線撮影のために−70℃で6時間にわたり露光した。ラットIL−8/GroホモログCINC(ImmunoLaboratories、日本)に対するポリクロナール抗体を用いた市販のELISA法によって、基準時間、LAD結紮後6、12、24、及び48時間目に、4匹のラットにおいて、ラットIL−8/Gro−αの血清量を測定した。ラットIL−8/Gro−αタンパク質の既知の量に対して作成された光学濃度(OD)値の標準曲線に従って、各血清試料中のタンパク質の量を計算した。
【0212】
実験手順及び結果
1.内皮前駆細胞の選択的輸送
98%を超える純度になるように、G−CSFによって動員されたヒトCD34細胞に免疫的選択を施した後、60〜80%が幹細胞因子受容体CD117を同時発現した。四重パラメーター解析によって(図7a)、CD117bright細胞の10〜15%が胚血管芽細胞の表現型特性を示し、転写因子GATA−2とGATA−3、及び内皮前駆細胞の指標となることが最近示された(79)AC133と同様に、VEGFR−2及びTie−2の低レベルの表面発現を伴うことが見出された。これらの細胞は、vWF、eNOS及びE−セレクチンなどの成熟内皮細胞のマーカーを発現していなかったが、CXCケモカイン受容体1、2、及び4に対しては陽性であった。DiI標識された2.0×106のヒトCD34+細胞(純度>98%、40%、及び2%)をLAD結紮したRowettヌードラットに静脈内注入すると、48時間でラット心筋の稠密な浸潤を伴った(図7b)。心臓の局所梗塞に影響されない部位にはDiI標識細胞はほとんど検出されず(図示せず)、DiI標識細胞は擬似手術したラットからの心筋には浸潤しなかった(図7b)ことから、これらのDiI標識細胞は特に梗塞部位に向けて輸送されていた。98%を超える純度のヒトCD34+細胞を与えたラットは、注入後2週間までに、梗塞床の微小血管分布が増加し、マトリックス沈着及び繊維症が減少することを示した(図7c)。98%を超える純度のCD34+を含む2.0×106の細胞を与えたラットの梗塞床においては、2%又は40%の何れかの純度のCD34+を含む2.0×106の細胞を与えたラットの類似の領域においてよりも、1高倍率視野当たりの第VIII因子陽性な毛細血管の数が3倍以上多くなった(p<0.01、図7c)。さらに、これらの毛細血管の大多数は、HLAクラスI分子を発現したことからヒト由来であった(図示せず)。このように、ヒト骨髄由来の様々な細胞集団が梗塞床に遊走するが、血管形成には臨界数の内皮前駆細胞が選択的に輸送されることが必要であると考えられる。
【0213】
2.梗塞された心筋によるCXCケモカインの産生に対する虚血の影響
ヒト白血球の化学走性及び組織の浸潤は、特異的なケモカインとCXC細胞表面受容体との相互作用によって制御されるため、本発明者らは次に、梗塞されたラット心筋によって産生されるCXCケモカインへの虚血の影響を検討した。図8a〜cに見られるように、梗塞された心筋では、CXCR1/2に結合したELR陽性のケモカインであるIL−8及びGro−αのmRNA発現が時間に依存した増加を示し、LAD結紮後6〜12時間で発現が最大となった。非梗塞の心筋と比較して、LAD結紮後の組織は、mRNAの総含量に対して規格化した後で、これらのELR陽性の血管新生誘発性ケモカインの7.2〜7.5倍高いmRNA量を発現した(p<0.001)。さらに、血清のIL−8のレベルは、LAD結紮後6〜12時間以内に8〜10倍増加し(p<0.001)、48時間でもなお増加したままであった(図8d)。IL−8又はGro−αの何れかに対する、又はこれらの血管新生誘発性ケモカインの表面受容体(CXCR1又はCXCR2)に対する遮断mAbを同時投与すると、ヒト血管芽細胞の心筋での輸送を対照抗体に比較して40〜60%低下させた(p<0.01、図8E)。
【0214】
3.ケモカインに対するヒト骨髄由来CD34+血管芽細胞の化学走性応答
その後の実験で、本発明者らはヒト骨髄由来CD34+血管芽細胞のIL−8に対するインビトロ及びインビボでの化学走性応答を直接測定した。図9aに示すように、ヒトCD34+細胞のインビトロでの化学走性は、濃度0.2〜5μ/mlのIL−8によって、用量に依存して誘導された。骨髄間質細胞によって恒常的に産生されるELR−ケモカインであるSDF−1は、IL−8と類似の濃度にて、ほぼ同程度のCD34+細胞の化学走性を誘導した(図9b)。これに対して、増殖因子VEGF又は幹細胞因子(SCF)によっては、化学走性は誘導されなかった。さらに、IL−8を1μg/mlだけ非梗塞の心臓内に注入すると、CD34+細胞のインビボでの化学走性を誘導したが(図9c)、対照として用いたVEGF及びSCFは何れもインビボでの化学走性に何ら効果がなかった(図9d)。これらの結果を総合すると、ELR陽性なケモカインの組織発現が亢進されると、骨髄由来の内皮前駆細胞の組織虚血部位への化学走性を誘導することによって、インビボでの血管形成が増強されることが示唆される。
【0215】
4.骨髄から心筋へのヒトCD34陽性細胞の輸送を再誘導するためのCXCR4/SDF−1相互作用の遮断
LAD結紮したラットからの虚血性の血清は、静脈内に注入したヒトCD34+血管芽細胞が損傷した心筋へ輸送されるのを増強するうえに、CD34+CD117bright血管芽細胞集団の循環を急速に増大させ、同時にこれらの細胞の骨髄への輸送を増加させた。図10aに示すように、VEGF又は虚血の血清の何れかを2日間培養すると、CD34+CD117bright血管芽細胞の増殖を、それぞれ2.8倍及び4.3倍に増加させた(p<0.01)。さらに、図10bに示すように、2×106のヒトCD34陽性細胞(純度>95%)を静脈内に注入後2〜14日で、LAD結紮後の虚血ラットから得た骨髄は、正常なラットの骨髄と比較して、5〜8倍高いレベルのヒトCD34+CD117bright血管芽細胞を含有していた(p<0.001)。SDF−1は骨髄間質細胞によって恒常的に発現され、活発に循環している循環CD34+細胞の骨髄での遊走を優先的に促進するため(80)、本発明者らは、ヒトCD34+CD117bright血管芽細胞の虚血ラット骨髄へのホーミングの増加が、SDF−1/CXCR4相互作用の増大のためかどうかを検討した。図10cに示すように、ヒトCXCR4又はラットSDF−1に対するmAbの同時投与は、抗CD34対照抗体と比較して、静脈内投与されたCD34+ヒト血管芽細胞の虚血ラット骨髄への遊走を著しく阻害した(いずれもp<0.001)。さらに、ヒトCXCR4又はラットSDF−1に対するmAbの同時投与は、CD34+ヒト血管芽細胞の虚血ラット心筋への輸送を、それぞれ平均24%及び17%増加させた(いずれもp<0.001、図10d)。2週までには、ヒトCD34+細胞を抗CXCR4mAbと組合せて与えたラットの心筋梗塞床は、CD34+細胞をアイソタイプの対照抗体と組合せて与えたラットの心筋梗塞床と比較して、微小血管分布が3倍超の増加を示した。これらの結果から、静脈内注入したCD34+血管芽細胞は梗塞心筋へ輸送され、ELR+のケモカインの産生が増大するのに応答して血管形成を誘導するが、このプロセスの効率は、血管芽細胞の骨髄へのSDF−1応答性の遊走が伴うことによって著しく低下することが示唆される。CXCR4/SDF−1相互作用を遮断すると、梗塞後に、拡大され循環するヒトCD34陽性細胞集団の骨髄から心筋への輸送を再誘導し、その結果、梗塞床の新血管新生を増強させる。
【0216】
5.心筋機能の改善
LAD結紮後、左心室機能は極度に低下するが、98%を超える純度のCD34+細胞を注入すると2週間以内に左心室の大きさと機能に顕著な回復を伴い、これらの効果は経過観察した15週の全期間にわたり持続した(図11a及びb)。98%を超える純度のCD34+細胞を与えたラットでは、左心室の収縮期末期の面積が梗塞直後に比べて15週までに平均37±6%だけ減少し(図11a)、左心室駆出率(LVEF)は、15週までに平均34±4%だけ回復した(図11b、p<0.001)。純度2%又は40%のCD34+を含む、G−CSFによって動員された2×106のヒト細胞の静脈内注入が、虚血心筋へのほぼ同程度の輸送にもかかわらず心筋機能を顕著には改善しなかったことから、これらのパラメーターの改善は、注入されたCD34+細胞の数に依存する(図11a及びb)。しかし、純度40%のCD34+を含む、G−CSFによって動員されたヒト骨髄由来の細胞と共に、抗CXCR4mAbを同時投与すると、98%を超える純度のCD34+で観察されるレベルまで、LVEFの回復が顕著に改善しLVAsが顕著に減少した。トリクローム染色によって、左心室の塊及びコラーゲン沈着における顕著な差異がこれらのグループ間に観察された(図11c)。純度2%のCD34を含む2×106のヒト細胞を与えたラットでは、左心室の前壁が繊維組織によって完全に入れ替わり、中隔の著しい代償性肥大が存在した。純度40%のCD34を含む2×106のヒト細胞を与えたラットの心臓でも同様の変化が見られた。これに対して、98%を超える純度のCD34+を含む2×106のヒト細胞を与えたラットの心臓では、これより著しく大きな前壁の塊が維持され、中隔の大きさは正常であり、コラーゲンの沈着は最小限であった。特に興味深いことには、純度40%のCD34を含む2×106のヒト細胞を抗CXCR4mAbと共に与えたラットの心臓も、同様に、心筋前壁の塊の増加、中隔肥大の減少、及びコラーゲン沈着の減少を示した。総合すると、繊維性瘢痕/正常左心室心筋の平均比率は、純度2%及び40%のCD34+細胞を与えたラットが36〜45%であるのに対して、98%を超える純度のCD34+細胞を与えたラット、又は純度40%のCD34+細胞を抗CXCR4mAbと共に注入したラットでは、それぞれ、13%及び21%であった(p<0.01、図11d)。したがって、臨界数の内皮前駆細胞の選択的な輸送を増大することによって梗塞床の血管形成が増強されると、組織修復のプロセスがさらに抑制され、生存可能な心筋を救出し、心臓機能を改善することになる。
【0217】
考察
本研究は、虚血組織によって産生されるELR+ケモカインが、骨髄由来の内皮細胞前駆細胞に対して化学遊走物質の濃度勾配を生じることによって、虚血部位での代償的な血管形成の進展を制御することを実証するものである。ELR+CXCケモカインであるIL−8とELR−CXCケモカインであるSDF−1は何れも、CD34+内皮前駆細胞の化学走性、並びに成熟内皮に対して同様の効果を示すが(73)、異なる血管外の部位で発現すると、内皮前駆細胞の方向性の出現に対してそれらは生物学的に相反する影響を及ぼし、その結果、組織の新血管新生に対してもそれらは生物学的に相反する影響を及ぼす。これらの相互作用を理解することで、本発明者らは、心筋での輸送を増加させ、梗塞床の血管形成を誘導し、虚血後の心室の組織修復を抑制し、心筋機能を改善するために、ヒト骨髄由来のCD34+細胞の特異的なサブセットの化学走性特性を操作し増強することができた。
【0218】
基底膜を通る骨髄由来の前駆細胞の遊走は、メタロプロテイナーゼ−9(MMP−9、ゼラチナーゼB)などのタンパク質分解酵素の分泌に依存するため(81)、心臓内のメタロプロテイナーゼの活性が、血管芽細胞の循環からの血管外遊出及び梗塞域内への経内皮遊走を決定する重要な要因かもしれない。IL−8は、好中球中の細胞内貯蔵顆粒から潜在型MMP−9を迅速に(20分以内)放出するように誘導し(82〜83)、非ヒト霊長類におけるインビボでの静脈内投与後、血清のMMP−9のレベルを最大1000倍増加させる(84)。IL−8は骨髄前駆細胞におけるMMP−9の発現を顕著に増加させ(81)、MCP−9に対する中和抗体は骨髄前駆細胞の遊走を抑制するため(85)、本発明者らの研究の結果は、血管芽細胞の浸潤とその後の梗塞床の血管形成が、IL−8に依存したMMP−9分泌の増加に起因するかもしれないことを示唆している。
【0219】
潜在型MMP−9の活性化及びこれと同時に起こるマウスの心筋梗塞瘢痕組織内の新血管新生は、梗塞床に浸潤する骨髄前駆細胞によって同時発現されるウロキナーゼタイプのプラスミノゲン活性化因子(u−PA)に依存することが示された(81)。ヒト細胞内のu−PAの転写及びタンパク質分解活性は、G−CSF及び他のコロニー刺激因子によって顕著に増加する(86〜88)。IL−8によって誘導される化学走性及び前駆細胞の遊走には、機能性G−CSF受容体を介して送達されるシグナルの存在がさらに必要とされるため(89)、IL−8によって媒介される虚血部位への血管芽細胞の輸送にはu−PA活性の増加が必要である可能性がある。このことは、IL−8が高レベルで産生されたにもかかわらず、心筋梗塞後に通常観察される梗塞床における新血管新生の程度が限られていたこと(62、63)を説明すると思われるし、さらに虚血組織の血行再建術治療に使用するためにコロニー刺激因子をインビボ投与又はエクソビボ投与してヒト骨髄由来の血管芽細胞を動員し分化させることに理論的根拠を与える。
【0220】
CXCケモカインであるSDF−1の骨髄間質細胞による恒常的な産生は、造血前駆細胞の骨髄へのホーミング及び移植にとって必須であると考えられる(76〜78)。さらに、SDF−1 −/−マウスは胃腸管の血管新生(50)及び心室中隔の形成(90)の両者に欠陥があるため、非造血組織におけるSDF−1の発現は発達中の血管系において役割を果たす。骨髄由来の内皮前駆細胞はCXCR4を発現し(80)、本明細書で示すようにSDF−1に対して化学走性の応答を示すため、胚形成中又は組織損傷後に、非造血部位でのSDF−1の発現が誘導されることは、組織の新血管新生のプロセスにおいて重要な要素であるかもしれない(91)。CXCR4/SDF−1相互作用の遮断によって、本発明者らがヒト骨髄由来の血管芽細胞の組織虚血部位への輸送を再誘導することができたということは、SDF−1がヒト内皮前駆細胞に対する生物学的に活性な化学走性因子であること、並びにそれが非造血部位で発現されると血管新生誘発活性を有するかもしれないことを強く主張するものである。虚血組織部位におけるSDF−1及び他のケモカインの発現の増大及び局在化が、血管形成を増強する上でELR+CXCケモカインと相乗的であり得るか否かは、将来の研究課題であろう。総合すると、本研究の結果は、IL−8、Gro−α、及びSDF−1を含むCXCケモカインがヒト骨髄に依存する血管形成を制御する際に中心的な役割を果たすこと、さらに、これらのケモカインとヒト骨髄由来の自家血管芽細胞上に存在するそれらの受容体との相互作用を操作することは、組織虚血後の治療的血管形成の潜在的な効力を増強できることを示唆するものである。
【0221】
第3群の実験
実験方法と結果
1.心筋虚血は、CCケモカイン産生を誘導してヒトCD34+血管芽細胞のCCケモカイン受容体の発現を増加させる
ヒト単核細胞の化学走性及び組織浸潤は細胞表面受容体と特定のケモカインリガンドとの相互作用によって制御されるので、血管芽細胞のCCケモカイン受容体発現及び梗塞ラット心筋によるCCケモカイン産生の速度論に対する虚血の影響を研究した。図12に示したように、LAD結紮ラットの血清と共にCD34+CD117bright血管芽細胞を培養すると、CCR1及びCCR2の表面発現は増加したが、CCR3及びCCR5の表面発現に変化はなかった。
【0222】
図13に示したように、梗塞した心筋では、時間に応じていくつかのCCR−結合ケモカインのmRNA発現が増加することが示された。梗塞した心筋では、総mRNA含量で正規化したところ(全てp<0.001)、CCR−2結合CCケモカインMCP−1が8倍を上回る濃度で発現し、MCP−3およびRANTESのmRNAが3〜3.5倍高い濃度で発現し、CCR−3結合ケモカインエオタキシンも発現することが発見された。研究した梗塞組織全てにおいて、これらのCCケモカイン類の発現増加が示されたが、CCR5−結合CCケモカインMIP−1アルファ又はMIP−1ベータの発現誘導は示されなかったので、この遺伝子発現パターンは比較的特異的なものと考えられる。
【0223】
2.虚血性心筋へのヒトCD34+血管芽細胞の輸送は、CC及びCXCケモカイン類の誘導発現によって調節される
次に、虚血性心筋へのヒトの血管芽細胞の輸送が、前記で確認したCCケモカインの誘導発現と関連するかどうかについて研究した。MCP−1、MCP−3、及びRANTESに対する遮断mAb、又はエオタキシンに対する遮断mAbを同時投与すると、ヒトの血管芽細胞の心筋への輸送が対照抗体と比較して40−60%減少した(p<0.01)、(図14)。CCケモカインが血管芽細胞の虚血性心筋への化学走性を媒介することを証明するために、エオタキシンに応じた血管芽細胞の化学走性をインビボで測定した。図15に示したように、心臓の非梗塞部にエオタキシンを心臓内注射すると、CD34+血管芽細胞輸送の1.5−1.7倍の増加が誘導されたが、成長因子VEGF及び幹細胞因子を注射しても、血管芽細胞増殖は増加するものの化学走性に何ら影響はなかった(図示せず)。
【0224】
第4群の実験
筋細胞の大きさの決定。正常なラットの心臓及び梗塞組織切片の梗塞部位並びに梗塞周囲縁、遠位部の筋細胞をトリクローム染色して、大きさを測定した。10−15倍の高倍率視野の各々について100〜200個の筋細胞の横径及び縦径(mm)をイメージプロプラスソフトウェアを用いて400xで測定した。
【0225】
パラフィン組織切片のDNA末端標識による筋細胞アポトーシスの測定
単一細胞レベルにおけるアポトーシスをin situで検出するために、デオキシヌクレオチジルトランスフェラーゼ(TdT)によるDNA末端標識、TUNEL法を用いた(ベーリンガーマンハイム、ドイツ・マンハイム)。生理食塩液又はCD34+ヒト細胞のいずれかを注射して2週間後のLAD結紮ラット及び陰性対照として健康なラットから、ラットの心筋組織切片を切除した。簡単に説明すると、組織を脱パラフィン化してプロティナーゼKで消化し、37℃で60分間、浸潤雰囲気中でTdT及びフルオレセイン標識dUTPとインキュベートした。フルオレセイン特異的抗体複合アルカリホスファターゼで30分間インキュベーションした後、DNAが断片化された核を青色に染色して、TUNEL染色を視覚化した。
【0226】
1.新血管新生は肥大した心筋をアポトーシスから保護する
新血管新生を誘導して心臓機能の改善をもたらす機構を研究した。LAD結紮後2週間経つと、生理食塩水対照群の梗塞周囲縁の筋細胞の外観はゆがんで形が不規則となり、筋細胞の直径は梗塞のないラットと同様となることが(0.020mm+/−0.002 対 0.019mm+/−0.001)結果から示された。一方、CD34+細胞を投与したラットの梗塞周囲縁の筋細胞の形は規則正しい楕円形をしており、対照群ラットの筋細胞より有意に大きかった(直径0.036mm+/−0.004 対 0.019mm+/−0.001、p<0.01)。筋細胞特異的マーカーであるデスミンの染色及びDNA末端標識を同時に行うと、CD34+細胞を注射したラットの梗塞した左心室では、生理食塩液対照群と比較すると、検出されたアポトーシスした筋細胞の数は6分の1であった(アポトーシス指数1.2+0.6対7.1+0.7、p<0.01)。これらの差は梗塞周囲縁内において特に顕著で、生理食塩水処理対照群の形状の不規則な小筋細胞は、アポトーシス核指数が最高であった。また、生理食塩水対照群では左心室壁の75〜80%に筋細胞のアポトーシスが広がっていたが、CD34+細胞を注射したラットでは、筋細胞のアポトーシスは梗塞部から遠位の左心室の最大20〜25%に検出されたにすぎなかった。これらの結果を総合すると、CD34+細胞注射によって誘導された梗塞領域の血管形成及び梗塞周囲の血管新生は、生理食塩水対照群において明らかな遠心性に広がるアポトーシス誘発プロセスを阻止して、梗塞周囲領域内の肥大した心筋の生存を可能にし、心筋機能を改善する。
【0227】
2.早期の新血管新生は後期の心筋組織修復を予防する
最後の実験群によって、対照群および実験群(生理食塩液対CD34+細胞)における2週間後の梗塞周囲縁の筋細胞のアポトーシスの程度と、その後4ヶ月にわたる心筋の組織修復の進行との比較を明らかにした。LVEFが初期に減少し、LVASが増加することは同じであるが、2週間後のコラーゲン沈着又は瘢痕組織/正常な左心室心筋の平均比率は、マッソントリクローム染色によって確認したところ、生理食塩水を投与したラットでは12%であったのに対しCD34+細胞を投与されたラットでは3%であった。梗塞して15週間後では、瘢痕/正常な左心室心筋の平均比率は、CD34+細胞を投与したラットでは13%であったのに対して、それ以外の実験群(生理食塩水、CD34−、SVEC)ではいずれも36−45%であった(p<0.01)。CD34+細胞を投与したラットでは、ラット由来の筋細胞のみを含む前自由壁内に、生存可能な心筋量の有意な増加が示され、ヒトではなくラットのMHC分子を発現していることから、筋細胞はヒト幹細胞前駆体から再分化するのではなく本来の筋細胞を再利用することが確かめられた。対照群では、左心室壁厚のほとんど全体に渡ってコラーゲン沈着および瘢痕形成が広がっており、動脈瘤性拡張および典型的なEKG異常をともなっているが、CD34+細胞を投与したラットでは、左心室壁の壁厚の20〜50%のみに広がっていた。さらに、非梗塞領域における病的なコラーゲン沈着は、CD34+細胞を投与したラットでは著しく減少した。これらの結果を考え合わせると、2週間後に梗塞周囲の筋細胞においてアポトーシスの減少が観察されたことによって、肥大しているが生存可能な筋細胞の生存が延長され、15週間後までに心筋とコラーゲン及び線維組織との置換が阻止されることが示された。
【0228】
考察
梗塞周囲縁および筋細胞間で増殖した毛細血管がラット由来であることが観察され、血管形成に加えて、ヒトの血管芽細胞又は同時投与したその他の骨髄由来因子は、血管新生誘発因子の豊富な発生源となることが可能で、既存の脈管系からさらに血管新生を誘導することを可能にすることを示している。
【参照文献】
【0229】
【特許請求の範囲】
【請求項1】
患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、
(c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。
【請求項2】
工程(b)の後であり且つ工程(c)の前に、前記内皮前駆細胞を増殖因子と接触させることによって、前記内皮前駆細胞を増殖させる請求項1の方法。
【請求項3】
前記増殖因子がサイトカインである請求項2の方法。
【請求項4】
前記サイトカインがVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである請求項3の方法。
【請求項5】
前記増殖因子がケモカインである請求項2の方法。
【請求項6】
前記ケモカインがインターロイキン−8である請求項2の方法。
【請求項7】
前記内皮前駆細胞が、増殖の前に、他の幹細胞から分離される請求項3の方法。
【請求項8】
前記虚血によって損傷を受けた組織が心筋である請求項1の方法。
【請求項9】
前記虚血によって損傷を受けた組織が神経系組織である請求項1の方法。
【請求項10】
前記幹細胞が前記患者の骨髄から採取される請求項1の方法。
【請求項11】
前記骨髄からの前記幹細胞の採取が、前記患者の骨髄からの吸引によって為される請求項10の方法。
【請求項12】
前記患者からの前記幹細胞の採取が、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員することと、
(b)前記幹細胞を含有する血液の試料を前記患者から採取することと
を備えた方法によって為される請求項1の方法。
【請求項13】
前記増殖因子が、皮下、経口、静脈内、又は筋肉内から患者に導入される請求項12の方法。
【請求項14】
前記増殖因子が動員を誘導するケモカインである請求項12の方法。
【請求項15】
前記ケモカインがインターロイキン−8である請求項14の方法。
【請求項16】
前記増殖因子がサイトカインである請求項12の方法。
【請求項17】
前記サイトカインがG−CSF、M−CSF、又はGM−CSFである請求項16の方法。
【請求項18】
前記内皮前駆細胞が、CD117の発現に基づいて回収される請求項1の方法。
【請求項19】
前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される請求項1の方法。
【請求項20】
前記内皮前駆細胞が、CD34、VEGF−R、Tie−2,GATA−3、又はAC133のうちの1以上の発現に基づいて回収される請求項1の方法。
【請求項21】
前記患者が、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している請求項1の方法。
【請求項22】
前記患者が虚血によって損傷を受けた組織を患う前に工程(a)が行われ、前記患者が虚血によって損傷を受けた組織を患った後に工程(c)が行われる請求項1の方法。
【請求項23】
工程(b)と(c)の間の一期間に、記内皮前駆細胞が凍結される請求項1の方法。
【請求項24】
増殖された後、工程(c)が行われる前の一期間に、前記内皮前駆細胞が凍結される請求項3の方法。
【請求項25】
末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記内皮前駆細胞が前記患者に導入される請求項1の方法。
【請求項26】
以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う請求項1の方法。
【請求項27】
患者の梗塞周囲組織における血管新生を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記患者の梗塞周囲組織における血管新生を前記内皮前駆細胞が刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。
【請求項28】
患者の虚血によって損傷を受けた組織における内皮前駆細胞を増加する方法であって、
(a)前記患者に内皮前駆細胞を投与することと、
(b)前記患者にケモカインを投与し、損傷を受けた組織における内皮前駆細胞を引き付けることとを備えた方法。
【請求項29】
前記ケモカインが、内皮前駆細胞を投与する前に患者に投与される請求項28の方法。
【請求項30】
前記ケモカインが、内皮前駆細胞と同時に患者に投与される請求項28の方法。
【請求項31】
前記ケモカインが、内皮前駆細胞を投与する後に患者に投与される請求項28の方法。
【請求項32】
前記ケモカインがCXCケモカインである請求項28の方法。
【請求項33】
前記CXCケモカインがインターロイキン−8、Gro−α、又は間質由来因子−1である請求項28の方法。
【請求項34】
前記ケモカインがCCケモカインである請求項28の方法。
【請求項35】
前記CCケモカインがRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4である請求項34の方法。
【請求項36】
患者の末梢循環、心筋、左心室、右心室、冠動脈、脊髄液、神経組織、虚血性組織、又は虚血後組織への注入によって、前記ケモカインが前記患者に投与される請求項28の方法。
【請求項37】
患者の虚血によって損傷を受けた組織への内皮前駆細胞又は血管芽細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法。
【請求項38】
間質由来因子−1(SDF−1)とCXCR4との相互作用が、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される請求項37の方法。
【請求項39】
アンギオテンシン変換酵素阻害剤、AT1−受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備えた請求項38の方法。
【請求項40】
患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法。
【請求項41】
SDF−1の産生が、患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される請求項40の方法。
【請求項42】
患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることによって前記患者の前記癌を治療することを備えた方法。
【請求項43】
患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることにより、前記患者の前記癌を治療することを備えた方法。
【請求項44】
患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法。
【請求項45】
患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法。
【請求項46】
前記受容体がCD117である請求項57又は58の方法。
【請求項47】
内皮前駆細胞又はマスト前駆細胞中に存在する目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法。
【請求項48】
前記ベクターがトランスフェクションによって前記細胞中に挿入される請求項47の方法。
【請求項49】
前記プロモーターがプレプロエンドセリン−1プロモーターである請求項62の方法。
【請求項50】
前記プロモーターが哺乳動物起源である請求項64の方法。
【請求項51】
前記プロモーターがヒト起源である請求項65の方法。
【請求項52】
癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させる効果を有する一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物。
【請求項53】
患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で採取した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法。
【請求項54】
患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で採取したマスト前駆細胞の中からGATA−2を発現しているものを回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法。
【請求項55】
前記異常が虚血によって損傷を受けた組織である請求項53又は54の方法。
【請求項56】
前記遺伝子産物がプロエンドセリンである請求項53又は54の方法。
【請求項57】
前記遺伝子産物がエンドセリンである請求項53又は54の方法。
【請求項58】
心筋梗塞に罹患した患者の心筋の機能を改善させる方法であって、
(a)前記患者中のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法。
【請求項59】
前記患者が哺乳動物起源である請求項42、43、44又は45の何れかに記載の方法。
【請求項60】
前記哺乳動物がヒト起源である請求項59の方法。
【請求項61】
患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法。
【請求項62】
前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として得られる請求項61の方法。
【請求項63】
患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、
(c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法。
【請求項64】
前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として得られる請求項63の方法。
【請求項65】
心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法。
【請求項66】
心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法。
【請求項67】
内皮前駆細胞を前記患者に導入することをさらに備えた請求項66の方法。
【請求項68】
前記患者にG−CSFを導入して内皮前駆細胞を動員することをさらに備えた請求項67の方法。
【請求項1】
患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、
(c)患者の虚血によって損傷を受けた前記組織における血管形成を前記内皮前駆細胞が刺激するように、工程(b)で得た前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。
【請求項2】
工程(b)の後であり且つ工程(c)の前に、前記内皮前駆細胞を増殖因子と接触させることによって、前記内皮前駆細胞を増殖させる請求項1の方法。
【請求項3】
前記増殖因子がサイトカインである請求項2の方法。
【請求項4】
前記サイトカインがVEGF、FGF、G−CSF、IGF、M−CSF、又はGM−CSFである請求項3の方法。
【請求項5】
前記増殖因子がケモカインである請求項2の方法。
【請求項6】
前記ケモカインがインターロイキン−8である請求項2の方法。
【請求項7】
前記内皮前駆細胞が、増殖の前に、他の幹細胞から分離される請求項3の方法。
【請求項8】
前記虚血によって損傷を受けた組織が心筋である請求項1の方法。
【請求項9】
前記虚血によって損傷を受けた組織が神経系組織である請求項1の方法。
【請求項10】
前記幹細胞が前記患者の骨髄から採取される請求項1の方法。
【請求項11】
前記骨髄からの前記幹細胞の採取が、前記患者の骨髄からの吸引によって為される請求項10の方法。
【請求項12】
前記患者からの前記幹細胞の採取が、
(a)前記患者に増殖因子を導入して、前記幹細胞を前記患者の血液中に動員することと、
(b)前記幹細胞を含有する血液の試料を前記患者から採取することと
を備えた方法によって為される請求項1の方法。
【請求項13】
前記増殖因子が、皮下、経口、静脈内、又は筋肉内から患者に導入される請求項12の方法。
【請求項14】
前記増殖因子が動員を誘導するケモカインである請求項12の方法。
【請求項15】
前記ケモカインがインターロイキン−8である請求項14の方法。
【請求項16】
前記増殖因子がサイトカインである請求項12の方法。
【請求項17】
前記サイトカインがG−CSF、M−CSF、又はGM−CSFである請求項16の方法。
【請求項18】
前記内皮前駆細胞が、CD117の発現に基づいて回収される請求項1の方法。
【請求項19】
前記内皮前駆細胞が、GATA−2被活性化遺伝子産物の発現に基づいて回収される請求項1の方法。
【請求項20】
前記内皮前駆細胞が、CD34、VEGF−R、Tie−2,GATA−3、又はAC133のうちの1以上の発現に基づいて回収される請求項1の方法。
【請求項21】
前記患者が、以下のうちの1以上:心筋梗塞、慢性心不全、虚血性心疾患、冠動脈疾患、糖尿病性心疾患、出血性卒中、血栓性卒中、塞栓性卒中、虚血肢、又は組織が虚血になるその他の疾病に罹患したことがある、又は罹患している請求項1の方法。
【請求項22】
前記患者が虚血によって損傷を受けた組織を患う前に工程(a)が行われ、前記患者が虚血によって損傷を受けた組織を患った後に工程(c)が行われる請求項1の方法。
【請求項23】
工程(b)と(c)の間の一期間に、記内皮前駆細胞が凍結される請求項1の方法。
【請求項24】
増殖された後、工程(c)が行われる前の一期間に、前記内皮前駆細胞が凍結される請求項3の方法。
【請求項25】
末梢循環、心筋、左心室、右心室、冠動脈、脳脊髄液、神経組織、虚血性組織、又は虚血後組織中に直接注入することによって、前記内皮前駆細胞が前記患者に導入される請求項1の方法。
【請求項26】
以下のもの:プラスミノーゲン活性化因子阻害剤の阻害剤、アンギオテンシン変換酵素阻害剤、又はβ−遮断薬のうちの1以上を患者に投与することをさらに備え、工程(c)の前、同時、又は後に、このような投与を行う請求項1の方法。
【請求項27】
患者の梗塞周囲組織における血管新生を刺激する方法であって、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記前駆細胞を増殖因子と接触させることによって、工程(b)で回収した前記内皮前駆細胞を増殖させることと、
(d)前記患者の梗塞周囲組織における血管新生を前記内皮前駆細胞が刺激するように、工程(c)で得た増殖した前記内皮前駆細胞を前記患者の別の部位に導入することとを備えた方法。
【請求項28】
患者の虚血によって損傷を受けた組織における内皮前駆細胞を増加する方法であって、
(a)前記患者に内皮前駆細胞を投与することと、
(b)前記患者にケモカインを投与し、損傷を受けた組織における内皮前駆細胞を引き付けることとを備えた方法。
【請求項29】
前記ケモカインが、内皮前駆細胞を投与する前に患者に投与される請求項28の方法。
【請求項30】
前記ケモカインが、内皮前駆細胞と同時に患者に投与される請求項28の方法。
【請求項31】
前記ケモカインが、内皮前駆細胞を投与する後に患者に投与される請求項28の方法。
【請求項32】
前記ケモカインがCXCケモカインである請求項28の方法。
【請求項33】
前記CXCケモカインがインターロイキン−8、Gro−α、又は間質由来因子−1である請求項28の方法。
【請求項34】
前記ケモカインがCCケモカインである請求項28の方法。
【請求項35】
前記CCケモカインがRANTES、EOTAXIN、MCP−1、MCP−2、MCP−3、又はMCP−4である請求項34の方法。
【請求項36】
患者の末梢循環、心筋、左心室、右心室、冠動脈、脊髄液、神経組織、虚血性組織、又は虚血後組織への注入によって、前記ケモカインが前記患者に投与される請求項28の方法。
【請求項37】
患者の虚血によって損傷を受けた組織への内皮前駆細胞又は血管芽細胞の輸送を増加させる方法であって、間質由来因子−1とCXCR4との何らかの相互作用を阻害することを備えた方法。
【請求項38】
間質由来因子−1(SDF−1)とCXCR4との相互作用が、前記患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される請求項37の方法。
【請求項39】
アンギオテンシン変換酵素阻害剤、AT1−受容体遮断薬、又はβ遮断薬を前記患者に投与することをさらに備えた請求項38の方法。
【請求項40】
患者の骨髄への内皮前駆細胞の輸送を減少させる方法であって、前記患者の骨髄での間質由来因子−1の産生を阻害することを備えた方法。
【請求項41】
SDF−1の産生が、患者に抗SDF−1又は抗CXCR4モノクローナル抗体を投与することによって阻害される請求項40の方法。
【請求項42】
患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインのエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることによって前記患者の前記癌を治療することを備えた方法。
【請求項43】
患者の癌を治療する方法であって、前記癌に伴って増殖している細胞によって産生された特異的ケモカインに対する特異的受容体であり、内皮前駆細胞上に位置する特異的受容体のエピトープに対して誘導されたモノクローナル抗体を前記患者に投与して、このような増殖している細胞への内皮前駆細胞の輸送を減少させることにより、前記患者の前記癌を治療することを備えた方法。
【請求項44】
患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管形成を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法。
【請求項45】
患者の腫瘍を治療する方法であって、内皮前駆細胞上の特異的な受容体に対するアンタゴニストを前記患者に投与して、前記患者の腫瘍における血管新生を誘導する前記前駆細胞の能力を減弱せしめることにより、前記腫瘍を治療することを備えた方法。
【請求項46】
前記受容体がCD117である請求項57又は58の方法。
【請求項47】
内皮前駆細胞又はマスト前駆細胞中に存在する目的の遺伝子を発現させる方法であって、GATA−2モチーフを含有するプロモーターと前記目的の遺伝子とを含むベクターを前記細胞中に挿入することを備えた方法。
【請求項48】
前記ベクターがトランスフェクションによって前記細胞中に挿入される請求項47の方法。
【請求項49】
前記プロモーターがプレプロエンドセリン−1プロモーターである請求項62の方法。
【請求項50】
前記プロモーターが哺乳動物起源である請求項64の方法。
【請求項51】
前記プロモーターがヒト起源である請求項65の方法。
【請求項52】
癌によって産生される特異的なケモカインのエピトープに対して誘導されたモノクローナル抗体であって、前記癌への内皮前駆細胞の輸送を低減させる効果を有する一定量のモノクローナル抗体と、薬学的に許容される担体とを備えた組成物。
【請求項53】
患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)工程(b)で採取した内皮前駆細胞の中からGATA−2を発現している内皮前駆細胞を回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法。
【請求項54】
患者の異常を治療する方法であって、
前記異常は、前記患者におけるGATA−2被活性化遺伝子産物の発現によって治療され、
(a)前記患者のある部位から幹細胞を採取することと、
(b)工程(a)で採取した前記幹細胞からマスト前駆細胞を回収することと、
(c)工程(b)で採取したマスト前駆細胞の中からGATA−2を発現しているものを回収することと、
(d)工程(c)でGATA−2を発現しているとして回収された前記細胞がGATA−2被活性化遺伝子産物を発現するように誘導することと、
(e)前記患者の別の部位に、GATA−2被活性化遺伝子産物を発現している工程(d)で得られた前記細胞を導入して前記異常を治療することとを備えた方法。
【請求項55】
前記異常が虚血によって損傷を受けた組織である請求項53又は54の方法。
【請求項56】
前記遺伝子産物がプロエンドセリンである請求項53又は54の方法。
【請求項57】
前記遺伝子産物がエンドセリンである請求項53又は54の方法。
【請求項58】
心筋梗塞に罹患した患者の心筋の機能を改善させる方法であって、
(a)前記患者中のある部位から幹細胞を採取することと、
(b)前記幹細胞からCD117を発現する細胞を回収することと、
(c)前記細胞が前記患者の心筋の機能を改善するように、前記患者の異なる部位に前記回収した細胞を導入することとを備えた方法。
【請求項59】
前記患者が哺乳動物起源である請求項42、43、44又は45の何れかに記載の方法。
【請求項60】
前記哺乳動物がヒト起源である請求項59の方法。
【請求項61】
患者の虚血によって損傷を受けた組織における血管形成を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、 (c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管形成を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法。
【請求項62】
前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として得られる請求項61の方法。
【請求項63】
患者の虚血によって損傷を受けた組織における血管新生を刺激する方法であって、
(a)同種異系の幹細胞を取得することと、
(b)工程(a)で採取した前記幹細胞から内皮前駆細胞を回収することと、
(c)前記内皮前駆細胞が患者の虚血によって損傷を受けた前記組織における血管新生を刺激するように、工程(b)で回収した前記内皮前駆細胞を前記患者に導入することとを備えた方法。
【請求項64】
前記同種異系の幹細胞が、胚、胎児、又は臍帯血を起源として得られる請求項63の方法。
【請求項65】
心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者にG−CSFを注入して内皮前駆細胞を動員することを備えた方法。
【請求項66】
心筋梗塞に罹患した患者の心筋の機能を改善する方法であって、前記患者に抗CXCR4抗体を注入することを備えた方法。
【請求項67】
内皮前駆細胞を前記患者に導入することをさらに備えた請求項66の方法。
【請求項68】
前記患者にG−CSFを導入して内皮前駆細胞を動員することをさらに備えた請求項67の方法。
【図1A】

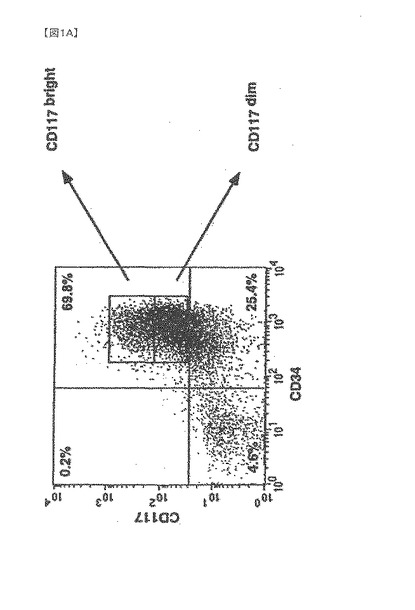
【図1B】
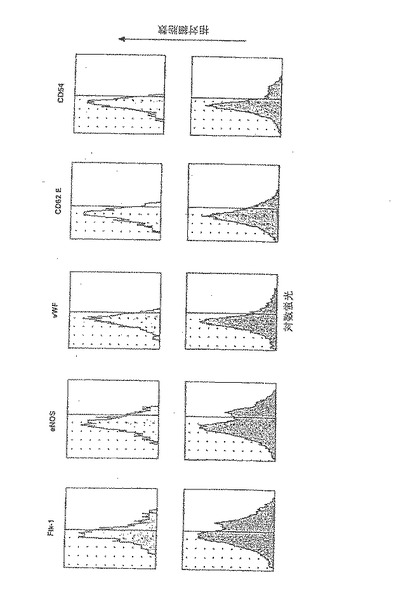
【図1C】
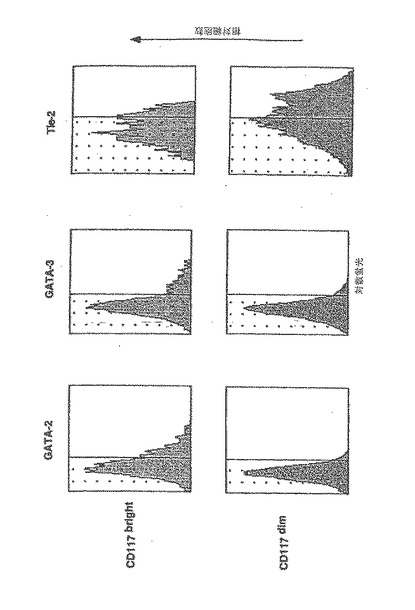
【図1D】
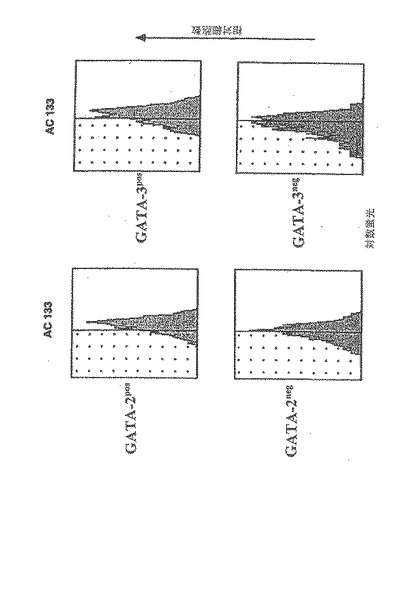
【図2A】
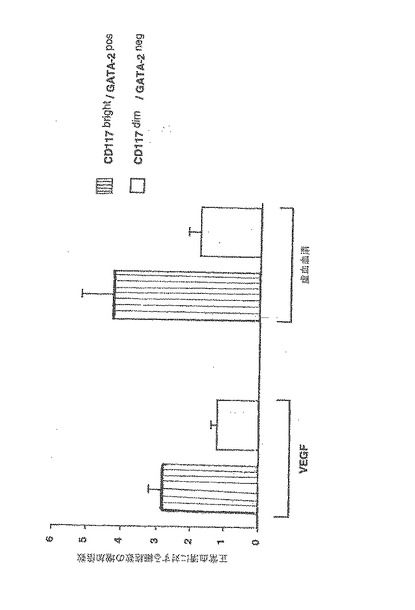
【図2B】

【図2C】
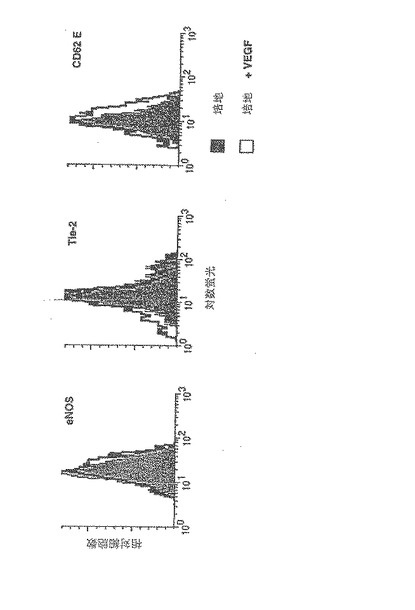
【図3A】
【図3B】

【図3C】
【図3D】

【図3E】

【図4−1】
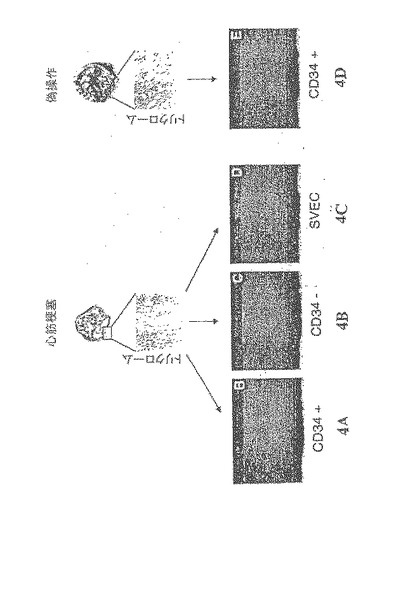
【図4−2】

【図4−3】
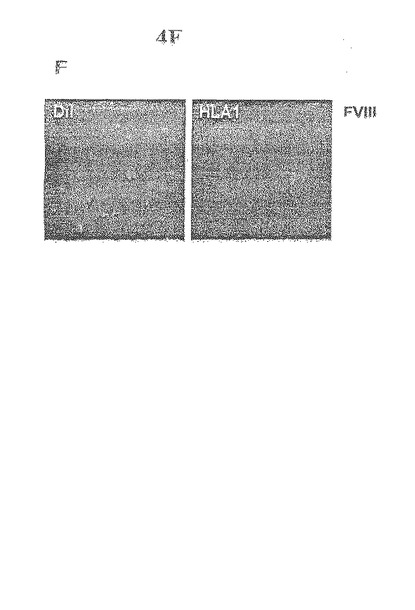
【図5A】
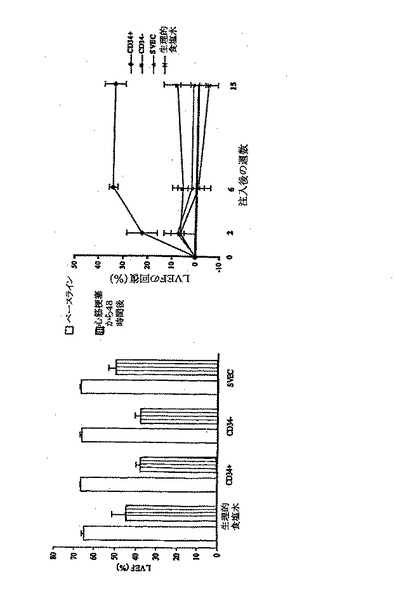
【図5B】
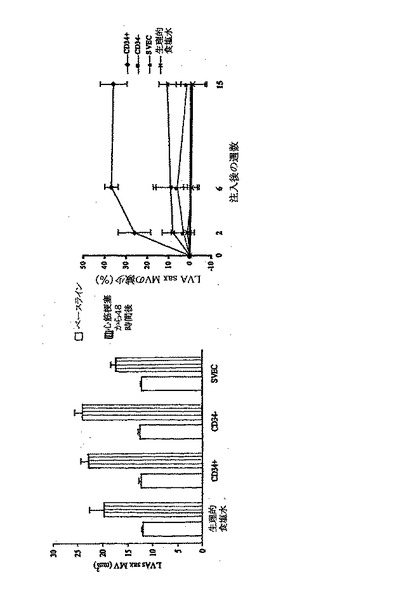
【図5C】

【図5D】
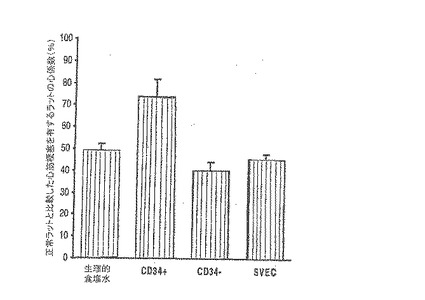
【図6A】

【図6B】

【図6C】

【図6D】

【図6E】
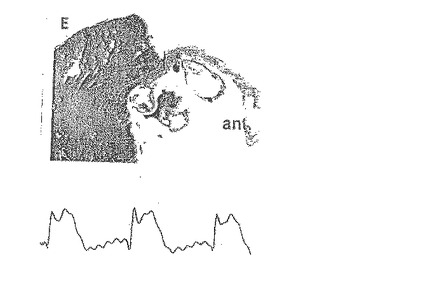
【図6F】
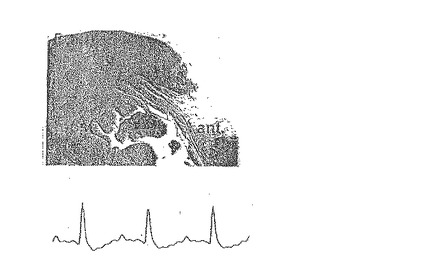
【図6G】

【図7A】
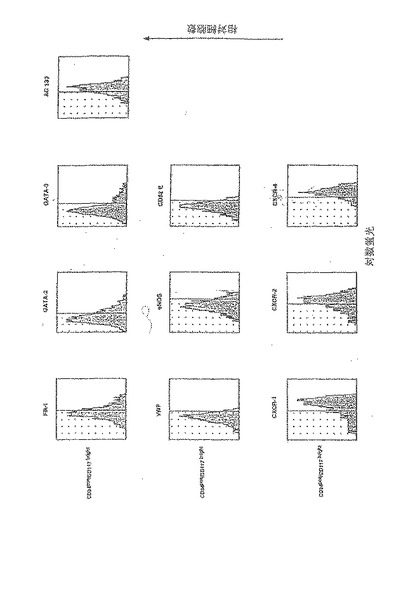
【図7B】

【図7C】

【図8A】

【図8B】
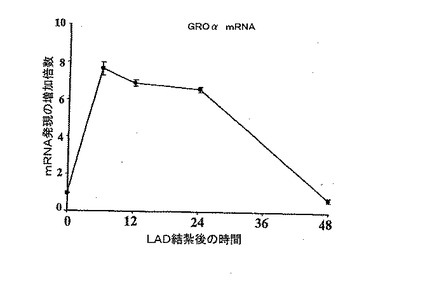
【図8C】

【図8D】
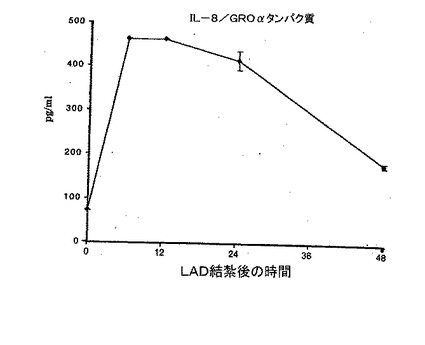
【図8E】

【図9A】

【図9B】
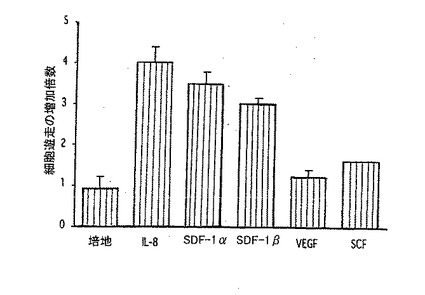
【図9C】
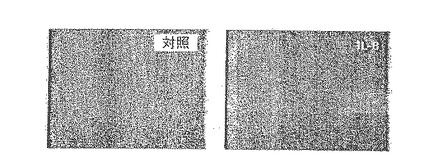
【図9D】
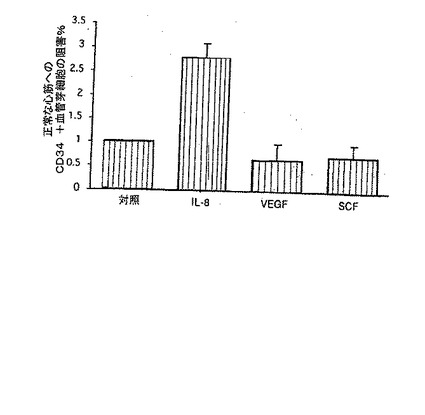
【図10A】
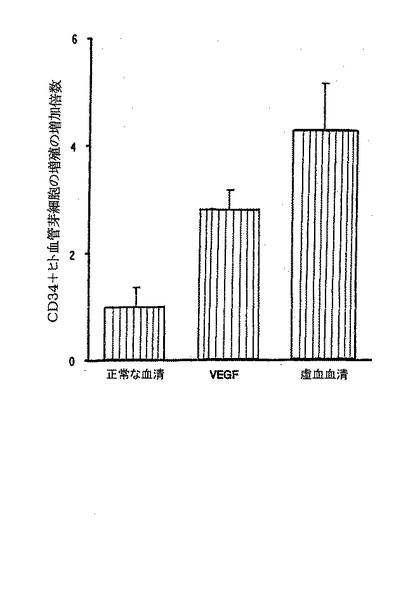
【図10B】
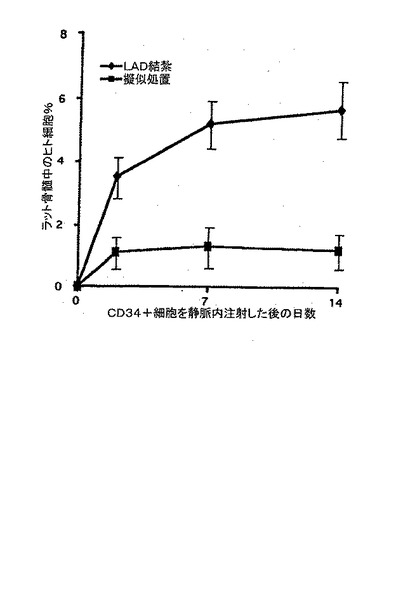
【図10C】

【図10D】
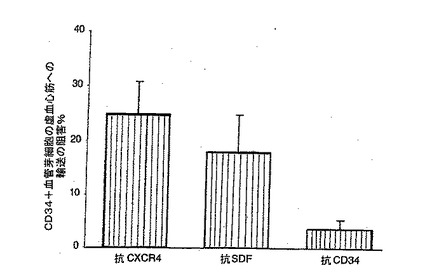
【図11A】
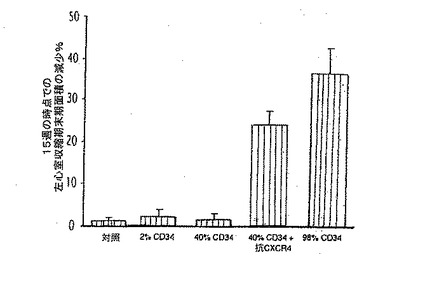
【図11B】
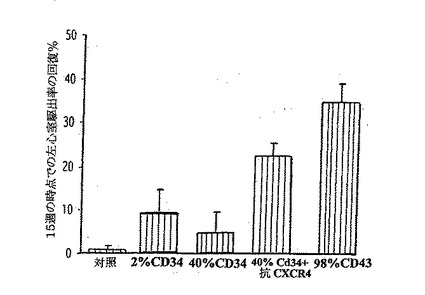
【図11C】
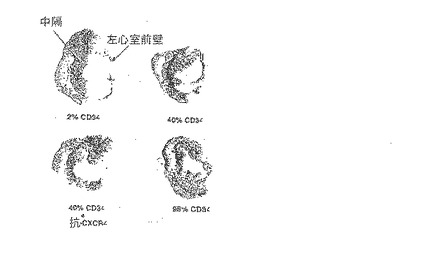
【図11D】
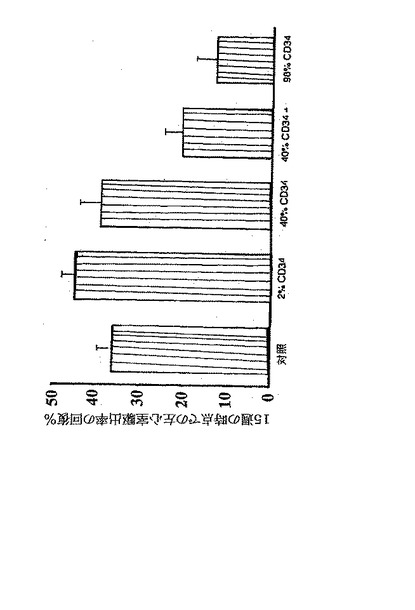
【図12】

【図13】
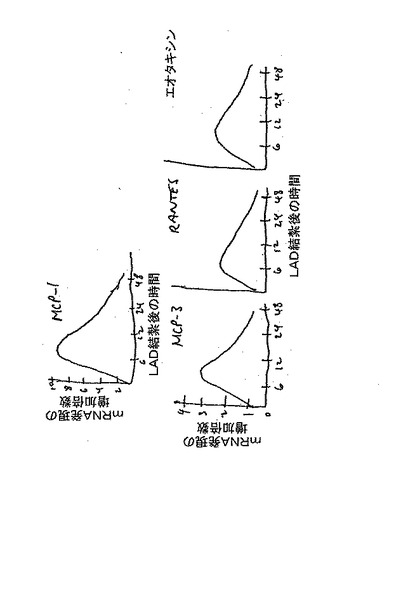
【図14】

【図15】
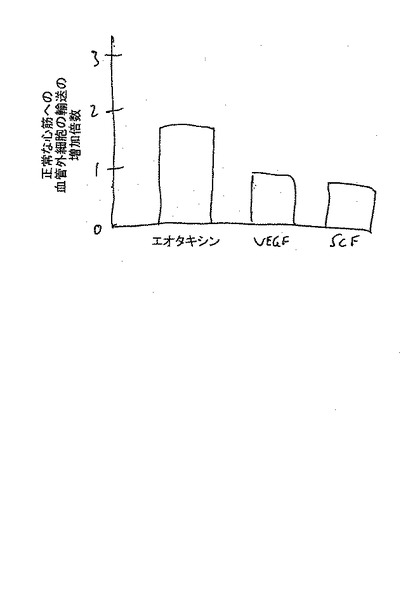
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図4−1】
【図4−2】
【図4−3】
【図5A】
【図5B】
【図5C】
【図5D】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図7A】
【図7B】
【図7C】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図10C】
【図10D】
【図11A】
【図11B】
【図11C】
【図11D】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2013−63979(P2013−63979A)
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−235072(P2012−235072)
【出願日】平成24年10月24日(2012.10.24)
【分割の表示】特願2002−501968(P2002−501968)の分割
【原出願日】平成13年6月5日(2001.6.5)
【出願人】(592104782)ザ・トラスティーズ・オブ・コランビア・ユニバーシティー・イン・ザ・シティー・オブ・ニューヨーク (21)
【氏名又は名称原語表記】THE TRUSTEES OF COLUMBIA UNIVERSITY IN THE CITY OF NEW YORK
【Fターム(参考)】
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−235072(P2012−235072)
【出願日】平成24年10月24日(2012.10.24)
【分割の表示】特願2002−501968(P2002−501968)の分割
【原出願日】平成13年6月5日(2001.6.5)
【出願人】(592104782)ザ・トラスティーズ・オブ・コランビア・ユニバーシティー・イン・ザ・シティー・オブ・ニューヨーク (21)
【氏名又は名称原語表記】THE TRUSTEES OF COLUMBIA UNIVERSITY IN THE CITY OF NEW YORK
【Fターム(参考)】
[ Back to top ]