
ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法
【課題】 HDL上昇、さらには動脈硬化性疾患の予防治療に有用な物質として期待される、ヒトABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングする方法。
【解決手段】 プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【解決手段】 プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト肝臓においてABCA1遺伝子の発現を制御するプロモーター・エンハンサー領域の発見に基づき、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法に関するものである。特に、プロモーター活性の増加、あるいは特定の非翻訳領域を含むmRNAの増加を指標として、肝臓におけるABCA1発現の増加により動脈硬化性疾患を予防または治療する作用を有する物質をスクリーニングする方法に関するものである。
【背景技術】
【0002】
血中高密度リポタンパク質(HDL)コレステロールの低下は動脈硬化の危険因子である。HDLコレステロールが10mg/dL上昇すれば冠動脈疾患のリスクが20〜30%低下すると予想されている。また人工HDL粒子が動脈硬化病巣を直接退縮する治療効果を持つことが実証され、HDL上昇薬が待望されているが、HDLを直接上昇する薬は実用化されていない(非特許文献1、Nat Rev Drug Discov. 2005 4, 193-205)。
【0003】
HDLは、ABC膜トランスポーターA1(ABCA1)が細胞内のリン脂質とコレステロールを、細胞外のアポリポプロティンA−Iに輸送して形成する。ABCA1はHDLの形成に必須の役割を持ち、機能不全をもたらす遺伝子変異はHDL欠損症の原因となる(非特許文献2、Nat. Genet. 1999; 22:352)。逆に、ABCA1過剰発現マウスではHDLが上昇する(非特許文献3、J. Biol. Chem. 2001; 276:33969)。ABCA1は各組織に発現するが、肝ABCA1が血中HDLの8割を産生することが肝特異的欠損マウスの研究により判明した(非特許文献4、J.Clin.Invest. 2005; 115, 1333)。したがって、HDL上昇には、肝臓においてABCA1の発現を増加させることが極めて有力な手段と考えられる。
【0004】
ABCA1の遺伝子転写制御に関しては、マクロファージなど末梢細胞のABCA1遺伝子の転写はエクソン1上流のプロモーター領域(末梢型プロモーター)により制御され、このプロモーターは核内受容体LXRが直接結合して活性化されることが知られている(非特許文献5、Biochem. Biophys. Res. Commun. 2000; 274, 794)。
【0005】
LXRアゴニストはABCA1発現促進を介してHDL上昇効果を示す。しかしLXR活性化は同時に、中性脂質合成のマスターレギュレーターである転写因子SREBP−1cの発現を促進する。その結果、脂肪酸生合成酵素系の遺伝子発現が増加し脂肪肝や血清トリグリセリド上昇が引き起こされ、このような副作用がLXRアゴニストの臨床薬としての開発の障害となっている(非特許文献6、Biochem. Pharmacol., 2009; 77, 1316-1327)。
【0006】
発明者らはラット肝臓のABCA1発現は、LXR依存の末梢型プロモーターとともに、肝臓に特異的な肝型プロモーターにより二重の制御を受けることを見いだした(非特許文献7、J. Biol. Chem. 2007; 282:21090)。すなわちラット肝臓にはエクソン2から転写が開始される肝特異的なABCA1 mRNA(L2)が存在しており、ABCA1遺伝子のエクソン2上流領域が肝臓に特異的なプロモーターとして機能する。肝型プロモーターにはステロール応答配列SREが存在しており、コレステロール減少により活性型に転換されて核内移行する転写因子SREBP−2が直接結合して活性化する機構を明らかにした。このプロモーター領域は、ラット・マウス・ヒトに共通することから、この肝型プロモーターを活性化する薬物は、LXRの活性化による副作用を示さずに肝ABCA1発現を促進できると考えられた。そして、発明者らは、この肝型プロモーターが、ヒトに有効なABCA1発現促進薬のスクリーニングに使用できることを考えた(特許文献1 特開2007−082445号)。
【0007】
マクロファージや繊維芽細胞などでは、コレステロール合成の律速酵素であるHMG−CoA還元酵素の阻害剤スタチン系薬剤がABCA1発現を低下させる。これは、コレステロール合成過程で生じるLXRの内因性リガンドがスタチン系薬剤の作用によって減少し、ABCA1遺伝子プロモーター活性が低下することによる(非特許文献8、Arterioscler. Thromb. Vasc. Biol. 2004; 24, 2365)。しかしながら、スタチン系薬剤には、血中LDL濃度の低下に加えて、血中HDL濃度の上昇効果も認められており、スタチン系薬剤はヒト肝がん由来細胞HepG2において、ABCA1遺伝子の発現促進作用を示すことも知られている(非特許文献9、Biochem. Biophys. Res. Commun. 2004; 324, 835)。発明者らは、スタチン系薬物は肝由来細胞において末梢型プロモーターの活性を低下させる一方、肝型プロモーターを活性化することによりABCA1発現増加をもたらす機構を明らかにしてきた(非特許文献7)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2007−082445号公報
【非特許文献】
【0009】
【非特許文献1】Nat Rev Drug Discov. 2005 4, 193-205
【非特許文献2】Nat. Genet. 1999; 22:352
【非特許文献3】J. Biol. Chem. 2001; 276:33969
【非特許文献4】J. Clin. Invest. 2005; 115, 1333
【非特許文献5】Biochem. Biophys. Res. Commun. 2000; 274, 794
【非特許文献6】Biochem. Pharmacol., 2009; 77, 1316-1327
【非特許文献7】J. Biol. Chem. 2007; 282:21090
【非特許文献8】Arterioscler. Thromb. Vasc. Biol. 2004; 24, 2365
【非特許文献9】Biochem. Biophys. Res. Commun. 2004; 324, 835
【発明の概要】
【発明が解決しようとする課題】
【0010】
ABCA1は各組織に発現するが、肝ABCA1が血中HDLの8割を産生することが肝特異的欠損マウスの研究により判明した(非特許文献4)。また、肝特異的なABCA1欠損マウスでは動脈硬化が増悪することがわかった(Arterioscler. Thromb. Vasc. Biol 2009; 29:548)。したがって、肝臓においてABCA1の発現を増加させることが、HDL上昇、さらには動脈硬化性疾患の予防治療に極めて重要と考えられ、肝臓におけるABCA1遺伝子発現の制御機構の解明、さらにはそれを利用した制御薬剤のスクリーニング方法が望まれていた。
【0011】
以上に鑑みて、本発明の課題は、ヒト肝細胞のABCA1の上昇作用を有し、動脈硬化に伴う疾患の予防および/または治療に有用な医薬に用いることができる物質をスクリーニングする新規な方法を提供することにある。
【0012】
なお、本発明の方法において用いられるヒトABCA1遺伝子は、Gene Bankデータベース上に「ATP結合カセット輸送体A1(ATP-binding cassette transporter A1)」として公開されている。また、国際公開第00/078972号パンフレットには、「ABC1」をコードするものとしてヒトABCA1遺伝子が、またそれと共に上記のエクソン1上流のプロモーターが開示されている。
【課題を解決するための手段】
【0013】
すなわち、本発明の一の側面によると、プロモーター活性を有するDNA、ならびにヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【0014】
前記プロモーター活性を有するDNAが、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAであることが好ましい。
【0015】
前記ヒトABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質であることが好ましい。
【発明の効果】
【0016】
本発明にかかるABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法によれば、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、動脈硬化性疾患の予防および/または治療剤を試験するための新規方法を確立することができ、HDL上昇薬の開発に有用である。また、本発明の方法は、従来のABCA1遺伝子発現促進物質のスクリーニング方法に比べ、ヒトHDL上昇作用に対して、より効果的な分子を検出できる可能性がある。
【図面の簡単な説明】
【0017】
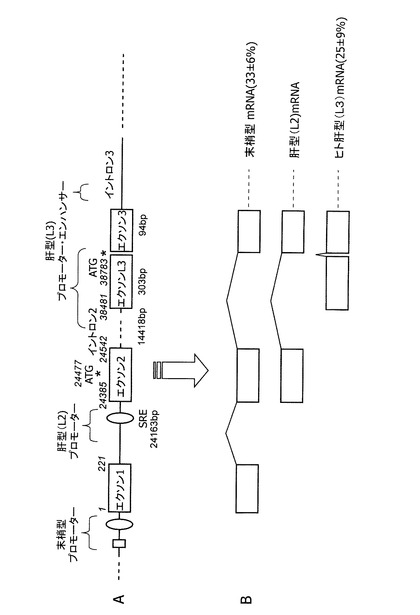
【図1】図1AはヒトABCA1遺伝子の模式図を示す。模式図中の数値は、ヒトABCA1遺伝子のDNAにおいて、末梢型ABCA1 mRNAの転写開始点を1とした塩基位置を示すものである。図1Bは、5’−RACE法により解析したヒト肝臓で発現するABCA1のmRNAの5’末端の模式図を示す。括弧内は、ヒト肝臓における、各mRNAの存在比を示す。
【図2】図2Aは、ヒト各組織(肝臓、筋肉、腎臓、膵臓、心臓、脳、胎盤および肺)で発現する総ABCA1 mRNA量を示したグラフであり、図2Bは、ヒト各組織で発現するヒト肝型(L3型)ABCA1 mRNA量を示したグラフである。
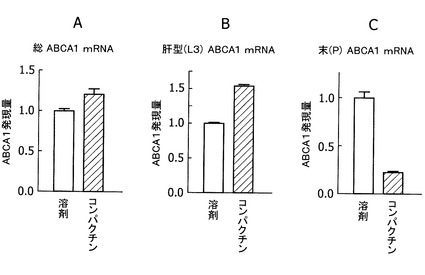
【図3】図3Aは、コンパクチンで処理したヒト肝由来培養細胞JHH5における総ABCA1 mRNAの発現量変化を示すグラフであり、図3Bは、コンパクチンで処理したヒト肝由来培養細胞JHH−5におけるヒト肝型(L3)ABCA1 mRNAの発現量変化を示すグラフであり、図3Cは、コンパクチンで処理したヒト肝由来培養細胞JHH−5における末梢型(P)ABCA1 mRNAの発現量変化を示すグラフである。
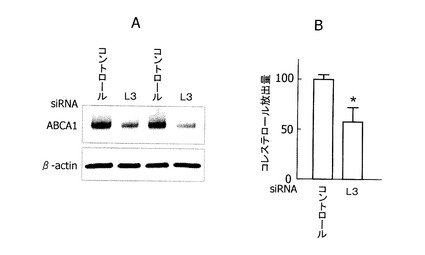
【図4】図4Aは、ヒト肝型(L3型)ABCA1 mRNAの発現をsiRNAを用いてノックダウンしたヒト肝由来培養細胞JHH−5における、ABCA1蛋白の発現量をウェスタンブロットで測定した写真であり、図4BはアポA−I依存のコレステロール細胞外排出量(HDL産生量)を示したグラフである。
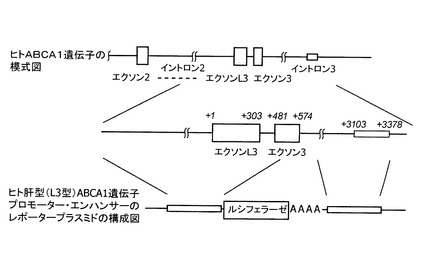
【図5】図5は、ヒトABCA1遺伝子の模式図、およびヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーのレポータープラスミドの構成図である。模式図中の数値は、ヒトABCA1遺伝子のDNAにおいて、L3型ABCA1 mRNAの転写開始点を1とした塩基位置を示し、転写開始点の下流をプラス、上流をマイナスで表したものである。
【図6】図6は、ヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーがコンパクチンにより活性化されることを示すグラフである。
【図7】図7は、L3型エンハンサー配列をSV−40プロモーターを導入したルシフェラーゼプラスミドに組み込んでも、コンパクチンによる活性化が認められること、しかしSREBP−2の発現増加により影響されないことを示すグラフである。
【図8】図8は、ヒト肝型(L3型)ABCA1遺伝子エンハンサーに含まれる配列を示す。上の行の左端が5’側であり、下の行の右端が3’側である。
【図9】図9は、ヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーの活性が、配列a、配列bあるいは配列cの欠失により消失したことを示すグラフである。
【図10】図10は、ヒト肝型(L3型)ABCA1遺伝子エンハンサーの活性が、転写調節因子HNF4alphaの発現ベクターを導入することにより上昇し、配列cの欠失により消失したことを示すグラフである。
【図11】図11は、コンパクチンで処理したヒト肝由来培養細胞JHH−5における、HNF4alpha蛋白の発現量をウェスタンブロットで測定した写真である。
【発明を実施するための形態】
【0018】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって、限定されるものではない。
【0019】
発明者らは、ヒト肝および肝由来細胞のABCA1発現制御をさらに詳細に解析したところ、ヒト肝には従来から知られているマウス・ラットに共通の肝型(L2型)ABCA1 mRNAに加えて、さらに別の肝型(L3型)ABCA1 mRNAが存在することを見いだした。このヒト肝型(L3型)ABCA1 mRNAは、ヒト肝に存在する総ABCA1 mRNAの25%に相当しており、ヒト肝由来細胞をスタチン系薬剤で処理すると発現が促進されることを見いだした。L3型の転写開始点はエクソン2の翻訳開始点より下流であるが、L3型mRNAは機能的ABCA1をコードしていることは、L3型ABCA1 mRNAをsiRNAを用いてノックダウンすると、ABCA1蛋白質の発現が減少し、HDL形成能も低下したという事実により確認された。
【0020】
引き続き、対応するL3型プロモーター・エンハンサー領域を同定したところ、スタチン系薬物による活性化を担うエンハンサー領域は、既報のL2型プロモーター(特許文献1、非特許文献7)とは異なりSREBP−2には応答せず、新規なメカニズムでヒトABCA1発現を制御することを見いだした。したがって、L3型プロモーター・エンハンサー領域はヒトで肝臓のABCA1発現を制御し、血中HDLレベルを調節する重要な役割をもつと考えることができる。L3型プロモーター・エンハンサー領域の活性化、あるいは新規非翻訳領域を含むL3型ABCA1 mRNAの発現増加を指標とする薬物スクリーニング法は、ヒトにおいて有効なHDL上昇薬の開発に極めて有用性が高いと考えられる。
【0021】
さらには、発明者らは、L3型エンハンサーは、L3型プロモーター以外のプロモーターを用いた場合でも、ヒトABCA1発現を促進することを見出した。
【0022】
上記のヒト肝臓におけるABCA1遺伝子の発現調節に新たな制御機構が存在するという発見に基づき、ABCA1遺伝子発現を調節する物質を選別または同定するための方法として、新規ヒト肝型(本明細書中、L3型とも表記する)ABCA1 mRNAをコードする遺伝子の、エクソン3上流(本明細書中、エクソンL3とも表記する)部分を用いる方法、およびイントロン2に存在する新規ヒト肝型ABCA1プロモーターとイントロン3に存在するエンハンサー(本明細書中、L3型ABCA1プロモーター等とも表記する)を用いる方法を開発し、本発明を完成させた。
【0023】
本発明が開示する新規ヒト肝型(L3型)プロモーターは、従来から知られている、ABCA1遺伝子のエクソン1上流のプロモーター(国際公開第00/078972号パンフレット参照)あるいはエクソン2上流のL2型プロモーターとは全く異なる領域に存在し、DNA配列も異なる。このL3型プロモーターに関しては、ABCA1遺伝子転写制御、HDL形成との関連は報告されておらず、動脈硬化性疾患の治療または予防剤の試験法としての有用性も知られていない。上記したように、ピタバスタチン、シンバスタチン、アトルバスタチンがヒト肝癌細胞HepG2においてABCA1のmRNAを増加させることが示されている(非特許文献9)。その作用機序については、発明者らが見いだした肝型(L2)プロモーターが想定されていた。しかし、本発明にかかる新規ヒト肝型(L3型)ABCA1プロモーターおよび当該プロモーターによる発現制御については、従来技術文献において、記載も示唆もしていない。同様に、本発明が開示する新規肝型ABCA1エンハンサーについても、これらの従来技術文献において何ら報告されたものではなく、発明者らによる今般の研究によりはじめて、明らかにされたものである。
【0024】
本発明は、一実施の形態によると、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法に関する。
【0025】
本発明はまた、別の実施の形態によると、このようなスクリーニング方法に用いることができる、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに関する。
【0026】
ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNA、およびヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAは、ヒトに由来する配列を有するものであれば、天然のものであってもよく、また人工的なものであってもよい。すなわち、本発明の別の側面によると、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAとを含んでなる組み換えDNAが提供される。
【0027】
ヒトABCA1遺伝子のイントロン2は、ヒトABCA1遺伝子のエクソン2の下流かつエクソン3の上流に存在する領域であり、イントロン3はエクソン3の下流かつエクソン4の上流に存在する領域である。具体的には、ヒトABCA1遺伝子のエクソン2として、配列番号1に記載するヒトABCA1遺伝子のDNA配列の24385位から24542位の配列を有するものが、イントロン2として、配列番号1に記載の配列の24543位から38960位の配列を有するものが、エクソン3として、配列番号1に記載の配列の38961位から39054位の配列を有するものが、イントロン3として、配列番号1に記載の配列の39055位から43587位の配列を有するものが、およびエクソン4として、配列番号1に記載の配列の43588位から43729位の配列を有するものが、それぞれ見出されている。また、本明細書中、エクソンL3と記す領域は、配列番号1に記載の配列の38481位から38783位の配列を有する領域である。配列番号2にエクソンL3の配列を示す。なお、配列番号1は、ヒトABCA1遺伝子のエクソン1から始まる配列である。また、ヒトにおける上記イントロン2、エクソンL3の相同体、変異体および誘導体の配列は、当業者であれば、ヒトにおける上記イントロン2および/またはエクソン3、4の配列との相同性に基づき決定することができる。
【0028】
ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、配列番号1に示すヒトABCA1遺伝子の塩基配列の38055位から38960位までの塩基を含むことが好ましい。ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、特には、(a)配列番号1に示すヒトABCA1遺伝子の塩基配列の38055位から38960位までの塩基(配列番号3)、配列38055位から配列39054位までの塩基(配列番号4)、配列38055位から配列39224位までの塩基(配列番号5)に記載の配列を有するDNAとすることができる。
【0029】
また、イントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(a)のDNAと実質的に同一の配列を有するDNAとすることができる。具体的には、イントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(b)配列番号3、配列番号4、配列番号5のいずれかに記載の配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。当該技術分野における通常の知識を有する者であれば、野生型プロモーターのヌクレオチド配列の部分的な置換、挿入または欠失などの改変により、野生型プロモーターと同等またはそれ以上のプロモーター活性を有するDNAを調製することが可能である。
【0030】
また、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(c)配列番号3、配列番号4、配列番号5のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。このようなDNAには、(a)のDNAの、自然界に存在する変異型遺伝子もしくは人為的に改変した変異型遺伝子または異種生物由来の相同遺伝子等が含まれる。
【0031】
相同性の比較は、通常、容易に入手できる配列比較プログラムを用いて行うことができる。商業的に入手可能なそのようなコンピュータープログラムは2以上の配列の間の%相同性を計算することができる。ほとんどの配列比較方法においては、全体としての相同性スコアに不当にペナルティを与えることなく可能な挿入および欠失を考慮に入れて、最適な整列を生じさせるよう設計される。これは、「ギャップ」を配列の整列に挿入して、局所的な相同性を最大化するよう努めることによって達成される。このような方法では、できる限り少数のギャップにおける配列の整列が、多くのギャップにおけるものより高いスコアを達成するように、各ギャップに「ギャップペナルティ」を割り当てて、相同性を計算する。ほとんどの整列プログラムはギャップペナルティを設定することができる。しかしながら、配列比較のためのそのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。このような配列比較を実行できるソフトウェアの例として、BLASTパッケージ(Ausubel et al.,1999 ibid-Chapter18参照)、FASTA(Atschul et al.,1990,J.Mol.Biol.,403-410)およびGENEWORKSスイートが挙げられる。
【0032】
また、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAの少なくとも一部とすることができる。ここで、DNAは通常、互いに相補的な二本鎖DNA構造を有している。このため、(a)のDNAとハイブリダイズするDNAとは、(a)のDNAのいずれかの鎖とハイブリダイズする鎖を有するDNAをいうことに留意すべきである。
【0033】
なお、ストリンジェントな条件およびハイブリダイゼーションの方法は、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に従って設定することができる。具体的には、ストリンジェントな条件は、「1xSSC、0.1%SDS、37℃」とすることができ、よりストリンジェントな条件は、「0.5xSSC、0.1%SDS、42℃」とすることができ、さらにストリンジェントな条件は、「0.2xSSC、0.1%SDS、65℃」とすることができる(1xSSC:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)。ただし、これらの条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記または他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0034】
なお、(b)〜(d)のDNAのプロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法等により測定することができる。また、(b)〜(d)のDNAは、(a)のDNAと「同様」のプロモーター活性を有することが好ましい。ここで、「同様」のプロモーター活性を有するとは、質的に類似のプロモーター活性を有することをいい、例えば、(b)〜(d)のDNAが、(a)のDNAと少なくとも1つの因子について同一の制御を受ける場合が含まれる。一方で、「同様」のプロモーター活性を有するとは、量的に同程度のプロモーター活性を有することを意図するものではない。また、当業者には明らかなように、プロモーター活性を発揮するために、DNAは、配列番号3、配列番号4、配列番号5のいずれかに記載の配列(またはこれと実質的に同一の配列)の全てを有する必要はなく、その一部であってもよい。上記したように、プロモーター活性を有するのに必要な領域は、当業者であれば、ルシフェラーゼアッセイ法等により適宜決定することができる。
【0035】
ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(e)配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置(配列番号1のヒトABCA1 DNAの38481位)下流3103位から下流3378位までの塩基(配列番号6)を含むことが好ましく、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から下流3172位から下流3378位までの塩基(配列番号7)を含むことがさらに好ましく、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から下流3172位から下流3259位までの塩基(配列番号8)を含むことが最も好ましい。
【0036】
さらには、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から3172位から3383位までの塩基(配列番号9)と、L3型mRNAの転写開始位置から3230位から3243位までの塩基(配列番号10)と、L3型mRNAの転写開始位置から3244位から3256位までの塩基(配列番号11)をすべて有するDNAであることが必要である。
【0037】
これらの3つの配列が保持される限り、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(f)配列番号6、配列番号7、配列番号8のいずれかに記載の好ましいDNAの配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列であってもよい。
また、これらの3つの配列が保持される限り、(g)配列番号6、配列番号7、配列番号8のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有するものであってもよい。相同性の比較に関しては、(c)のDNAについて述べたのと同様にして行うことができる。
さらに、これらの3つの配列が保持される限り、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(h)配列番号6、配列番号7、配列番号8のいずれかに記載の好ましいDNAとストリンジェントな条件でハイブリダイズし、エンハンサー活性を有するDNAの少なくとも一部とすることができる。
【0038】
上記に配列を示したもの以外であっても、本願発明において、エンハンサー活性を有するのに必要な領域は、当業者であれば適宜決定することができる。エンハンサー活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。
【0039】
次に、本発明の一実施の形態による、これらのDNAを用いた、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法において用いることができる、組換えDNAについて説明する。組換えDNAは、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを含んでなる。
【0040】
プロモーター活性を有するDNAとしては、上記において説明した、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAを用いることができる。さらには、プロモーター活性を有するDNAとしては、ヒトABCA1遺伝子に由来する配列を有するものに限定されず、任意のプロモーターの塩基配列を有するものを用いることができる。当業者には既知のプロモーターとしては、例えば、SV−40プロモーター、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、EFプロモーター、CAGプロモーターが挙げられるが、これらには限定されない。
いっぽう、エンハンサー活性を有するDNAとしては、上記において説明した、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAを用いることができる。
【0041】
これらのプロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとは、組換えDNAにおいては、任意の組み合わせで用いることができる。すなわち、(a)、(b)、(c)、(d)のいずれかから選択されるプロモーター活性を有するDNAもしくはその他の任意プロモーター活性を有するDNAと、(e)、(f)、(g)、(h)のいずれかから選択されるエンハンサー活性を有するDNAとを、任意の組み合わせで含む組換えDNAとすることができる。
【0042】
なお、本発明にかかる組換えDNAにおいて、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAを用いる場合でも、そのほかに転写最小プロモーターをさらに有することができる。最小プロモーターの例として、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、PGKプロモーター、SV40プロモーターなどが挙げられるが、これらに限定されるものではない。
【0043】
本発明にかかるDNAは、ゲノムライブラリーからの選別や、ゲノムDNAを鋳型としたポリメラーゼ連鎖反応(PCR)により取得できる。ゲノムライブラリーからの選別は、適当なベクターに組み込んだゲノムDNAと、目的のDNAの一部の配列を有する標識DNAとのハイブリダイゼーションによって行うことができる。ハイブリダイゼーションは、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、添付の使用説明書に記載の方法に従って行なうことができる。また、ゲノムDNAを鋳型としたPCRは、目的のDNAの一部の配列を有するプライマーを用いて行うことができる。ヌクレオチド配列の改変は、例えば、制限酵素またはDNAエキソヌクレアーゼによる欠失導入、部位特異的変異誘発法による変異導入、変異プライマーを用いたPCR法による改変、合成変異DNAの直接導入などの方法により行うことができる。
【0044】
本発明の他の側面によると、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを有する組換えベクターが提供される。組換えベクターは、当該プロモーター活性を有するDNA、及びエンハンサー活性を有するDNAに作動可能に連結した構造遺伝子をさらに有してもよい。以下に詳細に説明するように、構造遺伝子は、例えば、組換えベクターをスクリーニングに用いる場合、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。
【0045】
本発明にあっては、ベクターは、任意のベクター、例えば、バクテリオファージ、プラスミド、コスミド、ファージミドなどであってよい。より具体的には、本発明に用いることができるベクターの例として、動物細胞用のベクターであるpCDベクター、cDM8ベクター、レトロウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、アデノアソシエイトウイルスベクターなどや、細菌用のベクターであるpUCなどを挙げることができる。構造遺伝子をベクター内に組み込むためには、構造遺伝子が正しく転写される方向に、さらにプロモーターおよびエンハンサーが構造遺伝子に対して適正に機能する位置関係になる様に、プロモーター領域の下流であって、エンハンサー領域の上流に構造遺伝子を連結すればよい。プロモーターと構造遺伝子との連結、及びエンハンサーと構造遺伝子との連結は、制限酵素切断部位を利用して行うことができる。また、適当な制限酵素部位がなくてもリガーゼ反応を用いて行うことができる。
【0046】
なお、「作動可能に連結」とは、一の遺伝子要素が、他の遺伝子要素に対して予測されるように機能できるように配置されていることをいう。例えば、プロモーターが、構造遺伝子の転写の開始を助ける場合には、プロモーターは構造遺伝子に作動可能に結合している。同様に、エンハンサーが構造遺伝子の転写を促進する場合には、エンハンサーが構造遺伝子に作動可能に結合している。この機能的な関係が維持される限り、プロモーターと構造遺伝子との間、あるいは構造遺伝子とエンハンサーとの間に残基が介在してもよい。
【0047】
本発明の他の側面によると、プロモーター活性を有する外因性のDNA及びヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。本明細書において、形質転換体は、広く、外部からDNAが導入された結果、新しい形質を獲得した生物をいう。形質転換体は、任意の生物であってよく、また、任意の生物に由来する任意の細胞または組織であってよい。特に、本発明にかかる形質転換体は、上記組換えベクターで形質転換された形質転換体を含む。
【0048】
組換えベクターにより形質転換することができる宿主の例として、例えば、エシェリヒア属菌、バチルス属菌、酵母、昆虫細胞、昆虫、動物細胞などを挙げることができる。これらの宿主への形質転換の方法の例として、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、アデノウイルスベクターまたはレトロウイルスベクターによる遺伝子導入法、マイクロインジェクション法などを挙げることができる。
【0049】
本発明の他の側面によると、ヒトABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。本発明にかかるキットは、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。以下に詳細に説明するように、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることで、ヒトABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングすることができる。すなわち、本発明にかかるキットは、上記組換えベクターおよび/または上記形質転換体を有する。
【0050】
本発明にかかるキットには、スクリーニングの方法に応じて、酵素、酵素基質、緩衝液等の試薬、培地、道具等を組み合せることができる。酵素基質の例として、ルシフェリン、ATP、[14C]クロラムフェニコール、アセチルCoAを挙げることができる。緩衝液として、解析に用いる酵素の活性の維持に好適な緩衝液を適宜選択することができる。道具の例として、TLC用展開シート、メンブレン等を挙げることができる。
【0051】
本発明の他の側面によると、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供され、動脈硬化性疾患の予防または治療に有用な物質を探索するために用いることができる。本発明にかかるスクリーニング方法は、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする。
【0052】
具体的には、本発明にかかるスクリーニング方法は、
被検物質を供するステップと、
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0053】
被検物質として、単一の化合物または組成物、天然または合成化合物、有機または無機化合物、タンパク質、ペプチド、オリゴヌクレオチド、ポリヌクレオチド、細胞抽出物、細胞培養上清、その他の任意の物質を供することができる。
【0054】
スクリーニングは、インビトロおよびインビボのいずれでも行うことができる。本発明にかかるスクリーニング方法において用いられる細胞または生物として、細菌等の原核生物(大腸菌等)、酵母(サッカロマイセス(Saccharomyces)属、クライヴェロマイセス(Kluyveromyces)属等)、昆虫、植物もしくは動物またはこれらに由来する細胞が挙げられる。動物の例として、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物を挙げることができる。本発明で用いられる細胞は、実験室で培養することのできる培養細胞であることが好ましく、初代培養肝細胞であることがさらに好ましい。細胞は、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物に由来することができ、ヒトに由来することが好ましい。また、細胞は、肝臓に由来することが好ましい。
【0055】
本発明に用いられる細胞または生物は、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有することが好ましい。当該DNAは、内因性であっても、または外因性であってもよい。外因性である場合、当該DNAは、任意の手段により細胞または生物に導入されてもよい。例えば、スクリーニングに培養細胞を用いる場合、培養細胞への当該DNAの導入方法として、本発明の属する技術分野において汎用される任意の方法を採用することができる。例えば、DNAの導入方法の例として、リン酸カルシウム−DNA共沈法、リポフェクション法、DEAE−デキストラン法、電気パルス穿孔法等が挙げられるが、これらに限定されない。いずれの方法においても、用いる細胞に応じて至適化されたトランスフェクション条件を用いることが好ましい。
【0056】
構造遺伝子は、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。当業者に明らかなように、上記構造遺伝子は、その転写量および/または発現量の測定方法に応じて、適宜選択することができる。上記構造遺伝子の例として、ヒトABCA1、所定の活性を有するタンパク質(ルシフェラーゼ等)、所定の抗体が結合できるタンパク質、所定の物質が特異的に結合できるタンパク質(GST等)等をコードする遺伝子を挙げることができ、特に、ホタルルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼおよび緑色蛍光蛋白質からなる群より選択されるタンパク質をコードする遺伝子(いわゆるレポーター遺伝子)を挙げることができる。
【0057】
被検物質を細胞または生物に接触させることは、当業者に明らかなように、任意の方法により行うことができる。例えば、培養細胞を用いる場合、被検物質を培地中に添加することで、接触させることができる。すなわち、培養細胞を用いる場合、細胞を、被検物質の存在下または非存在下で培養することが好ましい。また、動物を用いる場合は、任意の手段による投与により、例えば、任意の経路からの注射により、接触させることができる。被検物質を細胞または生物へ接触させる時間は、被験物質の効果、濃度、細胞または生物の種類および技術的考慮に応じて、適宜設定することができる。また、被験物質の濃度は、その毒性、透過性、細胞数、処理時間等に応じて、適宜設定することができ、通常1nM〜1mMであることが好ましい。培養細胞を用いる場合、一般的にこのような処理は、多くの試験を同時に実施できるマルチウェルプレートで実施することが好ましい。培養細胞を用いる場合、被験化合物の処理によるレポーター遺伝子の発現が測定可能な条件は、細胞が生存してレポーター遺伝子の発現産物が生産可能な条件であれば良い。特に好ましい条件にあっては、好ましくは使用される細胞株に適した培地を使用し、4〜6%の炭酸ガス存在下、36〜38℃で5〜72時間培養する。
【0058】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定し、これらの転写量および/または発現量を比較することで、当該被検物質を、ヒトABCA1遺伝子の転写を調整する作用を有する物質の候補物質とすることができるか否か判断することができる。具体的には、ある被検物質を接触させた群で、接触させなかった群と比べて、上記遺伝子の転写量および/または発現量が変化した場合、特に増加した場合、当該被検物質を候補物質とすることができる。
【0059】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定するステップと、被検物質を接触させた群と接触させなかった群の転写量および/または発現量を比較するステップは、当業者に明らかなように、任意の方法により行うことができる。具体的には、ノーザンブロッティング法、DNAアレイ法等により、上記遺伝子の転写量を測定することができる。また、ルシフェラーゼアッセイ法やCAT法等により、上記遺伝子の発現量を測定することができる。さらに、上記所定の遺伝子に対する抗体が入手可能な場合は、ELISA法、免疫沈降法、ウエスタンブロッティング法により、当該遺伝子の発現量を測定することができる。また、上記所定の遺伝子として、GST等の所定の物質に特異的に結合するタンパク質の遺伝子を用いる場合は、当該タンパク質を沈降させ、沈降したタンパク質の量を測定することで、上記遺伝子の発現量を測定することができる。
【0060】
[ノーザンブロッティング法]
ノーザンブロッティング法は、変性したRNAを変性条件下のアガロース電気泳動で分離し、ニトロセルロースフィルターに移して、標識した特異的なプローブで検出するものである。RNAを変性させるのは、RNAは分子内に二次構造を持つため、そのままではサイズに従った正確な分離ができず、さらに、フィルターは一本鎖の核酸しか結合できないためである。
【0061】
[ルシフェラーゼアッセイ法、CAT法]
ルシフェラーゼアッセイ法およびCAT法は、目的とする遺伝子の転写調節領域の下流にレポーターとしてLUC(ルシフェラーゼ)またはCAT(クロラムフェニコールアセチルトランスフェラーゼ)遺伝子を組み込んだプラスミドを作成し、そのプラスミドを導入した細胞の酵素活性を測定するものである。具体的には、ルシフェラーゼアッセイ法は、マグネシウム存在下、ルシフェラーゼがルシフェリンとATPから酸化ルシフェリンとAMPを作る反応を触媒する際に発する波長560nmの光を、ルミノメーターを使って検出するものである。CAT法の場合、基質の[14C]クロラムフェニコールが、CATによりアセチル化されて、[14C]アセチルクロラムフェニコールが産生される。このサンプルを酢酸エチルで抽出し、薄層のシリカゲルプレート上に展開後、移動度の異なるスポットの放射活性を測定する。
【0062】
本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質が提供される。特に、上記したように、本発明にかかるスクリーニング方法によると、ヒトABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。このため、本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、動脈硬化性疾患の予防または治療に有用な物質が提供される。
【0063】
また、本発明のさらに他の側面によると、ヒトの新規肝型ABCA1 mRNAの5’側領域をコードする、ヒトABCA1遺伝子の新規エクソンL3中の配列に基づいて作成したプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とする、ヒトABCA1遺伝子の発現を促進する作用を有する物質のスクリーニング方法が提供される。
【0064】
ヒトABCA1遺伝子の新規エクソンL3は、配列番号2に示すものであって、配列番号1に示すヒトABCA1遺伝子の塩基配列の38481位から38783位までの303塩基に対応する。本実施形態において用いられるプライマーは、ヒトABCA1遺伝子において、新規エクソンL3の上流および下流の配列の一部と実質的に相補的なプライマー対であればよく、プライマー配列の長さおよびヒトABCA1遺伝子のどの部位と実質的に相補的であるかなどは特に限定されない。ここで、実質的に相補的とは、ヒトABCA1遺伝子の配列とアニールできる程度に相補的であればよいことを意味する。たとえば、これらのプライマーは、5’側又は3’側、又はその両方に、ヒトABCA1遺伝子の配列と一部に相補的ではない配列を含んでも、これらとアニールできる限り、本発明のプライマーに含まれるものとする。また、非特異的な増幅を防ぐためや、適当な制限酵素認識部位を導入するために、ヒトABCA1遺伝子と相補的でないミスマッチ配列を持つプライマーを使用することができる。また、プライマーは、検出のために標識されていてもよく、例えば、プライマーは、放射性同位体、蛍光物質、酵素、その他の化学物質等により標識されていてもよい。
【0065】
具体的には、好ましいヒトL3型ABCA1検出用フォワードプライマーとしては、配列番号12に記載されているものが挙げられるが、これらには限定されない。また、ヒトL3型ABCA1検出用リバースプライマーとしては、配列番号13に記載されているものが挙げられるが、これらには限定されない。また、これらのプライマーと組み合わせて使用しうる好ましいプローブとしては、配列番号14に記載されるものを使用することができる。ほかに、好ましいプローブ、フォワードプライマー、リバースプライマーの組み合わせとして、配列番号15、配列番号16、配列番号17に記載されたものが、さらに別の好ましいプローブ、フォワードプライマー、リバースプライマーの組み合わせとして、配列番号18、配列番号19、配列番号20に記載されたものが挙げられる。
【0066】
これらのPCRプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とする、ヒトABCA1遺伝子の発現を促進する作用を有する物質のスクリーニング方法は、
被検物質を供するステップと、
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップであって、PCRプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とするステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0067】
具体的には、かかる方法は、既に説明したプロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いる、スクリーニング方法に準じて行うことができる。PCRプライマーを用いてヒトABCA1のmRNA発現量を測定するステップにおいては、好ましくは、配列番号12、13に記載のプライマー対を用いて、ヒト新規肝型ABCA1のmRNA発現量を測定する。かかるスクリーニング方法によれば、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質、さらにはヒトにおいて動脈硬化性疾患の予防または治療に有用な物質を提供することができる。
【0068】
また、本発明の他の側面によると、転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤が提供される。転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質は、フォルスコリン、イソブチルメチルキサンチン(IBMX)、cAMPアナログなど、細胞内cAMPレベルを上昇させることでHNF4alphaの転写コアクチベーターとして働くPGC−1alphaの発現を上昇させる物質も含む。このような、転写調整因子HNF4alphaの発現量や活性を増加する物質は、L3型ABCA1 mRNAを増加することが期待される。
【0069】
本実施の形態による、転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤は、本発明の一実施の形態として説明した、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法により、同定することができる。さらに具体的には、スクリーニング方法において、特に、エンハンサーとして、配列番号1に示すヒトABCA1遺伝子のL3型mRNAの転写開始位置から3244位から3256位までの塩基(配列番号11)を含む配列を用いることにより同定することができる。かかる配列を含むものであれば、エンハンサー配列の長短は問わず、また、プロモーターも任意のものであってよい。
【0070】
本発明は以下の実施例においてより具体的に説明される。但し、本発明はこれらの実施例に限定されるものでない。
【実施例1】
【0071】
[ヒト肝臓で発現するABCA1 mRNAの検出]
5’−RACE法により、ヒト肝臓で発現しているABCA1のmRNAの5’末端領域を解析した。プライマーはABCA1のmRNAエクソン4およびエクソン5の配列を基に設計した。配列番号21に、用いた遺伝子特異的プライマーの配列を示す。これらのプライマーを用い、GeneRacerキット(Invitrogen社製)の添付プロトコールに従い、ヒト肝臓由来総RNA(Human Liver Total RNA;クロンテック社製)を鋳型として、cDNAを増幅した。
【0072】
ヒト肝臓において、3種類の異なる転写開始点を持つABCA1のmRNAを検出した。2種類は既報の末梢型(配列番号22)および肝型(L2型)(配列番号23)であり、新たに見いだしたヒト肝型(L3型)のABCA1 mRNAの最初のエクソンの配列を配列番号2に示す。また、L3型のABCA1 mRNAの塩基配列を、配列番号24に示す。図1Bは、検出したABCA1のmRNAの5’末端の模式図である。ここで末梢型と仮称したmRNAは、これまで5’−RACE法を用いてヒト繊維芽細胞やヒト単球由来THP−1細胞から同定されたABCA1のmRNA[NM_005502]であり、既報のプロモーター領域(エクソン1上流、Proc. Natl. Acad. Sci. 2000; 97, 7987)により転写制御を受けると考えられる。L2型と命名したmRNAは、ヒト、マウス、ラットに共通に発現しており、エクソン2から転写が開始されるmRNAであり、既報の肝型プロモーター領域(エクソン2上流、特許文献1:非特許文献7)により転写制御を受けると考えられる。ヒト肝型(L3型)と命名したmRNAは、イントロン3中の新たなエクソン(エクソンL3)から転写が開始されるmRNAである。ヒトのエクソンL3領域に相同な領域はマウス、ラットには存在しない。
【実施例2】
【0073】
[ヒト組織におけるABCA1のmRNA発現分布]
ヒト肝臓から得られたmRNA(Human Liver Total RNA;クロンテック社製)を用い、ヒト末梢型ABCA1 mRNAおよびヒト肝型(L3型)ABCA1 mRNAの存在比を調べた。ヒト肝型(L3型)ABCA1 mRNA量はエクソンL3領域(ヒトL3型特有の非翻訳領域)を認識するプライマー・プローブ、ヒト末梢型ABCA1 mRNA量はエクソン1領域(末梢型特有の非翻訳領域)を認識するプライマー・プローブ、また総ABCA1 mRNA量はタンパク質をコードするエクソン12領域の配列を基に作成したプライマー・プローブを用いて、ヒト肝型ABCA1 cDNAを標準品とした検量線を作成し、リアルタイムRT−PCRにより定量した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。以下に、ヒト肝型(L3型)ABCA1検出用プローブおよび総ABCA1検出用プローブの配列を示す。
【0074】
・ヒトL3型ABCA1検出用標識プローブ(配列番号14)
FAM-TGCCTGCTTTCAGAGTGCTCAATGTGC -TAMRA
・ヒトL3型ABCA1検出用フォワードプライマー(配列番号12)
GCACTATGCTAAGGCTGGGAAT
・ヒトL3型ABCA1検出用リバースプライマー(配列番号13)
TCGTGCTTTATCTGGTTCACTTCT
・ヒト末梢型ABCA1検出用標識プローブ(配列番号25)
FAM- TTCCCCGGTTCTGTTTTCTCCCCTT -TAMRA
・ヒト末梢型ABCA1検出用フォワードプライマー(配列番号26)
GGCGCTTTGCTCCTTGTTT
・ヒト末梢型ABCA1検出用リバースプライマー(配列番号27)
TCTCTTTCTCCTACCCCTTGACA
・ヒト総ABCA1検出用標識プローブ(配列番号28)
FAM-ACACCTGGAGAGAAGCTTTCAACGAGACTAACC -TAMRA
・ヒト総ABCA1検出用フォワードプライマー(配列番号29)
TGTCCAGTCCAGTAATGGTTCTGT
・ヒト総ABCA1検出用リバースプライマー(配列番号30)
AAGCGAGATATGGTCCGGATT
【0075】
ヒト各組織(肝臓、筋肉、腎臓、膵臓、心臓、脳、胎盤および肺)から得られたmRNA(ヒトMTCパネルI;クロンテック社製)を用い、各組織における総ABCA1 mRNAおよびヒト肝型(L3型)ABCA1 mRNAの分布をリアルタイムRT−PCRにより定量し、骨格筋の存在量に対する相対比で示した(図2)。総ABCA1 mRNAも肝臓は骨格筋の2倍程度発現していたが、L3型は、ほぼ肝臓に特異的に発現しており、ヒト肝型(L3型)と呼ぶことにした。
【実施例3】
【0076】
[ヒト肝臓における肝型(L3型)ABCA1 mRNAの存在比]
ヒト肝臓から得られたmRNA(クロンテック社製)4例について、ヒト末梢型およびL3型ABCA1 mRNAを定量して、総ABCA1 mRNA量と比較したところ、末梢型は33.4±5.5%存在するのに対し、L3型は26.3±9.4%存在していた。
【実施例4】
【0077】
[ヒト肝型(L3型)ABCA1 mRNA発現のスタチン系薬剤による誘導]
ヒト肝ガン由来細胞JHH−5は10%のFCSを含むWilliams培地で培養し、被験化合物を含む無血清培地(0.2%BSA)で24時間処理した。被験化合物として、HMG−CoA還元酵素阻害剤・スタチンの一種であるコンパクチン(最終濃度50μM)を添加し、溶剤のみを添加したものを対照とした。細胞からRNeasy(QIAGEN社製)を用いてRNAを調製した。リアルタイム定量RT−PCRにより、調製したRNA中における総ABCA1およびL3型のABCA1 mRNA量を測定した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。
【0078】
ヒト肝ガン由来細胞JHH−5細胞を50μMのコンパクチンで処理すると、総ABCA1mRNA量が増加した(図3A)。コンパクチン処理した細胞では、末梢型のABCA1 mRNAは低下していたが(図3C)、L3型のABCA1 mRNA量が増加していた(図3B)。したがって、L3型のABCA1 mRNA量を指標に、肝臓でのABCA1 mRNA発現を調整する作用を有する物質をスクリーニングすることの有用性が示された。
【実施例5】
【0079】
[ヒト肝型(L3型)ABCA1 mRNAの機能]
ヒト肝ガン由来細胞JHH−5細胞を、L3型ABCA1 mRNAに対するsiRNAで32時間処理し、次いで50μMのコンパクチンを含む無血清培地(0.2%BSA)で16時間処理した。対照としてStealth RNAi negative control high GC(インビトロゲン社製)を用いた。細胞を可溶化してABCA1蛋白質の発現量をウェスタンブロットで測定した。あるいはRNAi処理後に、細胞の培地に[3H]コレステロールを添加して24時間培養し、洗浄した後に50μMのコンパクチンで16時間処理した。次いでアポリポタンパクA−Iを添加あるいは非添加の培地で6時間培養し、アポリポタンパクA−Iに依存した培地へのコレステロール排出量を測定した。以下に、ヒト肝型(L3型)ABCA1に対するsiRNAの配列を示す。
【0080】
・ヒト肝型(L3型)ABCA1に対するsiRNA センス1(配列番号31)
AACCCAGGCACACUCCUCCUGUAGC
・ヒト肝型(L3型)ABCA1に対するsiRNA アンチセンス1(配列番号32)
GCUACAGGAGGAGUGUGCCUGGGUU
【0081】
JHH−5細胞のL3型ABCA1 mRNAをsiRNAを用いてノックダウンすると、ABCA1蛋白発現は図4Aに示すように低下した。またアポリポタンパクA−Iに依存した培地へのコレステロール排出(HDL産生)は、約50%低下した(図4)。したがって、L3型ABCA1 mRNAは、機能的でHDL産生に寄与しているABCA1蛋白をコードすることが判明した。
【実施例6】
【0082】
[コンパクチンによる新規なヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーの活性化]
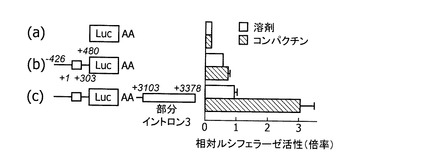
ヒト肝型(L3型)ABCA1 mRNAの発現調節領域の解析を行った。ヒトゲノムDNAから、L3型ABCA1 mRNAの転写開始点(配列番号1のヒトABCA1 DNAの38481位)の上流426塩基から下流480塩基まで(配列番号3)をクローニングし、得られた領域を、ホタルルシフェラーゼベクターpGL3(プロメガ社製)に組み込み、プロモーターのレポータープラスミドを構築した。プラスミドの構築の模式図を図5に、レポータープラスミドの構成を図6(b)に示す。以降の図面において、図中の数字は、L3型ABCA1 mRNAの転写開始点からの塩基の位置を示し、プラスは転写開始点から下流、マイナスは転写開始点から上流を示す。また、イントロン3に属する部分配列であるL3型mRNAの転写開始点の下流3103塩基から3378塩基まで(配列番号6)をクローニングし、上記プロモーターのレポータープラスミドのルシフェラーゼ遺伝子下流に組み込み、プロモーター・エンハンサーのレポータープラスミドを構築した。プロモーター・エンハンサーのレポータープラスミドの構成を図6(c)に示す。図6(a)は、コントロールとして用いた、プロモーターもエンハンサーも組み込まないプラスミドの構成である。
【0083】
また、上記L3型転写開始点の下流3103塩基から3378塩基(配列番号6)をSV−40プロモーターを有するホタルルシフェラーゼベクター(プロメガ社製)に組み込み、エンハンサーレポータープラスミドを構築した。レポータープラスミドの構成を図7(b)に示す。図7(a)は、コントロールとして用いた、SV−40プロモーターを有するが、エンハンサーを組み込まないプラスミドの構成である。
【0084】
JHH−5細胞への一過性トランスフェクションにより、レポータープラスミドおよび遺伝子導入効率補正用のウミシイタケルシフェラーゼレポータープラスミドphRL−TK(プロメガ社製)を形質導入し、50μMのコンパクチンを含む5%の脱脂血清培地で16時間処理した。対照として、同様に溶剤(ジメチルスルホキシド(DMSO))で処理した。その後、細胞を回収し、デュアルルシフェラーゼアッセイシステム(プロメガ社製)により細胞内ルシフェラーゼ活性を測定した。なお、転写活性は次のように定義して評価した。
(転写活性)=(ホタルルシフェラーゼ活性測定値)/(ウミシイタケルシフェラーゼ活性測定値)
【0085】
さらに、スタチン系薬剤で処理する代わりに、活性型SREBP2を過剰発現させた際の転写活性も測定した。活性型SREBP2は、エンハンサーレポータープラスミドに加えて、pME−SREBP2(1−480)(J. Biol. Chem. 2003; 278, 36176)を導入し、過剰発現した。pME−SREBP2(1−480)プラスミドは、活性型SREBP2であるSREBP2のアミノ酸残基1〜480を発現することができるベクターである。対照として空ベクターpME−18Sプラスミドを共導入した(図7中のmock)。
【0086】
JHH−5細胞をコンパクチン処理することにより、ヒト肝型(L3型)転写開始点上流領域を組み込んだレポータープラスミドの転写活性はわずかに上昇した(図6(b))。さらに転写開始点下流領域を組み込んだレポータープラスミドの転写活性は、コンパクチン処理により大きく増大した(図6(c))。この結果は、スタチン系薬剤によるL3型ABCA1 mRNAの応答(実施例4、図3C)とよく一致した。L3型下流領域(エンハンサー領域)は人工のSV−40プロモーターを有するレポータープラスミドに組み込んだ場合でも、コンパクチン処理により活性を大きく増大させるエンハンサー活性を示した(図7(b))。しかしながら、エンハンサー活性はSREBP2の過剰発現により影響されなかった。したがって、該肝臓型転写開始点上流領域(プロモーター領域)および下流領域(エンハンサー領域)が肝臓でのABCA1遺伝子発現を制御していることが示された。また、スタチン系薬物による活性化は、SREBP2が直接活性化するL2プロモーターとは異なるメカニズムによることが示された。
【実施例7】
【0087】
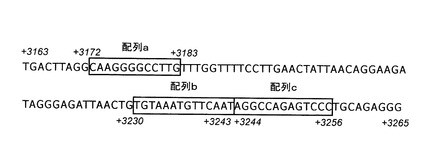
[ヒト肝型(L3型)ABCA1遺伝子エンハンサー活性を調節する配列]
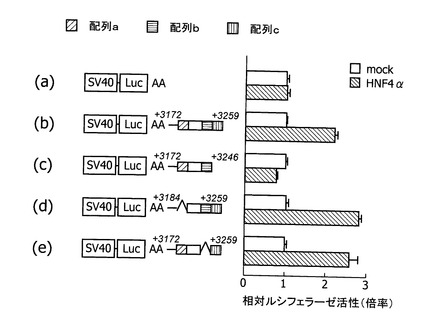
実施例6で得られたL3型ABCA1遺伝子エンハンサーの配列について、5’側ならびに3’側を短縮したレポータープラスミド、ならびにエンハンサー配列を部分的に欠失したプラスミドを作成した(図9)。図9(a)はエンハンサー配列を組み込まなかったプラスミドであり、図9(b)は図6(c)と同様に配列番号6に示すエンハンサー配列を組み込んだプラスミドである。図9(c)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3378塩基(配列番号7に示す)を、図9(d)はエンハンサー配列としてL3型転写開始点の下流3184塩基から3378塩基を、図9(e)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3259塩基(配列番号8)を、図9(f)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3246塩基を、図9(g)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3228塩基、及び3242塩基から3259塩基を、それぞれ組み込んだプラスミドである。実施例6に記載した方法で、これらのプラスミドをJHH−5細胞に形質導入し、50μMコンパクチンで処理し、転写活性を測定した。
【0088】
実施例6で得られたL3型ABCA1遺伝子エンハンサーの短縮(図9(c)〜(g)により、L3転写開始点下流3172塩基から3259塩基まで(配列番号8)がコンパクチン処理による活性化を担うことが判明した(図9(e))。さらに、図8に示す、ヒトABCA1遺伝子中の配列a(配列番号9)、配列b(配列番号10)、あるいは配列c(配列番号11)を欠失すると、コンパクチンによる活性化が著しく低下することがわかった(図9(d)、(f)、(g))。したがって、エンハンサー内に存在するこの三つの配列がヒトのABCA1遺伝子の転写調節に機能していると考えられる。
【実施例8】
【0089】
[転写調節因子HNF4alphaによるヒト肝型(L3型)ABCA1遺伝子エンハンサー活性の調節]
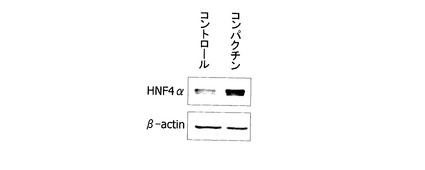
実施例6で得られたL3型ABCA1遺伝子エンハンサーの配列について、エンハンサー配列を部分的に欠失したプラスミドを作成した(図10)。プロモーターとしては、SV40を用いた。図10(a)はエンハンサー配列を組み込まなかったプラスミドであり、図10(b)はL3型転写開始点の下流3172塩基から3259塩基(配列番号8)を組み込んだプラスミドである。図10(c)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3246塩基を、図10(d)はエンハンサー配列としてL3型転写開始点の下流3184塩基から3259塩基を、図10(e)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3229塩基及び3244塩基から3259塩基を、それぞれ組み込んだプラスミドである。実施例6に記載した方法で、これらのプラスミドをHepG2細胞に形質導入し、エンハンサーレポータープラスミドに加えて、転写調節因子HNF4alphaの発現ベクターを導入し、転写活性を測定した。mockとして、空ベクターpCMV5プラスミドを共導入した。図11は、50μMのコンパクチンで処理したJHH−5細胞において、転写調節因子HNF4alpha蛋白質の発現量が増加していることを示すウエスタンブロットである。コントロールでは、溶剤(ジメチルスルホキシド(DMSO))で処理した。50μMコンパクチンが、HNF4alpha蛋白発現を増加させ、L3型ABCA1遺伝子プロモーター・エンハンサーを活性化し、L3型ABCA1 mRNAを増加させることが明らかになった。また、図10のグラフに示すように、転写調節因子HNF4alphaは、配列cに依存して、レポーター遺伝子の転写を増加することがわかった。
【産業上の利用可能性】
【0090】
本発明に係るスクリーニング方法によれば、動脈硬化性疾患の予防または治療に有用な物質を発見することができ、HDL上昇や動脈硬化性疾患の予防治療に極めて有用である。
【技術分野】
【0001】
本発明は、ヒト肝臓においてABCA1遺伝子の発現を制御するプロモーター・エンハンサー領域の発見に基づき、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法に関するものである。特に、プロモーター活性の増加、あるいは特定の非翻訳領域を含むmRNAの増加を指標として、肝臓におけるABCA1発現の増加により動脈硬化性疾患を予防または治療する作用を有する物質をスクリーニングする方法に関するものである。
【背景技術】
【0002】
血中高密度リポタンパク質(HDL)コレステロールの低下は動脈硬化の危険因子である。HDLコレステロールが10mg/dL上昇すれば冠動脈疾患のリスクが20〜30%低下すると予想されている。また人工HDL粒子が動脈硬化病巣を直接退縮する治療効果を持つことが実証され、HDL上昇薬が待望されているが、HDLを直接上昇する薬は実用化されていない(非特許文献1、Nat Rev Drug Discov. 2005 4, 193-205)。
【0003】
HDLは、ABC膜トランスポーターA1(ABCA1)が細胞内のリン脂質とコレステロールを、細胞外のアポリポプロティンA−Iに輸送して形成する。ABCA1はHDLの形成に必須の役割を持ち、機能不全をもたらす遺伝子変異はHDL欠損症の原因となる(非特許文献2、Nat. Genet. 1999; 22:352)。逆に、ABCA1過剰発現マウスではHDLが上昇する(非特許文献3、J. Biol. Chem. 2001; 276:33969)。ABCA1は各組織に発現するが、肝ABCA1が血中HDLの8割を産生することが肝特異的欠損マウスの研究により判明した(非特許文献4、J.Clin.Invest. 2005; 115, 1333)。したがって、HDL上昇には、肝臓においてABCA1の発現を増加させることが極めて有力な手段と考えられる。
【0004】
ABCA1の遺伝子転写制御に関しては、マクロファージなど末梢細胞のABCA1遺伝子の転写はエクソン1上流のプロモーター領域(末梢型プロモーター)により制御され、このプロモーターは核内受容体LXRが直接結合して活性化されることが知られている(非特許文献5、Biochem. Biophys. Res. Commun. 2000; 274, 794)。
【0005】
LXRアゴニストはABCA1発現促進を介してHDL上昇効果を示す。しかしLXR活性化は同時に、中性脂質合成のマスターレギュレーターである転写因子SREBP−1cの発現を促進する。その結果、脂肪酸生合成酵素系の遺伝子発現が増加し脂肪肝や血清トリグリセリド上昇が引き起こされ、このような副作用がLXRアゴニストの臨床薬としての開発の障害となっている(非特許文献6、Biochem. Pharmacol., 2009; 77, 1316-1327)。
【0006】
発明者らはラット肝臓のABCA1発現は、LXR依存の末梢型プロモーターとともに、肝臓に特異的な肝型プロモーターにより二重の制御を受けることを見いだした(非特許文献7、J. Biol. Chem. 2007; 282:21090)。すなわちラット肝臓にはエクソン2から転写が開始される肝特異的なABCA1 mRNA(L2)が存在しており、ABCA1遺伝子のエクソン2上流領域が肝臓に特異的なプロモーターとして機能する。肝型プロモーターにはステロール応答配列SREが存在しており、コレステロール減少により活性型に転換されて核内移行する転写因子SREBP−2が直接結合して活性化する機構を明らかにした。このプロモーター領域は、ラット・マウス・ヒトに共通することから、この肝型プロモーターを活性化する薬物は、LXRの活性化による副作用を示さずに肝ABCA1発現を促進できると考えられた。そして、発明者らは、この肝型プロモーターが、ヒトに有効なABCA1発現促進薬のスクリーニングに使用できることを考えた(特許文献1 特開2007−082445号)。
【0007】
マクロファージや繊維芽細胞などでは、コレステロール合成の律速酵素であるHMG−CoA還元酵素の阻害剤スタチン系薬剤がABCA1発現を低下させる。これは、コレステロール合成過程で生じるLXRの内因性リガンドがスタチン系薬剤の作用によって減少し、ABCA1遺伝子プロモーター活性が低下することによる(非特許文献8、Arterioscler. Thromb. Vasc. Biol. 2004; 24, 2365)。しかしながら、スタチン系薬剤には、血中LDL濃度の低下に加えて、血中HDL濃度の上昇効果も認められており、スタチン系薬剤はヒト肝がん由来細胞HepG2において、ABCA1遺伝子の発現促進作用を示すことも知られている(非特許文献9、Biochem. Biophys. Res. Commun. 2004; 324, 835)。発明者らは、スタチン系薬物は肝由来細胞において末梢型プロモーターの活性を低下させる一方、肝型プロモーターを活性化することによりABCA1発現増加をもたらす機構を明らかにしてきた(非特許文献7)。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2007−082445号公報
【非特許文献】
【0009】
【非特許文献1】Nat Rev Drug Discov. 2005 4, 193-205
【非特許文献2】Nat. Genet. 1999; 22:352
【非特許文献3】J. Biol. Chem. 2001; 276:33969
【非特許文献4】J. Clin. Invest. 2005; 115, 1333
【非特許文献5】Biochem. Biophys. Res. Commun. 2000; 274, 794
【非特許文献6】Biochem. Pharmacol., 2009; 77, 1316-1327
【非特許文献7】J. Biol. Chem. 2007; 282:21090
【非特許文献8】Arterioscler. Thromb. Vasc. Biol. 2004; 24, 2365
【非特許文献9】Biochem. Biophys. Res. Commun. 2004; 324, 835
【発明の概要】
【発明が解決しようとする課題】
【0010】
ABCA1は各組織に発現するが、肝ABCA1が血中HDLの8割を産生することが肝特異的欠損マウスの研究により判明した(非特許文献4)。また、肝特異的なABCA1欠損マウスでは動脈硬化が増悪することがわかった(Arterioscler. Thromb. Vasc. Biol 2009; 29:548)。したがって、肝臓においてABCA1の発現を増加させることが、HDL上昇、さらには動脈硬化性疾患の予防治療に極めて重要と考えられ、肝臓におけるABCA1遺伝子発現の制御機構の解明、さらにはそれを利用した制御薬剤のスクリーニング方法が望まれていた。
【0011】
以上に鑑みて、本発明の課題は、ヒト肝細胞のABCA1の上昇作用を有し、動脈硬化に伴う疾患の予防および/または治療に有用な医薬に用いることができる物質をスクリーニングする新規な方法を提供することにある。
【0012】
なお、本発明の方法において用いられるヒトABCA1遺伝子は、Gene Bankデータベース上に「ATP結合カセット輸送体A1(ATP-binding cassette transporter A1)」として公開されている。また、国際公開第00/078972号パンフレットには、「ABC1」をコードするものとしてヒトABCA1遺伝子が、またそれと共に上記のエクソン1上流のプロモーターが開示されている。
【課題を解決するための手段】
【0013】
すなわち、本発明の一の側面によると、プロモーター活性を有するDNA、ならびにヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【0014】
前記プロモーター活性を有するDNAが、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAであることが好ましい。
【0015】
前記ヒトABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質であることが好ましい。
【発明の効果】
【0016】
本発明にかかるABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法によれば、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、動脈硬化性疾患の予防および/または治療剤を試験するための新規方法を確立することができ、HDL上昇薬の開発に有用である。また、本発明の方法は、従来のABCA1遺伝子発現促進物質のスクリーニング方法に比べ、ヒトHDL上昇作用に対して、より効果的な分子を検出できる可能性がある。
【図面の簡単な説明】
【0017】
【図1】図1AはヒトABCA1遺伝子の模式図を示す。模式図中の数値は、ヒトABCA1遺伝子のDNAにおいて、末梢型ABCA1 mRNAの転写開始点を1とした塩基位置を示すものである。図1Bは、5’−RACE法により解析したヒト肝臓で発現するABCA1のmRNAの5’末端の模式図を示す。括弧内は、ヒト肝臓における、各mRNAの存在比を示す。
【図2】図2Aは、ヒト各組織(肝臓、筋肉、腎臓、膵臓、心臓、脳、胎盤および肺)で発現する総ABCA1 mRNA量を示したグラフであり、図2Bは、ヒト各組織で発現するヒト肝型(L3型)ABCA1 mRNA量を示したグラフである。
【図3】図3Aは、コンパクチンで処理したヒト肝由来培養細胞JHH5における総ABCA1 mRNAの発現量変化を示すグラフであり、図3Bは、コンパクチンで処理したヒト肝由来培養細胞JHH−5におけるヒト肝型(L3)ABCA1 mRNAの発現量変化を示すグラフであり、図3Cは、コンパクチンで処理したヒト肝由来培養細胞JHH−5における末梢型(P)ABCA1 mRNAの発現量変化を示すグラフである。
【図4】図4Aは、ヒト肝型(L3型)ABCA1 mRNAの発現をsiRNAを用いてノックダウンしたヒト肝由来培養細胞JHH−5における、ABCA1蛋白の発現量をウェスタンブロットで測定した写真であり、図4BはアポA−I依存のコレステロール細胞外排出量(HDL産生量)を示したグラフである。
【図5】図5は、ヒトABCA1遺伝子の模式図、およびヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーのレポータープラスミドの構成図である。模式図中の数値は、ヒトABCA1遺伝子のDNAにおいて、L3型ABCA1 mRNAの転写開始点を1とした塩基位置を示し、転写開始点の下流をプラス、上流をマイナスで表したものである。
【図6】図6は、ヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーがコンパクチンにより活性化されることを示すグラフである。
【図7】図7は、L3型エンハンサー配列をSV−40プロモーターを導入したルシフェラーゼプラスミドに組み込んでも、コンパクチンによる活性化が認められること、しかしSREBP−2の発現増加により影響されないことを示すグラフである。
【図8】図8は、ヒト肝型(L3型)ABCA1遺伝子エンハンサーに含まれる配列を示す。上の行の左端が5’側であり、下の行の右端が3’側である。
【図9】図9は、ヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーの活性が、配列a、配列bあるいは配列cの欠失により消失したことを示すグラフである。
【図10】図10は、ヒト肝型(L3型)ABCA1遺伝子エンハンサーの活性が、転写調節因子HNF4alphaの発現ベクターを導入することにより上昇し、配列cの欠失により消失したことを示すグラフである。
【図11】図11は、コンパクチンで処理したヒト肝由来培養細胞JHH−5における、HNF4alpha蛋白の発現量をウェスタンブロットで測定した写真である。
【発明を実施するための形態】
【0018】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって、限定されるものではない。
【0019】
発明者らは、ヒト肝および肝由来細胞のABCA1発現制御をさらに詳細に解析したところ、ヒト肝には従来から知られているマウス・ラットに共通の肝型(L2型)ABCA1 mRNAに加えて、さらに別の肝型(L3型)ABCA1 mRNAが存在することを見いだした。このヒト肝型(L3型)ABCA1 mRNAは、ヒト肝に存在する総ABCA1 mRNAの25%に相当しており、ヒト肝由来細胞をスタチン系薬剤で処理すると発現が促進されることを見いだした。L3型の転写開始点はエクソン2の翻訳開始点より下流であるが、L3型mRNAは機能的ABCA1をコードしていることは、L3型ABCA1 mRNAをsiRNAを用いてノックダウンすると、ABCA1蛋白質の発現が減少し、HDL形成能も低下したという事実により確認された。
【0020】
引き続き、対応するL3型プロモーター・エンハンサー領域を同定したところ、スタチン系薬物による活性化を担うエンハンサー領域は、既報のL2型プロモーター(特許文献1、非特許文献7)とは異なりSREBP−2には応答せず、新規なメカニズムでヒトABCA1発現を制御することを見いだした。したがって、L3型プロモーター・エンハンサー領域はヒトで肝臓のABCA1発現を制御し、血中HDLレベルを調節する重要な役割をもつと考えることができる。L3型プロモーター・エンハンサー領域の活性化、あるいは新規非翻訳領域を含むL3型ABCA1 mRNAの発現増加を指標とする薬物スクリーニング法は、ヒトにおいて有効なHDL上昇薬の開発に極めて有用性が高いと考えられる。
【0021】
さらには、発明者らは、L3型エンハンサーは、L3型プロモーター以外のプロモーターを用いた場合でも、ヒトABCA1発現を促進することを見出した。
【0022】
上記のヒト肝臓におけるABCA1遺伝子の発現調節に新たな制御機構が存在するという発見に基づき、ABCA1遺伝子発現を調節する物質を選別または同定するための方法として、新規ヒト肝型(本明細書中、L3型とも表記する)ABCA1 mRNAをコードする遺伝子の、エクソン3上流(本明細書中、エクソンL3とも表記する)部分を用いる方法、およびイントロン2に存在する新規ヒト肝型ABCA1プロモーターとイントロン3に存在するエンハンサー(本明細書中、L3型ABCA1プロモーター等とも表記する)を用いる方法を開発し、本発明を完成させた。
【0023】
本発明が開示する新規ヒト肝型(L3型)プロモーターは、従来から知られている、ABCA1遺伝子のエクソン1上流のプロモーター(国際公開第00/078972号パンフレット参照)あるいはエクソン2上流のL2型プロモーターとは全く異なる領域に存在し、DNA配列も異なる。このL3型プロモーターに関しては、ABCA1遺伝子転写制御、HDL形成との関連は報告されておらず、動脈硬化性疾患の治療または予防剤の試験法としての有用性も知られていない。上記したように、ピタバスタチン、シンバスタチン、アトルバスタチンがヒト肝癌細胞HepG2においてABCA1のmRNAを増加させることが示されている(非特許文献9)。その作用機序については、発明者らが見いだした肝型(L2)プロモーターが想定されていた。しかし、本発明にかかる新規ヒト肝型(L3型)ABCA1プロモーターおよび当該プロモーターによる発現制御については、従来技術文献において、記載も示唆もしていない。同様に、本発明が開示する新規肝型ABCA1エンハンサーについても、これらの従来技術文献において何ら報告されたものではなく、発明者らによる今般の研究によりはじめて、明らかにされたものである。
【0024】
本発明は、一実施の形態によると、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法に関する。
【0025】
本発明はまた、別の実施の形態によると、このようなスクリーニング方法に用いることができる、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに関する。
【0026】
ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNA、およびヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAは、ヒトに由来する配列を有するものであれば、天然のものであってもよく、また人工的なものであってもよい。すなわち、本発明の別の側面によると、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAとを含んでなる組み換えDNAが提供される。
【0027】
ヒトABCA1遺伝子のイントロン2は、ヒトABCA1遺伝子のエクソン2の下流かつエクソン3の上流に存在する領域であり、イントロン3はエクソン3の下流かつエクソン4の上流に存在する領域である。具体的には、ヒトABCA1遺伝子のエクソン2として、配列番号1に記載するヒトABCA1遺伝子のDNA配列の24385位から24542位の配列を有するものが、イントロン2として、配列番号1に記載の配列の24543位から38960位の配列を有するものが、エクソン3として、配列番号1に記載の配列の38961位から39054位の配列を有するものが、イントロン3として、配列番号1に記載の配列の39055位から43587位の配列を有するものが、およびエクソン4として、配列番号1に記載の配列の43588位から43729位の配列を有するものが、それぞれ見出されている。また、本明細書中、エクソンL3と記す領域は、配列番号1に記載の配列の38481位から38783位の配列を有する領域である。配列番号2にエクソンL3の配列を示す。なお、配列番号1は、ヒトABCA1遺伝子のエクソン1から始まる配列である。また、ヒトにおける上記イントロン2、エクソンL3の相同体、変異体および誘導体の配列は、当業者であれば、ヒトにおける上記イントロン2および/またはエクソン3、4の配列との相同性に基づき決定することができる。
【0028】
ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、配列番号1に示すヒトABCA1遺伝子の塩基配列の38055位から38960位までの塩基を含むことが好ましい。ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、特には、(a)配列番号1に示すヒトABCA1遺伝子の塩基配列の38055位から38960位までの塩基(配列番号3)、配列38055位から配列39054位までの塩基(配列番号4)、配列38055位から配列39224位までの塩基(配列番号5)に記載の配列を有するDNAとすることができる。
【0029】
また、イントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(a)のDNAと実質的に同一の配列を有するDNAとすることができる。具体的には、イントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(b)配列番号3、配列番号4、配列番号5のいずれかに記載の配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。当該技術分野における通常の知識を有する者であれば、野生型プロモーターのヌクレオチド配列の部分的な置換、挿入または欠失などの改変により、野生型プロモーターと同等またはそれ以上のプロモーター活性を有するDNAを調製することが可能である。
【0030】
また、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(c)配列番号3、配列番号4、配列番号5のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。このようなDNAには、(a)のDNAの、自然界に存在する変異型遺伝子もしくは人為的に改変した変異型遺伝子または異種生物由来の相同遺伝子等が含まれる。
【0031】
相同性の比較は、通常、容易に入手できる配列比較プログラムを用いて行うことができる。商業的に入手可能なそのようなコンピュータープログラムは2以上の配列の間の%相同性を計算することができる。ほとんどの配列比較方法においては、全体としての相同性スコアに不当にペナルティを与えることなく可能な挿入および欠失を考慮に入れて、最適な整列を生じさせるよう設計される。これは、「ギャップ」を配列の整列に挿入して、局所的な相同性を最大化するよう努めることによって達成される。このような方法では、できる限り少数のギャップにおける配列の整列が、多くのギャップにおけるものより高いスコアを達成するように、各ギャップに「ギャップペナルティ」を割り当てて、相同性を計算する。ほとんどの整列プログラムはギャップペナルティを設定することができる。しかしながら、配列比較のためのそのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。このような配列比較を実行できるソフトウェアの例として、BLASTパッケージ(Ausubel et al.,1999 ibid-Chapter18参照)、FASTA(Atschul et al.,1990,J.Mol.Biol.,403-410)およびGENEWORKSスイートが挙げられる。
【0032】
また、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAは、(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAの少なくとも一部とすることができる。ここで、DNAは通常、互いに相補的な二本鎖DNA構造を有している。このため、(a)のDNAとハイブリダイズするDNAとは、(a)のDNAのいずれかの鎖とハイブリダイズする鎖を有するDNAをいうことに留意すべきである。
【0033】
なお、ストリンジェントな条件およびハイブリダイゼーションの方法は、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に従って設定することができる。具体的には、ストリンジェントな条件は、「1xSSC、0.1%SDS、37℃」とすることができ、よりストリンジェントな条件は、「0.5xSSC、0.1%SDS、42℃」とすることができ、さらにストリンジェントな条件は、「0.2xSSC、0.1%SDS、65℃」とすることができる(1xSSC:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)。ただし、これらの条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記または他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0034】
なお、(b)〜(d)のDNAのプロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法等により測定することができる。また、(b)〜(d)のDNAは、(a)のDNAと「同様」のプロモーター活性を有することが好ましい。ここで、「同様」のプロモーター活性を有するとは、質的に類似のプロモーター活性を有することをいい、例えば、(b)〜(d)のDNAが、(a)のDNAと少なくとも1つの因子について同一の制御を受ける場合が含まれる。一方で、「同様」のプロモーター活性を有するとは、量的に同程度のプロモーター活性を有することを意図するものではない。また、当業者には明らかなように、プロモーター活性を発揮するために、DNAは、配列番号3、配列番号4、配列番号5のいずれかに記載の配列(またはこれと実質的に同一の配列)の全てを有する必要はなく、その一部であってもよい。上記したように、プロモーター活性を有するのに必要な領域は、当業者であれば、ルシフェラーゼアッセイ法等により適宜決定することができる。
【0035】
ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(e)配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置(配列番号1のヒトABCA1 DNAの38481位)下流3103位から下流3378位までの塩基(配列番号6)を含むことが好ましく、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から下流3172位から下流3378位までの塩基(配列番号7)を含むことがさらに好ましく、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から下流3172位から下流3259位までの塩基(配列番号8)を含むことが最も好ましい。
【0036】
さらには、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、配列番号1に示すヒトABCA1遺伝子において、L3型mRNAの転写開始位置から3172位から3383位までの塩基(配列番号9)と、L3型mRNAの転写開始位置から3230位から3243位までの塩基(配列番号10)と、L3型mRNAの転写開始位置から3244位から3256位までの塩基(配列番号11)をすべて有するDNAであることが必要である。
【0037】
これらの3つの配列が保持される限り、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(f)配列番号6、配列番号7、配列番号8のいずれかに記載の好ましいDNAの配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列であってもよい。
また、これらの3つの配列が保持される限り、(g)配列番号6、配列番号7、配列番号8のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有するものであってもよい。相同性の比較に関しては、(c)のDNAについて述べたのと同様にして行うことができる。
さらに、これらの3つの配列が保持される限り、ヒトABCA1遺伝子のイントロン3中の塩基配列を含み、エンハンサー活性を有するDNAは、(h)配列番号6、配列番号7、配列番号8のいずれかに記載の好ましいDNAとストリンジェントな条件でハイブリダイズし、エンハンサー活性を有するDNAの少なくとも一部とすることができる。
【0038】
上記に配列を示したもの以外であっても、本願発明において、エンハンサー活性を有するのに必要な領域は、当業者であれば適宜決定することができる。エンハンサー活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。
【0039】
次に、本発明の一実施の形態による、これらのDNAを用いた、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法において用いることができる、組換えDNAについて説明する。組換えDNAは、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを含んでなる。
【0040】
プロモーター活性を有するDNAとしては、上記において説明した、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAを用いることができる。さらには、プロモーター活性を有するDNAとしては、ヒトABCA1遺伝子に由来する配列を有するものに限定されず、任意のプロモーターの塩基配列を有するものを用いることができる。当業者には既知のプロモーターとしては、例えば、SV−40プロモーター、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、EFプロモーター、CAGプロモーターが挙げられるが、これらには限定されない。
いっぽう、エンハンサー活性を有するDNAとしては、上記において説明した、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAを用いることができる。
【0041】
これらのプロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとは、組換えDNAにおいては、任意の組み合わせで用いることができる。すなわち、(a)、(b)、(c)、(d)のいずれかから選択されるプロモーター活性を有するDNAもしくはその他の任意プロモーター活性を有するDNAと、(e)、(f)、(g)、(h)のいずれかから選択されるエンハンサー活性を有するDNAとを、任意の組み合わせで含む組換えDNAとすることができる。
【0042】
なお、本発明にかかる組換えDNAにおいて、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAを用いる場合でも、そのほかに転写最小プロモーターをさらに有することができる。最小プロモーターの例として、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、PGKプロモーター、SV40プロモーターなどが挙げられるが、これらに限定されるものではない。
【0043】
本発明にかかるDNAは、ゲノムライブラリーからの選別や、ゲノムDNAを鋳型としたポリメラーゼ連鎖反応(PCR)により取得できる。ゲノムライブラリーからの選別は、適当なベクターに組み込んだゲノムDNAと、目的のDNAの一部の配列を有する標識DNAとのハイブリダイゼーションによって行うことができる。ハイブリダイゼーションは、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、添付の使用説明書に記載の方法に従って行なうことができる。また、ゲノムDNAを鋳型としたPCRは、目的のDNAの一部の配列を有するプライマーを用いて行うことができる。ヌクレオチド配列の改変は、例えば、制限酵素またはDNAエキソヌクレアーゼによる欠失導入、部位特異的変異誘発法による変異導入、変異プライマーを用いたPCR法による改変、合成変異DNAの直接導入などの方法により行うことができる。
【0044】
本発明の他の側面によると、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを有する組換えベクターが提供される。組換えベクターは、当該プロモーター活性を有するDNA、及びエンハンサー活性を有するDNAに作動可能に連結した構造遺伝子をさらに有してもよい。以下に詳細に説明するように、構造遺伝子は、例えば、組換えベクターをスクリーニングに用いる場合、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。
【0045】
本発明にあっては、ベクターは、任意のベクター、例えば、バクテリオファージ、プラスミド、コスミド、ファージミドなどであってよい。より具体的には、本発明に用いることができるベクターの例として、動物細胞用のベクターであるpCDベクター、cDM8ベクター、レトロウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、アデノアソシエイトウイルスベクターなどや、細菌用のベクターであるpUCなどを挙げることができる。構造遺伝子をベクター内に組み込むためには、構造遺伝子が正しく転写される方向に、さらにプロモーターおよびエンハンサーが構造遺伝子に対して適正に機能する位置関係になる様に、プロモーター領域の下流であって、エンハンサー領域の上流に構造遺伝子を連結すればよい。プロモーターと構造遺伝子との連結、及びエンハンサーと構造遺伝子との連結は、制限酵素切断部位を利用して行うことができる。また、適当な制限酵素部位がなくてもリガーゼ反応を用いて行うことができる。
【0046】
なお、「作動可能に連結」とは、一の遺伝子要素が、他の遺伝子要素に対して予測されるように機能できるように配置されていることをいう。例えば、プロモーターが、構造遺伝子の転写の開始を助ける場合には、プロモーターは構造遺伝子に作動可能に結合している。同様に、エンハンサーが構造遺伝子の転写を促進する場合には、エンハンサーが構造遺伝子に作動可能に結合している。この機能的な関係が維持される限り、プロモーターと構造遺伝子との間、あるいは構造遺伝子とエンハンサーとの間に残基が介在してもよい。
【0047】
本発明の他の側面によると、プロモーター活性を有する外因性のDNA及びヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。本明細書において、形質転換体は、広く、外部からDNAが導入された結果、新しい形質を獲得した生物をいう。形質転換体は、任意の生物であってよく、また、任意の生物に由来する任意の細胞または組織であってよい。特に、本発明にかかる形質転換体は、上記組換えベクターで形質転換された形質転換体を含む。
【0048】
組換えベクターにより形質転換することができる宿主の例として、例えば、エシェリヒア属菌、バチルス属菌、酵母、昆虫細胞、昆虫、動物細胞などを挙げることができる。これらの宿主への形質転換の方法の例として、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、アデノウイルスベクターまたはレトロウイルスベクターによる遺伝子導入法、マイクロインジェクション法などを挙げることができる。
【0049】
本発明の他の側面によると、ヒトABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。本発明にかかるキットは、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。以下に詳細に説明するように、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることで、ヒトABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングすることができる。すなわち、本発明にかかるキットは、上記組換えベクターおよび/または上記形質転換体を有する。
【0050】
本発明にかかるキットには、スクリーニングの方法に応じて、酵素、酵素基質、緩衝液等の試薬、培地、道具等を組み合せることができる。酵素基質の例として、ルシフェリン、ATP、[14C]クロラムフェニコール、アセチルCoAを挙げることができる。緩衝液として、解析に用いる酵素の活性の維持に好適な緩衝液を適宜選択することができる。道具の例として、TLC用展開シート、メンブレン等を挙げることができる。
【0051】
本発明の他の側面によると、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供され、動脈硬化性疾患の予防または治療に有用な物質を探索するために用いることができる。本発明にかかるスクリーニング方法は、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする。
【0052】
具体的には、本発明にかかるスクリーニング方法は、
被検物質を供するステップと、
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0053】
被検物質として、単一の化合物または組成物、天然または合成化合物、有機または無機化合物、タンパク質、ペプチド、オリゴヌクレオチド、ポリヌクレオチド、細胞抽出物、細胞培養上清、その他の任意の物質を供することができる。
【0054】
スクリーニングは、インビトロおよびインビボのいずれでも行うことができる。本発明にかかるスクリーニング方法において用いられる細胞または生物として、細菌等の原核生物(大腸菌等)、酵母(サッカロマイセス(Saccharomyces)属、クライヴェロマイセス(Kluyveromyces)属等)、昆虫、植物もしくは動物またはこれらに由来する細胞が挙げられる。動物の例として、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物を挙げることができる。本発明で用いられる細胞は、実験室で培養することのできる培養細胞であることが好ましく、初代培養肝細胞であることがさらに好ましい。細胞は、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物に由来することができ、ヒトに由来することが好ましい。また、細胞は、肝臓に由来することが好ましい。
【0055】
本発明に用いられる細胞または生物は、プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有することが好ましい。当該DNAは、内因性であっても、または外因性であってもよい。外因性である場合、当該DNAは、任意の手段により細胞または生物に導入されてもよい。例えば、スクリーニングに培養細胞を用いる場合、培養細胞への当該DNAの導入方法として、本発明の属する技術分野において汎用される任意の方法を採用することができる。例えば、DNAの導入方法の例として、リン酸カルシウム−DNA共沈法、リポフェクション法、DEAE−デキストラン法、電気パルス穿孔法等が挙げられるが、これらに限定されない。いずれの方法においても、用いる細胞に応じて至適化されたトランスフェクション条件を用いることが好ましい。
【0056】
構造遺伝子は、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。当業者に明らかなように、上記構造遺伝子は、その転写量および/または発現量の測定方法に応じて、適宜選択することができる。上記構造遺伝子の例として、ヒトABCA1、所定の活性を有するタンパク質(ルシフェラーゼ等)、所定の抗体が結合できるタンパク質、所定の物質が特異的に結合できるタンパク質(GST等)等をコードする遺伝子を挙げることができ、特に、ホタルルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼおよび緑色蛍光蛋白質からなる群より選択されるタンパク質をコードする遺伝子(いわゆるレポーター遺伝子)を挙げることができる。
【0057】
被検物質を細胞または生物に接触させることは、当業者に明らかなように、任意の方法により行うことができる。例えば、培養細胞を用いる場合、被検物質を培地中に添加することで、接触させることができる。すなわち、培養細胞を用いる場合、細胞を、被検物質の存在下または非存在下で培養することが好ましい。また、動物を用いる場合は、任意の手段による投与により、例えば、任意の経路からの注射により、接触させることができる。被検物質を細胞または生物へ接触させる時間は、被験物質の効果、濃度、細胞または生物の種類および技術的考慮に応じて、適宜設定することができる。また、被験物質の濃度は、その毒性、透過性、細胞数、処理時間等に応じて、適宜設定することができ、通常1nM〜1mMであることが好ましい。培養細胞を用いる場合、一般的にこのような処理は、多くの試験を同時に実施できるマルチウェルプレートで実施することが好ましい。培養細胞を用いる場合、被験化合物の処理によるレポーター遺伝子の発現が測定可能な条件は、細胞が生存してレポーター遺伝子の発現産物が生産可能な条件であれば良い。特に好ましい条件にあっては、好ましくは使用される細胞株に適した培地を使用し、4〜6%の炭酸ガス存在下、36〜38℃で5〜72時間培養する。
【0058】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定し、これらの転写量および/または発現量を比較することで、当該被検物質を、ヒトABCA1遺伝子の転写を調整する作用を有する物質の候補物質とすることができるか否か判断することができる。具体的には、ある被検物質を接触させた群で、接触させなかった群と比べて、上記遺伝子の転写量および/または発現量が変化した場合、特に増加した場合、当該被検物質を候補物質とすることができる。
【0059】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定するステップと、被検物質を接触させた群と接触させなかった群の転写量および/または発現量を比較するステップは、当業者に明らかなように、任意の方法により行うことができる。具体的には、ノーザンブロッティング法、DNAアレイ法等により、上記遺伝子の転写量を測定することができる。また、ルシフェラーゼアッセイ法やCAT法等により、上記遺伝子の発現量を測定することができる。さらに、上記所定の遺伝子に対する抗体が入手可能な場合は、ELISA法、免疫沈降法、ウエスタンブロッティング法により、当該遺伝子の発現量を測定することができる。また、上記所定の遺伝子として、GST等の所定の物質に特異的に結合するタンパク質の遺伝子を用いる場合は、当該タンパク質を沈降させ、沈降したタンパク質の量を測定することで、上記遺伝子の発現量を測定することができる。
【0060】
[ノーザンブロッティング法]
ノーザンブロッティング法は、変性したRNAを変性条件下のアガロース電気泳動で分離し、ニトロセルロースフィルターに移して、標識した特異的なプローブで検出するものである。RNAを変性させるのは、RNAは分子内に二次構造を持つため、そのままではサイズに従った正確な分離ができず、さらに、フィルターは一本鎖の核酸しか結合できないためである。
【0061】
[ルシフェラーゼアッセイ法、CAT法]
ルシフェラーゼアッセイ法およびCAT法は、目的とする遺伝子の転写調節領域の下流にレポーターとしてLUC(ルシフェラーゼ)またはCAT(クロラムフェニコールアセチルトランスフェラーゼ)遺伝子を組み込んだプラスミドを作成し、そのプラスミドを導入した細胞の酵素活性を測定するものである。具体的には、ルシフェラーゼアッセイ法は、マグネシウム存在下、ルシフェラーゼがルシフェリンとATPから酸化ルシフェリンとAMPを作る反応を触媒する際に発する波長560nmの光を、ルミノメーターを使って検出するものである。CAT法の場合、基質の[14C]クロラムフェニコールが、CATによりアセチル化されて、[14C]アセチルクロラムフェニコールが産生される。このサンプルを酢酸エチルで抽出し、薄層のシリカゲルプレート上に展開後、移動度の異なるスポットの放射活性を測定する。
【0062】
本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、ヒトABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質が提供される。特に、上記したように、本発明にかかるスクリーニング方法によると、ヒトABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。このため、本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、動脈硬化性疾患の予防または治療に有用な物質が提供される。
【0063】
また、本発明のさらに他の側面によると、ヒトの新規肝型ABCA1 mRNAの5’側領域をコードする、ヒトABCA1遺伝子の新規エクソンL3中の配列に基づいて作成したプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とする、ヒトABCA1遺伝子の発現を促進する作用を有する物質のスクリーニング方法が提供される。
【0064】
ヒトABCA1遺伝子の新規エクソンL3は、配列番号2に示すものであって、配列番号1に示すヒトABCA1遺伝子の塩基配列の38481位から38783位までの303塩基に対応する。本実施形態において用いられるプライマーは、ヒトABCA1遺伝子において、新規エクソンL3の上流および下流の配列の一部と実質的に相補的なプライマー対であればよく、プライマー配列の長さおよびヒトABCA1遺伝子のどの部位と実質的に相補的であるかなどは特に限定されない。ここで、実質的に相補的とは、ヒトABCA1遺伝子の配列とアニールできる程度に相補的であればよいことを意味する。たとえば、これらのプライマーは、5’側又は3’側、又はその両方に、ヒトABCA1遺伝子の配列と一部に相補的ではない配列を含んでも、これらとアニールできる限り、本発明のプライマーに含まれるものとする。また、非特異的な増幅を防ぐためや、適当な制限酵素認識部位を導入するために、ヒトABCA1遺伝子と相補的でないミスマッチ配列を持つプライマーを使用することができる。また、プライマーは、検出のために標識されていてもよく、例えば、プライマーは、放射性同位体、蛍光物質、酵素、その他の化学物質等により標識されていてもよい。
【0065】
具体的には、好ましいヒトL3型ABCA1検出用フォワードプライマーとしては、配列番号12に記載されているものが挙げられるが、これらには限定されない。また、ヒトL3型ABCA1検出用リバースプライマーとしては、配列番号13に記載されているものが挙げられるが、これらには限定されない。また、これらのプライマーと組み合わせて使用しうる好ましいプローブとしては、配列番号14に記載されるものを使用することができる。ほかに、好ましいプローブ、フォワードプライマー、リバースプライマーの組み合わせとして、配列番号15、配列番号16、配列番号17に記載されたものが、さらに別の好ましいプローブ、フォワードプライマー、リバースプライマーの組み合わせとして、配列番号18、配列番号19、配列番号20に記載されたものが挙げられる。
【0066】
これらのPCRプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とする、ヒトABCA1遺伝子の発現を促進する作用を有する物質のスクリーニング方法は、
被検物質を供するステップと、
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップであって、PCRプライマーを用いてヒトABCA1のmRNA発現量を測定することを特徴とするステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0067】
具体的には、かかる方法は、既に説明したプロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いる、スクリーニング方法に準じて行うことができる。PCRプライマーを用いてヒトABCA1のmRNA発現量を測定するステップにおいては、好ましくは、配列番号12、13に記載のプライマー対を用いて、ヒト新規肝型ABCA1のmRNA発現量を測定する。かかるスクリーニング方法によれば、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質、さらにはヒトにおいて動脈硬化性疾患の予防または治療に有用な物質を提供することができる。
【0068】
また、本発明の他の側面によると、転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤が提供される。転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質は、フォルスコリン、イソブチルメチルキサンチン(IBMX)、cAMPアナログなど、細胞内cAMPレベルを上昇させることでHNF4alphaの転写コアクチベーターとして働くPGC−1alphaの発現を上昇させる物質も含む。このような、転写調整因子HNF4alphaの発現量や活性を増加する物質は、L3型ABCA1 mRNAを増加することが期待される。
【0069】
本実施の形態による、転写因子HNF4alphaの量あるいは活性を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤は、本発明の一実施の形態として説明した、肝臓においてヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法により、同定することができる。さらに具体的には、スクリーニング方法において、特に、エンハンサーとして、配列番号1に示すヒトABCA1遺伝子のL3型mRNAの転写開始位置から3244位から3256位までの塩基(配列番号11)を含む配列を用いることにより同定することができる。かかる配列を含むものであれば、エンハンサー配列の長短は問わず、また、プロモーターも任意のものであってよい。
【0070】
本発明は以下の実施例においてより具体的に説明される。但し、本発明はこれらの実施例に限定されるものでない。
【実施例1】
【0071】
[ヒト肝臓で発現するABCA1 mRNAの検出]
5’−RACE法により、ヒト肝臓で発現しているABCA1のmRNAの5’末端領域を解析した。プライマーはABCA1のmRNAエクソン4およびエクソン5の配列を基に設計した。配列番号21に、用いた遺伝子特異的プライマーの配列を示す。これらのプライマーを用い、GeneRacerキット(Invitrogen社製)の添付プロトコールに従い、ヒト肝臓由来総RNA(Human Liver Total RNA;クロンテック社製)を鋳型として、cDNAを増幅した。
【0072】
ヒト肝臓において、3種類の異なる転写開始点を持つABCA1のmRNAを検出した。2種類は既報の末梢型(配列番号22)および肝型(L2型)(配列番号23)であり、新たに見いだしたヒト肝型(L3型)のABCA1 mRNAの最初のエクソンの配列を配列番号2に示す。また、L3型のABCA1 mRNAの塩基配列を、配列番号24に示す。図1Bは、検出したABCA1のmRNAの5’末端の模式図である。ここで末梢型と仮称したmRNAは、これまで5’−RACE法を用いてヒト繊維芽細胞やヒト単球由来THP−1細胞から同定されたABCA1のmRNA[NM_005502]であり、既報のプロモーター領域(エクソン1上流、Proc. Natl. Acad. Sci. 2000; 97, 7987)により転写制御を受けると考えられる。L2型と命名したmRNAは、ヒト、マウス、ラットに共通に発現しており、エクソン2から転写が開始されるmRNAであり、既報の肝型プロモーター領域(エクソン2上流、特許文献1:非特許文献7)により転写制御を受けると考えられる。ヒト肝型(L3型)と命名したmRNAは、イントロン3中の新たなエクソン(エクソンL3)から転写が開始されるmRNAである。ヒトのエクソンL3領域に相同な領域はマウス、ラットには存在しない。
【実施例2】
【0073】
[ヒト組織におけるABCA1のmRNA発現分布]
ヒト肝臓から得られたmRNA(Human Liver Total RNA;クロンテック社製)を用い、ヒト末梢型ABCA1 mRNAおよびヒト肝型(L3型)ABCA1 mRNAの存在比を調べた。ヒト肝型(L3型)ABCA1 mRNA量はエクソンL3領域(ヒトL3型特有の非翻訳領域)を認識するプライマー・プローブ、ヒト末梢型ABCA1 mRNA量はエクソン1領域(末梢型特有の非翻訳領域)を認識するプライマー・プローブ、また総ABCA1 mRNA量はタンパク質をコードするエクソン12領域の配列を基に作成したプライマー・プローブを用いて、ヒト肝型ABCA1 cDNAを標準品とした検量線を作成し、リアルタイムRT−PCRにより定量した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。以下に、ヒト肝型(L3型)ABCA1検出用プローブおよび総ABCA1検出用プローブの配列を示す。
【0074】
・ヒトL3型ABCA1検出用標識プローブ(配列番号14)
FAM-TGCCTGCTTTCAGAGTGCTCAATGTGC -TAMRA
・ヒトL3型ABCA1検出用フォワードプライマー(配列番号12)
GCACTATGCTAAGGCTGGGAAT
・ヒトL3型ABCA1検出用リバースプライマー(配列番号13)
TCGTGCTTTATCTGGTTCACTTCT
・ヒト末梢型ABCA1検出用標識プローブ(配列番号25)
FAM- TTCCCCGGTTCTGTTTTCTCCCCTT -TAMRA
・ヒト末梢型ABCA1検出用フォワードプライマー(配列番号26)
GGCGCTTTGCTCCTTGTTT
・ヒト末梢型ABCA1検出用リバースプライマー(配列番号27)
TCTCTTTCTCCTACCCCTTGACA
・ヒト総ABCA1検出用標識プローブ(配列番号28)
FAM-ACACCTGGAGAGAAGCTTTCAACGAGACTAACC -TAMRA
・ヒト総ABCA1検出用フォワードプライマー(配列番号29)
TGTCCAGTCCAGTAATGGTTCTGT
・ヒト総ABCA1検出用リバースプライマー(配列番号30)
AAGCGAGATATGGTCCGGATT
【0075】
ヒト各組織(肝臓、筋肉、腎臓、膵臓、心臓、脳、胎盤および肺)から得られたmRNA(ヒトMTCパネルI;クロンテック社製)を用い、各組織における総ABCA1 mRNAおよびヒト肝型(L3型)ABCA1 mRNAの分布をリアルタイムRT−PCRにより定量し、骨格筋の存在量に対する相対比で示した(図2)。総ABCA1 mRNAも肝臓は骨格筋の2倍程度発現していたが、L3型は、ほぼ肝臓に特異的に発現しており、ヒト肝型(L3型)と呼ぶことにした。
【実施例3】
【0076】
[ヒト肝臓における肝型(L3型)ABCA1 mRNAの存在比]
ヒト肝臓から得られたmRNA(クロンテック社製)4例について、ヒト末梢型およびL3型ABCA1 mRNAを定量して、総ABCA1 mRNA量と比較したところ、末梢型は33.4±5.5%存在するのに対し、L3型は26.3±9.4%存在していた。
【実施例4】
【0077】
[ヒト肝型(L3型)ABCA1 mRNA発現のスタチン系薬剤による誘導]
ヒト肝ガン由来細胞JHH−5は10%のFCSを含むWilliams培地で培養し、被験化合物を含む無血清培地(0.2%BSA)で24時間処理した。被験化合物として、HMG−CoA還元酵素阻害剤・スタチンの一種であるコンパクチン(最終濃度50μM)を添加し、溶剤のみを添加したものを対照とした。細胞からRNeasy(QIAGEN社製)を用いてRNAを調製した。リアルタイム定量RT−PCRにより、調製したRNA中における総ABCA1およびL3型のABCA1 mRNA量を測定した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。
【0078】
ヒト肝ガン由来細胞JHH−5細胞を50μMのコンパクチンで処理すると、総ABCA1mRNA量が増加した(図3A)。コンパクチン処理した細胞では、末梢型のABCA1 mRNAは低下していたが(図3C)、L3型のABCA1 mRNA量が増加していた(図3B)。したがって、L3型のABCA1 mRNA量を指標に、肝臓でのABCA1 mRNA発現を調整する作用を有する物質をスクリーニングすることの有用性が示された。
【実施例5】
【0079】
[ヒト肝型(L3型)ABCA1 mRNAの機能]
ヒト肝ガン由来細胞JHH−5細胞を、L3型ABCA1 mRNAに対するsiRNAで32時間処理し、次いで50μMのコンパクチンを含む無血清培地(0.2%BSA)で16時間処理した。対照としてStealth RNAi negative control high GC(インビトロゲン社製)を用いた。細胞を可溶化してABCA1蛋白質の発現量をウェスタンブロットで測定した。あるいはRNAi処理後に、細胞の培地に[3H]コレステロールを添加して24時間培養し、洗浄した後に50μMのコンパクチンで16時間処理した。次いでアポリポタンパクA−Iを添加あるいは非添加の培地で6時間培養し、アポリポタンパクA−Iに依存した培地へのコレステロール排出量を測定した。以下に、ヒト肝型(L3型)ABCA1に対するsiRNAの配列を示す。
【0080】
・ヒト肝型(L3型)ABCA1に対するsiRNA センス1(配列番号31)
AACCCAGGCACACUCCUCCUGUAGC
・ヒト肝型(L3型)ABCA1に対するsiRNA アンチセンス1(配列番号32)
GCUACAGGAGGAGUGUGCCUGGGUU
【0081】
JHH−5細胞のL3型ABCA1 mRNAをsiRNAを用いてノックダウンすると、ABCA1蛋白発現は図4Aに示すように低下した。またアポリポタンパクA−Iに依存した培地へのコレステロール排出(HDL産生)は、約50%低下した(図4)。したがって、L3型ABCA1 mRNAは、機能的でHDL産生に寄与しているABCA1蛋白をコードすることが判明した。
【実施例6】
【0082】
[コンパクチンによる新規なヒト肝型(L3型)ABCA1遺伝子プロモーター・エンハンサーの活性化]
ヒト肝型(L3型)ABCA1 mRNAの発現調節領域の解析を行った。ヒトゲノムDNAから、L3型ABCA1 mRNAの転写開始点(配列番号1のヒトABCA1 DNAの38481位)の上流426塩基から下流480塩基まで(配列番号3)をクローニングし、得られた領域を、ホタルルシフェラーゼベクターpGL3(プロメガ社製)に組み込み、プロモーターのレポータープラスミドを構築した。プラスミドの構築の模式図を図5に、レポータープラスミドの構成を図6(b)に示す。以降の図面において、図中の数字は、L3型ABCA1 mRNAの転写開始点からの塩基の位置を示し、プラスは転写開始点から下流、マイナスは転写開始点から上流を示す。また、イントロン3に属する部分配列であるL3型mRNAの転写開始点の下流3103塩基から3378塩基まで(配列番号6)をクローニングし、上記プロモーターのレポータープラスミドのルシフェラーゼ遺伝子下流に組み込み、プロモーター・エンハンサーのレポータープラスミドを構築した。プロモーター・エンハンサーのレポータープラスミドの構成を図6(c)に示す。図6(a)は、コントロールとして用いた、プロモーターもエンハンサーも組み込まないプラスミドの構成である。
【0083】
また、上記L3型転写開始点の下流3103塩基から3378塩基(配列番号6)をSV−40プロモーターを有するホタルルシフェラーゼベクター(プロメガ社製)に組み込み、エンハンサーレポータープラスミドを構築した。レポータープラスミドの構成を図7(b)に示す。図7(a)は、コントロールとして用いた、SV−40プロモーターを有するが、エンハンサーを組み込まないプラスミドの構成である。
【0084】
JHH−5細胞への一過性トランスフェクションにより、レポータープラスミドおよび遺伝子導入効率補正用のウミシイタケルシフェラーゼレポータープラスミドphRL−TK(プロメガ社製)を形質導入し、50μMのコンパクチンを含む5%の脱脂血清培地で16時間処理した。対照として、同様に溶剤(ジメチルスルホキシド(DMSO))で処理した。その後、細胞を回収し、デュアルルシフェラーゼアッセイシステム(プロメガ社製)により細胞内ルシフェラーゼ活性を測定した。なお、転写活性は次のように定義して評価した。
(転写活性)=(ホタルルシフェラーゼ活性測定値)/(ウミシイタケルシフェラーゼ活性測定値)
【0085】
さらに、スタチン系薬剤で処理する代わりに、活性型SREBP2を過剰発現させた際の転写活性も測定した。活性型SREBP2は、エンハンサーレポータープラスミドに加えて、pME−SREBP2(1−480)(J. Biol. Chem. 2003; 278, 36176)を導入し、過剰発現した。pME−SREBP2(1−480)プラスミドは、活性型SREBP2であるSREBP2のアミノ酸残基1〜480を発現することができるベクターである。対照として空ベクターpME−18Sプラスミドを共導入した(図7中のmock)。
【0086】
JHH−5細胞をコンパクチン処理することにより、ヒト肝型(L3型)転写開始点上流領域を組み込んだレポータープラスミドの転写活性はわずかに上昇した(図6(b))。さらに転写開始点下流領域を組み込んだレポータープラスミドの転写活性は、コンパクチン処理により大きく増大した(図6(c))。この結果は、スタチン系薬剤によるL3型ABCA1 mRNAの応答(実施例4、図3C)とよく一致した。L3型下流領域(エンハンサー領域)は人工のSV−40プロモーターを有するレポータープラスミドに組み込んだ場合でも、コンパクチン処理により活性を大きく増大させるエンハンサー活性を示した(図7(b))。しかしながら、エンハンサー活性はSREBP2の過剰発現により影響されなかった。したがって、該肝臓型転写開始点上流領域(プロモーター領域)および下流領域(エンハンサー領域)が肝臓でのABCA1遺伝子発現を制御していることが示された。また、スタチン系薬物による活性化は、SREBP2が直接活性化するL2プロモーターとは異なるメカニズムによることが示された。
【実施例7】
【0087】
[ヒト肝型(L3型)ABCA1遺伝子エンハンサー活性を調節する配列]
実施例6で得られたL3型ABCA1遺伝子エンハンサーの配列について、5’側ならびに3’側を短縮したレポータープラスミド、ならびにエンハンサー配列を部分的に欠失したプラスミドを作成した(図9)。図9(a)はエンハンサー配列を組み込まなかったプラスミドであり、図9(b)は図6(c)と同様に配列番号6に示すエンハンサー配列を組み込んだプラスミドである。図9(c)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3378塩基(配列番号7に示す)を、図9(d)はエンハンサー配列としてL3型転写開始点の下流3184塩基から3378塩基を、図9(e)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3259塩基(配列番号8)を、図9(f)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3246塩基を、図9(g)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3228塩基、及び3242塩基から3259塩基を、それぞれ組み込んだプラスミドである。実施例6に記載した方法で、これらのプラスミドをJHH−5細胞に形質導入し、50μMコンパクチンで処理し、転写活性を測定した。
【0088】
実施例6で得られたL3型ABCA1遺伝子エンハンサーの短縮(図9(c)〜(g)により、L3転写開始点下流3172塩基から3259塩基まで(配列番号8)がコンパクチン処理による活性化を担うことが判明した(図9(e))。さらに、図8に示す、ヒトABCA1遺伝子中の配列a(配列番号9)、配列b(配列番号10)、あるいは配列c(配列番号11)を欠失すると、コンパクチンによる活性化が著しく低下することがわかった(図9(d)、(f)、(g))。したがって、エンハンサー内に存在するこの三つの配列がヒトのABCA1遺伝子の転写調節に機能していると考えられる。
【実施例8】
【0089】
[転写調節因子HNF4alphaによるヒト肝型(L3型)ABCA1遺伝子エンハンサー活性の調節]
実施例6で得られたL3型ABCA1遺伝子エンハンサーの配列について、エンハンサー配列を部分的に欠失したプラスミドを作成した(図10)。プロモーターとしては、SV40を用いた。図10(a)はエンハンサー配列を組み込まなかったプラスミドであり、図10(b)はL3型転写開始点の下流3172塩基から3259塩基(配列番号8)を組み込んだプラスミドである。図10(c)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3246塩基を、図10(d)はエンハンサー配列としてL3型転写開始点の下流3184塩基から3259塩基を、図10(e)はエンハンサー配列としてL3型転写開始点の下流3172塩基から3229塩基及び3244塩基から3259塩基を、それぞれ組み込んだプラスミドである。実施例6に記載した方法で、これらのプラスミドをHepG2細胞に形質導入し、エンハンサーレポータープラスミドに加えて、転写調節因子HNF4alphaの発現ベクターを導入し、転写活性を測定した。mockとして、空ベクターpCMV5プラスミドを共導入した。図11は、50μMのコンパクチンで処理したJHH−5細胞において、転写調節因子HNF4alpha蛋白質の発現量が増加していることを示すウエスタンブロットである。コントロールでは、溶剤(ジメチルスルホキシド(DMSO))で処理した。50μMコンパクチンが、HNF4alpha蛋白発現を増加させ、L3型ABCA1遺伝子プロモーター・エンハンサーを活性化し、L3型ABCA1 mRNAを増加させることが明らかになった。また、図10のグラフに示すように、転写調節因子HNF4alphaは、配列cに依存して、レポーター遺伝子の転写を増加することがわかった。
【産業上の利用可能性】
【0090】
本発明に係るスクリーニング方法によれば、動脈硬化性疾患の予防または治療に有用な物質を発見することができ、HDL上昇や動脈硬化性疾患の予防治療に極めて有用である。
【特許請求の範囲】
【請求項1】
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【請求項2】
前記プロモーター活性を有するDNAが、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAである、請求項1に記載の方法。
【請求項3】
前記ヒトABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項1または2に記載の方法。
【請求項1】
プロモーター活性を有するDNAと、ヒトABCA1遺伝子のイントロン3中の配列を含み、エンハンサー活性を有するDNAとを用いることを特徴とする、ヒトABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【請求項2】
前記プロモーター活性を有するDNAが、ヒトABCA1遺伝子のイントロン2中の塩基配列を含み、プロモーター活性を有するDNAである、請求項1に記載の方法。
【請求項3】
前記ヒトABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項1または2に記載の方法。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図7】

【図8】
【図9】

【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−19728(P2012−19728A)
【公開日】平成24年2月2日(2012.2.2)
【国際特許分類】
【出願番号】特願2010−159674(P2010−159674)
【出願日】平成22年7月14日(2010.7.14)
【出願人】(597128004)国立医薬品食品衛生研究所長 (22)
【Fターム(参考)】
【公開日】平成24年2月2日(2012.2.2)
【国際特許分類】
【出願日】平成22年7月14日(2010.7.14)
【出願人】(597128004)国立医薬品食品衛生研究所長 (22)
【Fターム(参考)】
[ Back to top ]