
ヒトANTのアイソフォームの発現の調節方法
【課題】 核酸塩基を転移させるあらゆる方法と組み合わせて、ヒトおよび動物の治療に使用することができる新規な産生物を提供する。
【解決手段】 ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とするiRNA。
【解決手段】 ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とするiRNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトANTのアイソフォームの発現の調節方法に関するものであり、特に二本鎖干渉RNA(iRNA)及び上記調節のためのその使用、及び該アイソフォームをコードするcDNAの使用に関する。
【背景技術】
【0002】
アデニンヌクレオチド輸送体(ANT)は、ミトコンドリアの内膜に最も豊富に存在するタンパク質である。ANTは、2つの異なる機能を有しており、1つは、ミトコンドリアの内膜を通過するアデニンヌクレオチドの輸送に関与していることである(酸化的リン酸化を目的とするADPの取り込み、一般的な代謝のためのATPの細胞質ゾルへの放出)。二つ目は、ミトコンドリアのアポトーシス過程で必須の役割を果たしていることである。これはANTが非特異的な細孔の立体配置を取ることができるからであり、これによりミトコンドリア膜の透過性及び細胞死を生じさせる(Kroemer & Reed 2000)。
【0003】
ANTをコードする遺伝子は、多くの種、たとえば、酵母、種々の植物、ウシ、ラット、マウス及びヒトにおいてクローン化されている。これら全ての種は、いくつかのアイソフォームをもっており、それらの遺伝子の構造は、多くは3イントロンで分離された4エキソンからなる構造となっている。ヒトANTは3つのアイソフォーム(ANT1、ANT2、ANT3)が存在し、それらは3つの異なる核酸遺伝子でコードされ、該遺伝子はクローン化されており、それらの配列は決定されている。ANT1(染色体4)は主として心臓及び骨格筋に発現する。ANT1の突然変異(アラニン114のプロリンへの置換)に関係したヒト遺伝病が知られている。この病気は進行性外眼筋麻痺(ミトコンドリアDNAの実質的欠損によって特徴付けられる稀な疾患)である。ANT2(X染色体)は成熟した組織では非常に弱く発現される。ANT2の最も高い発現レベルは、筋芽細胞及び腫瘍細胞のような増殖細胞で観察される。ANT2は、又、SV40ウイルスで形質転換した細胞、及びミトコンドリアDNAを欠損した系(rho°)で特異的に見出される。ANT3(X及びY染色体の擬似常染色体領域)は全ての分化した組織のいたるところで発現する。
【0004】
アポトーシスは、3つのフェーズで起こる細胞の自殺の過程である。それらのフェーズとは、プレ−ミトコンドリアフェーズ(不均一性)、ミトコンドリアフェーズ(死の決定)、及び退化フェーズ(細胞の腐敗)である。ミトコンドリア内膜に挿入されているタンパク質であるANTは、ミトコンドリアの役割を大幅に変化させる細孔を形成する能力がある。ANTがオープン細孔状態にあるとき、ミトコンドリアは細胞破壊器官となる。
【0005】
以下に述べる点は、今日では確立されている。
・ANTの細孔機能を誘導することにより、in vitroで細胞を死滅させることが可能である(Belzac、Jacototら、Cancer Res.2001 Feb 15.61(4):1260−4)。
・ANTの細孔機能をブロックすることにより、ex vivoで心臓細胞(単離した虚血性心臓)を保護することが可能である(Di Lisaら、J Biol Chem.2000 Nov 9)。
・ANTを失活させることにより、in vivoで、虚血性脳疾患に続く神経の死から保護することが可能である(Caoら、J Cereb Blood Flow Metab.2001 Apr.21(4):321−333)。
【0006】
したがって、ANTはアポトーシスの主要なコントロールポイントであり、Bax(プロアポトーシス)腫瘍サプレッサー及びBcl−2(アンチアポトーシス)オンコプロテインのような内因性タンパク質によって調節される。また、ANTはVpr(HIV由来のプロアポトーシス)及びvMIA(CMV由来のアンチアポトーシス)のようなウイルス内因性タンパク質によって調節される。それゆえ、ANTはアポトーシスの病理学的脱調節に対抗するための理想的な標的である。
【0007】
最近のデータにより、二本鎖RNA(dsRNA)は、それらを構成する二本鎖のいずれかの配列と非常に相同的な配列を有する遺伝子の発現の消失を誘導することが明らかになっている。この現象は、RNA干渉又はiRNAと呼ばれ、結局メッセンジャーRNAの分解をもたらす(Hammondら、2001、Sharp、2001)。Tuschlらは、21−ヌクレオチドの二本鎖(小さい干渉RNA、siRNA)の哺乳類細胞への導入によって、遺伝子の発現が特異的に抑制されることを証明した(Elbashirら、2001)。トランスフェクション後、siRNAは、標的遺伝子の発現を抑制するために、細胞成分(DICER酵素及びRISC複合体)と共に行動する。
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明者は、選択された方法で、ヒトANTのアイソフォームの発現レベルに作用することにより、治療を目的として、アポトーシスを調節することが可能であることを見出した。
【0009】
特に、各ANTのアイソフォームをコードする配列である、限定された21−ヌクレオチドの領域からデザインされたiRNAによって、トランスフェクション後、各アイソフォームの発現を選択的に抑制することができる二本鎖iRNAを得ることが可能であることを見出した。
【0010】
したがって、本発明の目的は、核酸塩基を転移させるあらゆる方法と組み合わせて、ヒトおよび動物の治療に使用することができる新規な産生物を提供することである。
【課題を解決するための手段】
【0011】
本発明は、ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とする。
【0012】
好ましくは、本発明のiRNAは、18〜25、より好ましくは21ヌクレオチドのsiRNA(小さい干渉RNA)である。
【0013】
好ましいiRNAは、SEQ IDNo.1及びSEQ IDNo.2、SEQ IDNo.3及びSEQ IDNo.4、SEQ IDNo.5及びSEQ IDNo.6の配列をもつ鎖からなる二本鎖より選択される。
SEQ IDNo.1:5’−acagaucagugcugagaagdTdT−3’
SEQ IDNo.2:5’−cuucucagcacugaucugudTdT−3’
SEQ IDNo.3:5’−gcagaucacugcagauaagdTdT−3’
SEQ IDNo.4:5’−cuuaccugcagugaucugcdTdT−3’
SEQ IDNo.5:5’−gggcaucguggacugcauudTdT−3’
SEQ IDNo.6:5’−aaugcaguccacgaugcccdTdT−3’
【0014】
本発明は、また、上記した少なくとも1つのiRNA、あるいは該iRNAの各鎖をコードするDNA配列を含む構造物である。
【0015】
本発明の1態様においては、上記構造物は、iRNAがペプチド、リポソーム、ナノ粒子(ナノスフィア、ナノチューブ)、又はウレアポリマーのような非天然オリゴマーからなるベクターと結合しており、該ベクターは、iRNAの導入、iRNAの膜、組織又は生物学的外皮、特に、細胞膜、ミトコンドリア膜、核膜、皮膚、粘膜、内皮細胞壁、血液脳関門の通過、iRNAのバイオアベイラビリティー、iRNAの安定性、及びiRNAの薬理学的分布を促進することを特徴とする。
【0016】
他の態様においては、該構造物は、iRNAが核酸を転移させるためのベクター、例えば、レトロウイルス(Barton and Medzhitov、PNAS、2002、vol.99(23):p14943−14945)、トランスポゾン、アデノウイルス(Xiaら、Nature Bidech、2002、vol.20、p1005−1010)又はプラスミド(Brummelkampら、Cancel Call、2002、p243−247)と結合していることを特徴とする。
【0017】
本発明は、また、薬剤学的に許容できる担体と組み合わせて、上記した少なくとも1つのiRNA又は上記構造物の有効量を含んでいることを特徴とする薬剤組成物である。
【0018】
好ましい薬剤組成物は、注射剤として投与できる形態のものである。
【0019】
他の形態としては、経口、非経口、直腸又は局所的投与が適している(Levisら、Nature Genetics、2002、vol.32、p107−108)。
【0020】
上記したiRNA、構造物及び薬剤組成物は、それらがミトコンドリア膜の透過性及びアポトーシス、ネクローシス及び自己分解型及び関連した機構の細胞死を調節(誘導又は抑制)する能力を有していることを特徴とする。
【0021】
本発明の組成物は、ヒトANTのアイソフォームの発現を調節することができ、この点で、特にアポトーシスの脱調節及び他の関連した機構の細胞死に関係する病気治療に有益である。
【0022】
したがって、本発明は、ミトコンドリア透過膜電位(ΔΨm)の低下及びアポトーシス、ネクローシスの脱調節及び他の関連した機構の細胞死を誘導/促進するための(siRNA−ANT2)、又は、逆に、抑制するための(siRNA−ANT及び/又はsiRNA−ANT3)、siRNA−ANT1、siRNA−ANT2及び/又はsiRNA−ANT3の使用に関する。
【0023】
したがって、本発明は、また、ミトコンドリア透過膜電位(ΔΨm)の低下及びアポトーシスを誘導/促進するための(hANT1 cDNA及び/又はhANT3 cDNA)、又は、逆に、抑制するための(hANT2 cDNA)、 hANT1、hANT2及び/又はhANT3 cDNAの使用に関する。
【0024】
特に、例えば、各種のガン、全身性エリテマトーデスや関節炎のような自己免疫疾患において、アポトーシスが起こらない状態を治療するために、上記使用がなされる。
【0025】
他の使用としては、上記組成物を、過剰のアポトーシス、例えば、神経退化疾患(アルツハイマー病、パーキンソン病、ハンティングトン病)及び虚血性脳疾患、虚血性心疾患を治療するために使用するというものである。
【0026】
例えば、ANT1又はANT3 siRNA、あるいは代わりにANT2 cDNAを、虚血性疾患の状態又は神経退化病の状態において神経死を抑制するために、あるいは、虚血性疾患の状態において心筋の死、又は肝細胞の死(ウイルス感染、薬物関連中毒)を抑制するために使用することができる。例えば、h−ANT2 siRNA及び/又はh−ANT1又はh−ANT3 cDNAを、腫瘍細胞アポトーシス又は自己反応性リンパ球アポトーシスを誘導するために使用することができる。
【0027】
上記薬剤組成物は、HIV感染症の治療に対しても非常に有益である。
【0028】
本発明の他の特徴及び長所は、図1〜6を参照しつつ、後述される。
【発明を実施するための最良の形態】
【0029】
トランスフェクション24時間後、細胞を溶解して、ANTのアイソフォームの発現を、抗V5モノクローナル抗体を使用するウエスタンブロッティング法で決定する。
【0030】
共トランスフェクション
HeLa細胞を、ウシ胎児血清10%を加えたDMEM/Glutamax−Iを分注した6ウェルプレート中で培養する。24時間後、3μlのlipofectamine 2000(Invitrogen社製)、3μgのsiRNA、血清フリーDMEM中の1μgのベクターpcDNA3.1V5−hANT1又は2(最終容量500μl)を加えて、上記細胞をトランスフェクトする。トランスフェクション後6時間、該細胞を6時間リンスし、24、48又は72時間培養を行う。
【0031】
細胞抽出物の調製及びウエスタンブロッティング
上記細胞を、100μlの溶解バッファー(25mM Tris−HCl、pH7.5、25mM NaCl、5mM EDTA、1%Triton X−100、プロテアーゼ阻害剤カクテル)に再懸濁し、4℃、13000rpmで10分間遠心する。10μlの上澄み液を集め、Bradfordテストを行う。その後、SDS−Laemmliバッファーの存在下、100℃で3分間変性させた後、該抽出物をSDS−PAGEゲルで解析する。移動後、タンパク質は、抗V5抗体(1/5000 Invitrogen社)を用いて検出する。
【0032】
ヒトANTのアイソフォームのクローニング、及び発現ベクターの産生
293T細胞及びHeLa細胞の全RNAを単離し(Trizol protocol)、オリゴdTタイププライマーと共に、逆転写/増幅実験に使用した。ヒトANTのアイソフォーム(hANT1、hANT2及びhANT3)に特異的なプライマーは、各アイソフォームの完全なcDNAを特異的に増幅するため、GenBankにある配列に基づき合成した(表1)。これらの産物は、その後、dアデノシン残基を末端に付加した後、ベクターpGEM−Tでサブクローン化した。各挿入物の配列を確認した(図1)。3つのアイソフォームをコードするcDNAは、その後、発現ベクター:pcDNA3.1(version+、Invitrogen社)及びpIRES−eGFP(Clontech)でクローン化した。3つのアイソフォームに相当するV5エピトープを有する融合タンパク質を産生するため、増幅アプローチ(表2)によって、3つのアイソフォームをコードするcDNAの末端を修飾(ストップコドンの変異及び制限酵素認識配列の付加)し、これらの産物をベクターpcDNA3.1−V5(versionA、Invitrogen社)でサブクローン化することが可能となった。最終構造物はシーケンシングで確認した。
【0033】
ヒトANTのアイソフォームのアポトーシスの可能性
トランスフェクションの実験を、何も含まないベクターpIRES−2−GFPをコントロールとして、また、3つのヒトANTのアイソフォームをコードするcDNAの配列を含むベクターpIRES−2−eGFPを用いて、293T細胞で行った。トランスフェクション後のある時間において、フローサイトメトリで細胞を解析した。
【0034】
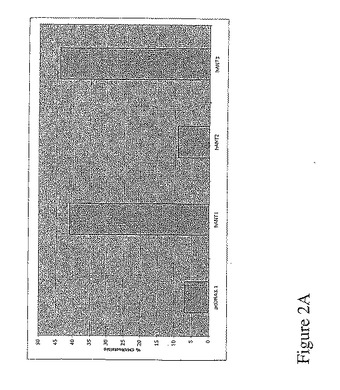
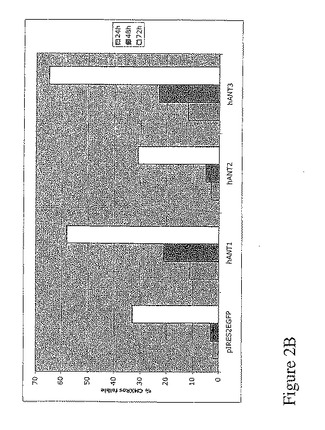
その結果、hANT1及びhANT3のアイソフォームの発現により、ミトコンドリア膜の電位が消失し、それによりアポトーシスを起し、一方、hANT2のアイソフォームの発現は、ミトコンドリアの統合性に影響を与えなかった(図2)。
【0035】
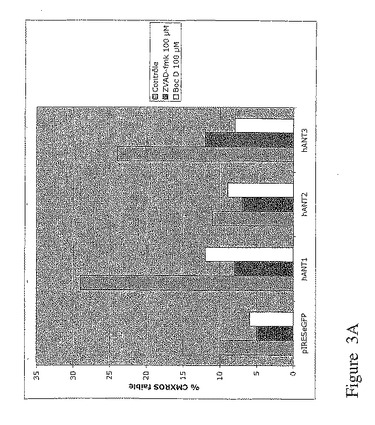
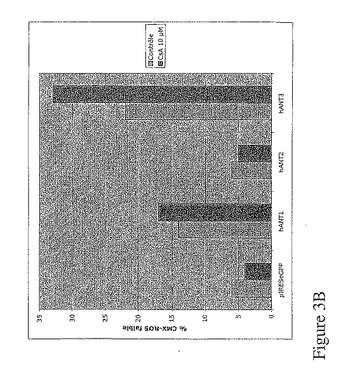
同様の実験アプローチを使用して、本発明者は、hANT1及びhANT3のアイソフォームの発現に関係するアポトーシスは、カスパーゼ阻害剤(ZVADとBoc D)で抑制されるが(図3A)、シクロスポリンA(CsA)では抑制されないことを証明した(図3B)。
【0036】
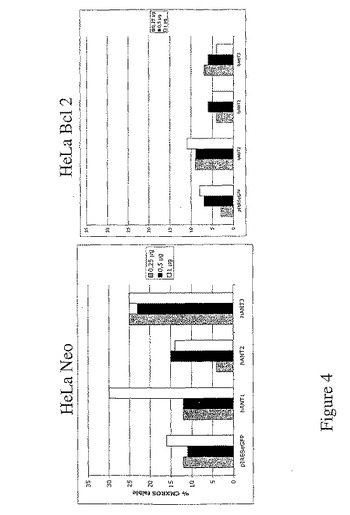
また、本発明者は、Bcl2タンパク質を過剰発現するHeLa細胞を用いて、Bcl2タンパク質がhANT1及びhANT2のアイソフォームで誘導されるアポトーシスを抑制することができることを証明した(図4)。
【0037】
hANT1及びhANT2のアイソフォームの細胞内局在性
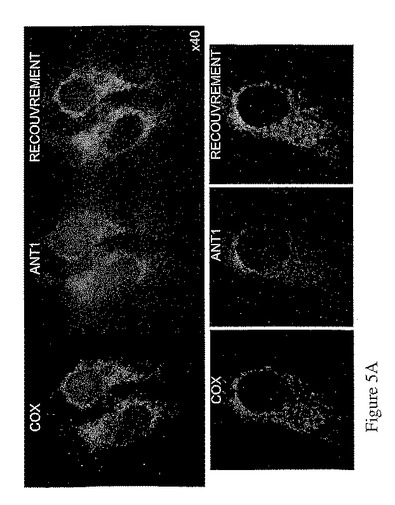
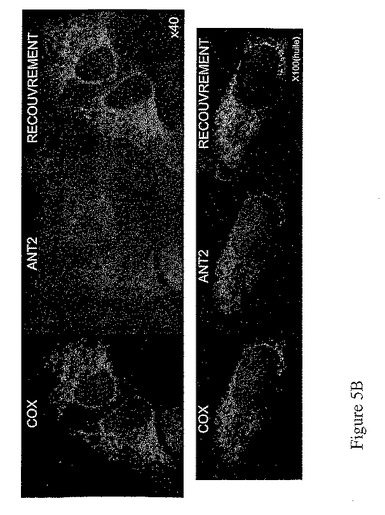
hANT1−V5及びhANT2−V5融合タンパク質をコードする構造物を用いたHeLa細胞のトランスフェクション後、本発明者は、hANT1及びhANT2のアイソフォームの細胞内局在性を決定するため、免疫標識を行った。抗V5抗体で得られたシグナル及びCOX(ミトコンドリアタンパク質)に対抗する抗体で得られたシグナルによる局在性の解析により、hANT1及びhANT2に対し、ミトコンドリアにおける局在性を証明した(図5)。
【0038】
ヒトANTのアイソフォームの二本鎖iRNA
iRNAの調製
ヒトANT1(AAACAGATCAGTGCTGAGAAG、ヌクレオチド 127−147)、ヒトANT2(AAGCAGATCACTGCAGATAAG、ヌクレオチド 127−147)、4つの突然変異をもつヒトANT2(AAGCGGATCGCTACAAATAAG、ヌクレオチド 127−147)及びヒトANT3(AAGGGCATCGTGGACTGCATT、ヌクレオチド 127−147)のcDNA配列に相当する二本鎖iRNAを、Elbashirら(2001)の方法に基づき設計した。該二本鎖は、Proligo(フランス)により調製した。
hANT1(127−147)
DNA 5’−aaacagatcagtgctgagaag−3’(SEQ ID No.7)
配列: 5’−acagaucagugcugagaagdTdT−3’(SEQ ID No.8)
二本鎖iRNA:5’−cuucucagcacugaucugudTdT−3’(SEQ ID No.9)
hANT2(127−147)
DNA 5’−aagcagatcactgcagataag−3’(SEQ ID No.10)
配列: 5’−gcagaucacugcagauaagdTdT−3’(SEQ ID No.11)
二本鎖iRNA:5’−cuuaucugcagugaucugcdTdT−3’(SEQ ID No.12)
hANT2mut(127−147)
DNA配列 5’−aagcggatcgctacaaataag−3’(SEQ ID No.13)
二本鎖iRNA:5’−gcggaucgcuacaaauaagdTdT−3’(SEQ ID No.14)
5’−cuuauuuguagcgauccgcdTdT−3’(SEQ ID No.15)
hANT3mut(154−174)
DNA配列 5’−aagggcatcgtggactgcatt−3’(SEQ ID No.16)
二本鎖iRNA:5’−gggcaucguggacugcauudTdT−3’(SEQ ID No.17)
5’−aaugcaguccacgaugcccdTdT−3’(SEQ ID No.18)
【0039】
各表は、使用されたプライマーの配列を示し、表1は、RT/PCR実験において、3つのヒトANTアイソフォームをコードするcDNAをクローンするためのもの、表2は、hANT−V5融合タンパク質をコードするcDNAを含む発現ベクターを作成するためのものである。
【0040】
【表1】

3つのヒトANTのアイソフォームをコードするcDNAをクローン化するためのRT/PCR実験
【0041】
【表2】
hANT−V5融合タンパク質をコードするcDNAを含む表現ベクターの作成
【0042】
参照:
Hammond, S.M., Caudy, A.A. and Hannon, G.J. (2001). Post-transcriptional gene
silencing by double-stranded RNA. Nat Rev Genet, 2, 110-119.
Sharp, P.A. (2001). RNA interference-2001. Genes Dev. 15, 485-490.
Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K. and Tuschl, T. (2001). Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells, Nature, 411, 494-498.
【図面の簡単な説明】
【0043】
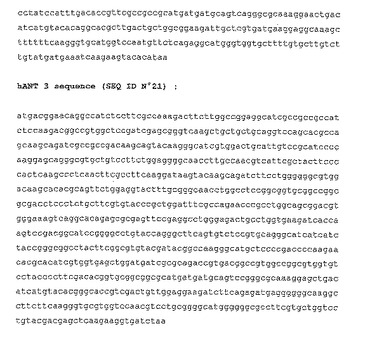
【図1】ヒトANTの3つのアイソフォームをコードする完全なcDNA配列。293T及びHeLa細胞由来のRNAを用いてRT/PCR後に単離。
【図2A】図2Aは、アポトーシスを誘導するhANT1アイソフォームとhANT3アイソフォームの発現。1μgのベクターpIRES−2−eGFPと1μgのベクターpcDNA3.1−hANTの共トランスフェクション24時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図2B】図2Bは、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション24、48、72時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図2C】図2Cは、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション24、48、72時間後における293T細胞の低倍数体の核の頻度についてのフローサイトメトリ解析。
【図3A】hANT1とhANT3の発現によって誘導されるアポトーシスは、ZVAD及びBoc Dで抑制されるが、CsAでは抑制されない。図3Aは、10μMのCsAの存在又は不存在下、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション48時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図3B】hANT1とhANT3の発現によって誘導されるアポトーシスは、ZVAD及びBoc Dで抑制されるが、CsAでは抑制されない。図3Bは、100μMのZVAD−fmk又は100μMのBoc Dの存在又は不存在下、1μgのpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション48時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図4】hANT1とhANT3のアイソフォームの発現によって誘導されるアポトーシスは、Bc12の発現で抑制される。HeLa Neo及びBc12細胞に1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTをトランスフェクトし、72時間後、GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図5A】図5Aは、hANT1とhANT2のアイソフォームの細胞内局在性。HeLa細胞に1μgのベクターpcDNA3.1V5−hANT1(A)又は1μgのベクターpcDNA3.1v5−hANT1(B)をトランスフェクトし、パラホルムアルデヒドで固定化する。HANT− V5融合タンパク質とCOXミトコンドリアタンパク質の共在性は、V5エピトープ(緑色蛍光)及びCOXタンパク質(赤色蛍光)の免疫蛍光検出によって決定する。混合色のイメージは、共在性を示す緑色蛍光と赤色蛍光のスーパーインポジションを表す。
【図5B】図5Bは、hANT1とhANT2のアイソフォームの細胞内局在性。HeLa細胞に1μgのベクターpcDNA3.1V5−hANT1(A)又は1μgのベクターpcDNA3.1v5−hANT1(B)をトランスフェクトし、パラホルムアルデヒドで固定化する。HANT− V5融合タンパク質とCOXミトコンドリアタンパク質の共在性は、V5エピトープ(緑色蛍光)及びCOXタンパク質(赤色蛍光)の免疫蛍光検出によって決定する。混合色のイメージは、共在性を示す緑色蛍光と赤色蛍光のスーパーインポジションを表す。
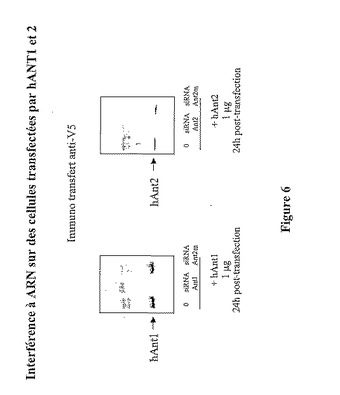
【図6】特異的なsiRNAを使用したヒトANTのアイソフォーム1及び2の発現の特異的抑制。(A)HeLa細胞に、最初、発現ベクターpcDNA3.1V5−hANT1、次いでhANT1又はhANT2mutに対して特異的なsiRNAを共トランスフェクトする。(B)HeLa細胞に、最初、発現ベクターpcDNA3.1V5−hANT2、次いでhANT2又はhANT2mutに対して特異的なsiRNAを共トランスフェクトする。
【技術分野】
【0001】
本発明は、ヒトANTのアイソフォームの発現の調節方法に関するものであり、特に二本鎖干渉RNA(iRNA)及び上記調節のためのその使用、及び該アイソフォームをコードするcDNAの使用に関する。
【背景技術】
【0002】
アデニンヌクレオチド輸送体(ANT)は、ミトコンドリアの内膜に最も豊富に存在するタンパク質である。ANTは、2つの異なる機能を有しており、1つは、ミトコンドリアの内膜を通過するアデニンヌクレオチドの輸送に関与していることである(酸化的リン酸化を目的とするADPの取り込み、一般的な代謝のためのATPの細胞質ゾルへの放出)。二つ目は、ミトコンドリアのアポトーシス過程で必須の役割を果たしていることである。これはANTが非特異的な細孔の立体配置を取ることができるからであり、これによりミトコンドリア膜の透過性及び細胞死を生じさせる(Kroemer & Reed 2000)。
【0003】
ANTをコードする遺伝子は、多くの種、たとえば、酵母、種々の植物、ウシ、ラット、マウス及びヒトにおいてクローン化されている。これら全ての種は、いくつかのアイソフォームをもっており、それらの遺伝子の構造は、多くは3イントロンで分離された4エキソンからなる構造となっている。ヒトANTは3つのアイソフォーム(ANT1、ANT2、ANT3)が存在し、それらは3つの異なる核酸遺伝子でコードされ、該遺伝子はクローン化されており、それらの配列は決定されている。ANT1(染色体4)は主として心臓及び骨格筋に発現する。ANT1の突然変異(アラニン114のプロリンへの置換)に関係したヒト遺伝病が知られている。この病気は進行性外眼筋麻痺(ミトコンドリアDNAの実質的欠損によって特徴付けられる稀な疾患)である。ANT2(X染色体)は成熟した組織では非常に弱く発現される。ANT2の最も高い発現レベルは、筋芽細胞及び腫瘍細胞のような増殖細胞で観察される。ANT2は、又、SV40ウイルスで形質転換した細胞、及びミトコンドリアDNAを欠損した系(rho°)で特異的に見出される。ANT3(X及びY染色体の擬似常染色体領域)は全ての分化した組織のいたるところで発現する。
【0004】
アポトーシスは、3つのフェーズで起こる細胞の自殺の過程である。それらのフェーズとは、プレ−ミトコンドリアフェーズ(不均一性)、ミトコンドリアフェーズ(死の決定)、及び退化フェーズ(細胞の腐敗)である。ミトコンドリア内膜に挿入されているタンパク質であるANTは、ミトコンドリアの役割を大幅に変化させる細孔を形成する能力がある。ANTがオープン細孔状態にあるとき、ミトコンドリアは細胞破壊器官となる。
【0005】
以下に述べる点は、今日では確立されている。
・ANTの細孔機能を誘導することにより、in vitroで細胞を死滅させることが可能である(Belzac、Jacototら、Cancer Res.2001 Feb 15.61(4):1260−4)。
・ANTの細孔機能をブロックすることにより、ex vivoで心臓細胞(単離した虚血性心臓)を保護することが可能である(Di Lisaら、J Biol Chem.2000 Nov 9)。
・ANTを失活させることにより、in vivoで、虚血性脳疾患に続く神経の死から保護することが可能である(Caoら、J Cereb Blood Flow Metab.2001 Apr.21(4):321−333)。
【0006】
したがって、ANTはアポトーシスの主要なコントロールポイントであり、Bax(プロアポトーシス)腫瘍サプレッサー及びBcl−2(アンチアポトーシス)オンコプロテインのような内因性タンパク質によって調節される。また、ANTはVpr(HIV由来のプロアポトーシス)及びvMIA(CMV由来のアンチアポトーシス)のようなウイルス内因性タンパク質によって調節される。それゆえ、ANTはアポトーシスの病理学的脱調節に対抗するための理想的な標的である。
【0007】
最近のデータにより、二本鎖RNA(dsRNA)は、それらを構成する二本鎖のいずれかの配列と非常に相同的な配列を有する遺伝子の発現の消失を誘導することが明らかになっている。この現象は、RNA干渉又はiRNAと呼ばれ、結局メッセンジャーRNAの分解をもたらす(Hammondら、2001、Sharp、2001)。Tuschlらは、21−ヌクレオチドの二本鎖(小さい干渉RNA、siRNA)の哺乳類細胞への導入によって、遺伝子の発現が特異的に抑制されることを証明した(Elbashirら、2001)。トランスフェクション後、siRNAは、標的遺伝子の発現を抑制するために、細胞成分(DICER酵素及びRISC複合体)と共に行動する。
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明者は、選択された方法で、ヒトANTのアイソフォームの発現レベルに作用することにより、治療を目的として、アポトーシスを調節することが可能であることを見出した。
【0009】
特に、各ANTのアイソフォームをコードする配列である、限定された21−ヌクレオチドの領域からデザインされたiRNAによって、トランスフェクション後、各アイソフォームの発現を選択的に抑制することができる二本鎖iRNAを得ることが可能であることを見出した。
【0010】
したがって、本発明の目的は、核酸塩基を転移させるあらゆる方法と組み合わせて、ヒトおよび動物の治療に使用することができる新規な産生物を提供することである。
【課題を解決するための手段】
【0011】
本発明は、ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とする。
【0012】
好ましくは、本発明のiRNAは、18〜25、より好ましくは21ヌクレオチドのsiRNA(小さい干渉RNA)である。
【0013】
好ましいiRNAは、SEQ IDNo.1及びSEQ IDNo.2、SEQ IDNo.3及びSEQ IDNo.4、SEQ IDNo.5及びSEQ IDNo.6の配列をもつ鎖からなる二本鎖より選択される。
SEQ IDNo.1:5’−acagaucagugcugagaagdTdT−3’
SEQ IDNo.2:5’−cuucucagcacugaucugudTdT−3’
SEQ IDNo.3:5’−gcagaucacugcagauaagdTdT−3’
SEQ IDNo.4:5’−cuuaccugcagugaucugcdTdT−3’
SEQ IDNo.5:5’−gggcaucguggacugcauudTdT−3’
SEQ IDNo.6:5’−aaugcaguccacgaugcccdTdT−3’
【0014】
本発明は、また、上記した少なくとも1つのiRNA、あるいは該iRNAの各鎖をコードするDNA配列を含む構造物である。
【0015】
本発明の1態様においては、上記構造物は、iRNAがペプチド、リポソーム、ナノ粒子(ナノスフィア、ナノチューブ)、又はウレアポリマーのような非天然オリゴマーからなるベクターと結合しており、該ベクターは、iRNAの導入、iRNAの膜、組織又は生物学的外皮、特に、細胞膜、ミトコンドリア膜、核膜、皮膚、粘膜、内皮細胞壁、血液脳関門の通過、iRNAのバイオアベイラビリティー、iRNAの安定性、及びiRNAの薬理学的分布を促進することを特徴とする。
【0016】
他の態様においては、該構造物は、iRNAが核酸を転移させるためのベクター、例えば、レトロウイルス(Barton and Medzhitov、PNAS、2002、vol.99(23):p14943−14945)、トランスポゾン、アデノウイルス(Xiaら、Nature Bidech、2002、vol.20、p1005−1010)又はプラスミド(Brummelkampら、Cancel Call、2002、p243−247)と結合していることを特徴とする。
【0017】
本発明は、また、薬剤学的に許容できる担体と組み合わせて、上記した少なくとも1つのiRNA又は上記構造物の有効量を含んでいることを特徴とする薬剤組成物である。
【0018】
好ましい薬剤組成物は、注射剤として投与できる形態のものである。
【0019】
他の形態としては、経口、非経口、直腸又は局所的投与が適している(Levisら、Nature Genetics、2002、vol.32、p107−108)。
【0020】
上記したiRNA、構造物及び薬剤組成物は、それらがミトコンドリア膜の透過性及びアポトーシス、ネクローシス及び自己分解型及び関連した機構の細胞死を調節(誘導又は抑制)する能力を有していることを特徴とする。
【0021】
本発明の組成物は、ヒトANTのアイソフォームの発現を調節することができ、この点で、特にアポトーシスの脱調節及び他の関連した機構の細胞死に関係する病気治療に有益である。
【0022】
したがって、本発明は、ミトコンドリア透過膜電位(ΔΨm)の低下及びアポトーシス、ネクローシスの脱調節及び他の関連した機構の細胞死を誘導/促進するための(siRNA−ANT2)、又は、逆に、抑制するための(siRNA−ANT及び/又はsiRNA−ANT3)、siRNA−ANT1、siRNA−ANT2及び/又はsiRNA−ANT3の使用に関する。
【0023】
したがって、本発明は、また、ミトコンドリア透過膜電位(ΔΨm)の低下及びアポトーシスを誘導/促進するための(hANT1 cDNA及び/又はhANT3 cDNA)、又は、逆に、抑制するための(hANT2 cDNA)、 hANT1、hANT2及び/又はhANT3 cDNAの使用に関する。
【0024】
特に、例えば、各種のガン、全身性エリテマトーデスや関節炎のような自己免疫疾患において、アポトーシスが起こらない状態を治療するために、上記使用がなされる。
【0025】
他の使用としては、上記組成物を、過剰のアポトーシス、例えば、神経退化疾患(アルツハイマー病、パーキンソン病、ハンティングトン病)及び虚血性脳疾患、虚血性心疾患を治療するために使用するというものである。
【0026】
例えば、ANT1又はANT3 siRNA、あるいは代わりにANT2 cDNAを、虚血性疾患の状態又は神経退化病の状態において神経死を抑制するために、あるいは、虚血性疾患の状態において心筋の死、又は肝細胞の死(ウイルス感染、薬物関連中毒)を抑制するために使用することができる。例えば、h−ANT2 siRNA及び/又はh−ANT1又はh−ANT3 cDNAを、腫瘍細胞アポトーシス又は自己反応性リンパ球アポトーシスを誘導するために使用することができる。
【0027】
上記薬剤組成物は、HIV感染症の治療に対しても非常に有益である。
【0028】
本発明の他の特徴及び長所は、図1〜6を参照しつつ、後述される。
【発明を実施するための最良の形態】
【0029】
トランスフェクション24時間後、細胞を溶解して、ANTのアイソフォームの発現を、抗V5モノクローナル抗体を使用するウエスタンブロッティング法で決定する。
【0030】
共トランスフェクション
HeLa細胞を、ウシ胎児血清10%を加えたDMEM/Glutamax−Iを分注した6ウェルプレート中で培養する。24時間後、3μlのlipofectamine 2000(Invitrogen社製)、3μgのsiRNA、血清フリーDMEM中の1μgのベクターpcDNA3.1V5−hANT1又は2(最終容量500μl)を加えて、上記細胞をトランスフェクトする。トランスフェクション後6時間、該細胞を6時間リンスし、24、48又は72時間培養を行う。
【0031】
細胞抽出物の調製及びウエスタンブロッティング
上記細胞を、100μlの溶解バッファー(25mM Tris−HCl、pH7.5、25mM NaCl、5mM EDTA、1%Triton X−100、プロテアーゼ阻害剤カクテル)に再懸濁し、4℃、13000rpmで10分間遠心する。10μlの上澄み液を集め、Bradfordテストを行う。その後、SDS−Laemmliバッファーの存在下、100℃で3分間変性させた後、該抽出物をSDS−PAGEゲルで解析する。移動後、タンパク質は、抗V5抗体(1/5000 Invitrogen社)を用いて検出する。
【0032】
ヒトANTのアイソフォームのクローニング、及び発現ベクターの産生
293T細胞及びHeLa細胞の全RNAを単離し(Trizol protocol)、オリゴdTタイププライマーと共に、逆転写/増幅実験に使用した。ヒトANTのアイソフォーム(hANT1、hANT2及びhANT3)に特異的なプライマーは、各アイソフォームの完全なcDNAを特異的に増幅するため、GenBankにある配列に基づき合成した(表1)。これらの産物は、その後、dアデノシン残基を末端に付加した後、ベクターpGEM−Tでサブクローン化した。各挿入物の配列を確認した(図1)。3つのアイソフォームをコードするcDNAは、その後、発現ベクター:pcDNA3.1(version+、Invitrogen社)及びpIRES−eGFP(Clontech)でクローン化した。3つのアイソフォームに相当するV5エピトープを有する融合タンパク質を産生するため、増幅アプローチ(表2)によって、3つのアイソフォームをコードするcDNAの末端を修飾(ストップコドンの変異及び制限酵素認識配列の付加)し、これらの産物をベクターpcDNA3.1−V5(versionA、Invitrogen社)でサブクローン化することが可能となった。最終構造物はシーケンシングで確認した。
【0033】
ヒトANTのアイソフォームのアポトーシスの可能性
トランスフェクションの実験を、何も含まないベクターpIRES−2−GFPをコントロールとして、また、3つのヒトANTのアイソフォームをコードするcDNAの配列を含むベクターpIRES−2−eGFPを用いて、293T細胞で行った。トランスフェクション後のある時間において、フローサイトメトリで細胞を解析した。
【0034】
その結果、hANT1及びhANT3のアイソフォームの発現により、ミトコンドリア膜の電位が消失し、それによりアポトーシスを起し、一方、hANT2のアイソフォームの発現は、ミトコンドリアの統合性に影響を与えなかった(図2)。
【0035】
同様の実験アプローチを使用して、本発明者は、hANT1及びhANT3のアイソフォームの発現に関係するアポトーシスは、カスパーゼ阻害剤(ZVADとBoc D)で抑制されるが(図3A)、シクロスポリンA(CsA)では抑制されないことを証明した(図3B)。
【0036】
また、本発明者は、Bcl2タンパク質を過剰発現するHeLa細胞を用いて、Bcl2タンパク質がhANT1及びhANT2のアイソフォームで誘導されるアポトーシスを抑制することができることを証明した(図4)。
【0037】
hANT1及びhANT2のアイソフォームの細胞内局在性
hANT1−V5及びhANT2−V5融合タンパク質をコードする構造物を用いたHeLa細胞のトランスフェクション後、本発明者は、hANT1及びhANT2のアイソフォームの細胞内局在性を決定するため、免疫標識を行った。抗V5抗体で得られたシグナル及びCOX(ミトコンドリアタンパク質)に対抗する抗体で得られたシグナルによる局在性の解析により、hANT1及びhANT2に対し、ミトコンドリアにおける局在性を証明した(図5)。
【0038】
ヒトANTのアイソフォームの二本鎖iRNA
iRNAの調製
ヒトANT1(AAACAGATCAGTGCTGAGAAG、ヌクレオチド 127−147)、ヒトANT2(AAGCAGATCACTGCAGATAAG、ヌクレオチド 127−147)、4つの突然変異をもつヒトANT2(AAGCGGATCGCTACAAATAAG、ヌクレオチド 127−147)及びヒトANT3(AAGGGCATCGTGGACTGCATT、ヌクレオチド 127−147)のcDNA配列に相当する二本鎖iRNAを、Elbashirら(2001)の方法に基づき設計した。該二本鎖は、Proligo(フランス)により調製した。
hANT1(127−147)
DNA 5’−aaacagatcagtgctgagaag−3’(SEQ ID No.7)
配列: 5’−acagaucagugcugagaagdTdT−3’(SEQ ID No.8)
二本鎖iRNA:5’−cuucucagcacugaucugudTdT−3’(SEQ ID No.9)
hANT2(127−147)
DNA 5’−aagcagatcactgcagataag−3’(SEQ ID No.10)
配列: 5’−gcagaucacugcagauaagdTdT−3’(SEQ ID No.11)
二本鎖iRNA:5’−cuuaucugcagugaucugcdTdT−3’(SEQ ID No.12)
hANT2mut(127−147)
DNA配列 5’−aagcggatcgctacaaataag−3’(SEQ ID No.13)
二本鎖iRNA:5’−gcggaucgcuacaaauaagdTdT−3’(SEQ ID No.14)
5’−cuuauuuguagcgauccgcdTdT−3’(SEQ ID No.15)
hANT3mut(154−174)
DNA配列 5’−aagggcatcgtggactgcatt−3’(SEQ ID No.16)
二本鎖iRNA:5’−gggcaucguggacugcauudTdT−3’(SEQ ID No.17)
5’−aaugcaguccacgaugcccdTdT−3’(SEQ ID No.18)
【0039】
各表は、使用されたプライマーの配列を示し、表1は、RT/PCR実験において、3つのヒトANTアイソフォームをコードするcDNAをクローンするためのもの、表2は、hANT−V5融合タンパク質をコードするcDNAを含む発現ベクターを作成するためのものである。
【0040】
【表1】
3つのヒトANTのアイソフォームをコードするcDNAをクローン化するためのRT/PCR実験
【0041】
【表2】
hANT−V5融合タンパク質をコードするcDNAを含む表現ベクターの作成
【0042】
参照:
Hammond, S.M., Caudy, A.A. and Hannon, G.J. (2001). Post-transcriptional gene
silencing by double-stranded RNA. Nat Rev Genet, 2, 110-119.
Sharp, P.A. (2001). RNA interference-2001. Genes Dev. 15, 485-490.
Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K. and Tuschl, T. (2001). Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells, Nature, 411, 494-498.
【図面の簡単な説明】
【0043】
【図1】ヒトANTの3つのアイソフォームをコードする完全なcDNA配列。293T及びHeLa細胞由来のRNAを用いてRT/PCR後に単離。
【図2A】図2Aは、アポトーシスを誘導するhANT1アイソフォームとhANT3アイソフォームの発現。1μgのベクターpIRES−2−eGFPと1μgのベクターpcDNA3.1−hANTの共トランスフェクション24時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図2B】図2Bは、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション24、48、72時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図2C】図2Cは、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション24、48、72時間後における293T細胞の低倍数体の核の頻度についてのフローサイトメトリ解析。
【図3A】hANT1とhANT3の発現によって誘導されるアポトーシスは、ZVAD及びBoc Dで抑制されるが、CsAでは抑制されない。図3Aは、10μMのCsAの存在又は不存在下、1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション48時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図3B】hANT1とhANT3の発現によって誘導されるアポトーシスは、ZVAD及びBoc Dで抑制されるが、CsAでは抑制されない。図3Bは、100μMのZVAD−fmk又は100μMのBoc Dの存在又は不存在下、1μgのpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTのトランスフェクション48時間後における293T細胞のフローサイトメトリ解析。GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図4】hANT1とhANT3のアイソフォームの発現によって誘導されるアポトーシスは、Bc12の発現で抑制される。HeLa Neo及びBc12細胞に1μgのベクターpIRES−2−eGFP又は1μgの各ベクターpIRES−eGFP−hANTをトランスフェクトし、72時間後、GFP陽性細胞のみのCMXRosレベルの強度を定性的に評価。
【図5A】図5Aは、hANT1とhANT2のアイソフォームの細胞内局在性。HeLa細胞に1μgのベクターpcDNA3.1V5−hANT1(A)又は1μgのベクターpcDNA3.1v5−hANT1(B)をトランスフェクトし、パラホルムアルデヒドで固定化する。HANT− V5融合タンパク質とCOXミトコンドリアタンパク質の共在性は、V5エピトープ(緑色蛍光)及びCOXタンパク質(赤色蛍光)の免疫蛍光検出によって決定する。混合色のイメージは、共在性を示す緑色蛍光と赤色蛍光のスーパーインポジションを表す。
【図5B】図5Bは、hANT1とhANT2のアイソフォームの細胞内局在性。HeLa細胞に1μgのベクターpcDNA3.1V5−hANT1(A)又は1μgのベクターpcDNA3.1v5−hANT1(B)をトランスフェクトし、パラホルムアルデヒドで固定化する。HANT− V5融合タンパク質とCOXミトコンドリアタンパク質の共在性は、V5エピトープ(緑色蛍光)及びCOXタンパク質(赤色蛍光)の免疫蛍光検出によって決定する。混合色のイメージは、共在性を示す緑色蛍光と赤色蛍光のスーパーインポジションを表す。
【図6】特異的なsiRNAを使用したヒトANTのアイソフォーム1及び2の発現の特異的抑制。(A)HeLa細胞に、最初、発現ベクターpcDNA3.1V5−hANT1、次いでhANT1又はhANT2mutに対して特異的なsiRNAを共トランスフェクトする。(B)HeLa細胞に、最初、発現ベクターpcDNA3.1V5−hANT2、次いでhANT2又はhANT2mutに対して特異的なsiRNAを共トランスフェクトする。
【特許請求の範囲】
【請求項1】
ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とするiRNA。
【請求項2】
18〜25、特には21ヌクレオチドのsiRNAであることを特徴とする請求項1記載のiRNA。
【請求項3】
SEQ IDNo.1、SEQ IDNo.2又はSEQ IDNo.3をもつことを特徴とする請求項2記載のiRNA。
【請求項4】
請求項1〜3のいずれかに記載の少なくとも1つのiRNA、あるいは該iRNAの各鎖をコードするDNA配列を含む構造物。
【請求項5】
iRNAがペプチド、リポソーム、ナノ粒子(ナノスフィア、ナノチューブ)、又はウレアポリマーのような非天然オリゴマーからなるベクターと結合しており、該ベクターは、iRNAの導入、iRNAの膜、組織又は生物学的外皮、特に、細胞膜、ミトコンドリア膜、核膜、皮膚、粘膜、内皮細胞壁、血液脳関門の通過、iRNAのバイオアベイラビリティー、iRNAの安定性、及びiRNAの薬理学的分布を促進することを特徴とする請求項4記載の構造物。
【請求項6】
ベクターが、核酸を転移させるためのベクター、例えば、レトロウイルス、トランスポゾン、アデノウイルス又はプラスミドであることを特徴とする請求項4記載の構造物。
【請求項7】
薬剤学的に許容できる担体と組み合わせて、請求項1〜3のいずれか1項に記載した少なくとも1つのiRNA又は請求項4〜6のいずれか1項に記載した構造物の有効量を含んでいることを特徴とする薬剤組成物。
【請求項8】
注射剤として投与できる形態、又は経口、非経口、直腸又は局所的投与できる形態であることを特徴とする請求項7記載の薬剤組成物。
【請求項9】
ミトコンドリア膜の透過性及びアポトーシス、ネクローシス及び自己分解型及び関連した機構の細胞死を調節(誘導又は抑制)する能力を有していることを特徴とする請求項1〜3のいずれか1項に記載したiRNA、又は請求項1〜6のいずれか1項に記載した構造物、又は請求項7又は8に記載した薬剤組成物。
【請求項1】
ANTのアイソフォームの発現を選択的に抑制することができるiRNAであって、該iRNAは二本鎖RNAであり、二本鎖の内の1つはANTのアイソフォームをコードしているmRNAの断片と非常に相同性が高いことを特徴とするiRNA。
【請求項2】
18〜25、特には21ヌクレオチドのsiRNAであることを特徴とする請求項1記載のiRNA。
【請求項3】
SEQ IDNo.1、SEQ IDNo.2又はSEQ IDNo.3をもつことを特徴とする請求項2記載のiRNA。
【請求項4】
請求項1〜3のいずれかに記載の少なくとも1つのiRNA、あるいは該iRNAの各鎖をコードするDNA配列を含む構造物。
【請求項5】
iRNAがペプチド、リポソーム、ナノ粒子(ナノスフィア、ナノチューブ)、又はウレアポリマーのような非天然オリゴマーからなるベクターと結合しており、該ベクターは、iRNAの導入、iRNAの膜、組織又は生物学的外皮、特に、細胞膜、ミトコンドリア膜、核膜、皮膚、粘膜、内皮細胞壁、血液脳関門の通過、iRNAのバイオアベイラビリティー、iRNAの安定性、及びiRNAの薬理学的分布を促進することを特徴とする請求項4記載の構造物。
【請求項6】
ベクターが、核酸を転移させるためのベクター、例えば、レトロウイルス、トランスポゾン、アデノウイルス又はプラスミドであることを特徴とする請求項4記載の構造物。
【請求項7】
薬剤学的に許容できる担体と組み合わせて、請求項1〜3のいずれか1項に記載した少なくとも1つのiRNA又は請求項4〜6のいずれか1項に記載した構造物の有効量を含んでいることを特徴とする薬剤組成物。
【請求項8】
注射剤として投与できる形態、又は経口、非経口、直腸又は局所的投与できる形態であることを特徴とする請求項7記載の薬剤組成物。
【請求項9】
ミトコンドリア膜の透過性及びアポトーシス、ネクローシス及び自己分解型及び関連した機構の細胞死を調節(誘導又は抑制)する能力を有していることを特徴とする請求項1〜3のいずれか1項に記載したiRNA、又は請求項1〜6のいずれか1項に記載した構造物、又は請求項7又は8に記載した薬剤組成物。
【図1】



【図4】
【図6】
【図4】
【図6】
【公表番号】特表2007−527200(P2007−527200A)
【公表日】平成19年9月27日(2007.9.27)
【国際特許分類】
【出願番号】特願2006−502111(P2006−502111)
【出願日】平成16年1月21日(2004.1.21)
【国際出願番号】PCT/FR2004/000127
【国際公開番号】WO2004/067558
【国際公開日】平成16年8月12日(2004.8.12)
【出願人】(505275815)
【Fターム(参考)】
【公表日】平成19年9月27日(2007.9.27)
【国際特許分類】
【出願日】平成16年1月21日(2004.1.21)
【国際出願番号】PCT/FR2004/000127
【国際公開番号】WO2004/067558
【国際公開日】平成16年8月12日(2004.8.12)
【出願人】(505275815)
【Fターム(参考)】
[ Back to top ]