
ヒトCD154−結合性の合成ペプチドおよびその使用
本発明は、7残基長のアミノ酸配列を含み、好ましくは両端に2個のシステイン残基が側面にあり、ヒトCD154を特異的に認識することができ、CD40:CD154相互作用を阻止でき、それにより該相互作用に依存する生物学的作用を阻害することができる合成ペプチドに関する。本発明のペプチドは、好ましくは環状であり、診断応用および治療応用に、特に腫瘍、炎症性疾患および移植による拒絶反応の診断および治療に、使用するのに適している。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトCD40リガンド(CD40LまたはCD154)に結合できる合成ペプチドに関する。特に、本発明は、CD154の活性部位と特異的に結合でき、それにより、CD154とその天然リガンドであるCD40との相互作用を阻害し、同様に該相互作用に依存する生物学的作用も阻害するアミノ酸配列を有する、合成ペプチドに関する。本発明はまた、炎症性、免疫性または血液学的起源を有する疾患の診断および治療における該ペプチドの使用に関する。
【背景技術】
【0002】
CD40は、B細胞表面で構成的に発現される分子であり、そこでCD40は成熟過程の間に決定的に重要な役割を果たす。CD40は、多くのさらなる細胞型、例えば単球、繊維芽細胞、樹状細胞、内皮細胞、平滑筋細胞および上皮細胞の表面でも発現される(van Kooten et al, J. Leukoc. Biol. 2000)。そのリガンドであるCD154(CD40Lとも称される)は、T細胞、マクロファージ、血小板、単球、NK細胞および内皮細胞の表面で活性化に伴って一過的に発現される。特に、B細胞の表面で発現されるCD40と活性型T細胞の表面で発現されるCD154との間の相互作用は、B細胞のクローン増殖およびアイソタイプスイッチを引き起こし、結果としてそれらの形質転換を導き形質細胞となる。
【0003】
有効な免疫応答の発達におけるCD40:CD154相互作用の重要性は、CD154のコード遺伝子における突然変異が、IgG、IgAおよびIgEを欠くがIgMの過剰産生により特徴づけられ、そして重篤な反復性感染を伴う重篤なヒト型の免疫不全、高IgM症候群(HIM)の原因となるという事実によって示される(Aruffo A. et al, Cell 1993)。
【0004】
ここ数年、多くの研究によりCD40:CD154相互作用が炎症性病因(inflammatory etiogenesis)を伴う多くの過程、例えばアテローム性動脈硬化症、自己免疫疾患、移植後の拒絶反応、移植片対宿主病などに関与することが実証された(Schonbeck U. et al, Circ. Res. 2001;Henn V. et al, Nature 1998;Biancone L. et al, Int. J. Mol. Med. 1999;Buchner K. et al, J. Pathol. 2003)。CD40の発現もしくはそのリガンドCD154の発現または両方の分子の共発現が、多くの腫瘍においても実際に示され、これはこのシグナル伝達経路が腫瘍発生に関与することを示唆するものである(Bussolati B. et al, Int. J. Cancer 2002;Biancone L. et al, J. Immunol. 1999;Cantaluppi V. et al, Int. J. Immunopathol. Pharmacol. 2006;Hill S.C. et al, J. Immunol. 2005;Melichar B. et al, Gynecol. Oncol. 2007;Eliopoulos A.G. et al, Curr. Opin. Pharmacol. 2004)。特に、CD40:CD154シグナル伝達経路が、慢性リンパ性白血病(CLL)の進行に関与すると考えられているのは、腫瘍細胞の生存と内皮細胞の血管新生誘導性の特徴の両方を促進することが示されたからである(Dicker F. et al, Blood 2006)。
【0005】
CD40:CD154相互作用を阻害すること目的とした多くのアプローチが、新規の治療機会を提供するために開発された。特に、有望な結果が、自己免疫疾患、アテローム性動脈硬化症および移植による拒絶反応の異なる実験モデルで得られた。抗CD154モノクローナル抗体に基づくアプローチによるCD40:CD154相互作用の阻害は、自己免疫疾患の多くのモデル、例えば全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、乾癬およびクローン病において臨床改善をもたらした(Boumpas D.T. et al, Arthritis Rheum. 2003; Liossis S.N. et al, Bio Drugs 2004; Daoussis D. et al, Clin. Diagn. Lab. Immun. 2004)。抗CD154抗体は、特許文献においても開示されている。国際公開第WO2005/003175号は、アグリコシル(aglycosyl)抗CD154抗体を開示する;国際公開第WO2006/033702号は、抗CD154抗体およびペプチド、特に抗体軽鎖および重鎖の可変領域を開示する。さらに、多数の書籍により、膵島、皮膚、骨髄、心臓および腎臓移植後のマウスにおけるモノクローナル抗体の使用が、その移植された臓器の延長された存続をもたらすことが実際に示された(Molano R.D. et al, Diabetes 2001; Quezada S.A. et al, Blood 2003; Elster E.A. et al, Transpl. Immunol. 2004; Xu H. et al, J. Clin. Investig. 2006; Snanoudj R. et al, Transpl. Int. 2006; Nanji S.A. et al, Diabetes 2006)。補助刺激の阻止は、補助刺激分子に対するモノクローナル抗体、Fab断片または融合タンパク質を用いて達成しうる。しかしながら、これらのアプローチの結果は、Fc部分の認識に拠る免疫原性作用および血栓形成促進性の副作用の危険性によって(Henn V. et al, Nature 1998)および脆弱な組織浸透性や免疫抑制により、しばしば損なわれる。内在する免疫原性を低下させるために、ヒト化抗体およびミニ抗体が開発されたが、残念ながら親和性が犠牲にされた。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第WO2005/003175号
【特許文献2】国際公開第WO2006/033702号
【非特許文献】
【0007】
【非特許文献1】van Kooten et al, J. Leukoc. Biol. 2000
【非特許文献2】Aruffo A. et al, Cell 1993
【非特許文献3】Schonbeck U. et al, Circ. Res. 2001
【非特許文献4】Henn V. et al, Nature 1998
【非特許文献5】Biancone L. et al, Int. J. Mol. Med. 1999
【非特許文献6】Buchner K. et al, J. Pathol. 2003
【非特許文献7】Bussolati B. et al, Int. J. Cancer 2002
【非特許文献8】Biancone L. et al, J. Immunol. 1999
【非特許文献9】Cantaluppi V. et al, Int. J. Immunopathol. Pharmacol. 2006
【非特許文献10】Hill S.C. et al, J. Immunol. 2005
【非特許文献11】Melichar B. et al, Gynecol. Oncol. 2007
【非特許文献12】Eliopoulos A.G. et al, Curr. Opin. Pharmacol. 2004
【非特許文献13】Dicker F. et al, Blood 2006
【非特許文献14】Boumpas D.T. et al, Arthritis Rheum. 2003
【非特許文献15】Liossis S.N. et al, Bio Drugs 2004
【非特許文献16】Daoussis D. et al, Clin. Diagn. Lab. Immun. 2004
【非特許文献17】Molano R.D. et al, Diabetes 2001
【非特許文献18】Quezada S.A. et al, Blood 2003
【非特許文献19】Elster E.A. et al, Transpl. Immunol. 2004
【非特許文献20】Xu H. et al, J. Clin. Investig. 2006
【非特許文献21】Snanoudj R. et al, Transpl. Int. 2006
【非特許文献22】Nanji S.A. et al, Diabetes 2006
【非特許文献23】Henn V. et al, Nature 1998
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の1つの目的は、CD40:CD154相互作用を阻害でき、該相互作用に依存する生物学的作用を阻止でき、および先行技術、特にモノクローナル抗CD154抗体の使用に関する技術の限界および欠点を克服する試薬を提供することである。
【課題を解決するための手段】
【0009】
この目的は、合成ペプチドによって達成され、それは好ましくは環状であり、ヒトCD154(ヒトCD40Lとも称される)の活性部位と特異的に結合できる。
【0010】
本発明のペプチドは、配列表中の配列番号:13で示されるエプタ(epta)−アミノ酸のCD154−結合性配列を含む。
【図面の簡単な説明】
【0011】
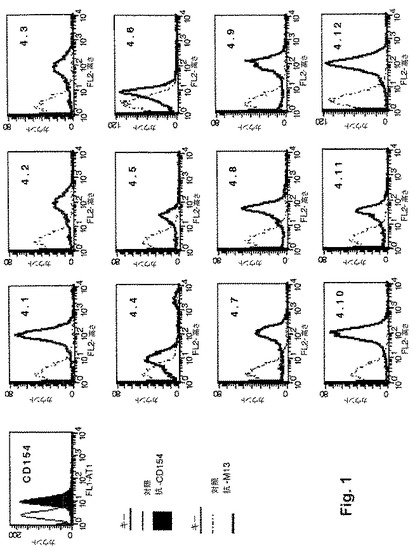
【図1】図1のヒストグラムは、細胞表面上でのヒトCD154発現のためにトランスフェクトされたネズミ骨髄腫細胞系のJ558L細胞に対する、選択されたファージクローンの結合性を示す。
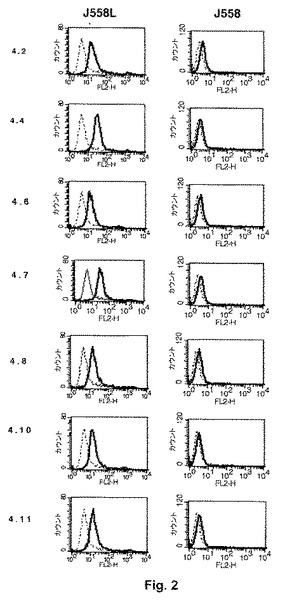
【図2】図2は、ヒトCD154結合性クローンから得られ6個のヒスチジン・テイル(−His6)がタグ付けされた7個のペプチドの、J558L CD154+細胞に対する結合性を示す。
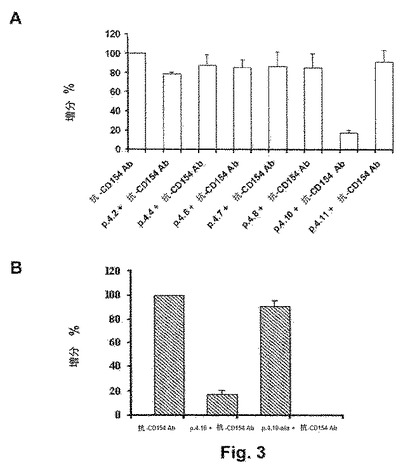
【図3】図3Aは、抗CD154ペプチドとCD154の活性部位に特異的な抗ヒトCD154フルオレセイン複合体化抗体との間の競合を示す。図3Bは、ペプチド4.10と、対照として用いられるペプチド4.10−ala(配列番号:8)との比較を示す。
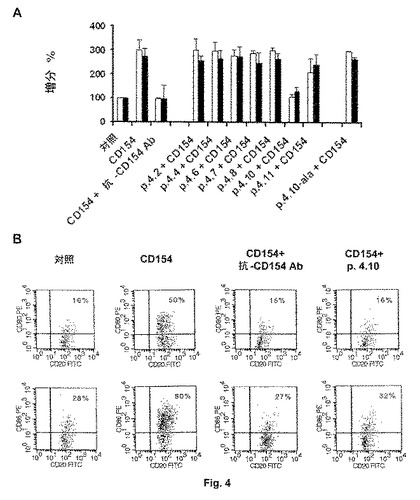
【図4】4Aのヒストグラムは、可溶性の組み換えヒトCD154分子(rhsCD154)を用いた刺激により誘導された活性B化細胞に対する抗CD154ペプチドおよび対照ペプチド4.10−alaの効果を示す。
【図5】図5Aは、刺激しない基底条件下での、IL4およびCD154を用いた72時間の刺激後、ブロッキング抗ヒトCD154抗体と組み合わせたIL4およびCD154を用いる72時間の刺激後、またはペプチド4.10(配列番号:6)と組み合わせたIL4およびCD154を用いる72時間の刺激後の、アイソタイプスイッチを受けてIgGを発現するナイーブB細胞の存在をそれぞれ示す。
【図6】図6は、ペプチド4.10(配列番号:6)の、対照ペプチドの4.10−ala(配列番号:8)の、またはブロッキング抗ヒトCD154抗体の非存在下または存在下での、HUVEC(HUVEC=ヒト臍静脈内皮細胞)の運動性に対するCD154誘導性の刺激効果を示す。
【図7】図7Aは、刺激なしの基底条件下での或いはブロッキング抗CD154抗体またはペプチド4.10(配列番号:6)または対照ペプチドの4.10−ala(配列番号:8)のいずれかの存在/非存在下での、rhsCD154による刺激後に血管様の細胞コードを形成するマトリゲルに撒かれたHUVECの能力を示す。図7B〜Fは、上記の様々な実験条件下、マトリゲル上での細胞コード形成能についての代表的な画像を示す。
【図8】図8は、250μlのマトリゲルと組み合わせ、SCIDマウスの臀部に皮下接種した内皮HUVEC細胞に対する、ペプチド4.10の存在/非存在下でのrhsCD154誘導刺激のインビボ効果を示す。
【図9】図9は、IL−4刺激により誘導されるCLL細胞の生存を阻害する、本発明のペプチド4.10(配列番号:6)の能力を示す。
【図10】図10は、0.5μMのADPを単独で用いて刺激することによる、あるいはrhsCD154を用いて又はペプチド4.10(配列番号:6)と組み合わせたrhsCD154を用いて又はrhsCD154および抗ヒトCD154抗体からなる免疫複合体と組み合わせたrhsCD154を用いて37℃で10分間刺激した後に0.5μMのADPを用いて刺激することによる、ヒト血小板凝集を示す。
【発明を実施するための形態】
【0012】
好ましい実施形態によれば、本発明のペプチドは、7アミノ酸長から30アミノ酸長である。
【0013】
さらにより好ましい実施形態に関して、本発明のペプチドは、配列番号:13で示されるエプタ−アミノ酸のCD154結合性配列の両端の側面にシステイン残基を付加することにより結果として生じる配列表中の配列番号:6で示される、ノナ−アミノ酸配列からなる。この実施形態は、配列番号:6がその末端の2個のシステイン間でジスルフィド結合を形成することによる環化を受けることができる点で特に有利である。
【0014】
本発明のペプチドは、それが投与される患者に望ましくない免疫応答を誘導することがないという利点を有する。
【0015】
さらなる態様に関して、本発明のペプチドは、複数のペプチドコピーを有する多量体ペプチド構造物の形式で提供され、該ペプチドコピーの各々は直鎖もしくは環状のいずれかである。多量体ペプチド構造物は、四量体構造物であることが好ましい。この多量体ペプチド構造物は、複数のペプチドコピーから成り、その各コピーは少なくとも1個のアミノ酸スペーサー、好ましくはG(Gly)残基により少なくとも1個の他のコピーと結合しており、各ペプチドコピーは好ましくは3個のK(Lys)残基からなるアミノ酸コアと直接もしくは間接的に結合している。
【0016】
特に好ましい本発明の多量体ペプチド構造物は、以下の式で表される:
【化1】
[式中、LはLeuであり、PはProであり、TはThrであり、RはArgであり、HはHisであり、MはMetであり、AはAlaであり、GはGlyであり、およびKはLysである]。
【0017】
以上で示される多量体構造物は、CD154−結合性の配列である配列番号:13を直鎖で含む場合でさえ、組み換え体の可溶性CD154によるCD40刺激に対して、60μMの濃度で、ペプチド4.10と同じ阻害活性を発揮することが示された。さらに、四量体構造物は、ペプチド4.10のように、サブリミナル用量(subliminar dose)(0.3U/ml)のトロンビンによるヒト血小板凝集を活性化することを結果として不活性にした。これらの結果は、CD154−結合性の配列(配列番号:13)は結果として環状および直鎖のいずれの場合でもCD154に結合することができ、生物学的機能を発揮することができることを示す。環状ペプチドはそのアミノ酸配列をタンパク質分解(例えば、血漿中での)から保護しうるという利点を有すると同時に、ペプチドの多量体型を設計しうるということは、分子サイズ、クリアランス速度および結果としての半減期を調節することを可能にする。
【0018】
これまでに言及したように、本発明の範囲には、診断および治療応用における本発明に関するペプチドまたは多量体構造物の使用もまた含まれる。
【0019】
さらに、本発明の範囲には、本発明のペプチドまたは多量体ペプチド構造物が診断剤(例えば、インビボもしくはインビトロ・アッセイのための検出可能な分子)または治療剤(例えば、活性化内皮細胞、腫瘍細胞または活性化の際に細胞表面にCD154を発現できる他のいずれかの細胞に対して、細胞傷害効果を発揮することができる抗炎症剤、抗腫瘍剤または細胞傷害剤)と連結され、インビボもしくはインビトロでの検出を可能にするコンジュゲート、ならびにCD40:CD154相互作用に関係する多くの疾患もしくは障害の処置のための医薬を製造するためのコンジュゲートが含まれる。そのような疾患または状態の例としては、炎症性疾患(例えばアテローム性動脈硬化症、自己免疫疾患、移植による拒絶反応、関節炎を伴う炎症、接触皮膚炎、高IgE症候群、炎症性腸疾患、アレルギー性喘息を含むアレルギー、特発性炎症性疾患)、腫瘍性疾患(例えば慢性リンパ性白血病)、移植片対宿主病が含まれる。自己免疫疾患の例としては、全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、クローン病、乾癬がある。本発明のペプチドまたは多量体構造物あるいはコンジュゲートは、免疫寛容の誘導のための使用にも適している。
【0020】
合成ペプチドは、分子間の相互作用を阻害するための、mAbの代替的アプローチを示す。合成ペプチドの使用を下支えする理論は、タンパク質はただ外部の狭い領域だけを介してその生物学的作用を発揮するのであり、そのわずか2個、3個のアミノ酸だけが特有の受容体機能に重要であるというものである。従って、必須のアミノ酸残基を保持するより短い立体構造的に正しい形式で、その領域に対応した配列を合成することができる。このように、標的とする受容体の活性部位を特異的に認識するが、生物学的に不活性であり立体的な障害を与え、それにより受容体とその天然タンパク質との間の相互作用を妨害する短いペプチドを得ることが可能である。アゴニストと受容体との間の相互作用を模倣することができる、そのようなペプチドの主な理論的な利点は、その限られた大きさであり、それは該ペプチドを水中で容易に可溶化させ、非免疫原性とさせ、該ペプチドが長期間投与されることを可能にする(Allen S.D. et al, J. Peptide Res. 2005; Ladner R.C. et al, DDT 2004)。
【0021】
しかしながら、CD40:CD154相互作用を効果的に阻害することができるアミノ酸配列を同定することは、自明な結果ではない。以下でさらに詳細に例示されるように、本発明のペプチド配列(配列番号:13または配列番号:16)の単一アミノ酸残基のモディファイでさえ、事実上、ヒトCD154の活性部位に対する結合能を失わせることとなり、結果としてペプチドの阻害能が無くなることとなる。
【0022】
上述のように、本発明に関するペプチドは、配列表中の配列番号:13で示されるアミノ酸配列または配列番号:16で示されるペプチド配列を含む。該ペプチドは、ヒトCD154の活性部位に対して非常に高い結合特異性を示す。該ペプチドは、CD154の活性部位を認識し結合することができ、結果としてインビボおよびインビトロの両方でその受容体を効果的に阻止でき、該相互作用に起因する生物学的作用を阻害することとなる。以下の説明において、配列番号:6からなる本発明のペプチドは、「ペプチド4.10」で示される。
【0023】
以下の実験の項で例示される結果は、本発明のペプチド4.10が、組み換え体の可溶型のヒトCD154(組み換えヒト可溶性CD154、またはrhsCD154)による刺激によってB細胞上のCD40の活性化を妨害できることを示す。CD40:CD154相互作用に対して発揮される干渉作用は、B細胞の活性化および結果として生じる免疫応答の両方を妨げる。本発明者らは、ペプチド4.10が、B細胞に対する他の補助刺激分子、例えばCD80およびCD86の誘導を阻害し、同様に免疫グロブリンのアイソタイプスイッチの誘導も阻害できることを実際に示した。さらに、ペプチド4.10は、遊走するおよび血管様構造物を形成する細胞の能力として評価される内皮細胞のrhsCD154誘導型の活性化を、インビトロで阻止することができる。ペプチド4.10のそのような抗血管形成活性は、SCIDマウスでのネズミ血管形成マトリゲル・モデルにおいて、インビボでさらにはっきりと確認でき、そのことを以下でより詳細に記載する。最後に、本発明者らにより得られた予備的なデータは、ペプチド4.10がCLL患者由来のB細胞に劇的なアポトーシスを誘導することができることを示す。
【0024】
短いペプチドを含むコンジュゲートおよび医薬組成物の調製および投与技術のさらなる詳細は、先行技術において知られており;それは本発明の理解に本質的なものではないため、その詳述は本明細書には必要ない。
【0025】
以下の実験の項で説明するように、本明細書は、ヒトCD154に特異的に向けられる7個のアミノ酸配列の同定を説明する。特に、ヒトCD154に結合することができる7個の環状エプタ−ペプチドが選択され、特徴づけられた。ヒトCD154のN末端ドメインを含む融合タンパク質を、ペプチドを選択するために用いた。7個の選択された及び特徴づけられたエプタ−ペプチドの中で、CD40:CD154相互作用を阻害できることが実際に確認された唯一のものが、本発明のペプチド4.10である。さらに、ペプチド4.10のアミノ酸配列を、ペプチド4.10の有用な代替形態を選りすぐる試みの中で、1個もしくは2個のアミノ酸残基を変異によりモディファイした。しかしながら、その代替形態はいずれも、CD40:CD154相互作用を阻害する能力を維持せず、その相互作用に関係する生物学的作用も維持しなかった。
【0026】
以下で示される添付図および表を含み、以下の詳細な記載は、単なる例示として提供される。
【実施例1】
【0027】
図1のヒストグラムは、細胞表面上でのヒトCD154発現のためにトランスフェクトされたネズミ骨髄腫細胞系のJ558L細胞に対する、選択されたファージクローンの結合性を示す。この細胞に対して選択されたファージの結合性は、J558L細胞をファージとインキュベートさせ、次いで抗M13モノクローナル抗体とインキュベートさせて、フローサイトメトリーにより実際に示された。各実験条件について10,000事象を分析し、その値は3回の別個の実験の平均値および標準偏差である。12個の選択したクローンはすべてJ558L細胞と結合する。
【0028】
表1は、ヒトCD154結合性クローンから得られたペプチド挿入体の7個のアミノ酸配列を示す。4.1、4.5および4.9と称するクローンの挿入体は組み換えにより失われ、一方で4.2および4.3と称するクローンの挿入体ならびに4.7および4.12と称するクローンの挿入体は同一であることが確認された。
【0029】
図2は、ヒトCD154結合性クローンから得られ6個のヒスチジン・テイル(−His6)がタグ付けされた7個のペプチドの、J558L CD154+細胞に対する結合性を示す。この細胞へのペプチドの結合性は、フローサイトメトリーにより実際に示された。この細胞を、−His6タグ付きペプチドの各々とインキュベートし、次いで−His6テイルに特異的なフィコエリトリン複合体化(conjugated)二次抗体とインキュベートした。点線のヒストグラムは、アイソトープ対照(isotopic control)であり、一方、黒線はCD154特異的ペプチドの結合強度である[4.2=4.3=CPSGHTKAC(配列番号:1)、4.4=CGTHSSRIC(配列番号:2)、4.6=CLGTQNKEC(配列番号:3)、4.7=4.12=CTPGKPHSC(配列番号:4)、4.8=CKAASANIC(配列番号:5)、4.10=CLPTRHMAC(配列番号:6)および4.11=CLSAVHNMC(配列番号:7)]。各実験条件について10,000事象を分析し、その値は3回の別個の実験の平均値および標準偏差である。ペプチドはすべてJ558L CD154+細胞上のCD154と特異的に結合することができ、対照体のJ558細胞と結合できない。
【0030】
図3Aは、抗CD154ペプチドとCD154の活性部位に特異的な抗ヒトCD154フルオレセイン複合体化抗体との間の競合を示す。ヒトCD154をトランスフェクトしたネズミ骨髄腫細胞であるJ558Lを、6個のヒスチジン・テイル(−His6)タグ付きの7個の抗CD154ペプチドと一緒にインキュベートし、次いで抗CD154フルオレセイン複合体化抗体と一緒にインキュベートした。J558L細胞表面に発現されたCD154との結合に関して抗CD154抗体と競合するペプチドの能力は、フローサイトメトリーにより、抗ヒトCD154フルオレセイン複合体化抗体を単独で用いてインキュベートした細胞により示される陽性対照と比較した蛍光強度の低下として評価した。ペプチド4.10(配列番号:6)が、抗ヒトCD154抗体と競合でき、その結合性を有意に示すことができた唯一のものであることは、図から明らかである。図3Bは、ペプチド4.10と、対照として用いられるペプチド4.10−ala(配列番号:8)との比較を示す。対照ペプチドのアミノ酸配列は、ペプチド4.10のアミノ酸配列とは、5番目のアミノ酸であるアルギニン残基がアラニン残基に置換されている点で異なっている。図3Bで明らかに示されるように、このアルギニン残基を欠くことで、ヒトCD154に対して抗CD154フルオレセイン複合体化抗体と競合する該ペプチドの能力が完全に不活化する結果となるため、そのアルギニン残基が必須であることは明らかである。
【0031】
4Aのヒストグラムは、可溶性の組み換えヒトCD154分子(rhsCD154)を用いた刺激により誘導された活性B化細胞に対する抗CD154ペプチドおよび対照ペプチド4.10−alaの効果を示す。ブロッキング抗ヒトCD154抗体または7個の選択されたペプチドのいずれか1個の非存在下もしくは存在下で、B細胞をrhsCD154を用いて48時間刺激した。末梢B細胞の活性化は、フローサイトメトリーにより、補助刺激分子CD80およびCD86の膜発現で評価した。図4Aで明らかに示されるように、ヒト可溶性CD154(rhsCD154)を用いた48時間のB細胞の刺激は、細胞表面でのCD80およびCD86の強い発現を誘導する。対照的に、rhsCD154をブロッキング抗ヒトCD154抗体と又はペプチド4.10(配列番号:6)と組み合わせて用いた場合、その活性化は抑制される。残りのペプチドはいずれも、rhsCD154誘導型のB細胞の活性化を有意に阻害することはできない。図4Bは、サイトメトリー分析の詳細を示す。
【0032】
図5Aは、刺激しない基底条件下での、IL4およびCD154を用いた72時間の刺激後、ブロッキング抗ヒトCD154抗体と組み合わせたIL4およびCD154を用いる72時間の刺激後、またはペプチド4.10(配列番号:6)と組み合わせたIL4およびCD154を用いる72時間の刺激後の、アイソタイプスイッチを受けてIgGを発現するナイーブB細胞の存在をそれぞれ示す。ペプチド4.10(配列番号:6)は、ちょうど抗CD154抗体のように、rhsCD154およびIL4による刺激によって誘導される免疫グロブリンのアイソタイプスイッチを強力に阻害することができる。図5Bは、サイトメトリー分析の詳細を示す。
【0033】
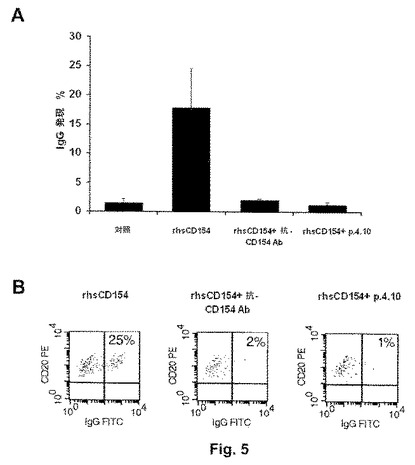
図6は、ペプチド4.10(配列番号:6)の、対照ペプチドの4.10−ala(配列番号:8)の、またはブロッキング抗ヒトCD154抗体の非存在下または存在下での、HUVEC(HUVEC=ヒト臍静脈内皮細胞)の運動性に対するCD154誘導性の刺激効果を示す。特に、図6Aは、様々な実験条件下で4時間の期間にわたり、細胞遊走のタイムラプス・キネティクスを示し;図6Bは、上記の実験条件下で刺激後30分間で細胞の遊走速度に観察された差異を示し;図6Cは、タイムラプス分析の代表的な画像を示す。対照ペプチドの4.10−ala(配列番号:8)とは反対に、ペプチド4.10(配列番号:6)は、内皮細胞のrhsCD154誘導性の運動性を阻害する。
【0034】
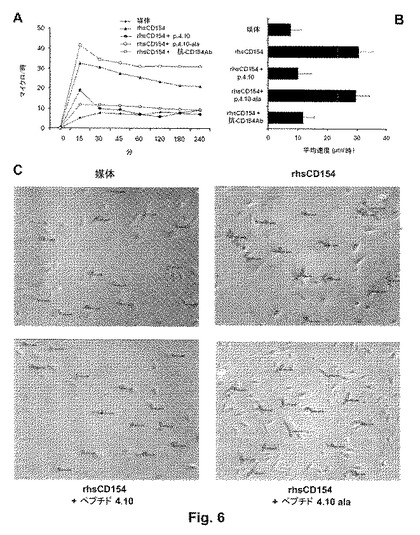
図7Aは、刺激なしの基底条件下での或いはブロッキング抗CD154抗体またはペプチド4.10(配列番号:6)または対照ペプチドの4.10−ala(配列番号:8)のいずれかの存在/非存在下での、rhsCD154による刺激後に血管様の細胞コードを形成するマトリゲルに撒かれたHUVECの能力を示す。図7B〜Fは、上記の様々な実験条件下、マトリゲル上での細胞コード形成能についての代表的な画像を示す(B=刺激なしの対照;C=rhsCD154;D=rhsCD154+4.10ペプチド;E=rhsCD154+4.10−alaペプチド;F=rhsCD154+ブロッキング抗CD154抗体)。本発明のペプチド4.10(配列番号:6)は、ペプチド4.10−ala(配列番号:8)と対照的に、細胞コードを形成する内皮細胞のrhsCD154誘導性の能力を阻害する。
【0035】
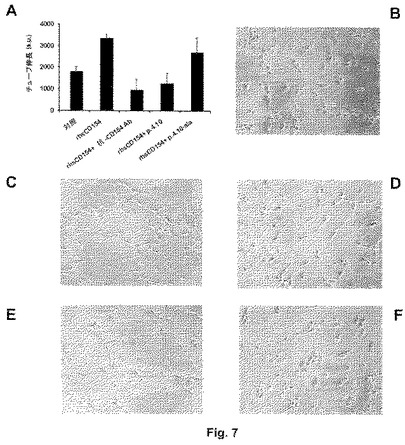
図8は、250μlのマトリゲルと組み合わせ、SCIDマウスの臀部に皮下接種した内皮HUVEC細胞に対するペプチド4.10の存在/非存在下でのrhsCD154誘導刺激のインビボ効果を示す。接種の7日後に、マウスを屠殺し、移植片を取り出し、10%ホルマリン溶液中で固定化し、パラフィンに包埋し、組織分析のために調製した。図8Aは、ヘマトキシリン−エオシン染色した組織切片において、基底条件下およびそれ以外の上記実験条件下での血管形成領域の量子化(quantization)を示す。図8Bおよび8Cは、rhsCD154を単独で用いた及びペプチド4.10(配列番号:6)と組み合わせてrhsCD154を用いた内皮細胞の刺激により得られた血管形成の代表的な2枚の画像をそれぞれ示す。各実験条件に対して、6匹のSCIDマウスを用いた。ペプチド4.10は、インビボでのrhsCD154誘導性の血管形成を完全に阻害する。
【0036】
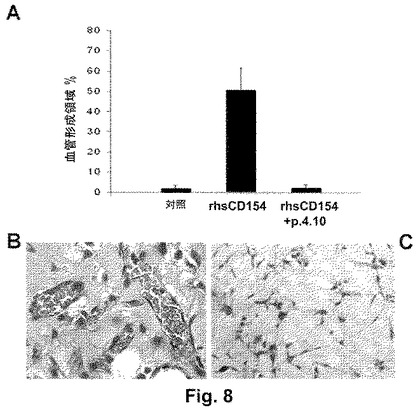
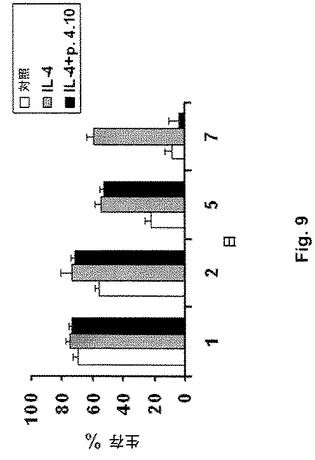
図9は、IL−4刺激により誘導されるCLL細胞の生存を阻害する、本発明のペプチド4.10(配列番号:6)の能力を示す。
【0037】
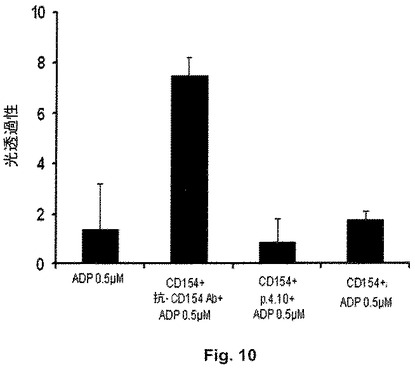
図10は、0.5μMのADPを単独で用いて刺激することによる、あるいはrhsCD154を用いて又はペプチド4.10(配列番号:6)と組み合わせたrhsCD154を用いて又はrhsCD154および抗ヒトCD154抗体からなる免疫複合体と組み合わせたrhsCD154を用いて37℃で10分間刺激した後に0.5μMのADPを用いて刺激することによる、ヒト血小板凝集を示す。血小板凝集は、血小板凝集計を用い、対照と比較した光透過性の増加として測定した。rhsCD154と組み合わせたヒトCD154抗体によるヒト血小板のプライミングは、0.5μMのADPを用いた刺激により血小板凝集を増大させる。反対に、単独のrhsCD154またはペプチド4.10と組み合わせたrhsCD154は、0.5μMのADPを用いた刺激により誘導される血小板凝集レベルに影響を及ぼさない。これらの結果は、本発明のペプチド4.10(配列番号:6)とCD154分子との相互作用は、血小板凝集を誘導することはないが、これとは対照的に、抗CD154抗体との相互作用では血小板凝集が誘導される。
【0038】
以下の実験の項は、単なる例示として提供され、添付する特許請求の範囲により規定される本発明の範囲を限定することは意図しない。
【0039】
実験の項
この研究を通して、7個のアミノ酸残基とジスルフィド結合による環化を可能にする両端で側面に位置する2個のシステインを含むペプチド配列パネルから成り、M13ファージで発現され、キャプシドpIIIファージタンパク質と遺伝子工学的に融合され、そのファージ表面に無作為に発現される、ペプチドライブラリー(ペプチドの多様性<109)を用いた。このファージライブラリーを、既知のモデル(Hetian L. et al, J. Biol. Chem. 2002)から本発明者らにより発展されたバイオパニング・インビトロ・モデルによってスクリーニングした。ファージ(200μlのTBS中、1×1011CFU)を、ヒト組み換えCD154と一緒に室温で1時間インキュベートして、その後プレートに撒いた。このファージをTBS−Tを用いて数回洗浄してCD154と結合していない非特異的なファージを取り除き、CD154と結合したファージを、100μlの0.2M グリシン、pH2.2を用いて溶出することにより回収し、15μlの1M Tris−HCl、pH9.1を用いて10分間中和した。ファージ数は、溶出液の段階希釈物をLB寒天培地(アガロース7g/リッター、MgCl2・6H2O 1g;Sigma)上の宿主のテトラサイクリン耐性の大腸菌ER2738細胞(New England Biolabs, Hitchin, U.K.)に加え、テトラサイクリン(Kramel Biotech, Cramlington, U.K.)存在下のIPTG/X−Gal LB寒天培地に撒いて、滴定することにより評価した。
【0040】
37℃で12時間のインキュベーション後に、青プラークとしてプレート上で目視できるファージコロニーの数を計数した。評価したファージを、テトラサイクリン存在下のIPTG/X−Gal LB寒天培地のプレート上でファージを培養することにより増幅し、3.3%のポリエチレングリコール8,000/0.4 NaCl(Sigma)中で遠心分離することにより沈殿させた。次いで、そのファージを上記のように滴定し、ヒト組み換えCD154で被覆した新しいプレート上で再度インキュベートした。スクリーニング過程および増幅過程を4回繰返して、ヒトCD154特異的なファージクローンのライブラリーを豊富化した。4回目の増幅ラウンドの後に、12個のファージクローンを無作為に選び出し、個々に増幅し、細胞表面で発現するCD154を認識するその能力を試験した。選択された個々のクローンの特異的な結合性は、参照細胞系としてJ558Lと称されるヒトCD154の発現のためにトランスフェクトされたネズミ骨髄腫細胞系を用いたフローサイトメトリーにより、インビトロで実際に示した。室温で30分間のインキュベーション後のファージ(1011cfu)のJ558L細胞に対する結合性は、その細胞をファージタンパク質M13に対するモノクローナル抗体(Pharmacia, Uppsala, Sweden)と一緒にインキュベートし、続いて抗マウス・フィコエリトリン複合体化抗体(Sigma)と一緒にインキュベートする間接免疫蛍光法によるフローサイトメトリーによって検出した。図1で示されるように、12個のファージクローンはすべて、ヒトCD154発現細胞(J558L)と特異的に結合できたが、その対照体であるCD154−陰性(J558)細胞とは結合しなかった。
【0041】
この12個のファージクローンのペプチド挿入体の配列決定により、表Iに列記される7個の異なる配列が得られた。ペプチドを合成し、インビトロおよびインビボでの局在を容易にするためにビオチン(−bio)または6−ヒスチジン・テイル(−his6)と複合体化させた。同様に、このペプチドのJ558L細胞に対する結合性は、細胞をhis6タグ付きペプチド(60μMの−his6ペプチド)と一緒に室温で30分間インキュベートし、次いで抗ポリヒスチジン−フィコエリトリン複合体化抗体(Sigma)と一緒に4℃で30分間インキュベートして、フローサイトメトリーにより実際に示した。7個の選択されたペプチドはすべて、J558L CD154+細胞に対して良好な親和性を示した。さらに、該ペプチドと、CD154の活性部位を特異的に認識する抗ヒトCD154フルオレセイン複合体化抗体(Serotec)との競合アッセイを行った。J558L細胞を、his6−ペプチドと一緒に室温で30分間インキュベートし、次いで、抗ヒトCD154フルオレセイン複合体化抗体と一緒にさらに30分間室温でインキュベートした。図3Aは、ペプチド4.10が、抗ヒトCD154フルオレセイン複合体化抗体とJ558L細胞との相互作用を有意に阻害できる唯一のものであることを示す。ペプチド4.10のアミノ酸配列中の、CD154との結合に必須のアミノ酸残基を特定するために、3種の異なるペプチドを合成し、その各々は、点突然変異を有しており、それらは4番目、5番目、6番目のアミノ酸配列がそれぞれアラニン残基に置換されることによりペプチド4.10のアミノ酸配列とは異なっている。
【0042】
変異ペプチドは、J558L細胞表面に発現されたヒトCD154を認識することができなかった(データは示さない)。最も低い反応性のペプチド、4.10−alaと称するアミノ酸配列CLPTAHMAC(配列番号:8)を有するペプチドを、対照ペプチドとして選んだ(図3B)。
【0043】
同様の結果が、表IIで列記されるアミノ酸配列を有する4個の改変されたペプチド4.101、4.102、4.103および4.104を用いて得られた。これらのペプチドはいずれも、CD154+ J558Lの細胞表面に発現されたヒトCD154と結合しないことが示された(データは示さない)。
【0044】
B細胞は、その細胞膜上でCD40分子を構成的に発現することが知られている(van Kooten C. et al, J. Leukoc. Biol. 2000)。CD154を発現する活性化T細胞によるCD40の刺激は、結果としてB細胞の活性化、増殖およびアイソタイプスイッチを引き起こす。本発明者らは、フィコール・グラジエント遠心分離法および免疫磁気分離法(MACSシステム−Milteniy)により、健常ドナーの末梢全血から分離したB細胞を用いた。次いで、それにより得られた休止期のB細胞を、可溶型のヒト組み換えCD154(rhsCD154)を用いて刺激し、細胞膜上での補助刺激分子CD80(B7−1)およびCD86(B7−2)の活性化および発現を誘導した。rhsCD154(100ng/ml)による48時間の刺激後に、末梢B細胞の細胞表面上にCD80およびCD86の膜発現において劇的な増加が観察された。この発現は、それぞれ、基底レベルと比較して300%および274%にまで増加した。対照的に、リンパ球刺激の前にペプチド4.10(配列番号:6)と一緒にrhsCD154(100ng/ml)で37℃にて15分間あらかじめインキュベートすることで、B細胞刺激およびその関連するCD80およびCD86の発現を阻止した。ペプチド4.10(配列番号:6)の阻害作用は、60μMの濃度でピークに達し、CD80およびCD86の発現はそれぞれ107%および130%にまで低下した(図4Aおよび4B)。その効果はより低い濃度(30μM)の場合でさえ観察することが可能であった。
【0045】
反対に、同じ濃度(60μM)の他のペプチドと一緒にrhsCD154(100ng/ml)を用いてあらかじめインキュベートしても、rhsCD154による刺激によって誘導されるCD80およびCD86の発現に有意な影響はなかった。より高い濃度(250μM)では、ペプチド4.6(配列番号:3)および4.11(配列番号:7)はまた、B細胞活性化の阻害を示した:特に、ペプチド4.6(配列番号:3)は、CD80の発現を117%に及びCD86の発現を122%に低下させたが、一方で、ペプチド4.11(配列番号:7)の効果は安定した再現性はなかった。反対に、対照ペプチドの4.10−ala(配列番号:8)は、250μMの濃度でさえrhsCD154(100ng/ml)と一緒にあらかじめインキュベートしても、rhsCD154による刺激によって誘導されるCD80およびCD86の発現には影響せず、またB細胞の活性化を有意に低下させることもなく、さらにCD80およびCD86の発現レベルを低下させることもなかった(図4A)。ブロッキング対照として、実施した全ての実験でB細胞の刺激前に、rhsCD154(100ng/ml)をブロッキング抗ヒトCD154抗体(Alexis)と一緒に37℃で15分間あらかじめインキュベートすることで、CD40媒介型のB細胞の活性化を完全に阻害し、CD80およびCD86の発現を基底レベルにまで低下させた(図4Aおよび4B)。この結果は、5回の別個の実験の平均値±標準偏差である。
【0046】
同様の結果はまた、ヒト脾臓フラグメントから単離されたB細胞を用いて得られた。さらに、ナイーブB細胞集団で行われた別の実験でも、ペプチド4.10がCD40媒介型のB細胞Igのアイソタイプスイッチを効果的に阻害することが示された。これらの実験は、免疫磁気分離法(MACSシステム Milteniy)によりB細胞プールから成熟リンパ球を失わせることによる陰性選択によって末梢血から得られたCD27−ナイーブB細胞で行った。本発明者らは、ナイーブB細胞を、IL−4(0.4ng/ml)およびrhsCD154(100ng/ml)の存在下の低グルコースDMEM(Sigma)中で96時間培養した場合に、膜IgGを発現するB細胞の数に有意な増加(25%)が誘導されたことを実際に示した。対照的に、ペプチド4.10(配列番号:6)と組み合わせたIL4(0.4ng/ml)およびrhsCD154(100ng/ml)を用いて刺激されたB細胞は非常に低い割合でアイソタイプスイッチを受けた(図5Aおよび5B)。同様に、ブロッキング抗ヒトCD154抗体と組み合わせたIL4(0.4ng/ml)およびrhsCD154(100ng/ml)によるナイーブB細胞の刺激では、アイソタイプスイッチは誘導されなかった(図5Aおよび5B)。
【0047】
炎症過程は、ひとたび誘発されると特定の受容体またはリンパ球補助刺激分子、例えばCD40およびそのリガンドCD154を発現する免疫系以外の細胞、例えば血管壁の内皮細胞により保存されうることが知られている。かくして、本発明者らは、ペプチド4.10(配列番号:6)が、リンパ球刺激および/または成熟を阻害することに加えて、内皮細胞の細胞表面に発現されるCD40分子の刺激を阻止して、炎症を制限できることを実際に示した。本発明者らは、rhsCD154(100ng/ml)の投与による内皮CD40刺激によって誘導される血管新生誘導効果は、単一細胞の遊走および基底マトリクス(マトリゲル)上で血管様の細胞コードを形成する細胞の能力の両方のようにインビトロ評価され、本発明の抗CD154ペプチド4.10(配列番号:6)を用いてrhsCD154(100ng/ml)を37℃で15分間あらかじめインキュベートすることにより阻害されたことが実際に示された。特に、細胞遊走実験は、20%血清存在下の完全内皮細胞培養液中4×103細胞密度でゼラチン上に撒かれたHUVEC細胞を用いて実施した。後日、その内皮培地は、rhsCD154(100ng/ml)存在下もしくは非存在下の減少した血清濃度(5%)を含む低グルコースDMEM(Sigma)に交換した。
【0048】
さらに、阻害効果を評価するために、rhsCD154(100ng/ml)を、ペプチド4.10(配列番号:6)と又は対照ペプチドの4.10−ala(配列番号:8)と又はブロッキング抗ヒトCD154抗体と一緒に37℃で15分間インキュベートした。次いで、フラスコを、恒温チャンバー(約37℃)を備えた位相差倒立顕微鏡下で4時間、10×倍率で観察した。細胞を、顕微鏡に繋がれたカメラを用いて規則的に15分間隔で4時間の全期間にわたって撮影した。各細胞の遊走速度は、画像分析ソフトウェアを用いて、各単一フレームにおいて各単独細胞それぞれの核の位置に基づいて計算し、結果として、平均の細胞遊走速度(±標準偏差)を各実験条件ごとに計算した。図6Aおよび6Bで示されるように、rhsCD154(100ng/ml)による内皮CD40の刺激は、細胞の基底の運動性(1時間あたり常に12μm未満)において約350%(1時間あたり42μm)の増加を誘導し、一方でrhsCD154(100ng/ml)をペプチド4.10(配列番号:6)と共に37℃で15分間あらかじめインキュベートすることで、ならびにブロッキング抗ヒトCD154抗体と一緒にあらかじめインキュベートすることで、細胞の運動性は基底レベルまで顕著に低下した。反対に、対照ペプチド4.10−ala(配列番号:8)は、内皮CD40の刺激に影響を及ぼさなかった(図6A、6B、6C)。
【0049】
rhsCD154(100ng/ml)によるヒト内皮CD40刺激を妨げるペプチド4.10の能力のさらなる確認として、HUVEC(3.5×104)をインビトロ血管新生アッセイにかけた。HUVECを、5%血清が補充された基底RPMI培地中、巨大な細胞の組織体の形成を促進することができる腫瘍マトリックス(tumor matrix)(マトリゲル、BD)上に撒いた。37℃、5%CO2で4時間インキュベーションして、細胞を位相差倒立顕微鏡下で観察した。図7Aおよび7Bは、rhsCD154(100ng/ml)による刺激が、4時間後にすでに複雑な内皮ネットワークの形成を誘導したが、一方でrhsCD154(100ng/ml)をペプチド4.10(配列番号:6)と一緒に、ならびにブロッキング抗ヒトCD154抗体と一緒に37℃で15分間あらかじめインキュベートすることで、刺激を完全に無効にした(図7A、7Dおよび7F)。反対に、rhsCD154(100ng/ml)を対照ペプチドの4.10−ala(配列番号:8)と一緒に37℃で15分間あらかじめインキュベートすることでは、血管様の細胞コードを形成する細胞の能力に影響を及ぼさなかった(図7E)。他の抗ヒトCD154ペプチド、例えば活性部位と特異的に結合しない4.6(配列番号:3)および4.11(配列番号:7)は、内皮のCD40刺激を有意に低下させることはできなかった(データは示さない)。
【0050】
最後に、ペプチド4.10の抗血管形成作用を、インビボ血管形成アッセイで試験した。200μlのHANK溶液中に再懸濁した内皮HUVEC細胞(2×106)を、rhsCD154(100ng/ml)を含む液状の500μlのマトリゲルと混合し、ペプチド4.10(配列番号:6)の存在下および非存在下でSCIDマウスの右側の臀部および左側の臀部にそれぞれ皮下接種した。6日後に、マウスを屠殺し、マトリゲルプラグを回収し、10%ホルマリン溶液中で少なくとも24時間固定化し、次いで免疫組織分析のために処理した。HUVECおよびrhsCD154移植片のヘマトキシリン−エオシン染色した組織切片は、強い血管形成を示し(図8Aおよび8B)、sCD154をペプチド4.10(配列番号:6)であらかじめインキュベートした接種では著しく阻害されたか又は完全に見られず、そこでは大部分の細胞がアポトーシスを受けていた(図8Aおよび8C)。
【0051】
ペプチド4.10(配列番号:6)は、インビボで得られたCLL腫瘍細胞のIL−4存在下での培養による生存を阻害できることもはっきり示した。CLL細胞のアポトーシスに対する耐性および増強された生存は、慢性リンパ性白血病患者におけるインビボでのこれらの腫瘍細胞の伸展(expansion)によるものと考えられる。これらの結果は、CD40とそのリガンドであるCD154との相互作用が、CLL細胞の生存を刺激するのに重要であること、およびCD40:CD154相互作用のCD154の活性部位を認識することができるペプチドによる阻害が、これらの細胞の生存を阻害しうることを示す(図9)。
【0052】
最後に、ペプチド4.10の主な利点の1つは、ヒト血小板に対する反応性を欠いていることである。これまで、有望な抗CD154モノクローナル抗体を試験するために実施された多くの臨床試験が、劇的な血栓形成促進性の副作用により突然中断されたことが知られている(Kelsoe G. et al, J. Clin. Invest. 2003)。仮説は、抗CD154抗体が、免疫グロブリンFc部分に対する受容体(FcR)を発現するヒト血小板表面において交差反応を誘発し、活性化後にCD154分子を誘発する、というものである。したがって、本発明者らは、血小板凝集アッセイにより、ペプチド4.10の血小板に対する作用を試験した。このような実験は、磁気攪拌器およびリアルタイム光学密度計を備えた恒温凝集計を使用して行った。血小板の凝集は、参照試料(ブランク)に対する光学密度の変化として評価した。この実験は、5U/mlのヘパリンを含む試験管に収集された健常ドナーの末梢全血から抽出された血小板で行った。特に、多血小板血漿(platelet enriched plasma:PRP)は900rpmで20分間の全血の遠心分離により得られ、一方で参照試料として用いられる少血小板血漿(platelet poor plasma:PPP)は3000rpmで10分間の別の遠心分離後の上清を回収することにより得られた。Langer(Langer F. et al, Thromb. Haemost. 2005)により既に示されたように、血小板を組み換えヒト可溶性CD154および抗CD154モノクローナル抗体からなる免疫複合体と一緒に37℃で10分間インキュベートすることによるプライミングは、0.5μMのADPによる次の刺激により誘導される血小板の凝集を増強した(図10)。対照的に、CD154単独による又はペプチド4.10(60μM)との組み合わせによるプライミングは、0.5μMのADPによる刺激により誘導される血小板の凝集を増強しなかった(図10)。同様の結果が、0.3U/mlのトロンビンを用いて血小板凝集を誘導することで得られた。
【0053】
内皮細胞は、特に炎症部位で、その表面に高頻度でCD40を発現するため、および血管形成は幾つかの炎症過程および炎症性疾患の発症において主要な役割を果たしうるため、本発明の4.10ペプチドの上記特性により、該ペプチドは見込みのある抗炎症アプローチの開発に関する非常に有望な分子である。さらに、4.10ペプチド(配列番号:6)により発揮されるリンパ球活性化における阻害作用は、移植または腫瘍細胞増殖がCD40による刺激に起因する抗アポトーシス作用によって支援される腫瘍性疾患、例えば慢性リンパ性白血病(CLL)に対する免疫抑制治療として、この分子の見込みのある使用を提供する。
【0054】
【表1】
(*)本発明に関するペプチド
【0055】
【表2】
【0056】
参考文献
1. van Kooten, C., and J. Banchereau. 2000. CD40-CD40 ligand. J. Leukoc. Biol. 67: 2-17.
2. Aruffo, A., M. Farrington, D. Hollenbaugh, X. Li, A. Milatovich, S. Nonoyama, J. Bajorath, L. S. Grosmaire, R. Stenkamp, M. Neubauer, et al. 1993. The CD40 ligand, gp39, is defective in activated T cells from patients with X-linked hyper-IgM syndrome. Cell. 72: 291-300.
3. Schonbeck, U., and P. Libby. 2001. CD40 signaling and plaque instability. Circ. Res. 89: 1092-1103.
4. Henn, V., J. R. Slupsky, M. Grafe, I. Anagnostopoulos, R. Forster, G. Muller-Berghaus, and R. A. Kroczek. 1998. CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells. Nature. 391: 591-594.
5. Biancone, L., V. Cantaluppi, and G. Camussi. 1999. CD40-CD154 interaction in experimental and human disease. Int. J. Mol. Med. 3: 343-353.
6. Buchner, K., V. Henn, M. Grafe, O. J. de Boer, A. E. Becker, and R. A. Kroczek. 2003. CD40 ligand is selectively expressed on CD4+ T cells and platelets: implications for CD40-CD40L signalling in atherosclerosis. J. Pathol. 201: 288-295.
7. Bussolati, B., S. Russo, I. Deambrosis, V. Cantaluppi, A. Volpe, U. Ferrando, and G. Camussi. 2002. Expression of CD154 on renal cell carcinomas and effect on cell proliferation, motility and platelet-activating factor synthesis. Int. J. Cancer. 100: 654-661.
8. Biancone, L., V. Cantaluppi, M. Boccellino, L. Del Sorbo, S. Russo, .A. Albini, I. Stamenkovic, and G. Camussi. 1999. Activation of CD40 favors the growth and vascularization of Kaposi's sarcoma. J. Immunol. 163: 6201-6208.
9. Cantaluppi, V., M. C. Deregibus, L. Biancone, I. Deambrosis, B. Bussolati, A. Albini, and G. Camussi. 2006. The expression of CD154 by Kaposi's sarcoma cells mediates the anti-apoptotic and migratory effects of HIV-1-TAT protein. Int. J. Immunopathol. Pharmacol. 19: 81-96.
10. Hill, S. C., S. J. Youde, S. Man, G. R. Teale, A. J. Baxendale, A. Hislop, C. C. Davies, D. M. Luesley, A. M. Blom, A. B. Rickinson, L. S. Young, and A. G. Eliopoulos. 2005. Activation of CD40 in cervical carcinoma cells facilitates CTL responses and augments chemotherapy-induced apoptosis. J. Immunol. 174: 41-50.
【0057】
11. Melichar, B., R. Patenia, S. Gallardo, K. Melicharova, W. Hu, and R. S. Freedman. 2007. Expression of CD40 and growth-inhibitory activity of CD40 ligand in ovarian cancer cell lines. Gynecol. Oncol. 104: 707-713.
12. Eliopoulos, A. G., and L. S. Young. The role of the CD40 pathway in the pathogenesis and treatment of cancer. 2004. Curr. Opin. Pharmacol. 4: 360-367-
13. Dicker F., Kater A.P., Prada C.E., Fukuda T., and Kipps T.J. 2006. CD154 induces p73 to oevercome the resistance to apoptosis of chronic lymphocytic leukemia cells lacking functional p53. Blood. 108: 450-3457.
14. Boumpas, D. T., R. Furie, S. Manzi, G. G. Illei, D. J. Wallace, J. E. Balow, and A. Waishnaw. 2003. A short course of BG 9588 (anti CD40 ligand antibody) improves serologic activity and decreases hematuria in patients with proliferative lupus glomerulonephritis. Arthritis Rheum. 46: 3251-3258.
15. Liossis, S. N., and P. P. Sfikakis. 2004. Costimulation blockade in the treatment of rheumatic diseases. BioDrugs. 18: 95-102.
16. Daoussis, D., A. P. Andonopoulos, S. N. Liossis. 2004. Targeting CD40L: a promising therapeutic approach. Clin. Diagn. Lab. Immunol. 11: 635-641.
17. Molano, R. D., T. Berney, H. Li, P. Cattan, A. Pileggi, C. Vizzardelli, N. S. Kenyon, C. Ricordi, L. C. Burkly, and L. Inverardi. 2001. Prolonged islet graft survival in NOD mice by blockade of the CD40-CD154 pathway of T-cell costimulation. Diabetes. 2001. 50: 270-276.
18. Quezada, S. A., B. Fuller, L. Z. Jarvinen, M. Gonzalez, B. R. Blazar, A. Y. Rudensky, T. B. Strom, and R. J. Noelle. 2003. Mechanisms of donor-specific transfusion tolerance: preempitive induction of clonal T-cell exhaustion via indirect presentation. Blood. 102: 1920-1926.
19. Elster, E. A., D. A. Hale, R. B. Mannon, L. C. Cendales, S. J. Swanson, and A. D. Kirk. 2004. The road to tolerance: renal transplant tolerance induction in nonhuman primate studies and clinical trials. Transpl. Immunol. 13: 87-99.
20. Xu, H., X. Zhang, R. B. Mannon, and A. D. Kirk. 2006. Platelet-derived or soluble CD154 induces vascularized allograft rejection independent of cell-bound CD154. J. Clin. Invest. 116: 769-774.
【0058】
21. Snanoudj, R., H. de Preneuf, C. Creput, N. Arzouk, B. Deroure, S. Beaudreuil, A. Durrbach, and B. Charpentier. 2006. Costimulation blockade and its possible future use in clinical transplantation. Transpl. Int. 19: 693-704.
22. Nanji, S. A., W. W. Hancock, B. Luo, C. D. Schur, R. L. Pawlick, L. F. Zhu, C. C. Anderson, and A. M. Shapiro. 2006. Costimulation blockade of both inducible costimulator and CD40 ligand induces dominant tolerance to islet allografts and prevents spontaneous autoimmune diabetes in the NOD mouse. Diabetes. 55: 27-33.
23. Allen, S. D., S. V. Rawale, C. C. Whitacre, and P. T. P. Kaumaya. 2005. Therapeutic peptidomimetic strategies for autoimmune diseases: costimulation blockade. J. Peptide Res. 65: 591-604.
24. Ladner, R.C., A.K. Sato, J. Gorzelany and M. de Souza. 2004. Phage display-derived peptides as therapeutic alternatives to antibodies. DDT. 12:525-529.
25. Hetian, L., A. Ping, S. Shumei, L. Xiaoying, H. Luowen, W. Jian , M. Lin, L. Meisheng, Y. Junshan and S.Chengchao. 2002. A novel peptide isolated from phage display library inhibits tumor growth and metastasis by blocking the binding of vascular endothelial growth factor to its kinase domain receptor. J. Biol. Chem. 277: 43137-43142.
26. Kelsoe, G. 2003. Therapeutic CD154 antibody for lupus: promise for the future? J. Clin. Invest. 112: 1480-1482.
27. Langer, F., S. B. Ingersoll, A. Amirkhosravi, T. Meyer, F. A. Siddiqui, S. Ahmad, J. M. Walker, M. Amaya, H., Desai, J. L. Francis. 2005. The role of CD40 in CD40L- and antibody-mediated platelet activation. Thromb. Haemost. 93:1137-1146.
【技術分野】
【0001】
本発明は、ヒトCD40リガンド(CD40LまたはCD154)に結合できる合成ペプチドに関する。特に、本発明は、CD154の活性部位と特異的に結合でき、それにより、CD154とその天然リガンドであるCD40との相互作用を阻害し、同様に該相互作用に依存する生物学的作用も阻害するアミノ酸配列を有する、合成ペプチドに関する。本発明はまた、炎症性、免疫性または血液学的起源を有する疾患の診断および治療における該ペプチドの使用に関する。
【背景技術】
【0002】
CD40は、B細胞表面で構成的に発現される分子であり、そこでCD40は成熟過程の間に決定的に重要な役割を果たす。CD40は、多くのさらなる細胞型、例えば単球、繊維芽細胞、樹状細胞、内皮細胞、平滑筋細胞および上皮細胞の表面でも発現される(van Kooten et al, J. Leukoc. Biol. 2000)。そのリガンドであるCD154(CD40Lとも称される)は、T細胞、マクロファージ、血小板、単球、NK細胞および内皮細胞の表面で活性化に伴って一過的に発現される。特に、B細胞の表面で発現されるCD40と活性型T細胞の表面で発現されるCD154との間の相互作用は、B細胞のクローン増殖およびアイソタイプスイッチを引き起こし、結果としてそれらの形質転換を導き形質細胞となる。
【0003】
有効な免疫応答の発達におけるCD40:CD154相互作用の重要性は、CD154のコード遺伝子における突然変異が、IgG、IgAおよびIgEを欠くがIgMの過剰産生により特徴づけられ、そして重篤な反復性感染を伴う重篤なヒト型の免疫不全、高IgM症候群(HIM)の原因となるという事実によって示される(Aruffo A. et al, Cell 1993)。
【0004】
ここ数年、多くの研究によりCD40:CD154相互作用が炎症性病因(inflammatory etiogenesis)を伴う多くの過程、例えばアテローム性動脈硬化症、自己免疫疾患、移植後の拒絶反応、移植片対宿主病などに関与することが実証された(Schonbeck U. et al, Circ. Res. 2001;Henn V. et al, Nature 1998;Biancone L. et al, Int. J. Mol. Med. 1999;Buchner K. et al, J. Pathol. 2003)。CD40の発現もしくはそのリガンドCD154の発現または両方の分子の共発現が、多くの腫瘍においても実際に示され、これはこのシグナル伝達経路が腫瘍発生に関与することを示唆するものである(Bussolati B. et al, Int. J. Cancer 2002;Biancone L. et al, J. Immunol. 1999;Cantaluppi V. et al, Int. J. Immunopathol. Pharmacol. 2006;Hill S.C. et al, J. Immunol. 2005;Melichar B. et al, Gynecol. Oncol. 2007;Eliopoulos A.G. et al, Curr. Opin. Pharmacol. 2004)。特に、CD40:CD154シグナル伝達経路が、慢性リンパ性白血病(CLL)の進行に関与すると考えられているのは、腫瘍細胞の生存と内皮細胞の血管新生誘導性の特徴の両方を促進することが示されたからである(Dicker F. et al, Blood 2006)。
【0005】
CD40:CD154相互作用を阻害すること目的とした多くのアプローチが、新規の治療機会を提供するために開発された。特に、有望な結果が、自己免疫疾患、アテローム性動脈硬化症および移植による拒絶反応の異なる実験モデルで得られた。抗CD154モノクローナル抗体に基づくアプローチによるCD40:CD154相互作用の阻害は、自己免疫疾患の多くのモデル、例えば全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、乾癬およびクローン病において臨床改善をもたらした(Boumpas D.T. et al, Arthritis Rheum. 2003; Liossis S.N. et al, Bio Drugs 2004; Daoussis D. et al, Clin. Diagn. Lab. Immun. 2004)。抗CD154抗体は、特許文献においても開示されている。国際公開第WO2005/003175号は、アグリコシル(aglycosyl)抗CD154抗体を開示する;国際公開第WO2006/033702号は、抗CD154抗体およびペプチド、特に抗体軽鎖および重鎖の可変領域を開示する。さらに、多数の書籍により、膵島、皮膚、骨髄、心臓および腎臓移植後のマウスにおけるモノクローナル抗体の使用が、その移植された臓器の延長された存続をもたらすことが実際に示された(Molano R.D. et al, Diabetes 2001; Quezada S.A. et al, Blood 2003; Elster E.A. et al, Transpl. Immunol. 2004; Xu H. et al, J. Clin. Investig. 2006; Snanoudj R. et al, Transpl. Int. 2006; Nanji S.A. et al, Diabetes 2006)。補助刺激の阻止は、補助刺激分子に対するモノクローナル抗体、Fab断片または融合タンパク質を用いて達成しうる。しかしながら、これらのアプローチの結果は、Fc部分の認識に拠る免疫原性作用および血栓形成促進性の副作用の危険性によって(Henn V. et al, Nature 1998)および脆弱な組織浸透性や免疫抑制により、しばしば損なわれる。内在する免疫原性を低下させるために、ヒト化抗体およびミニ抗体が開発されたが、残念ながら親和性が犠牲にされた。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第WO2005/003175号
【特許文献2】国際公開第WO2006/033702号
【非特許文献】
【0007】
【非特許文献1】van Kooten et al, J. Leukoc. Biol. 2000
【非特許文献2】Aruffo A. et al, Cell 1993
【非特許文献3】Schonbeck U. et al, Circ. Res. 2001
【非特許文献4】Henn V. et al, Nature 1998
【非特許文献5】Biancone L. et al, Int. J. Mol. Med. 1999
【非特許文献6】Buchner K. et al, J. Pathol. 2003
【非特許文献7】Bussolati B. et al, Int. J. Cancer 2002
【非特許文献8】Biancone L. et al, J. Immunol. 1999
【非特許文献9】Cantaluppi V. et al, Int. J. Immunopathol. Pharmacol. 2006
【非特許文献10】Hill S.C. et al, J. Immunol. 2005
【非特許文献11】Melichar B. et al, Gynecol. Oncol. 2007
【非特許文献12】Eliopoulos A.G. et al, Curr. Opin. Pharmacol. 2004
【非特許文献13】Dicker F. et al, Blood 2006
【非特許文献14】Boumpas D.T. et al, Arthritis Rheum. 2003
【非特許文献15】Liossis S.N. et al, Bio Drugs 2004
【非特許文献16】Daoussis D. et al, Clin. Diagn. Lab. Immun. 2004
【非特許文献17】Molano R.D. et al, Diabetes 2001
【非特許文献18】Quezada S.A. et al, Blood 2003
【非特許文献19】Elster E.A. et al, Transpl. Immunol. 2004
【非特許文献20】Xu H. et al, J. Clin. Investig. 2006
【非特許文献21】Snanoudj R. et al, Transpl. Int. 2006
【非特許文献22】Nanji S.A. et al, Diabetes 2006
【非特許文献23】Henn V. et al, Nature 1998
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の1つの目的は、CD40:CD154相互作用を阻害でき、該相互作用に依存する生物学的作用を阻止でき、および先行技術、特にモノクローナル抗CD154抗体の使用に関する技術の限界および欠点を克服する試薬を提供することである。
【課題を解決するための手段】
【0009】
この目的は、合成ペプチドによって達成され、それは好ましくは環状であり、ヒトCD154(ヒトCD40Lとも称される)の活性部位と特異的に結合できる。
【0010】
本発明のペプチドは、配列表中の配列番号:13で示されるエプタ(epta)−アミノ酸のCD154−結合性配列を含む。
【図面の簡単な説明】
【0011】
【図1】図1のヒストグラムは、細胞表面上でのヒトCD154発現のためにトランスフェクトされたネズミ骨髄腫細胞系のJ558L細胞に対する、選択されたファージクローンの結合性を示す。
【図2】図2は、ヒトCD154結合性クローンから得られ6個のヒスチジン・テイル(−His6)がタグ付けされた7個のペプチドの、J558L CD154+細胞に対する結合性を示す。
【図3】図3Aは、抗CD154ペプチドとCD154の活性部位に特異的な抗ヒトCD154フルオレセイン複合体化抗体との間の競合を示す。図3Bは、ペプチド4.10と、対照として用いられるペプチド4.10−ala(配列番号:8)との比較を示す。
【図4】4Aのヒストグラムは、可溶性の組み換えヒトCD154分子(rhsCD154)を用いた刺激により誘導された活性B化細胞に対する抗CD154ペプチドおよび対照ペプチド4.10−alaの効果を示す。
【図5】図5Aは、刺激しない基底条件下での、IL4およびCD154を用いた72時間の刺激後、ブロッキング抗ヒトCD154抗体と組み合わせたIL4およびCD154を用いる72時間の刺激後、またはペプチド4.10(配列番号:6)と組み合わせたIL4およびCD154を用いる72時間の刺激後の、アイソタイプスイッチを受けてIgGを発現するナイーブB細胞の存在をそれぞれ示す。
【図6】図6は、ペプチド4.10(配列番号:6)の、対照ペプチドの4.10−ala(配列番号:8)の、またはブロッキング抗ヒトCD154抗体の非存在下または存在下での、HUVEC(HUVEC=ヒト臍静脈内皮細胞)の運動性に対するCD154誘導性の刺激効果を示す。
【図7】図7Aは、刺激なしの基底条件下での或いはブロッキング抗CD154抗体またはペプチド4.10(配列番号:6)または対照ペプチドの4.10−ala(配列番号:8)のいずれかの存在/非存在下での、rhsCD154による刺激後に血管様の細胞コードを形成するマトリゲルに撒かれたHUVECの能力を示す。図7B〜Fは、上記の様々な実験条件下、マトリゲル上での細胞コード形成能についての代表的な画像を示す。
【図8】図8は、250μlのマトリゲルと組み合わせ、SCIDマウスの臀部に皮下接種した内皮HUVEC細胞に対する、ペプチド4.10の存在/非存在下でのrhsCD154誘導刺激のインビボ効果を示す。
【図9】図9は、IL−4刺激により誘導されるCLL細胞の生存を阻害する、本発明のペプチド4.10(配列番号:6)の能力を示す。
【図10】図10は、0.5μMのADPを単独で用いて刺激することによる、あるいはrhsCD154を用いて又はペプチド4.10(配列番号:6)と組み合わせたrhsCD154を用いて又はrhsCD154および抗ヒトCD154抗体からなる免疫複合体と組み合わせたrhsCD154を用いて37℃で10分間刺激した後に0.5μMのADPを用いて刺激することによる、ヒト血小板凝集を示す。
【発明を実施するための形態】
【0012】
好ましい実施形態によれば、本発明のペプチドは、7アミノ酸長から30アミノ酸長である。
【0013】
さらにより好ましい実施形態に関して、本発明のペプチドは、配列番号:13で示されるエプタ−アミノ酸のCD154結合性配列の両端の側面にシステイン残基を付加することにより結果として生じる配列表中の配列番号:6で示される、ノナ−アミノ酸配列からなる。この実施形態は、配列番号:6がその末端の2個のシステイン間でジスルフィド結合を形成することによる環化を受けることができる点で特に有利である。
【0014】
本発明のペプチドは、それが投与される患者に望ましくない免疫応答を誘導することがないという利点を有する。
【0015】
さらなる態様に関して、本発明のペプチドは、複数のペプチドコピーを有する多量体ペプチド構造物の形式で提供され、該ペプチドコピーの各々は直鎖もしくは環状のいずれかである。多量体ペプチド構造物は、四量体構造物であることが好ましい。この多量体ペプチド構造物は、複数のペプチドコピーから成り、その各コピーは少なくとも1個のアミノ酸スペーサー、好ましくはG(Gly)残基により少なくとも1個の他のコピーと結合しており、各ペプチドコピーは好ましくは3個のK(Lys)残基からなるアミノ酸コアと直接もしくは間接的に結合している。
【0016】
特に好ましい本発明の多量体ペプチド構造物は、以下の式で表される:
【化1】
[式中、LはLeuであり、PはProであり、TはThrであり、RはArgであり、HはHisであり、MはMetであり、AはAlaであり、GはGlyであり、およびKはLysである]。
【0017】
以上で示される多量体構造物は、CD154−結合性の配列である配列番号:13を直鎖で含む場合でさえ、組み換え体の可溶性CD154によるCD40刺激に対して、60μMの濃度で、ペプチド4.10と同じ阻害活性を発揮することが示された。さらに、四量体構造物は、ペプチド4.10のように、サブリミナル用量(subliminar dose)(0.3U/ml)のトロンビンによるヒト血小板凝集を活性化することを結果として不活性にした。これらの結果は、CD154−結合性の配列(配列番号:13)は結果として環状および直鎖のいずれの場合でもCD154に結合することができ、生物学的機能を発揮することができることを示す。環状ペプチドはそのアミノ酸配列をタンパク質分解(例えば、血漿中での)から保護しうるという利点を有すると同時に、ペプチドの多量体型を設計しうるということは、分子サイズ、クリアランス速度および結果としての半減期を調節することを可能にする。
【0018】
これまでに言及したように、本発明の範囲には、診断および治療応用における本発明に関するペプチドまたは多量体構造物の使用もまた含まれる。
【0019】
さらに、本発明の範囲には、本発明のペプチドまたは多量体ペプチド構造物が診断剤(例えば、インビボもしくはインビトロ・アッセイのための検出可能な分子)または治療剤(例えば、活性化内皮細胞、腫瘍細胞または活性化の際に細胞表面にCD154を発現できる他のいずれかの細胞に対して、細胞傷害効果を発揮することができる抗炎症剤、抗腫瘍剤または細胞傷害剤)と連結され、インビボもしくはインビトロでの検出を可能にするコンジュゲート、ならびにCD40:CD154相互作用に関係する多くの疾患もしくは障害の処置のための医薬を製造するためのコンジュゲートが含まれる。そのような疾患または状態の例としては、炎症性疾患(例えばアテローム性動脈硬化症、自己免疫疾患、移植による拒絶反応、関節炎を伴う炎症、接触皮膚炎、高IgE症候群、炎症性腸疾患、アレルギー性喘息を含むアレルギー、特発性炎症性疾患)、腫瘍性疾患(例えば慢性リンパ性白血病)、移植片対宿主病が含まれる。自己免疫疾患の例としては、全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、クローン病、乾癬がある。本発明のペプチドまたは多量体構造物あるいはコンジュゲートは、免疫寛容の誘導のための使用にも適している。
【0020】
合成ペプチドは、分子間の相互作用を阻害するための、mAbの代替的アプローチを示す。合成ペプチドの使用を下支えする理論は、タンパク質はただ外部の狭い領域だけを介してその生物学的作用を発揮するのであり、そのわずか2個、3個のアミノ酸だけが特有の受容体機能に重要であるというものである。従って、必須のアミノ酸残基を保持するより短い立体構造的に正しい形式で、その領域に対応した配列を合成することができる。このように、標的とする受容体の活性部位を特異的に認識するが、生物学的に不活性であり立体的な障害を与え、それにより受容体とその天然タンパク質との間の相互作用を妨害する短いペプチドを得ることが可能である。アゴニストと受容体との間の相互作用を模倣することができる、そのようなペプチドの主な理論的な利点は、その限られた大きさであり、それは該ペプチドを水中で容易に可溶化させ、非免疫原性とさせ、該ペプチドが長期間投与されることを可能にする(Allen S.D. et al, J. Peptide Res. 2005; Ladner R.C. et al, DDT 2004)。
【0021】
しかしながら、CD40:CD154相互作用を効果的に阻害することができるアミノ酸配列を同定することは、自明な結果ではない。以下でさらに詳細に例示されるように、本発明のペプチド配列(配列番号:13または配列番号:16)の単一アミノ酸残基のモディファイでさえ、事実上、ヒトCD154の活性部位に対する結合能を失わせることとなり、結果としてペプチドの阻害能が無くなることとなる。
【0022】
上述のように、本発明に関するペプチドは、配列表中の配列番号:13で示されるアミノ酸配列または配列番号:16で示されるペプチド配列を含む。該ペプチドは、ヒトCD154の活性部位に対して非常に高い結合特異性を示す。該ペプチドは、CD154の活性部位を認識し結合することができ、結果としてインビボおよびインビトロの両方でその受容体を効果的に阻止でき、該相互作用に起因する生物学的作用を阻害することとなる。以下の説明において、配列番号:6からなる本発明のペプチドは、「ペプチド4.10」で示される。
【0023】
以下の実験の項で例示される結果は、本発明のペプチド4.10が、組み換え体の可溶型のヒトCD154(組み換えヒト可溶性CD154、またはrhsCD154)による刺激によってB細胞上のCD40の活性化を妨害できることを示す。CD40:CD154相互作用に対して発揮される干渉作用は、B細胞の活性化および結果として生じる免疫応答の両方を妨げる。本発明者らは、ペプチド4.10が、B細胞に対する他の補助刺激分子、例えばCD80およびCD86の誘導を阻害し、同様に免疫グロブリンのアイソタイプスイッチの誘導も阻害できることを実際に示した。さらに、ペプチド4.10は、遊走するおよび血管様構造物を形成する細胞の能力として評価される内皮細胞のrhsCD154誘導型の活性化を、インビトロで阻止することができる。ペプチド4.10のそのような抗血管形成活性は、SCIDマウスでのネズミ血管形成マトリゲル・モデルにおいて、インビボでさらにはっきりと確認でき、そのことを以下でより詳細に記載する。最後に、本発明者らにより得られた予備的なデータは、ペプチド4.10がCLL患者由来のB細胞に劇的なアポトーシスを誘導することができることを示す。
【0024】
短いペプチドを含むコンジュゲートおよび医薬組成物の調製および投与技術のさらなる詳細は、先行技術において知られており;それは本発明の理解に本質的なものではないため、その詳述は本明細書には必要ない。
【0025】
以下の実験の項で説明するように、本明細書は、ヒトCD154に特異的に向けられる7個のアミノ酸配列の同定を説明する。特に、ヒトCD154に結合することができる7個の環状エプタ−ペプチドが選択され、特徴づけられた。ヒトCD154のN末端ドメインを含む融合タンパク質を、ペプチドを選択するために用いた。7個の選択された及び特徴づけられたエプタ−ペプチドの中で、CD40:CD154相互作用を阻害できることが実際に確認された唯一のものが、本発明のペプチド4.10である。さらに、ペプチド4.10のアミノ酸配列を、ペプチド4.10の有用な代替形態を選りすぐる試みの中で、1個もしくは2個のアミノ酸残基を変異によりモディファイした。しかしながら、その代替形態はいずれも、CD40:CD154相互作用を阻害する能力を維持せず、その相互作用に関係する生物学的作用も維持しなかった。
【0026】
以下で示される添付図および表を含み、以下の詳細な記載は、単なる例示として提供される。
【実施例1】
【0027】
図1のヒストグラムは、細胞表面上でのヒトCD154発現のためにトランスフェクトされたネズミ骨髄腫細胞系のJ558L細胞に対する、選択されたファージクローンの結合性を示す。この細胞に対して選択されたファージの結合性は、J558L細胞をファージとインキュベートさせ、次いで抗M13モノクローナル抗体とインキュベートさせて、フローサイトメトリーにより実際に示された。各実験条件について10,000事象を分析し、その値は3回の別個の実験の平均値および標準偏差である。12個の選択したクローンはすべてJ558L細胞と結合する。
【0028】
表1は、ヒトCD154結合性クローンから得られたペプチド挿入体の7個のアミノ酸配列を示す。4.1、4.5および4.9と称するクローンの挿入体は組み換えにより失われ、一方で4.2および4.3と称するクローンの挿入体ならびに4.7および4.12と称するクローンの挿入体は同一であることが確認された。
【0029】
図2は、ヒトCD154結合性クローンから得られ6個のヒスチジン・テイル(−His6)がタグ付けされた7個のペプチドの、J558L CD154+細胞に対する結合性を示す。この細胞へのペプチドの結合性は、フローサイトメトリーにより実際に示された。この細胞を、−His6タグ付きペプチドの各々とインキュベートし、次いで−His6テイルに特異的なフィコエリトリン複合体化(conjugated)二次抗体とインキュベートした。点線のヒストグラムは、アイソトープ対照(isotopic control)であり、一方、黒線はCD154特異的ペプチドの結合強度である[4.2=4.3=CPSGHTKAC(配列番号:1)、4.4=CGTHSSRIC(配列番号:2)、4.6=CLGTQNKEC(配列番号:3)、4.7=4.12=CTPGKPHSC(配列番号:4)、4.8=CKAASANIC(配列番号:5)、4.10=CLPTRHMAC(配列番号:6)および4.11=CLSAVHNMC(配列番号:7)]。各実験条件について10,000事象を分析し、その値は3回の別個の実験の平均値および標準偏差である。ペプチドはすべてJ558L CD154+細胞上のCD154と特異的に結合することができ、対照体のJ558細胞と結合できない。
【0030】
図3Aは、抗CD154ペプチドとCD154の活性部位に特異的な抗ヒトCD154フルオレセイン複合体化抗体との間の競合を示す。ヒトCD154をトランスフェクトしたネズミ骨髄腫細胞であるJ558Lを、6個のヒスチジン・テイル(−His6)タグ付きの7個の抗CD154ペプチドと一緒にインキュベートし、次いで抗CD154フルオレセイン複合体化抗体と一緒にインキュベートした。J558L細胞表面に発現されたCD154との結合に関して抗CD154抗体と競合するペプチドの能力は、フローサイトメトリーにより、抗ヒトCD154フルオレセイン複合体化抗体を単独で用いてインキュベートした細胞により示される陽性対照と比較した蛍光強度の低下として評価した。ペプチド4.10(配列番号:6)が、抗ヒトCD154抗体と競合でき、その結合性を有意に示すことができた唯一のものであることは、図から明らかである。図3Bは、ペプチド4.10と、対照として用いられるペプチド4.10−ala(配列番号:8)との比較を示す。対照ペプチドのアミノ酸配列は、ペプチド4.10のアミノ酸配列とは、5番目のアミノ酸であるアルギニン残基がアラニン残基に置換されている点で異なっている。図3Bで明らかに示されるように、このアルギニン残基を欠くことで、ヒトCD154に対して抗CD154フルオレセイン複合体化抗体と競合する該ペプチドの能力が完全に不活化する結果となるため、そのアルギニン残基が必須であることは明らかである。
【0031】
4Aのヒストグラムは、可溶性の組み換えヒトCD154分子(rhsCD154)を用いた刺激により誘導された活性B化細胞に対する抗CD154ペプチドおよび対照ペプチド4.10−alaの効果を示す。ブロッキング抗ヒトCD154抗体または7個の選択されたペプチドのいずれか1個の非存在下もしくは存在下で、B細胞をrhsCD154を用いて48時間刺激した。末梢B細胞の活性化は、フローサイトメトリーにより、補助刺激分子CD80およびCD86の膜発現で評価した。図4Aで明らかに示されるように、ヒト可溶性CD154(rhsCD154)を用いた48時間のB細胞の刺激は、細胞表面でのCD80およびCD86の強い発現を誘導する。対照的に、rhsCD154をブロッキング抗ヒトCD154抗体と又はペプチド4.10(配列番号:6)と組み合わせて用いた場合、その活性化は抑制される。残りのペプチドはいずれも、rhsCD154誘導型のB細胞の活性化を有意に阻害することはできない。図4Bは、サイトメトリー分析の詳細を示す。
【0032】
図5Aは、刺激しない基底条件下での、IL4およびCD154を用いた72時間の刺激後、ブロッキング抗ヒトCD154抗体と組み合わせたIL4およびCD154を用いる72時間の刺激後、またはペプチド4.10(配列番号:6)と組み合わせたIL4およびCD154を用いる72時間の刺激後の、アイソタイプスイッチを受けてIgGを発現するナイーブB細胞の存在をそれぞれ示す。ペプチド4.10(配列番号:6)は、ちょうど抗CD154抗体のように、rhsCD154およびIL4による刺激によって誘導される免疫グロブリンのアイソタイプスイッチを強力に阻害することができる。図5Bは、サイトメトリー分析の詳細を示す。
【0033】
図6は、ペプチド4.10(配列番号:6)の、対照ペプチドの4.10−ala(配列番号:8)の、またはブロッキング抗ヒトCD154抗体の非存在下または存在下での、HUVEC(HUVEC=ヒト臍静脈内皮細胞)の運動性に対するCD154誘導性の刺激効果を示す。特に、図6Aは、様々な実験条件下で4時間の期間にわたり、細胞遊走のタイムラプス・キネティクスを示し;図6Bは、上記の実験条件下で刺激後30分間で細胞の遊走速度に観察された差異を示し;図6Cは、タイムラプス分析の代表的な画像を示す。対照ペプチドの4.10−ala(配列番号:8)とは反対に、ペプチド4.10(配列番号:6)は、内皮細胞のrhsCD154誘導性の運動性を阻害する。
【0034】
図7Aは、刺激なしの基底条件下での或いはブロッキング抗CD154抗体またはペプチド4.10(配列番号:6)または対照ペプチドの4.10−ala(配列番号:8)のいずれかの存在/非存在下での、rhsCD154による刺激後に血管様の細胞コードを形成するマトリゲルに撒かれたHUVECの能力を示す。図7B〜Fは、上記の様々な実験条件下、マトリゲル上での細胞コード形成能についての代表的な画像を示す(B=刺激なしの対照;C=rhsCD154;D=rhsCD154+4.10ペプチド;E=rhsCD154+4.10−alaペプチド;F=rhsCD154+ブロッキング抗CD154抗体)。本発明のペプチド4.10(配列番号:6)は、ペプチド4.10−ala(配列番号:8)と対照的に、細胞コードを形成する内皮細胞のrhsCD154誘導性の能力を阻害する。
【0035】
図8は、250μlのマトリゲルと組み合わせ、SCIDマウスの臀部に皮下接種した内皮HUVEC細胞に対するペプチド4.10の存在/非存在下でのrhsCD154誘導刺激のインビボ効果を示す。接種の7日後に、マウスを屠殺し、移植片を取り出し、10%ホルマリン溶液中で固定化し、パラフィンに包埋し、組織分析のために調製した。図8Aは、ヘマトキシリン−エオシン染色した組織切片において、基底条件下およびそれ以外の上記実験条件下での血管形成領域の量子化(quantization)を示す。図8Bおよび8Cは、rhsCD154を単独で用いた及びペプチド4.10(配列番号:6)と組み合わせてrhsCD154を用いた内皮細胞の刺激により得られた血管形成の代表的な2枚の画像をそれぞれ示す。各実験条件に対して、6匹のSCIDマウスを用いた。ペプチド4.10は、インビボでのrhsCD154誘導性の血管形成を完全に阻害する。
【0036】
図9は、IL−4刺激により誘導されるCLL細胞の生存を阻害する、本発明のペプチド4.10(配列番号:6)の能力を示す。
【0037】
図10は、0.5μMのADPを単独で用いて刺激することによる、あるいはrhsCD154を用いて又はペプチド4.10(配列番号:6)と組み合わせたrhsCD154を用いて又はrhsCD154および抗ヒトCD154抗体からなる免疫複合体と組み合わせたrhsCD154を用いて37℃で10分間刺激した後に0.5μMのADPを用いて刺激することによる、ヒト血小板凝集を示す。血小板凝集は、血小板凝集計を用い、対照と比較した光透過性の増加として測定した。rhsCD154と組み合わせたヒトCD154抗体によるヒト血小板のプライミングは、0.5μMのADPを用いた刺激により血小板凝集を増大させる。反対に、単独のrhsCD154またはペプチド4.10と組み合わせたrhsCD154は、0.5μMのADPを用いた刺激により誘導される血小板凝集レベルに影響を及ぼさない。これらの結果は、本発明のペプチド4.10(配列番号:6)とCD154分子との相互作用は、血小板凝集を誘導することはないが、これとは対照的に、抗CD154抗体との相互作用では血小板凝集が誘導される。
【0038】
以下の実験の項は、単なる例示として提供され、添付する特許請求の範囲により規定される本発明の範囲を限定することは意図しない。
【0039】
実験の項
この研究を通して、7個のアミノ酸残基とジスルフィド結合による環化を可能にする両端で側面に位置する2個のシステインを含むペプチド配列パネルから成り、M13ファージで発現され、キャプシドpIIIファージタンパク質と遺伝子工学的に融合され、そのファージ表面に無作為に発現される、ペプチドライブラリー(ペプチドの多様性<109)を用いた。このファージライブラリーを、既知のモデル(Hetian L. et al, J. Biol. Chem. 2002)から本発明者らにより発展されたバイオパニング・インビトロ・モデルによってスクリーニングした。ファージ(200μlのTBS中、1×1011CFU)を、ヒト組み換えCD154と一緒に室温で1時間インキュベートして、その後プレートに撒いた。このファージをTBS−Tを用いて数回洗浄してCD154と結合していない非特異的なファージを取り除き、CD154と結合したファージを、100μlの0.2M グリシン、pH2.2を用いて溶出することにより回収し、15μlの1M Tris−HCl、pH9.1を用いて10分間中和した。ファージ数は、溶出液の段階希釈物をLB寒天培地(アガロース7g/リッター、MgCl2・6H2O 1g;Sigma)上の宿主のテトラサイクリン耐性の大腸菌ER2738細胞(New England Biolabs, Hitchin, U.K.)に加え、テトラサイクリン(Kramel Biotech, Cramlington, U.K.)存在下のIPTG/X−Gal LB寒天培地に撒いて、滴定することにより評価した。
【0040】
37℃で12時間のインキュベーション後に、青プラークとしてプレート上で目視できるファージコロニーの数を計数した。評価したファージを、テトラサイクリン存在下のIPTG/X−Gal LB寒天培地のプレート上でファージを培養することにより増幅し、3.3%のポリエチレングリコール8,000/0.4 NaCl(Sigma)中で遠心分離することにより沈殿させた。次いで、そのファージを上記のように滴定し、ヒト組み換えCD154で被覆した新しいプレート上で再度インキュベートした。スクリーニング過程および増幅過程を4回繰返して、ヒトCD154特異的なファージクローンのライブラリーを豊富化した。4回目の増幅ラウンドの後に、12個のファージクローンを無作為に選び出し、個々に増幅し、細胞表面で発現するCD154を認識するその能力を試験した。選択された個々のクローンの特異的な結合性は、参照細胞系としてJ558Lと称されるヒトCD154の発現のためにトランスフェクトされたネズミ骨髄腫細胞系を用いたフローサイトメトリーにより、インビトロで実際に示した。室温で30分間のインキュベーション後のファージ(1011cfu)のJ558L細胞に対する結合性は、その細胞をファージタンパク質M13に対するモノクローナル抗体(Pharmacia, Uppsala, Sweden)と一緒にインキュベートし、続いて抗マウス・フィコエリトリン複合体化抗体(Sigma)と一緒にインキュベートする間接免疫蛍光法によるフローサイトメトリーによって検出した。図1で示されるように、12個のファージクローンはすべて、ヒトCD154発現細胞(J558L)と特異的に結合できたが、その対照体であるCD154−陰性(J558)細胞とは結合しなかった。
【0041】
この12個のファージクローンのペプチド挿入体の配列決定により、表Iに列記される7個の異なる配列が得られた。ペプチドを合成し、インビトロおよびインビボでの局在を容易にするためにビオチン(−bio)または6−ヒスチジン・テイル(−his6)と複合体化させた。同様に、このペプチドのJ558L細胞に対する結合性は、細胞をhis6タグ付きペプチド(60μMの−his6ペプチド)と一緒に室温で30分間インキュベートし、次いで抗ポリヒスチジン−フィコエリトリン複合体化抗体(Sigma)と一緒に4℃で30分間インキュベートして、フローサイトメトリーにより実際に示した。7個の選択されたペプチドはすべて、J558L CD154+細胞に対して良好な親和性を示した。さらに、該ペプチドと、CD154の活性部位を特異的に認識する抗ヒトCD154フルオレセイン複合体化抗体(Serotec)との競合アッセイを行った。J558L細胞を、his6−ペプチドと一緒に室温で30分間インキュベートし、次いで、抗ヒトCD154フルオレセイン複合体化抗体と一緒にさらに30分間室温でインキュベートした。図3Aは、ペプチド4.10が、抗ヒトCD154フルオレセイン複合体化抗体とJ558L細胞との相互作用を有意に阻害できる唯一のものであることを示す。ペプチド4.10のアミノ酸配列中の、CD154との結合に必須のアミノ酸残基を特定するために、3種の異なるペプチドを合成し、その各々は、点突然変異を有しており、それらは4番目、5番目、6番目のアミノ酸配列がそれぞれアラニン残基に置換されることによりペプチド4.10のアミノ酸配列とは異なっている。
【0042】
変異ペプチドは、J558L細胞表面に発現されたヒトCD154を認識することができなかった(データは示さない)。最も低い反応性のペプチド、4.10−alaと称するアミノ酸配列CLPTAHMAC(配列番号:8)を有するペプチドを、対照ペプチドとして選んだ(図3B)。
【0043】
同様の結果が、表IIで列記されるアミノ酸配列を有する4個の改変されたペプチド4.101、4.102、4.103および4.104を用いて得られた。これらのペプチドはいずれも、CD154+ J558Lの細胞表面に発現されたヒトCD154と結合しないことが示された(データは示さない)。
【0044】
B細胞は、その細胞膜上でCD40分子を構成的に発現することが知られている(van Kooten C. et al, J. Leukoc. Biol. 2000)。CD154を発現する活性化T細胞によるCD40の刺激は、結果としてB細胞の活性化、増殖およびアイソタイプスイッチを引き起こす。本発明者らは、フィコール・グラジエント遠心分離法および免疫磁気分離法(MACSシステム−Milteniy)により、健常ドナーの末梢全血から分離したB細胞を用いた。次いで、それにより得られた休止期のB細胞を、可溶型のヒト組み換えCD154(rhsCD154)を用いて刺激し、細胞膜上での補助刺激分子CD80(B7−1)およびCD86(B7−2)の活性化および発現を誘導した。rhsCD154(100ng/ml)による48時間の刺激後に、末梢B細胞の細胞表面上にCD80およびCD86の膜発現において劇的な増加が観察された。この発現は、それぞれ、基底レベルと比較して300%および274%にまで増加した。対照的に、リンパ球刺激の前にペプチド4.10(配列番号:6)と一緒にrhsCD154(100ng/ml)で37℃にて15分間あらかじめインキュベートすることで、B細胞刺激およびその関連するCD80およびCD86の発現を阻止した。ペプチド4.10(配列番号:6)の阻害作用は、60μMの濃度でピークに達し、CD80およびCD86の発現はそれぞれ107%および130%にまで低下した(図4Aおよび4B)。その効果はより低い濃度(30μM)の場合でさえ観察することが可能であった。
【0045】
反対に、同じ濃度(60μM)の他のペプチドと一緒にrhsCD154(100ng/ml)を用いてあらかじめインキュベートしても、rhsCD154による刺激によって誘導されるCD80およびCD86の発現に有意な影響はなかった。より高い濃度(250μM)では、ペプチド4.6(配列番号:3)および4.11(配列番号:7)はまた、B細胞活性化の阻害を示した:特に、ペプチド4.6(配列番号:3)は、CD80の発現を117%に及びCD86の発現を122%に低下させたが、一方で、ペプチド4.11(配列番号:7)の効果は安定した再現性はなかった。反対に、対照ペプチドの4.10−ala(配列番号:8)は、250μMの濃度でさえrhsCD154(100ng/ml)と一緒にあらかじめインキュベートしても、rhsCD154による刺激によって誘導されるCD80およびCD86の発現には影響せず、またB細胞の活性化を有意に低下させることもなく、さらにCD80およびCD86の発現レベルを低下させることもなかった(図4A)。ブロッキング対照として、実施した全ての実験でB細胞の刺激前に、rhsCD154(100ng/ml)をブロッキング抗ヒトCD154抗体(Alexis)と一緒に37℃で15分間あらかじめインキュベートすることで、CD40媒介型のB細胞の活性化を完全に阻害し、CD80およびCD86の発現を基底レベルにまで低下させた(図4Aおよび4B)。この結果は、5回の別個の実験の平均値±標準偏差である。
【0046】
同様の結果はまた、ヒト脾臓フラグメントから単離されたB細胞を用いて得られた。さらに、ナイーブB細胞集団で行われた別の実験でも、ペプチド4.10がCD40媒介型のB細胞Igのアイソタイプスイッチを効果的に阻害することが示された。これらの実験は、免疫磁気分離法(MACSシステム Milteniy)によりB細胞プールから成熟リンパ球を失わせることによる陰性選択によって末梢血から得られたCD27−ナイーブB細胞で行った。本発明者らは、ナイーブB細胞を、IL−4(0.4ng/ml)およびrhsCD154(100ng/ml)の存在下の低グルコースDMEM(Sigma)中で96時間培養した場合に、膜IgGを発現するB細胞の数に有意な増加(25%)が誘導されたことを実際に示した。対照的に、ペプチド4.10(配列番号:6)と組み合わせたIL4(0.4ng/ml)およびrhsCD154(100ng/ml)を用いて刺激されたB細胞は非常に低い割合でアイソタイプスイッチを受けた(図5Aおよび5B)。同様に、ブロッキング抗ヒトCD154抗体と組み合わせたIL4(0.4ng/ml)およびrhsCD154(100ng/ml)によるナイーブB細胞の刺激では、アイソタイプスイッチは誘導されなかった(図5Aおよび5B)。
【0047】
炎症過程は、ひとたび誘発されると特定の受容体またはリンパ球補助刺激分子、例えばCD40およびそのリガンドCD154を発現する免疫系以外の細胞、例えば血管壁の内皮細胞により保存されうることが知られている。かくして、本発明者らは、ペプチド4.10(配列番号:6)が、リンパ球刺激および/または成熟を阻害することに加えて、内皮細胞の細胞表面に発現されるCD40分子の刺激を阻止して、炎症を制限できることを実際に示した。本発明者らは、rhsCD154(100ng/ml)の投与による内皮CD40刺激によって誘導される血管新生誘導効果は、単一細胞の遊走および基底マトリクス(マトリゲル)上で血管様の細胞コードを形成する細胞の能力の両方のようにインビトロ評価され、本発明の抗CD154ペプチド4.10(配列番号:6)を用いてrhsCD154(100ng/ml)を37℃で15分間あらかじめインキュベートすることにより阻害されたことが実際に示された。特に、細胞遊走実験は、20%血清存在下の完全内皮細胞培養液中4×103細胞密度でゼラチン上に撒かれたHUVEC細胞を用いて実施した。後日、その内皮培地は、rhsCD154(100ng/ml)存在下もしくは非存在下の減少した血清濃度(5%)を含む低グルコースDMEM(Sigma)に交換した。
【0048】
さらに、阻害効果を評価するために、rhsCD154(100ng/ml)を、ペプチド4.10(配列番号:6)と又は対照ペプチドの4.10−ala(配列番号:8)と又はブロッキング抗ヒトCD154抗体と一緒に37℃で15分間インキュベートした。次いで、フラスコを、恒温チャンバー(約37℃)を備えた位相差倒立顕微鏡下で4時間、10×倍率で観察した。細胞を、顕微鏡に繋がれたカメラを用いて規則的に15分間隔で4時間の全期間にわたって撮影した。各細胞の遊走速度は、画像分析ソフトウェアを用いて、各単一フレームにおいて各単独細胞それぞれの核の位置に基づいて計算し、結果として、平均の細胞遊走速度(±標準偏差)を各実験条件ごとに計算した。図6Aおよび6Bで示されるように、rhsCD154(100ng/ml)による内皮CD40の刺激は、細胞の基底の運動性(1時間あたり常に12μm未満)において約350%(1時間あたり42μm)の増加を誘導し、一方でrhsCD154(100ng/ml)をペプチド4.10(配列番号:6)と共に37℃で15分間あらかじめインキュベートすることで、ならびにブロッキング抗ヒトCD154抗体と一緒にあらかじめインキュベートすることで、細胞の運動性は基底レベルまで顕著に低下した。反対に、対照ペプチド4.10−ala(配列番号:8)は、内皮CD40の刺激に影響を及ぼさなかった(図6A、6B、6C)。
【0049】
rhsCD154(100ng/ml)によるヒト内皮CD40刺激を妨げるペプチド4.10の能力のさらなる確認として、HUVEC(3.5×104)をインビトロ血管新生アッセイにかけた。HUVECを、5%血清が補充された基底RPMI培地中、巨大な細胞の組織体の形成を促進することができる腫瘍マトリックス(tumor matrix)(マトリゲル、BD)上に撒いた。37℃、5%CO2で4時間インキュベーションして、細胞を位相差倒立顕微鏡下で観察した。図7Aおよび7Bは、rhsCD154(100ng/ml)による刺激が、4時間後にすでに複雑な内皮ネットワークの形成を誘導したが、一方でrhsCD154(100ng/ml)をペプチド4.10(配列番号:6)と一緒に、ならびにブロッキング抗ヒトCD154抗体と一緒に37℃で15分間あらかじめインキュベートすることで、刺激を完全に無効にした(図7A、7Dおよび7F)。反対に、rhsCD154(100ng/ml)を対照ペプチドの4.10−ala(配列番号:8)と一緒に37℃で15分間あらかじめインキュベートすることでは、血管様の細胞コードを形成する細胞の能力に影響を及ぼさなかった(図7E)。他の抗ヒトCD154ペプチド、例えば活性部位と特異的に結合しない4.6(配列番号:3)および4.11(配列番号:7)は、内皮のCD40刺激を有意に低下させることはできなかった(データは示さない)。
【0050】
最後に、ペプチド4.10の抗血管形成作用を、インビボ血管形成アッセイで試験した。200μlのHANK溶液中に再懸濁した内皮HUVEC細胞(2×106)を、rhsCD154(100ng/ml)を含む液状の500μlのマトリゲルと混合し、ペプチド4.10(配列番号:6)の存在下および非存在下でSCIDマウスの右側の臀部および左側の臀部にそれぞれ皮下接種した。6日後に、マウスを屠殺し、マトリゲルプラグを回収し、10%ホルマリン溶液中で少なくとも24時間固定化し、次いで免疫組織分析のために処理した。HUVECおよびrhsCD154移植片のヘマトキシリン−エオシン染色した組織切片は、強い血管形成を示し(図8Aおよび8B)、sCD154をペプチド4.10(配列番号:6)であらかじめインキュベートした接種では著しく阻害されたか又は完全に見られず、そこでは大部分の細胞がアポトーシスを受けていた(図8Aおよび8C)。
【0051】
ペプチド4.10(配列番号:6)は、インビボで得られたCLL腫瘍細胞のIL−4存在下での培養による生存を阻害できることもはっきり示した。CLL細胞のアポトーシスに対する耐性および増強された生存は、慢性リンパ性白血病患者におけるインビボでのこれらの腫瘍細胞の伸展(expansion)によるものと考えられる。これらの結果は、CD40とそのリガンドであるCD154との相互作用が、CLL細胞の生存を刺激するのに重要であること、およびCD40:CD154相互作用のCD154の活性部位を認識することができるペプチドによる阻害が、これらの細胞の生存を阻害しうることを示す(図9)。
【0052】
最後に、ペプチド4.10の主な利点の1つは、ヒト血小板に対する反応性を欠いていることである。これまで、有望な抗CD154モノクローナル抗体を試験するために実施された多くの臨床試験が、劇的な血栓形成促進性の副作用により突然中断されたことが知られている(Kelsoe G. et al, J. Clin. Invest. 2003)。仮説は、抗CD154抗体が、免疫グロブリンFc部分に対する受容体(FcR)を発現するヒト血小板表面において交差反応を誘発し、活性化後にCD154分子を誘発する、というものである。したがって、本発明者らは、血小板凝集アッセイにより、ペプチド4.10の血小板に対する作用を試験した。このような実験は、磁気攪拌器およびリアルタイム光学密度計を備えた恒温凝集計を使用して行った。血小板の凝集は、参照試料(ブランク)に対する光学密度の変化として評価した。この実験は、5U/mlのヘパリンを含む試験管に収集された健常ドナーの末梢全血から抽出された血小板で行った。特に、多血小板血漿(platelet enriched plasma:PRP)は900rpmで20分間の全血の遠心分離により得られ、一方で参照試料として用いられる少血小板血漿(platelet poor plasma:PPP)は3000rpmで10分間の別の遠心分離後の上清を回収することにより得られた。Langer(Langer F. et al, Thromb. Haemost. 2005)により既に示されたように、血小板を組み換えヒト可溶性CD154および抗CD154モノクローナル抗体からなる免疫複合体と一緒に37℃で10分間インキュベートすることによるプライミングは、0.5μMのADPによる次の刺激により誘導される血小板の凝集を増強した(図10)。対照的に、CD154単独による又はペプチド4.10(60μM)との組み合わせによるプライミングは、0.5μMのADPによる刺激により誘導される血小板の凝集を増強しなかった(図10)。同様の結果が、0.3U/mlのトロンビンを用いて血小板凝集を誘導することで得られた。
【0053】
内皮細胞は、特に炎症部位で、その表面に高頻度でCD40を発現するため、および血管形成は幾つかの炎症過程および炎症性疾患の発症において主要な役割を果たしうるため、本発明の4.10ペプチドの上記特性により、該ペプチドは見込みのある抗炎症アプローチの開発に関する非常に有望な分子である。さらに、4.10ペプチド(配列番号:6)により発揮されるリンパ球活性化における阻害作用は、移植または腫瘍細胞増殖がCD40による刺激に起因する抗アポトーシス作用によって支援される腫瘍性疾患、例えば慢性リンパ性白血病(CLL)に対する免疫抑制治療として、この分子の見込みのある使用を提供する。
【0054】
【表1】
(*)本発明に関するペプチド
【0055】
【表2】
【0056】
参考文献
1. van Kooten, C., and J. Banchereau. 2000. CD40-CD40 ligand. J. Leukoc. Biol. 67: 2-17.
2. Aruffo, A., M. Farrington, D. Hollenbaugh, X. Li, A. Milatovich, S. Nonoyama, J. Bajorath, L. S. Grosmaire, R. Stenkamp, M. Neubauer, et al. 1993. The CD40 ligand, gp39, is defective in activated T cells from patients with X-linked hyper-IgM syndrome. Cell. 72: 291-300.
3. Schonbeck, U., and P. Libby. 2001. CD40 signaling and plaque instability. Circ. Res. 89: 1092-1103.
4. Henn, V., J. R. Slupsky, M. Grafe, I. Anagnostopoulos, R. Forster, G. Muller-Berghaus, and R. A. Kroczek. 1998. CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells. Nature. 391: 591-594.
5. Biancone, L., V. Cantaluppi, and G. Camussi. 1999. CD40-CD154 interaction in experimental and human disease. Int. J. Mol. Med. 3: 343-353.
6. Buchner, K., V. Henn, M. Grafe, O. J. de Boer, A. E. Becker, and R. A. Kroczek. 2003. CD40 ligand is selectively expressed on CD4+ T cells and platelets: implications for CD40-CD40L signalling in atherosclerosis. J. Pathol. 201: 288-295.
7. Bussolati, B., S. Russo, I. Deambrosis, V. Cantaluppi, A. Volpe, U. Ferrando, and G. Camussi. 2002. Expression of CD154 on renal cell carcinomas and effect on cell proliferation, motility and platelet-activating factor synthesis. Int. J. Cancer. 100: 654-661.
8. Biancone, L., V. Cantaluppi, M. Boccellino, L. Del Sorbo, S. Russo, .A. Albini, I. Stamenkovic, and G. Camussi. 1999. Activation of CD40 favors the growth and vascularization of Kaposi's sarcoma. J. Immunol. 163: 6201-6208.
9. Cantaluppi, V., M. C. Deregibus, L. Biancone, I. Deambrosis, B. Bussolati, A. Albini, and G. Camussi. 2006. The expression of CD154 by Kaposi's sarcoma cells mediates the anti-apoptotic and migratory effects of HIV-1-TAT protein. Int. J. Immunopathol. Pharmacol. 19: 81-96.
10. Hill, S. C., S. J. Youde, S. Man, G. R. Teale, A. J. Baxendale, A. Hislop, C. C. Davies, D. M. Luesley, A. M. Blom, A. B. Rickinson, L. S. Young, and A. G. Eliopoulos. 2005. Activation of CD40 in cervical carcinoma cells facilitates CTL responses and augments chemotherapy-induced apoptosis. J. Immunol. 174: 41-50.
【0057】
11. Melichar, B., R. Patenia, S. Gallardo, K. Melicharova, W. Hu, and R. S. Freedman. 2007. Expression of CD40 and growth-inhibitory activity of CD40 ligand in ovarian cancer cell lines. Gynecol. Oncol. 104: 707-713.
12. Eliopoulos, A. G., and L. S. Young. The role of the CD40 pathway in the pathogenesis and treatment of cancer. 2004. Curr. Opin. Pharmacol. 4: 360-367-
13. Dicker F., Kater A.P., Prada C.E., Fukuda T., and Kipps T.J. 2006. CD154 induces p73 to oevercome the resistance to apoptosis of chronic lymphocytic leukemia cells lacking functional p53. Blood. 108: 450-3457.
14. Boumpas, D. T., R. Furie, S. Manzi, G. G. Illei, D. J. Wallace, J. E. Balow, and A. Waishnaw. 2003. A short course of BG 9588 (anti CD40 ligand antibody) improves serologic activity and decreases hematuria in patients with proliferative lupus glomerulonephritis. Arthritis Rheum. 46: 3251-3258.
15. Liossis, S. N., and P. P. Sfikakis. 2004. Costimulation blockade in the treatment of rheumatic diseases. BioDrugs. 18: 95-102.
16. Daoussis, D., A. P. Andonopoulos, S. N. Liossis. 2004. Targeting CD40L: a promising therapeutic approach. Clin. Diagn. Lab. Immunol. 11: 635-641.
17. Molano, R. D., T. Berney, H. Li, P. Cattan, A. Pileggi, C. Vizzardelli, N. S. Kenyon, C. Ricordi, L. C. Burkly, and L. Inverardi. 2001. Prolonged islet graft survival in NOD mice by blockade of the CD40-CD154 pathway of T-cell costimulation. Diabetes. 2001. 50: 270-276.
18. Quezada, S. A., B. Fuller, L. Z. Jarvinen, M. Gonzalez, B. R. Blazar, A. Y. Rudensky, T. B. Strom, and R. J. Noelle. 2003. Mechanisms of donor-specific transfusion tolerance: preempitive induction of clonal T-cell exhaustion via indirect presentation. Blood. 102: 1920-1926.
19. Elster, E. A., D. A. Hale, R. B. Mannon, L. C. Cendales, S. J. Swanson, and A. D. Kirk. 2004. The road to tolerance: renal transplant tolerance induction in nonhuman primate studies and clinical trials. Transpl. Immunol. 13: 87-99.
20. Xu, H., X. Zhang, R. B. Mannon, and A. D. Kirk. 2006. Platelet-derived or soluble CD154 induces vascularized allograft rejection independent of cell-bound CD154. J. Clin. Invest. 116: 769-774.
【0058】
21. Snanoudj, R., H. de Preneuf, C. Creput, N. Arzouk, B. Deroure, S. Beaudreuil, A. Durrbach, and B. Charpentier. 2006. Costimulation blockade and its possible future use in clinical transplantation. Transpl. Int. 19: 693-704.
22. Nanji, S. A., W. W. Hancock, B. Luo, C. D. Schur, R. L. Pawlick, L. F. Zhu, C. C. Anderson, and A. M. Shapiro. 2006. Costimulation blockade of both inducible costimulator and CD40 ligand induces dominant tolerance to islet allografts and prevents spontaneous autoimmune diabetes in the NOD mouse. Diabetes. 55: 27-33.
23. Allen, S. D., S. V. Rawale, C. C. Whitacre, and P. T. P. Kaumaya. 2005. Therapeutic peptidomimetic strategies for autoimmune diseases: costimulation blockade. J. Peptide Res. 65: 591-604.
24. Ladner, R.C., A.K. Sato, J. Gorzelany and M. de Souza. 2004. Phage display-derived peptides as therapeutic alternatives to antibodies. DDT. 12:525-529.
25. Hetian, L., A. Ping, S. Shumei, L. Xiaoying, H. Luowen, W. Jian , M. Lin, L. Meisheng, Y. Junshan and S.Chengchao. 2002. A novel peptide isolated from phage display library inhibits tumor growth and metastasis by blocking the binding of vascular endothelial growth factor to its kinase domain receptor. J. Biol. Chem. 277: 43137-43142.
26. Kelsoe, G. 2003. Therapeutic CD154 antibody for lupus: promise for the future? J. Clin. Invest. 112: 1480-1482.
27. Langer, F., S. B. Ingersoll, A. Amirkhosravi, T. Meyer, F. A. Siddiqui, S. Ahmad, J. M. Walker, M. Amaya, H., Desai, J. L. Francis. 2005. The role of CD40 in CD40L- and antibody-mediated platelet activation. Thromb. Haemost. 93:1137-1146.
【特許請求の範囲】
【請求項1】
CD154受容体の活性部位に選択的に結合することができ、CD40:CD154相互作用を阻害することができるペプチドであって、配列番号:13で示されるCD154結合性のエプタ−アミノ酸配列を含む、ペプチド。
【請求項2】
7アミノ酸長から30アミノ酸長である、請求項1記載のペプチド。
【請求項3】
配列番号:13で示されるCD154結合性のアミノ酸配列が、該配列の両端の側面に2個のシステインをさらに含み、それにより配列番号:6で示されるノナ−アミノ酸配列を提供することとなる、請求項1または2記載のペプチド。
【請求項4】
配列番号:6で示されるアミノ酸配列からなる、請求項3記載のペプチド。
【請求項5】
環状である、請求項3または4記載のペプチド。
【請求項6】
請求項1〜5のいずれか一項記載のペプチドの複数のコピーを含む、多量体構造物。
【請求項7】
請求項1〜5のいずれか一項記載のペプチドの各コピーが、直鎖または環状のいずれかである、請求項6記載の多量体構造物。
【請求項8】
請求項1〜5のいずれか一項記載のペプチドの各コピーが、少なくとも1つの他のコピーと少なくとも1つのアミノ酸スペーサーにより連結されている、請求項6または7記載の多量体構造物。
【請求項9】
アミノ酸スペーサーが、Gly(G)残基である、請求項8記載の多量体構造物。
【請求項10】
ペプチドの各コピーが、直接または間接的に連結されるアミノ酸コアを含む、請求項8または9記載の多量体構造物。
【請求項11】
アミノ酸コアが、複数のLys(K)残基からなる、請求項10記載の多量体構造物。
【請求項12】
ペプチドが、配列番号:13で示される直鎖のエプタ−アミノ酸配列からなる、請求項6〜11のいずれか一項記載の多量体構造物。
【請求項13】
以下の構造
【化1】
[式中、LはLeuであり、PはProであり、TはThrであり、RはArgであり、HはHisであり、MはMetであり、AはAlaであり、GはGlyであり、およびKはLysである]
からなる、請求項12記載の多量体構造物。
【請求項14】
請求項1〜5のいずれか一項記載の少なくとも1つのペプチド、または請求項6〜13のいずれか一項記載の少なくとも1つの多量体構造物を含み、生体分子、診断剤および治療剤からなる群より選択される分子と複合体化された、コンジュゲート。
【請求項15】
治療剤が、抗炎症性剤、免疫抑制剤、免疫調節剤または抗腫瘍剤である、請求項14記載のコンジュゲート。
【請求項16】
治療剤が、活性化内皮細胞、腫瘍細胞または活性化によりその表面にCD154を発現する細胞に対して細胞傷害効果を発揮することができる、請求項14記載のコンジュゲート。
【請求項17】
診断剤が、インビボで検出可能な分子である、請求項14記載のコンジュゲート。
【請求項18】
診断剤が、インビトロアッセイのための検出可能な分子である、請求項14記載のコンジュゲート。
【請求項19】
医薬としての、請求項1〜5のいずれか一項記載のペプチド、または請求項6〜13のいずれか一項記載の多量体構造物。
【請求項20】
CD40:CD154相互作用に関係する疾患または障害の処置のための医薬を製造するための、請求項1〜5のいずれか一項記載のペプチドまたは請求項6〜13のいずれか一項記載の多量体構造物の使用。
【請求項21】
疾患または障害が、炎症である、請求項20記載の使用。
【請求項22】
炎症が、アテローム性動脈硬化症、自己免疫疾患または移植による拒絶反応から選択される炎症性疾患である、請求項21記載の使用。
【請求項23】
自己免疫疾患が、全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、クローン病または乾癬である、請求項22記載の使用。
【請求項24】
炎症が、関節炎を伴う炎症、接触皮膚炎、高IgE症候群、炎症性腸疾患、アレルギー性喘息を含むアレルギーおよび特発性炎症性疾患からなる群より選択される、請求項21記載の使用。
【請求項25】
移植による拒絶反応が、移植された膵臓の膵島細胞、皮膚、骨髄、肝臓、心臓および腎臓を含む、請求項22記載の使用。
【請求項26】
疾患が、腫瘍性疾患である、請求項20記載の使用。
【請求項27】
腫瘍性疾患が、慢性リンパ性白血病(CLL)である、請求項26記載の使用。
【請求項28】
疾患が、移植片対宿主病である、請求項20記載の使用。
【請求項29】
請求項1〜5のいずれか一項記載のペプチドまたは請求項6〜13のいずれか一項記載の多量体構造物の有効量、および医薬上許容されるビヒクルおよび/または賦形剤を含む、医薬組成物。
【請求項1】
CD154受容体の活性部位に選択的に結合することができ、CD40:CD154相互作用を阻害することができるペプチドであって、配列番号:13で示されるCD154結合性のエプタ−アミノ酸配列を含む、ペプチド。
【請求項2】
7アミノ酸長から30アミノ酸長である、請求項1記載のペプチド。
【請求項3】
配列番号:13で示されるCD154結合性のアミノ酸配列が、該配列の両端の側面に2個のシステインをさらに含み、それにより配列番号:6で示されるノナ−アミノ酸配列を提供することとなる、請求項1または2記載のペプチド。
【請求項4】
配列番号:6で示されるアミノ酸配列からなる、請求項3記載のペプチド。
【請求項5】
環状である、請求項3または4記載のペプチド。
【請求項6】
請求項1〜5のいずれか一項記載のペプチドの複数のコピーを含む、多量体構造物。
【請求項7】
請求項1〜5のいずれか一項記載のペプチドの各コピーが、直鎖または環状のいずれかである、請求項6記載の多量体構造物。
【請求項8】
請求項1〜5のいずれか一項記載のペプチドの各コピーが、少なくとも1つの他のコピーと少なくとも1つのアミノ酸スペーサーにより連結されている、請求項6または7記載の多量体構造物。
【請求項9】
アミノ酸スペーサーが、Gly(G)残基である、請求項8記載の多量体構造物。
【請求項10】
ペプチドの各コピーが、直接または間接的に連結されるアミノ酸コアを含む、請求項8または9記載の多量体構造物。
【請求項11】
アミノ酸コアが、複数のLys(K)残基からなる、請求項10記載の多量体構造物。
【請求項12】
ペプチドが、配列番号:13で示される直鎖のエプタ−アミノ酸配列からなる、請求項6〜11のいずれか一項記載の多量体構造物。
【請求項13】
以下の構造
【化1】
[式中、LはLeuであり、PはProであり、TはThrであり、RはArgであり、HはHisであり、MはMetであり、AはAlaであり、GはGlyであり、およびKはLysである]
からなる、請求項12記載の多量体構造物。
【請求項14】
請求項1〜5のいずれか一項記載の少なくとも1つのペプチド、または請求項6〜13のいずれか一項記載の少なくとも1つの多量体構造物を含み、生体分子、診断剤および治療剤からなる群より選択される分子と複合体化された、コンジュゲート。
【請求項15】
治療剤が、抗炎症性剤、免疫抑制剤、免疫調節剤または抗腫瘍剤である、請求項14記載のコンジュゲート。
【請求項16】
治療剤が、活性化内皮細胞、腫瘍細胞または活性化によりその表面にCD154を発現する細胞に対して細胞傷害効果を発揮することができる、請求項14記載のコンジュゲート。
【請求項17】
診断剤が、インビボで検出可能な分子である、請求項14記載のコンジュゲート。
【請求項18】
診断剤が、インビトロアッセイのための検出可能な分子である、請求項14記載のコンジュゲート。
【請求項19】
医薬としての、請求項1〜5のいずれか一項記載のペプチド、または請求項6〜13のいずれか一項記載の多量体構造物。
【請求項20】
CD40:CD154相互作用に関係する疾患または障害の処置のための医薬を製造するための、請求項1〜5のいずれか一項記載のペプチドまたは請求項6〜13のいずれか一項記載の多量体構造物の使用。
【請求項21】
疾患または障害が、炎症である、請求項20記載の使用。
【請求項22】
炎症が、アテローム性動脈硬化症、自己免疫疾患または移植による拒絶反応から選択される炎症性疾患である、請求項21記載の使用。
【請求項23】
自己免疫疾患が、全身性エリテマトーデス(SLE)、リウマチ性関節炎(RA)、クローン病または乾癬である、請求項22記載の使用。
【請求項24】
炎症が、関節炎を伴う炎症、接触皮膚炎、高IgE症候群、炎症性腸疾患、アレルギー性喘息を含むアレルギーおよび特発性炎症性疾患からなる群より選択される、請求項21記載の使用。
【請求項25】
移植による拒絶反応が、移植された膵臓の膵島細胞、皮膚、骨髄、肝臓、心臓および腎臓を含む、請求項22記載の使用。
【請求項26】
疾患が、腫瘍性疾患である、請求項20記載の使用。
【請求項27】
腫瘍性疾患が、慢性リンパ性白血病(CLL)である、請求項26記載の使用。
【請求項28】
疾患が、移植片対宿主病である、請求項20記載の使用。
【請求項29】
請求項1〜5のいずれか一項記載のペプチドまたは請求項6〜13のいずれか一項記載の多量体構造物の有効量、および医薬上許容されるビヒクルおよび/または賦形剤を含む、医薬組成物。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2011−505404(P2011−505404A)
【公表日】平成23年2月24日(2011.2.24)
【国際特許分類】
【出願番号】特願2010−536413(P2010−536413)
【出願日】平成20年11月27日(2008.11.27)
【国際出願番号】PCT/EP2008/066349
【国際公開番号】WO2009/071486
【国際公開日】平成21年6月11日(2009.6.11)
【出願人】(597075904)フレゼニウス メディカル ケア ドイッチェランド ゲゼルシャフト ミット ベシュレンクテル ハフツング (55)
【Fターム(参考)】
【公表日】平成23年2月24日(2011.2.24)
【国際特許分類】
【出願日】平成20年11月27日(2008.11.27)
【国際出願番号】PCT/EP2008/066349
【国際公開番号】WO2009/071486
【国際公開日】平成21年6月11日(2009.6.11)
【出願人】(597075904)フレゼニウス メディカル ケア ドイッチェランド ゲゼルシャフト ミット ベシュレンクテル ハフツング (55)
【Fターム(参考)】
[ Back to top ]