
ヒトFATP5遺伝子のプロモーターおよび使用
本発明は、ヒトFATP5タンパク質のプロモーターに対応することを特徴とする単離されたヒト核酸、および、前記プロモーターの依存下に置かれたタンパク質、特にヒトFATP5タンパク質をコードする単離されたヒト核酸に関する。本発明はさらに、前記核酸を含むベクター、または前記核酸または前記ベクターが導入された宿主細胞に関する。最後に、本発明は前記核酸、前記ベクターまたは前記宿主細胞の、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質を同定する方法における使用、および当該同定方法に関する。発明は、最後に、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質、および、前記物質の、糖尿病を予防および/または処置するための、または肝臓における脂肪酸のβ酸化を誘導するための医薬の製造への使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、ヒトFATP5タンパク質のプロモーターに対応することを特徴とする単離ヒト核酸、および前記プロモーターの依存下に置かれた、タンパク質、特にヒトFATP5タンパク質をコードする単離ヒト核酸に関する。
本発明はさらに、前記核酸を含むベクター、または、前記核酸もしくは前記ベクターが導入された宿主細胞に関する。最後に、本発明は、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質を同定する方法における、前記核酸、前記ベクターまたは前記宿主細胞の使用、および前記同定方法に関する。
本発明は、最後に、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質、および、糖尿病を予防および/または処置するための、または、肝臓における脂肪酸のβ酸化を誘導するための医薬の製造における前記物質の使用に関する。
【0002】
II型糖尿病は、世界中において主要な公衆衛生上の問題である。現在、およそ1億2500万人が本症に罹患しており、このうち米国内に存在する者だけで1500万人を占める。予測は、2010年に、糖尿病に罹患している患者数が2倍になることを示している。現在米国で糖尿病を患っている90〜95%の人々がII型糖尿病に罹患している。II型は、インスリン耐性と高血糖症とを特徴とする代謝病であり、しばしば高血圧、脂質障害および肥満を伴う。成人において、この病的状態は一般に30才から現れ、心臓、血管、眼、腎臓および/または神経への損傷を含む合併症を特徴とする。
【0003】
今のところ、残念なことに、原因に対する治療的および/または予防的処置は存在せず、処置は、一般に対症療法的となる傾向がある。
したがって、糖尿病の原因を処置することが可能な物質が真に求められている。また、糖尿病の原因を処置することができる前記物質を特定するために、前記物質を特定する効果的方法も常に変わらず求められている。
本発明の目的の1つは、そのような方法を提供することである。
【0004】
長鎖脂肪酸(LCFA)は生物のための重要なエネルギー源であり、ミトコンドリアにおけるそのβ酸化を介してATPを提供する。これらはまた、多数の多様な細胞過程、例えば細胞シグナリング経路または遺伝子発現の調節のための基質でもある。例として、脂肪酸は心臓にエネルギーの恒常的で効果的な供給を提供するために不可欠である。
LCFAは、リポタンパク質リパーゼによる脂肪分解の後で粘膜の細胞に入り、そしてその後、小胞体でトリグリセリドにエステル化される。トリグリセリドはその後、アポリポタンパク質とともに脂肪粒子(lipoparticle)複合体内に組み込まれ、極低密度リポタンパク質(VLDL)またはキロミクロンを形成し、そしてエキソサイトーシスによって細胞から出て行き、全身循環中に廃棄される。リポタンパク質リパーゼは、これらの粒子を血管内皮細胞の表面で非エステル型脂肪酸に変換し、これは、その後アルブミンと結合することができる。
【0005】
ラット細胞における脂肪酸の捕獲の動態は、最初の30秒間にわたる急速な線形段階と、それに続く、初期捕獲速度の低減を伴う移行段階、および低い蓄積速度を有する後期に分けられる。最初の捕獲速度は、脂肪酸のベクトル的に一方向の流れに対応し、インキュベーション培地に適用される脂肪酸濃度の計算された作用によって特徴づけることができる。動態、LCFAによる競合、およびプロテアーゼでの細胞の処理による脂肪酸輸送の抑制は、輸送体に媒介された過程を支持するものである。
【0006】
最初の脂肪酸輸送タンパク質(FATP)は、ラット脂肪細胞において1994年に同定された。2つの対応する遺伝子がその後に同定され、そして現在はFATP1およびアシルコエンザイムAシンテターゼ(ACS)と呼ばれている。マウス遺伝子ライブラリーのスクリーニングにより、FATPファミリーの全ての成員における高度に保存された311個のアミノ酸からなるサイン配列(signature sequence)によって特徴づけられるタンパク質のファミリーが、その後明らかになった。これらはすべてAMP結合部位を含む。これまでに、6種のマウスFATPおよび6種のヒトFATPが同定された。コエンザイムAシンテターゼ活性は、現在FATP1、2、4および5に関連している。LCFA捕獲におけるこれらの役割は、これらの酵素活性から推論することができる。あるいは、FATPの発現は、遊離脂肪酸の細胞内レベルの増加を導くことができ、したがって、VLACS(超長鎖アシル−CoAシンテターゼ)活性を誘導することができる。実際、アシル化に関与する酵素、例えばアシルコエンザイムAシンテターゼ(ACS)は、PPAR(ペルオキシゾーム増殖因子活性化受容体)転写因子およびある種の長鎖脂肪酸によって正に調節されている。
【0007】
アシル化は、脂肪酸を活性化し、アシル−CoAの活性代謝誘導体を形成し、これはいかなる代謝経路にも入ることができるようになる。小胞体において、脂肪酸はトリグリセリドの合成に関与することができる。ミトコンドリアおよびペルオキシソームにおいて、FATPは脂肪酸の分解を誘導することができる。
FATPは、そのアミノ末端に応じて異なる細胞内位置を占めている。これらのN末端は、脂肪酸を異なる細胞内コンパートメントに分配し、その後別々の代謝経路へのこれらの組み込みを誘導するために、様々なパートナーと相互作用することができる。
【0008】
これらは少なくとも2つの相同サインユニットを保有しているが、異なるFATPは別々の染色体位置、特有のプロモーターおよび遺伝子調節、特有の分布プロファイル、異なる細胞位置、および専門化された機能を有する。
従来技術において、FATP5は、このタンパク質について検出可能な活性に応じて、例えば超長鎖アシル−CoAシンテターゼ関連物(VLACSR)、超長鎖アシル−CoAシンテターゼホモログ(VLCS−H2)、コリル−コエンザイムAリガーゼまたは胆汁酸コエンザイムAシンテターゼ(BACS)などの異なる名称で見出される。
【0009】
ヒトにおいて、タンパク質はもっぱら肝臓で発現される。
マウスにおいては、2.6kbのメッセンジャーRNAが肝臓で非常に豊富であることが知られている。より短いメッセンジャーRNAのより低いレベルの発現が、脳、肺、睾丸および脾臓(2.5kb)において、そして、骨格筋(2.2kb)において検出された。転写物の存在は、PCR技法により心臓において示されたが、腎臓においては示されなかった。
ヒトにおいて、転写物は、そのマウスホモログと対照的に、線維芽細胞、脳または心臓で検出することができなかった。
サザンブロット分析は、VLACSR遺伝子がヒトおよびマウスの双方で単一コピーで存在することを示す。
【0010】
例えば、FATP5が長鎖脂肪酸および超長鎖脂肪酸(例えばC18〜C26)を活性化できることが報告されている。FATP5は、ケノデオキシコレート、デオキシコレート、リトコレートおよびトリヒドロキシコレスタノン酸(trihydroxycholestanoic acid)を活性化することができる。
本発明者らの知る限りでは、脂肪酸分解におけるFATP5の関与は、これまで全く記載されたことはなかった。FATP5の役割は、腸肝再循環を介した脂肪酸の再活性化だけに限定されていた。
従来技術において、国際出願WO 01/21795は、FATP5が脂肪酸輸送体として役割を果たし、そうして細胞による脂肪酸捕獲の過程に関与することを示唆している。
【0011】
さらにまた、糖尿病に罹患した者、または糖尿病を発症しやすい者は、血流中に高レベルの遊離脂肪酸を有することが知られている。血漿中の遊離脂肪酸の過剰は、インスリン感受性組織、例えば骨格筋において、エネルギーを産生するためのブドウ糖の利用に対する競合者のように作用する。脂肪酸は、異なるレベルで糖代謝に干渉する。これらは、間接的に解糖を阻害し、グリコーゲンの形態でのブドウ糖の貯蔵を減らし、間接的にグルコース輸送を阻害する。これは、遊離脂肪酸が過剰である場合におけるその優先的な利用、および、高血糖症の発症を導く、循環中のブドウ糖濃度の増加を示す傾向がある。
【0012】
驚くべきことに、そして、予想を超えて、本発明者らは今回、FATP5が肝臓における脂肪酸の分解に関与することを見出した。同様に、本発明者らは、この特性が、結果として間違いなく血漿中のブドウ糖および脂質濃度を減少させることを示すことができた。特に、本発明者らは、FATP5のメッセンジャーRNAの発現レベルが、糖尿病の肥満ラット(Zucker糖尿病肥満型ラット(ZDFラット))において、非糖尿病ラット(Zucker痩せ型ラット(ZLCラット))におけるFATP5のメッセンジャーRNAの発現レベルに比べて大幅に減少していることを示すことができた。
したがって、FATP5の過剰発現が、結果として遊離脂肪酸のレベルを減らし得ることを想起することが可能となり、したがって、FATP5の発現レベルを増やす能力を有する任意の物質が、糖尿病の予防的または治療的処置用の医薬の製造のための良い候補であり得ることを十分に予測することができる。
本発明は、この応用分野に含まれる。
【0013】
本発明者らは、ヒトFATP5遺伝子のプロモーターである、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、ヒト核酸配列を単離した。この配列は、従来技術においてFATP5プロモーターの配列であると考えられていた配列と、わずかにしか相同ではない。
本発明は、したがって、第一に、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、単離されたヒト核酸配列に関する。前記配列は、ヒトFATP5遺伝子のプロモーターに対応する。
【0014】
本発明はさらに、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列からなることを特徴とする、単離されたヒト核酸配列に関する。
本発明の1つの特定の実施例において、配列番号1の番号で特定されるヌクレオチド配列を少なくとも含む核酸配列は、タンパク質、特にヒトFATP5タンパク質をコードする。特に好ましくは、配列番号1の番号で特定されるヌクレオチド配列を少なくとも含み、かつ、ヒトFATP5タンパク質をコードする前記核酸配列は、添付された配列表において配列番号2の番号で特定されるヌクレオチド配列に対応する。
【0015】
本発明はさらに、上記と相補的なセンスまたはアンチセンス核酸配列、および、本発明による核酸配列のうちの1つと、少なくとも80%、好ましくは少なくとも90%の同一性パーセントを有する任意の核酸配列に関する。
参照配列と少なくともX%の同一性パーセントを有する核酸配列は、本発明において、参照配列の100ヌクレオチドあたり100−X個までの変更を含み得るが、前記参照配列の機能的な特性を保持している核酸配列として定義される。本発明に関して、「変更」は、参照配列における連続的または散剤的なヌクレオチドの欠失、置換または挿入を含む。
本発明による少なくとも80%、好ましくは少なくとも90%の同一性パーセントを有する核酸配列は、アレル変異体、すなわち配列番号1または配列番号2の配列の個体変異に対応する全ての配列を含む。これらの自然な変異配列は、哺乳類、特にヒトに存在する多型に対応する。
【0016】
本発明はさらに、上記に定義した本発明の核酸配列が挿入されたクローニングおよび/または発現ベクターに関する。好ましくは、本発明のベクターは、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含む核酸配列を含む。
本発明の1つの特定の態様において、本発明のベクターはまた、上記に定義された、ヒトFATP5遺伝子のプロモーターとして同定された核酸配列の依存下に置かれた、検出可能なタンパク質をコードする核酸配列を含む。特に、このプロモーターは、配列番号1の配列に対応する。「検出可能なタンパク質」とは、一度発現されると、当業者に知られている任意の技術によって容易に同定される任意のタンパク質を意味すると理解される。FATP5自体、またはマーカーとして用いられるタンパク質、例えば緑色蛍光タンパク質(GFP)、レニラ(renilla)またはルシフェラーゼを、用いることができるタンパク質として挙げることができる。
【0017】
かかるベクターは、宿主細胞における表現、および任意にタンパク質の分泌のために必要な要素を含むことができる。
前記ベクターは、好ましくは、翻訳開始および終止シグナル、ならびに適切な転写調節領域を含む。これらは細胞中で安定して維持することができる必要があり、そして、翻訳されたタンパク質の分泌を指示する特定のシグナルをコードする配列を任意に含むことができる。
本発明による核酸配列は、選択された宿主内の自律複製型ベクター中に、または選択された宿主の組込型ベクター中に挿入することができる。
自律複製系のなかでは、宿主細胞にしたがって、プラスミドまたはウイルス型のシステムを用いることが好ましい。
本発明によって用いることができるプラスミドベクターは、核酸配列の発現を可能にする任意の既知のプラスミドであってもよい。
【0018】
「核酸配列の発現」とは、本発明によれば、本発明の核酸配列のRNAへの転写、および任意にこれに続く前記RNAのタンパク質への翻訳を可能にする本発明のベクターの能力を意味するものと理解される。
好ましくは、本発明によるプラスミドは、上記の単離されたヒト核酸配列の発現を可能にする。特に好ましくは、本発明によるプラスミドは、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列、特にFATP5をコードする核酸を少なくとも含むことを特徴とする核酸配列の発現を可能にする。
【0019】
以下のものを、本発明によって用いることができるプラスミドの例として挙げることができる:プラスミドpCMVTag(Stratagene, La Jolla, USA)、pcDNA3(Invitrogen, Cergy Pontoise, France)、pSG5(Stratagene, La Jolla, USA)またはpGL2およびpGL3(Promega, Mannheim, Germany)。
ウイルスベクターは、特にアデノウイルス、レトロウイルス、レンチウイルス、ポックスウイルスまたはヘルペスウイルスであってもよい。当業者は、これらの系の各々のために用いることができる技術に精通している。
核酸配列を宿主細胞の染色体に組み込むことが望ましい場合は、例えば、プラスミドまたはウイルス型のシステムを用いることが可能であり、このようなウイルスは、例えばレトロウイルスまたはアデノ随伴ウイルス(AAV)である。
【0020】
非ウイルスベクターの中では、好ましいのは、裸のDNAまたはRNAなどの裸のポリヌクレオチド、バクテリア人工染色体(BAC)、酵母における発現のための酵母人工染色体(YAC)、マウス細胞における発現のためのマウス人工染色体(MAC)、および、好ましくは、ヒト細胞における発現のためのヒト人工染色体(HAC)である。
本発明によると、ベクターは好ましくはプラスミドまたはアデノウイルスである。
かかるベクターは、当業者によって一般に用いられている方法によって調製し、そして、得られた組換えベクターは、標準的な方法、例えばリポフェクション、エレクトロポレーション、熱衝撃、膜の化学的透過化処理後の形質転換、または細胞融合によって適切な宿主に導入することができる。
【0021】
本発明はさらに、本発明による少なくとも1種の核酸配列または本発明による少なくとも1種のベクターが導入された、形質転換された宿主細胞、特に真核細胞および原核細胞に関する。
細菌細胞、酵母細胞および動物細胞、特に哺乳類細胞を、本発明に関して用いることができる細胞として挙げることができる。昆虫細胞、例えばバキュロウイルスを伴う方法を用いることができるものも挙げることができる。
【0022】
本発明はさらに、上記の宿主細胞を、該細胞中に導入された本発明による核酸配列の、タンパク質の形態での発現を可能にする条件下で培養することを特徴とする、タンパク質を産生する方法に関する。
本発明はさらに、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定のための、上記の本発明の核酸配列、ベクターまたは宿主細胞の使用に関する。
【0023】
本発明はさらに、上記の本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質を特定する方法であって、
a)上記の核酸配列、ベクターまたは宿主細胞を、ヌクレオチド配列の発現に適した媒体中で、上記の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質と接触させること、および
b)上記の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルを測定すること、
を特徴とする方法に関する。
【0024】
「ヌクレオチド配列の発現に適した媒体」は、本明細書中では、本発明の核酸配列からの、RNA、特にメッセンジャーRNA、および任意にタンパク質の合成を可能にする任意の発現系を意味すると理解される。
1つの特定の態様において、本発明の方法は、b)で決定された発現レベルと、同定する物質に接触させなかった対照における、上記の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルとの比較のための、追加の工程を含む。
比較は、本発明の核酸配列に対する被験物質の調節能力を評価することを可能にする。
【0025】
「調節する」または「調節能力」は、本発明によれば、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を刺激または阻害する被験物質の能力を意味すると理解される。
本発明の核酸配列の依存下に置かれたヌクレオチド配列の発現レベルは、自体公知の従来のmRNAまたはタンパク質分析技法で測定することができ、以下の技術を、非限定的な例として言及することができる:RT−PCR、ノザンブロッティング、ウェスタンブロッティング、RIA、ELISA、免疫沈降、および免疫細胞化学または免疫組織化学分析技術。細胞もしくは分子生物学、または免疫細胞化学もしくは免疫組織化学分析の実験プロトコルに関するすべてのことについては、本特許出願の各所において、当業者に完全に精通している多数の著作、特に「Current Protocols in Immunology; John Wiley and Sons, Teton Data System, Jackson, Wyoming (ISBN 0-471-30660-6, 2003)」を参照することができる。
前記測定は、プローブ、プライマーまたは抗体を用いて有利になされる。
【0026】
本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質は、生体高分子、例えば、核酸、脂質、糖、タンパク質、ペプチド、タンパク質−脂質、タンパク質−糖、ペプチド−脂質もしくはペプチド−砂糖ハイブリッド化合物、または化学的分枝もしくは化学分子が付加されたタンパク質またはペプチドであってもよい。
本発明はさらに、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質であって、本発明の方法により入手可能な前記物質に関する。
【0027】
本発明はさらに、本発明による核酸配列と相互作用することが可能な物質を同定する方法であって、
a)上記の核酸配列、ベクターまたは宿主細胞を、本発明による核酸配列と相互作用することが可能な物質に接触させること、および
b)前記核酸配列と前記被験物質との間の相互作用を評価すること、
を特徴とする方法に関する。
本発明を実行する1つの特定の方法では、前記同定方法の目的は、本発明による核酸配列と結合することが可能な物質を特定することであり、前記方法は、例えば、異種相互作用の場合は熱量測定であり、または、ペプチド−ペプチド相互作用の場合は、2ハイブリッド試験である。
【0028】
最後に、本発明は、本発明による核酸配列と相互作用すること、または結合することが可能な物質であって、上記の同定方法のうちの1つによって入手できる前記物質に関する。
本発明によれば、前記核酸配列と前記被験物質との間の相互作用および/または結合は、任意の既知の技法によって評価することができる。
【0029】
本発明者らは、FATP5が肝臓における脂肪酸の分解に関与していることを示した。同様に、本発明者らは、この特性が、結果として間違いなく血漿ブドウ糖および脂質濃度を減少させることを示すことができた。特に、本発明者らは、FATP5メッセンジャーRNAの発現レベルが、糖尿病ラット(Zucker糖尿病肥満型ラット(ZDFラット))において、非糖尿病ラット(ZLCラット)におけるFATP5メッセンジャーRNAの発現に比べて大幅に減少していることを示すことができた。
したがって、本発明はさらに、糖尿病を予防および/または処置するための医薬の製造のための、本発明による物質の使用に関する。
本発明はさらに、肝臓における脂肪酸のβ酸化を誘導するための医薬の製造のための、本発明による物質の使用に関する。
【0030】
上記の事項に加えて、本発明はまた、本発明をいかに実行するかについての例および添付の図面を参照した以下の説明から明らかになる他の事項を含む。
以下の例は、いかなる意味においても制限を意味することなく本発明を例証する。
【0031】
例1:脂肪酸のβ酸化におけるhFATP5の関与の証明
材料および方法
A)FATP5発現プロファイル
a)RNAの調製
全RNAは、細胞および組織から、ChomczynskiおよびSacchi(Chomczynski et al., 1987)により記載された技法を用いて調製した。RNAは260nmでの光学密度の測定によって定量し、そして、RNAの品質は260/280光学密度比の測定によってチェックした。
1μgの各サンプルを、酵素MMLV−RT(Gibco BRL, Paisley, UK)を用いた逆転写(RT)反応においてテンプレートとして用い、ポリメラーゼ連鎖反応を、2μlのRT産物について実行した。特異的プライマー(Eurogentec, Seraing, Belgium)を、hFATP5およびrFATP5について調製した。PCR産物の特異性を確保するために、熱変性温度(Tm)は各々の特定の標的について最適化し、サイクル数は、転写産物の量に応じて、25〜30の間に設定した。
【0032】
【表1】

【0033】
DNA PCR産物は、1%のアガロースゲル上で分析した。シグナルは、Gel Doc(登録商標)2000装置(Bio-rad, Marnes La Coquette, France)を用いて、密度測定値として定量化した。in vivo転写分析については、定量化は、特定の各標的の値の、内部対照(アクチン)の値に対する比として計算した。結果は、1に設定された対照と比較した誘導レベルとして表示した。
【0034】
定量分析は、32キャピラリーLight Cycler装置(Roche)にて、二本鎖DNAを標識するフルオロフォアであるサイバーグリーンを用いて、二段階PCR(Roche, Mannheim, Germany)によって行った。縦座標の増幅曲線の微分値(PCRの最中の蛍光の増大を表す)と、横座標のサイクル数との交点として表された各標的についての結果は、対照遺伝子で正規化した。
【表2】
【0035】
B)Zucker糖尿病肥満型ラットにおける糖尿病発症の動態
Genetic Model Inc.(Charles River Laboratories, Wilmington, MA, USA)に由来する5週齢の雄のZucker糖尿病肥満型ラット(ZDF/GMI−fa/fa)および同週齢の雄のZucker痩せ型ラットは、無制限量のPurina 5008(PMI Nutrition International, Wellingborough, Northamptonshire, UK)で飼育した。
6週齢、10週齢および20週齢において、48頭のラットを、16頭(8頭のZucker糖尿病肥満型ラットおよび8頭のZucker痩せ型ラット)の3群に分けた。動物の重量およびその摂食量は、実験の間中モニターした。
【0036】
血液サンプルは、日中、8時間の食餌療法期間の後、麻酔下にて眼窩採血により採取した。麻酔は、ペントバルビタールの腹腔内投与によって誘発した。動物は頚椎脱臼によって安楽死させ、肝臓を摘出し、重量を測定し、さらなる分析のために液体窒素中で直ちに凍結させた。
【表3】
【0037】
b)分析
血清グルコース、コレステロール、トリグリセリドおよび遊離脂肪酸は、酵素によるバイオアッセイを用いて決定した。
インスリンレベルは、放射免疫分析により決定した。
肝臓の一部は、組織学的研究のために保存した。
【0038】
C)アデノウイルス中へのhFATP5の完全な配列のクローニング
別途示さない限り、用いた技法は、供給者によって、または分子生物学に関する多数の古典的な著作に記載されたものである。
hFATP5の完全なコード配列(配列番号2の配列の1575位におけるATGから)は、Adeno-X(Clontech, Palo Alto, USA)のPT3414−1発現系にクローニングした。
ヒト肝細胞培養物の全RNAは、Qiagen(Qiagen, Courtaboeuf, France)からの、RNeasy miniprep kitで抽出した。
【0039】
次いで、第一DNA鎖を以下の通りに合成した。
【表4】
【0040】
次に、以下のプライマー:
配列h5_4As(Invitrogen, Cergy Pontoise, France):配列番号19:GCTCTAGATCAGAGCCTCCAGGTTCCCTCACACACAGCC
配列h5_1S(Invitrogen, Cergy Pontoise, France):配列番号20:GCTCTAGATGGTACCATGGGTGTCAGGCAACAG
を用い、以下の反応媒体:
センスオリゴ(10pM)1μl
アンチセンスオリゴ(10pM)1μl
TaqプラチナPfx+バッファー10×+MgSO4(Invitrogen, Cergy Pontoise, France)
dNTP(Promega, Williamsburg, IA)
最終容量は20μl
により、PCRを実行した。
【0041】
次に、以下のプログラムにしたがって増幅を行った:
94℃で5分間を1サイクル、次いで94℃での30秒、次に55℃での30秒、そして68℃での2分を含む25サイクル。
その後、産物を4℃で維持した。
得られたPCR断片を、QIAquick PCR精製キット(Qiagen, Courtaboeuf, France)を用いて精製した。
精製した断片を、次に、XbaI(NEB, Beverly, MA)で消化し、ClontechからのpAdenoXキットのベクターpShuttle中に、供給者の推奨にしたがってクローニングした。
【0042】
得られたプラスミドを、DH5d型の大腸菌に導入し、そして、クローンをこれらのカナマイシン耐性によって選択した。次に、陽性クローンのプラスミドDNAを精製し、制限分析によりチェックし、配列決定した。タンパク質の発現は、T7発現系を用いて、in vitroで確認した。pShuttleの発現カセットを、PI−Sce/I−CeuIを用いて切断し、Adeno-X(登録商標)ウイルスDNAにライゲーションした。ライゲーション産物をSwaIで消化し、そして大腸菌中に導入した。アンピシリン耐性クローンを選択した。
対象となる遺伝子を含んでいる組換えアデノウイルスDNAをPacIで消化し、ヒト胎児腎細胞(293細胞)中に導入した。アデノウイルス組換え体を回収し、3回の増幅サイクルにより再増幅した。ウイルスはCICs法によって精製し、次にSephadex G50カラムで洗浄した。ウイルス粒子は、96穴プレートにおけるプラーク形成アッセイにより力価測定した(Pfu)。
【0043】
D)ラット肝細胞の調製
ラット肝細胞は、コラゲナーゼ灌流によって雄ラットの肝臓から単離した。ラットを、ペントバルビタールの腹腔内投与によって麻酔した。次に、肝臓を門脈において、まず200mlの肝臓潅流培地で、そして次に、10mMのHepes、4mMのCaCl2および14mgのBlendzyme3が添加された200mlのHanksバッファーで灌流した。肝臓を切開し、肝細胞洗浄培地(Gibco BRL, Paisley, UK)中で細切し、70μmのフィルターで濾過した。
細胞は遠心分離し、肝細胞洗浄培地中で3回洗浄した後、細胞の生存率をトリパンブルー排除法により評価した(生存率>85%)。細胞は、UltroserSF(2容量%)、ペニシリン(100U/ml)、ストレプトマイシン(100μg/ml)、遊離脂肪酸/BSA(SIGMA, St. Louis, MO, USA)(0.2重量/容量%)、L−グルタミン(2mM)、デキサメタゾン(1μM)、トリヨードサイロニン(T3、SIGMA, St. Louis, MO, USA)(100nM)およびインスリン(100nM)が添加されたWilliams培地中に接種した。4時間後に、培養培地を、UltroserまたはBSAを含まない同様のウィリアムズ培地と交換した。
【0044】
E)糖尿病を発症中のZDFラットへのhFATP5アデノウイルスおよびLACZアデノウイルスの感染
Genetic Model Inc.(Charles River Laboratories, Wilmington, MA, USA)に由来する5週齢の雄のZucker糖尿病肥満型ラット(ZDF/GMI−fa/fa)および同週齢の雄のZucker痩せ型ラットは、無制限量のPurina 5008(PMI Nutrition International, Wellingborough, Northamptonshire, UK)で飼育した。
8週齢において、5.109PfuのアデノウイルスまたはPBS(対照用)を、ラットの尾静脈中に注射した。注射容量は、1ラットあたり2ml未満であった。3頭のラットをpAdhFATP5に、そして3頭をpAdLacZに感染させた。ラットは、10週で安楽死させた。ブドウ糖、遊離脂肪酸およびトリグリセリドレベルを、血漿中で測定した。
【0045】
F)機能試験
a)脂肪酸の捕獲
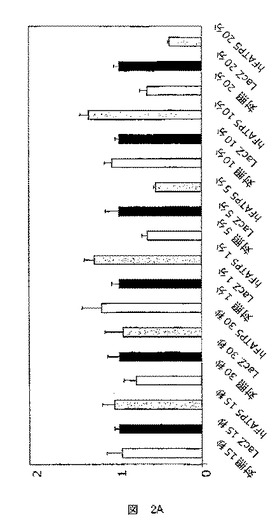
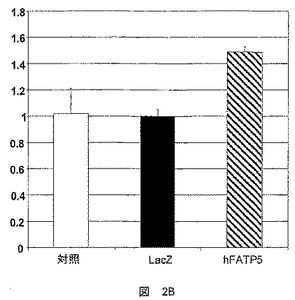
BSAと複合体化した最終20μMの14C標識オレイン酸(図2B)または最終78nMのトリチウム標識オレイン酸(図2A)を、輸送試験に用いた。
ラット肝細胞(106細胞/コンパートメント)を6穴プレートのウェルに接種し、37℃で4時間インキュベートした。3T3−L1細胞(ATCC No. CL-173、Manassas、VA、USA)(5.105細胞/コンパートメント)は、2%のウシ胎仔血清(FCS)を含むDMEM中でインキュベートした。感染は多重度が1であり、一晩継続した。
【0046】
細胞はトリプシンで処理し、回収し、遠心分離し、PBSで再懸濁して5.105細胞/mlの懸濁液を得た。200μlの細胞懸濁液(105細胞)は、5mlのポリプロピレンチューブ中に置いた。PBS中の細胞懸濁液は、水浴中37℃で5分間、振盪しながらプレインキュベートした。脂肪酸の捕獲のために、2倍に濃縮した脂肪酸/BSAの原液を各チューブに等量加えた。所定の間隔で、0.1%のBSAと200μMのフロレチン(SIGMA, St. Louis, MO, USA)を含むPBSの冷液(0℃)(洗浄液)5mlを各チューブに添加することにより捕獲を停止させた。細胞を遠心分離し、二回洗浄した。遠心分離残渣を、400μlの溶解緩衝液(水中の0.4%のSDS)で再懸濁した。次に4mlのシンチレーター溶液を加え、取り込まれた脂肪酸をTri-Carb 2100カウンター(Packard, Meriden, CT, USA)でカウントした。結果はLacZの誘導の割合として表し、1に対して正規化した。
【0047】
b)脂肪酸のアシル化
ラット肝細胞(106細胞/コンパートメント)を、6穴プレートのウェルに接種し、37℃で4時間インキュベートした。次に培地を回収し、2mlの伝染性混合物(DMEM、2%のFCS)を各ウェルに加えた。感染は多重度が1であり、一晩継続した。細胞をトリプシンで処理し、回収し、遠心分離した。遠心残渣を、300μlの緩衝液A(pH8.5の500mMのtris HCl、1mMのMgCl2、100mMのNaCl、1mMのATP、0.1%のTritonX 100)で再懸濁した。細胞を超音波処理し、氷中で30分間インキュベートした。タンパク質レベルは、ブラッドフォード法により定量化した。1試験あたり、20μgのタンパク質を含む20μlの抽出物を用い、ガラスチューブ中に置いた。180μlの反応混合物(pH8.5の50mMのtris HCl、150μMのコエンザイムA、300μMのDTT、10mMのATP、10mMのMgCl2、0.1%のTritonX 100、14Cで標識された10μMのパルミチン酸またはリグノセリン酸)を、試験を開始するために加えた。反応は、5〜30分後に800μlの1%の過塩素酸の添加により停止させた。
【0048】
標識されたオレイル−CoAのレベルを評価するために、反応混合物を2.25mlのイソプロピル/ヘプタン/硫酸(40/10/1)で抽出した。抽出物を混合し、有機相を回収した。2回の連続した抽出を実行した。4mlのシンチレーター溶液を1mlの水相に加え、Tri-Carb 2100TRカウンターでカウントした。アシルCoAの量を測定するために、沸騰させたタンパク質の対照抽出物で測定した放射活性を、対応する試験値から差し引いた。
結果は、1について正規化したLacZの誘導の割合として表した。
【0049】
c)脂肪酸の酸化
ラット肝細胞(6.105細胞/コンパートメント)を、12穴プレートのウェルに接種し、37℃で4時間インキュベートした。感染は多重度が100であり、一晩継続した。次に、培地を回収し、64nMのオレイン酸[9,10 3H]と2%のBSAとが添加された500μlのDMEMを各ウェルに加えた。37℃での12時間のインキュベーションの後、培地をマイクロチューブへ移し、過剰なオレイン酸[3H]を、50μlの10%のトリクロロ酢酸と50μlの20%のBSAとで沈殿させた。混合物を、12,400rpmで2分間遠心分離した。上清をマイクロチューブに移し、500μlの水を加えた。インキュベーションを、50℃で18時間継続した。4mlのシンチレーター溶液の添加の後、トリチウム水を測定し、Tri-Carb 2100カウンターでカウントした。
結果は、1について正規化したLacZの誘導の割合として表した。
【0050】
d)脂肪酸のエステル化
脂肪酸の酸化は培地中で測定したが、トリグリセリドは、標識された脂肪酸とともにインキュベートした細胞から抽出した。ラット肝細胞をトリプシンで処理し、全く同一のチューブ3本中で混合した。細胞は、18,500rpmで2分間遠心分離した。各ウェルを重炭酸塩ベースの溶液(クレブス−リンゲル重炭酸バッファー:1.2mMのKH2PO4、26mMのNaHCO3、1.3mMのMgCl2、124mMのNaCl、5mMのKCl、10mMのブドウ糖)で洗浄し、そしてこの緩衝液を、遠心分離残渣を再懸濁するのに用いた。
【0051】
細胞を二度目の遠心分離に供し、上清を回収した。この行程を繰り返した。次に遠心分離残渣を100μlの水中に再懸濁し、さらなる分析のために液体窒素で凍結することができる。その後、遠心分離残渣を、室温(25℃)に放置した。次に、500μlのアセトンを加え、混合物を濃縮器を用いて、真空下で乾燥させた。残渣を、基質として5μlの50nMのトリオレインを含む、300μlのクロロホルム/メタノール混合物(1/1)で再懸濁した。混合後、全体を遠心分離し、上清を除去した。2回の抽出を行った。遠心分離残渣は、30μlのクロロホルム/メタノール混合物(1/1)で再懸濁し、G60シリコーンゲルでプレコートされた薄層クロマトグラフィープレート上に堆積させた。移動は、200mlの緩衝液(H−ヘキサン/ジエチルエーテル/メタノール/酢酸 90/20/2/3)で、1.5時間行った。
【0052】
結果:
A)hFATP5の細胞および組織分布
PCRによる研究により、ヒトFATP5の、肝臓、初代肝細胞およびHepG2ヒト細胞系の細胞における排他的な発現が確認された。
これらの結果を図1に示す。
【0053】
B)FATP5の機能的特徴化
a)アデノウイルスの構築
肝臓での脂肪酸の代謝におけるFATP5の機能を特徴化するため、hFATP5遺伝子を担持するアデノウイルス(pAdeno−hFATP5)、対照アデノウイルスpAdeno−LacZ、およびアデノウイルスpAdeno−CD36を設計し、最後のものは脂肪酸輸送についての陽性対照としてのものである。
【0054】
b)脂肪酸の捕獲
オレイン酸の捕獲は、3T3−L1細胞(図2A)およびアデノウイルスに感染したウィスターラットから単離したラット肝細胞(RH)(図2B)において試験した。
FATP5の過剰発現は、3T3−L1細胞における脂肪酸輸送活性に影響を及ぼさなかったが(図2A)、ラット肝細胞において脂肪酸輸送を有意に増大させた。
この結果は、脂肪酸輸送活性が細胞系によって別様に行われていることを示唆するものである。これは、細胞系に固有の種々のコファクターの効果的な動員による可能性がある。以前の結果は、FATPがACSまたはFATなどのパートナーと協働して作用し得ることを示唆している。
ここで提示された結果は、FATP5が、タンパク質が発現された細胞によって別様に、脂肪酸輸送に作用することを示唆するものである。
さらにまた、前脂肪細胞系におけるFATP5の効果の欠如、およびラット肝細胞における脂肪酸輸送の急速な増大は、FATP5が、単なるありふれた輸送体であると言うよりは、むしろ、肝細胞における脂肪酸輸送に影響する特有の機能を有することを示唆する。
【0055】
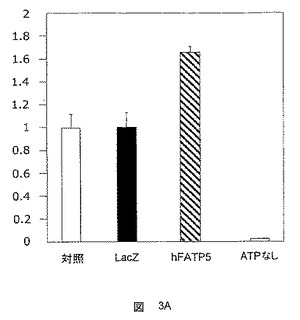
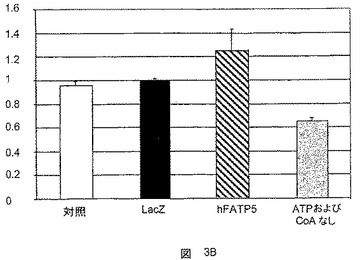
c)ACSおよびVLACS活性に関する試験
ACS試験は、パルミチン酸塩およびリグノセリン酸塩を用いて、アデノウイルスに感染した初代ラット肝細胞において行った。hFATP5の過剰発現による、リグノセリン酸塩とのインキュベーション後のVLACS活性の有意な増大は、30分のインキュベーション期間でのみ検出可能であった(図3A)。hFATP5の過剰発現によるACS活性の誘導が間接効果である可能性が示唆される。
超長鎖アシルCoAシンテターゼ活性はわずかに増大したが、CD36の過剰発現がリグノセリン酸塩のアシル化に影響を及ぼさないことから(図3B)、これはhFATP5に特有のことである。
FATPタンパク質のVLACS活性に関する先に発表された結果は、このようにして確認された。しかしながら、誘導動態には明確な差異がある。結果は、FATP5が、超長鎖脂肪酸より大きな脂肪酸のクラスを代表するパルミチン酸塩などのC16脂肪酸に対する有意な効果を示すことから、FATP5がより幅広い範囲の活性を有することを示唆している。
【0056】
d)脂肪酸のβ酸化に対するhFATP5発現の効果
hFATP5の過剰発現によって脂肪酸の分解の程度が調節されるかどうかを分析するために、トリチウム化オレイン酸の、β酸化の最終産物であるトリチウム水への変換を測定した。
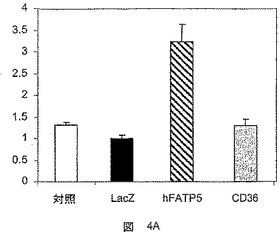
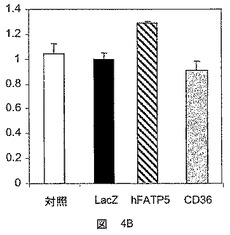
CD36対照遺伝子の過剰発現は、HepG2細胞(図4A)またはラット初代肝細胞(図4B)におけるβ酸化に影響を及ぼさず、これは、CD36が脂肪酸のエステル化を導くが、その分解は導かないとする仮説に対応するものである。
対照的に、hFATP5の過剰発現は、HepG2肝細胞におけるβ酸化の程度を顕著に増大させた(3倍超)(図4A)。
hFATP5の過剰発現は、初代肝細胞において脂肪酸の分解を誘導する。
【0057】
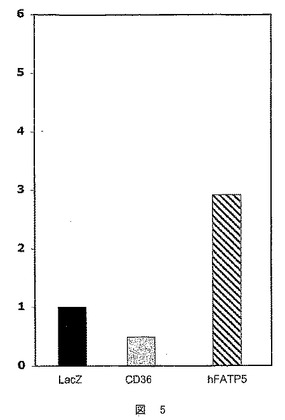
e)hFATP5が脂肪酸のエステル化よりはむしろそのβ酸化に関与することの確認
hFATP5の発現が脂肪酸の分解のみを導き、そのエステル化度に影響しないことを確認するために、HepG2細胞における同じ実験の中で、β酸化試験を、エステル化試験により補完した。
hFATP5の過剰発現は、エステル化度に影響を及ぼさなかった(図5)。
このことは、肝臓における脂肪酸の分解のみからなる、hFATP5の新規な機能的活性を明確に示すものであり、一方、FAT/CD36は、脂肪酸をエステル化過程に導く。hFATP5は、β酸化アクティベーターである。
【0058】
f)Zucker糖尿病肥満型ラットにおける糖尿病発症の最中のhFATP5の発現の調節
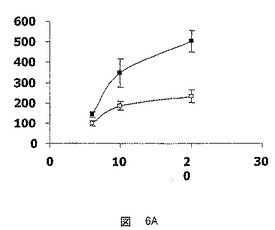
Zucker糖尿病肥満型ラットにおける、このモデルでのNIDDMの発症の最中のFATP5遺伝子の発現レベルの研究を行い、結果をインスリンおよびブドウ糖レベルにおける変動と相関させた。
Zucker糖尿病肥満型ラットは、ヒトの場合と同様の経過で、糖尿病を自然に発症し(図6Aおよび6B)、
−6週齢の雄のZDF(fa/fa)ラットについて観察される、高インスリン血症および正常血糖の状態、
−10週齢の雄のZDF(fa/fa)ラットについて観察される、高インスリン血症および高血糖の状態、および、
−20週齢の雄のZDFラットについて観察される、インスリン欠乏および高血糖の状態、
を有する。
hFATP5の発現の定量的および半定量的な分析による研究(表IIIおよび図7)は、hFATP5メッセンジャーRNAのレベルがZDFラットにおいて10週で低減することを示した。同じ期間にわたって、グリコーゲンレベルはZDFラットの方が、ZLCラットよりも明らかに高かった(表III)。
【0059】
【表5】
生化学的分析は、血漿中の遊離脂肪酸のレベルが10週齢および20週齢のZLCラットでより高いが、ZDFラットの肝臓中の遊離脂肪酸のレベルはZLCラットと比較して低かったことを明らかにした(図8A)。
この10週の期間は、ZDFラットおよびZLCラットの体重増加曲線の乖離によって特徴づけられ、これは糖尿病の発症を示す(図8B)。
【0060】
g)脂肪酸のアシル化へのFATP5の過剰発現の効果
ZDFおよびZLCラットの初代肝細胞におけるhFATP5の過剰発現の機能的効果を分析した。
FATP5の過剰発現は、ZDFラットの初代肝細胞において、早くには5分で、そして遅くには30分で、脂肪酸のアシル化を極めて有意に増大させた。
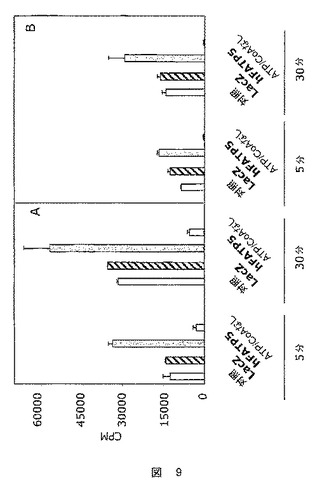
これらの結果を図9に示す。
【0061】
h)糖尿病肥満型ラットの肝臓における脂肪酸分解不足の解消
脂肪酸のベータ酸化は、ZDFラットの肝臓において、ZLC対照ラットの肝臓において生じるものと比べて不足している。FATP5の過剰発現は、この不足を解消する。FATP5の過剰発現は、対照ラットには影響を及ぼさない。これらの結果を図10に示す。
【0062】
i)FATP5の過剰発現は、ZDFラットの血中のブドウ糖および脂質のレベルを低減する
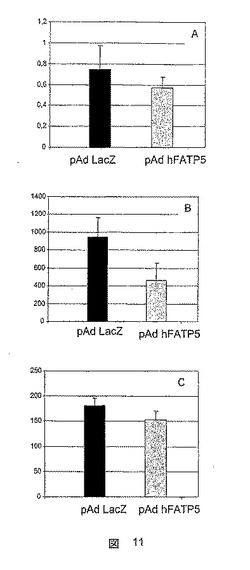
ブドウ糖レベルは、FATP5を担持しているアデノウイルスに感染したZDFラットにおいて、10週齢で、LacZ対照アデノウイルスに感染したラットと比較して、有意に低減した(図11C)。この低下は、遊離脂肪酸レベルの低下(図11A)、および血漿トリグリセリドレベルの大幅かつ急速な低下(図11B)と相関している。
これは、FATP5の発現が糖代謝に影響を及ぼし、in vivoで高血糖症を軽減することの証明である。
【0063】
例2:ヒトFATP5遺伝子の推定プロモーター領域の特徴化およびプロモーター活性の証明
FATP5遺伝子産物(AF064255またはNM_012254)は、脂質合成に関与するタンパク質のファミリーに属する。
FATP5遺伝子産物は、複合脂質の合成、そして特に脂肪酸の伸長に関与している。FATP5遺伝子は、第19染色体に位置している。
FATP5をコードする配列に関する一般的な情報は、以下の通りである(表IV)。
【表6】
【0064】
a)予測によるFATP5の転写開始点(TSS)の位置、および相補DNA末端の迅速増幅技術(cDNA末端の迅速増幅、RACE)による同定
FATP5遺伝子は、第19染色体に位置し、14kbにわたって分配された10個のエクソンで構成されている。FATP5遺伝子を含むコンティグNT_011104は、2002年8月に公開された。
mRNAおよびESTからマッピングした5’UTR領域は、34bpのサイズを示す(5’UTR1)。したがって、mRNAおよびGenBankによって公開されているEST、およびDBTSS(ヒト転写開始点データベース)に由来する情報に関連して決定される、最も確からしい「生物学的」転写開始部位は、図12Bで示したとおりに位置している。
b)予測によるFATP5プロモーターの位置
材料および方法
HepG2細胞のRNAの調製:
RNAは、Qiagen RNeasy mini kit protocol(Qiagen, Courtaboeuf, France)に従って、100万個のHepG2細胞から精製した。
5’RACE:
TSSは、供給者の推奨に従って、GeneRacerキット、カタログ番号L1500-01、L1500-02、L1502-01、L1502-02バージョンJ05300225-0355を用いて同定した。
結果:
上記のようにして決定した開始点の上流の10,000bpの領域をコンティグNT_011104から選択した。分析した領域は、10023位のコード領域の上流、すなわちATGの上流でちょうど終止する。
予測は、推定プロモーター領域同定用のソフトウェアを用いて行った。これらのソフトウェア、Promoter Inspector、First EFなどの各々は、PolII真核生物プロモーターの特徴(TATAシグナル、GC組成の偏り、階層的組織など)に基づいて構築された数理モデルを用いたものである。異なるアプローチ(二次判別分析、マルコフ連鎖など)を特徴とし、異なる特徴に基づくソフトウェアは、同じ位置に見出された予測を立証することを可能にする。
最も適切な予測の結果を、下表Vにまとめた。
【0065】
【表7】
【0066】
これらの予測は、ヒトFATP5遺伝子のプロモーターを、種々のソフトウェアによって同定された両極端の位置により決定された、7400〜10,000bpの間の領域内に位置づけることを可能にするものである(図12A)。
プロモーター領域は、別な遺伝子が8500位に終止し、そして10,000位がATPコドンの位置であることから、塩基8500と10,000との間であるはずである。実際、8500bpの位置から上流の領域は、機能が知られていないNM_032792遺伝子に対応し、そして10,000bpの位置から下流の領域は、FATP5遺伝子に対応する。この情報は、7400〜9100bpの間の領域内にプロモーターを位置づけた、生物情報ソフトウェアでなされたプロモーターの予測に匹敵するはずである。
【0067】
ヒトFATP5遺伝子のエクソン1の上流に位置するこの領域は、この遺伝子の調節にとって重要な推定プロモーターまたは調節エレメントを含むと推測される。Transfac(転写因子データベース)のテンプレートで分析され、中程度に保存されていることが示された領域は、境界7400〜10,023の間に広がる。ESTおよび全長型のmRNAの生物学的データは、転写開始点(TSS)を9990位に位置付けることを可能にした。
したがって、図13に記載された配列は、ヒトFATP5遺伝子の推定プロモーターに対応する。この配列は、添付された配列表中、配列番号1の番号で特定される。
9990位以外のTSSは、9928位において確認することができた。しかしながら、細胞種によって、別のTSSを代替物として用いることができる。
【0068】
c)ヒトFATP5遺伝子のプロモータ配列の、国際出願WO 0121795の先行技術との比較分析
本発明の対象となるヒトFATP5遺伝子の推定プロモーター配列と、種々のアクセス可能な配列バンク、例えば「Htgs」(終止していない(non-terminated)配列)、「Chromosome」(完全なゲノム、染色体および終止している(terminated)コンティグ)、および最後にnr(全ての公表された配列)などに含まれる配列との類似性について、一連の検索を行った。
ヒトFATP5遺伝子(コンティグNT_011104)のゲノム環境と、国際出願WO 01/21785に公開されているプロモーター領域との間の類似性の検索は、配列類似性がないことを示した。
GenBankの「Chromosome」部門に対する類似性検索は、このデータベースで参照されているヒトまたは他の種(酵母、ショウジョウバエなど)との有意な類似性を見出すことができなかった。
【0069】
Htgsデータベースに含まれる配列中での類似性検索は、国際出願WO 01/21785のプロモータ配列に対応し得るヒトコンティグを特徴づけることができなかった。
国際出願WO 01/21785のプロモータ配列がヒトFATP5遺伝子のゲノム環境との類似性を示さないことを確認するために、これらの2つの配列をアラインメントした。有意な類似性は見出されなかった。
国際出願WO 01/21785に公開されているFATP5遺伝子のプロモータ配列の分析は、この配列が、本発明の対象となるヒトFATP5遺伝子のプロモーターには対応しないが、マウスFATP5遺伝子のプロモーターに対応することを示した。
結論として、種々の検索により、本発明の対象となるヒトFATP5遺伝子のプロモーターの配列は、いかなる既知の配列とも有意な類似性を有しないと結論付けることができる。
【0070】
d)本発明の対象となるヌクレオチド配列のプロモーター活性の確認
材料および方法
FATP5の推定プロモータ配列の増幅:
テンプレート:ヒト胎盤のゲノムDNA(Sigma, St. Louis, MO)、
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号3:CGCTCGAGCTGTGAGCACCTGGATCAGTGCGTGCC、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号4:CCCAAGCTTGGTACCAGCTCCTCCCTAGG、
TaqプラチナPfx+バッファー10×+MgSO4(Invitrogen, Cergy Pontoise, France)、
dNTP(Promega, Williamsburg, IA)。
hFATP5プロモーターに対応するインサートの精製:インサートのサイズ=1573塩基。
増幅プログラム:
94℃で5分間のインキュベーションサイクルを1回、
その後、94℃で30秒間、55℃で30秒間、68℃で2分間のインキュベーションサイクルを25回。
【0071】
得られた物質は、QIAquick PCR精製キット(Qiagen, Courtaboeuf, France)で精製し、4℃に保った。
ヒトFATP5遺伝子のプロモーターを、ルシフェラーゼ遺伝子を含むベクターpGL3にクローニングするための制限:
【表8】
2時間、37℃でのインキュベーション。
Quick ligation kit(NEB, Beverly, MA)、cat#M2200Sの標準的プロトコルによるライゲーション。
30秒間、42℃の熱衝撃の標準的プロトコルによる形質転換。
細菌:DH5アルファ(Invitrogen, Cergy Pontoise, France)。
50μg/mlのアンピシリンを含むLB寒天(Invitrogen, Cergy Pontoise, France)のディッシュへのプレーティング。
37℃での一晩のインキュベーション。
スクリーニング:
各クローンを50μg/mlのアンピシリンを含むLB寒天上に継代し、20μlの水中で再懸濁した。
懸濁液を、95℃で10分間加熱した。
【0072】
pGL3クローン/hFATP5プロモーターの選択のためのPCR条件
テンプレート:細菌溶解物
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号5:TTCATTACATCTGTGTGTTGGTTTTTTGTGTG、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号6:TATGCAGTTGCTCTCCAGCGGTTCCATCTTCC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)、50μg/ml。
増幅プログラム:
94℃で5分間のインキュベーションサイクルを1回、
その後、94℃で30秒間、55℃で30秒間、72℃で2分間のインキュベーションサイクルを25回、
その後、72℃で7分間のサイクルを1回。
得られた物質は、4℃に保った。
プラスミドの作製は、EndoFree Plasmid maxi kit(Qiagen, Courtaboeuf, France, cat#12362)の標準的プロトコルに従った。
配列決定は、BigDye terminator V2.0 cycle sequencing kit(Applied Biosystems, Foster City, CA, cat#4314415)の標準的プロトコルに従った。
【0073】
結果:
ヒトFATP5プロモーターの1573塩基の断片をpGL3にクローニングし、配列決定した。断片の連続した欠失による構築物は、FATP5の肝での発現に特異的なプロモーター領域を同定することを可能にする。
HEP2、HELAおよびHEK293肝細胞の一過性トランスフェクション:
スクリーニングのための最小可能なプロモーター断片を特定するために、一過性にトランスフェクトした細胞を用いた。トランスフェクションは、jetPEIプロトコル(Polytransfection, Illkirch, France)により、供給者の推奨に従って行った。
第3日:照度計による測定:
トランスフェクション混合物を含む培地を除去した。
プレートを、250μlのPBSで2回洗浄し、そして、1/5に希釈した100μlの溶解バッファーを加えた。プレートを、振盪させながら室温で30分間インキュベートした。
各ウェルからの10μlの溶解液を、Greiner 96穴白色プレート(Greiner Bio-One, Frickenhausen, Germany)に分配した。
【0074】
ルシフェラーゼ活性は、Dual Luciferase kit(Promega, Madison, WI, USA)を用いて測定した。測定値は、TR717マイクロプレートルミノメーター(Applied Biosystems, Foster City, CA, USA)で取得した。
ルシフェラーゼ発光の結果はレニラに対して正規化し、プロモーターを含む構築物の、空ベクターpGL3と比較した誘導として表した。
結果:
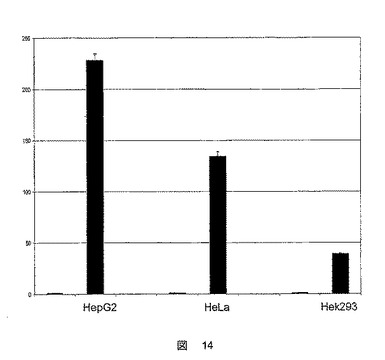
FATP5プロモーターの断片全体のプロモーター活性が確認された。図14に示したこれらの結果は、本核酸配列が実際にFATP5プロモーターであり、かつそれが機能的であることを確認するものである。ルシフェラーゼ活性の最も高い値は、HepG2ヒト肝細胞で観察された。
【0075】
例3:安定してヒトFATP5プロモーターを発現している細胞でのスクリーニング試験
a)ヒトFATP5遺伝子の推定プロモーターの、ハイグロマイシン耐性を有するベクターpGL3ハイグロマイシンへのクローニング
ベクターpGL3ハイグロマイシンの作製:
Tkプロモーターの制御下にあるハイグロマイシンをコードする遺伝子(サイズ:1671塩基)を、ベクターpREP4(Invitrogen, Cergy Pontoise, France)から、以下のプライマーを用いて増幅した。
材料と方法:
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号7:GAAGATCTCTGCTTCATCCCCGTGGC、
オリゴアンチセンス(Invitrogen, Cergy Pontoise, France):配列番号8:GAAGATCTACCAGACCCCACGCAACGC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)。
増幅プログラムおよび得られた物質の精製は、hFATP5プロモーターに対応するインサートの増幅および精製のために先に用いたのと同じプロトコルに従って行った。
得られた断片は、ベクターpGL3−Basic(Promega, Williamsburg, IA)のBglII部位にクローニングした。クローニングのための制限、ライゲーションおよび形質転換は、hFATP5プロモーターに対応するインサートの増幅および精製のために先に用いたのと同一のプロトコルに従って行った。
Tkハイグロマイシン断片を有するプラスミドpGL3が組み込まれたクローンは、pGL3クローン/hFATP5プロモーターの選択に用いたのと同じプロトコルによるPCRにより、テンプレートとして細菌溶解物を、および:
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号9:CTGCTTCATCCCCGTGGC、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号10:ACCAGACCCCACGCAACGC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)、
を用いて選択した。
hFATP5プロモーターを含むpGL3ハイグロマイシンクローンの選択、プラスミドの作製および配列決定は、pGL3クローン/hFATP5プロモーターの選択のために用いたのと同じプロトコルに従って行った。
【0076】
b)HepG2肝細胞の安定したトランスフェクション
用いた培地を下表に記載する。
【表9】
【0077】
c)細胞のプレーティング
HepG2細胞の培養物を調製した。細胞は、80%コンフルエントに達したときに剥離し、6穴プレート中の培地に、1ウェルあたり320,000細胞の割合で再接種した。細胞は、37℃、5%CO2で、24時間インキュベートした。
【0078】
d)トランスフェクションおよび活性化
トランスフェクションは、a)で調製した500ngのプラスミドを用いて、jetPEIプロトコル(Polytransfection, Illkirch, France)により、供給者の推奨に従って行った。
6穴プレートで培養した細胞の培地を吸引した。1mlの無血清DMEMと80μlのトランスフェクション混合物とを各ウェルに入れた。次に、細胞を、5%のCO2を含む雰囲気下、37℃で2時間放置した。この2時間が経過したときに、FCSを含まない培地1mlと、2%の終濃度のUltroser SF(USF, BioSepra, Cergy Pontoise, France)とを加えた。
【0079】
e)トランスフェクション細胞の選択
48時間の培養後に、各ウェルの培地を吸引した。10%のFCSと終濃度0.2mg/mlのハイグロマイシンとを含む2mlの培養培地を加えた。細胞は、80%コンフルエントに達したときに剥離した。
次に、2個のウェルからの細胞を混合し、150mmのペトリ皿中の同じ選択培地に接種した。選択培養は、細胞クローンが現れるまで維持した。
【0080】
f)クローンの単離
クローンが明確に区別されるようになったら、これらを、シリンダーを用いて残りの培養物から単離した。次に、シリンダーに含まれた細胞をPBSで洗浄し、培地の吸引後に取り出した。
細胞を96穴プレートに配置し、そして10%のFCSと終濃度0.25mg/mlのハイグロマイシンとを含む250μlのDMEMを加えた。培養は、80%コンフルエントになるまで継続した。
【0081】
g)安定してヒトFATP5プロモーターを発現するHepG2細胞のスクリーニング
ストックを用意するために、クローンを増幅した。
各クローンに由来する細胞は、24穴プレートにて、10%のFCS、ペニシリン/ストレプトマイシン混合物(0.125%)および0.25mg/mlのハイグロマイシンを含むDMEM中、80%コンフルエントまで培養した。
被験化合物による処置は、24時間の期間適用した。この24時間が経過したときに、ルシフェラーゼ活性を、細胞溶解物において、Applied Biosystems TR717発光リーダー(California, CA)を用いて測定した。
ルシフェラーゼ活性を示している細胞は、この活性が、これらの細胞が受け取ったプラスミドに含まれる推定プロモーターによって誘導されたことを示す。
この結果は、本発明の核酸が、実際に、その依存下に置かれた遺伝子の発現を導くことができるプロモーターであることを示すものである。
【0082】
例4:糖尿病表現型の解消およびインスリン増感剤によるZDFラットにおけるFATP5遺伝子の発現の調節
レキシノイド(rexinoid)と呼ばれるRXR核受容器のアゴニストは、インスリン増感剤であり、糖尿病肥満型動物モデル(2型糖尿病)において有益な効果を有する。ベキサロテンであるタルグレチン(登録商標)(CAS 1543559-49-0)は、核受容体ヘテロダイマー、RXR−PPAR(ペルオキシゾーム増殖因子活性化受容体)またはRXR−LXR(肝臓X受容体)を選択的に活性化する。
雄の糖尿病肥満型ラット(Zucker糖尿病肥満型ラット(ZDFラット))は、食物と水に自由にアクセスできる動物ケージ内に維持し、夜昼リズム(nyctohemeral rhythm)が観察された。
【0083】
化合物LGD 1069を、0.3、1.0、3.0、10または30mg/kg/日の用量で食餌に加え、7週齢より50日間投与した。4頭の動物は、LGD 1069の添加なしで給餌した:ZDF2頭とZLC2頭。動物を絶食させ、その翌日に安楽死させた。組織を回収し、液体窒素中で直ちに凍結した。
肝RNAは、ChomczynskiおよびSacchiに由来する従来の技法で抽出した。発現は、FATP5に特異的な放射性cDNAプローブを用いたハイブリダイゼーションによるノーザンブロッティングによって分析した。
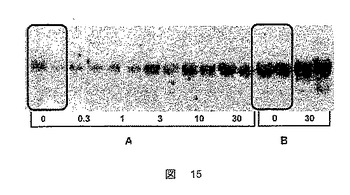
結果を図15に示す。
ZDFラットの肝臓におけるFATP5の発現の、ZLCラットに比した負の調節は、RXRアゴニストによる処置によって打ち消され、これは糖尿病の進行を逆転させた。
【0084】
例5:真核細胞(CHO:チャイニーズハムスター卵巣細胞)におけるFATP5受容体を介した脂肪酸の能動輸送を測定するための試験
蛍光標識された脂肪酸模倣分子の使用は、FATP受容体を過剰発現した細胞における脂肪酸の細胞内蓄積の、マイクロタイタープレート形式での迅速な測定を提供する。
試験の説明
【表10】
必要な専門器材
−CO2インキュベーター、
−顕微鏡、
−マルチドロップ、
−マイクロプレートウォッシャー、
−フルオレセイン用フィルターを有するマイクロプレート用蛍光計。
【0085】
プロトコル
第1日
96穴マイクロプレートへの細胞の分配
FATP受容体を過剰発現するCHO真核細胞を、96穴培養プレートに、1ウェルあたり約10,000細胞の密度で、以下の手順に従って分配する。
−層流フード下で、4mlの細胞剥離溶液を225cm3の培養フラスコに37℃で加える。
−溶液を、細胞単層上、手動による軌道回転運動で10〜15秒間振盪し、その後、10mlの血清用ピペットで吸引する。
−4mlの細胞剥離溶液を加え、フラスコを(最長で)10分間インキュベーター中に戻す。10分後に、細胞の剥離を顕微鏡下でチェックする。細胞がすべて剥離するまで操作を続ける。
−8mlの培地をフラスコに加え、細胞塊をほぐすために、内容物を何度か上から下へ混合する。この細胞混合物を、37℃に予熱した88mlの培地に加える。
【0086】
マルチドロップの準備
−マルチドロップを層流フードに配置する。マルチドロップのヘッドおよびチューブを、20〜30mlの70%エタノール、次いで20〜30mlの無菌水、そして最後に20〜30mlの培地をポンプ吸引することにより洗浄する。
−マルチドロップのチューブを細胞を含むフラスコに配置し、およそ5〜10mlの培地を除去する。1ウェルあたり100μlの細胞を加える(10,000細胞)。
−マイクロプレートを、37℃、10%CO2で48時間、インキュベーター内に置く。
【0087】
第2日:被験分子の添加
被験分子を、以下のプレートレイアウトに従って添加する:
【表11】
−マルチドロップを用いて、95μlのDMEM(フェノールレッド不含)を、96穴マイクロプレートの各ウェルに分配する。
−被験分子を、Tと表示されたウェルに終濃度5mg/ml(1%のDMSO)で同時に分配する。
−200mMのラウリン酸を、陰性対照ウェル(NC)に分配する。
−5μlのPBSを、陽性対照ウェル(PC)に分配する。
【0088】
第3日
−細胞を、PBSで2回洗浄する。
−細胞を、PBS中に0.1mMのBODIPY-FAと、脂肪酸を含まない0.1%のBSAとを含む脂肪酸模倣溶液中、2分間、37℃でインキュベートする。
−2分後に、細胞を事前に氷中に配置しておいたPBS/0.1%BSA混合物で4回洗浄する。
−プレートを、蛍光マイクロプレートリーダーにて、励起波長として485nmを、放出波長として530nmを用いて読み込む。
結果を、プローブなし(PC)の対照細胞のパーセンテージとして、任意の蛍光単位で表す。
この試験は、真核細胞内へのFATP5受容体を介した脂肪酸の能動輸送を測定することを可能にする。これはまた、FATP5受容体を介した脂肪酸の能動輸送に対する所定の分子の影響を測定することを可能にし、それによってヒトFATP5遺伝子のプロモーターの活性を刺激または阻害するその能力を決定することを可能にする。
【図面の簡単な説明】
【0089】
【図1】PCRによって分析された、ヒトFATP5遺伝子の発現の細胞および組織分布を示した図である。NCI−H295:副腎細胞系、THP−1:単球、THP1D:THP−1単球の分化したマクロファージ、JurkatおよびJurkat JR:T細胞、CEM:急性リンパ芽球性白血病細胞系の細胞、HH:ヒト初代肝細胞、HepG2、HuH7:ヒト肝細胞、CaCo2:結腸細胞、CaCo2D:分化したCaCo2細胞、CASMC:冠状動脈平滑筋細胞、ASMC:大動脈平滑筋細胞。
【図2A】3T3−L1細胞での脂肪酸捕獲試験で得られた結果を示した図である。結果は、各条件についての、時間の関数としての脂肪酸捕獲の誘導を表す。対照条件は、発現が細胞による脂肪酸捕獲に影響を及ぼさない遺伝子を担持するアデノウイルスに感染した細胞による捕獲の測定値を表す。
【図2B】ラット肝細胞(RH)での脂肪酸捕獲試験で得られた結果を示した図である(時間:1分)。結果は、各条件についての、時間の関数としての脂肪酸捕獲の誘導を表す。対照条件は、発現が細胞による脂肪酸捕獲に影響を及ぼさない遺伝子を担持するアデノウイルスに感染した細胞による捕獲の測定値を表す。
【図3A】hFATP5遺伝子を担持するアデノウイルスへの感染後における、ACS活性を決定するための試験において得られた結果を示した図である。結果は、誘導係数として表す。
【図3B】hFATP5遺伝子を担持するアデノウイルスへの感染後における、VLACS活性を決定するための試験において得られた結果を示した図である。結果は、誘導係数として表す。
【0090】
【図4A】hFATP5の発現の、脂肪酸のβ酸化への影響を決定するための試験において得られた結果を示した図である。4A:48時間におけるHepG2、4B:12時間におけるRH。結果は、誘導係数として表す。
【図4B】hFATP5の発現の、脂肪酸のβ酸化への影響を決定するための試験において得られた結果を示した図である。4A:48時間におけるHepG2、4B:12時間におけるRH。結果は、誘導係数として表す。
【図5】hFATP5の発現の、48時間におけるHepG2細胞での脂肪酸のエステル化度への影響を決定するための試験において得られた結果を示した図である。結果は、3H2O/3TG比として表す。
【図6A】hFATP5の発現の、Zucker糖尿病肥満型ラットでの糖尿病の発症の最中におけるhFATP5の発現の調節への影響を決定するための試験において得られた結果を示した図である。A:ブドウ糖レベルの測定値、B:インスリンレベルの測定値、−■−:雄のZDFラット(fa/fa)、−□−:雄のZLCラット(fa/+)。
【図6B】hFATP5の発現の、Zucker糖尿病肥満型ラットでの糖尿病の発症の最中におけるhFATP5の発現の調節への影響を決定するための試験において得られた結果を示した図である。A:ブドウ糖レベルの測定値、B:インスリンレベルの測定値、−■−:雄のZDFラット(fa/fa)、−□−:雄のZLCラット(fa/+)。
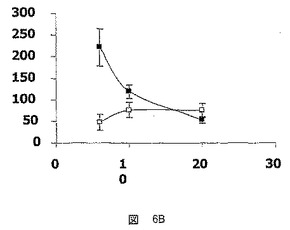
【図7】Zucker糖尿病肥満型ラット(ZDF:−●−)およびZLCラット(−○−)におけるhFATP5メッセンジャーRNAのレベルを測定するための試験において得られた結果を示した図である。
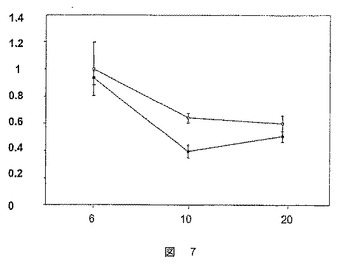
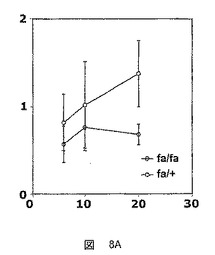
【図8A】生化学分析の結果を示した図である。A:肝臓における遊離脂肪酸レベルの測定値(−θ−:ZDFラット(fa/fa)、−○−:ZLCラット(fa/+))、B:体重増加曲線(−●−:ZDFラット(fa/fa)、−□−:ZLCラット(fa/+))。
【図8B】生化学分析の結果を示した図である。A:肝臓における遊離脂肪酸レベルの測定値(−θ−:ZDFラット(fa/fa)、−○−:ZLCラット(fa/+))、B:体重増加曲線(−●−:ZDFラット(fa/fa)、−□−:ZLCラット(fa/+))。
【0091】
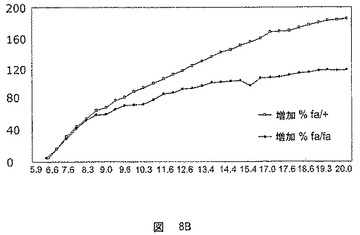
【図9】8週齢のZDFおよびZLCラットにおける、各条件につき5および30分でのオレイン酸のアシル化の程度を示した図である。A:ZDFラット、B:ZLCラット。
【図10】ZDFラットの初代肝細胞における、ZLC対照ラットと比較したβ酸化の程度の抑制の解消を示した図である。A:8週齢のラット、B:10週齢のラット。(C=対照、Z=LacZ、h5=hFATP5)。
【図11】LacZ対照アデノウイルスに感染したラットと比較した、FATP5を担時するアデノウイルスに感染したZDFラットにおける生化学パラメータの測定値を示した図である(遊離脂肪酸(A)、トリグリセリド(B)およびブドウ糖(C))。
【図12】FATP5遺伝子の転写開始点(TSS)の位置を示した図である。A:バイオインフォマティクスによる予測的な方法論、B:RACEによる実験的な方法論。
【図13】ヒトFATP5遺伝子の推定プロモーターの配列を示した図である。この配列は、添付された配列表において配列番号1の番号で特定される。
【図14】ヒトFATP5遺伝子のプロモータ配列の、HepG2、HeLaおよびHek293細胞における活性を示した図である。結果は、空プラスミドpGL3で形質転換した細胞で測定した対照と比較した誘導として表示してある。
【図15】ZDFラット(A)またはZLCラット(B)をインスリン増感剤として知られる化合物で処置した後の、FATP5メッセンジャーRNAのレベルの変動を示した図である。
【技術分野】
【0001】
本出願は、ヒトFATP5タンパク質のプロモーターに対応することを特徴とする単離ヒト核酸、および前記プロモーターの依存下に置かれた、タンパク質、特にヒトFATP5タンパク質をコードする単離ヒト核酸に関する。
本発明はさらに、前記核酸を含むベクター、または、前記核酸もしくは前記ベクターが導入された宿主細胞に関する。最後に、本発明は、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質を同定する方法における、前記核酸、前記ベクターまたは前記宿主細胞の使用、および前記同定方法に関する。
本発明は、最後に、ヒトFATP5タンパク質の前記プロモーターの依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質、および、糖尿病を予防および/または処置するための、または、肝臓における脂肪酸のβ酸化を誘導するための医薬の製造における前記物質の使用に関する。
【0002】
II型糖尿病は、世界中において主要な公衆衛生上の問題である。現在、およそ1億2500万人が本症に罹患しており、このうち米国内に存在する者だけで1500万人を占める。予測は、2010年に、糖尿病に罹患している患者数が2倍になることを示している。現在米国で糖尿病を患っている90〜95%の人々がII型糖尿病に罹患している。II型は、インスリン耐性と高血糖症とを特徴とする代謝病であり、しばしば高血圧、脂質障害および肥満を伴う。成人において、この病的状態は一般に30才から現れ、心臓、血管、眼、腎臓および/または神経への損傷を含む合併症を特徴とする。
【0003】
今のところ、残念なことに、原因に対する治療的および/または予防的処置は存在せず、処置は、一般に対症療法的となる傾向がある。
したがって、糖尿病の原因を処置することが可能な物質が真に求められている。また、糖尿病の原因を処置することができる前記物質を特定するために、前記物質を特定する効果的方法も常に変わらず求められている。
本発明の目的の1つは、そのような方法を提供することである。
【0004】
長鎖脂肪酸(LCFA)は生物のための重要なエネルギー源であり、ミトコンドリアにおけるそのβ酸化を介してATPを提供する。これらはまた、多数の多様な細胞過程、例えば細胞シグナリング経路または遺伝子発現の調節のための基質でもある。例として、脂肪酸は心臓にエネルギーの恒常的で効果的な供給を提供するために不可欠である。
LCFAは、リポタンパク質リパーゼによる脂肪分解の後で粘膜の細胞に入り、そしてその後、小胞体でトリグリセリドにエステル化される。トリグリセリドはその後、アポリポタンパク質とともに脂肪粒子(lipoparticle)複合体内に組み込まれ、極低密度リポタンパク質(VLDL)またはキロミクロンを形成し、そしてエキソサイトーシスによって細胞から出て行き、全身循環中に廃棄される。リポタンパク質リパーゼは、これらの粒子を血管内皮細胞の表面で非エステル型脂肪酸に変換し、これは、その後アルブミンと結合することができる。
【0005】
ラット細胞における脂肪酸の捕獲の動態は、最初の30秒間にわたる急速な線形段階と、それに続く、初期捕獲速度の低減を伴う移行段階、および低い蓄積速度を有する後期に分けられる。最初の捕獲速度は、脂肪酸のベクトル的に一方向の流れに対応し、インキュベーション培地に適用される脂肪酸濃度の計算された作用によって特徴づけることができる。動態、LCFAによる競合、およびプロテアーゼでの細胞の処理による脂肪酸輸送の抑制は、輸送体に媒介された過程を支持するものである。
【0006】
最初の脂肪酸輸送タンパク質(FATP)は、ラット脂肪細胞において1994年に同定された。2つの対応する遺伝子がその後に同定され、そして現在はFATP1およびアシルコエンザイムAシンテターゼ(ACS)と呼ばれている。マウス遺伝子ライブラリーのスクリーニングにより、FATPファミリーの全ての成員における高度に保存された311個のアミノ酸からなるサイン配列(signature sequence)によって特徴づけられるタンパク質のファミリーが、その後明らかになった。これらはすべてAMP結合部位を含む。これまでに、6種のマウスFATPおよび6種のヒトFATPが同定された。コエンザイムAシンテターゼ活性は、現在FATP1、2、4および5に関連している。LCFA捕獲におけるこれらの役割は、これらの酵素活性から推論することができる。あるいは、FATPの発現は、遊離脂肪酸の細胞内レベルの増加を導くことができ、したがって、VLACS(超長鎖アシル−CoAシンテターゼ)活性を誘導することができる。実際、アシル化に関与する酵素、例えばアシルコエンザイムAシンテターゼ(ACS)は、PPAR(ペルオキシゾーム増殖因子活性化受容体)転写因子およびある種の長鎖脂肪酸によって正に調節されている。
【0007】
アシル化は、脂肪酸を活性化し、アシル−CoAの活性代謝誘導体を形成し、これはいかなる代謝経路にも入ることができるようになる。小胞体において、脂肪酸はトリグリセリドの合成に関与することができる。ミトコンドリアおよびペルオキシソームにおいて、FATPは脂肪酸の分解を誘導することができる。
FATPは、そのアミノ末端に応じて異なる細胞内位置を占めている。これらのN末端は、脂肪酸を異なる細胞内コンパートメントに分配し、その後別々の代謝経路へのこれらの組み込みを誘導するために、様々なパートナーと相互作用することができる。
【0008】
これらは少なくとも2つの相同サインユニットを保有しているが、異なるFATPは別々の染色体位置、特有のプロモーターおよび遺伝子調節、特有の分布プロファイル、異なる細胞位置、および専門化された機能を有する。
従来技術において、FATP5は、このタンパク質について検出可能な活性に応じて、例えば超長鎖アシル−CoAシンテターゼ関連物(VLACSR)、超長鎖アシル−CoAシンテターゼホモログ(VLCS−H2)、コリル−コエンザイムAリガーゼまたは胆汁酸コエンザイムAシンテターゼ(BACS)などの異なる名称で見出される。
【0009】
ヒトにおいて、タンパク質はもっぱら肝臓で発現される。
マウスにおいては、2.6kbのメッセンジャーRNAが肝臓で非常に豊富であることが知られている。より短いメッセンジャーRNAのより低いレベルの発現が、脳、肺、睾丸および脾臓(2.5kb)において、そして、骨格筋(2.2kb)において検出された。転写物の存在は、PCR技法により心臓において示されたが、腎臓においては示されなかった。
ヒトにおいて、転写物は、そのマウスホモログと対照的に、線維芽細胞、脳または心臓で検出することができなかった。
サザンブロット分析は、VLACSR遺伝子がヒトおよびマウスの双方で単一コピーで存在することを示す。
【0010】
例えば、FATP5が長鎖脂肪酸および超長鎖脂肪酸(例えばC18〜C26)を活性化できることが報告されている。FATP5は、ケノデオキシコレート、デオキシコレート、リトコレートおよびトリヒドロキシコレスタノン酸(trihydroxycholestanoic acid)を活性化することができる。
本発明者らの知る限りでは、脂肪酸分解におけるFATP5の関与は、これまで全く記載されたことはなかった。FATP5の役割は、腸肝再循環を介した脂肪酸の再活性化だけに限定されていた。
従来技術において、国際出願WO 01/21795は、FATP5が脂肪酸輸送体として役割を果たし、そうして細胞による脂肪酸捕獲の過程に関与することを示唆している。
【0011】
さらにまた、糖尿病に罹患した者、または糖尿病を発症しやすい者は、血流中に高レベルの遊離脂肪酸を有することが知られている。血漿中の遊離脂肪酸の過剰は、インスリン感受性組織、例えば骨格筋において、エネルギーを産生するためのブドウ糖の利用に対する競合者のように作用する。脂肪酸は、異なるレベルで糖代謝に干渉する。これらは、間接的に解糖を阻害し、グリコーゲンの形態でのブドウ糖の貯蔵を減らし、間接的にグルコース輸送を阻害する。これは、遊離脂肪酸が過剰である場合におけるその優先的な利用、および、高血糖症の発症を導く、循環中のブドウ糖濃度の増加を示す傾向がある。
【0012】
驚くべきことに、そして、予想を超えて、本発明者らは今回、FATP5が肝臓における脂肪酸の分解に関与することを見出した。同様に、本発明者らは、この特性が、結果として間違いなく血漿中のブドウ糖および脂質濃度を減少させることを示すことができた。特に、本発明者らは、FATP5のメッセンジャーRNAの発現レベルが、糖尿病の肥満ラット(Zucker糖尿病肥満型ラット(ZDFラット))において、非糖尿病ラット(Zucker痩せ型ラット(ZLCラット))におけるFATP5のメッセンジャーRNAの発現レベルに比べて大幅に減少していることを示すことができた。
したがって、FATP5の過剰発現が、結果として遊離脂肪酸のレベルを減らし得ることを想起することが可能となり、したがって、FATP5の発現レベルを増やす能力を有する任意の物質が、糖尿病の予防的または治療的処置用の医薬の製造のための良い候補であり得ることを十分に予測することができる。
本発明は、この応用分野に含まれる。
【0013】
本発明者らは、ヒトFATP5遺伝子のプロモーターである、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、ヒト核酸配列を単離した。この配列は、従来技術においてFATP5プロモーターの配列であると考えられていた配列と、わずかにしか相同ではない。
本発明は、したがって、第一に、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、単離されたヒト核酸配列に関する。前記配列は、ヒトFATP5遺伝子のプロモーターに対応する。
【0014】
本発明はさらに、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列からなることを特徴とする、単離されたヒト核酸配列に関する。
本発明の1つの特定の実施例において、配列番号1の番号で特定されるヌクレオチド配列を少なくとも含む核酸配列は、タンパク質、特にヒトFATP5タンパク質をコードする。特に好ましくは、配列番号1の番号で特定されるヌクレオチド配列を少なくとも含み、かつ、ヒトFATP5タンパク質をコードする前記核酸配列は、添付された配列表において配列番号2の番号で特定されるヌクレオチド配列に対応する。
【0015】
本発明はさらに、上記と相補的なセンスまたはアンチセンス核酸配列、および、本発明による核酸配列のうちの1つと、少なくとも80%、好ましくは少なくとも90%の同一性パーセントを有する任意の核酸配列に関する。
参照配列と少なくともX%の同一性パーセントを有する核酸配列は、本発明において、参照配列の100ヌクレオチドあたり100−X個までの変更を含み得るが、前記参照配列の機能的な特性を保持している核酸配列として定義される。本発明に関して、「変更」は、参照配列における連続的または散剤的なヌクレオチドの欠失、置換または挿入を含む。
本発明による少なくとも80%、好ましくは少なくとも90%の同一性パーセントを有する核酸配列は、アレル変異体、すなわち配列番号1または配列番号2の配列の個体変異に対応する全ての配列を含む。これらの自然な変異配列は、哺乳類、特にヒトに存在する多型に対応する。
【0016】
本発明はさらに、上記に定義した本発明の核酸配列が挿入されたクローニングおよび/または発現ベクターに関する。好ましくは、本発明のベクターは、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含む核酸配列を含む。
本発明の1つの特定の態様において、本発明のベクターはまた、上記に定義された、ヒトFATP5遺伝子のプロモーターとして同定された核酸配列の依存下に置かれた、検出可能なタンパク質をコードする核酸配列を含む。特に、このプロモーターは、配列番号1の配列に対応する。「検出可能なタンパク質」とは、一度発現されると、当業者に知られている任意の技術によって容易に同定される任意のタンパク質を意味すると理解される。FATP5自体、またはマーカーとして用いられるタンパク質、例えば緑色蛍光タンパク質(GFP)、レニラ(renilla)またはルシフェラーゼを、用いることができるタンパク質として挙げることができる。
【0017】
かかるベクターは、宿主細胞における表現、および任意にタンパク質の分泌のために必要な要素を含むことができる。
前記ベクターは、好ましくは、翻訳開始および終止シグナル、ならびに適切な転写調節領域を含む。これらは細胞中で安定して維持することができる必要があり、そして、翻訳されたタンパク質の分泌を指示する特定のシグナルをコードする配列を任意に含むことができる。
本発明による核酸配列は、選択された宿主内の自律複製型ベクター中に、または選択された宿主の組込型ベクター中に挿入することができる。
自律複製系のなかでは、宿主細胞にしたがって、プラスミドまたはウイルス型のシステムを用いることが好ましい。
本発明によって用いることができるプラスミドベクターは、核酸配列の発現を可能にする任意の既知のプラスミドであってもよい。
【0018】
「核酸配列の発現」とは、本発明によれば、本発明の核酸配列のRNAへの転写、および任意にこれに続く前記RNAのタンパク質への翻訳を可能にする本発明のベクターの能力を意味するものと理解される。
好ましくは、本発明によるプラスミドは、上記の単離されたヒト核酸配列の発現を可能にする。特に好ましくは、本発明によるプラスミドは、添付された配列表において配列番号1の番号で特定されるヌクレオチド配列、特にFATP5をコードする核酸を少なくとも含むことを特徴とする核酸配列の発現を可能にする。
【0019】
以下のものを、本発明によって用いることができるプラスミドの例として挙げることができる:プラスミドpCMVTag(Stratagene, La Jolla, USA)、pcDNA3(Invitrogen, Cergy Pontoise, France)、pSG5(Stratagene, La Jolla, USA)またはpGL2およびpGL3(Promega, Mannheim, Germany)。
ウイルスベクターは、特にアデノウイルス、レトロウイルス、レンチウイルス、ポックスウイルスまたはヘルペスウイルスであってもよい。当業者は、これらの系の各々のために用いることができる技術に精通している。
核酸配列を宿主細胞の染色体に組み込むことが望ましい場合は、例えば、プラスミドまたはウイルス型のシステムを用いることが可能であり、このようなウイルスは、例えばレトロウイルスまたはアデノ随伴ウイルス(AAV)である。
【0020】
非ウイルスベクターの中では、好ましいのは、裸のDNAまたはRNAなどの裸のポリヌクレオチド、バクテリア人工染色体(BAC)、酵母における発現のための酵母人工染色体(YAC)、マウス細胞における発現のためのマウス人工染色体(MAC)、および、好ましくは、ヒト細胞における発現のためのヒト人工染色体(HAC)である。
本発明によると、ベクターは好ましくはプラスミドまたはアデノウイルスである。
かかるベクターは、当業者によって一般に用いられている方法によって調製し、そして、得られた組換えベクターは、標準的な方法、例えばリポフェクション、エレクトロポレーション、熱衝撃、膜の化学的透過化処理後の形質転換、または細胞融合によって適切な宿主に導入することができる。
【0021】
本発明はさらに、本発明による少なくとも1種の核酸配列または本発明による少なくとも1種のベクターが導入された、形質転換された宿主細胞、特に真核細胞および原核細胞に関する。
細菌細胞、酵母細胞および動物細胞、特に哺乳類細胞を、本発明に関して用いることができる細胞として挙げることができる。昆虫細胞、例えばバキュロウイルスを伴う方法を用いることができるものも挙げることができる。
【0022】
本発明はさらに、上記の宿主細胞を、該細胞中に導入された本発明による核酸配列の、タンパク質の形態での発現を可能にする条件下で培養することを特徴とする、タンパク質を産生する方法に関する。
本発明はさらに、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定のための、上記の本発明の核酸配列、ベクターまたは宿主細胞の使用に関する。
【0023】
本発明はさらに、上記の本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質を特定する方法であって、
a)上記の核酸配列、ベクターまたは宿主細胞を、ヌクレオチド配列の発現に適した媒体中で、上記の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質と接触させること、および
b)上記の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルを測定すること、
を特徴とする方法に関する。
【0024】
「ヌクレオチド配列の発現に適した媒体」は、本明細書中では、本発明の核酸配列からの、RNA、特にメッセンジャーRNA、および任意にタンパク質の合成を可能にする任意の発現系を意味すると理解される。
1つの特定の態様において、本発明の方法は、b)で決定された発現レベルと、同定する物質に接触させなかった対照における、上記の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルとの比較のための、追加の工程を含む。
比較は、本発明の核酸配列に対する被験物質の調節能力を評価することを可能にする。
【0025】
「調節する」または「調節能力」は、本発明によれば、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を刺激または阻害する被験物質の能力を意味すると理解される。
本発明の核酸配列の依存下に置かれたヌクレオチド配列の発現レベルは、自体公知の従来のmRNAまたはタンパク質分析技法で測定することができ、以下の技術を、非限定的な例として言及することができる:RT−PCR、ノザンブロッティング、ウェスタンブロッティング、RIA、ELISA、免疫沈降、および免疫細胞化学または免疫組織化学分析技術。細胞もしくは分子生物学、または免疫細胞化学もしくは免疫組織化学分析の実験プロトコルに関するすべてのことについては、本特許出願の各所において、当業者に完全に精通している多数の著作、特に「Current Protocols in Immunology; John Wiley and Sons, Teton Data System, Jackson, Wyoming (ISBN 0-471-30660-6, 2003)」を参照することができる。
前記測定は、プローブ、プライマーまたは抗体を用いて有利になされる。
【0026】
本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質は、生体高分子、例えば、核酸、脂質、糖、タンパク質、ペプチド、タンパク質−脂質、タンパク質−糖、ペプチド−脂質もしくはペプチド−砂糖ハイブリッド化合物、または化学的分枝もしくは化学分子が付加されたタンパク質またはペプチドであってもよい。
本発明はさらに、本発明による核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質であって、本発明の方法により入手可能な前記物質に関する。
【0027】
本発明はさらに、本発明による核酸配列と相互作用することが可能な物質を同定する方法であって、
a)上記の核酸配列、ベクターまたは宿主細胞を、本発明による核酸配列と相互作用することが可能な物質に接触させること、および
b)前記核酸配列と前記被験物質との間の相互作用を評価すること、
を特徴とする方法に関する。
本発明を実行する1つの特定の方法では、前記同定方法の目的は、本発明による核酸配列と結合することが可能な物質を特定することであり、前記方法は、例えば、異種相互作用の場合は熱量測定であり、または、ペプチド−ペプチド相互作用の場合は、2ハイブリッド試験である。
【0028】
最後に、本発明は、本発明による核酸配列と相互作用すること、または結合することが可能な物質であって、上記の同定方法のうちの1つによって入手できる前記物質に関する。
本発明によれば、前記核酸配列と前記被験物質との間の相互作用および/または結合は、任意の既知の技法によって評価することができる。
【0029】
本発明者らは、FATP5が肝臓における脂肪酸の分解に関与していることを示した。同様に、本発明者らは、この特性が、結果として間違いなく血漿ブドウ糖および脂質濃度を減少させることを示すことができた。特に、本発明者らは、FATP5メッセンジャーRNAの発現レベルが、糖尿病ラット(Zucker糖尿病肥満型ラット(ZDFラット))において、非糖尿病ラット(ZLCラット)におけるFATP5メッセンジャーRNAの発現に比べて大幅に減少していることを示すことができた。
したがって、本発明はさらに、糖尿病を予防および/または処置するための医薬の製造のための、本発明による物質の使用に関する。
本発明はさらに、肝臓における脂肪酸のβ酸化を誘導するための医薬の製造のための、本発明による物質の使用に関する。
【0030】
上記の事項に加えて、本発明はまた、本発明をいかに実行するかについての例および添付の図面を参照した以下の説明から明らかになる他の事項を含む。
以下の例は、いかなる意味においても制限を意味することなく本発明を例証する。
【0031】
例1:脂肪酸のβ酸化におけるhFATP5の関与の証明
材料および方法
A)FATP5発現プロファイル
a)RNAの調製
全RNAは、細胞および組織から、ChomczynskiおよびSacchi(Chomczynski et al., 1987)により記載された技法を用いて調製した。RNAは260nmでの光学密度の測定によって定量し、そして、RNAの品質は260/280光学密度比の測定によってチェックした。
1μgの各サンプルを、酵素MMLV−RT(Gibco BRL, Paisley, UK)を用いた逆転写(RT)反応においてテンプレートとして用い、ポリメラーゼ連鎖反応を、2μlのRT産物について実行した。特異的プライマー(Eurogentec, Seraing, Belgium)を、hFATP5およびrFATP5について調製した。PCR産物の特異性を確保するために、熱変性温度(Tm)は各々の特定の標的について最適化し、サイクル数は、転写産物の量に応じて、25〜30の間に設定した。
【0032】
【表1】
【0033】
DNA PCR産物は、1%のアガロースゲル上で分析した。シグナルは、Gel Doc(登録商標)2000装置(Bio-rad, Marnes La Coquette, France)を用いて、密度測定値として定量化した。in vivo転写分析については、定量化は、特定の各標的の値の、内部対照(アクチン)の値に対する比として計算した。結果は、1に設定された対照と比較した誘導レベルとして表示した。
【0034】
定量分析は、32キャピラリーLight Cycler装置(Roche)にて、二本鎖DNAを標識するフルオロフォアであるサイバーグリーンを用いて、二段階PCR(Roche, Mannheim, Germany)によって行った。縦座標の増幅曲線の微分値(PCRの最中の蛍光の増大を表す)と、横座標のサイクル数との交点として表された各標的についての結果は、対照遺伝子で正規化した。
【表2】
【0035】
B)Zucker糖尿病肥満型ラットにおける糖尿病発症の動態
Genetic Model Inc.(Charles River Laboratories, Wilmington, MA, USA)に由来する5週齢の雄のZucker糖尿病肥満型ラット(ZDF/GMI−fa/fa)および同週齢の雄のZucker痩せ型ラットは、無制限量のPurina 5008(PMI Nutrition International, Wellingborough, Northamptonshire, UK)で飼育した。
6週齢、10週齢および20週齢において、48頭のラットを、16頭(8頭のZucker糖尿病肥満型ラットおよび8頭のZucker痩せ型ラット)の3群に分けた。動物の重量およびその摂食量は、実験の間中モニターした。
【0036】
血液サンプルは、日中、8時間の食餌療法期間の後、麻酔下にて眼窩採血により採取した。麻酔は、ペントバルビタールの腹腔内投与によって誘発した。動物は頚椎脱臼によって安楽死させ、肝臓を摘出し、重量を測定し、さらなる分析のために液体窒素中で直ちに凍結させた。
【表3】
【0037】
b)分析
血清グルコース、コレステロール、トリグリセリドおよび遊離脂肪酸は、酵素によるバイオアッセイを用いて決定した。
インスリンレベルは、放射免疫分析により決定した。
肝臓の一部は、組織学的研究のために保存した。
【0038】
C)アデノウイルス中へのhFATP5の完全な配列のクローニング
別途示さない限り、用いた技法は、供給者によって、または分子生物学に関する多数の古典的な著作に記載されたものである。
hFATP5の完全なコード配列(配列番号2の配列の1575位におけるATGから)は、Adeno-X(Clontech, Palo Alto, USA)のPT3414−1発現系にクローニングした。
ヒト肝細胞培養物の全RNAは、Qiagen(Qiagen, Courtaboeuf, France)からの、RNeasy miniprep kitで抽出した。
【0039】
次いで、第一DNA鎖を以下の通りに合成した。
【表4】
【0040】
次に、以下のプライマー:
配列h5_4As(Invitrogen, Cergy Pontoise, France):配列番号19:GCTCTAGATCAGAGCCTCCAGGTTCCCTCACACACAGCC
配列h5_1S(Invitrogen, Cergy Pontoise, France):配列番号20:GCTCTAGATGGTACCATGGGTGTCAGGCAACAG
を用い、以下の反応媒体:
センスオリゴ(10pM)1μl
アンチセンスオリゴ(10pM)1μl
TaqプラチナPfx+バッファー10×+MgSO4(Invitrogen, Cergy Pontoise, France)
dNTP(Promega, Williamsburg, IA)
最終容量は20μl
により、PCRを実行した。
【0041】
次に、以下のプログラムにしたがって増幅を行った:
94℃で5分間を1サイクル、次いで94℃での30秒、次に55℃での30秒、そして68℃での2分を含む25サイクル。
その後、産物を4℃で維持した。
得られたPCR断片を、QIAquick PCR精製キット(Qiagen, Courtaboeuf, France)を用いて精製した。
精製した断片を、次に、XbaI(NEB, Beverly, MA)で消化し、ClontechからのpAdenoXキットのベクターpShuttle中に、供給者の推奨にしたがってクローニングした。
【0042】
得られたプラスミドを、DH5d型の大腸菌に導入し、そして、クローンをこれらのカナマイシン耐性によって選択した。次に、陽性クローンのプラスミドDNAを精製し、制限分析によりチェックし、配列決定した。タンパク質の発現は、T7発現系を用いて、in vitroで確認した。pShuttleの発現カセットを、PI−Sce/I−CeuIを用いて切断し、Adeno-X(登録商標)ウイルスDNAにライゲーションした。ライゲーション産物をSwaIで消化し、そして大腸菌中に導入した。アンピシリン耐性クローンを選択した。
対象となる遺伝子を含んでいる組換えアデノウイルスDNAをPacIで消化し、ヒト胎児腎細胞(293細胞)中に導入した。アデノウイルス組換え体を回収し、3回の増幅サイクルにより再増幅した。ウイルスはCICs法によって精製し、次にSephadex G50カラムで洗浄した。ウイルス粒子は、96穴プレートにおけるプラーク形成アッセイにより力価測定した(Pfu)。
【0043】
D)ラット肝細胞の調製
ラット肝細胞は、コラゲナーゼ灌流によって雄ラットの肝臓から単離した。ラットを、ペントバルビタールの腹腔内投与によって麻酔した。次に、肝臓を門脈において、まず200mlの肝臓潅流培地で、そして次に、10mMのHepes、4mMのCaCl2および14mgのBlendzyme3が添加された200mlのHanksバッファーで灌流した。肝臓を切開し、肝細胞洗浄培地(Gibco BRL, Paisley, UK)中で細切し、70μmのフィルターで濾過した。
細胞は遠心分離し、肝細胞洗浄培地中で3回洗浄した後、細胞の生存率をトリパンブルー排除法により評価した(生存率>85%)。細胞は、UltroserSF(2容量%)、ペニシリン(100U/ml)、ストレプトマイシン(100μg/ml)、遊離脂肪酸/BSA(SIGMA, St. Louis, MO, USA)(0.2重量/容量%)、L−グルタミン(2mM)、デキサメタゾン(1μM)、トリヨードサイロニン(T3、SIGMA, St. Louis, MO, USA)(100nM)およびインスリン(100nM)が添加されたWilliams培地中に接種した。4時間後に、培養培地を、UltroserまたはBSAを含まない同様のウィリアムズ培地と交換した。
【0044】
E)糖尿病を発症中のZDFラットへのhFATP5アデノウイルスおよびLACZアデノウイルスの感染
Genetic Model Inc.(Charles River Laboratories, Wilmington, MA, USA)に由来する5週齢の雄のZucker糖尿病肥満型ラット(ZDF/GMI−fa/fa)および同週齢の雄のZucker痩せ型ラットは、無制限量のPurina 5008(PMI Nutrition International, Wellingborough, Northamptonshire, UK)で飼育した。
8週齢において、5.109PfuのアデノウイルスまたはPBS(対照用)を、ラットの尾静脈中に注射した。注射容量は、1ラットあたり2ml未満であった。3頭のラットをpAdhFATP5に、そして3頭をpAdLacZに感染させた。ラットは、10週で安楽死させた。ブドウ糖、遊離脂肪酸およびトリグリセリドレベルを、血漿中で測定した。
【0045】
F)機能試験
a)脂肪酸の捕獲
BSAと複合体化した最終20μMの14C標識オレイン酸(図2B)または最終78nMのトリチウム標識オレイン酸(図2A)を、輸送試験に用いた。
ラット肝細胞(106細胞/コンパートメント)を6穴プレートのウェルに接種し、37℃で4時間インキュベートした。3T3−L1細胞(ATCC No. CL-173、Manassas、VA、USA)(5.105細胞/コンパートメント)は、2%のウシ胎仔血清(FCS)を含むDMEM中でインキュベートした。感染は多重度が1であり、一晩継続した。
【0046】
細胞はトリプシンで処理し、回収し、遠心分離し、PBSで再懸濁して5.105細胞/mlの懸濁液を得た。200μlの細胞懸濁液(105細胞)は、5mlのポリプロピレンチューブ中に置いた。PBS中の細胞懸濁液は、水浴中37℃で5分間、振盪しながらプレインキュベートした。脂肪酸の捕獲のために、2倍に濃縮した脂肪酸/BSAの原液を各チューブに等量加えた。所定の間隔で、0.1%のBSAと200μMのフロレチン(SIGMA, St. Louis, MO, USA)を含むPBSの冷液(0℃)(洗浄液)5mlを各チューブに添加することにより捕獲を停止させた。細胞を遠心分離し、二回洗浄した。遠心分離残渣を、400μlの溶解緩衝液(水中の0.4%のSDS)で再懸濁した。次に4mlのシンチレーター溶液を加え、取り込まれた脂肪酸をTri-Carb 2100カウンター(Packard, Meriden, CT, USA)でカウントした。結果はLacZの誘導の割合として表し、1に対して正規化した。
【0047】
b)脂肪酸のアシル化
ラット肝細胞(106細胞/コンパートメント)を、6穴プレートのウェルに接種し、37℃で4時間インキュベートした。次に培地を回収し、2mlの伝染性混合物(DMEM、2%のFCS)を各ウェルに加えた。感染は多重度が1であり、一晩継続した。細胞をトリプシンで処理し、回収し、遠心分離した。遠心残渣を、300μlの緩衝液A(pH8.5の500mMのtris HCl、1mMのMgCl2、100mMのNaCl、1mMのATP、0.1%のTritonX 100)で再懸濁した。細胞を超音波処理し、氷中で30分間インキュベートした。タンパク質レベルは、ブラッドフォード法により定量化した。1試験あたり、20μgのタンパク質を含む20μlの抽出物を用い、ガラスチューブ中に置いた。180μlの反応混合物(pH8.5の50mMのtris HCl、150μMのコエンザイムA、300μMのDTT、10mMのATP、10mMのMgCl2、0.1%のTritonX 100、14Cで標識された10μMのパルミチン酸またはリグノセリン酸)を、試験を開始するために加えた。反応は、5〜30分後に800μlの1%の過塩素酸の添加により停止させた。
【0048】
標識されたオレイル−CoAのレベルを評価するために、反応混合物を2.25mlのイソプロピル/ヘプタン/硫酸(40/10/1)で抽出した。抽出物を混合し、有機相を回収した。2回の連続した抽出を実行した。4mlのシンチレーター溶液を1mlの水相に加え、Tri-Carb 2100TRカウンターでカウントした。アシルCoAの量を測定するために、沸騰させたタンパク質の対照抽出物で測定した放射活性を、対応する試験値から差し引いた。
結果は、1について正規化したLacZの誘導の割合として表した。
【0049】
c)脂肪酸の酸化
ラット肝細胞(6.105細胞/コンパートメント)を、12穴プレートのウェルに接種し、37℃で4時間インキュベートした。感染は多重度が100であり、一晩継続した。次に、培地を回収し、64nMのオレイン酸[9,10 3H]と2%のBSAとが添加された500μlのDMEMを各ウェルに加えた。37℃での12時間のインキュベーションの後、培地をマイクロチューブへ移し、過剰なオレイン酸[3H]を、50μlの10%のトリクロロ酢酸と50μlの20%のBSAとで沈殿させた。混合物を、12,400rpmで2分間遠心分離した。上清をマイクロチューブに移し、500μlの水を加えた。インキュベーションを、50℃で18時間継続した。4mlのシンチレーター溶液の添加の後、トリチウム水を測定し、Tri-Carb 2100カウンターでカウントした。
結果は、1について正規化したLacZの誘導の割合として表した。
【0050】
d)脂肪酸のエステル化
脂肪酸の酸化は培地中で測定したが、トリグリセリドは、標識された脂肪酸とともにインキュベートした細胞から抽出した。ラット肝細胞をトリプシンで処理し、全く同一のチューブ3本中で混合した。細胞は、18,500rpmで2分間遠心分離した。各ウェルを重炭酸塩ベースの溶液(クレブス−リンゲル重炭酸バッファー:1.2mMのKH2PO4、26mMのNaHCO3、1.3mMのMgCl2、124mMのNaCl、5mMのKCl、10mMのブドウ糖)で洗浄し、そしてこの緩衝液を、遠心分離残渣を再懸濁するのに用いた。
【0051】
細胞を二度目の遠心分離に供し、上清を回収した。この行程を繰り返した。次に遠心分離残渣を100μlの水中に再懸濁し、さらなる分析のために液体窒素で凍結することができる。その後、遠心分離残渣を、室温(25℃)に放置した。次に、500μlのアセトンを加え、混合物を濃縮器を用いて、真空下で乾燥させた。残渣を、基質として5μlの50nMのトリオレインを含む、300μlのクロロホルム/メタノール混合物(1/1)で再懸濁した。混合後、全体を遠心分離し、上清を除去した。2回の抽出を行った。遠心分離残渣は、30μlのクロロホルム/メタノール混合物(1/1)で再懸濁し、G60シリコーンゲルでプレコートされた薄層クロマトグラフィープレート上に堆積させた。移動は、200mlの緩衝液(H−ヘキサン/ジエチルエーテル/メタノール/酢酸 90/20/2/3)で、1.5時間行った。
【0052】
結果:
A)hFATP5の細胞および組織分布
PCRによる研究により、ヒトFATP5の、肝臓、初代肝細胞およびHepG2ヒト細胞系の細胞における排他的な発現が確認された。
これらの結果を図1に示す。
【0053】
B)FATP5の機能的特徴化
a)アデノウイルスの構築
肝臓での脂肪酸の代謝におけるFATP5の機能を特徴化するため、hFATP5遺伝子を担持するアデノウイルス(pAdeno−hFATP5)、対照アデノウイルスpAdeno−LacZ、およびアデノウイルスpAdeno−CD36を設計し、最後のものは脂肪酸輸送についての陽性対照としてのものである。
【0054】
b)脂肪酸の捕獲
オレイン酸の捕獲は、3T3−L1細胞(図2A)およびアデノウイルスに感染したウィスターラットから単離したラット肝細胞(RH)(図2B)において試験した。
FATP5の過剰発現は、3T3−L1細胞における脂肪酸輸送活性に影響を及ぼさなかったが(図2A)、ラット肝細胞において脂肪酸輸送を有意に増大させた。
この結果は、脂肪酸輸送活性が細胞系によって別様に行われていることを示唆するものである。これは、細胞系に固有の種々のコファクターの効果的な動員による可能性がある。以前の結果は、FATPがACSまたはFATなどのパートナーと協働して作用し得ることを示唆している。
ここで提示された結果は、FATP5が、タンパク質が発現された細胞によって別様に、脂肪酸輸送に作用することを示唆するものである。
さらにまた、前脂肪細胞系におけるFATP5の効果の欠如、およびラット肝細胞における脂肪酸輸送の急速な増大は、FATP5が、単なるありふれた輸送体であると言うよりは、むしろ、肝細胞における脂肪酸輸送に影響する特有の機能を有することを示唆する。
【0055】
c)ACSおよびVLACS活性に関する試験
ACS試験は、パルミチン酸塩およびリグノセリン酸塩を用いて、アデノウイルスに感染した初代ラット肝細胞において行った。hFATP5の過剰発現による、リグノセリン酸塩とのインキュベーション後のVLACS活性の有意な増大は、30分のインキュベーション期間でのみ検出可能であった(図3A)。hFATP5の過剰発現によるACS活性の誘導が間接効果である可能性が示唆される。
超長鎖アシルCoAシンテターゼ活性はわずかに増大したが、CD36の過剰発現がリグノセリン酸塩のアシル化に影響を及ぼさないことから(図3B)、これはhFATP5に特有のことである。
FATPタンパク質のVLACS活性に関する先に発表された結果は、このようにして確認された。しかしながら、誘導動態には明確な差異がある。結果は、FATP5が、超長鎖脂肪酸より大きな脂肪酸のクラスを代表するパルミチン酸塩などのC16脂肪酸に対する有意な効果を示すことから、FATP5がより幅広い範囲の活性を有することを示唆している。
【0056】
d)脂肪酸のβ酸化に対するhFATP5発現の効果
hFATP5の過剰発現によって脂肪酸の分解の程度が調節されるかどうかを分析するために、トリチウム化オレイン酸の、β酸化の最終産物であるトリチウム水への変換を測定した。
CD36対照遺伝子の過剰発現は、HepG2細胞(図4A)またはラット初代肝細胞(図4B)におけるβ酸化に影響を及ぼさず、これは、CD36が脂肪酸のエステル化を導くが、その分解は導かないとする仮説に対応するものである。
対照的に、hFATP5の過剰発現は、HepG2肝細胞におけるβ酸化の程度を顕著に増大させた(3倍超)(図4A)。
hFATP5の過剰発現は、初代肝細胞において脂肪酸の分解を誘導する。
【0057】
e)hFATP5が脂肪酸のエステル化よりはむしろそのβ酸化に関与することの確認
hFATP5の発現が脂肪酸の分解のみを導き、そのエステル化度に影響しないことを確認するために、HepG2細胞における同じ実験の中で、β酸化試験を、エステル化試験により補完した。
hFATP5の過剰発現は、エステル化度に影響を及ぼさなかった(図5)。
このことは、肝臓における脂肪酸の分解のみからなる、hFATP5の新規な機能的活性を明確に示すものであり、一方、FAT/CD36は、脂肪酸をエステル化過程に導く。hFATP5は、β酸化アクティベーターである。
【0058】
f)Zucker糖尿病肥満型ラットにおける糖尿病発症の最中のhFATP5の発現の調節
Zucker糖尿病肥満型ラットにおける、このモデルでのNIDDMの発症の最中のFATP5遺伝子の発現レベルの研究を行い、結果をインスリンおよびブドウ糖レベルにおける変動と相関させた。
Zucker糖尿病肥満型ラットは、ヒトの場合と同様の経過で、糖尿病を自然に発症し(図6Aおよび6B)、
−6週齢の雄のZDF(fa/fa)ラットについて観察される、高インスリン血症および正常血糖の状態、
−10週齢の雄のZDF(fa/fa)ラットについて観察される、高インスリン血症および高血糖の状態、および、
−20週齢の雄のZDFラットについて観察される、インスリン欠乏および高血糖の状態、
を有する。
hFATP5の発現の定量的および半定量的な分析による研究(表IIIおよび図7)は、hFATP5メッセンジャーRNAのレベルがZDFラットにおいて10週で低減することを示した。同じ期間にわたって、グリコーゲンレベルはZDFラットの方が、ZLCラットよりも明らかに高かった(表III)。
【0059】
【表5】
生化学的分析は、血漿中の遊離脂肪酸のレベルが10週齢および20週齢のZLCラットでより高いが、ZDFラットの肝臓中の遊離脂肪酸のレベルはZLCラットと比較して低かったことを明らかにした(図8A)。
この10週の期間は、ZDFラットおよびZLCラットの体重増加曲線の乖離によって特徴づけられ、これは糖尿病の発症を示す(図8B)。
【0060】
g)脂肪酸のアシル化へのFATP5の過剰発現の効果
ZDFおよびZLCラットの初代肝細胞におけるhFATP5の過剰発現の機能的効果を分析した。
FATP5の過剰発現は、ZDFラットの初代肝細胞において、早くには5分で、そして遅くには30分で、脂肪酸のアシル化を極めて有意に増大させた。
これらの結果を図9に示す。
【0061】
h)糖尿病肥満型ラットの肝臓における脂肪酸分解不足の解消
脂肪酸のベータ酸化は、ZDFラットの肝臓において、ZLC対照ラットの肝臓において生じるものと比べて不足している。FATP5の過剰発現は、この不足を解消する。FATP5の過剰発現は、対照ラットには影響を及ぼさない。これらの結果を図10に示す。
【0062】
i)FATP5の過剰発現は、ZDFラットの血中のブドウ糖および脂質のレベルを低減する
ブドウ糖レベルは、FATP5を担持しているアデノウイルスに感染したZDFラットにおいて、10週齢で、LacZ対照アデノウイルスに感染したラットと比較して、有意に低減した(図11C)。この低下は、遊離脂肪酸レベルの低下(図11A)、および血漿トリグリセリドレベルの大幅かつ急速な低下(図11B)と相関している。
これは、FATP5の発現が糖代謝に影響を及ぼし、in vivoで高血糖症を軽減することの証明である。
【0063】
例2:ヒトFATP5遺伝子の推定プロモーター領域の特徴化およびプロモーター活性の証明
FATP5遺伝子産物(AF064255またはNM_012254)は、脂質合成に関与するタンパク質のファミリーに属する。
FATP5遺伝子産物は、複合脂質の合成、そして特に脂肪酸の伸長に関与している。FATP5遺伝子は、第19染色体に位置している。
FATP5をコードする配列に関する一般的な情報は、以下の通りである(表IV)。
【表6】
【0064】
a)予測によるFATP5の転写開始点(TSS)の位置、および相補DNA末端の迅速増幅技術(cDNA末端の迅速増幅、RACE)による同定
FATP5遺伝子は、第19染色体に位置し、14kbにわたって分配された10個のエクソンで構成されている。FATP5遺伝子を含むコンティグNT_011104は、2002年8月に公開された。
mRNAおよびESTからマッピングした5’UTR領域は、34bpのサイズを示す(5’UTR1)。したがって、mRNAおよびGenBankによって公開されているEST、およびDBTSS(ヒト転写開始点データベース)に由来する情報に関連して決定される、最も確からしい「生物学的」転写開始部位は、図12Bで示したとおりに位置している。
b)予測によるFATP5プロモーターの位置
材料および方法
HepG2細胞のRNAの調製:
RNAは、Qiagen RNeasy mini kit protocol(Qiagen, Courtaboeuf, France)に従って、100万個のHepG2細胞から精製した。
5’RACE:
TSSは、供給者の推奨に従って、GeneRacerキット、カタログ番号L1500-01、L1500-02、L1502-01、L1502-02バージョンJ05300225-0355を用いて同定した。
結果:
上記のようにして決定した開始点の上流の10,000bpの領域をコンティグNT_011104から選択した。分析した領域は、10023位のコード領域の上流、すなわちATGの上流でちょうど終止する。
予測は、推定プロモーター領域同定用のソフトウェアを用いて行った。これらのソフトウェア、Promoter Inspector、First EFなどの各々は、PolII真核生物プロモーターの特徴(TATAシグナル、GC組成の偏り、階層的組織など)に基づいて構築された数理モデルを用いたものである。異なるアプローチ(二次判別分析、マルコフ連鎖など)を特徴とし、異なる特徴に基づくソフトウェアは、同じ位置に見出された予測を立証することを可能にする。
最も適切な予測の結果を、下表Vにまとめた。
【0065】
【表7】
【0066】
これらの予測は、ヒトFATP5遺伝子のプロモーターを、種々のソフトウェアによって同定された両極端の位置により決定された、7400〜10,000bpの間の領域内に位置づけることを可能にするものである(図12A)。
プロモーター領域は、別な遺伝子が8500位に終止し、そして10,000位がATPコドンの位置であることから、塩基8500と10,000との間であるはずである。実際、8500bpの位置から上流の領域は、機能が知られていないNM_032792遺伝子に対応し、そして10,000bpの位置から下流の領域は、FATP5遺伝子に対応する。この情報は、7400〜9100bpの間の領域内にプロモーターを位置づけた、生物情報ソフトウェアでなされたプロモーターの予測に匹敵するはずである。
【0067】
ヒトFATP5遺伝子のエクソン1の上流に位置するこの領域は、この遺伝子の調節にとって重要な推定プロモーターまたは調節エレメントを含むと推測される。Transfac(転写因子データベース)のテンプレートで分析され、中程度に保存されていることが示された領域は、境界7400〜10,023の間に広がる。ESTおよび全長型のmRNAの生物学的データは、転写開始点(TSS)を9990位に位置付けることを可能にした。
したがって、図13に記載された配列は、ヒトFATP5遺伝子の推定プロモーターに対応する。この配列は、添付された配列表中、配列番号1の番号で特定される。
9990位以外のTSSは、9928位において確認することができた。しかしながら、細胞種によって、別のTSSを代替物として用いることができる。
【0068】
c)ヒトFATP5遺伝子のプロモータ配列の、国際出願WO 0121795の先行技術との比較分析
本発明の対象となるヒトFATP5遺伝子の推定プロモーター配列と、種々のアクセス可能な配列バンク、例えば「Htgs」(終止していない(non-terminated)配列)、「Chromosome」(完全なゲノム、染色体および終止している(terminated)コンティグ)、および最後にnr(全ての公表された配列)などに含まれる配列との類似性について、一連の検索を行った。
ヒトFATP5遺伝子(コンティグNT_011104)のゲノム環境と、国際出願WO 01/21785に公開されているプロモーター領域との間の類似性の検索は、配列類似性がないことを示した。
GenBankの「Chromosome」部門に対する類似性検索は、このデータベースで参照されているヒトまたは他の種(酵母、ショウジョウバエなど)との有意な類似性を見出すことができなかった。
【0069】
Htgsデータベースに含まれる配列中での類似性検索は、国際出願WO 01/21785のプロモータ配列に対応し得るヒトコンティグを特徴づけることができなかった。
国際出願WO 01/21785のプロモータ配列がヒトFATP5遺伝子のゲノム環境との類似性を示さないことを確認するために、これらの2つの配列をアラインメントした。有意な類似性は見出されなかった。
国際出願WO 01/21785に公開されているFATP5遺伝子のプロモータ配列の分析は、この配列が、本発明の対象となるヒトFATP5遺伝子のプロモーターには対応しないが、マウスFATP5遺伝子のプロモーターに対応することを示した。
結論として、種々の検索により、本発明の対象となるヒトFATP5遺伝子のプロモーターの配列は、いかなる既知の配列とも有意な類似性を有しないと結論付けることができる。
【0070】
d)本発明の対象となるヌクレオチド配列のプロモーター活性の確認
材料および方法
FATP5の推定プロモータ配列の増幅:
テンプレート:ヒト胎盤のゲノムDNA(Sigma, St. Louis, MO)、
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号3:CGCTCGAGCTGTGAGCACCTGGATCAGTGCGTGCC、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号4:CCCAAGCTTGGTACCAGCTCCTCCCTAGG、
TaqプラチナPfx+バッファー10×+MgSO4(Invitrogen, Cergy Pontoise, France)、
dNTP(Promega, Williamsburg, IA)。
hFATP5プロモーターに対応するインサートの精製:インサートのサイズ=1573塩基。
増幅プログラム:
94℃で5分間のインキュベーションサイクルを1回、
その後、94℃で30秒間、55℃で30秒間、68℃で2分間のインキュベーションサイクルを25回。
【0071】
得られた物質は、QIAquick PCR精製キット(Qiagen, Courtaboeuf, France)で精製し、4℃に保った。
ヒトFATP5遺伝子のプロモーターを、ルシフェラーゼ遺伝子を含むベクターpGL3にクローニングするための制限:
【表8】
2時間、37℃でのインキュベーション。
Quick ligation kit(NEB, Beverly, MA)、cat#M2200Sの標準的プロトコルによるライゲーション。
30秒間、42℃の熱衝撃の標準的プロトコルによる形質転換。
細菌:DH5アルファ(Invitrogen, Cergy Pontoise, France)。
50μg/mlのアンピシリンを含むLB寒天(Invitrogen, Cergy Pontoise, France)のディッシュへのプレーティング。
37℃での一晩のインキュベーション。
スクリーニング:
各クローンを50μg/mlのアンピシリンを含むLB寒天上に継代し、20μlの水中で再懸濁した。
懸濁液を、95℃で10分間加熱した。
【0072】
pGL3クローン/hFATP5プロモーターの選択のためのPCR条件
テンプレート:細菌溶解物
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号5:TTCATTACATCTGTGTGTTGGTTTTTTGTGTG、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号6:TATGCAGTTGCTCTCCAGCGGTTCCATCTTCC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)、50μg/ml。
増幅プログラム:
94℃で5分間のインキュベーションサイクルを1回、
その後、94℃で30秒間、55℃で30秒間、72℃で2分間のインキュベーションサイクルを25回、
その後、72℃で7分間のサイクルを1回。
得られた物質は、4℃に保った。
プラスミドの作製は、EndoFree Plasmid maxi kit(Qiagen, Courtaboeuf, France, cat#12362)の標準的プロトコルに従った。
配列決定は、BigDye terminator V2.0 cycle sequencing kit(Applied Biosystems, Foster City, CA, cat#4314415)の標準的プロトコルに従った。
【0073】
結果:
ヒトFATP5プロモーターの1573塩基の断片をpGL3にクローニングし、配列決定した。断片の連続した欠失による構築物は、FATP5の肝での発現に特異的なプロモーター領域を同定することを可能にする。
HEP2、HELAおよびHEK293肝細胞の一過性トランスフェクション:
スクリーニングのための最小可能なプロモーター断片を特定するために、一過性にトランスフェクトした細胞を用いた。トランスフェクションは、jetPEIプロトコル(Polytransfection, Illkirch, France)により、供給者の推奨に従って行った。
第3日:照度計による測定:
トランスフェクション混合物を含む培地を除去した。
プレートを、250μlのPBSで2回洗浄し、そして、1/5に希釈した100μlの溶解バッファーを加えた。プレートを、振盪させながら室温で30分間インキュベートした。
各ウェルからの10μlの溶解液を、Greiner 96穴白色プレート(Greiner Bio-One, Frickenhausen, Germany)に分配した。
【0074】
ルシフェラーゼ活性は、Dual Luciferase kit(Promega, Madison, WI, USA)を用いて測定した。測定値は、TR717マイクロプレートルミノメーター(Applied Biosystems, Foster City, CA, USA)で取得した。
ルシフェラーゼ発光の結果はレニラに対して正規化し、プロモーターを含む構築物の、空ベクターpGL3と比較した誘導として表した。
結果:
FATP5プロモーターの断片全体のプロモーター活性が確認された。図14に示したこれらの結果は、本核酸配列が実際にFATP5プロモーターであり、かつそれが機能的であることを確認するものである。ルシフェラーゼ活性の最も高い値は、HepG2ヒト肝細胞で観察された。
【0075】
例3:安定してヒトFATP5プロモーターを発現している細胞でのスクリーニング試験
a)ヒトFATP5遺伝子の推定プロモーターの、ハイグロマイシン耐性を有するベクターpGL3ハイグロマイシンへのクローニング
ベクターpGL3ハイグロマイシンの作製:
Tkプロモーターの制御下にあるハイグロマイシンをコードする遺伝子(サイズ:1671塩基)を、ベクターpREP4(Invitrogen, Cergy Pontoise, France)から、以下のプライマーを用いて増幅した。
材料と方法:
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号7:GAAGATCTCTGCTTCATCCCCGTGGC、
オリゴアンチセンス(Invitrogen, Cergy Pontoise, France):配列番号8:GAAGATCTACCAGACCCCACGCAACGC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)。
増幅プログラムおよび得られた物質の精製は、hFATP5プロモーターに対応するインサートの増幅および精製のために先に用いたのと同じプロトコルに従って行った。
得られた断片は、ベクターpGL3−Basic(Promega, Williamsburg, IA)のBglII部位にクローニングした。クローニングのための制限、ライゲーションおよび形質転換は、hFATP5プロモーターに対応するインサートの増幅および精製のために先に用いたのと同一のプロトコルに従って行った。
Tkハイグロマイシン断片を有するプラスミドpGL3が組み込まれたクローンは、pGL3クローン/hFATP5プロモーターの選択に用いたのと同じプロトコルによるPCRにより、テンプレートとして細菌溶解物を、および:
センスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号9:CTGCTTCATCCCCGTGGC、
アンチセンスオリゴ(Invitrogen, Cergy Pontoise, France):配列番号10:ACCAGACCCCACGCAACGC、
Goldstar+バッファー10×+MgCl2(Eurogentec, Seraing, Belgium)、
を用いて選択した。
hFATP5プロモーターを含むpGL3ハイグロマイシンクローンの選択、プラスミドの作製および配列決定は、pGL3クローン/hFATP5プロモーターの選択のために用いたのと同じプロトコルに従って行った。
【0076】
b)HepG2肝細胞の安定したトランスフェクション
用いた培地を下表に記載する。
【表9】
【0077】
c)細胞のプレーティング
HepG2細胞の培養物を調製した。細胞は、80%コンフルエントに達したときに剥離し、6穴プレート中の培地に、1ウェルあたり320,000細胞の割合で再接種した。細胞は、37℃、5%CO2で、24時間インキュベートした。
【0078】
d)トランスフェクションおよび活性化
トランスフェクションは、a)で調製した500ngのプラスミドを用いて、jetPEIプロトコル(Polytransfection, Illkirch, France)により、供給者の推奨に従って行った。
6穴プレートで培養した細胞の培地を吸引した。1mlの無血清DMEMと80μlのトランスフェクション混合物とを各ウェルに入れた。次に、細胞を、5%のCO2を含む雰囲気下、37℃で2時間放置した。この2時間が経過したときに、FCSを含まない培地1mlと、2%の終濃度のUltroser SF(USF, BioSepra, Cergy Pontoise, France)とを加えた。
【0079】
e)トランスフェクション細胞の選択
48時間の培養後に、各ウェルの培地を吸引した。10%のFCSと終濃度0.2mg/mlのハイグロマイシンとを含む2mlの培養培地を加えた。細胞は、80%コンフルエントに達したときに剥離した。
次に、2個のウェルからの細胞を混合し、150mmのペトリ皿中の同じ選択培地に接種した。選択培養は、細胞クローンが現れるまで維持した。
【0080】
f)クローンの単離
クローンが明確に区別されるようになったら、これらを、シリンダーを用いて残りの培養物から単離した。次に、シリンダーに含まれた細胞をPBSで洗浄し、培地の吸引後に取り出した。
細胞を96穴プレートに配置し、そして10%のFCSと終濃度0.25mg/mlのハイグロマイシンとを含む250μlのDMEMを加えた。培養は、80%コンフルエントになるまで継続した。
【0081】
g)安定してヒトFATP5プロモーターを発現するHepG2細胞のスクリーニング
ストックを用意するために、クローンを増幅した。
各クローンに由来する細胞は、24穴プレートにて、10%のFCS、ペニシリン/ストレプトマイシン混合物(0.125%)および0.25mg/mlのハイグロマイシンを含むDMEM中、80%コンフルエントまで培養した。
被験化合物による処置は、24時間の期間適用した。この24時間が経過したときに、ルシフェラーゼ活性を、細胞溶解物において、Applied Biosystems TR717発光リーダー(California, CA)を用いて測定した。
ルシフェラーゼ活性を示している細胞は、この活性が、これらの細胞が受け取ったプラスミドに含まれる推定プロモーターによって誘導されたことを示す。
この結果は、本発明の核酸が、実際に、その依存下に置かれた遺伝子の発現を導くことができるプロモーターであることを示すものである。
【0082】
例4:糖尿病表現型の解消およびインスリン増感剤によるZDFラットにおけるFATP5遺伝子の発現の調節
レキシノイド(rexinoid)と呼ばれるRXR核受容器のアゴニストは、インスリン増感剤であり、糖尿病肥満型動物モデル(2型糖尿病)において有益な効果を有する。ベキサロテンであるタルグレチン(登録商標)(CAS 1543559-49-0)は、核受容体ヘテロダイマー、RXR−PPAR(ペルオキシゾーム増殖因子活性化受容体)またはRXR−LXR(肝臓X受容体)を選択的に活性化する。
雄の糖尿病肥満型ラット(Zucker糖尿病肥満型ラット(ZDFラット))は、食物と水に自由にアクセスできる動物ケージ内に維持し、夜昼リズム(nyctohemeral rhythm)が観察された。
【0083】
化合物LGD 1069を、0.3、1.0、3.0、10または30mg/kg/日の用量で食餌に加え、7週齢より50日間投与した。4頭の動物は、LGD 1069の添加なしで給餌した:ZDF2頭とZLC2頭。動物を絶食させ、その翌日に安楽死させた。組織を回収し、液体窒素中で直ちに凍結した。
肝RNAは、ChomczynskiおよびSacchiに由来する従来の技法で抽出した。発現は、FATP5に特異的な放射性cDNAプローブを用いたハイブリダイゼーションによるノーザンブロッティングによって分析した。
結果を図15に示す。
ZDFラットの肝臓におけるFATP5の発現の、ZLCラットに比した負の調節は、RXRアゴニストによる処置によって打ち消され、これは糖尿病の進行を逆転させた。
【0084】
例5:真核細胞(CHO:チャイニーズハムスター卵巣細胞)におけるFATP5受容体を介した脂肪酸の能動輸送を測定するための試験
蛍光標識された脂肪酸模倣分子の使用は、FATP受容体を過剰発現した細胞における脂肪酸の細胞内蓄積の、マイクロタイタープレート形式での迅速な測定を提供する。
試験の説明
【表10】
必要な専門器材
−CO2インキュベーター、
−顕微鏡、
−マルチドロップ、
−マイクロプレートウォッシャー、
−フルオレセイン用フィルターを有するマイクロプレート用蛍光計。
【0085】
プロトコル
第1日
96穴マイクロプレートへの細胞の分配
FATP受容体を過剰発現するCHO真核細胞を、96穴培養プレートに、1ウェルあたり約10,000細胞の密度で、以下の手順に従って分配する。
−層流フード下で、4mlの細胞剥離溶液を225cm3の培養フラスコに37℃で加える。
−溶液を、細胞単層上、手動による軌道回転運動で10〜15秒間振盪し、その後、10mlの血清用ピペットで吸引する。
−4mlの細胞剥離溶液を加え、フラスコを(最長で)10分間インキュベーター中に戻す。10分後に、細胞の剥離を顕微鏡下でチェックする。細胞がすべて剥離するまで操作を続ける。
−8mlの培地をフラスコに加え、細胞塊をほぐすために、内容物を何度か上から下へ混合する。この細胞混合物を、37℃に予熱した88mlの培地に加える。
【0086】
マルチドロップの準備
−マルチドロップを層流フードに配置する。マルチドロップのヘッドおよびチューブを、20〜30mlの70%エタノール、次いで20〜30mlの無菌水、そして最後に20〜30mlの培地をポンプ吸引することにより洗浄する。
−マルチドロップのチューブを細胞を含むフラスコに配置し、およそ5〜10mlの培地を除去する。1ウェルあたり100μlの細胞を加える(10,000細胞)。
−マイクロプレートを、37℃、10%CO2で48時間、インキュベーター内に置く。
【0087】
第2日:被験分子の添加
被験分子を、以下のプレートレイアウトに従って添加する:
【表11】
−マルチドロップを用いて、95μlのDMEM(フェノールレッド不含)を、96穴マイクロプレートの各ウェルに分配する。
−被験分子を、Tと表示されたウェルに終濃度5mg/ml(1%のDMSO)で同時に分配する。
−200mMのラウリン酸を、陰性対照ウェル(NC)に分配する。
−5μlのPBSを、陽性対照ウェル(PC)に分配する。
【0088】
第3日
−細胞を、PBSで2回洗浄する。
−細胞を、PBS中に0.1mMのBODIPY-FAと、脂肪酸を含まない0.1%のBSAとを含む脂肪酸模倣溶液中、2分間、37℃でインキュベートする。
−2分後に、細胞を事前に氷中に配置しておいたPBS/0.1%BSA混合物で4回洗浄する。
−プレートを、蛍光マイクロプレートリーダーにて、励起波長として485nmを、放出波長として530nmを用いて読み込む。
結果を、プローブなし(PC)の対照細胞のパーセンテージとして、任意の蛍光単位で表す。
この試験は、真核細胞内へのFATP5受容体を介した脂肪酸の能動輸送を測定することを可能にする。これはまた、FATP5受容体を介した脂肪酸の能動輸送に対する所定の分子の影響を測定することを可能にし、それによってヒトFATP5遺伝子のプロモーターの活性を刺激または阻害するその能力を決定することを可能にする。
【図面の簡単な説明】
【0089】
【図1】PCRによって分析された、ヒトFATP5遺伝子の発現の細胞および組織分布を示した図である。NCI−H295:副腎細胞系、THP−1:単球、THP1D:THP−1単球の分化したマクロファージ、JurkatおよびJurkat JR:T細胞、CEM:急性リンパ芽球性白血病細胞系の細胞、HH:ヒト初代肝細胞、HepG2、HuH7:ヒト肝細胞、CaCo2:結腸細胞、CaCo2D:分化したCaCo2細胞、CASMC:冠状動脈平滑筋細胞、ASMC:大動脈平滑筋細胞。
【図2A】3T3−L1細胞での脂肪酸捕獲試験で得られた結果を示した図である。結果は、各条件についての、時間の関数としての脂肪酸捕獲の誘導を表す。対照条件は、発現が細胞による脂肪酸捕獲に影響を及ぼさない遺伝子を担持するアデノウイルスに感染した細胞による捕獲の測定値を表す。
【図2B】ラット肝細胞(RH)での脂肪酸捕獲試験で得られた結果を示した図である(時間:1分)。結果は、各条件についての、時間の関数としての脂肪酸捕獲の誘導を表す。対照条件は、発現が細胞による脂肪酸捕獲に影響を及ぼさない遺伝子を担持するアデノウイルスに感染した細胞による捕獲の測定値を表す。
【図3A】hFATP5遺伝子を担持するアデノウイルスへの感染後における、ACS活性を決定するための試験において得られた結果を示した図である。結果は、誘導係数として表す。
【図3B】hFATP5遺伝子を担持するアデノウイルスへの感染後における、VLACS活性を決定するための試験において得られた結果を示した図である。結果は、誘導係数として表す。
【0090】
【図4A】hFATP5の発現の、脂肪酸のβ酸化への影響を決定するための試験において得られた結果を示した図である。4A:48時間におけるHepG2、4B:12時間におけるRH。結果は、誘導係数として表す。
【図4B】hFATP5の発現の、脂肪酸のβ酸化への影響を決定するための試験において得られた結果を示した図である。4A:48時間におけるHepG2、4B:12時間におけるRH。結果は、誘導係数として表す。
【図5】hFATP5の発現の、48時間におけるHepG2細胞での脂肪酸のエステル化度への影響を決定するための試験において得られた結果を示した図である。結果は、3H2O/3TG比として表す。
【図6A】hFATP5の発現の、Zucker糖尿病肥満型ラットでの糖尿病の発症の最中におけるhFATP5の発現の調節への影響を決定するための試験において得られた結果を示した図である。A:ブドウ糖レベルの測定値、B:インスリンレベルの測定値、−■−:雄のZDFラット(fa/fa)、−□−:雄のZLCラット(fa/+)。
【図6B】hFATP5の発現の、Zucker糖尿病肥満型ラットでの糖尿病の発症の最中におけるhFATP5の発現の調節への影響を決定するための試験において得られた結果を示した図である。A:ブドウ糖レベルの測定値、B:インスリンレベルの測定値、−■−:雄のZDFラット(fa/fa)、−□−:雄のZLCラット(fa/+)。
【図7】Zucker糖尿病肥満型ラット(ZDF:−●−)およびZLCラット(−○−)におけるhFATP5メッセンジャーRNAのレベルを測定するための試験において得られた結果を示した図である。
【図8A】生化学分析の結果を示した図である。A:肝臓における遊離脂肪酸レベルの測定値(−θ−:ZDFラット(fa/fa)、−○−:ZLCラット(fa/+))、B:体重増加曲線(−●−:ZDFラット(fa/fa)、−□−:ZLCラット(fa/+))。
【図8B】生化学分析の結果を示した図である。A:肝臓における遊離脂肪酸レベルの測定値(−θ−:ZDFラット(fa/fa)、−○−:ZLCラット(fa/+))、B:体重増加曲線(−●−:ZDFラット(fa/fa)、−□−:ZLCラット(fa/+))。
【0091】
【図9】8週齢のZDFおよびZLCラットにおける、各条件につき5および30分でのオレイン酸のアシル化の程度を示した図である。A:ZDFラット、B:ZLCラット。
【図10】ZDFラットの初代肝細胞における、ZLC対照ラットと比較したβ酸化の程度の抑制の解消を示した図である。A:8週齢のラット、B:10週齢のラット。(C=対照、Z=LacZ、h5=hFATP5)。
【図11】LacZ対照アデノウイルスに感染したラットと比較した、FATP5を担時するアデノウイルスに感染したZDFラットにおける生化学パラメータの測定値を示した図である(遊離脂肪酸(A)、トリグリセリド(B)およびブドウ糖(C))。
【図12】FATP5遺伝子の転写開始点(TSS)の位置を示した図である。A:バイオインフォマティクスによる予測的な方法論、B:RACEによる実験的な方法論。
【図13】ヒトFATP5遺伝子の推定プロモーターの配列を示した図である。この配列は、添付された配列表において配列番号1の番号で特定される。
【図14】ヒトFATP5遺伝子のプロモータ配列の、HepG2、HeLaおよびHek293細胞における活性を示した図である。結果は、空プラスミドpGL3で形質転換した細胞で測定した対照と比較した誘導として表示してある。
【図15】ZDFラット(A)またはZLCラット(B)をインスリン増感剤として知られる化合物で処置した後の、FATP5メッセンジャーRNAのレベルの変動を示した図である。
【特許請求の範囲】
【請求項1】
添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、単離されたヒト核酸配列。
【請求項2】
添付された配列表において配列番号1の番号で特定されるヌクレオチド配列からなることを特徴とする、単離されたヒト核酸配列。
【請求項3】
ヒトFATP5タンパク質をコードすることを特徴とする、請求項1に記載の核酸配列。
【請求項4】
添付された配列表において配列番号2の番号で特定されるヌクレオチド配列と対応することを特徴とする、請求項3に記載の核酸配列。
【請求項5】
請求項1〜4のいずれかに記載の核酸配列を含むことを特徴とするベクター。
【請求項6】
請求項1に記載の核酸配列を含むことを特徴とする、請求項5に記載のベクター。
【請求項7】
請求項1に記載の核酸配列の依存下に置かれた検出可能なタンパク質をコードする核酸配列をも含むことを特徴とする、請求項5または6に記載のベクター。
【請求項8】
検出可能なタンパク質が、FATP5、緑色蛍光タンパク質(GFP)、レニラおよびルシフェラーゼから選択されるタンパク質であることを特徴とする、請求項7に記載のベクター。
【請求項9】
プラスミドであることを特徴とする、請求項5〜8のいずれかに記載のベクター。
【請求項10】
請求項1〜4のいずれかに記載の核酸配列または請求項5〜9のいずれかに記載のベクターが導入された宿主細胞。
【請求項11】
原核生物または真核生物細胞であることを特徴とする、請求項10に記載の細胞。
【請求項12】
細菌であることを特徴とする、請求項11に記載の原核細胞。
【請求項13】
タンパク質を製造する方法であって、請求項10〜12のいずれかに記載の宿主細胞を、前記細胞に導入されたヌクレオチド配列のタンパク質の形態での発現を可能にする条件下で培養することを特徴とする、前記方法。
【請求項14】
請求項1〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞の、請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定のための使用。
【請求項15】
請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定方法であって、
a)請求項2〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞を、ヌクレオチド配列の発現に適した媒体中で、請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質と接触させること、および、
b)請求項1に記載の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルを、任意の適切な手段により測定すること、
を特徴とする前記方法。
【請求項16】
b)で決定された発現レベルと、同定される物質に接触させなかった対照における、請求項1に記載の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルとの比較のための追加の工程を含むことを特徴とする、請求項15に記載の方法。
【請求項17】
請求項1に記載の核酸配列と相互作用することが可能な物質の同定方法であって、
a)請求項1〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞を、請求項1に記載の核酸配列と相互作用することが可能な物質に接触させること、および、
b)請求項1に記載の核酸配列と被験物質との間の相互作用を評価すること、
を特徴とする前記方法。
【請求項18】
請求項1に記載の核酸配列に結合することが可能な物質を同定することを目的とする、請求項17に記載の同定方法。
【請求項1】
添付された配列表において配列番号1の番号で特定されるヌクレオチド配列を少なくとも含むことを特徴とする、単離されたヒト核酸配列。
【請求項2】
添付された配列表において配列番号1の番号で特定されるヌクレオチド配列からなることを特徴とする、単離されたヒト核酸配列。
【請求項3】
ヒトFATP5タンパク質をコードすることを特徴とする、請求項1に記載の核酸配列。
【請求項4】
添付された配列表において配列番号2の番号で特定されるヌクレオチド配列と対応することを特徴とする、請求項3に記載の核酸配列。
【請求項5】
請求項1〜4のいずれかに記載の核酸配列を含むことを特徴とするベクター。
【請求項6】
請求項1に記載の核酸配列を含むことを特徴とする、請求項5に記載のベクター。
【請求項7】
請求項1に記載の核酸配列の依存下に置かれた検出可能なタンパク質をコードする核酸配列をも含むことを特徴とする、請求項5または6に記載のベクター。
【請求項8】
検出可能なタンパク質が、FATP5、緑色蛍光タンパク質(GFP)、レニラおよびルシフェラーゼから選択されるタンパク質であることを特徴とする、請求項7に記載のベクター。
【請求項9】
プラスミドであることを特徴とする、請求項5〜8のいずれかに記載のベクター。
【請求項10】
請求項1〜4のいずれかに記載の核酸配列または請求項5〜9のいずれかに記載のベクターが導入された宿主細胞。
【請求項11】
原核生物または真核生物細胞であることを特徴とする、請求項10に記載の細胞。
【請求項12】
細菌であることを特徴とする、請求項11に記載の原核細胞。
【請求項13】
タンパク質を製造する方法であって、請求項10〜12のいずれかに記載の宿主細胞を、前記細胞に導入されたヌクレオチド配列のタンパク質の形態での発現を可能にする条件下で培養することを特徴とする、前記方法。
【請求項14】
請求項1〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞の、請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定のための使用。
【請求項15】
請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質の同定方法であって、
a)請求項2〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞を、ヌクレオチド配列の発現に適した媒体中で、請求項1に記載の核酸配列の依存下に置かれたヌクレオチド配列の発現を調節することが可能な物質と接触させること、および、
b)請求項1に記載の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルを、任意の適切な手段により測定すること、
を特徴とする前記方法。
【請求項16】
b)で決定された発現レベルと、同定される物質に接触させなかった対照における、請求項1に記載の核酸配列の依存下に置かれた前記ヌクレオチド配列の発現のレベルとの比較のための追加の工程を含むことを特徴とする、請求項15に記載の方法。
【請求項17】
請求項1に記載の核酸配列と相互作用することが可能な物質の同定方法であって、
a)請求項1〜4のいずれかに記載の核酸配列、請求項5〜9のいずれかに記載のベクターまたは請求項10〜12のいずれかに記載の宿主細胞を、請求項1に記載の核酸配列と相互作用することが可能な物質に接触させること、および、
b)請求項1に記載の核酸配列と被験物質との間の相互作用を評価すること、
を特徴とする前記方法。
【請求項18】
請求項1に記載の核酸配列に結合することが可能な物質を同定することを目的とする、請求項17に記載の同定方法。
【図1】


【図2A】
【図2B】
【図3A】
【図3B】
【図4A】
【図4B】
【図5】
【図6A】
【図6B】
【図7】
【図8A】
【図8B】
【図9】
【図10】

【図11】
【図12】

【図13】

【図14】
【図15】
【図2A】
【図2B】
【図3A】
【図3B】
【図4A】
【図4B】
【図5】
【図6A】
【図6B】
【図7】
【図8A】
【図8B】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2007−533293(P2007−533293A)
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願番号】特願2006−524261(P2006−524261)
【出願日】平成16年8月4日(2004.8.4)
【国際出願番号】PCT/EP2004/008747
【国際公開番号】WO2005/021590
【国際公開日】平成17年3月10日(2005.3.10)
【出願人】(591032596)メルク パテント ゲゼルシャフト ミット ベシュレンクテル ハフトング (1,043)
【氏名又は名称原語表記】Merck Patent Gesellschaft mit beschraenkter Haftung
【住所又は居所原語表記】Frankfurter Str. 250,D−64293 Darmstadt,Federal Republic of Germany
【Fターム(参考)】
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願日】平成16年8月4日(2004.8.4)
【国際出願番号】PCT/EP2004/008747
【国際公開番号】WO2005/021590
【国際公開日】平成17年3月10日(2005.3.10)
【出願人】(591032596)メルク パテント ゲゼルシャフト ミット ベシュレンクテル ハフトング (1,043)
【氏名又は名称原語表記】Merck Patent Gesellschaft mit beschraenkter Haftung
【住所又は居所原語表記】Frankfurter Str. 250,D−64293 Darmstadt,Federal Republic of Germany
【Fターム(参考)】
[ Back to top ]