
ヒトSAA3核酸分子、タンパク質およびこれらの使用方法
ヒト乳房上皮細胞から単離、精製、およびキャラクタライズしたヒト血清アミロイドA3(SAA3)をコードする核酸配列を開示する。該核酸配列がコードするタンパク質、およびこうしたタンパク質の使用方法についても開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、免疫学および哺乳動物免疫系の分野に関する。特に本発明は、これまでその遺伝子が偽遺伝子としてキャラクタライズされていたので存在しないと考えられてきた血清アミロイドA(SAA3)の新規cDNAおよびアイソフォームを提供する。
【技術背景】
【0002】
いくつかの科学または特許刊行物を、本発明が属する技術の水準を説明するために、本明細書において参照する。これらの刊行物を全て、参照によりその全内容を本明細書に組み入れる。
【0003】
哺乳動物は、さらなる組織損傷を防ぐため、損傷を受けた組織の修復を開始するため、および病原性生物を隔離し破壊するために、複雑な一連の生物学的反応を実行することで、組織損傷、外傷、または感染症に応答する。この過程は炎症反応と呼ばれており、その炎症反応の初期および中間段階は急性期応答と呼ばれている。
【0004】
急性期応答は、広範な種類の仲介物質(サイトカイン、インターロイキンおよび腫瘍壊死因子など)を伴う。急性期応答はまた、肝臓の生合成プロファイルの過激な変更を伴う。通常の状況下では、肝臓は定常状態濃度で様々な血漿タンパク質を合成する。このうちのいくつかのタンパク質、「急性期」タンパク質は、炎症応答において、通常の状態でみられるレベルより何倍も大きいレベルへと誘導される。急性期タンパク質は、Steel および Whitehead (Immunology Today 15: 81-87, 1994)によって総評されている。
【0005】
大量に誘導される急性期タンパク質の1つは、血清アミロイドA(SAA)である。SAAは、炎症応答の急性期に高密度リポタンパク質3(HDL3)と急速に会合および蓄積する小さなアポリポタンパク質である。ほとんどのSAAアイソフォームは炎症に応答して誘導されるが、一方で特定のSAA(例えばヒトSAA4)は構成的に発現しているかまたは炎症応答で最小限しか誘導されないようである。
【0006】
血清アミロイドAタンパク質(SAA)は、今日までに調査された全ての脊椎動物において産生されるアポリポタンパク質のスーパーファミリーを含む(C.M. Uhlar, A.S. Whitehead, Serum amyloid A, the major vertebrate acute-phase reactant, Eur. J. Biochem. 265 (1999) 501-523)。生物種によっては、SAAをコードする3または4つの遺伝子座が特定されており、これらの遺伝子は肝臓からおよび/または肝臓外から区別されて発現されている(J.D. Sipe, Serum amyloid A: from fibril to function. Current status, Amyloid: Int. J. Exp. Clin. Invest. 7 (2000) 10-12)。急性期血清アミロイドAタンパク質(A-SAA)は、主に肝臓によって合成され、血漿の高密度リポタンパク質3(HDL3)画分と大部分が会合している(G.A. Coetzee, A.F. Strachan, D.R. Van Der Westhuyzen, H.C. Hoffe, M.S. Jeenah, F.C. De Beer, Serum amyloid A-containing human high density lipoprotein, J. Biol. Chem. 261 (1986) 9644-9651; N. Eriksen, E.P. Benditt, Isolation and characterization of the amyloid-related apoprotein (SAA) from human high density lipoprotein, Proc. Natl. Acad. Sci. USA 77 (1980) 6860-6864)。肝臓によって誘導されたA-SAAレベルは、炎症性サイトカインであるIL-1、IL-6およびTNF-αに応答して急性期に1000倍増加し得る(L.E. Jensen, A.S. Whitehead, Regulation of serum amyloid A protein expression during the acute-phase response, Biochem. J. 334 (1998) 489-503)。血液の循環におけるA-SAAの劇的な増加は、主に転写の増加によって達成される(C.M. Uhlar, A.S. Whitehead, Serum amyloid A, the major vertebrate acute-phase reactant, Eur. J. Biochem. 265 (1999) 501-523)。
【0007】
肝臓は、SAA産生の主要な部位と考えられてきた。しかしながら、限定的な証拠に基づくことではあるが、肝臓外でのSAA産生が見いだされた。例えば、SAA mRNAの発現は、ヒトのアテローム性動脈硬化の病巣、ならびにヒト培養平滑筋細胞および単核白血球/マクロファージ細胞系で報告されており(Meek ら, 1994; Urieli-Shoval ら, 1994; Yamada ら, 1996)、またウサギ骨膜繊維芽細胞からSAAの独特のアイソフォーム(SAA3)が産生されている(Mitchell ら, J. Clin. Invest. 87: 1177-1185, 1991)。さらに最近、SAA mRNAが多くの組織学的に正常な上皮組織(胃、腸、扁桃腺、乳房、前立腺、甲状腺、肺、膵臓、腎臓、皮膚および脳神経など)で広く発現していることが発見された(Urieli-Shoval ら, J. Histochem. Cytochem. 46: 1377-1384, 1998)。
【0008】
多数の研究者による実験的証拠は、SAAタンパク質の多様な機能を示唆しているが、そうした機能には以下のものが含まれる。免疫応答の抑制(M.A. Aldo-Benson, M.D. Benson, SAA suppression of immune response in vitro: evidence for an effect on T cell-macrophage interaction, J. Immunol. 128 (1982) 2390-2392)、血小板凝集の阻害(S. Zimlichman, A. Danaon, I. Nathan, G. Mozes, R. Shainkin-Kestenbaum, Serum amyloid A, an acute phase protein, inhibits platelet activation, J. Lab. Clin. Med. 116 (1990) 180-186)、コレステロール/脂質代謝への関与(R. Kisilevsky, L. Subrahmanyan, Serum amyloid A changes high density lipoprotein's cellular affinity. A clue to serum amyloid A's principal function, Lab. Invest. 66 (1992) 778-785; R.L. Meek, N. Eriksen, E.P. Benditt, Murine serum amyloid A3 is a high density apolipoprotein and is secreted by macrophages, Proc. Natl. Acad. Sci. USA 89 (1992) 7949-7952)、内毒素の解毒への関与(C. Baumberger, R.J. Ulevitch, J.M. Dayer, Modulation of endotoxic activity of lipopolysaccharide by high-density lipoprotein, Pathobiology 59 (1991) 378-383)、コラゲナーゼ活性の誘導(C.E. Brinckerhoff, T.I. Mitchell, M.J. Karmilowicz, B. Kluve-Beckerman, M.D. Benson, Autocrine induction of collagenase by serum amyloid A-like and β2-microglobulin-like proteins, Science 243 (1989) 655-657)、好中球のオキシダティブバーストの阻害(R.P. Linke, V. Bock, G. Valet, G. Tothe, Inhibition of the oxidative burst response of N-formyl peptide-stimulated neutrophils by serum amyloid A protein, Biochem. Biophys. Res. Commun. 176 (1991) 1100-1105; M.E. Gatt, S. Urieli-Shoval, L. Preciado-Patt, M. Fridkin, S. Calco, Y. Azar, Y. Matzner, Effect of serum amyloid A on selected in vitro functions of isolated human neutrophils, J. Lab. Clin. Med. 132 (1998) 414-420)、単核白血球、多形核白血球およびT細胞の移動の誘導(R. Badaloto, J.M. Wang, W.J. Murphy, A.R. Lloyd, D.F. Michiel, L.L. Bausserman, D.J. Kelvin, J.J. Oppenheim, Serum amyloid A is a chemoattractant: induction of migration, adhesion and tissue infiltration of monocytes and polymorphonuclear leukocytes, J. Exp. Med. 180 (1994) 203-209; L. Xu, R. Badolato, W.J. Murphy, D.L. Longo, M. Anver, S. Hale, J.J. Oppenheim, J.M. Wang, A novel biologic function of serum amyloid A. Induction of T lymphocyte migration and adhesion, J. Immunol. 155 (1995) 1184-1190)、ならびに細胞外マトリックス構成要素への細胞付着の阻害(L. Preciado-Patt, D. Levartowsky, M. Pras, R. Hershkoviz, O. Likder, M. Fridkin, Inhibition of cell adhesion to glycoproteins of the extracellular matrix by peptides corresponding to serum amyloid A. Toward understanding the physiological role of an enigmatic protein, Eur. J. Biochem. 223 (1994) 35-42)。しかしながら、正常状態および疾病状態におけるSAAの主要な生理学的役割はよく分かっていない。
【0009】
ヒトSAA mRNAおよびタンパク質の肝臓外発現は、マクロファージ細胞、脂肪細胞、平滑筋細胞、および内皮細胞で実証されており、これらの産生部位で機能している蓋然性が高いことを示唆する。Urieli-Shovalらは、ヒトSAAの肝臓外発現が、主として各種組織の上皮の構成要素に局所化されていることを明らかにした(S. Urieli-Shoval, P. Cohen, S. Eisenberg, Y. Matzner, Widespread expression of serum amyloid A in histologically normal human tissues: predominant localization to the epithelium, J. Histochem. Cytochem. 46 (1998) 1377-1384)。
【0010】
ヒトでは4つのSAA遺伝子が染色体11p15.1上にクラスター化されている(G.C. Sellar, S.A. Jordan, W.A. Bickmore, J.A. Fantes, V. van Heyningen, A.S. Whitehead, The human serum amyloid A protein (SAA) superfamily gene cluster: mapping to chromosome 11p15.1 by physical and genetic linkage analysis, Genomics 19 (1994) 221-227)。高誘導性SAA1およびSAA2遺伝子は、90%同一な104残基のA-SAAタンパク質をコードする。SAA1およびSAA2遺伝子は、それらのプロモーター領域、エクソンおよびイントロンの全体で約95%のヌクレオチド配列同一性を有する(J.C. Betts, M.R. Edbrooke, R.V. Thakker, P. Woo, The human acute-phase serum amyloid A gene family: structure, evolution and expression in hepatoma cells, Scand. J. Immunol. 34 (1991) 471-482; P. Woo, J. Sipe, C.A. Dinarello, H. R. Colten, Structure of a human serum amyloid A gene and modulation of its expression in transfected L cells, J. Biol. Chem. 262 (1987) 15790-15795)。配列が類似しているにもかかわらず、最近の研究によって、サイトカイン依存性誘導発現という状況で、SAA2と比較してSAA1の転写発現が、グルココルチコイドから異なる増強を受ける、ということが実証された(C.F. Thorn, A.S. Whitehead, Differential glucocorticoid enhancement of the cytokine-driven transcriptional activat
ion of human acute phase serum amyloid A genes, SAA1 and SAA2, J. Immunol. 169 (2002) 399-406)。最初にBettsらによって記述されたヒトSAA4(J.C. Betts, M.R. Edbrooke, R.V. Thakker, P. Woo, The human acute-phase serum amyloid A gene family: structure, evolution and expression in hepatoma cells, Scand. J. Immunol. 34 (1991) 471-482)は、構成的なSAA4(C-SAA4)をコードする(D.M. Steel, G.C. Sellar, C.M. Uhlar, S. Simon, F.C. DeBeer, A.S. Whitehead, A constitutively expressed serum amyloid A protein gene (SAA4) is closely linked to, and share structural similarities with, an acute-phase serum amyloid A protein gene (SAA2), Genomics 16 (1993) 447-454; A.S. Whitehead, M.C. de Beer, D.M. Steel, M. Rits, J.M. Lelias, W.S. Lane, F.C. de Beer, Identification of novel members of the serum amyloid A protein superfamily as constitutive apolipoproteins of high density lipoproteins, J. Biol. Chem. 267 (1992) 3862-3867)。A-SAA1およびA-SAA2とは対照的に、C-SAA4は急性期応答において著しく誘導されない。C-SAA4はA-SAA1およびA-SAA2よりも8残基長く、どちらのA-SSAタンパク質とも55%の同一性しか示さない。C-SAA4は、正常および急性期HDL3の両方において低レベルでしか存在せず、このタンパク質の蓋然性の高いハウスキーピング機能を示唆している。
【0011】
ヒトSAA3遺伝子は最初SackおよびTalbotによって同定され、A-SAAと71%の同一性を示す104残基のタンパク質をコードすると予測された(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)。しかしながら、後のゲノムに基づく研究により、予測されたエキソン3に存在する1ヌクレオチドの挿入によって結果的に末端切断型ヒトSAA3タンパク質が得られることが明らかになった(B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661)。今日まで、SAA3転写産物もタンパク質も調査されたヒト組織または細胞系で検出されておらず、SAA3のプロモーターと推測された領域が機能することも示されていない(S. Urieli-Shoval, P. Cohen, S. Eisenberg, Y. Matzner, Widespread expression of serum amyloid A in histologically normal human tissues: predominant localization to the epithelium, J. Histochem. Cytochem. 46 (1998) 1377-1384); B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661; S. Urieli-Shoval, R.L. Meek, R.H. Hanson, N. Eriksen, E.P. Benditt, huma
n serum amyloid A genes are expressed in monocyte/macrophage cell lines, Am. J. Pathol. 145 (1994) 650-660; G.C. Sellar, A.S. Whitehead, Localization of four human serum amyloid A (SAA) protein superfamily genes to Chromosome 11p: Characterization of a fifth SAA-related gene sequence, Genomics 16 (1993) 774-776)。これらの研究は、SAA3遺伝子が偽遺伝子であるか、またはSAA3発現に適当な誘導条件および/または細胞型が用いられなかったことを示唆している。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明者らは初めて、ヒト細胞において、SAA3遺伝子の誘導された発現を確認および実証し、SAA3遺伝子の転写産物(cDNA)を同定した。本発明の目的は、ヒトSAA3転写産物およびそれがコードするタンパク質を提供することである。
【0013】
また本発明の別の目的は、ヒトSAA3タンパク質をコードするヌクレオチド配列を提供することである。
【0014】
また本発明のもう1つ別の目的は、ヒトSAA3タンパク質を構成するアミノ酸配列を提供することである。
【0015】
また本発明のさらに別の目的は、組換えSAA3の産生、SAA3発現の役割をさらに明らかにするためのアッセイにおける使用、急性期免疫応答のさらなる理解などのために、本発明の配列を使用するための組換えDNAプロトコルを提供することである。
【0016】
さらに本発明の別の目的は、例えばリポ多糖類(LPS)またはプロラクチン(PRL)のようなSAA3発現の誘導因子を同定するためのアッセイを提供すること、およびこれらの化合物の存在下で、機能的に連結された配列をヒト細胞において発現誘導させることができるプロモーター領域を提供することである。
【課題を解決するための手段】
【0017】
本発明において、ヒト乳房上皮細胞から分離および精製したヒト血清アミロイドA3(SAA3)タンパク質を提供する。
【0018】
本発明の別の態様において、哺乳動物ヒトSAA3をコードする単離された核酸分子を提供する。核酸分子は遺伝子、cDNA、またはRNAであってよく、一本鎖または二本鎖であってよい。好ましい実施形態において、核酸分子は配列番号2またはその保存的に改変された変異体をコードする配列を含む。最も好ましい実施形態において、核酸分子は配列番号1またはその保存的に改変された変異体を含み、たとえば、発現された他のヒトSAA3配列、同じように同定された配列、または本明細書に記載するようにLPSまたはプロラクチンにより誘導されたSAA配列をコードする他のあらゆる核酸配列が含まれる。
【0019】
本発明において、ヒトSAA3遺伝子の配列(イントロン領域およびフランキング領域が含まれる)が決定された。このことから本発明にはまた、1以上の天然非コード領域もしくはイントロン領域またはその保存的に改変された変異体が含まれる、発現されたヒトSAA3をコードするヌクレオチド配列が含まれる。
【0020】
本発明の別の態様において、ヒトSAA3の1以上のエピトープに免疫学的に特異的な抗体を提供する。好ましくは、抗体は、ヒトSAA3を血清SAAから区別するヒトSAA3の少なくとも1つのエピトープに免疫学的に特異的である。
【0021】
また他の実施形態において、ヒトSAA3と天然で関連しているプロモーター領域は、遺伝子移植のプロトコルに、または、SAA産生を刺激するために使用して、動物の乳房もしくは他の乳房組織に関連する疾患の治療を補助することができる。例として、ヒトSAA3プロモーターはプロラクチンにより誘導される。すなわち、プロラクチンその他のヒトSAA3誘導因子を投与して、前記動物の乳房組織によるSAA3の産生を刺激し、増加させることができるであろう。プロモーター領域はまた、乳房細胞における異種性ヌクレオチド配列の発現のための乳房組織特異性を提供する。
【0022】
ウシと同様にヒトにおいて、乳房関連SAA3はムチン3(MUC3)産生を刺激する能力を有する。このことからSAA3は、腸の感染症または不充分なレベルのムチンと関連する他の疾患状態、例えば旅行者下痢、乳児下痢、壊死性腸炎、および尿路感染症などを治療および予防するために使用することができ、集団の動物における下痢を防止するための手段として獣医薬を提供する。従って本発明には、上記の疾患および類似の病状を有する他の疾患を治療するための、医薬として有効な量のSAA3ペプチドおよび担体を含む医薬組成物が含まれる。最後に、ムチンを産生する粘膜表面(例えば鼻咽頭、嚢、など)の他の上皮細胞(epithelial cell lining)もまた、ムチン産生を刺激するために本発明の医薬組成物で処置してそれらと関連する感染症を予防または治療することができる。
【0023】
また別の実施形態において、本発明は乳房および他の細胞におけるSAA発現の影響を調べるための、ならびに炎症または組織損傷におけるSAAの役割をさらに明らかにするためのアッセイからなる。本発明の配列はまた、他のSAAアイソフォームを同定するために、あるいは乳腺炎または乳房組織の炎症もしくは感染と関連する他の乳房組織疾患または損傷を診断するために使用することができる。
【0024】
本発明の他の特徴および効果は、以下の図面、詳細な説明および実施例を参照することによって、さらに理解されるであろう。
【発明を実施するための最良の形態】
【0025】
1.定義
本発明の組成物および方法に関する様々な用語を、本明細書全体にわたり、そして特許請求の範囲において用いる。
【0026】
単位、接頭語、および記号は、SI単位系で許された形式で示すことができる。特に断らない限り、核酸は左から右に5'から3'の向きで記述し、アミノ酸配列は左から右にアミノからカルボキシの向きで記述する。数値範囲は、範囲を定める数自体を含み、定義される範囲内の各々の整数が含まれる。アミノ酸は、本明細書において、一般に知られている3文字記号またはIUPAC-IUB Biochemical nomenclature Commisionにより推奨される1文字記号のいずれかにより表され得る。同様に、ヌクレオチドは一般に認められた1文字コードで表すことができる。特に規定しない限り、本明細書で用いるソフトウェア、電気および電子の用語は、The New IEEE Standard Dictionary of Electrical and Electronics Terms (第5版,1993)に定義されている通りである。以下に定義する用語は、明細書全体でより十分に定義される。
【0027】
「増幅する」とは、少なくとも1つの核酸配列を鋳型として用いて、ある核酸配列の複数のコピー、または核酸配列に相補的な複数のコピーを作成することを意味する。増幅システムには、ポリメラーゼ連鎖反応(PCR)システム、リガーゼ連鎖反応(LCR)システム、核酸配列に基づく増幅(NASBA, Canteen, Mississauga, Ontario)、Q-βレプリカーゼシステム、転写に基づく増幅システム(TAS)および鎖置換増幅(SDA)が含まれる。例えばDiagnostic Molecular Microbiology: Principles and Applications, D.H. Persingら,編, American Society for Microbiology, Washington, D.C. (1993)を参照のこと。増幅の産物は増幅産物(amplicon)と呼ぶ。
【0028】
「抗体」という用語には、抗体の抗原結合性形態(例えばFab、F(ab)2)が含まれる。「抗体」という用語はしばしば、検体(抗原)に特異的に結合し認識する1以上の免疫グロブリン遺伝子によって実質的にコードされるポリペプチドまたはそのフラグメントを言う。また一方、様々な抗体フラグメントは完全な抗体の消化の観点から定義することができるものの、当業者ならばそのようなフラグメントは化学的に、または組換えDNA手法を利用することにより、新規に合成することができると理解するであろう。従って、本明細書で用いる抗体という用語にはまた、一本鎖Fv、キメラ抗体(すなわち異なる種からの定常および可変領域を含むもの)、ヒト化抗体(すなわち非ヒト起源の相補性決定領域(CDR)を含むもの)およびヘテロコンジュゲート抗体(例えば二重特異性抗体)などの抗体フラグメントが含まれる。
【0029】
本明細書において用いる「アンチセンス配向」とは、アンチセンス鎖が転写される方向性で、プロモーターに機能的に連結している二本鎖ポリヌクレオチド配列をいう。アンチセンス鎖は、内因性転写産物の翻訳がしばしば阻害されるくらい、内因性転写産物に対して十分相補的である。
【0030】
本明細書において用いる「SAA3」および「M-SAA3」という用語は互換的に使用され、限定するものではないが、本明細書において開示する配列、起源を問わないそれらの保存的に改変された変異体、ならびに本明細書に開示されたアッセイで実証されるようにSAA3の生物学的性質を保持する他のあらゆる変異体が含まれる。
【0031】
本明細書において用いる「染色体領域」には、その染色体領域が含むDNAの線状部分に関して測定されうるある長さの染色体が含まれる。染色体領域は2つの固有なDNA配列、すなわちマーカーにより定義することができる。
【0032】
「保存的に改変された変異体」という用語は、アミノ酸および核酸配列の両方に適用されるものであり、特定の配列への言及がなされるあらゆる場合に含まれることを意図したものである。特定の核酸配列に関して、保存的に改変された変異体はアミノ酸配列の同一または保存的に改変された変異体をコードする核酸を意味する。遺伝暗号の縮重のために、多数の機能的に同一な核酸が、任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全てアミノ酸のアラニンをコードする。このことから、アラニンがコドンによって指定される各位置で、コードされるポリペプチドを変更することなくコドンを上述の対応するコドンのいずれにも変更することができる。このような核酸変異は「サイレント変異」であり、保存的に改変された変異の1つの種を表す。ポリペプチドをコードする本発明の各核酸配列はまた、遺伝暗号を参照して、核酸の可能なあらゆるサイレント変異を表す。当業者ならば、核酸における各コドンを、機能的に同一の分子を生じるために改変することが出来ると理解できるであろう(ただし、通常メチオニンの唯一のコドンであるAUG、および通常トリプトファンをコードする唯一のコドンであるUGGは除く)。それゆえ、本発明のポリペプチドをコードする核酸の各々のサイレント変異は、それぞれの記載されるポリペプチド配列に暗黙的に含まれており、本発明の範囲内にある。
【0033】
アミノ酸配列については、当業者ならば、コードされる配列中の単一のアミノ酸またはわずかな割合のアミノ酸を変更する、追加するまたは欠失させる核酸、ペプチド、ポリペプチドまたはタンパク質配列への個別の置換、欠失または追加は、その変更が結果的に1つのアミノ酸を化学的に類似するアミノ酸で置換するものである場合に、「保存的に改変された変異体」であると認識するであろう。このことから、1〜15からなる整数の群から選択される数のアミノ酸残基を、このように変更することができる。従って、例えば、1、2、3、4、5、7、または10の変更を行うことができる。保存的に改変された変異体は典型的には、それらが誘導された未改変のポリペプチド配列と類似の生物学的活性を示す。例えば、基質特異性、酵素活性、またはリガンド/受容体結合は、一般にその天然の基質について天然のタンパク質の少なくとも30%、40%、50%、60%、70%、80%または90%である。機能的に類似のアミノ酸を規定する保存的置換表は当技術分野でよく知られている。
【0034】
以下の6つの群はそれぞれ、互いに保存的な置換であるアミノ酸を含有する。
【0035】
1) アラニン(A),セリン(S),トレオニン(T);
2) アスパラギン酸(D),グルタミン酸(E);
3) アスパラギン(N),グルタミン(Q);
4) アルギニン(R), リジン(K);
5) イソロイシン(I),ロイシン(L),メチオニン(M),バリン(V);
6) フェニルアラニン(F),チロシン(Y),トリプトファン(W)。
【0036】
Creighton (1984) Proteins W.H. Freeman and Companyを参照のこと。
【0037】
所定の核酸に関して、「コードする」または「コードされる」とは、所定のタンパク質へと翻訳される情報を含むことを意味する。タンパク質をコードする核酸は、核酸の翻訳される領域内に非翻訳領域(例えばイントロン)を含んでもよく、またはそのような介在する非翻訳配列を欠いてもよい(例えばcDNAにおけるように)。タンパク質をコードする情報は、コドンの使用により特定される。典型的には、アミノ酸配列は「普遍的」遺伝暗号を用いて核酸によりコードされる。しかしながら、普遍的遺伝暗号の変形、例えばいくつかの植物、動物および真菌ミトコンドリア、細菌のMycoplasma capricolumn、または繊毛虫のマクロヌクレアスに存在するようなものをそれらにおいて核酸を発現させる場合に用いてもよい。
【0038】
核酸を合成的に作成または変更する場合、核酸を発現させようと意図する宿主の知られている優先コドンを利用することができる。
【0039】
本明細書において、特定のポリヌクレオチドまたはそれがコードするタンパク質に関して用いる「全長配列」は、その特定のタンパク質の天然(非合成)の内因性で生物学的に活性な形態の全アミノ酸配列を有することを意味する。ある配列が全長なのかを判別する方法は当技術分野でよく知られており、例えば、ノーザンまたはウェスタンブロット法、プライマー伸長法、S1プロテクション法、およびリボヌクレアーゼプロテクション法などの模範的な技術が含まれる。既知の全長相同(オーソロガスおよび/またはパラロガス)配列との比較もまた、本発明の全長配列を同定するために使用することができる。さらに、mRNAの5'および3'非翻訳領域に典型的に存在するコンセンサス配列は、ポリヌクレオチドが全長であるか識別するのに役立つ。例えば、コンセンサス配列ANNNNAUGG(ここで下線のコドンはN末端のメチオニンを表す)は、ポリヌクレオチドが完全な5'末端を有するか判別するのに役立つ。3'末端におけるコンセンサス配列、例えばポリアデニル化配列などは、ポリヌクレオチドが完全な3'末端を有するか判別するのに役立つ。
【0040】
タンパク質またはペプチドに関して、「分離(単離)したタンパク質(もしくはペプチド)」または「分離し精製したタンパク質(もしくはペプチド)」という用語を、本明細書において場合により用いる。この用語は、そのタンパク質が「実質的に純粋」な形態で存在するように、天然でそのタンパク質に結びついている他のタンパク質から十分に分離されたタンパク質を意味し得る。また、この用語は、単離された核酸分子の発現により産生されるタンパク質を意味し得る。
【0041】
核酸分子に関連して、「単離された核酸」という用語を場合により用いる。この用語はDNAに適用する場合、そのDNAが由来する生物の天然に存在するゲノムで(5'および3'方向に)直接隣接している配列から切り離されたDNA分子を意味する。例えば、「単離された核酸」は、プラスミドもしくはウィルスベクターなどのベクターに挿入した、または原核生物もしくは真核生物のゲノムDNAに組み込まれたDNA分子を含み得る。「単離された核酸分子」はまた、cDNA分子からなることもできる。
【0042】
RNA分子に関連して、「単離された核酸」という用語は、上記に定義した単離されたDNA分子がコードするRNA分子を主に意味する。また、この用語はそのRNA分子が「実質的に純粋」な形態(「実質的に純粋」という用語は以下で定義する)で存在するように、天然の状態(すなわち細胞または組織内)でそのRNA分子に結びついているRNA分子から十分に分離したRNA分子を意味し得る。
【0043】
核酸に関して本明細書において用いる「異種の」とは、外来種が起源であるか、または、同種が起源である場合には人間の意図的な介入により組成および/または遺伝子座において天然の形態から実質的に改変した核酸である。例えば異種構造遺伝子に機能的に連結したプロモーターは、その構造遺伝子の起源とは異なる種に由来し、または同種由来の場合にはその遺伝子およびプロモーターの片方または両方がそれらの元の形態から実質的に改変されたものである。異種タンパク質は外来種が起源であってもよく、または同種由来の場合には人間の意図的な介入によりその元の形態から実質的に改変されたものである。
【0044】
「宿主細胞」とは、ベクターを含有し、そのベクターの複製および/または発現を支持する細胞を意味する。宿主細胞は、原核細胞、例えば大腸菌(E. coli)などであってもよく、または真核細胞、例えば酵母、昆虫、両生類または哺乳動物細胞などであってもよい。「ハイブリダイゼーション複合体」という用語には、互いに選択的にハイブリダイズする、2つの一本鎖核酸配列が形成する二重核酸構造が含まれる。
【0045】
細胞に核酸を挿入するという文脈での「導入する」という用語は、「トランスフェクション」または「トランスフォーメーション(形質転換)」または「トランスダクション(形質導入)」を意味し、また、核酸を真核または原核細胞に組み入れることが含まれ、ここで核酸は細胞のゲノムに組み込まれてもよく(例えば染色体、プラスミド、色素体、もしくはミトコンドリアDNA)、自立したレプリコンに変換させてもよく、または一時的に発現(例えばトランスフェクトされたmRNA)させてもよい。
【0046】
特に断らない限り、用語「SAA3をコードする核酸」とは、本発明の核酸であり、SAA3をコードする本発明のポリヌクレオチドを含む核酸を意味する。「SAA3遺伝子」とは、本発明の遺伝子であり、全長SAA3ポリヌクレオチドの異種ゲノム形態を意味する。
【0047】
本明細書において特定のマーカーに関して用いる「規定され、それを含む染色体領域内に局在化され」とは、その記述されたマーカーによって範囲を定められまたそれを含む連続している一定の長さの染色体をいう。
【0048】
本明細書において用いる「マーカー」とは、ある染色体上の独特の位置を同定するために役立つその染色体上の座をいう。「多型マーカー」とは、異なる形態のマーカーが、相同なペアとして存在するとき、そのペアの両方の染色体の伝達が可能になるようにする複数の形態(対立遺伝子)で出現するマーカーをいう。遺伝子型は、1つまたは複数のマーカーを使用することにより定義することができる。
【0049】
本明細書において用いる「核酸」には、一本鎖または二本鎖のいずれの形態のデオキシリボヌクレオチドまたはリボヌクレオチドポリマーも含まれ、特に断らない限り、天然のヌクレオチドと似た様式で一本鎖核酸にハイブリダイズするという点で天然ヌクレオチドの本質的な性質を有する既知のアナログも包含する(例えばペプチド核酸)。
【0050】
「核酸ライブラリー」とは、特定の生物のゲノムの転写された全フラクションを含みかつこれらを実質的に表現する単離されたDNAまたはRNA分子のコレクションを意味する。模範的な核酸ライブラリー、例えばゲノミックまたはcDNAライブラリーなどの構築は、例えば、BergerおよびKimmel, Guide to Molecular Cloning Techniques, Methods in Enzymology, Vol. 152, Academic Press, Inc., San Diego, CA (Berger); Sambrook ら, Molecular Cloning - A Laboratory Manual, 2nd ed., Vol. 1-3 (1989);およびCurrent Protocols in Molecular Biology, F.M. Ausubel ら, Eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc. (1994)などの標準的な分子生物学文献に教示されている。
【0051】
本明細書において用いる「ポリヌクレオチド」には、天然に生ずるヌクレオチドと実質的に同じヌクレオチド配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズする、および/または天然に生ずるヌクレオチドと同じアミノ酸への翻訳を可能にする、という意味で天然リボヌクレオチドに必須の性質を有するデオキシリボヌクレオチド、リボポリヌクレオチド、またはそのアナログが含まれる。ポリヌクレオチドは全長であってもよく、または天然もしくは異種の構造または調節遺伝子のサブシークエンスであってもよい。特に断らない限り、この用語には、特定の配列のみならずその相補配列も含まれる。このことから、安定性のためまたは他の目的のために修飾したバックボーンを有するDNAまたはRNAもまた、本明細書において意図する「ポリヌクレオチド」である。その上、例えば2つだけ例を挙げると、普通ではない塩基(イノシン等)または修飾された塩基(例えばトリチル化塩基など)を含むDNAまたはRNAは、本明細書において用いる通り、ポリヌクレオチドである。当業者に知られている様々な有用な目的を果たす、DNAおよびRNAへの非常に多様な改変がなされることが理解される。本明細書において用いる通り、ポリヌクレオチドという用語は、そのような化学的、酵素的または代謝的に改変された形態のポリヌクレオチド、ならびに、ウィルスおよび細胞(他のものに加え、単純および複雑細胞が含まれる)に特徴的なDNAおよびRNAの化学的形態を含む。
【0052】
「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は、本明細書において相互に交換できるように使用され、アミノ酸残基のポリマーを意味する。これらの用語は、天然に生ずるアミノ酸ポリマーのみならず、1以上のアミノ酸残基が対応する天然に生ずるアミノ酸の人工的な化学的アナログであるアミノ酸ポリマーにも適用される。このような天然に生ずるアミノ酸のアナログに必須の性質は、タンパク質に組み込まれた場合、天然に生ずるアミノ酸のみからなるタンパク質によって誘導される抗体に、前記アナログを組み入れたタンパク質が特異的に反応することである。「ポリペプチド」、「ペプチド」、および「タンパク質」という用語はまた、限定するものではないが、リン酸化、グリコシル化、脂質結合、硫酸化、グルタミン酸残基のγ-カルボキシル化、ヒドロキシル化、およびADP-リボシル化が含まれる修飾を含む。広く知られており上記に記述した通り、ポリペプチドは完全に線形ではないと理解される。例えば、ポリペプチドは、ユビキチン化の結果分枝したものであってもよく、また、一般に翻訳後の事象の結果、環状であってもよく(それらは分枝していてもしていなくてもよい)、前記事象には天然のプロセス事象、および天然には起こらない人為的操作によってもたらされる事象が含まれる。環状、分枝状、および分枝した環状ポリペプチドは、非翻訳の天然のプロセスによっても、また完全に合成的な方法によっても、合成することができる。さらに本発明は、本発明のタンパク質のメチオニン含有およびメチオニン非含有アミノ末端変異体の両方の使用を含む。タンパク質に関して、「N末端領域」という用語は、タンパク質のアミノ末端に隣接する約50アミノ酸残基を含む。
【0053】
本明細書において用いる「TFLKモチーフ」は、SAA3のTFLK活性部位の構造的完全性および生物学的活性を維持するアミノ酸または他のものによる任意の構築物を含む。
【0054】
本明細書において用いる「組換え」には、異種核酸を導入することにより改変した細胞もしくはベクター、またはそのように改変した細胞から誘導される細胞が含まれる。したがって例えば、組換え細胞は、細胞の天然(非組換え)形態で同一の形態で見られない遺伝子を発現するか、または他の場合には異常発現するか、または低発現するかもしくは全く発現しない天然の遺伝子を、人為的な介入の結果として発現する。本明細書において用いる「組換え」という用語は、自然に起きる事象(例えば自発的突然変異、天然の形質転換/形質導入/転位)、例えば人為的な介入無しで起こる事象など、による細胞またはベクターの変更は包含しない。
【0055】
本明細書において用いる通り、「組換え発現カセット」は、宿主細胞内で特定の核酸の転写を可能にする一連の特定の核酸エレメントを有し、組換え的にまたは合成的に作り出された核酸構築物である。組換え発現カセットは、プラスミド、染色体、ミトコンドリアDNA、色素体DNA、ウィルスまたは核酸フラグメントに組み入れることができる。典型的には、発現ベクターの組換え発現カセット部分には、様々な配列の他に、転写されるべき核酸、およびプロモーターが含まれる。
【0056】
「残基」または「アミノ酸残基」、または「アミノ酸」という用語は、本明細書において互いに交換できるように用いられ、タンパク質、ポリペプチドまたはペプチド(これらをまとめて「タンパク質」と呼ぶ)に組み入れられたアミノ酸を意味する。アミノ酸は天然に生ずるアミノ酸であってもよく、他に限定が無ければ、天然に生ずるアミノ酸と似た様式で機能することができる天然アミノ酸の非天然アナログを包含してもよい。
【0057】
「ストリンジェント条件」または「ストリンジェントなハイブリダイゼーション条件」という用語には、プローブが、他の配列に対してハイブリダイズするのに比べて検出できるほど強い度合い(例えば少なくともバックグランドよりも2倍強く)で、標的配列にハイブリダイズする条件への言及が含まれる。ストリンジェント条件は配列依存的であり、異なる状況下では異なり得る。ハイブリダイゼーションおよび/または洗浄条件のストリンジェンシーを制御することにより、プローブと100%相補的な標的配列を同定することができる(相同プローブ)。これとは別に、より低い程度の類似性が検出できるよう配列にいくらかのミスマッチが許容されるようストリンジェンシー条件を調節することができる(非相同プローブ)。一般に、プローブは長さにして約1000ヌクレオチド未満であり、場合により長さにして500ヌクレオチド未満である。
【0058】
典型的には、ストリンジェント条件は、塩濃度が約1.5M Naイオン未満、典型的には約0.01〜1.0M Naイオン濃度(または他の塩)で、pHが7.0〜8.3であり、短いプローブ(例えば10〜50ヌクレオチド)については温度が少なくても約30℃であり、長いプローブ(例えば50ヌクレオチドより長いもの)については温度が少なくとも約60℃であるものである。ストリンジェント条件はまた、ホルムアミドなどの不安定化剤を添加することにより達成することができる。模範的な低ストリンジェンシー条件には、37℃で、30〜35%ホルムアミド、1M NaCl、1%SDS(ドデシル硫酸ナトリウム)を有するバッファー溶液でハイブリダイゼーションし、50〜55℃で、1×〜2×のSSC(20× SSC=3.0M NaCl/0.3Mクエン酸三ナトリウム)で洗浄することが含まれる。模範的な中程度のストリンジェンシー条件には、37℃で、40〜45%ホルムアミド、1M NaCl、1% SDSでのハイブリダイゼーション、および55℃〜50℃で、0.5×〜1× SSCでの洗浄が含まれる。模範的な高ストリンジェンシー条件には、37℃で、50%ホルムアミド、1M NaCl、1% SDSでのハイブリダイゼーション、および60℃〜65℃で、0.1× SSCでの洗浄が含まれる。
【0059】
特異性は典型的には、ハイブリダイゼーション後の洗浄の関数であり、重要な要素は最終洗浄溶液のイオン強度および温度である。DNA-DNAハイブリドについては、TmはMeinkoth および Wahl, Anal. Biochem., 138:267-284 (1984)の式:
Tm=81.5℃ + 16.6 (log M) + 0.41 (%GC) -0.61 (% form) - 500/L
から概算することができるが、ここでMは一価カチオンのモル濃度であり、%GCはDNA中のグアノシンおよびシトシンヌクレオチドの百分率であり、% formはハイブリダイゼーション溶液中のホルムアミドの百分率であり、Lは塩基対で表したハイブリッドの長さである。Tmは相補的な標的配列の50%が完全に一致するプローブにハイブリダイズする温度(所定のイオン強度およびpH下での)である。Tmは1%のミスマッチにつき約1℃減少する。したがって、Tm、ハイブリダイゼーションおよび/または洗浄条件は、所望の相同性の配列にハイブリダイズするよう調節することが出来る。例えば≧90%の相同性を有する配列を所望ならば、Tmは10℃低下させることができる。一般にストリンジェント条件は、所定のイオン強度およびpHで、その特定の配列およびその相補配列の熱融点(Tm)よりも約5℃低く選択される。しかしながら、非常にストリンジェントな条件は、熱融点(Tm)よりも1、2、3または4℃低いハイブリダイゼーションおよび/または洗浄を利用することができ、中程度にストリンジェントな条件は熱融点(Tm)よりも6、7、8、9または10℃低いハイブリダイゼーションおよび/または洗浄を利用することができ、低ストリンジェントな条件は熱融点(Tm)よりも11、12、13、14、15または20℃低いハイブリダイゼーションおよび/または洗浄を利用することができる。この式、ハイブリダイゼーションおよび洗浄組成、ならびに所望のTmを用いれば、当業者はハイブリダイゼーションおよび/または洗浄溶液のストリンジェンシーのバリエーションが本質的に記載されたと理解できるであろう。所望のミスマッチの度合いが45℃(水溶液)または32℃(ホルムアミド溶液)より低いTmを与えるならば、より高い温度を使用できるようSSC濃度を上げることが好ましい。核酸のハイブリダイゼーションの広範囲な指針は、Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acids Probes, Part I, Chapter 2, Ausubel, ら, Eds., Greene Publishing and Wiley-Interscience, New York (1995)に見られる。
【0060】
「実質的に純粋」という用語は、対象とする化合物(例えば核酸、オリゴヌクレオチド、タンパク質など)を重量で少なくとも50〜60%含む調製物を意味する。より好ましくは、調製物は対象とする化合物を重量で少なくとも75%、最も好ましくは90〜99%含む。純度は、対象とする化合物に適当な方法で測定する(例えばクロマトグラフィー法、アガロースまたはポリアクリルアミドゲル電気泳動、HPLC解析など)。
【0061】
核酸配列およびアミノ酸配列は、核酸またはアミノ酸の類似配列を並べ、このことにより相違を明確にするコンピュータプログラムを用いて比較することができる。BLASTプログラム(NCBI)およびそれに使われているパラメータは多くの実施者によりアミノ酸配列フラグメントを並べるために使用されている。しかし、これと同等のアライメントおよび類似性/相同性評価は、標準的なアライメントソフトウェアを使用して得ることができる。例えば、the Genetics Computer Group in Madison, Wisconsinから取得できるGCG Wisconsin Package version 9.1およびBest-Fit programにより使用されるデフォルトパラメータ(ギャップ生成ペナルティー=12、ギャップ延長ペナルティー=4)もまた、配列相同性および類似性を比較するために使用することが出来る。
【0062】
「実質的に同一」という用語は、タンパク質の性質(すなわちタンパク質の構造、安定性、基質特異性および/または生物学的活性)に実質的に影響を及ぼさない配列変化を有する核酸またはアミノ酸配列を意味する。とりわけ核酸配列への言及について、「実質的に同一」という用語は、コード領域および発現を左右する保存配列を意味し、コードされるポリペプチド中の、同じアミノ酸をコードする縮重コドンまたは保存的な置換アミノ酸をコードする異なるコドンを主に意味することを意図する。アミノ酸配列に関して、「実質的に同一」という用語は、一般に、保存的な置換および/またはポリペプチドの構造もしくは機能の決定に関与しない領域での変化を意味する。
【0063】
「%相同」および「%類似」という用語もまた、アミノ酸および核酸配列の間の比較において、本明細書で使用する。アミノ酸配列に関して、「%相同」とは、配列解析プログラムによって比較されるアミノ酸配列中の同一アミノ酸とマッチする対象アミノ酸配列中のアミノ酸の百分率を意味する。「%類似」とは、同一または保存アミノ酸とマッチする対象アミノ酸配列中のアミノ酸の百分率を意味する。保存アミノ酸とは、1つを別のものと入れ替えたとしても、結果的に得られるタンパク質の三次構造を認識できるほどには変化させないような、構造は異なるものの物理的性質は類似しているアミノ酸である。保存的な置換は、Taylor (1986, J. Theor. Biol. 119:205)に定義されている。核酸分子に関して、「%相同」とは、配列解析プログラムにより同一ヌクレオチドとマッチする対象核酸配列中のヌクレオチドの百分率を意味する。
【0064】
抗体に関して、用語「免疫学的に特異的」とは、対象とするタンパク質の1以上のエピトープに結合し、しかし抗原性の生物学的分子の混合集団を含有するサンプル中の他の分子を実質的に認識せずそれらに結合しない抗体を意味する。
【0065】
オリゴヌクレオチドまたは他の一本鎖核酸分子に関して、「特異的にハイブリダイズする」という用語は、当技術分野で一般に用いられる所定の条件、すなわちストリンジェンシーの条件下でのハイブリダイゼーションを可能にする、十分に相補的な配列である2つの一本鎖核酸分子の間の結合を意味する(「実質的に相補的」とも呼ばれる)。特に、この用語は、オリゴヌクレオチドと、一本鎖DNAまたはRNA分子内に含有される実質的に相補的な配列とのハイブリダイゼーションを意味し、かつ、該オリゴヌクレオチドと非相補的な配列の一本鎖核酸とのハイブリダイゼーションを実質的に除外する。
【0066】
「コード配列」または「コード領域」は、配列が発現する場合に遺伝子産物を産生させるのに必要な配列情報を有する核酸分子を意味する。
【0067】
「機能的に連結」または「機能的に挿入」という用語は、コード配列の発現に必要な制御配列が、コード配列を発現できるよう、コード配列に対して適切な位置で核酸分子内に配置されることを意味する。この同じ定義を場合により、発現ベクター内の転写制御因子(例えばエンハンサー)の配置に適用する。
【0068】
転写および翻訳制御配列は、宿主細胞におけるコード配列の発現を提供するDNA調節配列、例えばプロモーター、エンハンサー、ポリアデニル化シグナル、ターミネーター、などである。
【0069】
「プロモーター」、「プロモーター領域」または「プロモーター配列」という用語は、一般にコード領域の5'または3'側、またはコード領域内、またはイントロン内に見ることのできる、遺伝子の転写制御領域を意味する。典型的には、プロモーターは、細胞内のRNAポリメラーゼに結合し、下流(3'方向)にあるコード配列の転写を開始させることができるDNA調節領域である。典型的な5'プロモーター配列は、その3'末端で転写開始部位に接しており、上流(5'方向)に伸びてバックグラウンドを超える検出可能なレベルの転写を開始させるのに必要な最小の数の塩基またはエレメントを含む。プロモーター配列内には、転写開始部位(ヌクレアーゼS1でのマッピングにより都合よく定義される)のみならず、RNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が存在する。
【0070】
「ベクター」は、別の核酸断片の複製または発現をもたらすよう、その断片を機能的に挿入することのできるレプリコン、例えばプラスミド、ファージ、コスミド、またはウィルスである。
【0071】
「核酸構築物」または「DNA構築物」という用語は、適当な調節配列に機能的に連結され、細胞を形質転換するためにベクターに挿入された1以上のコード配列を指して用いられることがある。この用語は「形質転換用DNA」と互いに交換できるよう用いることができる。このような核酸構築物は、対象とする遺伝子産物をコードする配列とあわせて、選択マーカー遺伝子および/またはレポーター遺伝子を含有していてもよい。
【0072】
「選択(可能な)マーカー遺伝子」という用語は、発現させた場合、選択可能な表現型、例えば抗生物質耐性を形質転換細胞に附与する産物をコードする遺伝子を意味する。
【0073】
「レポーター遺伝子」という用語は、標準的手法により、直接または間接的に簡便に検出できる産物をコードする遺伝子を意味する。
【0074】
核酸構築物の「異種」領域は、より大きな分子中の核酸分子の特定可能な断片(複数であってもよい)であり、この核酸分子は天然では前記より大きな分子と結合していないものである。このことから、異種領域が哺乳動物遺伝子をコードする場合、その遺伝子は通常、起源生物のゲノム内でその哺乳動物ゲノムDNAの両端につながっていないDNAが両端につながっている。別の例では、異種領域はコード配列それ自身が天然には見いだされない構築物である(例えば、ゲノムコード配列がイントロンを含有するcDNA、または天然遺伝子とは異なるコドンを有する合成配列)。対立遺伝子のバリエーションや天然に起こる突然変異事象は、本明細書において定義するDNAの異種領域を生み出さない。上記に定義した用語「DNA構築物」はまた、異種領域、特に細胞の形質転換に使用するために構築したものに言及するために用いられる。
【0075】
外因性または異種DNAを細胞内に導入した場合に、細胞はそのDNAにより「形質転換」または「形質導入」される。形質転換用DNAは、細胞のゲノム内に組み入れられても(共有結合的連結)よく、また組み入れられていなくてもよい。例えば原核生物、酵母および哺乳動物細胞の場合、形質転換用DNAはエピソームエレメント、例えばプラスミドなどに維持されてもよい。真核細胞に関しては、安定的に形質転換された細胞は、染色体複製を通じて娘細胞により遺伝で受け継がれるように、その形質転換用DNAが染色体に組み入れられているものである。この安定性は、真核細胞が細胞系、すなわちその形質転換用DNAを含有する娘細胞の集団からなるクローンを樹立する能力によって実証される。
【0076】
「クローン」は、有糸分裂により単一細胞または共通の祖先から誘導される細胞の集団である。「細胞系」は、何世代にも渡ってin vitroで安定に増殖することができる一次細胞のクローンである。
【0077】
2.詳細な説明
血清アミロイドA(SAA)は、主に肝臓で産生され、組織損傷または感染に関連した炎症応答の一部として哺乳動物の血清に増加したレベルで生じる急性期タンパク質である。本発明者らはSAAの独特のアイソフォームであるSAA3を見いだした。このSAA3は、それまでその転写産物もタンパク質もヒト細胞で決して検出されず、その遺伝子が非発現、したがって偽遺伝子とみなされてきたために、存在しないと考えられてきた。
【0078】
以下の説明は、本発明の実施に伴う一般的な手法を示す。具体的な材料が記述されていることに関して、以下の説明は例示のみを目的としたものに過ぎず、本発明を制限することを意図したものではない。特に断らない限り、一般的なクローニング手法、例えばSambrook ら, Molecular Cloning, Cold Spring Harbor Laboratory (1989) (以下「Sambrookら」) またはAusubelら(eds) Current Protocols in Molecular Biology, John Wiley & Sons (1999) (以下「Ausubel ら」)に示されている手法を用いる。
【0079】
A. SAA3、SAA3特異的抗体、およびSAA3をコードする核酸分子の調製
1. タンパク質および抗体
ヒトSAA3は、血清からSAAを精製するために開発された種々の方法(参照により本明細書に組み入れた資料に詳述されている)により調製することができる。疎水性クロマトグラフィーマトリックス系および溶離液のバリエーション、例えばSmithらが記載したもの(Protein Expression & Purification 2: 158-161, 1991)などもまた利用可能である。
【0080】
これとは別に、アミノ酸配列情報が利用可能(例えば配列番号2)であることからヒトSAA3をコードする核酸分子を単離することが可能である。この方法は、当技術分野で広く知られた方法に従い、選択した生物種からのcDNA発現ライブラリーを抗ヒトSAA3抗体を用いてスクリーニングすることにより実現することが出来る。これとは別に、図2(配列番号1)の一部または全部をコードする一連の縮重オリゴヌクレオチドプローブを、以下に詳述する通り、cDNAまたはゲノミックライブラリーをスクリーニングするために用いることができる。
【0081】
一旦取得したならば、cDNAまたは遺伝子をin vitro転写のために適当なin vitro転写ベクター、例えばpSP64またはpSP65にクローンしてもよく、次いで適当な無細胞翻訳系、例えば小麦麦芽またはウサギ網状赤血球で無細胞翻訳をしてもよい。In vitro転写および翻訳系は、例えばPromega Biotech, Madison, Wisconsin or BRL, Rockville, Marylandから市販されている。pCITE in vitro翻訳システム(Novagen)もまた利用可能である。
【0082】
好適な実施形態において、適当な原核または真核細胞系での発現によって、より多くの量のタンパク質を産生することができる。例えば、ヒトSAA3をコードするDNA分子の一部または全部を、細菌細胞(例えば大腸菌)または酵母細胞(例えばSaccharomyces cerevisiae)または哺乳動物細胞での発現に合わせたベクターに挿入してもよい。これらのベクターは、宿主細胞内でDNAが発現できるようにする様式で配置された、宿主細胞内でのこのDNAの発現に必要な調節因子を含む。発現に必要なこのような調節因子には、プロモーター配列、転写開始配列および場合によりエンハンサー配列が含まれる。
【0083】
組換え原核または真核細胞系での遺伝子発現により産生されたヒトSAA3は、当技術分野で既知の方法により精製することが出来る。好ましい実施形態においては、市販の発現/分泌系が利用可能であり、ここでは組換えタンパク質が発現され、周囲の媒体から容易に精製できるように、その後宿主細胞から分泌される。発現/分泌ベクターを使用しない場合、これとは別のアプローチは、組換えタンパク質をアフィニティー分離、例えばその組換えタンパク質に特異的に結合する抗体との免疫学的相互作用により精製することを伴う。このような方法は当業者に一般的に利用されている。
【0084】
本発明はまた、ヒトSAA3に結合する能力のある、1以上の選択された種からの抗体を提供する。選択されたヒトSAA3の一部または全部に対するポリクローナルまたはモノクローナル抗体は、標準的な方法で調製することが出来る。モノクローナル抗体は、Kohler and Milsteinの一般的な方法に従い、標準的なプロトコルに沿って調製することが出来る。好適な実施形態において、ヒトSAA3の選択されたエピトープに免疫特異的に反応し、ヒトSAA3を他のSAAから区別する抗体を調製する。
【0085】
2.核酸分子
配列情報を取得したならば、ヒトSAA3をコードする核酸分子を2つの一般的方法によって用意することが出来る。ヒトSAA3は(1)適当なヌクレオチド三リン酸から合成してもよく、または(2)生物学的取得源から単離してもよい。いずれの方法も当技術分野で広く知られた方法を利用する。
【0086】
ヌクレオチド配列情報が利用可能であれば、本発明の単離された核酸分子をオリゴヌクレオチド合成により調製することができる。合成オリゴヌクレオチドは、Applied Biosystems 38A DNA Synthesizerまたは類似の装置により実施されるホスホルアミダイト(phosphoramadite)法により用意することができる。得られる構築物は、当技術分野で既知の方法、例えば高性能液体クロマトグラフィー(HPLC)により精製することが出来る。長い二本鎖ポリヌクレオチド、例えば本発明のDNA分子は、現在のオリゴヌクレオチド合成方法に固有の大きさの限界のために、段階的に合成しなければならない。このことから例えば、長い二本鎖分子を、適当な相補性を有する小さないくつかの断片として合成してもよい。このようにして製造された相補的断片を、各断片が隣接する断片との結合のために適当な結合性のある末端を有するようにアニールしてもよい。隣接断片は、DNAリガーゼの存在下で結合性のある末端をアニールさせることによりライゲートさせて、全長二重鎖分子を構築することができる。このようにして構築した合成DNA分子は次いで、適当なベクターにクローニングして増幅することができる。
【0087】
ヒトSAA3をコードする核酸分子はまた、当技術分野で広く知られた方法を用いて興味有る哺乳動物種から単離することができる。選択した生物種からの核酸分子は、cDNAまたはゲノミックライブラリーを、ヒトSAA3コード遺伝子に特異的な核酸配列にマッチするよう設計されたオリゴヌクレオチドでスクリーニングすることにより単離することができる。ある種からの遺伝子が所望ならば、ゲノミックライブラリーをスクリーニングする。これとは別に、タンパク質コード配列に特に興味があるのならば、cDNAライブラリーをスクリーニングする。適当なアミノ酸残基をコードするために1より多くの核酸残基が使用可能な縮重の位置では、全ての適当な核酸残基を組み入れて混合オリゴヌクレオチド集団を作成してもよく、またはイノシンなどの中性塩基を用いてもよい。オリゴヌクレオチド設計の方針は当技術分野で広く知られている(Sambrookら, Molecular Cloning, 1989, Cold Spring Harbor Press, Cold Spring Harbor NYも参照のこと)。
【0088】
これとは別に、ヒトSAA3タンパク質の一部をコードするように、PCR(ポリメラーゼ連鎖反応)プライマーを上述の方法により設計してもよく、これらのプライマーを用いて単離されたcDNAまたはゲノムDNAから核酸を増幅することができる。好適な実施形態において、ヒトSAA3をコードする核酸を単離するために用いるオリゴヌクレオチドは、血清SAAとは異なり、ヒトSAA3に独特の配列をコードするよう設計される。
【0089】
本発明において、ヒトSAA3をコードする核酸分子と適当な配列相同性を有する核酸は、適当なストリンジェンシーのハイブリダイゼーションおよび洗浄条件を用いて同定することができる。例えば、Sambrookら(1989, 上記に同じ)の方法に沿って、5× SSC、5× デンハート試薬(Denhardt's Reagent)、1.0% SDS、100μg/ml変性済み断片化サケ精子DNA、0.05%ピロリン酸ナトリウムおよび最大50%までのホルムアミドを含むハイブリダイゼーション溶液を用いてハイブリダイゼーションを行うことができる。ハイブリダイゼーションは少なくとも6時間にわたり37〜42℃で行う。ハイブリダイゼーションに次いで、フィルターを以下の通り洗浄する。 (1)2× SSCおよび1%SDS中で室温で5分間、(2)2× SSCおよび0.1%SDS中で室温で15分間、(3)1× SSCおよび1%SDS中で37℃で30分〜1時間、(4)溶液を30分毎に交換しながら、1× SSCおよび1%SDS中で42〜65℃で2時間。
【0090】
特定の配列相同性を有する核酸分子の間のハイブリダイゼーションを達成するのに必要なストリンジェンシー条件を計算するための一般式を挙げる(Sambrookら,1989,上記に同じ):
Tm = 81.5℃ + 16.6Log [Na+] + 0.41(% G+C) - 0.63 (% ホルムアミド) - 600/二本鎖中のbp数。
【0091】
上述の式の一例として、[N+] = [0.368]かつ50% ホルムアミドで,GC含量が42%でプローブの平均長さが200塩基の場合、Tmは57℃である。DNA二本鎖のTmは、相同性が1%減少するごとに1〜1.5℃低下する。したがって、約75%より高い配列相同性を有する標的は、42℃のハイブリダイゼーション温度を用いることにより観察する。上述のハイブリダイゼーションおよび洗浄条件を用いて、好適な実施形態において、ハイブリダイゼーション温度が37℃であり最終洗浄を42℃で行い、より好適な実施形態ではハイブリダイゼーションを42℃で行い最終洗浄を50℃で行い、最も好ましい実施形態ではハイブリダイゼーションを42℃で行い最終洗浄を65℃で行う。
【0092】
ハイブリダイゼーションおよび洗浄のストリンジェンシーは主に、溶液の塩濃度および温度に依存する。一般に、プローブの標的とのアニーリングの速度を最大化するために、ハイブリダイゼーションは通常、ハイブリッドの計算上のTmよりも20〜25℃低い塩および温度条件で行われる。洗浄条件は、プローブの標的に対する相同性の度合いについて可能な限りストリンジェントにするべきである。一般に、洗浄条件は、そのハイブリッドのTmより約12〜20℃低くなるように選択される。本発明の核酸に関しては、中程度のストリンジェンシーのハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および55℃で15分間、2× SSCおよび0.5%SDSでの洗浄として定義される。高度にストリンジェントなハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および65℃で15分間、1× SSCおよび0.5%SDSでの洗浄として定義される。非常に高度にストリンジェントなハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および65℃で15分間、0.1× SSCおよび0.5%SDSでの洗浄として定義される。
【0093】
本発明の核酸分子は、任意の適当なクローニングベクター中でDNAとして維持することができる。好適な実施形態において、クローンはプラスミドクローニング/発現ベクター、例えばpGEM-T(Promega Biotech, Madison, WI)またはpBluescript(Stratagene, La Jolla, CA)などで維持されるが、これらはどちらも適当な大腸菌宿主細胞中で増殖させることができる。
【0094】
本発明のヒトSAA3コード核酸分子には、一本または二本鎖であってもよい、cDNA、ゲノムDNA、RNA、およびその断片が含まれる。従って本発明は、本発明の核酸分子の配列少なくとも1つとハイブリダイズする能力のある配列を有するオリゴヌクレオチド(DNAまたはRNAのセンスまたはアンチセンス鎖)を提供する。このようなオリゴヌクレオチドは、例えばPCR増幅などによる、試験サンプル中のヒトSAA3コード遺伝子またはmRNAを検出するためのプローブとして有用である。
【0095】
B. ヒトSAA3タンパク質の使用
抗体および核酸
SAAレベルは、動物または患者から得られたサンプル中の疾患または他の炎症症状を診断または検出するために使用することができる。好適な実施形態において、これらのレベルを、乳房の感染症または症状を同定するためにアッセイする。増加したSAAまたはSAA3レベルは、急性期と関連する可能性があり、感染症または疾患の診断に用いることができる。SAA3はまた、ムチン産生を刺激することから医薬として使用してもよく、そしてムチンMUC3の不十分なレベルと関連する腸の不具合を軽減するために投与してもよい。
【0096】
1. タンパク質および抗体
精製したヒトSAA3またはその断片は、ポリクローナルまたはモノクローナル抗体を産生するために使用することができ、これらの抗体は培養細胞または組織中での、および無傷の生物中でのタンパク質の存在および蓄積に対する敏感な検出試薬としての機能を果たしうる。組換え技術は、選択されたヒトSAA3の一部または全部を含有する融合タンパク質の発現を可能にする。SAA3タンパク質の全長タンパク質または断片を、このタンパク質の様々なエピトープに特異的な一連のモノクローナルまたはポリクローナル抗体を生成するために好適に利用することができ、このことによりこのタンパク質のより高い検出感度を提供する。好適な実施形態においては、ヒトSAA3を血清SAAと識別するヒトSAA3の断片を、エピトープ特異的抗体を生成するために利用する。
【0097】
ヒトSAA3に免疫学的に特異的なポリクローナルまたはモノクローナル抗体は、そのタンパク質を検出および定量するために設計した種々のアッセイに使用することができる。このようなアッセイには、限定するものではないが(1)免疫沈降に次ぐタンパク質定量、(2)イムノブロット解析(例えばドットブロット、ウェスタンブロット)(3)放射免疫アッセイ(4)ネフェロメトリー、比濁または免疫クロマトグラフィー(横方向流動)アッセイ、(5)ELISAおよび種々の定量的迅速検定が含まれる酵素結合アッセイ(例えばディップ・スティックなどの検査)が含まれる。
【0098】
ヒトSAA3に免学特異的に相互作用するポリクローナルまたはモノクローナル抗体は、このタンパク質を同定し精製するために利用することができる。例えば、抗体は、それら抗体が免疫特異的に相互作用するタンパク質のアフィニティー分離に利用することができる。抗体はまた、タンパク質および他の生物学的分子の混合物を含有するサンプルから、タンパク質を免疫沈降するために使用することができる。
【0099】
2.核酸
ヒトSAA3をコードする核酸分子は、本発明における種々の目的のために使用することができる。DNA、RNAまたはそのフラグメントは、その遺伝子の存在をおよび/または発現を検出するためのプローブとして使用することができる。このようなアッセイのためにヒトSAA3をコードする核酸をプローブとして利用することができる方法には、限定するものではないが、(1)in situハイブリダイゼーション、(2)サザンハイブリダイゼーション、(3)ノーザンハイブリダイゼーション、(4)類別された(assorted)増幅反応、例えばポリメラーゼ連鎖反応(PCR)および逆転写酵素-PCR(RT-PCR)が含まれる。
【0100】
例示した本発明のヒトSAA3をコードする核酸はまた、ヒトを含む他の生物種由来の関連遺伝子を同定するためのプローブとして利用することができる。当技術分野で広く知られ上述した通り、ハイブリダイゼーションのストリンジェンシーは、核酸プローブと様々な程度の相同性を有する相補配列とのハイブリダイゼーションを可能にするために調節することができる。
【0101】
ヒトSAA3をコードする核酸の前述の使用に加え、これらは形質転換(トランスジェニック)細胞、組織および生物の創造において有用性が期待される。
【0102】
本発明はヒトSAA3タンパク質をコードする、新規な精製および単離された核酸配列を提供する。現在のところ好適な形態では、DNA配列は新規SAA3またはその保存的に改変された変異体をコードするcDNA配列を含み、これらはプロラクチンまたはLPSに応答して乳房上皮細胞で発現され、活性TFLK領域を含んでなり、そして本明細書において開示するタンパク質の生物学的活性を保有する。より好適な実施形態において、核酸配列は配列番号1と少なくとも約80%の相同性、またはコードされるアミノ酸配列と80%の相同性がある。具体的には、単離された配列を配列番号1に表す。これとは別のDNA形態、例えばゲノムDNAおよびヌクレオチドから部分または全化学合成により調製したDNAならびに欠失または突然変異を有するDNAもまた、本発明の範囲内にある。
【0103】
本発明においてさらにゲノムSAA3配列を同定しキャラクタライズした。ヒトSAA3配列とともにイントロンを含むゲノム領域が使用可能であり、ヌクレオチド配列の最も効率的な発現を達成するためにしばしば必要である。SAA3の5'領域もまた開示するが、これはプロモーター領域の一部を含み、異種遺伝子の一時的および空間的(spatial)発現に使用することができるSAA3プロモーターを単離するために使用することができる。このプロモーターは乳房上皮細胞への組織優先的発現(tissue preferred expression)を提供することができ、またプロラクチンまたはLPSの存在下での機能的に連結された配列の誘導発現も提供することができる。
【0104】
本発明により提供されたDNA配列と、同種または異種発現制御DNA配列、例えばプロモーター、オペレーター、およびレギュレーターなどとの結合は、mRNAからのin vivoおよびin vitro転写を可能にし、次いでこれは翻訳が可能であり、本発明の新規タンパク質および関連ポリおよびオリゴペプチドを大量に提供する。本発明の目下好適なDNA発現系において、SAA3をコードするDNAは、このタンパク質のin vitro転写および翻訳を可能にする制御プロモーターDNA配列に機能的に連結されている。
【0105】
DNA配列を、適当なウィルスおよび環状DNAプラスミドベクターを潜在的に伴う、標準的な形質転換および形質導入プロセスによって原核および真核宿主細胞に導入することはまた、本発明の意図の内にあり、有用なタンパク質をこれまで天然の取得源からは入手できなかったほど大量に提供すると予想される。哺乳動物宿主細胞の使用は、以下により詳しく記述する通り、本発明の組換え発現産物に最良の生物学的活性を附与するために必要であるかもしれない、翻訳後修飾(例えば末端切断化、グリコシル化、およびチロシン、セリンまたはトレオニンリン酸化)を提供すると予想される。
【0106】
細胞を形質転換し、ベクターを構築し、メッセンジャーRNAを抽出し、cDNAライブラリーを調製するなどのために用いるほとんどの技術は、当技術分野で広く実施されており、ほとんどの当業者は具体的な条件および手順を説明する標準的な資料文献に精通している。しかしながら、便宜のために、以下の段落はガイドラインとして役立つであろう。
【0107】
宿主および制御配列
ヒトSAA3をコードする配列を発現させるために、原核および真核細胞系の両方を使用することができる。むろん、原核細胞宿主がクローニング手順のためには最も好都合である。原核細胞は大腸菌の種々の株が最も頻出の代表であるが、一方、他の微生物株もまた使用可能である。宿主と両立できる種から誘導された複製サイト、選択マーカーおよび制御配列を含有するプラスミドベクターが使用される。例えば、大腸菌は典型的にはBolivarら(Gene (1977) 2:95)により大腸菌種から誘導されたプラスミドであるpBR322の誘導体を用いて形質転換する。pBR322はアンピシリンおよびテトラサイクリン耐性の遺伝子を含有し、従って所望のベクターを構築するにあたり、保持または破壊することができる複数の選択マーカーを提供する。転写開始のためのプロモーターを、場合によりオペレータを、リボソーム結合サイト配列とともに含むと本明細書において定義した、一般的に使われる原核細胞制御配列は、βラクターゼ(ペニシリナーゼ)およびラクトース(lac)プロモーター系(Changら, Nature (1977) 198:1056)、およびトリプトファン(trp)プロモーター系(Goeddel,ら, 核酸s Res (1980) 8:4057)ならびにラムダ誘導PLプロモーターおよびN遺伝子リボソーム結合サイト(Shimatakeら Nature (1981) 292:128)などの一般に使われるプロモーターを含む。
【0108】
細菌に加え、真核細胞微生物、例えば酵母もまた宿主として使用できる。いくつかの他の株または種が一般に入手可能であるが、Saccharomyces cerevisiaeの研究株、パン酵母が最もよく使われる。例えばBroach, J.R., Meth Enz (1983) 101:307の2μ複製起点、または他の酵母で両立できる複製起点(例えばStinchcomb,ら, Nature (1979) 282:39, Tschumper, G.,ら, Gene (1980) 10:157 および Clarke, L,ら, Meth Enx (1983) 101:300を参照のこと)を採用したベクターを用いることができる。酵母ベクターのための制御配列には、解糖酵素の合成のためのプロモーター(Hess,ら, J Adv Enzyme Reg (1968) 7:149; Holland,ら, Biochemistry (1978) 17:4900)が含まれる。当技術分野で知られているさらなるプロモーターには、3-ホスホグリセリン酸キナーゼのプロモーター(Hitzeman,ら J Biol Chem (1980) 255:2073)が含まれる。増殖条件および/または遺伝的バックグラウンドにより転写が制御されているというさらなる利点を有する他のプロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロームC、酸性フォスファターゼ、窒素代謝に関連している分解酵素、α因子システム、ならびにマルトースおよびガラクトース利用を担う酵素のプロモーター領域である。また、コード配列の3'末端にはターミネーター配列が望ましいと考えられている。このようなターミネーターは、酵母由来遺伝子のコード配列に続く3'非翻訳領域に見られる。
【0109】
また多細胞生物から誘導される真核宿主細胞培養で、ポリペプチドをコードする遺伝子を発現させることも、もちろん可能である。例えば、Axel,ら, 米国特許第4,399,216号を参照のこと。これらの系は、イントロンをスプライスで除く能力という追加の利点を有し、そのことからゲノム断片を直接発現させるために使用できる。有用な宿主細胞系にはVEROおよびHeLa細胞、ならびにチャイニーズハムスター卵巣(CHO)細胞が含まれる。このような細胞のための発現ベクターには通常、哺乳動物細胞と両立できるプロモーターおよび制御配列が含まれ、例えば一般に使われているシミアンウィルス40(SV40)からの初期および後期プロモーター(Fiers, ら, Nature (1978) 273:113)、またはポリオーマ、アデノウィルス2、ウシ乳頭腫ウィルス、またはトリ肉腫ウィルスに由来するものなどの他のウィルスプロモーターが挙げられる。制御可能なプロモーターであるhMT1I(Karin, M.,ら, Nature (1982) 299:797-802)もまた使用可能である。哺乳動物細胞宿主系形質転換の一般的な態様は、Axel(上記に同じ)により記述されている。「エンハンサー」領域もまた発現を最適化するのにいまや重要のようである。これらは一般に、非コードDNA領域内においてプロモーターの上流または下流に見られる配列である。複製起点は、必要ならば、ウィルス起源から取得してもよい。一方、染色体への組み入れは真核細胞におけるDNA複製の一般的な機構である。
【0110】
形質転換
用いる宿主細胞に依存して、形質転換をその細胞に適当な標準的な技術を用いて行う。Cohen, S.N., Proc Natl Acad Sci (USA) 1972) 69:2110によって記載された塩化カルシウムを用いるカルシウム処理法、またはManiatisら, Molecular Cloning: A Laboratory Manual (1982) Cold Spring Harbor Press, p. 254 and Hanahan, D., J Mol Biol (1983) 166:557-580に記載されている塩化ルビジウム法を、実質的に細胞壁を有する原核細胞または他の細胞に用いることができる。そのような細胞壁を有しない哺乳動物細胞については、Graham およびvan der Eb, Virology (1978) 52:546のリン酸カルシウム沈殿法を、場合によりWigler, M., ら, Cell (1979) 16:777-785によって改変して、使用してもよい。酵母への形質転換は、Beggs, J.D. Nature (1978) 275:104-109 or of Hinnen, A.,ら, Proc Natl Acad Sci (USA) (1978) 75:1929の方法により行うことが出来る。
【0111】
ベクター構築
所望のコードおよび制御配列を含有する適当なベクターの構築は、当技術分野でよく理解されている標準的なライゲーションおよび制限技術を用いる。単離したプラスミド、DNA配列、または合成オリゴヌクレオチドを、所望の形態に切断し、調整し、再度ライゲーションする。
【0112】
ベクターを形成するDNA配列は、いくつかの起源から入手可能である。バックボーンベクターおよび制御システムは、構築する配列の大部分に用いる、入手可能な「ホスト」ベクターに一般に見られる。典型的な配列は上記に記載した。適当なコード配列については、開始時の構築は、適切な配列をcDNAまたはゲノミックDNAライブラリーから取り出すことに関する。一方、一旦配列が開示されたならば、遺伝子配列全体をin vitroで個別のヌクレオシド誘導体から出発して合成することが可能である。遺伝子または相当な大きさのcDNA(例えば500〜1000bp)の全配列は、個別の重複する相補的オリゴヌクレオチドを合成し、一本鎖非重複部分をデオキシリボヌクレオチド三リン酸の存在下でDNAポリメラーゼを用いて埋めることで調製してもよい。このやり方は配列が既知のいくつかの遺伝子の構築について用いられ成功している。例えば、Edge, M.D., Nature (1981) 292:756; Nambair, K.P., ら, Science (1984) 223:1299; Jay, Ernest, J Biol Chem (1984) 259:6311を参照のこと。
【0113】
合成オリゴヌクレオチドは、Edgeら, Nature (上記に同じ) およびDuckworthら, 核酸s Res (1981) 9:1691によって記載されたホスホトリエステル法、またはBeaucage, S.L.,およびCaruthers, M.H., Tet Letts (1981) 22:1859 ならびにMatteucci, M.D.,およびCaruthers, M.H., J Am Chem Soc (1981) 103:3185によって記載されたホスホルアミダイト法により調製することができ、また市販の自動オリゴヌクレオチド合成機を用いて調製することが出来る。アニーリング前のまたは標識化のための一本鎖のキナーゼ処理は、過剰の、例えば1 nmole基質に対して約10単位のポリヌクレオチドキナーゼを、50mM Tris、pH7.6、10mM MgCl2、5mM ジチオトレイトール、1〜2mM ATP、1.7 pmole γ32P-ATP(2.9mCi/mmole)、0.1mM スペルミジン、0.1mM EDTAの存在下で用いて達成される。
【0114】
このようにして一度所望のベクターの構成要素を手に入れたならば、それらを標準的な制限およびライゲーション手順により切り取ってライゲートすることができる。
【0115】
部位特異的DNA切断は、当技術分野で一般に理解されている条件下で、適当な制限酵素(複数であってもよい)で処理することにより行うが、それらの条件は個別に市販の制限酵素の製造業者により特定されている。例えばNew England Biolabs製品カタログを参照のこと。一般にプラスミドまたはDNA配列約1μgを、約20μlのバッファー溶液中で1単位の酵素により切断する。本発明の実施例では、典型的には、過剰量の制限酵素を使用して基質DNAの完全消化を保証する。約37℃で約1時間〜2時間のインキュベーション時間が実行可能であるが、バリエーションも許容可能である。各インキュベーションの後、タンパク質をフェノール/クロロホルム抽出で除去し、次いでエーテル抽出を行ってもよく、核酸を水性画分からエタノールを用いて沈殿させて回収する。所望により、切断した断片の大きさによる分離を、標準技術を用いてポリアクリルアミドゲルまたはアガロースゲル電気泳動で行うことができる。大きさによる分離の一般的な記載は、Methods in Enzymology (1980) 65:499-560に見られる。
【0116】
制限切断断片は、大腸菌DNAポリメラーゼIのラージフラグメント(クレノウ)で、4種のデオキシヌクレオチド三リン酸(dNTP)の存在下で、20℃〜25℃で50mM Tris pH7.6、50mM NaCl、6mM MgCl2、6mM DTTおよび0.1〜1.0mM dNTPs約15〜25分のインキュベーション時間を用いて処理することにより平滑末端化することができる。クレノウフラグメントは、5'側一本鎖突出部を埋めていくが、突き出た3'側一本鎖をたとえ4種のdNTPが存在していても削り取る。所望ならば、突出部の性質により決定される限定的範囲内で、選択的修復をdNTPの1種または選択したもののみを供給することにより行うことができる。クレノウでの処理後、混合物をフェノール/クロロホルムで抽出しエタノールで沈殿させる。適当な条件下でのS1ヌクレアーゼまたはBAL-31を用いた処理は、あらゆる一本鎖部分の加水分解という結果を招く。
【0117】
ライゲーションは標準的な条件および温度に沿って15〜50μlの容量で行われる。標準的な条件および温度は例えば、20mM Tris-HCl pH 7.5、10mM MgCl2、10mM DTT、33μg/ml BSA、10mM〜50mM NaCl、および0℃で40μM ATP、0.01〜0.02(Weiss)単位T4 DNAリガーゼ(「粘着末端」ライゲーション用)、または14℃で1mM ATP、0.3〜0.6(Weiss)単位T4 DNAリガーゼ(「平滑末端」ライゲーション用)である。分子間での「粘着末端」ライゲーションは、通常33〜100μg/ml合計DNA濃度で行う(5〜100nM合計最終濃度)。分子間での平滑末端ライゲーションは1μM合計最終濃度で行う。
【0118】
「ベクター断片」を用いるベクター構築において、ベクター断片は一般に5'リン酸を除去しベクターの自己ライゲーションを防ぐために、細菌由来アルカリフォスファターゼ(BAP)または仔牛小腸由来アルカリフォスファターゼで処理する。処理はpH 8で約10mM Tris-HCl、1mM EDTAで、ベクター1μgあたり約1単位のBAPまたはCIPを用いて60℃で約1時間行う。核酸断片を回収するために、調製物をフェノール/クロロホルムで抽出しエタノールで沈殿させる。これとは別に、二重に消化したベクターの再ライゲーションを、追加の制限酵素消化により、および/または不要断片の分離により防止することができる。
【0119】
配列変更を要する、cDNAまたはゲノムDNAから誘導されたベクター中の部分については、部位特異的プライマー導入突然変異(Zoller, M.J.,およびSmith, M. 核酸s Res (1982) 10:6487-6500 and Adelman, J.P.,ら, DNA (1983) 2:183-193)を用いることができる。これは、所望の突然変異を表す限定的なミスマッチ部分以外は、突然変異を導入すべき一本鎖ファージDNAに相補的なプライマー合成オリゴヌクレオチドを用いることにより行う。手短に述べると、合成オリゴヌクレオチドをファージに相補的な鎖を合成するようし向けるプライマーとして使用し、結果的に得られる部分的または完全二本鎖DNAをファージ担持宿主細菌へと形質転換する。形質転換した細菌の培養物をトップアガーにプレートし、ファージを抱えている単一細胞からのプラーク形成を可能にする。
【0120】
理論的には、新しいプラークの50%が一本鎖として突然変異型のファージを含有し、50%が元の配列を保有する。結果的に得られるプラークは、完全なマッチの結合は可能であるものの、元の鎖とのミスマッチが結合を防止するのに十分な温度で、キナーゼ処理済み合成プライマーとハイブリダイゼーション後洗浄する。次いでプローブとハイブリダイズするプラークを選び出し、培養し、DNAを回収する。
【0121】
構築物の確認
プラスミド構築の正しいライゲーションは、まず大腸菌(E. coli)MC1061株(Casadaban, M., ら, J Mol Biol (1980) 138:179-207)または他の適当な宿主をライゲーション混合物で形質転換することにより確認できる。成功した形質転換体は、当技術分野で理解されている通り、アンピシリン、テトラサイクリン、または構築プラスミドの様式に依存する別のマーカーを用いた別の抗生物質耐性を用いることにより選択する。形質転換体のプラスミドは次いで、Clewell, D.B.,ら, Proc Natl Acad Sci (USA) (1969) 62:1159の方法に沿って調製し、場合によりクロラムフェニコール増幅(Clewell, D.B., J Bacteriol (1972) 110:667)を行う。複数の微少DNAプレップを一般的に用いる。例えばHolmes, D.S., ら, Anal Biochem Acids Res (1979) 7:1513-1523を参照のこと。単離したDNAは、制限処理により解析および/またはSanger, F., ら, Proc Natl Acad Sci (USA) (1977) 74:5463のジデオキシヌクレオチド法、さらにはMessing,ら, 核酸s Res (1981) 9:309, Maxam,ら, Methods in Enzymology (1980) 65:499が記載した方法により配列解析する。
【0122】
宿主細胞の例示
本発明におけるクローニングおよび原核細胞発現に用いる宿主細胞は以下の通りである。
【0123】
ほとんどの細菌プロモーターの制御下でのクローニングおよび配列解析、ならびに構築物の発現のために、例えばMC1061、DH1、RR1、C600hfl、K803、HB101、JA221、およびJM101などの大腸菌株を使用することができる。
【0124】
3.SAA3の発見に基づくさらなる使用
ヒトSAA3はまた、いくつかの他の目的のために使用することができる。これには、限定するものではないが、(1)腸または脈管構造をまたぐ分子の送達のための担体(2)新生児における腸粘膜の発達のための栄養的サプリメント(3)免疫応答の調節因子(注入または経口投与を通して)が含まれる。
【0125】
4.医薬製剤
本発明において、SAA3およびより具体的にはその活性部位(すなわちTFLKモチーフ)は、腸におけるムチン産生を刺激する。これは、ムチンが腸感染症の予防および治療において主要な役割を果たし、多くのプロバイオティック微生物(probiotics)がムチン産生を誘導することにより作用することが示されたことから、大変意義深い。Mackら,"Probiotics inhibit enteropathogenic Escherechia coli adherence in vitro by inducing intestinal mucin gene expression", 1999, Am J Physiol, 276:G941-950を参照のこと。そこに開示されている内容を参照により本明細書に組み入れる。このことから、本発明にはまたヒトSAA3に関連する、ヒトおよび動物のための医薬製剤が含まれる。医薬の分野に精通したものならば、医薬組成物の用量および投与計画が投与等ではなく、そのヒトおよび動物の年齢、健康状態、性別、大きさおよび体重によって変わることが容易に理解できるであろう。これらのパラメーターは、きちんと確立された手順と解析、例えばフェーズI、II、III臨床試験で、各システムについて求めることができる。
【0126】
投与については、SAA3は製薬上許容可能な担体、例えば適当な液体ビヒクルまたは賦形剤、および場合により補助剤または添加剤と組み合わせることができる。液体ビヒクルおよび賦形剤は常套的なものであり、市販されている。それらの実例としては、蒸留水、生理食塩水、デキストロース水溶液などが挙げられる。
【0127】
一般に、活性化合物に加えてさらに、本明細書の医薬組成物は、活性化合物を医薬的に使用できる製剤へと加工するのを促進する、適当な賦形剤および補助剤を含有してもよい。経口用剤形は錠剤、糖衣剤、およびカプセル剤を包含する。直腸投与できる製剤には坐剤が含まれる。他の剤形には、非経口または経口投与のための適当な溶液、および口腔または舌下投与することができる組成物が含まれる。
【0128】
本発明の医薬製剤は、当技術分野でよく知られた様式で製造する。例えば、医薬製剤は、慣用の混合、顆粒化、糖衣化、溶解、凍結乾燥処理などの手段により製造できる。用いる処理方法は、最終的には用いる活性成分の物理的性質に依存する。
【0129】
適当な賦形剤は、特に、糖、例えばラクトースまたはスクロース、マンニトールまたはソルビトール、セルロース調製物および/またはリン酸カルシウム、例えばリン酸三カルシウムまたはリン酸水素カルシウム、ならびに結合剤、例えばデンプン、糊、(例えばトウモロコシデンプン、小麦デンプン、コメデンプン、ジャガイモデンプン、ゼラチン、ガムトラガカント、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドンを使用)である。所望により、崩壊剤を添加することができ、その例としては上述のデンプンならびにカルボキシメチルデンプン、架橋ポリビニルピロリドン、寒天、もしくはアルギン酸またはその塩、例えばアルギン酸ナトリウムなどが挙げられる。補助剤は、流動調節剤および滑沢剤、例えばシリカ、タルク、ステアリン酸もしくはその塩、例えばステアリン酸マグネシウムまたはステアリン酸カルシウムおよび/またはポリエチレングリコールである。糖衣の芯は適当なコーティングとともに提供してもよく、そのコーティングは所望ならば胃液に耐性であってもよい。
【0130】
この目的のために濃縮した糖溶液を用いることができ、それは場合によりアラビアガム、タルク、ポリビニルピロリドン、ポリエチレングリコールおよび/または二酸化チタン、ラッカー溶液ならびに適当な有機溶媒または溶媒混合物を含有してもよい。胃液に耐性のコーティングを製造するためには、適当なセルロース調製物の溶液、例えばアセチルセルロースフタレート、またはヒドロキシプロピルメチルセルロースフタレート、染料および色素を、例えば同定または化合物用量の異なる組み合わせをキャラクタライズするために、糖衣剤コーティングの錠剤に加えることができる。
【0131】
経口に用いることのできる他の医薬製剤には、ゼラチンでできたプッシュフィット(push-fit)カプセル剤、ならびにゼラチンおよび例えばグリセロールまたはソルビトールなどの可塑剤でできた軟性密封カプセル剤が含まれる。プッシュフィットカプセル剤は顆粒の形態で活性化合物を含有してもよく、顆粒は充填剤例えばラクトース、結合剤(例えばデンプン)、および/または滑沢剤(例えばタルクまたはステアリン酸マグネシウム)ならびに場合により安定剤と混合してもよい。軟カプセル剤については、活性化合物を好ましくは適当な液体、例えば脂肪油、流動パラフィン、または液状ポリエチレングリコールに溶解または懸濁させる。さらに安定剤を添加してもよい。直腸に用いることができる医薬製剤には、例えば、活性化合物と坐剤用の基剤との組み合わせからなる坐剤が含まれる。適当な坐剤用の基剤は、例えば天然または合成トリグリセリド、パラフィン炭化水素、ポリエチレングリコールまたはより高級なアルコールである。さらに、活性化合物と基剤の組み合わせからなるゼラチン性直腸用カプセルを用いることも可能である。基剤の材料となりうるものには、液状トリグリセリド、ポリエチレングリコール、またはパラフィン炭化水素が含まれる。
【0132】
非経口投与の適当な製剤には、水に溶けうるまたは水に懸濁されうる形態の活性化合物の水溶液が含まれる。さらに、適切な油性注入懸濁物として、活性化合物の懸濁剤を投与してもよい。適当な親油性溶媒またはビヒクルには、脂肪油、例えばゴマ油、あるいは合成脂肪酸エステル、例えばオレイン酸エチルまたはトリグリセリドが含まれる。水性注入懸濁物は、懸濁物の粘性を増加させる物質を含有してもよく、そうした物質には例えば、カルボキシメチルセルロースナトリウム、ソルビトールおよび/またはデキストランが含まれ、場合により懸濁物はまた、安定剤を含有してもよい。
【0133】
慣用の担体を用いた投与の他に、活性成分を、当業者に知られているいくつかの特殊化された薬剤送達技術により投与してもよい。以下の例は、例証のみを目的として挙げるものであり、全く本発明を制限することを意図したものではない。
【0134】
本明細書において用いる「有効量」という用語は、本明細書において開示する方法および手順によって決定される通り、病原体の粘膜細胞への付着が減少するように、ムチン産生を増加させるのに十分な量のヒトSAA3を意味する。
【0135】
本発明において、新規ヒトSAA3およびさらに特定するとそのTFLKモチーフ活性部位は、ムチン産生、より具体的にはMUC3産生を刺激することが示された。ムチン産生は大腸菌の付着を阻害することが示されており、同じことをするプロバイオティック物質は、ムチンの刺激を通して機能することが示されている。ヒトSAA3および/またはペプチドは、プロバイオティックの代わりに使用することができる。
【0136】
腸の感染症におけるムチンの重要性は、感染性生物が疾患を引き起こすのに必要な事象を防ぐ能力にある。
【0137】
ムチンは腸の上皮細胞により産生され、その表面へと分泌される。このことから、ムチンは戦略上、腸の上皮細胞と腸管内に取り込まれた加害性因子(すなわち感染因子、有害物質)との間に位置する。
【0138】
ムチンはまた、細菌の腸管の上皮細胞への付着を阻害する。細菌による腸の内層細胞への結合が、侵襲、毒素送達および下痢疾患の罹患の最初の段階である。腸内病原体の結合が阻害されれば、疾患は発症しない。
【0139】
ムチンはウィルスの複製を阻害することが示されている。
【0140】
ムチンは、先天免疫の一部であり、腸の基本的な防御系である。したがって、抗体/T細胞によって駆動される後天性免疫系と比較して、ムチンが提供する利点には以下のものが含まれる:加害性因子に対して即時または急速に誘導可能な応答、広範な作用スペクトル、局所的効力、および動物界にわたる保存。
【0141】
感染因子に起因するムチン分泌の増加は、よく知られた臨床的現象である。ムチンは、病原性の腸内細菌が腸の細胞に付着するのを阻害し、このことから感染を防止する。これは、ムチンが、通常腸の細胞に付着するために使われる細菌の細胞壁の構造物に付着することにより成し遂げられる。プロバイオティック細菌(非病原性細菌)は、病原性腸内細菌による腸管内側の上皮細胞への付着を防ぐ。プロバイオティック細菌から分泌される物質は、腸の細胞により多くのムチンを産生させ、これはプロバイオティック物質が感染を防ぐ1つの機構である。
【0142】
上記のことは、数多くの腸内病理に対する本発明のペプチドの薬学的用途を実証している。例えば旅行者下痢の予防が挙げられる。多くの感染性生物は事実上地域的なものであり、自分たちの地域から出た旅行者は、通常これらの生物にそれまでに曝露されたことがなく、従ってそれらへの免疫を形成していない。多くの人が旅行前に抗生剤を使うものの、いくつかの抗生剤は有害な副作用を有し、また生物は多くの抗生剤への耐性を獲得しつつある。
【0143】
別の潜在的な用途は、とりわけ軍隊において、赤痢や他の感染性疾患を防止することである。ヒトSAA3は、腸関連感染症を減少または防止する迅速、安全、かつ効果的な手段である。
【0144】
別の例には、幼児下痢の予防または治療が含まれる。母乳で育てられた幼児は、調乳で育てられた幼児よりも感染がずっと少ない。初乳は幼児に有益な天然物質であり、ヒトSAA3は初乳の成分であることから、調乳のかけがえのない天然添加物となる。調乳は広く市販されており、例えばInfamel(商標), Similac(商標), Carnation Good Start(商標),およびGerber(商標)が挙げられる。プロバイオティックは、ウィルスによって引き起こされた下痢の重症度を減少させ、回復時間を短縮することが示されている。このことからヒトSAA3の別の用途は、上記症状を有する小児のためのものであり、この用途は病院での入院期間や経費を減少させることによる経済的影響をも有するであろう。
【0145】
別の例には壊死性腸炎(NEC)の予防または治療が含まれる。これは未熟児におこる深刻な合併症である。現在用いられている種々の生殖技術のために、未熟児の数は爆発的に増えた。NECのための治療法は、ここ数十年変わっていない。未熟児の腸の細菌がNECの罹患に主要な役割を果たすため、この症状の治療は、その未熟児に食事を与えず、強力な抗生剤を与え、そして腸に穴が開かないことを願うことからなる。
【0146】
ヒトSAA3の別の用途には、発生地域での下痢の予防が含まれる。大腸菌O-157:H7の発生は、溶血性尿毒症候群による死へとつながりうる。SAA3はムチン産生を誘導し、これは腸の上皮細胞に大腸菌が付着するのを防ぎ、したがってこの感染症を予防することができよう。
【0147】
別の例には、尿路感染症の治療または予防が含まれる。膀胱上皮細胞は、腸の上皮細胞によく似ており、ムチンを産生する能力を有する。したがって泌尿器の感染症の予防(導尿カテーテルを使っている入院患者も含まれる)もまた、本発明の医薬組成物の用途となる。
【0148】
本明細書には、腸のムチンのアップレギュレーションが含まれるものの、他の粘膜表面(例えば鼻咽頭、膀胱、など)内層の上皮細胞もまたムチンを産生する。これらのムチンは、腸のムチンに類似して感染症を防止する機能を果たし、また本発明における治療の効果的な標的にもなりうる。
【0149】
以下の実施例を、本発明をより詳細に説明するために提供する。これらは本発明を例証することを意図したものであり、限定することを意図したものではない。
【実施例】
【0150】
細胞系および培養
ヒト乳腺上皮細胞系MCF-7(ATCC)およびT47-D(ATCC)を本研究において用いた。これらのヒト乳房上皮細胞は両方ともプロラクチン(PRL)受容体を発現することが明らかにされている(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)ことから、これらの細胞を選択した。MCF-7細胞は、0.2U/ml インシュリン、2mMグルタミン、4.5g/Lグルコース、50μg/mLストレプトマイシン、50U/mLペニシリン、および10%FBSを添加したダルベッコ変法イーグル培地(DMEM)で増殖させた。T47-D細胞は、MCF-7細胞について記載したものと同じものを添加したRPMI 1640培地にさらに10mM HEPESおよび1mMピルビン酸ナトリウムを添加して培養した。細胞培養物は、37℃で5% CO2を含む加湿雰囲気でインキュベートした。細胞培養継代のために、付着細胞を、トリプシン-EDTA(Invitrogen Corp.)を用いて解放させた。
【0151】
細胞刺激アッセイ
M-SAA3発現を検出するために、これらのヒト乳腺細胞を、100ng/mLのヒト組換えプロラクチン(PRL)(Sigma)または20μg/mLの大腸菌リポポリサッカライド(LPS)(Difco)のいずれかで、0時間、0.5時間、4時間、24時間および48時間刺激させた。LPSで誘導する乳房細胞については、細胞をコンフルーエント状態まで増殖させ、次いで1μg/mLヒドロコルチゾンを含有しFBSを2%しか含有しない以外は上記と同じものを添加したDMEM中のLPSで刺激した。PRLで刺激する乳腺細胞については、細胞をMATRIGEL Matrix (BD Biosciences)でコーティングしたウェル上で85%コンフルーエント状態まで増殖させ、次いで以前に記載された通りのものを添加したDMEMおよびF12培地の無血清の1:1の混合物中のPRLで誘導した(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)。この研究においてもちいたPRL濃度は、ヒトの授乳におけるPRLの生理学的レベルに基づくものであり、それは典型的には100〜200ng/mLである。これに対して基底のレベルは2〜15ng/mLの範囲内である(M.C. Neville, J. Morton, Physiology and endocrine changes underlying human lactogenesis II, J. Nutr. 131 (2001) 3005S-3008S)。48時間刺激については、24時間後に培地を、適当な成分および刺激剤を含有する新鮮な培地で交換した。
【0152】
RNA単離およびRT-PCR
全RNAは、刺激していない、または刺激した乳腺上皮細胞から製造者が記載した手順に従いTRIZOL(Invitrogen Life Technologies)を用いて単離した。RNAはエタノール沈殿で濃縮し-80℃で保存した。RNA全体は1%(重量/容量)アガロースゲルで分画化し、次いでゲルを臭化エチジウムで染色することにより可視化した。第一のcDNA鎖合成は、全RNAを5μg、ポリd(T)14オリゴヌクレオチドを40μMおよび製造者の推奨に従ってSuperScriptII RNase H- Reverse Transcriptase(Invitrogen Life Technologies)を用いて行った。第二のcDNA鎖合成および二本鎖cDNAの増幅はAmpliTaq Gold DNA Polymerase (Applied Biosystems)を用いて行った。第二鎖cDNA合成に用いるフォワードF1およびリバースR1プライマー(表1)は、予測されるヒトSAA3 cDNA配列(GenBank登録番号X13895)の提唱されているエキソン2のアンチセンス鎖およびエキソン3のセンス鎖にそれぞれ相補的である。
【0153】
表1 ヒトM-SAA3 cDNAを取得するために用いたヒトSAA3特異的オリゴヌクレオチドプライマー
RT-PCRプライマー オリゴヌクレオチド配列 *bp♯
F1 5'-GATGAAGCTCTCCACTGGCATCA-3' 56 bp 配列番号5
R1 5'-TCAGAGTAGGCTCTCCACATGTCTT-3' 154 bp 配列番号6
F4 5'-AACTTGAAACAGAATGTGTATTATCCTTGGTTG-3' 1 bp 配列番号7
RACEプライマー オリゴヌクレオチド配列 *bp♯
R3 (1o 5'RACE) 5'-CAGCTGCCTTGAGGAATGTTAACCAT-3' 116 bp 配列番号8
R4 (2o 5'RACE) 5'-GCTGCTGACACCCAGGACCAG-3' 90 bp 配列番号9
F1b (1o 3'RACE) 5'-AGCAGGATGAAGCTCTCCTCTGGCATCA-3' 51 bp 配列番号10
F2c (2o 3'RACE) 5'-CAGCCAAGGATGGTTAACATTCCTCAAGGCA-3' 107 bp 配列番号11
*(bp♯は、本研究において得られたヒトM-SAA3 cDNA配列の5'ヌクレオチドに対する、図2の中のプライマーの位置に対応する。)
GeneAmp PCR System 2700 サーモサイクラー(Applied Biosystems)を用いるサイクリングパラメータは、95℃で10分を1サイクル、94℃で30秒、58℃で40秒、60℃で20秒を43サイクル、次いで60℃で10分を1サイクルであった。LPSまたはPRLで刺激した後に得られる123塩基対のRT-PCR産物は、pCRII-TOPO (Invitrogen Life Technologies)にクローニングした。クローニングしたインサートは同一性を確認するために両方向の配列を決定した。390塩基対のグリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)cDNA断片を増幅させるフォワードおよびリバースプライマーを含有する対照RT反応もまた、mRNA分解が生じなかったことを保証するために、各cDNA調製について行った。
【0154】
cDNA末端の5'および3'迅速増幅
全長M-SAA3 cDNAは、市販の5'RACE および GeneRacer System(Invitrogen Life Technologies)を製造業者の推奨に従って用いて、cDNA末端の5'および3'迅速増幅法(RACE)により取得した。簡潔には、刺激していない、および刺激した乳腺上皮細胞の両方由来の全RNA5μgを、5'RACE用遺伝子特異的リバースR1プライマー、および3'GeneRacer手順用ポリd(T)18GeneRacerアダプタープライマー(Invitrogen Life Technologies)を用いて逆転写した。ネステッド一次および二次PCR用の遺伝子特異的RACEプライマーは、製造業者の推奨に従って設計し、上述の123塩基対中央M-SAA3 cDNA断片の選択した領域に相補的であった。5'および3'RACE手順に用いたネステッド一次および二次PCRプライマーを表1に示す。一次PCRは第一鎖cDNAを1μL含有する50μLの容量中で行った。一次PCRに用いた熱サイクリングパラメータは、95℃で9分を1サイクル、94℃で30秒および72℃で1.5分を5サイクル、94℃で30秒および70℃で1.5分を5サイクル、94℃で30秒、65℃で1分、および72℃で1.5分を25サイクル、次いで72℃で10分を1サイクルであった。二次PCRは、適当な一次PCR混合物1μL、アダプタープライマー、およびフォワードまたはリバースネステッド二次遺伝子特異的プライマーのいずれか、を含有する50μL容量中で行った。熱サイクルパラメータは、一次PCRに用いたものと同じであり、ただし4つめのサイクリングパラメータとして25サイクルの代わりに33サイクルを用いた。結果的に得られる二次PCR増幅産物は、同一性を確認し、また全長ヒトM-SAA3 cDNA配列を取得するために配列決定した。
【0155】
M-SAA3のヌクレオチド配列解析
クローニングした123塩基対RT-PCR増幅産物は、ベクターに基づくSp6およびT7プライマーを用いて配列決定し、また二次5'および3'RACE産物は、遺伝子特異的ネステッド二次プライマーを用いて配列決定した。DNA配列は、the Wisconsin Genetics Computer Group (GCG) Package (バージョン10.2)の中のBESTFITおよびPILEUPプログラムを用いて解析した。M-SAA3 cDNAのヌクレオチド配列をGenBankデータベースに、登録番号AY209188で登録した。
【0156】
M-SAA3発現の誘導および検出
ヒト乳腺上皮細胞をPRLまたはLPSで刺激した後にSAA3が発現するかどうか判断するために、SAA3特異的F1およびR1プライマー(表1)を用いてRT-PCRを行った。GAPDH cDNAに相補的なフォワードおよびリバースプライマーを含有する対照RT反応は、各cDNA調製物に関して、予想される390塩基対のRT-PCR産物を増幅し、このことはcDNAを用意するために用いたmRNAが高い完全性を有していたことを保証する(図1)。SAA3特異的F1およびR1プライマーとPRLで0.5時間、4時間もしくは24時間、またはLPSで48時間刺激したヒトMCF-7乳腺細胞から調製したcDNAとのRT-PCRから、結果的に長さにして123塩基対の増幅産物が得られた。図1に、SAA3特異的F1およびR1プライマーとPRLで0.5時間、またはLPSで48時間刺激した乳房細胞から得られた123塩基対のRT-PCR産物を示す。また、図1に示されている通り、刺激していない乳腺細胞より調製したcDNAからは、SAA3特異的F1およびR1プライマーを用いても検出可能な増幅産物が得られなかった。この123塩基対産物の存在は、SAA3特異的F1およびR1プライマーとLPS刺激後わずか0.5時間のMCF-7細胞から調製したcDNAとのRT-PCRから検出され、またSAA3特異的F1およびR1プライマーとLPS刺激から48時間後の該細胞から調製したcDNAとのRT-PCRからも一貫して検出された。PRLで48時間刺激した後のマトリックス上のMCF-7細胞があまりにも非付着性になってしまったため、この時点以降のcDNAは調製しなかった。
【0157】
SAA3特異的F1およびR1プライマーとPRLまたはLPSで刺激したMCF-7細胞から調製したcDNAとから得られた123塩基対のRT-PCR産物の配列決定は、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によって当初推測されたヒトSAA3 cDNA配列のある領域と97%の相同性を示した。この乳房関連SAA3(M-SAA3)123塩基対RT-PCR産物のヌクレオチド配列を、ヒトM-SAA3の全長cDNA配列を含む両側のヌクレオチドと共に図2に示す。123塩基対のM-SAA3 RT-PCR断片は、スプライスジャンクションを含有し、そのスプライシングジャンクションでは、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)が推測した通り、2692ヌクレオチドのイントロンが正確にスプライシングで除かれた。このイントロンがヒトSAA3遺伝子中に存在することは、SAA3特異的F1およびR1プライマーと鋳型としてのヒトゲノムDNAとを用いてPCRを行うことにより確認した。このイントロンの不在は、このRT-PCR産物がM-SAA3 mRNAから生成されたものであって、ゲノムDNA混入物から生成されたものではないことを追認するものである。123塩基対のM-SAA3 cDNA産物はまた、刺激したT47-Dヒト乳腺細胞系から作成されたcDNA調製物からも検出された。
【0158】
全長ヒトM-SAA3 cDNAの単離および解析
全長M-SAA3 cDNAを取得するために、PRLまたはLPSで刺激した乳房MCF-7細胞から単離したRNAを用いて、cDNA末端の5'および3'迅速増幅(RACE)を行った。結果的に得られたM-SAA3特異的ネステッド二次プライマーを用いた二次PCRからの150塩基対の5'RACE産物、および、550塩基対の3'GeneRacer RACE産物を配列決定した。RACE手順により得られた二次PCR産物は両方とも、123塩基対のM-SAA3中央cDNA断片の配列と重複していた。最初の123塩基対のRT-PCR産物と同じように、これらの二次PCR増幅産物は、PRLまたはLPSで刺激したMCF-7乳房細胞からのcDNA調製物についてのみ検出された。5'RACE二次PCR産物が、M-SAA3 mRNAに由来するものであってゲノムDNAに由来するものではないことを確かめるために、M-SAA3特異的フォワードF4およびリバースR1プライマー(表1)をRT-PCRに用いた。結果的に得られた178塩基対のM-SAA3 cDNA断片は、予想通り第一イントロンを含有せず、この5'RACE二次PCR産物が確かにM-SAA3転写産物の5'領域の一部であるという証拠を提供した。同様に、二次PCRで得られた3'RACE産物は、M-SAA3 cDNA中の最後のイントロンを含有しなかった。これらの結果は一緒になって、RT-PCRおよび二次RACE産物がM-SAA3 mRNAに由来するものであって、ゲノムDNAに由来するものではないことをさらに検証した。さらに、これらの増幅産物はPRLまたはLPSで刺激した乳腺細胞に由来するcDNA調製物のみから検出することができ、このことは、これらの刺激物質がヒトM-SAA3遺伝子の転写発現を誘導することを追認した。
【0159】
PRLまたはLPSで刺激した後のヒトM-SAA3 cDNAについて得られた完全なヌクレオチド配列は長さにして655塩基対であり、これを図2に示す。また図2には、本研究で得られたヒトM-SAA3 cDNAと、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515) (GenBank 登録番号X13895)(配列番号3)が提唱した予測されるヒトSAA3 cDNA配列との比較を示す。他の3つのヒトSAA遺伝子の4エクソン構造と異なり、M-SAA3遺伝子は3エクソン構造を有する。SackおよびTalbotによって元々エクソン1と予測された領域はヒトM-SAA3 cDNAに存在せず、また当初予測されていたイントロン1全体がスプライシングで除かれることもない。そうではなく、5'側非翻訳領域(UTR)は、元々イントロン1の一部であろうと予測されていた最後の52ヌクレオチドを含有する。M-SAA3 cDNAの5'領域は妥当なKozakコンセンサス配列を含有し、最も有望な翻訳開始コドンはヌクレオチド57〜59に位置している(M. Kozak, Structural features in eukaryotic mRNAs that modulate the initiation of translation, J. Biol. Chem. 266 (1991) 19867-19870)。
【0160】
本発明者らの結果は、ヒトSAA3遺伝子の最初のイントロンが長さにして2692ヌクレオチドであり、GenBankに寄託されているいくつかのゲノムDNA配列(登録番号 AC055860.13およびAC108007.5)と一致していることを示した。このイントロンはそもそもSackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によってイントロン2として、そしてヒトSAA3遺伝子(GenBank登録番号X13895)中で長さにして2638ヌクレオチドであると予測されていた。本研究において得られたM-SAA3 cDNAは、イントロン1についてのスプライスサイト接合点が、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によってなされたイントロン2についての予測と一致することを示した。M-SAA3の最後のイントロンであるイントロン2は、長さにして321塩基対である。このイントロンのM-SAA3 mRNAからのプロセッシングは、イントロン3であろうと予測されていたこの領域に位置する、元々予測されていた273塩基対イントロンと類似していたものの、同一ではなかった。より詳しくは、M-SAA3の最後のまたは第3エキソンは、元々イントロン3の一部であろうと考えられていた、追加の71ヌクレオチドを含有していた。本研究において得られたヒトM-SAA3 cDNAと、GenBankに寄託されたヒトSAA3遺伝子配列(登録番号X13895)とのさらなる比較はまた、イントロン1およびイントロン2の両方が、スプライスのドナー/アクセプターサイト用のGT-AGコンセンサス配列と一致することを示した。
【0161】
ヒトM-SAA3 cDNAのヌクレオチド204における一塩基(T)挿入の存在は、フレームシフトを引き起こし、そして元々ヒトSAA3にコードされる遺伝子産物について予測されていたものよりも上流に翻訳停止コドンを生じさせた(配列番号3および4)(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515) (図2)。この一塩基(T)挿入は以前の研究で得られた結果を支持するものであり(B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661; (G.C. Sellar, A.S. Whitehead, Localization of four human serum amyloid A (SAA) protein superfamily genes to Chromosome 11p: Characterization of a fifth SAA-related gene sequence, Genomics 16 (1993) 774-776)、サンガーセンターヒトゲノム配列計画(www.ensemble.org/genome/central)より得られるヒトゲノムのこの領域のコンティグと一致した。それ故、結果的に得られる3'UTRは長さにして406塩基対であり、もともとヒトSAA3 cDNAについて予測されていた145塩基対3'UTRではない。図2に示すとおり、本研究において得られたヒトM-SAA3 cDNA配列と、予測されたSAA3 cDNAとの間にはまた、いくつかのさらなるヌクレオチドの相違が発見され、そのうちの4つは同族コドンがコードする予測される残基を変更する。一方、ポリ(A)尾部の21ヌクレオチド上流に位置する、M-SAA3転写産物のポリアデニル化シグナル(AAUAAA)は、ヒトSAA3遺伝子中のこのコンセンサス配列の位置について、SackおよびTalbotによってなされた予測と一致した(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)。
【0162】
ヒトM-SAA3と他のSAA3アイソフォームとの比較
ヒトM-SAA3 cDNAは、典型的に他のSAAタンパク質に見られる18残基のシグナルペプチドを含有する60アミノ酸の前駆体タンパク質に理論的に翻訳されるオープンリーディングフレームを含有した(図3)。成熟42アミノ酸タンパク質は、計算上4866.69ダルトンの分子量と理論上9.31のpIを有する。図3に示す通り、予測されるヒトM-SAA3タンパク質と、ウサギ、ハムスター、ウシ、およびマウスSAA3アイソフォームとの配列アライメントは、最初の48個のN末端残基について相当なアミノ酸相同性(〜94%)を示す。一方、ヒトM-SAA3転写産物の塩基204におけるヌクレオチド挿入は、下流で前駆タンパク質のコドン61で翻訳停止コドンを生じさせ、したがって他の哺乳動物SAA3アイソフォームよりも小さなタンパク質が結果的に得られる(図3)。コドン50におけるこの塩基挿入でコードされる予測残基、ならびにそれに続くヒトSAA3前駆体タンパク質のアミノ酸はまた、他の哺乳動物SAA3アイソフォームのこの領域のアミノ酸と著しくことなる(図3)。さらにヒトSAA3のこれらのC末端側の11個のアミノ酸、および前駆体タンパク質のコドン50のヌクレオチド挿入より前に位置する3つの残基は、以前に報告さたヒトSAA3の配列(GenBank登録番号X13895)(配列番号3および4)と異なる。
【0163】
ヒトM-SAA発現の調節
ホルモンであるPRLおよびグラム陰性細菌由来の外膜構成要素である内毒素LPSは、本研究においてヒトM-SAA3発現を刺激することが示された。ヒトおよび動物において、LPSはグラム陰性細菌感染症の存在を示す。内毒素ショックの主要なメディエーターであるLPS(E.T. Rietschel, T. Kirikae, F.U. Schade, U. Mamat, G. Schmidt, H. Loppnow, A.J. Ulmer, U. Zahringer, U. Seydel, F.D. Padowa, M. Schreier, H. Brade, Bacterial endotoxin: molecular relationships of structure to activity and function, FASEB J. 8 (1994) 217-225)は、授乳ウシ由来の乳房上皮細胞の一次培養物を、in vitroで用量依存的に炎症性サイトカインIL-1およびIL-6を産生するように刺激することが示されている(H. Okada, H. Ohtsuka, S. Kon-nai, R. Kirisawa, Y. Yokomizo, T. Yoshino, T.J. Rosol, Effects of lipopolysaccharide on production of interleukin-1 and interleukin-6 by bovine mammary epithelial cells in vitro, J. Vet. Med. Sci. 61 (1999) 33-35)。これらの乳房細胞はまた、炎症性サイトカインTNF-α転写産物を発現することが示されている(H. Okada, T. Ito, H. Ohtsuka, R. Kirisawa, H. Iwai, K. Yamashita, T. Yoshino, T.J. Rosol, Detection of interleukin-1 and interleukin-6 on cryopreserved bovine mammary epithelial cells in vitro, J. Vet. Med. Sci. 59 (1997) 503-507)。同様にヒト乳房上皮細胞の一次
培養物もまた、IL-6とTNF-αを産生することが示されている(F. Basolo, P.G. Conaldi, L. Fiore, S. Calvo, A. Toniolo, Normal breast epithelial cells produce interleukins 6 and 8 together with tumor-necrosis factor; defective IL6 expression in mammary carcinoma, Int. J. Cancer 55 (1993) 926-930)。炎症性サイトカインIL-1、IL-6、およびTNF-αは、特定の前後関係でSAA遺伝子の発現を活性化し、免疫応答の大きさを決定することが知られている。興味深いことに、ヒトSAA3プロモーターは、4つの推定上の急性期応答因子(APRF)コンセンサス結合サイト(CTGGGA)(配列番号12)を含有し、このことはLPSなどの内毒素が炎症性サイトカインの発現を誘導し、次いでそれらの炎症性サイトカインがM-SAA3発現を刺激することを示唆している。
【0164】
M-SAA3発現の別の誘導因子はPRLである。この多機能性ホルモンは、広範なスペクトルの生理学的プロセスを調節し、それには乳腺発達、授乳、ならびに内分泌、傍分泌および/または自己分泌機構による免疫機能が含まれる(M.C. Neville, J. Morton, Physiology and endocrine changes underlying human lactogenesis II, J. Nutr. 131 (2001) 3005S-3008S)。分子レベルでは、PRLは、多数の細胞(乳腺細胞も含まれる)に見られるヘマトポエチン/サイトカイン受容体スーパーファミリーのメンバーであるPRL受容体のホモダイマー化を誘導する(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236; C. Bole-Feysot, V. Goffin, M. Edery, N. Binart, P.A. Kelly, Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice, Endocr. Rev. 19 (1998) 225-268)。これは、受容体関連チロシン特異的キナーゼであるJAKの活性化を開始させ、それに続く情報変換因子(transducer)および転写活性化因子(STAT)の活性化を開始させる。STAT3は、炎症および組織損傷に応答して種々の急性期遺伝子の発現を誘導することがわかっている(J.N. Ihle, STATs: signal transducers and activators of transcription, Cell 84 (1996) 331-334)。in vitroおよびin vivoでのSTAT5の調節は、この因子が催乳性ホルモンシグナル経路において中心的で重要な役割を有し(B. Groner, F. Gouilleux, Prolactin-mediated gene activation in mammary epithelial cells, Curr. Opin. Genet. Dev. 5 (1995) 587-594)、またPRLに応答して乳房組織で母乳タンパク質の発現をアップレギュレートすることを示している(J.N. Ihle, STATs: signal transducers and activators of transcription, Cell 84 (1996) 331-334)。興味深いことに、ヒトSAA3のプロモーター領域(GenBank登録番号X13895)は、それぞれ5'UTRの1607塩基対(TTCCCGGAA)(配列番号15)および429塩基対(TTCCAAGGAA)(配列番号16)上流に位置する、推定上のSTAT3様 (TTCC(G=C)GGAA)(配列番号13)およびSTAT5様結合部位(TTCC(A>T)GGAA)(配列番号14)を含有する。これらのcis作用座因子は、LPSまたはPRL刺激に続くM-SAA3転写発現のアップレギュレーションを部分的に担っている可能性がある。PRLがヒト乳房上皮細胞系でSTAT5の活性化を誘導することは、以前に報告されている(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)。IL-6がヒト乳房上皮細胞系のJAK/STAT経路を活性化することが示されているものの(A. Badache, N.E. Hynes, Interleukin 6 inhibits proliferation and, in cooperation with an epidermal growth factor receptor autocrine loop, increases migration of T47D breast cancer cells, Cancer Res. 61 (2001) 383-391)、LPSが、ヒト乳房上皮細胞におけるSTAT因子活性化に与える影響は未だ不明である。
【0165】
SAAの正確な生物学的機能は解明されていないが、いくつかの研究は、種々のSAAアイソフォームのN末端領域が、それらの構造および機能の性質を部分的に担うことを示唆している。例えば、A-SAA1およびA-SAA2のN末端領域は、コレステロールに特異的に結合し、コレステロールをHepG2肝臓および大動脈平滑筋細胞へと輸送するが、このことは急性期応答中およびアテローム硬化症における、A-SAA1およびA-SAA2によるコレステロールフラックス調節のもっともらしい役割を示唆する(J. Liang, B.M. Schreiber, M. Salmona, G. Phillip, W.A. Gonnerman, F.C. de Beer, J.D. Sipe, Amino terminal region of acute phase, but not constitutive, serum amyloid A (apoSAA) specifically binds and transports cholesterol into aortic smooth muscule and HepG2 cells, J. Lipid Res. 37 (1996) 2109-2116)。別の研究は、A-SAAのN末端領域が、二次性アミロイド症に見られるアミロイドフィブリルの主要な構成要素であるアミロイドAの前駆体であることを実証した(A. Husebekk, B. Skogen, G. Husby, G. Marhaug, Transformation of amyloid precursor SAA to protein AA and incorporation in amyloid fibrils in vivo, Scand. J. Immunol. 21 (1985) 283-287; J.J. Liepnieks, B.K. Beckerman, M.D. Benson, Characterization of amyloid A protein in human secondary amyloidosis: the predominant deposition of serum amyloid A1, Biochim. Biophys. Acta 1270 (1995) 81-86)。本発明者らは以前に、いくつかの哺乳動物の初乳からM-SAA3アイソフォームを単離した(T.L. McDonald, M.A. Larson, D.R. Mack, A. Weber, Elevated extrahepatic expression and secretion of mammary-associated serum amyloid A 3 (M-SAA3) into colostrum, Vet. Immunol. Immunopathol. 83 (2001) 205-213)。さらに、ヒト腸上皮細胞を、合成したM-SAA3のN末端側の10アミノ酸で前処理することは、保護性腸ムチンMUC3の産生を刺激し、かつ未処理の細胞と比較して、腸病原性大腸菌(EPEC)がこれらの腸上皮細胞に付着するのを著しく減少させる(D.R. Mack, T.L. McDonald, M.A. Larson, S. Wei, A. Weber, The conserved TFLK motif of mammary-associated serum amyloid A3 is responsible for upregulation of intestinal MUC3 mucin expression in vitro, Pediatr. Res. 53 (2003) 137-142; M.A. Larson, S.H. Wei, A. Weber, D.R. Mack, T.L. McDonald, Human mammary-associated serum amyloid A3 peptide enhances intestinal MUC3 expression and inhibits EPEC adherence, Biochem. Biophys. Res. Comm. 300 (2003) 531-540)。これらの研究は一緒になって、種々のSAAアイソフォームに対して、組成、構造、および発現または局在化の場所に基づいて、機能を帰するが、これらの機能は全て、正または負の結果に寄与する。
【0166】
引用文献
本明細書において引用した全ての文献を、参照により明白にその全内容を本明細書に組み入れる。これには以下の公表された特許出願および特許も含まれる。米国特許第6509444、WO 01/31006、およびPCT/US99/19428;
M.A. Larson, A. Weber, A.T. Weber, T.L. McDonald, Differential expression of bovine mammary-associated serum amyloid A3 is induced by prolactin or lipopolysaccharide, (準備中);
G. Ramadori, J.D. Sipe, H.R. Colten, Expression and regulation of the murine serum amyloid A (SAA) gene in extrahepatic sites, J. Immunol. 135 (1985) 3645-3647;
R.L. Meek, E.P. Benditt, Amyloid A gene family expression in different mouse tissues, J. Exp. Med. 164 (1986) 2006-2017;
R.L. Meek, E.P. Benditt, Rat tissues express serum amyloid A protein-related mRNAs, Proc. Natl. Acad. Sci. USA 86(1989) 1890-1894。
【図面の簡単な説明】
【0167】
【図1】リポ多糖(LPS)またはプロラクチン(PRL)のいずれかで刺激する前および後のヒト乳腺上皮細胞系MCF-7から単離したM-SAA3 mRNAのRT-PCR解析を示すゲル。RT-PCRに用いたヒトSAA-3特異的F1およびR1プライマー(表1参照)は、予測エキソン2および3(GenBank 登録番号X13895)、またはこの研究で決定したエクソン1および2における残基をコードするヌクレオチドに対応する。レーン1はDNAマーカー、レーン2は逆転写酵素無しで調製したcDNAサンプルのRT-PCR増幅の代表例、レーン3はヒトGAPDH特異的プライマーを使用した刺激していない乳腺上皮細胞から調製したcDNAサンプルのRT-PCR増幅の代表例、レーン4は刺激していないMCF-7細胞、レーン5はPRLで刺激したMCF-7細胞、レーン6は刺激していないMCF-7細胞、レーン7はLPSで刺激したMCF-7細胞である。
【図2】本明細書に記載したヒトM-SAA3 cDNA配列と、ヒトSAA3遺伝子の最初に推測されたcDNA配列(GenBank登録番号X13895)(配列番号3および4)との比較を示す。GCGパッケージ(バージョン10.2)のBESTFITプログラムを、ヌクレオチド配列アライメントを得るために用いた。同一のヌクレオチドを縦の線で示し、ダッシュで類似性を最大化するために導入したギャップを示す。ヒトSAA3 cDNA配列の5'末端からのヌクレオチドの数を右に示す。最初の123 bp RT-PCR産物を生成するために用いたヒトSAA3特異的フォワードF1およびリバースR1プライマーの位置を、しかるべきヌクレオチド配列の上に示し、各プライマーの方向を矢印で示す。推定される翻訳開始コドンを太字で下線を引いて示す。M-SAA3転写産物におけるイントロン1と2のスプライス部位接合部を、下向きの矢印で接合部の上に示す。アスタリスクは、推定されるM-SAA3タンパク質が結果的にフレームシフトする(読み枠がずれる)1塩基挿入を示す。太字および二重の下線は、推定される翻訳停止コドンを示す。ポリ(A)シグナルは、下線を引いたポリアデニル化のためのコンセンサス配列の上に示されており、ポリA尾部は太字である。
【図3】推定されるヒトSAA3タンパク質と、ウサギ、ハムスター、ウシおよびマウス由来のSAA3アイソフォームとの比較を示す。アミノ酸配列のアライメントは、GCGパッケージのPILEUPプログラムを用いて得た。哺乳動物およびSAA3タンパク質の略記に続けて、対応する登録番号を以下に示す。rab-SAA3=ウサギSAA3(GenBank M64696)、ham-SAA3=ハムスターSAA3(GenBank M33431)、bov-SAA3=ウシSAA3(GenBank AF335552)、mou-SAA3=マウスSAA3(SWISS-PROT P04918)、およびhum-SAA3=ヒトSAA3。少なくとも3種のSAA3アイソフォームにおける同一または類似する残基を暗い背景で強調した。ダッシュは類似性を最大化するために導入したギャップを表す。適切なSAA3アイソフォームの後に前駆タンパク質のN末端からの残基の数を右に示し、シグナルペプチドの推定される切断部位を下向きの三角形(▽)で示す。
【技術分野】
【0001】
本発明は、免疫学および哺乳動物免疫系の分野に関する。特に本発明は、これまでその遺伝子が偽遺伝子としてキャラクタライズされていたので存在しないと考えられてきた血清アミロイドA(SAA3)の新規cDNAおよびアイソフォームを提供する。
【技術背景】
【0002】
いくつかの科学または特許刊行物を、本発明が属する技術の水準を説明するために、本明細書において参照する。これらの刊行物を全て、参照によりその全内容を本明細書に組み入れる。
【0003】
哺乳動物は、さらなる組織損傷を防ぐため、損傷を受けた組織の修復を開始するため、および病原性生物を隔離し破壊するために、複雑な一連の生物学的反応を実行することで、組織損傷、外傷、または感染症に応答する。この過程は炎症反応と呼ばれており、その炎症反応の初期および中間段階は急性期応答と呼ばれている。
【0004】
急性期応答は、広範な種類の仲介物質(サイトカイン、インターロイキンおよび腫瘍壊死因子など)を伴う。急性期応答はまた、肝臓の生合成プロファイルの過激な変更を伴う。通常の状況下では、肝臓は定常状態濃度で様々な血漿タンパク質を合成する。このうちのいくつかのタンパク質、「急性期」タンパク質は、炎症応答において、通常の状態でみられるレベルより何倍も大きいレベルへと誘導される。急性期タンパク質は、Steel および Whitehead (Immunology Today 15: 81-87, 1994)によって総評されている。
【0005】
大量に誘導される急性期タンパク質の1つは、血清アミロイドA(SAA)である。SAAは、炎症応答の急性期に高密度リポタンパク質3(HDL3)と急速に会合および蓄積する小さなアポリポタンパク質である。ほとんどのSAAアイソフォームは炎症に応答して誘導されるが、一方で特定のSAA(例えばヒトSAA4)は構成的に発現しているかまたは炎症応答で最小限しか誘導されないようである。
【0006】
血清アミロイドAタンパク質(SAA)は、今日までに調査された全ての脊椎動物において産生されるアポリポタンパク質のスーパーファミリーを含む(C.M. Uhlar, A.S. Whitehead, Serum amyloid A, the major vertebrate acute-phase reactant, Eur. J. Biochem. 265 (1999) 501-523)。生物種によっては、SAAをコードする3または4つの遺伝子座が特定されており、これらの遺伝子は肝臓からおよび/または肝臓外から区別されて発現されている(J.D. Sipe, Serum amyloid A: from fibril to function. Current status, Amyloid: Int. J. Exp. Clin. Invest. 7 (2000) 10-12)。急性期血清アミロイドAタンパク質(A-SAA)は、主に肝臓によって合成され、血漿の高密度リポタンパク質3(HDL3)画分と大部分が会合している(G.A. Coetzee, A.F. Strachan, D.R. Van Der Westhuyzen, H.C. Hoffe, M.S. Jeenah, F.C. De Beer, Serum amyloid A-containing human high density lipoprotein, J. Biol. Chem. 261 (1986) 9644-9651; N. Eriksen, E.P. Benditt, Isolation and characterization of the amyloid-related apoprotein (SAA) from human high density lipoprotein, Proc. Natl. Acad. Sci. USA 77 (1980) 6860-6864)。肝臓によって誘導されたA-SAAレベルは、炎症性サイトカインであるIL-1、IL-6およびTNF-αに応答して急性期に1000倍増加し得る(L.E. Jensen, A.S. Whitehead, Regulation of serum amyloid A protein expression during the acute-phase response, Biochem. J. 334 (1998) 489-503)。血液の循環におけるA-SAAの劇的な増加は、主に転写の増加によって達成される(C.M. Uhlar, A.S. Whitehead, Serum amyloid A, the major vertebrate acute-phase reactant, Eur. J. Biochem. 265 (1999) 501-523)。
【0007】
肝臓は、SAA産生の主要な部位と考えられてきた。しかしながら、限定的な証拠に基づくことではあるが、肝臓外でのSAA産生が見いだされた。例えば、SAA mRNAの発現は、ヒトのアテローム性動脈硬化の病巣、ならびにヒト培養平滑筋細胞および単核白血球/マクロファージ細胞系で報告されており(Meek ら, 1994; Urieli-Shoval ら, 1994; Yamada ら, 1996)、またウサギ骨膜繊維芽細胞からSAAの独特のアイソフォーム(SAA3)が産生されている(Mitchell ら, J. Clin. Invest. 87: 1177-1185, 1991)。さらに最近、SAA mRNAが多くの組織学的に正常な上皮組織(胃、腸、扁桃腺、乳房、前立腺、甲状腺、肺、膵臓、腎臓、皮膚および脳神経など)で広く発現していることが発見された(Urieli-Shoval ら, J. Histochem. Cytochem. 46: 1377-1384, 1998)。
【0008】
多数の研究者による実験的証拠は、SAAタンパク質の多様な機能を示唆しているが、そうした機能には以下のものが含まれる。免疫応答の抑制(M.A. Aldo-Benson, M.D. Benson, SAA suppression of immune response in vitro: evidence for an effect on T cell-macrophage interaction, J. Immunol. 128 (1982) 2390-2392)、血小板凝集の阻害(S. Zimlichman, A. Danaon, I. Nathan, G. Mozes, R. Shainkin-Kestenbaum, Serum amyloid A, an acute phase protein, inhibits platelet activation, J. Lab. Clin. Med. 116 (1990) 180-186)、コレステロール/脂質代謝への関与(R. Kisilevsky, L. Subrahmanyan, Serum amyloid A changes high density lipoprotein's cellular affinity. A clue to serum amyloid A's principal function, Lab. Invest. 66 (1992) 778-785; R.L. Meek, N. Eriksen, E.P. Benditt, Murine serum amyloid A3 is a high density apolipoprotein and is secreted by macrophages, Proc. Natl. Acad. Sci. USA 89 (1992) 7949-7952)、内毒素の解毒への関与(C. Baumberger, R.J. Ulevitch, J.M. Dayer, Modulation of endotoxic activity of lipopolysaccharide by high-density lipoprotein, Pathobiology 59 (1991) 378-383)、コラゲナーゼ活性の誘導(C.E. Brinckerhoff, T.I. Mitchell, M.J. Karmilowicz, B. Kluve-Beckerman, M.D. Benson, Autocrine induction of collagenase by serum amyloid A-like and β2-microglobulin-like proteins, Science 243 (1989) 655-657)、好中球のオキシダティブバーストの阻害(R.P. Linke, V. Bock, G. Valet, G. Tothe, Inhibition of the oxidative burst response of N-formyl peptide-stimulated neutrophils by serum amyloid A protein, Biochem. Biophys. Res. Commun. 176 (1991) 1100-1105; M.E. Gatt, S. Urieli-Shoval, L. Preciado-Patt, M. Fridkin, S. Calco, Y. Azar, Y. Matzner, Effect of serum amyloid A on selected in vitro functions of isolated human neutrophils, J. Lab. Clin. Med. 132 (1998) 414-420)、単核白血球、多形核白血球およびT細胞の移動の誘導(R. Badaloto, J.M. Wang, W.J. Murphy, A.R. Lloyd, D.F. Michiel, L.L. Bausserman, D.J. Kelvin, J.J. Oppenheim, Serum amyloid A is a chemoattractant: induction of migration, adhesion and tissue infiltration of monocytes and polymorphonuclear leukocytes, J. Exp. Med. 180 (1994) 203-209; L. Xu, R. Badolato, W.J. Murphy, D.L. Longo, M. Anver, S. Hale, J.J. Oppenheim, J.M. Wang, A novel biologic function of serum amyloid A. Induction of T lymphocyte migration and adhesion, J. Immunol. 155 (1995) 1184-1190)、ならびに細胞外マトリックス構成要素への細胞付着の阻害(L. Preciado-Patt, D. Levartowsky, M. Pras, R. Hershkoviz, O. Likder, M. Fridkin, Inhibition of cell adhesion to glycoproteins of the extracellular matrix by peptides corresponding to serum amyloid A. Toward understanding the physiological role of an enigmatic protein, Eur. J. Biochem. 223 (1994) 35-42)。しかしながら、正常状態および疾病状態におけるSAAの主要な生理学的役割はよく分かっていない。
【0009】
ヒトSAA mRNAおよびタンパク質の肝臓外発現は、マクロファージ細胞、脂肪細胞、平滑筋細胞、および内皮細胞で実証されており、これらの産生部位で機能している蓋然性が高いことを示唆する。Urieli-Shovalらは、ヒトSAAの肝臓外発現が、主として各種組織の上皮の構成要素に局所化されていることを明らかにした(S. Urieli-Shoval, P. Cohen, S. Eisenberg, Y. Matzner, Widespread expression of serum amyloid A in histologically normal human tissues: predominant localization to the epithelium, J. Histochem. Cytochem. 46 (1998) 1377-1384)。
【0010】
ヒトでは4つのSAA遺伝子が染色体11p15.1上にクラスター化されている(G.C. Sellar, S.A. Jordan, W.A. Bickmore, J.A. Fantes, V. van Heyningen, A.S. Whitehead, The human serum amyloid A protein (SAA) superfamily gene cluster: mapping to chromosome 11p15.1 by physical and genetic linkage analysis, Genomics 19 (1994) 221-227)。高誘導性SAA1およびSAA2遺伝子は、90%同一な104残基のA-SAAタンパク質をコードする。SAA1およびSAA2遺伝子は、それらのプロモーター領域、エクソンおよびイントロンの全体で約95%のヌクレオチド配列同一性を有する(J.C. Betts, M.R. Edbrooke, R.V. Thakker, P. Woo, The human acute-phase serum amyloid A gene family: structure, evolution and expression in hepatoma cells, Scand. J. Immunol. 34 (1991) 471-482; P. Woo, J. Sipe, C.A. Dinarello, H. R. Colten, Structure of a human serum amyloid A gene and modulation of its expression in transfected L cells, J. Biol. Chem. 262 (1987) 15790-15795)。配列が類似しているにもかかわらず、最近の研究によって、サイトカイン依存性誘導発現という状況で、SAA2と比較してSAA1の転写発現が、グルココルチコイドから異なる増強を受ける、ということが実証された(C.F. Thorn, A.S. Whitehead, Differential glucocorticoid enhancement of the cytokine-driven transcriptional activat
ion of human acute phase serum amyloid A genes, SAA1 and SAA2, J. Immunol. 169 (2002) 399-406)。最初にBettsらによって記述されたヒトSAA4(J.C. Betts, M.R. Edbrooke, R.V. Thakker, P. Woo, The human acute-phase serum amyloid A gene family: structure, evolution and expression in hepatoma cells, Scand. J. Immunol. 34 (1991) 471-482)は、構成的なSAA4(C-SAA4)をコードする(D.M. Steel, G.C. Sellar, C.M. Uhlar, S. Simon, F.C. DeBeer, A.S. Whitehead, A constitutively expressed serum amyloid A protein gene (SAA4) is closely linked to, and share structural similarities with, an acute-phase serum amyloid A protein gene (SAA2), Genomics 16 (1993) 447-454; A.S. Whitehead, M.C. de Beer, D.M. Steel, M. Rits, J.M. Lelias, W.S. Lane, F.C. de Beer, Identification of novel members of the serum amyloid A protein superfamily as constitutive apolipoproteins of high density lipoproteins, J. Biol. Chem. 267 (1992) 3862-3867)。A-SAA1およびA-SAA2とは対照的に、C-SAA4は急性期応答において著しく誘導されない。C-SAA4はA-SAA1およびA-SAA2よりも8残基長く、どちらのA-SSAタンパク質とも55%の同一性しか示さない。C-SAA4は、正常および急性期HDL3の両方において低レベルでしか存在せず、このタンパク質の蓋然性の高いハウスキーピング機能を示唆している。
【0011】
ヒトSAA3遺伝子は最初SackおよびTalbotによって同定され、A-SAAと71%の同一性を示す104残基のタンパク質をコードすると予測された(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)。しかしながら、後のゲノムに基づく研究により、予測されたエキソン3に存在する1ヌクレオチドの挿入によって結果的に末端切断型ヒトSAA3タンパク質が得られることが明らかになった(B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661)。今日まで、SAA3転写産物もタンパク質も調査されたヒト組織または細胞系で検出されておらず、SAA3のプロモーターと推測された領域が機能することも示されていない(S. Urieli-Shoval, P. Cohen, S. Eisenberg, Y. Matzner, Widespread expression of serum amyloid A in histologically normal human tissues: predominant localization to the epithelium, J. Histochem. Cytochem. 46 (1998) 1377-1384); B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661; S. Urieli-Shoval, R.L. Meek, R.H. Hanson, N. Eriksen, E.P. Benditt, huma
n serum amyloid A genes are expressed in monocyte/macrophage cell lines, Am. J. Pathol. 145 (1994) 650-660; G.C. Sellar, A.S. Whitehead, Localization of four human serum amyloid A (SAA) protein superfamily genes to Chromosome 11p: Characterization of a fifth SAA-related gene sequence, Genomics 16 (1993) 774-776)。これらの研究は、SAA3遺伝子が偽遺伝子であるか、またはSAA3発現に適当な誘導条件および/または細胞型が用いられなかったことを示唆している。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明者らは初めて、ヒト細胞において、SAA3遺伝子の誘導された発現を確認および実証し、SAA3遺伝子の転写産物(cDNA)を同定した。本発明の目的は、ヒトSAA3転写産物およびそれがコードするタンパク質を提供することである。
【0013】
また本発明の別の目的は、ヒトSAA3タンパク質をコードするヌクレオチド配列を提供することである。
【0014】
また本発明のもう1つ別の目的は、ヒトSAA3タンパク質を構成するアミノ酸配列を提供することである。
【0015】
また本発明のさらに別の目的は、組換えSAA3の産生、SAA3発現の役割をさらに明らかにするためのアッセイにおける使用、急性期免疫応答のさらなる理解などのために、本発明の配列を使用するための組換えDNAプロトコルを提供することである。
【0016】
さらに本発明の別の目的は、例えばリポ多糖類(LPS)またはプロラクチン(PRL)のようなSAA3発現の誘導因子を同定するためのアッセイを提供すること、およびこれらの化合物の存在下で、機能的に連結された配列をヒト細胞において発現誘導させることができるプロモーター領域を提供することである。
【課題を解決するための手段】
【0017】
本発明において、ヒト乳房上皮細胞から分離および精製したヒト血清アミロイドA3(SAA3)タンパク質を提供する。
【0018】
本発明の別の態様において、哺乳動物ヒトSAA3をコードする単離された核酸分子を提供する。核酸分子は遺伝子、cDNA、またはRNAであってよく、一本鎖または二本鎖であってよい。好ましい実施形態において、核酸分子は配列番号2またはその保存的に改変された変異体をコードする配列を含む。最も好ましい実施形態において、核酸分子は配列番号1またはその保存的に改変された変異体を含み、たとえば、発現された他のヒトSAA3配列、同じように同定された配列、または本明細書に記載するようにLPSまたはプロラクチンにより誘導されたSAA配列をコードする他のあらゆる核酸配列が含まれる。
【0019】
本発明において、ヒトSAA3遺伝子の配列(イントロン領域およびフランキング領域が含まれる)が決定された。このことから本発明にはまた、1以上の天然非コード領域もしくはイントロン領域またはその保存的に改変された変異体が含まれる、発現されたヒトSAA3をコードするヌクレオチド配列が含まれる。
【0020】
本発明の別の態様において、ヒトSAA3の1以上のエピトープに免疫学的に特異的な抗体を提供する。好ましくは、抗体は、ヒトSAA3を血清SAAから区別するヒトSAA3の少なくとも1つのエピトープに免疫学的に特異的である。
【0021】
また他の実施形態において、ヒトSAA3と天然で関連しているプロモーター領域は、遺伝子移植のプロトコルに、または、SAA産生を刺激するために使用して、動物の乳房もしくは他の乳房組織に関連する疾患の治療を補助することができる。例として、ヒトSAA3プロモーターはプロラクチンにより誘導される。すなわち、プロラクチンその他のヒトSAA3誘導因子を投与して、前記動物の乳房組織によるSAA3の産生を刺激し、増加させることができるであろう。プロモーター領域はまた、乳房細胞における異種性ヌクレオチド配列の発現のための乳房組織特異性を提供する。
【0022】
ウシと同様にヒトにおいて、乳房関連SAA3はムチン3(MUC3)産生を刺激する能力を有する。このことからSAA3は、腸の感染症または不充分なレベルのムチンと関連する他の疾患状態、例えば旅行者下痢、乳児下痢、壊死性腸炎、および尿路感染症などを治療および予防するために使用することができ、集団の動物における下痢を防止するための手段として獣医薬を提供する。従って本発明には、上記の疾患および類似の病状を有する他の疾患を治療するための、医薬として有効な量のSAA3ペプチドおよび担体を含む医薬組成物が含まれる。最後に、ムチンを産生する粘膜表面(例えば鼻咽頭、嚢、など)の他の上皮細胞(epithelial cell lining)もまた、ムチン産生を刺激するために本発明の医薬組成物で処置してそれらと関連する感染症を予防または治療することができる。
【0023】
また別の実施形態において、本発明は乳房および他の細胞におけるSAA発現の影響を調べるための、ならびに炎症または組織損傷におけるSAAの役割をさらに明らかにするためのアッセイからなる。本発明の配列はまた、他のSAAアイソフォームを同定するために、あるいは乳腺炎または乳房組織の炎症もしくは感染と関連する他の乳房組織疾患または損傷を診断するために使用することができる。
【0024】
本発明の他の特徴および効果は、以下の図面、詳細な説明および実施例を参照することによって、さらに理解されるであろう。
【発明を実施するための最良の形態】
【0025】
1.定義
本発明の組成物および方法に関する様々な用語を、本明細書全体にわたり、そして特許請求の範囲において用いる。
【0026】
単位、接頭語、および記号は、SI単位系で許された形式で示すことができる。特に断らない限り、核酸は左から右に5'から3'の向きで記述し、アミノ酸配列は左から右にアミノからカルボキシの向きで記述する。数値範囲は、範囲を定める数自体を含み、定義される範囲内の各々の整数が含まれる。アミノ酸は、本明細書において、一般に知られている3文字記号またはIUPAC-IUB Biochemical nomenclature Commisionにより推奨される1文字記号のいずれかにより表され得る。同様に、ヌクレオチドは一般に認められた1文字コードで表すことができる。特に規定しない限り、本明細書で用いるソフトウェア、電気および電子の用語は、The New IEEE Standard Dictionary of Electrical and Electronics Terms (第5版,1993)に定義されている通りである。以下に定義する用語は、明細書全体でより十分に定義される。
【0027】
「増幅する」とは、少なくとも1つの核酸配列を鋳型として用いて、ある核酸配列の複数のコピー、または核酸配列に相補的な複数のコピーを作成することを意味する。増幅システムには、ポリメラーゼ連鎖反応(PCR)システム、リガーゼ連鎖反応(LCR)システム、核酸配列に基づく増幅(NASBA, Canteen, Mississauga, Ontario)、Q-βレプリカーゼシステム、転写に基づく増幅システム(TAS)および鎖置換増幅(SDA)が含まれる。例えばDiagnostic Molecular Microbiology: Principles and Applications, D.H. Persingら,編, American Society for Microbiology, Washington, D.C. (1993)を参照のこと。増幅の産物は増幅産物(amplicon)と呼ぶ。
【0028】
「抗体」という用語には、抗体の抗原結合性形態(例えばFab、F(ab)2)が含まれる。「抗体」という用語はしばしば、検体(抗原)に特異的に結合し認識する1以上の免疫グロブリン遺伝子によって実質的にコードされるポリペプチドまたはそのフラグメントを言う。また一方、様々な抗体フラグメントは完全な抗体の消化の観点から定義することができるものの、当業者ならばそのようなフラグメントは化学的に、または組換えDNA手法を利用することにより、新規に合成することができると理解するであろう。従って、本明細書で用いる抗体という用語にはまた、一本鎖Fv、キメラ抗体(すなわち異なる種からの定常および可変領域を含むもの)、ヒト化抗体(すなわち非ヒト起源の相補性決定領域(CDR)を含むもの)およびヘテロコンジュゲート抗体(例えば二重特異性抗体)などの抗体フラグメントが含まれる。
【0029】
本明細書において用いる「アンチセンス配向」とは、アンチセンス鎖が転写される方向性で、プロモーターに機能的に連結している二本鎖ポリヌクレオチド配列をいう。アンチセンス鎖は、内因性転写産物の翻訳がしばしば阻害されるくらい、内因性転写産物に対して十分相補的である。
【0030】
本明細書において用いる「SAA3」および「M-SAA3」という用語は互換的に使用され、限定するものではないが、本明細書において開示する配列、起源を問わないそれらの保存的に改変された変異体、ならびに本明細書に開示されたアッセイで実証されるようにSAA3の生物学的性質を保持する他のあらゆる変異体が含まれる。
【0031】
本明細書において用いる「染色体領域」には、その染色体領域が含むDNAの線状部分に関して測定されうるある長さの染色体が含まれる。染色体領域は2つの固有なDNA配列、すなわちマーカーにより定義することができる。
【0032】
「保存的に改変された変異体」という用語は、アミノ酸および核酸配列の両方に適用されるものであり、特定の配列への言及がなされるあらゆる場合に含まれることを意図したものである。特定の核酸配列に関して、保存的に改変された変異体はアミノ酸配列の同一または保存的に改変された変異体をコードする核酸を意味する。遺伝暗号の縮重のために、多数の機能的に同一な核酸が、任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全てアミノ酸のアラニンをコードする。このことから、アラニンがコドンによって指定される各位置で、コードされるポリペプチドを変更することなくコドンを上述の対応するコドンのいずれにも変更することができる。このような核酸変異は「サイレント変異」であり、保存的に改変された変異の1つの種を表す。ポリペプチドをコードする本発明の各核酸配列はまた、遺伝暗号を参照して、核酸の可能なあらゆるサイレント変異を表す。当業者ならば、核酸における各コドンを、機能的に同一の分子を生じるために改変することが出来ると理解できるであろう(ただし、通常メチオニンの唯一のコドンであるAUG、および通常トリプトファンをコードする唯一のコドンであるUGGは除く)。それゆえ、本発明のポリペプチドをコードする核酸の各々のサイレント変異は、それぞれの記載されるポリペプチド配列に暗黙的に含まれており、本発明の範囲内にある。
【0033】
アミノ酸配列については、当業者ならば、コードされる配列中の単一のアミノ酸またはわずかな割合のアミノ酸を変更する、追加するまたは欠失させる核酸、ペプチド、ポリペプチドまたはタンパク質配列への個別の置換、欠失または追加は、その変更が結果的に1つのアミノ酸を化学的に類似するアミノ酸で置換するものである場合に、「保存的に改変された変異体」であると認識するであろう。このことから、1〜15からなる整数の群から選択される数のアミノ酸残基を、このように変更することができる。従って、例えば、1、2、3、4、5、7、または10の変更を行うことができる。保存的に改変された変異体は典型的には、それらが誘導された未改変のポリペプチド配列と類似の生物学的活性を示す。例えば、基質特異性、酵素活性、またはリガンド/受容体結合は、一般にその天然の基質について天然のタンパク質の少なくとも30%、40%、50%、60%、70%、80%または90%である。機能的に類似のアミノ酸を規定する保存的置換表は当技術分野でよく知られている。
【0034】
以下の6つの群はそれぞれ、互いに保存的な置換であるアミノ酸を含有する。
【0035】
1) アラニン(A),セリン(S),トレオニン(T);
2) アスパラギン酸(D),グルタミン酸(E);
3) アスパラギン(N),グルタミン(Q);
4) アルギニン(R), リジン(K);
5) イソロイシン(I),ロイシン(L),メチオニン(M),バリン(V);
6) フェニルアラニン(F),チロシン(Y),トリプトファン(W)。
【0036】
Creighton (1984) Proteins W.H. Freeman and Companyを参照のこと。
【0037】
所定の核酸に関して、「コードする」または「コードされる」とは、所定のタンパク質へと翻訳される情報を含むことを意味する。タンパク質をコードする核酸は、核酸の翻訳される領域内に非翻訳領域(例えばイントロン)を含んでもよく、またはそのような介在する非翻訳配列を欠いてもよい(例えばcDNAにおけるように)。タンパク質をコードする情報は、コドンの使用により特定される。典型的には、アミノ酸配列は「普遍的」遺伝暗号を用いて核酸によりコードされる。しかしながら、普遍的遺伝暗号の変形、例えばいくつかの植物、動物および真菌ミトコンドリア、細菌のMycoplasma capricolumn、または繊毛虫のマクロヌクレアスに存在するようなものをそれらにおいて核酸を発現させる場合に用いてもよい。
【0038】
核酸を合成的に作成または変更する場合、核酸を発現させようと意図する宿主の知られている優先コドンを利用することができる。
【0039】
本明細書において、特定のポリヌクレオチドまたはそれがコードするタンパク質に関して用いる「全長配列」は、その特定のタンパク質の天然(非合成)の内因性で生物学的に活性な形態の全アミノ酸配列を有することを意味する。ある配列が全長なのかを判別する方法は当技術分野でよく知られており、例えば、ノーザンまたはウェスタンブロット法、プライマー伸長法、S1プロテクション法、およびリボヌクレアーゼプロテクション法などの模範的な技術が含まれる。既知の全長相同(オーソロガスおよび/またはパラロガス)配列との比較もまた、本発明の全長配列を同定するために使用することができる。さらに、mRNAの5'および3'非翻訳領域に典型的に存在するコンセンサス配列は、ポリヌクレオチドが全長であるか識別するのに役立つ。例えば、コンセンサス配列ANNNNAUGG(ここで下線のコドンはN末端のメチオニンを表す)は、ポリヌクレオチドが完全な5'末端を有するか判別するのに役立つ。3'末端におけるコンセンサス配列、例えばポリアデニル化配列などは、ポリヌクレオチドが完全な3'末端を有するか判別するのに役立つ。
【0040】
タンパク質またはペプチドに関して、「分離(単離)したタンパク質(もしくはペプチド)」または「分離し精製したタンパク質(もしくはペプチド)」という用語を、本明細書において場合により用いる。この用語は、そのタンパク質が「実質的に純粋」な形態で存在するように、天然でそのタンパク質に結びついている他のタンパク質から十分に分離されたタンパク質を意味し得る。また、この用語は、単離された核酸分子の発現により産生されるタンパク質を意味し得る。
【0041】
核酸分子に関連して、「単離された核酸」という用語を場合により用いる。この用語はDNAに適用する場合、そのDNAが由来する生物の天然に存在するゲノムで(5'および3'方向に)直接隣接している配列から切り離されたDNA分子を意味する。例えば、「単離された核酸」は、プラスミドもしくはウィルスベクターなどのベクターに挿入した、または原核生物もしくは真核生物のゲノムDNAに組み込まれたDNA分子を含み得る。「単離された核酸分子」はまた、cDNA分子からなることもできる。
【0042】
RNA分子に関連して、「単離された核酸」という用語は、上記に定義した単離されたDNA分子がコードするRNA分子を主に意味する。また、この用語はそのRNA分子が「実質的に純粋」な形態(「実質的に純粋」という用語は以下で定義する)で存在するように、天然の状態(すなわち細胞または組織内)でそのRNA分子に結びついているRNA分子から十分に分離したRNA分子を意味し得る。
【0043】
核酸に関して本明細書において用いる「異種の」とは、外来種が起源であるか、または、同種が起源である場合には人間の意図的な介入により組成および/または遺伝子座において天然の形態から実質的に改変した核酸である。例えば異種構造遺伝子に機能的に連結したプロモーターは、その構造遺伝子の起源とは異なる種に由来し、または同種由来の場合にはその遺伝子およびプロモーターの片方または両方がそれらの元の形態から実質的に改変されたものである。異種タンパク質は外来種が起源であってもよく、または同種由来の場合には人間の意図的な介入によりその元の形態から実質的に改変されたものである。
【0044】
「宿主細胞」とは、ベクターを含有し、そのベクターの複製および/または発現を支持する細胞を意味する。宿主細胞は、原核細胞、例えば大腸菌(E. coli)などであってもよく、または真核細胞、例えば酵母、昆虫、両生類または哺乳動物細胞などであってもよい。「ハイブリダイゼーション複合体」という用語には、互いに選択的にハイブリダイズする、2つの一本鎖核酸配列が形成する二重核酸構造が含まれる。
【0045】
細胞に核酸を挿入するという文脈での「導入する」という用語は、「トランスフェクション」または「トランスフォーメーション(形質転換)」または「トランスダクション(形質導入)」を意味し、また、核酸を真核または原核細胞に組み入れることが含まれ、ここで核酸は細胞のゲノムに組み込まれてもよく(例えば染色体、プラスミド、色素体、もしくはミトコンドリアDNA)、自立したレプリコンに変換させてもよく、または一時的に発現(例えばトランスフェクトされたmRNA)させてもよい。
【0046】
特に断らない限り、用語「SAA3をコードする核酸」とは、本発明の核酸であり、SAA3をコードする本発明のポリヌクレオチドを含む核酸を意味する。「SAA3遺伝子」とは、本発明の遺伝子であり、全長SAA3ポリヌクレオチドの異種ゲノム形態を意味する。
【0047】
本明細書において特定のマーカーに関して用いる「規定され、それを含む染色体領域内に局在化され」とは、その記述されたマーカーによって範囲を定められまたそれを含む連続している一定の長さの染色体をいう。
【0048】
本明細書において用いる「マーカー」とは、ある染色体上の独特の位置を同定するために役立つその染色体上の座をいう。「多型マーカー」とは、異なる形態のマーカーが、相同なペアとして存在するとき、そのペアの両方の染色体の伝達が可能になるようにする複数の形態(対立遺伝子)で出現するマーカーをいう。遺伝子型は、1つまたは複数のマーカーを使用することにより定義することができる。
【0049】
本明細書において用いる「核酸」には、一本鎖または二本鎖のいずれの形態のデオキシリボヌクレオチドまたはリボヌクレオチドポリマーも含まれ、特に断らない限り、天然のヌクレオチドと似た様式で一本鎖核酸にハイブリダイズするという点で天然ヌクレオチドの本質的な性質を有する既知のアナログも包含する(例えばペプチド核酸)。
【0050】
「核酸ライブラリー」とは、特定の生物のゲノムの転写された全フラクションを含みかつこれらを実質的に表現する単離されたDNAまたはRNA分子のコレクションを意味する。模範的な核酸ライブラリー、例えばゲノミックまたはcDNAライブラリーなどの構築は、例えば、BergerおよびKimmel, Guide to Molecular Cloning Techniques, Methods in Enzymology, Vol. 152, Academic Press, Inc., San Diego, CA (Berger); Sambrook ら, Molecular Cloning - A Laboratory Manual, 2nd ed., Vol. 1-3 (1989);およびCurrent Protocols in Molecular Biology, F.M. Ausubel ら, Eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc. (1994)などの標準的な分子生物学文献に教示されている。
【0051】
本明細書において用いる「ポリヌクレオチド」には、天然に生ずるヌクレオチドと実質的に同じヌクレオチド配列にストリンジェントなハイブリダイゼーション条件下でハイブリダイズする、および/または天然に生ずるヌクレオチドと同じアミノ酸への翻訳を可能にする、という意味で天然リボヌクレオチドに必須の性質を有するデオキシリボヌクレオチド、リボポリヌクレオチド、またはそのアナログが含まれる。ポリヌクレオチドは全長であってもよく、または天然もしくは異種の構造または調節遺伝子のサブシークエンスであってもよい。特に断らない限り、この用語には、特定の配列のみならずその相補配列も含まれる。このことから、安定性のためまたは他の目的のために修飾したバックボーンを有するDNAまたはRNAもまた、本明細書において意図する「ポリヌクレオチド」である。その上、例えば2つだけ例を挙げると、普通ではない塩基(イノシン等)または修飾された塩基(例えばトリチル化塩基など)を含むDNAまたはRNAは、本明細書において用いる通り、ポリヌクレオチドである。当業者に知られている様々な有用な目的を果たす、DNAおよびRNAへの非常に多様な改変がなされることが理解される。本明細書において用いる通り、ポリヌクレオチドという用語は、そのような化学的、酵素的または代謝的に改変された形態のポリヌクレオチド、ならびに、ウィルスおよび細胞(他のものに加え、単純および複雑細胞が含まれる)に特徴的なDNAおよびRNAの化学的形態を含む。
【0052】
「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は、本明細書において相互に交換できるように使用され、アミノ酸残基のポリマーを意味する。これらの用語は、天然に生ずるアミノ酸ポリマーのみならず、1以上のアミノ酸残基が対応する天然に生ずるアミノ酸の人工的な化学的アナログであるアミノ酸ポリマーにも適用される。このような天然に生ずるアミノ酸のアナログに必須の性質は、タンパク質に組み込まれた場合、天然に生ずるアミノ酸のみからなるタンパク質によって誘導される抗体に、前記アナログを組み入れたタンパク質が特異的に反応することである。「ポリペプチド」、「ペプチド」、および「タンパク質」という用語はまた、限定するものではないが、リン酸化、グリコシル化、脂質結合、硫酸化、グルタミン酸残基のγ-カルボキシル化、ヒドロキシル化、およびADP-リボシル化が含まれる修飾を含む。広く知られており上記に記述した通り、ポリペプチドは完全に線形ではないと理解される。例えば、ポリペプチドは、ユビキチン化の結果分枝したものであってもよく、また、一般に翻訳後の事象の結果、環状であってもよく(それらは分枝していてもしていなくてもよい)、前記事象には天然のプロセス事象、および天然には起こらない人為的操作によってもたらされる事象が含まれる。環状、分枝状、および分枝した環状ポリペプチドは、非翻訳の天然のプロセスによっても、また完全に合成的な方法によっても、合成することができる。さらに本発明は、本発明のタンパク質のメチオニン含有およびメチオニン非含有アミノ末端変異体の両方の使用を含む。タンパク質に関して、「N末端領域」という用語は、タンパク質のアミノ末端に隣接する約50アミノ酸残基を含む。
【0053】
本明細書において用いる「TFLKモチーフ」は、SAA3のTFLK活性部位の構造的完全性および生物学的活性を維持するアミノ酸または他のものによる任意の構築物を含む。
【0054】
本明細書において用いる「組換え」には、異種核酸を導入することにより改変した細胞もしくはベクター、またはそのように改変した細胞から誘導される細胞が含まれる。したがって例えば、組換え細胞は、細胞の天然(非組換え)形態で同一の形態で見られない遺伝子を発現するか、または他の場合には異常発現するか、または低発現するかもしくは全く発現しない天然の遺伝子を、人為的な介入の結果として発現する。本明細書において用いる「組換え」という用語は、自然に起きる事象(例えば自発的突然変異、天然の形質転換/形質導入/転位)、例えば人為的な介入無しで起こる事象など、による細胞またはベクターの変更は包含しない。
【0055】
本明細書において用いる通り、「組換え発現カセット」は、宿主細胞内で特定の核酸の転写を可能にする一連の特定の核酸エレメントを有し、組換え的にまたは合成的に作り出された核酸構築物である。組換え発現カセットは、プラスミド、染色体、ミトコンドリアDNA、色素体DNA、ウィルスまたは核酸フラグメントに組み入れることができる。典型的には、発現ベクターの組換え発現カセット部分には、様々な配列の他に、転写されるべき核酸、およびプロモーターが含まれる。
【0056】
「残基」または「アミノ酸残基」、または「アミノ酸」という用語は、本明細書において互いに交換できるように用いられ、タンパク質、ポリペプチドまたはペプチド(これらをまとめて「タンパク質」と呼ぶ)に組み入れられたアミノ酸を意味する。アミノ酸は天然に生ずるアミノ酸であってもよく、他に限定が無ければ、天然に生ずるアミノ酸と似た様式で機能することができる天然アミノ酸の非天然アナログを包含してもよい。
【0057】
「ストリンジェント条件」または「ストリンジェントなハイブリダイゼーション条件」という用語には、プローブが、他の配列に対してハイブリダイズするのに比べて検出できるほど強い度合い(例えば少なくともバックグランドよりも2倍強く)で、標的配列にハイブリダイズする条件への言及が含まれる。ストリンジェント条件は配列依存的であり、異なる状況下では異なり得る。ハイブリダイゼーションおよび/または洗浄条件のストリンジェンシーを制御することにより、プローブと100%相補的な標的配列を同定することができる(相同プローブ)。これとは別に、より低い程度の類似性が検出できるよう配列にいくらかのミスマッチが許容されるようストリンジェンシー条件を調節することができる(非相同プローブ)。一般に、プローブは長さにして約1000ヌクレオチド未満であり、場合により長さにして500ヌクレオチド未満である。
【0058】
典型的には、ストリンジェント条件は、塩濃度が約1.5M Naイオン未満、典型的には約0.01〜1.0M Naイオン濃度(または他の塩)で、pHが7.0〜8.3であり、短いプローブ(例えば10〜50ヌクレオチド)については温度が少なくても約30℃であり、長いプローブ(例えば50ヌクレオチドより長いもの)については温度が少なくとも約60℃であるものである。ストリンジェント条件はまた、ホルムアミドなどの不安定化剤を添加することにより達成することができる。模範的な低ストリンジェンシー条件には、37℃で、30〜35%ホルムアミド、1M NaCl、1%SDS(ドデシル硫酸ナトリウム)を有するバッファー溶液でハイブリダイゼーションし、50〜55℃で、1×〜2×のSSC(20× SSC=3.0M NaCl/0.3Mクエン酸三ナトリウム)で洗浄することが含まれる。模範的な中程度のストリンジェンシー条件には、37℃で、40〜45%ホルムアミド、1M NaCl、1% SDSでのハイブリダイゼーション、および55℃〜50℃で、0.5×〜1× SSCでの洗浄が含まれる。模範的な高ストリンジェンシー条件には、37℃で、50%ホルムアミド、1M NaCl、1% SDSでのハイブリダイゼーション、および60℃〜65℃で、0.1× SSCでの洗浄が含まれる。
【0059】
特異性は典型的には、ハイブリダイゼーション後の洗浄の関数であり、重要な要素は最終洗浄溶液のイオン強度および温度である。DNA-DNAハイブリドについては、TmはMeinkoth および Wahl, Anal. Biochem., 138:267-284 (1984)の式:
Tm=81.5℃ + 16.6 (log M) + 0.41 (%GC) -0.61 (% form) - 500/L
から概算することができるが、ここでMは一価カチオンのモル濃度であり、%GCはDNA中のグアノシンおよびシトシンヌクレオチドの百分率であり、% formはハイブリダイゼーション溶液中のホルムアミドの百分率であり、Lは塩基対で表したハイブリッドの長さである。Tmは相補的な標的配列の50%が完全に一致するプローブにハイブリダイズする温度(所定のイオン強度およびpH下での)である。Tmは1%のミスマッチにつき約1℃減少する。したがって、Tm、ハイブリダイゼーションおよび/または洗浄条件は、所望の相同性の配列にハイブリダイズするよう調節することが出来る。例えば≧90%の相同性を有する配列を所望ならば、Tmは10℃低下させることができる。一般にストリンジェント条件は、所定のイオン強度およびpHで、その特定の配列およびその相補配列の熱融点(Tm)よりも約5℃低く選択される。しかしながら、非常にストリンジェントな条件は、熱融点(Tm)よりも1、2、3または4℃低いハイブリダイゼーションおよび/または洗浄を利用することができ、中程度にストリンジェントな条件は熱融点(Tm)よりも6、7、8、9または10℃低いハイブリダイゼーションおよび/または洗浄を利用することができ、低ストリンジェントな条件は熱融点(Tm)よりも11、12、13、14、15または20℃低いハイブリダイゼーションおよび/または洗浄を利用することができる。この式、ハイブリダイゼーションおよび洗浄組成、ならびに所望のTmを用いれば、当業者はハイブリダイゼーションおよび/または洗浄溶液のストリンジェンシーのバリエーションが本質的に記載されたと理解できるであろう。所望のミスマッチの度合いが45℃(水溶液)または32℃(ホルムアミド溶液)より低いTmを与えるならば、より高い温度を使用できるようSSC濃度を上げることが好ましい。核酸のハイブリダイゼーションの広範囲な指針は、Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acids Probes, Part I, Chapter 2, Ausubel, ら, Eds., Greene Publishing and Wiley-Interscience, New York (1995)に見られる。
【0060】
「実質的に純粋」という用語は、対象とする化合物(例えば核酸、オリゴヌクレオチド、タンパク質など)を重量で少なくとも50〜60%含む調製物を意味する。より好ましくは、調製物は対象とする化合物を重量で少なくとも75%、最も好ましくは90〜99%含む。純度は、対象とする化合物に適当な方法で測定する(例えばクロマトグラフィー法、アガロースまたはポリアクリルアミドゲル電気泳動、HPLC解析など)。
【0061】
核酸配列およびアミノ酸配列は、核酸またはアミノ酸の類似配列を並べ、このことにより相違を明確にするコンピュータプログラムを用いて比較することができる。BLASTプログラム(NCBI)およびそれに使われているパラメータは多くの実施者によりアミノ酸配列フラグメントを並べるために使用されている。しかし、これと同等のアライメントおよび類似性/相同性評価は、標準的なアライメントソフトウェアを使用して得ることができる。例えば、the Genetics Computer Group in Madison, Wisconsinから取得できるGCG Wisconsin Package version 9.1およびBest-Fit programにより使用されるデフォルトパラメータ(ギャップ生成ペナルティー=12、ギャップ延長ペナルティー=4)もまた、配列相同性および類似性を比較するために使用することが出来る。
【0062】
「実質的に同一」という用語は、タンパク質の性質(すなわちタンパク質の構造、安定性、基質特異性および/または生物学的活性)に実質的に影響を及ぼさない配列変化を有する核酸またはアミノ酸配列を意味する。とりわけ核酸配列への言及について、「実質的に同一」という用語は、コード領域および発現を左右する保存配列を意味し、コードされるポリペプチド中の、同じアミノ酸をコードする縮重コドンまたは保存的な置換アミノ酸をコードする異なるコドンを主に意味することを意図する。アミノ酸配列に関して、「実質的に同一」という用語は、一般に、保存的な置換および/またはポリペプチドの構造もしくは機能の決定に関与しない領域での変化を意味する。
【0063】
「%相同」および「%類似」という用語もまた、アミノ酸および核酸配列の間の比較において、本明細書で使用する。アミノ酸配列に関して、「%相同」とは、配列解析プログラムによって比較されるアミノ酸配列中の同一アミノ酸とマッチする対象アミノ酸配列中のアミノ酸の百分率を意味する。「%類似」とは、同一または保存アミノ酸とマッチする対象アミノ酸配列中のアミノ酸の百分率を意味する。保存アミノ酸とは、1つを別のものと入れ替えたとしても、結果的に得られるタンパク質の三次構造を認識できるほどには変化させないような、構造は異なるものの物理的性質は類似しているアミノ酸である。保存的な置換は、Taylor (1986, J. Theor. Biol. 119:205)に定義されている。核酸分子に関して、「%相同」とは、配列解析プログラムにより同一ヌクレオチドとマッチする対象核酸配列中のヌクレオチドの百分率を意味する。
【0064】
抗体に関して、用語「免疫学的に特異的」とは、対象とするタンパク質の1以上のエピトープに結合し、しかし抗原性の生物学的分子の混合集団を含有するサンプル中の他の分子を実質的に認識せずそれらに結合しない抗体を意味する。
【0065】
オリゴヌクレオチドまたは他の一本鎖核酸分子に関して、「特異的にハイブリダイズする」という用語は、当技術分野で一般に用いられる所定の条件、すなわちストリンジェンシーの条件下でのハイブリダイゼーションを可能にする、十分に相補的な配列である2つの一本鎖核酸分子の間の結合を意味する(「実質的に相補的」とも呼ばれる)。特に、この用語は、オリゴヌクレオチドと、一本鎖DNAまたはRNA分子内に含有される実質的に相補的な配列とのハイブリダイゼーションを意味し、かつ、該オリゴヌクレオチドと非相補的な配列の一本鎖核酸とのハイブリダイゼーションを実質的に除外する。
【0066】
「コード配列」または「コード領域」は、配列が発現する場合に遺伝子産物を産生させるのに必要な配列情報を有する核酸分子を意味する。
【0067】
「機能的に連結」または「機能的に挿入」という用語は、コード配列の発現に必要な制御配列が、コード配列を発現できるよう、コード配列に対して適切な位置で核酸分子内に配置されることを意味する。この同じ定義を場合により、発現ベクター内の転写制御因子(例えばエンハンサー)の配置に適用する。
【0068】
転写および翻訳制御配列は、宿主細胞におけるコード配列の発現を提供するDNA調節配列、例えばプロモーター、エンハンサー、ポリアデニル化シグナル、ターミネーター、などである。
【0069】
「プロモーター」、「プロモーター領域」または「プロモーター配列」という用語は、一般にコード領域の5'または3'側、またはコード領域内、またはイントロン内に見ることのできる、遺伝子の転写制御領域を意味する。典型的には、プロモーターは、細胞内のRNAポリメラーゼに結合し、下流(3'方向)にあるコード配列の転写を開始させることができるDNA調節領域である。典型的な5'プロモーター配列は、その3'末端で転写開始部位に接しており、上流(5'方向)に伸びてバックグラウンドを超える検出可能なレベルの転写を開始させるのに必要な最小の数の塩基またはエレメントを含む。プロモーター配列内には、転写開始部位(ヌクレアーゼS1でのマッピングにより都合よく定義される)のみならず、RNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が存在する。
【0070】
「ベクター」は、別の核酸断片の複製または発現をもたらすよう、その断片を機能的に挿入することのできるレプリコン、例えばプラスミド、ファージ、コスミド、またはウィルスである。
【0071】
「核酸構築物」または「DNA構築物」という用語は、適当な調節配列に機能的に連結され、細胞を形質転換するためにベクターに挿入された1以上のコード配列を指して用いられることがある。この用語は「形質転換用DNA」と互いに交換できるよう用いることができる。このような核酸構築物は、対象とする遺伝子産物をコードする配列とあわせて、選択マーカー遺伝子および/またはレポーター遺伝子を含有していてもよい。
【0072】
「選択(可能な)マーカー遺伝子」という用語は、発現させた場合、選択可能な表現型、例えば抗生物質耐性を形質転換細胞に附与する産物をコードする遺伝子を意味する。
【0073】
「レポーター遺伝子」という用語は、標準的手法により、直接または間接的に簡便に検出できる産物をコードする遺伝子を意味する。
【0074】
核酸構築物の「異種」領域は、より大きな分子中の核酸分子の特定可能な断片(複数であってもよい)であり、この核酸分子は天然では前記より大きな分子と結合していないものである。このことから、異種領域が哺乳動物遺伝子をコードする場合、その遺伝子は通常、起源生物のゲノム内でその哺乳動物ゲノムDNAの両端につながっていないDNAが両端につながっている。別の例では、異種領域はコード配列それ自身が天然には見いだされない構築物である(例えば、ゲノムコード配列がイントロンを含有するcDNA、または天然遺伝子とは異なるコドンを有する合成配列)。対立遺伝子のバリエーションや天然に起こる突然変異事象は、本明細書において定義するDNAの異種領域を生み出さない。上記に定義した用語「DNA構築物」はまた、異種領域、特に細胞の形質転換に使用するために構築したものに言及するために用いられる。
【0075】
外因性または異種DNAを細胞内に導入した場合に、細胞はそのDNAにより「形質転換」または「形質導入」される。形質転換用DNAは、細胞のゲノム内に組み入れられても(共有結合的連結)よく、また組み入れられていなくてもよい。例えば原核生物、酵母および哺乳動物細胞の場合、形質転換用DNAはエピソームエレメント、例えばプラスミドなどに維持されてもよい。真核細胞に関しては、安定的に形質転換された細胞は、染色体複製を通じて娘細胞により遺伝で受け継がれるように、その形質転換用DNAが染色体に組み入れられているものである。この安定性は、真核細胞が細胞系、すなわちその形質転換用DNAを含有する娘細胞の集団からなるクローンを樹立する能力によって実証される。
【0076】
「クローン」は、有糸分裂により単一細胞または共通の祖先から誘導される細胞の集団である。「細胞系」は、何世代にも渡ってin vitroで安定に増殖することができる一次細胞のクローンである。
【0077】
2.詳細な説明
血清アミロイドA(SAA)は、主に肝臓で産生され、組織損傷または感染に関連した炎症応答の一部として哺乳動物の血清に増加したレベルで生じる急性期タンパク質である。本発明者らはSAAの独特のアイソフォームであるSAA3を見いだした。このSAA3は、それまでその転写産物もタンパク質もヒト細胞で決して検出されず、その遺伝子が非発現、したがって偽遺伝子とみなされてきたために、存在しないと考えられてきた。
【0078】
以下の説明は、本発明の実施に伴う一般的な手法を示す。具体的な材料が記述されていることに関して、以下の説明は例示のみを目的としたものに過ぎず、本発明を制限することを意図したものではない。特に断らない限り、一般的なクローニング手法、例えばSambrook ら, Molecular Cloning, Cold Spring Harbor Laboratory (1989) (以下「Sambrookら」) またはAusubelら(eds) Current Protocols in Molecular Biology, John Wiley & Sons (1999) (以下「Ausubel ら」)に示されている手法を用いる。
【0079】
A. SAA3、SAA3特異的抗体、およびSAA3をコードする核酸分子の調製
1. タンパク質および抗体
ヒトSAA3は、血清からSAAを精製するために開発された種々の方法(参照により本明細書に組み入れた資料に詳述されている)により調製することができる。疎水性クロマトグラフィーマトリックス系および溶離液のバリエーション、例えばSmithらが記載したもの(Protein Expression & Purification 2: 158-161, 1991)などもまた利用可能である。
【0080】
これとは別に、アミノ酸配列情報が利用可能(例えば配列番号2)であることからヒトSAA3をコードする核酸分子を単離することが可能である。この方法は、当技術分野で広く知られた方法に従い、選択した生物種からのcDNA発現ライブラリーを抗ヒトSAA3抗体を用いてスクリーニングすることにより実現することが出来る。これとは別に、図2(配列番号1)の一部または全部をコードする一連の縮重オリゴヌクレオチドプローブを、以下に詳述する通り、cDNAまたはゲノミックライブラリーをスクリーニングするために用いることができる。
【0081】
一旦取得したならば、cDNAまたは遺伝子をin vitro転写のために適当なin vitro転写ベクター、例えばpSP64またはpSP65にクローンしてもよく、次いで適当な無細胞翻訳系、例えば小麦麦芽またはウサギ網状赤血球で無細胞翻訳をしてもよい。In vitro転写および翻訳系は、例えばPromega Biotech, Madison, Wisconsin or BRL, Rockville, Marylandから市販されている。pCITE in vitro翻訳システム(Novagen)もまた利用可能である。
【0082】
好適な実施形態において、適当な原核または真核細胞系での発現によって、より多くの量のタンパク質を産生することができる。例えば、ヒトSAA3をコードするDNA分子の一部または全部を、細菌細胞(例えば大腸菌)または酵母細胞(例えばSaccharomyces cerevisiae)または哺乳動物細胞での発現に合わせたベクターに挿入してもよい。これらのベクターは、宿主細胞内でDNAが発現できるようにする様式で配置された、宿主細胞内でのこのDNAの発現に必要な調節因子を含む。発現に必要なこのような調節因子には、プロモーター配列、転写開始配列および場合によりエンハンサー配列が含まれる。
【0083】
組換え原核または真核細胞系での遺伝子発現により産生されたヒトSAA3は、当技術分野で既知の方法により精製することが出来る。好ましい実施形態においては、市販の発現/分泌系が利用可能であり、ここでは組換えタンパク質が発現され、周囲の媒体から容易に精製できるように、その後宿主細胞から分泌される。発現/分泌ベクターを使用しない場合、これとは別のアプローチは、組換えタンパク質をアフィニティー分離、例えばその組換えタンパク質に特異的に結合する抗体との免疫学的相互作用により精製することを伴う。このような方法は当業者に一般的に利用されている。
【0084】
本発明はまた、ヒトSAA3に結合する能力のある、1以上の選択された種からの抗体を提供する。選択されたヒトSAA3の一部または全部に対するポリクローナルまたはモノクローナル抗体は、標準的な方法で調製することが出来る。モノクローナル抗体は、Kohler and Milsteinの一般的な方法に従い、標準的なプロトコルに沿って調製することが出来る。好適な実施形態において、ヒトSAA3の選択されたエピトープに免疫特異的に反応し、ヒトSAA3を他のSAAから区別する抗体を調製する。
【0085】
2.核酸分子
配列情報を取得したならば、ヒトSAA3をコードする核酸分子を2つの一般的方法によって用意することが出来る。ヒトSAA3は(1)適当なヌクレオチド三リン酸から合成してもよく、または(2)生物学的取得源から単離してもよい。いずれの方法も当技術分野で広く知られた方法を利用する。
【0086】
ヌクレオチド配列情報が利用可能であれば、本発明の単離された核酸分子をオリゴヌクレオチド合成により調製することができる。合成オリゴヌクレオチドは、Applied Biosystems 38A DNA Synthesizerまたは類似の装置により実施されるホスホルアミダイト(phosphoramadite)法により用意することができる。得られる構築物は、当技術分野で既知の方法、例えば高性能液体クロマトグラフィー(HPLC)により精製することが出来る。長い二本鎖ポリヌクレオチド、例えば本発明のDNA分子は、現在のオリゴヌクレオチド合成方法に固有の大きさの限界のために、段階的に合成しなければならない。このことから例えば、長い二本鎖分子を、適当な相補性を有する小さないくつかの断片として合成してもよい。このようにして製造された相補的断片を、各断片が隣接する断片との結合のために適当な結合性のある末端を有するようにアニールしてもよい。隣接断片は、DNAリガーゼの存在下で結合性のある末端をアニールさせることによりライゲートさせて、全長二重鎖分子を構築することができる。このようにして構築した合成DNA分子は次いで、適当なベクターにクローニングして増幅することができる。
【0087】
ヒトSAA3をコードする核酸分子はまた、当技術分野で広く知られた方法を用いて興味有る哺乳動物種から単離することができる。選択した生物種からの核酸分子は、cDNAまたはゲノミックライブラリーを、ヒトSAA3コード遺伝子に特異的な核酸配列にマッチするよう設計されたオリゴヌクレオチドでスクリーニングすることにより単離することができる。ある種からの遺伝子が所望ならば、ゲノミックライブラリーをスクリーニングする。これとは別に、タンパク質コード配列に特に興味があるのならば、cDNAライブラリーをスクリーニングする。適当なアミノ酸残基をコードするために1より多くの核酸残基が使用可能な縮重の位置では、全ての適当な核酸残基を組み入れて混合オリゴヌクレオチド集団を作成してもよく、またはイノシンなどの中性塩基を用いてもよい。オリゴヌクレオチド設計の方針は当技術分野で広く知られている(Sambrookら, Molecular Cloning, 1989, Cold Spring Harbor Press, Cold Spring Harbor NYも参照のこと)。
【0088】
これとは別に、ヒトSAA3タンパク質の一部をコードするように、PCR(ポリメラーゼ連鎖反応)プライマーを上述の方法により設計してもよく、これらのプライマーを用いて単離されたcDNAまたはゲノムDNAから核酸を増幅することができる。好適な実施形態において、ヒトSAA3をコードする核酸を単離するために用いるオリゴヌクレオチドは、血清SAAとは異なり、ヒトSAA3に独特の配列をコードするよう設計される。
【0089】
本発明において、ヒトSAA3をコードする核酸分子と適当な配列相同性を有する核酸は、適当なストリンジェンシーのハイブリダイゼーションおよび洗浄条件を用いて同定することができる。例えば、Sambrookら(1989, 上記に同じ)の方法に沿って、5× SSC、5× デンハート試薬(Denhardt's Reagent)、1.0% SDS、100μg/ml変性済み断片化サケ精子DNA、0.05%ピロリン酸ナトリウムおよび最大50%までのホルムアミドを含むハイブリダイゼーション溶液を用いてハイブリダイゼーションを行うことができる。ハイブリダイゼーションは少なくとも6時間にわたり37〜42℃で行う。ハイブリダイゼーションに次いで、フィルターを以下の通り洗浄する。 (1)2× SSCおよび1%SDS中で室温で5分間、(2)2× SSCおよび0.1%SDS中で室温で15分間、(3)1× SSCおよび1%SDS中で37℃で30分〜1時間、(4)溶液を30分毎に交換しながら、1× SSCおよび1%SDS中で42〜65℃で2時間。
【0090】
特定の配列相同性を有する核酸分子の間のハイブリダイゼーションを達成するのに必要なストリンジェンシー条件を計算するための一般式を挙げる(Sambrookら,1989,上記に同じ):
Tm = 81.5℃ + 16.6Log [Na+] + 0.41(% G+C) - 0.63 (% ホルムアミド) - 600/二本鎖中のbp数。
【0091】
上述の式の一例として、[N+] = [0.368]かつ50% ホルムアミドで,GC含量が42%でプローブの平均長さが200塩基の場合、Tmは57℃である。DNA二本鎖のTmは、相同性が1%減少するごとに1〜1.5℃低下する。したがって、約75%より高い配列相同性を有する標的は、42℃のハイブリダイゼーション温度を用いることにより観察する。上述のハイブリダイゼーションおよび洗浄条件を用いて、好適な実施形態において、ハイブリダイゼーション温度が37℃であり最終洗浄を42℃で行い、より好適な実施形態ではハイブリダイゼーションを42℃で行い最終洗浄を50℃で行い、最も好ましい実施形態ではハイブリダイゼーションを42℃で行い最終洗浄を65℃で行う。
【0092】
ハイブリダイゼーションおよび洗浄のストリンジェンシーは主に、溶液の塩濃度および温度に依存する。一般に、プローブの標的とのアニーリングの速度を最大化するために、ハイブリダイゼーションは通常、ハイブリッドの計算上のTmよりも20〜25℃低い塩および温度条件で行われる。洗浄条件は、プローブの標的に対する相同性の度合いについて可能な限りストリンジェントにするべきである。一般に、洗浄条件は、そのハイブリッドのTmより約12〜20℃低くなるように選択される。本発明の核酸に関しては、中程度のストリンジェンシーのハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および55℃で15分間、2× SSCおよび0.5%SDSでの洗浄として定義される。高度にストリンジェントなハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および65℃で15分間、1× SSCおよび0.5%SDSでの洗浄として定義される。非常に高度にストリンジェントなハイブリダイゼーションは、42℃で6×SSC、5×デンハート溶液、0.5% SDS、100μg/ml変性済みサケ精子DNAのハイブリダイゼーション、および65℃で15分間、0.1× SSCおよび0.5%SDSでの洗浄として定義される。
【0093】
本発明の核酸分子は、任意の適当なクローニングベクター中でDNAとして維持することができる。好適な実施形態において、クローンはプラスミドクローニング/発現ベクター、例えばpGEM-T(Promega Biotech, Madison, WI)またはpBluescript(Stratagene, La Jolla, CA)などで維持されるが、これらはどちらも適当な大腸菌宿主細胞中で増殖させることができる。
【0094】
本発明のヒトSAA3コード核酸分子には、一本または二本鎖であってもよい、cDNA、ゲノムDNA、RNA、およびその断片が含まれる。従って本発明は、本発明の核酸分子の配列少なくとも1つとハイブリダイズする能力のある配列を有するオリゴヌクレオチド(DNAまたはRNAのセンスまたはアンチセンス鎖)を提供する。このようなオリゴヌクレオチドは、例えばPCR増幅などによる、試験サンプル中のヒトSAA3コード遺伝子またはmRNAを検出するためのプローブとして有用である。
【0095】
B. ヒトSAA3タンパク質の使用
抗体および核酸
SAAレベルは、動物または患者から得られたサンプル中の疾患または他の炎症症状を診断または検出するために使用することができる。好適な実施形態において、これらのレベルを、乳房の感染症または症状を同定するためにアッセイする。増加したSAAまたはSAA3レベルは、急性期と関連する可能性があり、感染症または疾患の診断に用いることができる。SAA3はまた、ムチン産生を刺激することから医薬として使用してもよく、そしてムチンMUC3の不十分なレベルと関連する腸の不具合を軽減するために投与してもよい。
【0096】
1. タンパク質および抗体
精製したヒトSAA3またはその断片は、ポリクローナルまたはモノクローナル抗体を産生するために使用することができ、これらの抗体は培養細胞または組織中での、および無傷の生物中でのタンパク質の存在および蓄積に対する敏感な検出試薬としての機能を果たしうる。組換え技術は、選択されたヒトSAA3の一部または全部を含有する融合タンパク質の発現を可能にする。SAA3タンパク質の全長タンパク質または断片を、このタンパク質の様々なエピトープに特異的な一連のモノクローナルまたはポリクローナル抗体を生成するために好適に利用することができ、このことによりこのタンパク質のより高い検出感度を提供する。好適な実施形態においては、ヒトSAA3を血清SAAと識別するヒトSAA3の断片を、エピトープ特異的抗体を生成するために利用する。
【0097】
ヒトSAA3に免疫学的に特異的なポリクローナルまたはモノクローナル抗体は、そのタンパク質を検出および定量するために設計した種々のアッセイに使用することができる。このようなアッセイには、限定するものではないが(1)免疫沈降に次ぐタンパク質定量、(2)イムノブロット解析(例えばドットブロット、ウェスタンブロット)(3)放射免疫アッセイ(4)ネフェロメトリー、比濁または免疫クロマトグラフィー(横方向流動)アッセイ、(5)ELISAおよび種々の定量的迅速検定が含まれる酵素結合アッセイ(例えばディップ・スティックなどの検査)が含まれる。
【0098】
ヒトSAA3に免学特異的に相互作用するポリクローナルまたはモノクローナル抗体は、このタンパク質を同定し精製するために利用することができる。例えば、抗体は、それら抗体が免疫特異的に相互作用するタンパク質のアフィニティー分離に利用することができる。抗体はまた、タンパク質および他の生物学的分子の混合物を含有するサンプルから、タンパク質を免疫沈降するために使用することができる。
【0099】
2.核酸
ヒトSAA3をコードする核酸分子は、本発明における種々の目的のために使用することができる。DNA、RNAまたはそのフラグメントは、その遺伝子の存在をおよび/または発現を検出するためのプローブとして使用することができる。このようなアッセイのためにヒトSAA3をコードする核酸をプローブとして利用することができる方法には、限定するものではないが、(1)in situハイブリダイゼーション、(2)サザンハイブリダイゼーション、(3)ノーザンハイブリダイゼーション、(4)類別された(assorted)増幅反応、例えばポリメラーゼ連鎖反応(PCR)および逆転写酵素-PCR(RT-PCR)が含まれる。
【0100】
例示した本発明のヒトSAA3をコードする核酸はまた、ヒトを含む他の生物種由来の関連遺伝子を同定するためのプローブとして利用することができる。当技術分野で広く知られ上述した通り、ハイブリダイゼーションのストリンジェンシーは、核酸プローブと様々な程度の相同性を有する相補配列とのハイブリダイゼーションを可能にするために調節することができる。
【0101】
ヒトSAA3をコードする核酸の前述の使用に加え、これらは形質転換(トランスジェニック)細胞、組織および生物の創造において有用性が期待される。
【0102】
本発明はヒトSAA3タンパク質をコードする、新規な精製および単離された核酸配列を提供する。現在のところ好適な形態では、DNA配列は新規SAA3またはその保存的に改変された変異体をコードするcDNA配列を含み、これらはプロラクチンまたはLPSに応答して乳房上皮細胞で発現され、活性TFLK領域を含んでなり、そして本明細書において開示するタンパク質の生物学的活性を保有する。より好適な実施形態において、核酸配列は配列番号1と少なくとも約80%の相同性、またはコードされるアミノ酸配列と80%の相同性がある。具体的には、単離された配列を配列番号1に表す。これとは別のDNA形態、例えばゲノムDNAおよびヌクレオチドから部分または全化学合成により調製したDNAならびに欠失または突然変異を有するDNAもまた、本発明の範囲内にある。
【0103】
本発明においてさらにゲノムSAA3配列を同定しキャラクタライズした。ヒトSAA3配列とともにイントロンを含むゲノム領域が使用可能であり、ヌクレオチド配列の最も効率的な発現を達成するためにしばしば必要である。SAA3の5'領域もまた開示するが、これはプロモーター領域の一部を含み、異種遺伝子の一時的および空間的(spatial)発現に使用することができるSAA3プロモーターを単離するために使用することができる。このプロモーターは乳房上皮細胞への組織優先的発現(tissue preferred expression)を提供することができ、またプロラクチンまたはLPSの存在下での機能的に連結された配列の誘導発現も提供することができる。
【0104】
本発明により提供されたDNA配列と、同種または異種発現制御DNA配列、例えばプロモーター、オペレーター、およびレギュレーターなどとの結合は、mRNAからのin vivoおよびin vitro転写を可能にし、次いでこれは翻訳が可能であり、本発明の新規タンパク質および関連ポリおよびオリゴペプチドを大量に提供する。本発明の目下好適なDNA発現系において、SAA3をコードするDNAは、このタンパク質のin vitro転写および翻訳を可能にする制御プロモーターDNA配列に機能的に連結されている。
【0105】
DNA配列を、適当なウィルスおよび環状DNAプラスミドベクターを潜在的に伴う、標準的な形質転換および形質導入プロセスによって原核および真核宿主細胞に導入することはまた、本発明の意図の内にあり、有用なタンパク質をこれまで天然の取得源からは入手できなかったほど大量に提供すると予想される。哺乳動物宿主細胞の使用は、以下により詳しく記述する通り、本発明の組換え発現産物に最良の生物学的活性を附与するために必要であるかもしれない、翻訳後修飾(例えば末端切断化、グリコシル化、およびチロシン、セリンまたはトレオニンリン酸化)を提供すると予想される。
【0106】
細胞を形質転換し、ベクターを構築し、メッセンジャーRNAを抽出し、cDNAライブラリーを調製するなどのために用いるほとんどの技術は、当技術分野で広く実施されており、ほとんどの当業者は具体的な条件および手順を説明する標準的な資料文献に精通している。しかしながら、便宜のために、以下の段落はガイドラインとして役立つであろう。
【0107】
宿主および制御配列
ヒトSAA3をコードする配列を発現させるために、原核および真核細胞系の両方を使用することができる。むろん、原核細胞宿主がクローニング手順のためには最も好都合である。原核細胞は大腸菌の種々の株が最も頻出の代表であるが、一方、他の微生物株もまた使用可能である。宿主と両立できる種から誘導された複製サイト、選択マーカーおよび制御配列を含有するプラスミドベクターが使用される。例えば、大腸菌は典型的にはBolivarら(Gene (1977) 2:95)により大腸菌種から誘導されたプラスミドであるpBR322の誘導体を用いて形質転換する。pBR322はアンピシリンおよびテトラサイクリン耐性の遺伝子を含有し、従って所望のベクターを構築するにあたり、保持または破壊することができる複数の選択マーカーを提供する。転写開始のためのプロモーターを、場合によりオペレータを、リボソーム結合サイト配列とともに含むと本明細書において定義した、一般的に使われる原核細胞制御配列は、βラクターゼ(ペニシリナーゼ)およびラクトース(lac)プロモーター系(Changら, Nature (1977) 198:1056)、およびトリプトファン(trp)プロモーター系(Goeddel,ら, 核酸s Res (1980) 8:4057)ならびにラムダ誘導PLプロモーターおよびN遺伝子リボソーム結合サイト(Shimatakeら Nature (1981) 292:128)などの一般に使われるプロモーターを含む。
【0108】
細菌に加え、真核細胞微生物、例えば酵母もまた宿主として使用できる。いくつかの他の株または種が一般に入手可能であるが、Saccharomyces cerevisiaeの研究株、パン酵母が最もよく使われる。例えばBroach, J.R., Meth Enz (1983) 101:307の2μ複製起点、または他の酵母で両立できる複製起点(例えばStinchcomb,ら, Nature (1979) 282:39, Tschumper, G.,ら, Gene (1980) 10:157 および Clarke, L,ら, Meth Enx (1983) 101:300を参照のこと)を採用したベクターを用いることができる。酵母ベクターのための制御配列には、解糖酵素の合成のためのプロモーター(Hess,ら, J Adv Enzyme Reg (1968) 7:149; Holland,ら, Biochemistry (1978) 17:4900)が含まれる。当技術分野で知られているさらなるプロモーターには、3-ホスホグリセリン酸キナーゼのプロモーター(Hitzeman,ら J Biol Chem (1980) 255:2073)が含まれる。増殖条件および/または遺伝的バックグラウンドにより転写が制御されているというさらなる利点を有する他のプロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロームC、酸性フォスファターゼ、窒素代謝に関連している分解酵素、α因子システム、ならびにマルトースおよびガラクトース利用を担う酵素のプロモーター領域である。また、コード配列の3'末端にはターミネーター配列が望ましいと考えられている。このようなターミネーターは、酵母由来遺伝子のコード配列に続く3'非翻訳領域に見られる。
【0109】
また多細胞生物から誘導される真核宿主細胞培養で、ポリペプチドをコードする遺伝子を発現させることも、もちろん可能である。例えば、Axel,ら, 米国特許第4,399,216号を参照のこと。これらの系は、イントロンをスプライスで除く能力という追加の利点を有し、そのことからゲノム断片を直接発現させるために使用できる。有用な宿主細胞系にはVEROおよびHeLa細胞、ならびにチャイニーズハムスター卵巣(CHO)細胞が含まれる。このような細胞のための発現ベクターには通常、哺乳動物細胞と両立できるプロモーターおよび制御配列が含まれ、例えば一般に使われているシミアンウィルス40(SV40)からの初期および後期プロモーター(Fiers, ら, Nature (1978) 273:113)、またはポリオーマ、アデノウィルス2、ウシ乳頭腫ウィルス、またはトリ肉腫ウィルスに由来するものなどの他のウィルスプロモーターが挙げられる。制御可能なプロモーターであるhMT1I(Karin, M.,ら, Nature (1982) 299:797-802)もまた使用可能である。哺乳動物細胞宿主系形質転換の一般的な態様は、Axel(上記に同じ)により記述されている。「エンハンサー」領域もまた発現を最適化するのにいまや重要のようである。これらは一般に、非コードDNA領域内においてプロモーターの上流または下流に見られる配列である。複製起点は、必要ならば、ウィルス起源から取得してもよい。一方、染色体への組み入れは真核細胞におけるDNA複製の一般的な機構である。
【0110】
形質転換
用いる宿主細胞に依存して、形質転換をその細胞に適当な標準的な技術を用いて行う。Cohen, S.N., Proc Natl Acad Sci (USA) 1972) 69:2110によって記載された塩化カルシウムを用いるカルシウム処理法、またはManiatisら, Molecular Cloning: A Laboratory Manual (1982) Cold Spring Harbor Press, p. 254 and Hanahan, D., J Mol Biol (1983) 166:557-580に記載されている塩化ルビジウム法を、実質的に細胞壁を有する原核細胞または他の細胞に用いることができる。そのような細胞壁を有しない哺乳動物細胞については、Graham およびvan der Eb, Virology (1978) 52:546のリン酸カルシウム沈殿法を、場合によりWigler, M., ら, Cell (1979) 16:777-785によって改変して、使用してもよい。酵母への形質転換は、Beggs, J.D. Nature (1978) 275:104-109 or of Hinnen, A.,ら, Proc Natl Acad Sci (USA) (1978) 75:1929の方法により行うことが出来る。
【0111】
ベクター構築
所望のコードおよび制御配列を含有する適当なベクターの構築は、当技術分野でよく理解されている標準的なライゲーションおよび制限技術を用いる。単離したプラスミド、DNA配列、または合成オリゴヌクレオチドを、所望の形態に切断し、調整し、再度ライゲーションする。
【0112】
ベクターを形成するDNA配列は、いくつかの起源から入手可能である。バックボーンベクターおよび制御システムは、構築する配列の大部分に用いる、入手可能な「ホスト」ベクターに一般に見られる。典型的な配列は上記に記載した。適当なコード配列については、開始時の構築は、適切な配列をcDNAまたはゲノミックDNAライブラリーから取り出すことに関する。一方、一旦配列が開示されたならば、遺伝子配列全体をin vitroで個別のヌクレオシド誘導体から出発して合成することが可能である。遺伝子または相当な大きさのcDNA(例えば500〜1000bp)の全配列は、個別の重複する相補的オリゴヌクレオチドを合成し、一本鎖非重複部分をデオキシリボヌクレオチド三リン酸の存在下でDNAポリメラーゼを用いて埋めることで調製してもよい。このやり方は配列が既知のいくつかの遺伝子の構築について用いられ成功している。例えば、Edge, M.D., Nature (1981) 292:756; Nambair, K.P., ら, Science (1984) 223:1299; Jay, Ernest, J Biol Chem (1984) 259:6311を参照のこと。
【0113】
合成オリゴヌクレオチドは、Edgeら, Nature (上記に同じ) およびDuckworthら, 核酸s Res (1981) 9:1691によって記載されたホスホトリエステル法、またはBeaucage, S.L.,およびCaruthers, M.H., Tet Letts (1981) 22:1859 ならびにMatteucci, M.D.,およびCaruthers, M.H., J Am Chem Soc (1981) 103:3185によって記載されたホスホルアミダイト法により調製することができ、また市販の自動オリゴヌクレオチド合成機を用いて調製することが出来る。アニーリング前のまたは標識化のための一本鎖のキナーゼ処理は、過剰の、例えば1 nmole基質に対して約10単位のポリヌクレオチドキナーゼを、50mM Tris、pH7.6、10mM MgCl2、5mM ジチオトレイトール、1〜2mM ATP、1.7 pmole γ32P-ATP(2.9mCi/mmole)、0.1mM スペルミジン、0.1mM EDTAの存在下で用いて達成される。
【0114】
このようにして一度所望のベクターの構成要素を手に入れたならば、それらを標準的な制限およびライゲーション手順により切り取ってライゲートすることができる。
【0115】
部位特異的DNA切断は、当技術分野で一般に理解されている条件下で、適当な制限酵素(複数であってもよい)で処理することにより行うが、それらの条件は個別に市販の制限酵素の製造業者により特定されている。例えばNew England Biolabs製品カタログを参照のこと。一般にプラスミドまたはDNA配列約1μgを、約20μlのバッファー溶液中で1単位の酵素により切断する。本発明の実施例では、典型的には、過剰量の制限酵素を使用して基質DNAの完全消化を保証する。約37℃で約1時間〜2時間のインキュベーション時間が実行可能であるが、バリエーションも許容可能である。各インキュベーションの後、タンパク質をフェノール/クロロホルム抽出で除去し、次いでエーテル抽出を行ってもよく、核酸を水性画分からエタノールを用いて沈殿させて回収する。所望により、切断した断片の大きさによる分離を、標準技術を用いてポリアクリルアミドゲルまたはアガロースゲル電気泳動で行うことができる。大きさによる分離の一般的な記載は、Methods in Enzymology (1980) 65:499-560に見られる。
【0116】
制限切断断片は、大腸菌DNAポリメラーゼIのラージフラグメント(クレノウ)で、4種のデオキシヌクレオチド三リン酸(dNTP)の存在下で、20℃〜25℃で50mM Tris pH7.6、50mM NaCl、6mM MgCl2、6mM DTTおよび0.1〜1.0mM dNTPs約15〜25分のインキュベーション時間を用いて処理することにより平滑末端化することができる。クレノウフラグメントは、5'側一本鎖突出部を埋めていくが、突き出た3'側一本鎖をたとえ4種のdNTPが存在していても削り取る。所望ならば、突出部の性質により決定される限定的範囲内で、選択的修復をdNTPの1種または選択したもののみを供給することにより行うことができる。クレノウでの処理後、混合物をフェノール/クロロホルムで抽出しエタノールで沈殿させる。適当な条件下でのS1ヌクレアーゼまたはBAL-31を用いた処理は、あらゆる一本鎖部分の加水分解という結果を招く。
【0117】
ライゲーションは標準的な条件および温度に沿って15〜50μlの容量で行われる。標準的な条件および温度は例えば、20mM Tris-HCl pH 7.5、10mM MgCl2、10mM DTT、33μg/ml BSA、10mM〜50mM NaCl、および0℃で40μM ATP、0.01〜0.02(Weiss)単位T4 DNAリガーゼ(「粘着末端」ライゲーション用)、または14℃で1mM ATP、0.3〜0.6(Weiss)単位T4 DNAリガーゼ(「平滑末端」ライゲーション用)である。分子間での「粘着末端」ライゲーションは、通常33〜100μg/ml合計DNA濃度で行う(5〜100nM合計最終濃度)。分子間での平滑末端ライゲーションは1μM合計最終濃度で行う。
【0118】
「ベクター断片」を用いるベクター構築において、ベクター断片は一般に5'リン酸を除去しベクターの自己ライゲーションを防ぐために、細菌由来アルカリフォスファターゼ(BAP)または仔牛小腸由来アルカリフォスファターゼで処理する。処理はpH 8で約10mM Tris-HCl、1mM EDTAで、ベクター1μgあたり約1単位のBAPまたはCIPを用いて60℃で約1時間行う。核酸断片を回収するために、調製物をフェノール/クロロホルムで抽出しエタノールで沈殿させる。これとは別に、二重に消化したベクターの再ライゲーションを、追加の制限酵素消化により、および/または不要断片の分離により防止することができる。
【0119】
配列変更を要する、cDNAまたはゲノムDNAから誘導されたベクター中の部分については、部位特異的プライマー導入突然変異(Zoller, M.J.,およびSmith, M. 核酸s Res (1982) 10:6487-6500 and Adelman, J.P.,ら, DNA (1983) 2:183-193)を用いることができる。これは、所望の突然変異を表す限定的なミスマッチ部分以外は、突然変異を導入すべき一本鎖ファージDNAに相補的なプライマー合成オリゴヌクレオチドを用いることにより行う。手短に述べると、合成オリゴヌクレオチドをファージに相補的な鎖を合成するようし向けるプライマーとして使用し、結果的に得られる部分的または完全二本鎖DNAをファージ担持宿主細菌へと形質転換する。形質転換した細菌の培養物をトップアガーにプレートし、ファージを抱えている単一細胞からのプラーク形成を可能にする。
【0120】
理論的には、新しいプラークの50%が一本鎖として突然変異型のファージを含有し、50%が元の配列を保有する。結果的に得られるプラークは、完全なマッチの結合は可能であるものの、元の鎖とのミスマッチが結合を防止するのに十分な温度で、キナーゼ処理済み合成プライマーとハイブリダイゼーション後洗浄する。次いでプローブとハイブリダイズするプラークを選び出し、培養し、DNAを回収する。
【0121】
構築物の確認
プラスミド構築の正しいライゲーションは、まず大腸菌(E. coli)MC1061株(Casadaban, M., ら, J Mol Biol (1980) 138:179-207)または他の適当な宿主をライゲーション混合物で形質転換することにより確認できる。成功した形質転換体は、当技術分野で理解されている通り、アンピシリン、テトラサイクリン、または構築プラスミドの様式に依存する別のマーカーを用いた別の抗生物質耐性を用いることにより選択する。形質転換体のプラスミドは次いで、Clewell, D.B.,ら, Proc Natl Acad Sci (USA) (1969) 62:1159の方法に沿って調製し、場合によりクロラムフェニコール増幅(Clewell, D.B., J Bacteriol (1972) 110:667)を行う。複数の微少DNAプレップを一般的に用いる。例えばHolmes, D.S., ら, Anal Biochem Acids Res (1979) 7:1513-1523を参照のこと。単離したDNAは、制限処理により解析および/またはSanger, F., ら, Proc Natl Acad Sci (USA) (1977) 74:5463のジデオキシヌクレオチド法、さらにはMessing,ら, 核酸s Res (1981) 9:309, Maxam,ら, Methods in Enzymology (1980) 65:499が記載した方法により配列解析する。
【0122】
宿主細胞の例示
本発明におけるクローニングおよび原核細胞発現に用いる宿主細胞は以下の通りである。
【0123】
ほとんどの細菌プロモーターの制御下でのクローニングおよび配列解析、ならびに構築物の発現のために、例えばMC1061、DH1、RR1、C600hfl、K803、HB101、JA221、およびJM101などの大腸菌株を使用することができる。
【0124】
3.SAA3の発見に基づくさらなる使用
ヒトSAA3はまた、いくつかの他の目的のために使用することができる。これには、限定するものではないが、(1)腸または脈管構造をまたぐ分子の送達のための担体(2)新生児における腸粘膜の発達のための栄養的サプリメント(3)免疫応答の調節因子(注入または経口投与を通して)が含まれる。
【0125】
4.医薬製剤
本発明において、SAA3およびより具体的にはその活性部位(すなわちTFLKモチーフ)は、腸におけるムチン産生を刺激する。これは、ムチンが腸感染症の予防および治療において主要な役割を果たし、多くのプロバイオティック微生物(probiotics)がムチン産生を誘導することにより作用することが示されたことから、大変意義深い。Mackら,"Probiotics inhibit enteropathogenic Escherechia coli adherence in vitro by inducing intestinal mucin gene expression", 1999, Am J Physiol, 276:G941-950を参照のこと。そこに開示されている内容を参照により本明細書に組み入れる。このことから、本発明にはまたヒトSAA3に関連する、ヒトおよび動物のための医薬製剤が含まれる。医薬の分野に精通したものならば、医薬組成物の用量および投与計画が投与等ではなく、そのヒトおよび動物の年齢、健康状態、性別、大きさおよび体重によって変わることが容易に理解できるであろう。これらのパラメーターは、きちんと確立された手順と解析、例えばフェーズI、II、III臨床試験で、各システムについて求めることができる。
【0126】
投与については、SAA3は製薬上許容可能な担体、例えば適当な液体ビヒクルまたは賦形剤、および場合により補助剤または添加剤と組み合わせることができる。液体ビヒクルおよび賦形剤は常套的なものであり、市販されている。それらの実例としては、蒸留水、生理食塩水、デキストロース水溶液などが挙げられる。
【0127】
一般に、活性化合物に加えてさらに、本明細書の医薬組成物は、活性化合物を医薬的に使用できる製剤へと加工するのを促進する、適当な賦形剤および補助剤を含有してもよい。経口用剤形は錠剤、糖衣剤、およびカプセル剤を包含する。直腸投与できる製剤には坐剤が含まれる。他の剤形には、非経口または経口投与のための適当な溶液、および口腔または舌下投与することができる組成物が含まれる。
【0128】
本発明の医薬製剤は、当技術分野でよく知られた様式で製造する。例えば、医薬製剤は、慣用の混合、顆粒化、糖衣化、溶解、凍結乾燥処理などの手段により製造できる。用いる処理方法は、最終的には用いる活性成分の物理的性質に依存する。
【0129】
適当な賦形剤は、特に、糖、例えばラクトースまたはスクロース、マンニトールまたはソルビトール、セルロース調製物および/またはリン酸カルシウム、例えばリン酸三カルシウムまたはリン酸水素カルシウム、ならびに結合剤、例えばデンプン、糊、(例えばトウモロコシデンプン、小麦デンプン、コメデンプン、ジャガイモデンプン、ゼラチン、ガムトラガカント、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドンを使用)である。所望により、崩壊剤を添加することができ、その例としては上述のデンプンならびにカルボキシメチルデンプン、架橋ポリビニルピロリドン、寒天、もしくはアルギン酸またはその塩、例えばアルギン酸ナトリウムなどが挙げられる。補助剤は、流動調節剤および滑沢剤、例えばシリカ、タルク、ステアリン酸もしくはその塩、例えばステアリン酸マグネシウムまたはステアリン酸カルシウムおよび/またはポリエチレングリコールである。糖衣の芯は適当なコーティングとともに提供してもよく、そのコーティングは所望ならば胃液に耐性であってもよい。
【0130】
この目的のために濃縮した糖溶液を用いることができ、それは場合によりアラビアガム、タルク、ポリビニルピロリドン、ポリエチレングリコールおよび/または二酸化チタン、ラッカー溶液ならびに適当な有機溶媒または溶媒混合物を含有してもよい。胃液に耐性のコーティングを製造するためには、適当なセルロース調製物の溶液、例えばアセチルセルロースフタレート、またはヒドロキシプロピルメチルセルロースフタレート、染料および色素を、例えば同定または化合物用量の異なる組み合わせをキャラクタライズするために、糖衣剤コーティングの錠剤に加えることができる。
【0131】
経口に用いることのできる他の医薬製剤には、ゼラチンでできたプッシュフィット(push-fit)カプセル剤、ならびにゼラチンおよび例えばグリセロールまたはソルビトールなどの可塑剤でできた軟性密封カプセル剤が含まれる。プッシュフィットカプセル剤は顆粒の形態で活性化合物を含有してもよく、顆粒は充填剤例えばラクトース、結合剤(例えばデンプン)、および/または滑沢剤(例えばタルクまたはステアリン酸マグネシウム)ならびに場合により安定剤と混合してもよい。軟カプセル剤については、活性化合物を好ましくは適当な液体、例えば脂肪油、流動パラフィン、または液状ポリエチレングリコールに溶解または懸濁させる。さらに安定剤を添加してもよい。直腸に用いることができる医薬製剤には、例えば、活性化合物と坐剤用の基剤との組み合わせからなる坐剤が含まれる。適当な坐剤用の基剤は、例えば天然または合成トリグリセリド、パラフィン炭化水素、ポリエチレングリコールまたはより高級なアルコールである。さらに、活性化合物と基剤の組み合わせからなるゼラチン性直腸用カプセルを用いることも可能である。基剤の材料となりうるものには、液状トリグリセリド、ポリエチレングリコール、またはパラフィン炭化水素が含まれる。
【0132】
非経口投与の適当な製剤には、水に溶けうるまたは水に懸濁されうる形態の活性化合物の水溶液が含まれる。さらに、適切な油性注入懸濁物として、活性化合物の懸濁剤を投与してもよい。適当な親油性溶媒またはビヒクルには、脂肪油、例えばゴマ油、あるいは合成脂肪酸エステル、例えばオレイン酸エチルまたはトリグリセリドが含まれる。水性注入懸濁物は、懸濁物の粘性を増加させる物質を含有してもよく、そうした物質には例えば、カルボキシメチルセルロースナトリウム、ソルビトールおよび/またはデキストランが含まれ、場合により懸濁物はまた、安定剤を含有してもよい。
【0133】
慣用の担体を用いた投与の他に、活性成分を、当業者に知られているいくつかの特殊化された薬剤送達技術により投与してもよい。以下の例は、例証のみを目的として挙げるものであり、全く本発明を制限することを意図したものではない。
【0134】
本明細書において用いる「有効量」という用語は、本明細書において開示する方法および手順によって決定される通り、病原体の粘膜細胞への付着が減少するように、ムチン産生を増加させるのに十分な量のヒトSAA3を意味する。
【0135】
本発明において、新規ヒトSAA3およびさらに特定するとそのTFLKモチーフ活性部位は、ムチン産生、より具体的にはMUC3産生を刺激することが示された。ムチン産生は大腸菌の付着を阻害することが示されており、同じことをするプロバイオティック物質は、ムチンの刺激を通して機能することが示されている。ヒトSAA3および/またはペプチドは、プロバイオティックの代わりに使用することができる。
【0136】
腸の感染症におけるムチンの重要性は、感染性生物が疾患を引き起こすのに必要な事象を防ぐ能力にある。
【0137】
ムチンは腸の上皮細胞により産生され、その表面へと分泌される。このことから、ムチンは戦略上、腸の上皮細胞と腸管内に取り込まれた加害性因子(すなわち感染因子、有害物質)との間に位置する。
【0138】
ムチンはまた、細菌の腸管の上皮細胞への付着を阻害する。細菌による腸の内層細胞への結合が、侵襲、毒素送達および下痢疾患の罹患の最初の段階である。腸内病原体の結合が阻害されれば、疾患は発症しない。
【0139】
ムチンはウィルスの複製を阻害することが示されている。
【0140】
ムチンは、先天免疫の一部であり、腸の基本的な防御系である。したがって、抗体/T細胞によって駆動される後天性免疫系と比較して、ムチンが提供する利点には以下のものが含まれる:加害性因子に対して即時または急速に誘導可能な応答、広範な作用スペクトル、局所的効力、および動物界にわたる保存。
【0141】
感染因子に起因するムチン分泌の増加は、よく知られた臨床的現象である。ムチンは、病原性の腸内細菌が腸の細胞に付着するのを阻害し、このことから感染を防止する。これは、ムチンが、通常腸の細胞に付着するために使われる細菌の細胞壁の構造物に付着することにより成し遂げられる。プロバイオティック細菌(非病原性細菌)は、病原性腸内細菌による腸管内側の上皮細胞への付着を防ぐ。プロバイオティック細菌から分泌される物質は、腸の細胞により多くのムチンを産生させ、これはプロバイオティック物質が感染を防ぐ1つの機構である。
【0142】
上記のことは、数多くの腸内病理に対する本発明のペプチドの薬学的用途を実証している。例えば旅行者下痢の予防が挙げられる。多くの感染性生物は事実上地域的なものであり、自分たちの地域から出た旅行者は、通常これらの生物にそれまでに曝露されたことがなく、従ってそれらへの免疫を形成していない。多くの人が旅行前に抗生剤を使うものの、いくつかの抗生剤は有害な副作用を有し、また生物は多くの抗生剤への耐性を獲得しつつある。
【0143】
別の潜在的な用途は、とりわけ軍隊において、赤痢や他の感染性疾患を防止することである。ヒトSAA3は、腸関連感染症を減少または防止する迅速、安全、かつ効果的な手段である。
【0144】
別の例には、幼児下痢の予防または治療が含まれる。母乳で育てられた幼児は、調乳で育てられた幼児よりも感染がずっと少ない。初乳は幼児に有益な天然物質であり、ヒトSAA3は初乳の成分であることから、調乳のかけがえのない天然添加物となる。調乳は広く市販されており、例えばInfamel(商標), Similac(商標), Carnation Good Start(商標),およびGerber(商標)が挙げられる。プロバイオティックは、ウィルスによって引き起こされた下痢の重症度を減少させ、回復時間を短縮することが示されている。このことからヒトSAA3の別の用途は、上記症状を有する小児のためのものであり、この用途は病院での入院期間や経費を減少させることによる経済的影響をも有するであろう。
【0145】
別の例には壊死性腸炎(NEC)の予防または治療が含まれる。これは未熟児におこる深刻な合併症である。現在用いられている種々の生殖技術のために、未熟児の数は爆発的に増えた。NECのための治療法は、ここ数十年変わっていない。未熟児の腸の細菌がNECの罹患に主要な役割を果たすため、この症状の治療は、その未熟児に食事を与えず、強力な抗生剤を与え、そして腸に穴が開かないことを願うことからなる。
【0146】
ヒトSAA3の別の用途には、発生地域での下痢の予防が含まれる。大腸菌O-157:H7の発生は、溶血性尿毒症候群による死へとつながりうる。SAA3はムチン産生を誘導し、これは腸の上皮細胞に大腸菌が付着するのを防ぎ、したがってこの感染症を予防することができよう。
【0147】
別の例には、尿路感染症の治療または予防が含まれる。膀胱上皮細胞は、腸の上皮細胞によく似ており、ムチンを産生する能力を有する。したがって泌尿器の感染症の予防(導尿カテーテルを使っている入院患者も含まれる)もまた、本発明の医薬組成物の用途となる。
【0148】
本明細書には、腸のムチンのアップレギュレーションが含まれるものの、他の粘膜表面(例えば鼻咽頭、膀胱、など)内層の上皮細胞もまたムチンを産生する。これらのムチンは、腸のムチンに類似して感染症を防止する機能を果たし、また本発明における治療の効果的な標的にもなりうる。
【0149】
以下の実施例を、本発明をより詳細に説明するために提供する。これらは本発明を例証することを意図したものであり、限定することを意図したものではない。
【実施例】
【0150】
細胞系および培養
ヒト乳腺上皮細胞系MCF-7(ATCC)およびT47-D(ATCC)を本研究において用いた。これらのヒト乳房上皮細胞は両方ともプロラクチン(PRL)受容体を発現することが明らかにされている(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)ことから、これらの細胞を選択した。MCF-7細胞は、0.2U/ml インシュリン、2mMグルタミン、4.5g/Lグルコース、50μg/mLストレプトマイシン、50U/mLペニシリン、および10%FBSを添加したダルベッコ変法イーグル培地(DMEM)で増殖させた。T47-D細胞は、MCF-7細胞について記載したものと同じものを添加したRPMI 1640培地にさらに10mM HEPESおよび1mMピルビン酸ナトリウムを添加して培養した。細胞培養物は、37℃で5% CO2を含む加湿雰囲気でインキュベートした。細胞培養継代のために、付着細胞を、トリプシン-EDTA(Invitrogen Corp.)を用いて解放させた。
【0151】
細胞刺激アッセイ
M-SAA3発現を検出するために、これらのヒト乳腺細胞を、100ng/mLのヒト組換えプロラクチン(PRL)(Sigma)または20μg/mLの大腸菌リポポリサッカライド(LPS)(Difco)のいずれかで、0時間、0.5時間、4時間、24時間および48時間刺激させた。LPSで誘導する乳房細胞については、細胞をコンフルーエント状態まで増殖させ、次いで1μg/mLヒドロコルチゾンを含有しFBSを2%しか含有しない以外は上記と同じものを添加したDMEM中のLPSで刺激した。PRLで刺激する乳腺細胞については、細胞をMATRIGEL Matrix (BD Biosciences)でコーティングしたウェル上で85%コンフルーエント状態まで増殖させ、次いで以前に記載された通りのものを添加したDMEMおよびF12培地の無血清の1:1の混合物中のPRLで誘導した(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)。この研究においてもちいたPRL濃度は、ヒトの授乳におけるPRLの生理学的レベルに基づくものであり、それは典型的には100〜200ng/mLである。これに対して基底のレベルは2〜15ng/mLの範囲内である(M.C. Neville, J. Morton, Physiology and endocrine changes underlying human lactogenesis II, J. Nutr. 131 (2001) 3005S-3008S)。48時間刺激については、24時間後に培地を、適当な成分および刺激剤を含有する新鮮な培地で交換した。
【0152】
RNA単離およびRT-PCR
全RNAは、刺激していない、または刺激した乳腺上皮細胞から製造者が記載した手順に従いTRIZOL(Invitrogen Life Technologies)を用いて単離した。RNAはエタノール沈殿で濃縮し-80℃で保存した。RNA全体は1%(重量/容量)アガロースゲルで分画化し、次いでゲルを臭化エチジウムで染色することにより可視化した。第一のcDNA鎖合成は、全RNAを5μg、ポリd(T)14オリゴヌクレオチドを40μMおよび製造者の推奨に従ってSuperScriptII RNase H- Reverse Transcriptase(Invitrogen Life Technologies)を用いて行った。第二のcDNA鎖合成および二本鎖cDNAの増幅はAmpliTaq Gold DNA Polymerase (Applied Biosystems)を用いて行った。第二鎖cDNA合成に用いるフォワードF1およびリバースR1プライマー(表1)は、予測されるヒトSAA3 cDNA配列(GenBank登録番号X13895)の提唱されているエキソン2のアンチセンス鎖およびエキソン3のセンス鎖にそれぞれ相補的である。
【0153】
表1 ヒトM-SAA3 cDNAを取得するために用いたヒトSAA3特異的オリゴヌクレオチドプライマー
RT-PCRプライマー オリゴヌクレオチド配列 *bp♯
F1 5'-GATGAAGCTCTCCACTGGCATCA-3' 56 bp 配列番号5
R1 5'-TCAGAGTAGGCTCTCCACATGTCTT-3' 154 bp 配列番号6
F4 5'-AACTTGAAACAGAATGTGTATTATCCTTGGTTG-3' 1 bp 配列番号7
RACEプライマー オリゴヌクレオチド配列 *bp♯
R3 (1o 5'RACE) 5'-CAGCTGCCTTGAGGAATGTTAACCAT-3' 116 bp 配列番号8
R4 (2o 5'RACE) 5'-GCTGCTGACACCCAGGACCAG-3' 90 bp 配列番号9
F1b (1o 3'RACE) 5'-AGCAGGATGAAGCTCTCCTCTGGCATCA-3' 51 bp 配列番号10
F2c (2o 3'RACE) 5'-CAGCCAAGGATGGTTAACATTCCTCAAGGCA-3' 107 bp 配列番号11
*(bp♯は、本研究において得られたヒトM-SAA3 cDNA配列の5'ヌクレオチドに対する、図2の中のプライマーの位置に対応する。)
GeneAmp PCR System 2700 サーモサイクラー(Applied Biosystems)を用いるサイクリングパラメータは、95℃で10分を1サイクル、94℃で30秒、58℃で40秒、60℃で20秒を43サイクル、次いで60℃で10分を1サイクルであった。LPSまたはPRLで刺激した後に得られる123塩基対のRT-PCR産物は、pCRII-TOPO (Invitrogen Life Technologies)にクローニングした。クローニングしたインサートは同一性を確認するために両方向の配列を決定した。390塩基対のグリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)cDNA断片を増幅させるフォワードおよびリバースプライマーを含有する対照RT反応もまた、mRNA分解が生じなかったことを保証するために、各cDNA調製について行った。
【0154】
cDNA末端の5'および3'迅速増幅
全長M-SAA3 cDNAは、市販の5'RACE および GeneRacer System(Invitrogen Life Technologies)を製造業者の推奨に従って用いて、cDNA末端の5'および3'迅速増幅法(RACE)により取得した。簡潔には、刺激していない、および刺激した乳腺上皮細胞の両方由来の全RNA5μgを、5'RACE用遺伝子特異的リバースR1プライマー、および3'GeneRacer手順用ポリd(T)18GeneRacerアダプタープライマー(Invitrogen Life Technologies)を用いて逆転写した。ネステッド一次および二次PCR用の遺伝子特異的RACEプライマーは、製造業者の推奨に従って設計し、上述の123塩基対中央M-SAA3 cDNA断片の選択した領域に相補的であった。5'および3'RACE手順に用いたネステッド一次および二次PCRプライマーを表1に示す。一次PCRは第一鎖cDNAを1μL含有する50μLの容量中で行った。一次PCRに用いた熱サイクリングパラメータは、95℃で9分を1サイクル、94℃で30秒および72℃で1.5分を5サイクル、94℃で30秒および70℃で1.5分を5サイクル、94℃で30秒、65℃で1分、および72℃で1.5分を25サイクル、次いで72℃で10分を1サイクルであった。二次PCRは、適当な一次PCR混合物1μL、アダプタープライマー、およびフォワードまたはリバースネステッド二次遺伝子特異的プライマーのいずれか、を含有する50μL容量中で行った。熱サイクルパラメータは、一次PCRに用いたものと同じであり、ただし4つめのサイクリングパラメータとして25サイクルの代わりに33サイクルを用いた。結果的に得られる二次PCR増幅産物は、同一性を確認し、また全長ヒトM-SAA3 cDNA配列を取得するために配列決定した。
【0155】
M-SAA3のヌクレオチド配列解析
クローニングした123塩基対RT-PCR増幅産物は、ベクターに基づくSp6およびT7プライマーを用いて配列決定し、また二次5'および3'RACE産物は、遺伝子特異的ネステッド二次プライマーを用いて配列決定した。DNA配列は、the Wisconsin Genetics Computer Group (GCG) Package (バージョン10.2)の中のBESTFITおよびPILEUPプログラムを用いて解析した。M-SAA3 cDNAのヌクレオチド配列をGenBankデータベースに、登録番号AY209188で登録した。
【0156】
M-SAA3発現の誘導および検出
ヒト乳腺上皮細胞をPRLまたはLPSで刺激した後にSAA3が発現するかどうか判断するために、SAA3特異的F1およびR1プライマー(表1)を用いてRT-PCRを行った。GAPDH cDNAに相補的なフォワードおよびリバースプライマーを含有する対照RT反応は、各cDNA調製物に関して、予想される390塩基対のRT-PCR産物を増幅し、このことはcDNAを用意するために用いたmRNAが高い完全性を有していたことを保証する(図1)。SAA3特異的F1およびR1プライマーとPRLで0.5時間、4時間もしくは24時間、またはLPSで48時間刺激したヒトMCF-7乳腺細胞から調製したcDNAとのRT-PCRから、結果的に長さにして123塩基対の増幅産物が得られた。図1に、SAA3特異的F1およびR1プライマーとPRLで0.5時間、またはLPSで48時間刺激した乳房細胞から得られた123塩基対のRT-PCR産物を示す。また、図1に示されている通り、刺激していない乳腺細胞より調製したcDNAからは、SAA3特異的F1およびR1プライマーを用いても検出可能な増幅産物が得られなかった。この123塩基対産物の存在は、SAA3特異的F1およびR1プライマーとLPS刺激後わずか0.5時間のMCF-7細胞から調製したcDNAとのRT-PCRから検出され、またSAA3特異的F1およびR1プライマーとLPS刺激から48時間後の該細胞から調製したcDNAとのRT-PCRからも一貫して検出された。PRLで48時間刺激した後のマトリックス上のMCF-7細胞があまりにも非付着性になってしまったため、この時点以降のcDNAは調製しなかった。
【0157】
SAA3特異的F1およびR1プライマーとPRLまたはLPSで刺激したMCF-7細胞から調製したcDNAとから得られた123塩基対のRT-PCR産物の配列決定は、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によって当初推測されたヒトSAA3 cDNA配列のある領域と97%の相同性を示した。この乳房関連SAA3(M-SAA3)123塩基対RT-PCR産物のヌクレオチド配列を、ヒトM-SAA3の全長cDNA配列を含む両側のヌクレオチドと共に図2に示す。123塩基対のM-SAA3 RT-PCR断片は、スプライスジャンクションを含有し、そのスプライシングジャンクションでは、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)が推測した通り、2692ヌクレオチドのイントロンが正確にスプライシングで除かれた。このイントロンがヒトSAA3遺伝子中に存在することは、SAA3特異的F1およびR1プライマーと鋳型としてのヒトゲノムDNAとを用いてPCRを行うことにより確認した。このイントロンの不在は、このRT-PCR産物がM-SAA3 mRNAから生成されたものであって、ゲノムDNA混入物から生成されたものではないことを追認するものである。123塩基対のM-SAA3 cDNA産物はまた、刺激したT47-Dヒト乳腺細胞系から作成されたcDNA調製物からも検出された。
【0158】
全長ヒトM-SAA3 cDNAの単離および解析
全長M-SAA3 cDNAを取得するために、PRLまたはLPSで刺激した乳房MCF-7細胞から単離したRNAを用いて、cDNA末端の5'および3'迅速増幅(RACE)を行った。結果的に得られたM-SAA3特異的ネステッド二次プライマーを用いた二次PCRからの150塩基対の5'RACE産物、および、550塩基対の3'GeneRacer RACE産物を配列決定した。RACE手順により得られた二次PCR産物は両方とも、123塩基対のM-SAA3中央cDNA断片の配列と重複していた。最初の123塩基対のRT-PCR産物と同じように、これらの二次PCR増幅産物は、PRLまたはLPSで刺激したMCF-7乳房細胞からのcDNA調製物についてのみ検出された。5'RACE二次PCR産物が、M-SAA3 mRNAに由来するものであってゲノムDNAに由来するものではないことを確かめるために、M-SAA3特異的フォワードF4およびリバースR1プライマー(表1)をRT-PCRに用いた。結果的に得られた178塩基対のM-SAA3 cDNA断片は、予想通り第一イントロンを含有せず、この5'RACE二次PCR産物が確かにM-SAA3転写産物の5'領域の一部であるという証拠を提供した。同様に、二次PCRで得られた3'RACE産物は、M-SAA3 cDNA中の最後のイントロンを含有しなかった。これらの結果は一緒になって、RT-PCRおよび二次RACE産物がM-SAA3 mRNAに由来するものであって、ゲノムDNAに由来するものではないことをさらに検証した。さらに、これらの増幅産物はPRLまたはLPSで刺激した乳腺細胞に由来するcDNA調製物のみから検出することができ、このことは、これらの刺激物質がヒトM-SAA3遺伝子の転写発現を誘導することを追認した。
【0159】
PRLまたはLPSで刺激した後のヒトM-SAA3 cDNAについて得られた完全なヌクレオチド配列は長さにして655塩基対であり、これを図2に示す。また図2には、本研究で得られたヒトM-SAA3 cDNAと、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515) (GenBank 登録番号X13895)(配列番号3)が提唱した予測されるヒトSAA3 cDNA配列との比較を示す。他の3つのヒトSAA遺伝子の4エクソン構造と異なり、M-SAA3遺伝子は3エクソン構造を有する。SackおよびTalbotによって元々エクソン1と予測された領域はヒトM-SAA3 cDNAに存在せず、また当初予測されていたイントロン1全体がスプライシングで除かれることもない。そうではなく、5'側非翻訳領域(UTR)は、元々イントロン1の一部であろうと予測されていた最後の52ヌクレオチドを含有する。M-SAA3 cDNAの5'領域は妥当なKozakコンセンサス配列を含有し、最も有望な翻訳開始コドンはヌクレオチド57〜59に位置している(M. Kozak, Structural features in eukaryotic mRNAs that modulate the initiation of translation, J. Biol. Chem. 266 (1991) 19867-19870)。
【0160】
本発明者らの結果は、ヒトSAA3遺伝子の最初のイントロンが長さにして2692ヌクレオチドであり、GenBankに寄託されているいくつかのゲノムDNA配列(登録番号 AC055860.13およびAC108007.5)と一致していることを示した。このイントロンはそもそもSackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によってイントロン2として、そしてヒトSAA3遺伝子(GenBank登録番号X13895)中で長さにして2638ヌクレオチドであると予測されていた。本研究において得られたM-SAA3 cDNAは、イントロン1についてのスプライスサイト接合点が、SackおよびTalbot (G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)によってなされたイントロン2についての予測と一致することを示した。M-SAA3の最後のイントロンであるイントロン2は、長さにして321塩基対である。このイントロンのM-SAA3 mRNAからのプロセッシングは、イントロン3であろうと予測されていたこの領域に位置する、元々予測されていた273塩基対イントロンと類似していたものの、同一ではなかった。より詳しくは、M-SAA3の最後のまたは第3エキソンは、元々イントロン3の一部であろうと考えられていた、追加の71ヌクレオチドを含有していた。本研究において得られたヒトM-SAA3 cDNAと、GenBankに寄託されたヒトSAA3遺伝子配列(登録番号X13895)とのさらなる比較はまた、イントロン1およびイントロン2の両方が、スプライスのドナー/アクセプターサイト用のGT-AGコンセンサス配列と一致することを示した。
【0161】
ヒトM-SAA3 cDNAのヌクレオチド204における一塩基(T)挿入の存在は、フレームシフトを引き起こし、そして元々ヒトSAA3にコードされる遺伝子産物について予測されていたものよりも上流に翻訳停止コドンを生じさせた(配列番号3および4)(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515) (図2)。この一塩基(T)挿入は以前の研究で得られた結果を支持するものであり(B. Kluve-Beckerman, M.L. Drumm, M.D. Benson, Nonexpression of the human serum amyloid A three (SAA3) gene, DNA Cell Biol. 10 (1991) 651-661; (G.C. Sellar, A.S. Whitehead, Localization of four human serum amyloid A (SAA) protein superfamily genes to Chromosome 11p: Characterization of a fifth SAA-related gene sequence, Genomics 16 (1993) 774-776)、サンガーセンターヒトゲノム配列計画(www.ensemble.org/genome/central)より得られるヒトゲノムのこの領域のコンティグと一致した。それ故、結果的に得られる3'UTRは長さにして406塩基対であり、もともとヒトSAA3 cDNAについて予測されていた145塩基対3'UTRではない。図2に示すとおり、本研究において得られたヒトM-SAA3 cDNA配列と、予測されたSAA3 cDNAとの間にはまた、いくつかのさらなるヌクレオチドの相違が発見され、そのうちの4つは同族コドンがコードする予測される残基を変更する。一方、ポリ(A)尾部の21ヌクレオチド上流に位置する、M-SAA3転写産物のポリアデニル化シグナル(AAUAAA)は、ヒトSAA3遺伝子中のこのコンセンサス配列の位置について、SackおよびTalbotによってなされた予測と一致した(G.H. Sack, C.C. Talbot, Jr., The human serum amyloid A (SAA)-encoding gene GSAA1: nucleotide sequence and possible autocrine-collagenase-inducer function, Gene 84 (1989) 509-515)。
【0162】
ヒトM-SAA3と他のSAA3アイソフォームとの比較
ヒトM-SAA3 cDNAは、典型的に他のSAAタンパク質に見られる18残基のシグナルペプチドを含有する60アミノ酸の前駆体タンパク質に理論的に翻訳されるオープンリーディングフレームを含有した(図3)。成熟42アミノ酸タンパク質は、計算上4866.69ダルトンの分子量と理論上9.31のpIを有する。図3に示す通り、予測されるヒトM-SAA3タンパク質と、ウサギ、ハムスター、ウシ、およびマウスSAA3アイソフォームとの配列アライメントは、最初の48個のN末端残基について相当なアミノ酸相同性(〜94%)を示す。一方、ヒトM-SAA3転写産物の塩基204におけるヌクレオチド挿入は、下流で前駆タンパク質のコドン61で翻訳停止コドンを生じさせ、したがって他の哺乳動物SAA3アイソフォームよりも小さなタンパク質が結果的に得られる(図3)。コドン50におけるこの塩基挿入でコードされる予測残基、ならびにそれに続くヒトSAA3前駆体タンパク質のアミノ酸はまた、他の哺乳動物SAA3アイソフォームのこの領域のアミノ酸と著しくことなる(図3)。さらにヒトSAA3のこれらのC末端側の11個のアミノ酸、および前駆体タンパク質のコドン50のヌクレオチド挿入より前に位置する3つの残基は、以前に報告さたヒトSAA3の配列(GenBank登録番号X13895)(配列番号3および4)と異なる。
【0163】
ヒトM-SAA発現の調節
ホルモンであるPRLおよびグラム陰性細菌由来の外膜構成要素である内毒素LPSは、本研究においてヒトM-SAA3発現を刺激することが示された。ヒトおよび動物において、LPSはグラム陰性細菌感染症の存在を示す。内毒素ショックの主要なメディエーターであるLPS(E.T. Rietschel, T. Kirikae, F.U. Schade, U. Mamat, G. Schmidt, H. Loppnow, A.J. Ulmer, U. Zahringer, U. Seydel, F.D. Padowa, M. Schreier, H. Brade, Bacterial endotoxin: molecular relationships of structure to activity and function, FASEB J. 8 (1994) 217-225)は、授乳ウシ由来の乳房上皮細胞の一次培養物を、in vitroで用量依存的に炎症性サイトカインIL-1およびIL-6を産生するように刺激することが示されている(H. Okada, H. Ohtsuka, S. Kon-nai, R. Kirisawa, Y. Yokomizo, T. Yoshino, T.J. Rosol, Effects of lipopolysaccharide on production of interleukin-1 and interleukin-6 by bovine mammary epithelial cells in vitro, J. Vet. Med. Sci. 61 (1999) 33-35)。これらの乳房細胞はまた、炎症性サイトカインTNF-α転写産物を発現することが示されている(H. Okada, T. Ito, H. Ohtsuka, R. Kirisawa, H. Iwai, K. Yamashita, T. Yoshino, T.J. Rosol, Detection of interleukin-1 and interleukin-6 on cryopreserved bovine mammary epithelial cells in vitro, J. Vet. Med. Sci. 59 (1997) 503-507)。同様にヒト乳房上皮細胞の一次
培養物もまた、IL-6とTNF-αを産生することが示されている(F. Basolo, P.G. Conaldi, L. Fiore, S. Calvo, A. Toniolo, Normal breast epithelial cells produce interleukins 6 and 8 together with tumor-necrosis factor; defective IL6 expression in mammary carcinoma, Int. J. Cancer 55 (1993) 926-930)。炎症性サイトカインIL-1、IL-6、およびTNF-αは、特定の前後関係でSAA遺伝子の発現を活性化し、免疫応答の大きさを決定することが知られている。興味深いことに、ヒトSAA3プロモーターは、4つの推定上の急性期応答因子(APRF)コンセンサス結合サイト(CTGGGA)(配列番号12)を含有し、このことはLPSなどの内毒素が炎症性サイトカインの発現を誘導し、次いでそれらの炎症性サイトカインがM-SAA3発現を刺激することを示唆している。
【0164】
M-SAA3発現の別の誘導因子はPRLである。この多機能性ホルモンは、広範なスペクトルの生理学的プロセスを調節し、それには乳腺発達、授乳、ならびに内分泌、傍分泌および/または自己分泌機構による免疫機能が含まれる(M.C. Neville, J. Morton, Physiology and endocrine changes underlying human lactogenesis II, J. Nutr. 131 (2001) 3005S-3008S)。分子レベルでは、PRLは、多数の細胞(乳腺細胞も含まれる)に見られるヘマトポエチン/サイトカイン受容体スーパーファミリーのメンバーであるPRL受容体のホモダイマー化を誘導する(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236; C. Bole-Feysot, V. Goffin, M. Edery, N. Binart, P.A. Kelly, Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice, Endocr. Rev. 19 (1998) 225-268)。これは、受容体関連チロシン特異的キナーゼであるJAKの活性化を開始させ、それに続く情報変換因子(transducer)および転写活性化因子(STAT)の活性化を開始させる。STAT3は、炎症および組織損傷に応答して種々の急性期遺伝子の発現を誘導することがわかっている(J.N. Ihle, STATs: signal transducers and activators of transcription, Cell 84 (1996) 331-334)。in vitroおよびin vivoでのSTAT5の調節は、この因子が催乳性ホルモンシグナル経路において中心的で重要な役割を有し(B. Groner, F. Gouilleux, Prolactin-mediated gene activation in mammary epithelial cells, Curr. Opin. Genet. Dev. 5 (1995) 587-594)、またPRLに応答して乳房組織で母乳タンパク質の発現をアップレギュレートすることを示している(J.N. Ihle, STATs: signal transducers and activators of transcription, Cell 84 (1996) 331-334)。興味深いことに、ヒトSAA3のプロモーター領域(GenBank登録番号X13895)は、それぞれ5'UTRの1607塩基対(TTCCCGGAA)(配列番号15)および429塩基対(TTCCAAGGAA)(配列番号16)上流に位置する、推定上のSTAT3様 (TTCC(G=C)GGAA)(配列番号13)およびSTAT5様結合部位(TTCC(A>T)GGAA)(配列番号14)を含有する。これらのcis作用座因子は、LPSまたはPRL刺激に続くM-SAA3転写発現のアップレギュレーションを部分的に担っている可能性がある。PRLがヒト乳房上皮細胞系でSTAT5の活性化を誘導することは、以前に報告されている(E. Canbay, M. Norman, E. Kilic, V. Goffin, I. Zachary, Prolactin stimulates the JAK2 and focal adhesion kinase pathways in human breast carcinoma T47-D cells, Biochem. J. 324 (1991) 231-236)。IL-6がヒト乳房上皮細胞系のJAK/STAT経路を活性化することが示されているものの(A. Badache, N.E. Hynes, Interleukin 6 inhibits proliferation and, in cooperation with an epidermal growth factor receptor autocrine loop, increases migration of T47D breast cancer cells, Cancer Res. 61 (2001) 383-391)、LPSが、ヒト乳房上皮細胞におけるSTAT因子活性化に与える影響は未だ不明である。
【0165】
SAAの正確な生物学的機能は解明されていないが、いくつかの研究は、種々のSAAアイソフォームのN末端領域が、それらの構造および機能の性質を部分的に担うことを示唆している。例えば、A-SAA1およびA-SAA2のN末端領域は、コレステロールに特異的に結合し、コレステロールをHepG2肝臓および大動脈平滑筋細胞へと輸送するが、このことは急性期応答中およびアテローム硬化症における、A-SAA1およびA-SAA2によるコレステロールフラックス調節のもっともらしい役割を示唆する(J. Liang, B.M. Schreiber, M. Salmona, G. Phillip, W.A. Gonnerman, F.C. de Beer, J.D. Sipe, Amino terminal region of acute phase, but not constitutive, serum amyloid A (apoSAA) specifically binds and transports cholesterol into aortic smooth muscule and HepG2 cells, J. Lipid Res. 37 (1996) 2109-2116)。別の研究は、A-SAAのN末端領域が、二次性アミロイド症に見られるアミロイドフィブリルの主要な構成要素であるアミロイドAの前駆体であることを実証した(A. Husebekk, B. Skogen, G. Husby, G. Marhaug, Transformation of amyloid precursor SAA to protein AA and incorporation in amyloid fibrils in vivo, Scand. J. Immunol. 21 (1985) 283-287; J.J. Liepnieks, B.K. Beckerman, M.D. Benson, Characterization of amyloid A protein in human secondary amyloidosis: the predominant deposition of serum amyloid A1, Biochim. Biophys. Acta 1270 (1995) 81-86)。本発明者らは以前に、いくつかの哺乳動物の初乳からM-SAA3アイソフォームを単離した(T.L. McDonald, M.A. Larson, D.R. Mack, A. Weber, Elevated extrahepatic expression and secretion of mammary-associated serum amyloid A 3 (M-SAA3) into colostrum, Vet. Immunol. Immunopathol. 83 (2001) 205-213)。さらに、ヒト腸上皮細胞を、合成したM-SAA3のN末端側の10アミノ酸で前処理することは、保護性腸ムチンMUC3の産生を刺激し、かつ未処理の細胞と比較して、腸病原性大腸菌(EPEC)がこれらの腸上皮細胞に付着するのを著しく減少させる(D.R. Mack, T.L. McDonald, M.A. Larson, S. Wei, A. Weber, The conserved TFLK motif of mammary-associated serum amyloid A3 is responsible for upregulation of intestinal MUC3 mucin expression in vitro, Pediatr. Res. 53 (2003) 137-142; M.A. Larson, S.H. Wei, A. Weber, D.R. Mack, T.L. McDonald, Human mammary-associated serum amyloid A3 peptide enhances intestinal MUC3 expression and inhibits EPEC adherence, Biochem. Biophys. Res. Comm. 300 (2003) 531-540)。これらの研究は一緒になって、種々のSAAアイソフォームに対して、組成、構造、および発現または局在化の場所に基づいて、機能を帰するが、これらの機能は全て、正または負の結果に寄与する。
【0166】
引用文献
本明細書において引用した全ての文献を、参照により明白にその全内容を本明細書に組み入れる。これには以下の公表された特許出願および特許も含まれる。米国特許第6509444、WO 01/31006、およびPCT/US99/19428;
M.A. Larson, A. Weber, A.T. Weber, T.L. McDonald, Differential expression of bovine mammary-associated serum amyloid A3 is induced by prolactin or lipopolysaccharide, (準備中);
G. Ramadori, J.D. Sipe, H.R. Colten, Expression and regulation of the murine serum amyloid A (SAA) gene in extrahepatic sites, J. Immunol. 135 (1985) 3645-3647;
R.L. Meek, E.P. Benditt, Amyloid A gene family expression in different mouse tissues, J. Exp. Med. 164 (1986) 2006-2017;
R.L. Meek, E.P. Benditt, Rat tissues express serum amyloid A protein-related mRNAs, Proc. Natl. Acad. Sci. USA 86(1989) 1890-1894。
【図面の簡単な説明】
【0167】
【図1】リポ多糖(LPS)またはプロラクチン(PRL)のいずれかで刺激する前および後のヒト乳腺上皮細胞系MCF-7から単離したM-SAA3 mRNAのRT-PCR解析を示すゲル。RT-PCRに用いたヒトSAA-3特異的F1およびR1プライマー(表1参照)は、予測エキソン2および3(GenBank 登録番号X13895)、またはこの研究で決定したエクソン1および2における残基をコードするヌクレオチドに対応する。レーン1はDNAマーカー、レーン2は逆転写酵素無しで調製したcDNAサンプルのRT-PCR増幅の代表例、レーン3はヒトGAPDH特異的プライマーを使用した刺激していない乳腺上皮細胞から調製したcDNAサンプルのRT-PCR増幅の代表例、レーン4は刺激していないMCF-7細胞、レーン5はPRLで刺激したMCF-7細胞、レーン6は刺激していないMCF-7細胞、レーン7はLPSで刺激したMCF-7細胞である。
【図2】本明細書に記載したヒトM-SAA3 cDNA配列と、ヒトSAA3遺伝子の最初に推測されたcDNA配列(GenBank登録番号X13895)(配列番号3および4)との比較を示す。GCGパッケージ(バージョン10.2)のBESTFITプログラムを、ヌクレオチド配列アライメントを得るために用いた。同一のヌクレオチドを縦の線で示し、ダッシュで類似性を最大化するために導入したギャップを示す。ヒトSAA3 cDNA配列の5'末端からのヌクレオチドの数を右に示す。最初の123 bp RT-PCR産物を生成するために用いたヒトSAA3特異的フォワードF1およびリバースR1プライマーの位置を、しかるべきヌクレオチド配列の上に示し、各プライマーの方向を矢印で示す。推定される翻訳開始コドンを太字で下線を引いて示す。M-SAA3転写産物におけるイントロン1と2のスプライス部位接合部を、下向きの矢印で接合部の上に示す。アスタリスクは、推定されるM-SAA3タンパク質が結果的にフレームシフトする(読み枠がずれる)1塩基挿入を示す。太字および二重の下線は、推定される翻訳停止コドンを示す。ポリ(A)シグナルは、下線を引いたポリアデニル化のためのコンセンサス配列の上に示されており、ポリA尾部は太字である。
【図3】推定されるヒトSAA3タンパク質と、ウサギ、ハムスター、ウシおよびマウス由来のSAA3アイソフォームとの比較を示す。アミノ酸配列のアライメントは、GCGパッケージのPILEUPプログラムを用いて得た。哺乳動物およびSAA3タンパク質の略記に続けて、対応する登録番号を以下に示す。rab-SAA3=ウサギSAA3(GenBank M64696)、ham-SAA3=ハムスターSAA3(GenBank M33431)、bov-SAA3=ウシSAA3(GenBank AF335552)、mou-SAA3=マウスSAA3(SWISS-PROT P04918)、およびhum-SAA3=ヒトSAA3。少なくとも3種のSAA3アイソフォームにおける同一または類似する残基を暗い背景で強調した。ダッシュは類似性を最大化するために導入したギャップを表す。適切なSAA3アイソフォームの後に前駆タンパク質のN末端からの残基の数を右に示し、シグナルペプチドの推定される切断部位を下向きの三角形(▽)で示す。
【特許請求の範囲】
【請求項1】
SAA3タンパク質をコードし、以下からなる群より選択される核酸配列を含む、単離された核酸分子:
(a)配列番号1に記載の配列を含むヌクレオチド配列;
(b)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、配列番号1に記載の配列の少なくとも20の連続したヌクレオチドを含むヌクレオチド配列;
(c)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、配列番号1に記載の配列に対して少なくとも80%の配列相同性を有するヌクレオチド配列;
(d)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、高ストリンジェンシー条件下で配列番号1に記載の配列とハイブリダイズするヌクレオチド配列;
(e)配列番号2に記載のアミノ酸配列を有するタンパク質をコードするヌクレオチド配列。
【請求項2】
請求項1に記載の核酸分子を含む、ベクター。
【請求項3】
請求項2に記載のベクターで形質転換した、宿主細胞。
【請求項4】
宿主細胞が細菌細胞である、請求項3に記載の宿主細胞。
【請求項5】
宿主細胞が動物細胞である、請求項3に記載の宿主細胞。
【請求項6】
細胞がヒト由来である、請求項5に記載の動物細胞。
【請求項7】
細胞が乳房細胞である、請求項5に記載の動物細胞。
【請求項8】
プロラクチンまたはLPS刺激に応答して動物細胞における機能的に連結された核酸配列の発現を促進させる核酸分子であって、ここで前記核酸分子が以下からなる群より選択されるものである、前記核酸分子:
(a)配列番号1に記載の配列の5'側56ヌクレオチドのうちの少なくとも20の連続したヌクレオチドを含むヌクレオチド配列;
(b)SAA3をコードする配列と天然で関連しており、ヒト動物細胞における該SAA3コード配列の発現を促進させるヌクレオチド配列;
(c)配列番号1に記載の配列の最初の56ヌクレオチドに対して少なくとも80%の配列相同性を有するヌクレオチド配列;
(d)配列番号1に記載の配列の最初の56ヌクレオチドと、高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列。
【請求項9】
請求項8に記載の核酸分子を含む、ベクター。
【請求項10】
請求項9に記載のベクターで形質転換した、宿主細胞。
【請求項11】
ゲノムに安定に組み込まれている異種配列に機能的に連結した、請求項8に記載のヌクレオチド配列を有する動物細胞。
【請求項12】
動物細胞が哺乳動物由来である、請求項11に記載の動物細胞。
【請求項13】
細胞が乳腺細胞である、請求項12に記載の哺乳動物細胞。
【請求項14】
乳腺細胞が上皮乳腺細胞である、請求項13に記載の哺乳動物細胞。
【請求項15】
細胞において異種配列の転写を誘導する方法であって、プロラクチンまたはLPSなどの刺激に応答して細胞における異種配列の転写を誘導するプロモーターに機能的に連結された前記異種配列を含んでなる核酸分子で細胞を形質転換し、ここで前記プロモーターは配列番号1に記載の配列の最初の56ヌクレオチドを含んでなるヌクレオチド配列を含んでなるものであり;次いで前記細胞を前記刺激に曝露することを含んでなる前記方法。
【請求項16】
細胞が細菌細胞である、請求項15に記載の方法。
【請求項17】
細胞が動物細胞である、請求項15に記載の方法。
【請求項18】
細胞が乳房細胞である、請求項15に記載の方法。
【請求項19】
細胞が乳房上皮細胞である、請求項15に記載の方法。
【請求項20】
以下よりなる群から選択され、かつSAA3活性を有する単離されたポリペプチド:
(a)配列番号2に記載のアミノ酸配列を含んでなるポリペプチド;
(b)配列番号1に記載の配列を含んでなるヌクレオチド配列によりコードされるポリペプチド;
(c)GenBank寄託番号AY209188として寄託されたヌクレオチド配列によりコードされるアミノ酸配列を含んでなるポリペプチド;
(d)配列番号1に記載の配列と少なくとも80%の配列相同性を有するヌクレオチド配列がコードするポリペプチド;
(e)配列番号2に記載の配列と少なくとも80%の配列相同性を有するアミノ酸配列を含んでなるポリペプチド;
(f) (a)〜(e)のいずれかのアミノ酸配列の少なくとも30個の連続したアミノ酸を含んでなるポリペプチド。
【請求項21】
SAA3活性を有するポリペプチドをコードする単離された核酸分子であって、以下よりなる群から選択される核酸分子:
(a)配列番号1に記載の配列を含んでなる核酸分子;
(b)配列番号2に記載のアミノ酸配列をコードする配列を含んでなる核酸分子;
(c)GenBank寄託番号AY209188として寄託された配列を含んでなる核酸分子;
(d) (a)、(b)、または(c)の配列に対応するアンチセンス配列を含んでなる核酸分子;
(e)配列番号1に記載のヌクレオチド配列と、少なくとも80%の配列相同性を有する配列を含んでなる核酸分子;
(f)配列番号1に記載の配列と、高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列;
(g) (a)〜(e)のいずれかの配列の少なくとも50個の連続した核酸の配列を含んでなる核酸分子。
【請求項22】
宿主細胞で発現を促進するプロモーターに機能的に連結された、請求項21に記載の核酸分子を含んでなる、ヌクレオチド構築物。
【請求項1】
SAA3タンパク質をコードし、以下からなる群より選択される核酸配列を含む、単離された核酸分子:
(a)配列番号1に記載の配列を含むヌクレオチド配列;
(b)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、配列番号1に記載の配列の少なくとも20の連続したヌクレオチドを含むヌクレオチド配列;
(c)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、配列番号1に記載の配列に対して少なくとも80%の配列相同性を有するヌクレオチド配列;
(d)SAA3タンパク質の生物学的活性を有するタンパク質をコードする、高ストリンジェンシー条件下で配列番号1に記載の配列とハイブリダイズするヌクレオチド配列;
(e)配列番号2に記載のアミノ酸配列を有するタンパク質をコードするヌクレオチド配列。
【請求項2】
請求項1に記載の核酸分子を含む、ベクター。
【請求項3】
請求項2に記載のベクターで形質転換した、宿主細胞。
【請求項4】
宿主細胞が細菌細胞である、請求項3に記載の宿主細胞。
【請求項5】
宿主細胞が動物細胞である、請求項3に記載の宿主細胞。
【請求項6】
細胞がヒト由来である、請求項5に記載の動物細胞。
【請求項7】
細胞が乳房細胞である、請求項5に記載の動物細胞。
【請求項8】
プロラクチンまたはLPS刺激に応答して動物細胞における機能的に連結された核酸配列の発現を促進させる核酸分子であって、ここで前記核酸分子が以下からなる群より選択されるものである、前記核酸分子:
(a)配列番号1に記載の配列の5'側56ヌクレオチドのうちの少なくとも20の連続したヌクレオチドを含むヌクレオチド配列;
(b)SAA3をコードする配列と天然で関連しており、ヒト動物細胞における該SAA3コード配列の発現を促進させるヌクレオチド配列;
(c)配列番号1に記載の配列の最初の56ヌクレオチドに対して少なくとも80%の配列相同性を有するヌクレオチド配列;
(d)配列番号1に記載の配列の最初の56ヌクレオチドと、高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列。
【請求項9】
請求項8に記載の核酸分子を含む、ベクター。
【請求項10】
請求項9に記載のベクターで形質転換した、宿主細胞。
【請求項11】
ゲノムに安定に組み込まれている異種配列に機能的に連結した、請求項8に記載のヌクレオチド配列を有する動物細胞。
【請求項12】
動物細胞が哺乳動物由来である、請求項11に記載の動物細胞。
【請求項13】
細胞が乳腺細胞である、請求項12に記載の哺乳動物細胞。
【請求項14】
乳腺細胞が上皮乳腺細胞である、請求項13に記載の哺乳動物細胞。
【請求項15】
細胞において異種配列の転写を誘導する方法であって、プロラクチンまたはLPSなどの刺激に応答して細胞における異種配列の転写を誘導するプロモーターに機能的に連結された前記異種配列を含んでなる核酸分子で細胞を形質転換し、ここで前記プロモーターは配列番号1に記載の配列の最初の56ヌクレオチドを含んでなるヌクレオチド配列を含んでなるものであり;次いで前記細胞を前記刺激に曝露することを含んでなる前記方法。
【請求項16】
細胞が細菌細胞である、請求項15に記載の方法。
【請求項17】
細胞が動物細胞である、請求項15に記載の方法。
【請求項18】
細胞が乳房細胞である、請求項15に記載の方法。
【請求項19】
細胞が乳房上皮細胞である、請求項15に記載の方法。
【請求項20】
以下よりなる群から選択され、かつSAA3活性を有する単離されたポリペプチド:
(a)配列番号2に記載のアミノ酸配列を含んでなるポリペプチド;
(b)配列番号1に記載の配列を含んでなるヌクレオチド配列によりコードされるポリペプチド;
(c)GenBank寄託番号AY209188として寄託されたヌクレオチド配列によりコードされるアミノ酸配列を含んでなるポリペプチド;
(d)配列番号1に記載の配列と少なくとも80%の配列相同性を有するヌクレオチド配列がコードするポリペプチド;
(e)配列番号2に記載の配列と少なくとも80%の配列相同性を有するアミノ酸配列を含んでなるポリペプチド;
(f) (a)〜(e)のいずれかのアミノ酸配列の少なくとも30個の連続したアミノ酸を含んでなるポリペプチド。
【請求項21】
SAA3活性を有するポリペプチドをコードする単離された核酸分子であって、以下よりなる群から選択される核酸分子:
(a)配列番号1に記載の配列を含んでなる核酸分子;
(b)配列番号2に記載のアミノ酸配列をコードする配列を含んでなる核酸分子;
(c)GenBank寄託番号AY209188として寄託された配列を含んでなる核酸分子;
(d) (a)、(b)、または(c)の配列に対応するアンチセンス配列を含んでなる核酸分子;
(e)配列番号1に記載のヌクレオチド配列と、少なくとも80%の配列相同性を有する配列を含んでなる核酸分子;
(f)配列番号1に記載の配列と、高ストリンジェンシー条件下でハイブリダイズするヌクレオチド配列;
(g) (a)〜(e)のいずれかの配列の少なくとも50個の連続した核酸の配列を含んでなる核酸分子。
【請求項22】
宿主細胞で発現を促進するプロモーターに機能的に連結された、請求項21に記載の核酸分子を含んでなる、ヌクレオチド構築物。
【図1】


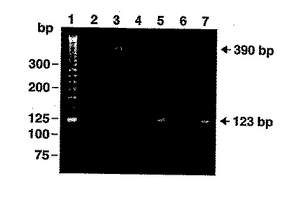
【図2−1】

【図2−2】
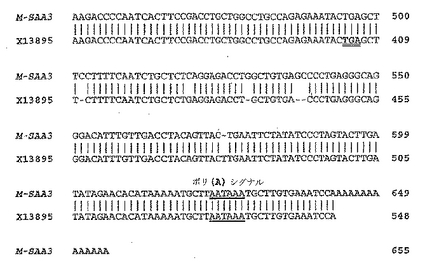
【図3】
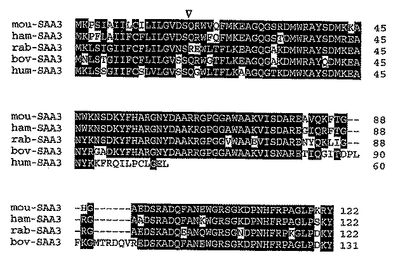
【図2−1】
【図2−2】
【図3】
【公表番号】特表2006−516194(P2006−516194A)
【公表日】平成18年6月29日(2006.6.29)
【国際特許分類】
【出願番号】特願2005−518834(P2005−518834)
【出願日】平成16年1月21日(2004.1.21)
【国際出願番号】PCT/US2004/001488
【国際公開番号】WO2004/065554
【国際公開日】平成16年8月5日(2004.8.5)
【出願人】(500525863)ボード・オブ・リージェンツ・オブ・ザ・ユニバーシティ・オブ・ネブラスカ (2)
【Fターム(参考)】
【公表日】平成18年6月29日(2006.6.29)
【国際特許分類】
【出願日】平成16年1月21日(2004.1.21)
【国際出願番号】PCT/US2004/001488
【国際公開番号】WO2004/065554
【国際公開日】平成16年8月5日(2004.8.5)
【出願人】(500525863)ボード・オブ・リージェンツ・オブ・ザ・ユニバーシティ・オブ・ネブラスカ (2)
【Fターム(参考)】
[ Back to top ]