
ヒドロゲナーゼ発現ベクター
【課題】エシェリヒア属等に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属に属する微生物内でNAD+の還元を行う可溶型ヒドロゲナーゼを誘導発現可能なベクターの提供を課題とする。
【解決手段】本発明者らは、大量発現系の構築が可能な大腸菌株(Esherichia coli)を宿主とする、ヒドロゲナーゼを発現する形質転換系の構築を行った。
その結果、本発明者らは、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子を導入することによって、エシェリヒア属等に属する微生物内でNAD+還元型ヒドロゲナーゼを誘導発現させることに成功した。
【解決手段】本発明者らは、大量発現系の構築が可能な大腸菌株(Esherichia coli)を宿主とする、ヒドロゲナーゼを発現する形質転換系の構築を行った。
その結果、本発明者らは、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子を導入することによって、エシェリヒア属等に属する微生物内でNAD+還元型ヒドロゲナーゼを誘導発現させることに成功した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、エシェリヒア属に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属等に属する微生物内でヒドロゲナーゼを誘導発現可能なベクター及びその利用に関する。
【背景技術】
【0002】
微生物を利用した工業原料等有用物質の製造プロセスは、一般にエネルギー消費が少なく、廃棄物も少ないという特徴がある。このため、近年、資源枯渇やCO2等排出物の環境への影響が懸念されている中、微生物機能を活用した有用物質の生産(バイオプロセス)技術の開発が環境調和型循環産業システムにおける製造技術基盤として必要とされている。
【0003】
微生物が持つ酵素は、原料から特定の光学活性を持つ物質を合成するという選択性を有するものが多い。そのため、酵素を用いて効率よく目的物質を合成、取得できるので無駄がないとされている。また、微生物による物質生産は高価な触媒や高温高圧などの厳しい処理条件を必要とせず、常温、常圧、pH中性付近などの比較的温和な条件で進むため、扱いやすくかつ消費するエネルギーも少ない点が利点である。
【0004】
物質生産に利用される微生物の酵素反応の多くは酸化還元反応であり、特に基質を還元する反応の場合、基質のほかに還元力となる捕酵素を必要とする。補酵素としてよく使用されるNADH、NADPHはともに高価であり、電気化学的な方法により再生が試みられているが実用的とは言えず、酵素によりこれらの再生を行う試みが行われている。
【0005】
微生物による物質生産における捕酵素再生酵素として、NAD+還元型ヒドロゲナーゼを利用するには、物質生産を行う菌にヒドロゲナーゼを異種発現させる必要がある。しかし、ヒドロゲナーゼの異種微生物を宿主とした遺伝子発現はきわめて困難であり、ヒドロゲナーゼを異種発現させた場合には、不活性型であり封入体となっている例が多く見られる。これはヒドロゲナーゼが複雑な翻訳後修飾を得るためだと思われる。多種多様なヒドロゲナーゼが存在する中で、現在のところ12個の発現例のみが報告されている(非特許文献1〜12)。
【0006】
Ralstonia eutropha(旧Alcaligenes eutrophus, Wautersia eutropha, ATCC 17697)は、土壌に生息するグラム陰性好気性菌である。本菌は有機物を炭素源として従属栄養的に生育するほかに、水素、酸素、二酸化炭素による独立栄養的な生育も行う。その場合、炭素固定はカルビンサイクルで行われ、電子伝達系を共役したヒドロゲナーゼでエネルギー生産が行われている。R. eutrophaは、エネルギー生産を担う膜結合型ヒドロゲナーゼ(Membrane-bound hydrogenase(MBH))、水素を基質としてNAD+の還元を行う可溶型ヒドロゲナーゼ(soluble hydrogenase(SH))、水素を感知しヒドロゲナーゼの遺伝子発現の制御に関わる制御ヒドロゲナーゼ(regulatory hydrogenase(RH))の3種類のヒドロゲナーゼを保有している(非特許文献13)。
【0007】
この中で微生物による物質生産における捕酵素再生酵素として、R. eutropha由来の水素を基質としてNAD+の還元を行う可溶型ヒドロゲナーゼの異種宿主での発現がこれまで試みられてきたが、成功例は報告されていなかった。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Karube I. et al, FEBS, 1993, 158-1, 119-122
【非特許文献2】Rousset M. et al, J Bacteriol., 1998, 180(18), 4982-4986
【非特許文献3】Asada Y. et al, Biochim Biophys Acta., 2000, 1490(3), 269-278
【非特許文献4】Porthun A. et al, Arch Microbiol., 2002, 177(2), 159-166
【非特許文献5】Posewitz MC. et al, J Biol Chem., 2004, 279(24), 25711-25720
【非特許文献6】Mishra J. et al, Biochem Biophys Res Commun., 2004, 324(2), 679-685
【非特許文献7】Girbal L. et al, Appl Environ Microbiol., 2005, 71(5), 2777-2781
【非特許文献8】Lenz O. et al, J Bacteriol., 2005, 187(18), 6590-6595
【非特許文献9】King PW. et al, J Bacteriol., 2006, 188(6), 2163-2172
【非特許文献10】Lenz O. et al, FEBS Lett., 2007, 10, 581(17), 3322-3326
【非特許文献11】Sybirna K. et al, BMC Biotechnol., 2008, 8, 73
【非特許文献12】Sun J. et al, PLoS ONE, 2010, 5, 5, e10526
【非特許文献13】Burgdorf T. et al, J Mol Microbiol Biotechnol, 2005, 10, 181-196
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は上記状況に鑑みてなされたものであり、本発明が解決しようとする課題は、エシェリヒア属等に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属に属する微生物内でNAD+の還元を行う可溶型ヒドロゲナーゼを誘導発現可能なベクターを提供することである。
【課題を解決するための手段】
【0010】
上記課題を解決すべく、本発明者らは、大量発現系の構築が可能な大腸菌株(Esherichia coli)を宿主とする、ヒドロゲナーゼを発現する形質転換系の構築を行った。
【0011】
まず、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3(配列番号:4)に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ(構造遺伝子hoxFUYHWI)をコードするポリヌクレオチドを導入した(pKV15358-4、配列番号:1)。
【0012】
プラスミドベクターpKV15358-3はコクリア バリアンス(Kocuria varians)NBRC15358由来のプラスミド(pKV15358-1)のレスキュープラスミドを小型化し、さらにコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株における翻訳において最適なSD前後配列を付加した、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株用ベクターである。
【0013】
次に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼを導入した該プラスミド(pKV15358-4)を、様々な培養条件下で大腸菌(Esherichia coli)JM109株により発現させ、無細胞抽出液の調製を行った後に、ヒドロゲナーゼの活性測定を行った。その結果、微好気条件において培養を行った場合に、ヒドロゲナーゼ活性が観測され、該プラスミドが大腸菌を宿主としてNAD+還元型ヒドロゲナーゼを発現することが明らかとなった。
【0014】
即ち、本発明者らは、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子を導入することによって、エシェリヒア属等に属する微生物内でNAD+還元型ヒドロゲナーゼを誘導発現させることに成功し、これにより本発明を完成するに至った。
【0015】
本発明は、より具体的には以下の〔1〕〜〔9〕を提供するものである。
〔1〕配列番号:4に記載の塩基配列からなるポリヌクレオチド、又は配列番号:4に記載の塩基配列からなるポリヌクレオチドの相補鎖とストリンジェントな条件化でハイブリダイズするポリヌクレオチド、及びヒドロゲナーゼをコードするポリヌクレオチドを含む、微生物内でヒドロゲナーゼを発現可能なベクター。
〔2〕前記微生物が、エシェリヒア属又はコクリア属に属する微生物であることを特徴とする、〔1〕に記載のベクター。
〔3〕前記ヒドロゲナーゼが、可溶型ヒドロゲナーゼであることを特徴とする、〔1〕又は〔2〕に記載のベクター。
〔4〕前記ヒドロゲナーゼが、NAD+還元能及び/又は酸素耐性を有することを特徴とする、〔1〕〜〔3〕のいずれかに記載のベクター。
〔5〕前記ヒドロゲナーゼが、下記(a)〜(d)のいずれかに記載のタンパク質複合体であることを特徴とする、〔1〕〜〔4〕のいずれかに記載のベクター;
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
〔6〕前記ヒドロゲナーゼを発現させる微生物がエシェリヒア属に属する微生物であることを特徴とする、〔1〕〜〔5〕のいずれかに記載のベクター。
〔7〕〔1〕〜〔6〕のいずれかに記載の組換えベクターにより、宿主を形質転換した形質転換体。
〔8〕 〔7〕に記載の形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法。
〔9〕微好気的条件で形質転換体を培養することを特徴とする、〔8〕に記載の製造方法。
【発明の効果】
【0016】
Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子が導入されたコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3を、大量発現系の構築が可能なエシェリヒア属に属する微生物に導入することにより、NAD+還元型ヒドロゲナーゼの大量発現系の構築が可能となる。また、本発明のプラスミドが導入された微生物を用いて、微生物発現系においてNAD+還元型ヒドロゲナーゼの活性(例えば触媒活性)を発現させることが可能となる。
【図面の簡単な説明】
【0017】
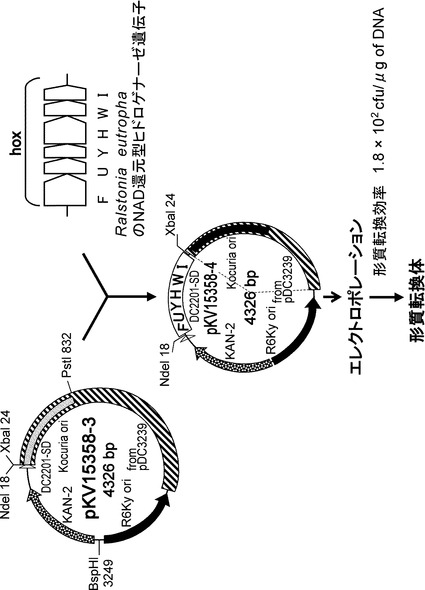
【図1】hoxFUYHWIを保持するkocuria rhizophila DC2201の調製方法を示す図である。
【発明を実施するための形態】
【0018】
本発明は、エシェリヒア属又はコクリア属に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属又はコクリア属に属する微生物内でヒドロゲナーゼを誘導発現可能なベクターを提供する。
【0019】
本発明のベクターは、エシェリヒア属又はコクリア属に属する微生物内で複製可能なDNA領域、及びエシェリヒア属に属する微生物に耐性を与える薬剤マーカーを有する。本発明のベクターは、エシェリヒア属又はコクリア属微生物内において複製可能なシャトルベクタープラスミドであり、ヒドロゲナーゼをエシェリヒア属又はコクリア属微生物内で発現させるのに有効なベクターである。
【0020】
本発明のシャトルベクターとしては、コクリア バリアンス(Kocuria varians)NBRC15358株から得られる環状プラスミドを挙げることができる。このプラスミドの複製に関与するDNA領域を明らかにし、当該DNA領域を含有させたプラスミドを作製すると、このようなプラスミドは、エシェリヒア属又はコクリア属微生物内で複製することが可能となる。
【0021】
エシェリヒア属又はコクリア属に属する微生物内で複製可能なDNA領域は、概領域を含むベクターをエシェリヒア属又はコクリア属微生物に導入したときに、ベクターが複製できる領域である限り特に制限されないが、例えば、コクリア バリアンス(Kocuria varians)株が保有する内因性プラスミドベクターpKV15358-1 (配列番号:2)に由来する自律複製領域を挙げることができる。pKV15358-1に由来する自律複製領域は、pKV15358-1の全長であってもよいが、遺伝子操作を容易にするために、pKV15358-1の一部であることが好ましい。さらに、コクリア属微生物で機能しうる限り、配列番号:2に記載された塩基配列において1もしくは数個の塩基が置換・欠失・挿入等された配列を有するDNA領域、又は配列番号:2に記載の塩基配列からなるポリヌクレオチドもしくは該配列から調製されうるプローブとストリンジェントな条件化でハイブリダイズするDNA領域であってもよい。ここで、数個とは例えば、2〜50個、好ましくは2〜10個、より好ましくは2〜5個である。また、ストリンジェントな条件としては、通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件が挙げられる。1もしくは数個の塩基の置換・欠失・挿入は、出願時に当業者に公知の技術により行うことができる。
【0022】
上述のようなベクターpKV15358-1の自律複製に必要な領域は、抗生物質感受性宿主細胞内での抗生物質耐性遺伝子の発現を指標とする欠失実験によって特定できる。即ち、ベクターpKV15358-1に、カナマイシン耐性遺伝子やアンピシリン耐性遺伝子などの薬剤耐性遺伝子を導入した後、該遺伝子を消化しない単独又は組み合わせの制限酵素によって消化し、DNA連結酵素により再環状化し、その後薬剤感受性の菌体を形質転換させる。この中から薬剤耐性を指標にして選択される薬剤体制菌株には、ベクターpKV15358-1の自律複製領域を含む派生ベクターを持ち、それらのベクターの制限酵素地図を比較することにより、ベクターpKV15358-1の自律複製領域が特定できる。
【0023】
本発明において、エシェリヒア属又はコクリア属微生物に耐性を与える薬剤マーカーとしては、該細菌に導入したときに薬剤耐性を付与する遺伝子であれば特に制限されないが、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、又はクロラムフェニコール耐性遺伝子等を挙げることができる。
【0024】
本発明のプラスミドベクターは、上記自律複製領域及び薬剤耐性マーカーを含むベクターであるが、このようなベクターとしては具体的には、pKV15358-2(配列番号:3)を挙げることができる。pKV15358-2は、上記pKV15358-1を制限酵素Xba1で切断して得られる自律複製領域を含む断片と、カナマイシン耐性遺伝子等により構成されるベクターである。尚、本発明のベクターはpKV15358-2が改変されたものであってもよい。改変としては、複製能や薬剤耐性に関与しない部位における変異、制限酵素部位のうちの一部の欠損や置換などを挙げることができる。
【0025】
本発明のベクターは、さらにコクリア属に属する微生物由来のSD配列を有していてもよい。SD配列は特に限定されないが、好ましくは配列番号:5に記載のSD配列を挙げることができる。SD配列をさらに含むベクターとしては、pKV15358-3(配列番号:4)を挙げることができる。
【0026】
また、コクリア属微生物において機能しうるプロモーターなどがさらに組み込まれていてもよい。プロモーターは特に限定されるものではないが、スーパーオキシドジスムターゼをコードする遺伝子を発現するプロモーターを挙げることができる。
【0027】
本発明において、エシェリヒア属に属する微生物は特に限定されるものではないが、好ましい例として、Esherichia coli JM109株を挙げることができる。
【0028】
本発明は、ヒドロゲナーゼをコードするDNAが挿入されたコクリア属微生物用誘導発現シャトルベクターにより、宿主を形質転換した形質転換体に関する。
【0029】
また、本発明は、当該形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法に関する。
【0030】
本発明のシャトルベクターを用いて発現をさせたい所望のヒドロゲナーゼをエシェリヒア属又はコクリア属微生物において発現させるためには、本発明のベクターにヒドロゲナーゼをコードするDNAを挿入した組換えベクターを作製する必要がある。
【0031】
本発明の組換えベクターは、本発明のベクター(例えば、pKV15358-3)にヒドロゲナーゼをコードするDNAを挿入して作製する。
【0032】
本発明においてヒドロゲナーゼの種類は特に制限させるものではないが、好ましくは可溶型ヒドロゲナーゼを、さらに好ましくはNAD+還元能及び/又は酸素耐性を有するヒドロゲナーゼを例示することができる。
【0033】
本発明においてヒドロゲナーゼとしては、より具体的には、下記(a)〜(d)のいずれかに記載のタンパク質複合体を挙げることができる。
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
【0034】
Ralstonia eutropha由来のNAD+還元型ヒドロゲナーゼ構造遺伝子hoxFの塩基配列を配列番号:6、アミン酸配列を配列番号:7に、hoxUの塩基配列を配列番号:8、アミノ酸配列を配列番号:9に、hoxYの塩基配列を配列番号:10、アミノ酸配列を配列番号:11に、hoxHの塩基配列を配列番号:12、アミノ酸配列を配列番号:13に、hoxWの塩基配列を配列番号:14に、アミノ酸配列を配列番号:15に、hoxIの塩基配列を配列番号:16に、アミノ酸配列を配列番号:17に示す。
【0035】
ここで、複数とは例えば、2〜50個、好ましくは2〜10個、より好ましくは2〜5個である。また、ストリンジェントな条件としては、通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件が挙げられる。1もしくは複数の塩基の置換・欠失・挿入は、出願時に当業者に公知の技術により行うことができる。
【0036】
該ヒドロゲナーゼを導入したベクターとしては、pKV15358-4(配列番号:1)を挙げることができる。
【0037】
本発明の形質転換体は、宿主を本発明のベクターまたは本発明の組換えベクターで形質転換することで作製できる。形質転換の方法は、例えばエレクトロポレーション法、リン酸カルシウム法、DEAEデキストラン法を用いることが出来る。ヒドロゲナーゼを発現させるための宿主及びベクタープラスミドの増殖、回収に用いる宿主として、エシェリヒア属又はコクリア属微生物を用いることができる。
【0038】
本発明は、上述の形質転換体を培養し、得られる培養物からヒドロゲナーゼを単離することにより、ヒドロゲナーゼを製造することができる。形質転換体の培養方法は、宿主に用いるエシェリヒア属又はコクリア属微生物に適した方法を適宜選択することができる。
【0039】
本発明のベクターは、誘導剤(例えば、ε−カプロラクタム、イソバレロニトリル)によってヒドロゲナーゼの発現を誘導することができる。
【0040】
本発明において「培養物」とは、菌体、培養物、無細胞抽出液、細胞膜などの培養により得られるものを意味する。無細胞抽出液は、培養後の菌体を、例えばリン酸ナトリウム緩衝液を加えてホモジナイザーなどで物理的に破砕した後、遠心分離し、破砕できない菌体(細胞)が存在しないように上清を回収して得ることができる。細胞膜は、上記遠心で得られたペレットを溶解バッファーで懸濁することにより得ることができる。
【0041】
発現されたヒドロゲナーゼは、培養液をそのまま用いてもよいし、透析や硫安沈殿などの公知の方法、あるいはゲルろ過、イオン交換、アフィニティー等の各種クロマトグラフィーなどの公知の方法を単独または適宜組み合わせることによって、濃縮、精製したものを用いてもよい。
【0042】
本発明において、ヒドロゲナーゼの発現を行う際、微好気条件で形質転換体を培養してもよい。
【0043】
本発明において微好気的条件とは、静置培養あるいは緩やかな振とう条件を与えることによって、完全な嫌気状態ではないものの、溶存酸素濃度が0.5ppm以下に抑えられた培養条件のことを指す。
【0044】
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0045】
〔実施例1〕pKV15358-3へのNAD+還元型ヒドロゲナーゼ遺伝子の導入
コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3(配列番号:4)に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ(構造遺伝子hoxFUYHWI)をコードするポリヌクレオチドを導入した(pKV15358-4、配列番号:1)。
【0046】
プラスミドベクターpKV15358-3(配列番号:4)はコクリア バリアンス(Kocuria varians)NBRC15358由来のプラスミド(pKV15358-1、配列番号:2)のレスキュープラスミドを小型化し、さらにコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株における翻訳において最適なSD前後配列(配列番号:5)を付加した、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株用ベクターである。
【0047】
〔実施例2〕Esherichia coli JM109株を用いたNAD+還元型ヒドロゲナーゼの生産
次に、ラルストニア・ユートロファ(Ralstonia eutropha)のNAD+還元型ヒドロゲナーゼを導入した該プラスミド(pKV15358-4)を、様々な培養条件[1]下で大腸菌(Esherichia coli)JM109株により発現させ、無細胞抽出液(CFE)の調製[2]を行った後に、ヒドロゲナーゼの活性測定[3]を行った。
【0048】
その結果、微好気条件において培養を行った場合に、ヒドロゲナーゼ活性が観測され、該プラスミドが大腸菌を宿主としてNAD+還元型ヒドロゲナーゼを発現することが明らかとなった。
【0049】
〔注釈〕
[1]培養条件
1)好気的条件:500 mL容三角フラスコにLB培地100 mLを分注し、オートクレーブ後にカナマイシン50 mg/mLを100μl加えた(終濃度50μg/mL)。前培養液を1 mL植菌して、シリコン栓で蓋をして37℃ 150 rpmで約6時間振盪培養した。
2)微好気的条件:300 mL容三角フラスコに5 mM(NH4)2HPO4を含むLB培地240 mLを分注し、オートクレーブ後に20%Glucoseを2.4 mL(終濃度0.2%)カナマイシン50 mg/mLを240μL加えた(終濃度50μg/ml)。微好気培養では培養においては生育が極端に遅くなるためにGlucoseを添加して生育を早めている。前培養液を2.4 mL植菌して、シリコン栓で蓋をして37℃ 90 rpmで約6時間振盪培養した。
【0050】
[2]無細胞抽出液(CFE)の調製
培養液を遠心(10,000×g、4℃、10 min)して得られた菌体を5 mLの50mM HEPES-KOH buffer(pH7.0)にボルテックスを用いて懸濁し、50 mL容チューブに移して再び遠心(8,000×g、4℃、10min)した後上清を捨てた。集菌体を4倍量の50mM HEPES−KOH(pH7.0)にボルテックスを用いて再度懸濁し、その懸濁液に超音波破砕機(SONIFIER 250(BRAND))を使用して40 Wの出力で5分間細胞破砕を行った。破砕を施した溶液を超遠心用チューブに移し、超遠心(100,000×g、4℃、1 hr)にかけた。得られた上清をCFEとし、1.5 mlチューブに1 mlずつ分注して−80℃で保存した。
【0051】
[3]ヒドロゲナーゼの活性測定
NAD+還元活性の測定は、NADHが340 nmの波長を特異的に吸収することを利用し、以下のように行った。
【0052】
ガラスセルに、200μLのCFEを入れ、50 mM HEPES-KOH buffer(pH 7.0)により2447μLにfill upした。ブチルゴム栓とアルミキャップで密閉した後、H2を1分通気した。これに活性化のため2 mM NADHを13μLシリンジで添加し(終濃度10μM)、30℃で10分間インキュベートした。インキュベート終了後セルを吸光度計に設置し、シリンジを用いて40μlの100 mM NAD+(終濃度1.6mM)を加え、反応開始後すぐにAbs340の変化を測定した。
【0053】
酵素活性1Uを、1分間に1μmolのNAD+を還元する量と定義し、比活性(U/mg-protein)を以下の式で算出した。
【0054】
【数1】

・NADHのモル吸光係数ε340=6.3(mM-1cm-1)とした。
・CFEのタンパク質濃度は、Bovine serum albumin をスタンダートとし、Bio-Rad Protein Assay Dye Reagentを使用して測定した。
【技術分野】
【0001】
本発明は、エシェリヒア属に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属等に属する微生物内でヒドロゲナーゼを誘導発現可能なベクター及びその利用に関する。
【背景技術】
【0002】
微生物を利用した工業原料等有用物質の製造プロセスは、一般にエネルギー消費が少なく、廃棄物も少ないという特徴がある。このため、近年、資源枯渇やCO2等排出物の環境への影響が懸念されている中、微生物機能を活用した有用物質の生産(バイオプロセス)技術の開発が環境調和型循環産業システムにおける製造技術基盤として必要とされている。
【0003】
微生物が持つ酵素は、原料から特定の光学活性を持つ物質を合成するという選択性を有するものが多い。そのため、酵素を用いて効率よく目的物質を合成、取得できるので無駄がないとされている。また、微生物による物質生産は高価な触媒や高温高圧などの厳しい処理条件を必要とせず、常温、常圧、pH中性付近などの比較的温和な条件で進むため、扱いやすくかつ消費するエネルギーも少ない点が利点である。
【0004】
物質生産に利用される微生物の酵素反応の多くは酸化還元反応であり、特に基質を還元する反応の場合、基質のほかに還元力となる捕酵素を必要とする。補酵素としてよく使用されるNADH、NADPHはともに高価であり、電気化学的な方法により再生が試みられているが実用的とは言えず、酵素によりこれらの再生を行う試みが行われている。
【0005】
微生物による物質生産における捕酵素再生酵素として、NAD+還元型ヒドロゲナーゼを利用するには、物質生産を行う菌にヒドロゲナーゼを異種発現させる必要がある。しかし、ヒドロゲナーゼの異種微生物を宿主とした遺伝子発現はきわめて困難であり、ヒドロゲナーゼを異種発現させた場合には、不活性型であり封入体となっている例が多く見られる。これはヒドロゲナーゼが複雑な翻訳後修飾を得るためだと思われる。多種多様なヒドロゲナーゼが存在する中で、現在のところ12個の発現例のみが報告されている(非特許文献1〜12)。
【0006】
Ralstonia eutropha(旧Alcaligenes eutrophus, Wautersia eutropha, ATCC 17697)は、土壌に生息するグラム陰性好気性菌である。本菌は有機物を炭素源として従属栄養的に生育するほかに、水素、酸素、二酸化炭素による独立栄養的な生育も行う。その場合、炭素固定はカルビンサイクルで行われ、電子伝達系を共役したヒドロゲナーゼでエネルギー生産が行われている。R. eutrophaは、エネルギー生産を担う膜結合型ヒドロゲナーゼ(Membrane-bound hydrogenase(MBH))、水素を基質としてNAD+の還元を行う可溶型ヒドロゲナーゼ(soluble hydrogenase(SH))、水素を感知しヒドロゲナーゼの遺伝子発現の制御に関わる制御ヒドロゲナーゼ(regulatory hydrogenase(RH))の3種類のヒドロゲナーゼを保有している(非特許文献13)。
【0007】
この中で微生物による物質生産における捕酵素再生酵素として、R. eutropha由来の水素を基質としてNAD+の還元を行う可溶型ヒドロゲナーゼの異種宿主での発現がこれまで試みられてきたが、成功例は報告されていなかった。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Karube I. et al, FEBS, 1993, 158-1, 119-122
【非特許文献2】Rousset M. et al, J Bacteriol., 1998, 180(18), 4982-4986
【非特許文献3】Asada Y. et al, Biochim Biophys Acta., 2000, 1490(3), 269-278
【非特許文献4】Porthun A. et al, Arch Microbiol., 2002, 177(2), 159-166
【非特許文献5】Posewitz MC. et al, J Biol Chem., 2004, 279(24), 25711-25720
【非特許文献6】Mishra J. et al, Biochem Biophys Res Commun., 2004, 324(2), 679-685
【非特許文献7】Girbal L. et al, Appl Environ Microbiol., 2005, 71(5), 2777-2781
【非特許文献8】Lenz O. et al, J Bacteriol., 2005, 187(18), 6590-6595
【非特許文献9】King PW. et al, J Bacteriol., 2006, 188(6), 2163-2172
【非特許文献10】Lenz O. et al, FEBS Lett., 2007, 10, 581(17), 3322-3326
【非特許文献11】Sybirna K. et al, BMC Biotechnol., 2008, 8, 73
【非特許文献12】Sun J. et al, PLoS ONE, 2010, 5, 5, e10526
【非特許文献13】Burgdorf T. et al, J Mol Microbiol Biotechnol, 2005, 10, 181-196
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は上記状況に鑑みてなされたものであり、本発明が解決しようとする課題は、エシェリヒア属等に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属に属する微生物内でNAD+の還元を行う可溶型ヒドロゲナーゼを誘導発現可能なベクターを提供することである。
【課題を解決するための手段】
【0010】
上記課題を解決すべく、本発明者らは、大量発現系の構築が可能な大腸菌株(Esherichia coli)を宿主とする、ヒドロゲナーゼを発現する形質転換系の構築を行った。
【0011】
まず、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3(配列番号:4)に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ(構造遺伝子hoxFUYHWI)をコードするポリヌクレオチドを導入した(pKV15358-4、配列番号:1)。
【0012】
プラスミドベクターpKV15358-3はコクリア バリアンス(Kocuria varians)NBRC15358由来のプラスミド(pKV15358-1)のレスキュープラスミドを小型化し、さらにコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株における翻訳において最適なSD前後配列を付加した、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株用ベクターである。
【0013】
次に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼを導入した該プラスミド(pKV15358-4)を、様々な培養条件下で大腸菌(Esherichia coli)JM109株により発現させ、無細胞抽出液の調製を行った後に、ヒドロゲナーゼの活性測定を行った。その結果、微好気条件において培養を行った場合に、ヒドロゲナーゼ活性が観測され、該プラスミドが大腸菌を宿主としてNAD+還元型ヒドロゲナーゼを発現することが明らかとなった。
【0014】
即ち、本発明者らは、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子を導入することによって、エシェリヒア属等に属する微生物内でNAD+還元型ヒドロゲナーゼを誘導発現させることに成功し、これにより本発明を完成するに至った。
【0015】
本発明は、より具体的には以下の〔1〕〜〔9〕を提供するものである。
〔1〕配列番号:4に記載の塩基配列からなるポリヌクレオチド、又は配列番号:4に記載の塩基配列からなるポリヌクレオチドの相補鎖とストリンジェントな条件化でハイブリダイズするポリヌクレオチド、及びヒドロゲナーゼをコードするポリヌクレオチドを含む、微生物内でヒドロゲナーゼを発現可能なベクター。
〔2〕前記微生物が、エシェリヒア属又はコクリア属に属する微生物であることを特徴とする、〔1〕に記載のベクター。
〔3〕前記ヒドロゲナーゼが、可溶型ヒドロゲナーゼであることを特徴とする、〔1〕又は〔2〕に記載のベクター。
〔4〕前記ヒドロゲナーゼが、NAD+還元能及び/又は酸素耐性を有することを特徴とする、〔1〕〜〔3〕のいずれかに記載のベクター。
〔5〕前記ヒドロゲナーゼが、下記(a)〜(d)のいずれかに記載のタンパク質複合体であることを特徴とする、〔1〕〜〔4〕のいずれかに記載のベクター;
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
〔6〕前記ヒドロゲナーゼを発現させる微生物がエシェリヒア属に属する微生物であることを特徴とする、〔1〕〜〔5〕のいずれかに記載のベクター。
〔7〕〔1〕〜〔6〕のいずれかに記載の組換えベクターにより、宿主を形質転換した形質転換体。
〔8〕 〔7〕に記載の形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法。
〔9〕微好気的条件で形質転換体を培養することを特徴とする、〔8〕に記載の製造方法。
【発明の効果】
【0016】
Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ遺伝子が導入されたコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3を、大量発現系の構築が可能なエシェリヒア属に属する微生物に導入することにより、NAD+還元型ヒドロゲナーゼの大量発現系の構築が可能となる。また、本発明のプラスミドが導入された微生物を用いて、微生物発現系においてNAD+還元型ヒドロゲナーゼの活性(例えば触媒活性)を発現させることが可能となる。
【図面の簡単な説明】
【0017】
【図1】hoxFUYHWIを保持するkocuria rhizophila DC2201の調製方法を示す図である。
【発明を実施するための形態】
【0018】
本発明は、エシェリヒア属又はコクリア属に属する微生物内で複製可能なシャトルベクターであって、エシェリヒア属又はコクリア属に属する微生物内でヒドロゲナーゼを誘導発現可能なベクターを提供する。
【0019】
本発明のベクターは、エシェリヒア属又はコクリア属に属する微生物内で複製可能なDNA領域、及びエシェリヒア属に属する微生物に耐性を与える薬剤マーカーを有する。本発明のベクターは、エシェリヒア属又はコクリア属微生物内において複製可能なシャトルベクタープラスミドであり、ヒドロゲナーゼをエシェリヒア属又はコクリア属微生物内で発現させるのに有効なベクターである。
【0020】
本発明のシャトルベクターとしては、コクリア バリアンス(Kocuria varians)NBRC15358株から得られる環状プラスミドを挙げることができる。このプラスミドの複製に関与するDNA領域を明らかにし、当該DNA領域を含有させたプラスミドを作製すると、このようなプラスミドは、エシェリヒア属又はコクリア属微生物内で複製することが可能となる。
【0021】
エシェリヒア属又はコクリア属に属する微生物内で複製可能なDNA領域は、概領域を含むベクターをエシェリヒア属又はコクリア属微生物に導入したときに、ベクターが複製できる領域である限り特に制限されないが、例えば、コクリア バリアンス(Kocuria varians)株が保有する内因性プラスミドベクターpKV15358-1 (配列番号:2)に由来する自律複製領域を挙げることができる。pKV15358-1に由来する自律複製領域は、pKV15358-1の全長であってもよいが、遺伝子操作を容易にするために、pKV15358-1の一部であることが好ましい。さらに、コクリア属微生物で機能しうる限り、配列番号:2に記載された塩基配列において1もしくは数個の塩基が置換・欠失・挿入等された配列を有するDNA領域、又は配列番号:2に記載の塩基配列からなるポリヌクレオチドもしくは該配列から調製されうるプローブとストリンジェントな条件化でハイブリダイズするDNA領域であってもよい。ここで、数個とは例えば、2〜50個、好ましくは2〜10個、より好ましくは2〜5個である。また、ストリンジェントな条件としては、通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件が挙げられる。1もしくは数個の塩基の置換・欠失・挿入は、出願時に当業者に公知の技術により行うことができる。
【0022】
上述のようなベクターpKV15358-1の自律複製に必要な領域は、抗生物質感受性宿主細胞内での抗生物質耐性遺伝子の発現を指標とする欠失実験によって特定できる。即ち、ベクターpKV15358-1に、カナマイシン耐性遺伝子やアンピシリン耐性遺伝子などの薬剤耐性遺伝子を導入した後、該遺伝子を消化しない単独又は組み合わせの制限酵素によって消化し、DNA連結酵素により再環状化し、その後薬剤感受性の菌体を形質転換させる。この中から薬剤耐性を指標にして選択される薬剤体制菌株には、ベクターpKV15358-1の自律複製領域を含む派生ベクターを持ち、それらのベクターの制限酵素地図を比較することにより、ベクターpKV15358-1の自律複製領域が特定できる。
【0023】
本発明において、エシェリヒア属又はコクリア属微生物に耐性を与える薬剤マーカーとしては、該細菌に導入したときに薬剤耐性を付与する遺伝子であれば特に制限されないが、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、又はクロラムフェニコール耐性遺伝子等を挙げることができる。
【0024】
本発明のプラスミドベクターは、上記自律複製領域及び薬剤耐性マーカーを含むベクターであるが、このようなベクターとしては具体的には、pKV15358-2(配列番号:3)を挙げることができる。pKV15358-2は、上記pKV15358-1を制限酵素Xba1で切断して得られる自律複製領域を含む断片と、カナマイシン耐性遺伝子等により構成されるベクターである。尚、本発明のベクターはpKV15358-2が改変されたものであってもよい。改変としては、複製能や薬剤耐性に関与しない部位における変異、制限酵素部位のうちの一部の欠損や置換などを挙げることができる。
【0025】
本発明のベクターは、さらにコクリア属に属する微生物由来のSD配列を有していてもよい。SD配列は特に限定されないが、好ましくは配列番号:5に記載のSD配列を挙げることができる。SD配列をさらに含むベクターとしては、pKV15358-3(配列番号:4)を挙げることができる。
【0026】
また、コクリア属微生物において機能しうるプロモーターなどがさらに組み込まれていてもよい。プロモーターは特に限定されるものではないが、スーパーオキシドジスムターゼをコードする遺伝子を発現するプロモーターを挙げることができる。
【0027】
本発明において、エシェリヒア属に属する微生物は特に限定されるものではないが、好ましい例として、Esherichia coli JM109株を挙げることができる。
【0028】
本発明は、ヒドロゲナーゼをコードするDNAが挿入されたコクリア属微生物用誘導発現シャトルベクターにより、宿主を形質転換した形質転換体に関する。
【0029】
また、本発明は、当該形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法に関する。
【0030】
本発明のシャトルベクターを用いて発現をさせたい所望のヒドロゲナーゼをエシェリヒア属又はコクリア属微生物において発現させるためには、本発明のベクターにヒドロゲナーゼをコードするDNAを挿入した組換えベクターを作製する必要がある。
【0031】
本発明の組換えベクターは、本発明のベクター(例えば、pKV15358-3)にヒドロゲナーゼをコードするDNAを挿入して作製する。
【0032】
本発明においてヒドロゲナーゼの種類は特に制限させるものではないが、好ましくは可溶型ヒドロゲナーゼを、さらに好ましくはNAD+還元能及び/又は酸素耐性を有するヒドロゲナーゼを例示することができる。
【0033】
本発明においてヒドロゲナーゼとしては、より具体的には、下記(a)〜(d)のいずれかに記載のタンパク質複合体を挙げることができる。
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
【0034】
Ralstonia eutropha由来のNAD+還元型ヒドロゲナーゼ構造遺伝子hoxFの塩基配列を配列番号:6、アミン酸配列を配列番号:7に、hoxUの塩基配列を配列番号:8、アミノ酸配列を配列番号:9に、hoxYの塩基配列を配列番号:10、アミノ酸配列を配列番号:11に、hoxHの塩基配列を配列番号:12、アミノ酸配列を配列番号:13に、hoxWの塩基配列を配列番号:14に、アミノ酸配列を配列番号:15に、hoxIの塩基配列を配列番号:16に、アミノ酸配列を配列番号:17に示す。
【0035】
ここで、複数とは例えば、2〜50個、好ましくは2〜10個、より好ましくは2〜5個である。また、ストリンジェントな条件としては、通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件が挙げられる。1もしくは複数の塩基の置換・欠失・挿入は、出願時に当業者に公知の技術により行うことができる。
【0036】
該ヒドロゲナーゼを導入したベクターとしては、pKV15358-4(配列番号:1)を挙げることができる。
【0037】
本発明の形質転換体は、宿主を本発明のベクターまたは本発明の組換えベクターで形質転換することで作製できる。形質転換の方法は、例えばエレクトロポレーション法、リン酸カルシウム法、DEAEデキストラン法を用いることが出来る。ヒドロゲナーゼを発現させるための宿主及びベクタープラスミドの増殖、回収に用いる宿主として、エシェリヒア属又はコクリア属微生物を用いることができる。
【0038】
本発明は、上述の形質転換体を培養し、得られる培養物からヒドロゲナーゼを単離することにより、ヒドロゲナーゼを製造することができる。形質転換体の培養方法は、宿主に用いるエシェリヒア属又はコクリア属微生物に適した方法を適宜選択することができる。
【0039】
本発明のベクターは、誘導剤(例えば、ε−カプロラクタム、イソバレロニトリル)によってヒドロゲナーゼの発現を誘導することができる。
【0040】
本発明において「培養物」とは、菌体、培養物、無細胞抽出液、細胞膜などの培養により得られるものを意味する。無細胞抽出液は、培養後の菌体を、例えばリン酸ナトリウム緩衝液を加えてホモジナイザーなどで物理的に破砕した後、遠心分離し、破砕できない菌体(細胞)が存在しないように上清を回収して得ることができる。細胞膜は、上記遠心で得られたペレットを溶解バッファーで懸濁することにより得ることができる。
【0041】
発現されたヒドロゲナーゼは、培養液をそのまま用いてもよいし、透析や硫安沈殿などの公知の方法、あるいはゲルろ過、イオン交換、アフィニティー等の各種クロマトグラフィーなどの公知の方法を単独または適宜組み合わせることによって、濃縮、精製したものを用いてもよい。
【0042】
本発明において、ヒドロゲナーゼの発現を行う際、微好気条件で形質転換体を培養してもよい。
【0043】
本発明において微好気的条件とは、静置培養あるいは緩やかな振とう条件を与えることによって、完全な嫌気状態ではないものの、溶存酸素濃度が0.5ppm以下に抑えられた培養条件のことを指す。
【0044】
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0045】
〔実施例1〕pKV15358-3へのNAD+還元型ヒドロゲナーゼ遺伝子の導入
コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株内で複製できるプラスミドベクターpKV15358-3(配列番号:4)に、Ralstonia eutrophaのNAD+還元型ヒドロゲナーゼ(構造遺伝子hoxFUYHWI)をコードするポリヌクレオチドを導入した(pKV15358-4、配列番号:1)。
【0046】
プラスミドベクターpKV15358-3(配列番号:4)はコクリア バリアンス(Kocuria varians)NBRC15358由来のプラスミド(pKV15358-1、配列番号:2)のレスキュープラスミドを小型化し、さらにコクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株における翻訳において最適なSD前後配列(配列番号:5)を付加した、コクリア リゾフィラ(Kocuria rhizophila)NBRC 103217株用ベクターである。
【0047】
〔実施例2〕Esherichia coli JM109株を用いたNAD+還元型ヒドロゲナーゼの生産
次に、ラルストニア・ユートロファ(Ralstonia eutropha)のNAD+還元型ヒドロゲナーゼを導入した該プラスミド(pKV15358-4)を、様々な培養条件[1]下で大腸菌(Esherichia coli)JM109株により発現させ、無細胞抽出液(CFE)の調製[2]を行った後に、ヒドロゲナーゼの活性測定[3]を行った。
【0048】
その結果、微好気条件において培養を行った場合に、ヒドロゲナーゼ活性が観測され、該プラスミドが大腸菌を宿主としてNAD+還元型ヒドロゲナーゼを発現することが明らかとなった。
【0049】
〔注釈〕
[1]培養条件
1)好気的条件:500 mL容三角フラスコにLB培地100 mLを分注し、オートクレーブ後にカナマイシン50 mg/mLを100μl加えた(終濃度50μg/mL)。前培養液を1 mL植菌して、シリコン栓で蓋をして37℃ 150 rpmで約6時間振盪培養した。
2)微好気的条件:300 mL容三角フラスコに5 mM(NH4)2HPO4を含むLB培地240 mLを分注し、オートクレーブ後に20%Glucoseを2.4 mL(終濃度0.2%)カナマイシン50 mg/mLを240μL加えた(終濃度50μg/ml)。微好気培養では培養においては生育が極端に遅くなるためにGlucoseを添加して生育を早めている。前培養液を2.4 mL植菌して、シリコン栓で蓋をして37℃ 90 rpmで約6時間振盪培養した。
【0050】
[2]無細胞抽出液(CFE)の調製
培養液を遠心(10,000×g、4℃、10 min)して得られた菌体を5 mLの50mM HEPES-KOH buffer(pH7.0)にボルテックスを用いて懸濁し、50 mL容チューブに移して再び遠心(8,000×g、4℃、10min)した後上清を捨てた。集菌体を4倍量の50mM HEPES−KOH(pH7.0)にボルテックスを用いて再度懸濁し、その懸濁液に超音波破砕機(SONIFIER 250(BRAND))を使用して40 Wの出力で5分間細胞破砕を行った。破砕を施した溶液を超遠心用チューブに移し、超遠心(100,000×g、4℃、1 hr)にかけた。得られた上清をCFEとし、1.5 mlチューブに1 mlずつ分注して−80℃で保存した。
【0051】
[3]ヒドロゲナーゼの活性測定
NAD+還元活性の測定は、NADHが340 nmの波長を特異的に吸収することを利用し、以下のように行った。
【0052】
ガラスセルに、200μLのCFEを入れ、50 mM HEPES-KOH buffer(pH 7.0)により2447μLにfill upした。ブチルゴム栓とアルミキャップで密閉した後、H2を1分通気した。これに活性化のため2 mM NADHを13μLシリンジで添加し(終濃度10μM)、30℃で10分間インキュベートした。インキュベート終了後セルを吸光度計に設置し、シリンジを用いて40μlの100 mM NAD+(終濃度1.6mM)を加え、反応開始後すぐにAbs340の変化を測定した。
【0053】
酵素活性1Uを、1分間に1μmolのNAD+を還元する量と定義し、比活性(U/mg-protein)を以下の式で算出した。
【0054】
【数1】
・NADHのモル吸光係数ε340=6.3(mM-1cm-1)とした。
・CFEのタンパク質濃度は、Bovine serum albumin をスタンダートとし、Bio-Rad Protein Assay Dye Reagentを使用して測定した。
【特許請求の範囲】
【請求項1】
配列番号:4に記載の塩基配列からなるポリヌクレオチド、又は配列番号:4に記載の塩基配列からなるポリヌクレオチドの相補鎖とストリンジェントな条件化でハイブリダイズするポリヌクレオチド、及びヒドロゲナーゼをコードするポリヌクレオチドを含む、微生物内でヒドロゲナーゼを発現可能なベクター。
【請求項2】
前記微生物が、エシェリヒア属又はコクリア属に属する微生物であることを特徴とする、請求項1に記載のベクター。
【請求項3】
前記ヒドロゲナーゼが、可溶型ヒドロゲナーゼであることを特徴とする、請求項1又は2に記載のベクター。
【請求項4】
前記ヒドロゲナーゼが、NAD+還元能及び/又は酸素耐性を有することを特徴とする、請求項1〜3のいずれかに記載のベクター。
【請求項5】
前記ヒドロゲナーゼが、下記(a)〜(d)のいずれかに記載のタンパク質複合体であることを特徴とする、請求項1〜4のいずれかに記載のベクター;
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
【請求項6】
前記ヒドロゲナーゼを発現させる微生物がエシェリヒア属に属する微生物であることを特徴とする、請求項1〜5のいずれかに記載のベクター。
【請求項7】
請求項1〜6のいずれかに記載の組換えベクターにより、宿主を形質転換した形質転換体。
【請求項8】
請求項7に記載の形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法。
【請求項9】
微好気的条件で形質転換体を培養することを特徴とする、請求項8に記載の製造方法。
【請求項1】
配列番号:4に記載の塩基配列からなるポリヌクレオチド、又は配列番号:4に記載の塩基配列からなるポリヌクレオチドの相補鎖とストリンジェントな条件化でハイブリダイズするポリヌクレオチド、及びヒドロゲナーゼをコードするポリヌクレオチドを含む、微生物内でヒドロゲナーゼを発現可能なベクター。
【請求項2】
前記微生物が、エシェリヒア属又はコクリア属に属する微生物であることを特徴とする、請求項1に記載のベクター。
【請求項3】
前記ヒドロゲナーゼが、可溶型ヒドロゲナーゼであることを特徴とする、請求項1又は2に記載のベクター。
【請求項4】
前記ヒドロゲナーゼが、NAD+還元能及び/又は酸素耐性を有することを特徴とする、請求項1〜3のいずれかに記載のベクター。
【請求項5】
前記ヒドロゲナーゼが、下記(a)〜(d)のいずれかに記載のタンパク質複合体であることを特徴とする、請求項1〜4のいずれかに記載のベクター;
(a)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列からなるタンパク質を含むタンパク質複合体、
(b)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体、
(c)配列番号:7、9、11、13、15及び17に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、及び/又は付加したアミノ酸配列からなるタンパク質を含むタンパク質複合体、又は
(d)配列番号:6、8、10、12、14及び16に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質を含むタンパク質複合体。
【請求項6】
前記ヒドロゲナーゼを発現させる微生物がエシェリヒア属に属する微生物であることを特徴とする、請求項1〜5のいずれかに記載のベクター。
【請求項7】
請求項1〜6のいずれかに記載の組換えベクターにより、宿主を形質転換した形質転換体。
【請求項8】
請求項7に記載の形質転換体を培養する工程、及び得られる培養物からヒドロゲナーゼを単離する工程を含む、ヒドロゲナーゼの製造方法。
【請求項9】
微好気的条件で形質転換体を培養することを特徴とする、請求項8に記載の製造方法。
【図1】

【公開番号】特開2013−32(P2013−32A)
【公開日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願番号】特願2011−132257(P2011−132257)
【出願日】平成23年6月14日(2011.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成23年3月5日、社団法人日本農芸化学会発行の刊行物「日本農芸化学会2011年度大会講演要旨集」に発表。
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構、微生物機能を活用した高度製造基盤技術開発事業、産業技術力強化法第19条の適用を受けるもの
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
【公開日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願日】平成23年6月14日(2011.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成23年3月5日、社団法人日本農芸化学会発行の刊行物「日本農芸化学会2011年度大会講演要旨集」に発表。
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構、微生物機能を活用した高度製造基盤技術開発事業、産業技術力強化法第19条の適用を受けるもの
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
[ Back to top ]