
ピロリ菌リポ多糖類の外殻エピトープ

もっとも一般的なヒトの病原体であるヘリコバクター・ピロリは、ヒトの慢性胃炎、消化性潰瘍、および、胃癌の進行に関連している。本発明は式I:Rがα1,3−DD−ヘプタンに結合したα1,6−グルカンで置換したα−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされる:の構造を含むα1,6−グルカン化合物を含むヘリコバクター・ピロリ化合物に関する。この化合物を含む組成物、化合物の使用、および、化合物に対して産生される抗体についても同様に記載される。
【化1】

【化1】
【発明の詳細な説明】
【技術分野】
【0001】
(先願の情報)
本出願は、2009年7月31日に出願され米国仮特許出願第61/230,315号の利益を主張し、その全体は引用によって本明細書に組み入れられる。
【0002】
(発明の分野)
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌(H.pylori)外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【背景技術】
【0003】
ヘリコバクター・ピロリは、ヒトの慢性胃炎、消化性潰瘍、および、胃癌に関連する最も一般的な細菌感染と認められる。ピロリ菌感染は、世界の人口の約半分に蔓延していると推測され、開発途上国では70%、先進工業国では20−30%にまで達している(Dunn et al., 1997)。大部分の人は小児期にピロリ菌にかかり、開発途上国の子供の間での感染の有病率は、社会経済的地位の低さと衛生状態の悪さに関連している(Castillo−Rojas et al., 2004)。ピロリ菌は胃癌の相対リスクを少なくとも6倍増加させるため、世界保健機構(WHO)によってクラスIの発癌物質であると確認されている。胃癌は世界で2番目に多い死亡原因であり、毎年700,000人が死亡している(Parkin et al., 2002)。
【0004】
ピロリ菌の現在の根絶方策はプロトンポンプインヒビターと抗生物質の使用に基づいている。しかしながら、化学療法の介入治療の有効性は、ピロリ菌分離物中の抗生物質耐性の頻度、および、再感染に対する免疫の欠如によって限定されてしまう。したがって、ピロリ菌感染の予防および根絶のためのグローバルな戦略を提供するために新しい治療が必要である。
【0005】
近年の調査がピロリ菌の病因におけるタンパク成分の役割と、防御免疫にタンパク質成分の役割に焦点が当てられている一方で、ワクチン製剤中にタンパク質以外の抗原を含めるという可能性を探った研究は比較的少ない。例えば、多糖類に基づいた複合体(conjugate)ワクチンは、全身感染を防ぎ、宿主のコロニー形成を阻害することが知られている(Anderson et al.,1986; Chu et al., 1991; Pon et al.,1997; Passwell et al.,2001; Passwell et al.,2003)。腸内病原菌の近年の研究は、人間に使用するための候補ワクチンとして、LPS複合体に基づいた手法を検査している(Gu et al.,1996; Mieszala et al., 2003; Cox et al.,2005; Yu and Gu, 2007)。リポ多糖類(LPS)はピロリ菌の主要な細胞表面成分である。多くのピロリ菌で行なわれた構造研究(Monteiro, 2001)は、O鎖と、脂質A部分に付けられたコアオリゴ糖とからなるLPSの構造モデルをもたらした。ほとんどのピロリ菌株のO鎖多糖類骨格(backbone)の構造は独特で、ヒト胃細胞と腫瘍細胞の細胞表面にある決定因子を模倣するII型および/またはI型のルイス(Le)式血液型決定因子を表示し(Wirth et al.,1996)、これらは、萎縮性胃炎につながる有害な自己免疫反応に関与していることもある(Appelmelk et al.,1996)。加えて、ピロリ菌LPSの分離外殻領域は、2つの異常な高分子成分:DD−ヘプトグリカン、および、α1,6−グルカンを含む(Monteiro, 2001)。外殻領域ピロリ菌LPS分離株中のα1,6−グルカンポリマーは、HP0159オープンリーディングフレームの生成物によって合成される。ピロリ菌中でのHP0159遺伝子の存在および発現は一般的である。
【0006】
多くのピロリ菌LPS生合成遺伝子が特徴付けられ、病因とコロニー形成におけるそれらの役割が測定されてきた(Logan et al., 2000; Logan et al., 2005; Hiratsuka et al., 2005; Chandan et al., 2007; Altman et al., 2008)。ピロリ菌HP0826変異株が構築され、この変異が、Le抗原を欠いた、切断されたLPSの形成をもたらすことが実証された。しかしながら、LPSの構造の十分な特徴づけは達成されなかった。
【0007】
当該技術分野での進展にもかかわらず、ピロリ菌の複数のタイプにわたって有効な免疫原性エピトープは捉えがたいままである。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【課題を解決するための手段】
【0009】
本発明は式Iの構造を含む、α1,6−グルカンを含むヘリコバクター・ピロリ化合物を提供し、
【0010】
【化1】
【0011】
Rは、α1,3−DD−ヘプタンに結合したα1,6−グルカンで置換したα−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされる。記載されたような化合物において、α1,6−グルカンは、約3から約12のα1,6−結合グルコース残基を含み、α1,3−DD−ヘプタンは、約2から約6のα1,3−結合ヘプトース残基を含んでもよい。
【0012】
上記の化合物のR基は、以下のとおりであってもよい。
【0013】
【化2】
【0014】
ここで、β−GlcNAc残基Lは、Hep GのO−2に結合する。記載されたような化合物では、グルカンの残基QおよびZは、α1,6−結合グルコース残基であり、nは約1乃至11の間の任意の値であってもよく、残基T、YおよびXは、α1,3−結合ヘプトース残基であり、mは約0から4の間の任意の値であってもよい。
【0015】
上記のような化合物は、ピロリ菌株HP0826::Kanから分離されるか、または、精製される。
【0016】
上記のような化合物では、式Iの構造は、Kdo残基Cに共有結合している脂質A部分をさらに含んでもよい。脂質A分子は、O−脱アシル化するか、または、Kdo残基のケトシド結合の加水分解によって開裂する。
【0017】
本発明は同様に、リンカー分子、タンパク質担体、または、その組み合わせに共役した、ほぼ線形のα1,6−グルカンを含む化合物を含む複合体を提供する。ほぼ線形のα1,6−グルカンを含む化合物は、本明細書に記載の化合物であり、ここで、式Iの構造は、リンカー分子、タンパク質担体、またはその組み合わせに共役される。ほぼ線形のα1,6−グルカンを含む化合物は、代替的に、デキストランT5などのデキストランであってもよい。タンパク質担体は、破傷風トキソイドまたはウシ血清アルブミンであってもよい。
【0018】
本発明は、上記のように、1つ以上の化合物または複合体を含む組成物も含む。
【0019】
本発明は、上記のようにα1,6−グルカンのエピトープを含む化合物に対する抗体をさらに含む。抗体はモノクローナル抗体1C4F9であってもよい。本発明はハイブリドーマ細胞株1C4F9をさらに包含し、これはモノクローナル抗体1C4F9を作り出す。
【0020】
上記のモノクローナル抗体は、そのような処置を必要としている個体において、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすために利用されてもよい。
【0021】
本発明は、同様に、個体内でのピロリ菌に対する免疫反応を誘発するために、上記の有効な量の組成物の使用を提供する。組成物中の化合物は適切な担体タンパク質に共役され、さらに、組成物中の化合物は、リポ多糖類の2−ケト−3−デオキシ−オクツロソン酸(Kdo)を介して、適切な担体タンパク質に共役してもよい。
【0022】
本発明は、本明細書に記載の免疫原性の組成物で哺乳動物を免疫化することによって作られた免疫抗血清をさらに提供する。免疫抗血清は、ピロリ菌の同種および異種の分類可能なおよび分類不可能な変異株および野生株における、α1,6−結合グルカンエピトープを認識するIgGを含む。IgGは、変異株(mutant)および野生株のa1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こす。
【0023】
本発明の追加の態様および利点は、以下の記載を考慮して明らかとなる。本発明の範囲内の様々な変化および修正は、この発明の教示を踏まえて当業者に明らかになるため、詳細な記載および実施例は、本発明の好ましい実施形態を示す一方で、たんなる説明として与えられるものである。
【図面の簡単な説明】
【0024】
本発明のこうした実施形態と他の実施形態は、添付の図面を参照しつつ、ほんの一例としてここで記載される。
【図1】陽イオンモードでピロリ菌株26695 HP0826::Kanから脱脂したLPSの分画1におけるLPSグリコフォームのCE−MS分析を示す。図1Aが抽出された質量スペクトル(m/z 1000−1500)を示す一方で、図1Bは、m/z 1266.3でのイオンの生成イオンスペクトルを示している。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図2】陰イオンモードでピロリ菌株26695 HP0826::KanからO−脱アシル化したLPSのLPSグリコフォームのCE−MS分析を示す。図2Aが抽出された質量スペクトル(m/z 600−2000)を示す一方で、図2Bは、m/z 1597.7でのイオンの生成イオンスペクトルを示している。脂質A部分に相当するフラグメントイオンは星印で印をつけている。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3A】ピロリ菌株26695 HP0826::Kanからの主要なLPSグリコフォームの構造を示す。アシル化は示されていない。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3B】図3AのLPSのKOH脱アシル化の生成物が、図3Bで示される(化合物1〜4)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3C】化合物4の脱アミノの生成物が、図3Cで示される(化合物5および6)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3D】化合物6の過ヨウ素酸酸化の生成物が図3Dで示される(化合物7および8)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3E】菌株26695(FIGURE 3E)からのピロリ菌LPS(図3E)と、菌株26695 HP0826::Kan変異株からのピロリ菌LPS(図3F)に関して当該技術分野で先に提案された構造も同様に示される(出典「Logan et al., 2000」)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3F】菌株26695(FIGURE 3E)からのピロリ菌LPS(図3E)と、菌株26695 HP0826::Kan変異株からのピロリ菌LPS(図3F)に関して当該技術分野で先に提案された構造も同様に示される(出典「Logan et al., 2000」)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図4】本発明のLPSベースの複合体の調製のための反応スキームを示す。
【図5】陰イオンモードで脱脂されたピロリ菌O:3 HP0826::Kan LPSの主要な分画のCE−MS分析を示す。●:側鎖に1つのHep残基を含むLPSグリコフォーム、R=Hex5−13、Hep、HexNAc、Fuc; ■:側鎖に2つのHep残基を含むLPSグリコフォーム、R=Hex8−13、Hep、Hep、HexNAc、Fuc
【図6】26695 HP0479::Kan(図6A)、および、26695 HP0826::Kan(図6B)による阻害ELISAによって、dLPS−TT複合体により誘発されたウサギ抗体の特異性を測定するためのグラフである。LPSのコーティングは以下のように描かれる:黒四角−26695;黒菱形−26695 HP0826::Kan;黒丸−26695 HP0159::Kan;黒三角−26695 HP0479::Kan;白丸−SS1;白四角−SS1 HP0826::Kan;白三角−SS1 HP0159::Kan;白逆三角−SS1 HP0479::Kan
【図7】CD−1マウス中のピロリ菌特異抗体反応を表すグラフを示す。マウスは、1μg/マウスのコレラ毒素で補助された(adjuvanted)25μg/マウスのdLPS−TT複合体(黒棒)、1μg/マウスのコレラ毒素で補助されたPJ2細胞を含まない超音波処理物(斜線棒)、または、食塩水(白棒)で、週1回の間隔で4度、ワクチン接種した。最初の免疫化から4週間後、血清、糞便、膣の洗浄サンプルを採取し、ピロリ菌に特異的なIgGおよびIgAについてアッセイした。個々のマウスは、群平均を示す(n=5/群)水平棒でグラフ上にプロットされる。片側マン・ホイットニー検定(one−tailed Mann Whitney test)によって、※ p<0.05、※※ p<0.01
【図8】CD−1マウスの胃におけるピロリ菌負荷(burden)を表す棒グラフである。1μg/マウスのコレラ毒素で補助された25μg/マウスのdLPS−TT複合体(黒棒)、1μg/マウスのコレラ毒素で補助されたPJ2細胞を含まない超音波処理物(斜線棒)、または、食塩水(白棒)で、マウスに週1回の間隔で4度、ワクチン接種した。第1の免疫から5週間後に、〜108cfu ピロリ菌、PJ2菌株によって、一日おきに3度、マウスに経口でチューブから栄養補給させた。4週間後にマウスを殺処分し、胃からの一般細菌を数えた。棒は、4−5のマウス±SEMの群を表わす(片側マン・ホイットニー検定によって、※p<0.05)。
【発明を実施するための形態】
【0025】
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【0026】
他に定義されない限り、本明細書で用いられる全ての技術的及び科学的用語は、本発明が属する技術分野の当業者によって一般に理解されるものと同じ意味を有する。本明細書に記載されているこれらのものと類似又は同等の任意の方法及び材料は、本発明の実施又は試験で用いることができるが、好ましい方法と材料がこれより記載される。以下に記載される全ての刊行物は、引用することによって本明細書に組み入れられる。
【0027】
本明細書で使用されているように、「精製された(purified)」とは、必ずしも絶対的な純度を意味するわけではないが、むしろ相対的な定義として意図されている。同様に、本明細書で使用されているように、「分離された(isolated)」は、その本来の環境から何かを除去することを言う。
【0028】
ヘリコバクター・ピロリは、ヒトの慢性胃炎、消化性潰瘍、および、胃癌に一般的に関連する細菌性病原菌である。ピロリ菌感染に関連する胃癌のリスクの増加の結果として、それはクラスI発癌物質として分類されている。リポ多糖類(LPS)はピロリ菌の主要な細胞表面成分である。ピロリ菌LPSの構造研究に関する従来の出版物は、O鎖多糖類がコアオリゴ糖に共有結合される提案されたモデルをもたらし、コアオリゴ糖は順に脂質A部分に付く。ほとんどのピロリ菌株のO鎖多糖類骨格は独特で、タイプ2および/またはタイプ1のルイス(Le)血液型決定子を表示することができ、この多糖類成分は抗原である。「分類可能な」ピロリ菌株は、抗ルイス抗体(抗Le)によって認識することができるルイスエピトープ(LeXおよび/またはLeY抗原)を有し、このような抗体は市販で入手可能であり、分類の助けとなる。「分類不可能な」菌株はルイス構造を含んでいない。
【0029】
さらなる構造研究は、ピロリ菌LPSの外殻領域が2つの異常な高分子成分、DD−ヘプトグリカン、および、α1,6−グルカン側鎖を含む(Monteiro, 2001)。Loganら(2000)はLPSの構造についても洞察した。具体的に、もし側鎖中のDD−ヘプトース(DD−Hep)が骨格中のDD−Hepに付けられていれば、その一方で、α1,6−グルカンがこの側鎖DD−Hepに付き、別の分岐を形成していると仮定した場合の提案された構造(図3Eおよび図3Fを参照)。具体的に、Loganら(2000)は、0826変異株LPS内のグルカン鎖の長さを、FAB−MSスペクトル(高速原子衝撃質量分析)に基づいた1乃至3のグルコースとなるように決定した。ピロリ菌株26695 LPS中でのα1,6−結合グルコースの存在も記載され、メチル化分析データに基づいているが、グルカンの長さは決められなかった。菌株26695 LPSにおいて、3−置換HepはO鎖のGlcNAc単位とコアとの間で結合を形成する。ピロリ菌HP0826::Kan LPSでの3−結合ヘプトースの存在も記載されているが、α1,3−ヘプタンの存在またはその長さについては記載されなかった。ヘプタンまたはグルカンの側鎖の構造または長さに関するさらなる情報は提供されなかった。ピロリ菌LPSの構造は、現在さらに定義されている。
【0030】
本発明は式Iの構造を含む、新規なα1,6−グルカンを含むヘリコバクター・ピロリ化合物を提供し、
【0031】
【化3】
【0032】
ここで、Rは、α1,6−グルカンで、その後、α1,3−DD−ヘプタンで置換された、α−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされる。
【0033】
上記のような構造では、三糖類(α−DDHep−3−α−L−Fuc−3−β−GlcNAc)のβ−GlcNAcは、α−DDHep Gに結合される。三糖類のα−DDHepは、α1,6−グルカンに結合され、これは順にα1,3−DD−ヘプタンに結合される。α1,3−DD−ヘプタンはその後、β−GlcNAc残基に結合され、後者はO鎖多糖類に付着点を提供する。「結合した(linked)」または「置換した(substituted)」との用語によって、2つの部分が共有結合によって連結することが意味される。
【0034】
本明細書で使用される「α1,6−グルカン」という用語は、「グルカン」、「α1,6−グルカン側鎖」、「グルカン側鎖」、「α1,6−グルカン部分」、および/または、「グルカン部分」と交互に呼ばれる。α1,6−グルカンは、α1,6−Oグリコシド結合によって結合したブドウ糖モノマーの線形の多糖類鎖である。任意の方法に限定することを意図していない1つの例では、α1,6−グルカンは線形の多糖類であってもよい。α1,6−グルカンが任意の適切な量のα1,6−グルコース残基を含んでもよい。例えば、任意の方法で限定することを望まず、グルカンは、約3乃至約12のα1,6−結合したグルコース残基を含んでもよく、具体的には、グルカン部分は約3、4、5、6、7、8、9、10、11、または、12のα1,6−結合グルコース残基、あるいは、ちょうど列挙された任意の2つの値によって定義される任意の範囲を含んでもよい。1つの非限定的な実施例では、α1,6−グルカンは、9−12のα1,6−グルコース残基を含んでもよい。別の非限定的な実施例では、α1,6−グルカンは、10のα1,6−グルコース残基を含んでもよい。
【0035】
本明細書で使用されるような「グリコフォーム」という用語は、同じLPS構造を含むが、α1,6−グルコースまたはα1,3−ヘプトース残基の数が異なる、さまざまな形態またはタイプの化合物を示す。例えば、および、理論に縛られることを望まずに、各々のグリコフォームは、特定の長さのグルカンおよび/またはヘプタン部分、あるいは、それらの組み合わせを含んでもよい。
【0036】
本明細書で使用されるような「α1,3−ヘプタン」という用語は、「α1,3−DD−ヘプタン」、「ヘプタン」、「α1,3−ヘプタン側鎖」、「ヘプタン側鎖」、「α1,3−ヘプタン部分」、「ヘプタン部分」、および/または、「DD−ヘプトグリカン」と交互に呼ばれる。α1,3−ヘプタンは、α1,3−Oグリコシド結合によって結合したヘプトースモノマーの多糖類鎖である。任意の方法に限定することを意図していない1つの例では、α1,3−ヘプタンは線形の多糖類であってもよい。α1,3−ヘプタンは、任意の適切な量のα1,3−ヘプトース残基を含んでもよい。例えば、任意の方法で限定することを望まずに、ヘプタンは、約2乃至約6のα1,3−結合ヘプトース残基を含んでもよく、具体的には、ヘプタン部分は、約2、3、4、5、または、6のα1,3−結合ヘプトース残基、または、ちょうど列挙された任意の2つの値によって定義される任意の範囲を含んでもよい。
【0037】
上記のα1,3−DDヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャップされる。「キャップされた(capped)」という用語によって、β−GlcNAc残基が側鎖の最後の残基であることを意味し、「末端の(terminated)」という用語が使用されてもよい。β−GlcNAcはヘプトースの位置O−2を介してDD−Hepに結合されてもよい。理論に縛られることを望まずに、β−GlcNAc残基はO鎖多糖類に付着点を与えてもよい。
【0038】
1つの非限定的な実施例において、Rは以下のとおりであり、
【0039】
【化4】
【0040】
ここで、β−GlcNAc残基Lは、Hep残基GのO−2に結合される。この実施例では、β−GlcNAc残基Wは、O鎖多糖類に付着点を与えてもよい。ちょうど記載された化合物では、グルカンの残基QおよびZは、α1,6−結合グルコース残基であり、nは、グルカンがα1,6−結合で約3乃至約12のグルコース残基を含むように、約1乃至11の間の任意の値でもよい。1つの具体的な非限定的な例において、主要なグリコフォームは、10の連続するα1,6−結合グルコース残基(n=9)を含んでもよい。ちょうど記載された化合物では、残基T、Y、および、Xは、α1,3−結合ヘプトース残基であり、mは、ヘプタンがα1,3−結合において約2乃至約6のヘプトース残基を含むように、約0乃至4の間の任意の値であってもよく、1つの具体的な非限定的な例において、主要なグリコフォームは4つの連続するα1,3−結合ヘプトース残基(m=2)を含んでもよい。
【0041】
構造は任意の適切なピロリ菌株から分離および/または精製され、例えば、任意の方法で限定することを望まずに、切断されたピロリ菌LPS分子は、限定されないが、O鎖多糖類を欠く同質遺伝子変異株をもたらすHP0826遺伝子中の変異株を有するピロリ菌株などの分類不可能なピロリ菌株(すなわち、ルイス抗原が欠けたもの)から分離される。非限定的な例において、本明細書に記載されている化合物はピロリ菌株26695 HP0826::Kanまたは菌株PJ2から分離および/または精製されてもよい。
【0042】
「内殻分子(inner core molecular)」とも本明細書で呼ばれる式Iの構造は、Kdo残基(例えば、Kdo残基C)に共有結合している脂質A部分をさらに含む。他の実施形態では、脂質A分子はO−脱アシル化しても、または、完全に脱アシル化してもよい。さらに他の実施形態では、脂質A部分は、Kdo残基のケトシド結合の加水分解を介して開裂してもよい。理論によって縛られることを望まず、脂質Aの開裂は、LPSの毒性を除去し、かつ、複合体の可能な凝集と不溶性を回避するために行われてもよい。当業者は、脂質A部分のケトシド結合のO−脱アシル化、脱アシル化、または、加水分解のための方法に精通しているであろう。(例を参照。「Holst et al.,1991; Altman et al., 2003」)
【0043】
本発明は、同様に、タンパク質担体に共役されたほぼ線形のα1,6−グルカンを含有する化合物を含む複合体を提供する。ほぼ線形のα1,6−グルカンを含有する化合物は、α1,6−結合グルコース残基で構成された任意の適切なほぼ線形の多糖類であってもよい。「ほぼ線形の」という用語によって、α1,6−グルカンが分岐をほとんど含まず、例えば、任意の方法で限定することを望まずに、α1,6−グルカンがα1,6−グルカン中に約0乃至約5%の分岐を含んでもよいことを意味する。具体的に、α1,6−グルカンは、約0、0.5、1、1.5、2、2.5、3、3.5、4、4.5、または、5%の分岐またはその間の任意の量を含んでもよく、化合物は混合物であってもよく、このとき、混合物内の分岐の量は、1つの化合物から別の化合物まで様々である。1つの非限定的な例では、α1,6−グルカンを含む化合物は上記のような、および、本明細書で記載されるような構造であってもよい。別の実施例では、α1,6−グルカンを含有する化合物はデキストランであってもよい。デキストランは上記のような、かつ、1〜10kDaの分子量を有する必要条件を満たす任意の適切なデキストランであってもよく、例えば、しかしながら任意の方法で限定することを望まずに、デキストランは、約1、3.5、5、6.5、8、または10kDa、または、その間の任意の値であってもよい。具体的な非限定的な例において、デキストランはデキストランT5であってもよい。さらに別の例では、α1,6−グルカンを含有する化合物は、約5〜8の間のα1,6−結合グルコース残基の直鎖であってもよく、例えば、任意の方法で限定することを望まずに、α1,6−グルカンを包含する化合物は、5、6、7、または、8のα1,6−結合グルコース残基の直鎖であってもよい。
【0044】
α1,6−グルカンを含有する化合物は、リンカー分子および/または担体タンパク質に共役し、当業者によって理解されるであろうように、本明細書に記載される構造は、担体タンパク質に直接共役されるか、または、順に担体タンパク質に共役されるリンカー分子(本明細書では「リンカー」とも呼ばれる)に変化してもよい。「共役した」という用語によって、構造がリンカー分子および/または担体タンパク質に共有結合している(covalently attached or linked)ことを意味する。リンカーおよび/または担体タンパク質の共有結合のための方法は当業者に周知であり、当業者によって認識されるように、共有結合のための方法(および、リンカーが存在しているかどうか)は、使用される担体タンパク質によって変化する。任意の方法で限定することを望まずに、このような1つの方法が図4に示され、これは「Fernandez−Santana et al. (1998)」からの出典であり、この方法では、脱アシル化したLPSまたは脱脂したLPSは、Kdo残基のカルボキシル基をリンカー分子に共有結合し、その後、マレイミド官能性を導入することによって活性化される。活性化されたLPSを、チオレート化した担体タンパク質と混ぜ合わせることで、共役した構造を生み出す。当業者によって評価されるように、本明細書に記載の方法は、一般的なものであり、従って、任意の他の適切な方法が使用されてもよい(例えば、限定されないが、Chu et al., 1991; Cox et al.,2005; Gu et al., 1996; Mieszala et al.,2003; Yu and Gu, 2007によって記載されたもの)。α1,6−グルカンを含有する化合物は、構造中の任意の適切なカルボキシル基を介して、リンカー/担体タンパク質に付けられてもよい。特定の非限定的な例では、α1,6−グルカンを含有する化合物は、本発明の構造であってもよく、内殻Kdo残基Cによって付けられてもよい。
【0045】
担体タンパク質は免疫原性の担体を含む、当該技術分野で知られている任意の適切な担体であってもよい。例えば、担体タンパク質は、破傷風トキソイド、ウシ血清アルブミン(BSA)、ジフテリアトキソイド(Diphteria toxoid)、変異株ジフテリアトキソイド、CRM、CRM197タンパク質、シュードモナスAタンパク質(Pseudomonas A protein)、コレラ毒素(CT)タンパク質、コレラ毒素変異株CT−E29Hタンパク質、および、例えば、限定されないが、鞭毛、線毛、および、他の毒素の一部などの当該技術分野で知られている他のものであってもよい。
【0046】
先に示されたように、複合体は、担体タンパク質を、自然発生の基を介して本発明の構造に直接結合させるか、または、一級アミノ基、ヒドラジド、チオール、カルボキシルなどに決して限定されないがこれらを含むスペーサーまたはリンカー分子の導入によって結合させることによって、複合体を調整することができる。
【0047】
本発明は、本明細書に記載の1以上の化合物、本明細書に記載の1以上の複合体、または、それらの組み合わせを含む組成物をさらに提供する。1つの実施形態では、組成物は、上記の化合物のグリコフォームの混合物を含む。例えば、任意の方法で限定することを望まずに、組成物は、グルカン部分中に10の連続するα1,6−結合グルコース残基(n=9)を含む主要なグリコフォームを含んでもよい。同様に、任意の方法で限定することを望まずに、組成物は、本明細書に記載の1以上のグリコフォームから調製された複合体を含むことができる。実施例で実証されたように、複合体の調製に使用されたピロリ菌株26695 HP0826::Kanによって合成されたLPS構造は、3つのグリコフォーム:主鎖(backbone)オリゴ糖I、GlcNAcと[GlcNAc、Fuc、Hep]でキャップした主鎖オリゴ糖I、および、もっとも長いグルカン鎖がCE−MS分析によって測定されるように約12のα−1,6−結合残基に対応する、α−1,6−結合グルカンを含むオリゴ糖の混合物であった(表2(図2))。
【0048】
ちょうど記載されたような組成物は免疫原性のものであってもよい。「免疫原性の」という用語によって、組成物がピロリ菌の野生株および/または変異株に対する免疫反応を誘発することを意味する。免疫反応は、ピロリ菌の分類可能な、および、分類不可能な菌株に対する広範な免疫原性反応を提供してもよい。
【0049】
本発明は、同様に、個体内でのピロリ菌に対する免疫反応を誘発するために、本明細書に記載の有効な量の組成物の使用を提供する。前に記載されたように、組成物は本発明に従って1以上の化合物を含んでもよい。1以上の化合物は、リンカーおよび/または適切な担体分子に共役してもよい。
【0050】
本発明は、上記の免疫原性の組成物で哺乳動物を免疫化することにより産生される免疫抗血清をさらに提供する。免疫抗血清は、ピロリ菌の同種および異種の、分類可能なおよび分類不可能な、変異株および野生型であり、α1,6−結合グルカンエピトープを認識する免疫後血清IgGを含むか、または、産生する。IgGは、変異株および野生株のα1,6−グルカンを発現するピロリ菌の補体を媒介とした溶菌を引き起こすこともある。
【0051】
本発明はさらに抗α1,6−グルカン抗体を提供する。抗体は、本明細書に記載の本発明の化合物に対して産生されるか、または、デキストラン(例えば、限定されないが、BSAデキストラン複合体などのデキストラン複合体)といった他のα1,6−グルカンを含む分子に対して産生されてもよい。1つの非限定的な例において、抗体は、当該技術分野で知られている既知の方法に従って産出されたモノクローナル抗体であってもよい(実施例10を参照。Altman et al.,2005)。例えば、任意の方法で限定することを望まずに、抗体は、ピロリ菌HP0826変異株LPSの外殻領域にあるα1,6−グルカンエピトープに対して産生されたモノクローナル抗体であってもよく、より具体的には、抗体は、図5で示す化合物に対して産生されてもよい。さらにもっと具体的な例において、モノクローナル抗体はハイブリドーマ細胞株1C4F9によって産生されたIgM 1C4F9であってもよい。この細胞株は、2009年7月30日にカナダの国際預託機関(international depositary authority)(国立微生物学研究所(カナダ マニトバ州 ウィニペグ アーリントン・ストリート 1015)に預けられ、受け入れ番号300709−01を割り当てられた。例で実証されたように、1C4F9は、LPS中のα1,6−グルカンエピトープと、分類可能な、および、分類不可能なピロリ菌のすべての細胞を認識し、生きている細菌の表面で容易に入手可能であり、さらに、LPS−OH−TTとdLPS−BSAまたはdLPS−TT複合体の両方で同じようによく反応した。
【0052】
本発明は、このような処置を必要としている個体において、変異株および野生株の、分類可能なおよび分類不可能なα1,6−グルカンを発現するピロリ菌の補体を媒介とした溶菌を引き起こすために、1C4F9抗体の使用を提供する。先に記載されたように、分類可能なピロリ菌株は抗Le抗体によって認識されるルイスの抗原を有し、その一方で、分類不可能なピロリ菌株はそのような抗原を有していない。しかしながら、分類可能なおよび分類不可能な菌株の両方がα1,6−グルカンを含んでおり、両方の菌株は1C4F9抗体によって認識される。
【0053】
現在、当該技術分野では、ピロリ菌株はルイスの抗原に対する市販の抗体を使用して分類されるが、しかしながら、分類不可能な株がルイスの構造を含んでいないため、この手法を用いてそれらを分類することはできない。ほとんどの分類可能なおよび分類不可能なピロリ菌株は、α1,6−グルカンエピトープを運ぶため、抗α1,6−グルカン抗体(mAb 1C4F9など)は、ピロリ菌分離物(isolate)を選別して特徴づける追加の方法を提供することができる。
【0054】
ピロリ菌株26695 HP0826::KanのLPSを精製し、その化学構造を、組成物、メチル化、徹底した核磁気共鳴(NMR)分析に加え、CE−MSデータによって測定した。ピロリ菌HP0826変異株LPSの外殻領域中のα1,6−結合グルカンの存在も実証された。この構造はα1,6−グルカンに特有のモノクローナル抗体、1C4F9によって認識された。後者の抗体は、ピロリ菌(O):3 HP0826::Kan突然変異株のホルマリンで固定した細胞を用いて精製した。これらの抗体は細胞表面接触可能であり殺菌性であった。以前は、ルイスの構造のみが抗原であると信じられていた。したがって、α1,6−グルカンに対する抗体の生成は予期しえなかった。
【0055】
ピロリ菌LPSのワクチンの可能性を調べるために、ピロリ菌26695 HP0826::Kan変異株の修飾LPSを共役して、破傷風トキソイド(TT)またはウシ血清アルブミン(BSA)にした。部分的に脱脂したLPSまたは脱脂したLPSの調製のための2つの手法:やかなヒドラジン分解によるLPSのO脱アシル化(LPS−OH)、または、穏やかな酸処理によるLPSの脱脂(dLPS)fを利用した。脱脂および/または部分的な脱脂のさらなる方法は技術分野において周知であり、このような適切な方法は本発明の範囲内で用いられてもよい。LPS−OHおよびdLPSの両方を、2−ケト−3−デオキシ−オクツロソン酸(Kdo)残基を介して、ジアミノ基を含むスペーサーに共有結合させ、その後、マレイミド官能性の導入と、チオール化したTTまたはBSAとの共役によって、それぞれ、複合体、LPS−OH−TT、dLPS−BSA、および、dLPS−TTを得た。個別の実験では、分類不可能な株PJ2を脱脂し、共役のために利用することによって、dLPS(PJ2)−TT複合体を得た。
【0056】
dLPS(PJ2)−TTと同様に、LPS−OH−TT、dLPS−BSA、dLPS−TT複合体は、IgM 1C4F9による間接的なELISAによって設定されるように、表面接触可能なα1,6−グルカン決定子の抗原を保持した。抗体はα1,6−グルカン決定子への優れた特異性を有することを示し、その結合特性が、ピロリ菌の分類可能なおよび分類不可能な菌株からの生成されたLPSと、イソマルト−系(isomalt−series)からのオリゴ糖とによる阻害ELISAによって測定された。これらの研究により、1C4F9が5または6の連続するα1,6−結合グルコース残基についての必要条件、Kabat(1993)が主張した部位を結合する抗−α−(1→6)デキストランの大きさと一致したパターンを有することが分かった。
【0057】
LPS−OH−TT、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TT複合体は、マウスとウサギにおいては免疫原性であり、ピロリ菌の異種の、同種の、および、野生型の菌株からのLPSに対する有意なIgG抗体反応を誘発した。免疫抗原に対する10倍強固なIgG免疫反応が、dLPSを含む複合体を受け取ったマウスおよびウサギで生じた。LPS−OH−TT、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TTのいずれかで免疫化したウサギの免疫後の血清は、ピロリ菌の26695 HP0826::Kan変異株および野生型26695菌株に対する殺菌活性を示した。
【0058】
要約すると、これらの結果は、Le抗原を欠いた、かつ、長いα1,6−グルカン鎖を保有する、脱脂したか部分的に脱脂したピロリ菌LPSベースのタンパク質複合体が、マウスとウサギの両方において免疫原性であり、殺菌性の抗体を誘発するということを示唆している。このエピトープが免疫原性であると同定されたことに留意することが重要である。このことは以前は確立されていなかった。なぜなら、抗原性であり、かつ、特定の抗体を産生すると知られていたのは、ルイスの構造だけだったからである。
【0059】
本発明は、以下の実施例においてさらに例証される。しかしながら、これらの実施例がたんなる説明目的のためのもので、任意の方法で本発明の範囲を制限するためには使用されてはならない。
【実施例】
【0060】
(実施例1:ピロリ菌26696 HP0826::Kan LPSの分離および構造分析)
ヘリコバクター・ピロリ菌株26695は、R.Alm博士(マイアミ州ボストンのアストラゼネカ)から入手し、ピロリ菌(O):3分離株はJ.Penner博士から、J99は、D.Taylor博士(カナダ・エドモントンのアルバータ大学)から入手し、SS1はA.Lee博士(オーストラリア・シドニーのニューサウスウェールズ大学)から、PJ1とPJ2の臨床分離株は、W.Conlan博士(IBS、NRC)からの新鮮な臨床分離株であり、および、M6はK.Eaton博士(ミシガン州のミシガン州立大学)から入手した。
【0061】
細菌株の増殖は、Hiratsukaら(2005)によって記載されたように実行された。手短にいえば、先に記載されたように(Hiratsuka et al)、微好気性環境下で、7%のウマ血液を含む、抗生物質を補ったコロンビア血液寒天培地(Difco)上で、細胞を37°Cで48時間増殖させた(Kan20μg/mL)。液体培養中の増殖のために、10%ウシ胎児血清を含む抗生物質を補ったブルセラ菌ブロスに、48時間のコロンビア血液寒天培地/ウマ血液プレートから採取したピロリ菌細胞を接種し、以前に記載されたように(Hiratsuka et al、2005)、微好気性条件下で(85%N2、10%CO2、5%O2)攪拌機内で48時間インキュベートした。
【0062】
ピロリ菌株を上記の液体培養において培養し、細菌増殖の遠心分離によって得られた浸潤細胞を、エタノール、アセトン、および、石油エーテル(light petroleum)によって2度連続して洗浄し、空気乾燥させた。WestphalとJann(1965)の暖かいフェノール水抽出手順(hot phenol−water extraction procedure)によって、空気乾燥した細胞塊からLPSを抽出した。広範な透析と凍結乾燥後に、水相からLPSを得た。超遠心分離(105,000×g,4°C、12時間)によってピロリ菌LPSをさらに精製し、ペレットを蒸留水で懸濁し、凍結乾燥させた。
【0063】
アルジトール酢酸方法(Sawardeker el al,1967)によって糖組成分析を行なった。加水分解を100°Cで4Mトリフルオロ酢酸で4時間行うか、または、100°Cで2Mトリフルオロ酢酸で16時間行い、その後、NaBH4によるH2Oの還元と、その後の無水酢酸/ピリジンによるアセチル化が続く。アルジトール酢酸誘導体を先に記載したように分析した(Altman et al,2003)。Ciucanu & Kerek(1984)の方法によって、および、先に記載したように(Altman et al,2003)、ガス液体クロマトグラフィー−質量分析法(GLC−MS)による完全メチル化されたアルジトール酢酸誘導体の特徴付けによって、メチル化分析を行った。
【0064】
アルジトール酢酸としてのピロリ菌26695 HP0826::Kanから精製されたLPSの糖分析は、L−フコース(L−Fuc)、D−グルコース(D−Glc)、D−ガラクトース(D−Gal)、N−アセチル−D−グルコサミン(D−GlcNAc)、D−グリセロ−D−マンノ−ヘプトース(DD−Hep)、および、L−グリセロ−D−マンノ−ヘプトース(LD−Hep)が、0.4:5.0:1.5:4.3:6.4:1.4の適切なモル比で存在していることを明らかにし、このことは、O鎖を欠く構造の存在を示している(Logan et al.,2000)。未処置の26695 HP0826::Kan LPS上で行なわれたメチル化分析は、これらの発見と一致しており、末端のL−Fuc、3−置換したL−Fuc、末端D−Glu、末端D−Gal、3−置換したグルコース、4−置換したD−Gal、6−置換したグルコース、末端DD−Hep、2−置換したDD−Hep、6−置換したDD−Hep、3−置換したDD−Hep、7−置換したDD−Hep、2,7−置換したDD−Hep、2−置換したLD−Hep、3−置換したLD−Hep、末端D−GlcNAc、および、3−置換D−GlcNAcが、0.1:1.0:1.1:0.1:1.2:1.0:6.0:0.4:1.2:1.1:3.1:0.5:1.6:1.2:0.1:0.3:0.4の近似モル比で存在していることを示す。O鎖を含んでいるLe抗原の特徴である3,4−置換したD−GlcNAcおよび2−結合D−Galは、まったく検知されなかった。LPSの純度はRNA由来のリビトールの欠如によって確認された。26695 LPSの組成物およびメチル化分析は、他で報告されている(Logan et al.,2005)。すべての糖がピラノース形態であった。
【0065】
(実施例2):キャピラリー電気泳動法−質量分析法(CE−MS)によって脱脂したピロリ菌26695 HP0826::Kan LPSによる特徴づけ
実施例1で得られた生成した26695 HP0826::Kan LPS(20mg)を、0.1Mの酢酸ナトリウム緩衝液(pH4.2)において100°Cで2時間加水分解し、先に記載したように(Altman et al,2003)Bio−GelP−2カラムでのゲルろ過によって分画することで、脱脂したLPS(dLPS)を生成した。3つの分画(分画1−3)を集めて、キャピラリー電気泳動質量分析法(CE−MS;表1)によって分析した。
【0066】
CE−MSに関して、Prince CEシステム(Prince Technologies,オランダ)を、4000QTRAP質量分析計(Applied Biosystems/MDS Sciex,カナダ)につないだ。シース溶液(イソプロパノール−メタノール、2:1)を、流速1.0μL/分で送達した。pH 9.0の脱イオン水中の15mM酢酸アンモニウムを用いて、約90cmの長さの剥き出しの石英ガラスの毛細管上で分離物を得た。5kVのエレクトロスプレーイオン化電圧は陽イオン検出モードに使用した。4000da/sの走査速度で増強された産生イオン走査モード(EPI)を用いて、タンデム質量スペクトルを得た。(12の値の)カーテン(curtain)および(「高い」縮尺に設定された)衝突ガスとして、窒素を使用した。
【0067】
陽イオンモードでの主要な分画1のCE−MS分析は、約11のα−1,6−結合残基に相当する最長のグルカン鎖である、Hex残基の連続的な追加に一致して、m/z 1212.3、m/z 1266.3、および、m/z 1320.4で一連の三価イオンの存在を確認した。イオンの信号強度に基づいて、m/z 1266.3での最も豊富なグリコフォームは、10のα−1,6−結合Hexを含む(図1)。陽イオン検出モードにおけるm/z 1266.3でのイオンの生成スペクトルは、以前観察された主鎖コア断片(core fragment)Hex2Hep3(PEtn)KdoとHexNAcHex2Hep3(PEtn)Kdoそれぞれに一致して、m/z 1244.5とm/z 1447.6での診断のイオンの存在を確認した(図1)。LPSコア中にα1,6−グルカンを保有する分画1および2を組み合わせて共役のために用いた。
【0068】
【表1】
【0069】
(実施例3):CE−MSによるO−脱アシル化した26695 HP0826::Kan LPSの特徴付け
(実施例1の)26695 HP0826::Kan LPSのO−脱アシル化は、幾つかの修正をしたHolst et al.に従って実行された(1991)。手短にいえば、LPS(4mg)を4時間、37°Cで無水ヒドラジン(0.2ml)中で撹拌した。反応混合物を冷却し、冷たいアセトン(2ml)をゆっくりと加えることで超過ヒドラジンを破壊した。30分後、沈殿したO−脱アシル化したLPS(LPS−OH)を、遠心分離(4°C、9300×g、10分)によって集めた。ペレットを冷たいアセトンで2度洗浄し、水に溶解させ、凍結乾燥させることでLPS−OH(3.5mg)を得た。
【0070】
陽イオンモードでのO−脱アシル化した26695 HP0826::Kan LPS(LPS−OH)のCE−MS分析を、実施例2に記載のように行った。CE−MSの結果は脱脂したLPSについて得られたMSデータと一致しており、主鎖オリゴ糖、および、HexNAc、および、[HexAc、Fuc、Hep]のそれぞれでキャップした主鎖オリゴ糖のそれぞれの存在と一致して、m/z 1137.2、m/z 1239.2、および、m/z 1408.0で3つの主要な二価イオンを提供したが(表2)、その一方で、m/z 1597.7、m/z 1651.4、および、m/z 1705.5での三価イオンは、約12のα−1,6−結合残基に相当する最長のグルカン鎖である、グルカンの存在と一致した(表2、図2)。m/z 1597.7のMS/MSスペクトルは、[Fuc,Hep]と[Fuc,Hep,HexNAc]のそれぞれの連続的な損失と一致して、m/z 1447.6とm/z 1244.5での診断のイオンの存在を示し(図2)、さらに、Kdo残基を介してO−脱アシル化された脂質A(脂質A−OH)に結合したコア断片FucHex2HexNAcHep4(PEtn)Kdoに一致して、m/z 1786.8で一価イオンをもたらし、その一方で、m/z 444.4およびm/z 887.8でのイオンは、2つのアミド結合した3−ヒドロキシオクタデカン[C18:0(3−OH)]脂肪酸鎖により置換されたジグルコサミン(diglucosamine)主鎖からなる脂質A−OH部分に相当した(図2)。
【0071】
【表2】
【0072】
(実施例4):ピロリ菌株26695 HP0826::Kan LPSのNMR分析
PEtN置換基のアルカリ加水分解がアグリコンを含まない還元末端を残すため(Holst et al.,1991)、(実施例1の)ピロリ菌株26695 HP0826::KanからのLPSの分解は、脂質A GlcNの即時還元のためのNaBH4の存在下で、4M KOHによる完全な脱アシル化で始まった。ゲルクロマトグラフィーによる生成物の分離は、クロマトグラムのオリゴ糖領域で溶出された、2つの分画をもたらした。更なる分析によって、これらの分画は、グルカン鎖の長さによって明らかに異なる、類似する化合物を含むことが示された。低分子量分画(Lower molecular mass fraction)は、質量分析法によって同定された化合物1−3を含み、両方の分画は、グルカン部分の異なる長さの化合物4を含んでいた(図3)。
【0073】
NMRスペクトル(DQCOSY、TOCSY、NOESY、1H−13C HSQC、および、HMBC)は、先に記載されたような(Brisson、2002)標準のソフトウェアを用いてVarian 500または600MHzの分光計で行われた。NMR実験はすべて、1Hスペクトルにはδ2.225ppmで、13Cスペクトルには31.45ppmで、内部基準としてアセトンを用いて25°Cで行われた。
【0074】
その複雑さゆえにスペクトルを完全に解釈することはできなかったが、両方の分画のNMRスペクトルは化合物4に相当した。α−1,6−グルカン、および、DD−ヘプタンの末端残基のH−1プロトンは、グルカンおよびヘプタンH−1の残りには重複せず、したがって、図3に示されるように、全体的な構造内でこれらのホモポリマーの位置を同定することができる。しかしながら、α−1,6−グルカン鎖の還元末端Glu残基Zの結合位置は測定されなかった。それは、同定されていない陽子(後に、HepNのH−6に割り当てられる)との強いNOE相関を示した。化合物4のNMRデータは、ヘプタン部分(X)の第1のHep残基がα−1,6−グルカンに属する非還元末端Glc残基VのO−3に結合されたため、α−1,6−グルカンがDD−ヘプタンと内殻との間に置かれたことを明確に示した。DD−ヘプタンの非還元末端は、O−3でβ−GlcN Wに置換された。小さなオリゴ糖1−3と同一の化合物は、α−1,6−グルカンがほとんどない変異株HP0159::Kanのピロリ菌LPSの主要な構成要素として先に発見され、それらのNMRデータが公表された(Altman et al.,2008)。化合物4に関して選択されたNMRデータが表3に示される。Kdoに結合した4つの近接するヘプトース残基の以前提案された配列は、調べた化合物のいずれにも存在しなかったことが容易にわかるであろう(図3)。β−GlcN残基Lは、HepGのO−2に直接結合し、LPS構造の任意の部分においてHepのO−7につなげられたGlcNはなかった。
【0075】
【表3】
【0076】
LPS構造のさらなる分析について、化合物1−4を脱アミノ化した。手短にいえば、サンプルを10%AcOHに溶解させ、過度のNaNO2を加えた。3時間後、Sephadex G−15カラムで、ゲルクロマトグラフィーによって生成物を分離した。その後、化合物のいくつかをNaBD4で還元し、脱塩した。結果として生じる生成物が化合物5および6であった。(上記のように行なわれた)化合物5のNMRスペクトルは、(アルカリ条件下のリン酸塩移動によって)HepEの位置6または7で、リン酸塩を含む2つの異性体を示した。再度、3つの近接するHep残基が存在し、以前に提案された構造が正確である場合に予測されるであろうように、4つではなかった。脱アミノ処置(上記の)による全側鎖の除去は、GlcN LがOS5の糖と分子の残部の間の接続を形成することを確認した。
【0077】
化合物6は、還元末端に、DD−ヘプタン、α−1,6−グルカン、および、三糖類DDHep−Fuc−anh−Man−ol(GlcN Lに由来するN−M−Lおよびanh−Man L)を含んでいた。この生成物のスペクトルはそれほど密集しておらず、(DDHep NのH−1とH−6の間のTOCSYからの)DDHep NのO−6に対するα−1,6−グルカンの付着点を同定することが可能になった。分岐糖を全く示さなかったメチル化分析によって確認されるように、構造は直線的であった。予測された生成物はすべて、提案された構造に一致して同定された(表4)。
【0078】
【表4】
【0079】
DD−ヘプタン−α−1,6−グルカン領域の構造のさらなる確認が、化合物6の過ヨウ素酸酸化の結果から得られた。過ヨウ素酸酸化は24時間、過度の0.1M NalO4で行なわれた。エチレングリコールを加え、Sephadex G−15カラムでのゲルクロマトグラフィーによって生成物を分離させた。それをNaBD4で還元し、脱塩し、2%AcOHによって、100°Cで3時間加水分解した。2つの主要な生成物7と8を得て、Sephadex G−15カラムでのゲルクロマトグラフィーによって分離した。
【0080】
NaBD4による酸化オリゴ糖の還元は、NMRスペクトル(APT−HSQC中のCH2OH信号に対するCHDOH信号の反転位相)での酸化炭素の同定を可能にした。化合物7および8の構造は、NMR分光法およびメチル化分析によって測定された。化合物7の形成は、DDHep Nが位置3では置換されなかったことを証明した。化合物6のメチル化が末端の3−および6−置換DD Hepのみを示したため、化合物6のNMRデータから提案されたように、DD−Hep Nは位置6で置換された。還元末端でグリセロール(Gro)を有する化合物8の形成は、酸化還元後にグリセロールを産生可能なオリゴ糖6の他の成分がなかったため、OS6中のGlc Vが位置6で次のGlc残基をグリコシル化したことを確認した。そのことは、同様に、末端グルコースの位置3でのα−1,6−グルカンの非還元末端へのDD−ヘプタン部分の付着を証明した。
【0081】
DD−ヘプタン領域のサイズを推定することができる。化合物8はDD−ヘプトースに由来する4つのマンノース残基を含んでいた。化合物8のスペクトルはよく分解し、H−1信号の統合はほぼ等モルの比率を与え、4つのマンノース残基の存在を示した。したがって、化合物6の完全なDD−ヘプタン領域は、ほとんど5つのDD−ヘプトース単位からなり、そのうちのひとつ(末端)は、過ヨウ素酸酸化によって取り除かれた。この結論は質量分析法によって確認された。ピロリ菌株26695 HP0826::Kanによって産生された主要なLPSグリコフォームの構造が図3に示される。
【0082】
(実施例5):LPS−OH−TT、dLPS−BSA、dLPS−TT、および、dLPS(PJ2)−TT複合体の調製および特徴付け
ウシ血清アルブミン(BSA)および/または破傷風トキソイド(TT)に対する、LPS−OH(実施例3を参照)およびdLPS(実施例2を参照)のKdo結合複合体を調製した。
【0083】
LPS−OH(4789Daの推定平均分子量に基づき、4mg、0.8μmol)またはdLPS(3779Daの推定平均分子量に基づき、4mg、1μmol)を、0.1M NaCl(0.4 mL)を含む0.1M 2−(N−モルホリノ)エタンスルホン酸酸(MES;Sigma−Aldrich、セントルイス、ミズーリ州)緩衝液(pH 4.8)に溶解させた。1−エチル−3−ジメチルアミノプロピルカルボジイミド(EDC;34.38mg、100:1モル比;Sigma−Aldrich)を加え、その後、1,8−ジアミノ−3,6−ジオキサオクタン(15μL、103μmol; Sigma−Aldrich)を加え、その反応を22°Cで4時間、pH 4.8で維持した。溶液をpH 7.0に調節し、蒸留水に対して透析し、Microsepの遠心分離装置1,000Da cutoff(Pall Life Sciences,Ann Arbor、ミシガン州)を用いて脱塩または凍結乾燥させた。
【0084】
基本的に、Fernandez−Santana et al. (1998)によってオリゴ糖について記載されたように、共役手順を行った。手短に言えば、乾燥したDMSO中で3−マレイミドプロピオン酸(maleimidopropionic acid)N−ヒドロキシスクシンイミドエステル(BMPS;2mg、7.5μmol;Sigma−Aldrich)と、スペーサーを含んでいるLPS−OH(2mg、0.4μmol)またはdLPS(2mg、0.5μmol)を22°Cで24時間、反応させた。溶液を蒸留水に対して透析し、凍結乾燥させた。
【0085】
BSAの活性化のために、乾燥したDMSO中で3,3’−ジチオジプロピオン酸ジ(N−ヒドロキシスクシンイミドエステル)(DTSP;0.63mg、1.6μmol;Sigma−Aldrich)を、6mM EDTA(最終濃度4mg/mL)を含む10mM PBSの緩衝液(pH 8.0)に事前に溶解しておいたウシ血清アルブミン(BSA)(分子量66,320Da)(8mg、0.12μmol)の溶液に、N2雰囲気下で加えた。その混合物を22°Cで2時間撹拌した。その後に、N2雰囲気下でジチオトレイトール(DTT; 7.12mg、46μmol; Sigma−Aldrich)を加え、その混合物を4°Cで1時間撹拌した。再生セルロース細胞膜30,000Da cutoff(YM30、Millipore)全体の圧力源としてN2を用いて、撹拌した限外濾過細胞(Millipore、Billerica、マサチューセッツ州)中に5mM EDTAを含む10mM PBSの緩衝液(pH7.2)に対して、結果として生じた溶液を4°Cで透析した。タンパク質とSHの含有量を、ビシンコニン酸タンパク質アッセイキット(BCA;Pierce、Rockford、イリノイ州)とEllman(1959)方法によってそれぞれ測定した。20−22SH基のモル置換が得られた。
【0086】
破傷風トキソイド(TT)(分子量、150,000Da)の活性化のために、6mM EDTA(最終濃度4mg/mL)を含む10mM PBS緩衝液(pH 8.0)中のTT溶液(4mg、0.03μmol)に、N2雰囲気下で、乾燥したDMSO(25μL)中のDTSP(0.316mg、0.8μmol)を加え、その反応をBSAについて上記したように処理を進めた。その後に、N2雰囲気下で、ジチオトレイトール(DTT;3.56mg、23μmol; Sigma−Aldrich)を加え、その混合物を4°Cで1時間撹拌した。活性化したTTを、撹拌した限外濾過細胞(Millipore)に移し、再生セルロース細胞膜、100,000Da cutoff(YM100、Millipore)にわたって、5mM EDTAを含む10mM PBSの緩衝液(pH7.2)に対して、4°Cで透析した。
【0087】
5mM EDTAを含む10mM PBS緩衝液(pH 7.2)中のBSA−SH21−22またはTT−SH21−22の溶液に、10mM PBSの緩衝液中のマレイミド官能基を有するLPS−OHまたはdLPS誘導体の溶液を、N2雰囲気下で加えた。その反応物を4°Cで24時間攪拌した。その後に、N−エチルマレイミド(1mg;Sigma−Aldrich)を加えた。その反応を22°Cで30分間進め、結果として生じる複合体を、4°Cで4時間、10mM PBS緩衝液(pH 7.2)に対して透析し、0.22μmポリフッ化ビニリデン(PVDF)膜(Millex−GV、Millipore、Cork、アイルランド)を用いて、フィルタを殺菌した。複合体は、その炭水化物とタンパク質含有量について、それぞれLPS−OHまたはdLPSおよびBSAを標準として、中性糖のためのフェノール硫酸方法(Dubois et al、1956)、および、BCAタンパク質アッセイキットを用いて分析された。複合体の効率は、10mM PBS緩衝液(pH 7.2)で平衡に保たれたSuperose 12 10/300 GLカラム(Amersham Biosciences、ウプサラ、スウェーデン)を用いて、高性能液体クロマトグラフィー(HPLC; Agilent 1200Series、Agilent Technologies、Waldbronn、ドイツ)によって確認された。クロマトグラフィーは、室温で、流速0.5 mL/分で行われた。溶出はダイオード・アレー検出器(diode array detector)(Agilent Technologies)で210nmおよび280nmでモニターされた。
【0088】
スペーサーの存在は、CH2NH2基に対応する3.22ppmでの新しい陽子共鳴の出現によって、1H−NMR分光法で確認された。β−マレイミドプロピオネート(maleimidopropionate)のCH2αおよびCH=CH基にそれぞれ対応する2.55ppmおよび6.9ppmでの陽子共鳴の存在によって確認されたように、スペーサー分子のアミン基が、3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステルとの反応によってさらに誘導体化されたことで、マレイミド官能基を有するLPS−OHまたはdLPSを得た。内殻のLD−Hepのホスホエタノールアミンの誘導体化は、化学量論量の試薬の使用により最小限に抑えられた。糖複合体は、担体タンパク質のチオ化(thiolation)とチオ化したタンパク質のマレイミド官能基を有するLPS−OHまたはdLPSへの追加によって得られた(図4)。共役の効率はHPLCによってモニターされた。得られた複合体はその炭水化物とタンパク質含有量について分析された。
【0089】
3つの複合体におけるTTに対するLPS−OHのモル比は、10:1から20:1まで及び、産出量は炭水化物含有量に基づいて13%から22%までで変動した(表5)。BSAまたはTTへのdLPSの共役は、著しく炭水化物含有量の多いdLPS−BSA−2、dLPS−TT、または、dLPS(PJ2)−TT複合体を産出した(表5)。LPS−OH−TTとdLPS−BSAまたはdLPS−TTの複合体の両方は、ELISAによってα1,6−グルカンに特異的なmAbと均等によく反応し、このことは、グルカンエピトープの形態が不変だったことを示唆している。
【0090】
【表5】
【0091】
(実施例6):マウスとウサギにおける、LPS−OH−TT、dLPS−BSA、dLPS−TTの複合体、および、dLPS(PJ2)−TT複合体の免疫原性
実施例5の複合体(LPS−OH−TT、dLPS−BSA、dLPS−TT、および、dLPS(PJ2)−TT)の免疫性をマウスとウサギで試験した。
【0092】
生後6−8週の5匹の雌BALB/cマウスを、適切な複合体を用いて腹腔内で免疫化した。各々のマウスは、1回の注入当たり0.2mL Ribiアジュバント中の2μgまたは10μgの炭水化物を受け取った。マウスを21日目と42日目にブーストして、血清を51日目に末端の心臓穿刺(terminal heart Puncture)から回収した。
【0093】
3匹のニュージーランドホワイトウサギを、適切な複合体を用いて皮下で免疫化した。各々のウサギは、0.5mLの不完全フロインドアジュバント中の10μgまたは50μgの炭水化物を受け取った。ウサギを28日目と56日目にブーストして、65日目に失血後から回収した。
【0094】
血清中の抗LPS抗体のレベルは、精製されたLPSを被膜抗原(1μg/ウェル)として用いるELISAによって測定された。PBSで洗浄した後に、PBSまたはMilk希釈液/ブロッキング溶液(blocking solution)(MDB)(KPL、Gaithersburg、MD)中の1%(w/v)のウシ血清アルブミン(BSA)によって、プレートを37°Cで1時間ブロック(block)した。希釈したマウスまたはウサギの免疫前後の血清を加え、プレートを37°Cで2時間インキュベートした。阻害ELISAについては、26695 HP0479::Kan LPSまたは26695 HP0826::Kan LPSを阻害する一連の希釈物を、OD450=0.6−0.8を与えたウサギ血清の事前に測定された希釈物と混合した。この混合物を22°Cで15分間インキュベートし、吸収したLPS抗原で遮断された元々のマイクロタイタープレートに移し、そこで37°Cで2時間、さらにインキュベートした。この工程の後は、間接的なELISA処置に従った。手短にいえば、プレートをPBSと第2抗体で洗浄し、ヤギ抗マウスIgG+IgMワサビダイコンペルオキシダーゼ複合体(Caltag、So.サンフランシスコ、カリフォルニア州)を室温で1時間加えた。最終的な洗浄工程の後、3,3’,5,5’−テトラメチルベンジデン(benzidene)(TMB)(KPL、Gaithersburg、MD)基質を加え、1Mのリン酸で反応を止めた。マイクロタイタープレートリーダー(Dynatech,Chantilly,VA)を用いて、450nmで吸光度を測定した。
【0095】
阻害割合を以下の式を用いて計算した。
%阻害=100×[(インヒビターを含むOD−インヒビターを含まないOD)/インヒビターを含まないOD]
【0096】
阻害対ログ濃度曲線が各インヒビターについてプロットされ、半最大阻害濃度(half maximal inhibitory concentration)(IC50)に必要な濃度を、外挿曲線(extrapolation Curve)から決定した。
【0097】
反応は、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TTのいずれかで免疫化した動物よりもLPS−OH−TTで免疫化したマウスとウサギの方で全般的に弱かったが、全ての複合体は、3度の注入後(表6乃至9)、ウサギとマウスの両方で同種(26695 HP0826::Kan)と対応する野生型(26695)株からのLPSに対するIgG反応を誘発した。LPS−OH、dLPS、または、dLPS(PJ2)のいずれかと、タンパク質担体(アジュバントを有する)との混合物で免疫化した対照ウサギは、3度の免疫化後に、ピロリ菌の同族の菌株(26695 HP0826::Kan変異株)、または、対応する野生型26695菌株からのLPSに対する特異的な反応は全くないか、低い値しか示さなかった。
【0098】
【表6】
【0099】
【表7】
【0100】
【表8】
【0101】
【表9】
【0102】
交差反応性研究は、様々なLPS グリコタイプ (Monteiro,2001)を代表するピロリ菌株、および、選択された突然変異株からのLPSに対して、LPS−OH−TT−2、LPS−OH−TT−3、LPS−OH−TT−4、dLPS−BSA−2、dLPS−TT、または、dLPS(PJ2)のいずれかで免疫化したウサギの免疫後血清を用いて行なわれた。結果を表10、11および12で示す。
【0103】
【表10】
【0104】
【表11】
【0105】
【表12】
【0106】
α1,6−グルカンを加えることができないピロリ菌株を代表するものである(Logan et al.,2005)SS1およびSS1 HP0826::Kan LPSによって、弱い交差反応性しか得られなかったため、LPS−OH−TT−2、LPS−OH−TT−3、またはLPS−OH−TT−4で免疫化したウサギから得た免疫後の血清の反応性は、α1,6−グルカンの存在の必要性を示した(表10)。驚くことに、dLPS−TTで免疫化したウサギから得た血清は同様に、任意の1,6−グルカンを含まなかった殻LPSエピトープ、即ち、株SS1、SS1 HP0826::Kan、M6およびJ99からのLPS(表11)を認識し、これはより広い殻の認識を示し、おそらくはTT担体タンパク質の存在により、dLPS−TT結合体がより免疫原性であったことを推測している(表11)。あるいは、結合体が3のグリコフォームの混合物から調製されるように、免疫反応は他の主要でないLPS構成要素へと生成されていたかもしれない。このことは、観察された交差反応について説明する。
【0107】
dLPS−TT結合体によって誘発されるウサギ血清の結合特異性を調べるため、阻害ELISAを、2つのグリコフォーム(およその比率1:1)、すなわち、線形の主鎖オリゴ糖構造および[GlcNAc,Fuc]にキャッピングされた線形の主鎖オリゴ糖(Hiratsuka et al.,2005)、ならびに、26695 HP0826::Kan LPSからなる、精製された26695 HP0479::Kan LPSを用いて実行した。26695 HP0479::Kan LPSがインヒビターとして使用された時、株SS1、SS1 HP0826::Kan HP0159::KanおよびSS1 HP0479::Kanからのグルカン−陰性LPSへのウサギ血清の結合(図6)が有意に阻害され、その一方で、26695 LPSがコーティング抗原(図6)として使用された時、26695 HP0826::Kan LPSが最も効果的なインヒビターであった。
【0108】
異種の分類可能な株および分類不可能な株を認識する、ウサギの免疫後血清の能力も同様に、ピロリ菌の分類可能な株および分類不可能な株両方のピロリ菌代表物(representatives)の選択された臨床分離株に対する細胞全体の間接的ELISA(WCE)において確認された。細胞全体の間接的ELISA(WCE)を、前述に記載の通りに実行した(Altman et al.,2008)。手短に言えば、マイクロタイタープレートのウェル(well)を、4°Cで一晩中、108細胞/mLで、100μLの細菌懸濁液で覆った。その後、ウェルをメタノールで固定し、37°Cで2時間、200μLのMilk希釈液/ブロッキング溶液(MDB)(KPL,Gaithersburg,MD)でブロックした。続いて、MDB中で1:500に希釈した100μLの1C4F9腹水を用いて、ウェルを37°Cで2時間インキュベートし、その後、MDB中で1:1,000に希釈した抗マウスIgG+IgMホースラディッシュペルオキシダーゼ(Caltag)を用いて、室温で1時間インキュベートした。間接的ELISAに関して記載されるように、基質TMBを加えた。非特異性のバックグラウンド値を、細菌細胞、第2の抗体結合体および基質を含む、陰性対照のウェルのOD450として決定した。これらOD450値は≦0.2であった。OD450<0.2の光学的濃度値を、陰性反応として分類し、OD450値≧0.2を陽性反応として分類した。プレート間の一貫性を保証するために、ピロリ菌株26695細胞を陽性対照として使用した。アッセイは10%以上は変わらなかった。
【0109】
dLPS−TT複合体で免疫化したウサギ由来の血清が、細胞全体の間接的ELISAを使用して試験されたすべての株(表13)に対し、もっとも強い交差反応性を示した。
【0110】
表13:ピロリ菌の臨床分離株に対する細胞全体の間接的ELISAbにおけるウサギの免疫後血清aの交差反応性。
【0111】
【表13】
【0112】
(実施例7:ウサギ抗血清の殺菌力)
ワクチン接種前またはワクチン接種後のウサギの血清(実施例6)を使用して、殺菌アッセイを行なった。
【0113】
プレートで増殖した(Plate−grown)ピロリ菌細胞を採取し、プレート1枚当たり5mLのPBSで洗浄した。遠心分離の後、ペレットを25mLのPBS中に懸濁した。最終的な細菌懸濁液を、4x106 CFU/mLでダルベッコ燐酸緩衝生理食塩水(DPBS)(Invitrogen,Grand Island,ニューヨーク)内で希釈した。殺菌アッセイを、平底のマイクロタイタープレート(ICN)内で行なった。補体除去した(de−complemented)免疫前または免疫後の血清(50μL)の10倍の連続希釈物を、各々のウェルに加えた。その後、細菌懸濁液(25μL)を加え、37°Cで15分間プレインキュベートした。仔ウサギの補体(Cedarlane Laboratories,Hornby,ON)を1:50に希釈し、25μLを適切なウェルに加えた。プレートを37°Cで45分間インキュベートし、次に、氷の上に置いた。10μLのアリコートを、コロンビア血液寒天(Columbia blood agar)上で3回播種し、4日間増殖させ、その後プレートを数えて、コロニー形成単位(CFU)の数を測定した。バクテリアと補体を含むが血清を含まない対照プレートを使用し、死滅率(percentage killing)を計算した。殺菌力は、50%を死滅させた血清のもっとも高い希釈として決定した。
【0114】
ワクチン接種前の血清またはタンパク質担体とLPS−OH、dLPSまたはdLPS(PJ2)のいずれかとの混合物(アジュバントを有する)により免疫化されたウサギから得た血清は、同種のピロリ菌26695 HP0826::Kanまたは野生型の26695菌株に対する殺菌力を示さないか、または、それらに対する低レベルの殺菌力を示した。LPS−OH−TT−4、dLPS−BSA−2、dLPS−TTおよびdLPS(PJ2)−TTで免疫化したウサギから得た免疫後血清は、26695 HP0826::Kan変異株および対応する野生型菌株の両方に対し、有意で機能的な活性を示し(表14)、dLPS−TT結合体で免疫化したウサギからのワクチン接種後の血清は、野生型菌株26695に対してもっとも高い機能的な活性を示した(表14)。
【0115】
【表14】
【0116】
更に、ウサギ血清の殺菌力も同様に、ピロリ菌の選択された臨床分離株、すなわち、002CL、0153CLおよび058CLに対して試験した(Altman et al.,2008)。これら分離株を、抗−α1,6−グルカンmAbsを用いるWCEアッセイにおけるそのOD450値に基づき、高度、中度および低度の結合剤の代表物として選択した:株002CL−O450 1.361、高度の結合剤;株153CL−OD450 0.29、中度の結合剤;および株058CL−OD450 0.162、低度の結合剤である。dLPS−TTまたはdLPS(PJ2)−TT複合体のいずれかで免疫化したウサギからの免疫後血清のみが、試験された3つの臨床分離株により機能的な活性を示したことを強調することが重要である(表15)。
【0117】
【表15】
【0118】
(実施例8:マウスにおけるdLPS−TT複合体による防御試験)
候補ワクチンとしてのdLPS−TT複合体の潜在性を、非近交系のCD−1マウスにて評価した。
【0119】
1μg/マウスのコレラ毒素(CT;Sigma)で補助された25μg/マウスのdLPS−TT複合体、1mg/マウスのコレラ毒素で補助された31.5μg/マウスのPJ2細胞を含まない音波処理物(PJ2 cell−free sonicate)(対照)、または食塩水(対照)を、グループ5のマウスに、週1回の間隔で4度、鼻腔内にワクチン接種した。最後の免疫の1週間後、血清、糞および膣の洗浄サンプルを、ピロリ菌特異的IgGおよびIgAのために採取し、分析した。血清IgGおよび血清、糞および膣のIgA抗体レベルを、標準のELISAによって測定した。プレートを本明細書に記載の通り、1μg/ウェルのピロリ菌株26695 HP0826::Kan LPSで覆った。血清サンプルを、IgGアッセイのため1:100に希釈し、IgAアッセイのため1:50に希釈した。糞のサンプルを1:2に希釈し、膣のサンプルを1:20に希釈した。ピロリ菌特異的抗体レベルを決定し、GraphPad software version 5.0を使用して、マン・ホイットニー検定によってデータを分析した。
【0120】
最初の免疫の5週間後、〜108 cfuピロリ菌株PJ2を一日おきに3回、マウスに経口的に強制投与した(Altman et al.,2003)。4週間後に、マウスを殺し、胃からのその一般細菌を数え、GraphPad software version 5.0を使用して、マン・ホイットニー検定によってデータを分析した。
【0121】
CTに加えてdLPS−TT複合体による鼻腔内の免疫化は、ELISAによって測定されるように、非常に強力なピロリ菌26695 HP0826::Kan LPS特異的IgG反応を誘発し、中度のピロリ菌26695 HP0826::Kan LPS特異血清IgAも同様に検出された(表16および17、図7)。加えて、膣または糞のサンプルにおけるピロリ菌26695 HP0826::Kan LPS特異的反応も同様に検知した(p<0.01)(表17、図7)。
【0122】
【表16】
【0123】
【表17】
【0124】
長いα1,6−グルカンを含有すると以前に示された分類不可能なピロリ菌マウス−コロニー形成株(colonizing strain)PJ2を、全てのマウスのグループにその後経口胃(orogastrically)接種した(Altman et al.,2003)。防御は、対照グループと比較して、ワクチン接種したグループにおける細菌の負荷の統計的に有意な減少として定義される。CTを加えたdLPS−TT複合体により鼻腔内で免疫化したマウスのグループに関するコロニー形成のデータは、統計的に有意であった(p<0.05)(図8)。さらに、これらのデータは、ピロリ菌26695 HP0826::Kan LPS特異血清IgA反応が、単独では細菌負荷に対する防御に十分ではないということ、および、LPS特異血清IgG反応も同様に、防御の増加に寄付するということを示唆する。
【0125】
〈実施例9:デキストラン−BSA複合体および免疫原性の調製および特性付け並びに機能的な活性の研究〉
250μLの0.2Mのホウ酸塩緩衝液(pH9.0)中に、10mgのデキストランT5(MW 5 KDa,Pharmacosmos A/S,Holbaek,Denmark)を溶解させることにより、デキストラン−BSA複合体を調製した。これを、0.2Mのホウ酸塩緩衝液(pH9.0)中の1,8−ジアミノ−3,6−ジオキサオクタン(25mL)およびシアノヒドリドほう酸ナトリウム(10mg)を含む溶液(250μl)に加えた(「Roy et al.,1984」に記載の通り)。反応を55°Cで5日間行なった。LPSベースの複合体の調製に関して上述されたように、反応生成物を精製した。
【0126】
この反応はスペーサーの導入をもたらす。LPSベースの複合体に関して記載されるように、3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステルとの反応によって、スペーサー分子のアミン基をさらに誘導体化した(実施例5を参照)。担体タンパク質のチオール化とマレイミドにより官能化されたデキストランT5のチオール化したタンパク質の付加とによって、糖複合体を得た。複合体においてBSAに対するデキストランのモル比は20:1であり、収量は炭水化物含有量に基づき、32%であった。
【0127】
様々なLPSグリコ型(glycotypes)のピロリ菌代表物および選択された突然変異株からのLPSに対するデキストラン−BSAにより免疫化されたウサギの免疫後血清を用いて、交差反応性研究を行なった(表18)。
【0128】
【表18】
【0129】
交差反応性がグルカン−陰性株SS1およびSS1 HP0826::Kanにより得られなかったため、デキストラン−BSAにより免疫化されたウサギから得た免疫後血清の反応は、α1,6−グルカンが存在するための必要条件を表した。デキストラン−BSAにより免疫化したウサギから得た免疫後血清は、26695 HP0826::Kan変異株および対応する野生型菌株の両方に対する機能的な活性を示した(表19)。
【0130】
【表19】
【0131】
異種の分類可能な株および分類不可能な株を認識し、殺菌性の抗体を誘発するといったウサギの免疫後血清の能力は、α1,6−結合グルコース反復単位の線形な主鎖を含むポリマーであるデキストランからなる複合体、または、連続するα1,6−結合グルコースの残基からなる最適化された線形のオリゴ糖を含む複合体からなる複合体、および、適切なタンパク質が、ピロリ菌感染に対する防御を与えるのに十分であるという可能性を示す。
【0132】
(実施例10:抗グルカンmAbsの特異性の研究)
ピロリ菌株O:3 HP0826::Kanに対して特異的なモノクローナル抗体を生成し、その特異性を研究した。
【0133】
前に記載されたように、ハイブリドーマを生成した(Altman et al.,2005)。82日にわたって5度、ピロリ菌株O:3 HP0826::Kanの108細胞(200μL)ホルマリン固定化細胞を、6匹のBALB/cマウス(Charles River Laboratories,St Constant,QC)に腹腔内に(i.p.)注入することで、有意な滴定濃度を達成した。最後の静脈内(i.v.)注入を行い、続いて3日後に融合させた。Kohler and Milstein(1975)に従い、2匹のマウスの脾臓細胞をSp2/Oプラズマ細胞腫細胞株と融合した。368のウェルからの最初の融合の上澄みを、間接的な酵素結合免疫測定法(ELISA)によって選別した。
【0134】
ELISAによるハイブリドーマの選別に関して、マイクロタイタープレート(ICN,Costa Mesa,CA)を、37°Cで3時間、50mMの炭酸塩バッファー(pH9.8)中の10μg/mLの対応するピロリ菌O:3 HP0826::Kan LPSで覆った。PBSでの洗浄後、プレートをPBS中の5%(w/v)のウシ血清アルブミン(BSA)でブロックした。使い切った上澄みを加え、プレートを室温で3時間インキュベートした。プレートをPBSと第2抗体で洗浄し、ヤギ抗マウスIgG+IgMホースラディッシュペルオキシダーゼ複合体(Caltag,So.San Francisco,CA)を室温で1時間加えた。最後の洗浄の工程後、3,3’,5,5’−テトラメチルベンジデン(tetramethylbenzidene)(TMB)(KPL,Gaithersburg,MD)基質を加え、反応を1Mのリン酸を用いて止めた。マイクロタイタープレートリーダー(Dynatech,Chantilly,VA)を使用し、吸光度を450nmで測定した。この工程の後に、実施例6に記載の通り、間接的ELISA手順を続けた。
【0135】
制限のある希釈クローニング(dilution cloning)に従って、安定した2つのハイブリドーマを得た。1つのクローン、1C4F9、IgMを、さらなる特徴付けのために選択した。腹水はBALB/cマウスにおいて上昇し、製造業者のプロトコルに従って、IgM−特異的アフィニティカラム(Pierce,Rockford,IL)を使用し、1C4F9モノクローナル抗体(mAb)を腹水から精製した。
【0136】
グルカンに特異的な抗体の特異性に関する研究は、連続するα1,6−結合グルコースの残基Glc→[α(1→6)Glc]n=1−6および選択的な精製されたピロリ菌LPSからなる、一連の線形のオリゴ糖を用いて行われた。阻害ELISAに関して、イソマルト系(isomalto−series)[Glcα1→(6Glcα1→)n=1−6](USBiologicals,Swampscott,MS)またはピロリ菌26695、O:3およびPJ2 LPSインヒビターからの線形のα1,6−結合グルコースを含むオリゴ糖の一連の希釈物を調製して、OD450=1を与えた精製1C4F9の以前に調製された希釈物と混合した。インキュベーション(22°C、15分)の後、この混合物を37°Cでもう2時間インキュベートした場合、吸着したLPS抗原を含む最初のブロックされたマイクロタイタープレートに混合物を移動させた。この工程の後、実施例6に記載の通り、間接的ELISAの手順を続けた。阻害の割合を、以下の式を使用して計算した:
【0137】
%阻害=100x[(インヒビターを含むOD−インヒビターを含まないOD)/インヒビターを含まないOD]
【0138】
阻害対ログの濃度の曲線を各々のインヒビターに関してプロットし、半分の最大阻害濃度に必要な濃度(IC50)を外挿曲線から決定した。
【0139】
イソマルトヘキサオース(n=5)およびイソマルトヘプタオース(isomaltoheptaose)(n=6)を使用すると、阻害特性が最適化した(表20)。グルカン鎖(Altman et al.,2003)に平均で6から8の残基を含むと以前に示されたピロリ菌PJ2 LPSは、最も有効なインヒビターであった(表20)。
【0140】
【表20】
【0141】
(実施例11:抗グルカンmAbsを使用する接触性の研究(accessibility study)および殺菌アッセイ)
抗グルカンmAbsは、間接型のIF顕微鏡観察法研究によって実証されるように、代表的なピロリ菌株から生存する細菌の表面に容易に接触可能であった(表21)。α1,6−グルカンおよびCagAの両方を、細菌の表面上で同時に識別する。
【0142】
モノクローナル抗体の殺菌力を決定するために、補体除去された腹水(50μL)からの1C4F9 mAbの10倍の連続希釈物を各々のウェルに加え、その後、細菌懸濁液(25μL)を加え、37°Cで15分間プレインキュベートした。この工程の後、実施例7に記載の通り、手順を続けた。1C4F9の細胞表面結合は、ピロリ菌の野生株および変異株に対する殺菌アッセイによって測定されるように、機能的な活性と相互に関連付けられた(表21)。
【0143】
【表21】
【0144】
本発明の好ましい実施形態が上述された一方で、様々な修正がその中で行われること、添付の請求項が、本発明の精神および範囲を逸脱することなく、全てのそのような修正を含むことを意図していることを認識し、かつ理解されたい。
【0145】
(引用文)
すべての特許、特許出願および出版物は、本明細書を指し、本明細書により出願の全体にわたって引用文に組み込まれる。
Altman E, Chandan V, Larocque S, Aubry A, Logan SM, Vinogradov E, Li J. Effect of the HP0159 ORF mutation on the lipopolysaccharide structure and colonizing ability of Helicobacter pylori. FEMS Immunol Med Microbiol 2008; 53: 204−213.
Altman E, Fernandez H, Chandan V, Harrison BA, Wilson Schuster M, Otth Rademacher L, Toledo C. Analysis of Helicobacter pylori isolates from Chile: occurrence of a selective type 1 Lewis b antigen expression in lipopolysaccharide. J Med Microbiol 2008; 57: 585−591.
Altman E, Harrison BA, Hirama T, Chandan V, To R, MacKenzie R. Characterization of murine monoclonal antibodies against Helicobacter pylori lipopolysaccharide specific for Lex and Ley blood group antigens. Biochem Cell Biol 2005; 83: 589−596.
Altman E, Smirnova N, Li J, Aubry A, Logan SM. Occurrence of a nontypable Helicobacter pylori strain lacking Lewis blood group O antigens and DD−heptoglycan: evidence for the role of the core alpha1,6−glucan in colonization. Glycobiology 2003; 13: 777−783.
Anderson PW, Pichichero ME, Insel RA, Betts R, Eby R, Smith DH. Vaccines consisting of periodate−cleaved oligosaccharides from the capsule of Haemophilus influenzae type b coupled to a protein carrier: structural and temporal requirements for priming in the human infant. J Immunol 1986; 137: 1181−1186.
Angelakopoulos H, Hohmann EL. Pilot study of phoP/phoQ−deleted Salmonella enterica serovar Typhimurium expressing Helicobacter pylori urease in adult volunteers. Infect Immun 2000; 68: 2135−2141.
Appelmelk BJ, Simmons−Smit L, Negrini R, Moran AP, Aspinall GO, Forte JG et al. Potential role of molecular mimicry between Helicobacter pylori lipopolysaccharide and host Lewis blood group antigens in autoimmunity. Infect Immun 1996; 64: 2031−2040.
Brisson J−R, Crawford E, Uhrin D, Khieu NH, Perry MB, Severn WB, Richards JC. The core oligosaccharide component from Mannheimia (Pasteurella) haemolytica serotype A1 lipopolysaccharide contains L−glycero−D−manno− and D−glycero−D−manno−heptoses: Analysis of the structure and conformation by high−resolution NMR spectroscopy. Can J Chem 2002; 80: 949−963.
Castillo−Rojas G, Mazari−Hiriart M & Lopez−Vidal Y (2004). Helicobacter pylori: focus on CagA and VacA major virulence factors. Salud publica de Mexico 46: 538−548.
Chandan V, Logan SM, Harrison BA, Vinogradov E, Aubry A, Stupak J, Li J, Altman, E. Characterization of a waaF mutant of Helicobacter pylori strains 26695 provides evidence that an extended lipopolysaccharide structure has a limited role in the invasion of gastric cancer cells. Biochem Cell Biol 2007; 85: 582−590.
Chu C, Liu B, Watson D, Szu S, Bryla D, Shiloach J, Schneerson R, Robbins JB. Preparation, characterization, and immunogenicity of conjugates composed of the O−specific polysaccharide of Shigella dysenteriae type 1 (Shiga’s bacillus) bound to tetanus toxoid. Infect Immun 1991; 59: 4450−4458.
Ciucanu I, Kerek F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr Res 2004; 131: 209−217.
Ciucanu I, Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr Res 2004, 131: 209−217.
Cox AD, Zou W, Gidney MAJ, Lacelle S, Plested JS, Makepeace K, Wright JC, Coull PA, Moxon ER, Richards JC. Candidacy of LPS−based glycoconjugates to prevent invasive meningococcal disease: Developmental chemistry and investigation of immunological responses following immunization of mice and rabbits. Vaccine 2005; 23: 5045−5054.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956; 28: 350−356.
Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clin Microbiol Rev 1997; 10: 720−741.
Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959; 82: 70−77.
Fernandez−Santana V, Gonzalez−Lio R, Sarracent−Perez J, Verez−Bencomo V. Conjugation of 5−azido−3−oxapentyl glycosides with thiolated proteins through the use of thiophilic derivatives. Glycoconjugate J 1998; 15: 549−553.
Gu X−X, Tsai C−M, Ueyama T, Barenkamp SJ, Robbins JB, Lim DJ. Synthesis, characterization, and immunological properties of detoxified lipooligosaccharide from nontypable Haemophilus influenzae conjugated to proteins. Infect Immun 1996; 64: 4047−4053.
Hiratsuka K, Logan SM, Conlan JW, Chandan V, Aubry A, Smirnova N, Ulrichsen H, Chan KHN, Griffith DW, Harrison BA, Li J, Altman E. Identification of a D−glycero−D−manno−heptosyltransferase gene from Helicobacter pylori. J Bacteriol 2005; 187: 5156−5165.
Holst O, Brade L, Kosma P, Brade, H. Structure, serological specificity, and synthesis of artificial glycoconjugates representing the genus−specific lipopolysaccharide epitope of Chlamidia spp. J Bacteriol 1991; 173:1862−1866.
Kabat EA. Chapter 11. Molecular Biology of Anti−・−(1・6)dextrans. Antibody responses to a Single−Site−Filling Antigenic Determinant. In: Carbohydrate Antigens (vol. 159). Garegg PJ, Lindberg AA, eds. ACS Symposium Series, Washington: American Chemical Society,1993; 146−58.
Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975; 256: 495−497.
Logan SM, Altman E, Mykytczuk O, Brisson J−R, Chandan V, St Michael F, Masson A, Leclerc S, Hiratsuka K, Smirnova N, Li J, Wu Y, Wakarchuk WW. Novel biosynthetic functions of lipopolysaccharide rfaJ homologs from Helicobacter pylori. Glycobiology 2005; 15: 721−733.
Logan SM, Conlan JW, Monteiro MA, Wakarchuk WW, Altman E. Functional genomics of Helicobacter pylori: identification of a ・−1,4 galactosyltransferase and generation of mutants with altered lipopolysaccharide. Mol Microbiol 2000; 35: 1156−1167.
Mieszala M, Kogan G, Jennings HJ. Conjugation of meningococcal lipooligosaccharides through their lipid A terminus conserves their epitopes and results in conjugate vaccines having improved immunological properties. Carbohydr Res 2003; 338: 167−175.
Monteiro MA. Helicobacter pylori: a wolf in sheep’s clothing: the glycotype families of Helicobacter pylori lipopolysaccharides expressing histo−blood groups: structure, biosynthesis, and role in pathogenesis. Adv Carbohydr Chem Biochem 2001; 57: 99−158.
Passwell JH, Ashkenazi S, Harlev E, Miron D, Ramon R, Farzam N, Lerner−Geva L, Levi Y, Chu C, Shiloach J, Robbins JB, Schneerson R, Israel Shigella Study Group. Safety and immunogenicity of Shigella sonnei−CRM9 and Shigella flexneri type 2a−rEPAsucc conjugate vaccines in one− to four−year old children. Pediatr Infect Dis J 2003; 22: 701−706.
Passwell JH, Harlev E, Ashkenazi S, Chu C, Miron D, Ramon R, Farzan N, Shiloach J, Bryla DA, Majadly F, Roberson R, Robbins JB, Schneerson R. Safety and immunogenicity of improved Shigella O−specific polysaccharide−protein conjugate vaccines in adults in Israel. Infect Immun 2000; 69: 1351−1357.
Pon RA, Lussier M, Yang Q−L, Jennings HJ. N−Propionylated group B meningococcal polysaccharide mimics a unique bactericidal capsular epitope in group B Neisseria meningitidis. J Exp Med 1997; 185: 1929−1938.
Rossi G, Ruggiero P, Peppoloni S, Pancotto L, Fortuna D, Lauretti L, Volpini G, Mancianti S, Corazza M, Taccini E, Di Pisa F, Rappuoli R, Del Giudice G. Therapeutic vaccination against Helicobacter pylori in the beagle dog experimental model: safety, immunogenicity, and efficacy. Infect Immun 2004; 72: 3252−3259.
Roy, R., Katzenellenbogen, E., and Jennings, H.J. 1984. Improved procedures for the conjugation of oligosaccharides to protein by reductive amination. Can J. Biochem. Cell Biol. 62, 270−275.
Ruggiero P, Peppoloni S, Rappuoli R, Del Giudice G. The quest for a vaccine against Helicobacter pylori: how to move from mouse to man? Microbes Infect 2003; 5: 749−756.
Sawardeker JH, Sloneker JH, Jeannes A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal Chem 1967; 39: 1602−1604.
Westphal, O Jann K. Bacterial polysaccharides. Extraction with phenol−water and further applications of the procedure. Meth Carbohydr Chem 1965; 5: 83−91.
Wirth HP, Yang M, Karita M, Blaser MJ. Expression of the human cell surface glycoconjugates Lewis X and Lewis Y by Helicobacter pylori isolates is related to cagA status. Infect Immun 1996; 64: 4598−4605.
Yokota, S., Amano, K., Fujii, N., Yokochi, T. Comparison of serum antibody titers to Helicobacter pylori lipopolysaccharides, CagA, VacA and partially purified cellular extracts in a Japanese population. FEMS Microbiol Lett 2000; 185: 193−198.
Yu S, Gu X−X. Biological and immunological characteristics of lipooligosaccharide−based conjugate vaccines for serotype C Moraxella catarrhalis. Infect Immun 2007; 75: 2974−2980.
【図1A】
【図1B】
【図2A】
【図2B】
【技術分野】
【0001】
(先願の情報)
本出願は、2009年7月31日に出願され米国仮特許出願第61/230,315号の利益を主張し、その全体は引用によって本明細書に組み入れられる。
【0002】
(発明の分野)
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌(H.pylori)外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【背景技術】
【0003】
ヘリコバクター・ピロリは、ヒトの慢性胃炎、消化性潰瘍、および、胃癌に関連する最も一般的な細菌感染と認められる。ピロリ菌感染は、世界の人口の約半分に蔓延していると推測され、開発途上国では70%、先進工業国では20−30%にまで達している(Dunn et al., 1997)。大部分の人は小児期にピロリ菌にかかり、開発途上国の子供の間での感染の有病率は、社会経済的地位の低さと衛生状態の悪さに関連している(Castillo−Rojas et al., 2004)。ピロリ菌は胃癌の相対リスクを少なくとも6倍増加させるため、世界保健機構(WHO)によってクラスIの発癌物質であると確認されている。胃癌は世界で2番目に多い死亡原因であり、毎年700,000人が死亡している(Parkin et al., 2002)。
【0004】
ピロリ菌の現在の根絶方策はプロトンポンプインヒビターと抗生物質の使用に基づいている。しかしながら、化学療法の介入治療の有効性は、ピロリ菌分離物中の抗生物質耐性の頻度、および、再感染に対する免疫の欠如によって限定されてしまう。したがって、ピロリ菌感染の予防および根絶のためのグローバルな戦略を提供するために新しい治療が必要である。
【0005】
近年の調査がピロリ菌の病因におけるタンパク成分の役割と、防御免疫にタンパク質成分の役割に焦点が当てられている一方で、ワクチン製剤中にタンパク質以外の抗原を含めるという可能性を探った研究は比較的少ない。例えば、多糖類に基づいた複合体(conjugate)ワクチンは、全身感染を防ぎ、宿主のコロニー形成を阻害することが知られている(Anderson et al.,1986; Chu et al., 1991; Pon et al.,1997; Passwell et al.,2001; Passwell et al.,2003)。腸内病原菌の近年の研究は、人間に使用するための候補ワクチンとして、LPS複合体に基づいた手法を検査している(Gu et al.,1996; Mieszala et al., 2003; Cox et al.,2005; Yu and Gu, 2007)。リポ多糖類(LPS)はピロリ菌の主要な細胞表面成分である。多くのピロリ菌で行なわれた構造研究(Monteiro, 2001)は、O鎖と、脂質A部分に付けられたコアオリゴ糖とからなるLPSの構造モデルをもたらした。ほとんどのピロリ菌株のO鎖多糖類骨格(backbone)の構造は独特で、ヒト胃細胞と腫瘍細胞の細胞表面にある決定因子を模倣するII型および/またはI型のルイス(Le)式血液型決定因子を表示し(Wirth et al.,1996)、これらは、萎縮性胃炎につながる有害な自己免疫反応に関与していることもある(Appelmelk et al.,1996)。加えて、ピロリ菌LPSの分離外殻領域は、2つの異常な高分子成分:DD−ヘプトグリカン、および、α1,6−グルカンを含む(Monteiro, 2001)。外殻領域ピロリ菌LPS分離株中のα1,6−グルカンポリマーは、HP0159オープンリーディングフレームの生成物によって合成される。ピロリ菌中でのHP0159遺伝子の存在および発現は一般的である。
【0006】
多くのピロリ菌LPS生合成遺伝子が特徴付けられ、病因とコロニー形成におけるそれらの役割が測定されてきた(Logan et al., 2000; Logan et al., 2005; Hiratsuka et al., 2005; Chandan et al., 2007; Altman et al., 2008)。ピロリ菌HP0826変異株が構築され、この変異が、Le抗原を欠いた、切断されたLPSの形成をもたらすことが実証された。しかしながら、LPSの構造の十分な特徴づけは達成されなかった。
【0007】
当該技術分野での進展にもかかわらず、ピロリ菌の複数のタイプにわたって有効な免疫原性エピトープは捉えがたいままである。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【課題を解決するための手段】
【0009】
本発明は式Iの構造を含む、α1,6−グルカンを含むヘリコバクター・ピロリ化合物を提供し、
【0010】
【化1】
【0011】
Rは、α1,3−DD−ヘプタンに結合したα1,6−グルカンで置換したα−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされる。記載されたような化合物において、α1,6−グルカンは、約3から約12のα1,6−結合グルコース残基を含み、α1,3−DD−ヘプタンは、約2から約6のα1,3−結合ヘプトース残基を含んでもよい。
【0012】
上記の化合物のR基は、以下のとおりであってもよい。
【0013】
【化2】
【0014】
ここで、β−GlcNAc残基Lは、Hep GのO−2に結合する。記載されたような化合物では、グルカンの残基QおよびZは、α1,6−結合グルコース残基であり、nは約1乃至11の間の任意の値であってもよく、残基T、YおよびXは、α1,3−結合ヘプトース残基であり、mは約0から4の間の任意の値であってもよい。
【0015】
上記のような化合物は、ピロリ菌株HP0826::Kanから分離されるか、または、精製される。
【0016】
上記のような化合物では、式Iの構造は、Kdo残基Cに共有結合している脂質A部分をさらに含んでもよい。脂質A分子は、O−脱アシル化するか、または、Kdo残基のケトシド結合の加水分解によって開裂する。
【0017】
本発明は同様に、リンカー分子、タンパク質担体、または、その組み合わせに共役した、ほぼ線形のα1,6−グルカンを含む化合物を含む複合体を提供する。ほぼ線形のα1,6−グルカンを含む化合物は、本明細書に記載の化合物であり、ここで、式Iの構造は、リンカー分子、タンパク質担体、またはその組み合わせに共役される。ほぼ線形のα1,6−グルカンを含む化合物は、代替的に、デキストランT5などのデキストランであってもよい。タンパク質担体は、破傷風トキソイドまたはウシ血清アルブミンであってもよい。
【0018】
本発明は、上記のように、1つ以上の化合物または複合体を含む組成物も含む。
【0019】
本発明は、上記のようにα1,6−グルカンのエピトープを含む化合物に対する抗体をさらに含む。抗体はモノクローナル抗体1C4F9であってもよい。本発明はハイブリドーマ細胞株1C4F9をさらに包含し、これはモノクローナル抗体1C4F9を作り出す。
【0020】
上記のモノクローナル抗体は、そのような処置を必要としている個体において、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすために利用されてもよい。
【0021】
本発明は、同様に、個体内でのピロリ菌に対する免疫反応を誘発するために、上記の有効な量の組成物の使用を提供する。組成物中の化合物は適切な担体タンパク質に共役され、さらに、組成物中の化合物は、リポ多糖類の2−ケト−3−デオキシ−オクツロソン酸(Kdo)を介して、適切な担体タンパク質に共役してもよい。
【0022】
本発明は、本明細書に記載の免疫原性の組成物で哺乳動物を免疫化することによって作られた免疫抗血清をさらに提供する。免疫抗血清は、ピロリ菌の同種および異種の分類可能なおよび分類不可能な変異株および野生株における、α1,6−結合グルカンエピトープを認識するIgGを含む。IgGは、変異株(mutant)および野生株のa1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こす。
【0023】
本発明の追加の態様および利点は、以下の記載を考慮して明らかとなる。本発明の範囲内の様々な変化および修正は、この発明の教示を踏まえて当業者に明らかになるため、詳細な記載および実施例は、本発明の好ましい実施形態を示す一方で、たんなる説明として与えられるものである。
【図面の簡単な説明】
【0024】
本発明のこうした実施形態と他の実施形態は、添付の図面を参照しつつ、ほんの一例としてここで記載される。
【図1】陽イオンモードでピロリ菌株26695 HP0826::Kanから脱脂したLPSの分画1におけるLPSグリコフォームのCE−MS分析を示す。図1Aが抽出された質量スペクトル(m/z 1000−1500)を示す一方で、図1Bは、m/z 1266.3でのイオンの生成イオンスペクトルを示している。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図2】陰イオンモードでピロリ菌株26695 HP0826::KanからO−脱アシル化したLPSのLPSグリコフォームのCE−MS分析を示す。図2Aが抽出された質量スペクトル(m/z 600−2000)を示す一方で、図2Bは、m/z 1597.7でのイオンの生成イオンスペクトルを示している。脂質A部分に相当するフラグメントイオンは星印で印をつけている。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3A】ピロリ菌株26695 HP0826::Kanからの主要なLPSグリコフォームの構造を示す。アシル化は示されていない。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3B】図3AのLPSのKOH脱アシル化の生成物が、図3Bで示される(化合物1〜4)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3C】化合物4の脱アミノの生成物が、図3Cで示される(化合物5および6)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3D】化合物6の過ヨウ素酸酸化の生成物が図3Dで示される(化合物7および8)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3E】菌株26695(FIGURE 3E)からのピロリ菌LPS(図3E)と、菌株26695 HP0826::Kan変異株からのピロリ菌LPS(図3F)に関して当該技術分野で先に提案された構造も同様に示される(出典「Logan et al., 2000」)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図3F】菌株26695(FIGURE 3E)からのピロリ菌LPS(図3E)と、菌株26695 HP0826::Kan変異株からのピロリ菌LPS(図3F)に関して当該技術分野で先に提案された構造も同様に示される(出典「Logan et al., 2000」)。PEtn=ホスホエタノールアミン;Glc=D−グルコピラノース;Gal=D−ガラクトピラノース;Kdo=2−ケト−3−デオキシ−オクツロソン酸;LDHep=L−グリセロ−D−マンノ−ヘプトース;DDHep=D−グリセロ−D−マンノ−ヘプトース;GlcNAc=2−アセタミド−2−デオキシ−D−グルコース;GlcN=2−アミノ−2−デオキシ−D−グルコース;Fuc=L−フコース;P=リン酸塩;および、Gro=グリセロール。
【図4】本発明のLPSベースの複合体の調製のための反応スキームを示す。
【図5】陰イオンモードで脱脂されたピロリ菌O:3 HP0826::Kan LPSの主要な分画のCE−MS分析を示す。●:側鎖に1つのHep残基を含むLPSグリコフォーム、R=Hex5−13、Hep、HexNAc、Fuc; ■:側鎖に2つのHep残基を含むLPSグリコフォーム、R=Hex8−13、Hep、Hep、HexNAc、Fuc
【図6】26695 HP0479::Kan(図6A)、および、26695 HP0826::Kan(図6B)による阻害ELISAによって、dLPS−TT複合体により誘発されたウサギ抗体の特異性を測定するためのグラフである。LPSのコーティングは以下のように描かれる:黒四角−26695;黒菱形−26695 HP0826::Kan;黒丸−26695 HP0159::Kan;黒三角−26695 HP0479::Kan;白丸−SS1;白四角−SS1 HP0826::Kan;白三角−SS1 HP0159::Kan;白逆三角−SS1 HP0479::Kan
【図7】CD−1マウス中のピロリ菌特異抗体反応を表すグラフを示す。マウスは、1μg/マウスのコレラ毒素で補助された(adjuvanted)25μg/マウスのdLPS−TT複合体(黒棒)、1μg/マウスのコレラ毒素で補助されたPJ2細胞を含まない超音波処理物(斜線棒)、または、食塩水(白棒)で、週1回の間隔で4度、ワクチン接種した。最初の免疫化から4週間後、血清、糞便、膣の洗浄サンプルを採取し、ピロリ菌に特異的なIgGおよびIgAについてアッセイした。個々のマウスは、群平均を示す(n=5/群)水平棒でグラフ上にプロットされる。片側マン・ホイットニー検定(one−tailed Mann Whitney test)によって、※ p<0.05、※※ p<0.01
【図8】CD−1マウスの胃におけるピロリ菌負荷(burden)を表す棒グラフである。1μg/マウスのコレラ毒素で補助された25μg/マウスのdLPS−TT複合体(黒棒)、1μg/マウスのコレラ毒素で補助されたPJ2細胞を含まない超音波処理物(斜線棒)、または、食塩水(白棒)で、マウスに週1回の間隔で4度、ワクチン接種した。第1の免疫から5週間後に、〜108cfu ピロリ菌、PJ2菌株によって、一日おきに3度、マウスに経口でチューブから栄養補給させた。4週間後にマウスを殺処分し、胃からの一般細菌を数えた。棒は、4−5のマウス±SEMの群を表わす(片側マン・ホイットニー検定によって、※p<0.05)。
【発明を実施するための形態】
【0025】
本発明は新しいヘリコバクター・ピロリLPS外殻エピトープに関する。より具体的には、本発明は、新しいピロリ菌外殻エピトープ、その合成、特徴づけ、および、複合体に関する。
【0026】
他に定義されない限り、本明細書で用いられる全ての技術的及び科学的用語は、本発明が属する技術分野の当業者によって一般に理解されるものと同じ意味を有する。本明細書に記載されているこれらのものと類似又は同等の任意の方法及び材料は、本発明の実施又は試験で用いることができるが、好ましい方法と材料がこれより記載される。以下に記載される全ての刊行物は、引用することによって本明細書に組み入れられる。
【0027】
本明細書で使用されているように、「精製された(purified)」とは、必ずしも絶対的な純度を意味するわけではないが、むしろ相対的な定義として意図されている。同様に、本明細書で使用されているように、「分離された(isolated)」は、その本来の環境から何かを除去することを言う。
【0028】
ヘリコバクター・ピロリは、ヒトの慢性胃炎、消化性潰瘍、および、胃癌に一般的に関連する細菌性病原菌である。ピロリ菌感染に関連する胃癌のリスクの増加の結果として、それはクラスI発癌物質として分類されている。リポ多糖類(LPS)はピロリ菌の主要な細胞表面成分である。ピロリ菌LPSの構造研究に関する従来の出版物は、O鎖多糖類がコアオリゴ糖に共有結合される提案されたモデルをもたらし、コアオリゴ糖は順に脂質A部分に付く。ほとんどのピロリ菌株のO鎖多糖類骨格は独特で、タイプ2および/またはタイプ1のルイス(Le)血液型決定子を表示することができ、この多糖類成分は抗原である。「分類可能な」ピロリ菌株は、抗ルイス抗体(抗Le)によって認識することができるルイスエピトープ(LeXおよび/またはLeY抗原)を有し、このような抗体は市販で入手可能であり、分類の助けとなる。「分類不可能な」菌株はルイス構造を含んでいない。
【0029】
さらなる構造研究は、ピロリ菌LPSの外殻領域が2つの異常な高分子成分、DD−ヘプトグリカン、および、α1,6−グルカン側鎖を含む(Monteiro, 2001)。Loganら(2000)はLPSの構造についても洞察した。具体的に、もし側鎖中のDD−ヘプトース(DD−Hep)が骨格中のDD−Hepに付けられていれば、その一方で、α1,6−グルカンがこの側鎖DD−Hepに付き、別の分岐を形成していると仮定した場合の提案された構造(図3Eおよび図3Fを参照)。具体的に、Loganら(2000)は、0826変異株LPS内のグルカン鎖の長さを、FAB−MSスペクトル(高速原子衝撃質量分析)に基づいた1乃至3のグルコースとなるように決定した。ピロリ菌株26695 LPS中でのα1,6−結合グルコースの存在も記載され、メチル化分析データに基づいているが、グルカンの長さは決められなかった。菌株26695 LPSにおいて、3−置換HepはO鎖のGlcNAc単位とコアとの間で結合を形成する。ピロリ菌HP0826::Kan LPSでの3−結合ヘプトースの存在も記載されているが、α1,3−ヘプタンの存在またはその長さについては記載されなかった。ヘプタンまたはグルカンの側鎖の構造または長さに関するさらなる情報は提供されなかった。ピロリ菌LPSの構造は、現在さらに定義されている。
【0030】
本発明は式Iの構造を含む、新規なα1,6−グルカンを含むヘリコバクター・ピロリ化合物を提供し、
【0031】
【化3】
【0032】
ここで、Rは、α1,6−グルカンで、その後、α1,3−DD−ヘプタンで置換された、α−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされる。
【0033】
上記のような構造では、三糖類(α−DDHep−3−α−L−Fuc−3−β−GlcNAc)のβ−GlcNAcは、α−DDHep Gに結合される。三糖類のα−DDHepは、α1,6−グルカンに結合され、これは順にα1,3−DD−ヘプタンに結合される。α1,3−DD−ヘプタンはその後、β−GlcNAc残基に結合され、後者はO鎖多糖類に付着点を提供する。「結合した(linked)」または「置換した(substituted)」との用語によって、2つの部分が共有結合によって連結することが意味される。
【0034】
本明細書で使用される「α1,6−グルカン」という用語は、「グルカン」、「α1,6−グルカン側鎖」、「グルカン側鎖」、「α1,6−グルカン部分」、および/または、「グルカン部分」と交互に呼ばれる。α1,6−グルカンは、α1,6−Oグリコシド結合によって結合したブドウ糖モノマーの線形の多糖類鎖である。任意の方法に限定することを意図していない1つの例では、α1,6−グルカンは線形の多糖類であってもよい。α1,6−グルカンが任意の適切な量のα1,6−グルコース残基を含んでもよい。例えば、任意の方法で限定することを望まず、グルカンは、約3乃至約12のα1,6−結合したグルコース残基を含んでもよく、具体的には、グルカン部分は約3、4、5、6、7、8、9、10、11、または、12のα1,6−結合グルコース残基、あるいは、ちょうど列挙された任意の2つの値によって定義される任意の範囲を含んでもよい。1つの非限定的な実施例では、α1,6−グルカンは、9−12のα1,6−グルコース残基を含んでもよい。別の非限定的な実施例では、α1,6−グルカンは、10のα1,6−グルコース残基を含んでもよい。
【0035】
本明細書で使用されるような「グリコフォーム」という用語は、同じLPS構造を含むが、α1,6−グルコースまたはα1,3−ヘプトース残基の数が異なる、さまざまな形態またはタイプの化合物を示す。例えば、および、理論に縛られることを望まずに、各々のグリコフォームは、特定の長さのグルカンおよび/またはヘプタン部分、あるいは、それらの組み合わせを含んでもよい。
【0036】
本明細書で使用されるような「α1,3−ヘプタン」という用語は、「α1,3−DD−ヘプタン」、「ヘプタン」、「α1,3−ヘプタン側鎖」、「ヘプタン側鎖」、「α1,3−ヘプタン部分」、「ヘプタン部分」、および/または、「DD−ヘプトグリカン」と交互に呼ばれる。α1,3−ヘプタンは、α1,3−Oグリコシド結合によって結合したヘプトースモノマーの多糖類鎖である。任意の方法に限定することを意図していない1つの例では、α1,3−ヘプタンは線形の多糖類であってもよい。α1,3−ヘプタンは、任意の適切な量のα1,3−ヘプトース残基を含んでもよい。例えば、任意の方法で限定することを望まずに、ヘプタンは、約2乃至約6のα1,3−結合ヘプトース残基を含んでもよく、具体的には、ヘプタン部分は、約2、3、4、5、または、6のα1,3−結合ヘプトース残基、または、ちょうど列挙された任意の2つの値によって定義される任意の範囲を含んでもよい。
【0037】
上記のα1,3−DDヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャップされる。「キャップされた(capped)」という用語によって、β−GlcNAc残基が側鎖の最後の残基であることを意味し、「末端の(terminated)」という用語が使用されてもよい。β−GlcNAcはヘプトースの位置O−2を介してDD−Hepに結合されてもよい。理論に縛られることを望まずに、β−GlcNAc残基はO鎖多糖類に付着点を与えてもよい。
【0038】
1つの非限定的な実施例において、Rは以下のとおりであり、
【0039】
【化4】
【0040】
ここで、β−GlcNAc残基Lは、Hep残基GのO−2に結合される。この実施例では、β−GlcNAc残基Wは、O鎖多糖類に付着点を与えてもよい。ちょうど記載された化合物では、グルカンの残基QおよびZは、α1,6−結合グルコース残基であり、nは、グルカンがα1,6−結合で約3乃至約12のグルコース残基を含むように、約1乃至11の間の任意の値でもよい。1つの具体的な非限定的な例において、主要なグリコフォームは、10の連続するα1,6−結合グルコース残基(n=9)を含んでもよい。ちょうど記載された化合物では、残基T、Y、および、Xは、α1,3−結合ヘプトース残基であり、mは、ヘプタンがα1,3−結合において約2乃至約6のヘプトース残基を含むように、約0乃至4の間の任意の値であってもよく、1つの具体的な非限定的な例において、主要なグリコフォームは4つの連続するα1,3−結合ヘプトース残基(m=2)を含んでもよい。
【0041】
構造は任意の適切なピロリ菌株から分離および/または精製され、例えば、任意の方法で限定することを望まずに、切断されたピロリ菌LPS分子は、限定されないが、O鎖多糖類を欠く同質遺伝子変異株をもたらすHP0826遺伝子中の変異株を有するピロリ菌株などの分類不可能なピロリ菌株(すなわち、ルイス抗原が欠けたもの)から分離される。非限定的な例において、本明細書に記載されている化合物はピロリ菌株26695 HP0826::Kanまたは菌株PJ2から分離および/または精製されてもよい。
【0042】
「内殻分子(inner core molecular)」とも本明細書で呼ばれる式Iの構造は、Kdo残基(例えば、Kdo残基C)に共有結合している脂質A部分をさらに含む。他の実施形態では、脂質A分子はO−脱アシル化しても、または、完全に脱アシル化してもよい。さらに他の実施形態では、脂質A部分は、Kdo残基のケトシド結合の加水分解を介して開裂してもよい。理論によって縛られることを望まず、脂質Aの開裂は、LPSの毒性を除去し、かつ、複合体の可能な凝集と不溶性を回避するために行われてもよい。当業者は、脂質A部分のケトシド結合のO−脱アシル化、脱アシル化、または、加水分解のための方法に精通しているであろう。(例を参照。「Holst et al.,1991; Altman et al., 2003」)
【0043】
本発明は、同様に、タンパク質担体に共役されたほぼ線形のα1,6−グルカンを含有する化合物を含む複合体を提供する。ほぼ線形のα1,6−グルカンを含有する化合物は、α1,6−結合グルコース残基で構成された任意の適切なほぼ線形の多糖類であってもよい。「ほぼ線形の」という用語によって、α1,6−グルカンが分岐をほとんど含まず、例えば、任意の方法で限定することを望まずに、α1,6−グルカンがα1,6−グルカン中に約0乃至約5%の分岐を含んでもよいことを意味する。具体的に、α1,6−グルカンは、約0、0.5、1、1.5、2、2.5、3、3.5、4、4.5、または、5%の分岐またはその間の任意の量を含んでもよく、化合物は混合物であってもよく、このとき、混合物内の分岐の量は、1つの化合物から別の化合物まで様々である。1つの非限定的な例では、α1,6−グルカンを含む化合物は上記のような、および、本明細書で記載されるような構造であってもよい。別の実施例では、α1,6−グルカンを含有する化合物はデキストランであってもよい。デキストランは上記のような、かつ、1〜10kDaの分子量を有する必要条件を満たす任意の適切なデキストランであってもよく、例えば、しかしながら任意の方法で限定することを望まずに、デキストランは、約1、3.5、5、6.5、8、または10kDa、または、その間の任意の値であってもよい。具体的な非限定的な例において、デキストランはデキストランT5であってもよい。さらに別の例では、α1,6−グルカンを含有する化合物は、約5〜8の間のα1,6−結合グルコース残基の直鎖であってもよく、例えば、任意の方法で限定することを望まずに、α1,6−グルカンを包含する化合物は、5、6、7、または、8のα1,6−結合グルコース残基の直鎖であってもよい。
【0044】
α1,6−グルカンを含有する化合物は、リンカー分子および/または担体タンパク質に共役し、当業者によって理解されるであろうように、本明細書に記載される構造は、担体タンパク質に直接共役されるか、または、順に担体タンパク質に共役されるリンカー分子(本明細書では「リンカー」とも呼ばれる)に変化してもよい。「共役した」という用語によって、構造がリンカー分子および/または担体タンパク質に共有結合している(covalently attached or linked)ことを意味する。リンカーおよび/または担体タンパク質の共有結合のための方法は当業者に周知であり、当業者によって認識されるように、共有結合のための方法(および、リンカーが存在しているかどうか)は、使用される担体タンパク質によって変化する。任意の方法で限定することを望まずに、このような1つの方法が図4に示され、これは「Fernandez−Santana et al. (1998)」からの出典であり、この方法では、脱アシル化したLPSまたは脱脂したLPSは、Kdo残基のカルボキシル基をリンカー分子に共有結合し、その後、マレイミド官能性を導入することによって活性化される。活性化されたLPSを、チオレート化した担体タンパク質と混ぜ合わせることで、共役した構造を生み出す。当業者によって評価されるように、本明細書に記載の方法は、一般的なものであり、従って、任意の他の適切な方法が使用されてもよい(例えば、限定されないが、Chu et al., 1991; Cox et al.,2005; Gu et al., 1996; Mieszala et al.,2003; Yu and Gu, 2007によって記載されたもの)。α1,6−グルカンを含有する化合物は、構造中の任意の適切なカルボキシル基を介して、リンカー/担体タンパク質に付けられてもよい。特定の非限定的な例では、α1,6−グルカンを含有する化合物は、本発明の構造であってもよく、内殻Kdo残基Cによって付けられてもよい。
【0045】
担体タンパク質は免疫原性の担体を含む、当該技術分野で知られている任意の適切な担体であってもよい。例えば、担体タンパク質は、破傷風トキソイド、ウシ血清アルブミン(BSA)、ジフテリアトキソイド(Diphteria toxoid)、変異株ジフテリアトキソイド、CRM、CRM197タンパク質、シュードモナスAタンパク質(Pseudomonas A protein)、コレラ毒素(CT)タンパク質、コレラ毒素変異株CT−E29Hタンパク質、および、例えば、限定されないが、鞭毛、線毛、および、他の毒素の一部などの当該技術分野で知られている他のものであってもよい。
【0046】
先に示されたように、複合体は、担体タンパク質を、自然発生の基を介して本発明の構造に直接結合させるか、または、一級アミノ基、ヒドラジド、チオール、カルボキシルなどに決して限定されないがこれらを含むスペーサーまたはリンカー分子の導入によって結合させることによって、複合体を調整することができる。
【0047】
本発明は、本明細書に記載の1以上の化合物、本明細書に記載の1以上の複合体、または、それらの組み合わせを含む組成物をさらに提供する。1つの実施形態では、組成物は、上記の化合物のグリコフォームの混合物を含む。例えば、任意の方法で限定することを望まずに、組成物は、グルカン部分中に10の連続するα1,6−結合グルコース残基(n=9)を含む主要なグリコフォームを含んでもよい。同様に、任意の方法で限定することを望まずに、組成物は、本明細書に記載の1以上のグリコフォームから調製された複合体を含むことができる。実施例で実証されたように、複合体の調製に使用されたピロリ菌株26695 HP0826::Kanによって合成されたLPS構造は、3つのグリコフォーム:主鎖(backbone)オリゴ糖I、GlcNAcと[GlcNAc、Fuc、Hep]でキャップした主鎖オリゴ糖I、および、もっとも長いグルカン鎖がCE−MS分析によって測定されるように約12のα−1,6−結合残基に対応する、α−1,6−結合グルカンを含むオリゴ糖の混合物であった(表2(図2))。
【0048】
ちょうど記載されたような組成物は免疫原性のものであってもよい。「免疫原性の」という用語によって、組成物がピロリ菌の野生株および/または変異株に対する免疫反応を誘発することを意味する。免疫反応は、ピロリ菌の分類可能な、および、分類不可能な菌株に対する広範な免疫原性反応を提供してもよい。
【0049】
本発明は、同様に、個体内でのピロリ菌に対する免疫反応を誘発するために、本明細書に記載の有効な量の組成物の使用を提供する。前に記載されたように、組成物は本発明に従って1以上の化合物を含んでもよい。1以上の化合物は、リンカーおよび/または適切な担体分子に共役してもよい。
【0050】
本発明は、上記の免疫原性の組成物で哺乳動物を免疫化することにより産生される免疫抗血清をさらに提供する。免疫抗血清は、ピロリ菌の同種および異種の、分類可能なおよび分類不可能な、変異株および野生型であり、α1,6−結合グルカンエピトープを認識する免疫後血清IgGを含むか、または、産生する。IgGは、変異株および野生株のα1,6−グルカンを発現するピロリ菌の補体を媒介とした溶菌を引き起こすこともある。
【0051】
本発明はさらに抗α1,6−グルカン抗体を提供する。抗体は、本明細書に記載の本発明の化合物に対して産生されるか、または、デキストラン(例えば、限定されないが、BSAデキストラン複合体などのデキストラン複合体)といった他のα1,6−グルカンを含む分子に対して産生されてもよい。1つの非限定的な例において、抗体は、当該技術分野で知られている既知の方法に従って産出されたモノクローナル抗体であってもよい(実施例10を参照。Altman et al.,2005)。例えば、任意の方法で限定することを望まずに、抗体は、ピロリ菌HP0826変異株LPSの外殻領域にあるα1,6−グルカンエピトープに対して産生されたモノクローナル抗体であってもよく、より具体的には、抗体は、図5で示す化合物に対して産生されてもよい。さらにもっと具体的な例において、モノクローナル抗体はハイブリドーマ細胞株1C4F9によって産生されたIgM 1C4F9であってもよい。この細胞株は、2009年7月30日にカナダの国際預託機関(international depositary authority)(国立微生物学研究所(カナダ マニトバ州 ウィニペグ アーリントン・ストリート 1015)に預けられ、受け入れ番号300709−01を割り当てられた。例で実証されたように、1C4F9は、LPS中のα1,6−グルカンエピトープと、分類可能な、および、分類不可能なピロリ菌のすべての細胞を認識し、生きている細菌の表面で容易に入手可能であり、さらに、LPS−OH−TTとdLPS−BSAまたはdLPS−TT複合体の両方で同じようによく反応した。
【0052】
本発明は、このような処置を必要としている個体において、変異株および野生株の、分類可能なおよび分類不可能なα1,6−グルカンを発現するピロリ菌の補体を媒介とした溶菌を引き起こすために、1C4F9抗体の使用を提供する。先に記載されたように、分類可能なピロリ菌株は抗Le抗体によって認識されるルイスの抗原を有し、その一方で、分類不可能なピロリ菌株はそのような抗原を有していない。しかしながら、分類可能なおよび分類不可能な菌株の両方がα1,6−グルカンを含んでおり、両方の菌株は1C4F9抗体によって認識される。
【0053】
現在、当該技術分野では、ピロリ菌株はルイスの抗原に対する市販の抗体を使用して分類されるが、しかしながら、分類不可能な株がルイスの構造を含んでいないため、この手法を用いてそれらを分類することはできない。ほとんどの分類可能なおよび分類不可能なピロリ菌株は、α1,6−グルカンエピトープを運ぶため、抗α1,6−グルカン抗体(mAb 1C4F9など)は、ピロリ菌分離物(isolate)を選別して特徴づける追加の方法を提供することができる。
【0054】
ピロリ菌株26695 HP0826::KanのLPSを精製し、その化学構造を、組成物、メチル化、徹底した核磁気共鳴(NMR)分析に加え、CE−MSデータによって測定した。ピロリ菌HP0826変異株LPSの外殻領域中のα1,6−結合グルカンの存在も実証された。この構造はα1,6−グルカンに特有のモノクローナル抗体、1C4F9によって認識された。後者の抗体は、ピロリ菌(O):3 HP0826::Kan突然変異株のホルマリンで固定した細胞を用いて精製した。これらの抗体は細胞表面接触可能であり殺菌性であった。以前は、ルイスの構造のみが抗原であると信じられていた。したがって、α1,6−グルカンに対する抗体の生成は予期しえなかった。
【0055】
ピロリ菌LPSのワクチンの可能性を調べるために、ピロリ菌26695 HP0826::Kan変異株の修飾LPSを共役して、破傷風トキソイド(TT)またはウシ血清アルブミン(BSA)にした。部分的に脱脂したLPSまたは脱脂したLPSの調製のための2つの手法:やかなヒドラジン分解によるLPSのO脱アシル化(LPS−OH)、または、穏やかな酸処理によるLPSの脱脂(dLPS)fを利用した。脱脂および/または部分的な脱脂のさらなる方法は技術分野において周知であり、このような適切な方法は本発明の範囲内で用いられてもよい。LPS−OHおよびdLPSの両方を、2−ケト−3−デオキシ−オクツロソン酸(Kdo)残基を介して、ジアミノ基を含むスペーサーに共有結合させ、その後、マレイミド官能性の導入と、チオール化したTTまたはBSAとの共役によって、それぞれ、複合体、LPS−OH−TT、dLPS−BSA、および、dLPS−TTを得た。個別の実験では、分類不可能な株PJ2を脱脂し、共役のために利用することによって、dLPS(PJ2)−TT複合体を得た。
【0056】
dLPS(PJ2)−TTと同様に、LPS−OH−TT、dLPS−BSA、dLPS−TT複合体は、IgM 1C4F9による間接的なELISAによって設定されるように、表面接触可能なα1,6−グルカン決定子の抗原を保持した。抗体はα1,6−グルカン決定子への優れた特異性を有することを示し、その結合特性が、ピロリ菌の分類可能なおよび分類不可能な菌株からの生成されたLPSと、イソマルト−系(isomalt−series)からのオリゴ糖とによる阻害ELISAによって測定された。これらの研究により、1C4F9が5または6の連続するα1,6−結合グルコース残基についての必要条件、Kabat(1993)が主張した部位を結合する抗−α−(1→6)デキストランの大きさと一致したパターンを有することが分かった。
【0057】
LPS−OH−TT、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TT複合体は、マウスとウサギにおいては免疫原性であり、ピロリ菌の異種の、同種の、および、野生型の菌株からのLPSに対する有意なIgG抗体反応を誘発した。免疫抗原に対する10倍強固なIgG免疫反応が、dLPSを含む複合体を受け取ったマウスおよびウサギで生じた。LPS−OH−TT、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TTのいずれかで免疫化したウサギの免疫後の血清は、ピロリ菌の26695 HP0826::Kan変異株および野生型26695菌株に対する殺菌活性を示した。
【0058】
要約すると、これらの結果は、Le抗原を欠いた、かつ、長いα1,6−グルカン鎖を保有する、脱脂したか部分的に脱脂したピロリ菌LPSベースのタンパク質複合体が、マウスとウサギの両方において免疫原性であり、殺菌性の抗体を誘発するということを示唆している。このエピトープが免疫原性であると同定されたことに留意することが重要である。このことは以前は確立されていなかった。なぜなら、抗原性であり、かつ、特定の抗体を産生すると知られていたのは、ルイスの構造だけだったからである。
【0059】
本発明は、以下の実施例においてさらに例証される。しかしながら、これらの実施例がたんなる説明目的のためのもので、任意の方法で本発明の範囲を制限するためには使用されてはならない。
【実施例】
【0060】
(実施例1:ピロリ菌26696 HP0826::Kan LPSの分離および構造分析)
ヘリコバクター・ピロリ菌株26695は、R.Alm博士(マイアミ州ボストンのアストラゼネカ)から入手し、ピロリ菌(O):3分離株はJ.Penner博士から、J99は、D.Taylor博士(カナダ・エドモントンのアルバータ大学)から入手し、SS1はA.Lee博士(オーストラリア・シドニーのニューサウスウェールズ大学)から、PJ1とPJ2の臨床分離株は、W.Conlan博士(IBS、NRC)からの新鮮な臨床分離株であり、および、M6はK.Eaton博士(ミシガン州のミシガン州立大学)から入手した。
【0061】
細菌株の増殖は、Hiratsukaら(2005)によって記載されたように実行された。手短にいえば、先に記載されたように(Hiratsuka et al)、微好気性環境下で、7%のウマ血液を含む、抗生物質を補ったコロンビア血液寒天培地(Difco)上で、細胞を37°Cで48時間増殖させた(Kan20μg/mL)。液体培養中の増殖のために、10%ウシ胎児血清を含む抗生物質を補ったブルセラ菌ブロスに、48時間のコロンビア血液寒天培地/ウマ血液プレートから採取したピロリ菌細胞を接種し、以前に記載されたように(Hiratsuka et al、2005)、微好気性条件下で(85%N2、10%CO2、5%O2)攪拌機内で48時間インキュベートした。
【0062】
ピロリ菌株を上記の液体培養において培養し、細菌増殖の遠心分離によって得られた浸潤細胞を、エタノール、アセトン、および、石油エーテル(light petroleum)によって2度連続して洗浄し、空気乾燥させた。WestphalとJann(1965)の暖かいフェノール水抽出手順(hot phenol−water extraction procedure)によって、空気乾燥した細胞塊からLPSを抽出した。広範な透析と凍結乾燥後に、水相からLPSを得た。超遠心分離(105,000×g,4°C、12時間)によってピロリ菌LPSをさらに精製し、ペレットを蒸留水で懸濁し、凍結乾燥させた。
【0063】
アルジトール酢酸方法(Sawardeker el al,1967)によって糖組成分析を行なった。加水分解を100°Cで4Mトリフルオロ酢酸で4時間行うか、または、100°Cで2Mトリフルオロ酢酸で16時間行い、その後、NaBH4によるH2Oの還元と、その後の無水酢酸/ピリジンによるアセチル化が続く。アルジトール酢酸誘導体を先に記載したように分析した(Altman et al,2003)。Ciucanu & Kerek(1984)の方法によって、および、先に記載したように(Altman et al,2003)、ガス液体クロマトグラフィー−質量分析法(GLC−MS)による完全メチル化されたアルジトール酢酸誘導体の特徴付けによって、メチル化分析を行った。
【0064】
アルジトール酢酸としてのピロリ菌26695 HP0826::Kanから精製されたLPSの糖分析は、L−フコース(L−Fuc)、D−グルコース(D−Glc)、D−ガラクトース(D−Gal)、N−アセチル−D−グルコサミン(D−GlcNAc)、D−グリセロ−D−マンノ−ヘプトース(DD−Hep)、および、L−グリセロ−D−マンノ−ヘプトース(LD−Hep)が、0.4:5.0:1.5:4.3:6.4:1.4の適切なモル比で存在していることを明らかにし、このことは、O鎖を欠く構造の存在を示している(Logan et al.,2000)。未処置の26695 HP0826::Kan LPS上で行なわれたメチル化分析は、これらの発見と一致しており、末端のL−Fuc、3−置換したL−Fuc、末端D−Glu、末端D−Gal、3−置換したグルコース、4−置換したD−Gal、6−置換したグルコース、末端DD−Hep、2−置換したDD−Hep、6−置換したDD−Hep、3−置換したDD−Hep、7−置換したDD−Hep、2,7−置換したDD−Hep、2−置換したLD−Hep、3−置換したLD−Hep、末端D−GlcNAc、および、3−置換D−GlcNAcが、0.1:1.0:1.1:0.1:1.2:1.0:6.0:0.4:1.2:1.1:3.1:0.5:1.6:1.2:0.1:0.3:0.4の近似モル比で存在していることを示す。O鎖を含んでいるLe抗原の特徴である3,4−置換したD−GlcNAcおよび2−結合D−Galは、まったく検知されなかった。LPSの純度はRNA由来のリビトールの欠如によって確認された。26695 LPSの組成物およびメチル化分析は、他で報告されている(Logan et al.,2005)。すべての糖がピラノース形態であった。
【0065】
(実施例2):キャピラリー電気泳動法−質量分析法(CE−MS)によって脱脂したピロリ菌26695 HP0826::Kan LPSによる特徴づけ
実施例1で得られた生成した26695 HP0826::Kan LPS(20mg)を、0.1Mの酢酸ナトリウム緩衝液(pH4.2)において100°Cで2時間加水分解し、先に記載したように(Altman et al,2003)Bio−GelP−2カラムでのゲルろ過によって分画することで、脱脂したLPS(dLPS)を生成した。3つの分画(分画1−3)を集めて、キャピラリー電気泳動質量分析法(CE−MS;表1)によって分析した。
【0066】
CE−MSに関して、Prince CEシステム(Prince Technologies,オランダ)を、4000QTRAP質量分析計(Applied Biosystems/MDS Sciex,カナダ)につないだ。シース溶液(イソプロパノール−メタノール、2:1)を、流速1.0μL/分で送達した。pH 9.0の脱イオン水中の15mM酢酸アンモニウムを用いて、約90cmの長さの剥き出しの石英ガラスの毛細管上で分離物を得た。5kVのエレクトロスプレーイオン化電圧は陽イオン検出モードに使用した。4000da/sの走査速度で増強された産生イオン走査モード(EPI)を用いて、タンデム質量スペクトルを得た。(12の値の)カーテン(curtain)および(「高い」縮尺に設定された)衝突ガスとして、窒素を使用した。
【0067】
陽イオンモードでの主要な分画1のCE−MS分析は、約11のα−1,6−結合残基に相当する最長のグルカン鎖である、Hex残基の連続的な追加に一致して、m/z 1212.3、m/z 1266.3、および、m/z 1320.4で一連の三価イオンの存在を確認した。イオンの信号強度に基づいて、m/z 1266.3での最も豊富なグリコフォームは、10のα−1,6−結合Hexを含む(図1)。陽イオン検出モードにおけるm/z 1266.3でのイオンの生成スペクトルは、以前観察された主鎖コア断片(core fragment)Hex2Hep3(PEtn)KdoとHexNAcHex2Hep3(PEtn)Kdoそれぞれに一致して、m/z 1244.5とm/z 1447.6での診断のイオンの存在を確認した(図1)。LPSコア中にα1,6−グルカンを保有する分画1および2を組み合わせて共役のために用いた。
【0068】
【表1】
【0069】
(実施例3):CE−MSによるO−脱アシル化した26695 HP0826::Kan LPSの特徴付け
(実施例1の)26695 HP0826::Kan LPSのO−脱アシル化は、幾つかの修正をしたHolst et al.に従って実行された(1991)。手短にいえば、LPS(4mg)を4時間、37°Cで無水ヒドラジン(0.2ml)中で撹拌した。反応混合物を冷却し、冷たいアセトン(2ml)をゆっくりと加えることで超過ヒドラジンを破壊した。30分後、沈殿したO−脱アシル化したLPS(LPS−OH)を、遠心分離(4°C、9300×g、10分)によって集めた。ペレットを冷たいアセトンで2度洗浄し、水に溶解させ、凍結乾燥させることでLPS−OH(3.5mg)を得た。
【0070】
陽イオンモードでのO−脱アシル化した26695 HP0826::Kan LPS(LPS−OH)のCE−MS分析を、実施例2に記載のように行った。CE−MSの結果は脱脂したLPSについて得られたMSデータと一致しており、主鎖オリゴ糖、および、HexNAc、および、[HexAc、Fuc、Hep]のそれぞれでキャップした主鎖オリゴ糖のそれぞれの存在と一致して、m/z 1137.2、m/z 1239.2、および、m/z 1408.0で3つの主要な二価イオンを提供したが(表2)、その一方で、m/z 1597.7、m/z 1651.4、および、m/z 1705.5での三価イオンは、約12のα−1,6−結合残基に相当する最長のグルカン鎖である、グルカンの存在と一致した(表2、図2)。m/z 1597.7のMS/MSスペクトルは、[Fuc,Hep]と[Fuc,Hep,HexNAc]のそれぞれの連続的な損失と一致して、m/z 1447.6とm/z 1244.5での診断のイオンの存在を示し(図2)、さらに、Kdo残基を介してO−脱アシル化された脂質A(脂質A−OH)に結合したコア断片FucHex2HexNAcHep4(PEtn)Kdoに一致して、m/z 1786.8で一価イオンをもたらし、その一方で、m/z 444.4およびm/z 887.8でのイオンは、2つのアミド結合した3−ヒドロキシオクタデカン[C18:0(3−OH)]脂肪酸鎖により置換されたジグルコサミン(diglucosamine)主鎖からなる脂質A−OH部分に相当した(図2)。
【0071】
【表2】
【0072】
(実施例4):ピロリ菌株26695 HP0826::Kan LPSのNMR分析
PEtN置換基のアルカリ加水分解がアグリコンを含まない還元末端を残すため(Holst et al.,1991)、(実施例1の)ピロリ菌株26695 HP0826::KanからのLPSの分解は、脂質A GlcNの即時還元のためのNaBH4の存在下で、4M KOHによる完全な脱アシル化で始まった。ゲルクロマトグラフィーによる生成物の分離は、クロマトグラムのオリゴ糖領域で溶出された、2つの分画をもたらした。更なる分析によって、これらの分画は、グルカン鎖の長さによって明らかに異なる、類似する化合物を含むことが示された。低分子量分画(Lower molecular mass fraction)は、質量分析法によって同定された化合物1−3を含み、両方の分画は、グルカン部分の異なる長さの化合物4を含んでいた(図3)。
【0073】
NMRスペクトル(DQCOSY、TOCSY、NOESY、1H−13C HSQC、および、HMBC)は、先に記載されたような(Brisson、2002)標準のソフトウェアを用いてVarian 500または600MHzの分光計で行われた。NMR実験はすべて、1Hスペクトルにはδ2.225ppmで、13Cスペクトルには31.45ppmで、内部基準としてアセトンを用いて25°Cで行われた。
【0074】
その複雑さゆえにスペクトルを完全に解釈することはできなかったが、両方の分画のNMRスペクトルは化合物4に相当した。α−1,6−グルカン、および、DD−ヘプタンの末端残基のH−1プロトンは、グルカンおよびヘプタンH−1の残りには重複せず、したがって、図3に示されるように、全体的な構造内でこれらのホモポリマーの位置を同定することができる。しかしながら、α−1,6−グルカン鎖の還元末端Glu残基Zの結合位置は測定されなかった。それは、同定されていない陽子(後に、HepNのH−6に割り当てられる)との強いNOE相関を示した。化合物4のNMRデータは、ヘプタン部分(X)の第1のHep残基がα−1,6−グルカンに属する非還元末端Glc残基VのO−3に結合されたため、α−1,6−グルカンがDD−ヘプタンと内殻との間に置かれたことを明確に示した。DD−ヘプタンの非還元末端は、O−3でβ−GlcN Wに置換された。小さなオリゴ糖1−3と同一の化合物は、α−1,6−グルカンがほとんどない変異株HP0159::Kanのピロリ菌LPSの主要な構成要素として先に発見され、それらのNMRデータが公表された(Altman et al.,2008)。化合物4に関して選択されたNMRデータが表3に示される。Kdoに結合した4つの近接するヘプトース残基の以前提案された配列は、調べた化合物のいずれにも存在しなかったことが容易にわかるであろう(図3)。β−GlcN残基Lは、HepGのO−2に直接結合し、LPS構造の任意の部分においてHepのO−7につなげられたGlcNはなかった。
【0075】
【表3】
【0076】
LPS構造のさらなる分析について、化合物1−4を脱アミノ化した。手短にいえば、サンプルを10%AcOHに溶解させ、過度のNaNO2を加えた。3時間後、Sephadex G−15カラムで、ゲルクロマトグラフィーによって生成物を分離した。その後、化合物のいくつかをNaBD4で還元し、脱塩した。結果として生じる生成物が化合物5および6であった。(上記のように行なわれた)化合物5のNMRスペクトルは、(アルカリ条件下のリン酸塩移動によって)HepEの位置6または7で、リン酸塩を含む2つの異性体を示した。再度、3つの近接するHep残基が存在し、以前に提案された構造が正確である場合に予測されるであろうように、4つではなかった。脱アミノ処置(上記の)による全側鎖の除去は、GlcN LがOS5の糖と分子の残部の間の接続を形成することを確認した。
【0077】
化合物6は、還元末端に、DD−ヘプタン、α−1,6−グルカン、および、三糖類DDHep−Fuc−anh−Man−ol(GlcN Lに由来するN−M−Lおよびanh−Man L)を含んでいた。この生成物のスペクトルはそれほど密集しておらず、(DDHep NのH−1とH−6の間のTOCSYからの)DDHep NのO−6に対するα−1,6−グルカンの付着点を同定することが可能になった。分岐糖を全く示さなかったメチル化分析によって確認されるように、構造は直線的であった。予測された生成物はすべて、提案された構造に一致して同定された(表4)。
【0078】
【表4】
【0079】
DD−ヘプタン−α−1,6−グルカン領域の構造のさらなる確認が、化合物6の過ヨウ素酸酸化の結果から得られた。過ヨウ素酸酸化は24時間、過度の0.1M NalO4で行なわれた。エチレングリコールを加え、Sephadex G−15カラムでのゲルクロマトグラフィーによって生成物を分離させた。それをNaBD4で還元し、脱塩し、2%AcOHによって、100°Cで3時間加水分解した。2つの主要な生成物7と8を得て、Sephadex G−15カラムでのゲルクロマトグラフィーによって分離した。
【0080】
NaBD4による酸化オリゴ糖の還元は、NMRスペクトル(APT−HSQC中のCH2OH信号に対するCHDOH信号の反転位相)での酸化炭素の同定を可能にした。化合物7および8の構造は、NMR分光法およびメチル化分析によって測定された。化合物7の形成は、DDHep Nが位置3では置換されなかったことを証明した。化合物6のメチル化が末端の3−および6−置換DD Hepのみを示したため、化合物6のNMRデータから提案されたように、DD−Hep Nは位置6で置換された。還元末端でグリセロール(Gro)を有する化合物8の形成は、酸化還元後にグリセロールを産生可能なオリゴ糖6の他の成分がなかったため、OS6中のGlc Vが位置6で次のGlc残基をグリコシル化したことを確認した。そのことは、同様に、末端グルコースの位置3でのα−1,6−グルカンの非還元末端へのDD−ヘプタン部分の付着を証明した。
【0081】
DD−ヘプタン領域のサイズを推定することができる。化合物8はDD−ヘプトースに由来する4つのマンノース残基を含んでいた。化合物8のスペクトルはよく分解し、H−1信号の統合はほぼ等モルの比率を与え、4つのマンノース残基の存在を示した。したがって、化合物6の完全なDD−ヘプタン領域は、ほとんど5つのDD−ヘプトース単位からなり、そのうちのひとつ(末端)は、過ヨウ素酸酸化によって取り除かれた。この結論は質量分析法によって確認された。ピロリ菌株26695 HP0826::Kanによって産生された主要なLPSグリコフォームの構造が図3に示される。
【0082】
(実施例5):LPS−OH−TT、dLPS−BSA、dLPS−TT、および、dLPS(PJ2)−TT複合体の調製および特徴付け
ウシ血清アルブミン(BSA)および/または破傷風トキソイド(TT)に対する、LPS−OH(実施例3を参照)およびdLPS(実施例2を参照)のKdo結合複合体を調製した。
【0083】
LPS−OH(4789Daの推定平均分子量に基づき、4mg、0.8μmol)またはdLPS(3779Daの推定平均分子量に基づき、4mg、1μmol)を、0.1M NaCl(0.4 mL)を含む0.1M 2−(N−モルホリノ)エタンスルホン酸酸(MES;Sigma−Aldrich、セントルイス、ミズーリ州)緩衝液(pH 4.8)に溶解させた。1−エチル−3−ジメチルアミノプロピルカルボジイミド(EDC;34.38mg、100:1モル比;Sigma−Aldrich)を加え、その後、1,8−ジアミノ−3,6−ジオキサオクタン(15μL、103μmol; Sigma−Aldrich)を加え、その反応を22°Cで4時間、pH 4.8で維持した。溶液をpH 7.0に調節し、蒸留水に対して透析し、Microsepの遠心分離装置1,000Da cutoff(Pall Life Sciences,Ann Arbor、ミシガン州)を用いて脱塩または凍結乾燥させた。
【0084】
基本的に、Fernandez−Santana et al. (1998)によってオリゴ糖について記載されたように、共役手順を行った。手短に言えば、乾燥したDMSO中で3−マレイミドプロピオン酸(maleimidopropionic acid)N−ヒドロキシスクシンイミドエステル(BMPS;2mg、7.5μmol;Sigma−Aldrich)と、スペーサーを含んでいるLPS−OH(2mg、0.4μmol)またはdLPS(2mg、0.5μmol)を22°Cで24時間、反応させた。溶液を蒸留水に対して透析し、凍結乾燥させた。
【0085】
BSAの活性化のために、乾燥したDMSO中で3,3’−ジチオジプロピオン酸ジ(N−ヒドロキシスクシンイミドエステル)(DTSP;0.63mg、1.6μmol;Sigma−Aldrich)を、6mM EDTA(最終濃度4mg/mL)を含む10mM PBSの緩衝液(pH 8.0)に事前に溶解しておいたウシ血清アルブミン(BSA)(分子量66,320Da)(8mg、0.12μmol)の溶液に、N2雰囲気下で加えた。その混合物を22°Cで2時間撹拌した。その後に、N2雰囲気下でジチオトレイトール(DTT; 7.12mg、46μmol; Sigma−Aldrich)を加え、その混合物を4°Cで1時間撹拌した。再生セルロース細胞膜30,000Da cutoff(YM30、Millipore)全体の圧力源としてN2を用いて、撹拌した限外濾過細胞(Millipore、Billerica、マサチューセッツ州)中に5mM EDTAを含む10mM PBSの緩衝液(pH7.2)に対して、結果として生じた溶液を4°Cで透析した。タンパク質とSHの含有量を、ビシンコニン酸タンパク質アッセイキット(BCA;Pierce、Rockford、イリノイ州)とEllman(1959)方法によってそれぞれ測定した。20−22SH基のモル置換が得られた。
【0086】
破傷風トキソイド(TT)(分子量、150,000Da)の活性化のために、6mM EDTA(最終濃度4mg/mL)を含む10mM PBS緩衝液(pH 8.0)中のTT溶液(4mg、0.03μmol)に、N2雰囲気下で、乾燥したDMSO(25μL)中のDTSP(0.316mg、0.8μmol)を加え、その反応をBSAについて上記したように処理を進めた。その後に、N2雰囲気下で、ジチオトレイトール(DTT;3.56mg、23μmol; Sigma−Aldrich)を加え、その混合物を4°Cで1時間撹拌した。活性化したTTを、撹拌した限外濾過細胞(Millipore)に移し、再生セルロース細胞膜、100,000Da cutoff(YM100、Millipore)にわたって、5mM EDTAを含む10mM PBSの緩衝液(pH7.2)に対して、4°Cで透析した。
【0087】
5mM EDTAを含む10mM PBS緩衝液(pH 7.2)中のBSA−SH21−22またはTT−SH21−22の溶液に、10mM PBSの緩衝液中のマレイミド官能基を有するLPS−OHまたはdLPS誘導体の溶液を、N2雰囲気下で加えた。その反応物を4°Cで24時間攪拌した。その後に、N−エチルマレイミド(1mg;Sigma−Aldrich)を加えた。その反応を22°Cで30分間進め、結果として生じる複合体を、4°Cで4時間、10mM PBS緩衝液(pH 7.2)に対して透析し、0.22μmポリフッ化ビニリデン(PVDF)膜(Millex−GV、Millipore、Cork、アイルランド)を用いて、フィルタを殺菌した。複合体は、その炭水化物とタンパク質含有量について、それぞれLPS−OHまたはdLPSおよびBSAを標準として、中性糖のためのフェノール硫酸方法(Dubois et al、1956)、および、BCAタンパク質アッセイキットを用いて分析された。複合体の効率は、10mM PBS緩衝液(pH 7.2)で平衡に保たれたSuperose 12 10/300 GLカラム(Amersham Biosciences、ウプサラ、スウェーデン)を用いて、高性能液体クロマトグラフィー(HPLC; Agilent 1200Series、Agilent Technologies、Waldbronn、ドイツ)によって確認された。クロマトグラフィーは、室温で、流速0.5 mL/分で行われた。溶出はダイオード・アレー検出器(diode array detector)(Agilent Technologies)で210nmおよび280nmでモニターされた。
【0088】
スペーサーの存在は、CH2NH2基に対応する3.22ppmでの新しい陽子共鳴の出現によって、1H−NMR分光法で確認された。β−マレイミドプロピオネート(maleimidopropionate)のCH2αおよびCH=CH基にそれぞれ対応する2.55ppmおよび6.9ppmでの陽子共鳴の存在によって確認されたように、スペーサー分子のアミン基が、3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステルとの反応によってさらに誘導体化されたことで、マレイミド官能基を有するLPS−OHまたはdLPSを得た。内殻のLD−Hepのホスホエタノールアミンの誘導体化は、化学量論量の試薬の使用により最小限に抑えられた。糖複合体は、担体タンパク質のチオ化(thiolation)とチオ化したタンパク質のマレイミド官能基を有するLPS−OHまたはdLPSへの追加によって得られた(図4)。共役の効率はHPLCによってモニターされた。得られた複合体はその炭水化物とタンパク質含有量について分析された。
【0089】
3つの複合体におけるTTに対するLPS−OHのモル比は、10:1から20:1まで及び、産出量は炭水化物含有量に基づいて13%から22%までで変動した(表5)。BSAまたはTTへのdLPSの共役は、著しく炭水化物含有量の多いdLPS−BSA−2、dLPS−TT、または、dLPS(PJ2)−TT複合体を産出した(表5)。LPS−OH−TTとdLPS−BSAまたはdLPS−TTの複合体の両方は、ELISAによってα1,6−グルカンに特異的なmAbと均等によく反応し、このことは、グルカンエピトープの形態が不変だったことを示唆している。
【0090】
【表5】
【0091】
(実施例6):マウスとウサギにおける、LPS−OH−TT、dLPS−BSA、dLPS−TTの複合体、および、dLPS(PJ2)−TT複合体の免疫原性
実施例5の複合体(LPS−OH−TT、dLPS−BSA、dLPS−TT、および、dLPS(PJ2)−TT)の免疫性をマウスとウサギで試験した。
【0092】
生後6−8週の5匹の雌BALB/cマウスを、適切な複合体を用いて腹腔内で免疫化した。各々のマウスは、1回の注入当たり0.2mL Ribiアジュバント中の2μgまたは10μgの炭水化物を受け取った。マウスを21日目と42日目にブーストして、血清を51日目に末端の心臓穿刺(terminal heart Puncture)から回収した。
【0093】
3匹のニュージーランドホワイトウサギを、適切な複合体を用いて皮下で免疫化した。各々のウサギは、0.5mLの不完全フロインドアジュバント中の10μgまたは50μgの炭水化物を受け取った。ウサギを28日目と56日目にブーストして、65日目に失血後から回収した。
【0094】
血清中の抗LPS抗体のレベルは、精製されたLPSを被膜抗原(1μg/ウェル)として用いるELISAによって測定された。PBSで洗浄した後に、PBSまたはMilk希釈液/ブロッキング溶液(blocking solution)(MDB)(KPL、Gaithersburg、MD)中の1%(w/v)のウシ血清アルブミン(BSA)によって、プレートを37°Cで1時間ブロック(block)した。希釈したマウスまたはウサギの免疫前後の血清を加え、プレートを37°Cで2時間インキュベートした。阻害ELISAについては、26695 HP0479::Kan LPSまたは26695 HP0826::Kan LPSを阻害する一連の希釈物を、OD450=0.6−0.8を与えたウサギ血清の事前に測定された希釈物と混合した。この混合物を22°Cで15分間インキュベートし、吸収したLPS抗原で遮断された元々のマイクロタイタープレートに移し、そこで37°Cで2時間、さらにインキュベートした。この工程の後は、間接的なELISA処置に従った。手短にいえば、プレートをPBSと第2抗体で洗浄し、ヤギ抗マウスIgG+IgMワサビダイコンペルオキシダーゼ複合体(Caltag、So.サンフランシスコ、カリフォルニア州)を室温で1時間加えた。最終的な洗浄工程の後、3,3’,5,5’−テトラメチルベンジデン(benzidene)(TMB)(KPL、Gaithersburg、MD)基質を加え、1Mのリン酸で反応を止めた。マイクロタイタープレートリーダー(Dynatech,Chantilly,VA)を用いて、450nmで吸光度を測定した。
【0095】
阻害割合を以下の式を用いて計算した。
%阻害=100×[(インヒビターを含むOD−インヒビターを含まないOD)/インヒビターを含まないOD]
【0096】
阻害対ログ濃度曲線が各インヒビターについてプロットされ、半最大阻害濃度(half maximal inhibitory concentration)(IC50)に必要な濃度を、外挿曲線(extrapolation Curve)から決定した。
【0097】
反応は、dLPS−BSA、dLPS−TT、または、dLPS(PJ2)−TTのいずれかで免疫化した動物よりもLPS−OH−TTで免疫化したマウスとウサギの方で全般的に弱かったが、全ての複合体は、3度の注入後(表6乃至9)、ウサギとマウスの両方で同種(26695 HP0826::Kan)と対応する野生型(26695)株からのLPSに対するIgG反応を誘発した。LPS−OH、dLPS、または、dLPS(PJ2)のいずれかと、タンパク質担体(アジュバントを有する)との混合物で免疫化した対照ウサギは、3度の免疫化後に、ピロリ菌の同族の菌株(26695 HP0826::Kan変異株)、または、対応する野生型26695菌株からのLPSに対する特異的な反応は全くないか、低い値しか示さなかった。
【0098】
【表6】
【0099】
【表7】
【0100】
【表8】
【0101】
【表9】
【0102】
交差反応性研究は、様々なLPS グリコタイプ (Monteiro,2001)を代表するピロリ菌株、および、選択された突然変異株からのLPSに対して、LPS−OH−TT−2、LPS−OH−TT−3、LPS−OH−TT−4、dLPS−BSA−2、dLPS−TT、または、dLPS(PJ2)のいずれかで免疫化したウサギの免疫後血清を用いて行なわれた。結果を表10、11および12で示す。
【0103】
【表10】
【0104】
【表11】
【0105】
【表12】
【0106】
α1,6−グルカンを加えることができないピロリ菌株を代表するものである(Logan et al.,2005)SS1およびSS1 HP0826::Kan LPSによって、弱い交差反応性しか得られなかったため、LPS−OH−TT−2、LPS−OH−TT−3、またはLPS−OH−TT−4で免疫化したウサギから得た免疫後の血清の反応性は、α1,6−グルカンの存在の必要性を示した(表10)。驚くことに、dLPS−TTで免疫化したウサギから得た血清は同様に、任意の1,6−グルカンを含まなかった殻LPSエピトープ、即ち、株SS1、SS1 HP0826::Kan、M6およびJ99からのLPS(表11)を認識し、これはより広い殻の認識を示し、おそらくはTT担体タンパク質の存在により、dLPS−TT結合体がより免疫原性であったことを推測している(表11)。あるいは、結合体が3のグリコフォームの混合物から調製されるように、免疫反応は他の主要でないLPS構成要素へと生成されていたかもしれない。このことは、観察された交差反応について説明する。
【0107】
dLPS−TT結合体によって誘発されるウサギ血清の結合特異性を調べるため、阻害ELISAを、2つのグリコフォーム(およその比率1:1)、すなわち、線形の主鎖オリゴ糖構造および[GlcNAc,Fuc]にキャッピングされた線形の主鎖オリゴ糖(Hiratsuka et al.,2005)、ならびに、26695 HP0826::Kan LPSからなる、精製された26695 HP0479::Kan LPSを用いて実行した。26695 HP0479::Kan LPSがインヒビターとして使用された時、株SS1、SS1 HP0826::Kan HP0159::KanおよびSS1 HP0479::Kanからのグルカン−陰性LPSへのウサギ血清の結合(図6)が有意に阻害され、その一方で、26695 LPSがコーティング抗原(図6)として使用された時、26695 HP0826::Kan LPSが最も効果的なインヒビターであった。
【0108】
異種の分類可能な株および分類不可能な株を認識する、ウサギの免疫後血清の能力も同様に、ピロリ菌の分類可能な株および分類不可能な株両方のピロリ菌代表物(representatives)の選択された臨床分離株に対する細胞全体の間接的ELISA(WCE)において確認された。細胞全体の間接的ELISA(WCE)を、前述に記載の通りに実行した(Altman et al.,2008)。手短に言えば、マイクロタイタープレートのウェル(well)を、4°Cで一晩中、108細胞/mLで、100μLの細菌懸濁液で覆った。その後、ウェルをメタノールで固定し、37°Cで2時間、200μLのMilk希釈液/ブロッキング溶液(MDB)(KPL,Gaithersburg,MD)でブロックした。続いて、MDB中で1:500に希釈した100μLの1C4F9腹水を用いて、ウェルを37°Cで2時間インキュベートし、その後、MDB中で1:1,000に希釈した抗マウスIgG+IgMホースラディッシュペルオキシダーゼ(Caltag)を用いて、室温で1時間インキュベートした。間接的ELISAに関して記載されるように、基質TMBを加えた。非特異性のバックグラウンド値を、細菌細胞、第2の抗体結合体および基質を含む、陰性対照のウェルのOD450として決定した。これらOD450値は≦0.2であった。OD450<0.2の光学的濃度値を、陰性反応として分類し、OD450値≧0.2を陽性反応として分類した。プレート間の一貫性を保証するために、ピロリ菌株26695細胞を陽性対照として使用した。アッセイは10%以上は変わらなかった。
【0109】
dLPS−TT複合体で免疫化したウサギ由来の血清が、細胞全体の間接的ELISAを使用して試験されたすべての株(表13)に対し、もっとも強い交差反応性を示した。
【0110】
表13:ピロリ菌の臨床分離株に対する細胞全体の間接的ELISAbにおけるウサギの免疫後血清aの交差反応性。
【0111】
【表13】
【0112】
(実施例7:ウサギ抗血清の殺菌力)
ワクチン接種前またはワクチン接種後のウサギの血清(実施例6)を使用して、殺菌アッセイを行なった。
【0113】
プレートで増殖した(Plate−grown)ピロリ菌細胞を採取し、プレート1枚当たり5mLのPBSで洗浄した。遠心分離の後、ペレットを25mLのPBS中に懸濁した。最終的な細菌懸濁液を、4x106 CFU/mLでダルベッコ燐酸緩衝生理食塩水(DPBS)(Invitrogen,Grand Island,ニューヨーク)内で希釈した。殺菌アッセイを、平底のマイクロタイタープレート(ICN)内で行なった。補体除去した(de−complemented)免疫前または免疫後の血清(50μL)の10倍の連続希釈物を、各々のウェルに加えた。その後、細菌懸濁液(25μL)を加え、37°Cで15分間プレインキュベートした。仔ウサギの補体(Cedarlane Laboratories,Hornby,ON)を1:50に希釈し、25μLを適切なウェルに加えた。プレートを37°Cで45分間インキュベートし、次に、氷の上に置いた。10μLのアリコートを、コロンビア血液寒天(Columbia blood agar)上で3回播種し、4日間増殖させ、その後プレートを数えて、コロニー形成単位(CFU)の数を測定した。バクテリアと補体を含むが血清を含まない対照プレートを使用し、死滅率(percentage killing)を計算した。殺菌力は、50%を死滅させた血清のもっとも高い希釈として決定した。
【0114】
ワクチン接種前の血清またはタンパク質担体とLPS−OH、dLPSまたはdLPS(PJ2)のいずれかとの混合物(アジュバントを有する)により免疫化されたウサギから得た血清は、同種のピロリ菌26695 HP0826::Kanまたは野生型の26695菌株に対する殺菌力を示さないか、または、それらに対する低レベルの殺菌力を示した。LPS−OH−TT−4、dLPS−BSA−2、dLPS−TTおよびdLPS(PJ2)−TTで免疫化したウサギから得た免疫後血清は、26695 HP0826::Kan変異株および対応する野生型菌株の両方に対し、有意で機能的な活性を示し(表14)、dLPS−TT結合体で免疫化したウサギからのワクチン接種後の血清は、野生型菌株26695に対してもっとも高い機能的な活性を示した(表14)。
【0115】
【表14】
【0116】
更に、ウサギ血清の殺菌力も同様に、ピロリ菌の選択された臨床分離株、すなわち、002CL、0153CLおよび058CLに対して試験した(Altman et al.,2008)。これら分離株を、抗−α1,6−グルカンmAbsを用いるWCEアッセイにおけるそのOD450値に基づき、高度、中度および低度の結合剤の代表物として選択した:株002CL−O450 1.361、高度の結合剤;株153CL−OD450 0.29、中度の結合剤;および株058CL−OD450 0.162、低度の結合剤である。dLPS−TTまたはdLPS(PJ2)−TT複合体のいずれかで免疫化したウサギからの免疫後血清のみが、試験された3つの臨床分離株により機能的な活性を示したことを強調することが重要である(表15)。
【0117】
【表15】
【0118】
(実施例8:マウスにおけるdLPS−TT複合体による防御試験)
候補ワクチンとしてのdLPS−TT複合体の潜在性を、非近交系のCD−1マウスにて評価した。
【0119】
1μg/マウスのコレラ毒素(CT;Sigma)で補助された25μg/マウスのdLPS−TT複合体、1mg/マウスのコレラ毒素で補助された31.5μg/マウスのPJ2細胞を含まない音波処理物(PJ2 cell−free sonicate)(対照)、または食塩水(対照)を、グループ5のマウスに、週1回の間隔で4度、鼻腔内にワクチン接種した。最後の免疫の1週間後、血清、糞および膣の洗浄サンプルを、ピロリ菌特異的IgGおよびIgAのために採取し、分析した。血清IgGおよび血清、糞および膣のIgA抗体レベルを、標準のELISAによって測定した。プレートを本明細書に記載の通り、1μg/ウェルのピロリ菌株26695 HP0826::Kan LPSで覆った。血清サンプルを、IgGアッセイのため1:100に希釈し、IgAアッセイのため1:50に希釈した。糞のサンプルを1:2に希釈し、膣のサンプルを1:20に希釈した。ピロリ菌特異的抗体レベルを決定し、GraphPad software version 5.0を使用して、マン・ホイットニー検定によってデータを分析した。
【0120】
最初の免疫の5週間後、〜108 cfuピロリ菌株PJ2を一日おきに3回、マウスに経口的に強制投与した(Altman et al.,2003)。4週間後に、マウスを殺し、胃からのその一般細菌を数え、GraphPad software version 5.0を使用して、マン・ホイットニー検定によってデータを分析した。
【0121】
CTに加えてdLPS−TT複合体による鼻腔内の免疫化は、ELISAによって測定されるように、非常に強力なピロリ菌26695 HP0826::Kan LPS特異的IgG反応を誘発し、中度のピロリ菌26695 HP0826::Kan LPS特異血清IgAも同様に検出された(表16および17、図7)。加えて、膣または糞のサンプルにおけるピロリ菌26695 HP0826::Kan LPS特異的反応も同様に検知した(p<0.01)(表17、図7)。
【0122】
【表16】
【0123】
【表17】
【0124】
長いα1,6−グルカンを含有すると以前に示された分類不可能なピロリ菌マウス−コロニー形成株(colonizing strain)PJ2を、全てのマウスのグループにその後経口胃(orogastrically)接種した(Altman et al.,2003)。防御は、対照グループと比較して、ワクチン接種したグループにおける細菌の負荷の統計的に有意な減少として定義される。CTを加えたdLPS−TT複合体により鼻腔内で免疫化したマウスのグループに関するコロニー形成のデータは、統計的に有意であった(p<0.05)(図8)。さらに、これらのデータは、ピロリ菌26695 HP0826::Kan LPS特異血清IgA反応が、単独では細菌負荷に対する防御に十分ではないということ、および、LPS特異血清IgG反応も同様に、防御の増加に寄付するということを示唆する。
【0125】
〈実施例9:デキストラン−BSA複合体および免疫原性の調製および特性付け並びに機能的な活性の研究〉
250μLの0.2Mのホウ酸塩緩衝液(pH9.0)中に、10mgのデキストランT5(MW 5 KDa,Pharmacosmos A/S,Holbaek,Denmark)を溶解させることにより、デキストラン−BSA複合体を調製した。これを、0.2Mのホウ酸塩緩衝液(pH9.0)中の1,8−ジアミノ−3,6−ジオキサオクタン(25mL)およびシアノヒドリドほう酸ナトリウム(10mg)を含む溶液(250μl)に加えた(「Roy et al.,1984」に記載の通り)。反応を55°Cで5日間行なった。LPSベースの複合体の調製に関して上述されたように、反応生成物を精製した。
【0126】
この反応はスペーサーの導入をもたらす。LPSベースの複合体に関して記載されるように、3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステルとの反応によって、スペーサー分子のアミン基をさらに誘導体化した(実施例5を参照)。担体タンパク質のチオール化とマレイミドにより官能化されたデキストランT5のチオール化したタンパク質の付加とによって、糖複合体を得た。複合体においてBSAに対するデキストランのモル比は20:1であり、収量は炭水化物含有量に基づき、32%であった。
【0127】
様々なLPSグリコ型(glycotypes)のピロリ菌代表物および選択された突然変異株からのLPSに対するデキストラン−BSAにより免疫化されたウサギの免疫後血清を用いて、交差反応性研究を行なった(表18)。
【0128】
【表18】
【0129】
交差反応性がグルカン−陰性株SS1およびSS1 HP0826::Kanにより得られなかったため、デキストラン−BSAにより免疫化されたウサギから得た免疫後血清の反応は、α1,6−グルカンが存在するための必要条件を表した。デキストラン−BSAにより免疫化したウサギから得た免疫後血清は、26695 HP0826::Kan変異株および対応する野生型菌株の両方に対する機能的な活性を示した(表19)。
【0130】
【表19】
【0131】
異種の分類可能な株および分類不可能な株を認識し、殺菌性の抗体を誘発するといったウサギの免疫後血清の能力は、α1,6−結合グルコース反復単位の線形な主鎖を含むポリマーであるデキストランからなる複合体、または、連続するα1,6−結合グルコースの残基からなる最適化された線形のオリゴ糖を含む複合体からなる複合体、および、適切なタンパク質が、ピロリ菌感染に対する防御を与えるのに十分であるという可能性を示す。
【0132】
(実施例10:抗グルカンmAbsの特異性の研究)
ピロリ菌株O:3 HP0826::Kanに対して特異的なモノクローナル抗体を生成し、その特異性を研究した。
【0133】
前に記載されたように、ハイブリドーマを生成した(Altman et al.,2005)。82日にわたって5度、ピロリ菌株O:3 HP0826::Kanの108細胞(200μL)ホルマリン固定化細胞を、6匹のBALB/cマウス(Charles River Laboratories,St Constant,QC)に腹腔内に(i.p.)注入することで、有意な滴定濃度を達成した。最後の静脈内(i.v.)注入を行い、続いて3日後に融合させた。Kohler and Milstein(1975)に従い、2匹のマウスの脾臓細胞をSp2/Oプラズマ細胞腫細胞株と融合した。368のウェルからの最初の融合の上澄みを、間接的な酵素結合免疫測定法(ELISA)によって選別した。
【0134】
ELISAによるハイブリドーマの選別に関して、マイクロタイタープレート(ICN,Costa Mesa,CA)を、37°Cで3時間、50mMの炭酸塩バッファー(pH9.8)中の10μg/mLの対応するピロリ菌O:3 HP0826::Kan LPSで覆った。PBSでの洗浄後、プレートをPBS中の5%(w/v)のウシ血清アルブミン(BSA)でブロックした。使い切った上澄みを加え、プレートを室温で3時間インキュベートした。プレートをPBSと第2抗体で洗浄し、ヤギ抗マウスIgG+IgMホースラディッシュペルオキシダーゼ複合体(Caltag,So.San Francisco,CA)を室温で1時間加えた。最後の洗浄の工程後、3,3’,5,5’−テトラメチルベンジデン(tetramethylbenzidene)(TMB)(KPL,Gaithersburg,MD)基質を加え、反応を1Mのリン酸を用いて止めた。マイクロタイタープレートリーダー(Dynatech,Chantilly,VA)を使用し、吸光度を450nmで測定した。この工程の後に、実施例6に記載の通り、間接的ELISA手順を続けた。
【0135】
制限のある希釈クローニング(dilution cloning)に従って、安定した2つのハイブリドーマを得た。1つのクローン、1C4F9、IgMを、さらなる特徴付けのために選択した。腹水はBALB/cマウスにおいて上昇し、製造業者のプロトコルに従って、IgM−特異的アフィニティカラム(Pierce,Rockford,IL)を使用し、1C4F9モノクローナル抗体(mAb)を腹水から精製した。
【0136】
グルカンに特異的な抗体の特異性に関する研究は、連続するα1,6−結合グルコースの残基Glc→[α(1→6)Glc]n=1−6および選択的な精製されたピロリ菌LPSからなる、一連の線形のオリゴ糖を用いて行われた。阻害ELISAに関して、イソマルト系(isomalto−series)[Glcα1→(6Glcα1→)n=1−6](USBiologicals,Swampscott,MS)またはピロリ菌26695、O:3およびPJ2 LPSインヒビターからの線形のα1,6−結合グルコースを含むオリゴ糖の一連の希釈物を調製して、OD450=1を与えた精製1C4F9の以前に調製された希釈物と混合した。インキュベーション(22°C、15分)の後、この混合物を37°Cでもう2時間インキュベートした場合、吸着したLPS抗原を含む最初のブロックされたマイクロタイタープレートに混合物を移動させた。この工程の後、実施例6に記載の通り、間接的ELISAの手順を続けた。阻害の割合を、以下の式を使用して計算した:
【0137】
%阻害=100x[(インヒビターを含むOD−インヒビターを含まないOD)/インヒビターを含まないOD]
【0138】
阻害対ログの濃度の曲線を各々のインヒビターに関してプロットし、半分の最大阻害濃度に必要な濃度(IC50)を外挿曲線から決定した。
【0139】
イソマルトヘキサオース(n=5)およびイソマルトヘプタオース(isomaltoheptaose)(n=6)を使用すると、阻害特性が最適化した(表20)。グルカン鎖(Altman et al.,2003)に平均で6から8の残基を含むと以前に示されたピロリ菌PJ2 LPSは、最も有効なインヒビターであった(表20)。
【0140】
【表20】
【0141】
(実施例11:抗グルカンmAbsを使用する接触性の研究(accessibility study)および殺菌アッセイ)
抗グルカンmAbsは、間接型のIF顕微鏡観察法研究によって実証されるように、代表的なピロリ菌株から生存する細菌の表面に容易に接触可能であった(表21)。α1,6−グルカンおよびCagAの両方を、細菌の表面上で同時に識別する。
【0142】
モノクローナル抗体の殺菌力を決定するために、補体除去された腹水(50μL)からの1C4F9 mAbの10倍の連続希釈物を各々のウェルに加え、その後、細菌懸濁液(25μL)を加え、37°Cで15分間プレインキュベートした。この工程の後、実施例7に記載の通り、手順を続けた。1C4F9の細胞表面結合は、ピロリ菌の野生株および変異株に対する殺菌アッセイによって測定されるように、機能的な活性と相互に関連付けられた(表21)。
【0143】
【表21】
【0144】
本発明の好ましい実施形態が上述された一方で、様々な修正がその中で行われること、添付の請求項が、本発明の精神および範囲を逸脱することなく、全てのそのような修正を含むことを意図していることを認識し、かつ理解されたい。
【0145】
(引用文)
すべての特許、特許出願および出版物は、本明細書を指し、本明細書により出願の全体にわたって引用文に組み込まれる。
Altman E, Chandan V, Larocque S, Aubry A, Logan SM, Vinogradov E, Li J. Effect of the HP0159 ORF mutation on the lipopolysaccharide structure and colonizing ability of Helicobacter pylori. FEMS Immunol Med Microbiol 2008; 53: 204−213.
Altman E, Fernandez H, Chandan V, Harrison BA, Wilson Schuster M, Otth Rademacher L, Toledo C. Analysis of Helicobacter pylori isolates from Chile: occurrence of a selective type 1 Lewis b antigen expression in lipopolysaccharide. J Med Microbiol 2008; 57: 585−591.
Altman E, Harrison BA, Hirama T, Chandan V, To R, MacKenzie R. Characterization of murine monoclonal antibodies against Helicobacter pylori lipopolysaccharide specific for Lex and Ley blood group antigens. Biochem Cell Biol 2005; 83: 589−596.
Altman E, Smirnova N, Li J, Aubry A, Logan SM. Occurrence of a nontypable Helicobacter pylori strain lacking Lewis blood group O antigens and DD−heptoglycan: evidence for the role of the core alpha1,6−glucan in colonization. Glycobiology 2003; 13: 777−783.
Anderson PW, Pichichero ME, Insel RA, Betts R, Eby R, Smith DH. Vaccines consisting of periodate−cleaved oligosaccharides from the capsule of Haemophilus influenzae type b coupled to a protein carrier: structural and temporal requirements for priming in the human infant. J Immunol 1986; 137: 1181−1186.
Angelakopoulos H, Hohmann EL. Pilot study of phoP/phoQ−deleted Salmonella enterica serovar Typhimurium expressing Helicobacter pylori urease in adult volunteers. Infect Immun 2000; 68: 2135−2141.
Appelmelk BJ, Simmons−Smit L, Negrini R, Moran AP, Aspinall GO, Forte JG et al. Potential role of molecular mimicry between Helicobacter pylori lipopolysaccharide and host Lewis blood group antigens in autoimmunity. Infect Immun 1996; 64: 2031−2040.
Brisson J−R, Crawford E, Uhrin D, Khieu NH, Perry MB, Severn WB, Richards JC. The core oligosaccharide component from Mannheimia (Pasteurella) haemolytica serotype A1 lipopolysaccharide contains L−glycero−D−manno− and D−glycero−D−manno−heptoses: Analysis of the structure and conformation by high−resolution NMR spectroscopy. Can J Chem 2002; 80: 949−963.
Castillo−Rojas G, Mazari−Hiriart M & Lopez−Vidal Y (2004). Helicobacter pylori: focus on CagA and VacA major virulence factors. Salud publica de Mexico 46: 538−548.
Chandan V, Logan SM, Harrison BA, Vinogradov E, Aubry A, Stupak J, Li J, Altman, E. Characterization of a waaF mutant of Helicobacter pylori strains 26695 provides evidence that an extended lipopolysaccharide structure has a limited role in the invasion of gastric cancer cells. Biochem Cell Biol 2007; 85: 582−590.
Chu C, Liu B, Watson D, Szu S, Bryla D, Shiloach J, Schneerson R, Robbins JB. Preparation, characterization, and immunogenicity of conjugates composed of the O−specific polysaccharide of Shigella dysenteriae type 1 (Shiga’s bacillus) bound to tetanus toxoid. Infect Immun 1991; 59: 4450−4458.
Ciucanu I, Kerek F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr Res 2004; 131: 209−217.
Ciucanu I, Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr Res 2004, 131: 209−217.
Cox AD, Zou W, Gidney MAJ, Lacelle S, Plested JS, Makepeace K, Wright JC, Coull PA, Moxon ER, Richards JC. Candidacy of LPS−based glycoconjugates to prevent invasive meningococcal disease: Developmental chemistry and investigation of immunological responses following immunization of mice and rabbits. Vaccine 2005; 23: 5045−5054.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956; 28: 350−356.
Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clin Microbiol Rev 1997; 10: 720−741.
Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys 1959; 82: 70−77.
Fernandez−Santana V, Gonzalez−Lio R, Sarracent−Perez J, Verez−Bencomo V. Conjugation of 5−azido−3−oxapentyl glycosides with thiolated proteins through the use of thiophilic derivatives. Glycoconjugate J 1998; 15: 549−553.
Gu X−X, Tsai C−M, Ueyama T, Barenkamp SJ, Robbins JB, Lim DJ. Synthesis, characterization, and immunological properties of detoxified lipooligosaccharide from nontypable Haemophilus influenzae conjugated to proteins. Infect Immun 1996; 64: 4047−4053.
Hiratsuka K, Logan SM, Conlan JW, Chandan V, Aubry A, Smirnova N, Ulrichsen H, Chan KHN, Griffith DW, Harrison BA, Li J, Altman E. Identification of a D−glycero−D−manno−heptosyltransferase gene from Helicobacter pylori. J Bacteriol 2005; 187: 5156−5165.
Holst O, Brade L, Kosma P, Brade, H. Structure, serological specificity, and synthesis of artificial glycoconjugates representing the genus−specific lipopolysaccharide epitope of Chlamidia spp. J Bacteriol 1991; 173:1862−1866.
Kabat EA. Chapter 11. Molecular Biology of Anti−・−(1・6)dextrans. Antibody responses to a Single−Site−Filling Antigenic Determinant. In: Carbohydrate Antigens (vol. 159). Garegg PJ, Lindberg AA, eds. ACS Symposium Series, Washington: American Chemical Society,1993; 146−58.
Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975; 256: 495−497.
Logan SM, Altman E, Mykytczuk O, Brisson J−R, Chandan V, St Michael F, Masson A, Leclerc S, Hiratsuka K, Smirnova N, Li J, Wu Y, Wakarchuk WW. Novel biosynthetic functions of lipopolysaccharide rfaJ homologs from Helicobacter pylori. Glycobiology 2005; 15: 721−733.
Logan SM, Conlan JW, Monteiro MA, Wakarchuk WW, Altman E. Functional genomics of Helicobacter pylori: identification of a ・−1,4 galactosyltransferase and generation of mutants with altered lipopolysaccharide. Mol Microbiol 2000; 35: 1156−1167.
Mieszala M, Kogan G, Jennings HJ. Conjugation of meningococcal lipooligosaccharides through their lipid A terminus conserves their epitopes and results in conjugate vaccines having improved immunological properties. Carbohydr Res 2003; 338: 167−175.
Monteiro MA. Helicobacter pylori: a wolf in sheep’s clothing: the glycotype families of Helicobacter pylori lipopolysaccharides expressing histo−blood groups: structure, biosynthesis, and role in pathogenesis. Adv Carbohydr Chem Biochem 2001; 57: 99−158.
Passwell JH, Ashkenazi S, Harlev E, Miron D, Ramon R, Farzam N, Lerner−Geva L, Levi Y, Chu C, Shiloach J, Robbins JB, Schneerson R, Israel Shigella Study Group. Safety and immunogenicity of Shigella sonnei−CRM9 and Shigella flexneri type 2a−rEPAsucc conjugate vaccines in one− to four−year old children. Pediatr Infect Dis J 2003; 22: 701−706.
Passwell JH, Harlev E, Ashkenazi S, Chu C, Miron D, Ramon R, Farzan N, Shiloach J, Bryla DA, Majadly F, Roberson R, Robbins JB, Schneerson R. Safety and immunogenicity of improved Shigella O−specific polysaccharide−protein conjugate vaccines in adults in Israel. Infect Immun 2000; 69: 1351−1357.
Pon RA, Lussier M, Yang Q−L, Jennings HJ. N−Propionylated group B meningococcal polysaccharide mimics a unique bactericidal capsular epitope in group B Neisseria meningitidis. J Exp Med 1997; 185: 1929−1938.
Rossi G, Ruggiero P, Peppoloni S, Pancotto L, Fortuna D, Lauretti L, Volpini G, Mancianti S, Corazza M, Taccini E, Di Pisa F, Rappuoli R, Del Giudice G. Therapeutic vaccination against Helicobacter pylori in the beagle dog experimental model: safety, immunogenicity, and efficacy. Infect Immun 2004; 72: 3252−3259.
Roy, R., Katzenellenbogen, E., and Jennings, H.J. 1984. Improved procedures for the conjugation of oligosaccharides to protein by reductive amination. Can J. Biochem. Cell Biol. 62, 270−275.
Ruggiero P, Peppoloni S, Rappuoli R, Del Giudice G. The quest for a vaccine against Helicobacter pylori: how to move from mouse to man? Microbes Infect 2003; 5: 749−756.
Sawardeker JH, Sloneker JH, Jeannes A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal Chem 1967; 39: 1602−1604.
Westphal, O Jann K. Bacterial polysaccharides. Extraction with phenol−water and further applications of the procedure. Meth Carbohydr Chem 1965; 5: 83−91.
Wirth HP, Yang M, Karita M, Blaser MJ. Expression of the human cell surface glycoconjugates Lewis X and Lewis Y by Helicobacter pylori isolates is related to cagA status. Infect Immun 1996; 64: 4598−4605.
Yokota, S., Amano, K., Fujii, N., Yokochi, T. Comparison of serum antibody titers to Helicobacter pylori lipopolysaccharides, CagA, VacA and partially purified cellular extracts in a Japanese population. FEMS Microbiol Lett 2000; 185: 193−198.
Yu S, Gu X−X. Biological and immunological characteristics of lipooligosaccharide−based conjugate vaccines for serotype C Moraxella catarrhalis. Infect Immun 2007; 75: 2974−2980.
【図1A】
【図1B】
【図2A】
【図2B】
【特許請求の範囲】
【請求項1】
式Iの構造を含む、α1,6−グルカンを含むヘリコバクター・ピロリ化合物であって、
【化1】
Rは、α1,3−DD−ヘプタンに結合したα1,6−グルカンで置換したα−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、
α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされることを特徴とする化合物。
【請求項2】
グルカンは、3から12のα1,6−結合グルコース残基を含むことを特徴とする請求項1に記載の化合物。
【請求項3】
ヘプタン部分が2から6のα1,3−結合ヘプトース残基を含むことを特徴とする請求項1に記載の化合物。
【請求項4】
Rが以下を含み、
【化2】
β−GlcNAc残基LがHep GのO−2に結合する場合、n=1乃至11であり、m=0乃至4であることを特徴とする請求項1に記載の化合物。
【請求項5】
n=9であることを特徴とする請求項4に記載の化合物。
【請求項6】
m=2であることを特徴とする請求項4または5に記載の化合物。
【請求項7】
式Iの構造がKdo残基Cに共有結合している脂質A部分をさらに含むことを特徴とする請求項1乃至6のいずれか1つに記載の化合物。
【請求項8】
脂質A分子は、O−脱アシル化するか、または、Kdo残基のケトシド結合の加水分解によって開裂することを特徴とする請求項7に記載の化合物。
【請求項9】
前記化合物はピロリ菌株HP0826::Kanから分離されるか、または、精製されることを特徴とする請求項1乃至6のいずれか1つに記載の化合物。
【請求項10】
リンカー分子、タンパク質担体、または、その組み合わせに共役した、ほぼ線形のα1,6−グルカンを含む化合物を含む複合体。
【請求項11】
ほぼ線形のα1,6−グルカンを含む化合物は、請求項1乃至9のいずれか1つに記載の化合物であり、
式Iの構造は、リンカー分子、タンパク質担体、またはその組み合わせに共役されることを特徴とする請求項10に記載の複合体。
【請求項12】
ほぼ線形のα1,6−グルカンを含む化合物がデキストランであることを特徴とする請求項10に記載の複合体。
【請求項13】
タンパク質担体が、破傷風トキソイドまたはウシ血清アルブミン、CRM、または、CRM197であることを特徴とする請求項9乃至12のいずれか1つに記載の複合体。
【請求項14】
請求項1乃至9のいずれか1つに記載の1以上の化合物、および/または、請求項10乃至13のいずれか1つに記載の複合体を含む組成物。
【請求項15】
請求項1のα1,6−グルカンエピトープを含む化合物を認識する抗体。
【請求項16】
抗体がモノクローナル抗体1C4F9であることを特徴とする請求項15に記載の抗体。
【請求項17】
処置を必要としている個体において、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすための、請求項15に記載のモノクローナル抗体の使用。
【請求項18】
請求項15に記載のモノクローナル抗体を、必要としている個体に投与する工程を含む、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を誘発する方法。
【請求項19】
個体内でピロリ菌に対する免疫反応を誘発するための、有効な量の請求項14に記載の組成物の使用。
【請求項20】
有効な量の請求項14に記載の組成物を個体に投与する工程を含む、前記個体でのピロリ菌に対する免疫反応を誘発する方法。
【請求項21】
化合物が適切な担体タンパク質に共役されることを特徴とする請求項19に記載の使用、または、請求項20に記載の方法。
【請求項22】
化合物が、リポ多糖類の2−ケト−3−デオキシ−オクツロソン酸(Kdo)を介して、適切な担体タンパク質に共役されることを特徴とする請求項19に記載の使用、または、請求項20に記載の方法。
【請求項23】
免疫原性の請求項14に記載の組成物で哺乳動物を免疫化することによって作られた免疫抗血清。
【請求項24】
免疫抗血清は、ピロリ菌の同種および異種の、分類可能なおよび分類不可能な、変異株および野生株における、α1,6−結合グルカンエピトープを認識するIgGを含むことを特徴とする請求項23に記載の免疫抗血清。
【請求項25】
IgGは、変異体および野生型のα1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすことを特徴とする請求項24に記載の免疫抗血清。
【請求項26】
ハイブリドーマ細胞株1C4F9。
【請求項1】
式Iの構造を含む、α1,6−グルカンを含むヘリコバクター・ピロリ化合物であって、
【化1】
Rは、α1,3−DD−ヘプタンに結合したα1,6−グルカンで置換したα−DDHep−3−α−L−Fuc−3−β−GlcNAc三糖類であり、
α1,3−DD−ヘプタンの最後のDD−Hep残基は、β−GlcNAc残基でキャッピングされることを特徴とする化合物。
【請求項2】
グルカンは、3から12のα1,6−結合グルコース残基を含むことを特徴とする請求項1に記載の化合物。
【請求項3】
ヘプタン部分が2から6のα1,3−結合ヘプトース残基を含むことを特徴とする請求項1に記載の化合物。
【請求項4】
Rが以下を含み、
【化2】
β−GlcNAc残基LがHep GのO−2に結合する場合、n=1乃至11であり、m=0乃至4であることを特徴とする請求項1に記載の化合物。
【請求項5】
n=9であることを特徴とする請求項4に記載の化合物。
【請求項6】
m=2であることを特徴とする請求項4または5に記載の化合物。
【請求項7】
式Iの構造がKdo残基Cに共有結合している脂質A部分をさらに含むことを特徴とする請求項1乃至6のいずれか1つに記載の化合物。
【請求項8】
脂質A分子は、O−脱アシル化するか、または、Kdo残基のケトシド結合の加水分解によって開裂することを特徴とする請求項7に記載の化合物。
【請求項9】
前記化合物はピロリ菌株HP0826::Kanから分離されるか、または、精製されることを特徴とする請求項1乃至6のいずれか1つに記載の化合物。
【請求項10】
リンカー分子、タンパク質担体、または、その組み合わせに共役した、ほぼ線形のα1,6−グルカンを含む化合物を含む複合体。
【請求項11】
ほぼ線形のα1,6−グルカンを含む化合物は、請求項1乃至9のいずれか1つに記載の化合物であり、
式Iの構造は、リンカー分子、タンパク質担体、またはその組み合わせに共役されることを特徴とする請求項10に記載の複合体。
【請求項12】
ほぼ線形のα1,6−グルカンを含む化合物がデキストランであることを特徴とする請求項10に記載の複合体。
【請求項13】
タンパク質担体が、破傷風トキソイドまたはウシ血清アルブミン、CRM、または、CRM197であることを特徴とする請求項9乃至12のいずれか1つに記載の複合体。
【請求項14】
請求項1乃至9のいずれか1つに記載の1以上の化合物、および/または、請求項10乃至13のいずれか1つに記載の複合体を含む組成物。
【請求項15】
請求項1のα1,6−グルカンエピトープを含む化合物を認識する抗体。
【請求項16】
抗体がモノクローナル抗体1C4F9であることを特徴とする請求項15に記載の抗体。
【請求項17】
処置を必要としている個体において、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすための、請求項15に記載のモノクローナル抗体の使用。
【請求項18】
請求項15に記載のモノクローナル抗体を、必要としている個体に投与する工程を含む、α1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を誘発する方法。
【請求項19】
個体内でピロリ菌に対する免疫反応を誘発するための、有効な量の請求項14に記載の組成物の使用。
【請求項20】
有効な量の請求項14に記載の組成物を個体に投与する工程を含む、前記個体でのピロリ菌に対する免疫反応を誘発する方法。
【請求項21】
化合物が適切な担体タンパク質に共役されることを特徴とする請求項19に記載の使用、または、請求項20に記載の方法。
【請求項22】
化合物が、リポ多糖類の2−ケト−3−デオキシ−オクツロソン酸(Kdo)を介して、適切な担体タンパク質に共役されることを特徴とする請求項19に記載の使用、または、請求項20に記載の方法。
【請求項23】
免疫原性の請求項14に記載の組成物で哺乳動物を免疫化することによって作られた免疫抗血清。
【請求項24】
免疫抗血清は、ピロリ菌の同種および異種の、分類可能なおよび分類不可能な、変異株および野生株における、α1,6−結合グルカンエピトープを認識するIgGを含むことを特徴とする請求項23に記載の免疫抗血清。
【請求項25】
IgGは、変異体および野生型のα1,6−グルカンを発現するピロリ菌株の補体を媒介とした溶菌を引き起こすことを特徴とする請求項24に記載の免疫抗血清。
【請求項26】
ハイブリドーマ細胞株1C4F9。
【図3D】

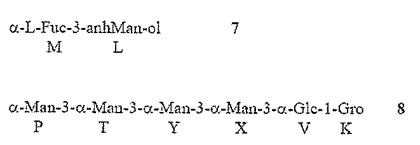
【図3E】
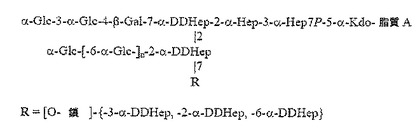
【図4】

【図5】
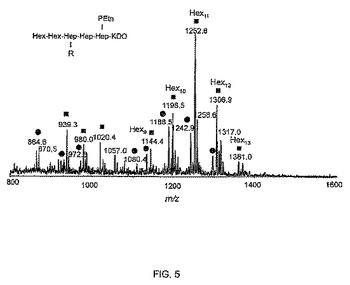
【図6】
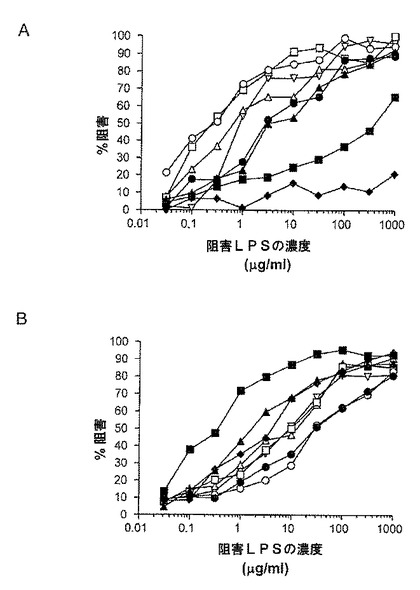
【図8】
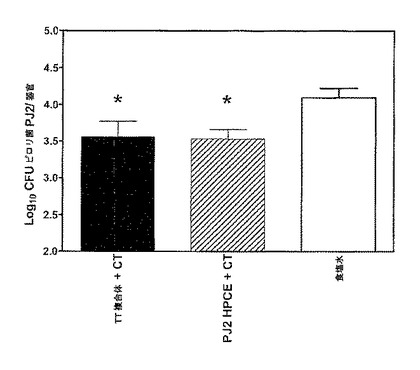
【図3A】
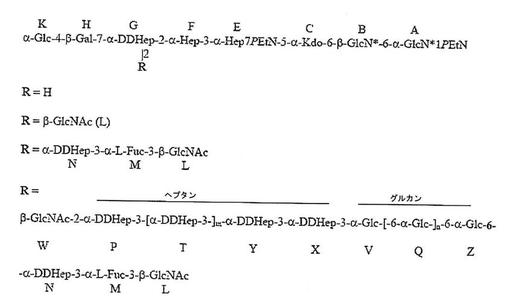
【図3B】
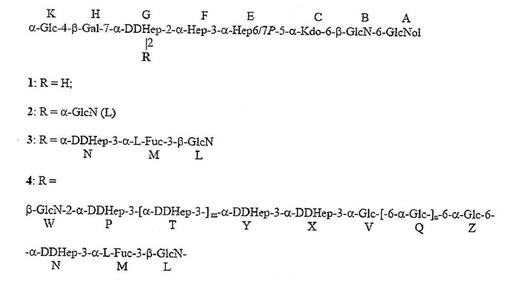
【図3C】
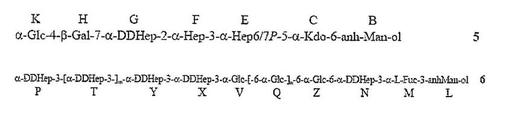
【図3F】
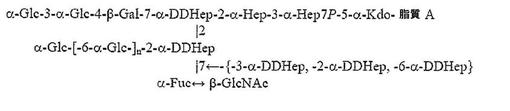
【図7】

【図3E】
【図4】
【図5】
【図6】
【図8】
【図3A】
【図3B】
【図3C】
【図3F】
【図7】
【公表番号】特表2013−501080(P2013−501080A)
【公表日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2012−521916(P2012−521916)
【出願日】平成22年7月30日(2010.7.30)
【国際出願番号】PCT/CA2010/001173
【国際公開番号】WO2011/011879
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(506175792)ナショナル リサーチ カウンシル オブ カナダ (9)
【Fターム(参考)】
【公表日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成22年7月30日(2010.7.30)
【国際出願番号】PCT/CA2010/001173
【国際公開番号】WO2011/011879
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(506175792)ナショナル リサーチ カウンシル オブ カナダ (9)
【Fターム(参考)】
[ Back to top ]