
ピロロキノリンキノン(QUININE)依存性可溶性グルコースデヒドロゲナーゼの突然変異体
本発明は、アシネトバクター・カルコアセチカスの野生型PQQ−sGDHのタンパク質配列(配列番号2)の428位にアミノ酸置換を含有する、PQQ−sGDHの新規の突然変異体に関する。本発明は、グルコースアッセイ、特に糖尿病の被験体における血中グルコースアッセイにおいて関心がもたれるグルコース電極の開発、及びグルコースを燃料として利用するバイオ燃料電池の実現のための上記PQQ−sGDH突然変異体の使用にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、グルコースアッセイ、特に糖尿病の被験体の血中グルコースアッセイにおいて関心がもたれるグルコース電極の開発の分野、及びグルコースを燃料として使用するバイオ燃料電池を応用するための分野に関する。
【0002】
本発明は、より具体的には、野生型酵素に比べて有利な特性を有する酵素ピロロキノリンキノン可溶性グルコースデヒドロゲナーゼ(PQQ s−GDHとも呼ばれる)の突然変異体に関する。
【背景技術】
【0003】
2型糖尿病はフランスにおいて200万人近くの人々に影響を与えているが、この数に疾患に気付いていない600000人を加える必要がある。米国においては状況は更に深刻である。先進国では、糖尿病は20歳〜65歳という年齢層での失明の主要な原因である。
【0004】
この疾患のモニタリング及び管理は、とりわけ血中グルコースの毎日の定量及びインシュリンの注射に基づく。様々な会社によって、患者が血糖値を自分で測定することを可能にするグルコースセンサーが提供されている。これらのセンサーは電流測定センサー、電位差測定センサー又は電量測定センサーであり得るが、全てグルコースを酸化することが可能な酵素の使用に基づいている。2つの主要な酵素はグルコースオキシダーゼ及びPQQ s−GDHである。
【0005】
ピロロキノリンキノングルコースデヒドロゲナーゼは、アシネトバクター・カルコアセチカス(Acinetobacter calcoaceticus)において同定されたキノプロテインファミリーに属し、2つのタイプがある(一方は膜酵素であり、他方は可溶性酵素である)。これらの酵素はD−グルコノラクトンへのグルコースの酸化を触媒し、電子供給体として使用することができる。
【0006】
アシネトバクター・カルコアセチカスのPQQ s−GDHは、約50kDaの2つのサブユニット(アポ酵素)とピロロキノリンキノン補因子(PQQ)とからなるホモ二量体から構成される(非特許文献1)。この酵素は活性部位を有し、PQQの存在下ではその内部でグルコースの酸化が触媒される。この活性部位は76位、144位、169位、343位、346位及び428位に位置するアミノ酸から構成される。
【0007】
PQQ s−GDHは容易に大量に得ることができるため、産業的にかなりの関心がもたれている。このため、PQQ s−GDHは血糖値の自己モニタリングのためのグルコースセンサーに使用される主要な酵素となっている。キノプロテインの利点は、電子受容体としてO2を利用するグルコースオキシダーゼとは対照的に、酸素に依存しないことである。
【0008】
その物理化学的特性に基づくと、PQQ s−GDHをバイオ燃料電池(その原理は電流を発生するグルコースの酸化に基づく)の製造に使用することも想定される。
【0009】
PQQ s−GDHの欠点は、その熱安定性の低さ、生理的pHでの安定性の低さ及び活性の穏やかさである。
【0010】
幾つかの著作物において、その特性を改善するためにPQQ s−GDHの変異体を開発することが試みられている。
【0011】
「突然変異体」又は「変異体」は、そのタンパク質配列が、野生型PQQ s−GDHのタンパク質配列と比べて少なくとも1つのアミノ酸の挿入、欠失及び/又は置換を含むPQQ s−GDHを意味する。以下、PQQ s−GDHの参照ヌクレオチド配列及び参照タンパク質配列は、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列とする(それぞれ配列番号1及び配列番号2)。
【0012】
PQQ s−GDHの安定性を改善することを目的とする突然変異
特許文献1は、2つのサブユニットがジスルフィド架橋によって結合するように突然変異させたPQQ s−GDHを記載している。この突然変異は、415位及び414位の少なくとも1つのアミノ酸、並びに/又は同時に340位及び418位に位置する2つのアミノ酸をシステイン残基で置き換えることからなる。これらの修飾は、より良好な熱安定性を酵素に与えている。
【0013】
PQQ s−GDHをその基質による阻害に対する感受性が低下したものにすることを目的とする突然変異
特許文献2は、領域349〜377内の少なくとも1つのアミノ酸を異なるアミノ酸で置き換えたPQQ s−GDHを記載している。この修飾は、基質による阻害に対する感受性が低下した酵素をもたらし、したがって高濃度のグルコースの存在下で使用することができる。
【0014】
PQQ s−GDHをその天然基質(グルコース)に対してより特異的なものにすることを目的とする突然変異
野生型PQQ s−GDHを修飾して、より基質特異的なものにすることも可能である。或る基質に対して特異的な酵素は該基質が関与する反応しか触媒しないが、逆に基質特異性の低い酵素は、その天然基質に構造的に類似した基質に基づく反応を触媒することが可能である。PQQ s−GDHの天然基質はグルコースであるが、野生型PQQ s−GDHの特異性は低く、他の単糖類及び二糖類の酸化を触媒することも可能である。
【0015】
幾つかの著作物が、野生型PQQ s−GDHをグルコースに対してより特異的にするために修飾することを提案している:
このため、特許文献3は49位、67位〜69位、76位、89位、129位〜131位、167位〜170位、174位、188位、189位、207位、215位、245位、249位、300位、341位〜343位、349位、351位及び429位に位置するアミノ酸の少なくとも1つを置換すること、並びに/又はアミノ酸を428位と429位との間に導入することによってPQQ s−GDHを修飾することを提案している;
特許文献4は、428位と429位との間にロイシン、リシン又はアラニンを挿入することによってPQQ s−GDHを修飾することを提案している;
特許文献5は、PQQ s−GDHの75位、326位〜354位、278位〜320位及び162位〜197位のうちの1つの位置でアミノ酸を置換することを記載している;
特許文献6は、PQQ s−GDHの428位と429位との間にアミノ酸を挿入し、任意で428位のアミノ酸をロイシン、プロリン又はバリンで置換することを提言している;
特許文献7は、125位、128位、142位、168位、169位、170位、224位、230位、236位、345位、351位、416位又は428位に1つ又は複数の置換を有するアシネトバクター属のPQQ s−GDHの突然変異体を記載している。この文献は更に、多数の突然変異部位を有する具体的な突然変異体を列挙している;
特許文献8は、428位と429位との間にロイシン、アラニン又はリシンの挿入を有するアシネトバクター・バウマンニ(Acinetobacter baumanii)のPQQ s−GDHの突然変異体、又は429位のアミノ酸がフェニルアラニン、プロリン、ロイシン又はチロシンで置換されている突然変異体を挙げている;
特許文献9は、348位のアミノ酸をアラニン、グリシン又はセリンで、428位のアミノ酸をロイシン、プロリン又はバリンで置換することを提案している。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第7,244,600号
【特許文献2】米国特許第7,244,581号
【特許文献3】米国特許出願公開第2007/0105173号
【特許文献4】欧州特許出願公開第1367120号
【特許文献5】米国特許第7,037,698号
【特許文献6】米国特許出願公開第2007/0243566号
【特許文献7】国際公開第2006/085509号
【特許文献8】欧州特許出願公開第1666586号
【特許文献9】米国特許第7,132,270号
【非特許文献】
【0017】
【非特許文献1】Oubrieet al., J. Mol. Biol; 289,319-333 (1999)
【発明の概要】
【発明が解決しようとする課題】
【0018】
グルコースセンサーで使用するためには、より活性の高い、すなわちD−グルコノラクトンへのグルコースのより速い変換反応を可能にするPQQ s−GDH突然変異体を得ることが必要とされるが、これは既存の突然変異体では可能ではない。
【0019】
したがって、十分な又は更には改善された熱安定性及びpH安定性を保持したうえで、野生型PQQ s−GDHよりも良好な活性を示すPQQ s−GDHを開発することが依然として必要とされている。
【課題を解決するための手段】
【0020】
この要求を満たすために、本発明者らは、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの新規の突然変異体を開発した。これらの突然変異体は、428位に位置するアスパラギンがシステイン、チロシン、アラニン、アスパラギン酸又はグルタミン酸で置換されているようなものである。
【0021】
したがって、本発明の第1の目的は、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDH突然変異体であって、428位に位置するそのアミノ酸が、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHのタンパク質配列(配列番号2)と比べてシステイン、チロシン、アラニン、アスパラギン酸又はグルタミン酸を含む群から選択されるアミノ酸で置換されていることを特徴とする、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDH突然変異体に関する。
【0022】
本発明に至った研究において、本発明者らは、PQQ s−GDH突然変異体であって、428位に位置するそのアミノ酸が、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHのタンパク質配列(配列番号2)と比べてリシンで置換されていることを特徴とする、PQQ s−GDH突然変異体も作製した(実施例第4部を参照されたい)。この突然変異体は配列番号10のタンパク質配列を有し、配列番号9の核酸分子によってコードされる。この突然変異体は、下記表Iに提示される配列番号17及び配列番号18のオリゴヌクレオチドを用いて作製することができる。
【0023】
アミノ酸のナンバリングは、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列を基準とする。しかしながら、本発明によるPQQ s−GDH突然変異体は、このアシネトバクター・カルコアセチカスの野生型PQQ s−GDHの突然変異体に限定されず、本発明はアシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも90%の同一率を有するPQQ s−GDHの突然変異体にも関する。
【0024】
参照配列であるアシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列(配列番号2)に対する配列の同一性は、2つの配列をそれらの間で最大の一致が得られるようにアラインメントした場合に同一であるアミノ酸残基のパーセンテージに応じて評価する。
【0025】
当業者は配列を比較するためのコンピュータプログラム、例えばBLAST suiteのプログラム(Altschul et al., NAR, 25, 3389-3402)を用いて同一率を算出することができる。BLASTプログラムは、参照配列として示した配列番号2全体からなる比較ウィンドウ上で採用される。
【0026】
参照配列と少なくともX%の同一性を有するアミノ酸配列を有するペプチドは、本発明においては、該参照ペプチドの機能的特性(本発明の場合、そのグルコース酸化の酵素活性)を保持したまま、参照配列の100個のアミノ酸に対して最大で100−X個の変化をその配列中に含むことのできるペプチドとして定義される。本発明の意味において、変化という用語は、参照配列におけるアミノ酸の欠失、置換、又は連続的若しくは分散した挿入を含む。
【0027】
アシネトバクター・カルコアセチカスのPQQ s−GDHの428位に位置するアミノ酸に対応するアミノ酸は、該相同酵素とアシネトバクター・カルコアセチカスのPQQ s−GDHとの配列アラインメントによって同定される。
【0028】
本発明は特に、配列番号4、配列番号6、配列番号8、配列番号12及び配列番号14(それぞれPQQ s−GDHのシステイン突然変異体、チロシン突然変異体、アラニン突然変異体、アスパラギン酸突然変異体及びグルタミン酸突然変異体のアミノ酸配列に対応する)から選択されるアミノ酸配列を有するPQQ s−GDH突然変異体に関する。これらの突然変異酵素は、好適なオリゴヌクレオチド対によるアシネトバクター・カルコアセチカスのPQQ s−GDH遺伝子の突然変異によって得られるヌクレオチド断片によってコードされる。
【0029】
これらの本発明による新規のPQQ s−GDH突然変異体は、市販のグルコースセンサーに使用される酵素であるアシネトバクター・カルコアセチカスの野生型酵素に比べて以下の改善された性能を示す:
これらの活性は、とりわけ生理的濃度、すなわち1mM〜10mMのグルコースで野生型酵素の活性よりも高い。酵素の活性は、PQQ s−GDHによるグルコースのグルコノラクトンへの酸化反応中に起こるレドックス試薬の着色をモニタリングすることによって定量化することができる。レドックス試薬は例えば、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム及びフェロセンである;
これらは高濃度でのグルコースの阻害効果に対する感受性が低い。
【0030】
PQQ s−GDH突然変異体の基質に対する特異性は、基質であるグルコース及び幾つかの他の糖類(二糖類等)に対する該突然変異体の酵素活性を比較することによって評価することができる。
【0031】
本発明によるPQQ s−GDH突然変異体の有利な特性によって、それらはグルコースをエネルギー源として用いるバイオ燃料電池等の生体電気システム及びグルコースバイオセンサーに特に好適に使用されるものとなる。
【0032】
本発明は、野生型PQQ s−GDH(例えばアシネトバクター・カルコアセチカスの野生型PQQ s−GDH)を、表Iに示されるオリゴヌクレオチド対からなる群から選択されるオリゴヌクレオチド対で修飾することによって得られる、本発明によるPQQ s−GDH突然変異体をコードする核酸分子にも関する。
【0033】
【表1】

【0034】
本発明によるPQQ s−GDH突然変異体をコードする核酸分子は特に、アシネトバクター・カルコアセチカスによって生成される配列番号1の配列の野生型酵素をコードする遺伝子のヌクレオチド配列を修飾することによって作製することができる。遺伝子の配列を修飾する幾つかの技法が当業者に既知である(Igarashi et al., Archives of Biochemistry and Biophysics 428 (2004) 52-63による概説を参照されたい)。特定の作製方法によると、本発明によるPQQ s−GDH突然変異体をコードする核酸分子は、導入すべき突然変異を有するオリゴヌクレオチドの存在下でのPCRによる突然変異誘発によって作製される(下記の実施例を参照されたい)。
【0035】
特定の実施の形態によると、本発明は、その配列が配列番号3、配列番号5、配列番号7、配列番号11及び配列番号13の配列からなる群から選択される、本発明によるPQQ s−GDH突然変異体をコードする核酸分子に関する。本発明によるPQQ s−GDH突然変異体をコードする核酸分子を、次いでプラスミド等の発現ベクターにクローニングした後、細菌、酵母又は細胞培養物等の好適な宿主において形質転換させることができる。
【0036】
発現ベクターとは、その発現に不可欠なシグナル、特にプロモーター(構成的又は誘導性)、リボソーム結合部位、転写終結シグナル、及び任意で抗生物質抵抗性遺伝子等の選択マーカーの間にヌクレオチドコード配列を挿入することを可能にする領域を有するベクターを意味する。
【0037】
本発明は、上記核酸分子を含む発現ベクター、及び該発現ベクターで形質転換され、本発明によるPQQ s−GDH突然変異体を発現する宿主細胞に更に関する。
【0038】
発現ベクターは当業者に既知の任意の方法、特に宿主細胞の膜透過性を例えばカルシウムイオンの存在下で変化させること、又はエレクトロポレーションによって宿主細胞内に導入することができる。
【0039】
本発明によるPQQ s−GDH突然変異体を発現するように形質転換した宿主細胞の培養の後、該細胞を遠心分離によって回収し、本発明による該PQQ s−GDH突然変異体を含む酵素を放出するように溶解することができる。
【0040】
大腸菌(Escherichia coli)が宿主微生物である場合、使用することのできるプラスミドは、特にプラスミドpBluescript、pUC18等である。
【0041】
例としては、使用することのできる宿主細胞は、大腸菌W3110、大腸菌C600、大腸菌JM109、大腸菌JM101、大腸菌DH5α等を含む。
【0042】
好ましくは、本発明によるPQQ s−GDH突然変異体は大腸菌JM101株において生成される。それらをコードする核酸分子は、アシネトバクター・カルコアセチカスのPQQ s−GDH遺伝子を修飾し、ベクターpUC18にクローニングすることによって得られる(Cleton-Jansen et al., Mol. Gen. Genet. 217 (1989) 430-436)。このように生成された突然変異体は、PQQ s−GDHのシグナル配列のために細菌のペリプラズムに輸送される。次いで、この突然変異体を超音波処理によって細菌を破壊した後に精製する。
【0043】
本発明は、溶液中のグルコースを定量するため、すなわちサンプル、特に生体サンプル、特に血液中のグルコース濃度を測定するための本発明によるPQQ s−GDH突然変異体の使用にも関する。
【0044】
所与の生体サンプルにおける溶液中のグルコースの定量は、レドックス試薬及び本発明によるPQQ s−GDH突然変異体を該サンプルに導入した後、得られる着色の強さをグルコース含量が既知の標準溶液と比較することによって行うことができる。
【0045】
本発明は、本発明によるPQQ s−GDH突然変異体を含むことを特徴とする、溶液中のグルコースを定量するためのキットにも関する。
【0046】
上記アッセイキットは通常、グルコースアッセイを行うために必要とされる試薬、特に緩衝液(任意の緩衝液を本発明によるキットに使用することができ、リン酸緩衝液及び酢酸緩衝液、トリスヒドロキシメチルアミノメタン(TRIS)を含む緩衝液、N−モルホリノ−3−プロパンスルホン酸(MPOS)を含む緩衝液、4−(2−ヒドロキシエチル)−1−ピペラジンエタンスルホン酸(HEPES)を含む緩衝液、TRIS−酢酸緩衝液等の緩衝液の混合物を含む緩衝液が挙げられるが、これらに限定されない)、レドックス試薬(PQQ s−GDH突然変異体を酸化する任意の試薬であり得るが、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム、フェロセン、並びにフェロセンメタノール、フェロセンカルボン酸等のフェロセンから誘導される錯体、並びにオスミウム及びルテニウムの錯体を含む群から選択することができる)、検量線を作成するための標準グルコース溶液、及びアッセイを行うために必要とされる使用説明書を更に含む。
【0047】
本発明は、導電性金属、特に白金、銅、銀、アルミニウム、金若しくは鋼、又はガラス状炭素、カーボンファイバー、カーボンナノチューブファイバー若しくはダイヤモンド等の炭素といった導電材料を含むグルコース電極に更に関する。該導電材料は、少なくとも1つの本発明によるPQQ s−GDH突然変異体を含む被着物で被覆されている。該被着物は、導電材料の導電性を改善するためのレドックスポリマーを更に含んでいてもよい。
【0048】
レドックスポリマーは、フェロセン、オスミウム及びルテニウムをベースとしたポリマー、並びにポリピロール及びポリアニリン等の導電性ポリマーから選択される。
【0049】
PQQ s−GDH突然変異体を上記導電材料上に固定化する方法は、特にポリマーマトリクスへのPQQ s−GDH突然変異体の封入、ポリマー膜の表面上へのPQQ s−GDH突然変異体の吸着、共有結合による固定、又は電着を含む、当業者が利用可能な従来の方法から選択することができる(Gao et al., Chem. Int. ED. 2002, 41, No. 5, 810-813)。
【0050】
固定化されたPQQ s−GDH突然変異体は、補因子PQQと会合したアポ酵素からなるのが好ましいが、アポ酵素を単独で固定化し、補因子PQQを、例えば反応混合物において溶液中に更に供給することも可能である。
【0051】
かかる電極は、グルコースベースのバイオ燃料電池又はグルコースバイオセンサー等の生体電気システムにおいて有利に使用される。
【0052】
本発明はしたがって、本発明による電極を含むグルコースバイオセンサーにも関する。
【0053】
グルコースバイオセンサーは、生物学的標的を認識することが可能なバイオレセプターが固定化された電極からなる。バイオレセプターへの生物学的標的の結合によって、膜の物理化学的変化、及び電極に取り付けられた電気化学的変換器(電流測定変換器、電位差測定変換器、電気伝導測定変換器等)による電気信号の生成がもたらされる。本発明の場合、バイオレセプターは本発明によるPQQ s−GDH突然変異体であり、生物学的標的はその基質(グルコース)である。
【0054】
1つの実施の形態によると、PQQ s−GDH突然変異体が固定化された電極はまた、電極からの該突然変異体の脱離を防止する膜で被覆されている。該膜はナフィオン、セルロース、又は任意の生体適合性材料、すなわち生理環境と適合する材料からなるものであり得る。
【0055】
本発明の1つの実施の形態によると、グルコースバイオセンサーは皮下に埋め込まれ、血中グルコース濃度を記録することを可能にする。
【0056】
本発明は、グルコースをエネルギー源として用い、アノードである本発明による第一電極と、カソードである第二電極とを備えるバイオ燃料電池にも関する。カソードは例えば、酸素を還元することを可能にし、銅をベースとする酵素群(マルチ銅オキシダーゼ)、特にビリルビンオキシダーゼ及びラッカーゼから選択される酵素を担持する酵素電極であり得る。カソードは例えば白金、金、又は白金合金若しくは金合金の金属電極であってもよい。
【0057】
本発明は、サンプルにおけるグルコースの溶液アッセイの方法であって、
a)その還元が色の変化をもたらすレドックス試薬、及び本発明によるPQQ s−GDH突然変異体を上記サンプル中に導入する工程、
b)酵素反応後にサンプルの着色の強さを測定する工程、
c)工程b)において測定された着色の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)上記サンプル中のグルコース濃度を決定する工程
を含む、サンプルにおけるグルコースの溶液アッセイの方法に更に関する。
【0058】
その還元が色の変化をもたらすレドックス試薬は、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム及びフェロセンから選択される。
【0059】
本発明は、サンプルのグルコースアッセイの方法であって、
a)本発明によるグルコース電極を上記サンプル中に導入する工程、
b)サンプル中の電流の強さを測定する工程、
c)工程b)において測定された電流の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)上記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルのグルコースアッセイの方法にも関する。
【0060】
本発明は、上記に提示された構成に加えて、他の構成を更に含むが、これらは本発明の適用例及び添付の図面を参照して、下記に与えられる説明から明らかとなる。
【図面の簡単な説明】
【0061】
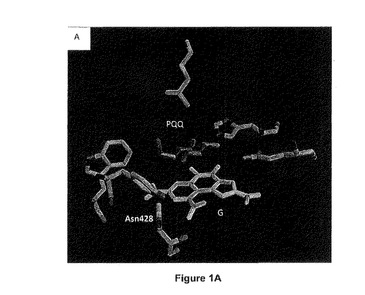
【図1A】野生型のアシネトバクター・カルコアセチカスのPQQ s−GDHの活性部位を示す概略図である。この図は非特許文献1に記載の座標pdb 1cq1に従って、PyMolソフトウェア(DeLano Scientific LLCにより公開、2006年版)を用いて作成した。「G」はグルコースを表し、「PQQ」は補酵素PQQを表す。活性部位のアミノ酸残基のナンバリングは配列番号2に対応する。
【図1B】Asn428Cysの位置で突然変異させたアシネトバクター・カルコアセチカスのPQQ s−GDHの活性部位を示す概略図である。この図は非特許文献1に記載の座標pdb 1cq1に従って、PyMolソフトウェア(DeLano Scientific LLCにより公開、2006年版)を用いて作成した。「G」はグルコースを表し、「PQQ」は補酵素PQQを表す。活性部位のアミノ酸残基のナンバリングは配列番号2に対応する。
【図2A】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2B】428位でチロシンに突然変異させた(N428Y)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
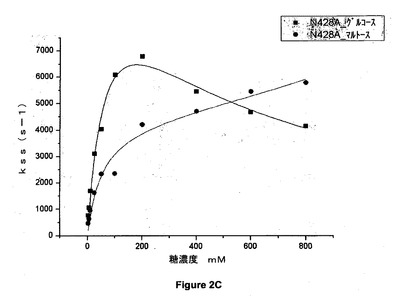
【図2C】428位でアラニンに突然変異させた(N428A)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
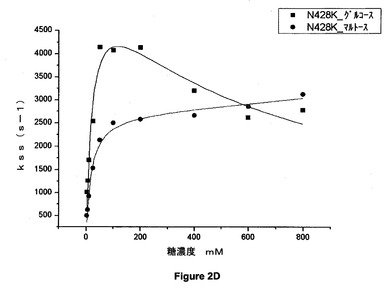
【図2D】428位でリシンに突然変異させた(N428K)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
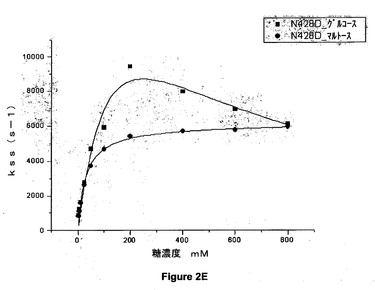
【図2E】428位でアスパラギン酸に突然変異させた(N428D)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
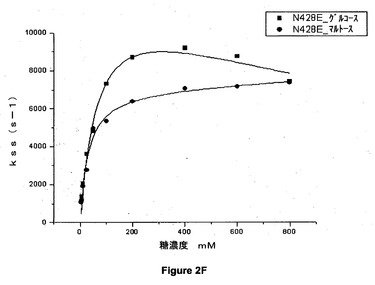
【図2F】428位でグルタミン酸に突然変異させた(N428E)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
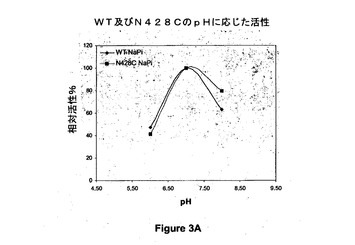
【図3A】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示す図である。
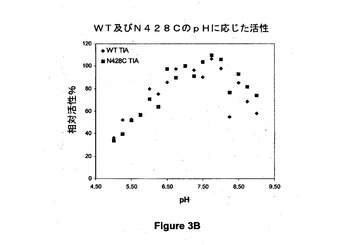
【図3B】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示す図である。
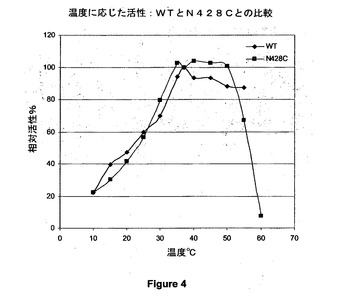
【図4】野生型(WT)PQQ s−GDH及び428位でシステインに突然変異させたPQQ s−GDH(N428C)の温度に応じた活性曲線を示す図である。
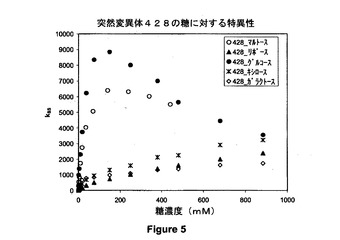
【図5】428位でシステインに突然変異させたPQQ s−GDH(N428C)の異なる基質に応じた活性を示すグラフである。
【実施例】
【0062】
1.材料
1.1.大腸菌の菌株
DH5α:supE44、ΔlacU169、(Φ80 lacZDM15)、hsdR17、recA1、endA1、gyrA96、thi−1、relA1(Hanahan, 1983)。この株はプラスミド作製及び定方向突然変異誘発に使用される。
【0063】
JM101:F’traD36 proA+B+ lacIq Δ(lacZ)M15/Δ(lac−proAB) glnV thi。この株は、三角フラスコ内でのアシネトバクター・カルコアセチカスのPQQグルコースデヒドロゲナーゼの生成に使用される。この株は、lacプロモーターの発現を阻害するプラスミドF−pro−lacIを保有する。
【0064】
1.2.プラスミド
pgp492:Nora Goosenから提供、アシネトバクター・カルコアセチカスの可溶性PQQグルコースデヒドロゲナーゼ遺伝子のコード配列をクローニングすることによって得られた組み換えプラスミド(Cleton-Jansen et al., Mol. Gen. Genet. 217 (1989) 430-436)。
【0065】
1.3.培養培地
LB富栄養培地:トリプトン10g/L;酵母エキス5g/L;NaCl 5g/L;蒸留水適量(全量1L);pH無調整、1バールで50分間オートクレーブ。
【0066】
2.遺伝子工学技法
2.1.スーパーコンピテント細菌の形質転換
スーパーコンピテント細菌DH5αは、Inoueの方法によって作製する(Sambrook and Russell (2001). Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press)。
【0067】
2.2.DNAの調製
プラスミドDNA精製キット(Quiagen)を少量及び大量のDNA調製に使用する。
【0068】
2.3.二本鎖DNAのシークエンシング
二本鎖DNAを、シークエンシングキットBigDyeターミネーターv1.1又はv3.1を用いてシークエンシングする。試薬は異なる蛍光マーカーを有する4つのddNTP(BigDyeターミネーター)、AmpliTaq DNAポリメラーゼ、及び反応に必要とされる他の全ての成分を含有する。伸長産物をシークエンサーABI 3130xlにかける前に精製して、取り込まれなかったマーカー、塩及び他の汚染物質を除去しなくてはならない。
【0069】
2.4.PCR−定方向突然変異誘発
突然変異誘発プロトコルでは、プラスミドの二本鎖の複製に関してポリメラーゼTaqよりも6倍信頼性の高いパイロコッカス・フリオサス(Pyrococcus furiosus)のDNAポリメラーゼPfuを用いてPCRを行う。表Iに列挙したオリゴデオキシリボヌクレオチド(各々がプラスミドの鎖に相補的である)は、DNAポリメラーゼPfuによるDNA合成のプライマーとして働き、非再結合(non-religated)末端で突然変異したプラスミドをもたらす。
【0070】
次いで、親DNAに特異的なエンドヌクレアーゼDpn I(標的配列5’−Gm6ATC−3’)で増幅産物を処理し、それにより突然変異を含有する新合成(neosynthesized)DNAを選択することを可能にする。大半の大腸菌株から単離されるDNAは「DAMメチル化(methylated DAM)」されており(DAM:DNAアデニンメチラーゼ;GATC配列のアデニン上のメチルの存在を示唆する)、したがってDpn Iによって消化することができる。消化混合物の一部を使用して、突然変異プラスミドの末端と再結合するスーパーコンピテントDH5α細菌を形質転換する。
【0071】
3.アシネトバクター・カルコアセチカスのPQQ s−GDHの生成、精製及び特徴決定
3.1.野生型PQQ s−GDH及び突然変異PQQ s−GDHの生成
アポ酵素s−GDHを大腸菌JM101株において、野生型s−GDH又は突然変異s−GDHをコードする配列を有する組み換えプラスミドpgp492によって生成させる。アンピシリン(200mg/L)を添加したLB培地(LBA)50mLの前培養物に、アンピシリン(100mg/mL)を添加したLB寒天皿上で単離されたクローンを播種し、37℃で一晩220rpmで攪拌する。培養物をLBA培地に100倍で播種する。これを37℃で攪拌しながら(220rpm)、OD600nmが0.4OD600nm/mL〜1OD600nm/mLとなるまでインキュベートする。次いで、培養物を400μMのIPTGによって誘導した後、25℃で20時間攪拌する(180rpm〜220rpm)。遠心分離(5285g、4℃)によって回収した細胞を、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)で洗浄し、−20℃又は添加剤を加えずに−80℃で保管する。
【0072】
3.2.野生型PQQ s−GDH酵素及び突然変異PQQ s−GDH酵素の精製
3.2.1.細胞破壊及びDNase Iでの処理
1リットルの培養物から得られた細胞ペレットを、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)20mL中に溶解させ、40Wの超音波処理出力で超音波1秒、停止1秒のサイクルで3分間超音波処理する。得られたサンプル(粗抽出物と呼ぶ)に最後に2mM MgCl2を添加し、室温で30分間DNase I(粗抽出物1mL当たり1U)で処理する。次いで、不溶性細胞残屑を20000gで45分間の遠心分離によって粗抽出物から除去する。
【0073】
3.2.2.陽イオン交換クロマトグラフィー
カットオフ値(cutoff threshold)が0.22μmのフィルター(Millex−GS 0.22μm、Millipore)で濾過して、OD280nmが10となるように希釈した超音波処理上清を、AKTA purifierシステム(GE Healthcare(登録商標))に接続し、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)で平衡化した陽イオン交換カラムsource 30S(GE Healthcare(登録商標))に投入する。溶出は同緩衝液中0Mから1MまでというNaCl勾配で、5mL/分の流速で行う。s−GDHタンパク質を含有する画分を合わせ、Amicon YM10膜上での遠心分離によって濾過することで濃縮する。この段階でs−GDHタンパク質は純粋であり、硫酸アンモニウム(90%飽和)の存在下、−20℃で沈殿形態で保管することができる。
【0074】
3.2.3.野生型s−GDH酵素及び突然変異s−GDH酵素の特徴決定
3.2.3.1.分子量の決定
精製後のs−GDH単量体の分子量を、MALDI質量分析によって決定する。タンパク質をPD10カラム(GE Healthcare)上で脱塩し、重炭酸アンモニウム50mMの緩衝液(pH7.5)で溶出する。次いで、凍結乾燥したサンプルをMALDI−TOFによって分析すると、分子量が50234.83Daであることが確認される。この分子量はシグナルペプチドを有しないPQQ s−GDHの理論分子量に一致する。
【0075】
3.2.3.2.補因子PQQによるs−GDHの再構成
精製(Tris 20mM、CaCl2 3mMの緩衝液(pH7.5))の終了時に、酵素溶液をPQQと共に室温で15分間プレインキュベートする。添加したPQQは、酵素のモル濃度(molarity of the concentration)の2倍に等しい最終濃度に相当する。次いで、Pipes 20mM、CaCl2 3mMの緩衝液(pH7)で平衡化したPD10カラム(GE Healthcare)上で脱塩することによって過剰のPQQを除去する。
【0076】
3.2.3.3.濃度の測定
溶液中の酵素濃度を、補因子PQQを有する又は有しない酵素(二量体又は単量体;吸光係数はそれぞれ1.28l・g−1・cm−1又は1.67l・g−1・cm−1である)を用いて280nmで測定したODから算出する(Olsthoorn et al., 1997)。
【0077】
3.2.3.4.酵素試験
酵素試験は、リン酸ナトリウム20mMの緩衝液(pH7)中、37℃、3mLの容量でVarianの分光光度計を用いて、電子受容体として働くPMSを用いたDCIPの還元による消失を600nmで時間に応じてモニタリングすることで行う。酵素の特異的活性は、タンパク質1mg当たり1分間に消失したDCIPのμmol量で表される。PMS及びDCIPの濃度はそれぞれ0.6mM及び0.06mMである。酵素を−0.05OD600nm/分と−0.2OD600nm/分との間の傾きが測定されるように希釈する。
【0078】
4.野生型PQQ s−GDH及び突然変異PQQ s−GDHの酵素特性を研究する技法
4.1.定常状態での速度定数(kcat)及びミカエリス定数(KM)の決定
実験はリン酸ナトリウム20mMの緩衝液(pH7)中37℃で、Varianの分光光度計上で行う。この試験における基質(グルコース及びマルトース)の濃度は、0mM〜800mMの間で変化させる。
【0079】
PMSを第1の電子受容体として使用し、酵素活性を第2の電子受容体として使用されるDCIPの還元によって600nmでモニタリングする。PMS及びDCIPの初期濃度はそれぞれ0.6mM及び0.06mMである。酵素を添加することによって試験を開始する。実験点は、以下の等式によるSigma−plot 6.0ソフトウェアを用いた等式1、等式2又は等式3による非線形回帰によって、分析する:
(1)ミカエリス−メンテンモデル
kss=kcat×[S]/(KM+[S])
(2)二重直角双曲線関数(Double hyperbole)
kss=kcat1×[S]/(KM1+[S])+kcat2×[S]/KM2+[S])
(3)競合的阻害によるミカエリス−メンテンモデル
v/vmax=[S]/KS×(1+[S]/KS+[I]/KI)
【0080】
結果
図2A、図2B、図2C、図2D、図2E及び図2Fのグラフは、野生型(WT)及び428位で突然変異させた(N428C、N428Y、N428A、N428K、N428D及びN428E)アシネトバクター・カルコアセチカスの可溶性PQQグルコースデヒドロゲナーゼのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示す。
【0081】
これらの結果から、マルトース及びグルコースという2つの基質について、本発明による突然変異酵素が野生型酵素の活性よりも高い活性を示すことが示される。これらの突然変異体の一部は野生型酵素の2倍の活性を有している。
【0082】
4.2.pHに応じた調査
4.2.1.pHに応じた活性
pHに応じた反応速度定数の変化の調査を、野生型酵素及び突然変異酵素をTris 120mM、イミダゾール30mM、酢酸30mMから構成される混合緩衝液(TIA)(NaClを用いてイオン強度を190mMに調整する)、又はpH範囲が6〜8のリン酸ナトリウム20mMの緩衝液(NaPi)中でインキュベートすることによって、5〜9のpH範囲にわたって行う。Varianの分光光度計を用いて37℃で実験を行う。PMSを第1の電子受容体として0.6mMの濃度で使用する。活性を、第2の電子受容体として0.06mMの濃度で使用されるDCIPの消失によって600nmでモニタリングする。酵素を添加することによって試験を開始する。野生型酵素又は突然変異酵素の最適活性は100%に対応し、各pHに対する活性を示す。
【0083】
結果
図3A及び図3Bは、野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示すグラフである。これらの結果から、PQQ s−GDHのシステイン突然変異体(N428C)が野生型酵素の活性に匹敵する相対活性を示すことが示され、この突然変異がpHに関わらず相対活性の喪失をもたらさないことが実証される。
【0084】
4.2.2.pHに応じた安定性
野生型PQQ s−GDH又は突然変異PQQ s−GDHのpHに応じた安定性を、精製した酵素を均質になるまでpH範囲5〜9の混合緩衝液で希釈することによって決定する。この混合緩衝液はTris 120mM、イミダゾール30mM、酢酸30mMから構成され、そのイオン強度をNaClを用いて190mMに調整する。希釈した酵素溶液(1μg/ml〜6μg/ml)を37℃でプレインキュベートする。時間に応じて様々なサンプルをとる。残存活性をリン酸ナトリウム20mMの緩衝液(pH7)中、0.06mM DCIP、0.6mM PMSの存在下でVarianの分光光度計を用いて37℃で測定する。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。
【0085】
結果
本発明による突然変異酵素は、その安定性が試験したpH範囲にわたって一定であるか否かを問わないが、pHに関わらず野生型酵素よりも良好な安定性を示す。特に、突然変異酵素はpHが7を超えても安定なままである。
【0086】
4.3.温度に応じた調査
4.3.1.温度に応じた活性
反応速度定数の変化を、リン酸ナトリウム20mMの緩衝液(pH7)中、0.06mMのDCIP及び0.6mMのPMSの存在下でpHに応じて調査する。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。温度は10℃〜60℃で変化させる。温度制御したVarianの分光光度計CARY UV Biomelt上で活性をモニタリングする。酵素を添加することによって試験を開始する。
【0087】
結果
図4は、野生型(WT)及び突然変異させた(N428)PQQ s−GDHの温度に応じた活性曲線を示す。
【0088】
この曲線から、30℃〜50℃で突然変異酵素が野生型酵素よりも良好な活性を示すことが示される。
【0089】
4.3.2.温度に応じた酵素の安定性
この研究は、リン酸ナトリウム20mMの緩衝液(pH7)中、0.06mMのDCIP及び0.6mMのPMSの存在下で行う。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。温度は10℃〜60℃で変化させる。温度制御したVarianの分光光度計CARY UV Biomelt上で37℃で活性をモニタリングする。酵素を添加することによって試験を開始する。
【0090】
結果
200分未満のインキュベーション時間では、突然変異酵素N428Cが40℃及び50℃で野生型酵素よりも良好な活性を示すことが分かり、したがって血中グルコースアッセイキットについてのその利点が実証される。
【0091】
4.4.基質に応じた調査
システイン突然変異体(N428C)の特異性を様々な基質に関して評価した。
【0092】
この特異性の測定に採用したプロトコルは第4.1.項に記載しているものと同一であるが、異なる基質を用いる。
【0093】
得られた結果を下記表II及び図5に提示する。
【0094】
【表2】
【0095】
結果
突然変異体N428Cのkssの生値は大半の基質について野生型酵素の値よりも高いが、相対活性(或る基質に対する酵素の特異性を別の基質と比較して特徴付ける)から、突然変異体N428Cがグルコースに対してより特異的であることが示される。したがって、4mMの基質では、マルトースを基質とすると、野生型酵素についてはグルコースと比べた相対活性は81%であることが測定されるのに対して、突然変異体N428Cの示すグルコースと比べたマルトースの相対活性は61%である。
【0096】
この同じプロトコルをアスパラギン酸(N428D)突然変異体、グルタミン酸(N428E)突然変異体、アラニン(N428A)突然変異体、チロシン(N428Y)突然変異体及びリシン(N428K)突然変異体のグルコース及びマルトースに対する特異性を評価するために採用した。
【0097】
結果を下記表IIIに提示する。
【0098】
【表3】
【技術分野】
【0001】
本発明は、グルコースアッセイ、特に糖尿病の被験体の血中グルコースアッセイにおいて関心がもたれるグルコース電極の開発の分野、及びグルコースを燃料として使用するバイオ燃料電池を応用するための分野に関する。
【0002】
本発明は、より具体的には、野生型酵素に比べて有利な特性を有する酵素ピロロキノリンキノン可溶性グルコースデヒドロゲナーゼ(PQQ s−GDHとも呼ばれる)の突然変異体に関する。
【背景技術】
【0003】
2型糖尿病はフランスにおいて200万人近くの人々に影響を与えているが、この数に疾患に気付いていない600000人を加える必要がある。米国においては状況は更に深刻である。先進国では、糖尿病は20歳〜65歳という年齢層での失明の主要な原因である。
【0004】
この疾患のモニタリング及び管理は、とりわけ血中グルコースの毎日の定量及びインシュリンの注射に基づく。様々な会社によって、患者が血糖値を自分で測定することを可能にするグルコースセンサーが提供されている。これらのセンサーは電流測定センサー、電位差測定センサー又は電量測定センサーであり得るが、全てグルコースを酸化することが可能な酵素の使用に基づいている。2つの主要な酵素はグルコースオキシダーゼ及びPQQ s−GDHである。
【0005】
ピロロキノリンキノングルコースデヒドロゲナーゼは、アシネトバクター・カルコアセチカス(Acinetobacter calcoaceticus)において同定されたキノプロテインファミリーに属し、2つのタイプがある(一方は膜酵素であり、他方は可溶性酵素である)。これらの酵素はD−グルコノラクトンへのグルコースの酸化を触媒し、電子供給体として使用することができる。
【0006】
アシネトバクター・カルコアセチカスのPQQ s−GDHは、約50kDaの2つのサブユニット(アポ酵素)とピロロキノリンキノン補因子(PQQ)とからなるホモ二量体から構成される(非特許文献1)。この酵素は活性部位を有し、PQQの存在下ではその内部でグルコースの酸化が触媒される。この活性部位は76位、144位、169位、343位、346位及び428位に位置するアミノ酸から構成される。
【0007】
PQQ s−GDHは容易に大量に得ることができるため、産業的にかなりの関心がもたれている。このため、PQQ s−GDHは血糖値の自己モニタリングのためのグルコースセンサーに使用される主要な酵素となっている。キノプロテインの利点は、電子受容体としてO2を利用するグルコースオキシダーゼとは対照的に、酸素に依存しないことである。
【0008】
その物理化学的特性に基づくと、PQQ s−GDHをバイオ燃料電池(その原理は電流を発生するグルコースの酸化に基づく)の製造に使用することも想定される。
【0009】
PQQ s−GDHの欠点は、その熱安定性の低さ、生理的pHでの安定性の低さ及び活性の穏やかさである。
【0010】
幾つかの著作物において、その特性を改善するためにPQQ s−GDHの変異体を開発することが試みられている。
【0011】
「突然変異体」又は「変異体」は、そのタンパク質配列が、野生型PQQ s−GDHのタンパク質配列と比べて少なくとも1つのアミノ酸の挿入、欠失及び/又は置換を含むPQQ s−GDHを意味する。以下、PQQ s−GDHの参照ヌクレオチド配列及び参照タンパク質配列は、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列とする(それぞれ配列番号1及び配列番号2)。
【0012】
PQQ s−GDHの安定性を改善することを目的とする突然変異
特許文献1は、2つのサブユニットがジスルフィド架橋によって結合するように突然変異させたPQQ s−GDHを記載している。この突然変異は、415位及び414位の少なくとも1つのアミノ酸、並びに/又は同時に340位及び418位に位置する2つのアミノ酸をシステイン残基で置き換えることからなる。これらの修飾は、より良好な熱安定性を酵素に与えている。
【0013】
PQQ s−GDHをその基質による阻害に対する感受性が低下したものにすることを目的とする突然変異
特許文献2は、領域349〜377内の少なくとも1つのアミノ酸を異なるアミノ酸で置き換えたPQQ s−GDHを記載している。この修飾は、基質による阻害に対する感受性が低下した酵素をもたらし、したがって高濃度のグルコースの存在下で使用することができる。
【0014】
PQQ s−GDHをその天然基質(グルコース)に対してより特異的なものにすることを目的とする突然変異
野生型PQQ s−GDHを修飾して、より基質特異的なものにすることも可能である。或る基質に対して特異的な酵素は該基質が関与する反応しか触媒しないが、逆に基質特異性の低い酵素は、その天然基質に構造的に類似した基質に基づく反応を触媒することが可能である。PQQ s−GDHの天然基質はグルコースであるが、野生型PQQ s−GDHの特異性は低く、他の単糖類及び二糖類の酸化を触媒することも可能である。
【0015】
幾つかの著作物が、野生型PQQ s−GDHをグルコースに対してより特異的にするために修飾することを提案している:
このため、特許文献3は49位、67位〜69位、76位、89位、129位〜131位、167位〜170位、174位、188位、189位、207位、215位、245位、249位、300位、341位〜343位、349位、351位及び429位に位置するアミノ酸の少なくとも1つを置換すること、並びに/又はアミノ酸を428位と429位との間に導入することによってPQQ s−GDHを修飾することを提案している;
特許文献4は、428位と429位との間にロイシン、リシン又はアラニンを挿入することによってPQQ s−GDHを修飾することを提案している;
特許文献5は、PQQ s−GDHの75位、326位〜354位、278位〜320位及び162位〜197位のうちの1つの位置でアミノ酸を置換することを記載している;
特許文献6は、PQQ s−GDHの428位と429位との間にアミノ酸を挿入し、任意で428位のアミノ酸をロイシン、プロリン又はバリンで置換することを提言している;
特許文献7は、125位、128位、142位、168位、169位、170位、224位、230位、236位、345位、351位、416位又は428位に1つ又は複数の置換を有するアシネトバクター属のPQQ s−GDHの突然変異体を記載している。この文献は更に、多数の突然変異部位を有する具体的な突然変異体を列挙している;
特許文献8は、428位と429位との間にロイシン、アラニン又はリシンの挿入を有するアシネトバクター・バウマンニ(Acinetobacter baumanii)のPQQ s−GDHの突然変異体、又は429位のアミノ酸がフェニルアラニン、プロリン、ロイシン又はチロシンで置換されている突然変異体を挙げている;
特許文献9は、348位のアミノ酸をアラニン、グリシン又はセリンで、428位のアミノ酸をロイシン、プロリン又はバリンで置換することを提案している。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第7,244,600号
【特許文献2】米国特許第7,244,581号
【特許文献3】米国特許出願公開第2007/0105173号
【特許文献4】欧州特許出願公開第1367120号
【特許文献5】米国特許第7,037,698号
【特許文献6】米国特許出願公開第2007/0243566号
【特許文献7】国際公開第2006/085509号
【特許文献8】欧州特許出願公開第1666586号
【特許文献9】米国特許第7,132,270号
【非特許文献】
【0017】
【非特許文献1】Oubrieet al., J. Mol. Biol; 289,319-333 (1999)
【発明の概要】
【発明が解決しようとする課題】
【0018】
グルコースセンサーで使用するためには、より活性の高い、すなわちD−グルコノラクトンへのグルコースのより速い変換反応を可能にするPQQ s−GDH突然変異体を得ることが必要とされるが、これは既存の突然変異体では可能ではない。
【0019】
したがって、十分な又は更には改善された熱安定性及びpH安定性を保持したうえで、野生型PQQ s−GDHよりも良好な活性を示すPQQ s−GDHを開発することが依然として必要とされている。
【課題を解決するための手段】
【0020】
この要求を満たすために、本発明者らは、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの新規の突然変異体を開発した。これらの突然変異体は、428位に位置するアスパラギンがシステイン、チロシン、アラニン、アスパラギン酸又はグルタミン酸で置換されているようなものである。
【0021】
したがって、本発明の第1の目的は、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDH突然変異体であって、428位に位置するそのアミノ酸が、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHのタンパク質配列(配列番号2)と比べてシステイン、チロシン、アラニン、アスパラギン酸又はグルタミン酸を含む群から選択されるアミノ酸で置換されていることを特徴とする、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDH突然変異体に関する。
【0022】
本発明に至った研究において、本発明者らは、PQQ s−GDH突然変異体であって、428位に位置するそのアミノ酸が、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHのタンパク質配列(配列番号2)と比べてリシンで置換されていることを特徴とする、PQQ s−GDH突然変異体も作製した(実施例第4部を参照されたい)。この突然変異体は配列番号10のタンパク質配列を有し、配列番号9の核酸分子によってコードされる。この突然変異体は、下記表Iに提示される配列番号17及び配列番号18のオリゴヌクレオチドを用いて作製することができる。
【0023】
アミノ酸のナンバリングは、アシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列を基準とする。しかしながら、本発明によるPQQ s−GDH突然変異体は、このアシネトバクター・カルコアセチカスの野生型PQQ s−GDHの突然変異体に限定されず、本発明はアシネトバクター・カルコアセチカスの野生型PQQ s−GDHに対して少なくとも90%の同一率を有するPQQ s−GDHの突然変異体にも関する。
【0024】
参照配列であるアシネトバクター・カルコアセチカスの野生型PQQ s−GDHの配列(配列番号2)に対する配列の同一性は、2つの配列をそれらの間で最大の一致が得られるようにアラインメントした場合に同一であるアミノ酸残基のパーセンテージに応じて評価する。
【0025】
当業者は配列を比較するためのコンピュータプログラム、例えばBLAST suiteのプログラム(Altschul et al., NAR, 25, 3389-3402)を用いて同一率を算出することができる。BLASTプログラムは、参照配列として示した配列番号2全体からなる比較ウィンドウ上で採用される。
【0026】
参照配列と少なくともX%の同一性を有するアミノ酸配列を有するペプチドは、本発明においては、該参照ペプチドの機能的特性(本発明の場合、そのグルコース酸化の酵素活性)を保持したまま、参照配列の100個のアミノ酸に対して最大で100−X個の変化をその配列中に含むことのできるペプチドとして定義される。本発明の意味において、変化という用語は、参照配列におけるアミノ酸の欠失、置換、又は連続的若しくは分散した挿入を含む。
【0027】
アシネトバクター・カルコアセチカスのPQQ s−GDHの428位に位置するアミノ酸に対応するアミノ酸は、該相同酵素とアシネトバクター・カルコアセチカスのPQQ s−GDHとの配列アラインメントによって同定される。
【0028】
本発明は特に、配列番号4、配列番号6、配列番号8、配列番号12及び配列番号14(それぞれPQQ s−GDHのシステイン突然変異体、チロシン突然変異体、アラニン突然変異体、アスパラギン酸突然変異体及びグルタミン酸突然変異体のアミノ酸配列に対応する)から選択されるアミノ酸配列を有するPQQ s−GDH突然変異体に関する。これらの突然変異酵素は、好適なオリゴヌクレオチド対によるアシネトバクター・カルコアセチカスのPQQ s−GDH遺伝子の突然変異によって得られるヌクレオチド断片によってコードされる。
【0029】
これらの本発明による新規のPQQ s−GDH突然変異体は、市販のグルコースセンサーに使用される酵素であるアシネトバクター・カルコアセチカスの野生型酵素に比べて以下の改善された性能を示す:
これらの活性は、とりわけ生理的濃度、すなわち1mM〜10mMのグルコースで野生型酵素の活性よりも高い。酵素の活性は、PQQ s−GDHによるグルコースのグルコノラクトンへの酸化反応中に起こるレドックス試薬の着色をモニタリングすることによって定量化することができる。レドックス試薬は例えば、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム及びフェロセンである;
これらは高濃度でのグルコースの阻害効果に対する感受性が低い。
【0030】
PQQ s−GDH突然変異体の基質に対する特異性は、基質であるグルコース及び幾つかの他の糖類(二糖類等)に対する該突然変異体の酵素活性を比較することによって評価することができる。
【0031】
本発明によるPQQ s−GDH突然変異体の有利な特性によって、それらはグルコースをエネルギー源として用いるバイオ燃料電池等の生体電気システム及びグルコースバイオセンサーに特に好適に使用されるものとなる。
【0032】
本発明は、野生型PQQ s−GDH(例えばアシネトバクター・カルコアセチカスの野生型PQQ s−GDH)を、表Iに示されるオリゴヌクレオチド対からなる群から選択されるオリゴヌクレオチド対で修飾することによって得られる、本発明によるPQQ s−GDH突然変異体をコードする核酸分子にも関する。
【0033】
【表1】
【0034】
本発明によるPQQ s−GDH突然変異体をコードする核酸分子は特に、アシネトバクター・カルコアセチカスによって生成される配列番号1の配列の野生型酵素をコードする遺伝子のヌクレオチド配列を修飾することによって作製することができる。遺伝子の配列を修飾する幾つかの技法が当業者に既知である(Igarashi et al., Archives of Biochemistry and Biophysics 428 (2004) 52-63による概説を参照されたい)。特定の作製方法によると、本発明によるPQQ s−GDH突然変異体をコードする核酸分子は、導入すべき突然変異を有するオリゴヌクレオチドの存在下でのPCRによる突然変異誘発によって作製される(下記の実施例を参照されたい)。
【0035】
特定の実施の形態によると、本発明は、その配列が配列番号3、配列番号5、配列番号7、配列番号11及び配列番号13の配列からなる群から選択される、本発明によるPQQ s−GDH突然変異体をコードする核酸分子に関する。本発明によるPQQ s−GDH突然変異体をコードする核酸分子を、次いでプラスミド等の発現ベクターにクローニングした後、細菌、酵母又は細胞培養物等の好適な宿主において形質転換させることができる。
【0036】
発現ベクターとは、その発現に不可欠なシグナル、特にプロモーター(構成的又は誘導性)、リボソーム結合部位、転写終結シグナル、及び任意で抗生物質抵抗性遺伝子等の選択マーカーの間にヌクレオチドコード配列を挿入することを可能にする領域を有するベクターを意味する。
【0037】
本発明は、上記核酸分子を含む発現ベクター、及び該発現ベクターで形質転換され、本発明によるPQQ s−GDH突然変異体を発現する宿主細胞に更に関する。
【0038】
発現ベクターは当業者に既知の任意の方法、特に宿主細胞の膜透過性を例えばカルシウムイオンの存在下で変化させること、又はエレクトロポレーションによって宿主細胞内に導入することができる。
【0039】
本発明によるPQQ s−GDH突然変異体を発現するように形質転換した宿主細胞の培養の後、該細胞を遠心分離によって回収し、本発明による該PQQ s−GDH突然変異体を含む酵素を放出するように溶解することができる。
【0040】
大腸菌(Escherichia coli)が宿主微生物である場合、使用することのできるプラスミドは、特にプラスミドpBluescript、pUC18等である。
【0041】
例としては、使用することのできる宿主細胞は、大腸菌W3110、大腸菌C600、大腸菌JM109、大腸菌JM101、大腸菌DH5α等を含む。
【0042】
好ましくは、本発明によるPQQ s−GDH突然変異体は大腸菌JM101株において生成される。それらをコードする核酸分子は、アシネトバクター・カルコアセチカスのPQQ s−GDH遺伝子を修飾し、ベクターpUC18にクローニングすることによって得られる(Cleton-Jansen et al., Mol. Gen. Genet. 217 (1989) 430-436)。このように生成された突然変異体は、PQQ s−GDHのシグナル配列のために細菌のペリプラズムに輸送される。次いで、この突然変異体を超音波処理によって細菌を破壊した後に精製する。
【0043】
本発明は、溶液中のグルコースを定量するため、すなわちサンプル、特に生体サンプル、特に血液中のグルコース濃度を測定するための本発明によるPQQ s−GDH突然変異体の使用にも関する。
【0044】
所与の生体サンプルにおける溶液中のグルコースの定量は、レドックス試薬及び本発明によるPQQ s−GDH突然変異体を該サンプルに導入した後、得られる着色の強さをグルコース含量が既知の標準溶液と比較することによって行うことができる。
【0045】
本発明は、本発明によるPQQ s−GDH突然変異体を含むことを特徴とする、溶液中のグルコースを定量するためのキットにも関する。
【0046】
上記アッセイキットは通常、グルコースアッセイを行うために必要とされる試薬、特に緩衝液(任意の緩衝液を本発明によるキットに使用することができ、リン酸緩衝液及び酢酸緩衝液、トリスヒドロキシメチルアミノメタン(TRIS)を含む緩衝液、N−モルホリノ−3−プロパンスルホン酸(MPOS)を含む緩衝液、4−(2−ヒドロキシエチル)−1−ピペラジンエタンスルホン酸(HEPES)を含む緩衝液、TRIS−酢酸緩衝液等の緩衝液の混合物を含む緩衝液が挙げられるが、これらに限定されない)、レドックス試薬(PQQ s−GDH突然変異体を酸化する任意の試薬であり得るが、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム、フェロセン、並びにフェロセンメタノール、フェロセンカルボン酸等のフェロセンから誘導される錯体、並びにオスミウム及びルテニウムの錯体を含む群から選択することができる)、検量線を作成するための標準グルコース溶液、及びアッセイを行うために必要とされる使用説明書を更に含む。
【0047】
本発明は、導電性金属、特に白金、銅、銀、アルミニウム、金若しくは鋼、又はガラス状炭素、カーボンファイバー、カーボンナノチューブファイバー若しくはダイヤモンド等の炭素といった導電材料を含むグルコース電極に更に関する。該導電材料は、少なくとも1つの本発明によるPQQ s−GDH突然変異体を含む被着物で被覆されている。該被着物は、導電材料の導電性を改善するためのレドックスポリマーを更に含んでいてもよい。
【0048】
レドックスポリマーは、フェロセン、オスミウム及びルテニウムをベースとしたポリマー、並びにポリピロール及びポリアニリン等の導電性ポリマーから選択される。
【0049】
PQQ s−GDH突然変異体を上記導電材料上に固定化する方法は、特にポリマーマトリクスへのPQQ s−GDH突然変異体の封入、ポリマー膜の表面上へのPQQ s−GDH突然変異体の吸着、共有結合による固定、又は電着を含む、当業者が利用可能な従来の方法から選択することができる(Gao et al., Chem. Int. ED. 2002, 41, No. 5, 810-813)。
【0050】
固定化されたPQQ s−GDH突然変異体は、補因子PQQと会合したアポ酵素からなるのが好ましいが、アポ酵素を単独で固定化し、補因子PQQを、例えば反応混合物において溶液中に更に供給することも可能である。
【0051】
かかる電極は、グルコースベースのバイオ燃料電池又はグルコースバイオセンサー等の生体電気システムにおいて有利に使用される。
【0052】
本発明はしたがって、本発明による電極を含むグルコースバイオセンサーにも関する。
【0053】
グルコースバイオセンサーは、生物学的標的を認識することが可能なバイオレセプターが固定化された電極からなる。バイオレセプターへの生物学的標的の結合によって、膜の物理化学的変化、及び電極に取り付けられた電気化学的変換器(電流測定変換器、電位差測定変換器、電気伝導測定変換器等)による電気信号の生成がもたらされる。本発明の場合、バイオレセプターは本発明によるPQQ s−GDH突然変異体であり、生物学的標的はその基質(グルコース)である。
【0054】
1つの実施の形態によると、PQQ s−GDH突然変異体が固定化された電極はまた、電極からの該突然変異体の脱離を防止する膜で被覆されている。該膜はナフィオン、セルロース、又は任意の生体適合性材料、すなわち生理環境と適合する材料からなるものであり得る。
【0055】
本発明の1つの実施の形態によると、グルコースバイオセンサーは皮下に埋め込まれ、血中グルコース濃度を記録することを可能にする。
【0056】
本発明は、グルコースをエネルギー源として用い、アノードである本発明による第一電極と、カソードである第二電極とを備えるバイオ燃料電池にも関する。カソードは例えば、酸素を還元することを可能にし、銅をベースとする酵素群(マルチ銅オキシダーゼ)、特にビリルビンオキシダーゼ及びラッカーゼから選択される酵素を担持する酵素電極であり得る。カソードは例えば白金、金、又は白金合金若しくは金合金の金属電極であってもよい。
【0057】
本発明は、サンプルにおけるグルコースの溶液アッセイの方法であって、
a)その還元が色の変化をもたらすレドックス試薬、及び本発明によるPQQ s−GDH突然変異体を上記サンプル中に導入する工程、
b)酵素反応後にサンプルの着色の強さを測定する工程、
c)工程b)において測定された着色の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)上記サンプル中のグルコース濃度を決定する工程
を含む、サンプルにおけるグルコースの溶液アッセイの方法に更に関する。
【0058】
その還元が色の変化をもたらすレドックス試薬は、2,6−ジクロロフェノールインドフェノール(DCIP)と組み合わせたフェナジンメトスルフェート(PMS)、フェリシアン化カリウム及びフェロセンから選択される。
【0059】
本発明は、サンプルのグルコースアッセイの方法であって、
a)本発明によるグルコース電極を上記サンプル中に導入する工程、
b)サンプル中の電流の強さを測定する工程、
c)工程b)において測定された電流の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)上記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルのグルコースアッセイの方法にも関する。
【0060】
本発明は、上記に提示された構成に加えて、他の構成を更に含むが、これらは本発明の適用例及び添付の図面を参照して、下記に与えられる説明から明らかとなる。
【図面の簡単な説明】
【0061】
【図1A】野生型のアシネトバクター・カルコアセチカスのPQQ s−GDHの活性部位を示す概略図である。この図は非特許文献1に記載の座標pdb 1cq1に従って、PyMolソフトウェア(DeLano Scientific LLCにより公開、2006年版)を用いて作成した。「G」はグルコースを表し、「PQQ」は補酵素PQQを表す。活性部位のアミノ酸残基のナンバリングは配列番号2に対応する。
【図1B】Asn428Cysの位置で突然変異させたアシネトバクター・カルコアセチカスのPQQ s−GDHの活性部位を示す概略図である。この図は非特許文献1に記載の座標pdb 1cq1に従って、PyMolソフトウェア(DeLano Scientific LLCにより公開、2006年版)を用いて作成した。「G」はグルコースを表し、「PQQ」は補酵素PQQを表す。活性部位のアミノ酸残基のナンバリングは配列番号2に対応する。
【図2A】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2B】428位でチロシンに突然変異させた(N428Y)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2C】428位でアラニンに突然変異させた(N428A)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2D】428位でリシンに突然変異させた(N428K)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2E】428位でアスパラギン酸に突然変異させた(N428D)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図2F】428位でグルタミン酸に突然変異させた(N428E)アシネトバクター・カルコアセチカスのPQQ s−GDHのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示すグラフである。
【図3A】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示す図である。
【図3B】野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示す図である。
【図4】野生型(WT)PQQ s−GDH及び428位でシステインに突然変異させたPQQ s−GDH(N428C)の温度に応じた活性曲線を示す図である。
【図5】428位でシステインに突然変異させたPQQ s−GDH(N428C)の異なる基質に応じた活性を示すグラフである。
【実施例】
【0062】
1.材料
1.1.大腸菌の菌株
DH5α:supE44、ΔlacU169、(Φ80 lacZDM15)、hsdR17、recA1、endA1、gyrA96、thi−1、relA1(Hanahan, 1983)。この株はプラスミド作製及び定方向突然変異誘発に使用される。
【0063】
JM101:F’traD36 proA+B+ lacIq Δ(lacZ)M15/Δ(lac−proAB) glnV thi。この株は、三角フラスコ内でのアシネトバクター・カルコアセチカスのPQQグルコースデヒドロゲナーゼの生成に使用される。この株は、lacプロモーターの発現を阻害するプラスミドF−pro−lacIを保有する。
【0064】
1.2.プラスミド
pgp492:Nora Goosenから提供、アシネトバクター・カルコアセチカスの可溶性PQQグルコースデヒドロゲナーゼ遺伝子のコード配列をクローニングすることによって得られた組み換えプラスミド(Cleton-Jansen et al., Mol. Gen. Genet. 217 (1989) 430-436)。
【0065】
1.3.培養培地
LB富栄養培地:トリプトン10g/L;酵母エキス5g/L;NaCl 5g/L;蒸留水適量(全量1L);pH無調整、1バールで50分間オートクレーブ。
【0066】
2.遺伝子工学技法
2.1.スーパーコンピテント細菌の形質転換
スーパーコンピテント細菌DH5αは、Inoueの方法によって作製する(Sambrook and Russell (2001). Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press)。
【0067】
2.2.DNAの調製
プラスミドDNA精製キット(Quiagen)を少量及び大量のDNA調製に使用する。
【0068】
2.3.二本鎖DNAのシークエンシング
二本鎖DNAを、シークエンシングキットBigDyeターミネーターv1.1又はv3.1を用いてシークエンシングする。試薬は異なる蛍光マーカーを有する4つのddNTP(BigDyeターミネーター)、AmpliTaq DNAポリメラーゼ、及び反応に必要とされる他の全ての成分を含有する。伸長産物をシークエンサーABI 3130xlにかける前に精製して、取り込まれなかったマーカー、塩及び他の汚染物質を除去しなくてはならない。
【0069】
2.4.PCR−定方向突然変異誘発
突然変異誘発プロトコルでは、プラスミドの二本鎖の複製に関してポリメラーゼTaqよりも6倍信頼性の高いパイロコッカス・フリオサス(Pyrococcus furiosus)のDNAポリメラーゼPfuを用いてPCRを行う。表Iに列挙したオリゴデオキシリボヌクレオチド(各々がプラスミドの鎖に相補的である)は、DNAポリメラーゼPfuによるDNA合成のプライマーとして働き、非再結合(non-religated)末端で突然変異したプラスミドをもたらす。
【0070】
次いで、親DNAに特異的なエンドヌクレアーゼDpn I(標的配列5’−Gm6ATC−3’)で増幅産物を処理し、それにより突然変異を含有する新合成(neosynthesized)DNAを選択することを可能にする。大半の大腸菌株から単離されるDNAは「DAMメチル化(methylated DAM)」されており(DAM:DNAアデニンメチラーゼ;GATC配列のアデニン上のメチルの存在を示唆する)、したがってDpn Iによって消化することができる。消化混合物の一部を使用して、突然変異プラスミドの末端と再結合するスーパーコンピテントDH5α細菌を形質転換する。
【0071】
3.アシネトバクター・カルコアセチカスのPQQ s−GDHの生成、精製及び特徴決定
3.1.野生型PQQ s−GDH及び突然変異PQQ s−GDHの生成
アポ酵素s−GDHを大腸菌JM101株において、野生型s−GDH又は突然変異s−GDHをコードする配列を有する組み換えプラスミドpgp492によって生成させる。アンピシリン(200mg/L)を添加したLB培地(LBA)50mLの前培養物に、アンピシリン(100mg/mL)を添加したLB寒天皿上で単離されたクローンを播種し、37℃で一晩220rpmで攪拌する。培養物をLBA培地に100倍で播種する。これを37℃で攪拌しながら(220rpm)、OD600nmが0.4OD600nm/mL〜1OD600nm/mLとなるまでインキュベートする。次いで、培養物を400μMのIPTGによって誘導した後、25℃で20時間攪拌する(180rpm〜220rpm)。遠心分離(5285g、4℃)によって回収した細胞を、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)で洗浄し、−20℃又は添加剤を加えずに−80℃で保管する。
【0072】
3.2.野生型PQQ s−GDH酵素及び突然変異PQQ s−GDH酵素の精製
3.2.1.細胞破壊及びDNase Iでの処理
1リットルの培養物から得られた細胞ペレットを、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)20mL中に溶解させ、40Wの超音波処理出力で超音波1秒、停止1秒のサイクルで3分間超音波処理する。得られたサンプル(粗抽出物と呼ぶ)に最後に2mM MgCl2を添加し、室温で30分間DNase I(粗抽出物1mL当たり1U)で処理する。次いで、不溶性細胞残屑を20000gで45分間の遠心分離によって粗抽出物から除去する。
【0073】
3.2.2.陽イオン交換クロマトグラフィー
カットオフ値(cutoff threshold)が0.22μmのフィルター(Millex−GS 0.22μm、Millipore)で濾過して、OD280nmが10となるように希釈した超音波処理上清を、AKTA purifierシステム(GE Healthcare(登録商標))に接続し、Tris 20mM、CaCl2 3mMの緩衝液(pH7.5)で平衡化した陽イオン交換カラムsource 30S(GE Healthcare(登録商標))に投入する。溶出は同緩衝液中0Mから1MまでというNaCl勾配で、5mL/分の流速で行う。s−GDHタンパク質を含有する画分を合わせ、Amicon YM10膜上での遠心分離によって濾過することで濃縮する。この段階でs−GDHタンパク質は純粋であり、硫酸アンモニウム(90%飽和)の存在下、−20℃で沈殿形態で保管することができる。
【0074】
3.2.3.野生型s−GDH酵素及び突然変異s−GDH酵素の特徴決定
3.2.3.1.分子量の決定
精製後のs−GDH単量体の分子量を、MALDI質量分析によって決定する。タンパク質をPD10カラム(GE Healthcare)上で脱塩し、重炭酸アンモニウム50mMの緩衝液(pH7.5)で溶出する。次いで、凍結乾燥したサンプルをMALDI−TOFによって分析すると、分子量が50234.83Daであることが確認される。この分子量はシグナルペプチドを有しないPQQ s−GDHの理論分子量に一致する。
【0075】
3.2.3.2.補因子PQQによるs−GDHの再構成
精製(Tris 20mM、CaCl2 3mMの緩衝液(pH7.5))の終了時に、酵素溶液をPQQと共に室温で15分間プレインキュベートする。添加したPQQは、酵素のモル濃度(molarity of the concentration)の2倍に等しい最終濃度に相当する。次いで、Pipes 20mM、CaCl2 3mMの緩衝液(pH7)で平衡化したPD10カラム(GE Healthcare)上で脱塩することによって過剰のPQQを除去する。
【0076】
3.2.3.3.濃度の測定
溶液中の酵素濃度を、補因子PQQを有する又は有しない酵素(二量体又は単量体;吸光係数はそれぞれ1.28l・g−1・cm−1又は1.67l・g−1・cm−1である)を用いて280nmで測定したODから算出する(Olsthoorn et al., 1997)。
【0077】
3.2.3.4.酵素試験
酵素試験は、リン酸ナトリウム20mMの緩衝液(pH7)中、37℃、3mLの容量でVarianの分光光度計を用いて、電子受容体として働くPMSを用いたDCIPの還元による消失を600nmで時間に応じてモニタリングすることで行う。酵素の特異的活性は、タンパク質1mg当たり1分間に消失したDCIPのμmol量で表される。PMS及びDCIPの濃度はそれぞれ0.6mM及び0.06mMである。酵素を−0.05OD600nm/分と−0.2OD600nm/分との間の傾きが測定されるように希釈する。
【0078】
4.野生型PQQ s−GDH及び突然変異PQQ s−GDHの酵素特性を研究する技法
4.1.定常状態での速度定数(kcat)及びミカエリス定数(KM)の決定
実験はリン酸ナトリウム20mMの緩衝液(pH7)中37℃で、Varianの分光光度計上で行う。この試験における基質(グルコース及びマルトース)の濃度は、0mM〜800mMの間で変化させる。
【0079】
PMSを第1の電子受容体として使用し、酵素活性を第2の電子受容体として使用されるDCIPの還元によって600nmでモニタリングする。PMS及びDCIPの初期濃度はそれぞれ0.6mM及び0.06mMである。酵素を添加することによって試験を開始する。実験点は、以下の等式によるSigma−plot 6.0ソフトウェアを用いた等式1、等式2又は等式3による非線形回帰によって、分析する:
(1)ミカエリス−メンテンモデル
kss=kcat×[S]/(KM+[S])
(2)二重直角双曲線関数(Double hyperbole)
kss=kcat1×[S]/(KM1+[S])+kcat2×[S]/KM2+[S])
(3)競合的阻害によるミカエリス−メンテンモデル
v/vmax=[S]/KS×(1+[S]/KS+[I]/KI)
【0080】
結果
図2A、図2B、図2C、図2D、図2E及び図2Fのグラフは、野生型(WT)及び428位で突然変異させた(N428C、N428Y、N428A、N428K、N428D及びN428E)アシネトバクター・カルコアセチカスの可溶性PQQグルコースデヒドロゲナーゼのグルコース又はマルトースの存在下での定常状態の速度論パラメータを示す。
【0081】
これらの結果から、マルトース及びグルコースという2つの基質について、本発明による突然変異酵素が野生型酵素の活性よりも高い活性を示すことが示される。これらの突然変異体の一部は野生型酵素の2倍の活性を有している。
【0082】
4.2.pHに応じた調査
4.2.1.pHに応じた活性
pHに応じた反応速度定数の変化の調査を、野生型酵素及び突然変異酵素をTris 120mM、イミダゾール30mM、酢酸30mMから構成される混合緩衝液(TIA)(NaClを用いてイオン強度を190mMに調整する)、又はpH範囲が6〜8のリン酸ナトリウム20mMの緩衝液(NaPi)中でインキュベートすることによって、5〜9のpH範囲にわたって行う。Varianの分光光度計を用いて37℃で実験を行う。PMSを第1の電子受容体として0.6mMの濃度で使用する。活性を、第2の電子受容体として0.06mMの濃度で使用されるDCIPの消失によって600nmでモニタリングする。酵素を添加することによって試験を開始する。野生型酵素又は突然変異酵素の最適活性は100%に対応し、各pHに対する活性を示す。
【0083】
結果
図3A及び図3Bは、野生型(WT)及び428位でシステインに突然変異させた(N428C)アシネトバクター・カルコアセチカスのPQQ s−GDHのpHに応じた活性を示すグラフである。これらの結果から、PQQ s−GDHのシステイン突然変異体(N428C)が野生型酵素の活性に匹敵する相対活性を示すことが示され、この突然変異がpHに関わらず相対活性の喪失をもたらさないことが実証される。
【0084】
4.2.2.pHに応じた安定性
野生型PQQ s−GDH又は突然変異PQQ s−GDHのpHに応じた安定性を、精製した酵素を均質になるまでpH範囲5〜9の混合緩衝液で希釈することによって決定する。この混合緩衝液はTris 120mM、イミダゾール30mM、酢酸30mMから構成され、そのイオン強度をNaClを用いて190mMに調整する。希釈した酵素溶液(1μg/ml〜6μg/ml)を37℃でプレインキュベートする。時間に応じて様々なサンプルをとる。残存活性をリン酸ナトリウム20mMの緩衝液(pH7)中、0.06mM DCIP、0.6mM PMSの存在下でVarianの分光光度計を用いて37℃で測定する。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。
【0085】
結果
本発明による突然変異酵素は、その安定性が試験したpH範囲にわたって一定であるか否かを問わないが、pHに関わらず野生型酵素よりも良好な安定性を示す。特に、突然変異酵素はpHが7を超えても安定なままである。
【0086】
4.3.温度に応じた調査
4.3.1.温度に応じた活性
反応速度定数の変化を、リン酸ナトリウム20mMの緩衝液(pH7)中、0.06mMのDCIP及び0.6mMのPMSの存在下でpHに応じて調査する。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。温度は10℃〜60℃で変化させる。温度制御したVarianの分光光度計CARY UV Biomelt上で活性をモニタリングする。酵素を添加することによって試験を開始する。
【0087】
結果
図4は、野生型(WT)及び突然変異させた(N428)PQQ s−GDHの温度に応じた活性曲線を示す。
【0088】
この曲線から、30℃〜50℃で突然変異酵素が野生型酵素よりも良好な活性を示すことが示される。
【0089】
4.3.2.温度に応じた酵素の安定性
この研究は、リン酸ナトリウム20mMの緩衝液(pH7)中、0.06mMのDCIP及び0.6mMのPMSの存在下で行う。グルコース濃度は、野生型酵素の試験については75mM、突然変異体N428Cの試験については150mMである。温度は10℃〜60℃で変化させる。温度制御したVarianの分光光度計CARY UV Biomelt上で37℃で活性をモニタリングする。酵素を添加することによって試験を開始する。
【0090】
結果
200分未満のインキュベーション時間では、突然変異酵素N428Cが40℃及び50℃で野生型酵素よりも良好な活性を示すことが分かり、したがって血中グルコースアッセイキットについてのその利点が実証される。
【0091】
4.4.基質に応じた調査
システイン突然変異体(N428C)の特異性を様々な基質に関して評価した。
【0092】
この特異性の測定に採用したプロトコルは第4.1.項に記載しているものと同一であるが、異なる基質を用いる。
【0093】
得られた結果を下記表II及び図5に提示する。
【0094】
【表2】
【0095】
結果
突然変異体N428Cのkssの生値は大半の基質について野生型酵素の値よりも高いが、相対活性(或る基質に対する酵素の特異性を別の基質と比較して特徴付ける)から、突然変異体N428Cがグルコースに対してより特異的であることが示される。したがって、4mMの基質では、マルトースを基質とすると、野生型酵素についてはグルコースと比べた相対活性は81%であることが測定されるのに対して、突然変異体N428Cの示すグルコースと比べたマルトースの相対活性は61%である。
【0096】
この同じプロトコルをアスパラギン酸(N428D)突然変異体、グルタミン酸(N428E)突然変異体、アラニン(N428A)突然変異体、チロシン(N428Y)突然変異体及びリシン(N428K)突然変異体のグルコース及びマルトースに対する特異性を評価するために採用した。
【0097】
結果を下記表IIIに提示する。
【0098】
【表3】
【特許請求の範囲】
【請求項1】
配列番号2のアシネトバクター・カルコアセチカスのPQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDHの突然変異体であって、428位に位置するそのアミノ酸が配列番号2と比べてシステイン、アラニン又はアスパラギン酸を含む群から選択されるアミノ酸で置換されていることを特徴とする、配列番号2のアシネトバクター・カルコアセチカスのPQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDHの突然変異体。
【請求項2】
PQQ s−GDHの突然変異体であって、428位に位置するそのアミノ酸が配列番号2と比べて置換されており、配列番号4、配列番号6、配列番号8、配列番号12及び配列番号14を含む群から選択されるアミノ酸配列を有することを特徴とする、PQQ s−GDHの突然変異体。
【請求項3】
請求項1又は2に記載の突然変異体PQQ s−GDHをコードすることを特徴とする核酸分子。
【請求項4】
配列番号15と配列番号16との対、配列番号19と配列番号20との対、配列番号21と配列番号22との対、配列番号23と配列番号24との対、及び配列番号25と配列番号26との対を含む群から選択されるオリゴヌクレオチド対による、配列番号1の配列を有する核酸分子の突然変異によって得られることを特徴とする、請求項3に記載の核酸分子。
【請求項5】
配列番号3、配列番号5、配列番号7、配列番号11及び配列番号13を含む群から選択される配列を有することを特徴とする、請求項3又は4に記載の核酸分子。
【請求項6】
請求項3〜5のいずれか一項に記載の核酸分子を含むことを特徴とする発現ベクター。
【請求項7】
請求項6に記載の発現ベクターによって形質転換されていることを特徴とする、請求項1又は2に記載の酵素を発現する宿主細胞。
【請求項8】
サンプル中のグルコース濃度を測定するための請求項1又は2に記載のPQQ s−GDH突然変異体の使用。
【請求項9】
前記サンプルが生体サンプル、特に血液であることを特徴とする、請求項8に記載の使用。
【請求項10】
請求項1又は2に記載のPQQ s−GDHの突然変異体を含むことを特徴とするグルコースアッセイキット。
【請求項11】
請求項1又は2に記載のPQQ s−GDHの突然変異体を少なくとも1つ含む被着物で被覆された導電材料を含むことを特徴とするグルコース電極。
【請求項12】
請求項11に記載の電極からなることを特徴とするグルコースセンサー。
【請求項13】
アノードである請求項11に記載の第一電極と、カソードである第二電極とを備えることを特徴とするグルコースバイオ燃料電池。
【請求項14】
サンプルにおいて溶液中のグルコースを定量する方法であって、
a)その還元が色の変化をもたらすレドックス試薬と、請求項1又は2に記載のPQQ s−GDH突然変異体とを前記サンプル中に導入する工程、
b)酵素反応後に前記サンプルの着色の強さを測定する工程、
c)工程b)において測定された着色の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)前記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルにおいて溶液中のグルコースを定量する方法。
【請求項15】
サンプルのグルコースアッセイを行う方法であって、
a)請求項11に記載のグルコース電極を前記サンプル中に導入する工程、
b)前記サンプル中の電流の強さを測定する工程、
c)工程b)において測定された電流の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)前記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルのグルコースアッセイを行う方法。
【請求項1】
配列番号2のアシネトバクター・カルコアセチカスのPQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDHの突然変異体であって、428位に位置するそのアミノ酸が配列番号2と比べてシステイン、アラニン又はアスパラギン酸を含む群から選択されるアミノ酸で置換されていることを特徴とする、配列番号2のアシネトバクター・カルコアセチカスのPQQ s−GDHに対して少なくとも80%の同一率を有するPQQ s−GDHの突然変異体。
【請求項2】
PQQ s−GDHの突然変異体であって、428位に位置するそのアミノ酸が配列番号2と比べて置換されており、配列番号4、配列番号6、配列番号8、配列番号12及び配列番号14を含む群から選択されるアミノ酸配列を有することを特徴とする、PQQ s−GDHの突然変異体。
【請求項3】
請求項1又は2に記載の突然変異体PQQ s−GDHをコードすることを特徴とする核酸分子。
【請求項4】
配列番号15と配列番号16との対、配列番号19と配列番号20との対、配列番号21と配列番号22との対、配列番号23と配列番号24との対、及び配列番号25と配列番号26との対を含む群から選択されるオリゴヌクレオチド対による、配列番号1の配列を有する核酸分子の突然変異によって得られることを特徴とする、請求項3に記載の核酸分子。
【請求項5】
配列番号3、配列番号5、配列番号7、配列番号11及び配列番号13を含む群から選択される配列を有することを特徴とする、請求項3又は4に記載の核酸分子。
【請求項6】
請求項3〜5のいずれか一項に記載の核酸分子を含むことを特徴とする発現ベクター。
【請求項7】
請求項6に記載の発現ベクターによって形質転換されていることを特徴とする、請求項1又は2に記載の酵素を発現する宿主細胞。
【請求項8】
サンプル中のグルコース濃度を測定するための請求項1又は2に記載のPQQ s−GDH突然変異体の使用。
【請求項9】
前記サンプルが生体サンプル、特に血液であることを特徴とする、請求項8に記載の使用。
【請求項10】
請求項1又は2に記載のPQQ s−GDHの突然変異体を含むことを特徴とするグルコースアッセイキット。
【請求項11】
請求項1又は2に記載のPQQ s−GDHの突然変異体を少なくとも1つ含む被着物で被覆された導電材料を含むことを特徴とするグルコース電極。
【請求項12】
請求項11に記載の電極からなることを特徴とするグルコースセンサー。
【請求項13】
アノードである請求項11に記載の第一電極と、カソードである第二電極とを備えることを特徴とするグルコースバイオ燃料電池。
【請求項14】
サンプルにおいて溶液中のグルコースを定量する方法であって、
a)その還元が色の変化をもたらすレドックス試薬と、請求項1又は2に記載のPQQ s−GDH突然変異体とを前記サンプル中に導入する工程、
b)酵素反応後に前記サンプルの着色の強さを測定する工程、
c)工程b)において測定された着色の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)前記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルにおいて溶液中のグルコースを定量する方法。
【請求項15】
サンプルのグルコースアッセイを行う方法であって、
a)請求項11に記載のグルコース電極を前記サンプル中に導入する工程、
b)前記サンプル中の電流の強さを測定する工程、
c)工程b)において測定された電流の強さと、グルコース含量が既知の標準溶液について測定された強さとを比較する工程、
d)前記サンプル中のグルコース濃度を決定する工程
を含むことを特徴とする、サンプルのグルコースアッセイを行う方法。
【図1A】

【図1B】

【図2A】

【図2B】

【図2C】
【図2D】
【図2E】
【図2F】
【図3A】
【図3B】
【図4】
【図5】
【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図3A】
【図3B】
【図4】
【図5】
【公表番号】特表2013−500033(P2013−500033A)
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願番号】特願2012−522198(P2012−522198)
【出願日】平成22年7月20日(2010.7.20)
【国際出願番号】PCT/FR2010/000522
【国際公開番号】WO2011/012779
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(502205846)サントル ナショナル ドゥ ラ ルシェルシュ シアンティフィク (154)
【Fターム(参考)】
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願日】平成22年7月20日(2010.7.20)
【国際出願番号】PCT/FR2010/000522
【国際公開番号】WO2011/012779
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(502205846)サントル ナショナル ドゥ ラ ルシェルシュ シアンティフィク (154)
【Fターム(参考)】
[ Back to top ]