
ファージ頭殻提示腸管免疫誘導型ワクチン
【課題】安価かつ効率的に腸管免疫を誘導できる腸管免疫誘導型ワクチンを提供する。
【解決手段】腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチン。
【解決手段】腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチン。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ファージ頭殻提示腸管免疫誘導型ワクチンに関するものである。より詳しくは、腸管感染病原体に感染するバクテリオファージ頭殻表層に抗原タンパク質が提示された腸管免疫誘導型ワクチンに関する。
【背景技術】
【0002】
近年、腸管出血性大腸菌O157:H7及びノロウイルスに代表される経口感染症が多く発生しており、発展途上国においては、経口感染による細菌性下痢は乳幼児の死亡原因の筆頭に挙げられる。
【0003】
しかし、現行のワクチンのほとんどは、皮下に接種する注射剤であり、少頻度で起こる副作用や投与時の疼痛が臨床上問題となっている。これを改良するための様々な投与方法が検討されているが、中でも、経口ワクチンは安全性及が高く、投与が簡便である剤形である。経口生ワクチンとしてはポリオワクチン(例えば特許文献1参照)が代表的であり、ポリオワクチンは、腸管内壁から感染し、体内でIgG、IgM等の抗体を産生する。
【0004】
一方、バクテリオファージは、一回の感染サイクルで、ファージが感染した一個の宿主細菌から100〜200の娘ファージ粒子を放出し、爆発的自己複製能力を有する。また、Tファージ等はその頭殻内に、自身の複製に必要な遺伝情報をその中に効率良く保持している。近年、バクテリオファージを利用した細菌感染症の治療・予防法が報告されている(特許文献2、3参照)。特許文献2は、ファージ表面タンパク質と細菌細胞への結合能を付与する抗体フラグメントとからなる融合タンパク質をファージ表面に提示するバクテリオファージを用いて、細菌細胞の増殖を阻害する方法である。しかし、この方法では免疫は誘導されない。また、特許文献3の方法では、抗原タンパク質をコードする遺伝子が組み込まれたファージゲノムを筋肉注射によって免疫細胞(動物細胞)に取り込み、抗原タンパク質を発現させることにより、抗体産生を誘導する。この場合、抗原タンパク質はファージ表面に直接は提示されない。
【0005】
ところで、腸管は、自己と非自己とを識別し、病原体の侵入を阻止する最前線の感染防御バリアーである。腸管の粘膜上皮は抗原が最初に生体に侵入する場であり、腸管のリンパ組織には最も多くのリンパ球が存在する。消化管関連リンパ組織(GALT)はバクテリアと食事抗原、常在細菌と病原菌とを区別し、GALTに存在するリンパ球は分泌型IgA抗体を産生する。しかしながら、これまで腸管免疫誘導型のワクチンは実用化された例がない。
【0006】
近年、本発明者らは、健康なヒトや動物の腸管内に、大腸菌JM109に感染するバクテリオファージが高濃度(105/g-糞便)で存在することを報告している(非特許文献1参照)。バクテリオファージが腸管内に共存することにより、細菌感染症の発症を抑制していることが考えられる。
【0007】
【特許文献1】特表2005-517471号公報
【特許文献2】特表2004-505648号公報
【特許文献3】特開2004-525931号公報
【非特許文献1】FEMS Microbiol. Letter, 211, 77-83 (2002)
【発明の開示】
【発明が解決しようとする課題】
【0008】
従って、本発明は、安価かつ効率的に腸管免疫を誘導できる腸管ワクチンを提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、斯かる現状に鑑み鋭意検討した結果、感染病原体が存在する腸管内で爆発的自己複製能力を有し、かつ抗原タンパク質を直接その表層に提示する組換えバクテリオファージを作製し、これを経口投与すれば、安全、安価かつ効率的に腸管免疫を誘導できることを見出し、本発明を完成した。
【0010】
すなわち、本発明は、腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチンを提供する。
【発明の効果】
【0011】
本発明のファージワクチンは、安全、簡便かつ効率的に、しかも安価に腸管免疫を誘導することができ、腸管からの感染症を防ぐことができる。
【発明を実施するための最良の形態】
【0012】
本発明のファージワクチンに用いる組換えバクテリオファージ(以下、「ファージ」とする)は、その頭殻内に、腸管感染病原体由来の抗原タンパク質をコードする遺伝子、及びファージ頭殻を構成するタンパク質の一つであるSOC(Small Outer Capsid)タンパク質をコードする遺伝子が組み込まれたファージゲノムを有する。ファージ頭殻を構成するタンパク質としては他にHOC(Highly antigenic Outer Capsid)タンパク質が知られており、soc遺伝子の代わりに、hoc遺伝子を用いることもできる。
【0013】
本発明で使用されるファージとしては、Tファージ(例えばT1〜T7)の他に、溶原性のp1ファージ、λファージ等も使用できるが、自己複製能の点からTファージが好ましく、特にT2ファージが好ましい。
【0014】
抗原タンパク質としては、腸管感染病原体に由来するものあれば特に限定されず、完全長タンパク質でも又はその断片でもよいが、分子量が数十〜数百Daのものが好ましい。
【0015】
当該腸管感染病原体としては、腸管に感染するものであれば特に制限されず、例えば、黄色ブドウ球菌(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)、ヘリコバクター・ピロリ(Helicobacter pylori)、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)、ストレプトコッカス・ムタンス(Streptococcus mutans)、ストレプトコッカス・オラリス(Streptococcus oralis)、ストレプトコッカス・パラサンギス(Streptococcus parasanguis)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、ストレプトコッカス・ビリダンス(Streptococcus viridans)、A群連鎖球菌及び嫌気性連鎖球菌、ヘモフィルス・インフルエンザ(Hemophilus influenzae)、赤痢菌(Shigella dysenteriae)、マイコバクテリウム・レプラエ(Mycobacterium leprae)、マイコバクテリウム・アシアチカム(Mycobacterium asiaticum)、マイコバクテリウム・イントラセルラレ(Mycobacterium intracellulare)、マイコプラズマ・ニューモニエ(Mycoplasma pneumoniae)、マイコプラズマ・ホミニス(Mycobacterium hominis)、クレブシエラ.ニューモニエ(Klebsiella pneumoniae)、緑膿菌(Pseudomonas aeruginosa)、プロピオニバクテリウム.アクネス(Propionibacterium acnes)、トレポネーマ・パリダム(Treponema pallidum)、トレポネーマ・ペルタヌエ(Treponema pertanue)、トレポネーマ・カラテウム(Treponema carateum)、大腸菌(Escherichia coli)、ネズミチフス菌(Salmonella typhimurium)、ボレリア・ブルグドルフェリ(Borrelia burgdorferi)、レプトスピレックス・ヘモラギア(Leptospirex hemoragia)、シトロバクター・フルエンジー(Citrobacter fruendii)等が挙げられ、この中で大腸菌、黄色ブドウ球菌又は赤痢菌が好ましく、特に大腸菌が好ましい。
【0016】
前記の2種の遺伝子が組み込まれた組換えファージは、具体的には、野生型T2ファージゲノム(Genbank accession number: AF374619)のsoc遺伝子とその上流に位置するsoc1遺伝子との間にβ-ガラクトシダーゼ遺伝子を挿入したT2-βファージを作製し、このファージを、大腸菌O157:H7の接着因子(インチミン:Int)をコードする遺伝子断片intを挿入したベクターpUC-socintによって形質転換した大腸菌に感染させ、β-ガラクトシダーゼ遺伝子を抗原ペプチド遺伝子と組換えることにより、容易に作製することができる。
【0017】
腸管出血性大腸菌O157:H7の接着因子インチミンのアミノ酸配列を配列番号6に、当該アミノ酸配列をコードする塩基配列を配列番号5に、本発明で実際に使用したインチミンポリペプチドのアミノ酸配列を配列番号4に、当該アミノ酸配列をコードする塩基配列を配列番号3にそれぞれ示す。また、soc1遺伝子の塩基配列を配列番号7、soc遺伝子の塩基配列を配列番号8にそれぞれ示す。
【0018】
上記の組換えファージを、ヒトを含む動物の腸管感染病原体に感染させると、ファージ頭殻表面に抗原タンパク質、詳細にはSOCと抗原タンパク質とから成る融合タンパク質が一定の頻度が提示され、この抗原タンパク質に対する抗体産生が誘導される。組換えファージの腸管感染病原体への感染は、経口投与による。この組換えファージは、野生型ファージと同様に、腸管感染病原体に感染することにより容易に複製することができる。
【0019】
本発明のファージワクチンによって予防又は治療可能な腸管感染症としては、例えば、NAGビブリオ感染症、アデノウイルス感染症、アメーバ赤痢、ウエルシュ菌感染症、エルシニア感染症、エロモナスハイドロフィラ/ソブリア感染症、感染性胃腸炎(ウイルス性胃腸炎・ノロウイルス感染症)、カンピロバクター腸炎、クリプトスポリジウム症、下痢原性大腸菌感染症、コレラ、細菌性赤痢、サルモネラ症、ジアルジア症、セレウス菌感染症、腸炎ビブリオ、腸管出血性大腸菌感染症、腸チフス・パラチフス、ビブリオ・フルビアリス/ファーニシ感染症、ブドウ球菌食中毒、プレシオモナスシゲロイデス感染症等が挙げられる。
【0020】
本発明のファージワクチンは、IgA、IgG、IgM等の抗体の産生を誘導するが、従来のポリオワクチン等の経口ワクチンに比べて、特にIgAの産生率が高い。抗体産生量の測定は、通常の蛍光抗体法、酵素免疫測定法(ELISA)、ラジオイムノアッセイ等によればよい。
【0021】
本発明のファージワクチンの用量は、経口投与される対象の免疫応答性、体重、年齢、性別、症状等に応じて変動するが、成人一人一回当り、通常、100万〜1兆PFU(Plaque Forming Unit)であり、好ましくは1億〜100億PFUである。
【0022】
本発明のファージワクチンには、アジュバントを更に添加することができる。アジュバントとしては、例えば、腸管免疫を誘導する際に、コレラ毒素や病原性大腸菌の産生する易熱分解性エンテロトキシン等が挙げられる。
【実施例】
【0023】
次に実施例を挙げて本発明を詳細に説明するが、本発明はこれら実施例に何ら限定されるものではない。
【0024】
<材料>
(1)大腸菌O157:H7濃縮液
LB液体培地(Bacto trypton 10g、Yeast extract 5g、NaCl 10gを脱イオン水1Lに溶解)に、大腸菌O157:H7(American Type Culture Collectionから入手)を1億CFU(Colony Forming Unit)/mlで播種し、37℃で終夜、振盪培養した。菌体培養液1.5 ml(約109 CFU/ml)を4℃、3,000rpmで15分間遠心し、ペレットを得た。このペレットをPBS(0.8 mlx2)で洗浄し、上清を捨てた後、再度PBS 0.15 mlを加えて、菌体濃縮液0.15 ml(約1010 CFU/ml)を得た。
【0025】
(2)インチミンポリペプチド
大腸菌O157:H7の外膜タンパク質である接着因子インチミン(Int)ポリペプチドを以下のようにして調製した。
(i)pETintの構築
pET14bベクター(Novagen社)のBamHIサイト及びNdeI及びサイトに、intexp+(Nde)(5'-ggaattccatatggatcaaaccaaggccagcattac-3':配列番号1)(下線は、制限酵素Nde認識サイトを示す)及びintexp-(Bam)(5'-cgggatccttattctacacaaaccgcatag-3':配列番号2)(下線は、制限酵素BamHI認識サイトを示す)をプライマーとしたPCRにより増幅したgene eae の3’側の約1/3に相当する配列(接着に関わる部分)(配列番号3)を挿入して、pETintを構築した。
(ii)Intポリペプチドの発現・精製
インチミンポリペプチドを以下の手順により発現・精製した。LB培地30 mlに(i)のpETintで形質転換した大腸菌BL21の前培養液300 μlを播種し、37℃で1時間培養した。対数増殖期の初期にさしかかったところで終濃度が1 mMとなるようにIPTG(イソプロピル-1-チオ-β-D-ガラクトシダーゼ)を添加し、28℃で4時間培養した。その培養液を遠心分離(4,000 xg、10分、4℃)にて集菌し、20 mM Tris-HCl(pH 8.0)3 mlに懸濁した。スピッツ管に1.5 mlの菌体液を入れ、on-1分、off-30秒で10分間の超音波破砕を行った。破砕後、遠心分離(15,000 rpm、15分、4℃)を行い、可溶画分と不溶画分とを分離し、得られた可溶画分を0.45 μmのフィルターでろ過した。この溶液を、ニッケルイオンを定着させた、His-trap(登録商標)Chelatingカラム(Amaersham Pharmacia Biotech社)に通した。その後、カラムを数回洗浄し、溶出バッファー(イミダゾール300 mM)によって、N-末端側にHisタグが融合されたIntポリペプチド(配列番号3及び4)を回収した。
【0026】
実施例1 組換えT2intファージの構築
(1)T2-βファージの構築
まず、pUC118ベクターのPstIサイト及びHindIIIサイトに、PCRにより増幅した野生型T2ファージのsoc1-soc断片を挿入し、pUC-Socを得た。大腸菌K12染色体DNAを鋳型としPCRを行うことにより、β-ガラクトシダーゼ遺伝子を得た。このβ-ガラクトシダーゼ遺伝子をpUC-Socのsoc1遺伝子(配列番号7)とsoc遺伝子(配列番号8)との間に挿入し、pUC-βSocを得た。β-ガラクトシダーゼ及びSocが独立に発現するように、β-ガラクトシダーゼのストップコドンは残した。尚、野生型T2ファージは、有坂文雄助教授(東京工業大学大学院生命理工学研究科)より頂戴した。
【0027】
次に、pUC-Socを用い、エレクトロポレーションによって大腸菌BEを形質転換した。この形質転換体を、アンピシリンを添加したLB培地で37℃で振盪培養(120 rpm)した。対数増殖期初期において、培養液にMOI(Multiplicity ofinfection)が0.01[PFU/CFU]となるようにT2ファージを添加した。ファージ添加後、培養液のOD660を経時的に測定し、溶菌に伴う濁度の現象が確認された後(ファージ添加後約4時間)、培養液にクロロホルムを添加し、未溶菌菌体の溶菌を行った。遠心分離(17,400 xg、10分、4℃)により菌体破砕物を除去し、ファージライセートを調製した。このファージライセートを104 PFU/mlとなるようにSM bufferで希釈した。この希釈液と大腸菌JM109の培養液とを混合し、X-gal溶液(40 mg/ml)を50 μl含む軟寒天を用いてプラークアッセイを行い、103 PFU/plateとなるようにプラークを形成させた。形成されたプラークについて、青色を呈するファージを選択することによって、β-ガラクトシダーゼを発現するファージ(T2-βファージ)を得た。
【0028】
(2)pUC-socintの構築
大腸菌O157:H7の染色体DNAを鋳型としPCRを行うことにより、大腸菌O157:H7の接着因子(Int)をコードする遺伝子断片intを得た。pUC-Socのsoc1遺伝子とsoc遺伝子との間にこのint遺伝子を挿入し、pUC-socintを構築した。接着因子(Int)及びSocが共発現するようにintのストップコドンは削除した。
【0029】
(3)組換え体のスクリーニング
上記(1)のT2-βファージはβ-ガラクトシダーゼを発現し、X-galを含むLB培地で大腸菌JM109とプラークアッセイを行うと、青いプラークを形成するが、soc1遺伝子下流で相同組換えが起こり、β-ガラクトシダーゼ遺伝子が別の遺伝子と組換わると、透明なプラークを形成する。この手法を用いることにより、相同組換え後の組換え体のスクリーニングを容易に行うことができる。
組換え体のスクリーニングは以下のようにして行った。まず、組換え用ベクターとしてpUC-socintを用い、エレクトロポレーションによって大腸菌BEを形質転換した。この形質転換体を、アンピシリンを添加したLB培地で37℃で振盪培養(120 rpm)した。対数増殖期初期において、培養液にMOI(Multiplicity of infection)が0.01[PFU/CFU]となるようにT2-βファージを添加した。ファージ添加後、培養液のOD660を経時的に測定し、溶菌に伴う濁度の現象を確認した後(ファージ添加後約4時間)、培養液にクロロホルムを添加し、未溶菌菌体の溶菌を行った。遠心分離(17,400 xg、10分、4℃)により菌体破砕物を除去し、ファージライセートを調製した。このファージライセートを104 PFU/mlとなるようにSM bufferで希釈した。次に、この希釈液と、大腸菌JM109の培養液とを混合し、X-gal溶液(40 mg/ml)を50 μl含む軟寒天を用いてプラークアッセイを行い、103 PFU/plateとなるようにプラークを形成させた。形成されたプラークについて、透明なプラークを選択することにより、インチミン組換え体ファージ(以下、「T2intファージ」とする)をスクリーニングした。
【0030】
実施例2 IgA抗体の検出
まず、以下の4溶液:
A:負対照群(材料(1)で得た大腸菌O157:H7 + 野生型T2ファージ(1010 PFU/マウス))
B:対照群(材料(2)で得たインチミンポリペプチド(0.5 μg/マウス))
C:対照群(実施例1で得たT2intファージ(1010 PFU/マウス))
D:ファージ投与群(材料(1)で得た大腸菌O157:H7 + T2intファージ(1010 PFU/マウス))
を調製した。尚、野生型T2ファージは、抗原(インチミン:int)は発現していない。T2intファージは組換えファージで、頭殻表層にintを発現している。
A〜Dの溶液0.1 mlずつを、4週齢の雌性BALB/cAnNCr1Cr1jマウス(体重約20 g、日本チャールズ・リバー社より購入)にゾンデにより胃内に投与した。投与は各群についてマウス6匹で行った。また、最初の投与から2週間後にブースター投与を行った。
【0031】
次に、産生されたIgA抗体を投与後のマウスの糞便から抽出するため、糞便20 mgをPBS 1 mlに溶解し、滅菌した割り箸でホモジナイズ後、この懸濁液を60分間かけて4〜6回、ボルテックスし、4℃、1,600 xgで15分遠心した。この上清を採り、更に4℃、7,200 xgで10分遠心し、得られた上清を糞便由来溶液とした。
【0032】
糞便由来溶液中のインチミン特異的IgA抗体量をELISAにより測定した。インチミン特異的IgA抗体量は、「インチミン特異的IgA抗体濃度」を「総IgA抗体濃度」で除して得られる値によって評価した。これらの「インチミン特異的IgA抗体濃度」及び「総IgA抗体濃度」は、以下のようにして別々に測定した:
(i)96穴プレートに、carbonate bufferで希釈した抗原(「特異的IgA抗体」についてはインチミン、「総IgA抗体」については抗マウスIgAを各々使用)を100 μl加え、4℃で一晩培養した;
(ii)次いで、PBS-Tween 200 μl(x3)でプレートを洗浄後、ブロックエース(雪印社製)を200 μl加え、ウェルをブロッキングした;
(iii)次いで、PBS-Tween 200 μl(x3)でプレートを洗浄し、適当に希釈したIgA抗体溶液を100 μl加え、1時間振盪した。濃度既知のIgA抗体を15.625〜1000 ng/mlの範囲で希釈し、同様に100 μl加えた;
(iv)次いで、PBS-Tween 200 μl(x5)でプレートを洗浄し、HRP標識二次抗体(抗マウスIgA-α)を加え、1時間振盪し、PBS-Tween 200 μl(x4)、PBS 200 μl(x1)で洗浄した;
(v)次いで、TMB substrate kit(フナコシ社)を各ウェルに100 μlずつ加えた後、反応を停止し、OD 450 nmの吸光度を測定した。
【0033】
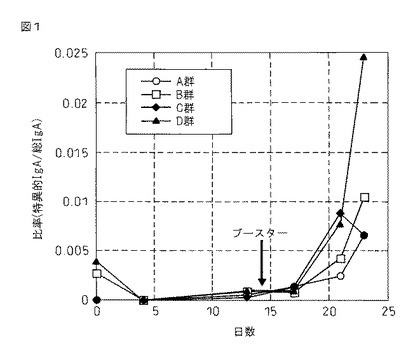
結果を図1に示す。図1から明らかなように、B〜Dの各投与群で抗インチミンIgA抗体の産生が見られ、特にD投与群では有意にその産生が認められた。
従って、本発明のファージワクチンは、抗体産生を効率的に誘導することが判った。
【図面の簡単な説明】
【0034】
【図1】図1は、A〜Dの各群投与後の抗インチミンIgA抗体の産生の結果を示す図である。
【技術分野】
【0001】
本発明は、ファージ頭殻提示腸管免疫誘導型ワクチンに関するものである。より詳しくは、腸管感染病原体に感染するバクテリオファージ頭殻表層に抗原タンパク質が提示された腸管免疫誘導型ワクチンに関する。
【背景技術】
【0002】
近年、腸管出血性大腸菌O157:H7及びノロウイルスに代表される経口感染症が多く発生しており、発展途上国においては、経口感染による細菌性下痢は乳幼児の死亡原因の筆頭に挙げられる。
【0003】
しかし、現行のワクチンのほとんどは、皮下に接種する注射剤であり、少頻度で起こる副作用や投与時の疼痛が臨床上問題となっている。これを改良するための様々な投与方法が検討されているが、中でも、経口ワクチンは安全性及が高く、投与が簡便である剤形である。経口生ワクチンとしてはポリオワクチン(例えば特許文献1参照)が代表的であり、ポリオワクチンは、腸管内壁から感染し、体内でIgG、IgM等の抗体を産生する。
【0004】
一方、バクテリオファージは、一回の感染サイクルで、ファージが感染した一個の宿主細菌から100〜200の娘ファージ粒子を放出し、爆発的自己複製能力を有する。また、Tファージ等はその頭殻内に、自身の複製に必要な遺伝情報をその中に効率良く保持している。近年、バクテリオファージを利用した細菌感染症の治療・予防法が報告されている(特許文献2、3参照)。特許文献2は、ファージ表面タンパク質と細菌細胞への結合能を付与する抗体フラグメントとからなる融合タンパク質をファージ表面に提示するバクテリオファージを用いて、細菌細胞の増殖を阻害する方法である。しかし、この方法では免疫は誘導されない。また、特許文献3の方法では、抗原タンパク質をコードする遺伝子が組み込まれたファージゲノムを筋肉注射によって免疫細胞(動物細胞)に取り込み、抗原タンパク質を発現させることにより、抗体産生を誘導する。この場合、抗原タンパク質はファージ表面に直接は提示されない。
【0005】
ところで、腸管は、自己と非自己とを識別し、病原体の侵入を阻止する最前線の感染防御バリアーである。腸管の粘膜上皮は抗原が最初に生体に侵入する場であり、腸管のリンパ組織には最も多くのリンパ球が存在する。消化管関連リンパ組織(GALT)はバクテリアと食事抗原、常在細菌と病原菌とを区別し、GALTに存在するリンパ球は分泌型IgA抗体を産生する。しかしながら、これまで腸管免疫誘導型のワクチンは実用化された例がない。
【0006】
近年、本発明者らは、健康なヒトや動物の腸管内に、大腸菌JM109に感染するバクテリオファージが高濃度(105/g-糞便)で存在することを報告している(非特許文献1参照)。バクテリオファージが腸管内に共存することにより、細菌感染症の発症を抑制していることが考えられる。
【0007】
【特許文献1】特表2005-517471号公報
【特許文献2】特表2004-505648号公報
【特許文献3】特開2004-525931号公報
【非特許文献1】FEMS Microbiol. Letter, 211, 77-83 (2002)
【発明の開示】
【発明が解決しようとする課題】
【0008】
従って、本発明は、安価かつ効率的に腸管免疫を誘導できる腸管ワクチンを提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、斯かる現状に鑑み鋭意検討した結果、感染病原体が存在する腸管内で爆発的自己複製能力を有し、かつ抗原タンパク質を直接その表層に提示する組換えバクテリオファージを作製し、これを経口投与すれば、安全、安価かつ効率的に腸管免疫を誘導できることを見出し、本発明を完成した。
【0010】
すなわち、本発明は、腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチンを提供する。
【発明の効果】
【0011】
本発明のファージワクチンは、安全、簡便かつ効率的に、しかも安価に腸管免疫を誘導することができ、腸管からの感染症を防ぐことができる。
【発明を実施するための最良の形態】
【0012】
本発明のファージワクチンに用いる組換えバクテリオファージ(以下、「ファージ」とする)は、その頭殻内に、腸管感染病原体由来の抗原タンパク質をコードする遺伝子、及びファージ頭殻を構成するタンパク質の一つであるSOC(Small Outer Capsid)タンパク質をコードする遺伝子が組み込まれたファージゲノムを有する。ファージ頭殻を構成するタンパク質としては他にHOC(Highly antigenic Outer Capsid)タンパク質が知られており、soc遺伝子の代わりに、hoc遺伝子を用いることもできる。
【0013】
本発明で使用されるファージとしては、Tファージ(例えばT1〜T7)の他に、溶原性のp1ファージ、λファージ等も使用できるが、自己複製能の点からTファージが好ましく、特にT2ファージが好ましい。
【0014】
抗原タンパク質としては、腸管感染病原体に由来するものあれば特に限定されず、完全長タンパク質でも又はその断片でもよいが、分子量が数十〜数百Daのものが好ましい。
【0015】
当該腸管感染病原体としては、腸管に感染するものであれば特に制限されず、例えば、黄色ブドウ球菌(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)、ヘリコバクター・ピロリ(Helicobacter pylori)、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)、ストレプトコッカス・ムタンス(Streptococcus mutans)、ストレプトコッカス・オラリス(Streptococcus oralis)、ストレプトコッカス・パラサンギス(Streptococcus parasanguis)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、ストレプトコッカス・ビリダンス(Streptococcus viridans)、A群連鎖球菌及び嫌気性連鎖球菌、ヘモフィルス・インフルエンザ(Hemophilus influenzae)、赤痢菌(Shigella dysenteriae)、マイコバクテリウム・レプラエ(Mycobacterium leprae)、マイコバクテリウム・アシアチカム(Mycobacterium asiaticum)、マイコバクテリウム・イントラセルラレ(Mycobacterium intracellulare)、マイコプラズマ・ニューモニエ(Mycoplasma pneumoniae)、マイコプラズマ・ホミニス(Mycobacterium hominis)、クレブシエラ.ニューモニエ(Klebsiella pneumoniae)、緑膿菌(Pseudomonas aeruginosa)、プロピオニバクテリウム.アクネス(Propionibacterium acnes)、トレポネーマ・パリダム(Treponema pallidum)、トレポネーマ・ペルタヌエ(Treponema pertanue)、トレポネーマ・カラテウム(Treponema carateum)、大腸菌(Escherichia coli)、ネズミチフス菌(Salmonella typhimurium)、ボレリア・ブルグドルフェリ(Borrelia burgdorferi)、レプトスピレックス・ヘモラギア(Leptospirex hemoragia)、シトロバクター・フルエンジー(Citrobacter fruendii)等が挙げられ、この中で大腸菌、黄色ブドウ球菌又は赤痢菌が好ましく、特に大腸菌が好ましい。
【0016】
前記の2種の遺伝子が組み込まれた組換えファージは、具体的には、野生型T2ファージゲノム(Genbank accession number: AF374619)のsoc遺伝子とその上流に位置するsoc1遺伝子との間にβ-ガラクトシダーゼ遺伝子を挿入したT2-βファージを作製し、このファージを、大腸菌O157:H7の接着因子(インチミン:Int)をコードする遺伝子断片intを挿入したベクターpUC-socintによって形質転換した大腸菌に感染させ、β-ガラクトシダーゼ遺伝子を抗原ペプチド遺伝子と組換えることにより、容易に作製することができる。
【0017】
腸管出血性大腸菌O157:H7の接着因子インチミンのアミノ酸配列を配列番号6に、当該アミノ酸配列をコードする塩基配列を配列番号5に、本発明で実際に使用したインチミンポリペプチドのアミノ酸配列を配列番号4に、当該アミノ酸配列をコードする塩基配列を配列番号3にそれぞれ示す。また、soc1遺伝子の塩基配列を配列番号7、soc遺伝子の塩基配列を配列番号8にそれぞれ示す。
【0018】
上記の組換えファージを、ヒトを含む動物の腸管感染病原体に感染させると、ファージ頭殻表面に抗原タンパク質、詳細にはSOCと抗原タンパク質とから成る融合タンパク質が一定の頻度が提示され、この抗原タンパク質に対する抗体産生が誘導される。組換えファージの腸管感染病原体への感染は、経口投与による。この組換えファージは、野生型ファージと同様に、腸管感染病原体に感染することにより容易に複製することができる。
【0019】
本発明のファージワクチンによって予防又は治療可能な腸管感染症としては、例えば、NAGビブリオ感染症、アデノウイルス感染症、アメーバ赤痢、ウエルシュ菌感染症、エルシニア感染症、エロモナスハイドロフィラ/ソブリア感染症、感染性胃腸炎(ウイルス性胃腸炎・ノロウイルス感染症)、カンピロバクター腸炎、クリプトスポリジウム症、下痢原性大腸菌感染症、コレラ、細菌性赤痢、サルモネラ症、ジアルジア症、セレウス菌感染症、腸炎ビブリオ、腸管出血性大腸菌感染症、腸チフス・パラチフス、ビブリオ・フルビアリス/ファーニシ感染症、ブドウ球菌食中毒、プレシオモナスシゲロイデス感染症等が挙げられる。
【0020】
本発明のファージワクチンは、IgA、IgG、IgM等の抗体の産生を誘導するが、従来のポリオワクチン等の経口ワクチンに比べて、特にIgAの産生率が高い。抗体産生量の測定は、通常の蛍光抗体法、酵素免疫測定法(ELISA)、ラジオイムノアッセイ等によればよい。
【0021】
本発明のファージワクチンの用量は、経口投与される対象の免疫応答性、体重、年齢、性別、症状等に応じて変動するが、成人一人一回当り、通常、100万〜1兆PFU(Plaque Forming Unit)であり、好ましくは1億〜100億PFUである。
【0022】
本発明のファージワクチンには、アジュバントを更に添加することができる。アジュバントとしては、例えば、腸管免疫を誘導する際に、コレラ毒素や病原性大腸菌の産生する易熱分解性エンテロトキシン等が挙げられる。
【実施例】
【0023】
次に実施例を挙げて本発明を詳細に説明するが、本発明はこれら実施例に何ら限定されるものではない。
【0024】
<材料>
(1)大腸菌O157:H7濃縮液
LB液体培地(Bacto trypton 10g、Yeast extract 5g、NaCl 10gを脱イオン水1Lに溶解)に、大腸菌O157:H7(American Type Culture Collectionから入手)を1億CFU(Colony Forming Unit)/mlで播種し、37℃で終夜、振盪培養した。菌体培養液1.5 ml(約109 CFU/ml)を4℃、3,000rpmで15分間遠心し、ペレットを得た。このペレットをPBS(0.8 mlx2)で洗浄し、上清を捨てた後、再度PBS 0.15 mlを加えて、菌体濃縮液0.15 ml(約1010 CFU/ml)を得た。
【0025】
(2)インチミンポリペプチド
大腸菌O157:H7の外膜タンパク質である接着因子インチミン(Int)ポリペプチドを以下のようにして調製した。
(i)pETintの構築
pET14bベクター(Novagen社)のBamHIサイト及びNdeI及びサイトに、intexp+(Nde)(5'-ggaattccatatggatcaaaccaaggccagcattac-3':配列番号1)(下線は、制限酵素Nde認識サイトを示す)及びintexp-(Bam)(5'-cgggatccttattctacacaaaccgcatag-3':配列番号2)(下線は、制限酵素BamHI認識サイトを示す)をプライマーとしたPCRにより増幅したgene eae の3’側の約1/3に相当する配列(接着に関わる部分)(配列番号3)を挿入して、pETintを構築した。
(ii)Intポリペプチドの発現・精製
インチミンポリペプチドを以下の手順により発現・精製した。LB培地30 mlに(i)のpETintで形質転換した大腸菌BL21の前培養液300 μlを播種し、37℃で1時間培養した。対数増殖期の初期にさしかかったところで終濃度が1 mMとなるようにIPTG(イソプロピル-1-チオ-β-D-ガラクトシダーゼ)を添加し、28℃で4時間培養した。その培養液を遠心分離(4,000 xg、10分、4℃)にて集菌し、20 mM Tris-HCl(pH 8.0)3 mlに懸濁した。スピッツ管に1.5 mlの菌体液を入れ、on-1分、off-30秒で10分間の超音波破砕を行った。破砕後、遠心分離(15,000 rpm、15分、4℃)を行い、可溶画分と不溶画分とを分離し、得られた可溶画分を0.45 μmのフィルターでろ過した。この溶液を、ニッケルイオンを定着させた、His-trap(登録商標)Chelatingカラム(Amaersham Pharmacia Biotech社)に通した。その後、カラムを数回洗浄し、溶出バッファー(イミダゾール300 mM)によって、N-末端側にHisタグが融合されたIntポリペプチド(配列番号3及び4)を回収した。
【0026】
実施例1 組換えT2intファージの構築
(1)T2-βファージの構築
まず、pUC118ベクターのPstIサイト及びHindIIIサイトに、PCRにより増幅した野生型T2ファージのsoc1-soc断片を挿入し、pUC-Socを得た。大腸菌K12染色体DNAを鋳型としPCRを行うことにより、β-ガラクトシダーゼ遺伝子を得た。このβ-ガラクトシダーゼ遺伝子をpUC-Socのsoc1遺伝子(配列番号7)とsoc遺伝子(配列番号8)との間に挿入し、pUC-βSocを得た。β-ガラクトシダーゼ及びSocが独立に発現するように、β-ガラクトシダーゼのストップコドンは残した。尚、野生型T2ファージは、有坂文雄助教授(東京工業大学大学院生命理工学研究科)より頂戴した。
【0027】
次に、pUC-Socを用い、エレクトロポレーションによって大腸菌BEを形質転換した。この形質転換体を、アンピシリンを添加したLB培地で37℃で振盪培養(120 rpm)した。対数増殖期初期において、培養液にMOI(Multiplicity ofinfection)が0.01[PFU/CFU]となるようにT2ファージを添加した。ファージ添加後、培養液のOD660を経時的に測定し、溶菌に伴う濁度の現象が確認された後(ファージ添加後約4時間)、培養液にクロロホルムを添加し、未溶菌菌体の溶菌を行った。遠心分離(17,400 xg、10分、4℃)により菌体破砕物を除去し、ファージライセートを調製した。このファージライセートを104 PFU/mlとなるようにSM bufferで希釈した。この希釈液と大腸菌JM109の培養液とを混合し、X-gal溶液(40 mg/ml)を50 μl含む軟寒天を用いてプラークアッセイを行い、103 PFU/plateとなるようにプラークを形成させた。形成されたプラークについて、青色を呈するファージを選択することによって、β-ガラクトシダーゼを発現するファージ(T2-βファージ)を得た。
【0028】
(2)pUC-socintの構築
大腸菌O157:H7の染色体DNAを鋳型としPCRを行うことにより、大腸菌O157:H7の接着因子(Int)をコードする遺伝子断片intを得た。pUC-Socのsoc1遺伝子とsoc遺伝子との間にこのint遺伝子を挿入し、pUC-socintを構築した。接着因子(Int)及びSocが共発現するようにintのストップコドンは削除した。
【0029】
(3)組換え体のスクリーニング
上記(1)のT2-βファージはβ-ガラクトシダーゼを発現し、X-galを含むLB培地で大腸菌JM109とプラークアッセイを行うと、青いプラークを形成するが、soc1遺伝子下流で相同組換えが起こり、β-ガラクトシダーゼ遺伝子が別の遺伝子と組換わると、透明なプラークを形成する。この手法を用いることにより、相同組換え後の組換え体のスクリーニングを容易に行うことができる。
組換え体のスクリーニングは以下のようにして行った。まず、組換え用ベクターとしてpUC-socintを用い、エレクトロポレーションによって大腸菌BEを形質転換した。この形質転換体を、アンピシリンを添加したLB培地で37℃で振盪培養(120 rpm)した。対数増殖期初期において、培養液にMOI(Multiplicity of infection)が0.01[PFU/CFU]となるようにT2-βファージを添加した。ファージ添加後、培養液のOD660を経時的に測定し、溶菌に伴う濁度の現象を確認した後(ファージ添加後約4時間)、培養液にクロロホルムを添加し、未溶菌菌体の溶菌を行った。遠心分離(17,400 xg、10分、4℃)により菌体破砕物を除去し、ファージライセートを調製した。このファージライセートを104 PFU/mlとなるようにSM bufferで希釈した。次に、この希釈液と、大腸菌JM109の培養液とを混合し、X-gal溶液(40 mg/ml)を50 μl含む軟寒天を用いてプラークアッセイを行い、103 PFU/plateとなるようにプラークを形成させた。形成されたプラークについて、透明なプラークを選択することにより、インチミン組換え体ファージ(以下、「T2intファージ」とする)をスクリーニングした。
【0030】
実施例2 IgA抗体の検出
まず、以下の4溶液:
A:負対照群(材料(1)で得た大腸菌O157:H7 + 野生型T2ファージ(1010 PFU/マウス))
B:対照群(材料(2)で得たインチミンポリペプチド(0.5 μg/マウス))
C:対照群(実施例1で得たT2intファージ(1010 PFU/マウス))
D:ファージ投与群(材料(1)で得た大腸菌O157:H7 + T2intファージ(1010 PFU/マウス))
を調製した。尚、野生型T2ファージは、抗原(インチミン:int)は発現していない。T2intファージは組換えファージで、頭殻表層にintを発現している。
A〜Dの溶液0.1 mlずつを、4週齢の雌性BALB/cAnNCr1Cr1jマウス(体重約20 g、日本チャールズ・リバー社より購入)にゾンデにより胃内に投与した。投与は各群についてマウス6匹で行った。また、最初の投与から2週間後にブースター投与を行った。
【0031】
次に、産生されたIgA抗体を投与後のマウスの糞便から抽出するため、糞便20 mgをPBS 1 mlに溶解し、滅菌した割り箸でホモジナイズ後、この懸濁液を60分間かけて4〜6回、ボルテックスし、4℃、1,600 xgで15分遠心した。この上清を採り、更に4℃、7,200 xgで10分遠心し、得られた上清を糞便由来溶液とした。
【0032】
糞便由来溶液中のインチミン特異的IgA抗体量をELISAにより測定した。インチミン特異的IgA抗体量は、「インチミン特異的IgA抗体濃度」を「総IgA抗体濃度」で除して得られる値によって評価した。これらの「インチミン特異的IgA抗体濃度」及び「総IgA抗体濃度」は、以下のようにして別々に測定した:
(i)96穴プレートに、carbonate bufferで希釈した抗原(「特異的IgA抗体」についてはインチミン、「総IgA抗体」については抗マウスIgAを各々使用)を100 μl加え、4℃で一晩培養した;
(ii)次いで、PBS-Tween 200 μl(x3)でプレートを洗浄後、ブロックエース(雪印社製)を200 μl加え、ウェルをブロッキングした;
(iii)次いで、PBS-Tween 200 μl(x3)でプレートを洗浄し、適当に希釈したIgA抗体溶液を100 μl加え、1時間振盪した。濃度既知のIgA抗体を15.625〜1000 ng/mlの範囲で希釈し、同様に100 μl加えた;
(iv)次いで、PBS-Tween 200 μl(x5)でプレートを洗浄し、HRP標識二次抗体(抗マウスIgA-α)を加え、1時間振盪し、PBS-Tween 200 μl(x4)、PBS 200 μl(x1)で洗浄した;
(v)次いで、TMB substrate kit(フナコシ社)を各ウェルに100 μlずつ加えた後、反応を停止し、OD 450 nmの吸光度を測定した。
【0033】
結果を図1に示す。図1から明らかなように、B〜Dの各投与群で抗インチミンIgA抗体の産生が見られ、特にD投与群では有意にその産生が認められた。
従って、本発明のファージワクチンは、抗体産生を効率的に誘導することが判った。
【図面の簡単な説明】
【0034】
【図1】図1は、A〜Dの各群投与後の抗インチミンIgA抗体の産生の結果を示す図である。
【特許請求の範囲】
【請求項1】
腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチン。
【請求項2】
前記バクテリオファージが、その頭殻内に、前記抗原タンパク質をコードする遺伝子、及びファージ頭殻構成SOC(Small Outer Capsid)タンパク質をコードする遺伝子を含む、請求項1記載の腸管免疫誘導型ワクチン。
【請求項3】
前記バクテリオファージがT2ファージであって、前記抗原タンパク質が腸管出血性大腸菌O157:H7の接着因子インチミンである、請求項1又は2記載の腸管免疫誘導型ワクチン。
【請求項4】
IgA抗体を産生する、請求項1〜3のいずれか1項記載の腸管免疫誘導型ワクチン。
【請求項1】
腸管感染病原体に感染するバクテリオファージを含む腸管免疫誘導型ワクチンであって、当該バクテリオファージの頭殻表面に腸管感染病原体由来の抗原タンパク質が提示される、前記ワクチン。
【請求項2】
前記バクテリオファージが、その頭殻内に、前記抗原タンパク質をコードする遺伝子、及びファージ頭殻構成SOC(Small Outer Capsid)タンパク質をコードする遺伝子を含む、請求項1記載の腸管免疫誘導型ワクチン。
【請求項3】
前記バクテリオファージがT2ファージであって、前記抗原タンパク質が腸管出血性大腸菌O157:H7の接着因子インチミンである、請求項1又は2記載の腸管免疫誘導型ワクチン。
【請求項4】
IgA抗体を産生する、請求項1〜3のいずれか1項記載の腸管免疫誘導型ワクチン。
【図1】


【公開番号】特開2007−204415(P2007−204415A)
【公開日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願番号】特願2006−24518(P2006−24518)
【出願日】平成18年2月1日(2006.2.1)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【Fターム(参考)】
【公開日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願日】平成18年2月1日(2006.2.1)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【Fターム(参考)】
[ Back to top ]