
フィブロネクチンのエキストラドメインBに対する特異的結合活性を有する修飾ユビキチンタンパク質
本発明は、フィブロネクチンのエキストラドメインB(ED−B)に結合可能な修飾ユビキチンから得られる新規な組換えタンパク質に関する。さらに、本発明は、薬学的活性成分および/または診断用活性成分と融合した前記組換えタンパク質を含む、融合タンパク質に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、フィブロネクチンのエキストラドメインB(ED−B)に結合可能な、新規なタンパク質、特に、ヘテロ多量体タンパク質に関する。さらに、本発明は、薬学的活性成分および/または診断用活性成分と融合した前記結合タンパク質を含む融合タンパク質に関する。さらに、本発明は、このような結合タンパク質または融合タンパク質の製造方法、ならびに、前記結合タンパク質または融合タンパク質を含む医薬組成物/診断用組成物を対象とする。加えて、本発明は、相互作用する結合決定領域(BDR)を少なくとも2個備えた直鎖状ポリユビキチン鎖を含む足場タンパク質をベースとするライブラリーに関する。
【0002】
別の実施形態では、本発明は、前記結合タンパク質または前記融合タンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを含むベクター、および、前記タンパク質、前記融合タンパク質、前記ベクターおよび/または前記ポリヌクレオチドを含む宿主細胞を対象とする。好ましい実施形態において、前記結合タンパク質または前記融合タンパク質は、医薬品または診断剤に含まれる。さらに、前記組換えタンパク質または前記融合タンパク質の製造方法、ならびに、医学的治療方法における前記タンパク質の使用についても記載する。
【背景技術】
【0003】
免疫グロブリンではないアミノ酸からなる結合分子の需要が高まりつつある。現在に至るまで、抗体は、最もよく確立された結合分子に類別されるが、免疫グロブリン分子にはいくつかの主要な欠点があることから、高い親和性と特異性でリガンドを標的とするために、新たな結合分子がなお求められている。免疫グロブリン分子は、非常に容易に製造でき、ほとんど全ての標的を対象とし得るが、その分子構造は極めて複雑である。そのため、容易に取扱い可能な、より小さな分子によって、抗体を代用することが絶えず求められている。これら代替結合剤は、例えば、疾病の診断、予防、および治療といった医療分野において、有用である。
【0004】
比較的明らかな三次元構造を有するタンパク質は、通常、タンパク質足場と呼ばれ、前記代替結合剤の設計の開始物質として用いられる。これら足場は、通常、特定またはランダムな配列変異を許容する1以上の領域を含んでおり、このような配列のランダム化が、タンパク質ライブラリーの製造のためにしばしば行われる。このようなライブラリーから、特異的結合分子を選択してもよい。抗体よりもサイズが小さく、かつ、標的抗原への親和性が、抗体に匹敵するか、あるいはより高い分子は、薬物動態的な性質および免疫原性の点から、抗体よりも優れていると予想される。
【0005】
従来の多くのアプローチにおいて、結合タンパク質の開始物質として、タンパク質足場が用いられている。例えば、国際公開第99/16873号パンフレット(特許文献1)において、特定のリガンドに結合活性を示すリポカリンファミリー(いわゆるアンチカリン)の修飾タンパク質が開発されている。リポカリンファミリーのペプチド構造は、遺伝子工学的方法を用いた、天然のリガンド結合ポケット内でのアミノ酸置換によって修飾されている。アンチカリンは、免疫グロブリンと同様に、分子構造の同定または結合に使用可能である。抗体と同様の方法で、可動性ループ構造が修飾され、これらの修飾により、天然のものとは異なるリガンドの認識が可能となる。
【0006】
国際公開第01/04144号パンフレット(特許文献2)には、それ自体には結合部分がないβシート構造タンパク質のタンパク質表面における、結合ドメインの人工的な生成が記載されている。この手法によれば、高い親和性と特異性でリガンドと相互作用する、新規に生成された人工結合ドメイン(例えば、眼水晶体構造タンパク質であるγクリスタンのバリエーション)が得られる。アンチカリンに関して先に述べたような可動性ループ構造から形成される、既存の結合部位の修飾とは対照的に、これらの結合ドメインは、βシートの表面に新規に生成される。しかしならが、国際公開第01/04144号パンフレットには、新規結合特性を生じるための、比較的大きなタンパク質の改変について記載されているのみである。国際公開第01/04144号パンフレットのタンパク質は、その大きさのため、ある程度の労力を必要とする方法でしか、遺伝子工学レベルで修飾できない。さらに、これまでに開示されたタンパク質においては、タンパク質の全体構造を維持するために、全アミノ酸のうち、比較的小さいパーセンテージのみが修飾されていた。したがって、従前は存在しない結合特性の発生に利用できるのは、タンパク質表面の比較的に小さい領域のみである。さらに、国際公開第01/04144号パンフレットには、γ−クリスタンに対する結合特性の生成のみが記載されている。
【0007】
国際公開第04/106368号パンフレット(特許文献3)には、ユビキチンタンパク質に基づく人工結合タンパク質の製造が記載されている。ユビキチンは、小型、単量体、細胞質性のタンパク質であり、配列が高度に保存され、原虫から脊椎動物にいたるまでのあらゆる既知の真核細胞に存在する。生物において、ユビキチンは、細胞タンパク質の制御分解の調節において重要な役割を担う。この目的のために、分解予定のタンパク質は、酵素カスケードを通過する間に、ユビキチンまたはポリユビキチン鎖と共有結合し、この標識のおかげで、選択的に分解される。最近の研究結果によると、ユビキチン、またはユビキチンによるタンパク質の標識は、それぞれ、数種のタンパク質の移入またはその遺伝子制御等の他の細胞プロセスにおいても、重要な役割を担っている。
【0008】
ユビキチンは、その生理的機能の解明に加えて、主としてその構造およびタンパク質の化学特性のために、研究対象となっている。ユビキチンのポリペプチド鎖は、並外れてコンパクトなα/β構造に折りたたまれた76個のアミノ酸からなる(Vijay−Kumar, 1987(非特許文献1))。前記ポリペプチド鎖のほぼ87%が、水素結合を介した二次構造エレメントの形成に関与する。二次構造は、3回転半のαヘリックス、ならびに、4つの鎖からなる逆平行βシートである。これらの要素の特徴的な配置(逆平行βシートがタンパク質表面に露出し、その裏側にαヘリックスが包まれ、これが前記逆平行βシート上に垂直に延びている)が、一般的に、いわゆるユビキチン様折り畳みモチーフと考えられている。さらなる構造的な特徴は、前記αヘリックスと前記βシート間のタンパク質内部における際立った疎水性領域である。
【0009】
その小さなサイズのため、ユビキチンの人工的な調製は、化学合成によっても、バイオテクノロジー手法によっても実施可能である。その有利な折り畳み特性のため、ユビキチンは、遺伝子工学により、大腸菌等の微生物を使用して比較的大量に、そのサイトゾルまたはペリプラズム空間に製造できる。ペリプラズムにおける優勢な酸化条件のため、後者の戦略は、一般的に、分泌タンパク質の製造に使用される。簡易かつ効率的な細菌調製により、ユビキチンは、その製造に問題がある他の異種タンパク質の調製のための融合パートナーとして使用できる。ユビキチンとの融合によって、溶解度の改善、およびそれによる製造収率の改善が達成できる。
【0010】
抗体または他の代替足場と比べ、ユビキチンタンパク質をベースとする人工結合タンパク質(Affilin(登録商標)ともいう)は、小さいサイズ、高い安定性、高い親和性、高い特異性、費用対効果の高い微生物による製造、血清半減期の調整という利点を有している。しかしながら、特定の標的に対する高い親和性を利用した新たな治療方法の観点から、それらのタンパク質をさらに開発する必要がある。国際公開第05/05730号パンフレット(特許文献4)には、人工結合タンパク質を得るためのユビキチン足場の使用が、概略的に記載されているが、フィブロネクチンのED−Bとの特異的かつ高親和性の結合を得るためにどのようにユビキチンタンパク質を修飾すべきかについては記載されていない。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】国際公開第99/16873号パンフレット
【特許文献2】国際公開第01/04144号パンフレット
【特許文献3】国際公開第04/106368号パンフレット
【特許文献4】国際公開第05/05730号パンフレット
【特許文献5】国際公開第2008/022759号パンフレット
【特許文献6】国際公開第97/45544号パンフレット
【特許文献7】国際公開第07/054120号パンフレット
【特許文献8】国際公開第99/58570号パンフレット
【特許文献9】国際公開第01/62800号パンフレット
【特許文献10】国際公開第06/119897号パンフレット
【特許文献11】国際公開第07/128563号パンフレット
【特許文献12】国際公開第01/62298号パンフレット
【特許文献13】国際公開第07/115837号パンフレット
【非特許文献】
【0012】
【非特許文献1】Vijay−Kumar, 1987
【非特許文献2】Menrad u. Menssen, 2005
【発明の概要】
【発明が解決しようとする課題】
【0013】
国際公開第2008/022759号パンフレット(特許文献5)には、組換え結合タンパク質が記載されており、新しい結合タンパク質を得るために、FYNキナーゼのSrcホモロジー3ドメイン(SH3)を使用している。タンパク質治療法および/またはタンパク質診断法を開発するために、RTループおよび/またはSrcループを変異させて標的特異性を設計できることが見出された。足場として利用されるリポカリンにおいてと同様に、変異誘発されるアミノ酸残基は、抗体/抗原結合機能の基礎をなす原理を擬態する可変性および可動性のループ領域内に存在している。相互作用部位の全体的な可動性により、抗体がエピトープに結合するが、これは主としてエンタルピーによって駆動されるプロセスである。しかしながら、このプロセスは、可動相補性決定領域の会合による移動度の欠失によって、不利なエントロピー寄与を招く。本発明者らは、それとは反対に、ユビキチンを足場として使用し、アミノ酸残基をそもそも可動性ループ領域内で改変するのではなく、βシート領域の強固で不動のβ鎖内、または前記β鎖に密接した範囲内で改変した。ED−Bとの結合領域として、ユビキチンの不動かつ強固なβ鎖または前記β鎖に密接した範囲内でアミノ酸残基を選択する利点は、特に、以下の通りである。結合パートナー同士は、強い結合に適した相補的形状をすでに示していると考えられる。したがって、これらの相互作用には、前記結合パートナーのより強固な構造における形状の相補性、電荷、および親水性/疎水性要素が関係する。これらの強固な本体の相互作用は、界面を最適化し、生物機能に対応する。
【0014】
フィブロネクチン(FN)は、健康な組織および体液において大量に発現する、重要なクラスの高分子量細胞外マトリックス糖タンパク質である。その主な役割は、多数の異なる細胞外マトリックスへの細胞の接着を促進することである。
【0015】
培養下の非形質転換細胞の表面にはフィブロネクチンが存在し、形質転換細胞にはフィブロネクチンが存在しないことから、フィブロネクチンは、重要な接着タンパク質として同定された。フィブロネクチンは、多数の様々な他の分子、例えば、コラーゲン、ヘパラン硫酸プロテオグリカンおよびフィブリン等と相互作用し、細胞の形状および細胞骨格の形成を制御する。さらに、フィブロネクチンは、胚形成時の細胞移入および細胞分化に関与する。フィブロネクチンはまた、創傷治癒においても重要な役割を担い、マクロファージおよび他の免疫細胞の移入を促進し、また、血餅の形成においても、血管の損傷領域への血小板の接着を可能とすることにより、重要な役割を担う。
【0016】
フィブロネクチンのエキストラドメインB(ED−B)は、一次RNA転写物の選択的スプライシングによってフィブロネクチン分子に挿入された、小さいドメインである。前記分子は、細胞外マトリックスのフィブロネクチン分子において、存在するかまたは取去されているかのいずれかであり、新しい血管の周辺に大量に発現するが、実質的に全ての正常成人の組織(子宮と卵巣を除く)において検出不能であるため、血管新生や組織の再構築と関連する最も選択的なマーカーの一つとなっている。ED−Bは、癌および乾癬に関与することが知られている。高レベルのED−B発現が、乳癌、結腸直腸癌、膵臓癌、非小細胞肺癌、肝細胞癌、頭蓋内髄膜腫、ヒトの皮膚癌、膠芽細胞腫を含む多くのヒトの固体癌のエンティティにおいて検出された(Menrad u. Menssen、2005(非特許文献2))。さらに、ED−Bは、診断剤と結合でき、診断ツールとして有利に使用できる。一例として、例えば、動脈硬化プラークの分子の画像化や、例えば、癌患者の免疫シンチグラフィーによる、癌検出おける使用があげられる。さらに多くの診断用途が考えられる。
【0017】
フィブロネクチンのヒトエキストラドメインB(ED−B)の91個のアミノ酸からなるアミノ酸配列を、配列番号2に示す。前記タンパク質を発現させるには、開始メチオニンを加えなければならない。ED−Bは、哺乳類、例えば、げっ歯類、ウシ、霊長類、肉食動物、ヒト等に保存されている。ヒトED−Bと100%の配列同一性を有する動物の例は、ラット(Rattus norvegicus)、ウシ(Bos taurus)、マウス(Mus musculus)、ウマ(Equus caballus)、アカゲザル(Macaca mulatta)、ハイイロオオカミ(Canis lupus familiaris)、チンパンジー(Pan troglodytes)である。
【0018】
ED−Bは、新生血管構造に特異的に蓄積し、癌における分子介入の標的となる。フィブロネクチンのED−Bドメインに対する抗体または抗体断片の多くは、当該技術分野において、癌およびその他の適応症の有望な治療薬として公知である(例えば、国際公開第97/45544号パンフレット(特許文献6)、国際公開第07/054120号パンフレット(特許文献7)、国際公開第99/58570号パンフレット(特許文献8)、国際公開第01/62800号パンフレット(特許文献9)参照)。ヒト一本鎖Fv抗体断片ScFvL19(L19ともいう)は、フィブロネクチンのED−Bドメインに特異的であり、実験腫瘍モデルおよび癌患者の両方において、選択的に新生脈管構造の腫瘍(tumor neovasculature)を標的とすることが確認されている。さらに、IL−12、IL−2、IL−10、IL−15、IL−24またはGM−CSF等のサイトカインと、抗ED−B抗体または抗ED−B抗体断片を含むコンジュゲートは、特に、癌、血管新生、または腫瘍の成長を阻害する医薬品の製造のための標的薬剤として記載されている(例えば、国際公開第06/119897号パンフレット(特許文献10)、国際公開第07/128563号パンフレット(特許文献11)、国際公開第01/62298号パンフレット(特許文献12)参照)。細胞傷害性薬剤または免疫刺激薬剤のような適切なエフェクター機能と抱合させた、抗ED−B抗体またはL19等抗ED−B抗体断片による固体腫瘍の新生脈管構造の選択的なターゲティングは、動物実験では成功することが判明している。膵臓癌の治療のため、インターロイキン−2部分(IL−2)と抗ED−B抗体部分とを含む融合タンパク質を、小分子ゲムシタビン(2’−デオキシ−2’、2’−デフルオロシチジン)と併用した(例えば、国際公開第07/115837号パンフレット(特許文献13)参照)。
【0019】
前記先行技術文献には、新しいED−B結合タンパク質の製造のため、抗体を含む様々なタンパク質足場を使用することが記載されている。
【0020】
現在入手可能な化合物よるED−Bのターゲティングには、ある種の不都合が伴う。ED−B抗原に対して、同等またはより高い親和性を有する、より小さい分子(本発明の、ユビキチンベースのED−B結合タンパク質等)は、抗体や他の結合タンパク質に対して、顕著な利点を有すると期待される。
【0021】
癌は、世界的に主要な死亡原因であるため、癌治療のための改善された薬剤の必要性が高まっている。現在の化学療法剤および放射線治療は、選択性が低いという欠点があり、多くの化学療法剤は、腫瘍部位に蓄積せず、よって腫瘍内において十分なレベルに到達できない。癌の効果的な治療に対する高い医学的ニーズが存在する。
【0022】
よって、本発明の目的は、フィブロネクチンの細胞外ドメイン(ED−B)に非常に高い親和性で特異的に結合できる、ユビキチンベースの新規結合タンパク質の提供である。本発明のさらに別の目的は、例えば、癌の治療に使用する、ED−Bに対して非常に高い結合特異性を有する新規結合タンパク質の同定および提供である。さらに、前記結合分子を製造するための方法も提供する。
【課題を解決するための手段】
【0023】
前記課題は、提出した独立請求項の主題によって解決される。本発明の好ましい実施形態は、従属請求項、ならびに下記の記載、実施例、および図面に含まれる。
【図面の簡単な説明】
【0024】
【図1】図1は、種々の腫瘍におけるED−Bの発生を列記した表を示す。
【図2】図2Aは、修飾ユビキチンED−Bバインダー5E1に関する濃度依存性ELISAの結果を示す。図2Bは、修飾ユビキチンED−Bバインダー1H4に関する濃度依存性ELISAの結果を示す。
【図3】図3は、修飾ユビキチン単量体からなる四量体と比較した、修飾ユビキチン単量体のKd値を、表に示す。
【図4A】図4Aは、単量体41B10について、結合親和性がKd=9.45μMであることを示す。
【図4B】図4Bは、41B10が異なる第2単量体と結合して得られた46H9について、結合親和性がKd=131nMであることを示す。
【図5A】図5Aは、修飾ユビキチンベースのED−B結合エフェクター融合タンパク質の模式図である。
【図5B】図5Bは、前記修飾ユビキチンエフェクター結合5E1−TNFコンジュゲートが、アポトースシス促進活性(L929アポトーシスアッセイで測定)を有すること示す。
【図5C】図5Cは、1H4−TNFα融合物のED−Bへの高結合親和性(Kd=15.1nM)を示す。
【図6】図6は、サイトカイン、例えば、TNFαに融合した、修飾ユビキチンベースのED−B結合へテロ二量体分子の親和性および活性を示す。
【図6A】図6Aは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd 50.7nM)。
【図6B】図6Bは、サイトカインTNFαと遺伝子融合し、前記ヘテロ二量体24H12が多量体化された、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd=5.6nM)。
【図6C】図6Cは、ヘテロ二量体修飾ユビキチンライブラリーから選択される候補例、例えば、ヘテロ二量体クローン9E12、22D1、24H12、41B10の分析結果を示す。
【図6D】図6Dは、Biacore(登録商標)を使用した、前記修飾ヘテロ二量体ユビキチン分子9E12の、無標識相互作用分析による分析結果を示す。
【図6E】図6Eは、Biacore(登録商標)を使用した、前記修飾ヘテロ二量体ユビキチン分子41B10の、無標識相互作用分析による分析結果を示す。
【図7】図7は、結合親和性および結合特異性に対する、異なる修飾がなされたユビキチン変異体の寄与を示す。
【図8】図8は、配列アライメントを示す。
【図9】図9は、「Ub2_TsX9」(両単量体の45位がトリプトファンに修飾され、2つの単量体の間にリンカーGIG(77〜79位;80位のメチオニンから第2単量体が開始)を有し、前記第2単量体の最後のC末端アミノ酸において、グリシンからアラニンに置換されているユビキチン)に対する、修飾ユビキチンへテロ二量体変異体1041−D11(1行目)の配列アライメントを示す。
【図10】図10は、ヒトED−Bへの前記ヘテロ二量体ユビキチン変異体1041−D11の結合についての、濃度依存性ELISAを示す。
【図11】図11は、フィブロネクチン断片を含む固定化されたED−B(67B89)に対するヘテロ二量体ユビキチン変異体1041−D11の結合を、遊離標的の量を増加させながら分析した競合濃度依存性ELISAを示す。
【図12】図12は、Biacore(登録商標)を使用した、前記修飾へテロ二量体ユビキチン分子1041−D11の、無標識相互作用分析における分析結果を示す。
【図13】図13は、結合活性の血清安定性を同時に分析する濃度依存性ELISAにおける、ED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。
【図14】図14は、ヘテロ二量体ユビキチン変異体1041−D11とフィブロネクチン断片との複合体形成の、SE−HPLCによる分析を示す。
【図15A】図15Aは、固定化されたヒト胎児肺繊維芽細胞(Wi38)への前記ヘテロ二量体ユビキチン変異体1041−D11の結合を示す。
【図15B】図15Bは、バイタル(vital)ヒト胎児肺繊維芽細胞(Wi38)への結合を示す。
【図15C】図15Cは、固定化されたマウスBalb 3T3細胞への結合を示す。
【図15D】図15Dは、固定化されたマウスST−2細胞への結合を示す。
【図16A】図16Aは、哺乳類組織切片における、ターゲットへのヘテロ二量体ユビキチン変異体1041−D11の特異性を示す。
【図16B】図16Bは、野生型ユビキチンとの比較における、腫瘍組織での1041−D11の蓄積を示す。
【図17A】図17Aは、1041−D11TNFα融合タンパク質におけるTNFαのアポトーシス誘導活性についての、細胞ベースアッセイ(cell based assay)(L929細胞)によるテストを示す。
【図17B】図17Bは、1041−D11TNFα融合タンパク質におけるTNFαのアポトーシス誘導活性についての、細胞ベースアッセイ(L929細胞)によるテストを示す。
【図17C】図17Cは、標的ED−Bに対するヘテロ二量体ユビキチン1041−D11TNFα融合タンパク質の高い選択性を証明している。
【図17D】図17Dは、修飾ユビキチンED−B結合1041−D11TNFα融合タンパク質についての、Biacoreアッセイによる結合分析を示す。
【図17E】図17Eは、培養細胞における変異体1041−D11で観察された高い結合特異性が、1041−D11がTNFαに融合されても維持されることを示す。
【図18】図18は、TNFαに融合した変異体1041−D11とメルファランとの併用による、7日間のマウスの処置期間中のin vivoでの相対的腫瘍成長を示す。
【図19】図19は、ED−Bに対して驚くほど強力な結合親和性を有することが見出されたさらなる16種類の配列について、コンセンサスな位置およびアミノ酸置換を示す。
【発明を実施するための形態】
【0025】
より具体的には、本発明者らは、配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有する修飾ユビキチンタンパク質を含む、ヒトフィブロネクチンのED−Bに結合可能なタンパク質であり、
前記フィブロネクチンのED−Bに対し、Kd=10−6〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも4個のアミノ酸が修飾されたタンパク質を提供する。
【0026】
好ましい実施形態において、前記タンパク質は、組換えタンパク質である。
【0027】
本発明のさらに別の実施形態では、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、4、5、6、7、8、9個、または全てのアミノ酸が修飾される。
【0028】
本願で使用されている重要な用語の定義
「フィブロネクチンのエキストラドメインB」という用語、またはその略語「ED−B」は、配列番号2との配列同一性が、少なくとも70%、任意に75%以上、さらに任意に80%、85%、90%、95%、96%、または97%以上、または100%を示し、先に定義したED−Bの機能を有する、全てのタンパク質を含む。
【0029】
「結合可能なタンパク質」または「結合タンパク質」という用語は、下記にさらに詳細に定義するように、ED−Bに対する結合ドメインを含むユビキチンタンパク質を指す。このようなユビキチンベースの結合タンパク質は、いずれも、例えば、多量体化部位(multimerization moieties)、ポリペプチドタグ、ポリペプチドリンカー等の、結合ドメインでない付加的なタンパク質ドメイン、および/または、非タンパク性のポリマー分子を含んでもよい。非タンパク性のポリマー分子は、例えば、ヒドロキシエチルでんぷん、ポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレン等である。
【0030】
抗体およびその断片は、当業者に周知である。本発明の結合タンパク質は、抗体またはその断片、例えば、FabもしくはscFv断片等ではない。また、本発明の結合ドメインは、抗体中に存在する免疫グロブリンフォールドを含まない。
【0031】
本明細書において、「リガンド」、「標的」および「結合パートナー」は、同義に用いられ、置き換え可能である。リガンドは、前述のヘテロ多量体修飾ユビキチンタンパク質に、本明細書中で定義される親和性をもって結合可能な任意の分子である。
【0032】
「ユビキチンタンパク質」という用語は、配列番号1のユビキチン、および、下記の定義によるその修飾物を包含する。ユビキチンは、真核生物において、高度に保存されている。例えば、今日まで研究されてきた全ての哺乳類において、ユビキチンは、同一のアミノ酸配列を有する。ヒト、げっ歯類、ブタ、および霊長類由来のユビキチン分子が、特に好ましい。また、任意の他の真核生物起源のユビキチンを使用することもできる。例えば、酵母由来のユビキチンは、配列番号1と3個のアミノ酸が異なるのみである。前記「ユビキチンタンパク質」という用語に包含される前記ユビキチンタンパク質は、通常、配列番号1に対し、70%を超える、好ましくは75%を超える、80%を超える、85%を超える、90%を超える、95%を超える、96%を超えるアミノ酸同一性を示すか、あるいは、97%までの配列同一性を示す。
【0033】
「修飾ユビキチンタンパク質」という用語は、アミノ酸の置換、挿入、または欠失のいずれか一つ、あるいはそれらの組み合わせによる、前記ユビキチンタンパク質の修飾物を指す。本発明の修飾ユビキチンタンパク質は、標的に対する新規な結合親和性を有する改変タンパク質である。
【0034】
配列番号1のアミノ酸配列に対する前記ユビキチン誘導体の配列同一性の程度の決定には、例えば、SIM Local similarity program(Xiaoquin Huang and Webb Miller, Advances in Applied Mathematics, vol. 12: 337-357, 1991)またはClustal,W.を使用できる(Thompson et al., Nucleic Acids Res., 22(22): 4673-4680, 1994.)。好ましくは、配列番号1に対する前記修飾タンパク質の配列同一性の程度は、配列番号1の完全配列に対して相対的に決定される。
【0035】
本発明の「ヘテロ二量体融合タンパク質」または「ヘテロ二量体タンパク質」は、異なる修飾がなされた2つの単量体ユビキチンタンパク質を含むタンパク質と考えられる。前記2つの単量体ユビキチンタンパク質は、特異的結合パートナーであるED−Bに対する一価の結合特性(結合ドメイン)を共同でもたらす、2つの相互作用結合ドメイン領域を有する。ヘテロ二量体は、2つの単量体ユビキチン分子の融合により得られるが、これらの両分子は、本明細書に記載のように、異なる修飾がなされている。
【0036】
異なる修飾がなされたユビキチン単量体を多量体化して一価の結合活性を有するヘテロ多量体結合タンパク質(ここでは、ヘテロ二量体タンパク質)を生成する利点は、ED−Bに対する新規な高親和性の結合特性を発生させるために修飾することが可能なアミノ酸残基の総数の増加にある。主な利点は、修飾されるアミノ酸が増加しても、ED−Bに対する前記新規に生成された結合タンパク質の足場の全体的な安定性を低下させることなく、タンパク質化学的完全性(protein−chemical integrity)が維持されることである。ED−Bに対する新規結合部位を発生させるために修飾可能な残基の総数は、前記修飾残基が2つの単量体ユビキチンタンパク質に割り当てられることで増加する。修飾数は、修飾単量体ユビキチン分子数に応じて、2倍にできる。ユビキチンベースのED−B結合タンパク質のモジュール構造により、修飾アミノ酸の総数を増加できる。これは、前記修飾アミノ酸が、2つの単量体ユビキチン分子に含まれるからである。本発明の方法は、ED−Bに対する一価の特異性を有するヘテロ二量体ユビキチン分子の同定を提供する。
【0037】
したがって、結合パートナーに対して共通の結合部位を有するヘテロ二量体の使用は、最終的な結合分子のタンパク質化学的完全性に過度に影響を与えない修飾残基数を増加させる可能性を切り開く。なぜなら、これらの修飾残基の全量が、前記二量体を形成する2つの単量体ユニットに分散するためである。ED−Bに結合する前記ヘテロ二量体修飾ユビキチンタンパク質は、タンパク質ライブラリーに存在する。
【0038】
「一価」とは、前記修飾二量体ユビキチンの第1および第2単量体ユニットにおいて生成された両結合領域が、共に、ED−Bに相乗的に組み合わせられるように結合する機能、すなわち、両結合領域が、共同で一価の結合活性を形成するように作用する能力と理解すべきである。前記へテロ二量体分子における前記第1および前記第2修飾ユビキチン両方の各結合領域を別々に見た場合、前記二量体分子と比較して、ED−Bへの結合の効率および親和性は、明らかに低いと考えられる。前記修飾ユビキチンが、各単量体タンパク質を単独で使用するよりも、より効率的にED−Bに結合可能となるように、両結合領域は、前記ヘテロ二量体修飾ユビキチンタンパク質表面に、アミノ酸の連続領域として形成される固有の結合部位を形成する。本発明によれば、最も有力な結合ユビキチン分子をスクリーニングした後に、前記2つの単量体タンパク質が互いに結合されるのではなく、既に前記ヘテロ二量体ユビキチンが存在する状態でスクリーニング工程を行うことが、特に重要である。最も有力な結合ユビキチン分子の配列情報を取得した後に、任意の他の方法、例えば、化学合成または遺伝子工学手法により、例えば、前記2つの同定した単量体ユビキチンユニット同士を結合することにより、これらの分子を得てもよい。
【0039】
本発明によれば、1つのリガンドに結合する異なる修飾がなされた前記少なくとも2つのユビキチン単量体は、例えば、遺伝学的手法を使用して、互いにヘッドトゥテイル融合で結合される。前記異なる修飾がなされた融合ユビキチン単量体は、一価で結合し、両方の「結合ドメイン領域」(「BDR」)が共に作用する場合にのみ効果的である。本明細書中において、「結合ドメイン領域」は、標的への結合に関与する、配列番号1の2、4、6、8、62、63、64、65、66および68位における、少なくとも4個、好ましくは6個のアミノ酸が修飾されたユビキチン単量体における領域と定義される。
【0040】
前記ヘテロ二量体タンパク質を形成する、前記修飾され結合されたユビキチン単量体は、一つの連続的な結合領域を介して同一のエピトープに結合する。前記ヘテロマーのこの連続的な領域は、異なる修飾がなされた2つのユビキチン単量体により形成される前記2つのモジュールの両結合決定領域により形成される。
【0041】
「ヘッドトゥテイル融合」は、2つのタンパク質を、前記二量体に含まれるユニット数に応じて、N−C−N−C方向に連結することにより、互いに融合させることと理解すべきである。このヘッドトゥテイル融合において、前記ユビキチン単量体は、リンカーを介さずに直接連結されてもよい。または、前記ユビキチン単量体の融合を、リンカーを介して行うこともできる。前記リンカーとしては、例えば、少なくともGIGのアミノ酸配列もしくは少なくともSGGGGのアミノ酸配列を有するリンカー、または、任意の他のリンカー、例えば、GIG、SGGGG、SGGGGIG、SGGGGSGGGGIGもしくはSGGGGSGGGGがあげられる。2つのユビキチン単量体の遺伝子融合用の他のリンカーも、当該技術分野では公知であり、使用可能である。
【0042】
本発明の修飾ユビキチンタンパク質は、標的またはリガンド(両用語は、同じ意味として使用される)としてのED−Bに対する新規な結合親和性を有する改変タンパク質である。「置換」という用語は、例えば、元のアミノ酸への化学基もしくは残基の置換または付加によるアミノ酸の化学的修飾も含む。βシート領域の少なくとも1つのβシート鎖に位置するアミノ酸または前記βシート鎖に隣接した3つ目までのアミノ酸を含む、タンパク質の少なくとも1つの表面露出領域におけるアミノ酸の置換が重要である。
【0043】
本発明によれば、ED−Bに特異的な新規の結合ドメインの生成のためのアミノ酸の置換を、任意の所望のアミノ酸を用いて行うことができる。すなわち、ED−Bに対する新規結合特性を発生させるための修飾において、前記アミノ酸が、置換するアミノ酸のものと類似する特定の化学的特性または側鎖を有するよう配慮する必要はなく、よって任意の所望のアミノ酸をこの目的に使用することができる。
【0044】
本発明によれば、選択されたアミノ酸の修飾工程は、ランダム突然変異誘発による遺伝子レベルでの突然変異誘発、すなわち、前記選択されたアミノ酸のランダム置換により行われることが好ましい。ユビキチンの修飾は、各タンパク質に属するDNAを改変する遺伝子工学的手法により行われることが好ましい。ユビキチンタンパク質の発現は、その後、原核生物または真核生物内で行われることが好ましい。
【0045】
置換は、特に、ユビキチンタンパク質のβシートの4つのβ鎖の表面露出アミノ酸、または、前記βシート鎖に隣接する3つ目までの表面露出アミノ酸において行われる。各β鎖は、通常、5〜7個のアミノ酸からなる。配列番号1を参照すると、例えば、前記β鎖は、通常、アミノ酸残基2〜7、12〜16、41〜45および65〜71を包含する。これに加えて好ましく修飾され得る領域として、前記βシート鎖に隣接する3つ目までのアミノ酸位置(すなわち、1番目、2番目または3番目)が挙げられる。これに加えて好ましく修飾され得る好ましい領域としては、特に、アミノ酸残基8〜11、62〜64および72〜75が挙げられる。前記好ましい領域は、2つのβ鎖を互いに連結するβターンを含む。好ましいβターンとして、アミノ酸残基62〜64があげられる。前記βシート鎖に密接する最も好ましいアミノ酸は、8位のアミノ酸である。また、アミノ酸置換のさらなる好ましい例は、36、44、70および/または71位である。例えば、これに加えて好ましく修飾され得る領域としては、62、63、および64位のアミノ酸(3個のアミノ酸)、72、73位のアミノ酸(2個のアミノ酸)、または8位のアミノ酸(1個のアミノ酸)が挙げられる。
【0046】
好ましい実施形態において、前記アミノ酸残基は、アミノ酸置換により改変される。欠失および挿入も可能である。付加または欠失され得るアミノ酸数は、単量体ユビキチンサブユニットにおいて、1、2、3、4、5、6、7、8、9、または10個のアミノ酸に限定され、それに応じて、前記二量体ユビキチンタンパク質については、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、18、20、22、24、26、または28個のアミノ酸に限定され、通常は、前記単量体タンパク質における修飾数のx倍である。一実施形態において、アミノ酸挿入はなされない。さらに別の実施形態において、欠失は行われない。
【0047】
本発明の修飾ユビキチンタンパク質が、1以上のアミノ酸の置換、欠失および/または付加を含む場合には、対応するタンパク質同士を割り当てるために、野生型ヒトユビキチン(配列番号1)のアミノ酸位置と、前記修飾ユビキチンとのアライメントを行わなければならない。融合タンパク質の場合(後述を参照)、各単量体ユビキチンサブユニットのナンバリング(およびアライメント)は、同様の方法によりなされる。すなわち、例えば、二量体のアライメントは、各サブユニットについて、1位のアミノ酸から開始される。
【0048】
本発明によれば、好ましくは哺乳類由来の、例えば、ヒトの単量体ユビキチンにおいて、β鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置に存在するアミノ酸の、少なくとも10%、好ましくは少なくとも20%、さらに好ましくは少なくとも25%を修飾、好ましくは置換することができ、これにより、以前は存在しなかった結合特性が発生する。β鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置に存在するアミノ酸の、好ましくは最大で約50%、さらに好ましくは最大で約40%もしくは約35%、または約30%もしくは約25%までが修飾、好ましくは置換される。1つのβ鎖内において、通常、1〜4個のアミノ酸が修飾される。一実施形態では、好ましくは、第1β鎖および第4β鎖、例えば、アミノ酸残基2〜7もしくは65〜71の領域において、6個のアミノ酸のうちの3個が修飾される。
【0049】
ヘテロ二量体の構成ユニットとして使用される本発明の修飾単量体ユビキチンは、合計でアミノ酸の20%までを占める。これを考慮すると、配列番号1に対する前記修飾ユビキチンタンパク質の配列同一性は、少なくとも60%である。本発明のさらに別の実施形態において、アミノ酸レベルでの配列同一性は、配列番号1のアミノ酸配列に対して、少なくとも60%、70%、少なくとも80%、さらに、少なくとも90%または少なくとも95%である。本発明は、配列番号1のアミノ酸配列と比較して、65%を超える、75%超える、85%超える、または97%超える前記修飾ユビキチンタンパク質のアミノ酸配列同一性も包含する。
【0050】
本発明のさらに別の実施形態において、ユビキチンは、配列番号1の2、4、6、8、62、63、64、65、66および/または68位における、3、4、5、6、または7個のアミノ酸が修飾される。他の実施形態では、これらの位置が修飾されるユビキチンを、既に前修飾(pre−modified)されていた。例えば、さらなる修飾は、74および75位のアミノ酸または45位のアミノ酸における修飾を含むことができ、この修飾により、より良好な安定性またはタンパク質化学特性が発生する。修飾ユビキチン単量体は、配列番号1のユビキチンにおける、合計9、10、11、12、13、14、および、最大で15個のアミノ酸が修飾、好ましくは置換されることで得られる。一例によれば、14個の置換および1個の欠失を有する修飾単量体ユビキチンを得ることができた。これは、ユビキチンの総アミノ酸数の約20%の割合に相当する。このことは、非常に驚くべきことであり、通常、もっと低い割合でも、タンパク質のフォールディングを阻害するのに十分であるため、予測できなかったことである。
【0051】
本発明の一実施形態において、これらのアミノ酸は、タンパク質表面上に連続的な領域を形成する、新規な結合特性を有する領域を生成するために修飾される。このようにして、ED−Bへの結合特性を有する連続的な領域を生成することができる。本発明において、「連続的な領域」は、以下の通りである。側鎖の電荷、空間的構造、および疎水性/親水性により、アミノ酸は、それらに応じた形で環境と相互作用する。前記環境は、溶媒、一般的には水、または、例えば、空間的に近いアミノ酸等の他の分子であり得る。タンパク質に関する構造情報および各種ソフトウェアにより、前記タンパク質の表面を特徴付けできる。例えば、タンパク質の原子と溶媒との界面領域について、この界面領域がどのような構造であるか、溶媒に接触しやすい表面領域はどれか、または、電荷が表面においてどのように分布しているかについての情報も含めて、この方法により可視化できる。連続的な領域は、例えば、適切なソフトウェアを使用したこの種の可視化により明らかにすることができる。このような方法は、当業者に公知である。本発明によれば、基本的に、表面露出領域全体を、新規結合特性の生成のために修飾される表面上の前記連続的な領域として使用することができる。一実施形態において、この目的のために、修飾は、αへリックス領域も含むことができる。ヘテロ二量体修飾ユビキチンタンパク質において、結合決定領域は、1つの連続的な領域を共同で形成する2つの前記表面露出領域を含み、前記連続的な領域は、1つの結合決定領域の2倍の長さを有する。
【0052】
βシート領域の少なくとも一本のβ鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置のいずれかを含む、前記タンパク質の少なくとも1つの表面露出領域においてアミノ酸を修飾することが重要である。前記「βシート構造」は、本質的にシート状であり、ほぼ完全に伸長している(stretched)ことにより定義される。ポリペプチド鎖の連続したセグメントから形成されるαへリックスとは対照的に、βシートは、ポリペプチド鎖の異なる領域により形成され得る。このようにして、一次構造では離れて位置する領域同士が、互いに近接することができる。β鎖は、一般的に、5〜10アミノ酸長(通常、ユビキチンにおいて5〜6残基)であって、ほぼ完全に伸長した立体構造である。前記β鎖は、互いに近接しており、一方の鎖のC=O基と他方の鎖のNH基との間で、水素結合が形成される。逆もまた同様である。βシートは、複数の鎖から形成され、シート状構造を有することができ、Cα原子の位置が、シート状平面の上方または下方の間を行ったり来たりする。アミノ酸側鎖は、このパターンに追随し、よって、二者択一的に、上の方向又は下の方向を指す。前記β鎖の方向により、前記シートは、平行シートおよび逆平行シートに分類される。本発明によれば、両方とも、変異可能であり、請求項記載のタンパク質の調製に使用可能である。
【0053】
前記β鎖および前記βシート構造の突然変異誘発のために、表面に近いβ鎖または前記β鎖(前記βシートの1つの鎖)に隣接する3つ目までのアミノ酸位置が、ユビキチンにおいて選択される。表面露出アミノ酸は、入手可能なX線結晶構造により同定できる。結晶構造が入手できない場合には、コンピューター解析を用いて、入手可能な一次構造について表面露出βシート領域および個々のアミノ酸位置の接触性(accessibility)を予測するか、あるいは3次元タンパク質構造をモデル化し、このようにして、潜在的な表面露出アミノ酸についての情報を入手することを試みることができる。さらなる開示は、例えば、J. Mol. Biol., 1987 Apr 5; 194(3):531−44. Vijay−Kumar S, Bugg C.E., Cook W.Jから得ることができる。
【0054】
しかしながら、突然変異させるアミノ酸位置の時間のかかる事前選択を省略して、前記βシートまたは前記β鎖に隣接する3つ目までのアミノ酸位置における修飾を行うことも可能である。前記βシート構造または前記βシート鎖に隣接する3つ目までのアミノ酸をコードするDNA領域を、DNA環境(DNA environment)から単離し、ランダム突然変異誘発に供し、その後、それらを予め除去したタンパク質をコードするDNAに再度組み込む。続いて、所望の結合特性を有する変異体の選択工程を行う。
【0055】
本発明の他の実施形態においては、表面に近い、前記β鎖または前記β鎖に隣接する3つ目までのアミノ酸位置は、前述のように選択され、これらの選択された領域内の突然変異誘発するアミノ酸位置が同定される。ついで、このようにして選択されたアミノ酸位置は、その後、部位特異的突然変異誘発により、DNAレベルで突然変異を誘発され得る。すなわち、特定のアミノ酸をコードするコドンが、予め選択された他の特定のアミノ酸をコードするコドンにより置換される。あるいは、この置換は、ランダム突然変異誘発により行われる。この場合、置換されるアミノ酸位置は規定されるが、新規な未決定のアミノ酸をコードするコドンは規定されない。
【0056】
「表面露出アミノ酸」は、周囲の溶媒に接触可能なアミノ酸である。タンパク質におけるアミノ酸の接触性が、モデルトリペプチドGly−X−Glyにおけるアミノ酸の接触性と比較して8%を超える場合、前記アミノ酸は、「表面露出」と呼ばれる。これらのタンパク質領域または個々のアミノ酸位置もまた、それぞれ、本発明により選択される、潜在的な結合パートナーに対する好ましい結合部位である。また、参考文献として、Caster et al., 1983 Science, 221, 709 − 713, and Shrake & Rupley, 1973 J. Mol. Biol. 79(2):351−371があげられ、開示全体を、引用により本願に取り込む。
【0057】
アミノ酸置換により、新規に生成された人工結合部位の領域が、その親タンパク質と比べて、ならびに互いに相違するユビキチンタンパク質足場のバリエーションを、それぞれの配列セグメントの標的突然変異誘発により生成できる。この場合、極性、電荷、溶解性、疎水性/親水性等の特定の特性を有するアミノ酸を、それぞれ、他の特性を有するアミノ酸と交換または置換できる。置換に加えて、「突然変異誘発」、「修飾」、および「交換」という用語は、挿入および欠失も含む。タンパク質レベルでは、前記修飾を、当業者に公知の方法によるアミノ酸側鎖の化学変性により行うこともできる。
【0058】
ユビキチンの突然変異誘発方法
各配列セグメントの突然変異誘発の開始点として、例えば、当業者に公知の方法により調製、改変、および増幅が可能なユビキチンのcDNAを使用できる。一次配列の比較的狭い領域(例えば、1〜3個のアミノ酸)におけるユビキチンの部位特異的改変には、市販の試薬および方法が利用可能である(「Quick Change」、Stratagene;「Mutagene ファージミド in vitro Mutagenesis Kit」、Biorad)。より大きい領域の部位特異的な突然変異誘発には、具体的な実施形態として、例えば、ポリメラーゼ連鎖反応(PCR)が、当業者に利用可能である。この目的のために、所望の位置に縮退した塩基対組成を有する合成オリゴデオキシヌクレオチドの混合物が、例えば、突然変異の導入に使用できる。これは、イノシン等の、ゲノムDNAには自然発生しない塩基対類似体の使用によっても達成できる。
【0059】
βシート領域の1つ以上のβ鎖または前記βシート鎖に隣接する3つ目までのアミノ酸位置の突然変異誘発の開始点は、例えば、ユビキチンのcDNAでもよいし、ゲノムDNAでもよい。また、ユビキチンタンパク質をコードする遺伝子は、合成によって調製されてもよい。
【0060】
突然変異誘発には、それ自体公知の様々な方法が利用可能である。例えば、部位特異的突然変異誘発法、ランダム突然変異誘発法、PCRを使用する突然変異誘発、または類似の方法が挙げられる。
【0061】
本発明の好ましい実施形態において、突然変異が誘発されるアミノ酸位置は予め定められている。修飾されるアミノ酸の選択は、修飾されるべきアミノ酸に関する請求項1の限定を満たすように行われる。いずれの場合にも、一般的に、様々な変異体のライブラリーが確立され、それ自体公知の方法を用いてスクリーニングされる。一般的に、修飾されるアミノ酸の事前選択は、修飾されるユビキチンタンパク質について、十分な構造情報が入手できる場合には、特に容易に実施できる。
【0062】
例えば、PCR、化学的突然変異誘発、またはバクテリアの突然変異誘発株の使用により、標的突然変異誘発およびより長い配列セグメントの突然変異誘発を行う方法もまた、従来技術に属しており、本発明において利用可能である。
【0063】
本発明の一実施形態において、突然変異誘発は、アミノ酸コドンNNKを有するDNAオリゴヌクレオチドのアセンブリにより行われる。ただし、他のコドン(トリプレット)も使用可能であることも理解すべきである。突然変異は、好ましくはβシート構造が維持されるように行われる。一般的に、突然変異誘発は、タンパク質表面に露出する安定なβシート領域の外側で行われる。それには、部位特異的突然変異誘発およびランダム突然変異誘発の両方が含まれる。一次構造の比較的狭い領域(例えば、3〜5個のアミノ酸)を含む部位特異的突然変異誘発は、市販のキットにより行える。前記市販のキットは、Stratagene(登録商標)(QuickChange(登録商標))またはBio−Rad(登録商標)(Mutagene(登録商標) ファージミド in vitro mutagenesis kit)(米国特許第5,789,166号明細書および米国特許第4,873,192号明細書参照)があげられる。
【0064】
より広い領域を部位特異的突然変異誘発に供する場合、DNAカセットを調製しなければならない。ここで、突然変異が誘発される領域は、突然変異させる位置および変化していない位置を含むオリゴヌクレオチドのアセンブリにより得られる(Nord et al., 1997 Nat. Biotechnol. 8, 772−777; McConell and Hoess, 1995 J. Mol. Biol. 250, 460−470.)。ランダム突然変異誘発は、突然変異誘発株におけるDNAの増殖、または、PCR増幅(エラープローンPCR)により導入できる(例えば、Pannekoek et al., 1993 Gene 128, 135 140)。この目的のため、エラー率が増大したポリメラーゼが使用される。導入される突然変異誘発の度合いを高めるため、あるいは、異なる変異同士をそれぞれ組み合わせるために、PCR断片における変異を、DNAシャッフリングにより組み合わせることができる(Stemmer,1994 Nature 370,389−391)。酵素に関するこれらの突然変異誘発戦略についての概説は、Kuchner and Arnold (1997) TIBTECH 15, 523−530の概説中に記載されている。このランダム突然変異誘発を選択されたDNA領域において実施するためにも、突然変異誘発に使用するDNAカセットの構築が必要である。
【0065】
ランダム修飾は、当該技術分野において、十分に確立された公知の方法により行われる。「ランダムに修飾されたヌクレオチド配列またはアミノ酸配列」は、多数の位置において、ヌクレオチドまたはアミノ酸が挿入、欠失、または置換されており、特性が予測できないヌクレオチド配列またはアミノ酸配列である。多くの場合、挿入されたランダムヌクレオチド(アミノ酸)配列またはヌクレオチド(アミノ酸)配列は、(例えば、ランダム化合成またはPCRを介した突然変異誘発の結果)「完全にランダム」となる。ただし、前記ランダム配列は、共通の機能的特性(例えば、発現産物のリガンドへの反応性)を有する配列も含むことができる。あるいは、前記ランダム配列は、最終的な発現産物が、例えば、異なるアミノ酸が均等に分布した完全にランダムな配列であるという意味で、ランダムとなり得る。
【0066】
ランダム化された断片をベクター中に適切に導入するために、本発明では、前記ランダムヌクレオチドは、部位特異的PCRを介した突然変異誘発の原理により、発現ベクター中に導入されるのが好ましい。ただし、他の選択肢も当業者に公知であり、例えば、合成ランダム配列ライブラリーをベクターに挿入することも可能である。
【0067】
融合PCRにより変異体またはライブラリーを発生させるために、例えば、3回のPCR反応を行ってもよい。2回のPRC反応は、部分的に重なった中間体断片を発生するために行われる。3回目のPCR反応は、前記中間体断片を融合するために行われる。
【0068】
ライブラリーまたは変異体株の構築方法は、所望の制限酵素認識部位周辺のプライマー(制限酵素認識部位プライマー)であるフォワード制限プライマーとリバース制限プライマーからなる第1のプライマーセット、および、例えば、対象のコドンの上流および下流周辺のプライマー(変異原性プライマー)であるフォワード変異原性プライマーおよびリバース変異原性プライマーからなる第2のプライマーセットを構築する工程を含んでもよい。一実施形態において、前記プライマーは、対象のコドンのすぐ上流およびすぐ下流に構築される。前記制限プライマーおよび前記変異原性プライマーを使用して、第1中間体断片および第2中間体断片が構築される。2回のPCR反応により、これらの直線状の中間体断片が生成される。これらの各直線状の中間体断片は、対象の変異コドンを少なくとも1つと、フランキングヌクレオチド配列と、切断部位とを含む。3回目のPCR反応では、前記2つの中間体断片、前記フォワード制限プライマー、および前記リバース制限プライマーを使用して、直線状の融合産物が生成される。一方、前記直線状産物の未結合の末端は、制限酵素で切断され、前記直線状産物に付着末端が生じる。前記直線状産物の前記付着末端は、DNAリガーゼの使用により融合され、環状産物、例えば、環状のポリヌクレオチド配列が生成される。
【0069】
前記中間体断片の構築のため、フォワードプライマーおよびリバースプライマーの2つのセットの設計および合成は、制限酵素切断部位およびそのフランキングヌクレオチド配列を含む第1のセット、ならびに、対象の変異コドンを少なくとも1つ含む第2のセット(変異原性プライマー)を用意することによって行われる。当業者であれば、前記変異体の数は、所望の変異アミノ酸修飾の数に対応することを認識するであろう。本発明者は、他の制限酵素を前記工程に使用する場合には、この切断部位の正確な位置、ならびに、前記フォワードプライマーおよび前記リバースプライマーの対応する配列を、これに応じて改変すればよいと企図している。その他の方法も当該技術分野において利用可能であり、これらを代わりに使用してもよい。
【0070】
本発明に従って発現産物のランダム化断片を足場に導入することとは別に、少なくとも1つの融合パートナーをコードするヌクレオチド配列にランダム化ヌクレオチド配列を融合させることにより、前記ランダム配列を融合パートナーに連結することがしばしば必要となる。このような融合パートナーは、例えば、前記発現産物の発現および/または精製/単離および/またはさらなる安定化を促進できる。
【0071】
本発明の一例によれば、単量体ユビキチンの2、4、6、8、62、63、64、65、66および/または68位における、少なくとも3または4個のアミノ酸のランダム置換を、PCRにより非常に簡易に行うことができる。これは、前述の位置が、前記タンパク質のアミノ末端またはカルボキシ末端の近くに局在しているためである。したがって、操作されるコドンは、対応するcDNA鎖の5’末端および3’末端に位置する。そこで、突然変異誘発PCR反応に使用される第1のオリゴデオキシヌクレオチドは、変異される2、4、6および/または8位のコドンから離れており、その配列は、ユビキチンのアミノ末端のコード鎖に対応する。したがって、第2のオリゴデオキシヌクレオチドは、変異される62、63、64、65、66および/または68位から離れており、カルボキシ末端のポリペプチド配列の非コード鎖に、少なくとも部分的に対応する。両オリゴデオキシヌクレオチドにより、単量体ユビキチンをコードするDNA配列を鋳型として用い、ポリメラーゼ連鎖反応を行うことができる。
【0072】
また、得られた増幅産物を、例えば、制限エンドヌクレアーゼの認識配列を導入するフランキングオリゴデオキシヌクレオチドを使用する、他のポリメラーゼ連鎖反応に添加することが可能である。本発明では、得られた遺伝子カセットを、所定のハプテンまたは抗原に対する結合特性を有するユビキチン変異体の単離のための次の選択工程での使用に適したベクター系中に導入するのが好ましい。
【0073】
ユビキチンの修飾領域
修飾領域は、基本的にが、結合パートナーであるED−Bに接触可能かどうか、ならびに、タンパク質の全体構造が、推測上修飾に耐性を示すかどうかに応じて選択できる。
【0074】
表面露出β鎖における修飾に加えて、前記タンパク質の他の表面露出領域における修飾も可能であり、前記修飾は、前記β鎖に隣接する3つ目までのアミノ酸位置において行われことが好ましい。これらの修飾領域は、新規に生成される、ED−Bに対する高結合親和性での結合に関与する。
【0075】
本発明の別の好ましい実施形態では、ユビキチン、好ましくは、哺乳類またはヒトのユビキチンにおける表面露出アミノ酸の少なくとも3、4、または6個、任意に、少なくとも8、10、12個、最大で15個を修飾することができ、ここで、前記修飾としては、置換が好ましい。これは、ユビキチンにおける3、4、5、6、7、8、9、10、11、12、13、14、または15個の表面露出アミノ酸の修飾を含む。これらの4個以上最大15個の表面露出修飾アミノ酸により、所定の結合パートナーに対して結合親和性を有する領域が形成される。本明細書中において、この領域を、「結合ドメイン領域」(BDR)と定義する。これに関しては、少なくとも2個、任意に少なくとも4個、さらに任意に少なくとも6、8、10、12個、最大で15個の表面露出アミノ酸が、βシート領域内に存在すること、すなわち、単一のβシート鎖に存在するか、もしくは複数のβ鎖に分散すること、または、βシート鎖に隣接する3つ目までのアミノ酸位置に存在することが特に好ましい。修飾された、好ましくは置換された全てのアミノ酸のうちの少なくとも3個が、一次配列において、互いに直接隣接していることがさらに好ましい。
【0076】
本発明の別の任意の実施形態において、前記タンパク質における4つのβ鎖の1つもしくは2つにおけるアミノ酸、好ましくは2つにおけるアミノ酸、または、前記4つのβ鎖のうちの好ましくは2つに隣接する3つ目までのアミノ酸位置におけるアミノ酸が、修飾されて、新規の結合特性を生成している。前記4つのβ鎖のうちの3つもしくは4つにおける修飾、または前記β鎖のうちの3つもしくは4つに隣接する3つ目までのアミノ酸位置における修飾も、ED−B結合発生のために、任意で行ってもよい。
【0077】
アミノ末端鎖およびカルボキシ末端鎖におけるアミノ酸、または、アミノ末端鎖およびカルボキシ末端鎖に隣接する3つ目までのアミノ酸位置におけるアミノ酸を修飾、好ましくは置換して、ED−Bへの新規の結合部位を生成することが特に好ましい。これに関し、前記カルボキシ末端βシート鎖に隣接する4つ目までのアミノ酸が修飾、好ましくは置換されること、および、前記アミノ末端βシート鎖に隣接する1つ目までのアミノ酸が修飾、好ましくは置換されることが特に好ましい。
【0078】
哺乳類のユビキチン、好ましくはヒトのユビキチンにおける下記の位置の表面露出アミノ酸の少なくとも3つが、修飾される、好ましくは置換されることが特に好ましい:2、4、6、8、62、63、64、65、66、68位。前記アミノ酸グループから選択されるこれら少なくとも4つのアミノ酸により、ユビキチンの表面に連続的な表面露出領域が形成されるが、前記領域は、結合パートナーであるED−Bに対し、以前は存在しなかった結合親和性を有する修飾タンパク質の生成に特に適していることが見出された。これらのアミノ酸残基のうち、少なくとも3つが修飾されなければならない。前記アミノ酸残基の3、4、5、6、7、8、9または10個が、任意にさらに別のアミノ酸残基と共に、任意に修飾される。
【0079】
上記修飾後、本発明者らは、実施例で述べるアミノ酸修飾ユビキチン配列が、非常に高い親和性(Kd値10−9以下)でED−Bと結合することを見出した。
【0080】
融合タンパク質
別の好ましい実施形態において、本発明は、薬学的活性成分および/または診断用活性成分と融合した本発明の結合タンパク質を含む融合タンパク質に関する。
【0081】
さらに別の態様において、本発明は、薬学的活性成分および/または診断用活性成分と融合した本発明のヘテロ二量体結合タンパク質を含む融合タンパク質に関する。本発明の融合タンパク質は、非ポリペプチド成分、例えば、非ペプチドリンカー、例えば、治療または診断に関連する放射線核種に対する非ペプチドリガンドを含んでもよい。前記融合タンパク質は、さらに、低分子の有機化合物または非アミノ酸化合物、例えば、糖、オリゴ糖、多糖、脂肪酸等を含んでもよい。本発明の好ましい一実施形態において、前記ユビキチンベースのED−B結合ヘテロマー分子は、治療または診断に関連する特性を有するタンパク質またはペプチドに、共有または非共有結合される。
【0082】
ED−B結合能を有するユビキチンベースの融合タンパク質を得るための方法について、いくつかの例を、以下に示す。
a)ユビキチンに存在するリジン残基を介した前記タンパク質の結合;
b)システイン残基を介した前記ヘテロ二量体ユビキチン結合タンパク質の結合
前記システイン残基は、C末端に位置するか、または任意の他の部位(例えば、24もしくは57位のアミノ酸残基)に位置し得る;マレイミド選択的成分による結合;
c)ペプチド性またはタンパク質性の結合−遺伝子融合(好ましくはC末端またはN末端)
d)タグに基づく融合−標的タンパク質ED−BのC末端またはN末端に位置するタンパク質またはペプチド。融合「タグ」、例えば、ポリヒスチジン(特に、放射性標識に関する)。
【0083】
対象のタンパク質を支持体に共有および非共有結合させるための、これらおよび他の方法は、当該技術分野において周知であるので、さらに詳細には説明しない。
【0084】
前記活性成分は、任意に、サイトカインであり、好ましくは、腫瘍壊死因子(例えば、TNFα、TNFβ)、インターロイキン(例えば、IL−2、IL−12、IL−10、IL−15、IL−24、IL−3、IL−4、IL−5、IL−6、IL−7、IL−9、IL−11、IL−13、IL−8、IL−1α、IL−1β)、インターフェロン(例えば、IFNα、IFNβ、IFNγ)、GM−CSF、GRO(GROα、GROβ、GROγ)、MIP(MIP−1−α、MIP−1β、MIP−3α、MIP−3β)、TGF−β LIF1 CD80、CD−40リガンド、B70、LT−β、Fas−リガンド、ENA−78、LDGF−PBP、GCP−2、PF4、Mig、IP−10、SDF−1α/β、BUNZO/STRC33、I−TAC、BLC/BCA−1、MDC、TECK、TARC、RANTES、HCC−1、HCC−4、DC−CK1、MCP−1−5、エオタキシン、エオタキシン−2、I−309、MPIF−1、6Ckine、CTACK、MEC、リンホタクチン、フラクタルカイン等からなる群から選択されるサイトカインである。
【0085】
本発明において使用される最も好ましいサイトカインの一つは、TNFである。炎症性サイトカインであるTNFは、哺乳類の体内において、抗腫瘍作用を含む複数の活性を有する。前記抗腫瘍作用は、ヒトにおいて有効投与量を用いた場合に許容できない毒性を示すことから、現在、臨床上活用されていない。現在、TNFは、メルファラン等の細胞増殖抑制物質との併用で、治療に使用される。
【0086】
さらに、前記ヘテロ多量体ユビキチン結合タンパク質に結合可能な前記活性成分は、任意に、毒性化合物、好ましくは、低分子有機化合物またはポリペプチドであり、任意に、例えば、サポリン(saporin)、トランケート緑膿菌外毒素A(truncated Pseudomonas exotoxin A)、組換えゲロニン(recombinant gelonin)、リシン−A鎖(Ricin−A chain)、カリケアマイシン(calicheamicin)、ネオカルチノスタチン(neocarzinostatin)、エスペラミシン(esperamicin)、ダイネミシン(dynemicin)、ケダルシジン(kedarcidin)、マデュロペプチン(maduropeptin)、ドキソルビシン(doxorubicin)、ダウノルビシン(daunorubicin)、アウリスタチン(auristatin)、コレラトキシン(cholera toxin)、モデシン(modeccin)、ジフテリア毒素(diphtheria toxin)からなる群から選択される毒性化合物である。
【0087】
本発明のさらなる実施形態において、前記本発明のユビキチン結合タンパク質は、人工アミノ酸を含んでもよい。
【0088】
前記本発明の融合タンパク質のさらなる実施形態において、前記活性成分は、蛍光色素である。好ましくは、前記活性成分は、ガンマ放射同位体のグループ、好ましくは、99Tc、123I、111In、陽電子放出体のグループ、好ましくは、18F、64Cu、68Ga、86Y、124I、ベータ放出体のグループ、好ましくは、131I、90Y、177Lu、67Cu、またはアルファ放射体のグループ、好ましくは、213Bi、211Atのいずれかのグループから選択される放射線核種;Alexa FluorまたはCy dyes(Berlier et al., J Histochem Cytochem. 51 (12): 1699−1712, 2003);光増感剤;プロコアグラント因子、好ましくは、組織因子(例えば、tTF(短縮化組織因子);プロドラッグ活性化のための酵素、好ましくは、カルボキシぺプチダーゼ、グルクロニダーゼおよびグルコシダーゼからなる群から選択される一つの酵素;および/または機能性Fcドメイン、好ましくは、ヒト機能性Fcドメインからなる群から選択される1つの成分である。
【0089】
本発明の融合タンパク質に関するさらなる実施形態は、さらに、血清半減期を調整する成分を含み、好ましくは、ポリエチレングリコールアルブミン結合ペプチドおよび免疫グロブリンからなる群から選択される成分を含む。
【0090】
結合特異性
本発明の融合タンパク質の結合特異性(解離定数)は、非融合タンパク質に関して先に定義した通りであり、Kdで示す。本発明によれば、「Kd」という用語は、特異的結合親和性を定義するものであり、本発明においては、特異的結合親和性は、10−5〜10−12Mの範囲である。10−5M以下の値であれば、定量化可能な結合親和性であると考えることができる。用途に応じ、Kdの値は、例えば、クロマトグラフィーへの適用の場合には、10−6M〜10−12Mが好ましく、10−6M〜10−11Mがさらに好ましく、または、診断もしくは治療への適用の場合には、10−9M〜10−12Mが好ましい。さらに好ましい結合親和性は、10−7〜10−10Mであり、好ましくは、10−11Mである。
【0091】
前記結合親和性を決定する方法は、それ自体公知であり、例えば、下記の方法から選択できる:ELISA、表面プラズモン共鳴(SPR)技術(例えば、Biacore(登録商標)により提供される)、蛍光分光法、等温滴定熱量測定(ITC)、超遠心分析法、FACS。当該技術分野において利用可能なその他の方法もまた、当業者の一般的な知識の範疇において使用可能である。
【0092】
上記修飾後、本発明者らは、実施例で述べるアミノ酸修飾ユビキチン配列が、非常に高い親和性(Kd値10−12M以下)で標的と結合することを見出した。
【0093】
ユビキチンの多量体化
本発明のさらなる実施形態では、ED−Bに結合可能な前記ユビキチンタンパク質を、ED−Bに結合可能な少なくとも1つの第2のユビキチンタンパク質と融合させ、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合される、前記ユビキチンタンパク質の多量体、任意に二量体または三量体を得る。あるいは、ED−Bに結合可能な前記ユビキチンタンパク質を、ED−Bに結合可能な少なくとも1つの第2のユビキチンタンパク質と融合させ、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成される、前記ユビキチンタンパク質の多量体、任意に二量体または三量体を得る。
【0094】
本明細書中において、「多量体」は、同一または1以上の異なる単量体ユビキチンタンパク質を含み得るタンパク質と考えられる。前記多量体が同一の単量体ユビキチンユニットを含有する場合には、ホモマーまたはホモ多量体(例えば、ホモ二量体またはホモ三量体)が形成される。二量体が異なる修飾がなされた2つの単量体を含む場合、前記二量体は、「異種二量体」または「ヘテロ二量体」と呼ばれる。2以上の異なる単量体ユビキチンユニットが存在する場合、ヘテロマーまたはヘテロ多量体が形成される。この発明の最も好ましい実施形態においては、ED−Bへの結合能を有する前記ヘテロマーは、少なくとも2つの異なる修飾ユビキチン単量体からなる。本発明の「ヘテロ二量体」は、特定の結合パートナーに対し、一価の結合特性をそれぞれ有する2つの異なる単量体ユビキチンタンパク質からなる、修飾ユビキチンタンパク質の融合物と考えられる。
【0095】
よって、本発明の「ヘテロ多量体」、任意に「ヘテロ二量体」は、特定の結合パートナーED−Bに対して共同で一価の結合特性を示す、異なる修飾がなされた少なくとも2つの(あるいは、少なくとも1つは修飾され、1つは非修飾である)単量体ユビキチンタンパク質の融合物と考えられる。本発明の修飾ヘテロ二量体ED−B結合ユビキチンタンパク質は、各単量体ユビキチンタンパク質を個々にスクリーニングし、その後、これら2つを結合して得るのではなく、前記ED−Bリガンドに対して共同での一価の結合活性を示す第1単量体ユニットおよび第2単量体ユニットからなるヘテロ二量体タンパク質をスクリーニングすることによって得られることを強調しておく。前記サブユニットのそれぞれは、ED−Bへの結合親和性がかなり制限され、結合した二量体修飾ユビキチンタンパク質のみが、本明細書に記載の優れた結合特性を有すると考えられる(例えば、図4参照)。
【0096】
本発明によれば、ヘッドトゥテイル融合により遺伝学的に結合された(genetically linked)、異なる修飾がなされた2つのユビキチン単量体(あるいは、少なくとも1つは修飾され、1つは非修飾である)は、ED−Bの同じエピトープに結合し、両方の結合ドメイン領域が共に作用する場合のみ効果を示す。前記単量体の前記BDRは、単一の連続的な結合領域を形成する。
【0097】
本発明の一実施形態においては、本発明により提供される修飾ユビキチンタンパク質を、異なる特異性のユビキチンタンパク質に、好ましくは部位特異的に結合させることができ、これにより、結合パートナーに対して一価の結合特性を有する、モジュール構成されたユビキチンが得られる。
【0098】
よって、例えば、上述の手順で得られたユビキチンの2つの変異体を、部位特異的に互いに結合させることができる。したがって、多量体を形成する場合、前記単量体ユビキチンユニットを生成し、これらをヘッドトゥテイルで結合させることによって形成可能である。換言すると、前記単量体ユニットは、前記多量体内に含まれるユニットの数に応じ、N−C−N−C−方向に結合される。前記多量体化は、リンカーを使用することなく直接行うか、またはリンカーを介して行うことができる。
【0099】
本発明の別の実施形態では、ED−Bに結合する修飾ユビキチン単量体の2つの変異体を、適切な方法で互いに遺伝学的に結合させることができ、これにより、極めて特異的な結合分子が得られる。ヘッドトゥテイル融合により遺伝学的に結合した修飾ユビキチン単量体の2つの変異体が、同じエピトープに結合し、両方の結合ドメイン領域が共に作用する場合のみ効果を示すことが特に好ましい。換言すると、これらは、前記2つのモジュールの両結合領域が共に作用することによって形成される単一の連続的な結合領域を介して、同一のエピトープに結合する。
【0100】
したがって、フィブロネクチンのED−Bに効果的に結合させるべく本発明により修飾されたユビキチンタンパク質は、多量体化されてもよく、特に、二量体化または三量体化されてもよい。ユビキチン単量体の融合は、リンカーを介して行うことができる。前記リンカーとしては、例えば、少なくともGIGの配列、または、任意の他のリンカー、例えば、SGGGGIGまたはSGGGGSGGGGIGがあげられる。2以上のユビキチン単量体の遺伝子融合用の他のリンカーを使用することもできる。好ましくは、このようなユビキチンタンパク質は、少なくとも2つの結合決定領域(BDR)からなり、各BDRにおいて、2、4、6、8、62、63、64、65、66、68位におけるアミノ酸の少なくとも1個が修飾され、前記BDRが、互いに遺伝子融合(genetically fused)されている。前記標的への結合は、両BDRが共同で媒介する。
【0101】
前記修飾ユビキチンタンパク質のさらなる多量体化を、例えば、前記修飾ユビキチンタンパク質を、サイトカインのような多量体化ドメインを有するエフェクター分子、例えば、TNFαに融合させることによって行うことができる。1以上の修飾ユビキチン単量体(二量体等、好ましくはヘテロ二量体)と、TNFαとの融合は、リンカーを介して行うことができる。さらに別の実施形態においては、前記多量体化は、2以上の修飾ユビキチン分子を遺伝子融合させるか、あるいはポリエチレングリコール(PEG)リンカーを使用することによって行われる。さらに別の実施形態においては、前記多量体化ドメインが、薬学的活性成分としても作用する。一例を挙げると、TNFαは、多量体化ドメインおよび医薬品成分の両方の役割を果たす。
【0102】
本発明のさらなる実施形態において、本発明の多量体化修飾ユビキチンタンパク質は、ヘテロマーからなる。「ヘテロマー」という用語は、異なる組成のユビキチン単量体が、多量体化分子に含まれることを意味する。遺伝子融合された2つのユビキチン単量体からなるヘテロ二量体であって、前記2つのユビキチン単量体が、結合ドメイン領域は異なるが、単一の結合部位を有しているものが特に好ましい。これらのヘテロ二量体は、例えば、TNF、最も好ましくは、TNFαを使用することによって多量体化できる。ユビキチンヘテロ二量体を使用する利点の1つは、高い結合特異性でED−Bを標的とすることができることである。
【0103】
二量体化、または一般的には多量体化のさらなる利点は、ED−Bに対する前記新たに生成された結合タンパク質の足場の全体的な安定性を低下させることなく、タンパク質化学的完全性を維持しながら、修飾可能なアミノ酸残基の数を増加できることにある。一方では、ED−Bに対する結合部位を生成するために修飾可能な残基の総数は、前記修飾残基を、2つまたは3つのユビキチンタンパク質の足場に割り当てることができるために増加する。他方では、単一のユビキチンタンパク質の足場上の残基の数は減少するが、前記修飾結合分子のタンパク質化学的安定性は維持される。要約すると、前記修飾アミノ酸は、2個または3個のユビキチン分子に含まれるため、前記ユビキチンベースのED−B結合タンパク質のモジュール構造により、修飾アミノ酸の総数を増加することが可能となる。先に述べた修飾数は、修飾ユビキチン一分子当たりの数である。
【0104】
さらに、多量体化により、標的物質に対する2つ(以上)の結合部位と、1つの一価の特異性(一つのシングルエピトープに対する)を有する修飾ユビキチン分子を提供し得るが、二特異性または多特異性(よって、2以上の異なるエピトープを同時に認識する)が付与される場合もある。
【0105】
2以上の修飾ユビキチン分子を、好ましくはヘッドトゥテイル遺伝子融合により融合する場合、修飾数は、修飾ユビキチン分子数に応じて、2倍またはx倍にできる。代替例として、前記融合を翻訳後に行うこともできる。
【0106】
修飾された状態の単量体ユビキチンタンパク質の使用に関し、二量体、特に、ホモ二量体またはヘテロ二量体等の多量体構造を使用する利点は、下層にある足場のタンパク質化学的完全性に影響を与えることなく修飾可能な残基の数を増加できることにある。
【0107】
(ED−Bに結合する修飾ユビキチンタンパク質)
本発明により、ED−Bに結合する修飾ユビキチン単量体が同定された。ED−Bに特異的に結合する好ましい修飾ユビキチンヘテロ二量体は、以下の通りである。
1.第1ユビキチン分子の結合決定領域(BDR1):2、4、6、62〜68位の修飾アミノ酸。好ましくは、第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
2.第1ユビキチン分子の結合決定領域(BDR1):6、8、62〜68位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
3.第1ユビキチン分子の結合決定領域(BDR1):2、4、6、8、62〜68位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:2、4、6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
4.第1ユビキチン分子の結合決定領域(BDR1):6、8、63〜66位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:2、6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
【0108】
実施形態において、前記融合タンパク質は、前記ユビキチンタンパク質の遺伝子融合による二量体であり、第1ユビキチン単量体の6、8、63〜66位のアミノ酸が置換され、第2ユビキチン単量体の6、8、62〜66位、および、任意に2位のアミノ酸が置換されている。好ましくは、
6位におけるリジン(K)のトリプトファン(W)またはフェニルアラニン(F)への置換、
8位におけるロイシン(L)のトリプトファンまたはフェニルアラニン(W、F)への置換、
63位におけるリジン(K)のアルギニン(R)またはヒスチジン(H)への置換、
64位におけるグルタミン酸(E)のリジン(K)、アルギニン(R)、またはヒスチジン(H)への置換、
65位におけるセリン(S)のフェニルアラニン(F)またはトリプトファン(W)への置換、
66位におけるトレオニン(T)のプロリン(P)への置換;
前記第2ユビキチン単量体において、
6位におけるリジン(K)のトレオニン(T)、アスパラギン(N)、セリン(S)、またはグルタミン(Q)への置換
8位におけるロイシン(L)のグルタミン(Q)、トレオニン(T)、アスパラギン(N)、またはセリン(S)への置換
62位におけるグルタミン(Q)のトリプトファン(W)またはフェニルアラニン(F)への置換
63位におけるリジン(K)のセリン(S)、トレオニン(T)、アスパラギン(N)、またはグルタミン(Q)への置換
64位におけるグルタミン酸(E)のアスパラギン(N)、セリン(S)、トレオニン(T)、またはグルタミン(Q)への置換
65位におけるセリン(S)のフェニルアラニン(F)またはトリプトファン(W)への置換
66位におけるトレオニン(T)のグルタミン酸(E)またはアスパラギン酸(D)への置換
任意に、2位におけるグルタミン(Q)のアルギニン(R)、ヒスチジン(H)、またはリジン(K)への置換が好ましい。
【0109】
最も好ましい置換は、下記の通りである。
(1)第1単量体ユニットにおける、少なくともK6W、L8W、K63R、E64K、S65F、およびT66Pの置換;
および、第2単量体ユニットにおける、少なくともK6T、L8Q、Q62W、K63S、E64N、S65WおよびT66Eの置換;任意にさらにQ2Rの置換、または
(2)第1単量体ユニットにおける、少なくともQ2T、F4W、K6H、Q62N、K63F、E64K、S65LおよびT66Sの置換;
および、第2単量体ユニットにおける、少なくともK6X、L8X、Q62X、K63X、E64X、S65XおよびT66Xの置換;任意にさらにQ2Xの置換、Xは、任意のアミノ酸。
【0110】
ED−Bへの結合タンパク質を生成するための前記第1ユビキチン単量体における特に好ましい置換は、下記の通りである。
2位:Q→T、4位:F→W、6位:K→H、62位:Q→N、63位:K→F、64位:E→K、65位:S→L、66位:T→S
【0111】
2つの単量体のヘッドトゥテイルでの結合には、リンカーを使用しなくてもよいし、どのようなリンカーを使用してもよい。好ましいリンカーは、配列番号32のリンカー、または、GIG配列、SGGGGIG配列、もしくはSGGGGSGGGGIG配列のリンカーである。
【0112】
好ましい実施形態において、ED−Bに対する結合のために共に作用する2つの結合決定領域(BDR)を備えたユビキチンヘテロ二量体は、配列番号33または34のアミノ酸配列を含む。薬学的活性成分としてTNFαを含む、本発明の好ましい融合タンパク質は、配列番号35または36の配列を有する。さらに好ましいタンパク質は、下記配列(配列番号47)を有するものであるが、下記配列中、XXXXは、どのようなアミノ酸でもよい。リンカーとして、SGGGGSGGGGIG配列が使用される(イタリック体で示す)。他の種類のリンカー、またはリンカーなしも、実施可能な代替例と理解すべきである。
【化1】

【0113】
これらの配列のタンパク質のコンセンサス配列の例を、図19に示す。
【0114】
さらに別の実施形態において、本発明の融合タンパク質は、TNFαに融合したユビキチンヘテロ二量体の融合タンパク質の三量体であり、前記融合タンパク質は、配列番号35または36の配列を有するか、あるいは、配列番号35または36に対して90%以上の配列同一性、好ましくは95%を超える配列同一性、96%を超える配列同一性、または97%までの配列同一性を有し、また同時に、フィブロネクチンのED−Bドメインに対する元の融合タンパク質の高親和性での結合能を、維持または向上することが好ましい。
【0115】
本発明のさらなる態様において、本発明は、前述のタンパク質または融合タンパク質をコードするポリヌクレオチドをも包含する。さらに、前記ポリヌクレオチドを含むベクターも、本発明に包含される。
【0116】
本発明のさらなる態様において、本明細書中に記載のタンパク質もしくは融合タンパク質、および/あるいは、本発明の組換えタンパク質もしくは組換え融合タンパク質をコードするポリヌクレオチド、または、前記ポリヌクレオチドを含むベクターを含む、宿主細胞が包含される。
【0117】
(本発明のタンパク質、例えば、TNFα等のエフェクターに融合したED−B特異性ヘテロ二量体ユビキチンベース結合タンパク質の使用)
本発明の修飾ユビキチンED−B結合タンパク質は、例えば、in vitroまたはin vivoで使用する診断手段の調製、ならびに治療手段の調製に使用される。前記本発明のタンパク質は、例えば、直接的なエフェクター分子(修飾物質、拮抗物質、作用物質)または抗原認識ドメインとして使用できる。ED−B抗原が大量に出現する腫瘍の例を、図1の表に示す。
【0118】
選択する融合パートナーに応じて、本発明の医薬組成物は、癌、例えば、乳癌および結腸直腸癌、またはED−Bが大量に出現する任意の他の腫瘍性疾患(図1に列記したこれらの例を参照)の治療を対象とするように適合される。
【0119】
前記組成物は、治療上の有効量を含むように適合される。投与量は、治療対象の組織、疾患の種類、患者の年齢および体重、ならびにそれ自体公知である他の要因によって決まる。
【0120】
前記組成物は、薬学的または診断的に許容されるキャリアを含み、さらに、それ自体公知の助剤を任意に含み得る。これらは、例えば、安定化剤、界面活性剤、塩類、緩衝剤、着色剤等を含むが、これらに限定されない。
【0121】
前記医薬組成物は、局所塗布用の液状製剤、クリーム、ローションの剤形;エアロゾル;粉末、細粒、錠剤、座剤、カプセル剤の剤形;エマルジョン、リポソーム製剤の剤形を取り得る。前記組成物は、無菌、非発熱原性、等張性であり、それ自体公知である、薬学的に既存かつ許容可能な添加剤を含むことが好ましい。加えて、米国薬局方またはRemington’s Pharmaceutical Sciences、Mac Publishing Company(1990)の規則も参照する。
【0122】
ヒトおよび獣医学の医薬療法および予防法の分野において、本発明により修飾されたED−B結合ユビキチンタンパク質を少なくとも1つ含む薬学的に有効な医薬品を、それ自体公知の方法により調製できる。ガレノス式製剤学(galenic preparation)に応じて、これらの組成物を、静脈内投与、腹腔内投与、筋肉内投与、皮下投与、経皮投与、または他の適用方法により投与できる。医薬調合物の種類は、治療対象の疾患の種類、疾患の重症度、治療対象の患者、および医学分野の当業者に公知の他の要因によって決定される。投与は、注射もしくは点滴による非経口的な投与、全身的、直腸的、経皮的な投与、または従来使用されている他の方法による投与のいずれかとすることができる。
【0123】
一実施形態において、前記医薬組成物は、本発明のタンパク質もしくは融合タンパク質、またはこれらの混合物を含み、さらに、1以上の化学療法剤、好ましくは、下記表に示すものから選択される化学療法剤を含む。
【0124】
【表1】
【0125】
好ましい実施形態において、前記化学療法剤は、メルファラン、ドキソルビシン、シクロフォスファミド、ダクチノマイシン、フルオロデオキシウラシル、シスプラチン、パクリタキセル、およびゲムシタビンから選択されるか、またはキナーゼ阻害剤群から選択される。
【0126】
本発明の「医薬組成物」は、種々の活性成分と希釈剤および/または担体が互いに混合された組成物の形態で存在してもよいし、あるいは、前記活性成分が、部分的にまたは全体的に別個に存在する組み合わせ製剤の形態を取ってもよい。このような組み合わせまたは組み合わせ製剤の一例は、複数の部品からなるキットである。
【0127】
本発明の「組成物」は、少なくとも2つの薬学的活性化合物を含む。これらの化合物は、同時に投与してもよいし、1分〜数日の時間間隔で個々に投与してもよい。これらの化合物は、同じ経路で投与してもよいし、異なる経路、例えば、活性化合物の1つを経口投与し、他の活性化合物を非経口投与することも可能である。また、前記活性化合物は、一つの医薬品、例えば、一つの点滴液中に調剤されてもよいし、別々に調剤された両化合物を含むキットとして提供されてもよい。また、両化合物を、2以上の包装内に存在させることも可能である。
【0128】
特に好ましい組み合わせは、本発明の融合タンパク質とメルファランおよび/またはドキソルビシン(リポソーム製剤)である。ATC L01分類の抗悪性腫瘍剤以外にも、本発明のTNF融合タンパク質は、サイトカインとその誘導体、放射性医薬品、細胞ベースの治療薬、およびナノ粒子を含む、他の抗悪性腫瘍物質と併用できる。
【0129】
腫瘍透過処理活性(tumor permeabilisation activity)により、本発明のTNF融合タンパク質は(さらに、本発明の他の組換えタンパク質/融合タンパク質も)、世界保健機関提供の解剖治療化学分類法(ATC)においてL01として列挙される全ての抗悪性腫瘍剤と併用できる。ここで、驚くべきことに、TNFαに融合したユビキチンヘテロ二量体の融合タンパク質、好ましくは、配列番号35または36の配列を有する融合タンパク質が、有利に治療に適用できることがわかった。TNFαは毒性が高く、よって、低用量でしか投与できないが、このような用量は、通常、最小治療閾値を下回る(よって、治療上不活性である)。TNFαのこの毒性のため、治療上有効な濃度を達成するために、現在、TNFαの使用に際しては、四肢分離かん流アプローチが選択されている。四肢かん流は、腕または足に直接抗癌剤を輸送するために使用できる医療技術である。四肢への血流および四肢からの血流を止血帯で一次的に停止させ、抗癌剤を直接四肢の血液に注入する。これにより、患者は、癌が発生している領域に高い投与量のTNFαを受けることができる。
【0130】
しかしながら、本発明のTNFα融合タンパク質を適用すれば、非毒性でありながら、治療的に有効な濃度でTNFαを投与することが可能となる。TNFαは、本発明の(結合)融合タンパク質に連結されるため、疾病部位(例えば、腫瘍部位)において直接活性を示し、よって、「遊離」TNFαの量を激減させることができる。
【0131】
TNFαの全身性副作用は、TNFαを本発明の融合タンパク質として投与することにより、著しく軽減できる。本発明のTNFα融合タンパク質を使用することにより、治療効果に達するためのTNFαの全投与量を大幅に低減でき、特に、化学療法剤(上記参照)を併用する全身的な腫瘍治療において、有利に使用できる(四肢かん流の必要性および制約がない)。
【0132】
さらなる実施形態において、前記医薬組成物は、複数の部品からなるキットの形態であり、本発明の組換えタンパク質/組換え融合タンパク質、および1以上の化学療法剤が別々に提供される。
【0133】
(本発明のED−B結合タンパク質の製造方法)
本発明のED−B結合タンパク質は、例えば、単純な有機合成戦略、固相支援合成技術等の多数の従来公知の技術のいずれかにより、または、市販の自動合成装置により、調製すればよい。また、本発明のED−B結合タンパク質は、従来の遺伝子組換え技術を単独で使用するか、または、従来の合成技術と併用することにより、調製してもよい。
【0134】
本発明の他の態様において、組換え修飾タンパク質の製造方法が提供される。前記方法は、少なくとも下記の工程を含む。
a)ユビキチンタンパク質を提供する工程
b)フィブロネクチンのED−Bを提供する工程
c)配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために前記ユビキチンタンパク質を修飾する工程であり、
2、4、6、62、63、64、65、66、および/または68位における、少なくとも4個のアミノ酸を、置換、欠失、または付加により修飾する工程
d)前記修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
e)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bと結合する修飾ユビキチンタンパク質をスクリーニングする工程
f)前記修飾ユビキチンタンパク質を単離する任意の工程
【0135】
前記修飾は、任意にDNAレベルでの遺伝子工学により行われてもよく、前記修飾タンパク質の発現は、任意に原核生物、真核生物、またはin vitroにおいて行われてもよい。
【0136】
さらなる実施形態において、前記修飾工程は、化学的合成工程を含む。
【0137】
さらに別の実施形態において、前記方法は、前記修飾ユビキチンタンパク質が、薬学的活性成分(任意にサイトカイン、好ましくはTNFα)または診断用成分と融合するように適合されている。あるいは、前記方法は、前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合させるか、または、前記組換えタンパク質を少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成させるように適合されている。
【0138】
本発明によれば、修飾タンパク質は、さらに、化学的合成により調製されてもよい。この実施形態において、請求項1の前記c)工程からd)工程は、一つの工程で行われる。
【0139】
本発明の別の態様においては、ヘテロ多量体融合タンパク質の製造方法が提供される。前記方法は、以下の工程を含む。
a)好適なリンカーを介して結合された2以上の修飾ユビキチン単量体を含む多量体ユビキチンタンパク質を提供する工程であり、
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために、前記多量体ユビキチンタンパク質における各単量体が修飾されており、
各単量体において、2、4、6、62、63、64、65、66、および/または68位における少なくとも4個のアミノ酸が、置換、欠失、または付加により修飾されている工程
b)フィブロネクチンのED−Bを提供する工程
c)前記ヘテロ多量体修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
d)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bに結合する修飾ユビキチンタンパク質をスクリーニングする工程
e)前記修飾ヘテロ多量体ユビキチンタンパク質を単離する任意の工程
【0140】
本発明の別の態様においては、組換え修飾ユビキチンタンパク質の製造方法が提供される。前記方法は、少なくとも以下の工程を含む。
a)単量体ユビキチンタンパク質から生じる、異なる修飾がなされた二量体ユビキチンタンパク質群を提供する工程であり、
前記群が、互いにヘッドトゥテイル配置で結合された2つの修飾ユビキチン単量体を含む二量体ユビキチンタンパク質を含み、
前記二量体タンパク質の各単量体は、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも6個のアミノ酸の置換により、異なる修飾がなされ、
前記置換は、下記(1)または(2)を含む、前記工程
(1)第1単量体ユニットにおける、少なくとも6、8、63、64、65および66位のアミノ酸の置換、および、
第2単量体ユニットにおける、少なくとも6、8、62、63、64、65および66位のアミノ酸の置換、任意にさらに2位のアミノ酸の置換、または
(2)第1単量体ユニットにおける、少なくとも2、4、6、62、63、64、65および66位のアミノ酸の置換、および、
第2単量体ユニットにおける、少なくとも6、8、62、63、64、65および66位のアミノ酸の置換、任意にさらに2位のアミノ酸の置換
b)潜在的なリガンドとして、フィブロネクチンのエキストラドメインB(ED−B)を提供する工程
c)前記異なる修飾がなされたタンパク質群を、前記フィブロネクチンのエキストラドメインB(ED−B)と接触させる工程
d)スクリーニング処理により、修飾二量体ユビキチンタンパク質を同定する工程であり、
前記修飾二量体ユビキチンタンパク質は、前記フィブロネクチンのエキストラドメインB(ED−B)と、Kd10−7〜10−12Mの範囲の特異的結合親和性で結合し、前記フィブロネクチンのエキストラドメインB(ED−B)に対して、一価の結合活性を示すタンパク質である、前記工程
e)前記修飾二量体ユビキチンタンパク質を、前記結合親和性を用いて単離する任意の工程
【0141】
前記好ましい実施形態において、前記ヘテロ多量体ユビキチンタンパク質は、ヘテロ二量体ユビキチンタンパク質である。
【0142】
さらに別の態様において、本発明は、上述の直線状ポリユビキチン鎖をベースとし、少なくとも2つのBDRにおいてランダム化されているライブラリーに関する。
【0143】
本発明のさらなる態様において、前述の2つのライブラリーの融合により得られるDNAを含む融合ライブラリーが提供される。ヘテロ二量体ユビキチン融合タンパク質を得るために、各ライブラリーは、異なる修飾がなされた単量体ユビキチンタンパク質ユニットをコードし、前記単量体ユニットは、互いにヘッドトゥテイル配置で結合される。前記ライブラリーは、前記フィブロネクチンのエキストラドメインB(ED−B)に対して一価の結合活性を示すユビキチンのヘテロ二量体融合タンパク質をコードする。前記相互結合は、当業者に公知のリンカーのいずれか1つ、または本明細書に記載のリンカーを用いて行われる。本発明の一実施形態において、TNFαは、同時に薬学的活性化合物としての役割も果たすリンカーとして使用される。
【0144】
複合ライブラリーの製造の概要を、実施例1で述べる。ただし、このようなライブラリーの品質に注意を払わなければならない。抗体または足場技術におけるライブラリーの品質は、そもそも、複雑性(個々の変異体の数)および機能性(得られた候補の構造的およびタンパク質化学的完全性)に左右される。しかしながら、両特性が互いに悪影響を及ぼし合う場合があり、前記足場における修飾位置の数を増加させてライブラリーの複雑性を向上しようとすれば、変異体のタンパク質化学的特性の低下を招くおそれがある。これにより、溶解度の低下、凝集、および/または低収量が起こることもある。これは、エネルギー的に良好なタンパク質パッケージを有する未変性の足場からの逸脱がより大きくなることによる。
【0145】
このため、このような足場ライブラリーの適切な構築は、標的への結合性を最適化するために、元の配列にできるだけ多くの変異を導入することと、一方で、タンパク質化学的な悪影響を避けるために、元の一次配列をできるだけ保存するという、両極端な姿勢の間でバランスをとる作業となる。
【0146】
本開示は、本発明の態様または実施形態を考慮して考え得る、本明細書に記載の特徴の各組み合わせも包含する。
【0147】
標的ED−Bに対して結合親和性を有する修飾ユビキチンタンパク質の選択、および、結合親和性を発生させる修飾アミノ酸の決定
例えば、ヘテロ二量体修飾ユビキチンタンパク質をコードする少なくとも2つのDNAライブラリーが、各単量体ユビキチンユニットにおいて、選択されたアミノ酸に異なる修飾をすることにより確立された後、これらのライブラリーを、例えば、リンカー技術によって遺伝子融合し、ヘテロ二量体修飾ユビキチンタンパク質をコードするDNA分子を得る。これらのライブラリーのDNAによりタンパク質を発現させ、得られた修飾二量体タンパク質を、本発明によりED−Bに接触させ、結合親和性が存在する場合には、パートナー同士の結合が任意に可能となる。
【0148】
接触工程およびスクリーニング工程が、ヘテロ二量体ユビキチンタンパク質について、既に行われていることが本発明の重要な態様である。この工程により、ED−Bへの一価の結合活性を提供するユビキチンタンパク質のスクリーニングが可能となる。
【0149】
本発明による接触は、適切な提示および選択方法、例えば、ファージディスプレイ法、リボソームディスプレイ法、mRNAディスプレイ法、細胞表面ディスプレイ法、酵母表面ディスプレイ法、または細菌表面ディスプレイ法等の方法により行うことが好ましく、ファージディスプレイ法により行うことが好ましい。完全な開示としては、下記の参考文献を参照できる;Hoess, Curr. Opin. Struct. Biol.. 3 (1993), 572−579;Wells and Lowmann, Curr. Opin. Struct. Biol. 2 (1992), 597−604;Kay et al., Phage Display of Peptides and Proteins−A Laboratory Manual (1996), Academic Press。前述の方法は、当業者に公知であり、それらを改変した方法も含めて、本発明において使用可能である。
【0150】
本発明において、前記修飾タンパク質が、所定の結合パートナーに対して、定量化可能な結合親和性を有するかどうかの決定は、好ましくは、下記の方法の1以上により、行うことができる:ELISA、表面プラズモン共鳴分光法、蛍光分光法、FACS、等温滴定熱量測定、および超遠心分析法。
【0151】
ファージディスプレイ選択法
本願に適合するファージディスプレイ法の一種を、結合特性を示すユビキチン変異体についての、本発明による選択手法の一例として後述する。同様に、例えば、細菌上(bacterial surface display;Daugherty et al., 1998, Protein Eng. 11(9):825−832)もしくは酵母細胞上に提示する方法(yeast surface display; Kieke et al., 1997 Protein Eng. 10(11):1303−10)、または、リボソームディスプレイ(Hanes and Pluckthun, 1997 Proc Natl Acad Sci U S A. 94(10):4937−4942; He and Taussig, 1997 Nucleic Acids Res. 25(24):5132−5134)、cisディスプレイ(Odegrip et al., 2004 Proc Natl Acad Sci U S A. 101(9):2806−2810)もしくはmRNAディスプレイ等の無細胞選択システムを適用できる。後者の場合、遺伝子型と表現型との一時的な物理的結合が、リボソームを介したタンパク質変異体の適切なmRNAへの結合により達成される。
【0152】
本明細書に記載のファージディスプレイ法において、ユビキチンの組換え変異体は、繊維状ファージ上に提示され、一方、提示される変異体をコードするDNAは、同時に、ファージエンベロープ中に、一本鎖の形で詰め込まれる。このように、親和性の向上という枠組みにおいて、特定の特性を有する変異体をライブラリーから選択することができ、それらの遺伝情報は、適切な細菌への感染により増幅できるか、または、異なる濃縮サイクルに添加できる。ファージ表面での変異したユビキチンの提示は、アミノ末端シグナル配列、好ましくは、PelBシグナル配列、および、前記ファージのカプシドまたは表面タンパク質への遺伝子融合により達成される。カプシドタンパク質pIIIまたはその断片へのカルボキシ末端融合が好ましい。また、コードされた融合タンパク質は、例えば、親和性クロマトグラフィーによる検出および/または精製のための親和性タグもしくは抗体エピトープ、または、親和性濃縮過程における融合タンパク質の特異的な切断のためのプロテアーゼ認識配列等の、さらなる機能性成分を含むことができる。また、例えば、ユビキチン変異体の遺伝子と、ファージカプシドタンパク質またはその断片のコード領域との間に、アンバー停止コドンを存在させることができる。前記アンバー停止コドンは、部分的には1つのアミノ酸の導入に起因して、適切なサプレッサー株内での翻訳中には認識されない。
【0153】
所定のハプテンまたは抗原への結合特性を有するユビキチン変異体の単離に関する選択方法に適した細菌ベクターであって、前述の融合タンパク質の遺伝子カセットが挿入される細菌ベクターは、ファージミドという。数ある中でも、それは、繊維状ファージの遺伝子間領域(例えば、M13もしくはf1)またはその一部分を含み、例えば、M13K07等のヘルパーファージにより前記ファージミドを担持する細菌細胞の重複感染の際、ファージカプシド中に、ファージミドDNAの閉鎖鎖がパッケージングされる。このようにして生成されたファージミドは、細菌により分泌され、コードされた各ユビキチン変異体を、前記カプシドタンパク質pIIIまたはその断片への融合により、細菌表面に提示する。天然型pIIIカプシドタンパク質は、ファージミド中に存在し、適合するバクテリア株に再感染する能力、およびそれゆえに同一DNAを増幅する可能性を保持する。このため、前記ユビキチン変異体の表現型、すなわち、その潜在的な結合特性と、その遺伝子型との間の物理的結合が確保される。
【0154】
得られたファージミドを、そこに提示されたユビキチン変異体の所定のハプテンまたは抗原への結合に関し、当業者に公知の手法により選択することができる。この目的のために、提示されたユビキチン変異体を、例えば、マイクロタイタープレート上に結合した標的物質に一時的に固定化することができ、非結合の変異体を分離した後に、特異的に溶出することができる。前記溶出は、例えば、100mMトリエチルアミン等の塩基性溶液により行うことが好ましい。あるいは、前記溶出は、酸性条件下で、タンパク質分解または感染細菌の直接添加により行うことができる。このようにして得られたファージミドは、所定のハプテンまたは抗原への結合特性を有するユビキチン変異体の選択と増幅の連続サイクルにより、再増幅と濃縮が可能である。
【0155】
このようにして得られた前記ユビキチン変異体のさらなる特性評価は、ファージミドの形態、すなわち、ファージに融合した形態で、または、対応する遺伝子カセットを好適な発現ベクターにクローニングした後に、可溶性タンパク質の形態で行うことができる。適切な方法は、当業者に公知であるか、または前記文献に記載されている。前記特性評価は、例えば、DNA配列の決定、よって、単離された変異体の一次配列の決定を含み得る。さらに、単離された変異体の親和性および特異性を、例えば、ELISAもしくは表面プラズモン共鳴分光法等の標準的な生化学的方法、蛍光分光法、FACS、等温滴定熱量測定、超遠心分析法、またはその他の方法により検出できる。安定性分析については、例えば、化学的または物理的アンフォールディングについての分光法が、当業者に公知である。
【0156】
リボソームディスプレイ選択法
本発明のさらなる実施形態であるリボソームディスプレイ法において、ユビキチン変異体は、無細胞転写/翻訳系により調製され、対応するmRNAおよびリボソームとの複合体として提示される。この目的のために、前述のDNAライブラリーが、基礎として使用され、前記DNAライブラリー中で、変異体の遺伝子は、発現およびタンパク質生合成のための対応する制御配列との融合物の形態で存在する。前記遺伝子ライブラリーの3’末端での前記停止コドンの欠失、および適切な実験条件(低温、高Mg2+濃度)により、新生タンパク質、mRNA、およびリボソームからなる三元複合体が、in vitroでの転写/翻訳の間、維持される。
【0157】
ヘテロ二量体修飾ユビキチンタンパク質を含むタンパク質ライブラリーが、各単量体ユビキチンユニットにおいて、選択されたアミノ酸に異なる修飾をすることにより確立された後、前記修飾二量体タンパク質を、本発明によりED−Bに接触させ、結合親和性が存在する場合には、パートナー同士の結合が可能となる。これらのタンパク質ライブラリーは、前記修飾タンパク質と前記ED−B標的タンパク質間の接触を可能とする方法でディスプレイを行うか、前記修飾タンパク質を提示する任意の他の方法を使用する、ディスプレイ法のライブラリーの形態であってもよい。前記ディスプレイ法は、任意に、ファージディスプレイ法、リボソームディスプレイ法、TATファージディスプレイ法、酵母ディスプレイ法、細菌ディスプレイ法、またはmRNAディスプレイ法である。
【0158】
Kd=10−7〜10−12Mの範囲の特異的結合親和性によるED−Bへの結合活性に関する修飾ユビキチン変異体の選択を、当業者に公知の方法により行うことができる。この目的のために、例えば、リボソーム複合体上に提示される前記ユビキチン変異体を、例えば、マイクロタイタープレート上に結合した標的物質に一時的に固定化することができ、または、溶液中での結合後に磁性粒子に結合することができる。非結合変異体を分離した後に、リボソーム複合体を破壊することにより、結合活性を有する変異体の遺伝子情報を、mRNAの形態で特異的に溶出できる。前記溶出は、50mM EDTAで行うことが好ましい。このようにして得られたmRNAは、単離して適切な方法を使用してDNAに逆転写でき(逆転写反応)、このようにして得られた前記DNAは、再増幅できる。
【0159】
in vitroでの転写/翻訳、選択および増幅の連続サイクルにより、所定のハプテンまたは抗原に対する結合特性を有するユビキチン変異体を濃縮できる。
【0160】
ED−B結合タンパク質の特性評価
このようにして得られたユビキチン変異体のさらなる特性評価は、上述の通り、対応する遺伝子カセットを好適な発現ベクターにクローニングした後に、可溶性タンパク質の形態で行うことができる。適切な方法は、当業者に公知であるか、または前記文献に記載されている。
【0161】
好ましくは、所定の結合パートナーに対して結合親和性を有するタンパク質の検出工程の後に、検出したタンパク質の単離および/または濃縮工程が続く。
【0162】
本発明により修飾したユビキチンタンパク質の発現に続いて、それ自体公知の方法により、これをさらに精製して濃縮することができる。選択される方法は、例えば、使用する発現ベクター、宿主生物、使用予定分野、タンパク質の大きさ、およびその他の要因等、それ自体当業者に公知の複数の要因により決定される。簡易に精製するために、本発明により修飾したタンパク質を、分離材料に対して高い親和性を有する他のペプチド配列に融合させることができる。好ましくは、ユビキチンタンパク質の機能性に有害な影響を及ぼさないか、あるいは、特定のプロテアーゼ切断部位の導入により精製後に分離可能な融合物が選択される。このような方法もまた、それ自体当業者に公知である。
【0163】
(図面の簡単な説明)
図1は、種々の腫瘍におけるED−Bの発生について列記した表を示す。
【0164】
図2Aは、修飾ユビキチンED−Bバインダー5E1に関する濃度依存性ELISAの結果を示す。前記修飾ユビキチンED−Bバインダー5E1は、23nMの親和性でヒトED−Bに結合し(黒丸)、一方、BSAへは結合しなかった(白丸)。
【0165】
図2Bは、修飾ユビキチンED−Bバインダー1H4に関する濃度依存性ELISAの結果を示す。前記修飾ユビキチンED−Bバインダー1H4は、7.8nMの親和性でヒトED−Bに結合した(黒丸)。コントロールとして、BSAへの結合をプロットしている(白丸)。
【0166】
図3は、四量体化により親和性が向上することを示している。図3の表に、修飾ユビキチン単量体のKd値を、修飾ユビキチン単量体からなる四量体のものと比較して示している。図中の例は、ユビキチン変異体5E1および1H4である。ED−B結合を、c−FN(細胞性フィブロネクチン)への結合と比較している。図3は、標的ED−Bへの四量体変異体の結合親和性(例えば、5E1は56nM、1H4は1.4nM)が、前記単量体の結合親和性(5E1は4.51μM、1H4は9.98μM)と比べて著しく高いことを示している。
【0167】
図4は、前方(第1)修飾ユビキチン単量体(BDR1を有する)と異なる修飾がされた後方(第2)ユビキチン単量体(BDR2を有する)との遺伝子組換えによるヘテロ二量体の生成により、親和性および特異性が有意に向上することを示している。前記修飾ユビキチン分子は、Biacore、蛍光偏光測定、細胞および組織切片への結合により分析した。複数の変異体のヒトED−Bへの結合に関する濃度依存性ELISA(conc−ELISA)の結果を示す。
【0168】
図4Aは、単量体41B10について、結合親和性がKd=9.45μMであることを示している。
【0169】
図4Bは、41B10が異なる第2単量体と結合して得られた46H9について、結合親和性がKd=131nMであることを示している。
【0170】
図5は、サイトカイン(例えば、TNFα)と融合した特異的変異体を示す。前記融合タンパク質は、前記修飾ユビキチン単量体を三量体化したものであり、生物学的に活性な分子である。
【0171】
図5Aは、修飾ユビキチンベースのED−B結合エフェクター融合タンパク質の模式図である。同図において、緑色(上部の構造)は、エフェクター、例えば、サイトカイン、好ましくは、TNFα;茶色、薄い茶色は、修飾ユビキチン単量体(Affilin(登録商標))である。
【0172】
図5Bは、前記修飾ユビキチンエフェクター結合5E1−TNFコンジュゲートが、アポトースシス促進活性(L929アポトーシスアッセイで測定)を有すること示す。
【0173】
図5Cは、1H4−TNFα融合物のED−Bへの高結合親和性(Kd=15.1nM)を示す(適合線により連結された黒丸)。コントロールとして、BSAへの結合をプロットしている(線により連結されていない黒丸)。
【0174】
図6は、サイトカイン、例えば、TNFαに融合した、修飾ユビキチンベースのED−B結合へテロ二量体分子の親和性および活性を示す。
・修飾ユビキチンベースのED−B結合サイトカイン融合物のアポトーシス誘導活性:EC50 0.78±0.24pM
・遊離サイトカインのアポトーシス誘導活性:EC50 3.14±3.59pM
【0175】
図6Aは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd 50.7nM)。
【0176】
図6Bは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12がサイトカインTNFαに遺伝子融合し、これにより多量体化した前記ヘテロ二量体24H12の親和性を示す(Kd=5.6nM)。
【0177】
図6Cは、ヘテロ二量体修飾ユビキチンライブラリーから選択される候補例、例えば、ヘテロ二量体クローン9E12、22D1、24H12、41B10の分析結果を示す。標的ED−Bに対するKd ELISA値は、コントロールとして用いた細胞基質のフィブロネクチンと比べて向上しており、前記標的への特異的結合が確認された。
【0178】
図6Dは、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾ヘテロ二量体ユビキチン分子9E12の分析結果を示す。種々の濃度(図中の凡例参照:0〜15μMの9E12)の前記ヘテロ二量体ユビキチン変異体について、チップ(Biacore)に固定化されたED−Bへの結合を分析し、前記ヘテロ二量体変異体9E12とED−B間の相互作用を分析した。会合解離曲線の分析からは、Kdを、決定できなかった。
【0179】
図6Eは、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾ヘテロ二量体ユビキチン分子41B10の分析結果を示す。種々の濃度(図中の凡例参照:0〜15μMの41B10)の前記ヘテロ二量体ユビキチン変異体について、チップ(Biacore)に固定化されたED−Bへの結合を分析し、前記ヘテロ二量体変異体41B10とED−Bとの間の相互作用を分析した。会合解離曲線を分析したところ、Kdは、623nM(623×10−9M、6.2×10−7M)であった。
【0180】
図7は、異なる修飾がなされたユビキチンベースの変異体の、結合親和性および結合特異性に対する寄与を示す。前記異なる変異体は、小文字で示す共通の配列モジュールを有する。前記変異体を、そのED−B結合に関して分析した。図3は、修飾ユビキチンへテロ二量体が得られる、単量体の種々の組み合わせを示す。ヘテロ二量体変異体46−A5、50−G11、および46−H4は全て、BDR1が同一の第1(前方)修飾単量体(同図において、「a」で示す)を有するが、BDR2の異なる位置が修飾された第2(後方)ユビキチン単量体を有する。変異体52−D10および52−B3は、46−H9と比べて、BDR1が異なる第1(前方)修飾単量体を有するが、BDR2が同一の第2(後方)ユビキチン単量体(同図において、「e」で示す)を有している。
【0181】
前記修飾ユビキチンヘテロ二量体は、下記配列を有する。
46−H4:配列番号25、45−H9:配列番号26、46−A5:配列番号27、50−G11:配列番号28、52−B3:配列番号29、52−D10:配列番号30
【0182】
上記配列は、実験過程において、配列LEHHHHHH(配列番号31)を有するHisタグの付加により修飾した。
【0183】
図7からわかるように、46−H4は、ED−Bへの優れた結合親和性を有する(Kd=189nM)。46−A5および52−D10は、結合活性を有さない。また、他の修飾ユビキチンタンパク質は、ED−Bへの結合活性が46−H4と比べて低い。したがって、ヘテロ二量体変異体における両単量体が、標的への高い結合親和性のために必要とされ、両単量体が標的に対する一価の結合を示すと結論付けることができる。
【0184】
高いED−B結合活性を有する修飾ユビキチンヘテロ二量体46−H9は、野生型ユビキチン単量体と比較して、前記2つの単量体の両結合ドメイン領域における下記アミノ酸の置換により同定される。
第1モジュール(BDR1)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64A、S65T、T66L
第2モジュール(BDR2)において、(e)K6H、L8M、Q62K、K63P、E64I、S65A、T66E
50G11
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(c)K6M、L8R、Q62M、K63N、E64A、S65R、T66L
46H4
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(d)K6G、L8W、Q62T、K63Q、E64Q、S65T、T66R
52B3
第1モジュールにおいて、(g)Q2R、F4P、K6Y、Q62P、K63P、E64F、S65A、T66R
第2モジュール(46H9)において、K6H、L8M、Q62K、K63P、E64I、S65A、T66E
52D10(ED−Bに結合しない)
第1モジュールにおいて、Q2V、F4C、K6R、Q62T、K63A、E64P、S65G、T66D
第2モジュール(46H9)において、(e)K6H、L8M、Q62K、K63P、E64I、S65A、T66E
46A5(ED−Bに結合しない)
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(b)K6L、L8M、Q62L、K63A、E64F、S65A
【0185】
図8は、配列アライメントを示す。1行目:野生型ユビキチンタンパク質の2つの単量体(1行目)は、77位から88位までに存在する、12個のアミノ酸からなるリンカーSGGGGSGGGGIGにより結合される。BDR2を有する第2単量体は、89位のメチオニンから始まる。この二量体野生型ユビキチンタンパク質を、第1単量体および第2単量体に異なる修飾を有し、これにより2つのBDRが生じた、前記修飾ユビキチンへテロ二量体変異体46−H9(2行目)とアライメントさせる。標的への一価の結合のために、両BDRは、標的との結合において共に作用する。
【0186】
図9は、「Ub2_TsX9」(両単量体の45位がトリプトファンに修飾され、2つの単量体の間にリンカーGIG(77〜79位;80位のメチオニンから第2単量体が開始)を有し、前記第2単量体の最後のC末端アミノ酸において、グリシンからアラニンに置換されているユビキチン)に対する、修飾ユビキチンへテロ二量体変異体1041−D11(1行目)の配列アライメントを示す。3行目は、二量体である野生型ユビキチンであり、リンカーアライメントを示さない(このため、77位のメチオニンから第2単量体が開始する)、「Ubi−Dimer wt」を示す。4行目は、ヒトの野生型ユビキチンである「Ubi−Monomer wt」を示す。
【0187】
図10は、ヒトED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合についての、濃度依存性ELISAを示す。変異体1041−D11は、非常に高いED−Bへの結合親和性を示す(Kd=6.9nM=6.9×10−9M)。黒丸は、ヘテロ二量体ユビキチン変異体1041−D11の、フィブロネクチン断片を含むED−B(67B89−t0という)に対する結合親和性を示す。対照的に、この変異体は、ネガティブコントロール(6789−t0という)には結合しない(白丸)。
【0188】
図11は、フィブロネクチン断片を含む固定化されたED−B(67B89)に対するヘテロ二量体ユビキチン変異体1041−D11の結合を、遊離標的の量を増加させながら分析した競合濃度依存性ELISAの結果を示す。ヘテロ二量体ユビキチン変異体1041−D11は、非常に高いED−Bへの結合親和性を示す(IC50=140nM)。
【0189】
図12は、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾へテロ二量体ユビキチン分子1041−D11の分析結果を示す。種々の濃度(図中の凡例参照:0〜200nM 1041−D11)の前記ヘテロ二量体ユビキチン変異体について、SAチップ(Biacore)に固定化された、フィブロネクチン断片を含むED−B(67B89という)への結合を分析した。会合解離曲線を分析したところ、Kdは1nM(1×10−9M)であり、koff率は7.7×10−4s−1であり、このkoff率により、1041−D11とED−Bとの複合体の半減期が長いことが示される。
【0190】
図13は、結合活性の血清安定性を同時に分析する濃度依存性ELISAにおける、ED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。同図には、種々の異なる条件、例えば、前記変異体の、マウス血清中、ラット血清中、またはコントロールであるPBST中での、37℃で1時間でのプレインキュベーション等が示されている。Kd値は、全て10〜20nMの範囲内である。したがって、ED−Bへの前記ヘテロ二量体1041−D11の結合は、血清による有意な影響を受けないと結論付けることができる。
【0191】
図14は、ヘテロ二量体ユビキチン変異体1041−D11とフィブロネクチン断片との複合体形成の、SE−HPLCによる分析を示す。
【0192】
図14Aは、1041−D11とED−Bとの複合体形成を示す。3回のHPLCのランを重ねている。保持時間21.651分の青色のピークは、純粋な1041−D11に由来する。保持時間26.289分の黒色のピークは、フィブロネクチン断片67B89を表す。1041−D11と67B89との混合物により、SE−HPLC後の保持時間21.407分の赤色のピークが得られた。1041−D11のピークの保持時間が短くなる側へのシフト、および、67B89のピークの消失は、1041−D11と可溶性ED−Bとの複合体が形成されたことを示す。
【0193】
図14Bは、1041−D11(青色、21.944分)、ED−Bを含まないフィブロネクチン断片6789(黒色、26.289分)、および1041−D11と6789との混合物(21.929分および26.289分にピークを有する赤色線)について、3回のSE−HPLCのランを重ねて示している。1041−D11のピークのシフトは、ほとんど観察されなかった。この事実は、6789のピークが消失しなかったことと併せて、ED−Bを含まないフィブロネクチン断片6789との有意な結合を示さないことを表す。
【0194】
図15は、ヘテロ二量体ユビキチン変異体1041−D11の培養細胞への結合を示す。
【0195】
図15Aは、固定されたヒト胎児肺繊維芽細胞(Wi38)へのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。図15において、1列目は、抗ED−B抗体を使用したコントロールを示し、2列目は、タンパク質濃度58.7nMでの変異体のインキュベーションを示し、3列目は、10倍濃度(587nM)の1041−D11タンパク質でのインキュベーションを示し、4列目は、PBSを用いたネガティブコントロールを示す。1行目において、ヒトWi38繊維芽細胞を、位相差で示し、2行目において、免疫蛍光法で示し、3行目において、核のDAPI染色を示す。変異体1041−D11は、ED−B含有細胞外マトリックスに対する高特異性で、Wi38に結合すると結論付けることができる。EDBを低レベルで発現するNHDF細胞を使用するコントロールを行った(データは示さず)。前記変異体は、これらの細胞には結合しなかった。
【0196】
図15Bは、バイタル(vital)ヒト胎児肺繊維芽細胞(Wi38)への結合を示す。前記ネガティブコントロールのNHDF細胞は、初代の正常繊維芽細胞であり、EDB−フィブロネクチンを低レベルで発現する。1行目および3行目は、種々のタンパク質濃度での変異体、ならびにネガティブコントロールを示す。2行目および4行目は、EDB抗体を用いたコントロールのインキュベーションを示す。1〜2行目は、Wi38細胞株における前記変異体とポジティブコントロールを示す。3〜4行目は、NHDF細胞のインキュベーションを示す。変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、Wi38に結合することがわかる。
【0197】
図15Cは、固定されたマウスBalb 3T3細胞への結合を示す。前記変異体について、3つの異なるタンパク質濃度(1、10、50nM)でテストした。1行目は、細胞における前記変異体(SPVF−28−1041−411−TsX9)を示し、2行目は、ポジティブコントロール(Fv28−EDB−抗体)を示し、3行目は、ネガティブコントロール(UB2_TsS9;配列番号1に相当する非修飾ユビキチン)を用いたインキュベーションを示す。変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、マウスBalb 3T3細胞に結合することがわかる。
【0198】
図15Dは、固定されたマウスST−2細胞への結合を示す。前記変異体について、3つの異なるタンパク質濃度(1、10、50nM)でテストした。1行目は、細胞における前記変異体(SPVF−28−1041−411−TsX9)を示し、2行目は、ポジティブコントロール(Fv28−EDB−抗体)を示し、3行目は、ネガティブコントロール(UB2_TsS9;配列番号1に相当する非修飾ユビキチン)を用いたインキュベーションを示す。前記ヘテロ二量体ユビキチン変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、マウスBalb ST−2細胞に結合することがわかる。
【0199】
図16Aは、哺乳類組織切片における、ヘテロ二量体ユビキチン変異体1041−D11の標的に対する特異性を示す。7つの検体から採取したF9腫瘍組織について評価を行った。10nM〜100nMの範囲の異なる濃度のヘテロ二量体ユビキチン変異体1041−D11を用いた免疫組織化学検査において、マウス由来のF9腫瘍でのED−B特異的血管染色(vascular staining)が見られた。ED−Bは、腫瘍血管系の高特異的マーカーである。標的タンパク質であるED−Bは、血管の反管腔側に局在する。変異体1041−D11は、F9腫瘍由来の組織切片において、特異的に血管系を装飾する。得られた結果は、抗体フラグメントL19に匹敵する。さらに、48組織をテストしたところ、FDA関連パネルにおける48組織のいずれにおいても、非特異的な染色は観察されなかった。
【0200】
図16Bは、腫瘍組織における1041−D11の蓄積を、野生型ユビキチン(同図において、Ub2(NCP2))と比較して示している。F9腫瘍組織について、30分から16時間の間の異なる時点で、1041−D11および野生型ユビキチンの存在を分析した。腫瘍組織における1041−D11の最も高い蓄積は、投与から30分後および16時間後に観察されている。一方、F9腫瘍組織における野生型ユビキチンの蓄積は低い。前記変異体は、ED−Bを発現する腫瘍において、野生型ユビキチンと比べて濃縮されている。このことは、1041−D11の腫瘍組織指向性のターゲッティングの証拠となる。また、癌モデルにおける1041−D11の腫瘍/血液比は、動物における1041−D11変異体in vivo活性を明らかに証明している(データは示さず)。
【0201】
図17は、1041−D11−TNFα融合タンパク質の、ED−Bに対する高い選択性および特異性を示す。
【0202】
図17Aおよび17B:1041−D11−TNFα融合タンパク質のアポトーシス誘導TNFα活性を、細胞ベースアッセイ(cell based assay)(L929細胞)によりテストした。前記両図は、培養細胞において、前記1041−D11−TNFα融合タンパク質(図17B)が、遊離TNFα(図17B)と同程度に活性であることを、明らかに示している。
【0203】
図17Cは、標的ED−Bに対する、ヘテロ二量体ユビキチン1041−D11−TNFα融合タンパク質の高い選択性を証明している。前記ヒトED−Bフィブロネクチンドメイン67B89は、見かけのKD値1.8nMで1041−D11変異体に結合し(黒丸)、標的への高い親和性を示す。ED−Bドメインを欠損するヒトフィブロネクチン(h6789)には、1041−D11−TNFαは結合しない(白丸)。
【0204】
図17Dは、修飾ユビキチンベースのED−B結合1041−D11−TNFα融合タンパク質についての、Biacoreアッセイによる結合分析を示す。結果から、1041−D11TNFα融合タンパク質が、KD値1.13nMの高い親和性を示すことが証明された。
【0205】
図17Eは、培養細胞における変異体1041−D11で観察された高い結合特異性が、1041−D11がTNFαに融合されても維持されることを示す。前記融合タンパク質は、EDB発現細胞に特異的に結合する。したがって、1041−D11TNFα融合タンパク質は、非常に高い親和性および特異性で標的ED−B(ターゲット(+))に結合する。ED−Bを含まない血清(ターゲット(−))では、交差反応は観察されなかった。
【0206】
図18は、TNFαに融合した変異体1041−D11とメルファランとの併用による、7日間のマウスの処置期間中のin vivoでの相対的腫瘍成長を示す。データから、1041−D11−TNFαを細胞増殖抑制剤であるメルファランと併用した場合には、mTNFαとメルファランとを併用した場合、またはメルファラン単独の場合と比べ、より効率的に相対的腫瘍成長が減少されることをはっきりと示している。7日間の処置後の前記腫瘍成長動態は、1041−D11−mTNFαによる高効率での減少を示す。このことは、融合タンパク質1041−D11−TNFαとメルファランとの併用による腫瘍治療の有効性の明らかな証拠である。ED−Bは、マウスおよびヒトを含む数種の哺乳類において同一である。このため、前記結果から、ヒトにおける変異体1041−D11−TNFαの性能が予測される。
【0207】
図19は、ED−Bに対して驚くほど強力な結合親和性を有することが見出されたさらなる16種類の配列について、コンセンサスな位置およびアミノ酸置換を示す。前記コンセンサスなアミノ酸位置は、第1単量体の結合決定領域2、4、6、62、63、64、65、66位にあり、一方、コンセンサスなアミノ酸置換は、Q2T、F4W、K6H、Q62N、K63F、E64K、S65L、およびT66Sである。図2からわかるように、4種類のファミリーの配列を濃縮できた(コンセンサス配列、文字サイズはアミノ酸の発生頻度に対応する)。85および87位は、前記ヘテロ二量体タンパク質において、前記第2単量体における6および8位に対応する位置であり、141〜145位は、62〜64位に対応する。暗青色で示したTWH NFKLSは、1071−C12由来である。赤色で示した残基は、前記4種類のファミリーの配列のうちの1種類に属する。赤色で示した残基が、主に濃縮され(178/457配列)、HIT ELISAによれば、最も強力な結合分子を含む。
【実施例】
【0208】
下記実施例は、本発明をさらに説明するために提供するものである。本発明は、特に、ユビキチンの修飾を例にあげて実証されている。しかしながら、本発明は、これに限定されるものではなく、以下の実施例は、単に、前述の記載に基づいた本発明の実施可能性を示しているにすぎない。本発明の完全な開示のため、本願および付属書類に引用されている文献についても言及しているが、これらの引用文献は全て、引用により、その開示全体が本願に取り込まれている。
【0209】
[実施例1]
修飾ユビキチンタンパク質ベースのED−B結合タンパク質の同定
【0210】
単量体結合タンパク質のライブラリーの構築およびクローニング
特に示さない限り、例えば、Sambrookらに記載されているような、確立された組換え遺伝学的手法を使用した。合計10の選択されたアミノ酸位置における協奏的な変異誘発によって、高度な複雑性を有するヒトユビキチン単量体のランダムライブラリーを用意した。NNKトリプレットによって置換された修飾アミノ酸は、ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸を含む。
【0211】
ヘテロ二量結合タンパク質のライブラリーの構築とクローニング
特に示さない限り、例えば、Sambrookらに記載されているような、確立された組換え遺伝学的手法を使用した。合計15の選択されたアミノ酸位置における協奏的な変異誘発によって、高度な複雑性を有するヒトユビキチンヘテロ二量体のランダムライブラリーを用意した。NNKトリプレットによって置換された修飾アミノ酸は、近位(第1)ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸、および遠位(第2)ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸を含む。両ユビキチン単量体は、少なくともGIG配列、または少なくともSGGGG配列を有するグリシン/セリンリンカーによって遺伝学的に結合(ヘッドトゥテイル配置)しており、リンカー配列の例としては、GIG、SGGGG、SGGGGIG、SGGGGSGGGGIG(配列番号32)またはSGGGGSGGGGがあげられるが、その他のリンカーでもよい。
【0212】
TATファージディスプレイ選択
ユビキチンライブラリーを、例えば、選択システムとしてTATファージディスプレイを使用し、標的に対して濃縮した。当該技術分野において公知である他の選択方法を用いることもできる。前記標的は、タンパク質結合表面上に、またはタンパク質に共有結合されたビオチン化残基を介して、非特異的に固定化され得る。ストレプトアビジンビーズまたはニュートラアビジンストリップ上へのビオチンを介した固定化が好ましい。標的結合ファージは、溶液中または固定化標的上のいずれかにおいて選択される。例えば、ビオチン化され固定化された標的とファージとを、インキュベートし、続いて、マトリックスに結合したファージの洗浄およびマトリックス結合ファージの溶出を行った。標的のインキュベーションに続く各サイクルにおいて、前記ビーズを磁力により溶液から分離し、数回洗浄した。最初の選択サイクルにおいて、ビオチン化標的を、ニュートラアビジンストリップに固定化し、一方、2回目から4回目のサイクルにおいて、溶液中での選択を行い、続いて、Streptavidin−coated Dynabeads(登録商標)(Invitrogen社製)上に、標的とファージとの複合体を固定化した。最初の2回の選択サイクルでの洗浄後、標的が結合した修飾ユビキチン分子のファージを、酸性溶液での溶出により遊離させた。選択サイクルにおいて、過剰量の標的を用いた競合的溶出により、3回目および4回目のファージ溶出を行った。溶出したファージを再増幅した。バインダーの特異性を誘導するため、選択に際し、標的に類似するタンパク質を含めてもよい。
【0213】
TATファージディスプレイ選択の代替:リボソームディスプレイ選択
ユビキチンライブラリーを、例えば、選択システムとしてリボソームディスプレイを使用して、標的に対して濃縮した(Zahnd et al., 2007、Ohashi et al., 2007)。当該技術分野において公知である他の選択方法を用いることもできる。前記標的を、標準的な方法によってビオチン化し、Streptavidin−coated Dynabeads(登録商標)(Invitrogen社製)に固定化した。リボソーム、mRNA、および新生ユビキチンポリペプチドを含む三元複合体を、PURExpres(商標) In Vitro Protein Synthesis Kit(NEB社製)を用いて構築した。選択の一次ラウンドを2回行い、三元複合体をインキュベートし、続いて、同様の選択ラウンドを2回行った。標的インキュベーションに続く各サイクルにおいて、ビーズを磁力により溶液から分離し、ストリンジェンシーを増加させながら、リボソームディスプレイバッファーで洗浄した。最初の2回の選択サイクルでの洗浄後、前記ビーズを再度磁力により溶液から分離し、50mM EDTAの添加により、標的が結合した修飾ユビキチン分子のmRNAをリボソームから遊離させた。選択サイクルにおいて、過剰量の標的を用いた競合的溶出により、3回目および4回目のmRNAの溶出を行った(Lipovsek and Pluckthun, 2004)。各サイクルの後、RNeasy MinElute Cleanup Kit(Qiagen社製、ドイツ)、Turbo DNA−free Kit(Applied Biosystems社製、アメリカ)、およびTranscriptor Reverse Transcriptase(Roche社製、ドイツ)を用いて、RNAの精製とcDNAの合成を行った。
【0214】
濃縮プールのクローニング
4回目の選択サイクルの後、合成cDNAを、F1プライマー(GGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATG)(配列番号9)およびWUBI(co)RD_xhoプライマー(AAAAAAAAACTCGAGACCGCCACGCAGACGCAGAACCAG)(配列番号10)を用いたPCRにより増幅し、制限ヌクレアーゼNdeIおよびXhoI(Promega社製、アメリカ)で切断し、適合性付着末端を介して発現ベクターpET−20b(+)(Merck社製、ドイツ)に連結した。
【0215】
単一コロニーのヒット解析
NovaBlue(DE3)細胞(Merck社製、ドイツ)への形質転換の後、アンピシリン耐性単一コロニーを、200μl SOBAG培地(100μg/ml アンピシリンおよび20g/l グルコースを含むSOB培地)において、37℃で6時間培養した。500μlの自己誘導培地ZYM−5052(Studier社製、2005)を用い、96ディープウェルプレート(Genetix社製、イギリス)において、37℃で16時間培養することにより、ED−B結合修飾ユビキチンを発現させた。4℃、3600×gで、15分間遠心分離して、細胞を回収した。その後、ウェルあたり300μlの溶解バッファーを用いて、37℃で30分間インキュベートし、前記細胞を溶解した。前記溶解バッファーは、0.2×BugBuster(登録商標)(Merck社製、ドイツ)、0.3mg/mlリゾチーム(VWR社製、ドイツ)、0.2mM PMSF(Roth社製、ドイツ)、3mM MgCl2および0.2U/ml Benzonase(VWR社製、ドイツ)を含む、50mM NaH2PO4、300mM NaCl(pH8)とした。4℃、3600×gで、30分間遠心分離した後、得られた上清を、4μg/ml ED−B、およびセイヨウワサビぺルオキシダーゼ(POD)とのユビキチン特異的FabフラグメントコンジュゲートでコートしたNunc MediSorp plates(Thermo Fisher Scientific社製、アメリカ)を用いて、ELISAによりスクリーニングした。検出試薬として、TMB−Plus(Biotrend社製、ドイツ)を用いた。ウェルあたり50μlの0.2M H2SO4溶液を用いて黄色を発色させ、プレートリーダーにおいて450nmと620nmを測定した。
【0216】
数サイクルの選択ディスプレイを、ED−Bに対して行った。最後の2サイクルの選択において、結合分子を、過剰量の遊離ED−Bを用いて溶出した。これらのED−B結合変異体を、他のものとの間で同定した。
【化2】
【0217】
野生型ユビキチン単量体(Ubi monomer wt)と、野生型ユビキチン二量体(ubi dimer wt)、野生型ユビキチンタンパク質(図9におけるUb2−TsX、各単量体の45位における置換およびC末端における2つの置換を有する)、および修飾ユビキチンヘテロ二量体変異体1041−D11との配列アライメントを、図9に示す。Ub2−TsXにおいて、前記単量体のC末端の置換(GGからAA)は、血清における安定性を増加させる。これは、デユビキチナーゼは、ユビキチンのGGの後ろを切断し、AAの後ろを切断しないためである。野生型ユビキチンの二次構造は、前記C末端における置換を有するユビキチンと比較して、ほぼ同一である。
【0218】
1041−D11(図9、配列番号36)または1045−D10と呼ばれる、優れたED−B結合活性を有する修飾ユビキチンは、野生型と比較して、後述のアミノ酸置換によって同定される。
第1モジュール
K6W、L8W、K63R、E64K、S65F、T66P
第2モジュール
K6T、L8Q、Q62W、K63S、E64N、S65W、T66E
任意に、Q2R(当該置換は、変異体1041−D11に存在し、変異体1045−D10には存在しない)
融合タンパク質に適した好ましいリンカーは、配列番号32のリンカーまたはGIG配列である。ただし、代わりに使用できる、多くのリンカーが考えられる。
【0219】
さらに好ましい例として、下記の配列のタンパク質(配列番号47)が提示され、前記配列において、Xはどのようなアミノ酸でもよい。リンカーとして、ここでは、SGGGGSGGGGIGが使用される(イタリック体で示す)。ただし、他の種類のリンカー、またはリンカーなしも、代替可能である。
【化3】
これらの配列を有するタンパク質の例を、図19に示す。
【0220】
[実施例2]
ED−B結合修飾ユビキチン変異体とTNFα(例えば、ヒトTNFα、TNFαという)からの融合タンパク質の生成
【0221】
前記変異体は、修飾ユビキチン、例えば、ヘテロ二量体変異体1041−D11と、哺乳類のTNFα、例えば、マウスまたはヒトのTNFαとの間の融合タンパク質として、大腸菌内で発現させることができる。融合タンパク質のタンパク質解析には、タンパク質発現と純度、非凝集能、細胞培養におけるTNFα活性、標的タンパク質ED−Bへの親和性、選択性、細胞培養における特異的結合が含まれる。F9腫瘍を有するマウスにおける腫瘍の縮小を誘導するための動物実験における必須条件は、マウスTNFαとの融合物である。
【0222】
工程1:融合タンパク質のクローニングのためのベクターの生成(pETSUMO−TNFα)
pETSUMOadaptは、修飾ベクターpETSUMO(Invitrogen社製)であり、付加マルチクローニングサイト(MCS)の挿入によって修飾されている。pETSUMOadaptへのTNFαのクローン化から開始し、ED−Bに結合する修飾ユビキチン変異体を挿入するための制限部位を導入した。得られた構築物は、下記DNA配列(配列番号11)の構造体His6−SUMO−TNFαの構造を有する。
ATGGGCAGCAGCCATCATCATCATCATCACGGCAGCGGCCTGGTGCCGCGCGGCAGCGCTAGCATGTCGGACTCAGAAGTCAATCAAGAAGCTAAGCCAGAGGTCAAGCCAGAAGTCAAGCCTGAGACTCACATCAATTTAAAGGTGTCCGATGGATCTTCAGAGATCTTCTTCAAGATCAAAAAGACCACTCCTTTAAGAAGGCTGATGGAAGCGTTCGCTAAAAGACAGGGTAAGGAAATGGACTCCTTAAGATTCTTGTACGACGGTATTAGAATTCAAGCTGATCAGACCCCTGAAGATTTGGACATGGAGGATAACGATATTATTGAGGCTCACAGAGAACAGATTGGTGGTGTGCGTAGCAGCAGCCGTACCCCGAGCGATAAACCGGTGGCGCATGTGGTGGCGAATCCGCAGGCGGAAGGCCAGCTGCAGTGGCTGAACCGTCGTGCGAATGCGCTGCTGGCCAACGGCGTGGAACTGCGTGATAATCAGCTGGTTGTGCCGAGCGAAGGCCTGTATCTGATTTATAGCCAGGTGCTGTTTAAAGGCCAGGGCTGCCCGAGCACCCATGTGCTGCTGACCCATACCATTAGCCGTATTGCGGTGAGCTATCAGACCAAAGTGAACCTGCTGTCTGCGATTAAAAGCCCGTGCCAGCGTGAAACCCCGGAAGGCGCGGAAGCGAAACCGTGGTATGAACCGATTTATCTGGGCGGCGTGTTTCAGCTGGAAAAAGGCGATCGTCTGAGCGCGGAAATTAACCGTCCGGATTATCTGGATTTTGCGGAAAGCGGCCAGGTGTATTTTGGCATTATTGCGCTGTAATAA
【0223】
TNFα配列を、BamHIおよびXhoI部位の導入により、PCRで増幅した。
使用したプライマーは、以下の通りである。
【化4】
前記fwプライマー(配列番号12)は、TNFαの最初の15塩基対(下線部)を認識し、BamHI配列(太字部)を有する。前記revプライマー(配列番号13)は、TNFαの最後の塩基対、終止コドン(下線部)、およびXhoI制限部位(太字部)を含む。
【0224】
<PCR反応ミックス(100μl)>
84.5μl H2O;
10μl 10×Pwoバッファー+Mg;
2μl 10mM dNTPs(=200μM);
各0.5μl 100μM プライマーfw/rev(=各0.5μM);
2μl DNA(=0.25μg);
0.5μl Pwoポリメラーゼ(=2.5U;Roche)
【0225】
<PCRプログラム>
94℃で3分、94℃で30秒、60℃で30秒、72℃で2分(工程2〜4:30サイクル)、72℃で5分、続いて4℃で処理し、続いて、Qiagen−MinElute−Kit(10μl EBに溶出)によるPCR産物の精製を行った。前記PCR産物を、BamHI−XhoI制限およびライゲーションにより、ベクターpETSUMOadaptのMCSに導入した。
【0226】
<制限ミックス(100μl)>
ベクター:83μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BamHI(=30U;NEB),
1.5μl XhoI(=30U;NEB);
1.65μl ベクター;
3h、37℃でインキュベーション
PCR産物:76.5μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BamHI(=30U;NEB),
1.5μl XhoI(=30U;NEB);
8μl 挿入断片;
3h、37℃でインキュベーション
1%アガロースゲル(100V、60分間ラン)において制限ミックスを分離;
ベクター断片(5659bp)と挿入断片(491bp)の切断;
Qiagen gel extraction kitを用いた精製(30μl EBに溶出)。
【0227】
<ライゲーション(20μl)>
15.2μl H2O;
2μl 10×T4−DNAリガーゼバッファー;
2.26μl ベクター(200ng);
0.54μl 挿入断片(40ng)
5分間、65℃でインキュベーション;
16℃に冷却;
1μl T4−DNAリガーゼ(=3U;NEB)添加;
16時間、16℃でインキュベーション
【0228】
<NaAc/イソプロパノール沈殿>
ライゲーションミクスチャー(20μl)+2.2μl 3M NaAc(pH5.0)+22.2μl イソプロパノール;
30分間、−20℃;
15分間、4℃、13000Upm;
500μlの70% EtOHにペレットを再懸濁;
スピン;
10μlのH2Oにペレットを再懸濁
【0229】
<形質転換>
エレクトロコンピテントNovablue(DE3)細胞(40μlアリコート)と10μlのライゲーション産物とを混合;
0.1cm エレクトロポレーションキュベットに移動;
エレクトロポレーターでパルスを印加(1.8kV、50μF、100 Ohm);
37℃、220Upm、45分間、1mlのSOC培地と、溶液とをインキュベート;
カナマイシン含有LBプレートに、100μlを播き、37℃、一晩インキュベーション
【0230】
工程2:修飾ユビキチンベースのEDB融合タンパク質のクローニング
EDB結合修飾ユビキチンベースの変異体とTNFαとの融合物の生成のために、PCRにより、対象であるEDB修飾ユビキチンベースの配列をpET20bベクターから増幅し、BsaIおよびBamHI制限部位を導入する。この方法は、単量体および二量体EDB修飾ユビキチンベースの変異体に好適である。単量体WTユビキチン(Wubi)用のプライマーは、下記の通りである。
【化5】
【0231】
前記fwプライマー(配列番号14)は、修飾ユビキチンの最初の15塩基対(下線部)を認識し、BsaI配列(太字部)を有する。前記revプライマー(配列番号15)は、修飾ユビキチンの最後の15塩基対を認識し、アミノ酸リンカー(SGGGG配列)およびBamHI制限部位(太字部分)を挿入する。修飾ユビキチンベースの変異体それぞれについて、特異的fwプライマーを使用する。単量体EDB修飾ユビキチンベースの変異体1H4、5E1および4B10のプライマーは、下記の通りである。
【化6】
【0232】
前記revプライマーは、全ての単量体修飾ユビキチンベースの変異体に使用する。二量体修飾ユビキチンベースの変異体用のrevプライマーは、下記の通りである。
【化7】
【0233】
二量体WTユビキチン(WubiHubi)のクローニング用および二量体EDB修飾ユビキチンベースの変異体用のfwプライマーは、下記の通りである。
【化8】
【0234】
<PCRミックス(100μl)>
84.5μl H2O;
10μl 10×Pwoバッファー+Mg;
2μl 10mM dNTPs(=200μM);
各0.5μl 100μM プライマーfw/rev(=0.5μM);
2μl DNA(変異体により決定);
0.5μl Pwoポリメラーゼ(=2.5U;Roche)
【0235】
<PCRプログラム>
1.94℃で3分
2.94℃で30秒
3.60℃で30秒
4.72℃で2分(工程2〜4:30サイクル)
5.72℃で5分、続いて4℃
【0236】
アガロースゲルでPCR産物を精製し、必要なバンドを切り出し、Qiagen−gel extraction kitで精製する。BsaI−BamHI制限により、前記PCR産物をクローニングする(pETSUMO−TNFα内に)。
【0237】
<制限ミックス(100μl)>
75μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BsaI(=30U;NEB);
8μl DNA(ベクターまたはPCR産物)2時間、50℃でインキュベーション、
10分間、65℃、
3μl BamHIの添加(=30U;NEB)、
2時間、37℃
1%アガロースゲルにおいて制限ミックスを分離;
ベクター断片および挿入断片の切断;
Qiagen−gel extraction kitを用いた精製(30μl EBに溶出)。
【0238】
<ライゲーション(20μl)>
12.5μl H2O;
2μl 10×T4−DNAリガーゼバッファー;
5μl ベクター(66ng);
0.5μl 挿入断片(変異体の種類により変わる)
5分間、65℃でインキュベーション;
16℃に冷却;
1μl T4−DNAリガーゼ(=3U;NEB)添加;
16時間、16℃でインキュベーション
【0239】
<NaAc/イソプロパノール沈殿>(工程1参照)
【0240】
<形質転換>
エレクトロコンピテントNovablue(DE3)細胞の形質転換は、前述の通りである。結果として、下記の融合構築物が得られた。すなわち、His6−SUMO−修飾ユビキチン−SGGGG−TNFαを有するpETSUMOadaptにおけるEDB修飾ユビキチンとTNFαとの融合構築物である(単量体修飾ユビキチンの場合、359個のアミノ酸からなり、二量体修飾ユビキチンの場合、447個のアミノ酸からなる)。
【0241】
[実施例3]
ユビキチンベースのTNFα融合タンパク質の発現と精製
【0242】
DNA配列解析により、SUMO−TNFα融合タンパク質の正確な配列が示された。変異体の発現のために、前培養物をLB/カナマイシンで1:100に希釈し、前記培養液を、600nmの光学密度(OD600)0.5まで、200rpm、37℃で攪拌することで、クローンを振とうフラスコで培養した。発現は、IPTG(最終濃度1mM)の添加で誘導した。30℃、4時間、200rpmで、培養を続けた。4℃、6000×gで20分間遠心分離し、細菌細胞を回収した。前記細胞ぺレットを、ベンゾナーゼおよびリゾチームを含むNPI20バッファー30mlに懸濁した。細胞を、氷上で超音波処理(3×20秒)によって破砕した。懸濁液を4℃、40000×gで30分間遠心分離した後、可溶性タンパク質を含む上清を得た。両タンパク質を、室温での親和性クロマトグラフィーにより、精製した。Ni−アガロース(5ml、GE Healthcare社製)の一本のカラムを、50mlのNPI−20で平衡化した。可溶性タンパク質を含む前記上清を、前記カラムにアプライし、続いて、NPI−20で洗浄した。前記結合したタンパク質を、NPI−20から50%NPI−500(100ml)の直線勾配で溶出した。各画分は、SDS−PAGEにより、その純度を解析した。好適な画分をプールし、SUMOヒドロラーゼ切断バッファー(50mM Tris,300mM NaCl、pH8.0)で平衡化したゲルろ過カラム(Superdex 75、1.6×60cm、GE Healthcare社製)に、流速1ml/分で、アプライした。
【0243】
切断反応は、製造元(Invitrogen社)の使用説明書にしたがって行った。切断後、前記タンパク質をNi−アガロースカラム(5ml、GE Healthcare社製)にアプライした。Hisタグ化SUMOヒドロラーゼおよびHisタグ化SUMOは、前記カラムに結合し、正しい融合タンパク質(Hisタグフリー)は、前記カラムを通過した。前記タンパク質の純度は、rpHPLC解析およびゲル電気泳動によって、証明された。前記三量体(TNFαを介する)が正しい分子量を有することは、分析用SEC解析(10/30 Superdex G75、GE Healthcare社製)を用いて確認した。
【0244】
[実施例4]
修飾ユビキチンベースのED−B結合変異体のヒトED−Bに対する結合分析
【0245】
実施例4A:濃度依存ELISAによる修飾ユビキチンベースのED−B結合変異体の結合分析
ユビキチンベースの変異体のヒトED−Bに対する結合を、濃度依存ELISAによって分析した。精製タンパク質を、ヒトED−B、BSAおよび細胞フィブロネクチン(cFN)でコーティングされた複数のNUNC−medisorpプレートに、量を増加させながらアプライした。ウェルあたり抗原50μl(10μg/ml)でのコーティングを、4℃で一晩行った。前記プレートを、0.1% Tween20を含むPBS(pH7.4;PBST)で洗浄した後、前記ウェルを、37℃で2時間、ブロッキング溶液(PBS pH7.4;3% BSA;0.5% Tween20)を用いてブロッキングした。前記ウェルを、PBSTでさらに3回洗浄した。
【0246】
前記ウェルにおいて、異なる濃度の修飾ユビキチンベースのED−B結合タンパク質(50μl量)を、室温で1時間インキュベートした(図10、開始濃度として、500nMの1041−D11タンパク質を使用した)。PBSTで前記ウェルを洗浄した後、抗ユビキチンfabフラグメント(AbyD)PODコンジュゲートを、PBSTに適切に希釈(例えば、1:2000または1:6500)して、アプライした。前記プレートを、ウェルあたり300μlのバッファーPBSTで、3回洗浄した。50μlのTMB基質溶液(KEM−EN−Tec)を各ウェルに加え、15分間インキュベートした。ウェルあたり50μlの0.2M H2SO4を加えて、反応を停止させ、前記ELISAプレートを、TECAN Sunrise ELISA−Readerを用いて読み取った。参照波長を620nmとして、450nmで光度計による吸光光度測定を行った。図1は、みかけKD値11nMという、ED−Bに対する1H4の特異的結合を明瞭に示している。変異体5E1は、みかけKD値7.7μM、4B10は、みかけKD値280nMを、それぞれ示している。図10は、変異体1041−D11のED−Bに対する非常に高い結合親和性を示す(KD=6.9nM)。このように、ユビキチン野生型のわずかな修飾(各単量体において8個の置換まで)により、ED−Bに対する非常に高い結合親和性がもたらされる。
【0247】
実施例4B:競合的濃度依存ELISAによる、修飾ユビキチンベースのED−B結合変異体の結合分析
競合的濃度依存ELISAにより、量が増加する遊離標的の存在下、フィブロネクチン断片(67B89)を含む固定化ED−Bに対するユビキチン変異体1041−D11の結合を分析した。ELISAの条件は、1041−D11タンパク質を、ED−B(67B89)(0μM〜10μM)、またはネガティブコントロール6789(0μM〜10μM)で、1時間プレインキュベートし、その後、その混合物を、Medisorp−plate上に配置した標的67B89に添加した以外は、実施例5Aで述べた通りである。これに続き、前記変異体を、対応する抗体によって検出した(抗ユビキチン−Fab−POD;希釈度1:6500)。図11は、変異体1041−D11が、ED−Bに対して非常に高い結合親和性を有することを示す(IC50=140nM)。図10に示した結果が裏付けられている。すなわち、ユビキチン野生型のわずかな修飾(各単量体において8個の置換まで)のみにより、ED−Bに対する非常に高い結合親和性がもたらされる。
【0248】
実施例4C:結合活性の血清安定性を同時に分析する濃度依存ELISAによる修飾ユビキチンベースのED−B結合変異体の結合分析
当該技術分野において周知の方法により、前述(実施例5Aおよび5B)と同様に、ELISAを行った。ED−B(ここでは、67B89という)を、マイクロタイタープレートにコーティングし、前記変異体を、ED−Bに結合させ、特異的ユビキチン抗体により検出する(抗ユビキチン−Fab−POD)。この分析において、前記変異体は、異なる方法で処理される。すなわち、前記変異体を、37℃で1時間、マウス血清中でインキュベートする処理(図13参照、青丸部分)、前記変異体を、37℃で1時間、ラット血清中でインキュベートする処理(図13、赤丸部分)、または、前記変異体を、37℃で1時間、PBSでインキュベートする処理(図13、黒丸部分)である。図13は、変異体1041−D11の全てのKDが、10.3nM(PBS)から20.74nM(マウス血清)の間にあることを示す。
【0249】
実施例4D:Biacore分析による修飾ユビキチンベースのED−B結合変異体の結合分析
当業者に公知の方法を用いて、CM5チップ(Biacore)に固定化したED−B含有フィブロネクチン断片(67B89という)に対する結合について、前記変異体を異なる濃度で分析した(例えば、0〜200nMの変異体、好ましくは1041−D11)。得られたデータは、BIA評価ソフトウェアおよび1:1−Langmuir−fittingにより処理した。図12に示すように、変異体1041−D11のKDは、1.0nMであった。結合速度定数は、kon=7.6×105M−1s−1、koff=7.7×10−4s−1であった。図17Dに示すように、融合タンパク質1041−D11−TNFαのKDは、1.13nMであった。結合速度定数は、kon=4.5×105M−1s−1、koff=5.0×10−4s−1であった。
【0250】
実施例4E:SE−HPLCによる修飾ユビキチンベースのED−B結合変異体の複合体構造の分析
複合体構造の分析のために、Tricorn Superdex75 5/150 GLカラム(GE−Healthcare社製)(V=3ml)を使用し、50μlのタンパク質をアプライした。さらなる条件は、バッファー:1×PBS(pH7.3)、流速:0.3ml/分、ラン:45分(サンプルの注入:15分後)とした。条件:0.72nmol 1041−D11タンパク質+0.72nmol ED−B(67B89またはネガティブコントロール6789ともいう)を、室温で1時間インキュベートし、複合体構造を分析するために、カラムにアプライした。図14では、前記変異体のみを黒で示し、標的ED−Bのみを青で示し、ED−Bとの複合体を構成する変異体結合をピンクで示す。図14Aは、前記変異体を有するED−Bを示す。図14Bは、ED−Bを有さない変異体を示す。同図は、変異体1041−D11が、ED−B(67B89)と複合体を構築するが、6789とは複合体を構築しないことを示す。
【0251】
[実施例5]
TNFαの生物学的分析
TNFα修飾ユビキチンベースのED−B結合融合体の生理学的TNFα活性を、L929アポトーシス分析によって決定した(Flick et al.、1984 J. Immunol. Methods. 68:167−175)。この分析では、TNFαが、ピコモル範囲のEC50値で、アクチノマイシンD感受性細胞の細胞死を効率的に促進する。
【0252】
細胞を、FBSと抗生物質を含む培地に再懸濁した。密度3.5×105細胞/mlの細胞懸濁液100μlを、96ウェル標準細胞培養プレートのウェルに播き、加湿したCO2インキュベーターで一晩インキュベートした。その後、前記培養培地を除去し、FBS、アクチノマイシンDおよび抗生物質を含む培地50μlを各ウェルに加え、その後、さらに30分間インキュベートした。その後、各テスト項目50μlと、10−7から10−18Mの適切な濃度範囲のTNFα修飾ユビキチンベースのED−B結合融合体またはヒト組換えTNFαコントロールを添加した。さらに48時間インキュベートした後、細胞生存の尺度である代謝活性を、WST−1試薬(Roche社製)を用いて決定した。
【0253】
1つのテスト項目あたり、少なくとも3通りの独立した実験を行い、各実験は3回行った。TNFα修飾ユビキチンベースのED−B結合融合タンパク質の各テストは、分析間のばらつきについての情報を得るために、ヒト組換えTNFαの投与量範囲のテストと並行して行った。
【0254】
定量評価は、EC50値、すなわち、半数の細胞の生存を促進するテスト項目濃度による値に基づく。
【表2】
mub:修飾ユビキチンベースのED−B結合
【0255】
修飾ユビキチンベースのED−B結合融合体の、1つのユビキチン単量体構築物(Wubi)および3つのユビキチン二量体構築物のTNFαを分析した。TNFα部分に結合した修飾ユビキチンベースのED−B結合変異体に依存して、TNFαに関連する活性は、約1桁分増加(SPWF−28_24−H12_TNF−alpha)または減少した(SPWF−28_22−D1_TNF−alpha、Wubi−Hubi−TNF−alpha)。変異体1041−D11 TNFα分析は、図17参照。
【0256】
[実施例6]
細胞培養分析におけるユビキチン変異体の結合分析
培養細胞に対する変異体1041−D11等の結合をテストした。ED−Bが高発現レベルである正常ヒト胎児肺線維芽細胞(Wi38細胞)、マウス胚性線維芽細胞株(Balb 3T3)、マウス骨髄(ST−2)単球/マクロファージ(RAW264.7)由来のストローマ細胞株、NHDF細胞、およびマウス線維芽細胞(LM)を含む、異なる培養細胞を分析した。
【0257】
変異体1041−D11(異なる濃度)またはED−B特異的抗体(500nM FV28 CH4/F1 1×PBS)を、Wi38細胞(60000細胞/ml、ATCC由来)と共にインキュベートし(1時間、37℃)、その後、メタノールで固定し(5分、−20℃)、ブロッキングした(5%ウマ/PBS、1時間)。その後、rabbit−a−Strep−Tag−IgG(GenScript社製 A00875、1:500)と共に1時間インキュベートし、a−rabbit−IgG*Alexa488−AK(Invitrogen社製 A11008、1:1000)と共に1時間インキュベートした。核を、DAPIで染色した。図15Aの1列目は、EDB抗体を用いたコントロールを示し、2列目は、58.7nMのタンパク質濃度での変異体のインキュベーションを示し、3列目は、10倍高濃度(587nM)の1041−D11タンパク質のインキュベーションを示し、4列目は、PBSでのネガティブコントロールである。1行目には、ヒトWi38線維芽細胞を位相差で示し、2行目には、免疫蛍光法で示し、3行目には、DAPI染色を示す。これらの写真から、変異体1041−D11は、ED−B含有細胞外マトリックスに対する高特異性で、固定Wi38細胞に結合すると結論づけられる。ネガティブコントロール細胞型NHDFは、EDBフィブロネクチンを低レベルで発現する初代の正常線維芽細胞である(データ示さず)。前記変異体は、これら細胞に結合しない。
【0258】
図15Bは、バイタル(vital)Wi38細胞での変異体1041−D11の分析を示す。ネガティブコントロール細胞型NHDFは、EDBフィブロネクチンを低レベルで発現する初代の正常線維芽細胞である。前記細胞を、チャンバースライドに播いた(NUNC、60000細胞/ml)。結合能の分析のため、前記細胞を、−20℃で5分間、100%MeOHで固定した。非特異的結合をブロックするために、前記細胞を、37℃で1時間、5%ウマ血清でインキュベートした。前記細胞を、異なる濃度の変異体1041−D11と、ポジティブコントロールであるED−B特異的抗体FV28 CH4/F1またはネガティブコントロールであるUB_2とで、室温で1時間、テストした。rabbit−a−Strep−Tag−IgG(GenScript社製 A00875、1:500)との1時間のインキュベーション、a−rabbit−IgG* Alexa488−AK(Invitrogen社製 A11008、1:1000)との1時間のインキュベーションにより、検証を行った。核を、DAPIで染色した。図15Bの1行目および3行目は、異なるタンパク質濃度の変異体およびネガティブコントロールを示す。2行目および4行目は、EDB抗体を用いたコントロールのインキュベーションを示す。最初の2行は、Wi38細胞株における前記変異体とポジティブコントロールを示す。3行目および4行目は、NHDF細胞のインキュベーションを示す。写真からわかるように、変異体1041−D11は、ED−B含有細胞外マトリックスに対する高い特異性で、バイタル(vital)Wi38細胞に結合することがわかる。低EDBを含まないNHDF細胞を用いた対照試験を行った(データ示さず)。前記変異体は、それらの細胞に結合しない。
【0259】
異なる細胞タイプ、例えば、Balb3T3(ATCC、Kat−Nr.30−2002)、Raw(Lonza社製、Kat−Nr.BE12−115F/U1)、ST−2(Lonza社製、Kat−Nr.BE12−115F/U1)を用いて、同様の実験を行った。図15CおよびDは、ED−Bの結合が、マウスBalb3T3とST−2細胞に高い特異性であることを示す。単球/マクロファージ(Raw)への結合は、見られなかった(データ示さず)。
【0260】
概要を前述したとおり、図16Aは、組織切片における1041−D11の特異性を示す。7つの検体から採取したF9腫瘍組織を評価した。500nM 1041−D11を用いた免疫組織化学から、マウス由来のF9腫瘍におけるED−B特異的血管が染色される結果が得られた。ED−Bは、腫瘍脈管系に対する高い特異性のマーカーである。標的タンパク質EDBは、血管の反管腔側に局在する。1041−D11は、F9腫瘍由来の組織切片において、血管系を特異的に装飾する。得られた結果は、抗体フラグメントL19の組織特異性に匹敵する。さらに、48組織をテストした。FDA関連パネルにおける48組織のいずれにおいても、非特異的な染色は観察されなかった。図16Bは、野生型ユビキチンと比較した、腫瘍細胞における1041−D11の蓄積を示す。このように、ED−Bに特異的に結合する修飾ユビキチンベースの融合タンパク質は、ED−Bに基づく癌の標的治療に好適である。
【0261】
[実施例7]
1041D11−TNFαのin vivoでの有効性
1041−D11−TNFαの治療上の有効性を確立するために、当該化合物をマウスモデルにおけるF9テラトーマ(Borsi et al.、2003 Blood 102, 4384−4392参照)に関してテストした。マウスにおけるED−B発現は、in vivoの状態でヒトと同等であり、1041−D11−mTNFα、好ましくはこれをメルファラン等の細胞傷害性化合物と併用した場合の癌に対する治療効果を評価する上で好適である。F9テラトーマは、高い血管密度を有する高悪性度の腫瘍である。Borsiらには、EDB抗体を介したマウスTNFαのターゲティングは、メルファランの有効性を向上させるものであり、このことは、腫瘍の成長の遅延によって実証されていると記載されている。有効性の検証のための実験計画は、Borsi、2003から採用した。
【0262】
ステージ1では、腫瘍対体重比率、体重減少、および生存率に関するエンドポイントにより、薬理的に活性かつ許容可能な投与量を規定した。本発明者らは、1041D11−TNFαが、最も高い投与量(6.75pmol/g)で許容されるが、腫瘍の成長に対して抑制効果を有さず(3、4、および8日後に体重の10%を上回る→動物死亡)、一方、最も低投与量(0.25pmol/g)の1041D11−TNFαが、腫瘍の成長を遅らせるようであることを見出した。使用したさらに別の投薬グループでは、投与量2.25pmol/g 1041D11−TNFαから、低下させた。
【0263】
研究のステージ2では、腫瘍成長の遅延をエンドポイントとするメルファランの投与量依存的な有効性を明らかにした(動物の体重減少>10%、腫瘍>体重の10%、腫瘍の潰瘍化)。この研究においては、メルファランと併用した1041D11/mTNFα(マウスTNFα)についてテストを行った。168匹の動物を使用し、14投薬グループ(各グループとも、F9腫瘍の大きさが300〜400mm3の時点で採用した8匹のマウスからなる)について、テストサンプルを静脈(i.v.)に投与し、24時間後にメルファランを腹腔内(i.p.)に注射した。表1は、投薬スケジュールを示す。
【表3】
【0264】
図18は、処置期間中(7日間)の相対的腫瘍成長を示す。図18aは、メルファランと併用した本発明の化合物1041−D11−TNFαが、メルファランと併用したmTNFαまたはメルファラン単独よりも効率的に、相対的腫瘍成長を減少させることを明確に示している。処置後7日間の腫瘍成長動態は、1041−D11−mTNFαによる腫瘍の有意な減少を示す。これは、メルファランとの併用の有効性の明らかな証拠である。
【0265】
刊行物
1. Birchler, M., F. Viti, L. Zardi, B. Spiess, and D. Neri. 1999. Selective targeting and photocoagulation of ocular angiogenesis mediated by a phage-derived human antibody fragment. Nat Biotechnol 17:984-8.
2. Brenmoehl, J., M. Lang, M. Hausmann, S. N. Leeb, W. Falk, J. Scholmerich, M. Goke, and G. Rogler. 2007. Evidence for a differential expression of fibronectin splice forms ED-A and ED-B in Crohn’s disease (CD) mucosa. Int J Colorectal Dis 22:611-23.
3. Dubin, D., J. H. Peters, L. F. Brown, B. Logan, K. C. Kent, B. Berse, S. Berven, B. Cercek, B. G. Sharifi, R. E. Pratt, and et al. 1995. Balloon catheterization induced arterial expression of embryonic fibronectins. Arterioscler Thromb Vasc Biol 15:1958-67.
4. Goodsell, D. S. 2001. FUNDAMENTALS OF CANCER MEDICINE: the Molecular Perspective: Antibodies. the Oncologist 6:547-548.
5. Kaczmarek, J., P. Castellani, G. Nicolo, B. Spina, G. Allemanni, and L. Zardi. 1994. Distribution of oncofetal fibronectin isoforms in normal, hyperplastic and neoplastic human breast tissues. Int J Cancer 59:11-6.
6. Menrad, A., and H. D. Menssen. 2005. ED-B fibronectin as a target for antibody-based cancer treatments. Expert Opin Ther Targets 9:491-500.
7. Pujuguet, P., A. Hammann, M. Moutet, J. L. Samuel, F. Martin, and M. Martin. 1996. Expression of fibronectin ED-A+ and ED-B+ isoforms by human and experimental colorectal cancer. Contribution of cancer cells and tumor-associated myofibroblasts. Am J Pathol 148:579-92.
8. Trachsel, E., M. Kaspar, F. Bootz, M. Detmar, and D. Neri. 2007. A human mAb specific to oncofetal fibronectin selectively targets chronic skin inflammation in vivo. J Invest Dermatol 127:881-6.
9. Van Vliet, A., H. J. Baelde, L. J. Vleming, E. de Heer, and J. A. Bruijn. 2001. Distribution of fibronectin isoforms in human renal disease. J Pathol 193:256-62.
10. Lipovsek, D., and Pluckthun, A. (2004). In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 290, 51-67.
11. Ohashi, H., Shimizu, Y., Ying, B.W., and Ueda, T. (2007). Efficient protein selection based on ribosome display system with purified components. Biochem Biophys. Res. Commun. 352, 270-276.
12. Studier, F.W. (2005). Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41, 207-234.
13. Zahnd, C., Amstutz, P., and Pluckthun, A. (2007). Ribosome display: selecting and evolving proteins in vitro that specifically bind to a target. Nat. Methods 4, 269-279.
【技術分野】
【0001】
本発明は、フィブロネクチンのエキストラドメインB(ED−B)に結合可能な、新規なタンパク質、特に、ヘテロ多量体タンパク質に関する。さらに、本発明は、薬学的活性成分および/または診断用活性成分と融合した前記結合タンパク質を含む融合タンパク質に関する。さらに、本発明は、このような結合タンパク質または融合タンパク質の製造方法、ならびに、前記結合タンパク質または融合タンパク質を含む医薬組成物/診断用組成物を対象とする。加えて、本発明は、相互作用する結合決定領域(BDR)を少なくとも2個備えた直鎖状ポリユビキチン鎖を含む足場タンパク質をベースとするライブラリーに関する。
【0002】
別の実施形態では、本発明は、前記結合タンパク質または前記融合タンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを含むベクター、および、前記タンパク質、前記融合タンパク質、前記ベクターおよび/または前記ポリヌクレオチドを含む宿主細胞を対象とする。好ましい実施形態において、前記結合タンパク質または前記融合タンパク質は、医薬品または診断剤に含まれる。さらに、前記組換えタンパク質または前記融合タンパク質の製造方法、ならびに、医学的治療方法における前記タンパク質の使用についても記載する。
【背景技術】
【0003】
免疫グロブリンではないアミノ酸からなる結合分子の需要が高まりつつある。現在に至るまで、抗体は、最もよく確立された結合分子に類別されるが、免疫グロブリン分子にはいくつかの主要な欠点があることから、高い親和性と特異性でリガンドを標的とするために、新たな結合分子がなお求められている。免疫グロブリン分子は、非常に容易に製造でき、ほとんど全ての標的を対象とし得るが、その分子構造は極めて複雑である。そのため、容易に取扱い可能な、より小さな分子によって、抗体を代用することが絶えず求められている。これら代替結合剤は、例えば、疾病の診断、予防、および治療といった医療分野において、有用である。
【0004】
比較的明らかな三次元構造を有するタンパク質は、通常、タンパク質足場と呼ばれ、前記代替結合剤の設計の開始物質として用いられる。これら足場は、通常、特定またはランダムな配列変異を許容する1以上の領域を含んでおり、このような配列のランダム化が、タンパク質ライブラリーの製造のためにしばしば行われる。このようなライブラリーから、特異的結合分子を選択してもよい。抗体よりもサイズが小さく、かつ、標的抗原への親和性が、抗体に匹敵するか、あるいはより高い分子は、薬物動態的な性質および免疫原性の点から、抗体よりも優れていると予想される。
【0005】
従来の多くのアプローチにおいて、結合タンパク質の開始物質として、タンパク質足場が用いられている。例えば、国際公開第99/16873号パンフレット(特許文献1)において、特定のリガンドに結合活性を示すリポカリンファミリー(いわゆるアンチカリン)の修飾タンパク質が開発されている。リポカリンファミリーのペプチド構造は、遺伝子工学的方法を用いた、天然のリガンド結合ポケット内でのアミノ酸置換によって修飾されている。アンチカリンは、免疫グロブリンと同様に、分子構造の同定または結合に使用可能である。抗体と同様の方法で、可動性ループ構造が修飾され、これらの修飾により、天然のものとは異なるリガンドの認識が可能となる。
【0006】
国際公開第01/04144号パンフレット(特許文献2)には、それ自体には結合部分がないβシート構造タンパク質のタンパク質表面における、結合ドメインの人工的な生成が記載されている。この手法によれば、高い親和性と特異性でリガンドと相互作用する、新規に生成された人工結合ドメイン(例えば、眼水晶体構造タンパク質であるγクリスタンのバリエーション)が得られる。アンチカリンに関して先に述べたような可動性ループ構造から形成される、既存の結合部位の修飾とは対照的に、これらの結合ドメインは、βシートの表面に新規に生成される。しかしならが、国際公開第01/04144号パンフレットには、新規結合特性を生じるための、比較的大きなタンパク質の改変について記載されているのみである。国際公開第01/04144号パンフレットのタンパク質は、その大きさのため、ある程度の労力を必要とする方法でしか、遺伝子工学レベルで修飾できない。さらに、これまでに開示されたタンパク質においては、タンパク質の全体構造を維持するために、全アミノ酸のうち、比較的小さいパーセンテージのみが修飾されていた。したがって、従前は存在しない結合特性の発生に利用できるのは、タンパク質表面の比較的に小さい領域のみである。さらに、国際公開第01/04144号パンフレットには、γ−クリスタンに対する結合特性の生成のみが記載されている。
【0007】
国際公開第04/106368号パンフレット(特許文献3)には、ユビキチンタンパク質に基づく人工結合タンパク質の製造が記載されている。ユビキチンは、小型、単量体、細胞質性のタンパク質であり、配列が高度に保存され、原虫から脊椎動物にいたるまでのあらゆる既知の真核細胞に存在する。生物において、ユビキチンは、細胞タンパク質の制御分解の調節において重要な役割を担う。この目的のために、分解予定のタンパク質は、酵素カスケードを通過する間に、ユビキチンまたはポリユビキチン鎖と共有結合し、この標識のおかげで、選択的に分解される。最近の研究結果によると、ユビキチン、またはユビキチンによるタンパク質の標識は、それぞれ、数種のタンパク質の移入またはその遺伝子制御等の他の細胞プロセスにおいても、重要な役割を担っている。
【0008】
ユビキチンは、その生理的機能の解明に加えて、主としてその構造およびタンパク質の化学特性のために、研究対象となっている。ユビキチンのポリペプチド鎖は、並外れてコンパクトなα/β構造に折りたたまれた76個のアミノ酸からなる(Vijay−Kumar, 1987(非特許文献1))。前記ポリペプチド鎖のほぼ87%が、水素結合を介した二次構造エレメントの形成に関与する。二次構造は、3回転半のαヘリックス、ならびに、4つの鎖からなる逆平行βシートである。これらの要素の特徴的な配置(逆平行βシートがタンパク質表面に露出し、その裏側にαヘリックスが包まれ、これが前記逆平行βシート上に垂直に延びている)が、一般的に、いわゆるユビキチン様折り畳みモチーフと考えられている。さらなる構造的な特徴は、前記αヘリックスと前記βシート間のタンパク質内部における際立った疎水性領域である。
【0009】
その小さなサイズのため、ユビキチンの人工的な調製は、化学合成によっても、バイオテクノロジー手法によっても実施可能である。その有利な折り畳み特性のため、ユビキチンは、遺伝子工学により、大腸菌等の微生物を使用して比較的大量に、そのサイトゾルまたはペリプラズム空間に製造できる。ペリプラズムにおける優勢な酸化条件のため、後者の戦略は、一般的に、分泌タンパク質の製造に使用される。簡易かつ効率的な細菌調製により、ユビキチンは、その製造に問題がある他の異種タンパク質の調製のための融合パートナーとして使用できる。ユビキチンとの融合によって、溶解度の改善、およびそれによる製造収率の改善が達成できる。
【0010】
抗体または他の代替足場と比べ、ユビキチンタンパク質をベースとする人工結合タンパク質(Affilin(登録商標)ともいう)は、小さいサイズ、高い安定性、高い親和性、高い特異性、費用対効果の高い微生物による製造、血清半減期の調整という利点を有している。しかしながら、特定の標的に対する高い親和性を利用した新たな治療方法の観点から、それらのタンパク質をさらに開発する必要がある。国際公開第05/05730号パンフレット(特許文献4)には、人工結合タンパク質を得るためのユビキチン足場の使用が、概略的に記載されているが、フィブロネクチンのED−Bとの特異的かつ高親和性の結合を得るためにどのようにユビキチンタンパク質を修飾すべきかについては記載されていない。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】国際公開第99/16873号パンフレット
【特許文献2】国際公開第01/04144号パンフレット
【特許文献3】国際公開第04/106368号パンフレット
【特許文献4】国際公開第05/05730号パンフレット
【特許文献5】国際公開第2008/022759号パンフレット
【特許文献6】国際公開第97/45544号パンフレット
【特許文献7】国際公開第07/054120号パンフレット
【特許文献8】国際公開第99/58570号パンフレット
【特許文献9】国際公開第01/62800号パンフレット
【特許文献10】国際公開第06/119897号パンフレット
【特許文献11】国際公開第07/128563号パンフレット
【特許文献12】国際公開第01/62298号パンフレット
【特許文献13】国際公開第07/115837号パンフレット
【非特許文献】
【0012】
【非特許文献1】Vijay−Kumar, 1987
【非特許文献2】Menrad u. Menssen, 2005
【発明の概要】
【発明が解決しようとする課題】
【0013】
国際公開第2008/022759号パンフレット(特許文献5)には、組換え結合タンパク質が記載されており、新しい結合タンパク質を得るために、FYNキナーゼのSrcホモロジー3ドメイン(SH3)を使用している。タンパク質治療法および/またはタンパク質診断法を開発するために、RTループおよび/またはSrcループを変異させて標的特異性を設計できることが見出された。足場として利用されるリポカリンにおいてと同様に、変異誘発されるアミノ酸残基は、抗体/抗原結合機能の基礎をなす原理を擬態する可変性および可動性のループ領域内に存在している。相互作用部位の全体的な可動性により、抗体がエピトープに結合するが、これは主としてエンタルピーによって駆動されるプロセスである。しかしながら、このプロセスは、可動相補性決定領域の会合による移動度の欠失によって、不利なエントロピー寄与を招く。本発明者らは、それとは反対に、ユビキチンを足場として使用し、アミノ酸残基をそもそも可動性ループ領域内で改変するのではなく、βシート領域の強固で不動のβ鎖内、または前記β鎖に密接した範囲内で改変した。ED−Bとの結合領域として、ユビキチンの不動かつ強固なβ鎖または前記β鎖に密接した範囲内でアミノ酸残基を選択する利点は、特に、以下の通りである。結合パートナー同士は、強い結合に適した相補的形状をすでに示していると考えられる。したがって、これらの相互作用には、前記結合パートナーのより強固な構造における形状の相補性、電荷、および親水性/疎水性要素が関係する。これらの強固な本体の相互作用は、界面を最適化し、生物機能に対応する。
【0014】
フィブロネクチン(FN)は、健康な組織および体液において大量に発現する、重要なクラスの高分子量細胞外マトリックス糖タンパク質である。その主な役割は、多数の異なる細胞外マトリックスへの細胞の接着を促進することである。
【0015】
培養下の非形質転換細胞の表面にはフィブロネクチンが存在し、形質転換細胞にはフィブロネクチンが存在しないことから、フィブロネクチンは、重要な接着タンパク質として同定された。フィブロネクチンは、多数の様々な他の分子、例えば、コラーゲン、ヘパラン硫酸プロテオグリカンおよびフィブリン等と相互作用し、細胞の形状および細胞骨格の形成を制御する。さらに、フィブロネクチンは、胚形成時の細胞移入および細胞分化に関与する。フィブロネクチンはまた、創傷治癒においても重要な役割を担い、マクロファージおよび他の免疫細胞の移入を促進し、また、血餅の形成においても、血管の損傷領域への血小板の接着を可能とすることにより、重要な役割を担う。
【0016】
フィブロネクチンのエキストラドメインB(ED−B)は、一次RNA転写物の選択的スプライシングによってフィブロネクチン分子に挿入された、小さいドメインである。前記分子は、細胞外マトリックスのフィブロネクチン分子において、存在するかまたは取去されているかのいずれかであり、新しい血管の周辺に大量に発現するが、実質的に全ての正常成人の組織(子宮と卵巣を除く)において検出不能であるため、血管新生や組織の再構築と関連する最も選択的なマーカーの一つとなっている。ED−Bは、癌および乾癬に関与することが知られている。高レベルのED−B発現が、乳癌、結腸直腸癌、膵臓癌、非小細胞肺癌、肝細胞癌、頭蓋内髄膜腫、ヒトの皮膚癌、膠芽細胞腫を含む多くのヒトの固体癌のエンティティにおいて検出された(Menrad u. Menssen、2005(非特許文献2))。さらに、ED−Bは、診断剤と結合でき、診断ツールとして有利に使用できる。一例として、例えば、動脈硬化プラークの分子の画像化や、例えば、癌患者の免疫シンチグラフィーによる、癌検出おける使用があげられる。さらに多くの診断用途が考えられる。
【0017】
フィブロネクチンのヒトエキストラドメインB(ED−B)の91個のアミノ酸からなるアミノ酸配列を、配列番号2に示す。前記タンパク質を発現させるには、開始メチオニンを加えなければならない。ED−Bは、哺乳類、例えば、げっ歯類、ウシ、霊長類、肉食動物、ヒト等に保存されている。ヒトED−Bと100%の配列同一性を有する動物の例は、ラット(Rattus norvegicus)、ウシ(Bos taurus)、マウス(Mus musculus)、ウマ(Equus caballus)、アカゲザル(Macaca mulatta)、ハイイロオオカミ(Canis lupus familiaris)、チンパンジー(Pan troglodytes)である。
【0018】
ED−Bは、新生血管構造に特異的に蓄積し、癌における分子介入の標的となる。フィブロネクチンのED−Bドメインに対する抗体または抗体断片の多くは、当該技術分野において、癌およびその他の適応症の有望な治療薬として公知である(例えば、国際公開第97/45544号パンフレット(特許文献6)、国際公開第07/054120号パンフレット(特許文献7)、国際公開第99/58570号パンフレット(特許文献8)、国際公開第01/62800号パンフレット(特許文献9)参照)。ヒト一本鎖Fv抗体断片ScFvL19(L19ともいう)は、フィブロネクチンのED−Bドメインに特異的であり、実験腫瘍モデルおよび癌患者の両方において、選択的に新生脈管構造の腫瘍(tumor neovasculature)を標的とすることが確認されている。さらに、IL−12、IL−2、IL−10、IL−15、IL−24またはGM−CSF等のサイトカインと、抗ED−B抗体または抗ED−B抗体断片を含むコンジュゲートは、特に、癌、血管新生、または腫瘍の成長を阻害する医薬品の製造のための標的薬剤として記載されている(例えば、国際公開第06/119897号パンフレット(特許文献10)、国際公開第07/128563号パンフレット(特許文献11)、国際公開第01/62298号パンフレット(特許文献12)参照)。細胞傷害性薬剤または免疫刺激薬剤のような適切なエフェクター機能と抱合させた、抗ED−B抗体またはL19等抗ED−B抗体断片による固体腫瘍の新生脈管構造の選択的なターゲティングは、動物実験では成功することが判明している。膵臓癌の治療のため、インターロイキン−2部分(IL−2)と抗ED−B抗体部分とを含む融合タンパク質を、小分子ゲムシタビン(2’−デオキシ−2’、2’−デフルオロシチジン)と併用した(例えば、国際公開第07/115837号パンフレット(特許文献13)参照)。
【0019】
前記先行技術文献には、新しいED−B結合タンパク質の製造のため、抗体を含む様々なタンパク質足場を使用することが記載されている。
【0020】
現在入手可能な化合物よるED−Bのターゲティングには、ある種の不都合が伴う。ED−B抗原に対して、同等またはより高い親和性を有する、より小さい分子(本発明の、ユビキチンベースのED−B結合タンパク質等)は、抗体や他の結合タンパク質に対して、顕著な利点を有すると期待される。
【0021】
癌は、世界的に主要な死亡原因であるため、癌治療のための改善された薬剤の必要性が高まっている。現在の化学療法剤および放射線治療は、選択性が低いという欠点があり、多くの化学療法剤は、腫瘍部位に蓄積せず、よって腫瘍内において十分なレベルに到達できない。癌の効果的な治療に対する高い医学的ニーズが存在する。
【0022】
よって、本発明の目的は、フィブロネクチンの細胞外ドメイン(ED−B)に非常に高い親和性で特異的に結合できる、ユビキチンベースの新規結合タンパク質の提供である。本発明のさらに別の目的は、例えば、癌の治療に使用する、ED−Bに対して非常に高い結合特異性を有する新規結合タンパク質の同定および提供である。さらに、前記結合分子を製造するための方法も提供する。
【課題を解決するための手段】
【0023】
前記課題は、提出した独立請求項の主題によって解決される。本発明の好ましい実施形態は、従属請求項、ならびに下記の記載、実施例、および図面に含まれる。
【図面の簡単な説明】
【0024】
【図1】図1は、種々の腫瘍におけるED−Bの発生を列記した表を示す。
【図2】図2Aは、修飾ユビキチンED−Bバインダー5E1に関する濃度依存性ELISAの結果を示す。図2Bは、修飾ユビキチンED−Bバインダー1H4に関する濃度依存性ELISAの結果を示す。
【図3】図3は、修飾ユビキチン単量体からなる四量体と比較した、修飾ユビキチン単量体のKd値を、表に示す。
【図4A】図4Aは、単量体41B10について、結合親和性がKd=9.45μMであることを示す。
【図4B】図4Bは、41B10が異なる第2単量体と結合して得られた46H9について、結合親和性がKd=131nMであることを示す。
【図5A】図5Aは、修飾ユビキチンベースのED−B結合エフェクター融合タンパク質の模式図である。
【図5B】図5Bは、前記修飾ユビキチンエフェクター結合5E1−TNFコンジュゲートが、アポトースシス促進活性(L929アポトーシスアッセイで測定)を有すること示す。
【図5C】図5Cは、1H4−TNFα融合物のED−Bへの高結合親和性(Kd=15.1nM)を示す。
【図6】図6は、サイトカイン、例えば、TNFαに融合した、修飾ユビキチンベースのED−B結合へテロ二量体分子の親和性および活性を示す。
【図6A】図6Aは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd 50.7nM)。
【図6B】図6Bは、サイトカインTNFαと遺伝子融合し、前記ヘテロ二量体24H12が多量体化された、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd=5.6nM)。
【図6C】図6Cは、ヘテロ二量体修飾ユビキチンライブラリーから選択される候補例、例えば、ヘテロ二量体クローン9E12、22D1、24H12、41B10の分析結果を示す。
【図6D】図6Dは、Biacore(登録商標)を使用した、前記修飾ヘテロ二量体ユビキチン分子9E12の、無標識相互作用分析による分析結果を示す。
【図6E】図6Eは、Biacore(登録商標)を使用した、前記修飾ヘテロ二量体ユビキチン分子41B10の、無標識相互作用分析による分析結果を示す。
【図7】図7は、結合親和性および結合特異性に対する、異なる修飾がなされたユビキチン変異体の寄与を示す。
【図8】図8は、配列アライメントを示す。
【図9】図9は、「Ub2_TsX9」(両単量体の45位がトリプトファンに修飾され、2つの単量体の間にリンカーGIG(77〜79位;80位のメチオニンから第2単量体が開始)を有し、前記第2単量体の最後のC末端アミノ酸において、グリシンからアラニンに置換されているユビキチン)に対する、修飾ユビキチンへテロ二量体変異体1041−D11(1行目)の配列アライメントを示す。
【図10】図10は、ヒトED−Bへの前記ヘテロ二量体ユビキチン変異体1041−D11の結合についての、濃度依存性ELISAを示す。
【図11】図11は、フィブロネクチン断片を含む固定化されたED−B(67B89)に対するヘテロ二量体ユビキチン変異体1041−D11の結合を、遊離標的の量を増加させながら分析した競合濃度依存性ELISAを示す。
【図12】図12は、Biacore(登録商標)を使用した、前記修飾へテロ二量体ユビキチン分子1041−D11の、無標識相互作用分析における分析結果を示す。
【図13】図13は、結合活性の血清安定性を同時に分析する濃度依存性ELISAにおける、ED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。
【図14】図14は、ヘテロ二量体ユビキチン変異体1041−D11とフィブロネクチン断片との複合体形成の、SE−HPLCによる分析を示す。
【図15A】図15Aは、固定化されたヒト胎児肺繊維芽細胞(Wi38)への前記ヘテロ二量体ユビキチン変異体1041−D11の結合を示す。
【図15B】図15Bは、バイタル(vital)ヒト胎児肺繊維芽細胞(Wi38)への結合を示す。
【図15C】図15Cは、固定化されたマウスBalb 3T3細胞への結合を示す。
【図15D】図15Dは、固定化されたマウスST−2細胞への結合を示す。
【図16A】図16Aは、哺乳類組織切片における、ターゲットへのヘテロ二量体ユビキチン変異体1041−D11の特異性を示す。
【図16B】図16Bは、野生型ユビキチンとの比較における、腫瘍組織での1041−D11の蓄積を示す。
【図17A】図17Aは、1041−D11TNFα融合タンパク質におけるTNFαのアポトーシス誘導活性についての、細胞ベースアッセイ(cell based assay)(L929細胞)によるテストを示す。
【図17B】図17Bは、1041−D11TNFα融合タンパク質におけるTNFαのアポトーシス誘導活性についての、細胞ベースアッセイ(L929細胞)によるテストを示す。
【図17C】図17Cは、標的ED−Bに対するヘテロ二量体ユビキチン1041−D11TNFα融合タンパク質の高い選択性を証明している。
【図17D】図17Dは、修飾ユビキチンED−B結合1041−D11TNFα融合タンパク質についての、Biacoreアッセイによる結合分析を示す。
【図17E】図17Eは、培養細胞における変異体1041−D11で観察された高い結合特異性が、1041−D11がTNFαに融合されても維持されることを示す。
【図18】図18は、TNFαに融合した変異体1041−D11とメルファランとの併用による、7日間のマウスの処置期間中のin vivoでの相対的腫瘍成長を示す。
【図19】図19は、ED−Bに対して驚くほど強力な結合親和性を有することが見出されたさらなる16種類の配列について、コンセンサスな位置およびアミノ酸置換を示す。
【発明を実施するための形態】
【0025】
より具体的には、本発明者らは、配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有する修飾ユビキチンタンパク質を含む、ヒトフィブロネクチンのED−Bに結合可能なタンパク質であり、
前記フィブロネクチンのED−Bに対し、Kd=10−6〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも4個のアミノ酸が修飾されたタンパク質を提供する。
【0026】
好ましい実施形態において、前記タンパク質は、組換えタンパク質である。
【0027】
本発明のさらに別の実施形態では、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、4、5、6、7、8、9個、または全てのアミノ酸が修飾される。
【0028】
本願で使用されている重要な用語の定義
「フィブロネクチンのエキストラドメインB」という用語、またはその略語「ED−B」は、配列番号2との配列同一性が、少なくとも70%、任意に75%以上、さらに任意に80%、85%、90%、95%、96%、または97%以上、または100%を示し、先に定義したED−Bの機能を有する、全てのタンパク質を含む。
【0029】
「結合可能なタンパク質」または「結合タンパク質」という用語は、下記にさらに詳細に定義するように、ED−Bに対する結合ドメインを含むユビキチンタンパク質を指す。このようなユビキチンベースの結合タンパク質は、いずれも、例えば、多量体化部位(multimerization moieties)、ポリペプチドタグ、ポリペプチドリンカー等の、結合ドメインでない付加的なタンパク質ドメイン、および/または、非タンパク性のポリマー分子を含んでもよい。非タンパク性のポリマー分子は、例えば、ヒドロキシエチルでんぷん、ポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレン等である。
【0030】
抗体およびその断片は、当業者に周知である。本発明の結合タンパク質は、抗体またはその断片、例えば、FabもしくはscFv断片等ではない。また、本発明の結合ドメインは、抗体中に存在する免疫グロブリンフォールドを含まない。
【0031】
本明細書において、「リガンド」、「標的」および「結合パートナー」は、同義に用いられ、置き換え可能である。リガンドは、前述のヘテロ多量体修飾ユビキチンタンパク質に、本明細書中で定義される親和性をもって結合可能な任意の分子である。
【0032】
「ユビキチンタンパク質」という用語は、配列番号1のユビキチン、および、下記の定義によるその修飾物を包含する。ユビキチンは、真核生物において、高度に保存されている。例えば、今日まで研究されてきた全ての哺乳類において、ユビキチンは、同一のアミノ酸配列を有する。ヒト、げっ歯類、ブタ、および霊長類由来のユビキチン分子が、特に好ましい。また、任意の他の真核生物起源のユビキチンを使用することもできる。例えば、酵母由来のユビキチンは、配列番号1と3個のアミノ酸が異なるのみである。前記「ユビキチンタンパク質」という用語に包含される前記ユビキチンタンパク質は、通常、配列番号1に対し、70%を超える、好ましくは75%を超える、80%を超える、85%を超える、90%を超える、95%を超える、96%を超えるアミノ酸同一性を示すか、あるいは、97%までの配列同一性を示す。
【0033】
「修飾ユビキチンタンパク質」という用語は、アミノ酸の置換、挿入、または欠失のいずれか一つ、あるいはそれらの組み合わせによる、前記ユビキチンタンパク質の修飾物を指す。本発明の修飾ユビキチンタンパク質は、標的に対する新規な結合親和性を有する改変タンパク質である。
【0034】
配列番号1のアミノ酸配列に対する前記ユビキチン誘導体の配列同一性の程度の決定には、例えば、SIM Local similarity program(Xiaoquin Huang and Webb Miller, Advances in Applied Mathematics, vol. 12: 337-357, 1991)またはClustal,W.を使用できる(Thompson et al., Nucleic Acids Res., 22(22): 4673-4680, 1994.)。好ましくは、配列番号1に対する前記修飾タンパク質の配列同一性の程度は、配列番号1の完全配列に対して相対的に決定される。
【0035】
本発明の「ヘテロ二量体融合タンパク質」または「ヘテロ二量体タンパク質」は、異なる修飾がなされた2つの単量体ユビキチンタンパク質を含むタンパク質と考えられる。前記2つの単量体ユビキチンタンパク質は、特異的結合パートナーであるED−Bに対する一価の結合特性(結合ドメイン)を共同でもたらす、2つの相互作用結合ドメイン領域を有する。ヘテロ二量体は、2つの単量体ユビキチン分子の融合により得られるが、これらの両分子は、本明細書に記載のように、異なる修飾がなされている。
【0036】
異なる修飾がなされたユビキチン単量体を多量体化して一価の結合活性を有するヘテロ多量体結合タンパク質(ここでは、ヘテロ二量体タンパク質)を生成する利点は、ED−Bに対する新規な高親和性の結合特性を発生させるために修飾することが可能なアミノ酸残基の総数の増加にある。主な利点は、修飾されるアミノ酸が増加しても、ED−Bに対する前記新規に生成された結合タンパク質の足場の全体的な安定性を低下させることなく、タンパク質化学的完全性(protein−chemical integrity)が維持されることである。ED−Bに対する新規結合部位を発生させるために修飾可能な残基の総数は、前記修飾残基が2つの単量体ユビキチンタンパク質に割り当てられることで増加する。修飾数は、修飾単量体ユビキチン分子数に応じて、2倍にできる。ユビキチンベースのED−B結合タンパク質のモジュール構造により、修飾アミノ酸の総数を増加できる。これは、前記修飾アミノ酸が、2つの単量体ユビキチン分子に含まれるからである。本発明の方法は、ED−Bに対する一価の特異性を有するヘテロ二量体ユビキチン分子の同定を提供する。
【0037】
したがって、結合パートナーに対して共通の結合部位を有するヘテロ二量体の使用は、最終的な結合分子のタンパク質化学的完全性に過度に影響を与えない修飾残基数を増加させる可能性を切り開く。なぜなら、これらの修飾残基の全量が、前記二量体を形成する2つの単量体ユニットに分散するためである。ED−Bに結合する前記ヘテロ二量体修飾ユビキチンタンパク質は、タンパク質ライブラリーに存在する。
【0038】
「一価」とは、前記修飾二量体ユビキチンの第1および第2単量体ユニットにおいて生成された両結合領域が、共に、ED−Bに相乗的に組み合わせられるように結合する機能、すなわち、両結合領域が、共同で一価の結合活性を形成するように作用する能力と理解すべきである。前記へテロ二量体分子における前記第1および前記第2修飾ユビキチン両方の各結合領域を別々に見た場合、前記二量体分子と比較して、ED−Bへの結合の効率および親和性は、明らかに低いと考えられる。前記修飾ユビキチンが、各単量体タンパク質を単独で使用するよりも、より効率的にED−Bに結合可能となるように、両結合領域は、前記ヘテロ二量体修飾ユビキチンタンパク質表面に、アミノ酸の連続領域として形成される固有の結合部位を形成する。本発明によれば、最も有力な結合ユビキチン分子をスクリーニングした後に、前記2つの単量体タンパク質が互いに結合されるのではなく、既に前記ヘテロ二量体ユビキチンが存在する状態でスクリーニング工程を行うことが、特に重要である。最も有力な結合ユビキチン分子の配列情報を取得した後に、任意の他の方法、例えば、化学合成または遺伝子工学手法により、例えば、前記2つの同定した単量体ユビキチンユニット同士を結合することにより、これらの分子を得てもよい。
【0039】
本発明によれば、1つのリガンドに結合する異なる修飾がなされた前記少なくとも2つのユビキチン単量体は、例えば、遺伝学的手法を使用して、互いにヘッドトゥテイル融合で結合される。前記異なる修飾がなされた融合ユビキチン単量体は、一価で結合し、両方の「結合ドメイン領域」(「BDR」)が共に作用する場合にのみ効果的である。本明細書中において、「結合ドメイン領域」は、標的への結合に関与する、配列番号1の2、4、6、8、62、63、64、65、66および68位における、少なくとも4個、好ましくは6個のアミノ酸が修飾されたユビキチン単量体における領域と定義される。
【0040】
前記ヘテロ二量体タンパク質を形成する、前記修飾され結合されたユビキチン単量体は、一つの連続的な結合領域を介して同一のエピトープに結合する。前記ヘテロマーのこの連続的な領域は、異なる修飾がなされた2つのユビキチン単量体により形成される前記2つのモジュールの両結合決定領域により形成される。
【0041】
「ヘッドトゥテイル融合」は、2つのタンパク質を、前記二量体に含まれるユニット数に応じて、N−C−N−C方向に連結することにより、互いに融合させることと理解すべきである。このヘッドトゥテイル融合において、前記ユビキチン単量体は、リンカーを介さずに直接連結されてもよい。または、前記ユビキチン単量体の融合を、リンカーを介して行うこともできる。前記リンカーとしては、例えば、少なくともGIGのアミノ酸配列もしくは少なくともSGGGGのアミノ酸配列を有するリンカー、または、任意の他のリンカー、例えば、GIG、SGGGG、SGGGGIG、SGGGGSGGGGIGもしくはSGGGGSGGGGがあげられる。2つのユビキチン単量体の遺伝子融合用の他のリンカーも、当該技術分野では公知であり、使用可能である。
【0042】
本発明の修飾ユビキチンタンパク質は、標的またはリガンド(両用語は、同じ意味として使用される)としてのED−Bに対する新規な結合親和性を有する改変タンパク質である。「置換」という用語は、例えば、元のアミノ酸への化学基もしくは残基の置換または付加によるアミノ酸の化学的修飾も含む。βシート領域の少なくとも1つのβシート鎖に位置するアミノ酸または前記βシート鎖に隣接した3つ目までのアミノ酸を含む、タンパク質の少なくとも1つの表面露出領域におけるアミノ酸の置換が重要である。
【0043】
本発明によれば、ED−Bに特異的な新規の結合ドメインの生成のためのアミノ酸の置換を、任意の所望のアミノ酸を用いて行うことができる。すなわち、ED−Bに対する新規結合特性を発生させるための修飾において、前記アミノ酸が、置換するアミノ酸のものと類似する特定の化学的特性または側鎖を有するよう配慮する必要はなく、よって任意の所望のアミノ酸をこの目的に使用することができる。
【0044】
本発明によれば、選択されたアミノ酸の修飾工程は、ランダム突然変異誘発による遺伝子レベルでの突然変異誘発、すなわち、前記選択されたアミノ酸のランダム置換により行われることが好ましい。ユビキチンの修飾は、各タンパク質に属するDNAを改変する遺伝子工学的手法により行われることが好ましい。ユビキチンタンパク質の発現は、その後、原核生物または真核生物内で行われることが好ましい。
【0045】
置換は、特に、ユビキチンタンパク質のβシートの4つのβ鎖の表面露出アミノ酸、または、前記βシート鎖に隣接する3つ目までの表面露出アミノ酸において行われる。各β鎖は、通常、5〜7個のアミノ酸からなる。配列番号1を参照すると、例えば、前記β鎖は、通常、アミノ酸残基2〜7、12〜16、41〜45および65〜71を包含する。これに加えて好ましく修飾され得る領域として、前記βシート鎖に隣接する3つ目までのアミノ酸位置(すなわち、1番目、2番目または3番目)が挙げられる。これに加えて好ましく修飾され得る好ましい領域としては、特に、アミノ酸残基8〜11、62〜64および72〜75が挙げられる。前記好ましい領域は、2つのβ鎖を互いに連結するβターンを含む。好ましいβターンとして、アミノ酸残基62〜64があげられる。前記βシート鎖に密接する最も好ましいアミノ酸は、8位のアミノ酸である。また、アミノ酸置換のさらなる好ましい例は、36、44、70および/または71位である。例えば、これに加えて好ましく修飾され得る領域としては、62、63、および64位のアミノ酸(3個のアミノ酸)、72、73位のアミノ酸(2個のアミノ酸)、または8位のアミノ酸(1個のアミノ酸)が挙げられる。
【0046】
好ましい実施形態において、前記アミノ酸残基は、アミノ酸置換により改変される。欠失および挿入も可能である。付加または欠失され得るアミノ酸数は、単量体ユビキチンサブユニットにおいて、1、2、3、4、5、6、7、8、9、または10個のアミノ酸に限定され、それに応じて、前記二量体ユビキチンタンパク質については、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、18、20、22、24、26、または28個のアミノ酸に限定され、通常は、前記単量体タンパク質における修飾数のx倍である。一実施形態において、アミノ酸挿入はなされない。さらに別の実施形態において、欠失は行われない。
【0047】
本発明の修飾ユビキチンタンパク質が、1以上のアミノ酸の置換、欠失および/または付加を含む場合には、対応するタンパク質同士を割り当てるために、野生型ヒトユビキチン(配列番号1)のアミノ酸位置と、前記修飾ユビキチンとのアライメントを行わなければならない。融合タンパク質の場合(後述を参照)、各単量体ユビキチンサブユニットのナンバリング(およびアライメント)は、同様の方法によりなされる。すなわち、例えば、二量体のアライメントは、各サブユニットについて、1位のアミノ酸から開始される。
【0048】
本発明によれば、好ましくは哺乳類由来の、例えば、ヒトの単量体ユビキチンにおいて、β鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置に存在するアミノ酸の、少なくとも10%、好ましくは少なくとも20%、さらに好ましくは少なくとも25%を修飾、好ましくは置換することができ、これにより、以前は存在しなかった結合特性が発生する。β鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置に存在するアミノ酸の、好ましくは最大で約50%、さらに好ましくは最大で約40%もしくは約35%、または約30%もしくは約25%までが修飾、好ましくは置換される。1つのβ鎖内において、通常、1〜4個のアミノ酸が修飾される。一実施形態では、好ましくは、第1β鎖および第4β鎖、例えば、アミノ酸残基2〜7もしくは65〜71の領域において、6個のアミノ酸のうちの3個が修飾される。
【0049】
ヘテロ二量体の構成ユニットとして使用される本発明の修飾単量体ユビキチンは、合計でアミノ酸の20%までを占める。これを考慮すると、配列番号1に対する前記修飾ユビキチンタンパク質の配列同一性は、少なくとも60%である。本発明のさらに別の実施形態において、アミノ酸レベルでの配列同一性は、配列番号1のアミノ酸配列に対して、少なくとも60%、70%、少なくとも80%、さらに、少なくとも90%または少なくとも95%である。本発明は、配列番号1のアミノ酸配列と比較して、65%を超える、75%超える、85%超える、または97%超える前記修飾ユビキチンタンパク質のアミノ酸配列同一性も包含する。
【0050】
本発明のさらに別の実施形態において、ユビキチンは、配列番号1の2、4、6、8、62、63、64、65、66および/または68位における、3、4、5、6、または7個のアミノ酸が修飾される。他の実施形態では、これらの位置が修飾されるユビキチンを、既に前修飾(pre−modified)されていた。例えば、さらなる修飾は、74および75位のアミノ酸または45位のアミノ酸における修飾を含むことができ、この修飾により、より良好な安定性またはタンパク質化学特性が発生する。修飾ユビキチン単量体は、配列番号1のユビキチンにおける、合計9、10、11、12、13、14、および、最大で15個のアミノ酸が修飾、好ましくは置換されることで得られる。一例によれば、14個の置換および1個の欠失を有する修飾単量体ユビキチンを得ることができた。これは、ユビキチンの総アミノ酸数の約20%の割合に相当する。このことは、非常に驚くべきことであり、通常、もっと低い割合でも、タンパク質のフォールディングを阻害するのに十分であるため、予測できなかったことである。
【0051】
本発明の一実施形態において、これらのアミノ酸は、タンパク質表面上に連続的な領域を形成する、新規な結合特性を有する領域を生成するために修飾される。このようにして、ED−Bへの結合特性を有する連続的な領域を生成することができる。本発明において、「連続的な領域」は、以下の通りである。側鎖の電荷、空間的構造、および疎水性/親水性により、アミノ酸は、それらに応じた形で環境と相互作用する。前記環境は、溶媒、一般的には水、または、例えば、空間的に近いアミノ酸等の他の分子であり得る。タンパク質に関する構造情報および各種ソフトウェアにより、前記タンパク質の表面を特徴付けできる。例えば、タンパク質の原子と溶媒との界面領域について、この界面領域がどのような構造であるか、溶媒に接触しやすい表面領域はどれか、または、電荷が表面においてどのように分布しているかについての情報も含めて、この方法により可視化できる。連続的な領域は、例えば、適切なソフトウェアを使用したこの種の可視化により明らかにすることができる。このような方法は、当業者に公知である。本発明によれば、基本的に、表面露出領域全体を、新規結合特性の生成のために修飾される表面上の前記連続的な領域として使用することができる。一実施形態において、この目的のために、修飾は、αへリックス領域も含むことができる。ヘテロ二量体修飾ユビキチンタンパク質において、結合決定領域は、1つの連続的な領域を共同で形成する2つの前記表面露出領域を含み、前記連続的な領域は、1つの結合決定領域の2倍の長さを有する。
【0052】
βシート領域の少なくとも一本のβ鎖内または前記βシート鎖に隣接する3つ目までのアミノ酸位置のいずれかを含む、前記タンパク質の少なくとも1つの表面露出領域においてアミノ酸を修飾することが重要である。前記「βシート構造」は、本質的にシート状であり、ほぼ完全に伸長している(stretched)ことにより定義される。ポリペプチド鎖の連続したセグメントから形成されるαへリックスとは対照的に、βシートは、ポリペプチド鎖の異なる領域により形成され得る。このようにして、一次構造では離れて位置する領域同士が、互いに近接することができる。β鎖は、一般的に、5〜10アミノ酸長(通常、ユビキチンにおいて5〜6残基)であって、ほぼ完全に伸長した立体構造である。前記β鎖は、互いに近接しており、一方の鎖のC=O基と他方の鎖のNH基との間で、水素結合が形成される。逆もまた同様である。βシートは、複数の鎖から形成され、シート状構造を有することができ、Cα原子の位置が、シート状平面の上方または下方の間を行ったり来たりする。アミノ酸側鎖は、このパターンに追随し、よって、二者択一的に、上の方向又は下の方向を指す。前記β鎖の方向により、前記シートは、平行シートおよび逆平行シートに分類される。本発明によれば、両方とも、変異可能であり、請求項記載のタンパク質の調製に使用可能である。
【0053】
前記β鎖および前記βシート構造の突然変異誘発のために、表面に近いβ鎖または前記β鎖(前記βシートの1つの鎖)に隣接する3つ目までのアミノ酸位置が、ユビキチンにおいて選択される。表面露出アミノ酸は、入手可能なX線結晶構造により同定できる。結晶構造が入手できない場合には、コンピューター解析を用いて、入手可能な一次構造について表面露出βシート領域および個々のアミノ酸位置の接触性(accessibility)を予測するか、あるいは3次元タンパク質構造をモデル化し、このようにして、潜在的な表面露出アミノ酸についての情報を入手することを試みることができる。さらなる開示は、例えば、J. Mol. Biol., 1987 Apr 5; 194(3):531−44. Vijay−Kumar S, Bugg C.E., Cook W.Jから得ることができる。
【0054】
しかしながら、突然変異させるアミノ酸位置の時間のかかる事前選択を省略して、前記βシートまたは前記β鎖に隣接する3つ目までのアミノ酸位置における修飾を行うことも可能である。前記βシート構造または前記βシート鎖に隣接する3つ目までのアミノ酸をコードするDNA領域を、DNA環境(DNA environment)から単離し、ランダム突然変異誘発に供し、その後、それらを予め除去したタンパク質をコードするDNAに再度組み込む。続いて、所望の結合特性を有する変異体の選択工程を行う。
【0055】
本発明の他の実施形態においては、表面に近い、前記β鎖または前記β鎖に隣接する3つ目までのアミノ酸位置は、前述のように選択され、これらの選択された領域内の突然変異誘発するアミノ酸位置が同定される。ついで、このようにして選択されたアミノ酸位置は、その後、部位特異的突然変異誘発により、DNAレベルで突然変異を誘発され得る。すなわち、特定のアミノ酸をコードするコドンが、予め選択された他の特定のアミノ酸をコードするコドンにより置換される。あるいは、この置換は、ランダム突然変異誘発により行われる。この場合、置換されるアミノ酸位置は規定されるが、新規な未決定のアミノ酸をコードするコドンは規定されない。
【0056】
「表面露出アミノ酸」は、周囲の溶媒に接触可能なアミノ酸である。タンパク質におけるアミノ酸の接触性が、モデルトリペプチドGly−X−Glyにおけるアミノ酸の接触性と比較して8%を超える場合、前記アミノ酸は、「表面露出」と呼ばれる。これらのタンパク質領域または個々のアミノ酸位置もまた、それぞれ、本発明により選択される、潜在的な結合パートナーに対する好ましい結合部位である。また、参考文献として、Caster et al., 1983 Science, 221, 709 − 713, and Shrake & Rupley, 1973 J. Mol. Biol. 79(2):351−371があげられ、開示全体を、引用により本願に取り込む。
【0057】
アミノ酸置換により、新規に生成された人工結合部位の領域が、その親タンパク質と比べて、ならびに互いに相違するユビキチンタンパク質足場のバリエーションを、それぞれの配列セグメントの標的突然変異誘発により生成できる。この場合、極性、電荷、溶解性、疎水性/親水性等の特定の特性を有するアミノ酸を、それぞれ、他の特性を有するアミノ酸と交換または置換できる。置換に加えて、「突然変異誘発」、「修飾」、および「交換」という用語は、挿入および欠失も含む。タンパク質レベルでは、前記修飾を、当業者に公知の方法によるアミノ酸側鎖の化学変性により行うこともできる。
【0058】
ユビキチンの突然変異誘発方法
各配列セグメントの突然変異誘発の開始点として、例えば、当業者に公知の方法により調製、改変、および増幅が可能なユビキチンのcDNAを使用できる。一次配列の比較的狭い領域(例えば、1〜3個のアミノ酸)におけるユビキチンの部位特異的改変には、市販の試薬および方法が利用可能である(「Quick Change」、Stratagene;「Mutagene ファージミド in vitro Mutagenesis Kit」、Biorad)。より大きい領域の部位特異的な突然変異誘発には、具体的な実施形態として、例えば、ポリメラーゼ連鎖反応(PCR)が、当業者に利用可能である。この目的のために、所望の位置に縮退した塩基対組成を有する合成オリゴデオキシヌクレオチドの混合物が、例えば、突然変異の導入に使用できる。これは、イノシン等の、ゲノムDNAには自然発生しない塩基対類似体の使用によっても達成できる。
【0059】
βシート領域の1つ以上のβ鎖または前記βシート鎖に隣接する3つ目までのアミノ酸位置の突然変異誘発の開始点は、例えば、ユビキチンのcDNAでもよいし、ゲノムDNAでもよい。また、ユビキチンタンパク質をコードする遺伝子は、合成によって調製されてもよい。
【0060】
突然変異誘発には、それ自体公知の様々な方法が利用可能である。例えば、部位特異的突然変異誘発法、ランダム突然変異誘発法、PCRを使用する突然変異誘発、または類似の方法が挙げられる。
【0061】
本発明の好ましい実施形態において、突然変異が誘発されるアミノ酸位置は予め定められている。修飾されるアミノ酸の選択は、修飾されるべきアミノ酸に関する請求項1の限定を満たすように行われる。いずれの場合にも、一般的に、様々な変異体のライブラリーが確立され、それ自体公知の方法を用いてスクリーニングされる。一般的に、修飾されるアミノ酸の事前選択は、修飾されるユビキチンタンパク質について、十分な構造情報が入手できる場合には、特に容易に実施できる。
【0062】
例えば、PCR、化学的突然変異誘発、またはバクテリアの突然変異誘発株の使用により、標的突然変異誘発およびより長い配列セグメントの突然変異誘発を行う方法もまた、従来技術に属しており、本発明において利用可能である。
【0063】
本発明の一実施形態において、突然変異誘発は、アミノ酸コドンNNKを有するDNAオリゴヌクレオチドのアセンブリにより行われる。ただし、他のコドン(トリプレット)も使用可能であることも理解すべきである。突然変異は、好ましくはβシート構造が維持されるように行われる。一般的に、突然変異誘発は、タンパク質表面に露出する安定なβシート領域の外側で行われる。それには、部位特異的突然変異誘発およびランダム突然変異誘発の両方が含まれる。一次構造の比較的狭い領域(例えば、3〜5個のアミノ酸)を含む部位特異的突然変異誘発は、市販のキットにより行える。前記市販のキットは、Stratagene(登録商標)(QuickChange(登録商標))またはBio−Rad(登録商標)(Mutagene(登録商標) ファージミド in vitro mutagenesis kit)(米国特許第5,789,166号明細書および米国特許第4,873,192号明細書参照)があげられる。
【0064】
より広い領域を部位特異的突然変異誘発に供する場合、DNAカセットを調製しなければならない。ここで、突然変異が誘発される領域は、突然変異させる位置および変化していない位置を含むオリゴヌクレオチドのアセンブリにより得られる(Nord et al., 1997 Nat. Biotechnol. 8, 772−777; McConell and Hoess, 1995 J. Mol. Biol. 250, 460−470.)。ランダム突然変異誘発は、突然変異誘発株におけるDNAの増殖、または、PCR増幅(エラープローンPCR)により導入できる(例えば、Pannekoek et al., 1993 Gene 128, 135 140)。この目的のため、エラー率が増大したポリメラーゼが使用される。導入される突然変異誘発の度合いを高めるため、あるいは、異なる変異同士をそれぞれ組み合わせるために、PCR断片における変異を、DNAシャッフリングにより組み合わせることができる(Stemmer,1994 Nature 370,389−391)。酵素に関するこれらの突然変異誘発戦略についての概説は、Kuchner and Arnold (1997) TIBTECH 15, 523−530の概説中に記載されている。このランダム突然変異誘発を選択されたDNA領域において実施するためにも、突然変異誘発に使用するDNAカセットの構築が必要である。
【0065】
ランダム修飾は、当該技術分野において、十分に確立された公知の方法により行われる。「ランダムに修飾されたヌクレオチド配列またはアミノ酸配列」は、多数の位置において、ヌクレオチドまたはアミノ酸が挿入、欠失、または置換されており、特性が予測できないヌクレオチド配列またはアミノ酸配列である。多くの場合、挿入されたランダムヌクレオチド(アミノ酸)配列またはヌクレオチド(アミノ酸)配列は、(例えば、ランダム化合成またはPCRを介した突然変異誘発の結果)「完全にランダム」となる。ただし、前記ランダム配列は、共通の機能的特性(例えば、発現産物のリガンドへの反応性)を有する配列も含むことができる。あるいは、前記ランダム配列は、最終的な発現産物が、例えば、異なるアミノ酸が均等に分布した完全にランダムな配列であるという意味で、ランダムとなり得る。
【0066】
ランダム化された断片をベクター中に適切に導入するために、本発明では、前記ランダムヌクレオチドは、部位特異的PCRを介した突然変異誘発の原理により、発現ベクター中に導入されるのが好ましい。ただし、他の選択肢も当業者に公知であり、例えば、合成ランダム配列ライブラリーをベクターに挿入することも可能である。
【0067】
融合PCRにより変異体またはライブラリーを発生させるために、例えば、3回のPCR反応を行ってもよい。2回のPRC反応は、部分的に重なった中間体断片を発生するために行われる。3回目のPCR反応は、前記中間体断片を融合するために行われる。
【0068】
ライブラリーまたは変異体株の構築方法は、所望の制限酵素認識部位周辺のプライマー(制限酵素認識部位プライマー)であるフォワード制限プライマーとリバース制限プライマーからなる第1のプライマーセット、および、例えば、対象のコドンの上流および下流周辺のプライマー(変異原性プライマー)であるフォワード変異原性プライマーおよびリバース変異原性プライマーからなる第2のプライマーセットを構築する工程を含んでもよい。一実施形態において、前記プライマーは、対象のコドンのすぐ上流およびすぐ下流に構築される。前記制限プライマーおよび前記変異原性プライマーを使用して、第1中間体断片および第2中間体断片が構築される。2回のPCR反応により、これらの直線状の中間体断片が生成される。これらの各直線状の中間体断片は、対象の変異コドンを少なくとも1つと、フランキングヌクレオチド配列と、切断部位とを含む。3回目のPCR反応では、前記2つの中間体断片、前記フォワード制限プライマー、および前記リバース制限プライマーを使用して、直線状の融合産物が生成される。一方、前記直線状産物の未結合の末端は、制限酵素で切断され、前記直線状産物に付着末端が生じる。前記直線状産物の前記付着末端は、DNAリガーゼの使用により融合され、環状産物、例えば、環状のポリヌクレオチド配列が生成される。
【0069】
前記中間体断片の構築のため、フォワードプライマーおよびリバースプライマーの2つのセットの設計および合成は、制限酵素切断部位およびそのフランキングヌクレオチド配列を含む第1のセット、ならびに、対象の変異コドンを少なくとも1つ含む第2のセット(変異原性プライマー)を用意することによって行われる。当業者であれば、前記変異体の数は、所望の変異アミノ酸修飾の数に対応することを認識するであろう。本発明者は、他の制限酵素を前記工程に使用する場合には、この切断部位の正確な位置、ならびに、前記フォワードプライマーおよび前記リバースプライマーの対応する配列を、これに応じて改変すればよいと企図している。その他の方法も当該技術分野において利用可能であり、これらを代わりに使用してもよい。
【0070】
本発明に従って発現産物のランダム化断片を足場に導入することとは別に、少なくとも1つの融合パートナーをコードするヌクレオチド配列にランダム化ヌクレオチド配列を融合させることにより、前記ランダム配列を融合パートナーに連結することがしばしば必要となる。このような融合パートナーは、例えば、前記発現産物の発現および/または精製/単離および/またはさらなる安定化を促進できる。
【0071】
本発明の一例によれば、単量体ユビキチンの2、4、6、8、62、63、64、65、66および/または68位における、少なくとも3または4個のアミノ酸のランダム置換を、PCRにより非常に簡易に行うことができる。これは、前述の位置が、前記タンパク質のアミノ末端またはカルボキシ末端の近くに局在しているためである。したがって、操作されるコドンは、対応するcDNA鎖の5’末端および3’末端に位置する。そこで、突然変異誘発PCR反応に使用される第1のオリゴデオキシヌクレオチドは、変異される2、4、6および/または8位のコドンから離れており、その配列は、ユビキチンのアミノ末端のコード鎖に対応する。したがって、第2のオリゴデオキシヌクレオチドは、変異される62、63、64、65、66および/または68位から離れており、カルボキシ末端のポリペプチド配列の非コード鎖に、少なくとも部分的に対応する。両オリゴデオキシヌクレオチドにより、単量体ユビキチンをコードするDNA配列を鋳型として用い、ポリメラーゼ連鎖反応を行うことができる。
【0072】
また、得られた増幅産物を、例えば、制限エンドヌクレアーゼの認識配列を導入するフランキングオリゴデオキシヌクレオチドを使用する、他のポリメラーゼ連鎖反応に添加することが可能である。本発明では、得られた遺伝子カセットを、所定のハプテンまたは抗原に対する結合特性を有するユビキチン変異体の単離のための次の選択工程での使用に適したベクター系中に導入するのが好ましい。
【0073】
ユビキチンの修飾領域
修飾領域は、基本的にが、結合パートナーであるED−Bに接触可能かどうか、ならびに、タンパク質の全体構造が、推測上修飾に耐性を示すかどうかに応じて選択できる。
【0074】
表面露出β鎖における修飾に加えて、前記タンパク質の他の表面露出領域における修飾も可能であり、前記修飾は、前記β鎖に隣接する3つ目までのアミノ酸位置において行われことが好ましい。これらの修飾領域は、新規に生成される、ED−Bに対する高結合親和性での結合に関与する。
【0075】
本発明の別の好ましい実施形態では、ユビキチン、好ましくは、哺乳類またはヒトのユビキチンにおける表面露出アミノ酸の少なくとも3、4、または6個、任意に、少なくとも8、10、12個、最大で15個を修飾することができ、ここで、前記修飾としては、置換が好ましい。これは、ユビキチンにおける3、4、5、6、7、8、9、10、11、12、13、14、または15個の表面露出アミノ酸の修飾を含む。これらの4個以上最大15個の表面露出修飾アミノ酸により、所定の結合パートナーに対して結合親和性を有する領域が形成される。本明細書中において、この領域を、「結合ドメイン領域」(BDR)と定義する。これに関しては、少なくとも2個、任意に少なくとも4個、さらに任意に少なくとも6、8、10、12個、最大で15個の表面露出アミノ酸が、βシート領域内に存在すること、すなわち、単一のβシート鎖に存在するか、もしくは複数のβ鎖に分散すること、または、βシート鎖に隣接する3つ目までのアミノ酸位置に存在することが特に好ましい。修飾された、好ましくは置換された全てのアミノ酸のうちの少なくとも3個が、一次配列において、互いに直接隣接していることがさらに好ましい。
【0076】
本発明の別の任意の実施形態において、前記タンパク質における4つのβ鎖の1つもしくは2つにおけるアミノ酸、好ましくは2つにおけるアミノ酸、または、前記4つのβ鎖のうちの好ましくは2つに隣接する3つ目までのアミノ酸位置におけるアミノ酸が、修飾されて、新規の結合特性を生成している。前記4つのβ鎖のうちの3つもしくは4つにおける修飾、または前記β鎖のうちの3つもしくは4つに隣接する3つ目までのアミノ酸位置における修飾も、ED−B結合発生のために、任意で行ってもよい。
【0077】
アミノ末端鎖およびカルボキシ末端鎖におけるアミノ酸、または、アミノ末端鎖およびカルボキシ末端鎖に隣接する3つ目までのアミノ酸位置におけるアミノ酸を修飾、好ましくは置換して、ED−Bへの新規の結合部位を生成することが特に好ましい。これに関し、前記カルボキシ末端βシート鎖に隣接する4つ目までのアミノ酸が修飾、好ましくは置換されること、および、前記アミノ末端βシート鎖に隣接する1つ目までのアミノ酸が修飾、好ましくは置換されることが特に好ましい。
【0078】
哺乳類のユビキチン、好ましくはヒトのユビキチンにおける下記の位置の表面露出アミノ酸の少なくとも3つが、修飾される、好ましくは置換されることが特に好ましい:2、4、6、8、62、63、64、65、66、68位。前記アミノ酸グループから選択されるこれら少なくとも4つのアミノ酸により、ユビキチンの表面に連続的な表面露出領域が形成されるが、前記領域は、結合パートナーであるED−Bに対し、以前は存在しなかった結合親和性を有する修飾タンパク質の生成に特に適していることが見出された。これらのアミノ酸残基のうち、少なくとも3つが修飾されなければならない。前記アミノ酸残基の3、4、5、6、7、8、9または10個が、任意にさらに別のアミノ酸残基と共に、任意に修飾される。
【0079】
上記修飾後、本発明者らは、実施例で述べるアミノ酸修飾ユビキチン配列が、非常に高い親和性(Kd値10−9以下)でED−Bと結合することを見出した。
【0080】
融合タンパク質
別の好ましい実施形態において、本発明は、薬学的活性成分および/または診断用活性成分と融合した本発明の結合タンパク質を含む融合タンパク質に関する。
【0081】
さらに別の態様において、本発明は、薬学的活性成分および/または診断用活性成分と融合した本発明のヘテロ二量体結合タンパク質を含む融合タンパク質に関する。本発明の融合タンパク質は、非ポリペプチド成分、例えば、非ペプチドリンカー、例えば、治療または診断に関連する放射線核種に対する非ペプチドリガンドを含んでもよい。前記融合タンパク質は、さらに、低分子の有機化合物または非アミノ酸化合物、例えば、糖、オリゴ糖、多糖、脂肪酸等を含んでもよい。本発明の好ましい一実施形態において、前記ユビキチンベースのED−B結合ヘテロマー分子は、治療または診断に関連する特性を有するタンパク質またはペプチドに、共有または非共有結合される。
【0082】
ED−B結合能を有するユビキチンベースの融合タンパク質を得るための方法について、いくつかの例を、以下に示す。
a)ユビキチンに存在するリジン残基を介した前記タンパク質の結合;
b)システイン残基を介した前記ヘテロ二量体ユビキチン結合タンパク質の結合
前記システイン残基は、C末端に位置するか、または任意の他の部位(例えば、24もしくは57位のアミノ酸残基)に位置し得る;マレイミド選択的成分による結合;
c)ペプチド性またはタンパク質性の結合−遺伝子融合(好ましくはC末端またはN末端)
d)タグに基づく融合−標的タンパク質ED−BのC末端またはN末端に位置するタンパク質またはペプチド。融合「タグ」、例えば、ポリヒスチジン(特に、放射性標識に関する)。
【0083】
対象のタンパク質を支持体に共有および非共有結合させるための、これらおよび他の方法は、当該技術分野において周知であるので、さらに詳細には説明しない。
【0084】
前記活性成分は、任意に、サイトカインであり、好ましくは、腫瘍壊死因子(例えば、TNFα、TNFβ)、インターロイキン(例えば、IL−2、IL−12、IL−10、IL−15、IL−24、IL−3、IL−4、IL−5、IL−6、IL−7、IL−9、IL−11、IL−13、IL−8、IL−1α、IL−1β)、インターフェロン(例えば、IFNα、IFNβ、IFNγ)、GM−CSF、GRO(GROα、GROβ、GROγ)、MIP(MIP−1−α、MIP−1β、MIP−3α、MIP−3β)、TGF−β LIF1 CD80、CD−40リガンド、B70、LT−β、Fas−リガンド、ENA−78、LDGF−PBP、GCP−2、PF4、Mig、IP−10、SDF−1α/β、BUNZO/STRC33、I−TAC、BLC/BCA−1、MDC、TECK、TARC、RANTES、HCC−1、HCC−4、DC−CK1、MCP−1−5、エオタキシン、エオタキシン−2、I−309、MPIF−1、6Ckine、CTACK、MEC、リンホタクチン、フラクタルカイン等からなる群から選択されるサイトカインである。
【0085】
本発明において使用される最も好ましいサイトカインの一つは、TNFである。炎症性サイトカインであるTNFは、哺乳類の体内において、抗腫瘍作用を含む複数の活性を有する。前記抗腫瘍作用は、ヒトにおいて有効投与量を用いた場合に許容できない毒性を示すことから、現在、臨床上活用されていない。現在、TNFは、メルファラン等の細胞増殖抑制物質との併用で、治療に使用される。
【0086】
さらに、前記ヘテロ多量体ユビキチン結合タンパク質に結合可能な前記活性成分は、任意に、毒性化合物、好ましくは、低分子有機化合物またはポリペプチドであり、任意に、例えば、サポリン(saporin)、トランケート緑膿菌外毒素A(truncated Pseudomonas exotoxin A)、組換えゲロニン(recombinant gelonin)、リシン−A鎖(Ricin−A chain)、カリケアマイシン(calicheamicin)、ネオカルチノスタチン(neocarzinostatin)、エスペラミシン(esperamicin)、ダイネミシン(dynemicin)、ケダルシジン(kedarcidin)、マデュロペプチン(maduropeptin)、ドキソルビシン(doxorubicin)、ダウノルビシン(daunorubicin)、アウリスタチン(auristatin)、コレラトキシン(cholera toxin)、モデシン(modeccin)、ジフテリア毒素(diphtheria toxin)からなる群から選択される毒性化合物である。
【0087】
本発明のさらなる実施形態において、前記本発明のユビキチン結合タンパク質は、人工アミノ酸を含んでもよい。
【0088】
前記本発明の融合タンパク質のさらなる実施形態において、前記活性成分は、蛍光色素である。好ましくは、前記活性成分は、ガンマ放射同位体のグループ、好ましくは、99Tc、123I、111In、陽電子放出体のグループ、好ましくは、18F、64Cu、68Ga、86Y、124I、ベータ放出体のグループ、好ましくは、131I、90Y、177Lu、67Cu、またはアルファ放射体のグループ、好ましくは、213Bi、211Atのいずれかのグループから選択される放射線核種;Alexa FluorまたはCy dyes(Berlier et al., J Histochem Cytochem. 51 (12): 1699−1712, 2003);光増感剤;プロコアグラント因子、好ましくは、組織因子(例えば、tTF(短縮化組織因子);プロドラッグ活性化のための酵素、好ましくは、カルボキシぺプチダーゼ、グルクロニダーゼおよびグルコシダーゼからなる群から選択される一つの酵素;および/または機能性Fcドメイン、好ましくは、ヒト機能性Fcドメインからなる群から選択される1つの成分である。
【0089】
本発明の融合タンパク質に関するさらなる実施形態は、さらに、血清半減期を調整する成分を含み、好ましくは、ポリエチレングリコールアルブミン結合ペプチドおよび免疫グロブリンからなる群から選択される成分を含む。
【0090】
結合特異性
本発明の融合タンパク質の結合特異性(解離定数)は、非融合タンパク質に関して先に定義した通りであり、Kdで示す。本発明によれば、「Kd」という用語は、特異的結合親和性を定義するものであり、本発明においては、特異的結合親和性は、10−5〜10−12Mの範囲である。10−5M以下の値であれば、定量化可能な結合親和性であると考えることができる。用途に応じ、Kdの値は、例えば、クロマトグラフィーへの適用の場合には、10−6M〜10−12Mが好ましく、10−6M〜10−11Mがさらに好ましく、または、診断もしくは治療への適用の場合には、10−9M〜10−12Mが好ましい。さらに好ましい結合親和性は、10−7〜10−10Mであり、好ましくは、10−11Mである。
【0091】
前記結合親和性を決定する方法は、それ自体公知であり、例えば、下記の方法から選択できる:ELISA、表面プラズモン共鳴(SPR)技術(例えば、Biacore(登録商標)により提供される)、蛍光分光法、等温滴定熱量測定(ITC)、超遠心分析法、FACS。当該技術分野において利用可能なその他の方法もまた、当業者の一般的な知識の範疇において使用可能である。
【0092】
上記修飾後、本発明者らは、実施例で述べるアミノ酸修飾ユビキチン配列が、非常に高い親和性(Kd値10−12M以下)で標的と結合することを見出した。
【0093】
ユビキチンの多量体化
本発明のさらなる実施形態では、ED−Bに結合可能な前記ユビキチンタンパク質を、ED−Bに結合可能な少なくとも1つの第2のユビキチンタンパク質と融合させ、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合される、前記ユビキチンタンパク質の多量体、任意に二量体または三量体を得る。あるいは、ED−Bに結合可能な前記ユビキチンタンパク質を、ED−Bに結合可能な少なくとも1つの第2のユビキチンタンパク質と融合させ、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成される、前記ユビキチンタンパク質の多量体、任意に二量体または三量体を得る。
【0094】
本明細書中において、「多量体」は、同一または1以上の異なる単量体ユビキチンタンパク質を含み得るタンパク質と考えられる。前記多量体が同一の単量体ユビキチンユニットを含有する場合には、ホモマーまたはホモ多量体(例えば、ホモ二量体またはホモ三量体)が形成される。二量体が異なる修飾がなされた2つの単量体を含む場合、前記二量体は、「異種二量体」または「ヘテロ二量体」と呼ばれる。2以上の異なる単量体ユビキチンユニットが存在する場合、ヘテロマーまたはヘテロ多量体が形成される。この発明の最も好ましい実施形態においては、ED−Bへの結合能を有する前記ヘテロマーは、少なくとも2つの異なる修飾ユビキチン単量体からなる。本発明の「ヘテロ二量体」は、特定の結合パートナーに対し、一価の結合特性をそれぞれ有する2つの異なる単量体ユビキチンタンパク質からなる、修飾ユビキチンタンパク質の融合物と考えられる。
【0095】
よって、本発明の「ヘテロ多量体」、任意に「ヘテロ二量体」は、特定の結合パートナーED−Bに対して共同で一価の結合特性を示す、異なる修飾がなされた少なくとも2つの(あるいは、少なくとも1つは修飾され、1つは非修飾である)単量体ユビキチンタンパク質の融合物と考えられる。本発明の修飾ヘテロ二量体ED−B結合ユビキチンタンパク質は、各単量体ユビキチンタンパク質を個々にスクリーニングし、その後、これら2つを結合して得るのではなく、前記ED−Bリガンドに対して共同での一価の結合活性を示す第1単量体ユニットおよび第2単量体ユニットからなるヘテロ二量体タンパク質をスクリーニングすることによって得られることを強調しておく。前記サブユニットのそれぞれは、ED−Bへの結合親和性がかなり制限され、結合した二量体修飾ユビキチンタンパク質のみが、本明細書に記載の優れた結合特性を有すると考えられる(例えば、図4参照)。
【0096】
本発明によれば、ヘッドトゥテイル融合により遺伝学的に結合された(genetically linked)、異なる修飾がなされた2つのユビキチン単量体(あるいは、少なくとも1つは修飾され、1つは非修飾である)は、ED−Bの同じエピトープに結合し、両方の結合ドメイン領域が共に作用する場合のみ効果を示す。前記単量体の前記BDRは、単一の連続的な結合領域を形成する。
【0097】
本発明の一実施形態においては、本発明により提供される修飾ユビキチンタンパク質を、異なる特異性のユビキチンタンパク質に、好ましくは部位特異的に結合させることができ、これにより、結合パートナーに対して一価の結合特性を有する、モジュール構成されたユビキチンが得られる。
【0098】
よって、例えば、上述の手順で得られたユビキチンの2つの変異体を、部位特異的に互いに結合させることができる。したがって、多量体を形成する場合、前記単量体ユビキチンユニットを生成し、これらをヘッドトゥテイルで結合させることによって形成可能である。換言すると、前記単量体ユニットは、前記多量体内に含まれるユニットの数に応じ、N−C−N−C−方向に結合される。前記多量体化は、リンカーを使用することなく直接行うか、またはリンカーを介して行うことができる。
【0099】
本発明の別の実施形態では、ED−Bに結合する修飾ユビキチン単量体の2つの変異体を、適切な方法で互いに遺伝学的に結合させることができ、これにより、極めて特異的な結合分子が得られる。ヘッドトゥテイル融合により遺伝学的に結合した修飾ユビキチン単量体の2つの変異体が、同じエピトープに結合し、両方の結合ドメイン領域が共に作用する場合のみ効果を示すことが特に好ましい。換言すると、これらは、前記2つのモジュールの両結合領域が共に作用することによって形成される単一の連続的な結合領域を介して、同一のエピトープに結合する。
【0100】
したがって、フィブロネクチンのED−Bに効果的に結合させるべく本発明により修飾されたユビキチンタンパク質は、多量体化されてもよく、特に、二量体化または三量体化されてもよい。ユビキチン単量体の融合は、リンカーを介して行うことができる。前記リンカーとしては、例えば、少なくともGIGの配列、または、任意の他のリンカー、例えば、SGGGGIGまたはSGGGGSGGGGIGがあげられる。2以上のユビキチン単量体の遺伝子融合用の他のリンカーを使用することもできる。好ましくは、このようなユビキチンタンパク質は、少なくとも2つの結合決定領域(BDR)からなり、各BDRにおいて、2、4、6、8、62、63、64、65、66、68位におけるアミノ酸の少なくとも1個が修飾され、前記BDRが、互いに遺伝子融合(genetically fused)されている。前記標的への結合は、両BDRが共同で媒介する。
【0101】
前記修飾ユビキチンタンパク質のさらなる多量体化を、例えば、前記修飾ユビキチンタンパク質を、サイトカインのような多量体化ドメインを有するエフェクター分子、例えば、TNFαに融合させることによって行うことができる。1以上の修飾ユビキチン単量体(二量体等、好ましくはヘテロ二量体)と、TNFαとの融合は、リンカーを介して行うことができる。さらに別の実施形態においては、前記多量体化は、2以上の修飾ユビキチン分子を遺伝子融合させるか、あるいはポリエチレングリコール(PEG)リンカーを使用することによって行われる。さらに別の実施形態においては、前記多量体化ドメインが、薬学的活性成分としても作用する。一例を挙げると、TNFαは、多量体化ドメインおよび医薬品成分の両方の役割を果たす。
【0102】
本発明のさらなる実施形態において、本発明の多量体化修飾ユビキチンタンパク質は、ヘテロマーからなる。「ヘテロマー」という用語は、異なる組成のユビキチン単量体が、多量体化分子に含まれることを意味する。遺伝子融合された2つのユビキチン単量体からなるヘテロ二量体であって、前記2つのユビキチン単量体が、結合ドメイン領域は異なるが、単一の結合部位を有しているものが特に好ましい。これらのヘテロ二量体は、例えば、TNF、最も好ましくは、TNFαを使用することによって多量体化できる。ユビキチンヘテロ二量体を使用する利点の1つは、高い結合特異性でED−Bを標的とすることができることである。
【0103】
二量体化、または一般的には多量体化のさらなる利点は、ED−Bに対する前記新たに生成された結合タンパク質の足場の全体的な安定性を低下させることなく、タンパク質化学的完全性を維持しながら、修飾可能なアミノ酸残基の数を増加できることにある。一方では、ED−Bに対する結合部位を生成するために修飾可能な残基の総数は、前記修飾残基を、2つまたは3つのユビキチンタンパク質の足場に割り当てることができるために増加する。他方では、単一のユビキチンタンパク質の足場上の残基の数は減少するが、前記修飾結合分子のタンパク質化学的安定性は維持される。要約すると、前記修飾アミノ酸は、2個または3個のユビキチン分子に含まれるため、前記ユビキチンベースのED−B結合タンパク質のモジュール構造により、修飾アミノ酸の総数を増加することが可能となる。先に述べた修飾数は、修飾ユビキチン一分子当たりの数である。
【0104】
さらに、多量体化により、標的物質に対する2つ(以上)の結合部位と、1つの一価の特異性(一つのシングルエピトープに対する)を有する修飾ユビキチン分子を提供し得るが、二特異性または多特異性(よって、2以上の異なるエピトープを同時に認識する)が付与される場合もある。
【0105】
2以上の修飾ユビキチン分子を、好ましくはヘッドトゥテイル遺伝子融合により融合する場合、修飾数は、修飾ユビキチン分子数に応じて、2倍またはx倍にできる。代替例として、前記融合を翻訳後に行うこともできる。
【0106】
修飾された状態の単量体ユビキチンタンパク質の使用に関し、二量体、特に、ホモ二量体またはヘテロ二量体等の多量体構造を使用する利点は、下層にある足場のタンパク質化学的完全性に影響を与えることなく修飾可能な残基の数を増加できることにある。
【0107】
(ED−Bに結合する修飾ユビキチンタンパク質)
本発明により、ED−Bに結合する修飾ユビキチン単量体が同定された。ED−Bに特異的に結合する好ましい修飾ユビキチンヘテロ二量体は、以下の通りである。
1.第1ユビキチン分子の結合決定領域(BDR1):2、4、6、62〜68位の修飾アミノ酸。好ましくは、第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
2.第1ユビキチン分子の結合決定領域(BDR1):6、8、62〜68位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
3.第1ユビキチン分子の結合決定領域(BDR1):2、4、6、8、62〜68位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:2、4、6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
4.第1ユビキチン分子の結合決定領域(BDR1):6、8、63〜66位の修飾アミノ酸。好ましくは第2ユビキチン分子に遺伝子融合されている。
第2ユビキチン分子:2、6、8、62〜68位の修飾アミノ酸によりBDR2が規定されている。
【0108】
実施形態において、前記融合タンパク質は、前記ユビキチンタンパク質の遺伝子融合による二量体であり、第1ユビキチン単量体の6、8、63〜66位のアミノ酸が置換され、第2ユビキチン単量体の6、8、62〜66位、および、任意に2位のアミノ酸が置換されている。好ましくは、
6位におけるリジン(K)のトリプトファン(W)またはフェニルアラニン(F)への置換、
8位におけるロイシン(L)のトリプトファンまたはフェニルアラニン(W、F)への置換、
63位におけるリジン(K)のアルギニン(R)またはヒスチジン(H)への置換、
64位におけるグルタミン酸(E)のリジン(K)、アルギニン(R)、またはヒスチジン(H)への置換、
65位におけるセリン(S)のフェニルアラニン(F)またはトリプトファン(W)への置換、
66位におけるトレオニン(T)のプロリン(P)への置換;
前記第2ユビキチン単量体において、
6位におけるリジン(K)のトレオニン(T)、アスパラギン(N)、セリン(S)、またはグルタミン(Q)への置換
8位におけるロイシン(L)のグルタミン(Q)、トレオニン(T)、アスパラギン(N)、またはセリン(S)への置換
62位におけるグルタミン(Q)のトリプトファン(W)またはフェニルアラニン(F)への置換
63位におけるリジン(K)のセリン(S)、トレオニン(T)、アスパラギン(N)、またはグルタミン(Q)への置換
64位におけるグルタミン酸(E)のアスパラギン(N)、セリン(S)、トレオニン(T)、またはグルタミン(Q)への置換
65位におけるセリン(S)のフェニルアラニン(F)またはトリプトファン(W)への置換
66位におけるトレオニン(T)のグルタミン酸(E)またはアスパラギン酸(D)への置換
任意に、2位におけるグルタミン(Q)のアルギニン(R)、ヒスチジン(H)、またはリジン(K)への置換が好ましい。
【0109】
最も好ましい置換は、下記の通りである。
(1)第1単量体ユニットにおける、少なくともK6W、L8W、K63R、E64K、S65F、およびT66Pの置換;
および、第2単量体ユニットにおける、少なくともK6T、L8Q、Q62W、K63S、E64N、S65WおよびT66Eの置換;任意にさらにQ2Rの置換、または
(2)第1単量体ユニットにおける、少なくともQ2T、F4W、K6H、Q62N、K63F、E64K、S65LおよびT66Sの置換;
および、第2単量体ユニットにおける、少なくともK6X、L8X、Q62X、K63X、E64X、S65XおよびT66Xの置換;任意にさらにQ2Xの置換、Xは、任意のアミノ酸。
【0110】
ED−Bへの結合タンパク質を生成するための前記第1ユビキチン単量体における特に好ましい置換は、下記の通りである。
2位:Q→T、4位:F→W、6位:K→H、62位:Q→N、63位:K→F、64位:E→K、65位:S→L、66位:T→S
【0111】
2つの単量体のヘッドトゥテイルでの結合には、リンカーを使用しなくてもよいし、どのようなリンカーを使用してもよい。好ましいリンカーは、配列番号32のリンカー、または、GIG配列、SGGGGIG配列、もしくはSGGGGSGGGGIG配列のリンカーである。
【0112】
好ましい実施形態において、ED−Bに対する結合のために共に作用する2つの結合決定領域(BDR)を備えたユビキチンヘテロ二量体は、配列番号33または34のアミノ酸配列を含む。薬学的活性成分としてTNFαを含む、本発明の好ましい融合タンパク質は、配列番号35または36の配列を有する。さらに好ましいタンパク質は、下記配列(配列番号47)を有するものであるが、下記配列中、XXXXは、どのようなアミノ酸でもよい。リンカーとして、SGGGGSGGGGIG配列が使用される(イタリック体で示す)。他の種類のリンカー、またはリンカーなしも、実施可能な代替例と理解すべきである。
【化1】
【0113】
これらの配列のタンパク質のコンセンサス配列の例を、図19に示す。
【0114】
さらに別の実施形態において、本発明の融合タンパク質は、TNFαに融合したユビキチンヘテロ二量体の融合タンパク質の三量体であり、前記融合タンパク質は、配列番号35または36の配列を有するか、あるいは、配列番号35または36に対して90%以上の配列同一性、好ましくは95%を超える配列同一性、96%を超える配列同一性、または97%までの配列同一性を有し、また同時に、フィブロネクチンのED−Bドメインに対する元の融合タンパク質の高親和性での結合能を、維持または向上することが好ましい。
【0115】
本発明のさらなる態様において、本発明は、前述のタンパク質または融合タンパク質をコードするポリヌクレオチドをも包含する。さらに、前記ポリヌクレオチドを含むベクターも、本発明に包含される。
【0116】
本発明のさらなる態様において、本明細書中に記載のタンパク質もしくは融合タンパク質、および/あるいは、本発明の組換えタンパク質もしくは組換え融合タンパク質をコードするポリヌクレオチド、または、前記ポリヌクレオチドを含むベクターを含む、宿主細胞が包含される。
【0117】
(本発明のタンパク質、例えば、TNFα等のエフェクターに融合したED−B特異性ヘテロ二量体ユビキチンベース結合タンパク質の使用)
本発明の修飾ユビキチンED−B結合タンパク質は、例えば、in vitroまたはin vivoで使用する診断手段の調製、ならびに治療手段の調製に使用される。前記本発明のタンパク質は、例えば、直接的なエフェクター分子(修飾物質、拮抗物質、作用物質)または抗原認識ドメインとして使用できる。ED−B抗原が大量に出現する腫瘍の例を、図1の表に示す。
【0118】
選択する融合パートナーに応じて、本発明の医薬組成物は、癌、例えば、乳癌および結腸直腸癌、またはED−Bが大量に出現する任意の他の腫瘍性疾患(図1に列記したこれらの例を参照)の治療を対象とするように適合される。
【0119】
前記組成物は、治療上の有効量を含むように適合される。投与量は、治療対象の組織、疾患の種類、患者の年齢および体重、ならびにそれ自体公知である他の要因によって決まる。
【0120】
前記組成物は、薬学的または診断的に許容されるキャリアを含み、さらに、それ自体公知の助剤を任意に含み得る。これらは、例えば、安定化剤、界面活性剤、塩類、緩衝剤、着色剤等を含むが、これらに限定されない。
【0121】
前記医薬組成物は、局所塗布用の液状製剤、クリーム、ローションの剤形;エアロゾル;粉末、細粒、錠剤、座剤、カプセル剤の剤形;エマルジョン、リポソーム製剤の剤形を取り得る。前記組成物は、無菌、非発熱原性、等張性であり、それ自体公知である、薬学的に既存かつ許容可能な添加剤を含むことが好ましい。加えて、米国薬局方またはRemington’s Pharmaceutical Sciences、Mac Publishing Company(1990)の規則も参照する。
【0122】
ヒトおよび獣医学の医薬療法および予防法の分野において、本発明により修飾されたED−B結合ユビキチンタンパク質を少なくとも1つ含む薬学的に有効な医薬品を、それ自体公知の方法により調製できる。ガレノス式製剤学(galenic preparation)に応じて、これらの組成物を、静脈内投与、腹腔内投与、筋肉内投与、皮下投与、経皮投与、または他の適用方法により投与できる。医薬調合物の種類は、治療対象の疾患の種類、疾患の重症度、治療対象の患者、および医学分野の当業者に公知の他の要因によって決定される。投与は、注射もしくは点滴による非経口的な投与、全身的、直腸的、経皮的な投与、または従来使用されている他の方法による投与のいずれかとすることができる。
【0123】
一実施形態において、前記医薬組成物は、本発明のタンパク質もしくは融合タンパク質、またはこれらの混合物を含み、さらに、1以上の化学療法剤、好ましくは、下記表に示すものから選択される化学療法剤を含む。
【0124】
【表1】
【0125】
好ましい実施形態において、前記化学療法剤は、メルファラン、ドキソルビシン、シクロフォスファミド、ダクチノマイシン、フルオロデオキシウラシル、シスプラチン、パクリタキセル、およびゲムシタビンから選択されるか、またはキナーゼ阻害剤群から選択される。
【0126】
本発明の「医薬組成物」は、種々の活性成分と希釈剤および/または担体が互いに混合された組成物の形態で存在してもよいし、あるいは、前記活性成分が、部分的にまたは全体的に別個に存在する組み合わせ製剤の形態を取ってもよい。このような組み合わせまたは組み合わせ製剤の一例は、複数の部品からなるキットである。
【0127】
本発明の「組成物」は、少なくとも2つの薬学的活性化合物を含む。これらの化合物は、同時に投与してもよいし、1分〜数日の時間間隔で個々に投与してもよい。これらの化合物は、同じ経路で投与してもよいし、異なる経路、例えば、活性化合物の1つを経口投与し、他の活性化合物を非経口投与することも可能である。また、前記活性化合物は、一つの医薬品、例えば、一つの点滴液中に調剤されてもよいし、別々に調剤された両化合物を含むキットとして提供されてもよい。また、両化合物を、2以上の包装内に存在させることも可能である。
【0128】
特に好ましい組み合わせは、本発明の融合タンパク質とメルファランおよび/またはドキソルビシン(リポソーム製剤)である。ATC L01分類の抗悪性腫瘍剤以外にも、本発明のTNF融合タンパク質は、サイトカインとその誘導体、放射性医薬品、細胞ベースの治療薬、およびナノ粒子を含む、他の抗悪性腫瘍物質と併用できる。
【0129】
腫瘍透過処理活性(tumor permeabilisation activity)により、本発明のTNF融合タンパク質は(さらに、本発明の他の組換えタンパク質/融合タンパク質も)、世界保健機関提供の解剖治療化学分類法(ATC)においてL01として列挙される全ての抗悪性腫瘍剤と併用できる。ここで、驚くべきことに、TNFαに融合したユビキチンヘテロ二量体の融合タンパク質、好ましくは、配列番号35または36の配列を有する融合タンパク質が、有利に治療に適用できることがわかった。TNFαは毒性が高く、よって、低用量でしか投与できないが、このような用量は、通常、最小治療閾値を下回る(よって、治療上不活性である)。TNFαのこの毒性のため、治療上有効な濃度を達成するために、現在、TNFαの使用に際しては、四肢分離かん流アプローチが選択されている。四肢かん流は、腕または足に直接抗癌剤を輸送するために使用できる医療技術である。四肢への血流および四肢からの血流を止血帯で一次的に停止させ、抗癌剤を直接四肢の血液に注入する。これにより、患者は、癌が発生している領域に高い投与量のTNFαを受けることができる。
【0130】
しかしながら、本発明のTNFα融合タンパク質を適用すれば、非毒性でありながら、治療的に有効な濃度でTNFαを投与することが可能となる。TNFαは、本発明の(結合)融合タンパク質に連結されるため、疾病部位(例えば、腫瘍部位)において直接活性を示し、よって、「遊離」TNFαの量を激減させることができる。
【0131】
TNFαの全身性副作用は、TNFαを本発明の融合タンパク質として投与することにより、著しく軽減できる。本発明のTNFα融合タンパク質を使用することにより、治療効果に達するためのTNFαの全投与量を大幅に低減でき、特に、化学療法剤(上記参照)を併用する全身的な腫瘍治療において、有利に使用できる(四肢かん流の必要性および制約がない)。
【0132】
さらなる実施形態において、前記医薬組成物は、複数の部品からなるキットの形態であり、本発明の組換えタンパク質/組換え融合タンパク質、および1以上の化学療法剤が別々に提供される。
【0133】
(本発明のED−B結合タンパク質の製造方法)
本発明のED−B結合タンパク質は、例えば、単純な有機合成戦略、固相支援合成技術等の多数の従来公知の技術のいずれかにより、または、市販の自動合成装置により、調製すればよい。また、本発明のED−B結合タンパク質は、従来の遺伝子組換え技術を単独で使用するか、または、従来の合成技術と併用することにより、調製してもよい。
【0134】
本発明の他の態様において、組換え修飾タンパク質の製造方法が提供される。前記方法は、少なくとも下記の工程を含む。
a)ユビキチンタンパク質を提供する工程
b)フィブロネクチンのED−Bを提供する工程
c)配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために前記ユビキチンタンパク質を修飾する工程であり、
2、4、6、62、63、64、65、66、および/または68位における、少なくとも4個のアミノ酸を、置換、欠失、または付加により修飾する工程
d)前記修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
e)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bと結合する修飾ユビキチンタンパク質をスクリーニングする工程
f)前記修飾ユビキチンタンパク質を単離する任意の工程
【0135】
前記修飾は、任意にDNAレベルでの遺伝子工学により行われてもよく、前記修飾タンパク質の発現は、任意に原核生物、真核生物、またはin vitroにおいて行われてもよい。
【0136】
さらなる実施形態において、前記修飾工程は、化学的合成工程を含む。
【0137】
さらに別の実施形態において、前記方法は、前記修飾ユビキチンタンパク質が、薬学的活性成分(任意にサイトカイン、好ましくはTNFα)または診断用成分と融合するように適合されている。あるいは、前記方法は、前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合させるか、または、前記組換えタンパク質を少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成させるように適合されている。
【0138】
本発明によれば、修飾タンパク質は、さらに、化学的合成により調製されてもよい。この実施形態において、請求項1の前記c)工程からd)工程は、一つの工程で行われる。
【0139】
本発明の別の態様においては、ヘテロ多量体融合タンパク質の製造方法が提供される。前記方法は、以下の工程を含む。
a)好適なリンカーを介して結合された2以上の修飾ユビキチン単量体を含む多量体ユビキチンタンパク質を提供する工程であり、
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために、前記多量体ユビキチンタンパク質における各単量体が修飾されており、
各単量体において、2、4、6、62、63、64、65、66、および/または68位における少なくとも4個のアミノ酸が、置換、欠失、または付加により修飾されている工程
b)フィブロネクチンのED−Bを提供する工程
c)前記ヘテロ多量体修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
d)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bに結合する修飾ユビキチンタンパク質をスクリーニングする工程
e)前記修飾ヘテロ多量体ユビキチンタンパク質を単離する任意の工程
【0140】
本発明の別の態様においては、組換え修飾ユビキチンタンパク質の製造方法が提供される。前記方法は、少なくとも以下の工程を含む。
a)単量体ユビキチンタンパク質から生じる、異なる修飾がなされた二量体ユビキチンタンパク質群を提供する工程であり、
前記群が、互いにヘッドトゥテイル配置で結合された2つの修飾ユビキチン単量体を含む二量体ユビキチンタンパク質を含み、
前記二量体タンパク質の各単量体は、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも6個のアミノ酸の置換により、異なる修飾がなされ、
前記置換は、下記(1)または(2)を含む、前記工程
(1)第1単量体ユニットにおける、少なくとも6、8、63、64、65および66位のアミノ酸の置換、および、
第2単量体ユニットにおける、少なくとも6、8、62、63、64、65および66位のアミノ酸の置換、任意にさらに2位のアミノ酸の置換、または
(2)第1単量体ユニットにおける、少なくとも2、4、6、62、63、64、65および66位のアミノ酸の置換、および、
第2単量体ユニットにおける、少なくとも6、8、62、63、64、65および66位のアミノ酸の置換、任意にさらに2位のアミノ酸の置換
b)潜在的なリガンドとして、フィブロネクチンのエキストラドメインB(ED−B)を提供する工程
c)前記異なる修飾がなされたタンパク質群を、前記フィブロネクチンのエキストラドメインB(ED−B)と接触させる工程
d)スクリーニング処理により、修飾二量体ユビキチンタンパク質を同定する工程であり、
前記修飾二量体ユビキチンタンパク質は、前記フィブロネクチンのエキストラドメインB(ED−B)と、Kd10−7〜10−12Mの範囲の特異的結合親和性で結合し、前記フィブロネクチンのエキストラドメインB(ED−B)に対して、一価の結合活性を示すタンパク質である、前記工程
e)前記修飾二量体ユビキチンタンパク質を、前記結合親和性を用いて単離する任意の工程
【0141】
前記好ましい実施形態において、前記ヘテロ多量体ユビキチンタンパク質は、ヘテロ二量体ユビキチンタンパク質である。
【0142】
さらに別の態様において、本発明は、上述の直線状ポリユビキチン鎖をベースとし、少なくとも2つのBDRにおいてランダム化されているライブラリーに関する。
【0143】
本発明のさらなる態様において、前述の2つのライブラリーの融合により得られるDNAを含む融合ライブラリーが提供される。ヘテロ二量体ユビキチン融合タンパク質を得るために、各ライブラリーは、異なる修飾がなされた単量体ユビキチンタンパク質ユニットをコードし、前記単量体ユニットは、互いにヘッドトゥテイル配置で結合される。前記ライブラリーは、前記フィブロネクチンのエキストラドメインB(ED−B)に対して一価の結合活性を示すユビキチンのヘテロ二量体融合タンパク質をコードする。前記相互結合は、当業者に公知のリンカーのいずれか1つ、または本明細書に記載のリンカーを用いて行われる。本発明の一実施形態において、TNFαは、同時に薬学的活性化合物としての役割も果たすリンカーとして使用される。
【0144】
複合ライブラリーの製造の概要を、実施例1で述べる。ただし、このようなライブラリーの品質に注意を払わなければならない。抗体または足場技術におけるライブラリーの品質は、そもそも、複雑性(個々の変異体の数)および機能性(得られた候補の構造的およびタンパク質化学的完全性)に左右される。しかしながら、両特性が互いに悪影響を及ぼし合う場合があり、前記足場における修飾位置の数を増加させてライブラリーの複雑性を向上しようとすれば、変異体のタンパク質化学的特性の低下を招くおそれがある。これにより、溶解度の低下、凝集、および/または低収量が起こることもある。これは、エネルギー的に良好なタンパク質パッケージを有する未変性の足場からの逸脱がより大きくなることによる。
【0145】
このため、このような足場ライブラリーの適切な構築は、標的への結合性を最適化するために、元の配列にできるだけ多くの変異を導入することと、一方で、タンパク質化学的な悪影響を避けるために、元の一次配列をできるだけ保存するという、両極端な姿勢の間でバランスをとる作業となる。
【0146】
本開示は、本発明の態様または実施形態を考慮して考え得る、本明細書に記載の特徴の各組み合わせも包含する。
【0147】
標的ED−Bに対して結合親和性を有する修飾ユビキチンタンパク質の選択、および、結合親和性を発生させる修飾アミノ酸の決定
例えば、ヘテロ二量体修飾ユビキチンタンパク質をコードする少なくとも2つのDNAライブラリーが、各単量体ユビキチンユニットにおいて、選択されたアミノ酸に異なる修飾をすることにより確立された後、これらのライブラリーを、例えば、リンカー技術によって遺伝子融合し、ヘテロ二量体修飾ユビキチンタンパク質をコードするDNA分子を得る。これらのライブラリーのDNAによりタンパク質を発現させ、得られた修飾二量体タンパク質を、本発明によりED−Bに接触させ、結合親和性が存在する場合には、パートナー同士の結合が任意に可能となる。
【0148】
接触工程およびスクリーニング工程が、ヘテロ二量体ユビキチンタンパク質について、既に行われていることが本発明の重要な態様である。この工程により、ED−Bへの一価の結合活性を提供するユビキチンタンパク質のスクリーニングが可能となる。
【0149】
本発明による接触は、適切な提示および選択方法、例えば、ファージディスプレイ法、リボソームディスプレイ法、mRNAディスプレイ法、細胞表面ディスプレイ法、酵母表面ディスプレイ法、または細菌表面ディスプレイ法等の方法により行うことが好ましく、ファージディスプレイ法により行うことが好ましい。完全な開示としては、下記の参考文献を参照できる;Hoess, Curr. Opin. Struct. Biol.. 3 (1993), 572−579;Wells and Lowmann, Curr. Opin. Struct. Biol. 2 (1992), 597−604;Kay et al., Phage Display of Peptides and Proteins−A Laboratory Manual (1996), Academic Press。前述の方法は、当業者に公知であり、それらを改変した方法も含めて、本発明において使用可能である。
【0150】
本発明において、前記修飾タンパク質が、所定の結合パートナーに対して、定量化可能な結合親和性を有するかどうかの決定は、好ましくは、下記の方法の1以上により、行うことができる:ELISA、表面プラズモン共鳴分光法、蛍光分光法、FACS、等温滴定熱量測定、および超遠心分析法。
【0151】
ファージディスプレイ選択法
本願に適合するファージディスプレイ法の一種を、結合特性を示すユビキチン変異体についての、本発明による選択手法の一例として後述する。同様に、例えば、細菌上(bacterial surface display;Daugherty et al., 1998, Protein Eng. 11(9):825−832)もしくは酵母細胞上に提示する方法(yeast surface display; Kieke et al., 1997 Protein Eng. 10(11):1303−10)、または、リボソームディスプレイ(Hanes and Pluckthun, 1997 Proc Natl Acad Sci U S A. 94(10):4937−4942; He and Taussig, 1997 Nucleic Acids Res. 25(24):5132−5134)、cisディスプレイ(Odegrip et al., 2004 Proc Natl Acad Sci U S A. 101(9):2806−2810)もしくはmRNAディスプレイ等の無細胞選択システムを適用できる。後者の場合、遺伝子型と表現型との一時的な物理的結合が、リボソームを介したタンパク質変異体の適切なmRNAへの結合により達成される。
【0152】
本明細書に記載のファージディスプレイ法において、ユビキチンの組換え変異体は、繊維状ファージ上に提示され、一方、提示される変異体をコードするDNAは、同時に、ファージエンベロープ中に、一本鎖の形で詰め込まれる。このように、親和性の向上という枠組みにおいて、特定の特性を有する変異体をライブラリーから選択することができ、それらの遺伝情報は、適切な細菌への感染により増幅できるか、または、異なる濃縮サイクルに添加できる。ファージ表面での変異したユビキチンの提示は、アミノ末端シグナル配列、好ましくは、PelBシグナル配列、および、前記ファージのカプシドまたは表面タンパク質への遺伝子融合により達成される。カプシドタンパク質pIIIまたはその断片へのカルボキシ末端融合が好ましい。また、コードされた融合タンパク質は、例えば、親和性クロマトグラフィーによる検出および/または精製のための親和性タグもしくは抗体エピトープ、または、親和性濃縮過程における融合タンパク質の特異的な切断のためのプロテアーゼ認識配列等の、さらなる機能性成分を含むことができる。また、例えば、ユビキチン変異体の遺伝子と、ファージカプシドタンパク質またはその断片のコード領域との間に、アンバー停止コドンを存在させることができる。前記アンバー停止コドンは、部分的には1つのアミノ酸の導入に起因して、適切なサプレッサー株内での翻訳中には認識されない。
【0153】
所定のハプテンまたは抗原への結合特性を有するユビキチン変異体の単離に関する選択方法に適した細菌ベクターであって、前述の融合タンパク質の遺伝子カセットが挿入される細菌ベクターは、ファージミドという。数ある中でも、それは、繊維状ファージの遺伝子間領域(例えば、M13もしくはf1)またはその一部分を含み、例えば、M13K07等のヘルパーファージにより前記ファージミドを担持する細菌細胞の重複感染の際、ファージカプシド中に、ファージミドDNAの閉鎖鎖がパッケージングされる。このようにして生成されたファージミドは、細菌により分泌され、コードされた各ユビキチン変異体を、前記カプシドタンパク質pIIIまたはその断片への融合により、細菌表面に提示する。天然型pIIIカプシドタンパク質は、ファージミド中に存在し、適合するバクテリア株に再感染する能力、およびそれゆえに同一DNAを増幅する可能性を保持する。このため、前記ユビキチン変異体の表現型、すなわち、その潜在的な結合特性と、その遺伝子型との間の物理的結合が確保される。
【0154】
得られたファージミドを、そこに提示されたユビキチン変異体の所定のハプテンまたは抗原への結合に関し、当業者に公知の手法により選択することができる。この目的のために、提示されたユビキチン変異体を、例えば、マイクロタイタープレート上に結合した標的物質に一時的に固定化することができ、非結合の変異体を分離した後に、特異的に溶出することができる。前記溶出は、例えば、100mMトリエチルアミン等の塩基性溶液により行うことが好ましい。あるいは、前記溶出は、酸性条件下で、タンパク質分解または感染細菌の直接添加により行うことができる。このようにして得られたファージミドは、所定のハプテンまたは抗原への結合特性を有するユビキチン変異体の選択と増幅の連続サイクルにより、再増幅と濃縮が可能である。
【0155】
このようにして得られた前記ユビキチン変異体のさらなる特性評価は、ファージミドの形態、すなわち、ファージに融合した形態で、または、対応する遺伝子カセットを好適な発現ベクターにクローニングした後に、可溶性タンパク質の形態で行うことができる。適切な方法は、当業者に公知であるか、または前記文献に記載されている。前記特性評価は、例えば、DNA配列の決定、よって、単離された変異体の一次配列の決定を含み得る。さらに、単離された変異体の親和性および特異性を、例えば、ELISAもしくは表面プラズモン共鳴分光法等の標準的な生化学的方法、蛍光分光法、FACS、等温滴定熱量測定、超遠心分析法、またはその他の方法により検出できる。安定性分析については、例えば、化学的または物理的アンフォールディングについての分光法が、当業者に公知である。
【0156】
リボソームディスプレイ選択法
本発明のさらなる実施形態であるリボソームディスプレイ法において、ユビキチン変異体は、無細胞転写/翻訳系により調製され、対応するmRNAおよびリボソームとの複合体として提示される。この目的のために、前述のDNAライブラリーが、基礎として使用され、前記DNAライブラリー中で、変異体の遺伝子は、発現およびタンパク質生合成のための対応する制御配列との融合物の形態で存在する。前記遺伝子ライブラリーの3’末端での前記停止コドンの欠失、および適切な実験条件(低温、高Mg2+濃度)により、新生タンパク質、mRNA、およびリボソームからなる三元複合体が、in vitroでの転写/翻訳の間、維持される。
【0157】
ヘテロ二量体修飾ユビキチンタンパク質を含むタンパク質ライブラリーが、各単量体ユビキチンユニットにおいて、選択されたアミノ酸に異なる修飾をすることにより確立された後、前記修飾二量体タンパク質を、本発明によりED−Bに接触させ、結合親和性が存在する場合には、パートナー同士の結合が可能となる。これらのタンパク質ライブラリーは、前記修飾タンパク質と前記ED−B標的タンパク質間の接触を可能とする方法でディスプレイを行うか、前記修飾タンパク質を提示する任意の他の方法を使用する、ディスプレイ法のライブラリーの形態であってもよい。前記ディスプレイ法は、任意に、ファージディスプレイ法、リボソームディスプレイ法、TATファージディスプレイ法、酵母ディスプレイ法、細菌ディスプレイ法、またはmRNAディスプレイ法である。
【0158】
Kd=10−7〜10−12Mの範囲の特異的結合親和性によるED−Bへの結合活性に関する修飾ユビキチン変異体の選択を、当業者に公知の方法により行うことができる。この目的のために、例えば、リボソーム複合体上に提示される前記ユビキチン変異体を、例えば、マイクロタイタープレート上に結合した標的物質に一時的に固定化することができ、または、溶液中での結合後に磁性粒子に結合することができる。非結合変異体を分離した後に、リボソーム複合体を破壊することにより、結合活性を有する変異体の遺伝子情報を、mRNAの形態で特異的に溶出できる。前記溶出は、50mM EDTAで行うことが好ましい。このようにして得られたmRNAは、単離して適切な方法を使用してDNAに逆転写でき(逆転写反応)、このようにして得られた前記DNAは、再増幅できる。
【0159】
in vitroでの転写/翻訳、選択および増幅の連続サイクルにより、所定のハプテンまたは抗原に対する結合特性を有するユビキチン変異体を濃縮できる。
【0160】
ED−B結合タンパク質の特性評価
このようにして得られたユビキチン変異体のさらなる特性評価は、上述の通り、対応する遺伝子カセットを好適な発現ベクターにクローニングした後に、可溶性タンパク質の形態で行うことができる。適切な方法は、当業者に公知であるか、または前記文献に記載されている。
【0161】
好ましくは、所定の結合パートナーに対して結合親和性を有するタンパク質の検出工程の後に、検出したタンパク質の単離および/または濃縮工程が続く。
【0162】
本発明により修飾したユビキチンタンパク質の発現に続いて、それ自体公知の方法により、これをさらに精製して濃縮することができる。選択される方法は、例えば、使用する発現ベクター、宿主生物、使用予定分野、タンパク質の大きさ、およびその他の要因等、それ自体当業者に公知の複数の要因により決定される。簡易に精製するために、本発明により修飾したタンパク質を、分離材料に対して高い親和性を有する他のペプチド配列に融合させることができる。好ましくは、ユビキチンタンパク質の機能性に有害な影響を及ぼさないか、あるいは、特定のプロテアーゼ切断部位の導入により精製後に分離可能な融合物が選択される。このような方法もまた、それ自体当業者に公知である。
【0163】
(図面の簡単な説明)
図1は、種々の腫瘍におけるED−Bの発生について列記した表を示す。
【0164】
図2Aは、修飾ユビキチンED−Bバインダー5E1に関する濃度依存性ELISAの結果を示す。前記修飾ユビキチンED−Bバインダー5E1は、23nMの親和性でヒトED−Bに結合し(黒丸)、一方、BSAへは結合しなかった(白丸)。
【0165】
図2Bは、修飾ユビキチンED−Bバインダー1H4に関する濃度依存性ELISAの結果を示す。前記修飾ユビキチンED−Bバインダー1H4は、7.8nMの親和性でヒトED−Bに結合した(黒丸)。コントロールとして、BSAへの結合をプロットしている(白丸)。
【0166】
図3は、四量体化により親和性が向上することを示している。図3の表に、修飾ユビキチン単量体のKd値を、修飾ユビキチン単量体からなる四量体のものと比較して示している。図中の例は、ユビキチン変異体5E1および1H4である。ED−B結合を、c−FN(細胞性フィブロネクチン)への結合と比較している。図3は、標的ED−Bへの四量体変異体の結合親和性(例えば、5E1は56nM、1H4は1.4nM)が、前記単量体の結合親和性(5E1は4.51μM、1H4は9.98μM)と比べて著しく高いことを示している。
【0167】
図4は、前方(第1)修飾ユビキチン単量体(BDR1を有する)と異なる修飾がされた後方(第2)ユビキチン単量体(BDR2を有する)との遺伝子組換えによるヘテロ二量体の生成により、親和性および特異性が有意に向上することを示している。前記修飾ユビキチン分子は、Biacore、蛍光偏光測定、細胞および組織切片への結合により分析した。複数の変異体のヒトED−Bへの結合に関する濃度依存性ELISA(conc−ELISA)の結果を示す。
【0168】
図4Aは、単量体41B10について、結合親和性がKd=9.45μMであることを示している。
【0169】
図4Bは、41B10が異なる第2単量体と結合して得られた46H9について、結合親和性がKd=131nMであることを示している。
【0170】
図5は、サイトカイン(例えば、TNFα)と融合した特異的変異体を示す。前記融合タンパク質は、前記修飾ユビキチン単量体を三量体化したものであり、生物学的に活性な分子である。
【0171】
図5Aは、修飾ユビキチンベースのED−B結合エフェクター融合タンパク質の模式図である。同図において、緑色(上部の構造)は、エフェクター、例えば、サイトカイン、好ましくは、TNFα;茶色、薄い茶色は、修飾ユビキチン単量体(Affilin(登録商標))である。
【0172】
図5Bは、前記修飾ユビキチンエフェクター結合5E1−TNFコンジュゲートが、アポトースシス促進活性(L929アポトーシスアッセイで測定)を有すること示す。
【0173】
図5Cは、1H4−TNFα融合物のED−Bへの高結合親和性(Kd=15.1nM)を示す(適合線により連結された黒丸)。コントロールとして、BSAへの結合をプロットしている(線により連結されていない黒丸)。
【0174】
図6は、サイトカイン、例えば、TNFαに融合した、修飾ユビキチンベースのED−B結合へテロ二量体分子の親和性および活性を示す。
・修飾ユビキチンベースのED−B結合サイトカイン融合物のアポトーシス誘導活性:EC50 0.78±0.24pM
・遊離サイトカインのアポトーシス誘導活性:EC50 3.14±3.59pM
【0175】
図6Aは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12の親和性を示す(Kd 50.7nM)。
【0176】
図6Bは、修飾ユビキチンベースのED−B結合へテロ二量体分子24H12がサイトカインTNFαに遺伝子融合し、これにより多量体化した前記ヘテロ二量体24H12の親和性を示す(Kd=5.6nM)。
【0177】
図6Cは、ヘテロ二量体修飾ユビキチンライブラリーから選択される候補例、例えば、ヘテロ二量体クローン9E12、22D1、24H12、41B10の分析結果を示す。標的ED−Bに対するKd ELISA値は、コントロールとして用いた細胞基質のフィブロネクチンと比べて向上しており、前記標的への特異的結合が確認された。
【0178】
図6Dは、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾ヘテロ二量体ユビキチン分子9E12の分析結果を示す。種々の濃度(図中の凡例参照:0〜15μMの9E12)の前記ヘテロ二量体ユビキチン変異体について、チップ(Biacore)に固定化されたED−Bへの結合を分析し、前記ヘテロ二量体変異体9E12とED−B間の相互作用を分析した。会合解離曲線の分析からは、Kdを、決定できなかった。
【0179】
図6Eは、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾ヘテロ二量体ユビキチン分子41B10の分析結果を示す。種々の濃度(図中の凡例参照:0〜15μMの41B10)の前記ヘテロ二量体ユビキチン変異体について、チップ(Biacore)に固定化されたED−Bへの結合を分析し、前記ヘテロ二量体変異体41B10とED−Bとの間の相互作用を分析した。会合解離曲線を分析したところ、Kdは、623nM(623×10−9M、6.2×10−7M)であった。
【0180】
図7は、異なる修飾がなされたユビキチンベースの変異体の、結合親和性および結合特異性に対する寄与を示す。前記異なる変異体は、小文字で示す共通の配列モジュールを有する。前記変異体を、そのED−B結合に関して分析した。図3は、修飾ユビキチンへテロ二量体が得られる、単量体の種々の組み合わせを示す。ヘテロ二量体変異体46−A5、50−G11、および46−H4は全て、BDR1が同一の第1(前方)修飾単量体(同図において、「a」で示す)を有するが、BDR2の異なる位置が修飾された第2(後方)ユビキチン単量体を有する。変異体52−D10および52−B3は、46−H9と比べて、BDR1が異なる第1(前方)修飾単量体を有するが、BDR2が同一の第2(後方)ユビキチン単量体(同図において、「e」で示す)を有している。
【0181】
前記修飾ユビキチンヘテロ二量体は、下記配列を有する。
46−H4:配列番号25、45−H9:配列番号26、46−A5:配列番号27、50−G11:配列番号28、52−B3:配列番号29、52−D10:配列番号30
【0182】
上記配列は、実験過程において、配列LEHHHHHH(配列番号31)を有するHisタグの付加により修飾した。
【0183】
図7からわかるように、46−H4は、ED−Bへの優れた結合親和性を有する(Kd=189nM)。46−A5および52−D10は、結合活性を有さない。また、他の修飾ユビキチンタンパク質は、ED−Bへの結合活性が46−H4と比べて低い。したがって、ヘテロ二量体変異体における両単量体が、標的への高い結合親和性のために必要とされ、両単量体が標的に対する一価の結合を示すと結論付けることができる。
【0184】
高いED−B結合活性を有する修飾ユビキチンヘテロ二量体46−H9は、野生型ユビキチン単量体と比較して、前記2つの単量体の両結合ドメイン領域における下記アミノ酸の置換により同定される。
第1モジュール(BDR1)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64A、S65T、T66L
第2モジュール(BDR2)において、(e)K6H、L8M、Q62K、K63P、E64I、S65A、T66E
50G11
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(c)K6M、L8R、Q62M、K63N、E64A、S65R、T66L
46H4
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(d)K6G、L8W、Q62T、K63Q、E64Q、S65T、T66R
52B3
第1モジュールにおいて、(g)Q2R、F4P、K6Y、Q62P、K63P、E64F、S65A、T66R
第2モジュール(46H9)において、K6H、L8M、Q62K、K63P、E64I、S65A、T66E
52D10(ED−Bに結合しない)
第1モジュールにおいて、Q2V、F4C、K6R、Q62T、K63A、E64P、S65G、T66D
第2モジュール(46H9)において、(e)K6H、L8M、Q62K、K63P、E64I、S65A、T66E
46A5(ED−Bに結合しない)
第1モジュール(46H9)において、(a)Q2G、F4V、K6R、Q62P、K63H、E64P、S65T、T66L
第2モジュールにおいて、(b)K6L、L8M、Q62L、K63A、E64F、S65A
【0185】
図8は、配列アライメントを示す。1行目:野生型ユビキチンタンパク質の2つの単量体(1行目)は、77位から88位までに存在する、12個のアミノ酸からなるリンカーSGGGGSGGGGIGにより結合される。BDR2を有する第2単量体は、89位のメチオニンから始まる。この二量体野生型ユビキチンタンパク質を、第1単量体および第2単量体に異なる修飾を有し、これにより2つのBDRが生じた、前記修飾ユビキチンへテロ二量体変異体46−H9(2行目)とアライメントさせる。標的への一価の結合のために、両BDRは、標的との結合において共に作用する。
【0186】
図9は、「Ub2_TsX9」(両単量体の45位がトリプトファンに修飾され、2つの単量体の間にリンカーGIG(77〜79位;80位のメチオニンから第2単量体が開始)を有し、前記第2単量体の最後のC末端アミノ酸において、グリシンからアラニンに置換されているユビキチン)に対する、修飾ユビキチンへテロ二量体変異体1041−D11(1行目)の配列アライメントを示す。3行目は、二量体である野生型ユビキチンであり、リンカーアライメントを示さない(このため、77位のメチオニンから第2単量体が開始する)、「Ubi−Dimer wt」を示す。4行目は、ヒトの野生型ユビキチンである「Ubi−Monomer wt」を示す。
【0187】
図10は、ヒトED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合についての、濃度依存性ELISAを示す。変異体1041−D11は、非常に高いED−Bへの結合親和性を示す(Kd=6.9nM=6.9×10−9M)。黒丸は、ヘテロ二量体ユビキチン変異体1041−D11の、フィブロネクチン断片を含むED−B(67B89−t0という)に対する結合親和性を示す。対照的に、この変異体は、ネガティブコントロール(6789−t0という)には結合しない(白丸)。
【0188】
図11は、フィブロネクチン断片を含む固定化されたED−B(67B89)に対するヘテロ二量体ユビキチン変異体1041−D11の結合を、遊離標的の量を増加させながら分析した競合濃度依存性ELISAの結果を示す。ヘテロ二量体ユビキチン変異体1041−D11は、非常に高いED−Bへの結合親和性を示す(IC50=140nM)。
【0189】
図12は、Biacore(登録商標)を使用した無標識相互作用分析による、前記修飾へテロ二量体ユビキチン分子1041−D11の分析結果を示す。種々の濃度(図中の凡例参照:0〜200nM 1041−D11)の前記ヘテロ二量体ユビキチン変異体について、SAチップ(Biacore)に固定化された、フィブロネクチン断片を含むED−B(67B89という)への結合を分析した。会合解離曲線を分析したところ、Kdは1nM(1×10−9M)であり、koff率は7.7×10−4s−1であり、このkoff率により、1041−D11とED−Bとの複合体の半減期が長いことが示される。
【0190】
図13は、結合活性の血清安定性を同時に分析する濃度依存性ELISAにおける、ED−Bへのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。同図には、種々の異なる条件、例えば、前記変異体の、マウス血清中、ラット血清中、またはコントロールであるPBST中での、37℃で1時間でのプレインキュベーション等が示されている。Kd値は、全て10〜20nMの範囲内である。したがって、ED−Bへの前記ヘテロ二量体1041−D11の結合は、血清による有意な影響を受けないと結論付けることができる。
【0191】
図14は、ヘテロ二量体ユビキチン変異体1041−D11とフィブロネクチン断片との複合体形成の、SE−HPLCによる分析を示す。
【0192】
図14Aは、1041−D11とED−Bとの複合体形成を示す。3回のHPLCのランを重ねている。保持時間21.651分の青色のピークは、純粋な1041−D11に由来する。保持時間26.289分の黒色のピークは、フィブロネクチン断片67B89を表す。1041−D11と67B89との混合物により、SE−HPLC後の保持時間21.407分の赤色のピークが得られた。1041−D11のピークの保持時間が短くなる側へのシフト、および、67B89のピークの消失は、1041−D11と可溶性ED−Bとの複合体が形成されたことを示す。
【0193】
図14Bは、1041−D11(青色、21.944分)、ED−Bを含まないフィブロネクチン断片6789(黒色、26.289分)、および1041−D11と6789との混合物(21.929分および26.289分にピークを有する赤色線)について、3回のSE−HPLCのランを重ねて示している。1041−D11のピークのシフトは、ほとんど観察されなかった。この事実は、6789のピークが消失しなかったことと併せて、ED−Bを含まないフィブロネクチン断片6789との有意な結合を示さないことを表す。
【0194】
図15は、ヘテロ二量体ユビキチン変異体1041−D11の培養細胞への結合を示す。
【0195】
図15Aは、固定されたヒト胎児肺繊維芽細胞(Wi38)へのヘテロ二量体ユビキチン変異体1041−D11の結合を示す。図15において、1列目は、抗ED−B抗体を使用したコントロールを示し、2列目は、タンパク質濃度58.7nMでの変異体のインキュベーションを示し、3列目は、10倍濃度(587nM)の1041−D11タンパク質でのインキュベーションを示し、4列目は、PBSを用いたネガティブコントロールを示す。1行目において、ヒトWi38繊維芽細胞を、位相差で示し、2行目において、免疫蛍光法で示し、3行目において、核のDAPI染色を示す。変異体1041−D11は、ED−B含有細胞外マトリックスに対する高特異性で、Wi38に結合すると結論付けることができる。EDBを低レベルで発現するNHDF細胞を使用するコントロールを行った(データは示さず)。前記変異体は、これらの細胞には結合しなかった。
【0196】
図15Bは、バイタル(vital)ヒト胎児肺繊維芽細胞(Wi38)への結合を示す。前記ネガティブコントロールのNHDF細胞は、初代の正常繊維芽細胞であり、EDB−フィブロネクチンを低レベルで発現する。1行目および3行目は、種々のタンパク質濃度での変異体、ならびにネガティブコントロールを示す。2行目および4行目は、EDB抗体を用いたコントロールのインキュベーションを示す。1〜2行目は、Wi38細胞株における前記変異体とポジティブコントロールを示す。3〜4行目は、NHDF細胞のインキュベーションを示す。変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、Wi38に結合することがわかる。
【0197】
図15Cは、固定されたマウスBalb 3T3細胞への結合を示す。前記変異体について、3つの異なるタンパク質濃度(1、10、50nM)でテストした。1行目は、細胞における前記変異体(SPVF−28−1041−411−TsX9)を示し、2行目は、ポジティブコントロール(Fv28−EDB−抗体)を示し、3行目は、ネガティブコントロール(UB2_TsS9;配列番号1に相当する非修飾ユビキチン)を用いたインキュベーションを示す。変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、マウスBalb 3T3細胞に結合することがわかる。
【0198】
図15Dは、固定されたマウスST−2細胞への結合を示す。前記変異体について、3つの異なるタンパク質濃度(1、10、50nM)でテストした。1行目は、細胞における前記変異体(SPVF−28−1041−411−TsX9)を示し、2行目は、ポジティブコントロール(Fv28−EDB−抗体)を示し、3行目は、ネガティブコントロール(UB2_TsS9;配列番号1に相当する非修飾ユビキチン)を用いたインキュベーションを示す。前記ヘテロ二量体ユビキチン変異体1041−D11が、ED−B含有細胞外マトリックスに対する高特異性で、マウスBalb ST−2細胞に結合することがわかる。
【0199】
図16Aは、哺乳類組織切片における、ヘテロ二量体ユビキチン変異体1041−D11の標的に対する特異性を示す。7つの検体から採取したF9腫瘍組織について評価を行った。10nM〜100nMの範囲の異なる濃度のヘテロ二量体ユビキチン変異体1041−D11を用いた免疫組織化学検査において、マウス由来のF9腫瘍でのED−B特異的血管染色(vascular staining)が見られた。ED−Bは、腫瘍血管系の高特異的マーカーである。標的タンパク質であるED−Bは、血管の反管腔側に局在する。変異体1041−D11は、F9腫瘍由来の組織切片において、特異的に血管系を装飾する。得られた結果は、抗体フラグメントL19に匹敵する。さらに、48組織をテストしたところ、FDA関連パネルにおける48組織のいずれにおいても、非特異的な染色は観察されなかった。
【0200】
図16Bは、腫瘍組織における1041−D11の蓄積を、野生型ユビキチン(同図において、Ub2(NCP2))と比較して示している。F9腫瘍組織について、30分から16時間の間の異なる時点で、1041−D11および野生型ユビキチンの存在を分析した。腫瘍組織における1041−D11の最も高い蓄積は、投与から30分後および16時間後に観察されている。一方、F9腫瘍組織における野生型ユビキチンの蓄積は低い。前記変異体は、ED−Bを発現する腫瘍において、野生型ユビキチンと比べて濃縮されている。このことは、1041−D11の腫瘍組織指向性のターゲッティングの証拠となる。また、癌モデルにおける1041−D11の腫瘍/血液比は、動物における1041−D11変異体in vivo活性を明らかに証明している(データは示さず)。
【0201】
図17は、1041−D11−TNFα融合タンパク質の、ED−Bに対する高い選択性および特異性を示す。
【0202】
図17Aおよび17B:1041−D11−TNFα融合タンパク質のアポトーシス誘導TNFα活性を、細胞ベースアッセイ(cell based assay)(L929細胞)によりテストした。前記両図は、培養細胞において、前記1041−D11−TNFα融合タンパク質(図17B)が、遊離TNFα(図17B)と同程度に活性であることを、明らかに示している。
【0203】
図17Cは、標的ED−Bに対する、ヘテロ二量体ユビキチン1041−D11−TNFα融合タンパク質の高い選択性を証明している。前記ヒトED−Bフィブロネクチンドメイン67B89は、見かけのKD値1.8nMで1041−D11変異体に結合し(黒丸)、標的への高い親和性を示す。ED−Bドメインを欠損するヒトフィブロネクチン(h6789)には、1041−D11−TNFαは結合しない(白丸)。
【0204】
図17Dは、修飾ユビキチンベースのED−B結合1041−D11−TNFα融合タンパク質についての、Biacoreアッセイによる結合分析を示す。結果から、1041−D11TNFα融合タンパク質が、KD値1.13nMの高い親和性を示すことが証明された。
【0205】
図17Eは、培養細胞における変異体1041−D11で観察された高い結合特異性が、1041−D11がTNFαに融合されても維持されることを示す。前記融合タンパク質は、EDB発現細胞に特異的に結合する。したがって、1041−D11TNFα融合タンパク質は、非常に高い親和性および特異性で標的ED−B(ターゲット(+))に結合する。ED−Bを含まない血清(ターゲット(−))では、交差反応は観察されなかった。
【0206】
図18は、TNFαに融合した変異体1041−D11とメルファランとの併用による、7日間のマウスの処置期間中のin vivoでの相対的腫瘍成長を示す。データから、1041−D11−TNFαを細胞増殖抑制剤であるメルファランと併用した場合には、mTNFαとメルファランとを併用した場合、またはメルファラン単独の場合と比べ、より効率的に相対的腫瘍成長が減少されることをはっきりと示している。7日間の処置後の前記腫瘍成長動態は、1041−D11−mTNFαによる高効率での減少を示す。このことは、融合タンパク質1041−D11−TNFαとメルファランとの併用による腫瘍治療の有効性の明らかな証拠である。ED−Bは、マウスおよびヒトを含む数種の哺乳類において同一である。このため、前記結果から、ヒトにおける変異体1041−D11−TNFαの性能が予測される。
【0207】
図19は、ED−Bに対して驚くほど強力な結合親和性を有することが見出されたさらなる16種類の配列について、コンセンサスな位置およびアミノ酸置換を示す。前記コンセンサスなアミノ酸位置は、第1単量体の結合決定領域2、4、6、62、63、64、65、66位にあり、一方、コンセンサスなアミノ酸置換は、Q2T、F4W、K6H、Q62N、K63F、E64K、S65L、およびT66Sである。図2からわかるように、4種類のファミリーの配列を濃縮できた(コンセンサス配列、文字サイズはアミノ酸の発生頻度に対応する)。85および87位は、前記ヘテロ二量体タンパク質において、前記第2単量体における6および8位に対応する位置であり、141〜145位は、62〜64位に対応する。暗青色で示したTWH NFKLSは、1071−C12由来である。赤色で示した残基は、前記4種類のファミリーの配列のうちの1種類に属する。赤色で示した残基が、主に濃縮され(178/457配列)、HIT ELISAによれば、最も強力な結合分子を含む。
【実施例】
【0208】
下記実施例は、本発明をさらに説明するために提供するものである。本発明は、特に、ユビキチンの修飾を例にあげて実証されている。しかしながら、本発明は、これに限定されるものではなく、以下の実施例は、単に、前述の記載に基づいた本発明の実施可能性を示しているにすぎない。本発明の完全な開示のため、本願および付属書類に引用されている文献についても言及しているが、これらの引用文献は全て、引用により、その開示全体が本願に取り込まれている。
【0209】
[実施例1]
修飾ユビキチンタンパク質ベースのED−B結合タンパク質の同定
【0210】
単量体結合タンパク質のライブラリーの構築およびクローニング
特に示さない限り、例えば、Sambrookらに記載されているような、確立された組換え遺伝学的手法を使用した。合計10の選択されたアミノ酸位置における協奏的な変異誘発によって、高度な複雑性を有するヒトユビキチン単量体のランダムライブラリーを用意した。NNKトリプレットによって置換された修飾アミノ酸は、ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸を含む。
【0211】
ヘテロ二量結合タンパク質のライブラリーの構築とクローニング
特に示さない限り、例えば、Sambrookらに記載されているような、確立された組換え遺伝学的手法を使用した。合計15の選択されたアミノ酸位置における協奏的な変異誘発によって、高度な複雑性を有するヒトユビキチンヘテロ二量体のランダムライブラリーを用意した。NNKトリプレットによって置換された修飾アミノ酸は、近位(第1)ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸、および遠位(第2)ユビキチン単量体内の2、4、6、8、62、63、64、65、66、68位から選択された少なくとも3個のアミノ酸を含む。両ユビキチン単量体は、少なくともGIG配列、または少なくともSGGGG配列を有するグリシン/セリンリンカーによって遺伝学的に結合(ヘッドトゥテイル配置)しており、リンカー配列の例としては、GIG、SGGGG、SGGGGIG、SGGGGSGGGGIG(配列番号32)またはSGGGGSGGGGがあげられるが、その他のリンカーでもよい。
【0212】
TATファージディスプレイ選択
ユビキチンライブラリーを、例えば、選択システムとしてTATファージディスプレイを使用し、標的に対して濃縮した。当該技術分野において公知である他の選択方法を用いることもできる。前記標的は、タンパク質結合表面上に、またはタンパク質に共有結合されたビオチン化残基を介して、非特異的に固定化され得る。ストレプトアビジンビーズまたはニュートラアビジンストリップ上へのビオチンを介した固定化が好ましい。標的結合ファージは、溶液中または固定化標的上のいずれかにおいて選択される。例えば、ビオチン化され固定化された標的とファージとを、インキュベートし、続いて、マトリックスに結合したファージの洗浄およびマトリックス結合ファージの溶出を行った。標的のインキュベーションに続く各サイクルにおいて、前記ビーズを磁力により溶液から分離し、数回洗浄した。最初の選択サイクルにおいて、ビオチン化標的を、ニュートラアビジンストリップに固定化し、一方、2回目から4回目のサイクルにおいて、溶液中での選択を行い、続いて、Streptavidin−coated Dynabeads(登録商標)(Invitrogen社製)上に、標的とファージとの複合体を固定化した。最初の2回の選択サイクルでの洗浄後、標的が結合した修飾ユビキチン分子のファージを、酸性溶液での溶出により遊離させた。選択サイクルにおいて、過剰量の標的を用いた競合的溶出により、3回目および4回目のファージ溶出を行った。溶出したファージを再増幅した。バインダーの特異性を誘導するため、選択に際し、標的に類似するタンパク質を含めてもよい。
【0213】
TATファージディスプレイ選択の代替:リボソームディスプレイ選択
ユビキチンライブラリーを、例えば、選択システムとしてリボソームディスプレイを使用して、標的に対して濃縮した(Zahnd et al., 2007、Ohashi et al., 2007)。当該技術分野において公知である他の選択方法を用いることもできる。前記標的を、標準的な方法によってビオチン化し、Streptavidin−coated Dynabeads(登録商標)(Invitrogen社製)に固定化した。リボソーム、mRNA、および新生ユビキチンポリペプチドを含む三元複合体を、PURExpres(商標) In Vitro Protein Synthesis Kit(NEB社製)を用いて構築した。選択の一次ラウンドを2回行い、三元複合体をインキュベートし、続いて、同様の選択ラウンドを2回行った。標的インキュベーションに続く各サイクルにおいて、ビーズを磁力により溶液から分離し、ストリンジェンシーを増加させながら、リボソームディスプレイバッファーで洗浄した。最初の2回の選択サイクルでの洗浄後、前記ビーズを再度磁力により溶液から分離し、50mM EDTAの添加により、標的が結合した修飾ユビキチン分子のmRNAをリボソームから遊離させた。選択サイクルにおいて、過剰量の標的を用いた競合的溶出により、3回目および4回目のmRNAの溶出を行った(Lipovsek and Pluckthun, 2004)。各サイクルの後、RNeasy MinElute Cleanup Kit(Qiagen社製、ドイツ)、Turbo DNA−free Kit(Applied Biosystems社製、アメリカ)、およびTranscriptor Reverse Transcriptase(Roche社製、ドイツ)を用いて、RNAの精製とcDNAの合成を行った。
【0214】
濃縮プールのクローニング
4回目の選択サイクルの後、合成cDNAを、F1プライマー(GGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATG)(配列番号9)およびWUBI(co)RD_xhoプライマー(AAAAAAAAACTCGAGACCGCCACGCAGACGCAGAACCAG)(配列番号10)を用いたPCRにより増幅し、制限ヌクレアーゼNdeIおよびXhoI(Promega社製、アメリカ)で切断し、適合性付着末端を介して発現ベクターpET−20b(+)(Merck社製、ドイツ)に連結した。
【0215】
単一コロニーのヒット解析
NovaBlue(DE3)細胞(Merck社製、ドイツ)への形質転換の後、アンピシリン耐性単一コロニーを、200μl SOBAG培地(100μg/ml アンピシリンおよび20g/l グルコースを含むSOB培地)において、37℃で6時間培養した。500μlの自己誘導培地ZYM−5052(Studier社製、2005)を用い、96ディープウェルプレート(Genetix社製、イギリス)において、37℃で16時間培養することにより、ED−B結合修飾ユビキチンを発現させた。4℃、3600×gで、15分間遠心分離して、細胞を回収した。その後、ウェルあたり300μlの溶解バッファーを用いて、37℃で30分間インキュベートし、前記細胞を溶解した。前記溶解バッファーは、0.2×BugBuster(登録商標)(Merck社製、ドイツ)、0.3mg/mlリゾチーム(VWR社製、ドイツ)、0.2mM PMSF(Roth社製、ドイツ)、3mM MgCl2および0.2U/ml Benzonase(VWR社製、ドイツ)を含む、50mM NaH2PO4、300mM NaCl(pH8)とした。4℃、3600×gで、30分間遠心分離した後、得られた上清を、4μg/ml ED−B、およびセイヨウワサビぺルオキシダーゼ(POD)とのユビキチン特異的FabフラグメントコンジュゲートでコートしたNunc MediSorp plates(Thermo Fisher Scientific社製、アメリカ)を用いて、ELISAによりスクリーニングした。検出試薬として、TMB−Plus(Biotrend社製、ドイツ)を用いた。ウェルあたり50μlの0.2M H2SO4溶液を用いて黄色を発色させ、プレートリーダーにおいて450nmと620nmを測定した。
【0216】
数サイクルの選択ディスプレイを、ED−Bに対して行った。最後の2サイクルの選択において、結合分子を、過剰量の遊離ED−Bを用いて溶出した。これらのED−B結合変異体を、他のものとの間で同定した。
【化2】
【0217】
野生型ユビキチン単量体(Ubi monomer wt)と、野生型ユビキチン二量体(ubi dimer wt)、野生型ユビキチンタンパク質(図9におけるUb2−TsX、各単量体の45位における置換およびC末端における2つの置換を有する)、および修飾ユビキチンヘテロ二量体変異体1041−D11との配列アライメントを、図9に示す。Ub2−TsXにおいて、前記単量体のC末端の置換(GGからAA)は、血清における安定性を増加させる。これは、デユビキチナーゼは、ユビキチンのGGの後ろを切断し、AAの後ろを切断しないためである。野生型ユビキチンの二次構造は、前記C末端における置換を有するユビキチンと比較して、ほぼ同一である。
【0218】
1041−D11(図9、配列番号36)または1045−D10と呼ばれる、優れたED−B結合活性を有する修飾ユビキチンは、野生型と比較して、後述のアミノ酸置換によって同定される。
第1モジュール
K6W、L8W、K63R、E64K、S65F、T66P
第2モジュール
K6T、L8Q、Q62W、K63S、E64N、S65W、T66E
任意に、Q2R(当該置換は、変異体1041−D11に存在し、変異体1045−D10には存在しない)
融合タンパク質に適した好ましいリンカーは、配列番号32のリンカーまたはGIG配列である。ただし、代わりに使用できる、多くのリンカーが考えられる。
【0219】
さらに好ましい例として、下記の配列のタンパク質(配列番号47)が提示され、前記配列において、Xはどのようなアミノ酸でもよい。リンカーとして、ここでは、SGGGGSGGGGIGが使用される(イタリック体で示す)。ただし、他の種類のリンカー、またはリンカーなしも、代替可能である。
【化3】
これらの配列を有するタンパク質の例を、図19に示す。
【0220】
[実施例2]
ED−B結合修飾ユビキチン変異体とTNFα(例えば、ヒトTNFα、TNFαという)からの融合タンパク質の生成
【0221】
前記変異体は、修飾ユビキチン、例えば、ヘテロ二量体変異体1041−D11と、哺乳類のTNFα、例えば、マウスまたはヒトのTNFαとの間の融合タンパク質として、大腸菌内で発現させることができる。融合タンパク質のタンパク質解析には、タンパク質発現と純度、非凝集能、細胞培養におけるTNFα活性、標的タンパク質ED−Bへの親和性、選択性、細胞培養における特異的結合が含まれる。F9腫瘍を有するマウスにおける腫瘍の縮小を誘導するための動物実験における必須条件は、マウスTNFαとの融合物である。
【0222】
工程1:融合タンパク質のクローニングのためのベクターの生成(pETSUMO−TNFα)
pETSUMOadaptは、修飾ベクターpETSUMO(Invitrogen社製)であり、付加マルチクローニングサイト(MCS)の挿入によって修飾されている。pETSUMOadaptへのTNFαのクローン化から開始し、ED−Bに結合する修飾ユビキチン変異体を挿入するための制限部位を導入した。得られた構築物は、下記DNA配列(配列番号11)の構造体His6−SUMO−TNFαの構造を有する。
ATGGGCAGCAGCCATCATCATCATCATCACGGCAGCGGCCTGGTGCCGCGCGGCAGCGCTAGCATGTCGGACTCAGAAGTCAATCAAGAAGCTAAGCCAGAGGTCAAGCCAGAAGTCAAGCCTGAGACTCACATCAATTTAAAGGTGTCCGATGGATCTTCAGAGATCTTCTTCAAGATCAAAAAGACCACTCCTTTAAGAAGGCTGATGGAAGCGTTCGCTAAAAGACAGGGTAAGGAAATGGACTCCTTAAGATTCTTGTACGACGGTATTAGAATTCAAGCTGATCAGACCCCTGAAGATTTGGACATGGAGGATAACGATATTATTGAGGCTCACAGAGAACAGATTGGTGGTGTGCGTAGCAGCAGCCGTACCCCGAGCGATAAACCGGTGGCGCATGTGGTGGCGAATCCGCAGGCGGAAGGCCAGCTGCAGTGGCTGAACCGTCGTGCGAATGCGCTGCTGGCCAACGGCGTGGAACTGCGTGATAATCAGCTGGTTGTGCCGAGCGAAGGCCTGTATCTGATTTATAGCCAGGTGCTGTTTAAAGGCCAGGGCTGCCCGAGCACCCATGTGCTGCTGACCCATACCATTAGCCGTATTGCGGTGAGCTATCAGACCAAAGTGAACCTGCTGTCTGCGATTAAAAGCCCGTGCCAGCGTGAAACCCCGGAAGGCGCGGAAGCGAAACCGTGGTATGAACCGATTTATCTGGGCGGCGTGTTTCAGCTGGAAAAAGGCGATCGTCTGAGCGCGGAAATTAACCGTCCGGATTATCTGGATTTTGCGGAAAGCGGCCAGGTGTATTTTGGCATTATTGCGCTGTAATAA
【0223】
TNFα配列を、BamHIおよびXhoI部位の導入により、PCRで増幅した。
使用したプライマーは、以下の通りである。
【化4】
前記fwプライマー(配列番号12)は、TNFαの最初の15塩基対(下線部)を認識し、BamHI配列(太字部)を有する。前記revプライマー(配列番号13)は、TNFαの最後の塩基対、終止コドン(下線部)、およびXhoI制限部位(太字部)を含む。
【0224】
<PCR反応ミックス(100μl)>
84.5μl H2O;
10μl 10×Pwoバッファー+Mg;
2μl 10mM dNTPs(=200μM);
各0.5μl 100μM プライマーfw/rev(=各0.5μM);
2μl DNA(=0.25μg);
0.5μl Pwoポリメラーゼ(=2.5U;Roche)
【0225】
<PCRプログラム>
94℃で3分、94℃で30秒、60℃で30秒、72℃で2分(工程2〜4:30サイクル)、72℃で5分、続いて4℃で処理し、続いて、Qiagen−MinElute−Kit(10μl EBに溶出)によるPCR産物の精製を行った。前記PCR産物を、BamHI−XhoI制限およびライゲーションにより、ベクターpETSUMOadaptのMCSに導入した。
【0226】
<制限ミックス(100μl)>
ベクター:83μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BamHI(=30U;NEB),
1.5μl XhoI(=30U;NEB);
1.65μl ベクター;
3h、37℃でインキュベーション
PCR産物:76.5μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BamHI(=30U;NEB),
1.5μl XhoI(=30U;NEB);
8μl 挿入断片;
3h、37℃でインキュベーション
1%アガロースゲル(100V、60分間ラン)において制限ミックスを分離;
ベクター断片(5659bp)と挿入断片(491bp)の切断;
Qiagen gel extraction kitを用いた精製(30μl EBに溶出)。
【0227】
<ライゲーション(20μl)>
15.2μl H2O;
2μl 10×T4−DNAリガーゼバッファー;
2.26μl ベクター(200ng);
0.54μl 挿入断片(40ng)
5分間、65℃でインキュベーション;
16℃に冷却;
1μl T4−DNAリガーゼ(=3U;NEB)添加;
16時間、16℃でインキュベーション
【0228】
<NaAc/イソプロパノール沈殿>
ライゲーションミクスチャー(20μl)+2.2μl 3M NaAc(pH5.0)+22.2μl イソプロパノール;
30分間、−20℃;
15分間、4℃、13000Upm;
500μlの70% EtOHにペレットを再懸濁;
スピン;
10μlのH2Oにペレットを再懸濁
【0229】
<形質転換>
エレクトロコンピテントNovablue(DE3)細胞(40μlアリコート)と10μlのライゲーション産物とを混合;
0.1cm エレクトロポレーションキュベットに移動;
エレクトロポレーターでパルスを印加(1.8kV、50μF、100 Ohm);
37℃、220Upm、45分間、1mlのSOC培地と、溶液とをインキュベート;
カナマイシン含有LBプレートに、100μlを播き、37℃、一晩インキュベーション
【0230】
工程2:修飾ユビキチンベースのEDB融合タンパク質のクローニング
EDB結合修飾ユビキチンベースの変異体とTNFαとの融合物の生成のために、PCRにより、対象であるEDB修飾ユビキチンベースの配列をpET20bベクターから増幅し、BsaIおよびBamHI制限部位を導入する。この方法は、単量体および二量体EDB修飾ユビキチンベースの変異体に好適である。単量体WTユビキチン(Wubi)用のプライマーは、下記の通りである。
【化5】
【0231】
前記fwプライマー(配列番号14)は、修飾ユビキチンの最初の15塩基対(下線部)を認識し、BsaI配列(太字部)を有する。前記revプライマー(配列番号15)は、修飾ユビキチンの最後の15塩基対を認識し、アミノ酸リンカー(SGGGG配列)およびBamHI制限部位(太字部分)を挿入する。修飾ユビキチンベースの変異体それぞれについて、特異的fwプライマーを使用する。単量体EDB修飾ユビキチンベースの変異体1H4、5E1および4B10のプライマーは、下記の通りである。
【化6】
【0232】
前記revプライマーは、全ての単量体修飾ユビキチンベースの変異体に使用する。二量体修飾ユビキチンベースの変異体用のrevプライマーは、下記の通りである。
【化7】
【0233】
二量体WTユビキチン(WubiHubi)のクローニング用および二量体EDB修飾ユビキチンベースの変異体用のfwプライマーは、下記の通りである。
【化8】
【0234】
<PCRミックス(100μl)>
84.5μl H2O;
10μl 10×Pwoバッファー+Mg;
2μl 10mM dNTPs(=200μM);
各0.5μl 100μM プライマーfw/rev(=0.5μM);
2μl DNA(変異体により決定);
0.5μl Pwoポリメラーゼ(=2.5U;Roche)
【0235】
<PCRプログラム>
1.94℃で3分
2.94℃で30秒
3.60℃で30秒
4.72℃で2分(工程2〜4:30サイクル)
5.72℃で5分、続いて4℃
【0236】
アガロースゲルでPCR産物を精製し、必要なバンドを切り出し、Qiagen−gel extraction kitで精製する。BsaI−BamHI制限により、前記PCR産物をクローニングする(pETSUMO−TNFα内に)。
【0237】
<制限ミックス(100μl)>
75μl H2O;
10μl 10×NEバッファー3;
1μl 100×BSA;
3μl BsaI(=30U;NEB);
8μl DNA(ベクターまたはPCR産物)2時間、50℃でインキュベーション、
10分間、65℃、
3μl BamHIの添加(=30U;NEB)、
2時間、37℃
1%アガロースゲルにおいて制限ミックスを分離;
ベクター断片および挿入断片の切断;
Qiagen−gel extraction kitを用いた精製(30μl EBに溶出)。
【0238】
<ライゲーション(20μl)>
12.5μl H2O;
2μl 10×T4−DNAリガーゼバッファー;
5μl ベクター(66ng);
0.5μl 挿入断片(変異体の種類により変わる)
5分間、65℃でインキュベーション;
16℃に冷却;
1μl T4−DNAリガーゼ(=3U;NEB)添加;
16時間、16℃でインキュベーション
【0239】
<NaAc/イソプロパノール沈殿>(工程1参照)
【0240】
<形質転換>
エレクトロコンピテントNovablue(DE3)細胞の形質転換は、前述の通りである。結果として、下記の融合構築物が得られた。すなわち、His6−SUMO−修飾ユビキチン−SGGGG−TNFαを有するpETSUMOadaptにおけるEDB修飾ユビキチンとTNFαとの融合構築物である(単量体修飾ユビキチンの場合、359個のアミノ酸からなり、二量体修飾ユビキチンの場合、447個のアミノ酸からなる)。
【0241】
[実施例3]
ユビキチンベースのTNFα融合タンパク質の発現と精製
【0242】
DNA配列解析により、SUMO−TNFα融合タンパク質の正確な配列が示された。変異体の発現のために、前培養物をLB/カナマイシンで1:100に希釈し、前記培養液を、600nmの光学密度(OD600)0.5まで、200rpm、37℃で攪拌することで、クローンを振とうフラスコで培養した。発現は、IPTG(最終濃度1mM)の添加で誘導した。30℃、4時間、200rpmで、培養を続けた。4℃、6000×gで20分間遠心分離し、細菌細胞を回収した。前記細胞ぺレットを、ベンゾナーゼおよびリゾチームを含むNPI20バッファー30mlに懸濁した。細胞を、氷上で超音波処理(3×20秒)によって破砕した。懸濁液を4℃、40000×gで30分間遠心分離した後、可溶性タンパク質を含む上清を得た。両タンパク質を、室温での親和性クロマトグラフィーにより、精製した。Ni−アガロース(5ml、GE Healthcare社製)の一本のカラムを、50mlのNPI−20で平衡化した。可溶性タンパク質を含む前記上清を、前記カラムにアプライし、続いて、NPI−20で洗浄した。前記結合したタンパク質を、NPI−20から50%NPI−500(100ml)の直線勾配で溶出した。各画分は、SDS−PAGEにより、その純度を解析した。好適な画分をプールし、SUMOヒドロラーゼ切断バッファー(50mM Tris,300mM NaCl、pH8.0)で平衡化したゲルろ過カラム(Superdex 75、1.6×60cm、GE Healthcare社製)に、流速1ml/分で、アプライした。
【0243】
切断反応は、製造元(Invitrogen社)の使用説明書にしたがって行った。切断後、前記タンパク質をNi−アガロースカラム(5ml、GE Healthcare社製)にアプライした。Hisタグ化SUMOヒドロラーゼおよびHisタグ化SUMOは、前記カラムに結合し、正しい融合タンパク質(Hisタグフリー)は、前記カラムを通過した。前記タンパク質の純度は、rpHPLC解析およびゲル電気泳動によって、証明された。前記三量体(TNFαを介する)が正しい分子量を有することは、分析用SEC解析(10/30 Superdex G75、GE Healthcare社製)を用いて確認した。
【0244】
[実施例4]
修飾ユビキチンベースのED−B結合変異体のヒトED−Bに対する結合分析
【0245】
実施例4A:濃度依存ELISAによる修飾ユビキチンベースのED−B結合変異体の結合分析
ユビキチンベースの変異体のヒトED−Bに対する結合を、濃度依存ELISAによって分析した。精製タンパク質を、ヒトED−B、BSAおよび細胞フィブロネクチン(cFN)でコーティングされた複数のNUNC−medisorpプレートに、量を増加させながらアプライした。ウェルあたり抗原50μl(10μg/ml)でのコーティングを、4℃で一晩行った。前記プレートを、0.1% Tween20を含むPBS(pH7.4;PBST)で洗浄した後、前記ウェルを、37℃で2時間、ブロッキング溶液(PBS pH7.4;3% BSA;0.5% Tween20)を用いてブロッキングした。前記ウェルを、PBSTでさらに3回洗浄した。
【0246】
前記ウェルにおいて、異なる濃度の修飾ユビキチンベースのED−B結合タンパク質(50μl量)を、室温で1時間インキュベートした(図10、開始濃度として、500nMの1041−D11タンパク質を使用した)。PBSTで前記ウェルを洗浄した後、抗ユビキチンfabフラグメント(AbyD)PODコンジュゲートを、PBSTに適切に希釈(例えば、1:2000または1:6500)して、アプライした。前記プレートを、ウェルあたり300μlのバッファーPBSTで、3回洗浄した。50μlのTMB基質溶液(KEM−EN−Tec)を各ウェルに加え、15分間インキュベートした。ウェルあたり50μlの0.2M H2SO4を加えて、反応を停止させ、前記ELISAプレートを、TECAN Sunrise ELISA−Readerを用いて読み取った。参照波長を620nmとして、450nmで光度計による吸光光度測定を行った。図1は、みかけKD値11nMという、ED−Bに対する1H4の特異的結合を明瞭に示している。変異体5E1は、みかけKD値7.7μM、4B10は、みかけKD値280nMを、それぞれ示している。図10は、変異体1041−D11のED−Bに対する非常に高い結合親和性を示す(KD=6.9nM)。このように、ユビキチン野生型のわずかな修飾(各単量体において8個の置換まで)により、ED−Bに対する非常に高い結合親和性がもたらされる。
【0247】
実施例4B:競合的濃度依存ELISAによる、修飾ユビキチンベースのED−B結合変異体の結合分析
競合的濃度依存ELISAにより、量が増加する遊離標的の存在下、フィブロネクチン断片(67B89)を含む固定化ED−Bに対するユビキチン変異体1041−D11の結合を分析した。ELISAの条件は、1041−D11タンパク質を、ED−B(67B89)(0μM〜10μM)、またはネガティブコントロール6789(0μM〜10μM)で、1時間プレインキュベートし、その後、その混合物を、Medisorp−plate上に配置した標的67B89に添加した以外は、実施例5Aで述べた通りである。これに続き、前記変異体を、対応する抗体によって検出した(抗ユビキチン−Fab−POD;希釈度1:6500)。図11は、変異体1041−D11が、ED−Bに対して非常に高い結合親和性を有することを示す(IC50=140nM)。図10に示した結果が裏付けられている。すなわち、ユビキチン野生型のわずかな修飾(各単量体において8個の置換まで)のみにより、ED−Bに対する非常に高い結合親和性がもたらされる。
【0248】
実施例4C:結合活性の血清安定性を同時に分析する濃度依存ELISAによる修飾ユビキチンベースのED−B結合変異体の結合分析
当該技術分野において周知の方法により、前述(実施例5Aおよび5B)と同様に、ELISAを行った。ED−B(ここでは、67B89という)を、マイクロタイタープレートにコーティングし、前記変異体を、ED−Bに結合させ、特異的ユビキチン抗体により検出する(抗ユビキチン−Fab−POD)。この分析において、前記変異体は、異なる方法で処理される。すなわち、前記変異体を、37℃で1時間、マウス血清中でインキュベートする処理(図13参照、青丸部分)、前記変異体を、37℃で1時間、ラット血清中でインキュベートする処理(図13、赤丸部分)、または、前記変異体を、37℃で1時間、PBSでインキュベートする処理(図13、黒丸部分)である。図13は、変異体1041−D11の全てのKDが、10.3nM(PBS)から20.74nM(マウス血清)の間にあることを示す。
【0249】
実施例4D:Biacore分析による修飾ユビキチンベースのED−B結合変異体の結合分析
当業者に公知の方法を用いて、CM5チップ(Biacore)に固定化したED−B含有フィブロネクチン断片(67B89という)に対する結合について、前記変異体を異なる濃度で分析した(例えば、0〜200nMの変異体、好ましくは1041−D11)。得られたデータは、BIA評価ソフトウェアおよび1:1−Langmuir−fittingにより処理した。図12に示すように、変異体1041−D11のKDは、1.0nMであった。結合速度定数は、kon=7.6×105M−1s−1、koff=7.7×10−4s−1であった。図17Dに示すように、融合タンパク質1041−D11−TNFαのKDは、1.13nMであった。結合速度定数は、kon=4.5×105M−1s−1、koff=5.0×10−4s−1であった。
【0250】
実施例4E:SE−HPLCによる修飾ユビキチンベースのED−B結合変異体の複合体構造の分析
複合体構造の分析のために、Tricorn Superdex75 5/150 GLカラム(GE−Healthcare社製)(V=3ml)を使用し、50μlのタンパク質をアプライした。さらなる条件は、バッファー:1×PBS(pH7.3)、流速:0.3ml/分、ラン:45分(サンプルの注入:15分後)とした。条件:0.72nmol 1041−D11タンパク質+0.72nmol ED−B(67B89またはネガティブコントロール6789ともいう)を、室温で1時間インキュベートし、複合体構造を分析するために、カラムにアプライした。図14では、前記変異体のみを黒で示し、標的ED−Bのみを青で示し、ED−Bとの複合体を構成する変異体結合をピンクで示す。図14Aは、前記変異体を有するED−Bを示す。図14Bは、ED−Bを有さない変異体を示す。同図は、変異体1041−D11が、ED−B(67B89)と複合体を構築するが、6789とは複合体を構築しないことを示す。
【0251】
[実施例5]
TNFαの生物学的分析
TNFα修飾ユビキチンベースのED−B結合融合体の生理学的TNFα活性を、L929アポトーシス分析によって決定した(Flick et al.、1984 J. Immunol. Methods. 68:167−175)。この分析では、TNFαが、ピコモル範囲のEC50値で、アクチノマイシンD感受性細胞の細胞死を効率的に促進する。
【0252】
細胞を、FBSと抗生物質を含む培地に再懸濁した。密度3.5×105細胞/mlの細胞懸濁液100μlを、96ウェル標準細胞培養プレートのウェルに播き、加湿したCO2インキュベーターで一晩インキュベートした。その後、前記培養培地を除去し、FBS、アクチノマイシンDおよび抗生物質を含む培地50μlを各ウェルに加え、その後、さらに30分間インキュベートした。その後、各テスト項目50μlと、10−7から10−18Mの適切な濃度範囲のTNFα修飾ユビキチンベースのED−B結合融合体またはヒト組換えTNFαコントロールを添加した。さらに48時間インキュベートした後、細胞生存の尺度である代謝活性を、WST−1試薬(Roche社製)を用いて決定した。
【0253】
1つのテスト項目あたり、少なくとも3通りの独立した実験を行い、各実験は3回行った。TNFα修飾ユビキチンベースのED−B結合融合タンパク質の各テストは、分析間のばらつきについての情報を得るために、ヒト組換えTNFαの投与量範囲のテストと並行して行った。
【0254】
定量評価は、EC50値、すなわち、半数の細胞の生存を促進するテスト項目濃度による値に基づく。
【表2】
mub:修飾ユビキチンベースのED−B結合
【0255】
修飾ユビキチンベースのED−B結合融合体の、1つのユビキチン単量体構築物(Wubi)および3つのユビキチン二量体構築物のTNFαを分析した。TNFα部分に結合した修飾ユビキチンベースのED−B結合変異体に依存して、TNFαに関連する活性は、約1桁分増加(SPWF−28_24−H12_TNF−alpha)または減少した(SPWF−28_22−D1_TNF−alpha、Wubi−Hubi−TNF−alpha)。変異体1041−D11 TNFα分析は、図17参照。
【0256】
[実施例6]
細胞培養分析におけるユビキチン変異体の結合分析
培養細胞に対する変異体1041−D11等の結合をテストした。ED−Bが高発現レベルである正常ヒト胎児肺線維芽細胞(Wi38細胞)、マウス胚性線維芽細胞株(Balb 3T3)、マウス骨髄(ST−2)単球/マクロファージ(RAW264.7)由来のストローマ細胞株、NHDF細胞、およびマウス線維芽細胞(LM)を含む、異なる培養細胞を分析した。
【0257】
変異体1041−D11(異なる濃度)またはED−B特異的抗体(500nM FV28 CH4/F1 1×PBS)を、Wi38細胞(60000細胞/ml、ATCC由来)と共にインキュベートし(1時間、37℃)、その後、メタノールで固定し(5分、−20℃)、ブロッキングした(5%ウマ/PBS、1時間)。その後、rabbit−a−Strep−Tag−IgG(GenScript社製 A00875、1:500)と共に1時間インキュベートし、a−rabbit−IgG*Alexa488−AK(Invitrogen社製 A11008、1:1000)と共に1時間インキュベートした。核を、DAPIで染色した。図15Aの1列目は、EDB抗体を用いたコントロールを示し、2列目は、58.7nMのタンパク質濃度での変異体のインキュベーションを示し、3列目は、10倍高濃度(587nM)の1041−D11タンパク質のインキュベーションを示し、4列目は、PBSでのネガティブコントロールである。1行目には、ヒトWi38線維芽細胞を位相差で示し、2行目には、免疫蛍光法で示し、3行目には、DAPI染色を示す。これらの写真から、変異体1041−D11は、ED−B含有細胞外マトリックスに対する高特異性で、固定Wi38細胞に結合すると結論づけられる。ネガティブコントロール細胞型NHDFは、EDBフィブロネクチンを低レベルで発現する初代の正常線維芽細胞である(データ示さず)。前記変異体は、これら細胞に結合しない。
【0258】
図15Bは、バイタル(vital)Wi38細胞での変異体1041−D11の分析を示す。ネガティブコントロール細胞型NHDFは、EDBフィブロネクチンを低レベルで発現する初代の正常線維芽細胞である。前記細胞を、チャンバースライドに播いた(NUNC、60000細胞/ml)。結合能の分析のため、前記細胞を、−20℃で5分間、100%MeOHで固定した。非特異的結合をブロックするために、前記細胞を、37℃で1時間、5%ウマ血清でインキュベートした。前記細胞を、異なる濃度の変異体1041−D11と、ポジティブコントロールであるED−B特異的抗体FV28 CH4/F1またはネガティブコントロールであるUB_2とで、室温で1時間、テストした。rabbit−a−Strep−Tag−IgG(GenScript社製 A00875、1:500)との1時間のインキュベーション、a−rabbit−IgG* Alexa488−AK(Invitrogen社製 A11008、1:1000)との1時間のインキュベーションにより、検証を行った。核を、DAPIで染色した。図15Bの1行目および3行目は、異なるタンパク質濃度の変異体およびネガティブコントロールを示す。2行目および4行目は、EDB抗体を用いたコントロールのインキュベーションを示す。最初の2行は、Wi38細胞株における前記変異体とポジティブコントロールを示す。3行目および4行目は、NHDF細胞のインキュベーションを示す。写真からわかるように、変異体1041−D11は、ED−B含有細胞外マトリックスに対する高い特異性で、バイタル(vital)Wi38細胞に結合することがわかる。低EDBを含まないNHDF細胞を用いた対照試験を行った(データ示さず)。前記変異体は、それらの細胞に結合しない。
【0259】
異なる細胞タイプ、例えば、Balb3T3(ATCC、Kat−Nr.30−2002)、Raw(Lonza社製、Kat−Nr.BE12−115F/U1)、ST−2(Lonza社製、Kat−Nr.BE12−115F/U1)を用いて、同様の実験を行った。図15CおよびDは、ED−Bの結合が、マウスBalb3T3とST−2細胞に高い特異性であることを示す。単球/マクロファージ(Raw)への結合は、見られなかった(データ示さず)。
【0260】
概要を前述したとおり、図16Aは、組織切片における1041−D11の特異性を示す。7つの検体から採取したF9腫瘍組織を評価した。500nM 1041−D11を用いた免疫組織化学から、マウス由来のF9腫瘍におけるED−B特異的血管が染色される結果が得られた。ED−Bは、腫瘍脈管系に対する高い特異性のマーカーである。標的タンパク質EDBは、血管の反管腔側に局在する。1041−D11は、F9腫瘍由来の組織切片において、血管系を特異的に装飾する。得られた結果は、抗体フラグメントL19の組織特異性に匹敵する。さらに、48組織をテストした。FDA関連パネルにおける48組織のいずれにおいても、非特異的な染色は観察されなかった。図16Bは、野生型ユビキチンと比較した、腫瘍細胞における1041−D11の蓄積を示す。このように、ED−Bに特異的に結合する修飾ユビキチンベースの融合タンパク質は、ED−Bに基づく癌の標的治療に好適である。
【0261】
[実施例7]
1041D11−TNFαのin vivoでの有効性
1041−D11−TNFαの治療上の有効性を確立するために、当該化合物をマウスモデルにおけるF9テラトーマ(Borsi et al.、2003 Blood 102, 4384−4392参照)に関してテストした。マウスにおけるED−B発現は、in vivoの状態でヒトと同等であり、1041−D11−mTNFα、好ましくはこれをメルファラン等の細胞傷害性化合物と併用した場合の癌に対する治療効果を評価する上で好適である。F9テラトーマは、高い血管密度を有する高悪性度の腫瘍である。Borsiらには、EDB抗体を介したマウスTNFαのターゲティングは、メルファランの有効性を向上させるものであり、このことは、腫瘍の成長の遅延によって実証されていると記載されている。有効性の検証のための実験計画は、Borsi、2003から採用した。
【0262】
ステージ1では、腫瘍対体重比率、体重減少、および生存率に関するエンドポイントにより、薬理的に活性かつ許容可能な投与量を規定した。本発明者らは、1041D11−TNFαが、最も高い投与量(6.75pmol/g)で許容されるが、腫瘍の成長に対して抑制効果を有さず(3、4、および8日後に体重の10%を上回る→動物死亡)、一方、最も低投与量(0.25pmol/g)の1041D11−TNFαが、腫瘍の成長を遅らせるようであることを見出した。使用したさらに別の投薬グループでは、投与量2.25pmol/g 1041D11−TNFαから、低下させた。
【0263】
研究のステージ2では、腫瘍成長の遅延をエンドポイントとするメルファランの投与量依存的な有効性を明らかにした(動物の体重減少>10%、腫瘍>体重の10%、腫瘍の潰瘍化)。この研究においては、メルファランと併用した1041D11/mTNFα(マウスTNFα)についてテストを行った。168匹の動物を使用し、14投薬グループ(各グループとも、F9腫瘍の大きさが300〜400mm3の時点で採用した8匹のマウスからなる)について、テストサンプルを静脈(i.v.)に投与し、24時間後にメルファランを腹腔内(i.p.)に注射した。表1は、投薬スケジュールを示す。
【表3】
【0264】
図18は、処置期間中(7日間)の相対的腫瘍成長を示す。図18aは、メルファランと併用した本発明の化合物1041−D11−TNFαが、メルファランと併用したmTNFαまたはメルファラン単独よりも効率的に、相対的腫瘍成長を減少させることを明確に示している。処置後7日間の腫瘍成長動態は、1041−D11−mTNFαによる腫瘍の有意な減少を示す。これは、メルファランとの併用の有効性の明らかな証拠である。
【0265】
刊行物
1. Birchler, M., F. Viti, L. Zardi, B. Spiess, and D. Neri. 1999. Selective targeting and photocoagulation of ocular angiogenesis mediated by a phage-derived human antibody fragment. Nat Biotechnol 17:984-8.
2. Brenmoehl, J., M. Lang, M. Hausmann, S. N. Leeb, W. Falk, J. Scholmerich, M. Goke, and G. Rogler. 2007. Evidence for a differential expression of fibronectin splice forms ED-A and ED-B in Crohn’s disease (CD) mucosa. Int J Colorectal Dis 22:611-23.
3. Dubin, D., J. H. Peters, L. F. Brown, B. Logan, K. C. Kent, B. Berse, S. Berven, B. Cercek, B. G. Sharifi, R. E. Pratt, and et al. 1995. Balloon catheterization induced arterial expression of embryonic fibronectins. Arterioscler Thromb Vasc Biol 15:1958-67.
4. Goodsell, D. S. 2001. FUNDAMENTALS OF CANCER MEDICINE: the Molecular Perspective: Antibodies. the Oncologist 6:547-548.
5. Kaczmarek, J., P. Castellani, G. Nicolo, B. Spina, G. Allemanni, and L. Zardi. 1994. Distribution of oncofetal fibronectin isoforms in normal, hyperplastic and neoplastic human breast tissues. Int J Cancer 59:11-6.
6. Menrad, A., and H. D. Menssen. 2005. ED-B fibronectin as a target for antibody-based cancer treatments. Expert Opin Ther Targets 9:491-500.
7. Pujuguet, P., A. Hammann, M. Moutet, J. L. Samuel, F. Martin, and M. Martin. 1996. Expression of fibronectin ED-A+ and ED-B+ isoforms by human and experimental colorectal cancer. Contribution of cancer cells and tumor-associated myofibroblasts. Am J Pathol 148:579-92.
8. Trachsel, E., M. Kaspar, F. Bootz, M. Detmar, and D. Neri. 2007. A human mAb specific to oncofetal fibronectin selectively targets chronic skin inflammation in vivo. J Invest Dermatol 127:881-6.
9. Van Vliet, A., H. J. Baelde, L. J. Vleming, E. de Heer, and J. A. Bruijn. 2001. Distribution of fibronectin isoforms in human renal disease. J Pathol 193:256-62.
10. Lipovsek, D., and Pluckthun, A. (2004). In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 290, 51-67.
11. Ohashi, H., Shimizu, Y., Ying, B.W., and Ueda, T. (2007). Efficient protein selection based on ribosome display system with purified components. Biochem Biophys. Res. Commun. 352, 270-276.
12. Studier, F.W. (2005). Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41, 207-234.
13. Zahnd, C., Amstutz, P., and Pluckthun, A. (2007). Ribosome display: selecting and evolving proteins in vitro that specifically bind to a target. Nat. Methods 4, 269-279.
【特許請求の範囲】
【請求項1】
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有する修飾ユビキチンタンパク質を少なくとも1つ含む、フィブロネクチンのED−Bドメインに結合可能なタンパク質であり、
前記フィブロネクチンのED−Bドメインに対し、Kd=10−5〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3個のアミノ酸が、置換、欠失、または付加によって修飾されたタンパク質。
【請求項2】
前記修飾により、前記タンパク質の表面に位置するアミノ酸の連続的なストレッチが形成され、
前記置換されたアミノ酸の少なくとも4個が、βシート内またはβシート鎖に隣接する3つ目までのアミノ酸位置に存在する、請求項1記載のタンパク質。
【請求項3】
前記修飾が、前記ユビキチンの2〜7位、12〜16位、41〜45位、および65〜71位の範囲にあるβシート鎖内、ならびに/または62〜64位もしくは8〜11位の範囲にあるβシート鎖に隣接する3つ目までのアミノ酸位置に存在し、
前記タンパク質が、修飾後、配列番号1で示す対応配列に対して70%以上のアミノ酸配列同一性を有し、
好ましくは、前記修飾が、前記ユビキチンタンパク質の前記βシート鎖内に位置するアミノ酸の少なくとも10%、最大で40%に対する修飾を含む、請求項1または2記載のタンパク質。
【請求項4】
2、4、6、8、62、63、64、65、66、および/または68位に位置する少なくとも4個のアミノ酸が修飾され、
さらに追加で1〜7個のアミノ酸が修飾され、
前記追加で修飾されるアミノ酸は、任意に、36、44、70、71、72、および73位の1以上のアミノ酸から選択される、請求項1から3のいずれか一項に記載のタンパク質。
【請求項5】
異なる修飾がなされた少なくとも2つのユビキチンタンパク質がヘッドトゥテイル配置で互いに結合されるか、あるいは、
少なくとも1つの修飾ユビキチンタンパク質と少なくとも1つの非修飾ユビキチンタンパク質がヘッドトゥテイル配置で互いに結合されたヘテロ多量体修飾ユビキチンタンパク質であり、
前記多量体タンパク質は、任意に二量体または三量体タンパク質である、請求項1から4のいずれか一項に記載のタンパク質。
【請求項6】
修飾多量体ユビキチンタンパク質、任意に、修飾二量体または三量体ユビキチンタンパク質を含む、フィブロネクチンのエキストラドメインB(ED−B)に結合可能なタンパク質であり、
少なくとも2つの単量体ユビキチンユニットがヘッドトゥテイル配置で互いに結合され、
前記多量体タンパク質の各単量体は、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3または4個のアミノ酸の置換により修飾され、
前記修飾単量体ユビキチンユニットが、配列番号1に対し、60%以上、85%以上、および90%以上の群から選択される少なくとも一つのアミノ酸同一性を有し、
前記タンパク質が、前記フィブロネクチンのED−Bドメインに対して、Kd=10−5〜10−12Mの特異的結合親和性を有し、前記フィブロネクチンのエキストラドメインB(ED−B)に対して一価の結合活性を示し、
前記フィブロネクチンのED−Bドメインに対し、Kd=10−5〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、各単量体ユビキチンにおいて、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3または4個のアミノ酸が、置換、欠失、または付加により修飾されているタンパク質。
【請求項7】
前記多量体が、同一または異なる修飾ユビキチンタンパク質により形成されている、請求項6記載のタンパク質。
【請求項8】
前記修飾ユビキチンタンパク質が、遺伝子融合または翻訳後融合されている、請求項7記載のタンパク質。
【請求項9】
前記ユビキチンタンパク質のヘテロ二量体である請求項6から8のいずれか一項に記載のタンパク質であり、
第1のユビキチン単量体の少なくとも6、8、63〜66位、ならびに、第2のユビキチン単量体の6、8、62〜66位、さらに任意に2位に修飾を有し、
好ましくは、K6W、L8W、K63R、E64K、S65F、T66P、ならびに、K6T、L8Q、Q62W、K63S、E64N、S65W、T66E、任意にQ2Rに修飾を有するタンパク質。
【請求項10】
配列番号47の配列を含む前記ユビキチンタンパク質のヘテロ二量体である、請求項5から8のいずれか一項に記載のタンパク質。
【請求項11】
ユビキチン単量体同士が、リンカー、好ましくは、配列番号32のリンカーにより結合されるか、またはGIG配列によって結合される、請求項7から10のいずれか一項に記載のタンパク質。
【請求項12】
配列番号33または34のユビキチンヘテロ二量体を含む、請求項10または11記載のタンパク質。
【請求項13】
薬学的活性成分および/または診断用活性成分と融合した請求項1から12のいずれか一項に記載のタンパク質を含む融合タンパク質であり、
前記薬学的活性成分が、任意にサイトカイン、ケモカイン、細胞傷害性化合物、または酵素であり、前記サイトカインは好ましくはTNFαであり、
前記診断用活性成分が、蛍光化合物、光増感剤、または放射性核種から選択される融合タンパク質。
【請求項14】
前記融合タンパク質が、多量体修飾ユビキチンタンパク質を含み、前記多量体修飾ユビキチンタンパク質が、任意にサイトカインである薬学的または診断用活性成分と任意に融合しているか、あるいは
前記タンパク質が、多量体修飾ユビキチンタンパク質を含み、前記多量体が、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成されている、請求項13記載の融合タンパク質。
【請求項15】
ユビキチンヘテロ二量体がTNFαに融合して得られる融合タンパク質の三量体である、請求項13または14記載の融合タンパク質であり、
前記融合タンパク質は、好ましくは、配列番号35、36、または47の配列を有するか、あるいは、配列番号35、36、または47の配列に対して90%以上の同一性を有する融合タンパク質。
【請求項16】
組換えタンパク質または組換え融合タンパク質である、請求項1から15のいずれか一項に記載のタンパク質または融合タンパク質。
【請求項17】
請求項1から16のいずれか一項に記載の組換えタンパク質もしくは組換え融合タンパク質、またはこれらの組み合わせと、薬学的に許容される担体とを含む医薬組成物。
【請求項18】
さらに、1以上の化学療法剤を含み、
前記化学療法剤が、好ましくは、メルファラン、ドキソルビシン、シクロフォスファミド、ダクチノマイシン、フルオロデオキシウラシル、シスプラチン、パクリタキセル、およびゲムシタビンから選択されるか、あるいはキナーゼ阻害剤群、または放射性医薬品から選択される、請求項17記載の医薬組成物。
【請求項19】
混合製剤の形態、好ましくは、複数の部品からなるキットの形態である、請求項18記載の医薬組成物。
【請求項20】
請求項1から16のいずれか一項に記載の組換えタンパク質または融合タンパク質をコードするポリヌクレオチド。
【請求項21】
請求項20記載のポリヌクレオチドを含むベクター。
【請求項22】
請求項1から4のいずれか一項に記載のタンパク質、
請求項5から12のいずれか一項に記載のタンパク質、
請求項13から16のいずれか一項に記載の融合タンパク質、
請求項21記載のベクター、および/または
請求項19記載のポリヌクレオチドを含む、宿主細胞。
【請求項23】
請求項1から4のいずれか一項に記載の組換えタンパク質、請求項5から12のいずれか一項に記載の多量体タンパク質、または請求項13から16のいずれか一項に記載の融合タンパク質と、
診断上許容される担体とを含む、診断剤。
【請求項24】
以下の工程を含む、請求項1から4のいずれか一項に記載の組換えタンパク質の製造方法。
a)ユビキチンタンパク質を提供する工程
b)フィブロネクチンのED−Bドメインを提供する工程
c)配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために前記ユビキチンタンパク質を修飾する工程であり、
2、4、6、8、62、63、64、65、66、および/または68位における、少なくとも3または4個のアミノ酸を、置換、欠失、または付加によって修飾する工程
d)前記修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bドメインと接触させる工程
e)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bドメインに結合する修飾ユビキチンタンパク質をスクリーニングする工程
f)前記修飾ユビキチンタンパク質を単離する工程
【請求項25】
前記修飾工程が化学的合成工程を含む、請求項24記載の方法。
【請求項26】
前記修飾が、DNAレベルの遺伝子操作によって行われ、
前記タンパク質の発現が、原核もしくは真核生物内、またはin vitroで行われる、請求項24または25記載の方法。
【請求項27】
請求項24から26のいずれか一項に記載の方法であり、
前記組換えタンパク質を、任意にサイトカイン、好ましくはTNFαである薬学的活性成分、または診断用成分と融合させるか、
前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合させるか、あるいは
前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体、二量体または三量体を、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成させる方法。
【請求項28】
以下の工程を含む、請求項5から14のいずれか一項に記載のヘテロ多量体融合タンパク質の製造方法。
a)直接または好適なリンカーを介して結合された2以上の修飾ユビキチン単量体を含む多量体ユビキチンタンパク質を提供する工程であり、
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために、前記多量体ユビキチンタンパク質の各単量体を修飾し、各単量体において、2、4、6、62、63、64、65、66、および/または68位における、少なくとも3または4個のアミノ酸を、置換、欠失、または付加によって修飾する工程
b)フィブロネクチンのED−Bを提供する工程
c)前記ヘテロ多量体修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
d)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bに結合する修飾ユビキチンタンパク質をスクリーニングする工程
e)前記修飾ヘテロ多量体ユビキチンタンパク質を単離する任意の工程
【請求項29】
前記ヘテロ多量体ユビキチンタンパク質がヘテロ二量体ユビキチンタンパク質である、請求項28記載の方法。
【請求項30】
医学的治療方法に使用するための、請求項1から のいずれか一項に記載の組換えタンパク質、請求項5から12のいずれか一項に記載の多量体タンパク質、請求項13から16のいずれか一項に記載の融合タンパク質。
【請求項31】
請求項5から16の一項以上に規定する修飾多量体ユビキチンタンパク質またはユビキチン融合タンパク質の直鎖を含み、少なくとも2つの相互作用する結合決定領域(BDR)においてランダム化されているライブラリー。
【請求項1】
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有する修飾ユビキチンタンパク質を少なくとも1つ含む、フィブロネクチンのED−Bドメインに結合可能なタンパク質であり、
前記フィブロネクチンのED−Bドメインに対し、Kd=10−5〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3個のアミノ酸が、置換、欠失、または付加によって修飾されたタンパク質。
【請求項2】
前記修飾により、前記タンパク質の表面に位置するアミノ酸の連続的なストレッチが形成され、
前記置換されたアミノ酸の少なくとも4個が、βシート内またはβシート鎖に隣接する3つ目までのアミノ酸位置に存在する、請求項1記載のタンパク質。
【請求項3】
前記修飾が、前記ユビキチンの2〜7位、12〜16位、41〜45位、および65〜71位の範囲にあるβシート鎖内、ならびに/または62〜64位もしくは8〜11位の範囲にあるβシート鎖に隣接する3つ目までのアミノ酸位置に存在し、
前記タンパク質が、修飾後、配列番号1で示す対応配列に対して70%以上のアミノ酸配列同一性を有し、
好ましくは、前記修飾が、前記ユビキチンタンパク質の前記βシート鎖内に位置するアミノ酸の少なくとも10%、最大で40%に対する修飾を含む、請求項1または2記載のタンパク質。
【請求項4】
2、4、6、8、62、63、64、65、66、および/または68位に位置する少なくとも4個のアミノ酸が修飾され、
さらに追加で1〜7個のアミノ酸が修飾され、
前記追加で修飾されるアミノ酸は、任意に、36、44、70、71、72、および73位の1以上のアミノ酸から選択される、請求項1から3のいずれか一項に記載のタンパク質。
【請求項5】
異なる修飾がなされた少なくとも2つのユビキチンタンパク質がヘッドトゥテイル配置で互いに結合されるか、あるいは、
少なくとも1つの修飾ユビキチンタンパク質と少なくとも1つの非修飾ユビキチンタンパク質がヘッドトゥテイル配置で互いに結合されたヘテロ多量体修飾ユビキチンタンパク質であり、
前記多量体タンパク質は、任意に二量体または三量体タンパク質である、請求項1から4のいずれか一項に記載のタンパク質。
【請求項6】
修飾多量体ユビキチンタンパク質、任意に、修飾二量体または三量体ユビキチンタンパク質を含む、フィブロネクチンのエキストラドメインB(ED−B)に結合可能なタンパク質であり、
少なくとも2つの単量体ユビキチンユニットがヘッドトゥテイル配置で互いに結合され、
前記多量体タンパク質の各単量体は、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3または4個のアミノ酸の置換により修飾され、
前記修飾単量体ユビキチンユニットが、配列番号1に対し、60%以上、85%以上、および90%以上の群から選択される少なくとも一つのアミノ酸同一性を有し、
前記タンパク質が、前記フィブロネクチンのED−Bドメインに対して、Kd=10−5〜10−12Mの特異的結合親和性を有し、前記フィブロネクチンのエキストラドメインB(ED−B)に対して一価の結合活性を示し、
前記フィブロネクチンのED−Bドメインに対し、Kd=10−5〜10−12Mの特異的結合親和性で検出可能な結合を示す修飾ユビキチンタンパク質を得るため、各単量体ユビキチンにおいて、配列番号1の2、4、6、8、62、63、64、65、66、および68位における、少なくとも3または4個のアミノ酸が、置換、欠失、または付加により修飾されているタンパク質。
【請求項7】
前記多量体が、同一または異なる修飾ユビキチンタンパク質により形成されている、請求項6記載のタンパク質。
【請求項8】
前記修飾ユビキチンタンパク質が、遺伝子融合または翻訳後融合されている、請求項7記載のタンパク質。
【請求項9】
前記ユビキチンタンパク質のヘテロ二量体である請求項6から8のいずれか一項に記載のタンパク質であり、
第1のユビキチン単量体の少なくとも6、8、63〜66位、ならびに、第2のユビキチン単量体の6、8、62〜66位、さらに任意に2位に修飾を有し、
好ましくは、K6W、L8W、K63R、E64K、S65F、T66P、ならびに、K6T、L8Q、Q62W、K63S、E64N、S65W、T66E、任意にQ2Rに修飾を有するタンパク質。
【請求項10】
配列番号47の配列を含む前記ユビキチンタンパク質のヘテロ二量体である、請求項5から8のいずれか一項に記載のタンパク質。
【請求項11】
ユビキチン単量体同士が、リンカー、好ましくは、配列番号32のリンカーにより結合されるか、またはGIG配列によって結合される、請求項7から10のいずれか一項に記載のタンパク質。
【請求項12】
配列番号33または34のユビキチンヘテロ二量体を含む、請求項10または11記載のタンパク質。
【請求項13】
薬学的活性成分および/または診断用活性成分と融合した請求項1から12のいずれか一項に記載のタンパク質を含む融合タンパク質であり、
前記薬学的活性成分が、任意にサイトカイン、ケモカイン、細胞傷害性化合物、または酵素であり、前記サイトカインは好ましくはTNFαであり、
前記診断用活性成分が、蛍光化合物、光増感剤、または放射性核種から選択される融合タンパク質。
【請求項14】
前記融合タンパク質が、多量体修飾ユビキチンタンパク質を含み、前記多量体修飾ユビキチンタンパク質が、任意にサイトカインである薬学的または診断用活性成分と任意に融合しているか、あるいは
前記タンパク質が、多量体修飾ユビキチンタンパク質を含み、前記多量体が、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成されている、請求項13記載の融合タンパク質。
【請求項15】
ユビキチンヘテロ二量体がTNFαに融合して得られる融合タンパク質の三量体である、請求項13または14記載の融合タンパク質であり、
前記融合タンパク質は、好ましくは、配列番号35、36、または47の配列を有するか、あるいは、配列番号35、36、または47の配列に対して90%以上の同一性を有する融合タンパク質。
【請求項16】
組換えタンパク質または組換え融合タンパク質である、請求項1から15のいずれか一項に記載のタンパク質または融合タンパク質。
【請求項17】
請求項1から16のいずれか一項に記載の組換えタンパク質もしくは組換え融合タンパク質、またはこれらの組み合わせと、薬学的に許容される担体とを含む医薬組成物。
【請求項18】
さらに、1以上の化学療法剤を含み、
前記化学療法剤が、好ましくは、メルファラン、ドキソルビシン、シクロフォスファミド、ダクチノマイシン、フルオロデオキシウラシル、シスプラチン、パクリタキセル、およびゲムシタビンから選択されるか、あるいはキナーゼ阻害剤群、または放射性医薬品から選択される、請求項17記載の医薬組成物。
【請求項19】
混合製剤の形態、好ましくは、複数の部品からなるキットの形態である、請求項18記載の医薬組成物。
【請求項20】
請求項1から16のいずれか一項に記載の組換えタンパク質または融合タンパク質をコードするポリヌクレオチド。
【請求項21】
請求項20記載のポリヌクレオチドを含むベクター。
【請求項22】
請求項1から4のいずれか一項に記載のタンパク質、
請求項5から12のいずれか一項に記載のタンパク質、
請求項13から16のいずれか一項に記載の融合タンパク質、
請求項21記載のベクター、および/または
請求項19記載のポリヌクレオチドを含む、宿主細胞。
【請求項23】
請求項1から4のいずれか一項に記載の組換えタンパク質、請求項5から12のいずれか一項に記載の多量体タンパク質、または請求項13から16のいずれか一項に記載の融合タンパク質と、
診断上許容される担体とを含む、診断剤。
【請求項24】
以下の工程を含む、請求項1から4のいずれか一項に記載の組換えタンパク質の製造方法。
a)ユビキチンタンパク質を提供する工程
b)フィブロネクチンのED−Bドメインを提供する工程
c)配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために前記ユビキチンタンパク質を修飾する工程であり、
2、4、6、8、62、63、64、65、66、および/または68位における、少なくとも3または4個のアミノ酸を、置換、欠失、または付加によって修飾する工程
d)前記修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bドメインと接触させる工程
e)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bドメインに結合する修飾ユビキチンタンパク質をスクリーニングする工程
f)前記修飾ユビキチンタンパク質を単離する工程
【請求項25】
前記修飾工程が化学的合成工程を含む、請求項24記載の方法。
【請求項26】
前記修飾が、DNAレベルの遺伝子操作によって行われ、
前記タンパク質の発現が、原核もしくは真核生物内、またはin vitroで行われる、請求項24または25記載の方法。
【請求項27】
請求項24から26のいずれか一項に記載の方法であり、
前記組換えタンパク質を、任意にサイトカイン、好ましくはTNFαである薬学的活性成分、または診断用成分と融合させるか、
前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体を、任意にサイトカインである薬学的活性成分、または診断用成分と任意に融合させるか、あるいは
前記組換えタンパク質を、少なくとも1つの第2の組換えユビキチンタンパク質と融合させて前記組換えユビキチンタンパク質の多量体、任意に二量体または三量体とし、この多量体、二量体または三量体を、任意にTNFαである前記薬学的活性成分、または前記診断用成分を介して形成させる方法。
【請求項28】
以下の工程を含む、請求項5から14のいずれか一項に記載のヘテロ多量体融合タンパク質の製造方法。
a)直接または好適なリンカーを介して結合された2以上の修飾ユビキチン単量体を含む多量体ユビキチンタンパク質を提供する工程であり、
配列番号1のアミノ酸配列に対して60%以上のアミノ酸配列同一性を有するタンパク質を得るために、前記多量体ユビキチンタンパク質の各単量体を修飾し、各単量体において、2、4、6、62、63、64、65、66、および/または68位における、少なくとも3または4個のアミノ酸を、置換、欠失、または付加によって修飾する工程
b)フィブロネクチンのED−Bを提供する工程
c)前記ヘテロ多量体修飾ユビキチンタンパク質を、前記フィブロネクチンのED−Bと接触させる工程
d)10−5〜10−12Mの特異的結合親和性で前記フィブロネクチンのED−Bに結合する修飾ユビキチンタンパク質をスクリーニングする工程
e)前記修飾ヘテロ多量体ユビキチンタンパク質を単離する任意の工程
【請求項29】
前記ヘテロ多量体ユビキチンタンパク質がヘテロ二量体ユビキチンタンパク質である、請求項28記載の方法。
【請求項30】
医学的治療方法に使用するための、請求項1から のいずれか一項に記載の組換えタンパク質、請求項5から12のいずれか一項に記載の多量体タンパク質、請求項13から16のいずれか一項に記載の融合タンパク質。
【請求項31】
請求項5から16の一項以上に規定する修飾多量体ユビキチンタンパク質またはユビキチン融合タンパク質の直鎖を含み、少なくとも2つの相互作用する結合決定領域(BDR)においてランダム化されているライブラリー。
【図1】

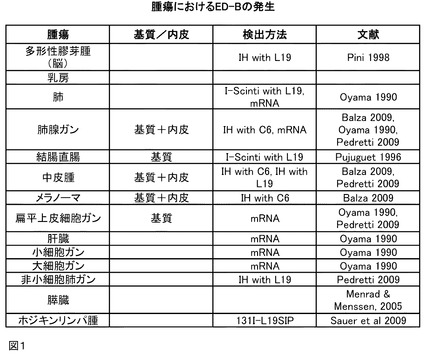
【図4A】
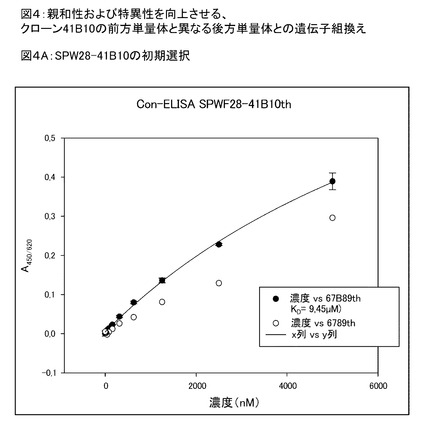
【図4B】
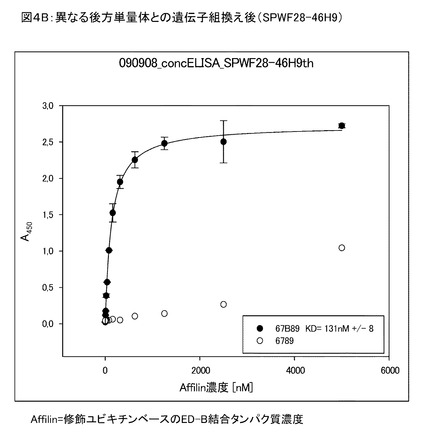
【図5B】
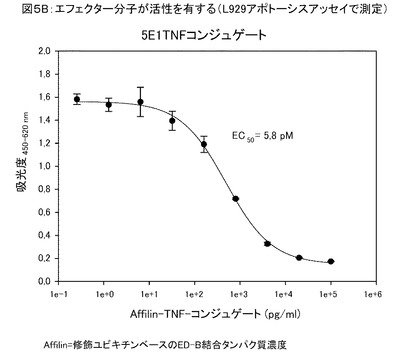
【図5C】
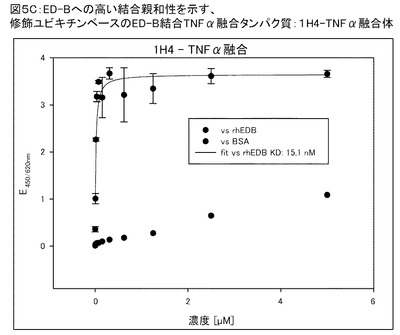
【図6】

【図6A】
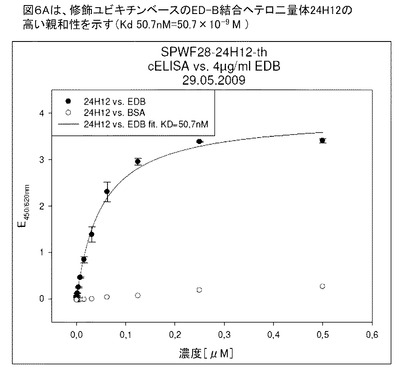
【図6B】
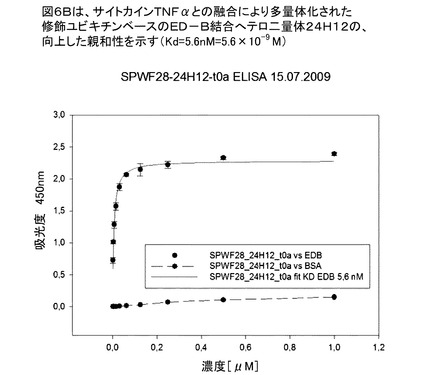
【図6C】
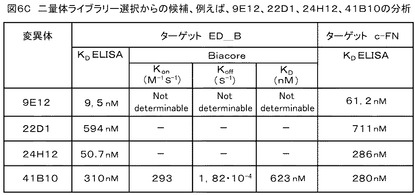
【図11】
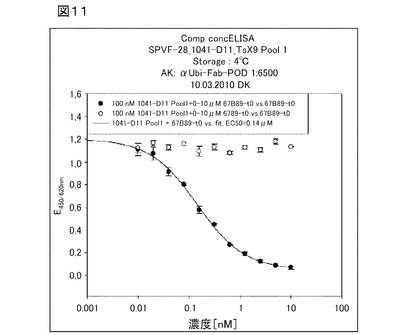
【図17A】
【図17B】
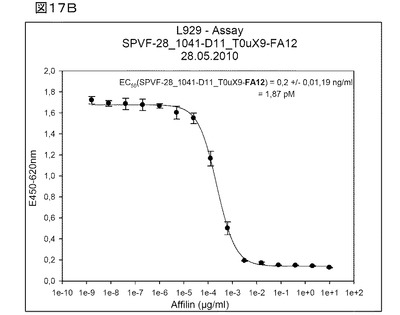
【図17C】
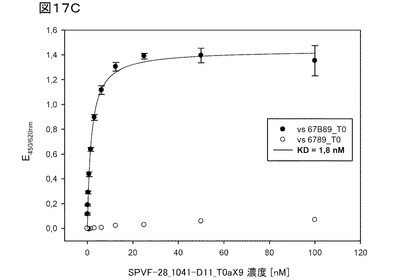
【図18】
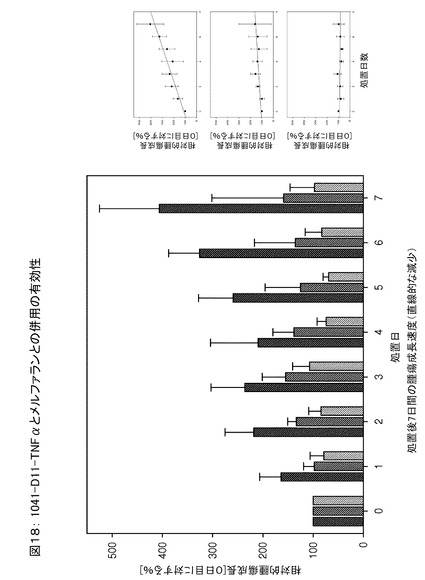
【図2】

【図3】
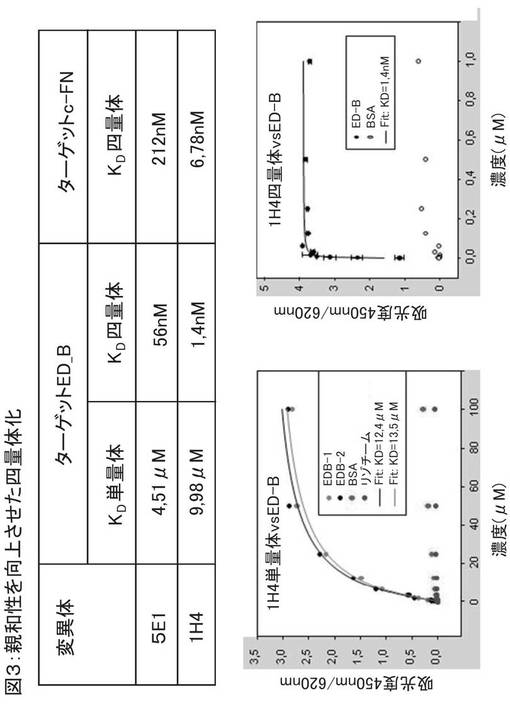
【図5A】

【図6D】
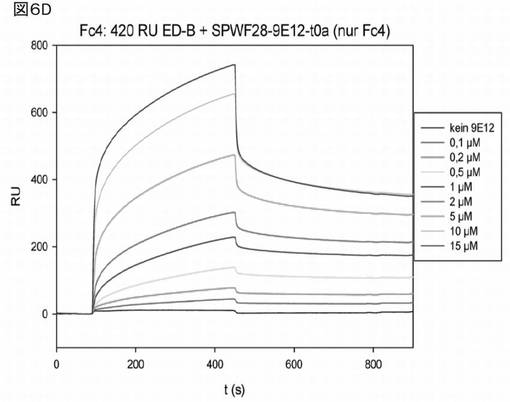
【図6E】
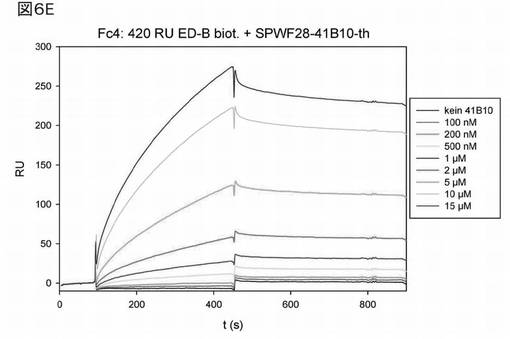
【図7】
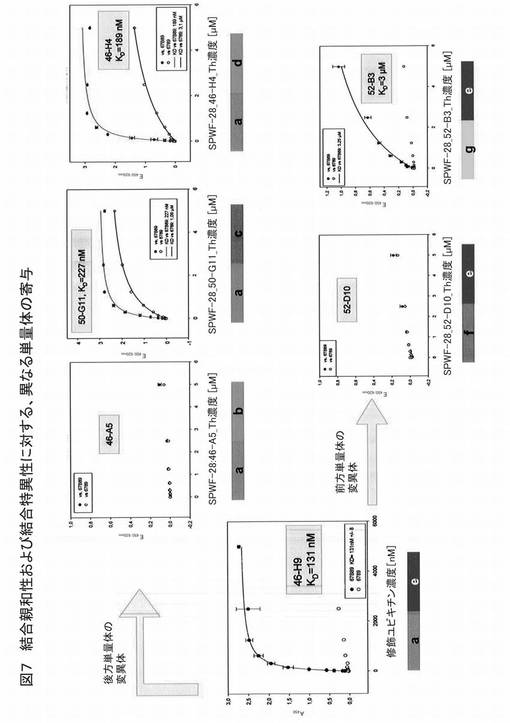
【図8】

【図9】

【図10】
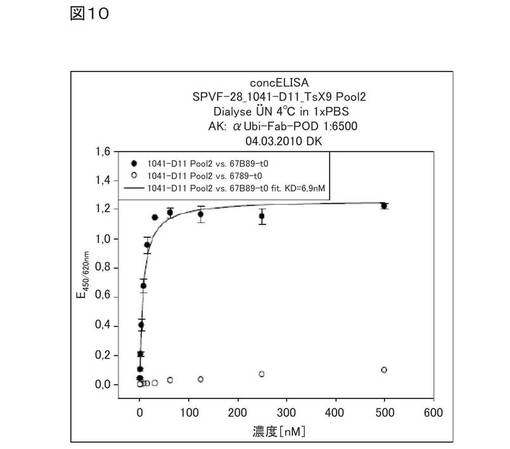
【図12】
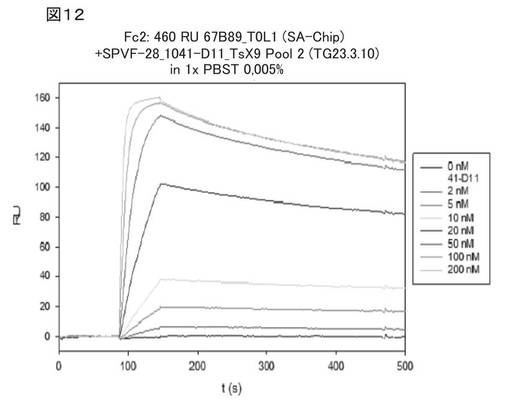
【図13】
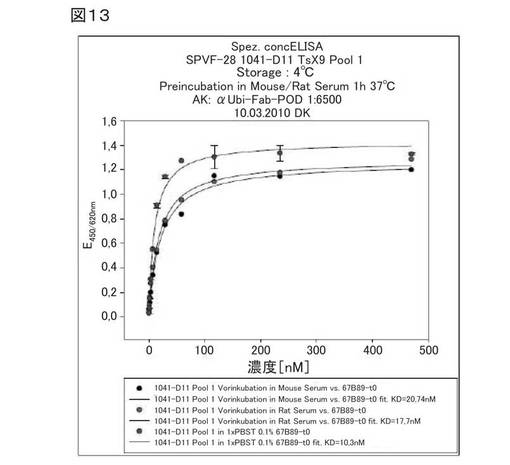
【図14】
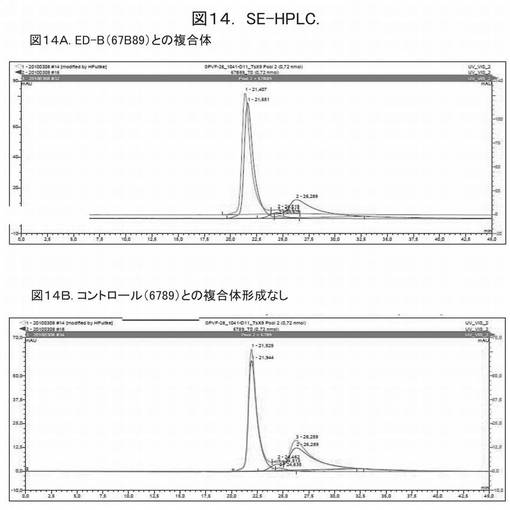
【図15A】

【図15B】
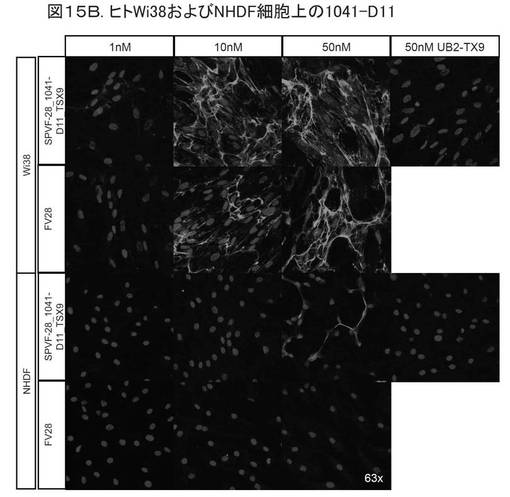
【図15C】
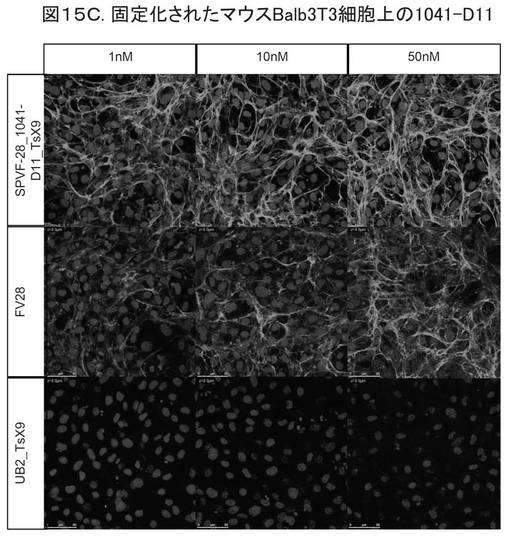
【図15D】
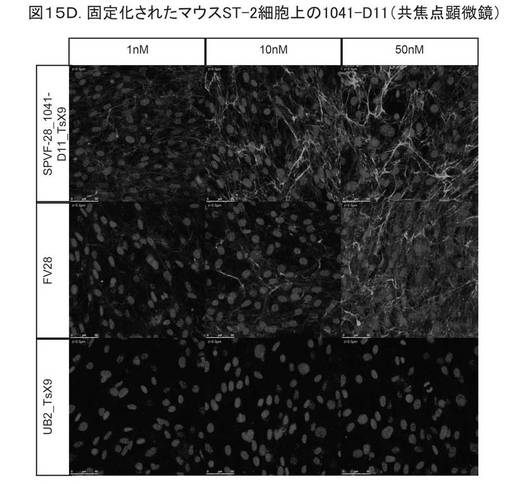
【図16A】

【図16B】
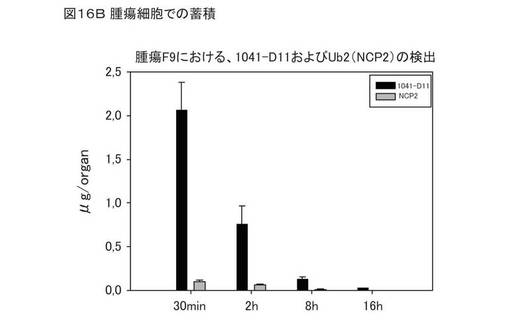
【図17D】
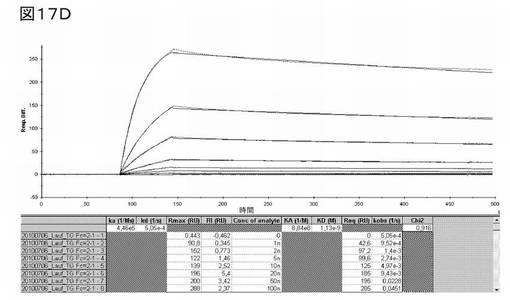
【図17E】
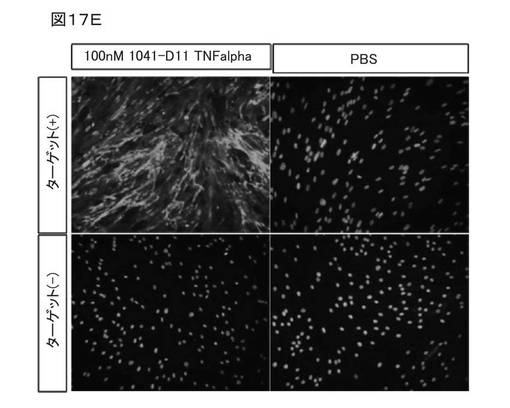
【図19】

【図4A】
【図4B】
【図5B】
【図5C】
【図6】
【図6A】
【図6B】
【図6C】
【図11】
【図17A】
【図17B】
【図17C】
【図18】
【図2】
【図3】
【図5A】
【図6D】
【図6E】
【図7】
【図8】
【図9】
【図10】
【図12】
【図13】
【図14】
【図15A】
【図15B】
【図15C】
【図15D】
【図16A】
【図16B】
【図17D】
【図17E】
【図19】
【公表番号】特表2013−513375(P2013−513375A)
【公表日】平成25年4月22日(2013.4.22)
【国際特許分類】
【出願番号】特願2012−542582(P2012−542582)
【出願日】平成22年12月14日(2010.12.14)
【国際出願番号】PCT/EP2010/069666
【国際公開番号】WO2011/073209
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(501139870)シル プロテインズ ゲーエムベーハー (7)
【氏名又は名称原語表記】Scil Proteins GmbH
【住所又は居所原語表記】Heinrich−Damerow−Str. 01, D−06120, Halle, Germany
【Fターム(参考)】
【公表日】平成25年4月22日(2013.4.22)
【国際特許分類】
【出願日】平成22年12月14日(2010.12.14)
【国際出願番号】PCT/EP2010/069666
【国際公開番号】WO2011/073209
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(501139870)シル プロテインズ ゲーエムベーハー (7)
【氏名又は名称原語表記】Scil Proteins GmbH
【住所又は居所原語表記】Heinrich−Damerow−Str. 01, D−06120, Halle, Germany
【Fターム(参考)】
[ Back to top ]