
フィブロネクチンのED―Bドメインに対する抗体、それらの構造及び用途
【課題】腫瘍関連抗原を標的にすることによって固形腫瘍を治療する抗体を提供する。
【解決手段】フィブロネクチン(FN)は、細胞外マトリックス及び体液の多機能性高分子量の糖蛋白成分である。正常細胞の定着(establishment)及び維持、細胞移動、うっ血、血栓症、創傷の治癒、腫瘍原性(oncogenic)転換等、多くの異なる生物過程に関与する、フィブロネクチン(FN)のED−B腫瘍胎児ドメインに対して特異的であり、且つその腫瘍胎児ドメインに直接結合する特異性結合抗体又は抗体断片でありそのような結合メンバを製造するための物質及び方法。
【解決手段】フィブロネクチン(FN)は、細胞外マトリックス及び体液の多機能性高分子量の糖蛋白成分である。正常細胞の定着(establishment)及び維持、細胞移動、うっ血、血栓症、創傷の治癒、腫瘍原性(oncogenic)転換等、多くの異なる生物過程に関与する、フィブロネクチン(FN)のED−B腫瘍胎児ドメインに対して特異的であり、且つその腫瘍胎児ドメインに直接結合する特異性結合抗体又は抗体断片でありそのような結合メンバを製造するための物質及び方法。
【発明の詳細な説明】
【技術分野】
【0001】
背景
本発明は、フィブロネクチン、ED−B(これはまた、免疫細胞学と生体内(in vivo)腫瘍を標的にすることとの両方によって実証されている通り、成長している腫瘍の新生血管形成(neovasculature)において滲出する)の胎児イソ型(foetal isoform)のための特殊な結合剤に関する。本発明はまた、かかる特殊な結合剤に係る物質及び方法にも関する。
【背景技術】
【0002】
腫瘍治療の現行技術の大抵のものの第一の狙いは、腫瘍のできるだけ多くの構成細胞を殺すことにある。化学療法及び放射線療法を用いて経験してきた成功が限られているのは、治療の特殊性(specificity,専門性)が比較的欠如していること、及び正常組織に及ぼす有毒性副作用の傾向があることと関係がある。腫瘍のための治療の選択性を改善し得る一つの方法は、結合タンパク質を通して、腫瘍へ物質(agent,活性物質)を運ぶことである。この結合タンパク質は通常、腫瘍の表面上に滲出した標識抗原(marker antigen)のための特殊性を有する、抗体結合ドメイン(binding domain,結合領域)から成る。俗に「マジック弾(magic bullets)」と呼ばれている、標的にされた腫瘍のかかる形態は、げっ歯類(rodents)からの単クローン性抗体(mAbs)によって、主に実証されてきた。mAbは、細胞表面上に滲出したいわゆる腫瘍関連抗原に対して特異性がある。そのようなmAbは、細胞毒部分(例えば、毒素又は薬物)へ化学的に結合していても良いし、或いは組換え融合蛋白質として産生しても良い。ここに、mAbを記号化した遺伝子と毒素とは一緒に連結され、縦列に滲出される。
【0003】
「マジック弾」のアプローチは、ヒト・癌の治療において重要であり有効であり、リンパ起始(lymphoid origin)の腫瘍を標的にする場合に最も注目されている(ここに、悪性細胞は循環中の治効量に最も影響を受けやすい)が、このアプローチには限界がある。しかし、固体腫瘍の治療は、全細胞塊の微小部分(腫瘍の最も外側の周辺の細胞が支配的である)のみが循環中の治療的免疫接合(therapeutic immunoconjugates)にさらされるという点で、重大な臨床的問題を残している;これらの周辺標的は、腫瘍内部に対していわゆる「結合部障壁(binding site barrier)」を形成する(非特許文献1)。腫瘍内部における組織の構造は、一般的に線維支質を有し、あまりにも稠密であり、腫瘍細胞に密に包み込まれているため、抗体の大きさ範囲の分子は浸透することができない。更に、腫瘍は、リンパ排液が欠如しているため、大きな間隙圧力を有することが知られている。腫瘍はまた、外因性分子の流入を妨げる。腫瘍内への治療薬の取込みに影響を及ぼす諸因子に関する最近の報告については、非特許文献2を参照。
【0004】
腫瘍関連抗原を標的にすることによって固形腫瘍を治療することには明らかに限界があるものの、これらの腫瘍は、抗体を治療するために代替的な抗原を標的にするという共通の特徴を有するのである。一旦、腫瘍がある大きさを越えて成長したとき、腫瘍は一般的に、成長を支えるための十分な酸素と養分とを、独立した血液供給に依存する。もし、この血液供給を妨げるか、又は遮ることができるのなら、過程において、現実的に何千もの腫瘍細胞を餓死させることができる。腫瘍が成長するにつれて、腫瘍は、隣接した毛管内皮細胞に作用する脈管由来諸因子の別種の配列を生じさせ、それらを誘導して(inducing)増殖し移動しながら、スウィッチを受けて脈管由来表現型になる。これらの新たに形成された血管の構造は、正常組織の秩序のある構造とは著しく対照的に、出口のない末端(blind endings)と漏出の増大に通じる穿孔とを有し、高度に崩壊している。血管形成の誘導は、ある細胞表面の抗原の表現のアップレギュレーション(upreglation)によって成し遂げられ、それらの多くは正常組織の脈管構造に共通している。
【0005】
腫瘍の新生血管形成に特有な抗原を識別することは、血管を標的にすることによる、固形腫瘍の一般的療法を開発する上での主要な限定因子(limiting factor)であった。本発明の主題である抗原は、かかる問題に直接向いている。
【0006】
腫瘍が進行している間、周囲組織の細胞外マトリックスは、二つの主要過程、即ち、(1)正常組織の細胞外マトリックス成分のタンパク分解の低下、及び(2)腫瘍細胞と、腫瘍誘導された(tumour−induced)シトキンによって活性化された支質細胞とによる、細胞外マトリックス成分のドゥノボ(de novo)合成によって、再構築される。これらの二つの過程は、安定な状態で、「腫瘍細胞外マトリックス(tumoral extracellular matrix)」を生じさせる。この腫瘍細胞外マトリックスは、腫瘍の進行にとって一層適した環境を与え、しかも定性的かつ定量的に正常組織のそれと全く異なっている。このマトリックスの諸成分の中には、テナシン及びフィブロネクチン(FN)のイソ型が多量存在する。これらの蛋白質のイソ型としての記述(description)から、それらの構造上の大きな異質性が認識される。この構造上の異質性は、転写レベル、転写後レベル及び翻訳後レベルでもたらされる。本発明の主題は、フィブロネクチンのイソ型の一種の、いわゆるB+イソ型(B−FN)である。
【0007】
フィブロネクチン(FN)は、細胞外マトリックス及び体液の多機能性高分子量の糖蛋白成分である。これらの成分は、正常細胞の定着(establishment)及び維持、細胞移動、うっ血、血栓症、創傷の治癒、腫瘍原性(oncogenic)転換等、多くの異なる生物過程に関与している(報告に関しては、非特許文献3〜6;ハイネス,1990;非特許文献7を参照)。FNsの構造上の相違は、FN一次転写物の三つの領域(ED−A、ED−B及びIIICS)によってもたらされ(非特許文献5及び8)、少なくとも20種の異なるイソ型が生じる。これらイソ型の幾つかは、腫瘍及び正常組織では区別して表現される。FN−プレ−mRNAのスプライシングパターンは、進行中のものに特異的な組織態様に調整されるだけではなく、移動済み細胞及び悪性疾患の中で調整解除されることが知られている(非特許文献8〜16)。実際、ED−A、ED−B及びIIICSの配列を有するFNイソ型は、大部分、正常細胞中よりも移動済み細胞及び悪性腫瘍細胞の中に表現される。特に、ED−B配列(B+イソ型)を有するFNイソ型は、創傷治癒の間だけではなく、胎児組織及び腫瘍組織の中に高度に表現される。しかし、成人の正常組織中では表現が制限される(非特許文献13及び17〜23)。B+FNの分子は、成熟した血管中では検知できないが、正常な発育(例えば、子宮内膜の発育)、病の進行(例えば、糖尿病性の網膜障害の)及び腫瘍の進行の状態にある脈管由来の血管中では、アップレギュレーションとなる(非特許文献24)。
【0008】
ED−B配列は、単エクソンによってコード化された完全型III−相同(homology)の繰り返しであり、91種のアミノ酸から成る。細胞型特異性結合部を有する、代替的スプライシングされたIIICSイソ型とは対照的に、A+イソ型及びB+イソ型の生物機能は、まだ推測の域を出ていない(非特許文献25)。
【0009】
B+イソ型の存在自体は、腫瘍誘導された新生抗原を構成する。しかし、加えるに、ED−B表現は、III型の繰り返し7以内の、正常に潜在する(normally cryptic)抗原を露出する(前のED−B);このエピトープはFN分子欠乏ED−Bの中に露出されないので、ED−B表現は直接的にも間接的にも新生抗原の表現を誘導することとなる。この潜在的抗原の部位は、BC−1と呼ばれる単クローン性抗体(mAb)の標的を形成する(非特許文献26)。このmAbの特異性と生物諸特性は、特許文献1に記載されており、イギリス(UK),ソールズベリー(Salisbury),ポートンダウン(Porton Down),動物細胞培養欧州共同体(the European Collection of Animal Cell Cultures)に委託されている融合細胞腫(番号88042101)から入手することができる。mAbは、成熟した血管内皮への交差反応性なしで、腫瘍の脈管由来の血管を一局部に止めるのに使用され成功してきており、抗体を使用して血管を標的にするためのFNイソ型の可能性を説明している。
【0010】
しかし、BC−1 mAbの特異性に対する幾つかの注意点が残っている。BC−1がB+イソ型を直接的に認識しないという事実により、幾つかの組織において、BC−1によって認識されるエピトープはED−Bが存在しなくても暴露され(unmasked)、その結果、不要なBC−1の交差反応性へ間接的に導き得るかどうかという疑問が生じた。また、BC−1は、ヒト・B+イソ型には厳密に特異的であり、生物分布(biodistribution)及びBC−1の腫瘍局在化に関する、動物の研究はできないということを意味する。B+イソ型を有する組換え融合蛋白に対する多クローン性抗体は生成されてきた(非特許文献27)が、それら抗体は、エピトープを暴露するためのN−グリカナーゼ(N−glycanase)で処理されたFNと反応するのみである。
【0011】
マウス・単クローン性抗体を使用する上での更なる一般的問題は、ヒト・抗マウス抗体(human anti−mouse antibody)(HAMA)の反応である(非特許文献28及び29)。HAMA反応は、治療量の減量に役立つ、投与済み抗体の中和から、アレルギー反応、血清病及び腎臓障害まで、ある範囲の効果を有する。
【0012】
組換えED−Bと反応する多クローン性抗血清は確認されてきた(上記を参照)が、マウスの免疫化の結果として生じるBC−1と同一の特異性を有するmAbの分離は、一般的に困難であることが分かった。なぜなら、ヒト及びマウスのED−B蛋白は、ほぼ100%の配列相同を示すからである。従って、ヒト・蛋白は、それへの免疫反応を備えていないマウスにとっては自己抗原であるように見えるかも知れない。実際、何十年にも渡るこの分野の徹底した研究において、ED−Bを直接的に認識するものは何もなく、単mAbのみがB+FNイソ型(BC−1)に対する間接的反応性を有するものとして認定されてきた。BC−1の特異性は、ED−B自体の部分のためというよりむしろ、潜在的エピトープのために、ED−Bの結果として暴露されるということは間違いなく意義がある。
ED−Bは、マウスFNが恐らく欠けていて、マウスの免疫系による「自己」と見られないものと思われる。
【0013】
本発明は、以前使用されたものに対する代替的方法を使用して実現された。ここでは、フィブロネクチン又はED−Bによる従来の免疫化は必要でない:ED−Bイソ型に対する特異性を有する抗体は、線維状バクテリオファージの表面に表示される、ヒト・抗体の種々の領域のライブラリ(libraries)から単鎖Fvs(scFvs)として得られてきた(非特許文献30及び特許文献2〜6をも参照)。
【0014】
ED−Bを有する組換えFN−断片(fragments)と、組換えED−B自体との両者の上に直接選択することにより、特異性scFvが分離され得ることを、我々は、抗体ファージライブラリ(antibody phage library)を使用することによって見いだした。このとき、これら抗体は固形表面の上に被覆される(「パンニング(panning)」)。抗原のこれら同一源もまた、「二代雑種第二子世代(second generation)」のscFvを生成するのに使用され成功し、「親和力成熟(affinity maturation)」の過程で、親クローンに関する諸特性が改善されてきた。分離されたscFvは、N−グリカナーゼで前処理をしないで、ヒト・FNのB+イソ型と強く且つ特異的に反応することを我々は見いだした。
【0015】
抗腫瘍の適用において、本発明によって提供されるヒト・抗体抗原結合ドメインは、HAMA反応を受けないという利点を有する。また、本明細書中に例証される通り、これら結合ドメインは、生体外でも生体内でも、腫瘍組織の免疫組織化学分析において有用である。これらの用途及び他の用途は、本明細書中で更に論じられ、当業者には容易に理解できる。
【0016】
術語
特異的結合メンバ(specific binding member)
特異性結合メンバとは、相互に対して結合特異性を有する一対の分子のメンバをいう。特異性結合ペアメンバは、自然界で得られるが、全部又は一部を合成によって生成することができる。一対の分子の一つのメンバは、その表面上に、領域又はキャビティ(cavity,空洞)を有する。この領域又はキャビティは、その一対の分子の他のメンバに特異的に結合し、それ故に、その極性空隙機構に対して相補的である。このように、ペアメンバは相互に特異的に結合する特性を有する。特異性結合メンバのタイプの例は、抗原−抗体、ビオチンーアビジン、ホルモン−ホルモン受容体、受容体−リガンド、酵素−基質である。
【0017】
抗体(antibody)
抗体とは、自然界に存在し、又は一部若しくは全部を合成によって生成される免疫グロブリンをいう。この術語には、結合ドメインを有するあらゆるポリペプチド又は蛋白質も含まれる。この結合ドメインは、抗体の結合ドメインであるか、又は抗体の結合ドメインと同種のものである。これらは、自然源から得ることができるが、一部又は全部を合成によって生成することができる。抗体の例は、免疫グロブリンのイソ型(isotypes)及びこれらイソ型の亜綱(subclasses);Fab,scFv,Fv,dAb,Fd等の抗原結合ドメインから成る断片;並びにダイヤボディ(diabodies)である。
【0018】
単クローン抗体及び他の抗体を利用し、組換えDNA技術の技法を使用して、原抗体の特異性を保持する、他の抗体又はキメラ(chimeric)分子を生成することは可能である。そのような技法には、免疫グロブリンの不変領域又は不変領域プラス外郭構造に対する抗体の、免疫グロブリン可変領域又は相補性決定領域(CDRs)をコード化するDNAを導入することを伴うこともある。例えば、特許文献7〜9を参照。抗体を生成する融合細胞腫又は他の細胞は、遺伝的変異(genetic mutation)又は他の変化を受けることもある。これらの変化によって、生成される諸抗体の結合特異性を変えても良いし変えなくても良い。
【0019】
抗体は多くの方法によって変性することができるのであるから、用語「抗体」は、必要とされる特異性を持つ結合ドメインを有する、あらゆる特異性結合メンバ又は物質と解釈されるべきである。このように、この用語は、抗体断片、誘導体、機能的に同等の物、及び抗体の同族体を意味し、自然界で得られようと全部又は一部を合成によって得られようといずれにせよ、免疫グロブリン結合ドメインから成るあらゆるポリペプチドを含んでいる。それ故に、もう一つのポリペプチドに融合した、免疫グロブリン結合ドメイン又は同等物から成るキメラ分子が含有される。キメラ抗体のクローン化及び表現は、特許文献10及び11に記載されている。
【0020】
抗体全体の断片は結合性抗原の機能を果たし得るということが示された。結合性断片の例は、(i)VL、VH、CL及びCH1のドメインから成るFab断片;(ii)VH及びCH1のドメインから成るFd断片;(iii)単抗体のVL及びVHのドメインから成るFv断片;(iv)VHドメインから成るdAb断片(非特許文献31);(v)分離されたCDR領域;(vi)F(ab’)2断片、二つの連結したFab断片から成る二価の断片;(vii)単鎖Fv分子(scFv)(ここに、VHドメイン及びVLドメインは、この二つのドメインを結合させて抗原結合部を形成させるペプチドリンカーによって連結される(非特許文献32及び33);(viii)二重特異性単鎖Fv二量体(特許文献12);並びに(ix)「ダイアボディ(diabodies)」、遺伝子融合によって構成される多価断片又は多重特異性断片(特許文献13及び非特許文献34)である。
【0021】
ダイアボディはポリペプチドの多量体(multimers)であり、各々ポリペプチドは、免疫グロブリンの軽鎖(light chain,L鎖)の結合性領域を含む第一ドメインと、免疫グロブリンの重鎖(heavy chain,H鎖)の結合性領域を含む第二ドメインとから成る。この二つのドメインは、(例えば、ペプチドリンカーによって)連結されているが、相互に結合して(associate)抗原結合部を形成することはできない。抗原結合部は、多量体内の一つのポリペプチドから成る第一ドメインと、多量体内のもう一つのポリペプチドから成る第二ドメインとの結合(association)によって形成される(特許文献13)。
【0022】
二重特異性抗体を使用する場合、これらは、従来の二重特異性抗体であっても良い。これらは、種々の方法で製造される(非特許文献35);例えば、化学的に調製又はハイブリッド融合細胞腫から調製することができる。或いは、これら二重特異性抗体は、上述の二重特異性抗体断片のいずれかであっても良い。完全な抗体よりむしろ、scFv二量体又はダイヤボディを使用するのが好ましい場合もある。ダイヤボディ及びscFvは、可変ドメインのみを使用し、抗イディオタイプ(anti−idiotypic,抗個特異的抗原)反応の影響を潜在的に減少させながら、Fc領域なしで構成し得る。二重特異性抗体の他の形態には、非特許文献36に記載の単鎖「ヤーヌシンズ(Janusins)」が含まれる。
【0023】
二重特異性の完全抗体とは対照的に、二重特異性抗体のダイヤボディは、特に有用である場合もある。なぜなら、これらダイヤボディは、大腸菌(E.coli.)中で容易に構築され表現されるからである。適切な結合特異性のダイヤボディ(及び抗体断片等の他の多くのポリペプチド)は、ライブラリからファージ表示(phage display)を使用しながら、容易に選択することができる(特許文献13)。もし、ダイヤボディの一つの腕(arm)が、例えば、抗原Xに向けられた特異性で一定に保持されるなら、そのとき、他方の腕は変化し、適切な特異性の抗体が選ばれる所にライブラリが造られる。
【0024】
抗原結合性ドメイン
抗原結合性ドメインとは、抗原の一部又は全部に特異的に結合して相補的である領域から成る抗体の一部をいう。抗原が大きい所では、抗体はその抗原の特定の部分(この部分はエピトープと呼ばれる)に結合するだけであろう。抗原結合性ドメインは、一種以上の抗体可変ドメインによって供給されるだろう。抗原結合性ドメインは、抗体軽鎖可変領域(VL)と、抗体重鎖可変領域(VH)とから成るのが好ましい。
【0025】
特異性
特異性とは、特異性結合ペアの一方のメンバが、その特異性結合のパートナに比べて、他の分子に対するいかなる有意な結合性をも示さない状況をいう。この用語はまた、例えば、抗原のひとつのメンバによって運ばれる特定のエピトープに対して、抗原結合性ドメインが特異的である場合にも適用できる。この場合、抗原結合性ドメインを運ぶ特異性結合メンバは、そのエピトープを運ぶ種々の抗原に結合することができるだろう。
【0026】
機能的同等変異体
機能的同等変異体とは、他の分子(親)とは構造的に相違しているが、幾つかの重要な相同を残しており、また、親分子の生物機能(例えば、特定の抗原又はエピトープと結合する能力)の少なくとも幾つかをも残している分子(変異体)をいう。変異体は、断片、誘導体又は突然変異体の形態で存在することがある。変異体、誘導体又は突然変異体は、一種以上のアミノ酸の付加、欠失(deletion)、置換若しくは挿入;又は他の分子の連結による親分子の変性によって得られることがある。これらの変化は、ヌクレオチド又は蛋白のレベルで行われることがある。例えば、コード化されたポリペプチドは、Fab断片であることがある。このFab断片は、そのとき、もう一つの源からのFc尾部に連結される。代替的には、酵素、フラワーセイン(flourescein)等のマーカーが連結されることがある。
【特許文献1】欧州特許第0,344,134B1号明細書
【特許文献2】国際公開第92/01047号パンフレット
【特許文献3】国際公開第92/20791号パンフレット
【特許文献4】国際公開第93/06213号パンフレット
【特許文献5】国際公開第93/11236号パンフレット
【特許文献6】国際公開第93/19172号パンフレット
【特許文献7】欧州特許出願公開第184187号明細書
【特許文献8】英国特許出願公開第2188638号明細書
【特許文献9】欧州特許出願公開第239400号明細書
【特許文献10】欧州特許出願公開第0120694号明細書
【特許文献11】欧州特許出願公開第0125023号明細書
【特許文献12】国際特許出願番号第PCT/US92/09965号
【特許文献13】国際公開第94/13804号パンフレット
【非特許文献1】Juweidら著、Cancer Res.、1992年、52巻、p.5144〜5153
【非特許文献2】Jain.R.著、Sci.Am.、1994年、271巻、p.58〜65
【非特許文献3】Alitaloら著、Adv.Cancer Res.、1982年、37巻、p.111〜158
【非特許文献4】Yamada著、Ann.Rev.Biochem.、1983年、52巻、p.761〜799
【非特許文献5】Hynes著、Ann.Rev.Cell Biol.、1985年、l巻、p.67〜90
【非特許文献6】Ruoslahti著、Ann.Rev.Biochem.、1988年、57巻、p.375〜413
【非特許文献7】Owensら著、Oxf.Surv.Eucaryot.Genes、1986年、3巻、p.141〜160
【非特許文献8】Zardiら著、EMBO J.、1987年、6巻、p.2337〜2342
【非特許文献9】Castellaniら著、J.Cell Biol.、1986年、103巻、p.1671〜1677
【非特許文献10】Borsiら著、J.Cell.Biol.、1987年、104巻、p.595〜600
【非特許文献11】Vartioら著、J.Cell Science、1987年、88巻、p.419〜430
【非特許文献12】Baroneら著、EMBO J.、1989年、8巻、p.1079〜1085
【非特許文献13】Carnemollaら著、J.Cell Biol.、1989年、108巻、p.1139〜1148
【非特許文献14】Oyamaら著、J.Biol.Chem.、1989年、p.10331〜10334
【非特許文献15】Oyamaら著、Cancer Res.、1990年、50巻、p.1075〜1078
【非特許文献16】Borsiら著、Exp.Cell Res.、1992年、199巻、p.98〜105
【非特許文献17】Norton及びHynes著、Mol.Cell.Biol.、1987年、7巻、p.4297〜4307
【非特許文献18】Schwarzbauerら著、EMBO J.、1987年、6巻、p.2573〜2580
【非特許文献19】Gutman及びKornblihtt著、Proc.Natl.Acad.Sci.(USA)、1987年、84巻、p.7179〜7182
【非特許文献20】Kornblihttら著、EMBO J.、1985年、4巻、p.1755
【非特許文献21】Ffrench−Constantら著、J.Cell Biol.、1989年、109巻、p.903〜914
【非特許文献22】Ffrench−Constantら著、Development、1989年、106巻、p.375〜388
【非特許文献23】Laitinenら著、Lab.Invest.、1991年、64巻、p.375〜388
【非特許文献24】Castellaniら著、Int.J.Cancer、1994年、59巻、p.612〜618
【非特許文献25】Humphriesら著、J.Cell Biol.、1986年、103巻、p.2637〜2647
【非特許文献26】Carnemollaら著、J.Biol.Chem.、1992年、p.24689〜24692
【非特許文献27】Petersら著、Cell Adhes.Commun.、1995年、3巻、p.67〜89
【非特許文献28】Schroffら著、Cancer Res.、1985年、45巻、p.879〜885
【非特許文献29】DeJagerら著、Proc.Am.Assoc.CancerRes.、1988年、29巻、p.377
【非特許文献30】Nissimら著、EMBO J.、1994年、13巻、p.692〜698
【非特許文献31】Ward,E.S.ら著、Nature、1989年、341巻、p.544〜546
【非特許文献32】Birdら著、Science、1988年、242巻、p.423〜426
【非特許文献33】Hustonら著、PNAS USA、1988年、85巻、p.5879〜5883
【非特許文献34】Holligerら著、Proc.Natl.Acad.Sci.USA、1993年、90巻、p.6444〜6448
【非特許文献35】Holliger,P.及びWinter,G.著、Current Opinion Biotechnol.、1993年、4巻、p.446〜449
【非特許文献36】Trauneckerら著、Embo Journal、1991年、10巻、p.3655〜3659
【非特許文献37】Winter,G.及びC.Milstein著、Nature、1991年、349巻、p.293〜299
【非特許文献38】Tomlinson I.M.ら著、J.Mol.Biol.、1992年、227巻、p.776〜798
【非特許文献39】Ledermann J.A.ら著、Int.J.Cancer、1991年、47巻、p.659〜664
【非特許文献40】Bagshawe K.D.ら著、Antibody,Immunoconjugates and Radiopharmaceuticals、1991年、4巻、p.915〜922
【非特許文献41】Pluckthun,A.著、Bio/Technology、1991年、9巻、p.545〜551
【非特許文献42】Reff,M.E.著、Curr.Opinion Biotech.、1993年、4巻、p.573〜576
【非特許文献43】Trill J.J.ら著、Curr.Opinion Biotech.、1995年、6巻、p.553〜560
【非特許文献44】分子クローニング(Molecular Cloning),実験マニュアル(Laboratory Manual):第2版,サンブルック(Sambrook)ら著、1989年、Cold Spring Harbor Laboratory 出版
【非特許文献45】分子生物学における短い実験記録(Short Protocols in Molecular Biology)、第2版,オスベル(Ausubel)等編集,ジョン・ウィリー(John Wiley)及びサンズ(Sons)、1992年
【非特許文献46】Pierschbacherら著、Cell、1981年、26巻、p.259〜267
【非特許文献47】Marksら著、J.Mol.Biol.、1991年、222巻、p.581〜597
【非特許文献48】Hoogenboomら著、Nucl.Acids Res.、1991年、19巻、p.4133〜4137
【非特許文献49】Gibson TJ、博士論文、1984年、University of Cambridge, Cambridge, UK
【非特許文献50】Hawkinsら著、J.Mol.Biol.、1992年、226巻、p.889〜896
【非特許文献51】Crothers及びMetzger著、Immunochemistry、1972年、9巻、p.341〜357
【非特許文献52】Marksら著、Bio/Technology、1992年、10巻、p.779〜783
【非特許文献53】Lowら著、J.Mol.Biol.、1996年、260巻、p.359〜368
【非特許文献54】Griffithsら著、EMBO J.、1994年、13巻、p.3245〜3260
【非特許文献55】Vaughanら著、Nature Biotechnol.、1996年、14巻、p.309〜314
【非特許文献56】Borsiら著、Anal.Biochem.、1991年、192巻、p.372〜379
【非特許文献57】Saginatiら著、Eur.J.Biochem.、1992年、205巻、p.545〜549
【非特許文献58】Siriら著、Nucl.Acids Res.、1991年、19巻、p.525〜531
【非特許文献59】Brownら著、Amer.J.Pathol.、1993年、142巻、p.793〜801
【非特許文献60】Kaczmarekら著、Int.J.Cancer、1994年、58巻、p.11〜16
【非特許文献61】Folliら著、Cancer Res.、1994年、54巻、p.2643〜2649
【非特許文献62】McCafferty,J.、Griffiths,A.D.、Winter,G.及びChiswell,D.J.、Nature (London)、1990年、”Phage antibodies: filamentous phage displaying antibody variable domains.”、348巻、p.552〜554
【非特許文献63】Packら著、Bio/Technology、1993年、11巻、p.1271〜1277
【非特許文献64】Neriら著、Nature Biotechnology、1996年、14巻、p.385〜390
【非特許文献65】Ausubelら編、John Wiley & Sons、1992年、Short Protocols in Molecular Biology(第2版)
【非特許文献66】Borsiら著、Int.J.Cancer、1992年、52巻、p.688〜692
【非特許文献67】Jonsson著、BioTechniques、1991年、11巻、p.620〜627
【非特許文献68】Neriら著、Bio/Techniques、1996年、20巻、p.708〜713
【発明の開示】
【発明が解決しようとする課題】
【0027】
本発明によると、フィブロネクチン(FN)のED−B胎児ドメインに対して特異性のある特異性結合メンバが提供される。
【課題を解決するための手段】
【0028】
本発明による特異性結合メンバは,ED−Bドメインに直接結合する。一つの具体例において、特異性結合メンバは、蛋白分解酵素サーモリシン(thermolysin)でFNを処理した後、ED−Bを含有するFNの一つに、又はいかなるものにも、又は全部に結合する。更なる具体例において、特異性結合メンバは、III型相同の繰返しを含有するFNの一つに、又はいかなるものにも、又は全部に結合する。これら繰返しには、ED−Bドメインが含まれる。既知のFNは、二つの論文(非特許文献13及び26)の中に確認される。「ED−Bを含有する全FN」への言及は、これらの論文中で、ED−Bを含有していると確認される全FNへの言及として理解されても良い。
【発明を実施するための最良の形態】
【0029】
特異性結合メンバは、ヒトED−Bと結合するのが好ましく、また、マウス、ラット及び/又はニワトリのような、他の少なくとも一種のB+FNと結合するのが好ましい。特異性結合ペアメンバは、ヒト・フィブロネクチンED−Bにも;動物モデル中のsbbメンバのテスト及び分析が可能な、マウス・フィブロネクチンED−B等の非ヒト・フィブロネクチンED−Bにも結合し得るのが好ましい。
【0030】
本発明による特異性結合ペアメンバは、本明細書のどこかで検討され公式に入手できる沈積済み抗体BC−1と競うことなく、フィブロネクチンED−Bに結合する。BC−1は、ヒトB+イソ型に対してまさに特異的である。本発明による特異性結合ペアメンバは、BC−1と同一のエピトープには結合しない。
【0031】
本発明による特異性結合メンバのB+FNへの結合は、ED−Bドメインによって抑制されることがある。
【0032】
本発明の一つの面において、結合ドメインは、精製済み単量体として測定されるとき、ED−B FNに対して6×10−8M以下の解離定数(Kd)を有する。
【0033】
本発明の一つの面において、結合ドメインは、フィブロネクチンED−BをN−グリカナーゼ(N−glycanase)で前処理しないで、フィブロネクチンED−Bと反応する(即ち、結合することができる)。
【0034】
本発明による特異性結合ペアメンバは、分離体として、又は精製された形で、即ち、他の特異性結合ペアメンバ(例えば、抗体又は抗体断片)を含有していない製剤の状態、又はフィブロネクチンED−Bと結合することのできる他の特異性結合ペアメンバを含有していない状態で提供されることがある。本発明による特異性結合メンバは、実質的に純粋な形で提供されるのが好ましい。それらは、伝統的な融合細胞腫技術を使って得られた抗体と縛りつけるよりむしろ、単一クローンからのものという意味で『単クローン』であるのかも知れない。検討した通り、本発明による特異性結合ペアメンバは、バクテリオファージ表示技術(bacteriophage display technology)及び/又は組換えによる表現(例えば、細菌細胞、ホスト細胞)を使用して得られることがある。フィブロネクチンED−Bに直接結合する、単クローンの特異性結合ペアメンバについては、従来全く開示されていない。
【0035】
この特異性結合メンバは、抗体から成るのが好ましい。特異性結合メンバは、単鎖Fv(scFv)等の抗体断片の形態のポリペプチド配列から成ることがある。Fab、Fab”、F(ab’)2、Fabc、Facb、ダイアボディ等の、抗体断片の他のタイプが利用されることもある(非特許文献37及び特許文献13)。特異性結合メンバは、完全抗体の形であることがある。完全抗体は、抗体イソ型(例えば、IgG、IgA、IgD、IgE及びIgM)の形のいずれか、及びイソ型亜綱(例えば、IgG1又はIgG4)の形のいずれかであることがある。
【0036】
抗体は、あらゆる起源、例えば、ヒト、マウス、ヒツジ、ウサギのものでよい。他の誘導は当業者には明らかである。抗体は、ヒト起源が好ましい。「ヒト」は、ヒトのcDNA、蛋白又はペプチドライブラリから部分的に又は完全に誘導される抗体を意味する。この用語は、抗体分子にヒト特性を与えて、その分子にヒト免疫系の防衛をバイパスさせるために変性された、非ヒト起源のヒト化されたペプチド及び蛋白を意味する。
【0037】
特異性結合メンバは、設計された抗体[例えば、明示されている(disclosed)フィブロネクチンED−Bに対して一つの結合腕(即ち、特異性ドメイン)、及び異なる特異性に対する他の腕、又は二価若しくは多価の分子を有する、二重特異性抗体分子(又は、F(ab’)2等の断片)]の形態であることもある。
【0038】
特異性結合メンバは、抗体配列の他に、他のアミノ酸(例えば、ペプチド若しくはポリペプチドを形成するもの、又は抗原に結合する能力の他にもう一つの機能特性をその分子に与えるもの)を含むことがある。例えば、特異性結合メンバは、標識、酵素又はそれらの断片等を含む。
【0039】
結合ドメインは、胚芽細胞系列分節(germ line segment)又は再配列された遺伝子分節(gene segment)によってコード化されたVHドメインの一部又は全部から成ることがある。結合ドメインは、VLカッパ(κ)ドメイン又はVLラムダ(λ)ドメインの一部又は全部から成ることがある。
【0040】
結合ドメインは、VH1、VH3又はVH4の胚芽細胞系列遺伝子配列、又はその再配列形から成ることがある。
【0041】
本発明による特異性結合メンバは、ヒト・胚芽細胞系列DP47由来の重鎖可変領域(「VH」ドメイン)(それの配列は、図1aに示す;残基は1〜98)から成ることがある。「DP」命名法は、非特許文献38に記載されている。CDR3のアミノ酸配列は、Ser Leu Pro Lysであってもよい。CDR3のアミノ酸配列は、Gly Val Gly Ala Phe Arg Pro Tyr Arg Lys His Gluであってもよい。このように、本発明による特異性結合メンバの結合ドメインは、CGS1及びCGS2について図1aに示されるアミノ酸配列から成るVHドメインを含有することがある。
【0042】
結合ドメインは、ヒト・胚芽細胞系列DP16由来の軽鎖可変領域(「VL」ドメイン)(それの配列は、コドン1〜90として図1bに示す)から成ることがある。
【0043】
VLドメインは、CDR3配列 Asn Ser Ser Pro Val Val Leu Asn Gly Val Valから成ることがある。VLドメインは、CDR3配列 Asn Ser Ser Pro Phe Glu His Asn Leu Val Valから成ることがある。
【0044】
本発明の特異性結合メンバは、図1に示される配列と機能的に同等の変異体から成ることがある。例えば、本明細書に開示される特性が保持されるという条件で、一種以上のアミノ酸が挿入され、削除され、置換され又は追加された。例えば、特異性結合メンバがED−Bに結合するという条件で、CDR3配列は、変性されることがある;或いは、一つ以上の変化が外郭構造領域に生じることがある;或いは、外郭構造がもう一つの外郭構造若しくは変性形態と置き換えられることがある。
【0045】
本明細書に開示される抗体の抗原結合ドメインのVLドメイン若しくはVHドメイン由来の一種以上のCDRのものは、いわゆる「CDR−移植」で使用されることがある。「CDR−移植」では、特許文献9に開示されるように、第一抗体の一種以上のCDR配列は、その抗体のものでない配列(例えば、もう一つの抗体の配列)の外郭構造の内部に置かれる。CGS1及びCGS2のためのCDR配列は、図1a及び図1bに示す。
【0046】
本発明による特異性結合メンバは、フィブロネクチンED−Bへの結合に関し、本明細書に記載のscFv又は抗体と競合するものであることがある。複数の結合メンバ間の競合は、生体外で容易に分析されることがある。例えば、報告者の特異性分子を一つの結合メンバ(これは、識別されていない他の結合メンバの存在下に検出され得る)と識別して、同一エピトープ又はオーバーラップしているエピトープと結合する特異性結合メンバを同定し得るようにすることによって、容易に分析されることがある。
【0047】
本発明による特異性結合メンバは、特異性結合メンバをそれのエピトープに結合させる又は結合するがままにしておく過程を含む方法で使用することがある。結合は、哺乳動物(例えば、ヒト、又はマウス等のげっ歯動物)に特異性結合メンバを投与した結果として生じることがある。
【0048】
本発明は、腫瘍に対する診断薬として使用するための、上記のように特異性結合メンバを使用する方法を提供する。以下に記載する動物をモデルとした実験的証拠によって、本発明による結合メンバは生体内位置決定を行う上で有効であることが示される。
【0049】
本発明による好ましい特異性結合メンバには、ヒト・腫瘍に結合するもの(例えば、クリオスタット分節で、浸潤性の脈管形成性現象型(phenotype)を示すもの);胚腫瘍に結合するもの(例えば、クリオスタット分節で)が含まれる。結合は、免疫細胞化学による染色法によって証明されることがある。
【0050】
好ましい具体例において、特異性結合メンバは、テナシン、細胞外マトリックス蛋白質には結合しない、又は有意には結合しない。
【0051】
もう一つの好ましい具体例において、特異性結合メンバは、正常なヒト皮膚(例えば、クリオスタット分節で、及び/又は免疫細胞化学による染色法を用いて実証されるように)には結合しない、又は有意には結合しない。
【0052】
本発明による特異性結合メンバの更なる具体例は、肝臓、脾臓、腎臓、胃、小腸、大腸、卵巣、子宮、膀胱、膵臓、副腎、骨格筋、心臓、肺、甲状腺及び脳から選ばれる一種以上の正常組織には結合しない、又は有意には結合しない(例えば、クリオスタット分節で、及び/又は免疫細胞化学による染色法を用いて実証されるように)。
【0053】
ED−Bのための特異性結合メンバは、生体内の標的剤(targeting agent)として使用されることがある。この標的剤は、フィブロネクチンED−Bを表現する 腫瘍又はフィブロネクチンED−Bと結合した腫瘍の存在又は位置を特異的に実証するのに使用されることがある。それは、造影剤(imaging agent)として使用されることがある。本発明は、フィブロネクチンED−B表現を表現する、又はフィブロネクチンED−B表現と結合した細胞又は腫瘍の存在を決定する方法を提供する。この方法は、供給された特異性結合メンバに細胞を接触させ、次いで、その細胞に対する特異性結合メンバの結合を決定する過程を含む。その方法は、本体から除去された細胞の試験サンプルについて、生体内又は生体外で行われることがある。
【0054】
細胞サンプルに関する抗体の反応性は、あらゆる適切な手段によって決定されることがある。個々のリポーター分子を用いて識別することも可能である。そのリポーター分子は、直接的又は間接的に検出可能な、好ましくは測定可能な信号を発生することがある。リポーター分子の連結は、直接的又は間接的、共有結合的(例えば、ペプチド結合による)、或いは非共有結合的であることがある。ペプチド結合による連結は、抗体とリポーター分子とをコード化する遺伝子融合の組換え表現の結果として行われることがある。
【0055】
一つの好ましい態様は、スペクトル的に分離された吸収特性又は放出特性を有する個々の蛍光色素、蛍光物質又はレーザー色素と、各抗体との共有結合によることがある。適した蛍光色素には、フルオレセイン、ローダミン、フィコエリトリン、及びテキサス・レッド(Texas Red)が含まれる。適した色素産生色素には、ジアミノベンジジンが含まれる。
【0056】
他のリポーターには、着色され、磁性若しくは常磁性であるラテックス・ビーズ(latex beads)等の粒状物質又は高分子コロイド粒子;及び直接的又は間接的に検出可能な信号を発生して、可視的に観察することが可能であり、電子化により検出することが可能であり、さもなければ記録することが可能である生物活性剤又は化学的活性剤が含まれる。これらの分子は、例えば、顕色若しくは変色し又は電気特性を変化させる反応に、触媒作用を及ぼす酵素であってもよい。これらは、分子的に励起され、エネルギー状態間における電子遷移が特徴的なスペクトル吸収又はスペクトル発光を生じることがある。これらには、バイオセンサーと連結して使用される化学的構成要素(chemical entities)が含まれる。ビオチン/アビジン又はビオチン/ストレプトアビジン、及びアルカリリン酸分解酵素の検出システムが採用されることがある。
【0057】
結合を決定する態様は、本発明の特徴ではなく、当業者は、彼等の好み及び通常の知識に従って、適切な態様を選ぶことができる。
【0058】
個々の抗体−リポーター連結によって発生する信号は、細胞サンプル中における、関連する抗体結合の定量的な絶対データ又は相対データを得るのに使用されることがある(標準及び試験)。加えて、プロピジウムヨウ化物(propidium iodide)等の一般的核染料は、サンプル中の全細胞の個体数を数えるのに使用されることがある。125I、111In、99mTc等の放射性ヌクレオチドが抗体に付着したとき、もし、その抗体が正常組織よりむしろ腫瘍の中に優先的に局在化していれば、腫瘍組織中の放射性ラベルの存在は、ガンマカメラを使用して、検知し定量化し得る。得られる腫瘍像の質(quality)は、信号対雑音比に直接相関している。
【0059】
抗体は、新たに血管が新生した腫瘍を追跡するための診断剤として利用されることがある。また、抗体は、細胞毒剤を運ぶため又は新たな血管内の凝固を引き起こすために、(例えば、変性された形で)採用されることもある。かくして、成長している腫瘍に対する酸素及び栄養素の補給を断って、腫瘍治療の間接的形態を形成する。
【0060】
本発明は、治療試薬として使用するための、上述の特異性結合メンバの使用方法をも提供する。そのとき、その特異性結合メンバは、例えば、融合蛋白として結合され、化合され又は設計されて、作用因子機能が与えられる。本発明の特異性結合メンバは、重要な抗体を表現する腫瘍又はその抗体と結合した腫瘍に至る毒素、放射能、T細胞、キラー細胞又は他の分子を標的とするのに使用される。
【0061】
従って、本発明の更なる面は、与えられた特異性結合メンバを投与することから成る処置方法;そのような特異性結合メンバから成る薬剤組成物;及び投与のための薬剤の製造過程において(例えば、薬学的に許容できる賦形剤を用いて特異性結合メンバを調製することから成る、薬剤組成物又は医薬品組成物の製法において)、特異性結合メンバを使用する方法を提供する。
【0062】
本発明によると、提供される組成物は、個体に投与されることがある。投与は、「治療的に有効な量」であるのが好ましい。これは、患者に効果が現れるのに十分な量である。そのような効果は、少なくとも一つの症状の少なくとも回復であることがある。実際の投与量と、投与の割合及び時間−クール(course)とは、治療されているものの性質(nature)及び激しさ(severity)に依存する。投与量の決定等の治療の処方箋は、一般の診療医及び他の医師の責任による。抗体の適切な投与量は、当業において周知である。非特許文献39及び40を参照。
【0063】
組成物は、単独で又は他の治療法と組合わせ、治療される容態によるが、同時に又は逐次的に投与することがある。
【0064】
本発明による薬剤組成物、及び本発明により使用するための薬剤組成物は、有効成分に加えて、薬学的に許容される賦形剤、キャリア、緩衝剤、安定剤又は当業者に周知の他の物質を含有することがある。そのような物質は、非毒性であるべきであり、また、有効成分の効能を妨げてはいけない。キャリア又は他の物質の正確な性質は、投与の経路に依存する。経路は、経口であることもあり、注射(例えば、静脈注射)によることもある。
【0065】
経口のための薬剤組成物、錠剤、カプセル、粉末又は液体の形態であることがある。錠剤は、ゼラチン、佐剤(adjuvant)等の固形キャリアを含有することがある。液体の薬剤組成物は、一般的には、水、石油、動物油、植物油、鉱油合成油等の液体キャリアを含有する。生理的塩類溶液;ブドウ糖若しくは他のサッカライド溶液;又はエチレングリコール、プロピレングリコール、ポリエチレングリコール等のグリコールが含まれることがある。
【0066】
静脈注射剤、又は苦痛の位置での注射剤のためには、有効成分は、非経口的に許容される水溶液の形態であろう。この水溶液は、発熱因子を含まず、適切なpH、等張性及び安定性を有する。当業者は、例えば、塩化ナトリウム注射剤、リンガー液、乳酸加リンガー液等の等張性媒体を使用して、適切な溶液をうまく調製することができる。保存剤、安定剤、緩衝剤、酸化防止剤及び/又は他の添加剤は、必要に応じ、含有させてもよい。
【0067】
本発明の特異性結合メンバは、核酸のコード化による表現によって造られることがある。特異性結合メンバの製法(この製法は、特異性結合メンバのために核酸をコード化することによる表現から成る)が本発明の一つの面であるように、与えられるあらゆる特異性結合メンバをコード化する核酸それ自体、本発明の一つの面である。表現は、適切な条件下、核酸を含有する組換えホスト細胞を培養することによって、都合よく達成されることがある。
【0068】
核酸は、本明細書に記載の抗体抗原結合ドメインのあらゆるアミノ酸配列、又は機能的に同等のあらゆる形態をコード化することがある。一種以上のヌクレオチドの添加、置換、削除又は挿入によって、ヌクレオチド・レベルでの変化が生じることがある。この変化は、遺伝子コードの縮重(degeneracy)に依存し、アミノ酸レベルに反映されるかも知れないし、反映されないかも知れない。
【0069】
異なるホスト細胞の一形態でのポリペプチドのクローン化及び表現のためのシステムは、周知である。適切なホスト細胞には、細菌系、哺乳動物の細胞系、酵母菌系及びバクロウィルス系が含まれる。非相同のポリペプチドを表現するために、当該技術において入手可能な哺乳動物・細胞系には、チャイニーズハムスターの子房細胞、ヒーラー細胞、ベビーハムスターの腎臓細胞、及びその他多数が含まれる。通常の好ましい細菌ホストは、大腸菌である。
【0070】
大腸菌等の原核細胞中の抗体断片及び抗体の表現は、当業ではうまく確立されている。報告としては、例えば非特許文献41を参照。培養における真核細胞中の表現もまた、特異性結合メンバを製造するための一つの選択肢として(最近の報告を参照(例えば、非特許文献42及び43)、当業者は入手することができる。
【0071】
適切な調節配列;プロモーター配列;ターミネーター配列;ポリアデニル化配列;エンハンサー配列;マーカー遺伝子;及び適切な他の配列を含有する適切なベクターを選択することができる、又は組み立てることができる。ベクターは、適切な、プラスミド、ウィルス性のもの(例えば、ファージ)又はファージミド(phagemid)であることがある。更なる詳細については、例えば、非特許文献44を参照。核酸を操作するための多数の技術及び実験記録(例えば、核酸構成体の調製;突然変異生成;配列すること;細胞中へのDNAの導入と遺伝子表現;及び蛋白の分析、に関するもの)は、非特許文献45に詳細に記載されている。サンブルック等の開示内容、及びオスベル等の開示内容は、言及することによって本明細書に取り入れる。
【0072】
本発明の更なる面によると、本明細書に開示される核酸を含有するホスト細胞が提供される。また、更なる面によると、そのような核酸をホスト細胞の中に導入することを含む方法が提供される。この導入のためには、利用可能なあらゆる技術が採用されることがある。真核細胞のための適切な技術には、リン酸カルシウムのトランスフェクション(transfection,移入);DEAE−デキストラン;エレクトロポレーション;リポソーム媒介(liposome−mediated)トランスフェクション;及びレトロウィルス又は他のウィルス(例えば、ワクシニア、若しくは昆虫細胞に対するバクロウィルス)を用いたトランスダクションが含まれることがある。細菌細胞のための適切な技術には、塩化カルシウム転換;エレクトロポレーション;及びバクテリオファージを用いたトランスフェクションが含まれることがある。
【0073】
導入によって、核酸からの表現が生じる又は許容される(例えば、遺伝子の表現のための条件下、ホスト細胞が培養される)結果となることがある。
【0074】
一つの具体例によると、本発明の核酸は、ホスト細胞のゲノム(例えば、染色体)の中に組込まれる。組込みは、標準的技術に従い、ゲノムとの組換えを促進する配列の封入(inclusion)によって促進されることがある。
【0075】
特異性結合メンバが生成された後、その特異性結合メンバは、検討された通り、例えば、本明細書に開示される方法のあらゆるもの(例えば、特異性結合メンバの他に、そのメンバの細胞への結合を決定するための一種以上の試薬を有するキット(kit)等の、薬剤生成物又は診断薬生成物の調製)において使用されることがある。
【0076】
本発明及び具体例の更なる面は、当業者には明白であろう。本発明が十分に理解されるべく、次の諸例を単なる例証として提供するが、これに限定されるものではない。次の諸図面が参考となる。
【0077】
図1は、scFv CGS−1及びCGS−2のVH並びにVLの、整列したアミノ酸配列を示す。図1aはVH配列を示し、図1bはVL配列を示す。CDRs(1、2及び3)を示す。両方のscFvへの最も相同のヒト・胚芽細胞系列VHは、VH3科のDP47分節である。両方のクローンのVL分節はDPL6、scFvライブラリを造るのに使用される軽鎖である(非特許文献30)。二つのクローンを相互に識別する残基は、下線が引いてある。
【0078】
図2:図2Aは、ヒトFNサブユニットのドメイン構造のモデルを示す。FN−プレ−mRNAの交互スプライシング(alternative splicing)による、IIICS、ED−A及びED−Bの変異領域を示す。この図はまた、ED−Bを含有する主要サーモリシン消化生成物の他に、内部相同をも示す(非特許文献8)。図2Bは、プラスマの4〜18%SDS−PAGE;WI38VA;それらのクーマシーブルー(Coomassie Blue)染色済みサーモリシン消化生成物;並びに、BC−1、IST−6、CGS−1及びCGS−2で調べた免疫ブロット分析を示す。FNの1μg/mg(レーン3)及び10μg/mg(レーン4)でサーモリシンを使用した、不消化(レーン1)及び消化済みプラスマFN。FNの1μg/mg(レーン5)、5μg/mg(レーン6)及び10μg/mg(レーン7)でサーモリシンを使用した、不消化(レーン2)及び消化済みWI38VA FN。右側の番号は、図2Aに示される主要サーモリシン消化生成物を示す。左側の値は、キロドルトン(kD)での分子量基準(molecular weight standards)を示す。
【0079】
図3:図3Aは、大腸菌に表現された融合蛋白及び組換え蛋白の中に含有されるFNIII型繰返し配列;これら蛋白とCGS−1及びCGS−2との反応性;並びに、これら蛋白とmAbs BC−1及びIST−6との反応性を示す。図3Bは、クーマシーブルー染色済みゲル;及び、CGS−1、CGS−2、BC−1、IST−6で調べた免疫ブロット分析を示す。レーンの番号は、図の上部のペプチド構造の番号と一致する。左側の値は、kDでの分子量基準を示す。
【0080】
図4: マウスの赤外線イメージャー;タングステン・ハロゲンランプを備えた、黒色の非蛍光性箱と;CY7赤外線蛍光物質に対して特異的な励起・放出フィルターと;コンピュータ制御の8−ビット単色CCD−カメラとから成る、標的実験のために使用したマウス・イメージャー(imager)。
【0081】
図5:B−FNに向けられた単量体のscFc(CGS−1)及び二量体のscFc(CGS−1)2を用いて、蛍光によって標識化された抗体断片を、標的としてのF9ネズミの奇形癌に向けられている。リゾチームに対して結合特異性を有する二量体scFc(D1.3)2は、ネガティブ対照として使用した。
【0082】
図6:B−FNの同一エピトープに向けられた、親和性成熟のscFv(CGS−2)及び低結合性scFv(28SI)を使用して、蛍光によって標識化された抗体断片が、F9ネズミの奇形癌に向けられている。標的化は、結像を部分的に覆い隠す黒色外皮によって48時間覆われた、大きい腫瘍(約0.6g)でも、小さい腫瘍(約0.2g)でも検出された。
【0083】
本明細書に記載の全資料は、言及することによってここに取り入れる。
【実施例】
【0084】
諸例の目録
例1・・・ヒトFNのED−Bドメインに特異的なヒトscFvの分離。
例2・・・ヒトFNのED−Bドメインに特異的なヒトscFvの親和性成熟。
例3・・・ED−B含有フィブロネクチンに対する親和性成熟済みscFvの特異性。
例4・・・ヒト及びマウスの腫瘍分節を免疫細胞化学的に染色する上で、親和性成熟済み抗ED−B scFvを使用する方法。
例5・・・ヒト腫瘍を生体内で標的化する上で、親和性成熟済み抗ED−B scFvを使用する方法。
例6・・・ヌードマウスに異種組織移植されたF9ネズミの奇形癌の標的化。
【0085】
例1・・・ヒトFNのED−Bドメインに特異的なヒトscFvの分離
ヒトscFvファージ・ライブラリ(非特許文献30)は、組換え抗体の選択のために使用した。ED−Bのイソ型の二つの異なる形態は、選択のための抗原の供給源として使用し、両方とも、イソ型は組換えヒト・蛋白であった。
【0086】
III型の繰返し2−11(B−)及び2−11(B+)を含有する組換えFNペプチドは、大腸菌中に表現された。
【0087】
クローンpFH154からのFN cDNA(非特許文献20)、λF10及びλF20(非特許文献13)を使用して、組み立てられた。ベース(bases)2229〜4787の中に広がるcDNA構成体(非特許文献20)は、キアゲン(Qiagen)からのQIA表現キットを用いて、ベクターpQE−3/5の中に挿入された(チャトワース,CA)。組換え体FN−III 2−11(B−)及び(B+)は、セファローズ(Sepharose)4Bに結合したmAb 3E3(非特許文献46)を使用した免疫親和性クロマトグラフィによって精製された。III型相同繰返し7B89、789、ED−B及びFN−6を含有する組換えFN断片を調製するためのDNA断片は、テンプレートとして、クローンFN2−11(B+)及びFN2−11(B−)からのcDNAを使用しながら、U1tMa DNAポリメラーゼを使用したポリメラーゼ鎖反応(polymerase chain reaction)(PCR)の増幅(パーキン・エルマー(Perkin Elmer)によって生成された。プライマーは、QIA表現キット(キアゲン)を使用して、pQE−12の中にPCR生成物がクローン化するように設計された。これらは、実質的に大腸菌の中に転換され、表現された。全cDNAクローンは、シーケナーゼ(Sequenase)2.0DNA配列キット(USB)を使用して配列された。
【0088】
組換え蛋白は、製造者の使用説明書(キアゲン)に従い、FN断片のカルボキシ末端のヘキサヒスチジン標識を使用して、Ni−NTAクロマトグラフィ(IMAC)によって精製した。ED−B−βGal融合蛋白は、ED−B cDNAをλgt11バクテリオファージ・ベクターの中にクローン化して、クローンλED−Bを与えることによって調製された。(ED−B配列の一部を含有する)クローンλchFN60は、クローン化されたニワトリ・FN pchFN60(ノートン(Norton)等,1987)からの融合蛋白として誘導された。
【0089】
ヒト・scFvファージライブラリを選択するために、3ラウンドのパニング(panning)が、二つの異なる組換え抗原(7B89及びED−B)の各々に対して行われた。これら抗原は両方とも、PBS(20mMリン酸塩緩衝剤、0.15M NaCl、pH7.2)中の50μg/mlで、一晩、免疫チューブ(ナンク(Nunc);Maxisorp,Roskilde,デンマーク)の上にコーティングされた。第一抗原は、組換えFN断片7B89(この中で、ED−Bドメインは、隣接するIII型相同繰返しによって、側面が接触する。)であり、これは、一晩、4℃でコーティングされた。使用された第二抗原は、カルボキシ末端のヘキサヒスチジン標識を有する組換えED−B(非特許文献8)であった。この蛋白は、リシン残基を含有していないので、第一アミノ酸の末端アミノ基は、サイト特異的な(site−specific)共有結合による、反応性ELSAプレートへのED−Bの固定に利用できる(ナンク(Nunc);コバリンク(Covalink)。コーティングは、一晩、室温で行った。
【0090】
3ラウンドのパニングの後、溶離されたファージは、HB2151大腸菌細胞の中に感染させ、記載した通りにプレートした(非特許文献30)。選択の各ラウンドの後、95のアンピシリン−抵抗性単一コロニーをスクリーニングし、ELISAによって抗原特異性scFvを同定した。パニングのために使用した抗体上に最も高いELISA信号を与えたクローンは、更なる分析のため、及び親和性成熟のために選定した。これらのクローンはまた、例4に一層詳しく記載される免疫細胞化学的染色によって、多種多様の膠芽腫及び胸部腫瘍の切断面に特異的な染色を与えることも実証された。
【0091】
例2・・・ヒトFNのED−Bドメインに特異的なヒトscFvの親和性成熟
(7B89と共に選択した)クローン35GE及び(ED−Bドメインのみと共に選択した)28SIは、親和性成熟のための候補抗体として選定した。親和性を改善するための手段として軽鎖を多種産生すべく、我々は、次いで、206=6.4×107の潜在的配列の多様性という条件で、縮重したオリゴヌクレオチド及びPCRを使用しながら、軽鎖CDR3の中央の6個の残基(DSSGNH)をランダム化することに基づき(図1)、単純な親和性成熟の方策を調べた。(重鎖CDR3に沿った)この領域は、抗原結合部位の中央に配置されている(パドラン(Padlan),1994)。我々は、また、その選択を支配する静電気の影響の可能性を回避すべく、6個の残基がセリンへ伸びる直前に、アルギニン残基を変異させた。
【0092】
「親」scFv断片を表現する単一細胞コロニーからのプラスミドは、プライマー LMB3(5’ CAG GAA ACA GCT ATG 3’)及びCDR3−6−VL−FOR(5’ CTT GGT CCC TCC GCC GAA TAC CAC MNN MNN MNN MNN MNN MNN AGA GGA GTT ACA GTA ATA GTC AGC CTC 3’)(94C[1’]−55C[1’]−72C[1’30”]、25サイクル)を用いて増幅されるPCRであった(緩衝剤及び条件については、非特許文献47を参照)。得られた生成物は、(オリジナルscFv遺伝子を含有するプラスミドの痕跡を除去するため)ゲル精製し、次いで、プライマー LMB3及びJI−Not−FOR(5’ ATT GCT TTT CTT TTT TGC GGC CGC GCC TAG GAC GGT CAG CTT GGT CCC TCC GCC 3’)(94C[1’]−55C[1’]−72C[1’30”]、25サイクル)を用いて、第二増幅段階のためのテンプレート(template,鋳型)として使用した。アガロース・ゲル上に正確な分子量の単結合として伝わる粗PCR生成物は、スピン結合(Spin−Bind)(FMC,ロックランド(Rockland),ME,USA)を用いて、Ncol/Notlで二重消化され(double−digested)結さつされて(ligated)、ゲル精製されたNcol/Notl消化済みファージミドpHEN1になる(非特許文献48)PCR混合物から直接精製した。このpHEN1は、単消化済みベクターから二重消化済みベクターを分離するのを促進するためにダミーのNcol/Not1挿入物(insert)を含有する。このベクターは、キアゲン(Qiagen)(チャッツワース(Chatsworth),CA,米国)のプラスミド・マキシプレ・キット(maxi−pre kit)で調製した。消化済みプラスミド及び挿入物の約5μgを、結さつ混合物(ligation mix)中で使用した。
【0093】
結さつ混合物はフェノールで一回抽出し、フェノール/クロロホルム/イソアミルアルコール(25:25:1)で一回抽出し、次いで、キャリアとしてグリコーゲンを用いてエタノール沈降(ボーリンガー(Boehringer),Mannheim,ドイツ)させ、次いで、高速真空で乾燥させた。ペレットは、水20μl中に再び懸濁させ、次いで、エレクトロコンピテント(electrocompetent)のTG1大腸菌細胞(非特許文献49)中でエレクトロポレーションさせた(electroporated)。我々は、グリセロールのストックを使用するときは、形質転換細胞109/μgのタイター(titre)を有するエレクトロコンピテント細胞を典型的に使用した。或いは、新たに調製したエレクトロコンピテントを有する形質転換細胞1010/μg)のタイター(titre)を有するエレクトロコンピテント細胞を典型的に使用した。これによって、本明細書に概説する手順により、107を越えるクローンが産生された。
【0094】
次いで、成熟ライブラリは、ニッシム・ライブラリ(非特許文献30)に関し、ファージ粒子を産生すべく処理した。このファージ粒子は、抗原として7B89(10μg/ml)を使用している免疫チューブ(immunotubes)上の選定1ラウンドのために使用し、次いで、動的選定(kinetic selection)(非特許文献50)の1ラウンドを行った。この選定過程は、選定の第一ラウンドからのミルク−PBS(2%MPBS)中で5分間、ビオチニル化7B89(10nM)をファージ懸濁液(約1012t.u.)で培養し、次いで、非ビオチニル化7B89(1μM)を添加し、次いで、30分間、競合を続行させることによって行った。次いで、2%MPBS中に予ブロックされた(preblocked)、ストレプトアビジンでコーティングした動力ビード(dynabeads)(Dynal:M480)100μlを、反応混合物に添加し、2分間混合し、次いで、磁石上で捕獲し、次いで、(PBS+0.1%Tween−20)とPBSとで交互に10回洗浄した。ファージは、トリエチルアミン0.5ml、100mMを用いてビードから溶離した。次いで、この溶液は、0.25ml、1MのTris、pH7.4で中和し、次いで、HB2151細胞(非特許文献30)を急激に成長させて感染させるのに使用した。95のアンピシリン−抵抗性単一コロニーは、scFV含有の上澄み液(非特許文献30)を生成するのに使用した。この上澄み液は、最良のバインダーを同定すべく、ELISA、免疫細胞化学及びBIAコア(BIAcore)によってスクリーニングを行った。次いで、この上澄み液は、pDN268表現ベクター(ネリ(Neri)等,1996)のSfil/Notlサイトの間でサブクローン化した。このベクターは、ScFVのC−末端に、リン酸化が可能な標識、FLAGエピトープ及びヘキサヒスチジン標識を付加する。
【0095】
pDN268中でサブクローン化した関連抗体の単クローンは、アンピシリン100mg/l及びグルコース0.1%を含有する2xTY中、37℃で成長した。細胞培養が、OD600=0.8に到達したとき、IPTGを最終濃度がmMになるまで添加し、成長は30℃で16〜20時間の間続けた。遠心分離(GS−3 Sorvall roter,7000rpm、30分間)を行った後、上澄み液を瀘過し、濃縮し、次いで、ミニセッテ(フィルトロン)接線流れ装置(Minisette(Filtron)tangential flow apparatus)を使用して、負荷緩衝液(loading buffer)(リン酸塩50mM、pH7.4、NaCl 500mM、イミダゾール20mM)の中で交換した。得られた溶液は、Ni−NTA樹脂(キアゲン)の上に負荷させ、負荷緩衝液50mlで洗浄し、次いで、溶離緩衝液(リン酸塩50mM、pH7.4、NaCl 500mM、イミダゾール20mM)で溶離した。精製した抗体は、SDS−PAGE(ラエムリ(Laemmli),1970)によって分析し、4℃のPBSに対して透析した。多価scFv断片は、結合活性効果に基づき、BIAコア上で人工的に優れた結合を示すことがあるということが知られている(非特許文献30及び51)ので、精製済みscFv製剤は、S−75コラム搭載のFPLC装置(ファルマシア(Pharmacia))を用い、ゲル濾過によって更に処理した。FPLC−精製済み単量体断片の抗体濃度は、分光測定器によって、scFv溶液1mg/mlについて1.4ユニットの280nmでの吸光度を想定して、決定した。
【0096】
PBSにおける0.1〜1μMの範囲の種々の濃度での、一価のscFvの結合を、BIAコア機器(ファルマシア・バイオセンサー)上で、次の抗原を使用しながら測定した。(i)ストレプトアビジンをコーティングしたチツプ上に固定された、ビオチニル化済み組換えFN断片7B89の1000レゾナンス・ユニット(Resonance Units)(RU)。これは、scFvの250RUによって特異的に結合する;(ii)N−末端アミノ基で化学的に固定された、組換えED−Bの200RU。これは、scFvの600RUによって特異的に結合する;(iii)ED−Bに富むフィブロネクチンWI38VAの3500RU(例3を参照)。これは、scFvの150RUによって特異的に結合する。データの動的分析は、製造者インストラクションに従って行った。抗体含有上澄み液のBIAコア定性分析に基づいて、各scFvクローンの一つの親和性成熟済みバージョン(version)、即ち、78B9断片を有する選定からのクローンCGS−1、及びED−B組換えFN断片を有する選定からのCGS−2を選定した。会合速度定数(kon)及び解離速度定数(koff)を、scFvとオリジナル・クローン28SI両方の、計算による平衡解離定数(Kd)と共に、表1に示す。クローンのCGS−1もCGS−2も、ナノモルの範囲のKdを有するが、クローンCGS−2は、センサー・チップ上で試験した3種の蛋白全てに関し、(110nMから改善されて)1nMのKdを与え(表1)、その親クローンを越えて最良の改善を示した。この改善は、主として、単量体抗体の製剤で測定された(示さず)一層緩慢な動的解離定数(10−4S−1以下)による。
【0097】
成熟方法は、大部分に共通しているように思われ、マルトース結合蛋白;シトクロムC;マウス・エンドグリン(endoglin)の細胞外ドメイン(D.N.,L.
Wyder,R.Klemenz);サイトメガロウィルス(A.P.,G.
Neri,R.Botti,P.N.);核腫瘍マーカーHMGI−C蛋白(A.P.,P.Soldani,V.Giancotti,P.N.);及び卵巣腫瘍マーカーの胎盤アルカリホスファターゼ(Deonarain及びA.A.Epenetos)に対する親和性改善済み抗体を産生した。従って、この成熟方法は、少なくとも他の成熟方法(非特許文献52及び53)と同程度に有効であるように思え、しかも、非常に大きいファージ抗体ライブラリ(非特許文献54及び55)から誘導されたものと類似の親和性を有する抗体を産生する。
【0098】
親和性成熟済みクローンCGS−1及びCGS−2は、ヒト・胚芽細胞系列の抗体V遺伝子のデータベース(V−BASE)に配列し整列し、次いで、マクベクター(MacVector)ソフトウェアを用いて翻訳した。両クローンのVH遺伝子は、ヒト・胚芽細胞系列DP47(VH3)にほとんど相同であり、各クローンの他に、異なるVH CDR3配列を有した(図1)。両クローンのVL遺伝子は、ニッシム等に記載のヒト・合成scFv全範囲(repertoire)の構造に使用されるDPL16胚芽細胞系列であった。VL CDR3配列は、任意の順序に配列された残基の6個から4個が、互いに相違した(図1b)。
【0099】
【表1】

【0100】
例3・・・ED−B含有フィブロネクチンに対する親和性成熟済みscFvsの特異性
二つの親和性成熟済みscFvs、CGS−1及びCGS−2の免疫反応性は、初期にELISAによって評価し、次いで、mAb BC−1、及びmAbIST−6と直接比較した。mAb BC−1は、B−FNイソ型を認識し、mAb IST−6は、ED−Bを欠いているFNイソ型を認識するのみである(非特許文献13及び26)。これらmAbの特性は、以前報告された(非特許文献13及び26)。その後、サーモリシン処理によって誘導されたFN断片と組換え融合蛋白との大きいパネルを使用して、特異性の正確な分析が行われた。
【0101】
ELISA及び免疫ブロット分析のために使用した抗原は、次の通りに調製した。FNは、ヒト・プラスマ;及び以前報告されたWI38VA13細胞系列の調整済み媒体(非特許文献8)から精製した。精製したFNは、非特許文献13によって報告されたように、サーモリシン(プロテアーゼ X型;シグマ・ケミカル社(Sigma Chemical Co.)を用いて消化した。天然のFN 110kD(B−)及び天然のFN 120kD(B+)の断片(図2参照)は、以前報告されたように(非特許文献56)、FN消化物から精製した。テナシン−Cの大きいイソ型は、非特許文献57によって報告されたように精製した。組換え蛋白は、例1に記載の通りに表現し、精製した。SDS−PAGE及びウェスタン・ブロッティング(Western blotting)は、非特許文献13によって報告されたように、実施した。
【0102】
ELISAに使用された全抗原は、PBSで50〜100μg/mlに希釈し、免疫−プレート・ウェル(Immuno−Platewells)(Nunc,Roskilde,デンマーク)の上に4℃で一晩コーティングした。非結合抗原は、PBSで除去し、次いで、プレートは、ウシ由来血清アルブミン(BSA)3%(w/v)含有するPBSを用いて、37℃で2時間の間、ブロックした。この後、0.05%トゥイーン(Tween)20含有PBS(PBST)で4回洗浄した。次いで、抗体は、37℃で1.5時間の間、結合させた:scFvsは、標識配列(FLAG標識に対してはmAb M2[Kodak,New Haven CT];又はmyc標識に対しては9E10[ATCC,Rockville, MD])に向けられた抗血清を用い、前もって温置した。試験を行った対照抗体は、mAbs BC−1及びIST−6であった。PBSTで4回洗浄した後、プレートは、ヤギの抗マウスIgG(Bio−SPA Division,Milan,イタリア)を(PBST+3%BSAで)1:2000に希釈し、ビオチニル化して、37℃で1時間の間、培養した。洗浄を繰り返し、次いで、ストレプトアビジンービオチニル化したアルカリ・ホスファターゼ複合体(Bio−SPA Division,Milan,イタリア)を、37℃で1時間の間、添加した(MgCl2 2mMを含有するPBSTに1:800で希釈された)。反応は、ジエタノールアミン10%、pH9.8中で、ホスファターゼ基質の錠剤(Sigma)を使用して進め、光学的濃度は、405nmで読み取った。結果は、次の表2に示す。
【0103】
【表2】
【0104】
scFv及び単クローン抗体と、フィブロネクチン誘導抗原との免疫反応性は、ELISAによって測定した。測定値は、バックグランド信号を減じた後の、405nmで測定したODを示す。データは、4回の実験の平均であり、これらデータは、最大10%の標準偏差を示す。
【0105】
実験に用いたフィブロネクチンの異なる形態の素性(identity)は、次の通りである:即ち、プラスマ FN=ヒト・プラスマフィブロネクチン;WI38−VA FN:SV4O−転換済み線維芽細胞の上澄み液からのプラスマフィブロネクチン(非特許文献8);n110kD=ED−Bを含有しない、FNドメイン4で処理したサーモリシン;n120kD=ED−Bを含有した、FNドメイン4で処理したサーモリシン;recFN7B89=隣接するIII型FN相同繰り返しによって側面に位置するED−Bドメイン;recFN789=ED−Bドメインを有する、III型FN相同の繰り返し;recED−B=組換えED−B単独;recFN6=組換えFNドメイン6。
【0106】
CGS−1もCGS−2も、ED−B配列を含有する、天然のFN断片の全て又は組換えFN断片の全ての他に、組換えED−Bペプチドを認識した。しかし、それらは、ED−Bを欠くFN断片のいずれにも結合しなかった。また、CGS−1及びCGS−2は、テナシン(これは、15個のIII型相同繰り返しを含む:非特許文献58);及びプラスマ FN(これは、サーモリシン消化生成物(非特許文献8)中に、検出可能なレベルのED−B配列を含有していない)と反応しなかった。対照的に、CGS−1及びCGS−2は、SV40−転換済み細胞系列WI38VAから精製したFNと強く反応した。精製済みFN及びその細胞系列から調製した全RNAを使用した、サーモリシン消化及びS1ヌクレアーゼの実験によって示されるように(非特許文献8;Borsi等,1992)、この細胞からのFN分子約70〜90%には、ED−Bが含まれる。可溶性組換えED−Bを使用して、WI38VA細胞上のFに対するCGS−1及び/又はCGS−2の結合を抑制することによって、FNのED−B成分に対するscFvの特異性を更に実証した(データは示していない)。
【0107】
データによると、CGS−1及びCGS−2のみが、ED−Bドメインを含有するFN誘導体と特異的に反応することが確認される。それらは双方とも、組換えED−Bの場合を除き、mAb BC−1と同様の反応性を示す。組換えED−Bは、BC−1によって認識されていない。mAb対照に関連して得られたELISA信号の強度は、ED−B含有抗原に対する二つのscFvの高い特異性を反映する。
【0108】
CGS−1及びCGS−2の特異性は、プラスマ及びWI38VAの細胞からのFNと、それらのサーモリシン消化物とを使用しながら、免疫ブロット分析により調べた。サーモリシン消化によると、WI38VA細胞からのFN(その大部分はED−Bを含有する)は、(ED−Bを含有する)120kD断片と、ED−Bを欠く僅かな110kD断片とを生成する(図2A;非特許文献8)。120kDドメインの更なる消化は、二つの断片:即ち、そのカルボキシ末端にほぼ完全なED−B配列を有する85kD断片;及び35kD配列を生成する(図2A;非特許文献8)。
【0109】
図2Bの左側には、免疫ブロット分析によって分析した蛋白断片のクーマシー染色済みゲルがある。プラスマFN(レーン1)と、蛋白のサーモリシン消化物(110kD蛋白を含有するレーン3,消化済み110kD蛋白を含有するレーン4)とは、CGS−1及びCGS−2によって認識されなかった。対照的に、WI38VA細胞からの、ED−Bに富むFNは、元のまま及びサーモリシン消化を増大した後の両方において、両方のscFv断片によって認識された。CGS−1によって特異的に認識し得た、最も小さいFN−誘導断片は、(ほとんど全てがIII型の繰り返し2〜11による)120kD蛋白であり、一方、CGS−2は、ED−BのN−末端の他に、繰り返し2〜7による85kD断片を認識し得た(図2B;非特許文献8)。これらの結果は、二つのscFvが反応性であり、ED−B配列内のエピトープを識別することを示す。85kDドメインに対するCGS−2の結合は、このクローンのためのエピトープがED−Bのアミノ末端に存在することを示す。対照的に、120kDドメインが85kDへ消化されるときの、CGS−1の結合の損失は、それによって、ED−B分子のカルボキシ末端の方に向いて一層多く配置されたエピトープが認識されることを実証する。
【0110】
CGS−1及びCGS−2の優れた特異性は、ED−B配列を持った又は持たない融合蛋白及び組換えFN断片を用いた免疫ブロット分析によって更に研究した。FN融合蛋白は、非特許文献13によって記述されている通りに調製した。これらの実験結果は図3に示す;ヒト・FNのドメインの構造の概略図関連は、非特許文献26を参照。得られた結合のプルフィルによると、精製済みFNと蛋白分解の分割生成物とに関する免疫ブロット分析及びELISAによって従来見いだされていたものが、本質的に確認される。即ち、CGS−1及びCGS−2は、ED−B含有FN断片(レーン2及び4)と強い反応性があったが、ED−Bを欠くFN配列(レーン1及び3)には反応性を全く示さなかった。CGS−1は、ヒト・ED−B融合蛋白(レーン5)ともニワトリ・ED−B融合蛋白(レーン6)とも反応しなかったが、CGS−2は、両方の断片と強く反応した(図3)。この結果は、CGS−1によって認識されたED−B含有FN中のエピトープの、ある立体配座上の制約を反映しているのかも知れない。例えば、エピトープが、変性に敏感であり、又はSDS−PAGEによって分断される(fractionated)とき、正確には現出されず、ニトロセルロース等の固形支持体に転写されるということは起こり得る。
【0111】
まとめて考えると、これらの結果によると、CGS−1及びCGS−2は、互いに異なり且つED−B構造(これは、mAb BC−1によって認識される)と異なる領域で、ED−B含有FNに強く且つ特異的に結合することが実証される。
【0112】
例4・・・ヒト及びマウスの腫瘍を免疫細胞化学的に染色する上で親和性成熟済み抗ED−B scFvsを使用する方法
CGS−1及びCGS−2はいずれも、種々の正常で新生のヒト・組織中に、ED−B含有FN分子を免疫局在化する(immunolocalise)ために使用されてきた。B−FNイソ型は、皮膚創傷の治癒の間、マクロファージ及び線維芽細胞の中に表現されることが知られていたので、正常組織には皮膚が選ばれた(非特許文献8及び非特許文献59)。選定された二つのヒト・腫瘍は、従来、抗フィブロネクチンmAbで染色する特異性のために分析されてきた。この腫瘍の血管内の内皮細胞は、B−FNイソ型の表現を含む血管形成過程が増加して、高度に増殖性の状態にあるため、膠芽腫(glioblastoma)の多形は詳細に研究されてきた(Carnemolla等,1994)。更に、正常で増殖性で新生のヒト・胸組織の多様のパネルを用いた研究によって、血管形成とB−FN表現の間の相互関係に関する一層の証拠が提供された(非特許文献60)。
【0113】
本明細書に記載の諸実験のために、CGS−1及びCGS−2の免疫組織化学的染色は、mAb BC−1(これは、B−FNイソ型を認識する)の免疫組織化学的染色;及び既知の全FNイソ型変異体(IST−4)に反応する、又はED−Bを欠くFNイソ型(IST−6)のみに反応することが知られている他のmAbの免疫組織化学的染色と比較した。これら対照抗体の全ての特徴は、従来報告されている(非特許文献13及び26)。
【0114】
正常で新生の組織は、手術中に採取されるサンプルから得た。組織の調製及び固定が、FN含有分子の正確で感度のよい検出のために重要であることは既に実証されていた(Carnemolla等,1994)。免疫組織化学のために、5μm厚さのクリオスタット分節(cryostat sections)は、空気乾燥され、冷アセトン中に10分間、固定された。免疫染色は、ストレプトアビジン−ビオチン・アルカリ・ホスファターゼ複合体染色キット(Bio−SPA Division,Milan,イタリア)と、ナフトール−AS−MX−リン酸塩と、ファースト・レッド(Fast Red)TR(Sigma)とを用いて行った。ギル・ヘマトキシリン(Gill’s haematoxylin)は、対比染色として使用した。次いで、従来、「Carnemolla等,1994」によって報告されているように、グリセロゲル(glycergel)(Dake,Carpenteria,CA)中に固定した。実験で特異性を更に分析するため、組織のポジティブ染色(positive staining)が得られる所で、抗体を組換えED−Bドメインで培養し、次いで、従来記述された通りに検出することによって、ED−Bに対する特異性を実証した。
【0115】
これらの全実験結果によって、CGS−1もCGS−2も、mAb BC−1と同一の組織構造と反応することが分かった。CGS−1、CGS−2及びBC−1を用い、皮膚で得られた染色パターンは、真皮(dermis)中に表現されたFNからのED−Bは存在しないことを反映している。浸潤性管の癌分節の染色において、CGS−1、CGS−2及びBC−1は、制限された染色分布を示し、新生細胞と基質(stroma)との間の境界に限定した。このことは、全FNは腫瘍基質全体に均一に分布しているが、B−FNの表現は、ある領域に限定されていて、しかも、従来、mAb BC−1を用いて(非特許文献60)、浸潤性管の癌の中に(ケースの95%で)うまく局在化させられたのはこれらの領域であるという事実と一致する。
【0116】
膠芽腫の多様な癌のBC−1を染色することにおける従来の発見が、確認された。非特許文献24は、糸球体様脈管構造の染色の典型的なパターンを観察していたが、我々の実験では、CGS−1及びCGS−2は、定性的には同一の結果を与えることが分かった。
【0117】
しかし、CGS−1とCGS−2とmAb BC−1との間には、重要な相違が存在する。即ち、BC−1は厳密にヒト・特異性であるのに、二つのヒト・scFvはニワトリ及びマウスのB−FNの両方に結合することが実証された。CGS−2はニワトリ・胚芽と反応し(データは示さず)、CGS−1及びCGS−2はマウス・腫瘍と反応した。
【0118】
マウスF9の奇形癌の分節上の血管構造のCGS−1染色も示した。対照的に、試験を行った全ての正常マウス組織(肝臓、脾臓、腎臓、胃、小腸、大腸、卵巣、子宮、膀胱、膵臓、副腎、骨格筋、心臓、肺、甲状腺及び脳)は、CGS−1及びCGS−2とネガティブ染色反応を示した(データは示さず)。そのF9奇形癌分節中で染色された構造は、組換えED−Bドメインを使用して、得られた染色を完全に抑制することによって、ED−B特異性であることが分かった(データは示さず)。
【0119】
例5・・・ヒト腫瘍を生体内で標的化する上で、親和性成熟済み抗ED−B scFvを使用する方法
ヒト・メラノーマの細胞系列SKMEL−28は、一つの側腹部の皮下に1×107の細胞/マウスを注射することにより、6〜10週間の老齢ヌードマウス(Balb−c又はMF−1;Harlan UK)中の異種組織移植済み腫瘍を成長させるのに使用した。腫瘍を有するマウスは、腫瘍が直径約1cmに達したとき、PBSのscFv1−Cy1溶液1mg/mlの100μlを用いて、尾部の静脈に注射した。
【0120】
CY7による組換え抗体の標識化は、PBS(1mg/ml)の抗体溶液1m1に「1M炭酸ナトリウム、pH=9.3」100μlと、「CY7−bis−Osu(Amersham;Cat.Nr.PA17000;DMSO中2mg/ml)」200μlを添加することによって達成した。室温で、30分後、「1MTris、pH=7.4」100μlをその混合物に添加し、次いで、標識化された抗体は、PBSと平衡状態に置いた、使い捨てPD10カラム(Pharmacia Biotech,Piscataway,NJ,米国)を使用して、未反応染料から分離した。溶離した生の抗体断片は、セントリコン(Centricon)−10チューブ(Amicon,Beverly,MA,米国)を用いて、約1mg/mlに濃縮した。達成された標識化比は、通常、1モルのCY7:1モルの抗体に近い。これは、抗体溶液1mg/mlは280nmで1.4単位の吸収を与え、747nmでのCY7のモル吸光係数は200,000(M−1cm−1)であると想定し、また、280nmでのCY7吸収を無視し、lcmキュベットを用い分光器によって評価した。標識化した後の、抗体サンプルの免疫反応性は、抗原カラム上の親和性−クロマトグラフ法によるバンド−シフト(Neri等,1996)によって;又はBIAコア分析によって確認した。マウスは、酸素/フルオロエタンの混合物の吸入による麻酔の下、一定時間間隔で、ホーム・ビルト(home−built)マウス−イメージャー(mouse−imager)を用いてイメージした。結果の再現性を確実なものとすべく、各サンプルに対し、2〜8匹の動物を研究した。操作手順は、D.ネリ(Neri)に発行されたUKプロジェクト・ライセンス(Project Licence)「腫瘍の標的化(Tumour Targeting)」(UK PPL 80/1056)に従って行った。
【0121】
非特許文献61の光検出装置の改造型として、赤外線蛍光物質CY7が使用できる赤外線マウス・イメージャーを造った。赤外線照射を選定し、一層優れた組織貫通を得た。CY7(>76nm)の蛍光は、人には見えず、コンピューター制御のCCD−カメラを使用する必要がある。このマウス・イメージャーは、黒に塗装され、光が入らないようにした箱を有した。この箱には、100Wタングステン・ハロゲン・ランプを備え、CY7(クローマ社(Chroma Corporation),Brattleboro,VT,米国;673〜748nm)のために特別に設計した直径50mmの励起フィルターを備えた。得られた照射線は、5×10cmの大きさの領域全体に渡り、ほぼ均一であった。マウスはイメージングのためにその中に置いた。蛍光は、C−マウント・レンズと50mm放出フィルター(クローマ社,Brattleboro,VT,米国;765〜855nm)を備えた、8ビットのモノクロのプルニックス(Pulnix)CCDカメラによって検出し、次いで、イメージ(Image)DOK装置(Kinetic Imaging Ltd.,リバプール、UK)に接続した。この装置は、フレーム・グラバー(frame−grabber)と、配列イメージを捕獲し集積化を行うためのソフトウェアとを備えたコンピューターから成る。それぞれ50msで捕捉された配列のイメージは、典型的には、平均化プロセスに使用した。これの番号は、一つの動物の一連の画像では一定に保持し、異なる時点で標的化されている腫瘍が直接比較できるようにした。次いで、TIFFフォーマットでの画像は、プログラム・グラフィック・コンバーターを使用して、PICTファイルに変換し、次いで、パワー・マッキントッシュ7100/66コンピューターを用いて、精巧に仕上げた。
【0122】
この装置のデザインの概略図は、図4に示す。
【0123】
これらの実験によって、両方のscFvは、巨視的レベルに視覚化されたとき、腫瘍に局在化していることが実証された。
【0124】
二つの抗EDB scFvを用いて、成長している腫瘍の新生血管形成を標的化する巨視的実証は、次のように詳述される。
【0125】
一つの側腹部に、異種組織移植されたSKMEL−28のヒト・メラノーマ、又はマウスF9奇形癌を有する、ヌードマウス及び/又はSCIDマウスは、FLAG標識(tag)を有する標識化されていないscFv断片、又はビオチニル化されたscFv断片を用いて注射した。
【0126】
マウスは、注射の後、異なる時点で犠牲となり、腫瘍及び非腫瘍の分節を得た。次いで、これら分節は、抗FLAG M2抗体(Kodak,181)又はストレプトアビジン−ベースの検出試薬を使用し、従来の免疫組織化学プロトコールで染色した。最適標的化は、一般に注射の12時間後に得られた。CGS1もCGS2も、異種組織移植されたヒト・腫瘍、及びマウスの奇形癌の両方の新生血管に結合することが実証された。
【0127】
例6・・・ヌードマウスに異種組織移植されたF9ネズミの奇形癌の標的化
我々は、ネズミF9の奇形癌細胞4×106を皮下注射することによって、ヌードマウスの側腹部で固形腫瘍を成長させた。この腫瘍は、マウス内で非常に急速に成長し、注射後、約1週間で1cmに達し、高度に血管が新生した。抗体の標的化をイメージすべく、我々は、非特許文献61の光検出手順の改造型を使用した。この改造型は、上記に詳述する通り(図4参照)、種々の時点でイメージされた同一動物に関する腫瘍標的化及び抗体の排除を可能にする。
【0128】
腫瘍に標的化し、抗体の検出を容易にするため、表現ベクターPGIN50のSfil/Notl部位における抗体をサブクローン化することによって、scFv(CGS−1)、scFv(CGS−2)及び抗リゾチーム scFv(D1.3)(非特許文献62)に、ホモ二量体化標識(非特許文献63)を追加した。このベクターは、pD268(非特許文献64)の誘導体である。この中で、標識のHis6配列は、配列:GGC LTD TLQ AFT DQL EDE KSA LQT EIA HLL KEK EKL EFI LAA Hによって置換される。これは、共有の抗体断片のホモ二量体化のためのFos蛋白の両親媒性のらせんと、システイン残基とを含む(Abate等,1990)。完全な共有の二量体化は、達成されなかった。抗体断片約30〜50%は、共有結合した二量体から成った。
【0129】
抗体断片は、雌鶏、卵のリゾチーム(D1.3)又は7B89(抗ED−B抗体;Carnemolla等,1996)をCNBr活性化されたセファローズ(Sepharose)(Pharmacia Biotech,Piscataway,NJ,米国)にカップリングすることによって得たカラムのアフィニティ・クロマトグラフィによって精製した。上澄み液は、アフィニティ支持体の上に供給し、次いで、PBSで洗浄し、PBS+0.5MNaClで洗浄し、次いで、100mMのEt3Nで溶離した。次いで、抗体は、PBSに対して透析して分離した。
【0130】
抗体は上述の通り、標識化し、次いで、PBSの1mg/ml scFv1−Cy71溶液100μlで、腫瘍を有するマウスの尾部の静脈に注射した。このとき、腫瘍は、直径約1cmに達していた。
【0131】
図5に示す通り、scFv(CGS−1)は、3日間以下の間、腫瘍上に局在化した(この期間に腫瘍からの急速なクリアランスもあったが)。しかし、大たい骨の、ある染色も存在した。CGS−1の腫瘍への標的化の実施は、C末端に、システイン残基を含有する両親媒性のらせんを導入して、抗体の二量体化(非特許文献63)を促進することによって、顕著に改善された。実際、二量体のscFv(CGS−2)2の局在化は、24〜72時間、有意には減少しないように思えた。対照的に、ネガティブ対照(二量体の抗体scFv(D1.3)2、抗リゾチーム抗体)は、急速なクリアランスを示し、腫瘍又は大たい骨の検出可能な局在化は全く見られなかった。
【0132】
scFv(28SI)は、6時間で腫瘍ターゲッティングをわずかに示した(図示せず)。しかし、24時間又はそれ以降において、全く検出できなかった(図6)。親和性の成熟は、ターゲッティングを著しく改善することとなった。このように、scFv(CGS−2)は、単量体(図6)としてであろうと、二量体(図示せず)としてであろうと、小さいF9腫瘍及び大きいF9腫瘍を有効に標的化した。2日後、腫瘍1g当り抗体の注射投与量%は、scFv(CGS−2)単量体では約2、scFv(CGS−2)二量体では3〜4であることが分かった。腫瘍に運ばれたscFv(CGS−2)による投与量もまた、scFv(CGS−1)によるものより大きく(図5及び6)、それらの各々親和性と相互関係がある(表1)。しかし、scFv(28SI)もscFv(CGS−2)も、蛋白質分割しないように思え、また肝臓の取り込みが大きいことを示す(図6)ところが、scFv(CGS−1)抗体は、有意に一層安定であり、肝臓の取り込みが一層小さいことを示す(図5)。
【0133】
【表3】
【0134】
【表4】
【0135】
【表5】
【0136】
【表6】
【0137】
【表7】
【0138】
【表8】
【0139】
【表9】
【図面の簡単な説明】
【0140】
【図1a】scFv CGS−1及びCGS−2のVHの、整列したアミノ酸配列であって、CDRs(1、2及び3)を示す。
【図1b】scFv CGS−1及びCGS−2のVLの、整列したアミノ酸配列であって、CDRs(1、2及び3)を示す。
【図2A】ヒトFNサブユニットのドメイン構造のモデルを示す。
【図2B】プラスマの4〜18%SDS−PAGE;WI38VA;それらのクーマシーブルー(Coomassie Blue)染色済みサーモリシン消化生成物;並びに、BC−1、IST−6、CGS−1及びCGS−2で調べた免疫ブロット分析を示す。
【図3A】大腸菌に表現された融合蛋白及び組換え蛋白の中に含有されるFNIII型繰返し配列;これら蛋白とCGS−1及びCGS−2との反応性;並びに、これら蛋白とmAbs BC−1及びIST−6との反応性を示す。
【図3B】クーマシーブルー染色済みゲル;及び、CGS−1、CGS−2、BC−1、IST−6で調べた免疫ブロット分析を示す。
【図4】マウスの赤外線イメージャー;タングステン・ハロゲンランプを備えた、黒色の非蛍光性箱と;CY7赤外線蛍光物質に対して特異的な励起・放出フィルターと;コンピュータ制御の8−ビット単色CCD−カメラとから成る、標的実験のために使用したマウス・イメージャー(imager)である。
【図5】B−FNに向けられた単量体のscFc(CGS−1)及び二量体のscFc(CGS−1)2を用いて、蛍光によって標識化された抗体断片を、標的としてのF9ネズミの奇形癌に向けられている。
【図6】B−FNの同一エピトープに向けられた、親和性成熟のscFv(CGS−2)及び低結合性scFv(28SI)を使用して、蛍光によって標識化された抗体断片が、F9ネズミの奇形癌に向けられている。
【技術分野】
【0001】
背景
本発明は、フィブロネクチン、ED−B(これはまた、免疫細胞学と生体内(in vivo)腫瘍を標的にすることとの両方によって実証されている通り、成長している腫瘍の新生血管形成(neovasculature)において滲出する)の胎児イソ型(foetal isoform)のための特殊な結合剤に関する。本発明はまた、かかる特殊な結合剤に係る物質及び方法にも関する。
【背景技術】
【0002】
腫瘍治療の現行技術の大抵のものの第一の狙いは、腫瘍のできるだけ多くの構成細胞を殺すことにある。化学療法及び放射線療法を用いて経験してきた成功が限られているのは、治療の特殊性(specificity,専門性)が比較的欠如していること、及び正常組織に及ぼす有毒性副作用の傾向があることと関係がある。腫瘍のための治療の選択性を改善し得る一つの方法は、結合タンパク質を通して、腫瘍へ物質(agent,活性物質)を運ぶことである。この結合タンパク質は通常、腫瘍の表面上に滲出した標識抗原(marker antigen)のための特殊性を有する、抗体結合ドメイン(binding domain,結合領域)から成る。俗に「マジック弾(magic bullets)」と呼ばれている、標的にされた腫瘍のかかる形態は、げっ歯類(rodents)からの単クローン性抗体(mAbs)によって、主に実証されてきた。mAbは、細胞表面上に滲出したいわゆる腫瘍関連抗原に対して特異性がある。そのようなmAbは、細胞毒部分(例えば、毒素又は薬物)へ化学的に結合していても良いし、或いは組換え融合蛋白質として産生しても良い。ここに、mAbを記号化した遺伝子と毒素とは一緒に連結され、縦列に滲出される。
【0003】
「マジック弾」のアプローチは、ヒト・癌の治療において重要であり有効であり、リンパ起始(lymphoid origin)の腫瘍を標的にする場合に最も注目されている(ここに、悪性細胞は循環中の治効量に最も影響を受けやすい)が、このアプローチには限界がある。しかし、固体腫瘍の治療は、全細胞塊の微小部分(腫瘍の最も外側の周辺の細胞が支配的である)のみが循環中の治療的免疫接合(therapeutic immunoconjugates)にさらされるという点で、重大な臨床的問題を残している;これらの周辺標的は、腫瘍内部に対していわゆる「結合部障壁(binding site barrier)」を形成する(非特許文献1)。腫瘍内部における組織の構造は、一般的に線維支質を有し、あまりにも稠密であり、腫瘍細胞に密に包み込まれているため、抗体の大きさ範囲の分子は浸透することができない。更に、腫瘍は、リンパ排液が欠如しているため、大きな間隙圧力を有することが知られている。腫瘍はまた、外因性分子の流入を妨げる。腫瘍内への治療薬の取込みに影響を及ぼす諸因子に関する最近の報告については、非特許文献2を参照。
【0004】
腫瘍関連抗原を標的にすることによって固形腫瘍を治療することには明らかに限界があるものの、これらの腫瘍は、抗体を治療するために代替的な抗原を標的にするという共通の特徴を有するのである。一旦、腫瘍がある大きさを越えて成長したとき、腫瘍は一般的に、成長を支えるための十分な酸素と養分とを、独立した血液供給に依存する。もし、この血液供給を妨げるか、又は遮ることができるのなら、過程において、現実的に何千もの腫瘍細胞を餓死させることができる。腫瘍が成長するにつれて、腫瘍は、隣接した毛管内皮細胞に作用する脈管由来諸因子の別種の配列を生じさせ、それらを誘導して(inducing)増殖し移動しながら、スウィッチを受けて脈管由来表現型になる。これらの新たに形成された血管の構造は、正常組織の秩序のある構造とは著しく対照的に、出口のない末端(blind endings)と漏出の増大に通じる穿孔とを有し、高度に崩壊している。血管形成の誘導は、ある細胞表面の抗原の表現のアップレギュレーション(upreglation)によって成し遂げられ、それらの多くは正常組織の脈管構造に共通している。
【0005】
腫瘍の新生血管形成に特有な抗原を識別することは、血管を標的にすることによる、固形腫瘍の一般的療法を開発する上での主要な限定因子(limiting factor)であった。本発明の主題である抗原は、かかる問題に直接向いている。
【0006】
腫瘍が進行している間、周囲組織の細胞外マトリックスは、二つの主要過程、即ち、(1)正常組織の細胞外マトリックス成分のタンパク分解の低下、及び(2)腫瘍細胞と、腫瘍誘導された(tumour−induced)シトキンによって活性化された支質細胞とによる、細胞外マトリックス成分のドゥノボ(de novo)合成によって、再構築される。これらの二つの過程は、安定な状態で、「腫瘍細胞外マトリックス(tumoral extracellular matrix)」を生じさせる。この腫瘍細胞外マトリックスは、腫瘍の進行にとって一層適した環境を与え、しかも定性的かつ定量的に正常組織のそれと全く異なっている。このマトリックスの諸成分の中には、テナシン及びフィブロネクチン(FN)のイソ型が多量存在する。これらの蛋白質のイソ型としての記述(description)から、それらの構造上の大きな異質性が認識される。この構造上の異質性は、転写レベル、転写後レベル及び翻訳後レベルでもたらされる。本発明の主題は、フィブロネクチンのイソ型の一種の、いわゆるB+イソ型(B−FN)である。
【0007】
フィブロネクチン(FN)は、細胞外マトリックス及び体液の多機能性高分子量の糖蛋白成分である。これらの成分は、正常細胞の定着(establishment)及び維持、細胞移動、うっ血、血栓症、創傷の治癒、腫瘍原性(oncogenic)転換等、多くの異なる生物過程に関与している(報告に関しては、非特許文献3〜6;ハイネス,1990;非特許文献7を参照)。FNsの構造上の相違は、FN一次転写物の三つの領域(ED−A、ED−B及びIIICS)によってもたらされ(非特許文献5及び8)、少なくとも20種の異なるイソ型が生じる。これらイソ型の幾つかは、腫瘍及び正常組織では区別して表現される。FN−プレ−mRNAのスプライシングパターンは、進行中のものに特異的な組織態様に調整されるだけではなく、移動済み細胞及び悪性疾患の中で調整解除されることが知られている(非特許文献8〜16)。実際、ED−A、ED−B及びIIICSの配列を有するFNイソ型は、大部分、正常細胞中よりも移動済み細胞及び悪性腫瘍細胞の中に表現される。特に、ED−B配列(B+イソ型)を有するFNイソ型は、創傷治癒の間だけではなく、胎児組織及び腫瘍組織の中に高度に表現される。しかし、成人の正常組織中では表現が制限される(非特許文献13及び17〜23)。B+FNの分子は、成熟した血管中では検知できないが、正常な発育(例えば、子宮内膜の発育)、病の進行(例えば、糖尿病性の網膜障害の)及び腫瘍の進行の状態にある脈管由来の血管中では、アップレギュレーションとなる(非特許文献24)。
【0008】
ED−B配列は、単エクソンによってコード化された完全型III−相同(homology)の繰り返しであり、91種のアミノ酸から成る。細胞型特異性結合部を有する、代替的スプライシングされたIIICSイソ型とは対照的に、A+イソ型及びB+イソ型の生物機能は、まだ推測の域を出ていない(非特許文献25)。
【0009】
B+イソ型の存在自体は、腫瘍誘導された新生抗原を構成する。しかし、加えるに、ED−B表現は、III型の繰り返し7以内の、正常に潜在する(normally cryptic)抗原を露出する(前のED−B);このエピトープはFN分子欠乏ED−Bの中に露出されないので、ED−B表現は直接的にも間接的にも新生抗原の表現を誘導することとなる。この潜在的抗原の部位は、BC−1と呼ばれる単クローン性抗体(mAb)の標的を形成する(非特許文献26)。このmAbの特異性と生物諸特性は、特許文献1に記載されており、イギリス(UK),ソールズベリー(Salisbury),ポートンダウン(Porton Down),動物細胞培養欧州共同体(the European Collection of Animal Cell Cultures)に委託されている融合細胞腫(番号88042101)から入手することができる。mAbは、成熟した血管内皮への交差反応性なしで、腫瘍の脈管由来の血管を一局部に止めるのに使用され成功してきており、抗体を使用して血管を標的にするためのFNイソ型の可能性を説明している。
【0010】
しかし、BC−1 mAbの特異性に対する幾つかの注意点が残っている。BC−1がB+イソ型を直接的に認識しないという事実により、幾つかの組織において、BC−1によって認識されるエピトープはED−Bが存在しなくても暴露され(unmasked)、その結果、不要なBC−1の交差反応性へ間接的に導き得るかどうかという疑問が生じた。また、BC−1は、ヒト・B+イソ型には厳密に特異的であり、生物分布(biodistribution)及びBC−1の腫瘍局在化に関する、動物の研究はできないということを意味する。B+イソ型を有する組換え融合蛋白に対する多クローン性抗体は生成されてきた(非特許文献27)が、それら抗体は、エピトープを暴露するためのN−グリカナーゼ(N−glycanase)で処理されたFNと反応するのみである。
【0011】
マウス・単クローン性抗体を使用する上での更なる一般的問題は、ヒト・抗マウス抗体(human anti−mouse antibody)(HAMA)の反応である(非特許文献28及び29)。HAMA反応は、治療量の減量に役立つ、投与済み抗体の中和から、アレルギー反応、血清病及び腎臓障害まで、ある範囲の効果を有する。
【0012】
組換えED−Bと反応する多クローン性抗血清は確認されてきた(上記を参照)が、マウスの免疫化の結果として生じるBC−1と同一の特異性を有するmAbの分離は、一般的に困難であることが分かった。なぜなら、ヒト及びマウスのED−B蛋白は、ほぼ100%の配列相同を示すからである。従って、ヒト・蛋白は、それへの免疫反応を備えていないマウスにとっては自己抗原であるように見えるかも知れない。実際、何十年にも渡るこの分野の徹底した研究において、ED−Bを直接的に認識するものは何もなく、単mAbのみがB+FNイソ型(BC−1)に対する間接的反応性を有するものとして認定されてきた。BC−1の特異性は、ED−B自体の部分のためというよりむしろ、潜在的エピトープのために、ED−Bの結果として暴露されるということは間違いなく意義がある。
ED−Bは、マウスFNが恐らく欠けていて、マウスの免疫系による「自己」と見られないものと思われる。
【0013】
本発明は、以前使用されたものに対する代替的方法を使用して実現された。ここでは、フィブロネクチン又はED−Bによる従来の免疫化は必要でない:ED−Bイソ型に対する特異性を有する抗体は、線維状バクテリオファージの表面に表示される、ヒト・抗体の種々の領域のライブラリ(libraries)から単鎖Fvs(scFvs)として得られてきた(非特許文献30及び特許文献2〜6をも参照)。
【0014】
ED−Bを有する組換えFN−断片(fragments)と、組換えED−B自体との両者の上に直接選択することにより、特異性scFvが分離され得ることを、我々は、抗体ファージライブラリ(antibody phage library)を使用することによって見いだした。このとき、これら抗体は固形表面の上に被覆される(「パンニング(panning)」)。抗原のこれら同一源もまた、「二代雑種第二子世代(second generation)」のscFvを生成するのに使用され成功し、「親和力成熟(affinity maturation)」の過程で、親クローンに関する諸特性が改善されてきた。分離されたscFvは、N−グリカナーゼで前処理をしないで、ヒト・FNのB+イソ型と強く且つ特異的に反応することを我々は見いだした。
【0015】
抗腫瘍の適用において、本発明によって提供されるヒト・抗体抗原結合ドメインは、HAMA反応を受けないという利点を有する。また、本明細書中に例証される通り、これら結合ドメインは、生体外でも生体内でも、腫瘍組織の免疫組織化学分析において有用である。これらの用途及び他の用途は、本明細書中で更に論じられ、当業者には容易に理解できる。
【0016】
術語
特異的結合メンバ(specific binding member)
特異性結合メンバとは、相互に対して結合特異性を有する一対の分子のメンバをいう。特異性結合ペアメンバは、自然界で得られるが、全部又は一部を合成によって生成することができる。一対の分子の一つのメンバは、その表面上に、領域又はキャビティ(cavity,空洞)を有する。この領域又はキャビティは、その一対の分子の他のメンバに特異的に結合し、それ故に、その極性空隙機構に対して相補的である。このように、ペアメンバは相互に特異的に結合する特性を有する。特異性結合メンバのタイプの例は、抗原−抗体、ビオチンーアビジン、ホルモン−ホルモン受容体、受容体−リガンド、酵素−基質である。
【0017】
抗体(antibody)
抗体とは、自然界に存在し、又は一部若しくは全部を合成によって生成される免疫グロブリンをいう。この術語には、結合ドメインを有するあらゆるポリペプチド又は蛋白質も含まれる。この結合ドメインは、抗体の結合ドメインであるか、又は抗体の結合ドメインと同種のものである。これらは、自然源から得ることができるが、一部又は全部を合成によって生成することができる。抗体の例は、免疫グロブリンのイソ型(isotypes)及びこれらイソ型の亜綱(subclasses);Fab,scFv,Fv,dAb,Fd等の抗原結合ドメインから成る断片;並びにダイヤボディ(diabodies)である。
【0018】
単クローン抗体及び他の抗体を利用し、組換えDNA技術の技法を使用して、原抗体の特異性を保持する、他の抗体又はキメラ(chimeric)分子を生成することは可能である。そのような技法には、免疫グロブリンの不変領域又は不変領域プラス外郭構造に対する抗体の、免疫グロブリン可変領域又は相補性決定領域(CDRs)をコード化するDNAを導入することを伴うこともある。例えば、特許文献7〜9を参照。抗体を生成する融合細胞腫又は他の細胞は、遺伝的変異(genetic mutation)又は他の変化を受けることもある。これらの変化によって、生成される諸抗体の結合特異性を変えても良いし変えなくても良い。
【0019】
抗体は多くの方法によって変性することができるのであるから、用語「抗体」は、必要とされる特異性を持つ結合ドメインを有する、あらゆる特異性結合メンバ又は物質と解釈されるべきである。このように、この用語は、抗体断片、誘導体、機能的に同等の物、及び抗体の同族体を意味し、自然界で得られようと全部又は一部を合成によって得られようといずれにせよ、免疫グロブリン結合ドメインから成るあらゆるポリペプチドを含んでいる。それ故に、もう一つのポリペプチドに融合した、免疫グロブリン結合ドメイン又は同等物から成るキメラ分子が含有される。キメラ抗体のクローン化及び表現は、特許文献10及び11に記載されている。
【0020】
抗体全体の断片は結合性抗原の機能を果たし得るということが示された。結合性断片の例は、(i)VL、VH、CL及びCH1のドメインから成るFab断片;(ii)VH及びCH1のドメインから成るFd断片;(iii)単抗体のVL及びVHのドメインから成るFv断片;(iv)VHドメインから成るdAb断片(非特許文献31);(v)分離されたCDR領域;(vi)F(ab’)2断片、二つの連結したFab断片から成る二価の断片;(vii)単鎖Fv分子(scFv)(ここに、VHドメイン及びVLドメインは、この二つのドメインを結合させて抗原結合部を形成させるペプチドリンカーによって連結される(非特許文献32及び33);(viii)二重特異性単鎖Fv二量体(特許文献12);並びに(ix)「ダイアボディ(diabodies)」、遺伝子融合によって構成される多価断片又は多重特異性断片(特許文献13及び非特許文献34)である。
【0021】
ダイアボディはポリペプチドの多量体(multimers)であり、各々ポリペプチドは、免疫グロブリンの軽鎖(light chain,L鎖)の結合性領域を含む第一ドメインと、免疫グロブリンの重鎖(heavy chain,H鎖)の結合性領域を含む第二ドメインとから成る。この二つのドメインは、(例えば、ペプチドリンカーによって)連結されているが、相互に結合して(associate)抗原結合部を形成することはできない。抗原結合部は、多量体内の一つのポリペプチドから成る第一ドメインと、多量体内のもう一つのポリペプチドから成る第二ドメインとの結合(association)によって形成される(特許文献13)。
【0022】
二重特異性抗体を使用する場合、これらは、従来の二重特異性抗体であっても良い。これらは、種々の方法で製造される(非特許文献35);例えば、化学的に調製又はハイブリッド融合細胞腫から調製することができる。或いは、これら二重特異性抗体は、上述の二重特異性抗体断片のいずれかであっても良い。完全な抗体よりむしろ、scFv二量体又はダイヤボディを使用するのが好ましい場合もある。ダイヤボディ及びscFvは、可変ドメインのみを使用し、抗イディオタイプ(anti−idiotypic,抗個特異的抗原)反応の影響を潜在的に減少させながら、Fc領域なしで構成し得る。二重特異性抗体の他の形態には、非特許文献36に記載の単鎖「ヤーヌシンズ(Janusins)」が含まれる。
【0023】
二重特異性の完全抗体とは対照的に、二重特異性抗体のダイヤボディは、特に有用である場合もある。なぜなら、これらダイヤボディは、大腸菌(E.coli.)中で容易に構築され表現されるからである。適切な結合特異性のダイヤボディ(及び抗体断片等の他の多くのポリペプチド)は、ライブラリからファージ表示(phage display)を使用しながら、容易に選択することができる(特許文献13)。もし、ダイヤボディの一つの腕(arm)が、例えば、抗原Xに向けられた特異性で一定に保持されるなら、そのとき、他方の腕は変化し、適切な特異性の抗体が選ばれる所にライブラリが造られる。
【0024】
抗原結合性ドメイン
抗原結合性ドメインとは、抗原の一部又は全部に特異的に結合して相補的である領域から成る抗体の一部をいう。抗原が大きい所では、抗体はその抗原の特定の部分(この部分はエピトープと呼ばれる)に結合するだけであろう。抗原結合性ドメインは、一種以上の抗体可変ドメインによって供給されるだろう。抗原結合性ドメインは、抗体軽鎖可変領域(VL)と、抗体重鎖可変領域(VH)とから成るのが好ましい。
【0025】
特異性
特異性とは、特異性結合ペアの一方のメンバが、その特異性結合のパートナに比べて、他の分子に対するいかなる有意な結合性をも示さない状況をいう。この用語はまた、例えば、抗原のひとつのメンバによって運ばれる特定のエピトープに対して、抗原結合性ドメインが特異的である場合にも適用できる。この場合、抗原結合性ドメインを運ぶ特異性結合メンバは、そのエピトープを運ぶ種々の抗原に結合することができるだろう。
【0026】
機能的同等変異体
機能的同等変異体とは、他の分子(親)とは構造的に相違しているが、幾つかの重要な相同を残しており、また、親分子の生物機能(例えば、特定の抗原又はエピトープと結合する能力)の少なくとも幾つかをも残している分子(変異体)をいう。変異体は、断片、誘導体又は突然変異体の形態で存在することがある。変異体、誘導体又は突然変異体は、一種以上のアミノ酸の付加、欠失(deletion)、置換若しくは挿入;又は他の分子の連結による親分子の変性によって得られることがある。これらの変化は、ヌクレオチド又は蛋白のレベルで行われることがある。例えば、コード化されたポリペプチドは、Fab断片であることがある。このFab断片は、そのとき、もう一つの源からのFc尾部に連結される。代替的には、酵素、フラワーセイン(flourescein)等のマーカーが連結されることがある。
【特許文献1】欧州特許第0,344,134B1号明細書
【特許文献2】国際公開第92/01047号パンフレット
【特許文献3】国際公開第92/20791号パンフレット
【特許文献4】国際公開第93/06213号パンフレット
【特許文献5】国際公開第93/11236号パンフレット
【特許文献6】国際公開第93/19172号パンフレット
【特許文献7】欧州特許出願公開第184187号明細書
【特許文献8】英国特許出願公開第2188638号明細書
【特許文献9】欧州特許出願公開第239400号明細書
【特許文献10】欧州特許出願公開第0120694号明細書
【特許文献11】欧州特許出願公開第0125023号明細書
【特許文献12】国際特許出願番号第PCT/US92/09965号
【特許文献13】国際公開第94/13804号パンフレット
【非特許文献1】Juweidら著、Cancer Res.、1992年、52巻、p.5144〜5153
【非特許文献2】Jain.R.著、Sci.Am.、1994年、271巻、p.58〜65
【非特許文献3】Alitaloら著、Adv.Cancer Res.、1982年、37巻、p.111〜158
【非特許文献4】Yamada著、Ann.Rev.Biochem.、1983年、52巻、p.761〜799
【非特許文献5】Hynes著、Ann.Rev.Cell Biol.、1985年、l巻、p.67〜90
【非特許文献6】Ruoslahti著、Ann.Rev.Biochem.、1988年、57巻、p.375〜413
【非特許文献7】Owensら著、Oxf.Surv.Eucaryot.Genes、1986年、3巻、p.141〜160
【非特許文献8】Zardiら著、EMBO J.、1987年、6巻、p.2337〜2342
【非特許文献9】Castellaniら著、J.Cell Biol.、1986年、103巻、p.1671〜1677
【非特許文献10】Borsiら著、J.Cell.Biol.、1987年、104巻、p.595〜600
【非特許文献11】Vartioら著、J.Cell Science、1987年、88巻、p.419〜430
【非特許文献12】Baroneら著、EMBO J.、1989年、8巻、p.1079〜1085
【非特許文献13】Carnemollaら著、J.Cell Biol.、1989年、108巻、p.1139〜1148
【非特許文献14】Oyamaら著、J.Biol.Chem.、1989年、p.10331〜10334
【非特許文献15】Oyamaら著、Cancer Res.、1990年、50巻、p.1075〜1078
【非特許文献16】Borsiら著、Exp.Cell Res.、1992年、199巻、p.98〜105
【非特許文献17】Norton及びHynes著、Mol.Cell.Biol.、1987年、7巻、p.4297〜4307
【非特許文献18】Schwarzbauerら著、EMBO J.、1987年、6巻、p.2573〜2580
【非特許文献19】Gutman及びKornblihtt著、Proc.Natl.Acad.Sci.(USA)、1987年、84巻、p.7179〜7182
【非特許文献20】Kornblihttら著、EMBO J.、1985年、4巻、p.1755
【非特許文献21】Ffrench−Constantら著、J.Cell Biol.、1989年、109巻、p.903〜914
【非特許文献22】Ffrench−Constantら著、Development、1989年、106巻、p.375〜388
【非特許文献23】Laitinenら著、Lab.Invest.、1991年、64巻、p.375〜388
【非特許文献24】Castellaniら著、Int.J.Cancer、1994年、59巻、p.612〜618
【非特許文献25】Humphriesら著、J.Cell Biol.、1986年、103巻、p.2637〜2647
【非特許文献26】Carnemollaら著、J.Biol.Chem.、1992年、p.24689〜24692
【非特許文献27】Petersら著、Cell Adhes.Commun.、1995年、3巻、p.67〜89
【非特許文献28】Schroffら著、Cancer Res.、1985年、45巻、p.879〜885
【非特許文献29】DeJagerら著、Proc.Am.Assoc.CancerRes.、1988年、29巻、p.377
【非特許文献30】Nissimら著、EMBO J.、1994年、13巻、p.692〜698
【非特許文献31】Ward,E.S.ら著、Nature、1989年、341巻、p.544〜546
【非特許文献32】Birdら著、Science、1988年、242巻、p.423〜426
【非特許文献33】Hustonら著、PNAS USA、1988年、85巻、p.5879〜5883
【非特許文献34】Holligerら著、Proc.Natl.Acad.Sci.USA、1993年、90巻、p.6444〜6448
【非特許文献35】Holliger,P.及びWinter,G.著、Current Opinion Biotechnol.、1993年、4巻、p.446〜449
【非特許文献36】Trauneckerら著、Embo Journal、1991年、10巻、p.3655〜3659
【非特許文献37】Winter,G.及びC.Milstein著、Nature、1991年、349巻、p.293〜299
【非特許文献38】Tomlinson I.M.ら著、J.Mol.Biol.、1992年、227巻、p.776〜798
【非特許文献39】Ledermann J.A.ら著、Int.J.Cancer、1991年、47巻、p.659〜664
【非特許文献40】Bagshawe K.D.ら著、Antibody,Immunoconjugates and Radiopharmaceuticals、1991年、4巻、p.915〜922
【非特許文献41】Pluckthun,A.著、Bio/Technology、1991年、9巻、p.545〜551
【非特許文献42】Reff,M.E.著、Curr.Opinion Biotech.、1993年、4巻、p.573〜576
【非特許文献43】Trill J.J.ら著、Curr.Opinion Biotech.、1995年、6巻、p.553〜560
【非特許文献44】分子クローニング(Molecular Cloning),実験マニュアル(Laboratory Manual):第2版,サンブルック(Sambrook)ら著、1989年、Cold Spring Harbor Laboratory 出版
【非特許文献45】分子生物学における短い実験記録(Short Protocols in Molecular Biology)、第2版,オスベル(Ausubel)等編集,ジョン・ウィリー(John Wiley)及びサンズ(Sons)、1992年
【非特許文献46】Pierschbacherら著、Cell、1981年、26巻、p.259〜267
【非特許文献47】Marksら著、J.Mol.Biol.、1991年、222巻、p.581〜597
【非特許文献48】Hoogenboomら著、Nucl.Acids Res.、1991年、19巻、p.4133〜4137
【非特許文献49】Gibson TJ、博士論文、1984年、University of Cambridge, Cambridge, UK
【非特許文献50】Hawkinsら著、J.Mol.Biol.、1992年、226巻、p.889〜896
【非特許文献51】Crothers及びMetzger著、Immunochemistry、1972年、9巻、p.341〜357
【非特許文献52】Marksら著、Bio/Technology、1992年、10巻、p.779〜783
【非特許文献53】Lowら著、J.Mol.Biol.、1996年、260巻、p.359〜368
【非特許文献54】Griffithsら著、EMBO J.、1994年、13巻、p.3245〜3260
【非特許文献55】Vaughanら著、Nature Biotechnol.、1996年、14巻、p.309〜314
【非特許文献56】Borsiら著、Anal.Biochem.、1991年、192巻、p.372〜379
【非特許文献57】Saginatiら著、Eur.J.Biochem.、1992年、205巻、p.545〜549
【非特許文献58】Siriら著、Nucl.Acids Res.、1991年、19巻、p.525〜531
【非特許文献59】Brownら著、Amer.J.Pathol.、1993年、142巻、p.793〜801
【非特許文献60】Kaczmarekら著、Int.J.Cancer、1994年、58巻、p.11〜16
【非特許文献61】Folliら著、Cancer Res.、1994年、54巻、p.2643〜2649
【非特許文献62】McCafferty,J.、Griffiths,A.D.、Winter,G.及びChiswell,D.J.、Nature (London)、1990年、”Phage antibodies: filamentous phage displaying antibody variable domains.”、348巻、p.552〜554
【非特許文献63】Packら著、Bio/Technology、1993年、11巻、p.1271〜1277
【非特許文献64】Neriら著、Nature Biotechnology、1996年、14巻、p.385〜390
【非特許文献65】Ausubelら編、John Wiley & Sons、1992年、Short Protocols in Molecular Biology(第2版)
【非特許文献66】Borsiら著、Int.J.Cancer、1992年、52巻、p.688〜692
【非特許文献67】Jonsson著、BioTechniques、1991年、11巻、p.620〜627
【非特許文献68】Neriら著、Bio/Techniques、1996年、20巻、p.708〜713
【発明の開示】
【発明が解決しようとする課題】
【0027】
本発明によると、フィブロネクチン(FN)のED−B胎児ドメインに対して特異性のある特異性結合メンバが提供される。
【課題を解決するための手段】
【0028】
本発明による特異性結合メンバは,ED−Bドメインに直接結合する。一つの具体例において、特異性結合メンバは、蛋白分解酵素サーモリシン(thermolysin)でFNを処理した後、ED−Bを含有するFNの一つに、又はいかなるものにも、又は全部に結合する。更なる具体例において、特異性結合メンバは、III型相同の繰返しを含有するFNの一つに、又はいかなるものにも、又は全部に結合する。これら繰返しには、ED−Bドメインが含まれる。既知のFNは、二つの論文(非特許文献13及び26)の中に確認される。「ED−Bを含有する全FN」への言及は、これらの論文中で、ED−Bを含有していると確認される全FNへの言及として理解されても良い。
【発明を実施するための最良の形態】
【0029】
特異性結合メンバは、ヒトED−Bと結合するのが好ましく、また、マウス、ラット及び/又はニワトリのような、他の少なくとも一種のB+FNと結合するのが好ましい。特異性結合ペアメンバは、ヒト・フィブロネクチンED−Bにも;動物モデル中のsbbメンバのテスト及び分析が可能な、マウス・フィブロネクチンED−B等の非ヒト・フィブロネクチンED−Bにも結合し得るのが好ましい。
【0030】
本発明による特異性結合ペアメンバは、本明細書のどこかで検討され公式に入手できる沈積済み抗体BC−1と競うことなく、フィブロネクチンED−Bに結合する。BC−1は、ヒトB+イソ型に対してまさに特異的である。本発明による特異性結合ペアメンバは、BC−1と同一のエピトープには結合しない。
【0031】
本発明による特異性結合メンバのB+FNへの結合は、ED−Bドメインによって抑制されることがある。
【0032】
本発明の一つの面において、結合ドメインは、精製済み単量体として測定されるとき、ED−B FNに対して6×10−8M以下の解離定数(Kd)を有する。
【0033】
本発明の一つの面において、結合ドメインは、フィブロネクチンED−BをN−グリカナーゼ(N−glycanase)で前処理しないで、フィブロネクチンED−Bと反応する(即ち、結合することができる)。
【0034】
本発明による特異性結合ペアメンバは、分離体として、又は精製された形で、即ち、他の特異性結合ペアメンバ(例えば、抗体又は抗体断片)を含有していない製剤の状態、又はフィブロネクチンED−Bと結合することのできる他の特異性結合ペアメンバを含有していない状態で提供されることがある。本発明による特異性結合メンバは、実質的に純粋な形で提供されるのが好ましい。それらは、伝統的な融合細胞腫技術を使って得られた抗体と縛りつけるよりむしろ、単一クローンからのものという意味で『単クローン』であるのかも知れない。検討した通り、本発明による特異性結合ペアメンバは、バクテリオファージ表示技術(bacteriophage display technology)及び/又は組換えによる表現(例えば、細菌細胞、ホスト細胞)を使用して得られることがある。フィブロネクチンED−Bに直接結合する、単クローンの特異性結合ペアメンバについては、従来全く開示されていない。
【0035】
この特異性結合メンバは、抗体から成るのが好ましい。特異性結合メンバは、単鎖Fv(scFv)等の抗体断片の形態のポリペプチド配列から成ることがある。Fab、Fab”、F(ab’)2、Fabc、Facb、ダイアボディ等の、抗体断片の他のタイプが利用されることもある(非特許文献37及び特許文献13)。特異性結合メンバは、完全抗体の形であることがある。完全抗体は、抗体イソ型(例えば、IgG、IgA、IgD、IgE及びIgM)の形のいずれか、及びイソ型亜綱(例えば、IgG1又はIgG4)の形のいずれかであることがある。
【0036】
抗体は、あらゆる起源、例えば、ヒト、マウス、ヒツジ、ウサギのものでよい。他の誘導は当業者には明らかである。抗体は、ヒト起源が好ましい。「ヒト」は、ヒトのcDNA、蛋白又はペプチドライブラリから部分的に又は完全に誘導される抗体を意味する。この用語は、抗体分子にヒト特性を与えて、その分子にヒト免疫系の防衛をバイパスさせるために変性された、非ヒト起源のヒト化されたペプチド及び蛋白を意味する。
【0037】
特異性結合メンバは、設計された抗体[例えば、明示されている(disclosed)フィブロネクチンED−Bに対して一つの結合腕(即ち、特異性ドメイン)、及び異なる特異性に対する他の腕、又は二価若しくは多価の分子を有する、二重特異性抗体分子(又は、F(ab’)2等の断片)]の形態であることもある。
【0038】
特異性結合メンバは、抗体配列の他に、他のアミノ酸(例えば、ペプチド若しくはポリペプチドを形成するもの、又は抗原に結合する能力の他にもう一つの機能特性をその分子に与えるもの)を含むことがある。例えば、特異性結合メンバは、標識、酵素又はそれらの断片等を含む。
【0039】
結合ドメインは、胚芽細胞系列分節(germ line segment)又は再配列された遺伝子分節(gene segment)によってコード化されたVHドメインの一部又は全部から成ることがある。結合ドメインは、VLカッパ(κ)ドメイン又はVLラムダ(λ)ドメインの一部又は全部から成ることがある。
【0040】
結合ドメインは、VH1、VH3又はVH4の胚芽細胞系列遺伝子配列、又はその再配列形から成ることがある。
【0041】
本発明による特異性結合メンバは、ヒト・胚芽細胞系列DP47由来の重鎖可変領域(「VH」ドメイン)(それの配列は、図1aに示す;残基は1〜98)から成ることがある。「DP」命名法は、非特許文献38に記載されている。CDR3のアミノ酸配列は、Ser Leu Pro Lysであってもよい。CDR3のアミノ酸配列は、Gly Val Gly Ala Phe Arg Pro Tyr Arg Lys His Gluであってもよい。このように、本発明による特異性結合メンバの結合ドメインは、CGS1及びCGS2について図1aに示されるアミノ酸配列から成るVHドメインを含有することがある。
【0042】
結合ドメインは、ヒト・胚芽細胞系列DP16由来の軽鎖可変領域(「VL」ドメイン)(それの配列は、コドン1〜90として図1bに示す)から成ることがある。
【0043】
VLドメインは、CDR3配列 Asn Ser Ser Pro Val Val Leu Asn Gly Val Valから成ることがある。VLドメインは、CDR3配列 Asn Ser Ser Pro Phe Glu His Asn Leu Val Valから成ることがある。
【0044】
本発明の特異性結合メンバは、図1に示される配列と機能的に同等の変異体から成ることがある。例えば、本明細書に開示される特性が保持されるという条件で、一種以上のアミノ酸が挿入され、削除され、置換され又は追加された。例えば、特異性結合メンバがED−Bに結合するという条件で、CDR3配列は、変性されることがある;或いは、一つ以上の変化が外郭構造領域に生じることがある;或いは、外郭構造がもう一つの外郭構造若しくは変性形態と置き換えられることがある。
【0045】
本明細書に開示される抗体の抗原結合ドメインのVLドメイン若しくはVHドメイン由来の一種以上のCDRのものは、いわゆる「CDR−移植」で使用されることがある。「CDR−移植」では、特許文献9に開示されるように、第一抗体の一種以上のCDR配列は、その抗体のものでない配列(例えば、もう一つの抗体の配列)の外郭構造の内部に置かれる。CGS1及びCGS2のためのCDR配列は、図1a及び図1bに示す。
【0046】
本発明による特異性結合メンバは、フィブロネクチンED−Bへの結合に関し、本明細書に記載のscFv又は抗体と競合するものであることがある。複数の結合メンバ間の競合は、生体外で容易に分析されることがある。例えば、報告者の特異性分子を一つの結合メンバ(これは、識別されていない他の結合メンバの存在下に検出され得る)と識別して、同一エピトープ又はオーバーラップしているエピトープと結合する特異性結合メンバを同定し得るようにすることによって、容易に分析されることがある。
【0047】
本発明による特異性結合メンバは、特異性結合メンバをそれのエピトープに結合させる又は結合するがままにしておく過程を含む方法で使用することがある。結合は、哺乳動物(例えば、ヒト、又はマウス等のげっ歯動物)に特異性結合メンバを投与した結果として生じることがある。
【0048】
本発明は、腫瘍に対する診断薬として使用するための、上記のように特異性結合メンバを使用する方法を提供する。以下に記載する動物をモデルとした実験的証拠によって、本発明による結合メンバは生体内位置決定を行う上で有効であることが示される。
【0049】
本発明による好ましい特異性結合メンバには、ヒト・腫瘍に結合するもの(例えば、クリオスタット分節で、浸潤性の脈管形成性現象型(phenotype)を示すもの);胚腫瘍に結合するもの(例えば、クリオスタット分節で)が含まれる。結合は、免疫細胞化学による染色法によって証明されることがある。
【0050】
好ましい具体例において、特異性結合メンバは、テナシン、細胞外マトリックス蛋白質には結合しない、又は有意には結合しない。
【0051】
もう一つの好ましい具体例において、特異性結合メンバは、正常なヒト皮膚(例えば、クリオスタット分節で、及び/又は免疫細胞化学による染色法を用いて実証されるように)には結合しない、又は有意には結合しない。
【0052】
本発明による特異性結合メンバの更なる具体例は、肝臓、脾臓、腎臓、胃、小腸、大腸、卵巣、子宮、膀胱、膵臓、副腎、骨格筋、心臓、肺、甲状腺及び脳から選ばれる一種以上の正常組織には結合しない、又は有意には結合しない(例えば、クリオスタット分節で、及び/又は免疫細胞化学による染色法を用いて実証されるように)。
【0053】
ED−Bのための特異性結合メンバは、生体内の標的剤(targeting agent)として使用されることがある。この標的剤は、フィブロネクチンED−Bを表現する 腫瘍又はフィブロネクチンED−Bと結合した腫瘍の存在又は位置を特異的に実証するのに使用されることがある。それは、造影剤(imaging agent)として使用されることがある。本発明は、フィブロネクチンED−B表現を表現する、又はフィブロネクチンED−B表現と結合した細胞又は腫瘍の存在を決定する方法を提供する。この方法は、供給された特異性結合メンバに細胞を接触させ、次いで、その細胞に対する特異性結合メンバの結合を決定する過程を含む。その方法は、本体から除去された細胞の試験サンプルについて、生体内又は生体外で行われることがある。
【0054】
細胞サンプルに関する抗体の反応性は、あらゆる適切な手段によって決定されることがある。個々のリポーター分子を用いて識別することも可能である。そのリポーター分子は、直接的又は間接的に検出可能な、好ましくは測定可能な信号を発生することがある。リポーター分子の連結は、直接的又は間接的、共有結合的(例えば、ペプチド結合による)、或いは非共有結合的であることがある。ペプチド結合による連結は、抗体とリポーター分子とをコード化する遺伝子融合の組換え表現の結果として行われることがある。
【0055】
一つの好ましい態様は、スペクトル的に分離された吸収特性又は放出特性を有する個々の蛍光色素、蛍光物質又はレーザー色素と、各抗体との共有結合によることがある。適した蛍光色素には、フルオレセイン、ローダミン、フィコエリトリン、及びテキサス・レッド(Texas Red)が含まれる。適した色素産生色素には、ジアミノベンジジンが含まれる。
【0056】
他のリポーターには、着色され、磁性若しくは常磁性であるラテックス・ビーズ(latex beads)等の粒状物質又は高分子コロイド粒子;及び直接的又は間接的に検出可能な信号を発生して、可視的に観察することが可能であり、電子化により検出することが可能であり、さもなければ記録することが可能である生物活性剤又は化学的活性剤が含まれる。これらの分子は、例えば、顕色若しくは変色し又は電気特性を変化させる反応に、触媒作用を及ぼす酵素であってもよい。これらは、分子的に励起され、エネルギー状態間における電子遷移が特徴的なスペクトル吸収又はスペクトル発光を生じることがある。これらには、バイオセンサーと連結して使用される化学的構成要素(chemical entities)が含まれる。ビオチン/アビジン又はビオチン/ストレプトアビジン、及びアルカリリン酸分解酵素の検出システムが採用されることがある。
【0057】
結合を決定する態様は、本発明の特徴ではなく、当業者は、彼等の好み及び通常の知識に従って、適切な態様を選ぶことができる。
【0058】
個々の抗体−リポーター連結によって発生する信号は、細胞サンプル中における、関連する抗体結合の定量的な絶対データ又は相対データを得るのに使用されることがある(標準及び試験)。加えて、プロピジウムヨウ化物(propidium iodide)等の一般的核染料は、サンプル中の全細胞の個体数を数えるのに使用されることがある。125I、111In、99mTc等の放射性ヌクレオチドが抗体に付着したとき、もし、その抗体が正常組織よりむしろ腫瘍の中に優先的に局在化していれば、腫瘍組織中の放射性ラベルの存在は、ガンマカメラを使用して、検知し定量化し得る。得られる腫瘍像の質(quality)は、信号対雑音比に直接相関している。
【0059】
抗体は、新たに血管が新生した腫瘍を追跡するための診断剤として利用されることがある。また、抗体は、細胞毒剤を運ぶため又は新たな血管内の凝固を引き起こすために、(例えば、変性された形で)採用されることもある。かくして、成長している腫瘍に対する酸素及び栄養素の補給を断って、腫瘍治療の間接的形態を形成する。
【0060】
本発明は、治療試薬として使用するための、上述の特異性結合メンバの使用方法をも提供する。そのとき、その特異性結合メンバは、例えば、融合蛋白として結合され、化合され又は設計されて、作用因子機能が与えられる。本発明の特異性結合メンバは、重要な抗体を表現する腫瘍又はその抗体と結合した腫瘍に至る毒素、放射能、T細胞、キラー細胞又は他の分子を標的とするのに使用される。
【0061】
従って、本発明の更なる面は、与えられた特異性結合メンバを投与することから成る処置方法;そのような特異性結合メンバから成る薬剤組成物;及び投与のための薬剤の製造過程において(例えば、薬学的に許容できる賦形剤を用いて特異性結合メンバを調製することから成る、薬剤組成物又は医薬品組成物の製法において)、特異性結合メンバを使用する方法を提供する。
【0062】
本発明によると、提供される組成物は、個体に投与されることがある。投与は、「治療的に有効な量」であるのが好ましい。これは、患者に効果が現れるのに十分な量である。そのような効果は、少なくとも一つの症状の少なくとも回復であることがある。実際の投与量と、投与の割合及び時間−クール(course)とは、治療されているものの性質(nature)及び激しさ(severity)に依存する。投与量の決定等の治療の処方箋は、一般の診療医及び他の医師の責任による。抗体の適切な投与量は、当業において周知である。非特許文献39及び40を参照。
【0063】
組成物は、単独で又は他の治療法と組合わせ、治療される容態によるが、同時に又は逐次的に投与することがある。
【0064】
本発明による薬剤組成物、及び本発明により使用するための薬剤組成物は、有効成分に加えて、薬学的に許容される賦形剤、キャリア、緩衝剤、安定剤又は当業者に周知の他の物質を含有することがある。そのような物質は、非毒性であるべきであり、また、有効成分の効能を妨げてはいけない。キャリア又は他の物質の正確な性質は、投与の経路に依存する。経路は、経口であることもあり、注射(例えば、静脈注射)によることもある。
【0065】
経口のための薬剤組成物、錠剤、カプセル、粉末又は液体の形態であることがある。錠剤は、ゼラチン、佐剤(adjuvant)等の固形キャリアを含有することがある。液体の薬剤組成物は、一般的には、水、石油、動物油、植物油、鉱油合成油等の液体キャリアを含有する。生理的塩類溶液;ブドウ糖若しくは他のサッカライド溶液;又はエチレングリコール、プロピレングリコール、ポリエチレングリコール等のグリコールが含まれることがある。
【0066】
静脈注射剤、又は苦痛の位置での注射剤のためには、有効成分は、非経口的に許容される水溶液の形態であろう。この水溶液は、発熱因子を含まず、適切なpH、等張性及び安定性を有する。当業者は、例えば、塩化ナトリウム注射剤、リンガー液、乳酸加リンガー液等の等張性媒体を使用して、適切な溶液をうまく調製することができる。保存剤、安定剤、緩衝剤、酸化防止剤及び/又は他の添加剤は、必要に応じ、含有させてもよい。
【0067】
本発明の特異性結合メンバは、核酸のコード化による表現によって造られることがある。特異性結合メンバの製法(この製法は、特異性結合メンバのために核酸をコード化することによる表現から成る)が本発明の一つの面であるように、与えられるあらゆる特異性結合メンバをコード化する核酸それ自体、本発明の一つの面である。表現は、適切な条件下、核酸を含有する組換えホスト細胞を培養することによって、都合よく達成されることがある。
【0068】
核酸は、本明細書に記載の抗体抗原結合ドメインのあらゆるアミノ酸配列、又は機能的に同等のあらゆる形態をコード化することがある。一種以上のヌクレオチドの添加、置換、削除又は挿入によって、ヌクレオチド・レベルでの変化が生じることがある。この変化は、遺伝子コードの縮重(degeneracy)に依存し、アミノ酸レベルに反映されるかも知れないし、反映されないかも知れない。
【0069】
異なるホスト細胞の一形態でのポリペプチドのクローン化及び表現のためのシステムは、周知である。適切なホスト細胞には、細菌系、哺乳動物の細胞系、酵母菌系及びバクロウィルス系が含まれる。非相同のポリペプチドを表現するために、当該技術において入手可能な哺乳動物・細胞系には、チャイニーズハムスターの子房細胞、ヒーラー細胞、ベビーハムスターの腎臓細胞、及びその他多数が含まれる。通常の好ましい細菌ホストは、大腸菌である。
【0070】
大腸菌等の原核細胞中の抗体断片及び抗体の表現は、当業ではうまく確立されている。報告としては、例えば非特許文献41を参照。培養における真核細胞中の表現もまた、特異性結合メンバを製造するための一つの選択肢として(最近の報告を参照(例えば、非特許文献42及び43)、当業者は入手することができる。
【0071】
適切な調節配列;プロモーター配列;ターミネーター配列;ポリアデニル化配列;エンハンサー配列;マーカー遺伝子;及び適切な他の配列を含有する適切なベクターを選択することができる、又は組み立てることができる。ベクターは、適切な、プラスミド、ウィルス性のもの(例えば、ファージ)又はファージミド(phagemid)であることがある。更なる詳細については、例えば、非特許文献44を参照。核酸を操作するための多数の技術及び実験記録(例えば、核酸構成体の調製;突然変異生成;配列すること;細胞中へのDNAの導入と遺伝子表現;及び蛋白の分析、に関するもの)は、非特許文献45に詳細に記載されている。サンブルック等の開示内容、及びオスベル等の開示内容は、言及することによって本明細書に取り入れる。
【0072】
本発明の更なる面によると、本明細書に開示される核酸を含有するホスト細胞が提供される。また、更なる面によると、そのような核酸をホスト細胞の中に導入することを含む方法が提供される。この導入のためには、利用可能なあらゆる技術が採用されることがある。真核細胞のための適切な技術には、リン酸カルシウムのトランスフェクション(transfection,移入);DEAE−デキストラン;エレクトロポレーション;リポソーム媒介(liposome−mediated)トランスフェクション;及びレトロウィルス又は他のウィルス(例えば、ワクシニア、若しくは昆虫細胞に対するバクロウィルス)を用いたトランスダクションが含まれることがある。細菌細胞のための適切な技術には、塩化カルシウム転換;エレクトロポレーション;及びバクテリオファージを用いたトランスフェクションが含まれることがある。
【0073】
導入によって、核酸からの表現が生じる又は許容される(例えば、遺伝子の表現のための条件下、ホスト細胞が培養される)結果となることがある。
【0074】
一つの具体例によると、本発明の核酸は、ホスト細胞のゲノム(例えば、染色体)の中に組込まれる。組込みは、標準的技術に従い、ゲノムとの組換えを促進する配列の封入(inclusion)によって促進されることがある。
【0075】
特異性結合メンバが生成された後、その特異性結合メンバは、検討された通り、例えば、本明細書に開示される方法のあらゆるもの(例えば、特異性結合メンバの他に、そのメンバの細胞への結合を決定するための一種以上の試薬を有するキット(kit)等の、薬剤生成物又は診断薬生成物の調製)において使用されることがある。
【0076】
本発明及び具体例の更なる面は、当業者には明白であろう。本発明が十分に理解されるべく、次の諸例を単なる例証として提供するが、これに限定されるものではない。次の諸図面が参考となる。
【0077】
図1は、scFv CGS−1及びCGS−2のVH並びにVLの、整列したアミノ酸配列を示す。図1aはVH配列を示し、図1bはVL配列を示す。CDRs(1、2及び3)を示す。両方のscFvへの最も相同のヒト・胚芽細胞系列VHは、VH3科のDP47分節である。両方のクローンのVL分節はDPL6、scFvライブラリを造るのに使用される軽鎖である(非特許文献30)。二つのクローンを相互に識別する残基は、下線が引いてある。
【0078】
図2:図2Aは、ヒトFNサブユニットのドメイン構造のモデルを示す。FN−プレ−mRNAの交互スプライシング(alternative splicing)による、IIICS、ED−A及びED−Bの変異領域を示す。この図はまた、ED−Bを含有する主要サーモリシン消化生成物の他に、内部相同をも示す(非特許文献8)。図2Bは、プラスマの4〜18%SDS−PAGE;WI38VA;それらのクーマシーブルー(Coomassie Blue)染色済みサーモリシン消化生成物;並びに、BC−1、IST−6、CGS−1及びCGS−2で調べた免疫ブロット分析を示す。FNの1μg/mg(レーン3)及び10μg/mg(レーン4)でサーモリシンを使用した、不消化(レーン1)及び消化済みプラスマFN。FNの1μg/mg(レーン5)、5μg/mg(レーン6)及び10μg/mg(レーン7)でサーモリシンを使用した、不消化(レーン2)及び消化済みWI38VA FN。右側の番号は、図2Aに示される主要サーモリシン消化生成物を示す。左側の値は、キロドルトン(kD)での分子量基準(molecular weight standards)を示す。
【0079】
図3:図3Aは、大腸菌に表現された融合蛋白及び組換え蛋白の中に含有されるFNIII型繰返し配列;これら蛋白とCGS−1及びCGS−2との反応性;並びに、これら蛋白とmAbs BC−1及びIST−6との反応性を示す。図3Bは、クーマシーブルー染色済みゲル;及び、CGS−1、CGS−2、BC−1、IST−6で調べた免疫ブロット分析を示す。レーンの番号は、図の上部のペプチド構造の番号と一致する。左側の値は、kDでの分子量基準を示す。
【0080】
図4: マウスの赤外線イメージャー;タングステン・ハロゲンランプを備えた、黒色の非蛍光性箱と;CY7赤外線蛍光物質に対して特異的な励起・放出フィルターと;コンピュータ制御の8−ビット単色CCD−カメラとから成る、標的実験のために使用したマウス・イメージャー(imager)。
【0081】
図5:B−FNに向けられた単量体のscFc(CGS−1)及び二量体のscFc(CGS−1)2を用いて、蛍光によって標識化された抗体断片を、標的としてのF9ネズミの奇形癌に向けられている。リゾチームに対して結合特異性を有する二量体scFc(D1.3)2は、ネガティブ対照として使用した。
【0082】
図6:B−FNの同一エピトープに向けられた、親和性成熟のscFv(CGS−2)及び低結合性scFv(28SI)を使用して、蛍光によって標識化された抗体断片が、F9ネズミの奇形癌に向けられている。標的化は、結像を部分的に覆い隠す黒色外皮によって48時間覆われた、大きい腫瘍(約0.6g)でも、小さい腫瘍(約0.2g)でも検出された。
【0083】
本明細書に記載の全資料は、言及することによってここに取り入れる。
【実施例】
【0084】
諸例の目録
例1・・・ヒトFNのED−Bドメインに特異的なヒトscFvの分離。
例2・・・ヒトFNのED−Bドメインに特異的なヒトscFvの親和性成熟。
例3・・・ED−B含有フィブロネクチンに対する親和性成熟済みscFvの特異性。
例4・・・ヒト及びマウスの腫瘍分節を免疫細胞化学的に染色する上で、親和性成熟済み抗ED−B scFvを使用する方法。
例5・・・ヒト腫瘍を生体内で標的化する上で、親和性成熟済み抗ED−B scFvを使用する方法。
例6・・・ヌードマウスに異種組織移植されたF9ネズミの奇形癌の標的化。
【0085】
例1・・・ヒトFNのED−Bドメインに特異的なヒトscFvの分離
ヒトscFvファージ・ライブラリ(非特許文献30)は、組換え抗体の選択のために使用した。ED−Bのイソ型の二つの異なる形態は、選択のための抗原の供給源として使用し、両方とも、イソ型は組換えヒト・蛋白であった。
【0086】
III型の繰返し2−11(B−)及び2−11(B+)を含有する組換えFNペプチドは、大腸菌中に表現された。
【0087】
クローンpFH154からのFN cDNA(非特許文献20)、λF10及びλF20(非特許文献13)を使用して、組み立てられた。ベース(bases)2229〜4787の中に広がるcDNA構成体(非特許文献20)は、キアゲン(Qiagen)からのQIA表現キットを用いて、ベクターpQE−3/5の中に挿入された(チャトワース,CA)。組換え体FN−III 2−11(B−)及び(B+)は、セファローズ(Sepharose)4Bに結合したmAb 3E3(非特許文献46)を使用した免疫親和性クロマトグラフィによって精製された。III型相同繰返し7B89、789、ED−B及びFN−6を含有する組換えFN断片を調製するためのDNA断片は、テンプレートとして、クローンFN2−11(B+)及びFN2−11(B−)からのcDNAを使用しながら、U1tMa DNAポリメラーゼを使用したポリメラーゼ鎖反応(polymerase chain reaction)(PCR)の増幅(パーキン・エルマー(Perkin Elmer)によって生成された。プライマーは、QIA表現キット(キアゲン)を使用して、pQE−12の中にPCR生成物がクローン化するように設計された。これらは、実質的に大腸菌の中に転換され、表現された。全cDNAクローンは、シーケナーゼ(Sequenase)2.0DNA配列キット(USB)を使用して配列された。
【0088】
組換え蛋白は、製造者の使用説明書(キアゲン)に従い、FN断片のカルボキシ末端のヘキサヒスチジン標識を使用して、Ni−NTAクロマトグラフィ(IMAC)によって精製した。ED−B−βGal融合蛋白は、ED−B cDNAをλgt11バクテリオファージ・ベクターの中にクローン化して、クローンλED−Bを与えることによって調製された。(ED−B配列の一部を含有する)クローンλchFN60は、クローン化されたニワトリ・FN pchFN60(ノートン(Norton)等,1987)からの融合蛋白として誘導された。
【0089】
ヒト・scFvファージライブラリを選択するために、3ラウンドのパニング(panning)が、二つの異なる組換え抗原(7B89及びED−B)の各々に対して行われた。これら抗原は両方とも、PBS(20mMリン酸塩緩衝剤、0.15M NaCl、pH7.2)中の50μg/mlで、一晩、免疫チューブ(ナンク(Nunc);Maxisorp,Roskilde,デンマーク)の上にコーティングされた。第一抗原は、組換えFN断片7B89(この中で、ED−Bドメインは、隣接するIII型相同繰返しによって、側面が接触する。)であり、これは、一晩、4℃でコーティングされた。使用された第二抗原は、カルボキシ末端のヘキサヒスチジン標識を有する組換えED−B(非特許文献8)であった。この蛋白は、リシン残基を含有していないので、第一アミノ酸の末端アミノ基は、サイト特異的な(site−specific)共有結合による、反応性ELSAプレートへのED−Bの固定に利用できる(ナンク(Nunc);コバリンク(Covalink)。コーティングは、一晩、室温で行った。
【0090】
3ラウンドのパニングの後、溶離されたファージは、HB2151大腸菌細胞の中に感染させ、記載した通りにプレートした(非特許文献30)。選択の各ラウンドの後、95のアンピシリン−抵抗性単一コロニーをスクリーニングし、ELISAによって抗原特異性scFvを同定した。パニングのために使用した抗体上に最も高いELISA信号を与えたクローンは、更なる分析のため、及び親和性成熟のために選定した。これらのクローンはまた、例4に一層詳しく記載される免疫細胞化学的染色によって、多種多様の膠芽腫及び胸部腫瘍の切断面に特異的な染色を与えることも実証された。
【0091】
例2・・・ヒトFNのED−Bドメインに特異的なヒトscFvの親和性成熟
(7B89と共に選択した)クローン35GE及び(ED−Bドメインのみと共に選択した)28SIは、親和性成熟のための候補抗体として選定した。親和性を改善するための手段として軽鎖を多種産生すべく、我々は、次いで、206=6.4×107の潜在的配列の多様性という条件で、縮重したオリゴヌクレオチド及びPCRを使用しながら、軽鎖CDR3の中央の6個の残基(DSSGNH)をランダム化することに基づき(図1)、単純な親和性成熟の方策を調べた。(重鎖CDR3に沿った)この領域は、抗原結合部位の中央に配置されている(パドラン(Padlan),1994)。我々は、また、その選択を支配する静電気の影響の可能性を回避すべく、6個の残基がセリンへ伸びる直前に、アルギニン残基を変異させた。
【0092】
「親」scFv断片を表現する単一細胞コロニーからのプラスミドは、プライマー LMB3(5’ CAG GAA ACA GCT ATG 3’)及びCDR3−6−VL−FOR(5’ CTT GGT CCC TCC GCC GAA TAC CAC MNN MNN MNN MNN MNN MNN AGA GGA GTT ACA GTA ATA GTC AGC CTC 3’)(94C[1’]−55C[1’]−72C[1’30”]、25サイクル)を用いて増幅されるPCRであった(緩衝剤及び条件については、非特許文献47を参照)。得られた生成物は、(オリジナルscFv遺伝子を含有するプラスミドの痕跡を除去するため)ゲル精製し、次いで、プライマー LMB3及びJI−Not−FOR(5’ ATT GCT TTT CTT TTT TGC GGC CGC GCC TAG GAC GGT CAG CTT GGT CCC TCC GCC 3’)(94C[1’]−55C[1’]−72C[1’30”]、25サイクル)を用いて、第二増幅段階のためのテンプレート(template,鋳型)として使用した。アガロース・ゲル上に正確な分子量の単結合として伝わる粗PCR生成物は、スピン結合(Spin−Bind)(FMC,ロックランド(Rockland),ME,USA)を用いて、Ncol/Notlで二重消化され(double−digested)結さつされて(ligated)、ゲル精製されたNcol/Notl消化済みファージミドpHEN1になる(非特許文献48)PCR混合物から直接精製した。このpHEN1は、単消化済みベクターから二重消化済みベクターを分離するのを促進するためにダミーのNcol/Not1挿入物(insert)を含有する。このベクターは、キアゲン(Qiagen)(チャッツワース(Chatsworth),CA,米国)のプラスミド・マキシプレ・キット(maxi−pre kit)で調製した。消化済みプラスミド及び挿入物の約5μgを、結さつ混合物(ligation mix)中で使用した。
【0093】
結さつ混合物はフェノールで一回抽出し、フェノール/クロロホルム/イソアミルアルコール(25:25:1)で一回抽出し、次いで、キャリアとしてグリコーゲンを用いてエタノール沈降(ボーリンガー(Boehringer),Mannheim,ドイツ)させ、次いで、高速真空で乾燥させた。ペレットは、水20μl中に再び懸濁させ、次いで、エレクトロコンピテント(electrocompetent)のTG1大腸菌細胞(非特許文献49)中でエレクトロポレーションさせた(electroporated)。我々は、グリセロールのストックを使用するときは、形質転換細胞109/μgのタイター(titre)を有するエレクトロコンピテント細胞を典型的に使用した。或いは、新たに調製したエレクトロコンピテントを有する形質転換細胞1010/μg)のタイター(titre)を有するエレクトロコンピテント細胞を典型的に使用した。これによって、本明細書に概説する手順により、107を越えるクローンが産生された。
【0094】
次いで、成熟ライブラリは、ニッシム・ライブラリ(非特許文献30)に関し、ファージ粒子を産生すべく処理した。このファージ粒子は、抗原として7B89(10μg/ml)を使用している免疫チューブ(immunotubes)上の選定1ラウンドのために使用し、次いで、動的選定(kinetic selection)(非特許文献50)の1ラウンドを行った。この選定過程は、選定の第一ラウンドからのミルク−PBS(2%MPBS)中で5分間、ビオチニル化7B89(10nM)をファージ懸濁液(約1012t.u.)で培養し、次いで、非ビオチニル化7B89(1μM)を添加し、次いで、30分間、競合を続行させることによって行った。次いで、2%MPBS中に予ブロックされた(preblocked)、ストレプトアビジンでコーティングした動力ビード(dynabeads)(Dynal:M480)100μlを、反応混合物に添加し、2分間混合し、次いで、磁石上で捕獲し、次いで、(PBS+0.1%Tween−20)とPBSとで交互に10回洗浄した。ファージは、トリエチルアミン0.5ml、100mMを用いてビードから溶離した。次いで、この溶液は、0.25ml、1MのTris、pH7.4で中和し、次いで、HB2151細胞(非特許文献30)を急激に成長させて感染させるのに使用した。95のアンピシリン−抵抗性単一コロニーは、scFV含有の上澄み液(非特許文献30)を生成するのに使用した。この上澄み液は、最良のバインダーを同定すべく、ELISA、免疫細胞化学及びBIAコア(BIAcore)によってスクリーニングを行った。次いで、この上澄み液は、pDN268表現ベクター(ネリ(Neri)等,1996)のSfil/Notlサイトの間でサブクローン化した。このベクターは、ScFVのC−末端に、リン酸化が可能な標識、FLAGエピトープ及びヘキサヒスチジン標識を付加する。
【0095】
pDN268中でサブクローン化した関連抗体の単クローンは、アンピシリン100mg/l及びグルコース0.1%を含有する2xTY中、37℃で成長した。細胞培養が、OD600=0.8に到達したとき、IPTGを最終濃度がmMになるまで添加し、成長は30℃で16〜20時間の間続けた。遠心分離(GS−3 Sorvall roter,7000rpm、30分間)を行った後、上澄み液を瀘過し、濃縮し、次いで、ミニセッテ(フィルトロン)接線流れ装置(Minisette(Filtron)tangential flow apparatus)を使用して、負荷緩衝液(loading buffer)(リン酸塩50mM、pH7.4、NaCl 500mM、イミダゾール20mM)の中で交換した。得られた溶液は、Ni−NTA樹脂(キアゲン)の上に負荷させ、負荷緩衝液50mlで洗浄し、次いで、溶離緩衝液(リン酸塩50mM、pH7.4、NaCl 500mM、イミダゾール20mM)で溶離した。精製した抗体は、SDS−PAGE(ラエムリ(Laemmli),1970)によって分析し、4℃のPBSに対して透析した。多価scFv断片は、結合活性効果に基づき、BIAコア上で人工的に優れた結合を示すことがあるということが知られている(非特許文献30及び51)ので、精製済みscFv製剤は、S−75コラム搭載のFPLC装置(ファルマシア(Pharmacia))を用い、ゲル濾過によって更に処理した。FPLC−精製済み単量体断片の抗体濃度は、分光測定器によって、scFv溶液1mg/mlについて1.4ユニットの280nmでの吸光度を想定して、決定した。
【0096】
PBSにおける0.1〜1μMの範囲の種々の濃度での、一価のscFvの結合を、BIAコア機器(ファルマシア・バイオセンサー)上で、次の抗原を使用しながら測定した。(i)ストレプトアビジンをコーティングしたチツプ上に固定された、ビオチニル化済み組換えFN断片7B89の1000レゾナンス・ユニット(Resonance Units)(RU)。これは、scFvの250RUによって特異的に結合する;(ii)N−末端アミノ基で化学的に固定された、組換えED−Bの200RU。これは、scFvの600RUによって特異的に結合する;(iii)ED−Bに富むフィブロネクチンWI38VAの3500RU(例3を参照)。これは、scFvの150RUによって特異的に結合する。データの動的分析は、製造者インストラクションに従って行った。抗体含有上澄み液のBIAコア定性分析に基づいて、各scFvクローンの一つの親和性成熟済みバージョン(version)、即ち、78B9断片を有する選定からのクローンCGS−1、及びED−B組換えFN断片を有する選定からのCGS−2を選定した。会合速度定数(kon)及び解離速度定数(koff)を、scFvとオリジナル・クローン28SI両方の、計算による平衡解離定数(Kd)と共に、表1に示す。クローンのCGS−1もCGS−2も、ナノモルの範囲のKdを有するが、クローンCGS−2は、センサー・チップ上で試験した3種の蛋白全てに関し、(110nMから改善されて)1nMのKdを与え(表1)、その親クローンを越えて最良の改善を示した。この改善は、主として、単量体抗体の製剤で測定された(示さず)一層緩慢な動的解離定数(10−4S−1以下)による。
【0097】
成熟方法は、大部分に共通しているように思われ、マルトース結合蛋白;シトクロムC;マウス・エンドグリン(endoglin)の細胞外ドメイン(D.N.,L.
Wyder,R.Klemenz);サイトメガロウィルス(A.P.,G.
Neri,R.Botti,P.N.);核腫瘍マーカーHMGI−C蛋白(A.P.,P.Soldani,V.Giancotti,P.N.);及び卵巣腫瘍マーカーの胎盤アルカリホスファターゼ(Deonarain及びA.A.Epenetos)に対する親和性改善済み抗体を産生した。従って、この成熟方法は、少なくとも他の成熟方法(非特許文献52及び53)と同程度に有効であるように思え、しかも、非常に大きいファージ抗体ライブラリ(非特許文献54及び55)から誘導されたものと類似の親和性を有する抗体を産生する。
【0098】
親和性成熟済みクローンCGS−1及びCGS−2は、ヒト・胚芽細胞系列の抗体V遺伝子のデータベース(V−BASE)に配列し整列し、次いで、マクベクター(MacVector)ソフトウェアを用いて翻訳した。両クローンのVH遺伝子は、ヒト・胚芽細胞系列DP47(VH3)にほとんど相同であり、各クローンの他に、異なるVH CDR3配列を有した(図1)。両クローンのVL遺伝子は、ニッシム等に記載のヒト・合成scFv全範囲(repertoire)の構造に使用されるDPL16胚芽細胞系列であった。VL CDR3配列は、任意の順序に配列された残基の6個から4個が、互いに相違した(図1b)。
【0099】
【表1】
【0100】
例3・・・ED−B含有フィブロネクチンに対する親和性成熟済みscFvsの特異性
二つの親和性成熟済みscFvs、CGS−1及びCGS−2の免疫反応性は、初期にELISAによって評価し、次いで、mAb BC−1、及びmAbIST−6と直接比較した。mAb BC−1は、B−FNイソ型を認識し、mAb IST−6は、ED−Bを欠いているFNイソ型を認識するのみである(非特許文献13及び26)。これらmAbの特性は、以前報告された(非特許文献13及び26)。その後、サーモリシン処理によって誘導されたFN断片と組換え融合蛋白との大きいパネルを使用して、特異性の正確な分析が行われた。
【0101】
ELISA及び免疫ブロット分析のために使用した抗原は、次の通りに調製した。FNは、ヒト・プラスマ;及び以前報告されたWI38VA13細胞系列の調整済み媒体(非特許文献8)から精製した。精製したFNは、非特許文献13によって報告されたように、サーモリシン(プロテアーゼ X型;シグマ・ケミカル社(Sigma Chemical Co.)を用いて消化した。天然のFN 110kD(B−)及び天然のFN 120kD(B+)の断片(図2参照)は、以前報告されたように(非特許文献56)、FN消化物から精製した。テナシン−Cの大きいイソ型は、非特許文献57によって報告されたように精製した。組換え蛋白は、例1に記載の通りに表現し、精製した。SDS−PAGE及びウェスタン・ブロッティング(Western blotting)は、非特許文献13によって報告されたように、実施した。
【0102】
ELISAに使用された全抗原は、PBSで50〜100μg/mlに希釈し、免疫−プレート・ウェル(Immuno−Platewells)(Nunc,Roskilde,デンマーク)の上に4℃で一晩コーティングした。非結合抗原は、PBSで除去し、次いで、プレートは、ウシ由来血清アルブミン(BSA)3%(w/v)含有するPBSを用いて、37℃で2時間の間、ブロックした。この後、0.05%トゥイーン(Tween)20含有PBS(PBST)で4回洗浄した。次いで、抗体は、37℃で1.5時間の間、結合させた:scFvsは、標識配列(FLAG標識に対してはmAb M2[Kodak,New Haven CT];又はmyc標識に対しては9E10[ATCC,Rockville, MD])に向けられた抗血清を用い、前もって温置した。試験を行った対照抗体は、mAbs BC−1及びIST−6であった。PBSTで4回洗浄した後、プレートは、ヤギの抗マウスIgG(Bio−SPA Division,Milan,イタリア)を(PBST+3%BSAで)1:2000に希釈し、ビオチニル化して、37℃で1時間の間、培養した。洗浄を繰り返し、次いで、ストレプトアビジンービオチニル化したアルカリ・ホスファターゼ複合体(Bio−SPA Division,Milan,イタリア)を、37℃で1時間の間、添加した(MgCl2 2mMを含有するPBSTに1:800で希釈された)。反応は、ジエタノールアミン10%、pH9.8中で、ホスファターゼ基質の錠剤(Sigma)を使用して進め、光学的濃度は、405nmで読み取った。結果は、次の表2に示す。
【0103】
【表2】
【0104】
scFv及び単クローン抗体と、フィブロネクチン誘導抗原との免疫反応性は、ELISAによって測定した。測定値は、バックグランド信号を減じた後の、405nmで測定したODを示す。データは、4回の実験の平均であり、これらデータは、最大10%の標準偏差を示す。
【0105】
実験に用いたフィブロネクチンの異なる形態の素性(identity)は、次の通りである:即ち、プラスマ FN=ヒト・プラスマフィブロネクチン;WI38−VA FN:SV4O−転換済み線維芽細胞の上澄み液からのプラスマフィブロネクチン(非特許文献8);n110kD=ED−Bを含有しない、FNドメイン4で処理したサーモリシン;n120kD=ED−Bを含有した、FNドメイン4で処理したサーモリシン;recFN7B89=隣接するIII型FN相同繰り返しによって側面に位置するED−Bドメイン;recFN789=ED−Bドメインを有する、III型FN相同の繰り返し;recED−B=組換えED−B単独;recFN6=組換えFNドメイン6。
【0106】
CGS−1もCGS−2も、ED−B配列を含有する、天然のFN断片の全て又は組換えFN断片の全ての他に、組換えED−Bペプチドを認識した。しかし、それらは、ED−Bを欠くFN断片のいずれにも結合しなかった。また、CGS−1及びCGS−2は、テナシン(これは、15個のIII型相同繰り返しを含む:非特許文献58);及びプラスマ FN(これは、サーモリシン消化生成物(非特許文献8)中に、検出可能なレベルのED−B配列を含有していない)と反応しなかった。対照的に、CGS−1及びCGS−2は、SV40−転換済み細胞系列WI38VAから精製したFNと強く反応した。精製済みFN及びその細胞系列から調製した全RNAを使用した、サーモリシン消化及びS1ヌクレアーゼの実験によって示されるように(非特許文献8;Borsi等,1992)、この細胞からのFN分子約70〜90%には、ED−Bが含まれる。可溶性組換えED−Bを使用して、WI38VA細胞上のFに対するCGS−1及び/又はCGS−2の結合を抑制することによって、FNのED−B成分に対するscFvの特異性を更に実証した(データは示していない)。
【0107】
データによると、CGS−1及びCGS−2のみが、ED−Bドメインを含有するFN誘導体と特異的に反応することが確認される。それらは双方とも、組換えED−Bの場合を除き、mAb BC−1と同様の反応性を示す。組換えED−Bは、BC−1によって認識されていない。mAb対照に関連して得られたELISA信号の強度は、ED−B含有抗原に対する二つのscFvの高い特異性を反映する。
【0108】
CGS−1及びCGS−2の特異性は、プラスマ及びWI38VAの細胞からのFNと、それらのサーモリシン消化物とを使用しながら、免疫ブロット分析により調べた。サーモリシン消化によると、WI38VA細胞からのFN(その大部分はED−Bを含有する)は、(ED−Bを含有する)120kD断片と、ED−Bを欠く僅かな110kD断片とを生成する(図2A;非特許文献8)。120kDドメインの更なる消化は、二つの断片:即ち、そのカルボキシ末端にほぼ完全なED−B配列を有する85kD断片;及び35kD配列を生成する(図2A;非特許文献8)。
【0109】
図2Bの左側には、免疫ブロット分析によって分析した蛋白断片のクーマシー染色済みゲルがある。プラスマFN(レーン1)と、蛋白のサーモリシン消化物(110kD蛋白を含有するレーン3,消化済み110kD蛋白を含有するレーン4)とは、CGS−1及びCGS−2によって認識されなかった。対照的に、WI38VA細胞からの、ED−Bに富むFNは、元のまま及びサーモリシン消化を増大した後の両方において、両方のscFv断片によって認識された。CGS−1によって特異的に認識し得た、最も小さいFN−誘導断片は、(ほとんど全てがIII型の繰り返し2〜11による)120kD蛋白であり、一方、CGS−2は、ED−BのN−末端の他に、繰り返し2〜7による85kD断片を認識し得た(図2B;非特許文献8)。これらの結果は、二つのscFvが反応性であり、ED−B配列内のエピトープを識別することを示す。85kDドメインに対するCGS−2の結合は、このクローンのためのエピトープがED−Bのアミノ末端に存在することを示す。対照的に、120kDドメインが85kDへ消化されるときの、CGS−1の結合の損失は、それによって、ED−B分子のカルボキシ末端の方に向いて一層多く配置されたエピトープが認識されることを実証する。
【0110】
CGS−1及びCGS−2の優れた特異性は、ED−B配列を持った又は持たない融合蛋白及び組換えFN断片を用いた免疫ブロット分析によって更に研究した。FN融合蛋白は、非特許文献13によって記述されている通りに調製した。これらの実験結果は図3に示す;ヒト・FNのドメインの構造の概略図関連は、非特許文献26を参照。得られた結合のプルフィルによると、精製済みFNと蛋白分解の分割生成物とに関する免疫ブロット分析及びELISAによって従来見いだされていたものが、本質的に確認される。即ち、CGS−1及びCGS−2は、ED−B含有FN断片(レーン2及び4)と強い反応性があったが、ED−Bを欠くFN配列(レーン1及び3)には反応性を全く示さなかった。CGS−1は、ヒト・ED−B融合蛋白(レーン5)ともニワトリ・ED−B融合蛋白(レーン6)とも反応しなかったが、CGS−2は、両方の断片と強く反応した(図3)。この結果は、CGS−1によって認識されたED−B含有FN中のエピトープの、ある立体配座上の制約を反映しているのかも知れない。例えば、エピトープが、変性に敏感であり、又はSDS−PAGEによって分断される(fractionated)とき、正確には現出されず、ニトロセルロース等の固形支持体に転写されるということは起こり得る。
【0111】
まとめて考えると、これらの結果によると、CGS−1及びCGS−2は、互いに異なり且つED−B構造(これは、mAb BC−1によって認識される)と異なる領域で、ED−B含有FNに強く且つ特異的に結合することが実証される。
【0112】
例4・・・ヒト及びマウスの腫瘍を免疫細胞化学的に染色する上で親和性成熟済み抗ED−B scFvsを使用する方法
CGS−1及びCGS−2はいずれも、種々の正常で新生のヒト・組織中に、ED−B含有FN分子を免疫局在化する(immunolocalise)ために使用されてきた。B−FNイソ型は、皮膚創傷の治癒の間、マクロファージ及び線維芽細胞の中に表現されることが知られていたので、正常組織には皮膚が選ばれた(非特許文献8及び非特許文献59)。選定された二つのヒト・腫瘍は、従来、抗フィブロネクチンmAbで染色する特異性のために分析されてきた。この腫瘍の血管内の内皮細胞は、B−FNイソ型の表現を含む血管形成過程が増加して、高度に増殖性の状態にあるため、膠芽腫(glioblastoma)の多形は詳細に研究されてきた(Carnemolla等,1994)。更に、正常で増殖性で新生のヒト・胸組織の多様のパネルを用いた研究によって、血管形成とB−FN表現の間の相互関係に関する一層の証拠が提供された(非特許文献60)。
【0113】
本明細書に記載の諸実験のために、CGS−1及びCGS−2の免疫組織化学的染色は、mAb BC−1(これは、B−FNイソ型を認識する)の免疫組織化学的染色;及び既知の全FNイソ型変異体(IST−4)に反応する、又はED−Bを欠くFNイソ型(IST−6)のみに反応することが知られている他のmAbの免疫組織化学的染色と比較した。これら対照抗体の全ての特徴は、従来報告されている(非特許文献13及び26)。
【0114】
正常で新生の組織は、手術中に採取されるサンプルから得た。組織の調製及び固定が、FN含有分子の正確で感度のよい検出のために重要であることは既に実証されていた(Carnemolla等,1994)。免疫組織化学のために、5μm厚さのクリオスタット分節(cryostat sections)は、空気乾燥され、冷アセトン中に10分間、固定された。免疫染色は、ストレプトアビジン−ビオチン・アルカリ・ホスファターゼ複合体染色キット(Bio−SPA Division,Milan,イタリア)と、ナフトール−AS−MX−リン酸塩と、ファースト・レッド(Fast Red)TR(Sigma)とを用いて行った。ギル・ヘマトキシリン(Gill’s haematoxylin)は、対比染色として使用した。次いで、従来、「Carnemolla等,1994」によって報告されているように、グリセロゲル(glycergel)(Dake,Carpenteria,CA)中に固定した。実験で特異性を更に分析するため、組織のポジティブ染色(positive staining)が得られる所で、抗体を組換えED−Bドメインで培養し、次いで、従来記述された通りに検出することによって、ED−Bに対する特異性を実証した。
【0115】
これらの全実験結果によって、CGS−1もCGS−2も、mAb BC−1と同一の組織構造と反応することが分かった。CGS−1、CGS−2及びBC−1を用い、皮膚で得られた染色パターンは、真皮(dermis)中に表現されたFNからのED−Bは存在しないことを反映している。浸潤性管の癌分節の染色において、CGS−1、CGS−2及びBC−1は、制限された染色分布を示し、新生細胞と基質(stroma)との間の境界に限定した。このことは、全FNは腫瘍基質全体に均一に分布しているが、B−FNの表現は、ある領域に限定されていて、しかも、従来、mAb BC−1を用いて(非特許文献60)、浸潤性管の癌の中に(ケースの95%で)うまく局在化させられたのはこれらの領域であるという事実と一致する。
【0116】
膠芽腫の多様な癌のBC−1を染色することにおける従来の発見が、確認された。非特許文献24は、糸球体様脈管構造の染色の典型的なパターンを観察していたが、我々の実験では、CGS−1及びCGS−2は、定性的には同一の結果を与えることが分かった。
【0117】
しかし、CGS−1とCGS−2とmAb BC−1との間には、重要な相違が存在する。即ち、BC−1は厳密にヒト・特異性であるのに、二つのヒト・scFvはニワトリ及びマウスのB−FNの両方に結合することが実証された。CGS−2はニワトリ・胚芽と反応し(データは示さず)、CGS−1及びCGS−2はマウス・腫瘍と反応した。
【0118】
マウスF9の奇形癌の分節上の血管構造のCGS−1染色も示した。対照的に、試験を行った全ての正常マウス組織(肝臓、脾臓、腎臓、胃、小腸、大腸、卵巣、子宮、膀胱、膵臓、副腎、骨格筋、心臓、肺、甲状腺及び脳)は、CGS−1及びCGS−2とネガティブ染色反応を示した(データは示さず)。そのF9奇形癌分節中で染色された構造は、組換えED−Bドメインを使用して、得られた染色を完全に抑制することによって、ED−B特異性であることが分かった(データは示さず)。
【0119】
例5・・・ヒト腫瘍を生体内で標的化する上で、親和性成熟済み抗ED−B scFvを使用する方法
ヒト・メラノーマの細胞系列SKMEL−28は、一つの側腹部の皮下に1×107の細胞/マウスを注射することにより、6〜10週間の老齢ヌードマウス(Balb−c又はMF−1;Harlan UK)中の異種組織移植済み腫瘍を成長させるのに使用した。腫瘍を有するマウスは、腫瘍が直径約1cmに達したとき、PBSのscFv1−Cy1溶液1mg/mlの100μlを用いて、尾部の静脈に注射した。
【0120】
CY7による組換え抗体の標識化は、PBS(1mg/ml)の抗体溶液1m1に「1M炭酸ナトリウム、pH=9.3」100μlと、「CY7−bis−Osu(Amersham;Cat.Nr.PA17000;DMSO中2mg/ml)」200μlを添加することによって達成した。室温で、30分後、「1MTris、pH=7.4」100μlをその混合物に添加し、次いで、標識化された抗体は、PBSと平衡状態に置いた、使い捨てPD10カラム(Pharmacia Biotech,Piscataway,NJ,米国)を使用して、未反応染料から分離した。溶離した生の抗体断片は、セントリコン(Centricon)−10チューブ(Amicon,Beverly,MA,米国)を用いて、約1mg/mlに濃縮した。達成された標識化比は、通常、1モルのCY7:1モルの抗体に近い。これは、抗体溶液1mg/mlは280nmで1.4単位の吸収を与え、747nmでのCY7のモル吸光係数は200,000(M−1cm−1)であると想定し、また、280nmでのCY7吸収を無視し、lcmキュベットを用い分光器によって評価した。標識化した後の、抗体サンプルの免疫反応性は、抗原カラム上の親和性−クロマトグラフ法によるバンド−シフト(Neri等,1996)によって;又はBIAコア分析によって確認した。マウスは、酸素/フルオロエタンの混合物の吸入による麻酔の下、一定時間間隔で、ホーム・ビルト(home−built)マウス−イメージャー(mouse−imager)を用いてイメージした。結果の再現性を確実なものとすべく、各サンプルに対し、2〜8匹の動物を研究した。操作手順は、D.ネリ(Neri)に発行されたUKプロジェクト・ライセンス(Project Licence)「腫瘍の標的化(Tumour Targeting)」(UK PPL 80/1056)に従って行った。
【0121】
非特許文献61の光検出装置の改造型として、赤外線蛍光物質CY7が使用できる赤外線マウス・イメージャーを造った。赤外線照射を選定し、一層優れた組織貫通を得た。CY7(>76nm)の蛍光は、人には見えず、コンピューター制御のCCD−カメラを使用する必要がある。このマウス・イメージャーは、黒に塗装され、光が入らないようにした箱を有した。この箱には、100Wタングステン・ハロゲン・ランプを備え、CY7(クローマ社(Chroma Corporation),Brattleboro,VT,米国;673〜748nm)のために特別に設計した直径50mmの励起フィルターを備えた。得られた照射線は、5×10cmの大きさの領域全体に渡り、ほぼ均一であった。マウスはイメージングのためにその中に置いた。蛍光は、C−マウント・レンズと50mm放出フィルター(クローマ社,Brattleboro,VT,米国;765〜855nm)を備えた、8ビットのモノクロのプルニックス(Pulnix)CCDカメラによって検出し、次いで、イメージ(Image)DOK装置(Kinetic Imaging Ltd.,リバプール、UK)に接続した。この装置は、フレーム・グラバー(frame−grabber)と、配列イメージを捕獲し集積化を行うためのソフトウェアとを備えたコンピューターから成る。それぞれ50msで捕捉された配列のイメージは、典型的には、平均化プロセスに使用した。これの番号は、一つの動物の一連の画像では一定に保持し、異なる時点で標的化されている腫瘍が直接比較できるようにした。次いで、TIFFフォーマットでの画像は、プログラム・グラフィック・コンバーターを使用して、PICTファイルに変換し、次いで、パワー・マッキントッシュ7100/66コンピューターを用いて、精巧に仕上げた。
【0122】
この装置のデザインの概略図は、図4に示す。
【0123】
これらの実験によって、両方のscFvは、巨視的レベルに視覚化されたとき、腫瘍に局在化していることが実証された。
【0124】
二つの抗EDB scFvを用いて、成長している腫瘍の新生血管形成を標的化する巨視的実証は、次のように詳述される。
【0125】
一つの側腹部に、異種組織移植されたSKMEL−28のヒト・メラノーマ、又はマウスF9奇形癌を有する、ヌードマウス及び/又はSCIDマウスは、FLAG標識(tag)を有する標識化されていないscFv断片、又はビオチニル化されたscFv断片を用いて注射した。
【0126】
マウスは、注射の後、異なる時点で犠牲となり、腫瘍及び非腫瘍の分節を得た。次いで、これら分節は、抗FLAG M2抗体(Kodak,181)又はストレプトアビジン−ベースの検出試薬を使用し、従来の免疫組織化学プロトコールで染色した。最適標的化は、一般に注射の12時間後に得られた。CGS1もCGS2も、異種組織移植されたヒト・腫瘍、及びマウスの奇形癌の両方の新生血管に結合することが実証された。
【0127】
例6・・・ヌードマウスに異種組織移植されたF9ネズミの奇形癌の標的化
我々は、ネズミF9の奇形癌細胞4×106を皮下注射することによって、ヌードマウスの側腹部で固形腫瘍を成長させた。この腫瘍は、マウス内で非常に急速に成長し、注射後、約1週間で1cmに達し、高度に血管が新生した。抗体の標的化をイメージすべく、我々は、非特許文献61の光検出手順の改造型を使用した。この改造型は、上記に詳述する通り(図4参照)、種々の時点でイメージされた同一動物に関する腫瘍標的化及び抗体の排除を可能にする。
【0128】
腫瘍に標的化し、抗体の検出を容易にするため、表現ベクターPGIN50のSfil/Notl部位における抗体をサブクローン化することによって、scFv(CGS−1)、scFv(CGS−2)及び抗リゾチーム scFv(D1.3)(非特許文献62)に、ホモ二量体化標識(非特許文献63)を追加した。このベクターは、pD268(非特許文献64)の誘導体である。この中で、標識のHis6配列は、配列:GGC LTD TLQ AFT DQL EDE KSA LQT EIA HLL KEK EKL EFI LAA Hによって置換される。これは、共有の抗体断片のホモ二量体化のためのFos蛋白の両親媒性のらせんと、システイン残基とを含む(Abate等,1990)。完全な共有の二量体化は、達成されなかった。抗体断片約30〜50%は、共有結合した二量体から成った。
【0129】
抗体断片は、雌鶏、卵のリゾチーム(D1.3)又は7B89(抗ED−B抗体;Carnemolla等,1996)をCNBr活性化されたセファローズ(Sepharose)(Pharmacia Biotech,Piscataway,NJ,米国)にカップリングすることによって得たカラムのアフィニティ・クロマトグラフィによって精製した。上澄み液は、アフィニティ支持体の上に供給し、次いで、PBSで洗浄し、PBS+0.5MNaClで洗浄し、次いで、100mMのEt3Nで溶離した。次いで、抗体は、PBSに対して透析して分離した。
【0130】
抗体は上述の通り、標識化し、次いで、PBSの1mg/ml scFv1−Cy71溶液100μlで、腫瘍を有するマウスの尾部の静脈に注射した。このとき、腫瘍は、直径約1cmに達していた。
【0131】
図5に示す通り、scFv(CGS−1)は、3日間以下の間、腫瘍上に局在化した(この期間に腫瘍からの急速なクリアランスもあったが)。しかし、大たい骨の、ある染色も存在した。CGS−1の腫瘍への標的化の実施は、C末端に、システイン残基を含有する両親媒性のらせんを導入して、抗体の二量体化(非特許文献63)を促進することによって、顕著に改善された。実際、二量体のscFv(CGS−2)2の局在化は、24〜72時間、有意には減少しないように思えた。対照的に、ネガティブ対照(二量体の抗体scFv(D1.3)2、抗リゾチーム抗体)は、急速なクリアランスを示し、腫瘍又は大たい骨の検出可能な局在化は全く見られなかった。
【0132】
scFv(28SI)は、6時間で腫瘍ターゲッティングをわずかに示した(図示せず)。しかし、24時間又はそれ以降において、全く検出できなかった(図6)。親和性の成熟は、ターゲッティングを著しく改善することとなった。このように、scFv(CGS−2)は、単量体(図6)としてであろうと、二量体(図示せず)としてであろうと、小さいF9腫瘍及び大きいF9腫瘍を有効に標的化した。2日後、腫瘍1g当り抗体の注射投与量%は、scFv(CGS−2)単量体では約2、scFv(CGS−2)二量体では3〜4であることが分かった。腫瘍に運ばれたscFv(CGS−2)による投与量もまた、scFv(CGS−1)によるものより大きく(図5及び6)、それらの各々親和性と相互関係がある(表1)。しかし、scFv(28SI)もscFv(CGS−2)も、蛋白質分割しないように思え、また肝臓の取り込みが大きいことを示す(図6)ところが、scFv(CGS−1)抗体は、有意に一層安定であり、肝臓の取り込みが一層小さいことを示す(図5)。
【0133】
【表3】
【0134】
【表4】
【0135】
【表5】
【0136】
【表6】
【0137】
【表7】
【0138】
【表8】
【0139】
【表9】
【図面の簡単な説明】
【0140】
【図1a】scFv CGS−1及びCGS−2のVHの、整列したアミノ酸配列であって、CDRs(1、2及び3)を示す。
【図1b】scFv CGS−1及びCGS−2のVLの、整列したアミノ酸配列であって、CDRs(1、2及び3)を示す。
【図2A】ヒトFNサブユニットのドメイン構造のモデルを示す。
【図2B】プラスマの4〜18%SDS−PAGE;WI38VA;それらのクーマシーブルー(Coomassie Blue)染色済みサーモリシン消化生成物;並びに、BC−1、IST−6、CGS−1及びCGS−2で調べた免疫ブロット分析を示す。
【図3A】大腸菌に表現された融合蛋白及び組換え蛋白の中に含有されるFNIII型繰返し配列;これら蛋白とCGS−1及びCGS−2との反応性;並びに、これら蛋白とmAbs BC−1及びIST−6との反応性を示す。
【図3B】クーマシーブルー染色済みゲル;及び、CGS−1、CGS−2、BC−1、IST−6で調べた免疫ブロット分析を示す。
【図4】マウスの赤外線イメージャー;タングステン・ハロゲンランプを備えた、黒色の非蛍光性箱と;CY7赤外線蛍光物質に対して特異的な励起・放出フィルターと;コンピュータ制御の8−ビット単色CCD−カメラとから成る、標的実験のために使用したマウス・イメージャー(imager)である。
【図5】B−FNに向けられた単量体のscFc(CGS−1)及び二量体のscFc(CGS−1)2を用いて、蛍光によって標識化された抗体断片を、標的としてのF9ネズミの奇形癌に向けられている。
【図6】B−FNの同一エピトープに向けられた、親和性成熟のscFv(CGS−2)及び低結合性scFv(28SI)を使用して、蛍光によって標識化された抗体断片が、F9ネズミの奇形癌に向けられている。
【特許請求の範囲】
【請求項1】
精製された単量体として測定されるとき、6×10−8M未満のED−Bに対する解離定数(Kd)を有する、フィブロネクチン(FN)の腫瘍胎児ED−Bドメインに対して特異的であり且つ該腫瘍胎児ドメインに直接結合するものである、分離された抗体又は抗体断片。
【請求項2】
ヒト起源の抗原結合ドメインを含む、請求項1に記載の分離された抗体又は抗体断片。
【請求項3】
FNを蛋白分解酵素サーモリシンで処理した後、ED−B含有FNの全てに結合する、請求項1に記載の分離された抗体又は抗体断片。
【請求項4】
B+イソ型のフィブロネクチンをN−グリカナーゼで処理しないで、B+イソ型のフィブロネクチンに結合する、請求項1に記載の分離された抗体又は抗体断片。
【請求項5】
CDR3配列 Ser Leu Pro Lys及びヒト胚芽細胞系列DP47(配列番号9に示す、1番目の残基であるGluから98番目の残基であるargまで)から誘導される変異性重(VH)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項6】
CDR3配列 Gly Val Gly Ala Phe Arg Pro Tyr Arg Lys His Glu及びヒト胚芽細胞系列DP47(配列番号10に示す、1番目の残基であるGluから98番目の残基であるargまで)から誘導される変異性重(VH)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項7】
Pro Val Val Leu Asn Gly Val ValのCDR3配列の残基及びヒト胚芽細胞系列DPL16(配列番号11に示す、1番目の残基であるSerから90番目の残基であるSerまで)から誘導される変異性軽(VL)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項8】
Pro Phe Glu His Asn Leu Val ValのCDR3配列の残基及びヒト胚芽細胞系列DPL16(配列番号12に示す、1番目の残基であるSerから90番目の残基であるSerまで)から誘導される変異性軽(VL)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項9】
抗体又は抗体断片は、scFv分子を含む、請求項1〜8のいずれか1項に記載の分離された抗体又は抗体断片。
【請求項10】
抗体又は抗体断片は、二量体のscFv分子を含む、請求項1〜8のいずれか1項に記載の分離された抗体又は抗体断片。
【請求項11】
請求項5〜8のいずれか1項に記載の分離された抗体断片。
【請求項12】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片をコード化する核酸。
【請求項13】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片をコードするファージ。
【請求項14】
請求項12に記載の核酸を用いて転換される又は移入される(transfected,トランスフェクションされる)試験管内のホスト細胞。
【請求項15】
腫瘍を標的化するための薬剤を調製するために、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片を使用する方法。
【請求項16】
腫瘍をイメージングするための薬剤を調製するために、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片を使用する方法。
【請求項17】
試験管内のホスト細胞中で請求項12に記載の核酸を発現することによって、分離された抗体又は抗体断片を製造することを含む、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片の製法。
【請求項18】
請求項1に記載の抗体又は抗体断片の製法であって、
a)ファージ中で発現された抗体又は抗体断片ライブラリを、フィブロネクチン蛋白から誘導したED−Bドメインを含む組換えフィブロネクチン断片でスクリ−ニングし、
b)ホスト細菌細胞をポジティブ・クローン(positive clones)に感染させ、
c)ポジティブ・ファージ・クローンを親和性熟成プロセスにかけ、
d)工程a)及びb)を繰り返して、抗原に対する改善された親和性を有するポジティブ・ファージ・クローンを選定し、
e)ホスト細胞をポジティブ・クローンに感染させ、次いで、該ホスト細胞からの抗体分子を精製する
諸工程から成り、
工程a)において、抗体断片がscFv分子である、抗体断片ライブラリをスクリーニングする上記製法。
【請求項19】
scFv分子がヒト起源の抗原結合ドメインを含む、請求項18に記載の製法。
【請求項20】
工程a)において、組換え抗原7B89又はED−Bを用いてスクリーニングする、請求項18記載の製法。
【請求項21】
生成抗体又は生成抗体断片は、フィブロネクチン(FN)の腫瘍胎児ED−Bドメインに対して特異的であり且つ該腫瘍胎児ドメインに直接結合する該抗原結合ドメインを含むものであり、且つ、ポジティブ・クローンの核酸が、該核酸によってコード化された該生成抗体または生成抗体断片の試験管内のホスト細胞中での発現によって製造に用いられる、請求項18〜20のいずれか1項に記載の製法。
【請求項22】
生成抗体が完全抗体の形をとる、請求項21に記載の製法。
【請求項23】
生成抗体断片がscFv分子の形をとる、請求項21に記載の製法。
【請求項24】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片、及び、細胞へ該分離された抗体又は抗体断片が結合するのを決定する一種以上の試薬を含む、診断薬キット。
【請求項1】
精製された単量体として測定されるとき、6×10−8M未満のED−Bに対する解離定数(Kd)を有する、フィブロネクチン(FN)の腫瘍胎児ED−Bドメインに対して特異的であり且つ該腫瘍胎児ドメインに直接結合するものである、分離された抗体又は抗体断片。
【請求項2】
ヒト起源の抗原結合ドメインを含む、請求項1に記載の分離された抗体又は抗体断片。
【請求項3】
FNを蛋白分解酵素サーモリシンで処理した後、ED−B含有FNの全てに結合する、請求項1に記載の分離された抗体又は抗体断片。
【請求項4】
B+イソ型のフィブロネクチンをN−グリカナーゼで処理しないで、B+イソ型のフィブロネクチンに結合する、請求項1に記載の分離された抗体又は抗体断片。
【請求項5】
CDR3配列 Ser Leu Pro Lys及びヒト胚芽細胞系列DP47(配列番号9に示す、1番目の残基であるGluから98番目の残基であるargまで)から誘導される変異性重(VH)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項6】
CDR3配列 Gly Val Gly Ala Phe Arg Pro Tyr Arg Lys His Glu及びヒト胚芽細胞系列DP47(配列番号10に示す、1番目の残基であるGluから98番目の残基であるargまで)から誘導される変異性重(VH)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項7】
Pro Val Val Leu Asn Gly Val ValのCDR3配列の残基及びヒト胚芽細胞系列DPL16(配列番号11に示す、1番目の残基であるSerから90番目の残基であるSerまで)から誘導される変異性軽(VL)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項8】
Pro Phe Glu His Asn Leu Val ValのCDR3配列の残基及びヒト胚芽細胞系列DPL16(配列番号12に示す、1番目の残基であるSerから90番目の残基であるSerまで)から誘導される変異性軽(VL)鎖領域を有する、請求項1に記載の分離された抗体又は抗体断片。
【請求項9】
抗体又は抗体断片は、scFv分子を含む、請求項1〜8のいずれか1項に記載の分離された抗体又は抗体断片。
【請求項10】
抗体又は抗体断片は、二量体のscFv分子を含む、請求項1〜8のいずれか1項に記載の分離された抗体又は抗体断片。
【請求項11】
請求項5〜8のいずれか1項に記載の分離された抗体断片。
【請求項12】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片をコード化する核酸。
【請求項13】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片をコードするファージ。
【請求項14】
請求項12に記載の核酸を用いて転換される又は移入される(transfected,トランスフェクションされる)試験管内のホスト細胞。
【請求項15】
腫瘍を標的化するための薬剤を調製するために、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片を使用する方法。
【請求項16】
腫瘍をイメージングするための薬剤を調製するために、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片を使用する方法。
【請求項17】
試験管内のホスト細胞中で請求項12に記載の核酸を発現することによって、分離された抗体又は抗体断片を製造することを含む、請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片の製法。
【請求項18】
請求項1に記載の抗体又は抗体断片の製法であって、
a)ファージ中で発現された抗体又は抗体断片ライブラリを、フィブロネクチン蛋白から誘導したED−Bドメインを含む組換えフィブロネクチン断片でスクリ−ニングし、
b)ホスト細菌細胞をポジティブ・クローン(positive clones)に感染させ、
c)ポジティブ・ファージ・クローンを親和性熟成プロセスにかけ、
d)工程a)及びb)を繰り返して、抗原に対する改善された親和性を有するポジティブ・ファージ・クローンを選定し、
e)ホスト細胞をポジティブ・クローンに感染させ、次いで、該ホスト細胞からの抗体分子を精製する
諸工程から成り、
工程a)において、抗体断片がscFv分子である、抗体断片ライブラリをスクリーニングする上記製法。
【請求項19】
scFv分子がヒト起源の抗原結合ドメインを含む、請求項18に記載の製法。
【請求項20】
工程a)において、組換え抗原7B89又はED−Bを用いてスクリーニングする、請求項18記載の製法。
【請求項21】
生成抗体又は生成抗体断片は、フィブロネクチン(FN)の腫瘍胎児ED−Bドメインに対して特異的であり且つ該腫瘍胎児ドメインに直接結合する該抗原結合ドメインを含むものであり、且つ、ポジティブ・クローンの核酸が、該核酸によってコード化された該生成抗体または生成抗体断片の試験管内のホスト細胞中での発現によって製造に用いられる、請求項18〜20のいずれか1項に記載の製法。
【請求項22】
生成抗体が完全抗体の形をとる、請求項21に記載の製法。
【請求項23】
生成抗体断片がscFv分子の形をとる、請求項21に記載の製法。
【請求項24】
請求項1〜11のいずれか1項に記載の分離された抗体又は抗体断片、及び、細胞へ該分離された抗体又は抗体断片が結合するのを決定する一種以上の試薬を含む、診断薬キット。
【図1a】


【図1b】

【図2A】

【図2B】
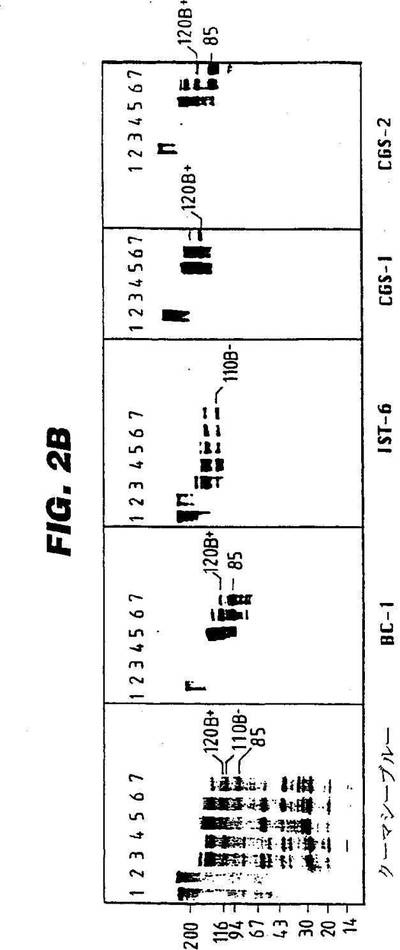
【図3A】
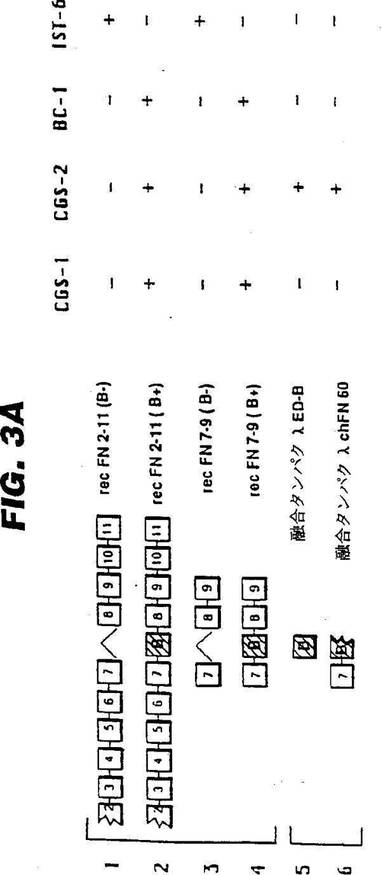
【図3B】
【図4】
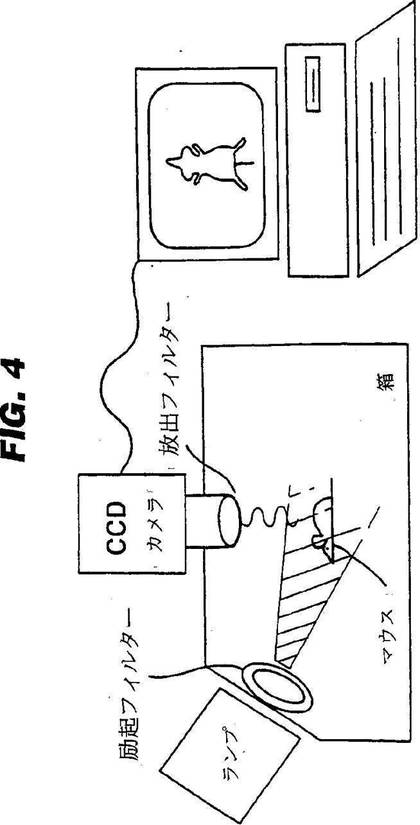
【図5】

【図6】

【図1b】
【図2A】
【図2B】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【公開番号】特開2009−50261(P2009−50261A)
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願番号】特願2008−204682(P2008−204682)
【出願日】平成20年8月7日(2008.8.7)
【分割の表示】特願平9−541828の分割
【原出願日】平成9年5月23日(1997.5.23)
【出願人】(508205729)ピロゲン ソシエタ ア レスポンサビリタ リミタータ (1)
【Fターム(参考)】
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願日】平成20年8月7日(2008.8.7)
【分割の表示】特願平9−541828の分割
【原出願日】平成9年5月23日(1997.5.23)
【出願人】(508205729)ピロゲン ソシエタ ア レスポンサビリタ リミタータ (1)
【Fターム(参考)】
[ Back to top ]