
フィブロネクチンのEDAドメインの使用を基にした剤および方法
本発明は、免疫賦活性剤の生成におけるポリペプチドの使用に関し、該ポリペプチドは、フィブロネクチンの該EDAドメイン、TLR4に結合できる該EDAドメインのフラグメントまたは、TLR4に結合でき、該EDAドメインのあらゆる形態または天然のフラグメントと70%以上のホモロジーを有する該EDAドメインの改変型またはそのフラグメントに対応する配列を含んでいる。本発明は、該剤の生成方法および用途にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
(本発明の技術分野)
本発明は、感染および腫瘍疾患の処置および予防のための医薬組成物、特に免疫治療用組成物の調製および使用に対する特定の範囲を有する、TLR4受容体(トル様受容体4)を発現する細胞への分子輸送のためのタンパク質性ベクター、該タンパク質性ベクターの調製およびその用途に関する。
【背景技術】
【0002】
(先行技術の説明)
病原体および癌は、依然として全世界で主な死亡要因である。ワクチンが現在存在しない疾患、例えばAIDSまたはマラリアを抑制するための、または慢性感染または癌を処置するための、さらに存在しているワクチンの効力および安全性改善のためのかかるワクチンの開発は、依然として高い優先事項である。多くの症例において、かかるワクチンの開発は、CD8+細胞傷害性Tリンパ球細胞(CTL)を特異的に刺激できるストラテジーを必要とする。
【0003】
CTLは、MHCクラスI分子と関連した短鎖ペプチドのT-細胞受容体(TCR)の提示により活性化される。これらのペプチド-MHCクラスI複合体は、至適CTL活性化を必要とする共刺激シグナルも提供できる抗原提示細胞(APC)の表面上に存在する。
【0004】
樹状細胞(DC)は、本来のTリンパ球と相互作用し、ヘルパーCD4+および細胞傷害性CD8+Tリンパ球を活性化する初期免疫応答を開始する特別な能力を有する最も強力なAPCである。DCによる抗原提示およびT細胞刺激はGuermonprezら("抗原提示およびDCによるT細胞刺激"Annu. Rev. Immunol. 2002, 20:621-627)に論説されており、出典明示によりこの文献を明細書の一部とする。
【0005】
継続する炎症および免疫応答がなければ、樹状細胞は、血液、末梢組織、リンパ球および二次リンパ組織中を巡回する。末梢組織において、樹状細胞は、自己および非自己抗原を取り込む。次いで、内在化された抗原はタンパク質分解ペプチドへとプロセシングされ、これらのペプチドは、MHCクラスIおよびII分子(各々CD8+またはCD4+Tリンパ球の活性化のため)上に担持される。抗原取込み、分解および担持に関するこの過程は、抗原提示と呼ばれる。しかしながら、刺激が無ければ、末梢樹状細胞は全く有効でない抗原を提示する。病原体または内因性シグナル由来の外因性シグナルは樹状細胞を誘導し、成熟と呼ばれる樹状細胞をAPCおよびTリンパ球アクチベーターに変換する発生プログラムに参加する。細菌およびウイルス産物、ならびに炎症性サイトカインおよび他の自己分子は、本来の樹状細胞表面レセプターと直接的な相互作用を介して樹状細胞成熟を誘導する。CD40独立経路およびCD40非独立経路を経るTリンパ球、および内皮細胞は、直接的な細胞対細胞接触およびサイトカインの分泌を通じて樹状細胞の最終的な成熟に関与する。危険を示すシグナルに遭遇した直後に、抗原摂取効率、細胞内輸送および分解およびMHC分子の細胞内輸送が修飾される。ペプチド担持、さらに半減期およびMHC分子の細胞表面への送達が増強される。T細胞共刺激分子の表面発現も増大する。こうして、樹状細胞は最も強力なAPCとなる、本来のTリンパ球を活性化でき、適応免疫応答を開始できる唯一のものである。その抗原提示能の修飾と同時に、成熟は末梢組織以外の樹状細胞の大量移動も誘導する。ケモカイン受容体および付着分子の発現における修飾、ならびに細胞骨格組織化の著しい変化は、リンパを介して二次リンパ組織にむかう樹状細胞の移動に関与する。
【0006】
樹状細胞成熟の誘導
樹状細胞は、2つのタイプのシグナルに応答する:病原体の直接的認識(特異的な型-認識受容体を介して)および感染の間接検出(炎症性サイトカイン、体内細胞化合物を介して、特異的免疫応答を継続する)。これらのシグナルに対する応答において、樹状細胞は活性化されて、樹状細胞を有効なT細胞刺激因子へと形質転換する成熟プログラムに入る。少なくとも5つの型の表面受容体が樹状細胞成熟を引き起こすことが報告されている:(i)トル様受容体(TLR)、(ii)サイトカイン受容体、(iii)TNF-受容体(TNF-R)ファミリー分子、(iv)FcRおよび(v)細胞死に対するセンサー。最も有効な成熟刺激のいくつかは、それらの各リガンドとTLR(TLR1-9)の相互作用により媒介される。KaishoおよびAkiraは、トル様受容体の知見("アジュバンド受容体としてのトル様受容体". Biochimica et Biophysica Acta, 2002, 1589: 1-13)を論じている。TLRは、マクロファージおよび樹状細胞上、ならびに他の細胞、例えばBリンパ球上で発現される。数種のTLRのリガンドもまた同定されている。大部分のこれらのリガンドは、その宿主には見出されない病原体から得られ、TLRは侵入微生物を検出するために重要であることが示唆されている。TLRによるリガンド認識は、炎症誘発性サイトカインの産生および共刺激分子の上方調節を誘導することによって先天性免疫の急速な活性化を誘起する。次に、活性化された先天性免疫は、有効な適応免疫を引き起こす。TLR4に関して、特異的に認識される分子パターンは、LPS(グラム陰性細菌)、リポタイコ酸(グラム陽性細菌)、タキソール、Fタンパク質(呼吸器合胞体ウイルス)、熱ショックタンパク質60、およびフィブロネクチンEDAドメインである。
【0007】
それ故に、至適T細胞応答を誘導出来る候補ワクチンは、いくつかの条件を満たさなけれならない。まず、抗原誘導性T細胞エピトープをMHCクラスIおよび/またはII分子に送達するためには、APCを標的としなければならない。従って、DCの標的化は、ワクチン開発のための新規送達系を設計する際に主な目的を示すことができる。さらに、該ベクターは、適切なシグナルをDCに送達して、その活性化を誘導するべきである。成熟シグナルがないとDCへの抗原送達は、Tヘルパーおよび細胞傷害性細胞の活性化よりもむしろ寛容を誘導できるであろう。さらに、その効果は、ベクターそれ自身に対して予め存在している免疫によって影響されるべきではない。
【0008】
抗原性ペプチドをMHCクラスIおよび/またはII分子に送達するための最初の方法は、合成ペプチドワクチンを基にしており、APCの表面上でこれらの分子に直接結合できる選択されたエピトープを含有している。ある場合では、これらのペプチドは、ネズミのモデルにおいては、腫瘍保護またはウイルス排除をもたらしたが、しかし他の場合において、それらは寛容誘導をもたらした。様々な型のペプチドを用いて行ったヒトの治験は、癌への適切な臨床応答を引き起こした。
【0009】
多くの様々なストラテジーは現在開発中である。基本的に、それらは2つのカテゴリーに分けることができる。第一のタイプは、APCによる抗原の合成またはこれらの細胞の細胞質へのその能動送達を基にしており、および"古典的"MHCI抗原プロセシング経路を利用する。第二のタイプは、APCの交差提示の能力に有利であり、遊離または細胞関連性の外因性抗原を基にしている。APCの細胞質への抗原の送達は、細菌毒素の手段によって行われてきた(Moron et al. "MHCクラスI経路への抗原送達のための新規ツール". TRENDS in Immunology, 2004; 25: 92-97)。例示としては、EP1188446A1は、細胞を発現するCD11bへの分子送達のための、百日咳菌(Bordetella pertussis)アデニルシクラーゼ毒素を基にしたタンパク質性ベクターに関する。
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、フィブロネクチン・エクストラ・ドメインA(EDA)、APCの適切な選択および成熟を誘導できる、最終的に効果的な特異的CTL応答をもたらすTLR4発現細胞へ抗原送達のための理論的手段として、TLR4に対する潜在的な天然リガンドに関する。フィブロネクチン分子は、1つの遺伝子産物であって、得られるタンパク質は、一つの前駆体mRNAの選択的スプライシングから生じる複数の形態で存在できる(Pankov R and Kenneth MY, "Fibronectin at a glance". Journal of Cell Science, 2002; 115:3861 3863)。スプライシングの主なタイプは、タイプIIIリピートの中心的な型において認められる。エキソン用法またはスキッピングは、2つのタイプIIIリピートのいずれかの包含または排除に導く:エクストラドメインB(EDB、EIIIBまたはEDIIとも呼ばれる)、およびエクストラドメインA(EDA、EIIIAまたはEDIとも呼ばれる)。選択的スプライシングによるEDAおよびEDBを含有する細胞性フィブロネクチンは組織損傷への応答において産生される。他の生物学機能のなかの一つは、EDAはプロテオグリカン放出およびメタロプロテイナーゼ(MMP1、3および9)および前炎症性サイトカインの発現を誘導することが示されている(Saito S et al. "フィブロネクチンエクストラドメインAはマトリックスメタロプロテイナーゼ遺伝子発現をインターロイキン-1依存機構により誘導する", J. Biol. Chem. 1999; 161:3071-3076を参照されたい)。EDAがTLR4を活性化でき、こうしてLPS様応答を誘導できることも示されている(Okamura Y et al.,"フィブロネクチンのエクストラドメインAはトル様受容体4を活性化する", J. Biol. Chem. 2001; 276:10229-10233)。
【0011】
前記したように、抗原に対する免疫応答を強化するためのストラテジーの開発は、癌または感染疾患の処置のためのワクチン開発に対する道をひらくものである。特に、C型肝炎ウイルス感染において、免疫応答は感染の排除において重要な役割を担っており、それ故、免疫的強化ストラテジーの使用はこの感染の処置および予防についての代替え案を構成すると説明されてきた。
【0012】
C型肝炎ウイルス(HCV)は、Flaviviridaeファミリーに属する一本鎖RNAウイルスである (Miller RH. and Purcell RH. 1990. PNAS. 87:2057)。このウイルスは、慢性肝炎および肝臓疾患の主な病因の一つとして認識されており、全世界で170万人に影響すると見積もられる(World-Health-Organisation. Hepatitis C. Wkly Epidemiol Rec 1997; 72:65)。HCV感染の主な態様の中の一つは、慢性化(感染者の70%)へと向かう高い傾向、および肝細胞癌の進行に関する高リスクを伴う肝硬変(20%)の進行である(Dienstag et al. Gastroenterology 1983; 85:439)。IFN-αによる処置は、HCV感染において最も一般的な治療であるが、処置患者のわずか20-30%においてのみ有効である(Camps et al. J Hepatol 1993; 17:390)。IFN-αおよびリバビリンの組合せは、これらの結果(患者の30-40%は持続的様式でウイルスを排除する)を改善したが、治療に対する抵抗性患者は依然として高い%を保持している(Poynard et al. Lancet 1998; 352:1426)。従って、慢性C型肝炎の処置のための新規治療ストラテジーの開発は極めて重要である。
【0013】
HCVゲノムは、9.6キロ塩基のゲノムであって、3つの構造タンパク質(コア、E1およびE2)および少なくとも6つの非構造タンパク質(NS2、NS3、NS4a、NS5aおよびNS5b)(主に、MEおよびFeinstone SM. (1997) Hepatology 25, 1527)をコードする十分なオープンリーディングフレームを挟んでいる5'および3'末端に高度に保存された非コード領域を含有する。
【0014】
HCVによる急性感染後またはIFN-αを用いる処置後のウイルス排除は、ウイルスタンパク質に対して強力な細胞性CD4およびCD8免疫応答の存在に関連している。特に、HCVの非構造のNS3タンパク質に対するCD4応答は、急性感染後ウイルス排除と関連しているが、一方でこのT細胞の非応答は、ウイルスの継続および慢性感染の確立に関する(Diepolder et al. Lancet 1995; 346:1006, Pape et al J Viral Hepat 1999; 6 Suppl 1:26-40)。また、様々な試験によりHCV感染患者のNS3タンパク質のなかで多様な細胞傷害性エピトープが同定されてきた。これらのデータから、NS3タンパク質は抗HCV細胞性応答の誘導のための良好な標的であり得ることが示唆された。
【課題を解決するための手段】
【0015】
本発明の詳細な説明
一態様において、本発明は、抗原刺激剤に対する免疫細胞応答の生成において、
-フィブロネクチンEDAドメイン(EDA)、または
-TLR4を結合できる該EDAドメインのフラグメント、または
-TLR4を結合でき、EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する、該EDAドメインまたはフラグメントの改変型、
に対応するアミノ酸配列を含むポリペプチドの使用に関する。
【0016】
本発明において、この剤はフィブロネクチンのEDAドメインおよび免疫応答を生じることが望まれる抗原の両方を包含し、これらの成分は個々の構成成分として存在するか、または共有結合している。
【0017】
本発明の具体的な実現において、エレメントc)で引用したTLR4を結合できる該EDAドメインまたはフラグメントの該改変型は、そのアミノ酸配列がエレメントa)およびb)に規定されたポリペプチドにおいて1以上のアミノ酸の置換、付加または削除から得られることを特徴とする。
【0018】
本発明の好ましい実現において、エレメントc)を結合できる該フラグメントは、EDAドメインまたはその対応するフラグメントの任意の天然形態と85%以上のホモロジーを有することを特徴とし、より好ましい実現において、フィブロネクチンEDAドメインの天然形態またはその対応するフラグメントと95%以上のホモロジーを有する。
【0019】
本発明に従って、特別な実現において、フィブロネクチンEDAドメインのアミノ酸配列は、TLR4を結合できるEDAのあらゆる天然形態のものである。このEDAドメインは、あらゆる動物種、特に哺乳動物、即ち齧歯類(マウス、ラット等)または霊長類(特にヒト)におけるドメインの天然形態の中から選択され得る。
【0020】
他の特定の実現において、免疫賦活剤は、該剤がTLR4を結合できることを特徴とするEDAドメインの部分アミノ酸配列を含む。
【0021】
本発明の他の別の特定の実現において、EDAドメインは、任意の該EDAドメインの天然形態またはフラグメントの修飾された改変型であり、またTLR4への結合特性を保持することを特徴とする。特別な実施形態において、かかる改変型EDAドメインは、EDAドメインのあらゆる天然形態と70%以上のホモロジーを有する。適切に修飾された改変型は、フィブロネクチンEDAドメイン天然形態の配列、またはそのもののフラグメントを他の候補ポリペプチドの配列をまず比較することによって選択され得る。任意のアラインメントアルゴリズム(例えばFASTA、Lipman DJ, Pearson WR. Rapid and sensitive protein similarity, Science. 1985 Mar 22;227(4693):1435-41)、またはコンピューターソフトウェア(即ち、Labvelocity Inc.からはJellifish または NCBIからはBlast software)は、ホモロジー分析に使用され得る。次に、70%以上のホモロジーを有する候補ポリペプチドの配列は、そのTLR4結合能力について試験される。TLR4結合特性は、任意の従来の結合アッセイ、例えばJohn Wiley & Sons (John E. Coligan, Ada M. Kruisbeek, David H. Margulies, Ethan M. Shevach, Warren Strober編)(定期的に更新される。May 1, 2005までに更新された)によって発表されたThe current protocols in Immunology およびThe current protocols in Protein Scienceに記述されているようなフローサイトメトリーを用いて評価され得る。
【0022】
本発明の一実施態様において、EDAドメインは、下記から選択された配列を含む:
a)マウスEDAドメイン(Entrez タンパク質:NM_010233、アミノ酸1721〜1810;配列番号:2、アミノ酸2-91)の完全なアミノ酸配列;
b)ヒトEDAドメイン(Entrez タンパク質 NM_002026、アミノ酸1631〜1716;配列番号:4)の完全なアミノ酸配列;および
c)TLR4発現細胞に結合し得る配列a)およびb)のフラグメント。
【0023】
別の特定の実現において、該EDAドメインは、下記から選択される配列を含む:
a)マウスフィブロネクチンEDAドメインの選択的にスプライシングされた形態に対応する配列番号:6のアミノ酸2-57;
b)ヒトEDAドメインの選択的にスプライシングされた形態である配列番号:8;および
c)TLR4発現細胞に結合できる配列a)およびb)のフラグメント。
【0024】
いくつかの実施形態において、免疫賦活性剤は1以上の目的とする分子をさらに包含し得る。免疫賦活性剤中に存在する場合、目的とする分子は、薬剤の他の成分との組合せにおいて該分子に対する免疫応答を生成するために効果的である量で投与され得る。
【0025】
好ましい実現において、EDAドメイン(またはそのフラグメントまたは改変型)および目的とする分子は、同ハイブリッド分子またはタンパク質性ベクター中に一緒に結合される。
【0026】
別の態様において、本発明は、上記したとおりのタンパク質性ベクターに関し、ここで目的とする分子は、ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質および化学物質からなる群から選択される。
【0027】
タンパク質性ベクターの特別な実現において、目的とする分子は、抗原またはエピトープである。本発明の一実施態様において、ベクターと結合される抗原は、ウイルス抗原、細菌抗原、真菌抗原または寄生抗原である。具体的な実現において、該ウイルス抗原は、C型肝炎ウイルス由来のウイルス抗原であって、好ましい実現において、該C型肝炎ウイルス抗原はNS3タンパク質またはその抗原性フラグメントである。NS3タンパク質は、N末端からの189個のアミノ酸を含むセリンプロテアーゼおよびC末端からの442個のアミノ酸を含むヘリカーゼ-ヌクレオシドトリホスファターゼを包含する2つのドメインを包含する67kDaタンパク質のC型肝炎ウイルスの非構造NS3タンパク質を指す。本発明のタンパク質ベクターに包含されるNS3タンパク質配列は、ヒトC型肝炎ウイルスの任意の株または単離株と対応可能である。
【0028】
他の実施形態において、該抗原は、腫瘍性抗原または腫瘍性抗原決定基である。本明細書で使用したとおりに、"エピトープ"は、MHCクラスIまたはクラスII分子と結合し、CD8+またはCD4+T細胞各々のT細胞受容体分子によって認識され、そして免疫応答を誘導し得るペプチド配列を指す。
【0029】
特定の実現において、目的とする該分子は、オボアルブミン(OVA 257-264)由来の抗原性細胞傷害性T決定基またはSIINFEKL(配列番号:2、アミノ酸95-102、これはエピトープ(QLE-SIINFEKL-TEW)のC末端およびN-末端で3つの付加アミノ酸により挟まれている)である。
【0030】
該抗原は、Th免疫応答を産生できる任意の材料、CD8+Tリンパ球応答、NK細胞応答、γ/δTリンパ球応答、または抗体応答であり得る。それらに限定するものではないが、適切な抗原は、ペプチド、ポリペプチド、脂質、糖脂質、ポリサッカリド、炭水化物、ポリヌクレオチド、プリオン、細菌、ウイルスまたは生存するか不活性化された真菌;および細菌、ウイルス、真菌、原生動物、腫瘍または微生物、毒素またはトキソイドから得られた抗原である。
【0031】
タンパク質性ベクターの特定の別の実現において、目的とする分子はアレルゲンである。
【0032】
この方法、EDAドメインは、TLR4発現細胞への抗原送達のためのベクターとしても作用する。
【0033】
本発明の目的とする他の特定の実現において、目的とする該分子は、タンパク質性ベクターと化学的または遺伝子的に結合された化学物質または薬物である。このように該タンパク質性ベクターは、TLR4発現細胞に標的化する特異的な薬物に有用である。
【0034】
特別な実現において、タンパク質性ベクターは、Tag配列も含む、例えばN末端ヒスチジンテイルも含むことを特徴とする。これは、タンパク質性ベクターが遺伝子設計方法によって生成される場合に、精製過程を単純化する。例示として、配列の配列番号:2および配列番号:6は、本発明のタンパク質性ベクターの特定の実現を示す。具体的に、本発明の実現を限定しないが、該タンパク質性ベクターは、配列番号:10を含み、NS3タンパク質のフラグメントを含む。
【0035】
タンパク質性ベクターに導入したEDAドメインは、TLR4に結合すること、さらに目的とする分子の転位をTLR4発現細胞のサイトゾルに利用することを特徴とする。
【0036】
本発明は、目的とする分子をTLR4発現細胞に標的化および輸送するためにタンパク質性ベクターの使用に関する。特定の実現において、TLR4発現細胞はあらゆる種類の抗原提示細胞(APC)である。好ましい実施形態において、かかるAPCは樹状細胞である。
【0037】
本発明の特定の実現において、タンパク質性ベクターは、目的とする抗原またはエピトープの輸送を促進して、プロセシングをさらに進めて、およびTリンパ球に対する抗原提示のためにMHC分子上に担持することを特徴とする。
【0038】
他の実現において、タンパク質性ベクターは、標的化したAPCの成熟を刺激できること、従ってMHC分子の発現および共刺激シグナルを増加することを特徴とする。特に有利な実施形態において、タンパク質性ベクターは、抗原提示の誘導とAPC成熟の促進を同時に行うことができ、こうして効果的な抗原特異的免疫応答を誘導できることを特徴とする。より好ましい実施形態において、この抗原特異的免疫応答はCTL応答である。
【0039】
本発明のタンパク質性ベクターは、組換えDNA技術によって得ることができる。従って、別の態様において、本発明は、本発明のタンパク質性ベクターをコードしている修飾された核酸に関する。この核酸は、タンパク質性ベクターのアミノ酸配列から容易に推定することができる。
【0040】
この修飾された核酸は、DNA構築体内に含有され得る。従って、本発明は、本発明のタンパク質性ベクターをコードする核酸を含むDNA構築体を提供する。このDNA構築体は、本発明のタンパク質性ベクターをコードしている核酸に作動可能に結合したコントロール配列を導入できる。"作動可能に結合した"とは、核酸を示す場合には、核酸が他の核酸配列と機能的な関係におかれる。"コントロール配列"は、特異的宿主細胞によって認識される発現シグナルであり、これは機能、例えば特定のコード配列(コントロール配列の例示は、プロモーター、エンハンサー、転写ターミネーター、リボソーム結合部位、タンパク質分泌または他の細胞内輸送のためのシグナルペプチド)の転写および翻訳を調節するものである。所望の配列を結合することは、従来の制限部位でのライゲーションによって達成される。かかる部位が存在しない場合に、合成オリゴヌクレオチドアダプターまたはリンカーは、従来方法に従って使用される。この意味においての利点は、このDNA構築体がDNA構築体によって、形質転換された宿主細胞の選択を可能にするモチーフまたは表現型をコードするマーカーまたは遺伝子も含むという事実により説明される。この発明によって提供された修飾された核酸およびDNA構築体は、数多くの実験手引き書(例えば、"Molecular Cloning: a Laboratory manual." Joseph Sambrook, David W. Russel Eds. 2001, 3rd ed. Cold Spring Harbor, New York)にまとめられている既知の従来方法によって得ることが出来る。
【0041】
特定の実現において、本発明により提供された修飾された核酸またはDNA構築体は、配列番号:1、配列番号:5、配列番号:9(EDA-NS3)または配列番号:11(EDA-OVA)を包含する。
【0042】
この発明によって提供された修飾された核酸またはDNA構築体は、適切なベクターに挿入され得る。従って、別の態様において、本発明は、上記修飾された核酸またはDNA構築体を含むベクター、例えば発現ベクターに関する。ベクターの選択は、ベクターが挿入されるべき宿主細胞に依存する。例示としては、核酸が挿入されるベクターは、プラスミドまたはウイルスであってよく、挿入の際に細胞ゲノム中に組み入れられていても、いなくてもよい。該ベクターは、従来方法(Sambrook et al., 2001, 上掲を引用する)により得られ得る。
【0043】
別の態様において、本発明は、この発明によって提供された修飾された核酸またはDNA構築体を含む宿主細胞、例えば形質転換宿主細胞に関する。本発明に従って、発現宿主細胞は、原核生物、即ちEscherichia coli、または真核宿主、即ち酵母(例えば、Saccharomyces cerevisiae、Pichia Pastoris)、昆虫細胞または哺乳動物細胞である。
【0044】
本発明の特定の実現において、本発明のタンパク質性ベクターをコードする修飾された核酸またはDNA構築体を含む発現ベクターは、イン・ビボの遺伝子移送または治療を目的としている。より具体的な実施態様において、発現ベクターはウイルスベクターである。この効果に対する適切なウイルスベクターは、次のものを包含する:アデノウイルス、アデノ関連ウイルス、レトロウイルス、レンチウイルス、アルファウイルス、ヘルペスウイルス、コロナウイルス誘導性ベクター等。
【0045】
別の態様において、本発明は、本発明のタンパク質性ベクターを作成するための方法に関するものであって、本発明の修飾された核酸またはDNA構築体を含有する宿主細胞を、タンパク質性ベクターの発現できる条件下で培養することを含む。宿主細胞の培養物を至適化するための条件は、用いた宿主細胞のタイプに依存する。所望により、本発明のタンパク質性ベクターを作成するための方法は、該ベクターの単離および精製を包含する。
【0046】
あるいは、本発明のタンパク質性ベクターは、他の従来方法によって得ることができる。かかる方法には、例えば固相化学合成;高速液体クロマトグラフィー(HPLC)を用いる精製;および、望ましくは従来技術、例えば配列決定または質量スペクトル分析法、アミノ酸分析、磁気共鳴技術等による分析が包含される。
【0047】
別の実現において、本発明のタンパク質性ベクターは、フィブロネクチンEDAドメイン(EDA)、またはTLR4を結合できるEDAドメインのフラグメント、または該EDAドメインの改変型に対応するアミノ酸配列を有するポリペプチドと、目的とする分子(例えば、ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質または他の化学物質)との共有結合によって得られる。これは、従来方法を用いて行うことができ、これはJohn Wiley & Sons により発表された実験手引き書、例えば"The current protocols in protein chemistry" (定期的に更新される。May 1, 2005までに更新されたもの)、またはGT Hermanson, AK Mallia および PK Smithによる"Immobilized affinity ligand Techniques"(Academic Press, Inc. Sandiego, CA, 1992)などにまとめられている。
【0048】
次に、本発明に従って、タンパク質性ベクターまたはこれをコードしている修飾された核酸およびDNA構築体、または該核酸または該DNA構築体を組み込んでいる発現ベクターおよび発現宿主細胞を、医薬組成物の調製において使用できる。
【0049】
他の実施形態において、本発明は、上記したとおり、免疫賦活性剤の調製において、該剤が医薬組成物であることを特徴とする、フィブロネクチンEDAドメイン(EDA)またはその同フラグメントまたは改変型に対応するアミノ酸配列を有するポリペプチドの使用を指す。
【0050】
特定の実施形態において、本発明の医薬組成物は、抗原提示細胞の成熟を刺激するため、または目的とする分子に特異的な有効な免疫応答を誘導するために使用され得る。特定の実施形態において、該医薬組成物は、免疫賦活性組成物が投与され得る対象におけるTh1免疫応答を誘導するために使用される。本明細書で使用したとおりに、"Th1免疫応答を誘導すること"とは、免疫賦活性組成物は混合したTh1/Th2応答を誘導する場合も包含できる。しかしながら、他の場合において、免疫賦活性組成物は、Th2免疫応答の誘導がほどんどないか、または実質的にないTh1免疫応答を誘導できる。特定の実現において、該医薬組成物はCTL応答を誘導するのに使用され得る。
【0051】
いくつかの実施形態において、免疫賦活性組成物は、免疫賦活性アジュバンドとして、例えば追加のアジュバンドと共に1以上の抗原と組み合わせて、または追加のアジュバンドを含まないで使用され得る。このように、いくつかの例において、免疫賦活性組成物はワクチンを形成できる。他の場合において、免疫賦活性組成物は、ワクチンとの結合において使用され得るアジュバンドとして機能し得る。
【0052】
フィブロネクチンEDAドメイン(またはそのフラグメントまたは改変型)を含むポリペプチドを包含する免疫賦活性組成物は、活性化されたCD8+Tリンパ球の拡大、メモリーCD8+Tリンパ球の生成、または両方を増強できる。このように、本発明の免疫賦活性組成物は、免疫賦活性組成物を受容する対象において抗原特異的細胞媒介性免疫を増強できる。
【0053】
特定の実現において、フィブロネクチンEDAドメイン(または、そのフラグメントまたは改変型)を含有する免疫賦活性組成物は、感染性疾患、腫瘍疾患またはアレルギー性疾患の処置および予防に有用である。本発明の具体的な実施形態において、該組成物はC型肝炎の処置および予防のために使用される。
【0054】
フィブロネクチンEDAドメイン(または、そのフラグメントまたは改変型)を含有する免疫賦活性組成物は、さらに担体、賦形剤および他の既知の医薬的に許容し得る成分を含有できる。
【0055】
本発明の免疫賦活性組成物は、当業者に既知の従来の方法(例えば、経口的に、皮下的に、局所的に)に従って、動物、例えば哺乳動物(ヒトまたは非ヒト)、鳥類などに投与され得る。
【0056】
本発明は、対象に対してフィブロネクチンEDAドメイン(またはそのフラグメントまたは改変型)を含む免疫賦活性組成物を投与することを含む、治療および/または予防方法も提供する。
【0057】
適切な投与経路は、経皮的または経粘膜的吸収、注射(例えば、皮下、腹腔内、筋肉内、静脈内など)、摂取、吸入などである。
【0058】
他のさらなる態様において、本発明は、少なくとも1つの許容し得る医薬担体および有効量のタンパク質性ベクターを、少なくとも1つのその発現形態または実施形態:
a)ポリペプチドの形態にあるタンパク質性ベクター;
b)該タンパク質性ベクターをコードする修飾された核酸;
c)該修飾された核酸を含む発現ベクター;または
d)該修飾された核酸も含む発現宿主細胞、
において包含する医薬組成物に関する。
【0059】
特定の実現において、医薬組成物は、有効量の樹状細胞を含んでいることを特徴とし、ここで該樹状細胞は、少なくとも1つのその発現形態または実施形態においてイン・ビトロでタンパク質性ベクターとインキュベートされている。より特定の実施態様において、該医薬組成物はワクチンまたは免疫治療用の組成物である。
【0060】
さらに、タンパク質性ベクターのいくつかの追加的使用は本明細書により提供される。本発明の一実施態様において、その発現形態のいずれかにおいてタンパク質性ベクターは、イン・ビトロまたはイン・ビボで樹状細胞成熟を誘導するために有効な医薬組成物の調製のために使用される。
【0061】
他の実施形態において、該タンパク質性ベクターは、タンパク質性ベクターと結合した目的とする分子(抗原またはエピトープ)に対して特異的な特異的免疫応答を誘導するための医薬組成物の調製のために使用される。この免疫応答は、体液性免疫応答(目的とする分子に対する抗体産生)、Tヘルパー応答、または細胞傷害性T細胞応答である。好ましい実施形態において、該免疫応答はCTL応答である。
【0062】
より特定の実現において、本発明は、感染疾患の処置および予防のための医薬組成物の調製におけるタンパク質性ベクターの使用に関する。該感染性疾患は、細菌、ウイルス、真菌または寄生感染疾患であり得る。
【0063】
別の特定の実現において、本発明は、腫瘍疾患の処置および予防のために有用な医薬組成物の調製において、本発明のタンパク質性ベクターの使用に関する。
【0064】
さらに別の特定の実現において、本発明は、アレルギー性疾患の処置および予防のために有用な医薬組成物の調製におけるタンパク質性ベクターの使用に関する。多くのアレルギー性疾患は、Th2免疫応答の活性化に関連する。従って、特異的なアレルゲンを担持するタンパク質性ベクターを使用することによって、Th2からTh1応答へのずれまたは切替えは、アレルギー性疾患に対して保護的または治療的効果をもち得る。
【0065】
本発明の特定の実施形態に従って、提案された医薬組成物は動物またはヒト宿主への投与に使用される。任意の適切な投与経路が使用され得る。特定の実現において、医薬組成物は、非経口経路(即ち、静脈、皮下、筋肉内)、経皮的経路または粘膜経路によって投与される。
【0066】
(図面の簡単な説明)
図1.産生および精製したEDAおよびEDA-SIINFEKLタンパク質のSDS-PAGE分析。EDAおよびEDA-SIINFEKLタンパク質の1つのアリコートを、15%ポリアクリルアミドゲル上におき、電気泳動に供した。分子量マーカー(MWM)をKDaで示す。EDAおよびEDA-SIINFEKLタンパク質(13-14 KDa)の推定分子量に対応するバンドを観察した。
【0067】
図2.タンパク質性ベクターEDA-SIINFEKLのTLR4への結合。2A.直接的な結合アッセイ。HEK293-LacZ細胞(HEK293-LacZ)およびHEK293-TLR4/MD2/CD14(HEK293 TLR4)細胞を、1μM EDA-SIINFEKLを用いてパルスし、パラホルムアルデヒドで固定し、抗Hisおよび抗EDA抗体で標識し、抗マウスIgG-FITCで顕色させ、フローサイトメトリーにより分析した。2B.TLR4発現細胞に対する抗TLR4抗体の結合を阻害するEDA能力。HEK-TLR4細胞を、500nM EDA-SIINFEKL タンパク質の存在または非存在において、2時間4℃でインキュベートした。細胞を洗浄し、FITCで染色した抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。2C.様々な濃度のEDA-SIINFEKLタンパク質を用いてTLR4発現細胞に結合する抗TLR4抗体の阻害%。2D.細胞付着アッセイ。トリチウム化したチミジンで染色したHEK-hTLR4またはHEK-LacZ細胞を、EDAタンパク質で予め被覆した前に96ウェルミクロプレートのウェルに分配し、2時間37℃の間インキュベートした。非付着細胞を除き、付着細胞を回収し、取り込んだ放射活性をTopcountシンチレーションカウンターで計測した。ウェルあたりの付着細胞数を、標準曲線を利用して計算した。
【0068】
図3.EDA-SIINFEKLはTLR4シグナル経路を活性化する。発現がNF-κB-誘導性ELAM-1プロモーターによって制御されるこのレポーター遺伝子でトランスフェクトしたHEK293/TLR4-MD2-CD14またはHEK293/LacZ発現細胞の培養物上清においてヒト分泌した胚アルカリホスファターゼ遺伝子の比色分析測定。トランスフェクションの24時間後に、細胞を様々な濃度のLPS、100 nM EDA-SIINFEKL タンパク質(EDA)またはプロテイナーゼKで予め消化した100 nM EDA-SIINFEKL タンパク質の存在または非存在下でインキュベートした。棒線は、NF-κBの誘導倍である(HEK293/TLR4-MD2-CD14由来の上清で得たODをHEK293/LacZからの上清で得たODによって割った)。
【0069】
図4.EDA-SIINFEKLは、イン・ビトロでDCによって炎症性サイトカインの分泌を誘導する。骨髄由来DCを、LPS(1μg/ml)、EDA-SIINFEKL(500 nM)、プロテイナーゼKで消化したEDA-SIINFEKL(500 nM)の存在下または生理食塩溶液で培養した。24時間後、IL-12(A)およびTNF-α(B)の存在をELISAにより培養物上清中で測定した。
【0070】
図5.EDA-SIINFEKLは、イン・ビボでCD11c DC 成熟を誘導する。樹状細胞の成熟は、T-細胞応答の最適な刺激のための要件である。成熟が起こる時に、APCは、表面分子、例えばクラスI(我々のモデルではH2Kb)およびクラスIIMHC(我々のモデルではIAb)、CD40、CD80およびCD86分子の発現を増加させる。従って、我々は、EDA-SIINFEKLがCD11c発現細胞のイン・ビボで成熟を誘導するかどうかを分析した。C57BL/6wtマウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いてi.v.で免疫化した。また、C57BL/6 TLR4 KOマウスをEDA-SIINFEKL(25μg)またはPBS単独を用いて免疫化した。15時間後にマウスを屠殺し、CD11c細胞をautoMACSにより精製した。細胞を標識し、H-2Kb、I-Ab、CD40、CD80およびCD86分子の発現についてフローサイトメトリーにより分析した。
【0071】
図6:EDA-SIINFEKLは、樹状細胞によりSIINFEKLエピトープに特異的なT細胞に対して効果的に提示される。我々は、このエピトープに特異的なOT-1 遺伝子組み換えマウス由来のT細胞に対して処理済CTLエピトープSIINFEKLの提示について、APCによって捕捉されるべきEDA-SIINFEKLの能力を特徴分析した。(A) OT-1 トランスジェニックマウスからの非付着細胞によるIFN-γ産生。骨髄由来DCを、培地単独、様々な濃度の合成SIINFEKLペプチド、SIINFEKL、およびEDA、EDA-SIINFEKL(融合タンパク質)またはEDA単独の存在下で培養した。24時間後、DCを回収し、105個の非付着性OT-1細胞の存在下でAPCとして使用した。さらなる24時間後に、上清を回収し、分泌したIFN-γを測定した。(B)TLR4分子依存性。B3Zハイブリドーマ細胞(105細胞/ウェル)を、C57BL/6 wt マウス由来の脾臓細胞の存在下、またはTLR4分子(105 細胞/ウェル)およびEDA-SIINFEKLタンパク質(100 nM)のためのノックアウトマウス由来の脾臓細胞と共にインキュベートした。(C)C57BL76wt マウスの脾臓由来の細胞を、抗TLR4抗体の存在または非存在(BおよびC)において、B3Zハイブリドーマ細胞およびEDA-SIINFEKLタンパク質と共に共培養した。培養物上清に分泌したIL-2の量を、CTLL 細胞系の使用を基にしたバイオアッセイによって測定した。(D)融合タンパク質EDA-SIINFEKLの抗原提示に対するクロロキン、モネンシン、ブレフェルジンまたはシクロヘキシミドの効果。骨髄由来樹状細胞を、30 mMのクロロキン、ブレフェルジン、モネンシン、または4μg/mlのシクロヘキシミドの非存在下または存在下で1時間インキュベートし、その後EDA-SIINFEKLまたは合成ペプチドSIINFEKL(白色の棒)を添加した。10時間の培養後に、DCをグルタルアルデヒド中で固定し、非付着性OT-1マウス細胞(105細胞/ウェル)との同時培養物中で抗原提示細胞(APC)(104 細胞/ウェル)として使用した。24時間後、分泌したIFN-γの量を市販のELISAにより測定した。
【0072】
APC上でのTLR4発現が提示過程を進行させ得るかどうかを試験するために、我々は、C57BL/6 wt マウスまたはTLR4 KO マウス由来の脾臓細胞および様々な濃度のEDA-SIINFEKL (nM)の存在において、B3Z ハイブリドーマ細胞(SIINFEKLエピトープに特異的)を培養した。培養上清に放出されたIL-2の量を、CTLLを基にしたバイオアッセイにより測定した(図6E)。
【0073】
図7.EDA-SIINFEKLによるマウスの免疫性付与は、SIINFEKLエピトープに特異的な細胞応答を誘導する。前記結果は、EDA-SIINFEKL組換えタンパク質が生物活性であり、APCを特異的に活性化することを示す。イン・ビボでの特異的なT細胞免疫応答の誘導は、ワクチンの開発のために重要である。このように、我々は、EDA-SIINFEKL融合タンパク質により免疫化されたマウスが、このエピトープに対して特異的な細胞応答を生じるかどうかを試験した。(A)IFN-γ産生細胞の誘導。0および10日目に、C57BL/6 マウスを、EDA-SIINFEKLの1.5 nmolまたはSIINFEKLペプチドの1.5 nmolを用いて免疫化した。20日目に、SIINFEKLの存在または非存在において脾臓細胞を48時間インキュベートし、そして分泌されたIFN-γの量をELISAにより測定した。(B)SIINFEKL-特異的CTL応答分析の誘導。EDA-SIINFEKLまたはSIINFEKLにより免疫化したマウス由来の脾臓細胞を、SIINFEKLペプチドの存在下で5日間再刺激した。このインキュベーション後に、従来のクロミウム51遊離アッセイにより、SIINFEKLペプチドの非存在または存在下でEL-4標的細胞に対するCTL活性を測定した。データは、3つのサンプルから正味の特異的分析物(SIINFEKLでパルスした標的細胞の%分解−非パルス化標的細胞の分解%)の値の平均%を示す。
【0074】
図8.EDA-SIINFEKLは、EG7 OVA発現細胞による腫瘍攻撃から守る。EG7OVA 腫瘍細胞の注射に対するマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを3 nmolのEDA-SIINFEKL、SIINFEKLまたは生理食塩溶液を用いて0および10日目にs.c.で免疫化した。20日後、二次免疫性付与マウスを105 EG7OVA 細胞を用いてs.c.攻撃した。腫瘍増殖を、カリパスを用いてモニターし、式 V=(L x w2)/2(式中、L, 長さ;w, 幅である)を用いてmm3で表した。腫瘍サイズが8cm3以上の体積に達した場合に、マウスを屠殺した。
【0075】
図9.EDAは、OVAタンパク質を用いる免疫性付与後に細胞傷害性応答の誘導においてアジュバンドとして作用する。EDAがイン・ビボで樹状細胞の成熟を促進することができれば、EDAはそれ自身によっては細胞傷害性応答を活性化できないが細胞傷害性エピトープを含有するタンパク質を用いる免疫性付与後にアジュバンドとして作用できるという可能性が存在する。この可能性を試験するために、我々は、その他のタイプのアジュバンドを用いないでEDA(50μg)とOVA タンパク質(500μg)により一群のマウス(A)、およびOVA(500μg)により別の群のマウス(B)を免疫化した。免疫性付与1週間後、マウスを屠殺し、脾臓細胞を、SIINFEKL合成ペプチドの存在下で培養した。5日後の培養物中で、SIINFEKLペプチドを用いてパルスしたEL-4 標的細胞に対する細胞傷害性応答を、従来のCr51遊離アッセイにおいて測定した。
【0076】
図10.EDAは、より大きな抗原についてのビヒクルとして作用できる。前記実験において、EDAタンパク質は、細胞傷害性エピトープを輸送するためのビヒクルとして作用でき、該エピトープに対するCTL応答の誘導を支持することが証明された。後者の工程において、EDAが比較的大きな抗原を輸送でき、抗原に対する細胞性応答の誘導を促進できるかどうかについての試験がのぞまれる。この目的により、我々は融合タンパク質EDA-OVAを構築し、イン・ビトロおよびイン・ビボでの下記実験を行った。(A)組換えEDA-OVAタンパク質のSDS-PAGE分析。組換えタンパク質EDA-OVAを、E. coliで発現し、アフィニティクロマトグラフィーにより精製した。脱塩し、中和し、濃縮し、SDS-PAGEにより分析した。該タンパク質の推定分子量に対応するおおよそ55kDaのバンドを観察した。(B)抗原提示実験、骨髄由来DCを、OVA、EDA-OVA(融合タンパク質)、EDA+OVAまたはEDA単独の存在下または非存在下で培養した。24時間後、DCをOT-1マウス由来の105非付着細胞の存在において抗原提示細胞として使用した。DCの存在下でOT-1マウス由来の非付着細胞によりIFN-γ産生を、市販のELISAにより定量化した。(C) EDA-OVAタンパク質は、イン・ビボでOVA特異的CTLを誘導する。C57BL/6 マウスを、1nmolのEDA-OVAを用いるか、または1 nmolのOVAを用いて免疫化した。免疫性付与7日後に、免疫化したマウスからの脾臓細胞を、SIINFEKLペプチドの存在下に5日間イン・ビトロで再刺激した。その後、SIINFEKLの非存在または存在下でインキュベートしたEL-4 細胞に対する特異的CTL活性を、従来のCr51 遊離アッセイにおいて測定した。データは、3つのサンプルから、正味の特異的溶菌値(SIINFEKLを用いてパルスした標的細胞の溶菌%−非パルス標的細胞の溶菌%)の平均%を示す。
【0077】
図11.EDA-NS3タンパク質は、NS3C型肝炎ウイルスタンパク質に対する特異的CTL応答を引き起こす。EDAタンパク質が大きな抗原についてのビヒクルとして作用できることが観察されると、C型肝炎ウイルスのNS3タンパク質(NS3タンパク質のプロテアーゼ領域からのアミノ酸1-196)に対する抗ウイルス応答を誘導するEDAの能力を、該ウイルスによる感染に対するワクチン接種ストラテジーとして分析した。(A)組換えEDA-NS3(1-196)タンパク質のSDS-PAGE分析。組換え融合タンパク質EDA-NS3を構築し、E. coliで発現し、SDS-PAGEにより分析した。(B)EDA-OVAタンパク質は、イン・ビボでOVA-特異的CTLを誘導する。HHDマウス(HLA-A2.1タンパク質についてトランスジェニック)を、生理食塩溶液に溶解したEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与1週間後、脾臓細胞を、イン・ビトロでNS3 1073ペプチド(HLA-A2拘束のためにNS3タンパク質由来の免疫優性の細胞傷害性T決定基を含有する)を用いて再刺激した。培養物中で5日後、ペプチド1073(V)(配列番号:20、CVNGVCWTV)と共に、または該ペプチド(配列番号:21、CLNGVCWTV)の1073(L)改変型と共に、またはペプチドの非存在下でインキュベートしたT2標的細胞に対して、細胞傷害性活性を、従来のCr51遊離アッセイを用いて測定した。(C)EDA-NS3タンパク質は、NS3タンパク質からの様々なエピトープに対して複数のエピトープ様応答を誘導する。EDA-NS3により免疫化したマウスから得た脾臓細胞を、ペプチド1038-1046、NS3 1073-1081またはNS3 1169-1177(NS3タンパク質のフラグメント1-196のなかで、HLA-A2拘束のために3つの細胞傷害性決定基を含有する)の存在下に、または組換えNS3タンパク質(Mikrogen)と共に、48時間イン・ビトロで再刺激した。上清に分泌されたIFN-γの量を市販のELISAにより測定した。(D)EDA-NS3タンパク質は、長期間持続する細胞傷害性応答を誘導する。HHDマウスを、生理食塩溶液中のEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与60日後、マウスを屠殺し、ペプチドNS3 1073に特異的なCTLの存在を、NS3 1073 ペプチドの非存在または存在中でインキュベートしたT2標的細胞を用いて、従来のクロミウム51遊離アッセイにおいて測定した。(E) EDA-NS3とインキュベートしたDCを有するC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現する組換えワクシニアウイルス vHCV(1-3011)による感染からマウスを保護する。EDA-NS3 タンパク質と予めインキュベートした 106DCを用いてマウスを免疫化し、7日後に、それらはi.p.経路による5x106 ワクシニアウイルスvHCV(1-3011)の攻撃を受けた。感染3日後にマウスを屠殺し、BSC-1細胞感染アッセイにより、卵巣組織mgあたりのウイルス負荷を定量した。
【0078】
実施例
実施例1.フィブロネクチンからのエクストラドメインAはTLR4と相互作用し、TLR4情報伝達経路を活性化する。
1.1 材料および方法
1.1.1 EDAおよび EDA-SIINFEKLタンパク質性ベクター組換えタンパク質の発現
組換えタンパク質性ベクターの調製
フィブロネクチンエクストラドメインA(EDA)を、肝臓損傷を誘導するために、特異的プライマーおよびコンカナバリン-Aにより処理したマウスから得た肝細胞由来のRNAを用いて、RT-PCRにより増幅した [Lasarte et al, Hepatology. 2003; 37(2):461-70.]。肝臓組織試験片を均質化し、Ultraturrax Driver T.25 (Janke & Kunkel, Ika-Labortechnik, Germany)を用いてUltraspec (Biotecx, Houston, TX, USA)において溶菌した。RNAを、Chomczynski および Sacchiらの方法に従って単離した(Chomczynski P および Sacchi N. グアニジン酸チオシアネート-フェノール-クロロホルム抽出によるRNA単離のシングル・ステップ方法. Anal Biochem 1987; 162: 156-159)。RNAを、5mMジチオスレイトール(DDT)、0.5mM デオキシヌクレオシド三リン酸(Boehringer Mannheim, Mannheim, Germany)、25U リボヌクレアーゼ阻害剤(Promega Corporation, Madison, WI, USA) および 200ng ランダムヘキサマー(Boehringer Mannheim)を加えた5xRT緩衝液(250mM トリス-HCl Ph 8.3, 375mM KCl, 15mM MgCl2)(20μL容量)中で200UのM-MuLV 逆トランスクリプターゼ(Gibco-BRL)を用いて逆転写した(60分、37℃で)。加熱(95℃、1分間)および氷上での急冷後、cDNAプール(0.3μg)を、0.08mM dNTP、上流および下流プライマー(各40ng)、1.5mM MgCl2およびTaq DNA ポリメラーゼの2U(Promega Corporation)を含有する10x緩衝溶液(100mM トリス-HCl pH9.3、500mM KCl、1% Triton X-100)(20μl)中で、PCR増幅に使用した。上流プライマーは(配列番号:13)
【化1】

(下線を引いた塩基を、制限酵素NdeIにより認識される配列を導入するためにプライマーに加えた。斜字体の配列はEDAの開始部分に対応する)であり、
下流プライマーは(配列番号:14)
【化2】
(下線を引いた塩基を、制限酵素NotIにより認識される配列を導入するためにプライマーに加えた。太字の配列は、両末端で3つのアミノ酸により挟まれたOVA CTL エピトープ(SIINFEKL)
【化3】
をコードしている配列に対応しており、斜字体の配列はEDAの末端に対応する)である。
【0079】
PCR増幅フラグメントを、TOPO TAクローニングキット(Invitrogen, Carlsbad, CA, USA)を用いてpCR2.1-TOPOにクローニングした。このプラスミドをNdeIおよびNotIで解裂させ、得られたDNAフラグメントをカルボキシ末端で6つのヒスチジン残基(6xHis tags)を担持する融合タンパク質の発現を可能にするNdeI/NotI消化したプラスミドpET20b (Novagen)でサブクローニングした。
【0080】
融合タンパク質EDA SIINFEKL-6xHisを発現する得られるプラスミドpET20b2-26を、組換えタンパク質性ベクターの発現のためにBL21(DE3)細胞にトランスフェクトした。トランスフェクトした細胞を、OD600が0.5-1に達するまで、37℃でLB(1l)中で生育させた。IPTGを、0.4 mMの終濃度まで該培養物に加えて、室温で終夜振盪しながらインキュベートした。細胞を遠心分離により回収し、0.1M トリス-HCl(pH=7,2)に再懸濁させ、リゾチームで処理し、French press (20.000 pstで2回通過)を使用して破砕し、遠心分離により清澄させ、濾過した。可溶性分画に存在する融合タンパク質をFPLCプラットホーム(AKTA, Pharmacia) を用いてアフィニティクロマトグラフィー(Histrap, Pharmacia)により精製した。溶出したタンパク質を、Hitrap脱塩カラム(Pharmacia)を用いて脱塩し、Amicon Ultra 4-5000 MWCO Centrifugal filter device (Millipore Carrighwahill, Ireland)を用いて濃縮した。組換えタンパク質性ベクターを、エンドトキシンのレベルが0.2EU/タンパク質μg以下(LAL アッセイによる評価, Cambrex)となるまで、Endotrap カラム(Profos Ag, Regensburg, Germany)を用いて、エンドトキシンから精製した。
【0081】
EDAタンパク質についての発現プラスミドを得るために、PCRをプライマー
【化4】
(配列番号:13)および
【化5】
(配列番号:15)、およびEDA-SIINFEKLプラスミドに記載したものと類似したクローニングストラテジーを用いて行い、プラスミドpET20bEDA1.2を得た。タンパク質(20μg)を、15% SDS-アクリルアミドゲルにおいて各試料に添加し、その後コマジー・ブルーにより染色した。推定分子量(13 kDa)に対応するバンドを観察した。
【0082】
1.1.2. TLR4に対するEDA-SIINFEKLの結合。フローサイトメトリーおよび付着アッセイ
組換えEDA-SIINFEKLタンパク質がTLR4発現細胞に結合できたかどうかを試験するために、我々は、HEK293発現ヒトTLR4-MD2-CD14 293 (Invivogenから)を使用した。我々は、負のコントロールとしてLacZ (Invivogen)で形質転換したHEK293細胞も使用した。細胞を、1 mM EDA-SIINFEKLと共に1時間4℃でパルス化し、PBSで洗浄し、PBS中で4%パラホルムアルデヒドを用いて10分間固定した。3回洗浄後、細胞を、1/100 抗His抗体(Qiagen)および1/200 抗CD16 (FcBlock, from Becton Dickinson)を用いて1時間30分間標識した。3回洗浄した後、細胞を、1/100希釈でフルオレセインにて標識した抗マウスIgGと共に30分間インキュベートし、フローサイトメトリーにより分析した。
【0083】
あるいは、EDA-SIINFEKLタンパク質がHEK-hTLR4 細胞に結合するFITC染色した抗ヒトTLR4抗体を阻害する能力を測定した。この目的のために、HEK TLR4 細胞を、EDA-SIINFEKLの様々な量の存在下または非存在のもとで、2時間4℃でインキュベートした。その後、細胞を洗浄し、抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。阻害%を、EDA-SIINFEKL の様々なアッセイ濃度に対して計算した。細胞付着アッセイを行った。HEK LacZまたはHEK hTLR4 細胞をトリチウム化したチミジンにより予め染色し、EDAタンパク質で予め被覆した96 ウェルプレートに分配した。2時間37℃でインキュベーションの後、非付着細胞を除去し、一方で付着細胞を回収し、取り込んだ放射活性を、Topcount シンチレーションカウンターで測定した。ウェルあたりの付着細胞数を、様々な染色細胞の濃度を用いて得た標準曲線を用いて計算した。
【0084】
1.1.3. TLR4情報伝達経路の活性化
HEK293/hTLR4-MD2-CD14またはHEK293/LacZ 発現細胞を、生成者指示書に従って(Invivogen)、ヒト分泌胚のアルカリホスファターゼ遺伝子(SEAP)を担持するプラスミドを用いてトランスフェクトした。SEAP発現をNF-κB-誘導性ELAM-1プロモーター(pNiFty-SEAP (Invivogen))によって制御した。トランスフェクション24時間後、細胞を、様々な濃度のLPS、100 nM EDA SIINFEKLタンパク質またはプロテイナーゼ-Kで予め消化した100 nM EDA-SIINFEKLタンパク質の存在または非存在においてインキュベートした。24時間後、レポーター遺伝子の発現を、比色分析アッセイ(Invivogen)により、培養物上清において測定した。図3において、棒線は、NF-γB誘導の倍数(HEK293/TLR4-MD2-CD14からの上清に対して得たODを、HEK293/LacZからの上清に対して得たODによって割る)を示す。このアッセイにおいてEDA調製物中のエンドトキシン汚染物質の量は0.0003μg/ml以下であった。
【0085】
1.2 結果
1.2.1. EDAの発現およびEDA-SIINFEKL組換え融合タンパク質
組換えEDA-SIINFEKLタンパク質をE. coliにおいて6xHis 融合タンパク質(配列番号:2)として発現させ、アフィニティクロマトグラフィーにより精製し、脱塩し、方法の章で説明したようにエンドトキシンから精製した。得られたタンパク質を、抗his抗体(図1)を用いてSDS-PAGEおよびウェスタンブロットにより分析した。推定分子量(13 kDa)に対応するバンドを、各タンパク質について観察した。
【0086】
1.2.2. EDA-SIINFEKL 融合タンパク質はTLR4を結合する
フィブロネクチンエクストラドメインAは、トル様受容体4を活性化することが記載されている(Okamura et al, JBC, 2001; 276:10229-10233)。しかしながら、TLR4に対するEDAの物理的結合に関する直接的な証拠はない。我々は、EDA-SIINFEKLタンパク質が、TLR4を発現する細胞に結合する能力を有するかどうかを最初に分析した。ヒトTLR4-MD2-CD14を発現するHEK293またはLacZ(Invivogen)でトランスフェクトしたHEK293細胞を、1μM EDA-SIINFEKLタンパク質を用いてパルスし、抗His抗体およびフルオレセインで標識した抗マウスIgG(方法を参照されたい)で標識し、フローサイトメトリーにより分析した。ヒトTLR4-MD2-CD14を発現するHEK 293 細胞は、LacZを発現するHEK 293 細胞よりわずかに高い蛍光強度を示した(図2A)ことがわかった。また、ヒトTLR4に結合するフルオレセイン染色抗体を阻害するEDA-SIINFEKLタンパク質の能力も測定した。この結果から、HEK-hTLR4細胞と500mM EDA-SIINFEKLタンパク質との前記インキュベーションにより、該抗体結合のおおよそ50%が阻害されたということが判った(図2B)。図2Cは、抗体結合に対するEDA-SIINFEKLの様々な用量の抑制効果を示す。一方、我々は、付着アッセイにおいてEDAタンパク質で前もって被覆したプラスチックウェルに対する、HEK hTLR4およびHEK LacZコントロール細胞の結合能力を測定した。HEK hTLR4 細胞は、EDAを含有するウェルに特異的に結合できることを示す。これらの全ての実験は、EDAおよびEDA-SIINFEKLタンパク質がTLR4を結合できることを示す。
【0087】
1.2.3. EDA-SIINFEKL 融合タンパク質はTLR4 情報伝達経路を活性化する
TLR4シグナル伝達はNF-κB輸送をもたらし、これは多様な遺伝子のプロモーターのなかのコンセンサスエレメントに結合する転写因子である。組換えEDA-SIINFEKLタンパク質がTLR4を活性化できるかどうかを決定するために、我々は、ヒト分泌胚のアルカリホスファターゼ遺伝子(SEAP)を担持するプラスミドでトランスフェクトしたHEK293/hTLR4-MD2-CD14またはHEK293/LacZ細胞を、NF-κB-誘導性ELAM-1プロモーター(pNiFty-SEAP(Invivogen))の制御の下で使用した。我々は、EDA-SIINFEKLタンパク質が、HEK293/hTLR4-MD2-CD14トランスフェクト細胞においてのみSEAPの発現を刺激でき、NF-κB誘導倍数が7に達し、細胞をLPS(0.01μg)とインキュベートした時に見出された値と類似していることを見出した(図3)。EDA-SIINFEKLタンパク質がプロテイナーゼKで予め消化されていれば、NF-κB核輸送を刺激するこの能力は完全に消失し(図3)、TLR4のEDA活性化は、組換えタンパク質調製において潜在的なLPSコンタミネーションによって説明できないことを示唆している。
【0088】
1.3 考察
我々は、E. coliにおいて組換え融合タンパク質EDA-SIINFEKL 6xHisを発現できる組換えプラスミドpET20b2-26を構築した。6つのヒスチジンの存在により、融合タンパク質の検出および精製が可能となる。こうして、我々は、十分量の融合タンパク質をE. coli培養物の細胞質分画から精製することが出来た。結合アッセイは、EDA-SIINFEKLタンパク質がTLR4に特異的に結合出来ることが示唆されたTLR4発現細胞で行った。さらに、我々は本明細書において、EDA-SIINFEKLがTLR4シグナル経路を活性化できることを示す。この活性化は、プロテイナーゼKによる先の消化によりNF-κB核輸送を刺激する能力が完全に消失するので、該タンパク質の潜在的なLPSコンタミネーションとは関連がない。さらに、このアッセイにおいて、EDA調製の際のエンドトキシンの汚染物質の量は、0.0003μg/ml以下であって(LALアッセイによって評価される)、このイン・ビトロのアッセイではTLR-4 シグナル経路を活性化することができない。これらの結果は、EDAタンパク質が、抗原をTLR4発現細胞に標的化するためにタンパク質性ビヒクルとして使用できたということを示唆する。樹状細胞は、トル様受容体および特定のTLR4を発現することが知られている。また、DCに対する最も強力な成熟刺激のいくつかは、TLR受容体とそれら各々のリガンドとの相互作用からおこる。こうして、EDAを含有する融合タンパク質および特定抗原の相互作用は、炎症性サイトカインの産生および共刺激分子の上方調節を誘導することによって先天性免疫の急速な活性化を促進できた。さらに、DCの表面に対するこの融合タンパク質の標的化は、DCにより抗原の捕捉およびエンドサイトーシスを増加でき、結果としてこの抗原に対する免疫応答が増強する。
【0089】
実施例2.融合タンパク質を含有するEDAはイン・ビトロおよびイン・ビボで樹状細胞成熟を誘導し、細胞傷害性Tリンパ球の誘導を可能にする
2.1 材料および方法
2.1.1. 骨髄由来樹状細胞産生
樹状細胞を骨髄細胞から成熟させた。ACK溶菌緩衝液をもちいて赤血球を溶菌した後に、細胞を洗浄し、次にCD4 (GK1; ATCC, Manassas, VA)、CD8(53.6.72; ATCC)、Ly-6G/Gr1(BD-Pharmingen; Sandiego CA)およびCD45R/B220 (BD-Pharmingen)に対する抗体の混合物とのインキュベーションにより、その後にウサギ補体によってリンパ球および顆粒球を枯渇させた。残存細胞を、mGM-CSF(20 ng/ml)およびmIL-4(20 ng/ml)(Peprotech; London, UK両方)を有する強化完全培地中で、12-ウェルプレートにおいて、106細胞/mlで生育させた。2日毎に、培地をサイトカイン含有する新規培地で置換した。非付着性樹状細胞を、7日で収穫し、37℃および5%CO2で、LPS(Sigma)(1μg/mlまたは15ng/ml)、EDA-SIINFEKL (500 nM)またはSIINFEKL (10μM)の存在または非存在下で培養した。幾つかの実験において、ポリミキシン(10μg/ml)を培養物に加え、エンドトキシン汚染物質の影響を阻害する。24時間培養後、上清を回収し、IL-12およびTNF-αを、生成者指示書に従ってELISA(BD-Pharmingen)によって測定した。
【0090】
2.1.2 EDA-SIINFEKLを用いる免疫性付与後のCD11c細胞のイン・ビボでの成熟の測定。プロテイナーゼKによる消化の効果
DCの成熟を、多様な表面マーカーの発現を測定するフローサイトメトリーによりイン・ビトロで評価した。C57BL6マウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独によりi.v.注射した。プロテイナーゼKによるEDA-SIINFEKLの消化を、アガロースプロテイナーゼK(Sigma, St Louis)を用いて実施した。つまり、洗浄緩衝液(20 mM トリス-HCl, pH 7.2, 1 mM EDTA, 1mM ClCa2)中で洗浄したプロテイナーゼアガロースビーズ(5 mg/ml)を使用して、EDA-SIINFEKLタンパク質またはLPSを20分30℃で消化した。アガロース-プロテイナーゼKビースを遠心分離により除去した。15時間の免疫性付与後、マウスを屠殺し、autoMACSによりCD11c細胞を精製した。細胞を標識し、フローサイトメトリーにより分析した。
【0091】
2.1.3. 抗原提示能力を分析するためのイン・ビトロ試験
我々は、このエピトープに特異的なOT-1トランスジェニックマウスからのT細胞またはTハイブリドーマB3Zの両方に対してプロセシングされたCTLエピトープSIINFEKLを提示するAPCにより捕捉されるべきEDA-SIINFEKLの能力を特徴づけた。骨髄由来DCを、様々な濃度のEDA-SIINFEKL、EDAおよびSIINFEKL(共有結合でない)またはSIINFEKLの存在下において培養した。培養24時間後、上清を回収し、IFN-γ産生をELISAにより測定した。あるいは、培養開始12時間後、DCを、回収し、0.05% グルタルアルデヒドで固定し、OT-1からの非付着細胞の様々な量の存在下またはB3ZハイブリドーマT細胞系の存在下に、APCとして使用される。いくつかの実験において、DCをクロロキン(3μM)、モネンシン(1μl Golgystop, Pharmingen)、ブレフェルジン(1μl Golgyplug, Pharmingen)、シクロヘキシミド(4μg/ml)の存在または非存在の下でインキュベートし、次いでEDA-SIINFEKLまたはSIINFEKLペプチドの存在下でインキュベートした。数例において、抗TLR4抗体を該培養物に添加した。
【0092】
処理されたAPCの存在下でB3Z 細胞の活性化を、IL-2産生を測定することによって行った。B3Z ハイブリドーマ細胞(105 細胞/ウェル)を、完全培地(10% FCS、2 mM グルタミン、100 U/ml ペニシリン、100μg/ml ストレプトマイシンおよび5x10-5M 2-メルカプトエタノールで強化したRPMI 1640)中で、18 時間C57BL/6 wtマウスまたはTLR4 KOマウス由来の脾臓細胞(105細胞/ウェル)、および様々な濃度のEDA-SIINFEKLの存在下に培養した。培養物上清に放出されたIL-2の量を、先に記載したとおりにCTLLを基にしたバイオアッセイにより測定した。
【0093】
2.1.4.イン・ビボでの免疫性付与後の細胞傷害性Tリンパ球(CTL)およびIFN-γ産生細胞の誘導の測定
C57BL6 マウスを、0日および10日目にEDA-SIINFEKL(50μg)またはSIINFEKL(50μg)を用いてi.v.により免疫化した。20日目で、SIINFEKLに対してCTL応答の分析のためにマウスを屠殺した。免疫化動物から脾臓細胞を、SIINFEKL の1mlあたり0.1μgの存在下に5 x 106細胞/ml(10 ml)を完全培地中で5日間培養した。5日目に、細胞をクロミウム放出アッセイのために回収した。分解活性を、様々な数のエフェクター細胞を、51Crを予め添加した1x 104 EL-4 標的細胞と共にSIINFEKLの存在下または非存在で4時間インキュベートすることによって測定した。溶菌比活性%を、式:(cpm 実験値 − cpm 自然発生値)/(cpm 最高値 − cpm 自然発生値) x 100、に従って計算した、式中自然発生値溶菌とは、エフェクター細胞の非存在下で培養した標的細胞に対応しており、最高値の溶菌は標的細胞を5% Triton x100とインキュベートすることによって得られる。
【0094】
SIINFEKLに対する応答においてIFN-γの産生を測定するために、免疫化マウス由来の脾臓細胞を、0.25 mlの最終容量で完全培地単独かまたはペプチド(30μM)と共に、8x105 細胞/ウェルで96-ウェルプレートに播種した。48 時間後に上清(50μl)を除去し、生成者指示書に従ってIFN-γをELISAにより測定した(Pharmingen, Sandiego, CA)。
【0095】
他の実験群において、タンパク質混合物中でアジュバンドとして作用するEDAタンパク質の能力を証明した。この場合において、C57BL/6 マウスの一群を、PBSにおいてOVA タンパク質(500μg)の存在下にEDA-SIINFEKL(50μg)を用いてi.v.により免疫化し、一方別の群はPBSにおいてOVAタンパク質(500μg)単独で免疫化した。1週間後、上記したとおりにマウスを屠殺し、両方の群でSIINFEKLに対するCTL応答を決定した。
【0096】
2.1.5. EG7OVA 腫瘍細胞攻撃に対する保護
マウスを、0および10日目にEDA-SIINFEKL(3 nmol)またはSIINFEKL(3 nmol)を用いてs.c.で免疫化した。二次免疫性付与後20日目のマウスを105 EG7OVA細胞を用いてs.c.により攻撃した。腫瘍成長を、カリパスで制御し、式V = (L x w2)/2を用いてmm3で表した。式中、Lは長さ;wは幅である。腫瘍サイズが8 cm3以上の大きさに達した時にマウスを屠殺した。
【0097】
2.2. 結果
2.2.1. EDA-SIINFEKL 融合タンパク質は、骨髄由来樹状細胞(BMDC)によるIL-12およびTNF-α産生を刺激する。
我々は、EDA-SIINFEKL組換えタンパク質がBMDCを刺激し、炎症性サイトカイン、例えばIL-12またはTNF-αを産生出来るかどうかを試験した。即ち、BMDCをSIINFEKL (10μM)、LPS(1μg/ml および 15ng/ml)またはEDA-SIINFEKL-6xHis (500nM)を用いて培養した。24時間後に、培養物上清中のIL-12またはTNF-αをELISAにより測定した。EDA-SIINFEKLが、BMDCにより非常に高レベルのIL-12またはTNF-α産生を刺激できたということが判った(図4)。該タンパク質がプロテイナーゼKで予め処理された時にこの免疫賦活性能力は消失し、該活性はタンパク質のサンプル中のエンドトキシンの潜在的痕跡を理由とするものではないことを示唆している。
【0098】
2.2.2. EDA-SIINFEKLはイン・ビボでCD11c発現DCのTLR4依存性の成熟を誘導する
樹状細胞(DC)は、ナイーブT細胞および抗原に対する二次応答を刺激する独特な能力を有する最も強力な抗原提示細胞である。DCは、抗原を捕捉する能力を持ち、それらをペプチドに分解して、細胞傷害性T細胞(CTL)またはTヘルパー細胞各々に対するMHCクラスIまたはII分子との会合において、このペプチドを提示する。未熟DCは、抗原を捕捉できるが、ナイーブT細胞を刺激出来るようになるためには分化または成熟しなければならない。従って、樹状細胞の成熟はT-細胞応答の至適刺激のための要件である。成熟がおこる時に、APCは、表面分子、例えばクラスIおよびクラスII MHC、CD40、CD80およびCD86分子の発現を増加させる。そのため、我々は、EDA-SIINFEKLがイン・ビボでCD11c発現細胞の成熟を誘導できるかどうかを分析した。C57BL6 マウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いて、i.v.で免疫化した。15時間後、マウスを屠殺し、CD11c細胞をautoMACSにより精製し、抗体を標識して、フローサイトメトリーにより分析した。我々は、MHCクラスIおよびクラスII分子、CD40およびCD86の発現を誘導できたことを見出した。タンパク質が免疫性付与前にプロテイナーゼ Kで消化した場合に、EDA-SIINFEKLのこの能力が完全に消失した(図 5)。プロテイナーゼ Kで消化されたLPSの消化物は、これらの成熟マーカーの発現を誘導するためのLPSの能力に対していずれの抑制効果も持たなかった(図示せず)。我々は、C57BL/6 TLR4 KO マウス由来のCD11c細胞の成熟を誘導するEDA-SIINFEKLの能力を試験し、そしてEDA-SIINFEKLがC57BL/6 wt マウスに見出された成熟マーカーの過剰発現を誘導できないことを見出した(図5)。
【0099】
2.2.3 EDA-SIINFEKLは、SIINFEKLエピトープに特異的なT細胞に対して樹状細胞により有効に提示される
我々は、このエピトープに特異的なOT-1 トランスジェニックマウス由来のT細胞に対して処理済みCTLエピトープSIINFEKLを提示するためにAPCにより捕捉されるべきEDA-SIINFEKLの能力を特徴決定した。骨髄由来DC(105細胞/ウェル)を、様々な濃度のEDA-SIINFEKL、EDA+SIINFEKL、EDAまたはSIINFEKLペプチドの存在において培養した。48時間後、105非付着性OT-1細胞を添加した。OT-1 非付着細胞によるIFN-γ産生を測定した(図 6A)。SIINFEKL ペプチドは、IFN-γ産生によって証明されたようなOT-1 マウス由来のT細胞に対して有効に提示された。EDA-SIINFEKLは、高レベルのIFN-γも誘導するが、しかしタンパク質の高用量は、IFN-γと同様のレベルか、または同等より高いレベルを得るために必要であり、EDAタンパク質がMHCクラスI分子に対するSIINFEKLエピトープを担持することを明確に示している。SIINFEKLペプチドとインキュベートしたDCへのEDAタンパク質の添加は、OT-1マウス由来のT細胞によりIFN-γ産生を増加しなかった。予測したように、EDA単独とインキュベートしたDCは、OT-1 マウス由来のT細胞を活性化しなかった。DC におけるTLR4分子発現が、EDA-SIINFEKL提示を増強できるかどうかを分析するために、SIINFEKL特異的B3Z細胞を、C57BL/6 wtまたはTLR4KOマウス由来の脾臓細胞の存在下に様々な濃度のEDA-SIINFEKLとインキュベートした。B3Z細胞に対するEDA-SIINFEKL提示は、TLR4を発現した抗原提示細胞においてより効率的であった(図6B)。また、B3Z 細胞に対するEDA-SIINFEKL提示は、抗TLR4抗体の添加によって完全に遮断され(図6C)、TLR4がEDA-SIINFEKL捕捉に関与することを示唆している。その後、我々は、EDA-SIINFEKLのプロセシングに対して様々な薬物の効果を試験し、そして我々は、この提示が、エンドソームおよび後期リソソーム(図 6D)における酸性化の既知阻害剤である、クロロキン以外のモネンシン、ブレフェルジンまたはシクロヘキシミドにより、完全に阻害されることを見出した。予測したように、SIINFEKL合成のペプチドの提示は、これらの薬物によって影響されなかった。これらのデータは、EDA-SIINFEKLの内在化は、マクロピノサイトーシスによって媒介されるのではなく、EDA-SIINFEKLはクラスI細胞質性プロセシング経路を介して処理されることを示す。
【0100】
2.2.4. EDA-SIINFEKLはイン・ビボにおいてSIINFEKLに特異的なCTLを誘導する。
前記結果は、EDA-SIINFEKLの組換えタンパク質は生物活性であり、およびAPCを特異的に活性化することを示す。イン・ビボで特異的なTリンパ球免疫応答の誘導はワクチンの開発に重要である。従って、我々は、EDA-SIINFEKL 融合タンパク質を用いて免疫化したマウスは、SIINFEKLエピトープによりパルス化した標的細胞に対して特異的CTL応答を進行させるかどうかを試験した。C57BL6マウスを、0および10日目に、PBS中のEDA-SIINFEKL(50μg)またはSIINFEKL(50μg)を用いてi.v.にて免疫化した。20日目に、マウスをSIINFEKLに対するCTL応答の分析のために屠殺した。EDA-SIINFEKLはSIINFEKLを用いてパルス化したEL-4標的細胞に対してCTLを誘導できたことが判った。一方で、CTL活性はマウスをSIINFEKL単独で免疫化した場合には見出されなかった(図7)。イン・ビボで樹状細胞の成熟を誘導するためのEDAの能力のために、アジュバンドとして作用するEDAの能力もまた、OVAタンパク質で免疫化した場合に分析された。この実験において、我々は、免疫性付与混合物中のEDAの存在により、免疫賦活性効果を有し、OVAタンパク質により誘導されたSIINFEKLに対するCTL応答誘導を増強できることを観察した(図9のパネルAおよびBで分解活性を比較する)。
【0101】
2.2.5. EDA-SIINFEKLはOVAタンパク質を発現する腫瘍細胞による攻撃からマウスを保護する。
EG7OVA腫瘍細胞に対してマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを、EDA-SIINFEKL(3 nmol)、SIINFEKL(3 nmol)または生理食塩を用いてs.c.にて免疫化した。二次免疫性付与20日後に、105 EG7OVA 細胞を用いてs.c.にてマウスを攻撃した。我々は、EDA-SIINFEKL 免疫性付与により、マウスを腫瘍成長から保護することが判った。SIINFEKLまたは生理食塩水を用いて免疫化した全てのマウスは腫瘍を進行させたが、EDA-SIINFEKLで免疫化したマウスの40%はが依然として腫瘍の存在はなく、残りの60%は腫瘍成長遅延を経験した(図8)。
【0102】
2.3. 考察
本試験において、組換えEDAタンパク質をエピトープ送達ベクターとして用いて、我々は、アジュバンドの必要性なく、イン・ビボでCTL応答を開始する免疫性付与のためのストラテジーを確立した。我々は、ベクターとしてEDA含有融合タンパク質の有効性に関与するメカニズムを同定し、抗原をTLR4発現細胞に送達し、そして抗原に対する細胞性免疫応答を誘導する。
【0103】
最初に、我々は、イン・ビトロでのEDAを用いるBMDCの刺激は炎症性サイトカイン、例えばIL-12およびTNF-αの産生を刺激できたことを見出した。これらのサイトカインは、抗原に対して強力な免疫応答の開始のために重要であることが知られている。さらに、イン・ビボでEDAによる免疫性付与によりDCの成熟を誘導でき、DCの表面に共刺激分子の発現を増加させ得ることを見出した。これらの共刺激分子の発現は、抗原に対する免疫応答の有効な誘導のために最も重要である。この効果は、TLR4の存在に依存しており、イン・ビボにおいてC57BL/6 TLR4 KO マウス由来の単離したCD11c細胞をEDAで予め免疫化したので、非免疫化動物において見出されたものと比較して、その成熟状態に対していずれの改善をも示さなかったということを見出した。
【0104】
我々は、APC上のTLR4分子の存在が、EDA-SIINFEKL融合タンパク質と共にインキュベーションした後に抗原提示を改善すること、およびこの提示がそのエンドリソソームタンパク質分解を阻害するクロロキンによって影響されないことを見出した。これらのデータは、EDA-SIINFEKLの内在化がマクロピノサイトーシスによって媒介されないことを示しており、組換えEDA-SIINFEKLタンパク質が細胞質性抗原として処理されることを示し、および該融合タンパク質がAPCプラズマ膜を介してサイトゾルに送達されなければならないことを言及する。
【0105】
最も重要なことには、我々は、組換え融合タンパク質EDA-SIINFEKLを有するマウスの免疫性付与は、SIINFEKLエピトープに対して特異的なイン・ビボCTL応答を誘導することができることを見出した。さらに、EDA-SIINFEKLを用いる免疫性付与は、EG7OVA腫瘍細胞による攻撃に対してマウスを保護できる。全てのこれらのデータから、EDAを含有するこのタンパク質性ベクターは、(i)抗原をTLR4発現細胞、特にプロフェッショナルAPCに標的化する;(ii)ベクター化(vectorised)抗原を古典的クラスI抗原プロセシング経路へ送達する;(iii)イン・ビボおよびイン・ビトロで樹状細胞成熟を誘導する;および (iv)アジュバンドの非存在下でベクター化抗原に対するCTLをイン・ビボで駆動し、そうして感染性剤または癌に対するワクチン接種ストラテジーにおいて使用することが出来ることが示された。また、EDA含有するこれらの融合タンパク質は、TLR4発現細胞に対して医薬関連分子の細胞質性送達のために働き得る。さらに、イン・ビボでの樹状細胞成熟を誘導するためのEDAの能力は、免疫性応答を与えることが望まれる抗原を含有する製剤におけるアジュバンドとしてのその使用を可能にし、EDAワクチン開発における使用のための可能性を開くものである。
【0106】
実施例3.EDAタンパク質を、ビヒクルとして少なくとも390個のアミノ酸の抗原を輸送するために使用できる
3.1 材料および方法
3.1.1 EDA-OVA および EDA-NS3組換えタンパク質の発現
EDA-OVA 発現プラスミドの構築のために、mRNAを、OVAタンパク質を発現するEG7OVA 腫瘍細胞から抽出した。プライマー
【化6】
(配列番号:16)および
【化7】
(配列番号:17) (下線の塩基を制限酵素NotIにより配列を導入したが、該配列の斜字体はオボアルブミンの開始および末端に属する)を用いるPCRによる逆転写および増幅の後に、該PCR生成物をTOPO TA kit (Invitrogen)を用いてpCR2.1-TOPOにクローニングし、NotIを用いて消化し、NotIで消化する前にプラスミドpET20EDA 1.2(EDA タンパク質を発現する)にサブクローニングした。構築物の正しい配向性を配列決定により評価した。プラスミド培養物により形質転換したE. coliの誘導後、可溶性画分中に存在する融合タンパク質を、FPLCプラトフォーム(AKTA, Pharmacia)を用いてアフィニティクロマトグラフィーにより精製した(Histrap, Pharmacia)。タンパク質を脱塩Hitrap columns(Pharmacia)を用いて脱塩し、フィルター装置を用いる遠心分離Amicon Ultra 4-5000 MWCO (Millipore Carrighwahill, Ireland)により濃縮した。組換えタンパク質ベクターを、エンドトキシンレベルがタンパク質(LAL アッセイによって測定した, Cambrex)の0.2 EU/μg以下となるまで、Endotrap endotoxin columns (Profos Ag, Regensburg, Germany)を用いて精製した。EDA-NS3 タンパク質構築体について、同様のストラテジーを、NS3タンパク質(アミノ酸1026-1221)のアミノ末端フラグメントの発現を可能にするプライマー
【化8】
(配列番号:18) および
【化9】
(配列番号:19)を用いて行った。E. coli画分の可溶性分画から抽出したEDA-OVA タンパク質とは異なり、EDA-NS3 タンパク質の場合には、該タンパク質を8M 尿素に予め溶解した封入体から精製した。Histrap columnsを用いるアフィニティクロマトグラフィーの後に、イオン交換クロマトグラフィー(DEAEセファロース)を行った。この第二クロマトグラフィーからの精製分画を、G25カラムでの再結合プロトコールに従って再結合させた。一旦再結合した後、EDA-NS3カラムを脱塩し、Endotrap columns (Profos, Germany)を用いてエンドトキシンを精製した。
このように精製されたタンパク質をSDS-PAGEにより分析した
【0107】
3.1.2 抗原提示能力を評価するためのイン・ビトロでの試験
APCにより捕捉されるべきEDA-OVAの能力およびOT-1トランスジェニックマウス由来のTリンパ球に対するCTLエピトープのプロセシングしたSIINFEKLのその後の提示を評価した。骨髄由来樹状細胞(105 細胞/ウェル)を様々な濃度のEDA-OVA、EDA +OVA(非共有結合)、OVAまたはEDAの存在において培養した。12時間後、OT-1 トランスジェニックマウス由来の105 細胞/ウェルの非付着細胞を添加した。培養の開始から24 時間に、該上清を市販のELISAアッセイにより分泌されたIFN-γ定量のために抽出した。
【0108】
3.1.3 イン・ビボT細胞傷害性リンパ球(CTL)誘導および免疫性付与後のIFN-γ産生細胞の測定
C57BL6またはHHD(HLA-A2.1 分子についてのトランスジェニック)マウスを、0および10日目に各々EDA-OVAまたはEDA-NS3(50μg)を用いてi.v.免疫化した。20日目に、マウスを、SIINFEKLまたはNS3 1073 ペプチドに対するCTL応答を決定するために屠殺した。免疫化動物由来の脾臓細胞を、5 x 106 細胞/ml(10 ml)でSIINFEKL(0.1μg/ml)またはNS3 1073(1μg/ml)の存在下に、完全培地中で5日間培養した。5日目に、細胞をクロミウム遊離分析のために回収した。分解活性を、51Crを予め添加した1 x 104 EL-4 標的細胞と様々な量のエフェクター細胞を、ペプチドを用いるかまたは用いないで4時間インキュベートして測定した。分解物比活性の%を、式(実験値 cpm− 自然発生値 cpm)/(最高値 cpm − 自然発生値 cpm) x 100に従って計算した、式中の自然発生値は、エフェクター細胞の非存在下でインキュベートした標的細胞に対応しており、最大の分解物は標的細胞を5%のTriton x100とインキュベートすることによって得られた。
【0109】
様々なペプチドに対する応答におけるIFN-γ産生を測定するために、免疫化マウス由来の脾臓細胞を、終濃度0.25 mlで、完全培地のみ、または30μM のペプチド NS3 1038-1046 (配列番号:23)、1073-1081(配列番号:20、CVNGVCWTV)、1169-1177(配列番号:22)と共に、または組換えNS3(1μg/ml)と共に、8 x 105 細胞/ウェルにて、96ウェルプレートに分配した。48h後に上清(50μl)を回収し、生成者指示書に従ってIFN-γをELISAにより測定した(Pharmingen, Sandiego, CA)。
【0110】
3.1.4 C型肝炎ウイルスポリプロテインを発現するvHCV1-1031ワクシニアウイルスによる感染に対する保護
C57BL/6を、EDA-NS3 タンパク質と共にパルス化された1 x 106の骨髄由来樹状細胞を用いて1日および10日目に免疫化した。二次免疫性付与後10日目に、マウスを、組換えvHCV 1-1031ワクシニアウイルスを5 x 106 pfuにてi.p.により感染させた。感染3日後、動物を屠殺し、ウイルス負荷/卵巣組織mgを、BSC-1 細胞系の使用を基にしたプラーク形成ユニット定量アッセイにより定量した。
【0111】
3.2 結果
3.2.1 EDA-OVAおよびEDA-NS3タンパク質の発現および精製
材料および方法の章に示したとおりに、EDA-OVAまたはEDA-NS3を、EDA-OVAの場合には形質転換したE. coli抽出物から可溶性分画を用いて精製し、EDA-NS3の場合には封入体を形成する。図10Aおよび11Aにおいて、両方のタンパク質についてSDS-PAGEの結果が示される。55kDaおよび32kDaサイズの各タンパク質に対応する一本の各バンドが得られた、これは各タンパク質について予測したサイズに対応している。
【0112】
3.2.2 OVAタンパク質に結合するEDAは、OT-1トランスジェニックマウス由来のTリンパ球に対するIVAの抗原提示を増強する
我々は、SIINFEKLエピトープに特異的なT細胞に対してSIINFEKLエピトープの抗原提示を増強するOVAタンパク質に対するEDA結合能力を分析した。我々は、EDA-OVAの存在下で培養された骨髄由来樹状細胞は、等モル量のOVAタンパク質単独(図9B)により誘導されたものよりも非付着性OT-1マウス細胞によるIFN-γ産生を強く刺激したことが観察された。我々は、IFN-γ産生はまた、OVAタンパク質単独と比較した場合に、EDAタンパク質およびOVAタンパク質を含んだ共培養物中で増強し、EDAにより誘導されたDC成熟は、T細胞活性化を増強できることを示唆していると考察した。
【0113】
3.2.3. OVAタンパク質またはNS3タンパク質に結合するEDAは、イン・ビボで特異的細胞傷害性T細胞誘導を増強する
SIINFEKLエピトープまたはNS31073エピトープ各々に対するCTL応答を誘導するEDA-OVAおよびEDA-NS3の能力を分析した。我々は、EDA-OVAタンパク質について、生理食塩溶液に溶解したこのタンパク質による免疫性付与は、SIINFEKLペプチドとインキュベートした標的細胞に対する細胞傷害性応答を誘導でき、OVAタンパク質単独による免疫性付与では誘導できないことを観察した(図9C)。同様の方法において、EDA-NS3タンパク質を用いるHHDマウス(HLA-A2.1に対するトランスジェニック)免疫性付与は、ペプチドNS31073(V)(配列番号:20、CVNGNCWTV)、このペプチド(配列番号:21、CLNGVCWTV)の改変型1073(L)と予めインキュベートした標的細胞に対して有効な細胞傷害性応答を誘導する(図11B)。我々は、EDA-NS3タンパク質によるHHDマウス免疫性付与が、ペプチドNS3 1038 (配列番号:23)、1073(配列番号:20、CVNGVCWTV)および1169(配列番号:22)(図11C)に特異的なIFN-γ産生細胞の活性化を誘導することを観察した。さらに、EDA-NS3 タンパク質によるマウス免疫性付与は、NS3 1073ペプチドに対して特異的な長時間持続する細胞傷害性細胞性応答を誘導する。実際に、EDA-NS3タンパク質により免疫化したマウスを免疫性付与60日後に屠殺した場合に、我々はこのペプチドに特異的なCTLの存在を検出した(図11D)。
【0114】
3.2.4 EDA-NS3タンパク質とインキュベートした樹状細胞によるC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現するワクシニアウイルスによる感染からマウスを保護する。
我々は、EDA-NS3タンパク質と共にイン・ビトロでインキュベートした樹状細胞を用いる免疫性付与により、C型肝炎ウイルスタンパク質を発現する組換えウイルスによる攻撃に対してマウスを保護できる細胞応答を誘導できたかどうかについて試験することを求めた。この理由から、我々は、EDA-NS3タンパクとインキュベートした骨髄由来樹状細胞または任意の抗原と予めインキュベートしていない樹状細胞を用いてC57/BL6マウスを免疫化した。二次免疫性付与10日後に、マウスを5 x 106 pfuの組換えvHCV 1-3011ワクシニアウイルスを用いて感染させた(Dr Rice, Washington University School of Medicine, St Louis, MOからのご厚意とGrakoui A, et al. J Virol. 1993; 67:1385による説明により)。3日後に、マウス両群においてウイルス負荷を測定した。この実験において、我々は、EDA-NS3 タンパク質とインキュベートしたDCによる免疫性付与により、組換えワクシニアウイルスによる感染に対して6%のマウスを保護できたことを観察した。
【0115】
3.3. 考察
前記結果に示したように、EDAタンパク質は、OVAタンパク質からTLR4分子発現細胞にSIINFEKLエピトープを標的化するための有用なベクターとして機能し、それらの免疫原性を増強出来る。より高分子のタンパク質の免疫原性を増強するEDAの能力を評価するために、我々は、完全なOVA(397個のアミノ酸)を含有するEDA-OVA 融合組換えタンパク質およびC型肝炎ウイルスからのNS3タンパク質のプロテアーゼ活性を有するフラグメントを含有するEDA-NS3タンパク質を構築した。
【0116】
これらの結果は、EDAタンパク質が、より大きな抗原を標的とする非常に有効なベクターとして作用できることを示す。このように、我々は、抗原提示アッセイにおいて、EDAに対するOVAの結合により、抗原提示細胞による抗原捕獲を増強させ、特異的T細胞活性化を増加させることを見出した。また、我々は、これらの融合タンパク質(EDA-OVAおよびEDA-NS3)を用いる免疫性付与が、これらの抗原に対する特異的細胞傷害性応答を誘導できることを観察した。これらの誘導性応答は長期間継続する。最終的に、我々は、樹状細胞と共にEDA-NS3タンパク質の投与は、C型肝炎ウイルスタンパク質を発現するワクシニアウイルスによる感染に対する保護的細胞性応答を誘導できることを観察した。
【0117】
これらのデータは、EDAタンパク質は、目的とする抗原に対して細胞性応答の誘導のたために非常に適切なタンパク質ベクターであり得る。EDAタンパク質を基にした融合タンパク質の構築は、腫瘍疾患または感染性剤によっておこる疾患に対するワクチン接種プロトコールにおいて適切なストラテジーである。
【図面の簡単な説明】
【0118】
【図1】図1.産生および精製したEDAおよびEDA-SIINFEKLタンパク質のSDS-PAGE分析。EDAおよびEDA-SIINFEKLタンパク質の1つのアリコートを、15%ポリアクリルアミドゲル上におき、電気泳動に供した。分子量マーカー(MWM)をKDaで示す。EDAおよびEDA-SIINFEKLタンパク質(13-14 KDa)の推定分子量に対応するバンドを観察した。
【図2】図2.タンパク質性ベクターEDA-SIINFEKLのTLR4への結合。2A.直接的な結合アッセイ。HEK293-LacZ細胞(HEK293-LacZ)およびHEK293-TLR4/MD2/CD14(HEK293 TLR4)細胞を、1μM EDA-SIINFEKLを用いてパルスし、パラホルムアルデヒドで固定し、抗Hisおよび抗EDA抗体で標識し、抗マウスIgG-FITCで顕色させ、フローサイトメトリーにより分析した。2B.TLR4発現細胞に対する抗TLR4抗体の結合を阻害するEDA能力。HEK-TLR4細胞を、500nM EDA-SIINFEKL タンパク質の存在または非存在において、2時間4℃でインキュベートした。細胞を洗浄し、FITCで染色した抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。2C.様々な濃度のEDA-SIINFEKLタンパク質を用いてTLR4発現細胞に結合する抗TLR4抗体の阻害%。2D.細胞付着アッセイ。トリチウム化したチミジンで染色したHEK-hTLR4またはHEK-LacZ細胞を、EDAタンパク質で予め被覆した前に96ウェルミクロプレートのウェルに分配し、2時間37℃の間インキュベートした。非付着細胞を除き、付着細胞を回収し、取り込んだ放射活性をTopcountシンチレーションカウンターで計測した。ウェルあたりの付着細胞数を、標準曲線を利用して計算した。
【図3】図3.EDA-SIINFEKLはTLR4シグナル経路を活性化する。発現がNF-κB-誘導性ELAM-1プロモーターによって制御されるこのレポーター遺伝子でトランスフェクトしたHEK293/TLR4-MD2-CD14またはHEK293/LacZ発現細胞の培養物上清においてヒト分泌した胚アルカリホスファターゼ遺伝子の比色分析測定。トランスフェクションの24時間後に、細胞を様々な濃度のLPS、100 nM EDA-SIINFEKL タンパク質(EDA)またはプロテイナーゼKで予め消化した100 nM EDA-SIINFEKL タンパク質の存在または非存在下でインキュベートした。棒線は、NF-κBの誘導倍である(HEK293/TLR4-MD2-CD14由来の上清で得たODをHEK293/LacZからの上清で得たODによって割った)。
【図4】図4.EDA-SIINFEKLは、イン・ビトロでDCによって炎症性サイトカインの分泌を誘導する。骨髄由来DCを、LPS(1μg/ml)、EDA-SIINFEKL(500 nM)、プロテイナーゼKで消化したEDA-SIINFEKL(500 nM)の存在下または生理食塩溶液で培養した。24時間後、IL-12(A)およびTNF-α(B)の存在をELISAにより培養物上清中で測定した。
【図5】図5.EDA-SIINFEKLは、イン・ビボでCD11c DC 成熟を誘導する。樹状細胞の成熟は、T-細胞応答の最適な刺激のための要件である。成熟が起こる時に、APCは、表面分子、例えばクラスI(我々のモデルではH2Kb)およびクラスIIMHC(我々のモデルではIAb)、CD40、CD80およびCD86分子の発現を増加させる。従って、我々は、EDA-SIINFEKLがCD11c発現細胞のイン・ビボで成熟を誘導するかどうかを分析した。C57BL/6wtマウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いてi.v.で免疫化した。また、C57BL/6 TLR4 KOマウスをEDA-SIINFEKL(25μg)またはPBS単独を用いて免疫化した。15時間後にマウスを屠殺し、CD11c細胞をautoMACSにより精製した。細胞を標識し、H-2Kb、I-Ab、CD40、CD80およびCD86分子の発現についてフローサイトメトリーにより分析した。
【図6】図6:EDA-SIINFEKLは、樹状細胞によりSIINFEKLエピトープに特異的なT細胞に対して効果的に提示される。我々は、このエピトープに特異的なOT-1 遺伝子組み換えマウス由来のT細胞に対して処理済CTLエピトープSIINFEKLの提示について、APCによって捕捉されるべきEDA-SIINFEKLの能力を特徴分析した。(A) OT-1 トランスジェニックマウスからの非付着細胞によるIFN-γ産生。骨髄由来DCを、培地単独、様々な濃度の合成SIINFEKLペプチド、SIINFEKL、およびEDA、EDA-SIINFEKL(融合タンパク質)またはEDA単独の存在下で培養した。24時間後、DCを回収し、105個の非付着性OT-1細胞の存在下でAPCとして使用した。さらなる24時間後に、上清を回収し、分泌したIFN-γを測定した。(B)TLR4分子依存性。B3Zハイブリドーマ細胞(105細胞/ウェル)を、C57BL/6 wt マウス由来の脾臓細胞の存在下、またはTLR4分子(105 細胞/ウェル)およびEDA-SIINFEKLタンパク質(100 nM)のためのノックアウトマウス由来の脾臓細胞と共にインキュベートした。(C)C57BL76wt マウスの脾臓由来の細胞を、抗TLR4抗体の存在または非存在(BおよびC)において、B3Zハイブリドーマ細胞およびEDA-SIINFEKLタンパク質と共に共培養した。培養物上清に分泌したIL-2の量を、CTLL 細胞系の使用を基にしたバイオアッセイによって測定した。(D)融合タンパク質EDA-SIINFEKLの抗原提示に対するクロロキン、モネンシン、ブレフェルジンまたはシクロヘキシミドの効果。骨髄由来樹状細胞を、30 mMのクロロキン、ブレフェルジン、モネンシン、または4μg/mlのシクロヘキシミドの非存在下または存在下で1時間インキュベートし、その後EDA-SIINFEKLまたは合成ペプチドSIINFEKL(白色の棒)を添加した。10時間の培養後に、DCをグルタルアルデヒド中で固定し、非付着性OT-1マウス細胞(105細胞/ウェル)との同時培養物中で抗原提示細胞(APC)(104 細胞/ウェル)として使用した。24時間後、分泌したIFN-γの量を市販のELISAにより測定した。した(図6E)。
【図7】図7.EDA-SIINFEKLによるマウスの免疫性付与は、SIINFEKLエピトープに特異的な細胞応答を誘導する。前記結果は、EDA-SIINFEKL組換えタンパク質が生物活性であり、APCを特異的に活性化することを示す。イン・ビボでの特異的なT細胞免疫応答の誘導は、ワクチンの開発のために重要である。このように、我々は、EDA-SIINFEKL融合タンパク質により免疫化されたマウスが、このエピトープに対して特異的な細胞応答を生じるかどうかを試験した。(A)IFN-γ産生細胞の誘導。0および10日目に、C57BL/6 マウスを、EDA-SIINFEKLの1.5 nmolまたはSIINFEKLペプチドの1.5 nmolを用いて免疫化した。20日目に、SIINFEKLの存在または非存在において脾臓細胞を48時間インキュベートし、そして分泌されたIFN-γの量をELISAにより測定した。(B)SIINFEKL-特異的CTL応答分析の誘導。EDA-SIINFEKLまたはSIINFEKLにより免疫化したマウス由来の脾臓細胞を、SIINFEKLペプチドの存在下で5日間再刺激した。このインキュベーション後に、従来のクロミウム51遊離アッセイにより、SIINFEKLペプチドの非存在または存在下でEL-4標的細胞に対するCTL活性を測定した。データは、3つのサンプルから正味の特異的分析物(SIINFEKLでパルスした標的細胞の%分解−非パルス化標的細胞の分解%)の値の平均%を示す。
【図8】図8.EDA-SIINFEKLは、EG7 OVA発現細胞による腫瘍攻撃から守る。EG7OVA 腫瘍細胞の注射に対するマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを3 nmolのEDA-SIINFEKL、SIINFEKLまたは生理食塩溶液を用いて0および10日目にs.c.で免疫化した。20日後、二次免疫性付与マウスを105 EG7OVA 細胞を用いてs.c.攻撃した。腫瘍増殖を、カリパスを用いてモニターし、式 V=(L x w2)/2(式中、L, 長さ;w, 幅である)を用いてmm3で表した。腫瘍サイズが8cm3以上の体積に達した場合に、マウスを屠殺した。
【図9】図9.EDAは、OVAタンパク質を用いる免疫性付与後に細胞傷害性応答の誘導においてアジュバンドとして作用する。EDAがイン・ビボで樹状細胞の成熟を促進することができれば、EDAはそれ自身によっては細胞傷害性応答を活性化できないが細胞傷害性エピトープを含有するタンパク質を用いる免疫性付与後にアジュバンドとして作用できるという可能性が存在する。この可能性を試験するために、我々は、その他のタイプのアジュバンドを用いないでEDA(50μg)とOVA タンパク質(500μg)により一群のマウス(A)、およびOVA(500μg)により別の群のマウス(B)を免疫化した。免疫性付与1週間後、マウスを屠殺し、脾臓細胞を、SIINFEKL合成ペプチドの存在下で培養した。5日後の培養物中で、SIINFEKLペプチドを用いてパルスしたEL-4 標的細胞に対する細胞傷害性応答を、従来のCr51遊離アッセイにおいて測定した。
【図10】図10.EDAは、より大きな抗原についてのビヒクルとして作用できる。前記実験において、EDAタンパク質は、細胞傷害性エピトープを輸送するためのビヒクルとして作用でき、該エピトープに対するCTL応答の誘導を支持することが証明された。後者の工程において、EDAが比較的大きな抗原を輸送でき、抗原に対する細胞性応答の誘導を促進できるかどうかについての試験がのぞまれる。この目的により、我々は融合タンパク質EDA-OVAを構築し、イン・ビトロおよびイン・ビボでの下記実験を行った。(A)組換えEDA-OVAタンパク質のSDS-PAGE分析。組換えタンパク質EDA-OVAを、E. coliで発現し、アフィニティクロマトグラフィーにより精製した。脱塩し、中和し、濃縮し、SDS-PAGEにより分析した。該タンパク質の推定分子量に対応するおおよそ55kDaのバンドを観察した。(B)抗原提示実験、骨髄由来DCを、OVA、EDA-OVA(融合タンパク質)、EDA+OVAまたはEDA単独の存在下または非存在下で培養した。24時間後、DCをOT-1マウス由来の105非付着細胞の存在において抗原提示細胞として使用した。DCの存在下でOT-1マウス由来の非付着細胞によりIFN-γ産生を、市販のELISAにより定量化した。(C) EDA-OVAタンパク質は、イン・ビボでOVA特異的CTLを誘導する。C57BL/6 マウスを、1nmolのEDA-OVAを用いるか、または1 nmolのOVAを用いて免疫化した。免疫性付与7日後に、免疫化したマウスからの脾臓細胞を、SIINFEKLペプチドの存在下に5日間イン・ビトロで再刺激した。その後、SIINFEKLの非存在または存在下でインキュベートしたEL-4 細胞に対する特異的CTL活性を、従来のCr51 遊離アッセイにおいて測定した。データは、3つのサンプルから、正味の特異的溶菌値(SIINFEKLを用いてパルスした標的細胞の溶菌%−非パルス標的細胞の溶菌%)の平均%を示す。
【図11】図11.EDA-NS3タンパク質は、NS3C型肝炎ウイルスタンパク質に対する特異的CTL応答を引き起こす。EDAタンパク質が大きな抗原についてのビヒクルとして作用できることが観察されると、C型肝炎ウイルスのNS3タンパク質(NS3タンパク質のプロテアーゼ領域からのアミノ酸1-196)に対する抗ウイルス応答を誘導するEDAの能力を、該ウイルスによる感染に対するワクチン接種ストラテジーとして分析した。(A)組換えEDA-NS3(1-196)タンパク質のSDS-PAGE分析。組換え融合タンパク質EDA-NS3を構築し、E. coliで発現し、SDS-PAGEにより分析した。(B)EDA-OVAタンパク質は、イン・ビボでOVA-特異的CTLを誘導する。HHDマウス(HLA-A2.1タンパク質についてトランスジェニック)を、生理食塩溶液に溶解したEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与1週間後、脾臓細胞を、イン・ビトロでNS3 1073ペプチド(HLA-A2拘束のためにNS3タンパク質由来の免疫優性の細胞傷害性T決定基を含有する)を用いて再刺激した。培養物中で5日後、ペプチド1073(V)(配列番号:20、CVNGVCWTV)と共に、または該ペプチド(配列番号:21、CLNGVCWTV)の1073(L)改変型と共に、またはペプチドの非存在下でインキュベートしたT2標的細胞に対して、細胞傷害性活性を、従来のCr51遊離アッセイを用いて測定した。(C)EDA-NS3タンパク質は、NS3タンパク質からの様々なエピトープに対して複数のエピトープ様応答を誘導する。EDA-NS3により免疫化したマウスから得た脾臓細胞を、ペプチド1038-1046、NS3 1073-1081またはNS3 1169-1177(NS3タンパク質のフラグメント1-196のなかで、HLA-A2拘束のために3つの細胞傷害性決定基を含有する)の存在下に、または組換えNS3タンパク質(Mikrogen)と共に、48時間イン・ビトロで再刺激した。上清に分泌されたIFN-γの量を市販のELISAにより測定した。(D)EDA-NS3タンパク質は、長期間持続する細胞傷害性応答を誘導する。HHDマウスを、生理食塩溶液中のEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与60日後、マウスを屠殺し、ペプチドNS3 1073に特異的なCTLの存在を、NS3 1073 ペプチドの非存在または存在中でインキュベートしたT2標的細胞を用いて、従来のクロミウム51遊離アッセイにおいて測定した。(E) EDA-NS3とインキュベートしたDCを有するC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現する組換えワクシニアウイルス vHCV(1-3011)による感染からマウスを保護する。EDA-NS3 タンパク質と予めインキュベートした 106DCを用いてマウスを免疫化し、7日後に、それらはi.p.経路による5x106 ワクシニアウイルスvHCV(1-3011)の攻撃を受けた。感染3日後にマウスを屠殺し、BSC-1細胞感染アッセイにより、卵巣組織mgあたりのウイルス負荷を定量した。
【技術分野】
【0001】
(本発明の技術分野)
本発明は、感染および腫瘍疾患の処置および予防のための医薬組成物、特に免疫治療用組成物の調製および使用に対する特定の範囲を有する、TLR4受容体(トル様受容体4)を発現する細胞への分子輸送のためのタンパク質性ベクター、該タンパク質性ベクターの調製およびその用途に関する。
【背景技術】
【0002】
(先行技術の説明)
病原体および癌は、依然として全世界で主な死亡要因である。ワクチンが現在存在しない疾患、例えばAIDSまたはマラリアを抑制するための、または慢性感染または癌を処置するための、さらに存在しているワクチンの効力および安全性改善のためのかかるワクチンの開発は、依然として高い優先事項である。多くの症例において、かかるワクチンの開発は、CD8+細胞傷害性Tリンパ球細胞(CTL)を特異的に刺激できるストラテジーを必要とする。
【0003】
CTLは、MHCクラスI分子と関連した短鎖ペプチドのT-細胞受容体(TCR)の提示により活性化される。これらのペプチド-MHCクラスI複合体は、至適CTL活性化を必要とする共刺激シグナルも提供できる抗原提示細胞(APC)の表面上に存在する。
【0004】
樹状細胞(DC)は、本来のTリンパ球と相互作用し、ヘルパーCD4+および細胞傷害性CD8+Tリンパ球を活性化する初期免疫応答を開始する特別な能力を有する最も強力なAPCである。DCによる抗原提示およびT細胞刺激はGuermonprezら("抗原提示およびDCによるT細胞刺激"Annu. Rev. Immunol. 2002, 20:621-627)に論説されており、出典明示によりこの文献を明細書の一部とする。
【0005】
継続する炎症および免疫応答がなければ、樹状細胞は、血液、末梢組織、リンパ球および二次リンパ組織中を巡回する。末梢組織において、樹状細胞は、自己および非自己抗原を取り込む。次いで、内在化された抗原はタンパク質分解ペプチドへとプロセシングされ、これらのペプチドは、MHCクラスIおよびII分子(各々CD8+またはCD4+Tリンパ球の活性化のため)上に担持される。抗原取込み、分解および担持に関するこの過程は、抗原提示と呼ばれる。しかしながら、刺激が無ければ、末梢樹状細胞は全く有効でない抗原を提示する。病原体または内因性シグナル由来の外因性シグナルは樹状細胞を誘導し、成熟と呼ばれる樹状細胞をAPCおよびTリンパ球アクチベーターに変換する発生プログラムに参加する。細菌およびウイルス産物、ならびに炎症性サイトカインおよび他の自己分子は、本来の樹状細胞表面レセプターと直接的な相互作用を介して樹状細胞成熟を誘導する。CD40独立経路およびCD40非独立経路を経るTリンパ球、および内皮細胞は、直接的な細胞対細胞接触およびサイトカインの分泌を通じて樹状細胞の最終的な成熟に関与する。危険を示すシグナルに遭遇した直後に、抗原摂取効率、細胞内輸送および分解およびMHC分子の細胞内輸送が修飾される。ペプチド担持、さらに半減期およびMHC分子の細胞表面への送達が増強される。T細胞共刺激分子の表面発現も増大する。こうして、樹状細胞は最も強力なAPCとなる、本来のTリンパ球を活性化でき、適応免疫応答を開始できる唯一のものである。その抗原提示能の修飾と同時に、成熟は末梢組織以外の樹状細胞の大量移動も誘導する。ケモカイン受容体および付着分子の発現における修飾、ならびに細胞骨格組織化の著しい変化は、リンパを介して二次リンパ組織にむかう樹状細胞の移動に関与する。
【0006】
樹状細胞成熟の誘導
樹状細胞は、2つのタイプのシグナルに応答する:病原体の直接的認識(特異的な型-認識受容体を介して)および感染の間接検出(炎症性サイトカイン、体内細胞化合物を介して、特異的免疫応答を継続する)。これらのシグナルに対する応答において、樹状細胞は活性化されて、樹状細胞を有効なT細胞刺激因子へと形質転換する成熟プログラムに入る。少なくとも5つの型の表面受容体が樹状細胞成熟を引き起こすことが報告されている:(i)トル様受容体(TLR)、(ii)サイトカイン受容体、(iii)TNF-受容体(TNF-R)ファミリー分子、(iv)FcRおよび(v)細胞死に対するセンサー。最も有効な成熟刺激のいくつかは、それらの各リガンドとTLR(TLR1-9)の相互作用により媒介される。KaishoおよびAkiraは、トル様受容体の知見("アジュバンド受容体としてのトル様受容体". Biochimica et Biophysica Acta, 2002, 1589: 1-13)を論じている。TLRは、マクロファージおよび樹状細胞上、ならびに他の細胞、例えばBリンパ球上で発現される。数種のTLRのリガンドもまた同定されている。大部分のこれらのリガンドは、その宿主には見出されない病原体から得られ、TLRは侵入微生物を検出するために重要であることが示唆されている。TLRによるリガンド認識は、炎症誘発性サイトカインの産生および共刺激分子の上方調節を誘導することによって先天性免疫の急速な活性化を誘起する。次に、活性化された先天性免疫は、有効な適応免疫を引き起こす。TLR4に関して、特異的に認識される分子パターンは、LPS(グラム陰性細菌)、リポタイコ酸(グラム陽性細菌)、タキソール、Fタンパク質(呼吸器合胞体ウイルス)、熱ショックタンパク質60、およびフィブロネクチンEDAドメインである。
【0007】
それ故に、至適T細胞応答を誘導出来る候補ワクチンは、いくつかの条件を満たさなけれならない。まず、抗原誘導性T細胞エピトープをMHCクラスIおよび/またはII分子に送達するためには、APCを標的としなければならない。従って、DCの標的化は、ワクチン開発のための新規送達系を設計する際に主な目的を示すことができる。さらに、該ベクターは、適切なシグナルをDCに送達して、その活性化を誘導するべきである。成熟シグナルがないとDCへの抗原送達は、Tヘルパーおよび細胞傷害性細胞の活性化よりもむしろ寛容を誘導できるであろう。さらに、その効果は、ベクターそれ自身に対して予め存在している免疫によって影響されるべきではない。
【0008】
抗原性ペプチドをMHCクラスIおよび/またはII分子に送達するための最初の方法は、合成ペプチドワクチンを基にしており、APCの表面上でこれらの分子に直接結合できる選択されたエピトープを含有している。ある場合では、これらのペプチドは、ネズミのモデルにおいては、腫瘍保護またはウイルス排除をもたらしたが、しかし他の場合において、それらは寛容誘導をもたらした。様々な型のペプチドを用いて行ったヒトの治験は、癌への適切な臨床応答を引き起こした。
【0009】
多くの様々なストラテジーは現在開発中である。基本的に、それらは2つのカテゴリーに分けることができる。第一のタイプは、APCによる抗原の合成またはこれらの細胞の細胞質へのその能動送達を基にしており、および"古典的"MHCI抗原プロセシング経路を利用する。第二のタイプは、APCの交差提示の能力に有利であり、遊離または細胞関連性の外因性抗原を基にしている。APCの細胞質への抗原の送達は、細菌毒素の手段によって行われてきた(Moron et al. "MHCクラスI経路への抗原送達のための新規ツール". TRENDS in Immunology, 2004; 25: 92-97)。例示としては、EP1188446A1は、細胞を発現するCD11bへの分子送達のための、百日咳菌(Bordetella pertussis)アデニルシクラーゼ毒素を基にしたタンパク質性ベクターに関する。
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、フィブロネクチン・エクストラ・ドメインA(EDA)、APCの適切な選択および成熟を誘導できる、最終的に効果的な特異的CTL応答をもたらすTLR4発現細胞へ抗原送達のための理論的手段として、TLR4に対する潜在的な天然リガンドに関する。フィブロネクチン分子は、1つの遺伝子産物であって、得られるタンパク質は、一つの前駆体mRNAの選択的スプライシングから生じる複数の形態で存在できる(Pankov R and Kenneth MY, "Fibronectin at a glance". Journal of Cell Science, 2002; 115:3861 3863)。スプライシングの主なタイプは、タイプIIIリピートの中心的な型において認められる。エキソン用法またはスキッピングは、2つのタイプIIIリピートのいずれかの包含または排除に導く:エクストラドメインB(EDB、EIIIBまたはEDIIとも呼ばれる)、およびエクストラドメインA(EDA、EIIIAまたはEDIとも呼ばれる)。選択的スプライシングによるEDAおよびEDBを含有する細胞性フィブロネクチンは組織損傷への応答において産生される。他の生物学機能のなかの一つは、EDAはプロテオグリカン放出およびメタロプロテイナーゼ(MMP1、3および9)および前炎症性サイトカインの発現を誘導することが示されている(Saito S et al. "フィブロネクチンエクストラドメインAはマトリックスメタロプロテイナーゼ遺伝子発現をインターロイキン-1依存機構により誘導する", J. Biol. Chem. 1999; 161:3071-3076を参照されたい)。EDAがTLR4を活性化でき、こうしてLPS様応答を誘導できることも示されている(Okamura Y et al.,"フィブロネクチンのエクストラドメインAはトル様受容体4を活性化する", J. Biol. Chem. 2001; 276:10229-10233)。
【0011】
前記したように、抗原に対する免疫応答を強化するためのストラテジーの開発は、癌または感染疾患の処置のためのワクチン開発に対する道をひらくものである。特に、C型肝炎ウイルス感染において、免疫応答は感染の排除において重要な役割を担っており、それ故、免疫的強化ストラテジーの使用はこの感染の処置および予防についての代替え案を構成すると説明されてきた。
【0012】
C型肝炎ウイルス(HCV)は、Flaviviridaeファミリーに属する一本鎖RNAウイルスである (Miller RH. and Purcell RH. 1990. PNAS. 87:2057)。このウイルスは、慢性肝炎および肝臓疾患の主な病因の一つとして認識されており、全世界で170万人に影響すると見積もられる(World-Health-Organisation. Hepatitis C. Wkly Epidemiol Rec 1997; 72:65)。HCV感染の主な態様の中の一つは、慢性化(感染者の70%)へと向かう高い傾向、および肝細胞癌の進行に関する高リスクを伴う肝硬変(20%)の進行である(Dienstag et al. Gastroenterology 1983; 85:439)。IFN-αによる処置は、HCV感染において最も一般的な治療であるが、処置患者のわずか20-30%においてのみ有効である(Camps et al. J Hepatol 1993; 17:390)。IFN-αおよびリバビリンの組合せは、これらの結果(患者の30-40%は持続的様式でウイルスを排除する)を改善したが、治療に対する抵抗性患者は依然として高い%を保持している(Poynard et al. Lancet 1998; 352:1426)。従って、慢性C型肝炎の処置のための新規治療ストラテジーの開発は極めて重要である。
【0013】
HCVゲノムは、9.6キロ塩基のゲノムであって、3つの構造タンパク質(コア、E1およびE2)および少なくとも6つの非構造タンパク質(NS2、NS3、NS4a、NS5aおよびNS5b)(主に、MEおよびFeinstone SM. (1997) Hepatology 25, 1527)をコードする十分なオープンリーディングフレームを挟んでいる5'および3'末端に高度に保存された非コード領域を含有する。
【0014】
HCVによる急性感染後またはIFN-αを用いる処置後のウイルス排除は、ウイルスタンパク質に対して強力な細胞性CD4およびCD8免疫応答の存在に関連している。特に、HCVの非構造のNS3タンパク質に対するCD4応答は、急性感染後ウイルス排除と関連しているが、一方でこのT細胞の非応答は、ウイルスの継続および慢性感染の確立に関する(Diepolder et al. Lancet 1995; 346:1006, Pape et al J Viral Hepat 1999; 6 Suppl 1:26-40)。また、様々な試験によりHCV感染患者のNS3タンパク質のなかで多様な細胞傷害性エピトープが同定されてきた。これらのデータから、NS3タンパク質は抗HCV細胞性応答の誘導のための良好な標的であり得ることが示唆された。
【課題を解決するための手段】
【0015】
本発明の詳細な説明
一態様において、本発明は、抗原刺激剤に対する免疫細胞応答の生成において、
-フィブロネクチンEDAドメイン(EDA)、または
-TLR4を結合できる該EDAドメインのフラグメント、または
-TLR4を結合でき、EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する、該EDAドメインまたはフラグメントの改変型、
に対応するアミノ酸配列を含むポリペプチドの使用に関する。
【0016】
本発明において、この剤はフィブロネクチンのEDAドメインおよび免疫応答を生じることが望まれる抗原の両方を包含し、これらの成分は個々の構成成分として存在するか、または共有結合している。
【0017】
本発明の具体的な実現において、エレメントc)で引用したTLR4を結合できる該EDAドメインまたはフラグメントの該改変型は、そのアミノ酸配列がエレメントa)およびb)に規定されたポリペプチドにおいて1以上のアミノ酸の置換、付加または削除から得られることを特徴とする。
【0018】
本発明の好ましい実現において、エレメントc)を結合できる該フラグメントは、EDAドメインまたはその対応するフラグメントの任意の天然形態と85%以上のホモロジーを有することを特徴とし、より好ましい実現において、フィブロネクチンEDAドメインの天然形態またはその対応するフラグメントと95%以上のホモロジーを有する。
【0019】
本発明に従って、特別な実現において、フィブロネクチンEDAドメインのアミノ酸配列は、TLR4を結合できるEDAのあらゆる天然形態のものである。このEDAドメインは、あらゆる動物種、特に哺乳動物、即ち齧歯類(マウス、ラット等)または霊長類(特にヒト)におけるドメインの天然形態の中から選択され得る。
【0020】
他の特定の実現において、免疫賦活剤は、該剤がTLR4を結合できることを特徴とするEDAドメインの部分アミノ酸配列を含む。
【0021】
本発明の他の別の特定の実現において、EDAドメインは、任意の該EDAドメインの天然形態またはフラグメントの修飾された改変型であり、またTLR4への結合特性を保持することを特徴とする。特別な実施形態において、かかる改変型EDAドメインは、EDAドメインのあらゆる天然形態と70%以上のホモロジーを有する。適切に修飾された改変型は、フィブロネクチンEDAドメイン天然形態の配列、またはそのもののフラグメントを他の候補ポリペプチドの配列をまず比較することによって選択され得る。任意のアラインメントアルゴリズム(例えばFASTA、Lipman DJ, Pearson WR. Rapid and sensitive protein similarity, Science. 1985 Mar 22;227(4693):1435-41)、またはコンピューターソフトウェア(即ち、Labvelocity Inc.からはJellifish または NCBIからはBlast software)は、ホモロジー分析に使用され得る。次に、70%以上のホモロジーを有する候補ポリペプチドの配列は、そのTLR4結合能力について試験される。TLR4結合特性は、任意の従来の結合アッセイ、例えばJohn Wiley & Sons (John E. Coligan, Ada M. Kruisbeek, David H. Margulies, Ethan M. Shevach, Warren Strober編)(定期的に更新される。May 1, 2005までに更新された)によって発表されたThe current protocols in Immunology およびThe current protocols in Protein Scienceに記述されているようなフローサイトメトリーを用いて評価され得る。
【0022】
本発明の一実施態様において、EDAドメインは、下記から選択された配列を含む:
a)マウスEDAドメイン(Entrez タンパク質:NM_010233、アミノ酸1721〜1810;配列番号:2、アミノ酸2-91)の完全なアミノ酸配列;
b)ヒトEDAドメイン(Entrez タンパク質 NM_002026、アミノ酸1631〜1716;配列番号:4)の完全なアミノ酸配列;および
c)TLR4発現細胞に結合し得る配列a)およびb)のフラグメント。
【0023】
別の特定の実現において、該EDAドメインは、下記から選択される配列を含む:
a)マウスフィブロネクチンEDAドメインの選択的にスプライシングされた形態に対応する配列番号:6のアミノ酸2-57;
b)ヒトEDAドメインの選択的にスプライシングされた形態である配列番号:8;および
c)TLR4発現細胞に結合できる配列a)およびb)のフラグメント。
【0024】
いくつかの実施形態において、免疫賦活性剤は1以上の目的とする分子をさらに包含し得る。免疫賦活性剤中に存在する場合、目的とする分子は、薬剤の他の成分との組合せにおいて該分子に対する免疫応答を生成するために効果的である量で投与され得る。
【0025】
好ましい実現において、EDAドメイン(またはそのフラグメントまたは改変型)および目的とする分子は、同ハイブリッド分子またはタンパク質性ベクター中に一緒に結合される。
【0026】
別の態様において、本発明は、上記したとおりのタンパク質性ベクターに関し、ここで目的とする分子は、ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質および化学物質からなる群から選択される。
【0027】
タンパク質性ベクターの特別な実現において、目的とする分子は、抗原またはエピトープである。本発明の一実施態様において、ベクターと結合される抗原は、ウイルス抗原、細菌抗原、真菌抗原または寄生抗原である。具体的な実現において、該ウイルス抗原は、C型肝炎ウイルス由来のウイルス抗原であって、好ましい実現において、該C型肝炎ウイルス抗原はNS3タンパク質またはその抗原性フラグメントである。NS3タンパク質は、N末端からの189個のアミノ酸を含むセリンプロテアーゼおよびC末端からの442個のアミノ酸を含むヘリカーゼ-ヌクレオシドトリホスファターゼを包含する2つのドメインを包含する67kDaタンパク質のC型肝炎ウイルスの非構造NS3タンパク質を指す。本発明のタンパク質ベクターに包含されるNS3タンパク質配列は、ヒトC型肝炎ウイルスの任意の株または単離株と対応可能である。
【0028】
他の実施形態において、該抗原は、腫瘍性抗原または腫瘍性抗原決定基である。本明細書で使用したとおりに、"エピトープ"は、MHCクラスIまたはクラスII分子と結合し、CD8+またはCD4+T細胞各々のT細胞受容体分子によって認識され、そして免疫応答を誘導し得るペプチド配列を指す。
【0029】
特定の実現において、目的とする該分子は、オボアルブミン(OVA 257-264)由来の抗原性細胞傷害性T決定基またはSIINFEKL(配列番号:2、アミノ酸95-102、これはエピトープ(QLE-SIINFEKL-TEW)のC末端およびN-末端で3つの付加アミノ酸により挟まれている)である。
【0030】
該抗原は、Th免疫応答を産生できる任意の材料、CD8+Tリンパ球応答、NK細胞応答、γ/δTリンパ球応答、または抗体応答であり得る。それらに限定するものではないが、適切な抗原は、ペプチド、ポリペプチド、脂質、糖脂質、ポリサッカリド、炭水化物、ポリヌクレオチド、プリオン、細菌、ウイルスまたは生存するか不活性化された真菌;および細菌、ウイルス、真菌、原生動物、腫瘍または微生物、毒素またはトキソイドから得られた抗原である。
【0031】
タンパク質性ベクターの特定の別の実現において、目的とする分子はアレルゲンである。
【0032】
この方法、EDAドメインは、TLR4発現細胞への抗原送達のためのベクターとしても作用する。
【0033】
本発明の目的とする他の特定の実現において、目的とする該分子は、タンパク質性ベクターと化学的または遺伝子的に結合された化学物質または薬物である。このように該タンパク質性ベクターは、TLR4発現細胞に標的化する特異的な薬物に有用である。
【0034】
特別な実現において、タンパク質性ベクターは、Tag配列も含む、例えばN末端ヒスチジンテイルも含むことを特徴とする。これは、タンパク質性ベクターが遺伝子設計方法によって生成される場合に、精製過程を単純化する。例示として、配列の配列番号:2および配列番号:6は、本発明のタンパク質性ベクターの特定の実現を示す。具体的に、本発明の実現を限定しないが、該タンパク質性ベクターは、配列番号:10を含み、NS3タンパク質のフラグメントを含む。
【0035】
タンパク質性ベクターに導入したEDAドメインは、TLR4に結合すること、さらに目的とする分子の転位をTLR4発現細胞のサイトゾルに利用することを特徴とする。
【0036】
本発明は、目的とする分子をTLR4発現細胞に標的化および輸送するためにタンパク質性ベクターの使用に関する。特定の実現において、TLR4発現細胞はあらゆる種類の抗原提示細胞(APC)である。好ましい実施形態において、かかるAPCは樹状細胞である。
【0037】
本発明の特定の実現において、タンパク質性ベクターは、目的とする抗原またはエピトープの輸送を促進して、プロセシングをさらに進めて、およびTリンパ球に対する抗原提示のためにMHC分子上に担持することを特徴とする。
【0038】
他の実現において、タンパク質性ベクターは、標的化したAPCの成熟を刺激できること、従ってMHC分子の発現および共刺激シグナルを増加することを特徴とする。特に有利な実施形態において、タンパク質性ベクターは、抗原提示の誘導とAPC成熟の促進を同時に行うことができ、こうして効果的な抗原特異的免疫応答を誘導できることを特徴とする。より好ましい実施形態において、この抗原特異的免疫応答はCTL応答である。
【0039】
本発明のタンパク質性ベクターは、組換えDNA技術によって得ることができる。従って、別の態様において、本発明は、本発明のタンパク質性ベクターをコードしている修飾された核酸に関する。この核酸は、タンパク質性ベクターのアミノ酸配列から容易に推定することができる。
【0040】
この修飾された核酸は、DNA構築体内に含有され得る。従って、本発明は、本発明のタンパク質性ベクターをコードする核酸を含むDNA構築体を提供する。このDNA構築体は、本発明のタンパク質性ベクターをコードしている核酸に作動可能に結合したコントロール配列を導入できる。"作動可能に結合した"とは、核酸を示す場合には、核酸が他の核酸配列と機能的な関係におかれる。"コントロール配列"は、特異的宿主細胞によって認識される発現シグナルであり、これは機能、例えば特定のコード配列(コントロール配列の例示は、プロモーター、エンハンサー、転写ターミネーター、リボソーム結合部位、タンパク質分泌または他の細胞内輸送のためのシグナルペプチド)の転写および翻訳を調節するものである。所望の配列を結合することは、従来の制限部位でのライゲーションによって達成される。かかる部位が存在しない場合に、合成オリゴヌクレオチドアダプターまたはリンカーは、従来方法に従って使用される。この意味においての利点は、このDNA構築体がDNA構築体によって、形質転換された宿主細胞の選択を可能にするモチーフまたは表現型をコードするマーカーまたは遺伝子も含むという事実により説明される。この発明によって提供された修飾された核酸およびDNA構築体は、数多くの実験手引き書(例えば、"Molecular Cloning: a Laboratory manual." Joseph Sambrook, David W. Russel Eds. 2001, 3rd ed. Cold Spring Harbor, New York)にまとめられている既知の従来方法によって得ることが出来る。
【0041】
特定の実現において、本発明により提供された修飾された核酸またはDNA構築体は、配列番号:1、配列番号:5、配列番号:9(EDA-NS3)または配列番号:11(EDA-OVA)を包含する。
【0042】
この発明によって提供された修飾された核酸またはDNA構築体は、適切なベクターに挿入され得る。従って、別の態様において、本発明は、上記修飾された核酸またはDNA構築体を含むベクター、例えば発現ベクターに関する。ベクターの選択は、ベクターが挿入されるべき宿主細胞に依存する。例示としては、核酸が挿入されるベクターは、プラスミドまたはウイルスであってよく、挿入の際に細胞ゲノム中に組み入れられていても、いなくてもよい。該ベクターは、従来方法(Sambrook et al., 2001, 上掲を引用する)により得られ得る。
【0043】
別の態様において、本発明は、この発明によって提供された修飾された核酸またはDNA構築体を含む宿主細胞、例えば形質転換宿主細胞に関する。本発明に従って、発現宿主細胞は、原核生物、即ちEscherichia coli、または真核宿主、即ち酵母(例えば、Saccharomyces cerevisiae、Pichia Pastoris)、昆虫細胞または哺乳動物細胞である。
【0044】
本発明の特定の実現において、本発明のタンパク質性ベクターをコードする修飾された核酸またはDNA構築体を含む発現ベクターは、イン・ビボの遺伝子移送または治療を目的としている。より具体的な実施態様において、発現ベクターはウイルスベクターである。この効果に対する適切なウイルスベクターは、次のものを包含する:アデノウイルス、アデノ関連ウイルス、レトロウイルス、レンチウイルス、アルファウイルス、ヘルペスウイルス、コロナウイルス誘導性ベクター等。
【0045】
別の態様において、本発明は、本発明のタンパク質性ベクターを作成するための方法に関するものであって、本発明の修飾された核酸またはDNA構築体を含有する宿主細胞を、タンパク質性ベクターの発現できる条件下で培養することを含む。宿主細胞の培養物を至適化するための条件は、用いた宿主細胞のタイプに依存する。所望により、本発明のタンパク質性ベクターを作成するための方法は、該ベクターの単離および精製を包含する。
【0046】
あるいは、本発明のタンパク質性ベクターは、他の従来方法によって得ることができる。かかる方法には、例えば固相化学合成;高速液体クロマトグラフィー(HPLC)を用いる精製;および、望ましくは従来技術、例えば配列決定または質量スペクトル分析法、アミノ酸分析、磁気共鳴技術等による分析が包含される。
【0047】
別の実現において、本発明のタンパク質性ベクターは、フィブロネクチンEDAドメイン(EDA)、またはTLR4を結合できるEDAドメインのフラグメント、または該EDAドメインの改変型に対応するアミノ酸配列を有するポリペプチドと、目的とする分子(例えば、ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質または他の化学物質)との共有結合によって得られる。これは、従来方法を用いて行うことができ、これはJohn Wiley & Sons により発表された実験手引き書、例えば"The current protocols in protein chemistry" (定期的に更新される。May 1, 2005までに更新されたもの)、またはGT Hermanson, AK Mallia および PK Smithによる"Immobilized affinity ligand Techniques"(Academic Press, Inc. Sandiego, CA, 1992)などにまとめられている。
【0048】
次に、本発明に従って、タンパク質性ベクターまたはこれをコードしている修飾された核酸およびDNA構築体、または該核酸または該DNA構築体を組み込んでいる発現ベクターおよび発現宿主細胞を、医薬組成物の調製において使用できる。
【0049】
他の実施形態において、本発明は、上記したとおり、免疫賦活性剤の調製において、該剤が医薬組成物であることを特徴とする、フィブロネクチンEDAドメイン(EDA)またはその同フラグメントまたは改変型に対応するアミノ酸配列を有するポリペプチドの使用を指す。
【0050】
特定の実施形態において、本発明の医薬組成物は、抗原提示細胞の成熟を刺激するため、または目的とする分子に特異的な有効な免疫応答を誘導するために使用され得る。特定の実施形態において、該医薬組成物は、免疫賦活性組成物が投与され得る対象におけるTh1免疫応答を誘導するために使用される。本明細書で使用したとおりに、"Th1免疫応答を誘導すること"とは、免疫賦活性組成物は混合したTh1/Th2応答を誘導する場合も包含できる。しかしながら、他の場合において、免疫賦活性組成物は、Th2免疫応答の誘導がほどんどないか、または実質的にないTh1免疫応答を誘導できる。特定の実現において、該医薬組成物はCTL応答を誘導するのに使用され得る。
【0051】
いくつかの実施形態において、免疫賦活性組成物は、免疫賦活性アジュバンドとして、例えば追加のアジュバンドと共に1以上の抗原と組み合わせて、または追加のアジュバンドを含まないで使用され得る。このように、いくつかの例において、免疫賦活性組成物はワクチンを形成できる。他の場合において、免疫賦活性組成物は、ワクチンとの結合において使用され得るアジュバンドとして機能し得る。
【0052】
フィブロネクチンEDAドメイン(またはそのフラグメントまたは改変型)を含むポリペプチドを包含する免疫賦活性組成物は、活性化されたCD8+Tリンパ球の拡大、メモリーCD8+Tリンパ球の生成、または両方を増強できる。このように、本発明の免疫賦活性組成物は、免疫賦活性組成物を受容する対象において抗原特異的細胞媒介性免疫を増強できる。
【0053】
特定の実現において、フィブロネクチンEDAドメイン(または、そのフラグメントまたは改変型)を含有する免疫賦活性組成物は、感染性疾患、腫瘍疾患またはアレルギー性疾患の処置および予防に有用である。本発明の具体的な実施形態において、該組成物はC型肝炎の処置および予防のために使用される。
【0054】
フィブロネクチンEDAドメイン(または、そのフラグメントまたは改変型)を含有する免疫賦活性組成物は、さらに担体、賦形剤および他の既知の医薬的に許容し得る成分を含有できる。
【0055】
本発明の免疫賦活性組成物は、当業者に既知の従来の方法(例えば、経口的に、皮下的に、局所的に)に従って、動物、例えば哺乳動物(ヒトまたは非ヒト)、鳥類などに投与され得る。
【0056】
本発明は、対象に対してフィブロネクチンEDAドメイン(またはそのフラグメントまたは改変型)を含む免疫賦活性組成物を投与することを含む、治療および/または予防方法も提供する。
【0057】
適切な投与経路は、経皮的または経粘膜的吸収、注射(例えば、皮下、腹腔内、筋肉内、静脈内など)、摂取、吸入などである。
【0058】
他のさらなる態様において、本発明は、少なくとも1つの許容し得る医薬担体および有効量のタンパク質性ベクターを、少なくとも1つのその発現形態または実施形態:
a)ポリペプチドの形態にあるタンパク質性ベクター;
b)該タンパク質性ベクターをコードする修飾された核酸;
c)該修飾された核酸を含む発現ベクター;または
d)該修飾された核酸も含む発現宿主細胞、
において包含する医薬組成物に関する。
【0059】
特定の実現において、医薬組成物は、有効量の樹状細胞を含んでいることを特徴とし、ここで該樹状細胞は、少なくとも1つのその発現形態または実施形態においてイン・ビトロでタンパク質性ベクターとインキュベートされている。より特定の実施態様において、該医薬組成物はワクチンまたは免疫治療用の組成物である。
【0060】
さらに、タンパク質性ベクターのいくつかの追加的使用は本明細書により提供される。本発明の一実施態様において、その発現形態のいずれかにおいてタンパク質性ベクターは、イン・ビトロまたはイン・ビボで樹状細胞成熟を誘導するために有効な医薬組成物の調製のために使用される。
【0061】
他の実施形態において、該タンパク質性ベクターは、タンパク質性ベクターと結合した目的とする分子(抗原またはエピトープ)に対して特異的な特異的免疫応答を誘導するための医薬組成物の調製のために使用される。この免疫応答は、体液性免疫応答(目的とする分子に対する抗体産生)、Tヘルパー応答、または細胞傷害性T細胞応答である。好ましい実施形態において、該免疫応答はCTL応答である。
【0062】
より特定の実現において、本発明は、感染疾患の処置および予防のための医薬組成物の調製におけるタンパク質性ベクターの使用に関する。該感染性疾患は、細菌、ウイルス、真菌または寄生感染疾患であり得る。
【0063】
別の特定の実現において、本発明は、腫瘍疾患の処置および予防のために有用な医薬組成物の調製において、本発明のタンパク質性ベクターの使用に関する。
【0064】
さらに別の特定の実現において、本発明は、アレルギー性疾患の処置および予防のために有用な医薬組成物の調製におけるタンパク質性ベクターの使用に関する。多くのアレルギー性疾患は、Th2免疫応答の活性化に関連する。従って、特異的なアレルゲンを担持するタンパク質性ベクターを使用することによって、Th2からTh1応答へのずれまたは切替えは、アレルギー性疾患に対して保護的または治療的効果をもち得る。
【0065】
本発明の特定の実施形態に従って、提案された医薬組成物は動物またはヒト宿主への投与に使用される。任意の適切な投与経路が使用され得る。特定の実現において、医薬組成物は、非経口経路(即ち、静脈、皮下、筋肉内)、経皮的経路または粘膜経路によって投与される。
【0066】
(図面の簡単な説明)
図1.産生および精製したEDAおよびEDA-SIINFEKLタンパク質のSDS-PAGE分析。EDAおよびEDA-SIINFEKLタンパク質の1つのアリコートを、15%ポリアクリルアミドゲル上におき、電気泳動に供した。分子量マーカー(MWM)をKDaで示す。EDAおよびEDA-SIINFEKLタンパク質(13-14 KDa)の推定分子量に対応するバンドを観察した。
【0067】
図2.タンパク質性ベクターEDA-SIINFEKLのTLR4への結合。2A.直接的な結合アッセイ。HEK293-LacZ細胞(HEK293-LacZ)およびHEK293-TLR4/MD2/CD14(HEK293 TLR4)細胞を、1μM EDA-SIINFEKLを用いてパルスし、パラホルムアルデヒドで固定し、抗Hisおよび抗EDA抗体で標識し、抗マウスIgG-FITCで顕色させ、フローサイトメトリーにより分析した。2B.TLR4発現細胞に対する抗TLR4抗体の結合を阻害するEDA能力。HEK-TLR4細胞を、500nM EDA-SIINFEKL タンパク質の存在または非存在において、2時間4℃でインキュベートした。細胞を洗浄し、FITCで染色した抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。2C.様々な濃度のEDA-SIINFEKLタンパク質を用いてTLR4発現細胞に結合する抗TLR4抗体の阻害%。2D.細胞付着アッセイ。トリチウム化したチミジンで染色したHEK-hTLR4またはHEK-LacZ細胞を、EDAタンパク質で予め被覆した前に96ウェルミクロプレートのウェルに分配し、2時間37℃の間インキュベートした。非付着細胞を除き、付着細胞を回収し、取り込んだ放射活性をTopcountシンチレーションカウンターで計測した。ウェルあたりの付着細胞数を、標準曲線を利用して計算した。
【0068】
図3.EDA-SIINFEKLはTLR4シグナル経路を活性化する。発現がNF-κB-誘導性ELAM-1プロモーターによって制御されるこのレポーター遺伝子でトランスフェクトしたHEK293/TLR4-MD2-CD14またはHEK293/LacZ発現細胞の培養物上清においてヒト分泌した胚アルカリホスファターゼ遺伝子の比色分析測定。トランスフェクションの24時間後に、細胞を様々な濃度のLPS、100 nM EDA-SIINFEKL タンパク質(EDA)またはプロテイナーゼKで予め消化した100 nM EDA-SIINFEKL タンパク質の存在または非存在下でインキュベートした。棒線は、NF-κBの誘導倍である(HEK293/TLR4-MD2-CD14由来の上清で得たODをHEK293/LacZからの上清で得たODによって割った)。
【0069】
図4.EDA-SIINFEKLは、イン・ビトロでDCによって炎症性サイトカインの分泌を誘導する。骨髄由来DCを、LPS(1μg/ml)、EDA-SIINFEKL(500 nM)、プロテイナーゼKで消化したEDA-SIINFEKL(500 nM)の存在下または生理食塩溶液で培養した。24時間後、IL-12(A)およびTNF-α(B)の存在をELISAにより培養物上清中で測定した。
【0070】
図5.EDA-SIINFEKLは、イン・ビボでCD11c DC 成熟を誘導する。樹状細胞の成熟は、T-細胞応答の最適な刺激のための要件である。成熟が起こる時に、APCは、表面分子、例えばクラスI(我々のモデルではH2Kb)およびクラスIIMHC(我々のモデルではIAb)、CD40、CD80およびCD86分子の発現を増加させる。従って、我々は、EDA-SIINFEKLがCD11c発現細胞のイン・ビボで成熟を誘導するかどうかを分析した。C57BL/6wtマウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いてi.v.で免疫化した。また、C57BL/6 TLR4 KOマウスをEDA-SIINFEKL(25μg)またはPBS単独を用いて免疫化した。15時間後にマウスを屠殺し、CD11c細胞をautoMACSにより精製した。細胞を標識し、H-2Kb、I-Ab、CD40、CD80およびCD86分子の発現についてフローサイトメトリーにより分析した。
【0071】
図6:EDA-SIINFEKLは、樹状細胞によりSIINFEKLエピトープに特異的なT細胞に対して効果的に提示される。我々は、このエピトープに特異的なOT-1 遺伝子組み換えマウス由来のT細胞に対して処理済CTLエピトープSIINFEKLの提示について、APCによって捕捉されるべきEDA-SIINFEKLの能力を特徴分析した。(A) OT-1 トランスジェニックマウスからの非付着細胞によるIFN-γ産生。骨髄由来DCを、培地単独、様々な濃度の合成SIINFEKLペプチド、SIINFEKL、およびEDA、EDA-SIINFEKL(融合タンパク質)またはEDA単独の存在下で培養した。24時間後、DCを回収し、105個の非付着性OT-1細胞の存在下でAPCとして使用した。さらなる24時間後に、上清を回収し、分泌したIFN-γを測定した。(B)TLR4分子依存性。B3Zハイブリドーマ細胞(105細胞/ウェル)を、C57BL/6 wt マウス由来の脾臓細胞の存在下、またはTLR4分子(105 細胞/ウェル)およびEDA-SIINFEKLタンパク質(100 nM)のためのノックアウトマウス由来の脾臓細胞と共にインキュベートした。(C)C57BL76wt マウスの脾臓由来の細胞を、抗TLR4抗体の存在または非存在(BおよびC)において、B3Zハイブリドーマ細胞およびEDA-SIINFEKLタンパク質と共に共培養した。培養物上清に分泌したIL-2の量を、CTLL 細胞系の使用を基にしたバイオアッセイによって測定した。(D)融合タンパク質EDA-SIINFEKLの抗原提示に対するクロロキン、モネンシン、ブレフェルジンまたはシクロヘキシミドの効果。骨髄由来樹状細胞を、30 mMのクロロキン、ブレフェルジン、モネンシン、または4μg/mlのシクロヘキシミドの非存在下または存在下で1時間インキュベートし、その後EDA-SIINFEKLまたは合成ペプチドSIINFEKL(白色の棒)を添加した。10時間の培養後に、DCをグルタルアルデヒド中で固定し、非付着性OT-1マウス細胞(105細胞/ウェル)との同時培養物中で抗原提示細胞(APC)(104 細胞/ウェル)として使用した。24時間後、分泌したIFN-γの量を市販のELISAにより測定した。
【0072】
APC上でのTLR4発現が提示過程を進行させ得るかどうかを試験するために、我々は、C57BL/6 wt マウスまたはTLR4 KO マウス由来の脾臓細胞および様々な濃度のEDA-SIINFEKL (nM)の存在において、B3Z ハイブリドーマ細胞(SIINFEKLエピトープに特異的)を培養した。培養上清に放出されたIL-2の量を、CTLLを基にしたバイオアッセイにより測定した(図6E)。
【0073】
図7.EDA-SIINFEKLによるマウスの免疫性付与は、SIINFEKLエピトープに特異的な細胞応答を誘導する。前記結果は、EDA-SIINFEKL組換えタンパク質が生物活性であり、APCを特異的に活性化することを示す。イン・ビボでの特異的なT細胞免疫応答の誘導は、ワクチンの開発のために重要である。このように、我々は、EDA-SIINFEKL融合タンパク質により免疫化されたマウスが、このエピトープに対して特異的な細胞応答を生じるかどうかを試験した。(A)IFN-γ産生細胞の誘導。0および10日目に、C57BL/6 マウスを、EDA-SIINFEKLの1.5 nmolまたはSIINFEKLペプチドの1.5 nmolを用いて免疫化した。20日目に、SIINFEKLの存在または非存在において脾臓細胞を48時間インキュベートし、そして分泌されたIFN-γの量をELISAにより測定した。(B)SIINFEKL-特異的CTL応答分析の誘導。EDA-SIINFEKLまたはSIINFEKLにより免疫化したマウス由来の脾臓細胞を、SIINFEKLペプチドの存在下で5日間再刺激した。このインキュベーション後に、従来のクロミウム51遊離アッセイにより、SIINFEKLペプチドの非存在または存在下でEL-4標的細胞に対するCTL活性を測定した。データは、3つのサンプルから正味の特異的分析物(SIINFEKLでパルスした標的細胞の%分解−非パルス化標的細胞の分解%)の値の平均%を示す。
【0074】
図8.EDA-SIINFEKLは、EG7 OVA発現細胞による腫瘍攻撃から守る。EG7OVA 腫瘍細胞の注射に対するマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを3 nmolのEDA-SIINFEKL、SIINFEKLまたは生理食塩溶液を用いて0および10日目にs.c.で免疫化した。20日後、二次免疫性付与マウスを105 EG7OVA 細胞を用いてs.c.攻撃した。腫瘍増殖を、カリパスを用いてモニターし、式 V=(L x w2)/2(式中、L, 長さ;w, 幅である)を用いてmm3で表した。腫瘍サイズが8cm3以上の体積に達した場合に、マウスを屠殺した。
【0075】
図9.EDAは、OVAタンパク質を用いる免疫性付与後に細胞傷害性応答の誘導においてアジュバンドとして作用する。EDAがイン・ビボで樹状細胞の成熟を促進することができれば、EDAはそれ自身によっては細胞傷害性応答を活性化できないが細胞傷害性エピトープを含有するタンパク質を用いる免疫性付与後にアジュバンドとして作用できるという可能性が存在する。この可能性を試験するために、我々は、その他のタイプのアジュバンドを用いないでEDA(50μg)とOVA タンパク質(500μg)により一群のマウス(A)、およびOVA(500μg)により別の群のマウス(B)を免疫化した。免疫性付与1週間後、マウスを屠殺し、脾臓細胞を、SIINFEKL合成ペプチドの存在下で培養した。5日後の培養物中で、SIINFEKLペプチドを用いてパルスしたEL-4 標的細胞に対する細胞傷害性応答を、従来のCr51遊離アッセイにおいて測定した。
【0076】
図10.EDAは、より大きな抗原についてのビヒクルとして作用できる。前記実験において、EDAタンパク質は、細胞傷害性エピトープを輸送するためのビヒクルとして作用でき、該エピトープに対するCTL応答の誘導を支持することが証明された。後者の工程において、EDAが比較的大きな抗原を輸送でき、抗原に対する細胞性応答の誘導を促進できるかどうかについての試験がのぞまれる。この目的により、我々は融合タンパク質EDA-OVAを構築し、イン・ビトロおよびイン・ビボでの下記実験を行った。(A)組換えEDA-OVAタンパク質のSDS-PAGE分析。組換えタンパク質EDA-OVAを、E. coliで発現し、アフィニティクロマトグラフィーにより精製した。脱塩し、中和し、濃縮し、SDS-PAGEにより分析した。該タンパク質の推定分子量に対応するおおよそ55kDaのバンドを観察した。(B)抗原提示実験、骨髄由来DCを、OVA、EDA-OVA(融合タンパク質)、EDA+OVAまたはEDA単独の存在下または非存在下で培養した。24時間後、DCをOT-1マウス由来の105非付着細胞の存在において抗原提示細胞として使用した。DCの存在下でOT-1マウス由来の非付着細胞によりIFN-γ産生を、市販のELISAにより定量化した。(C) EDA-OVAタンパク質は、イン・ビボでOVA特異的CTLを誘導する。C57BL/6 マウスを、1nmolのEDA-OVAを用いるか、または1 nmolのOVAを用いて免疫化した。免疫性付与7日後に、免疫化したマウスからの脾臓細胞を、SIINFEKLペプチドの存在下に5日間イン・ビトロで再刺激した。その後、SIINFEKLの非存在または存在下でインキュベートしたEL-4 細胞に対する特異的CTL活性を、従来のCr51 遊離アッセイにおいて測定した。データは、3つのサンプルから、正味の特異的溶菌値(SIINFEKLを用いてパルスした標的細胞の溶菌%−非パルス標的細胞の溶菌%)の平均%を示す。
【0077】
図11.EDA-NS3タンパク質は、NS3C型肝炎ウイルスタンパク質に対する特異的CTL応答を引き起こす。EDAタンパク質が大きな抗原についてのビヒクルとして作用できることが観察されると、C型肝炎ウイルスのNS3タンパク質(NS3タンパク質のプロテアーゼ領域からのアミノ酸1-196)に対する抗ウイルス応答を誘導するEDAの能力を、該ウイルスによる感染に対するワクチン接種ストラテジーとして分析した。(A)組換えEDA-NS3(1-196)タンパク質のSDS-PAGE分析。組換え融合タンパク質EDA-NS3を構築し、E. coliで発現し、SDS-PAGEにより分析した。(B)EDA-OVAタンパク質は、イン・ビボでOVA-特異的CTLを誘導する。HHDマウス(HLA-A2.1タンパク質についてトランスジェニック)を、生理食塩溶液に溶解したEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与1週間後、脾臓細胞を、イン・ビトロでNS3 1073ペプチド(HLA-A2拘束のためにNS3タンパク質由来の免疫優性の細胞傷害性T決定基を含有する)を用いて再刺激した。培養物中で5日後、ペプチド1073(V)(配列番号:20、CVNGVCWTV)と共に、または該ペプチド(配列番号:21、CLNGVCWTV)の1073(L)改変型と共に、またはペプチドの非存在下でインキュベートしたT2標的細胞に対して、細胞傷害性活性を、従来のCr51遊離アッセイを用いて測定した。(C)EDA-NS3タンパク質は、NS3タンパク質からの様々なエピトープに対して複数のエピトープ様応答を誘導する。EDA-NS3により免疫化したマウスから得た脾臓細胞を、ペプチド1038-1046、NS3 1073-1081またはNS3 1169-1177(NS3タンパク質のフラグメント1-196のなかで、HLA-A2拘束のために3つの細胞傷害性決定基を含有する)の存在下に、または組換えNS3タンパク質(Mikrogen)と共に、48時間イン・ビトロで再刺激した。上清に分泌されたIFN-γの量を市販のELISAにより測定した。(D)EDA-NS3タンパク質は、長期間持続する細胞傷害性応答を誘導する。HHDマウスを、生理食塩溶液中のEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与60日後、マウスを屠殺し、ペプチドNS3 1073に特異的なCTLの存在を、NS3 1073 ペプチドの非存在または存在中でインキュベートしたT2標的細胞を用いて、従来のクロミウム51遊離アッセイにおいて測定した。(E) EDA-NS3とインキュベートしたDCを有するC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現する組換えワクシニアウイルス vHCV(1-3011)による感染からマウスを保護する。EDA-NS3 タンパク質と予めインキュベートした 106DCを用いてマウスを免疫化し、7日後に、それらはi.p.経路による5x106 ワクシニアウイルスvHCV(1-3011)の攻撃を受けた。感染3日後にマウスを屠殺し、BSC-1細胞感染アッセイにより、卵巣組織mgあたりのウイルス負荷を定量した。
【0078】
実施例
実施例1.フィブロネクチンからのエクストラドメインAはTLR4と相互作用し、TLR4情報伝達経路を活性化する。
1.1 材料および方法
1.1.1 EDAおよび EDA-SIINFEKLタンパク質性ベクター組換えタンパク質の発現
組換えタンパク質性ベクターの調製
フィブロネクチンエクストラドメインA(EDA)を、肝臓損傷を誘導するために、特異的プライマーおよびコンカナバリン-Aにより処理したマウスから得た肝細胞由来のRNAを用いて、RT-PCRにより増幅した [Lasarte et al, Hepatology. 2003; 37(2):461-70.]。肝臓組織試験片を均質化し、Ultraturrax Driver T.25 (Janke & Kunkel, Ika-Labortechnik, Germany)を用いてUltraspec (Biotecx, Houston, TX, USA)において溶菌した。RNAを、Chomczynski および Sacchiらの方法に従って単離した(Chomczynski P および Sacchi N. グアニジン酸チオシアネート-フェノール-クロロホルム抽出によるRNA単離のシングル・ステップ方法. Anal Biochem 1987; 162: 156-159)。RNAを、5mMジチオスレイトール(DDT)、0.5mM デオキシヌクレオシド三リン酸(Boehringer Mannheim, Mannheim, Germany)、25U リボヌクレアーゼ阻害剤(Promega Corporation, Madison, WI, USA) および 200ng ランダムヘキサマー(Boehringer Mannheim)を加えた5xRT緩衝液(250mM トリス-HCl Ph 8.3, 375mM KCl, 15mM MgCl2)(20μL容量)中で200UのM-MuLV 逆トランスクリプターゼ(Gibco-BRL)を用いて逆転写した(60分、37℃で)。加熱(95℃、1分間)および氷上での急冷後、cDNAプール(0.3μg)を、0.08mM dNTP、上流および下流プライマー(各40ng)、1.5mM MgCl2およびTaq DNA ポリメラーゼの2U(Promega Corporation)を含有する10x緩衝溶液(100mM トリス-HCl pH9.3、500mM KCl、1% Triton X-100)(20μl)中で、PCR増幅に使用した。上流プライマーは(配列番号:13)
【化1】
(下線を引いた塩基を、制限酵素NdeIにより認識される配列を導入するためにプライマーに加えた。斜字体の配列はEDAの開始部分に対応する)であり、
下流プライマーは(配列番号:14)
【化2】
(下線を引いた塩基を、制限酵素NotIにより認識される配列を導入するためにプライマーに加えた。太字の配列は、両末端で3つのアミノ酸により挟まれたOVA CTL エピトープ(SIINFEKL)
【化3】
をコードしている配列に対応しており、斜字体の配列はEDAの末端に対応する)である。
【0079】
PCR増幅フラグメントを、TOPO TAクローニングキット(Invitrogen, Carlsbad, CA, USA)を用いてpCR2.1-TOPOにクローニングした。このプラスミドをNdeIおよびNotIで解裂させ、得られたDNAフラグメントをカルボキシ末端で6つのヒスチジン残基(6xHis tags)を担持する融合タンパク質の発現を可能にするNdeI/NotI消化したプラスミドpET20b (Novagen)でサブクローニングした。
【0080】
融合タンパク質EDA SIINFEKL-6xHisを発現する得られるプラスミドpET20b2-26を、組換えタンパク質性ベクターの発現のためにBL21(DE3)細胞にトランスフェクトした。トランスフェクトした細胞を、OD600が0.5-1に達するまで、37℃でLB(1l)中で生育させた。IPTGを、0.4 mMの終濃度まで該培養物に加えて、室温で終夜振盪しながらインキュベートした。細胞を遠心分離により回収し、0.1M トリス-HCl(pH=7,2)に再懸濁させ、リゾチームで処理し、French press (20.000 pstで2回通過)を使用して破砕し、遠心分離により清澄させ、濾過した。可溶性分画に存在する融合タンパク質をFPLCプラットホーム(AKTA, Pharmacia) を用いてアフィニティクロマトグラフィー(Histrap, Pharmacia)により精製した。溶出したタンパク質を、Hitrap脱塩カラム(Pharmacia)を用いて脱塩し、Amicon Ultra 4-5000 MWCO Centrifugal filter device (Millipore Carrighwahill, Ireland)を用いて濃縮した。組換えタンパク質性ベクターを、エンドトキシンのレベルが0.2EU/タンパク質μg以下(LAL アッセイによる評価, Cambrex)となるまで、Endotrap カラム(Profos Ag, Regensburg, Germany)を用いて、エンドトキシンから精製した。
【0081】
EDAタンパク質についての発現プラスミドを得るために、PCRをプライマー
【化4】
(配列番号:13)および
【化5】
(配列番号:15)、およびEDA-SIINFEKLプラスミドに記載したものと類似したクローニングストラテジーを用いて行い、プラスミドpET20bEDA1.2を得た。タンパク質(20μg)を、15% SDS-アクリルアミドゲルにおいて各試料に添加し、その後コマジー・ブルーにより染色した。推定分子量(13 kDa)に対応するバンドを観察した。
【0082】
1.1.2. TLR4に対するEDA-SIINFEKLの結合。フローサイトメトリーおよび付着アッセイ
組換えEDA-SIINFEKLタンパク質がTLR4発現細胞に結合できたかどうかを試験するために、我々は、HEK293発現ヒトTLR4-MD2-CD14 293 (Invivogenから)を使用した。我々は、負のコントロールとしてLacZ (Invivogen)で形質転換したHEK293細胞も使用した。細胞を、1 mM EDA-SIINFEKLと共に1時間4℃でパルス化し、PBSで洗浄し、PBS中で4%パラホルムアルデヒドを用いて10分間固定した。3回洗浄後、細胞を、1/100 抗His抗体(Qiagen)および1/200 抗CD16 (FcBlock, from Becton Dickinson)を用いて1時間30分間標識した。3回洗浄した後、細胞を、1/100希釈でフルオレセインにて標識した抗マウスIgGと共に30分間インキュベートし、フローサイトメトリーにより分析した。
【0083】
あるいは、EDA-SIINFEKLタンパク質がHEK-hTLR4 細胞に結合するFITC染色した抗ヒトTLR4抗体を阻害する能力を測定した。この目的のために、HEK TLR4 細胞を、EDA-SIINFEKLの様々な量の存在下または非存在のもとで、2時間4℃でインキュベートした。その後、細胞を洗浄し、抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。阻害%を、EDA-SIINFEKL の様々なアッセイ濃度に対して計算した。細胞付着アッセイを行った。HEK LacZまたはHEK hTLR4 細胞をトリチウム化したチミジンにより予め染色し、EDAタンパク質で予め被覆した96 ウェルプレートに分配した。2時間37℃でインキュベーションの後、非付着細胞を除去し、一方で付着細胞を回収し、取り込んだ放射活性を、Topcount シンチレーションカウンターで測定した。ウェルあたりの付着細胞数を、様々な染色細胞の濃度を用いて得た標準曲線を用いて計算した。
【0084】
1.1.3. TLR4情報伝達経路の活性化
HEK293/hTLR4-MD2-CD14またはHEK293/LacZ 発現細胞を、生成者指示書に従って(Invivogen)、ヒト分泌胚のアルカリホスファターゼ遺伝子(SEAP)を担持するプラスミドを用いてトランスフェクトした。SEAP発現をNF-κB-誘導性ELAM-1プロモーター(pNiFty-SEAP (Invivogen))によって制御した。トランスフェクション24時間後、細胞を、様々な濃度のLPS、100 nM EDA SIINFEKLタンパク質またはプロテイナーゼ-Kで予め消化した100 nM EDA-SIINFEKLタンパク質の存在または非存在においてインキュベートした。24時間後、レポーター遺伝子の発現を、比色分析アッセイ(Invivogen)により、培養物上清において測定した。図3において、棒線は、NF-γB誘導の倍数(HEK293/TLR4-MD2-CD14からの上清に対して得たODを、HEK293/LacZからの上清に対して得たODによって割る)を示す。このアッセイにおいてEDA調製物中のエンドトキシン汚染物質の量は0.0003μg/ml以下であった。
【0085】
1.2 結果
1.2.1. EDAの発現およびEDA-SIINFEKL組換え融合タンパク質
組換えEDA-SIINFEKLタンパク質をE. coliにおいて6xHis 融合タンパク質(配列番号:2)として発現させ、アフィニティクロマトグラフィーにより精製し、脱塩し、方法の章で説明したようにエンドトキシンから精製した。得られたタンパク質を、抗his抗体(図1)を用いてSDS-PAGEおよびウェスタンブロットにより分析した。推定分子量(13 kDa)に対応するバンドを、各タンパク質について観察した。
【0086】
1.2.2. EDA-SIINFEKL 融合タンパク質はTLR4を結合する
フィブロネクチンエクストラドメインAは、トル様受容体4を活性化することが記載されている(Okamura et al, JBC, 2001; 276:10229-10233)。しかしながら、TLR4に対するEDAの物理的結合に関する直接的な証拠はない。我々は、EDA-SIINFEKLタンパク質が、TLR4を発現する細胞に結合する能力を有するかどうかを最初に分析した。ヒトTLR4-MD2-CD14を発現するHEK293またはLacZ(Invivogen)でトランスフェクトしたHEK293細胞を、1μM EDA-SIINFEKLタンパク質を用いてパルスし、抗His抗体およびフルオレセインで標識した抗マウスIgG(方法を参照されたい)で標識し、フローサイトメトリーにより分析した。ヒトTLR4-MD2-CD14を発現するHEK 293 細胞は、LacZを発現するHEK 293 細胞よりわずかに高い蛍光強度を示した(図2A)ことがわかった。また、ヒトTLR4に結合するフルオレセイン染色抗体を阻害するEDA-SIINFEKLタンパク質の能力も測定した。この結果から、HEK-hTLR4細胞と500mM EDA-SIINFEKLタンパク質との前記インキュベーションにより、該抗体結合のおおよそ50%が阻害されたということが判った(図2B)。図2Cは、抗体結合に対するEDA-SIINFEKLの様々な用量の抑制効果を示す。一方、我々は、付着アッセイにおいてEDAタンパク質で前もって被覆したプラスチックウェルに対する、HEK hTLR4およびHEK LacZコントロール細胞の結合能力を測定した。HEK hTLR4 細胞は、EDAを含有するウェルに特異的に結合できることを示す。これらの全ての実験は、EDAおよびEDA-SIINFEKLタンパク質がTLR4を結合できることを示す。
【0087】
1.2.3. EDA-SIINFEKL 融合タンパク質はTLR4 情報伝達経路を活性化する
TLR4シグナル伝達はNF-κB輸送をもたらし、これは多様な遺伝子のプロモーターのなかのコンセンサスエレメントに結合する転写因子である。組換えEDA-SIINFEKLタンパク質がTLR4を活性化できるかどうかを決定するために、我々は、ヒト分泌胚のアルカリホスファターゼ遺伝子(SEAP)を担持するプラスミドでトランスフェクトしたHEK293/hTLR4-MD2-CD14またはHEK293/LacZ細胞を、NF-κB-誘導性ELAM-1プロモーター(pNiFty-SEAP(Invivogen))の制御の下で使用した。我々は、EDA-SIINFEKLタンパク質が、HEK293/hTLR4-MD2-CD14トランスフェクト細胞においてのみSEAPの発現を刺激でき、NF-κB誘導倍数が7に達し、細胞をLPS(0.01μg)とインキュベートした時に見出された値と類似していることを見出した(図3)。EDA-SIINFEKLタンパク質がプロテイナーゼKで予め消化されていれば、NF-κB核輸送を刺激するこの能力は完全に消失し(図3)、TLR4のEDA活性化は、組換えタンパク質調製において潜在的なLPSコンタミネーションによって説明できないことを示唆している。
【0088】
1.3 考察
我々は、E. coliにおいて組換え融合タンパク質EDA-SIINFEKL 6xHisを発現できる組換えプラスミドpET20b2-26を構築した。6つのヒスチジンの存在により、融合タンパク質の検出および精製が可能となる。こうして、我々は、十分量の融合タンパク質をE. coli培養物の細胞質分画から精製することが出来た。結合アッセイは、EDA-SIINFEKLタンパク質がTLR4に特異的に結合出来ることが示唆されたTLR4発現細胞で行った。さらに、我々は本明細書において、EDA-SIINFEKLがTLR4シグナル経路を活性化できることを示す。この活性化は、プロテイナーゼKによる先の消化によりNF-κB核輸送を刺激する能力が完全に消失するので、該タンパク質の潜在的なLPSコンタミネーションとは関連がない。さらに、このアッセイにおいて、EDA調製の際のエンドトキシンの汚染物質の量は、0.0003μg/ml以下であって(LALアッセイによって評価される)、このイン・ビトロのアッセイではTLR-4 シグナル経路を活性化することができない。これらの結果は、EDAタンパク質が、抗原をTLR4発現細胞に標的化するためにタンパク質性ビヒクルとして使用できたということを示唆する。樹状細胞は、トル様受容体および特定のTLR4を発現することが知られている。また、DCに対する最も強力な成熟刺激のいくつかは、TLR受容体とそれら各々のリガンドとの相互作用からおこる。こうして、EDAを含有する融合タンパク質および特定抗原の相互作用は、炎症性サイトカインの産生および共刺激分子の上方調節を誘導することによって先天性免疫の急速な活性化を促進できた。さらに、DCの表面に対するこの融合タンパク質の標的化は、DCにより抗原の捕捉およびエンドサイトーシスを増加でき、結果としてこの抗原に対する免疫応答が増強する。
【0089】
実施例2.融合タンパク質を含有するEDAはイン・ビトロおよびイン・ビボで樹状細胞成熟を誘導し、細胞傷害性Tリンパ球の誘導を可能にする
2.1 材料および方法
2.1.1. 骨髄由来樹状細胞産生
樹状細胞を骨髄細胞から成熟させた。ACK溶菌緩衝液をもちいて赤血球を溶菌した後に、細胞を洗浄し、次にCD4 (GK1; ATCC, Manassas, VA)、CD8(53.6.72; ATCC)、Ly-6G/Gr1(BD-Pharmingen; Sandiego CA)およびCD45R/B220 (BD-Pharmingen)に対する抗体の混合物とのインキュベーションにより、その後にウサギ補体によってリンパ球および顆粒球を枯渇させた。残存細胞を、mGM-CSF(20 ng/ml)およびmIL-4(20 ng/ml)(Peprotech; London, UK両方)を有する強化完全培地中で、12-ウェルプレートにおいて、106細胞/mlで生育させた。2日毎に、培地をサイトカイン含有する新規培地で置換した。非付着性樹状細胞を、7日で収穫し、37℃および5%CO2で、LPS(Sigma)(1μg/mlまたは15ng/ml)、EDA-SIINFEKL (500 nM)またはSIINFEKL (10μM)の存在または非存在下で培養した。幾つかの実験において、ポリミキシン(10μg/ml)を培養物に加え、エンドトキシン汚染物質の影響を阻害する。24時間培養後、上清を回収し、IL-12およびTNF-αを、生成者指示書に従ってELISA(BD-Pharmingen)によって測定した。
【0090】
2.1.2 EDA-SIINFEKLを用いる免疫性付与後のCD11c細胞のイン・ビボでの成熟の測定。プロテイナーゼKによる消化の効果
DCの成熟を、多様な表面マーカーの発現を測定するフローサイトメトリーによりイン・ビトロで評価した。C57BL6マウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独によりi.v.注射した。プロテイナーゼKによるEDA-SIINFEKLの消化を、アガロースプロテイナーゼK(Sigma, St Louis)を用いて実施した。つまり、洗浄緩衝液(20 mM トリス-HCl, pH 7.2, 1 mM EDTA, 1mM ClCa2)中で洗浄したプロテイナーゼアガロースビーズ(5 mg/ml)を使用して、EDA-SIINFEKLタンパク質またはLPSを20分30℃で消化した。アガロース-プロテイナーゼKビースを遠心分離により除去した。15時間の免疫性付与後、マウスを屠殺し、autoMACSによりCD11c細胞を精製した。細胞を標識し、フローサイトメトリーにより分析した。
【0091】
2.1.3. 抗原提示能力を分析するためのイン・ビトロ試験
我々は、このエピトープに特異的なOT-1トランスジェニックマウスからのT細胞またはTハイブリドーマB3Zの両方に対してプロセシングされたCTLエピトープSIINFEKLを提示するAPCにより捕捉されるべきEDA-SIINFEKLの能力を特徴づけた。骨髄由来DCを、様々な濃度のEDA-SIINFEKL、EDAおよびSIINFEKL(共有結合でない)またはSIINFEKLの存在下において培養した。培養24時間後、上清を回収し、IFN-γ産生をELISAにより測定した。あるいは、培養開始12時間後、DCを、回収し、0.05% グルタルアルデヒドで固定し、OT-1からの非付着細胞の様々な量の存在下またはB3ZハイブリドーマT細胞系の存在下に、APCとして使用される。いくつかの実験において、DCをクロロキン(3μM)、モネンシン(1μl Golgystop, Pharmingen)、ブレフェルジン(1μl Golgyplug, Pharmingen)、シクロヘキシミド(4μg/ml)の存在または非存在の下でインキュベートし、次いでEDA-SIINFEKLまたはSIINFEKLペプチドの存在下でインキュベートした。数例において、抗TLR4抗体を該培養物に添加した。
【0092】
処理されたAPCの存在下でB3Z 細胞の活性化を、IL-2産生を測定することによって行った。B3Z ハイブリドーマ細胞(105 細胞/ウェル)を、完全培地(10% FCS、2 mM グルタミン、100 U/ml ペニシリン、100μg/ml ストレプトマイシンおよび5x10-5M 2-メルカプトエタノールで強化したRPMI 1640)中で、18 時間C57BL/6 wtマウスまたはTLR4 KOマウス由来の脾臓細胞(105細胞/ウェル)、および様々な濃度のEDA-SIINFEKLの存在下に培養した。培養物上清に放出されたIL-2の量を、先に記載したとおりにCTLLを基にしたバイオアッセイにより測定した。
【0093】
2.1.4.イン・ビボでの免疫性付与後の細胞傷害性Tリンパ球(CTL)およびIFN-γ産生細胞の誘導の測定
C57BL6 マウスを、0日および10日目にEDA-SIINFEKL(50μg)またはSIINFEKL(50μg)を用いてi.v.により免疫化した。20日目で、SIINFEKLに対してCTL応答の分析のためにマウスを屠殺した。免疫化動物から脾臓細胞を、SIINFEKL の1mlあたり0.1μgの存在下に5 x 106細胞/ml(10 ml)を完全培地中で5日間培養した。5日目に、細胞をクロミウム放出アッセイのために回収した。分解活性を、様々な数のエフェクター細胞を、51Crを予め添加した1x 104 EL-4 標的細胞と共にSIINFEKLの存在下または非存在で4時間インキュベートすることによって測定した。溶菌比活性%を、式:(cpm 実験値 − cpm 自然発生値)/(cpm 最高値 − cpm 自然発生値) x 100、に従って計算した、式中自然発生値溶菌とは、エフェクター細胞の非存在下で培養した標的細胞に対応しており、最高値の溶菌は標的細胞を5% Triton x100とインキュベートすることによって得られる。
【0094】
SIINFEKLに対する応答においてIFN-γの産生を測定するために、免疫化マウス由来の脾臓細胞を、0.25 mlの最終容量で完全培地単独かまたはペプチド(30μM)と共に、8x105 細胞/ウェルで96-ウェルプレートに播種した。48 時間後に上清(50μl)を除去し、生成者指示書に従ってIFN-γをELISAにより測定した(Pharmingen, Sandiego, CA)。
【0095】
他の実験群において、タンパク質混合物中でアジュバンドとして作用するEDAタンパク質の能力を証明した。この場合において、C57BL/6 マウスの一群を、PBSにおいてOVA タンパク質(500μg)の存在下にEDA-SIINFEKL(50μg)を用いてi.v.により免疫化し、一方別の群はPBSにおいてOVAタンパク質(500μg)単独で免疫化した。1週間後、上記したとおりにマウスを屠殺し、両方の群でSIINFEKLに対するCTL応答を決定した。
【0096】
2.1.5. EG7OVA 腫瘍細胞攻撃に対する保護
マウスを、0および10日目にEDA-SIINFEKL(3 nmol)またはSIINFEKL(3 nmol)を用いてs.c.で免疫化した。二次免疫性付与後20日目のマウスを105 EG7OVA細胞を用いてs.c.により攻撃した。腫瘍成長を、カリパスで制御し、式V = (L x w2)/2を用いてmm3で表した。式中、Lは長さ;wは幅である。腫瘍サイズが8 cm3以上の大きさに達した時にマウスを屠殺した。
【0097】
2.2. 結果
2.2.1. EDA-SIINFEKL 融合タンパク質は、骨髄由来樹状細胞(BMDC)によるIL-12およびTNF-α産生を刺激する。
我々は、EDA-SIINFEKL組換えタンパク質がBMDCを刺激し、炎症性サイトカイン、例えばIL-12またはTNF-αを産生出来るかどうかを試験した。即ち、BMDCをSIINFEKL (10μM)、LPS(1μg/ml および 15ng/ml)またはEDA-SIINFEKL-6xHis (500nM)を用いて培養した。24時間後に、培養物上清中のIL-12またはTNF-αをELISAにより測定した。EDA-SIINFEKLが、BMDCにより非常に高レベルのIL-12またはTNF-α産生を刺激できたということが判った(図4)。該タンパク質がプロテイナーゼKで予め処理された時にこの免疫賦活性能力は消失し、該活性はタンパク質のサンプル中のエンドトキシンの潜在的痕跡を理由とするものではないことを示唆している。
【0098】
2.2.2. EDA-SIINFEKLはイン・ビボでCD11c発現DCのTLR4依存性の成熟を誘導する
樹状細胞(DC)は、ナイーブT細胞および抗原に対する二次応答を刺激する独特な能力を有する最も強力な抗原提示細胞である。DCは、抗原を捕捉する能力を持ち、それらをペプチドに分解して、細胞傷害性T細胞(CTL)またはTヘルパー細胞各々に対するMHCクラスIまたはII分子との会合において、このペプチドを提示する。未熟DCは、抗原を捕捉できるが、ナイーブT細胞を刺激出来るようになるためには分化または成熟しなければならない。従って、樹状細胞の成熟はT-細胞応答の至適刺激のための要件である。成熟がおこる時に、APCは、表面分子、例えばクラスIおよびクラスII MHC、CD40、CD80およびCD86分子の発現を増加させる。そのため、我々は、EDA-SIINFEKLがイン・ビボでCD11c発現細胞の成熟を誘導できるかどうかを分析した。C57BL6 マウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いて、i.v.で免疫化した。15時間後、マウスを屠殺し、CD11c細胞をautoMACSにより精製し、抗体を標識して、フローサイトメトリーにより分析した。我々は、MHCクラスIおよびクラスII分子、CD40およびCD86の発現を誘導できたことを見出した。タンパク質が免疫性付与前にプロテイナーゼ Kで消化した場合に、EDA-SIINFEKLのこの能力が完全に消失した(図 5)。プロテイナーゼ Kで消化されたLPSの消化物は、これらの成熟マーカーの発現を誘導するためのLPSの能力に対していずれの抑制効果も持たなかった(図示せず)。我々は、C57BL/6 TLR4 KO マウス由来のCD11c細胞の成熟を誘導するEDA-SIINFEKLの能力を試験し、そしてEDA-SIINFEKLがC57BL/6 wt マウスに見出された成熟マーカーの過剰発現を誘導できないことを見出した(図5)。
【0099】
2.2.3 EDA-SIINFEKLは、SIINFEKLエピトープに特異的なT細胞に対して樹状細胞により有効に提示される
我々は、このエピトープに特異的なOT-1 トランスジェニックマウス由来のT細胞に対して処理済みCTLエピトープSIINFEKLを提示するためにAPCにより捕捉されるべきEDA-SIINFEKLの能力を特徴決定した。骨髄由来DC(105細胞/ウェル)を、様々な濃度のEDA-SIINFEKL、EDA+SIINFEKL、EDAまたはSIINFEKLペプチドの存在において培養した。48時間後、105非付着性OT-1細胞を添加した。OT-1 非付着細胞によるIFN-γ産生を測定した(図 6A)。SIINFEKL ペプチドは、IFN-γ産生によって証明されたようなOT-1 マウス由来のT細胞に対して有効に提示された。EDA-SIINFEKLは、高レベルのIFN-γも誘導するが、しかしタンパク質の高用量は、IFN-γと同様のレベルか、または同等より高いレベルを得るために必要であり、EDAタンパク質がMHCクラスI分子に対するSIINFEKLエピトープを担持することを明確に示している。SIINFEKLペプチドとインキュベートしたDCへのEDAタンパク質の添加は、OT-1マウス由来のT細胞によりIFN-γ産生を増加しなかった。予測したように、EDA単独とインキュベートしたDCは、OT-1 マウス由来のT細胞を活性化しなかった。DC におけるTLR4分子発現が、EDA-SIINFEKL提示を増強できるかどうかを分析するために、SIINFEKL特異的B3Z細胞を、C57BL/6 wtまたはTLR4KOマウス由来の脾臓細胞の存在下に様々な濃度のEDA-SIINFEKLとインキュベートした。B3Z細胞に対するEDA-SIINFEKL提示は、TLR4を発現した抗原提示細胞においてより効率的であった(図6B)。また、B3Z 細胞に対するEDA-SIINFEKL提示は、抗TLR4抗体の添加によって完全に遮断され(図6C)、TLR4がEDA-SIINFEKL捕捉に関与することを示唆している。その後、我々は、EDA-SIINFEKLのプロセシングに対して様々な薬物の効果を試験し、そして我々は、この提示が、エンドソームおよび後期リソソーム(図 6D)における酸性化の既知阻害剤である、クロロキン以外のモネンシン、ブレフェルジンまたはシクロヘキシミドにより、完全に阻害されることを見出した。予測したように、SIINFEKL合成のペプチドの提示は、これらの薬物によって影響されなかった。これらのデータは、EDA-SIINFEKLの内在化は、マクロピノサイトーシスによって媒介されるのではなく、EDA-SIINFEKLはクラスI細胞質性プロセシング経路を介して処理されることを示す。
【0100】
2.2.4. EDA-SIINFEKLはイン・ビボにおいてSIINFEKLに特異的なCTLを誘導する。
前記結果は、EDA-SIINFEKLの組換えタンパク質は生物活性であり、およびAPCを特異的に活性化することを示す。イン・ビボで特異的なTリンパ球免疫応答の誘導はワクチンの開発に重要である。従って、我々は、EDA-SIINFEKL 融合タンパク質を用いて免疫化したマウスは、SIINFEKLエピトープによりパルス化した標的細胞に対して特異的CTL応答を進行させるかどうかを試験した。C57BL6マウスを、0および10日目に、PBS中のEDA-SIINFEKL(50μg)またはSIINFEKL(50μg)を用いてi.v.にて免疫化した。20日目に、マウスをSIINFEKLに対するCTL応答の分析のために屠殺した。EDA-SIINFEKLはSIINFEKLを用いてパルス化したEL-4標的細胞に対してCTLを誘導できたことが判った。一方で、CTL活性はマウスをSIINFEKL単独で免疫化した場合には見出されなかった(図7)。イン・ビボで樹状細胞の成熟を誘導するためのEDAの能力のために、アジュバンドとして作用するEDAの能力もまた、OVAタンパク質で免疫化した場合に分析された。この実験において、我々は、免疫性付与混合物中のEDAの存在により、免疫賦活性効果を有し、OVAタンパク質により誘導されたSIINFEKLに対するCTL応答誘導を増強できることを観察した(図9のパネルAおよびBで分解活性を比較する)。
【0101】
2.2.5. EDA-SIINFEKLはOVAタンパク質を発現する腫瘍細胞による攻撃からマウスを保護する。
EG7OVA腫瘍細胞に対してマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを、EDA-SIINFEKL(3 nmol)、SIINFEKL(3 nmol)または生理食塩を用いてs.c.にて免疫化した。二次免疫性付与20日後に、105 EG7OVA 細胞を用いてs.c.にてマウスを攻撃した。我々は、EDA-SIINFEKL 免疫性付与により、マウスを腫瘍成長から保護することが判った。SIINFEKLまたは生理食塩水を用いて免疫化した全てのマウスは腫瘍を進行させたが、EDA-SIINFEKLで免疫化したマウスの40%はが依然として腫瘍の存在はなく、残りの60%は腫瘍成長遅延を経験した(図8)。
【0102】
2.3. 考察
本試験において、組換えEDAタンパク質をエピトープ送達ベクターとして用いて、我々は、アジュバンドの必要性なく、イン・ビボでCTL応答を開始する免疫性付与のためのストラテジーを確立した。我々は、ベクターとしてEDA含有融合タンパク質の有効性に関与するメカニズムを同定し、抗原をTLR4発現細胞に送達し、そして抗原に対する細胞性免疫応答を誘導する。
【0103】
最初に、我々は、イン・ビトロでのEDAを用いるBMDCの刺激は炎症性サイトカイン、例えばIL-12およびTNF-αの産生を刺激できたことを見出した。これらのサイトカインは、抗原に対して強力な免疫応答の開始のために重要であることが知られている。さらに、イン・ビボでEDAによる免疫性付与によりDCの成熟を誘導でき、DCの表面に共刺激分子の発現を増加させ得ることを見出した。これらの共刺激分子の発現は、抗原に対する免疫応答の有効な誘導のために最も重要である。この効果は、TLR4の存在に依存しており、イン・ビボにおいてC57BL/6 TLR4 KO マウス由来の単離したCD11c細胞をEDAで予め免疫化したので、非免疫化動物において見出されたものと比較して、その成熟状態に対していずれの改善をも示さなかったということを見出した。
【0104】
我々は、APC上のTLR4分子の存在が、EDA-SIINFEKL融合タンパク質と共にインキュベーションした後に抗原提示を改善すること、およびこの提示がそのエンドリソソームタンパク質分解を阻害するクロロキンによって影響されないことを見出した。これらのデータは、EDA-SIINFEKLの内在化がマクロピノサイトーシスによって媒介されないことを示しており、組換えEDA-SIINFEKLタンパク質が細胞質性抗原として処理されることを示し、および該融合タンパク質がAPCプラズマ膜を介してサイトゾルに送達されなければならないことを言及する。
【0105】
最も重要なことには、我々は、組換え融合タンパク質EDA-SIINFEKLを有するマウスの免疫性付与は、SIINFEKLエピトープに対して特異的なイン・ビボCTL応答を誘導することができることを見出した。さらに、EDA-SIINFEKLを用いる免疫性付与は、EG7OVA腫瘍細胞による攻撃に対してマウスを保護できる。全てのこれらのデータから、EDAを含有するこのタンパク質性ベクターは、(i)抗原をTLR4発現細胞、特にプロフェッショナルAPCに標的化する;(ii)ベクター化(vectorised)抗原を古典的クラスI抗原プロセシング経路へ送達する;(iii)イン・ビボおよびイン・ビトロで樹状細胞成熟を誘導する;および (iv)アジュバンドの非存在下でベクター化抗原に対するCTLをイン・ビボで駆動し、そうして感染性剤または癌に対するワクチン接種ストラテジーにおいて使用することが出来ることが示された。また、EDA含有するこれらの融合タンパク質は、TLR4発現細胞に対して医薬関連分子の細胞質性送達のために働き得る。さらに、イン・ビボでの樹状細胞成熟を誘導するためのEDAの能力は、免疫性応答を与えることが望まれる抗原を含有する製剤におけるアジュバンドとしてのその使用を可能にし、EDAワクチン開発における使用のための可能性を開くものである。
【0106】
実施例3.EDAタンパク質を、ビヒクルとして少なくとも390個のアミノ酸の抗原を輸送するために使用できる
3.1 材料および方法
3.1.1 EDA-OVA および EDA-NS3組換えタンパク質の発現
EDA-OVA 発現プラスミドの構築のために、mRNAを、OVAタンパク質を発現するEG7OVA 腫瘍細胞から抽出した。プライマー
【化6】
(配列番号:16)および
【化7】
(配列番号:17) (下線の塩基を制限酵素NotIにより配列を導入したが、該配列の斜字体はオボアルブミンの開始および末端に属する)を用いるPCRによる逆転写および増幅の後に、該PCR生成物をTOPO TA kit (Invitrogen)を用いてpCR2.1-TOPOにクローニングし、NotIを用いて消化し、NotIで消化する前にプラスミドpET20EDA 1.2(EDA タンパク質を発現する)にサブクローニングした。構築物の正しい配向性を配列決定により評価した。プラスミド培養物により形質転換したE. coliの誘導後、可溶性画分中に存在する融合タンパク質を、FPLCプラトフォーム(AKTA, Pharmacia)を用いてアフィニティクロマトグラフィーにより精製した(Histrap, Pharmacia)。タンパク質を脱塩Hitrap columns(Pharmacia)を用いて脱塩し、フィルター装置を用いる遠心分離Amicon Ultra 4-5000 MWCO (Millipore Carrighwahill, Ireland)により濃縮した。組換えタンパク質ベクターを、エンドトキシンレベルがタンパク質(LAL アッセイによって測定した, Cambrex)の0.2 EU/μg以下となるまで、Endotrap endotoxin columns (Profos Ag, Regensburg, Germany)を用いて精製した。EDA-NS3 タンパク質構築体について、同様のストラテジーを、NS3タンパク質(アミノ酸1026-1221)のアミノ末端フラグメントの発現を可能にするプライマー
【化8】
(配列番号:18) および
【化9】
(配列番号:19)を用いて行った。E. coli画分の可溶性分画から抽出したEDA-OVA タンパク質とは異なり、EDA-NS3 タンパク質の場合には、該タンパク質を8M 尿素に予め溶解した封入体から精製した。Histrap columnsを用いるアフィニティクロマトグラフィーの後に、イオン交換クロマトグラフィー(DEAEセファロース)を行った。この第二クロマトグラフィーからの精製分画を、G25カラムでの再結合プロトコールに従って再結合させた。一旦再結合した後、EDA-NS3カラムを脱塩し、Endotrap columns (Profos, Germany)を用いてエンドトキシンを精製した。
このように精製されたタンパク質をSDS-PAGEにより分析した
【0107】
3.1.2 抗原提示能力を評価するためのイン・ビトロでの試験
APCにより捕捉されるべきEDA-OVAの能力およびOT-1トランスジェニックマウス由来のTリンパ球に対するCTLエピトープのプロセシングしたSIINFEKLのその後の提示を評価した。骨髄由来樹状細胞(105 細胞/ウェル)を様々な濃度のEDA-OVA、EDA +OVA(非共有結合)、OVAまたはEDAの存在において培養した。12時間後、OT-1 トランスジェニックマウス由来の105 細胞/ウェルの非付着細胞を添加した。培養の開始から24 時間に、該上清を市販のELISAアッセイにより分泌されたIFN-γ定量のために抽出した。
【0108】
3.1.3 イン・ビボT細胞傷害性リンパ球(CTL)誘導および免疫性付与後のIFN-γ産生細胞の測定
C57BL6またはHHD(HLA-A2.1 分子についてのトランスジェニック)マウスを、0および10日目に各々EDA-OVAまたはEDA-NS3(50μg)を用いてi.v.免疫化した。20日目に、マウスを、SIINFEKLまたはNS3 1073 ペプチドに対するCTL応答を決定するために屠殺した。免疫化動物由来の脾臓細胞を、5 x 106 細胞/ml(10 ml)でSIINFEKL(0.1μg/ml)またはNS3 1073(1μg/ml)の存在下に、完全培地中で5日間培養した。5日目に、細胞をクロミウム遊離分析のために回収した。分解活性を、51Crを予め添加した1 x 104 EL-4 標的細胞と様々な量のエフェクター細胞を、ペプチドを用いるかまたは用いないで4時間インキュベートして測定した。分解物比活性の%を、式(実験値 cpm− 自然発生値 cpm)/(最高値 cpm − 自然発生値 cpm) x 100に従って計算した、式中の自然発生値は、エフェクター細胞の非存在下でインキュベートした標的細胞に対応しており、最大の分解物は標的細胞を5%のTriton x100とインキュベートすることによって得られた。
【0109】
様々なペプチドに対する応答におけるIFN-γ産生を測定するために、免疫化マウス由来の脾臓細胞を、終濃度0.25 mlで、完全培地のみ、または30μM のペプチド NS3 1038-1046 (配列番号:23)、1073-1081(配列番号:20、CVNGVCWTV)、1169-1177(配列番号:22)と共に、または組換えNS3(1μg/ml)と共に、8 x 105 細胞/ウェルにて、96ウェルプレートに分配した。48h後に上清(50μl)を回収し、生成者指示書に従ってIFN-γをELISAにより測定した(Pharmingen, Sandiego, CA)。
【0110】
3.1.4 C型肝炎ウイルスポリプロテインを発現するvHCV1-1031ワクシニアウイルスによる感染に対する保護
C57BL/6を、EDA-NS3 タンパク質と共にパルス化された1 x 106の骨髄由来樹状細胞を用いて1日および10日目に免疫化した。二次免疫性付与後10日目に、マウスを、組換えvHCV 1-1031ワクシニアウイルスを5 x 106 pfuにてi.p.により感染させた。感染3日後、動物を屠殺し、ウイルス負荷/卵巣組織mgを、BSC-1 細胞系の使用を基にしたプラーク形成ユニット定量アッセイにより定量した。
【0111】
3.2 結果
3.2.1 EDA-OVAおよびEDA-NS3タンパク質の発現および精製
材料および方法の章に示したとおりに、EDA-OVAまたはEDA-NS3を、EDA-OVAの場合には形質転換したE. coli抽出物から可溶性分画を用いて精製し、EDA-NS3の場合には封入体を形成する。図10Aおよび11Aにおいて、両方のタンパク質についてSDS-PAGEの結果が示される。55kDaおよび32kDaサイズの各タンパク質に対応する一本の各バンドが得られた、これは各タンパク質について予測したサイズに対応している。
【0112】
3.2.2 OVAタンパク質に結合するEDAは、OT-1トランスジェニックマウス由来のTリンパ球に対するIVAの抗原提示を増強する
我々は、SIINFEKLエピトープに特異的なT細胞に対してSIINFEKLエピトープの抗原提示を増強するOVAタンパク質に対するEDA結合能力を分析した。我々は、EDA-OVAの存在下で培養された骨髄由来樹状細胞は、等モル量のOVAタンパク質単独(図9B)により誘導されたものよりも非付着性OT-1マウス細胞によるIFN-γ産生を強く刺激したことが観察された。我々は、IFN-γ産生はまた、OVAタンパク質単独と比較した場合に、EDAタンパク質およびOVAタンパク質を含んだ共培養物中で増強し、EDAにより誘導されたDC成熟は、T細胞活性化を増強できることを示唆していると考察した。
【0113】
3.2.3. OVAタンパク質またはNS3タンパク質に結合するEDAは、イン・ビボで特異的細胞傷害性T細胞誘導を増強する
SIINFEKLエピトープまたはNS31073エピトープ各々に対するCTL応答を誘導するEDA-OVAおよびEDA-NS3の能力を分析した。我々は、EDA-OVAタンパク質について、生理食塩溶液に溶解したこのタンパク質による免疫性付与は、SIINFEKLペプチドとインキュベートした標的細胞に対する細胞傷害性応答を誘導でき、OVAタンパク質単独による免疫性付与では誘導できないことを観察した(図9C)。同様の方法において、EDA-NS3タンパク質を用いるHHDマウス(HLA-A2.1に対するトランスジェニック)免疫性付与は、ペプチドNS31073(V)(配列番号:20、CVNGNCWTV)、このペプチド(配列番号:21、CLNGVCWTV)の改変型1073(L)と予めインキュベートした標的細胞に対して有効な細胞傷害性応答を誘導する(図11B)。我々は、EDA-NS3タンパク質によるHHDマウス免疫性付与が、ペプチドNS3 1038 (配列番号:23)、1073(配列番号:20、CVNGVCWTV)および1169(配列番号:22)(図11C)に特異的なIFN-γ産生細胞の活性化を誘導することを観察した。さらに、EDA-NS3 タンパク質によるマウス免疫性付与は、NS3 1073ペプチドに対して特異的な長時間持続する細胞傷害性細胞性応答を誘導する。実際に、EDA-NS3タンパク質により免疫化したマウスを免疫性付与60日後に屠殺した場合に、我々はこのペプチドに特異的なCTLの存在を検出した(図11D)。
【0114】
3.2.4 EDA-NS3タンパク質とインキュベートした樹状細胞によるC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現するワクシニアウイルスによる感染からマウスを保護する。
我々は、EDA-NS3タンパク質と共にイン・ビトロでインキュベートした樹状細胞を用いる免疫性付与により、C型肝炎ウイルスタンパク質を発現する組換えウイルスによる攻撃に対してマウスを保護できる細胞応答を誘導できたかどうかについて試験することを求めた。この理由から、我々は、EDA-NS3タンパクとインキュベートした骨髄由来樹状細胞または任意の抗原と予めインキュベートしていない樹状細胞を用いてC57/BL6マウスを免疫化した。二次免疫性付与10日後に、マウスを5 x 106 pfuの組換えvHCV 1-3011ワクシニアウイルスを用いて感染させた(Dr Rice, Washington University School of Medicine, St Louis, MOからのご厚意とGrakoui A, et al. J Virol. 1993; 67:1385による説明により)。3日後に、マウス両群においてウイルス負荷を測定した。この実験において、我々は、EDA-NS3 タンパク質とインキュベートしたDCによる免疫性付与により、組換えワクシニアウイルスによる感染に対して6%のマウスを保護できたことを観察した。
【0115】
3.3. 考察
前記結果に示したように、EDAタンパク質は、OVAタンパク質からTLR4分子発現細胞にSIINFEKLエピトープを標的化するための有用なベクターとして機能し、それらの免疫原性を増強出来る。より高分子のタンパク質の免疫原性を増強するEDAの能力を評価するために、我々は、完全なOVA(397個のアミノ酸)を含有するEDA-OVA 融合組換えタンパク質およびC型肝炎ウイルスからのNS3タンパク質のプロテアーゼ活性を有するフラグメントを含有するEDA-NS3タンパク質を構築した。
【0116】
これらの結果は、EDAタンパク質が、より大きな抗原を標的とする非常に有効なベクターとして作用できることを示す。このように、我々は、抗原提示アッセイにおいて、EDAに対するOVAの結合により、抗原提示細胞による抗原捕獲を増強させ、特異的T細胞活性化を増加させることを見出した。また、我々は、これらの融合タンパク質(EDA-OVAおよびEDA-NS3)を用いる免疫性付与が、これらの抗原に対する特異的細胞傷害性応答を誘導できることを観察した。これらの誘導性応答は長期間継続する。最終的に、我々は、樹状細胞と共にEDA-NS3タンパク質の投与は、C型肝炎ウイルスタンパク質を発現するワクシニアウイルスによる感染に対する保護的細胞性応答を誘導できることを観察した。
【0117】
これらのデータは、EDAタンパク質は、目的とする抗原に対して細胞性応答の誘導のたために非常に適切なタンパク質ベクターであり得る。EDAタンパク質を基にした融合タンパク質の構築は、腫瘍疾患または感染性剤によっておこる疾患に対するワクチン接種プロトコールにおいて適切なストラテジーである。
【図面の簡単な説明】
【0118】
【図1】図1.産生および精製したEDAおよびEDA-SIINFEKLタンパク質のSDS-PAGE分析。EDAおよびEDA-SIINFEKLタンパク質の1つのアリコートを、15%ポリアクリルアミドゲル上におき、電気泳動に供した。分子量マーカー(MWM)をKDaで示す。EDAおよびEDA-SIINFEKLタンパク質(13-14 KDa)の推定分子量に対応するバンドを観察した。
【図2】図2.タンパク質性ベクターEDA-SIINFEKLのTLR4への結合。2A.直接的な結合アッセイ。HEK293-LacZ細胞(HEK293-LacZ)およびHEK293-TLR4/MD2/CD14(HEK293 TLR4)細胞を、1μM EDA-SIINFEKLを用いてパルスし、パラホルムアルデヒドで固定し、抗Hisおよび抗EDA抗体で標識し、抗マウスIgG-FITCで顕色させ、フローサイトメトリーにより分析した。2B.TLR4発現細胞に対する抗TLR4抗体の結合を阻害するEDA能力。HEK-TLR4細胞を、500nM EDA-SIINFEKL タンパク質の存在または非存在において、2時間4℃でインキュベートした。細胞を洗浄し、FITCで染色した抗TLR4抗体とインキュベートし、フローサイトメトリーによって分析した。2C.様々な濃度のEDA-SIINFEKLタンパク質を用いてTLR4発現細胞に結合する抗TLR4抗体の阻害%。2D.細胞付着アッセイ。トリチウム化したチミジンで染色したHEK-hTLR4またはHEK-LacZ細胞を、EDAタンパク質で予め被覆した前に96ウェルミクロプレートのウェルに分配し、2時間37℃の間インキュベートした。非付着細胞を除き、付着細胞を回収し、取り込んだ放射活性をTopcountシンチレーションカウンターで計測した。ウェルあたりの付着細胞数を、標準曲線を利用して計算した。
【図3】図3.EDA-SIINFEKLはTLR4シグナル経路を活性化する。発現がNF-κB-誘導性ELAM-1プロモーターによって制御されるこのレポーター遺伝子でトランスフェクトしたHEK293/TLR4-MD2-CD14またはHEK293/LacZ発現細胞の培養物上清においてヒト分泌した胚アルカリホスファターゼ遺伝子の比色分析測定。トランスフェクションの24時間後に、細胞を様々な濃度のLPS、100 nM EDA-SIINFEKL タンパク質(EDA)またはプロテイナーゼKで予め消化した100 nM EDA-SIINFEKL タンパク質の存在または非存在下でインキュベートした。棒線は、NF-κBの誘導倍である(HEK293/TLR4-MD2-CD14由来の上清で得たODをHEK293/LacZからの上清で得たODによって割った)。
【図4】図4.EDA-SIINFEKLは、イン・ビトロでDCによって炎症性サイトカインの分泌を誘導する。骨髄由来DCを、LPS(1μg/ml)、EDA-SIINFEKL(500 nM)、プロテイナーゼKで消化したEDA-SIINFEKL(500 nM)の存在下または生理食塩溶液で培養した。24時間後、IL-12(A)およびTNF-α(B)の存在をELISAにより培養物上清中で測定した。
【図5】図5.EDA-SIINFEKLは、イン・ビボでCD11c DC 成熟を誘導する。樹状細胞の成熟は、T-細胞応答の最適な刺激のための要件である。成熟が起こる時に、APCは、表面分子、例えばクラスI(我々のモデルではH2Kb)およびクラスIIMHC(我々のモデルではIAb)、CD40、CD80およびCD86分子の発現を増加させる。従って、我々は、EDA-SIINFEKLがCD11c発現細胞のイン・ビボで成熟を誘導するかどうかを分析した。C57BL/6wtマウスを、EDA-SIINFEKL(25μg)、プロテイナーゼKで消化したEDA-SIINFEKL(25μg)、LPS(25μg)またはPBS単独を用いてi.v.で免疫化した。また、C57BL/6 TLR4 KOマウスをEDA-SIINFEKL(25μg)またはPBS単独を用いて免疫化した。15時間後にマウスを屠殺し、CD11c細胞をautoMACSにより精製した。細胞を標識し、H-2Kb、I-Ab、CD40、CD80およびCD86分子の発現についてフローサイトメトリーにより分析した。
【図6】図6:EDA-SIINFEKLは、樹状細胞によりSIINFEKLエピトープに特異的なT細胞に対して効果的に提示される。我々は、このエピトープに特異的なOT-1 遺伝子組み換えマウス由来のT細胞に対して処理済CTLエピトープSIINFEKLの提示について、APCによって捕捉されるべきEDA-SIINFEKLの能力を特徴分析した。(A) OT-1 トランスジェニックマウスからの非付着細胞によるIFN-γ産生。骨髄由来DCを、培地単独、様々な濃度の合成SIINFEKLペプチド、SIINFEKL、およびEDA、EDA-SIINFEKL(融合タンパク質)またはEDA単独の存在下で培養した。24時間後、DCを回収し、105個の非付着性OT-1細胞の存在下でAPCとして使用した。さらなる24時間後に、上清を回収し、分泌したIFN-γを測定した。(B)TLR4分子依存性。B3Zハイブリドーマ細胞(105細胞/ウェル)を、C57BL/6 wt マウス由来の脾臓細胞の存在下、またはTLR4分子(105 細胞/ウェル)およびEDA-SIINFEKLタンパク質(100 nM)のためのノックアウトマウス由来の脾臓細胞と共にインキュベートした。(C)C57BL76wt マウスの脾臓由来の細胞を、抗TLR4抗体の存在または非存在(BおよびC)において、B3Zハイブリドーマ細胞およびEDA-SIINFEKLタンパク質と共に共培養した。培養物上清に分泌したIL-2の量を、CTLL 細胞系の使用を基にしたバイオアッセイによって測定した。(D)融合タンパク質EDA-SIINFEKLの抗原提示に対するクロロキン、モネンシン、ブレフェルジンまたはシクロヘキシミドの効果。骨髄由来樹状細胞を、30 mMのクロロキン、ブレフェルジン、モネンシン、または4μg/mlのシクロヘキシミドの非存在下または存在下で1時間インキュベートし、その後EDA-SIINFEKLまたは合成ペプチドSIINFEKL(白色の棒)を添加した。10時間の培養後に、DCをグルタルアルデヒド中で固定し、非付着性OT-1マウス細胞(105細胞/ウェル)との同時培養物中で抗原提示細胞(APC)(104 細胞/ウェル)として使用した。24時間後、分泌したIFN-γの量を市販のELISAにより測定した。した(図6E)。
【図7】図7.EDA-SIINFEKLによるマウスの免疫性付与は、SIINFEKLエピトープに特異的な細胞応答を誘導する。前記結果は、EDA-SIINFEKL組換えタンパク質が生物活性であり、APCを特異的に活性化することを示す。イン・ビボでの特異的なT細胞免疫応答の誘導は、ワクチンの開発のために重要である。このように、我々は、EDA-SIINFEKL融合タンパク質により免疫化されたマウスが、このエピトープに対して特異的な細胞応答を生じるかどうかを試験した。(A)IFN-γ産生細胞の誘導。0および10日目に、C57BL/6 マウスを、EDA-SIINFEKLの1.5 nmolまたはSIINFEKLペプチドの1.5 nmolを用いて免疫化した。20日目に、SIINFEKLの存在または非存在において脾臓細胞を48時間インキュベートし、そして分泌されたIFN-γの量をELISAにより測定した。(B)SIINFEKL-特異的CTL応答分析の誘導。EDA-SIINFEKLまたはSIINFEKLにより免疫化したマウス由来の脾臓細胞を、SIINFEKLペプチドの存在下で5日間再刺激した。このインキュベーション後に、従来のクロミウム51遊離アッセイにより、SIINFEKLペプチドの非存在または存在下でEL-4標的細胞に対するCTL活性を測定した。データは、3つのサンプルから正味の特異的分析物(SIINFEKLでパルスした標的細胞の%分解−非パルス化標的細胞の分解%)の値の平均%を示す。
【図8】図8.EDA-SIINFEKLは、EG7 OVA発現細胞による腫瘍攻撃から守る。EG7OVA 腫瘍細胞の注射に対するマウスを保護するためのEDA-SIINFEKL融合タンパク質の能力を試験するために、マウスを3 nmolのEDA-SIINFEKL、SIINFEKLまたは生理食塩溶液を用いて0および10日目にs.c.で免疫化した。20日後、二次免疫性付与マウスを105 EG7OVA 細胞を用いてs.c.攻撃した。腫瘍増殖を、カリパスを用いてモニターし、式 V=(L x w2)/2(式中、L, 長さ;w, 幅である)を用いてmm3で表した。腫瘍サイズが8cm3以上の体積に達した場合に、マウスを屠殺した。
【図9】図9.EDAは、OVAタンパク質を用いる免疫性付与後に細胞傷害性応答の誘導においてアジュバンドとして作用する。EDAがイン・ビボで樹状細胞の成熟を促進することができれば、EDAはそれ自身によっては細胞傷害性応答を活性化できないが細胞傷害性エピトープを含有するタンパク質を用いる免疫性付与後にアジュバンドとして作用できるという可能性が存在する。この可能性を試験するために、我々は、その他のタイプのアジュバンドを用いないでEDA(50μg)とOVA タンパク質(500μg)により一群のマウス(A)、およびOVA(500μg)により別の群のマウス(B)を免疫化した。免疫性付与1週間後、マウスを屠殺し、脾臓細胞を、SIINFEKL合成ペプチドの存在下で培養した。5日後の培養物中で、SIINFEKLペプチドを用いてパルスしたEL-4 標的細胞に対する細胞傷害性応答を、従来のCr51遊離アッセイにおいて測定した。
【図10】図10.EDAは、より大きな抗原についてのビヒクルとして作用できる。前記実験において、EDAタンパク質は、細胞傷害性エピトープを輸送するためのビヒクルとして作用でき、該エピトープに対するCTL応答の誘導を支持することが証明された。後者の工程において、EDAが比較的大きな抗原を輸送でき、抗原に対する細胞性応答の誘導を促進できるかどうかについての試験がのぞまれる。この目的により、我々は融合タンパク質EDA-OVAを構築し、イン・ビトロおよびイン・ビボでの下記実験を行った。(A)組換えEDA-OVAタンパク質のSDS-PAGE分析。組換えタンパク質EDA-OVAを、E. coliで発現し、アフィニティクロマトグラフィーにより精製した。脱塩し、中和し、濃縮し、SDS-PAGEにより分析した。該タンパク質の推定分子量に対応するおおよそ55kDaのバンドを観察した。(B)抗原提示実験、骨髄由来DCを、OVA、EDA-OVA(融合タンパク質)、EDA+OVAまたはEDA単独の存在下または非存在下で培養した。24時間後、DCをOT-1マウス由来の105非付着細胞の存在において抗原提示細胞として使用した。DCの存在下でOT-1マウス由来の非付着細胞によりIFN-γ産生を、市販のELISAにより定量化した。(C) EDA-OVAタンパク質は、イン・ビボでOVA特異的CTLを誘導する。C57BL/6 マウスを、1nmolのEDA-OVAを用いるか、または1 nmolのOVAを用いて免疫化した。免疫性付与7日後に、免疫化したマウスからの脾臓細胞を、SIINFEKLペプチドの存在下に5日間イン・ビトロで再刺激した。その後、SIINFEKLの非存在または存在下でインキュベートしたEL-4 細胞に対する特異的CTL活性を、従来のCr51 遊離アッセイにおいて測定した。データは、3つのサンプルから、正味の特異的溶菌値(SIINFEKLを用いてパルスした標的細胞の溶菌%−非パルス標的細胞の溶菌%)の平均%を示す。
【図11】図11.EDA-NS3タンパク質は、NS3C型肝炎ウイルスタンパク質に対する特異的CTL応答を引き起こす。EDAタンパク質が大きな抗原についてのビヒクルとして作用できることが観察されると、C型肝炎ウイルスのNS3タンパク質(NS3タンパク質のプロテアーゼ領域からのアミノ酸1-196)に対する抗ウイルス応答を誘導するEDAの能力を、該ウイルスによる感染に対するワクチン接種ストラテジーとして分析した。(A)組換えEDA-NS3(1-196)タンパク質のSDS-PAGE分析。組換え融合タンパク質EDA-NS3を構築し、E. coliで発現し、SDS-PAGEにより分析した。(B)EDA-OVAタンパク質は、イン・ビボでOVA-特異的CTLを誘導する。HHDマウス(HLA-A2.1タンパク質についてトランスジェニック)を、生理食塩溶液に溶解したEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与1週間後、脾臓細胞を、イン・ビトロでNS3 1073ペプチド(HLA-A2拘束のためにNS3タンパク質由来の免疫優性の細胞傷害性T決定基を含有する)を用いて再刺激した。培養物中で5日後、ペプチド1073(V)(配列番号:20、CVNGVCWTV)と共に、または該ペプチド(配列番号:21、CLNGVCWTV)の1073(L)改変型と共に、またはペプチドの非存在下でインキュベートしたT2標的細胞に対して、細胞傷害性活性を、従来のCr51遊離アッセイを用いて測定した。(C)EDA-NS3タンパク質は、NS3タンパク質からの様々なエピトープに対して複数のエピトープ様応答を誘導する。EDA-NS3により免疫化したマウスから得た脾臓細胞を、ペプチド1038-1046、NS3 1073-1081またはNS3 1169-1177(NS3タンパク質のフラグメント1-196のなかで、HLA-A2拘束のために3つの細胞傷害性決定基を含有する)の存在下に、または組換えNS3タンパク質(Mikrogen)と共に、48時間イン・ビトロで再刺激した。上清に分泌されたIFN-γの量を市販のELISAにより測定した。(D)EDA-NS3タンパク質は、長期間持続する細胞傷害性応答を誘導する。HHDマウスを、生理食塩溶液中のEDA-NS3タンパク質の100μg/マウスを用いてi.v.経路により免疫化した。免疫性付与60日後、マウスを屠殺し、ペプチドNS3 1073に特異的なCTLの存在を、NS3 1073 ペプチドの非存在または存在中でインキュベートしたT2標的細胞を用いて、従来のクロミウム51遊離アッセイにおいて測定した。(E) EDA-NS3とインキュベートしたDCを有するC57BL/6 マウスの免疫性付与は、C型肝炎ウイルスタンパク質を発現する組換えワクシニアウイルス vHCV(1-3011)による感染からマウスを保護する。EDA-NS3 タンパク質と予めインキュベートした 106DCを用いてマウスを免疫化し、7日後に、それらはi.p.経路による5x106 ワクシニアウイルスvHCV(1-3011)の攻撃を受けた。感染3日後にマウスを屠殺し、BSC-1細胞感染アッセイにより、卵巣組織mgあたりのウイルス負荷を定量した。
【特許請求の範囲】
【請求項1】
抗原刺激剤に対する免疫細胞応答の生成において、
−フィブロネクチンEDAドメイン(EDA)、または
−TLR4を結合できる該EDAドメインのフラグメント、または
−TLR4を結合でき、該EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する該EDAドメインまたはフラグメントの改変型、
から選択されるアミノ酸配列を含むポリペプチドの使用。
【請求項2】
EDAドメインに対応するフラグメントが、
a)配列番号:2のアミノ酸2-91;
b)配列番号:4;および
c)TLR4発現細胞を結合できる配列a)およびb)のフラグメント、
から選択される配列を含むことを特徴とする、請求項1記載のポリペプチドの使用。
【請求項3】
EDAドメインに対応するフラグメントが、
a)配列番号:6のアミノ酸2-57;
b)配列番号:8;および
c)TLR4発現細胞を結合できる配列a)およびb)のフラグメント、
から選択される配列を含むことを特徴とする、請求項1記載のポリペプチドの使用。
【請求項4】
免疫賦活性剤が1以上の目的とする分子をさらに含むことを特徴とする、請求項1〜3のいずれかに記載のポリペプチドの使用。
【請求項5】
免疫賦活性剤がタンパク質性ベクターであり、ここで該ポリペプチドが目的とする該分子と結合されていることを特徴とする、請求項1〜4のいずれかに記載のポリペプチドの使用。
【請求項6】
アミノ酸配列が、下記のもの:
a)フィブロネクチンEDAドメイン(EDA)、
b)TLR4を結合できる該EDAドメインのフラグメント、または
c)TLR4を結合でき、該EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する該EDAドメインまたはそのフラグメントの改変型;
から選択されるポリペプチドを含むタンパク質性ベクターであって、ポリペプチドが次のもの;ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質および化学物質から選択される目的とする分子と結合することを特徴とする、タンパク質性ベクター。
【請求項7】
目的とする分子が抗原およびエピトープから選択されることを特徴とする、請求項6に記載のタンパク質性ベクター。
【請求項8】
目的とする分子がウイルス抗原、細菌抗原および寄生抗原から選択されることを特徴とする、請求項6または7のいずれかに記載のタンパク質性ベクター。
【請求項9】
目的とする分子がC型肝炎ウイルス由来のウイルス抗原であることを特徴とする、請求項6〜8のいずれかに記載のタンパク質性ベクター。
【請求項10】
C型肝炎ウイルス抗原がNS3タンパク質または該タンパク質の抗原性フラグメントであることを特徴とする、請求項9記載のタンパク質性ベクター。
【請求項11】
アミノ酸配列が配列番号:10であることを特徴とする、請求項10記載のタンパク質性ベクター。
【請求項12】
目的とする分子が腫瘍性抗原および腫瘍性抗原性決定基から選択されることを特徴とする、請求項6または7のいずれかに記載のタンパク質性ベクター。
【請求項13】
目的とする分子が、オボアルブミン(OVA 257-264)由来の抗原性細胞傷害性T決定基またはSIINFEKLであることを特徴とする、請求項12に記載のタンパク質性ベクター。
【請求項14】
アミノ酸配列が配列番号:2または配列番号:6から選択されることを特徴とする、請求項13記載のタンパク質性ベクター。
【請求項15】
目的とする分子がアレルゲンであることを特徴とする、請求項6記載のタンパク質性ベクター。
【請求項16】
目的とする分子が化学的または遺伝的にEDAドメインと結合された薬物であることを特徴とする、請求項6記載のタンパク質性ベクター。
【請求項17】
目的とする分子をTLR4発現細胞のサイトゾルに特異的に標的化および輸送するための、請求項6〜16いずれかに記載のタンパク質性ベクターの使用。
【請求項18】
TLR4発現細胞が抗原提示細胞であることを特徴とする、請求項17記載のタンパク質性ベクターの使用。
【請求項19】
抗原提示細胞が樹状細胞であることを特徴とする、請求項18記載のタンパク質性ベクターの使用。
【請求項20】
請求項6〜16のいずれかに規定されたタンパク質性ベクターをコードしていることを特徴とする、修飾された核酸。
【請求項21】
タンパク質性ベクターの発現を調節する作動可能に結合したコントロール配列も含むことを特徴とする、請求項20に記載の修飾された核酸。
【請求項22】
配列番号:1または配列番号:5を含むことを特徴とする、請求項20または21に記載の修飾された核酸。
【請求項23】
配列番号:9を含むことを特徴とする、請求項20または21に記載の修飾された核酸。
【請求項24】
発現ベクターが、請求項20〜23のいずれかに規定された修飾された核酸を含むことを特徴とする、請求項6〜16のいずれかに規定されたタンパク質性ベクターの遺伝子発現のための発現ベクター。
【請求項25】
ベクターがウイルスベクターであることを特徴とする、請求項24記載の発現ベクター。
【請求項26】
請求項20〜23のいずれかに規定された修飾された核酸、または請求項24または25に規定された発現ベクターを含むことを特徴とする、発現宿主細胞。
【請求項27】
宿主発現細胞が大腸菌であることを特徴とする、請求項26に記載の発現宿主細胞。
【請求項28】
タンパク質性ベクターの産生およびそのものの回収が可能となる条件下で、請求項26または27に規定された宿主発現細胞を培養することを含むことを特徴とする、請求項6〜16のいずれかに規定されたタンパク質性ベクターを産生するための方法。
【請求項29】
医薬組成物の調製における、請求項20〜23のいずれかに規定された修飾された核酸、請求項24または25に規定された発現ベクター、または請求項26または27に規定された宿主細胞の使用。
【請求項30】
免疫細胞応答促進剤が医薬組成物であることを特徴とする、請求項1〜5のいずれかに記載のポリペプチドの使用。
【請求項31】
医薬組成物が、抗原提示細胞の成熟を刺激することを特徴とする、請求項30に記載のポリペプチドの使用。
【請求項32】
医薬組成物が、目的とする分子に対して有効な免疫応答を誘導することを特徴とする、請求項30または31に記載のポリペプチドの使用。
【請求項33】
免疫応答がCTL応答であることを特徴とする、請求項32に記載のポリペプチドの使用。
【請求項34】
医薬組成物が感染性疾患の処置および予防に有用であることを特徴とする、請求項30〜33のいずれかに記載のポリペプチドの使用。
【請求項35】
医薬組成物がC型肝炎の処置および予防のための組成物であることを特徴とする、請求項34に記載のポリペプチドの使用。
【請求項36】
医薬組成物が腫瘍疾患の処置および予防のための組成物であることを特徴とする、請求項34に記載のポリペプチドの使用。
【請求項37】
医薬組成物がアレルギー性疾患の処置および予防のための組成物であることを特徴とする、請求項30〜33のいずれかに記載のポリペプチドの使用。
【請求項38】
少なくとも1つの許容し得る医薬担体、および少なくとも1つの下記成分:
i)請求項6〜16のいずれかに規定されたタンパク質性ベクター;
ii)請求項20〜23のいずれかに規定された修飾された核酸;
iii)請求項24または25に規定された修飾された核酸を含む発現ベクター;または
iv)請求項26または27に規定された該修飾された核酸も含む発現宿主細胞、
を含むことを特徴とする医薬組成物。
【請求項39】
ある量の樹状細胞を含むことを特徴とし、該樹状細胞がイン・ビトロで下記の少なくとも1つの成分;
i)請求項6〜16のいずれかに規定したタンパク質性ベクター;
ii)請求項20〜23のいずれかに規定した核酸;または
iii)請求項24または25に規定した該修飾された核酸を含む発現ベクター、
とインキュベートされている、医薬組成物。
【請求項40】
組成物がワクチンまたは免疫治療用の組成物であることを特徴とする、請求項38または39に記載の医薬組成物。
【請求項1】
抗原刺激剤に対する免疫細胞応答の生成において、
−フィブロネクチンEDAドメイン(EDA)、または
−TLR4を結合できる該EDAドメインのフラグメント、または
−TLR4を結合でき、該EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する該EDAドメインまたはフラグメントの改変型、
から選択されるアミノ酸配列を含むポリペプチドの使用。
【請求項2】
EDAドメインに対応するフラグメントが、
a)配列番号:2のアミノ酸2-91;
b)配列番号:4;および
c)TLR4発現細胞を結合できる配列a)およびb)のフラグメント、
から選択される配列を含むことを特徴とする、請求項1記載のポリペプチドの使用。
【請求項3】
EDAドメインに対応するフラグメントが、
a)配列番号:6のアミノ酸2-57;
b)配列番号:8;および
c)TLR4発現細胞を結合できる配列a)およびb)のフラグメント、
から選択される配列を含むことを特徴とする、請求項1記載のポリペプチドの使用。
【請求項4】
免疫賦活性剤が1以上の目的とする分子をさらに含むことを特徴とする、請求項1〜3のいずれかに記載のポリペプチドの使用。
【請求項5】
免疫賦活性剤がタンパク質性ベクターであり、ここで該ポリペプチドが目的とする該分子と結合されていることを特徴とする、請求項1〜4のいずれかに記載のポリペプチドの使用。
【請求項6】
アミノ酸配列が、下記のもの:
a)フィブロネクチンEDAドメイン(EDA)、
b)TLR4を結合できる該EDAドメインのフラグメント、または
c)TLR4を結合でき、該EDAドメインのあらゆる天然形態またはフラグメントと70%以上のホモロジーを有する該EDAドメインまたはそのフラグメントの改変型;
から選択されるポリペプチドを含むタンパク質性ベクターであって、ポリペプチドが次のもの;ポリペプチド、リポペプチド、オリゴサッカリド、ポリサッカリド、核酸、脂質および化学物質から選択される目的とする分子と結合することを特徴とする、タンパク質性ベクター。
【請求項7】
目的とする分子が抗原およびエピトープから選択されることを特徴とする、請求項6に記載のタンパク質性ベクター。
【請求項8】
目的とする分子がウイルス抗原、細菌抗原および寄生抗原から選択されることを特徴とする、請求項6または7のいずれかに記載のタンパク質性ベクター。
【請求項9】
目的とする分子がC型肝炎ウイルス由来のウイルス抗原であることを特徴とする、請求項6〜8のいずれかに記載のタンパク質性ベクター。
【請求項10】
C型肝炎ウイルス抗原がNS3タンパク質または該タンパク質の抗原性フラグメントであることを特徴とする、請求項9記載のタンパク質性ベクター。
【請求項11】
アミノ酸配列が配列番号:10であることを特徴とする、請求項10記載のタンパク質性ベクター。
【請求項12】
目的とする分子が腫瘍性抗原および腫瘍性抗原性決定基から選択されることを特徴とする、請求項6または7のいずれかに記載のタンパク質性ベクター。
【請求項13】
目的とする分子が、オボアルブミン(OVA 257-264)由来の抗原性細胞傷害性T決定基またはSIINFEKLであることを特徴とする、請求項12に記載のタンパク質性ベクター。
【請求項14】
アミノ酸配列が配列番号:2または配列番号:6から選択されることを特徴とする、請求項13記載のタンパク質性ベクター。
【請求項15】
目的とする分子がアレルゲンであることを特徴とする、請求項6記載のタンパク質性ベクター。
【請求項16】
目的とする分子が化学的または遺伝的にEDAドメインと結合された薬物であることを特徴とする、請求項6記載のタンパク質性ベクター。
【請求項17】
目的とする分子をTLR4発現細胞のサイトゾルに特異的に標的化および輸送するための、請求項6〜16いずれかに記載のタンパク質性ベクターの使用。
【請求項18】
TLR4発現細胞が抗原提示細胞であることを特徴とする、請求項17記載のタンパク質性ベクターの使用。
【請求項19】
抗原提示細胞が樹状細胞であることを特徴とする、請求項18記載のタンパク質性ベクターの使用。
【請求項20】
請求項6〜16のいずれかに規定されたタンパク質性ベクターをコードしていることを特徴とする、修飾された核酸。
【請求項21】
タンパク質性ベクターの発現を調節する作動可能に結合したコントロール配列も含むことを特徴とする、請求項20に記載の修飾された核酸。
【請求項22】
配列番号:1または配列番号:5を含むことを特徴とする、請求項20または21に記載の修飾された核酸。
【請求項23】
配列番号:9を含むことを特徴とする、請求項20または21に記載の修飾された核酸。
【請求項24】
発現ベクターが、請求項20〜23のいずれかに規定された修飾された核酸を含むことを特徴とする、請求項6〜16のいずれかに規定されたタンパク質性ベクターの遺伝子発現のための発現ベクター。
【請求項25】
ベクターがウイルスベクターであることを特徴とする、請求項24記載の発現ベクター。
【請求項26】
請求項20〜23のいずれかに規定された修飾された核酸、または請求項24または25に規定された発現ベクターを含むことを特徴とする、発現宿主細胞。
【請求項27】
宿主発現細胞が大腸菌であることを特徴とする、請求項26に記載の発現宿主細胞。
【請求項28】
タンパク質性ベクターの産生およびそのものの回収が可能となる条件下で、請求項26または27に規定された宿主発現細胞を培養することを含むことを特徴とする、請求項6〜16のいずれかに規定されたタンパク質性ベクターを産生するための方法。
【請求項29】
医薬組成物の調製における、請求項20〜23のいずれかに規定された修飾された核酸、請求項24または25に規定された発現ベクター、または請求項26または27に規定された宿主細胞の使用。
【請求項30】
免疫細胞応答促進剤が医薬組成物であることを特徴とする、請求項1〜5のいずれかに記載のポリペプチドの使用。
【請求項31】
医薬組成物が、抗原提示細胞の成熟を刺激することを特徴とする、請求項30に記載のポリペプチドの使用。
【請求項32】
医薬組成物が、目的とする分子に対して有効な免疫応答を誘導することを特徴とする、請求項30または31に記載のポリペプチドの使用。
【請求項33】
免疫応答がCTL応答であることを特徴とする、請求項32に記載のポリペプチドの使用。
【請求項34】
医薬組成物が感染性疾患の処置および予防に有用であることを特徴とする、請求項30〜33のいずれかに記載のポリペプチドの使用。
【請求項35】
医薬組成物がC型肝炎の処置および予防のための組成物であることを特徴とする、請求項34に記載のポリペプチドの使用。
【請求項36】
医薬組成物が腫瘍疾患の処置および予防のための組成物であることを特徴とする、請求項34に記載のポリペプチドの使用。
【請求項37】
医薬組成物がアレルギー性疾患の処置および予防のための組成物であることを特徴とする、請求項30〜33のいずれかに記載のポリペプチドの使用。
【請求項38】
少なくとも1つの許容し得る医薬担体、および少なくとも1つの下記成分:
i)請求項6〜16のいずれかに規定されたタンパク質性ベクター;
ii)請求項20〜23のいずれかに規定された修飾された核酸;
iii)請求項24または25に規定された修飾された核酸を含む発現ベクター;または
iv)請求項26または27に規定された該修飾された核酸も含む発現宿主細胞、
を含むことを特徴とする医薬組成物。
【請求項39】
ある量の樹状細胞を含むことを特徴とし、該樹状細胞がイン・ビトロで下記の少なくとも1つの成分;
i)請求項6〜16のいずれかに規定したタンパク質性ベクター;
ii)請求項20〜23のいずれかに規定した核酸;または
iii)請求項24または25に規定した該修飾された核酸を含む発現ベクター、
とインキュベートされている、医薬組成物。
【請求項40】
組成物がワクチンまたは免疫治療用の組成物であることを特徴とする、請求項38または39に記載の医薬組成物。
【図1】

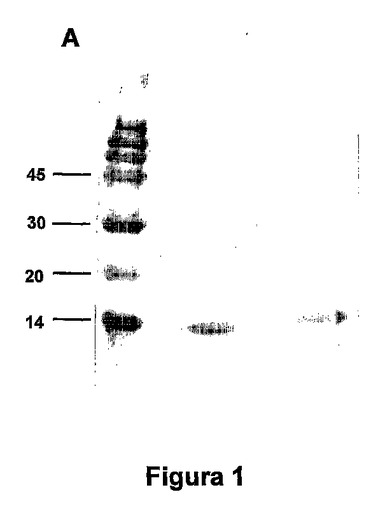
【図2】

【図3】

【図4】
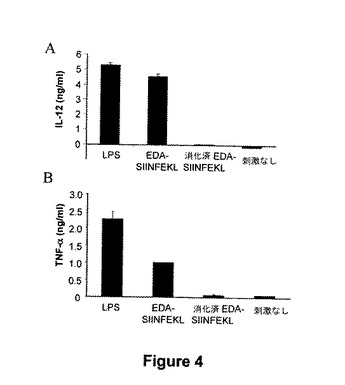
【図5−1】
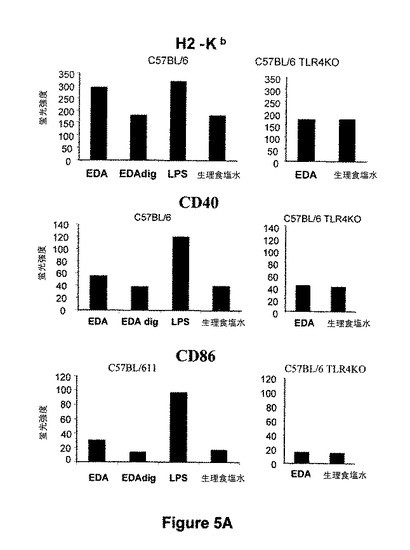
【図5−2】
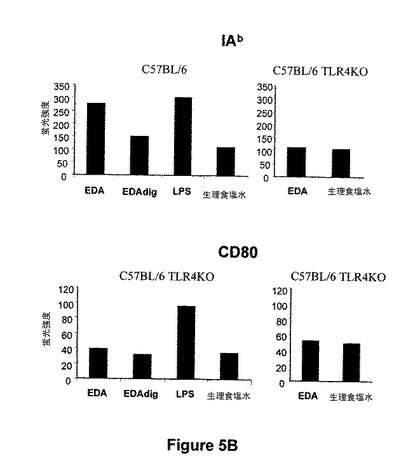
【図6】
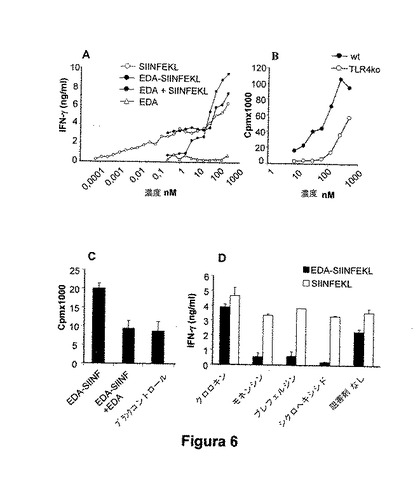
【図7】
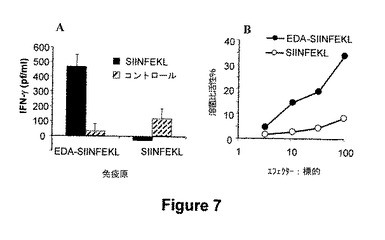
【図8】
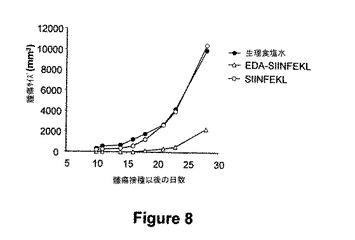
【図9】
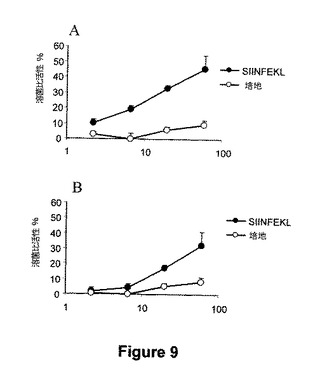
【図10】
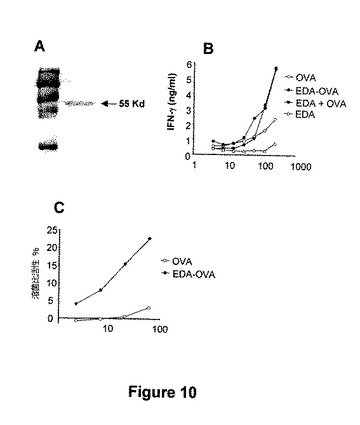
【図11】
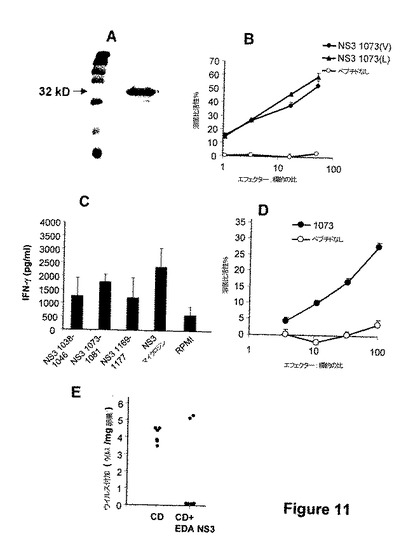
【図2】
【図3】
【図4】
【図5−1】
【図5−2】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2008−543301(P2008−543301A)
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願番号】特願2008−516349(P2008−516349)
【出願日】平成18年6月13日(2006.6.13)
【国際出願番号】PCT/ES2006/000343
【国際公開番号】WO2006/134190
【国際公開日】平成18年12月21日(2006.12.21)
【出願人】(506061716)プロイェクト、デ、ビオメディシナ、シーマ、ソシエダッド、リミターダ (34)
【氏名又は名称原語表記】PROYECTO DE BIOMEDICINA CIMA, S.L.
【Fターム(参考)】
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願日】平成18年6月13日(2006.6.13)
【国際出願番号】PCT/ES2006/000343
【国際公開番号】WO2006/134190
【国際公開日】平成18年12月21日(2006.12.21)
【出願人】(506061716)プロイェクト、デ、ビオメディシナ、シーマ、ソシエダッド、リミターダ (34)
【氏名又は名称原語表記】PROYECTO DE BIOMEDICINA CIMA, S.L.
【Fターム(参考)】
[ Back to top ]