
フルクトシルアミノ酸オキシダーゼ
プロトンリレー系に関与するアミノ酸残基で改変された変異体フルクトシルアミノ酸オキシダーゼが開示される。この変異体フルクトシルアミノ酸オキシダーゼは、オキシダーゼ活性が低下し、その一方そのデヒドロゲナーゼ活性は実質的に維持される。本発明はまた、糖化タンパク質を測定するためのアッセイ装置やアッセイ法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、糖化アルブミン、フルクトシルペプチド、HbA1cおよびフルクトシルバリン(FV)などの糖化タンパク質の測定用キットおよびセンサーに使用するためのフルクトシルアミノ酸オキシダーゼ(フルクトシルアミンオキシダーゼとも呼ばれる)に関する。より詳細には、本発明はオキシダーゼ活性の減少した変異体フルクトシルアミノ酸オキシダーゼに関する。
【背景技術】
【0002】
糖化タンパク質は、タンパク質上のアミノ基および糖の還元末端との間の共有結合を介して非酵素的に生成され、アマドリ化合物とも言われる。血中において、グルコースはヘモグロビンのβ−鎖のN−末端でバリンと結合し、糖化ヘモグロビン(グリコヘモグロビン;HbAlc)を生成する。ヘモグロビンに対するHbAlcの存在比は、正常な健常個体と比較して糖尿病患者においてより高く、HbAlcの血中濃度は、過去数週間の血糖値を反映することが知られている。したがって、HbAlcの血中濃度は、糖尿病の診断のための臨床試験において、また糖尿病患者の血糖コントロールにおいて非常に重要である。血中のHbAlc濃度は、フルクトシルバリンまたはフルクトシルバリルヒスチジンに特異性を有する酵素を用いて測定することが可能である。
【0003】
フルクトシルアミノ酸オキシダーゼは、様々な種類の株から単離されており、糖化アルブミン、HbAlcおよびフルクトシルアミノ酸などの糖化タンパク質がそのような酵素を用いて分析できることが示唆されている(特開昭61−268178号公報;特開昭61−280297号公報;特開平03−155780号公報;特開平05−192193号公報;特開平07−289253号公報;特開平08−154672号公報;特開2001−95598号公報;特開2003−79386号公報;特開2003−235585号公報;Agric. Biol. Chem., 53(1), 103-110, 1989;Agric. Biol. Chem., 55(2), 333-338, 1991;J. Biol. Chem., 269(44), 27297-27301, 1994;Appl. Environ. Microbiol., 61(12), 4487-4489, 1995;Biosci. Biotech. Biochem., 59(3), 487-491, 1995;J. Biol. Chem., 270(1), 218-224, 1995;J. Biol. Chem., 271(51), 32803-32809, 1996;およびJ. Biol. Chem., 272(6), 3437-3443, 1997)。
【0004】
フルクトシルアミノ酸オキシダーゼは、フルクトシルアミノ酸が酸化され、2−ケト−D−グルコースおよび対応するアミノ酸を生成する反応を触媒するFAD依存性酵素であり、FADの還元型(FADH2)の生成を伴う。FADH2は、次に電子アクセプターに電子を伝達し、その酸化型に変換される。酸素の存在下で、FADH2は人工電子アクセプター(アソメディエーター(asmediators)または電子メディエーターとも言う)に対してよりもむしろ酸素分子に優先的に電子を伝達する。したがって、フルクトシルアミノ酸を、メディエーターを用いてフルクトシルアミノ酸オキシダーゼによりアッセイする場合、アッセイの結果は、反応系における溶存酸素レベルに大きく影響を受ける。そのような不利益は、人工電子アクセプターを利用するポイント・オブ・ケア検査装置による血液試料の臨床検査において特に指摘される。人工電子メディエーターを用いる酵素センサーストリップに使用される酵素は、酸素に対して低い活性を有することが望まれる。
【0005】
したがって、本発明の目的は、酵素、特にその活性が溶存酸素レベルによりほとんど影響を受けないフルクトシルアミノ酸オキシダーゼを提供することである。
【発明の概要】
【0006】
一般的に言えば、本願の発明者らは、酵素、より詳細にはその活性が溶存酸素レベルによりほとんど影響を受けないフルクトシルアミノ酸オキシダーゼを提供することができる。より具体的には、これは、その野生型において優先的にオキシダーゼ活性を示す酵素のオキシダーゼ活性を減少させることにより、好ましくは同時に酵素のデヒドロゲナーゼ活性を増加させることにより達成される。以下により詳細に説明するように、これは野生型酵素を変異させることにより達成されている。
【0007】
本発明者らは、フルクトシルアミノ酸オキシダーゼの種々の変異体を製造し、驚くべきことに、特定の種類の変異体が、デヒドロゲナーゼ活性、詳細には、色素媒介(dye-mediated)デヒドロゲナーゼ活性を実質的に保持しながら減少したオキシダーゼ活性を示すことを見出した。
【0008】
本発明は、配列番号1記載のアミノ酸配列の56位に対応する位置で、アミノ酸残基Asnを、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼを提供する。
【0009】
好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有し、好ましくは、野生型フルクトシルアミノ酸オキシダーゼと比較して増加されたデヒドロゲナーゼ活性を有する。好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの30%以下のオキシダーゼ活性を有し、そしてまた、好ましくは野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する。
【0010】
別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1〜16からなる群より選択されるアミノ酸配列であって、配列番号1記載のアミノ酸配列の56位に対応する位置のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基により置換されているアミノ酸配列を有する。好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1記載のアミノ酸配列であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。より好ましくは、配列番号1の位置109−124のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられている。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号2記載のアミノ酸配列であって、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号3記載のアミノ酸配列であって、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号7記載のアミノ酸配列であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。より好ましくは、配列番号7の位置61−72のアミノ酸配列Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられている。
【0011】
別の側面では、本発明は、本発明の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチドを提供する。
【0012】
また別の側面では、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。
【0013】
さらに別の側面では、本発明は、本発明のベクターで形質転換された宿主細胞を提供する。
【0014】
別の側面において、本発明は、試料中の糖化タンパク質をアッセイするための方法であって、試料を本発明のフルクトシルアミノ酸オキシダーゼと接触させること、およびフルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む方法を提供する。
【0015】
なお別の側面において、本発明は、HbA1cをアッセイするための方法であって、試料中のHbA1cを消化してフルクトシルバリンまたはフルクトシルバリルヒスチジンを生成すること、該フルクトシルバリンまたはフルクトシルバリルヒスチジンを本発明のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンまたはフルクトシルバリルヒスチジンの量を測定することを含む方法を提供する。
【0016】
別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための装置であって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置を提供する。
【0017】
また別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキットを提供する。
【0018】
別の側面において、本発明は、電極に固定化されている本発明のフルクトシルアミノ酸オキシダーゼを有する酵素電極を提供する。
【0019】
別の側面において、本発明は、作用電極として本発明の酵素電極を含むフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーを提供する。
【0020】
また別の側面において、本発明は、配列番号1記載のアミノ酸配列であって、配列番号1の61−72位のアミノ酸配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaが、配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。なお別の側面において、本発明は、配列番号1記載のアミノ酸配列であって、配列番号1の109−124位のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。また別の側面において、本発明は、配列番号7記載のアミノ酸配列であって、配列番号7の61−72位のアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。また別の側面において、本発明は、配列番号7記載のアミノ酸配列であって、配列番号1の109−126位のアミノ酸配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaが、配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。
【発明の効果】
【0021】
本発明の変異されたフルクトシルアミノ酸オキシダーゼは、糖尿病状態の診断やコントロールにおいて臨床的に有用なヘモグロビン(HbA1c)などの糖化タンパク質の測定に使用することが可能である。
【図面の簡単な説明】
【0022】
【図1】いくつかの真核性FAODのN−末端付近のアミノ酸配列のアラインメントを示す。
【図2】フルクトシルアミノ酸オキシダーゼの反応系を示す。
【図3】精製されたN1−1 FAOD N47A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図4】粗PnFPOX野生型およびAsn56変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図5】PnFPOX Asn56変異体の速度論的パラメーターを示す。
【図6】精製されたPnFPOX N56A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図7】精製されたアマドリアーゼII野生型およびN52A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図8】精製されたアマドリアーゼII野生型およびN52A変異体の速度論的パラメーターを示す。
【図9】PnFPOX N56Aまたは野生型FAODを有する電極を用いるFV濃度のアッセイを示す。
【発明を実施するための形態】
【0023】
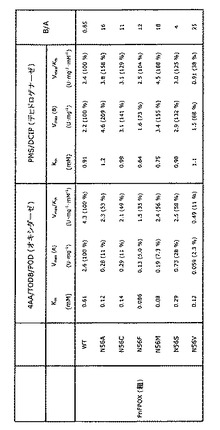
フルクトシルアミノ酸オキシダーゼ(FAOD)は、様々な真核性および原核性株に存在するということが知られている。表1は、今までに単離された、または予測されたFAODのいくつかの例を示す。
【0024】
【表1】
【0025】
真核性FAODは、基質特異性により、(i)α−フルクトシルアミノ酸優先(FAOD−P、FAOD−U、FAOX−E、FPOX−C、およびPnFPOX)、(ii)ε−フルクトシルアミノ酸優先(FAOD−F、FAOD−A、アマドリアーゼI、およびFAOD−Ao1)、および(iii)α−およびε−糖化アミノ酸両方に類似の活性(アマドリアーゼII、FAOD−Ao2、およびN1−1 FAOD)に分類することができる。一般に、α−フルクトシルアミノ酸に活性なFAODは、α−フルクトシルバリンおよびHbA1cのアッセイにおいて有用となり、ε−フルクトシルアミノ酸に活性なFAODは、ε−フルクトシルリジンおよび糖化アルブミンのアッセイにおいて有用となる。当業者は、酵素に変異を導入することにより、α−またはε−フルクトシルアミノ酸に対するその優先性を変えることができるということを理解するものである。
【0026】
本発明は、配列番号1記載のアミノ酸配列(PnFPOX)の56位に対応する位置で、アミノ酸残基AsnをAla、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼを提供する。
【0027】
本明細書において使用される用語、タンパク質の「変異体」は、表示される位置でタンパク質上の1つまたは複数のアミノ酸残基に置換を含む変異型タンパク質を意味する。用語変異体はまた、そのような変異体タンパク質をコードするポリヌクレオチドにも使用される。
【0028】
本明細書において使用される語句「対応する位置」は、デフォルトパラメーターを有するVector NTIのソフトウェアAlignX(インビトロジェン製;Lu, G., and Moriyama, E.N. (2004) Vector NTI, a balanced all-in-one sequence analysis suite. Brief Bioinform 5, 378-88参照)を用いて、参照アミノ酸配列のアミノ酸残基と位置合わせされる照会(query)アミノ酸配列におけるアミノ酸残基の位置を意味する。したがって、「配列番号1記載のアミノ酸配列の56位に対応する位置のAsn残基」は、照会アミノ酸配列が、デフォルトパラメーターを有するVector NTIのAlignXを用いて配列番号1とアラインメントされる場合に、配列番号1のAsn56と位置合わせされる照会アミノ酸配列におけるAsn残基を意味する。配列番号1自体のAsn47もこの用語により包含されることに留意すべきである。
【0029】
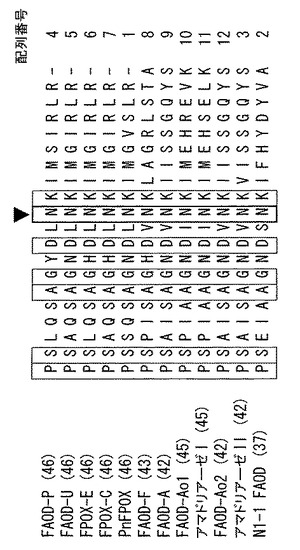
図1は、いくつかの真核性FAOD(ペニシリウム・ヤンシネルム(Penicillum janthinelum)AKU3413由来のFAOD−P、ウロクラディウム・エスピー(Ulocladium sp.)JS−103由来のFAOD−U、ユーペニシリウム・テレヌム(Eupenicillum terrenum)ATCC 18547由来のFPOX−E、コニオカエタ・エスピー(Coniochaeta sp.)NISL 9330由来のFPOX−C、ファエオスフェリア・ノドラム(Phaeosphaeria nodorum)由来のPnFPOX、フザリウム・オキシスポラム(Fusarium oxysporum)NBRC 9972由来のFAOD−F、アスペルギルス・テレウス(Aspergillus terreus)GP1由来のFAOD−A、アスペルギルス・フミガーツス(Aspergillus fumigatus)由来のアマドリアーゼI、アスペルギルス・オリゼー(Aspergillus oryzae)由来のFAOD−Ao1、アスペルギルス・フミガーツス(Aspergillus fumigatus)由来のアマドリアーゼII、アスペルギルス・オリゼー(Aspergillus oryzae)由来のFAOD−Ao2およびピチア・エスピー(Pichia sp.)N1−1由来のN1−1 FAOD)のN−末端付近の保存された領域内のアミノ酸配列のアラインメントを示す。これらのFAODの全配列は、配列番号1〜12に記載されている。アラインメントは、Vector NTI suite6.0のAlignXアプリケーションを用いて作製された。矢印はN1−1 FAODのAsn47に対応する残基を示す。括弧中の数字は、配列番号により表される各アミノ酸配列における、図1の一番左の「P」の位置を示す。当業者は、Blastなどの別のアラインメントソフトウェアプログラムが、同じまたは実質的に同じアラインメントを提供するであろう事を理解するものである。
【0030】
図1から、N1−1 FAODのAsn47を含む領域におけるアミノ酸配列が、真核性FAODのあいだで高度に保存されているということがわかる。したがって、当業者は、配列アラインメント用の任意の市販のソフトウェアプログラムを用いて保存された領域内のN1−1 FAODのAsn47に対応するAsn残基を容易に同定することができ、また、変異体フルクトシルアミノ酸オキシダーゼが、そのAsn残基上に修飾を導入することにより容易に製造されることを理解することができる。
【0031】
別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1〜15からなる群より選択されるアミノ酸配列であって、配列番号1記載のアミノ酸配列の56位に対応する位置でアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。
【0032】
好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1記載のアミノ酸配列(PnFPOX)であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。好ましくは、Asn残基はAlaで置換される。別の好ましい実施態様においては、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号2記載のアミノ酸配列(N1−1)であって、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。なお別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号3記載のアミノ酸配列(アマドリアーゼII)であって、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。
【0033】
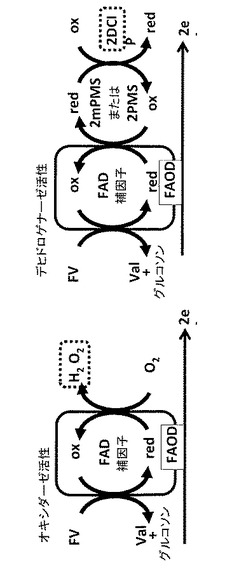
本発明の変異体フルクトシルアミノ酸オキシダーゼは、デヒドロゲナーゼ活性を実質的に保持しながら減少されたオキシダーゼ活性を示す。図2は、フルクトシルアミノ酸オキシダーゼの反応スキームを説明する。
【0034】
本明細書において、「オキシダーゼ活性」は、酸素を電子アクセプターとして利用しながらフルクトシルアミノ酸の酸化を触媒し、2−ケト−D−グルコースと対応するアミノ酸を生成する、フルクトシルアミノ酸オキシダーゼの酵素活性(Vmax)である。オキシダーゼ活性は、本技術分野において既知の任意の方法、例えば、4AA/TODB/POD(4−アミノアンチピリン/N,N−ビス(4−スルホブチル)−3−メチルアニリン二ナトリウム塩/西洋ワサビペルオキシダーゼ)などのH2O2検出用試薬やPt電極により、生成したH2O2の量を測定することによりアッセイすることができる。本明細書において、相対的または定量的活性という文脈では、オキシダーゼ活性は、具体的には、25℃で、10mM PPB、pH7.0、1.5mM TODB、2U/ml西洋ワサビペルオキシダーゼ(POD)、および1.5mM 4−アミノアンチピリン(4AA)において生成されたH2O2の量により測定される、単位時間当たり酸化された基質(フルクトシルアミノ酸)のモル量と定義される。キノンイミン色素の生成は、546nmで分光光度法により測定することができる。
【0035】
本明細書において、「デヒドロゲナーゼ活性」は、酸素以外の電子メディエーターを電子アクセプターとして利用しながらフルクトシルアミノ酸の酸化を触媒し、2−ケト−D−グルコースと対応するアミノ酸を生成する、フルクトシルアミノ酸オキシダーゼの酵素活性(Vmax)である。デヒドロゲナーゼ活性は、たとえば、mPMS/DCIP(メチル硫酸1−メトキシ−5−メチルフェナジニウム/2,6−ジクロロインドフェノール)、cPES(トリフルオロ−酢酸−1−(3−カルボキシ−プロポキシ)−5−エチル−フェナジニウム、NA BM31_1144(N,N−ビス−(ヒドロキシエチル)3−メトキシ−ニトロソアニリン塩酸塩、NA BM31_1008(N,N−ビス−ヒドロキシエチル−4−ニトロソアニリンおよびN−N−4−ジメチル−ニトロソアニリン)などを用いてメディエーターに移動した電子の量を測定することによりアッセイすることができる。
【0036】
本明細書において、相対的または定量的活性という文脈では、デヒドロゲナーゼ活性は、具体的には、25℃で、10mM PPB(pH7.0)、0.6mM DCIP、および6mM メトキシPMS(mPMS)においてメディエーターに移動された電子の量により測定される、単位時間当たり酸化された基質(フルクトシルアミノ酸)のモル量と定義される。
【0037】
本発明の変異体フルクトシルアミノ酸オキシダーゼは、デヒドロゲナーゼ活性を実質的に保持しながら、野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有する。
【0038】
好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型のフルクトシルアミノ酸オキシダーゼの50%以下のオキシダーゼ活性を有する。より好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの40%以下、より好ましくは30%以下、さらにより好ましくは20%以下、最も好ましくは15%以下のオキシダーゼ活性を有する。また、好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する。より好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの70%以上、より好ましくは90%以上、さらにより好ましくは100%以上、最も好ましくは100%より高いデヒドロゲナーゼ活性を有する。
【0039】
野生型フルクトシルアミノ酸オキシダーゼにおいて、オキシダーゼ活性とデヒドロゲナーゼ活性は、比率、デヒドロゲナーゼ活性/オキシダーゼ活性が、約0.8〜1.2の間と同程度にある。アッセイ系に溶存酸素が存在する場合、基質の酸化により生じた電子は、酸素に優先的に移動することになる。したがって、電子メディエーターの存在下で測定される酵素活性は、溶存酸素濃度によって大きな影響を受けることになる。対照的に、本発明の変異体フルクトシルアミノ酸オキシダーゼは、約2.0以上、好ましくは4.0以上、より好ましくは10以上、さらにより好ましくは20以上、最も好ましくは40以上の比率、デヒドロゲナーゼ活性/オキシダーゼ活性を有する。デヒドロゲナーゼ活性がオキシダーゼ活性を上回るので、本発明のフルクトシルアミノ酸オキシダーゼの酵素活性は、溶存酸素濃度の影響をほとんど受けないものとなる。これは、血液試料による臨床診断におけるフルクトシルアミノ酸オキシダーゼの利用において好都合なものである。
【0040】
別の側面において、本発明は、1つまたは複数のループ領域が改変または変異されているフルクトシルアミノ酸オキシダーゼを提供する。PnFPOXは、フルクトシルヘキサペプチドに対してFVHと比較して1%未満の活性レベルのオキシダーゼ活性を示す。一方、FPOX−Cは、活性を示さない。他方、FPOX−Cは、P.nFPOXよりもFVHに対してより高い活性を示す。両オキシダーゼのアミノ酸配列の比較から、FAODとFPOXとのあいだの有意差は、それらのループ領域に見られる。
【0041】
ループ1領域
PnFPOX (61)Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Ala(72)(配列番号:13)
FPOX−C (61)Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Ser(72)(配列番号:14)
【0042】
ループ2領域
PnFPOX (109)Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Glu(124)(配列番号:15)
FPOX−C (109)Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Ala(126)(配列番号:16)
【0043】
ループ1領域(12aa)では、5つのアミノ酸残基がPnFPOXとFPOX−Cとで異なる。ループ2領域(16aa)では、2つのアミノ酸挿入と7つのアミノ酸置換が見られる。
【0044】
驚くべきことに、PnFPOXとFPOX−Cとのループ領域が互に交換されているいくつかのキメラループ変異体が改善された活性を示すことが見出された。下記の実施例8に示されているように、ループ2領域がFPOX−Cループ2領域により置き換えられているPnFPOXは、野生型PnFPOXよりもFV、FVHおよびフルクトシルヘキサペプチド(F−HP)に対してより高いオキシダーゼ活性を示す。ループ1領域がPnFPOXループ1領域により置き換えられているFPOX−Cは、野生型FPOX−Cにおいては検出されない、F−HPに対してオキシダーゼ活性を示す。加えて、N56A変異を有するキメラループ変異体も、PnFPOXおよびFPOX−C由来のN56A変異体において観察されるように、減少したオキシダーゼ活性と増加したデヒドロゲナーゼ活性を示す。特に好ましいキメラループ変異体は、ループ2領域がFPOX−Cのループ2領域で置換されており、かつAsn56がAlaで置換されているPnFPOXである。別の特に好ましいキメラループ変異体は、ループ1領域がPnFPOXのループ1領域で置換されており、かつAsn56がAlaで置換されているFPOX−Cである。これらの結果は、PnFPOX由来のループ1領域とFPOX−C由来のループ2領域の組合せが、フルクトシルバリンおよびフルクトシルバリルヒスチジンならびにフルクトシルヘキサペプチドに対してより高い活性を提供するということを示唆する。
【0045】
別の側面において、本発明は、本発明の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチドを提供する。フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドのヌクレオチド配列は、公共のデータベースから容易に得ることができる。野生型フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドは、PCRまたは他の既知の技術を用いて各生物のゲノムからクローニングすることができる。変異は、部位特異的突然変異誘発法、PCR突然変異誘発法または本技術分野において周知の任意の他の技術により導入することができる。変異されるべきAsn残基は、本技術分野において利用可能な配列アラインメントのための任意のソフトウェアを用いて同定することができる。あるいは、変異体フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドは、一連の化学的に合成されたオリゴヌクレオチドを用いてPCRにより製造してもよいし、または完全に合成してもよい。
【0046】
変異されたフルクトシルアミノ酸オキシダーゼは、変異体遺伝子を適切な発現ベクターに挿入し、そのベクターを大腸菌(E. coli)細胞などの適切な宿主細胞に導入することにより製造することができる。形質転換体を培養し、その形質転換体において発現されたフルクトシルアミノ酸オキシダーゼを、細胞または培養培地から回収することができる。
【0047】
かくして得られた組換えフルクトシルアミノ酸オキシダーゼは、本技術分野において公知の任意の精製技術、たとえばイオン交換カラムクロマトグラフィー、アフィニティークロマトグラフィー、液体クロマトグラフィー、ろ過、限外ろ過、塩析沈殿、溶媒沈殿、免疫沈降、ゲル電気泳動、等電電気泳動および透析などにより精製することができる。
【0048】
したがって、本発明は、変異体フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドを含むベクター、そのようなベクターで形質転換された宿主細胞、ならびに該形質転換体を培養し、変異体フルクトシルアミノ酸オキシダーゼを培養物から回収および精製することにより、本発明の変異体フルクトシルアミノ酸オキシダーゼを製造する方法も包含する。
【0049】
本発明はまた、試料中の糖化タンパク質をアッセイするための方法も包含する。その方法は、試料を本発明のフルクトシルアミノ酸オキシダーゼと接触させること、およびフルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む。本発明によりアッセイされる糖化タンパク質は、たとえば、フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1c、フルクトシルヘキサペプチド、糖化アルブミンおよび他のフルクトシルアミノ酸である。なお別の側面において、本発明は、試料中のHbA1cを消化してフルクトシルバリンを生成すること、そのフルクトシルバリンを本発明のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンの量を測定することを含むHbA1cをアッセイするための方法を提供する。
【0050】
別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1c、フルクトシルヘキサペプチドまたは糖化アルブミンをアッセイするための装置であって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置を提供する。
【0051】
アッセイ装置は、任意の従来の市販されている血糖値をモニターするための電流測定バイオセンサーのテストストリップと同様の構造を有することができる。そのような装置の1つの例は、絶縁基材上に配置された2つの電極(作用電極と参照または対電極)、試薬ポートおよび試料受け(receiver)を有する。試薬ポートは、本発明の変異されたフルクトシルアミノ酸オキシダーゼおよびメディエーターを含む。血液試料などの試料が試料受けに添加されると、試料に含まれるフルクトシルアミノ酸がフルクトシルアミノ酸オキシダーゼと反応し、電流が生じ、試料中のフルクトシルアミノ酸の量を示す。酵素基質の測定に適した電気化学センサーの典型的な例は、たとえば国際公開第2004/113900号および米国特許第5,997,817号明細書から公知である。電気化学センサーの代わりとして、光学的検出技術も使用可能なはずである。典型的には、そのような光学装置は、酵素、電子移動メディエーターおよび指示薬を含む試薬系において起こる色の変化にもとづくものである。色の変化は、蛍光、吸光、緩和(remission)測定を用いて定量することができる。酵素基質の測定に適した光学装置の典型的な例が、たとえば米国特許第7,008,799号明細書、米国特許第6,036,919号明細書および米国特許第5,334,508号明細書から公知である。
【0052】
なお別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキットを提供する。
【0053】
フルクトシルバリンまたはフルクトシルバリルヒスチジンの測定のためのキットは、本発明の酵素を用いて構築することが可能である。本発明のフルクトシルアミノ酸オキシダーゼに加えて、キットは、測定に必要な緩衝液、適切なメディエーター、および必要ならばペルオキシダーゼなどの酵素、較正曲線の作製のためのフルクトシルバリンまたはフルクトシルバリルヒスチジンまたはそれらの誘導体の標準溶液、および使用説明書を含有する。本発明のフルクトシルアミノ酸オキシダーゼは、多様な形態、たとえば凍結乾燥試薬として、または適切なストレージソリューションの溶液として提供してもよい。
【0054】
本発明の酵素を用いて、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドアッセイキットを構築することも可能である。糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドは、酵素的にまたは化学的に消化され、フルクトシルバリン、フルクトシルバリルヒスチジンおよびフルクトシルヘキサペプチドといったフルクトシルアミノ酸またはフルクトシルペプチドを生じ、それが今度は本発明のフルクトシルアミノ酸オキシダーゼを用いて定量される。このようにして、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドがアッセイ可能である。したがって、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドのための本発明のアッセイキットは、フルクトシルバリンまたはフルクトシルバリルヒスチジンの測定のための前記キットにさらに加水分解するための試薬またはプロテアーゼを含有することができる。
【0055】
別の側面において、本発明は、電極上に固定化された本発明のフルクトシルアミノ酸オキシダーゼを有する酵素電極を提供する。
【0056】
別の側面において、本発明は、作用電極として本発明の酵素電極を含む、フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーを提供する。
【0057】
試料中のフルクトシルアミノ酸の濃度は、酵素反応により生じた電子の量を測定することにより決定することができる。本技術分野においては、炭素電極、金属電極、および白金電極などの様々なセンサー系が知られている。本発明の変異されたフルクトシルアミノ酸オキシダーゼは、電極上に固定される。固定化のための手段の例としては、架橋、高分子マトリックスへの封入、透析膜による被覆、光架橋性ポリマー、電気伝導性ポリマー、酸化−還元ポリマー、およびそれらの任意の組合せなどが挙げられる。
【0058】
測定が電流測定系において行なわれる場合、固定化された酵素を備えた炭素電極、金電極または白金電極が作用電極として、対電極(白金電極など)および参照電極(Ag/AgCl電極など)と共に使用される。電極は、メディエーターを含有する緩衝液中に挿入され、所定の温度に保たれる。所定の電圧を作用電極に適用し、その後試料を添加、そして電流の増加した値を測定する。アッセイに使用されるメディエーターの例としては、フェリシアン化カリウム、フェロセン、オスミウム誘導体、ルテニウム誘導体、フェナジンメトサルフェートなどが挙げられる。通常、1つの作用電極と1つの対電極または偽−参照電極とのいわゆる二電極系も使用することが可能である。
【0059】
さらに、フルクトシルアミノ酸は、炭素電極、金電極、または白金電極を用いる電流測定系において、固定化された電子メディエーターを用いてアッセイすることもできる。酵素は、作用電極を作製するために吸着または共有結合により高分子マトリックスにおいてフェリシアン化カリウム、フェロセン、オスミウム誘導体、フェナジンメトサルフェートなどの電子メディエーターと一緒に電極上に固定化される。この作用電極が対電極(白金電極など)および参照電極(Ag/AgCl電極など)と一緒に緩衝液中に挿入され、所定の温度に保たれる。所定の電圧を作用電極に適用し、その後試料を添加、そして電流の増加した値を測定する。
【0060】
糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドの測定のためのセンサーを作製するために、フルクトシルアミノ酸の測定用の前記センサーは、複合センサーを構築するためにさらに固定化されたプロテイナーゼ(プロテアーゼなど)を含む膜と組み合わせられる。複数の酵素の組合せによる連続反応にもとづくそのような複合センサーの構造は、本技術分野において周知である。たとえば、Anthony P. F. Tuner, Isao Karube and George S. Wilsonによる「バイオセンサー−基礎と応用」(“Biosensor-Fundamental and Applications”)、オクスフォード大学出版、1987年を参照。
【0061】
本明細書において引用した全ての特許文献および参考文献の内容は、参照により本明細書中に完全に組み込まれる。
【実施例】
【0062】
本発明は、以下の実施例により詳細に説明されるが、本発明はそれらの実施例に限定されるべきものではない。
【0063】
実施例1
N1−1 FAOD変異体の製造および特徴付
【0064】
単量体サルコシンオキシダーゼ(MSOX)の構造機能相関の研究により、FADから酸素への電子移動に関与するプロトンリレー系(PRS)が、Thr48およびLys265と4つのH2O分子とを含んでなるということを明らかにした(Trickey et al, Structure, 7, 331-345, 1999)。ピチア・エスピー(Pichia sp.)N1−1株由来のフルクトシルアミノ酸オキシダーゼ(N1−1 FAOD、配列番号:2)の活性部位は、MSOXと、特にプロトンリレー系を担うと予測される保存された残基において高い相同性を有する。
【0065】
N1−1 FAODの予測構造モデルを、MSOX構造を用いて構築し、アミノ酸残基Asn44、Ser46、Asn47、Lys48およびLys269がFADからのプロトンと電子の移動に関与していると予測した。単一または二重突然変異をN1−1 FAODの電子アクセプターアベイラビリティーを改変する目的でそれらのアミノ酸残基に導入した。
【0066】
N1−1 FAOD単一変異体の発現ベクターを、N1−1 FAOD野生型遺伝子を鋳型として、QuickChange(登録商標)法(ストラタジーン(stratagene))を用いて部位特異的突然変異誘発法により作製した。Asn44、Ser46、Asn47、Lys48またはLys269で変異を導入するためにプライマーを設計した。増幅産物は、DpnIにより消化し、大腸菌(E. coli)DH5αに形質転換し、LB寒天培地(50μg/ml カナマイシン)において37℃で一晩インキュベートした。クローンの変異された配列を、ABI Prism 3130 Genetic Analyzerにおいて、ABI Prism BigDye Terminator cycle sequencing kit v3.0を用いて確認した。次に、抽出したプラスミドを、NcoIおよびSalIで消化し、NcoIおよびSalIで消化したpET28(a)ベクターにライゲートした。ライゲーション混合物で形質転換した大腸菌(E. Coli)BL21(DE3)細胞を、LB寒天培地(50μg/ml カナマイシン)において37℃で一晩インキュベートした。二重変異体は、同じ方法を用いてプライマーの組合せにより作製した。
【0067】
N1−1 FAOD変異体の発現ベクターをもつ大腸菌(Escherichia coli)BL21(DE3)細胞を、50μg/mlのカナマイシンを含有するLB培地150mlにおいて37℃で培養した。A660値が0.6に達した後、0.3mM IPTGを添加し、25℃でさらに5時間インキュベートした。細胞を遠心分離により回収し、0.85% NaCl水溶液で二回洗浄した。その後、細胞を10mMのリン酸カリウム緩衝液(PPB)pH7.0に再懸濁し、超音波ホモジナイザーにより溶菌した。ライセートを10,000×g、4℃で10分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離し、その後上清を10mM PPB(pH7.0)に対して透析した。オキシダーゼ活性は、25℃、10mM PPB、pH7.0中で、基質(0.2、0.5、0.75、1、2、5mM f−αValまたは5mM f−αVal−His)の存在下で、1.5mM TODB、2U/ml西洋ワサビペルオキシダーゼ(POD)、および1.5mM 4−アミノアンチピリン(4AA)でアッセイした。キノンイミン色素の生成を分光光度計により546nmで測定した。デヒドロゲナーゼ活性は、25℃、10mM PPB(pH7.0)において、基質(0.2、0.5、1、2、5mM f−αValまたは5mM f−αVal−His)の存在下で、0.6mM DCIPおよび6mM メトキシPMS(mPMS)でアッセイした。
【0068】
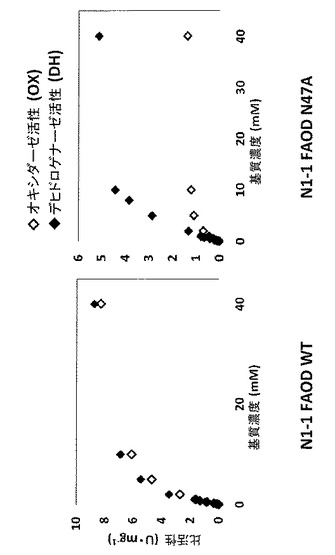
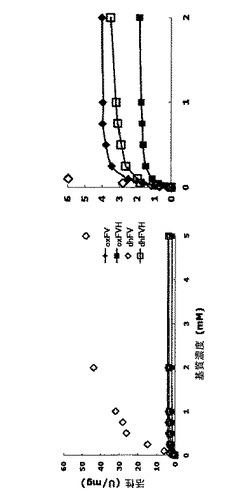
ほとんどのケースで、オキシダーゼ活性は完全にまたは実質的に消失したが、N47A、S46A、K48AおよびN44Aなどのいくつかの変異体は、相対的に高い活性を示した。なかでもN47Aは、最も目立った特徴を示した。N47Aのオキシダーゼ活性は野生型の活性の約20%に減少したが、N47Aのデヒドロゲナーゼ活性は野生型の活性の60%よりも高いままであった。結果として、N47Aは、オキシダーゼ活性よりも約4倍高いデヒドロゲナーゼ活性を示した。N47Aのこの特徴は、人工電子アクセプターと共にフルクトシルアミノ酸オキシダーゼを用いるセンサーストリップに有益なはずである。野生型N1−1 FAODおよびN47A変異体の粗調製物の速度論的パラメーターを表2に示す。
【0069】
【表2】
【0070】
実施例2
N1−1野生型およびN47A変異体FAODの精製
【0071】
組換えFAODは、次のように精製した。最初に、酵素を含有する水溶性画分を組換え大腸菌(Escherichia coli.)から調製した。発現ベクターを含む大腸菌を7LのLB培地(37℃、10L醗酵槽で、50μg/ml アンピシリン)において培養し、その後発現をIPTG(最終濃度:0.3mM)により約OD660=0.7で誘導し、培養温度を30℃まで下げた。細胞を100mM PPb(pH7.0)中に懸濁し、フレンチプレスを用いて4回破壊した。上清液を超遠心分離(40,000g、90分)に付し、上清を10mMのPPb(pH7.0)に対して4℃で一晩透析し水溶性画分を調製した。
【0072】
この水溶性画分を、さらに液体クロマトグラフィーに付し、精製酵素を調製した。酵素をさらに陰イオン交換クロマトグラフィー(DEAE−5PW)で精製した。水溶性画分を、10mM PPb(pH7.0)で平衡化した陰イオン交換クロマトグラフィーカラムDEAE−5PW(5.0mmI.D.×5cm、東ソー(Tosoh))に吸着させた。カラム体積の3倍量の10mM PPb(pH7.0)による平衡化の後、FAODを、0.7MのNaClを含有する10mM PPb(pH7.0)で溶出した。流速は1ml/分に設定し、溶出液を1分毎に回収した。280nmの吸収波長を溶出液のモニターに使用した。
【0073】
活性画分を、35%硫酸アンモニウムを用いて分離し、上清液を疎水性クロマトグラフィーに付した。活性画分を、35%硫酸アンモニウムを含有する10mM PPb(pH6.5)で平衡化した疎水性クロマトグラフィーカラム、Resource Phe(1ml、ファルマシア(Pharmacia))に吸着させた。カラム体積の3倍量の、35%硫酸アンモニウムを含有する10mM PPb(pH6.5)による平衡化の後、FAODを、10mM PPb(pH6.5)で溶出した。流速は2ml/分に設定し、溶出液を1分毎に回収した。活性画分を、45%硫酸アンモニウムで分離し、その後沈殿を、1%マンノースおよび100μM FADを含有する10mM PPb(pH7.0)に溶解し、同緩衝液に対して4℃で6時間透析した。それをさらに、100μM FADを含有する10mM PPb(pH8.0)に対して4℃で6時間透析した。透析した試料を次の陰イオン交換クロマトグラフィーに用いた。
【0074】
その試料を、10mM PPb(pH8.0)で平衡化した陰イオン交換クロマトグラフィーカラム、Bioasit Q(4.6mmI.D.×5cm、東ソー)に吸着させた。カラム体積の3倍量の10mM PPb(pH8.0)による平衡化の後、FAODを、0.3MのNaClを含有する10mM PPb(pH7.0)で溶出した。流速は1ml/分に設定し、溶出液を1分毎に回収した。活性画分を10mM PPb(pH7.0)に対して4℃で一晩透析した。試料の精製度をSDS/PAGEにより調べた。試料をPhast Gel 8−25を用いる電気泳動に付し、ゲルを銀で染色した。試料調製、電気泳動および染色は、Phast System(登録商標)に添付のマニュアルにしたがって行なった。
【0075】
精製した酵素のオキシダーゼおよびデヒドロゲナーゼ活性は、実施例1のように測定し、図3に示した。速度論的パラメーターは、表3に示す。
【0076】
【表3】
【0077】
実施例3
PnFPOX変異体の製造
【0078】
ファエオスフェリア・ノドラム(Phaeosphaeria nodorum)由来のフルクトシルアミノ酸オキシダーゼ(PnFPOX、配列番号:1)由来の変異体を製造し、特徴付けした。
【0079】
N1−1 FAODとPnFPOXとのアラインメント(図1)および実施例1および2の結果にもとづき、Asn56がFADからのプロトンおよび電子の移動に関与すると予測した。PnFAODの電子アクセプターのアベイラビリティーを改変することを目的にAsn56に種々の変異が導入された。
【0080】
変異は、実施例1のように部位特異的突然変異誘発法を用いて導入され、BL21(DE3)細胞を、野生型または変異体PnFPOXを含有する発現ベクターで形質転換した。培養した細胞を10mM PPB、pH7.0に再懸濁し、超音波処理により溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。オキシダーゼおよびデヒドロゲナーゼ活性を実施例1のように測定した。
【0081】
粗PnFPOX野生型およびAsn56変異体の活性は、図4にまとめる。比較的高いオキシダーゼ活性(野生型の41%)を示したAsn56Serを除き、Asn56変異体のほとんどは、それらのオキシダーゼ活性を劇的に減少させた(<野生型の20%)。なかでも、いくつかの変異体(Asn56Cys、Phe、Met、Val)は、比較的高いデヒドロゲナーゼ活性を示し(>野生型の60%)、Asn56CysおよびAsn56Metでは野生型より高かった。Asn56変異体の速度論的パラメーターは、図5にまとめる。Asn56MetおよびAsn56Valは、Asn56Alaよりも高い、Vmaxデヒドロゲナーゼ(B)/Vmaxオキシダーゼ(A)を示した。Asn56Asp、Glu、およびProは、オキシダーゼおよびデヒドロゲナーゼの両方の活性を失った。
【0082】
実施例4
PnFPOXN56A変異体の精製
【0083】
pEPN(pET28a−PnFPOX)−N56Aで形質転換されたBL21(DE3)を、50μg/ml カナマイシンを含有するLB培地(2L)において37℃で好気的に培養した。A660nm値が0.6に達した後、細胞を0.3mM IPTGにより誘発し、25℃で5.5時間インキュベーションを続けた。細胞を遠心分離により回収し、10mM PPB、pH7.0に再懸濁し、フレンチプレス(1,000kg/cm2)に3回かけて溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。その後上清を、25μM FADを含有する10mM PPB、pH8.0に対して透析し、粗酵素溶液をさらに精製した。
【0084】
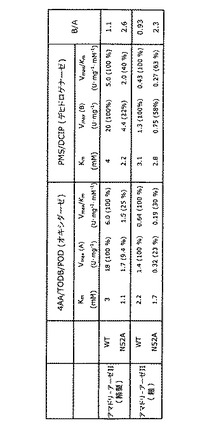
硫酸アンモニウムを透析した上清に35%飽和まで添加し、その後形成した沈殿を15,000gで20分間遠心分離によりペレットとした。硫酸アンモニウムを65%飽和まで添加した上清を、15,000gで20分間遠心分離した。得られた沈殿を、25μM FADおよび1%マンノースを含有する10mM PPB、pH8.0に溶解し、同緩衝液に対して4℃で透析し、その後、25μM FADを含有する10mM PPB、pH8.0に対して透析した。透析した酵素溶液を、10mM PPB、pH8.0で平衡化した、RESOURCE Q カラム(GE ヘルスケア(Healthcare))に適用した。活性な素通り(flow-through)画分を回収し、FAOD活性を示さない吸着されたタンパク質を、1M NaClで溶出した。活性な素通り画分を回収し、10mM PPB、pH7.0に対して透析した。透析した酵素溶液を、10mM PPB、pH7.0で平衡化したHiLoad 16/60 Superdex 75 pg カラム(GE ヘルスケア)に適用した。ゲルろ過クロマトグラフィーを同緩衝液で行なった。活性画分を回収し、精製した酵素溶液を100μM FADを含有する10mM PPB、pH7.0に対して透析し、4℃で保管した。その精製酵素の純度は、SDS−PAGEにより確認し、濃度は、DC Protein Assay Kit(バイオ−ラッド(Bio-Rad)、CA、米国)を用いて測定した。結果を表4に示す。
【0085】
【表4】
【0086】
N56A変異体FAODのオキシダーゼ活性およびデヒドロゲナーゼ活性を測定した。変異体FAODのSV曲線および速度論的パラメーターは、図6および表5にそれぞれ示す。図6において、左のグラフは0〜5mMの基質濃度における活性を示し、右のグラフは基質濃度0〜2mMの拡大図である。N56A変異体は、両基質に対して有意に増加したデヒドロゲナーゼ活性を示す。
【0087】
【表5】
【0088】
実施例5
アマドリアーゼII FAOD変異体の製造と特徴付
【0089】
アマドリアーゼII(配列番号:3)由来の変異体を製造し特徴付けを行なった。N1−1 FAODとアマドリアーゼIIとのアラインメント(図1)および実施例1〜4の結果にもとづき、Asn52がFADからのプロトンおよび電子の移動に関与すると予測した。Asn52Ala変異を有する変異体を、実施例1に記載された方法にしたがって製造した。
【0090】
BL21(DE3)細胞を、アマドリアーゼII野生型またはN52A変異体を含有する発現ベクターで形質転換した。培養した細胞を回収し、10mlの10mM PPB、pH7.0に再懸濁し、フレンチプレス(1,000kg/cm2)に2回かけて溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。その後、上清を25μM FADを含有する10mM PPB、pH8.0に対して透析し、粗酵素溶液を製造した。粗酵素溶液を、硫酸アンモニウム沈殿、陰イオン交換クロマトグラフィー(RESOURCE Q カラム(GE ヘルスケア))、およびゲルろ過クロマトグラフィー(HiLoad 16/60 Superdex 75 pg カラム(GE ヘルスケア))により精製した。
【0091】
FAOD活性を、基質としてのf−αValと共に4AA/TODB/POD(オキシダーゼ活性)およびPMS/DCIP(デヒドロゲナーゼ活性)を用いてアッセイした。結果を図7に示す。
【0092】
精製したアマドリアーゼII野生型およびAsn52Alaの速度論的パラメーターは図8にまとめる。変異体N52Aは、オキシダーゼ活性よりも約2倍高いデヒドロゲナーゼ活性を示した。
【0093】
実施例6
バイオセンサーの構築
【0094】
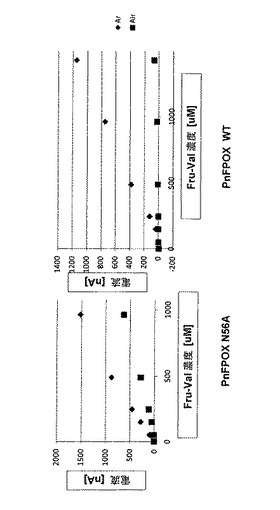
PnFPOX野生型またはN56A PnFPOXいずれかの40mU/5μl溶液を、2%AWP溶液と混合した。混合物5μlを再利用可能な金電極(表面積:7mm2)に適用し、30℃で30分間乾燥させた。電極を、UV−光で1分間照射し、固定化された野生型またはN56A FPOXを有する電極を製造した。電極を、2mMニトロソアニリン(NA;国際公開第2004/113900号のp.33ff.より公知)のPBS(pH7.4)溶液2mlに浸漬した。その後、+200mV対Ag/AgClを適用し、電流をモニターした。定常状態電流が観測された場合、異なる濃度のFVを含む試料溶液を反応混合物に添加し、電流の増加をモニターした。酸素の非存在下での酵素活性を測定するために、Arガスで反応チャンバー内を連続的に不活化した。
【0095】
図9は、酸素存在下または非存在下(Arガス下)における、FV濃度とPnFPOX N56Aまたは野生型酵素を有する電極の電流増加との相関関係を示す。電気化学反応に対する酸素の影響が観察されるが、酸素の存在の影響は野生型を用いた場合よりも少なかった。これらの結果は、本発明のフルクトシルアミノ酸オキシダーゼの活性は、溶存酸素レベルによる影響を受けにくいということを示唆した。
【0096】
実施例7
FPOX−C変異体の製造および特徴付
【0097】
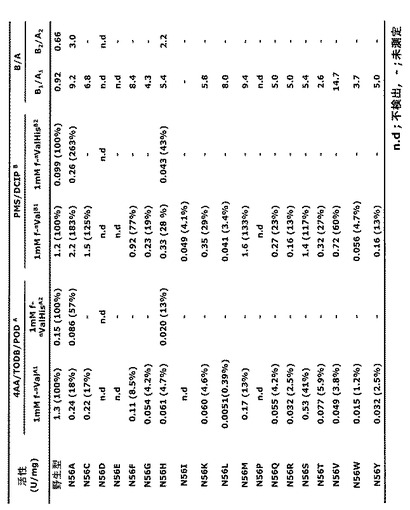
Asn52Ala変異を有するFPOX−C(配列番号:7)由来の変異体を、実施例5と同様の方法で製造した。その粗酵素調製物を、さらに精製することなく、酵素活性アッセイに使用した。FAOD活性を、基質としての1mM f−αValまたは1mM f−αVal−Hisのいずれかと共に4AA/TODB/POD(Ox:オキシダーゼ活性)およびPMS/DCIP(DH:デヒドロゲナーゼ活性)を用いてアッセイした。結果を以下の表6に示す。
【0098】
【表6】
【0099】
変異体FPOX−C N52Aは、f−αValおよびf−αVal−Hisの両方に対してオキシダーゼ活性よりも非常に高いデヒドロゲナーゼ活性を示した。
【0100】
実施例8
ループ変異体の製造および特徴付
【0101】
PnFPOXは、フルクトシルヘキサペプチドに対して、FVHと比較して1%未満のレベルでオキシダーゼ活性を示すが、FPOX−Cは活性を示さない。他方、FPOX−Cは、FVHに対してP.n FPOXよりもより高い活性を示す。両オキシダーゼのアミノ酸配列の比較から、FAODとFPOXとの間の有意な相違は、それらのループ領域に見られる。
【0102】
ループ1領域
PnFPOX (61)Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Ala(72)(配列番号:13)
FPOX−C (61)Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Ser(72)(配列番号:14)
【0103】
ループ2領域
PnFPOX (109)Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Glu(124)(配列番号:15)
FPOX−C (109)Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Ala(126)(配列番号:16)
【0104】
ループ1領域(12aa)では、5つのアミノ酸残基がPnFPOXとFPOX−Cとで異なる。ループ2領域(16aa)では、2つのアミノ酸挿入と7つのアミノ酸置換が見られる。
【0105】
1PnFPOXとFPOX−Cとのループ領域が互に交換されているキメラループ変異体、ならびにそのN56A変異体を製造した。上記に示した部分アミノ酸配列にもとづき、8種類の変異体を製造した。
【0106】
Pn FPOX/L1ExC(すなわち、ループ1領域がFPOX−C由来のループ1領域に置換されているPn FPOX)
Pn FPOX/L1ExC/N56A(すなわち、ループ1領域がFPOX−C由来のループ1領域に置換されているPn FPOXのN56A変異体)
Pn FPOX/L2ExC(すなわち、ループ2領域がFPOX−C由来のループ2領域に置換されているPn FPOX)
Pn FPOX/L2ExC/N56A(すなわち、ループ2領域がFPOX−C由来のループ2領域に置換されているPn FPOXのN56A変異体)
FPOX−C/L1ExPn(すなわち、ループ1領域がPn FPOX由来のループ1領域に置換されているFPOX−C)
FPOX−C/L1ExPn/N56A(すなわち、ループ1領域がPn FPOX由来のループ1領域に置換されているFPOX−CのN56A変異体)
FPOX−C/L2ExPn(すなわち、ループ2領域がPn FPOX由来のループ2領域に置換されているFPOX−C)
FPOX−C/L2ExPn/N56A(すなわち、ループ2領域がPn FPOX由来のループ2領域に置換されているFPOX−CのN56A変異体)
【0107】
変異の導入および粗酵素の製造は、実施例1および5に記載されているように行なった。FAOD活性を、基質としての1mM f−αVal、1mM f−αVal−Hisまたは5mMフルクトシルヘキサペプチド(F−HP)のいずれかと共に4AA/TODB/POD(Ox:オキシダーゼ活性)およびPMS/DCIP(DH:デヒドロゲナーゼ活性)を用いてアッセイした。
【0108】
Pn FPOX/L1ExCおよびPn FPOX/L2ExCは、野生型P.n.FPOXと比較してf−αVal−Hisに対して高い酸化活性を示した。FPOX−C/L1ExPnは、野生型FPOX−Cにおいては観察されないF−HPへの活性を示した。興味深いことに、Pn FPOX/L2ExCは、F−HPに対して活性を増加させたが、Pn FPOX/L1ExCは、F−HPに対して活性をほとんど失った。これらの結果は、F−HPに対するP.n.FPOX活性の改善が、ループ領域の改変によりさらに改善できたことを示した。
【0109】
これらのループ変異体へのN56A変異の導入は、オキシダーゼ活性の減少とデヒドロゲナーゼ活性の増加をもたらした。Pn FPOX/L1ExC/N56AおよびPn FPOX/L2ExC/N56Aの両方が、f−αValに対してそれらのオキシダーゼ活性と比較してより高いデヒドロゲナーゼ活性を示した。Pn FPOX/L2ExC/N56Aのf−αValに対するデヒドロゲナーゼ活性は、Pn FPOX/N56Aと比較して際立った増加を示し、FPOX−C/N56Aのものよりもより高かった。
【0110】
Pn FPOX/L1ExC/N56AおよびPn FPOX/L2ExC/N56Aの両方が、f−αVal−Hisに対してそれらのオキシダーゼ活性と比較してより高いデヒドロゲナーゼ活性を示した。最も高いデヒドロゲナーゼ活性は、Pn FPOX/L2ExC/N56Aにおいて観察され、これはFPOX−C/N56Aのものとほとんど同じである。
【0111】
代表的な結果を以下の表7に示す。
【0112】
【表7】
【技術分野】
【0001】
本発明は、糖化アルブミン、フルクトシルペプチド、HbA1cおよびフルクトシルバリン(FV)などの糖化タンパク質の測定用キットおよびセンサーに使用するためのフルクトシルアミノ酸オキシダーゼ(フルクトシルアミンオキシダーゼとも呼ばれる)に関する。より詳細には、本発明はオキシダーゼ活性の減少した変異体フルクトシルアミノ酸オキシダーゼに関する。
【背景技術】
【0002】
糖化タンパク質は、タンパク質上のアミノ基および糖の還元末端との間の共有結合を介して非酵素的に生成され、アマドリ化合物とも言われる。血中において、グルコースはヘモグロビンのβ−鎖のN−末端でバリンと結合し、糖化ヘモグロビン(グリコヘモグロビン;HbAlc)を生成する。ヘモグロビンに対するHbAlcの存在比は、正常な健常個体と比較して糖尿病患者においてより高く、HbAlcの血中濃度は、過去数週間の血糖値を反映することが知られている。したがって、HbAlcの血中濃度は、糖尿病の診断のための臨床試験において、また糖尿病患者の血糖コントロールにおいて非常に重要である。血中のHbAlc濃度は、フルクトシルバリンまたはフルクトシルバリルヒスチジンに特異性を有する酵素を用いて測定することが可能である。
【0003】
フルクトシルアミノ酸オキシダーゼは、様々な種類の株から単離されており、糖化アルブミン、HbAlcおよびフルクトシルアミノ酸などの糖化タンパク質がそのような酵素を用いて分析できることが示唆されている(特開昭61−268178号公報;特開昭61−280297号公報;特開平03−155780号公報;特開平05−192193号公報;特開平07−289253号公報;特開平08−154672号公報;特開2001−95598号公報;特開2003−79386号公報;特開2003−235585号公報;Agric. Biol. Chem., 53(1), 103-110, 1989;Agric. Biol. Chem., 55(2), 333-338, 1991;J. Biol. Chem., 269(44), 27297-27301, 1994;Appl. Environ. Microbiol., 61(12), 4487-4489, 1995;Biosci. Biotech. Biochem., 59(3), 487-491, 1995;J. Biol. Chem., 270(1), 218-224, 1995;J. Biol. Chem., 271(51), 32803-32809, 1996;およびJ. Biol. Chem., 272(6), 3437-3443, 1997)。
【0004】
フルクトシルアミノ酸オキシダーゼは、フルクトシルアミノ酸が酸化され、2−ケト−D−グルコースおよび対応するアミノ酸を生成する反応を触媒するFAD依存性酵素であり、FADの還元型(FADH2)の生成を伴う。FADH2は、次に電子アクセプターに電子を伝達し、その酸化型に変換される。酸素の存在下で、FADH2は人工電子アクセプター(アソメディエーター(asmediators)または電子メディエーターとも言う)に対してよりもむしろ酸素分子に優先的に電子を伝達する。したがって、フルクトシルアミノ酸を、メディエーターを用いてフルクトシルアミノ酸オキシダーゼによりアッセイする場合、アッセイの結果は、反応系における溶存酸素レベルに大きく影響を受ける。そのような不利益は、人工電子アクセプターを利用するポイント・オブ・ケア検査装置による血液試料の臨床検査において特に指摘される。人工電子メディエーターを用いる酵素センサーストリップに使用される酵素は、酸素に対して低い活性を有することが望まれる。
【0005】
したがって、本発明の目的は、酵素、特にその活性が溶存酸素レベルによりほとんど影響を受けないフルクトシルアミノ酸オキシダーゼを提供することである。
【発明の概要】
【0006】
一般的に言えば、本願の発明者らは、酵素、より詳細にはその活性が溶存酸素レベルによりほとんど影響を受けないフルクトシルアミノ酸オキシダーゼを提供することができる。より具体的には、これは、その野生型において優先的にオキシダーゼ活性を示す酵素のオキシダーゼ活性を減少させることにより、好ましくは同時に酵素のデヒドロゲナーゼ活性を増加させることにより達成される。以下により詳細に説明するように、これは野生型酵素を変異させることにより達成されている。
【0007】
本発明者らは、フルクトシルアミノ酸オキシダーゼの種々の変異体を製造し、驚くべきことに、特定の種類の変異体が、デヒドロゲナーゼ活性、詳細には、色素媒介(dye-mediated)デヒドロゲナーゼ活性を実質的に保持しながら減少したオキシダーゼ活性を示すことを見出した。
【0008】
本発明は、配列番号1記載のアミノ酸配列の56位に対応する位置で、アミノ酸残基Asnを、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼを提供する。
【0009】
好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有し、好ましくは、野生型フルクトシルアミノ酸オキシダーゼと比較して増加されたデヒドロゲナーゼ活性を有する。好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの30%以下のオキシダーゼ活性を有し、そしてまた、好ましくは野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する。
【0010】
別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1〜16からなる群より選択されるアミノ酸配列であって、配列番号1記載のアミノ酸配列の56位に対応する位置のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基により置換されているアミノ酸配列を有する。好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1記載のアミノ酸配列であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。より好ましくは、配列番号1の位置109−124のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられている。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号2記載のアミノ酸配列であって、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号3記載のアミノ酸配列であって、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。また好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号7記載のアミノ酸配列であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。より好ましくは、配列番号7の位置61−72のアミノ酸配列Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられている。
【0011】
別の側面では、本発明は、本発明の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチドを提供する。
【0012】
また別の側面では、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。
【0013】
さらに別の側面では、本発明は、本発明のベクターで形質転換された宿主細胞を提供する。
【0014】
別の側面において、本発明は、試料中の糖化タンパク質をアッセイするための方法であって、試料を本発明のフルクトシルアミノ酸オキシダーゼと接触させること、およびフルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む方法を提供する。
【0015】
なお別の側面において、本発明は、HbA1cをアッセイするための方法であって、試料中のHbA1cを消化してフルクトシルバリンまたはフルクトシルバリルヒスチジンを生成すること、該フルクトシルバリンまたはフルクトシルバリルヒスチジンを本発明のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンまたはフルクトシルバリルヒスチジンの量を測定することを含む方法を提供する。
【0016】
別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための装置であって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置を提供する。
【0017】
また別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキットを提供する。
【0018】
別の側面において、本発明は、電極に固定化されている本発明のフルクトシルアミノ酸オキシダーゼを有する酵素電極を提供する。
【0019】
別の側面において、本発明は、作用電極として本発明の酵素電極を含むフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーを提供する。
【0020】
また別の側面において、本発明は、配列番号1記載のアミノ酸配列であって、配列番号1の61−72位のアミノ酸配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaが、配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。なお別の側面において、本発明は、配列番号1記載のアミノ酸配列であって、配列番号1の109−124位のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。また別の側面において、本発明は、配列番号7記載のアミノ酸配列であって、配列番号7の61−72位のアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。また別の側面において、本発明は、配列番号7記載のアミノ酸配列であって、配列番号1の109−126位のアミノ酸配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaが、配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼを提供する。
【発明の効果】
【0021】
本発明の変異されたフルクトシルアミノ酸オキシダーゼは、糖尿病状態の診断やコントロールにおいて臨床的に有用なヘモグロビン(HbA1c)などの糖化タンパク質の測定に使用することが可能である。
【図面の簡単な説明】
【0022】
【図1】いくつかの真核性FAODのN−末端付近のアミノ酸配列のアラインメントを示す。
【図2】フルクトシルアミノ酸オキシダーゼの反応系を示す。
【図3】精製されたN1−1 FAOD N47A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図4】粗PnFPOX野生型およびAsn56変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図5】PnFPOX Asn56変異体の速度論的パラメーターを示す。
【図6】精製されたPnFPOX N56A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図7】精製されたアマドリアーゼII野生型およびN52A変異体のオキシダーゼ活性およびデヒドロゲナーゼ活性を示す。
【図8】精製されたアマドリアーゼII野生型およびN52A変異体の速度論的パラメーターを示す。
【図9】PnFPOX N56Aまたは野生型FAODを有する電極を用いるFV濃度のアッセイを示す。
【発明を実施するための形態】
【0023】
フルクトシルアミノ酸オキシダーゼ(FAOD)は、様々な真核性および原核性株に存在するということが知られている。表1は、今までに単離された、または予測されたFAODのいくつかの例を示す。
【0024】
【表1】
【0025】
真核性FAODは、基質特異性により、(i)α−フルクトシルアミノ酸優先(FAOD−P、FAOD−U、FAOX−E、FPOX−C、およびPnFPOX)、(ii)ε−フルクトシルアミノ酸優先(FAOD−F、FAOD−A、アマドリアーゼI、およびFAOD−Ao1)、および(iii)α−およびε−糖化アミノ酸両方に類似の活性(アマドリアーゼII、FAOD−Ao2、およびN1−1 FAOD)に分類することができる。一般に、α−フルクトシルアミノ酸に活性なFAODは、α−フルクトシルバリンおよびHbA1cのアッセイにおいて有用となり、ε−フルクトシルアミノ酸に活性なFAODは、ε−フルクトシルリジンおよび糖化アルブミンのアッセイにおいて有用となる。当業者は、酵素に変異を導入することにより、α−またはε−フルクトシルアミノ酸に対するその優先性を変えることができるということを理解するものである。
【0026】
本発明は、配列番号1記載のアミノ酸配列(PnFPOX)の56位に対応する位置で、アミノ酸残基AsnをAla、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼを提供する。
【0027】
本明細書において使用される用語、タンパク質の「変異体」は、表示される位置でタンパク質上の1つまたは複数のアミノ酸残基に置換を含む変異型タンパク質を意味する。用語変異体はまた、そのような変異体タンパク質をコードするポリヌクレオチドにも使用される。
【0028】
本明細書において使用される語句「対応する位置」は、デフォルトパラメーターを有するVector NTIのソフトウェアAlignX(インビトロジェン製;Lu, G., and Moriyama, E.N. (2004) Vector NTI, a balanced all-in-one sequence analysis suite. Brief Bioinform 5, 378-88参照)を用いて、参照アミノ酸配列のアミノ酸残基と位置合わせされる照会(query)アミノ酸配列におけるアミノ酸残基の位置を意味する。したがって、「配列番号1記載のアミノ酸配列の56位に対応する位置のAsn残基」は、照会アミノ酸配列が、デフォルトパラメーターを有するVector NTIのAlignXを用いて配列番号1とアラインメントされる場合に、配列番号1のAsn56と位置合わせされる照会アミノ酸配列におけるAsn残基を意味する。配列番号1自体のAsn47もこの用語により包含されることに留意すべきである。
【0029】
図1は、いくつかの真核性FAOD(ペニシリウム・ヤンシネルム(Penicillum janthinelum)AKU3413由来のFAOD−P、ウロクラディウム・エスピー(Ulocladium sp.)JS−103由来のFAOD−U、ユーペニシリウム・テレヌム(Eupenicillum terrenum)ATCC 18547由来のFPOX−E、コニオカエタ・エスピー(Coniochaeta sp.)NISL 9330由来のFPOX−C、ファエオスフェリア・ノドラム(Phaeosphaeria nodorum)由来のPnFPOX、フザリウム・オキシスポラム(Fusarium oxysporum)NBRC 9972由来のFAOD−F、アスペルギルス・テレウス(Aspergillus terreus)GP1由来のFAOD−A、アスペルギルス・フミガーツス(Aspergillus fumigatus)由来のアマドリアーゼI、アスペルギルス・オリゼー(Aspergillus oryzae)由来のFAOD−Ao1、アスペルギルス・フミガーツス(Aspergillus fumigatus)由来のアマドリアーゼII、アスペルギルス・オリゼー(Aspergillus oryzae)由来のFAOD−Ao2およびピチア・エスピー(Pichia sp.)N1−1由来のN1−1 FAOD)のN−末端付近の保存された領域内のアミノ酸配列のアラインメントを示す。これらのFAODの全配列は、配列番号1〜12に記載されている。アラインメントは、Vector NTI suite6.0のAlignXアプリケーションを用いて作製された。矢印はN1−1 FAODのAsn47に対応する残基を示す。括弧中の数字は、配列番号により表される各アミノ酸配列における、図1の一番左の「P」の位置を示す。当業者は、Blastなどの別のアラインメントソフトウェアプログラムが、同じまたは実質的に同じアラインメントを提供するであろう事を理解するものである。
【0030】
図1から、N1−1 FAODのAsn47を含む領域におけるアミノ酸配列が、真核性FAODのあいだで高度に保存されているということがわかる。したがって、当業者は、配列アラインメント用の任意の市販のソフトウェアプログラムを用いて保存された領域内のN1−1 FAODのAsn47に対応するAsn残基を容易に同定することができ、また、変異体フルクトシルアミノ酸オキシダーゼが、そのAsn残基上に修飾を導入することにより容易に製造されることを理解することができる。
【0031】
別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1〜15からなる群より選択されるアミノ酸配列であって、配列番号1記載のアミノ酸配列の56位に対応する位置でアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。
【0032】
好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号1記載のアミノ酸配列(PnFPOX)であって、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。好ましくは、Asn残基はAlaで置換される。別の好ましい実施態様においては、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号2記載のアミノ酸配列(N1−1)であって、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。なお別の好ましい実施態様において、本発明の変異体フルクトシルアミノ酸オキシダーゼは、配列番号3記載のアミノ酸配列(アマドリアーゼII)であって、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する。
【0033】
本発明の変異体フルクトシルアミノ酸オキシダーゼは、デヒドロゲナーゼ活性を実質的に保持しながら減少されたオキシダーゼ活性を示す。図2は、フルクトシルアミノ酸オキシダーゼの反応スキームを説明する。
【0034】
本明細書において、「オキシダーゼ活性」は、酸素を電子アクセプターとして利用しながらフルクトシルアミノ酸の酸化を触媒し、2−ケト−D−グルコースと対応するアミノ酸を生成する、フルクトシルアミノ酸オキシダーゼの酵素活性(Vmax)である。オキシダーゼ活性は、本技術分野において既知の任意の方法、例えば、4AA/TODB/POD(4−アミノアンチピリン/N,N−ビス(4−スルホブチル)−3−メチルアニリン二ナトリウム塩/西洋ワサビペルオキシダーゼ)などのH2O2検出用試薬やPt電極により、生成したH2O2の量を測定することによりアッセイすることができる。本明細書において、相対的または定量的活性という文脈では、オキシダーゼ活性は、具体的には、25℃で、10mM PPB、pH7.0、1.5mM TODB、2U/ml西洋ワサビペルオキシダーゼ(POD)、および1.5mM 4−アミノアンチピリン(4AA)において生成されたH2O2の量により測定される、単位時間当たり酸化された基質(フルクトシルアミノ酸)のモル量と定義される。キノンイミン色素の生成は、546nmで分光光度法により測定することができる。
【0035】
本明細書において、「デヒドロゲナーゼ活性」は、酸素以外の電子メディエーターを電子アクセプターとして利用しながらフルクトシルアミノ酸の酸化を触媒し、2−ケト−D−グルコースと対応するアミノ酸を生成する、フルクトシルアミノ酸オキシダーゼの酵素活性(Vmax)である。デヒドロゲナーゼ活性は、たとえば、mPMS/DCIP(メチル硫酸1−メトキシ−5−メチルフェナジニウム/2,6−ジクロロインドフェノール)、cPES(トリフルオロ−酢酸−1−(3−カルボキシ−プロポキシ)−5−エチル−フェナジニウム、NA BM31_1144(N,N−ビス−(ヒドロキシエチル)3−メトキシ−ニトロソアニリン塩酸塩、NA BM31_1008(N,N−ビス−ヒドロキシエチル−4−ニトロソアニリンおよびN−N−4−ジメチル−ニトロソアニリン)などを用いてメディエーターに移動した電子の量を測定することによりアッセイすることができる。
【0036】
本明細書において、相対的または定量的活性という文脈では、デヒドロゲナーゼ活性は、具体的には、25℃で、10mM PPB(pH7.0)、0.6mM DCIP、および6mM メトキシPMS(mPMS)においてメディエーターに移動された電子の量により測定される、単位時間当たり酸化された基質(フルクトシルアミノ酸)のモル量と定義される。
【0037】
本発明の変異体フルクトシルアミノ酸オキシダーゼは、デヒドロゲナーゼ活性を実質的に保持しながら、野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有する。
【0038】
好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型のフルクトシルアミノ酸オキシダーゼの50%以下のオキシダーゼ活性を有する。より好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの40%以下、より好ましくは30%以下、さらにより好ましくは20%以下、最も好ましくは15%以下のオキシダーゼ活性を有する。また、好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する。より好ましくは、本発明の変異体フルクトシルアミノ酸オキシダーゼは、野生型フルクトシルアミノ酸オキシダーゼの70%以上、より好ましくは90%以上、さらにより好ましくは100%以上、最も好ましくは100%より高いデヒドロゲナーゼ活性を有する。
【0039】
野生型フルクトシルアミノ酸オキシダーゼにおいて、オキシダーゼ活性とデヒドロゲナーゼ活性は、比率、デヒドロゲナーゼ活性/オキシダーゼ活性が、約0.8〜1.2の間と同程度にある。アッセイ系に溶存酸素が存在する場合、基質の酸化により生じた電子は、酸素に優先的に移動することになる。したがって、電子メディエーターの存在下で測定される酵素活性は、溶存酸素濃度によって大きな影響を受けることになる。対照的に、本発明の変異体フルクトシルアミノ酸オキシダーゼは、約2.0以上、好ましくは4.0以上、より好ましくは10以上、さらにより好ましくは20以上、最も好ましくは40以上の比率、デヒドロゲナーゼ活性/オキシダーゼ活性を有する。デヒドロゲナーゼ活性がオキシダーゼ活性を上回るので、本発明のフルクトシルアミノ酸オキシダーゼの酵素活性は、溶存酸素濃度の影響をほとんど受けないものとなる。これは、血液試料による臨床診断におけるフルクトシルアミノ酸オキシダーゼの利用において好都合なものである。
【0040】
別の側面において、本発明は、1つまたは複数のループ領域が改変または変異されているフルクトシルアミノ酸オキシダーゼを提供する。PnFPOXは、フルクトシルヘキサペプチドに対してFVHと比較して1%未満の活性レベルのオキシダーゼ活性を示す。一方、FPOX−Cは、活性を示さない。他方、FPOX−Cは、P.nFPOXよりもFVHに対してより高い活性を示す。両オキシダーゼのアミノ酸配列の比較から、FAODとFPOXとのあいだの有意差は、それらのループ領域に見られる。
【0041】
ループ1領域
PnFPOX (61)Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Ala(72)(配列番号:13)
FPOX−C (61)Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Ser(72)(配列番号:14)
【0042】
ループ2領域
PnFPOX (109)Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Glu(124)(配列番号:15)
FPOX−C (109)Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Ala(126)(配列番号:16)
【0043】
ループ1領域(12aa)では、5つのアミノ酸残基がPnFPOXとFPOX−Cとで異なる。ループ2領域(16aa)では、2つのアミノ酸挿入と7つのアミノ酸置換が見られる。
【0044】
驚くべきことに、PnFPOXとFPOX−Cとのループ領域が互に交換されているいくつかのキメラループ変異体が改善された活性を示すことが見出された。下記の実施例8に示されているように、ループ2領域がFPOX−Cループ2領域により置き換えられているPnFPOXは、野生型PnFPOXよりもFV、FVHおよびフルクトシルヘキサペプチド(F−HP)に対してより高いオキシダーゼ活性を示す。ループ1領域がPnFPOXループ1領域により置き換えられているFPOX−Cは、野生型FPOX−Cにおいては検出されない、F−HPに対してオキシダーゼ活性を示す。加えて、N56A変異を有するキメラループ変異体も、PnFPOXおよびFPOX−C由来のN56A変異体において観察されるように、減少したオキシダーゼ活性と増加したデヒドロゲナーゼ活性を示す。特に好ましいキメラループ変異体は、ループ2領域がFPOX−Cのループ2領域で置換されており、かつAsn56がAlaで置換されているPnFPOXである。別の特に好ましいキメラループ変異体は、ループ1領域がPnFPOXのループ1領域で置換されており、かつAsn56がAlaで置換されているFPOX−Cである。これらの結果は、PnFPOX由来のループ1領域とFPOX−C由来のループ2領域の組合せが、フルクトシルバリンおよびフルクトシルバリルヒスチジンならびにフルクトシルヘキサペプチドに対してより高い活性を提供するということを示唆する。
【0045】
別の側面において、本発明は、本発明の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチドを提供する。フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドのヌクレオチド配列は、公共のデータベースから容易に得ることができる。野生型フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドは、PCRまたは他の既知の技術を用いて各生物のゲノムからクローニングすることができる。変異は、部位特異的突然変異誘発法、PCR突然変異誘発法または本技術分野において周知の任意の他の技術により導入することができる。変異されるべきAsn残基は、本技術分野において利用可能な配列アラインメントのための任意のソフトウェアを用いて同定することができる。あるいは、変異体フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドは、一連の化学的に合成されたオリゴヌクレオチドを用いてPCRにより製造してもよいし、または完全に合成してもよい。
【0046】
変異されたフルクトシルアミノ酸オキシダーゼは、変異体遺伝子を適切な発現ベクターに挿入し、そのベクターを大腸菌(E. coli)細胞などの適切な宿主細胞に導入することにより製造することができる。形質転換体を培養し、その形質転換体において発現されたフルクトシルアミノ酸オキシダーゼを、細胞または培養培地から回収することができる。
【0047】
かくして得られた組換えフルクトシルアミノ酸オキシダーゼは、本技術分野において公知の任意の精製技術、たとえばイオン交換カラムクロマトグラフィー、アフィニティークロマトグラフィー、液体クロマトグラフィー、ろ過、限外ろ過、塩析沈殿、溶媒沈殿、免疫沈降、ゲル電気泳動、等電電気泳動および透析などにより精製することができる。
【0048】
したがって、本発明は、変異体フルクトシルアミノ酸オキシダーゼをコードするポリヌクレオチドを含むベクター、そのようなベクターで形質転換された宿主細胞、ならびに該形質転換体を培養し、変異体フルクトシルアミノ酸オキシダーゼを培養物から回収および精製することにより、本発明の変異体フルクトシルアミノ酸オキシダーゼを製造する方法も包含する。
【0049】
本発明はまた、試料中の糖化タンパク質をアッセイするための方法も包含する。その方法は、試料を本発明のフルクトシルアミノ酸オキシダーゼと接触させること、およびフルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む。本発明によりアッセイされる糖化タンパク質は、たとえば、フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1c、フルクトシルヘキサペプチド、糖化アルブミンおよび他のフルクトシルアミノ酸である。なお別の側面において、本発明は、試料中のHbA1cを消化してフルクトシルバリンを生成すること、そのフルクトシルバリンを本発明のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンの量を測定することを含むHbA1cをアッセイするための方法を提供する。
【0050】
別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1c、フルクトシルヘキサペプチドまたは糖化アルブミンをアッセイするための装置であって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置を提供する。
【0051】
アッセイ装置は、任意の従来の市販されている血糖値をモニターするための電流測定バイオセンサーのテストストリップと同様の構造を有することができる。そのような装置の1つの例は、絶縁基材上に配置された2つの電極(作用電極と参照または対電極)、試薬ポートおよび試料受け(receiver)を有する。試薬ポートは、本発明の変異されたフルクトシルアミノ酸オキシダーゼおよびメディエーターを含む。血液試料などの試料が試料受けに添加されると、試料に含まれるフルクトシルアミノ酸がフルクトシルアミノ酸オキシダーゼと反応し、電流が生じ、試料中のフルクトシルアミノ酸の量を示す。酵素基質の測定に適した電気化学センサーの典型的な例は、たとえば国際公開第2004/113900号および米国特許第5,997,817号明細書から公知である。電気化学センサーの代わりとして、光学的検出技術も使用可能なはずである。典型的には、そのような光学装置は、酵素、電子移動メディエーターおよび指示薬を含む試薬系において起こる色の変化にもとづくものである。色の変化は、蛍光、吸光、緩和(remission)測定を用いて定量することができる。酵素基質の測定に適した光学装置の典型的な例が、たとえば米国特許第7,008,799号明細書、米国特許第6,036,919号明細書および米国特許第5,334,508号明細書から公知である。
【0052】
なお別の側面において、本発明は、試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、本発明のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキットを提供する。
【0053】
フルクトシルバリンまたはフルクトシルバリルヒスチジンの測定のためのキットは、本発明の酵素を用いて構築することが可能である。本発明のフルクトシルアミノ酸オキシダーゼに加えて、キットは、測定に必要な緩衝液、適切なメディエーター、および必要ならばペルオキシダーゼなどの酵素、較正曲線の作製のためのフルクトシルバリンまたはフルクトシルバリルヒスチジンまたはそれらの誘導体の標準溶液、および使用説明書を含有する。本発明のフルクトシルアミノ酸オキシダーゼは、多様な形態、たとえば凍結乾燥試薬として、または適切なストレージソリューションの溶液として提供してもよい。
【0054】
本発明の酵素を用いて、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドアッセイキットを構築することも可能である。糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドは、酵素的にまたは化学的に消化され、フルクトシルバリン、フルクトシルバリルヒスチジンおよびフルクトシルヘキサペプチドといったフルクトシルアミノ酸またはフルクトシルペプチドを生じ、それが今度は本発明のフルクトシルアミノ酸オキシダーゼを用いて定量される。このようにして、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドがアッセイ可能である。したがって、糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドのための本発明のアッセイキットは、フルクトシルバリンまたはフルクトシルバリルヒスチジンの測定のための前記キットにさらに加水分解するための試薬またはプロテアーゼを含有することができる。
【0055】
別の側面において、本発明は、電極上に固定化された本発明のフルクトシルアミノ酸オキシダーゼを有する酵素電極を提供する。
【0056】
別の側面において、本発明は、作用電極として本発明の酵素電極を含む、フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーを提供する。
【0057】
試料中のフルクトシルアミノ酸の濃度は、酵素反応により生じた電子の量を測定することにより決定することができる。本技術分野においては、炭素電極、金属電極、および白金電極などの様々なセンサー系が知られている。本発明の変異されたフルクトシルアミノ酸オキシダーゼは、電極上に固定される。固定化のための手段の例としては、架橋、高分子マトリックスへの封入、透析膜による被覆、光架橋性ポリマー、電気伝導性ポリマー、酸化−還元ポリマー、およびそれらの任意の組合せなどが挙げられる。
【0058】
測定が電流測定系において行なわれる場合、固定化された酵素を備えた炭素電極、金電極または白金電極が作用電極として、対電極(白金電極など)および参照電極(Ag/AgCl電極など)と共に使用される。電極は、メディエーターを含有する緩衝液中に挿入され、所定の温度に保たれる。所定の電圧を作用電極に適用し、その後試料を添加、そして電流の増加した値を測定する。アッセイに使用されるメディエーターの例としては、フェリシアン化カリウム、フェロセン、オスミウム誘導体、ルテニウム誘導体、フェナジンメトサルフェートなどが挙げられる。通常、1つの作用電極と1つの対電極または偽−参照電極とのいわゆる二電極系も使用することが可能である。
【0059】
さらに、フルクトシルアミノ酸は、炭素電極、金電極、または白金電極を用いる電流測定系において、固定化された電子メディエーターを用いてアッセイすることもできる。酵素は、作用電極を作製するために吸着または共有結合により高分子マトリックスにおいてフェリシアン化カリウム、フェロセン、オスミウム誘導体、フェナジンメトサルフェートなどの電子メディエーターと一緒に電極上に固定化される。この作用電極が対電極(白金電極など)および参照電極(Ag/AgCl電極など)と一緒に緩衝液中に挿入され、所定の温度に保たれる。所定の電圧を作用電極に適用し、その後試料を添加、そして電流の増加した値を測定する。
【0060】
糖化アルブミン、HbA1cまたはフルクトシルヘキサペプチドの測定のためのセンサーを作製するために、フルクトシルアミノ酸の測定用の前記センサーは、複合センサーを構築するためにさらに固定化されたプロテイナーゼ(プロテアーゼなど)を含む膜と組み合わせられる。複数の酵素の組合せによる連続反応にもとづくそのような複合センサーの構造は、本技術分野において周知である。たとえば、Anthony P. F. Tuner, Isao Karube and George S. Wilsonによる「バイオセンサー−基礎と応用」(“Biosensor-Fundamental and Applications”)、オクスフォード大学出版、1987年を参照。
【0061】
本明細書において引用した全ての特許文献および参考文献の内容は、参照により本明細書中に完全に組み込まれる。
【実施例】
【0062】
本発明は、以下の実施例により詳細に説明されるが、本発明はそれらの実施例に限定されるべきものではない。
【0063】
実施例1
N1−1 FAOD変異体の製造および特徴付
【0064】
単量体サルコシンオキシダーゼ(MSOX)の構造機能相関の研究により、FADから酸素への電子移動に関与するプロトンリレー系(PRS)が、Thr48およびLys265と4つのH2O分子とを含んでなるということを明らかにした(Trickey et al, Structure, 7, 331-345, 1999)。ピチア・エスピー(Pichia sp.)N1−1株由来のフルクトシルアミノ酸オキシダーゼ(N1−1 FAOD、配列番号:2)の活性部位は、MSOXと、特にプロトンリレー系を担うと予測される保存された残基において高い相同性を有する。
【0065】
N1−1 FAODの予測構造モデルを、MSOX構造を用いて構築し、アミノ酸残基Asn44、Ser46、Asn47、Lys48およびLys269がFADからのプロトンと電子の移動に関与していると予測した。単一または二重突然変異をN1−1 FAODの電子アクセプターアベイラビリティーを改変する目的でそれらのアミノ酸残基に導入した。
【0066】
N1−1 FAOD単一変異体の発現ベクターを、N1−1 FAOD野生型遺伝子を鋳型として、QuickChange(登録商標)法(ストラタジーン(stratagene))を用いて部位特異的突然変異誘発法により作製した。Asn44、Ser46、Asn47、Lys48またはLys269で変異を導入するためにプライマーを設計した。増幅産物は、DpnIにより消化し、大腸菌(E. coli)DH5αに形質転換し、LB寒天培地(50μg/ml カナマイシン)において37℃で一晩インキュベートした。クローンの変異された配列を、ABI Prism 3130 Genetic Analyzerにおいて、ABI Prism BigDye Terminator cycle sequencing kit v3.0を用いて確認した。次に、抽出したプラスミドを、NcoIおよびSalIで消化し、NcoIおよびSalIで消化したpET28(a)ベクターにライゲートした。ライゲーション混合物で形質転換した大腸菌(E. Coli)BL21(DE3)細胞を、LB寒天培地(50μg/ml カナマイシン)において37℃で一晩インキュベートした。二重変異体は、同じ方法を用いてプライマーの組合せにより作製した。
【0067】
N1−1 FAOD変異体の発現ベクターをもつ大腸菌(Escherichia coli)BL21(DE3)細胞を、50μg/mlのカナマイシンを含有するLB培地150mlにおいて37℃で培養した。A660値が0.6に達した後、0.3mM IPTGを添加し、25℃でさらに5時間インキュベートした。細胞を遠心分離により回収し、0.85% NaCl水溶液で二回洗浄した。その後、細胞を10mMのリン酸カリウム緩衝液(PPB)pH7.0に再懸濁し、超音波ホモジナイザーにより溶菌した。ライセートを10,000×g、4℃で10分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離し、その後上清を10mM PPB(pH7.0)に対して透析した。オキシダーゼ活性は、25℃、10mM PPB、pH7.0中で、基質(0.2、0.5、0.75、1、2、5mM f−αValまたは5mM f−αVal−His)の存在下で、1.5mM TODB、2U/ml西洋ワサビペルオキシダーゼ(POD)、および1.5mM 4−アミノアンチピリン(4AA)でアッセイした。キノンイミン色素の生成を分光光度計により546nmで測定した。デヒドロゲナーゼ活性は、25℃、10mM PPB(pH7.0)において、基質(0.2、0.5、1、2、5mM f−αValまたは5mM f−αVal−His)の存在下で、0.6mM DCIPおよび6mM メトキシPMS(mPMS)でアッセイした。
【0068】
ほとんどのケースで、オキシダーゼ活性は完全にまたは実質的に消失したが、N47A、S46A、K48AおよびN44Aなどのいくつかの変異体は、相対的に高い活性を示した。なかでもN47Aは、最も目立った特徴を示した。N47Aのオキシダーゼ活性は野生型の活性の約20%に減少したが、N47Aのデヒドロゲナーゼ活性は野生型の活性の60%よりも高いままであった。結果として、N47Aは、オキシダーゼ活性よりも約4倍高いデヒドロゲナーゼ活性を示した。N47Aのこの特徴は、人工電子アクセプターと共にフルクトシルアミノ酸オキシダーゼを用いるセンサーストリップに有益なはずである。野生型N1−1 FAODおよびN47A変異体の粗調製物の速度論的パラメーターを表2に示す。
【0069】
【表2】
【0070】
実施例2
N1−1野生型およびN47A変異体FAODの精製
【0071】
組換えFAODは、次のように精製した。最初に、酵素を含有する水溶性画分を組換え大腸菌(Escherichia coli.)から調製した。発現ベクターを含む大腸菌を7LのLB培地(37℃、10L醗酵槽で、50μg/ml アンピシリン)において培養し、その後発現をIPTG(最終濃度:0.3mM)により約OD660=0.7で誘導し、培養温度を30℃まで下げた。細胞を100mM PPb(pH7.0)中に懸濁し、フレンチプレスを用いて4回破壊した。上清液を超遠心分離(40,000g、90分)に付し、上清を10mMのPPb(pH7.0)に対して4℃で一晩透析し水溶性画分を調製した。
【0072】
この水溶性画分を、さらに液体クロマトグラフィーに付し、精製酵素を調製した。酵素をさらに陰イオン交換クロマトグラフィー(DEAE−5PW)で精製した。水溶性画分を、10mM PPb(pH7.0)で平衡化した陰イオン交換クロマトグラフィーカラムDEAE−5PW(5.0mmI.D.×5cm、東ソー(Tosoh))に吸着させた。カラム体積の3倍量の10mM PPb(pH7.0)による平衡化の後、FAODを、0.7MのNaClを含有する10mM PPb(pH7.0)で溶出した。流速は1ml/分に設定し、溶出液を1分毎に回収した。280nmの吸収波長を溶出液のモニターに使用した。
【0073】
活性画分を、35%硫酸アンモニウムを用いて分離し、上清液を疎水性クロマトグラフィーに付した。活性画分を、35%硫酸アンモニウムを含有する10mM PPb(pH6.5)で平衡化した疎水性クロマトグラフィーカラム、Resource Phe(1ml、ファルマシア(Pharmacia))に吸着させた。カラム体積の3倍量の、35%硫酸アンモニウムを含有する10mM PPb(pH6.5)による平衡化の後、FAODを、10mM PPb(pH6.5)で溶出した。流速は2ml/分に設定し、溶出液を1分毎に回収した。活性画分を、45%硫酸アンモニウムで分離し、その後沈殿を、1%マンノースおよび100μM FADを含有する10mM PPb(pH7.0)に溶解し、同緩衝液に対して4℃で6時間透析した。それをさらに、100μM FADを含有する10mM PPb(pH8.0)に対して4℃で6時間透析した。透析した試料を次の陰イオン交換クロマトグラフィーに用いた。
【0074】
その試料を、10mM PPb(pH8.0)で平衡化した陰イオン交換クロマトグラフィーカラム、Bioasit Q(4.6mmI.D.×5cm、東ソー)に吸着させた。カラム体積の3倍量の10mM PPb(pH8.0)による平衡化の後、FAODを、0.3MのNaClを含有する10mM PPb(pH7.0)で溶出した。流速は1ml/分に設定し、溶出液を1分毎に回収した。活性画分を10mM PPb(pH7.0)に対して4℃で一晩透析した。試料の精製度をSDS/PAGEにより調べた。試料をPhast Gel 8−25を用いる電気泳動に付し、ゲルを銀で染色した。試料調製、電気泳動および染色は、Phast System(登録商標)に添付のマニュアルにしたがって行なった。
【0075】
精製した酵素のオキシダーゼおよびデヒドロゲナーゼ活性は、実施例1のように測定し、図3に示した。速度論的パラメーターは、表3に示す。
【0076】
【表3】
【0077】
実施例3
PnFPOX変異体の製造
【0078】
ファエオスフェリア・ノドラム(Phaeosphaeria nodorum)由来のフルクトシルアミノ酸オキシダーゼ(PnFPOX、配列番号:1)由来の変異体を製造し、特徴付けした。
【0079】
N1−1 FAODとPnFPOXとのアラインメント(図1)および実施例1および2の結果にもとづき、Asn56がFADからのプロトンおよび電子の移動に関与すると予測した。PnFAODの電子アクセプターのアベイラビリティーを改変することを目的にAsn56に種々の変異が導入された。
【0080】
変異は、実施例1のように部位特異的突然変異誘発法を用いて導入され、BL21(DE3)細胞を、野生型または変異体PnFPOXを含有する発現ベクターで形質転換した。培養した細胞を10mM PPB、pH7.0に再懸濁し、超音波処理により溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。オキシダーゼおよびデヒドロゲナーゼ活性を実施例1のように測定した。
【0081】
粗PnFPOX野生型およびAsn56変異体の活性は、図4にまとめる。比較的高いオキシダーゼ活性(野生型の41%)を示したAsn56Serを除き、Asn56変異体のほとんどは、それらのオキシダーゼ活性を劇的に減少させた(<野生型の20%)。なかでも、いくつかの変異体(Asn56Cys、Phe、Met、Val)は、比較的高いデヒドロゲナーゼ活性を示し(>野生型の60%)、Asn56CysおよびAsn56Metでは野生型より高かった。Asn56変異体の速度論的パラメーターは、図5にまとめる。Asn56MetおよびAsn56Valは、Asn56Alaよりも高い、Vmaxデヒドロゲナーゼ(B)/Vmaxオキシダーゼ(A)を示した。Asn56Asp、Glu、およびProは、オキシダーゼおよびデヒドロゲナーゼの両方の活性を失った。
【0082】
実施例4
PnFPOXN56A変異体の精製
【0083】
pEPN(pET28a−PnFPOX)−N56Aで形質転換されたBL21(DE3)を、50μg/ml カナマイシンを含有するLB培地(2L)において37℃で好気的に培養した。A660nm値が0.6に達した後、細胞を0.3mM IPTGにより誘発し、25℃で5.5時間インキュベーションを続けた。細胞を遠心分離により回収し、10mM PPB、pH7.0に再懸濁し、フレンチプレス(1,000kg/cm2)に3回かけて溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。その後上清を、25μM FADを含有する10mM PPB、pH8.0に対して透析し、粗酵素溶液をさらに精製した。
【0084】
硫酸アンモニウムを透析した上清に35%飽和まで添加し、その後形成した沈殿を15,000gで20分間遠心分離によりペレットとした。硫酸アンモニウムを65%飽和まで添加した上清を、15,000gで20分間遠心分離した。得られた沈殿を、25μM FADおよび1%マンノースを含有する10mM PPB、pH8.0に溶解し、同緩衝液に対して4℃で透析し、その後、25μM FADを含有する10mM PPB、pH8.0に対して透析した。透析した酵素溶液を、10mM PPB、pH8.0で平衡化した、RESOURCE Q カラム(GE ヘルスケア(Healthcare))に適用した。活性な素通り(flow-through)画分を回収し、FAOD活性を示さない吸着されたタンパク質を、1M NaClで溶出した。活性な素通り画分を回収し、10mM PPB、pH7.0に対して透析した。透析した酵素溶液を、10mM PPB、pH7.0で平衡化したHiLoad 16/60 Superdex 75 pg カラム(GE ヘルスケア)に適用した。ゲルろ過クロマトグラフィーを同緩衝液で行なった。活性画分を回収し、精製した酵素溶液を100μM FADを含有する10mM PPB、pH7.0に対して透析し、4℃で保管した。その精製酵素の純度は、SDS−PAGEにより確認し、濃度は、DC Protein Assay Kit(バイオ−ラッド(Bio-Rad)、CA、米国)を用いて測定した。結果を表4に示す。
【0085】
【表4】
【0086】
N56A変異体FAODのオキシダーゼ活性およびデヒドロゲナーゼ活性を測定した。変異体FAODのSV曲線および速度論的パラメーターは、図6および表5にそれぞれ示す。図6において、左のグラフは0〜5mMの基質濃度における活性を示し、右のグラフは基質濃度0〜2mMの拡大図である。N56A変異体は、両基質に対して有意に増加したデヒドロゲナーゼ活性を示す。
【0087】
【表5】
【0088】
実施例5
アマドリアーゼII FAOD変異体の製造と特徴付
【0089】
アマドリアーゼII(配列番号:3)由来の変異体を製造し特徴付けを行なった。N1−1 FAODとアマドリアーゼIIとのアラインメント(図1)および実施例1〜4の結果にもとづき、Asn52がFADからのプロトンおよび電子の移動に関与すると予測した。Asn52Ala変異を有する変異体を、実施例1に記載された方法にしたがって製造した。
【0090】
BL21(DE3)細胞を、アマドリアーゼII野生型またはN52A変異体を含有する発現ベクターで形質転換した。培養した細胞を回収し、10mlの10mM PPB、pH7.0に再懸濁し、フレンチプレス(1,000kg/cm2)に2回かけて溶菌した。ライセートを10,000g、4℃で20分間遠心分離し、上清を50,000rpm、4℃で60分間遠心分離した。その後、上清を25μM FADを含有する10mM PPB、pH8.0に対して透析し、粗酵素溶液を製造した。粗酵素溶液を、硫酸アンモニウム沈殿、陰イオン交換クロマトグラフィー(RESOURCE Q カラム(GE ヘルスケア))、およびゲルろ過クロマトグラフィー(HiLoad 16/60 Superdex 75 pg カラム(GE ヘルスケア))により精製した。
【0091】
FAOD活性を、基質としてのf−αValと共に4AA/TODB/POD(オキシダーゼ活性)およびPMS/DCIP(デヒドロゲナーゼ活性)を用いてアッセイした。結果を図7に示す。
【0092】
精製したアマドリアーゼII野生型およびAsn52Alaの速度論的パラメーターは図8にまとめる。変異体N52Aは、オキシダーゼ活性よりも約2倍高いデヒドロゲナーゼ活性を示した。
【0093】
実施例6
バイオセンサーの構築
【0094】
PnFPOX野生型またはN56A PnFPOXいずれかの40mU/5μl溶液を、2%AWP溶液と混合した。混合物5μlを再利用可能な金電極(表面積:7mm2)に適用し、30℃で30分間乾燥させた。電極を、UV−光で1分間照射し、固定化された野生型またはN56A FPOXを有する電極を製造した。電極を、2mMニトロソアニリン(NA;国際公開第2004/113900号のp.33ff.より公知)のPBS(pH7.4)溶液2mlに浸漬した。その後、+200mV対Ag/AgClを適用し、電流をモニターした。定常状態電流が観測された場合、異なる濃度のFVを含む試料溶液を反応混合物に添加し、電流の増加をモニターした。酸素の非存在下での酵素活性を測定するために、Arガスで反応チャンバー内を連続的に不活化した。
【0095】
図9は、酸素存在下または非存在下(Arガス下)における、FV濃度とPnFPOX N56Aまたは野生型酵素を有する電極の電流増加との相関関係を示す。電気化学反応に対する酸素の影響が観察されるが、酸素の存在の影響は野生型を用いた場合よりも少なかった。これらの結果は、本発明のフルクトシルアミノ酸オキシダーゼの活性は、溶存酸素レベルによる影響を受けにくいということを示唆した。
【0096】
実施例7
FPOX−C変異体の製造および特徴付
【0097】
Asn52Ala変異を有するFPOX−C(配列番号:7)由来の変異体を、実施例5と同様の方法で製造した。その粗酵素調製物を、さらに精製することなく、酵素活性アッセイに使用した。FAOD活性を、基質としての1mM f−αValまたは1mM f−αVal−Hisのいずれかと共に4AA/TODB/POD(Ox:オキシダーゼ活性)およびPMS/DCIP(DH:デヒドロゲナーゼ活性)を用いてアッセイした。結果を以下の表6に示す。
【0098】
【表6】
【0099】
変異体FPOX−C N52Aは、f−αValおよびf−αVal−Hisの両方に対してオキシダーゼ活性よりも非常に高いデヒドロゲナーゼ活性を示した。
【0100】
実施例8
ループ変異体の製造および特徴付
【0101】
PnFPOXは、フルクトシルヘキサペプチドに対して、FVHと比較して1%未満のレベルでオキシダーゼ活性を示すが、FPOX−Cは活性を示さない。他方、FPOX−Cは、FVHに対してP.n FPOXよりもより高い活性を示す。両オキシダーゼのアミノ酸配列の比較から、FAODとFPOXとの間の有意な相違は、それらのループ領域に見られる。
【0102】
ループ1領域
PnFPOX (61)Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Ala(72)(配列番号:13)
FPOX−C (61)Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Ser(72)(配列番号:14)
【0103】
ループ2領域
PnFPOX (109)Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Glu(124)(配列番号:15)
FPOX−C (109)Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Ala(126)(配列番号:16)
【0104】
ループ1領域(12aa)では、5つのアミノ酸残基がPnFPOXとFPOX−Cとで異なる。ループ2領域(16aa)では、2つのアミノ酸挿入と7つのアミノ酸置換が見られる。
【0105】
1PnFPOXとFPOX−Cとのループ領域が互に交換されているキメラループ変異体、ならびにそのN56A変異体を製造した。上記に示した部分アミノ酸配列にもとづき、8種類の変異体を製造した。
【0106】
Pn FPOX/L1ExC(すなわち、ループ1領域がFPOX−C由来のループ1領域に置換されているPn FPOX)
Pn FPOX/L1ExC/N56A(すなわち、ループ1領域がFPOX−C由来のループ1領域に置換されているPn FPOXのN56A変異体)
Pn FPOX/L2ExC(すなわち、ループ2領域がFPOX−C由来のループ2領域に置換されているPn FPOX)
Pn FPOX/L2ExC/N56A(すなわち、ループ2領域がFPOX−C由来のループ2領域に置換されているPn FPOXのN56A変異体)
FPOX−C/L1ExPn(すなわち、ループ1領域がPn FPOX由来のループ1領域に置換されているFPOX−C)
FPOX−C/L1ExPn/N56A(すなわち、ループ1領域がPn FPOX由来のループ1領域に置換されているFPOX−CのN56A変異体)
FPOX−C/L2ExPn(すなわち、ループ2領域がPn FPOX由来のループ2領域に置換されているFPOX−C)
FPOX−C/L2ExPn/N56A(すなわち、ループ2領域がPn FPOX由来のループ2領域に置換されているFPOX−CのN56A変異体)
【0107】
変異の導入および粗酵素の製造は、実施例1および5に記載されているように行なった。FAOD活性を、基質としての1mM f−αVal、1mM f−αVal−Hisまたは5mMフルクトシルヘキサペプチド(F−HP)のいずれかと共に4AA/TODB/POD(Ox:オキシダーゼ活性)およびPMS/DCIP(DH:デヒドロゲナーゼ活性)を用いてアッセイした。
【0108】
Pn FPOX/L1ExCおよびPn FPOX/L2ExCは、野生型P.n.FPOXと比較してf−αVal−Hisに対して高い酸化活性を示した。FPOX−C/L1ExPnは、野生型FPOX−Cにおいては観察されないF−HPへの活性を示した。興味深いことに、Pn FPOX/L2ExCは、F−HPに対して活性を増加させたが、Pn FPOX/L1ExCは、F−HPに対して活性をほとんど失った。これらの結果は、F−HPに対するP.n.FPOX活性の改善が、ループ領域の改変によりさらに改善できたことを示した。
【0109】
これらのループ変異体へのN56A変異の導入は、オキシダーゼ活性の減少とデヒドロゲナーゼ活性の増加をもたらした。Pn FPOX/L1ExC/N56AおよびPn FPOX/L2ExC/N56Aの両方が、f−αValに対してそれらのオキシダーゼ活性と比較してより高いデヒドロゲナーゼ活性を示した。Pn FPOX/L2ExC/N56Aのf−αValに対するデヒドロゲナーゼ活性は、Pn FPOX/N56Aと比較して際立った増加を示し、FPOX−C/N56Aのものよりもより高かった。
【0110】
Pn FPOX/L1ExC/N56AおよびPn FPOX/L2ExC/N56Aの両方が、f−αVal−Hisに対してそれらのオキシダーゼ活性と比較してより高いデヒドロゲナーゼ活性を示した。最も高いデヒドロゲナーゼ活性は、Pn FPOX/L2ExC/N56Aにおいて観察され、これはFPOX−C/N56Aのものとほとんど同じである。
【0111】
代表的な結果を以下の表7に示す。
【0112】
【表7】
【特許請求の範囲】
【請求項1】
配列番号1記載のアミノ酸配列の56位に対応する位置でアミノ酸残基AsnをAla、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼ。
【請求項2】
野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有し、かつ野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項3】
野生型フルクトシルアミノ酸オキシダーゼの30%以下のオキシダーゼ活性を有し、かつ野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項4】
野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項5】
配列番号1〜12からなる群より選択されるアミノ酸配列であり、配列番号1記載のアミノ酸配列の56位に対応する位置のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項6】
配列番号1記載のアミノ酸配列であり、56位のアミノ酸残基Ansが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項7】
配列番号1の109〜124位でアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaに置き換えられている請求項6記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項8】
配列番号2記載のアミノ酸配列であり、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項9】
配列番号3記載のアミノ酸配列であり、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項10】
配列番号7記載のアミノ酸配列であり、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項11】
配列番号7の61〜72位でアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaに置き換えられている請求項10記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項12】
請求項1〜11のいずれか1項に記載の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチド。
【請求項13】
請求項12記載のポリヌクレオチドを含むベクター。
【請求項14】
請求項13記載のベクターにより形質転換された宿主細胞。
【請求項15】
試料中の糖化タンパク質をアッセイする方法であって、該試料を請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼと接触させること、および該フルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む方法。
【請求項16】
試料中のHbA1cを消化してフルクトシルバリンまたはフルクトシルバリルヒスチジンを生成すること、該フルクトシルバリンまたはフルクトシルバリルヒスチジンを請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンまたはフルクトシルバリルヒスチジンの量を測定することを含むHbA1cをアッセイする方法。
【請求項17】
試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための装置であって、請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置。
【請求項18】
試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキット。
【請求項19】
電極上に固定された請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼを有する酵素電極。
【請求項20】
フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーであって、請求項19記載の酵素電極を作用電極として含む酵素センサー。
【請求項21】
配列番号1記載のアミノ酸配列であり、配列番号1の61〜72位のアミノ酸配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaが、配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項22】
配列番号1記載のアミノ酸配列であり、配列番号1の109〜124位のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項23】
配列番号7記載のアミノ酸配列であり、配列番号7の61〜72位のアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項24】
配列番号7記載のアミノ酸配列であり、配列番号1の109〜126位のアミノ酸配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaが、配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項1】
配列番号1記載のアミノ酸配列の56位に対応する位置でアミノ酸残基AsnをAla、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換することにより改変された変異体フルクトシルアミノ酸オキシダーゼ。
【請求項2】
野生型フルクトシルアミノ酸オキシダーゼと比較して減少されたオキシダーゼ活性を有し、かつ野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項3】
野生型フルクトシルアミノ酸オキシダーゼの30%以下のオキシダーゼ活性を有し、かつ野生型フルクトシルアミノ酸オキシダーゼの50%以上のデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項4】
野生型フルクトシルアミノ酸オキシダーゼと比較して増加したデヒドロゲナーゼ活性を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項5】
配列番号1〜12からなる群より選択されるアミノ酸配列であり、配列番号1記載のアミノ酸配列の56位に対応する位置のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項6】
配列番号1記載のアミノ酸配列であり、56位のアミノ酸残基Ansが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されているアミノ酸配列を有する請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項7】
配列番号1の109〜124位でアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaに置き換えられている請求項6記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項8】
配列番号2記載のアミノ酸配列であり、47位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項9】
配列番号3記載のアミノ酸配列であり、52位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項10】
配列番号7記載のアミノ酸配列であり、56位のアミノ酸残基Asnが、Ala、Cys、Phe、Met、SerおよびValから選択されるアミノ酸残基で置換されている請求項1記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項11】
配列番号7の61〜72位でアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaに置き換えられている請求項10記載の変異体フルクトシルアミノ酸オキシダーゼ。
【請求項12】
請求項1〜11のいずれか1項に記載の変異体フルクトシルアミノ酸オキシダーゼをコードする単離されたポリヌクレオチド。
【請求項13】
請求項12記載のポリヌクレオチドを含むベクター。
【請求項14】
請求項13記載のベクターにより形質転換された宿主細胞。
【請求項15】
試料中の糖化タンパク質をアッセイする方法であって、該試料を請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼと接触させること、および該フルクトシルアミノ酸オキシダーゼにより酸化された糖化タンパク質の量を測定することを含む方法。
【請求項16】
試料中のHbA1cを消化してフルクトシルバリンまたはフルクトシルバリルヒスチジンを生成すること、該フルクトシルバリンまたはフルクトシルバリルヒスチジンを請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼと接触させること、および酸化されたフルクトシルバリンまたはフルクトシルバリルヒスチジンの量を測定することを含むHbA1cをアッセイする方法。
【請求項17】
試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための装置であって、請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含む装置。
【請求項18】
試料中のフルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするためのキットであって、請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼおよび電子移動メディエーターを含むキット。
【請求項19】
電極上に固定された請求項1〜11のいずれか1項に記載のフルクトシルアミノ酸オキシダーゼを有する酵素電極。
【請求項20】
フルクトシルバリン、フルクトシルバリルヒスチジン、HbA1cまたはフルクトシルヘキサペプチドをアッセイするための酵素センサーであって、請求項19記載の酵素電極を作用電極として含む酵素センサー。
【請求項21】
配列番号1記載のアミノ酸配列であり、配列番号1の61〜72位のアミノ酸配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaが、配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項22】
配列番号1記載のアミノ酸配列であり、配列番号1の109〜124位のアミノ酸配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluが、配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項23】
配列番号7記載のアミノ酸配列であり、配列番号7の61〜72位のアミノ酸配列:Ile Arg Leu Arg Asn Lys Val Asp Leu Gln Met Serが、配列:Val Ser Leu Arg Asn Pro Val Asp Leu Gln Leu Alaで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【請求項24】
配列番号7記載のアミノ酸配列であり、配列番号1の109〜126位のアミノ酸配列:Lys Gln Tyr Gln Ala Leu His Asp Ala Gly Ala Gly Leu Glu Lys Thr His Alaが、配列:Ser Gly Tyr Gln Ala Leu Val Asp Ala Gly Leu Asp Ala Thr Asn Gluで置き換えられているアミノ酸配列を有する変異体フルクトシルアミノ酸オキシダーゼ。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−500729(P2013−500729A)
【公表日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2012−523236(P2012−523236)
【出願日】平成22年8月2日(2010.8.2)
【国際出願番号】PCT/EP2010/004708
【国際公開番号】WO2011/015326
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(501205108)エフ ホフマン−ラ ロッシュ アクチェン ゲゼルシャフト (285)
【出願人】(503195850)有限会社アルティザイム・インターナショナル (31)
【Fターム(参考)】
【公表日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成22年8月2日(2010.8.2)
【国際出願番号】PCT/EP2010/004708
【国際公開番号】WO2011/015326
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(501205108)エフ ホフマン−ラ ロッシュ アクチェン ゲゼルシャフト (285)
【出願人】(503195850)有限会社アルティザイム・インターナショナル (31)
【Fターム(参考)】
[ Back to top ]