
ブタサーコウイルス、ワクチンおよび診断試薬
【課題】生理標本または組織標本、特にPMWS症候群のブタの病変部の標本から単離可能なブタのサーコウイルス(CIRCOVIRUS)、特にII型サーコウイルスを提供する。
【解決手段】離乳後多機能萎縮症候群(post−weaning multisystemic Wasting syndrome:PMWS)の農場から得られた肺または神経節標本から単離した新規なブタサーコウイルス(CIRCOVIRUS)株と、この株の精製物、その無毒化または不活性化ワクチン、組換え生ワクチン、プラスミドワクチンおよびサブユニットワクチンと、診断試薬および診断方法。さらに、インビトロ発現ベクターのサブユニットの製造に用いられるDNA断片またはウイルスまたはプラスミド型のインビトロ発現ベクターに組み込まれる配列としてのDNA断片。
【解決手段】離乳後多機能萎縮症候群(post−weaning multisystemic Wasting syndrome:PMWS)の農場から得られた肺または神経節標本から単離した新規なブタサーコウイルス(CIRCOVIRUS)株と、この株の精製物、その無毒化または不活性化ワクチン、組換え生ワクチン、プラスミドワクチンおよびサブユニットワクチンと、診断試薬および診断方法。さらに、インビトロ発現ベクターのサブユニットの製造に用いられるDNA断片またはウイルスまたはプラスミド型のインビトロ発現ベクターに組み込まれる配列としてのDNA断片。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、PMWS症候群(ブタ多機能萎縮症候群:Porcine Multisystemic Wasting Syndrome)(離乳後多機能萎縮症候群:Post-weaning Multisystemic Wasting Syndromeともいわれる)の原因となる新規なブタサーコウイルス(PCV,Porcine Circo Virus)株と、その検出試薬および検出方法と、ワクチン接種方法と、ワクチンと、これらワクチンおよび試薬の製造方法とに関するものである。
【背景技術】
【0002】
ブタサーコウイルス(PCV)はブタ腎臓細胞株PK/15中に無毒病原性汚染物質として最初に見い出された。このウイルスPCVはチキン貧血症ウイルス(CAV、Chicken Anaemia Virus)および嘴・羽症候群ウイルス(PBFDV、Pscittacine Beak and Feather Disease Virus)とともにサーコウイルス群(Circoviridae)に分類された。これらのウイルスはエンベロプのない小さなウイルス(15〜24nm)で、その共通な特徴は1.76〜2.31kbの環状一本鎖のDNAからなるゲノムを含む点にある。このゲノムは約30kDaのポリペプチドをコード化すると考えられていた(非特許文献1)が、最近の研究で、複雑な転写がわかってきた(非特許文献2)。さらに、上記3種類のサーコウイルス(circovirus)はそのヌクレオチド配列に有意な相同性はなく、また、共通の抗原決定基もない。
【非特許文献1】Todd et al., Arch Virol 1991, 117; 129-135
【非特許文献2】Meehan B. M. et al., 1997,78; 221-227
【0003】
PK/15細胞由来のPCVは病原性ではないと考えられる。この配列は「非特許文献3」が明らかにしている。PCV株が病原性で、PMWS症候群と関連があるかもしれないと考えられるようになったのはほんの最近のことである(非特許文献4、5)。Nayer達はブタPCVのDNAがPMWS症候群を示すことをPCR法を用いて見い出した。しかし、野生型PCV株は単離または精製されていない。
【非特許文献3】"B. M. Meehan et al., J. Gen. Virol 1997 (78) 221-227"
【非特許文献4】Gupi P. S. Nayar et al., Can. Vet. J, Vol. 38, 1997: 385-387
【非特許文献5】Clark E. G., Proc. Am. Assoc. Swine Prac. 1997; 499-501
【0004】
カナダ、米国およびフランスで見られるPMWS症候群の臨床的な特徴は、体重を徐々に失い、頻呼吸症、呼吸困難および黄疸等を発症することである。病理的な面では、リンパ性または肉芽種性浸潤、リンパ節腫脹、まれには肝炎およびリンパ性または肉芽種性腎炎を発症する(非特許文献6〜10)。
【非特許文献6】Clark E. G., Proc. Am. Assoc. Swine Prac. 1997; 499-501;
【非特許文献7】La Semaine Veterinaire 1996 (834)、
【非特許文献8】La Semaine Veterinaire No. 26;
【非特許文献9】La Semaine Veterinaire 1997 (857):54;
【非特許文献10】Gupi P. S. Nayar et al., Can. Vet. J, Vol. 38, 1997: 385-387
【発明の開示】
【発明が解決しようとする課題】
【0005】
本出願人は、カナダ、米国(カリフォルニア)およびフランス(ブリタニー)の農場から得た肺または神経節の標本から5つの新規なPCV株(以下、本発明のサーコウイルス(circovirus)という)を単離することに成功した。これらのウイルスはPMWS症候群のブタの病変部に検出されるが、健康なブタには検出されない。
【0006】
本出願人はさらに、これらの4種の株(カナダおよび米国の株)と2種のフランスの株のゲノムの配列決定した。これらの株はヌクレオチドレベルで互いに非常に強い相同性(96%以上)超をを示す、一方、PK/15株ははるかに弱い相同性(約76%)しか示さない。従って、新規のタイプのブタサーコウイルス (circovirus)を代表するものと考えられ、PK/15に代表されるタイプI(第I群)に対して、本発明の新規な株はタイプII(第II群)とよぶことにする。
【課題を解決するための手段】
【0007】
本発明の対象は単離または精製物の形をした上記定義の第II群のブタサーコウイルス(circovirus)にある。
本発明はさらに、生理標本または組織標本、特にPMWS症候群のブタの病変部の標本から実施例に記載した方法で単離可能なブタのサーコウイルス(circovirus)、特にII型サーコウイルスを提供する。
【0008】
本発明の対象はECACC(European Collection of Cell Cultures, Centre for Applied Microbiology & Research, Porton Down, Salisbury, Wiltshire SP4 0JG, United Kingdom)に寄託された下記の5種の株の精製物にある:
1997年10月2日木曜日寄託:
寄託番号第V97100219号(以下、Imp.1008PCVという)
寄託番号第V97100218号(以下、Imp.1010PCVという)
寄託番号第V97100217号(以下、Imp.999PCVという)
1998年1月16日金曜日寄託:
寄託番号第V98011608号(以下、Imp.1011−48285という)
寄託番号第V98011609号(以下、Imp.1011−48121という)
【0009】
本発明はさらに、病気のブタから単離したブタサーコウイルス(circovirus)および/または本発明株と有意な血清学的類似性を有するサーコウイルスおよび/またはPK/15のPCVとハイブリダイゼーションが全く無く、厳密な条件下で本発明株とのクロスハイブリダイゼーションを有するサーコウイルスに関するものである。
【発明を実施するための最良の形態】
【0010】
ウイルス性株の増殖、特に全抗原(例えばウイルス)および/またはサブユニット(例えばポリペプチド)の製造は、PMWS症候群のブタの病変部の生理学標本または組織標本から単離されたウイルス株、特にブタ腎臓株細胞を汚染されていない細胞株(特にPCV、ペスチウイルス、ブタアデノウイルスおよびブタパルボウイルスに汚染されていないPK/15細胞)で増殖するのが有利である。
これらの単離株がPK/15細胞での培養で非常に生産的であり、ウイルスまたは抗原の製造、特に不活性化ワクチンの製造に有利であるということは驚くべきことであり、予想できなかったことである。
【0011】
従って、本発明の他の対象は本発明のサーコウイルス(circovirus)または病変部細胞、特にPMWS症候群のブタの細胞株の生理学標本または組織標本から単離される任意のブタサーコウイルスの少なくとも1つに感染された細胞をインビトロで培養した細胞株、特にPK/15細胞で継代した後に単離されるサーコウイルス(circovirus)の調製方法にある。
本発明のさらに別の対象は上記の培養抽出物または上澄み(必要に応じて一般的な方法でさらに精製されていてもよい)と、一般にインビトロの培養から得られる任意の抗原製剤にある。
本発明のさらに別の対象は免疫抗原性の有効成分と、少なくとも1つの上記定義の抗原を含むワクチンにある。
【0012】
本発明のさらに別の対象は、無毒化した生全ワクチンをベースとする免疫抗原性の有効成分またはこの有効成分を精製して得られるワクチンにある。この無毒化は通常の方法、例えば細胞での継代、好ましくはブタ細胞、特にPK/15細胞等の株細胞での継代(特に50〜150、特に100回の継代)で行なわれる。これらのワクチンは一般に獣医学上許容される賦形剤または希釈液と、さらには必要に応じて添加される獣医学上許容される補助剤およびフリーズドライ安定剤とを含むことができる。
上記の抗原製剤およびワクチンは103〜106のTCID50を有するのが好ましい。
【0013】
本発明のさらに別の対象は、不活化した本発明のサーコウイルス抗原をベースにした免疫抗原性有効成分またはワクチンにある。このワクチンは獣医学上許容される賦形剤または希釈液、さらには必要に応じて添加される獣医学上許容される補助剤を含むことができる。
【0014】
本発明のサーコウイルス(断片の形でもよい)は当業者に公知の方法で不活性化される。この不活性化は化学的方法、例えば抗原をホルムアルデヒド(ホルマリン)、パラフォルムアルデヒド、竈−プロピオラクトン、エチレンイミンまたはその誘導体等の試薬で処理して行なうことができる。好ましい不活性化法は試薬、特にエチレンイミンまたは竈−プロピオラクトンで処理する方法である。
本発明の不活性化ワクチンにはアジュバンドを加え、当業者に公知の方法でエマルジョンの形、例えば油中水エマルジョンまたは水中油エマルジョン等の乳剤の形にするのが好ましい。一般的なアジュバンド化合物を有効成分に添加してアジュバンド処理することもできる。
【0015】
使用可能な補剤(アジュバンド)としては例えば水酸化アルミニウム、サポニン(例えば非特許文献11参照)、Avridine(商標登録)(非特許文献12参照)、DDA(ジメチルジオクタデシルアンモニウムブロミド、非特許文献13参照)、ポリホスファゼン(非特許文献14参照)、さらには有機油、スクアラン(例えばSPT乳剤、非特許文献15参照)、スクアレーン(例えばMF59、非特許文献16参照)をベースとする水中油乳剤または代謝可能な油をベースとする油中水乳剤(好ましくは特許文献1、2に記載の乳剤を挙げることができる。補剤の組み合わせ、例えば乳剤AvrIDine(商標登録)またはDDAとの組み合せた選択することもできる。
これらのワクチンは106〜108のTCID50を有するのが好ましい。
【非特許文献11】Quillja saponinまたはQuil A;"Vaccin Design, The Subunit and Adjuvant Approach, 1995, Michael F. Powel と Mark J. Newman編、Plennum Press, New-York and London, p.210"
【非特許文献12】Vaccine Design p. 148
【非特許文献13】Vaccine Design p. 157
【非特許文献14】Vaccine Design p. 204
【非特許文献15】Vaccine Design p. 147
【非特許文献16】Vaccine Design p. 183
【特許文献1】国際特許第WO-A-94−20071号公報
【特許文献2】米国特許第5,422,109号明細書
【0016】
生ワクチン補剤は不活性化ワクチン用のものから選択するこができる。乳剤が好ましい。不活性化ワクチン用のものには特許文献3に記載のものも加えることができる。
凍結安定剤としては例えばSPGA(非特許文献17)、ソルビトール、マンニトール、スターチ、スクロース、デキストランまたはグルコース等の炭水化物、アルブミンまたはカゼイン等のプロテイン、これらの化合物の誘導体またはアルカリ金属燐酸塩等の緩衝剤を挙げることができる。
【特許文献3】国際特許第WO-A-94−16681号公報
【非特許文献17】Bovarnik et al., J. Bacteriology 59, 509, 950
【0017】
本件出願人は配列ID番号1〜4(必要な場合にはさらに配列ID番号6)で識別される4つの単離株のゲノムを得た。
本発明のさらに他の対象はこれらの配列の一部または全てを含むDNA断片にある。本発明がこれと均等な配列すなわち上記配列の株の特徴または上記配列によりコードされるポリペプチドの機能を変更しない配列も含むものであるということは当然である。また、コードの欠失(degenerescence)により変化した配列も含むということは理解てきよう。
本発明はさらに、厳密な条件下で上記配列と混成可能であり、および/または、上記定義の第II群に属する本発明株と高い相同性を有するという意味で均等な配列も含むものである。
【0018】
これらの配列およびその断片は適当なベクターを用いてインビトロまたはインビボでポリペプチドの発現に有利に用いることができる。
特に、これに使用可能な本発明のDNA断片を形成する読み取り枠はタイプIIのサーコウイルスのゲノム配列で同定されている。本発明は、少なくともこれらの読み取り枠(対応するアミノ酸配列)の一つを含む全てのポリペプチドに関するものである。特に、本発明は主としてORF4、ORF7、ORF10またはORF13で構成されるプロテインに関するものである。
インビトロのサブユニットを発現するための発現手段としてはE. coliまたはバキュロウイルス(特許文献3)を用いるのが好ましい。
【特許文献4】米国特許第4,745,051号明細書
【0019】
上記の単数または複数のコード配列またはその断片をバキュロウイルスゲノム(例えばAutographa californica Nuclear Polyhedrosis Virus AcNPV)に組み込み、次いで、それを昆虫細胞、例えばSpodoptera fruigiperda Sf9(寄託番号ATCC CRL 1711)で増殖される。また、イースト(例えば酵母)またはほ乳類細胞(例えばCHO、BHK)等の真核細胞でサブユニットを製造することもできる。
【0020】
本発明のさらに他の対象は、これらの発現手段によってインビトロで製造され、必要に応じて一般的な方法で精製されたポリペプチドにある。本発明のさらに他の対象は、このようにして得られた少なくとも1つのポリペプチドまたはその断片を獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤をさらに含むことができる)中に含むサブユニットワクチンにある。
【0021】
組み換え生ワクチンの製造時にインビボ発現させるには単数または複数のコード化配列またはその断片を単数または複数のポリペプチドを発現できる条件下で適当な発現ベクターに挿入する。適切なベクターとしては好ましくはブタで増殖可能でブタに対しては非病原性の生ウイルスを当業者に周知の方法で使用することができる。特に、アウジェスキー病ウイルス、ブタアデノウイルス等のブタヘルペスウイルス、ポックスウイルス、特にワクシニアウイルス、トリポックスウイルス、カナリアポックスウイルス、ブタポックスウイルス等を用いることができる(特許文献4〜7)。
【特許文献5】国際特許第WO-A-90−11092号公報
【特許文献6】国際特許第WO-A-93−19813号公報
【特許文献7】国際特許第WO-A-94−21797号公報
【特許文献8】国際特許第WO-A-95−20660号公報
【0022】
従って、本発明の他の対象は、このようにして精製されたベクターと、組み換え生ワクチンまたはプラスミドワクチン(ポリヌクレオチドまたはDNAワクチン)にある。これらのワクチンは獣医学上許容される賦形剤または希釈液を含むことができる。
本発明のワクチン(無毒化、不活性化、サブユニット、組み換えおよびプラスミド生ワクチン)は本発明の一つまたは複数(2、3つ)のサーコウイルスから成る一つまたは複数の有効成分(抗原)を含むことができる。
【0023】
本発明の上記各種ワクチンはブタの他の病原体に対するワクチンと組み合わせることができる。特にPMSW症候群に関連する病原体に対するワクチンと組み合わせることができる。本発明のワクチン、特に不活性化ワクチンはブタの他の病原体に対応する他のワクチン価を含むことができる。これらのブタの他の病原体の中では好ましくはPRRS(Porcine Reproductory and Respiratory Syndrome :ブタ生殖器および呼吸器症候群)(特許文献8〜10および非特許文献18を参照)および/またはMycoplasma hyopneumonia(特許文献11〜13および非特許文献19を参照)を挙げることができる。他の重要なワクチン価の中ではアクチノバシラス胸膜肺炎菌、E. coli、ブタ萎縮鼻炎(porcine Atrophic Rhinitis)、仮性狂犬病(アウジェスキー病)、ブタコレラ、ブタインフルエンザを挙げることができる。
【特許文献9】国際特許第WO-A-93−07898号公報
【特許文献10】国際特許第WO-A-94−18311号公報
【特許文献11】フランス国特許第2,709,966号公報
【非特許文献18】"C. Chareyre 他、IPVS Congress,Birmingham, England, 5-9 July 1998, p. 139の議事録"
【特許文献12】欧州特許第597,852号
【特許文献13】第550,477号、
【特許文献14】第571,648号
【非特許文献19】"第15回IPVS CongressのO. Martinon 達、p. 157, 284, 285およびG. Reynaud達、p. 150"
【0024】
本発明のさらに他の対象は、本発明のサーコウイルスに対してブタの免疫反応を誘発する方法、特にブタに有効なワクチン接種方法にある。
このワクチン接種方法では上記ワクチンを一回または複数回にわたってブタに投与する。同じワクチン接種プロトコルで複数種の前記ワクチンを組み合わせることも可能である。
このワクチンは大人のブタだけでなく、若いブタまたは妊娠中の雌に対して投与することができる。妊娠中の雌にワクチン接種することによって新生児に受動免疫(母親抗体:maternal antibody)を与えることができる。
【0025】
本発明はさらに、本発明のブタサーコウイルスの存在を診断する方法を提供する。従って、本発明の他の対象は診断テストと下記試薬を用いたテスト方法にある。
互いに異なるサーコウイルスの配列が分かると共通の配列を定義でき、公知の全てのブタサーコウイルスを識別できる試薬を製造することができる。
当業者は特定の診断を実施するために対応するPK/15サーコウイルスの配列とほとんどまたは全く相同性を示さない領域に対応する配列の断片を選択することができる。
【0026】
配列を並べることによって当業者は所望の試薬を選択することができる。
第1の試薬は上記DNA配列およびその断片で構成され、それは周知のハイブリッド法またはPCR(ポリメラーゼ鎖反応)法によってプローブまたはプライマーとして用いられる。
第2の試薬は上記ウイルスの配列によってコードされたポリペプチドまたはベクター(前記参照)を用いて発現されたまたは公知のペプチド合成法によって化学的に合成されたポリペプチドで構成される。
第3および第4の試薬はウイルス、ポリペプチドまたは断片、抽出物またはDNA配列でコードされたものから一般的な方法で製造可能なポリクローナル抗体およびモノクローナル抗体からなる。
【0027】
上記の第2、第3および第4の試薬は本発明の対象である診断法で用いることができる。この診断法では検査するブタから得られた生理学的液体標本(血液、血漿、血清等)または組織標本(神経節、肝臓、肺、腎臓等)中に本発明のサーコウイルスに特有な抗原の存在を調べて抗原自体を検出するか、この抗原に対する抗体を検出する。
【0028】
本発明の抗原および抗体は公知の任意の臨床診断法で用いることができるが、
上記の抗原および抗体を獣医、ブリーダーまたは動物の飼い主が直接現場で使用可能な方法で用いるのが好ましい。臨床技術および現場技術を有する当業者は上記の抗原および/または抗体を試薬として用いることができる。
【0029】
本発明で用いるのに好ましい診断法はウェスタンブロット法、免疫抗体法、ELISA(酵素結合免疫測定法)および免疫クロマトグラフィーである。
免疫クロマトグラフィー法に関しては特に非特許文献20と特許文献14〜29を参照することができる。
【非特許文献20】"Robert F. Zulk et al., Clin. Chem. 31/7, 1144-1150 (1985) "
【特許文献15】国際特許第WO-A-88−08534号公報
【特許文献16】国際特許第WO-A-91−12528号公報
【特許文献17】欧州特許第291,176号公報
【特許文献18】欧州特許第299,428号公報
【特許文献19】欧州特許第291,194号公報
【特許文献20】欧州特許第284,232号公報
【特許文献21】米国特許第5,120,643号明細書
【特許文献22】米国特許第5,030,558号明細書
【特許文献23】米国特許第5,266,497号明細書
【特許文献24】米国特許第4,740,468号明細書
【特許文献25】米国特許第5,266,497号明細書
【特許文献26】米国特許第4,855,240号明細書
【特許文献27】米国特許第5,451,504号明細書
【特許文献28】米国特許第5,141,850号明細書
【特許文献29】米国特許第5,232,835号明細書
【特許文献30】米国特許第5,238,652号明細書
【0030】
標本中に特有な抗体は間接法、競合法または置換法によって検出するのが好ましい。そのためには抗原自体を診断試薬として用いるか、抗体の認識を保持する抗原断片を診断試薬として用いる。標識化はペルオキシダーゼを用いた標識化または特殊な標識化、好ましくはコロイド性金を用いた標識化が有利である。
標本中の抗原自体をこの抗原に特有な標識化した抗体を用いて検出することもできる。標識化は上記のものが有利である。
競合法または置換法または抗原自体の検出に用いられる抗原に特有な抗体は、モノクローナル抗体またはポリクローナール抗体、これら抗体の断片、好ましくはFabまたはF(ab)'2断片であるである。
【0031】
本発明はさらに、上記抗体を生理学的液体標本または組織標本中の抗原の検出またはこれら標本または試験片中に存在する抗体の検出のための診断試薬として使用可能な本発明抗原に特有のモノクローナル抗体またはポリクローナル抗体の製造方法に関するものである。本発明はものでさらに、これらの抗体の免疫機能を有する断片、特にF(ab)およびF(ab)'2断片は本発明に含まれる。
抗体は一般的な方法で製造することができる。特に非特許文献21または非特許文献22を参照することができる。これらの内容は本件明細書の一部を成す。
特に、抗原またはその断片の少なくとも1つで免疫化されたネズミの脾臓細胞を適当な骨髄腫細胞と融合(この方法自体は公知)することができる。
【非特許文献21】"Antibodies, A Laboratory Manual, 1988, Cold Spring Harbor Laboratory, USA"
【非特許文献22】"J. W. Goding, Monoclonal Antibodies: Principles and Practice, Academic Press Inc.,"
【0032】
本発明の他の対象は上記抗原に特有なモノクローナル抗体またはポリクローナル抗体、特にネズミまたはウサギの抗体の純粋または部分的に純粋な製剤或いは粗原料からなる製剤にある。
本発明では上記のDNA配列を基にした所望のエピトープ(ワクチン用エピトープまたは診断用エピトープ)を決めることができる。本発明のサーコウイルスのゲノムのDNA配列から、エピトープを決めることができる。当業者は公知の方法、例えば適切なコンピュータプログラムまたはPEPSCANを用いてエピトープを求めることができる。このエピトープはプロテインの免疫優性領域であり、プロテインの表面に露出した領域である。このエピトープは抗体によって認識され、診断の分野において診断目的の抗体の調製または診断試薬として用いられる対応するペプチドの製造に用いられる。
少なくともエピトープは8〜9個のアミノ酸を有するペプチドである。一般には最小でも13〜25個のアミノ酸が好ましい。
【0033】
当業者は一つまたは複数の上記技術および利用可能なその他の技術を用いて診断用のペプチドまたは抗体で用いるエピトープを見付けることができる。
従って、本発明の他の対象はこの抗原および/またはこの抗原に特有なモノクローナル抗体またはポリクローナル抗体を含む診断キットにある。これらは特に、前記の診断技術による診断キットである。
【0034】
以下、下記の図を参照して本発明の実施例をさらに詳細に説明する。しかし、本発明が下記実施例に限定されるものではない。
配列IDリスト
配列ID番号1 = Imp.1011−48121株のゲノムのDNA配列
配列ID番号2 = Imp.1011−48285株のゲノムのDNA配列
配列ID番号3 = Imp.999株のゲノムのDNA配列
配列ID番号4 = Imp.1010株のゲノムのDNA配列
配列ID番号5 = PK/15株のゲノムのDNA配列
配列ID番号6 = 1997年10月3日のフランスの最初の出願におけるImp.999株のゲノムのDNA配列
【実施例】
【0035】
実施例1
ブタサーコウイルス(circovirus)株の培養および分離
組織標本はフランス、カナダおよび米国で子豚の肺およびリンパ節から集めた。これらの子豚は離乳後多機能萎縮症候群に典型的な臨床的徴候を示したものである。ウイルスを単離するため組織標本は死体解剖直後に−70℃で冷凍した。
ウイルス単離のために無菌の乳鉢と乳棒を用いて無菌の砂で組織を粉砕し、イール(Earl)塩 (EMEM, BioWhittaker UK Ltd., Workingham, UK)、ペニシリン(100IU/ml)およびストレプトマイシン(100μm/ml)を含む最小培地(MEM−SA培地)中に約15%の組織標本を含む懸濁液を調製した。粉砕物をMEM−SAに溶解し、+4℃、3000gで30分間、遠心分離にかけて上澄みを回収した。
【0036】
細胞培地の接種前に、100μlのクロロホルムを2mlの各上澄みに添加し、室温で10分間混合した。この混合物を超遠心分離管に移し、10分間3000gで遠心分離して上澄みを回収した。次いで、この上澄みをウイルス分離実験で種菌として用いた。
全てのウイルス単離研究はブタサーコウイルス(PCV)、ペスチウイルス、ブタアデノウイルスおよびブタパルヴォウイルスに感染していないことが分かっているPK/15細胞培地で実施した(非特許文献23)。
【非特許文献23】Allan G. et al., Pathogenesis of porcine circovirus experimental infections of colostrum-deprived piglets and examination of pig foetal material. Vet. Microbiol. 1995, 44, 49-64
【0037】
ブタサーコウイルスの単離は下記方法によって実施した:
PK/15細胞の単一層をトリプシン処理(トリプシン−versene混合物)によって集密的な培養から解離し、最終濃度が約400,000細胞/1mlであるペスチウイルス(=MEM−G培地)に感染していない15%のウシ胎児血清を含むMEM−SA培地に溶解した。次に、この細胞懸濁液の10mlの部分標本を2mlの前記接種材料の部分標本と混合し、最終混合物を2つの25cm2ファルコンフラスコに6mlの容積に分取した。+37℃で18時間、10%のCo2を含む雰囲気下で培養した。
【0038】
培養後、半集密的な単一層を300mMのD−グルコサミン(Cat # G48175、Sigma-Aldrich株式会社、Poole, UK)で処理し(非特許文献24)、さらに37℃で48〜72時間培養を続けた。この最終培養に続いて、2つのファルコンの1方を3回凍結/融解サイクルにかけた。残りのファルコンのPK/15細胞はトリプシン−versene溶液で処理し、20mlのMEM−G培地中に再懸濁し、400,000細胞/ml濃度で75cm2ファルコン中に接種した。新しく接種されたフラスコは凍結/融解サイクル後に得られた対応する可溶化液5mlを添加して「超感染」させた。
【非特許文献24】Tischr I. et al., Arch. Virol., 1987 96 39-57
【0039】
実施例2
免疫抗体法または切片上ハイブリッド形成法によるブタサーコウイルスを検出するためのするための細胞培地標本の調製
5mlの容積の「超感染」した懸濁液を回収し、無菌で脱脂したカバーガラスを含む直径55mmのペトリ皿中に接種した。フラスコ中およびカバーガラス上で+37℃で培養し、実施例1に記載のグルコサミンで処理した。カバーガラス上での培養はグルコサミン処理後24〜48時間で回収し、室温で10分間アセトンを用いて、または4時間緩衝剤処理した10%のホルムアルデヒドを用いて固定した。この固定後、全てのカバーガラスをシリカゲル上で−70℃で貯蔵してから、ハイブリッド形成の研究および細胞免疫の研究に使用した。
【0040】
実施例3
ハイブリッド形成法(in situ)によるPCV配列の検出法
ハイブリッド形成法(in situ)は病気のブタから集めた組織に対して実施し、ホルムアルデヒドで固定し、ウイルス単離(実施例2参照)用に接種した細胞培地の調整で用い、カバーガラスに固定した。
PK/15ブタサーコウイルス(PCV)および感染性ニワトリ貧血症ウイルス(CAV)に対応する完全なゲノムプローブを用いた。1.7キロの塩基ペア(kbp)の挿入断片の形にクローン化されたPCVゲノムの複製型を含むプラスミドpPCVI(Meehan B. et al. Sequence of porcine circovirus DNA: affinities with plant circovirus, J. Gen. Virol. 1997, 78, 221-227)をPCV固有DNA源として用いた。2.3kbpの鳥サーコウイルスCAVの複製型を含む鳥プラスミドpCAAIを負対照として用いた。2つのプラスミドのグリセロール源をアルカリ法(非特許文献25)によってプラスミドの製造および精製で用い、これらはプローブ調製用のテンプレートとして用いた。PCVおよびCAVの完全なゲノムを表すサーコウイルスプローブはメーカ推薦の方法で市販の非放射性ラベリングキット("DIG DNA Labelling Kit"、Boehringer Mannheim, Lewis, UK)を用いて上記精製プラスミド(各プローブに1μg)およびヘキサヌクレオチドプライマーから製造した。
【非特許文献25】Sambrook J. et al. Molecular cloning: A Laboratory Manual. 2nd Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1988
【0041】
ハイブリッド形成法(in situ)で使用する前に、プローブをジゴキシジェニンで標識化し、容積50〜100μlの純水に溶解した。
パラフィンに包み、ホルムアルデヒドで固定した病気のブタの組織標本と、ホルムアルデヒドで固定した感染細胞培地の製剤を下記方法でPCV核酸検出のために調製した:
5μm厚の切片をパラフィンに包まれた組織塊から切り出し、濃度を次第に薄くしたアルコール溶液中で再水和した。ホルムアルデヒドで固定した組織切片および細胞培地を15分間および5分間それぞれ5mMのEDTAを含む0.05MのTris−HCL緩衝剤(pH7.6)を用いた0.5%のプロテナーゼK溶液中で+37℃で培養した。次に、スライドガラスを加圧滅菌した蒸留水中の1%グリシン溶液中に30秒配置し、0.01MのPBS緩衝剤(燐酸緩衝食塩水)(pH7.2)で2度洗浄し、最後に無菌蒸留水で5分間洗浄した。スライドガラスを最後に外気で乾燥し、プローブと接触させた。
【0042】
各組織/プローブ製剤を清潔な脱脂カバーガラスで被覆し、+90℃の炉内に10分間配置した後、氷塊と1分間接触させ、最後に+37℃で18時間培養した。次に、調整物を2Xクエン酸ナトリウム塩(SSC)緩衝剤(pH7.0)に短時間含浸した後、2XのSSC緩衝剤中で5分間、2回洗浄し、最後にPBS緩衝剤中で5分間、2回洗浄した。
洗浄後、調整物を0.1Mのマレイン酸、0.15MのNaCL(pH7.5)(マレイン緩衝剤)の溶液中に10分間含浸し、マレイン緩衝剤中の遮断試薬の1%溶液(Cat # 1096176, Boehringer Mannheim UK, Lewis, East Sussex, UK)中で+37℃で20分間培養した。
【0043】
次に、調整物を遮断緩衝剤で希釈された抗ジゴキシジェニンモノクローナル性抗体(Boehringer Mannheim)の1/250溶液中で+37℃で1時間培養し、PBS中で洗浄し、最後に抗ネズミ免疫グロブリン抗体を用いて+37℃で30分間培養した。調整物をPBSで洗浄し、内因性ペルオキシダーゼ作用を室温で20分間、PBS中の0.5%水酸化ペルオキシド溶液で処理して遮断した。調整物を再度PBS中で洗浄し、使用直前に調製した3−アミノ−9−ジエチルカルバゾル(AEC)基質(CambrIDge Bioscience, CambrIDge, UK)で処理した。
最後に水道水で洗浄した後、調整物をヘマトキシリンで逆染色し、水道水で"青色化"して固定液(GVA Mount, CambrIDge Bioscience, CambrIDge, UK)で顕微鏡のカバーガラスに固定した。対照実験には無関係な負プローブ(CAV)と正プローブ(PCV)を病気のブタおよび病気ではないブタから得られた標本で使用することが含まれる。
【0044】
実施例4
免疫抗体法によるPCVの検出法
アセトンを用いて固定した全ての細胞倍地の最初のスクリーニングを貯蔵した大人のブタ血清の1/100希釈を用いた間接免疫抗体法(IIF)で実施した。この貯蔵血清は北アイルランドの25匹の大人の雌ブタからの血清を含み、PCV:ブタパルボウイルス、ブタアデノウイルスおよびPRRウイルスを含む広範囲のブタのウイルスに対する抗体を含むことで知られている。血清(PBSで希釈)を細胞培地と+37℃で1時間接触させた後、PBS中で2回洗浄してIIFを実施した。細胞培地をイソチオシアン酸フルオレセインに結合したPBSで1/80に希釈したウサギの抗ブタ免疫グロブリン抗体を用いて1時間染色し、PBSで洗浄し、グリセロール緩衝剤に固定し、紫外線下で顕微鏡観察した。
【0045】
実施例5
病気のブタの組織における切片上ハイブリッド形成結果
フランス、カナダおよびカリフォルニアの多機能性萎縮病変を有する子豚から集めた組織から調製したPCVゲノムプローブを用いた切片上ハイブリッド形成法から、研究された複数の病変部に病変部に関連するPCV核酸の存在が示された。これに対してPCVゲノムプローブを病気でないブタから集めた組織に用い時またはCAVプローブを病気のブタの組織に用いた時には何の徴候も見られなかった。PCV核酸の存在はサイトプラズムおよびカリフォルニアの子豚の肺の病変部を浸潤する多数の単核細胞の核に認められた。PCV核酸の存在は肺細胞、気管支および細気管支の上皮細胞および細動脈、細静脈およびリンパ管の内皮細胞にも見られた。
【0046】
病気のフランスのブタではPCV核酸の存在は多数の濾胞性リンパ細胞のサイトプラズマおよびリンパ節の同様な毛細血管内の単核細胞に検出された。また、PCV核酸は一次的合胞体にも検出された。これらの検出結果から、カリフォルニアのブタの肺、フランスのブタの腸間膜リンパ節およびカナダのブタの器官の標本を新規なブタサーコウイルス株の単離用に選択した。
【0047】
実施例6
新規なブタサーコウイルス株の細胞培養結果と、免疫抗体法による検出
多機能性萎縮症候群の臨床的徴候を示すフランスの子豚(Imp. 1008株)、カリフォルニアの子豚(Imp. 999株)およびカナダの子豚(Imp. 1010株)から集めた標本を培養した細胞培地には細胞変性作用(CPE)は全く見られなかった。しかし、アセトンを用いて固定した後、貯蔵されたブタのポリクローン化した血清を用いて培養した細胞培地から得られた製剤を免疫標識化することによって、カリフォルニアの子豚(Imp. 999株)の肺、フランスの子豚(Imp. 1008株)の縦隔リンパ節およびカナダの子豚(Imp. 1010株)の器官を用いて培養した培地中の多数の細胞に核蛍光が現れた。
【0048】
実施例7
ブタサーコウイルスのゲノムDNAの抽出
ブタサーコウイルス(PCV)の新規な株の複製型を72〜76時間の培養後に回収した感染したPK/15細胞培地(実施例1参照)(75cm2ファルコン10個)を用いて調製し、CAVの複製型のクローン化で記載したグルコサミンで処理した(非特許文献26)。この複製型の2本鎖DNAはHirt法(非特許文献27)の変形法で抽出した(非特許文献28に記載)。
【非特許文献26】Todd. D. et al., Dot blot hybrIDization assay for chicken anaemia agent using a cloned DNA probe. J. Clin. Microbiol. 1991, 29, 933-939
【非特許文献27】Hirt B. Selective extraction of polyoma virus DNA from infected cell cultures, J. Mol. Biol. 1967, 36, 365-369
【非特許文献28】Molitor T. W. et al. Porcine parvovirus DNA: characterization of the genomic and replicative form DNA of two virus isolates, Virology, 1984, 137, 241-254
【0049】
実施例8
ブタサーコウイルス Imp.999株のゲノムの複製型の制限地図
Hirt法で抽出されたDNA(1〜5μg)をメーカの推薦する方法に従ってS1ヌクレアーゼ(Amersham)で処理し、各種制限酵素(Boehringer Mannheim UK, Lewis, East Sussex, UK)で消化し、消化物をTodd達が記載の方法(非特許文献29)で、臭化エチジウムの存在下、1.5%のアガロースゲル上で電気泳動で分離した。Imp.999株の培地から抽出したDNAは独特のEcoRI部位、2つのSacI部位を有し、PstI部位は全く持たない。従って、この制限プロフィールは、逆にPstI部位を有し、EcoRI部位を全く持たないPCVのPK/15株(非特許文献30)が示す制限プロフィールとは異なっている。
【非特許文献29】Purification and biochemical characterization of chicken anemia agent. J. Gen. Virol. 1990, 71, 819-823
【非特許文献30】Meehan B. et al. Sequence of porcine circovirus) DNA: affinities with plant circovirus, 1997, 78, 221-227
【0050】
実施例9
ブタサーコウイルス Imp.999株のゲノムのクローン化
制限酵素EcoRIを有するPCV Imp.999株の2本鎖複製型の消化で生じた約1.8kbpの制限断片を市販のQiagen kit(QIAEXII Gel Extraction Kit, Cat # 20021, QIAGEN Ltd., Crawley, West Sussex, UK)を用いて1.5%アガロースゲル(実施例3参照)上で電気泳動で分離した。次に、このEcoRI−EcoRI制限断片を標準クローン化法(非特許文献31)によって予め同じ制限酵素を用いて消化し、脱リン酸したベクターpGEM-7(Promega, Medical Supply Company, Dublin, Ireland)と結合した。得られたプラスミドを標準的方法によってEscherichia coli JM109宿主株(Stratagene, La Jolla, USA)に変換した。同じく、PCV Imp.999株のEcoRI−EcoRI制限断片をベクターpBlueScript SK+(Stratagene, La Jolla, USA)のEcoRI部位にクローン化した。各宿主株に対して得られたクローンの中から所望の大きさの断片を有する少なくとも2つのクローンを選択した。次に、得られたクローンを培養し、Imp.999株の完全なゲノムを含むプラスミドを標準的なプラスミド調製および精製法によって少容量(2ml)または大容量(250ml)に精製した。
【非特許文献31】Sambrook J. et al. Molecular cloning: A Laboratory Manual. 2nd Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1988
【0051】
実施例10
PCV Imp.999株のゲノムDNAの配列(2本鎖複製型)
2つのEcoRI Imp.999のクローン配列(クローンpGEM-7/2およびpGEM-7/8)をメーカー推薦の方法で配列キット"AmpliTaq DNA polymerase FS"(Cat # 402079 PE Applied Biosystems, Warrington, UK)および"Applied BioSystems AB1373A automatic sequencing apparatus"を用いてSangerのジデオキシヌクレオチド法で決定した。最初の配列反応はM13「前方」および「後方」自在プライマーを用いて実施した。続く配列反応は「DNA歩行」法によって実施した。その後の配列に必要なオリゴヌクレオチドはLife Technology(Inchinnan Business Park, Paisley, UK)によって合成された。
【0052】
得られた配列は集め、MacDNASIS version 3.2のソフトウェア(Cat # 22020101, Appligene, Durham, UK)を用いて分析した。各種の読み取り枠を"National Center for Biotechnology Information"(NCBI, Bethesda, MD, USA)サーバーで入手可能なBLAST algorithmを用いて解析した。
クローンpGEM-7/8(配列ID番号6)から最初に得られた完全な配列(EcoRI-EcoRI)は図13、14に表してある。この配列はEcoRI部位のGの後に任意に始まり、ヌクレオチドに関してはいくつか不安定な部分を有している。
次いで、この配列が最適化された。配列ID番号3(図9〜14)はこの株の配列を全て表しており、EcoRI部位の始めすなわち最初のヌクレオチドとしてGから任意に始まる。
【0053】
同様の方法本発明の他の3つの分離株の配列を得た(配列ID番号1、2、4および図1〜4および図7、8参照)。これら4つの株のゲノムの大きさは下記の通りである:
Imp.1011−48121 1767個のヌクレオチド
Imp.1011−48285 1767個のヌクレオチド
Imp.999 1768個のヌクレオチド
Imp.1010 1768個のヌクレオチド
【0054】
実施例11
PCV Imp.999株の配列の分析
Imp.999株から得られた配列をGenBankのデータバンクに収容された配列との相同性テストで用いた結果、検出された有意な相同性はPK/15株の配列(寄託番号Y09921)(図9〜12参照)と約76%の相同性であった。
データバンク(NCBIサーバーのBLAST X algorithm)を用いた6つのフェーズでの配列の翻訳の相同性テストでは、アミノ酸レベルで、GenBank U49186配列でコードされるプラント(GenBank番号1841515)のサーコウイルスに類似のBBTBウイルスの理論的レプリカーゼに対応する読み取り枠と94%の相同性を示すということが分かった。
PCV Imp.999株から得られた配列と有意な相同性を示すものはデータバンクに収容された他の配列にはなかった。
多機能萎縮症候群の臨床的徴候を示すカリフォルニアの子豚から得られた病変部を用いて培養されたImp.999株から得られた配列のには解析から、この単分離ウイルスが新規なブタサーコウイルス株であることが明らかになった。
【0055】
実施例12
配列の比較分析
新規な4つのPCV株のヌクレオチド配列とPCVのPK/15株の配列(図9〜12)とを整列比較した。新規な4つの株と公知のPK/15株とに関する相同性マトリクスを作成した。結果は下記の通りである:
1:Imp.1011−48121
2:Imp.1011−48285
3:Imp.999
4:Imp.1010
5:PK/15
【0056】
【表1】

【0057】
2つのフランス株Imp. 1011-48121とImp. 1011-48285の相同性は99%以上(0.9977)である。2つの北米株Imp. 999とImp. 1010の相同性も99%以上(0.9949)以上である。フランス株と北米株との相同性はわずかに96%以上である。
これら全ての株とPK/15株との相同性は75〜76%の値に落ちる。
このことから、本発明の株はPK/15株に代表される種類とは異なる新規なブタサーコウイルスの代表であると言える。PK/15がタイプIを表すのに対し、PMWS症候群を示すブタから単離されたこの新規な株をタイプIIのブタサーコウイルスとよぶことにする。このタイプIIに属する株は実際には非常に離れた地理的領域から単離されたにもかかわらず、驚くべきヌクレオチド配列の相同性を示す。
【0058】
実施例13
新規なPCV株のゲノムによってコードされるプロテインの分析
Imp.1010単離株のヌクレオチド配列は多機能性萎縮症候群に関する他のサーコウイルス株を代表するものと考えられる。この配列をBLASTX algorithm(非特許文献32)と、ソフトウェアMacVector 6.0(Oxford Molecular Group, Oxford OX4 4GA, UK)のプログラムとを組み合わせを用いて詳細に分析した。この配列(環状ゲノム)上には20個のアミノ酸より大きいサイズの読み取り枠(またはORF)が13個検出できた。この13個のORFは下記の通りである:
【非特許文献32】Altschl et al. J. Mol. Biol. 1990. 215. 403-410
【0059】
【表2】
【0060】
各ORFの開始位置および終了位置は株1010のゲノムの図7、8に表された配列(配列ID番号4)を参照する。ORF1〜13の端部は株999と同一である。これらは下記ORF3と13を除き、株1011−48121および1011−48285とも同一である:
ORF3 1432−1539、センス、108nt、35aa
ORF3 314−1377、アンチセンス、705nt、234aa
これら13のORFの内で4つがクローン化ウイルスPCV PK/15のゲノム上に位置する類似のORFと有意な相同性を有する。多機能性萎縮症候群に関連するサーコウイルス単離株全てのゲノム上に存在する読み取り枠をそれぞれ分析した。これら4つのORFは下記の通り:
【0061】
【表3】
【0062】
各ORFの始めと終わりの位置は図7、8に表された配列(配列ID番号4)を参照する。ORFの大きさ(ヌクレオチド=ntで)は終止コドンを含む。
PCV Imp. 1010とPCV PK−15の単離株のゲノム機構の比較からから、2つのウイルスのゲノムに保存された4つのORFが同定できた。下記の表は観察された相同性の程度を表す:
【0063】
【表4】
【0064】
最大の相同性はORF4 Imp. 1010とORF1 PK−15との間に見られた(86%相同性)。このプロテインはおそらくウイルスのDNA複製に含まれ、ウイルスの複製に必須であるため、このことは予想されることである(非特許文献33)。
ORF13 Imp. 1010とORF2 PK−15との間の配列同一性は余り強くない(66.4%の同一性)が、この2つのORFはそれぞれ、鳥サーコウイルスCAV(非特許文献34)の主構造プロテインのN−末端領域と同一な良く保存されたN−末端基本領域を有している。さらに、ORF7 Imp. 1010とORF3 PK−15との間と、ORF10 Imp. 1010とORF4 PK−15との間に大きな違いが見られた。PCV PK−15のORF3とORF4と比較すると、それぞれにImp. 1010分離株のORF7とORF10のC−末端領域に欠失がある。最大の配列相同性はORF7/ORF3(重複部分の水準において61.5%の相同性)とORF10/ORF4(重複部分の水準において83%の相同性)のN−末端領域の水準において見られる。
【非特許文献33】Meehan et al., Gen. Virol. 1997,78; 221-227;Mankertz et al. J. Gen. Virol. 1998. 79. 381-384
【非特許文献34】Meehan et al., Arch. Virol. 1992, 124. 301-319
【0065】
ブタサーコウイルスのゲノム構造はゲノムが極端に高密度であるため非常に複雑であるように見える。主構造プロテインはおそらくブタサーコウイルスゲノムの同じ鎖上に位置する複数の読み取り枠間のスプライシングに由来するものである。従って、上記の表に記載の全ての読み取り枠(ORF1〜ORF13)でタイプIIのブタサーコウイルスにコードされた抗原プロテインの全てまたは一部を表すことができ、これらは特有の病気および/またはワクチン接種に使用可能な抗原である。本発明はこれらのORFを少なくとも1つ含む全てのプロテインに関するものである。本発明は主としてORF4、ORF7、ORF10またはORF13で構成されるプロテインに関するものである。
【0066】
実施例14
新規な株からクローンされたPCVゲノムの感染特性
Imp. 999単離株の完全なゲノム(複製型)を含むプラスミドpGEM-7/8をMeehan B. 達に記載の方法によってPK/15細胞に形質移入した(非特許文献35)。未感染のPK/15細胞に形質移入した後の最初の継代に対して実施された免疫抗体法の分析(実施例4参照)から、クローンpGEM-7/8のプラスミドは感染性PCVウイルスを誘導することが示された。感染性PCVの遺伝物質を含むクローンが利用可能になったことによってウイルス性ゲノムに任意の有用な処置をして無毒化ワクチンまたは組み換えワクチンを製造し、或いは診断キット用抗原を製造することができ、変成PCVウイルス(ブタで無毒化または欠失)を製造することができるようになった。
【非特許文献35】Characterization of viral DNAs from cells infected with chicken anemia agent: sequence analysis of the cloned replicative form and transfection capabilities of cloned genome gragments. Arch. Virol. 1992, 124. 301-319
【0067】
実施例15
インビトロ培養によるPCV抗原の製造
感染していないPK/15細胞の培養およびウイルス処置を実施例1と同じ方法で実施した。感染細胞を37℃で4日間培養後、トリプシン処理し、回収し、命名した。次の継代は1mlにつき400,000個の感染細胞を接種した。
【0068】
実施例16
ウイルス性抗原の不活化
ウイルス培養の最後に、感染した細胞を回収し、超音波(Branson Sonifier)またはロータースタータータイプのコロイドミル(UltraTurrax, IKA)を用いて溶解した。次に、懸濁液を3700gで30分間、遠心分離した。このウイルス懸濁液を+37℃で18時間、0.1%エチレンイミンで、或いは、+28℃で24時間、竈−プロピオラクトンで不活化した。ウイルス価が不十分な場合にはウイルス懸濁液を300kDaのメンブレン(Millipore PTMK300)を用いた限外濾過で濃縮した。不活化ウイルス懸濁液は+5℃で貯蔵した。
【0069】
実施例17
有機油をベースにしたワクチン乳剤の製造
下記処方でワクチンを調製した:
不活化ブタサーコウイルス懸濁液 : 250ml
MondanIDe(商標登録)ISA 70(SEPPIC) : 750ml
水相および油相は濾過によって別々に滅菌した。この乳剤はシルバーソン・タービン乳化機を用いて各成分を混合し、均質化して調製した。
一回のワクチン投与量は約107.5のTCID50を含む。皮膚経路の一回のワクチン投与容量は0.5ml、筋肉経路の投与量は2mlである。
【0070】
実施例18
代謝可能な油ベースのワクチン乳剤の製造
下記処方でワクチンを調製した:
不活化ブタサーコウイルス懸濁液: 200ml
Dehymuls HRE 7(Henkel) : 60ml
Radia 7204(Oleofina) : 740ml
水相および油相は濾過によって別々に滅菌した。この乳剤はシルバーソン・タービン乳化機を用いて各成分を混合し、均質化して調製した。
一回のワクチン投与量は約107.5のTCID50を含む。皮膚経路の一回のワクチン投与容量は0.5ml、筋肉内経路の投与量は2mlである。
【0071】
実施例19
過免疫血清(PCV−T)、PK/15から調製されたモノクローナル抗体F99およびカナダ株から調製された過免疫血清(PCV−C)で汚染されたアメリカおよびフランスのPCVウイルス株およびPK/15の間接免疫抗体法の結果
【0072】
【表5】
* 間接的免疫抗体法で正の反応を示した血清またはモノクローナル抗体の最後の希釈液の逆数。
【図面の簡単な説明】
【0073】
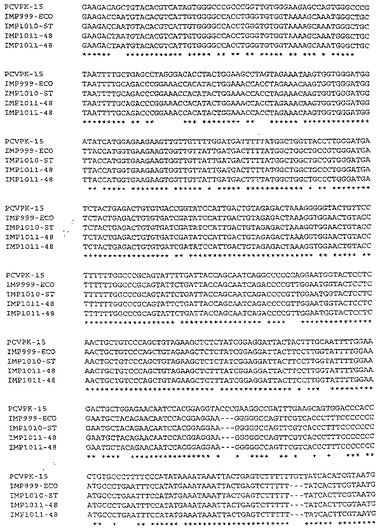
【図1】Imp.1011−48121株のゲノムのDNA配列
【図2】図1の続き
【図3】Imp.1011−48285株のゲノムのDNA配列
【図4】図3の続き
【図5】Imp.999株のゲノムのDNA配列
【図6】図5の続き
【図7】Imp.1010株のゲノムのDNA配列
【図8】図7の続き
【図9】PCV PK/15株の配列と図1〜8の4つの配列との整列化
【図10】図9の続き
【図11】図10の続き
【図12】図11の続き
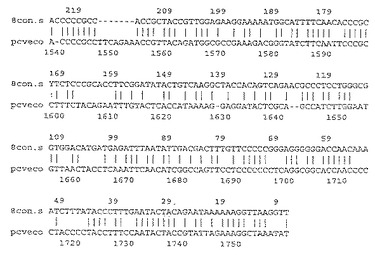
【図13】1997年10月3日のフランスの最初の出願におけるImp.999株のゲノムのDNA配列
【図14】図13の続き
【図15】PK/15株の配列と図13、14の配列との整列化
【図16】図15の続き
【図17】図16の続き
【図18】図17の続き
【技術分野】
【0001】
本発明は、PMWS症候群(ブタ多機能萎縮症候群:Porcine Multisystemic Wasting Syndrome)(離乳後多機能萎縮症候群:Post-weaning Multisystemic Wasting Syndromeともいわれる)の原因となる新規なブタサーコウイルス(PCV,Porcine Circo Virus)株と、その検出試薬および検出方法と、ワクチン接種方法と、ワクチンと、これらワクチンおよび試薬の製造方法とに関するものである。
【背景技術】
【0002】
ブタサーコウイルス(PCV)はブタ腎臓細胞株PK/15中に無毒病原性汚染物質として最初に見い出された。このウイルスPCVはチキン貧血症ウイルス(CAV、Chicken Anaemia Virus)および嘴・羽症候群ウイルス(PBFDV、Pscittacine Beak and Feather Disease Virus)とともにサーコウイルス群(Circoviridae)に分類された。これらのウイルスはエンベロプのない小さなウイルス(15〜24nm)で、その共通な特徴は1.76〜2.31kbの環状一本鎖のDNAからなるゲノムを含む点にある。このゲノムは約30kDaのポリペプチドをコード化すると考えられていた(非特許文献1)が、最近の研究で、複雑な転写がわかってきた(非特許文献2)。さらに、上記3種類のサーコウイルス(circovirus)はそのヌクレオチド配列に有意な相同性はなく、また、共通の抗原決定基もない。
【非特許文献1】Todd et al., Arch Virol 1991, 117; 129-135
【非特許文献2】Meehan B. M. et al., 1997,78; 221-227
【0003】
PK/15細胞由来のPCVは病原性ではないと考えられる。この配列は「非特許文献3」が明らかにしている。PCV株が病原性で、PMWS症候群と関連があるかもしれないと考えられるようになったのはほんの最近のことである(非特許文献4、5)。Nayer達はブタPCVのDNAがPMWS症候群を示すことをPCR法を用いて見い出した。しかし、野生型PCV株は単離または精製されていない。
【非特許文献3】"B. M. Meehan et al., J. Gen. Virol 1997 (78) 221-227"
【非特許文献4】Gupi P. S. Nayar et al., Can. Vet. J, Vol. 38, 1997: 385-387
【非特許文献5】Clark E. G., Proc. Am. Assoc. Swine Prac. 1997; 499-501
【0004】
カナダ、米国およびフランスで見られるPMWS症候群の臨床的な特徴は、体重を徐々に失い、頻呼吸症、呼吸困難および黄疸等を発症することである。病理的な面では、リンパ性または肉芽種性浸潤、リンパ節腫脹、まれには肝炎およびリンパ性または肉芽種性腎炎を発症する(非特許文献6〜10)。
【非特許文献6】Clark E. G., Proc. Am. Assoc. Swine Prac. 1997; 499-501;
【非特許文献7】La Semaine Veterinaire 1996 (834)、
【非特許文献8】La Semaine Veterinaire No. 26;
【非特許文献9】La Semaine Veterinaire 1997 (857):54;
【非特許文献10】Gupi P. S. Nayar et al., Can. Vet. J, Vol. 38, 1997: 385-387
【発明の開示】
【発明が解決しようとする課題】
【0005】
本出願人は、カナダ、米国(カリフォルニア)およびフランス(ブリタニー)の農場から得た肺または神経節の標本から5つの新規なPCV株(以下、本発明のサーコウイルス(circovirus)という)を単離することに成功した。これらのウイルスはPMWS症候群のブタの病変部に検出されるが、健康なブタには検出されない。
【0006】
本出願人はさらに、これらの4種の株(カナダおよび米国の株)と2種のフランスの株のゲノムの配列決定した。これらの株はヌクレオチドレベルで互いに非常に強い相同性(96%以上)超をを示す、一方、PK/15株ははるかに弱い相同性(約76%)しか示さない。従って、新規のタイプのブタサーコウイルス (circovirus)を代表するものと考えられ、PK/15に代表されるタイプI(第I群)に対して、本発明の新規な株はタイプII(第II群)とよぶことにする。
【課題を解決するための手段】
【0007】
本発明の対象は単離または精製物の形をした上記定義の第II群のブタサーコウイルス(circovirus)にある。
本発明はさらに、生理標本または組織標本、特にPMWS症候群のブタの病変部の標本から実施例に記載した方法で単離可能なブタのサーコウイルス(circovirus)、特にII型サーコウイルスを提供する。
【0008】
本発明の対象はECACC(European Collection of Cell Cultures, Centre for Applied Microbiology & Research, Porton Down, Salisbury, Wiltshire SP4 0JG, United Kingdom)に寄託された下記の5種の株の精製物にある:
1997年10月2日木曜日寄託:
寄託番号第V97100219号(以下、Imp.1008PCVという)
寄託番号第V97100218号(以下、Imp.1010PCVという)
寄託番号第V97100217号(以下、Imp.999PCVという)
1998年1月16日金曜日寄託:
寄託番号第V98011608号(以下、Imp.1011−48285という)
寄託番号第V98011609号(以下、Imp.1011−48121という)
【0009】
本発明はさらに、病気のブタから単離したブタサーコウイルス(circovirus)および/または本発明株と有意な血清学的類似性を有するサーコウイルスおよび/またはPK/15のPCVとハイブリダイゼーションが全く無く、厳密な条件下で本発明株とのクロスハイブリダイゼーションを有するサーコウイルスに関するものである。
【発明を実施するための最良の形態】
【0010】
ウイルス性株の増殖、特に全抗原(例えばウイルス)および/またはサブユニット(例えばポリペプチド)の製造は、PMWS症候群のブタの病変部の生理学標本または組織標本から単離されたウイルス株、特にブタ腎臓株細胞を汚染されていない細胞株(特にPCV、ペスチウイルス、ブタアデノウイルスおよびブタパルボウイルスに汚染されていないPK/15細胞)で増殖するのが有利である。
これらの単離株がPK/15細胞での培養で非常に生産的であり、ウイルスまたは抗原の製造、特に不活性化ワクチンの製造に有利であるということは驚くべきことであり、予想できなかったことである。
【0011】
従って、本発明の他の対象は本発明のサーコウイルス(circovirus)または病変部細胞、特にPMWS症候群のブタの細胞株の生理学標本または組織標本から単離される任意のブタサーコウイルスの少なくとも1つに感染された細胞をインビトロで培養した細胞株、特にPK/15細胞で継代した後に単離されるサーコウイルス(circovirus)の調製方法にある。
本発明のさらに別の対象は上記の培養抽出物または上澄み(必要に応じて一般的な方法でさらに精製されていてもよい)と、一般にインビトロの培養から得られる任意の抗原製剤にある。
本発明のさらに別の対象は免疫抗原性の有効成分と、少なくとも1つの上記定義の抗原を含むワクチンにある。
【0012】
本発明のさらに別の対象は、無毒化した生全ワクチンをベースとする免疫抗原性の有効成分またはこの有効成分を精製して得られるワクチンにある。この無毒化は通常の方法、例えば細胞での継代、好ましくはブタ細胞、特にPK/15細胞等の株細胞での継代(特に50〜150、特に100回の継代)で行なわれる。これらのワクチンは一般に獣医学上許容される賦形剤または希釈液と、さらには必要に応じて添加される獣医学上許容される補助剤およびフリーズドライ安定剤とを含むことができる。
上記の抗原製剤およびワクチンは103〜106のTCID50を有するのが好ましい。
【0013】
本発明のさらに別の対象は、不活化した本発明のサーコウイルス抗原をベースにした免疫抗原性有効成分またはワクチンにある。このワクチンは獣医学上許容される賦形剤または希釈液、さらには必要に応じて添加される獣医学上許容される補助剤を含むことができる。
【0014】
本発明のサーコウイルス(断片の形でもよい)は当業者に公知の方法で不活性化される。この不活性化は化学的方法、例えば抗原をホルムアルデヒド(ホルマリン)、パラフォルムアルデヒド、竈−プロピオラクトン、エチレンイミンまたはその誘導体等の試薬で処理して行なうことができる。好ましい不活性化法は試薬、特にエチレンイミンまたは竈−プロピオラクトンで処理する方法である。
本発明の不活性化ワクチンにはアジュバンドを加え、当業者に公知の方法でエマルジョンの形、例えば油中水エマルジョンまたは水中油エマルジョン等の乳剤の形にするのが好ましい。一般的なアジュバンド化合物を有効成分に添加してアジュバンド処理することもできる。
【0015】
使用可能な補剤(アジュバンド)としては例えば水酸化アルミニウム、サポニン(例えば非特許文献11参照)、Avridine(商標登録)(非特許文献12参照)、DDA(ジメチルジオクタデシルアンモニウムブロミド、非特許文献13参照)、ポリホスファゼン(非特許文献14参照)、さらには有機油、スクアラン(例えばSPT乳剤、非特許文献15参照)、スクアレーン(例えばMF59、非特許文献16参照)をベースとする水中油乳剤または代謝可能な油をベースとする油中水乳剤(好ましくは特許文献1、2に記載の乳剤を挙げることができる。補剤の組み合わせ、例えば乳剤AvrIDine(商標登録)またはDDAとの組み合せた選択することもできる。
これらのワクチンは106〜108のTCID50を有するのが好ましい。
【非特許文献11】Quillja saponinまたはQuil A;"Vaccin Design, The Subunit and Adjuvant Approach, 1995, Michael F. Powel と Mark J. Newman編、Plennum Press, New-York and London, p.210"
【非特許文献12】Vaccine Design p. 148
【非特許文献13】Vaccine Design p. 157
【非特許文献14】Vaccine Design p. 204
【非特許文献15】Vaccine Design p. 147
【非特許文献16】Vaccine Design p. 183
【特許文献1】国際特許第WO-A-94−20071号公報
【特許文献2】米国特許第5,422,109号明細書
【0016】
生ワクチン補剤は不活性化ワクチン用のものから選択するこができる。乳剤が好ましい。不活性化ワクチン用のものには特許文献3に記載のものも加えることができる。
凍結安定剤としては例えばSPGA(非特許文献17)、ソルビトール、マンニトール、スターチ、スクロース、デキストランまたはグルコース等の炭水化物、アルブミンまたはカゼイン等のプロテイン、これらの化合物の誘導体またはアルカリ金属燐酸塩等の緩衝剤を挙げることができる。
【特許文献3】国際特許第WO-A-94−16681号公報
【非特許文献17】Bovarnik et al., J. Bacteriology 59, 509, 950
【0017】
本件出願人は配列ID番号1〜4(必要な場合にはさらに配列ID番号6)で識別される4つの単離株のゲノムを得た。
本発明のさらに他の対象はこれらの配列の一部または全てを含むDNA断片にある。本発明がこれと均等な配列すなわち上記配列の株の特徴または上記配列によりコードされるポリペプチドの機能を変更しない配列も含むものであるということは当然である。また、コードの欠失(degenerescence)により変化した配列も含むということは理解てきよう。
本発明はさらに、厳密な条件下で上記配列と混成可能であり、および/または、上記定義の第II群に属する本発明株と高い相同性を有するという意味で均等な配列も含むものである。
【0018】
これらの配列およびその断片は適当なベクターを用いてインビトロまたはインビボでポリペプチドの発現に有利に用いることができる。
特に、これに使用可能な本発明のDNA断片を形成する読み取り枠はタイプIIのサーコウイルスのゲノム配列で同定されている。本発明は、少なくともこれらの読み取り枠(対応するアミノ酸配列)の一つを含む全てのポリペプチドに関するものである。特に、本発明は主としてORF4、ORF7、ORF10またはORF13で構成されるプロテインに関するものである。
インビトロのサブユニットを発現するための発現手段としてはE. coliまたはバキュロウイルス(特許文献3)を用いるのが好ましい。
【特許文献4】米国特許第4,745,051号明細書
【0019】
上記の単数または複数のコード配列またはその断片をバキュロウイルスゲノム(例えばAutographa californica Nuclear Polyhedrosis Virus AcNPV)に組み込み、次いで、それを昆虫細胞、例えばSpodoptera fruigiperda Sf9(寄託番号ATCC CRL 1711)で増殖される。また、イースト(例えば酵母)またはほ乳類細胞(例えばCHO、BHK)等の真核細胞でサブユニットを製造することもできる。
【0020】
本発明のさらに他の対象は、これらの発現手段によってインビトロで製造され、必要に応じて一般的な方法で精製されたポリペプチドにある。本発明のさらに他の対象は、このようにして得られた少なくとも1つのポリペプチドまたはその断片を獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤をさらに含むことができる)中に含むサブユニットワクチンにある。
【0021】
組み換え生ワクチンの製造時にインビボ発現させるには単数または複数のコード化配列またはその断片を単数または複数のポリペプチドを発現できる条件下で適当な発現ベクターに挿入する。適切なベクターとしては好ましくはブタで増殖可能でブタに対しては非病原性の生ウイルスを当業者に周知の方法で使用することができる。特に、アウジェスキー病ウイルス、ブタアデノウイルス等のブタヘルペスウイルス、ポックスウイルス、特にワクシニアウイルス、トリポックスウイルス、カナリアポックスウイルス、ブタポックスウイルス等を用いることができる(特許文献4〜7)。
【特許文献5】国際特許第WO-A-90−11092号公報
【特許文献6】国際特許第WO-A-93−19813号公報
【特許文献7】国際特許第WO-A-94−21797号公報
【特許文献8】国際特許第WO-A-95−20660号公報
【0022】
従って、本発明の他の対象は、このようにして精製されたベクターと、組み換え生ワクチンまたはプラスミドワクチン(ポリヌクレオチドまたはDNAワクチン)にある。これらのワクチンは獣医学上許容される賦形剤または希釈液を含むことができる。
本発明のワクチン(無毒化、不活性化、サブユニット、組み換えおよびプラスミド生ワクチン)は本発明の一つまたは複数(2、3つ)のサーコウイルスから成る一つまたは複数の有効成分(抗原)を含むことができる。
【0023】
本発明の上記各種ワクチンはブタの他の病原体に対するワクチンと組み合わせることができる。特にPMSW症候群に関連する病原体に対するワクチンと組み合わせることができる。本発明のワクチン、特に不活性化ワクチンはブタの他の病原体に対応する他のワクチン価を含むことができる。これらのブタの他の病原体の中では好ましくはPRRS(Porcine Reproductory and Respiratory Syndrome :ブタ生殖器および呼吸器症候群)(特許文献8〜10および非特許文献18を参照)および/またはMycoplasma hyopneumonia(特許文献11〜13および非特許文献19を参照)を挙げることができる。他の重要なワクチン価の中ではアクチノバシラス胸膜肺炎菌、E. coli、ブタ萎縮鼻炎(porcine Atrophic Rhinitis)、仮性狂犬病(アウジェスキー病)、ブタコレラ、ブタインフルエンザを挙げることができる。
【特許文献9】国際特許第WO-A-93−07898号公報
【特許文献10】国際特許第WO-A-94−18311号公報
【特許文献11】フランス国特許第2,709,966号公報
【非特許文献18】"C. Chareyre 他、IPVS Congress,Birmingham, England, 5-9 July 1998, p. 139の議事録"
【特許文献12】欧州特許第597,852号
【特許文献13】第550,477号、
【特許文献14】第571,648号
【非特許文献19】"第15回IPVS CongressのO. Martinon 達、p. 157, 284, 285およびG. Reynaud達、p. 150"
【0024】
本発明のさらに他の対象は、本発明のサーコウイルスに対してブタの免疫反応を誘発する方法、特にブタに有効なワクチン接種方法にある。
このワクチン接種方法では上記ワクチンを一回または複数回にわたってブタに投与する。同じワクチン接種プロトコルで複数種の前記ワクチンを組み合わせることも可能である。
このワクチンは大人のブタだけでなく、若いブタまたは妊娠中の雌に対して投与することができる。妊娠中の雌にワクチン接種することによって新生児に受動免疫(母親抗体:maternal antibody)を与えることができる。
【0025】
本発明はさらに、本発明のブタサーコウイルスの存在を診断する方法を提供する。従って、本発明の他の対象は診断テストと下記試薬を用いたテスト方法にある。
互いに異なるサーコウイルスの配列が分かると共通の配列を定義でき、公知の全てのブタサーコウイルスを識別できる試薬を製造することができる。
当業者は特定の診断を実施するために対応するPK/15サーコウイルスの配列とほとんどまたは全く相同性を示さない領域に対応する配列の断片を選択することができる。
【0026】
配列を並べることによって当業者は所望の試薬を選択することができる。
第1の試薬は上記DNA配列およびその断片で構成され、それは周知のハイブリッド法またはPCR(ポリメラーゼ鎖反応)法によってプローブまたはプライマーとして用いられる。
第2の試薬は上記ウイルスの配列によってコードされたポリペプチドまたはベクター(前記参照)を用いて発現されたまたは公知のペプチド合成法によって化学的に合成されたポリペプチドで構成される。
第3および第4の試薬はウイルス、ポリペプチドまたは断片、抽出物またはDNA配列でコードされたものから一般的な方法で製造可能なポリクローナル抗体およびモノクローナル抗体からなる。
【0027】
上記の第2、第3および第4の試薬は本発明の対象である診断法で用いることができる。この診断法では検査するブタから得られた生理学的液体標本(血液、血漿、血清等)または組織標本(神経節、肝臓、肺、腎臓等)中に本発明のサーコウイルスに特有な抗原の存在を調べて抗原自体を検出するか、この抗原に対する抗体を検出する。
【0028】
本発明の抗原および抗体は公知の任意の臨床診断法で用いることができるが、
上記の抗原および抗体を獣医、ブリーダーまたは動物の飼い主が直接現場で使用可能な方法で用いるのが好ましい。臨床技術および現場技術を有する当業者は上記の抗原および/または抗体を試薬として用いることができる。
【0029】
本発明で用いるのに好ましい診断法はウェスタンブロット法、免疫抗体法、ELISA(酵素結合免疫測定法)および免疫クロマトグラフィーである。
免疫クロマトグラフィー法に関しては特に非特許文献20と特許文献14〜29を参照することができる。
【非特許文献20】"Robert F. Zulk et al., Clin. Chem. 31/7, 1144-1150 (1985) "
【特許文献15】国際特許第WO-A-88−08534号公報
【特許文献16】国際特許第WO-A-91−12528号公報
【特許文献17】欧州特許第291,176号公報
【特許文献18】欧州特許第299,428号公報
【特許文献19】欧州特許第291,194号公報
【特許文献20】欧州特許第284,232号公報
【特許文献21】米国特許第5,120,643号明細書
【特許文献22】米国特許第5,030,558号明細書
【特許文献23】米国特許第5,266,497号明細書
【特許文献24】米国特許第4,740,468号明細書
【特許文献25】米国特許第5,266,497号明細書
【特許文献26】米国特許第4,855,240号明細書
【特許文献27】米国特許第5,451,504号明細書
【特許文献28】米国特許第5,141,850号明細書
【特許文献29】米国特許第5,232,835号明細書
【特許文献30】米国特許第5,238,652号明細書
【0030】
標本中に特有な抗体は間接法、競合法または置換法によって検出するのが好ましい。そのためには抗原自体を診断試薬として用いるか、抗体の認識を保持する抗原断片を診断試薬として用いる。標識化はペルオキシダーゼを用いた標識化または特殊な標識化、好ましくはコロイド性金を用いた標識化が有利である。
標本中の抗原自体をこの抗原に特有な標識化した抗体を用いて検出することもできる。標識化は上記のものが有利である。
競合法または置換法または抗原自体の検出に用いられる抗原に特有な抗体は、モノクローナル抗体またはポリクローナール抗体、これら抗体の断片、好ましくはFabまたはF(ab)'2断片であるである。
【0031】
本発明はさらに、上記抗体を生理学的液体標本または組織標本中の抗原の検出またはこれら標本または試験片中に存在する抗体の検出のための診断試薬として使用可能な本発明抗原に特有のモノクローナル抗体またはポリクローナル抗体の製造方法に関するものである。本発明はものでさらに、これらの抗体の免疫機能を有する断片、特にF(ab)およびF(ab)'2断片は本発明に含まれる。
抗体は一般的な方法で製造することができる。特に非特許文献21または非特許文献22を参照することができる。これらの内容は本件明細書の一部を成す。
特に、抗原またはその断片の少なくとも1つで免疫化されたネズミの脾臓細胞を適当な骨髄腫細胞と融合(この方法自体は公知)することができる。
【非特許文献21】"Antibodies, A Laboratory Manual, 1988, Cold Spring Harbor Laboratory, USA"
【非特許文献22】"J. W. Goding, Monoclonal Antibodies: Principles and Practice, Academic Press Inc.,"
【0032】
本発明の他の対象は上記抗原に特有なモノクローナル抗体またはポリクローナル抗体、特にネズミまたはウサギの抗体の純粋または部分的に純粋な製剤或いは粗原料からなる製剤にある。
本発明では上記のDNA配列を基にした所望のエピトープ(ワクチン用エピトープまたは診断用エピトープ)を決めることができる。本発明のサーコウイルスのゲノムのDNA配列から、エピトープを決めることができる。当業者は公知の方法、例えば適切なコンピュータプログラムまたはPEPSCANを用いてエピトープを求めることができる。このエピトープはプロテインの免疫優性領域であり、プロテインの表面に露出した領域である。このエピトープは抗体によって認識され、診断の分野において診断目的の抗体の調製または診断試薬として用いられる対応するペプチドの製造に用いられる。
少なくともエピトープは8〜9個のアミノ酸を有するペプチドである。一般には最小でも13〜25個のアミノ酸が好ましい。
【0033】
当業者は一つまたは複数の上記技術および利用可能なその他の技術を用いて診断用のペプチドまたは抗体で用いるエピトープを見付けることができる。
従って、本発明の他の対象はこの抗原および/またはこの抗原に特有なモノクローナル抗体またはポリクローナル抗体を含む診断キットにある。これらは特に、前記の診断技術による診断キットである。
【0034】
以下、下記の図を参照して本発明の実施例をさらに詳細に説明する。しかし、本発明が下記実施例に限定されるものではない。
配列IDリスト
配列ID番号1 = Imp.1011−48121株のゲノムのDNA配列
配列ID番号2 = Imp.1011−48285株のゲノムのDNA配列
配列ID番号3 = Imp.999株のゲノムのDNA配列
配列ID番号4 = Imp.1010株のゲノムのDNA配列
配列ID番号5 = PK/15株のゲノムのDNA配列
配列ID番号6 = 1997年10月3日のフランスの最初の出願におけるImp.999株のゲノムのDNA配列
【実施例】
【0035】
実施例1
ブタサーコウイルス(circovirus)株の培養および分離
組織標本はフランス、カナダおよび米国で子豚の肺およびリンパ節から集めた。これらの子豚は離乳後多機能萎縮症候群に典型的な臨床的徴候を示したものである。ウイルスを単離するため組織標本は死体解剖直後に−70℃で冷凍した。
ウイルス単離のために無菌の乳鉢と乳棒を用いて無菌の砂で組織を粉砕し、イール(Earl)塩 (EMEM, BioWhittaker UK Ltd., Workingham, UK)、ペニシリン(100IU/ml)およびストレプトマイシン(100μm/ml)を含む最小培地(MEM−SA培地)中に約15%の組織標本を含む懸濁液を調製した。粉砕物をMEM−SAに溶解し、+4℃、3000gで30分間、遠心分離にかけて上澄みを回収した。
【0036】
細胞培地の接種前に、100μlのクロロホルムを2mlの各上澄みに添加し、室温で10分間混合した。この混合物を超遠心分離管に移し、10分間3000gで遠心分離して上澄みを回収した。次いで、この上澄みをウイルス分離実験で種菌として用いた。
全てのウイルス単離研究はブタサーコウイルス(PCV)、ペスチウイルス、ブタアデノウイルスおよびブタパルヴォウイルスに感染していないことが分かっているPK/15細胞培地で実施した(非特許文献23)。
【非特許文献23】Allan G. et al., Pathogenesis of porcine circovirus experimental infections of colostrum-deprived piglets and examination of pig foetal material. Vet. Microbiol. 1995, 44, 49-64
【0037】
ブタサーコウイルスの単離は下記方法によって実施した:
PK/15細胞の単一層をトリプシン処理(トリプシン−versene混合物)によって集密的な培養から解離し、最終濃度が約400,000細胞/1mlであるペスチウイルス(=MEM−G培地)に感染していない15%のウシ胎児血清を含むMEM−SA培地に溶解した。次に、この細胞懸濁液の10mlの部分標本を2mlの前記接種材料の部分標本と混合し、最終混合物を2つの25cm2ファルコンフラスコに6mlの容積に分取した。+37℃で18時間、10%のCo2を含む雰囲気下で培養した。
【0038】
培養後、半集密的な単一層を300mMのD−グルコサミン(Cat # G48175、Sigma-Aldrich株式会社、Poole, UK)で処理し(非特許文献24)、さらに37℃で48〜72時間培養を続けた。この最終培養に続いて、2つのファルコンの1方を3回凍結/融解サイクルにかけた。残りのファルコンのPK/15細胞はトリプシン−versene溶液で処理し、20mlのMEM−G培地中に再懸濁し、400,000細胞/ml濃度で75cm2ファルコン中に接種した。新しく接種されたフラスコは凍結/融解サイクル後に得られた対応する可溶化液5mlを添加して「超感染」させた。
【非特許文献24】Tischr I. et al., Arch. Virol., 1987 96 39-57
【0039】
実施例2
免疫抗体法または切片上ハイブリッド形成法によるブタサーコウイルスを検出するためのするための細胞培地標本の調製
5mlの容積の「超感染」した懸濁液を回収し、無菌で脱脂したカバーガラスを含む直径55mmのペトリ皿中に接種した。フラスコ中およびカバーガラス上で+37℃で培養し、実施例1に記載のグルコサミンで処理した。カバーガラス上での培養はグルコサミン処理後24〜48時間で回収し、室温で10分間アセトンを用いて、または4時間緩衝剤処理した10%のホルムアルデヒドを用いて固定した。この固定後、全てのカバーガラスをシリカゲル上で−70℃で貯蔵してから、ハイブリッド形成の研究および細胞免疫の研究に使用した。
【0040】
実施例3
ハイブリッド形成法(in situ)によるPCV配列の検出法
ハイブリッド形成法(in situ)は病気のブタから集めた組織に対して実施し、ホルムアルデヒドで固定し、ウイルス単離(実施例2参照)用に接種した細胞培地の調整で用い、カバーガラスに固定した。
PK/15ブタサーコウイルス(PCV)および感染性ニワトリ貧血症ウイルス(CAV)に対応する完全なゲノムプローブを用いた。1.7キロの塩基ペア(kbp)の挿入断片の形にクローン化されたPCVゲノムの複製型を含むプラスミドpPCVI(Meehan B. et al. Sequence of porcine circovirus DNA: affinities with plant circovirus, J. Gen. Virol. 1997, 78, 221-227)をPCV固有DNA源として用いた。2.3kbpの鳥サーコウイルスCAVの複製型を含む鳥プラスミドpCAAIを負対照として用いた。2つのプラスミドのグリセロール源をアルカリ法(非特許文献25)によってプラスミドの製造および精製で用い、これらはプローブ調製用のテンプレートとして用いた。PCVおよびCAVの完全なゲノムを表すサーコウイルスプローブはメーカ推薦の方法で市販の非放射性ラベリングキット("DIG DNA Labelling Kit"、Boehringer Mannheim, Lewis, UK)を用いて上記精製プラスミド(各プローブに1μg)およびヘキサヌクレオチドプライマーから製造した。
【非特許文献25】Sambrook J. et al. Molecular cloning: A Laboratory Manual. 2nd Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1988
【0041】
ハイブリッド形成法(in situ)で使用する前に、プローブをジゴキシジェニンで標識化し、容積50〜100μlの純水に溶解した。
パラフィンに包み、ホルムアルデヒドで固定した病気のブタの組織標本と、ホルムアルデヒドで固定した感染細胞培地の製剤を下記方法でPCV核酸検出のために調製した:
5μm厚の切片をパラフィンに包まれた組織塊から切り出し、濃度を次第に薄くしたアルコール溶液中で再水和した。ホルムアルデヒドで固定した組織切片および細胞培地を15分間および5分間それぞれ5mMのEDTAを含む0.05MのTris−HCL緩衝剤(pH7.6)を用いた0.5%のプロテナーゼK溶液中で+37℃で培養した。次に、スライドガラスを加圧滅菌した蒸留水中の1%グリシン溶液中に30秒配置し、0.01MのPBS緩衝剤(燐酸緩衝食塩水)(pH7.2)で2度洗浄し、最後に無菌蒸留水で5分間洗浄した。スライドガラスを最後に外気で乾燥し、プローブと接触させた。
【0042】
各組織/プローブ製剤を清潔な脱脂カバーガラスで被覆し、+90℃の炉内に10分間配置した後、氷塊と1分間接触させ、最後に+37℃で18時間培養した。次に、調整物を2Xクエン酸ナトリウム塩(SSC)緩衝剤(pH7.0)に短時間含浸した後、2XのSSC緩衝剤中で5分間、2回洗浄し、最後にPBS緩衝剤中で5分間、2回洗浄した。
洗浄後、調整物を0.1Mのマレイン酸、0.15MのNaCL(pH7.5)(マレイン緩衝剤)の溶液中に10分間含浸し、マレイン緩衝剤中の遮断試薬の1%溶液(Cat # 1096176, Boehringer Mannheim UK, Lewis, East Sussex, UK)中で+37℃で20分間培養した。
【0043】
次に、調整物を遮断緩衝剤で希釈された抗ジゴキシジェニンモノクローナル性抗体(Boehringer Mannheim)の1/250溶液中で+37℃で1時間培養し、PBS中で洗浄し、最後に抗ネズミ免疫グロブリン抗体を用いて+37℃で30分間培養した。調整物をPBSで洗浄し、内因性ペルオキシダーゼ作用を室温で20分間、PBS中の0.5%水酸化ペルオキシド溶液で処理して遮断した。調整物を再度PBS中で洗浄し、使用直前に調製した3−アミノ−9−ジエチルカルバゾル(AEC)基質(CambrIDge Bioscience, CambrIDge, UK)で処理した。
最後に水道水で洗浄した後、調整物をヘマトキシリンで逆染色し、水道水で"青色化"して固定液(GVA Mount, CambrIDge Bioscience, CambrIDge, UK)で顕微鏡のカバーガラスに固定した。対照実験には無関係な負プローブ(CAV)と正プローブ(PCV)を病気のブタおよび病気ではないブタから得られた標本で使用することが含まれる。
【0044】
実施例4
免疫抗体法によるPCVの検出法
アセトンを用いて固定した全ての細胞倍地の最初のスクリーニングを貯蔵した大人のブタ血清の1/100希釈を用いた間接免疫抗体法(IIF)で実施した。この貯蔵血清は北アイルランドの25匹の大人の雌ブタからの血清を含み、PCV:ブタパルボウイルス、ブタアデノウイルスおよびPRRウイルスを含む広範囲のブタのウイルスに対する抗体を含むことで知られている。血清(PBSで希釈)を細胞培地と+37℃で1時間接触させた後、PBS中で2回洗浄してIIFを実施した。細胞培地をイソチオシアン酸フルオレセインに結合したPBSで1/80に希釈したウサギの抗ブタ免疫グロブリン抗体を用いて1時間染色し、PBSで洗浄し、グリセロール緩衝剤に固定し、紫外線下で顕微鏡観察した。
【0045】
実施例5
病気のブタの組織における切片上ハイブリッド形成結果
フランス、カナダおよびカリフォルニアの多機能性萎縮病変を有する子豚から集めた組織から調製したPCVゲノムプローブを用いた切片上ハイブリッド形成法から、研究された複数の病変部に病変部に関連するPCV核酸の存在が示された。これに対してPCVゲノムプローブを病気でないブタから集めた組織に用い時またはCAVプローブを病気のブタの組織に用いた時には何の徴候も見られなかった。PCV核酸の存在はサイトプラズムおよびカリフォルニアの子豚の肺の病変部を浸潤する多数の単核細胞の核に認められた。PCV核酸の存在は肺細胞、気管支および細気管支の上皮細胞および細動脈、細静脈およびリンパ管の内皮細胞にも見られた。
【0046】
病気のフランスのブタではPCV核酸の存在は多数の濾胞性リンパ細胞のサイトプラズマおよびリンパ節の同様な毛細血管内の単核細胞に検出された。また、PCV核酸は一次的合胞体にも検出された。これらの検出結果から、カリフォルニアのブタの肺、フランスのブタの腸間膜リンパ節およびカナダのブタの器官の標本を新規なブタサーコウイルス株の単離用に選択した。
【0047】
実施例6
新規なブタサーコウイルス株の細胞培養結果と、免疫抗体法による検出
多機能性萎縮症候群の臨床的徴候を示すフランスの子豚(Imp. 1008株)、カリフォルニアの子豚(Imp. 999株)およびカナダの子豚(Imp. 1010株)から集めた標本を培養した細胞培地には細胞変性作用(CPE)は全く見られなかった。しかし、アセトンを用いて固定した後、貯蔵されたブタのポリクローン化した血清を用いて培養した細胞培地から得られた製剤を免疫標識化することによって、カリフォルニアの子豚(Imp. 999株)の肺、フランスの子豚(Imp. 1008株)の縦隔リンパ節およびカナダの子豚(Imp. 1010株)の器官を用いて培養した培地中の多数の細胞に核蛍光が現れた。
【0048】
実施例7
ブタサーコウイルスのゲノムDNAの抽出
ブタサーコウイルス(PCV)の新規な株の複製型を72〜76時間の培養後に回収した感染したPK/15細胞培地(実施例1参照)(75cm2ファルコン10個)を用いて調製し、CAVの複製型のクローン化で記載したグルコサミンで処理した(非特許文献26)。この複製型の2本鎖DNAはHirt法(非特許文献27)の変形法で抽出した(非特許文献28に記載)。
【非特許文献26】Todd. D. et al., Dot blot hybrIDization assay for chicken anaemia agent using a cloned DNA probe. J. Clin. Microbiol. 1991, 29, 933-939
【非特許文献27】Hirt B. Selective extraction of polyoma virus DNA from infected cell cultures, J. Mol. Biol. 1967, 36, 365-369
【非特許文献28】Molitor T. W. et al. Porcine parvovirus DNA: characterization of the genomic and replicative form DNA of two virus isolates, Virology, 1984, 137, 241-254
【0049】
実施例8
ブタサーコウイルス Imp.999株のゲノムの複製型の制限地図
Hirt法で抽出されたDNA(1〜5μg)をメーカの推薦する方法に従ってS1ヌクレアーゼ(Amersham)で処理し、各種制限酵素(Boehringer Mannheim UK, Lewis, East Sussex, UK)で消化し、消化物をTodd達が記載の方法(非特許文献29)で、臭化エチジウムの存在下、1.5%のアガロースゲル上で電気泳動で分離した。Imp.999株の培地から抽出したDNAは独特のEcoRI部位、2つのSacI部位を有し、PstI部位は全く持たない。従って、この制限プロフィールは、逆にPstI部位を有し、EcoRI部位を全く持たないPCVのPK/15株(非特許文献30)が示す制限プロフィールとは異なっている。
【非特許文献29】Purification and biochemical characterization of chicken anemia agent. J. Gen. Virol. 1990, 71, 819-823
【非特許文献30】Meehan B. et al. Sequence of porcine circovirus) DNA: affinities with plant circovirus, 1997, 78, 221-227
【0050】
実施例9
ブタサーコウイルス Imp.999株のゲノムのクローン化
制限酵素EcoRIを有するPCV Imp.999株の2本鎖複製型の消化で生じた約1.8kbpの制限断片を市販のQiagen kit(QIAEXII Gel Extraction Kit, Cat # 20021, QIAGEN Ltd., Crawley, West Sussex, UK)を用いて1.5%アガロースゲル(実施例3参照)上で電気泳動で分離した。次に、このEcoRI−EcoRI制限断片を標準クローン化法(非特許文献31)によって予め同じ制限酵素を用いて消化し、脱リン酸したベクターpGEM-7(Promega, Medical Supply Company, Dublin, Ireland)と結合した。得られたプラスミドを標準的方法によってEscherichia coli JM109宿主株(Stratagene, La Jolla, USA)に変換した。同じく、PCV Imp.999株のEcoRI−EcoRI制限断片をベクターpBlueScript SK+(Stratagene, La Jolla, USA)のEcoRI部位にクローン化した。各宿主株に対して得られたクローンの中から所望の大きさの断片を有する少なくとも2つのクローンを選択した。次に、得られたクローンを培養し、Imp.999株の完全なゲノムを含むプラスミドを標準的なプラスミド調製および精製法によって少容量(2ml)または大容量(250ml)に精製した。
【非特許文献31】Sambrook J. et al. Molecular cloning: A Laboratory Manual. 2nd Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1988
【0051】
実施例10
PCV Imp.999株のゲノムDNAの配列(2本鎖複製型)
2つのEcoRI Imp.999のクローン配列(クローンpGEM-7/2およびpGEM-7/8)をメーカー推薦の方法で配列キット"AmpliTaq DNA polymerase FS"(Cat # 402079 PE Applied Biosystems, Warrington, UK)および"Applied BioSystems AB1373A automatic sequencing apparatus"を用いてSangerのジデオキシヌクレオチド法で決定した。最初の配列反応はM13「前方」および「後方」自在プライマーを用いて実施した。続く配列反応は「DNA歩行」法によって実施した。その後の配列に必要なオリゴヌクレオチドはLife Technology(Inchinnan Business Park, Paisley, UK)によって合成された。
【0052】
得られた配列は集め、MacDNASIS version 3.2のソフトウェア(Cat # 22020101, Appligene, Durham, UK)を用いて分析した。各種の読み取り枠を"National Center for Biotechnology Information"(NCBI, Bethesda, MD, USA)サーバーで入手可能なBLAST algorithmを用いて解析した。
クローンpGEM-7/8(配列ID番号6)から最初に得られた完全な配列(EcoRI-EcoRI)は図13、14に表してある。この配列はEcoRI部位のGの後に任意に始まり、ヌクレオチドに関してはいくつか不安定な部分を有している。
次いで、この配列が最適化された。配列ID番号3(図9〜14)はこの株の配列を全て表しており、EcoRI部位の始めすなわち最初のヌクレオチドとしてGから任意に始まる。
【0053】
同様の方法本発明の他の3つの分離株の配列を得た(配列ID番号1、2、4および図1〜4および図7、8参照)。これら4つの株のゲノムの大きさは下記の通りである:
Imp.1011−48121 1767個のヌクレオチド
Imp.1011−48285 1767個のヌクレオチド
Imp.999 1768個のヌクレオチド
Imp.1010 1768個のヌクレオチド
【0054】
実施例11
PCV Imp.999株の配列の分析
Imp.999株から得られた配列をGenBankのデータバンクに収容された配列との相同性テストで用いた結果、検出された有意な相同性はPK/15株の配列(寄託番号Y09921)(図9〜12参照)と約76%の相同性であった。
データバンク(NCBIサーバーのBLAST X algorithm)を用いた6つのフェーズでの配列の翻訳の相同性テストでは、アミノ酸レベルで、GenBank U49186配列でコードされるプラント(GenBank番号1841515)のサーコウイルスに類似のBBTBウイルスの理論的レプリカーゼに対応する読み取り枠と94%の相同性を示すということが分かった。
PCV Imp.999株から得られた配列と有意な相同性を示すものはデータバンクに収容された他の配列にはなかった。
多機能萎縮症候群の臨床的徴候を示すカリフォルニアの子豚から得られた病変部を用いて培養されたImp.999株から得られた配列のには解析から、この単分離ウイルスが新規なブタサーコウイルス株であることが明らかになった。
【0055】
実施例12
配列の比較分析
新規な4つのPCV株のヌクレオチド配列とPCVのPK/15株の配列(図9〜12)とを整列比較した。新規な4つの株と公知のPK/15株とに関する相同性マトリクスを作成した。結果は下記の通りである:
1:Imp.1011−48121
2:Imp.1011−48285
3:Imp.999
4:Imp.1010
5:PK/15
【0056】
【表1】
【0057】
2つのフランス株Imp. 1011-48121とImp. 1011-48285の相同性は99%以上(0.9977)である。2つの北米株Imp. 999とImp. 1010の相同性も99%以上(0.9949)以上である。フランス株と北米株との相同性はわずかに96%以上である。
これら全ての株とPK/15株との相同性は75〜76%の値に落ちる。
このことから、本発明の株はPK/15株に代表される種類とは異なる新規なブタサーコウイルスの代表であると言える。PK/15がタイプIを表すのに対し、PMWS症候群を示すブタから単離されたこの新規な株をタイプIIのブタサーコウイルスとよぶことにする。このタイプIIに属する株は実際には非常に離れた地理的領域から単離されたにもかかわらず、驚くべきヌクレオチド配列の相同性を示す。
【0058】
実施例13
新規なPCV株のゲノムによってコードされるプロテインの分析
Imp.1010単離株のヌクレオチド配列は多機能性萎縮症候群に関する他のサーコウイルス株を代表するものと考えられる。この配列をBLASTX algorithm(非特許文献32)と、ソフトウェアMacVector 6.0(Oxford Molecular Group, Oxford OX4 4GA, UK)のプログラムとを組み合わせを用いて詳細に分析した。この配列(環状ゲノム)上には20個のアミノ酸より大きいサイズの読み取り枠(またはORF)が13個検出できた。この13個のORFは下記の通りである:
【非特許文献32】Altschl et al. J. Mol. Biol. 1990. 215. 403-410
【0059】
【表2】
【0060】
各ORFの開始位置および終了位置は株1010のゲノムの図7、8に表された配列(配列ID番号4)を参照する。ORF1〜13の端部は株999と同一である。これらは下記ORF3と13を除き、株1011−48121および1011−48285とも同一である:
ORF3 1432−1539、センス、108nt、35aa
ORF3 314−1377、アンチセンス、705nt、234aa
これら13のORFの内で4つがクローン化ウイルスPCV PK/15のゲノム上に位置する類似のORFと有意な相同性を有する。多機能性萎縮症候群に関連するサーコウイルス単離株全てのゲノム上に存在する読み取り枠をそれぞれ分析した。これら4つのORFは下記の通り:
【0061】
【表3】
【0062】
各ORFの始めと終わりの位置は図7、8に表された配列(配列ID番号4)を参照する。ORFの大きさ(ヌクレオチド=ntで)は終止コドンを含む。
PCV Imp. 1010とPCV PK−15の単離株のゲノム機構の比較からから、2つのウイルスのゲノムに保存された4つのORFが同定できた。下記の表は観察された相同性の程度を表す:
【0063】
【表4】
【0064】
最大の相同性はORF4 Imp. 1010とORF1 PK−15との間に見られた(86%相同性)。このプロテインはおそらくウイルスのDNA複製に含まれ、ウイルスの複製に必須であるため、このことは予想されることである(非特許文献33)。
ORF13 Imp. 1010とORF2 PK−15との間の配列同一性は余り強くない(66.4%の同一性)が、この2つのORFはそれぞれ、鳥サーコウイルスCAV(非特許文献34)の主構造プロテインのN−末端領域と同一な良く保存されたN−末端基本領域を有している。さらに、ORF7 Imp. 1010とORF3 PK−15との間と、ORF10 Imp. 1010とORF4 PK−15との間に大きな違いが見られた。PCV PK−15のORF3とORF4と比較すると、それぞれにImp. 1010分離株のORF7とORF10のC−末端領域に欠失がある。最大の配列相同性はORF7/ORF3(重複部分の水準において61.5%の相同性)とORF10/ORF4(重複部分の水準において83%の相同性)のN−末端領域の水準において見られる。
【非特許文献33】Meehan et al., Gen. Virol. 1997,78; 221-227;Mankertz et al. J. Gen. Virol. 1998. 79. 381-384
【非特許文献34】Meehan et al., Arch. Virol. 1992, 124. 301-319
【0065】
ブタサーコウイルスのゲノム構造はゲノムが極端に高密度であるため非常に複雑であるように見える。主構造プロテインはおそらくブタサーコウイルスゲノムの同じ鎖上に位置する複数の読み取り枠間のスプライシングに由来するものである。従って、上記の表に記載の全ての読み取り枠(ORF1〜ORF13)でタイプIIのブタサーコウイルスにコードされた抗原プロテインの全てまたは一部を表すことができ、これらは特有の病気および/またはワクチン接種に使用可能な抗原である。本発明はこれらのORFを少なくとも1つ含む全てのプロテインに関するものである。本発明は主としてORF4、ORF7、ORF10またはORF13で構成されるプロテインに関するものである。
【0066】
実施例14
新規な株からクローンされたPCVゲノムの感染特性
Imp. 999単離株の完全なゲノム(複製型)を含むプラスミドpGEM-7/8をMeehan B. 達に記載の方法によってPK/15細胞に形質移入した(非特許文献35)。未感染のPK/15細胞に形質移入した後の最初の継代に対して実施された免疫抗体法の分析(実施例4参照)から、クローンpGEM-7/8のプラスミドは感染性PCVウイルスを誘導することが示された。感染性PCVの遺伝物質を含むクローンが利用可能になったことによってウイルス性ゲノムに任意の有用な処置をして無毒化ワクチンまたは組み換えワクチンを製造し、或いは診断キット用抗原を製造することができ、変成PCVウイルス(ブタで無毒化または欠失)を製造することができるようになった。
【非特許文献35】Characterization of viral DNAs from cells infected with chicken anemia agent: sequence analysis of the cloned replicative form and transfection capabilities of cloned genome gragments. Arch. Virol. 1992, 124. 301-319
【0067】
実施例15
インビトロ培養によるPCV抗原の製造
感染していないPK/15細胞の培養およびウイルス処置を実施例1と同じ方法で実施した。感染細胞を37℃で4日間培養後、トリプシン処理し、回収し、命名した。次の継代は1mlにつき400,000個の感染細胞を接種した。
【0068】
実施例16
ウイルス性抗原の不活化
ウイルス培養の最後に、感染した細胞を回収し、超音波(Branson Sonifier)またはロータースタータータイプのコロイドミル(UltraTurrax, IKA)を用いて溶解した。次に、懸濁液を3700gで30分間、遠心分離した。このウイルス懸濁液を+37℃で18時間、0.1%エチレンイミンで、或いは、+28℃で24時間、竈−プロピオラクトンで不活化した。ウイルス価が不十分な場合にはウイルス懸濁液を300kDaのメンブレン(Millipore PTMK300)を用いた限外濾過で濃縮した。不活化ウイルス懸濁液は+5℃で貯蔵した。
【0069】
実施例17
有機油をベースにしたワクチン乳剤の製造
下記処方でワクチンを調製した:
不活化ブタサーコウイルス懸濁液 : 250ml
MondanIDe(商標登録)ISA 70(SEPPIC) : 750ml
水相および油相は濾過によって別々に滅菌した。この乳剤はシルバーソン・タービン乳化機を用いて各成分を混合し、均質化して調製した。
一回のワクチン投与量は約107.5のTCID50を含む。皮膚経路の一回のワクチン投与容量は0.5ml、筋肉経路の投与量は2mlである。
【0070】
実施例18
代謝可能な油ベースのワクチン乳剤の製造
下記処方でワクチンを調製した:
不活化ブタサーコウイルス懸濁液: 200ml
Dehymuls HRE 7(Henkel) : 60ml
Radia 7204(Oleofina) : 740ml
水相および油相は濾過によって別々に滅菌した。この乳剤はシルバーソン・タービン乳化機を用いて各成分を混合し、均質化して調製した。
一回のワクチン投与量は約107.5のTCID50を含む。皮膚経路の一回のワクチン投与容量は0.5ml、筋肉内経路の投与量は2mlである。
【0071】
実施例19
過免疫血清(PCV−T)、PK/15から調製されたモノクローナル抗体F99およびカナダ株から調製された過免疫血清(PCV−C)で汚染されたアメリカおよびフランスのPCVウイルス株およびPK/15の間接免疫抗体法の結果
【0072】
【表5】
* 間接的免疫抗体法で正の反応を示した血清またはモノクローナル抗体の最後の希釈液の逆数。
【図面の簡単な説明】
【0073】
【図1】Imp.1011−48121株のゲノムのDNA配列
【図2】図1の続き
【図3】Imp.1011−48285株のゲノムのDNA配列
【図4】図3の続き
【図5】Imp.999株のゲノムのDNA配列
【図6】図5の続き
【図7】Imp.1010株のゲノムのDNA配列
【図8】図7の続き
【図9】PCV PK/15株の配列と図1〜8の4つの配列との整列化
【図10】図9の続き
【図11】図10の続き
【図12】図11の続き
【図13】1997年10月3日のフランスの最初の出願におけるImp.999株のゲノムのDNA配列
【図14】図13の続き
【図15】PK/15株の配列と図13、14の配列との整列化
【図16】図15の続き
【図17】図16の続き
【図18】図17の続き
【特許請求の範囲】
【請求項1】
タイプIIのブタサーコウイルス(circovirus)の精製物。
【請求項2】
ECACCに下記寄託番号で寄託された精製物から成る群の中から選択されるブタサーコウイルスの精製物:
寄託番号第V97100219号
寄託番号第V97100218号
寄託番号第V97100217号
寄託番号第V98011608号
寄託番号第V98011609号。
【請求項3】
PMWS症候群のブタの生理標本または組織標本、特に病変部から分離されるブタサーコウイルスに感染したインビトロ細胞培地の細胞で産成され、単離されたブタサーコウイルスの精製物。
【請求項4】
ブタの腎臓株細胞で産成され、単離される請求項3に記載のブタサーコウイルスの精製物。
【請求項5】
PCVに汚染されていないPK/15細胞で産成され、単離される請求項4に記載のブタサーコウイルスの精製物。
【請求項6】
請求項1のサーコウイルスに感染した細胞のインビトロ細胞培地から集められた培養抽出物または上澄み。
【請求項7】
請求項1に記載のサーコウイルスに感染した細胞のインビトロの細胞培地から集められた抗原製剤。
【請求項8】
抗原としてブタサーコウイルスを含む請求項7に記載の抗原製剤を含むワクチンまたは請求項6に記載の培養の上澄みまたは抽出物。
【請求項9】
無毒化した生の全抗原を獣医学上許容される賦形剤または希釈液中(必要に応じて獣医学上許容される補助剤およびフリーズドライ安定剤をさらに含むことができる)に含む請求項8に記載のワクチン。
【請求項10】
抗原が不活性化されており、ワクチンが獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤を含むことができる)を含む請求項9に記載のワクチン。
【請求項11】
下記配列群の中から選択される配列を含むDNA断片:
配列ID番号:1
配列ID番号:2
配列ID番号:3
配列ID番号:4
配列ID番号:6
【請求項12】
ORF1〜13から成る群の中から選択されるORFを含むDNA断片。
【請求項13】
ORF4,7,10および13から成る群の中から選択されるORFを含む請求項12に記載のDNA断片。
【請求項14】
請求項11〜13のいずれか一項に記載のDNA断片でコード化されたポリペプチド。
【請求項15】
請求項11〜13のいずれか一項に記載のDNA配列または断片がゲノム中に組み込まれ、インビトロで発現可能なインビトロ発現ベクター。
【請求項16】
E. coliおよびバキュロウイルスの中から選択される請求項15に記載の発現ベクター。
【請求項17】
必要に応じて精製された、請求項15または16に記載の発現ベクターによって製造されたポリペプチド。
【請求項18】
獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤をさらに含むことができる)中に請求項14または17に記載のポリペプチドを少なくとも1つ含むサブユニットワクチン。
【請求項19】
請求項11〜13のいずれか一項に記載DNA配列または断片がゲノム中に組み込まれ、インビボで発現可能なインビボ発現ベクター。
【請求項20】
ブタには病原性ではなく、ブタで増殖可能な生ウイルスとプラスミドの中から選択される請求項19に記載の発現ベクター。
【請求項21】
ウイルス性ベクターがアウジェスキー病ウイルス、ブタアデノウイルス等のブタヘルペスウイルス、特にワクシニアウイルス、トリポックスウイルス、カナリアポックスウイルス、ブタポックスウイルス等のポックスウイルスから選択される請求項20に記載の発現ベクター。
【請求項22】
獣医学上許容される賦形剤または希釈液中に請求項19〜21のいずれか一項に記載の発現ベクターを含む生ワクチンまたはプラスミド。
【請求項23】
請求項1〜4に定義の複数のブタサーコウイルスの抗原を含む請求項8〜10、18および22のいずれか一項に記載のワクチン。
【請求項24】
ブタの他の病原体に対応する少なくとも1つの他のワクチン価を有する請求項8〜10、18、22、23のいずれか一項に記載のワクチン。
【請求項25】
PRRS、Mycoplasma hyopneumonia、アクチノバシラス胸膜肺炎菌、E. coli、ブタ萎縮鼻炎、仮性狂犬病、ブタコレラ、ブタインフルエンザから成る群の中から選択される少なくとも1つの他のワクチン価を有する請求項24に記載のワクチン。
【請求項26】
PRRSおよびMycoplasma hyopneumoniaから成る群の中から選択される少なくとも1つの他のワクチン価を有する請求項24に記載のワクチン。
【請求項27】
請求項11〜13のいずれか一項に記載の配列の全てまたは一部を含むプローブまたはプライマー。
【請求項28】
請求項1〜5のいずれか一項に記載のサーコウイルス、請求項14または17に記載のポリペプチド、またはこれらの断片から調製されるモノクローナル抗原またはポリクローナル抗原。
【請求項29】
テストするブタの生理学的液体標本または組織標本で抗原の存在をテストし、抗原自体またはこの抗原に対する抗体を検出するブタサーコウイルスの検出方法。
【請求項1】
タイプIIのブタサーコウイルス(circovirus)の精製物。
【請求項2】
ECACCに下記寄託番号で寄託された精製物から成る群の中から選択されるブタサーコウイルスの精製物:
寄託番号第V97100219号
寄託番号第V97100218号
寄託番号第V97100217号
寄託番号第V98011608号
寄託番号第V98011609号。
【請求項3】
PMWS症候群のブタの生理標本または組織標本、特に病変部から分離されるブタサーコウイルスに感染したインビトロ細胞培地の細胞で産成され、単離されたブタサーコウイルスの精製物。
【請求項4】
ブタの腎臓株細胞で産成され、単離される請求項3に記載のブタサーコウイルスの精製物。
【請求項5】
PCVに汚染されていないPK/15細胞で産成され、単離される請求項4に記載のブタサーコウイルスの精製物。
【請求項6】
請求項1のサーコウイルスに感染した細胞のインビトロ細胞培地から集められた培養抽出物または上澄み。
【請求項7】
請求項1に記載のサーコウイルスに感染した細胞のインビトロの細胞培地から集められた抗原製剤。
【請求項8】
抗原としてブタサーコウイルスを含む請求項7に記載の抗原製剤を含むワクチンまたは請求項6に記載の培養の上澄みまたは抽出物。
【請求項9】
無毒化した生の全抗原を獣医学上許容される賦形剤または希釈液中(必要に応じて獣医学上許容される補助剤およびフリーズドライ安定剤をさらに含むことができる)に含む請求項8に記載のワクチン。
【請求項10】
抗原が不活性化されており、ワクチンが獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤を含むことができる)を含む請求項9に記載のワクチン。
【請求項11】
下記配列群の中から選択される配列を含むDNA断片:
配列ID番号:1
配列ID番号:2
配列ID番号:3
配列ID番号:4
配列ID番号:6
【請求項12】
ORF1〜13から成る群の中から選択されるORFを含むDNA断片。
【請求項13】
ORF4,7,10および13から成る群の中から選択されるORFを含む請求項12に記載のDNA断片。
【請求項14】
請求項11〜13のいずれか一項に記載のDNA断片でコード化されたポリペプチド。
【請求項15】
請求項11〜13のいずれか一項に記載のDNA配列または断片がゲノム中に組み込まれ、インビトロで発現可能なインビトロ発現ベクター。
【請求項16】
E. coliおよびバキュロウイルスの中から選択される請求項15に記載の発現ベクター。
【請求項17】
必要に応じて精製された、請求項15または16に記載の発現ベクターによって製造されたポリペプチド。
【請求項18】
獣医学上許容される賦形剤または希釈液(必要に応じて獣医学上許容される補助剤をさらに含むことができる)中に請求項14または17に記載のポリペプチドを少なくとも1つ含むサブユニットワクチン。
【請求項19】
請求項11〜13のいずれか一項に記載DNA配列または断片がゲノム中に組み込まれ、インビボで発現可能なインビボ発現ベクター。
【請求項20】
ブタには病原性ではなく、ブタで増殖可能な生ウイルスとプラスミドの中から選択される請求項19に記載の発現ベクター。
【請求項21】
ウイルス性ベクターがアウジェスキー病ウイルス、ブタアデノウイルス等のブタヘルペスウイルス、特にワクシニアウイルス、トリポックスウイルス、カナリアポックスウイルス、ブタポックスウイルス等のポックスウイルスから選択される請求項20に記載の発現ベクター。
【請求項22】
獣医学上許容される賦形剤または希釈液中に請求項19〜21のいずれか一項に記載の発現ベクターを含む生ワクチンまたはプラスミド。
【請求項23】
請求項1〜4に定義の複数のブタサーコウイルスの抗原を含む請求項8〜10、18および22のいずれか一項に記載のワクチン。
【請求項24】
ブタの他の病原体に対応する少なくとも1つの他のワクチン価を有する請求項8〜10、18、22、23のいずれか一項に記載のワクチン。
【請求項25】
PRRS、Mycoplasma hyopneumonia、アクチノバシラス胸膜肺炎菌、E. coli、ブタ萎縮鼻炎、仮性狂犬病、ブタコレラ、ブタインフルエンザから成る群の中から選択される少なくとも1つの他のワクチン価を有する請求項24に記載のワクチン。
【請求項26】
PRRSおよびMycoplasma hyopneumoniaから成る群の中から選択される少なくとも1つの他のワクチン価を有する請求項24に記載のワクチン。
【請求項27】
請求項11〜13のいずれか一項に記載の配列の全てまたは一部を含むプローブまたはプライマー。
【請求項28】
請求項1〜5のいずれか一項に記載のサーコウイルス、請求項14または17に記載のポリペプチド、またはこれらの断片から調製されるモノクローナル抗原またはポリクローナル抗原。
【請求項29】
テストするブタの生理学的液体標本または組織標本で抗原の存在をテストし、抗原自体またはこの抗原に対する抗体を検出するブタサーコウイルスの検出方法。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】
【図12】

【図13】

【図14】

【図15】

【図16】

【図17】

【図18】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公開番号】特開2008−86317(P2008−86317A)
【公開日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願番号】特願2007−254101(P2007−254101)
【出願日】平成19年9月28日(2007.9.28)
【分割の表示】特願2000−515010(P2000−515010)の分割
【原出願日】平成10年10月1日(1998.10.1)
【出願人】(503365659)
【出願人】(500155350)クィーンズ ユニバーシティ オブ ベルファスト (5)
【出願人】(500155361)ユニバーシティ オブ サスカチュワン (5)
【Fターム(参考)】
【公開日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願日】平成19年9月28日(2007.9.28)
【分割の表示】特願2000−515010(P2000−515010)の分割
【原出願日】平成10年10月1日(1998.10.1)
【出願人】(503365659)
【出願人】(500155350)クィーンズ ユニバーシティ オブ ベルファスト (5)
【出願人】(500155361)ユニバーシティ オブ サスカチュワン (5)
【Fターム(参考)】
[ Back to top ]