
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子
本発明は、ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ(CMP−Neu5Acヒドロキシラーゼ)のタンパク質、cDNA、およびゲノムDNA調節配列を提供する。さらに、本発明は、ブタ動物、組織、および臓器、ならびに機能的CMP−Neu5Acヒドロキシラーゼの発現を欠くこのような動物、組織、および臓器由来の細胞および細胞株を含む。このような動物、組織、臓器、および細胞を、研究および医学的治療法(異種移植が含まれる)ならびに産業上の家畜飼育操作で使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2003年6月6日提出の米国特許出願番号60/476,396号の優先権を主張する。
【0002】
(発明の分野)
本発明は、ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ(CMP−Neu5Acヒドロキシラーゼ)のタンパク質、cDNA、およびゲノムDNA調節配列を提供する。さらに、本発明は、ブタ動物、組織、および臓器、ならびに機能的CMP−Neu5Acヒドロキシラーゼの発現を欠くこのような動物、組織、および臓器由来の細胞および細胞株を含む。このような動物、組織、臓器、および細胞を、研究および医学的治療法(異種移植が含まれる)ならびに産業上の家畜飼育操作で使用することができる。さらに、異種移植で使用するためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く臓器、組織、および細胞の調製方法を提供する。
【背景技術】
【0003】
(発明の背景)
許容可能なヒトドナー臓器が利用不可能であること、宿主対移植片拒絶による長期成功率の低さ、ならびに感染症および癌の深刻なリスクは、組織および臓器移植分野が現在直面している主な課題である。許容可能な臓器に対する需要が供給を超えているので、多くの患者は毎年利用可能になる臓器を待っている間に死亡している。この需要を満たす一助とするために、研究は同種間移植の代替法の開発に焦点が当てられている。透析は腎不全患者が利用可能であり、人工心臓モデルが試験されており、臓器不全を補助または置換するための他の機械系が開発されている。しかし、このようなアプローチは非常に高価である。透析装置を頻繁且つ定期的に利用することが必要であるので、このような治療を受ける患者の自由および生活の質が非常に制限される。
【0004】
異種移植片移植は、ヒト移植のための人工臓器に代わる潜在的に魅力的なものである。非ヒト臓器の潜在的なプールは事実上限られている。ブタは、最も可能性が高い異種移植臓器の供給源と考えられている。ブタの供給は豊富にあり、交配プログラムが十分に確立されており、その臓器のサイズおよび生理学的性質はヒトに適合可能である。したがって、ブタ臓器での異種移植は、臨床的移植に利用可能な臓器の不足に対する潜在的な解決法を提供する。
【0005】
このような異種間組織の宿主拒絶は、依然としてこの分野における主な懸念である。異種移植に対する免疫学的障壁は、恐れられており、且つ依然として恐れられている。第1の免疫学的ハードルは、「超急性拒絶」(HAR)である。HARは、外来組織に結合する予め形成された高力価の天然の抗体の遍在と定義される。ドナー臓器内皮上の標的エピトープに対するこれらの天然の抗体の結合は、HARにおける開始事象と考えられる。この結合の後、レシピエントの血液でのドナー臓器の灌流から数分以内に、血小板およびフィブリンが沈殿し、最終的にドナー臓器が間質の浮腫および出血を起こし、その全てによってレシピエントで臓器不全が起こる(Strahanら(1996)Frontiers in Bioscience 1,pp.34−41)。
【0006】
類人猿または旧世界ザル(例えば、ヒヒ)からヒトへのいくつかの著名な異種移植は、拒絶することなく何ヵ月間も許容される。しかし、このような試みは、多数の免疫学的要因によって最終的に失敗している。HARを抑制するための強い免疫抑制を使用した場合でさえも、移植片内皮上の異種補体の活性化を制御するための移植片組織内の補体調節タンパク質(CRP)の不全に一部起因する軽度の先天性免疫応答により、最終的に移植臓器が破壊される(Starzl,Immunol.Rev.,141,213−44(1994))。ヒトへの異種移植に許容可能な臓器のプールを開発する目的で、研究者は、ヒトCRPを産生する動物を操作している(霊長類における異種移植片破壊を遅延させるが消失しないことが証明されているアプローチ)(McCurryら,Nat.Med.,1,423−27(1995);Bachら,Immunol.Today,17,379−84(1996))。
【0007】
補体媒介攻撃に加えて、調和しない異種移植片のヒトによる拒絶は、共通の抗原(多数の糖タンパク質および糖脂質のガラクトース−α(1,3)−ガラクトース(gal−α−gal)末端残基)によって媒介されるようである(Galiliら,Proc.Nat.Acad.Sci.(USA),84,1369−73(1987);Cooperら,Immunol.Rev.,141,31−58(1994);Galiliら,Springer Sem.Immunopathol,15,155−171(1993);Sandrinら,Transplant Rev.,8,134(1994))。この抗原は、ヒトA、B、およびO型血液抗原と化学的に関連し、多数の寄生虫ならびに細菌およびウイルスなどの感染因子に存在する。ほとんどの哺乳動物組織もこの抗原を含むが、旧世界ザル、類人猿、およびヒトは注目すべき例外である(Joziasseら,J.Biol.Chem.,264,14290−97(1989)を参照のこと)。このような炭水化物エピトープを持たない個体は、エピトープに特異的な天然に存在する抗体(IgMおよびIgG)を豊富に産生する。多くのヒトは、gal−α−gal炭水化物決定基に対して特異性を有する有意なレベルの循環IgGを示す(Galiliら,J.Exp.Med.,162,573−82(1985);Galiliら,Proc.Nat.Acad.Sci.(USA),84,1369−73(1987))。α−ガラクトシルトランスフェラーゼ(α−GT)酵素は、gal−α−gal部分の形成を触媒する。研究は、異種組織の細胞表面上でのgal−α−gal部分の発現を減少または消失するためのこの酵素の調整または消失に焦点が当てられている。
【0008】
ブタ由来のα−ガラクトシルトランスフェラーゼ遺伝子の消失は、ブタからヒトへの異種移植に対する最も有意なハードルの1つと長い間考えられている。ブタゲノム中の2つの対立遺伝子はα−GT遺伝子をコードする。ブタにおけるα−GT遺伝子の1対立遺伝子ノックアウトが2002年に報告された(DaiらNature Biotechnol.,20:251(2002);Laiら,Science,295:1089(2002))。
【0009】
最近、α−GT遺伝子の二重対立遺伝子ノックアウト(double allelic knockout)が行われた(Phelpsら,Science,299:pp.411−414(2003))。Revivicor Inc.のWO 2004/028243は、α1,3ガラクトシルトランスフェラーゼ(α1,3−GT)の全発現を欠くブタ動物、組織、臓器、細胞、および細胞株を記載している。したがって、α1,3−GTの機能的発現を欠く動物、組織、臓器、細胞、および細胞を、異種移植および他の医学的目的のために使用することができる。
【0010】
Immerge Biotherapeutics,Inc.のPCT特許出願WO2004/016742号は、α(1,3)−ガラクトシルトランスフェラーゼヌル細胞、GGTA−1ヌル細胞の選択方法、それから産生されたα(1,3)−ガラクトシルトランスフェラーゼヌルブタ(生存可能なGGTA−1ヌルブタと呼ばれる)、このようなブタの作製方法、および異種移植のための細胞、組織、および臓器の使用方法を記載している。
【0011】
gal−α−gal以外の最も初期の公知の異種抗原の1つは、Hnaganutiu Deicher抗体が認識し、且つ血清疾患と長期的に関連するエピトープである。エピトープは、N−グリコリルノイラミン酸(Neu5Gc)(炭水化物のシアル酸ファミリーのメンバー)として同定された。炭水化物のうち、シアル酸は豊富且つ遍在する。シアル酸は、N−アセチルノイラミン酸(Neu5Acyl)およびその誘導体に使用される総称である。N−アセチルノイラミン酸(Neu5Ac)およびN−グリコリルノイラミン酸(neu5Gc)は、シアル酸の最も豊富な誘導体のうちの2つである。
【0012】
Neu5Gcエピトープは、複合糖質のグリカン鎖中の末端に存在する。この露呈した位置のために、例えば、炎症反応、免疫細胞の成熟、分化プロセス、ホルモン結合、病原体結合、および毒素結合の場合の細胞認識で重要な役割を果たす(Varki,A.,Glycobiology,2,pp.25−40(1992))。
【0013】
Neu5Gcを含む複合糖質はヒトで免疫原性を示す。健常なヒトでは、Neu5Gcは検出可能でないが、Neu5Gcはほとんどの哺乳動物で豊富に存在する。人におけるNeu5Gcの欠如は、機能酵素の形成を防止するヒト遺伝子におけるエクソンの欠失に起因する(Chou,H.H.らProc.Natl.Acad.Sci.(USA),95,pp.11751−11756(1998);Irie,A.らJ.Biol.Chem.,273,pp.15866−15871(1998))。したがって、Neu5Gc含有複合糖質は抗原として作用し、抗体の形成を誘導することができる。歴史的に、この抗体はHanganutziu−Deicher(HD)抗原および抗体と呼ばれる(Hanganutziu,M.,CR Soc.Biol.(Paris),91,p.1457(1924);Deicher,H.,Z.Hyg.,106,p.561(1926))。Hanganutziu−Deicher抗原は、多数のヒト腫瘍(結腸癌、網膜芽腫、黒色腫、および乳癌)およびニワトリ腫瘍組織で検出可能である(Higashi,H.らCancer Res.,45,pp.3796−3802(1985))。腫瘍中の抗原量は非常に少ないにもかかわらず(通常、全量が1%未満のシアル酸、しばしば0.01から0.1%までの範囲)、Hanganutziu−Deicher抗体の形成を誘導することができる(Higashihara,T.ら,Int Arch Allergy Appl Immunol.,95,pp.231−235(1991))。この免疫反応は、ヒトへのNeu5Gc含有ブタ臓器の異種移植に対する潜在的な障壁である。
【0014】
Neu5Gcエピトープは、Neu5AcのN−アセチル部分への水酸基の付加によって形成される。ヒドロキシル化を触媒する酵素は、CMP−Neu5Acヒドロキシラーゼである。したがって、CMP−Neu5Acヒドロキシラーゼ遺伝子の発現は、細胞表面上のNeu5Gcエピトープの存在を決定する。哺乳動物におけるCMP−Neu5Acヒドロキシラーゼの精製研究により、シトクロムb5およびシトクロムb5レダクターゼに依存する可溶性細胞質オキシゲナーゼであることが示された(Kawano,T.ら,J.Biol.Chem.,269,pp.9024−9029(1994);Schneckenburger,P.ら,Glycoconj.J.,11,pp.194−203(1994);Schlenzka,W.ら,Glycobiology,4,pp.675−683(1994);Kozutsumi,Y.ら,J.Biochem.(Tokyo),108,pp.704−706(1990);およびShaw,L.ら,Eur.J.Biochem.,219,pp.1001−1011(1994))。
【0015】
Neu5Gcの別の重要な特徴は、細胞に侵入させる病原体の接着分子として作用することである(Kelm,S.およびSchauer,R.,Int.Rev.Cytol,179,pp.137−240(1997))。これにより、一定の家畜で疾患および経済的損失を被る。詳細には、K99線毛を有する腸管毒素原性大腸菌は、腸内吸収細胞および粘膜分泌細胞に結合したNue5Gcα2→3Galβ1→4Glcβ1→1’セラミド[GM3(Neu5Gc)]などのガングリオシド中のNeu5Gc、N−グリコシルシアロパラグロボシド、およびGM2(Neu5Gc)への結合によって新生仔ブタに感染し、潜在的に致死的下痢を引き起こす(Malykh,Y.ら,Biochem.J.,370,pp.601−607(2003);Kyogashima,M.ら(1993);Teneberg,S.ら,FEBS Leaers,263,pp.10−14(1990);Isobe,T.ら,Anal.Biochem.,236,pp.35−40(1996);Lindahl,M.およびCaristedt,I.,Gen.Microbiol.,136,pp.1609−1614(1990);King,T.P.ら,Proceedings of the 6th International Symposium on Digestive Physiology in Pigs,pp.290−293(1994))。ブタロタウイルスは、GM3(Neu5Gc)への結合によって新生ブタに感染して下痢を引き起こす。ブタ伝染性胃腸炎コロナウイルスは、α2,3結合Neu5Gcを含む複合糖質への侵入によってブタに感染する(Schultz,Bら,.L Virol.,70,pp.5634−5637(1996))。
【0016】
マウスの肝臓およびブタの顎下腺からCMP−Neu5Acヒドロキシラーゼが均一に単離され、特徴づけられている(Kawano,T.ら,J.Biol.Chem.,269,pp.9024−9029(1994);Schneckenburger,P.ら,Glycoconj.J.,11,pp.194−203(1994);and,Schlenzka,W.ら,Glycobiology,4,pp.675−683(1994))。
【0017】
Schlenzkaら(Glycobiology,Vol.4,pp.675−683(1994))は、イオン交換クロマトグラフィ、固定化トリアジン色素を使用したクロマトグラフィ、疎水性相互作用クロマトグラフィ、およびゲル濾過を使用して、ブタ顎下腺から酵素を精製した。Schneckenburgerら(Glycoconj.J.,Vol.11,pp.194−203(1994))は、マウス肝臓からCMP−Neu5Acヒドロキシラーゼを単離した。ブタ顎下腺およびマウス肝臓由来のCMP−Neu5Acヒドロキシラーゼは共に分子量65kDaの可溶性単量体である。CMP−Neu5Acおよびシトクロムb5とのその触媒的相互作用は、互いに類似している。これらの酵素の活性は、イオン含有補欠分子団に依存するようである。
【0018】
JP−A06113838は、マウスCMP−Neu5Acヒドロキシラーゼのタンパク質およびDNA配列ならびにこのヒドロキシラーゼに特異的に結合するモノクローナル抗体を記載している。
【0019】
Boehringer ManheimGMBHのPCT公開番号WO97/03200A1は、ブタCMP−Neu5Acヒドロキシラーゼの部分cDNAを開示している。この出願は、CMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン8の中央から始まるcDNA配列を開示している(GenBankアクセッション番号Y15010としてさらに開示されている)。
【0020】
Martensen,L.ら(Eur.J.Biochem.,Vol.268,pp.5157−5166(2001))は、ブタCMP−Neu5Acヒドロキシラーゼの全長アミノ酸配列を開示している。
【0021】
RBC BiotechnologyのPCT公開番号WO02/088351号は、ブタCMP−Neu5Acヒドロキシラーゼの部分cDNA配列およびゲノム配列(エクソン7〜11および各エクソン周囲の部分ゲノム配列)を開示している。さらに、任意選択的に、α−1,3−ガラクトシルトランスフェラーゼ遺伝子および/または補体タンパク質の挿入などの他のゲノム修飾と組み合わせたCMP−Neu5Acヒドロキシラーゼエピトープを欠くブタ細胞および動物の作製方法を提供する。
【発明の開示】
【発明が解決しようとする課題】
【0022】
本発明の目的は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列および調節配列を提供することである。
【0023】
本発明の目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子の全長cDNAおよび新規の変異型を提供することである。
【0024】
本発明の別の目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子をコードする新規の核酸配列およびアミノ酸配列を提供することである。
【0025】
本発明のさらなる目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子を欠損した細胞、組織、および/または臓器を提供することである。
【0026】
本発明の別の目的は、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物(特にブタ)を作製することである。
【0027】
本発明のなおさらなる目的は、必要とするヒトレシピエントへの非ヒト臓器の異種移植で使用するためのCMP−Neu5Acヒドロキシラーゼ遺伝子を欠損した細胞、組織、および/または臓器を提供することである。
【課題を解決するための手段】
【0028】
(発明の要旨)
ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の全長cDNA配列、ペプチド配列、およびゲノム機構を決定した。今日まで、部分cDNAおよびゲノム配列のみが同定されている。本発明は、新規のブタCMP−Neu5Acヒドロキシラーゼのタンパク質、cDNA、cDNA変異型、およびゲノムDNA配列を提供する。さらに、本発明は、ブタ動物、組織、および臓器、ならびに機能的CMP−Neu5Acヒドロキシラーゼの発現を欠くこのような動物、組織、および臓器由来の細胞および細胞株を含む。このような動物、組織、臓器、および細胞を、研究および医学的治療法(異種移植が含まれる)で使用することができる。さらに、異種移植で使用するためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く臓器、組織、および細胞の調製方法を提供する。
【0029】
本発明の1つの態様は、ブタCMP−Neu5Acヒドロキシラーゼの全長cDNAを提供する。全長cDNAを、表1(配列番号1)に示し、全長ペプチド配列を表2(配列番号2)で提供する。全長cDNAの開始コドンはエクソン4の3’部分に存在し、終止コドンはエクソン17の3’部分で見出される。配列番号1および2と少なくとも80%、85%、90%、95%、98%、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号1または2の少なくとも10、15、17、20、25、または30個のヌクレオチドまたはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号1とハイブリッド形成する任意のヌクレオチド配列およびそれに相同なヌクレオチドをさらに提供する。
【0030】
1つの実施形態では、CMP−Neu5Acヒドロキシラーゼの新規の変異型をコードする核酸配列およびペプチド配列を提供する(表3〜8、図2)。配列番号3は、エクソン1、4、5、6、7、8、9、10、11、12、13、15a、16、17、および18を含む遺伝子の変異型(変異型1)のcDNAを示す。配列番号5は、エクソン1、4、5、6、7、8、9、10、11、12、および12aを含む遺伝子の変異型(変異型2)のcDNAを示す。配列番号7は、エクソン1、4、5、6、7、8、9、10、11、および11aを含む遺伝子の変異型(変異型3)のcDNAを示す。配列番号4、6、および8は、それぞれ変異型1、変異型2、および変異型3のアミノ酸配列を示す。配列番号3〜8と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号3〜8の少なくとも10、15、17、20、25、または30個のヌクレオチドまたはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号3、5、および7とハイブリッド形成する任意のヌクレオチド配列ならびにそれに相同なヌクレオチドをさらに提供する。
【0031】
さらなる実施形態は、CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA配列を示す核酸配列を提供する(表9、図1)。配列番号10〜28は、それぞれエクソン1、4〜11、11a、12、12a、13〜15、15a、16〜18を示し、配列番号29〜45は、それぞれイントロン1a、1b、4〜15、15a、16、および17を示す。配列番号9は、CMP−Neu5Acヒドロキシラーゼ遺伝子の5’非翻訳領域を示す。配列番号46(表10)は、CMP−Neu5AcヒドロキシラーゼのゲノムDNAおよび調節配列を示す。
【0032】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号47に示す。配列番号47は、5’UTR、エクソン1、およびエクソン1(表11)3’に存在するイントロン配列の一部を含む5’連続ゲノム配列を示す。
【0033】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号48に示す。配列番号48は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、エクソン8、イントロン8、エクソン9、イントロン9、エクソン10、イントロン10、エクソン11、イントロン11、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18でイントロン配列を含む連続ゲノム配列を示す(表12)。さらに、配列番号48の少なくとも2775、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、または10,000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号48と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0034】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号49に示す。配列番号49は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、およびエクソン8でイントロン配列を含む連続ゲノム配列を示す。さらに、配列番号49の少なくとも1750、1800、1900、2000、2100、2200、2300、2400、2500、2600、2700、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4100、4200、4300、4400、4500、4600、4700、4800、4900、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号49と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0035】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号50に示す。配列番号50は、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18を含む連続ゲノム配列を示す。配列番号50の少なくとも150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1500、2000、2500、3000、3500、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20、000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0036】
さらなる実施形態では、配列番号9〜45、46、47、48、49、および50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号9〜45、46、47、および48の少なくとも10、15、17、20、25、30、50、100、150、200、300、400、500、または1000個の連続ヌクレオチドを含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号9〜45、46、47、48、49、および50とハイブリッド形成する任意のヌクレオチド配列ならびにこれらに相同なヌクレオチドをさらに提供する。
【0037】
本発明の別の態様は、CMP−Neu5AcヒドロキシラーゼをコードするcDNAまたはその変異型を含む核酸構築物を提供する。これらのcDNA配列は、配列番号1〜8またはその任意のフラグメントに由来し得る。構築物は、1つまたは1つを超える内部リボゾーム侵入部位(IRES)を含み得る。構築物はまた、CMP−Neu5Acヒドロキシラーゼをコードする核酸配列に作動可能に連結されたプロモーターを含み得るか、構築物はプロモーターを含まなくてよい。別の実施形態では、宿主ゲノムへの挿入がランダムまたはターゲティングされている核酸配列を含む核酸構築物を提供する。核酸配列に加えて、発現ベクターは、選択マーカー(例えば、増強緑色蛍光タンパク質(eGFP)遺伝子配列、開始配列および/またはエンハンサー配列、ポリAテール配列、および/または原核生物および/または真核生物の宿主細胞で構築物を発現させる核酸配列など)を含み得る。
【0038】
別の実施形態では、体細胞中の相同組換えを行うことができる核酸ターゲティングベクター構築物も提供する。これらのターゲティングベクターを、相同組換えを介してCMP−Neu5Acヒドロキシラーゼ遺伝子をターゲティングするために哺乳動物細胞に形質転換することができる。1つの実施形態では、ターゲティングベクターは、CMP−Neu5Acヒドロキシラーゼのゲノム配列に相同な3’組換えアームおよび5’組換えアームを含み得る。相同なDNA配列には、CMP−Neu5Acヒドロキシラーゼ配列と少なくとも15bp、20bp、25bp、50bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbp、または50kbpが相同な配列が含まれ得る。別の実施形態では、相同DNA配列には、1つまたは複数のイントロンおよび/またはエクソン配列が含まれ得る。特定の実施形態では、DNA配列は、CMP−Neu5Acヒドロキシラーゼ遺伝子のイントロン5およびイントロン6に相同であり得る(例えば、図6〜8を参照のこと)。別の特定の実施形態では、DNA配列は、CMP−Neu5Acのイントロン5、エクソン6の55bp部分、およびイントロン6と相同であることができ、エクソン6の55bp部分に対して3’方向にてインフレームで増強緑色蛍光タンパク質配列を含む(例えば、図10〜11を参照のこと)。
【0039】
本発明の別の実施形態は、ブタCMP−Neu5AcヒドロキシラーゼのcDNAまたはゲノム配列(配列番号1、3、5、7、9〜45、47、または48など)とハイブリッド形成することができるオリゴヌクレオチドプライマーを提供する。好ましい実施形態では、プライマーは、ストリンジェントな条件下で配列番号1、3、5、7、9〜45、46、47、または48とハイブリッド形成する。別の実施形態は、配列番号1、3、5、7、9〜45、46、47、または48などのブタCMP−Neu5Acヒドロキシラーゼ核酸配列とハイブリッド形成することができるオリゴヌクレオチドプローブを提供する。ポリヌクレオチドプライマーまたはプローブは、本発明のポリヌクレオチドとハイブリッド形成する少なくとも14塩基、20塩基、好ましくは30塩基、または50塩基を有し得る。プローブまたはプライマーは、少なくとも14ヌクレオチド長であり得るが、好ましい実施形態では、少なくとも15、20、25、28、または30ヌクレオチド長である。
【0040】
本発明の別の態様では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子を欠く哺乳動物細胞を提供する。相同組換えの結果としてこれらの細胞を得ることができる。特に、CMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子の不活化により、機能的Hanganutziu−Deicher抗原の発現能力が低下した細胞を産生することができる。
【0041】
本発明の実施形態では、得られたCMP−Neu5Acヒドロキシラーゼ酵素がもはやHanganutziu−Deicher抗原を作製することができないように本明細書中に記載のプロセス、配列、および/または構築物にしたがってCMP−Neu5Acヒドロキシラーゼ遺伝子の対立遺伝子を不活化する。1つの実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子をRNAに転写することができるが、タンパク質に翻訳することができない。別の実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を、不活性な短縮形態に転写することができる。このような短縮RNAは、翻訳することができないか、非機能的タンパク質に翻訳することができる。別の実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を遺伝子が転写しないような方法で不活化することができる。さらなる実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を転写し、その後非機能的タンパク質に翻訳することができる。
【0042】
本発明のさらなる態様では、CMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたゲノムターゲティング事象を介して不活化されたブタ動物を提供する。本発明の別の態様では、CMP−Neu5Acヒドロキシラーゼ遺伝子の両対立遺伝子が遺伝子ターゲティング事象を介して不活化されたブタ動物を提供する。相同組換えを介して遺伝子をターゲティングすることができる。他の実施形態では、遺伝子を破壊することができる(すなわち、遺伝コードの一部を変化させ、それにより遺伝子のそのセグメントの転写および/または翻訳に影響を与えることができる)。例えば、欠失(「ノックアウト」)技術または挿入(「ノックイン」)技術によって遺伝子を破壊することができる。既存配列の転写を調整する所望のタンパク質または調節配列のさらなる遺伝子を挿入することができる。
【0043】
本発明の別の態様では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の1つの対立遺伝子、任意選択的に両対立遺伝子を欠くブタ細胞を、クローン化トランスジェニック動物を産生するための除核卵母細胞への核導入のためのドナー細胞として使用することができる。あるいは、胚幹細胞中でブタCMP−Neu5Acヒドロキシラーゼノックアウトを作製し、その後子孫の産生に使用することができる。本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生した機能的CMP−Neu5Acヒドロキシラーゼ遺伝子の1つの対立遺伝子を欠く子孫を交配して、メンデル型遺伝子によって両対立遺伝子の機能性を欠く子孫をさらに産生することができる。異種移植ストラテジーで使用するために、細胞、組織、および/または臓器をこれらの動物から回収することができる。Hanganutziu−Deicher抗原は、Neu5Gcエピトープに起因する移植細胞、組織、または臓器の免疫拒絶を軽減させることができる。
【0044】
あるいは、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生したCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子を欠く動物は、腸管毒素原性感染および疾患(例えば、E.coli感染、ロタウイルス感染、および胃腸炎コロナウイルスなど)に対する感受性が低いか耐性を示し得る。このような動物を、例えば、商業的飼育で使用することができる。
【0045】
本発明の1つの態様では、本発明の任意の態様にしたがった方法によってブタを調製することができる。遺伝子操作ブタを、移植療法のための組織および/または臓器供給源として使用することができる。この様式で調製したブタ胚またはそれから発生した細胞株も細胞移植療法で使用することができる。したがって、本発明のさらなる態様では、CMP−Neu5Acヒドロキシラーゼを欠く胚または動物から調製されたブタCMP−Neu5Acヒドロキシラーゼを欠く遺伝子操作細胞のブタへの投与を含む治療方法を提供する。本発明のこの態様は、薬物におけるこのような細胞の使用(例えば、細胞移植療法)および移植のための細胞または組織移植片の調製におけるこのような胚由来の細胞の使用にまで及ぶ。細胞を、組織または臓器(例えば、心臓、肺、腎臓、膵臓、角膜、神経(例えば、脳、中枢神経系、脊髄)、皮膚)に組織化することができるか、細胞は、島細胞、血球(例えば、血球(すなわち、赤血球、白血球))または造血幹細胞もしくは他の幹細胞(例えば、骨髄)であり得る。
【0046】
本発明の別の態様では、CMP−Neu5Acヒドロキシラーゼ欠損ブタは他の異種抗原(例えば、ブタiGb3シンターゼ(例えば、を米国特許出願60/517,524号を参照のこと)および/またはブタForssmanシンターゼ(例えば、米国特許出願60/568,922号を参照のこと)など)をコードする遺伝子も欠く。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたα1,3ガラクトシルトランスフェラーゼ遺伝子およびCMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタ細胞を提供する。別の実施形態では、ブタ1,3ガラクトシルトランスフェラーゼ遺伝子ノックアウト細胞を、本明細書中に記載のプロセス、配列、および/または構築物によって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子をノックアウトするようにさらに修飾する。さらに、任意選択的に不都合な免疫応答に関連する1つまたは複数のさらなる遺伝子を欠く本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ欠損ブタを補体阻害タンパク質(例えば、CD59、DAF、および/またはMCPなど)を発現するように修飾することができ、MCPをCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子の発現を消失するようにさらに修飾することができる。これらの動物を、移植療法のための組織および/または臓器の供給源として使用することができる。これらの動物を、移植療法のための組織および/または臓器の供給源として使用することができる。この様式で調製したブタ胚またはそれから発生した細胞株も細胞移植療法で使用することができる。
【0047】
(発明の説明)
本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子の消失により、Neu5Gcエピトープに対するヒトの免疫応答を減少し、異種移植に対する免疫学的障壁を除去することができる。本発明は、全長cDNAおよびペプチドをコードする新規の核酸配列に関する。遺伝子のゲノム機構、イントロン配列、および調節領域に関する情報も提供する。1つの態様では、本発明は、配列番号1、3、5、または7の1つをコードする単離および実質的に精製されたcDNA分子またはそのフラグメントを提供する。本発明の別の態様では、CMP−Neu5Acヒドロラーゼ遺伝子の全長ゲノムを含むDNA配列を、配列番号9〜45、46、47、48、49、もしくは50またはそのフラグメントに提供する。別の態様では、配列番号1、3、5、7、9〜45、46、47、48、49、または50由来のブタCMP−Neu5AcヒドロキシラーゼcDNAまたはゲノム配列の増幅のためのプライマーを提供する。さらに、配列番号1、3、5、7、9〜45、46、47、48、49、もしくは50またはそのフラグメント由来のCMP−Neu5Acヒドロキシラーゼ核酸配列を同定するためのプローブを提供する。配列番号9〜45、46、47、48、49、もしくは50またはそのフラグメントによって示されるDNAを使用して、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタを構築物ことができる。したがって、本発明はまた、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタ染色体および本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生された機能的CMP−Neu5Acヒドロキシラーゼタンパク質を欠くトランスジェニックブタを提供する。このようなブタを、ヒトへの異種移植のための組織供給源として使用することができる。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物によって産生されたCMP−Neu5Acヒドロキシラーゼ欠損ブタは、異種移植における不都合な免疫応答に関連する他の遺伝子(例えば、α1,3ガラクトシルトランスフェラーゼ遺伝子、iGb3シンターゼ遺伝子、またはFSMシンターゼ遺伝子など)も欠く。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼおよび/または異種移植における不都合な免疫応答に関連する他の遺伝子を欠くブタは、補体阻害因子(例えば、CD59、DAF、および/またはMCPなど)を発現する。
【0048】
(定義)
「標的DNA配列」は、相同組換えによって修飾されるDNA配列である。標的DNAは、動物細胞の任意のオルガネラ(核およびミトコンドリアが含まれる)中に存在することができ、インタクトな遺伝子、エクソンもしくはイントロン、調節配列、または遺伝子間の任意の領域であり得る。
【0049】
「ターゲティングDNA配列」は、所望の配列修飾を含むDNA配列である。ターゲティングDNA配列は、標的DNAと実質的に同質遺伝子的であり得る。
【0050】
「相同DNA配列または相同DNA」は、基準DNA配列と少なくとも80%、85%、90%、95%、98%、または99%同一のDNA配列である。相同配列は、ストリンジェントな条件下で標的配列とハイブリッド形成し、ストリンジェントな条件には、配列が少なくとも85%、好ましくは少なくとも95%または98%同一である場合にハイブリッド形成が起こる条件が含まれる。
【0051】
「同質遺伝子または実質的に同質遺伝子のDNA配列」は、基準DNA配列と同一であるかほぼ同一であるDNA配列である。用語「実質的に同質遺伝子的な」は、基準DNA配列と少なくとも約97〜99%同一、好ましくは基準DNA配列と少なくとも約99.5〜99.9%同一、一定の用途では基準DNA配列と100%同一のDNAをいう。
【0052】
「相同組換え」は、配列相同性に基づいたDNA組換えプロセスをいう。
【0053】
「遺伝子ターゲティング」は、一方が染色体上に存在し、他方が染色体上に存在しない2つのDNA配列間の相同組換えをいう。
【0054】
「非相同または無作為な組み込み」は、DNAが相同組換えに関与しないゲノムに組み込まれる任意のプロセスをいう。
【0055】
「選択マーカー遺伝子」は、その発現により遺伝子を含む細胞が同定される遺伝子である。選択マーカーは、この遺伝子を含まない細胞の成長を防止または遅延させる培地上で細胞を増殖させるものであり得る。例には、抗生物質耐性遺伝子および生物を選択された代謝産物に対して成長させる遺伝子が含まれる。あるいは、この遺伝子は、細胞への容易に同定される表現型の付与によって形質転換体の目視によるスクリーニングを容易にすることができる。このような同定可能な表現型は、例えば、発光もしくは着色化合物の産生または細胞周囲の培地中の検出可能な変化であり得る。
【0056】
用語「連続する」は、本明細書中でその標準的な意味(すなわち、途切れないまたは連続的な)で使用される。
【0057】
用語「ブタ」は、任意のブタ種(Large White、Landrace、Meishan、Minipigが含まれる)をいう。
【0058】
用語「卵母細胞」は、卵形成の最終産物であり、且つ前駆体形態がそれぞれ卵原細胞、一次卵母細胞および二次卵母細胞でもある成熟動物の卵を説明する。
【0059】
用語「フラグメント」は、ヌクレオチドまたはペプチド配列の一部または部分配列を意味する。
【0060】
用語「誘導体」および「アナログ」は、このようなヌクレオチドまたはペプチドと本質的に同一の生物機能または生物活性を保持するヌクレオチド配列またはペプチド配列を意味する。例えば、アナログには、活性成熟ポリペプチドを産生するためのプロタンパク質部分の切断によって活性化することができるプロタンパク質が含まれる。
【0061】
本明細書中で提供したDNA(デオキシリボ核酸)配列を、塩基アデニン(A)、チミン(T)、シトシン(C)、およびグアニン(G)によって示す。
【0062】
本明細書中で提供したアミノ酸配列を、以下の略語で示す。
【0063】
【表1】

「トランスフェクション」は、宿主細胞へのDNAの移入をいう。細胞は、自然にはDNAを取り込まない。したがって、種々の技術的「トリック」を使用して、遺伝子導入を容易にする。多数のトランスフェクション法(例えば、CaPO4およびエレクトロポレーション)が当業者に公知である(J.Sambrook,E.Fritsch,T.Maniatis,Molecular Cloning:A Laboratory Manual,Cold Spring Laboratory Press,1989)。宿主細胞のトランスフェクションは、首尾の良いトランスフェクションの指標である。
【0064】
(I.ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の完全なcDNA配列および変異型)
本発明の1つの態様は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の新規の全長核酸cDNA配列を提供する(図2、表1、配列番号1)。本発明の別の態様は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の推定アミノ酸ペプチド配列を提供する(表2、配列番号2)。全長cDNAのATG開始コドンはエクソン4の3’部分に存在し、終止コドンTAGはエクソン17の3’部分で見出される。配列番号1または2と少なくとも90、95、98、または99%相同な核酸配列およびアミノ酸配列を提供する。さらに、配列番号1または2の少なくとも10、15、17、20、または25個の連続核酸またはアミノ酸を含むヌクレオチド配列およびペプチド配列も提供する。配列番号1〜2のフラグメント、誘導体、およびアナログをさらに提供する。配列番号1〜2のフラグメントには、少なくとも約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個のヌクレオチドを含む任意の連続する核酸配列またはペプチド配列が含まれ得る。
【0065】
【表1A−1】
【0066】
【表1A−2】
【0067】
【表1A−3】
【0068】
【表1A−4】
【0069】
【表2−1】
【0070】
【表2−2】
(変異型)
本発明の別の態様は、CMP−Neu5Acヒドロキシラーゼ遺伝子転写物の3つの新規の変異型の新規の核酸cDNA配列を提供する(図2、表3、5、および7、配列番号3、5、および7)。配列番号3は、エクソン1、4、5、6、7、8、9、10、11、12、13、15a、16、17、および18を含む遺伝子の変異型(変異型1)のcDNAを示す。エクソン15aは、通常はイントロン15(エクソン16の約460bp上流)中で認められる潜在性のエクソンである。変異型1の開始コドンはエクソン4中に存在し、終止コドンはエクソン17中に存在する。配列番号5は、エクソン1、4、5、6、7、8、9、10、11、12、および12aを含む遺伝子の変異型(変異型2)のcDNAを示す。エクソン12aは、イントロン12の部分配列から保持された潜在性のエクソンである(配列番号21を参照のこと)。変異型2の開始コドンはエクソン4中に存在し、終止コドンはエクソン12a中に存在する。配列番号7は、エクソン1、4、5、6、7、8、9、10、11、および11aを含む遺伝子の変異型(変異型3)のcDNAを示す。エクソン11aは、イントロン11の部分配列から保持された潜在性のエクソンである(配列番号19を参照のこと)。変異型3の開始コドンはエクソン4中に存在し、終止コドンはエクソン11a中に存在する。本発明の別の態様は、CMP−Neu5Acヒドロキシラーゼ遺伝子転写物の3つの新規の変異型の新規の推定アミノ酸ペプチド配列を提供する。配列番号4、6、および8は、それぞれ変異型1、変異型2、および変異型3のアミノ酸配列を示す。配列番号3〜8と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号3〜8の少なくとも10、15、17、20、25、30、50、100、150、200、300、400、500、または1000個の連続するヌクレオチド配列またはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。配列番号3〜8のフラグメント、誘導体、およびアナログをさらに提供する。配列番号3〜8のフラグメントには、少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、または10kpbを含む任意の連続核酸配列またはペプチド配列が含まれ得る。
【0071】
【表3−1】
【0072】
【表3−2】
【0073】
【表3−3】
【0074】
【表3−4】
【0075】
【表4】
【0076】
【表5−1】
【0077】
【表5−2】
【0078】
【表6】
【0079】
【表7−1】
【0080】
【表7−2】
【0081】
【表7−3】
【0082】
【表8】
本発明の他の態様では、CMP−Neu5AcヒドロキシラーゼをコードするcDNAまたはその変異型を含む核酸構築物を提供する。これらのcDNA配列は、配列番号1、3、5、または7であり得るか、配列番号2、4、6、もしくは8またはその任意のフラグメントに由来し得る。構築物は、1つまたは1つを超える内部リボゾーム侵入部位(IRES)を含み得る。構築物はまた、CMP−Neu5Acヒドロキシラーゼをコードする核酸配列に作動可能に連結されたプロモーターを含み得るか、構築物はプロモーターを含まなくてよい。別の実施形態では、宿主ゲノムへの挿入がランダムまたはターゲティングされている核酸配列を含む核酸構築物を提供する。核酸配列に加えて、発現ベクターは、選択マーカー(例えば、増強緑色蛍光タンパク質(eGFP)遺伝子配列、開始配列および/またはエンハンサー配列、ポリAテール配列、および/または原核生物および/または真核生物の宿主細胞で構築物を発現させる核酸配列など)を含み得る。適切なベクターおよび選択マーカーを以下に記載する。発現構築物は、転写開始、終結、および/またはリボゾーム結合部位のための部位をさらに含み得る。構築物は、任意の原核細胞または真核細胞(酵母細胞、E.coliなどの細菌細胞、CHO細胞などの哺乳動物細胞、および/または植物細胞が含まれるが、これらに限定されない)で発現することができる。
【0083】
このような構築物で使用するためのプロモーターには、λファージPLプロモーター、E.coli lacプロモーター、E.coli trpプロモーター、E.coli phoAプロモーター、E.coli tacプロモーター、SV40初期プロモーター、SV40後期プロモーター、レトロウイルスLTRプロモーター、PGKIプロモーター、GALIプロモーター、GALIO遺伝子プロモーター、CYCiプロモーター、PH05プロモーター、TRPIプロモーター、ADHIプロモーター、ADH2プロモーター、ホルグリムアルデヒド(forglymaldehyde)リン酸デヒドロゲナーゼプロモーター、ヘキソキナーゼプロモーター、ピルビン酸デカルボキシラーゼプロモーター、ホスホフルクロキナーゼプロモーター、トリオースリン酸イソメラーゼプロモーター、ホスホグルコースイソメラーゼプロモーター、グルコキナーゼα接合因子フェロモンプロモーター、PRBIプロモーター、GUT2プロモーター、GPDIプロモーター、メタロチオネインプロモーター、および/またはアデノウイルスおよびワクシニアウイルス由来などの哺乳動物ウイルスプロモーターが含まれるが、これらに限定されない。他のプロモーターが当業者に公知である。
【0084】
(II.CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列)
CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA機構を示す核酸配列(図1、表9)も提供する。配列番号10〜28は、それぞれエクソン1、4〜11、11a、12、12a、13〜15、15a、および16〜18を示す。エクソン11a、12a、および15aは、CMP−Neu5Acヒドロキシラーゼの一定の変異転写物で保持される潜在的エクソンである。配列番号29〜45は、エクソン1とエクソン4との間のイントロン配列(以後それぞれイントロン1aおよびイントロン1b)、4〜15、15a、16、および17を示す。イントロン15aは、潜在的エクソン15aに続くイントロン15の3’下流部分である。配列番号9は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の5’非翻訳領域を示す。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA配列を示す核酸配列(表10、配列番号46)も提供する。さらに、5’UTR、エクソン1、およびエクソン1とエクソン4との間に存在するイントロン配列の一部(イントロン1a)を含む5’連続ゲノム配列を示す連続ゲノム配列(配列番号47、表11)を提供する。エクソン1とエクソン4からエクソン18との間に存在するイントロン配列(イントロン1b)を含む連続ゲノム配列(配列番号48、表12)も提供する。配列番号9〜45、46、47、48、49、および50と少なくとも80%、85%、90%、95%、98%、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号9〜45、46、47、48、49、および50の少なくとも10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個の連続ヌクレオチド配列またはアミノ酸配列を含むヌクレオチド配列およびペプチド配列ならびにこれらと少なくとも80、85、90、95、98、または99%相同な任意のヌクレオチド配列も提供する。配列番号9〜45、46、47、48、49、および50のフラグメント、誘導体、およびアナログをさらに提供する。配列番号9〜45、46、47、48、49、および50のフラグメントには、任意の連続核酸配列もしくはペプチド配列または少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、もしくは10kpbが含まれ得る。
【0085】
さらに、ゲノム配列の推定転写因子結合部位の形態の調節領域を同定した(図4を参照のこと)。これらの結合部位は、ブタCMP−Neu5Acヒドロキシラーゼゲノムの5’UTRおよびエクソン1に存在し、転写因子(例えば、ETSF、MZF1、SF1、CMYB、MEF2、TATA、MEF2、NMP4、CAAT、AP1、BRN2、SATB1、ATF、GAT1、USF、WHN、NMP4、ZF5、NFKB、ZBP89、MOK2、ZF5、NFY、およびMYCMAXなど)の結合部位が含まれる。
【0086】
【表9−1】
【0087】
【表9−2】
【0088】
【表9−3】
【0089】
【表9−4】
【0090】
【表9−5】
【0091】
【表9−6】
【0092】
【表9−7】
【0093】
【表9−8】
【0094】
【表9−9】
【0095】
【表9−10】
【0096】
【表9−11】
【0097】
【表9−12】
【0098】
【表9−13】
【0099】
【表9−14】
【0100】
【表9−15】
【0101】
【表9−16】
【0102】
【表9−17】
【0103】
【表9−18】
【0104】
【表9−19】
【0105】
【表9−20】
【0106】
【表9−21】
【0107】
【表9−22】
【0108】
【表9−23】
【0109】
【表9−24】
【0110】
【表9−25】
【0111】
【表9−26】
【0112】
【表9−27】
【0113】
【表9−28】
【0114】
【表10−1】
【0115】
【表10−2】
【0116】
【表10−3】
【0117】
【表10−4】
【0118】
【表10−5】
【0119】
【表10−6】
【0120】
【表10−7】
【0121】
【表10−8】
【0122】
【表10−9】
【0123】
【表10−10】
【0124】
【表10−11】
【0125】
【表10−12】
【0126】
【表10−13】
【0127】
【表10−14】
【0128】
【表10−15】
【0129】
【表10−16】
【0130】
【表10−17】
【0131】
【表10−18】
【0132】
【表10−19】
【0133】
【表10−20】
【0134】
【表10−21】
【0135】
【表10−22】
【0136】
【表11】
【0137】
【表12−1】
【0138】
【表12−2】
【0139】
【表12−3】
【0140】
【表12−4】
【0141】
【表12−5】
【0142】
【表12−6】
【0143】
【表12−7】
【0144】
【表12−8】
【0145】
【表12−9】
【0146】
【表12−10】
【0147】
【表12−11】
【0148】
【表12−12】
【0149】
【表12−13】
【0150】
【表12−14】
【0151】
【表12−15】
【0152】
【表12−16】
【0153】
【表12−17】
【0154】
【表12−18】
【0155】
【表12−19】
【0156】
【表12−20】
【0157】
【表12−21】
【0158】
【表13−1】
【0159】
【表13−2】
【0160】
【表13−3】
【0161】
【表13−4】
【0162】
【表13−5】
【0163】
【表13−6】
【0164】
【表13−7】
【0165】
【表13−8】
【0166】
【表13−9】
【0167】
【表13−10】
配列番号49は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、およびエクソン8を含む連続ゲノム配列を示す(表13)。さらに、配列番号49の少なくとも1750、1800、1900、2000、2100、2200、2300、2400、2500、2600、2700、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4100、4200、4300、4400、4500、4600、4700、4800、4900、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号49と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0168】
【表14−1】
【0169】
【表14−2】
【0170】
【表14−3】
【0171】
【表14−4】
【0172】
【表14−5】
【0173】
【表14−6】
【0174】
【表14−7】
【0175】
【表14−8】
【0176】
【表14−9】
【0177】
【表14−10】
【0178】
【表14−11】
【0179】
【表14−12】
配列番号50は、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18を含む連続ゲノム配列を示す。さらに、配列番号50の少なくとも150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1500、2000、2500、3000、3500、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20、000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0180】
(VIII.オリゴヌクレオチドプローブおよびプライマー)
本発明は、さらに、上記配列(配列番号1、3、5、7、9〜45、46、47、48、49、および50)とハイブリッド形成するオリゴヌクレオチドプローブおよびプライマーを提供する。配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントと相同であり得るオリゴヌクレオチドを提供する。ストリンジェントな条件下で配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントとハイブリッド形成するオリゴヌクレオチドも提供する。ストリンジェントな条件は、配列が少なくとも約85%、約90%、約95%、または少なくとも約98%相同である場合のみハイブリッド形成する条件を記載する。あるいは、オリゴヌクレオチドは、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントとハイブリッド形成する少なくとも20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、45、46、47、48、49、50、75、または100塩基を有し得る。このようなオリゴヌクレオチドを、本明細書中に提供した配列を検出するためのプライマーおよびプローブとして使用することができる。プローブまたはプライマーは、少なくとも14ヌクレオチド長であり得るが、好ましい実施形態では、少なくとも15、20、25、28、30、または35ヌクレオチド長である。
【0181】
上記配列を考えると、当業者は、標準的なアルゴリズムを使用して、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメント中に含まれる配列と相同なオリゴヌクレオチドプローブおよびプライマーを構築することができる。相補的対合の規則は周知である:シトシン(「C」)は常にグアニン(「G」)と対合し、チミン(「T」)またはウラシル(「U」)は常にアデニン(「A」)と対合する。プライマーまたはプローブと十分にハイブリッド形成して対応する相補配列を認識することができる限り、プライマーまたはプローブは標的核酸配列と100%相補的である必要はないと認識される。一般に、一定の程度の対ミスマッチを許容することができる。
【0182】
プライマーのハイブリッド形成領域として使用されるオリゴヌクレオチド配列を、プローブのハイブリッド形成領域として使用することもできる。プローブとして使用するためのプライマー配列の適合性は、プライマーのハイブリッド形成特性に依存する。同様に、プローブとして使用されるオリゴヌクレオチドをプライマーとして使用することができる。
【0183】
これらの特定の実施形態では、例えば、5’または3’末端のいずれかへの標的配列に相補的であるか標的配列に相補的ではないヌクレオチドの付加によって特定のプライマーおよびプローブを調製することができることが当業者に明らかである。プライマー組成物を標的配列上の伸長開始点として作用する限り、およびプライマーおよびプローブが上記配列番号内に含まれる少なくとも14個の連続ヌクレオチドを含む限り、このような組成物は本発明の範囲内である。
【0184】
本明細書中のプローブおよびプライマーを、考慮すべき要因であるが、排他的でも決定的でもない以下の基準によって選択することができる。プローブおよびプライマーを、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントで同定されたCMP−Neu5Acヒドロキシラーゼ核酸配列領域から選択する。プローブおよびプライマーは、試験に悪影響を与えると予想される他の遺伝子配列との相同性を欠く。プローブまたはプライマーは、E.coli DNAポリメラーゼ(好ましくは、そのDNAポリメラーゼの一部はKlenowフラグメントと呼ばれる)などの増幅酵素による伸長を妨害することができる増幅核酸中での二次構造形成を欠く。増幅培地中での約15重量%まで、好ましくは5〜10重量%のジメチルスルホキシド(DMSO)および/または30〜40℃の漸増増幅温度の使用によってこれを行うことができる。
【0185】
好ましくは、プローブまたはプライマーは、式アデニン(A)+チミン(T)+シトシン(C)+グアニン(G)/シトシン(C)+グアニン(G)によって測定したところ、約50%のグアニンおよびシトシンヌクレオチドを含むべきである。好ましくは、プローブまたはプライマーは、ハイブリッドの安定性が減少し得るプライマーの3’末端に複数の連続したアデニン残基およびチミン残基を含まない。
【0186】
本発明のプローブおよびプライマーは、約10〜30ヌクレオチド長、好ましくは少なくとも10、11、12、13、14、15、20、25、または28ヌクレオチド長(詳細には、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、または30ヌクレオチドが含まれる)であり得る。本発明で使用されるヌクレオチドは、リボヌクレオチド、デオキシリボヌクレオチド、および修飾ヌクレオチド(そのハイブリッド形成特性が本質的に変化しない修飾基を含むイノシンまたはヌクレオチドなど)であり得る。プローブおよびプライマー配列を、本明細書を通して5’末端から3’末端の方向で一本鎖DNAオリゴヌクレオチドとして示す。このようなまたはその相補的形態またはそのRNA形態(TがUに置換されている)で任意のプローブを使用することができる。
【0187】
本発明のプローブおよびプライマーを、対応するヌクレオチド配列を含むインサートを含む組換えプラスミドのクローニング、任意選択的に適切なヌクレアーゼの使用時のクローン化プラスミドからの後者の切断およびその回収(例えば、分子量による分画)によって調製することができる。本発明のプローブおよびプライマーを、例えば、従来のホスホトリエステル法もしくはホスホジエステル法またはその自動化実施形態によって化学合成することも出きる。1つのこのような自動化実施形態では、出発材料としてジエチルホスホラミダイトを使用し、Beaucageら,Tetrahedron Letters 22:1859−1862(1981)に記載のように合成することができる。修飾固相上での1つのオリゴヌクレオチド合成方法は、米国特許第4,458,066号に記載されている。生物学的供給源(制限エンドヌクレアーゼ消化物など)から単離したプローブまたはプライマーを使用することも可能である。
【0188】
プライマーまたはプローブとして使用されるオリゴヌクレオチドは、ホスホロチオエート(Matsukura S.,Naibunpi Gakkai Zasshi.43(6):527−32(1967))、アルキルホスホロチオエート(Miller P.ら,Biochemistry 18(23):5134−43(1979)、ペプチド核酸(Nielsen P.ら,Science 254(5037):1497−500(1991);Nielsen P.ら,Nucleic−Acids−Res.21(2):197−200(1993))、モルホリノ核酸、固定核酸、偽環状オリゴ核酸塩基、2’−0,4’−C−エチレン架橋核酸などのヌクレオチドアナログも含み得るか、挿入剤(Asseline J.ら,Proc.Natl.Acad.Sci.USA 81(11):3297−301(1984))を含み得る。
【0189】
所望の特徴を有するプローブおよびプライマーのデザインのために、当業者に公知の以下の有用なガイドラインを適用することができる。ハイブリッド形成反応の範囲および特異性は多数の要因に影響を受け、1つまたは複数のこれらの要因の操作により、特定のプローブの正確な感度および特異性(その標的に完全に相補的であるかどうか)が決定される。本明細書中でさらに説明される種々のアッセイ条件の重要性および影響は、当業者に公知である。
【0190】
標的核酸ハイブリッドに対するプローブおよびプライマーの安定性は、アッセイ条件に適合するように選択すべきである。長いATリッチな配列の回避、GC塩基対でのハイブリッドの終結、および/または適切なTmでのプローブのデザインによってこれを行うことができる。プローブの開始点および終点を、その長さおよびGC率によってTmが最終アッセイが行われる温度よりも約2〜10℃高くなるように選択すべきである。G−C塩基対がさらなる水素結合によってA−T塩基対と比較して熱安定性が高いので、プローブの塩基組成は有意である。したがって、より高いG−C含量の相補核酸を含むハイブリッド形成は、より高い温度で安定である。プローブデザイン時にプローブが使用されるイオン強度およびインキュベーション温度などの条件も考慮すべきである。反応混合物のイオン強度が増加するにつれてハイブリッド形成が増加し、イオン強度の増加と共にハイブリッドの熱安定性が増加することが公知である。水素結合を破壊するホルムアミド、尿素、DIVISO、およびアルコールなどの化学試薬は、ハイブリッド形成のストリンジェンシーを増加させる。このような試薬による水素結合の不安定化によりTmが非常に減少し得る。一般に、所与の二重鎖の融解温度より5℃低い温度で約10〜50塩基長の合成オリゴヌクレオチドプローブの最適なハイブリッド形成が起こる。至適温度未満でのインキュベーションによりミスマッチ塩基配列がハイブリッド形成するので、特異性が減少し得る。高ストリンジェンシー条件下のみでハイブリッド形成するプローブを有することが望ましい。高ストリンジェンシー条件下のみで高相補核酸ハイブリッドが形成され、相補性が十分でないハイブリッドは形成されない。したがって、アッセイ条件のストリンジェンシーにより、ハイブリッドを形成する2つの核酸鎖間に必要な相補性が決定される。標的核酸と形成されたハイブリッドと非標的核酸と形成されたハイブリッドとの間の安定性の相違を最大にするなどのためにストリンジェンシーの程度を選択する。この場合、1塩基対の変化を検出する必要があり、非常に高いストリンジェンシー条件が必要である。
【0191】
標的核酸配列の長さ(したがって、プローブ配列の長さ)も重要であり得る。いくつかの場合、所望のハイブリッド形成特性を有するプローブおよびプライマーが得られる特定の領域由来の種々の位置および長さのいくつかの配列が存在し得る。他の場合、ある配列が1塩基しか異ならない別の配列よりも有意に良好であり得る。
【0192】
完全に相補的ではない核酸をハイブリッド形成することが可能であるが、主に完全に相補的な塩基の最も長いストレッチによりハイブリッドの安定性が決定される。異なる長さおよび塩基組成のオリゴヌクレオチドプローブおよびプライマーを使用することができるが、本発明の好ましいオリゴヌクレオチドプローブおよびプライマーは約14塩基長と30塩基長との間であり、標的核酸配列と完全に相補的な配列で十分なストレッチを有する。
【0193】
ハイブリッド形成に対して抑制的である強固な内部構造を形成することが公知の標的DNAまたはRNA中の領域はあまり好ましくない。同様に、広範な自己相補性を有するプローブは回避すべきである。上記で説明するように、ハイブリッド形成は、水素結合した二本鎖を形成する相補核酸の2つの一本鎖の会合である。2つの鎖のうちの1つの全体または一部がハイブリッドに関与する場合、新規のハイブリッドの形成への関与が低下し得ると予想される。十分な自己相補性が存在する場合、1つのプローブ型の分子内で形成された分子内および分子間ハイブリッドが存在し得る。このような構造を、慎重なプローブデザインによって回避することができる。目的の配列の大部分が一本鎖であるようなプローブのデザインにより、ハイブリッド形成の比率および範囲が非常に増加し得る。この相互作用型を検索するためにコンピュータプログラムを利用可能である。しかし、一定の例では、この相互作用型を回避することができない。
【0194】
特異的プライマーおよびオリゴヌクレオチドプローブに特異的な配列を、CMP−Neu5Acヒドロキシラーゼ核酸配列を増幅および検出することができるポリメラーゼ連鎖反応で使用することができる。
【0195】
(IV.CMP−Neu5Acヒドロキシラーゼ遺伝子の遺伝子ターゲティング)
遺伝子ターゲティングにより、動物細胞ゲノムの選択的操作が可能である。この技術を使用して、部位特異的様式および正確な様式で特定のDNA配列をターゲティングおよび修飾することができる。修飾のために異なるDNA配列型(調節領域、コード領域、および遺伝子間のDNA領域が含まれる)をターゲティングすることができる。調節領域の例には、プロモーター領域、エンハンサー領域、ターミネーター領域、およびイントロンが含まれる。これらの調節領域の修飾により、遺伝子発現のタイミングおよびレベルを変化させることができる。細胞内のタンパク質を変化、増強、または消失させるためにコード領域を修飾することができる。イントロンおよびエクソンならびに遺伝子間領域は、修飾に適切な標的である。
【0196】
DNA配列の修飾には、いくつかの型(挿入、欠失、置換、またはその任意の組み合わせが含まれる)が存在し得る。修飾の特定の例は、遺伝子産物の発現を破壊するヌクレオチド配列の部位特異的に相互作用による遺伝子の不活化(すなわち、「ノックアウト」)である。例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子の1つの破壊アプローチは、ターゲティングDNAと標的DNAとの間の相同組換えによって選択マーカーを標的遺伝子のコード領域に挿入することができるように選択マーカーをターゲティングDNAに挿入することである。例えば、図3、12、および13を参照のこと。この方法では、例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子配列が破壊され、コードされた酵素が機能しなくなる。
【0197】
(相同組換え)
相同組換えによって内因性遺伝子が部位特異的に修飾されるので、ゲノムを新規に変化するように操作することができる。最初の相同組換え工程は、ヘテロ二重鎖DNAを含む中間体組換え構造を形成するための相補配列を含む少なくとも1つのDNA鎖とのDNA二重鎖の対合を含むDNA鎖交換である(例えば、Radding,C.M.(1982)Ann.Rev.Genet.16:405;米国特許第4,888,274号を参照のこと)。ヘテロ二重鎖DNAは、いくつかの形態(1つの相補鎖がDNA二重鎖に侵入した三重鎖形態を含む3つのDNA鎖が含まれる)を取ることができ(Hsiehら,Genes and Development 4:1951(1990);Raoら(1991)PNAS 88:2984))、2つの相補DNA鎖がDNA二重鎖と対合する場合、古典的なHolliday組換え連結構造およびΧ構造(Holliday,R.,Genet.Res.5:282(1964))または二重Dループ(“Diagnostic Applications of Double−D Loop Formation”U.S.Ser.No.07/755,462,filed Sep.4,1991)を形成することができる。一旦形成されると、ヘテロ二重鎖構造を、侵入したDNA鎖の全部または一部がレシピエントDNA二重鎖にスプライシングされる鎖の破壊および交換、レシピエントDNA二重鎖の付加または置換によって解明することができる。あるいは、ヘテロ二重鎖構造によって遺伝子を変換することができ、テンプレートとして侵入鎖を使用したミスマッチ塩基の修復によって侵入鎖の配列がレシピエントDNA二重鎖に移行される(Genes,第3版(1987)Lewin,B.,John Wiley,New York,N.Y.;Lopezら,Nucleic Acids Res.15:5643(1987))。破壊および再連結機構または遺伝子変換機構のいずれかにより、相同対合連結部でのヘテロ二重鎖DNAの形成は、あるDNAから別のDNAへの遺伝子配列情報を伝達するように作用することができる。
【0198】
DNA分子間の遺伝子配列情報を伝達する相同組換え(遺伝子変換および古典的な鎖の破壊/再連結)の能力により、ターゲティングされた相同組換えが遺伝子工学および遺伝子操作における強力な方法となる。
【0199】
相同組換えでは、組み込まれるDNAが実質的に相同なDNA配列を含むゲノム中の部位と相互作用して組み込まれる。非相同性の(「無作為」または「不正の」)組み込みでは、組み込まれるDNAはゲノム中の相同配列で見出されないが、他の場所(多数の潜在的な位置)に組み込まれる。一般に、高等真核細胞を使用した研究によって、相同組換えの頻度は無作為な組み込みの頻度よりもはるかに低いことが明らかとなった。これらの頻度の比は、相同組換えを介した相互作用に依存する「遺伝子ターゲティング」(すなわち、ゲノム中での外因性「ターゲティングDNA」と対応する「標的DNA」との間の組換え)と直接関連する。
【0200】
多数の論文に哺乳動物細胞における相同組換えの使用が記載されている。これらの論文の例は、Kucherlapatiら,Proc.Natl.Acad.Sci.(USA)81:3153−3157,1984;Kucherlapatiら,MoL Cell.Bio.5:714−720,1985;Smithiesら,Nature 317:230−234,1985;Wakeら,Mol.Cell.Bio.8:2080−2089,1985;Ayaresら,Genetics 111:375−388,1985;Ayaresら,Mol.Cell.Bio.7:1656−1662,1986;Songら,Proc.Natl.Acad.Sci.USA 84:6820−6824,1987;ThomasらCell 44:419−428,1986;ThomasおよびCapecchi,Cell 51:503−512,1987;Nandiら,Proc.Natl.Acad Sci.USA 85:3845−3849,1988;およびMansourら,Nature 336:348−352,1988;EvansおよびKaufman,Nature 294:146−154,1981;Doetschmanら,Nature 330:576−578,1987;ThomaおよびCapecchi,Cell 51:503−512,4987;Thompsonら,Cell 56:316−321,1989である。
【0201】
本発明は、線維芽細胞などの細胞中でブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するために相同組換えを使用する。DNAは、機能的酵素の発現およびHanganutziu−Deicher抗原分子の産生を防止するために、天然の遺伝子の少なくとも一方、任意選択的には両方のコピーが変化した特定の対立遺伝子に遺伝子の少なくとも一部を含み得る。変化は、挿入、欠失、置換、またはその組み合せであり得る。変化がたった1つの遺伝子のコピーが不活化する導入である場合、標的遺伝子の1つの非変異コピーを有する細胞が増幅され、変化が第1の変化と同一でも異なっていてもよく(通常異なっている)、且つ欠失または置換が含まれる第2のターゲティング工程に供することができ、最初に移入された変化の少なくとも一部が重複し得る。この第2のターゲティング工程では、同一の相同性アームを有するが、異なる哺乳動物選択マーカーを含むターゲティングベクターを使用することができる。得られた形質転換体を、機能的標的抗原の非存在についてスクリーニングし、野生型標的遺伝子の非存在を確認するために細胞のDNAをさらにスクリーニングすることができる。あるいは、変異にヘテロ接合性を示す宿主の交配によって表現型に関するホモ接合性を達成することができる。
【0202】
遺伝子操作することができるブタ細胞を、種々の異なる臓器および組織(脳、心臓、肺、腺、脳、目、胃、脾臓、すい臓、腎臓、肝臓、腸、子宮、膀胱、皮膚、毛髪、爪、耳、鼻、口、唇、歯茎、歯、舌、唾液腺、扁桃腺、咽頭、食道、大腸、小腸、直腸、肛門、幽門、甲状腺、胸腺、腎上体、骨、軟骨、腱、靭帯、骨格筋、平滑筋、血管、血液、脊髄、気管、尿管、尿道、視床下部、下垂体、副腎、卵巣、輸卵管、子宮、膣、乳腺、精巣、精嚢、陰茎、リンパ、リンパ節、およびリンパ管などであるが、これらに限定されない)から得ることができる。本発明の1つの実施形態では、ブタ細胞は、上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋肉細胞、リン酸細胞(□hosphate cell)、卵丘細胞、上皮細胞、内皮細胞、ランゲルハンス島細胞、血球、血液前駆細胞、骨細胞、骨前駆細胞、神経幹細胞、始原幹細胞、肝細胞、ケラチノサイト、臍静脈内皮細胞、大動脈内皮細胞、微小血管内皮細胞、線維芽細胞、肝臓星細胞、大動脈平滑筋細胞、心筋細胞、ニューロン、クップフェル細胞、平滑筋細胞、シュワン細胞、上皮細胞、赤血球、血小板、好中球、リンパ球、単球、好酸球、好塩基球、脂肪細胞、軟骨細胞、膵島細胞、甲状腺細胞、副甲状腺細胞、耳下腺細胞、腫瘍細胞、グリア細胞、星状細胞、赤血球、白血球、マクロファージ、上皮細胞、体細胞、下垂体細胞、副腎細胞、有毛細胞、気泡細胞、腎細胞、網膜細胞、桿状体細胞、錐状体細胞、心臓細胞、ペースメーカー細胞、脾臓細胞、抗原提示細胞、記憶細胞、T細胞、B細胞、形質細胞、筋細胞、卵巣細胞、子宮細胞、前立腺細胞、腟上皮細胞、精子細胞、精巣細胞、生殖細胞、卵細胞、ライディッヒ細胞、尿細管周囲細胞、セルトリ細胞、ルテイン細胞、頸部細胞、子宮内膜細胞、乳房細胞、濾胞細胞、粘液細胞、繊毛細胞、非角化上皮細胞、角化上皮細胞、肺細胞、杯細胞、円柱上皮細胞、扁平上皮細胞、骨細胞、骨芽細胞、および破骨細胞からなる群から選択され得るが、これらに限定されない。
【0203】
1つの別の実施形態では、胚幹細胞を使用することができる。胚幹細胞株を使用することができるか、胚幹細胞株をブタ動物などの宿主から新たに得ることができる。細胞を、適切な線維芽細胞支持細胞層で成長させるか、白血病阻害因子(LIF)の存在下で成長させることができる。好ましい実施形態では、ブタ細胞は線維芽細胞であり得るが、1つの特定の実施形態では、ブタ細胞は胎児線維芽細胞であり得る。線維芽細胞は、発育中の胎児および成体動物から大量に得ることができので、好ましい体細胞型である。
【0204】
これらの細胞を急速な倍加時間で容易に増殖することができ、遺伝子ターゲティング手順で使用するためにクローンで増殖することができる。
【0205】
(ターゲティングベクター)
ターゲティングした対立遺伝子でホモ接合性の細胞を、標的対立遺伝子に相同であり、且つマーカー遺伝子を含むDNAの細胞への移入および組み込まれた構築物を含む細胞の選択によって産生することができる。標的対立遺伝子で標的ベクター中の相同DNAを染色体DNAで組換える(例えば、図3、12、および13を参照のこと)。マーカー遺伝子の両端を、相同DNA配列、3’組換えアーム、および5’組換えアームと隣接させることができる(例えば、図11を参照のこと)。ターゲティンベクターの構築方法は当該分野で記載されており、例えば、Daiら,Nature Biotechnology 20:251−255,2002;WO 00/51424を参照のこと。
【0206】
標的対立遺伝子での相同組換えのために種々の構築物を調製することができる。通常、構築物には、50bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbp、または50kbpの標的対立遺伝子と相同な配列が含まれ得る。配列には、任意のブタCMP−Neu5Acヒドロキシラーゼ遺伝子の任意の連続配列が含まれ得る(配列番号9〜45、46、47、48、49、もしくは50またはその任意の組み合わせもしくはフラグメントの少なくとも約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個の連続ヌクレオチドが含まれる)。配列番号9〜45、46、47、48、49、および50のフラグメントには、少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、または10kpbを含む任意の連続核酸配列またはペプチド配列が含まれ得る。
【0207】
標的DNA配列の相同性の範囲の決定に種々の検討材料(例えば、標的対立遺伝子のサイズ、配列の利用可能性、標的対立遺伝子における二重重複事象の相対的効率、および標的配列の他の配列との類似性など)が関与し得る。
【0208】
ターゲティングDNAには、実質的に同質遺伝子のDNAが修飾すべきゲノム中の対応する標的配列での所望の配列修飾物と隣接した配列が含まれ得る。実質的に同質遺伝子の配列は、対応する標的配列(所望の配列修飾を除く)を少なくとも約95%、97〜98%、99.0〜99.5%、99.6〜99.9%、または100%同一であり得る。ターゲティングDNAおよび標的DNAは、好ましくは、100%同一である少なくとも75、150、または500塩基対のDNAストレッチを共有することができる。したがって、ターゲティングDNAは、ターゲティングされる細胞株と密接に関連する細胞に由来し得るか、ターゲティングDNAはターゲティングされる細胞と同一の細胞株または動物の細胞に由来し得る。
【0209】
内因性標的CMP−Neu5Acヒドロキシラーゼを修飾するようにDNA構築物をデザインすることができる。構築物のターゲティングのための相同配列は、得られる遺伝子産物の機能を破壊するようにデザインされた1つまたは複数の欠失、挿入、置換、またはその組み合わせを有し得る。1つの実施形態では、変化は、読み取り枠中で標的遺伝子の上流配列と融合した選択マーカー遺伝子の挿入であり得る。
【0210】
適切な選択マーカーには、tk遺伝子(チミジンキナーゼ)などの一定の培地基質で成長する能力を付与する遺伝子またはHAT培地(ヒポキサンチン、アミノプテリン、およびチミジン)で成長する能力を付与するhprt遺伝子(ヒポキサンチンホスホリボシルトランスフェラーゼ);MAX培地(ミコフェノール酸)で成長させる細菌gpt遺伝子(グアニン/キサンチンホスホリボシルトランスフェラーゼ)が含まれるが、これらに限定されない。例えば、Song,K−Y.らProc.Net’l Acad.Sci.U.S.A.84:6820−6824(1987);Sambrook,J.ら,Molecular Cloning−A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.(1989),Chapter 16を参照のこと。選択マーカーの他の例には、抗生物質などの化合物に体する耐性を付与する遺伝子、選択された基質で成長する能力を付与する遺伝子、緑色蛍光タンパク質、増強緑色蛍光タンパク質(eGFP)などの発光物質などの検出可能なシグナルを発するタンパク質をコードする遺伝子が含まれる。広範な種々のこのようなマーカーが公知且つ利用可能であり、例えば、ネオマイシン耐性遺伝子(neo)(Southern,P.,およびP.Berg,J.Mol.Appl.Genet.1:327−341(1982));およびハイグロマイシン耐性遺伝子(hyg)(Nucleic Acids Research 11:6895−6911(1983),およびTe Riele,H.ら,Nature 348:649−651(1990))などの抗生物質耐性遺伝子が含まれる。他の選択マーカー遺伝子には、アセトヒドロキシ酸シンターゼ(AHAS)、アルカリホスファターゼ(AP)、βガラクトシダーゼ(LacZ)、βグルクロニダーゼ(GUS)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、緑色蛍光タンパク質(GFP)、赤色蛍光タンパク質(RFP)、黄色蛍光タンパク質(YFP)、シアン蛍光タンパク質(CFP)、西洋ワサビペルオキシダーゼ(HRP)、ルシフェラーゼ(Luc)、ノパリンシンターゼ(NOS)、オクトピンシンターゼ(OCS)、およびその誘導体が含まれる。アンピシリン、ブレオマイシン、クロラムフェニコール、ゲンタマイシン、ハイグロマイシン、カナマイシン、リンコマイシン、メトトレキセート、フォスフィノスリシン、ピューロマイシン、およびテトラサイクリンに耐性を付与する複数の選択マーカーを利用可能である。
【0211】
抗生物質耐性遺伝子およびネガティブ選択因子の組み込み方法は、当業者に熟知されている(例えば、WO 99/15650;米国特許第6,080,576号;米国特許第6.136,566号;Niwaら,J.Biochem.113:343−349(1993);およびYoshidaら,Transgenic Research,4:277−287(1995)を参照のこと)。
【0212】
本発明で有用なさらなる選択マーカー遺伝子は、例えば、以下の米国特許に記載されている:6,319,669;6,316,181;6,303,373;6,291,177;6,284,519;6,284,496;6,280,934;6,274,354;6,270,958;6,268,201;6,265,548;6,261,760;6,255,558;6,255,071;6,251,677;6,251,602;6,251,582;6,251,384;6,248,558;6,248,550;6,248,543;6,232,107;6,228,639;6,225,082;6,221,612;6,218,185;6,214,567;6,214,563;6,210,922;6,210,910;6,203,986;6,197,928;6,180,343;6,172,188;6,153,409;6,150,176;6,146,826;6,140,132;6,136,539;6,136,538;6,133,429;6,130,313;6,124,128;6,110,711;6,096,865;6,096,717;6,093,808;6,090,919;6,083,690;6,077,707;6,066,476;6,060,247;6,054,321;6,037,133;6,027,881;6,025,192;6,020,192;6,013,447;6,001,557;5,994,077;5,994,071;5,993,778;5,989,808;5,985,577;5,968,773;5,968,738;5,958,713;5,952,236;5,948,889;5,948,681;5,942,387;5,932,435;5,922,576;5,919,445;および5,914,233。
【0213】
選択マーカーの組み合わせも使用することができる。例えば、CMP−Neu5Acヒドロキシラーゼをターゲティングするために、neo遺伝子(上記で考察のようにそれ自体のプロモーターを含むか含まない)を、CMP−Neu5Acヒドロキシラーゼ遺伝子に相同なDNA配列にクローン化することができる。マーカーの組み合わせを使用するために、ターゲティングDNAの外側に存在するようにHSV−tk遺伝子をクローン化することができる(所望ならば、別の選択マーカーを反対側に位置づけることができる)。ターゲティングされるべき細胞へのDNA構築物の移入後、適切な抗生物質に対して細胞を選択することができる。この特定の例では、G418およびガンシクロビルに耐性を示す細胞は、neo遺伝子がCMP−Neu5Acヒドロキシラーゼ遺伝子に組換えられているが、二重重複領域の外側に存在することによりtk遺伝子が失われている相同組換えによって惹起されている可能性が最も高い。
【0214】
欠失は、少なくとも約50bp、より通常には少なくとも約100bp、一般に、せいぜい約20kbpであり得るが、欠失は、通常、少なくとも1つまたは複数のエクソンの一部を含むコード領域の一部、1つまたは複数のイントロンの一部を含むことができ、隣接非コード領域(特に、5’非コード領域(転写調節領域))の一部を含んでいても含んでいなくても良い。したがって、相同領域は、コード領域を超えて5’非コード領域または3’非コード領域に及び得る。挿入は、一般に10kbpを超えず、通常5kbpを超えず、一般に少なくとも50bp、より通常には少なくとも200bpであり得る。
【0215】
相同領域には、フレームシフトが起こる場合または鍵となるアミノ酸が変化した場合に標的遺伝子をさらに不活化することができるか、機能障害対立遺伝子などを修正することができる変異が含まれ得る。通常、変異は、約5%を超えない相同隣接配列のわずかな変化であり得る。
【0216】
当該分野で公知の方法にしたがって構築物を調製することができ、種々のフラグメントを1つにし、適切なベクターに移入し、クローン化し、分析し、その後所望の構築物を達成するまでさらに操作することができる(例えば、図5〜11を参照のこと)。制限分析、切り出し、プローブの同定を行うために配列に種々の修飾を行うことができる。所望ならばサイレント変異を移入することができる。種々の段階で、制限分析、配列決定、ポリメラーゼ連鎖反応を使用した増幅、プライマー修復、in vitro変異誘発などを使用することができる。
【0217】
原核生物複製系(例えば、E.coliによって認識可能な供給源)を含む細菌ベクターを使用して構築物を調製することができ、各段階で、構築物をクローン化および分析することができる。標的細胞への移入前に除去することができるマーカー(挿入のために使用されるマーカーと同一または異なる)を使用することができる。一旦構築物を含むベクターが完成すると、細菌配列の欠失、線状化、相同配列への短い欠失の移入などによってさらに操作することができる。最終的操作後、構築物を細胞に移入することができる。
【0218】
DNA構築物を宿主細胞に侵入させるために使用することができる技術には、リン酸カルシウム/DNA共沈、核へのDNAの微量注入、エレクトロポレーション、インタクトな細胞との細菌プロトプラストの融合、トランスフェクション、または当業者に公知の任意の他の技術が含まれる。DNAは、一本鎖もしくは二本鎖、線状もしくは環状、弛緩もしくはスーパーコイル状のDNAであり得る。哺乳動物細胞の種々のトランスフェクション技術については、例えば、Keownら,Methods in Enzymology Vol.185,pp.527−537(1990)を参照のこと。
【0219】
本発明は、さらに、上記で広範に記載されている1つまたは複数の配列を含む組換え構築物を含む(例えば、表9〜12)。構築物は、本発明の配列が正方向または逆方向で挿入されたプラスミドベクターまたはウイルスベクターなどのベクターを含む。構築物は、配列に作動可能に連結された調節配列(例えば、プロモーターが含まれる)も含み得る。多数の適切なベクターおよびプロモーターは当業者に公知であり、市販されている。例として以下のベクターを提供する:pBs、pQE−9(Qiagen)、phagescript、PsiX174、pBluescript SK、pBsKS、pNH8a、pNH16a、pNHl8a、pNH46a(Stratagene);pTrc99A、pKK223−3、pKK233−3、pDR540、pRIT5(Pharmacia);Eukaryotic:pWLneo、pSv2cat、pOG44、pXTI、pSG(Stratagene)pSVK3、pBPv、pMSG、pSVL(Pharmiacia)。また、宿主中で複製可能且つ生存可能な限り、任意の他のプラスミドおよびベクターを使用することができる。当該分野で公知且つ市販されているベクター(およびその変異型または誘導体)を、本発明の方法で使用するために本発明にしたがって1つまたは複数の組換え部位を含むように操作することができる。このようなベクターを、例えば、Vector Laboratories Inc.、Invitrogen、Promega、Novagen、NEB、Clontech、Boehringer Mannheim、Pharmacia、EpiCenter、OriGenes Technologies Inc.、Stratagene、PerkinElmer、Pharmingen、およびResearch Geneticsから入手することができる。目的の他のベクターには、pFastBac、pFastBacHT、pFastBacDUAL、pSFV、およびpTet−Splice(Invitrogen)、pEUK−C1、pPUR、pMAM、pMAMneo、pBI101、pBI121、pDR2、pCMVEBNA、およびpYACneo(Clontech)、pSVK3、pSVL、pMSG、pCHI 10、およびpKK232−8(Pharmacia、Inc.)、p3’SS、pXTI、pSG5、pPbac、pMbac、pMClneo、およびpOG44(Stratagene、Inc.)、およびpYES2、pAC360、pBlueBacHis A、B、およびC、pVL1392、pBlueBacIII、pCDM8、pcDNAI、pZeoSV、pcDNA3、pREP4、pCEP4、およびpEBVHis(Invitrogen、Corp.)、ならびにその変異型または誘導体などの真核生物発現ベクターが含まれる。
【0220】
本発明での使用に適切な他のベクターには、pUC18、pUC19、pBlueScript、pSPORT、コスミド、ファージミド、YAC’s(酵母人工染色体)、BAC’s(細菌人工染色体)、PI(Escherichia coliファージ)、pQE70、pQE60、pQE9(quagan)、pBSベクター、PhageScriptベクター、BlueScriptベクター、pNH8A、pNH16A、pNH18A、pNH46A(Stratagene)、pcDNA3(Invitrogen)、pGEX、pTrsfus、pTrc99A、pET−5、pET−9、pKK223−3、pKK233−3、pDR540、pRIT5(Pharmacia)、pSPORT1、pSPORT2、pCMVSPORT2.0、およびpSV−SPORT1(Invitrogen)、ならびにその変異型または誘導体が含まれる。レンチウイルスベクター(例えば、WO 03/059923;TiscorniaらPNAS100:1844−1848(2003)を参照のこと)などのウイルスベクターを使用することもできる。
【0221】
さらなる目的のベクターには、pTrxFus、pThioHis、pLEX、pTrcHis、pTrcHis2、pRSET、pBlueBacHis2、pcDNA3.1/His、pcDNA3.1(−)/Myc−His、pSecTag、pEBVHis、pPIC9K、pPIC3.5K、pA0815、pPICZ、pPICZA、pPICZB、pPICZC、pGAPZA、pGAPZB、pGAPZC、pBlueBac4.5、pBlueBacHis2、pMelBac、pSinRep5、pSinHis、pIND、pIND(SP1)、pVgRXR、pcDNA2.1、pYES2、pZErO1.1、pZErO−2.1、pCR−Blunt、pSE280、pSE380、pSE420、pVL1392、pVL1393、pCDM8、pcDNA1.1、pcDNA1.1/Amp、pcDNA3.1、pcDNA3.1/Zeo、pSe、SV2、pRc/CMV2、pRc/RSV、pREP4、pREP7、pREP8、pREP9、pREP 10、pCEP4、pEBVHis、pCR3.1、pCR2.1、pCR3.1−Uni、およびpCRBac(Invitrogen);λExCell、λgtll、pTrc99A、pKK223−3、pGEX−1λT、pGEX−2T、pGEX−2TK、pGEX−4T−1、pGEX−4T−2、pGEX−4T−3、pGEX−3X、pGEX−5X−1、pGEX−5X−2、pGEX−5X−3、pEZZ18、pRIT2T、pMC1871、pSVK3、pSVL、pMSG、pCH110、pKK232−8、pSL1180、pNEO、およびpUC4K(Pharmacia);pSCREEN−1b(+)、pT7Blue(R)、pT7Blue−2、pCITE−4abc(+)、pOCUS−2、pTAg、pET−32LIC、pET−30LIC、pBAC−2cp LIC、pBACgus−2cp LIC、pT7Blue−2 LIC、pT7Blue−2、λSCREEN−1、λBlueSTAR、pET−3abcd、pET−7abc、pET9abcd、pETllabcd、pET12abc、pET−14b、pET−15b、pET−16b、pET−17b−pET−17xb、pET−19b、pET−20b(+)、pET−21 abcd(+)、pET−22b(+)、pET−23abcd(+)、pET−24abcd(+)、pET−25b(+)、pET−26b(+)、pET−27b(+)、pET−28abc(+)、pET−29abc(+)、pET−30abc(+)、pET−31b(+)、pET− 32abc(+)、pET−33b(+)、pBAC−1、pBACgus−1、pBAC4x−1、pBACgus4x−1、pBAC−3cp、pBACgus−2cp、pBACsurf−1、plg、Signalplg、pYX、Selecta Vecta−Neo、Selecta Vecta−Hyg、およびSelecta Vecta−Gpt(Novagen);pLexA、pB42AD、pGBT9、pAS2−1、pGAD424、pACT2、pGAD GL、pGAD GH、pGAD10、pGilda、pEZM3、pEGFP、pEGFP−1、pEGFP−N、pEGFP−C、pEBFP、pGFPuv、pGFP、p6xHis−GFP、pSEAP2−Basic、pSEAP2−Contral、pSEAP2−プロモーター、pSEAP2−エンハンサー、pβgal−Basic、pβgal−Control、pβgal−プロモーター、pβgal−エンハンサー、pCMV、pTet−Off、pTet−On、pTK−Hyg、pRetro−Off、pRetro−On、pIRES1neo、pIRESi1hyg、pLXSN、pLNCX、pLAPSN、pMAMneo、pMAMneo−CAT、pMAMneo−LUC、pPUR、pSV2neo、pYEX4T−1/2/3、pYEX−S1、pBacPAK−His、pBacPAK8/9、pAcUW31、BacPAK6、pTriplEx、λgt10、λgt11、pWE15、およびλTriplEx(Clontech);Lambda ZAP II、pBK−CMV、pBK−RSV、pBluescript II KS +/−、pBlucscript II SK +/−、pAD−GAL4、pBD−GAL4 Cam、pSurfscript、Lambda FIX II、Lambda DASH、Lambda EMBL3、Lambda EMBL4、SuperCos、pCR−Scrigt Amp、pCR−Script Cam、pCR−Script Direct pBS +/−、pBC KS +/−、pBC 5K +/−、Phagescript、pCAL−n−EK、pCAL−n、pCAL−c、pCAL−kc、pET−3abcd、pET−11abcd、pSPUTK、pESP−I、pCMVLacI、pOPRSVI/MCS、pOPI3 CAT、pXT1、pSG5、pPbac、pMbac、pMClneo、pMC1neo Poly A、pOG44、pOG45、pFRTβGAL、pNEOβGAL、pRS403、pRS404、pRS405、pRS406、pRS413、pRS414、pRS415、およびpRS416(Stratagene)が含まれる。
【0222】
さらなるベクターには、例えば、pPC86、pDBLeu、pDBTrp、pPC97、p2.5、pGADI−3、pGAD10、pACt、pACT2、pGADGL、pOADGH、pAS2−1、pGAD424、pGBT8、pGBT9、pGAD−GAL4、pLexA.、pBD−GAL4、pHISi、pHISi−1、placZi、pB42AD、pDG202、pJK202、pJG4−5、pNLexA、pYESTrp、およびその変異型または誘導体が含まれる。
【0223】
また、宿主中で複製可能且つ生存可能である限り、当該分野で公知の任意の他のプラスミドおよびベクターを使用することができる。
【0224】
(相同組換えされた細胞の選択)
次いで、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の発現をノックアウトするために相同組換えを行った細胞を、適切に組み込まれた細胞を同定するために適切に選択された培地中で成長させることができる。次いで、所望の表現型を示す細胞を、制限分析、エレクトロポレーション、サザン分析、ポリメラーゼ連鎖反応、または当該分野で公知の別の技術によってさらに分析することができる。標的遺伝子部位で適切な挿入を示すフラグメントの同定によって、標的遺伝子を不活化するか修飾するために相同組換えを行った細胞を同定することができる。
【0225】
CMP−Neu5Acヒドロキシラーゼ遺伝子に挿入した選択マーカー遺伝子の存在により、宿主ゲノムへの標的構築物の組み込みが確立される。次いで、所望の表現型を示す細胞を、制限分析、エレクトロポレーション、サザン分析、ポリメラーゼ連鎖反応、モノクローナル抗体アッセイ、蛍光標示式細胞分取器(FACS)、または相同または非相同組換えが起こるかどうかを確立するためにDNAを分析するための当該分野で公知の任意の他の技術もしくは方法によってさらに分析することができる。挿入のためのプローブの使用およびその後の構築物の隣接領域を超えて及ぶCMP−Neu5Acヒドロキシラーゼ遺伝子の存在のためのインサートに隣接する5’領域および3’領域の配列決定または欠失が組み込まれている場合の欠失の存在の同定によってこれを決定することができる。構築物内の配列に相補的であり、且つ構築物の外側および標的対立遺伝子の配列と相補的であるプライマーを使用することもできる。この方法では、相同組換えが起こった場合に相補鎖中に存在する両プライマーを有するDNA二重鎖のみを得ることができる。プライマー配列または予想されるサイズの配列の存在の証明によって、相同組換えの発生が支持される。
【0226】
相同組換え事象のスクリーニングのために使用されるポリメラーゼ連鎖反応は、例えば、KimおよびSmithies,Nucleic Acids Res.16:8887−8903,1988;およびJoynerら,Nature 338:153−156,1989に記載されている。
【0227】
相同組換え事象の別のスクリーニング方法には、例えば、Malykhら,European Journal of Cell Biology 80,48−58(2001),Malykhら,Glycoconjugate J.15,885−893(1998)に記載のブタCMP−Neu5Acヒドロキシラーゼおよび/またはNeu5Gcに特異的なモノクローナル抗体またはポリクローナル抗体の使用が含まれる。
【0228】
相同組換え事象による機能的CMP−Neu5Acヒドロキシラーゼ発現を欠くブタ細胞のさらなる特徴づけには、サザン分析、ノーザンブロット分析、特異的レクチン結合アッセイ、および/または配列分析、例えば、Y.Malykhら,Biochem J.370:601−607(2003);Y.MalykhらEuropean Journal of Cell Biology 80:48−58(2001);Y.MalykhらGlycoconjugate J.15:885−893(1998)に記載の抗Neu5Gcまたは抗CMP−Neu5Acヒドロキシラーゼ抗体アッセイの使用が含まれるが、これらに限定されない。一般に、A.SharmaらTransplantation 75(4):430−436(2003)を参照のこと。
【0229】
第1ラウンドのターゲティングから得た細胞株は、ターゲティングされた対立遺伝子とヘテロ接合性である可能性がある。両対立遺伝子が修飾されたホモ接合性を多数の方法で達成することができる。1つのアプローチは、1つのコピーが修飾された多数の細胞を成長させ、その後これらの細胞を異なる選択マーカーを使用して残存するブタCMP−Neu5Acヒドロキシラーゼ対立遺伝子の別ラウンドのターゲティングに供することである。あるいは、伝統的なメンデルの遺伝的特徴にしたがった修飾対立遺伝子に対してヘテロ接合性の動物の交配によってホモ接合体を得ることができる。いくつかの状況では、2つの異なる修飾対立遺伝子を有することが望ましい。連続ラウンドの遺伝子ターゲティングまたはヘテロ接合体(それぞれ1つの所望の修飾対立遺伝子を保有する)の交配によってこれを行うことができる。
【0230】
(VIII.異種移植の免疫学的障壁を克服するためのさらなる遺伝子の遺伝子操作)
本発明の1つの態様では、非機能的CMP−Neu5Acヒドロキシラーゼ遺伝子に対してホモ接合性の細胞を、さらなる遺伝子操作に供することができる。例えば、宿主に異なる遺伝子の能力を置換、交換、または提供するために、内因性CMP−Neu5Acヒドロキシラーゼ対立遺伝子が非機能的になっているホモ接合性宿主にさらなる遺伝子の能力を移入することができる。同系遺伝子接合体化(homogenotization)後にマーカー遺伝子を除去することができる。場合によって拡大された配列を有し、元の構築物のマーカー遺伝子部分が欠失された実質的に相同なDNAを含む構築物の移入により、標的対立遺伝子との相同組換えを得ることができる。組み込みのためのマーカー遺伝子の組み合わせの使用により、ポジティブ選択および他のネガティブ選択を行い、除去工程でマーカー遺伝子が保持された細胞を選択することができる。
【0231】
1つの実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子およびα(1,3)GT遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的CMP−Neu5Acヒドロキシラーゼを欠く動物を産生し、その後この動物由来の細胞を使用してα(1,3)GT遺伝子をノックアウトすることができる。ホモ接合性α(1,3)GTネガティブブタが最近報告されている(Phelpsら,Science 2003;WO 04/028243)。あるいは、これらのα(1,3)GTノックアウト動物由来の細胞を使用して、CMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0232】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠き、且つヒト補体阻害タンパク質を産生するブタ細胞も提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、この動物由来の細胞をヒト補体阻害タンパク質(CD59(Philbrick,W.M.ら(1990)Eur.J.Immunol.20:87−92で報告されたcDNA)、ヒト崩壊促進因子(human decay accelerating factor)(DAF)(Medofら(1987)Proc.Natl.Acad.Sci.USA 84:2007で報告されたcDNA)、およびヒト膜補因子タンパク質(MCP)(Lublin,D.ら(1988)J.Exp.Med.168:181−194で報告されたcDNA)などであるが、これらに限定されない)を発現するようにさらに修飾することができる。
【0233】
別の実施形態では、ヒト補体阻害タンパク質を産生するトランスジェニックブタ由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。ヒト補体阻害タンパク質を産生するトランスジェニックブタは当該分野で公知である(例えば、米国特許第6,166,288号を参照のこと)。
【0234】
さらなる実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子およびブタForssmanシンターゼ(FSM)遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、gal−α−galエピトープの産生に関与し、且つ異種移植拒絶で役割を果たすブタFSMシンターゼ遺伝子をノックアウトするためにさらに修飾することができる。ブタFSMシンターゼ遺伝子は最近同定された(米国特許出願60/568,922号を参照のこと)。あるいは、これらのFSMシンターゼ遺伝子ノックアウト動物由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0235】
なおさらなる実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子およびブタイソグロボシド(isogloboside)3シンターゼ遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、ブタiGb3シンターゼ遺伝子をノックアウトするために使用することができる。ブタiGb3シンターゼ遺伝子は最近報告されている(米国特許出願第60/517,524号)。あるいは、これらのブタiGb3シンターゼ遺伝子ノックアウト動物由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0236】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子、α(1,3)GT遺伝子、FSMシンターゼ遺伝子、およびブタiGb3シンターゼ遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、α(1,3)GT遺伝子、FSMシンターゼ遺伝子、およびブタiGb3シンターゼ遺伝子をノックアウトするために使用することができる。ホモ接合性α(1,3)GTネガティブブタは最近報告されている(Phelpsらsupra,Science 2003;WO 04/028243)。あるいは、これらのα(1,3)GTノックアウト動物由来の細胞を使用して、ブタiGb3シンターゼ遺伝子、ブタFSMシンターゼ遺伝子、およびCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化し、さらに、ヒト補チア阻害タンパク質(CD59、ヒト崩壊促進因子(DAF)、およびヒト膜補因子タンパク質(MCP)などが含まれるが、これらに限定されない)を発現するようにさらに修飾することができる。
【0237】
(VIII.遺伝子操作動物の産生)
本発明は、ブタ全能性幹細胞の遺伝子操作によるCMP−Neu5Acヒドロキシラーゼ発現を欠くトランスジェニックブタの産生方法を提供する。1つの実施形態では、(a)動物中の1つまたは複数の標的CMP−Neu5Acヒドロキシラーゼ核酸ゲノム配列の同定、(b)CMP−Neu5Acヒドロキシラーゼ核酸ゲノム配列をターゲティングする1つまたは複数の相同組換えベクターの調製、(c)動物の複数の全能性細胞のゲノムへの1つまたは複数のターゲティングベクターの挿入およびそれによる複数のトランスジェニック全能性細胞の産生、(d)動物の四倍体胚盤胞を得ること、(e)四倍体胚盤胞への複数の全能性細胞の挿入およびそれによるトランスジェニック胚の産生、(f)レシピエント雌動物への胚の導入、および(g)雌動物中で胚を成長させて出産させることによって動物を産生することができる。トランスジェニックブタを作製するための本明細書中に記載のトランスジェニック動物の産生方法は、一般に、米国特許第6,492,575号に記載されている。
【0238】
別の実施形態では、全能性細胞は、胚幹(ES)細胞であり得る。胚盤胞からのES細胞の単離、ES細胞株の確立、およびその後のその培養を、例えば、Doetchmannら,J.Embryo.Exp.Morph.87:27−45(1985);Liら,Cell 69:915−926(1992);Robertson,E.J.“Tetracarcinomas and Embryonic Stem Cells:A Practical Approach,”E.J.Robertson編,IRL Press,Oxford,England(1987);WurstおよびJoyner,”Gene Targeting:A Practical Approach,”A.L.Joyner編,IRL Press,Oxford,England(1993);Hogenら,“Manipulating the Mouse Embryo:A Laboratory Manual,”Hogan,Beddington,CostantiniおよびLacy編,Cold Spring Harbor Laboratory Press,New York(1994);およびWangら,Nature 336:741−744(1992)に記載の従来の方法によって行う。例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子を変化させるためのターゲティングベクターでの胚幹細胞の形質転換後、細胞を適切な培地(例えば、ウシ胎児血清増強DMEMなど)中の支持細胞層にプレートすることができる。構築物を含む細胞を、選択培地の使用によって検出することができ、十分なコロニー成長期間後、コロニーを選別し、相同組換えの発生について分析することができる。構築物の配列内および配列外であるが標的対立遺伝子に存在するプライマーを使用したポリメラーゼ連鎖反応を使用することができる。次いで、相同組換えを示すコロニーを、胚操作および胚盤胞注射のために使用することができる。胚盤胞を、過剰排卵雌から得ることができる。次いで、胚幹細胞をトリプシン処理し、修飾細胞を胚盤胞を含む液滴に添加することができる。少なくとも1つの修飾胚幹細胞を、胚盤胞の胞胚腔に注射することができる。注射後、少なくとも1つの胚盤胞を、偽妊娠雌の各子宮角に戻すことができる。次いで、雌を出産させ、得られた同腹子を構築物を含む変異細胞についてスクリーニングした。形質転換ES細胞と異なる比率の胚盤胞を選択する。胚盤胞およびES細胞の異なる表現型の提供により、キメラ子孫を容易に検出することができ、次いで、修飾CMP−Neu5Acヒドロキシラーゼ遺伝子の存在を探索するために遺伝子型同定を行うことができる。
【0239】
本発明のさらなる実施形態では、全能性細胞は、胚生殖(EG)細胞であり得る。胚生殖細胞は、in vitroで培養およびトランスフェクトすることができ、その後にキメラの体細胞系列および生殖細胞系列に寄与する、ES細胞と機能的に等価な未分化の細胞である(Stewartら,Dev.Biol.161:626−628(1994))。EG細胞は、成長因子(白血病抑制因子、スチール因子(steel factor)、塩基性線維芽細胞成長因子)と組み合わせた始原生殖細胞(配偶子の先祖)の培養によって誘導される(Matsuiら,Cell 70:841−847(1992);Resnickら,Nature 359:550−551(1992))。Donovanら,“Transgenic Animals,Generation and Use,”L.M.Houdebine編,Harwood Academic Publishers(1997)などに記載の当業者に公知の方法を使用して、EG細胞を培養することができる。
【0240】
例えば、Jamesら,Genet.Res.Camb.60:185−194(1992);NagyおよびRossant,“Gene Targeting:A Practical Approach,”A.L.Joyner編,IRL Press,Oxford,England(1993);またはKubiakおよびTarkowski,Exp.Cell Res.157:561−566(1985)に記載の天然の接合子の産生および発生または2つの細胞胚の電気融合およびその後の培養によって本発明で使用するための四倍体胚盤胞を得ることができる。
【0241】
例えば、Wangら,EMBO J.10:2437−2450(1991)に記載の当該分野で公知の任意の方法によって、胚盤胞へのES細胞またはEG細胞の移入を行うことができる。
【0242】
「複数」の全能性細胞は、1つを超える任意の数の細胞を含み得る。例えば、本発明で使用される全能性細胞数は、約2〜約30細胞、約5〜約20細胞、または約5〜約10細胞であり得る。1つの実施形態では、1つの細胞懸濁液から採取した約5〜10個のES細胞を、顕微操作装置中の保持ピペットによって固定された胚盤胞に注射する。次いで、当該分野で公知の方法(例えば、Robertson,E.J.“Teratocarcinomas and Embryonic Stem Cells:A Practical Approach”IRL Press,Oxford,England(1987)を参照のこと)によって、胚を少なくとも3時間(場合により一晩)インキュベート後、雌レシピエント動物に移入する。次いで、雌動物中で胚を成長させて出産させることができる。
【0243】
(クローン化トランスジェニック子孫を産生するための体細胞核導入)
本発明は、体細胞核導入による機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタのクローニング方法を提供する。一般に、広範な種々の哺乳動物クローニング方法が現在急速に開発および報告されており、所望の結果を達成する任意の方法を本発明で使用することができる。このような方法の非限定的な例を以下に記載する。例えば、ドナー核の供給源として使用すべき所望の分化ブタ細胞を得る工程と、ブタから卵母細胞を得る工程と、卵母細胞を除核する工程と、除核卵母細胞に所望の分化細胞または細胞核を導入する工程と(例えば、NT単位を形成するための融合または注射による)、得られたNT単位を活性化する工程と、NT単位が胎児に成長するように宿主ブタに培養したNT単位を導入する工程とを含む核導入プロセスによってブタを産生することができる。
【0244】
核導入技術または核移植技術は当該分野で公知である(Campbellら,Theriogenology,43:181(1995);Collasら,Mol.Report Dev.,38:264−267(1994);Keeferら,Biol.Reprod.,50:935−939(1994);Simsら,Proc.Natl.Acad.Sci.,USA,90:6143−6147(1993);WO 94/26884;WO 94/24274,WO 90/03432,米国特許第4,944,384号および同第5,057,420号)。1つの非限定的な例では、除核卵母細胞への挿入前にドナー染色体またはドナー細胞が再プログラミングされた哺乳動物のクローニング方法を記載したCollasらの米国特許出願番号2003/0046722号などに記載の方法などの方法を提供する。本発明は、染色体、核、または細胞の卵母細胞の挿入または融合方法も記載する。
【0245】
CMP−Neu5Acヒドロキシラーゼ遺伝子を変化させるように修飾したドナー細胞核を、レシピエントブタ卵母細胞に導入する。この方法の使用は、特定のドナー細胞型に制限されない。ドナー細胞は、Wilmutら,Nature 385 810(1997);Campbellら,Nature 380 64−66(1996);またはCibelliら,Science 280 1256−1258(1998)などに記載のものであり得る。正常な核型の全細胞(核導入で首尾よく使用することができる胚細胞、胎児および成体の体細胞が含まれる)を、原理上は使用することができる。胎児線維芽細胞は、特に有用なドナー細胞クラスである。一般に適切な核導入方法は、Campbellら,Theriogenology 43 181(1995),Collasら,Mol.Reprod.Dev.38 264−267(1994),Keeferら,Biol.Reprod.50 935−939(1994),Simsら,Proc.Nat’l.Acad.Sci.USA 90 6143−6147(1993),WO−A−9426884,WO−A−9424274,WO−A−9807841,WO−A−9003432,米国特許第4,994,384号および同第5,057,420号に記載されている。分化または少なくとも部分的に分化したドナー細胞も使用することができる。ドナー細胞は培養中および静止期でもあり得るが、必ずしもそうではない。休止期の核ドナー細胞は、静止期に入るように誘導されるかin vivoで休止状態で存在し得る細胞である。先行技術の方法は、クローニング手順で胚細胞型も使用している(Campbellら(Nature,380:64−68,1996)およびSticeら(Biol.Reprod.,20 54:100−110,1996)。
【0246】
体細胞核ドナー細胞を、種々の異なる臓器および組織(皮膚、間葉、肺、膵臓、心臓、腸、胃、膀胱、血管、腎臓、尿道、生殖器、および胚、胎児、または成体動物の全部または一部の脱凝集調製物)から得ることができる。本発明の適切な実施形態では、核ドナー細胞は、上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋細胞、グラヌロース細胞、卵丘細胞、上皮細胞、または内皮細胞からなる群から選択される。別の実施形態では、核細胞は、胚幹細胞である。好ましい実施形態では、ドナー細胞として線維芽細胞を使用することができる。
【0247】
本発明の別の実施形態では、本発明の核ドナー細胞は、動物の生殖細胞である。胚、胎児、または成体期の動物種の任意の生殖細胞を、核ドナー細胞として使用することができる。適切な実施形態では、核ドナー細胞は胚生殖細胞である。
【0248】
受容細胞との連携を確実にするために、細胞周期の任意の時期(GO、GI、G2、S、M)で核ドナー細胞を停止することができる。当該分野で公知の任意の方法を使用して、細胞周期を操作することができる。細胞周期を制御する方法には、培養細胞の接触阻害によって誘導されたGO期静止、血清または他の必須栄養素の除去によって誘導されたGO期静止、老化によって誘導されたGO期静止、特定の成長因子の添加によって誘導されたGO期静止、熱ショック、高圧、もしくは化学物質、ホルモン、成長因子、もしくは他の物質での他の処理などの物理的もしくは化学的手段によって誘導されたGOもしくはGI期静止;任意の複製点手順で妨害する化学物質での処理によるS期の制御;蛍光標示式細胞分取、有糸分裂の除去、微小管破壊剤、有糸分裂の進行を破壊する任意の化学物質での処理を使用した選択によるM期の制御が含まれるが、これらに限定されない(Freshney,R.I,.“Culture of Animal Cells:A Manual of Basic Technique,”Alan R.Liss,Inc,New York(1983)も参照のこと)。
【0249】
卵母細胞の単離方法は当該分野で周知である。本質的に、これは、ブタの卵巣または生殖器官からの卵母細胞の単離を含み得る。ブタ卵母細胞の容易に利用可能な供給源は、食肉処理場の材料である。遺伝子工学、核導入、およびクローニングなどの技術の組み合わせのために、卵母細胞は遺伝学的に成熟していなければならず、その後にこれらの細胞を核導入のためのレシピエント細胞として使用することができ、これらの細胞を精子細胞による受精によって胚に発生することができる。このプロセスは、一般に、哺乳動物の卵巣(例えば、食肉処理場で得たウシ卵巣)からの未熟な(第I減数分裂前期(prophase I))卵母細胞の回収、卵母細胞が第II減数分裂中期に達するまで(この場合、ウシ卵母細胞は一般に吸引後18〜24時間で起こる)の受精または除核前の成熟培地での卵母細胞の成熟を必要とする。この期間は、「成熟期」として公知である。
【0250】
第II減数分裂中期の卵母細胞は、レシピエント卵母細胞であり得るが、この段階で卵母細胞は移入された核をそのままで受精精子で処理するために「活性化する」ことができるか、十分に活性化することができると考えられる。in vivoで成熟した第II減数分裂中期の卵母細胞は、核導入技術で首尾よく使用されている。本質的に第II減数分裂中期の卵母細胞を、発情期またはヒト絨毛性ゴナドトロピン(hCG)もしくは類似のホルモンの注射から35〜48時間後または39〜41時間後に非過剰排卵または過剰排卵ブタのいずれかから外科的に回収することができる。
【0251】
約10〜40時間、好ましくは約16〜18時間の範囲の定期成熟後、卵母細胞を除核することができる。除核前に卵母細胞を除去し、卵丘細胞の除去前に適切な培地(1mg/mlのヒアルロニダーゼを含むHECMなど)に入れることができる。次いで、裸の卵母細胞を極体についてスクリーニングし、次いで、極体の存在によって決定した選択された第II減数分裂中期の卵母細胞を核導入に使用する。除核を以下に示す。
【0252】
米国特許第4,994,384号などに記載の公知の方法によって除核を行うことができる。例えば、第II減数分裂中期の卵母細胞を、即時除核のために任意選択的に7.5mg/mlのサイトカラシンBを含むHECMに入れるか、適切な培地(例えば、CR1aa+10%発情期ウシ精子などの胚培養培地)に入れ、その後、好ましくはせいぜい24時間後、より好ましくは16〜18時間後に除核することができる。
【0253】
極体および隣接細胞質を除去するために、マイクロピペットを使用した顕微鏡手術で除核することができる。次いで、首尾よく除核された卵母細胞を同定するために、卵母細胞をスクリーニングすることができる。卵母細胞の1つのスクリーニング方法は、卵母細胞を1mg/mlの33342Hoechst色素を含むHECMで染色し、その後に10秒未満の紫外線照射下で卵母細胞を視覚化することである。次いで、首尾よく除核された卵母細胞を、適切な培養培地(CR1aa+10%精子)に入れることができる。
【0254】
次いで、除核卵母細胞と同種の1つの哺乳動物細胞を、NT単位を産生するために使用される除核卵母細胞の卵黄周囲腔に導入することができる。当該分野で公知の方法にしたがって、哺乳動物の細胞および除核卵母細胞を使用してNT単位を産生することができる。例えば、エレクトロポレーションによって細胞を融合することができる。原形質膜の一過性破壊に十分な電気パルスの印加によって電気融合を行う。膜が急速に再形成されるので、原形質膜のこの破壊は非常に短時間である。したがって、2つの隣接する膜の破壊が誘導され、再形成時に脂質二重層が混合する場合、2つの細胞間に小さなチャネルを開けることができる。このような小さな開口部の熱力学的不安定性のために、2つの細胞が1つになるまで拡大する。例えば、Pratherらの米国特許第4,997,384号を参照のこと。種々の電気融合培地(例えば、スクロース、マンニトール、ソルビトール、およびリン酸緩衝化溶液)を使用することができる。融合誘導因子としてSendaiウイルスを使用して融合を行うこともできる(Graham,Wister Inot.Symp.Monogr.,9,19,1969)。また、エレクトロポレーション融合を使用するよりもむしろ核を卵母細胞に直接注射することができる。例えば、CollasおよびBarnes,Mol.Reprod.Dev.,38:264−267(1994)を参照のこと。融合後、得られた融合NT単位を、活性化まで適切な培地(例えば、CR1aa培地)に入れる。典型的には、その後短時間で(例えば、24時間後または約4〜9時間後)活性化を行うことができる。
【0255】
所望の結果が達成される任意の方法によってNT単位を活性化することができる。このような方法には、例えば、本質的にNT単位への冷温または実質的に冷温ショックの印加による生物学的温度未満でのNT単位の培養が含まれる。胚が通常曝露される生理学的温度条件と比較して低温である室温でのNT単位の培養によってこれを最も都合良く行うことができる。あるいは、公知の活性化剤の適用によって活性化することができる。例えば、受精時の精子による卵母細胞の貫通は、融合前の卵母細胞を活性化して、核導入後により多数の生存可能な認識および複数の遺伝的に同一のブタが得られることを示した。また、電気的および化学的ショックなどの処理を使用して、融合後にNT胚を活性化することができる。例えば、Susko−Parrishらの米国特許第5,496,720号を参照のこと。さらに、卵母細胞中の2価の陽イオンレベルの増加および卵母細胞中の細胞タンパク質のリン酸化の減少によって同時または連続的に活性化を行うことができる。一般に、卵母細胞の細胞質への2価の陽イオン(例えば、イオノフォアの形態のマグネシウム、ストロンチウム、バリウム、またはカルシウム)の移入によってこれを行うことができる。2価の陽イオンレベルの他の増加方法には、電気ショック、エタノールでの処理、およびケージ化キレート剤での処理が含まれる。公知の方法(例えば、キナーゼインヒビター(例えば、6−ジメチル−アミノプリン、スタウロスポリン、2−アミノプリン、およびスフィンゴシンなどのセリン−トレオニンキナーゼインヒビター)の添加)によってリン酸化を減少させることができる。あるいは、卵母細胞へのリン酸塩(例えば、リン酸塩2Aおよびリン酸塩2B)の移入によって細胞タンパク質のリン酸化を阻害することができる。
【0256】
次いで、細胞コロニーの作製まで適切なin vitro培養培地で活性化NT単位を培養することができる。胚の培養および成熟に適切な培養培地は、当該分野で周知である。胚の培養および維持に使用することができる公知の培地の例には、Ham’s F−10+10%ウシ胎児血清(FCS)、組織培養培地−199(TCM−199)+10%ウシ胎児血清、タイロード−アルブミン−乳酸−ピルビン酸(TALP)、Dulbeccoリン酸緩衝化生理食塩水(PBS)、Eagle培地、およびWhitten培地が含まれる。
【0257】
その後、培養NT単位を洗浄し、好ましくは適切なコンフルエントの支持細胞層を含むウェルプレート中に含まれる適切な培地に入れることができる。適切な支持細胞層には、例として、線維芽細胞および上皮細胞が含まれる。NT単位がレシピエント雌への導入に適切なサイズに到達するまで、または細胞コロニーを産生するために使用することができる細胞を得るために、NT単位を支持細胞層上で培養する。好ましくは、少なくとも約2〜400個の細胞、より好ましくは約4〜128個の細胞、最も好ましくは少なくとも約50個の細胞までこれらのNT単位を培養することができる。
【0258】
次いで、活性化NT単位を、雌ブタの卵管に導入することができる(胚移植)。1つの実施形態では、雌ブタは、発情期同調レシピエント未経産ブタ(gilt)であり得る。交配未経産ブタ(large white/Duroc/Landrace)(280〜480lbs)を使用することができる。未経産ブタを、飼料と混合した18〜20mgのReguMate(Altrenogest,Hoechst,Warren,NJ)の経口投与によってレシピエント動物と同調させることができる。Regu−Mateを、14日間連続して与えることができる。次いで、最後のRegu−Mate処置から約150時間後に1000単位のヒト絨毛性ゴナドトロピン(hCG,Intervet America,Millsboro,DE)をi.m.投与することができる。hCG注射から約22〜26時間後に胚移植を行うことができる。1つの実施形態では、妊娠出産させて生きた子孫を誕生させることができる。別の実施形態では、早期に出産を終了させて、胚細胞を採取することができる。
【0259】
本発明における胚移植方法およびレシピエント動物の管理は、胚移植産業で使用される標準的な手順である。同期導入は、本発明の成功で重要である(すなわち、NT胚の時期をレシピエント雌の発情周期と同調させる)。例えば、Siedel,G.E.,Jr.“Critical review of embryo transfer procedures with cattle”in Fertilization and Embryonic Development in Vitro(1981)L.Mastroianni,Jr.およびJ.D.Biggers編,Plenum Press,New York,N.Y.,page 323を参照のこと。
【0260】
(VIII.ブタ動物、臓器、組織、細胞、および細胞株)
本発明は、CMP−Neu5Acヒドロキシラーゼ遺伝子の両対立遺伝子が不活化された生きたブタを提供する。本発明はまた、異種移植に有用なこのようなブタ由来の臓器、組織、および細胞を提供する。
【0261】
1つの実施形態では、機能的CMP−Neu5Acヒドロキシラーゼの任意の発現を欠くブタから得たブタ臓器、組織、および/または精製もしくは実質的に純粋な細胞もしくは細胞株を提供する。
【0262】
1つの実施形態では、本発明は、異種移植に有用な臓器を提供する。任意のブタ臓器(脳、心臓、肺、腺、脳、目、胃、脾臓、すい臓、腎臓、肝臓、腸、子宮、膀胱、皮膚、毛髪、爪、耳、鼻、口、唇、歯茎、歯、舌、唾液腺、扁桃腺、咽頭、食道、大腸、小腸、直腸、肛門、幽門、甲状腺、胸腺、腎上体、骨、軟骨、腱、靭帯、骨格筋、平滑筋、血管、血液、脊髄、気管、尿管、尿道、視床下部、下垂体、副腎、卵巣、輸卵管、子宮、膣、乳腺、精巣、精嚢、陰茎、リンパ、リンパ節、およびリンパ管が含まれるが、これらに限定されない)を使用することができる。
【0263】
別の実施形態では、本発明は、異種移植に有用な組織を提供する。任意のブタ組織(上皮組織、結合組織、血液組織、骨組織、軟骨組織、筋肉組織、神経組織、腺様組織、脂肪組織、疎性結合組織、骨組織、褐色脂肪組織、海綿状様骨組織、筋肉組織、軟骨細胞(cartaginous)、海綿組織、軟骨様組織、クロム親和組織、肉様組織、弾性組織、上皮組織、脂肪組織、軟骨様組織、Gaingee組織、膠様組織、肉芽組織、腸関連リンパ組織、ハーラー脈管組織、硬質(hard)造血組織、未分化組織、間質細胞、被覆組織、島組織、リンパ組織、リンパ様組織、間葉組織、中腎性組織、粘液様結合細胞、多房性脂肪細胞、骨髄組織、軟組織鼻点、造腎細胞、結節組織、骨細胞、造骨組織、類骨組織、根尖周囲組織、細網組織、細毛組織、外科用ゴム敷布、骨格筋組織、平滑筋組織、および皮下組織)を使用することができる。
【0264】
さらなる実施形態では、本発明は、機能的α1,3GTの発現を欠くブタ動物由来の細胞および細胞株を提供する。1つの実施形態では、これらの細胞または細胞株を異種移植のために使用することができる。任意のブタ組織または臓器由来の細胞(上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋細胞、リン酸細胞(□hosphate cell)、卵丘細胞、上皮細胞;内皮細胞、ランゲルハンス島細胞、膵臓インスリンを分泌する細胞、膵臓α−2細胞、膵臓β細胞、膵臓α−1細胞、血球、血液前駆細胞、骨細胞、骨前駆細胞、ニューロン幹細胞、始原幹細胞、肝細胞、ケラチノサイト、臍帯静脈内皮細胞、大動脈内皮細胞、微小血管系内皮細胞、線維芽細胞、肝臓星細胞、大動脈平滑筋細胞、心筋細胞、ニューロン、クップフェル細胞、平滑筋細胞、シュワン細胞、上皮細胞、赤血球、血小板、好中球、リンパ球、単球、好酸球、好塩基球、脂肪細胞、軟骨細胞、膵島細胞、甲状腺細胞、副甲状腺細胞、耳下腺細胞、腫瘍細胞、グリア細胞、星状細胞、赤血球、白血球、マクロファージ、上皮細胞、体細胞、下垂体細胞、副腎細胞、有毛細胞、気泡細胞、腎細胞、網膜細胞、桿状体細胞、錐状体細胞、心臓細胞、ペースメーカー細胞、脾臓細胞、抗原提示細胞、記憶細胞、T細胞、B細胞、形質細胞、筋細胞、卵巣細胞、子宮細胞、前立腺細胞、腟上皮細胞、精子細胞、精巣細胞、生殖細胞、卵細胞、ライディッヒ細胞、尿細管周囲細胞、セルトリ細胞、ルテイン細胞、頸部細胞、子宮内膜細胞、乳房細胞、濾胞細胞、粘液細胞、繊毛細胞、非角化上皮細胞、角化上皮細胞、肺細胞、杯細胞、円柱上皮細胞、ドーパミン作動性細胞、扁平上皮細胞、骨細胞、骨芽細胞、破骨細胞、胚幹細胞、線維芽細胞、および胎児線維芽細胞が含まれるが、これらに限定されない)を使用することができる。特定の実施形態では、機能的α−1,3−GT発現を欠くブタ由来の膵臓細胞(ランゲルハンス島細胞、インスリン分泌細胞、48のα−2細胞、β細胞、α−1細胞が含まれる)を提供する。
【0265】
非生存誘導体には、これらの組織誘導体を移植で使用する前に架橋または他の化学的処理によってさらに処理することができる、酵素または化学的処理によって生存細胞を剥ぎ取られた組織が含まれる。好ましい実施形態では、誘導体には、種々の組織(皮膚、泌尿器組織、膀胱組織、または臓器粘膜下組織が含まれる)由来の細胞外基質が含まれる。また、医療機器として心臓弁および他の非生存組織を含むために生存組織を剥ぎ取られた腱、関節、および骨を提供する。
【0266】
(治療上の使用)
広範な種々の方法にしたがって宿主に細胞を投与することができる。好ましい投与様式は、非経口、腹腔内、静脈内、皮内、硬膜外、脊髄内、胸骨内、関節内、滑液内、くも膜下腔内、動脈内、心臓内、筋肉内、鼻腔内、皮下、眼窩内、嚢内、局所、経皮パッチ、座剤を含む直腸、膣、もしくは尿道投与、経皮、鼻噴霧、外科用インプラント、内部外科用塗布剤(internal surgical paint)、注入ポンプ、またはカテーテルである。1つの実施形態では、薬剤およびキャリアを、直接組織注射もしくはボーラス、インプラント、微粒子、ミクロスフェア、ナノ粒子、またはナノスフェアなどの持続放出処方物で投与する。
【0267】
開示の細胞の注入によって治療することができる障害には、正常な血球の産生および成熟の機能障害の不全に起因する疾患(すなわち、再生不良性貧血および低増殖性幹細胞障害);造血臓器における新生物悪性疾患(例えば、白血病およびリンパ腫);非造血臓器の広範な悪性固形腫瘍;自己免疫疾患;および遺伝病が含まれるが、これらに限定されない。このような障害には、正常な血球の産生および成熟の不全または機能障害に起因する疾患(超増殖性幹細胞障害)(薬物、照射、または感染による特発性の再生不良性貧血、汎血球減少症、顆粒球減少症、血小板減少症、赤血球欠如症、ブラックファン−ダイヤモンド症候群が含まれる);造血性悪性腫瘍(急性リンパ芽球性(リンパ球性)白血病、慢性リンパ球性白血病、急性骨髄性白血病、慢性骨髄性白血病、急性悪性脊髄硬化症、多発性骨髄腫、真性多血症、原因不明の骨髄化生、ワルデンストレームマクログロブリン血症、ホジキンリンパ腫、非ホジキンリンパ腫が含まれる);悪性固形腫瘍患者における免疫抑制(悪性黒色腫、胃癌、卵巣癌、乳癌、小細胞肺癌、網膜芽細胞種、睾丸癌、膠芽細胞腫、横紋筋肉腫、神経芽腫、ユーイング肉腫、リンホニア(lymphonia)が含まれる);自己免疫疾患(関節リウマチ、1型糖尿病、慢性肝炎、多発性軟化症、全身性紅斑性狼瘡が含まれる);遺伝性(先天性)障害(貧血(家族性、再生不良性)、ファンコーニ貧血、ジヒドロ葉酸還元酵素欠損症、ホルムアミド転移酵素欠損症、レッシ−ナイハン症候群、先天性異常赤血球産生症症候群IIV、シバッハマン−ダイヤモンド症候群、ジヒドロ葉酸還元酵素欠損、ホリナミノ(forinamino)転移酵素欠損症、レッシ−ナイハン症候群、先天性球状赤血球症、先天的性楕円赤血球症、先天性ストマトサイト増加症、先天性Rh陰性疾患、発作性夜間血色素尿症、G6PD(グルコースリン酸デヒドロゲナーゼ)変異型1、2、3、ピルビン酸キナーゼ欠損症、先天的性エリトロポイエチン感受性、欠損症、鎌状赤血球症および鎌状赤血球傾向、サラセミアα、β、γ、メトヘモグロビン血症、先天性免疫障害、重症複合型免疫不全症(SCID)、ベアリンパ球症候群、イオノフォア応答性複合型免役不全、キャップ形成異常を伴う複合型免疫不全、ヌクレオシドホスホリラーゼ欠損症、顆粒球アクチン欠損症、乳児顆粒球減少症、ゴーシェ病、アデノシンデアミナーゼ欠損症、コストマン症候群、細網系異形成、先天性白血球機能障害症候群が含まれる);その他(骨粗鬆症、脊髄硬化症、後天性溶血性貧血、後天性免疫不全、原発性または続発性免疫不全を発症する感染症、細菌感染(例えば、ブルセラ病、リステリア病、結核、ハンセン病)、寄生虫感染(例えば、マラリア、リーシュマニア症)、真菌感染、加齢によるリンパ系細胞組の不均衡(disproportionsin)および免疫機能障害(impaired immune fimctions)を含む障害、貪食細胞障害、コストマン顆粒球減少症、慢性肉芽腫疾患、チェジアック−東症候群、好中球アクチン欠損症、好中球膜GP−180欠損症、代謝性貯蔵病、ムコ多糖沈着症、ムコリピドーシス、免疫機構に関与するその他の障害、ヴィスコット−オールドリッチ症候群、αランチリプシン(lantirypsin)欠損症など)が含まれるが、これらに限定されない。
【0268】
疾患または病変には、神経変性疾患、肝臓変性疾患病、腎臓変性疾患、脊髄損傷、頭部の外傷または手術、組織、臓器、または腺の変性を引き起こすウイルス感染などが含まれる。このような神経変性疾患には、AIDS痴呆;脱髄性疾患(多発性軟化症および急性トランスフェラーゼ脊髄炎など);錐体外路および小脳障害(皮質脊髄系(ecorticospinal system)の損傷など);大脳基底核障害または小脳障害;ハンティングトン舞踏病と老年舞踏病などの多動性障害;薬物油土迂生運動障害(CNSドーパミン受容体を遮断する薬物によって誘導されるものなど);低運動性障害(パーキンソン病など);進行性過剰核形成(progressive supra−nucleo palsy);小脳の構造損傷;脊髄小脳変性(脊髄性運動失調、フリートライヒ運動失調)、小脳皮質変性、多系統変性(multiple systems degenerations)(Mencel、Dejerine Thomas、Shi−Drager、およびMachado−Joseph)、全身性障害(systermioc disorders)(Rufsum疾患など)、βリポタンパク質欠損血症、失調症、毛細血管拡張;ミトコンドリア多システム障害;脱髄性コア障害(多発性軟化症、急性横断脊髄炎など);運動単位障害(神経原性筋萎縮(前角細胞萎縮(筋萎縮性側索硬化症など)、幼児型脊髄性筋萎縮、および若年性脊髄性筋萎縮)など);アルツハイマー病;中年のダウン症;びまん性レビー小体病;レビー小体型老人性痴呆;パーキンソン病、ヴェルニッケ−コルサコフ症候群;慢性アルコール中毒;クロイツフェルト・ヤコブ病;亜急性硬化性汎脳炎、ハレルホルデン−スパッツ疾患;ならびに拳闘家痴呆が含まれるが、これらに限定されない。例えば、Berkowら(編)(1987),The Merck Manual(15’)版),Merck and Co.,Rahway,NJ.を参照のこと。
【0269】
(産業的飼育での使用)
本発明は、CMP−Neu5Acヒドロキシラーゼ遺伝位の一方または両方の対立遺伝子が不活化された飼育適用を目的とする生きたブタを提供する。CMP−Neu5Acヒドロキシラーゼの一方または両方の対立遺伝子の不活化により、人畜共通疾患、ブタにおける感染症(例えば、E.coli、ブタロタウイルス、およびブタ伝染性胃腸炎コロナウイルス)、ならびに宿主動物においてNeu5Gcを使用する任意の他の人畜共通生物または腸管毒素原性生物に対するブタ動物の感受性を減少させることができる。疾患感受性の減少により、より多数の健康な動物の得る能力によって飼育操作がより経済的に実現し、腸管毒素原性生物に起因する動物の死亡が減少する。
【0270】
以下の実施例は例示目的で提供し、本発明を決して制限しない。
【実施例】
【0271】
(核酸の単離)
PCRベースの方法の組み合わせストラテジーを使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を同定した。このようなPCR方は、当該分野で周知であり、例えば、PCR Technology,H.A.Erlich編,Stockton Press,London,1989;PCR Protocols:A Guide to Methods and Applications,M.A.Innis,D.H.Gelfand,J.J.Sninsky,およびT.J.White編,Academic Pres,Inc.,New York,1990に記載されている。
【0272】
Trizol試薬(Gibco,Grand Island,NY)を使用して、成体ブタ(グレートヨークシャー種)から総RNAを抽出した。DNアーゼI(Ambion,Inc.,Austin,TX)での処理後、ポリA+RNAを、Dynabeads mRNA精製キット(Dynal,Oslo,Norway)を使用して分離した。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の5’末端または3’末端を同定するために、Marathon(登録商標)cDNA増幅キット(Clontech)を使用して、5’または3’−RACE(cDNA末端の迅速な複製)手順を行った。転写物のエクソン−イントロン境界または5’もしくは3’隣接領域を同定するために、UniversalGenomeWalkeerライブラリーキット(Clontech)を使用して、ブタGenomeWalker(登録商標)ライブラリーを構築した。GenBankアクセッションA59058によって提供された部分的cDNA配列から、遺伝子特異的およびネスト化プライマー対をデザインした。
【0273】
(cDNAおよびゲノムCMP−Neu5Acヒドロキシラーゼ配列の決定)
5’または3’RACE分析:ブタCMP−Neu5Acヒドロキシラーゼ遺伝子構築物の5’末端および3’末端を同定するために、テンプレートとして成体ブタ脾臓から単離したポリA+RNAと共にMarathon cDNA増幅キット(Clontech)を使用して、5’および3’−RACE手順を行った。20UのAMV−RTおよび1pmolの市販のcDNA合成プライマーを使用して、48℃で2時間のインキュベーションによって1μgのポリA+RNAから第1の鎖cDNAを合成した。第2の鎖cDNA合成は、RNアーゼH、E.coli DNAポリメラーゼI、E.coliDNAポリメラーゼI、およびE.coliDNAリガーゼから構成される市販の酵素カクテルでの16℃で1.5時間の全第1の鎖の反応を含んでいた。T4DNAポリメラーゼによる二本鎖cDNA末端の平滑末端化後、市販のMarathoncDNA Adapterを、精製された二本鎖cDNAのアリコートにライゲーションした。キット中の10mM ticme−KOH/0.1M EDTA緩衝液でのアダプターライゲーション産物の希釈により、PCR増幅のためのcDNAを用意した。
【0274】
ブタCMP−Neu5Acヒドロキシラーゼ遺伝子転写物の5’および3’のほとんどの配列を得るために、提供されたMarathon cDNA増幅プライマー組を、GenBankアクセッション番号A59058によって提供された配列に基づいて、遺伝子特異的プライマーおよびネスト化遺伝子特異的プライマーと対合した。これらのプライマー組を、表13に示す。この方法により、GenBankアクセッション番号A59058中に含まれる配列に基づいたオリゴヌクレオチドプライマーを、3’方向および5’方向に配向させ、これを使用して重複PCRフラグメントを作製する。これらの重複3’および5’産物を組み合わせて、インタクトな全長cDNAを産生する。この方法は、例えば、Innisら,supra;およびFrohmanら,Proc.Natl.Acad.Sci.,85:8998,1988に記載されており、さらに、例えば、米国特許第4,683,195号に記載されている。
【0275】
ゲノムウォーキング分析:ブタCMP−Neu5Acヒドロキシラーゼ転写物のエクソン−イントロン境界または5’もしくは3’隣接領域を同定するために、UniversalGenomeWalker(登録商標)ライブラリーキット(Clontech,Palo Alto,CA)を使用して、ブタGenomeWalker(登録商標)ライブラリーを構築した。
【0276】
簡単に述べれば、ブタゲノムDNAの5つのアリコートを、1平滑末端切断制限エンドヌクレアーゼ(Dral、EcoRV、PvulI、ScaI、またはStuI)で個別に消化した。フェノール−クロロホルム抽出、エタノール沈殿、および制限消化フラグメントの再懸濁後、各消化アリコートの一部を、キット中にGenomuWalkerアダプターとの個別のライゲーション反応で使用した。このプロセスにより、PCRベースのクローニングストラテジーで使用するための5つのライブラリーが作製された。表13で同定したプライマー対を、ゲノムウォーキングストラテジーで使用した。eLON−GaseまたはTaKaRa LATaq(Takara Shuzo Co.,Ltd.,Shiga,Japan)酵素のいずれかを、全GenomeWalker実験およびゲノムDNAの直接長(direct long)PCRのために使用した。PerkinElmer Gene Amp System 9600または9700サーモサイクラーによる全GenomeWalker実験で、製造者が推奨する熱サイクリング条件を使用した。
【0277】
【表15】
増幅産物のサブクローニングおよび配列決定:ゲノムDNAから増幅したPCR産物、、GeneWalker−PCR(Clontech)、および5’−3’−RACEを、Qiagenゲル抽出キット(Qiagen,Valencia,CA)を使用してゲル精製し、必要に応じて、Original TAクローニングキット(Invitrogen,Carlsbad,CA)中のpCRIIベクターにサブクローン化した。pCRIIライゲーションインサートのプラスミドDNAミニプレップを、説明書にしたがってQIAprepスピンミニプレップキット(Qiagen)を使用して調製した。ABI 377自動化配列分析器(Applied Biosystems,Inc.,Foster City,CA)をT7およびSP6プロモータープライマーまたは内部インサート配列からデザインしたプライマーでプライミングしたローダミンまたはBigDyeターミネーターサイクル配列決定キット(Applied Biosystems)のいずれかと共に使用して、クローン化インサートの自動化蛍光配列決定を行った。
【0278】
プライマー合成:他で記載しない限り、種々のPCRベースの方法でプライマーとして使用する全てのオリゴヌクレオチドを、固相合成およびホスホラミダイトヌクレオシド化学を使用したABI 394 DNA Synthesizer(Applied Biosystems,Inc.,Foster City CA)を使用して合成した。
【0279】
(転写因子結合部位の分析)
エクソン配列の228bpおよびエクソン1の601bp上流を使用して、可能な転写因子結合部位の分析を行った。www.genomatix.deで利用可能な「MatInspector」ソフトウェアを使用して、配列をスクリーニングした。配列は、以下の転写因子の結合部位を含む:MZF1、ETSF、SF1、CMYB、MEF2、NMP4、BRN2、AP1、GAT1、SATB1、ATF、USF、WHN、ZF5、NFκB、MOK2、NFY、MYCMAX、ZF5。図4を参照のこと。
【0280】
ブタCMP−Neu5Acヒドロキシラーゼ相同組換えターゲティングベクターの構築CMP−Neu5Acヒドロキシラーゼノックアウト標的ベクター。ノックアウトのためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6をターゲティングするベクターを構築することができる。第1の工程では、
【0281】
【化1】
などのプライマーを使用して、ターゲティングベクターの3’アームとして使用するためのイントロン6部分をPCRによって増幅する(図5を参照のこと)。約3.7kbの増幅されたPCR産物を、EcoRIおよびApaIを使用した制限酵素消化後にpCRIIベクターに挿入することができる。図6を参照のこと。
【0282】
3’アームの挿入後、
【0283】
【化2】
などのプライマーを使用して、ターゲティングベクターの5’アームとして使用するためのイントロン5部分をPCRによって増幅することができる(図5を参照のこと)。この最初の増幅後、それぞれ
【0284】
【化3】
などのプライマーと組み合わせてpDH1などのプライマーを使用してEcoNI制限部位を5’アームの3’部分に挿入するために連続的PCR増幅を行うことができる(図5を参照のこと)。操作EcoNI部位を含む約2.6kbの増幅PCR産物はApaIおよびEcoNIを使用して消化した制限酵素であり、これを予め挿入した3’アームを含むpCRIIベクターに挿入して(図7を参照のこと)、約6.3kbのブタCMP−Neu5Acヒドロキシラーゼターゲティング配列を含むターゲティングベクター(pDHΔex6)を作製することができる(図8を参照のこと)。
【0285】
EGFPノックイン標的ベクター。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6の3’末端への増強緑色蛍光タンパク質配列のインフレーム挿入によって、pDHΔex6をさらに修飾することができる。第1の工程では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のイントロン5部分およびエクソン6部分を、
【0286】
【化4】
などのプライマーを使用したPCRによって増幅し(図9を参照のこと)、MunIおよびEcoRvでの制限酵素消化後にEGFPおよびポリAテールを含むベクター(pIRES−EGFP)に挿入することができる。挿入後、
【0287】
【化5】
などのプライマーを使用した挿入を含むpIRES−EGFPベクターでPCR増幅を行うことができる(図9を参照のこと)。PCR増幅産物はEcoNIおよびMunIを使用して消化した制限酵素であり、これを予め構築したpDHΔex6ターゲティングベクターに挿入することができる(図10を参照のこと)。得られたターゲティングベクター(pDHΔex6)を図11に例示する。
【0288】
(ブタCMP−Neu5Acヒドロキシラーゼ欠損胎児線維芽細胞の産生)
妊娠33日の同一妊娠の10体の胎児から胎児線維芽細胞を単離する。頭部および内蔵の除去後、胎児をハンクス平衡塩類溶液(HBSS;Gibco−BRL,1 5 Rockville,MD)で洗浄し、20mlのHBSSに入れ、小さな手術用鋏でサイコロ状にする。組織をペレット化し、50ml試験管中で胎児あたり40mlのDMEMおよび100U/mlコラゲナーゼ(gbco−BRL)で再懸濁する。チューブを37℃の水浴中で撹拌しながら40分間インキュベートする。消化された組織を3〜4分間静置し、細胞豊富な上清を新たな50mlチューブに移し、ペレット化する。次いで、細胞を、10%ウシ胎児血清(FCS)、1×非必須アミノ酸、1mMピルビン酸ナトリウム、および2ng/ml bFGFを含む40mlのDMEMで再懸濁し、10cmの皿に播種する。トランスフェクションのために、10μgの線状化pDHΔex6EGFPベクターを、製造者のガイドラインにしたがってリポフェクタミン2000(Carlsbad,CA)を使用して200万個の細胞に移入する。48時間のトランスフェクション後、トランスフェクトした細胞を、2,000細胞/ウェルの密度で48ウェルプレートに播種し、コンフルエンスまで成長させる。コンフルエンス後、細胞を、相同組換えが起こってEGFPを発現する細胞のみが選択される蛍光標示式細胞分取(FACS)(FACSCalibur,Becton Dickenson,San Jose,CA)によって分取する(例えば、図13を参照のこと)。
【0289】
選択された細胞を、再播種し、コンフルエントまで成長させる。一旦コンフルエンスに到達すると、いくつかの小アリコートをその後の使用のために凍結させ、残りを相同組換えについてのPCRおよびサザンブロットによる検証のために使用する。推定ターゲティングクローンを、EGFPに相補的なプライマーおよびアンチセンスプライマーとしてベクターの外側の配列に相補的なプライマーを使用したエクソン6/EGFPインサートを介したPCRによってスクリーニングすることができる。ターゲティングされた対立遺伝子由来の推定バンドの存在によってポジティブクローンを同定するために、PCR産物を、EGFPプローブを使用したサザンブロッティングによって分析することができる。
【0290】
(核ドナーとして異種CMP−Neu5Acヒドロキシラーゼ欠損胎児線維芽細胞を使用したクローン化ブタの作製)
核導入のための細胞の調製:一般的に上記のようにブタCMP−Neu5Acヒドロキシラーゼにヘテロ接合性を示す細胞を産生するために、ドナー細胞を遺伝子操作する。上記に詳述のように産生した核ドナーとしてEGFP選択ブタ線維芽細胞を使用して、当該分野で周知の方法(Daiら,Nature Biotechnology 20:251255,2002;およびPolejaevaら,Nature 407:86−90,2000)によって核導入を行うことができる。
【0291】
例えば、I.Polejaevaら,Nature 407:505(2000)に記載のように、同期化された過剰排卵する性的に成熟したLarge−WhiteX Landacre外部交配未経産ブタから卵母細胞を単離することができる。ドナー細胞を、血清枯渇(0.5%)によって24時間と120時間との間の推定G0/G1期に同調させる。例えば、本質的にA.C.Boquestら,Biol.Reproduction 68:1283(2002)に記載のように、卵母細胞の除核、核導入、電気融合、および電気活性化を行うことができる。再構築した胚を一晩培養し、非同期(−1日)レシピエントの卵管に導入することができる。妊娠を確認し、実時間超音波によってモニタリングすることができる。
【0292】
ヘテロ接合性CMP−Neu5Acヒドロキシラーゼ1ノックアウト(SKO)雄ブタと雌ブタとの交配を行い、二重ノックアウト(DKO)ブタの小集団を確立することができる。
【0293】
(CMP−Neu5Acヒドロキシラーゼ欠損ブタの検証)
1ノックアウト雄ブタおよび雌ブタの交配後、二重ノックアウトブタの検証を行う。子孫由来の線維芽細胞を、1μgの抗N−グリコシルGM2モノクローナル抗体MK2−34(Seikagaku Kogyo,JP)と氷上で30分間インキュベートする。FITC抱合ヤギ抗マウスIgGを細胞および結合がNeu5GCの有無を示す抗体に添加し、それにより活性なCMP−Neu5Acヒドロキシラーゼの有無をフローサイトメトリー(FACSCalibur,Becton Dickenson,San Jose,CA)によって検出する。
【図面の簡単な説明】
【0294】
【図1】ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム機構を示す。黒塗りのバーは、それぞれ番号をつけたエクソンを示す。エクソン間のイントロンの長さは、相対距離を示す。(白抜きのボックスもいくつかの変異型で認められるエクソンを示す(図2を参照のこと);「開始」および「終止」は、それぞれ開始コドンおよび終止コドンを示す)。図の下におおよそのスケールを示す。
【図2】CMP−Neu5Acヒドロキシラーゼ遺伝子のcDNA配列を示す。変異型1は、エクソン14および15の代わりにエクソン15aを含む。変異型2はエクソン12aを含み、変異型3はエクソン11aを含む。「開始」および「終止」は、それぞれ開始コドンおよび終止コドンを示す。
【図3】その対応するゲノム機構と共にターゲティングベクターの4つの非限定的な例を示す。この特定の非限定的な例における選択マーカー遺伝子は、eGFP(増強緑色蛍光タンパク質)である。ブタCMP−Neu5Aceヒドロキシラーゼ遺伝子を不活化するためにGFPをDNA構築物に挿入することができる。
【図4】エクソン1(228bp)内に存在する転写因子結合部位および601bpにわたるその5’隣接領域を示す。
【図5】ブタCMP−Neu5Acヒドロキシラーゼ遺伝子ターゲティングベクターのDNA構築のために使用することができるオリゴヌクレオチド配列を示す。
【図6】プライマーpDH3およびpDH4を使用したブタCMP−Neu5Acヒドロキシラーゼ遺伝子からの3’アームセグメントの産生およびベクター(pCRII)へのその挿入を示す略図である。
【図7】プライマーpDH1およびpDH2、その後pDH2a、pDH2b、およびpDH2cを使用したブタCMP−Neu5Acヒドロキシラーゼ遺伝子からの5’アームセグメントの産生および3’アームが予め挿入されたベクター(pCRII)への挿入を示す略図である。
【図8】相同組換えによってブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6を欠失するために使用することができるターゲティングベクターを例示する図の非限定的な例である。
【図9】ノックインストラテジーで使用するための増強緑色蛍光タンパク質発現ベクターの作製で使用されるオリゴヌクレオチド配列を示す。
【図10】ポリAシグナルを有するEGFPフラグメントのターゲティングベクターpDHΔex6への挿入を例示する図である。
【図11】eGFP発現のためのノックインベクターを例示する図である。
【図12】ターゲティングカセットDNA構築物(pDHΔex6)とゲノムDNAとの間でフレームシフトが起こる相同組換えを例示する図である。
【図13】ターゲティングカセットDNA構築物(pDHΔex6)とゲノムDNAとの間でフレームシフトが起こる相同組換えを例示する図である。
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2003年6月6日提出の米国特許出願番号60/476,396号の優先権を主張する。
【0002】
(発明の分野)
本発明は、ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ(CMP−Neu5Acヒドロキシラーゼ)のタンパク質、cDNA、およびゲノムDNA調節配列を提供する。さらに、本発明は、ブタ動物、組織、および臓器、ならびに機能的CMP−Neu5Acヒドロキシラーゼの発現を欠くこのような動物、組織、および臓器由来の細胞および細胞株を含む。このような動物、組織、臓器、および細胞を、研究および医学的治療法(異種移植が含まれる)ならびに産業上の家畜飼育操作で使用することができる。さらに、異種移植で使用するためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く臓器、組織、および細胞の調製方法を提供する。
【背景技術】
【0003】
(発明の背景)
許容可能なヒトドナー臓器が利用不可能であること、宿主対移植片拒絶による長期成功率の低さ、ならびに感染症および癌の深刻なリスクは、組織および臓器移植分野が現在直面している主な課題である。許容可能な臓器に対する需要が供給を超えているので、多くの患者は毎年利用可能になる臓器を待っている間に死亡している。この需要を満たす一助とするために、研究は同種間移植の代替法の開発に焦点が当てられている。透析は腎不全患者が利用可能であり、人工心臓モデルが試験されており、臓器不全を補助または置換するための他の機械系が開発されている。しかし、このようなアプローチは非常に高価である。透析装置を頻繁且つ定期的に利用することが必要であるので、このような治療を受ける患者の自由および生活の質が非常に制限される。
【0004】
異種移植片移植は、ヒト移植のための人工臓器に代わる潜在的に魅力的なものである。非ヒト臓器の潜在的なプールは事実上限られている。ブタは、最も可能性が高い異種移植臓器の供給源と考えられている。ブタの供給は豊富にあり、交配プログラムが十分に確立されており、その臓器のサイズおよび生理学的性質はヒトに適合可能である。したがって、ブタ臓器での異種移植は、臨床的移植に利用可能な臓器の不足に対する潜在的な解決法を提供する。
【0005】
このような異種間組織の宿主拒絶は、依然としてこの分野における主な懸念である。異種移植に対する免疫学的障壁は、恐れられており、且つ依然として恐れられている。第1の免疫学的ハードルは、「超急性拒絶」(HAR)である。HARは、外来組織に結合する予め形成された高力価の天然の抗体の遍在と定義される。ドナー臓器内皮上の標的エピトープに対するこれらの天然の抗体の結合は、HARにおける開始事象と考えられる。この結合の後、レシピエントの血液でのドナー臓器の灌流から数分以内に、血小板およびフィブリンが沈殿し、最終的にドナー臓器が間質の浮腫および出血を起こし、その全てによってレシピエントで臓器不全が起こる(Strahanら(1996)Frontiers in Bioscience 1,pp.34−41)。
【0006】
類人猿または旧世界ザル(例えば、ヒヒ)からヒトへのいくつかの著名な異種移植は、拒絶することなく何ヵ月間も許容される。しかし、このような試みは、多数の免疫学的要因によって最終的に失敗している。HARを抑制するための強い免疫抑制を使用した場合でさえも、移植片内皮上の異種補体の活性化を制御するための移植片組織内の補体調節タンパク質(CRP)の不全に一部起因する軽度の先天性免疫応答により、最終的に移植臓器が破壊される(Starzl,Immunol.Rev.,141,213−44(1994))。ヒトへの異種移植に許容可能な臓器のプールを開発する目的で、研究者は、ヒトCRPを産生する動物を操作している(霊長類における異種移植片破壊を遅延させるが消失しないことが証明されているアプローチ)(McCurryら,Nat.Med.,1,423−27(1995);Bachら,Immunol.Today,17,379−84(1996))。
【0007】
補体媒介攻撃に加えて、調和しない異種移植片のヒトによる拒絶は、共通の抗原(多数の糖タンパク質および糖脂質のガラクトース−α(1,3)−ガラクトース(gal−α−gal)末端残基)によって媒介されるようである(Galiliら,Proc.Nat.Acad.Sci.(USA),84,1369−73(1987);Cooperら,Immunol.Rev.,141,31−58(1994);Galiliら,Springer Sem.Immunopathol,15,155−171(1993);Sandrinら,Transplant Rev.,8,134(1994))。この抗原は、ヒトA、B、およびO型血液抗原と化学的に関連し、多数の寄生虫ならびに細菌およびウイルスなどの感染因子に存在する。ほとんどの哺乳動物組織もこの抗原を含むが、旧世界ザル、類人猿、およびヒトは注目すべき例外である(Joziasseら,J.Biol.Chem.,264,14290−97(1989)を参照のこと)。このような炭水化物エピトープを持たない個体は、エピトープに特異的な天然に存在する抗体(IgMおよびIgG)を豊富に産生する。多くのヒトは、gal−α−gal炭水化物決定基に対して特異性を有する有意なレベルの循環IgGを示す(Galiliら,J.Exp.Med.,162,573−82(1985);Galiliら,Proc.Nat.Acad.Sci.(USA),84,1369−73(1987))。α−ガラクトシルトランスフェラーゼ(α−GT)酵素は、gal−α−gal部分の形成を触媒する。研究は、異種組織の細胞表面上でのgal−α−gal部分の発現を減少または消失するためのこの酵素の調整または消失に焦点が当てられている。
【0008】
ブタ由来のα−ガラクトシルトランスフェラーゼ遺伝子の消失は、ブタからヒトへの異種移植に対する最も有意なハードルの1つと長い間考えられている。ブタゲノム中の2つの対立遺伝子はα−GT遺伝子をコードする。ブタにおけるα−GT遺伝子の1対立遺伝子ノックアウトが2002年に報告された(DaiらNature Biotechnol.,20:251(2002);Laiら,Science,295:1089(2002))。
【0009】
最近、α−GT遺伝子の二重対立遺伝子ノックアウト(double allelic knockout)が行われた(Phelpsら,Science,299:pp.411−414(2003))。Revivicor Inc.のWO 2004/028243は、α1,3ガラクトシルトランスフェラーゼ(α1,3−GT)の全発現を欠くブタ動物、組織、臓器、細胞、および細胞株を記載している。したがって、α1,3−GTの機能的発現を欠く動物、組織、臓器、細胞、および細胞を、異種移植および他の医学的目的のために使用することができる。
【0010】
Immerge Biotherapeutics,Inc.のPCT特許出願WO2004/016742号は、α(1,3)−ガラクトシルトランスフェラーゼヌル細胞、GGTA−1ヌル細胞の選択方法、それから産生されたα(1,3)−ガラクトシルトランスフェラーゼヌルブタ(生存可能なGGTA−1ヌルブタと呼ばれる)、このようなブタの作製方法、および異種移植のための細胞、組織、および臓器の使用方法を記載している。
【0011】
gal−α−gal以外の最も初期の公知の異種抗原の1つは、Hnaganutiu Deicher抗体が認識し、且つ血清疾患と長期的に関連するエピトープである。エピトープは、N−グリコリルノイラミン酸(Neu5Gc)(炭水化物のシアル酸ファミリーのメンバー)として同定された。炭水化物のうち、シアル酸は豊富且つ遍在する。シアル酸は、N−アセチルノイラミン酸(Neu5Acyl)およびその誘導体に使用される総称である。N−アセチルノイラミン酸(Neu5Ac)およびN−グリコリルノイラミン酸(neu5Gc)は、シアル酸の最も豊富な誘導体のうちの2つである。
【0012】
Neu5Gcエピトープは、複合糖質のグリカン鎖中の末端に存在する。この露呈した位置のために、例えば、炎症反応、免疫細胞の成熟、分化プロセス、ホルモン結合、病原体結合、および毒素結合の場合の細胞認識で重要な役割を果たす(Varki,A.,Glycobiology,2,pp.25−40(1992))。
【0013】
Neu5Gcを含む複合糖質はヒトで免疫原性を示す。健常なヒトでは、Neu5Gcは検出可能でないが、Neu5Gcはほとんどの哺乳動物で豊富に存在する。人におけるNeu5Gcの欠如は、機能酵素の形成を防止するヒト遺伝子におけるエクソンの欠失に起因する(Chou,H.H.らProc.Natl.Acad.Sci.(USA),95,pp.11751−11756(1998);Irie,A.らJ.Biol.Chem.,273,pp.15866−15871(1998))。したがって、Neu5Gc含有複合糖質は抗原として作用し、抗体の形成を誘導することができる。歴史的に、この抗体はHanganutziu−Deicher(HD)抗原および抗体と呼ばれる(Hanganutziu,M.,CR Soc.Biol.(Paris),91,p.1457(1924);Deicher,H.,Z.Hyg.,106,p.561(1926))。Hanganutziu−Deicher抗原は、多数のヒト腫瘍(結腸癌、網膜芽腫、黒色腫、および乳癌)およびニワトリ腫瘍組織で検出可能である(Higashi,H.らCancer Res.,45,pp.3796−3802(1985))。腫瘍中の抗原量は非常に少ないにもかかわらず(通常、全量が1%未満のシアル酸、しばしば0.01から0.1%までの範囲)、Hanganutziu−Deicher抗体の形成を誘導することができる(Higashihara,T.ら,Int Arch Allergy Appl Immunol.,95,pp.231−235(1991))。この免疫反応は、ヒトへのNeu5Gc含有ブタ臓器の異種移植に対する潜在的な障壁である。
【0014】
Neu5Gcエピトープは、Neu5AcのN−アセチル部分への水酸基の付加によって形成される。ヒドロキシル化を触媒する酵素は、CMP−Neu5Acヒドロキシラーゼである。したがって、CMP−Neu5Acヒドロキシラーゼ遺伝子の発現は、細胞表面上のNeu5Gcエピトープの存在を決定する。哺乳動物におけるCMP−Neu5Acヒドロキシラーゼの精製研究により、シトクロムb5およびシトクロムb5レダクターゼに依存する可溶性細胞質オキシゲナーゼであることが示された(Kawano,T.ら,J.Biol.Chem.,269,pp.9024−9029(1994);Schneckenburger,P.ら,Glycoconj.J.,11,pp.194−203(1994);Schlenzka,W.ら,Glycobiology,4,pp.675−683(1994);Kozutsumi,Y.ら,J.Biochem.(Tokyo),108,pp.704−706(1990);およびShaw,L.ら,Eur.J.Biochem.,219,pp.1001−1011(1994))。
【0015】
Neu5Gcの別の重要な特徴は、細胞に侵入させる病原体の接着分子として作用することである(Kelm,S.およびSchauer,R.,Int.Rev.Cytol,179,pp.137−240(1997))。これにより、一定の家畜で疾患および経済的損失を被る。詳細には、K99線毛を有する腸管毒素原性大腸菌は、腸内吸収細胞および粘膜分泌細胞に結合したNue5Gcα2→3Galβ1→4Glcβ1→1’セラミド[GM3(Neu5Gc)]などのガングリオシド中のNeu5Gc、N−グリコシルシアロパラグロボシド、およびGM2(Neu5Gc)への結合によって新生仔ブタに感染し、潜在的に致死的下痢を引き起こす(Malykh,Y.ら,Biochem.J.,370,pp.601−607(2003);Kyogashima,M.ら(1993);Teneberg,S.ら,FEBS Leaers,263,pp.10−14(1990);Isobe,T.ら,Anal.Biochem.,236,pp.35−40(1996);Lindahl,M.およびCaristedt,I.,Gen.Microbiol.,136,pp.1609−1614(1990);King,T.P.ら,Proceedings of the 6th International Symposium on Digestive Physiology in Pigs,pp.290−293(1994))。ブタロタウイルスは、GM3(Neu5Gc)への結合によって新生ブタに感染して下痢を引き起こす。ブタ伝染性胃腸炎コロナウイルスは、α2,3結合Neu5Gcを含む複合糖質への侵入によってブタに感染する(Schultz,Bら,.L Virol.,70,pp.5634−5637(1996))。
【0016】
マウスの肝臓およびブタの顎下腺からCMP−Neu5Acヒドロキシラーゼが均一に単離され、特徴づけられている(Kawano,T.ら,J.Biol.Chem.,269,pp.9024−9029(1994);Schneckenburger,P.ら,Glycoconj.J.,11,pp.194−203(1994);and,Schlenzka,W.ら,Glycobiology,4,pp.675−683(1994))。
【0017】
Schlenzkaら(Glycobiology,Vol.4,pp.675−683(1994))は、イオン交換クロマトグラフィ、固定化トリアジン色素を使用したクロマトグラフィ、疎水性相互作用クロマトグラフィ、およびゲル濾過を使用して、ブタ顎下腺から酵素を精製した。Schneckenburgerら(Glycoconj.J.,Vol.11,pp.194−203(1994))は、マウス肝臓からCMP−Neu5Acヒドロキシラーゼを単離した。ブタ顎下腺およびマウス肝臓由来のCMP−Neu5Acヒドロキシラーゼは共に分子量65kDaの可溶性単量体である。CMP−Neu5Acおよびシトクロムb5とのその触媒的相互作用は、互いに類似している。これらの酵素の活性は、イオン含有補欠分子団に依存するようである。
【0018】
JP−A06113838は、マウスCMP−Neu5Acヒドロキシラーゼのタンパク質およびDNA配列ならびにこのヒドロキシラーゼに特異的に結合するモノクローナル抗体を記載している。
【0019】
Boehringer ManheimGMBHのPCT公開番号WO97/03200A1は、ブタCMP−Neu5Acヒドロキシラーゼの部分cDNAを開示している。この出願は、CMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン8の中央から始まるcDNA配列を開示している(GenBankアクセッション番号Y15010としてさらに開示されている)。
【0020】
Martensen,L.ら(Eur.J.Biochem.,Vol.268,pp.5157−5166(2001))は、ブタCMP−Neu5Acヒドロキシラーゼの全長アミノ酸配列を開示している。
【0021】
RBC BiotechnologyのPCT公開番号WO02/088351号は、ブタCMP−Neu5Acヒドロキシラーゼの部分cDNA配列およびゲノム配列(エクソン7〜11および各エクソン周囲の部分ゲノム配列)を開示している。さらに、任意選択的に、α−1,3−ガラクトシルトランスフェラーゼ遺伝子および/または補体タンパク質の挿入などの他のゲノム修飾と組み合わせたCMP−Neu5Acヒドロキシラーゼエピトープを欠くブタ細胞および動物の作製方法を提供する。
【発明の開示】
【発明が解決しようとする課題】
【0022】
本発明の目的は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列および調節配列を提供することである。
【0023】
本発明の目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子の全長cDNAおよび新規の変異型を提供することである。
【0024】
本発明の別の目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子をコードする新規の核酸配列およびアミノ酸配列を提供することである。
【0025】
本発明のさらなる目的は、CMP−Neu5Acヒドロキシラーゼ遺伝子を欠損した細胞、組織、および/または臓器を提供することである。
【0026】
本発明の別の目的は、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物(特にブタ)を作製することである。
【0027】
本発明のなおさらなる目的は、必要とするヒトレシピエントへの非ヒト臓器の異種移植で使用するためのCMP−Neu5Acヒドロキシラーゼ遺伝子を欠損した細胞、組織、および/または臓器を提供することである。
【課題を解決するための手段】
【0028】
(発明の要旨)
ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の全長cDNA配列、ペプチド配列、およびゲノム機構を決定した。今日まで、部分cDNAおよびゲノム配列のみが同定されている。本発明は、新規のブタCMP−Neu5Acヒドロキシラーゼのタンパク質、cDNA、cDNA変異型、およびゲノムDNA配列を提供する。さらに、本発明は、ブタ動物、組織、および臓器、ならびに機能的CMP−Neu5Acヒドロキシラーゼの発現を欠くこのような動物、組織、および臓器由来の細胞および細胞株を含む。このような動物、組織、臓器、および細胞を、研究および医学的治療法(異種移植が含まれる)で使用することができる。さらに、異種移植で使用するためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く臓器、組織、および細胞の調製方法を提供する。
【0029】
本発明の1つの態様は、ブタCMP−Neu5Acヒドロキシラーゼの全長cDNAを提供する。全長cDNAを、表1(配列番号1)に示し、全長ペプチド配列を表2(配列番号2)で提供する。全長cDNAの開始コドンはエクソン4の3’部分に存在し、終止コドンはエクソン17の3’部分で見出される。配列番号1および2と少なくとも80%、85%、90%、95%、98%、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号1または2の少なくとも10、15、17、20、25、または30個のヌクレオチドまたはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号1とハイブリッド形成する任意のヌクレオチド配列およびそれに相同なヌクレオチドをさらに提供する。
【0030】
1つの実施形態では、CMP−Neu5Acヒドロキシラーゼの新規の変異型をコードする核酸配列およびペプチド配列を提供する(表3〜8、図2)。配列番号3は、エクソン1、4、5、6、7、8、9、10、11、12、13、15a、16、17、および18を含む遺伝子の変異型(変異型1)のcDNAを示す。配列番号5は、エクソン1、4、5、6、7、8、9、10、11、12、および12aを含む遺伝子の変異型(変異型2)のcDNAを示す。配列番号7は、エクソン1、4、5、6、7、8、9、10、11、および11aを含む遺伝子の変異型(変異型3)のcDNAを示す。配列番号4、6、および8は、それぞれ変異型1、変異型2、および変異型3のアミノ酸配列を示す。配列番号3〜8と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号3〜8の少なくとも10、15、17、20、25、または30個のヌクレオチドまたはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号3、5、および7とハイブリッド形成する任意のヌクレオチド配列ならびにそれに相同なヌクレオチドをさらに提供する。
【0031】
さらなる実施形態は、CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA配列を示す核酸配列を提供する(表9、図1)。配列番号10〜28は、それぞれエクソン1、4〜11、11a、12、12a、13〜15、15a、16〜18を示し、配列番号29〜45は、それぞれイントロン1a、1b、4〜15、15a、16、および17を示す。配列番号9は、CMP−Neu5Acヒドロキシラーゼ遺伝子の5’非翻訳領域を示す。配列番号46(表10)は、CMP−Neu5AcヒドロキシラーゼのゲノムDNAおよび調節配列を示す。
【0032】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号47に示す。配列番号47は、5’UTR、エクソン1、およびエクソン1(表11)3’に存在するイントロン配列の一部を含む5’連続ゲノム配列を示す。
【0033】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号48に示す。配列番号48は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、エクソン8、イントロン8、エクソン9、イントロン9、エクソン10、イントロン10、エクソン11、イントロン11、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18でイントロン配列を含む連続ゲノム配列を示す(表12)。さらに、配列番号48の少なくとも2775、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、または10,000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号48と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0034】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号49に示す。配列番号49は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、およびエクソン8でイントロン配列を含む連続ゲノム配列を示す。さらに、配列番号49の少なくとも1750、1800、1900、2000、2100、2200、2300、2400、2500、2600、2700、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4100、4200、4300、4400、4500、4600、4700、4800、4900、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号49と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0035】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列を、配列番号50に示す。配列番号50は、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18を含む連続ゲノム配列を示す。配列番号50の少なくとも150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1500、2000、2500、3000、3500、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20、000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0036】
さらなる実施形態では、配列番号9〜45、46、47、48、49、および50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号9〜45、46、47、および48の少なくとも10、15、17、20、25、30、50、100、150、200、300、400、500、または1000個の連続ヌクレオチドを含むヌクレオチド配列およびペプチド配列も提供する。任意選択的にストリンジェントな条件下で配列番号9〜45、46、47、48、49、および50とハイブリッド形成する任意のヌクレオチド配列ならびにこれらに相同なヌクレオチドをさらに提供する。
【0037】
本発明の別の態様は、CMP−Neu5AcヒドロキシラーゼをコードするcDNAまたはその変異型を含む核酸構築物を提供する。これらのcDNA配列は、配列番号1〜8またはその任意のフラグメントに由来し得る。構築物は、1つまたは1つを超える内部リボゾーム侵入部位(IRES)を含み得る。構築物はまた、CMP−Neu5Acヒドロキシラーゼをコードする核酸配列に作動可能に連結されたプロモーターを含み得るか、構築物はプロモーターを含まなくてよい。別の実施形態では、宿主ゲノムへの挿入がランダムまたはターゲティングされている核酸配列を含む核酸構築物を提供する。核酸配列に加えて、発現ベクターは、選択マーカー(例えば、増強緑色蛍光タンパク質(eGFP)遺伝子配列、開始配列および/またはエンハンサー配列、ポリAテール配列、および/または原核生物および/または真核生物の宿主細胞で構築物を発現させる核酸配列など)を含み得る。
【0038】
別の実施形態では、体細胞中の相同組換えを行うことができる核酸ターゲティングベクター構築物も提供する。これらのターゲティングベクターを、相同組換えを介してCMP−Neu5Acヒドロキシラーゼ遺伝子をターゲティングするために哺乳動物細胞に形質転換することができる。1つの実施形態では、ターゲティングベクターは、CMP−Neu5Acヒドロキシラーゼのゲノム配列に相同な3’組換えアームおよび5’組換えアームを含み得る。相同なDNA配列には、CMP−Neu5Acヒドロキシラーゼ配列と少なくとも15bp、20bp、25bp、50bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbp、または50kbpが相同な配列が含まれ得る。別の実施形態では、相同DNA配列には、1つまたは複数のイントロンおよび/またはエクソン配列が含まれ得る。特定の実施形態では、DNA配列は、CMP−Neu5Acヒドロキシラーゼ遺伝子のイントロン5およびイントロン6に相同であり得る(例えば、図6〜8を参照のこと)。別の特定の実施形態では、DNA配列は、CMP−Neu5Acのイントロン5、エクソン6の55bp部分、およびイントロン6と相同であることができ、エクソン6の55bp部分に対して3’方向にてインフレームで増強緑色蛍光タンパク質配列を含む(例えば、図10〜11を参照のこと)。
【0039】
本発明の別の実施形態は、ブタCMP−Neu5AcヒドロキシラーゼのcDNAまたはゲノム配列(配列番号1、3、5、7、9〜45、47、または48など)とハイブリッド形成することができるオリゴヌクレオチドプライマーを提供する。好ましい実施形態では、プライマーは、ストリンジェントな条件下で配列番号1、3、5、7、9〜45、46、47、または48とハイブリッド形成する。別の実施形態は、配列番号1、3、5、7、9〜45、46、47、または48などのブタCMP−Neu5Acヒドロキシラーゼ核酸配列とハイブリッド形成することができるオリゴヌクレオチドプローブを提供する。ポリヌクレオチドプライマーまたはプローブは、本発明のポリヌクレオチドとハイブリッド形成する少なくとも14塩基、20塩基、好ましくは30塩基、または50塩基を有し得る。プローブまたはプライマーは、少なくとも14ヌクレオチド長であり得るが、好ましい実施形態では、少なくとも15、20、25、28、または30ヌクレオチド長である。
【0040】
本発明の別の態様では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子を欠く哺乳動物細胞を提供する。相同組換えの結果としてこれらの細胞を得ることができる。特に、CMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子の不活化により、機能的Hanganutziu−Deicher抗原の発現能力が低下した細胞を産生することができる。
【0041】
本発明の実施形態では、得られたCMP−Neu5Acヒドロキシラーゼ酵素がもはやHanganutziu−Deicher抗原を作製することができないように本明細書中に記載のプロセス、配列、および/または構築物にしたがってCMP−Neu5Acヒドロキシラーゼ遺伝子の対立遺伝子を不活化する。1つの実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子をRNAに転写することができるが、タンパク質に翻訳することができない。別の実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を、不活性な短縮形態に転写することができる。このような短縮RNAは、翻訳することができないか、非機能的タンパク質に翻訳することができる。別の実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を遺伝子が転写しないような方法で不活化することができる。さらなる実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子を転写し、その後非機能的タンパク質に翻訳することができる。
【0042】
本発明のさらなる態様では、CMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたゲノムターゲティング事象を介して不活化されたブタ動物を提供する。本発明の別の態様では、CMP−Neu5Acヒドロキシラーゼ遺伝子の両対立遺伝子が遺伝子ターゲティング事象を介して不活化されたブタ動物を提供する。相同組換えを介して遺伝子をターゲティングすることができる。他の実施形態では、遺伝子を破壊することができる(すなわち、遺伝コードの一部を変化させ、それにより遺伝子のそのセグメントの転写および/または翻訳に影響を与えることができる)。例えば、欠失(「ノックアウト」)技術または挿入(「ノックイン」)技術によって遺伝子を破壊することができる。既存配列の転写を調整する所望のタンパク質または調節配列のさらなる遺伝子を挿入することができる。
【0043】
本発明の別の態様では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の1つの対立遺伝子、任意選択的に両対立遺伝子を欠くブタ細胞を、クローン化トランスジェニック動物を産生するための除核卵母細胞への核導入のためのドナー細胞として使用することができる。あるいは、胚幹細胞中でブタCMP−Neu5Acヒドロキシラーゼノックアウトを作製し、その後子孫の産生に使用することができる。本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生した機能的CMP−Neu5Acヒドロキシラーゼ遺伝子の1つの対立遺伝子を欠く子孫を交配して、メンデル型遺伝子によって両対立遺伝子の機能性を欠く子孫をさらに産生することができる。異種移植ストラテジーで使用するために、細胞、組織、および/または臓器をこれらの動物から回収することができる。Hanganutziu−Deicher抗原は、Neu5Gcエピトープに起因する移植細胞、組織、または臓器の免疫拒絶を軽減させることができる。
【0044】
あるいは、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生したCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子を欠く動物は、腸管毒素原性感染および疾患(例えば、E.coli感染、ロタウイルス感染、および胃腸炎コロナウイルスなど)に対する感受性が低いか耐性を示し得る。このような動物を、例えば、商業的飼育で使用することができる。
【0045】
本発明の1つの態様では、本発明の任意の態様にしたがった方法によってブタを調製することができる。遺伝子操作ブタを、移植療法のための組織および/または臓器供給源として使用することができる。この様式で調製したブタ胚またはそれから発生した細胞株も細胞移植療法で使用することができる。したがって、本発明のさらなる態様では、CMP−Neu5Acヒドロキシラーゼを欠く胚または動物から調製されたブタCMP−Neu5Acヒドロキシラーゼを欠く遺伝子操作細胞のブタへの投与を含む治療方法を提供する。本発明のこの態様は、薬物におけるこのような細胞の使用(例えば、細胞移植療法)および移植のための細胞または組織移植片の調製におけるこのような胚由来の細胞の使用にまで及ぶ。細胞を、組織または臓器(例えば、心臓、肺、腎臓、膵臓、角膜、神経(例えば、脳、中枢神経系、脊髄)、皮膚)に組織化することができるか、細胞は、島細胞、血球(例えば、血球(すなわち、赤血球、白血球))または造血幹細胞もしくは他の幹細胞(例えば、骨髄)であり得る。
【0046】
本発明の別の態様では、CMP−Neu5Acヒドロキシラーゼ欠損ブタは他の異種抗原(例えば、ブタiGb3シンターゼ(例えば、を米国特許出願60/517,524号を参照のこと)および/またはブタForssmanシンターゼ(例えば、米国特許出願60/568,922号を参照のこと)など)をコードする遺伝子も欠く。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたα1,3ガラクトシルトランスフェラーゼ遺伝子およびCMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタ細胞を提供する。別の実施形態では、ブタ1,3ガラクトシルトランスフェラーゼ遺伝子ノックアウト細胞を、本明細書中に記載のプロセス、配列、および/または構築物によって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子をノックアウトするようにさらに修飾する。さらに、任意選択的に不都合な免疫応答に関連する1つまたは複数のさらなる遺伝子を欠く本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ欠損ブタを補体阻害タンパク質(例えば、CD59、DAF、および/またはMCPなど)を発現するように修飾することができ、MCPをCMP−Neu5Acヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子の発現を消失するようにさらに修飾することができる。これらの動物を、移植療法のための組織および/または臓器の供給源として使用することができる。これらの動物を、移植療法のための組織および/または臓器の供給源として使用することができる。この様式で調製したブタ胚またはそれから発生した細胞株も細胞移植療法で使用することができる。
【0047】
(発明の説明)
本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼ遺伝子の消失により、Neu5Gcエピトープに対するヒトの免疫応答を減少し、異種移植に対する免疫学的障壁を除去することができる。本発明は、全長cDNAおよびペプチドをコードする新規の核酸配列に関する。遺伝子のゲノム機構、イントロン配列、および調節領域に関する情報も提供する。1つの態様では、本発明は、配列番号1、3、5、または7の1つをコードする単離および実質的に精製されたcDNA分子またはそのフラグメントを提供する。本発明の別の態様では、CMP−Neu5Acヒドロラーゼ遺伝子の全長ゲノムを含むDNA配列を、配列番号9〜45、46、47、48、49、もしくは50またはそのフラグメントに提供する。別の態様では、配列番号1、3、5、7、9〜45、46、47、48、49、または50由来のブタCMP−Neu5AcヒドロキシラーゼcDNAまたはゲノム配列の増幅のためのプライマーを提供する。さらに、配列番号1、3、5、7、9〜45、46、47、48、49、もしくは50またはそのフラグメント由来のCMP−Neu5Acヒドロキシラーゼ核酸配列を同定するためのプローブを提供する。配列番号9〜45、46、47、48、49、もしくは50またはそのフラグメントによって示されるDNAを使用して、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタを構築物ことができる。したがって、本発明はまた、機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタ染色体および本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生された機能的CMP−Neu5Acヒドロキシラーゼタンパク質を欠くトランスジェニックブタを提供する。このようなブタを、ヒトへの異種移植のための組織供給源として使用することができる。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物によって産生されたCMP−Neu5Acヒドロキシラーゼ欠損ブタは、異種移植における不都合な免疫応答に関連する他の遺伝子(例えば、α1,3ガラクトシルトランスフェラーゼ遺伝子、iGb3シンターゼ遺伝子、またはFSMシンターゼ遺伝子など)も欠く。別の実施形態では、本明細書中に記載のプロセス、配列、および/または構築物にしたがって産生されたCMP−Neu5Acヒドロキシラーゼおよび/または異種移植における不都合な免疫応答に関連する他の遺伝子を欠くブタは、補体阻害因子(例えば、CD59、DAF、および/またはMCPなど)を発現する。
【0048】
(定義)
「標的DNA配列」は、相同組換えによって修飾されるDNA配列である。標的DNAは、動物細胞の任意のオルガネラ(核およびミトコンドリアが含まれる)中に存在することができ、インタクトな遺伝子、エクソンもしくはイントロン、調節配列、または遺伝子間の任意の領域であり得る。
【0049】
「ターゲティングDNA配列」は、所望の配列修飾を含むDNA配列である。ターゲティングDNA配列は、標的DNAと実質的に同質遺伝子的であり得る。
【0050】
「相同DNA配列または相同DNA」は、基準DNA配列と少なくとも80%、85%、90%、95%、98%、または99%同一のDNA配列である。相同配列は、ストリンジェントな条件下で標的配列とハイブリッド形成し、ストリンジェントな条件には、配列が少なくとも85%、好ましくは少なくとも95%または98%同一である場合にハイブリッド形成が起こる条件が含まれる。
【0051】
「同質遺伝子または実質的に同質遺伝子のDNA配列」は、基準DNA配列と同一であるかほぼ同一であるDNA配列である。用語「実質的に同質遺伝子的な」は、基準DNA配列と少なくとも約97〜99%同一、好ましくは基準DNA配列と少なくとも約99.5〜99.9%同一、一定の用途では基準DNA配列と100%同一のDNAをいう。
【0052】
「相同組換え」は、配列相同性に基づいたDNA組換えプロセスをいう。
【0053】
「遺伝子ターゲティング」は、一方が染色体上に存在し、他方が染色体上に存在しない2つのDNA配列間の相同組換えをいう。
【0054】
「非相同または無作為な組み込み」は、DNAが相同組換えに関与しないゲノムに組み込まれる任意のプロセスをいう。
【0055】
「選択マーカー遺伝子」は、その発現により遺伝子を含む細胞が同定される遺伝子である。選択マーカーは、この遺伝子を含まない細胞の成長を防止または遅延させる培地上で細胞を増殖させるものであり得る。例には、抗生物質耐性遺伝子および生物を選択された代謝産物に対して成長させる遺伝子が含まれる。あるいは、この遺伝子は、細胞への容易に同定される表現型の付与によって形質転換体の目視によるスクリーニングを容易にすることができる。このような同定可能な表現型は、例えば、発光もしくは着色化合物の産生または細胞周囲の培地中の検出可能な変化であり得る。
【0056】
用語「連続する」は、本明細書中でその標準的な意味(すなわち、途切れないまたは連続的な)で使用される。
【0057】
用語「ブタ」は、任意のブタ種(Large White、Landrace、Meishan、Minipigが含まれる)をいう。
【0058】
用語「卵母細胞」は、卵形成の最終産物であり、且つ前駆体形態がそれぞれ卵原細胞、一次卵母細胞および二次卵母細胞でもある成熟動物の卵を説明する。
【0059】
用語「フラグメント」は、ヌクレオチドまたはペプチド配列の一部または部分配列を意味する。
【0060】
用語「誘導体」および「アナログ」は、このようなヌクレオチドまたはペプチドと本質的に同一の生物機能または生物活性を保持するヌクレオチド配列またはペプチド配列を意味する。例えば、アナログには、活性成熟ポリペプチドを産生するためのプロタンパク質部分の切断によって活性化することができるプロタンパク質が含まれる。
【0061】
本明細書中で提供したDNA(デオキシリボ核酸)配列を、塩基アデニン(A)、チミン(T)、シトシン(C)、およびグアニン(G)によって示す。
【0062】
本明細書中で提供したアミノ酸配列を、以下の略語で示す。
【0063】
【表1】
「トランスフェクション」は、宿主細胞へのDNAの移入をいう。細胞は、自然にはDNAを取り込まない。したがって、種々の技術的「トリック」を使用して、遺伝子導入を容易にする。多数のトランスフェクション法(例えば、CaPO4およびエレクトロポレーション)が当業者に公知である(J.Sambrook,E.Fritsch,T.Maniatis,Molecular Cloning:A Laboratory Manual,Cold Spring Laboratory Press,1989)。宿主細胞のトランスフェクションは、首尾の良いトランスフェクションの指標である。
【0064】
(I.ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の完全なcDNA配列および変異型)
本発明の1つの態様は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の新規の全長核酸cDNA配列を提供する(図2、表1、配列番号1)。本発明の別の態様は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の推定アミノ酸ペプチド配列を提供する(表2、配列番号2)。全長cDNAのATG開始コドンはエクソン4の3’部分に存在し、終止コドンTAGはエクソン17の3’部分で見出される。配列番号1または2と少なくとも90、95、98、または99%相同な核酸配列およびアミノ酸配列を提供する。さらに、配列番号1または2の少なくとも10、15、17、20、または25個の連続核酸またはアミノ酸を含むヌクレオチド配列およびペプチド配列も提供する。配列番号1〜2のフラグメント、誘導体、およびアナログをさらに提供する。配列番号1〜2のフラグメントには、少なくとも約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個のヌクレオチドを含む任意の連続する核酸配列またはペプチド配列が含まれ得る。
【0065】
【表1A−1】
【0066】
【表1A−2】
【0067】
【表1A−3】
【0068】
【表1A−4】
【0069】
【表2−1】
【0070】
【表2−2】
(変異型)
本発明の別の態様は、CMP−Neu5Acヒドロキシラーゼ遺伝子転写物の3つの新規の変異型の新規の核酸cDNA配列を提供する(図2、表3、5、および7、配列番号3、5、および7)。配列番号3は、エクソン1、4、5、6、7、8、9、10、11、12、13、15a、16、17、および18を含む遺伝子の変異型(変異型1)のcDNAを示す。エクソン15aは、通常はイントロン15(エクソン16の約460bp上流)中で認められる潜在性のエクソンである。変異型1の開始コドンはエクソン4中に存在し、終止コドンはエクソン17中に存在する。配列番号5は、エクソン1、4、5、6、7、8、9、10、11、12、および12aを含む遺伝子の変異型(変異型2)のcDNAを示す。エクソン12aは、イントロン12の部分配列から保持された潜在性のエクソンである(配列番号21を参照のこと)。変異型2の開始コドンはエクソン4中に存在し、終止コドンはエクソン12a中に存在する。配列番号7は、エクソン1、4、5、6、7、8、9、10、11、および11aを含む遺伝子の変異型(変異型3)のcDNAを示す。エクソン11aは、イントロン11の部分配列から保持された潜在性のエクソンである(配列番号19を参照のこと)。変異型3の開始コドンはエクソン4中に存在し、終止コドンはエクソン11a中に存在する。本発明の別の態様は、CMP−Neu5Acヒドロキシラーゼ遺伝子転写物の3つの新規の変異型の新規の推定アミノ酸ペプチド配列を提供する。配列番号4、6、および8は、それぞれ変異型1、変異型2、および変異型3のアミノ酸配列を示す。配列番号3〜8と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号3〜8の少なくとも10、15、17、20、25、30、50、100、150、200、300、400、500、または1000個の連続するヌクレオチド配列またはアミノ酸配列を含むヌクレオチド配列およびペプチド配列も提供する。配列番号3〜8のフラグメント、誘導体、およびアナログをさらに提供する。配列番号3〜8のフラグメントには、少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、または10kpbを含む任意の連続核酸配列またはペプチド配列が含まれ得る。
【0071】
【表3−1】
【0072】
【表3−2】
【0073】
【表3−3】
【0074】
【表3−4】
【0075】
【表4】
【0076】
【表5−1】
【0077】
【表5−2】
【0078】
【表6】
【0079】
【表7−1】
【0080】
【表7−2】
【0081】
【表7−3】
【0082】
【表8】
本発明の他の態様では、CMP−Neu5AcヒドロキシラーゼをコードするcDNAまたはその変異型を含む核酸構築物を提供する。これらのcDNA配列は、配列番号1、3、5、または7であり得るか、配列番号2、4、6、もしくは8またはその任意のフラグメントに由来し得る。構築物は、1つまたは1つを超える内部リボゾーム侵入部位(IRES)を含み得る。構築物はまた、CMP−Neu5Acヒドロキシラーゼをコードする核酸配列に作動可能に連結されたプロモーターを含み得るか、構築物はプロモーターを含まなくてよい。別の実施形態では、宿主ゲノムへの挿入がランダムまたはターゲティングされている核酸配列を含む核酸構築物を提供する。核酸配列に加えて、発現ベクターは、選択マーカー(例えば、増強緑色蛍光タンパク質(eGFP)遺伝子配列、開始配列および/またはエンハンサー配列、ポリAテール配列、および/または原核生物および/または真核生物の宿主細胞で構築物を発現させる核酸配列など)を含み得る。適切なベクターおよび選択マーカーを以下に記載する。発現構築物は、転写開始、終結、および/またはリボゾーム結合部位のための部位をさらに含み得る。構築物は、任意の原核細胞または真核細胞(酵母細胞、E.coliなどの細菌細胞、CHO細胞などの哺乳動物細胞、および/または植物細胞が含まれるが、これらに限定されない)で発現することができる。
【0083】
このような構築物で使用するためのプロモーターには、λファージPLプロモーター、E.coli lacプロモーター、E.coli trpプロモーター、E.coli phoAプロモーター、E.coli tacプロモーター、SV40初期プロモーター、SV40後期プロモーター、レトロウイルスLTRプロモーター、PGKIプロモーター、GALIプロモーター、GALIO遺伝子プロモーター、CYCiプロモーター、PH05プロモーター、TRPIプロモーター、ADHIプロモーター、ADH2プロモーター、ホルグリムアルデヒド(forglymaldehyde)リン酸デヒドロゲナーゼプロモーター、ヘキソキナーゼプロモーター、ピルビン酸デカルボキシラーゼプロモーター、ホスホフルクロキナーゼプロモーター、トリオースリン酸イソメラーゼプロモーター、ホスホグルコースイソメラーゼプロモーター、グルコキナーゼα接合因子フェロモンプロモーター、PRBIプロモーター、GUT2プロモーター、GPDIプロモーター、メタロチオネインプロモーター、および/またはアデノウイルスおよびワクシニアウイルス由来などの哺乳動物ウイルスプロモーターが含まれるが、これらに限定されない。他のプロモーターが当業者に公知である。
【0084】
(II.CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム配列)
CMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA機構を示す核酸配列(図1、表9)も提供する。配列番号10〜28は、それぞれエクソン1、4〜11、11a、12、12a、13〜15、15a、および16〜18を示す。エクソン11a、12a、および15aは、CMP−Neu5Acヒドロキシラーゼの一定の変異転写物で保持される潜在的エクソンである。配列番号29〜45は、エクソン1とエクソン4との間のイントロン配列(以後それぞれイントロン1aおよびイントロン1b)、4〜15、15a、16、および17を示す。イントロン15aは、潜在的エクソン15aに続くイントロン15の3’下流部分である。配列番号9は、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の5’非翻訳領域を示す。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノムDNA配列を示す核酸配列(表10、配列番号46)も提供する。さらに、5’UTR、エクソン1、およびエクソン1とエクソン4との間に存在するイントロン配列の一部(イントロン1a)を含む5’連続ゲノム配列を示す連続ゲノム配列(配列番号47、表11)を提供する。エクソン1とエクソン4からエクソン18との間に存在するイントロン配列(イントロン1b)を含む連続ゲノム配列(配列番号48、表12)も提供する。配列番号9〜45、46、47、48、49、および50と少なくとも80%、85%、90%、95%、98%、または99%相同なヌクレオチド配列およびアミノ酸配列を提供する。さらに、配列番号9〜45、46、47、48、49、および50の少なくとも10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個の連続ヌクレオチド配列またはアミノ酸配列を含むヌクレオチド配列およびペプチド配列ならびにこれらと少なくとも80、85、90、95、98、または99%相同な任意のヌクレオチド配列も提供する。配列番号9〜45、46、47、48、49、および50のフラグメント、誘導体、およびアナログをさらに提供する。配列番号9〜45、46、47、48、49、および50のフラグメントには、任意の連続核酸配列もしくはペプチド配列または少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、もしくは10kpbが含まれ得る。
【0085】
さらに、ゲノム配列の推定転写因子結合部位の形態の調節領域を同定した(図4を参照のこと)。これらの結合部位は、ブタCMP−Neu5Acヒドロキシラーゼゲノムの5’UTRおよびエクソン1に存在し、転写因子(例えば、ETSF、MZF1、SF1、CMYB、MEF2、TATA、MEF2、NMP4、CAAT、AP1、BRN2、SATB1、ATF、GAT1、USF、WHN、NMP4、ZF5、NFKB、ZBP89、MOK2、ZF5、NFY、およびMYCMAXなど)の結合部位が含まれる。
【0086】
【表9−1】
【0087】
【表9−2】
【0088】
【表9−3】
【0089】
【表9−4】
【0090】
【表9−5】
【0091】
【表9−6】
【0092】
【表9−7】
【0093】
【表9−8】
【0094】
【表9−9】
【0095】
【表9−10】
【0096】
【表9−11】
【0097】
【表9−12】
【0098】
【表9−13】
【0099】
【表9−14】
【0100】
【表9−15】
【0101】
【表9−16】
【0102】
【表9−17】
【0103】
【表9−18】
【0104】
【表9−19】
【0105】
【表9−20】
【0106】
【表9−21】
【0107】
【表9−22】
【0108】
【表9−23】
【0109】
【表9−24】
【0110】
【表9−25】
【0111】
【表9−26】
【0112】
【表9−27】
【0113】
【表9−28】
【0114】
【表10−1】
【0115】
【表10−2】
【0116】
【表10−3】
【0117】
【表10−4】
【0118】
【表10−5】
【0119】
【表10−6】
【0120】
【表10−7】
【0121】
【表10−8】
【0122】
【表10−9】
【0123】
【表10−10】
【0124】
【表10−11】
【0125】
【表10−12】
【0126】
【表10−13】
【0127】
【表10−14】
【0128】
【表10−15】
【0129】
【表10−16】
【0130】
【表10−17】
【0131】
【表10−18】
【0132】
【表10−19】
【0133】
【表10−20】
【0134】
【表10−21】
【0135】
【表10−22】
【0136】
【表11】
【0137】
【表12−1】
【0138】
【表12−2】
【0139】
【表12−3】
【0140】
【表12−4】
【0141】
【表12−5】
【0142】
【表12−6】
【0143】
【表12−7】
【0144】
【表12−8】
【0145】
【表12−9】
【0146】
【表12−10】
【0147】
【表12−11】
【0148】
【表12−12】
【0149】
【表12−13】
【0150】
【表12−14】
【0151】
【表12−15】
【0152】
【表12−16】
【0153】
【表12−17】
【0154】
【表12−18】
【0155】
【表12−19】
【0156】
【表12−20】
【0157】
【表12−21】
【0158】
【表13−1】
【0159】
【表13−2】
【0160】
【表13−3】
【0161】
【表13−4】
【0162】
【表13−5】
【0163】
【表13−6】
【0164】
【表13−7】
【0165】
【表13−8】
【0166】
【表13−9】
【0167】
【表13−10】
配列番号49は、エクソン4に対して5’方向に、エクソン4、イントロン4、エクソン5、イントロン5、エクソン6、イントロン6、エクソン7、イントロン7、およびエクソン8を含む連続ゲノム配列を示す(表13)。さらに、配列番号49の少なくとも1750、1800、1900、2000、2100、2200、2300、2400、2500、2600、2700、2800、2900、3000、3100、3200、3300、3400、3500、3600、3700、3800、3900、4000、4100、4200、4300、4400、4500、4600、4700、4800、4900、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号49と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0168】
【表14−1】
【0169】
【表14−2】
【0170】
【表14−3】
【0171】
【表14−4】
【0172】
【表14−5】
【0173】
【表14−6】
【0174】
【表14−7】
【0175】
【表14−8】
【0176】
【表14−9】
【0177】
【表14−10】
【0178】
【表14−11】
【0179】
【表14−12】
配列番号50は、エクソン12、イントロン12、エクソン13、イントロン13、エクソン14、イントロン14、エクソン15、イントロン15、エクソン16、イントロン16、エクソン17、イントロン17、およびエクソン18を含む連続ゲノム配列を示す。さらに、配列番号50の少なくとも150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1500、2000、2500、3000、3500、4000、4500、5000、5500、6000、6500、7000、7500、8000、8500、9000、9500、10000、11000、12000、13000、14000、15000、16000、17000、18000、19000、または20、000個の連続ヌクレオチドを含むヌクレオチド配列ならびに配列番号50と少なくとも80、85、90、95、98、または99%相同なヌクレオチド配列を提供する。
【0180】
(VIII.オリゴヌクレオチドプローブおよびプライマー)
本発明は、さらに、上記配列(配列番号1、3、5、7、9〜45、46、47、48、49、および50)とハイブリッド形成するオリゴヌクレオチドプローブおよびプライマーを提供する。配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントと相同であり得るオリゴヌクレオチドを提供する。ストリンジェントな条件下で配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントとハイブリッド形成するオリゴヌクレオチドも提供する。ストリンジェントな条件は、配列が少なくとも約85%、約90%、約95%、または少なくとも約98%相同である場合のみハイブリッド形成する条件を記載する。あるいは、オリゴヌクレオチドは、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントとハイブリッド形成する少なくとも20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、45、46、47、48、49、50、75、または100塩基を有し得る。このようなオリゴヌクレオチドを、本明細書中に提供した配列を検出するためのプライマーおよびプローブとして使用することができる。プローブまたはプライマーは、少なくとも14ヌクレオチド長であり得るが、好ましい実施形態では、少なくとも15、20、25、28、30、または35ヌクレオチド長である。
【0181】
上記配列を考えると、当業者は、標準的なアルゴリズムを使用して、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメント中に含まれる配列と相同なオリゴヌクレオチドプローブおよびプライマーを構築することができる。相補的対合の規則は周知である:シトシン(「C」)は常にグアニン(「G」)と対合し、チミン(「T」)またはウラシル(「U」)は常にアデニン(「A」)と対合する。プライマーまたはプローブと十分にハイブリッド形成して対応する相補配列を認識することができる限り、プライマーまたはプローブは標的核酸配列と100%相補的である必要はないと認識される。一般に、一定の程度の対ミスマッチを許容することができる。
【0182】
プライマーのハイブリッド形成領域として使用されるオリゴヌクレオチド配列を、プローブのハイブリッド形成領域として使用することもできる。プローブとして使用するためのプライマー配列の適合性は、プライマーのハイブリッド形成特性に依存する。同様に、プローブとして使用されるオリゴヌクレオチドをプライマーとして使用することができる。
【0183】
これらの特定の実施形態では、例えば、5’または3’末端のいずれかへの標的配列に相補的であるか標的配列に相補的ではないヌクレオチドの付加によって特定のプライマーおよびプローブを調製することができることが当業者に明らかである。プライマー組成物を標的配列上の伸長開始点として作用する限り、およびプライマーおよびプローブが上記配列番号内に含まれる少なくとも14個の連続ヌクレオチドを含む限り、このような組成物は本発明の範囲内である。
【0184】
本明細書中のプローブおよびプライマーを、考慮すべき要因であるが、排他的でも決定的でもない以下の基準によって選択することができる。プローブおよびプライマーを、配列番号1、3、5、7、9〜45、46、47、48、49、および50ならびにそのフラグメントで同定されたCMP−Neu5Acヒドロキシラーゼ核酸配列領域から選択する。プローブおよびプライマーは、試験に悪影響を与えると予想される他の遺伝子配列との相同性を欠く。プローブまたはプライマーは、E.coli DNAポリメラーゼ(好ましくは、そのDNAポリメラーゼの一部はKlenowフラグメントと呼ばれる)などの増幅酵素による伸長を妨害することができる増幅核酸中での二次構造形成を欠く。増幅培地中での約15重量%まで、好ましくは5〜10重量%のジメチルスルホキシド(DMSO)および/または30〜40℃の漸増増幅温度の使用によってこれを行うことができる。
【0185】
好ましくは、プローブまたはプライマーは、式アデニン(A)+チミン(T)+シトシン(C)+グアニン(G)/シトシン(C)+グアニン(G)によって測定したところ、約50%のグアニンおよびシトシンヌクレオチドを含むべきである。好ましくは、プローブまたはプライマーは、ハイブリッドの安定性が減少し得るプライマーの3’末端に複数の連続したアデニン残基およびチミン残基を含まない。
【0186】
本発明のプローブおよびプライマーは、約10〜30ヌクレオチド長、好ましくは少なくとも10、11、12、13、14、15、20、25、または28ヌクレオチド長(詳細には、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、または30ヌクレオチドが含まれる)であり得る。本発明で使用されるヌクレオチドは、リボヌクレオチド、デオキシリボヌクレオチド、および修飾ヌクレオチド(そのハイブリッド形成特性が本質的に変化しない修飾基を含むイノシンまたはヌクレオチドなど)であり得る。プローブおよびプライマー配列を、本明細書を通して5’末端から3’末端の方向で一本鎖DNAオリゴヌクレオチドとして示す。このようなまたはその相補的形態またはそのRNA形態(TがUに置換されている)で任意のプローブを使用することができる。
【0187】
本発明のプローブおよびプライマーを、対応するヌクレオチド配列を含むインサートを含む組換えプラスミドのクローニング、任意選択的に適切なヌクレアーゼの使用時のクローン化プラスミドからの後者の切断およびその回収(例えば、分子量による分画)によって調製することができる。本発明のプローブおよびプライマーを、例えば、従来のホスホトリエステル法もしくはホスホジエステル法またはその自動化実施形態によって化学合成することも出きる。1つのこのような自動化実施形態では、出発材料としてジエチルホスホラミダイトを使用し、Beaucageら,Tetrahedron Letters 22:1859−1862(1981)に記載のように合成することができる。修飾固相上での1つのオリゴヌクレオチド合成方法は、米国特許第4,458,066号に記載されている。生物学的供給源(制限エンドヌクレアーゼ消化物など)から単離したプローブまたはプライマーを使用することも可能である。
【0188】
プライマーまたはプローブとして使用されるオリゴヌクレオチドは、ホスホロチオエート(Matsukura S.,Naibunpi Gakkai Zasshi.43(6):527−32(1967))、アルキルホスホロチオエート(Miller P.ら,Biochemistry 18(23):5134−43(1979)、ペプチド核酸(Nielsen P.ら,Science 254(5037):1497−500(1991);Nielsen P.ら,Nucleic−Acids−Res.21(2):197−200(1993))、モルホリノ核酸、固定核酸、偽環状オリゴ核酸塩基、2’−0,4’−C−エチレン架橋核酸などのヌクレオチドアナログも含み得るか、挿入剤(Asseline J.ら,Proc.Natl.Acad.Sci.USA 81(11):3297−301(1984))を含み得る。
【0189】
所望の特徴を有するプローブおよびプライマーのデザインのために、当業者に公知の以下の有用なガイドラインを適用することができる。ハイブリッド形成反応の範囲および特異性は多数の要因に影響を受け、1つまたは複数のこれらの要因の操作により、特定のプローブの正確な感度および特異性(その標的に完全に相補的であるかどうか)が決定される。本明細書中でさらに説明される種々のアッセイ条件の重要性および影響は、当業者に公知である。
【0190】
標的核酸ハイブリッドに対するプローブおよびプライマーの安定性は、アッセイ条件に適合するように選択すべきである。長いATリッチな配列の回避、GC塩基対でのハイブリッドの終結、および/または適切なTmでのプローブのデザインによってこれを行うことができる。プローブの開始点および終点を、その長さおよびGC率によってTmが最終アッセイが行われる温度よりも約2〜10℃高くなるように選択すべきである。G−C塩基対がさらなる水素結合によってA−T塩基対と比較して熱安定性が高いので、プローブの塩基組成は有意である。したがって、より高いG−C含量の相補核酸を含むハイブリッド形成は、より高い温度で安定である。プローブデザイン時にプローブが使用されるイオン強度およびインキュベーション温度などの条件も考慮すべきである。反応混合物のイオン強度が増加するにつれてハイブリッド形成が増加し、イオン強度の増加と共にハイブリッドの熱安定性が増加することが公知である。水素結合を破壊するホルムアミド、尿素、DIVISO、およびアルコールなどの化学試薬は、ハイブリッド形成のストリンジェンシーを増加させる。このような試薬による水素結合の不安定化によりTmが非常に減少し得る。一般に、所与の二重鎖の融解温度より5℃低い温度で約10〜50塩基長の合成オリゴヌクレオチドプローブの最適なハイブリッド形成が起こる。至適温度未満でのインキュベーションによりミスマッチ塩基配列がハイブリッド形成するので、特異性が減少し得る。高ストリンジェンシー条件下のみでハイブリッド形成するプローブを有することが望ましい。高ストリンジェンシー条件下のみで高相補核酸ハイブリッドが形成され、相補性が十分でないハイブリッドは形成されない。したがって、アッセイ条件のストリンジェンシーにより、ハイブリッドを形成する2つの核酸鎖間に必要な相補性が決定される。標的核酸と形成されたハイブリッドと非標的核酸と形成されたハイブリッドとの間の安定性の相違を最大にするなどのためにストリンジェンシーの程度を選択する。この場合、1塩基対の変化を検出する必要があり、非常に高いストリンジェンシー条件が必要である。
【0191】
標的核酸配列の長さ(したがって、プローブ配列の長さ)も重要であり得る。いくつかの場合、所望のハイブリッド形成特性を有するプローブおよびプライマーが得られる特定の領域由来の種々の位置および長さのいくつかの配列が存在し得る。他の場合、ある配列が1塩基しか異ならない別の配列よりも有意に良好であり得る。
【0192】
完全に相補的ではない核酸をハイブリッド形成することが可能であるが、主に完全に相補的な塩基の最も長いストレッチによりハイブリッドの安定性が決定される。異なる長さおよび塩基組成のオリゴヌクレオチドプローブおよびプライマーを使用することができるが、本発明の好ましいオリゴヌクレオチドプローブおよびプライマーは約14塩基長と30塩基長との間であり、標的核酸配列と完全に相補的な配列で十分なストレッチを有する。
【0193】
ハイブリッド形成に対して抑制的である強固な内部構造を形成することが公知の標的DNAまたはRNA中の領域はあまり好ましくない。同様に、広範な自己相補性を有するプローブは回避すべきである。上記で説明するように、ハイブリッド形成は、水素結合した二本鎖を形成する相補核酸の2つの一本鎖の会合である。2つの鎖のうちの1つの全体または一部がハイブリッドに関与する場合、新規のハイブリッドの形成への関与が低下し得ると予想される。十分な自己相補性が存在する場合、1つのプローブ型の分子内で形成された分子内および分子間ハイブリッドが存在し得る。このような構造を、慎重なプローブデザインによって回避することができる。目的の配列の大部分が一本鎖であるようなプローブのデザインにより、ハイブリッド形成の比率および範囲が非常に増加し得る。この相互作用型を検索するためにコンピュータプログラムを利用可能である。しかし、一定の例では、この相互作用型を回避することができない。
【0194】
特異的プライマーおよびオリゴヌクレオチドプローブに特異的な配列を、CMP−Neu5Acヒドロキシラーゼ核酸配列を増幅および検出することができるポリメラーゼ連鎖反応で使用することができる。
【0195】
(IV.CMP−Neu5Acヒドロキシラーゼ遺伝子の遺伝子ターゲティング)
遺伝子ターゲティングにより、動物細胞ゲノムの選択的操作が可能である。この技術を使用して、部位特異的様式および正確な様式で特定のDNA配列をターゲティングおよび修飾することができる。修飾のために異なるDNA配列型(調節領域、コード領域、および遺伝子間のDNA領域が含まれる)をターゲティングすることができる。調節領域の例には、プロモーター領域、エンハンサー領域、ターミネーター領域、およびイントロンが含まれる。これらの調節領域の修飾により、遺伝子発現のタイミングおよびレベルを変化させることができる。細胞内のタンパク質を変化、増強、または消失させるためにコード領域を修飾することができる。イントロンおよびエクソンならびに遺伝子間領域は、修飾に適切な標的である。
【0196】
DNA配列の修飾には、いくつかの型(挿入、欠失、置換、またはその任意の組み合わせが含まれる)が存在し得る。修飾の特定の例は、遺伝子産物の発現を破壊するヌクレオチド配列の部位特異的に相互作用による遺伝子の不活化(すなわち、「ノックアウト」)である。例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子の1つの破壊アプローチは、ターゲティングDNAと標的DNAとの間の相同組換えによって選択マーカーを標的遺伝子のコード領域に挿入することができるように選択マーカーをターゲティングDNAに挿入することである。例えば、図3、12、および13を参照のこと。この方法では、例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子配列が破壊され、コードされた酵素が機能しなくなる。
【0197】
(相同組換え)
相同組換えによって内因性遺伝子が部位特異的に修飾されるので、ゲノムを新規に変化するように操作することができる。最初の相同組換え工程は、ヘテロ二重鎖DNAを含む中間体組換え構造を形成するための相補配列を含む少なくとも1つのDNA鎖とのDNA二重鎖の対合を含むDNA鎖交換である(例えば、Radding,C.M.(1982)Ann.Rev.Genet.16:405;米国特許第4,888,274号を参照のこと)。ヘテロ二重鎖DNAは、いくつかの形態(1つの相補鎖がDNA二重鎖に侵入した三重鎖形態を含む3つのDNA鎖が含まれる)を取ることができ(Hsiehら,Genes and Development 4:1951(1990);Raoら(1991)PNAS 88:2984))、2つの相補DNA鎖がDNA二重鎖と対合する場合、古典的なHolliday組換え連結構造およびΧ構造(Holliday,R.,Genet.Res.5:282(1964))または二重Dループ(“Diagnostic Applications of Double−D Loop Formation”U.S.Ser.No.07/755,462,filed Sep.4,1991)を形成することができる。一旦形成されると、ヘテロ二重鎖構造を、侵入したDNA鎖の全部または一部がレシピエントDNA二重鎖にスプライシングされる鎖の破壊および交換、レシピエントDNA二重鎖の付加または置換によって解明することができる。あるいは、ヘテロ二重鎖構造によって遺伝子を変換することができ、テンプレートとして侵入鎖を使用したミスマッチ塩基の修復によって侵入鎖の配列がレシピエントDNA二重鎖に移行される(Genes,第3版(1987)Lewin,B.,John Wiley,New York,N.Y.;Lopezら,Nucleic Acids Res.15:5643(1987))。破壊および再連結機構または遺伝子変換機構のいずれかにより、相同対合連結部でのヘテロ二重鎖DNAの形成は、あるDNAから別のDNAへの遺伝子配列情報を伝達するように作用することができる。
【0198】
DNA分子間の遺伝子配列情報を伝達する相同組換え(遺伝子変換および古典的な鎖の破壊/再連結)の能力により、ターゲティングされた相同組換えが遺伝子工学および遺伝子操作における強力な方法となる。
【0199】
相同組換えでは、組み込まれるDNAが実質的に相同なDNA配列を含むゲノム中の部位と相互作用して組み込まれる。非相同性の(「無作為」または「不正の」)組み込みでは、組み込まれるDNAはゲノム中の相同配列で見出されないが、他の場所(多数の潜在的な位置)に組み込まれる。一般に、高等真核細胞を使用した研究によって、相同組換えの頻度は無作為な組み込みの頻度よりもはるかに低いことが明らかとなった。これらの頻度の比は、相同組換えを介した相互作用に依存する「遺伝子ターゲティング」(すなわち、ゲノム中での外因性「ターゲティングDNA」と対応する「標的DNA」との間の組換え)と直接関連する。
【0200】
多数の論文に哺乳動物細胞における相同組換えの使用が記載されている。これらの論文の例は、Kucherlapatiら,Proc.Natl.Acad.Sci.(USA)81:3153−3157,1984;Kucherlapatiら,MoL Cell.Bio.5:714−720,1985;Smithiesら,Nature 317:230−234,1985;Wakeら,Mol.Cell.Bio.8:2080−2089,1985;Ayaresら,Genetics 111:375−388,1985;Ayaresら,Mol.Cell.Bio.7:1656−1662,1986;Songら,Proc.Natl.Acad.Sci.USA 84:6820−6824,1987;ThomasらCell 44:419−428,1986;ThomasおよびCapecchi,Cell 51:503−512,1987;Nandiら,Proc.Natl.Acad Sci.USA 85:3845−3849,1988;およびMansourら,Nature 336:348−352,1988;EvansおよびKaufman,Nature 294:146−154,1981;Doetschmanら,Nature 330:576−578,1987;ThomaおよびCapecchi,Cell 51:503−512,4987;Thompsonら,Cell 56:316−321,1989である。
【0201】
本発明は、線維芽細胞などの細胞中でブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するために相同組換えを使用する。DNAは、機能的酵素の発現およびHanganutziu−Deicher抗原分子の産生を防止するために、天然の遺伝子の少なくとも一方、任意選択的には両方のコピーが変化した特定の対立遺伝子に遺伝子の少なくとも一部を含み得る。変化は、挿入、欠失、置換、またはその組み合せであり得る。変化がたった1つの遺伝子のコピーが不活化する導入である場合、標的遺伝子の1つの非変異コピーを有する細胞が増幅され、変化が第1の変化と同一でも異なっていてもよく(通常異なっている)、且つ欠失または置換が含まれる第2のターゲティング工程に供することができ、最初に移入された変化の少なくとも一部が重複し得る。この第2のターゲティング工程では、同一の相同性アームを有するが、異なる哺乳動物選択マーカーを含むターゲティングベクターを使用することができる。得られた形質転換体を、機能的標的抗原の非存在についてスクリーニングし、野生型標的遺伝子の非存在を確認するために細胞のDNAをさらにスクリーニングすることができる。あるいは、変異にヘテロ接合性を示す宿主の交配によって表現型に関するホモ接合性を達成することができる。
【0202】
遺伝子操作することができるブタ細胞を、種々の異なる臓器および組織(脳、心臓、肺、腺、脳、目、胃、脾臓、すい臓、腎臓、肝臓、腸、子宮、膀胱、皮膚、毛髪、爪、耳、鼻、口、唇、歯茎、歯、舌、唾液腺、扁桃腺、咽頭、食道、大腸、小腸、直腸、肛門、幽門、甲状腺、胸腺、腎上体、骨、軟骨、腱、靭帯、骨格筋、平滑筋、血管、血液、脊髄、気管、尿管、尿道、視床下部、下垂体、副腎、卵巣、輸卵管、子宮、膣、乳腺、精巣、精嚢、陰茎、リンパ、リンパ節、およびリンパ管などであるが、これらに限定されない)から得ることができる。本発明の1つの実施形態では、ブタ細胞は、上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋肉細胞、リン酸細胞(□hosphate cell)、卵丘細胞、上皮細胞、内皮細胞、ランゲルハンス島細胞、血球、血液前駆細胞、骨細胞、骨前駆細胞、神経幹細胞、始原幹細胞、肝細胞、ケラチノサイト、臍静脈内皮細胞、大動脈内皮細胞、微小血管内皮細胞、線維芽細胞、肝臓星細胞、大動脈平滑筋細胞、心筋細胞、ニューロン、クップフェル細胞、平滑筋細胞、シュワン細胞、上皮細胞、赤血球、血小板、好中球、リンパ球、単球、好酸球、好塩基球、脂肪細胞、軟骨細胞、膵島細胞、甲状腺細胞、副甲状腺細胞、耳下腺細胞、腫瘍細胞、グリア細胞、星状細胞、赤血球、白血球、マクロファージ、上皮細胞、体細胞、下垂体細胞、副腎細胞、有毛細胞、気泡細胞、腎細胞、網膜細胞、桿状体細胞、錐状体細胞、心臓細胞、ペースメーカー細胞、脾臓細胞、抗原提示細胞、記憶細胞、T細胞、B細胞、形質細胞、筋細胞、卵巣細胞、子宮細胞、前立腺細胞、腟上皮細胞、精子細胞、精巣細胞、生殖細胞、卵細胞、ライディッヒ細胞、尿細管周囲細胞、セルトリ細胞、ルテイン細胞、頸部細胞、子宮内膜細胞、乳房細胞、濾胞細胞、粘液細胞、繊毛細胞、非角化上皮細胞、角化上皮細胞、肺細胞、杯細胞、円柱上皮細胞、扁平上皮細胞、骨細胞、骨芽細胞、および破骨細胞からなる群から選択され得るが、これらに限定されない。
【0203】
1つの別の実施形態では、胚幹細胞を使用することができる。胚幹細胞株を使用することができるか、胚幹細胞株をブタ動物などの宿主から新たに得ることができる。細胞を、適切な線維芽細胞支持細胞層で成長させるか、白血病阻害因子(LIF)の存在下で成長させることができる。好ましい実施形態では、ブタ細胞は線維芽細胞であり得るが、1つの特定の実施形態では、ブタ細胞は胎児線維芽細胞であり得る。線維芽細胞は、発育中の胎児および成体動物から大量に得ることができので、好ましい体細胞型である。
【0204】
これらの細胞を急速な倍加時間で容易に増殖することができ、遺伝子ターゲティング手順で使用するためにクローンで増殖することができる。
【0205】
(ターゲティングベクター)
ターゲティングした対立遺伝子でホモ接合性の細胞を、標的対立遺伝子に相同であり、且つマーカー遺伝子を含むDNAの細胞への移入および組み込まれた構築物を含む細胞の選択によって産生することができる。標的対立遺伝子で標的ベクター中の相同DNAを染色体DNAで組換える(例えば、図3、12、および13を参照のこと)。マーカー遺伝子の両端を、相同DNA配列、3’組換えアーム、および5’組換えアームと隣接させることができる(例えば、図11を参照のこと)。ターゲティンベクターの構築方法は当該分野で記載されており、例えば、Daiら,Nature Biotechnology 20:251−255,2002;WO 00/51424を参照のこと。
【0206】
標的対立遺伝子での相同組換えのために種々の構築物を調製することができる。通常、構築物には、50bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbp、または50kbpの標的対立遺伝子と相同な配列が含まれ得る。配列には、任意のブタCMP−Neu5Acヒドロキシラーゼ遺伝子の任意の連続配列が含まれ得る(配列番号9〜45、46、47、48、49、もしくは50またはその任意の組み合わせもしくはフラグメントの少なくとも約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、110、120、130、140、150、160、170、180、190、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、700、750、800、850、900、1000、5000、または10,000個の連続ヌクレオチドが含まれる)。配列番号9〜45、46、47、48、49、および50のフラグメントには、少なくとも約10bp、15bp、17bp、20bp、50bp、100bp、500bp、1kbp、5kbp、または10kpbを含む任意の連続核酸配列またはペプチド配列が含まれ得る。
【0207】
標的DNA配列の相同性の範囲の決定に種々の検討材料(例えば、標的対立遺伝子のサイズ、配列の利用可能性、標的対立遺伝子における二重重複事象の相対的効率、および標的配列の他の配列との類似性など)が関与し得る。
【0208】
ターゲティングDNAには、実質的に同質遺伝子のDNAが修飾すべきゲノム中の対応する標的配列での所望の配列修飾物と隣接した配列が含まれ得る。実質的に同質遺伝子の配列は、対応する標的配列(所望の配列修飾を除く)を少なくとも約95%、97〜98%、99.0〜99.5%、99.6〜99.9%、または100%同一であり得る。ターゲティングDNAおよび標的DNAは、好ましくは、100%同一である少なくとも75、150、または500塩基対のDNAストレッチを共有することができる。したがって、ターゲティングDNAは、ターゲティングされる細胞株と密接に関連する細胞に由来し得るか、ターゲティングDNAはターゲティングされる細胞と同一の細胞株または動物の細胞に由来し得る。
【0209】
内因性標的CMP−Neu5Acヒドロキシラーゼを修飾するようにDNA構築物をデザインすることができる。構築物のターゲティングのための相同配列は、得られる遺伝子産物の機能を破壊するようにデザインされた1つまたは複数の欠失、挿入、置換、またはその組み合わせを有し得る。1つの実施形態では、変化は、読み取り枠中で標的遺伝子の上流配列と融合した選択マーカー遺伝子の挿入であり得る。
【0210】
適切な選択マーカーには、tk遺伝子(チミジンキナーゼ)などの一定の培地基質で成長する能力を付与する遺伝子またはHAT培地(ヒポキサンチン、アミノプテリン、およびチミジン)で成長する能力を付与するhprt遺伝子(ヒポキサンチンホスホリボシルトランスフェラーゼ);MAX培地(ミコフェノール酸)で成長させる細菌gpt遺伝子(グアニン/キサンチンホスホリボシルトランスフェラーゼ)が含まれるが、これらに限定されない。例えば、Song,K−Y.らProc.Net’l Acad.Sci.U.S.A.84:6820−6824(1987);Sambrook,J.ら,Molecular Cloning−A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.(1989),Chapter 16を参照のこと。選択マーカーの他の例には、抗生物質などの化合物に体する耐性を付与する遺伝子、選択された基質で成長する能力を付与する遺伝子、緑色蛍光タンパク質、増強緑色蛍光タンパク質(eGFP)などの発光物質などの検出可能なシグナルを発するタンパク質をコードする遺伝子が含まれる。広範な種々のこのようなマーカーが公知且つ利用可能であり、例えば、ネオマイシン耐性遺伝子(neo)(Southern,P.,およびP.Berg,J.Mol.Appl.Genet.1:327−341(1982));およびハイグロマイシン耐性遺伝子(hyg)(Nucleic Acids Research 11:6895−6911(1983),およびTe Riele,H.ら,Nature 348:649−651(1990))などの抗生物質耐性遺伝子が含まれる。他の選択マーカー遺伝子には、アセトヒドロキシ酸シンターゼ(AHAS)、アルカリホスファターゼ(AP)、βガラクトシダーゼ(LacZ)、βグルクロニダーゼ(GUS)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、緑色蛍光タンパク質(GFP)、赤色蛍光タンパク質(RFP)、黄色蛍光タンパク質(YFP)、シアン蛍光タンパク質(CFP)、西洋ワサビペルオキシダーゼ(HRP)、ルシフェラーゼ(Luc)、ノパリンシンターゼ(NOS)、オクトピンシンターゼ(OCS)、およびその誘導体が含まれる。アンピシリン、ブレオマイシン、クロラムフェニコール、ゲンタマイシン、ハイグロマイシン、カナマイシン、リンコマイシン、メトトレキセート、フォスフィノスリシン、ピューロマイシン、およびテトラサイクリンに耐性を付与する複数の選択マーカーを利用可能である。
【0211】
抗生物質耐性遺伝子およびネガティブ選択因子の組み込み方法は、当業者に熟知されている(例えば、WO 99/15650;米国特許第6,080,576号;米国特許第6.136,566号;Niwaら,J.Biochem.113:343−349(1993);およびYoshidaら,Transgenic Research,4:277−287(1995)を参照のこと)。
【0212】
本発明で有用なさらなる選択マーカー遺伝子は、例えば、以下の米国特許に記載されている:6,319,669;6,316,181;6,303,373;6,291,177;6,284,519;6,284,496;6,280,934;6,274,354;6,270,958;6,268,201;6,265,548;6,261,760;6,255,558;6,255,071;6,251,677;6,251,602;6,251,582;6,251,384;6,248,558;6,248,550;6,248,543;6,232,107;6,228,639;6,225,082;6,221,612;6,218,185;6,214,567;6,214,563;6,210,922;6,210,910;6,203,986;6,197,928;6,180,343;6,172,188;6,153,409;6,150,176;6,146,826;6,140,132;6,136,539;6,136,538;6,133,429;6,130,313;6,124,128;6,110,711;6,096,865;6,096,717;6,093,808;6,090,919;6,083,690;6,077,707;6,066,476;6,060,247;6,054,321;6,037,133;6,027,881;6,025,192;6,020,192;6,013,447;6,001,557;5,994,077;5,994,071;5,993,778;5,989,808;5,985,577;5,968,773;5,968,738;5,958,713;5,952,236;5,948,889;5,948,681;5,942,387;5,932,435;5,922,576;5,919,445;および5,914,233。
【0213】
選択マーカーの組み合わせも使用することができる。例えば、CMP−Neu5Acヒドロキシラーゼをターゲティングするために、neo遺伝子(上記で考察のようにそれ自体のプロモーターを含むか含まない)を、CMP−Neu5Acヒドロキシラーゼ遺伝子に相同なDNA配列にクローン化することができる。マーカーの組み合わせを使用するために、ターゲティングDNAの外側に存在するようにHSV−tk遺伝子をクローン化することができる(所望ならば、別の選択マーカーを反対側に位置づけることができる)。ターゲティングされるべき細胞へのDNA構築物の移入後、適切な抗生物質に対して細胞を選択することができる。この特定の例では、G418およびガンシクロビルに耐性を示す細胞は、neo遺伝子がCMP−Neu5Acヒドロキシラーゼ遺伝子に組換えられているが、二重重複領域の外側に存在することによりtk遺伝子が失われている相同組換えによって惹起されている可能性が最も高い。
【0214】
欠失は、少なくとも約50bp、より通常には少なくとも約100bp、一般に、せいぜい約20kbpであり得るが、欠失は、通常、少なくとも1つまたは複数のエクソンの一部を含むコード領域の一部、1つまたは複数のイントロンの一部を含むことができ、隣接非コード領域(特に、5’非コード領域(転写調節領域))の一部を含んでいても含んでいなくても良い。したがって、相同領域は、コード領域を超えて5’非コード領域または3’非コード領域に及び得る。挿入は、一般に10kbpを超えず、通常5kbpを超えず、一般に少なくとも50bp、より通常には少なくとも200bpであり得る。
【0215】
相同領域には、フレームシフトが起こる場合または鍵となるアミノ酸が変化した場合に標的遺伝子をさらに不活化することができるか、機能障害対立遺伝子などを修正することができる変異が含まれ得る。通常、変異は、約5%を超えない相同隣接配列のわずかな変化であり得る。
【0216】
当該分野で公知の方法にしたがって構築物を調製することができ、種々のフラグメントを1つにし、適切なベクターに移入し、クローン化し、分析し、その後所望の構築物を達成するまでさらに操作することができる(例えば、図5〜11を参照のこと)。制限分析、切り出し、プローブの同定を行うために配列に種々の修飾を行うことができる。所望ならばサイレント変異を移入することができる。種々の段階で、制限分析、配列決定、ポリメラーゼ連鎖反応を使用した増幅、プライマー修復、in vitro変異誘発などを使用することができる。
【0217】
原核生物複製系(例えば、E.coliによって認識可能な供給源)を含む細菌ベクターを使用して構築物を調製することができ、各段階で、構築物をクローン化および分析することができる。標的細胞への移入前に除去することができるマーカー(挿入のために使用されるマーカーと同一または異なる)を使用することができる。一旦構築物を含むベクターが完成すると、細菌配列の欠失、線状化、相同配列への短い欠失の移入などによってさらに操作することができる。最終的操作後、構築物を細胞に移入することができる。
【0218】
DNA構築物を宿主細胞に侵入させるために使用することができる技術には、リン酸カルシウム/DNA共沈、核へのDNAの微量注入、エレクトロポレーション、インタクトな細胞との細菌プロトプラストの融合、トランスフェクション、または当業者に公知の任意の他の技術が含まれる。DNAは、一本鎖もしくは二本鎖、線状もしくは環状、弛緩もしくはスーパーコイル状のDNAであり得る。哺乳動物細胞の種々のトランスフェクション技術については、例えば、Keownら,Methods in Enzymology Vol.185,pp.527−537(1990)を参照のこと。
【0219】
本発明は、さらに、上記で広範に記載されている1つまたは複数の配列を含む組換え構築物を含む(例えば、表9〜12)。構築物は、本発明の配列が正方向または逆方向で挿入されたプラスミドベクターまたはウイルスベクターなどのベクターを含む。構築物は、配列に作動可能に連結された調節配列(例えば、プロモーターが含まれる)も含み得る。多数の適切なベクターおよびプロモーターは当業者に公知であり、市販されている。例として以下のベクターを提供する:pBs、pQE−9(Qiagen)、phagescript、PsiX174、pBluescript SK、pBsKS、pNH8a、pNH16a、pNHl8a、pNH46a(Stratagene);pTrc99A、pKK223−3、pKK233−3、pDR540、pRIT5(Pharmacia);Eukaryotic:pWLneo、pSv2cat、pOG44、pXTI、pSG(Stratagene)pSVK3、pBPv、pMSG、pSVL(Pharmiacia)。また、宿主中で複製可能且つ生存可能な限り、任意の他のプラスミドおよびベクターを使用することができる。当該分野で公知且つ市販されているベクター(およびその変異型または誘導体)を、本発明の方法で使用するために本発明にしたがって1つまたは複数の組換え部位を含むように操作することができる。このようなベクターを、例えば、Vector Laboratories Inc.、Invitrogen、Promega、Novagen、NEB、Clontech、Boehringer Mannheim、Pharmacia、EpiCenter、OriGenes Technologies Inc.、Stratagene、PerkinElmer、Pharmingen、およびResearch Geneticsから入手することができる。目的の他のベクターには、pFastBac、pFastBacHT、pFastBacDUAL、pSFV、およびpTet−Splice(Invitrogen)、pEUK−C1、pPUR、pMAM、pMAMneo、pBI101、pBI121、pDR2、pCMVEBNA、およびpYACneo(Clontech)、pSVK3、pSVL、pMSG、pCHI 10、およびpKK232−8(Pharmacia、Inc.)、p3’SS、pXTI、pSG5、pPbac、pMbac、pMClneo、およびpOG44(Stratagene、Inc.)、およびpYES2、pAC360、pBlueBacHis A、B、およびC、pVL1392、pBlueBacIII、pCDM8、pcDNAI、pZeoSV、pcDNA3、pREP4、pCEP4、およびpEBVHis(Invitrogen、Corp.)、ならびにその変異型または誘導体などの真核生物発現ベクターが含まれる。
【0220】
本発明での使用に適切な他のベクターには、pUC18、pUC19、pBlueScript、pSPORT、コスミド、ファージミド、YAC’s(酵母人工染色体)、BAC’s(細菌人工染色体)、PI(Escherichia coliファージ)、pQE70、pQE60、pQE9(quagan)、pBSベクター、PhageScriptベクター、BlueScriptベクター、pNH8A、pNH16A、pNH18A、pNH46A(Stratagene)、pcDNA3(Invitrogen)、pGEX、pTrsfus、pTrc99A、pET−5、pET−9、pKK223−3、pKK233−3、pDR540、pRIT5(Pharmacia)、pSPORT1、pSPORT2、pCMVSPORT2.0、およびpSV−SPORT1(Invitrogen)、ならびにその変異型または誘導体が含まれる。レンチウイルスベクター(例えば、WO 03/059923;TiscorniaらPNAS100:1844−1848(2003)を参照のこと)などのウイルスベクターを使用することもできる。
【0221】
さらなる目的のベクターには、pTrxFus、pThioHis、pLEX、pTrcHis、pTrcHis2、pRSET、pBlueBacHis2、pcDNA3.1/His、pcDNA3.1(−)/Myc−His、pSecTag、pEBVHis、pPIC9K、pPIC3.5K、pA0815、pPICZ、pPICZA、pPICZB、pPICZC、pGAPZA、pGAPZB、pGAPZC、pBlueBac4.5、pBlueBacHis2、pMelBac、pSinRep5、pSinHis、pIND、pIND(SP1)、pVgRXR、pcDNA2.1、pYES2、pZErO1.1、pZErO−2.1、pCR−Blunt、pSE280、pSE380、pSE420、pVL1392、pVL1393、pCDM8、pcDNA1.1、pcDNA1.1/Amp、pcDNA3.1、pcDNA3.1/Zeo、pSe、SV2、pRc/CMV2、pRc/RSV、pREP4、pREP7、pREP8、pREP9、pREP 10、pCEP4、pEBVHis、pCR3.1、pCR2.1、pCR3.1−Uni、およびpCRBac(Invitrogen);λExCell、λgtll、pTrc99A、pKK223−3、pGEX−1λT、pGEX−2T、pGEX−2TK、pGEX−4T−1、pGEX−4T−2、pGEX−4T−3、pGEX−3X、pGEX−5X−1、pGEX−5X−2、pGEX−5X−3、pEZZ18、pRIT2T、pMC1871、pSVK3、pSVL、pMSG、pCH110、pKK232−8、pSL1180、pNEO、およびpUC4K(Pharmacia);pSCREEN−1b(+)、pT7Blue(R)、pT7Blue−2、pCITE−4abc(+)、pOCUS−2、pTAg、pET−32LIC、pET−30LIC、pBAC−2cp LIC、pBACgus−2cp LIC、pT7Blue−2 LIC、pT7Blue−2、λSCREEN−1、λBlueSTAR、pET−3abcd、pET−7abc、pET9abcd、pETllabcd、pET12abc、pET−14b、pET−15b、pET−16b、pET−17b−pET−17xb、pET−19b、pET−20b(+)、pET−21 abcd(+)、pET−22b(+)、pET−23abcd(+)、pET−24abcd(+)、pET−25b(+)、pET−26b(+)、pET−27b(+)、pET−28abc(+)、pET−29abc(+)、pET−30abc(+)、pET−31b(+)、pET− 32abc(+)、pET−33b(+)、pBAC−1、pBACgus−1、pBAC4x−1、pBACgus4x−1、pBAC−3cp、pBACgus−2cp、pBACsurf−1、plg、Signalplg、pYX、Selecta Vecta−Neo、Selecta Vecta−Hyg、およびSelecta Vecta−Gpt(Novagen);pLexA、pB42AD、pGBT9、pAS2−1、pGAD424、pACT2、pGAD GL、pGAD GH、pGAD10、pGilda、pEZM3、pEGFP、pEGFP−1、pEGFP−N、pEGFP−C、pEBFP、pGFPuv、pGFP、p6xHis−GFP、pSEAP2−Basic、pSEAP2−Contral、pSEAP2−プロモーター、pSEAP2−エンハンサー、pβgal−Basic、pβgal−Control、pβgal−プロモーター、pβgal−エンハンサー、pCMV、pTet−Off、pTet−On、pTK−Hyg、pRetro−Off、pRetro−On、pIRES1neo、pIRESi1hyg、pLXSN、pLNCX、pLAPSN、pMAMneo、pMAMneo−CAT、pMAMneo−LUC、pPUR、pSV2neo、pYEX4T−1/2/3、pYEX−S1、pBacPAK−His、pBacPAK8/9、pAcUW31、BacPAK6、pTriplEx、λgt10、λgt11、pWE15、およびλTriplEx(Clontech);Lambda ZAP II、pBK−CMV、pBK−RSV、pBluescript II KS +/−、pBlucscript II SK +/−、pAD−GAL4、pBD−GAL4 Cam、pSurfscript、Lambda FIX II、Lambda DASH、Lambda EMBL3、Lambda EMBL4、SuperCos、pCR−Scrigt Amp、pCR−Script Cam、pCR−Script Direct pBS +/−、pBC KS +/−、pBC 5K +/−、Phagescript、pCAL−n−EK、pCAL−n、pCAL−c、pCAL−kc、pET−3abcd、pET−11abcd、pSPUTK、pESP−I、pCMVLacI、pOPRSVI/MCS、pOPI3 CAT、pXT1、pSG5、pPbac、pMbac、pMClneo、pMC1neo Poly A、pOG44、pOG45、pFRTβGAL、pNEOβGAL、pRS403、pRS404、pRS405、pRS406、pRS413、pRS414、pRS415、およびpRS416(Stratagene)が含まれる。
【0222】
さらなるベクターには、例えば、pPC86、pDBLeu、pDBTrp、pPC97、p2.5、pGADI−3、pGAD10、pACt、pACT2、pGADGL、pOADGH、pAS2−1、pGAD424、pGBT8、pGBT9、pGAD−GAL4、pLexA.、pBD−GAL4、pHISi、pHISi−1、placZi、pB42AD、pDG202、pJK202、pJG4−5、pNLexA、pYESTrp、およびその変異型または誘導体が含まれる。
【0223】
また、宿主中で複製可能且つ生存可能である限り、当該分野で公知の任意の他のプラスミドおよびベクターを使用することができる。
【0224】
(相同組換えされた細胞の選択)
次いで、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の発現をノックアウトするために相同組換えを行った細胞を、適切に組み込まれた細胞を同定するために適切に選択された培地中で成長させることができる。次いで、所望の表現型を示す細胞を、制限分析、エレクトロポレーション、サザン分析、ポリメラーゼ連鎖反応、または当該分野で公知の別の技術によってさらに分析することができる。標的遺伝子部位で適切な挿入を示すフラグメントの同定によって、標的遺伝子を不活化するか修飾するために相同組換えを行った細胞を同定することができる。
【0225】
CMP−Neu5Acヒドロキシラーゼ遺伝子に挿入した選択マーカー遺伝子の存在により、宿主ゲノムへの標的構築物の組み込みが確立される。次いで、所望の表現型を示す細胞を、制限分析、エレクトロポレーション、サザン分析、ポリメラーゼ連鎖反応、モノクローナル抗体アッセイ、蛍光標示式細胞分取器(FACS)、または相同または非相同組換えが起こるかどうかを確立するためにDNAを分析するための当該分野で公知の任意の他の技術もしくは方法によってさらに分析することができる。挿入のためのプローブの使用およびその後の構築物の隣接領域を超えて及ぶCMP−Neu5Acヒドロキシラーゼ遺伝子の存在のためのインサートに隣接する5’領域および3’領域の配列決定または欠失が組み込まれている場合の欠失の存在の同定によってこれを決定することができる。構築物内の配列に相補的であり、且つ構築物の外側および標的対立遺伝子の配列と相補的であるプライマーを使用することもできる。この方法では、相同組換えが起こった場合に相補鎖中に存在する両プライマーを有するDNA二重鎖のみを得ることができる。プライマー配列または予想されるサイズの配列の存在の証明によって、相同組換えの発生が支持される。
【0226】
相同組換え事象のスクリーニングのために使用されるポリメラーゼ連鎖反応は、例えば、KimおよびSmithies,Nucleic Acids Res.16:8887−8903,1988;およびJoynerら,Nature 338:153−156,1989に記載されている。
【0227】
相同組換え事象の別のスクリーニング方法には、例えば、Malykhら,European Journal of Cell Biology 80,48−58(2001),Malykhら,Glycoconjugate J.15,885−893(1998)に記載のブタCMP−Neu5Acヒドロキシラーゼおよび/またはNeu5Gcに特異的なモノクローナル抗体またはポリクローナル抗体の使用が含まれる。
【0228】
相同組換え事象による機能的CMP−Neu5Acヒドロキシラーゼ発現を欠くブタ細胞のさらなる特徴づけには、サザン分析、ノーザンブロット分析、特異的レクチン結合アッセイ、および/または配列分析、例えば、Y.Malykhら,Biochem J.370:601−607(2003);Y.MalykhらEuropean Journal of Cell Biology 80:48−58(2001);Y.MalykhらGlycoconjugate J.15:885−893(1998)に記載の抗Neu5Gcまたは抗CMP−Neu5Acヒドロキシラーゼ抗体アッセイの使用が含まれるが、これらに限定されない。一般に、A.SharmaらTransplantation 75(4):430−436(2003)を参照のこと。
【0229】
第1ラウンドのターゲティングから得た細胞株は、ターゲティングされた対立遺伝子とヘテロ接合性である可能性がある。両対立遺伝子が修飾されたホモ接合性を多数の方法で達成することができる。1つのアプローチは、1つのコピーが修飾された多数の細胞を成長させ、その後これらの細胞を異なる選択マーカーを使用して残存するブタCMP−Neu5Acヒドロキシラーゼ対立遺伝子の別ラウンドのターゲティングに供することである。あるいは、伝統的なメンデルの遺伝的特徴にしたがった修飾対立遺伝子に対してヘテロ接合性の動物の交配によってホモ接合体を得ることができる。いくつかの状況では、2つの異なる修飾対立遺伝子を有することが望ましい。連続ラウンドの遺伝子ターゲティングまたはヘテロ接合体(それぞれ1つの所望の修飾対立遺伝子を保有する)の交配によってこれを行うことができる。
【0230】
(VIII.異種移植の免疫学的障壁を克服するためのさらなる遺伝子の遺伝子操作)
本発明の1つの態様では、非機能的CMP−Neu5Acヒドロキシラーゼ遺伝子に対してホモ接合性の細胞を、さらなる遺伝子操作に供することができる。例えば、宿主に異なる遺伝子の能力を置換、交換、または提供するために、内因性CMP−Neu5Acヒドロキシラーゼ対立遺伝子が非機能的になっているホモ接合性宿主にさらなる遺伝子の能力を移入することができる。同系遺伝子接合体化(homogenotization)後にマーカー遺伝子を除去することができる。場合によって拡大された配列を有し、元の構築物のマーカー遺伝子部分が欠失された実質的に相同なDNAを含む構築物の移入により、標的対立遺伝子との相同組換えを得ることができる。組み込みのためのマーカー遺伝子の組み合わせの使用により、ポジティブ選択および他のネガティブ選択を行い、除去工程でマーカー遺伝子が保持された細胞を選択することができる。
【0231】
1つの実施形態では、CMP−Neu5Acヒドロキシラーゼ遺伝子およびα(1,3)GT遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的CMP−Neu5Acヒドロキシラーゼを欠く動物を産生し、その後この動物由来の細胞を使用してα(1,3)GT遺伝子をノックアウトすることができる。ホモ接合性α(1,3)GTネガティブブタが最近報告されている(Phelpsら,Science 2003;WO 04/028243)。あるいは、これらのα(1,3)GTノックアウト動物由来の細胞を使用して、CMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0232】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠き、且つヒト補体阻害タンパク質を産生するブタ細胞も提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、この動物由来の細胞をヒト補体阻害タンパク質(CD59(Philbrick,W.M.ら(1990)Eur.J.Immunol.20:87−92で報告されたcDNA)、ヒト崩壊促進因子(human decay accelerating factor)(DAF)(Medofら(1987)Proc.Natl.Acad.Sci.USA 84:2007で報告されたcDNA)、およびヒト膜補因子タンパク質(MCP)(Lublin,D.ら(1988)J.Exp.Med.168:181−194で報告されたcDNA)などであるが、これらに限定されない)を発現するようにさらに修飾することができる。
【0233】
別の実施形態では、ヒト補体阻害タンパク質を産生するトランスジェニックブタ由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。ヒト補体阻害タンパク質を産生するトランスジェニックブタは当該分野で公知である(例えば、米国特許第6,166,288号を参照のこと)。
【0234】
さらなる実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子およびブタForssmanシンターゼ(FSM)遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、gal−α−galエピトープの産生に関与し、且つ異種移植拒絶で役割を果たすブタFSMシンターゼ遺伝子をノックアウトするためにさらに修飾することができる。ブタFSMシンターゼ遺伝子は最近同定された(米国特許出願60/568,922号を参照のこと)。あるいは、これらのFSMシンターゼ遺伝子ノックアウト動物由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0235】
なおさらなる実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子およびブタイソグロボシド(isogloboside)3シンターゼ遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、ブタiGb3シンターゼ遺伝子をノックアウトするために使用することができる。ブタiGb3シンターゼ遺伝子は最近報告されている(米国特許出願第60/517,524号)。あるいは、これらのブタiGb3シンターゼ遺伝子ノックアウト動物由来の細胞を使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化するようにさらに修飾することができる。
【0236】
別の実施形態では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子、α(1,3)GT遺伝子、FSMシンターゼ遺伝子、およびブタiGb3シンターゼ遺伝子を欠くブタ細胞を提供する。本発明にしたがって機能的ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を欠く動物を産生し、その後この動物由来の細胞を、α(1,3)GT遺伝子、FSMシンターゼ遺伝子、およびブタiGb3シンターゼ遺伝子をノックアウトするために使用することができる。ホモ接合性α(1,3)GTネガティブブタは最近報告されている(Phelpsらsupra,Science 2003;WO 04/028243)。あるいは、これらのα(1,3)GTノックアウト動物由来の細胞を使用して、ブタiGb3シンターゼ遺伝子、ブタFSMシンターゼ遺伝子、およびCMP−Neu5Acヒドロキシラーゼ遺伝子を不活化し、さらに、ヒト補チア阻害タンパク質(CD59、ヒト崩壊促進因子(DAF)、およびヒト膜補因子タンパク質(MCP)などが含まれるが、これらに限定されない)を発現するようにさらに修飾することができる。
【0237】
(VIII.遺伝子操作動物の産生)
本発明は、ブタ全能性幹細胞の遺伝子操作によるCMP−Neu5Acヒドロキシラーゼ発現を欠くトランスジェニックブタの産生方法を提供する。1つの実施形態では、(a)動物中の1つまたは複数の標的CMP−Neu5Acヒドロキシラーゼ核酸ゲノム配列の同定、(b)CMP−Neu5Acヒドロキシラーゼ核酸ゲノム配列をターゲティングする1つまたは複数の相同組換えベクターの調製、(c)動物の複数の全能性細胞のゲノムへの1つまたは複数のターゲティングベクターの挿入およびそれによる複数のトランスジェニック全能性細胞の産生、(d)動物の四倍体胚盤胞を得ること、(e)四倍体胚盤胞への複数の全能性細胞の挿入およびそれによるトランスジェニック胚の産生、(f)レシピエント雌動物への胚の導入、および(g)雌動物中で胚を成長させて出産させることによって動物を産生することができる。トランスジェニックブタを作製するための本明細書中に記載のトランスジェニック動物の産生方法は、一般に、米国特許第6,492,575号に記載されている。
【0238】
別の実施形態では、全能性細胞は、胚幹(ES)細胞であり得る。胚盤胞からのES細胞の単離、ES細胞株の確立、およびその後のその培養を、例えば、Doetchmannら,J.Embryo.Exp.Morph.87:27−45(1985);Liら,Cell 69:915−926(1992);Robertson,E.J.“Tetracarcinomas and Embryonic Stem Cells:A Practical Approach,”E.J.Robertson編,IRL Press,Oxford,England(1987);WurstおよびJoyner,”Gene Targeting:A Practical Approach,”A.L.Joyner編,IRL Press,Oxford,England(1993);Hogenら,“Manipulating the Mouse Embryo:A Laboratory Manual,”Hogan,Beddington,CostantiniおよびLacy編,Cold Spring Harbor Laboratory Press,New York(1994);およびWangら,Nature 336:741−744(1992)に記載の従来の方法によって行う。例えば、CMP−Neu5Acヒドロキシラーゼ遺伝子を変化させるためのターゲティングベクターでの胚幹細胞の形質転換後、細胞を適切な培地(例えば、ウシ胎児血清増強DMEMなど)中の支持細胞層にプレートすることができる。構築物を含む細胞を、選択培地の使用によって検出することができ、十分なコロニー成長期間後、コロニーを選別し、相同組換えの発生について分析することができる。構築物の配列内および配列外であるが標的対立遺伝子に存在するプライマーを使用したポリメラーゼ連鎖反応を使用することができる。次いで、相同組換えを示すコロニーを、胚操作および胚盤胞注射のために使用することができる。胚盤胞を、過剰排卵雌から得ることができる。次いで、胚幹細胞をトリプシン処理し、修飾細胞を胚盤胞を含む液滴に添加することができる。少なくとも1つの修飾胚幹細胞を、胚盤胞の胞胚腔に注射することができる。注射後、少なくとも1つの胚盤胞を、偽妊娠雌の各子宮角に戻すことができる。次いで、雌を出産させ、得られた同腹子を構築物を含む変異細胞についてスクリーニングした。形質転換ES細胞と異なる比率の胚盤胞を選択する。胚盤胞およびES細胞の異なる表現型の提供により、キメラ子孫を容易に検出することができ、次いで、修飾CMP−Neu5Acヒドロキシラーゼ遺伝子の存在を探索するために遺伝子型同定を行うことができる。
【0239】
本発明のさらなる実施形態では、全能性細胞は、胚生殖(EG)細胞であり得る。胚生殖細胞は、in vitroで培養およびトランスフェクトすることができ、その後にキメラの体細胞系列および生殖細胞系列に寄与する、ES細胞と機能的に等価な未分化の細胞である(Stewartら,Dev.Biol.161:626−628(1994))。EG細胞は、成長因子(白血病抑制因子、スチール因子(steel factor)、塩基性線維芽細胞成長因子)と組み合わせた始原生殖細胞(配偶子の先祖)の培養によって誘導される(Matsuiら,Cell 70:841−847(1992);Resnickら,Nature 359:550−551(1992))。Donovanら,“Transgenic Animals,Generation and Use,”L.M.Houdebine編,Harwood Academic Publishers(1997)などに記載の当業者に公知の方法を使用して、EG細胞を培養することができる。
【0240】
例えば、Jamesら,Genet.Res.Camb.60:185−194(1992);NagyおよびRossant,“Gene Targeting:A Practical Approach,”A.L.Joyner編,IRL Press,Oxford,England(1993);またはKubiakおよびTarkowski,Exp.Cell Res.157:561−566(1985)に記載の天然の接合子の産生および発生または2つの細胞胚の電気融合およびその後の培養によって本発明で使用するための四倍体胚盤胞を得ることができる。
【0241】
例えば、Wangら,EMBO J.10:2437−2450(1991)に記載の当該分野で公知の任意の方法によって、胚盤胞へのES細胞またはEG細胞の移入を行うことができる。
【0242】
「複数」の全能性細胞は、1つを超える任意の数の細胞を含み得る。例えば、本発明で使用される全能性細胞数は、約2〜約30細胞、約5〜約20細胞、または約5〜約10細胞であり得る。1つの実施形態では、1つの細胞懸濁液から採取した約5〜10個のES細胞を、顕微操作装置中の保持ピペットによって固定された胚盤胞に注射する。次いで、当該分野で公知の方法(例えば、Robertson,E.J.“Teratocarcinomas and Embryonic Stem Cells:A Practical Approach”IRL Press,Oxford,England(1987)を参照のこと)によって、胚を少なくとも3時間(場合により一晩)インキュベート後、雌レシピエント動物に移入する。次いで、雌動物中で胚を成長させて出産させることができる。
【0243】
(クローン化トランスジェニック子孫を産生するための体細胞核導入)
本発明は、体細胞核導入による機能的CMP−Neu5Acヒドロキシラーゼ遺伝子を欠くブタのクローニング方法を提供する。一般に、広範な種々の哺乳動物クローニング方法が現在急速に開発および報告されており、所望の結果を達成する任意の方法を本発明で使用することができる。このような方法の非限定的な例を以下に記載する。例えば、ドナー核の供給源として使用すべき所望の分化ブタ細胞を得る工程と、ブタから卵母細胞を得る工程と、卵母細胞を除核する工程と、除核卵母細胞に所望の分化細胞または細胞核を導入する工程と(例えば、NT単位を形成するための融合または注射による)、得られたNT単位を活性化する工程と、NT単位が胎児に成長するように宿主ブタに培養したNT単位を導入する工程とを含む核導入プロセスによってブタを産生することができる。
【0244】
核導入技術または核移植技術は当該分野で公知である(Campbellら,Theriogenology,43:181(1995);Collasら,Mol.Report Dev.,38:264−267(1994);Keeferら,Biol.Reprod.,50:935−939(1994);Simsら,Proc.Natl.Acad.Sci.,USA,90:6143−6147(1993);WO 94/26884;WO 94/24274,WO 90/03432,米国特許第4,944,384号および同第5,057,420号)。1つの非限定的な例では、除核卵母細胞への挿入前にドナー染色体またはドナー細胞が再プログラミングされた哺乳動物のクローニング方法を記載したCollasらの米国特許出願番号2003/0046722号などに記載の方法などの方法を提供する。本発明は、染色体、核、または細胞の卵母細胞の挿入または融合方法も記載する。
【0245】
CMP−Neu5Acヒドロキシラーゼ遺伝子を変化させるように修飾したドナー細胞核を、レシピエントブタ卵母細胞に導入する。この方法の使用は、特定のドナー細胞型に制限されない。ドナー細胞は、Wilmutら,Nature 385 810(1997);Campbellら,Nature 380 64−66(1996);またはCibelliら,Science 280 1256−1258(1998)などに記載のものであり得る。正常な核型の全細胞(核導入で首尾よく使用することができる胚細胞、胎児および成体の体細胞が含まれる)を、原理上は使用することができる。胎児線維芽細胞は、特に有用なドナー細胞クラスである。一般に適切な核導入方法は、Campbellら,Theriogenology 43 181(1995),Collasら,Mol.Reprod.Dev.38 264−267(1994),Keeferら,Biol.Reprod.50 935−939(1994),Simsら,Proc.Nat’l.Acad.Sci.USA 90 6143−6147(1993),WO−A−9426884,WO−A−9424274,WO−A−9807841,WO−A−9003432,米国特許第4,994,384号および同第5,057,420号に記載されている。分化または少なくとも部分的に分化したドナー細胞も使用することができる。ドナー細胞は培養中および静止期でもあり得るが、必ずしもそうではない。休止期の核ドナー細胞は、静止期に入るように誘導されるかin vivoで休止状態で存在し得る細胞である。先行技術の方法は、クローニング手順で胚細胞型も使用している(Campbellら(Nature,380:64−68,1996)およびSticeら(Biol.Reprod.,20 54:100−110,1996)。
【0246】
体細胞核ドナー細胞を、種々の異なる臓器および組織(皮膚、間葉、肺、膵臓、心臓、腸、胃、膀胱、血管、腎臓、尿道、生殖器、および胚、胎児、または成体動物の全部または一部の脱凝集調製物)から得ることができる。本発明の適切な実施形態では、核ドナー細胞は、上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋細胞、グラヌロース細胞、卵丘細胞、上皮細胞、または内皮細胞からなる群から選択される。別の実施形態では、核細胞は、胚幹細胞である。好ましい実施形態では、ドナー細胞として線維芽細胞を使用することができる。
【0247】
本発明の別の実施形態では、本発明の核ドナー細胞は、動物の生殖細胞である。胚、胎児、または成体期の動物種の任意の生殖細胞を、核ドナー細胞として使用することができる。適切な実施形態では、核ドナー細胞は胚生殖細胞である。
【0248】
受容細胞との連携を確実にするために、細胞周期の任意の時期(GO、GI、G2、S、M)で核ドナー細胞を停止することができる。当該分野で公知の任意の方法を使用して、細胞周期を操作することができる。細胞周期を制御する方法には、培養細胞の接触阻害によって誘導されたGO期静止、血清または他の必須栄養素の除去によって誘導されたGO期静止、老化によって誘導されたGO期静止、特定の成長因子の添加によって誘導されたGO期静止、熱ショック、高圧、もしくは化学物質、ホルモン、成長因子、もしくは他の物質での他の処理などの物理的もしくは化学的手段によって誘導されたGOもしくはGI期静止;任意の複製点手順で妨害する化学物質での処理によるS期の制御;蛍光標示式細胞分取、有糸分裂の除去、微小管破壊剤、有糸分裂の進行を破壊する任意の化学物質での処理を使用した選択によるM期の制御が含まれるが、これらに限定されない(Freshney,R.I,.“Culture of Animal Cells:A Manual of Basic Technique,”Alan R.Liss,Inc,New York(1983)も参照のこと)。
【0249】
卵母細胞の単離方法は当該分野で周知である。本質的に、これは、ブタの卵巣または生殖器官からの卵母細胞の単離を含み得る。ブタ卵母細胞の容易に利用可能な供給源は、食肉処理場の材料である。遺伝子工学、核導入、およびクローニングなどの技術の組み合わせのために、卵母細胞は遺伝学的に成熟していなければならず、その後にこれらの細胞を核導入のためのレシピエント細胞として使用することができ、これらの細胞を精子細胞による受精によって胚に発生することができる。このプロセスは、一般に、哺乳動物の卵巣(例えば、食肉処理場で得たウシ卵巣)からの未熟な(第I減数分裂前期(prophase I))卵母細胞の回収、卵母細胞が第II減数分裂中期に達するまで(この場合、ウシ卵母細胞は一般に吸引後18〜24時間で起こる)の受精または除核前の成熟培地での卵母細胞の成熟を必要とする。この期間は、「成熟期」として公知である。
【0250】
第II減数分裂中期の卵母細胞は、レシピエント卵母細胞であり得るが、この段階で卵母細胞は移入された核をそのままで受精精子で処理するために「活性化する」ことができるか、十分に活性化することができると考えられる。in vivoで成熟した第II減数分裂中期の卵母細胞は、核導入技術で首尾よく使用されている。本質的に第II減数分裂中期の卵母細胞を、発情期またはヒト絨毛性ゴナドトロピン(hCG)もしくは類似のホルモンの注射から35〜48時間後または39〜41時間後に非過剰排卵または過剰排卵ブタのいずれかから外科的に回収することができる。
【0251】
約10〜40時間、好ましくは約16〜18時間の範囲の定期成熟後、卵母細胞を除核することができる。除核前に卵母細胞を除去し、卵丘細胞の除去前に適切な培地(1mg/mlのヒアルロニダーゼを含むHECMなど)に入れることができる。次いで、裸の卵母細胞を極体についてスクリーニングし、次いで、極体の存在によって決定した選択された第II減数分裂中期の卵母細胞を核導入に使用する。除核を以下に示す。
【0252】
米国特許第4,994,384号などに記載の公知の方法によって除核を行うことができる。例えば、第II減数分裂中期の卵母細胞を、即時除核のために任意選択的に7.5mg/mlのサイトカラシンBを含むHECMに入れるか、適切な培地(例えば、CR1aa+10%発情期ウシ精子などの胚培養培地)に入れ、その後、好ましくはせいぜい24時間後、より好ましくは16〜18時間後に除核することができる。
【0253】
極体および隣接細胞質を除去するために、マイクロピペットを使用した顕微鏡手術で除核することができる。次いで、首尾よく除核された卵母細胞を同定するために、卵母細胞をスクリーニングすることができる。卵母細胞の1つのスクリーニング方法は、卵母細胞を1mg/mlの33342Hoechst色素を含むHECMで染色し、その後に10秒未満の紫外線照射下で卵母細胞を視覚化することである。次いで、首尾よく除核された卵母細胞を、適切な培養培地(CR1aa+10%精子)に入れることができる。
【0254】
次いで、除核卵母細胞と同種の1つの哺乳動物細胞を、NT単位を産生するために使用される除核卵母細胞の卵黄周囲腔に導入することができる。当該分野で公知の方法にしたがって、哺乳動物の細胞および除核卵母細胞を使用してNT単位を産生することができる。例えば、エレクトロポレーションによって細胞を融合することができる。原形質膜の一過性破壊に十分な電気パルスの印加によって電気融合を行う。膜が急速に再形成されるので、原形質膜のこの破壊は非常に短時間である。したがって、2つの隣接する膜の破壊が誘導され、再形成時に脂質二重層が混合する場合、2つの細胞間に小さなチャネルを開けることができる。このような小さな開口部の熱力学的不安定性のために、2つの細胞が1つになるまで拡大する。例えば、Pratherらの米国特許第4,997,384号を参照のこと。種々の電気融合培地(例えば、スクロース、マンニトール、ソルビトール、およびリン酸緩衝化溶液)を使用することができる。融合誘導因子としてSendaiウイルスを使用して融合を行うこともできる(Graham,Wister Inot.Symp.Monogr.,9,19,1969)。また、エレクトロポレーション融合を使用するよりもむしろ核を卵母細胞に直接注射することができる。例えば、CollasおよびBarnes,Mol.Reprod.Dev.,38:264−267(1994)を参照のこと。融合後、得られた融合NT単位を、活性化まで適切な培地(例えば、CR1aa培地)に入れる。典型的には、その後短時間で(例えば、24時間後または約4〜9時間後)活性化を行うことができる。
【0255】
所望の結果が達成される任意の方法によってNT単位を活性化することができる。このような方法には、例えば、本質的にNT単位への冷温または実質的に冷温ショックの印加による生物学的温度未満でのNT単位の培養が含まれる。胚が通常曝露される生理学的温度条件と比較して低温である室温でのNT単位の培養によってこれを最も都合良く行うことができる。あるいは、公知の活性化剤の適用によって活性化することができる。例えば、受精時の精子による卵母細胞の貫通は、融合前の卵母細胞を活性化して、核導入後により多数の生存可能な認識および複数の遺伝的に同一のブタが得られることを示した。また、電気的および化学的ショックなどの処理を使用して、融合後にNT胚を活性化することができる。例えば、Susko−Parrishらの米国特許第5,496,720号を参照のこと。さらに、卵母細胞中の2価の陽イオンレベルの増加および卵母細胞中の細胞タンパク質のリン酸化の減少によって同時または連続的に活性化を行うことができる。一般に、卵母細胞の細胞質への2価の陽イオン(例えば、イオノフォアの形態のマグネシウム、ストロンチウム、バリウム、またはカルシウム)の移入によってこれを行うことができる。2価の陽イオンレベルの他の増加方法には、電気ショック、エタノールでの処理、およびケージ化キレート剤での処理が含まれる。公知の方法(例えば、キナーゼインヒビター(例えば、6−ジメチル−アミノプリン、スタウロスポリン、2−アミノプリン、およびスフィンゴシンなどのセリン−トレオニンキナーゼインヒビター)の添加)によってリン酸化を減少させることができる。あるいは、卵母細胞へのリン酸塩(例えば、リン酸塩2Aおよびリン酸塩2B)の移入によって細胞タンパク質のリン酸化を阻害することができる。
【0256】
次いで、細胞コロニーの作製まで適切なin vitro培養培地で活性化NT単位を培養することができる。胚の培養および成熟に適切な培養培地は、当該分野で周知である。胚の培養および維持に使用することができる公知の培地の例には、Ham’s F−10+10%ウシ胎児血清(FCS)、組織培養培地−199(TCM−199)+10%ウシ胎児血清、タイロード−アルブミン−乳酸−ピルビン酸(TALP)、Dulbeccoリン酸緩衝化生理食塩水(PBS)、Eagle培地、およびWhitten培地が含まれる。
【0257】
その後、培養NT単位を洗浄し、好ましくは適切なコンフルエントの支持細胞層を含むウェルプレート中に含まれる適切な培地に入れることができる。適切な支持細胞層には、例として、線維芽細胞および上皮細胞が含まれる。NT単位がレシピエント雌への導入に適切なサイズに到達するまで、または細胞コロニーを産生するために使用することができる細胞を得るために、NT単位を支持細胞層上で培養する。好ましくは、少なくとも約2〜400個の細胞、より好ましくは約4〜128個の細胞、最も好ましくは少なくとも約50個の細胞までこれらのNT単位を培養することができる。
【0258】
次いで、活性化NT単位を、雌ブタの卵管に導入することができる(胚移植)。1つの実施形態では、雌ブタは、発情期同調レシピエント未経産ブタ(gilt)であり得る。交配未経産ブタ(large white/Duroc/Landrace)(280〜480lbs)を使用することができる。未経産ブタを、飼料と混合した18〜20mgのReguMate(Altrenogest,Hoechst,Warren,NJ)の経口投与によってレシピエント動物と同調させることができる。Regu−Mateを、14日間連続して与えることができる。次いで、最後のRegu−Mate処置から約150時間後に1000単位のヒト絨毛性ゴナドトロピン(hCG,Intervet America,Millsboro,DE)をi.m.投与することができる。hCG注射から約22〜26時間後に胚移植を行うことができる。1つの実施形態では、妊娠出産させて生きた子孫を誕生させることができる。別の実施形態では、早期に出産を終了させて、胚細胞を採取することができる。
【0259】
本発明における胚移植方法およびレシピエント動物の管理は、胚移植産業で使用される標準的な手順である。同期導入は、本発明の成功で重要である(すなわち、NT胚の時期をレシピエント雌の発情周期と同調させる)。例えば、Siedel,G.E.,Jr.“Critical review of embryo transfer procedures with cattle”in Fertilization and Embryonic Development in Vitro(1981)L.Mastroianni,Jr.およびJ.D.Biggers編,Plenum Press,New York,N.Y.,page 323を参照のこと。
【0260】
(VIII.ブタ動物、臓器、組織、細胞、および細胞株)
本発明は、CMP−Neu5Acヒドロキシラーゼ遺伝子の両対立遺伝子が不活化された生きたブタを提供する。本発明はまた、異種移植に有用なこのようなブタ由来の臓器、組織、および細胞を提供する。
【0261】
1つの実施形態では、機能的CMP−Neu5Acヒドロキシラーゼの任意の発現を欠くブタから得たブタ臓器、組織、および/または精製もしくは実質的に純粋な細胞もしくは細胞株を提供する。
【0262】
1つの実施形態では、本発明は、異種移植に有用な臓器を提供する。任意のブタ臓器(脳、心臓、肺、腺、脳、目、胃、脾臓、すい臓、腎臓、肝臓、腸、子宮、膀胱、皮膚、毛髪、爪、耳、鼻、口、唇、歯茎、歯、舌、唾液腺、扁桃腺、咽頭、食道、大腸、小腸、直腸、肛門、幽門、甲状腺、胸腺、腎上体、骨、軟骨、腱、靭帯、骨格筋、平滑筋、血管、血液、脊髄、気管、尿管、尿道、視床下部、下垂体、副腎、卵巣、輸卵管、子宮、膣、乳腺、精巣、精嚢、陰茎、リンパ、リンパ節、およびリンパ管が含まれるが、これらに限定されない)を使用することができる。
【0263】
別の実施形態では、本発明は、異種移植に有用な組織を提供する。任意のブタ組織(上皮組織、結合組織、血液組織、骨組織、軟骨組織、筋肉組織、神経組織、腺様組織、脂肪組織、疎性結合組織、骨組織、褐色脂肪組織、海綿状様骨組織、筋肉組織、軟骨細胞(cartaginous)、海綿組織、軟骨様組織、クロム親和組織、肉様組織、弾性組織、上皮組織、脂肪組織、軟骨様組織、Gaingee組織、膠様組織、肉芽組織、腸関連リンパ組織、ハーラー脈管組織、硬質(hard)造血組織、未分化組織、間質細胞、被覆組織、島組織、リンパ組織、リンパ様組織、間葉組織、中腎性組織、粘液様結合細胞、多房性脂肪細胞、骨髄組織、軟組織鼻点、造腎細胞、結節組織、骨細胞、造骨組織、類骨組織、根尖周囲組織、細網組織、細毛組織、外科用ゴム敷布、骨格筋組織、平滑筋組織、および皮下組織)を使用することができる。
【0264】
さらなる実施形態では、本発明は、機能的α1,3GTの発現を欠くブタ動物由来の細胞および細胞株を提供する。1つの実施形態では、これらの細胞または細胞株を異種移植のために使用することができる。任意のブタ組織または臓器由来の細胞(上皮細胞、線維芽細胞、神経細胞、ケラチノサイト、造血細胞、メラノサイト、軟骨細胞、リンパ球(BおよびT)、マクロファージ、単球、単核細胞、心筋細胞、他の筋細胞、リン酸細胞(□hosphate cell)、卵丘細胞、上皮細胞;内皮細胞、ランゲルハンス島細胞、膵臓インスリンを分泌する細胞、膵臓α−2細胞、膵臓β細胞、膵臓α−1細胞、血球、血液前駆細胞、骨細胞、骨前駆細胞、ニューロン幹細胞、始原幹細胞、肝細胞、ケラチノサイト、臍帯静脈内皮細胞、大動脈内皮細胞、微小血管系内皮細胞、線維芽細胞、肝臓星細胞、大動脈平滑筋細胞、心筋細胞、ニューロン、クップフェル細胞、平滑筋細胞、シュワン細胞、上皮細胞、赤血球、血小板、好中球、リンパ球、単球、好酸球、好塩基球、脂肪細胞、軟骨細胞、膵島細胞、甲状腺細胞、副甲状腺細胞、耳下腺細胞、腫瘍細胞、グリア細胞、星状細胞、赤血球、白血球、マクロファージ、上皮細胞、体細胞、下垂体細胞、副腎細胞、有毛細胞、気泡細胞、腎細胞、網膜細胞、桿状体細胞、錐状体細胞、心臓細胞、ペースメーカー細胞、脾臓細胞、抗原提示細胞、記憶細胞、T細胞、B細胞、形質細胞、筋細胞、卵巣細胞、子宮細胞、前立腺細胞、腟上皮細胞、精子細胞、精巣細胞、生殖細胞、卵細胞、ライディッヒ細胞、尿細管周囲細胞、セルトリ細胞、ルテイン細胞、頸部細胞、子宮内膜細胞、乳房細胞、濾胞細胞、粘液細胞、繊毛細胞、非角化上皮細胞、角化上皮細胞、肺細胞、杯細胞、円柱上皮細胞、ドーパミン作動性細胞、扁平上皮細胞、骨細胞、骨芽細胞、破骨細胞、胚幹細胞、線維芽細胞、および胎児線維芽細胞が含まれるが、これらに限定されない)を使用することができる。特定の実施形態では、機能的α−1,3−GT発現を欠くブタ由来の膵臓細胞(ランゲルハンス島細胞、インスリン分泌細胞、48のα−2細胞、β細胞、α−1細胞が含まれる)を提供する。
【0265】
非生存誘導体には、これらの組織誘導体を移植で使用する前に架橋または他の化学的処理によってさらに処理することができる、酵素または化学的処理によって生存細胞を剥ぎ取られた組織が含まれる。好ましい実施形態では、誘導体には、種々の組織(皮膚、泌尿器組織、膀胱組織、または臓器粘膜下組織が含まれる)由来の細胞外基質が含まれる。また、医療機器として心臓弁および他の非生存組織を含むために生存組織を剥ぎ取られた腱、関節、および骨を提供する。
【0266】
(治療上の使用)
広範な種々の方法にしたがって宿主に細胞を投与することができる。好ましい投与様式は、非経口、腹腔内、静脈内、皮内、硬膜外、脊髄内、胸骨内、関節内、滑液内、くも膜下腔内、動脈内、心臓内、筋肉内、鼻腔内、皮下、眼窩内、嚢内、局所、経皮パッチ、座剤を含む直腸、膣、もしくは尿道投与、経皮、鼻噴霧、外科用インプラント、内部外科用塗布剤(internal surgical paint)、注入ポンプ、またはカテーテルである。1つの実施形態では、薬剤およびキャリアを、直接組織注射もしくはボーラス、インプラント、微粒子、ミクロスフェア、ナノ粒子、またはナノスフェアなどの持続放出処方物で投与する。
【0267】
開示の細胞の注入によって治療することができる障害には、正常な血球の産生および成熟の機能障害の不全に起因する疾患(すなわち、再生不良性貧血および低増殖性幹細胞障害);造血臓器における新生物悪性疾患(例えば、白血病およびリンパ腫);非造血臓器の広範な悪性固形腫瘍;自己免疫疾患;および遺伝病が含まれるが、これらに限定されない。このような障害には、正常な血球の産生および成熟の不全または機能障害に起因する疾患(超増殖性幹細胞障害)(薬物、照射、または感染による特発性の再生不良性貧血、汎血球減少症、顆粒球減少症、血小板減少症、赤血球欠如症、ブラックファン−ダイヤモンド症候群が含まれる);造血性悪性腫瘍(急性リンパ芽球性(リンパ球性)白血病、慢性リンパ球性白血病、急性骨髄性白血病、慢性骨髄性白血病、急性悪性脊髄硬化症、多発性骨髄腫、真性多血症、原因不明の骨髄化生、ワルデンストレームマクログロブリン血症、ホジキンリンパ腫、非ホジキンリンパ腫が含まれる);悪性固形腫瘍患者における免疫抑制(悪性黒色腫、胃癌、卵巣癌、乳癌、小細胞肺癌、網膜芽細胞種、睾丸癌、膠芽細胞腫、横紋筋肉腫、神経芽腫、ユーイング肉腫、リンホニア(lymphonia)が含まれる);自己免疫疾患(関節リウマチ、1型糖尿病、慢性肝炎、多発性軟化症、全身性紅斑性狼瘡が含まれる);遺伝性(先天性)障害(貧血(家族性、再生不良性)、ファンコーニ貧血、ジヒドロ葉酸還元酵素欠損症、ホルムアミド転移酵素欠損症、レッシ−ナイハン症候群、先天性異常赤血球産生症症候群IIV、シバッハマン−ダイヤモンド症候群、ジヒドロ葉酸還元酵素欠損、ホリナミノ(forinamino)転移酵素欠損症、レッシ−ナイハン症候群、先天性球状赤血球症、先天的性楕円赤血球症、先天性ストマトサイト増加症、先天性Rh陰性疾患、発作性夜間血色素尿症、G6PD(グルコースリン酸デヒドロゲナーゼ)変異型1、2、3、ピルビン酸キナーゼ欠損症、先天的性エリトロポイエチン感受性、欠損症、鎌状赤血球症および鎌状赤血球傾向、サラセミアα、β、γ、メトヘモグロビン血症、先天性免疫障害、重症複合型免疫不全症(SCID)、ベアリンパ球症候群、イオノフォア応答性複合型免役不全、キャップ形成異常を伴う複合型免疫不全、ヌクレオシドホスホリラーゼ欠損症、顆粒球アクチン欠損症、乳児顆粒球減少症、ゴーシェ病、アデノシンデアミナーゼ欠損症、コストマン症候群、細網系異形成、先天性白血球機能障害症候群が含まれる);その他(骨粗鬆症、脊髄硬化症、後天性溶血性貧血、後天性免疫不全、原発性または続発性免疫不全を発症する感染症、細菌感染(例えば、ブルセラ病、リステリア病、結核、ハンセン病)、寄生虫感染(例えば、マラリア、リーシュマニア症)、真菌感染、加齢によるリンパ系細胞組の不均衡(disproportionsin)および免疫機能障害(impaired immune fimctions)を含む障害、貪食細胞障害、コストマン顆粒球減少症、慢性肉芽腫疾患、チェジアック−東症候群、好中球アクチン欠損症、好中球膜GP−180欠損症、代謝性貯蔵病、ムコ多糖沈着症、ムコリピドーシス、免疫機構に関与するその他の障害、ヴィスコット−オールドリッチ症候群、αランチリプシン(lantirypsin)欠損症など)が含まれるが、これらに限定されない。
【0268】
疾患または病変には、神経変性疾患、肝臓変性疾患病、腎臓変性疾患、脊髄損傷、頭部の外傷または手術、組織、臓器、または腺の変性を引き起こすウイルス感染などが含まれる。このような神経変性疾患には、AIDS痴呆;脱髄性疾患(多発性軟化症および急性トランスフェラーゼ脊髄炎など);錐体外路および小脳障害(皮質脊髄系(ecorticospinal system)の損傷など);大脳基底核障害または小脳障害;ハンティングトン舞踏病と老年舞踏病などの多動性障害;薬物油土迂生運動障害(CNSドーパミン受容体を遮断する薬物によって誘導されるものなど);低運動性障害(パーキンソン病など);進行性過剰核形成(progressive supra−nucleo palsy);小脳の構造損傷;脊髄小脳変性(脊髄性運動失調、フリートライヒ運動失調)、小脳皮質変性、多系統変性(multiple systems degenerations)(Mencel、Dejerine Thomas、Shi−Drager、およびMachado−Joseph)、全身性障害(systermioc disorders)(Rufsum疾患など)、βリポタンパク質欠損血症、失調症、毛細血管拡張;ミトコンドリア多システム障害;脱髄性コア障害(多発性軟化症、急性横断脊髄炎など);運動単位障害(神経原性筋萎縮(前角細胞萎縮(筋萎縮性側索硬化症など)、幼児型脊髄性筋萎縮、および若年性脊髄性筋萎縮)など);アルツハイマー病;中年のダウン症;びまん性レビー小体病;レビー小体型老人性痴呆;パーキンソン病、ヴェルニッケ−コルサコフ症候群;慢性アルコール中毒;クロイツフェルト・ヤコブ病;亜急性硬化性汎脳炎、ハレルホルデン−スパッツ疾患;ならびに拳闘家痴呆が含まれるが、これらに限定されない。例えば、Berkowら(編)(1987),The Merck Manual(15’)版),Merck and Co.,Rahway,NJ.を参照のこと。
【0269】
(産業的飼育での使用)
本発明は、CMP−Neu5Acヒドロキシラーゼ遺伝位の一方または両方の対立遺伝子が不活化された飼育適用を目的とする生きたブタを提供する。CMP−Neu5Acヒドロキシラーゼの一方または両方の対立遺伝子の不活化により、人畜共通疾患、ブタにおける感染症(例えば、E.coli、ブタロタウイルス、およびブタ伝染性胃腸炎コロナウイルス)、ならびに宿主動物においてNeu5Gcを使用する任意の他の人畜共通生物または腸管毒素原性生物に対するブタ動物の感受性を減少させることができる。疾患感受性の減少により、より多数の健康な動物の得る能力によって飼育操作がより経済的に実現し、腸管毒素原性生物に起因する動物の死亡が減少する。
【0270】
以下の実施例は例示目的で提供し、本発明を決して制限しない。
【実施例】
【0271】
(核酸の単離)
PCRベースの方法の組み合わせストラテジーを使用して、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子を同定した。このようなPCR方は、当該分野で周知であり、例えば、PCR Technology,H.A.Erlich編,Stockton Press,London,1989;PCR Protocols:A Guide to Methods and Applications,M.A.Innis,D.H.Gelfand,J.J.Sninsky,およびT.J.White編,Academic Pres,Inc.,New York,1990に記載されている。
【0272】
Trizol試薬(Gibco,Grand Island,NY)を使用して、成体ブタ(グレートヨークシャー種)から総RNAを抽出した。DNアーゼI(Ambion,Inc.,Austin,TX)での処理後、ポリA+RNAを、Dynabeads mRNA精製キット(Dynal,Oslo,Norway)を使用して分離した。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子の5’末端または3’末端を同定するために、Marathon(登録商標)cDNA増幅キット(Clontech)を使用して、5’または3’−RACE(cDNA末端の迅速な複製)手順を行った。転写物のエクソン−イントロン境界または5’もしくは3’隣接領域を同定するために、UniversalGenomeWalkeerライブラリーキット(Clontech)を使用して、ブタGenomeWalker(登録商標)ライブラリーを構築した。GenBankアクセッションA59058によって提供された部分的cDNA配列から、遺伝子特異的およびネスト化プライマー対をデザインした。
【0273】
(cDNAおよびゲノムCMP−Neu5Acヒドロキシラーゼ配列の決定)
5’または3’RACE分析:ブタCMP−Neu5Acヒドロキシラーゼ遺伝子構築物の5’末端および3’末端を同定するために、テンプレートとして成体ブタ脾臓から単離したポリA+RNAと共にMarathon cDNA増幅キット(Clontech)を使用して、5’および3’−RACE手順を行った。20UのAMV−RTおよび1pmolの市販のcDNA合成プライマーを使用して、48℃で2時間のインキュベーションによって1μgのポリA+RNAから第1の鎖cDNAを合成した。第2の鎖cDNA合成は、RNアーゼH、E.coli DNAポリメラーゼI、E.coliDNAポリメラーゼI、およびE.coliDNAリガーゼから構成される市販の酵素カクテルでの16℃で1.5時間の全第1の鎖の反応を含んでいた。T4DNAポリメラーゼによる二本鎖cDNA末端の平滑末端化後、市販のMarathoncDNA Adapterを、精製された二本鎖cDNAのアリコートにライゲーションした。キット中の10mM ticme−KOH/0.1M EDTA緩衝液でのアダプターライゲーション産物の希釈により、PCR増幅のためのcDNAを用意した。
【0274】
ブタCMP−Neu5Acヒドロキシラーゼ遺伝子転写物の5’および3’のほとんどの配列を得るために、提供されたMarathon cDNA増幅プライマー組を、GenBankアクセッション番号A59058によって提供された配列に基づいて、遺伝子特異的プライマーおよびネスト化遺伝子特異的プライマーと対合した。これらのプライマー組を、表13に示す。この方法により、GenBankアクセッション番号A59058中に含まれる配列に基づいたオリゴヌクレオチドプライマーを、3’方向および5’方向に配向させ、これを使用して重複PCRフラグメントを作製する。これらの重複3’および5’産物を組み合わせて、インタクトな全長cDNAを産生する。この方法は、例えば、Innisら,supra;およびFrohmanら,Proc.Natl.Acad.Sci.,85:8998,1988に記載されており、さらに、例えば、米国特許第4,683,195号に記載されている。
【0275】
ゲノムウォーキング分析:ブタCMP−Neu5Acヒドロキシラーゼ転写物のエクソン−イントロン境界または5’もしくは3’隣接領域を同定するために、UniversalGenomeWalker(登録商標)ライブラリーキット(Clontech,Palo Alto,CA)を使用して、ブタGenomeWalker(登録商標)ライブラリーを構築した。
【0276】
簡単に述べれば、ブタゲノムDNAの5つのアリコートを、1平滑末端切断制限エンドヌクレアーゼ(Dral、EcoRV、PvulI、ScaI、またはStuI)で個別に消化した。フェノール−クロロホルム抽出、エタノール沈殿、および制限消化フラグメントの再懸濁後、各消化アリコートの一部を、キット中にGenomuWalkerアダプターとの個別のライゲーション反応で使用した。このプロセスにより、PCRベースのクローニングストラテジーで使用するための5つのライブラリーが作製された。表13で同定したプライマー対を、ゲノムウォーキングストラテジーで使用した。eLON−GaseまたはTaKaRa LATaq(Takara Shuzo Co.,Ltd.,Shiga,Japan)酵素のいずれかを、全GenomeWalker実験およびゲノムDNAの直接長(direct long)PCRのために使用した。PerkinElmer Gene Amp System 9600または9700サーモサイクラーによる全GenomeWalker実験で、製造者が推奨する熱サイクリング条件を使用した。
【0277】
【表15】
増幅産物のサブクローニングおよび配列決定:ゲノムDNAから増幅したPCR産物、、GeneWalker−PCR(Clontech)、および5’−3’−RACEを、Qiagenゲル抽出キット(Qiagen,Valencia,CA)を使用してゲル精製し、必要に応じて、Original TAクローニングキット(Invitrogen,Carlsbad,CA)中のpCRIIベクターにサブクローン化した。pCRIIライゲーションインサートのプラスミドDNAミニプレップを、説明書にしたがってQIAprepスピンミニプレップキット(Qiagen)を使用して調製した。ABI 377自動化配列分析器(Applied Biosystems,Inc.,Foster City,CA)をT7およびSP6プロモータープライマーまたは内部インサート配列からデザインしたプライマーでプライミングしたローダミンまたはBigDyeターミネーターサイクル配列決定キット(Applied Biosystems)のいずれかと共に使用して、クローン化インサートの自動化蛍光配列決定を行った。
【0278】
プライマー合成:他で記載しない限り、種々のPCRベースの方法でプライマーとして使用する全てのオリゴヌクレオチドを、固相合成およびホスホラミダイトヌクレオシド化学を使用したABI 394 DNA Synthesizer(Applied Biosystems,Inc.,Foster City CA)を使用して合成した。
【0279】
(転写因子結合部位の分析)
エクソン配列の228bpおよびエクソン1の601bp上流を使用して、可能な転写因子結合部位の分析を行った。www.genomatix.deで利用可能な「MatInspector」ソフトウェアを使用して、配列をスクリーニングした。配列は、以下の転写因子の結合部位を含む:MZF1、ETSF、SF1、CMYB、MEF2、NMP4、BRN2、AP1、GAT1、SATB1、ATF、USF、WHN、ZF5、NFκB、MOK2、NFY、MYCMAX、ZF5。図4を参照のこと。
【0280】
ブタCMP−Neu5Acヒドロキシラーゼ相同組換えターゲティングベクターの構築CMP−Neu5Acヒドロキシラーゼノックアウト標的ベクター。ノックアウトのためのブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6をターゲティングするベクターを構築することができる。第1の工程では、
【0281】
【化1】
などのプライマーを使用して、ターゲティングベクターの3’アームとして使用するためのイントロン6部分をPCRによって増幅する(図5を参照のこと)。約3.7kbの増幅されたPCR産物を、EcoRIおよびApaIを使用した制限酵素消化後にpCRIIベクターに挿入することができる。図6を参照のこと。
【0282】
3’アームの挿入後、
【0283】
【化2】
などのプライマーを使用して、ターゲティングベクターの5’アームとして使用するためのイントロン5部分をPCRによって増幅することができる(図5を参照のこと)。この最初の増幅後、それぞれ
【0284】
【化3】
などのプライマーと組み合わせてpDH1などのプライマーを使用してEcoNI制限部位を5’アームの3’部分に挿入するために連続的PCR増幅を行うことができる(図5を参照のこと)。操作EcoNI部位を含む約2.6kbの増幅PCR産物はApaIおよびEcoNIを使用して消化した制限酵素であり、これを予め挿入した3’アームを含むpCRIIベクターに挿入して(図7を参照のこと)、約6.3kbのブタCMP−Neu5Acヒドロキシラーゼターゲティング配列を含むターゲティングベクター(pDHΔex6)を作製することができる(図8を参照のこと)。
【0285】
EGFPノックイン標的ベクター。ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6の3’末端への増強緑色蛍光タンパク質配列のインフレーム挿入によって、pDHΔex6をさらに修飾することができる。第1の工程では、ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のイントロン5部分およびエクソン6部分を、
【0286】
【化4】
などのプライマーを使用したPCRによって増幅し(図9を参照のこと)、MunIおよびEcoRvでの制限酵素消化後にEGFPおよびポリAテールを含むベクター(pIRES−EGFP)に挿入することができる。挿入後、
【0287】
【化5】
などのプライマーを使用した挿入を含むpIRES−EGFPベクターでPCR増幅を行うことができる(図9を参照のこと)。PCR増幅産物はEcoNIおよびMunIを使用して消化した制限酵素であり、これを予め構築したpDHΔex6ターゲティングベクターに挿入することができる(図10を参照のこと)。得られたターゲティングベクター(pDHΔex6)を図11に例示する。
【0288】
(ブタCMP−Neu5Acヒドロキシラーゼ欠損胎児線維芽細胞の産生)
妊娠33日の同一妊娠の10体の胎児から胎児線維芽細胞を単離する。頭部および内蔵の除去後、胎児をハンクス平衡塩類溶液(HBSS;Gibco−BRL,1 5 Rockville,MD)で洗浄し、20mlのHBSSに入れ、小さな手術用鋏でサイコロ状にする。組織をペレット化し、50ml試験管中で胎児あたり40mlのDMEMおよび100U/mlコラゲナーゼ(gbco−BRL)で再懸濁する。チューブを37℃の水浴中で撹拌しながら40分間インキュベートする。消化された組織を3〜4分間静置し、細胞豊富な上清を新たな50mlチューブに移し、ペレット化する。次いで、細胞を、10%ウシ胎児血清(FCS)、1×非必須アミノ酸、1mMピルビン酸ナトリウム、および2ng/ml bFGFを含む40mlのDMEMで再懸濁し、10cmの皿に播種する。トランスフェクションのために、10μgの線状化pDHΔex6EGFPベクターを、製造者のガイドラインにしたがってリポフェクタミン2000(Carlsbad,CA)を使用して200万個の細胞に移入する。48時間のトランスフェクション後、トランスフェクトした細胞を、2,000細胞/ウェルの密度で48ウェルプレートに播種し、コンフルエンスまで成長させる。コンフルエンス後、細胞を、相同組換えが起こってEGFPを発現する細胞のみが選択される蛍光標示式細胞分取(FACS)(FACSCalibur,Becton Dickenson,San Jose,CA)によって分取する(例えば、図13を参照のこと)。
【0289】
選択された細胞を、再播種し、コンフルエントまで成長させる。一旦コンフルエンスに到達すると、いくつかの小アリコートをその後の使用のために凍結させ、残りを相同組換えについてのPCRおよびサザンブロットによる検証のために使用する。推定ターゲティングクローンを、EGFPに相補的なプライマーおよびアンチセンスプライマーとしてベクターの外側の配列に相補的なプライマーを使用したエクソン6/EGFPインサートを介したPCRによってスクリーニングすることができる。ターゲティングされた対立遺伝子由来の推定バンドの存在によってポジティブクローンを同定するために、PCR産物を、EGFPプローブを使用したサザンブロッティングによって分析することができる。
【0290】
(核ドナーとして異種CMP−Neu5Acヒドロキシラーゼ欠損胎児線維芽細胞を使用したクローン化ブタの作製)
核導入のための細胞の調製:一般的に上記のようにブタCMP−Neu5Acヒドロキシラーゼにヘテロ接合性を示す細胞を産生するために、ドナー細胞を遺伝子操作する。上記に詳述のように産生した核ドナーとしてEGFP選択ブタ線維芽細胞を使用して、当該分野で周知の方法(Daiら,Nature Biotechnology 20:251255,2002;およびPolejaevaら,Nature 407:86−90,2000)によって核導入を行うことができる。
【0291】
例えば、I.Polejaevaら,Nature 407:505(2000)に記載のように、同期化された過剰排卵する性的に成熟したLarge−WhiteX Landacre外部交配未経産ブタから卵母細胞を単離することができる。ドナー細胞を、血清枯渇(0.5%)によって24時間と120時間との間の推定G0/G1期に同調させる。例えば、本質的にA.C.Boquestら,Biol.Reproduction 68:1283(2002)に記載のように、卵母細胞の除核、核導入、電気融合、および電気活性化を行うことができる。再構築した胚を一晩培養し、非同期(−1日)レシピエントの卵管に導入することができる。妊娠を確認し、実時間超音波によってモニタリングすることができる。
【0292】
ヘテロ接合性CMP−Neu5Acヒドロキシラーゼ1ノックアウト(SKO)雄ブタと雌ブタとの交配を行い、二重ノックアウト(DKO)ブタの小集団を確立することができる。
【0293】
(CMP−Neu5Acヒドロキシラーゼ欠損ブタの検証)
1ノックアウト雄ブタおよび雌ブタの交配後、二重ノックアウトブタの検証を行う。子孫由来の線維芽細胞を、1μgの抗N−グリコシルGM2モノクローナル抗体MK2−34(Seikagaku Kogyo,JP)と氷上で30分間インキュベートする。FITC抱合ヤギ抗マウスIgGを細胞および結合がNeu5GCの有無を示す抗体に添加し、それにより活性なCMP−Neu5Acヒドロキシラーゼの有無をフローサイトメトリー(FACSCalibur,Becton Dickenson,San Jose,CA)によって検出する。
【図面の簡単な説明】
【0294】
【図1】ブタCMP−Neu5Acヒドロキシラーゼ遺伝子のゲノム機構を示す。黒塗りのバーは、それぞれ番号をつけたエクソンを示す。エクソン間のイントロンの長さは、相対距離を示す。(白抜きのボックスもいくつかの変異型で認められるエクソンを示す(図2を参照のこと);「開始」および「終止」は、それぞれ開始コドンおよび終止コドンを示す)。図の下におおよそのスケールを示す。
【図2】CMP−Neu5Acヒドロキシラーゼ遺伝子のcDNA配列を示す。変異型1は、エクソン14および15の代わりにエクソン15aを含む。変異型2はエクソン12aを含み、変異型3はエクソン11aを含む。「開始」および「終止」は、それぞれ開始コドンおよび終止コドンを示す。
【図3】その対応するゲノム機構と共にターゲティングベクターの4つの非限定的な例を示す。この特定の非限定的な例における選択マーカー遺伝子は、eGFP(増強緑色蛍光タンパク質)である。ブタCMP−Neu5Aceヒドロキシラーゼ遺伝子を不活化するためにGFPをDNA構築物に挿入することができる。
【図4】エクソン1(228bp)内に存在する転写因子結合部位および601bpにわたるその5’隣接領域を示す。
【図5】ブタCMP−Neu5Acヒドロキシラーゼ遺伝子ターゲティングベクターのDNA構築のために使用することができるオリゴヌクレオチド配列を示す。
【図6】プライマーpDH3およびpDH4を使用したブタCMP−Neu5Acヒドロキシラーゼ遺伝子からの3’アームセグメントの産生およびベクター(pCRII)へのその挿入を示す略図である。
【図7】プライマーpDH1およびpDH2、その後pDH2a、pDH2b、およびpDH2cを使用したブタCMP−Neu5Acヒドロキシラーゼ遺伝子からの5’アームセグメントの産生および3’アームが予め挿入されたベクター(pCRII)への挿入を示す略図である。
【図8】相同組換えによってブタCMP−Neu5Acヒドロキシラーゼ遺伝子のエクソン6を欠失するために使用することができるターゲティングベクターを例示する図の非限定的な例である。
【図9】ノックインストラテジーで使用するための増強緑色蛍光タンパク質発現ベクターの作製で使用されるオリゴヌクレオチド配列を示す。
【図10】ポリAシグナルを有するEGFPフラグメントのターゲティングベクターpDHΔex6への挿入を例示する図である。
【図11】eGFP発現のためのノックインベクターを例示する図である。
【図12】ターゲティングカセットDNA構築物(pDHΔex6)とゲノムDNAとの間でフレームシフトが起こる相同組換えを例示する図である。
【図13】ターゲティングカセットDNA構築物(pDHΔex6)とゲノムDNAとの間でフレームシフトが起こる相同組換えを例示する図である。
【特許請求の範囲】
【請求項1】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離全長cDNA配列。
【請求項2】
前記配列が配列番号1である、請求項1に記載のcDNA。
【請求項3】
前記配列が配列番号3である、請求項1に記載のcDNA。
【請求項4】
前記配列が配列番号5である、請求項1に記載のcDNA。
【請求項5】
前記配列が配列番号7である、請求項1に記載のcDNA。
【請求項6】
前記配列が配列番号2である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項7】
前記配列が配列番号4である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項8】
前記配列が配列番号6である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項9】
前記配列が配列番号8である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項10】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする全長cDNA配列を含む、核酸構築物。
【請求項11】
前記cDNAが配列番号1である、請求項10に記載の構築物。
【請求項12】
前記cDNAが配列番号3である、請求項10に記載の構築物。
【請求項13】
前記cDNAが配列番号5である、請求項10に記載の構築物。
【請求項14】
前記cDNAが配列番号7である、請求項10に記載の構築物。
【請求項15】
プロモーターをさらに含む、請求項10〜請求項15のいずれか1項に記載の構築物。
【請求項16】
選択マーカーをさらに含む、請求項10〜請求項15のいずれか1項に記載の構築物。
【請求項17】
前記選択マーカーが緑色蛍光タンパク質である、請求項16に記載の構築物。
【請求項18】
請求項10〜請求項17のいずれか1項に記載の構築物を含む、トランスフェクトされた細胞。
【請求項19】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質を発現する、細胞。
【請求項20】
前記タンパク質配列が、配列番号2、配列番号4、配列番号6、および配列番号8からなる群から選択される、請求項19に記載の細胞。
【請求項21】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に相同な単離ヌクレオチド配列。
【請求項22】
前記配列が、配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に少なくとも80%相同である、請求項21に記載のヌクレオチド配列。
【請求項23】
前記配列が、配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に少なくとも90%相同である、請求項21に記載のヌクレオチド配列。
【請求項24】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に相同なヌクレオチド配列を含む核酸構築物。
【請求項25】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列とハイブリッド形成する単離ヌクレオチド配列。
【請求項26】
ストリンジェントな条件下でハイブリッド形成する、請求項25に記載のヌクレオチド配列。
【請求項27】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列を含むベクター。
【請求項28】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列を含むプラスミド。
【請求項29】
配列番号46のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項30】
配列番号47のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項31】
配列番号48のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項32】
配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28からなる群から選択される、単離ヌクレオチド配列。
【請求項33】
配列番号9、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、および配列番号45からなる群から選択される、単離ヌクレオチド配列。
【請求項34】
請求項31〜請求項33のいずれか1項に記載の配列を含む、ベクター。
【請求項35】
請求項31〜請求項33のいずれか1項に記載の配列を含む、核酸構築物。
【請求項36】
請求項31〜請求項33のいずれか1項に記載の配列を含む、プラスミド。
【請求項37】
配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号19、配列番号21、配列番号25、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、および配列番号45からなる群から選択される配列の少なくとも17個の連続核酸を含む、核酸構築物。
【請求項38】
配列番号47の少なくとも17個の連続核酸を含む、核酸構築物。
【請求項39】
配列番号48の少なくとも2,755個の連続核酸を含む、核酸構築物。
【請求項40】
配列番号49の少なくとも1,750個の連続核酸を含む、核酸構築物。
【請求項41】
配列番号50の少なくとも150個の連続核酸を含む、核酸構築物。
【請求項42】
請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に相同な配列を含む、単離ヌクレオチド。
【請求項43】
前記配列が請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に少なくとも80%相同である、請求項41に記載のヌクレオチド。
【請求項44】
前記配列が請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に少なくとも90%相同である、請求項21に記載のヌクレオチド。
【請求項45】
請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列とハイブリッド形成する、単離ヌクレオチド。
【請求項46】
ストリンジェントな条件下でハイブリッド形成する、請求項45に記載のヌクレオチド。
【請求項47】
(a)配列番号47に相同な少なくとも17個の連続核酸を含む第1のヌクレオチド配列と、
(b)選択マーカー遺伝子と、
(c)配列番号48に相同な少なくとも17個の連続核酸を含む第2のヌクレオチド配列とを含む、ターゲティングベクター。
【請求項48】
(a)配列番号48に相同な少なくとも17個の連続核酸を含む第1のヌクレオチド配列と、
(b)選択マーカー遺伝子と、
(c)配列番号48に相同な少なくとも17個の連続核酸を含む第2のヌクレオチド配列とを含み、前記第1および第2のヌクレオチド配列が重複しない、ターゲティングベクター。
【請求項49】
前記選択マーカーが緑色蛍光タンパク質である、請求項47または請求項48に記載のターゲティングベクター。
【請求項50】
前記第1のヌクレオチド配列が5’組換えアームを示す、請求項47または請求項48に記載のターゲティングベクター。
【請求項51】
前記第2のヌクレオチド配列が3’組換えアームを示す、請求項47または請求項48に記載のターゲティングベクター。
【請求項52】
前記選択マーカーが緑色蛍光タンパク質である、請求項49に記載のターゲティングベクター。
【請求項53】
前記第1のヌクレオチド配列が、イントロン5の少なくとも17個の連続ヌクレオチドを含むヌクレオチド配列に相同である、請求項48に記載のターゲティングベクター。
【請求項54】
前記第2のヌクレオチド配列が、イントロン6の少なくとも17個の連続ヌクレオチドを含むヌクレオチド配列に相同である、請求項48に記載のターゲティングベクター。
【請求項55】
請求項47〜請求項54のいずれか1項に記載のターゲティングベクターでトランスフェクトした、細胞。
【請求項56】
前記ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が相同組換えによって不活化されている、請求項55に記載の細胞。
【請求項57】
請求項55に記載の細胞を含む、ブタ動物。
【請求項58】
前記ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が相同組換えによって不活化されている、請求項57に記載の動物。
【請求項59】
請求項58に記載の動物から得た、臓器。
【請求項60】
請求項58に記載の動物から得た、組織。
【請求項61】
前記臓器が、心臓、肺、腎臓、および肝臓からなる群から選択される、請求項59に記載の臓器。
【請求項62】
(a)請求項47または請求項48に記載のターゲティングベクターでブタ細胞をトランスフェクトする工程と、
(b)ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が不活化されている、トランスフェクトされた細胞を選択する工程とを含む、遺伝子操作細胞を産生する方法。
【請求項63】
(a)請求項47または請求項48に記載のターゲティングベクターでブタ細胞をトランスフェクトする工程と、
(b)ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が不活化されている、トランスフェクトされた細胞(核ドナー細胞)を選択する工程と、
(c)前記核ドナー細胞の核を除核卵母細胞に導入して胚を産生する工程と、
(d)前記胚を動物に成長させる工程とを含む、遺伝子操作動物を産生する方法。
【請求項64】
請求項63に記載の動物に由来する、臓器。
【請求項65】
前記臓器が、心臓、肺、腎臓、および肝臓からなる群から選択される、請求項64に記載の臓器。
【請求項66】
請求項63に記載の動物に由来する、組織。
【請求項1】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離全長cDNA配列。
【請求項2】
前記配列が配列番号1である、請求項1に記載のcDNA。
【請求項3】
前記配列が配列番号3である、請求項1に記載のcDNA。
【請求項4】
前記配列が配列番号5である、請求項1に記載のcDNA。
【請求項5】
前記配列が配列番号7である、請求項1に記載のcDNA。
【請求項6】
前記配列が配列番号2である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項7】
前記配列が配列番号4である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項8】
前記配列が配列番号6である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項9】
前記配列が配列番号8である、CMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする単離アミノ酸配列。
【請求項10】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質をコードする全長cDNA配列を含む、核酸構築物。
【請求項11】
前記cDNAが配列番号1である、請求項10に記載の構築物。
【請求項12】
前記cDNAが配列番号3である、請求項10に記載の構築物。
【請求項13】
前記cDNAが配列番号5である、請求項10に記載の構築物。
【請求項14】
前記cDNAが配列番号7である、請求項10に記載の構築物。
【請求項15】
プロモーターをさらに含む、請求項10〜請求項15のいずれか1項に記載の構築物。
【請求項16】
選択マーカーをさらに含む、請求項10〜請求項15のいずれか1項に記載の構築物。
【請求項17】
前記選択マーカーが緑色蛍光タンパク質である、請求項16に記載の構築物。
【請求項18】
請求項10〜請求項17のいずれか1項に記載の構築物を含む、トランスフェクトされた細胞。
【請求項19】
ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼタンパク質を発現する、細胞。
【請求項20】
前記タンパク質配列が、配列番号2、配列番号4、配列番号6、および配列番号8からなる群から選択される、請求項19に記載の細胞。
【請求項21】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に相同な単離ヌクレオチド配列。
【請求項22】
前記配列が、配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に少なくとも80%相同である、請求項21に記載のヌクレオチド配列。
【請求項23】
前記配列が、配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に少なくとも90%相同である、請求項21に記載のヌクレオチド配列。
【請求項24】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列に相同なヌクレオチド配列を含む核酸構築物。
【請求項25】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列とハイブリッド形成する単離ヌクレオチド配列。
【請求項26】
ストリンジェントな条件下でハイブリッド形成する、請求項25に記載のヌクレオチド配列。
【請求項27】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列を含むベクター。
【請求項28】
配列番号1、配列番号3、配列番号5、および配列番号7からなる群から選択されるヌクレオチド配列を含むプラスミド。
【請求項29】
配列番号46のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項30】
配列番号47のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項31】
配列番号48のヌクレオチド配列を含む、単離ヌクレオチド。
【請求項32】
配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28からなる群から選択される、単離ヌクレオチド配列。
【請求項33】
配列番号9、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、および配列番号45からなる群から選択される、単離ヌクレオチド配列。
【請求項34】
請求項31〜請求項33のいずれか1項に記載の配列を含む、ベクター。
【請求項35】
請求項31〜請求項33のいずれか1項に記載の配列を含む、核酸構築物。
【請求項36】
請求項31〜請求項33のいずれか1項に記載の配列を含む、プラスミド。
【請求項37】
配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号19、配列番号21、配列番号25、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、および配列番号45からなる群から選択される配列の少なくとも17個の連続核酸を含む、核酸構築物。
【請求項38】
配列番号47の少なくとも17個の連続核酸を含む、核酸構築物。
【請求項39】
配列番号48の少なくとも2,755個の連続核酸を含む、核酸構築物。
【請求項40】
配列番号49の少なくとも1,750個の連続核酸を含む、核酸構築物。
【請求項41】
配列番号50の少なくとも150個の連続核酸を含む、核酸構築物。
【請求項42】
請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に相同な配列を含む、単離ヌクレオチド。
【請求項43】
前記配列が請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に少なくとも80%相同である、請求項41に記載のヌクレオチド。
【請求項44】
前記配列が請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列に少なくとも90%相同である、請求項21に記載のヌクレオチド。
【請求項45】
請求項29〜請求項33のいずれか1項に記載のヌクレオチド配列とハイブリッド形成する、単離ヌクレオチド。
【請求項46】
ストリンジェントな条件下でハイブリッド形成する、請求項45に記載のヌクレオチド。
【請求項47】
(a)配列番号47に相同な少なくとも17個の連続核酸を含む第1のヌクレオチド配列と、
(b)選択マーカー遺伝子と、
(c)配列番号48に相同な少なくとも17個の連続核酸を含む第2のヌクレオチド配列とを含む、ターゲティングベクター。
【請求項48】
(a)配列番号48に相同な少なくとも17個の連続核酸を含む第1のヌクレオチド配列と、
(b)選択マーカー遺伝子と、
(c)配列番号48に相同な少なくとも17個の連続核酸を含む第2のヌクレオチド配列とを含み、前記第1および第2のヌクレオチド配列が重複しない、ターゲティングベクター。
【請求項49】
前記選択マーカーが緑色蛍光タンパク質である、請求項47または請求項48に記載のターゲティングベクター。
【請求項50】
前記第1のヌクレオチド配列が5’組換えアームを示す、請求項47または請求項48に記載のターゲティングベクター。
【請求項51】
前記第2のヌクレオチド配列が3’組換えアームを示す、請求項47または請求項48に記載のターゲティングベクター。
【請求項52】
前記選択マーカーが緑色蛍光タンパク質である、請求項49に記載のターゲティングベクター。
【請求項53】
前記第1のヌクレオチド配列が、イントロン5の少なくとも17個の連続ヌクレオチドを含むヌクレオチド配列に相同である、請求項48に記載のターゲティングベクター。
【請求項54】
前記第2のヌクレオチド配列が、イントロン6の少なくとも17個の連続ヌクレオチドを含むヌクレオチド配列に相同である、請求項48に記載のターゲティングベクター。
【請求項55】
請求項47〜請求項54のいずれか1項に記載のターゲティングベクターでトランスフェクトした、細胞。
【請求項56】
前記ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が相同組換えによって不活化されている、請求項55に記載の細胞。
【請求項57】
請求項55に記載の細胞を含む、ブタ動物。
【請求項58】
前記ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が相同組換えによって不活化されている、請求項57に記載の動物。
【請求項59】
請求項58に記載の動物から得た、臓器。
【請求項60】
請求項58に記載の動物から得た、組織。
【請求項61】
前記臓器が、心臓、肺、腎臓、および肝臓からなる群から選択される、請求項59に記載の臓器。
【請求項62】
(a)請求項47または請求項48に記載のターゲティングベクターでブタ細胞をトランスフェクトする工程と、
(b)ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が不活化されている、トランスフェクトされた細胞を選択する工程とを含む、遺伝子操作細胞を産生する方法。
【請求項63】
(a)請求項47または請求項48に記載のターゲティングベクターでブタ細胞をトランスフェクトする工程と、
(b)ブタCMP−N−アセチルノイラミン酸ヒドロキシラーゼ遺伝子の少なくとも1つの対立遺伝子が不活化されている、トランスフェクトされた細胞(核ドナー細胞)を選択する工程と、
(c)前記核ドナー細胞の核を除核卵母細胞に導入して胚を産生する工程と、
(d)前記胚を動物に成長させる工程とを含む、遺伝子操作動物を産生する方法。
【請求項64】
請求項63に記載の動物に由来する、臓器。
【請求項65】
前記臓器が、心臓、肺、腎臓、および肝臓からなる群から選択される、請求項64に記載の臓器。
【請求項66】
請求項63に記載の動物に由来する、組織。
【図1】

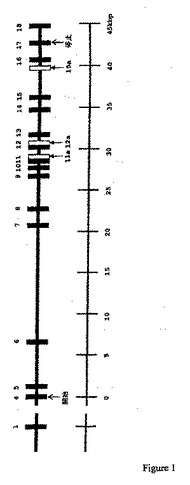
【図2】
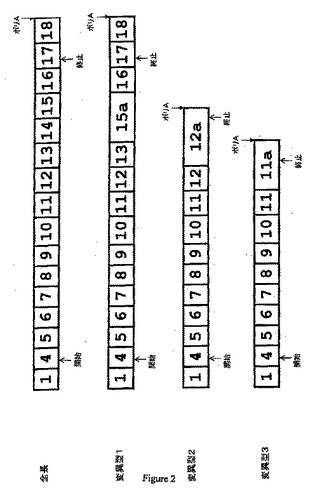
【図3】
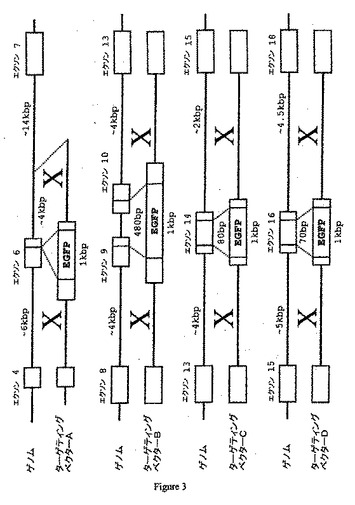
【図4】

【図5】

【図6】
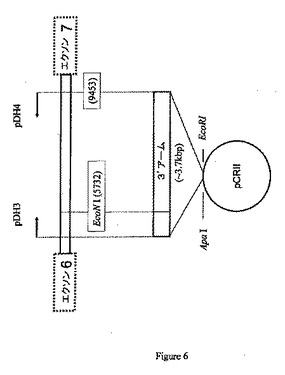
【図7】
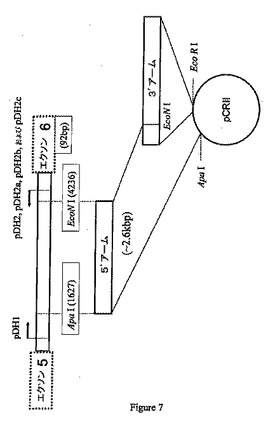
【図8】
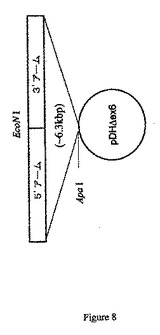
【図9】

【図10】
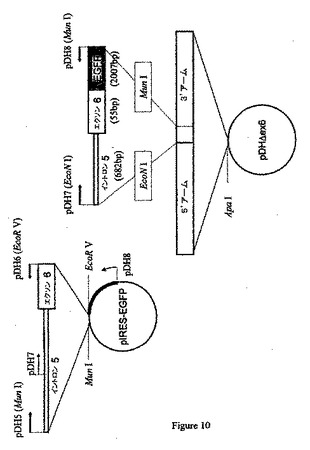
【図11】
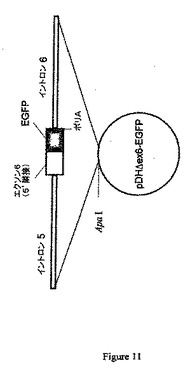
【図12】
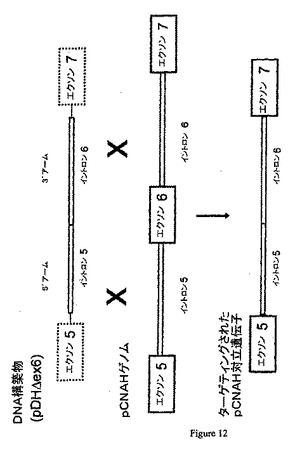
【図13】
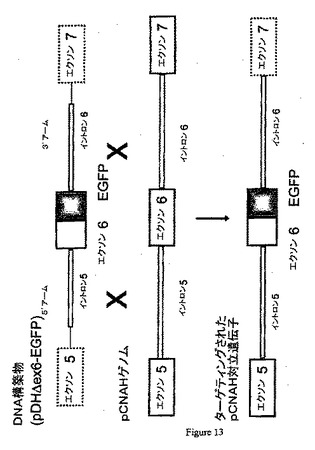
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2007−525955(P2007−525955A)
【公表日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願番号】特願2006−515268(P2006−515268)
【出願日】平成16年6月7日(2004.6.7)
【国際出願番号】PCT/US2004/018106
【国際公開番号】WO2004/108904
【国際公開日】平成16年12月16日(2004.12.16)
【出願人】(504279968)ユニバーシティー オブ ピッツバーグ (24)
【Fターム(参考)】
【公表日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願日】平成16年6月7日(2004.6.7)
【国際出願番号】PCT/US2004/018106
【国際公開番号】WO2004/108904
【国際公開日】平成16年12月16日(2004.12.16)
【出願人】(504279968)ユニバーシティー オブ ピッツバーグ (24)
【Fターム(参考)】
[ Back to top ]