
プライマーセット、アゾスピリラム属菌の検出キット及び検出方法
【課題】アゾスピリラム属菌を精度よく、迅速かつシンプルに検出・識別することのできるプライマーセットを提供する。
【解決手段】特定の塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、特定の塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。
【解決手段】特定の塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、特定の塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はプライマーセット、アゾスピリラム属菌の検出キット及び検出方法に係るものである。詳しくは、アゾスピリラム属菌16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅し得るプライマーセット、当該プライマーセットを用いたアゾスピリラム属菌の検出キット及び検出方法に係るものである。
【背景技術】
【0002】
アゾスピリラム属菌は、窒素固定能を有する好気性の桿状α−プロテオバクテリアとして知られている。アゾスピリラム属菌は世界中において、根圏土壌及び多種の植物根部から分離されている。多くのアゾスピリラム属菌株は、植物成長促進細菌(plant growth promoting bacteria,PGPB)として報告されている。植物成長促進細菌とは、作物の生長を促進させることができる細菌である。この成長促進は、アゾスピリラム属菌株による植物ホルモンの生成、窒素固定、植物免疫によってもたらされるものとして考えられている。従って、アゾスピリラム属の細菌は、バイオ肥料及びバイオ殺虫剤として利用され、近年の農業用の化学薬品を減らす需要により、多くの注目を集めている。
【0003】
しかしながら、根圏土壌及び植物体からのアゾスピリラム属菌の識別及び分離は、選択培地及び形態生物学的評価による従来的な方法に頼るしかなかった。従来の方法では、複数の選択培地(例えばNFb、RC培地など)を利用しながら、菌の形態または窒素固定能などを評価して、候補となる菌株を絞り込んだ上で、さらにrDNA配列の解読を行う必要があった。これらの従来の方法は、アゾスピリラム属菌に対する専門知識と実験経験、1〜2ヶ月の時間と、多大な労力とコストを必要とする。さらに、選択培地の利用と菌種特性の評価を繰り返すため、培地適応性と評価値の高い菌株が分離されやすく、分離した菌株には意図していない選択バイアスが生じることがある。
【0004】
最近のDNA配列解析技術の発展によって、ターゲットとなる微生物の検出及び識別において分子遺伝学的な識別法が考えられるようになった。非特許文献1〜非特許文献23は、アゾスピリラム属菌の検出と識別にかかわる情報を開示しており、これらのうち、特に非特許文献1〜3,7,10,11,13〜17は分子遺伝学的な識別法について開示しているものである。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Baldani JI, Krieg NR, Baldani VLD, Hartmann A, Dobereiner J (2005) Genus II. Azospirillum. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s manual of systematic bacteriology 2nd edn vol 2 part C. Springer, NY, pp 16−17
【非特許文献2】Bashan Y, Holguin G, Lifshitz R (1993) Isolation and characterization of plant growth−promoting rhizobacteria. In: Glick BR, Thompson JE (eds) Methods in plant molecular biology and biotechnology. CRC press, FL, pp 331−345
【非特許文献3】Baudoin E, Couillerot O, Spaepen S, Moenne−Loccoz Y, Nazaret S (2010) Applicability of the 16S−23S rDNA internal spacer for PCR detection of the phytostimulatory PGPR inoculants Azospirillum lipoferum CRT1 in field soil. J Appl Microbiol 108:25−38. doi:10.1111/j.1365−2672.2009.04393.x
【非特許文献4】Boddey RM, Baldani VLD, Baldani JI, Dobereiner J (1986) Effect of inoculation of Azospirillum spp. on nitrogen accumulation by field−grown wheat. Plant Soil 95:109−121. doi:10.1007/BF02378857
【非特許文献5】Burris RH (1972) Nitrogen fixing−assay methods and techniques. Methods Enzymol 24:415−431
【非特許文献6】Caceres EA (1982) Improved medium for isolation of Azospirillum spp. Appl Environ Microbiol 44:990−991
【非特許文献7】Cole JR, Chai B, Farris RJ, Wang Q, Kulam SA, McGarrell DM, Garrity GM, Tiedje JM (2005) The ribosomal database project (RDP−II): sequences and tools for high−throughput rRNA analysis. Nucleic Acids Res 33:294−296 doi:10.1093/nar/gki038
【非特許文献8】Dobbelaere S, Croonenborghs A, Thys A, Broek AV, Vanderleyden J (1999) Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil 212:155−164. doi:10.1007/BF02378857
【非特許文献9】Eckert B, Weber OB, Kirchh G, Halbritter A, Stoffels M, Hartmann A (2001) Azospirillum doebereinerae sp. nov., nitrogen−fixing bacterium associated with the C4−grass Miscanthus. Int J Syst Bacteriol 33:300−308
【非特許文献10】Fancelli S, Castaldini M, Ceccherini MT, Serio CD, Fani R, Gallori E, Marangolo M, Miclaus N, Bazzicalupo M (1998) Use of random amplified polymorphic DNA markers for the detection of Azospirillum strains in soil microcosms. Appl Microbiol Biotechnol 49:221−225. doi:10.1007/s002530051162
【非特許文献11】Grifoni A, Bazzicalupo M, Serio CD, Fancelli S, Fani R (1995)Identification of Azospirillum strains by restriction fragment length polymorphism of the 16S rDNA and of the histidine operon. FEMS Microbiol Lett 127:85−91. doi:10.1111/j.1574−6968.1995.tb07454.x
【非特許文献12】Glickmann E, Dessaux Y (1995) A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl Environ Microbiol 61:793−796
【非特許文献13】Jacoud C, Faure D, Wadoux P, Balley R (1998) Development of a strain−specific probe to follow inoculated Azospirillum lipoferum CRT1 under field conditions and enhancement of maize root development by inoculation. FEMS Microbial Ecol 27:43−51 doi:10.1111/j.1574−6941.1998.tb00524.x
【非特許文献14】Kabir MM, Faure D, Haurat J, Normand P, Jacoud C, Wadoux P, Bally R (1995) Oligonucleotide probes based on 16S rRNA sequences for the identification of four Azospirillum species. Can J Microbiol 41: 1081−1087. doi:10.1139/m95−151
【非特許文献15】Mehhaz S, Mirza MS, Haurat J, Bally R, Normand P, Bano A, Malik KA (2001) Isolation and 16S rRNA sequence analysis of the beneficial bacteria from the rhizosphere of rice. Can J Microbiol 47:110−117. doi:10.1139/cjm−47−2−110
【非特許文献16】Stoffels M, Castellanos T, Hartmann A (2001) Design and application of new 16S rRNA−targeted oligonucleotide probes for the Azospirillum−Skermanella−Rhodocista−cluster. Syst Appl Microbiol 24:83−97. doi:101078/0723−2020−00011
【非特許文献17】Xia Y, Embley TM, O’donnell AG (1994) Phylogenetic analysis of Azospirillum by direct sequencing of PCR amplified 16S rDNA. System Appl Microbiol 17:197−201
【非特許文献18】Lin SY,Young CC,Hupfer H,Siering C,Arun AB,Chen WM,Lai WA,Shen FT,Rekha PD, Yasshin AF.(2009)Azospirillum picis sp.nov.,isolated from discarded tar.Int J Syst Evol Microbiol 59:761−765.doi:10.1099/ijs.0.65837−0
【非特許文献19】Zhou Y,Wei W,Wang X,Xu L, Lai R(2009)Azospirillum palatum sp.nov.,isolated from forest soil in Zhejiang province,China.J Gen Appl Microbiol 55:1−7.
【非特許文献20】Peng G, Wang H,Zhang G,Hou W,Liu Y,Wang ET, Tan Z(2006)Azospirillum melinis sp.nov.,a group of diazotrophs isolated from tropical molasses grass.Int J Syst Evol Microbiol 56:1263−1271.doi:10.1099/ijs.0.64025−0
【非特許文献21】Mehnaz S,Weselowski B, Lazarovits G(2007a) Azospirillum canadense sp.nov.,a nitrogen−fixing bacterium isolated from corn rhizosphere.Int J Syst Evol Microbiol 57:620−624.doi:10.1099/ijs.0.64804−0
【非特許文献22】Mehnaz S,Weselowski B, Lazarovits G (2007b) Azospirillum zeae sp.nov.,a diazotrophic bacterium isolated from rhizosphere soil of Zea mays. Int J Syst Evol Microbiol 57:2805−2809.doi:10.1099/ijs.0.65128−0
【非特許文献23】Young CC,Hupfer H,Siering C,Ho MJ,Arun AB,Lai WA,Rekha PD,Shen FT, Hung MH, Chen WM, Yasshin AF(2008)Azospirillum rugosum sp.nov.,isolated from oil−contaminated soil.Int J Syst Evol Microbiol 58,959−963.doi:10.1099/ijs.0.65065−0
【非特許文献24】Han, S.O. and New, P.B. (1998) Isolation of Azospirillum spp. from natural soils by immunomagnetic separation. Soil Biol Biochem 30, 975−981.
【非特許文献25】Mcgilloway, R.L., Weaver, R.W., Ming, D.W. and Pillai, S.D. (2002) A PCR−MPN based quantitative approach to enumerate nitrifying bacteria in zeoponic substrates. J Rapid Meth Autom Microbiol 10, 49−58.
【非特許文献26】Mirza, M.S., Mehnaz, S., Normand, P., Combaret, C.P., Loccoz, Y.M., Bally, R. and Malik, K.A. (2006) Molecular characterization and PCR detection of a nitrogen−fixing Pseudomonas strain promoting rice growth. Biol Fertil Soils 43, 163−170.
【発明の概要】
【発明が解決しようとする課題】
【0006】
分子遺伝学的な識別法において、例えば、アゾスピリラム属菌については、ランダムに増幅される多型DNAの分析、及び増幅されたリボソームDNA断片の制限酵素分析をすることによって、アゾスピリラム属菌を他菌属と識別することが試みられた(非特許文献10、非特許文献11参照)。しかしながら、この方法は特定のアゾスピリラム属菌種、菌株のみを識別可能にすぎない。
【0007】
また、アゾスピリラム株の16S rDNAにハイブリダイズするオリゴヌクレオチドプローブも開発され、アゾスピリラム属に属する菌の所在をつきとめ、その動的特徴を知るために使用された(非特許文献13、非特許文献14、非特許文献16参照)。しかしながら、プローブを使用するこの方法は煩雑な実験操作を必要とし、精密な操作及び熟練性を必要とする。
【0008】
制限酵素及びプローブを用いた方法に比べ、プライマーセットを用いた方法は煩雑な実験操作を必要とせず、迅速かつシンプルな方法である。非特許文献3では、16S−23S rDNA遺伝子間スペーサ領域(ISR:internal spacer region)に基づき設計されたPCRプライマーセットfAZO/rAZOについて開示している。しかしこのプライマーセットは、菌株によってはアゾスピリラム属菌を認識できなかったり(偽陰性)、非アゾスピリラム属菌を誤認識したりする(偽陽性)ことがしばしばあり、検出精度に問題点がある。
【0009】
上記事情を鑑みて、本発明は、アゾスピリラム属菌をより精度よく、迅速かつシンプルに検出・識別することのできるプライマーセット、検出キット及び検出方法を提供することを目的とする。
【課題を解決するための手段】
【0010】
すなわち、本発明は以下の発明を包含する。
(1)アゾスピリラム属菌の16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅可能なプライマーセットであって、配列番号1で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。
(2)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる上記(1)に記載のプライマーセット。
(3)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなる上記(2)に記載のプライマーセット。
(4)上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加している上記(2)に記載のプライマーセット。
(5)上記(1)〜(4)の何れか一項に記載のプライマーセットを備えるアゾスピリラム属菌の検出キット。
(6)以下の工程:(a)対象サンプルからDNAを含有する試料を調製する工程、(b)上記試料に対して、上記(1)〜(4)の何れか一項に記載のプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、(c)上記工程(b)で得られる増幅断片の有無を検出する工程、を含むアゾスピリラム属菌の検出方法。
(7)上記対象サンプルが、土壌サンプル又は植物由来サンプルである(6)に記載の検出方法。
【発明の効果】
【0011】
本発明は、PCR法でアゾスピリラム属菌を検出することができるという効果を奏する。
【図面の簡単な説明】
【0012】
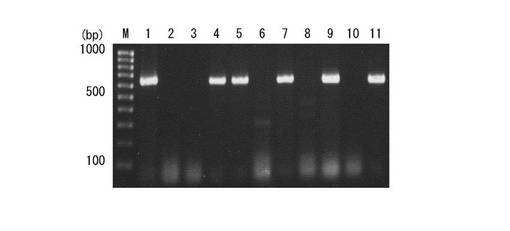
【図1】プライマーセットAz16S−Aを用いた、未知分離株に対するPCR実験の結果を示した図である。
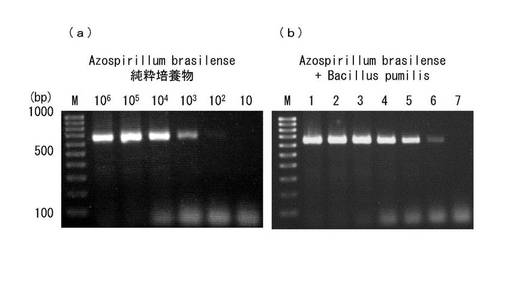
【図2】純粋培養物及び混合培養物における、プライマーセットAz16S−Aを用いたアゾスピリラム菌のPCR検出を示した図である。
【発明を実施するための形態】
【0013】
以下、本発明の実施の形態について、詳細に説明する。
【0014】
(プライマーセット)
本発明に係るプライマーセットは、アゾスピリラム属に属する細菌(アゾスピリラム属菌)の16S rDNAに由来するDNA断片を、ポリメラーゼ連鎖反応(PCR)によって増幅可能なものである。すなわち、本発明に係るプライマーセットは、アゾスピリラム属菌の16S rDNA及び16S rDNAの断片にハイブリダイズして、PCRにより所定サイズの上記DNA断片を増幅させる。
【0015】
上記プライマーセットは、アゾスピリラム属菌の16S rDNAに由来するDNA断片を、PCRによって選択的に増幅可能という特性を有する。ここで、「選択的に増幅可能」とは、当該プライマーセットを用いたPCRを行った場合に、アゾスピリラム属菌由来のDNA断片が専ら増幅産物として得られるが、他の多くの生物由来のDNA断片は増幅産物として実質的に得られないことを指す。実施例にも示すように、本発明に係るプライマーセットを用いた増幅では、アゾスピリラム属に属する複数種の細菌において共通のPCR増幅産物が得られる反面、アゾスピリラム属に近縁な細菌を含む多くの細菌ではPCR増幅産物が得られない。また、土壌サンプル及び植物含有サンプル(植物の根部等)に対するPCR増幅の結果から、本発明に係るプライマーは、植物、及びアゾスピリラム属菌ではない多くの土壌微生物由来のPCR増幅産物が実質的に得られないことが示されている(実施例参照)。従って、本発明に係るプライマーは、アゾスピリラム属菌の検出に極めて有用である。
【0016】
本発明に係るプライマーセットは、具体的には以下の(1)〜(4)の何れかに示すヌクレオチド(より具体的にはDNA)である。
(1)配列番号1で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。なお、配列番号1で表される塩基配列中のkはグアニン又はチミンを示す。なお、PCR増幅の選択性をより向上させる観点では、第一のプライマーは、配列番号1で示される塩基配列のうち連続する少なくとも18塩基部分を有するヌクレオチドからなることがより好ましい。また、同様に、第二のプライマーは、配列番号2で示される塩基配列のうち連続する少なくとも18塩基部分を有するヌクレオチドからなることがより好ましく、少なくとも20塩基部分を有するヌクレオチドからなることがさらに好ましい。また、後述するように第一のプライマー、及び第二のプライマーともに、上記条件を満たす複数種のプライマーの混合物であってもよく、単独(1種類)のプライマーであってもよい。
(2)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる上記(1)に記載のプライマーセット。
(3)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなる上記(2)に記載のプライマーセット。第一のプライマーとして当該混合物を用いれば、アゾスピリラム属菌に対する特異性がより高まり、より多種類にわたるアゾスピリラム属菌を増幅可能となる。
【0017】
上記第一のプライマーとして上記混合物を用いる場合、kがグアニンである上記ヌクレオチドとkがチミンである上記ヌクレオチドとの混合比率は特に限定されず、例えば、1:9〜9:1の範囲内、より好ましくは4:6〜6:4の範囲内とすることができる。
(4)上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加している上記(2)又は(3)に記載のプライマーセット。なお、基本となるプライマーに対して1〜2個の塩基に変異がある場合でも、所望のPCR増幅断片は得られる。また、上記(3)に記載のプライマーセットを構成する第一のプライマー(混合物)において、1〜2個の塩基が欠失又は置換する場合は、配列番号1で示される塩基配列におけるk以外の塩基が欠失又は置換しているものとする。
【0018】
また、特に限定されないが各プライマーのGC含量は、例えば、50%〜70%の範囲とするのが好ましく、50%〜65%の範囲とすることがより好ましい。プライマーのTm値も特に限定されないが、例えば、50℃〜65℃とすることが好ましく、50℃〜60℃とすることがより好ましい。
【0019】
本発明に係るプライマーセットは常法に従って合成することができる。また、本発明に係るプライマーセットを用いたPCRの反応条件は特に限定されない。
【0020】
(アゾスピリラム属菌の検出キット)
本発明に係るアゾスピリラム属菌の検出キットは、上記説明した本発明に係るプライマーセットを備える。この検出キットは、アゾスピリラム属菌の有無の判別を含む検出に用いる。さらに、必要に応じて、1)PCRに用いる各種試薬及び器具(ポリメラーゼ、PCRバッファー、各dNTP、ピペット等)、2)PCRに供するDNAを含有する試料を調製するための各種試薬及び器具(試験管、バッファー等)、3)PCR増幅断片を解析するための各種試薬及び器具(電気泳動ゲル材料、ピペット等)、4)さらなる詳細解析のため、アゾスピリラム属に属する特定種のみを増幅可能なPCRプライマーセット、5)検出キットの使用説明書、等の少なくとも1つを備えていてもよい。
【0021】
(アゾスピリラム属菌の検出方法)
本発明に係るアゾスピリラム属菌の検出方法は、以下の工程:
(a)対象サンプルからDNAを含有する試料を調製する工程、
(b)上記試料に対して、本発明に係るプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、
(c)上記工程(b)で得られる増幅断片の有無を検出する工程、
を含む。
【0022】
以下、各工程を説明する。
【0023】
(1)工程(a)
工程(a)は、対象サンプルからDNAを含有する試料を調製する工程である。
ここで、「対象サンプル」とは、アゾスピリラム属菌の有無を判別しようとするサンプルを意味する。対象サンプルは、アゾスピリラム属菌を含む可能性のあるものであればよく、限り特に限定されない。例えば、土壌サンプル、植物の根部等の植物由来サンプル、培地上の微生物コロニー、微生物を含む培養液等が挙げられる。
【0024】
DNAを含有する試料は、例えば、土壌サンプル等の対象サンプルから常法に従って調製することができる。例えば、ボイル法、CTAB法等が挙げられる。これらの中では、操作の簡便さの観点では、ボイル法が好ましい。なお、ボイル法とは、対象サンプルを液体中(例えば、滅菌蒸留水又はバッファー)に懸濁して短時間(例えば5分程度以下)加熱して、DNAを取り出す方法である。
【0025】
なお、調製されたDNAを含有する試料には、DNAの他に、各種RNA、タンパク質、細胞破砕物、等が含まれる。従って、該試料を常法に従って精製してもよい。但し、実施例にも示す通り、本発明の方法では、当該試料に対して一切の精製を行わなくとも、一度のPCRにより非常に高感度な検出を行うことができる。
【0026】
(2)工程(b)
工程(b)は、上記試料に対して、上記した本発明に係るプライマーセットを用いたPCRを行う工程である。工程(b)におけるPCRは、常法に従って行うことができる。工程(b)により、試料中にアゾスピリラム属菌が含まれている場合には、使用したプライマーの塩基配列を両端に有するアゾスピリラム16S rDNA由来のPCR増幅断片が得られる。
【0027】
(3)工程(c)
工程(c)は、工程(b)で得られるPCR増幅断片の有無を検出する工程である。PCR増幅断片(アンプリコン)の有無の確認は、常法に従って行うことができる。例えば、電気泳動によりPCR増幅断片の有無及びサイズを確認することができる。
【0028】
PCR増幅断片の有無の確認の結果、PCR増幅断片が存在する場合には、対象サンプルにアゾスピリラム属菌が存在すると判別することができる。また、PCR増幅断片の有無の確認の結果、PCR増幅断片が存在しない場合には、対象サンプルにアゾスピリラム属菌が存在していないと判別することができる。
【0029】
従来法による農業サンプルからのアゾスピリラム属菌の分離プロセスでは、選択培地の使用と菌の生理形態学的評価を繰り返し行うため、一定の特徴を持った菌株のみを分離する選択バイアスが生じる。例えば、非特許文献24では、窒素源を含まない培地を使用した従来の分離プロセスにおいて、高い窒素固定能を有するアゾスピリラム属菌が分離され、低い窒素固定能の菌株は分離できなかったことを開示している。後述する実施例にも示す通り、本発明の一形態としてのAz16S−Aプライマーセットを使用したアゾスピリラム属菌のPCR検出により、選択培地と生理形態学的評価の使用を減らすことができるため、結果的に選択バイアスを低減させた分離プロセスを実現できる。また、本発明の一形態であるこのPCR検出法は、農業サンプルから多様なアゾスピリラム属菌を分離し、新規の植物成長促進細菌株を探索する上で有用な手法、またはアゾスピリラム属菌集団の多様性と生態とを分析するための有用な手法として利用できる。多数のアゾスピリラム属菌及び分離株を対象にしてAz16S−AプライマーセットによるPCR増幅産物(アンプリコン)のDNA配列を決定したところ、少なくとも15の一塩基多型(SNP)が見出され、そのうち12の一塩基多型は購入可能な制限酵素の認識サイトである。さらに、この12のSNPのうちの一つは、アゾスピリラム属菌とR.centenariaとの識別に有用であると見込まれる。一塩基多型について更なる分析と確認が必要であるが、おそらく、Az16S−Aプライマーセットの利用により、一回のPCRと制限酵素処理(例えば、PCR−RFLP分析法)によって、アゾスピリラム属分離株またはアゾスピリラム属菌集団の精密な同定及び多様性分析の両方が可能である。さらに、発明者らの実験では、根圏土壌と植物体部位に対してAz16S−Aプライマーセットを使用し、アゾスピリラム属菌の局在性を予測している。このような微生物動態の分析では、非特許文献16で開示された、蛍光標識されたオリゴヌクレオチドプローブも使用できるが、しかし我々の方法は、アゾスピリラム属菌の動態分析後の分離プロセスにおいても利用できる点で優位性がある。PCR法は最確数(MPN)法と併用することによって、アゾスピリラム属菌集団の定量にも利用できる(非特許文献25、26参照)。本発明のPCR法を組み込んだ迅速、容易且つ選択バイアスが最小となったアゾスピリラム分離技術、及びAz16S−Aプライマー配列に基づくリアルタイムPCR定量技術に対するさらなる応用が考えられる。
【実施例】
【0030】
〔実施例1〕
<プライマーセットの設計及び作製>
アメリカ国立バイオテクノロジーインフォメーションセンター (NCBI, USA, http://www.ncbi.nlm.nih.gov/genbank/GenBankFtp.html)ジーンバンクヌクレオチドデータベースから合計30個のアゾスピリラム16S rDNA配列(14の種から)を入手し、バチルス属菌、Pseudomonas属菌、Roseomonas属菌を含む関連する類縁細菌群の16S rDNA配列と比較しながら、16S rDNAの開始コドンの下流300bp〜1200bpにおける、推測上の属特異的且つ保存的領域について同定した。
【0031】
推測上のアゾスピリラム属特異的領域に基づき、属特異的なプライマーセットを設計するために、Geneious Pro 4.8.5ソフトウエア(Biomatters Ltd社, Auckland,New Zealand)を使用して、アゾスピリラム及びその類縁細菌群に属する菌に対して、DNA配列アライメント解析を実施した。同様に、ジーンバンクから6つのipdC遺伝子配列(3つのアゾスピリラム属菌種から)、3つのnifA遺伝子配列(2つのアゾスピリラム属菌種から)、12のnifH遺伝子配列(3つのアゾスピリラム属菌種から)を入手し、プライマーセットを設計した。ipdC、nifA及びnifH遺伝子は何れも細菌による植物成長促進において重要な遺伝子である。具体的には、ipdC遺伝子は、インドール−3−酢酸生合成酵素をコード化するものであり、nifA遺伝子及びnifH遺伝子はそれぞれ、窒素固定のためのニトロゲナーゼ酵素の制御遺伝子及び構造遺伝子をコード化するものである。
【0032】
各プライマーセットの設計に際して、次の条件(1)〜(3)を適用した。(1)原則として20個前後、好ましくは20個を超えるヌクレオチド塩基を有するプライマーセットを設計し、(2)上記プライマーセットは200−800bpのDNA断片(アンプリコン)を生成させると予測されるものを選択し、(3)上記プライマーセットのヌクレオチドコードはアゾスピリラム属菌の間で同じかまたは最大で2塩基程度異なるものを許容するように設計した。表1は、Baudoinらによって報告されたfAZO/rAZOプライマー(非特許文献3参照)と共に、今回設計されたプライマーを表示したものである。設計されたが、簡易PCRを伴う予備分析(preliminary assays)によって排除されたものは、表1には示されていない。
【0033】
<結果>
表1は、今回設計した9つのプライマーセットと共に、Baudoinらによって報告されたプライマーセットfAZO/rAZO(非特許文献3参照)を表示したものである。プライマー名に16Sが含まれるもの、ipdが含まれるもの、nifが含まれるものは、順に、16S rDNAの塩基配列、ipdの塩基配列、nifの塩基配列に基づき設計されたものである。設計されたが予備分析(preliminary assays)によって排除されたものは、表1には示されていない。表において、KはG及びTの二種の塩基の混合(すなわち、Kがグアニンであるプライマーと、Kがチミンであるプライマーとの混合物。両者の混合比率は約1:1。)であることを意味している。アンプリコンサイズはアゾスピリラム・ブラシレンセ JCM 1224TSのDNA配列により推測されたものである。
【0034】
【表1】

【0035】
〔実施例2〕
<プライマーセットの属特異性評価>
12のアゾスピリラム標準株(reference strains)及び15の非アゾスピリラム標準株について、実施例1で作製したプライマーセットの属特異性について調べた。
【0036】
使用した12のアゾスピリラム標準株及び15の非アゾスピリラム標準株、合計27の標準株を以下に示した。TSは基準株であることを示している。
【0037】
(アゾスピリラム属の12の標準株)
(1)A. brasilense JCM 1224TS
(2)A. brasilense JCM 1225
(3)A. brasilense JCM 1226
(4)A. lipoferum JCM 1227
(5)A. lipoferum JCM 1228
(6)A. lipoferum JCM 1247TS
(7)A. lipoferum JCM 1270
(8)A. amazonense DSM 2787TS
(9)A. halopraeferens DSM 3675TS
(10)A. irakense DSM 11586TS
(11)A. picis DSM 19922TS
(12)A. rugosum DSM 19657TS
(非アゾスピリラム属の15の標準株)
アゾスピリラム属の類縁細菌群に属するもの:
(1)Rhodocista centenaria DSM 9894TS
(2)Magnetospirillum magnetotacticum DSM 3856TS
(3)Paeospirillum fulvum DSM 113TS
(4)Rhodospirillum rubrum DSM 467TS
(5)R. salinarum DSM 9154TS
(6)Thalassospira lucentensis DSM 14000TS
そのほかのプロテオバクテリア:
(7)Agrobacterium tumefaciens MAFF 301224
(8)Agrobacterium tumefaciens MAFF 301540
(9)Rhizobium leguminosarum MAFF 210039
(10)Alcaligenaceae sp. RERT 020126
(11)Stenotrophomonas maltophilia MAFF 301690
(12)Stenotrophomonas maltophilia RERT 020018
Firmicutesに属するもの:
(13)Bacillus pumilus MAFF 118256
(14)Bacillus pumilus RERT 020111
(15)Microbacterium esteraromaticum RERT 020020
12のアゾスピリラム及び15の非アゾスピリラム、合計27の標準株(表2を参照)は、ドイツ微生物・培養細胞収集有限会社(Deutsche Sammlung von Mikroorganismen und Zellkulturen、DSMZ)から(11のDSM菌株)、日本独立行政法人理化学研究所バイオリソースセンター微生物材料開発室(Japan Collection of Microorganisms、JCM)から(7つのJCM菌株)、日本独立行政法人農業生物資源研究所(National Institute of Agrobiological Sciences、NIAS)から(5つのMAFF菌株)、および発明者の細菌コレクションから(上記(10)、(12)、(14)、(15)のRERT菌株4つ)入手したものである。12のアゾスピリラム属菌株は、PCR実験の時点で、上記機関において入手できるアゾスピリラム属7種すべてを含む。International Journal of Systematic and Evolutionary Microbiology及びNCBIで報告された14のアゾスピリラム属菌種のうち(非特許文献18、非特許文献19参照)、その種の菌株の取得または培養の困難さのために7つの種は本実施例での考慮対象外とした(例えばA.doebereinerae DSMZ 13131TS)。
【0038】
これまでの系統分類学的解析では、種間の関係が分析間で多少異なる点はあるが、考慮対象外とした上記7つの種は何れも本研究で使用された7つのアゾスピリラム属菌種の間に位置するという結果を示す。考慮対象外とした上記7つの種は、
(1)A.doebereinerae(非特許文献9、非特許文献19)、
(2)A.largimobile(非特許文献9、非特許文献20)、
(3)A.canadense(非特許文献21、非特許文献23、非特許文献19)、
(4)A. melinis(非特許文献21、非特許文献23)、
(5)A. zeae(非特許文献22、非特許文献23)、
(6)A. oryzae(非特許文献20、非特許文献24)、
(7)A.palatum(非特許文献19参照)である。
【0039】
15の非アゾスピリラム菌株は、類縁細菌群(非特許文献17参照)とBaudoinら(非特許文献3参照)が研究において使用した土壌バクテリアの菌株から選択された。
【0040】
プライマーセットの属特異性の分析実験は、次のように行われた。27の標準株の定常期培養物(stationary−phase cultures)の一定分量を、遠心し、滅菌蒸留水に再懸濁させた溶液を95°Cにおいて5分間放置した後、遠心を行いPCRの鋳型となる細胞溶解物上清を得た。
【0041】
PCR反応混合物は、10xPCRバッファー2.5μl、1.5mM MgCl2、各dNTPを0.25mMずつ、各プライマーを0.02μMずつ、Taq DNAポリメラーゼ(BIOTAQ,BioLine,London,UK)を2.5ユニット、粗製のバクテリア溶解物10μlを含み、総量は25μlである。
【0042】
PCRはVeritiサーマルサイクラー(Applied Biosystems,CA,US)によって、下記条件に基づいて実施された。まず94°Cにおいて3分間放置し、初期変性させる。次に、下記操作を35サイクル繰り返す。各サイクルは、94°Cで30秒間変性させた後、各プライマーセットの最適温度で30秒アニーリングさせ、72°Cで1分間伸長させる。上記最適温度として、Az16S-A(配列番号1及び2として示す),Az16S-B、Az16S‐Cは58°Cである。AzipdC、AznifA−A、AznifA−Bは52°Cである。AznifH‐A,AznifH‐B,AznifH‐Cは55°Cである(塩基配列に関しては表1を参照)。上記35サイクルが終了した後、72°Cで5分間放置し最終的な伸長をさせた。Baudoinらによって報告されたfAZO/rAZOプライマーを使用したPCRは、彼らの報告した手順に従って実施した。PCR産物は、100−bpDNAラダー(HyperLadderIV,BioLine)と共に2%アガロースゲル中に展開され、臭化エチジウム溶液によって染色され、AE‐6905H Image Saver HR(ATTO,Tokyo,Japan)によって撮影された。
【0043】
このPCR実験によって選ばれたAz16S‐Aプライマーセットの属特異性は、さらに、35のアゾスピリラム様分離株及び70の未知の分離株に対しても、同様のPCR及び電気泳動の手順によって分析された(後述の実施例3を参照)。
【0044】
<結果>
上記属特異性分析実験の結果を、表2に示した。表2において+及び−はそれぞれ、予測アンプリコンの有無を示している。すなわち、+は予測アンプリコンが得られたことを指し、−は予測アンプリコンが得られなかったことを指す。
【0045】
【表2】
【0046】
実験が行われた10のプライマーセット(表1を参照)において、16S rDNA配列に基づき設計されたプライマーセットAz16S-Aは、12のアゾスピリラム標準株(7の種)すべて及び1の非アゾスピリラム属菌株Rhodocista centenaria DSM9894(表2参照)に対して、単一のアンプリコンを生成させることに成功した。アンプリコンは単一のDNAバンドとして観察され、上記DNAバンドの長さはおおよそ640bpであって(図1を参照)、この長さはA.brasilense 16S rDNA配列から予測された長さ(表1を参照)に一致するものである。なお、非アゾスピリラム属菌株Rhodocista centenariaは、自然界では温泉などの特殊環境に棲息することが報告されており、アゾスピリラム属菌とは棲息環境が異なる。
【0047】
プライマーセットAz16S-Bを用いたPCRは、12のアゾスピリラム標準株すべてから予測アンプリコンを生成させただけでなく、α−プロテオバクテリア及びβ−プロテオバクテリアにわたる10の非アゾスピリラム属菌株からも、予測アンプリコンを生成させた。
【0048】
プライマーセットAz16S-C、AznifH-A、及びAznifH-Bは、Baudoinらによって報告されたプライマーセットfAZO/rAZOと同じように、12のアゾスピリラム属菌株のうち、1−3の株に対してはアンプリコンを生成させることができず、17の非アゾスピリラム属菌株のうち、2-5の株に対しては複数のアンプリコンを生成させた。これらの4のプライマーセットにおいて、プライマーセットAznifH-A及びAznifH-Bは、他の2のプライマーセットに比べ比較的良い属特異性を示した。残りの4つのプライマーセットからは良くない結果が得られた。従って、プライマーセットAz16S-Aはアゾスピリラム属菌に対して最も優れた特異性を示した。
【0049】
ここで、図1について説明すると、図1は、プライマーセットAz16S−Aを用いた、未知分離株に対するPCR実験の結果を示している。PCR産物は2%アガロースに溶解され、臭化エチジウム溶液によって染色された。識別された分離株は、16SrDNA配列決定解析によって、下記のように確認された。レーン1はAchromobacter及びRahnellaによって汚染された、アゾスピリラム属菌培養物であり、レーン2はAchromobacter、レーン3はAchromobacter、レーン4はアゾスピリラム、レーン5はアゾスピリラム、レーン6はAchromobacter、レーン7はアゾスピリラム、レーン8はStenotrophomonas、レーン9はアゾスピリラム、レーン10はAchromobacter、レーン11は参照物としてのA. brasilense JCM 1224TS、レーンMは100−bpDNAラダーである。
【0050】
〔実施例3〕
<作製されたプライマーセットの利用可能性>
さらに、35のアゾスピリラム様分離株と、農業サンプルから得られた70の未知の分離株を用いて、選択されたプライマー候補の特異性、検出限界及び利用可能性について調べた。アゾスピリラム属菌の分離において検出限界は重要な面であって、根圏土壌及び植物体から分離するプロセスにおいては、アゾスピリラム属の菌落及び培養物はしばしば非アゾスピリラム微生物によって汚染されているからである。
【0051】
(使用された35のアゾスピリラム様分離株と70の未知の分離株)
アゾスピリラム様分離株は、根圏土壌並びに作物及び草の根部から収集されたものである。上記収集は、独立行政法人農業生物資源研究所(NIAS)と十勝農業協同組合連合会農産化学研究所(ARITFAC)によって、選択培地及び形態生理学評価の両方を用いて行われた。選択培地は例えば、窒素不含有ブロモチモールブルー(NFb)培地、Rojo Congo(コンゴレッド)培地である。形態生理学評価は、例えば、アセチレン減少分析(ARA)(非特許文献5、非特許文献9)である。これらの分離株のrDNA配列データは入手できないものであった。全部で35のアゾスピリラム様分離株を、上記収集されたコレクションからランダムに選出した。さらに、根圏土壌、穀物及び飼草の根部から、ARAとrDNA配列決定を用いずに、選択培地のみによって収集した我々のストックから、70の未知の分離株をランダムに選出した。これらの分離株の「属」及び「種」は未知であった。使用された35のアゾスピリラム様分離株は多数の、70の未知の分離株は少数の、アゾスピリラム属菌を含むものと考えられる。
【0052】
(作製したプライマーセットによる検出実験)
すべての菌株及び分離株に対して、16S rDNA配列決定解析を行った。ボイル法によって抽出された粗製DNAは、アゾスピリラム16S rDNA配列に基づき設計されたAz16S-D配列決定プライマーによって、PCR増幅された。Az16S-D配列決定プライマーセットのフォワードプライマーは5’CCGCGGTAATACGAAGGGGGCであり、リバースプライマーは5’GCCTTCCTCCGGCTTGTCACCGGCである。当該プライマーセットによるアンプリコンサイズは、おおよそ650bpと推測され、それは属レベルで系統発生的分類をするのには充分である。PCRはアニーリング温度60°Cで、前記のPCR実験で記載されているのと同じ手順によって実施された。PCR産物はMin−Elute PCR精製キット(QIAGEN,Hilden,Germany)によって精製され、Az16S-Dフォワードプライマー、BigDye Terminator v3.1サイクル配列決定キット,3730xlDNAアナライザー(Applied biosystems)によって配列決定された。配列データは、SEQ MATCH program ver.3.0(非特許文献7参照)を使用し、リボソームデータベースプロジェクト(リリース10、ミシガン州立大学微生物生態学センター[http://rdp.cme.msu.edu/index.jsp])で得られたデータと比較し、カットオフ類似性(Sab)値を>0.7に設定しマッチする微生物種を探索し上位20マッチに基づき菌株及び分離株の属を決定した。
【0053】
<結果>
上記作製したプライマーセットによる検出実験の結果を、表3に示した。
【0054】
【表3】
【0055】
表3は、35のアゾスピリラム様分離株及び70の未知分離株に対するPCR実験及びrDNA配列決定解析の結果を比較したものである。表3において、NIASは独立行政法人農業生物資源研究所であり、ARITFACは十勝農業協同組合連合会農産化学研究所である。+と−はそれぞれ、予測されたアンプリコンの有無を示している。
【0056】
表3によれば、プライマーセットAz16S-Aを用いたPCRは、NIASから入手した20のアゾスピリラム様分離株のうち17の菌株から、予測された単一のアンプリコンを得た。PCR検出においてアンプリコンが得られた17の分離株は、16S rDNA配列決定解析によって、アゾスピリラム属に分類され、残りの3つの株は、Williamsia、Bacillus、Rhizobiumに分類された。PCR実験及びrDNA配列決定解析の結果における完全な一致は、ARITFACから入手した15のアゾスピリラム様分離株及び我々のストックから入手した70の未知分離株においても見られた(表3及び図1を参照)。アンプリコンが得られた5つの未知分離株のうちの一つ(図1レーン1参照)のrDNA配列決定解析は、この培養物はアゾスピリラムのほかにも二つの種、Achromobacter及びRahnellaを含んでいることを示した。従って、プライマーセットAz16S-Aを用いたPCRは、混合培養物におけるアゾスピリラムを成功裏に検出することができた。Baudoinらによって報告されたプライマーセットfAZO/rAZOを用いたPCRでは、アゾスピリラム疑似分離株に対して、PCR実験及びrDNA配列決定解析の結果における完全な一致が見られた。しかし、70の未知分離株のうち(詳細は示していない)、2つの非アゾスピリラム分離物(BacillusとPantoea)からも、5つのアゾスピリラムと同様に、アンプリコンを生成させた。
【0057】
NIASから入手した分離株のうち、3つの非アゾスピリラム分離株は、Williamsia,Bacillus,Rhizobium種である。同じように、我々のストックからの67の非アゾスピリラム分離株のうち、34はBacillusとして同定され、8はMicrobacteriumとして、6はPaenibacillusとして、6はStenotrophomonasとして、6はAcromobacterとして、2はAlcaligenesとして、2はLysinibacillusとして、1はHerbaspirilluとして、1はPantoraとして、1はYersiniaとして同定された。
【0058】
〔実施例4〕
<アンプリコンの存在を示した分離株のキャラクタリゼーション>
上記実施例3において、70の未知分離株のうち、Az16S-Aプライマーを用いたPCRによって得られたアンプリコンの存在が確認されたのは5つの分離株である。この5つの分離株は、16S rDNA配列決定解析によって、アゾスピリラム属に分類された。これらの属種及び生理学的特性をさらに調べるため、この5つの分離株における炭素源利用、IAA生成、窒素固定化について、A. brasilense JCM 1224TS及びA. brasilense JCM 1226と比較しながら評価した。炭素源利用の評価についてはAPI20NEシステム(bioMERIEUX,Lyon,France)を用いて、製造元のプロトコルに従って実施した。IAA生成については、修飾されたSalkowski試薬(非特許文献12を参照)を用いて比色法によって計測された。分離株は1mMトリプトファンを補充したLuria−Bertani(LB)ブロスにおいて、30°C条件下において2日間、定常期になるまで培養され、それからIAAの計測に供された(非特許文献8を参照)。窒素固定能は、水素炎イオン検出器を備えたガスクロマトグラフ装置(GC-4000 GL Science,Tokyo,Japan)を使用して、ARA法(非特許文献9、非特許文献15を参照)によって見積もられた。ARAを行うため、バクテリア培養物は、2mlの濃度1%(v/v)の半固体NFb培地に添加され、9−ml密封バイアル瓶中において、30°C条件下で24時間培養された。上記培養後に10体積%(気体体積)のアセチレンをバイアル瓶中に導入し、さらに24時間培養し、それからガスクロマトグラフィーを実施した。それぞれの分離株のIAA生成及びアセチレン減少については、それぞれ3〜4のバイアル瓶で重複して実験された。
【0059】
<結果>
上記アンプリコンの存在を示した分離株のキャラクタリゼーション実験の結果を、表4に示した。
【0060】
【表4】
【0061】
表4は、いわゆる農業サンプルからPCR実験によって分離されたアゾスピリラム属菌の炭素源利用、インドール−3−酢酸(IAA)生成及びアセチレン減少について示した表である。炭素源利用タイプはAPI20NEシステムによって評価されたものであり、ABタイプはA. brasilenseに特有の利用型であって、N−アセチルグルコサミン(−)、L−アラビノース(+)、D−グルコース(−)、D−マンニトール(−)、D−マンノース(−)を利用するものである。
【0062】
我々のストックから入手の分離株のうち、アンプリコンが得られた5つの分離株は、A .brasilense(表4を参照)に特有的な炭素源利用型と同様の炭素源利用型を示した。IAA生成能及びアセチレン減少(窒素固定)能力は、5つの分離株すべてにおいて見られた。分離株におけるこれら二つの能力は、A. brasilense JCM 1224TSより優れていた。アセチレン減少に見られる分離株同士の大きな差異は、IAA生成に見られる分離株同士より顕著であった。分離株のIAA生成とアセチレン減少との間には、顕著な相関性は観察されなかった。これらの結果から、プライマーセットAz16S‐Aは、アゾスピリラム独特の生理学特性を示す分離株を正確に識別する能力を有することがさらに確認された。
【0063】
〔実施例5〕
<プライマーセットの検出限界>
Az16S-Aプライマーを用いたPCRを、異なる濃度(10‐106CFUml−1)のA. brasilense(JCM 1224TS)に対して、上記と同じ手順にて実施した。さらに、Bacillus pumilus(MAFF 118256)が混入されたA. brasilense(JCM 1224TS)培養物に対しても、PCRを実施した。バチルス属菌がモデル汚染菌として選ばれた理由は、我々の実験系において、アゾスピリラム選択培地に観察される非アゾスピリラム属菌の大部分においてバチルス属菌が存在するからである。混合培養物を得るために、アゾスピリラム及びバチルス培養物はそれぞれ、LB培養液において37℃条件下において48時間及び24時間培養された。それから、最終菌濃度が107CFUml−1となるように、A. brasilense:B. pumilusを1:1〜1:104の異なる比率で混合した。プライマーの検出限界とは、検出可能なPCRアンプリコンをもたらすことのできる、最小のアゾスピリラム濃度または比率である。
【0064】
<結果>
プライマーセットAz16S-Aを用いるPCRは、103CFUml−1の濃度を有するA. brasilense純粋培養物から、予測サイズに該当する検出可能な量のアンプリコンを生成させることが出来た(図2中の(a)を参照)。同様に、A. brasilense:B. pumilusを1:104またはそれ以上の比率で混合して得られた混合培養物からも、PCRでは同じように予想アンプリコンを生成することが出来た(図2中の(b)を参照)。A.brasilense:B. pumilusを1:104の比率で混合した混合培養物は、おおよそ103CFUml−1のA. brasilenseを含んでいる。したがって、プライマーセットの検出限界は、純粋培養物及び混合培養物において同じである。さらに、PCRの汚染菌存在時におけるアゾスピリラム菌の検出能力は、農業サンプルから得られた70の未知分離株においても確認された(図1レーン1参照)。
【0065】
なお、図2は純粋培養物及び混合培養物におけるプライマーセットAz16S−Aを用いたアゾスピリラム属菌のPCR検出を示した図である。図中(a)のPCR産物はA. brasilense JCM 1224TS純粋培養物から得られ、図中(b)のPCR産物はBacillus pumilus MAFF 118256が混合されたA. brasilense 培養物から得られた。PCR産物は2%アガロースに展開され、臭化エチジウム溶液によって染色された。図中(a)における、レーン上部の数字は、A. brasilense濃度(CFU ml−1)である。レーンMは100−bpDNAラダーである。図中(b)において、レーン1はA. brasilenseのみ、レーン2は、A. brasilense:B. pumilusを1:1で混合したもの、レーン3は、1:10で混合したもの、レーン4は、1:102で混合したもの、レーン5は、1:103で混合したもの、レーン6は1:104で混合したもの、レーン7は、B. pumilusのみのもの、を夫々示す。PCR産物は2%アガロースゲルに展開され、臭化エチジウム溶液によって染色された。
【産業上の利用可能性】
【0066】
本発明は、土壌サンプルを始め、各種サンプルからアゾスピリラム属菌を検出・同定する際に好適に利用することができる。
【技術分野】
【0001】
本発明はプライマーセット、アゾスピリラム属菌の検出キット及び検出方法に係るものである。詳しくは、アゾスピリラム属菌16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅し得るプライマーセット、当該プライマーセットを用いたアゾスピリラム属菌の検出キット及び検出方法に係るものである。
【背景技術】
【0002】
アゾスピリラム属菌は、窒素固定能を有する好気性の桿状α−プロテオバクテリアとして知られている。アゾスピリラム属菌は世界中において、根圏土壌及び多種の植物根部から分離されている。多くのアゾスピリラム属菌株は、植物成長促進細菌(plant growth promoting bacteria,PGPB)として報告されている。植物成長促進細菌とは、作物の生長を促進させることができる細菌である。この成長促進は、アゾスピリラム属菌株による植物ホルモンの生成、窒素固定、植物免疫によってもたらされるものとして考えられている。従って、アゾスピリラム属の細菌は、バイオ肥料及びバイオ殺虫剤として利用され、近年の農業用の化学薬品を減らす需要により、多くの注目を集めている。
【0003】
しかしながら、根圏土壌及び植物体からのアゾスピリラム属菌の識別及び分離は、選択培地及び形態生物学的評価による従来的な方法に頼るしかなかった。従来の方法では、複数の選択培地(例えばNFb、RC培地など)を利用しながら、菌の形態または窒素固定能などを評価して、候補となる菌株を絞り込んだ上で、さらにrDNA配列の解読を行う必要があった。これらの従来の方法は、アゾスピリラム属菌に対する専門知識と実験経験、1〜2ヶ月の時間と、多大な労力とコストを必要とする。さらに、選択培地の利用と菌種特性の評価を繰り返すため、培地適応性と評価値の高い菌株が分離されやすく、分離した菌株には意図していない選択バイアスが生じることがある。
【0004】
最近のDNA配列解析技術の発展によって、ターゲットとなる微生物の検出及び識別において分子遺伝学的な識別法が考えられるようになった。非特許文献1〜非特許文献23は、アゾスピリラム属菌の検出と識別にかかわる情報を開示しており、これらのうち、特に非特許文献1〜3,7,10,11,13〜17は分子遺伝学的な識別法について開示しているものである。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Baldani JI, Krieg NR, Baldani VLD, Hartmann A, Dobereiner J (2005) Genus II. Azospirillum. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s manual of systematic bacteriology 2nd edn vol 2 part C. Springer, NY, pp 16−17
【非特許文献2】Bashan Y, Holguin G, Lifshitz R (1993) Isolation and characterization of plant growth−promoting rhizobacteria. In: Glick BR, Thompson JE (eds) Methods in plant molecular biology and biotechnology. CRC press, FL, pp 331−345
【非特許文献3】Baudoin E, Couillerot O, Spaepen S, Moenne−Loccoz Y, Nazaret S (2010) Applicability of the 16S−23S rDNA internal spacer for PCR detection of the phytostimulatory PGPR inoculants Azospirillum lipoferum CRT1 in field soil. J Appl Microbiol 108:25−38. doi:10.1111/j.1365−2672.2009.04393.x
【非特許文献4】Boddey RM, Baldani VLD, Baldani JI, Dobereiner J (1986) Effect of inoculation of Azospirillum spp. on nitrogen accumulation by field−grown wheat. Plant Soil 95:109−121. doi:10.1007/BF02378857
【非特許文献5】Burris RH (1972) Nitrogen fixing−assay methods and techniques. Methods Enzymol 24:415−431
【非特許文献6】Caceres EA (1982) Improved medium for isolation of Azospirillum spp. Appl Environ Microbiol 44:990−991
【非特許文献7】Cole JR, Chai B, Farris RJ, Wang Q, Kulam SA, McGarrell DM, Garrity GM, Tiedje JM (2005) The ribosomal database project (RDP−II): sequences and tools for high−throughput rRNA analysis. Nucleic Acids Res 33:294−296 doi:10.1093/nar/gki038
【非特許文献8】Dobbelaere S, Croonenborghs A, Thys A, Broek AV, Vanderleyden J (1999) Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil 212:155−164. doi:10.1007/BF02378857
【非特許文献9】Eckert B, Weber OB, Kirchh G, Halbritter A, Stoffels M, Hartmann A (2001) Azospirillum doebereinerae sp. nov., nitrogen−fixing bacterium associated with the C4−grass Miscanthus. Int J Syst Bacteriol 33:300−308
【非特許文献10】Fancelli S, Castaldini M, Ceccherini MT, Serio CD, Fani R, Gallori E, Marangolo M, Miclaus N, Bazzicalupo M (1998) Use of random amplified polymorphic DNA markers for the detection of Azospirillum strains in soil microcosms. Appl Microbiol Biotechnol 49:221−225. doi:10.1007/s002530051162
【非特許文献11】Grifoni A, Bazzicalupo M, Serio CD, Fancelli S, Fani R (1995)Identification of Azospirillum strains by restriction fragment length polymorphism of the 16S rDNA and of the histidine operon. FEMS Microbiol Lett 127:85−91. doi:10.1111/j.1574−6968.1995.tb07454.x
【非特許文献12】Glickmann E, Dessaux Y (1995) A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl Environ Microbiol 61:793−796
【非特許文献13】Jacoud C, Faure D, Wadoux P, Balley R (1998) Development of a strain−specific probe to follow inoculated Azospirillum lipoferum CRT1 under field conditions and enhancement of maize root development by inoculation. FEMS Microbial Ecol 27:43−51 doi:10.1111/j.1574−6941.1998.tb00524.x
【非特許文献14】Kabir MM, Faure D, Haurat J, Normand P, Jacoud C, Wadoux P, Bally R (1995) Oligonucleotide probes based on 16S rRNA sequences for the identification of four Azospirillum species. Can J Microbiol 41: 1081−1087. doi:10.1139/m95−151
【非特許文献15】Mehhaz S, Mirza MS, Haurat J, Bally R, Normand P, Bano A, Malik KA (2001) Isolation and 16S rRNA sequence analysis of the beneficial bacteria from the rhizosphere of rice. Can J Microbiol 47:110−117. doi:10.1139/cjm−47−2−110
【非特許文献16】Stoffels M, Castellanos T, Hartmann A (2001) Design and application of new 16S rRNA−targeted oligonucleotide probes for the Azospirillum−Skermanella−Rhodocista−cluster. Syst Appl Microbiol 24:83−97. doi:101078/0723−2020−00011
【非特許文献17】Xia Y, Embley TM, O’donnell AG (1994) Phylogenetic analysis of Azospirillum by direct sequencing of PCR amplified 16S rDNA. System Appl Microbiol 17:197−201
【非特許文献18】Lin SY,Young CC,Hupfer H,Siering C,Arun AB,Chen WM,Lai WA,Shen FT,Rekha PD, Yasshin AF.(2009)Azospirillum picis sp.nov.,isolated from discarded tar.Int J Syst Evol Microbiol 59:761−765.doi:10.1099/ijs.0.65837−0
【非特許文献19】Zhou Y,Wei W,Wang X,Xu L, Lai R(2009)Azospirillum palatum sp.nov.,isolated from forest soil in Zhejiang province,China.J Gen Appl Microbiol 55:1−7.
【非特許文献20】Peng G, Wang H,Zhang G,Hou W,Liu Y,Wang ET, Tan Z(2006)Azospirillum melinis sp.nov.,a group of diazotrophs isolated from tropical molasses grass.Int J Syst Evol Microbiol 56:1263−1271.doi:10.1099/ijs.0.64025−0
【非特許文献21】Mehnaz S,Weselowski B, Lazarovits G(2007a) Azospirillum canadense sp.nov.,a nitrogen−fixing bacterium isolated from corn rhizosphere.Int J Syst Evol Microbiol 57:620−624.doi:10.1099/ijs.0.64804−0
【非特許文献22】Mehnaz S,Weselowski B, Lazarovits G (2007b) Azospirillum zeae sp.nov.,a diazotrophic bacterium isolated from rhizosphere soil of Zea mays. Int J Syst Evol Microbiol 57:2805−2809.doi:10.1099/ijs.0.65128−0
【非特許文献23】Young CC,Hupfer H,Siering C,Ho MJ,Arun AB,Lai WA,Rekha PD,Shen FT, Hung MH, Chen WM, Yasshin AF(2008)Azospirillum rugosum sp.nov.,isolated from oil−contaminated soil.Int J Syst Evol Microbiol 58,959−963.doi:10.1099/ijs.0.65065−0
【非特許文献24】Han, S.O. and New, P.B. (1998) Isolation of Azospirillum spp. from natural soils by immunomagnetic separation. Soil Biol Biochem 30, 975−981.
【非特許文献25】Mcgilloway, R.L., Weaver, R.W., Ming, D.W. and Pillai, S.D. (2002) A PCR−MPN based quantitative approach to enumerate nitrifying bacteria in zeoponic substrates. J Rapid Meth Autom Microbiol 10, 49−58.
【非特許文献26】Mirza, M.S., Mehnaz, S., Normand, P., Combaret, C.P., Loccoz, Y.M., Bally, R. and Malik, K.A. (2006) Molecular characterization and PCR detection of a nitrogen−fixing Pseudomonas strain promoting rice growth. Biol Fertil Soils 43, 163−170.
【発明の概要】
【発明が解決しようとする課題】
【0006】
分子遺伝学的な識別法において、例えば、アゾスピリラム属菌については、ランダムに増幅される多型DNAの分析、及び増幅されたリボソームDNA断片の制限酵素分析をすることによって、アゾスピリラム属菌を他菌属と識別することが試みられた(非特許文献10、非特許文献11参照)。しかしながら、この方法は特定のアゾスピリラム属菌種、菌株のみを識別可能にすぎない。
【0007】
また、アゾスピリラム株の16S rDNAにハイブリダイズするオリゴヌクレオチドプローブも開発され、アゾスピリラム属に属する菌の所在をつきとめ、その動的特徴を知るために使用された(非特許文献13、非特許文献14、非特許文献16参照)。しかしながら、プローブを使用するこの方法は煩雑な実験操作を必要とし、精密な操作及び熟練性を必要とする。
【0008】
制限酵素及びプローブを用いた方法に比べ、プライマーセットを用いた方法は煩雑な実験操作を必要とせず、迅速かつシンプルな方法である。非特許文献3では、16S−23S rDNA遺伝子間スペーサ領域(ISR:internal spacer region)に基づき設計されたPCRプライマーセットfAZO/rAZOについて開示している。しかしこのプライマーセットは、菌株によってはアゾスピリラム属菌を認識できなかったり(偽陰性)、非アゾスピリラム属菌を誤認識したりする(偽陽性)ことがしばしばあり、検出精度に問題点がある。
【0009】
上記事情を鑑みて、本発明は、アゾスピリラム属菌をより精度よく、迅速かつシンプルに検出・識別することのできるプライマーセット、検出キット及び検出方法を提供することを目的とする。
【課題を解決するための手段】
【0010】
すなわち、本発明は以下の発明を包含する。
(1)アゾスピリラム属菌の16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅可能なプライマーセットであって、配列番号1で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。
(2)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる上記(1)に記載のプライマーセット。
(3)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなる上記(2)に記載のプライマーセット。
(4)上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加している上記(2)に記載のプライマーセット。
(5)上記(1)〜(4)の何れか一項に記載のプライマーセットを備えるアゾスピリラム属菌の検出キット。
(6)以下の工程:(a)対象サンプルからDNAを含有する試料を調製する工程、(b)上記試料に対して、上記(1)〜(4)の何れか一項に記載のプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、(c)上記工程(b)で得られる増幅断片の有無を検出する工程、を含むアゾスピリラム属菌の検出方法。
(7)上記対象サンプルが、土壌サンプル又は植物由来サンプルである(6)に記載の検出方法。
【発明の効果】
【0011】
本発明は、PCR法でアゾスピリラム属菌を検出することができるという効果を奏する。
【図面の簡単な説明】
【0012】
【図1】プライマーセットAz16S−Aを用いた、未知分離株に対するPCR実験の結果を示した図である。
【図2】純粋培養物及び混合培養物における、プライマーセットAz16S−Aを用いたアゾスピリラム菌のPCR検出を示した図である。
【発明を実施するための形態】
【0013】
以下、本発明の実施の形態について、詳細に説明する。
【0014】
(プライマーセット)
本発明に係るプライマーセットは、アゾスピリラム属に属する細菌(アゾスピリラム属菌)の16S rDNAに由来するDNA断片を、ポリメラーゼ連鎖反応(PCR)によって増幅可能なものである。すなわち、本発明に係るプライマーセットは、アゾスピリラム属菌の16S rDNA及び16S rDNAの断片にハイブリダイズして、PCRにより所定サイズの上記DNA断片を増幅させる。
【0015】
上記プライマーセットは、アゾスピリラム属菌の16S rDNAに由来するDNA断片を、PCRによって選択的に増幅可能という特性を有する。ここで、「選択的に増幅可能」とは、当該プライマーセットを用いたPCRを行った場合に、アゾスピリラム属菌由来のDNA断片が専ら増幅産物として得られるが、他の多くの生物由来のDNA断片は増幅産物として実質的に得られないことを指す。実施例にも示すように、本発明に係るプライマーセットを用いた増幅では、アゾスピリラム属に属する複数種の細菌において共通のPCR増幅産物が得られる反面、アゾスピリラム属に近縁な細菌を含む多くの細菌ではPCR増幅産物が得られない。また、土壌サンプル及び植物含有サンプル(植物の根部等)に対するPCR増幅の結果から、本発明に係るプライマーは、植物、及びアゾスピリラム属菌ではない多くの土壌微生物由来のPCR増幅産物が実質的に得られないことが示されている(実施例参照)。従って、本発明に係るプライマーは、アゾスピリラム属菌の検出に極めて有用である。
【0016】
本発明に係るプライマーセットは、具体的には以下の(1)〜(4)の何れかに示すヌクレオチド(より具体的にはDNA)である。
(1)配列番号1で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなるプライマーセット。なお、配列番号1で表される塩基配列中のkはグアニン又はチミンを示す。なお、PCR増幅の選択性をより向上させる観点では、第一のプライマーは、配列番号1で示される塩基配列のうち連続する少なくとも18塩基部分を有するヌクレオチドからなることがより好ましい。また、同様に、第二のプライマーは、配列番号2で示される塩基配列のうち連続する少なくとも18塩基部分を有するヌクレオチドからなることがより好ましく、少なくとも20塩基部分を有するヌクレオチドからなることがさらに好ましい。また、後述するように第一のプライマー、及び第二のプライマーともに、上記条件を満たす複数種のプライマーの混合物であってもよく、単独(1種類)のプライマーであってもよい。
(2)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる上記(1)に記載のプライマーセット。
(3)上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなる上記(2)に記載のプライマーセット。第一のプライマーとして当該混合物を用いれば、アゾスピリラム属菌に対する特異性がより高まり、より多種類にわたるアゾスピリラム属菌を増幅可能となる。
【0017】
上記第一のプライマーとして上記混合物を用いる場合、kがグアニンである上記ヌクレオチドとkがチミンである上記ヌクレオチドとの混合比率は特に限定されず、例えば、1:9〜9:1の範囲内、より好ましくは4:6〜6:4の範囲内とすることができる。
(4)上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加している上記(2)又は(3)に記載のプライマーセット。なお、基本となるプライマーに対して1〜2個の塩基に変異がある場合でも、所望のPCR増幅断片は得られる。また、上記(3)に記載のプライマーセットを構成する第一のプライマー(混合物)において、1〜2個の塩基が欠失又は置換する場合は、配列番号1で示される塩基配列におけるk以外の塩基が欠失又は置換しているものとする。
【0018】
また、特に限定されないが各プライマーのGC含量は、例えば、50%〜70%の範囲とするのが好ましく、50%〜65%の範囲とすることがより好ましい。プライマーのTm値も特に限定されないが、例えば、50℃〜65℃とすることが好ましく、50℃〜60℃とすることがより好ましい。
【0019】
本発明に係るプライマーセットは常法に従って合成することができる。また、本発明に係るプライマーセットを用いたPCRの反応条件は特に限定されない。
【0020】
(アゾスピリラム属菌の検出キット)
本発明に係るアゾスピリラム属菌の検出キットは、上記説明した本発明に係るプライマーセットを備える。この検出キットは、アゾスピリラム属菌の有無の判別を含む検出に用いる。さらに、必要に応じて、1)PCRに用いる各種試薬及び器具(ポリメラーゼ、PCRバッファー、各dNTP、ピペット等)、2)PCRに供するDNAを含有する試料を調製するための各種試薬及び器具(試験管、バッファー等)、3)PCR増幅断片を解析するための各種試薬及び器具(電気泳動ゲル材料、ピペット等)、4)さらなる詳細解析のため、アゾスピリラム属に属する特定種のみを増幅可能なPCRプライマーセット、5)検出キットの使用説明書、等の少なくとも1つを備えていてもよい。
【0021】
(アゾスピリラム属菌の検出方法)
本発明に係るアゾスピリラム属菌の検出方法は、以下の工程:
(a)対象サンプルからDNAを含有する試料を調製する工程、
(b)上記試料に対して、本発明に係るプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、
(c)上記工程(b)で得られる増幅断片の有無を検出する工程、
を含む。
【0022】
以下、各工程を説明する。
【0023】
(1)工程(a)
工程(a)は、対象サンプルからDNAを含有する試料を調製する工程である。
ここで、「対象サンプル」とは、アゾスピリラム属菌の有無を判別しようとするサンプルを意味する。対象サンプルは、アゾスピリラム属菌を含む可能性のあるものであればよく、限り特に限定されない。例えば、土壌サンプル、植物の根部等の植物由来サンプル、培地上の微生物コロニー、微生物を含む培養液等が挙げられる。
【0024】
DNAを含有する試料は、例えば、土壌サンプル等の対象サンプルから常法に従って調製することができる。例えば、ボイル法、CTAB法等が挙げられる。これらの中では、操作の簡便さの観点では、ボイル法が好ましい。なお、ボイル法とは、対象サンプルを液体中(例えば、滅菌蒸留水又はバッファー)に懸濁して短時間(例えば5分程度以下)加熱して、DNAを取り出す方法である。
【0025】
なお、調製されたDNAを含有する試料には、DNAの他に、各種RNA、タンパク質、細胞破砕物、等が含まれる。従って、該試料を常法に従って精製してもよい。但し、実施例にも示す通り、本発明の方法では、当該試料に対して一切の精製を行わなくとも、一度のPCRにより非常に高感度な検出を行うことができる。
【0026】
(2)工程(b)
工程(b)は、上記試料に対して、上記した本発明に係るプライマーセットを用いたPCRを行う工程である。工程(b)におけるPCRは、常法に従って行うことができる。工程(b)により、試料中にアゾスピリラム属菌が含まれている場合には、使用したプライマーの塩基配列を両端に有するアゾスピリラム16S rDNA由来のPCR増幅断片が得られる。
【0027】
(3)工程(c)
工程(c)は、工程(b)で得られるPCR増幅断片の有無を検出する工程である。PCR増幅断片(アンプリコン)の有無の確認は、常法に従って行うことができる。例えば、電気泳動によりPCR増幅断片の有無及びサイズを確認することができる。
【0028】
PCR増幅断片の有無の確認の結果、PCR増幅断片が存在する場合には、対象サンプルにアゾスピリラム属菌が存在すると判別することができる。また、PCR増幅断片の有無の確認の結果、PCR増幅断片が存在しない場合には、対象サンプルにアゾスピリラム属菌が存在していないと判別することができる。
【0029】
従来法による農業サンプルからのアゾスピリラム属菌の分離プロセスでは、選択培地の使用と菌の生理形態学的評価を繰り返し行うため、一定の特徴を持った菌株のみを分離する選択バイアスが生じる。例えば、非特許文献24では、窒素源を含まない培地を使用した従来の分離プロセスにおいて、高い窒素固定能を有するアゾスピリラム属菌が分離され、低い窒素固定能の菌株は分離できなかったことを開示している。後述する実施例にも示す通り、本発明の一形態としてのAz16S−Aプライマーセットを使用したアゾスピリラム属菌のPCR検出により、選択培地と生理形態学的評価の使用を減らすことができるため、結果的に選択バイアスを低減させた分離プロセスを実現できる。また、本発明の一形態であるこのPCR検出法は、農業サンプルから多様なアゾスピリラム属菌を分離し、新規の植物成長促進細菌株を探索する上で有用な手法、またはアゾスピリラム属菌集団の多様性と生態とを分析するための有用な手法として利用できる。多数のアゾスピリラム属菌及び分離株を対象にしてAz16S−AプライマーセットによるPCR増幅産物(アンプリコン)のDNA配列を決定したところ、少なくとも15の一塩基多型(SNP)が見出され、そのうち12の一塩基多型は購入可能な制限酵素の認識サイトである。さらに、この12のSNPのうちの一つは、アゾスピリラム属菌とR.centenariaとの識別に有用であると見込まれる。一塩基多型について更なる分析と確認が必要であるが、おそらく、Az16S−Aプライマーセットの利用により、一回のPCRと制限酵素処理(例えば、PCR−RFLP分析法)によって、アゾスピリラム属分離株またはアゾスピリラム属菌集団の精密な同定及び多様性分析の両方が可能である。さらに、発明者らの実験では、根圏土壌と植物体部位に対してAz16S−Aプライマーセットを使用し、アゾスピリラム属菌の局在性を予測している。このような微生物動態の分析では、非特許文献16で開示された、蛍光標識されたオリゴヌクレオチドプローブも使用できるが、しかし我々の方法は、アゾスピリラム属菌の動態分析後の分離プロセスにおいても利用できる点で優位性がある。PCR法は最確数(MPN)法と併用することによって、アゾスピリラム属菌集団の定量にも利用できる(非特許文献25、26参照)。本発明のPCR法を組み込んだ迅速、容易且つ選択バイアスが最小となったアゾスピリラム分離技術、及びAz16S−Aプライマー配列に基づくリアルタイムPCR定量技術に対するさらなる応用が考えられる。
【実施例】
【0030】
〔実施例1〕
<プライマーセットの設計及び作製>
アメリカ国立バイオテクノロジーインフォメーションセンター (NCBI, USA, http://www.ncbi.nlm.nih.gov/genbank/GenBankFtp.html)ジーンバンクヌクレオチドデータベースから合計30個のアゾスピリラム16S rDNA配列(14の種から)を入手し、バチルス属菌、Pseudomonas属菌、Roseomonas属菌を含む関連する類縁細菌群の16S rDNA配列と比較しながら、16S rDNAの開始コドンの下流300bp〜1200bpにおける、推測上の属特異的且つ保存的領域について同定した。
【0031】
推測上のアゾスピリラム属特異的領域に基づき、属特異的なプライマーセットを設計するために、Geneious Pro 4.8.5ソフトウエア(Biomatters Ltd社, Auckland,New Zealand)を使用して、アゾスピリラム及びその類縁細菌群に属する菌に対して、DNA配列アライメント解析を実施した。同様に、ジーンバンクから6つのipdC遺伝子配列(3つのアゾスピリラム属菌種から)、3つのnifA遺伝子配列(2つのアゾスピリラム属菌種から)、12のnifH遺伝子配列(3つのアゾスピリラム属菌種から)を入手し、プライマーセットを設計した。ipdC、nifA及びnifH遺伝子は何れも細菌による植物成長促進において重要な遺伝子である。具体的には、ipdC遺伝子は、インドール−3−酢酸生合成酵素をコード化するものであり、nifA遺伝子及びnifH遺伝子はそれぞれ、窒素固定のためのニトロゲナーゼ酵素の制御遺伝子及び構造遺伝子をコード化するものである。
【0032】
各プライマーセットの設計に際して、次の条件(1)〜(3)を適用した。(1)原則として20個前後、好ましくは20個を超えるヌクレオチド塩基を有するプライマーセットを設計し、(2)上記プライマーセットは200−800bpのDNA断片(アンプリコン)を生成させると予測されるものを選択し、(3)上記プライマーセットのヌクレオチドコードはアゾスピリラム属菌の間で同じかまたは最大で2塩基程度異なるものを許容するように設計した。表1は、Baudoinらによって報告されたfAZO/rAZOプライマー(非特許文献3参照)と共に、今回設計されたプライマーを表示したものである。設計されたが、簡易PCRを伴う予備分析(preliminary assays)によって排除されたものは、表1には示されていない。
【0033】
<結果>
表1は、今回設計した9つのプライマーセットと共に、Baudoinらによって報告されたプライマーセットfAZO/rAZO(非特許文献3参照)を表示したものである。プライマー名に16Sが含まれるもの、ipdが含まれるもの、nifが含まれるものは、順に、16S rDNAの塩基配列、ipdの塩基配列、nifの塩基配列に基づき設計されたものである。設計されたが予備分析(preliminary assays)によって排除されたものは、表1には示されていない。表において、KはG及びTの二種の塩基の混合(すなわち、Kがグアニンであるプライマーと、Kがチミンであるプライマーとの混合物。両者の混合比率は約1:1。)であることを意味している。アンプリコンサイズはアゾスピリラム・ブラシレンセ JCM 1224TSのDNA配列により推測されたものである。
【0034】
【表1】
【0035】
〔実施例2〕
<プライマーセットの属特異性評価>
12のアゾスピリラム標準株(reference strains)及び15の非アゾスピリラム標準株について、実施例1で作製したプライマーセットの属特異性について調べた。
【0036】
使用した12のアゾスピリラム標準株及び15の非アゾスピリラム標準株、合計27の標準株を以下に示した。TSは基準株であることを示している。
【0037】
(アゾスピリラム属の12の標準株)
(1)A. brasilense JCM 1224TS
(2)A. brasilense JCM 1225
(3)A. brasilense JCM 1226
(4)A. lipoferum JCM 1227
(5)A. lipoferum JCM 1228
(6)A. lipoferum JCM 1247TS
(7)A. lipoferum JCM 1270
(8)A. amazonense DSM 2787TS
(9)A. halopraeferens DSM 3675TS
(10)A. irakense DSM 11586TS
(11)A. picis DSM 19922TS
(12)A. rugosum DSM 19657TS
(非アゾスピリラム属の15の標準株)
アゾスピリラム属の類縁細菌群に属するもの:
(1)Rhodocista centenaria DSM 9894TS
(2)Magnetospirillum magnetotacticum DSM 3856TS
(3)Paeospirillum fulvum DSM 113TS
(4)Rhodospirillum rubrum DSM 467TS
(5)R. salinarum DSM 9154TS
(6)Thalassospira lucentensis DSM 14000TS
そのほかのプロテオバクテリア:
(7)Agrobacterium tumefaciens MAFF 301224
(8)Agrobacterium tumefaciens MAFF 301540
(9)Rhizobium leguminosarum MAFF 210039
(10)Alcaligenaceae sp. RERT 020126
(11)Stenotrophomonas maltophilia MAFF 301690
(12)Stenotrophomonas maltophilia RERT 020018
Firmicutesに属するもの:
(13)Bacillus pumilus MAFF 118256
(14)Bacillus pumilus RERT 020111
(15)Microbacterium esteraromaticum RERT 020020
12のアゾスピリラム及び15の非アゾスピリラム、合計27の標準株(表2を参照)は、ドイツ微生物・培養細胞収集有限会社(Deutsche Sammlung von Mikroorganismen und Zellkulturen、DSMZ)から(11のDSM菌株)、日本独立行政法人理化学研究所バイオリソースセンター微生物材料開発室(Japan Collection of Microorganisms、JCM)から(7つのJCM菌株)、日本独立行政法人農業生物資源研究所(National Institute of Agrobiological Sciences、NIAS)から(5つのMAFF菌株)、および発明者の細菌コレクションから(上記(10)、(12)、(14)、(15)のRERT菌株4つ)入手したものである。12のアゾスピリラム属菌株は、PCR実験の時点で、上記機関において入手できるアゾスピリラム属7種すべてを含む。International Journal of Systematic and Evolutionary Microbiology及びNCBIで報告された14のアゾスピリラム属菌種のうち(非特許文献18、非特許文献19参照)、その種の菌株の取得または培養の困難さのために7つの種は本実施例での考慮対象外とした(例えばA.doebereinerae DSMZ 13131TS)。
【0038】
これまでの系統分類学的解析では、種間の関係が分析間で多少異なる点はあるが、考慮対象外とした上記7つの種は何れも本研究で使用された7つのアゾスピリラム属菌種の間に位置するという結果を示す。考慮対象外とした上記7つの種は、
(1)A.doebereinerae(非特許文献9、非特許文献19)、
(2)A.largimobile(非特許文献9、非特許文献20)、
(3)A.canadense(非特許文献21、非特許文献23、非特許文献19)、
(4)A. melinis(非特許文献21、非特許文献23)、
(5)A. zeae(非特許文献22、非特許文献23)、
(6)A. oryzae(非特許文献20、非特許文献24)、
(7)A.palatum(非特許文献19参照)である。
【0039】
15の非アゾスピリラム菌株は、類縁細菌群(非特許文献17参照)とBaudoinら(非特許文献3参照)が研究において使用した土壌バクテリアの菌株から選択された。
【0040】
プライマーセットの属特異性の分析実験は、次のように行われた。27の標準株の定常期培養物(stationary−phase cultures)の一定分量を、遠心し、滅菌蒸留水に再懸濁させた溶液を95°Cにおいて5分間放置した後、遠心を行いPCRの鋳型となる細胞溶解物上清を得た。
【0041】
PCR反応混合物は、10xPCRバッファー2.5μl、1.5mM MgCl2、各dNTPを0.25mMずつ、各プライマーを0.02μMずつ、Taq DNAポリメラーゼ(BIOTAQ,BioLine,London,UK)を2.5ユニット、粗製のバクテリア溶解物10μlを含み、総量は25μlである。
【0042】
PCRはVeritiサーマルサイクラー(Applied Biosystems,CA,US)によって、下記条件に基づいて実施された。まず94°Cにおいて3分間放置し、初期変性させる。次に、下記操作を35サイクル繰り返す。各サイクルは、94°Cで30秒間変性させた後、各プライマーセットの最適温度で30秒アニーリングさせ、72°Cで1分間伸長させる。上記最適温度として、Az16S-A(配列番号1及び2として示す),Az16S-B、Az16S‐Cは58°Cである。AzipdC、AznifA−A、AznifA−Bは52°Cである。AznifH‐A,AznifH‐B,AznifH‐Cは55°Cである(塩基配列に関しては表1を参照)。上記35サイクルが終了した後、72°Cで5分間放置し最終的な伸長をさせた。Baudoinらによって報告されたfAZO/rAZOプライマーを使用したPCRは、彼らの報告した手順に従って実施した。PCR産物は、100−bpDNAラダー(HyperLadderIV,BioLine)と共に2%アガロースゲル中に展開され、臭化エチジウム溶液によって染色され、AE‐6905H Image Saver HR(ATTO,Tokyo,Japan)によって撮影された。
【0043】
このPCR実験によって選ばれたAz16S‐Aプライマーセットの属特異性は、さらに、35のアゾスピリラム様分離株及び70の未知の分離株に対しても、同様のPCR及び電気泳動の手順によって分析された(後述の実施例3を参照)。
【0044】
<結果>
上記属特異性分析実験の結果を、表2に示した。表2において+及び−はそれぞれ、予測アンプリコンの有無を示している。すなわち、+は予測アンプリコンが得られたことを指し、−は予測アンプリコンが得られなかったことを指す。
【0045】
【表2】
【0046】
実験が行われた10のプライマーセット(表1を参照)において、16S rDNA配列に基づき設計されたプライマーセットAz16S-Aは、12のアゾスピリラム標準株(7の種)すべて及び1の非アゾスピリラム属菌株Rhodocista centenaria DSM9894(表2参照)に対して、単一のアンプリコンを生成させることに成功した。アンプリコンは単一のDNAバンドとして観察され、上記DNAバンドの長さはおおよそ640bpであって(図1を参照)、この長さはA.brasilense 16S rDNA配列から予測された長さ(表1を参照)に一致するものである。なお、非アゾスピリラム属菌株Rhodocista centenariaは、自然界では温泉などの特殊環境に棲息することが報告されており、アゾスピリラム属菌とは棲息環境が異なる。
【0047】
プライマーセットAz16S-Bを用いたPCRは、12のアゾスピリラム標準株すべてから予測アンプリコンを生成させただけでなく、α−プロテオバクテリア及びβ−プロテオバクテリアにわたる10の非アゾスピリラム属菌株からも、予測アンプリコンを生成させた。
【0048】
プライマーセットAz16S-C、AznifH-A、及びAznifH-Bは、Baudoinらによって報告されたプライマーセットfAZO/rAZOと同じように、12のアゾスピリラム属菌株のうち、1−3の株に対してはアンプリコンを生成させることができず、17の非アゾスピリラム属菌株のうち、2-5の株に対しては複数のアンプリコンを生成させた。これらの4のプライマーセットにおいて、プライマーセットAznifH-A及びAznifH-Bは、他の2のプライマーセットに比べ比較的良い属特異性を示した。残りの4つのプライマーセットからは良くない結果が得られた。従って、プライマーセットAz16S-Aはアゾスピリラム属菌に対して最も優れた特異性を示した。
【0049】
ここで、図1について説明すると、図1は、プライマーセットAz16S−Aを用いた、未知分離株に対するPCR実験の結果を示している。PCR産物は2%アガロースに溶解され、臭化エチジウム溶液によって染色された。識別された分離株は、16SrDNA配列決定解析によって、下記のように確認された。レーン1はAchromobacter及びRahnellaによって汚染された、アゾスピリラム属菌培養物であり、レーン2はAchromobacter、レーン3はAchromobacter、レーン4はアゾスピリラム、レーン5はアゾスピリラム、レーン6はAchromobacter、レーン7はアゾスピリラム、レーン8はStenotrophomonas、レーン9はアゾスピリラム、レーン10はAchromobacter、レーン11は参照物としてのA. brasilense JCM 1224TS、レーンMは100−bpDNAラダーである。
【0050】
〔実施例3〕
<作製されたプライマーセットの利用可能性>
さらに、35のアゾスピリラム様分離株と、農業サンプルから得られた70の未知の分離株を用いて、選択されたプライマー候補の特異性、検出限界及び利用可能性について調べた。アゾスピリラム属菌の分離において検出限界は重要な面であって、根圏土壌及び植物体から分離するプロセスにおいては、アゾスピリラム属の菌落及び培養物はしばしば非アゾスピリラム微生物によって汚染されているからである。
【0051】
(使用された35のアゾスピリラム様分離株と70の未知の分離株)
アゾスピリラム様分離株は、根圏土壌並びに作物及び草の根部から収集されたものである。上記収集は、独立行政法人農業生物資源研究所(NIAS)と十勝農業協同組合連合会農産化学研究所(ARITFAC)によって、選択培地及び形態生理学評価の両方を用いて行われた。選択培地は例えば、窒素不含有ブロモチモールブルー(NFb)培地、Rojo Congo(コンゴレッド)培地である。形態生理学評価は、例えば、アセチレン減少分析(ARA)(非特許文献5、非特許文献9)である。これらの分離株のrDNA配列データは入手できないものであった。全部で35のアゾスピリラム様分離株を、上記収集されたコレクションからランダムに選出した。さらに、根圏土壌、穀物及び飼草の根部から、ARAとrDNA配列決定を用いずに、選択培地のみによって収集した我々のストックから、70の未知の分離株をランダムに選出した。これらの分離株の「属」及び「種」は未知であった。使用された35のアゾスピリラム様分離株は多数の、70の未知の分離株は少数の、アゾスピリラム属菌を含むものと考えられる。
【0052】
(作製したプライマーセットによる検出実験)
すべての菌株及び分離株に対して、16S rDNA配列決定解析を行った。ボイル法によって抽出された粗製DNAは、アゾスピリラム16S rDNA配列に基づき設計されたAz16S-D配列決定プライマーによって、PCR増幅された。Az16S-D配列決定プライマーセットのフォワードプライマーは5’CCGCGGTAATACGAAGGGGGCであり、リバースプライマーは5’GCCTTCCTCCGGCTTGTCACCGGCである。当該プライマーセットによるアンプリコンサイズは、おおよそ650bpと推測され、それは属レベルで系統発生的分類をするのには充分である。PCRはアニーリング温度60°Cで、前記のPCR実験で記載されているのと同じ手順によって実施された。PCR産物はMin−Elute PCR精製キット(QIAGEN,Hilden,Germany)によって精製され、Az16S-Dフォワードプライマー、BigDye Terminator v3.1サイクル配列決定キット,3730xlDNAアナライザー(Applied biosystems)によって配列決定された。配列データは、SEQ MATCH program ver.3.0(非特許文献7参照)を使用し、リボソームデータベースプロジェクト(リリース10、ミシガン州立大学微生物生態学センター[http://rdp.cme.msu.edu/index.jsp])で得られたデータと比較し、カットオフ類似性(Sab)値を>0.7に設定しマッチする微生物種を探索し上位20マッチに基づき菌株及び分離株の属を決定した。
【0053】
<結果>
上記作製したプライマーセットによる検出実験の結果を、表3に示した。
【0054】
【表3】
【0055】
表3は、35のアゾスピリラム様分離株及び70の未知分離株に対するPCR実験及びrDNA配列決定解析の結果を比較したものである。表3において、NIASは独立行政法人農業生物資源研究所であり、ARITFACは十勝農業協同組合連合会農産化学研究所である。+と−はそれぞれ、予測されたアンプリコンの有無を示している。
【0056】
表3によれば、プライマーセットAz16S-Aを用いたPCRは、NIASから入手した20のアゾスピリラム様分離株のうち17の菌株から、予測された単一のアンプリコンを得た。PCR検出においてアンプリコンが得られた17の分離株は、16S rDNA配列決定解析によって、アゾスピリラム属に分類され、残りの3つの株は、Williamsia、Bacillus、Rhizobiumに分類された。PCR実験及びrDNA配列決定解析の結果における完全な一致は、ARITFACから入手した15のアゾスピリラム様分離株及び我々のストックから入手した70の未知分離株においても見られた(表3及び図1を参照)。アンプリコンが得られた5つの未知分離株のうちの一つ(図1レーン1参照)のrDNA配列決定解析は、この培養物はアゾスピリラムのほかにも二つの種、Achromobacter及びRahnellaを含んでいることを示した。従って、プライマーセットAz16S-Aを用いたPCRは、混合培養物におけるアゾスピリラムを成功裏に検出することができた。Baudoinらによって報告されたプライマーセットfAZO/rAZOを用いたPCRでは、アゾスピリラム疑似分離株に対して、PCR実験及びrDNA配列決定解析の結果における完全な一致が見られた。しかし、70の未知分離株のうち(詳細は示していない)、2つの非アゾスピリラム分離物(BacillusとPantoea)からも、5つのアゾスピリラムと同様に、アンプリコンを生成させた。
【0057】
NIASから入手した分離株のうち、3つの非アゾスピリラム分離株は、Williamsia,Bacillus,Rhizobium種である。同じように、我々のストックからの67の非アゾスピリラム分離株のうち、34はBacillusとして同定され、8はMicrobacteriumとして、6はPaenibacillusとして、6はStenotrophomonasとして、6はAcromobacterとして、2はAlcaligenesとして、2はLysinibacillusとして、1はHerbaspirilluとして、1はPantoraとして、1はYersiniaとして同定された。
【0058】
〔実施例4〕
<アンプリコンの存在を示した分離株のキャラクタリゼーション>
上記実施例3において、70の未知分離株のうち、Az16S-Aプライマーを用いたPCRによって得られたアンプリコンの存在が確認されたのは5つの分離株である。この5つの分離株は、16S rDNA配列決定解析によって、アゾスピリラム属に分類された。これらの属種及び生理学的特性をさらに調べるため、この5つの分離株における炭素源利用、IAA生成、窒素固定化について、A. brasilense JCM 1224TS及びA. brasilense JCM 1226と比較しながら評価した。炭素源利用の評価についてはAPI20NEシステム(bioMERIEUX,Lyon,France)を用いて、製造元のプロトコルに従って実施した。IAA生成については、修飾されたSalkowski試薬(非特許文献12を参照)を用いて比色法によって計測された。分離株は1mMトリプトファンを補充したLuria−Bertani(LB)ブロスにおいて、30°C条件下において2日間、定常期になるまで培養され、それからIAAの計測に供された(非特許文献8を参照)。窒素固定能は、水素炎イオン検出器を備えたガスクロマトグラフ装置(GC-4000 GL Science,Tokyo,Japan)を使用して、ARA法(非特許文献9、非特許文献15を参照)によって見積もられた。ARAを行うため、バクテリア培養物は、2mlの濃度1%(v/v)の半固体NFb培地に添加され、9−ml密封バイアル瓶中において、30°C条件下で24時間培養された。上記培養後に10体積%(気体体積)のアセチレンをバイアル瓶中に導入し、さらに24時間培養し、それからガスクロマトグラフィーを実施した。それぞれの分離株のIAA生成及びアセチレン減少については、それぞれ3〜4のバイアル瓶で重複して実験された。
【0059】
<結果>
上記アンプリコンの存在を示した分離株のキャラクタリゼーション実験の結果を、表4に示した。
【0060】
【表4】
【0061】
表4は、いわゆる農業サンプルからPCR実験によって分離されたアゾスピリラム属菌の炭素源利用、インドール−3−酢酸(IAA)生成及びアセチレン減少について示した表である。炭素源利用タイプはAPI20NEシステムによって評価されたものであり、ABタイプはA. brasilenseに特有の利用型であって、N−アセチルグルコサミン(−)、L−アラビノース(+)、D−グルコース(−)、D−マンニトール(−)、D−マンノース(−)を利用するものである。
【0062】
我々のストックから入手の分離株のうち、アンプリコンが得られた5つの分離株は、A .brasilense(表4を参照)に特有的な炭素源利用型と同様の炭素源利用型を示した。IAA生成能及びアセチレン減少(窒素固定)能力は、5つの分離株すべてにおいて見られた。分離株におけるこれら二つの能力は、A. brasilense JCM 1224TSより優れていた。アセチレン減少に見られる分離株同士の大きな差異は、IAA生成に見られる分離株同士より顕著であった。分離株のIAA生成とアセチレン減少との間には、顕著な相関性は観察されなかった。これらの結果から、プライマーセットAz16S‐Aは、アゾスピリラム独特の生理学特性を示す分離株を正確に識別する能力を有することがさらに確認された。
【0063】
〔実施例5〕
<プライマーセットの検出限界>
Az16S-Aプライマーを用いたPCRを、異なる濃度(10‐106CFUml−1)のA. brasilense(JCM 1224TS)に対して、上記と同じ手順にて実施した。さらに、Bacillus pumilus(MAFF 118256)が混入されたA. brasilense(JCM 1224TS)培養物に対しても、PCRを実施した。バチルス属菌がモデル汚染菌として選ばれた理由は、我々の実験系において、アゾスピリラム選択培地に観察される非アゾスピリラム属菌の大部分においてバチルス属菌が存在するからである。混合培養物を得るために、アゾスピリラム及びバチルス培養物はそれぞれ、LB培養液において37℃条件下において48時間及び24時間培養された。それから、最終菌濃度が107CFUml−1となるように、A. brasilense:B. pumilusを1:1〜1:104の異なる比率で混合した。プライマーの検出限界とは、検出可能なPCRアンプリコンをもたらすことのできる、最小のアゾスピリラム濃度または比率である。
【0064】
<結果>
プライマーセットAz16S-Aを用いるPCRは、103CFUml−1の濃度を有するA. brasilense純粋培養物から、予測サイズに該当する検出可能な量のアンプリコンを生成させることが出来た(図2中の(a)を参照)。同様に、A. brasilense:B. pumilusを1:104またはそれ以上の比率で混合して得られた混合培養物からも、PCRでは同じように予想アンプリコンを生成することが出来た(図2中の(b)を参照)。A.brasilense:B. pumilusを1:104の比率で混合した混合培養物は、おおよそ103CFUml−1のA. brasilenseを含んでいる。したがって、プライマーセットの検出限界は、純粋培養物及び混合培養物において同じである。さらに、PCRの汚染菌存在時におけるアゾスピリラム菌の検出能力は、農業サンプルから得られた70の未知分離株においても確認された(図1レーン1参照)。
【0065】
なお、図2は純粋培養物及び混合培養物におけるプライマーセットAz16S−Aを用いたアゾスピリラム属菌のPCR検出を示した図である。図中(a)のPCR産物はA. brasilense JCM 1224TS純粋培養物から得られ、図中(b)のPCR産物はBacillus pumilus MAFF 118256が混合されたA. brasilense 培養物から得られた。PCR産物は2%アガロースに展開され、臭化エチジウム溶液によって染色された。図中(a)における、レーン上部の数字は、A. brasilense濃度(CFU ml−1)である。レーンMは100−bpDNAラダーである。図中(b)において、レーン1はA. brasilenseのみ、レーン2は、A. brasilense:B. pumilusを1:1で混合したもの、レーン3は、1:10で混合したもの、レーン4は、1:102で混合したもの、レーン5は、1:103で混合したもの、レーン6は1:104で混合したもの、レーン7は、B. pumilusのみのもの、を夫々示す。PCR産物は2%アガロースゲルに展開され、臭化エチジウム溶液によって染色された。
【産業上の利用可能性】
【0066】
本発明は、土壌サンプルを始め、各種サンプルからアゾスピリラム属菌を検出・同定する際に好適に利用することができる。
【特許請求の範囲】
【請求項1】
アゾスピリラム属菌の16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅可能なプライマーセットであって、
配列番号1で示される塩基配列(kはグアニン又はチミンを表す)のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、
配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなることを特徴とするプライマーセット。
【請求項2】
上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、
上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる、ことを特徴とする請求項1に記載のプライマーセット。
【請求項3】
上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなることを特徴とする請求項2に記載のプライマーセット。
【請求項4】
上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加していることを特徴とする請求項2に記載のプライマーセット。
【請求項5】
請求項1〜4の何れか一項に記載のプライマーセットを備えることを特徴とするアゾスピリラム属菌の検出キット。
【請求項6】
以下の工程:
(a)対象サンプルからDNAを含有する試料を調製する工程、
(b)上記試料に対して、請求項1〜4の何れか一項に記載のプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、
(c)上記工程(b)で得られる増幅断片の有無を検出する工程、
を含むことを特徴とする、アゾスピリラム属菌の検出方法。
【請求項7】
上記対象サンプルが、土壌サンプル又は植物由来サンプルであることを特徴とする請求項6に記載の検出方法。
【請求項1】
アゾスピリラム属菌の16S rDNAに由来するDNA断片をポリメラーゼ連鎖反応によって増幅可能なプライマーセットであって、
配列番号1で示される塩基配列(kはグアニン又はチミンを表す)のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第一のプライマーと、
配列番号2で示される塩基配列のうち連続する少なくとも15塩基部分を有するヌクレオチドからなる第二のプライマーとからなることを特徴とするプライマーセット。
【請求項2】
上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンであるヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンであるヌクレオチドの少なくとも一方からなり、
上記第二のプライマーが配列番号2で示される塩基配列を有するヌクレオチドからなる、ことを特徴とする請求項1に記載のプライマーセット。
【請求項3】
上記第一のプライマーが、配列番号1で示される塩基配列においてkがグアニンである上記ヌクレオチド、及び配列番号1で示される塩基配列においてkがチミンである上記ヌクレオチドの混合物からなることを特徴とする請求項2に記載のプライマーセット。
【請求項4】
上記第一のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加されており、かつ上記第二のプライマーにおいて1〜2個の塩基が欠失、置換、又は付加していることを特徴とする請求項2に記載のプライマーセット。
【請求項5】
請求項1〜4の何れか一項に記載のプライマーセットを備えることを特徴とするアゾスピリラム属菌の検出キット。
【請求項6】
以下の工程:
(a)対象サンプルからDNAを含有する試料を調製する工程、
(b)上記試料に対して、請求項1〜4の何れか一項に記載のプライマーセットを用いてポリメラーゼ連鎖反応を行う工程、
(c)上記工程(b)で得られる増幅断片の有無を検出する工程、
を含むことを特徴とする、アゾスピリラム属菌の検出方法。
【請求項7】
上記対象サンプルが、土壌サンプル又は植物由来サンプルであることを特徴とする請求項6に記載の検出方法。
【図1】

【図2】
【図2】
【公開番号】特開2012−170448(P2012−170448A)
【公開日】平成24年9月10日(2012.9.10)
【国際特許分類】
【出願番号】特願2011−38657(P2011−38657)
【出願日】平成23年2月24日(2011.2.24)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成24年9月10日(2012.9.10)
【国際特許分類】
【出願日】平成23年2月24日(2011.2.24)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]